/












































































































































































































































































































































































































































































































































































































































































































































































































Author: Билич Г.Л.
Tags: общее школьное образование общеобразовательная школа биологические науки в целом биологические науки биология
ISBN: 978-5-699-98702-3
Year: 2018




Text
БИОЛОГИЯ
для поступающих в вузы
9-е издание
(переработанное и дополненное)
Г. Л. БИЛИЧ • Е. Ю. ЗИГАЛОВА
Москва
2018
373:57
28721
61
:
Giovanni Cancemi, Chilmay / Shutterstock.com
Shutterstock.com
:
magnetix, Jose Luis Calvo, Zuzanae, Timonina, Designua, NoPainNoGain,
chromatos, extender_01 / Shutterstock.com
Shutterstock.com
,
.
!" / #.$ .
%, &.' . (
. –
)
:
«*», 2018. – 7 8 4 .
ISBN 978-5 -699-98702-3
,
"
%"
"
,
%
!
-
"
+
,
;
"
!
<
<. = +
+
+
+!
< +
-
,
>
%
"
%
"
,
< . ?
%
,
<
!
<,
+ ,
;
<
,
+
%" -
%"
".
%
+
%
!"
;"
;
, +
!"
,
+
,
-
,
",
>
.
+
%
"
+
%
"
.
= +
%
%
,
+"
% &#* +
.
373:57
28721
ISBN 978-5-699-98702-3
© .
., . ., , 2017
©
«
« », 2018
61
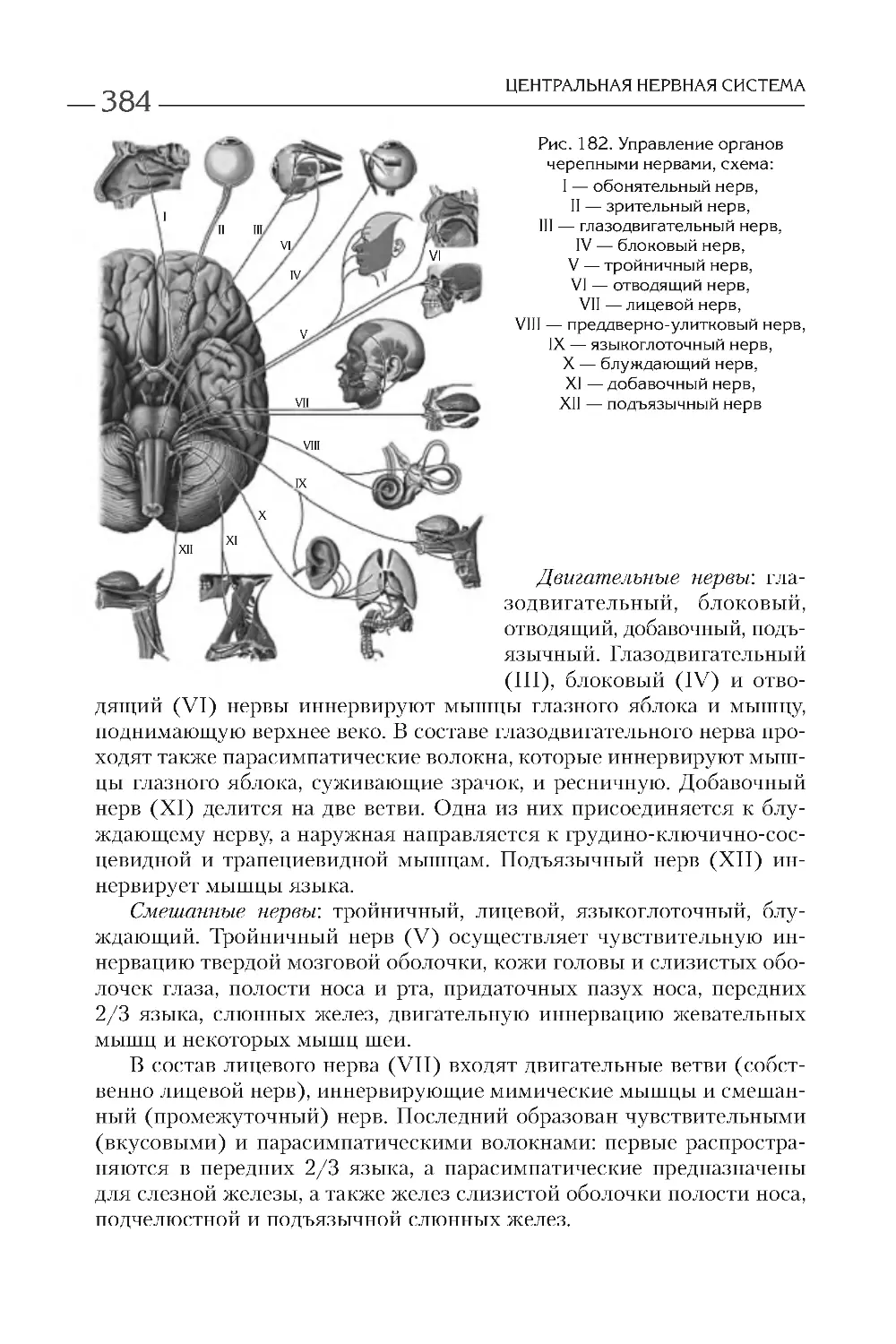
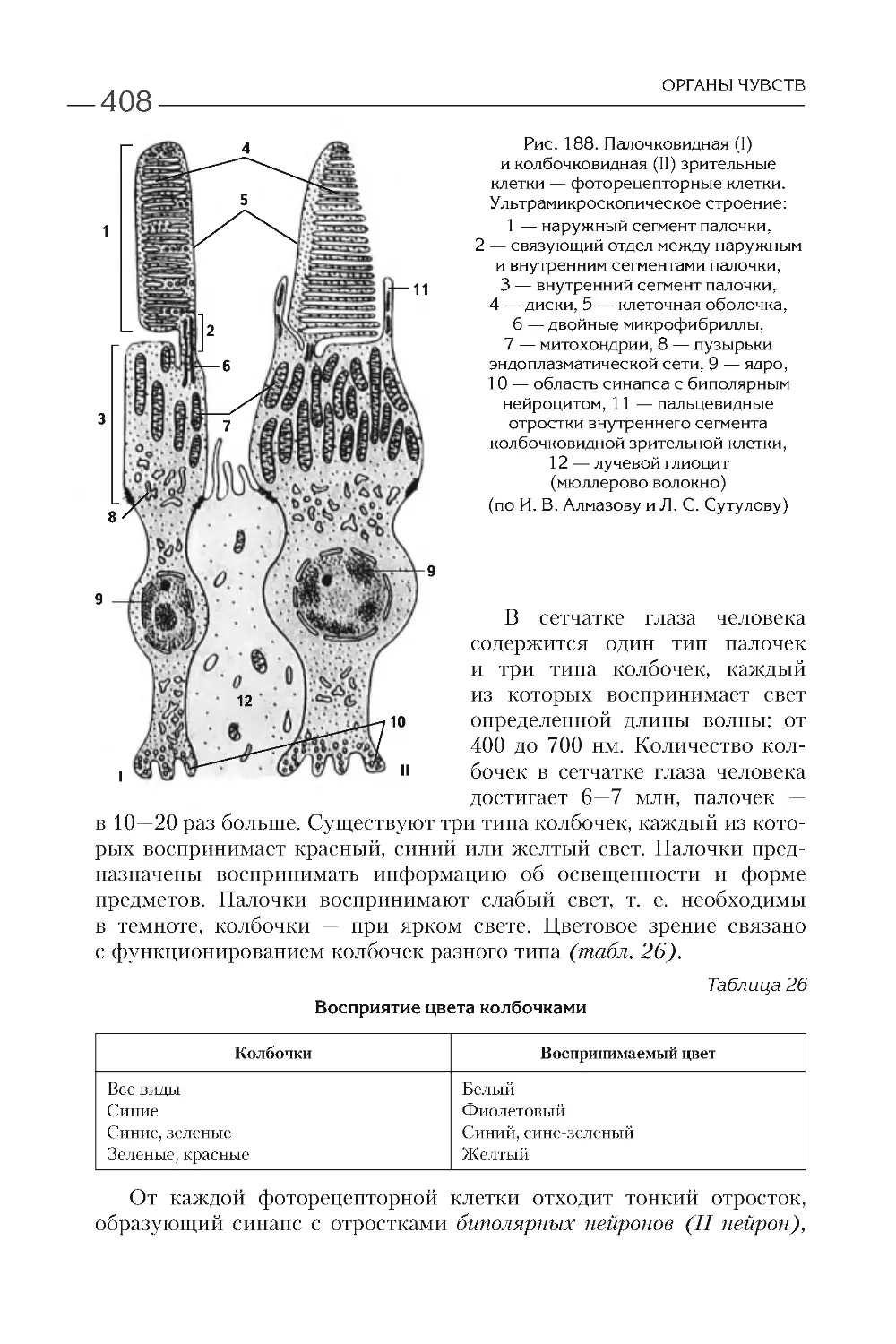
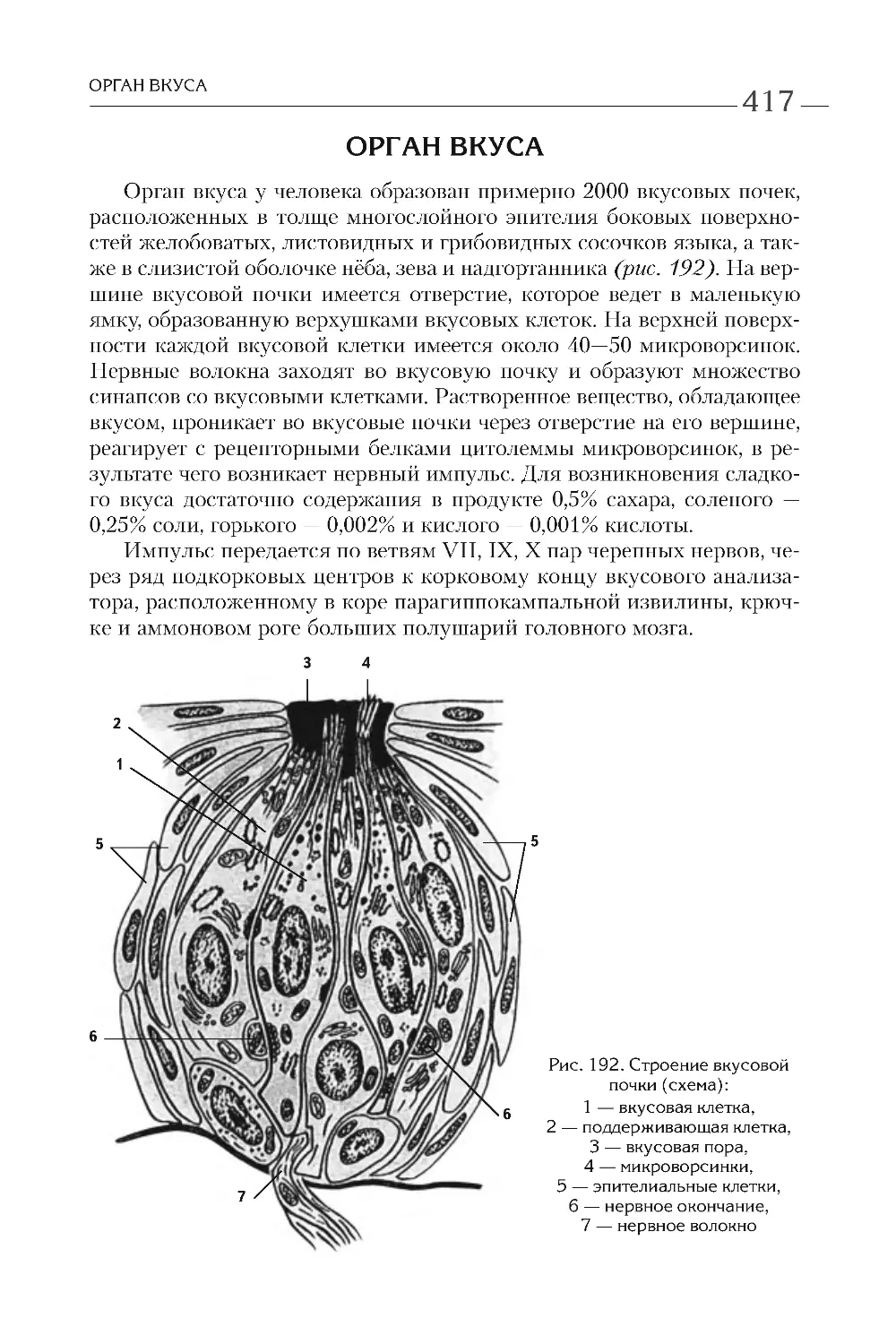
КЛЕТОЧНАЯ ТЕОРИЯ
Развитие цитологии связано с созданием
и усовершенствованием оптических уст-
ройств, позволяющих рассмотреть и изучить клетки. В 1609—1610 гг.
Галилео Галилей сконструировал первый микроскоп, однако лишь
в 1624 г. он его усовершенствовал так, что им можно было пользовать-
ся. Этот микроскоп увеличивал в 35–40 раз. Через год И. Фабер дал
прибору название «микроскоп».
В 1665 г. Роберт Гук впервые увидел в пробке ячейки, которым
дал название сеll — «клетка». В 70-х гг. XVII в. Марчелло Мальпиги
описал микроскопическое строение некоторых органов растений.
Благодаря усовершенствованию микроскопа Антоном ван Левен-
гуком появилась возможность изучать клетки и детальное строение
органов и тканей. В 1696 г. была опубликована его книга «Тайны при-
роды, открытые с помощью совершеннейших микроскопов». Левен-
гук впервые рассмотрел и описал эритроциты, сперматозоиды, открыл
дотоле неведомый и таинственный мир микроорганизмов, которые он
назвал инфузориями. Левенгук по праву считается основоположни-
ком научной микроскопии.
В 1715 г. X. Г. Гертель впервые использовал зеркало для освещения
микроскопических объектов, однако лишь через полтора столетия
Э. Аббе создал систему осветительных линз для микроскопа. В 1781 г.
Ф. Фонтана первый увидел и зарисовал животные клетки с их ядрами.
В первой половине XIX в. Ян Пуркинье усовершенствовал микроско-
пическую технику, что позволило ему описать клеточное ядро («заро-
дышевый пузырек») и клетки в различных органах животных. Ян Пур-
кинье впервые употребил термин «протоплазма». Р. Браун описал ядро
как постоянную структуру и предложил термин nucleus — «ядро».
В 1838 г. М . Шлейден создал теорию цитогенеза (клеткообразова-
ния). Его основная заслуга — постановка вопроса о возникновении
клеток в организме. Основываясь на работах Шлейдена, Теодор
Шванн создал клеточную теорию. В 1839 г. была опубликована его
бессмертная книга «Микроскопические исследования о соответствии
в структуре и росте животных и растений».
Основными исходными положениями клеточной теории были
следующие:
—
все ткани состоят из клеток;
—
клетки растений и животных имеют общие принципы строения,
так как возникают одинаковыми путями;
—
каждая отдельная клетка самостоятельна, а деятельность ор-
ганизма представляет собой сумму жизнедеятельности отдельных
клеток.
КЛЕТОЧНАЯ ТЕОРИЯ
4
Большое влияние на дальнейшее развитие клеточной теории ока-
зал Рудольф Вирхов. Он не только свел воедино все многочисленные
разрозненные факты, но и убедительно показал, что клетки являются
постоянной структурой и возникают только путем размножения себе
подобных — «каждая клетка из клетки» (omnia cellula e cellulae).
Во второй половине XIX в. возникло представление о клетке как
элементарном организме (Э. Брюкке, 1861). В 1874 г. Ж . Карнуа
ввел понятие «биология клетки», тем самым положив начало цитоло-
гии как науке о строении, функции и происхождении клеток.
В 1879—1882 гг. В. Флемминг описал митоз, в 1883 г. В. Вальдей-
ер ввел понятие «хромосомы», через год О. Гертвиг и Э. Страсбургер
одновременно и независимо друг от друга высказали гипотезу о том,
что наследственные признаки заключены в ядре. Конец XIX в. озна-
меновался открытием фагоцитоза Ильей Мечниковым (1892).
В начале XX в. Р. Гаррисон и А. Каррель разработали методы
культивирования клеток в пробирке наподобие одноклеточных орга-
низмов.
В 1928—1931 гг. Е. Руска, М. Кнолль и Б. Боррие сконструирова-
ли электронный микроскоп, благодаря которому было описано под-
линное строение клетки и открыты многие ранее неизвестные струк-
туры. А . Клод в 1929—1949 гг. впервые использовал для изучения
клеток электронный микроскоп и разработал методы фракционирова-
ния клеток с помощью ультрацентрифугирования. Все это позволило
по-новому увидеть клетку и интерпретировать собранные сведения.
Клетка является элементарной единицей всего живого, потому что
ей присущи все свойства живых организмов: высокоупорядоченное
строение, получение энергии извне и ее использование для выполне-
ния работы и поддержания упорядоченности (преодоление энтро-
пии), обмен веществ, активная реакция на раздражения, рост, разви-
тие, размножение, удвоение и передача биологической информации
потомкам, регенерация, адаптация к окружающей среде.
Клеточная теория в современной интерпретации включа-
ет следующие главные положения:
—
клетка является универсальной элементарной единицей живого;
—
клетки всех организмов принципиально сходны по своему стро-
ению, функции и химическому составу;
—
клетки размножаются только путем деления исходной клетки;
—
клетки хранят, перерабатывают и реализуют генетическую ин-
формацию;
—
многоклеточные организмы являются сложными клеточными
ансамблями, образующими целостные системы;
—
именно благодаря деятельности клеток в сложных организмах
осуществляются рост, развитие, обмен веществ и энергии.
5
ПРОКАРИОТИЧЕСКИЕ И ЭУКАРИОТИЧЕСКИЕ КЛЕТКИ
В XX в. за открытия в области цитологии и смежных наук были
присуждены Нобелевские премии. Среди лауреатов были:
в 1906 г. Камилло Гольджи и Сантьяго Рамон-и -Кахаль за откры-
тия в области структуры нейронов;
в 1908 г. Илья Мечников и Пауль Эрлих за открытия фагоцитоза
(Мечников) и антител (Эрлих);
в 1930 г. Карл Ландштейнер за открытие групп крови;
в 1931 г. Отто Варбург за открытие природы и механизмов дейст-
вия дыхательных ферментов цитохромоксидаз;
в 1946 г. Герман Меллер за открытие мутаций;
в 1953 г. Ханс Кребс за открытие цикла лимонной кислоты;
в 1959 г. Артур Корнберг и Северо Очоа за открытие механизмов
синтеза ДНК и РНК;
в 1962 г. Френсис Крик, Морис Уилкинсон и Джеймс Уотсон за
открытие молекулярной структуры нуклеиновых кислот и их значе-
ния для передачи информации в живых системах;
в 1963 г. Франсуа Жакоб, Андре Львов и Жак Моно за открытие
механизма синтеза белка;
в 1968 г. Хар Гобинд Корана, Маршалл Ниренберг и Роберт Хол-
ли за расшифровку генетического кода и его роли в синтезе белка;
в 1970 г. Джулиус Аксельрод, Бернард Кац и Ульф фон Эйлер за
открытие гуморальных медиаторов нервных окончаний и механизма
их хранения, выделения и инактивации;
в 1971 г. Эрл Сазерленд за открытие вторичного посредника
цАМФ и его роли в механизме действия гормонов;
в 1974 г. Кристиан де Дюв, Альберт Клод и Джордж Паладе за
открытия, касающиеся структурной и функциональной организации
клетки (ультраструктура и функция лизосом, комплекса Гольджи,
эндоплазматического ретикулума).
ПРОКАРИОТИЧЕСКИЕ
И ЭУКАРИОТИЧЕСКИЕ КЛЕТКИ
В настоящее время различают прокариотические и эукариотиче-
ские организмы. К первым принадлежат сине-зеленые водоросли, ак-
тиномицеты, бактерии, спирохеты, микоплазмы, риккетсии и хлами-
дии, ко вторым — большинство водорослей, грибы и лишайники,
растения и животные. В отличие от прокариотической, эукариотиче-
ская клетка имеет ядро, ограниченное оболочкой из двух мембран,
и большое количество мембранных органелл. Более детальные разли-
чия представлены в табл. 1 .
6
ХИМИЧЕСКАЯ ОРГАНИЗАЦИЯ КЛЕТКИ
Таблица 1
Признаки прокариотов и эукариотов
Признаки
Прокариоты
Эукариоты
Размер клеток
1—10 мкм
10—100 мкм
Вид метаболизма
Анаэробный
или аэробный
Аэробный
ДНК
Кольцевая в цитоплазме
Не кольцевая, очень длинная,
окружена ядерной оболочкой
Синтез РНК и белка
Итоидругое—
в цитоплазме
Синтез и процессинг РНК —
в ядре, белка — в цитоплазме
Органеллы
Нет или мало
Многочисленны и разно-
образны
Цитоскелет
Нет
Есть
Эндо- и экзоцитоз
Нет
Есть
Митохондрии
Нет
Есть
Эндоплазматическая сеть Нет
Есть
Комплекс Гольджи
Нет
Есть
Рибосомы
Есть: 70 S
Есть: 70 S — в митохондри-
ях, 80 S — в цитоплазме
Лизосомы
Нет
Есть
Внутриклеточное перева-
ривание
Нет
Есть
Деление клеток
Бинарное
Митоз (у половых клеток —
мейоз)
ХИМИЧЕСКАЯ ОРГАНИЗАЦИЯ КЛЕТКИ
Из всех элементов периодической системы Д. И. Менделеева
в организме человека обнаружено 86 постоянно присутствующих, из
них 25 необходимы для нормальной жизнедеятельности, 18 из кото-
рых необходимы абсолютно, а 7 полезны. Профессор Д. Р. Вильямc
назвал их «элементами жизни».
В состав веществ, участвующих в реакциях, связанных с жизнедея-
тельностью клетки, входят почти все известные химические элементы,
причем на долю четырех из них приходится около 98% массы клетки.
Это кислород (65—75%), углерод (15—18%), водород (8—10%) и азот
(1,5—3,0%). Остальные элементы подразделяются на две группы: ма-
7
ХИМИЧЕСКАЯ ОРГАНИЗАЦИЯ КЛЕТКИ
кроэлементы (около 1,9%) и микроэлементы (около 0,1%). К макро-
элементам относятся сера, фосфор, хлор, калий, натрий, магний, каль-
ций и железо, к микроэлементам — цинк, медь, йод, фтор, марганец,
селен, кобальт, молибден, стронций, никель, хром, ванадий и др. Несмо-
тря на очень малое содержание, микроэлементы играют важную роль.
Они влияют на обмен веществ. Без них невозможна нормальная жиз-
недеятельность каждой клетки в отдельности и организма как целого.
Клетка состоит из неорганических и органических веществ. Среди
неорганических преобладает вода, ее относительное количество со-
ставляет от 70 до 80%. Вода — универсальный растворитель, в ней
происходят все биохимические реакции в клетке, при участии воды
осуществляется ее теплорегуляция. Вещества, растворяющиеся в воде
(соли, основания, кислоты, белки, углеводы, спирты и др.), называют-
ся гидрофильными. Гидрофобные вещества (жиры и жироподобные) не
растворяются в воде. Есть органические вещества с вытянутыми мо-
лекулами, у которых один конец гидрофилен, другой же гидрофобен;
их называют амфипатическими. Примером амфипатических веществ
могут служить фосфолипиды, участвующие в образовании биологи-
ческих мембран.
Неорганические вещества (соли, кислоты, основания, положи-
тельные и отрицательные ионы) составляют от 1,0 до 1,5% массы
клетки. Среди органических веществ преобладают белки (10—20%),
жиры, или липиды (1—5%), углеводы (0,2—2,0%), нуклеиновые кис-
лоты (1—2%). Содержание низкомолекулярных веществ в клетке не
превышает 0,5%.
Молекула белка является полимером, который состоит из боль-
шого количества повторяющихся единиц (мономеров). Мономеры
белка — аминокислоты (их 20) соединены между собой пептидными
связями, образуя полипептидную цепь (первичную структуру белка).
Она закручивается в спираль, представляющую, в свою очередь, вто-
ричную структуру белка. Благодаря определенной пространственной
ориентации полипептидной цепи возникает третичная структура бел-
ка, которая определяет специфичность и биологическую активность
молекулы белка. Несколько третичных структур, объединяясь между
собой, образуют четвертичную структуру.
Белки выполняют важнейшие функции. Ферменты — биологиче-
ские катализаторы, увеличивающие скорость химических реакций
в клетке в сотни тысяч — миллионы раз, являются белками. Белки,
входя в состав всех клеточных структур, выполняют пластическую
(строительную) функцию. Они образуют клеточный скелет. Движе-
ния клеток также осуществляют специальные белки (актин, миозин,
динеин). Белки обеспечивают транспорт веществ в клетку, из клетки
и внутри клетки. Антитела, которые наряду с регуляторными выпол-
8
ХИМИЧЕСКАЯ ОРГАНИЗАЦИЯ КЛЕТКИ
няют и защитные функции, также являются белками. И, наконец, бел-
ки являются одним из источников энергии.
Углеводы подразделяются на моносахариды и полисахариды. Поли-
сахариды, подобно белкам, построены из мономеров — моносахаридов.
Среди моносахаридов в клетке наиболее важны глюкоза (содержит
шесть атомов углерода) и пентоза (пять атомов углерода). Пентозы вхо-
дят в состав нуклеиновых кислот. Моносахариды хорошо растворяются
в воде, полисахариды — плохо. В животных клетках полисахариды пред-
ставлены гликогеном, в растительных — в основном, растворимым крах-
малом и нерастворимыми целлюлозой, гемицеллюлозой, пектином и др.
Углеводы являются источником энергии. Сложные углеводы, соединен-
ные с белками (гликопротеины) и (или) жирами (гликолипиды), участ-
вуют в образовании клеточных поверхностей и взаимодействиях клеток.
К липидам относятся жиры и жироподобные вещества. Молекулы
жиров построены из глицерина и жирных кислот. К жироподобным
веществам относятся холестерин, некоторые гормоны, лецитин. Ли-
пиды, являющиеся основным компонентом клеточных мембран (они
описаны ниже), выполняют тем самым строительную функцию. Они
являются важнейшим источником энергии. Так, если при полном
окислении 1 г белка или углеводов освобождается 17,6 кДж энергии,
то при полном окислении 1 г жира — 38,9 кДж.
Нуклеиновые кислоты являются полимерными молекулами,
образованными мономерами — нуклеотидами, каждый из которых со-
стоит из пуринового или пиримидинового основания, сахара пентозы
и остатка фосфорной кислоты. Во всех клетках существует два типа
нуклеиновых кислот: дезоксирибонуклеиновая (ДНК) и рибонуклеи-
новая (РНК), которые отличаются по составу оснований и сахаров
(табл. 2, рис. 1).
Таблица 2
Состав нуклеиновых кислот
Кислота
Сахар
Азотистые основания
пуриновые
пиримидиновые
РНК
Рибоза
Аденин (А)
Гуанин (G)
Цитозин (С)
Урацил (U)
ДНК
Дезоксирибоза
Аденин (А)
Гуанин (G)
Цитозин (С)
Тимин (Т)
Молекула РНК образована одной полинуклеотидной цепью
(рис. 2).
Молекула ДНК состоит из двух разнонаправленных полинуклеотид-
ных цепей, закрученных одна вокруг другой в виде двойной спирали.
9
ХИМИЧЕСКАЯ ОРГАНИЗАЦИЯ КЛЕТКИ
Каждый нуклеотид состоит из азотистого основания, сахара и остатка
фосфорной кислоты. При этом основания расположены внутри двойной
спирали, а сахарофосфатный скелет — снаружи. Азотистые основания
обеих цепей соединены между собой комплементарно водородными свя-
зями, при этом аденин соединяется только с тимином, а цитозин с гуа-
нином (А=Т, G ≡С). В зависимости от номера атома по отношению
к связи с основанием концы цепи обозначают как 5 и 3 (см. рис. 1 и 2).
Рис. 1. Строение молекул нуклеиновых кислот:
I — РНК, II — нумерация атомов углерода в цикле пентозы, III — ДНК
Звездочкой (*) отмечены различия в строении ДНК и РНК. Валентные связи показаны
упрощенно. А — аденин, Т — тимин, С — цитозин, G — гуанин, U — урацил
O–
O–
O
O
O
OO
O
O
O
O
O
O
G
C
A
U*
O
O
O
O
O
O
O
CH2
CH2
CH2
CH2
OH
OH
OH
OH
*
OH
P
P
P
P
O
C5
C4
C3 C2
C1
O–
O–
O
O
O
OO
O
O
O
O
O
O
G
C
A
T*
O
O
O
O
O
O
O
CH2
CH2
CH2
CH2
OH
OH
OH
H
*
OH
P
P
P
P
5' конец
3' конец
I
II
III
10
ХИМИЧЕСКАЯ ОРГАНИЗАЦИЯ КЛЕТКИ
ДНК несет в себе генетическую информацию, закодированную по-
следовательностью азотистых оснований. Она определяет специфич-
ность синтезируемых клеткой белков, т. е. последовательность амино-
кислот в полипептидной цепи. Вместе с ДНК дочерним клеткам
передается генетическая информация, определяющая (во взаимодей-
ствии с условиями среды) все свойства клетки. ДНК содержится
в ядре и митохондриях, а у растений и в хлоропластах.
Все биохимические реакции в клетке строго структурированы и
осуществляются при участии высокоспецифических биокатализато-
ров — ферментов, или энзимов (от греч. en
—
в, zyme — брожение,
закваска) — белков, которые, соединяясь с биологическими молеку-
лами — субстратами, снижают энергию активации, необходимую для
осуществления той или иной реакции (энергия активации — это ми-
нимальное количество энергии, необходимое молекуле для вступле-
ния в химическую реакцию). Ферменты ускоряют реакцию на 10 по-
рядков (в 1010 раз).
Рис. 2 . Пространственная структура нуклеиновых кислот:
I — РНК, II — ДНК; ленты — сахарофосфатные остовы, А, С, G, Т, U —
азотистые основания, решетки между ними — водородные связи
(по Б. Албертсу и соавт., с изменениями)
5°
3°
5°
I
II
3°
11
СТРОЕНИЕ КЛЕТКИ
Названия всех ферментов складываются из двух частей. Первая
содержит указание либо на субстрат, либо на действие, либо на то
и другое. Вторая часть — окончание, оно всегда представлено буквами
«аза». Так, название фермента «сукцинатдегидрогеназа» означает, что
он воздействует на соединения янтарной кислоты («сукцинат-»), от-
нимая от них водород («-дегидроген-»).
По общему типу воздействия ферменты подразделяются на шесть
классов. Оксиредуктазы катализируют окислительно-восстанови-
тельные реакции, трансферазы участвуют в переносе функциональ-
ных групп, гидролазы обеспечивают реакции гидролиза, лиазы — при-
соединение групп по двойным связям, изомеразы осуществляют
перевод соединений в другую изомерную форму, а лигазы (не путать
с лиазами!) связывают молекулярные группировки в цепи.
Основа любого фермента — белок. Вместе с тем есть ферменты,
которые не обладают каталитической активностью, пока к белковой
основе (апоферменту) не присоединится более простая по строению
небелковая группировка — кофермент. Иногда коферменты имеют
собственные названия, иногда их обозначают буквами. Нередко в со-
став коферментов входят вещества, называемые теперь витаминами.
Многие витамины не синтезируются в организме и должны поэтому
поступать с пищей. При их недостатке возникают заболевания (ави-
таминозы), симптомы которых, по сути дела, это проявления недоста-
точной активности соответствующих ферментов.
Некоторые коферменты играют ключевую роль во многих важ-
нейших биохимических реакциях. В качестве примера можно приве-
сти кофермент А (КоА), который обеспечивает перенос группировок
уксусной кислоты. Кофермент никотинамидадениндинуклеотид
(сокращенно — НАД) обеспечивает перенос ионов водорода в окис-
лительно-восстановительных реакциях; таковы же и никотинами-
дадениндинуклеотидфосфат (НАДФ), флавинадениндинуклеотид
(ФАД) и ряд других. Кстати, никотинамид — один из витаминов.
СТРОЕНИЕ КЛЕТКИ
Клетка является основной структурной и функциональной едини-
цей живых организмов, осуществляющей рост, развитие, обмен ве-
ществ и энергии, хранящей, перерабатывающей и реализующей гене-
тическую информацию. Клетка представляет собой сложную систему
биополимеров, отделенную от внешней среды плазматической мем-
браной (цитолеммой, плазмалеммой) и состоящую из ядра и цито-
плазмы, в которой располагаются органеллы и включения.
Французский ученый, лауреат Нобелевской премии А. Львов,
основываясь на достижениях современной цитологии, писал: «Рас-
12
СТРОЕНИЕ КЛЕТКИ
сматривая живой мир на клеточном уровне, мы обнаруживаем его
единство: единство строения — каждая клетка содержит ядро, погру-
женное в цитоплазму; единство функции — обмен веществ, в основ-
ном, сходен во всех клетках; единство состава — главные макромоле-
кулы у всех живых существ состоят из одних и тех же малых молекул.
Для построения огромного разнообразия живых систем природа ис-
пользует ограниченное число строительных блоков». Вместе с тем
различные клетки имеют и специфические структуры. Это связано
с выполнением ими специальных функций.
Размеры клеток человека варьируют от нескольких микрометров
(например, малые лимфоциты — около 7) до 200 мкм (яйцеклетка).
Напомним, что один микрометр (мкм) = 10–6 м, 1 нанометр (нм) =
= 10–9 м, 1 ангстрем (Е) = 10–10 м. Форма клеток разнообразна. Они
могут быть шаровидными, овоидными, веретенообразными, плос-
кими, кубическими, призматическими, полигональными, пирами-
дальными, звездчатыми, чешуйчатыми, отростчатыми, амебовидны-
миидр.
Основными функциональными структурами клетки являются ее
поверхностный комплекс, цитоплазма и ядро (рис. 3).
Поверхностный комплекс включает в себя гликокаликс, плазма-
тическую мембрану (плазмалемму) и кортикальный слой цитоплаз-
мы. Нетрудно видеть, что резкого отграничения поверхностного ком-
плекса от цитоплазмы нет.
В цитоплазме выделяют гиалоплазму (матрикс, цитозоль), орга-
неллы и включения.
Основными структурными компонентами ядра являются карио-
лемма (кариотека), нуклеоплазма и хромосомы; петли некоторых хро-
мосом могут переплетаться, и в этой области образуется ядрышко.
Нередко к структурным элементам ядра относят хроматин. Однако,
по определению, хроматин — это вещество хромосом.
Плазмалемма, кариолемма и часть органелл образованы биологи-
ческими мембранами.
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Наиболее полно строение биологических мембран отражает жид-
костно-мозаичная модель, первоначальный вариант которой был
предложен в 1972 г. Г. Николсоном и С. Сингером. Мембрана состоит
из двух слоев амфипатических молекул липидов (билипидный слой,
или бислой). Каждая такая молекула имеет две части — головку
и хвост. Хвосты гидрофобны и обращены друг к другу. Головки, на-
13
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Рис. 3 . Ультрамикроскопическое строение клетки:
1 — цитолемма (плазматическая мембрана), 2 — пиноцитозные пузырьки,
3 — центросома клеточный центр (цитоцентр), 4 — гиалоплазма, 5 — эндоплазматиче-
ская сеть: А — мембрана зернистой сети, Б — рибосомы, 6 — связь перинуклеарного
пространства с полостями эндоплазматической сети, 7 — ядро, 8 — ядерные поры,
9 — незернистая (гладкая) эндоплазматическая сеть, 10 — ядрышко, 11 — внутренний
сетчатый аппарат (комплекс Гольджи), 12 — секреторные вакуоли, 13 — митохондрия,
14 — липосомы, 15 — три последовательные стадии фагоцитоза, 16 — связь клеточной
оболочки (цитолеммы) с мембранами эндоплазматической сети
1
7
13
2
8
14
3
9
15
4
10
16
Б
5
11
6
А
12
14
СТРОЕНИЕ КЛЕТКИ
против, гидрофильны и направлены кнаружи и внутрь клетки. В би-
липидный слой погружены молекулы белка (рис. 4).
На рис. 5 схематически представлена молекула фосфолипида фос-
фатидилхолина. Одна из жирных кислот — насыщенная, другая — не-
насыщенная. Молекулы липидов способны быстро диффундировать
в боковом направлении в пределах одного монослоя и крайне редко
переходят из одного монослоя в другой.
Химический состав цитолеммы эу- и прокариотических клеток су-
щественно отличается (таблица 3).
Рис. 4. Структура биологической мембраны:
1 — внешние белки, 2 — белок в толще мембраны, 3 — внутренние белки,
4 — интегральный (трансмембранный) белок, 5 — фосфолипиды билипидного слоя
Рис. 5. Молекула фосфолипида фосфатидилхолина:
А — полярная (гидрофильная) головка: 1 — холин, 2 — фосфат, 3 — глицерол;
Б — неполярный (гидрофобный) хвост: 4 — насыщенная жирная кислота,
5 — ненасыщенная жирная кислота, СН=СН — цисдвойная связь
1
A
Б
2
3
4
5
CH
CH
5
4
1
2
2
3
15
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Билипидный слой ведет себя как жидкость, обладающая значи-
тельным поверхностным натяжением. Вследствие этого он образует
замкнутые полости, которые не спадаются.
Таблица 3
Состав липидов клеточных мембран эукариот и прокариот
Липид
Эукариоты
Прокариоты
Холестерол
Фосфатидилэтаноламин
Фосфатидилсерин
Фосфатидилхолин
Сфингомиелин
Гликолипиды
Жирные кислоты
Имеется
Имеется
Имеется
Имеется
Имеется
Имеются2
Насыщенные и ненасы-
щенные (моно- и поли-)
Отсутствует1
Имеется в больших количествах
Имеется в следовых количествах
Отсутствует
Отсутствует
Отсутствуют
Только насыщенные или моно-
ненасыщенные3
1 У микоплазм в состав мембран входят экзогенные стерины.
2 У растений преобладают гликолипиды из глицерола.
3 У некоторых цианобактерий имеются полиненасыщенные жирные кислоты.
Некоторые белки проходят через всю толщу мембраны, так что
один конец молекулы обращен в пространство по одну сторону мем-
браны, другой — по другую. Их называют интегральными (трансмем-
бранными). Другие белки расположены так, что в околомембранное
пространство обращен лишь один конец молекулы, второй же конец
лежит во внутреннем или в наружном монослое мембраны. Такие бел-
ки называют внутренними или, соответственно, внешними (иногда те
и другие называют полуинтегральными). Некоторые белки (обычно
переносимые через мембрану и временно находящиеся в ней) могут
лежать между фосфолипидными слоями.
Концы белковых молекул, обращенные в околомембранное про-
странство, могут связываться с различными веществами, находя-
щимися в этом пространстве, поэтому интегральные белки играют
большую роль в организации трансмембранных процессов. С полуин-
тегральными белками всегда связаны молекулы, осуществляющие ре-
акции по восприятию сигналов из среды (молекулярные рецепторы)
или по передаче сигналов от мембраны в среду. Многие белки облада-
ют ферментативными свойствами.
Бислой асимметричен: в каждом монослое располагаются различ-
ные липиды, гликолипиды обнаруживаются только в наружном моно-
слое так, что их углеводные цепи направлены кнаружи. Молекулы хо-
лестерина в мембранах эукариот лежат во внутренней, обращенной
к цитоплазме половине мембраны. Цитохромы располагаются в наруж-
ном монослое, а АТФ-синтетазы — на внутренней стороне мембраны.
16
СТРОЕНИЕ КЛЕТКИ
Подобно липидам, белки также способны к латеральной диффу-
зии, однако скорость ее меньше, чем у липидных молекул. Переход из
одного монослоя в другой практически невозможен.
Из мембранных белков прокариот лучше всего изучен транспорт-
ный белок бактериородопсин, который содержится в «пурпурной
мембране» (Halobacterium halobium). L-спираль бактериородопсина
пересекает липидный бислой 7 раз (рис. 6).
Бактериородопсин представляет собой полипептидную цепь, со-
стоящую из 248 аминокислотных остатков и простетической груп-
пы — хромофора, поглощающего кванты света и ковалентно связан-
ного с лизином. Под влиянием кванта света хромофор возбуждается,
что приводит к конформационным изменениям полипептидной цепи.
Это вызывает перенос двух протонов с цитоплазматической поверх-
ности мембраны на ее внешнюю поверхность, в результате чего в мем-
бране возникает электрический потенциал, вызывающий синтез АТФ.
Среди мембранных белков прокариот различают пермеазы — пере-
носчики, ферменты, осуществляющие различные синтетические про-
цессы, в том числе синтез АТФ.
Концентрация веществ, в частности ионов, по обе стороны мем-
браны неодинакова, поэтому каждая сторона несет свой электриче-
ский заряд. Различия в концентрации ионов создают соответственно
и разность электрических потенциалов.
Поверхностный комплекс
Поверхностный комплекс обеспечивает взаимодействие клетки
с окружающей ее средой. В связи с этим он выполняет следующие
основные функции: разграничительную (барьерную), транспортную,
рецепторную (восприятие сигналов из внешней для клетки среды),
а также функцию передачи информации, воспринятой рецепторами,
глубоким структурам цитоплазмы.
Рис. 6 . Структура молекулы бактериородопсина
и ее расположение в липидном бислое:
1 — цитоплазма, 2 — спираль, 3 — липидный бислой (по Б. Албертсу и соавт.)
1
2
3
COOH
NH2
17
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Основой поверхностного комплекса является биологическая мем-
брана, называемая наружной клеточной мембраной (иначе — плаз -
малеммой). Ее толщина около 10 нм, так что в световом микроскопе
она неразличима. О строении и роли биологических мембран как та-
ковых сказано ранее, плазмалемма же обеспечивает, в первую очередь,
разграничительную функцию по отношению к внешней для клетки
среде. Естественно, она выполняет при этом и другие функции: транс-
портную и рецепторную (восприятия сигналов из внешней для клет-
ки среды). Плазмалемма, таким образом, обеспечивает поверхностные
свойства клетки (рис. 7).
Наружный и внутренний электроноплотные слои плазмалеммы
имеют толщину около 2—5 нм, средний электронопрозрачный слой —
около 3 нм. При замораживании — скалывании мембрана разделяется
на два слоя: слой А, содержащий многочисленные, иногда располо-
женные группами крупные частички размерами 8—9,5 нм, и слой В,
содержащий примерно такие же частички (но в меньшем количестве)
и мелкие углубления. Слой А — это скол внутренней (цитоплазмати-
ческой) половины мембраны, слой В — наружной.
В билипидный слой плазмалеммы погружены молекулы белка. Не-
которые из них (интегральные, или трансмембранные) проходят через
всю толщину мембраны, другие (периферические или внешние) лежат
во внутреннем или наружном монослоях мембраны. Некоторые интег-
ральные белки связаны нековалентными связями с белками цитоплаз-
мы. Подобно липидам, белковые молекулы также являются амфипати-
ческими: их гидрофобные участки окружены аналогичными «хвостам»
липидов, а гидрофильные обращены наружу или внутрь клетки.
Белки осуществляют бо
льшую часть мембранных функций: мно-
гие из них являются рецепторами, другие — ферментами, третьи —
Рис. 7 . Строение клеточной мембраны, схема: 1 — углеводные цепи,
2 — гликолипид, 3 — гликопротеид, 4 — углеводородный «хвост», 5 — полярная
«головка», 6 — белок, 7 — холестерин, 8 — микротрубочки (по А. Хэму и Д. Кормаку)
1
2
3
4
5
6
7
8
18
СТРОЕНИЕ КЛЕТКИ
переносчиками. Подобно липидам, белки также способны к латераль-
ной диффузии, однако скорость ее меньшая, чем у липидных молекул.
Переход молекул белка из одного монослоя в другой практически не-
возможен. Так как в каждом монослое содержатся свои белки, бислой
асимметричен. Несколько белковых молекул могут образовать канал,
через который проходят определенные ионы или молекулы.
Одной из важнейших функций биологических и, в том числе, плаз-
матической мембраны является транспорт. Напомним, что обра-
щенные друг к другу «хвосты» липидов образуют гидрофобный слой,
препятствующий проникновению полярных водорастворимых моле-
кул. Как правило, внутренняя цитоплазматическая поверхность плаз-
малеммы несет отрицательный заряд, что облегчает проникновение
в клетку положительно заряженных ионов.
Малые (18 Да) незаряженные молекулы воды быстро диффунди-
руют через мембраны, также быстро диффундируют малые полярные
молекулы (например, мочевина, СO2, глицерол), гидрофобные моле-
кулы (O2, N3, бензол), крупные незаряженные полярные молекулы
вообще не способны диффундировать (глюкоза, сахароза). В то же
время через цитолемму указанные вещества диффундируют легко
благодаря наличию в ней мембранных транспортных белков, специ-
фических для каждого химического соединения. Эти белки могут
функционировать по принципу унипорта (перенос одного вещества
через мембрану) или котранспорта (перенос двух веществ). Послед-
ний может быть в виде симпорта (перенос двух веществ в одном на-
правлении), либо антипорта (перенос двух веществ в противополож-
ных направлениях (рис. 8). При транспорте вторым веществом
Рис. 8. Схема функционирования транспортных белков:
1 — транспортируемая молекула, 2 — котранспортируемая молекула,
3 — липидный бислой, 4 — белок-переносчик, 5 — антипорт, 6 — симпорт,
7 — котранспорт, 8 — унипорт (по Б. Албертсу и соавт.)
8
5
4
3
2
1
6
1
7
5
19
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
является Н+
. Унипорт и симпорт являются основными способами пе-
реноса в прокариотическую клетку большей части веществ, необходи-
мых для ее жизнедеятельности.
Различают два типа транспорта: пассивный и активный. Первый не
требует затрат энергии, второй — энергозависимый (рис. 9). Пассивный
транспорт незаряженных молекул осуществляется по градиенту кон-
центрации, транспорт заряженных молекул зависит от градиента кон-
центрации Н+ и трансмембранной разности потенциалов, которые объ-
единяются в трансмембранный градиент Н+
, или электрохимический
протонный градиент (рис. 10). Как правило, внутренняя цитоплазмати-
ческая поверхность мембраны несет отрицательный заряд, что облегча-
ет проникновение в клетку положительно заряженных ионов.
Рис. 9. Схема пассивного транспорта по электрохимическому градиенту
и активного транспорта против электрохимического градиента:
1 — транспортируемая молекула, 2 — каналообразуюший белок,
3 — белок-переносчик, 4 — электрохимический градиент, 5 — энергия,
6 — активный транспорт, 7 — пассивный транспорт, 8 — облегченная диффузия,
9 — простая диффузия, 10 — липидный бислой
(по Б. Албертсу и соавт.)
Рис. 10. Электрохимический
протонный градиент.
Составляющие градиента:
1 — внутренняя митохондриальная
мембрана, 2 — матрикс,
3 — протондвижущая сила,
обусловленная мембранным
потенциалом, 4 — протондвижущая
сила, обусловленная градиентом
концентрации протонов
(по Б. Албертсу и соавт.)
7
8
9
10
11
5
4
3
2
1
6
1
Âíåêëåòî÷íîå
ïðîñòðàíñòâî
Öèòîïëàçìà
4
3
2
1
1
Êèñëàÿ ñðåäà
ΔΔΔΔΔV
ΔΔΔΔΔpH
Ùåëî÷íàÿ
ñðåäà
nH+
nH+
nH+
nH+
H+H
+
H+H
+
H+
H+
H+
2
20
СТРОЕНИЕ КЛЕТКИ
Диффузия (от лат. diffusio — распространение, растекание) — это
переход ионов или молекул, вызванный их броуновским движением,
через мембраны из зоны, где эти вещества находятся в более высокой
концентрации, в зону с более низкой концентрацией до тех пор, пока
концентрации по обе стороны мембраны выровняются. Диффузия
может быть нейтральной (незаряженные вещества проходят между
липидными молекулами или через белок, формирующий канал) или
облегченной (специфические белки-переносчики связывают вещест-
во и переносят его через мембрану). Облегченная диффузия протека-
ет быстрее, чем нейтральная. На рис. 11 показана гипотетическая
модель функционирования белков-переносчиков при облегченной
диффузии.
Вода поступает в клетку путем осмоса (от греч. osmos — толчок,
давление). В настоящее время математически доказывается наличие
в цитолемме мельчайших временны
х пор, возникающих по мере необ-
ходимости.
Активный транспорт осуществляют белки-переносчики, при этом
расходуется энергия, получаемая вследствие гидролиза АТФ или про-
тонного потенциала. Активный транспорт происходит против гради-
ента концентрации.
В транспортных процессах прокариотической клетки основную
роль играет электрохимический протонный градиент, при этом пере-
нос идет против градиента концентрации веществ. На цитолемме эука-
риотических клеток с помощью натриево-калиевого насоса поддержи-
вается мембранный потенциал. Этот насос, функционирующий как
антипорт, накачивающий против градиентов концентрации К+ в клет-
ку, а Nа+ во внеклеточную среду, является ферментом АТФ-азой
(рис. 12). При этом в АТФ-азе происходят конформационные измене-
ния, в результате которых Nа+ переносится через мембрану и выводит-
Рис. 11. Схема функционирования белков-переносчиков:
1 — транспортируемое вещество, 2 — градиент концентрации, 3 — транспортный
белок, осуществляющий облегченную диффузию, 4 — липидный бислой
(по Б. Албертсу и соавт.)
4
3
2
1
«Ïîí㻫Ïèíã»
À
À
À
À
À
À
À
À
À
À
21
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
ся во внеклеточную среду, а К+ переносится внутрь клетки. Процесс
напоминает модель облегченной диффузии, изображенной на рис. 11 .
АТФ-аза осуществляет также активный транспорт аминокислот
и сахаров. Аналогичный механизм присутствует в цитолемме аэробных
бактерий. Однако у них фермент вместо гидролиза АТФ осуществляет
его синтез из АДФ и фосфата, используя протонный градиент. Таким
же образом функционирует описанный выше бактериородопсин. Ины-
ми словами, один и тот же фермент осуществляет и синтез и гидролиз
АТФ.
В связи с наличием суммарного отрицательного заряда в цито-
плазме прокариотической клетки ряд незаряженных молекул перено-
сится по принципу симпорта с Н+
, источником энергии является
трансмембранный электрохимический градиент Н+ (например, гли-
цин, галактоза, глюкоза), отрицательно заряженные вещества перено-
сятся по принципу симпорта также с Н+ за счет градиента концентра-
ции Н+
, транспорт Nа+ осуществляется по принципу антипорта с Н+
,
который переносится в клетку также за счет градиента концентрации
Н+; механизм аналогичен (Nа+ К
+
) насосу эукариот. Положительно за-
ряженные вещества поступают в клетку по принципу унипорта за
счет трансмембранной разности электрических потенциалов.
Внешняя поверхность плазмалеммы покрыта гликокаликсом
(рис. 13). Толщина его различна и колеблется даже в разных участках
поверхности одной клетки от 7,5 до 200 нм. Гликокаликс представля-
Рис. 12. (Nа+К
+
)АТФ-аза:
1 — градиент концентрации ионов натрия, 2 — участок связывания калия,
3 — градиент концентрации ионов калия, 4 — участок связывания натрия.
I — внеклеточное пространство, II — внутриклеточное пространство
(по Б. Албертсу и соавт.)
I
II
4
3
2
1
Na+
Na+
Na+
K+
K+
ADP+Pi
ATP
22
СТРОЕНИЕ КЛЕТКИ
ет собой совокупность молекул, связанных с белками мембраны. По
составу эти молекулы могут представлять собой цепочки полисахари-
дов, гликолипидов и гликопротеинов.
Многие из молекул гликокаликса функционируют в качестве спе-
цифических молекулярных рецепторов. Концевой свободный отдел
рецептора обладает уникальной пространственной конфигурацией.
Поэтому с ним могут объединяться только те молекулы, находящиеся
вне клетки, которые обладают также уникальной конфигурацией, но
зеркально симметричной по отношению к рецептору. Именно благо-
даря существованию специфических рецепторов на поверхности
клетки могут закрепляться так называемые сигнальные молекулы,
в частности молекулы гормонов.
Чем больше конкретных специфических рецепторов находится
в гликокаликсе, тем активнее клетка реагирует на соответствующие
сигнальные вещества. Если в гликокаликсе нет молекул, специфиче-
ски связывающихся с внешними веществами, клетка на последние не
реагирует. Таким образом, гликокаликс наряду с самой плазмалеммой
обеспечивает и барьерную функцию поверхностного комплекса.
К глубокой поверхности плазмалеммы примыкают поверхностные
структуры цитоплазмы. Они связываются с белками плазмалеммы
и осуществляют передачу информации глубинным структурам, запу-
ская сложные цепи биохимических реакций. Они же, изменяя свое
взаимоположение, меняют конфигурацию плазмалеммы.
Межклеточные соединения
При контакте клеток друг с другом их плазмалеммы вступают во
взаимодействия. При этом образуются особые объединяющие струк-
1
2
3
45
Рис. 13. Гликокаликс:
1 — гликокаликс, выявленный специальным красителем (рутениевым красным),
2 — плазмалемма (часть гликокаликса на этом участке удалена), 3 — цитоплазма,
4 — кариотека, 5 — хроматин (по Б. Албертсу и соавт., с изменениями)
23
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
туры — межклеточные соединения (рис. 14). Они формируются при
образовании многоклеточного организма во время эмбрионального
развития и при образовании тканей. Межклеточные соединения по-
дразделяются на простые и сложные. В простых соединениях плаз-
малеммы соседних клеток формируют выросты наподобие зубцов, так
Рис. 14. Межклеточные соединения:
I — плотное соединение, II — десмосома, III — полудесмосома, IV — нексус
(щелевидное соединение); 1 — плазмалеммы смежных клеток, 2 — зоны слипания,
3 — электроноплотные пластинки, 4 — промежуточные филаменты (тонофиламенты),
закрепленные в пластинке, 5 — межклеточные филаменты, 6 — базальная мембрана,
7 — подлежащая соединительная ткань, 8 — коннексоны,
каждый из которых состоит из шести субъединиц с цилиндрическим каналом
(по А. Хэму и Д. Кормаку и по Б. Албертсу и соавт., с изменениями)
1
2
3
À
Ä
å
ñ
ì
î
ñ
î
ì
à
Á
1
2
3
Ïîëóäåñìîñîìà
Â
1
2
3
4
5
Ñîåäèíèòåëüíàÿ
òêàíü
1
1
2
Ã
24
СТРОЕНИЕ КЛЕТКИ
что зубец одной клетки внедряется между двумя зубцами другой (зуб-
чатое соединение) или переплетающихся между собой интердигита-
ций (пальцевидное соединение). Между плазмалеммами соседних кле-
ток всегда сохраняется межклеточная щель шириной 15—20 нм.
Сложные соединения, в свою очередь, подразделяются на адгезион-
ные, замыкающие и проводящие. К адгезионным соединениям отно-
сятся десмосома, полудесмосома и поясок сцепления (лентовидная дес-
мосома). Десмосома состоит из двух электроноплотных половин,
принадлежащих плазмалеммам соседних клеток, разделенных межкле-
точным пространством размерами около 25 нм, заполненным тонкофи-
бриллярным веществом гликопротеинной природы. К обращенным
к цитоплазме сторонам обеих пластинок десмосомы прикрепляются
кератиновые тонофиламенты, напоминающие по форме головные
шпильки. Помимо этого, через межклеточное пространство проходят
межклеточные волокна, соединяющие обе пластинки.
Полудесмосома, образованная лишь одной пластинкой с входя-
щими в нее тонофиламентами, прикрепляет клетку к базальной мем-
бране. Поясок сцепления, или лентовидная десмосома, представля-
ет собой «ленту», которая огибает всю поверхность клетки вблизи ее
апикального отдела. Ширина межклеточного пространства, заполнен-
ного волокнистым веществом, не превышает 15—20 нм. Цитоплазма-
тическая поверхность «ленты» уплотнена и укреплена сократитель-
ным пучком актиновых филаментов.
Плотные соединения, или запирающие зоны, проходят через
апикальные поверхности клеток в виде поясков шириной 0,5—0,6 мкм.
В плотных контактах между плазмалеммами соседних клеток практи-
чески нет межклеточного пространства и гликокаликса. Белковые мо-
лекулы обеих мембран контактируют между собой, поэтому через
плотные контакты молекулы не проходят. На плазмалемме одной
клетки имеется сеть гребешков, образованных цепочками белковых
частиц эллиптической формы, расположенных во внутреннем моно-
слое мембраны, которым на плазмалемме соседней клетки соответст-
вуют углубления, бороздки.
К проводящим соединениям относятся нексус, или щелевидный
контакт, и синапс. Через них из одной клетки в другую проходят во-
дорастворимые малые молекулы с молекулярной массой не более
1500 Да. Такими контактами соединены очень многие клетки челове-
ка (и животных). В нексусе между плазмалеммами соседних клеток
имеется пространство шириной 2—4 нм. Обе плазмалеммы соединены
между собой коннексонами — полыми гексагональными белковыми
структурами размерами около 9 нм, каждая из которых образована
шестью белковыми субъединицами. Методом замораживания и ска-
лывания показано, что на внутренней части мембраны имеются гекса-
25
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
гональные частички размерами 8—9 нм, а на наружной — соответ-
ствующие им ямки. Щелевые контакты играют важную роль в
осуществлении функции клеток, обладающих выраженной электри-
ческой активностью (например, кардиомиоциты). Синапсы играют
важную роль в осуществлении функций нервной системы.
Микроворсинки
Микроворсинки обеспечивают увеличение клеточной поверхно-
сти. Это, как правило, связано с осуществлением функции всасывания
веществ из внешней для клетки среды. Микроворсинки (рис. 15) явля-
ются производными поверхностного комплекса клетки. Они представ-
ляют собой выпячивания плазмалеммы длиной 1—2 и диаметром до
0,1 мкм. В гиалоплазме проходят продольные пучки актиновых микро-
филаментов, поэтому длина микроворсинок может изменяться. Это
один из способов регуляции активности поступления в клетку веществ.
У основания микроворсинки в поверхностном комплексе клетки про-
исходит объединение ее микрофиламентов с элементами цитоскелета.
Поверхность микроворсинок покрыта гликокаликсом. При особой
активности всасывания микроворсинки так близко располагаются
друг к другу, что их гликокаликс сливается. Такой комплекс называют
щеточной каймой. В щеточной кайме многие молекулы гликокаликса
обладают ферментативной активностью.
Особо крупные микроворсинки длиной до 7 мкм называют стерео-
цилиями (см. рис. 15). Они имеются у некоторых специализированных
клеток (например, у сенсорных клеток в органах равновесия и слуха).
Их роль связана не со всасыванием, а с тем, что они могут отклоняться
от своего первоначального положения. Такое изменение конфигурации
Рис. 15. Микроворсинки и реснички:
1 — микроворсинки, 2 — митохондрии, 3 — базальная мембрана,
4 — реснички, 5 — митохондрии
1
4
2
5
3
26
СТРОЕНИЕ КЛЕТКИ
поверхности клетки вызывает ее возбуждение, последнее воспринима-
ется нервными окончаниями, и сигналы поступают в центральную
нервную систему. Стереоцилии можно рассматривать как специальные
органеллы, развившиеся путем модификации микроворсинок.
Биологические мембраны разделяют клетку на отдельные обла-
сти, имеющие свои структурные и функциональные особенности —
компартменты, а также отграничивают клетку от окружающей ее
среды. Соответственно и мембраны, связанные с этими компартмен-
тами, имеют свои характерные черты.
Ядро
Оформленное ядро клетки (рис. 16) имеется только у эукариот.
У прокариот тоже имеются такие ядерные структуры, как хромосомы,
но они не заключены в особом компартменте. У большинства клеток
форма ядра шаровидная или овоидная, однако встречаются ядра и дру-
гой формы (кольцевидные, палочковидные, веретеновидные, бобовид-
ные, сегментированные и др.) . Размеры ядер колеблются в широких
пределах — от 3 до 25 мкм. Наиболее крупным ядром обладает яйце-
клетка. Большинство клеток человека имеет одно ядро, однако имеют-
ся двухъядерные (например, некоторые нейроны, клетки печени, кар-
диомиоциты). Двух-, а иногда и многоядерность иногда связана с
полиплоидией (от греч. polyploos — многократный, eidos — вид). Поли-
плоидия — это увеличение числа хромосомных наборов в ядрах клеток.
Иногда многоядерными клетками называют структуры, которые
образовались не вследствие полиплоидизации исходной клетки, а в ре-
зультате слияния нескольких одноядерных клеток. Такие структуры
Рис. 16. Ядро клетки:
1 — наружная мембрана
кариотеки (наружная ядерная
мембрана), 2 — перинуклеарное
пространство, 3 — внутренняя
мембрана кариотеки (внутренняя
ядерная мембрана), 4 — ядерная
ламина, 5 — поровый комплекс,
6 — рибосомы, 7 — нуклеоплазма
(ядерный сок), 8 — хроматин,
9 — цистерна гранулярной
эндоплазматической сети,
10 — ядрышко
(по Б. Албертсу и соавт.,
с изменениями)
1
5
9
2
3
4
6
7
8
10
27
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
имеют специальное название — симпласты; они встречаются, в частно-
сти, в составе скелетных поперечнополосатых мышечных волокон.
У эукариот хромосомы сосредоточены внутри ядра и отделены от
цитоплазмы ядерной оболочкой, или кариотекой. Кариотека образу-
ется за счет расширения и слияния друг с другом цистерн эндоплаз-
матической сети. Поэтому кариотека образована двумя мембрана-
ми — внутренней и наружной. Пространство между ними называют
перинуклеарным пространством. Оно имеет ширину 20—50 нм и со-
храняет сообщение с полостями эндоплазматической сети. Со сторо-
ны цитоплазмы наружная мембрана нередко покрыта рибосомами.
Местами внутренняя и наружная мембраны кариотеки сливаются,
а в месте слияния образуется по
ра. По
ра не зияет: между ее краями
упорядоченно располагаются белковые молекулы, так что в целом
формируется поровый комплекс.
Комплекс по
ры (рис. 17) представляет собой сложную структуру,
которая состоит из двух рядов связанных между собой белковых гра-
нул, каждая из которых содержит по 8 гранул, располагающихся на
равном расстоянии друг от друга по обе стороны ядерной оболочки.
Эти гранулы по размерам превосходят рибосомы. Гранулы, располо-
женные на цитоплазматической стороне поры, обусловливают осмио-
фильный материал, окружающий пору. В центре отверстия поры
иногда имеется крупная центральная гранула, связанная с гранулами,
описанными выше (возможно, это частицы, транспортирующиеся из
ядра в цитоплазму). Отверстие поры закрыто тонкой диафрагмой.
По-видимому, в поровых комплексах имеются цилиндрические кана-
лы диаметром около 9 нм и длиной около 15 нм.
Рис. 17. Поровый комплекс:
А — пространственная реконструкция, Б — схема основных структур,
В — схема молекулярной организации; 1 — периферические гранулы,
2 — центральная гранула, 3 — диафрагма поры (по Б. Албертсу и соавт., с изменениями)
À
Á
Â
3
1
2
2
1
28
СТРОЕНИЕ КЛЕТКИ
Через поровые комплексы осуществляется избирательный транс-
порт молекул и частиц из ядра в цитоплазму и обратно. Поры могут
занимать до 25% поверхности ядра. Количество пор у одного ядра до-
стигает 3000—4000, а их плотность составляет около 11 на 1 мкм2
ядерной оболочки. Из ядра в цитоплазму транспортируются в основ-
ном разные виды РНК. Из цитоплазмы в ядро поступают все фермен-
ты, необходимые для синтеза РНК, для регуляции интенсивности
этих синтезов и, в ряде клеток, молекулы гормонов, которые тоже ре-
гулируют активность синтезов РНК.
Внутренняя поверхность кариотеки связана с многочисленными
промежуточными филаментами (см. раздел «Цитоскелет»). В сово-
купности они образуют здесь тонкую пластинку, называемую ядер-
ной ламиной (рис. 18 и 19). К ней прикреплены хромосомы. Ядерная
пластинка связана с поровыми комплексами и играет главную роль
Рис. 18. Поверхностные структуры ядра:
1 — внутренняя ядерная мембрана, 2 — интегральные белки,
3 — белки ядерной ламины, 4 — хроматиновая фибрилла (часть хромосомы)
(по Б. Албертсу и соавт., с изменениями)
1
2
3
4
Рис. 19. Ядро и околоядерная область цитоплазмы:
1 — гранулярная эндоплазматическая сеть, 2 — поровые комплексы, 3 — внутренняя
ядерная мембрана, 4 — наружная ядерная мембрана, 5 — ядерная ламина
и субмембранный хроматин (по Б. Албертсу и соавт., с изменениями)
12345
29
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
в поддержании формы ядра. Она построена из промежуточных фила-
ментов особой структуры.
Нуклеоплазма представляет собой коллоид (обычно в форме
геля). По ней транспортируются различные молекулы, она содержит
множество разнообразных ферментов, в нее поступают с хромосом
РНК.
В живых клетках нуклеоплазма (кариоплазма) внешне гомогенна
(кроме ядрышка). После фиксации и обработки тканей для световой
или электронной микроскопии в кариоплазме становятся видны два
типа хроматина (от греч. chroma — краска): хорошо окрашивающийся
электронноплотный гетерохроматин, образованный осмиофильными
гранулами размером 10—15 нм и фибриллярными структурами тол-
щиной около 5 нм, и светлый эухроматин.
Гетерохроматин расположен в основном вблизи внутренней ядер-
ной мембраны, контактируя с ядерной пластинкой и оставляя свобод-
ными поры, и вокруг ядрышка. Эухроматин находится между скопле-
ниями гетерохроматина. По сути дела, хроматин — это комплексы
веществ, которыми образованы хромосомы — ДНК, белок и РНК
в соотношении 1 : 1,3 : 2. Основа каждой хромосомы образована ДНК,
молекула которой имеет вид спирали. Она упакована различными
белками, среди которых различают гистоновые и негистоновые. В ре-
зультате ассоциации ДНК с белками образуются дезоксинуклеопро-
теиды (ДНП).
Хромосомы и ядрышки
В хромосоме (рис. 20) молекула ДНК (см. рис. 1 и 2) упакована
компактно. Так, информация, заложенная в последовательности 1 млн
нуклеотидов при линейном расположении заняла бы отрезок длиной
0,34 мм. В результате компактизации она занимает объем 10–16 см3.
Длина одной хромосомы человека в растянутом виде около 5 см, дли-
на всех хромосом около 170 см, а их масса 6·10–12 г.
ДНК ассоциирована с белками-гистонами, в результате чего обра-
зуются нуклеосомы, являющиеся структурными единицами хромати-
на. Нуклеосомы, напоминающие бусины диаметром 10 нм, состоят из
8 молекул гистонов (по две молекулы гистонов Н2А, Н2Б, Н3 и Н4),
вокруг которых закручен участок ДНК, включающий 146 пар нуклео-
тидов. Между нуклеосомами располагаются линкерные участки ДНК,
состоящие из 60 пар нуклеотидов, а гистон Н1 обеспечивает взаим-
ный контакт соседних нуклеосом. Нуклеосомы — это лишь первый
уровень укладки ДНК.
Хроматин представлен в виде фибрилл толщиной около 30 нм, ко-
торые образуют петли длиной около 0,4 мкм каждая, содержащие от
30
СТРОЕНИЕ КЛЕТКИ
20 000 до 30 000 пар нуклеотидов, которые, в свою очередь, еще боль-
ше компактизируются, так что метафазная хромосома имеет средние
размеры 5 × 1,4 мкм.
В результате суперспирализации ДНК в делящемся ядре хромосо-
мы (от греч. chroma — краска, zoma — тело) становятся видимыми при
увеличении светового микроскопа. Каждая хромосома образована
одной длинной молекулой ДНК. Они представляют собой удлинен-
ные палочковидные структуры, имеющие два плеча, разделенные цен-
тромерой. В зависимости от ее расположения и взаимного расположе-
ния плеч выделяют три типа хромосом: метацентрические, имеющие
примерно одинаковые плечи; акроцентрические, имеющие одно очень
короткое и одно длинное плечо; субметацентрические, у которых одно
Рис. 20. Уровни упаковки ДНК в хромосоме:
I — двойная спираль молекулы ДНК, II — нуклеосомная нить, III — хроматиновая
фибрилла, IV — серия петельных доменов, V — конденсированный хроматин в составе
петельного домена, VI — метафазная хромосома; 1 — гистон Н1, 2 — ДНК,
3 — прочие гистоны, 4 — центромера, 5 — сестринские хроматиды
(по Б. Албертсу и др., с изменениями)
5'
3'
5'
3'
2нм
11 нм
30 нм
300 нм
700 нм
1400 нм
II
III
IV
V
VI
I
11
22
33
44
55
31
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
длинное и одно более короткое плечо. Некоторые акроцентрические
хромосомы имеют спутники (сателлиты) — мелкие участки короткого
плеча, соединенные с ним тонким неокрашивающимся фрагментом
(вторичная перетяжка). В хромосоме имеются эу- и гетерохроматино-
вые участки. Последние в неделящемся ядре (вне митоза) остаются
компактными. Чередование эу- и гетерохроматиновых участков ис-
пользуют для идентификации хромосом.
Метафазная хромосома состоит из двух соединенных центромерой
сестринских хроматид, каждая из которых содержит одну молекулу
ДНК, уложенную в виде суперспирали. При спирализации участки эу-
и гетерохроматина укладываются закономерным образом, так что по
протяжению хроматид образуются чередующиеся поперечные полосы.
Их выявляют при помощи специальных окрасок. Поверхность хромо-
сом покрыта различными молекулами, главным образом рибонуклео-
протеинами (РНП). В соматических клетках имеются по две копии ка-
ждой хромосомы, их называют гомологичными. Они одинаковы по
длине, форме, строению, расположению полос, несут одни и те же гены,
которые локализованы одинаково. Гомологичные хромосомы могут
различаться аллелями генов, содержащихся в них. Ген — это участок
молекулы ДНК, на котором синтезируется активная молекула РНК
(см. раздел «Синтез белков»). Гены, входящие в состав хромосом чело-
века, могут содержать до двух миллионов пар нуклеотидов.
Итак, хромосомы представляют собой двойные цепи ДНК, окру-
женные сложной системой белков. С одними участками ДНК связаны
гистоны. Они могут прикрывать их или освобождать. В первом слу-
чае данная область хромосомы не способна синтезировать РНК, во
втором же синтез происходит. Это один из способов регуляции функ-
циональной активности клетки путем дерепрессии и репрессии генов.
Существуют и иные способы такого управления.
Некоторые участки хромосом остаются окруженными белками
постоянно и в данной клетке никогда не участвуют в синтезе РНК.
Их можно называть блокированными. Механизмы блокирования раз-
нообразны. Обычно такие участки очень сильно спирализуются и по-
крываются не только гистонами, но и другими белками с более круп-
ными молекулами.
Деспирализованные активные участки хромосом не видны под
микроскопом. Лишь слабая гомогенная базофилия нуклеоплазмы
указывает на присутствие ДНК; их можно выявить также гистохими-
ческими методами. Такие участки относят к эухроматину. Неактив-
ные сильно спирализованные комплексы ДНК и высокомолекуляр-
ных белков выделяются при окрасках в виде глыбок гетерохроматина.
Хромосомы фиксированы на внутренней поверхности кариотеки
к ядерной ламине.
32
СТРОЕНИЕ КЛЕТКИ
В целом хромосомы в функционирующей клетке обеспечивают син-
тез РНК, необходимой для последующего синтеза белков. При этом осу-
ществляется считывание генетической информации — ее транскрипция.
Не вся хромосома принимает в ней непосредственное участие.
Разные участки хромосом обеспечивают синтез различных РНК.
Особенно выделяются участки, синтезирующие рибосомные РНК
(рРНК); ими обладают не все хромосомы. Эти участки называют
ядрышковыми организаторами. Ядрышковые организаторы образуют
петли. Верхушки петель разных хромосом тяготеют друг к другу и
встречаются вместе. Таким образом формируется структура ядра,
именуемая ядрышком (рис. 21). В нем различают три компонента.
Слабоокрашенный компонент соответствует петлям хромосом, фи-
бриллярный — транскрибированной рРНК и глобулярный — предше-
ственникам рибосом.
Ядрышки видны и в световом микроскопе. В зависимости от функ-
циональной активности клетки в образование ядрышка включаются то
меньшие, то большие участки организаторов. Иногда их группировка
может совершаться не в одном, а в нескольких местах. В этих случаях
в клетке обнаруживается несколько ядрышек. Области, в которых
ядрышковые организаторы активны, выявляют не только на электрон-
номикроскопическом уровне, но и светооптически при специальной
обработке препаратов (особые методы импрегнации серебром).
От ядрышка предшественники рибосом перемещаются к поровым
комплексам. При прохождении пор происходит дальнейшее форми-
рование рибосом.
Хромосомы являются ведущими компонентами клетки в регуляции
всех обменных процессов: любые метаболические реакции возможны
Рис. 21. Строение ядрышка:
I — схема, II — ядрышко в ядре клетки (электронно-микроскопическая фотография);
1 — кариотека, 2 — ядерная ламина, 3 — ядрышковые организаторы хромосом,
4 — концы хромосом, связанные с ядерной ламиной
(по Б. Албертсу и соавт., с изменениями)
11
22
33
44
II
I
33
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
только с участием ферментов, ферменты же всегда белки, белки син-
тезируются только с участием РНК.
Вместе с тем хромосомы являются и хранителями наследственных
свойств организма. Именно последовательность нуклеотидов в цепях
ДНК определяет генетический код. Совокупность всей генетической
информации, хранящейся в хромосомах, называют геномом. При
подготовке клетки к делению геном удваивается, а при самом делении
поровну распределяется между дочерними клетками. Все проблемы,
связанные с организацией генома и закономерностями передачи на-
следственной информации, излагаются в курсе генетики.
Кариотип
Метафазное ядро можно выделить из клетки, раздвинуть хромосо-
мы, сосчитать их и изучить их форму. Клетки особей каждого биоло-
гического вида имеют одинаковое количество хромосом. Каждая хро-
мосома во время метафазы имеет свои особенности строения.
Совокупность этих особенностей обозначается понятием «кариотип»
(рис. 22). Знание нормального кариотипа необходимо, чтобы выяв-
лять возможные отклонения. Такие отклонения всегда служат источ-
ником наследственных заболеваний.
Нормальный кариотип (набор хромосом) (от греч. karyon — ядро
ореха, typos — образец) человека включает 22 пары аутосом и одну
пару половых хромосом (либо XX у женщин, либо XY у мужчин).
В 1949 г. М . Барр обнаружил в ядрах нейронов кошек особые
плотные тельца, которые отсутствовали у самцов. Эти тельца имеют-
1
I
II
234
567891
01
1
12131415161718
19202122XY
Рис. 22 . Кариотип человека
(здорового мужчины)
I — кариотип, общий вид,
II — метафазные хромосомы
(по Б. Албертсу и соавт.)
34
СТРОЕНИЕ КЛЕТКИ
ся и в интерфазных ядрах других соматических клеток особей жен-
ского пола. Они были названы тельцами полового хроматина (тельца-
ми Барра). У человека они имеют диаметр около 1 мкм и лучше всего
идентифицируются в нейтрофильных сегментоядерных лейкоцитах,
где выглядят в виде «барабанной палочки», связанной с ядром. Раз-
личимы они хорошо и в эпителиоцитах слизистой оболочки щеки,
взятых путем соскоба. Тельца Барра представляют собой одну инак-
тивированную конденсированную Х-хромосому.
ЦИТОПЛАЗМА
Основными структурами цитоплазмы являются гиалоплазма (ма-
трикс), органеллы и включения.
Гиалоплазма
В физико-химическом отношении гиалоплазма (от греч. hyalos —
стекло) представляет собой коллоид, состоящий из воды, ионов и мно-
гих молекул органических веществ. Последние принадлежат ко всем
классам — и к углеводам, и к липидам, и к белкам, а также к комплекс-
ным соединениям типа гликолипидов, гликопротеинов и липопротеи-
нов. Многие из протеинов обладают ферментативной активностью.
В гиалоплазме протекает ряд важнейших биохимических реакций,
в частности, осуществляется гликолиз — филогенетически наиболее
древний процесс выделения энергии (от греч. glykys — сладкий и lysis —
распад), в результате чего шестиуглеродная молекула глюкозы распада-
ется на две трехуглеродные молекулы пировиноградной кислоты с
образованием АТФ (см. раздел «Основные реакции тканевого обмена»).
Молекулы гиалоплазмы, конечно, взаимодействуют между собой
весьма упорядоченно, но характер ее пространственной организации
пока недостаточно ясен, поэтому можно говорить лишь в общих чер-
тах, что гиалоплазма структурирована на молекулярном уровне.
Именно в гиалоплазме взвешены органеллы и включения.
Органеллы
Органеллами называют элементы цитоплазмы, структурирован-
ные на ультрамикроскопическом уровне и выполняющие конкретные
функции клетки; органеллы участвуют в осуществлении тех функций
клетки, которые необходимы для поддержания ее жизнедеятельно-
сти. Сюда относятся обеспечение ее энергетического обмена, синте-
тических процессов, обеспечение транспорта веществ и т. п .
Органеллы, присущие всем клеткам, называют органеллами
общего назначения, присущие же некоторым специализированным
35
ЦИТОПЛАЗМА
видам клеток — специальными. В зависимости от того, включает
структура органеллы биологическую мембрану или нет, различают
органеллы мембранные и немембранные.
Органеллы общего значения
Немембранные органеллы
К немембранным органеллам относятся цитоскелет, клеточный
центр и рибосомы.
Цитоскелет
Цитоскелет (клеточный скелет), в свою очередь, образован тремя
компонентами: микротрубочками, микрофиламентами и промежу-
точными филаментами.
Микротрубочки (рис. 23) пронизывают всю цитоплазму клетки.
Каждая из них представляют собой полый цилиндр диаметром 20—
30 нм. Стенка микротрубочки имеет толщину 6—8 нм. Она образова-
на 13 нитями (протофиламентами), скрученными по спирали одна
над другой. Каждая нить, в свою очередь, слагается из димеров белка
тубулина. Каждый димер представлен а- и b-тубулином. Синтез тубу-
линов происходит на мембранах гранулярной эндоплазматической
сети, а сборка в спирали — в клеточном центре.
Рис. 23. Строение микротрубочки:
1 — тубулиновые субъединицы,
2 — ассоциированные белки,
3 — перемещаемые частицы
11
22
33
36
СТРОЕНИЕ КЛЕТКИ
Соответственно многие микротрубочки имеют радиальное направ-
ление по отношению к центриолям. Отсюда они распространяются по
всей цитоплазме. Часть из них расположена под плазмалеммой, где
они вместе с пучками микрофиламентов участвуют в образовании
терминальной сети.
Микротрубочки прочны и образуют опорные структуры цитоске-
лета. Часть микротрубочек располагается в соответствии с силами
сжатия и натяжения, которые испытывает клетка. Особенно хорошо
это заметно в клетках эпителиальных тканей, которые разграничива-
ют разные среды организма.
Микротрубочки участвуют в транспорте веществ внутри клетки.
Со стенкой микротрубочки одним из своих концов связаны (ассоции-
рованы) белковые молекулы в виде коротких цепочек, которые спо-
собны в соответствующих условиях изменять свою пространственную
конфигурацию (конформация белка). В нейтральном положении це-
почка лежит параллельно поверхности стенки. При этом свободный
конец цепи может связываться с частицами, которые находятся
в окружающем гликокаликсе.
После связывания частицы белок изменяет конфигурацию и от-
клоняется от стенки, тем самым перемещая за собой и блокирован-
ную частицу. Отклоненная цепочка передает частицу той, что свисает
над ней, та тоже отклоняется и передает частицу далее. В связи с на-
личием конформируемых наружных цепей микротрубочки обеспечива-
ют основные потоки внутриклеточного активного транспорта.
Структура стенки микротрубочек может меняться при различных
воздействиях на них. В подобных случаях может нарушаться внутри-
клеточный транспорт. К числу блокаторов микротрубочек и, соответ-
ственно, внутриклеточного транспорта относится, в частности, алка-
лоид колхицин.
Промежуточные филаменты толщиной 8—10 нм представлены в
клетке длинными белковыми молекулами. Они тоньше микротрубочек,
но толще микрофиламентов, за что и получили свое название (рис. 24).
Рис. 24. Промежуточные филаменты в клетке (по К. де Дюву, с изменениями)
37
ЦИТОПЛАЗМА
Белки промежуточных филаментов принадлежат к четырем ос-
новным группам. Некоторые их характеристики приведены в табли-
це 4. Каждая группа, в свою очередь, включает в себя по нескольку
белков (так, известно более 20 видов кератинов).
Таблица 4
Виды промежуточных филаментов (по Б. Албертсу и соавт.)
Тип
филаментов
Образующие полипептиды
и их молекулярная масса (кД)
Некоторые структуры, в которых
встречаются данные филаменты
I
Кислые, нейтральные и основные
кератины (40—70)
Эпителиальные клетки и их про-
изводные (волосы, ногти и др.)
II
Виментин (53)
Клетки мезенхимного происхож-
дения
Десмин (52)
Мышечные клетки
Глиальный фибриллярный кислый
белок (45)
Астроциты и леммоциты
(шванновские клетки)
III
Белки нейрофиламентов (60, 100, 130) Нейроны
IV
Ядерные ламины А, В и С (65—75) Ядерная ламина во всех клетках
Каждый белок является антигеном, так что к нему можно создать
соответствующее антитело. Если каким-либо образом маркировать
антитело (например, прикрепив к нему флуоресцирующую метку),
то, вводя его в организм, можно обнаружить локализацию данного
белка. Белки промежуточных филаментов сохраняют свою специфич-
ность даже при значительных изменениях клетки, в том числе при ее
малигнизации. Поэтому, используя специфические меченые антитела
к белкам промежуточных филаментов, можно установить, какие клет-
ки были первичным источником опухоли.
Микрофиламенты — это белковые нити толщиной около 4 нм.
Большинство из них образовано молекулами актинов, которых выяв-
лено около 10 видов. Кроме того, актиновые филаменты могут груп-
пироваться в пучки, образующие собственно опорные структуры ци-
тоскелета.
Актин в клетке существует в двух формах: мономерной (глобуляр-
ный актин) и полимеризованной (фибриллярный актин). Кроме не-
посредственно актина, в построении микрофиламентов могут прини-
мать участие и другие пептиды: тропонины и тропомиозин (рис. 25).
Рис. 25 . Актиновый
микрофиламент:
1 — глобулы актина,
2 — тропомиозин,
3 — тропонины
(по Б. Албертсу и соавт.,
с изменениями)
38
СТРОЕНИЕ КЛЕТКИ
Филаменты актина способны образовывать комплексы с полимер-
ными же молекулами белка миозина. Когда миозин присутствует
в гиалоплазме в виде мономеров, он не вступает в комплекс с акти-
ном. Для полимеризации миозина необходимы ионы кальция. Связы-
вание его происходит с участием тропонина С (по названию элемента
кальция), освобождение — с участием тропонина I (ингибиторная мо-
лекула), комплексирование с тропомиозином — с участием тропони-
на Т. После того как возникает актино-миозиновый комплекс, актин
и миозин становятся способными смещаться в нем продольно относи-
тельно друг друга. Если концы комплекса скреплены с какими-либо
другими внутриклеточными структурами, последние сближаются.
Это лежит в основе мышечного сокращения.
Микрофиламентов особенно много в области цитоплазмы, относя-
щейся к поверхностному комплексу. Будучи соединенными с плазма-
леммой, они способны менять ее конфигурацию. Это важно для обес-
печения поступления веществ в клетку посредством пиноцитоза
и фагоцитоза. Этот же механизм используется клеткой при образова-
нии выростов ее поверхности — ламеллоподий. Клетка может закре-
питься ламеллоподией за окружающий субстрат и переместиться на
новое место.
Клеточный центр
Клеточный центр (рис. 26) образован двумя центриолями (дипло-
сома) и центросферой. Свое название органелла получила, так как
она обычно находится в глубоких отделах цитоплазмы, нередко вбли-
зи ядра или около формирующейся поверхности комплекса Гольджи.
Обе центриоли диплосомы расположены под углом друг к другу.
Основная функция клеточного центра — сборка микротрубочек.
Каждая центриоль представляет собой цилиндр, стенка которого,
в свою очередь, состоит из девяти комплексов микротрубочек длиной
около 0,5 и диаметром около 0,25 мкм. Каждый комплекс состоит из
трех микротрубочек и поэтому называется триплетом. Триплеты, рас-
положенные по отношению друг к другу под углом около 50°, состоят из
трех микротрубочек (изнутри кнаружи): полной А и неполных В и С
диаметром около 20 нм каждая. От трубочки А отходят две ручки. Одна
из них направлена к трубочке С соседнего триплета, другая — к центру
цилиндра, где внутренние ручки образуют фигуру звезды или спиц
колеса. Каждая микротрубочка имеет типичное строение (см. ранее).
Центриоли расположены взаимно перпендикулярно. Одна из них
упирается концом в боковую поверхность другой. Первая называется
дочерней, вторая — материнской. Дочерняя центриоль возникает
вследствие удвоения материнской. Материнская центриоль окружена
39
ЦИТОПЛАЗМА
электроноплотным ободком, образованным шаровидными сателли-
тами, соединенными плотным материалом с наружной стороной
каждого триплета. Средняя часть материнской центриоли может быть
также окружена комплексом фибриллярных структур, называемым
гало. Триплеты микротрубочек объединяются у основания материн-
ской центриоли электроноплотными скоплениями — корешками
(придатками).
К концу сателлитов и к области гало по цитоплазме транспортиру-
ются тубулины, и именно здесь происходит сборка микротрубочек.
Будучи собранными, они отделяются и направляются в разные участ-
12
3
4
5
9
8
7
6
Рис. 26. Клеточный центр и другие структуры цитоплазмы:
1 — центросфера, 2 — центриоль на поперечном срезе (триплеты микротрубочек,
радиальные спицы, центральная структура «колеса телеги»), 3 — центриоль
(продольный разрез), 4 — сателлиты, 5 — окаймленные пузырьки, 6 — зернистая
эндоплазматическая сеть, 7 — митохондрия, 8 — внутренний сетчатый аппарат
(комплекс Гольджи), 9 — микротрубочки
(по Р. Крстичу, с изменениями)
40
СТРОЕНИЕ КЛЕТКИ
ки цитоплазмы, чтобы занять свое место в структурах цитоскелета.
Возможно, сателлиты являются и источником материала для образо-
вания новых центриолей при их репликации. Область гиалоплазмы
вокруг центриолей и сателлита называется центросферой.
Центриоли являются саморегулирующимися структурами, кото-
рые удваиваются в клеточном цикле (см. раздел «Клеточный цикл»).
При удвоении вначале обе центриоли расходятся, и перпендикулярно
к базальному концу материнской возникает мелкая процентриоль,
образованная девятью одиночными микротрубочками. Затем к ка-
ждой из них путем самосборки из тубулина присоединяются еще две.
Центриоли участвуют в образовании базальных телец ресничек и жгу-
тиков и в образовании митотического веретена.
Рибосомы
Рибосомы (рис. 27) представляют собой тельца размерами
20 ×30 нм (константа седиментации 80). Рибосома состоит из двух
субъединиц — большой и малой. Каждая субъединица представляет
собой комплекс рибосомной РНК (рРНК) с белками. Большая субъе-
диница (константа седиментации 60) содержит три различные моле-
кулы рРНК, связанных с 40 молекулами белков; малая содержит одну
молекулу рРНК и 33 молекулы белков. Синтез рРНК осуществляется
на петлях хромосом — ядрышковых организаторах (в области ядрыш-
ка). Сборка рибосом осуществляется в области пор кариотеки.
Основная функция рибосом — сборка белковых молекул из ами-
нокислот, доставляемых к ним транспортными РНК (тРНК). Между
субъединицами рибосомы имеется щель, в которой проходит молеку-
ла информационной РНК (мРНК), а на большой субъединице — бо-
роздка, в которой располагается и по которой сползает формирующа-
яся белковая цепь. Сборка аминокислот производится в соответствии
с чередованием нуклеотидов в цепи мРНК. Таким способом осуществ-
ляется трансляция генетической информации.
Рис. 27. Рибосома:
I — малая субъединица,
II — большая субъединица,
III — объединение субъединиц;
верхний и нижний ряды —
изображения в разных проекциях
(по Б. Албертсу и соавт.,
с изменениями)
41
ЦИТОПЛАЗМА
Рибосомы могут находиться в гиалоплазме поодиночке либо груп-
пами в виде розеток, спиралей, завитков. Такие группы называют
полирибосомами (полисомами). Таким образом, молекула мРНК мо-
жет протягиваться по поверхности не только одной, но и нескольких
рядом лежащих рибосом. Значительная часть рибосом прикреплена
к мембранам: к поверхности эндоплазматической сети и к наружной
мембране кариотеки. Свободные рибосомы синтезируют белок, необ-
ходимый для жизнедеятельности самой клетки, прикрепленные —
белок, подлежащий выведению из клетки.
Количество рибосом в клетке может достигать десятков миллионов.
Мембранные органеллы
Каждая мембранная органелла представляет структуру цитоплаз-
мы, ограниченную мембраной. Вследствие этого внутри нее образует-
ся пространство, отграниченное от гиалоплазмы. Цитоплазма оказы-
вается таким образом разделенной на отдельные отсеки со своими
свойствами — компартменты. Наличие компартментов — одна из
важных особенностей эукариотических клеток.
К мембранным органеллам относятся митохондрии, эндоплазма-
тическая сеть (ЭПС), комплекс Гольджи, лизосомы и пероксисомы.
Некоторые авторы относят к общим органеллам также и микровор-
синки. Последние иногда причисляют к органеллам специальным, но
фактически они встречаются на поверхности любой клетки и будут
описаны вместе с поверхностным комплексом цитоплазмы. К. де Дюв
объединил ЭПС, комплекс Гольджи, лизосомы и пероксисомы поня-
тием вакуом (см. раздел «Комплекс Гольджи»).
Митохондрии
Митохондрии участвуют в процессах клеточного дыхания и пре-
образуют энергию, которая при этом освобождается, в форму, доступ-
ную для использования другими структурами клетки, поэтому за
ними закрепилось ставшее тривиальным образное название «энерге-
тические станции клетки».
Митохондрии, в отличие от других органелл, обладают собствен-
ной генетической системой, необходимой для их самовоспроизведе-
ния и синтеза белков. Они имеют свои ДНК, РНК и рибосомы, отли-
чающиеся от таковых в ядре и в других отделах цитоплазмы
собственной клетки. В то же время митохондриальные ДНК, РНК
и рибосомы весьма сходны с прокариотическими. Это послужило
толчком для разработки симбиотической гипотезы, согласно которой
митохондрии (и хлоропласты) возникли из симбиотических бакте-
42
СТРОЕНИЕ КЛЕТКИ
рий. Митохондриальная ДНК, кольцевидная (как у бактерий), на нее
приходится около 2% ДНК клетки.
Митохондрии (и хлоропласты) способны размножаться в клетке
путем бинарного деления. Таким образом, они являются самовоспро-
изводящимися органеллами. Вместе с тем генетическая информация,
содержащаяся в их ДНК, не обеспечивает их всеми необходимыми для
полного самовоспроизведения белками; часть этих белков кодируется
ядерными генами и поступает в митохондрии из гиалоплазмы, поэто-
му митохондрии в отношении их самовоспроизведения называют по-
луавтономными структурами. У человека и других млекопитающих ми-
тохондриальный геном наследуется от матери: при оплодотворении
митохондрии спермия в яйцеклетку не проникают. Такое, казалось бы,
отвлеченное чисто теоретическое положение в последние годы нашло
сугубо практическое применение: исследование последовательности
компонентов ДНК в митохондриях помогает выявлять генеалогиче-
ские связи по женской линии. Это бывает существенным для иденти-
фикации личности. Любопытными оказались и историко-этнографи-
ческие сопоставления. Так, в древних монгольских сказаниях
утверждалось, что три ветви этого народа произошли от трех матерей;
исследования митохондриальных ДНК действительно подтвердили,
что у представителей каждой ветви они обладают такими особыми
чертами, которых нет у других. Основные свойства митохондрий
и функции их структурных компонентов обобщены в табл. 5.
Таблица 5
Морфофункциональная организация митохондрий
Структуры
митохондрий
Состав
Функция
Наружная
мембрана
Около 20% всего белка мито-
хондрий. Ферменты липидного
обмена
Транспорт. Превращение
липидов в промежуточные
метаболиты
Межмембранное
пространство
Ферменты, использующие
АТФ для фосфорилирования
других нуклеотидов
Внутренняя
мембрана
Ферменты дыхательной цепи,
цитохромы, сукцинатдегидро-
геназа. Трансбелки
Создание электрохимического
протонного градиента. Перенос
метаболитов в матрикс и из него
Субмитохондри-
альные частицы
АТФ-синтетаза
Синтез и гидролиз АТФ
Матрикс
Ферменты (кроме сукцинат-
дегидрогеназы). ДНК, РНК,
рибосомы, ферменты, участву-
ющие в экспрессии генома
митохондрий
Цикл лимонной кислоты, прев-
ращение пирувата, аминокислот
и жирных кислот в ацетилкоэн-
зим А. Репликация, транскрип-
ция, трансляция
43
ЦИТОПЛАЗМА
В световом микроскопе митохондрии выглядят в виде округлых,
удлиненных или палочковидных структур длиной 0,3—5 и шириной
0,2—1 мкм. Каждая митохондрия образована двумя мембранами —
внешней и внутренней (рис. 28).
Между ними расположено межмембранное пространство шири-
ной 10—20 нм. Внешняя мембрана ровная, внутренняя же образует
многочисленные кристы, которые могут иметь вид складок и гребней.
Иногда кристы имеют вид трубочек диаметром 20—60 нм. Это наблю-
дается в клетках, которые синтезируют стероиды (здесь митохондрии
не только обеспечивают процессы дыхания, но и участвуют в синтезе
этих веществ). Благодаря кристам площадь внутренней мембраны су-
щественно увеличивается.
Пространство, ограниченное внутренней мембраной, заполнено
коллоидным митохондриальным матриксом. Он имеет мелкозерни-
стую структуру и содержит множество различных ферментов. В мат-
риксе также заключен собственный генетический аппарат митохон-
дрий (у растений кроме митохондрий ДНК содержится также и
в хлоропластах).
Со стороны матрикса к поверхности крист прикреплено множество
электроноплотных субмитохондриальных элементарных частиц (до
4000 на 1 мкм2 мембраны). Каждая из них имеет форму гриба (см.
рис. 28). Круглая головка диаметром 9—10 нм посредством тонкой
Рис. 28. Митохондрия:
I — общая схема строения: 1 — наружная мембрана, 2 — внутренняя мембрана,
3 — кристы, 4 — матрикс; II — схема строения кристы: 5 — складка внутренней
мембраны, 6 — грибовидные тельца (по Б. Албертсу и др.; по К. де Дюву, с изм.)
1
5
2
6
3
I
4
II
44
СТРОЕНИЕ КЛЕТКИ
ножки диаметром 3—4 нм прикрепляется к внутренней мембране.
В этих частицах сосредоточены АТФазы — ферменты, непосредственно
обеспечивающие синтез и распад АТФ. Эти процессы неразрывно свя-
заны с циклом трикарбоновых кислот (циклом лимонной кислоты, или
циклом Кребса, см. раздел «Основные реакции тканевого обмена»).
Количество, размеры и расположение митохондрий зависят от
функции клетки, в частности от ее потребности в энергии и от места,
где энергия расходуется. Так, в одной печеночной клетке их количест-
во достигает 2500. Множество крупных митохондрий содержится
в кардиомиоцитах и миосимпластах мышечных волокон. В спермиях
богатые кристами митохондрии окружают аксонему промежуточной
части жгутика. Есть клетки, в которых митохондрии имеют чрезвы-
чайно большие размеры. Такая митохондрия может ветвиться и обра-
зовывать трехмерную сеть. Это показано путем реконструкции струк-
туры клетки по отдельным последовательным срезам. На плоском
срезе видны лишь части этой митохондрии, что и создает впечатление
их множественности (рис. 29).
Эндоплазматическая сеть
Эндоплазматическая сеть (ЭПС), или, как ее нередко называют,
эндоплазматический ретикулум (ЭР), представляет собой единый не-
прерывный компартмент, ограниченный мембраной, образующей
множество инвагинаций и складок (рис. 30). Поэтому на электронно-
микроскопических фотографиях эндоплазматическая сеть выглядит
в виде множества трубочек, плоских или округлых цистерн, мембран-
ных пузырьков. На мембранах ЭПС совершаются многообразные
первичные синтезы веществ, необходимых для жизнедеятельности
Рис. 29. Гигантская митохондрия.
Реконструкция по серийным
электронно-микроскопическим
фотографиям срезов мышечного волокна
(по Ю. С . Ченцову, с изменениями)
45
ЦИТОПЛАЗМА
клетки. Первичными их можно условно назвать потому, что молеку-
лы этих веществ будут подвергаться дальнейшим химическим прев-
ращениям в других компартментах клетки.
Большинство веществ синтезируется на наружной поверхности
мембран ЭПС. Затем эти вещества переносятся через мембрану
внутрь компартмента и там транспортируются к местам дальнейших
биохимических превращений, в частности, к комплексу Гольджи. На
концах трубочек ЭПС они накапливаются и затем отделяются от них
в виде транспортных пузырьков. Каждый пузырек окружен, таким
образом, мембраной и перемещается в гиалоплазме к месту назначе-
ния. Как всегда, в транспорте принимают участие микротрубочки.
Среди продуктов, синтезируемых на мембранах ЭПС, особо отметим
те вещества, которые служат материалом для сборки мембран клетки
(окончательная сборка мембран осуществляется в комплексе Гольджи).
Различают два типа ЭПС: гранулярную (зернистую, шероховатую)
и агранулярную (гладкую). Обе они представляют собой единую
структуру.
Наружная, обращенная к гиалоплазме, сторона мембраны грану-
лярной ЭПС покрыта рибосомами. Поэтому при световой микроско-
пии гранулярная эндоплазматическая сеть выглядит в виде базофиль-
Рис. 30. Эндоплазматическая сеть: 1 — трубочки гладкой (агранулярной)
эндоплазматической сети, 2 — цистерны шероховатой гранулярной сети,
3 — поровый комплекс, 4 — наружная ядерная мембрана, покрытая рибосомами,
5 — внутренняя ядерная мембрана (по Крстичу, с изменениями)
11
22
33
44
55
46
СТРОЕНИЕ КЛЕТКИ
ного вещества, дающего положительную окраску на РНК. Здесь
осуществляется синтез белков. В клетках, специализирующихся на
синтезе белков, гранулярная эндоплазматическая сеть выглядит
в виде параллельных окончатых (фенестрированных), сообщающихся
между собой и с перинуклеарным пространством ламеллярных струк-
тур, между которыми лежит множество свободных рибосом.
Поверхность гладкой ЭПС лишена рибосом. Сама сеть представ-
ляет собой множество мелких трубочек диаметром около 50 нм ка-
ждая. Между трубочками часто расположены гранулы гликогена.
В некоторых клетках гладкая сеть образует выраженный лабиринт
(например, в гепатоцитах, в клетках Лейдига), в других — циркуляр-
ные пластинки (например, в ооцитах).
На мембранах гладкой сети синтезируются углеводы и липиды, сре-
ди них — гликоген и холестерин. Она принимает участие и в синтезе
стероидных гормонов (в клетках Лейдига, в корковых эндокриноцитах
надпочечника). Гладкая ЭПС принимает участие также в выделении ио-
нов хлора в париетальных клетках эпителия желез желудка. Будучи
депо ионов кальция, гладкая эндоплазматическая сеть участвует в со-
кращении кардиомиоцитов и волокон скелетной мышечной ткани. Она
же разграничивает будущие тромбоциты в мегакариоцитах. Чрезвычай-
но важна ее роль в детоксикации гепатоцитами веществ, которые посту-
пают из полости кишки по воротной вене в печеночные капилляры.
По просветам эндоплазматической сети синтезированные вещест-
ва транспортируются к комплексу Гольджи (но просветы сети не со-
общаются с просветами цистерн последнего). К комплексу Гольджи
вещества поступают в пузырьках, которые сначала отшнуровываются
от сети, транспортируются к комплексу и, наконец, сливаются с ним.
От комплекса Гольджи вещества транспортируются к местам своего
использования также в мембранных пузырьках. Следует подчеркнуть,
что одной из важнейших функций эндоплазматической сети является
синтез белков и липидов для всех клеточных органелл.
Комплекс Гольджи
Комплекс Гольджи (аппарат Гольджи, внутриклеточный сетчатый
аппарат, КГ) представляет собой совокупность цистерн, пузырьков,
пластинок, трубочек, мешочков. В световом микроскопе он выглядит
в виде сеточки, реально же представляет собой систему цистерн, ка-
нальцев и вакуолей.
Чаще всего в КГ выявляются три мембранных элемента: уплощен-
ные мешочки (цистерны), пузырьки и вакуоли (рис. 31). Основные
элементы комплекса Гольджи — диктиосомы (от греч. dyction — сеть).
Число их колеблется в разных клетках от одной до нескольких сотен.
47
ЦИТОПЛАЗМА
Диктиосомы связаны между собой каналами. Отдельная диктиосома
чаще всего имеет чашеобразную форму. Она имеет диаметр около
1 мкм и содержит 4—8 (в среднем 6) лежащих параллельно уплощен-
ных цистерн, пронизанных порами. Концы цистерн расширены. От
них отщепляются пузырьки и вакуоли, окруженные мембраной и со-
держащие различные вещества.
Множество мембранных пузырьков (в том числе и окаймленных)
имеет диаметр 50—65 нм. Более крупные секреторные гранулы имеют
диаметр от 66 до 100 нм. Часть вакуолей содержит гидролитические
ферменты, это предшественники лизосом.
Наиболее широкие уплощенные цистерны обращены в сторону
ЭПС. Транспортные пузырьки, несущие вещества — продукты пер-
вичных синтезов, присоединяются к этим цистернам. В цистернах
продолжается синтез полисахаридов, образуются комплексы белков,
углеводов и липидов, иначе говоря, приносимые макромолекулы мо-
дифицируются. Здесь происходит синтез полисахаридов, модифика-
ция олигосахаридов, образование белково-углеводных комплексов
и ковалентная модификация переносимых макромолекул.
По мере модификации вещества переходят из одних цистерн в
другие. На боковых поверхностях цистерн возникают выросты, куда
перемещаются вещества. Выросты отщепляются в виде пузырьков,
которые удаляются от КГ в различных направлениях по гиалоплазме.
Сторону КГ, куда поступают вещества от ЭПС, называют цисполю-
сом (формирующаяся поверхность), противоположную — транс-полю-
Рис. 31. Различные формы комплекса Гольджи
(по Б. Албертсу и соавт. и по Р. Крстичу, с изменениями)
48
СТРОЕНИЕ КЛЕТКИ
сом (зрелая поверхность). Таким образом, комплекс Гольджи структур-
но и биохимически поляризован. По направлению от цис-поверхности
к транс-поверхности увеличивается толщина мембран (от 6 до 8 нм),
а также содержание в них холестерина и углеводных компонентов
в мембранных гликопротеинах. Активность кислой фосфатазы, актив-
ность тиаминпирофосфатазы уменьшается по направлению от форми-
рующейся поверхности к зрелой. В последней цистерне транс-стороны
и окружающих ее окаймленных пузырьках имеется кислая фосфатаза.
Это особенно интересно в связи с вопросом о происхождении лизосом.
Судьба пузырьков, отщепляющихся от КГ, различна. Одни из них
направляются к поверхности клетки и выводят синтезированные ве-
щества в межклеточный матрикс. Часть этих веществ представляет
собой продукты метаболизма, часть же — специально синтезирован-
ные продукты, обладающие биологической активностью (секреты).
Чаще всего в таких случаях мембрана пузырька сливается с плазма-
леммой (есть и другие способы секреции, см. раздел «Экзоцитоз»).
В связи с такой функцией КГ часто располагается на той стороне
клетки, где происходит выведение веществ. Если оно осуществляется
равномерно со всех сторон, КГ представлен множественными диктио-
сомами, соединенными между собой каналами.
В процессе упаковки веществ в пузырьки расходуется значитель-
ное количество материала мембран. Он должен восполняться. Сборка
мембран — еще одна из функций КГ. Эта сборка совершается из ве-
ществ, поступающих, как обычно, от ЭПС. Элементы блоков мембран
создаются в полостях диктиосом, затем встраиваются в их мембраны
и наконец отделяются с пузырьками. Конкретная структура мембра-
ны зависит от того, куда она будет доставлена и где будет использо-
ваться.
Мембраны комплекса Гольджи образуются и поддерживаются гра-
нулярной эндоплазматической сетью — именно на ней синтезируются
мембранные компоненты. Эти компоненты переносятся транспорт-
ными пузырьками, отпочковывающимися от промежуточных зон сети
(транс-слияние), к формирующейся поверхности диктиосомы и сли-
ваются с ней (цис-слияние). От транс-стороны постоянно отпочковы-
ваются пузырьки, а мембраны цистерн постоянно обновляются. Они
поставляют клеточную мембрану, гликокаликс и синтезированные ве-
щества к плазмалемме. Таким образом обеспечивается обновление
плазматической мембраны.
Секреторный путь и обновление мембран представлены на рис. 32.
«Мембраны никогда не образуются de novo. Они всегда возника-
ют из предсуществующих мембран путем добавления дополнитель-
ных составных частей. Каждое поколение передает последующему,
в основном через яйцеклетку, запас заранее сформированных (пред-
49
ЦИТОПЛАЗМА
существующих) мембран, из которых путем прироста, прямо или опос-
редованно, образуются все мембраны организма» (К. де Дюв, 1987).
А. Новиков (1971) разработал концепцию ГЭРЛ (Г — Гольджи, ЭР —
эндоплазматический ретикулум (сеть), Л — лизосомы). ГЭРЛ (рис. 33)
включает в себя последний, зрелый мешочек диктиосомы, неправильной
формы, с многочисленными утолщениями (просекреторные гранулы,
или конденсирующие вакуоли), которые, отпочковываясь, превращают-
ся в секреторные гранулы. К нему прилежат лишенные рибосом цистер-
ны гранулярной эндоплазматической сети. Между ГЭРЛ и лежащей под
ним цистерной имеются каналы. От ГЭРЛ, содержащего кислую фосфа-
тазу, отпочковываются лизосомы, также содержащие этот фермент.
Рис. 32. Схема секреторного пути и обновления мембран:
1 — гранулярная эндоплазматическая сеть, 1.1 — область, где происходит синтез
белков, предназначенных для экспорта из клетки, 1.2 — область, где происходит
синтез белков, предназначенных для обновления мембран, 1.3 — область,
где происходит гликозилирование, 2 — транспортные пузырьки, где происходит
образование дисульфидных мостиков, 3 — комплекс Гольджи, где происходит
добавление липидов, сульфатирование, удаление боковых цепей, терминальное
гликозилирование, 4 — просекреторная гранула, где происходит протеолитическая
доработка, 5 — секреторная гранула, где происходит концентрация секрета,
6 — плазмалемма, 7 — экзоцитоз, 8 — встраивание в мембрану,
9 — сборка элементов мембраны (по К. де Дюву, с изменениями)
1.1
1.1
1.2
1.2
11
99
22
33
44
55
77
88
1.3
1.3
66
50
СТРОЕНИЕ КЛЕТКИ
Возможно, в ГЭРЛ поступают вещества из нижележащих цистерн
комплекса Гольджи и непосредственно из прилежащих к нему ци-
стерн эндоплазматической сети. Р. Крстич указал на наличие прямых
каналов между ГЭРЛ и прилежащими цистернами эндоплазматиче-
ской сети. Кроме того, в поры ГЭРЛ внедряются удлиненные пальце-
видные отростки цистерн эндоплазматической сети. От ГЭРЛ отходят
пальцевидные отростки, которые внедряются в поры предпоследней
цистерны диктиосомы.
Из сказанного ясно, что в КГ не только завершаются многообраз-
ные синтезы, но и происходит разделение синтезированных продук-
тов, сортировка в зависимости от их дальнейшего предназначения.
Такая функция КГ называется сегрегационной. Одним из важнейших
проявлений сегрегационной функции комплекса Гольджи является
сортировка веществ и их передвижение, которые осуществляются
с помощью окаймленных пузырьков. Главную роль в этом процессе
играют мембранные «адресные метки» — рецепторы, распознающие
специфические маркеры по принципу «замок — ключ».
Так, лизосомные ферменты сортируются в комплексе Гольджи
связанным с мембраной белком-рецептором, который «узнает» ман-
нозо-6 -фосфат, отбирает ферменты, способствует их упаковке в пу-
зырьки, окаймленные клатрином. Последние отпочковываются в виде
транспортных пузырьков, содержащих в мембране указанный рецеп-
тор. Таким образом, они функционируют как челноки, которые до-
ставляют рецептор маннозо-6-фосфата от транс-поверхности ком-
Рис. 33 . Схема комплекса ГЭРЛ (Гольджи, эндоплазматический ретикулум, лизосомы):
1 — цистерны гранулярной эндоплазматической сети, 2 — транспортные пузырьки,
3 — цис-цистерны комплекса Гольджи, 4 — лизосомы, 5 — соединительные канальцы,
6 — транс-цистерны комплекса Гольджи, 7 — конденсационные секреторные вакуоли
(по Р. Крстичу, с изменениями)
77
66
55
44
11
22
33
51
ЦИТОПЛАЗМА
плекса Гольджи к лизосомам и обратно; иными словами, рецептор
курсирует между строго специализированными мембранами.
Комплекс Гольджи является основной структурой вакуома, разде-
ляет его на эндоплазматический и экзоплазматический домены и в то
же время объединяет их функционально. Мембраны эндоплазматиче-
ского домена отличаются от мембран экзоплазматического. Послед-
ние сходны с плазмалеммой. В настоящее время вакуомом называют
вакуолярный аппарат и включают в него, кроме комплекса Гольджи и
ассоциированных с ним вакуолей, лизосом и пероксисом, также фаго-
сомы с эндосомами и саму плазмалемму.
Вещества циркулируют в клетке, будучи упакованными в мембра-
ны (перемещение содержимого клетки «в контейнерах», рис. 34).
Комплекс Гольджи (именно ГЭРЛ) является также центром циркуля-
ции мембран. При этом перед возвращением мембраны, отпочковав-
шейся от плазмалеммы в процессе эндоцитоза, эндосома освобожда-
ется от транспортированных в клетку веществ.
Рис. 34. Схема передвижения содержимого клетки в контейнерах («челноках»):
А — эндоплазматический домен, Б — экзоплазматический домен;
1 — эндоплазматическая сеть, 2 — комплекс Гольджи, 3 — плазмалемма,
4 — лизосомы, 5 — эндосомы, 6 — челнок Гольджи-лизосома через плазмалемму
и эндосому, 7 — челнок Гольджи-плазмалемма, 7а — кринофагическое отклонение,
8а, 8б — пути возвращения мембран плазмалеммы, 8в — челнок эндосома-лизосома,
9 — аутофагическая сегрегация, 10 — челнок плазмалемма-лизосома
(в обход эндосомы), 11 — челнок эндосома-лизосома,
12 — челнок плазмалемма-эндосома, 13 — прямой челнок Гольджи-лизосома.
Стрелки со светлыми концами — пути перемещений (по К. де Дюву, с изменениями)
11
Б
22
33
44
55
66
77
7а
7а
44
8а
8а
44
99
10
10
11
11
12
12
8б
8б
А
8в
8в
13
13
Б
55
52
СТРОЕНИЕ КЛЕТКИ
Положение комплекса Гольджи в клетке обусловлено ее функцио-
нальной специализацией. В секретирующих клетках он находится
между ядром и поверхностью выведения. Так, в бокаловидных клет-
ках ядро смещено к базальному концу, а комплекс Гольджи находится
между ним и апикальной поверхностью. В клетках эндокринных же-
лез, из которых секрет выводится в кровеносные капилляры, со всех
сторон окружающие клетку, комплекс Гольджи представлен многими
поверхностно лежащими диктиосомами. В гепатоцитах диктиосомы
располагаются группами: одни около билиарных участков, другие
около сосудистых. В плазматических клетках при изучении в свето-
вом микроскопе комплекс занимает светлую зону около ядра; он
окружен гранулярной эндоплазматической сетью и на ее базофиль-
ном фоне выглядит как «светлый дворик».
Во всех случаях вблизи комплекса Гольджи концентрируются ми-
тохондрии. Это связано с происходящими в нем энергозависимыми
реакциями.
Лизосомы
Каждая лизосома (рис. 35) представляет собой мембранный пузы-
рек диаметром 0,4—0,5 мкм. Его содержимое представляет собой гомо-
генный осмиофильный мелкозернистый материал. В нем содержится
около 50 видов различных гидролитических ферментов в дезактиви-
рованном состоянии (протеазы, липазы, фосфолипазы, нуклеазы, гли-
козидазы, фосфатазы, в том числе кислая фосфатаза; последняя явля-
ется маркером лизосом). Молекулы этих ферментов, как всегда,
синтезируются на рибосомах гранулярной ЭПС, откуда переносятся
транспортными пузырьками в КГ, где модифицируются. От зрелой по-
верхности цистерн КГ отпочковываются первичные лизосомы.
Все лизосомы клетки формируют лизосомное пространство, в кото-
ром с помощью протонного насоса постоянно поддерживается кислая
среда — рН колеблется в пределах 3,5—5,0. Мембраны лизосом устойчи-
вы к заключенным в них ферментам и предохраняют цитоплазму от их
действия. Это связано с особой конформацией молекул лизосомной
мембраны, при которой их химические связи скрыты. Повреждение или
нарушение проницаемости лизосомной мембраны приводит к актива-
ции ферментов и тяжелым повреждениям клетки вплоть до ее гибели.
Функция лизосом — внутриклеточный лизис («переварива-
ние») высокомолекулярных соединений и частиц. Последними могут
быть собственные органеллы и включения или частицы, поступившие
в клетку извне в ходе эндоцитоза (см. раздел «Эндоцитоз»). Захва-
ченные частицы обычно окружены мембраной. Такой комплекс назы-
вают фагосомой.
53
ЦИТОПЛАЗМА
Процесс внутриклеточного лизиса осуществляется в несколько эта-
пов. Сначала первичная лизосома сливается с фагосомой. Их комплекс
называют вторичной лизосомой (фаголизосомой). Во вторичной лизо-
соме ферменты активируются и расщепляют поступившие в клетку по-
лимеры до мономеров. Это происходит постепенно, поэтому вторичные
лизосомы идентифицируются благодаря наличию в них осмиофильного
материала различной электронной плотности. Продукты расщепления
транспортируются через лизосомную мембрану в цитозоль. Неперева-
7
8
9
10
11
5
4
3
2
1
6
12
13
14
15
16
H
+
À
Ò
Ð
H+
17
Рис. 35. Схема строения и функционирования лизосом (возможные пути
формирования вторичных лизосом путем слияния мишеней с первичными
лизосомами, содержащими новосинтезированные гидролитические ферменты):
1 — фагоцитоз, 2 — вторичная лизосома, 3 — фагосома, 4 — остаточное тельце,
5 — мультивезикулярное тельце, 6 — очистка лизосом от мономеров, 7 — пиноцитоз,
8 — аутофагосома, 9 — начало аутофагии, 10 — участок агранулярной
эндоплазматической сети, 11 — гранулярная эндоплазматическая сеть,
12 — протонный насос, 13 — первичные лизосомы, 14 — комплекс Гольджи,
15 — рециклирование мембран, 16 — плазмалемма, 17 — кринофагия;
пунктирные стрелки — направления движения
(по К. де Дюву и по Б. Албертсу и соавт., с изменениями)
54
СТРОЕНИЕ КЛЕТКИ
ренные вещества остаются в лизосоме и могут сохраняться в клетке,
окруженные мембраной, очень долго в виде остаточного тельца.
Остаточные тельца относят уже не к органеллам, а к включениям.
Возможен и другой путь превращений: вещества в фагосоме расщепля-
ются полностью, после чего мембрана фагосомы распадается. Фрагмен-
ты мембран направляются к КГ и используются в нем для сборки но-
вых. Вторичные лизосомы могут сливаться между собой, а также
с другими первичными лизосомами. При этом иногда образуются свое-
образные вторичные лизосомы — мультивезикулярные тельца.
В процессе жизнедеятельности клетки на разных иерархических
уровнях ее организации, начиная от молекул и кончая органеллами,
постоянно происходит перестройка структур. Вблизи поврежденных
или требующих замены участков цитоплазмы, обычно по соседству
с комплексом Гольджи, образуется полулунная двойная мембрана, ко-
торая растет, окружая со всех сторон поврежденные зоны (см. рис. 24).
Затем эта структура сливается с лизосомами. В такой аутофагосоме
(аутосоме) совершается лизис структур органеллы.
В других случаях в процессе макро- или микроаутофагии подлежа-
щие перевариванию структуры (например, гранулы секрета) впячивают-
ся в лизосомную мембрану, окружаются ею и подвергаются переварива-
нию. Образуется аутофагическая вакуоль. В результате множественной
микроаутофагии тоже формируются мультивезикулярные тельца (на-
пример, в нейронах мозга и кардиомиоцитах). Наряду с аутофагией в
некоторых клетках происходит и кринофагия (от греч. krinein — просеи-
вать, отделять) — слияние первичных лизосом с секреторными гранула-
ми. В лизосомах необновляющихся клеток в результате многократного
аутофагирования накапливается липофусцин — пигмент старения.
Таким образом, аутофагия представляет собой один из механиз-
мов обновления внутриклеточных структур — внутриклеточной фи-
зиологической регенерации. Путем аутофагии устраняются органеллы,
утратившие свою активность в процессе естественного их старения.
Устраняются также органеллы, ставшие избыточными, если в процес-
се нормальной жизнедеятельности снижается интенсивность физио-
логических процессов в клетке. Аутофагия — один из способов регуля-
ции функциональной активности. Поскольку изменения последней
цикличны, то аутофагия — один из механизмов реализации биологиче-
ских ритмов на клеточном уровне.
В некоторых случаях непереваренные остатки накапливаются
в лизосомах, что приводит к их перегрузке («хронический запор»).
Выделение непереваренных остатков путем экзоцитоза и их накопле-
ние во внеклеточной среде может вызвать повреждение внеклеточных
структур. Поэтому этот механизм реализуется редко. Наиболее часто
встречаются три типа пищеварительных расстройств клетки: внутри-
клеточный выброс, внеклеточный выброс и перегрузка (К. де Дюв).
55
ЦИТОПЛАЗМА
Пероксисомы
Пероксисомы (рис. 36) представляют собой мембранные пузырь-
ки диаметром от 0,2 до 0,5 мкм. Как и лизосомы, они отщепляются от
цистерн транс-полюса КГ. Есть также точка зрения, что мембраны пе-
роксисом образуются путем отпочкования от гладкой эндоплазмати-
ческой сети, а ферменты синтезируются полирибосомами цитозоля,
откуда и поступают в пероксисому. Под мембраной пузырька разли-
чают центральную более плотную часть и периферическую область.
Различают две формы пероксисом. Мелкие пероксисомы (диаме-
тром 0,15—0,25 мкм) имеются практически во всех клетках млекопи-
тающих (и человека), содержат мелкозернистый осмиофильный ма-
териал и морфологически мало отличаются от первичных лизосом.
Крупные пероксисомы (диаметром более 0,25 мкм) присутствуют
лишь в некоторых тканях (печень, почки). В них имеется кристалло-
видная сердцевина, в которой находятся ферменты в концентриро-
ванном виде. Наряду с пероксисомами встречаются и другие мем-
бранные микротельца диаметром от 0,5 до 10 мкм, содержащие
различные ферменты.
Пероксисомы содержат ферменты (пероксидазу, каталазу и окси-
дазу D-аминокислот). Пероксидаза участвует в обмене перекисных
соединений, в частности перекиси водорода, которая токсична для
клетки. Для биохимических реакций в пероксисомах используется
молекулярный кислород. Пероксисомы принимают также участие
Рис. 36. Пероксисома:
1 — мембрана пероксисомы, 2 — кристаллоид,
3 — включения гликогена около пероксисомы (по К. де Дюву, с изменениями)
3
2
1
56
СТРОЕНИЕ КЛЕТКИ
в нейтрализации многих других токсических соединений, например
этанола. Каталаза составляет среди ферментов пероксисом около 40%
всех белков. Пероксисомы участвуют также в обмене липидов, холе-
стерина и пуринов.
Специальные органеллы
Напомним, что органеллы называют специальными, если они есть
только у клеток, выполняющих особые специализированные функ-
ции. Таковы щеточная кайма, стереоцилии, базальный лабиринт, рес-
нички, кинетоцилии, жгутики, миофибриллы.
Среди специальных органелл в настоящем издании будут описаны
мерцательные реснички и жгутики, как наиболее распространенные.
Описание остальных специальных органелл дано в соответствующих
разделах курсов эмбриологии, общей и частной гистологии.
Реснички и жгутики
Ресничка (рис. 37) представляет собой вырост клетки, окружен-
ный плазмалеммой. У основания реснички на уровне кортикального
слоя цитоплазмы находится базальное тельце (кинетосома), которое
образовано девятью периферическими триплетами коротких микро-
трубочек, окружающих один белковый центральный цилиндр. Каж-
дый периферический триплет соединен с ним посредством белковых
«спиц».
От каждой наружной микротрубочки отходит отросток, напоми-
нающий флажок. Над базальным тельцем в названный цитоплазмати-
ческий вырост направляется осевая нить (осевой филамент, аксоне-
ма), тоже образованная микротрубочками (строение последних
описано ранее). От каждого базального тельца к соседнему (к основа-
нию другой реснички) отходит короткий боковой отросток, а в глубь
цитоплазмы — поперечно исчерченный базальный корешок (перио-
дичность исчерченности равна 65 нм).
Сразу над базальным тельцем микротрубочки аксонемы тоже
образуют девять периферических триплетов (см. рис. 26), но над
уровнем плазмалеммы в периферических комплексах одна из микро-
трубочек редуцируется; в центральной группе появляется пара ми-
кротрубочек, окруженная белковой оболочкой, имеющей вид цилин-
дра, поэтому на большей части протяжения реснички тянутся дуплеты
микротрубочек. Девять дуплетов располагаются на периферии,
один — в центре.
В результате на поперечном сечении реснички ее возвышающаяся
часть напоминает колесо с девятью спицами, в центре которого лежит
57
ЦИТОПЛАЗМА
нечетко контурирующаяся центральная капсула, окружающая две
центральные одиночные микротрубочки, с которыми она связана бел-
ковыми отростками. Периферические дуплеты состоят из двух ми-
кротрубочек (А и В) и окружают центральную капсулу (см. рис. 37).
От микротрубочки А к центральной капсуле отходит радиальная
«спица» с периодичностью расположения вдоль аксонемы, равной
29 нм. По направлению к соседней трубочке В отходит пара «ручек»,
Рис. 37. Ресничка:
I — ресничка на продольно-поперечном разрезе, II — внеклеточная часть,
III — поперечный срез через внеклеточную часть, IV — поперечный срез через
промежуточную область, соединяющую аксонему с базальным тельцем, V — срез
через базальное тельце, VI — внутриклеточная часть; 1— ресничка, 2 — периферические
дуплеты, 3 — центральный дуплет, 4 — центральная капсула, 5 — микроворсинки,
6 — наружные микротрубочки, 7 — базальное тельце, 8 — базальный корешок,
9 — цитолемма, 10 — периферические дуплеты, 11 — центральный дуплет,
12 — центральная капсула, 13 — спица, 14 — субфибрилла а, 15 — субфибрилла б,
16 — динеин, 17 — триплеты микротрубочек, 18 — триплеты, 19 — спицы,
20 — центральный цилиндр (по Р. Крстичу и по Ю. С . Ченцову, с изменениями)
I
11
22
33
44
55
66
77
88
99
10
10
11
11
12
12
13
13
14
14
15
15
16
16
20
20
17
17
18
18
19
19
II
III
IV
V
VI
58
СТРОЕНИЕ КЛЕТКИ
образованных, как и «спицы», белком динеином с периодичностью
24 нм. Соседние дуплеты микротрубочки соединены между собой
с периодичностью 86 нм поперечными мостиками, образованными
белком нексином.
Реснички являются производными не только поверхностного ком-
плекса клетки, но и клеточного центра. В начале их развития проис-
ходит многократная редупликация центриолей. Новые центриоли
парами мигрируют к поверхности клетки. Здесь происходит их моди-
фикация.
Одна из центриолей (проксимальная) ложится в поверхностном
комплексе вблизи от плазмалеммы у основания будущей реснички.
Другая центриоль (дистальная) ложится между ней и плазмалеммой.
Триплеты микротрубочек дистальной центриоли становятся дуплета-
ми, в проксимальной центриоли триплеты сохраняются. В клеточном
центре идет сборка микротрубочек, они направляются к дистальной
центриоли и наращивают ее длину. Дистальная центриоль растет
и влечет за собой плазмалемму. Рост аксонемы в длину происходит за
счет полимеризации тубулина у дистальных концов двух внутренних
трубочек каждого триплета базального тельца.
Жгутик эукариотической клетки (например, спермия) напомина-
ет ресничку, но он длиннее.
Реснички и жгутики выполняют функцию движения. Все реснички
клетки совершают координированные колебания. Это достигается по-
средством скольжения дуплетов микротрубочек относительно друг
друга. Оно обусловлено изменениями конфигурации молекул белка
динеина. Динеин обладает аденозинтрифосфатазной активностью.
При гидролизе АТФ выделяется свободная энергия, за счет которой
динеиновые ручки выпрямляются, контактируют с соседним дупле-
том микротрубочек и сдвигают его по направлению к верхушке рес-
нички. При регенерации АТФ ручки отделяются от соседнего дуплета
и опускаются вниз к основанию реснички.
Все реснички клетки совершают координированные колебатель-
ные движения. Они похожи на движения рук пловца брассом. Снача-
ла ресничка резко наклоняется над поверхностью клетки. При этом
слизь, которой обычно здесь покрыта поверхность, прогоняется в на-
правлении наклона. Далее наклоненная ресничка совершает поворот
на 180°. Поскольку она скользит параллельно поверхности клетки,
слизь над ней назад не возвращается. Затем ресничка снова выпрям-
ляется и перемещает слизь. Потом начинается новый цикл.
Число ресничек достигает нескольких сотен. Так, до 250 ресничек
длиной 5—15 мкм и диаметром 0,15—0,25 мкм покрывают апикаль-
ную поверхность реснитчатых эпителиоцитов верхних дыхательных
путей, маточных труб, семявыводящих канальцев.
59
ЦЕЛОСТНЫЕ РЕАКЦИИ КЛЕТКИ
Включения
Включениями целесообразно называть структурированные на уль-
трамикроскопическом уровне скопления веществ в клетке, возникающие
как продукты ее метаболизма. Будучи продуктами жизнедеятельности
клетки, сами включения почти не участвуют в осуществлении тех ее
активных функций, которые необходимы для поддержания этой жиз-
недеятельности. Включения могут активно использоваться клеткой,
но это осуществляется благодаря ферментным системам, которыми
обладают гиалоплазма и органеллы. Сами включения ферментатив-
ной активностью, как правило, не обладают.
В зависимости от состава и от способа использования клеткой
среди включений довольно условно различают трофические, пиг-
ментные и секреторные. Среди трофических включений упомянем
капли жира, гранулы гликогена (см. рис. 36), белковые гранулы. Эти
вещества накапливаются в клетке, а затем расходуются ею при воз-
никновении соответствующих функциональных потребностей. Боль-
шинство трофических включений лежит в гиалоплазме свободно.
Пигментные включения могут лежать и свободно, но могут быть
окружены мембраной. Часто мембраной окружены гранулы мелани-
на. Свободно в гиалоплазме предшественников эритроцитов распо-
лагается гемоглобин (у низших позвоночных и у птиц также и в са-
мих эритроцитах). Секреторные гранулы отделяются от комплекса
Гольджи и несут к плазмалемме синтезированные клеткой вещества;
они весьма разнообразны.
Нередко включениями называют структуры, присутствующие
в клетке временно. Это неточно. Гемоглобин, например, присутствует
в эритроцитах постоянно, столь же постоянны гранулы меланина
в пигментных клетках. В качестве включений рассматривают и оста-
точные тельца, возникающие после активных процессов фагоцитоза
и аутофагии, которые сохраняются в клетке иногда вплоть до ее гибе-
ли, но не принимают участия в обеспечении жизнедеятельности.
Совершенно резкую границу между органеллами и включениями
провести невозможно, поэтому в Международной гистологической
номенклатуре (Nomina Histologica, London, 1985) перечень их поме-
щен в одном разделе.
ЦЕЛОСТНЫЕ РЕАКЦИИ КЛЕТКИ
Все изменения состояния клетки и превращения веществ в ней
в конечном счете слагаются из отдельных звеньев биохимических
процессов. В свою очередь, каждый биохимический процесс представ-
ляет собой последовательную цепь многих единичных химических
60
СТРОЕНИЕ КЛЕТКИ
реакций. Эти цепи связаны между собой и совершаются в различных
компартментах клетки. Все биохимические реакции осуществляются
с помощью ферментов.
Детально процессы обмена веществ, в том числе и внутриклеточ-
ные, изучаются в курсе биохимии. В данной книге будут рассмотрены
не столько эти процессы, сколько участие в них клеточных структур.
Для осуществления биохимических реакций необходимо посту-
пление веществ в клетку — эндоцитоз, обмен веществ в клетке —
метаболизм и выведение продуктов метаболизма — экзоцитоз.
Эндоцитоз
Существует несколько способов эндоцитоза (от греч. endon — внутри,
kytos — клетка). Некоторые из них, такие как пассивный и активный
трансмембранный транспорт, описаны в разделе «Биологические мем-
браны». При эндоцитозе этот транспорт осуществляется плазмалеммой.
Кроме того, существуют более сложные способы — пиноцитоз (от греч.
pino — пью) и фагоцитоз (от греч. phagos — пожирающий). Обычно под
пиноцитозом понимают захват клеткой жидких коллоидных частиц, под
фагоцитозом — захват корпускул (более плотных и крупных частиц
вплоть до других клеток). Механизм пино- и фагоцитоза различен.
В общем виде поступление в клетку твердых частиц или капель
жидкости извне называется гетерофагией (от греч. heteros — иной).
Этот процесс наиболее широко распространен у простейших, но очень
важен и у человека (равно как и у других млекопитающих). Гетерофа-
гия играет существенную роль в защите организма (сегментоядерные
нейтрофильные гранулоциты, макрофагоциты), перестройке костной
ткани (остеокласты), образовании тироксина фолликулами щитовид-
ной железы, реабсорбции белка и других макромолекул в прокси-
мальном отделе нефрона и других процессах.
Пиноцитоз
Для того чтобы внешние молекулы поступили в клетку, они долж-
ны быть сначала связаны рецепторами гликокаликса (рис. 38).
В месте такого связывания под плазмалеммой обнаруживаются
молекулы белка клатрина. Плазмалемма вместе с присоединенными
извне молекулами и подстилаемая со стороны цитоплазмы клатри-
ном начинает впячиваться. Впячивание становится все глубже, его
края сближаются и затем смыкаются. В результате от плазмалеммы
отщепляется пузырек, несущий в себе захваченные молекулы. Клат-
рин на его поверхности выглядит на электронных микрофотографиях
как неровная каемка, поэтому такие пузырьки получили название
окаймленных.
61
ЦЕЛОСТНЫЕ РЕАКЦИИ КЛЕТКИ
Клатрин не дает возможности пузырькам присоединяться к вну-
триклеточным мембранам, поэтому окаймленные пузырьки могут
беспрепятственно транспортироваться в клетке именно к тем участ-
кам цитоплазмы, где должно использоваться их содержимое. Так
к ядру доставляются, в частности, стероидные гормоны. Однако обыч-
но окаймленные пузырьки сбрасывают кайму вскоре после отщепле-
ния от плазмалеммы. Клатрин переносится к плазмалемме и снова
может участвовать в реакциях эндоцитоза.
У поверхности клетки в цитоплазме имеются более постоянные
пузырьки — эндосомы. Окаймленные пузырьки сбрасывают клатрин
и сливаются с эндосомами, при этом объем и поверхность эндосом
увеличиваются. Затем «избыточная» часть эндосомы отщепляется
в виде нового пузырька, в котором нет поступивших в клетку веществ,
они остаются в эндосоме. Новый пузырек направляется к поверхно-
сти клетки и сливается с плазмалеммой. В результате убыль мембра-
ны, возникшая при отщеплении окаймленного пузырька, восстанав-
ливается, при этом в плазмалемму возвращаются и ее рецепторы.
Эндосомы погружаются в цитоплазму и встречаются с лизосома-
ми. Их мембраны сливаются. В возникшей таким образом вторичной
лизосоме вещества, поступившие в клетку, подвергаются разнообраз-
ным биохимическим превращениям. По завершении процесса мем-
брана лизосомы может распадаться на фрагменты, а продукты распа-
да вторичной лизосомы и ее содержимого становятся доступными
для внутриклеточных метаболических реакций. Например, аминокис-
лоты связываются тРНК и доставляются к рибосомам, а глюкоза мо-
жет поступать в комплекс Гольджи либо в канальцы агранулярной
ЭПС.
I
II
III
11
22
33
Рис. 38. Рецепторно-опосредованный эндоцитоз:
I — окаймленная ямка, II — промежуточная стадия, III — окаймленный пузырек;
1 — лиганды, 2 — мембранные рецепторы, 3 — клатрин
(по Б. Албертсу и соавт., с изменениями)
62
СТРОЕНИЕ КЛЕТКИ
Хотя эндосомы и не обладают клатриновой каймой, не все они
сливаются с лизосомами. Часть из них направляется от одной поверх-
ности клетки к другой (так происходит, если клетки образуют эпите-
лиальный пласт). Там мембрана эндосомы сливается с плазмалеммой
и содержимое выводится вовне. В результате вещества переносятся
через клетку из одной среды в другую без изменений. Этот процесс
называют трансцитозом. Существенно, что путем трансцитоза могут
переноситься и белковые молекулы, в частности иммуноглобулины.
Фагоцитоз
Если крупная частица имеет на поверхности молекулярные груп-
пировки, которые могут распознаваться рецепторами клетки, она свя-
зывается. Далеко не всегда чужеродные частицы сами обладают таки-
ми группировками. Однако, попадая в организм, они окружаются
молекулами иммуноглобулинов (опсонинами), которые всегда со-
держатся и в крови, и в межклеточной среде. Иммуноглобулины всег-
да распознаются клетками-фагоцитами.
После того как покрывающие чужеродную частицу опсонины свя-
зались с рецепторами фагоцита, активируется весь его поверхностный
комплекс. Актиновые микрофиламенты начинают взаимодействовать
с миозином, и конфигурация поверхности клетки изменяется. Вокруг
частицы вытягиваются выросты цитоплазмы фагоцита. Они охваты-
вают поверхность частицы и объединяются над ней. Наружные лист-
ки выростов сливаются, замыкая поверхность клетки (рис. 39).
Глубокие листки выростов образуют мембрану вокруг поглощен-
ной частицы — формируется фагосома. Фагосома сливается с лизосо-
мами, в результате чего возникает их комплекс — гетеролизосома
(гетеросома, или фаголизосома). В ней происходит лизис захвачен-
ных компонентов частицы. Часть продуктов лизиса выводится из ге-
теросомы и утилизируется клеткой, часть же может оказаться не под-
дающейся действию лизосомных ферментов.
Эти остатки, как уже сказано, образуют оста-
точные тельца.
Потенциально все клетки обладают способ-
ностью к фагоцитозу, но в организме лишь
некоторые специализируются в этом направле-
нии. Таковы нейтрофильные лейкоциты и мак-
рофаги.
Рис. 39 . Фагоцитоз:
1 — бактерия, 2 — псевдоподии фагоцита
(по Б. Албертсу и соавт., с изменениями)
22
11
63
ВНУТРИКЛЕТОЧНЫЕ БИОХИМИЧЕСКИЕ РЕАКЦИИ
ВНУТРИКЛЕТОЧНЫЕ
БИОХИМИЧЕСКИЕ РЕАКЦИИ
Управление внутриклеточными синтезами осуществляется из ядра
клетки. На активных участках хромосом синтезируются молекулы
РНК. Они транспортируются к поровым комплексам и поступают
в цитоплазму. На рибосомах из аминокислот происходит сборка бел-
ков, которые в соответствии с их назначением можно отнести к трем
группам. Одна группа — это структурные белки, которые используют-
ся клеткой для построения собственных органелл, другая — белки,
выделяемые клеткой вовне, это ее секреты; третья группа — фермен-
ты, которые обеспечивают все внутриклеточные биохимические прев-
ращения как катализаторы. Часть ферментов остается в цитоплазме.
Одни из них функционируют в гиалоплазме, другие же встраиваются
в органеллы. Часть ферментов направляется в ядро и там регулирует
считывание генетической информации с ДНК и матричный синтез
РНК. Также возвращаются в ядро те белки, которые участвуют в по-
строении самих хромосом.
Вещества, которые синтезируются на мембранах ЭПС, поступа-
ют в транспортные пузырьки и доставляются к комплексу Гольджи,
где происходит их модификация. В комплексе Гольджи осуществля-
ется сегрегация веществ, в результате чего они затем распределяют-
ся по клетке в зависимости от дальнейших путей использования (см.
рис. 34).
Синтез белка
Синтез каждого определенного специфического белка опреде-
ляется геном. Ген — это участок ДНК, являющийся элементарной
частицей генетической информации. Он характеризуется опреде-
ленной последовательностью нуклеотидов.
Центральная догма современной биологии характеризует одну из
основ жизни следующим образом:
Иными словами, наследственная информация, заключенная
в ДНК, передается по наследству благодаря ее самоудвоению (ре-
пликации). Генетическая информация, записанная в виде последо-
вательности нуклеотидов ДНК, в процессе транскрипции переписы-
64
СТРОЕНИЕ КЛЕТКИ
вается в нуклеотидную последовательность РНК, которая, в свою
очередь, определяет последовательность аминокислот соответству-
ющей белковой молекулы.
Благодаря наличию ядерной оболочки в клетках человека (и дру-
гих эукариот) процессы транскрипции и трансляции проходят в раз-
ных структурах и разделены во времени.
Синтез белка (трансляция) связан с процессом транскрипции —
переписывания информации, хранящейся в ДНК, поэтому мы начи-
наем описание с последнего.
Транскрипция осуществляется в ядре (рис. 40). Информация
о структуре белка, заключенная в ДНК, «переписывается» на инфор-
мационную РНК (мРНК). При этом с одного гена может «переписы-
ваться» множество молекул мРНК. Они подвергаются в ядре процес-
сингу, после чего транспортируются из ядра в цитоплазму, где
и выполняют свои функции. Процессинг (от англ. processing — обра-
ботка) — совокупность реакций, ведущих к превращению первичных
неактивных транскриптов в функционирующие молекулы.
В клетках существует три типа РНК. Среди них информационная
(мРНК) переносит информацию о нуклеотидной последовательности
ДНК к рибосомам. В образовании рибосом участвует рибосомная
РНК (рРНК). Небольшие транспортные РНК (тРНК) выполняют
двойную функцию: они присоединяют молекулу аминокислоты, тран-
спортируют ее к рибосоме и узнают триплет, соответствующий этой
аминокислоте в молекуле мРНК.
Рис. 40. Схема синтеза белка. Объяснения в тексте
Рибосома
РНК-полимераза
ДНК
Ядро
Аминокис лоты
Белок
Цитоплазма
ТРНК
МРНК
МРНК
65
ВНУТРИКЛЕТОЧНЫЕ БИОХИМИЧЕСКИЕ РЕАКЦИИ
В середине молекулы тРНК имеется группировка из трех азоти-
стых оснований, называемая антикодоном. Антикодон может связать-
ся с определенной группировкой трех оснований на мРНК — с кодо-
ном. Действительно, после сближения молекул антикодон тРНК
узнает кодон мРНК и спаривается с ним.
Генетический код, расшифрованный в 60-х годах XX в. М . Нирен-
бергом, У. Холлом и X. Кораной, основан на триплетах, или кодо-
нах, — три нуклеотида определяют присоединение к полипептидной
цепи одной аминокислоты (табл. 6).
Таблица 6
Генетический код
2-e положение
UСA G
1-е
п
о
л
о
ж
е
н
и
е
U
UUU Phe UCU Ser UAU Tyr UGU Cys U
3-е
п
о
л
о
ж
е
н
и
е
UUC Phe UCC Ser UAC Туг UGC Cys С
UUA Leu UCA Ser UAA
ochr UGA oral A
UUG Leu UCG Ser UAG amb UGG Try G
C
CUU Leu ECU Pro CAU His CGU Arg U
CUC Leu CCC Pro СAC His CGC Arg С
CUA Leu CCA Pro CAA Gki CGA Arg A
CUG Leu CCG Pro CAG Gki CGG Arg G
A
AUU Ilе ACU Thr AAU Asn AGU Ser U
AUC Ilе ACC Thr AAC Asn AGC Ser С
AUA Ilе АСА Thr AAA Lys AGA Arg A
AUG Met ACG Thr AAG Lys AGG Arg G
G
GUU Val GCU Ala GAU Asp GGU Gly U
GUC Val GCC Ala GAC Asp GGC Gly С
GUA Val GCA Ala GAA Gki GGA Gly A
GUGValGCGAlaGAGGkiGGGlyG
Триплетные комбинации азотистых оснований мРНК (U, А, С, G) определяют следую-
щие аминокислоты: Рhе — фенилаланин, Leu — лейцин, Ilе — изолейцин, Меt — метио-
нин, Val — валин, Ser — серин, Pro — пролин, Thr — треонин, Ala — аланин, Tyr — тирозин,
His — гистидин, Glh — глутамин, Asn — аспарагин, Lys — лизин, Asp — аспарагиновая
кислота, Glu — глутаминовая кислота, Cys — цистеин, Try — триптофан, Arg — аргинин,
Gly — глицин. Звездочкой обозначены стартовые кодоны; триплеты ochre, amber и oral
действуют как «стоп-кодоны» (по F. Crick).
66
СТРОЕНИЕ КЛЕТКИ
Генетический код отличается рядом важных свойств. Он трипле-
тен: именно три нуклеотида определяют присоединение к полипеп-
тидной цепи одной аминокислоты. Генетический код вырожден, т. е .
большинство аминокислот кодируются более чем одним триплетом.
При этом одна и та же аминокислота может кодироваться разными
триплетами, однако первые два нуклеотида для них всегда одинако-
вы. Например, триплет –С–С–С – кодирует пролин. Кроме того,
включение пролина может кодироваться триплетами ССU, ССА,
ССG. Триплет AUG кодирует первую аминокислоту — метилметио-
нин, с которой начинается синтез полипептидной цепи. Всего в гене-
тическом коде имеется 64 кодона, три из которых (UАА, UGA и UAG)
являются стоп-кодонами, завершающими синтез полипептидной
цепи.
Генетический код не перекрывается, хотя в нем отсутствуют зна-
ки, отделяющие один триплет от другого. Например, в последователь-
ности оснований UUCAUUGUU первые три основания кодируют
одну аминокислоту, вторые три — другую и т. д . Не может быть такой
ситуации в приведенном примере, когда основание UUC кодирует
одну аминокислоту, UCA — другую, а CAU — третью и т. д.
Код универсален, т. е . все живые организмы на планете Земля
(включая вирусы) имеют один и тот же код. Рамка считывания опре-
деляет положение первого нуклеотида кодона гена (или мРНК).
Рибосомная и транспортная РНК (рРНК и тРНК) синтезируются
на идентичных генах, которые (в отличие от генов мРНК) в каждой
клетке имеются в виде множества копий. При этом рРНК синтезиру-
ется на описанных ядрышковых организаторах — участках ДНК, име-
ющих форму петель, которые находятся в ядрышке. Предшественник
рРНК, синтезированный на ядрышковом организаторе, в ядрышке
соединяется с рибосомными белками, синтезированными в цитоплаз-
ме и транспортированными в ядро, образуя крупные рибонуклеопро-
теидные частицы. Последние претерпевают процессинг, в результате
которого в ядре образуются большая и малая субъединицы рибосом.
Предшественники рибосом транспортируются в цитоплазму, где
в ходе синтеза белка и происходит сборка самих рибосом.
Синтез самой молекулы белка (см. рис. 40) начинается с того, что
молекула тРНК связывается с соответствующей аминокислотой, в ре-
зультате чего образуется аминоацил-тРНК. Малая субъединица рибо-
сомы связывается с инициаторной тРНК, несущей молекулу метил-
метионина. Этот комплекс присоединяется к инициаторному кодону
мРНК (АИС). После этого к малой присоединяется большая субъ-
единица рибосомы. Реакции синтеза белка осуществляют рибосомы,
которые считывают информацию, заложенную в мРНК, продвигаясь
вдоль нее в направлении 5' ␚ 3'.
67
ВНУТРИКЛЕТОЧНЫЕ БИОХИМИЧЕСКИЕ РЕАКЦИИ
Рибосома связывает две молекулы тРНК: участок А рибосомы
связывает аминоацил-тРНК, участок Р рибосомы — аминоацил —
тРНК, связанную с растущей полипептидной цепью. Обе тРНК свя-
зываются с соседними кодонами мРНК. К рибосоме подходит следу-
ющая аминоацил-тРНК, и образуется первая пептидная связь.
Перемещаясь по цепи мРНК, рибосома присоединяет следующие
аминокислоты, которые связываются между собой, а молекулы тРНК
отделяются, чтобы вскоре присоединить новую аминокислоту. При
достижении рибосомой стоп-кодона синтез прекращается, потому что
к стоп-кодонам нет соответствующих антикодонов ни у одной т-РНК.
Полипептидная цепь отделяется от рибосомы.
Следует отметить, что в приведенном изложении схема белкового
синтеза сильно упрощена.
Основные реакции тканевого обмена
В клетках постоянно осуществляется метаболизм (от греч. meta-
bole — перемена, превращение), или обмен веществ, который пред-
ставляет собой совокупность процессов ассимиляции (реакций био-
синтеза сложных биологических молекул из более простых) и
диссимиляции (реакций расщепления). В результате диссимиляции
освобождается энергия, заключенная в химических связях пищевых
веществ. Эта энергия используется клеткой для осуществления раз-
личной работы, в том числе ассимиляции.
Первый закон термодинамики гласит: энергия не возникает и не
уничтожается, она лишь переходит из одного вида в другой, пригод-
ный для выполнения работы. Клетка использует энергию, заключен-
ную в химических связях аминокислот, моносахаридов и жирных
кислот, которые образуются в результате пищеварения из белков,
углеводов и жиров и поступают в клетку.
Схематично катаболизм пищевых веществ можно представить
следующим образом. В первой стадии происходит их расщепление до
мономеров. Во второй стадии, независимо от природы пищевого про-
дукта, образуется ацетилкоэнзим А (ацетил-КоА). Он, а также другие
ферменты, включающие в себя КоА, являются ключевыми звеньями
множества разнообразных биохимических реакций (рис. 41).
Рассмотрим более детально энергетический обмен на примере рас-
щепления глюкозы (рис. 42). Сначала глюкоза транспортируется
через плазматическую мембрану в цитоплазму клетки. В матриксе
цитоплазмы происходит ее бескислородное расщепление, или глико-
лиз, — многоступенчатый ферментативный процесс, в результате
которого из одной молекулы глюкозы образуются две молекулы
пирувата (пировиноградной кислоты — ПВК) и две молекулы адено-
зинтрифосфорной кислоты (АТФ).
68
СТРОЕНИЕ КЛЕТКИ
АТФ — нуклеотид, состоящий из аденина, рибозы и трех остатков
фосфорной кислоты. АТФ является универсальным переносчиком
и основным аккумулятором энергии в клетке, которая заключена
в высокоэнергетических связях между тремя остатками фосфорной
кислоты. При отщеплении от АТФ одной фосфатной группы образу-
ется АДФ (аденозиндифосфорная кислота) и фосфат и выделяется
свободная энергия, которая используется клеткой для осуществления
работы.
В результате гликолиза освобождается лишь около 5% энергии, за-
ключенной в химических связях молекулы глюкозы, остальная же ос-
вобождается в митохондриях в процессе аэробного окисления и тоже
запасается в АТФ. В митохондриях же АДФ, соединяясь с остатком
фосфорной кислоты, превращается в АТФ: АТФ АДФ + Pi (Pi — ор-
ганический фосфат). В расчете на один моль глюкозы образуется
36 молекул АТФ.
Рис. 41. Общая схема обмена веществ в клетке и роль СоА в нем
(по А. Ленинджеру, с изменениями)
Углеводы
Ацетил-CoA
Ацетил-CoA
CO2
O2
Ацетил-CoA
Окисление
через цикл
трикарбоно-
вых кислот
Синтез
жирных
кислот
Образование
катоновых
тел
Синтез
холестерина
Ацетоацетил-CoA
Малонил-CoA
Цитрат
Оксиметил-глуатирил-CoA
Жирные кислоты
Аминокислоты
69
ВНУТРИКЛЕТОЧНЫЕ БИОХИМИЧЕСКИЕ РЕАКЦИИ
Реакции окисления, приводящие
к освобождению энергии, осуществ-
ляются путем отнятия у окисляе-
мой молекулы отрицательно заря-
женного электрона. Этот электрон
связан с атомом водорода (Н+). Ак-
цептором электронов служат моле-
кулы никотинамидадениндинуклео-
тида (НАД+). Они и присоединяют
к себе этот ион водорода (реакция
восстановления). Восстановленная
молекула
никотинамидаденинди-
нуклеотида обозначается как НАДН.
Дальнейшие этапы окисления
происходят в митохондриях.
Следует подчеркнуть, что наибо-
лее важным источником энергии
в клетке являются жиры; их энерге-
тическая ценность выше, чем ценность гликогена, более чем в 6 раз,
а запасы жира в организме человека примерно в 30 раз больше, чем
запасы гликогена.
В митохондриях, точнее, в их матриксе, в цикле Кребса не проис-
ходит непосредственного синтеза АТФ. В цикле идет окисление моле-
кул, отделение четырех пар Н+
, которые используются для восстанов-
ления и фосфорилирования НАД с образованием НАДН и НАДФ,
перенос четырех пар высокоэнергетических электронов в дыхатель-
ную цепь, где они передаются на молекулярный кислород — конеч-
ный акцептор электронов, в результате чего образуется Н2O.
Основная часть АТФ синтезируется в процессе окислительного
фосфорилирования. Дыхательная цепь, или цепь переноса электро-
нов, является главной системой превращения энергии. Синтез АТФ
катализируется ферментом АТФ-синтетазой. П . Митчелл предложил
хемиоосмотическую гипотезу окислительного фосфорилирования
применительно к митохондриям. Согласно этой гипотезе, при тран-
спорте электронов по дыхательной цепи протоны «откачиваются» из
матрикса на наружную поверхность внутренней мембраны митохон-
Рис. 42. Этапы окисления глюкозы:
1 — гликолиз в цитозоле,
2 — окислительное фосфорилирование
в митохондриях; Пл — плазмалемма,
Вм и Нм — внутренняя и наружная
мембраны митохондрии
(по А. Ленинджеру, с изменениями)
Глюкозо-6 -фосфат
Глюкоза
CO2
CO2
H2O
O2
e–
Пл
Пл
1
2
Вм
Нм
Пируват
Ацетил-СoA
Цикл
трикарбоновых
кислот
(цикл Кребса)
70
СТРОЕНИЕ КЛЕТКИ
дрий, что вызывает возникновение электрохимического протонного
градиента по обеим сторонам внутренней митохондриальной мембра-
ны. При возникновении большого протонного градиента протоны на-
чинают перемещаться через АТФ-синтетазу в матрикс, их энергия
расходуется для синтеза АТФ.
По существу в дыхательной цепи происходит окисление водорода:
Н2+1⁄2O2␚Н2O.
Однако этот процесс происходит многоступенчато, причем атомы
водорода расщепляются на протоны, которые поступают в водную
среду, и высокоэнергетические электроны, которые транспортируют-
ся по дыхательной цепи; выделяемая ими энергия (порциями) расхо-
дуется для синтеза АТФ из АДФ и фосфата. Лишь на завершающем
этапе в конце дыхательной цепи протоны соединяются с электронами.
В состав дыхательной цепи входят два флавопротеидных фермен-
та (сукцинатдегидрогеназа и НАД-дегидрогеназа), четыре цитохрома,
негеминовое железо, медь и кофермент Q (убихинон). Согласно сов-
ременным представлениям, дыхательная цепь состоит из трех основ-
ных мембраносвязанных ферментных комплексов.
1. НАДН-дегидрогеназный комплекс, который передает электроны
от НАДН на переносчик электронов убихинон либо на нафтохиноны.
2. Убихинон переносит электроны на комплекс цитохромов и пе-
редает их на переносчик электронов цитохром С.
3. Цитохром С переносит электроны на цитохромоксидазный ком-
плекс, который передает их конечному акцептору электронов — кис-
лороду.
При переходе электронов от одного переносчика к другому их сво-
бодная энергия убывает, а освобождающаяся энергия используется для
«откачивания» протонов на наружную сторону мембраны, в результате
чего и создается электрохимический протонный градиент. Иными сло-
вами, энергия, освобождаемая в процессе переноса электронов по дыха-
тельной цепи, запасается в форме электрохимического протонного гра-
диента на мембране, в которую встроена дыхательная цепь.
АТФ-синтетаза представляет собой мембранный белковый ком-
плекс, который имеется во всех мембранах, осуществляющих окисли-
тельное фосфорилирование. Согласно хемиоосмотической гипотезе,
энергия перемещения протонов через АТФ-синтетазу в обратном на-
правлении (с наружной стороны мембраны на внутреннюю) использу-
ется для синтеза АТФ. Однако АТФ-синтетаза осуществляет не только
синтез, но и гидролиз АТФ. И тот и другой процесс сопряжены с пере-
движением протонов.
АДФ+Р1⇔АТФ
␘
АТФ-синтетаза
71
ВНУТРИКЛЕТОЧНЫЕ БИОХИМИЧЕСКИЕ РЕАКЦИИ
Экзоцитоз
Выведение веществ из клетки осуществляется благодаря существо-
ванию нескольких механизмов. Один из них — пассивный транспорт
вследствие разности концентраций внутри и вне плазмалеммы. Дру-
гой — это активный транспорт. Таким путем выводятся из клетки ионы
и мелкие молекулы (см. раздел «Биологические мембраны»). Третий
механизм обеспечивает выведение крупномолекулярных соединений.
Сначала крупномолекулярные соединения сегрегируются в ком-
плексе Гольджи в виде транспортных пузырьков. Последние с участи-
ем микротрубочек направляются к клеточной поверхности. Мембрана
пузырька встраивается в плазмалемму, и содержимое пузырька ока-
зывается за пределами клетки (рис. 43).
Слияние пузырька с плазмалеммой может совершаться без каких-
либо дополнительных сигналов. Такой экзоцитоз называют конститу-
тивным. Так выводится из клетки большинство продуктов ее собст-
венного метаболизма. Ряд клеток, однако, предназначен для синтеза
специальных соединений — секретов, которые используются в орга-
низме в других его частях. Для того чтобы транспортный пузырек
с секретом слился с плазмалеммой, необходимы сигналы извне. Толь-
ко тогда произойдет слияние и секрет освободится. Такой экзоцитоз
называют регулируемым. Сигнальные молекулы, способствующие
выведению секретов, называют либеринами (рилизинг-факторами),
а препятствующие выведению — статинами.
Мембрана транспортного пузырька встраивается в плазмалемму
и становится ее частью. И экзоцитоз, и возврат мембран эндосом
в нормально функционирующей клетке уравновешены с поглощением
мембран в ходе пино- и фагоцитоза.
Пути восприятия и передачи информации клеткой
Регуляция сложных многогранных упорядоченных функций клет-
ки осуществляется посредством сигналов, приводящих в действие
Рис. 43. Экзоцитоз. Объяснения в тексте (по Б. Албертсу и соавт., с изменениями)
72
СТРОЕНИЕ КЛЕТКИ
внутриклеточные пути передачи информации. Эти сигналы — моле-
кулы, которые распознаются рецепторами, встроенными в плазма-
лемму. Здесь расположены цепи мембранных белков, взаимодейству-
ющих друг с другом. В результате меняется их пространственная
конфигурация, и, как следствие этих конформаций, сигналы, воздей-
ствующие на клетку извне, преобразуются во внутриклеточные.
Известны два пути передачи сигналов. Принципиально они сход-
ны между собой и включают ряд звеньев: сигнальная молекула (пер-
вичный посредник) связывается с рецептором, что приводит к акти-
вации белка-преобразователя, передающего сигнал белку-усилителю.
Все три указанных белка являются внутримембранными. Фермент-
усилитель вызывает образование из фосфорилированного предшест-
венника вторичного посредника (мессенджера), который связывается
с протеинкиназой, активирующей клеточные реакции.
В первом пути вторичным посредником является циклический
аденозинмонофосфат (цАМФ). В осуществлении этого пути участву-
ют два типа рецепторов: стимулирующие (Rs) и ингибирующие (Ri).
Взаимодействие химического стимула с рецептором вызывает кон-
формационные изменения последнего, что передается через мембрану
соответственно Gs-белку или Gi-белку, который связывает внутри-
клеточный гуанозинтрифосфат (ГТФ). Это, в свою очередь, вызывает
новые конформационные изменения Gs-белка, который приобретает
способность активизировать фермент аденилатциклазу (Ац). В ре-
зультате воздействия фермента из АТФ образуется циклический аде-
нозинмонофосфат.
Во втором пути передачи информации имеются лишь стимулиру-
ющие рецепторы и Gs-белок, а в роли усилителя выступают фермен-
ты гуанилатциклаза (Гц) и фосфодиэстераза (ФДЕ). Последователь-
ность реакций в этом случае такая же, как и в первом пути. ФДЕ
расщепляет фосфатидилинозитол-4,5 дифосфат (ФИФ2) на диацил-
глицерол (ДГ) и инозитолтрифосфат (ИФ3). Гуанилатциклаза прев-
ращает ГТФ в циклический гуанозинмонофосфат (цГМФ).
ИФ3 освобождает ионы Са2+ из их внутриклеточных хранилищ,
тем самым повышая их концентрацию в цитозоле. Са2+ связывается
с определенными белками (например, кальмодулином, тропони-
ном С). Первый активирует протеинкиназу С, второй вызывает мы-
шечные сокращения. ДГ остается в плазмалемме и активирует проте-
инкиназу, которая фосфорилирует клеточные белки. Далее цГМФ
активирует специфическую протеинкиназу (G-киназу), которая так-
же фосфорилирует определенные белки. Так, цГМФ воздействует на
Nа+
-каналы плазмалеммы палочковых клеток сетчатки глаза. Три ука-
занные молекулы (ДГ, ИФ3
, цГМФ) являются вторичными посредни-
ками второго пути.
73
ЖИЗНЕННЫЙ ПУТЬ КЛЕТОК
В обоих путях передачи сигналов вторичные посредники связыва-
ются с регуляторными субъединицами фермента протеинкиназы
(ПК). Это приводит к освобождению ее каталитической субъедини-
цы, которая фосфорилирует определенные белки и благодаря этому
активизирует различные клеточные процессы, т. е . вызывает ответ
клетки на сигнал (например, сокращение, секрецию, репликацию,
транскрипцию, распад гликогена или жиров и т. д .).
Приводим некоторые клеточные ответы на гормоны, опосредуе-
мые цАМФ: синтез и секреция тироксина клетками щитовидной же-
лезы в ответ на воздействие тиреотропного гормона; секреция корти-
зола клетками коркового вещества надпочечников в ответ на
воздействие АКТГ; секреция прогестерона клетками желтого тела
яичника в ответ на воздействие лютеотропного гормона; распад гли-
когена в мышцах и печени; увеличение частоты и силы сердечных со-
кращений под влиянием адреналина; резорбция костной ткани под
влиянием гормона паращитовидных желез; реабсорбция воды в ка-
нальцах нефрона в ответ на воздействие вазопрессина.
Следующие клеточные реакции осуществляются через фосфоино-
зитидный путь: сокращение гладких миоцитов и секреция инсулина
b-клетками островков Лангерганса поджелудочной железы под влия-
нием ацетилхолина; секреция гистамина тучными клетками при воз-
действии антигена; секреция тромбоцитами серотонина и фактора ро-
ста под влиянием тромбина.
ЖИЗНЕННЫЙ ПУТЬ КЛЕТОК
Как известно, клетки не возникают сами по себе, а образуются только
при делении других. После деления во вновь образованной клетке не
всегда сразу существуют все системы, обеспечивающие ее специфиче-
скую функцию. Должно пройти некоторое время, чтобы сформирова-
лись все органеллы и были бы синтезированы все необходимые фермен-
ты. Этот отрезок времени называется созреванием. Созревание клетки
осуществляется на основе уже сложившейся ее полной детерминации.
Зрелая клетка может функционировать различное время. Некото-
рые клетки сохраняются в течение всей жизни особи (например, ней-
роны). Таких видов клеток немного. Большинство клеток гибнет и по
мере убыли замещается новыми. Скорость замещения у разных кле-
ток неодинакова.
Конечно, клетка может погибнуть в результате многих внешних
случайных причин, например от травмы, химического или радиаци-
онного поражения. В таком случае разрушение клетки происходит ха-
отично, а продукты распада ее сами оказывают раздражающее дейст-
вие на окружение. Развивается воспалительная реакция. Подобная
74
СТРОЕНИЕ КЛЕТКИ
случайная гибель клеток называется некрозом и служит предметом
изучения патологической анатомии.
Большинство клеток, однако, погибает тогда, когда проявляются
особые естественные генетические механизмы. Генетически запро-
граммированную клеточную гибель называют апоптозом. Механизм
возникновения апоптоза весьма сложен. Каждая клетка несет в хро-
мосомах гены, которые могут запускать синтез ферментов, стимули-
рующих ее к делению. Есть также гены, которые обеспечивают синтез
ферментов, препятствующих делению. Пока клетка функционирует,
эти синтезы уравновешены.
Для поддержания жизненного равновесия клетка должна также
получать сигналы от других клеток, нередко другого вида. Обычно
в качестве сигнальных выступают специфические молекулы олиго-
пептидов. Поскольку они поддерживают жизнь клеток, их назвали
цитокинами. Известно несколько десятков цитокинов. Действие их
разнообразно: на одни виды клеток более сильное, на другие — слабое
или даже может и не проявляться. Сейчас при описании межклеточ-
ных взаимодействий все чаще применяют термин «цитокинная сеть».
В жизненном пути многих видов клеток наступает момент, когда
функциональные их возможности исчерпываются. У таких клеток на-
рушается чувствительность к цитокинам и изменяется соотношение
активности генов, обеспечивающих внутреннее равновесие. Гены,
обеспечивающие размножение клетки, блокируются. Напротив, гены,
обеспечивающие синтез литических ферментов, стимулируются. По-
следние поступают в ядро и лизируют хроматин.
Хромосомы распадаются, синтезы в клетке прекращаются. Внешние
проявления такой гибели клеток разнообразны и известны давно. Их
называли пикнозом (сморщивание ядра), хроматолизисом (снижение
окрашиваемости ядра), кариорексисом (распад ядра на части). Лишь
недавно было показано, что это лишь частные проявления апоптоза.
Вслед за гибелью ядра разрушается и цитоплазма. Остатки фаго-
цитируются макрофагами. Материал погибших клеток перерабатыва-
ется макрофагами и может выводиться ими на поверхность. В таком
случае этот материал может опять использоваться другими клетками.
Вокруг клеток, подвергшихся апоптозу, воспалительный процесс не
возникает, и жизнедеятельность ткани, часть которой составляли по-
гибшие клетки, продолжается без нарушений.
КЛЕТОЧНЫЙ ЦИКЛ
События, изложенные в предыдущем разделе, описывают как бы
линейный путь жизни клетки. В результате таких событий числен-
ность клеточной популяции должна снижаться. Это действительно
75
КЛЕТОЧНЫЙ ЦИКЛ
происходит в некоторых тканях. В нервной ткани в течение жизни ор-
ганизма нейроны постоянно гибнут, но не восстанавливаются. Коли-
чество их при рождении, однако, настолько велико, что до наступле-
ния смерти способно обеспечить все необходимые связи и реакции.
Такие клеточные популяции называют стационарными. Ранее счита-
ли, что не восстанавливается численность мышечных клеток сердца —
кардиомиоцитов. Однако П. П . Румянцев доказал, что кардиомиоци-
ты также восстанавливаются.
В большинстве клеточных сообществ популяция способна восста-
навливаться. Естественно, восстановление ее численности совершает-
ся за счет новых клеточных делений. В сообществах, способных под-
держивать свою численность, кроме клеток, функционирующих,
линейно развивающихся и следующих к своей гибели, всегда есть и
такие, которые регулярно претерпевают циклические изменения. По-
добные клеточные популяции называют обновляющимися.
Клеточный цикл (рис. 44) представляет собой совокупность про-
цессов, происходящих в клетке при подготовке ее к делению и во вре-
мя собственно деления, в результате чего материнская клетка делится
на две дочерние. В цикле выделяют две фазы: автосинтетическую,
Рис. 44. Клеточный цикл:
I — последовательность фаз
клеточного цикла, II — клетка
в интерфазе после репликации
хромосом и центриолей,
III — профаза, IV — метафаза,
V — метафазная хромосома,
VI — анафаза, VII — телофаза,
VIII — сестринские клетки;
1 — ядро, 2 — сформировавшееся
митотическое веретено,
3 — хромосомы, 4 — астральные
микротрубочки, 5 — полюсные
микротрубочки, 6 — полюс
веретена, 7 — хроматиды,
8 — центромера, 9 — центромерные
микротрубочки, 10 — сестринские
хроматиды, 11 — центромерные
(кинетохорные) микротрубочки,
12 — раздвижение полюсов
в анафазе, 13 — вновь образованные
ядра, 14 —восстановление
интерфазных микротрубочек,
растущих от центросомы,
15 — остатки полюсных
микротрубочек, 16 — борозда
деления (по Б. Албертсу и соавт.,
с изменениями)
S
II
I
III
IV
VII
VIII
VI
V
22
77
88
99
66
10
10 11
11 10
10
12
12
14
14
15
15
16
16
13
13
66
55
44
33
11
M
G2
G1
76
СТРОЕНИЕ КЛЕТКИ
или интерфазу (подготовка клетки к делению), включающую пресин-
тетический (G1, от англ. gap — промежуток), синтетический (S) и
постсинтетический (G2) периоды, и деление клетки — митоз.
Хайфлик высказал точку зрения, согласно которой клетки от начала
своего возникновения после первого деления могут проходить несколь-
ко десятков клеточных циклов. После этого они погибают. Полагали, что
утрата клетками способности вступать в новые циклы и делиться — одна
из причин старения организма. Гипотеза эта была высказана в качестве
экстраполяции наблюдений в клеточных культурах фибробластов
in vitro; полагали, что и в живом организме дело должно обстоять так же.
Реальными наблюдениями in vivo гипотеза пока не подтверждена.
Интерфаза
Интерфаза — последовательность событий, подготавливающих
митоз. Весьма важным в интерфазе является матричный синтез ДНК
и удвоение хромосом — S-фаза. Промежуток между делением и на-
ступлением S-фазы называется фазой G1 (постмитотическая, или
пресинтетическая, фаза), а между S-фазой и митозом — фазой G2
(постсинтетическая, или премитотическая, фаза). В течение фазы G1
клетка диплоидна, в течение фазы S плоидность возрастает до четы-
рех, в фазе G2 клетка тетраплоидна.
В интерфазе скорость биосинтетических процессов возрастает в на-
правлении G1 ␚ S ␚ G2. В это время удваивается масса клетки и всех
ее компонентов, а также происходит удвоение центриолей (рис. 45).
В течение пресинтетической фазы G1 в клетке уже усилены био-
синтетические процессы и происходит подготовка к удвоению ДНК.
При этом развиваются преимущественно те органеллы, которые необ-
ходимы для синтеза ферментов, обеспечивающих, в свою очередь,
предстоящее удвоение ДНК (прежде всего это рибосомы). На мате-
ринской центриоли клеточного центра увеличивается количество са-
теллитов. Фаза G1 длится от нескольких часов до суток и более. Это
общая сущность S-фазы.
Само удвоение (репликация) хромосом весьма сложно и протека-
ет постепенно. Суть удвоения состоит в том, что на цепочке ДНК син-
тезируется точно такая же параллельная цепочка. Репликация (от
лат. replicatio — повторение) — это процесс передачи генетической
информации, хранящейся в родительской ДНК, путем точного ее
воспроизведения в дочерней клетке. При этом каждая родительская
цепь ДНК является матрицей для синтеза дочерней (матричный син-
тез ДНК).
Хромосома имеет структуру, обеспечивающую этот процесс. На
хромосоме находится небольшая область, которая не участвует в ма-
77
КЛЕТОЧНЫЙ ЦИКЛ
тричном синтезе — центромера (или центромер). Она подразделяет
хромосому на два плеча. На концах хромосомы находятся тоже обла-
сти, не участвующие в синтезе — теломеры.
Репликация основана на комплементарном спаривании основа-
ний. Вначале в одной точке ДНК обе ее цепи расходятся, образуя
асимметричную репликационную «вилку». Фермент ДНК-полимера-
за катализирует процесс полимеризации нуклеотидов только в на-
правлении 5' ␚ 3'. Напомним, что обе цепи ДНК антипараллельны,
поэтому синтез одной из дочерних цепей происходит непрерывно
(лидирующая цепь), другой (отстающей) — в виде отдельных фраг-
ментов размерами 10—200 нуклеотидов (фрагменты Оказаки). Впо-
следствии эти фрагменты соединяются под действием фермента
ДНК-лигазы.
Репликация начинается от середины каждого плеча, от участка, на-
зываемого сайтом инициации репликации. Распространяясь к теломе-
рам, репликация доходит до них и останавливается. Двигаясь к сере-
дине хромосомы, репликация доходит до центромеры и тоже
останавливается, однако центромерная область не удваивается. В ре-
S
G2M
G0
G1
1
3
3
2.1
1.4
3
1
2
1.1
1.2
1.3
1
2
1
2.1
3
Рис. 45. Изменения клеточного центра по ходу клеточного цикла:
G0, G1, S, G2 — периоды интерфазы клеточного цикла, М — митоз; 1 — материнская
центриоль, 1.1 — сателлит (область образования микротрубочек), 1.2 — придатки
материнской центриоли, 1.3 — микротрубочки, отходящие от клеточного центра,
1.4 — фибриллярное гало (область образования микротрубочек), 2 — дочерняя
центриоль первого поколения, в 3-периоде она становится материнской (2.1),
3 — дочерняя центриоль второго поколения. В периоде митоза изображена только
одна пара центриолей у одного из полюсов; центриоль 3 в новом цикле
рассматривается как дочерняя 2 (по Ю. С . Ченцову, с изменениями)
78
СТРОЕНИЕ КЛЕТКИ
зультате каждая хромосома имеет теперь две цепи ДНК. Каждая цепь
с окружающими их белками образует сестринские хроматиды. S -фаза
длится 8—12 часов.
В каждой хромосоме во время S-периода образуются группы ре-
пликационных «вилок» (20—80), которые возникают одновременно
у всех хромосом. При этом «вилки» расположены парами, которые
движутся в противоположных направлениях до тех пор, пока не
встретят соседнюю «вилку», так что образуются две дочерние спира-
ли. В результате репликации каждая из двух дочерних молекул ДНК
состоит из одной старой и одной новой цепи.
Мы привели лишь схематическое описание процесса репликации.
Он очень сложен, в нем участвует множество белков, способствую-
щих расплетанию ДНК, предотвращающих ее спутывание, а также
сшивающих фрагменты ДНК, инициаторных, корректирующих и др.
В S-периоде наиболее интенсивно синтезируется РНК и белки, свя-
занные с ДНК, и удваиваются центриоли.
В цитоплазме в течение S-фазы удваиваются не только цепи ДНК,
но и каждая из центриолей клеточного центра. Материнская центри-
оль строит свою новую дочернюю. Центриоль, бывшая до этого до-
черней, тоже строит свою пару, так что сама становится материнской.
Из этих четырех центриолей лишь исходная материнская участвует
в сборке микротрубочек. На мембранах ЭПС одновременно синтези-
руются белки (в том числе гистоны), необходимые для включения
в состав новой хроматиды.
В течение премитотической фазы G2 совершаются синтезы, необ-
ходимые для обеспечения непосредственно процесса деления. Коли-
чество ДНК и центриолей в клетке уже удвоено. Обе материнские
центриоли окутаны фибриллярным гало и осуществляют сборку ми-
кротрубочек. В этом периоде усиливается формирование лизосом, де-
лятся митохондрии и синтезируются новые белки, абсолютно необхо-
димые для осуществления митоза. К концу интерфазы хроматин
конденсирован, ядрышко хорошо видно, ядерная оболочка не нару-
шена, органеллы не изменены. Фаза G2 продолжается до 6 часов.
На протяжении каждой из названных фаз имеются так называе-
мые критические точки (регуляторные точки). Они были выявлены
при изучении действия повреждающих факторов на клеточное деле-
ние. Отрезки времени до и после такой точки различаются отношени-
ем к возможным повреждающим воздействиям. Если повреждающий
импульс (например, радиационный) действует перед прохождением
критической точки, процессы продолжаются до нее, и дальнейшая
подготовка к делению останавливается (наступает блок). Если им-
пульс совпадает со второй половиной фазы, процессы продолжаются
(при условии, что этот импульс не разрушает клетку как таковую).
79
МИТОЗ
Вместе с тем прохождение критических точек существенно и для
нормальной жизнедеятельности клетки. В ходе межклеточных взаимо-
действий под влиянием соответствующих цитокинов при прохождении
критической точки цикл может прерваться. В ядре после этого начина-
ются синтезы РНК, и клетка обретает специфическую функцию. Она
может выполнять ее до естественной гибели, но может и прекратить ее,
вернуться к точке прерванного цикла и продолжить деление.
В ходе последовательных делений свойства клеток могут изме-
няться. Это происходит за счет того, что у дочерних клеток в геноме
начинают экспрессироваться новые гены (существовавшие, конечно,
и ранее, но до сих пор блокированные), поэтому клетки-потомки
обычно более разнообразны, чем исходные формы. Клетки-предшест-
венники менее разнообразны, причем часто можно выявить клетки,
лежащие в основании всего дерева. Их называют стволовыми. И ство-
ловые, и клетки-предшественники образуют камбий клеточной попу-
ляции. Именно за счет деления клеток камбия и происходит замена
гибнущих элементов в обновляющихся популяциях, например фор-
менных клеток крови или клеток кожного эпителия.
Клетки, вышедшие из цикла в точке регуляции, не являются ни
стволовыми, ни клетками-предшественниками. Между тем они тоже
могут играть роль камбия, поскольку способны возвращаться в цикл.
Обычно это происходит при возрастании потребностей организма
и стимулируется гормонами и цитокинами. Этот способ увеличения
численности популяции обычен для многих внутренних органов, на-
пример наблюдается в паренхиме печени, почек и ряда других вну-
тренних органов. За счет этого, в частности, увеличивается масса ор-
ганов в детском возрасте. Такие клеточные популяции получили
название растущих. У взрослых организмов растущие популяции мо-
билизуются для пополнения убыли численности клеток после повре-
ждения части органа. Иначе говоря, они служат резервом не только
для физиологической, но и для репаративной регенерации ткани.
МИТОЗ
Когда подготовка к делению заканчивается, начинается непосред-
ственно митоз (от греч. mitos — нить). В нем различают четыре основ-
ные фазы: профазу, метафазу, анафазу и телофазу (см. рис. 44).
Иногда выделяют шесть фаз: профазу, прометафазу, метафазу, анафа-
зу, телофазу и цитокинез.
В течение профазы основные события происходят в ядре. На
участках эухроматина прекращается транскрипция. Они покрывают-
ся белками и по плотности становятся неотличимыми от гетерохро-
матина. Даже при разрешении светового микроскопа в ядре становят-
80
СТРОЕНИЕ КЛЕТКИ
ся видимыми многочисленные плотные базофильные скопления.
Затем начинается спирализация хромосом. Вследствие этого они ста-
новятся индивидуально различимыми. Спирализация, естественно,
захватывает и области ядрышковых организаторов, так что ядрышко
в результате распадается. Итак, к началу профазы хроматин конден-
сируется, в результате чего в ядре образуется плотный клубок. К кон-
цу профазы этот клубок разрыхляется (рыхлый клубок), становятся
видимыми d-хромосомы, каждая из которых состоит из двух хрома-
тид (s-хромосом), лежащих параллельно друг другу и связанных меж-
ду собой в области центромеры. В цитоплазме активизируется обра-
зование лизосом. Центриоли попарно расходятся к противоположным
концам клетки, которые теперь называют полюсами. Одновременно
на сателлитах центриолей идет интенсивная сборка микротрубочек.
События метафазы начинаются в цитоплазме. Лизосомы раство-
ряют ядерную оболочку, так что спирализованные хромосомы и кле-
точные центры оказываются в общем компартменте. Этому предшест-
вует фосфорилирование белков ядерной пластинки (ламины),
происходящее еще в профазе, что приводит к распаду пластинки,
а затем и самой нуклеолеммы. Фрагменты распавшейся ядерной обо-
лочки формируют мелкие мембранные пузырьки, цитоплазма клетки
смешивается с кариоплазмой. Комплекс Гольджи и ЭПС распадаются
на мелкие фрагменты в виде пузырьков.
На каждой центромере выявляется скопление специальных бел-
ков — кинетохор (от греч. kineo — подвижный и choreo — иду впе-
ред). Эти белки существуют и у хромосом неделящихся клеток, но
в этих условиях они выявляются лишь с помощью специального ме-
чения особыми антителами к ним.
Сборка микротрубочек на материнских центриолях продолжается,
так что в результате возникает биполярное митотическое веретено,
состоящее из этих микротрубочек и ассоциированных с ними белков.
Различают несколько видов микротрубочек. Многие нити расходятся
от центриолей (как от полюсов) во все стороны. Часть их образует
направленную к поверхности клетки астральную лучистость, другая
их часть направлена к экватору клетки — это полярные микротрубоч-
ки. У экватора полярные микротрубочки, связанные с разными полю-
сами, перекрывают друг друга. Кроме астральных и полярных микро-
трубочек от полюсов отходят кинетохорные — те, которые в области
экватора прикрепляются к кинетохорам хромосом. В клетках челове-
ка каждый кинетохор связан с 20—40 микротрубочками. Прикрепле-
ния микротрубочек к сестринским хроматидам гомологичных хромо-
сом происходят в случайном порядке.
Вся система микротрубочек и ассоциированного с ними тубулина
находится в динамическом равновесии. Иными словами, происходит
81
МИТОЗ
постоянная полимеризация тубулина и его деполимеризация. По обеим
сторонам d-хромосомы около ее центромеры расположены неболь-
шие участки материала умеренной электронной плотности, аналогич-
ные перицентриолярному материалу. Именно они и являются центра-
ми организации хромосомных микротрубочек из тубулина, тубулин
же синтезируется только в цитоплазме. Поэтому лишь после разру-
шения нуклеолеммы может произойти взаимодействие кинетохора
с тубулином и организация микротрубочек веретена. При более де-
тальном описании митоза эта стадия выделяется в качестве промета-
фазы. Она длится 10—20 мин.
В ходе собственно метафазы хромосомы перемещаются и распола-
гаются в одной плоскости перпендикулярно к оси между полюсами.
Образуется фигура, называемая материнской звездой. При этом все
хромосомы располагаются так, что их центромеры находятся в эква-
ториальной плоскости, пересекающей продольную ось веретена под
прямым углом (метафазная пластинка), причем каждый кинетохор
одной d-хромосомы обращен к одному из полюсов клетки.
В результате упорядочения положения хромосом система микро-
трубочек тоже упорядочивается. Они теперь образуют веретено деле-
ния (митотическое веретено). Хроматиды прочно присоединяются
к веретену благодаря взаимодействию кинетохорных трубочек с пе-
рицентриолярным веществом.
Каждая из метафазных хромосом состоит из двух фибрилл диаме-
тром 20—50 нм, которые уложены в плотный складчатый клубок. Фи-
бриллы имеют зернистый вид, так как срез препарата проходит через
этот клубок множество раз. При этом ДНК имеет более высокую
электронную плотность, чем связанный с ней белок. Напомним, что
именно в метафазе митоза определяют кариотип (см. ранее).
В S-периоде удваивается не вся ДНК одной хромосомы, а оста-
ется нереплицированным центромерный участок. В начале анафа-
зы происходит быстрая репликация ДНК в области центромеры,
что и служит сигналом к началу анафазы. Анафаза начинается вне-
запно с резкого разделения общей центромеры d-хромосомы, в ре-
зультате чего сестринские хроматиды становятся самостоятельны-
ми s-хромосомами.
Микротрубочки начинают укорачиваться: у кинетохоров происхо-
дит их разборка. В результате этого хроматиды подтягиваются к цен-
триолям. В это время s-хромосомы начинают передвигаться и с оди-
наковой скоростью (около 1 мкм в минуту) направляются к полюсам
клетки. Сами центриоли удаляются друг от друга в сторону полюсов
клетки. Образуется две дочерних звезды.
На хромосомы воздействуют две силы: тянущие, возникающие
вследствие деполимеризации хромосомных трубочек около полюсов
82
СТРОЕНИЕ КЛЕТКИ
веретена, и расталкивающие — в связи с полимеризацией тубулина
на концах непрерывных микротрубочек вблизи экватора. При этом
по мере расхождения хромосом веретено удлиняется, а степень пере-
крывания друг друга непрерывных трубочек уменьшается. Возмож-
но, источником сил, раздвигающих полюсы, является динеин, в то
время как движение хромосом к полюсам обусловлено микротру-
бочками.
В конце анафазы плазматическая мембрана как бы инвагинирует-
ся перпендикулярно к продольной оси митотического веретена, обра-
зуя борозду. В этой области под плазмалеммой появляется сократи-
мое кольцо, состоящее из актин- и миозинсодержащих нитей, которое
распадается после разделения клетки.
Телофаза завершает деление. Под плазмалеммой кольцом по про-
екции бывшей материнской звезды активируются элементы цитоске-
лета — актиновые микрофиламенты. Рядом с ними полимеризуется
миозин. Актино-миозиновое кольцо сжимается, и возникает перетяж-
ка плазмалеммы.
В телофазе разделившиеся группы хромосом подходят к полю-
сам, теряют хромосомные микротрубочки, разрыхляются, деконден-
сируются, переходя в хроматин, и начинают транскрибировать РНК.
Примерно в середине телофазы начинается образование нитчатой,
а затем гранулярной частей нуклеолонемы. К концу телофазы (по-
сле восстановления ядерной оболочки!) ядрышко полностью сфор-
мировано. Из мембранных пузырьков собираются комплекс Гольд-
жи и ЭПС.
Ядерная оболочка образуется из мембранных фрагментов внача-
ле в виде небольших шапочек, расположенных на поверхности фор-
мирующихся глыбок хроматина. Фрагменты оболочки растут, сли-
ваются между собой, окружая все ядро к концу телофазы. При этом
восстанавливаются ядерные поры и поровые комплексы, дефосфо-
рилируются белки ядерной пластинки, что приводит к ее восстанов-
лению.
В телофазе перед цитокинезом увеличивается биосинтез мем-
бран, которые необходимы для того, чтобы покрыть обе дочерние
клетки. Вновь синтезированные мембраны до момента разделения
клетки образуют на ее поверхности пузырьки, которые затем встра-
иваются в плазмалеммы дочерних клеток. Перетяжка становится все
более глубокой, и в результате в конце концов одна клетка разделя-
ется на две (цитокинез). Обе дочерние клетки диплоидны. Однако
не всегда деление ядра сопровождается разделением клетки, поэто-
му помимо телофазы (при полном делении клетки) и выделяют ци-
токинез.
83
МЕЙОЗ
После митоза в течение нескольких часов дочерние клетки связаны
между собой небольшим остаточным тельцем, образованным непрерыв-
ными микротрубочками и электроноплотным материалом матрикса.
Остаточное тельце покрыто плазмалеммой. Есть все основания считать,
что сила, необходимая для разделения клеток, возникает в результате
взаимного скольжения актиновых и миозиновых филаментов.
МЕЙОЗ
У организмов, размножающихся половым путем, имеется две ка-
тегории клеток: диплоидные и гаплоидные. К первым относятся сома-
тические и предшественницы половых клеток, ко вторым — зрелые
половые (гаметы). Уменьшение количества хромосом в два раза до-
стигается благодаря мейозу. Он включает в себя два последователь-
ных деления. После слияния гамет возникает новый одноклеточный
диплоидный организм (зигота), который несет не просто сумму при-
знаков своих родителей, а является индивидуумом с присущими
только ему свойствами.
При дальнейшем митотическом делении зиготы образуются ди-
плоидные же клетки, содержащие по два экземпляра каждой хромосо-
мы, которые называются гомологичными. Гомологичные хромосомы,
имеющие одинаковую длину и одинаковое расположение центромер,
содержат одинаковое количество генов, а эти гены имеют одну и ту же
линейную последовательность. Каждая из пары гомологичных хромо-
сом диплоидного организма происходит либо из ядра спермия, либо
из ядра яйцеклетки.
При образовании гамет в зрелом организме в результате мейоза
в каждую дочернюю клетку от всех пар гомологичных хромосом по-
падает лишь по одной из них. Это становится возможным потому, что
при мейозе происходит лишь одна репликация ДНК, за которой сле-
дуют два последовательных деления ядер (мейоз I и II) без повторно-
го синтеза ДНК. В результате из одной диплоидной образуются четы-
ре гаплоидные клетки.
Напомним, что перед началом мейоза в интерфазе клетка прошла
обычные фазы G1, S и G2, так что стала тетраплоидной. Иначе говоря,
произошла репликация ДНК и белков-гистонов хромосом, а сестрин-
ские хроматиды при этом остались связанными своими центромера-
ми, так что в ядре имеется по четыре набора каждой хромосомы. Уве-
личена масса клетки и ее органелл.
Каждое из двух делений мейоза (деления I и II) имеет свои отли-
чительные черты. Особенность деления I состоит в необычном
и сложном прохождении профазы (профаза I). Она подразделяется
84
СТРОЕНИЕ КЛЕТКИ
на несколько стадий: пролептонему, лептонему, зигонему, пахинему,
диплонему и диакинез (рис. 46, 47).
Во время пролептонемы (от греч. pro — период, leptos — тонкий,
nema — нить) происходит значительная, но не полная спирализация
хромосом. Ядерная оболочка сохраняется, ядрышко не распадается.
поэтому во время профазы мейоза возможны синтезы некоторых
РНК и белков. За счет этих синтезов в половых клетках (особенно
в женской) создаются запасы веществ, которые будут необходимы
для оплодотворения и ранних стадий развития зародыша.
Во время лептонемы хромосомы еще больше спирализуются, и
в ядре становятся видными тонкие нитевидные d-хромосомы (их 46,
т. е . два набора). Подчеркнем, что каждая гомологичная хромосома
Рис. 46. Основные стадии мейоза (по Мамонтову):
А — протофаза I, Б — метафаза I, В — анафаза I, Г — телофаза I,
Д — профаза II, Е — метафаза II, Ж — анафаза II, З — телофаза II
А
Б
В
Г
Д
Е
Ж
З
85
МЕЙОЗ
уже реплицирована и состоит из двух сестринских хроматид. Каждая
хромосома представляет собой тонкую фибриллу, состоящую из осе-
вой белковой нити, к которой прикрепляется хроматин сестринских
хроматид (петли ДНК). Хромосомы с помощью белковых скопле-
ний — прикрепительных дисков — закреплены обоими своими конца-
ми на внутренней мембране ядерной оболочки (ядерная оболочка со-
храняется, ядрышко хорошо видно).
Во время зигонемы (от греч. zygon — парный) гомологичные ди-
плоидные хромосомы выстраиваются рядом, обвивают друг друга,
укорачиваются и сцепляются между собой (конъюгация). Образуются
так называемые тетраплоидные биваленты (от лат. bi — двойной, va-
lens — сильный). Напомним, что каждая диплоидная хромосома из
одного бивалента происходит либо от отца, либо от матери. Половые
хромосомы располагаются около внутренней ядерной мембраны. Об-
ласть, занятая ими, называется половым пузырьком.
11
22
33
44
Рис. 47. Поведение хромосом при мейозе:
I и II — хроматиды в профазе мейоза, III — спаренные гомологичные хромосомы
при переходе к первой метафазе мейоза, IV — расхождение гомологичных хромосом
в первой анафазе мейоза, V — вторая метафаза, VI — расхождение сестринских
хроматид во второй анафазе; 1, 2, 3, 4 — хроматиды, 5 — соединяющиеся
кинетохорные нити, 6 — кинетохор, 7, 8 — плечи сестринских хроматид
(по Б. Албертсу и соавт., с изменениями)
11
22
33
44
11223344
77
77
88
88
88
I
II
III
IV
V
VI
55
66
86
СТРОЕНИЕ КЛЕТКИ
В зигонеме гомологичные d-хромосомы выстраиваются рядом,
сближаются, между ними образуются специализированные синапто-
немальные комплексы (от греч. synapsis — связь, соединение), кото-
рые представляют собой белковые структуры. При небольшом элек-
тронно-микроскопическом увеличении синаптонемальный комплекс
выглядит в виде двух электронно-плотных полос, разделенных свет-
лой полосой. При большом увеличении в комплексе видны две парал-
лельные боковые белковые нити длиной 120—150 нм и толщиной
10 нм каждая, соединенные тонкими поперечными полосами размера-
ми около 7 нм, по обе стороны от них лежат d-хромосомы. Их ДНК
формирует множество петель.
В центре комплекса проходит осевой элемент толщиной 20—40 нм.
Синаптонемальный комплекс удачно сравнивают с веревочной лест-
ницей, стороны которой образованы гомологичными хромосомами.
Именно в результате этого гомологичные хромосомы сцепляются
между собой и образуют биваленты, 46 d-хромосом образуют 23 бива-
лента. Каждый бивалент состоит из двух d-хромосом, т. е . из четы -
рех хроматид.
К концу зиготены каждая пара гомологичных хромосом связана
между собой с помощью синаптонемальных комплексов. Лишь поло-
вые хромосомы X и Y конъюгируют не полностью, т. к. они не полно-
стью гомологичны.
Пахинема (от греч. pahys — толстый) продолжается не менее чем
несколько суток. Процессы развертываются постепенно. Хромосо-
мы несколько укорачиваются и утолщаются. Между хроматидами
материнского и отцовского происхождения в нескольких местах
возникают соединения — хиазмы (от греч. chiasma — перекрест),
или рекомбинантные узелки. Они представляют собой белковые
комплексы размерами около 90 нм. В области каждой хиазмы про-
исходит обмен соответствующих участков гомологичных хромо-
сом — от отцовской к материнской, и наоборот. Этот процесс назы-
вают кроссинговером (от англ. crossing-over — перекрест). Таким
образом, кроссинговер обеспечивает многочисленные генетические
рекомбинации (рис. 48).
В каждом биваленте человека в профазе I кроссинговер происхо-
дит в среднем на двух-трех участках. Количество рекомбинантных
узелков равно количеству перекрестков.
По окончании кроссинговера хроматиды разъединяются, но оста-
ются связанными в области хиазм. Наступает стадия диплонемы.
В диплонеме (от греч. diploos — двойной) синаптонемальные ком-
плексы распадаются, конъюгировавшие хромосомы раздвигаются,
87
МЕЙОЗ
и гомологичные хромосомы каждого бивалента отодвигаются друг от
друга, но связь между ними сохраняется в зонах хиазм.
Между диплонемой и диакинезом нет четкой морфологической
границы, равно как и разграничений во времени. В диакинезе продол-
жается конденсация хромосом, они отделяются от нуклеолеммы,
но гомологичные d-хромосомы продолжают еще оставаться связан-
ными между собой хиазмами, а сестринские хроматиды каждой
d-хромосомы — центромерами. Благодаря наличию нескольких хиазм
биваленты образуют петли. В это время разрушаются ядерная обо-
лочка и ядрышки. Реплицированные центриоли направляются к по-
люсам, образуется веретено деления.
Вследствие сильно затянутой диплонемы профаза мейоза очень
длительна. При развитии спермиев она может длиться несколько су-
ток, а при развитии яйцеклеток — в течение многих лет. Половые
клетки в профазе мейоза называются гаметоцитами первого порядка
(первичными гаметоцитами, гаметоцитами I).
Метафаза I напоминает аналогичную стадию митоза. Хромосомы
устанавливаются в экваториальной плоскости, образуя метафазную
пластинку. В отличие от митоза, хромосомные микротрубочки при-
крепляются к центромере лишь с одной стороны (со стороны полю-
са), а центромеры гомологичных d-хромосом расположены по обеим
сторонам экватора. Связь между хромосомами с помощью хиазм про-
должает сохраняться.
В анафазе I хиазмы распадаются, гомологичные d-хромосомы от-
деляются друг от друга и расходятся к полюсам. Центромеры этих
хромосом, однако, в отличие от анафазы митоза, не реплицируются,
а значит, сестринские хроматиды не расходятся.
АА аа
ВВ bb
АА аа
ВВ
b
b
АА аа
ВВ
b
b
Биваленты
Биваленты
Кроссинговер
Кроссинговер
Генетическая
Генетическая
рекомбинация
рекомбинация
Рис. 48. Кроссинговер
88
СТРОЕНИЕ КЛЕТКИ
В телофазе I формируются ядерная оболочка и ядрышко, образу-
ется и углубляется борозда деления, происходит кариокинез. Сначала
наборы гомологичных d-хромосом находятся у полюсов. Хотя их чи-
сло уменьшилось вдвое, каждая из них состоит из двух генетически
различных хроматид. В результате цитокинеза в каждой дочерней
клетке сосредоточивается по 23-й d-хромосомы. Образовавшиеся
клетки называют гаметоцитами второго порядка (вторичными гаме-
тоцитами, гаметоцитами II).
Интерфаза II очень короткая. Ее важнейшая особенность состоит
в том, что не редуплицируется ДНК, то есть отсутствует S-фаза. Деле-
ние гаметоцита второго порядка совершается через профазу II, мета-
фазу II, анафазу II и телофазу II. Профаза II не длительна, и конъю-
гации хромосом при этом не наступает. В метафазе II 23 хромосомы
выстраиваются в плоскости экватора. В анафазе II ДНК в области
центромеры реплицируется, как это происходит и в анафазе митоза,
хромосомы расходятся к полюсам. В телофазе II образуются две до-
черние клетки.
Напомним, что в деление вступали не тетраплоидные клетки, как
при обычном митозе, а диплоидные. Поэтому каждая из новых клеток
гаплоидна. Восстановление диплоидности произойдет лишь в резуль-
тате слияния мужской и женской гамет, то есть при оплодотворе-
нии — образовании нового организма.
Итак, в результате двух последовательных делений мейоза II обра-
зуются четыре клетки, каждая из которых несет гаплоидный набор
s-хромосом. В таблице 7 приведены основные сходства и различия
между митозом и мейозом.
Таблица 7
Сравнительная характеристика митоза и мейоза
Этапы
Показатель
Митоз
Мейоз
Весь процесс Длительность
Короткий (при
образовании сома-
тических клеток)
Длительный
(при образовании гамет)
Интерфаза Расхождение хро-
мосом
Хроматиды (имеет-
ся длительная)
Гомологичные (имеется
длительная перед мейо-
зом I, короткая между
мейозом I и мейозом II)
S-фаза
Предшествует ка-
ждому делению
Только перед мейозом I,
отсутствует в интерфазе II
Рост клетки
Репликация орга-
нелл
Происходит
Происходит
Происходит
Происходит
89
МЕЙОЗ
Этапы
Показатель
Митоз
Мейоз
Профаза
Длительность
Одна короткая
Профаза I длительная (до
90% времени), профаза II
короткая
Хромосомы
Состоят из двух се-
стринских хрома-
тид, соединенных
центромерой
Состоят из двух сестрин-
ских хроматид, соединен-
ных центромерой
Взаимоотношения
гомологичных хро-
мосом
Обособлены
Конъюгируют с образова-
нием синаптонемальных
комплексов
Биваленты
Отсутствуют
Имеются
Хиазмы
Отсутствуют
Образуются
Кроссинговер
Отсутствует
Происходит
Метафаза Образование мета-
фазной пластинки
Происходит
В метафазе I отсутствует.
Только в метафазе II
Расположение цен-
тромер
В одной плоскости,
перпендикулярной
оси веретена на его
экваторе
В метафазе I над и под
экватором симметрично.
В метафазе II на экваторе
веретена
Хромосомные ми-
кротрубочки се-
стринских хрома-
тид
Направлены в раз-
ные стороны к про-
тивоположным по-
люсам
В метафазе I направлены
в одну сторону. В мета-
фазе II направлены в раз-
ные стороны
Анафаза
Репликация ДНК
в области центро-
мер и разделение
S-хромосом
Происходит
В анафазе I отсутствует,
происходит в анафазе II
Расхождение
d-хромосом
—
В анафазе I вследствие
распада хиазм
Расхождение
s-хромосом
Происходит вслед-
ствие разделения
центромер
В анафазе I не происходит.
Происходит в анафазе II
Генетическая иден-
тичность
Хроматиды иден-
тичны
Вследствие кроссинговера
хроматиды не идентичны
Телофаза
Количество хромо-
сом
Аналогично мате-
ринской клетке
(s-хромосомы)
Вдвое меньше, чем в роди-
тельской клетке (в телофа-
зе I s-хромосомы, в тело-
фазе II s-хромосомы)
Гомологичные хро-
мосомы в дочерних
клетках
Две разделившиеся
хроматиды попада-
ют в каждую клетку
Мейоз I — в каждую клет-
ку попадают две сестрин-
ские хроматиды, соединен-
ные в области центромеры.
Мейоз II — в каждую
клетку попадает одна
хроматида
Окончание табл. 7
90
ОСОБЕННОСТИ СТРОЕНИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
ОСОБЕННОСТИ СТРОЕНИЯ
РАСТИТЕЛЬНОЙ КЛЕТКИ
В клетках растений присутствуют все основные компоненты эука-
риотической клетки. Однако некоторые особенности, непосредствен-
но связанные с условиями функционирования растений, принципи-
ально отличают их от клеток животных. Из присущих животной
клетке органелл в растительной отсутствуют только центриоли (их
могут иметь одноклеточные растения), но имеются жесткая клеточ-
ная стенка, пластиды и вакуоли (рис. 49).
Клеточной стенкой обладают не только растения, но и грибы,
а также многие прокариоты. У растений формируются сначала пер-
вичная, а затем вторичная клеточная стенка (рис. 50). В клеточных
стенках присутствуют каркас из нерастяжимых пучков молекул цел-
люлозы и связующее вещество — гемицеллюлоза и пектины, которые
образуют матрикс стенки. Все эти вещества транспортируются в пу-
7
8
9
10
11
5
4
3
2
6
12
13
14
15
16
17
18
19
20
1
6
11
20
20
Рис. 49. Современная (обобщенная)
схема строения растительной
клетки, составленная по данным
электронно-микроскопического
исследования разных растительных
клеток:
1 — аппарат Гольджи,
2 — свободно расположенные
рибосомы, 3 — хлоропласты,
4 — межклеточные пространства,
5 — полисомы (несколько связанных
между собой рибосом),
6 — митохондрии, 7 — лизосомы,
8 — гранулярная (шероховатая)
эндоплазматическая сеть,
9 — агранулярная (гладкая)
эндоплазматическая сеть,
10 — микротрубочки,
11 — пластиды, 12 — плазмодесмы,
проходящие сквозь оболочку,
13 — клеточная оболочка,
14 — ядрышко, 15 — ядро
(в ядре спирали ДНК),
16 — поры в ядерной оболочке,
17 — плазмалемма,
18 — гиалоплазма (цитозоль),
19 — тонопласт, 20 — вакуоли
(по Арронет)
91
ОСОБЕННОСТИ СТРОЕНИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
зырьках комплекса Гольджи к плазматической мембране, где пузырь-
ки сливаются с ней и посредством экзоцитоза выбрасывают свое со-
держимое наружу. Эти вещества, попадая в пространство между
плазматической мембраной и срединной пластинкой, служат матери-
алом для образования клеточной стенки.
Первичная клеточная стенка характерна главным образом для ме-
ристемных и малодифференцированных клеток. Такие клетки спо-
собны значительно увеличивать свой объем и, соответственно, разме-
ры. Необходимо учитывать, что целлюлозные фибриллы нерастяжимы,
а увеличение линейных размеров осуществляется за счет смещения
относительно друг друга упомянутых фибрилл. Некоторые клетки
в течение всей жизни имеют только первичную оболочку, но у боль-
шинства клеток между плазматической мембраной и первичной стен-
кой откладывается вторичная. Ее строение в принципе сходно с пер-
вичной стенкой, но соотношение компонентов различно. Вторичная
стенка содержит значительно больше целлюлозы и меньше воды. Во
вторичной стенке имеется большое количество углублений — пор
(простых и окаймленных), по которым осуществляется транспорт ве-
ществ из одной клетки в другую.
Транспорт также осуществляется через мелкие сквозные отверстия,
которые пронизывают клеточные стенки — плазмодесмы (рис. 51).
5
6
À
Á
1
2
3
4
7
5
Â
7
8
Рис. 50. Схема строения
клеточной стенки:
А — общий вид,
Б — часть оболочки
при большом увеличении,
В — вид сверху;
1 — срединная пластинка,
2, 3, 4 — соответственно
внешний, средний и внутренний
слои вторичной оболочки,
5 — пора, 6 — слепая пора,
7 — плазмодесменные канальцы,
8 — поровое поле
(по Гуляеву)
92
ОСОБЕННОСТИ СТРОЕНИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
При формировании вторичной клеточной стенки линейный рост кле-
ток становится невозможен, поэтому этот процесс всегда сопровожда-
ется уменьшением объема протопласта (протопласт — содержимое
живой клетки, за исключением клеточной оболочки). Некоторые
клетки, имеющие вторичные утолщения оболочек, сохраняют живой
функционирующий протопласт (например, клетки колленхимы), но
очень часто утолщение приводит к нарушению транспорта веществ,
в результате чего протопласт отмирает, а главную функцию выполня-
ет мощная оболочка (например, склеренхима). В этом случае оболоч-
ки одревесневают, т.е. пропитываются лигнином. В результате повы-
шается механическая прочность и понижается водопроницаемость.
Кроме лигнина, в оболочке клеток могут накапливаться гидрофобные
вещества: растительные воски, кутин и суберин.
Клеточная стенка выполняет множество функций, но наиболее
важными являются две: роль наружного скелета и обеспечение воз-
можности тургора. Особенно важна клеточная стенка у высших на-
земных растений — она поддерживает тело над землей. Значительная
часть клеток живого растения мертва, а функционируют у них имен-
но толстые оболочки. Тургор — давление, которое оказывает прото-
пласт на клеточную стенку; если бы ее не было, клетка разорвалась
бы. Тургор выполняет функцию опоры у живых клеток, стенки кото-
6
4
3
1
5
2
Рис. 51. Плазмодесмы. Участок оболочек
трех смежных клеток при средних
увеличениях электронного микроскопа
(схематично):
1 — эндоплазматический ретикулум
смежных клеток сообщается между собой
через десмотубулы (каналы плазмодесм),
2 — плазмалемма выстилает каналы,
отграничивая цитоплазму от оболочки,
3 — элементы эндоплазматического
ретикулума, 4 — срединная пластинка,
5 — первичная оболочка,
6 — гиалоплазма
44
55
11
22
33
66
93
ОСОБЕННОСТИ СТРОЕНИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
рых не имеют сильно выраженного вторичного утолщения. Это осо-
бенно важно для травянистых растений.
Клеточные стенки разделяют организм растения на два простран-
ства. То, которое объединяет между собой протопласты, связанные
плазмодесмами, называется симпластом. Пространство, которое от-
граничено клеточными стенками и включает в себя межклетники, на-
зывается апопластом. Соответственно транспорт через плазмодесмы
называется симпластическим, а транспорт по оболочкам и межклет-
никам — апопластическим. Растительные клетки прочно связаны
между собой за счет срединной пластинки.
Пластиды. Пластиды являются органеллами, присущими исклю-
чительно растениям. В различных количествах они присутствуют
в любой живой растительной клетке и в своей совокупности образуют
пластид клетки. Выделяют три основных типа пластид: хлоропласты
(зеленые), хромопласты (от желтого до красного цветов) и лейкопла-
сты (бесцветные). Всех их объединяет общее происхождение, наличие
внутренних мембран, а также собственного генома и аппарата биосин-
теза белка, что говорит о некоторой автономности этих органелл.
Обычно в каждой клетке можно обнаружить только один тип пла-
стид.
Хлоропласты наиболее важны
для растения. Они встречаются
у большинства живых клеток зеле-
ных органов растения и часто зани-
мают бо
льшую часть объема про-
топласта. Хлоропласты — очень
крупные структуры (рис. 52). Не-
редко их размер превышает размер
ядра, но обычно ширина 2—4 мкм
и длина 5—10 мкм. Хроматофоры
(единственный хлоропласт водоро-
À
Á
1
2
3
4
5
6
7
8
9
10
1
2
Б
А
66
11
22
Рис. 52 . Строение хлоропласта:
А — продольный разрез через хлоропласт:
1 — наружная мембрана, 2 — внутренняя
мембрана, 3 — межмембранное пространство,
строма (матрикс), 4 — строма, 5 — граны
(стопки тилакоидов), 6 — ламеллы стромы
(межгранный тилакоид), 7 — капли жира,
8 — крахмальное зерно, 9 — ДНК,
10 — увеличенный участок стромы хлоропласта
(видны рибосомы); Б — трехмерная схема
расположения и взаимосвязи ламелл и гран
внутри хлоропласта: 1 — граны, 2 — ламеллы
94
ОСОБЕННОСТИ СТРОЕНИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
слей) еще больше — до 50 мкм в длину. Хлоропласты особенно много-
численны в тех клетках, которые хорошо освещены, клетки корня их
не имеют. Общая численность хлоропластов дерева может доходить
до ста миллиардов. У растущих в тени растений хлоропласты стано-
вятся крупнее и богаче хлорофиллом. Положение этих органелл
в клетке также непостоянно и зависит от интенсивности освещения.
Хлоропласты во многом сходны с митохондриями (см. рис. 52).
Они имеют оболочку, образованную двумя мембранами, между кото-
рыми находится межмембранное пространство. Оболочка ограничи-
вает содержимое хлоропласта, заполненное стромой (матриксом).
В отличие от митохондрий, хлоропласты имеют еще и третью мем-
бранную систему — ламеллярную. Она происходит от внутренней
мембраны оболочки.
Внутренние мембраны образуют мешочки с межмембранным про-
странством — тилакоиды. Они образуют стопки — граны, которые ле-
жат очень тесно. Межгранные тилакоиды простираются от одной гра-
ны до другой. Наличие и количество тилакоидов свидетельствует об
интенсивности фотосинтетических реакций, потому что именно в их
мембранах находятся фотосинтетические пигменты. Подобно мито-
хондриям, хлоропласты имеют собственную кольцевую ДНК, которая
находится в строме, и рибосомы. Хлоропласты самостоятельно синте-
зируют около сотни белков, которые входят в состав рибосом и мем-
бран тилакоидов. Однако органеллы нуждаются в белках, синтез ко-
торых определяется геномом ядра.
Хромопласты — это видоизмененные хлоропласты. При общем
типе строения хромопласты меньше, v них отсутствуют внутренняя
мембранная система и хлорофилл. Красный цвет обусловлен пигмен-
тами, которые относят к числу каротиноидов. Хромопласты имеются
в клетках лепестков цветов (что придает цветам яркость и способст-
вует привлечению опылителей), зрелых плодов, реже в вегетативных
органах (свекла, морковь, листья в период опадения).
Лейкопласты не содержат пигментов. Они имеют строение, общее
для всех пластид, но внутренняя мембранная система развита слабо.
Лейкопласты, запасающие крахмал, называют амилопластами, бел-
ки — протеинопластами, жиры — элайопластами (или олеопласта-
ми). Бесцветные пластиды растений, которых выращивали без осве-
щения, называют этиопластами (при наличии света они легко
превращаются в хлоропласты).
Хлоропласты способны делиться. У высших растений все пласти-
ды происходят от общего предшественника — пропластид, которые,
в свою очередь, развиваются из двухмембранных инициальных
частиц. Эти частицы наследуются только по материнской линии
через яйцеклетку, спермин их не содержат.
95
ДЕЛЕНИЕ РАСТИТЕЛЬНОЙ КЛЕТКИ
Вакуоли содержит каждая растительная клетка. В отличие от жи-
вотных клеток, вакуоли растений выполняют гораздо больше функ-
ций. В клетках растений крупные вакуоли рассматривают как четвер-
тый компонент (наряду с цитолеммой, цитоплазмой и ядром), т. к. их
нельзя отнести к органеллам цитоплазмы.
Вакуоли образуются из провакуолей — мембранных пузырьков,
которые отшнуровываются от элементов ЭПС или от мешочков ком-
плекса Гольджи. Затем пузырьки сливаются и образуют вакуоли зна-
чительных размеров. В старых клетках все вакуоли могут сливаться
в одну гигантскую центральную вакуоль, занимающую до 90% объема
протопласта. В процессе роста клетки ее объем увеличивается за счет
увеличения объема вакуоли, а не цитоплазмы.
От цитоплазмы вакуоль отделена собственной мембраной (тоно-
пластом), обеспечивающей транспорт веществ. Содержимое вакуолей
называется клеточным соком. Основную часть его составляет вода,
в которой растворены различные вещества. В вакуолях зрелых пло-
дов в большом количестве накапливаются глюкоза и фруктоза, а в ва-
куолях сахарных тростника и свеклы — сахароза. В вакуолях могут
также накапливаться и токсичные соединения.
Функции вакуолей разнообразны, наиболее существенными явля-
ются четыре: создание тургора, запасание необходимых клетке ве-
ществ и отложение веществ, вредных для клетки, а также фермента-
тивное расщепление органических соединений.
Включения растительной клетки. Трофические включения харак-
терны для всех клеток, но особенно многочисленны в органах, кото-
рые запасают питательные вещества (например, в корнях, паренхиме
побегов, семенах и др.). Они бывают трех типов: крахмальные зерна,
белковые гранулы, или кристаллы, и липидные капли. Включения мо-
гут быть отходами жизнедеятельности клетки.
ДЕЛЕНИЕ РАСТИТЕЛЬНОЙ КЛЕТКИ (ЦИТОКИНЕЗ)
Процесс начинается с появления в экваториальной плоскости деля-
щейся клетки структуры цилиндрической формы, образованной из
микротрубочек, — фрагмопласта (рис. 53). Затем вдоль этих микро-
трубочек транспортируются мембранные пузырьки, которые отшну-
ровываются от мешочков комплексом Гольджи. Эти пузырьки слива-
ются, образуя окруженный мембраной диск. Такой диск является
ранней клеточной пластинкой, с ней постоянно сливаются все новые
пузырьки. В итоге ранняя клеточная пластинка достигает плазматиче-
ской мембраны и сливается с ней, разделяя дочерние клетки. Раннюю
клеточную пластинку пронизывают элементы эндоплазматической
сети, которые образуют плазмодесмы, специфичные для растительных
96
ОСОБЕННОСТИ СТРОЕНИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
клеток. Пузырьки комплекса Гольджи, из которого образовалась ран-
няя клеточная стенка, содержат различные полисахариды, основные из
которых пектины и гемицеллюлоза. Связываясь между собой, эти ве-
щества образуют срединную пластинку, которая, в основном, состоит из
пектина. Позже в ее состав входят более плотные вещества — целлюло-
за и лигнин. Каждая клетка со всех сторон окружена срединной пла-
стинкой.
Àïïàðàò
Ãîëüäæè
Îñòàòêè ïîëþñíûõ
ìèêðîòðóáî÷åê âåðåòåíà
Ñòåíêà
ìàòåðèíñêîé êëåòêè
Ïóçûðüêè ñ ïðåäøåñò-
âåííèêàìè êë åòî÷íî é
ñòåíêè íàêàïëèâàþò-
ñÿ â ýêâàòîðèàëüíîé
çîíå ï åðåêðûâàíèÿ
ìèêðîòðóáî÷åê è
ñëèâàþòñÿ, îáðàçóÿ
ðàííþþ êëåòî÷íóþ
ïëàñòèíêó
ßäðî â ïîçäíåé
òåëîôàçå
Ïóçûðüêè êîìïëåêñà
Ãîëüäæè ïîäõîäÿò
ê ìèêðîòðóáî÷êàì
è äâèæóòñÿ ê ýêâàòîðó
Ïî ìåðå âêëþ÷åíèÿ
ìàòåðèàëà â êëåòî÷-
íóþ ñòåíêó, îáðàçóþ-
ùóþñ ÿ âíóòðè êëåòî÷-
íîé ïëàñòèíêè, ó
êðàåâ êëåòî÷í îé
ïëàñòèíêè íàêàïëè-
âàþ òñÿ íîâûå
ïóçûðüêè êîìïëåêñà
Ãîëüäæè; îíè ñëèâà-
þòñÿ ñ íåé è ðàñøè-
ðÿþò åå
Ñðåäèííàÿ
ïëàñòèíêà
Ðàííÿÿ
êëåòî÷íàÿ
ïëàñòèíêà
Ìèêðîòðóáî÷êè ôðàãìîïëàñòà
ðàñïàäàþòñÿ è ôîðìèðóþòñÿ
çàíîâî íà ïåðèôåðèè ðàííåé
êëåòî÷íîé ïëàñòèíêè
Ðàñøèðÿþùàÿñÿ
ìåìáðàíà êëåòî÷-
íîé ïëàñòèíêè
ñëèâàåòñÿ ñ
ïëàçìàòè÷åñêîé
ìåìáðàíîé;
ìàòåðèàë êëåòî ÷íîé
ñ ò å í êè ïðîäîëæàåò
îòêëàäûâàòüñÿ, ïîêà
íå áóäåò ãîòîâà
íîâàÿ ñòåíêà
Ïëàçìîäåñìû ìåæäó
ñîñåäíèìè êëåòêàìè
Âîññòàíîâèâøååñÿ
èíòåðôàçíîå ðàñïîëîæåíèå
êîðòèêàëüíûõ ìèêðîòðóáî÷åê
Рис. 53. Ход цитокинеза в клетках высших растений,
имеющих жесткую клеточную структуру
97
ФОТОСИНТЕЗ
На следующих этапах формируются сначала первичная, а затем
вторичная клеточная стенка. В клеточных стенках растений (как
в первичных, так и во вторичных) роль нерастяжимых элементов кар-
каса выполняют пучки молекул целлюлозы, а роль связующего ком-
понента принадлежит гемицеллюлозам и пектинам, которые образу-
ют матрикс клеточной стенки. Все эти вещества транспортируются
в пузырьках комплекса Гольджи к плазматической мембране, где пу-
зырьки сливаются с ней и посредством экзоцитоза выбрасывают со-
держащиеся в ней вещества наружу. Эти вещества, попадая в про-
странство между плазматической мембраной и срединной пластинкой,
служат материалом для образования клеточной стенки.
ФОТОСИНТЕЗ
Жизнь на Земле возможна благодаря световой (главным образом
солнечной) энергии, которая преобразуется в энергию химических
связей органических веществ, образующихся в процессе фотосинтеза.
Фотосинтезом обладают все растения и некоторые прокариоты (фо-
тосинтезирующие бактерии и сине-зеленые водоросли — цианобакте-
рии). Организмы, способные к фотосинтезу, называются фототрофа-
ми. Этот процесс представляет собой сложную цепь реакций,
в результате которых из углерода и водорода, полученных из неорга-
нических источников, образуются органические соединения.
Энергию для фотосинтеза дает свет, который улавливается особы-
ми молекулами — фотосинтетическими пигментами. Они представ-
лены молекулами, способными поглощать кванты света. Поскольку
при этом поглощается свет лишь определенной длины волны, часть
световых волн не поглощается, а отражается. В зависимости от спект-
рального состава отраженного света пигменты приобретают окраску —
зеленую, желтую, красную и др. Различают три класса фотосинтетиче-
ских пигментов — хлорофиллы, каротиноиды и фикобилины. Наиболее
важным пигментом является хлорофилл. Насчитывается свыше де-
сятка зеленых пигментов, отличающихся друг от друга атомными
группами, присоединенными к пиррольным структурам порфирино-
вого кольца, а также по некоторым другим характеристикам. Хлоро-
филл представляет собой сложный эфир дикарбоновой кислоты хло-
рофиллина с двумя спиртами — фитолом и метанолом. Структура
молекулы определяет свойства хлорофилла. Основой является пло-
ское порфириновое ядро, образованное четырьмя пиррольными коль-
цами, соединенными между собой метиновыми мостиками, с атомом
магния в центре (рис. 54). В ядре, кроме пиррола, содержатся также
его изомер — пирроленин и продукт неполного восстановления пирро-
ла — пирролин. Так как в ядре имеются многочисленные двойные свя-
98
ОСОБЕННОСТИ СТРОЕНИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
зи, там присутствуют делокализованные
π-электроны, которых в ядре насчитывает-
ся 18. Они очень важны для фотосинтеза.
Гидрофобный фитольный «хвост» удержи-
вает молекулу в гидрофобной части мем-
браны тилакоида хлоропласта, а гидро-
фильное порфириновое ядро обращено к
строме хлоропласта.
Растения и цианобактерии содержат
различные хлорофиллы типа а. У высших
растений, зеленых и эвгленовых водоро-
слей имеется хлорофилл b (он образуется
из хлорофилла а). Бурые и диатомовые
водоросли вместо хлорофилла b содержат
хлорофилл с, а красные водоросли — хлоро-
филл d. Хлорофиллы бактерий имеют не-
которые специфические особенности и на-
зываются бактериохлорофиллами.
Молекулы хлорофиллов способны, вза-
имодействуя друг с другом и молекулами
белков, создавать агрегаты, различающие-
ся по длине волн поглощенного света.
Хлорофилл а имеет два четко выраженных максимума поглощения:
660—663 нм и 428—430 нм. Хлорофилл b поглощает более короткие
волны в красной части спектра и более длинные в синей. Его макси-
мумы поглощения будут 642—644 нм и 452—455 нм соответственно.
Все хлорофиллы слабо поглощают желтый и оранжевый свет, а зеле-
ный они отражают, что и определяет зеленую окраску этого класса
пигментов (рис. 55).
Другую группу пигментов образуют каротиноиды. Эти жирораст-
воримые пигменты имеют различную окраску — от желтой до крас-
ной. Они содержатся во всех окрашенных пластидах (хлоропластах
и хромопластах) растений. Причем в зеленых частях растений хлоро-
филл маскирует каротиноиды, делая их незаметными до наступления
холодов. Осенью зеленые пигменты разрушаются, и каротиноиды ста-
новятся хорошо заметными, определяя окраску осенних листьев. Кро-
ме растений, каротиноиды синтезируют фототрофные бактерии
Рис. 54. Строение хлорофилла a:
1, 2, 3, 4 — пиррольные кольца; серым выделено
порфириновое ядро с делокализованными
π-электронами (пояснение в тексте)
Г
и
д
р
о
ф
о
б
н
а
я
ч
а
с
т
ь
(
о
с
т
а
т
о
к
ф
и
т
о
л
а
—
ф
и
т
и
л
)
12
3
99
ФОТОСИНТЕЗ
и грибы. Каротиноиды способны поглощать свет, недоступный для
других пигментов, и передавать его хлорофиллам.
Третьей группой пигментов являются фикобилины, присутствую-
щие у красных водорослей и цианобактерий. Фикобилины образуют
комплексы с белками — фикобилипротеиды (хромопротеиды).
Электроны молекул пигментов, поглотив энергию, переходят на
более высокий энергетический уровень, т. е. становятся возбужден-
ными. Однако рано или поздно они возвращаются на свой исходный
(стационарный) уровень, выделив энергию, полученную ранее при
возбуждении. Эта энергия используется для синтеза органических ве-
ществ из неорганических, запасаясь при этом в химических связях
синтезированных веществ.
В мембранах тилакоидов хлоропластов обнаружены комплексы
молекул, названные фотосистемой I и фотосистемой II. Они совмес-
тно обеспечивают трансформацию световой энергии. Каждая из фото-
систем имеет реакционный центр, который образован белками, ассо-
циированными с хлорофиллом. Пигменты реакционного центра
способны поглощать энергию света, которая переводит электроны
в возбужденное состояние, в результате чего они покидают молекулу
хлорофилла и переходят на расположенные поблизости молекулы-
переносчики. Вторым компонентом фотосистемы является антенный
комплекс. Кроме хлорофилла а, здесь присутствуют еще и дополни-
тельные пигменты — хлорофилл в, каротиноиды и фикобилины. Их
роль заключается в улавливании света с длиной волн, недоступной
для хлорофилла а. Молекулы пигментов антенных комплексов не осу-
ществляют фотохимических реакций, а передают полученную энер-
гию по цепи хлорофиллу реакционного центра. Направление переноса
Рис. 55. Спектры поглощения пигментов хлоропластов:
1 — хлорофилл а, 2 — хлорофилл b, 3 — b-каротин, 4 — фикобилины
100
ОСОБЕННОСТИ СТРОЕНИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
энергии (сами электроны здесь не передаются) в антенных комплексах
всегда ориентировано от пигментов, поглощающих самую короткую
часть спектра (каротиноидов), к более «длинноволновым» пигментам.
Такой процесс называется резонансной передачей энергии.
Наличие света необходимо не для всех этапов, а лишь вначале, по-
этому в фотосинтезе выделяют световую и темновую стадии.
Световая стадия фотосинтеза
На этом этапе происходит поглощение световой энергии пигмента-
ми, молекулы которых возбуждаются. Непосредственно фотохимиче-
ские реакции осуществляют реакционные центры фотосистем. Для
лучшего понимания всего процесса удобно начать с фотосистемы II.
Первичным донором электронов в фотосистеме II является хлорофилл
П680
.
В дальнейшем заряженный электрон передается по цепи пере-
носчиков, каждый раз теряя часть своей энергии. В итоге электрон
передается хлорофиллу фотосистемы I, которой для максимальной про-
дуктивности фотосинтеза необходимо работать согласовано с фотосис-
темой II (рис. 56), поэтому для последовательного продолжения цепи
реакций должна включиться фотосистема I. Содержащийся в ее реакци-
Рис. 56. Перенос электронов в процессе фотосинтеза в тилакоидной мембране.
Подвижными переносчиками электронов в этой цепи служат пластохинон (очень
сходный с убихиноном митохондрий), пластоцианин (небольшой медьсодержащий
белок) и ферредоксин (небольшой белок, содержащий железосерный центр).
Комплекс b6-f очень похож на комплекс b-с1 митохондрий и комплекс b-с бактерий:
все три комплекса принимают электроны от хинонов и перекачивают протоны.
Протоны, высвобождаемые при окислении воды, и протоны, захватываемые при
образовании NADPH, тоже участвуют в создании электрохимического протонного
градиента, доставляющего энергию для синтеза ATP; I — строма, II — тилакоидное
пространство; А — фотосистема II, Б — комплекс b6-f, В — фотосистема I,
Г — NADP -редуктаза; 1 — свет, 2 — антенный комплекс, 3 — ферредоксин,
4 — пластоцианин, 5 — пластохинон, 6 — фермент, расщепляющий воду,
7 — тилакоидная мембрана (по Албертсу и соавт., с изменениями)
22
66
77
55
44
33
11
11
I
A
Б
В
Г
II
NADP
NADP+
NADP
NADPH
HH+
nH+
4H++O2
2H2O
101
ФОТОСИНТЕЗ
онном центре хлорофилл П700
, два кванта света с несколько большей
длиной волны, чем хлорофилл фотосистемы II, возбуждается и соответ-
ственно теряет два электрона. После этого электроны перемещаются по
цепи переносчиков на NADP+
, который восстанавливается до NADPH.
В фотосистеме I утраченные электроны поступают от фотосисте-
мы II. В фотосистеме II электронные «дыры» заполняются электрона-
ми, которые добываются из ионов ОН– при разложении молекул воды
в результате фотолиза под действием света. Образовавшийся при
этом кислород не нужен для фотосинтетических реакций, поэтому он
выводится сначала из хлоропласта, а затем через межклетники и усть-
ица из растения.
ЭТО ВАЖНО!
Весь кислород воздуха на планете Земля был образован в результате
фотосинтетической деятельности зеленых растений.
Описанный путь переноса электрона по двум фотосистемам из-за
своей зигзагообразности (в силу того, что молекулы — переносчики
электрона погружены в мембрану тилакоида на разную глубину) на-
зывается Z-схемой, или нециклическим транспортом электронов
(рис. 57). Его результатом будет появление NADPH, разложение воды
и выделение кислорода, а также фотофосфорилирование, при кото-
ром энергия возбужденного электрона используется для присоедине-
ния неорганического фосфата к ADP, т. е. синтезу АТР.
В мембранах тилакоидов осуществляется еще и циклический
транспорт электронов. Он идет лишь в фотосистеме I. Его результа-
том будет синтез ATP, a NADPH при этом не образуется. Механизм
синтеза АТР в результате фосфорилирования ADP идет в соответст-
вии с хемиосмотической теорией. Согласно ей электрохимический
градиент в полости тилакоида создают расположенные в мембране
тилакоида многочисленные переносчики электронов, которые тран-
спортируют с одной поверхности мембраны на другую электроны
и протоны (последние необходимы для уравновешивания заряда),
обратно же переносится только электрон, а протоны остаются по дру-
гую сторону, потому что мембрана здесь для них непроницаема. В ре-
зультате избыточного накопления на одной стороне ионов водорода
формируется электрохимический протонный градиент. После того
как его значение достигнет определенной величины, протоны начина-
ют «протекать» через мембрану, но не во всех местах, а лишь через
особые белки, пронизывающие мембрану насквозь. Эти белки явля-
ются АТР-синтетазами, протоны проходят через них пассивно, одна-
ко, как и любой поток, они при этом совершают некоторую работу,
102
ОСОБЕННОСТИ СТРОЕНИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
которая используется для присоединения к ADP неорганического
фосфата, в результате чего образуется АТР.
Применительно к фотосинтезу этот процесс весьма упрощенно
выглядит следующим образом. После поглощения хлорофиллом
квантов света их электроны возбуждаются и передаются акцепторам,
расположенным на наружной поверхности мембраны. Электроны
с хлорофилла П680 поступают на жирорастворимый пластохинон. По-
лучая два электрона, он захватывает из стромы два протона, после
чего электроны передаются на железосерный белок Риске, затем на
цитохром f, пластоцианин и на длинноволновый хлорофилл П700.
В результате этого протоны поступают в полость тилакоида, где к ним
присоединяются еще два протона, которые образовались в результате
фотолиза воды. В конечном итоге в полости тилакоида формируется
избыток протонов, а затем происходят события, описанные выше.
Рис. 57. Нециклический транспорт электронов (нециклическое фосфорилирование)
в хлоропластах (Z-схема) и изменение редокс-потенциала, схема:
I — фотосистема II, II — фотосистема I; 1 — фермент, расщепляющий воду,
2 — хлорофилл П680, 3 — хлорофилл П680 в возбужденном состоянии, 4 — феофетин,
5 — первичный пластохинон фотосистемы II (QA), 6 — вторичный пластохинон
фотосистемы II (QB), 7 — жирорастворимый пластохинон (PQ), 8 — комплекс b6-f,
9 — железосерный белок Риске, 10 — цитохром f комплекса b6-f, 11 — пластоцианин,
12 — хлорофилл П700, 13 — хлорофилл П700 в возбужденном состоянии,
14 — первичный акцептор электронов фотосистемы I, 15, 16 — вторичные акцепторы
электронов фотосистемы I (железосерные белки), 17 — ферредоксин,
18 — NADP -редуктаза с FAD в качестве кофермента
Образуется электрохимический
градиент, генерирующий АТР
Направление потока электронов
Р
е
д
о
к
с
-
п
о
т
е
н
ц
и
а
л
,
м
В
т
I
2
3
13
14
15
16
12
11
7
6
5
4
17
1000
800
600
400
200
0
–200
– 400
–600
–800
Свет (2hν)
Свет (2hν)
2H2OO
2+4H+
nH+
2e–
2e–
H+ + NADP+ NADPH
II
103
ФОТОСИНТЕЗ
Темновая стадия фотосинтеза
Следующие реакции протекают в строме хлоропласта. Поскольку
для них не нужен свет, эту стадию фотосинтеза назвали темновой.
Именно в ней происходит биологическая фиксация углерода, в ре-
зультате чего синтезируются органические вещества, в химических
связях которых запасена энергия, первоначально полученная при воз-
буждении электрона хлорофилла квантом света.
Углекислый газ способен реагировать с пятиуглеродным соедине-
нием — рибулёзо-1,5-бифосфатом, которое образуется в строме в ре-
зультате фосфорилирования с помощью АТР молекул рибулёзо-
5-фосфата. Фермент присоединяет СO2 к рибулёзо-1,5-бифосфату,
и полученный в результате этого шестиуглеродный промежуточный
продукт быстро распадается на две молекулы 3-фосфоглицериновой
кислоты.
Следующим этапом является восстановление 3-фосфоглицерино-
вой кислоты до 3-фосфоглицеринового альдегида. Это достигается
путем фосфорилирования исходных триоз с помощью АТР, в резуль-
тате чего образуются молекулы 1,3-бифосфоглицериновой кислоты,
а они, в свою очередь, взаимодействуя с NADPH, восстанавливаются
до 3-фосфоглицеринового альдегида.
Возникает вопрос: из каких источников пополняются запасы
первичного акцептора углекислого газа? Регенерация рибулёзо-1,5-
бифосфата осуществляется по замкнутому циклу, который по имени
исследователей, открывших его, был назван циклом Кальвина — Бен-
сона (рис. 58). В этой цепи реакций в результате фиксации трех мо-
лекул СO2 образуется шесть молекул 3-фосфоглицериновой кисло-
ты, содержащих совместно восемнадцать атомов углерода (три из
них получены от трех фиксированных молекул углекислого газа).
Затем они проходят сложный цикл реакций, в результате которых
регенерируют три молекулы рибулёзо-1,5-бифосфата (совместно со-
держащих пятнадцать атомов углерода) и образуется одна трехугле-
родная молекула 3-фосфоглицеринового альдегида. Иными слова-
ми, в цикл вступают органические соединения (молекулы
рибулёзо-1,5-бифосфата) и неорганические (молекулы углекислого
газа), а в итоге образуются только органические — регенерирует то
же количество молекул рибулёзо-1,5-бифосфата и дополнительно
появляется органическая молекула 3-фосфоглицеринового альде-
гида.
Образовавшееся трехуглеродное соединение (молекула 3-фосфо-
глицеринового альдегида) уже само по себе является одним из проме-
жуточных продуктов гликолиза. Некоторая часть 3-фосфоглицерино-
вого альдегида остается в строме хлоропласта, где затем превращается
104
ОСОБЕННОСТИ СТРОЕНИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
в глюкозо-1 -фосфат, который, в свою очередь, сначала превращается
в ADP-глюкозу, а затем в крахмал. Особенно много крахмала откла-
дывается в хлоропластах днем при интенсивном течении фотосинте-
тических процессов; ночью под действием соответствующих фермен-
тов крахмал расщепляется до растворимых форм и используется
растением. Однако бо
льшая часть 3-фосфоглицеринового альдегида
переходит в гиалоплазму и вступает в реакции, которые представля-
ют собой гликолиз, но идущий в обратном порядке. При этом образу-
Рис. 58. Цикл фиксации углерода в темновой стадии фотосинтеза
(цикл Кальвина — Бенсона).
В ходе этого цикла из СО2 и Н2О образуются органические молекулы.
Для упрощения схемы многие промежуточные продукты на пути
от глицеральдегид-3-фосфата к рубулёзо-5 -фосфату опущены.
Участие воды в цикле также не показано
(по Б. Албертсу, с изменениями и дополнениями)
1C
3C
3C
6PI
2PI
3C
3C
3C
В результате фиксации трех молекул CO2 в конечном итоге
образуется одна молекула глицеральдегид-3 -фосфата,
при этом затрачивается 9 молекул АТР и 6 молекул NADPH
Сахара, жиры, аминокислоты
Одна молекула
Пять молекул
Шесть молекул
Шесть молекул
Шесть молекул
Три молекулы
Три молекулы
Три молекулы
5C
5C
3ADP
6ADP
6NADP
6NADP+
HCO
HCOH
HCOPOH
H
OH
O
105
ФОТОСИНТЕЗ
ется глюкозо-1 -фосфат и фруктозо-6-фосфат. В конечном итоге полу-
чается сахароза, которая загружается во флоэму и транспортируется
по ней к органам, нуждающимся в энергии, или к органам и тканям,
запасающим питательные вещества впрок.
ЭТО ВАЖНО!
Все органические вещества живых организмов (за исключением ве-
ществ, полученных в результате хемосинтеза) образуются в результате
биохимических превращений продуктов фотосинтеза.
СТРОЕНИЕ И ФУНКЦИОНИРОВАНИЕ
ПРОКАРИОТИЧЕСКОЙ КЛЕТКИ
Несмотря на единство строения, химического состава и функций,
присущее всем клеткам, между эу- и прокариотическими клетками
имеются существенные различия.
Прокариотическая клетка организована проще. Большинство из
них — это мелкие (как правило, не более 10 мкм) округлые, овальные
или удлиненные клетки, которые, в отличие от эукариот, не разделе-
ны на клеточные компартменты (рис. 59). Типичными прокариотами
являются бактерии, которые по форме подразделяют на сфериче-
ские — кокки, прямые или изогнутые палочки, спиральные или изви-
тые клетки (вибрионы и спириллы). Некоторые клетки, разделив-
шись, не расходятся, в результате чего образуются пары (диплококки,
диплобактерии), цепочки (стрептококки, стрептобактерии) или паке-
ты кокков (сарцины). В то же время прокариоты широко различаются
по своим физиологическим свойствам и очень быстро делятся. Так,
в течение 10—11 часов потомство одной-единственной клетки в бла-
гоприятных условиях может достичь 4 млрд особей.
ЭТО ВАЖНО!
Прокариоты легко адаптируются к условиям окружающей среды, у них
очень часты спонтанные мутации, а необычайное биохимическое мно-
гообразие способствует их повсеместному распространению на Земле.
Плазматическая мембрана (цитолемма) прокариот выполняет все
свойственные функции: транспортную, защитную, разграничитель-
ную, рецепции, восприятия сигналов внешней (для клетки) среды,
участия в иммунных процессах, обеспечения поверхностных свойств
клетки. Кроме того, плазматические мембраны у них выполняют еще
106
СТРОЕНИЕ И ФУНКЦИОНИРОВАНИЕ ПРОКАРИОТИЧЕСКОЙ КЛЕТКИ
ряд важнейших функций: в них локализуются ферменты цепи пере-
носа электронов и окислительного фосфорилирования, осуществля-
ется синтез компонентов клеточной стенки и капсулы, выведение
внеклеточных ферментов. У фотосинтезирующих организмов фото-
синтез также осуществляется на мембране; на внутренней стороне
мембраны расположены сайты связывания ДНК, каждая из дочерних
молекул ДНК после репликации прикрепляется к одному из сайтов,
À
7
8
9
10
11
5
4
3
2
12
13
14
15
16
17
18
19
20
1
6
Á
Â
1
2
3
4
5
1
2
3
Рис. 59. Строение бактериальной клетки:
А — бактериальная клетка: 1 — гранулы
поли-β-оксимасляной кислоты,
2 — жировые капельки, 3 — включения
серы, 4 — трубчатые тилакоиды,
5 — пластинчатые тилакоиды,
6 — пузырьки, 7 — хроматофоры,
8 — ядро (нуклеоиды), 9 — рибосомы,
10 — цитоплазма, 11 — базальное тельце,
12 — жгутики, 13 — капсула,
14 — клеточная стенка, 15 — цитоплазма-
тическая мембрана, 16 — мезосома,
17 — газовые вакуоли, 18 — ламеллярные
структуры, 19 — гранулы полисахарида,
20 — гранулы полифосфата.
Основные структуры бактериальной
клетки представлены в верхней части
рисунка, дополнительные мембранные
структуры, имеющиеся у фототрофных
и нефототрофных бактерий, — в средней
части, а включения запасных веществ —
в нижней части; Б — мембранные
структуры: 1 — везикулярные тилакоиды,
2 — тубулярные тилакоиды,
3 — ламеллярные тилакоиды,
4 — мезосомы, 5 — ламеллы; В — иные
структуры: 1 — хлоросомы, 2 — гозовые
вакуоли, 3 — карбоксисомы
(по Шлегелю, с изменениями)
107
СТРОЕНИЕ И ФУНКЦИОНИРОВАНИЕ ПРОКАРИОТИЧЕСКОЙ КЛЕТКИ
в результате роста мембраны молекулы ДНК расходятся, после чего
формируется перемычка, разделяющая клетку на две. Общий прин-
цип устройства мембран прокариот не отличается от эукариот, но
в химическом составе имеются различия (отсутствуют молекулы хо-
лестерина и некоторых других липидов, присущих мембранам эука-
риот). У некоторых микроорганизмов плазматическая мембрана впя-
чивается внутрь клетки, образуя стопки плоских мешочков, связанных
с цитолеммой. У цианобактерий и некоторых пурпурных бактерий
имеется множество мембранных тилакоидов, связанных с мембраной
и осуществляющих фотосинтез. Тилакоиды также являются произ-
водными цитолеммы, которые образуются в результате ее впячива-
ний. Некоторые прокариоты содержат газовые вакуоли (аэросомы).
Все мембранные структуры прокариотической клетки непрерывны.
Клеточная стенка прокариот выполняет многочисленные функ-
ции, в первую очередь, формообразование и защиту от осмотического
шока. В различных клетках клеточная стенка составляет от 5 до 50%
сухой массы клетки.
В соответствии с окраской бактерии делят на две группы — грам-
положительные и грамотрицательные, что связано с особенностями
строения их клеточных стенок. У грамположительных клеток цито-
лемма окружена толстой (20—80 нм) клеточной стенкой, состоящей
из пептидогликана муреина, тейхоевых кислот и полисахаридов;
у грамотрицательных — тонким (2—3 нм) слоем пептидогликана, по-
крытым наружной плазматической мембраной (рис. 60). Если обра-
ботать клетку лизоцимом или пенициллином, клеточная стенка раз-
рушается. Грамположительная клетка превращается в протопласт,
который весьма подвержен осмотическому шоку; грамотрицатель-
ная — в сферопласт, покрытый двумя мембранами и вследствие этого
менее чувствительный к осмотическому шоку.
Капсулы, слизь, влагалища. Многие бактерии (например, пнев-
мококки, клебсиеллы, некоторые клостридии и др.) и некоторые си-
не-зеленые водоросли кнаружи от клеточной стенки имеют слой
сильно оводненного материала, образующего капсулу. Капсулы связа-
ны с клеточной стенкой, имеют толщину до 10 мкм. Капсулы обеспе-
чивают устойчивость бактерий к ряду воздействий, например к фаго-
цитозу (повышая вирулентность бактерий), придают бактериям
специфичность (в ней могут находиться антигены), предохраняют
клетку от высыхания, механических повреждений, от действия виру-
сов, служат источником запасных питательных веществ, а также осу-
ществляют связь между клетками и способствуют прикреплению
к каким-либо поверхностям.
Подвижность прокариот. Прокариоты обладают жгутиками, рас-
положение которых может быть монополярным, биполярным и пери-
108
СТРОЕНИЕ И ФУНКЦИОНИРОВАНИЕ ПРОКАРИОТИЧЕСКОЙ КЛЕТКИ
трихиальным. По количеству жгутиков различают монотрихи (одна
нить) и политрихи (пучок нитей). Скорость движения прокариот
очень велика — от 1,6 до 12 мм/мин. Структура жгутика сложна
и совершенно отлична от строения ресничек и жгутиков эукариот
(рис. 61). Подвижные прокариоты совершают направленные движе-
ния по градиенту концентрации некоторые веществ — аттрактантов
или против градиента концентрации других веществ — репеллентов.
Такое поведение называется хемотаксисом. При движении жгутиков
против часовой стрелки клетка движется в одном направлении, при
движении жгутиков по часовой стрелке клетка кувыркается. Под вли-
янием аттрактантов клетка большую часть пути проделывает прямо-
линейно, под влиянием репеллентов клетка чаще кувыркается и уда-
ляется. Помимо хемотаксиса, существуют и другие виды таксиса.
À
Á
1
5
4
2
3
Êèøå÷íàÿ
ïàëî÷êà
(E.coli)
Ïîëîæåíèå
4 D-ala
3 Dpm
2 D-isoglu
1 L-ala
Çîëîòèñòûé
ñòàôèëîêîêê
(E.aureus)
Ïîëîæåíèå
4 D-ala
3 L-Lys
2 D-isoglu
1 L-ala
Ïîëîæåíèå
Ïîëîæåíèå
4
3
2
1
ÑÎ
ÑÍ(ÑÍ3)
4
3
2
1
ÑÎ
ÑÍ(ÑÍ3)
4
3
2
1
ÑÎ
ÑÍ(ÑÍ3)
GLY5
O
NAG
O
O
O
O
O
O
O
O
O
NAM
NAM
NAG
è ò.ä.
èò.ä.
Рис. 60 . Клеточная стенка
бактерий:
А — грамотрицательная,
Б — грамположительная;
1 — липопротеид,
2 — полисахаридные цепи,
3 — поперечные пептидные
связи, 4 — белок,
5 — тейховвая кислота
(по Рису и Стернбергу,
с изменениями)
109
СТРОЕНИЕ И ФУНКЦИОНИРОВАНИЕ ПРОКАРИОТИЧЕСКОЙ КЛЕТКИ
Аэротаксис — аэробные бактерии — устремляются к воздушной среде,
а анаэробные, наоборот, скапливаются в центре культуры. Фототак-
сис — пурпурные бактерии — перемещаются в зоны с наибольшей ос-
вещенностью, что связано с потребностью фототрофных бактерий в
квантах света для получения энергии. Магнитотаксис — железосодер-
жащие бактерии — движутся в магнитном поле в направлении линий.
Фимбрии, или пили, — это полые нити диаметром до 0,0015 мкм
и длиной от 0,3—5 мкм, образованные белком пилином. Они распола-
гаются по периферии клетки в количестве 100—250. Фимбрии участ-
вуют в процессах прикрепления бактерий к клеткам млекопитающих
(например, к слизистой оболочке кишечника). У некоторых бактерий
есть специализированные фимбрии, в частности F-фимбрии у кишеч-
ной палочки, участвующие в процессе конъюгации и могущие слу-
жить органом прикрепления бактериофагов.
Другие органеллы прокариот. Важными отличительными особен-
ностями прокариот являются малое количество мембранных орга-
нелл и наличие множества рибосом (от 5000 до 50 000 в клетке).
Внутриклеточные запасные вещества. В прокариотических клет-
ках могут откладываться полисахариды, липиды, полифосфаты. Каж-
дый вид прокариот накапливает один тип запасных веществ.
À
1
2
Á
8
9
10
7
5
4
6
3
2
1
Рис. 61. Схема вращения жгутика:
А — кишечная палочка: 1 — жгутик, 2 — направление вращения
жгутика; Б — схема «мотора», вращающего жгутик: 1 — крючок,
2 — волокно жгутика, 3 — кольцо L, 4 — наружная мембрана,
5 — кольцо P, 6 — пептидогликановый слой,
7 — цитоплазматическая мембрана, 8 — кольцо S («статор»),
9 — кольцо М («ротор»), 10 — стержень
(по Б. Албертсу и соавт., с изменениями)
110
СТРОЕНИЕ И ФУНКЦИОНИРОВАНИЕ ПРОКАРИОТИЧЕСКОЙ КЛЕТКИ
Покоящиеся формы. При неблагоприятных условиях внешней
среды многие прокариоты образуют покоящиеся формы, способные
сохранять жизнеспособность в течение длительного времени. К таким
формам относят эндо- и экзоспоры, цисты, бактероиды, гетероцисты
и др. Зрелая спора устойчива к различным неблагоприятным услови-
ям среды, включая высокую температуру, радиацию, ультрафиолето-
вые лучи, химические агенты.
ЭТО ВАЖНО!
Споры могут сохраняться в течение очень длительного времени (до
1000 лет) при нормальных условиях.
Однако при температуре 100 °С 90% спор бактерий гибнет через
11 минут; 99% спор актиномицетов погибает при температуре 75 °С
через 70 минут, а при высушивании они сохраняются до 15 лет. При
улучшении условий внешней среды спора прорастает. Прорастание
протекает в три этапа: активация, инициация и собственно прора-
стание. При прорастании спора теряет устойчивость к внешним
факторам.
Генетический аппарат прокариот. Одним из отличий прокариот
является отсутствие у них оформленного ядра. Вместо него имеется
нуклеоид. Величина генома у бактерий составляет от 0,8 × 106
до 8 × 106 пар нуклеотидов. Помимо хромосомы, у бактерий имеются
плазмиды — суперспирализованные молекулы двухнитчатой ДНК,
ковалентно замкнутые в кольцевую структуру. Бактериальные клетки
могут содержать несколько плазмид. Плазмиды способны реплициро-
ваться независимо от хромосомы. Плазмиды могут находиться в бак-
териальной клетке в двух состояниях — автономном (в цитоплазме)
и интегрированном (встраиваются в структуру бактериальной хромо-
сомы и реплицируются вместе с ней). Плазмиды способны перено-
ситься из клетки в клетку. Часто они контролируют у бактерий опре-
деленные свойства. К плазмидам относят также профаги — стадия
существования умеренных бактериофагов. С плазмидами связывают
патогенность ряда бактерий и их отдельных штаммов.
Метаболические процессы в микробной клетке. Большинство
микроорганизмов используют энергию, заключенную в различных
химических соединениях. Они подразделяются на две группы: хемо-
литотрофы (если используются неорганические соединения) и хемо-
органотрофы (если используются органические).
Автотрофы. К фотосинтезу способны немногие микроорганизмы
(фототрофы). Это цианобактерии и фотосинтезирующие бактерии.
Хемосинтезирующие автотрофные бактерии получают энергию за
111
ВИРУСЫ
счет окисления неорганических соединений. Их сравнительно немно-
го, но они оказали весьма существенное воздействие на течение гео-
химических процессов. В зависимости от используемых субстратов
хемосинтезирующие организмы подразделяют на: нитрифицирующие
бактерии, железобактерии, водородные бактерии, серобактерии и
бактерии, окисляющие сурьму.
Энергетический обмен. У микроорганизмов, как и у эукариот,
различают два типа дыхания: аэробное и анаэробное. Различают не-
сколько групп микроорганизмов.
Строгие (облигатные) аэробы растут только при наличии воздуха,
обладают набором ферментов для аэробного дыхания, производят
полное окисление углеводов до СО2 и Н2О.
Строгие анаэробы могут развиваться лишь в бескислородной сре-
де, обладают анаэробным дыханием.
Факультативные анаэробы способны развиваться как в бескисло-
родных, так и в кислородных условиях. Процесс дыхания у них про-
текает в две фазы: сначала фаза анаэробного роста, а затем потребле-
ние кислорода и более глубокое расщепление углеводов. Они
обладают обоими наборами ферментов.
Микроаэрофилы развиваются при низких концентрациях кислоро-
да — до 1%. К ним относят спирохеты, актиномицеты.
Аэротолерантные микробы могут развиваться при пониженных
концентрациях кислорода — до 10%. Примером могут служить кло-
стридии.
Капнические микробы требуют повышенного содержания углекис-
лоты (например, Brucella abortus).
Энергетический обмен у прокариот в основном включает в себя те
же этапы, что и у эукариот.
ВИРУСЫ
Вирусы представляют собой неклеточную форму жизни. Они не
способны к самостоятельному размножению и обмену веществ, по-
этому для реализаций этих функций вирусам необходима клетка-хо-
зяин. Вирусы были обнаружены русским ученым Д. И. Ивановским
в 1892 г. при изучении причин, вызывающих заболевание табака, ког-
да на листьях появлялась мозаика.
Строение вирусов. Вирусы не имеют клеточного строения
(рис. 62). Каждая вирусная частица устроена очень просто: она состо-
ит из расположенного в центре носителя генетической информации
и оболочки. Генетический материал представляет собой короткую мо-
лекулу нуклеиновой кислоты, это образует сердцевину вируса. Нукле-
112
ВИРУСЫ
иновая кислота у разных вирусов может быть представлена ДНК или
РНК, причем эти молекулы могут иметь необычное строение: наряду
с двунитчатой ДНК и однонитчатой РНК встречается однонитчатая
ДНК и двунитчатая РНК. Оболочка называется капсид. Она образо-
вана субъединицами — капсомерами, каждый из которых состоит из
одной или двух белковых молекул. Число капсомеров для каждого
вируса постоянно. Если вирусная частица, кроме капсида, больше не
имеет оболочки, ее называют простым вирусом, если имеется еще
одна — наружная, вирус называется сложным. У каждого вируса кап-
сомеры капсида располагаются в строго определенном порядке, бла-
годаря чему возникает определенный тип симметрии.
Жизненный цикл вирусов. Вирусы не могут самостоятельно раз-
множаться и осуществлять обмен веществ. В соответствии с этим
у них различают две жизненные формы: покоящаяся внеклеточная —
вирион и активно репродуцирующаяся внутриклеточная — вегета-
тивная. Вирионы демонстрируют отменную жизнеспособность.
В частности, они выдерживают давление до 6000 атм и переносят вы-
сокие дозы радиации, однако погибают при высокой температуре, об-
лучении ультрафиолетовыми лучами, а также воздействии кислот
и дезинфицирующих веществ. Взаимоотношения вируса с клеткой
последовательно проходят несколько стадий (рис. 63).
Первая стадия представляет собой адсорбцию вирионов на поверх-
ности клетки-мишени, которая для этого должна обладать соответст-
вующими поверхностными рецепторами. Именно с ними специфиче-
ски взаимодействует вирусная частица, после чего происходит их
3
2
4
1
5
Рис. 62. Сферический вирус:
1 — структурная единица
(субъединица), 2 — капсомер
(морфологическая единица),
3 — капсид, 4 — нуклеиновая
кислота, 5 — оболочка
113
ВИРУСЫ
прочное связывание. По этой причине клетки восприимчивы не ко
всем вирусам. Именно этим объясняется строгая определенность
путей проникновения вирусов. Например, клетки слизистой оболоч-
ки верхних дыхательных путей имеют рецепторы к вирусу гриппа,
а у клеток кожи их нет.
Вторая стадия состоит в проникновении целого вириона или его
нуклеиновой кислоты внутрь клетки-хозяина.
Третья стадия называется депротеинизация. В ходе ее происходит
освобождение носителя генетической информации вируса — его ну-
клеиновой кислоты от белковой оболочки (капсида). Именно она су-
щественным образом преобразует деятельность клетки-хозяина, под-
чиняя ее метаболизм своим потребностям и вынуждая ее синтезировать
определенные вещества. Сам вирус не обладает необходимыми для
этого механизмами, поэтому для синтеза нужных молекул он исполь-
зует клеточные ферменты (например, протеазы, РНК-полимеразы и
др.) и структуры (например, рибосомы).
В ходе четвертой стадии на основе вирусной нуклеиновой кисло-
ты происходит синтез необходимых для вируса соединений.
В пятой стадии происходит синтез компонентов вирусной части-
цы — нуклеиновой кислоты и белков капсида, причем все компонен-
ты синтезируются многократно.
7
5
4
3
2
6
1
Öèòîïëàçìà
Öèòîïëàçìà
1à
2à
3à
Ñö
Ñö
ÏÎ
Îò
ÎÂ
Рис. 63 . Проникновение
онковирусов в клетку
в процессе виропексиса (схема):
1 — внеклеточный онковирус
типа С, 2 — адсорбция вируса
на свободной клеточной
поверхности, 3 — рофеоцитоз
вируса, 1а, 2а, 3а — последова-
тельные этапы проникновения
онковирусов типа А,
почкующихся на поверхности
соседней клетки, 4 — локализация
вируса в фагосоме,
5 — проникновение сердцевины
вируса в цитоплазму,
6 — деструкция онковирусов
в фаголизосоме, 7 — лизосомы,
ПО — плазматическая оболочка
клетки, ОВ — оболочка вируса,
Сц — сердцевина, От — отростки
оболочки вируса
114
ВИРУСЫ
В ходе шестой стадии из синтезированных ранее многочисленных
копий нуклеиновой кислоты и белков формируются новые вирионы
путем самосборки.
Последняя — седьмая — стадия представляет собой выход вновь
собранных вирусных частиц из клетки-хозяина. У разных вирусов
этот процесс проходит неодинаково. У некоторых вирусов это сопро-
вождается гибелью клетки — лизис клетки. У других вирионы выхо-
дят из живой клетки путем отпочковывания, однако и в этом случае
клетка со временем погибнет, поскольку при отпочковывании повре-
ждается плазматическая мембрана.
Время, прошедшее с момента проникновения вируса в клетку до
выхода новых вирионов, называется скрытым, или латентным, пери-
одом. Оно может широко варьировать: от нескольких часов (пяти-ше-
сти у вирусов оспы и гриппа) до нескольких суток (вирусы кори, аде-
новирусы и др.).
У некоторых бактериофагов наряду с вирулентными (быстро раз-
вивающимися вирусами) имеются умеренные фаги. Их нуклеиновая
кислота после проникновения в бактериальную клетку интегрируется
в ДНК клетки и становится профагом. Профаг не оказывает литиче-
ского воздействия на клетку-хозяина и при делении реплицируется
вместе с клеточной ДНК. Бактерии, содержащие профаг, называют
лизогенными. Они проявляют устойчивость к содержащемуся в них
фагу, а также к близким к нему другим фагам. Лизогенные бактерии
могут менять свойства (например, выделять новые токсины).
Классификация вирусов. На систематическое положение вирусов
указывают разные показатели: тип нуклеиновой кислоты и количест-
во нитей (одно- или двунитчатая), ее масса и относительная доля
в вирусной частице. Кроме того, вирусы подразделяются в зависимо-
сти от формы капсида и строения оболочки, природы хозяина и мно-
гих других факторов. При обозначении конкретного вируса также не-
обходимо указывать переносчика, если он есть. По форме вириона
вирусы делят на: сферические (вирусы кори, гриппа, арбовирусы
и др.), палочковидные (вирусы мозаичной болезни табака, картофеля
и др.), кубоидальные (аденовирусы, реовирусы, вирусы оспы и др.)
и сперматозоидные (бактериофаги). В зависимости от поражаемой
клетки-мишени вирусы делят на вирусы животных, растений, грибов
и бактерий (бактериофаги, или фаги).
Происхождение вирусов. Природа вирусов пока до конца неиз-
вестна. Существует несколько гипотез. Согласно одной, вирусы пред-
ставляют собой результат морфофункционального регресса, связан-
ного с паразитическим образом жизни. Сторонники этой гипотезы
полагают, что предки вирусов имели клеточное строение. Несколько
отличается от этого другая гипотеза, постулирующая происхождение
115
ЭПИТЕЛИАЛЬНЫЕ ТКАНИ
вирусов из первобытных доклеточных организмов. По этой версии
предшественники вирусов еще тогда избрали паразитический образ
жизни, и, таким образом, они являются наиболее древними паразита-
ми. Согласно гипотезе об эндогенном происхождении, вирусы пред-
ставляют собой фрагмент когда-то клеточной нуклеиновой кислоты,
который приспособился к сепаратной репликации. Эту версию под-
тверждает существование в бактериальных клетках плазмид, поведе-
ние которых во многом сходно с вирусами. Наряду с этим существует
и «космическая» гипотеза, согласно которой вирусы были занесены
из Вселенной посредством каких-либо космических тел.
Значение вирусов. Все вирусы являются паразитами и поража-
ют все известные организмы. Многие из них (грипп, полиомиелит,
ВИЧ и др.) вызывают у людей тяжелые заболевания, нередко с ле-
тальным исходом. Однако вирусы могут быть полезными. Как и лю-
бые другие паразиты, они стимулируют деятельность защитных сил
организмов. Многие вирусы, поражающие бактерии, чрезвычайно
важны для медицины и ветеринарии, поскольку позволяют естест-
венным путем и без химических реагентов побеждать многие бакте-
риальные инфекции.
ТКАНИ
Ткань — это исторически сложившаяся общность клеток и меж-
клеточного вещества, объединенных единством происхождения, стро-
ения и функции. В организме человека выделяют четыре типа тканей:
эпителиальные, соединительные, мышечные и нервную.
ЭПИТЕЛИАЛЬНЫЕ ТКАНИ
Эпителиальные ткани покрывают поверхность тела и выстилают
слизистые оболочки, отделяя организм от внешней среды (покровный
эпителий), а также образуют железы (железистый эпителий). Эпите-
лий образует слой клеток, лежащих на тонкой базальной мембране,
лишенный кровеносных сосудов, его питание осуществляется за счет
подлежащей соединительной ткани. Базальная мембрана — слой
межклеточного вещества (белков и полисахаридов), располагающий-
ся на границе между различными тканями, например между эпители-
альным пластом и подлежащей соединительной тканью.
В зависимости от количества слоев клеток поверхностный эпите-
лий подразделяют на однослойный и многослойный (рис. 64). Одно-
слойный эпителий покрывает серозные оболочки (брюшина, плевра,
перикард), выстилает большинство слизистых оболочек, а многослой-
116
ТКАНИ
ный покрывает кожу и выстилает некоторые слизистые оболочки (на-
пример, конъюнктиву глаза, ротовую полость, глотку, пищевод, влага-
лище).
Железистый эпителий (железа) представляет собой орган, парен-
хима которого сформирована из железистых клеток. Железы под-
разделяются на экзокринные, имеющие выводные протоки; эндокрин-
ные, не имеющие выводных протоков и выделяющие синтезируемые
ими продукты непосредственно в межклеточное пространство, откуда
они поступают в кровь и лимфу; смешанные, состоящие из экзо-
и эндокринных отделов (например, поджелудочная железа). Кроме
того, имеется множество одноклеточных желез — бокаловидных кле-
ток, лежащих среди других эпителиальных клеток, покрывающих
слизистые оболочки полых органов пищеварительной, дыхательной
и половой систем, которые вырабатывают слизь (рис. 65).
Экзокринная железа состоит из начального (секреторного) отдела,
сформированного железистыми клетками, которые вырабатывают
различные секреты, и протоков. В зависимости от строения секретор-
ного отдела различают трубчатые (наподобие трубки), ацинозные
(напоминают грушу или удлиненную виноградину) и альвеолярные
(напоминают шарик), а также трубчато-ацинозные и трубчато-альве-
олярные железы, секреторные отделы которых имеют и ту и другую
форму. В зависимости от строения протоков железы подразделяются
Рис. 64. Строение эпителиальной ткани:
А — однослойный плоский эпителий, Б — однослойный кубический эпителий,
В — однослойный цилиндрический эпителий, Г — многослойный плоский
неороговевающий эпителий, Д — переходный эпителий,
Е — псевдомногослойный реснитчатый эпителий
АБ
В
ГДЕ
117
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
на простые, имеющие один проток, и сложные, в главные выводные
протоки которых вливается множество протоков, в каждый из кото-
рых, в свою очередь, открывается несколько секреторных отделов.
Железы вырабатывают различные секреты: белковый (серозные же-
лезы), слизь (слизистые) и смешанный.
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
Соединительные ткани представляют обширную группу, вклю-
чающую собственно соединительные ткани (рыхлая волокнистая
и плотная волокнистая неоформленная и оформленная), ткани со
специальными свойствами (ретикулярная, пигментная, жировая),
твердые скелетные (костная, хрящевая) и жидкие (кровь и лимфа).
Соединительные ткани выполняют различные функции: опорную
(или механическую), трофическую (или питательную), защитную.
В отличие от других тканей, соединительные сформированы из
многочисленных клеток и вырабатываемого ими межклеточного ве-
щества. Последнее состоит из аморфного вещества и различных воло-
кон (коллагеновых, эластических, ретикулярных). Межклеточное ве-
щество имеет различную консистенцию — от твердого у кости до
жидкого у крови и лимфы.
Многие клетки крови являются одновременно и клетками соеди-
нительной ткани, а другие — их предшественниками, поэтому целесо-
образно начать описание соединительных тканей с крови.
Рис. 65. Схема строения экзокринных и эндокринных желез:
А — экзокринная железа, Б — эндокринная железа; 1 — начальный отдел,
2 — секреторные гранулы, 3 — выводной проток экзокринной железы,
4 — покровный эпителий, 5 — соединительная ткань, 6 — кровеносный сосуд
5
А
4
Б
3
6
1
1
5
4
2
2
6
118
ТКАНИ
Кровь (рис. 66). «Кровь — особый сок», — восклицает Мефисто-
фель («Фауст» И. В . Гете). И действительно, жизнь человека связана
с кровью, которая выполняет следующие функции: транспортную,
трофическую (питательную), защитную, гемостатическую (крово-
останавливающую). Кроме того, кровь участвует в сохранении посто-
янного состава и свойств внутренней среды организма — гомеостаза
(от греч. homoios — одинаковый и stasis — состояние, неподвижность).
Общее количество крови у взрослого человека 4—6 л, что составляет
6—8% массы тела (у мужчин в среднем около 5,4 л, у женщин — около
4,5 л). Потеря 10% крови допустима, 30% — опасна, а 50% — смер-
тельна.
Кровь состоит из клеток (44% объема крови), взвешенных в жид-
ком межклеточном веществе сложного состава (плазма — 54% объе-
ма). Плазма — это жидкая часть крови, в которой содержится до 91%
воды, 6,5—8,0% белков, около 2% низкомолекулярных соединений,
рН плазмы колеблется в пределах от 7,37 до 7,43, а удельный вес
1,025—1,029. Плазма богата как электролитами, так и неэлектролита-
Рис. 66. Кровь:
А — мазок периферической крови взрослого человека (общий вид): 1 — эритроциты,
2 — лимфоциты, 3 — моноцит, 4 — нейтрофильные гранулоциты, 5 — эозинофильные
гранулоциты, 6 — базофильные гранулоциты, 7 — тромбоциты;
Б — клетки крови: I — базофильный гранулоцит, II — ацидофильный гранулоцит,
III — сегментоядерный нейтрофильный гранулоцит, IV — эритроцит, V — моноцит,
VI — тромбоциты, VII — лимфоцит
1
5
А
I
Б
II
III
IV
V
VI
VII
2
6
3
7
1
4
2
119
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
ми. Белки плазмы крови (6,5—8,0 г/л, альбумины и глобулины) вы-
полняют трофическую, транспортную, защитную, буферную функ-
ции; они также участвуют в свертывании крови и создании
коллоидно-осмотического давления. В крови содержатся безъядер-
ные клетки эритроциты — в среднем (4,0—5,0) × 1012/л (муж.
—
4,0×1012 — 5,6 × 1012/л; жен.
—
3,4 × 1012 – 5,0 × 1012/л), лейкоциты —
в среднем (4,0—6,0) × 109 (муж.
—
4,3×109 — 11,3×109 /л; жен.
—
3,2×109
—
10,2 × 109 /л), среди которых выделяют зернистые, или гра-
нулоциты, и незернистые, или агранулоциты (моноциты). В крови
имеются также кровяные пластинки (тромбоциты), число которых
составляет (180,0—320,0) × 109/л. В крови постоянно присутствуют
также клетки лимфоидного ряда (лимфоциты), которые являются
структурными элементами лимфоидной (иммунной) системы.
Эритроциты (от греч. erythros — красный), или красные кровя-
ные тельца, имеют форму двояковогнутых дисков диаметром от 7 до
10 мкм (рис. 67). Эритроцит — единственная клетка человека, лишен-
ная ядра. Эритроцит заполнен гемоглобином, осуществляющим пере-
нос кислорода и углекислого газа. Эритроциты выполняют свои
функции, только находясь в просвете сосудов. Общее количество эри-
троцитов взрослого человека достигает 25 × 1012
. Длительность жизни
эритроцитов около 120 дней, после чего они разрушаются и поглоща-
ются макрофагоцитами в селезенке, костном мозге и печени.
ЭТО ВАЖНО!
Общая площадь поверхности всех эритроцитов около 3800 м2. Если
сложить все эритроциты человека в один ряд, длина цепочки составит
175 000 км, ею можно было бы опоясать земной шар более четырех
раз.
Рис. 67. Нормальные эритроциты в форме двояковогнутого диска
120
ТКАНИ
В 1900—1901 гг. венский ученый К. Ландштейнер совершил одно
из величайших открытий в медицине, и в 1930 г. ему была присужде-
на Нобелевская премия «За открытие групп крови человека». Эри-
троцит покрыт цитолеммой толщиной около 7 нм, в которую встрое-
ны антигены систем АВО и резус. Антиген — любое вещество (обычно
в его состав входит белок), которое способно вызвать иммунную реак-
цию. Иммунная реакция — это ответ организма на внедрение чужого
агента. В плазме крови каждого человека имеются антитела против
антигенов эритроцитов, которые не содержатся в его собственной
крови. Антитело — это молекула белка, вырабатываемая одной из
клеток иммунной системы в ответ на внедрение антигена. Ландштей-
нер описал четыре группы крови (табл. 8).
Таблица 8
Группы крови человека
Группа крови
O (I)
А (II)
В (III)
АВ (ІV)
Частота в популяции
46%
42%
9%
3%
Агглютиногены
—
А
В
А+В
Агглютинины
а+b
b
а
—
Ученый обнаружил, что при смешивании плазмы крови одного че-
ловека и эритроцитов другого часто происходит их склеивание (агглю-
тинация). Это приводит к закупориванию мелких сосудов, что может
привести к смертельному исходу. Для разделения крови на группы
смешивали эритроциты с пробными сыворотками — так называемы-
ми сыворотками анти-А и анти-В . Ландштейнер выявил, что эритро-
циты группы О не агглютинируются ни одной из сывороток; эритро-
циты группы АВ агглютинируются обеими сыворотками; эритроциты
группы А агглютинируются сывороткой анти-А, но не агглютиниру-
ются сывороткой анти-В; наконец, эритроциты группы В агглютини-
руются сывороткой анти-В, но не агглютинируются сывороткой
анти А. В сыворотке крови группы О содержатся групповые антитела
анти-А и анти-В; в сыворотке группы А имеются только антитела ан-
ти-В, в сыворотке группы В — антитела анти-A, а в сыворотке АВ
групповые антитела отсутствуют.
ЭТО ВАЖНО!
В сыворотке крови содержатся только те антитела (изоагглютинины),
которые не агглютинируют эритроциты этой группы, поэтому при необ-
ходимости человеку следует переливать кровь только той же группы.
121
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
В 1940 г. Ландштейнер открыл еще один фактор крови — резус
(Rh-фактор), который впервые был обнаружен у обезьяны макаки
резус. У 85% людей эритроциты несут на своей поверхности
Rh-антиген, это Rh-положительные (Rh+), у других он отсутствует,
их называют резус-отрицательными (Rh-). Если человеку Rh- пере-
лить кровь от Rh+ донора, то у первого в течение двух-четырех меся-
цев будут продуцироваться Rh-антитела, и если ему перелить еще раз
Rh+ кровь, то произойдет агглютинация Rh+ эритроцитов. К . Ланд-
штейнер обнаружил связь между Rh-фактором и желтухой новоро-
жденных. Если Rh- женщина беременна от Rh+ мужчины, плод мо-
жет оказаться Rh+. Тогда при первой беременности в организме
матери вырабатываются Rh-антитела. При последующей беременно-
сти, если эта женщина вынашивает Rh+ плод, ее Rh-антитела прони-
кают через плаценту в кровь плода и вызывают у него агглютинацию
эритроцитов, что приводит к желтухе новорожденного.
Группы крови детей чаще всего зависят от группы крови одного из
родителей (табл. 9).
В настоящее время широко проводится переливание крови, как вы-
сокоэффективное средство лечения многих заболеваний. Это основ-
ное средство для спасения жизни человека при кровотечениях, выра-
женной анемии (малокровии), шоке, заражении крови (сепсис);
сложных операциях, сопровождающихся большой кровопотерей.
В 1914 г. А . Юстен открыл способность лимоннокислого натрия
предотвращать свертывание крови. Это создало предпосылки для за-
готовки, длительного хранения крови и возможность использовать ее
по мере необходимости. Человек, который сдает кровь, называется
донором (дающим).
Таблица 9
Группы крови родителей и детей
Мама
Папа
I
II
III
IV
I
I
I, II
I, III
II, III
II
I, II
II
I, II, III, IV
II, IV
III
I, III
I, II, III, IV
III
III, IV
IV
II, III
II, IV
III, IV
II, III, IV
Лейкоциты (от греч. leukos — белый) представляют собой ядро-
содержащие клетки, обладающие амебоидной подвижностью. В отли-
чие от эритроцитов, которые выполняют присущие им функции
в просвете кровеносных сосудов, лейкоциты осуществляют свои
122
ТКАНИ
в тканях, куда они мигрируют посредством диапедеза (от греч. dia —
сквозь, pedesis — прыжок). В одном мкл крови здорового человека со-
держится 4000—8000 лейкоцитов.
ЭТО ВАЖНО!
Если сложить все лейкоциты человека в один ряд, он вытянется на рас-
стояние около 525 км.
К зернистым лейкоцитам (гранулоцитам) относятся нейтрофиль-
ные, или полиморфноядерные, которые составляют от 93 до 96% всех
гранулоцитов (в среднем 4150 в 1 мкл крови). Время их циркуляции
в крови не превышает 8—12 ч, затем посредством диапедеза они миг-
рируют в соединительную ткань. Зрелый нейтрофильный гранулоцит
представляет собой сферическую клетку диаметром 10—12 мкм
с дольчатым ядром. В ядрах нейтрофильных гранулоцитов женщин
имеются тельца полового хроматина (тельца Барра)1 диаметром до
1,5—2,0 мкм. Цитоплазма гранулоцита богата гранулами двух типов:
нейтрофильными и азурофильными, которые участвуют в фагоцитозе
и инактивации фагоцитированного материала. Нейтрофилы — пер-
вая линия обороны внутренней среды организма. Осуществляя фаго-
цитоз продуктов распада и микроорганизмов, нейтрофильные грану-
лоциты погибают, а освобождающиеся при этом лизосомальные
ферменты разрушают окружающие ткани, способствуя формирова-
нию гнойника. В состав гноя обычно входят разрушенные нейтро-
фильные гранулоциты и продукты распада ткани. Количество
нейтрофильных гранулоцитов резко возрастает при острых воспали-
тельных и инфекционных заболеваниях.
Эозинофильные (ацидофильные) гранулоциты диаметром 10—
15 мкм составляют 0,5—5,0% циркулирующих лейкоцитов. В 1 мкл
крови их число колеблется в пределах от 120 до 300 (0,02—0,3 × 109/л).
Они циркулируют в крови не более 8 дней, после чего покидают кро-
веносное русло через мелкие венулы и проникают в рыхлую соедини-
тельную ткань. Особенно много их в слизистой оболочке кишечника
и дыхательных путей. Их двухлопастное ядро напоминает по форме
гантель. В цитоплазме имеется множество крупных красных или
оранжевых светопреломляющих несколько удлиненных гранул, кото-
рые представляют собой лизосомы. Эозинофильные гранулоциты
осуществляют фагоцитоз, однако менее активно, чем нейтрофильные.
Их подвижность также менее выражена. Эозинофильные гранулоци-
1 Тельце Барра — одна из двух Х-хромосом женских соматических клеток, которая
в интерфазе остается в конденсированном состоянии.
123
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
ты участвуют в иммунных реакциях. Количество эозинофильных гра-
нулоцитов в циркулирующей крови (эозинофилия) увеличивается
при паразитарных заболеваниях, аллергических и аутоиммунных
процессах.
Количество базофильных гранулоцитов в циркулирующей крови
невелико — около 0,5% всех лейкоцитов (40—50 клеток в 1 мкл крови,
0—0,65 × 109/л), а время их циркуляции не превышает 12—15 ч. Диа-
метр клетки 10—12 мкм, в световом микроскопе в клетке видно мно-
жество крупных темно-синих округлых или овальных гранул. Коли-
чество их столь велико, что они маскируют крупное ядро. Гранулы
содержат гистамин и гепарин. Базофилы также осуществляют фаго-
цитоз и участвуют в аллергических реакциях.
Лимфоциты, которые являются структурными элементами лим-
фоидной (иммунной) системы, составляют 25—40% всех лейкоцитов
(1000—4000 в 1 мкл), они преобладают в лимфе. Все лимфоциты име-
ют сферическую форму, но отличаются друг от друга размерами. Диа-
метр большей части лимфоцитов около 8 мкм (малые лимфоциты).
Лимфоциты подразделяются на две категории: тимус-зависимые
(Т-лимфоциты) осуществляют в основном клеточный иммунитет,
и бурсо-зависимые (В-лимфоциты) — гуморальный иммунитет (см.
раздел «Лимфоидные органы»). Морфологически они не отличаются
друг от друга (даже по своей ультраструктуре).
Моноциты составляют от 3 до 11% циркулирующих лейкоцитов
крови (90—600 в 1 мкл, 0,09—0,6 × 109/л). Время их пребывания в кро-
веносной системе 2—3 дня, после чего они мигрируют в ткани, где
превращаются в макрофаги и выполняют свою главную функцию —
защиту организма. Моноцит — клетка овальной формы, диаметром
около 15 мкм с крупным почкообразным, богатым хроматином ядром
и большим количеством цитоплазмы, в которой имеется множество
лизосом.
Тромбоциты, или кровяные пластинки, — уплощенные
овальные двояковыпуклые безъядерные фрагменты крупных клеток
мегакриоцитов диаметром 2—4 и толщиной 0,5—0,75 мкм (рис. 68).
Количество их достигает 250—350 тыс. в 1 мкл крови. Время их цир-
куляции в крови не превышает семи дней, после чего они попадают
в селезенку и легкие, где разрушаются.
ЭТО ВАЖНО!
Тромбоциты участвуют в свертывании крови, остановке кровотечений,
восстановительных процессах и в защите организма благодаря способ-
ности фагоцитировать вирусы, иммунные комплексы и неорганические
частички.
124
ТКАНИ
Остановка кровотечения. У здорового человека кровотечение
в результате ранений мелких сосудов прекращается в течение
1—3 мин. Это первичный гемостаз (от греч. haima — кровь, stasis— не-
подвижность), связанный с сужением сосудов и склеиванием тромбо-
цитов. При повреждении стенки кровеносного сосуда тромбоциты
прилипают к краям раны, из тромбоцитов высвобождаются биологи-
чески активные вещества, которые вызывают сужение сосудов. При
более значительных повреждениях благодаря сложному процессу
вторичного гомеостаза происходит остановка кровотечения. Под дей-
ствием ферментативной активности крови, которая получила назва-
ние «тромбокиназа», белок плазмы протромбин, образующийся в пе-
чени, превращается в тромбин, который вызывает переход
растворимого плазменного белка фибриногена, также образующегося
в печени, в нерастворимый фибрин. Последний и формирует основ-
ную часть тромба.
Рыхлая волокнистая соединительная ткань
Рыхлая волокнистая соединительная ткань (РВСТ) располагается
преимущественно по ходу кровеносных и лимфатических сосудов,
нервов, покрывает мышцы, образует строму (от греч. stroma — под-
1
2
3
Рис. 68. Тромбоцит:
1 — пучок микротрубочек,
2 — грануломер, 3 — гиаломер
(по Б. Албертсу и соавт., с изменениями)
125
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
стилка) — каркас органов, собственную пластинку слизистой оболоч-
ки, наружную оболочку внутренних органов. РВСТ состоит из много-
численных собственных и пришлых клеток: фибробласты, фиброциты,
ретикулярные, перициты, макрофагоциты, тканевые базофилы, плаз-
моциты, жировые клетки, пигментные, лимфоциты, гранулярные лей-
коциты, которые располагаются в межклеточном веществе, представ-
ленном коллагеновыми, эластическими, ретикулярными волокнами,
которые погружены в основное (аморфное) вещество (рис. 69).
Фибробласты (от греч. fibra — волокно, blastos — зародыш) — ос-
новные специализированные фиксированные клетки соединительной
ткани, богатые рибосомами, элементами зернистой (гранулярной) эн-
доплазмической сети и комплекса Гольджи. Фибробласты синтезиру-
ют и секретируют основные компоненты межклеточного вещества со-
единительной ткани: полисахариды, предшественники коллагена и
эластина. Фибробласты по мере старения превращаются в фиброци-
Рис. 69. Рыхлая волокнистая соединительная ткань:
1 — коллагеновое волокно, 2 — макрофагоцит, 3 — тканевый базофил, 4 — аморфное
межклеточное (основное) вещество, 5 — плазмоцит (плазматическая клетка),
6 — адипоцит (жировая клетка), 7 — эластическое волокно, 8 — фибробласт,
9 — эндотелиоцит, 10 — перицит, 11 — миоцит, 12 — кровеносный сосуд
1
2
34
5
6
7
89
10
11
12
126
ТКАНИ
ты, которые весьма слабо синтезируют компоненты межклеточного
вещества РВСТ. Фиброциты — многоотростчатые клетки веретено-
образной формы, бедные органеллами, образуют трехмерную сеть,
в пространствах которой располагаются различные клетки. Коллагено-
вые волокна образованы белком коллагеном. Три полипептидные цепи,
скручиваясь, образуют молекулу тропоколлагена. Молекулы тропо-
коллагена, объединяясь между собой, формируют коллагеновые пучки
толщиной до 150 мкм. Коллаген имеет спиральное строение, что обес-
печивает создание весьма прочных малорастяжимых структур.
Эластические волокна толщиной от 3 до 10 мкм образованы бел-
ком эластином, который также синтезируется фибробластами. В от-
личие от коллагеновых, эластические волокна способны растягивать-
ся в 1,5 раза, после чего возвращаются в исходное состояние.
Эластические волокна анастомозируют и переплетаются между со-
бой, образуя сети, окончатые пластины и мембраны.
Тонкие (от 100 нм до 1 мкм), разветвленные, малорастяжимые ре-
тикулярные волокна, переплетаясь между собой, образуют мелкопет-
листую сеть, в ячейках которой расположены клетки. Ретикулярные
волокна образуют каркасы органов кроветворения и иммунной систе-
мы, печени, поджелудочной железы и других паренхиматозных орга-
нов, окружают капилляры, кровеносные и лимфатические сосуды,
а также связаны с ретикулярными клетками.
Макрофаг (макрофагоцит). В 1882 г. И . И . Мечников впер-
вые описал фагоцитоз. Вонзая в прозрачное тело личинки морской
звезды шип розы, он наблюдал, что через несколько часов шип был
окутан слоем «подвижных клеток... Если заноза была предварительно
обмазана порошком кармина или краски индиго, то надвинувшиеся
клетки оказывались наполненными этими красками... Клетки эти
очень прожорливы и вбирают в себя все, что только могут захва-
тить», — писал И. И. Мечников. Он назвал эти клетки макрофагами
и указал на их возможную связь с моноцитами крови. В 70-х годах
XX века сформировалось представление о системе мононуклеарных
фагоцитов (СМФ), включающей группу клеток, объединенных общ-
ностью происхождения (из моноцитов крови), строения и функции
(активный фагоцит и пиноцитоз). Особенностью структуры макрофа-
гов является большое количество лизосом в их цитоплазме.
ЭТО ВАЖНО!
Основные функции макрофагов — это участие в естественном, специ-
фическом, противоопухолевом иммунитете и секреция различных био-
логически активных веществ. Макрофаги — вторая важнейшая, весьма
эффективная линия обороны.
127
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
Плазмоциты, или плазматические клетки, происходящие
из В-лимфоцитов, — это белоксинтезирующие клетки, богатые эле-
ментами шероховатой эндоплазматической сети, располагающиеся
вблизи мелких кровеносных сосудов в лимфоидных органах, в слизи-
стой оболочке пищеварительной и дыхательной систем. Они выраба-
тывают антитела (иммуноглобулины), чем определяется их важней-
шая роль в защите организма. Это третья линия обороны.
Тучные клетки, или тканевые базофилы, очень богаты круп-
ными (до 2 мкм) гранулами, содержащими биологически активные
вещества гистамин и гепарин, влияющие на кровеносные сосуды.
Ретикулярные клетки — удлиненные многоотростчатые клетки,
которые, соединяясь своими отростками, формируют сеть. При неблаго-
приятных условиях (инфекция, внедрение инородных частиц и т. д .) ре-
тикулярные клетки округляются, отделяются от ретикулярных волокон
и становятся способными к фагоцитозу. Ретикулярные клетки и волокна
образуют строму органов иммунной системы и кроветворения.
Жировые клетки, или адипоциты. Различают два типа жировых
клеток: белые и бурые. Зрелый однокапельный адипоцит белой жировой
ткани — крупная (50—120 мкм в диаметре) шаровидная клетка, почти
полностью занятая каплей жира. Однокапельный адипоцит осуществля-
ет синтез и внутриклеточное накопление липидов в качестве резервного
материала. Многокапельный адипоцит бурой жировой ткани содержит
множество капель жира и большое количество митохондрий.
Перициты окружают кровеносные капилляры, располагаясь кнару-
жи от эндотелии. Перициты — это отросчатые клетки, соприкасающиеся
отростками с каждым эндотелиоцитом. Они передают последним нерв-
ный импульс, который способствует накоплению или потере клеткой
жидкости. Это приводит к расширению или сужению просвета капилляра.
Пигментные клетки, содержащие пигмент меланин, залегают
в эпидермисе, особенно наружных половых органов и околососкового
поля, в радужке и собственно сосудистой оболочке глазного яблока,
в мягкой мозговой оболочке. На 1 мм2 поверхности кожи приходится
1200—1500 пигментных клеток. У представителей черной и желтой рас
количество их значительно больше. Цвет глаз зависит от генетически
обусловленного количества пигментных клеток в радужке глаза.
В рыхлой волокнистой соединительной ткани находятся также
клетки крови (моноциты, зернистые лейкоциты) и лимфоциты.
Плотная волокнистая соединительная ткань
Плотная волокнистая соединительная ткань (ПВСТ) характеризу-
ется сильным развитием волокнистых структур межклеточного веще-
ства, имеющих в основном веществе упорядоченное направление
128
ТКАНИ
(оформленная ткань) либо переплетающихся в разных направлениях
(неоформленная ткань). Плотная соединительная ткань выполняет
в основном опорную функцию.
Плотная оформленная волокнистая соединительная ткань фор-
мирует сухожилия, связки, фасции, пластины, эластический конус
гортани и ее голосовые связки, желтые связки, выйную связку копыт-
ных, входит в состав стенок артерий эластического типа. Главными
элементами ее являются тесно прилежащие друг к другу пучки колла-
геновых или эластических волокон, между которыми залегают много-
численные фиброциты.
ТКАНИ СО СПЕЦИАЛЬНЫМИ СВОЙСТВАМИ
Ткани со специальными свойствами расположены лишь в опреде-
ленных органах и участках тела и характеризуются особым строением
и своеобразной функцией (жировая, ретикулярная, пигментная).
Жировая ткань выполняет трофическую, депонирующую, фор-
мообразующую и терморегулирующую функцию. Жировая ткань
подразделяется на два типа: белую, образованную однокапельными
жировыми клетками, и бурую, образованную многокапельными.
У человека преобладает белая жировая ткань. Большая часть ее яв-
ляется резервной, это подкожная жировая клетчатка, сальники и др.
Она запасает жир — источник энергии, который расходуется по мере
необходимости. Количество бурой жировой ткани у человека невели-
ко, она имеется, главным образом, у новорожденного ребенка и рас-
положена в области шеи, в подмышечной ямке, под кожей спины
и боковых поверхностей туловища. Бурый цвет обусловлен множест-
вом кровеносных капилляров в ткани и митохондрий в клетках. Глав-
ная ее функция — теплопродукция. Бурая жировая ткань поддер-
живает температуру тела многих животных во время спячки
и температуру новорожденных детей.
Хрящевая ткань
К соединительным тканям относятся также хрящевая и костная
ткани.
Хрящевая ткань, содержащая 70—80% воды, 10—15% органиче-
ских и 4—7% неорганических веществ, состоит из хрящевых клеток
(хондробластов и хондроцитов) и основного (хрящевого межклеточ-
ного) вещества, находящегося в состоянии геля, в котором соедини-
тельнотканные волокна, в основном коллагеновые. Хондроциты рас-
полагаются в полостях — лакунах, окруженные межклеточным
веществом (рис. 70). Различают три типа хрящевой ткани.
129
ТКАНИ СО СПЕЦИАЛЬНЫМИ СВОЙСТВАМИ
1. Гиалиновый хрящ, из ко-
торого построены суставные,
реберные, эпифизарные хрящи
и ряд хрящей гортани; гладкий,
блестящий, голубовато-белого
цвета.
2. Эластический хрящ со-
держит в хрящевом основном
веществе
многочисленные
сложно переплетающиеся эла-
стические волокна. Он менее
прозрачен, желтоватого цвета,
очень упругий. Эластический
хрящ образует голосовые от-
ростки черпаловидных хрящей
гортани, надгортанник, хрящ
ушной раковины, хрящевую часть слуховой трубы и наружного слу-
хового прохода. В отличие от гиалинового, эластический хрящ не око-
стеневает с возрастом.
3. Волокнистый хрящ, в основном хрящевом веществе которого со-
держится большое количество коллагеновых волокон, придающих
хрящу большую прочность. Из него построены фиброзные кольца
межпозвоночных дисков, суставные диски и мениски, этим хрящом
покрыты суставные поверхности в височно-нижнечелюстном и гру-
дино-ключичном суставах.
Костная ткань
Костная ткань, отличающаяся особой механической прочностью,
состоит из костных клеток, замурованных в костное основное вещест-
во, содержащее коллагеновые волокна и пропитанное солями каль-
ция. Содержание воды в кости достигает 50%. В сухом остатке кост-
ной ткани около 33% органических веществ и 67% неорганических,
в основном это кристаллы гидроксиапатита.
Рис. 70. Изогенная группа (хондрон):
1 — хондроциты,
2 — капсула,
3 — межтерриториальный матрикс
2
3
1
130
ТКАНИ
Подобно хрящу, кость состоит из клеток и межклеточного вещест-
ва. Различают костные клетки двух типов: остеобласты и остеоциты.
Остеобласты — это многоугольные кубические отростчатые молодые
клетки, богатые элементами шероховатой эндоплазматической сети,
рибосомами (рис. 71). В них хорошо развит комплекс Гольджи. Их
многочисленные отростки контактируют между собой и с отростками
остеоцитов. Остеобласты синтезируют органические компоненты
межклеточного вещества (матрикс) и выделяют их из клетки через
всю поверхность в различных направлениях, что и приводит к образо-
ванию пещер (лакун), в которых они залегают, превращаясь в остео-
циты. Органический матрикс кости импрегнируется кристаллами ги-
дроксиапатита Са10(РО4)6(ОН)2 и аморфным фосфатом кальция
Са3(PO4)2, которые поступают в костную ткань из крови. Кристаллы
гидроксиапатита окутывают коллагеновые фибриллы и аморфное ве-
щество, а также расположены внутри фибрилл.
Остеоциты — зрелые, многоотростчатые веретенообразные клетки
с крупным округлым ядром и малым количеством органелл. Остеоциты
Рис. 71 . Остеобласт:
1 — матрикс кости,
2 — микроворсинки,
3 — комплекс Гольджи,
4 — цистерны гранулярной
эндоплазматической сети,
5 — митохондрии, 6 — ядро,
7 — ядрышко, 8 — остеоид,
9 — матриксные гранулы,
10 — остеоцит,
11 — оссеиновые волокна,
пропитанные кристаллами
кальция (оссифицированные)
(по Крстичу, с изменениями)
1
2
3
4
5
6
7
8
9
10
11
131
ТКАНИ СО СПЕЦИАЛЬНЫМИ СВОЙСТВАМИ
располагаются между костными пластинками в лакунах, однако тела
клеток не соприкасаются непосредственно с кальцинированным матрик-
сом, будучи окаймленными тонким слоем (1—2 мкм) неминерализован-
ной ткани. Очень длинные (до 50 мкм) отростки остеоцитов проходят
в канальцах, причем они отделены от кальцифицированного матрикса
пространством шириной около 0,1 мкм, в котором циркулирует тканевая
жидкость, осуществляющая питание клеток. Расстояние между каждым
остеоцитом и ближайшим капилляром не превышает 0,1—0,2 мм.
В костной ткани имеется еще одна категория клеток — остеокла-
сты, которые не являются костными, а имеют моноцитарное проис-
хождение и относятся к системе макрофагов. Остеокласты — это
крупные многоядерные (5—100 ядер) клетки размерами до 190 мкм,
которые разрушают кость и хрящ.
Различают два типа костной ткани ретикулофиброзную (грубово-
локнистую) и пластинчатую. Первая имеется у зародыша человека;
у взрослого она располагается в зонах прикрепления сухожилий к ко-
стям, в швах черепа после их зарастания.
Пластинчатая кость наиболее распространена в организме. Она
образована костными пластинками толщиной от 4 до 15 мкм, которые
состоят из остеоцитов и тонковолокнистого костного матрикса. Во-
локна, образующие пластинки, лежат параллельно друг другу и ори-
ентированы в определенном направлении. При этом волокна сосед-
них пластинок разнонаправлены и перекрещиваются почти под
прямым углом, формируя сводчатые арки, что обеспечивает большую
прочность кости.
ЭТО ВАЖНО!
Арка — наиболее прочная конструкция, которая повсеместно использу-
ется не только в архитектуре, но и в живом организме.
В зависимости от расположения костных пластинок различают
плотное (компактное) и губчатое костное вещество (трабекулярная
кость) (рис. 72). В компактном веществе костные пластинки распола-
гаются в определенном порядке, образуя сложные системы — остео-
ны. Остеон — структурная единица кости. Он состоит из 5—20 ци-
линдрических пластинок, вставленных одна в другую. В центре
каждого остеона расположен центральный канал (Гаверсов), в кото-
ром проходят кровеносные сосуды (рис. 73). Диаметр остеона —
0,3—0,4 мм. Каналы остеонов сообщаются между собой с помощью
коротких поперечных каналов. Между остеонами залегают интерсти-
циальные (вставочные, промежуточные) пластинки, кнаружи от них
находятся наружные окружающие (генеральные) пластинки, кнутри
внутренние окружающие (генеральные) пластинки.
132
ТКАНИ
Губчатое костное вещество состоит из костных пластинок и пере-
кладин (трабекул), перекрещивающихся между собой и образующих
множество ячеек. Направление перекладин совпадает с кривыми сжа-
тия и растяжения, образуя конструкции сводчатых арок (рис. 74). Та-
кое расположение костных трабекул под углом друг к другу обеспечи-
вает равномерную передачу давления или тяги мышц на кость. Внутри
5
3
6
7
1
4
2
Рис. 72. Строение
трубчатой кости:
1 — надкостница,
2 — компактное вещество
кости, 3 — слой наружных
окружающих пластинок,
4 — остеоны, 5 — слой
внутренних окружающих
пластинок,
6 — костномозговая
полость, 7 — костные
перекладины губчатой
кости
(по В. Баргману)
11
22
33
Рис. 73. Строение остеона:
1 — центральный канал
(канал остеона),
2 — пластинки остеона,
3 — костная клетка (остеоцит)
133
МЫШЕЧНЫЕ ТКАНИ
костей в костномозговых полостях и ячейках губчатого вещества на-
ходится костный мозг.
МЫШЕЧНЫЕ ТКАНИ
Мышечные ткани осуществляют функцию движения, способны
сокращаться. Существуют две разновидности мышечной ткани: ис-
черченная (скелетная и сердечная) — поперечнополосатая и неисчер-
ченная (гладкая).
Поперечнополосатая скелетная мышечная ткань образо-
вана цилиндрическими волокнами длиной от 1 до 40 мм и толщиной
до 0,1 мкм (рис. 75). Под плазматической мембраной (сарколеммой)
Рис. 75. Исчерченная (поперечнополосатая, скелетная) мышечная ткань:
1 — мышечное волокно, 2 — сарколемма, 3 — миофибриллы, 4 — ядра
11
22 33
44
11
5
22
6
33
II
44
II
1
55
2
66
3
I
4
IIII
Рис. 74. Митохондрия: I — общая схема строения:
1 — наружная мембрана, 2 — внутренняя
мембрана, 3 — кристы, 4 — матрикс; II — схема
строения кристы: 5 — складка внутренней
мембраны, 6 — грибовидные тельца
(по Б. Албертсу и др.; по К. де Дюву, с изм.)
134
ТКАНИ
располагается множество эллипсоидных ядер. Примерно две трети
объема волокна занимают цилиндрические миофибриллы, между ко-
торыми залегают многочисленные митохондрии. Волокна отличают-
ся поперечной исчерченностью: темные полосы (диск А) чередуются
со светлыми (диск I) (рис. 76). Диск А разделен светлой зоной (поло-
са Н), диск I — темной линией Z (телофрагма). Миофибриллы содер-
жат сократительные элементы — миофиламенты, среди которых раз-
личают толстые (миозиновые), занимающие диск А, и тонкие
(актиновые), лежащие в диске I и прикрепляющиеся к телофрагмам,
причем концы их проникают в диск А между толстыми филаментами.
Участок миофибриллы, расположенный между двумя телофрагмами,
представляет собой саркомер — сократительную единицу. На границе
Рис. 76. Строение двух миофибрилл поперечнополосатого мышечного волокна, схема:
1 — саркомер, 2 — полоска А (диск А), 3 — полоска H, 4 — линия М (мезофрагма)
в середине диска А, 5 — полоска I (диск I), 6 — линия (телофрагма) в середине диска I,
7 — митохондрия, 8 — конечная цистерна, 9 — саркоплазматический ретикулум,
10 — поперечные трубочки (по В. Г. Елисееву и др.)
11
55
99
22
66
10
10
33
77
44
88
135
МЫШЕЧНЫЕ ТКАНИ
между дисками А и I мембрана волокна впячивается, образуя Т-тру-
бочки, которые разветвляются внутри волокна. В поперечнополоса-
тых мышечных волокнах хорошо развита незернистая цитоплазмати-
ческая сеть, которая окружает саркомеры.
Скелетные мышцы иннервируются спинномозговыми и черепны-
ми нервами. Каждое мышечное волокно иннервируется аксоном или
его ветвью. Двигательный аксон несет импульс к сокращению мыш-
цы, при этом он контактирует с сарколеммой, образуя синапсоподоб-
ное нервно-мышечное окончание. Нервный импульс передается по
Т-трубочкам, а с них на конечные цистерны саркоплазматической
сети, вызывая изменение проницаемости последних, что ведет к выхо-
ду ионов кальция в цитоплазму. Это приводит к взаимодействию
актина с миозином и мышечному сокращению. Согласно теории
X. Хэксли и Т. Хэнсона, мышечное сокращение — это результат
скольжения тонких (актиновых) филаментов относительно толстых
(миозиновых), благодаря чему длина филаментов диска А изменяет-
ся, в то время как диск I уменьшается в размерах и исчезает.
В осуществлении мышечного сокращения принимают участие не-
сколько белков: актин, миозин, тропомиозин и тропонин. Актиновые
филаменты (F-актин) образованы двумя скрученными полимерными
волокнами, каждое из которых состоит из мономеров глобулярного
белка G-актина. Вокруг F-актина обвивается молекула тропомиозина,
залегающая в его спиральных желобках. Вдоль F-актина расположе-
ны молекулы тропонина, прикрепляющиеся и к тропомиозину. Тро-
нонин состоит из субъединиц Т (связывающей тропомиозин), I (свя-
зывающей актин и ингибирующей связывание актина с миозином)
и соединенной с ними С (связывающей Са2+).
Толстые филаменты состоят из молекул миозина, представляю-
щих собой нити, имеющие две шаровидные головки. В молекуле
миозина имеется два «шарнира», первый — между гидрофобным
«стволом» и гидрофильной «шейкой», другой — между «шейкой»
и «головками». Миозиновые молекулы, соединяясь своими гидро-
фобными «стволами», образуют стержень толстого миофиламента, из
которого выступают «шейки» и «головки», формирующие шесть спи-
ральных рядов. На головке миозина имеется специальный участок,
связывающий АТФ. Два стержня соединены между собой стволами,
образуя участок, лишенный «шеек» и «головок». Каждый миозино-
вый филамент окружен шестью актиновыми (рис. 77).
В основе мышечного сокращения лежит взаимодействие между
актином и миозином. Источником движущей силы мышечного сокра-
щения является освобождение энергии в результате гидролиза АТФ,
катализируемого миозином, который является актинзависимой
АТФ-азой. Этим свойством обладают миозиновые головки только
136
ТКАНИ
при условии их активации Са2+
. Напомним, что благодаря наличию в
молекуле миозина двух «шарнирных» устройств головки могут сги-
баться, прикрепляясь к актину и подтягивая актиновые филаменты
на 10 нм. Это возможно благодаря тому, что белок α-актинин, распо-
ложенный в области линии Z, закрепляет концы тонких (актиновых)
миофиламентов.
Поперечнополосатая сердечная мышечная ткань, которая
по своему строению и функции отличается от скелетных мышц, со-
стоит из кардиомиоцитов, образующих соединяющиеся друг с другом
комплексы. По своему строению сердечная мышечная ткань похожа
на скелетную (поперечнополосатая исчерченность), однако сокраще-
ния сердечной мышцы неподконтрольны сознанию человека, она ин-
нервируется вегетативной нервной системой.
Кардиомиоциты — клетки неправильной цилиндрической формы
длиной 100—150 мкм и диаметром 10—20 мкм (рис. 78). В световом
микроскопе видны многочисленные анастомозы, ветвления пучков
кардиомиоцитов, формирующих сети. Это связано с тем, что отдель-
ные клетки соединяются между собой нерегулярно. Каждый кардио-
миоцит имеет 1—2 овальных удлиненных ядра, лежащих в центре
и окруженных миофибриллами, расположенными по периферии
строго прямолинейно. На обоих полюсах ядра видны удлиненные
зоны цитоплазмы, лишенной миофибрилл. Весьма характерны кон-
такты двух соседних кардиомиоцитов, имеющих вид извилистых
темных полосок, вставочных дисков, которые активно участвуют
в передаче возбуждения от клетки к клетке. С помощью дисков кар-
диомиоциты соединяются друг с другом. Клетки богаты митохондри-
ями. Сарколемма кардиомиоцитов толщиной около 9 нм имеет мно-
жество микропиноцитозных инвагинаций, пузырьков.
Рис. 77. Строение саркомера, схема
137
МЫШЕЧНЫЕ ТКАНИ
Строение миофибрилл аналогично таковому скелетных мышц.
Однако в отличие от последних между миофибриллами кардиомио-
цитов нет столь четких границ. По периферии клетки и между мито-
хондриями находится множество частичек гликогена и элементов
гладкого эндоплазматического ретикулума. В кардиомиоцитах имеет-
ся очень большое количество крупных митохондрий с хорошо разви-
тыми кристами, которые располагаются группами между миофибрил-
лами. На уровне Z-линий плазмалемма кардиомиоцитов также
формирует Т-трубочки, вблизи которых сосредоточены скопления
цистерн гладкого эндоплазматического ретикулума. Однако триады
выражены менее четко, чем в скелетных мышцах.
Кардиомиоциты соединены между собой вставочными дисками,
которые на продольном разрезе имеют вид ступенек. Поперечные пе-
рекладины этих ступенек имеют отростки различной длины, между
которыми находятся инвагинации. В этих участках кардиомиоциты
соединяются между собой наподобие зубчатых швов черепа, а плазма-
леммы соседних клеток соединены между собой с помощью десмосом,
лентовидных поясков или пятен сцепления, к которым с обеих сторон
Рис. 78. Кардиомиоцит:
1 — базальная мембрана,
2 — окончание миопротофибрилл
на цитолемме кардиомиоцита,
3 — вставочный диск между кардио-
миоцитами, 4 — саркоплазматическая
сеть, 5 — саркосомы (митохондрии),
6 — миопротофибриллы, 7 — диск A
(анизотропный диск),
8 — диск I (изотропный диск),
9 — саркоплазма
(по В. Г. Елисееву и др.)
1
2
3
4
5
6
9
7
8
138
ТКАНИ
прикрепляются актиновые филаменты. Поперечные участки располо-
жены на месте Z-линий. На участках вставочного диска, лежащих па-
раллельно продольной оси кардиомиоцита (вертикальные линии сту-
пенек), находятся лентовидные десмосомы (пояски сцепления, к ним,
возможно, прикрепляются актиновые филаменты) и щелевидные
контакты, не связанные с миофиламентами. Через нексусы (щелевид-
ные контакты) осуществляются передача нервного возбуждения и об-
мен ионами между клетками.
НЕРВНАЯ ТКАНЬ
Нервная ткань образует центральную нервную систему (головной
и спинной мозг) и периферическую (нервы с их концевыми прибора-
ми, нервные узлы). Нервная ткань состоит из нейронов и нейроглии.
ЭТО ВАЖНО!
Нервная ткань обеспечивает анализ и синтез сигналов (импульсов), по-
ступающих в мозг. Она устанавливает взаимосвязь организма с внеш-
ней средой и участвует в координации функции внутри организма,
обеспечивая его целостность (вместе с гуморальной системой кровью,
лимфой).
Нейрон с отходящими от него отростками является структурно-
функциональной единицей нервной системы.
ЭТО ВАЖНО!
Основная функция нейрона — это получение, переработка, проведение
и передача информации, закодированной в виде электрических или хи-
мических сигналов.
В нейроне различают его тело (перикарион), где информация обра-
батывается, и отходящие от тела отростки, проводящие информацию
иногда на дальние расстояния. Один или несколько отростков, по кото-
рым нервный импульс приносится к телу нейрона, называется ден-
дритом. Единственный отросток, по которому нервный импульс
направляется от клетки — это аксон. Нервная клетка динамически по-
ляризована, т. е. способна пропускать импульс только в одном направ-
лении, от дендрита к телу клетки, где информация обрабатывается,
и далее к аксону. В зависимости от количества отростков различают
униполярные, или одноотростчатые (они имеются в эмбриональном
периоде), биполярные, или двухотростчатые, и мультиполярные, или
многоотростчатые, нейроны. Последние преобладают.
Как правило, нейроны — одноядерные клетки; два ядра имеют неко-
торые нейроны ганглиев вегетативной нервной системы. Сферическое
139
НЕРВНАЯ ТКАНЬ
ядро диаметром около 18 мкм в большинстве нейронов расположено
центрально (рис. 79). Основными особенностями строения нейронов
является наличие многочисленных нитей (нейрофибрилл) и скоплений
хроматофильного вещества (вещество Ниссля), богатого РНК, которое
представляет собой группы параллельных цистерн зернистой цитоплаз-
матической сети и полирибосомы, располагающиеся по всей цитоплаз-
ме клетки и в дендритах (отсутствуют в аксоне). Нейрофибриллы фор-
мируют в клетке густую трехмерную сеть, они пронизывают и отростки.
Вещество Ниссля и свободные рибосомы располагаются по всей цито-
плазме клетки и в дендритах, они отсутствуют в аксоне. Нейрофибрил-
лы обеспечивают прочность перикариона и отростков и осуществляют
химическую интеграцию клетки. Макромолекулы, синтезируемые в пе-
рикарионе, направляются в самые отдаленные участки отростков.
Нейроны воспринимают, проводят и передают информацию, зако-
дированную в виде электрических и химических сигналов. Заряжен-
ные молекулы или атомы называют ионами. Натрий, калий, кальций
и магнийположительные ионы; хлор, фосфат, остатки некоторых кис-
лот (например, угольной), крупные ионы белков отрицательные. Во
внеклеточной жидкости положительные и отрицательные ионы нахо-
дятся в равных соотношениях. Внутри клеток преобладают отрица-
тельно заряженные ионы, чем обусловлен общий отрицательный за-
ряд клетки. Калий — внутриклеточный ион, его концентрация
в нервных и мышечных клетках в 20—100 раз выше, чем вне клетки,
натрий — внеклеточный ион, его внутриклеточная концентрация
1
5
99
2
6
10
10
3
7
11
11
4
8
12
Рис. 79. Строение нервной клетки:
1 — аксонодендритический синапс,
2 — аксоносоматический синапс,
3 — пресинаптические пузырьки,
4 — пресинаптическая мембрана,
5 — синаптическая щель,
6 — постсинаптическая мембрана,
7 — эндоплазматическая сеть,
8 — митохондрия, 9 — внутренний
сетчатый аппарат (комплекс Гольджи),
10 — нейрофибриллы, 11 — ядро,
12 — ядрышко
140
ТКАНИ
в клетке в 5—15 раз ниже внеклеточной. И наоборот, внутриклеточ-
ная концентрация Cl — в 20—100 раз ниже внеклеточной.
По обе стороны мембраны нервных и мышечных клеток, между
внеклеточной и внутриклеточной жидкостями существует мембран-
ный потенциал — разность потенциалов, его величина 80 мВ. Это свя-
зано с избирательной проницаемостью плазматической мембраны для
различных ионов. К
+
легко диффундирует через мембрану. В связи
с его высоким содержанием в клетке он выходит из нее, вынося поло-
жительный заряд. Возникает мембранный потенциал. Мембранный
потенциал клетки, находящейся в состоянии покоя, называется
потенциалом покоя (рис. 80).
Мембрана
в покое
Ток натрия
внутрь
Аксон
Аксон
Пресинаптические
нервные
окончания
Перехват Ранье
Полная деполяризация
мембраны
Потенциал
действия
Плазматическая
мембрана
Медленно
Медленно
Медленно
Быстро
Быстро
Быстро
Высокая
Низкая
Невозбужденное
состояние
Невозбужденное
состояние
Медленно
Медленно
Медленно
+25
0
0123
м
с
Ф, мВ
Ф(мВ)
мВ
%75
%70
(К+)
(К+)
(Na+)
Высокая
Низкая
(К+)
(Na+)
(Na+)
Na+
Na+
Na+
Na+
Cl%
Cl%
K+
++
++
++
++
++
++
+
+
+
+
++
%
%%
%%
%%
%%
%%
%%
%%
%%
%
%
+
K+
K+
(К+)
(Na+)(
К
+)
(Na+)
%70 +20
+20
%75
%75
%70
%70
Пороговый потенциал
(изменение на % 10 мВ)
Тело клетки
Ядро
Дендриты
НЕЙРОН
Возможные
постсинаптические
участки
Шванновская клетка
Ток натрия
наружу
Возвращение
в состояние
покоя
Рис. 80. Ионные токи через мембрану аксона
при прохождении потенциала действия,
сопровождающееся изменением электрического напряжения (от –70 до +40 мВ),
обусловлено восстановлением равновесия между положительными и отрицательными
ионами по обе стороны мембраны, проницаемость которой на короткое время
увеличивается (по Стернбергу и соавт., с изменениями)
141
НЕРВНАЯ ТКАНЬ
Когда нервная или мышечная клетка активизируется, в ней возни-
кает потенциал действия — быстрый сдвиг мембранного потенциала
в положительную сторону. При этом в определенном участке мембра-
ны в ответ на раздражение клетка начинает терять свой отрицательный
заряд и Na+ устремляется в клетку, в результате чего на 1/1000 сек. на
этом участке возникает деполяризация, внутри клетки генерируется
положительный заряд — потенциал действия, или нервный импульс
(см. рис. 80). Таким образом, потенциал действия — это проникновение
потока ионов Na+ через мембрану в клетку. К
+
, содержащийся в боль-
шом количестве внутри клетки и обладающий высокой проницаемо-
стью, начинает покидать клетку. Это приводит к восстановлению в ней
отрицательного заряда. Движение ионов, возникающее вблизи деполя-
ризированного участка, приводит к деполяризации следующего участ-
ка мембраны, поэтому нервный импульс распространяется по нейрону.
Нейроны, которые передают возбуждение от точки восприятия
раздражения в центральную нервную систему и далее к рабочему ор-
гану, связаны между собой с помощью множества межклеточных кон-
тактов синапсов (от греч. synapsis — связь), передающими нервный
импульс от одного нейрона к другому (рис. 81, 82).
Рис. 81. Химический синапс:
1 — митохондрия, 2 — гладкий
эндоплазматический ретикулум,
3 — нейротрубочка,
4 — синаптические пузырьки,
5 — пресинаптическая мембрана
с гексагональной сетью,
6 — синаптическая щель,
7 — постсинаптическая мембрана,
8 — зернистая эндоплазматическая сеть,
9 — нейрофиламенты
1
2
3
4
5
6
7
8
9
П
Р
Е
С
И
Н
А
П
Т
И
Ч
Е
С
К
А
Я
Ч
А
С
Т
Ь
П
О
С
Т
С
И
Н
А
П
Т
И
Ч
Е
С
К
А
Я
Ч
А
С
Т
Ь
142
ТКАНИ
ЭТО ВАЖНО!
В синапсах происходит преобразование электрических сигналов в хи-
мические и обратное — химических в электрические.
Различают синапсы аксосоматические, в которых окончания аксо-
на одного нейрона образуют контакты с телом другого, аксодендрити-
ческие аксоны вступают в контакт с дендритами, а также аксоаксо-
нальные и дендродендритические, когда контактируют одноименные
отростки. Это создает возможность для проведения возбуждения по
одной из множества цепочек нейронов благодаря наличию физиоло-
гических контактов в определенных синапсах и физиологическому
разъединению в других.
Синапсы, в которых передача осуществляется с помощью биоло-
гически активных веществ, называются химическими, а вещества,
осуществляющие передачу, нейромедиаторами (от лат. mediator —
Рис. 82. Синаптическая передача, схема:
1а. Медиатор А, молекулы которого освобождаются из пресинаптического полюса,
связываются специфическими рецепторами постсинаптической мембраны.
Молекулы X, которые по своей конфигурации не подходят к этим рецепторам,
занять их не могут и потому не вызывают каких-либо синаптических эффектов.
1б. Молекулы М (например, молекулы некоторых психотропных препаратов) сходны
по своей конфигурации с молекулами нейромедиатора А и поэтому могут связываться
с рецепторами для этого медиатора, таким образом мешая ему выполнять свои функции.
2а и 2б. Некоторые вещества, называемые нейромодуляторами, способны воздейст-
вовать на окончание аксона, облегчая или подавляя высвобождение нейромедиатора
(по Ж. Годфруа)
Нервные
импульсы
Молекулы
медиатора А
Молек улы
модулятора R
Молекулы X
Нейрон II
Нейрон I
1а
1б
2а
2б
Рецепторные
учас тки
«Подходящие»
молекулы
психотропного
препарата
«Подходящие»
молекулы
психотропного
препарата
143
НЕРВНАЯ ТКАНЬ
посредник). Роль медиаторов выполняют норадреналин, ацетилхо-
лин, серотонин, дофамин и др. Медиатор поступает в синапс по пре-
синаптическому окончанию, которое ограничено пресинаптической
мембраной (пресинаптическая часть) и воспринимается постсинапти-
ческой мембраной (постсинаптическая часть). Между мембранами
расположена синаптическая щель. В пресинаптическом окончании
множество митохондрий и пресинаптических пузырьков, содержащих
медиатор. Нервный импульс, поступающий в пресинаптическое окон-
чание, вызывает освобождение в щель медиатора. Молекулы медиато-
ров реагируют со специфическими рецепторными белками клеточной
мембраны, меняя ее проницаемость для определенных ионов, что
приводит к возникновению потенциала действия (см. рис. 81). Наря-
ду с химическими имеются электротонические синапсы, в которых
передача импульсов происходит непосредственно биоэлектрическим
путем между контактирующими клетками.
ЭТО ВАЖНО!
В каждый нейрон поступает множество нервных импульсов от множе-
ства других нервных клеток, эти импульсы объединяются (интегриру-
ются) в теле нейрона.
В нервной системе существуют два вида синапсов: возбуждающие
и тормозящие. В возбуждающих синапсах одна клетка вызывает акти-
визацию другой. При этом возбуждающий медиатор вызывает депо-
ляризацию — поток ионов Na+ устремляется в клетку. В тормозящих
синапсах одна клетка тормозит активизацию другой. Это связано
с тем, что тормозящий медиатор вызывает устремление потока отри-
цательно заряженных ионов в клетки, поэтому деполяризации не про-
исходит.
Прежде чем рассказать о строении нервных волокон, необходимо
упомянуть еще об одной разновидности клеток нервной ткани. Кроме
нейронов, в нервной системе имеются клетки нейроглии (глиоциты),
выполняющие многообразные функции: опорную, трофическую, за-
щитную и секреторную. Среди них различают две группы: макро-
глию (эпендимоциты, олигодендроциты и астроциты) и микро-
г л и ю. Клетки глии выполняют опорную, трофическую, защитную,
разграничительную функции по отношению к нейронам. Количество
клеток глии в мозге человека примерно в 10 раз больше числа нейро-
нов. В центральной нервной системе олигодендроциты образуют мие-
линовую оболочку нервных волокон, в периферической нервной сис-
теме шванновские клетки образуют миелиновую оболочку нервных
волокон.
144
ТКАНИ
Нервные волокна представляют собой отростки нервных клеток
вместе с покрывающими их оболочками. Они подразделяются на ми-
елиновые и безмиелиновые волокна (рис. 83). Безмиелиновые нервные
волокна образованы одними или несколькими отростками нервных
клеток (осевыми цилиндрами), каждый из которых погружен в тело
шванновской клетки (клетка глии), прогибая ее цитоплазматическую
мембрану так, что между мембранами осевого цилиндра и шваннов-
ской клетки остается пространство. Скорость проведения нервного
импульса по безмиелиновому волокну менее 1 м/сек.
Миелиновые нервные волокна образованы одним осевым цилин-
дром, окруженным муфтой из шванновских клеток. Миелиновый
слой представляет собой многократно спирально закрученную вокруг
осевого цилиндра шванновскую клетку. Скорость проведения им-
пульса по миелиновому волокну — 70—100 м/сек.
Рис. 83. Строение нервных волокон, схема:
А — безмиелиновые волокна: 1 — шванновская клетка,
2 — нервные волокна, 3 — цитоплазма, 4 — ядро;
Б — образование миелина: 1 — ядро, 2 — цитоплазма,
3 — аксон, 4 — ядро шванновской клетки,
5 — плазматическия мембрана шванновской клетки;
В — строение миелинового волокна: 1 — нейрофибриллы,
2 — ядро шванновской клетки, 3 — миелин,
4 — цитоплазма шванновской клетки,
5 — плазматическия мембрана шванновской клетки,
6 — перехват Ранвье (граница между двумя
шванновскими клетками), 7 — аксон
1
1
1
5
5
А
Б
В
2
2
2
6
3
3
3
7
4
4
44
Миелин
145
НЕРВНАЯ ТКАНЬ
В зависимости от функции выделяют три основных типа нейронов.
1. Чувствительные, рецепторные, или афферентные, нейроны (от
лат. afferens — приносящий). Как правило, эти клетки имеют два вида
отростков. Дендрит следует на периферию и заканчивается чувстви-
тельными окончаниями-рецепторами, которые воспринимают внеш-
нее раздражение и трансформируют его энергию в энергию нервного
импульса; второй аксон направляется в головной или спинной мозг.
В зависимости от локализации различают несколько типов рецепто-
ров: 1) экстерорецепторы, воспринимающие раздражения внешней
среды, расположены в коже, слизистых оболочках и органах чувств;
2) интерорецепторы, получающие раздражение главным образом при
изменениях химического состава внутренней среды и давления, рас-
положены в сосудах, тканях и органах; 3) проприорецепторы, зало-
женные в мышцах, сухожилиях, связках, фасциях, надкостнице, су-
ставных капсулах.
2. Эфферентные. Тела эфферентных (эффекторных, двигательных
или секреторных) нейронов (от лат. efferens — выносящий) находят-
ся в ЦНС (или в симпатических и парасимпатических узлах). Их ак-
соны идут к рабочим органам (мышцам или железам). Различают два
вида рабочих, или исполнительных, органов: анимальные поперечно-
полосатые (скелетные) мышцы и вегетативные гладкие мышцы и же-
лезы. Соответственно этому имеются нервные окончания аксонов эф-
ферентных нейронов двух типов: двигательные и секреторные.
Первые (моторные) оканчиваются на мышечных волокнах, образуя
бляшки, которые в поперечнополосатых мышцах представляют аксо-
мышечные синапсы. Нервные окончания гладкой мышечной ткани
образуют вздутия, в которых также содержатся синаптические пу-
зырьки. Секреторные окончания контактируют с железистыми клет-
ками. Аксоны двигательных нейронов разветвляются, и каждый из
них иннервирует большое количество мышечных волокон. Оконча-
ние одного двигательного нейрона и иннервируемое им поперечнопо-
лосатое мышечное волокно образуют двигательную единицу.
3. Вставочные нейроны передают возбуждение с афферентного на
эфферентный нейрон.
ЭТО ВАЖНО!
Бо
льшая часть клеток человека постоянно обновляется. Так, продолжи-
тельность жизни эритроцитов 120 дней, печеночных клеток — 480 дней,
кишечного эпителия — 3 –5 дней. Последние обновляются со скоростью
1 млн клеток в минуту. Нервные клетки и мышечные волокна не делятся
после рождения.
Нервная, мышечная ткани и железистый эпителий относятся
к возбудимым тканям, которые в ответ на воздействие раздражителя
146
ОРГАНЫ, СИСТЕМЫ И АППАРАТЫ ОРГАНОВ
переходят из состояния покоя в состояние возбуждения. При этом
возбуждение, возникающее на одном участке мышечного или нервно-
го волокна, быстро передается на соседние участки этого волокна,
а также с нервного волокна на другие через синапс или с нервного
волокна на иннервируемую ими структуру. Возбудимость — это спо-
собность клеток воспринимать изменения внешней среды и отвечать
на них реакцией возбуждения. Проводимость — способность тканей
проводить возбуждение. Мышечные ткани обладают сократимостью,
т. е . способностью отвечать сокращением на раздражение.
ОРГАНЫ, СИСТЕМЫ И АППАРАТЫ ОРГАНОВ
Ткани образуют органы. Каждый орган отличается свойственной
лишь ему формой и строением, приспособленными к выполнению
определенной функции, содержит все виды тканей, однако одна из
них является основной, «рабочей», выполняющей главную функцию
органа. Так, в печени, легких, почках, железах — это эпителиальная.
В кости основная ткань соединительная (костная), в мозге — нервная.
Соединительная ткань выполняет опорную, механическую, трофиче-
скую функции, образует соединительнотканный каркас органа, его
строму. Мышечная ткань участвует в образовании стенок кровенос-
ных, лимфатических сосудов, пищеварительной системы, воздухонос-
ных и мочевыводящих путей. Нервная ткань представлена в виде не-
рвов (и их разветвлений), иннервирующих орган, нервных узлов,
лежащих в стенках органов или возле них.
Органы анатомически и функционально объединяются в систе-
мы органов. Система — это ряд органов, имеющих общий план стро-
ения, единство происхождения и выполняющих одну большую функ-
цию (например, пищеварения, дыхания). В организме человека
выделяют следующие системы органов: скелет, соединения костей,
мышечная, пищеварительная, дыхательная, мочевыделительная, по-
ловые (мужская и женская), нервная, лимфоидная и эндокринная, сер-
дечно-сосудистая, органы чувств (сенсорные системы).
Некоторые системы объединяются по функциональному принци-
пу в аппараты: они зачастую имеют различное строение и происхо-
ждение, могут быть не связаны анатомически, но их объединяет учас-
тие в выполнении общей функции (например, опорно-двигательный,
эндокринный аппараты), либо эти органы различны по своим функ-
циональным задачам, но связаны единым происхождением (напри-
мер, мочеполовой аппарат).
В организме человека выделяют сому (от греч. soma
—
тело),
включающую кости, соединения костей, кожу, мышцы, образующие
147
ОСОБЕННОСТИ СТРОЕНИЯ, РОСТА И РАЗВИТИЯ ЧЕЛОВЕКА
вместилища, полости и внутренности, расположенные внутри по-
лостей. К соме и внутренностям подходят и разветвляются в них кро-
веносные сосуды и нервы.
Основные принципы строения тела человека — это полярность
(различное строение и функция полюсов), сегментарность (более
четко сохранилась у человека лишь в области туловища), двусторон-
няя симметрия (сходство сторон, однако, не абсолютное) и корреля-
ция (соотношение между отдельными частями).
Анатомия человека традиционно называется нормальной анато-
мией. Каждый человек неповторим и отличается от другого только
ему присущими особенностями, и вместе с тем все люди принадлежат
к одному виду и обладают одинаковым планом строения.
ЭТО ВАЖНО!
Понятие «норма» отражает здоровое, реальное состояние человека.
Нормальным следует считать такое строение (состояние) организма,
органа, при котором функция не нарушается.
Норма характеризуется наличием индивидуальной изменчивости
(варианты нормы). Приведем пример. Положение слепой кишки
у взрослого человека варьирует: она лежит на различной высоте меж-
ду двумя полярными типами в правой подвздошной ямке или под пе-
ченью. Все это варианты нормы. А вот обратное расположение вну-
тренностей, при котором слепая кишка и печень расположены слева,
а желудок, селезенка и сигмовидная ободочная кишка справа, — явля-
ется аномалией. Аномалии — это отклонение от общей закономерно-
сти, выходящие за границы нормы.
ОСОБЕННОСТИ СТРОЕНИЯ, РОСТА
И РАЗВИТИЯ ЧЕЛОВЕКА
В индивидуальном развитии человека различают два основных пе-
риода: внутриутробный, или пренатальный, и внеутробный, или пост-
натальный.
Зародыш, или эмбрион, — организм, развивающийся в теле ма-
тери на ранних стадиях развития, начинающихся оплодотворением и
завершающихся рождением. В акушерстве зародышем называют ор-
ганизм лишь в течение первых восьми недель, когда происходят ос-
новные изменения его строения (эмбриональный период). Остальная,
бо
льшая часть внутриутробного развития (с 9-й по 38—39 -ю неделю)
называется плодным, или фетальным, периодом (от лат. fetus —
плод), а сам организм — плодом.
148
ОСОБЕННОСТИ СТРОЕНИЯ, РОСТА И РАЗВИТИЯ ЧЕЛОВЕКА
Беременность длится 280 дней, или 9 месяцев (10 лунных меся-
цев). В течение первого месяца развития происходит закладка основ-
ных органов зародыша. К концу четвертой недели эмбрион имеет
длину около 6 мм. У него уже есть недоразвитая сердечная мышца,
которая способна биться. Начинает развиваться головной мозг, пери-
ферическая нервная система, глотка, лицо. На голове, очень большой
по отношению к телу, формируются глаза. Тело имеет удлиненную
форму и заканчивается маленьким остроконечным хвостиком. На
25-й день начинает биться его сердце со скоростью 140 уд./мин. В те-
чение 5—8 -й недель у зародыша развиваются органы, появляются за-
чатки вначале верхних, а затем нижних конечностей в виде кожных
складок, в которые позднее врастают закладки костей, мышц, сосудов
и нервов. К концу второго месяца эмбрион достигает 2 см в длину.
Начиная с третьего месяца развития (плодный период) происхо-
дит дальнейший рост и дифференцировка органов и тканей.
В конце второго — начале третьего лунного месяца развития пло-
да можно различить относительно большую голову, на которой видны
рот, глаза и уши, туловище и конечности, на которых начинают разви-
ваться пальцы. В конце третьего лунного месяца возникают движения
конечностей, начинают образовываться ногти и индифферентные на-
ружные половые органы, которые возможно дифференцировать на
четвертом лунном месяце. К концу первого триместра плод достигает
7,5 см в длину. На четвертом месяце тело плода формируется оконча-
тельно. По мере развития физических черт становятся хорошо видны
ногти, брови и ресницы. Кожа покрывается тонкими, как пух, воло-
сками. Появляются сосательные движения. Плод достигает длины
около 12 см и веса 100—120 г.
К пятому месяцу плод свободно движется внутри матки, прослу-
шивается его сердцебиение. На пятом месяце развивается кожа, по-
крытая пушком, начинают функционировать сальные железы, а также
развиваться подкожная основа. К концу второго триметра плод до-
стигает 30 см длины, вес его — 500—600 г. К концу седьмого месяца
мозг и нервная система полностью развиты, на теле почти исчезают
волосы, похожие на пух, под кожей начинается отложение жира, кожа
более гладкая, открываются веки, яички у мальчиков опускаются
в мошонку. В течение девятого месяца интенсивно формируется под-
кожная основа, пушок выпадает, его замещают настоящие волосы,
уплотняются хрящи носа и ушной раковины, удлиняются ногти, вы-
ходя за пределы кончиков пальцев.
В конце первого лунного месяца беременности длина зародыша
составляет около 10 мм, в конце второго — 20–30 мм, а масса тела
равна 35 г, в конце шестого длина тела составляет 30 см, а его масса —
600–700 г, девятого — длина 47 см, масса — 2000–2500 г.
149
ОСОБЕННОСТИ СТРОЕНИЯ, РОСТА И РАЗВИТИЯ ЧЕЛОВЕКА
В течение первого года жизни ребенка происходит наибольший
прирост длины тела (на 21–25 см), в периоды раннего и первого дет-
ства скорость роста быстро уменьшается, в начале периода второго
детства скорость роста стабилизируется (4,5–5,5 см в год), а в кон-
це — резко возрастает.
В подростковом возрасте годичная прибавка длины тела у мальчи-
ков составляет в среднем 5,8 см, у девочек 5–5,7 см. При этом у дево-
чек наибольшее ускорение роста имеет место в возрасте от 10 до
13 лет, а у мальчиков — в подростковом, после чего оно замедляется.
Приводим общепринятую в настоящее время периодизацию пер-
вого (внутриутробного) и второго (внеутробного) периода жизни че-
ловека (табл. 10).
Таблица 10
Периоды жизни человека
Периоды
Возраст
Эмбриональный
0—8 недель
Переходный
8—16 недель
Плодный (фетальный)
4—10 месяцев
Новорожденный
1—10 дней
Грудной возраст
10дней—1год
Раннее детство
1—3 года
Первое детство
4—7 лет
Второе детство
8—12 лет (мальчики), 8—11 лет (девочки)
Подростковый возраст
13—16 лет (мальчики), 12—15 лет (девочки)
Юношеский возраст
17—21 (юноши), 16—20 (девушки)
Зрелый возраст, I период
22—35 (мужчины), 21—35 (женщины)
Зрелый возраст, II период
36—60 (мужчины), 36—55 (женщины)
Пожилой возраст
61—74 (мужчины), 56—74 (женщины)
Старческий возраст
75—90 лет (мужчины и женщины)
Долгожители
90 лет и выше
Однако некоторые возрастные периоды слишком растянуты во
времени, в течение которого происходят весьма существенные мор-
фологические изменения в организме. Правильнее было бы плодный
период жизни человека рассматривать по месяцам, а эмбриональ-
ный — даже по неделям.
После рождения длительность некоторых возрастных периодов
также требует пересмотра. Период новорожденности, адаптации орга-
низма к новым условиям существования, лучше рассматривать
150
ОСОБЕННОСТИ СТРОЕНИЯ, РОСТА И РАЗВИТИЯ ЧЕЛОВЕКА
по дням, грудной возраст — по месяцам, детские годы — по годам, зре-
лый возраст — десятилетиям, пожилой и старческий — по пятилетиям
или даже по трехлетиям.
Большинство антропометрических показателей имеют значитель-
ные индивидуальные колебания. Площадь поверхности тела и его от-
дельных фрагментов, пропорции зависят от возраста человека
(табл. 11). При оценке площади поверхности отдельных участков тела
взрослого человека можно применять «правило девятки», согласно
которому голова и шея составляют 9% площади поверхности тела,
верхние конечности (каждая 9%) — 18%, нижние (каждая 18%) —
36%, передняя часть туловища — 18%, задняя часть — 18%, промеж-
ность 1%, ладонь и пальцы — 1%.
Таблица 11
Длина, масса тела и площадь поверхности тела в различные
возрастные периоды постнатального онтогенеза
Показатели
Возрастные периоды (пол: м — мужской, ж — женский)
Новорож-
денный
8 лет
10 лет
12 лет
14 лет
мжмжмжмж м ж
Длина тела,
см
50,8 55,0 126,3 126,4 136,3 137,3 143,3 147,8 157,0 157,3
Масса тела, кг
3,5 3,4 26,1 25,6 32,9 31,8 35,8 38,5 46,1 49,1
Площадь
поверхности
тела, см2
2200
8690
9610
10 750
12 290
Показатели
Возрастные периоды (пол: м — мужской, ж — женский)
16 лет
18 лет
20 лет
24 года
25—60 лет
мжмжмжмж м ж
Длина тела,
см
169,8 160,0 172,3 161,8 173,6 162,8 174,7 162,7 174,5 162,6
Масса тела, кг 59,1 56,8 67,6 56,8 70,2 57,1 71,8 57,5 71,7 56,7
Площадь
поверхности
тела, см2
14 300
15 850
16 800
17255 18000 16000
Развитие человека происходит в течение всей его жизни, начиная
от образования зиготы и кончая смертью. Рост же (увеличение мас-
сы) заканчивается к 20—25 годам. Рост и развитие человека имеют
целый ряд закономерностей.
1. Генетическая детерминированность. Рост и развитие зависят
от генома человека, однако взаимодействие совокупности генов друг
151
ОСОБЕННОСТИ СТРОЕНИЯ, РОСТА И РАЗВИТИЯ ЧЕЛОВЕКА
с другом и с различными факторами внешней среды может в той или
иной мере влиять на фенотип.
2. Стадийность. Рост и развитие индивидуума протекают стадий-
но. При этом последовательность стадий также детерминирована. Од-
нако временны
е границы между отдельными стадиями варьируют.
Активность процесса различная в разных стадиях, что дает основание
говорить о цикличности. В каждой стадии в организме происходят
количественные и качественные изменения, что обусловливает не-
обратимость процесса.
3. Каждый период онтогенеза человека проявляется характер-
ными анатомо-физиологическими особенностями. Длина тела и его
масса являются интегральными показателями, позволяющими судить
о физическом развитии человека.
Рост человека продолжается в течение первых 20 лет его жизни.
Как правило, увеличение длины тела у мужчин заканчивается в воз-
расте 18—20 лет, у женщин — 16—20 лет. Впоследствии до 60—65 лет
длина тела не изменяется, а после этого в связи с уплощением меж-
позвоночных дисков, изменением осанки тела и уплощением сводов
стопы длина тела уменьшается примерно на 1—1,5 мм в год.
Масса тела ребенка удваивается к 6 месяцам, утраивается к году
и увеличивается примерно в 4 раза к двум годам. Увеличение длины
и массы тела идет примерно одинаковыми темпами. Максимальная
годичная прибавка массы тела имеет место в подростковом возрасте,
при этом максимум у девочек приходится на 13-й, у мальчиков на
15-й год жизни. Масса тела увеличивается до 20—25 лет. Обычно ста-
бильная масса тела сохраняется до 40—46 лет. Следует стремиться
к тому, чтобы в течение всей жизни масса тела человека сохранялась
в пределах цифр 19—20-летнего возраста.
За последние 100—150 лет наблюдается акселерация — ускорение
морфофункционального развития и созревания всего организма де-
тей и подростков, которая в большей степени проявляется в экономи-
чески развитых странах. У мужчин акселерация выражена в большей
степени. Так, масса тела новорожденных детей возросла в среднем на
100—300 г, годовалых на 1500—2000 г, длина тела на 5 см. Длина тела
детей в периодах второго детства и в подростковом увеличилась на
10—15 см, а взрослых мужчин — на 6—8 см. Сократился период уве-
личения длины тела человека в конце прошлого века — рост продол-
жался до 23—26 лет, в настоящее время у мужчин до 18—19, у женщин
до 16—17 лет. Ускорились прорезывание молочных и постоянных
зубов, психическое развитие, половое созревание. Средний возраст на-
чала менструаций у девочек уменьшился с 16,5 года в начале XIX века
до 12—13 лет в настоящее время, а наступление менопаузы — с 43—
152
ОСОБЕННОСТИ СТРОЕНИЯ, РОСТА И РАЗВИТИЯ ЧЕЛОВЕКА
45 лет в начале XX века до 48—50 лет. Комплекс изменений у взросло-
го человека называют «секулярным трендом» (вековая традиция).
Физическое развитие ребенка происходит не постепенно, а скач-
кообразно. Периоды роста не соответствуют описанным периодам
жизни человека (табл. 12).
У новорожденного голова округлая, большая (1/4 всей длины
тела, у взрослого 1/8), грудь и шея короткие, живот длинный, ноги
короткие, руки длинные. Окружность головы на 1—2 см больше
окружности груди, мозговой отдел черепа относительно больше лице-
вого. Форма грудной клетки бочкообразная. Позвоночник лишен из-
гибов, лишь незначительно выражен крестцовый мыс. Таз весьма под-
вижен, и кости, составляющие тазовую кость, не сращены между
собой. Внутренние органы относительно крупнее, чем у взрослого
человека. Так, масса печени новорожденного ребенка составляет
1/20 массы тела, в то время как у взрослого 1/50; длина кишечника
в 2 раза больше длины тела, у взрослого — в 4—4,5 раза; масса мозга
составляет 13—14% массы тела, у взрослого — лишь около 2%. Велики
надпочечники и вилочковая железа.
Таблица 12
Периоды роста человека
(по Г. Фанкони и А. Вальгреву, с изменениями)
Период
Возраст, годы
Характеристика
Грудной
1-й
Длина головы
в 4 раза меньше длины тела
Первое округление
(первый рост в ширину)
2—4
Длина головы
в 5 раз меньше длины тела
Первое вытягивание (в длину)
5—7
Длина головы
в 6 раз меньше длины тела
Второе округление
(второй рост в ширину)
8—10
Длина головы
в 6,5 раз меньше длины тела
Второе вытягивание (в длину)
11—15
Длина головы
в 7 раз меньше длины тела
Созревание
16—20
Длина головы
в 8 раз меньше длины тела
Зрелый возраст
20—24
То же
В грудном возрасте тело ребенка наиболее быстро растет. При-
мерно с шести месяцев начинается прорезывание молочных зубов. За
первый год жизни ряд органов и систем достигает величины взросло-
го (глаз, внутреннее ухо, центральная нервная система). В течение
первых лет жизни и в подростковом возрасте (мальчики — 13—16 лет,
девочки — 12 —15), когда происходит половое созревание, быстро рас-
153
ОСОБЕННОСТИ СТРОЕНИЯ, РОСТА И РАЗВИТИЯ ЧЕЛОВЕКА
тут и развиваются опорно-двигательный аппарат, пищеварительная,
дыхательная системы, мочеполовой аппарат.
В периоде раннего детства прорезываются все молочные зубы и
происходит первое «округление», т. е. увеличение массы тела опере-
жает рост тела в длину, быстро прогрессирует психическое развитие
ребенка и — что самое главное — речь, память. Ребенок начинает ори-
ентироваться в пространстве. В течение второго-третьего года разви-
вается воля («первый возраст упрямства»). В периоде первого детст-
ва рост в длину превалирует над увеличением массы тела. В конце
периода начинается прорезывание постоянных зубов. В связи с быст-
рым развитием мозга, масса которого к концу периода достигает уже
1100—1200 г, быстро развиваются умственные способности, казуаль-
ное мышление, длительно сохраняется способность узнавания, ориен-
тация во времени, в днях недели. В раннем и в первом детстве поло-
вые отличия (кроме первичных половых признаков) почти не
выражены.
В периоде второго детства вновь преобладает рост в ширину, одна-
ко в это время начинается половое созревание, более раннее у девочек,
что связано с усилением секреции женских половых гормонов, у них
в 8—9 лет расширяется таз и округляются бедра, увеличивается секре-
ция сальных желез и оволосение лобка. В том же возрасте (10—11 лет)
у мальчиков начинается рост гортани, яичек и полового члена, который
к 12 годам увеличивается на 0,5—0,7 см. К концу периода усиливается
рост тела в длину, темпы которого больше у девочек. В 10 лет происхо-
дит первый перекрест — длина и масса тела девочек превышает тако-
вую мальчиков. Прогрессирует психическое развитие детей. Развивает-
ся ориентация в отношении месяцев года и календарных лет.
В подростковом периоде быстро растут и развиваются половые
органы, развиваются вторичные половые признаки. У девочек увели-
чивается количество волос на коже половой области, появляются во-
лосы в подмышечных впадинах, увеличиваются размеры половых ор-
ганов, молочных желез; щелочная реакция влагалищного секрета
становится кислой, появляются менструации, меняется форма таза.
У мальчиков быстро увеличиваются яички и половой член, вначале
оволосение лобка развивается по женскому типу, набухают грудные
железы, к концу периода (15—16 лет) начинается рост волос на теле,
лице, в подмышечных впадинах, на лобке по мужскому типу, пигмен-
тируется кожа мошонки, еще больше увеличиваются половые органы,
возникают первые эякуляции. В подростковом возрасте рост тела
в длину превалирует над ростом в ширину, в 13—14 лет происходит
второй перекрест кривых роста мальчиков и девочек, после чего маль-
чики растут быстрее, чем девочки. В подростковом возрасте развива-
ется механическая и словесно-логическая память.
154
ОСОБЕННОСТИ СТРОЕНИЯ, РОСТА И РАЗВИТИЯ ЧЕЛОВЕКА
Юношеский возраст совпадает с периодом созревания. В юноше-
ском возрасте рост и развитие организма в основном завершаются,
и практически достигают морфофункциональной зрелости все аппа-
раты и системы органов.
Строение тела в зрелом возрасте изменяется мало, а в пожилом
и старческом прослеживаются характерные для этих возрастов пере-
стройки, которые изучает специальная наука геронтология (от греч.
geron — старик). Временны
е границы старения варьируют в широких
пределах у различных индивидуумов. В старческом возрасте проис-
ходит снижение адаптационных возможностей организма, изменяют-
ся морфофункциональные показатели всех аппаратов и систем орга-
нов, в этом важнейшая роль принадлежит лимфоидной, нервной
и кровеносной системам.
ВАЖНО!
Старение — генетически детерминированный процесс, но активный
образ жизни, регулярные занятия физической культурой замедляют
процесс старения, однако это возможно в пределах, обусловленных на-
следственными факторами.
Уже при первом взгляде на человека видны особенности его инди-
видуального строения. В анатомии имеется понятие о типах телосло-
жения. Телосложение определяется генетическими наследственными
факторами, влиянием внешней среды, социальными условиями. Вы-
деляют три типа телосложения человека: мезоморфный, брахиморф-
ный и долихоморфный. В табл. 13 приведены пропорции тела у лю-
дей разных типов телосложения.
Таблица 13
Характеристика пропорций тела человека
(по П. Н . Башкирову)
Типы
Относительные размеры частей тела к длине тела, %
Длина
Ширина
туловища
ноги
руки
плеч
таза
Долихоморфный
29,5
55,0
46,5
21,5
16,0
Мезоморфный
31,0
53,0
44,5
23,0
16,5
Брахиморфный
33,5
51,0
42,5
24,5
17,5
К мезоморфному (от греч. mesos — средний, morphe — вид, фор-
ма) типу телосложения (нормостеники) были отнесены те люди, чьи
анатомические особенности приближаются к усредненным параме-
трам нормы (с учетом возраста, пола и т. д .) Лица брахиморфного
155
БИОЛОГИЧЕСКАЯ И ПСИХОСОЦИАЛЬНАЯ СУЩНОСТЬ ЧЕЛОВЕКА
телосложения (от греч. brachys — короткий), или гиперстеники, отли-
чаются преобладанием поперечных размеров, упитанностью, имеют
не очень высокий рост. Сердце относительно больших размеров,
расположено поперечно благодаря высоко стоящей диафрагме. Это
приводит к укорочению легких, петли тонкой кишки расположены
преимущественно горизонтально. Люди долихоморфного типа тело-
сложения (от греч. dolichos — длинный) (астеники) отличаются строй-
ностью, легкостью, преобладанием продольных размеров над попереч-
ными, относительно более длинными конечностями, слабым развитием
мышц и жира, узкими костями. Их внутренности опущены, диафрагма
ниже, легкие длиннее, а сердце расположено почти вертикально.
Половые признаки отличают мужчину от женщины. Они делятся
на первичные (половые органы) и вторичные (например, развитие во-
лос на лобке, развитие молочных желез, изменение голоса и др.) . Об
этом подробно — в разделе «Половые системы».
БИОЛОГИЧЕСКАЯ И ПСИХОСОЦИАЛЬНАЯ
СУЩНОСТЬ ЧЕЛОВЕКА
Человек разумный (Homo sapiens sapiens) — уникальное существо.
Помимо анатомического строения, человека отличают от всех других
животных мышление, членораздельная речь, сознание, самосознание
(понимание своей индивидуальности), нравственность, способность к
творчеству, умозаключениям и предвидению, особая, присущая толь-
ко ему (!) человеческая сексуальность и свобода морального выбора.
Человек — микрокосмос, в нем заключена суть всех вещей. «Чело-
век — средоточение земли и неба» — утверждает конфуцианство (Ли
Цзи. Книга ритуалов, 7.3 .1 —7). Об этом же говорит и Талмуд: «Все,
что Всевышний создал в мире, Он создал и в человеке» (Авот, 31).
Одно из важнейших свойств человеческой личности — ответст-
венность и свобода выбора. Человек волен идти тем путем, по которо-
му он хочет идти. «Жизнь и смерть предложил Я тебе, благословение
и проклятие. Избери же жизнь, дабы жить тебе и потомству твое-
му» — сказано в Библии (Второзак., 30:19). Жизнь — это процесс ро-
ста и развития — физического и духовного. Лишь человеку присущи
самовоспитание и духовное совершенствование. Об этом прекрасно
говорит буддизм: «Постепенно, мало-помалу, время от времени, му-
дрец должен стряхивать с себя грязь, как серебряных дел мастер —
с серебра» (Дхаммапада, 23а).
Кто я? Зачем я здесь? Что я? Откуда я? Куда я иду? Вот основные
вопросы, волнующие каждого человека и все человечество.
156
БИОЛОГИЧЕСКАЯ И ПСИХОСОЦИАЛЬНАЯ СУЩНОСТЬ ЧЕЛОВЕКА
Кто я? Homo sapiens sapiens, человек разумный разумный. Обрати-
те внимание — слово «разумный» употребляется дважды в названии
человека. Великий поэт Фирдоуси писал о человеке:
Умом одаренный и мыслью богат,
Вместилище духа и разума он,
И мир бессловесных ему подчинен.
Зачем я здесь? Миссия человека — несение добра и исправление мира.
Поиски и обретение счастья и радости — пожалуй, основная цель каждо-
го человека. Это подчеркивают многие мировые религии. В Библии:
«Святой Дух обретает только в том, у кого сердце наполнено радостью»
(Иерусалимский талмуд, Сукка 5.1), в синтоизме: «Я сотворил людей,
поскольку желал видеть жизнь радостную» (Тереке, Офидэсаки, 14:25).
Что я? Сущность человека выражена в его нравственности и под-
линности. Будучи существом биологическим, человек занимает осо-
бое положение благодаря разуму, интеллекту и свободе выбора. Все
это можно объединить термином «дух». Именно дух — главное свой-
ство человеческой личности. Дух включает многочисленные сложные
и разнообразные эмоциональные и волевые качества (любовь, добро-
та, самопожертвование, раскаяние, благодарность и т. д .) .
Одна из важнейших особенностей человека — это его мораль (от
фр. morale — нравственность). Нравственность — совокупность норм
и принципов поведения людей по отношению к обществу и другим
людям. Нравственность является важной составляющей душевного
здоровья, душевного благополучия. Мораль связана со свободой вы-
бора между добром и злом в каждой конкретной жизненной ситуа-
ции. Чтобы сделать этот выбор, необходимо знать универсальную мо-
ральную систему и неукоснительно следовать ей. И здесь возникает,
пожалуй, главный, концептуальный для каждого человека и для всего
человечества вопрос: а существует ли такая система и является ли она
объективной? Существует ли абсолютная мораль?
Несмотря на культурные, этнические, образовательные, социаль-
ные и другие различия, у всех людей есть нечто общее (здесь я не имею
в виду анатомо-физиологические особенности вида Homo sapiens).
Следует обратить внимание на тот факт, что врачи лечат, а психоана-
литики анализируют любого человека, независимо от его религиозных
убеждений, национальности или социального положения.
Существуют объективные законы морали, столь же незыблемые,
как законы физики, химии, как обращение Земли вокруг Солнца или
закон сохранения и превращения энергии. Они существуют изначаль-
но, сколько существует человек, так как объективно отражают не би-
ологическую, а социальную сущность человека — личности, индиви-
дуума. И именно эти законы объединяют всех нас, людей, живущих
157
ПАССИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
на планете Земля в единую общность — человечество. Эти законы со-
зданы не людьми. Но только человек свободен в выборе, следовать
или не следовать этим законам. На них основаны взаимоотношения
людей, народов, государств, международное право.
В основе современной западной цивилизации лежит этический
монотеизм. Универсальная объективная моральная система — это де-
сять заповедей и семь законов сыновей Ноя, данных в Библии.
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
ПАССИВНАЯ ЧАСТЬ
ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
Одной из важнейших функций организма человека является пере-
движение в пространстве. Ее выполняет опорно-двигательный аппа-
рат, состоящий из двух частей: пассивной и активной. К первой отно-
сятся соединяющиеся между собой кости, ко второй — приводящие
их в движение мышцы.
Скелет (от греч. skeleton — высохший, высушенный) выполняет
множество функций: опорную, защитную, локомоторную, формоо-
бразующую, преодоление силы тяжести. Скелет всегда вызывал осо-
бый интерес ученых, философов, художников и поэтов. В «Послании
к Дельвигу» А. Пушкин писал:
.. . ему предмет
Необходимый был... скелет,
Предмет философам любезный,
Предмет приятный и полезный
Для глаз и сердца, слова нет...
Форма тела человека обусловлена скелетом, имеющим билате-
ральную симметрию и сегментарное строение (рис. 84). Общая масса
скелета составляет от 1/7 до 1/5 массы тела человека. В его состав
входит более 200 костей. Однако эта цифра весьма относительна.
Многочисленны индивидуальные вариации числа костей, у детей
в силу неполного окостенения костных фрагментов их больше. В лю-
бом учебнике можно прочитать о количестве костей следующее: по-
звоночный столб (26 костей), череп (29 костей), грудная клетка
(25 костей), кости верхних (64) и нижних (62) конечностей.
Кости скелета являются рычагами, приводимыми в движение
мышцами. В результате этого части тела изменяют положение по от-
ношению друг к другу и передвигают его в пространстве. К костям
прикрепляются связки, мышцы, сухожилия, фасции. Скелет образует
вместилище для органов, защищая их от внешних воздействий: в по-
158
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
лости черепа расположен го-
ловной мозг, в позвоночном ка-
нале — спинной, в грудной
клетке — сердце и крупные со-
суды, легкие, пищевод и др.,
в полости таза — мочеполовые
органы. Кости участвуют в ми-
неральном обмене, они являют-
ся депо кальция, фосфора и т. д .
Живая кость содержит витами-
ныA,D,Сидр.
Скелет образован разновид-
ностями соединительной тка-
ни — костной и хрящевой, кото-
рые состоят из клеток и плотного межклеточного вещества. Кость
и хрящ тесно связаны между собой общностью строения, происхож-
дения и функции. У зародыша человека и других позвоночных
животных скелет составляет около 50% массы всего тела. Однако
постепенно хрящ заменяется костью, и у взрослого человека масса хря-
ща составляет около 2% массы тела. Это суставные хрящи, межпозво-
ночные диски, хрящи носа, уха, гортани, трахеи, бронхов и ребер.
Хрящ образован хрящевой тканью. Хрящи выполняют следующие
функции: покрывают сочленовные поверхности, обладающие благода-
ря этому высокой устойчивостью к износу; суставные хрящи и межпоз-
воночные диски, являющиеся объектами приложения сил сжатия
и растяжения, осуществляют их передачу и амортизацию; хрящи воз-
духоносных путей и наружного уха формируют стенки полостей; к дру-
Рис. 84. Скелет человека, вид спереди:
1 — череп, 2 — позвоночный столб,
3 — ключица, 4 — ребро, 5 — грудина,
6 — плечевая кость, 7 — лучевая кость,
8 — локтевая кость, 9 — кости запястья,
10 — пястные кости, 11 — фаланги
пальцев кисти, 12 — подвздошная кость,
13 — крестец, 14 — лобковая кость,
15 — седалищная кость, 16 — бедренная
кость, 17 — надколенник,
18 — большеберцовая кость,
19 — малоберцовая кость,
20 — кости предплюсны,
21 — плюсневые кости,
22 — фаланги пальцев стоп
11
55
99
13
13
17
17
21
21
22
66
10
10
14
14
18
18
22
22
33
77
11
11
15
15
19
19
44
88
12
12
16
16
20
20
159
ПАССИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
гим хрящам прикрепляются мышцы, связки, сухожилия. Хрящи лише-
ны кровеносных сосудов, их питание осуществляется за счет диффузии
из окружающих тканей, снаружи их покрывает надхрящница.
Кости образованы костной тканью.
ВНИМАНИЕ!
Сопротивление свежей кости на излом такое же, как меди, и в девять
раз больше, чем свинца. Кость выдерживает сжатие 10 кг/мм2 (анало-
гично чугуну). А предел прочности ребер на излом — 110 кг/см2.
Кость как орган, кроме сочленовных поверхностей, покрыта снару-
жи надкостницей, которая прочно сращена с костью. Наружный слой
надкостницы — волокнистый, внутренний — остеогенный (костеобра-
зующий), прилежит непосредственно к костной ткани. В нем располо-
жены покоящиеся остеогенные клетки, за счет которых происходит
развитие, рост в толщину и восстановление костей после повреждения.
Кости отличаются друг от друга, при этом их форма и выполняе-
мая функция взаимосвязаны и взаимообусловлены. Различают кости
трубчатые (например, бедренная, плечевая, пястные, плюсневые
и др.); губчатые (например, тела позвонков, кости запястья и др.),
плоские (кости крыши черепа, грудина), смешанные (например, по-
звонки, кости основания черепа), воздухоносные (например, лобная,
клиновидная, верхняя челюсть) (рис. 85, табл. 14).
Рис. 85. Различные виды костей:
I — воздухоносная кость (решетчатая
кость), II — длинная (трубчатая) кость,
III — плоская кость, IV — губчатые
(короткие) кости, V — смешанная кость
ичные виды костей:
ная кость (решетчатая
нная (трубчатая) кость,
кость, IV — губчатые
и, V — смешанная кость
I
II
III
IV
V
160
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
У трубчатой кости различают ее удлиненную среднюю часть —
тело кости, или диафиз, обычно цилиндрической или близкой к трех-
гранной формы, и утолщенные концы — эпифизы (рис. 86). На них
располагаются суставные поверхности, покрытые суставным хрящом,
служащие для соединения с соседними костями. Участок кости, рас-
положенный между диафизом и эпифизом, называется метафизом.
В детском и юношеском возрасте рост костей в длину происходит за
счет гиалинового эпифизарного (метаэпифизарного) хряща, который
находится между диафизом и эпифизом трубчатой кости.
Рис. 86. Макрофаг:
1 — ядро, 2 — фаголизосомы (вторичные лизосомы), 3 — псевдоподии,
4 — митохондрия, 5 — остаточное тельце,
6 — цистерны гранулярной эндоплазматической сети (по Крстичу, с изм.)
11
22
33
44
55
66
161
ПАССИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
Таблица 14
Классификация костей
Вид костей
Части костей
Трубчатая (длинная, короткая)
Тело (диафиз), концы (эпифизы), метафизы,
отростки, выступы-бугры (апофизы)
Короткая (губчатая)
—
Плоская
Края, углы
Смешанная (ненормальная)
—
Воздухоносная
Воздухоносная полость
Кровоснабжение метаэпифизарного хряща осуществляется сосу-
дами эпифиза и диафиза. Кальцификация метаэпифизарного хряща
идет в направлении продольной оси кости по ходу коллагеновых во-
локон, расположенных между колонками. В этом хряще имеется зона
активно пролиферирующих клеток, расположенных в виде столбиков
или колонок.
Среди трубчатых костей выделяются длинные трубчатые кости
(плечевая, бедренная, кости предплечья и голени) и короткие (кости
пясти, плюсны, фаланги пальцев). Диафизы построены из компактной,
эпифизы — из губчатой кости, покрытой тонким слоем компактной.
Губчатые кости, так же как и эпифизы трубчатых костей, состоят
из губчатого вещества, покрытого тонким слоем компактного вещест-
ва. К этим костям также следует отнести кости, развивающиеся
в сухожилиях, — сесамовидные (гороховидная, надколенник). Губча-
тые кости имеют форму неправильного куба или многогранника с за-
кругленными углами и гранями. Такие
кости располагаются в местах, где боль-
шая нагрузка сочетается с большой под-
вижностью (рис. 87).
Плоские кости участвуют в образова-
нии полостей, поясов конечностей, вы-
полняют функцию защиты (кости крыши
черепа, грудина). К их поверхности при-
крепляются мышцы.
Рис. 87. Взаимоотношение компактного (1)
и губчатого (2) вещества в проксимальном
и дистальном эпифизах бедренной кости
2
1
162
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Смешанные кости имеют сложную форму. Они состоят из несколь-
ких частей, имеющих различное строение, очертание и происхожде-
ние, например позвонки, кости основания черепа.
Воздухоносные кости имеют в своем теле полость, выстланную
слизистой оболочкой и заполненную воздухом, например некоторые
кости черепа — лобная, клиновидная, решетчатая, верхняя челюсть.
Внутри костей в костномозговых полостях и ячейках губчатого ве-
щества, выстланных эндостом (слоем плоских остеогенных клеток,
лежащих на тонкой соединительнотканной пластинке), находится
костный мозг. В период внутриутробного развития, а также у новоро-
жденных во всех костных полостях находится красный костный мозг,
он выполняет кроветворную и защитную функцию. У взрослого чело-
века красный костный мозг содержится только в ячейках губчатого
вещества плоских костей (грудина, крылья подвздошных костей),
в губчатых костях и эпифизах трубчатых костей. В диафизах, т. е .
в костномозговых полостях, находится желтый костный мозг.
Кость живого человека — динамическая структура, в которой про-
исходят постоянный обмен веществ, анаболические и катаболические
процессы, разрушение старых и создание новых костных трабекул
и остеонов.
В зависимости от характера выполняемой работы меняются фор-
ма, ширина и длина костей, толщина компактного слоя, размеры кост-
номозговой полости и т. д . Весьма существенна формообразователь-
ная роль трудовых процессов, физкультуры и спорта. Все это
подтверждает правильность положения П. Ф. Лесгафта о том, что
рост и прочность костей определяются интенсивностью деятельнос-
ти прикрепляющихся к ним мышц.
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
Скелет человека состоит из позвоночного столба, ребер с груди-
ной, скелета конечностей и черепа (см. рис. 84). Скелет человека —
уникальное творение! Он существенно отличается от скелета живот-
ных. В первую очередь это относится к его пропорциям; черепу,
вмещающему головной мозг и органы чувств; свободным верхним ко-
нечностям, осуществляющим трудовые процессы; нижним конечно-
стям, служащим опорой при прямохождении. Скелет человека обла-
дает рядом характерных особенностей, наиболее важными из которых
являются вертикально расположенный позвоночный столб с изгиба-
ми; уплощенная широкая грудная клетка; череп, в котором преобла-
дает мозговой отдел, с округлой выпуклой крышей, возвышающейся
над лицевым отделом.
163
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
Соединения костей. Кости, соеди-
няясь между собой, образуют подвиж-
ные соединения или прочные непо-
движные конструкции. Все соединения
костей делятся на три большие группы:
непрерывные; полусуставы, или сим-
физы, и прерывные, или синовиальные
(суставы).
В непрерывных соединениях кости
связаны между собой с помощью раз-
личных видов соединительной ткани
(собственно соединительной ткани,
хряща, кости), в которых отсутствует
щель или полость между костями. Не-
прерывные соединения весьма прочны,
но неподвижны. К непрерывным сое-
динениям относятся связки, мембраны,
швы (например, черепа), соединения
диафизов костей с их эпифизами, которые с возрастом окостеневают,
превращаясь в костные (рис. 88).
Симфизы (от греч. symphisis — срастание) — это полуподвижные
хрящевые соединения. В толще хряща имеется небольшая щелевид-
ная полость, например межпозвоночные симфизы, лобковый и сим-
физ рукоятки грудины (рис. 89).
Суставы представляют собой прерывные соединения, у которых
между соединяющимися костями всегда имеется суставная щель
(рис. 90). Каждый сустав имеет сустав-
ные поверхности костей, покрытые,
как правило, гиалиновым суставным
хрящом, суставную капсулу и узкую
суставную полость, заполненную си-
новиальной жидкостью. Толщина су-
ставного хряща колеблется в пределах
от 0,2 до 6,0 мм и находится в прямой
зависимости от функциональной на-
грузки, испытываемой суставом. Чем
больше нагрузка, тем толще хрящ и
тем меньше он снашивается. Сустав-
ной хрящ лишен кровеносных сосудов
и надхрящницы. Основную роль в пи-
тании суставного хряща играет сино-
виальная жидкость. Наряду с этим его
питание осуществляется за счет диф-
Рис. 88. Швы черепа (вид сзади):
1 — теменно-сосцевидный шов,
2 — ламбдовидный шов,
3 — сагиттальный шов,
4 — шовные кости
Рис. 89. Симфиз:
1 — надкостница, 2 — кость,
3 — межлобковый диск,
4 — щель в межлобковом диске
4
3
2
1
4
1
2
3
164
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
фузии из капилляров подлежа-
щей костной пластинки. Сустав-
ной хрящ защищает суставные
концы кости от механических
воздействий, уменьшая давление
и равномерно распределяя его по
поверхности. Деформация хря-
ща, возникающая при движениях
в суставе, обратима.
Суставная капсула, прикре-
пляющаяся вблизи краев сустав-
ных поверхностей сочленяю-
щихся костей или отступая на
некоторое расстояние от них,
прочно срастается с надкостни-
цей, образуя замкнутую сустав-
ную полость. Капсула состоит из двух слоев. Наружный слой пред-
ставлен толстой прочной фиброзной мембраной. Внутренний слой
суставной капсулы образован тонкой гладкой блестящей синовиаль-
ной мембраной, которая выстилает изнутри фиброзную мембрану,
сумки и продолжается на поверхности кости, не покрытые суставным
хрящом. Синовиальная мембрана имеет множество небольших выро-
стов, обращенных в полость сустава, синовиальные ворсинки, очень
богатые кровеносными сосудами. Через ворсинки осуществляется
ультрафильтрация из кровеносного русла в полость сустава, в резуль-
тате чего образуется синовиальная жидкость и происходит всасыва-
ние веществ из нее.
В норме у живого человека суставная полость представляет собой
узкую щель, расположенную между покрытыми хрящом суставными
поверхностями и ограниченную синовиальной мембраной. Форма су-
ставной полости зависит от форм сочленяющихся поверхностей, на-
личия или отсутствия внутри сустава вспомогательных образований
(синовиальные складки, суставной диск или мениск) либо внутри-
капсульных связок.
Суставные поверхности редко полностью соответствуют друг дру-
гу по форме. Для достижения конгруэнтности (от лат. congruens —
Рис. 90. Схема строения сустава:
1 — надкостница, 2 — кость,
3 — суставная капсула,
4 — суставной хрящ,
5 — суставная полость
1
2
3
4
5
165
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
согласный между собой, соответствующий) в суставах имеется ряд
вспомогательных образований: хрящевых дисков, менисков, губ. Так,
в височно-нижнечелюстном суставе имеется хрящевой диск. Мени-
ски — это несплошные хрящевые или соединительнотканные пла-
стинки полулунной формы, расположенные между суставными
поверхностями. В коленном суставе имеются полукольцевые меди-
альный и латеральный мениски, которые расположены между сустав-
ными поверхностями бедренной и большеберцовой костей.
Движения в суставах совершаются вокруг трех осей: вокруг фрон-
тальной — сгибание и разгибание, при которых угол между сочленяю-
щимися костями уменьшается или увеличивается; вокруг сагитталь-
ной — приведение, при котором одна из сочленяющихся костей
приближается в срединной плоскости, и отведение, при котором кость
удаляется от нее; при вращении кость двигается вокруг своей про-
дольной оси. Круговое движение является сложным, благодаря по-
следовательному движению вокруг всех осей свободный конец дви-
жущейся кости (конечности, туловище, голова) описывает окружность.
Форма сочленяющихся поверхностей обусловливает количество осей,
вокруг которых может совершаться движение. В зависимости от этого
суставы делятся на одно-, двух- и многоосные.
Для удобства форму суставной поверхности сравнивают с отрез-
ком тела вращения. При этом каждая форма сустава содержит опре-
деленное число осей движения (рис. 91). Так, цилиндрический и блоко-
видный суставы являются одноосными. При вращении прямой линии
вокруг параллельной ей прямой оси возникает цилиндрическое тело
вращения. Цилиндрические суставы — это срединный атлантоосевой,
проксимальный лучелоктевой. Блок представляет собой цилиндр
с бороздой или гребнем, расположенными перпендикулярно оси ци-
линдра, на другой суставной поверхности имеется соответствующее
углубление или выступ. Примерами блоковидных суставов являются
межфаланговые суставы кисти. Разновидностью блоковидных суста-
вов является винтообразный сустав. Отличие винта от блока в том,
что борозда расположена не перпендикулярно оси, а по спирали. При-
мером винтообразного сустава может служить плечелоктевой сустав.
Эллипсовидный, мыщелковый и седловидный суставы являются
двухосными (см. рис. 91). При вращении половины эллипса вокруг
его диаметра образуется тело вращения — эллипс. Лучезапястный
сустав является эллипсовидным. Мыщелковый сустав по форме бли-
зок к блоковидному и эллипсовидному, его суставная головка — по-
добие эллипса, однако в отличие от первого суставная поверхность
располагается на мыщелке. Например, коленный и атлантозатылоч-
ный суставы — мыщелковые (первый комплексный, второй — комби-
нированный).
166
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Суставные поверхности седловидного сустава по форме напоми-
нают два седла с пересекающимися под прямым углом осями. Седло-
видный запястно-пястный сустав большого пальца характерен только
для человека и обусловливает противопоставление большого пальца
кисти остальным.
ВНИМАНИЕ!
Преобразование сустава в типично седловидный связано с трудовой де-
ятельностью.
Шаровидный и плоский суставы — многоосные. При вращении по-
ловины круга вокруг его диаметра образуется шар. Кроме движения
Б
В
Г
А
Рис. 91. Различные виды
суставных поверхностей, схема:
А — блоковидный,
Б — эллипсоидный,
В — седловидный,
Г — шаровидный
167
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
вокруг трех осей в этих суставах совершается еще и круговое движе-
ние (например, плечевой и тазобедренный суставы). Последний счи-
тают чашеобразным благодаря значительной глубине суставной ямки.
Плоские суставы также многоосные. Движения в них хотя и могут
производиться вокруг трех осей, отличаются малым объемом. Объем
движения в любом суставе зависит от его строения, разности угловых
размеров суставных поверхностей, а в плоских суставах величина
дуги движения незначительна, К плоским относятся, например, меж-
запястные, предплюсне-плюсневые суставы.
Простые суставы имеют две суставные поверхности, сложные —
более двух (например, локтевой сустав). Комбинированные суставы
представляют собой два анатомически изолированных сустава, кото-
рые функционируют совместно (например, височно-нижнечелюстные
суставы). В комплексных суставах между сочленяющимися суставны-
ми концами имеются диски или мениски (например, коленный сустав).
Скелет туловища
Скелет туловища образован позвоночным столбом и грудной
клеткой (см. рис. 84).
ВАЖНО!
Наличие позвоночного столба служит важнейшим отличительным при-
знаком всех представителей позвоночных животных.
Позвоночник связывает части тела, выполняет защитную и опор-
ную функции для спинного мозга и выходящих из позвоночного ка-
нала корешков спинномозговых нервов. Верхний конец позвоночника
поддерживает голову. Кости свободных конечностей прикрепляются
к скелету туловища посредством поясов. Позвоночник передает тя-
жесть тела поясу нижних конечностей.
Положение и форма позвоночника человека обусловливает воз-
можность прямохождения. Позвоночный столб выдерживает значи-
тельную часть тяжести человеческого тела.
ВНИМАНИЕ!
В строении позвонков четко выражена одна из самых важных законо-
мерностей строения костной системы: там, где при незначительном
объеме необходимо обеспечить прочность конструкции, сохраняя
ее легкость, имеется губчатое вещество, строго определенное
расположение перекладин которого, согласно линиям сил сжатия
и растяжения, обеспечивает прочность.
168
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Кроме того, прочность позвоночного столба как целого зависит
и от мощного связочного аппарата позвоночника. Будучи весьма
прочным, позвоночный столб удивительно подвижен.
Позвоночник человека представляет длинный изогнутый столб,
состоящий из 33—34 лежащих один на другом и постепенно увеличи-
вающихся в размерах сверху вниз позвонков; наиболее типично сле-
дующее их количество: шейных — 7, грудных — 12, поясничных — 5,
крестцовых — 5, копчиковых — 4 (рис. 92). Позвонки разных отделов
отличаются по форме и величине. Однако все они имеют ряд общих
признаков — они гомологичны.
Каждый позвонок состоит из тела, расположенного спереди, и дуги
(сзади), ограничивающих широкое позвоночное отверстие (рис. 93).
Накладываясь свободно одно на другое, отверстия образуют длинный
позвоночный канал, в котором залегает спинной мозг, надежно защи-
щенный стенками канала. От дуги позвонка отходят семь отростков.
Кзади направляется непарный остистый отросток. Вершины многих из
них легко прощупываются у человека по средней линии со спины.
Рис. 92. Позвоночный столб:
А — вид сбоку, Б — вид спереди, В — вид сзади. Отделы: I — шейный, II — грудной,
III — поясничный, IV — крестцовый, V — копчиковый; 1, 3 — шейный и поясничный
лордозы, 2, 4 — грудной и крестцовый кифозы, 5 — мыс
11
5
А
I
Б
II
В
III
IV
V
22
3
4
169
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
Во фронтальной плоскости справа и слева располагаются парные попе-
речные отростки. Вверх и вниз от дуги направлены верхние и нижние
суставные отростки. Основания суставных отростков ограничивают
верхнюю и нижнюю позвоночные вырезки. При соединении позвонков
друг с другом нижняя вырезка вышележащего позвонка и верхняя ни-
жележащего образуют справа и слева межпозвоночные отверстия, че-
рез которые проходят спинномозговые нервы и кровеносные сосуды.
Благодаря прямохождению человека существенно отличаются от
остальных I и II шейные позвонки, сочленяющиеся с черепом и несу-
щие на себе его тяжесть. Первый шейный позвонок, или атлант, ли-
шен остистого отростка (рис. 94). В древности Атласа, или Атланта,
Рис. 93. Позвонок:
А — вид сверху, Б — вид сбоку; 1 — тело, 2 — позвоночное отверстие,
3 — остистый отросток, 4 — верхний суставный отросток,
5 — поперечный отросток, 6 — нижний суставной отросток
1
1
5
А
Б
2
6
3
3
4
Рис. 94. Первый шейный
позвонок, или атлант,
вид сверху:
1 — задний бугорок,
2 — борозда позвоночной
артерии, 3 — отверстие
поперечного отростка,
4 — ямка зуба,
5 — передняя дуга,
6 — передний бугорок,
7 — поперечный
отросток, 8 — верхняя
суставная ямка,
9 — задняя дуга,
10 — позвоночное
отверстие
11
22
33
44
55
66
77
88
99 10
10
170
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
считали титаном, который за участие в борьбе против богов был при-
говорен навеки держать небесный свод. Средняя часть тела Атланта,
отделившись, приросла к телу II позвонка, образовав его зуб. Атлант
лишен и суставных отростков. Вместо них на верхней и нижней по-
верхностях латеральных масс находятся суставные ямки, верхние со-
членяются с мыщелками затылочной кости, образуя атлантозаты-
лочные суставы; нижние — с верхними суставными поверхностями
II позвонка, образуя боковые атлантоосевые суставы. Второй шей-
ный позвонок носит название «эпистрофей» (возвращение, оборачива-
ние) (рис. 95). Вокруг вышеупомянутого его зуба и происходят такие
вращения атланта вместе с черепом в срединном атлантоосевом
суставе. Шейные позвонки отличаются от других малыми размерами
и наличием небольшого отверстия в каждом поперечном отростке
(через отверстия проходит позвоночная артерия, направляющаяся
в полость черепа).
На боковых поверхностях тел грудных позвонков имеются ребер-
ные ямки для сочленения с головками ребер, а на утолщенных концах
поперечных отростков десяти верхних грудных позвонков имеются
реберные ямки, с которыми сочленяются соответствующие им по сче-
ту ребра. Пять крупных поясничных позвонков обеспечивают большую
подвижность этой части позвоночного столба.
Пять крестцовых позвонков у взрослого человека, срастаясь, обра-
зуют массивный крестец треугольной формы (рис. 96). Направлен-
ное вверх основание и суставные отростки I крестцового позвонка
сочленяются с V поясничным позвонком. В области соединения обра-
зуется закругленный направленный вперед мыс. На боковых частях
крестца находятся ушковидные поверхности для сочленения с тазо-
1
2
3
4
5
6
7
8
9
10
1
2
3
4
5
6
7
8
9
10
Рис. 95. Второй шейный позвонок,
или осевой позвонок, вид сбоку:
1 — передняя суставная
поверхность, 2 — зуб, 3 — задняя
суставная поверхность,
4 — верхняя суставная
поверхность, 5 — отверстие
поперечного отростка, 6 — дуга
позвонка, 7 — остистый отросток,
8 — нижний суставной отросток,
9 — поперечный отросток,
10 — тело позвонка
171
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
выми костями (крестцово-подвздошные суставы). Книзу крестец су-
живается, и его канал заканчивается крестцовой щелью. В крестцовом
канале находятся конечные нити спинного мозга и корешки пояснич-
ных и крестцовых спинномозговых нервов. По-латыни крестец —
os sacrum. «Сакральный» дословно — это «священный», но и «скры-
тый». Эта кость действительно в виде клина вставлена между под-
вздошными костями таза. Вместе с ними крестец образует свод, опи-
рающийся на головки бедренных костей. Форма крестца не позволяет
ему устремиться вниз. Не может крестец и вдавиться назад, а вперед
он не двигается из-за того, что боковые сочленения его с костями таза
не гладкие и не плоские, а бугристые. Копчик обычно срастается с вер-
хушкой крестца. У взрослого человека копчик образован двумя — пя-
тью (чаще четырьмя) рудиментарными копчиковыми позвонками.
Позвонки связаны между собой с помощью различного вида сое-
динений. Тела их соединены межпозвоночными дисками, толщина ко-
торых в грудном отделе 3—4 мм, в шейном 5—6 мм, в поясничном
10—12 мм. Межпозвоночные диски построены из волокнистого хря-
ща. Передняя и задняя продольные связки укрепляют соединения тел
позвонков. Суставные отростки выше- и нижележащего позвонков
сочленены между собой дугоотростчатыми суставами, а остистые
и поперечные — связками.
Рис. 96. Крестец и копчик:
А — вид спереди: 1 — основание крестца, 2 — верхний суставный отросток,
3 — латеральная часть, 4 — поперечные линии, 5 — передние крестцовые отверстия,
6 — верхушка крестца, 7 — копчик; Б — вид сзади: 1 — верхний суставный отросток,
2 — крестцовая бугристость, 3 — ушковидная поверхность, 4 — латеральный
крестцовый гребень, 5 — промежуточный крестцовый гребень, 6 — крестцовый рог,
7 — копчик, 8 — крестцовая щель, 9 — задние крестцовые отверстия,
10 — срединный крестцовый гребень, 11 — крестцовый канал
4
3
2
5
6
7
1
8
9
10
11
1
2
3
4
5
6
7
А
Б
172
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Позвоночник человека имеет несколько изгибов. Выпуклости по-
звоночного столба, обращенные дугой вперед, называются лордозами,
назад — кифозами (см. рис. 92). Шейный лордоз переходит в грудной
кифоз, который, в свою очередь, сменяется поясничным лордозом,
а затем крестцово-копчиковым кифозом.
ВНИМАНИЕ!
Функциональная роль изгибов очень велика. Благодаря им удары, тол-
чки и сотрясения, передающиеся позвоночнику при различных движе-
ниях, падении, ослабляются, и содержимое черепа как бы амортизиро-
вано.
Позвоночный столб человеческого зародыша и плода имеет форму
дуги, обращенной выпуклостью кзади. У новорожденного позвоноч-
ник почти прямой, кривизны развиваются постепенно в связи с тягой
мышц. Когда ребенок начинает держать голову, возникает шейный
лордоз (около 6 мес.); когда он садится — грудной кифоз (около
6 мес.), когда начинает стоять — поясничный лордоз (9—12 мес.),
а вместе с ним и крестцовый кифоз. Окончательное развитие изгибов
позвоночника происходит к шести-семи годам, в это время устанавли-
вается и центр тяжести — на уровне второго крестцового позвонка.
Отвесная линия этого центра проходит на 5 см кзади от поперечной
линии, соединяющей тазобедренные суставы, и на 3 см кпереди от по-
перечной оси голеностопных суставов.
Позвоночный столб человека весьма подвижен. Движения позво-
ночника осуществляются вокруг трех осей: поперечной — сгибание
вперед и разгибание назад, амплитуда этих движений 170—245°; са-
гиттальной — боковое сгибание вправо и влево, общий размах дви-
жений около 165°; продольной (вертикальной) — вращательные дви-
жения, общий размах — около 120° и как бы объединяющее их
круговое движение.
В шейном и поясничном отделах размах движений наибольший.
Объем движений в шейном отделе: сгибание — 70—79°, разгиба-
ние — 95—105°, вращение — 80—85°. В грудном отделе подвижность
ограничена наличием ребер и грудины, тонкостью межпозвоночных
дисков и частично направленными вертикально вниз остистыми от-
ростками. Сгибание, разгибание и боковые сгибания здесь относи-
тельно невелики: сгибание — до 35°, разгибание — до 50°, враще-
ние — до 20°. В поясничном отделе толстые межпозвоночные диски
способствуют большей подвижности (сгибание — до 60°, разгиба-
ние — до 40—45°), суставные отростки ограничивают ротацию и бо-
ковые движения позвоночника.
173
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
Грудная клетка образована соединенными между собой 12 парами
ребер, грудиной и 12 грудными позвонками (рис. 97). Ребро представ-
ляет собой длинную плоскую костную пластинку, переходящую спе-
реди в реберный хрящ (рис. 98). Костная часть ребра состоит из го-
ловки, на которой находится суставная поверхность для сочленения
с телами позвонков, шейки и тела. На телах десяти верхних ребер
имеются бугорки, снабженные суставными поверхностями для сочле-
нения с поперечными отростками позвонков. I —VII ребра называ-
ются истинными, хрящ каждого из них соединяется с грудиной,
5
6
7
8
4
3
2
1
Рис. 97. Грудная клетка, вид спереди:
1 — тело грудины,
2 — рукоятка грудины,
3 — верхняя апертура
грудной клетки, 4 — ключица,
5 — лопатка, 6 — ребра,
7 — мечевидный отросток грудины,
8 — реберная дуга
А
Б
2
3
4
5
6
7
1
8
Рис. 98. Ребра правые, вид сверху:
А — первое ребро, Б — второе ребро,
1 — угол ребра, 2 — бугорок ребра,
3 — шейка ребра, 4 — головка ребра,
5 — борозда подключичной вены,
6 — бугорок передней лестничной мышцы,
7 — борозда подключичной артерии,
8 — тело ребра
174
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
VIII—X ребра называются ложными, кон-
цы их хрящей срастаются между собой
и с хрящами нижних ребер, образуя ре-
берные дуги; XI—XII ребра — колеблю-
щиеся, так как их передние концы не до-
ходят до грудины и заканчиваются в
мышцах передней брюшной стенки.
Плоская грудина состоит из трех ча-
стей: широкой рукоятки вверху, удлинен-
ного тела и мечевидного отростка внизу
(рис. 99). На середине верхнего края ру-
коятки находится яремная вырезка, кото-
рая легко прощупывается у живого чело-
века. По бокам от нее имеются ключичные
вырезки для соединения с ключицами,
в результате чего образуются грудино-ключичные суставы, а ниже, на
боковых сторонах грудины, — реберные вырезки для прикрепления
хрящей I—VII ребер. Благодаря соединениям составляющих ее ко-
стей грудная клетка весьма подвижна. При вдохе и выдохе происхо-
дит вращение задних концов ребер в реберно-позвоночных суставах,
одновременно смещаются и ребра, и грудина. При вдохе передние
концы ребер и грудина поднимаются, межреберные промежутки рас-
ширяются, размеры грудной полости увеличиваются, при выдохе все
происходит наоборот.
Грудная клетка человека по форме напоминает бочку неправиль-
ной формы, расширенную в поперечном и уплощенную в передне-
заднем направлении. Через верхнее отверстие грудной клетки, кото-
рое ограничено первым грудным позвонком, первой парой ребер и
верхним краем грудины, проходят трахея, пищевод, сосуды и нервы.
Нижнее отверстие грудной клетки, ограниченное XII грудным по-
звонком, нижними ребрами, реберными хрящами и нижним концом
грудины, закрыто диафрагмой. Межреберные промежутки на всем
протяжении между позвоночником и грудиной заполнены межребер-
ными мышцами, в них проходят сосуды и нервы.
2
3
4
5
6
7
8
9
11
12
10
1
Рис. 99. Грудина, вид спереди:
1 — рукоятка грудины, 2 — тело грудины,
3 — мечевидный отросток, 4 — реберная вырезка
VII ребра; 5 — реберная вырезка VI ребра,
6 — реберная вырезка V ребра,
7 — реберная вырезка IV ребра, 8 — реберная
вырезка III ребра, 9 — реберная вырезка II ребра,
10 — реберная вырезка I ребра,
11 — ключичная вырезка, 12 — яремная вырезка
175
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
Череп
Череп — настолько типичный пример сквозной темы в литературе,
что среди литературоведов даже фигурирует термин «черепизм». Из
черепов победители воздвигали ужасающие своим видом пирамиды,
изображение черепа помещается на всевозможных предупреждающих
этикетках. Много информации о культе черепов. По форме черепа
пытались, да и сейчас кое-где пытаются, предсказать судьбу челове-
ка, его умственные способности, склонности (взгляды Ф. Галла,
Ч. Ломброзо, френологов). У многих народов череп подвергался
искусственной деформации. Будучи вместилищем головного мозга,
череп никогда и никем не воспринимался как простое собрание скре-
пленных между собой костей. Скорее как символ разума, человече-
ской мысли, чего-то таинственного, фетиш. Глядя на череп, человек
ощущает трагизм человеческого бытия. Надмирность сливается в этой
анатомической структуре с повседневностью. Поэтому-то зачастую
череп не просто хранили, но разукрашивали его, инкрустировали ме-
таллами, драгоценными камнями.
Череп условно подразделяется на мозговой и лицевой отделы.
Мозговой отдел (его объем 1400—1500 см3) является вместилищем
для головного мозга. В пределах висцерального, или лицевого, отдела
начинаются системы органов пищеварения, дыхания, лежат органы
чувств. Только у человека разумного округлый мозговой череп распо-
лагается над лицевым. Череп человека состоит из 23 костей, из них
8 парных и 7 непарных (рис. 100, 101).
Семь костей черепа воздухоносные, т. е . они имеют внутри поло-
сти, заполненные воздухом и сообщающиеся с полостью носа. Это
лобная, клиновидная, решетчатая, парные височная и верхняя челю-
сти. Наличие полостей уменьшает массу черепа при сохранении его
прочности. Кроме того, воздухоносные кости располагаются только
по периферии органов чувств, обеспечивая благодаря плохо проводя-
щей тепло воздушной среде как бы своеобразный термостат — тепло-
вую изоляцию органов зрения, обоняния и слуха. Последние же, как
оказалось, нормально функционируют лишь при температуре тела.
Воздухоносные полости участвуют в фонации.
Кости крыши черепа человека плоские, они состоят из более тол-
стой наружной и тонкой внутренней пластинок компактного костного
вещества; между ними заключено губчатое вещество (диплое), в ячей-
ках которого находится красный костный мозг и многочисленные
кровеносные сосуды (преобладают вены). Неровности на внутренней
поверхности костей черепа обусловлены давлением мозга: ямки соот-
ветствуют мозговым извилинам, а возвышения между ними — бо-
роздам.
176
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Мозговой отдел черепа взрослого человека составляют следую-
щие кости: лобная, затылочная, клиновидная, решетчатая, две височ-
ные и две теменные. Непарная воздухоносная лобная кость состоит
из чешуи, двух горизонтальных глазничных частей и носовой части.
Лобная кость формирует лоб и несет на себе лобные бугры, являющи-
еся характерной особенностью человека разумного, а также образует
верхние стенки глазниц, полости носа, височных ямок, нижнюю и пе-
реднюю стенки передней черепной ямки.
Парная теменная кость участвует в формировании крыши (сво-
да) черепа, в центре кости находится теменной бугор. Теменные кости
участвуют в образовании свода черепа. Затылочная кость, участвую-
щая в формировании свода и основания черепа, задней черепной
ямки, состоит из четырех частей (базилярной впереди, двух боковых
и чешуи), расположенных вокруг большого (затылочного) отверстия.
Рис. 100. Череп человека, вид сбоку:
1 — теменная кость, 2 — венечный шов, 3 — лобный бугор, 4 — височная поверхность
большого крыла клиновидной кости, 5 — глазничная пластинка решетчатой кости,
6 — слезная кость, 7 — носовая кость, 8 — височная ямка, 9 — передняя носовая ость,
10 — тело верхнечелюстной кости, 11 — нижняя челюсть, 12 — скуловая кость,
13 — скуловая дуга, 14 — шиловидный отросток, 15 — мыщелковый отросток
нижней челюсти, 16 — сосцевидный отросток, 17 — наружный слуховой проход,
18 — ламбдовидный шов, 19 — чешуя затылочной кости,
20 — верхняя височная линия , 21 — чешуйчатая часть височной кости
11
17
17
22
18
18
33
19
19
44
20
20
55
21
21
66
77
88
99
10
10
11
11
12
12
13
13
14
14
15
15
16
16
177
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
Два овальных затылочных мыщелка, расположенных на наружной
поверхности боковых частей, сочленяются с атлантом, образуя ат-
лантозатылочные суставы, в которых совершаются кивательные
движения и боковые наклоны головы.
Непарная воздухоносная клиновидная кость имеет тело, на верх-
ней поверхности которого находится гипофизарная ямка, где залегает
гипофиз. От тела в стороны отходят большие крылья, вверх и лате-
рально — малые крылья, вниз — крыловидные отростки. Клиновид-
ная кость участвует в формировании глазницы, передней черепной,
подвисочных и крыловидно-небных ямок.
Наиболее сложно устроена парная воздухоносная височная кость,
которая участвует в формировании свода и основания черепа и явля-
ется вместилищем органа слуха и равновесия. Она состоит из пира-
Рис. 101. Череп человека, вид спереди:
1 — венечный шов, 2 — теменная кость, 3 — глазничная часть лобной кости,
4 — глазничная поверхность большого крыла клиновидной кости, 5 — скуловая кость,
6 — нижняя носовая раковина, 7 — верхнечелюстная кость, 8 — подбородочный
выступ нижней челюсти, 9 — полость носа, 10 — сошник, 11 — перпендикулярная
пластинка решетчатой кости, 12 — глазничная поверхность верхнечелюстной кости,
13 — нижняя глазничная щель , 14 — слезная кость, 15 — глазничная пластина
решетчатой кости, 16 — верхняя глазничная щель, 17 — чешуйчатая часть
височной кости, 18 — скуловой отросток лобной кости, 19 — зрительный канал,
20 — носовая кость
Р 101Ч
11
17
17
22
18
18
33
19
19
44
20
20
55
66
77
88
99
10
10
11
11
12
12
13
13
14
14
14
14
15
15
16
16
178
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
миды, в которой расположена барабанная полость и внутреннее ухо;
барабанной части, в которой находится наружное слуховое отверстие
и наружный слуховой проход; чешуйчатой части, на наружной по-
верхности которой располагается глубокая нижнечелюстная ямка,
куда входит мыщелковый отросток нижней челюсти, образуя височ-
но-нижнечелюстной сустав. Скуловой отросток, соединяясь со скуло-
вой костью, формирует скуловую дугу.
Непарная воздухоносная решетчатая кость состоит из множест-
ва ячеек (решетчатые лабиринты), как бы подвешенных к решетчатой
пластинке, через отверстия которой в полость черепа входят обоня-
тельные нервы. От медиальных поверхностей лабиринтов с обеих сто-
рон отходят тонкие изогнутые верхняя и средняя носовые раковины,
свешивающиеся в полость носа.
Лицевой отдел черепа образован парными костями: верхними челю-
стями, небными, скуловыми, носовыми, слезными, нижними носовыми
раковинами, а также непарными: сошником и нижней челюстью. К лице-
вому черепу относят и подъязычную кость. Основную массу скелета ли-
цевого черепа образуют челюсти: две верхние и нижняя; другие мелкие
кости участвуют в формировании стенок глазниц, носовой и ротовой по-
лостей, определяя вместе с челюстями конфигурацию лицевого черепа.
Развитие головного мозга, характер питания и членораздельная речь на-
кладывают существенный отпечаток на строение лицевого черепа.
Основную роль в этом играет небольшой (по сравнению с животными)
скелет жевательного аппарата — челюстей и слабое развитие жеватель-
ных мышц, обусловленное характером питания человека разумного.
Верхняя челюсть — парная воздухоносная кость, которая состоит из
тела, содержащего гайморову пазуху, и четырех отростков: лобного, ску-
лового, небного и альвеолярного. Дуга последнего несет зубные альвео-
лы, отделенные друг от друга межальвеолярными перегородками. Неб-
ный отросток, соединяясь с одноименным отростком противоположной
кости срединным швом, участвует в образовании твердого нёба. Верхняя
челюсть участвует в формировании нижней стенки глазницы, боковой
стенки полости носа, подвисочной и крыловидно-небной ямок.
Парная небная кость, примыкающая сзади к верхней челюсти, уча-
ствует в образовании полости носа, передней части твердого нёба, носо-
вой перегородки, глазницы и крыловидно-небной ямки. Небная кость
состоит из соединенных под прямым углом горизонтальной и перпен-
дикулярной пластинок. Четырехугольные пластинки обеих небных ко-
стей, соединяясь между собой, образуют заднюю часть твердого нёба.
Нижняя носовая раковина — парная самостоятельная кость, кото-
рая расположена в носовой полости, отделяет средний носовой ход от
нижнего. Парная, очень тонкая и хрупкая слезная кость участвует
в образовании медиальной стенки глазницы. Парная скуловая кость
179
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
играет важную роль в создании рельефа лица, она укрепляет лицевой
череп. Располагающийся на задней поверхности носа сошник образует
большую часть носовой перегородки. Край сошника разделяет выход
из полости носа на две хоаны.
Нижняя челюсть имеет подковообразную форму. Это единственная
подвижная кость черепа, к которой прикрепляются жевательные мыш-
цы. Она состоит из тела и соединенных с ним под углом в 110—130°
двух ветвей. По средней линии виден обращенный кпереди подборо-
дочный выступ, являющийся отличительной чертой черепа человека
разумного. На задней поверхности челюсти по средней линии у челове-
ка имеется подбородочная ость (острие, шип), к которой прикрепляют-
ся мышцы. Верхний край тела нижней челюсти образует альвеолярную
дугу, несущую 16 зубных альвеол. Передний отдел дуги закруглен,
а сама альвеолярная часть тонкая, что также является одной из отличи-
тельных особенностей нижней челюсти человека. Ветви нижней челю-
сти направляются вверх и оканчиваются двумя отростками — венеч-
ным, к которому прикрепляется височная мышца, и мыщелковым,
участвующим в образовании комбинированного височно-нижнече -
люстного сустава, в котором осуществляются поднимание и опускание
нижней челюсти, смещение ее вперед и назад, боковые движения.
Особое место среди костей занимает дугообразная подъязычная
кость, расположенная в передней области шеи между гортанью
и нижней челюстью и соединенная с костями черепа лишь связками
и мышцами. Кость состоит из тела и двух пар рогов — больших и ма-
лых. От последних к шиловидным отросткам височных костей тянут-
ся связки, которые как бы подвешивают кость к черепу.
Соединения костей черепа преимущественно фиброзные (швы).
В области лицевого черепа швы ровные, гладкие, плоские (гармонич-
ные), у мозгового черепа — зубчатые, а между теменной и чешуей ви-
сочной кости — чешуйчатый. На основании черепа у ребенка имеются
хрящевые соединения (синхондрозы), которые с возрастом окостенева-
ют, превращаясь в синостозы. Лишь нижняя челюсть образует с чере-
пом подвижное соединение, височно-нижнечелюстной сустав.
Позвоночный столб соединяется с черепом атлантозатылочными,
срединным и латеральными атлантоосевыми суставами, в которых
осуществляются кивательные, вращательные движения, а также боко-
вые наклоны головы.
Череп как целое. Отдельные кости, соединяясь между собой, фор-
мируют сложный и весьма совершенный череп человека, анатомия
которого идеально соответствует выполняемым функциям. Свод че-
репа образован чешуей лобной и височной костей, латеральными ча-
стями больших крыльев клиновидной кости, теменными костями,
верхней частью чешуи затылочной кости. Прочие кости и их части
180
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
формируют основание черепа, которое прочно соединено с костями
лицевого черепа. Очертание крыши черепа у взрослого человека
овальное (у новорожденного — пятиугольное), при этом длина черепа
больше его ширины. На крыше черепа видны швы: по срединной ли-
нии сагиттальный (стреловидный), образованный при соединении со-
ответствующих краев теменных костей. Перпендикулярно к нему, т. е.
во фронтальной плоскости, проходит венечный шов, соединяющий
чешую лобной кости с теменными костями. Благодаря соединению
теменных костей с затылочной чешуей формируется ламбдовидный
шов, напоминающий соответствующую греческую букву лямбда. На
верхнебоковой поверхности с каждой стороны видны теменные буг-
ры. Задняя часть свода черепа образована большей частью затылоч-
ной и прилегающими к ней частями теменных и височных костей.
Снаружи в центре затылочной кости виден наружный затылочный
выступ (он прощупывается у живого человека).
При рассмотрении черепа сбоку видны лобная, теменная, височ-
ная, затылочная и клиновидная кости, а также скуловая дуга, образо-
ванная височным отростком скуловой кости и скуловым отростком
височной кости, верхняя и нижняя челюсти и две ямки: височная
и крыловидно-небная.
При изучении черепа спереди видны лобная область, глазницы,
вход в полость носа, верхние и нижние челюсти с зубами и скуловые
кости. Выпуклый лоб человека разумного образован чешуей лобной
кости, на которой по бокам расположены лобные бугры. Надбровные
дуги проходят над глазницами, а в надпереносье видна маленькая
площадка — глабелла.
На лицевом черепе имеется ряд весьма важных образований. Пар-
ная глазница представляет собой полость, напоминающую по форме
четырехгранную пирамиду с закругленными углами. Основание этой
пирамиды обращено вперед и образует вход в глазницу, а верхушка
направлена назад и медиально, в ней проходит канал зрительного
нерва. В полости глазницы расположены глазное яблоко и вспомога-
тельные образования органа зрения. У латерального края верхней
стенки глазницы расположена ямка слезной железы.
Полость носа занимает центральное положение в лицевом черепе.
На нижнем крае грушевидного отверстия выступает передняя носовая
ость, продолжающаяся кзади в костную перегородку носа, которая де-
лит его полость на две половины. Через парные хоаны полость носа
сообщается с полостью глотки. На боковой стенке полости носа, как
уже указывалось, выступают три носовые раковины, а также отростки
лабиринтов решетчатой кости. Раковины делят боковой отдел полости
носа на три носовых хода: нижний, средний и верхний, в которые от-
крываются носослезный канал и полости воздухоносных костей.
181
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
Полость рта впереди и с боков ограничена (из костных структур)
верхней и нижней альвеолярными дугами с зубами, частично телом и
ветвью нижней челюсти, а сверху — твердым нёбом.
Наружное основание черепа образовано нижними поверхностями
мозгового и лицевого черепа и простирается от зубов верхней челюсти
спереди до верхней выйной линии сзади, а также от нижнего края ску-
ловой дуги до противоположной (рис. 102). На наружном основании
черепа спереди видно костное небо, сзади — большое затылочное отвер-
Рис. 102. Наружное основание черепа:
1 — небный отросток верхней челюсти, 2 — срединный небный шов, 3 — резцовые от-
верстия, 4 — поперечный небный шов, 5 — хоаны, 6 — крыло сошника, 7 — крыловидная
ямка, 8 — латеральная пластинка крыловидного отростка, 9 — нижняя глазничная щель,
10 — скуловая дуга, 11 — крыловидный отросток, 12 — овальное отверстие,
13 — рваное отверстие, 14 — остистое отверстие, 15 — шиловидный отросток,
16 — глоточный бугорок, 17 — наружный слуховой проход, 18 — сосцевидный отросток,
19 — сосцевидная вырезка, 20 — нижняя выйная линия, 21 — большое (затылочное)
отверстие, 22 — наружный затылочный выступ, 23 — затылочный сосцевидный шов,
24 — мыщелковая ямка, 25 — мыщелковый канал, 26 — затылочный мыщелок,
27 — шилососцевидное отверстие, 28 — яремное отверстие, 29 — каменисто-
барабанная щель, 30 — наружное сонное отверстие, 31 — нижнечелюстная ямка,
32 — суставный бугорок, 33 — клиновидно-чешуйчатый шов, 34 — крыловидный
крючок, 35 — скуловерхнечелюстной шов, 36 — большое небное отверстие
11
17
17
22
18
18
33
19
19
44
20
20
55
21
21
66
22
22
77
23
23
88
24
24
99
25
25
10
10
26
26
11
11
27
27
12
12
28
28
13
13
29
29
14
14
30
30
15
15
31
31
16
16
32
32
33
33
34
34
35
35
36
36
182
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
стие с лежащими по бокам от него затылочными мыщелками, которые
сочленяются с атлантом. По бокам расположены сосцевидные отростки,
которые у человека развиты хорошо в связи с функцией грудино-клю-
чично-сосцевидных мышц и вертикальным положением головы. Строе-
нием нижней поверхности мозга обусловлен сложный рельеф внутрен-
него основания черепа (рис. 103). Здесь различают три ямки. В передней
из них лежат лобные доли полушарий большого мозга. Задние края ма-
лых крыльев клиновидной кости отделяют переднюю от более глубокой
средней черепной ямки, в которой располагаются височные доли боль-
ших полушарий головного мозга, а в гипофизарной ямке турецкого сед-
ла — гипофиз. В центре самой глубокой задней черепной ямки видно
Рис. 103. Внутреннее основание черепа:
1 — петушиный гребень, 2 — решетчатая пластинка, 3 — зрительный канал,
4 — гипофизарная ямка, 5 — круглое отверстие, 6 — спинка седла, 7 — овальное
отверстие, 8 — остистое отверстие, 9 — внутреннее слуховое отверстие, 10 — скат,
11 — яремное отверстие, 12 — подъязычный канал, 13 — ламбдовидный шов,
14 — борозда поперечного синуса, 15 — внутренний затылочный выступ,
16 — затылочная чешуя, 17 — большое (затылочное) отверстие, 18 — пирамида
(каменистая часть) височной кости, 19 — рваное отверстие, 20 — чешуйчатая часть
височной кости, 21 — большое крыло клиновидной кости, 22 — малое крыло
клиновидной кости, 23 — глазничная часть лобной кости
11
17
17
22
18
18
33
19
19
44
20
20
55
21
21
66
22
22
77
23
23
88
99
10
10
11
11
12
12
13
13
14
14
15
15
16
16
183
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
большое (затылочное) отверстие. В задней черепной ямке расположены
мозжечок, варолиев мост и продолговатый мозг. Основная роль в разви-
тии и формировании черепа принадлежит головному мозгу, жеватель-
ным мышцам, зубам, органам чувств.
Возрастные особенности строения черепа. В процессе индивиду-
ального развития череп претерпевает существенные изменения. У но-
ворожденного швов нет, пространства между костями заполнены сое-
динительной тканью (рис. 104). В участках, где сходятся несколько
костей, имеется шесть родничков, закрытых соединительнотканными
Рис. 104. Череп новорожденного;
вид сбоку (А) и сверху (Б):
1 — передний родничок,
2 — клиновидный родничок,
3 — большое крыло клиновидной
кости, 4 — лобный бугор,
5 — носовая кость, 6 — слезная кость,
7 — скуловая кость, 8 — верхняя
челюсть, 9 — нижняя челюсть,
10 — барабанное кольцо
височной кости, 11 — чешуйчатая
часть височной кости,
12 — латеральная часть затылочной
кости, 13 — сосцевидный родничок,
14 — затылочная чешуя, 15 — задний
родничок, 16 — теменной бугор,
17 — лобный шов
12
13
14
15
4
5
6
7
8
9
10
11
3
2
1
16
Б
17
15
4
1
16
А
Б
184
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
пластинками: два непарных (передний и задний) и два парных (кли-
новидный и сосцевидный). Самый крупный — передний, или лобный,
родничок ромбовидной формы — расположен там, где сближаются
правая и левая половины лобной и теменной костей. Благодаря род-
ничкам череп новорожденного очень эластичен, его форма может из-
меняться. Во время прохождения головки плода через родовые пути
края костной крыши черепа накладываются черепицеобразно один на
другой, что и приводит к уменьшению его размеров. Формирование
швов черепа заканчивается в основном к двум годам жизни, к этому
времени закрываются и роднички.
Именно в области родничков прикладывали медики средневеко-
вья горячее железо при болезнях мозга и глаз. После того как форми-
ровался рубец, они еще долго вызывали раздражающими средствами
нагноение, открывая тем самым, как они считали, путь для скаплива-
ющихся вредных веществ. Отсюда и происхождение термина fonta-
nella — фонтанчик. А в конфигурации швов пытались видеть различ-
ные буквы, символы, якобы информирующие о судьбе человека.
Соотношения мозгового и лицевого черепа у взрослого и новоро-
жденного различны. Лицо новорожденного короткое и широкое. Со-
отношение площади лицевого отдела к мозговому у новорожденного
равно 1 : 8, у двухлетнего ребенка — 1:6, у пятилетнего — 1:4, у де-
сятилетнего — 1 : 3, у взрослой женщины — 1 : 2,5, у взрослого мужчи-
ны — 1:2.
После рождения рост черепа происходит неравномерно.
1. Период энергичного роста — от рождения до семи лет. В течение
первого года жизни череп растет более или менее равномерно. От года до
трех лет особенно активно увеличивается его задняя часть, что связано
с переходом ребенка на втором году жизни к прямохождению. На вто-
ром-третьем годах жизни в связи с окончанием прорезывания молочных
зубов и усилением функции жевательных мышц значительно усиливает-
ся рост лицевого черепа, особенно его основания. К семи годам основа-
ние черепа достигает почти такой же длины, как у взрослого человека.
2. Период замедленного роста — от 7 до 12—13 лет (начало поло-
вого созревания). В это время в основном растет свод мозгового чере-
па, объем его полости достигает 1200—1300 см3.
3. В третий период — после 13 лет — активно растут лобный от-
дел мозгового и лицевой череп. Проявляются половые различия:
у мальчиков лицевой череп растет в длину сильнее, чем у девочек,
лицо удлиняется, поэтому если до периода половой зрелости у детей
лица округлые, то после ее наступления у юношей, как правило, лицо
вытягивается в длину, у девушек округлость сохраняется. Мужской
череп в связи с большими общими размерами тела крупнее женского.
Вместимость черепа у мужчин в среднем 1559 см3, у женщин 1347 см3,
185
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
но относительная вместимость черепа на 1 см длины тела у женщин
даже больше, чем у мужчин. Мозговой череп относительно сильнее
развит у женщин, а лицевой — у мужчин. Как правило, мужской че-
реп отличается выраженным рельефом в связи с более сильным раз-
витием прикрепляющихся к нему мышц, у женщин рельеф черепа бо-
лее сглаженный. Зарастание швов начинается в возрасте 20—30 лет,
причем у мужчин несколько раньше, чем у женщин.
4. Четвертый период — преобразования черепа в пожилом и стар-
ческом возрасте. В связи с выпадением зубов альвеолярные отрост-
ки верхней и альвеолярная часть нижней челюстей уменьшаются,
жевательная функция ослабевает, мышцы частично атрофируются,
изменяются. Челюсти утрачивают массивность, однако, если зубы
сохраняются, эти изменения не происходят. В силу уменьшения эла-
стичности череп становится более хрупким и легким.
Скелет конечностей
Конечности образованы поясами и свободными элементами, кото-
рые, в свою очередь, подразделены на три сегмента: верхний (прокси-
мальный) имеет одну кость, средний — две и нижний — множество ко-
стей. Кости образуют систему рычагов. Скелеты верхней и нижней
конечностей гомологичны. Пояс верхней конечности сформирован с ка-
ждой стороны двумя костями — лопаткой и ключицей, которые прикре-
плены к грудной клетке с помощью мышц и связок, а спереди посредст-
вом ключицы сочленяются с грудиной. Проксимальный отдел скелета
свободной верхней конечности образован плечевой костью, средний —
лучевой и локтевой костями и дистальный — 27 костями, которые,
в свою очередь, подразделяются на три части: запястье, пясть и фаланги
пальцев. Пояс нижних конечностей образован двумя тазовыми костями,
каждая из которых сзади сочленяется с крестцом, а спереди — друг
с другом. Скелет свободной нижней конечности также состоит из прок-
симального (бедро), среднего (большеберцовая и малоберцовая кости)
и дистального сегментов — 26 костей стопы, также подразделяющейся
на три части: предплюсну, плюсну и фаланги пальцев.
Только у человека разумного функция конечностей четко разгра-
ничена: верхние являются органом труда, нижние — опоры и передви-
жения. Особенности анатомического строения руки являются морфо-
логической основой ее функции — труда, специфического только для
человека разумного. Длинная ключица, соединяющая свободную
верхнюю конечность с костями туловища и отодвигающая руку от
него, способствует увеличению объема движений в плечевом суставе.
Соединения свободной верхней конечности, особенно в области пред-
плечья и кисти (в первую очередь, седловидный запястно-пястный
186
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
сустав большого пальца), и высокодифференцированные мышцы по-
зволяют выполнять многообразные тонкие и сложные трудовые про-
цессы. Скелет нижней конечности, являющейся у человека органом
опоры и перемещения тела в пространстве, состоит из более толстых
и массивных костей, соединенных между собой менее подвижными,
чем у верхних конечностей, сочленениями.
Кости верхней конечности
Кости пояса верхней конечнос-
ти (рис. 105). Ключица — парная,
S-образно изогнутая трубчатая кость,
которая сочленяется с грудиной и
с акромиальным отростком лопатки,
образуя грудино-ключичный и акро-
миально-ключичный суставы. В этих
суставах осуществляются опускание
и поднимание ключицы, ее движение
вперед и назад, а также вращение
и круговые движения.
Лопатка — плоская кость тре-
угольной формы, прилегающая к зад-
ней поверхности грудной клетки
своей реберной поверхностью. Ее
дорсальная поверхность разделена
костной остью на две ямки — надост-
ную и подостную, в которой залегают
одноименные мышцы. Ость продол-
жается латерально и кпереди в акро-
мион, сочленяющийся с ключицей.
4
3
2
5
6
7
8
9
1
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24 25
Рис. 105. Кости верхней конечности, правой, вид спереди:
1 — ключица, 2 — грудинный конец ключицы, 3 — лопатка,
4 — клювовидный отросток лопатки, 5 — суставная
впадина лопатки, 6 — плечевая кость, 7 — венечная ямка
плечевой кости, 8 — медиальный надмыщелок, 9 — блок
плечевой кости, 10 — венечный отросток локтевой кости,
11 — бугристость локтевой кости, 12 — локтевая кость,
13 — головка локтевой кости, 14 — кости запястья,
15 — I—V пястные кости, 16 — фаланги пальцев,
17 — шиловидный отросток лучевой кости, 18 — лучевая
кость, 19 — головка лучевой кости, 20 — гребень большого
бугорка, 21 — межбугорковая борозда, 22 — большой
бугорок, 23 — малый бугорок, 24 — головка
плечевой кости, 25 — акромион
187
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
Латеральный угол лопатки заканчивается утолщением с углубленной
суставной впадиной, участвующей в образовании плечевого сустава.
С глубокой древности лопатке уделяли очень большое внимание.
Гадание по лопатке — скапулимантия — было широко распространено
у древних греков и арабов. Для предсказаний также использовали ло-
патку оленей, баранов. Однако насечки на найденных лопатках ма-
монтов интерпретируют ныне как фиксацию фаз луны.
Кости свободной верхней конечности. Плечевая кость — длинная
трубчатая кость, состоящая из цилиндрического тела, которое внизу
приобретает трехгранную форму, вверху имеет шаровидную головку, со-
членяющуюся с лопаткой, образуя шаровидный многоосный плечевой
сустав, который у человека, в связи с прямохождением, наиболее свобо-
ден. В плечевом суставе осуществляется сгибание и разгибание руки, ее
отведение и приведение, вращение кнаружи и кнутри, а также круговое
движение. Движение руки выше плеча происходит благодаря тому, что
вся конечность движется вместе с плечевым поясом. Внизу плечевая
кость заканчивается сложно устроенным мыщелком, образованным
блоком и головкой, сочленяющимися с обеими костями предплечья.
Локтевая кость — длинная, трубчатая, ее тело напоминает трех-
гранную призму. Более массивный верхний эпифиз сочленяется
с плечевой и локтевой костями. Нижний эпифиз локтевой кости (ее
головка) сочленяется с лучевой костью. Длинная трубчатая лучевая
кость имеет головку, которая сочленяется с плечевой и локтевой ко-
стями. Сложный локтевой сустав образован тремя суставами: плече-
локтевым, плечелучевым и верхним лучелоктевым. В этом суставе
осуществляется сгибание и разгибание предплечья, его вращение
(пронация и супинация). Нижний эпифиз лучевой кости сочленяется
с верхним рядом костей запястья и локтевой костью.
Кисть делится на три отдела: запястье, пясть и пальцы (рис. 106).
Восемь костей запястья располагаются в два ряда. В проксимальном
лежат (начиная от лучевого края) ладьевидная, полулунная, трехгран-
ная, гороховидная (сесамовидная кость); в дистальном — кость-трапе-
ция (большая многоугольная), трапециевидная, головчатая и крючко-
видная. Кости запястья сочленяются между собой, кости верхнего
ряда — с запястной суставной поверхностью лучевой кости, образуя эл-
липсоидный лучезапястный сустав, в котором осуществляется сгиба-
ние, разгибание, приведение и отведение кисти. Кости, расположенные
в дистальном ряду запястья, соединяясь между собой и со второй-пятой
пястными костями, образуют суставы, укрепленные связками. Они
формируют твердую основу кисти, которая отличается большой проч-
ностью. Кости запястья образуют костный свод, обращенный выпукло-
стью к тылу кисти, а вогнутостью в сторону ладони. Благодаря этому
формируется борозда запястья, в которой проходят сухожилия пальцев.
188
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Пясть состоит из пяти костей, каждая из которых представляет со-
бой короткую трубчатую кость, сочленяющуюся с проксимальной фа-
лангой соответствующего пальца, образуя пястно-фаланговый сустав,
и с костями запястья, образуя запястно-пястные суставы. Особенно
важен седловидный запястно-пястный сустав большого пальца.
В нем совершаются разнообразные движения, среди которых важную
роль в трудовой деятельности играет противопоставление большого
пальца остальным. В древности пленным отрубали этот палец, дабы
Рис. 106. Кости кисти, правой, ладонная поверхность:
1 — локтевая кость, 2 — головка локтевой кости, 3 — шиловидный отросток локтевой
кости, 4 — полулунная кость, 5 — трехгранная кость, 6 — гороховидная кость,
7 — крючковидная кость, 8 — крючок крючковидной кости, 9 — кости запястья,
10 — основание пястной кости, 11 — тело пястной кости, 12 — головка пястной
кости, 13 — пястные кости (I–V), 14 — основание фаланги, 15 — тело фаланги,
16 — головка фаланги, 17 — кости пальцев, фаланги, 18 — бугристость дистальной
фаланги, 19 — дистальная фаланга (II), 20 — средняя фаланга (II), 21 — проксимальная
фаланга (II), 22 — дистальная фаланга (I), 23 — проксимальная фаланга (I),
24 — сесамовидные кости, 25 — пястная кость (I), 26 — трапециевидная кость,
27 — кость-трапеция, 28 — бугорок кости-трапеции, 29 — бугорок ладьевидной
кости, 30 — головчатая кость, 31 — ладьевидная кость, 32 — лучевая кость
11
1
7
1
7
22
18
18
33
19
19
44
20
20
55
21
21
66
22
22
77
23
23
88
24
24
9
9
25
25
10
10
26
26
11
11
27
27
12
12
28
28
1
3
1
3
29
29
14
14
30
30
15
15
31
31
16
16
32
32
189
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
они не могли взять в руки оружие; аналогично поступали и с гребца-
ми на галерах. В пястно-фаланговых суставах осуществляется сгиба-
ние и разгибание, приведение и отведение пальца.
Скелет пальцев образован короткими трубчатыми костями — фалан-
гами. У первого пальца — две фаланги, у второго-четвертого — по три.
Фаланги сочленяются между собой, образуя блоковидные межфаланго-
вые суставы, в которых совершается сгибание и разгибание фаланг.
Со времен палеолита известны изображения кисти, найденные на
камнях, всевозможных предметах, дверях и стенах домов, надгробных
памятниках. Существует большая литерату-
ра как о каждом пальце кисти, их предназна-
чении, символике, так и о всей кисти. Весьма
сложны не только функции, но и теории
происхождения асимметрии, а также мифо-
логические представления о правой руке и ее
отличиях от левой.
Кости нижней конечности
Нижняя конечность человека является
органом опоры и передвижения, и ее строе-
ние наилучшим образом приспособлено
к выполнению этих важных функций. Ко-
нечность состоит из пояса — это тазовые ко-
сти, между которыми сзади как бы вклини-
вается крестец, и свободной нижней
конечности (рис. 107).
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
Рис. 107. Кости нижней конечности, вид спереди:
1 — крестец, 2 — крестцово-подвздошный сустав,
3 — верхняя ветвь лобковой кости, 4 — симфизиальная
поверхность лобковой кости, 5 — нижняя ветвь лобковой
кости, 6 — ветвь седалищной кости, 7 — седалищный бугор,
8 — тело седалищной кости, 9 — медиальный надмыщелок
бедренной кости, 10 — медиальный мыщелок большеберцовой
кости, 11 — бугристость большеберцовой кости, 12 — тело
большеберцовой кости, 13 — медиальная лодыжка,
14 — фаланги пальцев, 15 — кости плюсны, 16 — кости
предплюсны, 17 — латеральная лодыжка, 18 — малоберцовая
кость, 19 — передний край большеберцовой кости,
20 — головка малоберцовой кости, 21 — латеральный мыщелок
большеберцовой кости, 22 — латеральный надмыщелок
бедренной кости, 23 — надколенник, 24 — бедренная кость,
25 — большой вертел бедренной кости, 26 — шейка
бедренной кости, 27 — головка бедренной кости,
28 — крыло подвздошной кости, 29 — подвздошный гребень
190
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Кости пояса нижней конечности. Тазовая кость — парная плоская
кость, образована подвздошной, лобковой и седалищной костями,
срастающимися между собой ко времени полового созревания в обла-
сти вертлужной впадины — глубокой ямки, сочленяющейся с голов-
кой бедренной кости. Подвздошная кость расположена над впадиной,
лобковая — кпереди и книзу, седалищная — книзу и сзади от нее. Се-
далищная и лобковая кости ограничивают крупное запирательное от-
верстие овальной формы, затянутое соединительнотканной запира-
тельной мембраной (рис. 108).
Седалищная кость имеет тело, которое участвует в образовании вер-
тлужной впадины, а ее ветвь ограничивает запирательное отверстие
и образует мощный седалищный бугор, хорошо выраженный у челове-
ка разумного. Лобковая кость также имеет тело, участвующее в форми-
ровании вертлужной впадины, и две ветви — верхнюю и нижнюю, сое-
Рис. 108. Таз:
А — вид сверху: 1 — расстояние между гребнями
подвздошных костей, 2 — поперечный размер
входа в малый таз, 3 — расстояние между
передними верхними остями, 4 — косой размер
входа в малый таз, 5 — прямой размер (истинная
конъюгата), 6 — лобковый симфиз,
7 — подвздошно-лобковое возвышение,
8 — верхняя передняя подвздошная ость,
9 — крыло подвздошной кости, 10 — крестцово-
подвздошное сочленение; Б — сагиттальный
распил, вид изнутри (со стороны таза):
1 — наружная конъюгата, 2 — прямой
размер (полости таза), 3 — ось таза,
4 — прямой размер (выхода из полости таза),
5 — диагональная конъюгата, 6 — истинная
(гинекологическая) конъюгата
4
3
2
5
6
1
4
3
2
5
6
7
8
9
10
1
Б
А
191
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
диняющиеся между собой под углом. На медиальной поверхности угла
имеется симфизиальная поверхность, которая, соединяясь с такой же
поверхностью противоположной кости, образует лобковый симфиз.
Подвздошная кость состоит из массивного тела и тонкого крыла,
оканчивающегося подвздошным гребнем. Вогнутая внутренняя по-
верхность крыла подвздошной кости формирует подвздошную ямку.
Крестцово-тазовая поверхность несет на себе ушковидную поверх-
ность, сочленяющуюся с одноименной поверхностью крестца, обра-
зуя плоский крестцово-подвздошный сустав, укрепленный мощными
связками, в котором движения практически отсутствуют. Благодаря
этому образуется прочное тазовое кольцо, имеющее арочное строение,
несущее на себе тяжесть туловища и передающее ее массивным ко-
стям свободной нижней конечности.
Таз состоит из двух отделов — большого таза и малого таза, которые
отделены один от другого пограничной линией, образованной дугообраз-
ной линией (правой и левой) подвздошных костей и гребнями лобко-
вых, сзади — мысом крестца, впереди — верхним краем лобкового сим-
физа. Большой таз образован крыльями подвздошных костей и телом
V поясничного позвонка. Малый таз ограничен ветвями лобковых и се-
далищных костей, седалищными буграми, крестцово-бугорными связка-
ми, крестцом и копчиком. Передняя стенка таза короткая — это описан-
ный выше симфиз. Длинная задняя стенка таза образована крестцом и
копчиком, боковые стенки — внутренними поверхностями тазовых ко-
стей и мощными связками (крестцово-бугорной и крестцово-остистой).
У женщин таз шире и короче, объем его и все размеры больше, чем
у мужчин. Кости женского таза тоньше, чем у мужчин. Крестец мужчи-
ны более узкий и вогнутый, а мыс выдается вперед, у женщин крестец
широкий и уплощенный, а мыс выражен в меньшей степени. Угол, под
которым соединяются нижние ветви лобковых костей (подлобковый
угол), у мужчин острый (около 70—75°), у женщин приближается
к прямому и даже тупому (90—100°). Седалищные бугры и крылья под-
вздошных костей у женщин расположены дальше друг от друга. Ниж-
нее отверстие женского таза шире, чем у мужчин, оно имеет форму по-
перечного овала. Итак, половые отличия женского таза сводятся
в основном к его большим размерам, большему объему и увеличению
нижнего отверстия. Это связано с выполнением основной функции: таз
женщины является вместилищем развивающегося в матке плода, кото-
рый во время родов покидает полость таза через нижнюю апертуру. По-
ловые различия таза начинают проявляться в возрасте 8—10 лет.
Кости свободной нижней конечности. Бедренная кость — наиболее
крупная, массивная трубчатая кость скелета человека. Существует
прямая корреляция между ее длиной и ростом человека. Шаровидная
головка бедренной кости сочленяется с вертлужной впадиной тазовой
192
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
кости, образуя шаровидный многоостный тазобедренный сустав, в ко-
тором осуществляются сгибание и разгибание бедра, его отведение
и приведение, вращение внутрь и кнаружи, круговые движения. Длин-
ная шейка, соединяющая головку с телом бедренной кости, располо-
жена под углом к последнему. У мужчин этот угол тупой (около 130°),
у женщин почти прямой. Тотчас под шейкой латерально расположен
большой вертел, с медиальной стороны находится малый вертел. Изо-
гнутое вперед тело бедренной кости цилиндрической формы характер-
но только для человека разумного. Наиболее сложно устроен нижний
эпифиз бедренной кости, на котором находятся два мощных мыщелка,
разделенные глубокой межмыщелковой ямкой, переходящей впереди
в надколенниковую поверхность. Надколенник представляет собой се-
самовидную кость, лежащую в толще сухожилия четырехглавой мыш-
цы бедра, легко прощупывающуюся у живого человека.
Массивная длинная трубчатая большеберцовая кость — единст-
венная из двух костей голени, которая сочленяется с бедренной. Мощ-
ный широкий верхний эпифиз имеет два мыщелка, несущие на своих
верхних концах суставные поверхности. Для достижения соответст-
вия формы мыщелков бедра суставным поверхностям большеберцо-
вой кости между ними находятся два хрящевых мениска. На боковой
поверхности латерального мыщелка большеберцовой кости имеется
суставная поверхность, сочленяющаяся с головкой малоберцовой ко-
сти. В образовании двухостного сложного комплексного коленного су-
става участвуют нижний эпифиз бедренной кости, надколенник и
верхний эпифиз большеберцовой кости. В коленном суставе осу-
ществляется сгибание и разгибание голени, а при ее полусогнутом по-
ложении — и вращение. Трехгранное тело большеберцовой кости пе-
реходит в ее нижний эпифиз примерно четырехугольной формы,
который несет на себе нижнюю суставную поверхность для сочлене-
ния с таранной костью стопы. Медиальный конец его оттянут и обра-
зует медиальную лодыжку.
Тонкая длинная трубчатая малоберцовая кость имеет головку, на
которой расположена суставная поверхность для сочленения с верх-
ним эпифизом большеберцовой кости. Трехгранное тело внизу окан-
чивается утолщенной латеральной лодыжкой, снабженной суставной
поверхностью.
В стопе различают предплюсну, плюсну и пальцы (рис. 109). Кости
предплюсны, испытывающие большую нагрузку, массивные, прочные.
Это семь коротких костей, расположенных в два ряда. В проксималь-
ном (заднем) — таранная и пяточная кости, в дистальном (переднем)
латерально располагается кубовидная кость, медиально — узкая ладье-
видная и впереди нее — три клиновидные кости. Нижняя суставная по-
верхность большеберцовой кости и суставные поверхности лодыжек
193
СКЕЛЕТ И ЕГО СОЕДИНЕНИЯ
образуют вилку, которая охватывает блок таранной кости сверху
и с боков, образуя сложный блоковидный голеностопный сустав, в ко-
тором осуществляется тыльное и подошвенное сгибание стопы.
Кости предплюсны соединены между собой множеством суставов.
Наиболее крупная пяточная кость соединяется с таранной костью
вверху и кубовидной спереди. Ладьевидная, кубовидная и три клино-
видные кости соединяются между собой, а первые две, кроме того, —
с пяточной и таранной; клиновидная и кубовидная — с плюсневыми
костями. Многочисленные соединения костей стопы укреплены проч-
ными связками. В межплюсневых суставах движения чаще всего соче-
танные: вращение пяточной кости вместе с ладьевидной и передним
концом стопы вокруг косой сагиттальной оси. При вращении стопы
внутрь (пронация) латеральный край ее поднимается, при вращении
кнаружи (супинация) медиальный край приподнимается, тыльная по-
верхность стопы поворачивается в латеральную сторону.
Рис. 109. Кости стопы, правой,
вид сверху:
1 — бугор пяточной кости,
2 — тело таранной кости,
3 — шейка таранной кости,
4 — головка таранной кости,
5 — таранная кость,
6 — ладьевидная кость,
7 — промежуточная клиновидная
кость, 8 — медиальная
клиновидная кость,
9 — основание плюсневой кости,
10 — тело плюсневой кости,
11 — головка плюсневой кости,
12 — плюсневая кость (I),
13 — основание фаланги,
14 — тело фаланги,
15 — головка фаланги,
16 — проксимальная фаланга (I),
17 — дистальная фаланга (I),
18 — дистальная фаланга (V),
19 — средняя фаланга (V),
20 — проксимальная фаланга (V),
21 — латеральная клиновидная
кость, 22 — бугористость
пятой (V) плюсневой кости,
23 — кубовидная кость,
24 — пяточная кость, 25 — задний
отросток таранной кости
11
17
17
22
18
18
33
19
19
44
20
20
55
21
21
66
22
22
77
23
23
88
24
24
99
25
25
10
10
11
11
12
12
13
13
14
14
15
15
16
16
194
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Пять коротких трубчатых плюсневых костей своими основаниями
сочленяются с клиновидными и кубовидной костями, образуя мало-
подвижные предплюсне-плюсневые суставы, а своими головками —
с основаниями соответствующих проксимальных фаланг.
Скелет пальцев образован короткими трубчатыми костями — фа-
лангами. Количество их соответствует фалангам пальцев кисти, однако
они отличаются небольшими размерами. Каждая проксимальная фа-
ланга своим основанием сочленяется с соответствующей плюсневой
костью. В эллипсоидных двухостных плюсне-фаланговых суставах
производится сгибание, разгибание, приведение и отведение пальцев.
Головка проксимальной фаланги сочленяется со средней фалангой, по-
следние — с основаниями дистальных фаланг. В блоковидных одноост-
ных межфаланговых суставах осуществляется сгибание и разгибание.
Стопа человека — орган опоры и передвижения — несет на себе
всю тяжесть человеческого тела. Это накладывает существенный от-
печаток на ее строение и характер соединения костей. Стопа постро-
ена по типу прочной и упругой сводчатой арки с короткими пальцами.
Основные особенности строения стопы современного человека раз-
умного — это наличие сводов, прочность, пронированное положение,
укрепление медиального края, укорочение пальцев, укрепление
и приведение первого пальца, который, в отличие от большого пальца
кисти, не противопоставляется остальным, и расширение его дисталь-
ной фаланги. Формирование сводов обусловлено тем, что кости меди-
ального края предплюсны лежат выше, чем кости латерального края.
Следует подчеркнуть, что лишь у человека разумного имеется сводча-
тая стопа. Она представлена пятью продольными и одним попереч-
ным сводами (дугами), которые обращены выпуклостью кверху. Сво-
ды образованы сочленяющимися между собой костями предплюсны
и плюсны. Каждый продольный свод начинается от одной и той же
точки пяточной кости, включает кости предплюсны и соответствую-
щую плюсневую кость. В образовании первого свода (медиального)
участвует таранная кость. Стопа в целом имеет три точки опоры: пя-
точный бугор и головки первой и пятой плюсневых костей. Продоль-
ные своды имеют неодинаковую высоту, наиболее высокий из них
второй свод (вторая дуга). В результате формируется поперечный свод
стопы, в образовании которого принимают участие ладьевидная, кли-
новидные и кубовидные кости. Конструкция стопы в виде сводчатой
арки у живого человека поддерживается благодаря форме костей,
прочности связок (пассивные «затяжки» стопы) и тонусу мышц (ак-
тивные «затяжки»). В соединениях костей стопы особенно ярко про-
является диалектическая зависимость структуры и функции. С помо-
щью специальных упражнений можно добиться изумительной
гибкости позвоночника, увеличить объем движений во всех суставах
и предотвратить их возрастные изменения.
195
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
АКТИВНАЯ ЧАСТЬ
ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
Скелетные мышцы
И. М. Сеченов в книге «Рефлексы головного мозга» пишет: «Все
бесконечное разнообразие внешних проявлений мозговой деятель-
ности сводится окончательно к одному лишь явлению — мышечному
движению. Смеется ли ребенок при виде игрушки, улыбается ли Га-
рибальди, когда его гонят за излишнюю любовь к Родине, дрожит ли
девушка при первой мысли о любви, создает ли Ньютон мировые за-
коны и пишет их на бумаге — везде окончательным фактом является
мышечное движение».
Скелетные мышцы приводят в движение кости, активно изменяют
положение тела человека, участвуют в образовании стенок ротовой,
брюшной полостей, таза, входят в состав стенок глотки, верхней части
пищевода, гортани, осуществляют движения глазного яблока и слухо-
вых косточек, дыхательные и глотательные движения. Скелетные
мышцы удерживают тело человека в равновесии, перемещают его
в пространстве. Общая масса скелетной мускулатуры у новорожден-
ного 20—22% массы тела; у взрослого человека достигает 40%; у по-
жилых и старых людей уменьшается до 25—30%. У человека около
400 поперечнополосатых мышц, сокращающихся произвольно под
воздействием импульсов, поступающих по нервам из центральной
нервной системы (рис. 110, 111).
Пучки поперечнополосатых мышечных волокон образуют скелет-
ные мышцы, которые иннервируются мотонейронами — двигательны-
ми нейронами передних рогов спинного мозга (см. «Спинной мозг»).
В зависимости от ряда морфофизиологических показателей (толщи-
ны волокон, содержания в них миоглобина, количества митохондрий,
активности окислительных ферментов) различают красные, белые
и промежуточные поперечнополосатые мышечные волокна. Красные
волокна богаты саркоплазмой, миоглобином и митохондриями, актив-
ность окислительных ферментов в них высокая, однако они самые
тонкие, количество миофибрилл в них невелико, и они расположены
группами. Более толстые промежуточные волокна беднее миоглоби-
ном и митохондриями. И, наконец, самые толстые белые волокна со-
держат меньше всего саркоплазмы, миоглобина и митохондрий, но
количество миофибрилл в них больше и располагаются они равно-
мерно, в них ниже активность окислительных ферментов. Структура
и функция волокон неразрывно связаны между собой. С функцио-
нальной точки зрения мышца состоит из двигательных единиц.
Каждая двигательная единица — это группа мышечных волокон
196
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
11
17
17
33
33
22
18
18
34
34
33
19
19
35
35
44
20
20
36
36
55
21
21
37
37
66
22
22
38
38
77
23
23
39
39
88
24
24
40
40
99
25
25
41
41
10
10
26
26
42
42
11
11
27
27
43
43
12
12
28
28
44
44
13
13
29
29
45
45
14
14
30
30
46
46
15
15
31
31
47
47
16
16
32
32
48
48
49
49
Рис. 110. Мышцы тела человека,
вид спереди:
1 — височная мышца, 2 — лобное
брюшко (затылочно-лобная мышца),
3 — круговая мышца рта,
4 — подбородочная мышца,
5 — грудино-подъязычная
мышца, 6 — локтевой
разгибатель запястья,
7 — разгибатель мизинца,
8 — разгибатель пальцев,
9 — локтевой сгибатель
запястья, 10 — локтевая
мышца, 11 — большая
грудная мышца, 12 — прямая
мышца живота, 13 — наружная
косая мышца живота,
14 — пирамидальная мышца,
15 — напрягатель широкой фасции
бедра, 16 — подвздошно-поясничная
мышца, 17 — гребенчатая мышца,
18 — длинная приводящая мышца,
19 — портняжная мышца,
20 — тонкая мышца, 21 — прямая
мышца бедра, 22 — латеральная
широкая мышца, 23 — медиальная
широкая мышца, 24 — икроножная
мышца, 25 — камбаловидная мышца,
26 — длинный сгибатель пальцев,
27 — длинный разгибатель пальцев
(сухожилие), 28 — мышца,
отводящая большой палец стопы,
29 — длинный разгибатель пальцев,
30 — передняя большеберцовая
мышца, 31 — короткий разгибатель
большого пальца кисти,
32 — длинная мышца, отводящая
большой палец кисти, 33 — короткий
лучевой разгибатель запястья,
35 — разгибатель пальцев,
36 — лучевой сгибатель запястья,
37 — длинный лучевой разгибатель
запястья, 38 — плечелучевая мышца,
39 — плечевая мышца,
40 — трехглавая мышца плеча,
41 — двуглавая мышца плеча,
42 — передняя зубчатая мышца,
43 — дельтовидная мышца,
44 — трапециевидная мышца,
45 — грудино-ключично-
сосцевидная мышца, 46 — мышца,
опускающая угол рта,
47 — жевательная мышца,
48 — большая скуловая мышца,
49 — круговая мышца глаза
197
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
11
17
17
22
18
18
33
19
19
44
20
20
55
21
21
66
22
22
77
23
23
88
24
24
99
25
25
10
10
26
26
11
11
27
27
12
12
28
28
13
13
29
29
14
14
30
30
15
15
31
31
16
16
32
32
33
33
34
34
Рис. 111. Мышцы тела человека,
вид сзади:
1 — височная мышца, 2 — затылочное
брюшко (затылочно-лобная мышца),
3 — полуостистая мышца головы,
4 — ременная мышца головы,
5 — грудино-ключично-сосцевидная
мышца, 6 — трапециевидная мышца,
7 — дельтовидная мышца,
8 — трехглавая мышца плеча,
9 — двуглавая мышца плеча,
10 — плечевая мышца, 11 — круглый
пронатор, 12 — плечелучевая мышца,
13 — лучевой сгибатель запястья,
14 — длинная ладонная мышца,
15 — локтевой сгибатель запястья,
16 — поверхностный сгибатель пальцев
(сухожилие), 17 — полусухожильная
мышца, 18 — полуперепончатая
мышца, 19 — двуглавая мышца бедра,
20 — икроножная мышца,
21 — камбаловидная мышца,
22 — длинная малоберцовая мышца
(сухожилие), 23 — короткая
малоберцовая мышца,
24 — подошвенная мышца,
25 — большая ягодичная мышца,
26 — средняя ягодичная мышца,
27 — наружная косая мышца живота,
28 — широчайшая мышца спины,
29 — передняя зубчатая мышца,
30 — большая круглая мышца,
31 — подостная мышца, 32 — малая
круглая мышца, 33 — плечелучевая
мышца, 34 — жевательная мышца
198
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
(миосимпластов), иннервируемых одним двигательным нейроном пе-
редних рогов спинного мозга, которые сокращаются одновременно.
У человека двигательная единица, как правило, состоит из 150 (и бо-
лее) мышечных волокон, причем в различных мышцах число волокон,
входящих в состав двигательной единицы, различно. Так, в наружной
прямой мышце глаза человека двигательная единица включает 13—
20 мышечных волокон, в двуглавой мышце плеча — 750—1000, в ме-
диальной головке икроножной мышцы — 1500—2000. Будучи интер-
нируемыми одним двигательным нейроном, все мышечные волокна
одной двигательной единицы сокращаются одновременно, но различ-
ные двигательные единицы могут сокращаться как одновременно, так
и последовательно.
Поперечнополосатые мышечные волокна одной двигательной еди-
ницы идентичны по своему строению и функциональным особенно-
стям. Различают две разновидности двигательных единиц: медленные
и быстрые. Медленные двигательные единицы состоят из небольшого
числа богатых митохондриями и окислительными ферментами крас-
ных мышечных волокон, которые хорошо кровоснабжаются (4—6 ка-
пилляров на одно мышечное волокно). Такие двигательные единицы
развивают небольшую силу, сокращаются медленно, выполняют дли-
тельную работу умеренной мощности, практически не утомляясь.
Быстрые двигательные единицы, в свою очередь, подразделяются
на две группы: легкоутомляемые и устойчивые к утомлению. Легко-
утомляемые образованы большим количеством белых мышечных во-
локон, они сокращаются с большой скоростью, развивая при этом
большую силу, однако быстро утомляются. Эти двигательные едини-
цы способны выполнять большую работу в течение короткого време-
ни. Быстрые, устойчивые к утомлению двигательные единицы силь-
ные и сокращаются быстро. Они образованы промежуточными
волокнами, которые по своим морфофункциональным особенностям
занимают положение между медленными неутомляемыми (красные)
и быстрыми утомляемыми (белые).
Скелетные мышцы человека содержат мышечные волокна всех ти-
пов, однако в зависимости от функции мышцы в ней преобладает тот
или иной тип волокон. Например, в четырехглавой мышце бедра че-
ловека относительное количество «красных» волокон (медленных
единиц) колеблется в пределах от 40 до 98%. В то же время соотноше-
ние тех и других в каждой мышце строго индивидуально и детерми-
нировано генетически. Чем больше в мышцах «белых» (быстрых) во-
локон, тем человек лучше приспособлен к выполнению физической
работы, требующей большой силы и скорости; чем больше «красных»
(медленных) волокон, тем выносливее человек. Аналогичные отноше-
ния и у животных. У длительно летающих птиц, например, в грудных
199
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
мышцах преобладают красные во-
локна, в то время как у нелетающих
кур — белые.
Мышца как орган состоит из пуч-
ков поперечнополосатых мышечных
волокон, каждое из которых покры-
то соединительнотканной оболочкой
(эндомизий). Пучки волокон различ-
ной величины отделены друг от дру-
га прослойками соединительной тка-
ни, которые образуют перимизий.
Мышца в целом покрыта наружным
перимизием (эпимизий), который
переходит на сухожилие (рис. 112).
Из эпимизия в мышцу проникают
кровеносные сосуды, разветвляю-
щиеся во внутренний перимизий и
эндомизий, в последнем располага-
ются капилляры и нервные волокна.
Мышцы и сухожилия богаты чувст-
вительными нервными окончаниями,
воспринимающими «мышечное и су-
хожильное чувство» — информацию
о тонусе мышечных волокон, степени
их сокращения, растяжении сухожи-
лий — и передающими ее по нервам
в мозг. Эти рецепторы образуют
нервно-мышечные и нервно-сухо-
жильные веретена, окруженные сое-
динительнотканной капсулой. Двига-
тельные окончания аксонов образуют
моторные бляшки (аксомышечные
синапсы), напоминающие по своему
строению синапсы.
Мышечные пучки формируют брюшко, переходящее в сухожиль-
ную часть. Проксимальный отдел мышцы — ее головка — начинается
от кости; дистальный конец — хвост (сухожилие) — прикрепляется
к другой кости. Исключением из этого правила являются мимические
мышцы, мышцы дна полости рта и промежности, которые не прикре-
пляются к костям. Начало мышцы находится проксимальнее, чем
точка ее прикрепления, которая располагается дистальнее. Начало со-
кращающейся мышцы остается неподвижным, это ее фиксированная
точка. На другой кости, к которой прикрепляются мышцы, находится
Рис. 112. Скелетная мышца:
1 — перимизий,
2 — мышечный пучок,
3 — эпимизий, 4 — фасция,
5 — артерия, 6 — кость,
7 — сухожилие, 8 — нерв,
9 — миофибриллы
11
99
22
22
33
44
55
66
77
88
44
33
200
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
АБ
В
Г
Д
Е
Ж
З
И
К
ЛМ
1
2
3
5
1
2
1
1
1
1
2
2
4
1
2
2
5
2
2
1
Рис. 113. Скелетные мышцы, разные по форме и строению:
А — веретенообразная мышца, Б — одноперистая мышца, В — двуперистая мышца,
Г — двуглавая мышца, Д — многоперистая мышца, Е — треугольная мышца,
Ж — двубрюшная мышца, З — круговая мышца, И — широкая мышца, имеющая
апоневроз, К — зубчатая мышца, Л — квадратная мышца, М — мышца, имеющая
сухожильные перемычки; 1 — сухожилие, 2 — брюшко, 3 — апоневроз, 4 — головка,
5 — сухожильная перемычка
201
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
подвижная точка. При сокращении мышцы она изменяет свое поло-
жение. При некоторых движениях положения фиксированной и под-
вижной точек меняются.
Сухожилия различных мышц отличаются друг от друга. Так, мыш-
цы конечностей имеют узкие и длинные сухожилия. Широкое и пло-
ское сухожилие, сухожильное растяжение, или апоневроз, характерно
для мышц, участвующих в формировании стенок полостей тела.
Брюшко некоторых мышц разделено промежуточным сухожилием,
например двубрюшная мышца. На протяжении некоторых мышц име-
ются промежуточные сухожилия, называемые сухожильными пере-
мычками, например прямая мышца живота. Будучи относительно
тонким, сухожилие мало растяжимо, обладает значительной прочно-
стью и выдерживает огромные нагрузки. Например, сухожилие четы-
рехглавой мышцы бедра способно выдержать растяжение силой
600 кг, ахиллово сухожилие — 400 кг.
Форма мышцы связана с ее функцией (рис. 113). На конечностях
чаще всего встречаются мышцы веретенообразной формы, так как
они прикрепляются общими своими концами к длинным костям, вы-
полняющим роль рычагов (например, двуглавая мышца плеча). Мыш-
цы лентовидной формы, либо в виде пластин, участвуют в образова-
нии стенок туловища (например, косые и поперечные мышцы живота).
Пучки некоторых мышц расположены циркулярно (например, круго-
вая мышца рта). Эти мышцы — сжиматели, они окружают ротовое,
заднепроходное и другие естественные отверстия тела человека.
Мышцы имеют ряд вспомогательных образований. Каждая мышца
или группа сходных по функциям мышц окружены собственной фас-
цией. Если мышцы лежат в несколько слоев, то между соседними
мышцами располагаются листки фасции: между поверхностными —
поверхностный, между глубокими — глубокий. Поверхностная фас-
ция отделяет мышцы от подкожной клетчатки. Она целиком окутыва-
ет все мышцы какой-нибудь области (например, конечности).
Мышечные перегородки разделяют группы мышц, выполняющих раз-
личную функцию.
Синовиальное влагалище отделяет движущееся сухожилие от не-
подвижных стенок фиброзного влагалища и устраняет их трение. Си-
новиальное влагалище представляет собой заполненную небольшим
количеством жидкости полость, ограниченную висцеральным и пари-
етальным листками.
Элементы биомеханики
При сокращении концы мышцы, прикрепленные к костям, при-
ближаются друг к другу. Кости, соединенные суставами, действуют
202
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
как рычаги. В биомеханике выделяют два типа рычагов: рычаг перво-
го рода — точки приложения действующих на него сил (сопротивле-
ния и приложения силы) находятся по разные стороны от точки опо-
ры; рычаг второго рода — обе силы прилагаются по одну сторону от
точки опоры (рис. 114).
Изменяя положение костных рычагов, мышцы действуют на суста-
вы. При этом каждая мышца влияет на сустав только в одном направ-
лении. У одноосного сустава (цилиндрический, блоковидный) имеют-
ся две действующие на него мышцы, являющиеся антагонистами; одна
мышца — сгибатель, другая — разгибатель. В то же время на каждый
сустав в одном направлении действуют, как правило, две и более мыш-
цы, являющиеся синергистами. У двуосного сустава (эллипсоидный,
мыщелковый, седловидный) мышцы группируются соответственно
двум его осям, вокруг которых совершаются движения. К шаровидно-
му суставу, имеющему три оси движения (многоостный сустав), мыш-
цы прилегают со всех сторон. Так, в плечевом суставе имеются мышцы
сгибатели и разгибатели (движения вокруг фронтальной оси), отводя-
щие и приводящие (движения вокруг сагиттальной оси) и вращатели,
осуществляющие движения вокруг продольной оси: вовнутрь (прона-
торы) и кнаружи (супинаторы).
В группе мышц синерги-
стов или антагонистов имеют-
ся главные, которые осуществ-
ляют конкретное движение,
и вспомогательные, которые
как бы моделируют движения
и создают его индивидуаль-
ный характер. При сокраще-
нии соответствующих мышц
тело человека, не совершая тех
или иных движений, удержи-
вается в определенном поло-
жении. Исходя из этого, раз-
личают три вида работы
мышц: преодолевающую, усту-
пающую и удерживающую.
À
Á
Â
I
À
Á
Â
II
À
Á
Â
III
Рис. 114. Действие мышц на костные рычаги:
I — рычаг равновесия, II — рычаг силы, III — рычаг
скорости; А — точка опоры, Б — точка приложения
силы, В — точка сопротивления
203
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
Точка приложения равнодействующих всех сил по отношению
к телу человека — это центр его тяжести. Общий центр тяжести у муж-
чин расположен на уровне II крестцового позвонка, у женщин — не-
сколько ниже, у детей — выше; у новорожденного — на уровне VI груд-
ного, у двухлетнего — I поясничного, у шестилетнего — III поясничного
позвонка.
Мышцы различных областей
Мышцы головы делятся на две группы: мимические и жеватель-
ные. Однако следует подчеркнуть, что в ряде случаев они функциони-
руют совместно (членораздельная речь, жевание, глотание, зевота).
Мимические мышцы располагаются под кожей лица, в основном ра-
диально или циркулярно вокруг ротового, носового отверстий, глаз-
ницы, наружного слухового прохода. Они начинаются от костей или
фасций и вплетаются в кожу, осуществляя мимические движения.
К мимическим относятся мышцы свода черепа, ушной раковины,
окружности глазной и ротовой щелей, носовых отверстий (рис. 115).
Жевательные мышцы располагаются на боковых отделах черепа
по четыре с каждой стороны, две из них (жевательная и височная) —
более поверхностно, две (крыловидные) — в нижне-височной ямке.
Все они начинаются на костях лица и прикрепляются к нижней челю-
сти, приводя ее в движение (рис. 116). Жевательные мышцы человека
менее развиты, чем у человекообразных обезьян.
12
13
14
15
4
5
6
3
2
1
7
8
9
10
11
16
Рис. 115. Мимические мышцы,
вид справа:
1 — сухожильный шлем (надчерепной
апоневроз), 2 — лобное брюшко
затылочно-лобной мышцы,
3 — круговая мышца глаза, 4 — мышца,
поднимающая верхнюю губу,
5 — мышца, поднимающая угол рта,
6 — круговая мышца рта, 7 — большая
скуловая мышца, 8 — мышца,
опускающая нижнюю губу,
9 — мышца, опускающая угол рта,
10 — мышца смеха, 11 — подкожная
мышца шеи, 12 — грудино-ключично-
сосцевидная мышца,
13 — трапециевидная мышца,
14 — задняя ушная мышца,
15 — затылочное брюшко
затылочно-лобной мышцы,
16 — верхняя ушная мышца
204
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Рис. 116. Жевательные мышцы,
вид справа:
А — скуловая дуга отпилена
и отвернута вместе с жевательной
мышцей: 1 — височная мышца,
2 — венечный отросток нижней
челюсти, 3 — жевательная мышца,
4 — латеральная крыловидная мышца;
Б — скуловая дуга и часть ветви
нижней челюсти удалены:
1 — скуловая дуга (отпилена),
2 — медиальная крыловидная мышца,
3 — угол нижней челюсти,
4 — ветвь нижней челюсти,
5 — латеральная крыловидная мышца
4
5
32
1
4
3
2
Б
А
1
4
3
2
5
6
7
8
9
10
11
12
13
14
15
16
17
18
1
Рис. 117. Поверхностные мышцы
спины (первый слой):
1 — трапециевидная мышца,
2 — ость лопатки, 3 — дельтовидная
мышца, 4 — подостная мышца,
5 — малая круглая мышца,
6 — большая круглая мышца,
7 — большая ромбовидная мышца,
8 — широчайшая мышца спины,
9 — поясничный треугольник,
10 — наружная косая мышца
живота, 11 — подвздошный
гребень, 12 — средняя ягодичная
мышца, 13 — большая ягодичная
мышца, 14 — апоневроз широчай-
шей мышцы спины, 15 — остистый
отросток XII грудного позвонка,
16 — трехглавая мышца плеча,
17 — грудино-ключично-
сосцевидная мышца,
18 — ременная мышца головы
205
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
Мышцы спины. Спина занимает заднюю поверхность туловища от
наружного затылочного выступа и верхней выйной линии наверху до
крестцово-подвздошных сочленений, задних отделов гребней под-
вздошных костей и копчика внизу. Спереди область спины ограниче-
на задними подмышечными линиями. Мышцы спины располагаются
послойно. Различают поверхностные и глубокие мышцы.
Поверхностные мышцы (рис. 117). Это трапециевидная, широчай-
шая спины, большая и малая ромбовидные, поднимающая лопатку,
верхняя и нижняя задние ромбовидные. Они прикрепляются к лопат-
ке, ключице, плечевой кости и осуществляют их движение. Послед-
ние две прикрепляются к ребрам, участвуют в акте вдоха.
Глубокие мышцы (рис. 118). К глубоким относятся ременная
мышца шеи и головы, мышца, выпрямляющая позвоночник, которые
особо развиты у человека в связи с прямохождением, а также попереч-
но-остистая, межостистые и межпоперечные. Эти мышцы разгиба-
ют позвоночник и удерживают тело человека в вертикальном положе-
нии. Четыре подзатылочные мышцы
(большая и малая прямые, верхняя
и нижняя косые) осуществляют дви-
жения головы.
Мышцы шеи. Область шеи навер-
ху ограничена линией, идущей по
нижнему краю тела и ветви нижней
челюсти до височно-нижнечелюстно-
го сустава и вершины сосцевидного
отростка височной кости, верхней
выйной линии, наружному затылоч-
ному выступу, внизу — яремной вы-
4
3
2
5
6
7
8
9
10
11
12
13
14
15
1
Рис. 118. Глубокие мышцы спины,
второй слой:
1 — полуостистая мышца головы, 2 — задняя
малая прямая мышца головы, 3 — верхняя
косая мышца головы, 4 — нижняя косая мышца
головы, 5 — полуостистая мышца головы,
6 — полуостистая мышца шеи,
7 — полуостистая мышца груди, 8 — мышцы,
поднимающие ребра, 9 — медиальные
межпоперечные мышцы поясницы,
10 — грудопоясничная фасция (глубокая
пластинка), 11 — подвздошно-реберная
мышца поясницы, 12 — подвздошно-реберная
мышца груди, 13 — наружные межреберные
мышцы, 14 — подвздошно-реберная мышца
шеи, 15 — длиннейшая мышца
206
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
резкой грудины, верхними краями ключицы и далее линией, соединя-
ющей последние с остистым отростком VII шейного позвонка. Шея
разделена позвоночником на два отдела: меньший задний (мышцы
этого отдела, относящиеся к мышцам спины, описаны выше) и перед-
ний — собственно область шеи. Движения шеи чаще всего комбини-
рованные, их совершает большое количество мышц, которые делятся
на две большие группы: мышцы, лежащие поверх гортани и кровенос-
ных сосудов, и глубокие (рис. 119).
Мышцы, лежащие поверх гортани и кровеносных сосудов, делятся
на три группы: поверхностные, надподъязычные и подподъязычные.
Последние две группы осуществляют движения подъязычной кости.
К поверхностным мышцам относятся подкожная мышца шеи, кото-
рая у человека сильно редуцирована и является остатком мышцы,
окутывающей все тело многих млекопитающих, и грудино-ключично-
сосцевидная, наиболее развитая у человека в связи с прямохождени-
ем. При двустороннем сокращении они запрокидывают голову, при
одностороннем — наклоняют ее.
Надподъязычные мышцы расположены между нижней челюстью и
подъязычной костью. Это двубрюшная, шилоподъязычная, челюстно-
подъязычная и подбородочно-подъязычная. Они поднимают подъязыч-
ную кость.
Подподъязычные мышцы расположены под кожей впереди гортани,
трахеи и щитовидной железы. Это грудино-подъязычная, грудино-щи-
4
5
6
3
2
1
7
8
9
10
Рис. 119. Мышцы шеи,
вид спереди:
1 — двубрюшная мышца
(переднее брюшко),
2 — челюстно-подъязычная
мышца, 3 — грудино-
ключично-сосцевидная мышца,
4 — грудино-подъязычные
мышцы, 5 — грудино-
щитовидная мышца,
6 — лопаточно-подъязычная
мышца, 7 — лестничные
мышцы (передняя, средняя,
задняя), 8 — подъязычная
кость, 9 — двубрюшная
мышца (заднее брюшко),
10 — шилоподъязычная мышца
207
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
товидная, лопаточно-подъязычная и щитоподъязычная. Они осу-
ществляют движения гортани и опускают подъязычную кость.
Глубокие мышцы расположены на шейном отделе позвоночника
(спереди и сбоку). К ним относятся лестничные, которые поднимают
I и II ребра, длинные мышцы головы и шеи, участвующие в движении
головы и шеи.
Мышцы груди. Область груди, расположенная на передней по-
верхности туловища, вверху ограничена яремной вырезкой грудины и
верхними краями ключиц, внизу — линией, проходящей через основа-
ние мечевидного отростка, реберные дуги по направлению к XII груд-
ному позвонку, с боков задними подмышечными линиями. Мышцы
груди также располагаются послойно.
Поверхностные мышцы (большая и малая грудные, подключичная
и передняя зубчатая) прикрепляются к лопатке, ключице и плечевой
кости, они осуществляют их движения (рис. 120).
К глубоким мышцам груди относятся наружные и внутренние
межреберные, подреберные, поднимающие ребра и поперечные. Они
4
3
2
5
6
7
8
9
1
10
11
Рис. 120. Поверхностные мышцы груди
и живота:
1 — большая грудная мышца,
2 — подмышечная полость, 3 — широчайшая
мышца спины, 4 — передняя зубчатая мышца,
5 — наружная косая мышца живота,
6 — апоневроз наружной косой мышцы
живота, 7 — пупочное кольцо, 8 — белая
линия живота, 9 — паховая связка,
10 — поверхностное паховое кольцо,
11 — семенной канатик
208
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
расположены целиком на груди и осуществляют движения ребер.
В усиленном вдохе участвуют также диафрагма, лестничные, груди-
но-ключично-сосцевидная, большие и малые грудные и другие мыш-
цы; в усиленном выдохе — подреберные, поперечная груди, мышцы
живота (рис. 121).
1
2
3
4
5
6
7
8
9
10
11
17
12
13
14
15
16
Рис. 121. Мышцы груди и живота. Большая грудная мышца (на обеих сторонах)
и наружная косая мышца (слева) отрезаны и удалены. На правой стороне рисунка
передняя стенка влагалища прямой мышцы живота удалена:
1 — грудино-ключично-сосцевидная мышца, 2 — трапециевидная мышца, 3 — большая
грудная мышца (отрезана), 4 — малая грудная мышца, 5 — передняя зубчатая мышца,
6 — прямая мышца живота, 7 — наружная косая мышца живота, 8 — пупочное кольцо,
9 — белая линия живота, 10 — апоневроз наружной косой мышцы живота,
11 — пирамидальная мышца, 12 — внутренняя косая мышца живота, 13 — наружная
косая мышца живота (отрезана и частично удалена), 14 — передняя зубчатая мышца,
15 — большая грудная мышца (отрезана), 16 — дельтовидная мышца,
17 — подключичная мышца
209
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
Мышцы живота. Область живота ограничена сверху линией, про-
ходящей через основание мечевидного отростка и реберные дуги, сни-
зу — подвздошными гребнями, паховыми складками, спереди — верх-
ними ветвями лобковых костей между лобковыми бугорками,
с боков — задними подмышечными линиями (см. рис. 120, 121). Под
диафрагмой расположена брюшная полость, в которой залегают
брюшные органы. Передняя боковая стенка брюшной полости обра-
зована тремя широкими мышцами живота, их сухожильными растя-
жениями и прямыми мышцами живота, задняя — поясничным отде-
лом позвоночника, большой поясничной и квадратной мышцей
поясницы, нижняя — подвздошными костями, мышцами диафрагмы
таза и мочеполовой диафрагмы. Выделяют собственно брюшную по-
лость и полость таза. Стенками последней служат сзади передняя
поверхность крестца с грушевидными мышцами, спереди и с боков —
тазовые кости с внутренними запирательными мышцами, снизу — ди-
афрагма таза и мочеполовая диафрагма. Брюшная полость выстлана
изнутри париетальным листком брюшины, который переходит на
внутренности (брюшина описана ниже).
В связи с прямохождением брюшная стенка человека не несет тя-
жести внутренностей. Она лишена костного скелета, мощные мышцы,
образующие брюшной пресс, компенсируют отсутствие скелета.
Мышцы живота расположены послойно. Различают три группы:
мышцы боковых стенок (наружная и внутренняя косые, поперечные),
мышцы передней стенки (прямая, пирамидальная) и мышцы задней
стенки (квадратная мышца поясницы). Мышцы живота — брюшной
пресс — предохраняют внутренности, оказывают на них давление
и удерживают в определенном положении, а также участвуют в дви-
жениях позвоночника и ребер.
Мышцы боковых стенок переходят в обширные апоневрозы. В ре-
зультате перекреста волокон апоневрозов обеих сторон образуется бе-
лая линия живота, расположенная по его средней линии от мечевид-
ного отростка грудины до лобкового симфиза. Почти на середине ее
находится пупочное кольцо. Нижний край апоневроза наружной ко-
сой мышцы перекидывается между верхней передней подвздошной
остью и лобковым бугорком, подворачиваясь внутрь в виде желоба.
Этот край называется паховой (пупартовой) связкой.
Щелевидный парный паховый канал длиной 4—5 см расположен
в паховой области в толще мышц живота, направлен косо книзу и ме-
диально, у мужчин через него проходит семенной канатик, у жен-
щин — круглая связка матки (рис. 122).
Диафрагма служит верхней стенкой брюшной полости, она участ-
вует в акте дыхания и вместе с мышцами живота в осуществлении
функции брюшного пресса (рис. 123). Диафрагма, которая имеется
210
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
4
3
2
5
6
7
8
9
1
Рис. 122. Паховый канал, вид спере-
ди. На правой стороне наружная
и внутренняя косые мышцы живота
разрезаны и отвернуты в сторону.
На левой стороне удалена передняя
стенка влагалища прямой мышцы
живота:
1 — паховый канал (вскрыт),
2 — семенной канатик, 3 — прямая
мышца живота, 4 — глубокое кольцо
пахового канала, 5 — поверхностное
кольцо пахового канала,
6 — апоневроз наружной косой
мышцы живота, 7 — поперечная
фасция живота, 8 — внутренняя
косая мышца живота,
9 — поперечная мышца живота
7
8
9
11
12
13
14
4
3
2
5
6
1
10
Рис. 123. Диафрагма, вид снизу,
со стороны брюшной полости:
1 — квадратная мышца поясницы,
2 — малая поясничная мышца,
3 — большая поясничная мышца,
4 — подвздошная фасция,
5 — поперечная фасция, 6 — большая
поясничная мышца (частично
удалена), 7 — подвздошная мышца,
8 — межпоперечные мышцы,
9 — латеральная дугообразная связка,
10 — медиальная дугообразная
связка, 11 — поясничная часть
диафрагмы, 12 — пищеводное
отверстие, 13 — отверстие
нижней полой вены,
14 — сухожильный центр
211
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
только у млекопитающих, представляет собой тонкую мышцу, изогну-
тую в виде купола, обращенного в грудную полость. Правая часть ку-
пола расположена несколько выше, чем левая. Мышечные пучки, кон-
вергируя от периферии к середине диафрагмы, переходят в ее
сухожильный центр. Через диафрагму проходят аорта, пищевод, сим-
патические стволы, вены, нервы и т. д .
Дно малого таза (нижняя стенка брюшной полости) сформирова-
но двумя группами мышц, образующих диафрагмы таза и мочеполо-
вую. Диафрагма таза образована мощной мышцей, поднимающей зад-
ний проход. Мышца каждой стороны треугольной формы состоит из
множества пучков, начинающихся на внутренней поверхности таза.
Обе мышцы спускаются вниз наподобие воронки, окружают конеч-
ный отдел прямой кишки и прикрепляются к копчику. Мышца фор-
мирует дно полости таза и укрепляет его, а также часть стенки моче-
вого пузыря, влагалища, мышца поднимает задний проход; у женщин,
кроме того, суживает влагалище. В поверхностном слое лежит наруж-
ный сфинктер заднего прохода, расположенный непосредственно под
кожей.
В мочеполовой диафрагме различают глубокое и поверхностное
пространство промежности. В первом находятся парная глубокая по-
перечная мышца промежности, укрепляющая диафрагму, и сфинктер
мочеиспускательного канала, являющийся, по существу, частью пре-
дыдущей мышцы. В поверхностном пространстве лежат парная луко-
вично-губчатая мышца, которая окружает у мужчин луковицу поло-
вого члена и его губчатое тело, у женщин — наружное отверстие
влагалища; седалищно-пещеристая мышца, способствующая эрекции
полового члена или клитора; поперечная мышца промежности. Мыш-
цы обеих диафрагм окутаны фасциальными листками. Большинство
мышц промежности вплетаются в сухожильный центр, который обра-
зован пучками плотной волокнистой соединительной ткани.
Мышцы верхней конечности. Рука как орган труда выполняет
многочисленные и разнообразные движения, которые осуществляет
большое количество мышц. Многие из них начинаются на ребрах,
грудине и позвоночнике и прикрепляются к костям пояса верхней ко-
нечности и плечевой кости. Они описаны выше. Мышцы верхней ко-
нечности подразделяются на мышцы плечевого пояса и мышцы сво-
бодной верхней конечности (рис. 124).
Мышцы плечевого пояса со всех сторон окружают плечевой су-
став. Поверхностный слой образован дельтовидной мышцей, глубо-
кий — над- и подостной, большой и малой круглыми, подлопаточной
и клювовидно-плечевой мышцами.
Мышцы свободной верхней конечности. Мышцы плеча делятся
на две группы: передние являются сгибателями (двуглавая и луче-
212
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
19
25
27
28
29
30
26
31
32
33
34
35
36
37
38
39
40
7
4
2
41
42
43
2
5
6
7
1
Б
А
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
Рис. 124. Мышцы верхней конечности
(А — вид спереди, Б — вид сзади): 1 — подлопаточная мышца, 2 — большая круглая
мышца, 3 — широчайшая мышца спины, 4 — трехглавая мышца плеча, 5 — локтевая
ямка, 6 — круглый пронатор, 7 — локтевой сгибатель запястья, 8 — длинная ладонная
мышца, 9 — поверхностный сгибатель пальцев, 10 — часть фасции предплечья,
11 — короткая ладонная мышца, 12 — возвышение мизинца, 13 — ладонный апоневроз,
14 — возвышение большого пальца, 15 — сухожилие длинной мышцы, отводящей
большой палец кисти, 16 — длинный сгибатель большого пальца кисти,
17 — поверхностный сгибатель пальцев, 18 — лучевой сгибатель запястья,
19 — плечелучевая мышца, 20 — апоневроз двуглавой мышцы плеча, 21 — сухожилие
двуглавой мышцы плеча, 22 — плечевая мышца, 23 — двуглавая мышца плеча,
24 — клювовидно-плечевая мышца, 25 — дельтовидная мышца, 26 — надостная мышца,
27 — длинный лучевой разгибатель запястья, 28 — локтевая мышца, 29 — короткий
лучевой разгибатель запястья, 30 — разгибатель пальцев, 31 — длинная мышца,
отводящая большой палец кисти, 32 — короткий разгибатель большого пальца кисти,
33 — сухожилие длинного разгибателя большого пальца кисти, 34 — первая тыльная
межкостная мышца, 35 — сухожилия разгибателя пальцев, 36 — сухожилие
разгибателя мизинца, 37 — сухожилие разгибателя указательного пальца,
38 — удерживатель разгибателей, 39 — локтевой разгибатель запястья,
40 — разгибатель мизинца, 41 — малая круглая мышца, 42 — подостная мышца
А
Б
213
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
вая), задние — разгибатели (трехглавая и локтевая). Мышцы пред-
плечья также делятся на две группы: переднюю и заднюю. К первой
относятся семь сгибателей кисти и пальцев: плечелучевая, лучевой
и локтевой сгибатели запястья, длинная ладонная (часто отсутству-
ет), поверхностный и глубокий сгибатели пальцев, длинный сгибатель
большого пальца и два пронатора: круглый и квадратный. Сгибатели
пальцев осуществляют чрезвычайно тонкие и высокодифференци-
рованные движения, которые свойственны лишь человеку разумно-
му. Благодаря специальным упражнениям можно достигнуть необы-
чайной точности и сложности движений. Во вторую группу входит
девять разгибателей кисти и пальцев: длинный и короткий лучевые
разгибатели запястья, локтевой разгибатель запястья, разгибатель
пальцев, разгибатели мизинцев и указательного пальца, длинный и
короткий разгибатели большого пальца, длинная мышца, отводящая
большой палец, и один супинатор. Все они располагаются в несколь-
ко слоев.
Мышцы кисти. В жизнедеятельности каждого человека кисть
в целом, и особенно пальцы, имеют первостепенное значение, так как
именно они непосредственно выполняют движения и соприкасаются
с предметами. Все эти движения осуществляются большим количест-
вом мышц предплечья и кисти. Последние (18) располагаются только
на ладонной поверхности, на тыльной лишь проходят сухожилия опи-
санных выше разгибателей, лежащих на предплечье. Мышцы кисти
делятся на три группы: мышца возвышения большого пальца (корот-
кая отводящая, короткий сгибатель, приводящая и противопоставля-
ющая большой палец); возвышения V пальца (короткая ладонная, от-
водящая, короткий сгибатель и противопоставляющая мизинец);
средняя группа (четыре червеобразные, три ладонные и четыре тыль-
ные межкостные). Мышцы большого пальца наиболее развиты у че-
ловека разумного по сравнению с другими приматами, так как они
изначально приспособлены для трудовых процессов.
Мышцы нижней конечности. Нижняя конечность человека, явля-
ясь органом опоры и передвижения, имеет наиболее мощную муску-
латуру, на долю которой приходится более 50% всей массы мышц. Со-
гласно делению конечности на сегменты, различают мышцы таза
и свободной нижней конечности (бедра, голени и стопы). Из всех
мышц нижней конечности у человека наиболее развиты большая яго-
дичная, выполняющая функции разгибателя бедра и поддержания
тела в вертикальном положении, четырехглавая мышца бедра, разги-
бающая голень и поддерживающая тело в вертикальном положении,
и камбаловидная, которая, и это особенно важно (!), осуществляет по-
дошвенное сгибание стопы, начальные этапы движения (предотвра-
щает наклон тела вперед) (рис. 125).
214
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Мышцы таза окружают со всех сторон тазобедренный сустав. Все
они начинаются от костей таза и прикрепляются к верхней трети бед-
ренной кости. Мышцы таза делятся на две группы: внутреннюю, кото-
рая расположена в полости таза (подвздошная, большая и малая пояс-
ничные, грушевидная, внутренняя запирательная), и наружную,
расположенную на боковой поверхности таза и в области ягодицы
(большая, средняя и малая ягодичные, квадратная бедра, напрягатель
широкой фасции, наружная запирательная и две близнечные). Мыш-
цы наружной группы лежат в несколько слоев. Они очень хорошо
развиты у человека в связи с прямохождением (особенно большая
ягодичная). Ягодичные мышцы регулируют равновесие тела при сто-
янии и ходьбе, у новорожденных и грудных детей они развиты слабо.
4
3
2
5
6
7
8
9
1
10
11
12
13
14
15
16
20
19
18
21
22
23
24
17
Рис. 125. Мышцы нижней
конечности, правой
(А — вид спереди, Б — вид сзади):
1 — портняжная мышца,
2 — подвздошно-поясничная
мышца, 3 — гребенчатая мышца,
4 — длинная приводящая мышца,
5 — тонкая мышца, 6 — икроножная
мышца (медиальная головка),
7 — камбаловидная мышца,
8 — сухожилие длинного
разгибателя большого пальца
стопы, 9 — нижний удерживатель
сухожилий-разгибателей,
10 — верхний удерживатель
сухожилий-разгибателей,
11 — длинный разгибатель пальцев,
12 — короткая малоберцовая
мышца, 13 — передняя
большеберцовая мышца,
14 — длинная малоберцовая мышца,
15 — четырехглавая мышца бедра,
16 — напрягатель широкой фасции,
17 — большая ягодичная мышца,
18 — подвздошно-большеберцовый
тракт, 19 — двуглавая мышца бедра,
20 — подколенная ямка,
21 — пяточное (ахиллово)
сухожилие, 22 — икроножная
мышца, 23 — полусухожильная
мышца, 24 — полуперепончатая
мышца
А
Б
215
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
По мере того как дети начинают ходить, развиваются ягодичные
мышцы.
Мышцы свободной нижней конечности. Мышцы бедра развиты
очень хорошо в связи с прямохождением. Они не только участвуют в
передвижении тела, но и удерживают тело в вертикальном положении.
Мышцы бедра делятся на три группы: передняя (разгибатели) — че -
тырехглавая и портняжная; задняя (сгибатели) — полусухожильная,
полуперепончатая, двуглавая; медиальная (приводящие) — гребенча-
тая, тонкая, длинная, короткая и большая приводящие. В связи с пря-
мохождением сгибание и разгибание в коленном суставе у человека
облегчено, поэтому мышцы-сгибатели (задняя группа) развиты сла-
бее, а четырехглавая лучше, чем у человекообразных обезьян.
Мышцы голени также участвуют в прямохождении и удержании
тела в вертикальном положении. Подобно предплечью, утолщенные
мышечные части лежат в проксимальном отделе, по направлению
к стопе они переходят в сухожилия. На голени отсутствуют вращате-
ли. Мышцы голени делятся на три группы: переднюю (тыльное сгиба-
ние стопы и разгибание пальцев) — передняя большеберцовая, длин-
ный разгибатель пальцев, длинный разгибатель большого пальца;
заднюю (подошвенное сгибание стопы и пальцев) — трехглавая, по-
дошвенная, подколенная, длинные сгибатели пальцев и большого паль-
ца стопы и задняя большеберцовая; латеральную (пронация и лате-
ральное сгибание стопы) — две малоберцовые: короткая и длинная. На
задней поверхности коленного сустава мышцы двуглавая и полупере-
пончатая (сверху) и обе головки икроножной (снизу) ограничивают
подколенную ямку, имеющую форму ромба, в которой проходят сосу-
ды и нервы.
В связи с прямохождением человеку постоянно приходится прео-
долевать силу тяжести, которая стремится согнуть нижние конечно-
сти в голеностопном суставе так, чтобы тело упало вперед. Поэтому
у человека имеется большое количество мышц, осуществляющих по-
дошвенное сгибание стопы (8), а малоберцовые мышцы являются
сгибателями. Этому способствует также развитие наружной лодыжки
у человека и наличие мощного ахиллова сухожилия.
Мышцы стопы. Движения пальцев стопы незначительны. Их осу-
ществляют, помимо мышц голени, собственные мышцы, расположен-
ные на тыле стопы (короткие разгибатели пальцев и большого паль-
ца) и на подошве, последние преобладают. Подошвенные мышцы
укрепляют свод стопы. Они делятся на три группы: медиальную, кото-
рая осуществляет движения большого пальца (отводящая, приводя-
щая и короткий сгибатель), латеральную, приводящую в движение
мизинец (отводящая и короткий сгибатель), среднюю (четыре черве-
образные, короткий сгибатель пальцев, квадратная мышца подошвы,
216
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
семь межкостных — три подошвенные и четыре тыльные). Фасция
подошвы резко утолщается и образует подошвенный апоневроз, иду-
щий от пяточного бугра к основаниям пальцев, от которого идут две
перегородки вглубь, разделяющие описанные группы мышц.
РАБОТОСПОСОБНОСТЬ, РАБОТА,
УТОМЛЕНИЕ И ОТДЫХ
Работа — это осуществление клеткой, органом, системой органов
или организмом свойственных им функций. Человек разумный вы-
полняет, как правило, общественно полезную работу. Научно-техни-
ческий прогресс изменил характер работы человека. На смену тяже-
лому физическому труду пришел труд умственный. «Большинство
современных рабочих выполняют задачи, требующие распознавания
образов, быстрого получения и переработки информации, а также
способности разрабатывать планы и принимать решения», — пишет
известный физиолог труда Г. Ульмер (1997). И это накладывает серь-
езный отпечаток на здоровье человека.
Работоспособность — это способность человека выполнять мак-
симально возможное количество работы на протяжении определенно-
го (заданного) времени и с определенной эффективностью. Работо-
способность, подобно работе, подразделяется на умственную и
физическую. Исходя из приведенного выше определения, умственная
работоспособность — это способность выполнять определенное ко-
личество работы, требующей значительной активации нервно-психи-
ческой сферы; физическая работоспособность — это способность вы-
полнять максимально возможное количество физической работы за
счет активации опорно-двигательного аппарата. Естественно, физиче-
ская работоспособность зависит и от состояния нервной системы, ин-
нервирующий опорно-двигательный аппарат.
Большое количество факторов влияет на работоспособность чело-
века. Это, в первую очередь, состояние его здоровья, уровень трениро-
ванности, опыт, физическое и психическое состояние, склонность
к данной работе (т. е . талант), мотивация к труду и эмоции, состояние
окружающей среды; организация труда. Важную роль играет опти-
мальная организация рабочего места, которая позволяет поддержать
необходимое положение тела и его сегментов для выполнения рабо-
ты. Выполнение работы требует затрат энергии. Общая потребность
в энергии — это сумма основного и рабочего обмена. Основной об-
мен — количество энергии, затрачиваемое организмом в условиях
полного покоя для поддержания жизни. У мужчин эта величина в
среднем составляет 1 ккал на 1 кг массы тела в 1 ч (4,2 кДж). У жен-
217
РАБОТОСПОСОБНОСТЬ, РАБОТА, УТОМЛЕНИЕ И ОТДЫХ
щин — 0,9 ккал (3,8 кДж). Рабочий обмен — это затрата энергии для
выполнения внешней работы. Общая потребность в энергии при ум-
ственном труде равна 2500—3200 ккал (10 475—13 410 кДж), при ме-
ханизированном труде или легкой немеханизированной работе —
3200—3500 ккал (13 410—14 665 кДж), при частично механизированном
труде или немеханизированном труде умеренной тяжести — 3500—
4500 ккал (14 665—18 855 кДж), при тяжелом немеханизированном
физическом труде — 4500—5000 ккал (18 855—20 950 кДж).
Умственная работа — это мышление. Умственная работа включает
прием и переработку информации, ее сравнение с информацией, хра-
нящейся в памяти, преобразование информации, определение про-
блем и путей их решения, формирование цели.
Умственная работа тесно связана с членораздельной речью, так
как человек имеет дело не с конкретными предметами, явлениями
или живыми организмами, а с определяющими их символами или по-
нятиями.
Умственная работа включает мыслительный и эмоциональный
компоненты. Мыслительный компонент связан с интеллектуальными
способностями человека, он требует обдумывания, концентрации
внимания. Эмоциональный компонент включает самооценку челове-
ка как субъекта умственного труда, оценку значимости цели и средств.
Эмоциональный компонент проявляется четкими реакциями вегета-
тивной нервной системы и изменениями настроения человека, вызы-
вая возникновение многочисленных эмоций.
Эмоциональные нагрузки, нервно-психическая перегрузка вызы-
вают стимуляцию симпатической части вегетативной нервной систе-
мы, что проявляется увеличением частоты пульса, минутного объема
сердца и дыхания, потоотделением. Физическая работа связана с дея-
тельностью опорно-двигательного аппарата, основную роль в этом
играют скелетные мышцы.
Если благодаря сокращению мышцы меняется положение части
тела, то преодолевается сила сопротивления, т. е. выполняется прео-
долевающая работа. Работа, при которой сила мышцы уступает дей-
ствию силы тяжести и удерживаемого груза, называется уступающей.
В этом случае мышца функционирует, однако она не укорачивается,
а удлиняется, например, когда невозможно поднять или удержать на
весу тело, имеющее большую массу. При большем усилии мышц при-
ходится опустить это тело на какую-нибудь поверхность.
Удерживающая работа выполняется, если благодаря сокращению
мышц тело или груз удерживается в определенном положении без пере-
мещения в пространстве, когда, например, человек держит груз, не дви-
гаясь. При этом мышцы сокращаются изометрически без изменения их
длины. Сила сокращения мышц уравновешивает массу тела и груза.
218
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Когда мышцы, сокращаясь, перемещают тело или его части в про-
странстве, они выполняют преодолевающую или уступающую работу,
которая является динамической. Статической является удерживающая
работа, при которой не происходит движений всего тела или его части.
При статической работе мышцы сокращаются изометрически, при
этом расстояние не преодолевается, но работа осуществляется.
Анатомический и физиологический поперечники характеризуют
величину или функцию той или иной мышцы. Анатомический попе-
речник — это площадь перпендикулярного длинной оси поперечного
сечения мышцы в определенном ее участке. Физиологический попе-
речник — это сумма площадей поперечных сечений всех мышечных
волокон, образующих мышцу. Первый показатель характеризует ве-
личину мышцы, второй — ее силу.
Абсолютная сила мышц вычисляется путем деления массы макси-
мального груза (кг), который может поднять мышца, на площадь ее
физиологического поперечника (см2). Этот показатель у человека для
разных мышц составляет от 6,24 до 16,8 кг/см2. Так, абсолютная сила
икроножной мышцы 5,9 кг/см2, трехглавой мышцы плеча 16,8 кг/см2,
двуглавой мышцы плеча 11,4 кг/см2. Напряжение, развиваемое
при сокращении одним мышечным волокном, колеблется в пределах
0,1—0,2 г.
Размах сокращения (амплитуда) зависит от длины мышечных во-
локон. В веретенообразных и лентовидных мышцах волокна длиннее,
а анатомический и физиологический поперечники совпадают. Поэто-
му сила этих мышц не очень большая, а амплитуда сокращения вели-
ка. В перистых мышцах физиологический поперечник значительно
больше анатомического, и соответственно их сила больше. В связи
с тем что мышечные волокна этих мышц короткие, амплитуда их со-
кращения невелика.
Одним из показателей эффективности физической работы являет-
ся коэффициент полезного действия, который показывает, какая
часть затраченной энергии превращается в энергию, осуществляю-
щую полезную внешнюю работу.
КПД (%) =
Энергия, затрачиваемая на внешнюю работу
× 100
Вырабатываемая энергия
КПД изолированной мышцы может достигнуть 35%. КПД в целом
организма при различных видах мышечной деятельности варьирует
в пределах от 3 до 25%.
При частом повторении одной и той же работы развивается рабо-
чий динамический стереотип — система рефлекторных действий, ко-
торые формируются при постоянном повторении одних и тех же раз-
219
РАБОТОСПОСОБНОСТЬ, РАБОТА, УТОМЛЕНИЕ И ОТДЫХ
дражителей. Рефлекторные реакции становятся автоматическими,
поэтому работа становится более энергетически экономичной и менее
утомительной, не требует постоянного внимания и сосредоточения.
Физическая нагрузка вызывает реакцию всех органов и систем.
В активно сокращающейся мышце увеличивается кровоток более чем
в 20 раз, усиливается обмен веществ. При умеренной физической на-
грузке обмен веществ в мышце осуществляется аэробно, во время тя-
желой работы часть энергии освобождается анаэробно. В результате
в мышцах образуется и накапливается молочная кислота. При нако-
плении значительных количеств молочной кислоты в мышечных во-
локнах развивается мышечное утомление.
При физической работе возрастают частота сердечных сокраще-
ний, ударный объем сердца, артериальное давление, потребление ор-
ганизмом кислорода. При легкой и умеренной физической работе
с постоянной нагрузкой в течение 5—10 мин. частота сердечных со-
кращений увеличивается, после чего достигает постоянного уровня,
или стационарного состояния, которое не приводит к утомлению че-
ловека в течение нескольких часов. Через 3—5 мин. после завершения
такой работы частота сердечных сокращений нормализуется. При тя-
желой работе не наступает стационарное состояние, развивается
утомление, частота сердечных сокращений увеличивается, а после
прекращения тяжелой работы период восстановления нормальной ча-
стоты сердечных сокращений длится несколько часов. У каждого че-
ловека есть свой, индивидуальный предел утомительной работы. Он
разделяет два уровня работоспособности. Работа, которую человек
может выполнять в течение 8 ч без развития признаков мышечного
утомления, считается легкой, она ниже предела.
Выше него находится область максимальной работоспособности,
выполнение которой существенно ограничено по времени. Макси-
мальная работоспособность снижается по мере увеличения длитель-
ности работы. Тренировка повышает работоспособность человека.
Как же определить предел утомительной динамической работы?
Одним из важных показателей является частота пульса, которая со-
храняется постоянно во время работы, не увеличиваясь в связи с
утомлением. У нетренированных людей в возрасте от 20 до 30 лет она
не превышает 130 ударов в 1 мин., менее чем через 5 минут после пре-
кращения работы частота пульса становится менее 100.
Восстановление — это процесс постепенного возвращения функ-
ций организма к исходному состоянию после прекращения работы. По
мере восстановления степень утомления уменьшается, а работоспособ-
ность увеличивается. Если человек выполняет работу, лежащую выше
пределов его утомления, необходимо периодически отдыхать. Лучше
много кратковременных перерывов для отдыха, чем один-два длинных.
220
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Даже в состоянии полного покоя скелетная мышца сохраняет
свою эластичность и определенную степень напряжения. Это назы-
вается мышечным тонусом. Мышечный тонус не сопровождается
утомлением.
Утомление — это физиологическое состояние человека, наступаю-
щее вследствие напряженной или длительной работы, которое выра-
жается во временном снижении работоспособности. Различают мы-
шечное (физическое) и центральное (нервно-психическое) утомление.
При тяжелой работе они сочетаются. Утомление характеризуется
уменьшением силы и выносливости мышц, нарушением координации
движений, увеличением энергозатрат для выполнения одной и той же
работы, нарушением памяти, скорости переработки информации, сос-
редоточения и т. д . Утомление субъективно ощущается человеком
в виде усталости, которая, кроме того, обусловлена потребностью во
сне. Усталость вызывает у человека желание прекратить работу или
уменьшить нагрузки. Утомление при тяжелой физической работе свя-
зано с накоплением в мышечных волокнах некоторых продуктов об-
мена (например, молочной кислоты). Отдых, особенно активный,
приводит к восстановлению работоспособности мышцы. Это связано
с удалением молочной кислоты и возобновлением запасов энергии
в мышце. Нервно-психическое (центральное) утомление вызвано
длительной напряженной умственной работой, однообразной моно-
тонной работой, шумом, плохими условиями труда, эмоциональными
факторами, заболеваниями.
Если же, несмотря на утомление, работа продолжается, возникает
истощение. Напомним, что тяжелые физические и нервно-психиче-
ские нагрузки вызывают стресс (вернее, дистресс). Различают острое
и хроническое истощение. Первое представляет собой резкое сниже-
ние работоспособности во время тяжелой работы, второе возникает
вследствие длительной напряженной или слишком часто повторяе-
мой тяжелой работы. Острое и хроническое истощение часто возни-
кает у профессиональных спортсменов, при спортивных соревновани-
ях и тренировках.
Отдых — это состояние покоя или особый, специально организо-
ванный вид деятельности, который снимает утомление и способству-
ет восстановлению работоспособности. И . М . Сеченов во второй по-
ловине XIX в. установил, что работа одних групп мышц конечностей
способствует устранению утомления других мышечных групп, выз-
ванного их работой. Это положение легло в основу определения двух
типов отдыха: активного и пассивного. Активный отдых — это отдых,
во время которого человек выполняет другой вид работы, отличный
от обычного выполняемого труда. Восстановление при активном от-
дыхе происходит быстрее и эффективнее, чем при пассивном отдыхе,
221
РАБОТОСПОСОБНОСТЬ, РАБОТА, УТОМЛЕНИЕ И ОТДЫХ
когда организм находится в условиях относительного покоя. Так, на-
пример, интенсивную умственную деятельность следует сменить фи-
зической активностью. И наоборот, интенсивную физическую — ум-
ственной.
Режим дня. Древнегреческое слово «диета» (diaita) означает
«образ жизни, режим». Это уже позже под диетой стали понимать ха-
рактер питания. Режим важен для человека любого возраста. Пере-
фразируя Пушкина, можно сказать: «Режиму все возрасты покорны».
Особенно это касается фундамента жизни детского, подросткового
и юношеского возрастов. Правильный режим дня школьника включа-
ет продолжительность учебного времени в школе и дома, сон, отдых,
прогулки, занятия физической культурой, режим питания (регуляр-
ное питание способствует хорошему аппетиту). Очень важен сон,
причем человек должен ложиться спать в одно и то же время, тогда он
быстро засыпает и спокойно спит. Продолжительность сна меняется
с возрастом: в 7—8 лет — 11 —11,5 ч, в 9—10 лет — 10—10,5,
в11—12лет—10ч,в13—15лет—9—9,5ч,в16—18лет—8—8,5ч.
Взрослому человеку необходимо спать 7,5—8 ч.
Работа и отдых должны чередоваться. Еще в XIX в. великий отече-
ственный физиолог И. М. Сеченов доказал, что отдых должен быть
активным. Для полноценного восстановления необходим активный
отдых. Особенно эффективно чередование физического и умственно-
го труда.
Пять основных условий успешного высокопроизводительного
труда:
1) усилия следует увеличивать постепенно;
2) следует соблюдать правильный ритм работы;
3) работа должна быть системной;
4) труд необходимо чередовать с активным отдыхом;
5) труд должен быть постоянным, ежедневным. Только в этом
случае вырабатываются и закрепляются навыки (Н. Е. Введенский).
Режим учебных заведений должен строиться с учетом анатомо-
физиологических особенностей детей соответствующих возрастных
групп. Однако есть и несколько общих требований:
не следует начинать занятия ранее 8 ч (в первом классе — не ра-
нее 8:30);
длительность урока не должна превышать 45 мин. (в первом клас-
се — 35 мин.);
после каждого урока следует делать десятиминутные перерывы,
а во второй половине учебного дня целесообразно делать два пе-
рерыва по 20 мин. каждый. В группах продленного дня после
окончания последнего урока необходим перерыв не менее 45 мин.
222
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Физическая активность
Великий врач древности Гиппократ назвал движение пищей для
жизни, а Плутарх — кладовой здоровья. Для сохранения и укрепле-
ния здоровья мышцы должны систематически работать.
Гиподинамия — одна из бед XX века во всем мире, особенно в нашей
стране. Около 70% населения России не занимается физкультурой.
Хронический дефицит двигательной активности детей тормозит их
нормальное физическое развитие. Физическая нагрузка — это величи-
на и интенсивность всей производимой человеком мышечной работы
связанной со всеми видами деятельности. Функция всех аппаратов
и систем организма взаимосвязана и зависит от состояния двигатель-
ного аппарата. Организм функционирует оптимально только при усло-
вии высокой физической активности. Двигательная активность явля-
ется наиболее естественным способом улучшения вегетативных
функций человека, обмена веществ. При низкой двигательной активно-
сти снижается сопротивляемость организма к разнообразным стрессо-
вым воздействиям, уменьшаются функциональные резервы различных
систем, ограничиваются рабочие возможности организма. Работа сер-
дца становится менее экономной, ограничиваются его потенциальные
резервы, угнетается функция желез внутренней секреции и в первую
очередь половых желез. Резко страдает сексуальность человека.
При высокой физической активности все органы и системы рабо-
тают весьма экономично, адаптационные резервы велики, сопротив-
ляемость организма к неблагоприятным условиям высокая. Чем боль-
ше привычная физическая активность, тем больше масса мышц
и выше максимальная способность к поглощению кислорода и мень-
ше масса жировой ткани. Чем выше максимальное поглощение кисло-
рода, тем интенсивнее снабжение им органов и тканей и уровень об-
мена веществ. В любом возрасте средний уровень максимального
поглощения кислорода на 10—20% выше у лиц, ведущих активный
образ жизни, чем у занятых умственной («сидячей») работой.
Из всех видов физической активности человеку (особенно не за-
нятому физическим трудом) необходима такая нагрузка, при которой
увеличивается снабжение организма кислородом и его потребление
(аэробная нагрузка). Наиболее эффективны (аэробические) пять ви-
дов физической активности — это ходьба (особенно скандинавская
ходьба), бег, плавание, езда на велосипеде и лыжи. Аэробные упражне-
ния требуют большого количества кислорода.
ВНИМАНИЕ!
Длительность и темп необходимо увеличивать постепенно, не следует
сразу испытывать сильные нагрузки. При физической нагрузке важна
постепенность.
223
ВНУТРЕННИЕ ОРГАНЫ
ВНУТРЕННИЕ ОРГАНЫ
В полостях тела человека расположены внутренние органы, или
внутренности. К ним относятся пищеварительная, дыхательная, моче-
выделительная и половая системы. Большинство органов этих систем
имеет трубчатое строение и построено по единому плану. Стенки
трубчатых органов на всем протяжении состоят из четырех слоев:
внутренней слизистой оболочки, подслизистой основы, мышечной
и наружной оболочек (рис. 126).
Подвижная складчатая слизистая оболочка состоит из трех пла-
стинок: эпителия, собственной и мышечной пластинок. Слизистая
оболочка покрыта слоем слизи, которая защищает ее от действия мно-
Рис. 126. Строение пищеварительной трубки (поперечное сечение):
1 — брыжейка, 2 — сложная железа, 3 — проток железы, 4 — просвет органа,
5 — подслизистое нервное сплетение (Мейснера), 6 — мышечно-кишечное нервное
сплетение (Ауэрбаха), 7 — продольный слой мышечной оболочки, 8 — круговой слой
мышечной оболочки, 9 — собственная пластинка слизистой оболочки,
10 — одиночный лимфатический фолликул, 11 — подслизистая основа,
12 — серозная оболочка, 13 — слизистая оболочка, 14 — складки слизистой оболочки
11
11
11
22
12
12
33
13
13
44
14
14
55
66
77
88
99
10
10
224
ВНУТРЕННИЕ ОРГАНЫ
гочисленных ферментов. Слизь вырабатывается одноклеточными
и многоклеточными железами, в изобилии имеющимися на протяже-
нии всей трубки. Эпителий на большем протяжении — однослойный
цилиндрический, но там, где происходит постоянная травматизация,
он — многослойный плоский неороговевающий (ротовая и носовая
полости, глотка, пищевод, заднепроходной канал, влагалище). Собст-
венная пластинка слизистой оболочки образована рыхлой волокни-
стой неоформленной соединительной тканью, в которой располага-
ются скопления лимфоидной ткани, выполняющие защитную
функцию, нервные элементы и сосуды (артериальные, венозные, лим-
фатические). Мышечная пластинка слизистой оболочки состоит из
гладких мышечных клеток.
Подслизистая основа образована рыхлой волокнистой неофор-
мленной соединительной тканью, в которой располагаются скопле-
ния лимфоидной ткани, подслизистое нервное сплетение, кровенос-
ные и лимфатические сосуды.
Мышечная оболочка чаще всего состоит из двух слоев: внутреннего
кругового и наружного продольного, разделенных прослойкой рых-
лой неоформленной соединительной ткани, в которой расположены
межмышечное нервное сплетение, кровеносные и лимфатические со-
суды. В большей части внутренних органов мышцы гладкие. Лишь
стенка ряда органов (глотки, верхней трети пищевода, наружный
сфинктер заднего прохода, мышцы гортани, наружный сфинктер мо-
чевого пузыря) образованы поперечнополосатой мышечной тканью,
которая сокращается произвольно. Благодаря сокращению мышц осу-
ществляются движения внутренних органов, например пищевые мас-
сы перемещаются по желудочно-кишечному тракту, а моча — по моче-
выводящим путям.
Наружная адвентициальная оболочка образована рыхлой волок-
нистой соединительной тканью, которая покрывает внутренние орга-
ны. Серозная оболочка (висцеральные листки брюшины или плевры)
окутывает бо
льшую часть органов пищеварительной системы (напри-
мер, желудок, кишечник, кроме части прямой кишки) и легкие.
Внутренние органы на всем протяжении обильно снабжены же-
лезами, которые являются производными эпителия. Железы выра-
батывают ферменты, необходимые для пищеварения, биологически
активные вещества и слизь, которая защищает слизистую оболочку
от травм и действия ферментов. Слизистая оболочка снабжена ог-
ромным количеством одноклеточных желез (бокаловидные клетки,
вырабатывающие слизь), расположенных внутри эпителиального
пласта, а также множеством многоклеточных желез, расположенных
как в самой слизистой оболочке, так и за пределами трубчатых
органов.
225
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
Сложная и многогранная жизнь человека связана с расходованием
веществ и энергии, поэтому человек (как и другие животные) нужда-
ется в постоянном введении в организм веществ, которые обеспечива-
ют его энергетические и пластические потребности.
Поиски пищи — основная забота человека как индивидуума и все-
го вида «человек разумный» от его возникновения и до наших дней.
И сегодня, в эпоху научно-технической революции она остается акту-
альной и очень далека от решения. В конце XX в. на земном шаре го-
лодают более миллиарда человек. И в то же время вопросы рацио-
нального питания, профилактики ожирения и других заболеваний,
связанных с неправильным питанием, являются одной из десяти ос-
новных проблем ООН.
Потребность в пищевых веществах — одна из основных потребно-
стей человека, которая зависит в первую очередь от энергетических
затрат организма. Эта потребность меняется в зависимости от пред-
шествующего питания. При избыточном поступлении пищевых
веществ происходит их депонирование в организме, при недоста-
точном — их мобилизация из депо и перестройка уровня функциони-
рования различных систем.
Пищеварительная система осуществляет механическую и химиче-
скую обработку пищи, расщепление пищевых веществ до мономеров,
всасывание переработанных и выделение непереработанных ингреди-
ентов. Вместе с тем органы пищеварения выполняют важнейшую за-
щитную функцию, предупреждая проникновение во внутреннюю сре-
ду организма чужеродных веществ. Голод и жажда, наряду с половым
влечением, усталостью и некоторыми другими, относятся к общим
ощущениям, которые вызваны стимулами внутренней среды организ-
ма. Эти ощущения приводят к возникновению мотиваций, направлен-
ных на их устранение. Так, дефицит пищи или воды вызывает раздра-
жение различных рецепторов, в результате чего человек ощущает
голод или жажду, возникает мотивация к поиску пищи или воды
и устранению их дефицита в процессе еды или питья. Иными слова-
ми, удовлетворение мотиваций устраняет причину, вызвавшую общее
ощущение голода или жажды. Следует подчеркнуть, что эти мотива-
ции врожденные.
Голод и аппетит. Потребление человеком пищи зависит от его
энергетических затрат в данный конкретный промежуток времени,
климатических условий и калорийности пищи. Это кратковременная
регуляция потребления пищи, которая накладывается на долговре-
менную регуляцию, направленную на возмещение нехватки пищи
в предшествующее время. Человек, который длительно голодал, воз-
226
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
вращаясь в нормальные условия, ест значительно больше. Выжившие
узники нацистских концлагерей описывают, как они после освобо-
ждения набрасывались на пищу и ели ее, не переставая, с радостью и
благодарностью, хотя это и было смертельно опасно. По мере восста-
новления массы тела потребность в пище медленно снижается. Дефи-
цит пищи вызывает сокращения стенки пустого желудка и снижение
уровня глюкозы в крови. Эти изменения возбуждают рецепторы стен-
ки желудка, гипоталамуса, печени, желудка, тонкой кишки и др. Сиг-
налы поступают в мозг, вызывая общее ощущение голода. Этот меха-
низм лежит в основе кратковременной регуляции потребления пищи.
Изменения жирового обмена и снижение теплопродукции, возбуждая
внутренние терморецепторы и липорецепторы, служат для долго-
срочной регуляции потребления пищи. К сожалению, человек — одно
из немногих животных, который ест и не испытывая голода. Человек
начинает ощущать насыщение во время еды еще до того, как происхо-
дит всасывание. Это связано со стимуляцией рецепторов полости
носа, рта, глотки, пищевода, желудка во время еды, затем в связи
со всасыванием пищевых веществ возбуждаются центральные глюко-,
термо- и липорецепторы. Информация обрабатывается главным обра-
зом в гипоталамусе, в результате чего и возникает ощущение насыще-
ния. Именно в гипоталамусе находятся «центры голода и насыще-
ния», связанные с лимбической системой.
Аппетит также относится к общим ощущениям. Причем аппетит
не всегда связан с голодом, он может возникать и самостоятельно, но
может быть составляющей голода. «В процессе эволюции аппетит
формируется не как реакция на уже возникшее истощение, но как ме-
ханизм, задолго предупреждающий такое истощение» (А. М. Уголев,
В. Г. Кассиль, 1961). Различают два вида аппетита: общий и избира-
тельный. Первый представляет собой эмоциональную реакцию на
пищу вообще, второй — на определенные виды пищи.
Жажда. Напомним, что 70—75% нашего тела составляет вода, при-
чем содержание ее весьма стабильно, а колебания у здорового челове-
ка в нормальных условиях происходят в пределах ±22% массы тела.
Потеря воды более 0,5% массы тела приводит к жажде. Человек теря-
ет воду с мочой, по
том, выдыхаемым воздухом. Это приводит к умень-
шению объема клеток, что в сочетании с уменьшением объема
внеклеточной жидкости вызывает ощущение жажды, которая сопро-
вождается сухостью во рту. Информация о внутриклеточном содер-
жании солей при потере воды воспринимается специальными рецеп-
торами (осморецепторами) гипоталамуса. Сухость во рту вызывает
раздражение соответствующих рецепторов слизистой оболочки поло-
сти рта и глотки, от которых нервные импульсы также передаются
в гипоталамус. Подобно чувству насыщения, ощущение утоления
227
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
жажды происходит до всасывания воды и нормализации ее внутри- и
внеклеточного содержания.
Пищеварительная система состоит из пищеварительной трубки,
длина которой у взрослого человека достигает 8—10 м, и ряда распо-
ложенных вне ее стенок крупных желез. Трубка образует множество
изгибов, петель, поэтому расстояние по прямой от рта до заднепро-
ходного отверстия составляет 70—90 см (рис. 127).
В переднем отделе пищеварительной системы (ротовая полость,
пищевод, желудок) происходит механическое измельчение пищи,
пища пережевывается, частично обрабатывается химически, всасыва-
ется вода, алкоголь и некоторые другие вещества и передвигается
в средний отдел — тонкую кишку, где пища подвергается химической
обработке, в результате чего образуются простые соединения (амино-
кислоты, жирные кислоты, моносахариды), которые всасываются
Рис. 127. Строение пищеварительной системы:
1 — околоушная (слюнная) железа, 2 — мягкое
нёбо, 3 — глотка, 4 — язык, 5 — пищевод,
6 — желудок, 7 — поджелудочная железа,
8 — проток поджелудочной железы, 9 — тощая
кишка, 10 — нисходящая ободочная кишка,
11 — поперечная ободочная кишка,
12 — сигмовидная ободочная кишка,
13 — наружный сфинктер заднего прохода,
14 — прямая кишка, 15 — подвздошная кишка,
16 — червеобразный отросток (аппендикс),
17 — слепая кишка, 18 — подвздошно-
слепокишечный клапан, 19 — восходящая
ободочная кишка, 20 — правый (печеночный)
изгиб ободочной кишки, 21 — двенадцати-
перстная кишка, 22 — желчный пузырь,
23 — печень, 24 — общий желчный проток,
25 — сфинктер привратника желудка,
26 — поднижнечелюстная (слюнная) железа,
27 — подъязычная (слюнная) железа,
28 — нижняя губа, 29 — полость рта,
30 — верхняя губа, 31 — зубы верхней
челюсти, 32 — твердое нёбо
11
11
11
21
21
31
31
22
12
12
22
22
32
32
33
13
13
23
23
44
14
14
24
24
55
15
15
25
25
66
16
16
26
26
77
17
17
27
27
88
18
18
28
28
99
19
19
29
29
10
10
20
20
30
30
228
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
в кровь и лимфу. В заднем отделе (толстая кишка) формируются ка-
ловые массы, непереваренные и непригодные к всасыванию вещества
удаляются наружу, интенсивно всасывается вода, с участием бактерий
происходит частичное переваривание некоторых видов клетчатки.
Толстая кишка обладает и выделительной функцией.
В эпителии желудочно-кишечного тракта на всем его протяжении
разбросано огромное количество эндокринных клеток, объединяю-
щихся в гастроэнтеропанкреатическую эндокринную систему, кото-
рые вырабатывают гормоны и биологически активные вещества.
ПОЛОСТЬ РТА
Полость рта подразделяется на два отдела: преддверие рта и собст-
венно полость рта (рис. 128). Преддверие рта ограничено губами и
щеками снаружи, зубами и деснами изнутри. Узкая ротовая щель че-
ловека ограничена губами, представляющими собой волокна круго-
вой мышцы рта, покрытые снаружи кожей, выстланные изнутри сли-
зистой оболочкой. Одним из отличительных признаков верхней губы
человека является желобок, расположенный посередине верхней
губы — фильтр. В толще щек находится щечная мышца (мышца тру-
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
Рис. 128. Полость рта
(вид спереди):
1 — уздечка нижней губы,
2 — подъязычный сосочек,
3 — дно полости рта,
4 — подъязычная складка,
5 — нижняя поверхность
языка,
6 — губная спайка,
7 — бахромчатая складка,
8 — спинка языка,
9 — уздечка верхней губы,
10 — десна,
11 — край языка,
12 — передняя язычная
железа, 13 — язычный нерв,
14 — нижняя продольная
мышца, 15 — уздечка языка,
16 — подъязычная железа,
17 — поднижнечелюстной
проток
229
ПОЛОСТЬ РТА
бачей). Слизистая оболочка сращена с альвеолярными дугами челю-
стей, образуя десны, покрывающие шейки зубов и тем самым охраня-
ющие их. В преддверии рта открывается множество мелких слюнных
желез, а также протоки околоушных желез. Собственно ротовая по-
лость ограничена сверху нёбом, которое разделяется на твердое и
мягкое. Твердое нёбо образовано небными отростками верхних челю-
стей и четырехугольными горизонтальными пластинками обеих неб-
ных костей, которые соединяются между собой. Задний отдел — мяг-
кое нёбо — небная занавеска — заканчивается удлиненным язычком.
Небная занавеска переходит по бокам в две пары дужек (задняя —
небно-глоточная, передняя — небноязычная), между которыми распо-
лагается небная миндалина. Дном полости рта является диафрагма
рта, образованная парной челюстно-подъязычной мышцей. Полость
рта сообщается с полостью глотки через зев, ограниченный мягким
нёбом, небными складками и корнем языка.
У новорожденных и детей первых трех месяцев жизни полость рта
чрезвычайно мала, а костное нёбо широкое и сплющенное. У детей
щеки выпуклые за счет жирового тела, которое с возрастом сплющи-
вается и отодвигается назад.
Язык человека образован поперечнополосатой мышечной тканью
и покрыт слизистой оболочкой (рис. 129А). Язык выполняет множе-
ство разнообразных функций: участвует в процессе жевания, члено-
раздельной речи, является органом вкуса. Чрезвычайно важна роль
языка при сосании молока матери новорожденным и грудным ребен-
ком. Губы ребенка захватывают сосок, фиксируя его; мягкое нёбо,
поднимаясь, закрывает зев, язык действует как поршень, отодвигаясь
назад, он создает отрицательное давление вместе с опускающейся
нижней челюстью. Затем челюсть поднимается, а альвеолярные дуги
сдавливают сосок, молоко заглатывается. Поэтому язык новорожден-
ного и грудного ребенка относительно большой, толстый, широкий
и занимает всю полость рта, в то же время он малоподвижен.
Уже древние греки понимали важную роль языка для человека.
Интересна характеристика языка, данная легендарным философом
и баснописцем Эзопом: «Есть ли что на свете лучше и прекраснее,
чем язык? Разве не языком держится вся философия и вся ученость?
Без языка нельзя сделать... порядок в государстве, законы, постанов-
ления — все это существует лишь благодаря языку. Всей нашей жизни
основа — язык; нет ничего лучше на свете... Что же на свете хуже язы-
ка? Язык несет нам раздоры, заговоры, обманы, побоища, зависть,
распри, войну; разве может быть что-то еще хуже, еще презреннее,
чем язык».
Слизистая оболочка языка покрыта многослойным плоским не-
ороговевающим эпителием. Передний отдел спинки языка усеян мно-
230
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
жеством сосочков, являющихся выростами собственной пластинки
слизистой оболочки, покрытых эпителием. У человека четыре вида
сосочков: нитевидные, грибовидные, желобовидные (окруженные ва-
лом) и листовидные (рис. 129Б). Больше всего на языке нитевидных
сосочков, которые придают языку бархатистый вид. Это высокие, уз-
кие выросты длиной 0,3 мм. Нитевидные сосочки имеют специализи-
рованные нервные окончания, которые воспринимают ощущения
прикосновения.
Количество грибовидных сосочков меньше, они закругленные, вы-
ступают над поверхностью языка, длина их 0,7—0,8 мм, диаметр 0,4—
1,0 мм, основание сужено, по форме своей сосочки напоминают гриб.
Сосочки, окруженные валом, или желобовидные (их 7—12), диаметром
2—3 мм образуют фигуру в виде римской цифры V, обращенной назад
острием, на границе между спинкой и корнем языка. По форме они
напоминают грибовидные, однако верхняя поверхность их уплощена,
а вокруг сосочка идет узкий глубокий желоб, заполненный слюной,
в него открываются протоки желез. Желоб снаружи обнесен валиком
слизистой оболочки. На поверхности грибовидных сосочков и боко-
вых поверхностях желобовидных сосочков в толще эпителия распо-
лагаются вкусовые почки — комплекс специализированных рецептор-
ных вкусовых клеток, образующих орган вкуса. Листовидные сосочки,
хорошо развитые у новорожденных и грудных детей, расположены по
краям языка в виде поперечно-вертикальных складок или листиков,
по 4—5 с каждой стороны, длиной 2—5 мм каждый. На листовидных
Рис. 129. Схема строения языка:
А: 1 — срединная бороздка языка, 2 — грибовидные сосочки,
3 — листовидные сосочки, 4 — желобововые сосочки, 5 — пограничная борозда,
6 — язычная миндалина, 7 — надгортанник;
Б: 1 — грибовидные сосочки, 2 — нитевидные сосочки, 3 — желобовые сосочки,
4 — лимфоидные узелки, 5 — конические сосочки
11
11
22
22
33
33
44
44
55
55
А
66
Б
77
231
ПОЛОСТЬ РТА
сосочках огромное количество вкусовых почек. У взрослых людей ли-
стовидные сосочки отсутствуют. На слизистой оболочке корня языка
сосочков нет. Переходя на нижнюю поверхность языка, слизистая
оболочка образует его уздечку.
Язык — мышечный орган. Мышцы языка подразделяются на две
группы: 1) наружные, начинающиеся на костях и оканчивающиеся в
языке, осуществляют движения языка и сохраняют его тонус; 2) соб-
ственные мышцы языка, не связанные с костями, состоят из пучков
продольных, поперечных и вертикальных волокон, переплетающихся
между собой и с наружными мышцами. При сокращении этих мышц
изменяется форма языка.
Зубы. У большинства млекопитающих, в том числе и человека,
последовательно сменяются два типа зубов: молочные и постоянные.
У взрослого человека 32 постоянных зуба (рис. 130). Основная функ-
ция зубов — механическое измельчение пищи. Различают три типа
зубов: резцы, служащие для захватывания и откусывания пищи; клы-
ки, которые дробят, разрывают пищу; коренные, которые растирают,
перемалывают пищу.
Рис. 130. Зубы верхней челюсти:
А, постоянные зубы: 1 — медиальный резец, 2 — латеральный резец, 3 — клык,
4 — первый малый коренной зуб, 5 — второй малый коренной зуб, 6 — первый
большой коренной зуб, 7 — второй большой коренной зуб, 8 — третий большой
коренной зуб; Б, молочные (сменные) зубы ребенка 4 лет: а — резцы, б — клык,
в — коренные зубы, г — альвеола III постоянного зуба
11
22
33
44
55
66
77
88
аа
бб
вв
гг
АБ
232
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
Форма зубов и их функция тесно связаны между собой. Кроме
указанных функций, у человека зубы участвуют в членораздельной
речи, придавая своеобразную окраску отдельным звукам. Зубы игра-
ют важную формообразующую роль. Напомним, что форма, рельеф
и структура костей зависят от деятельности прикрепляющихся к ним
мышц. Здоровые зубы обеспечивают нормальное функционирование
всего жевательного аппарата, это предотвращает развитие возрастных
изменений костей лицевого черепа. На каждой верхней и половине
нижней челюсти, начиная от средней вертикальной линии вправо
и влево расположены два резца, один клык, два малых коренных и три
больших коренных зуба.
Каждый зуб состоит из трех частей (рис. 131). Коронка — это бо-
лее массивный отдел зуба, выступающий над уровнем входа в альвео-
лу, несколько суженная шейка находится на границе между корнем и
коронкой, в этом месте с зубом соприкасается слизистая оболочка де-
сен. Каждый зуб имеет один, два или три корня. Корень, расположен-
ный в зубной альвеоле, оканчивается верхушкой, на которой располо-
жено маленькое отверстие, через которое в полость зуба входят сосуды
и нервы. Внутри зуба имеется полость, заполненная зубной пульпой,
Рис. 131. Строение зуба:
1 — эмаль, 2 — дентин,
3 — пульпа зуба, 4 — десна,
5 — цемент, 6 — периодонт,
7 — кость;
I — коронка зуба,
II — шейка зуба,
III — корень зуба,
IV — канал корня зуба
11
22
33
44
55
66
77
I
II
III
IV
233
ПОЛОСТЬ РТА
богатой сосудами и нервами. Зубы укреплены в зубных альвеолах че-
люстей. Корни зубов плотно срастаются с поверхностью зубных ячеек
посредством периодонта — пучков соединительнотканных волокон,
которые проникают с одной стороны в кость альвеолы, с другой —
в цемент корня зуба. Зуб построен главным образом из дентина, кото-
рый в области корня покрыт цементом, а в области коронки — эма-
лью. Будучи весьма прочными, зубы сохраняются в течение сотен ты-
сяч лет, что играет важную роль для палеонтологии и антропологии.
Прорезывание молочных зубов начинается на шестом-седьмом
месяце жизни ребенка и заканчивается к трем годам жизни. У ребен-
ка 20 молочных зубов. На каждой верхней и половине нижней челю-
сти ребенка расположены два резца, один клык, и два больших ко-
ренных зуба. С 6—7 лет начинается смена молочных зубов
постоянными. Сроки прорезывания молочных и постоянных зубов
представлены в табл. 15.
Таблица 15
Средние сроки прорезывания зубов
Зуб
Молочные
Постоянные
Внутренний резец
6—8 мес.
7—7,5 года
Наружный резец
7—9 мес.
8—8,5 года
Клык
15—20 мес.
11—11,5 года
Первый малый коренной
—
10—10,5 года
Второй малый коренной
—
11—11,5 года
Первый большой коренной
12—15 мес.
7—7,5 года
Второй большой коренной
20—24 мес.
12—12,5 года
Третий большой коренной
—
18—25 лет и позднее
Железы рта. В слизистой оболочке, подслизистой основе, толще
мышц, а также между слизистой оболочкой и надкостницей твердого
нёба находится множество мелких слюнных желез. В ротовую по-
лость открываются протоки трех пар больших слюнных желез: около-
ушных, подчелюстных и подъязычных и множества мелких, выделя-
ющих белковый секрет (околоушные и железы языка, расположенные
в области желобовидных сосочков); слизь (небные и задние язычные);
или смешанный секрет (губные, щечные, передние язычные, подъя-
зычные, поднижнечелюстные).
Самая крупная околоушная железа (масса 20—30 г) расположена
на боковой поверхности лица спереди и ниже ушной раковины. Вы-
водной околоушный проток (стеннонов) открывается в преддверии
234
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
рта на вершине сосочка, расположенного на уровне второго верхнего
большого коренного зуба.
Поднижнечелюстная железа (масса 13—16 г) располагается в под-
нижнечелюстном треугольнике поверхностно, ее выводной проток
(вартонов) открывается на сосочке сбоку от уздечки языка. Самая
маленькая подъязычная железа (масса около 5 г) расположена на
верхней поверхности диафрагмы рта. Ее главный — большой подъя-
зычный — проток открывается одним общим отверстием с протоком
поднижнечелюстной железы или рядом с ним.
Слюнные железы выделяют в сутки от 500 мл до 2 л слюны, состо-
ящей преимущественно из воды (до 99,5%), солей, ферментов (амила-
зы и глюкозидазы), слизи, электролитов, а также бактерицидного ве-
щества лизоцима и иммуноглобулинов. Наличие крупных слюнных
желез характерно только для млекопитающих (и человека). Основная
функция слюны — смачивание и частичная переработка пищи. У но-
ворожденных детей слюнные железы развиты слабо, их быстрый рост
происходит в период от 4 мес. до 2 лет.
В полости рта пища измельчается, увлажняется и смешивается со
слюной. Очень важно тщательное пережевывание пищи. Жевание
у здорового человека — это рефлекторный акт, который регулируется
корой больших полушарий головного мозга. Жевание осуществляет-
ся благодаря координированной деятельности челюстей, зубов, жева-
тельных и мимических мышц, языка, щек, нёба, дна полости рта.
Слюна смачивает пищу, а слизь, содержащаяся в слюне, облегчает
глотание. Уже в полости рта начинается расщепление углеводов.
Слюна растворяет молекулы вкусовых веществ, которые попадают во
вкусовые почки в растворенном виде. Кроме того, благодаря содержа-
нию лизоцима слюна дезинфицирует полость рта. Слюноотделение
является рефлекторным актом, слюноотделение усиливается уже при
виде пищи, в ответ на запах пищи и при мыслях о еде. В начале XX в.
И. П . Павлов обнаружил, что, как только пища попадает в рот собаки,
начинает рефлекторно вырабатываться слюна. Когда собака просто
видит пищу, то так же автоматически начинается слюноотделение, но
в этом случае рефлекс значительно менее постоянен и зависит от до-
полнительных факторов, таких как голод или переедание. Если соче-
тать пищу со звуковым или зрительным сигналом, то после несколь-
ких повторений слюна начинает вырабатываться в ответ на сигнал
даже без пищи. Павлов назвал это явление условным рефлексом. Та -
ким образом, один только вид или запах пищи действует как сигнал
для образования слюны. «Любое явление во внешнем мире может
быть превращено во временный сигнал объекта, стимулирующий
слюнные железы, — писал Павлов, — если стимуляция этим объектом
слизистой оболочки ротовой полости будет связана повторно... с воз-
235
ГЛОТКА И ПИЩЕВОД
действием определенного внешнего явления на другие чувствитель-
ные поверхности тела». Открытие И.П. Павловым условного рефлек-
са привело к созданию новой отрасли науки — физиологии высшей
нервной деятельности. В основе условного рефлекса лежит формиро-
вание новых или модификация существующих нервных связей, про-
исходящие в индивидуальной жизни животных и человека под влия-
нием изменений внешней среды. Эти временные связи тормозятся
при отмене подкрепления.
ГЛОТКА И ПИЩЕВОД
Глотка представляет собой воронкообразный канал длиной у взро-
слого человека 11—12 см (рис. 132). Верхняя стенка глотки сращена
с основанием черепа; на границе между VI и VII шейными позвонка-
ми она, суживаясь, переходит в пищевод. Полость глотки делится на
три части: верхнюю — носовую, среднюю — ротовую и нижнюю —
гортанную. На боковых стенках глотки с обеих сторон расположены
глоточные отверстия слуховой (евстахиевой) трубы, которая соеди-
няет глотку с полостью среднего уха и способствует сохранению в нем
постоянной величины атмосферного давления.
Поперечнополосатые мышцы глотки располагаются в двух на-
правлениях — продольном (подниматели) и поперечном, циркуляр-
ном (сжиматели). В полости глотки имеется важный защитный аппа-
рат — лимфоэпителиальное кольцо, названное по имени Н. И . Пирогова,
который впервые обратил на него внимание. Сюда входят небные,
язычная, глоточная и трубные миндалины (см. «Органы кроветворе-
ния и иммунной системы»).
Рис. 132. Строение глотки, схема:
1 — твердое нёбо, 2 — небный язычок,
3 — мягкое нёбо (небная занавеска),
4 — свод глотки, 5 — глоточная
(аденоидная) миндалина, 6 — носовая
часть глотки, 7 — гортанная часть
глотки, 8 — голосовая щель,
9 — надгортанник, 10 — ротовая часть
глотки, 11 — зев, 12 — язык
11
11
11
22
12
12
33
44
55
66
77
88
99
10
10
236
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
Глотание — рефлекторный акт, который начинается в тот момент,
когда пищевой комок соприкасается с нёбом, корнем языка или зад-
ней стенкой глотки. При глотании продольные мышцы глотки, сокра-
щаясь, поднимают ее, а циркулярные сокращаются сверху вниз, тем
самым продвигая пищу в направлении к пищеводу. В акте глотания
участвует также небная занавеска и мышцы языка. В глотке человека
происходит перекрест дыхательного и пищеварительного путей. При
глотании мягкое нёбо обособляет носоглотку, гортань поднимается,
надгортанник опускается и прикрывает вход в нее, язык отодвигается
назад, пища поступает в пищевод. При дыхании корень языка прижи-
мается к нёбу, закрывая выход из по-
лости рта, а надгортанник поднимает-
ся, открывая вход в гортань, куда
устремляется струя воздуха.
Пищевод человека — цилиндриче-
ская трубка длиной у взрослого чело-
века 22—30 см, сплющенная спереди
назад, которая в спокойном состоя-
нии имеет щелевидный просвет. Пи-
щевод начинается на уровне границы
между VI и VII шейными позвонками
и оканчивается на уровне XI грудного
позвонка. В пищеводе различают три
части: шейную, грудную и брюшную,
последняя, самая короткая (1—
1,5 см), находится под диафрагмой,
через пищеводное отверстие которой
пищевод проходит в брюшную по-
лость (рис. 133). Окруженный рых-
лой волокнистой неоформленной
соединительной тканью, пищевод
подвижен. Мышечная оболочка верх-
ней трети пищевода образована попе-
речнополосатыми мышечными во-
локнами, в средней они постепенно
замещаются гладкими, в нижней пол-
ностью состоят из гладких мышечных
волокон. Мышечная оболочка об-
условливает движения пищевода и
его постоянный тонус. Мышечные
волокна располагаются в два слоя:
внутренний кольцевой и наружный
продольный. Мышцы пищевода, по-
Рис. 133. Пищевод и желудок, схема:
1 — шейная часть, 2 — пищевод,
3 — грудная часть, 4 — тело
желудка, 5 — брюшная часть
пищевода, 6 — двенадцатиперстная
кишка, 7 — привратниковая часть
(пилорическая) желудка,
8 — диафрагмальное сужение
пищевода
11
22
33
55
44
88
66
77
237
ЖЕЛУДОК
следовательно сокращаясь сверху вниз, проталкивают пищевой комок
в желудок. При этом плотная пища проходит по пищеводу за 3—9 с,
жидкая — за 1—2 с.
ЖЕЛУДОК
Желудок, расположенный в левом подреберье и надчревной обла-
сти, напоминает реторту или грушу, однако форма его постоянно из-
меняется в зависимости от количества съеденной пищи, положения
тела и т. д . Вход в желудок — его кардиальная часть, слева от нее же-
лудок расширяется, образуя дно, которое переходит в тело. Левый вы-
пуклый край желудка формирует большую кривизну, правый, вогну-
тый, — малую кривизну. Выход из желудка называется привратником
(пилорус), он снабжен кольцевой мышцей — сфинктером. Суженная
часть желудка, примыкающая к привратнику, называется пилориче-
ской. Емкость желудка взрослого человека варьирует в зависимости
от принятой пищи и жидкости от 1,5 до 4 л (см. рис. 133).
Слизистая оболочка желудка покрыта однослойным цилиндриче-
ским железистым эпителием, выделяющим слизь, которая располагает-
ся в виде нескольких слоев, лежащих друг за другом, и выполняет за-
щитную функцию. Многочисленные (около 40 млн) желудочные
железы залегают в собственной пластинке слизистой оболочки почти
вплотную друг к другу (рис. 134). Различают три группы желез: пре-
обладают собственные (фундальные), у человека их около 35 млн, дли-
1
11
2
12
3
13
4
14
5
6
7
8
9
10
1
11
2
12
3
13
4444444444444444
14
5
6
7
8
9
10
Рис. 134. Желудок
(вскрыта его передняя стенка):
1 — кардиальная вырезка,
2 — дно желудка, 3 — передняя
стенка, 4 — тело желудка,
5 — большая кривизна желудка,
6 — желудок,
7 — привратниковая пещера,
8 — канал привратника,
9 — привратниковая
(пилорическая) часть,
10 — угловая вырезка,
11 — канал желудка,
12 — малая кривизна желудка,
13 — кардиальное отверстие,
14 — кардиальная часть
238
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
на каждой около 0,65 мм, диаметр 30–50 мкм; пилорические (около
3,5 млн) и кардиальные. В собственных железах выделяют четыре типа
клеток: главные вырабатывают пепсиноген и реннин; париетальные
(обкладочные) — компоненты соляной кислоты и внутренний антиане-
мический фактор; слизистые (добавочные и шеечные) — слизь; эндо-
кринные — серотонин, эндорфины и другие биологически активные
вещества. В пилорических железах имеется большое количество эндо-
кринных клеток, вырабатывающих серотонин, эндорфины, соматоста-
тин, гастрин (последний стимулирует секрецию соляной кислоты па-
риетальными клетками) и другие биологически активные вещества.
Желудок растет очень быстрыми темпами. Так, если масса вну-
тренних органов увеличивается от рождения до полового созревания
примерно в 12 раз, тело в целом в 20 и более раз, то желудок в 24 раза.
Объем желудка новорожденного составляет 30—35 см3
, через две не-
дели уже 90 см3, в три года 575—680 см3, у взрослого он равен 1200—
1600 см3, т. е. превосходит в 50 раз первоначальную величину. Желу-
дочные железы у новорожденного развиты слабо, количество их
составляет 120—123 на 1 мм2 (у взрослого 260—270 на 1 мм2), всего
около 500 000 желез. Их количество быстро увеличивается. В двухме-
сячном возрасте их насчитывается около 1,8 млн, в двухлетнем —
8 млн, в шестилетнем — 10 млн, в 15-летнем — 18 млн, а у взрослого —
около 40 млн.
Мышечная оболочка сформирована гладкой мышечной тканью,
образующей три слоя: наружный — продольный, средний — цирку-
лярный, внутренний — косой. Циркулярный слой наиболее развит
в пилорическом отделе, где образует упомянутый сжиматель при-
вратника (толщиной 3—5 мм), при сокращении которого закрывается
выход из желудка. Деятельность мышц желудка у живого человека
обусловливает его моторику, поддерживает тонус, почти стабильное
давление в просвете желудка и осуществляет перемешивание и опо-
рожнение.
Однокамерный желудок служит резервуаром для проглоченной
пищи, в котором пища интенсивно перемешивается и передвигается,
и, что самое важное, благодаря выделению желудочного сока (в со-
став которого входят пепсин, реннин, липаза, соляная кислота и слизь)
осуществляется частичная химическая переработка пищи. Кроме
того, желудок выполняет выделительную, эндокринную и всасыва-
тельную функции (всасываются сахара, спирт, вода, соли), в стенке
желудка образуется внутренний антианемический фактор, который
способствует поглощению поступающего с пищей витамина В12, что
предотвращает развитие анемии.
В желудке продолжается расщепление углеводов амилазой слю-
ны, осуществляется частичное расщепление белковых молекул, в том
239
ТОНКАЯ КИШКА
числе коллагена, а также жиров молока. У детей расщепляется в же-
лудке до 60% жира молока.
Железы желудка выделяют за сутки 1,5—2,5 л кислого желудочного
сока (рН 0,8—1,5), в котором содержится около 99% воды, соляная кис-
лота (0,3—0,5%), ферменты, слизь, соли и другие вещества. Под влия-
нием соляной кислоты пепсиноген превращается в активный пепсин.
Соляная кислота образуется уже в полости желудка из выделяемых
клетками желез ионов Н+ и Сl–
.
Слизь предохраняет слизистую обо-
лочку желудка от ее повреждения соляной кислотой и пепсином.
Количество желудочного сока, его состав, кислотность, содержание
ферментов зависят от количества, качества и консистенции пищи.
Пища, смешанная с желудочным соком, называется химусом. Желу-
дочный сок начинает вырабатываться условно-рефлекторно. Как толь-
ко пища попадает в рот, возбуждаются рецепторы органов вкуса и обо-
няния, благодаря этому усиливается выделение желудочного сока
(безусловный рефлекс). При попадании пищи в желудок выделение
желудочного сока усиливается. Белковая пища является наиболее эф-
фективным возбудителем желудочной секреции, максимум достигает-
ся во время второго часа. Самым слабым возбудителем желудочной
секреции является углеводная пища (в том числе хлеб). Жиры вначале
тормозят, а затем возбуждают желудочную секрецию. Переваривающая
активность желудочного сока наиболее высокая в ответ на поступление
белковой пищи, она ниже — при жировой и самая низкая — при угле-
водной. Стресс и сильные эмоции усиливают желудочную секрецию;
страх и депрессии угнетают ее. После приема смешанной пищи вначале
эвакуация происходит быстро, а затем постепенно замедляется. Эваку-
ация пищи из желудка в двенадцатиперстную кишку происходит от-
дельными порциями и обусловлена главным образом сокращениями
мышечной оболочки желудка, а пилорический жом препятствует ее
обратному забрасыванию в желудок. Пища находится в желудке раз-
личное время. Скорость эвакуации связана с количеством, составом
и степенью измельчения пищи в ротовой полости. Так, плохо переже-
ванная пища дольше задерживается в желудке, чем кашицеобразная
или жидкая. Эвакуация содержимого желудка начинается, когда оно
становится жидким (или полужидким). Быстрее всего удаляются из
желудка углеводы (через 1,5—2 ч), медленнее — белки, дольше всего
задерживаются в желудке жиры (4 ч и более).
ТОНКАЯ КИШКА
Тонкая кишка человека подразделяется на двенадцатиперстную
длиной 25—30 см, тощую длиной 2—2,5 м и подвздошную длиной
2,5—3,5 м кишки (см. рис. 127). Тощая кишка короче, а подвздошная
240
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
длиннее. Диаметр тонкой кишки не превышает 6 см. Тонкая кишка
образует множество петель, слизистая оболочка — многочисленные
круговые складки, благодаря чему увеличивается всасывательная по-
верхность слизистой оболочки, размер и количество складок умень-
шается по направлению к толстой кишке. У дистального конца под-
вздошной кишки складки исчезают. Поверхность слизистой оболочки
усеяна кишечными ворсинками и криптами (рис. 135). Ворсинки яв-
ляются выростами собственной пластинки слизистой оболочки.
В центре ворсинки проходит лимфатический капилляр, слепо начи-
нающийся на ее вершине. В каждую ворсинку входит по 1—2 артери-
олы, которые распадаются на капиллярные сети, расположенные
вблизи эпителиальных клеток. Из капилляров кровь собирается в ве-
нулу, проходящую вдоль оси ворсинки. Поверхность ворсинок покры-
та однослойным цилиндрическим эпителием, в котором имеются
клетки трех видов: выделяющие слизь бокаловидные, кишечные эпи-
телиоциты с исчерченной каемкой и небольшое количество эндо-
кринных клеток. Больше всего кишечных эпителиоцитов с исчерчен-
ной каемкой, на их обращенной в просвет кишечника (апикальной)
поверхности имеется каемка, образованная огромным количеством
микроворсинок (1500—3000 на поверхности каждой клетки), которые
увеличивают еще в 30—40 раз всасывающую поверхность.
Кишечные крипты (крипты Либеркюна) — углубления собствен-
ной пластинки слизистой оболочки в виде трубочек (длиной 0,25—
Рис. 135. Строение ворсинок
тонкой кишки:
1 — ворсинки,
2 — сеть лимфатических
и кровеносных сосудов
слизистой оболочки,
3 — одиночный лимфоидный
узелок, 4 — центральный
лимфатический (млечный)
капилляр, 5 — эпителий
11
22
33
44
55
241
ПЕЧЕНЬ И ЖЕЛЧНЫЙ ПУЗЫРЬ
0,5 мм, диаметром до 0,07 мм), устья которых открываются в просве-
тах между ворсинками. Количество их достигает 80—100 на 1 мм2
.
Крипты выстланы эпителиальными клетками пяти видов. Это кишеч-
ные эпителиоциты с исчерченной каемкой, бокаловидные, эндокрин-
ные, бескаемчатые и клетки Панета. Клетки первых трех типов ана-
логичны клеткам ворсинок. Эндокринные клетки вырабатывают
серотонин и кишечные гормоны (например, секретин, холецистоки-
нин и др.). На дне крипт располагаются клетки Панета, которые по
современным воззрениям вырабатывают лизоцим и фермент эрепсин,
участвующий в расщеплении дипептидов. Мелкие цилиндрические
бескаемчатые клетки, расположенные на дне крипт между клетками
Панета, активно делятся митотически и являются источником восста-
новления эпителия ворсинок и крипт.
В собственной пластинке слизистой оболочки имеется множество
одиночных лимфоидных узелков диаметром 0,5—3 мм, а также ско-
пления лимфоидных узелков, называемые пейеровыми бляшками.
Они расположены в основном в подвздошной кишке. Лимфоидная
ткань лучше выражена у детей. Количество одиночных узелков и их
агрегатов с возрастом уменьшается. У взрослых число пейеровых
бляшек достигает 30—40, у стариков их еще меньше. Лимфоидная
ткань, расположенная в стенке кишки, выполняет защитную и кро-
ветворную функции.
Мышечная оболочка тонкой кишки, состоящая из более мощного
внутреннего циркулярного и наружного продольного слоев, осу-
ществляет маятникообразные и перистальтические движения кишки
и обеспечивает постоянное тоническое сокращение ее мускулатуры.
Двенадцатиперстная кишка имеет форму подковы, огибающей го-
ловку поджелудочной железы. Слизистая оболочка двенадцатиперст-
ной кишки образует множество широких и коротких ворсинок (22—
40 на 1 мм2). Слизистая оболочка формирует, кроме круговых, и одну
продольную складку, которая заканчивается большим двенадцати-
перстным сосочком (фатеров), на вершине которого открываются об-
щий желчный проток и главный проток поджелудочной железы.
В подслизистой основе встречаются сложные разветвленные трубча-
тые железы, которые вырабатывают секрет, участвующий в перевари-
вании белков и углеводов, слизь, а также многочисленные гормоны
и биологически активные вещества.
ПЕЧЕНЬ И ЖЕЛЧНЫЙ ПУЗЫРЬ
Печень — самая крупная железа человека. Масса печени взрослого
человека составляет около 1/36 массы тела (1,5—2 кг), у новорожден-
ного — 1/20 (около 135 г), и она занимает бо
льшую часть брюшной
242
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
полости. Печень участвует в обмене белков, углеводов, жиров, вита-
минов и т. д . Среди многочисленных функций печени весьма важны
защитная, обезвреживающая, желчеобразовательная и др. В утробном
периоде печень является важным кроветворным органом. Печень рас-
положена справа под диафрагмой, лишь небольшая часть ее заходит у
взрослого человека влево от средней линии. Край печени острый, пе-
редне-верхняя (диафрагмальная) поверхность выпуклая соответст-
венно вогнутости диафрагмы (рис. 136).
Печень покрыта соединительнотканной оболочкой (глиссонова
капсула). Прослойки соединительной ткани внутри печени разделя-
ют ее перенхиму на шестиугольные дольки призматической формы,
около 1,5 мм в диаметре (классические дольки). Сложной и много-
78910
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
4
3
2
56
1
Рис. 136. Печень, вид снизу. Висцеральная поверхность:
1 — левая доля, 2 — левая треугольная связка, 3 — пищеводное вдавление, 4 — венозная
связка, 5 — сосочковый отросток, 6 — хвостатая доля, 7 — хвостатый отросток,
8 — воротная вена, 9 — нижняя полая вена, 10 — надпочечниковое вдавление,
11 — задняя часть диафрагмальной поверхности, 12 — место перехода брюшины,
13 — почечное вдавление, 14 — правая треугольная связка, 15 — двенадцатиперстно-
кишечное вдавление, 16 — правая доля, 17 — ободочнокишечное вдавление,
18 — желчный пузырь, 19 — квадратная доля, 20 — пузырный проток,
21 — круглая связка печени, 22 — общий желный проток,
23 — общий печеночный проток, 24 — собственная печеночная артерия,
25 — сальниковый бугор, 26 — желудочное вдавление
243
ПЕЧЕНЬ И ЖЕЛЧНЫЙ ПУЗЫРЬ
гранной функции печени соответствует характер ее сосудистой систе-
мы, строение и функция клеток, образующих печеночную ткань. Пе-
чень, в отличие от всех других органов, получает кровь из двух
источников: артериальную — из печеночной артерии и венозную — из
воротной вены. И та и другая кровь проходит через синусоидальные
кровеносные капилляры, по которым кровь течет очень медленно. Во-
ротная вена собирает кровь от желудка, тонкой кишки, поджелудоч-
ной железы, селезенки и большого сальника (рис. 137). Войдя в воро-
та печени, оба сосуда (печеночная артерия и воротная вена)
разделяются на долевые, сегментарные и т. д ., вплоть до междолько-
вых вен и артерий, которые проходят в междольковых соединитель-
нотканных прослойках вдоль боковых поверхностей классических
печеночных долек, между ними вместе с желчными протоками и лим-
фатическими сосудами, образуя так называемые триады. От междоль-
ковых сосудов под прямым углом отходят вокругдольковые, которые
окружают дольку наподобие кольца. От них начинаются синусои-
дальные капилляры, которые на периферии долек соединяются меж-
ду собой, образуя один капилляр, следующий к центру дольки, где
вливаются в центральную вену дольки. Последняя, в свою очередь,
впадает в поддольковую вену. Поддольковые вены являются началь-
ными сосудами системы печеночных вен, которые, укрупняясь, соби-
Рис. 137. Строение
(и кровоснабжение)
печеночной дольки, схема:
1 — воротная вена,
2 — печеночная артерия,
3 — сегментная вена
и артерия, 4 — междольковые
вена и артерия,
5 — вокругдольковые
вена и артерия,
6 — внутридольковые
гемокапилляры
(синусоидные сосуды),
7 — классическая
печеночная долька,
8 — центральная вена,
9 — поддольковая
(собирательная) вена,
10 — печеночная вена
Рис
(и кр
печено
1—
2—пе
3—
и артери
ве
5—
ве
6—в
ге
(сину
7—
печ
8—ц
9—
(соби
10—
11
22
33
44
55
66
77
88
99
10
10
244
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
раются в три-четыре печеночные вены, впадающие в нижнюю полую
вену. Через 1 г печеночной ткани в минуту проходит около 0,85 мл
крови, в течение часа вся кровь человека несколько раз проходит че-
рез синусоидальные капилляры печени. Это дало основание старым
анатомам назвать печень «самой нагруженной гаванью во всей реке
жизни».
Кровеносные капилляры имеют собственную стенку, образован-
ную двумя типами клеток: эндотелиальными клетками и расположен-
ными между ними звездчатыми клетками Купфера, имеющими длин-
ные отростки, свободно свисающие в просвет капилляра (рис. 138).
Эти клетки, относящиеся к системе макрофагов, обладают выражен-
ной фагоцитарной активностью. Желчные капилляры не имеют соб-
ственной стенки, плазматические мембраны соседних печеночных
7
8
9
10
11
12
4
3
2
5
6
1
Рис. 138. Строение печеночной балки (схема):
1 — вокругдольковая вена, 2 — гепатоцит, 3 — печеночная балка,
4 — синусоидный сосуд, 5 — вокругсинусоидное пространство
(пространство Диссе), 6 — центральная вена, 7 — вокругсинусоидный липоцит,
8 — желчный капилляр, 9 — звездчатый макрофагоцит, 10 — эндотелиальная клетка,
11 — вокругдольковый желчный проток, 12 — вокругдольковая артерия
245
ПЕЧЕНЬ И ЖЕЛЧНЫЙ ПУЗЫРЬ
клеток образуют стенку желчного капилляра. Иными словами, желч-
ные капилляры, по существу, являются расширенными зонами меж-
клеточных щелей. В то же время в местах перехода межклеточных
щелей в желчные капилляры участки плазматических мембран кле-
ток утолщены, благодаря этому желчные капилляры не сообщаются
с другими межклеточными щелями и у здорового человека желчь не
проникает в кровь. Желчные капилляры начинаются слепо вблизи
центральной вены и направляются к периферии дольки, где перехо-
дят в междольковые желчные протоки. У ворот печени путем слия-
ния правой и левой ветвей, приносящих желчь из соответствующих
долей печени, образуется общий печеночный проток.
Печеночные клетки (гепатоциты) богаты митохондриями (до
1000 и более в одной клетке), элементами зернистой и незернистой
цитоплазматической сети и комплекса Гольджи, рибосомами и осо-
бенно отложениями гликогена. Клетки располагаются в виде тяжей
(печеночные трабекулы) между капиллярами двумя рядами так, что-
бы плазматическая мембрана каждой из них обязательно контактиро-
вала одной своей стороной с просветом желчного капилляра, другой
соприкасалась со стенкой кровеносного капилляра. Для этого трабе-
кулы не могут одновременно в двух направлениях состоять более чем
из двух клеток, это возможно лишь в одном направлении: только
в толщину или в ширину. Такое строение способствует осуществле-
нию секреции гепатоцитов в двух направлениях: в желчные прото-
ки — желчь, в кровеносные капилляры — глюкозу, мочевину, белки,
жиры, витамины и т. д .
Желчь вырабатывается постоянно, однако есть основание считать,
что в печени существует суточный ритм: ночью преобладает синтез
гликогена, днем — желчи. В течение суток у человека образуется от
500 до 1000 мл желчи, ее рН = 7,8 — 8,6; содержание воды достигает
95—98%. В желчи имеются соли желчных кислот, билирубин, холе-
стерин, жирные кислоты, лецитин, минеральные элементы. Однако
в связи с ритмом питания нет необходимости в постоянном поступле-
нии желчи в двенадцатиперстную кишку. Этот процесс регулируется
гуморальными и нервно-рефлекторными механизмами.
Желчный пузырь, являющийся резервуаром для хранения желчи,
представляет собой удлиненный мешок длиной 8—12 см, шириной
4—5 см, с расширенным дном, напоминающим по своей форме грушу
емкостью около 40 см3 (см. рис. 136). Складчатая слизистая оболочка
выстлана однослойным цилиндрическим эпителием, поверхность ко-
торого покрыта исчерченной каемкой из микроворсинок, интенсивно
всасывающим воду, поэтому пузырная желчь сгущается в три-пять раз
по сравнению с желчью из общего печеночного протока. Пузырный
проток, соединяясь с общим печеночным, образует общий желчный
246
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
проток, который направляется вниз, прободает нисходящую часть
двенадцатиперстной кишки, сливаясь с протоком поджелудочной же-
лезы, и открывается на вершине большого сосочка двенадцатиперст-
ной кишки. Пучки мышечных волокон окружают конец общего желч-
ного протока в толще стенки кишки, образуя сфинктер Одди, который
препятствует затеканию содержимого двенадцатиперстной кишки
в желчный и панкреатический протоки. Выше него, над местом слия-
ния протока поджелудочной железы с общим желчным, располагается
еще один сфинктер общего желчного протока (Бойдена), который ре-
гулирует собственно приток желчи в кишку или желчный пузырь.
Одно из наиболее распространенных заболеваний желчных путей —
желчнокаменная болезнь. Камни образуются у тех людей, у которых
желчь перенасыщена холестерином, выпадающим в осадок в виде кри-
сталлов. Около 90% желчных камней состоит из холестерина.
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
Поджелудочная железа — вторая по величине железа пищевари-
тельного тракта (масса 60—100 г, длина 15—22 см), перекидывается
в поперечном направлении через тело первого поясничного позвонка.
Ее широкая головка располагается внутри подковы двенадцатиперст-
ной кишки и незаметно переходит в тело, которое заканчивается су-
женным хвостом, достигающим ворот селезенки (рис. 139).
Поджелудочная железа по существу состоит из двух желез. Экзо-
кринная часть вырабатывает у человека в течение суток 1500—
2000 мл водянистого панкреатического сока (рН = 8 —8,5), со-
держащего ферменты трипсин и химотрипсин, участвующие в
переваривании белков; амилазу, гликозидазу и галактозидазу, пере-
варивающие углеводы; липолитическую субстанцию, липазу, участ-
вующие в переваривании жиров; ферменты, расщепляющие нуклеи-
новые кислоты. Экзокринная часть поджелудочной железы
представляет собой сложную альвеолярно-трубчатую железу, разде-
ленную очень тонкими перегородками на дольки, в которых тесно
лежат ацинусы, образованные одним слоем железистых ацинозных
клеток, богатых элементами зернистой цитоплазматической сети
и гранулами, содержащими ферменты. Главный проток поджелудоч-
ной железы (вирсунгов) проходит слева направо через железу и от-
крывается на вершине большого сосочка двенадцатиперстной киш-
ки после слияния с общим желчным протоком.
Эндокринная часть, продуцирующая гормоны, регулирующие
углеводный и жировой обмен (инсулин, глюкагон), соматостатин
и др., образована группами клеток, которые располагаются в виде
островков диаметром 0,1—0,3 мм в толще железистых долек (остров-
247
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
ки Лангергаиса). Количество островков у взрослого человека коле-
блется от 200 тыс. до 1800 тыс. (см. «Эндокринный аппарат»).
Поджелудочная железа новорожденного очень мала, она весит
около 2—3 г. У новорожденных и детей раннего возраста железа отли-
чается большим абсолютным и относительным количеством панкреа-
тических островков.
Благодаря движениям тонкой кишки кислый химус, поступающий
из желудка, перемешивается со щелочными соками поджелудочной
железы, печени и кишечных желез, причем кишечное содержимое
постоянно контактирует со слизистой оболочкой тонкой кишки. В те-
чение суток у человека выделяется до 2,5 л кишечного сока. Его мно-
гочисленные ферменты, расщепляющие белки, жиры, углеводы, про-
исходят из разрушенных слущенных эпителиальных клеток слизистой
оболочки кишечника. В результате непрерывного процесса регенера-
ции клетки восстанавливаются.
7
8
9
10
11
12
13
14
15
16
17
18
4
3
2
5
6
1
Рис. 139. Желчный пузырь, общий желчный проток, поджелудочная железа
и двенадцатиперстная кишка, вид сзади. Часть стенки двенадцатиперстной кишки
и конечная часть общего желчного протока вскрыты:
1 — тело поджелудочной железы, 2 — селезеночная вена, 3 — воротная вена,
4 — общий печеночный проток, 5 — пузырный проток, 6 — шейка желчного пузыря,
7 — общий желчный проток, 8 — тело желчного пузыря, 9 — дно желчного пузыря,
10 — двенадцатиперстная кишка, 11 — сфинктер печеночно-поджелудочной ампулы
(сфинктер ампулы, сфинктер Одди), 12 — брюшина, 13 — проток поджелудочной
железы и его сфинктер, 14 — сфинктер общего желчного протока, 15 — головка
поджелудочной железы, 16 — верхняя брыжеечная артерия, 17 — верхняя
брыжеечная вена, 18 — хвост поджелудочной железы
248
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
В тонкой кишке продолжается химическая переработка пищи
и всасывание продуктов расщепления, а также механическое переме-
шивание и продвижение ее в направлении толстой кишки. Эндокрин-
ные клетки вырабатывают различные гормоны и биологически актив-
ные вещества. У человека всасывающая поверхность тонкой кишки,
благодаря наличию складок слизистой оболочки, ворсинок и микро-
ворсинок клеток кишечного эпителия, достигает 200 м2.
Во время пищеварения секреция желчи и ее выделение в просвет
кишечника резко увеличивается; вне его сфинктер Одди закрыт,
и желчь накапливается в желчном пузыре, где она концентрируется.
Уже через 3—12 мин после начала еды рефлекторно усиливается се-
креция желчи. Особенно интенсивно выделяется желчь при поедании
смешанной пищи, большое количество выделяется в ответ на потре-
бление жира, молока, мяса, яичных желтков. Из желчного пузыря
желчь выделяется в двенадцатиперстную кишку благодаря сокраще-
нию мышц желчного пузыря и расслаблению сфинктера общего желч-
ного протока. Желчь нейтрализует кислую реакцию химуса, инакти-
вирует пепсин, компоненты желчи эмульгируют жиры, содержащиеся
в пищевых продуктах, тем самым облегчая действие ферментов, рас-
щепляющих жиры, и стимулируют всасывание продуктов переработ-
ки жиров.
Секреция сока поджелудочной железы начинается рефлекторно
уже при мыслях о пище, взгляде на нее, обстановке еды. Поступление
пищи в ротовую полость, раздражение рецепторов органа вкуса и обо-
няния, поступление пищи в желудок и раздражение его рецепторов
рефлекторно усиливают секрецию сока поджелудочной железы, кото-
рая достигает максимума при поступлении химуса в двенадцатиперст-
ную кишку. Белковая и углеводная пища увеличивает секрецию сока
поджелудочной железы в первые два часа с максимумом во время
второго часа после еды, причем секреция длится от 4—5 ч (белковая
пища) до 9—10 ч (хлеб); при приеме жирной пищи (включая молоко)
секреция длится около 5 ч (максимум — в течение третьего часа).
В тонкой кишке происходят переваривание пищи и всасывание ее
компонентов. При этом в полости кишки под влиянием ферментов
кишечного и панкреатического соков, желчи происходит лишь расще-
пление молекул пищевых веществ до отдельных фрагментов. Оконча-
тельное расщепление происходит благодаря мембранному пищеваре-
нию, которое осуществляется на поверхности микроворсинок клеток
кишечного эпителия. В них обнаружено большое количество актив-
ных ферментов, участвующих в расщеплении и всасывании пищевых
продуктов. А . М. Уголев (1967) открыл пристеночное пищеварение,
которое, в отличие от полостного, происходящего в просвете кишки,
совершается на поверхности микроворсинок. Последние вырабатыва-
249
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
ют ряд собственных пищеварительных ферментов и адсорбируют на
своей поверхности некоторые ферменты из просвета кишечника и пи-
щевые вещества, которые наиболее интенсивно расщепляются и вса-
сываются. В результате расщепления белков образуются аминокисло-
ты, жиров — глицерин и жирные кислоты, углеводов — моносахариды.
При распаде пищевых веществ утрачиваются многие их свойства,
в том числе и вредные. Таким образом предотвращается попадание
в организм чужеродного белка.
Ритмичные сокращения ворсинок способствуют всасыванию ве-
ществ, которое осуществляется по направлению от наружной поверх-
ности эпителиальных клеток (обращенной в просвет кишки) к вну-
тренней (обращенной к кровеносным и лимфатическим капиллярам).
Аминокислоты и моносахариды всасываются в кровь; вода, минераль-
ные соли, витамины, жирные кислоты и глицерин — в лимфу. Актив-
ному пищеварению и всасыванию способствует высокий кровоток
в тонкой кишке, который во время еды составляет 400 мл/мин., а на
высоте пищеварения — 750—800 мл/мин.
Еще в начале XX в. И. П . Павлов показал, что в каждом отделе пи-
щеварительной системы вырабатываются различные ферменты, кото-
рые участвуют в расщеплении белков, жиров и углеводов. Он изучил
их взаимодействие и регуляцию выделения, совместную деятельность
органов пищеварения и влияние одного отдела на другой. В 1904 г.
Павлов был награжден Нобелевской премией по физиологии и меди-
цине «за работу по физиологии пищеварения, благодаря которой было
сформировано более ясное понимание жизненно важных аспектов это-
го вопроса».
Толстая кишка подразделяется на слепую с червеобразным от-
ростком, восходящую ободочную, поперечную ободочную, нисходя-
щую ободочную, сигмовидную ободочную и прямую. Длина толстой
кишки колеблется от 1,5 до 2 м, диаметр слепой кишки достигает
7—8 см, он постепенно уменьшается до 4 см в нисходящей ободочной
кишке (см. рис. 127).
Уже по своему внешнему виду толстая кишка отличается от тонкой
диаметром, наличием отростков брюшины, заполненных жиром; ти-
пичных вздутий (гаустр) и трех продольных мышечных лент (тений),
образованных наружным продольным слоем мускулатуры, который на
толстой кишке не создает сплошного покрытия. Ленты идут от основа-
ния червеобразного отростка до начала прямой кишки. В толстой киш-
ке отсутствуют пейеровы бляшки. Слизистая оболочка толстой кишки
лишена ворсинок, но в ней много складок полулунной формы и значи-
тельно большее число крипт, чем в тонкой, они крупнее и шире.
Тонкая кишка впадает в стенку толстой кишки, ниже впадения
слепая кишка образует мешок. Подвздошная кишка как бы вдается
250
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
своим концом внутрь толстой, где имеется сложное анатомическое
устройство — илеоцекальный клапан, снабженный мышечным сфин-
ктером и двумя губами. Этот клапан замыкает выход из тонкой киш-
ки, периодически он открывается, пропуская содержимое небольши-
ми порциями в толстую кишку; кроме того, он препятствует обратному
затеканию содержимого толстой кишки в тонкую (рис. 140). Слепая
кишка расположена в правой подвздошной ямке, ее длина и ширина
примерно равны (7—8 см); от задней стенки слепой кишки отходит
червеобразный отросток длиной 6—8 см, размеры его непостоянны
и варьируют в широких пределах.
Слепая кишка непосредственно переходит в восходящую ободоч-
ную кишку длиной 14—18 см, которая направляется вверх. У нижней
поверхности печени, изогнувшись примерно под прямым углом, она
переходит в поперечноободочную длиной 25—30 см, которая пересека-
ет брюшную полость справа налево. В левой части брюшной полости
у нижнего конца селезенки ободочная кишка вновь изгибается, пово-
рачивает вниз и переходит в нисходящую ободочную длиной около
10 см. В левой подвздошной ямке она образует петлю — сигмовидную
ободочную кишку — и опускается в малый таз, где загибается и, на-
правляясь вниз и влево, переходит на уровне мыса крестца в прямую
кишку, которая тянется до заднего прохода. Верхний тазовый отдел
прямой кишки длиной 12—15 см расположен в полости таза; книзу
кишка расширяется, образуя ампулу, диаметр которой при наполне-
7
8
9
10
11
4
3
2
5
6
1
Рис. 140. Подвздошно-слепокишечный
переход. Передняя стенка подвздошной
и слепой кишок удалена:
1 — сальниковые отростки,
2 — полулунные складки ободочной
кишки, 3 — подвздошная кишка,
4 — брыжейка аппендикса, 5 — аппендикс
(червеобразный отросток слепой кишки),
6 — отверстие аппендикса, 7 — уздечка
подвздошно-слепокишечного клапана,
8 — слепая кишка, 9 — подвздошно-
слепокишечный клапан (верхняя и нижняя
губы), 10 — гаустры ободочной кишки,
11 — свободная лента
251
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
нии может увеличиваться до 30—
40 см. Конечный отдел длиной 2,5—
4 см, который направляется назад
и вниз, называется заднепроходным
каналом. Он проходит сквозь тазо-
вое дно и заканчивается задним
проходом (рис. 141).
Слизистая оболочка прямой
кишки образует множество непо-
стоянных продольных и попереч-
ных складок. Продольные мы-
шечные волокна расположены на
прямой кишке сплошным слоем, который, утолщаясь в области задне-
проходного канала, образует внутренний сфинктер заднего прохода,
состоящий из гладких мышечных волокон. Непосредственно под ко-
жей лежит кольцеобразный наружный сфинктер (поперечнополоса-
тая мышечная ткань). Оба сфинктера, в обычном состоянии замыкаю-
щие задний проход, открываются при акте дефекации. Прямая кишка
до начала акта дефекации не содержит кала. Резервуаром кала явля-
ется тазовый отдел толстой кишки.
В любом возрасте длина толстой кишки равна длине тела. Длина
кишечника новорожденного 340—460 см, в течение первого года жиз-
ни увеличивается на 50%. Соотношение между длиной кишечника
и тела у новорожденного составляет 8,3 : 1, на первом году жизни —
6,6 : 1, в 16 лет — 7,6 :1, у взрослого — 5,4 :1. Соотношение между тол-
стой и тонкой кишкой у новорожденного составляет 1 : 5, оно не изме-
няется у грудного ребенка, у взрослого достигает 1 : 4.
В микрофлоре толстой кишки преобладают анаэробные палочки
(90%), остальные аэробные: кишечная палочка, молочнокислые бак-
терии и др. Микроорганизмы, населяющие толстую кишку, играют
важную роль в жизнедеятельности человека. Они участвуют в сбра-
живании углеводов, гнилостном разложении белков, расщеплении
желчных пигментов. Особенно важно равновесие между процессами
брожения и гниения: в результате брожения в кишечнике создается
кислая среда, препятствующая избыточному гниению. Нормальная
кишечная микрофлора способствует выработке организмом естест-
33
44
22
55
11
22
Рис. 141. Прямая кишка:
1 — общий покров, 2 — мышца, поднимаю-
щая задний проход, 3 — мышечная оболочка,
4 — ампула прямой кишки, 5 — наружный
сфинктер заднего прохода
252
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
венных защитных факторов; подавляет жизнедеятельность патоген-
ных микробов; синтезирует некоторые витамины (К, Е, В6, В12); рас-
щепляет небольшое количество клетчатки.
Из тонкой кишки через илеоцекальный клапан в толстую поступа-
ет 0,5—1 л химуса, который подвергается воздействию бактерий, насе-
ляющих толстую кишку. Здесь частично расщепляется растительная
клетчатка; разрушаются многие вещества, в результате чего образуют-
ся токсические соединения, обезвреживающиеся в печени; всасывает-
ся большое количество воды, минеральных веществ, водораствори-
мых витаминов и, в конечном итоге, образуется кал. За сутки при
правильном питании взрослый человек выделяет 150—200 г кала, со-
стоящего из 75—80% воды и 20—25% сухого остатка, в котором содер-
жатся клетчатка и другие неперевариваемые вещества, бактерии, не-
растворимые соли кальция и железа, небольшое количество жира,
продукты брожения и гниения. Кроме того, у здорового человека в те-
чение суток образуется и выводится 300—350 см3 газа.
Благодаря движениям толстой кишки каловые массы передвига-
ются в направлении прямой, ее заполнение, повышение давления до
40—50 мм рт. ст. и растяжение вызывают позыв к дефекации, которая
осуществляется благодаря сокращению мышечной оболочки прямой
кишки, непроизвольному расслаблению внутреннего сфинктера и
произвольному расслаблению наружного сфинктера заднего прохода.
Контроль над центром дефекации со стороны коры головного мозга
устанавливается у ребенка примерно с двух лет.
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
Пищеварительная система выполняет ряд сложных функций.
Главные из них:
1) секреторная выработка железистыми клетками различных пи-
щеварительных соков (слюна, желудочный, кишечный соки, сок под-
желудочной железы и желчь), состоящих из воды, ферментов, неорга-
нических веществ;
2) моторно-эвакуаторная, которую осуществляют мышцы пище-
варительного тракта, благодаря их деятельности пища измельчается,
продвигается в направлении от ротовой полости к прямой кишке
и смешивается с пищеварительными соками;
3) всасывательная — перенос продуктов пищеварения, а также
воды, витаминов и минеральных веществ во внутреннюю среду орга-
низма;
4) экскреторная (выделительная) — удаление непереваренных
остатков, а также продуктов метаболизма, солей тяжелых металлов,
253
ПИЩА И ХАРАКТЕР ПИТАНИЯ
некоторых токсинов и лекарственных веществ, а также микроорга-
низмов;
5) эндокринная — выработка эндокринными клетками и некото-
рыми железами (слюнные, поджелудочная) гормонов и других биоло-
гически активных веществ;
6) защитная — защита организма от вредных веществ, микроорга-
низмов;
7) гемопоэтическая — выработка желудочными клетками внутрен-
него антианемического фактора, способствующего всасыванию витами-
на В12 (цианокобаламина), без которого невозможно созревание эритро-
цитов; депонирование в слизистой оболочке желудка и тонкой кишки,
а также в печени ферритина, участвующего в синтезе гемоглобина;
8) рецепторная — слизистая оболочка желудочно-кишечного
тракта богата рецепторами, воспринимающими раздражения.
ПИЩА И ХАРАКТЕР ПИТАНИЯ
Пища, поступающая в желудочно-кишечный тракт человека, со-
стоит из нутриентов собственно питательных веществ, которые вса-
сываются, и балластных веществ, которые не всасываются, но играют
важную роль в жизни. В состав продуктов питания входят белки,
жиры, углеводы, клетчатка (или целлюлоза), витамины, минеральные
вещества, вода.
Одним из наиболее важных вопросов является калорийность
пищи. Каждое пищевое вещество обладает определенной энергетиче-
ской ценностью, или калорийностью (выражается в ккал на 100 г или
по системе СИ в кДж/100 г). При окислении 1 г глюкозы выделяется
3,75 ккал (16,5 кДж) энергии, сахарозы — 4 (16,7 кДж), крахмала —
4,1 (17,1), животного жира — 9,3 (37,0), белка — 5,6 (23,4) этилового
спирта — 7,1 (29,7).
В табл. 16 приведены примерные данные энергозатрат взрослых
людей в возрасте от 18 до 40 лет со средней массой тела 70 кг (мужчи-
ны) и 60 кг (женщины).
Таблица 16
Примерные энергозатраты в зависимости от характера нагрузки
Нагрузка
Энергозатраты
М (кДж) Ж (кДж) М (ккал) Ж (ккал)
Легкая физическая нагрузка
8800
7500
2100
1800
Средняя (большинство населения) 11 300
8400
2700
2000
Тяжелая
12 500
9200
3000
2200
Очень тяжелая
14 600
—
3500
—
254
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
После 40 лет энергозатраты, как правило, уменьшаются: от 40 до
49 лет — на 5%, от 50 до 59 лет — еще на 5%, каждые последующие
10лет—на10%.
Предельная интенсивность обменных процессов у мужчин состав-
ляет около 20 000 кДж/сут. (4800 ккал), у женщин 15 500 кДж
(3700 ккал). Этот предел не следует превышать при условии постоян-
ного выполнения чрезвычайно тяжелой работы в течение ряда лет.
Для видов спорта, требующих выносливости, эти цифры не должны
превышать 4200 кДж/ч (1000 ккал/ч).
В первые 6 мес. беременности ежедневно следует добавлять
400 кДж (96 ккал), в последние 3 мес.
—
900—1000 кДж (215—
239 ккал) ежедневно, при кормлении грудью — 2100 кДж (500 ккал)
ежедневно. В детском, подростковом и юношеском возрасте энергоза-
траты зависят и от возраста. В период второго детства энергозатраты
около 2200—2500 ккал, в период энергичного роста у подростков они
возрастают до 3000 ккал.
Увеличение энергозатрат при умственной работе связано с реф-
лекторным увеличением мышечного тонуса.
Подавляющее большинство людей в наше время малоактивны, их
энергетические затраты составляют не более 2300—2700 (м), 1800—
2000 (ж) ккал/сут.
Современные исследования позволяют считать, что 12—15% ка-
лорийности пищи должны составлять белки (из них 50% животные),
30—35% жиры и 50—55% углеводы. Суточные потребности взросло-
го человека, детей и подростков в нутриентах представлены
в табл. 17, 18.
Таблица 17
Физиологические потребности детей и подростков
в пищевых веществах и энергии
Вещества
Потребность в зависимости от возраста (года)
11—13
14—17
Белки (г),
из них животные
95
60
105
65
Жиры (г),
из них растительные
95
20
105
25
Углеводы (г)
380
420
Энергия (ккал)
2850
3000
255
ПИЩА И ХАРАКТЕР ПИТАНИЯ
Таблица 18
Средняя суточная потребность взрослого человека
в пищевых веществах и энергии
Пищевые вещества Потребность
Пищевые вещества
Потребность
Белки (г),
из них животные
80—100
50
Жиры (г),
из них растительные
80—100
20—25
Углеводы (г)
400—500 Фосфолипиды (г)
5
Клетчатка (г)
25
Холестерин (г)
Вода (г)
0,3—0,6
1750—2200
Кислоты (лимонная,
молочная и др.) (г)
2
В том числе питьевая (включая
чай, кофе и т. д.)
800—1000
Незаменимые
жирные кислоты (г)
3—6
В супах
В продуктах питания
250—500
700
Полноценное питание предусматривает наличие в пище всего на-
бора необходимых человеку веществ в соответствии с научно обосно-
ванными потребностями. Подчеркнем: научно обоснованными.
Зачастую данные о потребности в тех или иных веществах базиру-
ются на традиции или авторитете того или иного ученого и кочуют из
одной книги в другую. Совершенно правы американские специали-
сты по питанию, говоря: «Одним из основных источников путаницы
в вопросах современной диеты является избыток дезинформации
(подчеркнуто нами.
—
Г. Б . , Е. З .) о питании, которая значительно
превосходит объем действительных знаний по вопросу о питании...
многие... пренебрегают наукой о питании в пользу последней моды»
(«Безвредность пищевых продуктов»).
Если исходить только из калорийности пищи, то одни пищевые
продукты вполне можно заменить другими. Однако это не так, ибо,
помимо энергетической, пища выполняет и важнейшую пластиче-
скую функцию, влияет на все функции организма.
Соотношение белков, жиров и углеводов в полноценном рационе
должно доставлять 1:1,2:4,6, при этом 1000 ккал (4184 кДж) пищевые
продукты должны включать 30 г белка, 37 г жиров и 137 г углеводов.
Недостаточность питательных веществ приводит к уменьшению массы
тела, снижению работоспособности, нарушению роста и развития орга-
низма, угнетению восстановительных процессов, многим заболеваниям.
Белки
Белки, поступающие в организм с пищей, используются для:
роста (построения клеточных структур, клеток и тканей);
восстановления (регенерации) структур на всех иерархических
уровнях;
256
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
синтеза ферментов, белков, гормонов, гемоглобина, миоглобина;
энергетических нужд;
создания буферных систем, участвующих в поддержании постоян-
ства рН внутренней среды.
Для организма очень важен состав его аминокислот, среди кото-
рых имеются заменимые и незаменимые. Человек должен получать
необходимое количество незаменимых аминокислот (триптофан,
лейцин, изолейцин, валин, треонин, лизин, метионин, фенилаланин).
Детям наряду с ними необходим и гистидин. Белки, содержащие все
незаменимые аминокислоты в оптимальных количествах и соотноше-
ниях, являются полноценными и сбалансированными. Это белки яиц,
печени, молока и молочных продуктов (кроме сливок, сметаны и сли-
вочного масла), рыбы, мяса, птицы. Некоторые белки содержат все
аминокислоты, но одни из них в избытке, а другие в недостаточном
количестве; эти белки называют полноценными, но несбалансирован-
ными. Это белки зерновых культур (кроме кукурузы), сои, овощей,
мяса, богатого сухожилиями и фасциями. К неполноценным белкам,
в которых отсутствуют те или иные незаменимые аминокислоты, от-
носят белки бобовых растений (кроме сои), кукурузы и желатины.
Содержание белка в различных продуктах существенно варьиру-
ется.
Основными источниками белков являются следующие продукты:
яйца, молоко и молочные продукты, мясо и рыба, птица и зернобобо-
вые растения (фасоль, горох, чечевица, соя). Избыточный прием
естественных источников белка не приводит к патологическим явле-
ниям, кроме случаев повышенной чувствительности или аллергии.
Растительные белки имеют более низкую биологическую ценность по
сравнению с животными. Большинство растительных белков бедны
многими незаменимыми аминокислотами.
ВНИМАНИЕ!
Эталонный белок — яичный.
Правильная комбинация растительных и животных продуктов мо-
жет обеспечить требуемый уровень незаменимых аминокислот. Бел-
ковая недостаточность возникает в тех случаях, когда пища содержит
недостаточное количество отдельных аминокислот.
Одним из важнейших вопросов современной науки о питании яв-
ляется нормирование белка. Современная норма составляет 1,01 г
белка на 1 кг массы тела. В рационе детей и подростков количество
белка должно составлять 1,2—1,5 г/кг в сутки. Для взрослого (усред-
ненного) человека надежный (безопасный) уровень белка составляет
257
ПИЩА И ХАРАКТЕР ПИТАНИЯ
55—60 г в сутки. Однако это количество должно быть увеличено при
физической нагрузке, стрессе и т. д. Так, следует получать дополни-
тельно 10% белка при легких инфекционных заболеваниях, травмах,
огорчениях, заботах, бессоннице). При воздействии высокой темпера-
туры окружающей среды человек дополнительно теряет с потом боль-
шие количества азота вследствие разрушения белка. Тяжелая физиче-
ская работа требует потребления свыше 1 г белка на 1 кг массы тела
в сутки. При беременности и особенно при кормлении у женщин рез-
ко возрастает потребность в белке.
Жиры (липиды)
Жиры (липиды) входят в состав большинства пищевых продук-
тов. Ими особенно богаты мясо, птица, молоко и молочные продукты,
растительные масла. Жиры являются важным источником энергии,
в одном грамме жира запасено в два раза больше энергии, чем в грам-
ме углеводов. Жир защищает организм от холода. Именно в жирах
содержатся жирорастворимые витамины (A, D, Е, К). Жиры и жиро-
растворимые вещества придают вкус и аромат пищевым продуктам.
Жиры замедляют процесс пищеварения, что предотвращает ощуще-
ние голода.
Основными компонентами жиров являются жирные кислоты —
молекулы с длинной цепью, образованной атомами углерода и водо-
рода. Жирные кислоты отличаются длиной цепи и насыщенностью
водородом. Различают насыщенные и ненасыщенные жирные кисло-
ты. Насыщенные жирные кислоты, которые содержатся в основном
в животных продуктах, при комнатной температуре твердые, ненасы-
щенные жидкие, причем они тем более жидкие, чем они более нена-
сыщенные. Насыщенные жирные кислоты не содержат двойных свя-
зей, ненасыщенные содержат от одной (мононенасыщенные) до шести
(полиненасыщенные) двойных связей.
–С–С–С
–С=Сс–С
С=С–С=С=С –С=С
Насыщенная
Мононенасыщенная
Полиненасыщенная
Нормальная жизнедеятельность организма невозможна без посту-
пления с пищей незаменимых жирных кислот, которые не синтезиру-
ются в организме человека. Это олеиновая, линолевая, линоленовая и
арахидоновая кислоты. Они содержатся в растительных маслах. Ра-
цион человека должен включать от 80 до 100 г жиров в сутки (1,2—
1,3 г на 1 кг массы тела), в том числе 30—35 г растительного масла,
содержащего около 10 г полиненасыщенных жирных кислот.
Полиненасыщенные жирные кислоты входят в состав клеточных
мембран, регулируют их свойства, в состав миелиновых оболочек
258
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
нервных волокон, участвуют в образовании простагландинов, стаби-
лизируют стенки кровеносных сосудов, образуют с холестерином
соединения, которые легко выводятся из организма. Недостаточное
потребление полиненасыщенных жирных кислот способствует раз-
витию атеросклероза, гипертонической болезни, преждевременному
старению организма, сексуальным нарушениям у мужчин. Пищевая
ценность жиров связана, помимо энергетической, с содержанием
полиненасыщенных жирных кислот, фосфолипидов, витамина F
и жирорастворимых витаминов. В животных жирах преобладают на-
сыщенные жирные кислоты, в растительных — моно- и полиненасы-
щенные. Исключением являются пальмовое и кокосовое масла,
в которых мало ненасыщенных жирных кислот и очень много насы-
щенных.
В развитых странах сердечно-сосудистые заболевания являются
причиной половины всех случаев смертности: основной причиной
смерти мужчин и второй или третьей — женщин. Сердечно-сосудистые
заболевания (кроме врожденных) связаны с атеросклерозом. Основная
причина атеросклероза — избыток жиров в пище, особенно насыщен-
ных и богатых холестерином. Сокращение количества потребляемых
жиров с разумной заменой насыщенных жиров полиненасыщенны-
ми — возможное средство предупреждения атеросклероза и снижения
опасности возникновения инфаркта миокарда и инсульта.
Холестерин — важный компонент клеточных мембран и цитоплаз-
мы, участвует в обмене желчных кислот, в синтезе гормонов коры надпо-
чечников и гормонов половых желез. В обмене холестерина участвуют
некоторые витамины, полиненасыщенные жирные кислоты. Нормаль-
ное содержание холестерина в крови <200 мг% (<5,18 ммоль/л).
Предельно допустимым считается содержание холестерина >240 мг%
(>6,2 ммоль/л) крови. Более высокий уровень холестерина в крови по-
вышает опасность возникновения атеросклероза, ишемической болезни
сердца и ее осложнений (инфаркта и инсульта).
Холестерин переносится в организме кровью в составе белково-
жировых комплексов (липопротеинов). Различают липопротеины
низкой плотности (ЛПНП), очень низкой плотности (ЛПОНП) и вы-
сокой плотности (ЛПВП). Первые два типа («плохой» холестерин)
переносят холестерин от пищеварительной системы и могут отклады-
вать его на стенках кровеносных сосудов, приводя к развитию атеро-
склероза, сужению сосудов, возникновению инфарктов миокарда,
если это происходит в коронарных сосудах. ЛПВП («хороший» холе-
стерин), наоборот, выводит холестерин из тканей и кровеносных со-
судов и возвращает в печень, где он и уничтожается. Уровень ЛПНП
не должен превышать 350 мг% (3,5 г/л), а уровень ЛПВП не должен
быть ниже 220 мг% (2,2 г/л). Важно соотношение холестерина
259
ПИЩА И ХАРАКТЕР ПИТАНИЯ
и ЛПВП, если это отношение более 4, у человека имеется риск воз-
никновения ишемической болезни сердца.
Холестерин находится в животных жирах, печени, яйцах, молоч-
ном жире и т. д . Растения бедны холестерином, однако богаты фито-
стеринами, среди которых преобладают ситостерины. Особенно мно-
го их в растительных маслах. Потребление фитостеринов уменьшает
накопление холестерина в организме и тем самым снижает риск воз-
никновения ишемической болезни сердца. Не вызывает сомнений тот
факт, что по мере увеличения количества насыщенных жирных кис-
лот в пище возрастает уровень ЛПНП, ЛПОНП и холестерина в кро-
ви, по мере увеличения количества ненасыщенных жирных кислот
уровень холестерина в крови падает.
ВНИМАНИЕ!
В последние годы расширяется производство и употребление маргари-
на. Однако он содержит гидрогенизированные масла с очень высоким
содержанием насыщенных жирных кислот. Это определяет вредность
любого маргарина!
Углеводы
Углеводы широко используются в питании человека, они обеспе-
чивают 50—55% калорийности пищевого рациона, в некоторых стра-
нах эта цифра достигает 70%. Наиболее высокое содержание углево-
дов — в растительных продуктах, которые длительно сохраняются.
В питании человека используются все типы углеводов: моносахариды
(галактоза, фруктоза, глюкоза); дисахариды (сахароза, лактоза, маль-
тоза); полисахариды (усваивающийся крахмал и декстрины, неусваи-
вающаяся целлюлоза, пектины).
Углеводы — важнейший источник энергии для мышечной деятель-
ности, при недостатке углеводов распадаются жиры и белки, в крови
накапливаются токсичные кетоны. Для нормальной функции печени
необходимо достаточное количество сложного углевода гликогена в
печеночных клетках. Углеводы регулируют обмен белков и жиров.
Нормальное функционирование центральной нервной системы воз-
можно только при условии постоянного поступления в нее глюкозы.
Основными источниками углеводов являются растительные про-
дукты, в которых преобладает крахмал, расщепляющийся в организме
до глюкозы, которая всасывается в кровь и транспортируется к клет-
кам, использующим ее для получения энергии или превращающим ее
в гликоген или жир. Из растений, широко потребляемых в нашей
стране, наиболее богат крахмалом картофель. Плоды и овощи содер-
жат большое количество сахаров.
260
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
При рафинировании сахара из него удаляются все соли, витамины
и другие биологически активные вещества. В коричневом сахаре со-
храняется несколько большее количество важных биологически ак-
тивных веществ.
Углеводы хорошо усваиваются из различных продуктов, в преде-
лах 85–98%. К сожалению, избыток углеводов, особенно рафиниро-
ванных (сахар и кондитерские изделия), широко распространен во
всем мире, особенно в нашей стране.
ВНИМАНИЕ!
Сахар — это белый яд! Он не менее вреден, чем алкоголь и табак. По-
требление сахара более 6 кг в год на одного человека токсично.
Сахар ускоряет развитие диабета у предрасположенных людей,
увеличивает вероятность ожирения, которое в социальном плане куда
опаснее и серьезнее рака, способствует преждевременному (раннему)
развитию атеросклероза и его осложнений, импотенции у мужчин и
аноргазмии у женщин, увеличивает вероятность и степень кариеса зубов.
Уровень глюкозы натощак в крови составляет 1 г/л (3,9–
6,2 ммоль/л). При содержании сахара (глюкозы) в крови выше
6,7 ммоль/л (1,2 г/л) говорят о гипергликемии, ниже 2,2 ммоль/л —
о гипогликемии. Уже в первые минуты после еды повышается уро-
вень глюкозы в крови, через определенное время в ответ на резкое
увеличение ее содержания в крови достигается гликемический пик.
Это связано с тем, что эндокринная часть поджелудочной железы на-
чинает вырабатывать гормон инсулин, который способствует исполь-
зованию глюкозы, тем самым уровень ее в крови уменьшается, после
чего возвращается к норме (1 г/л). Мы рассмотрим регуляцию уровня
сахара в крови в разделе «Эндокринные железы».
В 70-е гг. XX в. Крапо и Женкинс в США и Канаде показали, что
влияние углеводов на организм можно оценить по гликемическому ин-
дексу (ГИ). Гликемический индекс (ГИ) определяется площадью тре-
угольника сахарной кривой после потребления конкретного продукта,
содержащего углеводы (рис. 142). ГИ чистой глюкозы составляет 100.
Гликемический индекс =
Площадь треугольника продукта
× 100
Площадь треугольника глюкозы
В зависимости от ГИ различают «хорошие» углеводы, которые не
повышают значительно уровень сахара в крови (зеленые овощи, по-
мидоры, баклажаны, чеснок, лук, молочные продукты, цельные злаки,
необработанный рис, овощи, многие фрукты, капуста, брокколи, чече-
вица, сухая фасоль и др.), и «плохие» углеводы, которые быстро и
261
ПИЩА И ХАРАКТЕР ПИТАНИЯ
резко увеличивают содержание сахара в крови (сахар, мед, картофель,
сладости, варенье, джемы, изделия из белой рафинированной муки,
сладкие напитки, пиво и др.). ГИ первых ниже 50, ГИ вторых выше.
Потребление «плохих» углеводов приводит к выделению большо-
го количества инсулина и постепенному нарушению функции эндо-
кринной части поджелудочной железы, что может привести к разви-
тию диабета второго типа.
Глюкоза является важным источником энергии, который исполь-
зуется экстренно или откладывается в печени и мышцах. Избыточная
неиспользованная глюкоза в составе углеводов с высоким ГИ превра-
щается в жир.
ВНИМАНИЕ!
Потребление углеводов с низким ГИ (ниже 50) снижает уровень холе-
стерина в крови и риск возникновения сердечно-сосудистых заболева-
ний у мужчин на 7—10% и у женщин на 25—30%.
Содержание глюкозы в крови, г/л
Время,
мин.
0
0,60
0,80
1
1,20
1,40
120
60
180
1
0
120
60
Высокий гликемический
индекс продукта
Содержание глюкозы в крови, г/л
Содержание глюкозы в крови, г/л
Низкий гликемический
индекс продукта
Время,
мин.
Время,
мин.
180
0
120
60
180
30
50
Рис. 142А. Сахарная кривая здорового человека
Рис. 142Б. Гликемический индекс
262
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
Балластные вещества
Необходимыми компонентами пищи являются не только собст-
венно питательные вещества, но и балластные вещества (пищевые во-
локна) — клетчатка, пектины, гемицеллюлоза, лигнин, кутин, воск.
Клетчатка (целлюлоза) представляет собой сложный углевод (поли-
сахарид), являющийся главной составной частью клеточных стенок
растительных клеток. Пектины (от греч. pektos — «свернувшийся, за-
мерзший») — также полисахариды, которые содержатся во всех на-
земных растениях, особенно много их в плодах, а также в некоторых
водорослях. Балластные вещества не перевариваются в кишечнике
человека. Они связывают воду, набухают (например, 100 г отрубей
связывает 400—500 мл воды), стимулируют пищеварение, способст-
вуют выведению из организма многих токсических веществ. Пище-
вые волокна, попадая в желудочно-кишечный тракт, стимулируют
двигательную активность кишечника, ускоряют всасывание веществ
в тонкой кишке, нормализуют внутрикишечное давление, увеличива-
ют массу кала.
Балластные вещества (особенно пектины) нормализуют обмен хо-
лестерина, замедляют процесс старения. Увеличение их количества
в составе пищи приводит к снижению уровня холестерина в крови.
Пищевые волокна обладают антитоксическим действием. Они снижа-
ют содержание глюкозы в крови и, что не менее важно, уменьшают
выработку инсулина. Пищевые волокна — эффективное средство для
профилактики и лечения ожирения, нарушений жирового и углевод-
ного обмена, гипертонической болезни, ишемической болезни сердца,
злокачественных заболеваний желудочно-кишечного тракта. Отруби,
особенно овсяные, способствуют выведению из крови ЛПНП и
ЛПОНП, не влияя на уровень ЛПВП.
Пектины усиливают двигательную активность желудочно-кишеч-
ного тракта, улучшают пищеварение и всасывание, оказывают защит-
ное действие благодаря способности связывать токсины, ионы тяже-
лых металлов (например, лития, алюминия, свинца, циркония и др.)
радионуклиды и, что не менее важно, холестерин, тем самым пектины
оказывают противоатеросклеротическое действие. При этом пектины
совершенно безвредны. Особенно богаты пектинами яблоки, свекло-
вичные, цитрусовые, некоторые водоросли. Следует подчеркнуть, что
витамин С улучшает связывание пектинами ионов тяжелых метал-
лов, токсинов, холестерина.
Потребность в пищевых волокнах взрослого человека, который не
занят тяжелым физическим трудом, составляет 25 г/сут. Один грамм
пищевых волокон содержится в 60 г цельного хлеба, 300 г хлеба из
муки высшего сорта, 10 г пшеничных или 24 г кукурузных отрубей,
263
ВИТАМИНЫ
85 г овсяных хлопьев, 250 г теста из муки высших сортов, 350 г фрук-
тов, 50 г орехов, 70—120 г лиственных овощей, 120—180 г бобовых,
125 г картофеля.
СВОБОДНЫЕ РАДИКАЛЫ
Молекулы, в которых присутствуют непарные электроны, называ-
ют свободными радикалами. Это весьма реактивные, заряженные
электрически фрагменты молекул, которые либо отбирают электроны
у других молекул, либо присоединяются к другим молекулам, что
приводит к разрушению биологических мембран, хромосомным нару-
шениям, мутациям и гибели клеток. Свободные радикалы являются
причиной возникновения очень многих заболеваний. Это сердечно-
сосудистые (в том числе атеросклероз) и злокачественные заболева-
ния, сексуальные расстройства у мужчин, снижение иммунитета.
Свободные радикалы ведут к преждевременному старению.
Человек обладает механизмами обезвреживания свободных ради-
калов. Им противостоят естественные мощные антиоксиданты, конт-
ролирующие возникновение свободных радикалов и защищающие от
них клетки и, в первую очередь, генетический аппарат — молекулу
ДНК. С возрастом эти естественные механизмы ослабевают, а нере-
дко ломаются. И тогда на помощь приходят антиоксиданты, способ-
ные нейтрализовать свободные радикалы. Антиоксиданты разруша-
ют свободные радикалы.
ВНИМАНИЕ!
Наиболее известные антиоксиданты — витамины С (аскорбиновая кис-
лота), Е (токоферол), бета-каротин и селен. Они способны снизить ве-
роятность развития ряда серьезных заболеваний у человека (злокачест-
венные опухоли, атеросклероз и его последствия, сексуальные
нарушения и др.), а также предотвратить преждевременное старение
и замедлить процесс естественного старения.
ВИТАМИНЫ
В 1880 г. русский врач Н. И . Лунин предположил, что «в молоке
помимо казеина, жира, молочного сахара и солей содержатся еще дру-
гие вещества, незаменимые для питания». В 1912 г. биохимик Кази-
мир Функ выделил из отрубей риса высокоактивное вещество, кото-
рое излечивало больных полиневритом голубей, и высказал мысль
о том, что этот препарат и другие ему подобные «являются жизненно
необходимыми аминами», и назвал их витаминами (от лат. vita —
«жизнь»). Витамины регулируют обмен веществ и энергии, клеточное
264
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
дыхание, окислительно-восстановительные реакции, участвуют в
синтезе гормонов, нуклеиновых кислот, свертывании крови.
Все витамины подразделяются на жирорастворимые (A, D, Е, К),
водорастворимые (С, Р, B1, B2, PP, В6, В12, фолиевая кислота) и вита-
миноподобные вещества (липоевая кислота, пангамовая кислота, хо-
лин, инозит, карнитин, s-метилметионин).
ВНИМАНИЕ!
Избыток водорастворимых витаминов легко выделяется с мочой, в то
время как жирорастворимые накапливаются в основном в печени, что
может вызвать нарушение ее функции.
Жирорастворимые витамины
Витамин А (ретинолы) обеспечивает нормальное зрение (входит
в состав зрительных ферментов), нормальный рост и развитие, оказы-
вает антиоксидантное и гипохолестериновое действие. Основные
источники — продукты животного происхождения: печень говяжья
и куриная, печень трески и других рыб, молоко и молочные продук-
ты, яйца. Предшественники ретинола — каратиноиды растений
(например, морковь) — в организме превращаются в ретинол. Су-
точная потребность взрослого человека составляет 4000—5000 ME
(1 ME = 0,3 мкг витамина А или ретинола). Мужчины старше 50 лет
и кормящие женщины должны получать в 1,5—2 раза больше. При-
знаки недостаточности: куриная (ночная) слепота, кожные высыпа-
ния, бледность и сухость кожи, ороговение волосяных фолликулов,
гнойничковые поражения кожи, сухость волос, ломкость ногтей, по-
вышенная утомляемость, снижение работоспособности.
Витамин D (кальциферолы). Один из важных факторов, регули-
рующих обмен кальция и фосфора, активирует всасывание кальция
и фосфора в тонкой кишке, участвует в механизме минерализации
костей. Витамин D образуется в организме из 7-дегидрохолестерина
при действии на кожу ультрафиолетовых лучей. Суточная потреб-
ность взрослого человека составляет 100 ME. Начиная с 3 лет —
100 ME, до 3 лет — 400 ME, беременные и кормящие женщины долж-
ны получать 500 ME (1 ME = 0,025 мкг холекальциферола).
Признаки недостаточности: повышенная раздражительность, беспо-
койство, общая слабость, потливость, запоздалое развитие зубов,
предрасположенность к заболеваниям дыхательных путей; наруше-
ние нормального процесса окостенения костей. У взрослых — общая
слабость, утомляемость, разрушаются зубы, у детей развивается
рахит.
265
ВИТАМИНЫ
Витамин Е (токоферолы). Активные антиоксиданты; они регули-
руют репродуктивную функцию и сексуальность человека, нормали-
зуют функции мышечной и эндокринной систем; оказывают антикан-
церогенное действие; замедляют процессы старения, уменьшают
вредные последствия стресса, курения и употребления алкоголя. Ос-
новные источники: зародыши злаков и зеленые овощи, растительные
масла, яйца, орехи. Суточная потребность взрослого человека: 10—
12 мг. Признаки недостаточности: мышечная гипотония, мышечная
слабость, мышечная дистрофия, склеродермия; нарушения структуры
и функции половых органов: у мужчин — сексуальные расстройства,
уменьшение либидо и потенции, нарушение сперматогенеза, дегене-
ративные изменения семенных канальцев, бесплодие, у женщин — на-
рушение либидо, бесплодие, предрасположенность к абортам.
Витамин К (филохиноны). Участие в процессах свертывания кро-
ви, синтезе протромбина. Основные источники: животные продукты,
цветная капуста, зеленый горошек, морковь, шпинат, томаты, мясо,
листья каштана, листья крапивы. У взрослого в сутки до 1,5 мг вита-
мина К2 синтезируется кишечной флорой. Суточная потребность:
0,2—0,3 мг. Признаки недостаточности: у новорожденных снижение
уровня протромбина, у взрослых — различные кровотечения.
Водорастворимые витамины
Витамин С (аскорбиновая кислота). Активное антиоксидантное
действие, участие в окислительно-восстановительных процессах, регу-
ляция обмена веществ; стимуляция синтеза коллагена; участие в вос-
становительных процессах; стабилизация стенок капилляров; повыше-
ние защитных механизмов и сопротивляемости организма; защитное
действие в отношении токсических веществ; антиканцерогенное дейст-
вие, замедление процессов старения; уменьшение вредных последствий
стресса, курения и употребления алкоголя. Основные источники —
фрукты, овощи: шиповник сухой, черная смородина, цитрусовые, брус-
ника, клюква, черноплодная рябина, яблоки, персики, черешня, земля-
ника, все виды капусты, малина, крыжовник, щавель, шпинат, салат,
зелень петрушки, свекла. Суточная потребность: 60–100 мг, беремен-
ные и кормящие женщины, люди старше 50 лет — в 2–3 раза больше.
ВНИМАНИЕ!
Эти количества позволяют человеку выжить. Американские специали-
сты по питанию рекомендуют до 1 г в сутки. Мы придерживаемся этой
точки зрения и рекомендуем для взрослых 500–1000 мг в сутки посто-
янно, кроме осени и лета, когда человек получает с пищей достаточное
количество витамина.
266
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
Признаки недостаточности: цинга, увеличение проницаемости
стенок сосудов; кровоточивость, множественные кровоизлияния; раз-
рыхленность и синюшность десен.
Витамины группы Р (биофлавоноиды) являются синергистами
витамина С. Функция: стабилизация капиллярной стенки и снижение
проницаемости сосудистой стенки, активизация окислительных про-
цессов в клетке, антигистаминное действие. Основные источники:
фрукты, овощи — шиповник сухой, черная смородина, цитрусовые,
черноплодная рябина, вишня, груша, айва, гранат, черешня темная,
щавель, крыжовник. Суточная потребность: 30–50 мг. Признаки не-
достаточности: общая слабость, склонность к геморрагиям, ломкость
капилляров и повышение их проницаемости.
Витамины группы В
Витамин B1 (тиамин). Функция: участие в обмене белков, азота,
жиров, углеводов, усиление превращения углеводов в жир; нормали-
зация деятельности нервной системы. Основные источники: цельные
зерновые продукты, из которых не удалены зародыши, дрожжи (осо-
бенно сухие), отруби, хлеб из цельного зерна, молоко и молочные
продукты, мясо, печень, почки, яйцо, горошек зеленый. Суточная по-
требность: 1,5—2,5 мг. Необходимость возрастает при потреблении
большого количества углеводов, при хроническом алкоголизме, са-
харном диабете. Признаки недостаточности: быстрая утомляемость
(психическая и физическая), потеря аппетита, запор, мышечная сла-
бость, боли в ногах, одышка, болезненность икроножных мышц, по-
вышенная чувствительность. В тяжелых случаях возникает болезнь
бери-бери.
Витамин В2 (рибофлавин). Участвует в процессах тканевого дыха-
ния, в окислительно-восстановительных процессах, в процессах роста,
в обмене белков, углеводов и жиров. Основные источники: дрожжи, го-
рох, гречневая крупа, макаронные изделия, хлеб, молоко и молочные
продукты, сыры, мясо, печень, почки, яйца, бобовые. Суточная потреб-
ность: 1,5—3,0 мг (0,07 мг на 1000 ккал). Чем больше потребление угле-
водов и жиров, тем выше потребность в витамине B2. Признаки недо-
статочности: сухость и синюшность губ, красная кайма и рубцы на
них, трещины и корочки в углах рта, сухой язык ярко-красного цвета,
дерматит носогубных складок, светобоязнь, конъюнктивит, блефарит.
Витамин РР (никотиновая кислота). Участвует в клеточном дыха-
нии, обмене белков, жиров и углеводов, влияет на функцию пищевари-
тельной системы, двигательную и секреторную функции желудка, функ-
ции печени и (экскреторной) поджелудочной железы, улучшает
использование растительных белков, осуществляет защиту организма от
267
ВИТАМИНЫ
радиации. Основные источники: растительные и животные продукты,
в которых витамин широко распространен. Мясо и мясные продукты,
печень, почки, рыба, молоко, сухофрукты, овощи (горошек, соя, чечеви-
ца), хлеб, крупы, рисовые и пшеничные отруби. Большое количество
легкоусвояемой никотиновой кислоты содержится в кофе. Молоко, бо-
бовые, мясо и мясные продукты, яйца, рыба очень богаты предшествен-
ником витамина PP триптофаном. Суточная потребность: новорожден-
ные и грудные дети — 5 —6 мг, дети — 10—15 мг, взрослые — 17 —20 мг.
В организме человека возможен эндогенный синтез из триптофана, по-
лучаемого с пищей (1 мг витамина РР из 60 мг триптофана).
ВНИМАНИЕ!
В кукурузе витамин РР находится в связанной форме и не освобождает-
ся в процессе пищеварения.
Признаки недостаточности: пеллагра (диарея, дерматит, демен-
ция), неврастения (раздражительность, бессонница, подавленность
и заторможенность), поносы, сухость и бледность кожных покровов,
сухой (или отечный) обложенный язык с трещинами, эритема кожи
шеи, тыльных поверхностей кистей.
Витамин В5 (пантотеновая кислота). Участвует в синтезе белков,
обмене жиров, регуляции функции нервной системы, в синтезе гор-
монов надпочечников и половых желез, оказывает гипохолестерине-
мическое действие. Основные источники: печень говяжья, дрожжи,
яйца, рыба, овощи, фрукты, мясо, хлеб ржаной, молоко. Суточная по-
требность: 5—10 мг. Признаки недостаточности: наблюдаются край-
не редко, возникает слабость, утомляемость, поражения кожи и сли-
зистых оболочек, невриты, сексуальные расстройства.
Витамин В6 (пиридоксин). Участвует в обмене белков и жиров,
в регуляции деятельности нервной системы, кроветворении, норма-
лизует обмен холестерина, оказывает антисклеротическое действие.
Основные источники: печень, мясо, рыба, фрукты и овощи, яйца, мо-
локо и молочные продукты, хлеб. Установлен синтез витамина B6 ки-
шечными бактериями у человека. Суточная потребность: 1,5—3,0 мг.
Потребность резко вырастает у беременных, кормящих женщин, у лиц
старше 70 лет, а также при алкоголизме. Признаки недостаточности:
при сбалансированном питании недостаточность не возникает. В ран-
нем детском возрасте — задержка роста, нарушение функции желу-
дочно-кишечного тракта, повышенная возбудимость. У взрослых —
потеря аппетита, тошнота, глоссит, жировая инфильтрация печени.
У беременных — бессонница, раздражительность, депрессия, тошнота,
рвота, стоматит, дерматит.
268
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
Витамин В9 (фолиевая кислота, витамин Вс). Участвует в крове-
творении (формировании нормальных эритроцитов), в синтезе ну-
клеиновых кислот, в синтезе белка, клеточном делении, обеспечивает
нормальное развитие и функцию мозга, нормализует сексуальную
функцию, способствует снижению уровня холестерина в крови, тем
самым оказывая антисклеротическое действие. Действие фолиевой
кислоты зависит от наличия в организме витамина В12
.
Основные
источники: дрожжи, печень, почки, мясо, яйца, зелень петрушки, шпи-
нат, молоко. Суточная потребность: 300—500 мг. Признаки недоста-
точности: анемия, повышение температуры, запоры или поносы,
глоссит (сухой ярко-красный язык), пониженная кислотность желу-
дочного сока.
Витамин В12 (цианокоболамин). Участвует в кроветворении, в
синтезе миелина в нервной системе, нуклеиновых кислот и белка, ли-
пидном обмене, оказывает антианемическое и липотропное действие.
Может усваиваться только при наличии внутреннего антианемиче-
ского фактора, вырабатываемого железами желудка (фактор Касла).
Основные источники: печень, почки, сельдь, скумбрия, сардины, мясо,
куры, яйца. Суточная потребность: 3—5 мкг. Признаки недостаточ-
ности: анемия; у детей, кроме того, замедление роста.
Витамин В8 (витамин ВН, биотин). Регуляция нервной трофики,
ключевая роль в жировом обмене (образовании жирных кислот),
участие в углеводном обмене, обмене аминокислот, гипохолестерине-
мическое действие. Основные источники: пивные дрожжи, почки,
яйца, яичный желток, капуста белокочанная, другие овощи, мясо,
рыба, пшеничная мука, кукуруза, овсяная крупа, горох, сыр, рис поли-
рованный. Суточная потребность: 150—200 мкг. Признаки недоста-
точности: дерматит, тошнота, отсутствие аппетита, гипотония, глос-
сит, кератоконъюнктивит, ацидоз, анемия, гилерхолестеринемия.
Авидин сырого яичного белка связывает биотин, образуя нерасще-
пляемое соединение.
Витаминоподобные вещества
К витаминоподобным веществам относятся липоевая кислота,
пангамовая кислота (витамин В15), холин, инозит, карнитин, S-метил-
метионин (витамин U). Все они оказывают липотропное действие,
нормализуют обмен холестерина. При сбалансированном питании не
возникает недостаточность этих веществ. Они содержатся в большин-
стве пищевых продуктов. S-метилметионин, оказывающий также про-
тивоязвенное действие, содержится в сырой белокочанной капусте,
кольраби, свекле, но при термической обработке витамин U полно-
стью разрушается.
269
ПОТРЕБНОСТЬ В ВИТАМИНАХ
ВНИМАНИЕ!
Большинство населения нашей страны страдает от неправильного пита-
ния и недостатка витаминов. Конечно, сегодня вряд ли можно встретить
больного, страдающего цингой или бери-бери. Но это совсем не значит,
что мы получаем витамины в достаточном количестве. Для оптимального
функционирования организма, особенно в современных неблагоприят-
ных условиях, необходимо значительно увеличить дозы витаминов.
ПОТРЕБНОСТЬ В ВИТАМИНАХ
Потребность в витаминах зависит от многих факторов. Главные
из них.
1. Пол. У мужчин потребность в витаминах выше, чем у женщин,
однако у последних она возрастает во время менструации, беременно-
сти и кормления грудью. Чем выше сексуальная активность мужчи-
ны, тем больше его потребность в витаминах.
2. Возраст. У людей старше 70 лет всасываемость витаминов из
кишечника и их усвояемость нарушены, поэтому потребность в вита-
минах возрастает на 25—30%.
3. Характер труда и физическая активность. Чем тяжелее труд
и выше физическая активность, тем выше потребность в витаминах.
4. Эмоциональное состояние и психические перегрузки. Нервно-
психическое напряжение, стресс требуют увеличения количества ви-
таминов, поступающих в организм.
5. Жизнь в зонах с холодным или жарким климатом требует уве-
личения количества витаминов на 30—60%.
6. Общее состояние организма. Практически при всех заболевани-
ях и травмах возрастает потребность в витаминах. Особенно ярко это
проявляется при гнойно-воспалительных и инфекционных заболева-
ниях, ожогах, травмах, после операций.
Витамины А, С и Е, а также селен являются мощными антиокси-
дантами. (Витамины А, С, Е являются синергистами.) Антиоксидан-
ты — вещества, препятствующие образованию свободных радикалов,
оказывающих патологическое воздействие на ткани, или способствую-
щие нейтрализации свободных радикалов. Антиоксиданты разрушают
свободные радикалы и предохраняют клетки от повреждающего дейст-
вия кислорода, продуктов распада липидов и других промежуточных
продуктов окисления, предотвращая тем самым их губительное действие
на живые клетки. Именно поэтому витамины А, С и Е защищают орга-
низм от злокачественных и сердечно-сосудистых заболеваний, и в пер-
вую очередь атеросклероза, являющегося основной причиной инфар-
ктов, инсультов, эндартерита и других грозных заболеваний, замедляют
процессы старения, защищают мозг, уменьшают вредное воздействие
стресса и загрязнений окружающей среды, курения и алкоголя. С возра-
270
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
стом функция иммунной системы медленно, но верно ослабевает. Вита-
мин А, так же как витамины С и Е, предотвращает старение иммунной
системы.
ВНИМАНИЕ!
Различные вредные воздействия, и в первую очередь табакокурение,
употребление алкогольных напитков, применение многих противозача-
точных средств, заболевания, травмы, стресс, загрязнение окружаю-
щей среды, интоксикации резко увеличивают образование в организме
свободных радикалов и, соответственно, потребность во многих вита-
минах, особенно витаминах-антиоксидантах А, С и Е.
МИНЕРАЛЬНЫЕ ВЕЩЕСТВА
Минеральные вещества играют важную роль в жизни организма, со-
хранении здоровья, недостаток или избыток многих из них может вызвать
ряд серьезных нарушений. Минеральные вещества участвуют в созда-
нии и поддержании постоянства внутренней среды организма (гомео-
стаза). Минеральные вещества поступают в организм с пищей и водой.
В табл. 19 представлены сведения об основных минеральных эле-
ментах, участвующих в биологических процессах в организме человека.
Таблица 19
Физиологическая роль, суточная потребность организма
и источники поступления основных минеральных веществ
Элементы
Физиологическая роль и суточная потребность
Источники
Натрий Содержится преимущественно во внеклеточной
жидкости и плазме крови. Играет роль в процессах
возбуждения, создании осмотического давления
жидкостей внутренней среды, распределении и вы-
ведении воды из организма, участвует в функции
бикарбонатной буферной системы. Суточная по-
требность—2—3г,аввидеNaCl—2—4г
Поваренная соль,
сыры, рыба и рыб-
ные консервы,
хлеб, грибы, мясо,
молоко, творог,
яйца
Калий
Содержится преимущественно внутри клеток,
а также в жидкостях внутренней среды. Играет
важную роль в процессах реполяризации после
возбуждения в нервных волокнах, сокращении
мышц, в том числе миокарда. Суточная потреб-
ность — 2—3 г
Овощи (карто-
фель), горох, чече-
вица, соя, мясо,
сухофрукты (абри-
косы, изюм), оре-
хи, мясо, молоко,
творог, рыба, хлеб
Кальций Структурный компонент тканей зубов и костей (до
99% общего количества кальция в организме). Уча-
ствует в регуляции функций и метаболизма клеток,
процессов возбуждения клеток, синаптической пе-
редаче, свертывании крови, сокращении мышц. Су-
точная потребность — 0,8—1,0 г
Молоко и молоч-
ные продукты,
сыры, рыба, щи,
зеленые овощи,
шпроты, сардины,
чеснок, кресс-
салат, яйца
271
МИНЕРАЛЬНЫЕ ВЕЩЕСТВА
Элементы
Физиологическая роль и суточная потребность
Источники
Хлор
Содержится во внеклеточной и внутриклеточной
жидкостях. Участвует в процессах возбуждения и
торможения, в проведении нервных импульсов, в си-
наптической передаче, образовании соляной кислоты
желудочного сока. Суточная потребность — 2 –4 г
Поваренная соль,
растительная
и животная пища,
жидкости, потреб-
ляемые при питье
Фосфор Содержание в клетках в 40 раз выше, чем во внекле-
точной среде, до 80% содержится в костях и зубах.
В составе фосфолипидов содержится в клеточных
мембранах. Необходимый элемент макроэргических
соединений (АТР) и их производных, циклических
нуклеотидов, коферментов, играющих важнейшую
роль в метаболизме и регуляции физиологических
функций. Суточная потребность — 0,8—1,2 г
Молоко, свежая
рыба, сыры, соя,
мясо, яйца, орехи,
горох, злаки, море-
продукты
Железо
Около 66% содержится в гемоглобине крови. Со-
держится в скелетных мышцах, печени, селезенке,
костном мозге, в составе ферментов. Основная
функция — связывание кислорода. Суточная
потребность — 10—15 мг
Икра паюсная, ке-
товая, мясо, печень,
свежая рыба, яйца,
сухофрукты, орехи,
крупа гречневая,
горох, пшено
Йод
Входит в состав гормонов щитовидной железы.
Суточная потребность — 125—150 мкг
Морепродукты,
рыбий жир, йоди-
рованная пищевая
соль
Магний Содержится в костной ткани, скелетных мышцах
и нервной системе. Входит в состав многих фер-
ментов и коферментов. Необходим для функции
клеточных мембран, деятельности миокарда и глад-
ких мышц. Суточная потребность — 250—350 мг
Какао, шоколад,
миндаль, овес, ку-
куруза, горох, соя,
хлеб из цельного
зерна, гречневая
крупа, мясо, моло-
ко, творог
Медь
Содержится в печени, селезенке, играет роль в про-
цессах всасывания железа, синтеза гемоглобина,
входит в состав ряда ферментов и пигментов.
Суточная потребность — 2 —5 мг
Яйца, печень, по-
чки, рыба, шпинат,
виноград, сухие
овощи
Фтор
Содержится в зубных тканях и необходим для их
сохранения. Входит в состав некоторых ферментов.
Суточная потребность — около 2 мг. При передози-
ровке токсичен
Пищевые продук-
ты, фторирован-
ные зубные пасты
и NaCl
Сера
Входит в состав аминокислот, белков (инсулин)
и витаминов (B1), участвует в обезвреживании ток-
синов в печени. Суточная потребность — 1 г
Мясо, печень,
рыба, яйца
Цинк
Важный компонент ряда мужских половых гормо-
нов. Необходим для процессов роста. Суточная
потребность — 12 —15 мг
Мясо (особенно
говядина, индейка),
бобы, крабы, яич-
ный желток, цель-
ное зерно пшени-
цы, отруби, семеч-
ки тыквы и подсол-
нечника, устрицы
Продолжение табл. 19
272
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
Элементы
Физиологическая роль и суточная потребность
Источники
Кобальт
Входит в состав витамина В12, необходим для нор-
мального эритропоэза. Содержится в печени, кост-
ной ткани. Суточная потребность точно неизвестна,
предположительно — 100–200 мкг
Печень
Марга-
нец
Входит в состав некоторых ферментных систем,
способствует нормальному функционированию
половой системы и кровеносных сосудов. Суточная
потребность — 2 –5 мг
Орехи, пряности
(имбирь, корица,
лавровые листья,
тимьян), чай, яч-
мень, рожь, гречи-
ха, пшеница
Селен
Способствует нормальному функционированию
мужской половой системы. Мощный антиоксидант.
Стабилизирует нуклеиновые кислоты, стимулирует
функцию лимфоидных органов, увеличивает выра-
ботку антител, повышает сопротивляемость орга-
низма, оказывает противоопухолевое действие.
Суточная потребность — 50–70 мкг
Каменная и мор-
ская соль, цельное
неочищенное зер-
но, отруби, чеснок,
кукуруза, дрожжи,
грибы, морепро-
дукты, почки, пе-
чень, сердце, яйца
Примечание. Всюду приведены суточные потребности взрослого чело-
века.
ВОДА
Около 60% массы тела взрослого человека приходится на долю
воды (у мужчин 61%, у женщин 54%). Разница связана с бо
льшим ко-
личеством жира в теле женщины. У новорожденного содержание
воды достигает 77%, после 70—80 лет снижается до 50%. Вода облада-
ет уникальными свойствами, которые чрезвычайно важны для самого
существования жизни. К ним относятся: способность молекул воды
слипаться между собой (сливаться с другими веществами); способ-
ность притягиваться к электрически заряженной поверхности и под-
ниматься по мелким порам; вода — растворитель. Она обладает высо-
кой теплопроводностью, высокой температурой кипения и замерзания
и максимальной плотностью при +40 °С; вода охлаждает тело при ис-
парении, тем самым регулирует температуру тела.
Вода — универсальная среда, участвующая во всех биохимиче-
ских и физиологических реакциях в организме. Пищеварение и вса-
сывание, выделение происходят в водной среде. Вода поступает в
организм человека через желудочно-кишечный тракт, его слизистая
оболочка всасывает воду. Выделение воды осуществляется почками
Окончание табл. 19
273
ВОДА
(800–1300 мл в сутки), с выдыхаемым воздухом (около 400 мл),
с потом (около 600 мл). Часть воды (300—400 мл в сутки) образует-
ся в процессе биохимических реакций. Так, при окислении 100 г
жира образуется 107 мл воды, углеводов — 55 мл, белков — 41 мл.
В обычных условиях потребность взрослого человека в воде состав-
ляет 40 г/кг массы тела, грудного ребенка — 120–150 г/кг. Суточная
потребность взрослого при умеренной физической нагрузке и нор-
мальной температуре составляет 1750–2200 мл, однако в виде
воды и напитков — лишь 800–1000. Потеря 6–8% воды приводит
к существенному нарушению обмена веществ, потеря 10% воды —
к необратимым патологическим изменениям в организме, 21% —
к смерти.
ПИЩЕВАРЕНИЕ
Пища в том виде, в каком она поступает в организм, не может всо-
саться в кровь и лимфу и не может быть использована для выполне-
ния различных жизненных функций. Для усвоения пищи она должна
подвергнуться механической и химической обработке в органах пи-
щеварительной системы. Пища измельчается в полости рта, переме-
шивается в желудке и тонкой кишке с пищеварительными соками,
ферменты которых расщепляют питательные вещества. Аминокисло-
ты, моносахариды и эмульгированные жиры всасываются и усваива-
ются организмом. Вода, минеральные вещества (соли), витамины ус-
ваиваются в натуральном виде. Механическая и химическая обработка
пищи и превращение ее в усваиваемые организмом вещества называ-
ется пищеварением. Оно осуществляется в пищеварительной трубке
при участии ферментов, выделяемых пищеварительными железами.
Поэтому пищеварение в желудке, тонкой кишке называют полостным
пищеварением. Переваривание пищи происходит также непосредст-
венно на поверхности микроворсинок эпителиальных клеток тонкой
кишки. Такое пищеварение называют контактным, или мембранным,
пищеварением. На поверхности микроворсинок и клеточной мембра-
ны эпителиоцитов имеется наиболее высокая концентрация пищева-
рительных ферментов. Мембранное пищеварение является как бы за-
ключительной фазой переваривания пищи, после чего расщепленные
белки и углеводы, эмульгированные жиры всасываются в кровенос-
ные и лимфатические капилляры.
Расщепление (переваривание) белков, жиров, углеводов про-
исходит с помощью пищеварительных ферментов. Ферменты содер-
жатся в слюне, желудочном соке, кишечном соке, в желчи и панкреа-
274
ПИЩЕВАРЕНИЕ
тическом соке, которые являются, соответственно, продуктами
секреции слюнных, желудочных, тонкокишечных и толстокишечных
желез, а также печени и поджелудочной железы.
ВНИМАНИЕ!
В течение суток в пищеварительную систему поступает примерно до
1,5—2 л слюны, 2,5 л желудочного сока, 2,5 л кишечного сока, 1,0 л
желчи, 1,5—2 л сока поджелудочной железы.
Ферменты являются важнейшими составляющими секретов пи-
щеварительных желез. Благодаря пищеварительным ферментам
белки расщепляются до аминокислот, жиры — до глицерина и жир-
ных кислот, углеводы — до моносахаридов. Пищеварительные фер-
менты представляют собой белки. Ферменты служат ускорителями
(катализаторами) реакций расщепления пищевых веществ. Фермен-
ты, расщепляющие белки — протеазы, жиры — липазы, углеводы —
α-амилазы. Для расщепления необходимы определенные условия,
температура тела и реакция среды (кислая или щелочная).
ПИЩЕВАРЕНИЕ В ПОЛОСТИ РТА
Механическая и химическая обработка пищи начинается в поло-
сти рта. Здесь зубы измельчают пищу, анализируются ее вкусовые ка-
чества. В ответ на раздражения вкусовых, тактильных и температур-
ных рецепторов, которые расположены в слизистой оболочке языка
и стенок полости рта, крупные и мелкие железы выделяют слюну. Вы-
деление слюны происходит рефлекторно. В полости рта начинается
переваривание углеводов, формируется пищевой комок. Средняя
длительность пребывания пищи в полости рта составляет 15–20 сек.
Слюна выделяется не только при непосредственном воздействии
пищи на нервные окончания в стенках полости рта (безусловный
рефлекс), а также в ответ на обонятельные, зрительные, слуховые и
другие воздействия (запах, цвет, разговор о еде) — условный рефлекс.
Один из важнейших физиологических процессов — жевание
—
ме-
ханическое измельчение пищи, ее смешивание со слюной, а также
рефлекторное воздействие на секреторную и двигательную функции
пищеварительной системы. В акте жевания участвуют челюсти, зубы,
жевательные и мимические мышцы, некоторые мышцы шеи, язык
и мягкое нёбо. Жевание регулируется рефлекторно с участием коры
полушарий большого мозга.
Глотание пережеванной и смоченной слюной пищи — это слож-
ный рефлекторный акт. Вход в носовую полость закрывается мягким
275
ПИЩЕВАРЕНИЕ В ЖЕЛУДКЕ
нёбом, надгортанник закрывает вход в гортань, задерживается дыха-
ние. Если человек во время еды разговаривает, то вход из глотки в
гортань не закрывается, пища может попасть в просвет гортани и
в дыхательные пути. Вот почему во время еды нельзя разговаривать.
Из ротовой полости пищевой комок движением корня языка через
зев попадает вначале в ротовую часть глотки. В это время продольные
мышцы глотки поднимают глотку, как бы натягивают ее на пищевой
комок. Одновременно круговые мышцы, сокращаясь, проталкивают
пищу из глотки в пищевод. Сокращения круговых и продольных мышц
пищевода продвигают пищу в желудок. Твердая пища проходит весь
путь от ротовой полости до желудка за 6–8 с, а жидкая — за 2–3 с.
ПИЩЕВАРЕНИЕ В ЖЕЛУДКЕ
Пища, поступившая из пищевода в желудок, находится в нем до
4—6 ч. В желудке пища перемешивается с желудочным соком и под
его действием переваривается. Пища в желудке превращается в жид-
кую кашицу. Ферменты пепсин и гастриксин переваривают (расще-
пляют) белки до крупных частиц полипептидов, еще не способных
всосаться в капилляры слизистой оболочки желудка. Пепсин также
створаживает казеин молока, который в желудке подвергается гидро-
лизу. Эмульгированные жирные компоненты молока расщепляет ли-
паза. Слизь (муцин) предохраняет слизистую оболочку желудка от
самопереваривания. При попадании в желудок алкоголя (спирта)
действие слизи ослабляется, и тогда создаются благоприятные усло-
вия для образования язв слизистой оболочки, для возникновения
воспалительных явлений гастрита.
Выделение желудочного сока начинается уже через 5—10 мин. по-
сле начала еды. Секреция желудочных желез продолжается все время,
пока пища находится в желудке. Состав желудочного сока и скорость
его выделения зависят от количества и качества пищи. Жир, крепкие
растворы сахара, а также отрицательные эмоции (гнев, печаль) тормо-
зят образование желудочного сока. Ускоряют образование и выделе-
ние желудочного сока экстракты мяса и овощей (бульоны из мяса
и овощей). Выделение желудочного сока происходит не только во
время еды, но уже при ощущении запаха пищи, ее виде, даже при раз-
говоре о еде. В этих случаях желудочный сок выделяется в результате
условно-рефлекторной деятельности организма.
Деятельность мышц желудка обусловливает его моторику, поддер-
живает тонус, почти стабильное давление в просвете желудка и осу-
ществляет перемешивание и опорожнение. В результате перемешива-
ния пищевых масс с желудочным соком образуется химус — жидкая
кашица, которая удаляется отдельными порциями из желудка. Быст-
276
ПИЩЕВАРЕНИЕ
рее всего из желудка выводятся углеводы, несколько медленнее —
белки, дольше всего (около 4 ч) — жиры. При этом жидкая и хорошо
переработанная пища эвакуируется быстрее, чем плотная и плохо пе-
режеванная.
Если пилорический сфинктер расслаблен и отверстие в двенадца-
типерстную кишку открыто, то пища поступает из желудка в двенад-
цатиперстную кишку. Если отверстие закрыто, то пищевая кашица
снова отбрасывается из привратника в глубь желудка и продолжает
перевариваться. После поступления порции пищи в двенадцатиперст-
ную кишку ее слизистая оболочка раздражается кислым содержимым
и механическим воздействием пищи. Пилорический сфинктер при
этом рефлекторно закрывает отверстие, ведущее из желудка в кишку.
После изменения рН в двенадцатиперстной кишке до щелочного
в связи с выделением в нее желчи и панкреатического сока в кишку
из желудка поступает новая порция кислот пищевой кашицы. Таким
образом, химус порциями из желудка выбрасывается в двенадцати-
перстную кишку.
ПИЩЕВАРЕНИЕ В ТОНКОЙ КИШКЕ
Кишечное пищеварение начинается уже в двенадцатиперстной
кишке, которая играет особую роль в пищеварении. В просвет двенад-
цатиперстной кишки выделяются не только секреты ее собственных
желез, но и желчь, а также панкреатический сок. Секрет желез две-
надцатиперстной кишки содержит муцин, защищающий слизистую
оболочку, а также ферменты, расщепляющие белок, и энтерокиназу,
превращающую неактивный фермент поджелудочного сока трипси-
ноген в активный трипсин. Панкреатический сок (секрет поджелудоч-
ной железы) бесцветный, имеет щелочную реакцию (рН 7,3—8,7). Он
содержит различные ферменты, переваривающие белки, жиры и угле-
воды. Под влиянием ферментов трипсина и химотрипсина белки рас-
щепляются до аминокислот; липаза расщепляет жиры до глицерина и
жирных кислот; амилаза и мальтаза расщепляют углеводы до моноса-
харидов. Секреция сока поджелудочной железы происходит рефлек-
торно в ответ на сигналы, идущие от рецепторов слизистой оболочки
полости рта, а также на раздражение слизистой оболочки двенадцати-
перстной кишки кислой пищевой кашицей, поступающей из желудка.
Желчь, образующаяся в печени в промежутке между приемами
пищи, поступает в желчный пузырь в жидком виде, концентрируется
там в 7—8 раз (вода всасывается в стенки желчного пузыря). Поступа-
ющая в двенадцатиперстную кишку желчь выделяется из желчного
пузыря. Желчь, имеющая золотисто-желтый цвет, содержит желчные
кислоты, желчные пигменты, холестерин и другие вещества. В тече-
277
ПИЩЕВАРЕНИЕ В ТОЛСТОЙ КИШКЕ
ние суток образуется 0,5—1,2 л желчи. Желчь эмульгирует жиры до
мельчайших капель и способствует их всасыванию, активирует пище-
варительные ферменты, замедляет гнилостные процессы, усиливает
перистальтику тонкой кишки. Желчеобразование и поступление жел-
чи в двенадцатиперстную кишку стимулируются присутствием пищи
в желудке и двенадцатиперстной кишке, а также видом и запахом
и регулируется нервным и гуморальным путями.
Из двенадцатиперстной кишки (благодаря ее перистальтике) пи-
щевая кашица продвигается в тощую, а затем в подвздошную кишку.
Выделяемый кишечными железами в ответ на механические и хими-
ческие раздражения кишечный сок (до 2,5 л в сутки) расщепляет пеп-
тиды (белки) до аминокислот, сахара
—
до глюкозы и фруктозы. В ки-
шечном соке содержится более 22 пищеварительных ферментов, в том
числе энтерокиназа (активатор трипсиногена поджелудочной желе-
зы), пептидаза, липаза, амилаза и фосфатаза, сахараза и др.
Различные участки тонкой кишки по-разному участвуют во вса-
сывании: жиры всасываются преимущественно в верхней половине
тонкой кишки, белки — в средней трети, вода — в подвздошной киш-
ке. Окончательное переваривание пищи (до аминокислот, моносаха-
ридов, эмульгированных жиров) и всасывание продуктов переварива-
ния происходит по мере продвижения пищевых масс в направлении
от двенадцатиперстной кишки в подвздошную кишку и далее в сле-
пую кишку. Движение пищевых масс происходит в результате сокра-
щения циркулярного и продольного мышечных слоев стенок тонкой
кишки. Выделяют два вида движений тонкой кишки: перистальтиче-
ские и маятникообразные.
ПИЩЕВАРЕНИЕ В ТОЛСТОЙ КИШКЕ
Из тонкой кишки непереваренные и невсосавшиеся в кровеносные
и лимфатические капилляры остатки пищи через подвздошно-слепо-
кишечное отверстие поступают в толстую кишку, где формируются
каловые массы, происходит реабсорбция воды, электролитов и водо-
растворимых витаминов. Толстая кишка обильно заселена микрофло-
рой, которая сбраживает углеводы и осуществляет гнилостное разло-
жение белков. Мышечная оболочка толстой кишки осуществляет
движения, благодаря чему содержимое перемешивается, от слепой
кишки в анальном направлении распространяются мощные волны со-
кращения, благодаря которым каловые массы перемещаются в сигмо-
видную кишку. Дефекация — опорожнение толстой кишки и удаление
кала — происходит произвольно и регулируется корой головного моз-
га. Напомним, что в дистальном отделе прямой кишки имеется два
сфинктера: поперечнополосатый произвольный наружный и гладко-
278
ДЫХАТЕЛЬНАЯ СИСТЕМА
мышечный непроизвольный внутренний. Оба сфинктера в состоянии
покоя сокращены, и задний проход замкнут.
Волнообразные сокращения сигмовидной ободочной кишки пере-
мещают каловые массы в прямую кишку, растяжение которой вызы-
вает нервные импульсы, в результате чего появляются осознанные
позывы к дефекации. Однако здоровый человек может сдержать по-
зыв к дефекации. Если количество кала превышает два литра, позыв
сдержать невозможно. При дефекации расслабляются непроизволь-
ный гладкомышечный внутренний сфинктер и поперечнополосатый
наружный сфинктер прямой кишки.
ВНИМАНИЕ!
Парасимпатические нервы усиливают, а симпатические тормозят мото-
рику толстой кишки.
ДЫХАТЕЛЬНАЯ СИСТЕМА
Дыхание — это процесс вентиляции легких и газообмена, сопрово-
ждающийся поглощением кислорода и выделением углекислого газа.
Дыхательная система выполняет важнейшую функцию — газообмен,
без которого невозможна жизнь, ибо превращение энергии в организ-
ме происходит в результате окислительного распада питательных ве-
ществ с участием кислорода. Удаление углекислого газа — одного из
конечных продуктов дыхательного обмена — еще одна важнейшая
функция газообмена. Перенос кислорода в организме включает сле-
дующие этапы: легочное (вне ее) дыхание — поступление кислорода
в альвеолы (легочная вентиляция) и диффузия кислорода из альвеол
в кровь капилляров малого круга; транспорт кислорода кровеносной
системой; тканевое дыхание — диффузия кислорода из капилляров
в ткани. Удаление и выведение углекислого газа происходит в обрат-
ном порядке.
Полость носа, носовая часть глотки, гортань, трахея, бронхи раз-
личных калибров, включая бронхиолы, являются воздухоносными
путями. В них воздух согревается, очищается от различных частиц и
увлажняется. Альвеолярные ходы и альвеолы являются собственно
респираторными отделами, в которых и происходит газообмен. У че-
ловека один из органов дыхательной системы — гортань — выполняет
две функции: воздухоносную и голосообразовательную (рис. 143).
Нормальное дыхание происходит через полость носа, где располо-
жен орган обоняния, который анализирует качество вдыхаемого воз-
духа. Полость носа выстлана изнутри слизистой оболочкой, которую
279
ДЫХАТЕЛЬНАЯ СИСТЕМА
можно разделить на две резко отличающиеся друг от друга по строе-
нию и функции части: дыхательную и обонятельную. Обонятельная
область, занимающая верхние носовые раковины и соответствующие
ей части перегородки и задний отдел верхней стенки полости носа,
покрыта цилиндрическим эпителием, в состав которого входят специ-
альные обонятельные нейросенсорные биполярные клетки, воспри-
нимающие запахи. Дыхательная область покрыта цилиндрическим
реснитчатым эпителием с большим количеством бокаловидных кле-
ток, вырабатывающих слизь. Эпителий покрыт слизью, которая бла-
годаря движению ресничек передвигается в направлении носоглотки,
куда и удаляется слизь. В полость носа выделяется секрет многочи-
сленных желез, которые в течение суток вырабатывают около 500 мл
жидкости. Слизь не только обволакивает частицы, но и увлажняет
Рис. 143. Дыхательная система:
1 — диафрагма, 2 — ребро, 3 — хрящевое кольцо не позволяет трахее опадать во
время дыхания, 4 — правое легкое, 5 — пищевод, по нему пища попадает в желудок,
6 — гортань, в ней находятся голосовые связки, 7 — носовая полость, здесь воздух
согревается, очищается и увлажняется, 8 — глотка, 9 — надгортанник, он закрывает
вход в гортань во время глотания, 10 — трахея, 11 — бронхи, 12 — левое легкое,
13 — межреберная мышца, она двигает ребра, 14 — сердце
11
11
11
22
12
12
33
13
13
44
14
14
55
66
77
88
99
10
10
280
ДЫХАТЕЛЬНАЯ СИСТЕМА
вдыхаемый воздух. Слизистая оболочка носа выполняет еще одну
функцию — она согревает воздух. Собственная пластинка слизистой
оболочки и подслизистая основа очень богаты артериями и венами.
Три носовые раковины увеличивают общую поверхность полости
носа. В области средней и нижней носовых раковин имеется пещери-
стая ткань, содержащая множество тонких вен, которые в обычных
условиях находятся в спавшемся состоянии, при наполнении их кро-
вью слизистая оболочка набухает. Воздух из полости носа поступает
через хоаны в носовую, затем ротовую части глотки и в гортань. Стро-
ение глотки описано выше.
ГОРТАНЬ
Сложное строение гортани связано именно с голосообразованием.
Устройство остальных воздухоносных путей гораздо более простое.
Человеческая гортань — самый прекрасный музыкальный инстру-
мент — расположена на уровне IV—VI шейных позвонков и связана с
подъязычной костью. Вверху гортань переходит в полость глотки,
внизу — в трахею. Снаружи ее положение заметно по выступу, назы-
ваемому «кадыком» («адамово яблоко»), более развитому у мужчин
и образованному соединением обеих пластинок щитовидного хряща.
Имеются значительные возрастные и половые особенности горта-
ни. Рост и функция гортани связаны с развитием половых желез.
У детей гортань расположена выше, чем у взрослых (нормальное по-
ложение устанавливается к 13—14 годам жизни), у стариков еще
ниже; у женщин несколько выше, чем у мужчин, причем гортань муж-
чины в среднем на 1/3 больше женской. У новорожденного гортань
относительно велика. В течение первых 4—5 лет жизни ребенка она
растет несколько медленнее трахеи. После шести лет рост гортани за-
медляется, но перед наступлением половой зрелости у мальчиков
рост ее быстро ускоряется и размеры стремительно увеличиваются.
В это время изменяется голос мальчиков.
Скелет гортани образован несколькими подвижно соединенными
между собой гиалиновыми и эластическими хрящами. Самый круп-
ный из гортанных хрящей — гиалиновый щитовидный, в котором раз-
личают две четырехугольные пластинки, соединяющиеся между со-
бой (упомянутый выступ гортани — «адамово яблоко») под прямым
(или почти прямым) углом у мужчин и тупым углом (около 120°)
у женщин. От задних краев отходят две пары рогов — верхние и ниж-
ние. Наиболее важны в функциональном отношении гиалиновые чер-
паловидные хрящи, от основания которых вперед отходит голосовой
отросток, состоящий из эластического хряща; назад и кнаружи — мы-
шечный отросток. К последнему прикрепляются мышцы, двигающие
281
ГОРТАНЬ
черпаловидный хрящ в перстнечерпаловидном суставе. При этом из-
меняется положение голосового отростка, к которому прикрепляются
голосовые связки. Сверху гортань покрыта надгортанником, состоя-
щим из эластического хряща. Надгортанник расположен впереди вхо-
да в гортань и прикреплен к щитовидному хрящу с помощью щито-
надгортанной связки. В основании гортани лежит гиалиновый
перстневидный хрящ, его дуга обращена вперед, а пластинка — назад.
Соединительнотканная связка соединяет нижний край хряща с пер-
вым хрящом трахеи (рис. 144).
Рис. 144. Хрящи, связки и суставы гортани:
А, вид спереди: 1 — тело подъязычной кости, 2 — зерновидный хрящ, 3 — верхний
рог щитовидного хряща, 4 — пластинка щитовидного хряща, 5 — нижний рог щито-
видного хряща, 6 — дуга перстневидного хряща, 7 — хрящи трахеи, 8 — кольцевые
связки, 9 — перстнещитовидный сустав, 10 — перстнещитовидная связка, 11 —
верхняя щитовидная вырезка, 12 — щитоподъязычная мембрана, 13 — срединная
щитоподъязычная связка, 14 — латеральная щитоподъязычная связка;
Б, вид сзади: 1 — надгортанник, 2 — большой рог подъязычной кости, 3 — зерновидный
хрящ, 4 — верхний рог щитовидного хряща, 5 — пластинка щитовидного хряща,
6 — черпаловидный хрящ, 7 — правый перстнечерпаловидный сустав, 8 — правый
перстнещитовидный сустав, 9 — хрящи трахеи, 10 — перепончатая стенка,
11 — пластинка перстневидного хряща, 12 — левый перстнещитовидный сустав трахеи,
13 — нижний рог щитовидного хряща, 14 — левый перстнечерпаловидный сустав,
15 — мышечный отросток черпаловидного хряща, 16 — голосовой отросток
черпаловидного хряща, 17 — щитонадгортанная связка, 18 — рожковидный хрящ,
19 — латеральная щитоподъязычная связка, 20 — щитоподъязычная мембрана
11
11
11
11
11
11
22
22
12
12
12
12
33
33
13
13
13
13
44
44
14
14
14
14
55
55
А
15
15
66
66
Б
16
16
77
77
17
17
88
88
18
18
99
99
19
19
10
10
10
10
20
20
282
ДЫХАТЕЛЬНАЯ СИСТЕМА
Хрящи соединяются между собой посредством связок и суставов.
Важнейший из них — перстнечерпаловидный сустав — расположен
между основанием черпаловидного хряща и соответствующей по-
верхностью перстневидного. Черпаловидный хрящ в этом суставе
вращается вокруг вертикальной оси, а также немного в стороны.
Перстнещитовидный сустав образован суставными поверхностями
нижних рогов щитовидного хряща и соответствующими площадками
перстневидного. Правый и левый суставы объединяются в один ком-
бинированный, в котором щитовидный хрящ наклоняется вперед,
удаляясь своей вырезкой от пластинки перстневидного и черпаловид-
ных хрящей, или выпрямляется, приближаясь к последним.
Полость гортани выстлана слизистой оболочкой, покрытой рес-
нитчатым эпителием с большим количеством бокаловидных клеток,
кроме голосовых связок, покрытых многослойным плоским неоро-
говевающим эпителием. Передняя и верхняя части задней поверх-
ности надгортанника также покрыты многослойным плоским не-
ороговевающим эпителием, бо
льшая часть задней — реснитчатым
эпителием.
Под слизистой оболочкой гортани лежит фиброзно-эластическая
мембрана. Часть ее, расположенная между щитовидным, черпаловид-
ным и перстневидным хрящами, называется эластическим конусом.
От внутренней поверхности угла щитовидного хряща к голосовым от-
росткам черпаловидных хрящей идут более плотные края конуса,
образующие голосовые связки, состоящие главным образом из эласти-
ческих волокон. Голосовые складки образованы слизистой оболочкой,
голосовой мышцей и описанной голосовой связкой.
Колебания голосовых связок при прохождении через них выдыха-
емого воздуха и создают звук, который в зависимости от натяжения
связок и ширины голосовой щели может меняться. Человек созна-
тельно регулирует этот процесс. Несмотря на сходство в строении
гортани человека и человекообразных обезьян, они не в состоянии
этого делать, поэтому и не могут издавать музыкальных звуков. Лишь
гиббоны способны воспроизводить звуки, напоминающие музыкаль-
ные. Следует подчеркнуть, что в гортани происходит лишь голосо-
образование. В членораздельной же речи принимают участие около-
носовые пазухи, губы, язык, мягкое нёбо, мимические мышцы.
Изменение положения хрящей гортани, натяжение голосовых свя-
зок, ширина голосовой щели регулируется работой поперечнополоса-
тых мышц гортани. Основная роль в этом принадлежит мышцам, на-
чинающимся на перстневидном хряще и прикрепляющимся к
мышечным отросткам черпаловидных.
Трахея, связанная с гортанью перстнетрахеальной связкой, начи-
нается на уровне верхнего края VII шейного и заканчивается на уров-
283
ГОРТАНЬ
не верхнего края V грудного позвонка, где и разделяется на два брон-
ха, образуя бифуркацию (рис. 145). В просвете трахеи на месте
бифуркации имеется полулунный выступ — киль. Начало трахеи
у грудных детей лежит высоко, на уровне IV—V шейных позвонков,
у взрослых — на уровне VI, у стариков опускается до VII шейного
позвонка. У женщин начало трахеи лежит несколько выше, чем у муж-
чин. При глотании или движениях головы положение верхнего конца
трахеи изменяется.
Стенка трахеи состоит из слизистой оболочки, подслизистой осно-
вы, волокнисто-мышечно-хрящевой и адвентициальной оболочек.
Слизистая оболочка трахеи выстлана реснитчатым псевдомногослой-
ным эпителием, содержащим большое количество бокаловидных
клеток. Волокнисто-мышечнохрящевая оболочка трахеи образована
7
8
9
10
11
12
13
14
15
4
3
2
5
6
1
2
3
Рис. 145. Трахея и бронхи:
1 — пищевод, 2 — трахея, 3 — аорта,
4 — левый главный бронх, 5 — левая
легочная артерия, 6 — левый верхний
долевой бронх, 7 — сегментарные бронхи
верхней доли левого легкого, 8 — левый
нижний долевой бронх, 9 — непарная
вена, 10 — сегментарные бронхи нижней
и средней долей правого легкого,
11 — правый нижний долевой бронх,
12 — правый средний долевой бронх,
13 — правый верхний долевой бронх,
14 — правый главный бронх,
15 — бифуркация трахеи
284
ДЫХАТЕЛЬНАЯ СИСТЕМА
16—20 гиалиновыми хрящами, каждый из которых представляет со-
бой дугу, открытую кзади, занимающую приблизительно две трети
окружности трахеи. Хрящи, покрытые надхрящницей, соединены
между собой кольцевыми связками. Отсутствие хрящей на задней
стенке весьма важно: благодаря этому пищевой комок, проходящий
по пищеводу, лежащему непосредственно позади трахеи, не испыты-
вает сопротивления с ее стороны. Вместе с тем трахея благодаря на-
личию в ее стенке хрящей, связанных плотной фиброзной тканью пе-
репончатой части, очень упруга и эластична. Трахея противостоит
значительному давлению извне, сохраняя просвет постоянно откры-
тым, и может растягиваться, изменяя свои продольные и поперечные
размеры. Трахея растет равномерно вместе с туловищем, однако наи-
более интенсивно в течение первого года жизни и в период полового
созревания.
Бронхи. «Бронхиальное дерево» состоит из ветвящихся бронхов,
просвет которых постепенно уменьшается. Главные бронхи обладают
выраженной упругостью, которая, как и способность к сопротивле-
нию, с возрастом увеличивается, у детей же хрящи тонкие, мягкие,
эластичные и обладают малой упругостью.
Главные бронхи не делятся дихотомически, от них отходят вторич-
ные, или долевые, бронхи. От правого — три: верхний, средний и ниж-
ний долевые, от левого — два: верхний и нижний долевые, делящиеся
на более мелкие третичные, или сегментарные, бронхи (справа — 10,
слева — 9), которые уже разделяются дихотомически (см. рис. 145).
При этом площадь сечения вышележащего бронха меньше, чем сумма
площадей сечений его ветвей. Главные бронхи являются бронхами
первого порядка, долевые — второго, а сегментарные — бронхами тре-
тьего порядка. В дальнейшем бронхи делятся на субсегментарные
(первой, второй, третьей генерации — всего 9—10), междольковые,
внутридольковые (рис. 146).
Бронхи выстланы цилиндрическим реснитчатым эпителием с
большим количеством бокаловидных клеток. В собственной пла-
стинке слизистой оболочки имеются непрерывные полоски эласти-
ческой ткани, разветвляющиеся по мере ветвления бронхов. Строе-
ние главных бронхов во многом напоминает строение трахеи.
Гиалиновые хрящи бронхов представляют собой дуги, открытые кза-
ди, где концы их соединяются перепончатой частью. Хрящи соеди-
няются между собой кольцевыми связками, аналогичными трахе-
альным. По мере уменьшения калибра бронха хрящи постепенно
меняют форму, образуя в начале полукольца, затем хрящевые пла-
стинки неодинаковой величины, которые совершенно исчезают
285
ГОРТАНЬ
в бронхиолах диаметром около 1 мм. В стенке главных бронхов мы-
шечная ткань располагается так же, как и в трахее. В стенке внутри-
легочных бронхов имеется круговой слой гладких мышечных воло-
кон, располагающихся между слизистой оболочкой и хрящами.
В мелких бронхах (диаметром до 1—2 мм) постепенно исчезают хря-
щевые пластинки и железы, а мышечная пластинка слизистой обо-
лочки становится относительно более толстой.
Диаметр самых мелких разветвлений воздухопроводящих путей —
бронхиол — от 0,5 до 1 мм. Имеется около 20 их генераций, послед-
няя — терминальные бронхиолы — делится на 14—16 дыхательных
(респираторных) бронхиол каждая. В стенках бронхиол, в отличие от
бронхов, отсутствуют хрящи. Слизистая оболочка бронхиол выстлана
однослойным реснитчатым эпителием, между клетками которого рас-
полагаются отдельные клетки Клара, которые, по современным дан-
ным, являются источником восстановления эпителия концевых брон-
хиол.
Рис. 146. Бронхиальное дерево, схемы:
А — ветвление бронхов в правом и левом легких: 1, 2 — главные бронхи,
3, 4 — долевые и сегментарные бронхи, 5–15 — ветви сегментарных бронхов,
дольковый бронх и его разветвления (не показаны) , 16 — конечная бронхиола,
17–19 — дыхательные бронхиолы (три порядка ветвлений), 20–22
—
альвеолярные
ходы (три порядка ветвлений), 23 — альвеолярные мешочки;
Б — строение ацинуса легкого: 1 — терминальная бронхиола, 2 — дыхательная
бронхиола первого порядка, 3 — дыхательные бронхиолы второго порядка,
4 — дыхательные бронхиолы третьего порядка, 5 — альвеолярные ходы,
6 — альвеолярные мешочки, 7 — альвеолы
11
11
21
21
22
22
22
22
33
33
23
23
44
44
23
23
55
55 15
15
66
66
16
16
77
17
17
18
18
19
19
20
20
А
Б
286
ДЫХАТЕЛЬНАЯ СИСТЕМА
ЛЕГКИЕ
Легкие по форме напоминают конус с закругленной верхушкой,
выступающей над первым ребром. Консистенция легкого мягкая,
упругая, напоминает губку; благодаря содержащемуся воздуху лег-
кие и их кусочки плавают в воде. Цвет легких у детей, особенно ран-
него возраста, бледно-розовый, у взрослых ткань постепенно темнеет,
появляются черные пятна ближе к поверхности за счет частиц угля,
которые откладываются в соединительнотканной основе легкого. На
медиальной (средостенной) поверхности каждого легкого располо-
жены ворота легкого, через которые проходят бронх, сосуды и нервы,
окруженные соединительной тканью, образующие корень легкого.
Каждое легкое разделяется глубокими щелями на доли: правое — на
три, левое — на две. Доли легких — это отдельные, до известной сте-
пени изолированные, обособленные анатомически и физиологически
участки легкого с вентилирующим их бронхом и собственной сосу-
дисто-нервной системой. Каждому сегментарному бронху соответст-
вует бронхолегочный сегмент. Сегмент — участок легочной ткани, до
известной степени изолированный в анатомическом и физиологиче-
ском отношении, имеющий свою сосудисто-нервную систему и вен-
тилируемый сегментарным бронхом. Сегменты образованы легочны-
ми дольками (в одном сегменте их число достигает примерно 80
долек), разделенными междольковыми соединительнотканными пе-
регородками. Долька представляет собой участок легочной ткани,
вентилируемый претерминальной (дольковой) бронхиолой, сопрово-
ждаемой конечными ветвлениями легочных артериол и венул, лим-
фатических сосудов и нервов. В верхушку каждой дольки входит
претерминальная дольковая бронхиола, которая разветвляется на
3—7 мельчайших концевых (терминальных) бронхиол диаметром
около 0,5—0,15 мм каждая.
Функциональной единицей легкого является ацинус. Это систе-
ма разветвлений одной концевой бронхиолы, делящейся на 14—16
дыхательных (респираторных) бронхиол первого порядка, которые
дихотомически делятся на респираторные бронхиолы второго по-
рядка. Последние, в свою очередь, также дихотомически разветвля-
ются на респираторные бронхиолы третьего порядка, образующие
2—3 генерации альвеолярных ходов (до 1500), несущих на себе
до 20 000 альвеолярных мешочков и альвеол (О. Вейбель). В одной
легочной дольке насчитывается около 50 ацинусов. Стенки терми-
нальных и дыхательных бронхиол окружены густой сетью эластиче-
ских волокон и связаны с эластическими волокнами легочной па-
ренхимы. Между спиральными пучками эластических волокон
287
ЛЕГКИЕ
имеются пучки гладких мышечных клеток. Благодаря этому при
вдохе бронхиолы не спадаются. Уже на стенках дыхательных брон-
хиол имеются бухтообразные выпячивания — альвеолы. Диаметр
альвеолярных ходов около 100 мкм. Вход в каждую альвеолу альве-
олярного хода окружен пучками гладких мышечных клеток. Следу-
ет особо подчеркнуть, что альвеолы, альвеолярные мешочки и ходы
являются не морфологическими структурами, а пространствами,
содержащими воздух.
Альвеолы напоминают пузырьки неправильной формы, они раз-
деляются межальвеолярными перегородками толщиной 2—8 мкм.
В каждой перегородке, обычно являющейся одновременно стенкой
двух (иногда и более) альвеол, расположена густая сеть кровенос-
ных капилляров, эластических, ретикулярных и коллагеновых воло-
кон и клеток соединительной ткани. Форма альвеол многоугольная,
11
22
33
44
55
55
66
66
77
88
88
Рис. 147. Строение
межальвеолярной перегородки:
1 — респираторный альвеолоцит, 2 — просвет
кровеносного капилляра, 3 — эндотелиальная
клетка, 4 — альвеолярный макрофаг,
5 — большой альвеолоцит, 6 — осмиофильные
тельца, 7 — эластическое волокно,
8 — просвет альвеол
288
ДЫХАТЕЛЬНАЯ СИСТЕМА
вход в альвеолу округлый. Количество альвеол в обоих легких чело-
века достигает 600—700 млн, а общая их поверхность колеблется
в пределах от 40 м2 при выдохе до 120 м2 при вдохе. Диаметр альвеол
новорожденного в среднем равен 150 мкм, взрослого — 280 мкм,
в старости объем альвеол увеличивается за счет исчезновения неко-
торых межальвеолярных перегородок. Их диаметр достигает 300—
350 мкм.
Альвеолы выстланы изнутри клетками двух типов: респираторны-
ми (дыхательными, чешуйчатыми) альвеолоцитами и большими (гра-
нулярными) альвеолоцитами (рис. 147). Преобладают дыхательные
альвеолоциты, которые выстилают около 87,5% поверхности альвеол.
Это уплощенные клетки толщиной 0,1—0,2 мкм. Лишь в области зале-
гания ядра, которое выбухает в просвет альвеолы, они утолщены. Та-
кое строение в наибольшей степени способствует газообмену. Боль-
шие (гранулярные) альвеолоциты — крупные округлые клетки
с большим округлым ядром, лежащие на базальной мембране и вы-
ступающие в просвет альвеолы. В каждой клетке находится от 2 до 10
окруженных мембраной слоистых округлых осмиофильных пластин-
чатых телец, богатых фосфолипидами. Тельца, выделяющиеся из
больших альвеолоцитов, по современным воззрениям вырабатывают
основную часть сурфактанта — вещества липопротеиновой природы,
выстилающего изнутри альвеолы в виде пленки. Основная функция
сурфактанта — поддержание поверхностного натяжения альвеолы,
ее способности к раздуванию при вдохе и противодействие спадению
при выдохе. Особенно важна роль сурфактанта при первом вдохе
у новорожденного. Сурфактант препятствует пропотеванию жидко-
сти в просвет альвеол и обладает бактерицидностью. Обычно боль-
шие альвеолоциты располагаются группами по две-три клетки. Счи-
тается, что большие альвеолоциты являются также источником
восстановления клеточной выстилки альвеол. В выстилке альвеол об-
наруживается еще один вид клеток — альвеолярные макрофаги, имею-
щие моноцитарное происхождение, относящиеся к фагоцитарной си-
стеме и активно фагоцитирующие частицы и сурфактант. Макрофаги
могут мигрировать в просвет альвеол и в ткань межальвеолярных пе-
регородок.
Воздушно-кровяной барьер (аэрогематический), через который
происходит газообмен, очень тонок (в среднем 0,2—0,5 мкм). Он обра-
зован тонкой (90—95 нм) цитоплазмой дыхательных альвеолоцитов,
базальной мембраной, на которой они лежат, сливающейся с базаль-
ной мембраной кровеносных капилляров (толщина общей мембраны
90—100 нм), и цитоплазмой эндотелиоцитов (толщиной 20—30 нм),
289
ФУНКЦИЯ ДЫХАТЕЛЬНОЙ СИСТЕМЫ
образующих стенку капилляра. Каждый капилляр граничит с одной
или несколькими альвеолами (см. рис. 147).
Плевра
Подобно брюшине, плевра состоит из двух листков: париетального
и висцерального. Висцеральный листок плотно срастается с легочной
тканью, покрывает легкое со всех сторон, заходит в щели между его
долями. Париетальная (пристеночная) плевра представляет собой
сплошной листок, который срастается с внутренней поверхностью
грудной полости и средостением, образуя замкнутый мешок, содержа-
щий легкое, покрытое висцеральной плеврой.
Полость плевры — узкая замкнутая щель между париетальной
и висцеральной плеврой, в которой находится небольшое количест-
во серозной жидкости, увлажняющей листки, тем самым облегчая
их движение при дыхании. На тех участках, где реберная плевра пе-
реходит в диафрагмальную и медиастинальную, образуются плев-
ральные синусы: реберно-диафрагмальный, диафрагмо-медиасти-
нальный и реберно-медиастинальный.
СРЕДОСТЕНИЕ
Между правой и левой плевральными полостями располагается
комплекс органов, называемых средостением. Спереди оно ограниче-
но грудиной, сзади — грудным отделом позвоночного столба, верхней
границей является верхняя апертура грудной клетки, нижней — ди-
афрагма. В средостении располагаются сердце, аорта, легочные арте-
рии и вены, вилочковая железа, пищевод, трахея, главные бронхи,
кровеносные и лимфатические сосуды, лимфатические узлы, симпа-
тические стволы, нервы и др.
ФУНКЦИЯ ДЫХАТЕЛЬНОЙ СИСТЕМЫ
Легочное дыхание осуществляется путем чередования вдоха, во
время которого атмосферный воздух, насыщенный кислородом, по-
ступает в альвеолы, и выдоха, при котором воздух, обогащенный угле-
кислым газом, удаляется в окружающую среду. Вдох осуществляется
благодаря сокращению главных (наружных межреберных мышц и ди-
афрагмы) и вспомогательных дыхательных мышц (грудино-ключич-
но-сосцевидная, большая и малая грудные и лестничные). В акте вы-
290
ДЫХАТЕЛЬНАЯ СИСТЕМА
доха участвуют внутренние межреберные мышцы и диафрагма
(главные), а также мышцы брюшного пресса. Выдох осуществляется
при расслаблении мышц вдоха и сокращении мышц выдоха. Припод-
нятая и расширенная при вдохе грудная клетка в силу своей тяжести
и при сокращении ряда мышц живота опускается. Мышцы воздейст-
вуют на реберно-позвоночные суставы, поднимая и опуская ребра.
Диафрагма уплощается во время вдоха и поднимается во время выдо-
ха (куполы выдаются в грудную клетку). В зависимости от преобла-
дания при дыхании поднимания ребер или уплощения диафрагмы
различают грудной (реберный) и брюшной (диафрагмальный) тип
дыхания. Первый тип преобладает у мужчин, второй — у женщин.
Однако с возрастом в связи с уменьшением подвижности грудной
клетки увеличивается роль брюшного дыхания. Брюшное дыхание
преобладает у работников физического труда, певцов. У беременных
женщин по мере увеличения срока беременности возрастает роль
грудного дыхания.
Легочная вентиляция меняется в зависимости от функциональ-
ного состояния организма. Интенсивность легочной вентиляции
определяется глубиной вдоха и частотой дыхательных движений.
Для суждения о морфофункциональном состоянии дыхательной
системы определяют ряд показателей. Одним из наиболее инфор-
мативных показателей легочной вентиляции служит минутный
объем воздуха (MOB), который оценивается по объему воздуха,
вдыхаемого или выдыхаемого за одну минуту. У взрослого здорово-
го человека частота дыхания в покое составляет 12—16 в 1 мин.,
MOB—6—10л·мин
-1
, при работе он возрастает до 30—100 л · мин
-1
.
В течение жизни человек делает около 700 млн вдохов и вдыхает
300—350 млн л воздуха.
Дыхательный объем — количество воздуха, которое человек вды-
хает и выдыхает при спокойном дыхании.
Резервный объем вдоха — количество воздуха, которое человек мо-
жет дополнительно вдохнуть после нормального вдоха.
Резервный объем выдоха — количество воздуха, которое человек
может дополнительно выдохнуть после спокойного выдоха.
Остаточный объем — количество воздуха, остающееся в легких
после максимального выдоха.
Жизненная емкость легких (ЖЕЛ) — наибольшее количество воз-
духа, которое можно выдохнуть после максимального вдоха. ЖЕЛ
складывается из дыхательного объема и резервных объемов вдоха
и выдоха. ЖЕЛ — один из важнейших показателей, позволяющих су-
дить о подвижности легких и грудной клетки. ЖЕЛ зависит от возра-
291
ФУНКЦИЯ ДЫХАТЕЛЬНОЙ СИСТЕМЫ
ста, пола, физической активности, размеров тела и т. д . После 40 лет
ЖЕЛ уменьшается тем больше, чем меньше физическая активность
человека. Как правило, у женщин ЖЕЛ на 20—25% меньше, чем
у мужчин. Так, у «среднего» взрослого здорового мужчины в возрасте
20—30 лет ЖЕЛ составляет 4,8 л, у женщины — 3,6 л; в возрасте 50—
60 лет соответственно 3,8 и 3,0 л.
У молодого мужчины ЖЕЛ в норме можно определить по фор-
муле:
ЖЕЛ(л)=рост(м)—2,5;уженщиныЖЕЛ(л)=рост(м)—2.
Резерв вдоха — максимальное количество воздуха, которое можно
вдохнуть после спокойного выдоха.
Функциональная остаточная емкость — количество воздуха, ос-
тающееся в легких после спокойного выдоха.
Общая емкость легких — количество воздуха, содержащееся в лег-
ких на высоте максимального вдоха. Общая емкость легких равна
сумме ЖЕЛ и остаточного объема.
Легочные объемы можно измерить с помощью спирометра. Наи-
более распространен водный спирометр, который представляет со-
бой цилиндр, помещенный дном кверху в резервуар с водой. Воздух,
попавший в этот цилиндр, не сообщается с внешней средой, а сам
цилиндр уравновешивается противовесом. Исследуемый берет в рот
широкую трубку с загубником, которая соединена с пространством
внутри цилиндра. Во время выдоха объем воздуха в цилиндре уве-
личивается, и он всплывает; при вдохе же цилиндр погружается. Эти
перемещения могут быть измерены при помощи калиброванной
шкалы или зарегистрированы посредством писчика на барабане ки-
мографа (в последнем случае получают так называемую спирограм-
му). В настоящее время существует множество модификаций спиро-
метра.
Альвеолярный воздух — это воздух, находящийся в альвеолах, он
отличается от атмосферного по концентрации содержащихся в нем
газов. В покое поглощение организмом «среднего» взрослого челове-
ка кислорода из альвеолярного воздуха составляет от 250 до 300 мл ·
мин-1
, а выделение углекислого газа — от 200 до 250 мл · мин
-1
. Кисло-
род в процессе диффузии проходит из просвета альвеолы в кровенос-
ные капилляры через аэрогематический барьер, плазму крови и мем-
брану эритроцита. Общее расстояние не превышает 5 мкм. СО2
диффундирует в обратном направлении. Диффузия осуществляется
благодаря градиенту парциальных давлений О2 и СО2 в альвеолярном
воздухе и в крови.
292
ДЫХАТЕЛЬНАЯ СИСТЕМА
В табл. 20 представлены парциальное давление и концентрация
газов в различных средах организма.
Таблица 20
Парциальное давление и концентрация газов в различных средах
(мм рт. ст.)
Среда
О2
СО2
Вдыхаемый воздух
160 (21%)
0,3 (0,04%)
Выдыхаемый воздух
(16%)
(4,5%)
Альвеолы
105 (14%)
40 (5,5%)
Артериальная кровь
105
40
Венозная кровь
40
45
Клетки
40
45
Гемоглобин (Hb) представляет собой хромопротеид, молекула кото-
рого состоит из четырех полипептидных цепей, в состав каждой из них
входит гем — протопорфирин, в центре которого находится ион двух-
валентного железа. Содержание Hb в крови здорового взрослого чело-
века составляет в среднем 158 г/л у мужчин и 140 г/л у женщин. Одна-
ко этот показатель колеблется в зависимости от возраста, состояния
здоровья, географических условий (высота над уровнем моря) и т. д .
Уменьшение содержания Hb ниже 130 г/л (мужчины) и 120 г/л (жен-
щины) называется анемией (от греч. a, an — начальная часть слова со
значением отрицания, haima — кровь). У здорового человека среднее
содержание Hb в одном эритроците составляет около 31 · 10-12
г (31 пг).
Сразу после диффузии в эритроциты О2 связывается с гемоглоби-
ном, в результате чего образуется оксигемоглобин (HbО2), который
диффундирует к центру эритроцита, при этом валентность железа не
меняется. 1 г гемоглобина связывает 1,34 мл О2, СО2 в эритроцитах
также связан с гемоглобином. СО2 диффундирует из эритроцитов
только после его освобождения из химической связи. Во время прохо-
ждения через легочные капилляры эритроциты захватывают кисло-
род, и в них увеличивается напряжение О2, в то же время напряжение
СО2 в крови снижается (рис. 148). Следует подчеркнуть, что у здоро-
вого человека напряжение дыхательных газов в крови становится
практически таким же, как их парциальные давления в альвеолах.
Окись углерода (СО) обладает гораздо большим сродством
к гемоглобину, чем кислород. Нb + СО ␝ НbСО (карбоксигемогло-
бин), распад которого происходит в 200 раз медленнее, чем оксиге-
293
ФУНКЦИЯ ДЫХАТЕЛЬНОЙ СИСТЕМЫ
моглобина. Нb, связанный с СО2
, называется карбогемоглобином
(НbСО2).
рН артериальной крови человека колеблется в узких пределах —
от 7,37 до 7,43. В регуляции кислотно-щелочного равновесия участву-
ет ряд механизмов: буферные свойства крови, газообмен в легких
и выделительная функция почек.
Буферные системы (от англ. to buff — смягчать толчки) — это со-
вокупность веществ, сохраняющих постоянство рН крови. В первую
очередь к ним относится бикарбонатная система, которая состоит из
относительно слабой угольной кислоты, образующейся при гидрата-
ции СО2, и сопряженного основания — бикарбоната:
СО2+Н2О␝Н2СО3␝Н++НСО3
–
.
Эта система тесно связана с дыхательной, которая, поддерживая
постоянное напряжение СО2 в крови, обеспечивает высокое содержа-
ние буферных систем. Кроме того, буферную функцию выполняют
фосфатная система, белки плазмы, буферные основания.
4
5
63
2
1
12
34
СО2
О2
СО2
Рис. 148. Аэрогематический барьер в легком:
1 — просвет альвеол, 2 — сурфактант, 3 — альвеолоцит, 4 — эндотелиоцит,
5 — просвет капилляра, 6 — эритроцит в просвете капилляра, стрелками показан
путь кислорода и углекислого газа через аэрогематический барьер
(между кровью и воздухом)
294
МОЧЕПОЛОВОЙ АППАРАТ
Напряжение О2 и СО2 в артериальной крови является основным,
конечным результатом внешнего дыхания. Сложная работа дыхатель-
ной системы призвана приспосабливать внешнее дыхание к постоян-
но меняющимся условиям внешней и внутренней среды организма.
Эта деятельность регулируется нервной системой. В продолговатом
мозге расположены центры вдоха и выдоха. Попеременные раздраже-
ния нейронов этих центров обусловливают ритмичные чередования
вдоха и выдоха. К дыхательным центрам постоянно поступают сигна-
лы о степени растяжения легких. Вдох и выдох запускаются по прин-
ципу отрицательной обратной связи.
Важную роль в регуляции дыхания (также по принципу обратной
связи) играют рН артериальной крови, напряжение в ней СО2 и О2.
Так, например, увеличение напряжения СО2 в артериальной крови
(гиперкапния) приводит к повышению минутного объема дыхания.
Как правило, при этом возрастают как дыхательный объем, так и ча-
стота дыхательных движений. Если снижается рН артериальной кро-
ви по сравнению с нормальным уровнем, вентиляция легких увеличи-
вается. Снижение напряжения О2 в артериальной крови (гипоксия)
сопровождается увеличением вентиляции легких. При этом газы кро-
ви и рН могут воздействовать на нейроны дыхательных центров как
непосредственно, так и путем возбуждения особых рецепторов — хе-
морецепторов, которые расположены в стенках некоторых крупных
сосудов (общей сонной артерии, дуги аорты).
Физическая активность приводит к увеличению вентиляции лег-
ких, т. к. сокращающиеся мышцы используют больше кислорода.
Кроме того, на дыхательные центры действуют сильные температур-
ные воздействия, температура тела, различные гормоны, боль.
МОЧЕПОЛОВОЙ АППАРАТ
Мочеполовой аппарат объединяет две системы органов, анатоми-
чески и физиологически различных, однако тесно связанных между
собой топографически и по своему происхождению (рис. 149, 150).
МОЧЕВЫЕ ОРГАНЫ
Почка человека и других млекопитающих имеет бобовидную фор-
му с закругленными верхним и нижним полюсами. Масса почки —120–
200 г. На вогнутом медиальном крае почки находится углубление — по-
чечные ворота, которые ведут в небольшую почечную пазуху. Это место
295
МОЧЕВЫЕ ОРГАНЫ
расположения нервов, кровеносных сосудов, почечной лоханки, чашек,
начала мочеточника и жировой ткани. Почки прилежат к задней стенке
брюшной полости и расположены вне брюшины.
После удаления содержимого пазухи на внутренней поверхности
почечной пазухи можно различить почечные сосочки. Число их колеб-
Рис. 149. Мочеполовой аппарат мужчины, вид спереди и справа:
1 — почка, 2 — корковое вещество почки, 3 — почечные пирамиды, 4 — почечная
лоханка, 5 — мочеточник, 6 — верхушка мочевого пузыря, 7 — срединная пупочная
связка, 8 — тело мочевого пузыря, 9 — тело полового члена, 10 — спинка полового
члена, 11 — головка полового члена, 12 — дольки яичка, 13 — яичко, 14 — придаток
яичка, 15 — семявыносящие протоки, 16 — корень полового члена, 17 — бульбоуре-
тральная железа, 18 — перепончатая часть мочеиспускательного канала, 19 — простата,
20 — семенной пузырек, 21 — ампула семявыносящего протока, 22 — дно мочевого
пузыря, 23 — почечные ворота, 24 — почечная артерия, 25 — почечная вена
11
11
11
22
12
12
33
13
13
44
14
14
55
15
15
66
16
16
77
17
17
88
18
18
99
19
19
10
10
20
20
21
21
22
22
23
23
24
24
25
25
296
МОЧЕПОЛОВОЙ АППАРАТ
Рис. 150. Мочеполовой аппарат женщины, вид спереди и справа:
1 — почка, 2 — мочеточник, 3 — дно матки, 4 — полость матки, 5 — тело матки,
6 — брыжейка маточной трубы, 7 — ампула маточной трубы, 8 — бахромки трубы,
9 — брыжейка матки (широкая связка матки), 10 — мочевой пузырь, 11 — слизистая
оболочка мочевого пузыря, 12 — устье мочеточника, 13 — ножка клитора, 14 — тело
клитора, 15 — головка клитора, 16 — наружное отверстие мочеиспускательного
канала (уретры), 17 — отверстие влагалища, 18 — большая железа преддверия
(бартолинова железа), 19 — луковица преддверия, 20 — женский мочеиспускательный
канал (женская уретра), 21 — влагалище, 22 — влагалищные складки, 23 — отверстие
матки, 24 — канал шейки матки, 26 — круглая связка матки, 26 — яичник,
27 — фолликул яичника, 28 — везикулярный привесок, 29 — придаток яичника
(надъяичник), 30 — трубные складки
11
11
11
21
21
22
12
12
22
22
33
13
13
23
23
44
14
14
24
24
55
15
15
25
25
66
16
16
26
26
77
17
17
27
27
88
18
18
28
28
99
19
19
29
29
10
10
20
20
30
30
297
МОЧЕВЫЕ ОРГАНЫ
лется от 5 до 15 (чаще 7 или 8). На вершине каждого сосочка от 10 до
20 и более сосочковых отверстий, с трудом различаемых невооружен-
ным глазом. Это устья мочевых канальцев, выразительно названных
старыми анатомами почечным, или «благословенным» ситом, в насто-
ящее время называемых решетчатым полем. Каждый сосочек обра-
щен внутрь полости малой почечной чашки. Иногда в одну чашку об-
ращены два или три сосочка, соединенных вместе; количество малых
чашек чаще всего 7—8 . Несколько малых составляют одну большую
чашку, которых у человека 2—3 . Большие чашки, сливаясь друг с дру-
гом, образуют одну общую полость — почечную лоханку, которая, по-
степенно суживаясь, переходит в мочеточник.
На фронтальном разрезе почки различают наружное, более свет-
лое, корковое и внутреннее, более темное, мозговое вещество. На све-
жих препаратах в корковом веществе видны две части: свернутая —
мелкие зерна и красные точки, представляющие собой почечные
тельца, а также радиальная исчерченность (лучистая часть) — это от-
ростки мозгового вещества, проникающие в корковое. У человека
мозговое вещество расположено в виде 7—10 пирамид, исчерченных
продольно благодаря наличию канальцев. Основание каждой пирами-
ды направлено к поверхности почки, а почечный сосочек — к лоханке.
Между пирамидами заходят прослойки коркового вещества, это по-
чечные столбы. Одна пирамида с прилежащим участком коркового
вещества образует одну почечную долю.
Невозможно понять строение и функцию почек, не зная их микро-
скопической структуры. Основной морфологической и функциональ-
ной единицей строения почки является нефрон (рис. 151).
ВНИМАНИЕ!
Нефрон — это почечное тельце и система канальцев, длина которых
в каждом нефроне 50–55 мм, а всех нефронов — около 100 км. В ка-
ждой почке более 1 млн нефронов, которые функционально связаны
с кровеносными сосудами.
Почечное тельце (мальпигиево) образовано клубочком капил-
ляров, окруженным капсулой клубочка, которая имеет форму двустен-
ной чаши. В эту чашу как бы вдавлен клубочек капилляра. Кровь по-
ступает в капилляры клубочка по приносящей артериоле, вытекает из
клубочка по выносящей артериоле. Иными словами, капиллярный
клубочек соединяет два артериальных сосуда, поэтому он назван «чу-
десной сетью». Выносящие артериолы вновь распадаются на капил-
298
МОЧЕПОЛОВОЙ АППАРАТ
ляры (вторичная капиллярная сеть), которые питают ткань почки
очищенной кровью и сокращаются в вены.
В течение суток через почки человека проходит около 1500 л крови.
Кровь, текущая в капиллярах клубочка, отделена от полости капсу-
лы лишь двумя слоями клеток фенестрированной капиллярной стенки
(цитоплазма эндотелиальных клеток, образующих стенку капилляров,
имеет множество пор и напоминает сито) и интимно сросшимся с ней
эпителием внутреннего листка капсулы. Из крови в просвет капилляра
через этот барьер поступают вещества мочи. Внутренний листок капсу-
лы образован эпителиальными клетками подоцитами, напоминающи-
ми спрутов. Это крупные клетки неправильной формы, имеющие не-
сколько больших широких отростков (цитотрабекулы), от которых
отходит множество мелких отростков цитоподий. Щели, разделяющие
цитоподии, соединяются с просветом капсулы.
В течение суток в просвет капсул фильтруется около 100 л первич-
ной мочи. Ее путь таков: кровь ␚ эндотелий капилляров ␚ базальная
мембрана ␚ щели между цитоподиями ␚ полость капсулы.
Рис. 151. Строение
и кровоснабжение нефрона, схема:
1 — почечное тельце,
2 — клубочек почечного тельца
(капилляры), 3 — капсула
клубочка (капсула Шумлянского –
Боумена), 4 — приносящая
клубочковая артериола,
5 — междольковая артерия,
6 — дуговая артерия, 7 — дуговая
вена, 8 — проксимальная часть
канальца нефрона, 9 — петля
нефрона, 10 — сосочковые
протоки, 11 — околоканальцевые
капилляры, 12 — дистальная
часть канальца нефрона,
13 — выносящая клубочковая
артериола
11
11
11
22
12
12
33
13
13
44
55
66
77
88
99
10
10
299
МОЧЕВЫЕ ОРГАНЫ
Проксимальный каналец нефрона, куда поступает моча из полости
капсулы, длиной около 14 мм и диаметром 50–60 мкм, образован од-
ним слоем высоких цилиндрических каемчатых клеток, на апикаль-
ной поверхности которых имеется щеточная каемка, состоящая из
множества микроворсинок. Около 85% натрия и воды, а также белок,
глюкоза, аминокислоты, кальций, фосфор из первичной мочи всасы-
ваются именно в проксимальных отделах. Проксимальный отдел пе-
реходит в тонкую нисходящую часть петли Генле (около 15 мкм в ди-
аметре). Через выстилающие ее плоские клетки всасывается вода;
восходящая часть — толстая (диаметр около 30 мкм), в ней происхо-
дит дальнейшая потеря натрия и накопление воды.
В коротком дистальном отделе происходит дальнейшее выделе-
ние натрия в тканевую жидкость и всасывание большого количества
воды. Процесс всасывания воды продолжается и в собирательных
трубочках. Всасывание воды в дистальной части и собирательных
трубочках регулируется антидиуретическим гормоном задней доли
гипофиза. В результате этого количество окончательной мочи по
сравнению с количеством первичной резко уменьшается (до 1,5 л
в сутки), в то же время возрастает концентрация веществ, не подвер-
гающихся обратному всасыванию.
Корковое вещество составляют почечные тельца и дистальные от-
делы нефронов. Мозговые лучи и мозговое вещество образованы пря-
мыми канальцами, мозговые лучи — нисходящими и восходящими
отделами петель корковых нефронов и начальными отделами собира-
тельных трубочек, а мозговое вещество почки — нисходящими и вос-
ходящими отделами и коленами петель нефронов, конечными отдела-
ми собирательных трубочек и сосочковыми протоками.
Почки являются не только органами выделения, но и своеобраз-
ной железой внутренней секреции. Между приносящей и выносящей
артериолами клубочка в зоне перехода восходящего колена петли
Генле в дистальный отдел находится участок, называемый плотным
пятном. В стенке канальца обнаруживается скопление множеств ядер.
В участках стенок приносящей и реже выносящей артериол, прилежа-
щих к плотному пятну, под эндотелиальными клетками находятся
особые, богатые гранулами юкстагломерулярные клетки, которые вы-
рабатывают белок — ренин, участвующий в регуляции кровяного дав-
ления, а также почечный эритропоэтический фактор, который стиму-
лирует образование эритроцитов.
Моча из сосочковых отверстий поступает в малые, затем в большие
почечные чашки и лоханку, переходящую в мочеточник. Стенки почеч-
ных чашек, лоханки, мочеточников и мочевого пузыря в основном по-
строены одинаково: они состоят из слизистой оболочки, покрытой пе-
реходным эпителием, мышечной и адвентициальной оболочек.
300
МОЧЕПОЛОВОЙ АППАРАТ
Мочеточники человека — цилиндрические трубки диаметром
6—8 мм — располагаются забрюшинно. Длина мочеточника взрослого
достигает 25—30 см, новорожденного — 5 —7 см. Они растут быстро,
и в течение первых двух лет их длина удваивается. У человека (как
и у других млекопитающих) мочеточники входят в мочевой пузырь,
косо прободая его стенку. Моча передвигается по мочеточникам бла-
годаря ритмическим перистальтическим сокращениям его толстой
мышечной оболочки, которая у детей развита слабо. Слизистая обо-
лочка мочеточника складчатая (поэтому его просвет на поперечном
разрезе имеет звездчатую форму) и выстлана переходным эпителием.
Мочевой пузырь взрослого человека лежит позади лобкового
симфиза. У новорожденных и детей первого года жизни его емкость
не превышает 50–80 см3, у взрослого — до 1 л. Основу стенки пузыря
составляют гладкие мышцы, которые располагаются в три слоя, пере-
плетающихся между собой, что способствует равномерному сокраще-
нию его стенок при мочеиспускании. Наиболее развит круговой слой,
который в области внутреннего отверстия мочеиспускательного кана-
ла образует сжиматель мочевого пузыря. Кроме него, у человека име-
ется поперечнополосатый наружный сжиматель мочеиспускательно-
го канала.
Слизистая оболочка при пустом пузыре складчатая, лежит на хо-
рошо развитой подслизистой основе. Слизистая оболочка выстлана
переходным эпителием. Клетки его поверхностного слоя в пустом мо-
чевом пузыре округлые, при наполнении пузыря и растяжении стен-
ки они уплощаются и истончаются. Однако эпителий остается непро-
ницаемым для мочи и надежно предохраняет мочевой пузырь от ее
всасывания. В нижней части пузыря расположено внутреннее отвер-
стие мочеиспускательного канала. Волокна внутреннего мышечного
слоя окружают устья мочеточников.
Моча накапливается в мочевом пузыре со скоростью около 50 мл
в 1 ч. Первые короткие позывы к мочеиспусканию появляются при
накоплении в пузыре 150—250 мл мочи, после накопления 250—
500 мл начинается его опорожнение. При наполненном пузыре возбу-
ждаются рецепторы растяжения, лежащие в его стенке. Это приводит
к активации парасимпатических крестцовых нейронов, иннервирую-
щих гладкие мышцы пузыря: они сокращаются, а под влиянием сим-
патических (внутренний сфинктер) и соматических (наружный сфин-
ктер) импульсов они расслабляются. Сокращение мышц брюшного
пресса также способствует увеличению давления внутри пузыря.
Способность контролировать мочеиспускание появляется в конце
первого года жизни и закрепляется в течение второго, приобретая
устойчивый характер к концу второго года.
301
ФУНКЦИЯ ПОЧЕК
Мочеиспускательный канал женщины представляет собой корот-
кую щелевидную трубку длиной 3—6 см, которая расположена позади
лобкового симфиза. Гладкие мышечные волокна стенки образуют два
слоя. Наружное отверстие находится в преддверии влагалища, впере-
ди и выше отверстия последнего и окружено поперечнополосатым на-
ружным сфинктером.
Мужской мочеиспускательный канал будет описан в разделе
«Мужские половые органы».
Функция почек
В течение суток человек потребляет примерно 2,5 л воды, в том чи-
сле 1500 мл в жидком виде и около 650 мл с твердой пищей. Кроме
того, в процессе распада белков, жиров и углеводов образуется еще
около 400 мл воды. Из организма вода выводится главным образом че-
рез почки 1,5 л в сутки, а также через легкие, кожу и частично с калом.
Почки очищают кровь от многих вредных веществ и выводят их
наружу. Так, например, с мочой выводятся конечные продукты обме-
на (мочевина, мочевая кислота, креатинин), многие лекарства, ионы
натрия, кальция, неорганический фосфат, вода. Так, содержание моче-
вины в плазме крови составляет 4,5 ммоль/л, в моче оно выше почти
в 70 раз (292—300 ммоль/л), содержание мочевой кислоты соответст-
венно 0,27 и 3 ммоль/л, содержание креатина соответственно 0,075
и 12 ммоль/л. Почки участвуют в поддержании кислотно-щелочного,
водного и электролитного состава, осмотического давления, постоян-
ства ионного состава и рН внутренней среды организма. Иными сло-
вами, почки поддерживают относительное постоянство состава крови
и жидкостей организма.
На деятельность почек влияют гормоны коры надпочечников (ми-
нералокортикоиды и глюкокортикоиды); антидиуретический гормон
(вазопрессин), выделяемый клетками гипоталамуса, он усиливает
обратное всасывание воды из первичной мочи в канальцах нефрона;
гормон паращитовидных желез и тиреокальцитонин.
Физические и химические свойства мочи
Моча представляет собой жидкость светло-желтого цвета. В моче
содержится 95% воды и 5% твердых веществ. Это мочевина (2%),
мочевая кислота (0,05%), креатинин (0,075%) и другие вещества,
в том числе соли калия, натрия. В течение суток из организма с мочой
выводится 25–30 г мочевины и до 25 г неорганических веществ. При
заболеваниях почек, при кратковременных физических нагрузках
в моче может появиться белок, которого в моче не должно быть. Реак-
302
МОЧЕПОЛОВОЙ АППАРАТ
ция мочи зависит от пищи. При употреблении преимущественно мяс-
ной пищи моча имеет кислую реакцию, при употреблении главным
образом растительной пищи реакция мочи щелочная.
Почки очищают кровь от многих вредных веществ и некоторых не-
обходимых для жизнедеятельности организма. Так, например, с мочой
выводятся конечные продукты обмена (мочевина, мочевая кислота,
креатинин), многие лекарства, ионы натрия, кальция, неорганический
фосфат, вода. Почки участвуют в поддержании кислотно-щелочного,
водного и электролитного состава, осмотического давления, постоянст-
ва ионного состава и рН внутренней среды организма. Иными словами,
почки поддерживают относительное постоянство состава крови и жид-
костей организма. Сказанное иллюстрирует табл. 21.
Таблица 21
Содержание некоторых веществ в плазме и моче
(по О. Гарту)
Вещество
Содержание, ммоль/л
Плазма
Моча
Na+
142
128
К+
4,5
54
Cl–
103
134
Глюкоза
5
Следы
Мочевина
4,5
292
Мочевая кислота
0,27
3,2
Креатинин
0,075
12
На деятельность почек влияют гормоны коры надпочечников (ми-
нералокортикоиды и глюкокортикоиды); антидиуретический гормон
(вазопрессин), выделяемый клетками гипоталамуса, он усиливает
обратное всасывание воды из первичной мочи в канальцах нефрона;
гормон паращитовидных желез и тиреокальцитонин.
Многие факторы влияют на мочеобразование и мочевыделение:
количество выпитой жидкости, содержание в пище поваренной соли,
температура окружающей среды, потоотделение, физическая актив-
ность. Чай, особенно зеленый, стимулирует образование мочи и ее
выделение, способствует улучшению функции почек, предотвращает
возникновение конкрементов.
303
БИОЛОГИЧЕСКАЯ И СОЦИАЛЬНАЯ СУЩНОСТЬ ПОЛА ЧЕЛОВЕКА
ПОЛОВЫЕ СИСТЕМЫ
БИОЛОГИЧЕСКАЯ И СОЦИАЛЬНАЯ СУЩНОСТЬ
ПОЛА ЧЕЛОВЕКА
Пол человека — сложное понятие, которое включает в себя: собст-
венно биологический пол (генетический, гонадный, гаметный и гормо-
нальный); род (психологические и культурные характеристики инди-
видуума); ядро родовой сущности (убежденность человека в его
принадлежности именно к этому, а не другому полу) складывается в ран-
нем детстве (до двух лет) и сохраняется в течение всей жизни человека.
Генетический пол зависит от наличия или отсутствия Y-хромосомы.
Напомним: во всех соматических клетках человека имеется по 46 хро-
мосом (диплоидный набор), среди которых 44 соматические и две по-
ловые: клетки женского организма имеют две половые Х-хромосомы,
мужского — одну X- и одну Y-хромосому Половые клетки имеют по
одной половой хромосоме: яйцеклетки только X, сперматозоиды X
или Y. Именно Y-хромосома, которой обладают только мужчины, на-
правляет в эмбриональном периоде развитие индифферентных поло-
вых желез по мужскому пути, превращая их в яички. Генетический
пол определяет истинный, или гонадный, пол (от греч. gone — семя),
т. е . пол, обусловленный строением половой железы (яичка или яич-
ника). В свою очередь, истинный пол представляет гаметный пол, т. е .
способность желез образовывать сперматозоиды или яйцеклетки,
и гормональный пол — способность вырабатывать мужские (андроге-
ны) или женские (эстрогены) половые гормоны. Половые гормоны
определяют морфологический пол, т. е. развитие мужских или жен-
ских половых органов и вторичных половых признаков. Они же (по-
ловые гормоны) определяют мозговой, или церебральный, пол, т. е .
половые различия головного мозга. Гражданский пол — это пол, офи-
циально зарегистрированный при рождении. Каждый человек ощу-
щает свою половую принадлежность, половую самоидентификацию,
идентичность. Гендер (синоним понятия «пол») подчиняется законо-
мерности осознания человеком своей сущности вообще и включает
в себя положительное свойство принадлежности к определенной
мужской или женской половине человечества и отрицательное свойст-
во исключения из той или иной группы. Гендерная роль — это внешнее
проявление и демонстрация поведения, которое позволяет окружаю-
щим судить о тендерной идентичности. Иными словами, гендерная
роль — это социальное выражение гендерной идентичности. Генетиче-
ский и гормональный пол определяет биологическую, психологическую
и социальную жизнь человека, его половое воспитание, половое само-
сознание, половое поведение, половую роль и выбор полового партнера.
304
ПОЛОВЫЕ СИСТЕМЫ
В организме и мужчин и женщин вырабатываются одни и те же по-
ловые гормоны: мужские андрогены (тестостерон) и женские (эстроге-
ны и прогестерон), но у мужчин преобладают первые (их уровень
в шесть раз выше, чем у женщин), а у женщин — вторые.
В таблице 22 приведены основные признаки пола человека.
Таблица 22
Пол человека
Пол
Признаки
Характеристика
Мужской пол
Женский пол
Генетический
Хромосомы
XY
XX
Гонадный
Половая железа
Яичко (семенник) Яичник
Гаметный
Половая клетка
Сперматозоид
Яйцеклетка
Гормональный
Гормоны
Андрогены
Эстрогены
Морфологический
(телесный)
Морфологические
Длина тела
Больше
Меньше
Масса тела
Больше
Меньше
Туловище
Короче
Длиннее
Конечности
Длиннее
Короче
Плечи
Шире
У
же
Таз
У
же
Шире
Грудная клетка
Длиннее, шире
Короче, у
же
Живот
Короче
Длиннее
Масса мышц
Больше
Меньше
Подкожная жиро-
вая клетчатка
Меньше
Больше
Кожа
Толще
Тоньше
Волосы
Больше на лице,
туловище, конечно-
стях, обильные на
лобке и животе до
пупка
На лице отсутству-
ют, меньше на ту-
ловище, отсутству-
ют на животе
Молочные железы
Не развиты
Развиты
Церебральный
(мозговой)
Половой димор-
физм головного
мозга
Гражданский
Официальная ре-
гистрация при ро-
ждении
Мужчина
Женщина
305
ВНУТРЕННИЕ МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ
Имеются определенные половые различия головного мозга, кото-
рые также обусловлены действием половых гормонов, в частности те-
стостерона во внеутробном периоде. Тестостерон влияет на скорость
роста развивающегося мозга и ответствен за различия в строении
мозга женщин и мужчин.
МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ
Мужские половые органы состоят из половой железы — яичка
с его оболочками и придатками, расположенными в мошонке, семя-
выносящих путей, вспомогательных половых желез и полового члена.
ВНУТРЕННИЕ МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ
Мужские половые железы человека (яички) выполняют две важ-
нейшие функции: в них образуются сперматозоиды и половые гормо-
ны, влияющие на развитие первичных и вторичных половых призна-
ков. Железа яйцевидной формы, плотная, размерами около 3×4×2 см.
До периода полового созревания яички и придатки развиваются мед-
ленно, затем рост их резко ускоряется. Так, у новорожденного маль-
чика масса яичка около 0,2 г, у годовалого — 1, в 14 лет — 2, в 15–
16лет—8,аувзрослого—15—25г.
Яичко покрыто плотной соединительнотканной белочной оболоч-
кой. От нее внутрь органа радиально отходят перегородочки, которые
своими противоположными краями прикрепляются к утолщению
оболочки в области заднего края яичка — средостению. Перегородоч-
ки делят яичко на множество долек (100—300), в которых располага-
Рис. 152. Схема строения яичка
и его придатка:
1 — семявыносящий проток,
2 — выносящие канальцы яичка,
3 — проток придатка, 4 — средостение
яичка, 5 — сеть яичка, 6 — прямые
семенные канальцы, 7–9
—
сообщения
между семенными канальцами
различных долек,
10 — белочная оболочка,
11 — долька придатка яичка
(по И.В. Алмазову и Л.С . Сутулову)
7
8
9
10
11
4
3
2
5
6
1
306
МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ
ется по одному-два извитых семенных канальца (рис. 152). Длина
каждого канальца 50—80 см. Общая длина всех канальцев одного яич-
ка достигает 300—400 м. У половозрелого мужчины стенка извитого
семенного канальца яичка выстлана слоем сперматогенного эпителия,
лежащего на базальной мембране. Эпителиосперматогенный слой со-
стоит из сперматогенных клеток, находящихся на разных стадиях раз-
вития, и поддерживающих эпителиоцитов (сустентоцитов, или кле-
ток Сертоли). Соседние клетки Сертоли соединены между собой
плотными контактами, благодаря чему сперматогенные клетки распо-
лагаются в двух ярусах. В наружном базальном слое залегают сперма-
тогонии. Близкое расположение их возле кровеносных капилляров
обеспечивает благоприятное поступление питательных веществ. Во
внутреннем адлюминальном слое находятся сперматоциты, спермати-
ды и сперматозоиды. Клетки Сертоли выполняют трофическую функ-
цию по отношению к сперматогенным элементам адлюминального
слоя и могут выполнять функции фагоцитов. Клетки Сертоли вместе
с другими структурами стенки канальца формируют гематотестику-
лярный барьер, препятствующий проникновению токсических ве-
ществ и антител из крови к сперматогенному эпителию, тем самым
способствуя сохранению микроокружения развивающихся половых
клеток. Являясь микроокружением сперматогенных клеток, клетки
Сертоли участвуют в сперматогенезе. Под влиянием фолликулости-
мулирующего гормона гипофиза клетки Сертоли синтезируют андро-
генсвязывающий белок (АСБ), который переносит мужские половые
гормоны к сперматогенным клеткам. Периодически контакты гемато-
тестикулярного барьера открываются, пропуская дифференцирую-
щие клетки в адлюминальную область.
В извитых семенных канальцах постоянно происходит образова-
ние огромного количества мужских половых клеток — сперматозои-
дов. У здорового взрослого мужчины в 1 мл3 спермы содержится око-
ло 100 млн сперматозоидов, а во время одного семяизвержения
выделяется 300—400 млн.
Вблизи средостения канальцы постепенно выпрямляются, перехо-
дят в прямые и впадают в сеть яичка, расположенную в его средосте-
нии. Канальцы сети открываются в 15—20 выносящих канальцев яич-
ка, которые прободают белочную оболочку и, извиваясь, входят
в придаток яичка.
Сперматозоид человека имеет головку, шейку и хвост (рис. 153).
Головка яйцевидной формы содержит ядро, обладающее, как и ядро
яйцеклетки, одним (гаплоидным) набором хромосом (23 хромосомы).
На переднем полюсе головки под плазматической мембраной распо-
ложена акросома. Содержащиеся в ней ферменты при оплодотворе-
нии растворяют плотную оболочку яйцеклетки и способствуют про-
307
ВНУТРЕННИЕ МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ
никновению сперматозоида в яйцеклетку. В шейке сперматозоида
расположено множество митохондрий, обеспечивающих сперматозо-
ид энергией для движения.
В яичке имеется еще одна разновидность клеток — интерстици-
альные эндокриноциты (клетки Лейдига). Это крупные клетки, кото-
рые располагаются между семенными канальцами вблизи капилля-
ров. В настоящее время считают, что клетки Лейдига активно
синтезируют мужской половой гормон — тестостерон (андроген), ко-
торый оказывает разностороннее действие на различные чувствитель-
ные к нему клетки мужского организма, стимулируя их рост и функ-
циональную активность. К ним относятся клетки простаты, семенных
пузырьков, желез крайней плоти, почек, кожи и др. Под влиянием ан-
дрогенов во внутриутробном периоде происходит дифференцировка
и последующее развитие половых органов, а затем половое созрева-
ние и возникновение вторичных половых признаков. Мужские поло-
вые гормоны влияют на половое поведение (либидо и потенцию). Ан-
дрогены стимулируют синтез белка и ускоряют рост тканей. Очень
важным является их воздействие на сперматогенез: низкая концент-
рация гормона активирует этот процесс, высокая — тормозит. Следу-
ет указать, что в яичках синтезируется и небольшое количество жен-
ских половых гормонов — эстрогенов. Андрогены и эстрогены вместе
с другими гормонами регулируют рост и развитие опорно-двигатель-
ного аппарата, они в определенном возрасте останавливают рост ко-
стей в длину.
Рис. 153. Строение сперматозоида:
I — головка, II — шейка, III — хвост; 1 — акросома, 2, 7 — цитолемма, 3 — ядро,
4 — проксимальная центриоль, 5 — митохондрии, 6 — дистальная центриоль,
8 — осевая нить (по В. Г. Елисееву и др.)
11
I
22
II
33
III
44
55
66
77
88
308
МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ
По мере старения мужчины в стенках извитых семенных каналь-
цев уменьшается количество половых клеток и увеличивается содер-
жание соединительной ткани, которая образует плотные оболочки
вокруг запустевших канальцев. Однако высокая сексуальная актив-
ность замедляет и тормозит этот процесс.
К яичку по заднему краю плотно прирастает придаток яичка, ко-
торый представляет собой систему канальцев, заполненных созрев-
шими сперматозоидами. Выносящие канальцы яичка, извиваясь, на-
правляются из сети яичка к придатку, образуя его головку. Каждый
выносящий каналец формирует дольку придатка. Все выносящие ка-
нальцы впадают в единственный очень длинный штопорообразно за-
крученный проток придатка. Он достигает 4—6 м в длину, его диаметр
около 5 мм. Придаток является не только хранилищем сперматозои-
дов, здесь они становятся способными к оплодотворению. Проток
придатка переходит в семявыносящий проток, который входит в со-
став семенного канатика.
Семявыносящий проток проходит через паховый канал, а далее по
боковой стенке таза вниз и назад, направляясь ко дну мочевого пузы-
ря, где оба протока сближаются. Благодаря мощной мышечной обо-
лочке семявыносящего протока его легко прощупать в паховом кана-
ле. Конечный отдел семявыносящего протока у человека расширяется,
образуя ампулу. В конечном отделе каждого семявыносящего протока
его стенка как бы выпячивается, образуя боковые выросты, семенные
пузырьки размерами 5×2×1 см. Их складчатая слизистая оболочка
вырабатывает густой желтоватый секрет, который смешивается со
спермой и разжижает ее, питает и активирует сперматозоиды. За-
остренный выделительный проток каждого семенного пузырька сое-
диняется с конечным отделом семявыносящего протока и образует
семявыбрасывающий проток длиной около 2 см, который прободает
простату и открывается в предстательную часть мужского мочеиспу-
скательного канала узким отверстием, расположенным у основания
семенного холмика.
Простата
Форма простаты напоминает каштан, своим основанием связан-
ный с мочевым пузырем, масса простаты взрослого мужчины — 18–
22 г. Простата развивается параллельно с яичками. Кастрация приво-
дит к атрофии простаты. Простата — это железисто-мышечный орган,
железистое вещество которого состоит из 30–60 простатических же-
лез. Пучки гладких мышечных клеток образуют вместе с прослойка-
ми соединительной ткани широкие толстые перегородки, отделяющие
309
ВНУТРЕННИЕ МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ
друг от друга простатические железы. Простата окружает начальную
часть мочеиспускательного канала (рис. 154).
Устья многочисленных проточков простатических желез открыва-
ются в мочеиспускательный канал. Эпителий желез и их проточков
вырабатывает жидкий беловатый секрет. Сокращение мышечного ап-
парата в момент семяизвержения (эякуляции) способствует выбрасы-
ванию секрета из простатических желез в мочеиспускательный канал.
Простата обладает эндокринной функцией. Она вырабатывает про-
стагландины и другие биологически активные вещества. Простата —
очень важный орган. Старые анатомы, подчеркивая ее роль, называли
простату «вторым сердцем мужчины».
Бульбоуретральные железы (куперовы) — парные округлые желе-
зы величиной с горошину каждая, расположенные между пучками
мышц мочеполовой диафрагмы. Выводной проток железы очень тон-
кий, длиной около 3–4 см, открывается в просвет мочеиспускательного
канала. Железы вырабатывают вязкий секрет, который предохраняет
слизистую оболочку мочеиспускательного канала от раздражающего
действия мочи.
Сперма — густая, беловатая или сероватая вязкая жидкость со
специфическим запахом, напоминающим запах свежих каштанов. Ве-
щества, секретируемые простатой, куперовыми железами и семенны-
ми пузырьками, добавляются к сперме во время ее продвижения по
Рис. 154. Конечные отделы
(ампулы) семявыносящих протоков,
семенные пузырьки и простата,
вид спереди
(ампулы семявыносящих протоков
и семенные пузырьки вскрыты
фронтальным (продольным)
разрезом. Передняя часть
простаты удалена, вскрыта пред-
стательная часть
мочеиспускательного канала):
1 — семявыносящий проток,
2 — семенной пузырек,
3 — выделительный проток,
4 — семявыбрасывающий проток,
5 — предстательная часть
мочеиспускательного канала,
6 — простата,
7 — предстательная маточка,
8 — семенной холмик,
9 — перепончатая часть
мочеиспускательного канала,
10 — устье семявыбрасывающего
протока, 11 — ампула
семявыносящего протока
11
11
11
22
33
44
55
66
77
88
99
10
10
310
МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ
семявыбрасывающему протоку и мочеиспускательному каналу, раз-
жижают ее, повышают жизнеспособность сперматозоидов и активи-
зируют их.
В состав спермы входят вода, слизь, сахар (фруктоза), основания,
огромное количество биологически активных веществ, включая про-
стагландины. Последние вызывают сокращение гладких мышц матки
и маточных труб. Около 30% семенной жидкости — это секрет проста-
ты, около 65—70% — семенных пузырьков. Во время одного семяиз-
вержения выбрасывается 3—5 мл спермы.
Семенной канатик представляет собой мягкий шнур длиной 15—
20 см, расположенный в паховом канале и начинающийся от верхнего
конца яичка. Он как бы подвешивает яичко. Канатик образован семя-
выносящим протоком, артериями и венами яичка и протока, лимфа-
тическими сосудами, нервными сплетениями, рудиментом влага-
лищного отростка брюшины, гладкими мышечными клетками и
соединительной тканью.
НАРУЖНЫЕ МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ
Мошонка — это отвисающее книзу выпячивание брюшной стенки,
расположенное между корнем полового члена и промежностью.
У здорового мужчины мошонка благодаря наличию гладких мышц
сокращена и приподнята. При половом возбуждении, понижении окру-
жающей температуры сокращение усиливается, и яички приподнима-
ются. При повышении температуры тела мошонка расслабляется. Это
один из первых признаков повышенной температуры у мужчины. Мо-
шонка — это как бы «физиологический термостат», поддерживающий
температуру яичек более низкой, чем температура тела, что является
необходимым условием нормального сперматогенеза.
Мужской половой член (penis, fallos) с глубокой древности явля-
ется символом силы, превосходства, господства. Во всех архаических
культурах фаллическая символика играла важную роль. И в раннем
детстве ребенок придает огромное значение половому члену. Ребенок
«приписывает всем людям, в том числе и женщинам, наличие пениса»
(З. Фрейд). Более того, у многих девочек возникает желание иметь
пенис, у мальчиков — страх его потерять («угроза кастрации»).
Половой член выполняет две функции. Он служит для выведения
мочи и для совокупления (введения в женское влагалище). Его ко-
рень прикреплен к лобковым костям и скрыт под кожей; подвижная
часть — тело — оканчивается утолщенной головкой, на вершине кото-
рой располагается наружное отверстие мочеиспускательного канала
(рис. 155). Тонкая, нежная, подвижная, растяжимая кожа полового
члена лежит на лишенной жировых клеток подкожной клетчатке, ко-
311
НАРУЖНЫЕ МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ
торая отсутствует в области го-
ловки. У основания головки
кожа образует циркулярную
свободную складку — крайнюю
плоть, скрывающую головку.
Уздечка, расположенная на
нижней поверхности головки,
соединяет крайнюю плоть с ко-
жей головки. Между крайней
плотью и кожей головки име-
ется небольшое пространство
(мешок), куда выделяется секрет многочисленных желез крайней
плоти, образующих беловатую смегму. Это пространство открывается
отверстием, через которое при отодвигании крайней плоти проходит
головка полового члена.
Половой член сформирован двумя пещеристыми и одним губча-
тым телами. Два пещеристых тела полового члена цилиндрической
формы с несколько заостренными концами прикрепляются к нижним
ветвям лобковых костей. Оба тела сходятся под лобковым симфизом
и далее срастаются, образуя на нижней поверхности желобок, где за-
легает губчатое тело полового члена, заканчивающееся впереди го-
ловкой, задний конец губчатого тела образует луковицу, расположен-
ную в толще мышц промежности.
Пещеристые и губчатое тела покрыты лишенной мышечных воло-
кон плотной соединительнотканной белочной оболочкой, которая от-
сутствует лишь на головке полового члена. От внутренней поверхно-
сти оболочки отходят отростки (трабекулы), которые сформированы
Рис. 155. Строение полового члена:
1 — головка полового члена, 2 — наружное
отверстие мочеиспускательного канала,
3 — венец головки, 4 — губчатое тело
полового члена, 5 — луковица полового
члена, 6 — седалищный бугор,
7 — артерия и вена полового члена,
8 — глубокая поперечная мышца,
9 — бульбоуретральная железа,
10 — сфинктер мочеиспускательного
канала, 11 — нижняя фасция мочеполовой
диафрагмы, 12 — ножка полового члена,
13 — пещеристые тела полового члена
(покрыты фасцией)
1
о
ка
д
11
11
11
22
12
12
33
13
13
44
55
66
77
88
99
10
10
312
МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ
плотной волокнистой соединительной тканью, содержащей множест-
во гладких мышечных клеток и эластических волокон. Трабекулы
разветвляются в ткани губчатого и пещеристых тел и переплетаются
между собой. Между ними образуется система ячеек (лакун, каверн),
которые представляют собой широкие кровеносные капилляры.
Головка полового члена образована плотной волокнистой соеди-
нительной тканью, обильно пронизанной сетью соединяющихся меж-
ду собой вен. Кожа полового члена, слизистая оболочка мочеиспуска-
тельного канала, белочная оболочка и трабекулы очень богаты
чувствительными нервными окончаниями.
МЕХАНИЗМ ЭРЕКЦИИ
Кровь к пещеристым телам доставляет в основном глубокая арте-
рия полового члена, которая распадается на ветви, идущие по трабеку-
лам. При спокойном состоянии полового члена они извитые, что дало
повод называть их завитковыми или улитковыми. Артерии открывают-
ся непосредственно в ячейки (каверны). Просвет этих артерий широ-
кий, а их стенки имеют толстую мышечную оболочку, помимо этого,
внутренняя оболочка артерий утолщена за счет добавочных пучков
гладких мышечных волокон, которые закрывают просвет при сокраще-
нии сосудистой стенки. В стенках вен также хорошо развит мышечный
слой. Основную роль в эрекции играют гладкие мышечные клетки ар-
терий, артериол и синусоидных капилляров. В покое при отсутствии
эрогенной стимуляции гладкие мышцы сокращены, поэтому стенки за-
витых артерий полового члена умеренно сокращены, и сами артерии
извитые. Межкавернозные вены и вены, проходящие под белочной
оболочкой пещеристых тел, открыты, и кровь легко оттекает по ним.
В результате психогенных влияний, идущих из коры головного мозга
в спинной, а также рефлекторных — вследствие раздражения чувстви-
тельных нервных окончаний, расположенных в коже половых органов
и эрогенных зон, возбуждаются парасимпатические нейроны крестцо-
вого отдела спинного мозга (центр эрекции). Нервный импульс, возни-
кающий в них, передается по тазовым нервам к половым органам. В ре-
зультате этого расслабляются гладкие мышцы трабекул пещеристых
тел полового члена и губчатого тела и завитковых артерий. Последние
выпрямляются, и кровь устремляется в ячейки (каверны) — они рас-
ширяются. Во время эрекции благодаря кровенаполнению пещер стен-
ки вен сдавлены, что препятствует оттоку крови из сосудистых поло-
стей. Эрекция усиливается благодаря одновременному рефлекторному
сокращению седалищно-пещеристых мышц и луковично-губчатых
мышц (это мышцы мочеполовой диафрагмы; первые начинаются от
ветвей обеих седалищных костей и вплетаются в белочную оболочку
313
ВНУТРЕННИЕ ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
пещеристых тел полового члена, вторые начинаются от луковицы по-
лового члена, окружая ее и губчатое тело, и вплетаются в его белочную
оболочку). При эрекции резко увеличиваются размеры полового члена,
он выпрямляется, становится плотным.
Мужской мочеиспускательный канал — это узкая трубка длиной
у новорожденного 5—6 см, у взрослого 16—22 см. В нем различают три
части: предстательную, прободающую простату; самую короткую пере-
пончатую, проходящую через диафрагму таза; и самую длинную губча-
тую, залегающую в губчатом теле полового члена. На задней стенке
предстательной части мочеиспускательного канала расположен неболь-
шой продолговатый гребень, выступающий в просвет канала, вершина
его образует семенной холмик, по бокам от которого открываются устья
семявыбрасывающих протоков. Семенной холмик образован пещери-
стой тканью, богатой гладкими мышечными клетками. В семенном хол-
мике имеется огромное количество нервных волокон и их чувствитель-
ных окончаний, которые являются точками наибольшей половой
чувствительности. Семенной холмик при эрекции набухает, увеличива-
ется, что препятствует затеканию спермы в мочевой пузырь. Вокруг
предстательной части мочеиспускательного канала поперечнополосатые
мышцы мочеполовой диафрагмы образуют его произвольный сфинктер.
Быстрый рост мужского полового члена (как и других половых
органов) происходит в период полового созревания.
ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
Женские половые органы, как и мужские, подразделяются на вну-
тренние (яичники, маточные трубы, матка и влагалище), расположен-
ные в полости малого таза, и наружные (женская половая область
и клитор), видимые снаружи (см. рис. 150).
ВНУТРЕННИЕ ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
Яичник — важнейшая парная железа, которая подобно яичку вы-
полняет две функции: образование яйцеклеток и выработку женских
половых гормонов, которые выделяются в кровь. Яичник, располо-
женный непосредственно ниже входа в малый таз, имеет овальную
форму. У новорожденной девочки масса яичника не превышает 0,15 г,
у нерожавшей женщины его масса 5—6 г, в возрасте 40—50 лет начи-
нается атрофия яичников, их масса уменьшается почти в два раза.
Один край яичника свободный, другой прикреплен к брыжейке.
Здесь в орган входят сосуды и нервы, поэтому он называется ворота-
ми яичника. Яичник покрыт соединительнотканной оболочкой, под
314
ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
ней располагается корковое вещество, состоящее из плотной волок-
нистой соединительной ткани, в которой находятся многочисленные
фолликулы — первичные (яйцеклетка — овогония, окруженная одним
слоем яичниковых фолликулоцитов), растущие (созревающие), атре-
тические, а также желтые тела и рубцы. Мозговая часть яичника обра-
зована соединительной тканью, в которой проходят сосуды и нервы.
В эмбриональном периоде первичные половые клетки мигрируют
в формирующуюся половую железу, где они превращаются в овогонии.
В отличие от мужских половых клеток, размножение женских проис-
ходит во внутриутробном периоде. Образуются первичные (приморди-
альные) фолликулы — овоцит первого порядка — яйцеклетка, окружен-
ная одним слоем фолликулярных клеток. У новорожденной девочки в
обоих яичниках имеется до 2 млн женских половых клеток. Количест-
во яйцеклеток после рождения не только не увеличивается, но быстро
уменьшается благодаря рассасыванию. До рождения девочки неболь-
шое количество примордиальных фолликулов растет, фолликулярные
клетки размножаются, эти фолликулы называются развивающимися.
Ко времени наступления половой зрелости в корковом веществе сохра-
няется около 300 000 первичных яйцеклеток. С наступлением половой
зрелости раз в месяц начинается созревание одного овоцита первого
порядка, которое заканчивается образованием зрелой гаплоидной яй-
цеклетки. Развивающиеся фолликулы преобразуются в зрелые пузыр-
чатые фолликулы яичника (граафовы пузырьки) в результате сложных
процессов овогенеза, которые происходят циклически каждые 28 дней.
При этом первичный фолликул растет, клетки фолликулярного эпите-
лия интенсивно размножаются, располагаются в виде многих слоев, во-
круг фолликула развивается соединительнотканная оболочка (тека),
клетки начинают вырабатывать жидкость фолликула, содержащую
гормоны эстрогена, которая раздвигает их. Одновременно растет и яй-
цеклетка, вокруг нее образуется блестящая оболочка (рис. 156).
Яйцеклетка (овоцит, ооцит) человека (и других млекопитающих)
относится к маложелтковым с равномерным распределением желточ-
ных включений. Яйцеклетка покрыта блестящей оболочкой, которая
окружена слоем питающих их фолликулярных клеток, вырабатываю-
щих женские половые гормоны, выполняющих по отношению к ово-
цит утрофическую, защитную и барьерную функции. Яйцеклетка,
окруженная слоем фолликулярных клеток, оттесняется к одному из
полюсов фолликула (яйценосный холмик), в связи с накоплением
жидкости образуется быстро увеличивающаяся в размерах полость —
граафов пузырек, стенка которого из-за избытка жидкости разрывает-
ся, и яйцеклетка, окруженная блестящей оболочкой и тремя-четырь-
мя тысячами фолликулярных клеток, выходит в свободную брюшную
полость, откуда попадает в маточную трубу, где и созревает.
315
ВНУТРЕННИЕ ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
В полость лопнувшего пузырька изливается кровь, сгусток крови
быстро замещается соединительной тканью, здесь развивается жел-
тое тело. Клетки фолликулярного эпителия размножаются, в них на-
капливается пигмент. Они превращаются в лютеоциты, продуцирую-
щие гормон прогестерон. Если яйцеклетка не была оплодотворена,
желтое тело функционирует 12—14 дней. Оно называется цикличе-
ским (менструальным) желтым телом. Если наступает беременность,
желтое тело сохраняется в течение всей беременности (желтое тело
беременности). Как только прекращается функционирование желтого
тела (менструального или беременности), оно атрофируется, в нем
разрастается соединительная ткань. На месте желтого тела остается
соединительнотканный рубец — беловатое тело. Итак, в течение при-
мерно 40 лет половой зрелости у женщины созревает и выделяется
около 400—500 яйцеклеток. Все прочие дегенерируют.
В яичниках образуются женские половые гормоны: эстрогены
и прогестерон, а также небольшое количество мужских половых
гормонов — андрогенов. Эстрогены обеспечивают развитие наруж-
ных женских половых органов, вторичных половых признаков, рост
Рис. 156. Строение пузырчатого (везикулярного) фолликула яичника
(граафова пузырька):
1 — наружная оболочка фолликула, 2 — внутренняя оболочка фолликула, 3 — полость
фолликула с фолликулярной жидкостью, 4 — яйценосный холмик, 5 — яйцеклетка,
6 — блестящая оболочка, 7 — лучистый венец, 8 — фолликулярные клетки
11
22
33
44
55
66
77
88
316
ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
и развитие опорно-двигательного аппарата, развитие тела по женско-
му типу, влияют на психику и поведение. Прогестерон оказывает вли-
яние на слизистую оболочку матки, готовя ее к внедрению (имплан-
тации) оплодотворенного яйца, росту и развитию плода, развитию
плаценты, молочных желез.
Маточная труба — парная, цилиндрической формы, расположена
в верхнем крае широкой связки матки. Длина ее у половозрелой жен-
щины 8—18 см, диаметр просвета 2—4 мм. В трубе различают четыре
части: маточную, которая проходит через стенку матки и открывается
в полость маточным отверстием; короткий перешеек, лежащий вбли-
зи матки; длинную ампулу и ее расширенную воронку, открывающу-
юся в брюшную полость вблизи яичника — брюшное отверстие. По-
следнее ограничено бахромкой трубы, одна из которых — яичниковая
бахромка — длиннее других.
Стенка трубы построена из складчатой слизистой оболочки, мышеч-
ной оболочки, состоящей из кругового и продольного слоев, и серозной
оболочки. Благодаря перистальтическим сокращениям мышечной обо-
лочки и движению ресничек эпителиальных клеток, выстилающих прос-
вет, яйцеклетка передвигается по трубе. Оплодотворение яйцеклетки
происходит в маточной трубе, откуда она переходит в полость матки.
Матка
Матка — полый толстостенный орган грушевидной формы. Узкая
полость матки, примерно треугольной формы, вверху сообщается с
трубами, а внизу через канал шейки матки — с влагалищем. Масса
матки новорожденной девочки 2—2,5 г, у нерожавшей женщины
40—50 г, у много раз рожавшей — в 1,5—2 раза больше. Интенсивный
рост матки начинается в начале периода полового созревания.
Стенка матки состоит из трех слоев: эндометрия (слизистая обо-
лочка), миометрия (мышечная оболочка) и периметрия (серозная
оболочка, или брюшина). Листки брюшины, покрывающие матку со
всех сторон, переходят в правую и левую широкие связки матки.
Шейка матки, кроме того, окружена околоматочной клетчаткой — па-
раметрием. Слизистая оболочка (эндометрий) в межменструальном
периоде гладкая, не имеет складок и непосредственно сращена с мыш-
цами. Она покрыта однослойным цилиндрическим эпителием, богата
капиллярными сетями и простыми трубчатыми маточными железа-
ми, пронизывающими всю толщину слизистой. В слизистой оболочке
различают два слоя: толстый поверхностный функциональный, кото-
рый отторгается во время менструации, и глубокий — базальный.
Миометрий образован гладкой мышечной тканью, в нем залегают
мощные венозные сплетения. При беременности гладкие мышечные
317
НАРУЖНЫЕ ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
клетки гипертрофируются, их размеры увеличиваются в 5–10 раз
в длину и в 3—4 раза в ширину, соответственно увеличиваются и раз-
меры матки. Резко возрастает количество кровеносных сосудов и ка-
пилляров. Непосредственно после родов масса матки достигает
1–1,5 кг, постепенно происходит ее обратное развитие, которое закан-
чивается через 6—8 недель после родов. Кровоснабжение матки
обильное, артерии мышечной оболочки спирально закручены, от них
отходят артерии, направляющиеся в эндометрий, где они разветвля-
ются на спиральные, питающие поверхностный функциональный
слой, и прямые, питающие базальный слой; и те и другие разветвля-
ются на артериолы и затем на капилляры.
Влагалище
Влагалище представляет собой уплощенную трубку длиной 7–9 см,
которая соединяет полость матки с наружными половыми органами
женщины. Влагалище направлено косо вверх примерно под углом
в 45°. Наружное отверстие влагалища открывается в его преддверие.
У девственниц оно закрыто девственной плевой, которая является
складкой слизистой оболочки, обычно кольцевидной формы. Стенка
влагалища состоит из складчатой слизистой оболочки, покрытой
неороговевающим многослойным эпителием и лишенной желез; мы-
шечной (круговые пучки вплетаются в продольный слой) и соедини-
тельнотканной адвентициальной оболочек. В эпителии влагалища
происходят циклические изменения в соответствии с фазами овариаль-
но-менструального цикла. Бо
льшая часть слизистой оболочки влагали-
ща относительно слабо иннервирована и имеет мало чувствительных
окончаний. Лишь вход во влагалище обладает огромным количеством
нервных окончаний. Клетки поверхностного слоя эпителия богаты
гликогеном, который у здоровой женщины под влиянием обитающих
во влагалище микробов распадается с образованием молочной кисло-
ты. Это придает влагалищной слизи кислую реакцию и обусловливает
ее бактерицидность по отношению к патогенным микробам.
Стенки влагалища весьма эластичны, они легко расслабляются и
сокращаются, поэтому полость меняет размеры и форму. Во время ро-
дов влагалище пропускает новорожденного, при половом акте влага-
лище легко приспосабливается к различным размерам мужского по-
лового члена.
НАРУЖНЫЕ ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
Женская половая область, или вульва, включает лобок («холм Ве-
неры»), большие и малые половые губы, клитор и преддверие влага-
318
ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
лища (рис. 157). У женщин в области лобка и больших половых губ
хорошо выражена подкожная жировая клетчатка. Кожа лобка богата
чувствительными нервными окончаниями. Большие половые губы, ог-
раничивающие половую щель, представляют собой складки кожи, по
краям которых обильно растут волосы. В обычном состоянии боль-
шие половые губы сомкнуты, при половом возбуждении они расхо-
дятся, открывая половую щель, которая увлажнена слизью. Малые
половые губы, лежащие кнутри от больших и отделенные от них боро-
здами, представляют собой складки кожи, лишенные жировой клет-
чатки и покрытые умеренно ороговевающим многослойным эпители-
ем, базальные клетки которого содержат большое количество
пигментных включений. Цвет малых половых губ у разных женщин
варьирует от розового до пурпурного. Передние верхние края малых
губ раздваиваются, окаймляя клитор, верхние части образуют край-
Рис. 157. Наружные женские половые органы, вид спереди:
1 — лобок, 2 — передняя спайка половых губ, 3 — крайняя плоть клитора, 4 — головка
клитора, 5 — большая половая губа, 6 — малая половая губа, 7 — устье большой
железы преддверия (бартолиниевой железы), 8 — уздечка малых половых губ,
9 — задняя спайка половых губ, 10 — задний проход, 11 — промежность,
12 — девственная плева, 13 — отверстие влагалища, 14 — преддверие влагалища,
15 — наружное отверстие мочеиспускательного канала, 16 — уздечка клитора
11
11
11
22
12
12
33
13
13
44
14
14
55
15
15
66
16
16
77
88
99
10
10
319
ОВАРИАЛЬНО-МЕНСТРУАЛЬНЫЙ ЦИКЛ
нюю плоть клитора, нижние — его уздечку. У многих женщин малые
половые губы выступают из-под больших.
Клитор, длиной 2,5—3,5 см, подобно мужскому половому члену,
состоит из двух пещеристых тел, разделенных перегородкой, и голов-
ки. Строение пещеристых тел клитора аналогично таковому полового
члена. Ножки клитора прикрепляются к нижним ветвям лобковых
костей. Чтобы увидеть клитор, следует слегка отодвинуть вверх его
крайнюю плоть.
Клитор — наиболее важная эрогенная зона женщины. Мастерс и
Джонсон называют клитор «уникальным органом, единственной
функцией которого является фокусирование и аккумуляция сексу-
альных ощущений и эротической радости».
Преддверие влагалища — это щель между малыми половыми губа-
ми, куда открываются наружные отверстия мочеиспускательного ка-
нала и влагалище, протоки множества малых и двух больших желез
преддверия (бартолиниевы). Луковица преддверия также состоит из
пещеристой ткани, которая расположена симметрично по бокам от
нижнего конца влагалища (аналог губчатого тела мужского полового
члена).
Наружные половые органы, особенно клитор и преддверие влага-
лища, имеют обильную иннервацию и множество различных чувстви-
тельных нервных окончаний. Наружные половые органы вместе с
влагалищем составляют единый совокупительный аппарат, предназ-
наченный для введения мужского полового члена и спермы и выведе-
ния плода.
ОВАРИАЛЬНО-МЕНСТРУАЛЬНЫЙ ЦИКЛ
У женщин созревание и выделение яйцеклетки из везикулярного
фолликула яичника (граафова пузырька) происходит циклически
(рис. 158). Этот процесс называется овуляцией. Овуляция сопрово-
ждается значительными изменениями всей половой системы. Крово-
течения связаны с отторжением поверхностного функционального
слоя слизистой оболочки матки. Овариально-менструальный цикл
находится под гормональным контролем: фолликулостимулирующий
(ФСГ) и лютеинизирующий (ЛГ) гормоны аденогипофиза, которые
регулируются единым гонадотропин-рилизинг-гормоном (ГТР-РГ)
гипоталамуса; женские половые гормоны эстрогены и прогестерон
(см. «Эндокринные железы»).
Обычно длительность цикла 28 дней (возможны колебания от 21
до 30 дней). В менструальном цикле различают три фазы. При
28-дневном цикле менструальная фаза длится около четырех дней.
Функциональный слой слизистой оболочки матки отторгается, на-
320
ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
ступает кровотечение. После прекращения менструации остается ба-
зальный слой слизистой оболочки, в котором сохраняются участки
маточных желез. В постменструальной фазе под влиянием женского
полового гормона эстрогена функциональный слой эндометрия реге-
нерирует, он утолщается, железы восстанавливаются. Эта фаза длит-
ся с 5-го по 14—15-й день, а в это время растет новый везикулярный
фолликул, который достигает зрелости к 14-му дню, стенка его лопа-
ется, и яйцеклетка выпадает в брюшную полость. Матка становится
способной к восприятию оплодотворенной яйцеклетки. В предмен-
струальной фазе (15—28-й дни) железы и слизистая оболочка матки
растут, набухают, секреция желез резко возрастает, толщина функ-
ционального слоя эндометрия быстро увеличивается, растут и его со-
суды.
Если яйцеклетка не оплодотворяется, начинается обратное разви-
тие желтого тела, продукция прогестерона резко уменьшается,
функциональный слой эндометрия начинает сморщиваться, спираль-
4
5
6
7
8
9
10
11
2
3
1
Э
ЭЭП
г
ЭЭ
Пг
Э
ФСГ ФСГ
ФСГ
ЛТГ
ЛГ
ЛГ
ЛГ
Дни47142128
59131721252933
2
ФСГ
Рис. 158. Овариально-менструальный цикл:
I — менструальная фаза, II — постменструальная фаза, III — предменструальная фаза;
1 — примордиальный фолликул в яичнике, 2 — первичные (растущие) фолликулы,
3 — вторичный фолликул (граафов пузырек), 4 — овуляция, 5 — желтое тело в стадии
расцвета, 6 — обратное развитие желтого тела, 7 — передняя доля гипофиза,
8 — задняя доля гипофиза, 9 — желтое тело беременности, 10 — оплодотворение,
11 — имплантированный зародыш. Стрелками показано действие фоллитропина
(ФСГ) на растущие фолликулы, лютропина (ЛГ) — на овуляцию и образование
желтых тел, лактотропина (пролактина, ЛТГ) — на сформированное желтое тело,
действие эстрогена (Э), стимулирующее рост эндометрия, — на матку
(постменструальная, или пролиферативная, фаза), прогестерона (Пг) —
на эндометрий (предменструальная фаза)
321
ОВАРИАЛЬНО-МЕНСТРУАЛЬНЫЙ ЦИКЛ
ные артерии все больше закручиваются, и кровоток через них умень-
шается, наступает их спазм. В результате этого возникает ишемия эн-
дометрия и его дегенеративные изменения. Вслед за спазмом
происходит расслабление стенки и восстановление кровообращения,
но сосуды становятся ломкими, рвутся, функциональный слой оттор-
гается, при этом повреждаются и вены, начинается кровотечение. На-
ступает очередная менструация.
Молочная железа
Женская грудь является символом женственности, сексуальности,
привлекательности. Грудь — предмет особой заботы женщин и восхи-
щения мужчин. Ч . Дарвин писал: «В зрелые годы, когда мы видим
предмет, похожий на женскую грудь, мы ощущаем прилив восторга,
который, по-видимому, влияет на все наши чувства». Размер и форма
груди совершенно не влияют на женскую сексуальность. Размеры
груди — скорее дело культуры и моды. Так, в Японии традиционно
было принято перевязывать грудь, чтобы она не росла.
Молочная железа, являющаяся видоизмененной потовой железой,
расположена на передней поверхности большой грудной мышцы.
У девственниц масса ее около 150—200 г, у кормящей женщины —
300—400 г, у мужчин она недоразвита. У новорожденной девочки
имеется лишь недоразвитая система протоков. В предпубертатном пе-
риоде у девочек быстро растет жировая ткань, к моменту половой зре-
лости железа становится округлой, но увеличение ее происходит
в основном за счет жировой ткани.
На передней поверхности железы в центре находится пигментиро-
ванный сосок (на его поверхности открываются 10—15 млечных пор),
окруженный также пигментированным околососковым кружком, диа-
метр ареолы 2—4 см. Кожа околососкового поля и особенно соска бо-
гато иннервируется. Здесь огромное количество чувствительных
нервных окончаний, поэтому у многих женщин вся грудь и особенно
сосок с околососковым полем являются важными эрогенными зона-
ми. В коже соска и околососкового кружка множество гладких мы-
шечных волокон. При половом возбуждении сосок резко набухает
и становится упругим, что связано с сокращением гладких мышечных
волокон.
У взрослой женщины железа состоит из 15–20 долей, между кото-
рыми располагается жировая и рыхлая волокнистая соединительная
ткань. Каждая доля — это сложная альвеолярная железа, выводной
проток которой направляется радиально к соску. Не доходя до соска,
проток, расширяясь, образует млечный синус. Однако концевые отде-
лы железы не кормящей женщины представляют собой лишь млечные
322
ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
альвеолярные протоки. Под влиянием эстрогена и прогестерона с кон-
ца пяти месяцев и до конца шести месяцев беременности на их концах
формируются альвеолы, образованные одним слоем цилиндрических
клеток. В дальнейшем до родов образование альвеол резко замедляет-
ся, но клетки секретируют, и образуемая ими жидкость (молозиво)
растягивает альвеолы, в результате чего железа продолжает набухать.
Этот процесс продолжается до и в первые 1–2 дня после родов.
Во время кормления грудью альвеолы молочных желез продуци-
руют молоко. Секреция молока стимулируется лактотропным гормо-
ном гипофиза. После окончания периода кормления ребенка посте-
пенно происходит обратное развитие молочной железы, лишь
сохраняются некоторые альвеолы.
ПОЛОВОЕ СОЗРЕВАНИЕ
При половом созревании, которое у девушек протекает в возрасте
9—13, у юношей — в возрасте 11–15 лет, в организме происходят важ-
нейшие морфофизиологические и психологические изменения, раз-
виваются вторичные половые признаки, начинаются менструации
(у девушек) и семяизвержения (у мальчиков). Все эти преобразова-
ния связаны с резким усилением синтеза и секреции гипоталамусом
гонадотропин-рилизинг-гормона, который стимулирует выработку
передней долей гипофиза фолликулостимулирующего и лютеинизи-
рующего гормонов (см. раздел «Эндокринные железы»). Последние
влияют на синтез и выделение половыми железами половых гормо-
нов. Уровень тестостерона у юношей повышается в 10—20 раз. В это
время половые гормоны стимулируют рост костей, который особенно
выражен в 12 лет у девушек и в 14 у юношей.
У девушек в период полового созревания развиваются молочные
железы. Это начинается с появления небольших бугорков, затем уве-
личиваются соски и околососковые поля, молочные железы растут
и округляются. Вслед за формированием молочных желез начинается
оволосение лобка по женскому типу, появляются волосы в подмы-
шечных впадинах, растут внутренние половые органы, щелочная сре-
да во влагалище сменяется кислой, меняется форма таза. В связи
с акселерацией средние сроки начала менструации 11,5—12,5 года, сто
лет тому назад эти сроки колебались в пределах 16—17 лет. У боль-
шинства девушек появляются выделения из влагалища.
У юношей процесс полового созревания начинается с увеличения
яичек и их придатков, полового члена, простаты, семявыносящих
протоков, семенных пузырьков. Очень важным проявлением полово-
го созревания являются поллюции (ночные семяизвержения), кото-
рые оказывают и серьезное психологическое влияние. Вначале оволо-
323
ГАМЕТОГЕНЕЗ
сение лобка начинается по женскому типу, набухают грудные железы,
к концу периода (15—16 лет) начинается рост волос на лице, теле,
в подмышечных впадинах, на лобке (по мужскому типу), пигментиру-
ется кожа мошонки, еще больше увеличиваются половые органы. Все
это хорошо видимые доказательства превращения юноши в мужчину.
Появляются волосы на конечностях, затем на животе и позже всего на
груди.
ВНИМАНИЕ!
Степень оволосения тела мужчины зависит от уровня мужского полового
гормона тестостерона в организме. Увы, это не касается волос на голове.
Наоборот, высокое содержание гормона способствует облысению.
Тестостерон оказывает серьезное влияние на рост гортани, в связи
с чем меняется тембр голоса. Это происходит в 13—14 лет. В связи
с акселерацией ломка голоса наступает на 4—5 лет раньше, чем 100
лет тому назад. Именно повышение уровня гормонов в крови оказы-
вает влияние на эротическое воображение, половое поведение, поло-
вую чувствительность. Чем выше уровень гормонов, тем чаще у юно-
шей возникает эрекция полового члена, у девушек — эрекция клитора,
любрикация. Половые гормоны «эротизируют» кору головного мозга.
Те юноши и девушки, у которых раньше происходит половое созрева-
ние, ведут более активную сексуальную жизнь. Однако в половом по-
ведении подростков важную роль играют и психосоциальные факто-
ры, а именно установки, принятые в определенной общественной
среде, личное отношение подростка к сексу, насколько важное значе-
ние конкретный индивидуум придает сексуальным проблемам.
ГАМЕТОГЕНЕЗ
Половые клетки, слияние которых дает новый организм, объеди-
няют термином гаметы. Женская гамета называется яйцеклеткой,
мужская — сперматозоидом. Все остальные клетки, не принимающие
непосредственного участия в образовании гамет, получили название
соматических клеток. Гаметогенез — поэтапное «сотворение» высо-
коспециализированных клеток, способных дать начало новому орга-
низму. Гаметогенез состоит из четырех стадий:
1) образование первичных половых клеток (ППК) и миграция их
в гонады;
2) размножение половых клеток в гонадах путем митоза;
3) уменьшение числа хромосом в каждой клетке в два раза в ре-
зультате мейоза;
324
ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
4) окончательное созревание и дифференцировка гамет, превра-
щение их в сперматозоиды и яйцеклетки, которые способны оплодо-
творять или быть оплодотворенными.
Гонады зародыша вначале содержат относительно небольшое чи-
сло заселивших их первичных половых клеток. Но попав в гонады,
половые клетки начинают энергично делиться, и их численность рез-
ко увеличивается. Клетки делятся митотически. Митоз обеспечивает
передачу двум дочерним клеткам совершенно одинаковых наборов
хромосом, содержащих наследственную информацию. Митотически
делящиеся женские половые клетки называют оогониями, а соответст-
вующие мужские — сперматогониями. В результате сперматогенеза
из одной сперматогонии образуется четыре сперматозоида, в резуль-
тате оогенеза – одна яйцеклетка.
ПОЛОВОЙ ЦИКЛ ЧЕЛОВЕКА
Половой цикл человека в связи с особенностями человеческой сек-
суальности тесно связан с психикой и зависит от многих психологи-
ческих, личностных и межличностных факторов. И у мужчины, и
у женщины половой цикл состоит из четырех фаз, последовательно
сменяющих друг друга: нарастающего полового возбуждения, сохране-
ния полового возбуждения на одном уровне (плато), оргазма и рассла-
бления (разрешения) (рис. 159). Эти фазы не всегда имеют четкие
временны
е границы, они различны по интенсивности и длительности
не только у разных людей, но даже у одного и того же человека. Как
у мужчины, так и у женщины при половом возбуждении обязательно
возникают две главные физиологические реакции: резкий прилив крови
к половым органам (а у женщин и к молочным железам) и мышечное
(вернее, нервно-мышечное) напряжение.
Возбуждение у мужчины проявляется эрекцией. Эрекция полового
члена — главное условие мужского полового цикла. Механизм эрекции
описан выше (см. раздел «Половой член»). Эрекция связана с возбу-
ждением парасимпатической нервной системы. Сексуальное возбу-
ждение у женщины через 10—40 сек. приводит к изменениям женских
половых органов, и в первую очередь к любрикации, увлажнению вла-
галища. Вначале на слизистой оболочке влагалища появляются от-
дельные капли, которые затем сливаются, увлажняя всю слизистую
оболочку. Любрикация свидетельствует о готовности женских поло-
вых путей к половому акту, она продолжается на протяжении всего
полового цикла, увеличивается объем клитора и его головки (эрек-
ция). Как правило, на высоте эрекции при условии готовности женщи-
ны половой член вводится во влагалище (интроитус) и начинается
собственно копулятивная, или фрикционная, стадия (от лат. copula-
325
ПОЛОВОЙ ЦИКЛ ЧЕЛОВЕКА
Оргазм
Плато
123
1
2
3
А
Р
а
з
р
е
ш
е
н
и
е
Р
а
з
р
е
ш
е
н
и
е
Р
а
з
р
е
ш
е
н
и
е
Возбуж-
дение
Оргазм
Рефрактерный
период
Сенсорно-
ирритативная
стадия
В
о
з
н
и
к
н
о
в
е
н
и
е
п
о
л
о
в
о
й
д
о
м
и
н
а
н
т
ы
П
о
я
в
л
е
н
и
е
э
р
е
к
ц
и
и
И
н
т
р
о
и
т
у
с
Н
а
ч
а
л
о
э
я
к
у
л
я
ц
и
и
О
т
н
о
с
и
т
е
л
ь
н
а
я
п
о
л
о
в
а
я
н
е
в
о
з
б
у
д
и
м
о
с
т
ь
А
б
с
о
л
ю
т
н
а
я
п
о
л
о
в
а
я
н
е
в
о
з
б
у
д
и
м
о
с
т
ь
Состо-
яние
нейрогу-
моральной
готовности
Психи-
ческая
стадия
Эрекци-
онная
стадия
Копуля-
тивная
(фрикци-
онная
стадия
Эяку-
ля-
тор-
ная
стадия
Рефрак-
терная
стадия
Рефрактерный
период
Плато
Б
В
Р
а
з
р
е
ш
е
н
и
е
Р
а
з
р
е
ш
е
н
и
е
Возбуж-
дение
Рис. 159. Половой цикл
(А, Б — по У. Мастерсу и В. Джонсон, В — по Г. С . Васильченко):
А — три варианта женского полового цикла: 1 — множественный оргазм,
2 — возбуждение, переходящее в фазу плато без достижения оргазма, в этом случае
фаза разрешения протекает очень медленно, 3 — резкое, прерывистое нарастание
возбуждения, достижение оргазма с быстрой фазой разрешения.
Б — наиболее типичные варианты мужского полового цикла, штриховой линией
показан один из возможных вариантов повторного оргазма
с эякуляцией после завершения рефракторного периода, возможны
и другие варианты развития мужского полового цикла, которые во многих случаях
совпадают по форме с кривыми 2 и 3, характеризующими женский половой цикл.
В — типичная кривая полового цикла мужчины
326
ЖЕНСКИЕ ПОЛОВЫЕ ОРГАНЫ
tion — «соединение»; frictio — «трение»). С точки зрения физиологии
в это время наблюдается плато, во время которого возбуждение сохра-
няется примерно на одном уровне.
Введение полового члена во влагалище и последующие фрикции
усиливают половое возбуждение женщины до определенного уров-
ня, после чего начинается фаза плато, во время которой сексуаль-
ное напряжение определенное время не нарастает и не снижается.
Длительность этой фазы широко варьирует. Если фрикции продол-
жаются, половое возбуждение усиливается и развивается следую-
щая фаза оргазм (от греч. orgao — «пылаю страстью»). Оргазм —
кульминация полового акта, в котором участвует весь организм
человека. Оргазм — это самое высокое наслаждение, какое может
испытывать человек. У мужчины оргазм достигается благодаря
эмиссии (от лат. emissio — «выпуск, испускание») и эякуляции (от
лат. ejaculatio — «извержение»).
Благодаря ритмичным сокращениям полового члена (пульсации),
мышц промежности, простаты с интервалом в 0,7—0,8 сек. семенная
жидкость перемещается в передний отдел мочеиспускательного ка-
нала, и начинается семяизвержение. Как правило, у мужчин оргазм
и эякуляция происходят одновременно, но это различные процессы.
Эякуляция — это выброс спермы. Причем в ряде случаев она не со-
провождается оргазмом.
Анатомо-физиологической основой женского оргазма являются рит-
мичные сокращения половых органов. Оргазм у женщин характеризует-
ся одновременными ритмичными сокращениями мускулатуры матки,
нижней трети влагалища («оргастической манжетки») и сфинктера
прямой кишки. В начале оргазма мышцы сокращаются с большими
интенсивностью и быстротой (интервал между сокращениями состав-
ляет примерно 0,8 сек.). Затем сила, длительность и периодичность
сокращений снижаются. «Мягкий» оргазм может сопровождаться
всего тремя-пятью сокращениями, тогда как сильный 10—15. Однако
у разных женщин и даже у одной и той же женщины некоторые оргаз-
мы происходят мягко, с ощущением слабого трепета или тепла, а дру-
гие можно сравнить со «взрывом авиабомбы». Изменения при оргазме
охватывают не только половые органы, но и все органы и системы. Во
время оргазма резко возрастает частота сердечных сокращений, кото-
рая может достигнуть 160—180 в 1 минуту; повышается артериальное
давление (систолическое — на 30—90 мм рт. ст., диастолическое — на
20—40 мм рт. ст.); частота дыхания увеличивается до 36—42 в 1 мину-
ту; сокращаются скелетные мышцы, особенно мимические, межребер-
ные и мышцы живота.
Между мужским и женским оргазмом много различий, однако
есть и важные анатомо-физиологические сходства: оргазм сопрово-
327
ПОЛОСТЬ ЖИВОТА. БРЮШИНА
ждается ритмическими сокращениями мышц половых органов, про-
межности, сфинктера прямой кишки; одни и те же нервы вызывают
сокращение мышц у мужчин и у женщин.
Женский оргазм — одна из важных отличительных особенностей
человека разумного. Современные исследователи единодушны во
мнении, что женщина — единственное существо женского пола, кото-
рое способно испытывать оргазм.
Фаза разрешения. После эякуляции половое возбуждение спадает.
Наступает рефрактерный период, длительность которого различна
не только у различных мужчин, но и у одного и того же мужчины.
У большинства мужчин в зрелом возрасте после эякуляции наступает
абсолютная рефрактерность, т. е . абсолютная половая невозбуди-
мость, когда практически эрекцию вызвать невозможно. Как правило,
мужчина засыпает. Женщина должна знать об этом и не обижаться.
На смену абсолютной приходит относительная рефрактерность, во
время которой может наступить повторная эрекция и последующий
половой акт.
Независимо от того, испытала ли женщина один или несколько
оргазмов, вслед за оргазмом наступает фаза разрешения. Если после
резкого возбуждения в фазе плато оргазм не наступил, фаза разреше-
ния может затянуться; у многих женщин развивается фрустрация —
тягостное психоэмоциональное состояние, вызванное острым чувст-
вом неудовлетворенности.
ПОЛОСТЬ ЖИВОТА. БРЮШИНА
Полость живота ограничена сверху диафрагмой, внизу продолжа-
ется в полость таза, выход из которой закрыт диафрагмой таза, задняя
стенка образована поясничным отделом позвоночника и мышцами
(квадратными поясницы и подвздошно-поясничными), передняя —
мышцами живота. Изнутри брюшная полость выстлана забрюшинной
фасцией, на которой лежит жировая ткань, и брюшиной. Пространст-
во, ограниченное брюшиной и забрюшинной фасцией, называется за-
брюшинным, в нем располагаются некоторые органы и жировая клет-
чатка (рис. 160).
Брюшина выстилает брюшную полость и покрывает расположен-
ные в ней внутренние органы. Брюшина образована соединитель-
нотканной пластинкой, которая покрыта однослойным плоским
эпителием мезотелием. В брюшине различают два листка: парие-
тальная брюшина выстилает стенки брюшной полости, висцераль-
ная брюшина покрывает органы. Общая площадь брюшины у взро-
328
ПОЛОСТЬ ЖИВОТА. БРЮШИНА
слого человека 1,6—1,75 м2. Оба листка брюшины переходят
непрерывно со стенок брюшной полости на органы и с органов на
стенки брюшной полости, ограничивая брюшинную полость. У жен-
щин полость брюшины открытая, она сообщается с внешней средой
через маточные трубы, полость матки и влагалище, у мужчин по-
лость замкнутая. Полость брюшины увлажнена небольшим количе-
ством серозной жидкости, что облегчает движение органов и предот-
вращает их трение.
Париетальная брюшина покрывает переднюю стенку брюшной
полости, вверху переходит на нижнюю поверхность диафрагмы,
а затем на заднюю и боковые стенки брюшной полости и на вну-
тренние органы, а внизу — на стенки и органы полости таза. Брю-
шина, переходя со стенок брюшной полости на органы или с одно-
Рис. 160. Отношение внутренних органов к брюшине.
Срединный (сагиттальный) разрез туловища:
1 — печень, 2 — печеночно-желудочная связка, 3 — сальниковая сумка,
4 — поджелудочная железа, 5 — двенадцатиперстная кишка, 6 — брыжейка тонкой
кишки, 7 — прямая кишка, 8 — мочевой пузырь, 9 — тонкая кишка, 10 — полость
большого сальника, 11 — поперечная ободочная кишка, 12 — брыжейка поперечной
ободочной кишки, 13 — желудок
11
11
11
22
12
12
33
13
13
44
55
66
77
88
99
10
10
329
ПОЛОСТЬ ЖИВОТА. БРЮШИНА
го органа на другой, образует складки, ямки, связки, брыжейки
и удвоения. На задней стенке брюшной полости брюшина покры-
вает органы, лежащие забрюшинно, и переходит на другие органы.
Органы, покрытые брюшиной только с одной стороны (поджелу-
дочная железа, бо
льшая часть двенадцатиперстной кишки, почки,
надпочечники и др.), лежат вне брюшины, забрюшинно (экстрапе-
ритонеально). Другие органы покрыты брюшиной только с трех
сторон и называются мезоперитонеально лежащими органами (вос-
ходящая и нисходящая ободочные кишки). Третья группа органов
покрыта брюшиной со всех сторон и занимает внутрибрюшинное
(интраперитонеальное) положение (желудок, тонкая кишка, сле-
пая кишка с червеобразным отростком, который имеет брыжейку,
поперечная и сигмовидная ободочные кишки, селезенка, печень).
Два листка брюшины, покрывающей сигмовидную ободочную киш-
ку со всех сторон, образуют ее брыжейку. К поперечной ободочной
кишке от задней стенки брюшной полости идут два листка брюши-
ны, которые образуют поперечно лежащую брыжейку поперечной
ободочной кишки. Непосредственно под брыжейкой поперечной
ободочной кишки от задней брюшной стенки начинается образо-
ванная двумя листками париетальной брюшины брыжейка тонкой
кишки. Между листками брыжеек расположены сосуды, нервы,
лимфатические узлы, клетчатка.
С нижней поверхности диафрагмы брюшина переходит на диаф-
рагмальную поверхность печени, а затем — на ее висцеральную по-
верхность. С задней брюшной стенки париетальная брюшина также
переходит на висцеральную поверхность печени, оба листка «встре-
чаются» у ворот печени, откуда направляются к малой кривизне же-
лудка, где расходятся, покрывая со всех сторон желудок и верхнюю
часть двенадцатиперстной кишки. При этом между воротами пече-
ни, с одной стороны, малой кривизной желудка и верхней частью
двенадцатиперстной кишки — с другой, образуется дупликатура
брюшины — малый сальник. В его правом крае между листками брю-
шины расположены справа налево общий желчный проток, воротная
вена и собственная печеночная артерия.
У большой кривизны желудка оба листка брюшины вновь сходят-
ся и спускаются вниз впереди поперечной ободочной и петель тонкой
кишки. Дойдя до лобкового симфиза, эти два листка брюшины заги-
баются кзади и поднимаются вверх позади нисходящих листков так-
же впереди поперечной ободочной и петель тонкой кишки. Эта длин-
ная складка, свисающая впереди поперечной ободочной кишки
и петель тонкой кишки в виде фартука и образованная четырьмя
330
ПОЛОСТЬ ЖИВОТА. БРЮШИНА
листками брюшины, называется большим сальником. Между листками
большого сальника находится жировая клетчатка. Выше брыжейки
поперечной ободочной кишки эти листки переходят в париетальную
брюшину задней брюшной стенки. Верхний листок направляется
вверх, впереди передней поверхности поджелудочной железы, и пере-
ходит на заднюю стенку брюшной полости и на диафрагму. Нижний
листок идет вниз и переходит в верхний листок брыжейки попереч-
ной ободочной кишки.
В полости малого таза брюшина покрывает верхний и частично
средний отделы прямой кишки, мочевые и половые органы. У муж-
чин брюшина переходит с передней поверхности прямой кишки на
заднюю, затем верхнюю стенку мочевого пузыря и продолжается в па-
риетальную брюшину, выстилающую переднюю брюшную стенку.
Между мочевым пузырем и прямой кишкой образуется прямокишеч-
но-пузырное углубление. У женщин брюшина с передней поверхно-
сти прямой кишки переходит на заднюю стенку верхней части влага-
лища, затем поднимается вверх, покрывая сзади, а затем спереди
матку и переходит на мочевой пузырь. Парная дупликатура брюши-
ны, широкая связка матки, соединяет матку с боковой стенкой малого
таза, маточная труба проходит внутри верхнего края широкой связки,
к широкой связке матки сзади прикрепляется яичник. Между маткой
и прямой кишкой образуется прямокишечно-маточное углубление
(дугласово пространство), ограниченное по бокам прямокишечно-ма-
точными складками. Между маткой и мочевым пузырем образуется
пузырно-маточное углубление.
331
СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
Жизнедеятельность организма возможна лишь при условии до-
ставки каждой клетке питательных веществ, кислорода, воды и удале-
ния выделяемых клеткой продуктов обмена веществ. Эту задачу вы-
полняет сосудистая система, представляющая собой систему трубок,
содержащих кровь и лимфу, и сердце — центральный орган, обуслов-
ливающий движение этой жидкости.
КРОВЕНОСНАЯ СИСТЕМА
Сердце и кровеносные сосуды образуют замкнутую систему, по ко-
торой кровь движется благодаря сокращениям сердечной мышцы
и гладких миоцитов стенок сосудов. Кровеносные сосуды представле-
ны артериями, несущими кровь от сердца; венами, по которым кровь
течет к сердцу; и микроциркуляторным руслом, состоящим из арте-
риол, капилляров и венул. Кровеносные сосуды отсутствуют лишь
в эпителиальном покрове кожи и слизистых оболочек, в волосах, ног-
тях, роговице глаз и суставных хрящах.
Стенка артерии состоит из трех оболочек: внутренней, средней и на-
ружной (рис. 161). Внутренняя оболочка образована эндотелием, кото-
рый выстилает просвет сосуда, подэндотелиальным слоем и внутренней
эластической мембраной. Средняя оболочка артерии состоит из распо-
ложенных спирально гладких миоцитов, между которыми проходит не-
большое количество коллагеновых и эластических волокон, и наружной
эластической мембраны, образованной продольными толстыми пере-
плетающимися волокнами. Наружная оболочка образована рыхлой во-
локнистой соединительной тканью, содержащей эластические и колла-
геновые волокна, в ней проходят кровеносные сосуды и нервы.
В зависимости от развития различных слоев стенки артерии по-
дразделяются на сосуды мышечного (преобладают), смешанного (мы-
шечно-эластического) и эластического типов. В стенке артерий мы-
шечного типа хорошо развита средняя оболочка. Миоциты и
эластические волокна располагаются в ней по типу пружины. Миоци-
ты средней оболочки стенки артерий мышечного типа своими сокра-
щениями регулируют приток крови к органам и тканям. По мере
уменьшения диаметра артерий все оболочки стенки артерий истонча-
ются. Наиболее тонкие артерии мышечного типа — артериолы, имею-
щие диаметр менее 100 мкм, переходят в капилляры.
К артериям смешанного типа относятся такие артерии, как сон-
ная и подключичная. В средней оболочке их стенки примерно равное
количество эластических волокон и миоцитов, появляются окончатые
эластические мембраны. К артериям эластического типа относятся
332
КРОВЕНОСНАЯ СИСТЕМА
аорта и легочный ствол, в которые кровь поступает под большим дав-
лением и с большой скоростью из сердца. Средняя оболочка образо-
вана концентрическими эластическими окончатыми мембранами,
между которыми залегают миоциты.
Дистальная часть сердечно-сосудистой системы — микроциркуля-
торное русло (рис. 162). Оно является путем местного кровотока, где
обеспечивается взаимодействие крови и тканей. Микроциркулятор-
ное русло начинается самым мелким артериальным сосудом — арте-
риолой и заканчивается венулой. Стенка артериолы содержит лишь
один ряд миоцитов. От артериол отходят прекапилляры и истинные
капилляры, у начала которых находятся гладкомышечные прекапил-
лярные сфинктеры, регулирующие кровоток. Истинные капилляры
вливаются в посткапилляры (посткапиллярные венулы). Посткапил-
ляры образуются из слияния двух или нескольких капилляров. По
мере слияния посткапилляров образуются венулы. Их калибр широ-
ко варьирует и в обычных условиях равен 25—50 мкм. Венулы влива-
ются в вены. К микроциркуляторному руслу относятся также и лим-
фатические капилляры.
Рис. 161. Строения стенки
артерии (А) и вены (Б)
мышечного типа среднего
калибра, схема:
I — внутренняя оболочка:
1 — эндотелий, 2 — базальная
мембрана, 3 — подэндотелиальный
слой, 4 — внутренняя эластическая
мембрана; II — средняя оболочка:
5 — миоциты, 6 — эластические
волокна, 7 — коллагеновые волокна;
III — наружная оболочка:
8 — наружная эластическая
мембрана, 9 — волокнистая
(рыхлая) соединительная ткань,
10 — кровеносные сосуды
(по В. Г. Елисееву и др.)
11
11
I
I
АБ
22
22
II
II
33
33
III
III
44
55
55
66
66
77
77
88
99
99
10
10
10
10
333
КРОВЕНОСНАЯ СИСТЕМА
Наиболее важный отдел кровеносной системы — это капилляры,
именно они осуществляют обмен веществ и газообмен. Общая обмен-
ная поверхность капилляров взрослого человека достигает 1000 м2
.
Кровеносные капилляры имеют стенки, образованные одним слоем
уплощенных эндотелиальных клеток — эндотелиоцитов, сплошной
или прерывистой базальной мембраной и редкими удлиненными пе-
рикапиллярными клетками, перицитами. Перицит передает эндоте-
лиальной клетке импульс, в результате чего она набухает или теряет
жидкость. Это и приводит к периодическим изменениям просвета ка-
пилляра. Посткапиллярные венулы диаметром 8—30 мкм, являющие-
ся конечным звеном микроциркуляторного русла, впадают в собира-
тельные венулы (диаметром 50—100 мкм), от них кровь оттекает
в мелкие собирательные вены (диаметром 100—300 мкм), которые,
сливаясь между собой, укрупняются.
Стенка вены также состоит из трех оболочек (см. рис. 161). Разли-
чают два типа вен — мышечный и безмышечный. В стенках безмы-
шечных вен отсутствуют гладкие мышечные клетки (например, вены
твердой и мягкой мозговой оболочек, сетчатки глаз, костей, селезенки
и плаценты). Они плотно сращены со стенками органов и поэтому не
спадаются. В стенках вен мышечного типа имеются гладкие мышеч-
ные клетки. На внутренней оболочке большинства средних и некото-
рых крупных вен имеются клапаны, которые пропускают кровь лишь
в направлении к сердцу, препятствуя обратному току крови в венах
и тем самым предохраняя сердце от излишней затраты энергии на
преодоление колебательных движений крови, постоянно возникаю-
щих в венах. Вены верхней половины тела не имеют клапанов.
Общее количество вен больше, чем артерий, а общая величина ве-
нозного русла превосходит артериальное. Скорость кровотока в венах
меньше, чем в артериях, в венах туловища и нижних конечностей
кровь течет против силы тяжести.
4
3
2
5
6
1
Рис. 162. Схема строения
микроциркуляторного русла:
1 — капиллярная сеть
(капилляры), 2 — посткапилляр
(посткапиллярная венула),
3 — артериоло-венулярный
анастомоз, 4 — венула,
5 — артериола, 6 — прекапилляр
(прекапиллярная артериола).
Стрелки слева — поступление
в ткани питательных веществ,
стрелки справа — выведение
из тканей продуктов обмена
334
КРОВЕНОСНАЯ СИСТЕМА
Сердце
Сердце, расположенное асимметрично в средостении, представля-
ет собой полый мышечный орган, разделенный внутри на четыре по-
лости: правое и левое предсердия и правый и левый желудочки. Пред-
сердия разделены межпредсердной, желудочки — межжелудочковой
перегородками (рис. 163). Размеры сердца здорового человека корре-
лируют с величиной его тела, а также зависят от интенсивности физи-
ческой нагрузки и обмена веществ. Средняя масса сердца у женщин
250 г, у мужчин 300 г.
Правое предсердие — кубической формы, в него впадают верх-
няя и нижняя полые вены и венечный синус сердца. Кпереди и впра-
во полость предсердия продолжается в правое ушко. Кровь из правого
предсердия при его сокращении поступает в правый желудочек че-
рез правое предсердно-желудочное отверстие, по краю которого рас-
положен предсердно-желудочковый (трехстворчатый) клапан, состо-
ящий из трех створок, образованных складками эндокарда и покрытых
эндотелием. От свободных краев створок начинаются сухожильные
хорды, прикрепленные концами к трем сосочковым мышцам, располо-
женным на внутренней поверхности правого желудочка. Эти мышцы
вместе с сухожильными хордами удерживают клапаны и при сокра-
щении (систоле) желудочка препятствуют обратному току крови
Рис. 163. Сердце.
Предсердия и желудочки сердца на его
продольном разрезе, вид спереди:
1 — устья правых легочных вен,
2 — левое предсердие, 3 — левая
легочная вена, 4 — межпредсердная
перегородка, 5 — левое предсердно-
желудочковое отверстие, 6 — передняя
створка и задняя створка левого
предсердно-желудочкового клапана:
7 — сухожильные хорды, 8 — левый
желудочек, 9 — миокард левого
желудочка, 10 — межжелудочковая
перегородка (мышечная часть),
11 — верхушка сердца, 12 — правый
желудочек, 13 — миокард правого
желудочка, 14 — перепончатая часть
межжелудочковой перегородки,
15 — створки правого предсердно-
желудочкового клапана, 16 — правое
предсердно-желудочковое отверстие,
17 — отверстие венечного синуса,
18 — правое предсердие,
19 — гребенчатые мышцы,
20 — устье нижней полой вены,
21 — овальная ямка
11
11
11
22
12
12
33
13
13
44
14
14
55
15
15
66
16
16
77
17
17
88
18
18
99
19
19
10
10
20
20
21
21
335
КРОВЕНОСНАЯ СИСТЕМА
в предсердие. Передневерхний отдел желудочка продолжается в ле-
гочный ствол.
При сокращении желудочка кровь выталкивается в легочный
ствол через отверстие легочного ствола, в области которого находит-
ся одноименный клапан. Клапан состоит из трех полулунных засло-
нок, свободно пропускающих кровь из желудочка в легочный ствол.
Соприкасаясь своими концами, они, подобно наполненным карманам,
закрывают отверстие и препятствуют обратному току крови. Это про-
исходит после опорожнения желудочка.
В левое предсердие открываются четыре легочные вены (по две с ка-
ждой стороны), кпереди и влево выпячивается левое ушко. Левый же-
лудочек имеет форму конуса, его миокард в 2—3 раза толще, чем у пра-
вого желудочка. Это связано с большой работой, производимой левым
желудочком. Из полости левого предсердия в левый желудочек ведет
левое предсердно-желудочковое отверстие овальной формы, снабжен-
ное левым предсердно-желудочковым двустворчатым клапаном (ми-
тральным). Из желудочка кровь направляется в отверстие аорты,
снабженное клапаном, состоящим из трех полулунных заслонок, имею-
щих такое же строение, как и клапан легочного ствола. На внутренней
поверхности левого желудочка, подобно правому, имеются две сосочко-
вые мышцы, от которых отходят тонкие сухожильные хорды, прикре-
пляющиеся к створкам левого предсердно-желудочкового клапана.
Стенка сердца состоит из трех слоев: наружного, или эпикарда,
среднего — миокарда, внутреннего — эндокарда. Эпикард, представ-
ляющий собой висцеральную пластину серозного перикарда, окуты-
вает сердце, начальные отделы легочного ствола и аорты, конечные
отделы полых вен. Эндокард, покрытый эндотелием, выстилает из-
нутри камеры сердца и его клапаны. Преобладающая часть сердечной
стенки — миокард, т. е. мышечный слой, образованный сердечной ис-
черченной (поперечнополосатой) мышечной тканью. Контакты двух
соседних кардиомиоцитов — в виде темных полосок, вставочных дис-
ков, которые активно участвуют в передаче возбуждения от клетки
к клетке. Миокард предсердий и желудочков разобщен, что создает
возможность отдельного их сокращения.
Последовательное сокращение и расслабление различных отде-
лов сердца связано с его строением и наличием проводящей систе-
мы, по которой распространяется импульс (рис. 164). Проводящая
предсердно-желудочковая система сердца состоит из синусно-пред-
сердного узла (Киса — Флака), который является водителем ритма
(пейсмейкером), предсердно-желудочкового узла (Ашоффа — Тава-
ра), предсердно-желудочкового пучка (пучка Гиса), его ножек и раз-
ветвлений (волокна Пуркинье). Проводящая система образована
сердечными проводящими волокнами, богато иннервируемыми
336
КРОВЕНОСНАЯ СИСТЕМА
нервами вегетативной нервной системы. Предсердия связаны между
собой синусно-предсердным узлом, а предсердия и желудочки —
предсердно-желудочковым пучком.
Две артерии, правая и левая венечные, ветви которых широко ана-
стомозируют между собой, снабжают сердце кровью. Они разветвля-
ются до капилляров во всех трех оболочках стенки сердца. Кровь
собирается в сердечные вены, далее — венозный синус, который непо-
средственно вливается в правое предсердие.
Перикард — это замкнутый мешок, в котором различают два
слоя: наружный — фиброзный перикард и внутренний — серозный пе-
рикард, который, в свою очередь, делится на два листка: висцераль-
ный, или эпикард, и париетальный, сращенный с внутренней поверх-
ностью серозного перикарда, выстилающий его изнутри. Между
висцеральным и париетальным листками находится щелевидная пе-
рикардиальная полость, содержащая небольшое количество серозной
жидкости, которая смачивает обращенные друг к другу поверхности
серозных листков, покрытых мезотелием. На крупных сосудах вблизи
сердца висцеральный и периетальный листки переходят непосредст-
венно один в другой.
В возрасте 30—40 лет в миокарде обычно начинается некоторое уве-
личение количества соединительной ткани, в ней появляются жировые
клетки. По мере старения человека жировая ткань накапливается под
эпикардом, происходит утолщение эндокарда. Эти изменения могут
быть в значительной мере замедлены или даже предотвращены благо-
даря регулярной физической нагрузке и правильному питанию.
Развитие мускулатуры тела влияет на величину сердца. Так, вели-
чина и масса сердца у лиц, занятых физическим трудом, и у спортсме-
нов больше, чем у представителей умственного труда. Причем виды
спорта, при которых физическое напряжение носит продолжитель-
Рис. 164. Схема проводящей системы сердца:
1 — синусно-предсердный узел, 2 — левое
предсердие, 3 — межпредсердная
перегородка, 4 — предсердно-желудочковый
узел, 5 — предсердно-желудочковый пучок,
6 — левая ножка предсердно-желудочкового
пучка, 7 — правая ножка предсердно-
желудочкового пучка, 8 — левый желудочек,
9 — проводящие мышечные волокна,
10 — межжелудочковая перегородка,
11 — правый желудочек, 12 — нижняя полая
вена, 13 — правое предсердие,
14 — верхняя полая вена
11
11
11
22
12
12
33
13
13
44
14
14
55
66
77
88
99
10
10
337
КРОВЕНОСНАЯ СИСТЕМА
ный характер (например, велосипед, гребля, марафонский бег, лыжи),
приводят к гипертрофии миокарда и увеличению размеров сердца.
Бег, плавание, бег на небольшие дистанции, бокс, легкая атлетика,
футбол и некоторые другие виды спорта приводят к менее выражен-
ному увеличению мышц сердца.
Функции сердца
Автоматизм (от греч. automatos — самодействующий, самопроиз-
вольный) сердца. Миокард, являясь мышечной тканью, обладает
свойствами возбудимости, проводимости и сократимости. Как мы пи-
сали выше, проводящая система сердца обеспечивает последователь-
ные сокращения и расслабления его отделов. Причем это происходит
автоматически. Автоматизм сердца — это его способность ритмически
сокращаться под влиянием возникающих в нем самом (в клетках его
проводящей системы) импульсов. Генератором этих импульсов явля-
ется синусно-предсердный узел, в клетках которого возникает потен-
циал действия (около 90—100 мВ), передающийся соседним клеткам
проводящей системы, а с них — через вставочные диски на рабочие
кардиомиоциты. Возбуждение распространяется по миокарду. Внача-
ле сокращаются предсердия, а затем желудочки. При этом миокард
сокращается, когда сила импульса достигает пороговой величины по
закону «всё или ничего». Согласно этому закону возбудимая ткань
дает максимальную ответную реакцию («всё») при пороговом или
надпороговым раздражении, но, если сила раздражения ниже порого-
вой, ответа нет («ничего»). Начав сокращаться, миокард уже отвечает
на другие стимулы, пока в нем не начнется процесс расслабления.
Здоровый миокард сокращается в течение всей жизни человека и не
испытывает при этом утомления. Это связано с рефрактерностью
(от фр. refractaire — невосприимчивость). Период абсолютной реф-
рактерности — это интервал времени, во время которого миокард не
отвечает ни на какие импульсы.
Миокард является возбудимой тканью. Его клетки обладают по-
тенциалом покоя, генерируют потенциал действия. Возбуждение,
которое возникло на любом участке миокарда, передается всем его
волокнам. Поэтому в ответ на адекватное раздражение происходит
возбуждение всех его волокон. Проводящая система обеспечивает ге-
нерацию возбуждения и его проведение к кардиомиоцитам. Деятель-
ностью сердца управляют сердечные центры, расположенные в про-
долговатом мозге и мосте, которые действуют через вегетативную
нервную систему. Симпатические нервы оказывают положительное
влияние (учащение сердечных сокращений и увеличение их силы),
парасимпатические — отрицательное (урежение сердечных сокраще-
338
КРОВЕНОСНАЯ СИСТЕМА
ний и уменьшение их силы). Кора головного мозга регулирует дея-
тельность сердечных центров через гипоталамус.
Движение крови по сосудам происходит главным образом благода-
ря нагнетательной функции сердца и сокращению мышц. Сердце — это
насос, нагнетающий кровь в сосуды. Физическая активность облегчает
работу сердца, а гиподинамия требует усиленной работы сердца, что
является одним из важных факторов нарушения его функции.
Кровь течет из аорты, в которой давление высокое (в среднем
100 мм рт. ст.), через капилляры, где давление очень низкое (15–25 мм
рт. ст.), через систему сосудов, в которых давление прогрессивно
уменьшается. Из капилляров кровь поступает в венулы (давление
12–15 мм рт. ст.), затем в вены (давление 3–5 мм рт. ст.) . В полых ве-
нах, по которым венозная кровь оттекает в правое предсердие, давле-
ние всего 1–3 мм рт. ст., а в самом предсердии — около 0 мм рт. ст.
Соответственно, уменьшается с 50 см/сек. в аорте до 0,07 см/сек.
в капиллярах и венулах.
В работе сердца чередуются сокращение (систола) и расслабление
(диастола). Во время общего расслабления сердца (диастола) кровь из
полых и легочных вен поступает, соответственно, в правое и левое пред-
сердия. После этого наступает сокращение (систола) предсердий, в ре-
зультате чего кровь из предсердий через предсердно-желудочковые от-
верстия нагнетается в желудочки. Затем начинается волна сокращений
желудочков, которая распространяется на оба желудочка, и кровь на-
гнетается в отверстия легочного ствола и аорты, в это время предсерд-
но-желудочковые клапаны закрываются. После этого наступает пауза.
Систола предсердий длится 0,1 сек., систола желудочков — 0,3 сек.,
общая пауза — 0,4 с. Эти три фазы составляют сердечный цикл. Во
время одного сердечного цикла предсердия сокращаются 0,1 сек. и от-
дыхают 0,7 сек.; желудочки, соответственно, — 0,3 сек. и 0,5 сек. В те-
чение суток сердце сокращается 8 часов и 16 часов отдыхает.
В связи с изменением давления в полостях сердца клапаны сердца,
легочной артерии и аорты открываются или закрываются. В начале
систолы желудочков предсердно-желудочковые клапаны закрывают-
ся, а полулунные клапаны аорты и легочной артерии открываются.
В период диастолы желудочков происходит систола предсердий,
предсердно-желудочковые клапаны открываются, и желудочки за-
полняются кровью. Возвращению крови из аорты и легочного ствола
препятствуют полулунные клапаны.
Во время систолы и диастолы возникают тоны сердца: первый —
систолический, более низкий и продолжительный, второй — диасто-
лический, короткий, высокий. Систолический тон, возникающий при
закрывании левого предсердного клапана, прослушивают в пятом
межреберье слева от грудины в области верхушки сердца; систоличе-
339
КРОВЕНОСНАЯ СИСТЕМА
ский тон, возникающий при закрытии правого предсердно-желудоч-
кового клапана, в месте соединения тела и мечевидного отростка гру-
дины. Диастолический тон аортального клапана прослушивают во
втором межреберье справа от грудины; диастолический тон клапана
легочного ствола — во втором межреберье слева от грудины. Сердеч-
ный толчок, возникающий вследствие изменения положения сердца
при систоле, благодаря чему левый желудочек ударяется о грудную
стенку, определяется в пятом межреберье слева от грудины (рис. 165).
Частота сердечных сокращений в минуту составляет в возрасте
одного года около 125 ударов в 1 мин., в два года — 105, в три года —
100,вчетыре—97,ввозрастеотпятидодесяти—90,с10до15—
75—78,с15до50—70,с50до60—74,с60до80—80.Несколько
любопытных цифр: в течение суток сердце бьется около 108 000 раз,
в течение жизни — 2 800 000 000—3 100 000 000 раз; через сердце про-
ходит 225—250 млн л крови.
Сердце приспосабливается к постоянно изменяющимся условиям
жизни человека. В покое желудочки взрослого человека выталкивают
в сосудистую систему около 5 л крови в минуту. Этот показатель —
минутный объем кровообращения (МОК) — при тяжелой физической
работе возрастает в 5—6 раз. Соотношение между МОК в покое и при
максимально напряженной мышечной работе говорит о функцио-
нальных резервах сердца, а значит, о функциональных резервах здо-
ровья. В то же время кровоток через сосуды самого сердца достигает
5% общего МОК. При интенсивной физической работе этот показа-
тель возрастает в 3—4 раза. Количество крови, выбрасываемое ка-
ждым желудочком во время систолы, составляет от 70 до 100 мл — это
ударный, или систолический, объем крови. Этот показатель также
увеличивается при физической нагрузке.
Средняя масса сердца взрослого человека составляет 300—320 г
(0,5% массы тела), в то же время в покое сердце потребляет около
25—30 мл O2 в минуту — около 10% общего потребления О2 в покое.
Рис. 165. Проекции точек
прослушивания тонов сердца:
1 — области прослушивания I тона,
2 — области прослушивания II тона
(по Коробкову и Чесноковой)
11
22
340
КРОВЕНОСНАЯ СИСТЕМА
При интенсивной мышечной деятельности потребление О2 сердцем
возрастает в 3—4 раза. В зависимости от нагрузки коэффициент по-
лезного действия (КПД) сердца составляет от 15 до 40%. Напомним,
что КПД современного тепловоза достигает 14—15%.
Биоэлектрическая активность сердца регистрируется с помощью
электрокардиографии, полученная кривая называется электрокардио-
граммой (ЭКГ). Впервые ЭКГ была записана в 1887 г. А . Уоллером.
В начале XX в. В. Эйнтховен разработал прибор для точной регистра-
ции небольших колебаний электрических потенциалов — струнный
гальванометр. Эйнтховен предложил также три точки тела, на кото-
рые следует накладывать электроды. При положении электродов на
правой и левой руках образуется отведение I, на правой руке и левой
ноге — отведение II, а на левой руке и левой ноге — отведение III. Эти
три отведения образуют равносторонний треугольник, и по их пара-
метрам можно определить угол, под которым сердце расположено
в грудной клетке. Согласно закону Эйнтховена, сумма потенциалов
в отведении I и III равна потенциалу в отведении II. В 1924 г. Эйнтхо-
вену была присуждена Нобелевская премия «за открытие механизма
электрокардиограммы». Нормальная ЭКГ состоит из нескольких зуб-
цов и комплекса колебаний, которые Эйнтховен назвал Р, QRS и Т.
Небольшой зубец Р отражает электрическую активность предсердий,
а быстрый высокоамплитудный комплекс QRS и более медленный
зубец Т — электрическую активность желудочков (рис. 166).
R
P
Q
T
(U)
S
<0,1с
Èíòеðâаë
,с
PQ<02
Дëèòеëüíîсòü
Êаëèбð.
1–
ìВ
+
0
–
Ç
ó
б
е
ö
Ð
С
е
г
ì
е
í
ò
P
Q
Ê
î
ì
ï
ë
е
ê
с
Q
R
S
С
е
г
ì
е
í
ò
S
T
Ç
ó
б
е
ö
Т
Ç
ó
б
е
ö
U
(
ï
î
с
ò
î
я
í
е
í
)
Èíòеðâаë Т
Q
çаâèсèò îò ÷асòîòû:
ïðè 70/ìèí
0,32 0 ,39
–
<0,12с
Рис. 166. Нормальная ЭКГ
человека, полученная путем
биполярного отведения
от поверхности тела
в направлении длинной
оси сердца (по Г. Антони)
341
КРОВЕНОСНАЯ СИСТЕМА
Кровоснабжение тела человека
Большой и малый круги кровообращения человека разобщены
(рис. 167).
Малый, или легочный, круг кровообращения начинается в пра-
вом желудочке сердца, из которого выходит легочный ствол, разделя-
ющийся на правую и левую легочные артерии, а последние развет-
вляются в легких соответственно ветвлению бронхов на артерии,
переходящие в капилляры. В капиллярных сетях, оплетающих альве-
олы, кровь отдает углекислый газ и обогащается кислородом. Артери-
альная кровь поступает из капилляров в вены, которые укрупняются
и по две с каждой стороны впадают в левое предсердие, где и заканчи-
вается малый круг кровообращения.
Большой, или телесный, круг кровообращения служит для до-
ставки всем органам и тканям тела питательных веществ и кислорода.
Он начинается в левом желудочке сердца, куда из левого предсердия
поступает артериальная кровь. Из левого желудочка выходит аорта,
от которой отходят артерии, идущие ко всем органам и тканям тела
и разветвляющиеся в их толще вплоть до артериол и капилляров —
последние переходят в венулы и далее в вены (рис. 168). Вены слива-
ются в два крупных ствола — верхнюю и нижнюю полые вены, кото-
рые впадают в правое предсердие сердца, где и заканчивается большой
круг кровообращения. Дополнением большого круга является третий
11
11
11
22
12
12
33
13
13
44
14
14
55
15
15
66
16
16
77
17
17
88
18
18
99
19
19
10
10
Рис. 167. Кровоснабжение тела человека.
Схема большого и малого кругов кровообращения:
1 — капилляры головы, верхних отделов туловища
и верхних конечностей, 2 — общая сонная артерия,
3 — легочные вены, 4 — дуга аорты, 5 — левое
предсердие, 6 — левый желудочек, 7 — аорта,
8 — печеночная артерия, 9 — капилляры печени,
10 — капилляры нижних отделов туловища и нижних
конечностей, 11 — верхняя брыжеечная артерия,
12 — нижняя полая вена, 13 — воротная вена,
14 — печеночные вены, 15 — правый желудочек,
16 — правое предсердие, 17 — верхняя полая вена,
18 — легочный ствол, 19 — капилляры легких
342
КРОВЕНОСНАЯ СИСТЕМА
(сердечный) круг кровообращения, обслуживающий само сердце.
Он начинается выходящими из аорты венечными артериями сердца
и заканчивается венами сердца. Последние сливаются в венечный
синус, впадающий в правое предсердие.
Аорта, расположенная слева от средней линии тела, своими ветвя-
ми кровоснабжает все органы и ткани. Она начинается расширени-
ем — луковицей аорты, от которой отходят правая и левая венечные
артерии. Луковица переходит в восходящую часть аорты. Изгибаясь
влево, дуга аорты переходит в нисходящую часть аорты. От вогнутой
стороны дуги аорты начинаются ветви к трахее, бронхам и к тимусу,
от выпуклой стороны дуги отходят три крупных сосуда: справа — пле-
чеголовной ствол, слева — левые общая сонная и подключичная
артерии.
Плечеголовной ствол длиной около 3 см направляется вверх, назад
и вправо, впереди трахеи, и на уровне правого грудино-ключичного
сустава делится на правые общую сонную и подключичную артерии.
1
2
3
4
56
7
8
9
10
11
12
13
14
15
16
17
18
Á
19
20
21
22
23
24
25
26
27
28
29
30 31
32
11
21
21
11
11
22
22
22
12
12
33
23
23
13
13
44
24
24
14
14
55
25
25
15
15
66
26
26
16
16
77
27
27
17
17
88
28
28
18
18
99
29
29
19
19
10
10
30
30
31
31
32
32
20
20
Рис. 168. Кровеносная система человека,
вид спереди:
1 — левая общая сонная артерия,
2 — внутренняя яремная вена,
3 — дуга аорты, 4 — подключичная вена,
5 — легочная артерия (левая),
6 — легочный ствол, 7 — левая легочная
вена, 8 — левый желудочек (сердца),
9 — нисходящая часть аорты,
10 — плечевая артерия, 11 — левая
желудочная артерия, 12 — нижняя полая
вена, 13 — общие подвздошные артерия
и вена, 14 — бедренная артерия,
15 — подколенная артерия, 16 — задняя
большеберцовая артерия, 17 — передняя
большеберцовая артерия, 18 — тыльные
артерия и вены и стопы, 19 — задние
большеберцовые артерия и вены,
20 — бедренная вена, 21 — внутренняя
подвздошная вена, 22 — наружные
подвздошные артерия и вена,
23 — поверхностная ладонная дуга
(артериальная), 24 — лучевые артерия
и вены, 25 — локтевые артерия и вены,
26 — воротная вена печени,
27 — плечевые артерия и вены,
28 — подмышечные артерия и вена,
29 — верхняя полая вена, 30 — правая
плечеголовная вена, 31 — плечеголовной
ствол, 32 — левая плечеголовная вена
343
КРОВЕНОСНАЯ СИСТЕМА
Общая сонная артерия (правая и левая) на уровне верхнего края
щитовидного хряща делится на наружную сонную артерию, развет-
вляющуюся вне полости черепа, и внутреннюю сонную артерию, про-
ходящую внутрь черепа и направляющуюся к мозгу. Наружная сон-
ная артерия снабжает кровью наружные части головы и шеи, полости
рта и носа, щитовидную железу, гортань, язык, нёбо, миндалины, гру-
дино-ключично-сосцевидную и затылочные мышцы, поднижнече-
люстную, подъязычную и околоушную слюнные железы, кожу, кости
и мышцы головы (мимические и жевательные), зубы верхней и ниж-
ней челюстей, твердую мозговую оболочку, наружное и среднее ухо.
Внутренняя сонная артерия направляется вверх к основанию черепа,
не отдавая ветвей, входит в полость, где делится на ряд ветвей, кото-
рые кровоснабжают мозг и глаза.
Подключичная артерия и ее ветви кровоснабжают шейный отдел
спинного мозга с его оболочками, ствол головного мозга, затылочные
и частично височные доли полушарий большого мозга, глубокие и от-
части поверхностные мышцы шеи, шейные позвонки, межреберные
мышцы первого, второго промежутков, часть мышц затылка, спины
и лопатки, диафрагму, кожу груди и верхней части живота, прямую
мышцу живота, молочную железу, гортань, трахею, пищевод, щито-
видную и вилочковую железы. Подключичная артерия переходит
в подмышечную артерию, которая кровоснабжает мышцы плечевого
пояса, кожу и мышцы боковой стенки грудной стенки, плечевой
и ключично-акромиальный суставы, содержимое подмышечной ямки.
Плечевая артерия является продолжением подмышечной; в локтевой
ямке делится на лучевую и локтевую артерии. Плечевая артерия кро-
воснабжает кожу и мышцы плеча, плечевую кость и локтевой сустав.
Лучевая артерия легко прощупывается в нижнем отделе вблизи ее
шиловидного отростка, будучи прикрытой лишь кожей и фасцией.
Здесь исследуют пульс. Лучевая артерия проходит на кисть и крово-
снабжает кожу и мышцы предплечья и кисти, лучевую кость, локте-
вой и лучезапястный суставы. Локтевая артерия располагается на
предплечье медиально в локтевой борозде параллельно локтевой ко-
сти, проходит на ладонную поверхность кисти. Она кровоснабжает
кожу и мышцы предплечья и кисти, локтевую кость, локтевой и луче-
запястный суставы. Локтевая и лучевая артерии образуют на кисти
две артериальные сети запястья: тыльную и ладонную, питающие
связки и суставы запястья, второй, третий, четвертый пальцы и две
артериальные ладонные дуги — глубокую и поверхностную.
Нисходящая часть аорты делится на две части: грудную и
брюшную. Грудная часть аорты снабжает кровью внутренние органы,
находящиеся в грудной полости, и ее стенки. Из грудной полости аор-
344
КРОВЕНОСНАЯ СИСТЕМА
та переходит в брюшную через аортальное отверстие диафрагмы. На
уровне IV поясничного позвонка аорта делится на две общие под-
вздошные артерии, после чего продолжается в виде срединной крест-
цовой артерии.
Брюшная часть аорты кровоснабжает брюшные внутренности
и стенки живота. Наиболее крупная ветвь — короткий толстый чрев-
ный ствол, который кровоснабжает желудок, двенадцатиперстную
кишку, поджелудочную железу, печень с желчным пузырем, селезен-
ку, малый и большой сальники. Верхняя и нижняя брыжеечные арте-
рии, отходящие от брюшной аорты, кровоснабжают поджелудочную
железу, тонкую кишку, толстую кишку.
Общая подвздошная артерия — это самая крупная артерия челове-
ка (за исключением аорты). Каждая из них делится на две артерии.
Внутренняя подвздошная артерия питает тазовую кость, крестец
и всю массу мышц малого и большого таза, ягодичной области, отча-
сти мускулы бедра, а также внутренности, расположенные в малом
тазу: прямую кишку, мочевой пузырь; у мужчин — семенные пузырь-
ки, семявыносящий проток, предстательную железу; у женщин —
матку и влагалище; наружные половые органы и промежность. На-
ружная подвздошная артерия кровоснабжает мышцы бедра, у муж-
чин — мошонку, у женщин — лобок и большие половые губы. Бед-
ренная артерия, являющаяся непосредственным продолжением
наружной подвздошной артерии, кровоснабжает бедренную кость,
кожу и мышцы бедра, кожу передней брюшной стенки, наружные по-
ловые органы, тазобедренный сустав.
Подколенная артерия является продолжением бедренной. Она ле-
жит в одноименной ямке, переходит на голень, где сразу же делится
на переднюю и заднюю большеберцовые артерии. Артерия кровоснаб-
жает кожу и близлежащие мышцы бедра и задней поверхности голе-
ни, коленный сустав. Задняя большеберцовая артерия кровоснабжает
кожу задней поверхности голени, кости, мышцы голени, коленный
и голеностопный суставы, мышцы стопы. Передняя большеберцовая
артерия кровоснабжает кожу и мышцы передней поверхности голени
и тыла стопы, коленный и голеностопный суставы, на стопе перехо-
дит в тыльную артерию стопы. Обе большеберцовые артерии образу-
ют на стопе подошвенную артерию-дугу, которая лежит на уровне
оснований плюсневых костей. От дуги отходят артерии, питающие
кожу и мышцы стопы и пальцев.
Вены большого круга кровообращения разделяются на три сис-
темы: система верхней полой вены; система нижней полой вены,
включающая систему воротной вены печени; система вен сердца,
образующих венечный синус сердца (рис. 169). Главный ствол ка-
345
КРОВЕНОСНАЯ СИСТЕМА
7
8
9
10
11
12
13
14
15
16
17
4
3
2
5
6
1
31
18
19
20
21
22
23
24
25
26
27
28
29
30
Рис. 169. Верхняя и нижняя полые вены и их притоки, вид спереди:
1 — яремная венозная дуга, 2 — внутренняя яремная вена, 3 — подключичная вена,
4 — левая плечеголовная вена, 5 — дуга аорты, 6 — латеральная подкожная вена
руки, 7 — медиальная подкожная вена руки, 8 — плечевая вена, 9 — верхняя
надчревная вена, 10 — нижняя полая вена, 11 — левая почечная вена, 12 — левая
яичниковая (яичковая) вена, 13 — левая нижняя надчревная вена, 14 — левая общая
подвздошная вена, 15 — внутренняя подвздошная вена, 16 — наружная подвздошная
вена, 17 — бедренная вена, 18 — глубокая вена бедра, 19 — поверхностная вена,
огибающая подвздошную кость, 20 — срединная крестцовая вена,
21 — поверхностная надчревная вена, 22 — правая яичниковая (яичковая) вена,
23 — правая почечная вена, 24 — задние межреберные вены, 25 — внутренняя
грудная вена, 26 — верхняя полая вена, 27 — правая плечеголовная вена,
28 — правая наружная яремная вена, 29 — правая внутренняя яремная вена,
30 — передняя яремная вена, 31 — правая позвоночная вена
346
КРОВЕНОСНАЯ СИСТЕМА
ждой из этих вен открывается самостоятельным отверстием в по-
лость правого предсердия. Вены системы верхней и нижней полых
вен аностомозируют между собой. Верхняя полая вена собирает
кровь из верхней половины тела, головы, шеи, верхней конечности
и грудной полости. Нижняя полая вена собирает кровь из нижних
конечностей, стенок и внутренностей таза и живота. Протоки ниж-
ней полой вены соответствуют парным ветвям аорты (за исключе-
нием печеночных).
Воротная вена собирает кровь из непарных органов брюшной по-
лости: селезенки, поджелудочной железы, большого сальника, желч-
ного пузыря и пищеварительного тракта, начиная с кардиального от-
дела желудка и кончая верхним отделом прямой кишки. В отличие от
всех прочих вен, воротная вена, войдя в ворота печени, вновь распада-
ется на все более мелкие ветви, вплоть до синусоидальных капилля-
ров печени, которые впадают в центральную вену в дольке. Из цент-
ральных вен образуются поддольковые вены, которые, укрупняясь,
собираются в печеночные вены, впадающие в нижнюю полую вену
(см. раздел «Органы пищеварения»).
Функция сердечно-сосудистой
системы
Кровь непрерывно движется по замкнутой сосудистой системе
в определенном направлении благодаря ритмичным сокращениям
сердца — мышечного насоса, перекачивающего кровь из вен в арте-
рии. У здорового человека количество притекающей к сердцу крови
равно количеству оттекающей. Скорость тока крови по артериям, ка-
пиллярам, венам различная и зависит от ширины просвета этих сосу-
дов. По капиллярам большого круга кровообращения кровь течет
медленно со скоростью 0,5 мм в 1 секунду. Медленное движение кро-
ви по капиллярам способствует взаимодействию между кровью и
прилежащими к капиллярам тканям. Наиболее быстро кровь движет-
ся в аорте (50 см в секунду), что в 1000 раз превышает скорость дви-
жения в капиллярах. Скорость тока крови в венах в два раза меньше,
чем в артериях, поскольку суммарная ширина просвета вен в два раза
больше, чем у артерий. Эти обменные процессы совершаются на ог-
ромной площади — 6300 м2. Такова общая поверхность стенок капил-
ляров в теле человека.
Из крови в ткани диффундирует кислород, питательные вещества,
гормоны. Из тканей в кровь через тонкие стенки капилляров выво-
дятся продукты обмена веществ. Поступление кислорода и других пи-
347
КРОВЕНОСНАЯ СИСТЕМА
тательных веществ в ткани происходит благодаря высокому давлению
крови в начальных отделах капилляров (до 30 мм рт. ст.). В венозном
отделе капилляров давление крови низкое (около 15 мм рт. ст.) и про-
дукты, подлежащие выведению из организма, поступают из тканей
в кровь (углекислый газ, мочевина и другие вещества).
Давление крови в сосудах (кровяное давление) — давление, которое
кровь оказывает на стенки кровеносных сосудов. Артериальное дав-
ление (АД) — это важнейший показатель, отражающий деятельность
сердечно-сосудистой системы в целом. Стабильность АД поддержи-
вается механизмами гомеостаза. Максимальное давление во время си-
столы называется систолическим давлением, минимальное во время
диастолы — диастолическим, разница между ними составляет пульсо-
вое давление.
У человека традиционно АД исследуют с помощью ртутного мано-
метра, поэтому его выражают в миллиметрах ртутного столба (в на-
стоящее время существует множество модификаций аппаратов для
определения АД). Для измерения АД по методу Рива — Роччи ниж-
нюю треть плеча оборачивают надувной резиновой манжетой аппара-
та, в которую с помощью ручной резиновой груши накачивают воз-
дух. Для выпуска воздуха служит клапан, поэтому можно установить
давление на любом уровне и измерить его с помощью манометра, сое-
диненного с манжетой. Стетоскоп накладывают на кожу передней
локтевой области в зоне прохождения плечевой артерии. В результате
нагнетания воздуха в манжету плечевая артерия сдавливается. Затем
медленно открывают клапан, воздух начинает выходить, поэтому дав-
ление в манжете уменьшается. Когда оно становится ниже систоличе-
ского, кровь поступает в артерию, начинают прослушиваться корот-
кие четкие звуки, или пульсовые удары, определяется систолическое
давление (рис. 170).
Давление в манжете, при котором звуки пульсовых ударов вновь
быстро исчезают, соответствует диастолическому. У человека в возра-
сте от 20 до 40 лет систолическое давление составляет 100–120 мм рт.
ст., диастолическое 70–80 мм рт. ст. Разницу между систолическим
и диастолическим давлением называют пульсовым давлением. Его ве-
личина в норме составляет 40–50 мм рт. ст. Индивидуальные неболь-
шие отклонения от этих цифр вполне обычны. Мышечное напряже-
ние, некоторые эмоциональные факторы (испуг, стресс, возбуждение)
приводят к повышению систолического давления. Обычно наимень-
шее систолическое давление наблюдается у человека во время сна.
Сильное потрясение может вызывать аномальное снижение давления
и привести к развитию недостаточности кровообращения.
348
КРОВЕНОСНАЯ СИСТЕМА
Пульс — последовательное ритмичное колебание стенок артерий,
возникающее в результате сокращений левого желудочка и соответст-
вующее частоте сердечных сокращений (обычно она характеризуется
числом сердечных сокращений за одну минуту). Пульс легко опреде-
ляется на таких поверхностных артериях, как лучевая (вблизи запя-
стья) и сонная (в области шеи). В среднем пульс взрослого человека в
покое составляет 60—80 ударов в минуту, однако физическая нагруз-
ка, травмы, заболевания и эмоциональное возбуждение могут приво-
дить к его значительному учащению. Пульс исследуется путем про-
стого прощупывания лучевой артерии в области лучезапястного
сустава, при этом следует обращать внимание на частоту пульса, его
ритм (ритмичный, аритмичный), высоту (высокий, низкий), напря-
жение (твердый, мягкий). Частота пульса зависит от физической ра-
боты и эмоционального состояния, высота — от ударного объема кро-
ви, напряжение — от артериального давления.
Общий объем крови в сосудах мужчин составляет в среднем 75—
77 мл/кг массы тела (около 5,4 л), женщин — 65 мл/кг (около 4,5 л).
Лишь около 9% всей крови взрослого человека находится в сосудах
малого круга кровообращения, около 84% — в сосудах большого круга
кровообращения и около 7% — в полостях сердца.
Ïëå÷åâàÿ àðòåðèÿ
Ìàíîìåòð
Ãðóøà
Êëàïàí
Äàâëåíèå êðîâè (â ïëå÷åâîé àðòåðèè)
Äàâëåíèå â ìàíæåòêå
Ñèñòîëè÷åñêîå
äàâëåíèå
Äèàñòîëè÷åñêîå
äàâëåíèå
Çâóêè Êîðîòêîâà
Âðåìÿ
êÏà ìì ðò. ñò.
20
15
10
0
150
125
75
100
0
Ìàíæåòêà Ïëå÷î
Рис. 170. Измерение кровяного давления непрямым методом
(по Зильбернаглю и Лангу)
349
КРОВЕНОСНАЯ СИСТЕМА
Движение крови по венам. Кровь по венам возвращается к серд-
цу. Движение крови по венам обеспечивается не силой сердечных
сокращений, а другими факторами. Давление крови, создаваемое со-
кращениями сердца, в начальных отделах вен (в венулах) низкое,
всего 10—15 мм рт. ст. Движению крови по тонкостенным венам
в сторону сердца способствуют: сокращение близлежащих к венам
скелетных мышц, которые сдавливают вены и тем самым проталки-
вают кровь к сердцу; наличие клапанов, которые препятствуют
обратному току крови и пропускают ее только в направлении серд-
ца; при дыхательных движениях отрицательное давление в грудной
полости оказывает присасывающее действие и помогает движению
крови по венам к сердцу.
Регуляция функций
сердечно-сосудистой системы
Деятельность сердца, тонус стенок кровеносных сосудов и поддер-
жание постоянства кровяного давления регулируются вегетативной
нервной системой, неподконтрольной нашему сознанию. Нервные
импульсы о состоянии сосудов, их тонуса поступают по сердечным
нервам в сосудодвигательный центр, расположенный в продолгова-
том мозге. При снижении давления крови в сосудах импульсы из со-
судодвигательных центров усиливают сокращения сердца, повышают
тонус сосудистых стенок, сосуды суживаются, давление крови в них
выравнивается. При повышении давления сила и частота сердечных
сокращений уменьшаются, тонус сосудов также уменьшается, сосуды
расширяются, давление нормализуется. Благодаря рефлекторным ме-
ханизмам осуществляется саморегуляция сосудистого тонуса и уров-
ня давления крови в сосудах.
В регуляции сосудистого тонуса (и, соответственно, давления
крови в сосудах) участвуют также гуморальные механизмы. Измене-
ния химического состава крови влияют на возбудимость и проводи-
мость нервных импульсов в сердце, на силу и частоту сердечных со-
кращений. При всплеске эмоций (радость, страх, гнев) в кровь
выбрасываются гормоны надпочечников (адреналин и норадрена-
лин), усиливающие работу сердца и суживающие сосуды. Гормон
гипоталамуса вазопрессин также суживает сосуды. Сосудорасширя-
ющее действие оказывают ацетилхолин, гистамин и другие биологи-
чески активные вещества. В экстремальных ситуациях, например
при больших кровопотерях, тонус сосудов поддерживается благода-
ря выбросу крови из так называемых кровяных депо (кожа, печень
350
КРОВЕНОСНАЯ СИСТЕМА
и др.). В то же время при потере более 30% крови биологические
механизмы не в состоянии обеспечить непрерывный ток крови, и че-
ловек может погибнуть.
ЛИМФАТИЧЕСКАЯ СИСТЕМА
Лимфатическая система, осуществляющая дренаж, по которому
тканевая жидкость оттекает в кровеносное русло, состоит из развет-
вленных в органах и тканях лимфатических капилляров (лимфока-
пилляров), лимфокапиллярных сетей, лимфатических сосудов, ство-
лов и протоков. По пути следования лимфатических сосудов лежат
лимфатические узлы, относящиеся к органам иммунной системы.
Тканевая жидкость (жидкость в межклеточных пространствах раз-
личных тканей) образуется в результате фильтрации из капилляров.
Она содержит воду, вещества, поступающие из крови, и продукты об-
мена. Лимфа (от греч. lympha — чистая вода) образуется из тканевой
жидкости. В норме у взрослого человека за сутки вырабатывается
около 2 л лимфы, в которой содержится около 20 г/л белка и огром-
ное количество лимфоцитов. Движение лимфы осуществляется бла-
годаря мышечным сокращениям; в тех лимфатических сосудах, где
имеются гладкие миоциты, лимфа движется благодаря их сокращени-
ям. Клапаны препятствуют обратному току лимфы.
Лимфатические капилляры осуществляют вместе с венами дренаж
тканей — всасывание воды и растворенных в ней кристаллоидов,
а также удаляют из тканей инородные частицы (обломки разрушен-
ных клеток, микробные тела). Капилляры образуют лимфокапилляр-
ные сети. По лимфатическим сосудам лимфа из капилляров течет
к региональным лимфатическим узлам и крупным коллекторным
лимфатическим стволам. По крупным лимфатическим коллекторам
—
к стволам (яремные, кишечные, бронхосредостенные, подключич-
ные, поясничные) и протокам (грудной, правый лимфатический), по
которым лимфа оттекает в вены. Стволы и протоки впадают в веноз-
ный угол справа и слева, образованный слиянием внутренней ярем-
ной и подключичной вен, или в одну из этих вен у места соединения
их друг с другом. Лежащие по пути тока лимфы лимфатические узлы
выполняют барьерно-фильтрационную, лимфоцитопоэтическую, им-
мунопоэтическую функции.
Лимфа образуется из тканевой жидкости, в ней содержится около
20 г/л белка. Скорость тока лимфы мала, однако она возрастает в 10—
15 раз при физической нагрузке, так как именно мышечные сокраще-
ния в основном способствуют движению лимфы.
351
ЛИМФОИДНАЯ СИСТЕМА
ЛИМФОИДНАЯ СИСТЕМА
(органы кроветворения и иммунной системы)
Иммунитет (от лат. immunitas — освобождение от чего-либо) — за-
щита организма от генетически чужеродных организмов и веществ,
к которым относятся микроорганизмы, вирусы, черви, различные бел-
ки, клетки, в том числе и измененные собственные. Известно, что при
клеточном делении, которое постоянно происходит в организме чело-
века, одна из миллиона образовавшихся клеток мутантная, т. е . генети-
чески чужеродна. В организме человека благодаря мутациям в каждый
конкретный момент должно быть около 10—20 млн генетически чуже-
родных клеток. Их совместное неправильное функционирование
должно было бы быстро привести к гибели организма. Почему же это-
го не происходит? Ответ на этот вопрос дали лауреаты Нобелевской
премии П. Медавар и Ф. Вернет. П . Медавар доказал, что механизмы
иммунитета удивительно точны. Они способны отличить чужеродную
клетку, содержащую всего один нуклеотид, отличающийся от генома
собственного организма. Ф. Вернет постулировал положение (назван-
ное «аксиомой Бернета»), что центральным биологическим механиз-
мом иммунитета является распознавание своего и чужого.
ВНИМАНИЕ!
Благодаря иммунитету уничтожаются и собственные клетки организма,
которые изменились генетически. А это происходит постоянно. Меха-
низмы иммунитета удивительно точные и целесообразные. Основной
биологический принцип иммунитета — распознание своего и чужого.
Основоположниками иммунологии — науки об иммунитете яв-
ляются Луи Пастер, Илья Мечников и Пауль Эрлих. В 1881 г.
Л. Пастер разработал принципы создания вакцин из ослабленных
микроорганизмов с целью предупреждения развития инфекционных
заболеваний. И. Мечников создал клеточную (фагоцитарную) тео-
рию иммунитета. П . Эрлих открыл антитела и создал гуморальную
теорию иммунитета, установив, что антитела передаются ребенку
с грудным молоком, создавая пассивный иммунитет. Эрлих разрабо-
тал метод изготовления дифтерийного антитоксина, благодаря чему
были спасены миллионы детских жизней. В 1908 г. И. Мечников
и П. Эрлих были удостоены Нобелевской премии за работы по тео-
рии иммунитета.
Выше мы писали об открытии в 1900 г. групп крови К. Ландштей-
нером. Он первым доказал наличие иммунологических различий ин-
дивидуумов в пределах одного вида. Известно, что организм отторга-
352
ЛИМФОИДНАЯ СИСТЕМА
ет пересаженные чужеродные ткани. В 40-х гг. XX в. было доказано,
что этот процесс опосредован иммунологическими механизмами. Од-
нако отторжение происходит не сразу и зависит от другого феноме-
на — иммунологической толерантности, открытой в 1953 г. одновре-
менно и независимо друг от друга П. Медаваром и М. Гашеком.
Иммунологическая толерантность (от лат. tolerantia — терпение) —
это распознавание и специфическая терпимость (напомним, что меха-
низмы иммунитета, распознавая чужеродное, нетерпимы к нему).
ВНИМАНИЕ!
Кроветворным органом у человека после его рождения является только
костный мозг.
Лимфоидная система объединяет органы и ткани, обеспечиваю-
щие защиту организма от генетически чужеродных клеток или ве-
ществ, поступающих извне или образующихся в организме. В состав
лимфоидной системы, по современным данным, входят все органы,
которые участвуют в образовании клеток, осуществляющих защит-
ные реакции организма: лимфоцитов, плазматических клеток. Лим-
фоидные органы построены из лимфоидной ткани, которая представ-
ляет собой ретикулярную строму и расположенные в ее петлях клетки
лимфоидного ряда: лимфоциты различной степени зрелости, моло-
дые и зрелые плазматические клетки, а также макрофаги и другие
клеточные элементы.
К лимфоидным органам относят: костный мозг, тимус, скопления
лимфоидной ткани, расположенные в стенках полых органов пищева-
рительной, дыхательной систем и мочеполового аппарата (миндалины,
лимфоидные (пейеровы) бляшки тонкой кишки, одиночные лимфоид-
ные узелки в слизистых оболочках внутренних органов), лимфатиче-
ские узлы, селезенку (рис. 171). Костный мозг и тимус, в которых из
стволовых клеток дифференцируются лимфоциты, являются цент-
ральными органами лимфоидной системы, остальные являются пери-
ферическими органами иммуногенеза — в эти органы лимфоциты вы-
селяются из центральных органов. Центральные органы лимфоидной
системы расположены в хорошо защищенных от внешних воздействий
местах. Периферические расположены на путях возможного внедрения
в организм генетически чужеродных веществ или на путях следования
таких веществ, образовавшихся в самом организме.
Органы лимфоидной системы вырабатывают иммунокомпонент-
ные клетки, в первую очередь лимфоциты, а также плазмоциты (плаз-
матические клетки), включают их в иммунный процесс, распознают
и уничтожают проникшие в организм или образовавшиеся в нем
353
ЛИМФОИДНАЯ СИСТЕМА
клетки и другие чужеродные
вещества. Антигены (от греч.
anti — приставка, обозна-
чающая противоположность,
genos — род, происхождение) —
это вещества, которые несут
признаки генетической чуже-
родности. При их введении в
организм развиваются специ-
фические иммунологические
реакции, в нем образуются
нейтрализующие их защитные
вещества — антитела, являю-
щиеся
иммуноглобулинами
(гуморальный
иммунитет),
или специфически реагирующие лимфоциты (клеточный иммуни-
тет). Т-лимфоциты обеспечивают осуществление клеточного (в основ-
ном) и гуморального иммунитета: они уничтожают чужеродные,
а также собственные измененные или погибшие клетки. В-лимфоци-
ты выполняют функции гуморального иммунитета. Производные
В-лимфоцитов — плазматические клетки — синтезируют и выделяют
в кровь и в секреты желез антитела, которые способны вступать в со-
единение с соответствующими антигенами и нейтрализовать их.
Антитела связываются с антигенами, что дает возможность поглощать
их фагоцитами. Антитела специфичны. Известно, что после некото-
рых инфекционных заболеваний, например дифтерии, человек не за-
болевает повторно. У него возникает активный иммунитет. Но если
перелить ребенку, не болевшему дифтерией, сыворотку крови челове-
ка, переболевшего ею, то первый становится невосприимчивым к диф-
терии, т. е . у него возникает пассивный иммунитет.
11
22
33
44
55
66
77
88
Рис. 171 . Схема расположения
центральных и периферических
органов иммунной системы человека:
1 — костный мозг, 2 — миндалины
лимфоидно-глоточного кольца,
3 — тимус, 4 — лимфатические узлы
(подмышечные), 5 — селезенка,
6 — лимфоидная (пейерова) бляшка,
7 — аппендикс, 8 — лимфоидные
узелки
354
ЛИМФОИДНАЯ СИСТЕМА
Большинство из имеющихся в организме лимфоцитов многократ-
но циркулируют между различными средами обитания: органы лим-
фоидной системы, где эти клетки образуются, лимфатические сосуды,
кровь, вновь органы лимфоидной системы и т. д . При этом считают,
что в костный мозг и тимус лимфоциты повторно не попадают. Об-
щая масса лимфоцитов в теле взрослого человека равна примерно
1500 г (6 × 1012 клеток). У новорожденного общая масса лимфоцитов
в среднем составляет 150 г. Большинство циркулирующих лимфоци-
тов — это малые лимфоциты диаметром около 8 мкм. Именно малый
лимфоцит является главной иммунокомпетентной клеткой. Основ-
ное свойство клеток лимфоидной системы — их способность взаимо-
действовать с огромным количеством антигенов.
Одним из важнейших свойств лимфоидной системы является им-
мунологическая память. В результате первой встречи запрограммиро-
ванного лимфоцита с определенным антигеном образуются две кате-
гории клеток: эффекторные, которые немедленно выполняют
специфическую функцию — секретируют антитела или реализуют
клеточные реакции, и клетки памяти, которые циркулируют длитель-
ное время. При повторном поступлении данного антигена они быстро
превращаются в лимфоциты-эффекторы, которые вступают в реак-
цию с антигеном. При каждом делении запрограммированного лим-
фоцита после его встречи с антигеном количество клеток памяти уве-
личивается.
Для осуществления иммунного ответа совершенно необходима
кооперация Т- и В-лимфоцитов. Однако для реализации иммунного
ответа недостаточно лишь Т- и В-лимфоцитов. Согласно современ-
ной трехклеточной схеме кооперации, образование антител осу-
ществляется благодаря совместной функции макрофага, Т- и В-лим -
фоцитов.
Один В-лимфоцит производит сотни плазматических клеток.
Плазматические клетки — это микроскопические фабрики, каждая из
которых вырабатывает огромное количество антител, готовых сра-
зиться со строго определенным антигеном. Антитела — это иммуно-
глобулины (Ig). Известно пять типов Ig: IgA, IgD, IgE, IgG и IgM.
Около 75% всех Ig — это IgG, которые вместе с IgM воздействуют на
бактерии и вирусы, IgA защищают слизистые оболочки пищевари-
тельной, дыхательной, мочевой и половой систем, IgE участвуют в ал-
лергических реакциях. Увеличение содержания IgM в организме сви-
детельствует об остром, недавно начавшемся заболевании; увеличение
IgG — о хроническом процессе.
Помимо этого, лимфоциты вырабатывают лимфокины, среди них
интерферон, который образуется под воздействием вируса. Интер-
ферон, в свою очередь, стимулирует клетки к выработке противови-
355
КОСТНЫЙ МОЗГ
русных белков. Интерферон активен не только против вируса, выз-
вавшего его образование, но и против других вирусов. Кроме того,
интерферон способствует увеличению числа Т-лимфоцитов.
Итак, основная функция лимфоидной системы — это нейтрализа-
ция, разрушение или удаление генетически чужеродных веществ, по-
падание которых в организм и вызывает развитие иммунного ответа.
Иммунитет специфичен.
КОСТНЫЙ МОЗГ
Костный мозг является одновременно органом кроветворения
и иммунной системы. Выделяют красный костный мозг, который
у взрослого человека располагается в ячейках губчатого вещества
плоских и коротких костей, эпифизов длинных костей, и желтый
костный мозг, заполняющий костномозговые полости диафизов длин-
ных (трубчатых) костей. Общая масса костного мозга у взрослого че-
ловека равна примерно 2,5—8 кг (4,5—4,7% массы тела). Около поло-
вины его составляет красный костный мозг, остальное — желтый.
Красный костный мозг состоит из стромы и кроветворных клеток
на разных стадиях развития. В нем содержатся стволовые кровет-
ворные клетки — предшественницы всех клеток крови и лимфоци-
тов. Ретикулярная ткань в виде ретикулярных клеток и волокон
образует трехмерный каркас костного мозга. В его петлях находятся
молодые и зрелые клетки крови, макрофаги, тучные и другие клет-
ки. Костный мозг располагается в виде шнуров цилиндрической
формы вокруг артериол. Шнуры отделены друг от друга синусоид-
ными капиллярами. Созревшие клетки крови проникают в просветы
синусоидов через временные миграционные поры, образующиеся
в цитоплазме эндотелиальных клеток только в момент прохождения
клеток.
У новорожденного ребенка красный костный мозг занимает все
костномозговые полости. Жировые клетки в красном костном мозге
впервые появляются после рождения (1—6 месяцев). После 4—5 лет
красный костный мозг в диафизах трубчатых костей постепенно на-
чинает замещаться желтым костным мозгом. К 20—25 годам желтый
костный мозг полностью заполняет костномозговые полости диафи-
зов трубчатых костей. Желтый костный мозг представлен в основном
жировой тканью, которая заместила ретикулярную. Что касается
костномозговых полостей плоских костей, то в них жировые клетки
составляют до 50% объема костного мозга. В старческом возрасте
костный мозг приобретает слизеподобную консистенцию (желатино-
вый костный мозг).
356
ЛИМФОИДНАЯ СИСТЕМА
ТИМУС
Тимус — центральный орган иммунной системы, в котором из кро-
ветворных стволовых клеток созревают и дифференцируются Т-лим-
фоциты, ответственные за реакции клеточного и гуморального имму-
нитета. Поступающие в тимус из костного мозга с током крови
стволовые клетки, пройдя ряд промежуточных стадий, превращаются
в конечном итоге в Т-лимфоциты. В дальнейшем Т-лимфоциты по-
ступают в кровь и лимфу, покидают с их током тимус и заселяют ти-
мусзависимые зоны периферических органов иммунной системы (се-
лезенки, лимфатических узлов). Тимус секретирует также вещества,
влияющие на дифференцировку Т-лимфоцитов.
Тимус, располагающийся за грудиной в передней части верхнего
средостения, состоит из двух вытянутых в длину асимметричных по
величине долей — правой и левой, сросшихся друг с другом в средней
их части или тесно соприкасающихся на уровне их середины. Тимус
достигает максимальных размеров к периоду полового созревания.
Его масса в 10—15 лет составляет в среднем 37,5 г. После 16 лет масса
тимуса постепенно уменьшается и в 16—20 лет равняется в среднем
25,5 г, а в 21—35 лет — 22,3 г. Лимфоидная ткань тимуса не исчезает
полностью даже в старческом возрасте. Она сохраняется, но в значи-
тельно меньшем количестве, чем в детском и подростковом возрасте.
Масса тимуса в 50—90 лет равна 13,4 г. В паренхиме тимуса рано по-
является жировая ткань.
Паренхима тимуса состоит из более темного, расположенного по
периферии долек коркового вещества и более светлого мозгового, за-
нимающего центральную часть долек. С возрастом зона коркового ве-
щества становится тоньше, преобладающим постепенно становится
мозговое вещество. В корковом веществе лимфоциты лежат более
плотно, чем в мозговом. Характерным для мозгового вещества явля-
ется наличие в нем слоистых телец тимуса (телец Гассаля). После
30–50 лет редко встречаются мелкие тельца.
ЛИМФОИДНАЯ ТКАНЬ СТЕНОК ОРГАНОВ
ПИЩЕВАРИТЕЛЬНОЙ И ДЫХАТЕЛЬНОЙ СИСТЕМ
Миндалины — небная и трубная (парные), язычная и глоточная
(непарные), расположенные в области зева, корня языка и носовой
части глотки, образуют защитное кольцо в области глотки. Они пред-
ставляют собой скопления диффузной лимфоидной ткани, содержа-
щие небольшие плотные лимфоидные узелки (фолликулы), располо-
женные в собственной пластинке слизистой оболочки.
357
ЛИМФОИДНАЯ ТКАНЬ
Язычная миндалина залегает в собственной пластинке слизистой
оболочки корня языка в виде скопления лимфоидной ткани. Она
достигает наиболее крупных размеров к 14—20 годам и состоит
из 80—90 лимфоидных узелков (фолликулов), число которых наи-
более велико в детском, подростковом и юношеском возрасте.
Основными клеточными элементами узелков являются лимфоциты
(до 95—98%).
Парная небная миндалина неправильной овоидной формы распо-
лагается в миндаликовой ямке (бухте), которая представляет собой
углубление между небно-язычной и небно-глоточной дужками. В соб-
ственной пластинке слизистой оболочки миндалины располагаются
округлые лимфоидные узелки. Наибольшее количество их наблюда-
ется в возрасте от 2 до 16 лет. К 8 —13 годам миндалины достигают
наибольших размеров, которые сохраняются примерно до 30 лет. Раз-
растание соединительной ткани внутри небной миндалины особенно
интенсивно происходит после 25—30 лет наряду с уменьшением ко-
личества лимфоидной ткани. После 40 лет в лимфоидной ткани редко
встречаются мелкие лимфоидные узелки.
Непарная глоточная миндалина располагается в области свода
и отчасти задней стенки глотки, между отверстиями правой и ле-
вой слуховых труб. Глоточная миндалина достигает наибольших
размеров в 8—20 лет, после 30 лет величина ее постепенно умень-
шается.
Парная трубная миндалина находится в области трубного валика,
отграничивающего сзади глоточное отверстие слуховой трубы. Труб-
ная миндалина достигает наибольшего развития в возрасте 4—7 лет.
Возрастная инволюция ее начинается в подростковом и юношеском
возрасте.
В толще слизистой оболочки и подслизистой основы органов пи-
щеварительной системы (глотки и пищевода, желудка, тонкой и тол-
стой кишки, желчного пузыря), а также органов дыхания (гортани,
трахеи, крупных бронхов) имеются одиночные лимфоидные узелки.
Они располагаются как сторожевые посты на протяжении всей длины
указанных органов, на различном расстоянии друг от друга и на раз-
личной глубине.
Групповые лимфоидные узелки червеобразного отростка (аппен-
дикса) в период их максимального развития (после рождения и до
16—17 лет) располагаются в слизистой оболочке и в подслизистой ос-
нове на всем его протяжении. Групповые узелки состоят из одиноч-
ных узелков, общее количество которых в стенке аппендикса у детей
и подростков достигает 450—550 штук. После 30—40 лет число узел-
358
ЛИМФОИДНАЯ СИСТЕМА
ков заметно уменьшается. У людей старше 60 лет лимфоидные узелки
в стенке червеобразного отростка встречаются редко.
Групповые (обобщенные) лимфоидные узелки, или — как их называ-
ли ранее — пейеровы бляшки, располагающиеся главным образом в
стенке подвздошной кишки, имеют вид плоских образований преиму-
щественно овальной или круглой формы, чуть-чуть выступающих в
просвет кишки. Количество их в детском возрасте достигает 50, в 16–
17 лет составляет 33–37. После 40 лет оно не превышает 20, а после
60 лет — 16. Групповые лимфоидные узелки построены из одиночных
узелков, между которыми располагаются тонкие пучки соединитель-
нотканных волокон.
ЛИМФАТИЧЕСКИЕ УЗЛЫ
Лимфатические узлы величиной от 0,5–1 мм до 7,5 см лежат на
пути следования лимфы от органов и тканей к лимфатическим прото-
кам и лимфатическим стволам. К выпуклой стороне каждого лимфа-
тического узла подходит 4–6 и более приносящих лимфатических со-
судов. После прохождения через лимфатический узел лимфа выходит
из него через 2–4 выносящих лимфатических сосуда, которые направ-
ляются или к следующему лимфатическому узлу этой же или сосед-
ней группы узлов, или к крупному коллекторному сосуду — протоку
или стволу. Лимфатические узлы располагаются группами, состоящи-
ми из двух узлов и более.
Каждый лимфатический узел покрыт соединительнотканной кап-
сулой, от которой внутрь узла отходят ответвления соединительной
ткани различной длины — капсулярные трабекулы (перекладины).
В том месте, где из лимфатического узла выходят выносящие лимфа-
тические сосуды, узел имеет небольшое вдавление — ворота, от кото-
рого в паренхиму лимфатического узла отходят воротные (хиларные)
трабекулы.
Внутри лимфатического узла между трабекулами находится стро-
ма, содержащая ретикулярные волокна и ретикулярные клетки, обра-
зующие трехмерную сеть, в петлях которой располагаются клеточные
элементы лимфоидного ряда. В паренхиме различают корковое и моз-
говое вещество. Корковое вещество занимает периферические отделы
узла, более светлое мозговое вещество лежит ближе к воротам узла в
его центральной части. В корковом веществе располагаются лимфо-
идные узелки округлой формы диаметром 0,5—1 мм, представляющие
собой скопления лимфоидных клеток, главным образом В-лимфоци-
тов. Вокруг лимфоидных узелков располагается диффузная лимфо-
359
СЕЛЕЗЕНКА
идная ткань. Кнутри от узелков, непосредственно на границе с мозго-
вым веществом, находится полоса лимфоидной ткани, получившая
название тимусзависимой паракортикальной зоны (околокорковое
вещество), содержащей преимущественно Т-лимфоциты.
Паренхима мозгового вещества представлена мякотными тяжа-
ми, которые соединяются друг с другом, образуя сложные перепле-
тения. Между мякотными тяжами располагаются мозговые проме-
жуточные синусы. Мякотные тяжи являются зоной скопления
В-лимфоцитов (как и лимфоидные узелки); здесь находятся также
плазматические клетки, макрофаги. Паренхима лимфатического
узла пронизана густой сетью каналов — лимфатическими синусами,
по которым поступающая в узел лимфа течет от подкапсульного
(краевого) синуса к воротному. Непосредственно под капсулой узла,
между капсулой и паренхимой, находится подкапсульный (краевой)
синус. В него впадают приносящие лимфатические сосуды, несущие
лимфу или от органа, для которого этот узел является регионарным,
или от предыдущего лимфатического узла. От подкапсульного сину-
са в паренхиму узла вдоль капсульных трабекул уходят промежу-
точные синусы коркового и мозгового вещества. Последние достига-
ют ворот лимфатического узла и впадают в воротный синус, из
которого берут начало выносящие лимфатические сосуды. В ворот-
ный синус впадает также подкапсульный (краевой) синус, охватыва-
ющий паренхиму органа по периферии и заканчивающийся в обла-
сти ворот узла.
СЕЛЕЗЕНКА
Селезенка находится на пути тока крови от кишечника к печени.
Она располагается в брюшной полости, в левом подреберье, на
уровне между IX—XI ребрами. Масса селезенки у взрослого мужчи-
ны составляет 192 г, у женщины — 153. Селезенка выполняет
многочисленные функции. Во внутриутробном периоде в ней обра-
зуются эритроциты и лимфоциты. После рождения селезенка не яв-
ляется кроветворным органом. В селезенке осуществляются важ-
ные иммунологические реакции. Антигены, циркулирующие в
крови, попадают в паренхиму селезенки, активируют лимфоциты,
способствуя их превращению в плазматические клетки, продуциру-
ющие антитела. Макрофаги селезенки фагоцитируют форменные
элементы крови, и в первую очередь эритроциты. При переварива-
нии эритроцитов освобождающееся из гемоглобина железо всасы-
360
ЛИМФОИДНАЯ СИСТЕМА
вается в кровь и повторно используется в костном мозге. Часть раз-
рушенного гемоглобина превращается макрофагами в билирубин.
В селезенке депонируется кровь и накапливаются клетки крови,
включая тромбоциты.
Строма селезенки образована ретикулярной тканью, в петлях ко-
торой расположены клетки крови, образующие паренхиму селезен-
ки — ее пульпу. Различают белую и красную пульпы. Белая пульпа
представляет собой типичную лимфоидную ткань, из которой состоят
лимфоидные узелки (фолликулы) селезенки и лимфатические пери-
артериальные влагалища, располагающиеся внутри красной пульпы,
которые в виде муфт окружают пульпарные артерии или начальные
отделы центральных артерий селезенки.
НЕСПЕЦИФИЧЕСКАЯ
СОПРОТИВЛЯЕМОСТЬ ОРГАНИЗМА
Наряду с иммунитетом организм человека обладает неспецифиче-
ской сопротивляемостью, которая зависит от многочисленных факто-
ров. К ним относится непроницаемость здоровой кожи и слизистых
оболочек для микроорганизмов; непроницаемость гистогематических
барьеров; бактерицидные вещества в биологических жидкостях орга-
низма (слюна, слеза, спинномозговая жидкость, кровь); выделение
вирусов почками; фагоцитарная система (макрофаги и микрофаги —
нейтрофильные гранулоциты); гидролитические ферменты; интерфе-
рон; лимфокины; система комплемента и др. Неспецифические за-
щитные факторы обеззараживают даже вещества, с которыми
организм ранее не встречался. Специфические начинают действовать
после первичного контакта с антигеном.
Барьеры играют очень важную роль в осуществлении защиты орга-
низма. Внешние барьеры (кожа, пищеварительная, дыхательная систе-
мы) препятствуют проникновению вредных веществ извне, печень
обезвреживает токсины, почки выводят токсичные продукты обмена
веществ. Внутренние барьеры (гистогематические) обладают избира-
тельной проницаемостью, они препятствуют поступлению во внутрен-
нюю среду вредных веществ, однако внутренние барьеры обеспечивают
необходимое микроокружение, при этом кровь непосредственно не
контактирует с клетками. Учение о гистогематических барьерах разра-
ботала советский ученый, академик Л.С . Штерн.
361
ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
В организме человека имеются три сложные системы управления
функциями — нервная, гуморальная и эндокринная, которые тесно
связаны между собой и осуществляют единую нейрогуморально-гор-
мональную регуляцию. Центральная нервная система, в том числе ее
высший отдел — кора головного мозга, регулирует функции желез
внутренней секреции. Это осуществляется как путем непосредствен-
ной иннервации желез, так и благодаря регуляции гипоталамусом де-
ятельности гипофиза.
Эндокринные железы не имеют выводных протоков, они выделя-
ют вырабатываемые ими гормоны в кровь или лимфу. Гормоны — би-
ологически активные вещества, которые отличаются рядом важных
свойств.
1. Действие гормонов носит дистантный характер. Иными словами,
органы, на которые гормоны действуют, расположены далеко от железы.
2. Действие гормонов строго специфично. Некоторые гормоны
действуют лишь на определенные клетки-мишени, другие — на мно-
жество различных клеток.
3. Гормоны обладают высокой биологической активностью.
4. Гормоны действуют только на живые клетки.
Гормоны участвуют в регуляции гомеостаза (постоянства внутрен-
ней среды), в обмене веществ, влияют на рост, дифференцировку, раз-
множение; обеспечивают ответную реакцию организма на изменения
внешней среды. Эндокринные железы анатомически и топографиче-
ски разобщены.
В настоящее время принято деление эндокринных желез на зави-
симые и независимые от передней доли гипофиза. К первым относятся
щитовидная железа, надпочечники (корковое вещество) и половые
железы. Взаимоотношения между передней долей гипофиза и завися-
щими от нее железами строятся по типу прямых и обратных связей.
Тропные (от греч. tropos — направление) гормоны передней доли ги-
пофиза активируют деятельность указанных желез, а их гормоны,
в свою очередь, воздействуют на нее, угнетая образование и выделе-
ние соответствующего гормона. Прочие железы (паращитовидная,
эпифиз, панкреатические островки, мозговое вещество надпочечни-
ков, параганглии) не подчинены непосредственному влиянию перед-
ней доли гипофиза (рис. 172).
ВНИМАНИЕ!
Гипоталамус является высшим центром регуляции эндокринных функ-
ций, он объединяет нервные и эндокринные регуляторные механизмы в
общую нейроэндокринную систему, координирует нервные и гормо-
нальные механизмы регуляции функций внутренних органов.
362
ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
Гипоталамус образует с гипофизом
единый функциональный комплекс, в кото-
ром первый играет регулирующую, а вто-
рой эффекторную роль. Гипоталамус вы-
деляет две группы веществ, которые
воздействуют на клетки передней доли
гипофиза: рилизинг-факторы, или либе-
рины, стимулирующие синтез и выделе-
ние ими гормонов (кортиколиберин, лю-
либерин, соматолиберин, тиреолиберин
и фоллиберин), статины тормозят синтез
и выделение гормонов (дофамин и сома-
тостатин). Гипофиз отвечает на поступаю-
щие в него из гипоталамуса сигналы выра-
боткой своих тропных гормонов, которые
направляются к периферическим эндо-
кринным железам. Кроме того, супраопти-
ческое и паравентрикулярное ядра гипо-
таламуса вырабатывают вазопрессин и
окситоцин, которые по разветвлениям аксонов нейросекреторных кле-
ток поступают в заднюю долю гипофиза, откуда разносятся кровью.
В табл. 23 приведены сведения об эндокринных железах и их гормонах.
Таблица 23
Эндокринные железы и их гормоны
Железы
внутренней
секреции
Название гормонов,
состав
Характер действия
12
3
Гипофиз
а) передняя
доля
Соматотропный, СТГ
(регулируется сомато-
статином и соматолибе-
рином)
Рост костей, мышц, органов. Анаболическое
действие. Увеличение относительного
содержания в организме белка и воды,
снижение жиров
Лактотропный, пролак-
тин, ЛТГ
Пролиферация роста молочных желез
и секреции молока
Тиреотропный, ТТГ (ре-
гулируется тиреолибе-
рином)
Стимуляция выработки тироксина щито-
видной железой
4
5
3
2
1
6
7
Рис. 172. Положение эндокринных желез
в теле человека
1 — гипофиз и эпифиз, 2 — паращитовидные
железы, 3 — щитовидная железа,
4 — надпочечники, 5 — панкреатические
островки, 6 — яичник, 7 — яичко
363
ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
12
3
Меланостимулирую-
щий, МСГ
Синтез меланина, распределение гранул
пигмента, пигментация кожи
Фолликулостимулиру-
ющий, ФСГ (регулиру-
ется фоллиберином)
У женщин: стимуляция роста фолликулов,
секреции эстрогенов и овуляции. У муж-
чин: опосредованная стимуляция сперма-
тогенеза, стимуляция развития семявыно-
сящих канальцев
Лютеинезирующий, ЛГ
(регулируется люлибе-
рином)
У женщин: стимуляция овуляции, образова-
ние желтого тела. Развитие и созревание поло-
вых клеток, секреция половых гормонов
Гормон, стимулирую-
щий интерстициальные
эндокриноциты
У мужчин: стимуляция синтеза и выделе-
ния тестостерона
Адренокортикотропный,
АКТГ (регулируется
кортиколиберином)
Регуляция образования и секреции глико-
кортикоидов коры надпочечников, мобили-
зация жира из жировой ткани
б) задняя
доля
Окситоцин
Сокращение матки. Сокращение миоэпители-
альных клеток ацинусов молочных желез,
выделение молока; усиление тонуса гладких
мышц желудочно-кишечного тракта
Антидиуретический
(АДГ), вазопрессин
Реабсорбция воды в почечных канальцах
(антидиуретическое действие). Сосудосу-
живающее действие (повышение кровяно-
го давления)
Щитовид-
ная железа
Тироксин (тетрайодти-
ронин, трийодтиронин)
(регулируется ТТГ)
Обеспечение роста, умственного и физиче-
ского развития; стимуляция энергетическо-
го обмена, синтеза белка и окислительного
катаболизма жиров и углеводов, поглоще-
ния кислорода и метаболизма всех клеток;
повышение чувствительности клеток к ка-
техоламинам; активация натриевого насоса
Тиреокальцитонин
Регуляция метаболизма кальция и фосфо-
ра, снижение уровня кальция в крови
Паращито-
видная
железа
Паратгормон
Регуляция метаболизма кальция и фосфо-
ра, повышение уровня кальция в крови
Поджелу-
дочная
железа
Инсулин
Регуляция обмена углеводов, белков,
жиров, снижение уровня сахара в крови
Глюкагон
Стимуляция распада гликогена, повышение
уровня сахара в крови
Соматостатин
Регуляция выделения инсулина и глюкаго-
на клетками островков поджелудочной же-
лезы; торможение двигательной активности
и секреции двенадцатиперстной кишки
Продолжение табл. 23
364
ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
12
3
Надпочеч-
ники
а) корковый
слой
Гидрокортизон (корти-
зол) (регулируется
АКТГ)
Регуляция обмена углеводов, белков,
жиров, катаболическое действие, противо-
воспалительное действие, повышение
устойчивости к инфекции
Альдостерон
Регуляция минерального обмена и водно-
солевого равновесия, увеличение активного
транспорта натрия через клеточные мем-
браны, повышение реабсорбции натрия
и воды в канальцах нефрона. Аналогичное
влияние на клетки потовых, слюнных
и кишечных желез. Участие в адаптации
организма к повышенной температуре
окружающей среды
Андрогены: см. «Гормоны половых желез»
б) мозговой
слой
(не зависит
от гипофиза)
Адреналин и норадрена-
лин (катехоламины)
Симпатическая стимуляция. Увеличение
частоты силы сердечных сокращений и кро-
вотока в мышцах, мозге, миокарде. Увеличе-
ние вентиляции легких, доставки кислорода
к мышцам, сердцу и мозгу.
Повышение содержания глюкозы и жирных
кислот в крови
Яичники
Эстрадиол, эстрон
(регулируется ФСГ)
Половая дифференцировка, развитие
половых органов, вторичных половых при-
знаков, половое поведение. Обеспечение
пролиферативной фазы эпителия слизи-
стой оболочки матки. Анаболическое
действие
Прогестерон
(регулируется ЛГ)
Подготовка слизистой оболочки матки
к имплантации зародыша. Нормальное
протекание беременности. Катаболический
эффект
Семенники
(яички)
Тестостерон
(регулируется ЛГ)
Половая дифференцировка, развитие
половых органов, вторичных половых
признаков, половое поведение. Анаболиче-
ское действие
Эпифиз
Мелатонин
Выработка и концентрация пигмента
в пигментных клетках, влияние на репро-
дуктивную функцию
Серотонин
Регуляция двигательной активности желу-
дочно-кишечного тракта, выделение слизи.
Серотонинергические нейроны головного
мозга участвуют в регуляции поведения,
сна, терморегуляции
Окончание табл. 23
365
ЩИТОВИДНАЯ ЖЕЛЕЗА
ГИПОФИЗ
Передняя доля гипофиза образована эпителиальными перекла-
динами, между ними располагаются тонкие соединительноткан-
ные прослойки, в которых лежат синусоидные капилляры. Задняя
доля образована мелкими многоотростчатыми клетками питуици-
тами и аксонами клеток ядер гипоталамуса, которые вырабатыва-
ют гормоны вазопрессин и окситоцин. По этим аксонам гормоны
транспортируются в заднюю долю гипофиза, откуда разносятся
кровью.
Секреция гормонов гипофизозависимых желез регулируется по
принципу обратной связи: при снижении концентрации определен-
ного гормона в крови соответствующие клетки гипоталамуса выра-
батывают либерин, который поступает по разветвлениям аксонов
в переднюю долю гипофиза. В ответ на это клетки передней доли
гипофиза выделяют тропный гормон, стимулирующий образование
гормона клетками конкретной железы. И наоборот, повышение со-
держания гормона в крови является сигналом для клеток гипотала-
муса прекратить выработку либерина. Клетки гипофиза отвечают на
это замедлением секреции и освобождением тройного гормона, что
приводит к подавлению секреции гормона эндокринной железой.
Иными словами, уровень гормона в крови регулируется по принципу
обратной связи.
Гипофиз является важнейшей железой внутренней секреции, ко-
торая регулирует деятельность целого ряда эндокринных желез. Он
расположен в гипофизарной ямке турецкого седла клиновидной ко-
сти. Воронка соединяет гипофиз с гипоталамусом. Масса гипофиза
у мужчин 0,5—0,6 г, у женщин 0,6—0,7 г. Будучи анатомически еди-
ным, гипофиз делится на две доли, имеющие различное происхожде-
ние: переднюю долю (аденогипофиз), которая крупнее (70—80% от
всей массы), и заднюю долю (нейрогипофиз).
ЩИТОВИДНАЯ ЖЕЛЕЗА
Щитовидная железа расположена на шее впереди гортани и верх-
него отдела трахеи. В ней различают две доли и перешеек. Масса же-
лезы взрослого человека составляет 20—30 г, в старческом возрасте —
меньше. Железа покрыта снаружи фиброзной капсулой, от которой
внутрь железы отходят перегородки трабекулы, которые, разветвля-
ясь, разделяют ее на дольки. Паренхима железы состоит из пузырь-
366
ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
ков-фолликулов, являющихся основными структурными и функцио-
нальными единицами. Стенка фолликула образована одним слоем
тироцитов. В полости фолликула содержится густой вязкий коллоид
щитовидной железы. Тироциты вырабатывают белковый компонент
тиреоглобулина, кроме того, они захватывают йод. Йодирование мо-
лекул происходит в полости фолликула, в коллоиде. Под влиянием
тиреотропного гормона передней доли гипофиза усиливается синтез
гормона, поглощение клетками йода и йодирование, а также расще-
пление тиреоглобулина, в результате чего освобождаются три- и тет-
райодтиронин, которые проходят через тироциты и поступают в ка-
пилляры.
В стенках фолликулов наряду с тироцитами между ними в базаль-
ной мембране имеются более крупные, светлые околофолликулярные
клетки (их верхушка не достигает просвета фолликула), продуциру-
ющие гормон тиреокальцитонин, участвующий в регуляции обмена
кальция и фосфора. Этот гормон является антагонистом гормона па-
ращитовидной железы. Он тормозит всасывание кальция из костей
и уменьшает содержание кальция в крови.
НАДПОЧЕЧНИК
Надпочечники располагаются забрюшинно в толще околопочеч-
ного жирового тела на уровне XI—XII грудных позвонков. Масса од-
ного надпочечника взрослого человека около 8—13 г. Будучи анато-
мически единым, надпочечник по существу состоит из двух желез,
представленных корковым и мозговым веществами. В корковом веще-
стве различают три зоны: клубочковую (наружную), пучковую (сред-
нюю) и сетчатую (на границе с мозговым слоем). Указанные зоны до-
статочно четко отделены друг от друга анатомически и, по
современным воззрениям, вырабатывают различные гормоны: клу-
бочковая — минералокортикоиды (альдостерон), пучковая — глюко-
кортикоиды (гидрокортизон, кортизон и кортикостерон), сетчатая —
андрогены, эстрогены и прогестерон (последние в небольшом
количестве). Передняя доля гипофиза регулирует глюкокортикоид-
ную и андрогенную функции (АКТГ).
Мозговое вещество продуцирует небольшое количество адренали-
на и норадреналина, и лишь при воздействии на организм сильных
раздражителей секреция этих гормонов резко усиливается.
367
НЕРВНАЯ СИСТЕМА
ПАРАЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Две пары мелких желез располагаются на задней поверхности до-
лей щитовидной железы. Железы образованы паратироцитами, кото-
рые продуцируют белковый паратиреоидный гормон, регулирующий
уровень кальция и опосредованно фосфора в крови, тем самым оказы-
вая влияние на возбудимость нервной и мышечной систем. Гормон
действует на кости, вызывая усиление функции остеокластов, кото-
рые производят деминерализацию костной ткани, выделение Са2+
в кровь. Тем самым поддерживается необходимый уровень Са2+
в кро-
ви. Избыток фосфора, также выделяющийся при этом из кости, уда-
ляется почками. Антагонистом паратиреоидного гормона является
тиреокальцитонин, вырабатываемый околофолликулярными клетка-
ми щитовидной железы. Секреция обоих гормонов контролируется
уровнем кальция в крови. Его снижение приводит к возбуждению се-
креции паратиреоидного гормона и, соответственно, к выделению
кальция в кровь. Повышенный уровень кальция приводит к активиза-
ции околофолликулярных клеток щитовидной железы, они выделяют
тиреокальцитонин, который усиливает фиксацию кальция, тем самым
снижая его уровень.
ПАНКРЕАТИЧЕСКИЕ ОСТРОВКИ
Поджелудочная железа состоит из экзокринной и эндокринной
частей. Экзокринная часть описана в разделе «Органы пищеварения».
Эндокринная часть образована группами панкреатических островков
(островки Лангерганса). Общее количество островков колеблется
в пределах 1—2 млн, а диаметр каждого 0,1—0,3 мм.
НЕРВНАЯ СИСТЕМА
Нервная система обеспечивает интеграцию всех частей организ-
ма в единое целое, осуществляет умственную деятельность, связь
организма с внешней средой (ощущения), управляет движениями,
регулирует все функции, включая человеческую сексуальность и ре-
продукцию (продолжение рода). Нервная система человека являет-
ся морфофизиологическим субстратом мышления, творчества, чле-
нораздельной речи, трудовой деятельности. Все функции, включая
умственную деятельность, осуществляют группы нервных клеток,
связанных между собой многочисленными синапсами.
368
НЕРВНАЯ СИСТЕМА
Нервную систему человека подразделяют на центральную
и периферическую. К центральной нервной системе относят спинной
и головной мозг, к периферической — отходящие от них парные спин-
номозговые и черепные нервы с корешками, их ветви, нервные окон-
чания и ганглии (нервные узлы, образованные телами нейронов) —
рис. 173.
Существует еще одна классификация, согласно которой единую
нервную систему также условно подразделяют на две части: сомати-
ческую (анимальную) и вегетативную (автономную). Соматическая
8
9
11
12
13
14
15
10
2
3
4
5
6
7
1
16
17
18
19
Рис. 173. Нервная система
(полусхематично):
1 — подкожный нерв, 2 — запирательный
нерв, 3 — бедренный нерв, 4 — локтевой
нерв, 5 — поясничное сплетение,
6 — срединный нерв, 7 — симпатический
ствол, 8 — плечевое сплетение,
9 — головной мозг, 10 — шейное сплетение,
11 — передние ветви (межреберные нервы),
12 — спинной мозг, 13 — лучевой нерв,
14 — конский хвост, 15 — крестцовое
сплетение, 16 — копчиковый нерв,
17 — седалищный нерв,
18 — большеберцовый нерв,
19 — общий малоберцовый нерв
369
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
нервная система иннервирует главным образом тело (кости, скелет-
ные мышцы, кожу) и обеспечивает связь организма с внешней средой.
Вегетативная (автономная) нервная система иннервирует все вну-
тренности, железы (в том числе и эндокринные), гладкие мышцы ор-
ганов и кожи, сосуды и сердце, а также обеспечивает обменные про-
цессы во всех органах и тканях.
ЦЕНТРАЛЬНАЯ НЕРВНАЯ
СИСТЕМА (ЦНС)
СПИННОЙ МОЗГ
Спинной мозг, расположенный в позвоночном канале, разделен на
две половины. На его боковых поверхностях симметрично входят зад-
ние (афферентные) и выходят передние (эфферентные) корешки
спинномозговых нервов. Участок спинного мозга, соответствующий
каждой паре корешков, называется сегментом (рис. 174). В пределах
Рис. 174. Спинной мозг (поперечный разрез) и рефлекторная дуга:
А — задняя срединная борозда, Б — белое вещество, В — задний рог, Г — задний
корешок, Д — спинномозговой узел, Е — боковой рог, Ж — передний корешок,
З — передний рог, И — передняя срединная щель; 1 — вставочный нейрон,
2 — афферентное нервное волокно, 3 — эфферентное нервное волокно,
4 — серая ветвь, 5 — белая ветвь, 6 — узел симпатического ствола,
7 — нервно-секреторное окончание
11
А
22
Б
33
В
44
Г
55
Д
66
Е
77
Ж
З
И
370
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
спинного мозга выделяют сегменты шейные (CI
–C
VII
), грудные (TI
–
TXII), поясничные (LI–LV), крестцовые (SI–SV) и копчиковые (CоI–
CоIV). Длина спинного мозга в среднем 45 см у мужчин и 41—42
у женщин, масса 34—88 г.
На поперечном разрезе спинного мозга видно расположенное вну-
три серое вещество и окружающее его со всех сторон белое. Серое ве-
щество образовано телами нервных клеток, которых в спинном мозге
насчитывают около 13 млн, началом их отростков, клетками глии.
Клетки, имеющие одинаковое строение и выполняющие одинаковые
функции, образуют ядра серого вещества. В сером веществе различа-
ют передние, задние, а в грудном (от I грудного до II—III поясничного
сегментов) и боковые столбы. На поперечном разрезе видны однои-
менные рога. В передних столбах (передних рогах) серого вещества
залегают двигательные нейроны, образующие ядра, являющиеся дви-
гательными соматическими центрами. Их аксоны выходят в составе
передних корешков, а затем спинномозговых нервов и направляются
на периферию, иннервируя скелетные мышцы. В задних — залегают
ядра, образованные мелкими вставочными нейронами, к которым в
составе задних, или чувствительных, корешков направляются аксоны
клеток, расположенных в спинномозговых узлах. Отростки вставоч-
ных нейронов осуществляют связь с нервными центрами головного
мозга, а также с несколькими соседними сегментами, с нейронами,
расположенными в передних рогах своего сегмента, выше и ниже ле-
жащих сегментов, т. е . связывают афферентные нейроны спинномоз-
говых узлов с нейронами передних рогов. В боковых рогах располо-
жены центры симпатической части вегетативной нервной системы.
Белое вещество спинного мозга представлено отростками нервных
клеток. Совокупность этих отростков составляет системы пучков
(тракты, или проводящие пути) спинного мозга.
ГОЛОВНОЙ МОЗГ
Головной мозг располагается в полости мозгового черепа, форма
которого определяется формой мозга, некоторыми этническими осо-
бенностями, полом и возрастом. Масса мозга взрослого человека око-
ло 1500 г (от 1100 до 2000, т. е. диапазон крайних индивидуальных
значений очень велик). В свою очередь, это давало и дает основания
выдвигать представления о зависимости от массы мозга гениально-
сти; о предначертанном поведении преступников; об умственном пре-
обладании мужчин над женщинами. Однако это не соответствует дей-
ствительности. Абсолютная масса мозга не позволяет судить об
интеллекте человека. Любопытные цифры приводит М. А . Гремяц-
371
ГОЛОВНОЙ МОЗГ
кий: масса мозга Тургенева была равна 2012 г, Кромвеля — 2000, Бай-
рона — 2238, Кювье — 1830, Шиллера —1871, Теккерея — 1294, поэта
Уитмена — 1282, врача Деллингера — 1207, Анатоля Франса — 1017 г.
Несмотря на то что масса мозга А. Франса была почти в два раза
меньше массы мозга И. Тургенева, оба они были гениальными писате-
лями и мыслителями.
ВНИМАНИЕ!
Интегративная деятельность головного мозга обеспечивает целенаправ-
ленное поведение человека, его умственную деятельность.
Головной мозг подразделяют на три основных отдела: ствол, моз-
жечок и конечный мозг (полушария большого мозга). Ствол включа-
ет продолговатый мозг, мост, средний и промежуточный мозг. Именно
отсюда выходят черепные нервы. Самая развитая, крупная и функци-
онально значимая часть мозга — это полушария большого мозга. В го-
ловном мозге выделяют передний мозг, который подразделяют на ко-
нечный и промежуточный, средний, задний и продолговатый.
Передний мозг. Конечный мозг, управляющий всей деятельностью
организма, состоит из двух полушарий, которые очень хорошо развиты
у человека разумного (рис. 175). Масса полушарий составляет около
78% общей массы головного мозга, а площадь поверхности коры обоих
полушарий человека достигает около 450 тыс. мм2, что зависит от нали-
чия большого количества борозд и извилин. Особенного развития у че-
ловека достигают лобные доли: их поверхность составляет около 29%
всей поверхности коры, а масса — более 50% массы головного мозга.
Полушария большого мозга отделены друг от друга продольной ще-
лью, в глубине которой видно соединяющее их мозолистое тело, обра-
зованное белым веществом, т. е. волокнами. Каждое полушарие состоит
из пяти долей: лобной, теменной, височной, затылочной и островковой.
Поперечная щель большого мозга отделяет затылочные доли полуша-
рий от мозжечка. Сзади и книзу от затылочных долей расположены
мозжечок и продолговатый мозг, переходящий в спинной.
Кора полушарий большого мозга образована серым веществом, ко-
торое лежит по периферии (на поверхности) полушарий. Толщина
коры различных участков полушарий колеблется от 1,3 до 5 мм. Коли-
чество нейронов в шестислойной коре у человека достигает 10—14 млрд.
Каждый из них связан с помощью синапсов с тысячами других нейро-
нов. Располагаются они правильно ориентированными «колонками».
Различные рецепторы воспринимают энергию раздражения и пе-
редают ее в виде нервного импульса в кору головного мозга, где про-
исходит анализ всех раздражений, которые поступают из внешней и
внутренней среды. В коре головного мозга располагаются центры
372
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
(корковые концы анализаторов, которые не имеют строго очерчен-
ных границ), регулирующие выполнение определенных функций.
В коре постцентральной извилины и верхней теменной дольки зале-
гают ядра коркового анализатора чувствительности (температур-
ной, болевой, осязательной, мышечного и сухожильного чувства)
противоположной половины тела. Причем вверху расположены про-
екции нижних конечностей и нижних отделов туловища, а внизу
проецируются рецепторные поля верхних частей тела и головы.
Пропорции тела весьма искажены, ибо на представительство в коре
кистей, языка, лица и губ приходится значительно большая пло-
щадь, чем на туловище и ноги, что соответствует их физиологиче-
ской значимости (рис. 176А).
Рис. 175. Головной мозг, вид сверху:
1 — лобный полюс, 2 — продольная щель большого мозга, 3 — верхняя лобная
борозда, 4 — нижняя лобная борозда, 5 — предцентральная борозда,
6 — предцентральная извилина, 7 — центральная борозда, 8 — постцентральная извилина,
9 — постцентральная борозда, 10 — межтеменная борозда, 11 — затылочный полюс,
12 — затылочная доля, 13 — теменно-затылочная борозда, 14 — угловая извилина,
15 — теменная доля, 16 — надкраевая извилина, 17 — латеральная борозда,
18 — средняя лобная извилина, 19 — лобная доля, 20 — верхняя лобная извилина
7
8
9
10
11
12
13
14
15
16
17
18
19
20
4
3
2
5
6
1
373
ГОЛОВНОЙ МОЗГ
Ядро двигательного анализатора находится главным образом
в предцентральной извилине («двигательная область коры»), и здесь
пропорции частей тела человека, как и в чувствительной зоне, весьма
искажены (рис. 176Б). Размеры проекционных зон различных частей
тела зависят не от их действительной величины, а от функционально-
го значения. Так, зоны кисти в коре полушарий большого мозга зна-
чительно больше, чем зоны туловища и нижней конечности, вместе
взятые. Двигательные области каждого из полушарий, весьма специа-
лизированные у человека, связаны со скелетными мышцами противо-
положной стороны тела. Если мышцы конечностей изолированно
связаны с одним из полушарий, то мышцы туловища, гортани и глот-
ки — с двигательными областями обоих полушарий. От двигательной
коры нервные импульсы направляются к нейронам спинного мозга,
а от них — к скелетным мышцам.
В коре височной доли находится ядро слухового анализатора. К ка-
ждому из полушарий подходят проводящие пути от рецепторов органа
слуха как левой, так и правой стороны. Ядро зрительного анализатора
располагается на медиальной поверхности затылочной доли. Причем
ядро правого полушария связано проводящими путями с латеральной
(височной) половиной сетчатки правого глаза и медиальной (носовой)
половиной сетчатки левого глаза; левого — с латеральной половиной
сетчатки левого и медиальной половиной сетчатки правого глаза.
Рис. 176. Расположение корковых центров:
А —корковый центр общей чувствительности (чувствительный «гомункулус»)
(из В. Пенфилда и И. Расмуссена). Изображения на поперечном срезе мозга (на уровне
постцентральной извилины) и относящиеся к ним обозначения показывают
пространственное представительство поверхности тела в коре большого мозга.
Б —двигательная область коры (двигательный «гомункулус»;
(из В. Пенфилда и И. Расмуссена). Изображение двигательного «гомункулуса»
отражает относительные размеры областей представительства отдельных участков
тела в коре предцентральной извилины большого мозга
Ïàëüöû
Ñ
ò
î
ï
à
Ñ
ò
î
ï
à
Ò
å
ë
î
Ò
å
ë
î
Ð
ó
ê
à
Ê
è
ñ
ò
ü
Ê
è
ñ
ò
ü
Ë
î
ê
î
ò
ü
Ë
è
ö
î
Ê
î
ë
å
í
î
Ï
à
ë
å
ö
Ï
à
ë
ü
ö
û
Í
î
ñ
Ø
å
ÿ
Ãóáû
Ãóáû
Ç
ó
á
û
Ã
ë
î
ò
ê
à
ß
ç
û
ê
×åëþñòü
АБ
374
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Благодаря близкому расположению ядер обонятельного (лимбиче-
ская система, крючок) и вкусового анализаторов (самые нижние отде-
лы коры постцентральной извилины) чувства обоняния и вкуса тесно
связаны между собой. Ядра вкусового и обонятельного анализаторов
обоих полушарий связаны проводящими путями с рецепторами как
левой, так и правой стороны.
Корковые концы анализаторов осуществляют анализ и синтез
сигналов, поступающих из внешней и внутренней среды организма,
составляющих первую сигнальную систему действительности
(И.П . Павлов). В отличие от первой, вторая сигнальная система име-
ется только у человека и тесно связана с членораздельной речью.
Речь и мышление человека осуществляются при участии всей коры
полушарий большого мозга. В то же время в коре полушарий большого
мозга человека имеются зоны, являющиеся центрами целого ряда спе-
циальных функций, связанных с речью. Двигательные анализаторы
устной и письменной речи располагаются в областях коры лобной доли
вблизи ядра двигательного анализатора. Центры зрительного и слухо-
вого восприятия речи находятся вблизи ядер анализаторов зрения и
слуха. При этом речевые анализаторы у правшей локализируются лишь
в левом полушарии, а у левшей — в большинстве случаев тоже слева.
Однако они могут располагаться справа или в обоих полушариях.
ВНИМАНИЕ!
Лобные доли являются морфологической основой психических функ-
ций человека и его разума.
В 1982 г. Р. Сперри был удостоен Нобелевской премии «за откры-
тия, касающиеся функциональной специализации полушарий мозга».
Он пишет: «Каждое полушарие... имеет как бы отдельное собственное
мышление». В то же время следует подчеркнуть, что оба полушария
здорового мозга работают вместе, образуя единый мозг.
ВНИМАНИЕ!
Исследования Сперри показали, что кора левого полушария отвечает за
вербальные (от лат. verbalis — словесный) операции и речь. Левое по-
лушарие ответственно за понимание речи, а также за выполнение дви-
жений и жестов, связанных с языком; за математические расчеты, аб-
страктное мышление, интерпретацию символических понятий. Кора
правого полушария контролирует выполнение невербальных функций,
она управляет интерпретацией зрительных образов, пространственных
взаимоотношений. Кора правого полушария дает возможность распоз-
навать предметы, но не позволяет выразить это словами. Кроме того,
правое полушарие распознает звуковые образы и воспринимает музы-
ку. Оба полушария ответственны за сознание и самосознание человека,
его социальные функции.
375
ГОЛОВНОЙ МОЗГ
Базальные (подкорковые, центральные) ядра и белое вещество
конечного мозга. В толще образованного нервными волокнами бело-
го вещества имеются скопления серого, складывающегося в отдельно
лежащие ядра, которые залегают ближе к основанию мозга. Они по-
лучают информацию об активности двигательной системы (полоса-
тое тело, ограда и миндалевидное тело), управляют движениями и
регулируют мышечный тонус. В толще белого вещества под мозоли-
стым телом располагаются боковые желудочки, которые являются по-
лостями полушарий большого мозга.
Промежуточный мозг, расположенный под мозолистым телом, со-
стоит из таламуса, эпиталамуса, метаталамуса и гипоталамуса
(рис. 177). Таламус (зрительный бугор) — парный, образованный
главным образом серым веществом, является подкорковым центром
всех видов чувствительности, в нем насчитывают несколько десятков
ядер, которые получают информацию от всех органов чувств и пере-
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
4
3
2
5
6
1
31
32
33
Рис. 177. Промежуточный и задний мозг. Вид со стороны полости третьего
желудочка мозга. Сагиттальный разрез ствола мозга:
1 — столб свода, 2 — межжелудочковое отверстие, 3 — межталамическое сращение,
4 — таламус, 5 — сосудистое сплетение третьего желудочка, 6 — гипоталамическая
борозда, 7 — треугольник поводка, 8 — шишковидное углубление, 9 — валик
мозолистого тела, 10 — шишковидная железа, 11 — крыша среднего мозга,
12 — водопровод среднего мозга, 13 — верхний мозговой парус, 14 — четвертый
желудочек, 15 — мозжечок, 16 — нижний мозговой парус, 17 — продолговатый мозг,
18 — задняя спайка, 19 — мост, 20 — корешок глазодвигательного нерва, 21 — заднее
продырявленное вещество, 22 — сосцевидное тело, 23 — углубление воронки,
24 — гипофиз, 25 — воронка, 26 — перекрест зрительных нервов, 27 — супраоптическое
углубление, 28 — терминальная пластинка, 29 — передняя спайка, 30 — колено
мозолистого тела, 31 — клюв мозолистого тела, 32 — прозрачная перегородка,
33 — ствол мозолистого тела
376
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
дают ее в кору головного мозга. Таламус связан с лимбической сис-
темой, ретикулярной формацией, гипоталамусом, мозжечком, ба-
зальными ганглиями. Через таламус сознание контролирует автома-
тические движения. Обращенные друг к другу медиальные поверхности
обоих таламусов образуют боковые стенки полости промежуточного
мозга — III желудочек.
Эпиталамус включает эпифиз (шишковидное тело), являющийся
железой внутренней секреции. Его гормоны влияют на развитие по-
ловых желез, тормозя их деятельность. Именно этот орган ранее рас-
сматривался в качестве «третьего глаза», а иногда и локализации
души. Метаталамус образован парными медиальным и латеральным
коленчатыми телами, лежащими позади каждого зрительного бугра.
Медиальное коленчатое тело наряду с нижними холмиками пластин-
ки крыши среднего мозга (четверохолмия) является подкорковым
центром слухового анализатора, а латеральное вместе с верхними
холмиками — подкорковым центром зрительного анализатора.
Гипоталамус располагается кпереди от ножек мозга и включает
в себя ряд структур: расположенную кпереди зрительную и обоня-
тельную части. К последней относится собственно подбугорье, или
гипоталамус, в котором расположены центры вегетативной части
нервной системы. Гипоталамус контролирует деятельность эндокрин-
ной системы человека благодаря тому, что его нейроны секретируют
нейрогормоны (вазопрессин и окситоцин), а также факторы, стиму-
лирующие или угнетающие выработку гормонов гипофизом.
ВНИМАНИЕ!
Гипоталамус, масса которого не превышает 5% мозга, является центром
регуляции эндокринных функций, он объединяет нервные и эндокрин-
ные регуляторные механизмы в общую нейроэндокринную систему.
В гипоталамусе залегают также нейроны, которые воспринимают
все изменения, происходящие в крови и спинномозговой жидкости
(температуру, состав, содержание гормонов и т. д .). Гипоталамус свя-
зан с корой большого мозга и лимбической системой. В гипоталамус
поступает информация из центров, регулирующих деятельность ды-
хательной и сердечно-сосудистой систем. В гипоталамусе расположе-
ны центры жажды, голода, центры, регулирующие эмоции и пове-
дение человека, сон и бодрствование, температуру тела и т. д. Из
гипоталамуса выделены обладающие морфиноподобным действием
энкефалины и эндорфины. Считают, что они влияют на поведение
(оборонительные, пищевые, половые реакции) и вегетативные про-
цессы, обеспечивающие выживание человека.
377
ГОЛОВНОЙ МОЗГ
ВНИМАНИЕ!
Гипоталамус регулирует все функции организма, кроме ритма сердца,
кровяного давления и спонтанных дыхательных движений, которые ре-
гулируются продолговатым мозгом.
Сосцевидные тела, образованные серым веществом, являются под-
корковыми центрами обонятельного анализатора. Кпереди от них
расположен серый бугор, в котором залегают ядра вегетативной нерв-
ной системы. Они оказывают влияние на эмоциональные реакции че-
ловека (агрессия, ярость), а также учащение или замедление сердце-
биений, повышение или понижение кровяного давления.
К среднему мозгу относят ножки мозга и крышу. Ножки мозга —
это белые округлые довольно толстые тяжи, выходящие из моста
и направляющиеся вперед в полушария большого мозга. В ножках за-
легают группы нейронов, богатые пигментом меланином, выделяю-
щиеся своим черным цветом (черное вещество). Другие пигментные
образования в ножках — это красные ядра. Черное вещество и крас-
ные ядра участвуют в регуляции мышечного тонуса и подсознатель-
ных автоматических движений.
В крыше среднего мозга различают пластинку в виде четверохол-
мия. Два верхних холмика являются подкорковыми центрами зри-
тельного анализатора, а нижние — слухового анализатора. В углубле-
нии между верхними холмиками лежит шишковидное тело (см.
«Эндокринная система»). Сильвиев водопровод — узкий канал дли-
ной 2 см, который соединяет III и IV желудочки. Вокруг водопровода
располагается центральное серое вещество, в котором заложены рети-
кулярная формация, ядра III и IV пар черепных нервов и др.
К заднему мозгу относятся мост, расположенный центрально,
и лежащий позади него мозжечок. У человека мост (варолиев мост)
достигает наибольшего развития, он выглядит в виде лежащего попе-
речно утолщенного валика. Мост состоит из множества нервных во-
локон, связывающих кору большого мозга со спинным мозгом и с ко-
рой полушарий мозжечка. Между волокнами залегают ретикулярная
формация, ядра V, VI, VII, VIII пар черепных нервов.
Мозжечок очень хорошо развит у человека в связи с прямохожде-
нием и трудовой деятельностью (рис. 178). Его масса у взрослого че-
ловека 120—160 г и составляет 8—12% массы головного мозга. В моз-
жечке различают два полушария и непарную срединную часть — червь.
Поверхности полушарий и червя разделяют поперечные параллель-
ные борозды, между которыми расположены узкие длинные листки
мозжечка. Благодаря этому его поверхность у взрослого человека со-
ставляет в среднем 850 см2. Мозжечок состоит из серого и белого ве-
щества. Белое вещество, проникая между серым, как бы ветвится,
378
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
образуя белые полоски, напоминая на срединном разрезе фигуру вет-
вящегося дерева — «древо жизни». Это понятие является издревле
символом всего существующего на Земле. Человек расположен в цен-
тре мироздания, а на «листочках этого дерева» в мозжечке якобы за-
писаны судьбы людей. Кора мозжечка состоит из серого вещества
толщиной 1—1,5 мм. Кроме того, в толще белого вещества имеются
скопления серого — четыре пары ядер. Волокна, связывающие мозже-
чок с другими отделами мозга, образуют три пары мозжечковых но-
жек: нижние направляются к продолговатому мозгу, средние — к мо-
сту, верхние — к четверохолмию.
Мозжечок играет основную роль в поддержании равновесия тела,
мышечного тонуса и координации движений. Из спинного мозга моз-
жечок получает информацию о положении частей тела и глаз. Мозже-
чок как бы согласовывает деятельность спинного мозга и двига-
тельной коры по осуществлению как тонких, так и быстрых и
последовательных движений. Он постоянно координирует сигналы,
4
5
6
3
2
1
А
Б
В
Г
Д
Е
И
Ж
З
Рис. 178. Мозжечок, срединный разрез через червя:
А — вершина, Б — скат, В — листок червя, Г — бугор, Д — пирамида червя,
Е — язычок червя, Ж — узелок, З — язычок мозжечка, И — центральная долька;
1 — червь, 2 — белые пластинки, 3 — полушарие мозжечка, 4 — сосудистая основа
IV желудочка, 5 — нижний мозговой парус, 6 — верхний мозговой парус
ГОЛОВНОЙ МОЗГ
379
идущие к мышцам от двигательных зон коры, на основании информа-
ции, получаемой мозгом от органов зрения, слуха и проприорецепто-
ров. Кора большого мозга управляет функциями мозжечка.
Продолговатый мозг является непосредственным продолжением
спинного мозга. Он построен из белого и серого вещества (рис. 179).
В последнем расположены многочисленные ядра, в том числе IX—XII
пар черепных нервов, ядра олив, центры дыхания и кровообращения,
ретикулярная формация. Белое вещество образовано нервными во-
локнами, которые составляют все чувствительные и двигательные
проводящие пути. Бо
льшая часть их перекрещивается в продолгова-
том мозге, так что левое полушарие связано с правой половиной тела,
и наоборот. Центры продолговатого мозга регулируют кровяное дав-
ление, сердечный ритм и спонтанные дыхательные движения.
На медиальной и нижней поверхностях полушарий выделяют ряд
образований, относящихся к лимбической системе, которая распола-
гается по краям полушарий (рис. 180). К ним относят в основном
структуры обонятельного мозга — наиболее древней части полуша-
рий. В описаниях морфологов так называемую лимбическую систему
представляют в виде «анатомического эмоционального кольца», в со-
став которого входят различные образования мозга. Это корковые
структуры: гиппокамп, парагиппокампальная, поясная извилины,
структуры обонятельного мозга (обонятельные луковицы, обонятель-
7
8
9
10
11
12
13
14
15
4
3
2
5
6
1
Рис. 179. Задняя сторона
продолговатого мозга, вид сверху:
1 — верхняя мозжечковая ножка,
2 — голубоватое место,
3 — срединная борозда
ромбовидной ямки, 4 — лицевой
бугорок, 5 — пограничная
борозда, 6 — мозговые полоски,
7 — нижняя мозжечковая ножка,
8 — треугольник подъязычного
нерва, 9 — задвижка,
10 — треугольник блуждающего
нерва, 11 — вестибулярное поле,
12 — средняя мозжечковая ножка,
13 — медиальное возвышение,
14 — верхний мозговой парус,
15 — верхние и нижние холмики
(среднего мозга)
380
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
ные бугорки, области коры над миндалиной, а также частично кора
лобной, островковой и височной долей; подкорковые структуры (мин-
далина, ядра перегородки, передние ядра таламуса), гипоталамус, сос-
цевидные тела. Все лимбические структуры связаны между собой и
с другими отделами мозга. Особенно богаты связи с гипоталамусом.
Кора лобных долей регулирует деятельность лимбической системы.
Через лимбическую систему проходят сигналы, направляющиеся от
всех органов чувств в кору полушарий, а также в обратном направле-
нии. Она обусловливает эмоциональный настрой человека и мотива-
ции, т. е . побуждение к действию, поведение, процессы научения и
памяти, а также обеспечивает общее улучшение приспособления ор-
ганизма к постоянно изменяющимся условиям внешней среды.
Ретикулярная формация (от лат. rete — сеть) представляет собой
совокупность клеток, клеточных скоплений и нервных волокон, рас-
положенных на всем протяжении ствола мозга (продолговатый мозг,
мост, средний и промежуточный мозг) и в центральных отделах спин-
ного мозга. Ретикулярная формация получает информацию от всех
органов чувств, внутренних и других органов, оценивает ее, фильтру-
ет и передает в лимбическую систему и кору большого мозга. Она ре-
гулирует уровень возбудимости и тонуса различных отделов цент-
ральной нервной системы, включая кору большого мозга, играет
Рис. 180. Структуры лимбической системы головного мозга:
1 — обонятельный треугольник, 2 — паратерминальная извилина, 3 — обонятельный
тракт, 4 — обонятельная луковица, 5 — поясная извилина, 6 — серый покров,
7 — свод, 8 — перешеек поясной извилины, 9 — терминальная полоска,
10 — парагиппокампальная извилина, 11 — мозговая полоска таламуса,
12 — гиппокамп, 13 — сосцевидное тело, 14 — миндалевидное тело, 15 — крючок
1
2
3
4
567
8
9
10
11
12
13
14
15
ГОЛОВНОЙ МОЗГ
381
важную роль в сознании, мышлении, памяти, восприятии, эмоциях,
сне, бодрствовании, вегетативных функциях, целенаправленных дви-
жениях, а также в механизмах формирования целостных реакций ор-
ганизма. Ретикулярная формация прежде всего выполняет функцию
фильтра, который позволяет важным для организма сенсорным сиг-
налам активировать кору мозга, но не пропускает привычные для него
или повторяющиеся сигналы.
Оболочки спинного и головного мозга
Спинной и головной мозг покрыты тремя оболочками (рис. 181).
Оболочки головного мозга в области большого затылочного отверстия
продолжаются в одноименные оболочки спинного мозга: наружная
твердая оболочка мозга, средняя паутинная и внутренняя мягкая обо-
лочка мозга. Непосредственно к наружной поверхности головного и
спинного мозга прилежит мягкая (сосудистая) оболочка, которая за-
ходит во все щели и борозды. От нее отходят соединительнотканные
волокна, которые вместе с кровеносными сосудами проникают в веще-
ство мозга. Кнаружи от сосудистой оболочки располагается паутинная
оболочка. Между веществом мозга, покрытым мягкой оболочкой и па-
4
5
6
7
8
9
10
11
3
2
1
Рис. 181. Взаимоотношение оболочек головного мозга и верхнего сагиттального
синуса со сводом черепа и поверхностью мозга; фронтальный разрез (схема):
1 — твердая оболочка головного мозга, 2 — свод черепа, 3 — грануляции паутинной
оболочки, 4 — верхний сагиттальный синус, 5 — кожа, 6 — эмиссарная вена,
7 — паутинная оболочка головного мозга, 8 — подпаутинное пространство,
9 — мягкая оболочка головного мозга, 10 — головной мозг, 11 — серп большого мозга
382
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
утинной оболочкой, находится подпаутинное (субарахноидальное)
пространство, заполненное спинномозговой жидкостью. В нижней ча-
сти позвоночного канала в подпаутинном пространстве свободно пла-
вают корешки спинномозговых нервов. Кверху подпаутинное про-
странство продолжается в одноименное пространство головного мозга.
Над крупными щелями и бороздами подпаутинное пространство ши-
рокое, образует вместилища, получившие название цистерн.
Подпаутинные пространства головного и спинного мозга сообща-
ются между собой в месте перехода спинного мозга в головной. В под-
паутинное пространство оттекает спинномозговая жидкость, образу-
ющаяся в желудочках головного мозга. Из боковых желудочков
жидкость оттекает в третий желудочек, из третьего — в четвертый,
а из него — в подпаутинное пространство. Обратное всасывание спин-
номозговой жидкости осуществляется через арахноидальные грануля-
ции — отростки паутинной оболочки, проникающие в просветы сину-
сов твердой оболочки головного мозга, а также в кровеносные и
лимфатические капилляры в местах выхода корешков черепных
и спинномозговых нервов из полости черепа и позвоночного канала.
Благодаря этому механизму спинномозговая жидкость постоянно
образуется и всасывается в кровь с одинаковой скоростью.
Снаружи от паутинной оболочки находится твердая оболочка
мозга, которая образована плотной волокнистой соединительной тка-
нью и отличается прочностью. От паутинной оболочки спинного моз-
га твердая отделена субдуральным пространством. Вверху субдураль-
ное пространство спинного мозга свободно сообщается с аналогичным
пространством полости черепа; внизу оно слепо заканчивается на
уровне II крестцового позвонка. Твердая оболочка спинного мозга
прочно срастается с краями большого (затылочного) отверстия и
вверху переходит в твердую оболочку головного мозга. Твердая обо-
лочка головного мозга срастается с надкостницей внутренней поверх-
ности костей основания мозгового черепа, особенно в местах их сое-
динения между собой и выхода черепных нервов из полости черепа,
с костями свода черепа она связана непрочно. Поверхность твердой
оболочки, обращенная в сторону мозга, гладкая; между ней и паутин-
ной оболочкой образуется узкое субдуральное пространство, в кото-
ром имеется небольшое количество жидкости.
На некоторых участках твердая оболочка головного мозга глубоко
впячивается в виде отростков в щели, отделяющие друг от друга ча-
сти мозга. В местах отхождения отростков оболочка расщепляется,
образуя каналы треугольной формы, выстланные эндотелием синусы
твердой мозговой оболочки; листки, образующие их стенки, туго натя-
нуты и не спадаются. В синусы из мозга по венам оттекает венозная
кровь, которая поступает затем во внутренние яремные вены.
383
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
Периферическая нервная система образована узлами (спинномоз-
говыми, черепными и вегетативными), нервами (31 пара спинномоз-
говых и 12 пар черепных) и нервными окончаниями (см. рис. 173).
Последние представлены рецепторами, воспринимающими раздраже-
ния внешней и внутренней среды, а также эффекторами, передающи-
ми нервные импульсы исполнительным органам. Каждый нерв состо-
ит из миелинизированных и немиелинизированных нервных волокон.
Снаружи нерв окружен соединительнотканной оболочкой — эпинев-
рием, в который входят питающие его сосуды. Нерв состоит из пуч-
ков, которые, в свою очередь, покрыты периневрием, а отдельные во-
локна — эндоневрием.
В зависимости от выполняемой функции различают нервы чувст-
вительные, двигательные и преимущественно смешанные. В перифе-
рической нервной системе человека преобладают смешанные нервы,
содержащие те и другие, а также симпатические волокна. Чувстви-
тельные нервы сформированы отростками (дендритами) нервных
клеток чувствительных узлов черепных нервов или спинномозговых
узлов. Двигательные нервы состоят из отростков (аксонов) нервных
клеток, лежащих в двигательных ядрах черепных нервов или в ядрах
передних столбов спинного мозга. Вегетативные нервы образованы
отростками клеток вегетативных ядер черепных нервов или боковых
столбов спинного мозга.
Черепные нервы. От ствола головного мозга отходит 12 пар че-
репных нервов (рис. 182). В их состав входят афферентные, т. е . «при-
ходящие», эфферентные, т. е . «уходящие», а также вегетативные во-
локна. Черепные нервы имеют собственные названия и порядковые
номера, обозначаемые римскими цифрами. Чувствительные нервы:
обонятельный, зрительный, преддверно-улитковый. Обонятельные
нервы (I) состоят из отростков рецепторных клеток, располагающих-
ся в слизистой оболочке обонятельной области полости носа, а зри-
тельные (II) — из отростков ганглиозных клеток сетчатой оболочки
глаза. В отличие от обонятельных нервов, которые образуют 15—20
нитей (нервов), зрительный нерв представлен единым стволом. Вой-
дя в полость черепа, правый и левый зрительные нервы перекрещива-
ются и продолжаются в зрительные тракты. Преддверно-улитковый
нерв (VIII) образован центральными отростками нейронов, залегаю-
щими в преддверном и улитковом узлах. Периферические отростки
клеток последних формируют нервы, заканчивающиеся, соответст-
венно, в вестибулярной части перепончатого лабиринта внутреннего
уха (орган равновесия) и в спиральном органе улиткового протока
(орган слуха).
384
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Двигательные нервы: гла-
зодвигательный, блоковый,
отводящий, добавочный, подъ-
язычный. Глазодвигательный
(III), блоковый (IV) и отво-
дящий (VI) нервы иннервируют мышцы глазного яблока и мышцу,
поднимающую верхнее веко. В составе глазодвигательного нерва про-
ходят также парасимпатические волокна, которые иннервируют мыш-
цы глазного яблока, суживающие зрачок, и ресничную. Добавочный
нерв (XI) делится на две ветви. Одна из них присоединяется к блу-
ждающему нерву, а наружная направляется к грудино-ключично-сос-
цевидной и трапециевидной мышцам. Подъязычный нерв (XII) ин-
нервирует мышцы языка.
Смешанные нервы: тройничный, лицевой, языкоглоточный, блу-
ждающий. Тройничный нерв (V) осуществляет чувствительную ин-
нервацию твердой мозговой оболочки, кожи головы и слизистых обо-
лочек глаза, полости носа и рта, придаточных пазух носа, передних
2/3 языка, слюнных желез, двигательную иннервацию жевательных
мышц и некоторых мышц шеи.
В состав лицевого нерва (VII) входят двигательные ветви (собст-
венно лицевой нерв), иннервирующие мимические мышцы и смешан-
ный (промежуточный) нерв. Последний образован чувствительными
(вкусовыми) и парасимпатическими волокнами: первые распростра-
няются в передних 2/3 языка, а парасимпатические предназначены
для слезной железы, а также желез слизистой оболочки полости носа,
подчелюстной и подъязычной слюнных желез.
I
II
III
IV
V
VI
VII
VIII
IX
X
XI
XII
Рис. 182. Управление органов
черепными нервами, схема:
I — обонятельный нерв,
II — зрительный нерв,
III — глазодвигательный нерв,
IV — блоковый нерв,
V — тройничный нерв,
VI — отводящий нерв,
VII — лицевой нерв,
VIII — преддверно-улитковый нерв,
IX — языкоглоточный нерв,
X — блуждающий нерв,
XI — добавочный нерв,
XII — подъязычный нерв
VIVI
385
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
В составе языкоглоточного нерва (IX) проходят двигательные,
чувствительные и парасимпатические волокна. Нерв осуществляет
чувствительную иннервацию слизистой оболочки задней трети язы-
ка, глотки, среднего уха, а также иннервирует мышцы глотки и около-
ушную слюнную железу.
Блуждающий нерв (X) осуществляет парасимпатическую иннер-
вацию органов шеи, грудной и брюшной полостей (до сигмовидной
ободочной кишки), а также содержит чувствительные и двигательные
волокна, которые иннервируют часть твердой оболочки головного
мозга, кожу наружного слухового прохода и ушной раковины, слизи-
стую оболочку и мышцы-сжиматели глотки, мышцы мягкого нёба,
слизистую оболочку и мышцы гортани, трахею, бронхи, пищевод,
сердце. В брюшной полости от ствола нерва отходят желудочные, пе-
ченочные и чревные ветви.
Спинномозговые нервы. Спинномозговые нервы (31 пара) фор-
мируются из двух корешков, отходящих от спинного мозга, — перед-
него (эфферентного) и заднего (афферентного), которые, соединяясь
между собой в межпозвоночном отверстии, образуют ствол спинно-
мозгового нерва (см. рис. 174). Это 8 шейных, 12 грудных, 5 пояснич-
ных, 5 крестцовых и 1 копчиковый нерв. Спинномозговые нервы со-
ответствуют сегментам спинного мозга. К заднему корешку прилежит
чувствительный спинномозговой узел, образованный телами круп-
ных афферентных Т-образных нейронов. Длинный отросток (ден-
дрит) направляется на периферию, где заканчивается рецептором,
а короткий аксон в составе заднего корешка входит в задние рога
спинного мозга. Волокна обоих корешков (переднего и заднего) обра-
зуют смешанные спинномозговые нервы, содержащие чувствитель-
ные, двигательные и вегетативные (симпатические) волокна. Послед-
ние имеются не во всех боковых рогах спинного мозга, а только в
VIII шейном, всех грудных и I—II поясничных нервах. В грудном от-
деле нервы сохраняют сегментарное строение (межреберные нервы),
а в остальных соединяются друг с другом петлями, образуя сплете-
ния: шейное, плечевое, поясничное, крестцовое и копчиковое, от кото-
рых отходят периферические нервы, иннервирующие кожу и скелет-
ные мышцы (рис. 183).
От шейного сплетения отходят чувствительные (кожные) нервы,
иннервирующие кожу затылочной области, ушной раковины, наруж-
ного слухового прохода, шеи; двигательные (мышечные) ветви к
близлежащим мышцам шеи и смешанный диафрагмальный нерв, ин-
нервирующий диафрагму.
Нервы плечевого сплетения иннервируют часть мышц шеи, мышцы
плечевого пояса, плечевой сустав, кожу и мышцы верхней конечности.
12 пар передних ветвей грудных нервов — это смешанные межребер-
386
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
ные нервы, иннервирующие все вентраль-
ные мышцы стенок грудной и брюшной
полостей, кожу передней и боковой по-
верхности груди и живота, молочную же-
лезу и осуществляющие чувствительную
иннервацию кожи туловища.
Нервы, выходящие из поясничного
сплетения, иннервируют кожу нижнего отдела передней брюшной
стенки и частично бедра, голени и стопы, наружных половых органов.
Мышечные нервы иннервируют мышцы стенок живота, передней
и медиальной групп мышц бедра. Крестцовое сплетение иннервирует
мышцы и частично кожу ягодичной области и промежности, кожу на-
ружных половых органов, кожу и мышцы задней поверхности бедра,
кости, суставы, мышцы и кожу голени и стопы. Ветви копчикового
сплетения иннервируют кожу в области копчика и в окружности зад-
него прохода.
ВЕГЕТАТИВНАЯ (АВТОНОМНАЯ)
НЕРВНАЯ СИСТЕМА (ВНС)
Вегетативная (от лат. vegeto — возбуждаю, оживляю) нервная си-
стема поддерживает постоянство внутренней среды организма, коор-
динирует и регулирует деятельность внутренних органов, обмен ве-
ществ, функциональную активность тканей. Пожалуй, наиболее
важна универсальность вегетативной нервной системы, которая ин-
нервирует весь без исключения организм, все органы, ткани. Вегета-
Рис. 183. Спинномозговые нервы:
1 — головной мозг в полости черепа,
2 — шейное сплетение (СI–VIII),
3 — диафрагмальный нерв, 4 — спинной мозг
в позвоночном канале, 5 — диафрагма,
6 — поясничное сплетение (LI–IV),
7 — бедренный нерв, 8 — крестцовое
сплетение (LIV–V; SI–III), 9 — мышечные ветви
седалищного нерва, 10 — общий
малоберцовый нерв, 11 — поверхностный
малоберцовый нерв, 12 — подкожный нерв,
13 — глубокий малоберцовый нерв,
14 — большеберцовый нерв,
15 — седалищный нерв, 16 — срединный нерв,
17 — локтевой нерв, 18 — лучевой нерв,
19 — мышечно-кожный нерв,
20 — подмышечный нерв,
21 — плечевое сплетение (СV–VIII; ТI)
11
11
11
22
12
12
33
13
13
44
14
14
55
15
15
66
16
16
77
17
17
88
18
18
99
19
19
10
10
20
20
21
21
387
ВЕГЕТАТИВНАЯ (АВТОНОМНАЯ) НЕРВНАЯ СИСТЕМА
тивная нервная система не находится под непосредственным конт-
ролем сознания, однако ее функция регулируется высшими отделами
мозга. ВНС действует совместно с соматической нервной системой.
Вегетативная нервная система подразделяется на две части: сим-
патическую (от греч. sympathes — чувствительный, восприимчивый
к влиянию) и парасимпатическую (от греч. para — возле, при). Каждая
из них имеет центральную и периферическую части (рис. 184). Цент-
ры вегетативной нервной системы расположены в четырех отделах
головного и спинного мозга, три из них — парасимпатические —
в стволе головного мозга и один — в крестцовом отделе спинного моз-
га. Единственный центр симпатической части расположен в правом и
левом боковых столбах (боковых рогах) VIII шейного, всех грудных
и I—II поясничных сегментов спинного мозга.
Рис. 184. Вегетативная (автономная) нервная система:
справа — симпатическая часть, слева — парасимпатическая часть. 1 — узлы
симпатического ствола, 2 — боковые рога спинного мозга, 3 — крестцовый отдел
парасимпатической части, 4 — блуждающий нерв, 5 — вегетативные нервные узлы
парасимпатической части, 6 — головной отдел парасимпатической части
11
22
33
44
55
66
388
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Периферическая часть вегетативной нервной системы образована
выходящими из головного и спинного мозга вегетативными нервами,
ветвями и нервными волокнами, вегетативными сплетениями и их уз-
лами (ганглиями), лежащими кпереди от позвоночника (предпозво-
ночные — превертебральные) и рядом с позвоночником (околопозво-
ночные — паравертебральные), а также расположенными вблизи
крупных сосудов возле органов и в их толще.
Рефлекторная дуга вегетативной нервной системы может быть
представлена следующим образом (см. рис. 174). От рецепторов пе-
редается возбуждение по волокнам афферентных нейронов, распо-
ложенных в спинномозговых узлах либо в узлах черепных нервов
или в узлах вегетативных сплетений. Аксоны этих нейронов в соста-
ве задних корешков вступают в спинной мозг (направляясь в боко-
вые рога) или в составе черепных нервов — в вегетативные ядра
мезенцефалического или бульварного отдела головного мозга. В бо-
ковых рогах, а также в указанных ядрах ствола головного мозга за-
легают ассоциативные мультиполярные нейроны. Их аксоны выхо-
дят из мозга в составе передних корешков спинномозговых или
черепных нервов. Это преганглионарные (предузловые) волокна, ко-
торые обычно миелинизированы. Они следуют к узлам внеорганных
или внутриорганных вегетативных сплетений, где образуют синапсы
с их клетками. В узлах лежат мультиполярные (вторые) нейроны
эфферентного вегетативного пути, аксоны которых, выйдя из ган-
глиев, образуют постганглионарные волокна (чаще всего немиели-
низированные), которые направляются к органам и тканям. Веге-
тативные волокна идут в составе соматических нервов или
самостоятельно в виде вегетативных нервов в оболочках стенок кро-
веносных сосудов.
Важной особенностью вегетативной нервной системы является
образование сплетений, к которым подходят симпатические или пара-
симпатические преганглионарные волокна. Околопозвоночные узлы
симпатической части вегетативной нервной системы, соединяясь
между собой с каждой стороны позвоночника межузловыми ветвями,
образуют правый и левый симпатические стволы, расположенные по
бокам от позвоночника. В каждом стволе различают 3 шейных, 10–
12 грудных, 4 поясничных и 4 крестцовых узла. На передней поверх-
ности крестца оба ствола сходятся, образуя непарный узел.
Аксоны нейронов боковых рогов спинного мозга (симпатиче-
ские) вначале идут в составе передних корешков спинномозговых
нервов, затем в составе этих нервов и отходящих от них белых сое-
динительных ветвей вступают в симпатический ствол (преганглио-
нарные волокна). Часть этих волокон заканчивается синапсами на
клетках узлов симпатического ствола. Аксоны этих клеток в виде
389
ВЕГЕТАТИВНАЯ (АВТОНОМНАЯ) НЕРВНАЯ СИСТЕМА
постганглионарных волокон выходят из симпатического ствола (па-
равертебральных узлов) в составе серых соединительных ветвей
(немиелинизированные), присоединяются к спинномозговым нер-
вам и иннервируют все органы и ткани, где эти нервы разветвляют-
ся, в том числе кровеносные сосуды, волосяные луковицы и потовые
железы кожи. Другая часть преганглионарных волокон (отростки
клеток первого нейрона эфферентного пути) не прерывается в узлах
симпатического ствола, а проходит через них транзитом и в составе
ветвей симпатического ствола (внутренностных нервов) входит в
узлы симпатических сплетений брюшной полости и таза (чревное,
аортальное, брыжеечные, верхнее и нижнее подчревные). В узлах
(превертебральных) этих сплетений преганглионарные волокна за-
канчиваются синапсами на нейронах узлов. Нервные клетки, распо-
ложенные в превертебральных узлах сплетений, являются вторыми
нейронами эфферентного пути симпатической иннервации внутрен-
них органов брюшной полости, таза, кровеносных и лимфатических
сосудов.
Аксоны эфферентных нейронов, расположенных в узлах симпа-
тических сплетений брюшной полости и таза, идут по двум направ-
лениям: в составе вегетативных нервов, содержащих постганглио-
нарные волокна, к внутренним органам, в виде постганглионарных
волокон, расположенных в оболочках кровеносных сосудов, также
к внутренним органам и другим органам, где эти сосуды разветвля-
ются.
Преганглионарные волокна парасимпатической части обычно бо-
лее длинные, чем преганглионарные симпатические, они идут в соста-
ве III, VII, IX и X пар черепных и II—IV крестцовых нервов. Аксоны
парасимпатических нейронов доходят до околоорганных вегетатив-
ных узлов или органных узлов вегетативных сплетений (сердечного,
легочного, пищеводного, желудочных, кишечного и др.), в которых
залегают тела клеток вторых эфферентных парасимпатических ней-
ронов, чьи аксоны направляются к рабочим органам. Парасимпатиче-
ская часть глазодвигательного нерва иннервирует ресничную мышцу
и мышцу-сфинктер зрачка. Парасимпатическая часть лицевого нерва
иннервирует слезную железу, а также железы слизистой оболочки по-
лости носа, нёба, поднижнечелюстную и подъязычную слюнные же-
лезы. Парасимпатическая часть языкоглоточного нерва иннервирует
околоушную слюнную железу. Парасимпатическая часть блуждаю-
щего нерва осуществляет парасимпатическую иннервацию гладкой
мускулатуры и желез органов шеи, груди и живота. Крестцовый от-
дел парасимпатической части вегетативной нервной системы осу-
ществляет парасимпатическую иннервацию гладких мышц и желез
органов таза.
390
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
ВНИМАНИЕ!
Большинство внутренних органов иннервируется обеими частями веге-
тативной нервной системы, которые оказывают на них различное, иног-
да противоположное влияние (табл. 24), обусловленное действиями
медиаторов. Основным медиатором симпатической нервной системы
является норадреналин, парасимпатической — ацетилхолин.
Таблица 24
Влияние симпатической и парасимпатической части
вегетативной нервной системы на органы
Орган или система
Стимуляция
парасимпатических нервов
симпатических нервов
Сердце
Уменьшение частоты
сокращений
Увеличение частоты
сокращений
Уменьшение силы сокра-
щений (предсердий)
Увеличение силы
сокращений
Кровеносные сосуды
Артерии кожи и слизи-
стых
Сужение
Артерии брюшной
полости
Сужение
Артерии скелетных
мышц
Сужение
Расширение (только под
действием адреналина крови)
Расширение (холинергическое)
Коронарные артерии
Расширение
Сужение
Расширение (только под дей-
ствием адреналина крови)
Артерии полового чле-
на, а также, возможно,
клитора и малых поло-
вых губ
Расширение
Вены
Сужение
Сосуды мозга
Расширение
Сужение
Желудочно-кишечный тракт
Продольные и цирку-
лярные мышцы
Усиление моторики
Ослабление моторики
Сфинктеры
Расслабление
Сокращение
Мочевой пузырь
Детрузор (мышца,
выталкивающая мочу)
Сокращение
Расслабление
Внутренний сфинктер
Сокращение
391
ВЕГЕТАТИВНАЯ (АВТОНОМНАЯ) НЕРВНАЯ СИСТЕМА
Орган или система
Стимуляция
парасимпатических нервов
симпатических нервов
Половые органы
Семенные пузырьки
Сокращение
Семявыносящий
проток
Сокращение
Матка
Сокращение
Расслабление (в зависимости
от вида животного и гормо-
нального фона)
Мышца, расширяющая
зрачок
Сокращение (мидриаз)
Сфинктер зрачка
Сокращение (миоз)
Цилиарная мышца
Сокращение (аккомода-
ция)
Незначительное расслабление
Трахеобронхиальные
мышцы
Сокращение
Расслабление (в основном
под действием адреналина)
Мышцы, поднимаю-
щие волосы
Сокращение
Экзокринные железы
Слюнные железы
Обильное выделение се-
розного секрета
Небольшое выделение слизи-
стого секрета (из подчелюст-
ной железы)
Слезные железы
Секреция
Пищеварительные
железы
Секреция
Снижение секреции
Железы носоглотки
Секреция
Бронхиальные железы
Секреция
Потовые железы
Секреция (холинергическая)
Метаболизм
Печень
Гликогенолиз
Гликонеогенез
Жировые клетки
Липолиз (повышение уровня
свободных жирных кислот
в крови)
Секреция инсулина
(β-клетками островка
Лангерганса)
Снижение
Окончание табл. 24
392
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Симпатическая нервная система опосредует реакцию организма
типа «борьбы или бегства». Расширение бронхов и увеличение легоч-
ной вентиляции, увеличение частоты и силы сердечных сокращений,
сужение артерий кожи, желудочно-кишечного тракта, почек и расши-
рение артерий мышц, миокарда приводит к увеличению доставки кис-
лорода мышцам и сердцу, благодаря чему они усиливают сокращения.
Этому способствует усиление распада гликогена в печени и жира
в жировой ткани, что улучшает снабжение мышц, сердца и мозга глю-
козой и жирными кислотами.
Преобладание активности парасимпатической системы обеспечи-
вает реакции типа «отдыха и восстановления», что приводит к восста-
новлению сил организма. При этом сила, частота сердечных сокраще-
ний и просвет воздухоносных путей уменьшаются, артерии скелетных
мышц суживаются, а желудочно-кишечного тракта — расширяются.
Это приводит к уменьшению кровотока в мышцах, миокарде и его
увеличению в пищеварительном тракте, что усиливает пищеварение.
ВНИМАНИЕ!
Эрекция полового члена и клитора, возбуждение женских половых орга-
нов связаны с возбуждением парасимпатических нейронов крестцового
отдела спинного мозга; во время оргазма происходит активация симпати-
ческих нейронов нижних грудных и верхних поясничных сегментов.
Центральная регуляция функций вегетативной нервной системы
осуществляется корой больших полушарий через гипоталамус и ствол
мозга (главным образом продолговатый мозг). Из этих структур вы-
ходят основные проводящие пути, которые направляются к преган-
глионарным нейронам.
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ
Высшая нервная деятельность (ВНД) — это интегративная функ-
ция коры полушарий большого мозга и подкорковых центров, обеспе-
чивающая наиболее оптимальное приспособление человека (и живот-
ных) к окружающей среде. ВНД основана на взаимодействии
врожденных безусловных рефлексов и приобретенных условных реф-
лексов, в котором у человека важную роль играет вторая сигнальная
система. И .М . Сеченов (1829—1905) в книге «Рефлексы головного
мозга» распространил рефлекторный принцип на все виды сознатель-
ной и бессознательной деятельности организма, включая психику.
И.П. Павлов (1849—1935) разработал лабораторные методы объек-
тивного изучения приспособительной деятельности человека и жи-
вотных, в первую очередь — метод условных рефлексов.
393
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ
ВНИМАНИЕ!
И.П . Павлов первым установил, что в основе психической деятельности
человека лежат физиологические процессы, происходящие в коре по-
лушарий большого мозга.
Особенности высшей нервной деятельности человека
Человек воспринимает внешний мир органами чувств, т. е . первой
сигнальной системой. Общими для животных и человека являются ана-
лиз и синтез конкретных сигналов, предметов и явлений внешнего мира,
составляющих первую сигнальную систему. Однако только человек
обладает второй сигнальной системой действительности, специфиче-
ским раздражителем которой является слово с заложенным в него смы-
слом, слово, которое обозначает предметы и явления окружающего мира.
ВНИМАНИЕ!
И.П . Павлов называет второй сигнальной системой действительности
нервные процессы, возникающие в полушариях большого мозга в ре-
зультате восприятия сигналов окружающего мира в виде речевых обо-
значений предметов и явлений.
Слово воспринимается человеком как услышанное (слуховой ана-
лизатор), как написанное (зрительный анализатор) или как произне-
сенное (двигательный анализатор). Во всех случаях данные раздра-
жители объединяются смыслом слова. Слова приобретают смысл
в результате возникновения прочной связи в коре полушарий боль-
шого мозга между центрами возбуждения, возникающими под дейст-
вием конкретных объектов окружающего мира, и центрами возбужде-
ния, возникающими при произнесении слов, обозначающих эти
предметы или действия. В результате образования таких связей слова
могут заменить конкретный раздражитель окружающей среды и
сделаться его символом. Наличие второй сигнальной системы вно-
сит новый принцип в деятельность мозга человека. Слово как сигнал
сигналов дает возможность отвлечься от конкретных предметов и яв-
лений. Развитие словесной сигнализации сделало возможным обо-
бщение и отвлечение, что находит свое выражение в характерных для
человека явлениях — мышлении и понятиях. Способность мыслить
путем абстрактных (отвлеченных) образов, понятий, выражаемых
произнесенными или написанными словами, сделало возможным воз-
никновение абстрактно-обобщенного мышления.
Вторая сигнальная система человека является основой сугубо че-
ловеческого словесно-логического мышления, основой формирова-
ния знаний об окружающем мире посредством словесных абстракций
и основой человеческого сознания.
394
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Типы нервной деятельности
Нервные реакции у разных людей различают по силе и подвижно-
сти. Эти индивидуальные особенности обусловлены взаимоотноше-
ниями процессов возбуждения и торможения. На основании разли-
чий нервных реакций, в первую очередь по силе нервных процессов,
И.П . Павлов выделил сильный и слабый типы нервной системы.
Сильный тип нервной системы может быть неуравновешенным или
уравновешенным. Неуравновешенный тип (безудержный, холерик)
отличается повышенной возбудимостью, взрывчатостью, процессы
возбуждения преобладают над процессами торможения. Уравнове-
шенный тип нервной системы может различаться по подвижности
нервных процессов, по быстроте реагирования, перестройке поведе-
ния. При подвижном типе (сангвиник) возможна быстрая переориен-
тация нервных процессов в ответ на смену жизненных обстоятельств.
При инертном типе нервной системы (флегматик) переориентация
деятельности дается с трудом, протекает медленно.
Типы нервной системы, выделенные И.П. Павловым, соответству-
ют классификации темпераментов человека, предложенной почти
2500 лет назад великим врачом древнего мира Гиппократом. Он по-
дразделял людей по темпераменту на холериков (неуравновешенных,
легковозбудимых), сангвиников (уравновешенных, с живой, подвиж-
ной нервной системой оптимистов), флегматиков (уравновешенных,
спокойных, рассудительных, инертных) и меланхоликов (слабый тип
нервной системы: мрачные, подавленные, вечные скептики).
Тип нервной системы наследуется, однако впоследствии существен-
ное влияние на него оказывает окружающая среда. Особенности харак-
тера формируются в индивидуальной жизни человека. Слабый тип (ме-
ланхолик) формируется при воспитании в тепличных условиях, когда за
ребенка все и всегда решают взрослые, когда ему шагу не дают сделать
самостоятельно, лишают его инициативы. Изоляция ребенка от трудно-
стей, от влияния внешней среды, даже при врожденном сильном типе
нервной системы, может сформировать лишь пассивно-защитные реак-
ции. Постановка слишком трудных, непосильных задач может вызвать
перенапряжение корковых процессов возбуждения или торможения, что
приводит к срывам нервной деятельности, неврозам. Психические функ-
ции у человека нарушаются при действии алкоголя, наркотиков, при
этом серьезно страдают механизмы нервных процессов.
В зависимости от преобладания у человека функции первой или
второй сигнальной систем различают три типа высшей нервной дея-
тельности. Художественный тип. Это люди, у которых восприятие
внешней среды отличается яркой образностью. Первая сигнальная
система доминирует. Мыслительный тип. Доминирует вторая сиг-
395
ИНТЕГРАТИВНАЯ ФУНКЦИЯ НЕРВНОЙ СИСТЕМЫ
нальная система, преобладает способность к анализу и синтезу, уста-
новлению причинно-следственных связей. Средний тип. Роль обеих
сигнальных систем уравновешена.
ИНТЕГРАТИВНАЯ ФУНКЦИЯ НЕРВНОЙ СИСТЕМЫ
Интегративные функции лежат в основе ритмов мозга, в т. ч . ци-
кла «сон — бодрствование», сознания и мышления, членораздельной
речи, научения и памяти, мотиваций и эмоций, интеллекта и творче-
ства. Морфологическим субстратом интегративных функций являет-
ся кора полушарий большого мозга, а именно новая кора и лим-
бическая система. Интегративные функции ЦНС не связаны с
управлением движениями и вегетативными функциями, а также с об-
работкой сигналов, поступающих от органов чувств.
Ритмы мозга
Жизнедеятельность организма человека происходит ритмично,
и эта ритмичность также координируется и регулируется нервной
системой. Ритмы подчиняются общему принципу иерархичности
строения живого. Так, существуют клеточные, тканевые, органные,
системные и организменные ритмы. Различают несколько типов био-
логических ритмов: околосуточные, или циркадианные (от лат. cir-
ca — «около», dies — «день»), это ритмы с периодичностью, примерно
равной суткам. У человека их около 300. Основной из них — это цикл
сна и бодрствования, с которым синхронизированы многие другие,
например суточные колебания температуры тела, выделения многих
гормонов, мочеотделение, подъем и спад умственной и физической
работоспособности. Инфрадианные (от лат. infra — «меньше») ритмы
более продолжительные, они повторяются реже одного раза в сутки.
Например, репродуктивный цикл у женщины. Хорошо изучены се-
зонные и годовые ритмы животных.
Большинство людей ощущают сезонные ритмы. В последние годы
изучены и описаны трехлетние и семилетние циклы у людей. Так, на-
пример, у многих спортсменов достижения подчиняются трехлетним
ритмам, у многих художников, поэтов, композиторов, писателей твор-
ческие подъемы и спады подчиняются семилетним ритмам. Все они
начинаются с рождения человека и протекают в течение всей жизни.
Ультрадианные (от лат. ultra — «сверх») ритмы повторяются чаще
одного раза в сутки, например фазы сна и бодрствования. У человека
наблюдаются циклические колебания познавательной деятельности
и внимания с периодом 90—100 мин. Цикличность электрической ак-
тивности мозга, независимо от сна или бодрствования, составляет
396
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
около 90 мин. В основе ритмов лежат взаимоотношения между Сол-
нцем, Землей и Луной. Все циркадианные ритмы связаны с главным
ритмом природы — вращением Земли.
Сон и бодрствование
Сон — это не перерыв в деятельности мозга, а просто иное состоя-
ние сознания. Во время сна активно функционируют нейроны, причем
во сне спонтанная активность многих нейронов даже усиливается. Сон
является одной из важных биологических потребностей вида «человек
разумный». Назначение его до конца не известно. Длительность сна
у большинства людей колеблется в пределах от 6 до 8 часов в сутки.
Сознание и мышление
Сознание — это, пожалуй, главный признак существования челове-
ка, которое характеризуется в первую очередь осознанием своего «я»,
собственной индивидуальности и индивидуальности других людей.
Морфологической основой сознания являются функционирующие
нейроны коры больших полушарий головного мозга. Сознание поро-
ждается в результате определенной функциональной активности ЦНС
в процессе взаимодействия коры больших полушарий и подкорковых
структур. Важнейшую роль в этом играет ретикулярная формация.
Мышление — это понимание и способность манипулировать сло-
весными символами, или понятиями. Мышление представляет собой
процесс познавательной деятельности человека, который предполага-
ет мысленную манипуляцию любыми словесными символами, идея-
ми, образами, суждениями, воспоминаниями, понятиями, образами
восприятия, убеждениями или намерениями. Мышление включает
решение задач, интеллектуальные функции, творчество, научение, па-
мять, воображение и т. д. В основе мышления лежат память и члено-
раздельная речь; иными словами, мышление вербально. Мышление
является функцией мозга. Главную роль в этом процессе играют лоб-
ные доли больших полушарий головного мозга. Сущность мышле-
ния — это мысленное моделирование человеком различных событий.
Членораздельная речь
Сознание человека тесно связано с членораздельной речью и зави-
сит от нее. Человек осознаёт только те предметы и явления, которые
превращаются в словесные символы. Большинство животных обща-
ются между собой с помощью различных сигналов. Членораздельная
речь уникальна и отличается от этих сигналов, которые, кстати, суще-
397
ИНТЕГРАТИВНАЯ ФУНКЦИЯ НЕРВНОЙ СИСТЕМЫ
ствуют и у человека (например, гримаса или вскрик от боли), тем, что
передает не только то, что имеется в настоящее время в данной кон-
кретной ситуации, но и то, что было в прошлом, что может произойти
в будущем. Речь передает сведения об опыте, которым говорящий не
обладает.
ВНИМАНИЕ!
И.П . Павлов назвал речь второй сигнальной системой, которая отли-
чается от первой (органы чувств) отвлечением и обобщением.
В коре полушарий большого мозга находятся три важнейших цен-
тра, связанных с речью: зрительный — на медиальной поверхности
затылочных долей в области шпорной борозды; слуховой — в обеих
височных долях; сомато-сенсорный — в прецентральных извилинах
обоих полушарий, именно его нейроны управляют мышцами лица,
туловища и конечностей. Следует знать о межполушарных различиях,
открытых Р. Сперри, который показал, что изолированное левое по-
лушарие столь же эффективно обеспечивает сознание человека и чле-
нораздельную речь, как оба полушария, в то время как изолированное
правое полушарие не может обеспечить устную и письменную речь.
В 1982 году Р. Сперри был удостоен Нобелевской премии «за откры-
тия, касающиеся функциональной специализации полушарий мозга».
Левое полушарие не только воспринимает речь, но и генерирует
звуки речи. Левое полушарие доминирует и в двигательных актах
речи. Правое полушарие способствует выделению сигналов из шума,
опознаёт интонации и музыкальные мелодии, оно обеспечивает пони-
мание устной и письменной речи, участвует в модуляции голоса. Од-
нако правильнее говорить не о доминировании левого полушария,
а о взаимодополняющей специализации обоих полушарий с преобла-
данием речевых функций, как правило, у левого.
Именно левое полушарие определяет главным образом лингви-
стические (вербальные) способности человека. Корковые центры ле-
вого полушария играют основную (специфическую) роль в восприя-
тии, запоминании и воспроизведении речи. Только их нормальное
функционирование приводит к полноценной речевой функции.
Научение и память
Научение — это процесс усвоения знаний или овладения ими. Са-
мая простая и самая распространенная форма научения — это привы-
кание. Память — это психическая функция сохранения информации
о каких-либо раздражениях, а также идеях, образах, событиях после
398
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
того, как они уже не действуют на человека, и способность сохранять
и впоследствии использовать (извлекать) эту информацию. Память
характеризуется пятью процессами: запоминание информации, ее со-
хранение, извлечение, воспроизведение и забывание.
ВНИМАНИЕ!
Основная отличительная особенность памяти человека — способность
формулировать идеи в виде словесных символов и сохранять их в аб-
страктной форме.
Различают три вида памяти. Сенсорная память. Информация, полу-
ченная рецепторами, сохраняется около 1/4 сек., в течение которых
в головном мозге решается вопрос о необходимости ее сохранения.
Кратковременная (первичная) память. Если сенсорная информация
необходима мозгу, она обрабатывается, интегрируется и сохраняется
около 20 сек. Емкость кратковременной памяти ограничивается семью
элементами (от 5 до 9). Если необходимо сохранить в кратковременной
памяти большее число элементов, мозг группирует их в соответствую-
щее количество. Кратковременная память является основой выполне-
ния текущих поведенческих и мыслительных операций. Долговременная
память. Если информация, хранящаяся в кратковременной памяти, не
нужна или не используется, она быстро стирается. Если она необходи-
ма, то переводится в долговременную память. Чем чаще человек извле-
кает эту информацию, тем лучше она фиксируется. Долговременная па-
мять зависит от частоты повторения, от контекста, уровня мотивации,
т. е . необходимости запомнить ту или иную информацию. Долговремен-
ную память подразделяют на вторичную и третичную. Долгосрочная
память является основным видом памяти человека, благодаря ей чело-
век может существовать как индивидуум. В этой памяти хранятся все
без исключения образы, события, знания, умения, навыки. Долговре-
менная память является основой условно-рефлекторной деятельности
человека. Забывание. Способность забывать имеет огромное значение
в выживании людей. Мозг должен освобождаться от ненужного груза
впечатлений и сведений. Память как бы сама регулирует нагрузку, гото-
вится к приему новой информации. При этом старая информация не
исчезает бесследно, но переходит из активной памяти в пассивную, от-
куда иногда ее удается извлечь. Это замечательное свойство спасает
многих людей в трагических ситуациях. Забывание зависит от многих
причин: если информация, хранящаяся в памяти, не используется, она
забывается; помехи, связанные с наслоением какой-то предшествующей
или последующей информации, также способствуют забыванию преды-
дущих сведений; часто происходит подавление или активное забывание,
связанное с бессознательной психической защитой.
399
ИНТЕГРАТИВНАЯ ФУНКЦИЯ НЕРВНОЙ СИСТЕМЫ
ВНИМАНИЕ!
Распространено заблуждение, что с возрастом память ухудшается. Это
не совсем так. Память ухудшается, если ее не тренировать. Безделье —
худший враг памяти. При высокой умственной активности память со-
храняется независимо от возраста. Особенно важно это для людей
старше 70—80 лет. Несколько примеров. Великий немецкий поэт
И.В. Гете закончил «Фауста» в 83 года. А Владимир Зельдин сыграл свой
последний спектакль в 101 год и несколько месяцев.
Поведение
Поведение включает различные действия, деятельность, реакции,
движения, процессы, операции, совершаемые индивидуумом. Поведе-
ние человека связано с его умственной деятельностью, мышлением,
способностью к умозаключениям и членораздельной речью. Формы
поведения человека принято подразделять на врожденные и приобре-
тенные. Наследственно закрепленные рефлексы лежат в основе при-
способительных поведенческих актов, проявляющихся без предвари-
тельного обучения. И.П. Павлов рассматривал врожденное поведение
как совокупность сложнейших безусловных рефлексов (инстинктов).
Индивидуальный опыт приобретается различными путями, в ос-
нове которых лежит общая способность живых организмов к обуче-
нию. Поведение человека — это, во-первых, генетически детермини-
рованная видоспецифическая программа, а во-вторых, гибкая система
конкретных приспособлений к изменяющимся условиям внешней
среды. Среди факторов, ответственных за организацию конкретного
поведения, выделяют генетические, субъективные (мотивации и эмо-
ции) и объективные внешние (пространство и время). Важную роль
играет пищевое поведение (поиск пищи, слюноотделение, усиление
двигательной активности и кровообращения желудочно-кишечного
тракта и т. д .); половое поведение, направленное на реализацию чело-
веческой сексуальности, включая репродукцию; социальное поведение
(взаимоотношения в социальной группе).
Поведенческие реакции человека могут быть вызваны воздействием
на организм физиологически значимых раздражителей внешней или
внутренней среды и возникновением потребностей. В настоящее время
общепризнанно, что все формы поведения имеют определенный мотив,
т. е. направлены на удовлетворение потребностей организма. И.П. Пав-
лов считал потребности фундаментом поведения и психики, «рефлексом
цели». Различают три вида потребностей: биологические, социальные и
идеальные. Биологические, или жизненные, потребности направлены на
сохранение целостности организма и вида (потребности в еде, питье, сне
и т. д .) . Социальные — это потребности принадлежать к определенной
группе и следовать поведенческим нормам данной группы, следовать
нравственным и эстетическим нормам общества. Идеальные потребно-
400
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
сти, или потребности познания и творчества, — это потребности челове-
ка в познании мира и своего места в нем, смысла жизни, потребности
приобретения знаний. Биологические потребности являются преимуще-
ственно врожденными, в то время как социальные и идеальные приобре-
таются в процессе индивидуального развития на основе биологических
потребностей. Согласно иерархической теории потребностей человека,
созданной американским психологом А. Маслоу, самореализация явля-
ется высшей потребностью человека. Путь к ней идет через последова-
тельное удовлетворение нижележащих потребностей.
ВНИМАНИЕ!
Самореализация человека — это любовь и творчество (рис. 185).
Мотивация
Мотивация (от лат. movere — «двигаться» или франц. motif —
«побудительная причина»). «Мотивация — это совокупность факто-
ров, определяющих поведение» (Ж. Годфруа, 1992). Мотивации свя-
заны, в первую очередь, с потребностями человека и его влечениями.
Мотивация всегда порождается потребностью. Мотивация — это тол-
чок к целенаправленному действию, вызванный потребностью. Вле-
чение — это внутреннее состояние человека, заставляющее его дейст-
Саìîðеаëèçаöèя
(ëюбîâü, òâîð÷есòâî)
Êîгíèòèâíûе
è ýсòеòè÷есêèе ïîòðебíîсòè
(â ïîðядêе, сïðаâедëèâîсòè,
êðасîòе, сèììеòðèè)
Пîòðебíîсòè â óâажеíèè
(â îдîбðеíèè, бëагîдаðíîсòè,
ïðèçíаíèè, êîìïеòеíöèè)
Пîòðебíîсòè â ïðèâяçаííîсòяõ,
õîðîшеì îòíîшеíèè, â ïðè÷асòíîсòè ê гðóïïе è дð.
Пîòðебíîсòè â беçîïасíîсòè
(фèçè÷есêîй è ïсèõîëîгè÷есêîй)
Фèçèîëîгè÷есêèе ïîòðебíîсòè
(â ïèще, âîде, êèсëîðîде, сíе,
îïòèìаëüíîй òеìïеðаòóðе, èçбежаòü бîëè è дð.)
Рис. 185. Иерархия
потребностей человека
(по Ж. Годфруа,
с изменениями)
401
ИНТЕГРАТИВНАЯ ФУНКЦИЯ НЕРВНОЙ СИСТЕМЫ
вовать определенным образом. Склонность к добру и склонность
к злу — два врожденных инстинкта, свойственные каждому человеку
и оказывающие влияние на принимаемые им решения.
Мотивации человека, лежащие в основе его поведения, неотдели-
мы от эмоций. Эмоции (от лат. emovere — «возбуждать, волновать») —
это психические переживания, душевные волнения, возникающие в
результате воздействия тех или иных стимулов внешней или внутрен-
ней среды. Различают десять фундаментальных эмоций: интерес, ра-
дость, удивление, горе, гнев, отвращение, презрение, страх, стыд, вина.
Эти функциональные эмоции, взаимодействуя между собой, могут
создавать устойчивые комплексы (например, любовь, враждебность,
тревожность, депрессия и др.). Эмоция — психическое переживание,
душевное волнение, которое может быть приятным или неприятным
для человека. Эмоции имеют обычно три составляющие: субъектив-
ную, физиологическую и поведенческую. Например, страх может до-
ставлять человеку неприятные субъективные ощущения, приводить к
изменению некоторых физиологических характеристик (например, к
учащению сердцебиения, усиленному потоотделению и т. д .) и застав-
лять его избегать таких ситуаций, которые провоцируют у него страх.
Эмоции проявляются в виде субъективных переживаний человека,
в его поведении и реакциях вегетативной нервной системы. Эмоции
связаны и с удовлетворением потребностей. Анатомическим субстра-
том эмоции являются лимбическая система, ретикулярная формация,
гипоталамус, кора лобных долей больших полушарий головного мозга
(см. раздел «Головной мозг»). Реализация эмоций осуществляется лим-
бической системой мозга, все структуры которой связаны между собой
многочисленными прямыми и обратными нервными связями.
Эмоции могут быть положительными и отрицательными: чувство
безопасности и страх, любовь и ненависть, радость и горе, симпатия и
антипатия, расположение и гнев и др. Положительные эмоции спо-
собствуют сохранению и укреплению здоровья и долголетию, отрица-
тельные ухудшают здоровье, угнетают защитные силы организма. Ин-
тенсивность эмоций связана с уровнем активации организма, но
направленность эмоций, их положительная или отрицательная окра-
ска зависят от того, как человек воспринимает данную ситуацию; оп-
тимисты воспринимают эмоции иначе, чем пессимисты. Первые легче
достигают самоуважения и самореализации, чем вторые.
ВНИМАНИЕ!
Оптимисты менее подвержены заболеваниям сердечно-сосудистой си-
стемы, злокачественным заболеваниям, стрессу. Оптимисты живут
дольше и быстрее выздоравливают после различных болезней и опера-
ций. Оптимисты реже страдают сексуальными расстройствами.
402
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Интеллект
Память и членораздельная речь являются основой мышления.
Мышление — это процесс познавательной деятельности, характеризу-
ющийся обобщенным и опосредованным отражением действитель-
ности. Интеллект человека связан с мышлением (от лат. intellectus —
«ум, рассудок, разум») человека.
Интеллект — это глобальная способность разумно действовать,
рационально мыслить и хорошо справляться с жизненными обстоя-
тельствами, т. е. адаптироваться к постоянно меняющимся условиям
внешней среды. Различают конкретный интеллект, который позво-
ляет решать ежедневные практические проблемы; абстрактный
интеллект, с помощью которого человек способен оперировать
абстрактными символами. Абстрактный интеллект связан с разго-
ворной речью. Человек рождается, имея генетически детерминиро-
ванные определенные врожденные способности (потенциальный
интеллект). Использование их, решение различных проблем спо-
собствует развитию кристаллического интеллекта, который включа-
ет в себя знания в различных областях. Иными словами, потенци-
альный интеллект является основой развития кристаллического,
который формируется в результате взаимодействий первого с внеш-
ней средой.
Физическое и психическое состояние матери во время беременно-
сти оказывает большое влияние на потенциальный интеллект ребен-
ка. После его рождения важную роль играет питание, психическая
стимуляция, количество детей в семье, социальное положение, тип
воспитания в семье и многие другие факторы.
В начале XX в. немецкий психолог Штерн разработал концепцию
коэффициента интеллектуальности (IQ), согласно которой у нор-
мального ребенка интеллектуальный (умственный) и хронологиче-
ский возраст совпадают. Сегодня существуют специальные таблицы,
позволяющие оценить IQ у людей различных возрастных групп.
Серьезные исследования, проведенные уже во второй половине XX в.,
показали, что примерно у 70% людей IQ лежит в пределах от 84 до
116 баллов, около 2% страдают умственной отсталостью (IQ < 70),
у 13% интеллект снижен (70—84), у 13% — повышен (116—132) и
примерно 2% имеют очень высокий интеллект — сверходаренные
люди. Однако многочисленные исследования показали, что сверхода-
ренные люди далеко не всегда гениальные художники, ученые, музы-
канты, писатели. Они удачливы, имеют прекрасное социальное поло-
жение, довольны жизнью. И этого вполне достаточно.
403
ОРГАНЫ ЧУВСТВ
ОРГАНЫ ЧУВСТВ
Взаимодействие организма с внешней средой осуществляется ор-
ганами чувств, или анализаторами. Выделяют органы зрения, слуха,
равновесия, вкуса, обоняния и осязания (кожного чувства). С помо-
щью органов чувств человек не только «ощущает» внешний мир бла-
годаря труду и членораздельной речи, на основе ощущений человек
обладает особыми, присущими только ему социальными формами от-
ражения — сознанием, самосознанием, способностью предвидения
(прогнозирования), творчеством и др.
И. П. Павлов разработал принципиально новое учение об анализа-
торах. Согласно ему каждый анализатор является комплексным «ме-
ханизмом», который не только воспринимает сигналы внешней среды
и преобразует их энергию в нервный импульс, но и производит выс-
ший анализ и синтез.
Каждый анализатор состоит из трех частей. Периферическая часть
(рецепторная клетка) воспринимает энергию внешнего раздражения и
перерабатывает ее в нервный импульс. При этом каждая рецепторная
клетка воспринимает раздражения с определенной зоны — рецептивно-
го поля, представляющего собой все точки периферического отдела
анализатора, возбуждение которых влияет на данный нейрон. Чем
больше число рецепторов, воспринимающих стимул, и частота нерв-
ных импульсов, тем больше размеры воспринимаемого раздражения
и его сила. Следующее звено — проводящие пути, по которым нервный
импульс следует к нервному центру. Проводящие пути проходят через
несколько уровней переключения (в спинном мозге, стволе, головном
мозге и таламусе) и достигают коркового конца анализатора (сенсорно-
го центра), который расположен в соответствующих участках коры го-
ловного мозга. В корковом центре сигналы внешнего мира реконструи-
руются, сличаются. Сигналы, поступающие от различных анализаторов,
интегрируются с информацией, которая хранится (накапливается)
в памяти для создания определенного восприятия внешнего мира, т. е.
имеет место высший анализ.
Любое ощущение имеет четыре параметра: пространственный,
временной, интенсивность (или количество) и качество (или модаль-
ность) — см. табл. 25. Каждый анализатор реагирует оптимально
только на определенные адекватные стимулы. Однако специфичность
анализаторов связана со специфичностью рецепторов и характером
центральной обработки информации.
Для восприятия важную роль играет предшествующий опыт. Раздра-
жения внешней среды интерпретируются мозгом с учетом накопленной
информации. Если же информация новая или неполная, мозг выдвигает
некую гипотезу (Ж. Годфруа, 1992). Восприятие человека осуществляет-
404
ОРГАНЫ ЧУВСТВ
ся по определенным принципам. Самый важный из этих принципов со-
стоит в том, что любой образ или предмет воспринимается как фигура,
выделяющаяся на каком-то фоне. Наш мозг действительно обладает тен-
денцией (по-видимому, врожденной) структурировать сигналы таким
образом, что все, что меньше или имеет более правильную конфигура-
цию, а главное — то, что имеет для нас какой-то смысл, воспринимается
как фигура; она выступает на некотором фоне, а сам фон воспринимает-
ся гораздо менее структурированным. Это относится прежде всего к зре-
нию, но также и к другим чувствам. Второй принцип — это принцип за-
полнения пробелов, проявляющийся в том, что наш мозг всегда старается
свести фрагментарное изображение в фигуру с простым и полным кон-
туром. Поэтому, когда предмет, образ, мелодия, слово или фраза пред-
ставлены лишь разрозненными элементами, мозг будет систематически
пытаться собрать их воедино и добавить недостающие части.
Таблица 25
Основные категории в области сенсорных процессов —
модальность и качество (по Ф. Блуму и соавт., с изменениями)
Модальность
Чувствительный
орган
Качество
Рецепторы
Зрение
Сетчатка глаза
Контрастность
Движение
Размеры
Цвет
Яркость
Палочки и колбоч-
ки — отростки
фоторецепторных
клеток
Слух
Улитка
Высота
Тембр
Волосковые
рецепторные клетки
Равновесие
Вестибулярный
орган
Сила тяжести
Вращение
Волосковые рецеп-
торные клетки
Осязание
Кожа
Давление
Вибрация
Тепло
Холод
Рецепторы кожи
Вкус
Язык (вкусовые
почки)
Вкус: сладкий, кислый,
горький, соленый
Вкусовые сосочки
языка (вкусовые
клетки)
Обоняние
Обонятельная
область слизистой
оболочки полости
носа
Запахи: цветочные
(душистые), фруктовые
(эфирные), мускусные
(амброзиевые), камфар-
ные или миндальные
(ароматные), чесночные,
хлорные или серные
(чесночные), горелые,
потовые, зловонные
(отталкивающие), гни-
лостные (тошнотворные)
Обонятельные
клетки
405
ОРГАН ЗРЕНИЯ
Объединение (группировка) элементов — это еще один аспект ор-
ганизации восприятия. Элементы могут объединяться по разным
признакам, например таким, как близость, сходство, непрерывность
(воображаемая) или симметрия. Принцип сходства состоит в том, что
нам легче объединять схожие элементы. Элементы будут также орга-
низовываться в единую форму, если они сохраняют одно направле-
ние. Это принцип непрерывности. Окружающий мир воспринимается
в трех измерениях благодаря наличию парных, симметрично располо-
женных органов чувств. Кроме того, человек воспринимает движение
и время, последнее имеет пределы от 1/18 до 2 сек.
В центральной нервной системе, куда поступают нервные им-
пульсы, вся информация обрабатывается в структурах мозга, ответ-
ственных за членораздельную речь. В результате и возникает вос-
приятие — способность видеть, слышать, осязать, ощущать вкусы,
запахи и положение тела в пространстве.
ОРГАН ЗРЕНИЯ
Орган зрения состоит из глазного яблока со зрительным нервом
и вспомогательных органов глаза. Глазное яблоко человека шаровид-
ной формы, относительно велико, его объем у взрослого человека
в среднем 7,5 см3. Глазное яблоко состоит из ядра, образованного тре-
мя оболочками: фиброзной, сосудистой и внутренней, или сетчаткой
(рис. 186). Наружная фиброзная оболочка подразделяется на задний
отдел — склеру (плотная соединительнотканная оболочка) и прозрач-
ную выпуклую роговицу, лишенную кровеносных сосудов.
Сосудистая оболочка глазного яблока (хориоидеа) расположена
под склерой, имеет толщину 0,1—0,22 мм. Она богата кровеносными
Рис. 186. Строение глазного яблока.
Горизонтальный разрез:
1 — фиброзная оболочка (склера),
2 — собственно сосудистая оболочка,
3 — сетчатка, 4 — радужка,
5 — зрачок, 6 — роговица,
7 — хрусталик, 8 — передняя камера
глазного яблока, 9 — задняя камера
глазного яблока, 10 — ресничный
поясок, 11 — ресничное тело,
12 — стекловидное тело,
13 — центральная ямка,
14 — зрительный нерв,
15 — диск зрительного нерва.
Сплошной линией показана наружная
ось глаза, пунктирной — зрительная
1
11
2
12
13
14
15
3
4
5
6
7
8
9
10
1
11
2
12
13
14
1
15
3
4
5
6
7
8
9
10
406
ОРГАНЫ ЧУВСТВ
сосудами и состоит из трех частей: собственно сосудистой оболочки,
ресничного тела и радужки. Основу собственно сосудистой оболочки
составляет густая сеть переплетающихся между собой артерий и вен,
между которыми располагается рыхлая волокнистая соединительная
ткань, богатая крупными пигментными клетками.
Кпереди сосудистая оболочка переходит в утолщенное ресничное
тело кольцевидной формы, которое предназначено для аккомодации
глаза и поддерживает, фиксирует и растягивает хрусталик. На разре-
зах, проведенных через меридианы глазного яблока, ресничное тело
выглядит как треугольник, обращенный своим основанием в перед-
нюю камеру глаза, а кзади вершиной, переходящей в собственно сосу-
дистую оболочку. Ресничное тело делится на две части: внутрен-
нюю — ресничный венец и наружную — ресничный кружок. От
поверхности последнего по направлению к хрусталику отходят 70—75
ресничных отростков длиной около 2 мм каждый, к которым прикре-
пляются волокна ресничного пояска (цинновой связки), идущие к
хрусталику. Сзади ресничное тело и его отростки покрыты ресничной
частью сетчатки. Бо
льшая часть ресничного тела — это ресничная
мышца. При ее сокращении хрусталик расправляется, округляется,
вследствие этого выпуклость и преломляющая сила его увеличивают-
ся, происходит аккомодация на близлежащие предметы. Гладкие мы-
шечные клетки в старческом возрасте частично атрофируются, на их
месте появляются участки соединительной ткани, что приводит к на-
рушению аккомодации и возникновению дальнозоркости.
Ресничное тело кпереди продолжается в радужку, которая, распо-
лагаясь между роговицей и хрусталиком, представляет собой круглый
диск с отверстием в центре (зрачок). В толще сосудистого слоя ра-
дужки проходят две мышцы. Волокна одной, расположенные цирку-
лярно, образуют сфинктер (суживатель) зрачка; другой, расширяю-
щей зрачок (его дилататор), имеют радиальное направление и лежат
в задней части сосудистого слоя глазного яблока. Расширитель зрачка
иннервируется симпатическими, сфинктер — парасимпатическими
волокнами. Различное количество и качество пигмента меланина
в радужке обусловливает цвет глаз — карий, черный (при наличии
большого количества пигмента) или голубой, зеленоватый (если мало
пигмента), альбиносов.
Многие издревле старались выявить связь между жизнью, наклон-
ностями к различным психическим, эмоциональным состояниям и
зрачком. В наше время все большее внимание привлекается к иридо-
диагностике, когда радужку приравнивают к своеобразному табло,
информирующему о локализации в теле различных заболеваний.
Внутренняя светочувствительная оболочка глазного яблока —
сетчатка — на всем протяжении прилежит к сосудистой оболочке
407
ОРГАН ЗРЕНИЯ
(рис. 187). Она состоит из двух листков: внутреннего — светочувст-
вительного (нервная часть) и наружного — пигментного. В десяти-
слойной сетчатке выделяют радиально ориентированные трехнейрон-
ные цепи, представленные наружным фоторецепторным слоем,
средним — ассоциативным и внутренним — ганглионарным.
К сосудистой оболочке снаружи прилежит слой, состоящий из
пигментных эпителиальных клеток, которые соприкасаются со слоем
палочек и колбочек (рис. 188). И те и другие представляют собой пе-
риферические отростки фоторецепторных клеток (I нейрон). Каждая
палочка состоит из наружного и внутреннего сегментов. Первый —
светочувствительный — образован сдвоенными мембранными диска-
ми, которые представляют собой складки плазматической мембраны
(отделенные от нее), в которую встроен зрительный пурпур — родоп-
син. Во внутреннем сегменте залегают органеллы. Колбочки отлича-
ются от палочек большей величиной и характером дисков. В наруж-
ном сегменте колбочек впячивания плазматической мембраны
образуют полудиски, которые сохраняют связь с мембраной. Зритель-
ный пигмент поглощает часть падающего на него света и отражает
остальную часть. Каждая палочка или колбочка содержит пигмент,
который поглощает лучи с определенной длиной световой волны.
Поглощая фотон света, зрительный пигмент меняет свою конфигура-
цию, при этом освобождается энергия, которая используется для
осуществления цепи химических реакций, что и приводит к возник-
новению нервного импульса.
4
3
2
5
6
1
Рис. 187. Расположение нейронов
в сетчатке глаза (схема):
1 — колбочки, 2 — палочки,
3 — пигментные клетки,
4 — биполярные клетки,
5 — ганглиозные клетки,
6 — нервные волокна.
Cтрелкой показано направление
пучка света
408
ОРГАНЫ ЧУВСТВ
В сетчатке глаза человека
содержится один тип палочек
и три типа колбочек, каждый
из которых воспринимает свет
определенной длины волны: от
400 до 700 нм. Количество кол-
бочек в сетчатке глаза человека
достигает 6—7 млн, палочек —
в 10—20 раз больше. Существуют три типа колбочек, каждый из кото-
рых воспринимает красный, синий или желтый свет. Палочки пред-
назначены воспринимать информацию об освещенности и форме
предметов. Палочки воспринимают слабый свет, т. е. необходимы
в темноте, колбочки — при ярком свете. Цветовое зрение связано
с функционированием колбочек разного типа (табл. 26).
Таблица 26
Восприятие цвета колбочками
Колбочки
Воспринимаемый цвет
Все виды
Синие
Синие, зеленые
Зеленые, красные
Белый
Фиолетовый
Синий, сине-зеленый
Желтый
От каждой фоторецепторной клетки отходит тонкий отросток,
образующий синапс с отростками биполярных нейронов (II нейрон),
Рис. 188. Палочковидная (I)
и колбочковидная (II) зрительные
клетки — фоторецепторные клетки.
Ультрамикроскопическое строение:
1 — наружный сегмент палочки,
2 — связующий отдел между наружным
и внутренним сегментами палочки,
3 — внутренний сегмент палочки,
4 — диски, 5 — клеточная оболочка,
6 — двойные микрофибриллы,
7 — митохондрии, 8 — пузырьки
эндоплазматической сети, 9 — ядро,
10 — область синапса с биполярным
нейроцитом, 11 — пальцевидные
отростки внутреннего сегмента
колбочковидной зрительной клетки,
12 — лучевой глиоцит
(мюллерово волокно)
(по И. В . Алмазову и Л. С. Сутулову)
I
II
7
8
9
10
4
3
2
5
6
1
11
12
9
409
ОРГАН ЗРЕНИЯ
которые, в свою очередь, передают возбуждение крупным ганглиоз-
ным клеткам (III нейрон). Их аксоны (500 тыс. — 1 млн) и образуют
зрительный нерв, который направляется в полость черепа через канал
зрительного нерва.
Хрусталик — абсолютно прозрачная двояковыпуклая линза диа-
метром около 9 мм. Коэффициент преломления хрусталика в поверх-
ностных слоях равен 1,32, в центральных — 1,42. Хрусталик как бы
подвешен на цинновой связке, волокна которой также прозрачны, они
сливаются с веществом хрусталика и передают ему движения реснич-
ной мышцы. При натяжении связки (расслабление ресничной мыш-
цы) хрусталик уплощается (установка на дальнее видение), при рас-
слаблении связки (сокращение ресничной мышцы) выпуклость
хрусталика увеличивается (установка на ближнее видение). Это и на-
зывается аккомодацией глаза. При нормальном зрении человек спосо-
бен четко видеть предметы на расстоянии 60 м. Минимальный предел
четкого видения меняется с возрастом: в 10 лет — 7 см, в 20 лет —
15см,в40лет—25см,в50лет—40см,чтосвязаносвозрастными
изменениями хрусталика и развитием дальнозоркости. Глаз человека
значительно менее совершенен, чем у многих животных. Так, напри-
мер, беркут замечает зайца с высоты более 3 км, а сокол — голубя на
расстоянии более 8 км.
Стекловидное тело, заполняющее пространство между сетчаткой
и хрусталиком, представляет собой прозрачное аморфное межкле-
точное вещество желеобразной консистенции, его индекс светопре-
ломления (1,334) близок к индексу светопреломления хрусталика
(1,32—1,42).
Принцип устройства фотоаппарата аналогичен строению глаза.
Роль диафрагмы в глазу выполняет зрачок, который в зависимости от
освещенности суживается (при ярком свете) или расширяется (при
тусклом свете). Объективом служат хрусталик и стекловидное тело.
Световые лучи в глазу попадают на сетчатку, при этом изображение
перевернутое. Пучок света благодаря светопреломляющим средам
(и в первую очередь аккомодации хрусталика) попадает на желтое
пятно сетчатки, являющееся зоной наилучшего видения. Световые
волны достигают колбочек и палочек лишь после того, как пройдут
почти всю толщу сетчатки.
На нижней поверхности мозга зрительные нервы перекрещивают-
ся, причем перекрещиваются лишь волокна, идущие от медиальной
(носовой) половины сетчатки. В каждом зрительном тракте проходят
волокна, несущие импульсы от клеток медиальной половины сетчат-
ки противоположного глаза и латеральной (височной) половины сво-
ей стороны. Часть волокон зрительного тракта после «переключения»
в одном из подкорковых центров несут импульсы к клеткам коры за-
410
ОРГАНЫ ЧУВСТВ
тылочной доли больших полушарий возле шпорной борозды, где и
заканчиваются (корковый конец зрительного анализатора). Другая
часть волокон образует синапсы с нейронами верхнего холмика чет-
верохолмия, откуда нервные импульсы следуют в ядра глазодвига-
тельного нерва, иннервирующие мышцы глаза, мышцу, суживающую
зрачок, и ресничную. Таким образом, в ответ на попадание световых
волн в глаз зрачок суживается, а глазные яблоки поворачиваются
в направлении пучка света.
Вспомогательные органы глаза. Глазное яблоко может вращаться
так, чтобы на рассматриваемом предмете сходились обе зрительные
оси. Этому, кроме того, способствуют аккомодация, а также движения
тела и головы. Четыре прямые (верхняя, нижняя, медиальная, лате-
ральная) и две косые (верхняя и нижняя) поперечнополосатые мыш-
цы составляют двигательный аппарат глаза. Прямые мышцы повора-
чивают глазное яблоко в соответствующем направлении, косые —
вокруг сагиттальной оси: верхняя — вниз и кнаружи, нижняя — вверх
и кнаружи. Благодаря содружественному действию указанных мышц
движения обоих глазных яблок — синхронные.
Веки защищают глазное яблоко спереди. Они представляют собой
кожные складки, ограничивающие глазную щель и закрывающие ее
при смыкании. В толще хрящей века заложены открывающиеся по их
краям разветвленные сальные (мейбомиевы) железы. Задняя поверх-
ность век покрыта конъюнктивой, которая продолжается в конъюн-
ктиву глаза. Конъюнктива — тонкая слизистая оболочка, которая ог-
раничивает конъюнктивальный мешок. По краям век располагаются
в 2—3 ряда ресницы, в их волосяные сумки открываются выводные
протоки сальных и потовых ресничных желез. На каждом веке около
80 ресниц, которые защищают глаза от попадания инородных частиц.
Ресницы обновляются примерно в течение 100 дней. Человек регу-
лярно моргает, примерно один раз за пять секунд.
Слезный аппарат включает одноименную железу и систему слез-
ных путей (рис. 189). От 5 до 12 выводных канальцев открываются
в конъюнктивальный мешок. У медиального угла глаза, на краях век,
там, где они расходятся, окружая слезное озеро, расположен слезный
сосочек, на котором имеются узкие отверстия — слезные точки, даю-
щие начало слезным канальцам, впадающим в слезный мешок, ниж-
ний конец которого переходит в носослезный проток, открывающий-
ся в полость носа. Вот почему при обильном слезовыделении человек
начинает сморкаться.
Слезы увлажняют конъюнктиву глаза и обезвреживают микроор-
ганизмы. Без слез конъюнктива и роговица высохли бы, и человек
ослеп. Ежедневно слезные железы вырабатывают около 100 мл слезы.
Со слезами выделяются из организма химические вещества, образую-
411
ПРЕДДВЕРНО-УЛИТКОВЫЙ ОРГАН
щиеся при нервном напряжении или эмоциональном стрессе. Гормон
пролактин способствует выработке и выделению слезной жидкости,
поэтому женщины плачут значительно чаще, чем мужчины. Слеза
имеет слабощелочную реакцию, в основном состоит из воды, в кото-
рой содержится около 1,5% NaCl, 0,5% альбумина и слизь. Кроме того,
в слезе имеется лизоцим, обладающий бактерицидным действием.
Глаз издревле наделялся священными функциями. Поэтому умер-
шему глаза сразу стараются закрыть, дабы он не сглазил. В любом
языке имеется много эмоциональных эпитетов, связанных с органом
зрения. Весьма сложна символика, касающаяся левого и правого гла-
за, «дурного глаза», количества глаз (циклопы, «третий глаз» и т. п .).
ПРЕДДВЕРНО-УЛИТКОВЫЙ ОРГАН
(ОРГАН СЛУХА И РАВНОВЕСИЯ)
Органы слуха и равновесия (статического чувства) у человека
объединены между собой в сложную систему, разделенную на три от-
дела: наружное ухо, среднее ухо и внутреннее ухо (рис. 190).
Наружное ухо. Ушная раковина — эластический хрящ сложной
формы, на дне которого находится наружное слуховое отверстие.
Считают, что расстояние между ушами помогает определению на-
правления источника звука. Одновременно человек получает инфор-
мацию о времени, фазе и силе звука. Рисунок ушной раковины чело-
века индивидуален, не изменяется с возрастом. Мифологичны
представления о форме мочки уха, его оттопыренности, символике
4
5
6
3
2
1
Рис. 189. Слезный аппарат глаза,
правого:
1 — слезная железа; 2 — верхнее
веко; 3 — слезный каналец,
4 — слезное озеро, 5 — слезный
мешок, 6 — носослезный проток
412
ОРГАНЫ ЧУВСТВ
ношения сережек. Мышцы ушной раковины почти не развиты, поэто-
му мы не в состоянии «навострить уши», «держать ушки на макуш-
ке», «держать ухо востро», «хлопать ушами» и т. п . Ушная раковина
отнюдь не массивный рупор. Здесь на маленькой площади встречают-
ся пять нервов различной природы (тройничный, веточки шейного
сплетения, промежуточного, языкоглоточного, блуждающего нервов).
Отсюда не только обусловленность рефлекторных связей с внутрен-
ними органами, но и представительство последних на ушной ракови-
не. Иными словами, на ушной раковине представлена вся чувстви-
тельность тела и внутренних органов. Поэтому можно использовать
ушную раковину для получения биологически важной информации
о состоянии любого органа.
Наружный слуховой проход длиной 33—35 мм закрыт барабанной
перепонкой, которая отделяет наружное ухо от среднего. Она пред-
ставляет собой пластинку толщиной 0,1 мм, имеющую форму эллип-
са, размерами 9×11 мм. В эпителии, выстилающем наружный слухо-
вой проход, наряду с большим количеством сальных имеются особые
трубчатые серные железы (видоизмененные потовые), вырабатываю-
щие вязкий, желтоватый секрет — «ушную серу». Вот почему даже
Рис. 190. Орган слуха и
равновесия и его части:
А — орган слуха и равновесия
на продольном разрезе,
Б — слуховые косточки,
В — улитка (улитковые
протоки «лестницы» вскрыты),
Г — улитковый проток
на поперечном разрезе;
1 — ушная раковина,
2 — наружный слуховой
проход, 3 — барабанная
перепонка, 4 — молоточек,
5 — наковальня, 6 — стремя,
7 — слуховая труба, 8 — улитка,
9 — лестница преддверия,
10 — барабанная лестница,
11 — улитковый проток,
12 — спиральный (кортиев)
орган, 13 — покровная
мембрана
11
11
11
22
12
12
33
13
13
44
А
55
Б
66
В
77
Г
88
99 10
10
413
ПРЕДДВЕРНО-УЛИТКОВЫЙ ОРГАН
при самой сильной жаре у человека из наружного слухового прохода
не выделяется пот.
Среднее ухо представляет собой воздухоносную барабанную по-
лость объемом около 1 см, расположенную в толще пирамиды височ-
ной кости. В барабанной полости находятся три слуховые косточки
и сухожилия мышц. Барабанная полость продолжается в слуховую
(евстахиеву) трубу, которая открывается в носовой части глотки.
Труба выполняет очень важную функцию — способствует выравнива-
нию давления воздуха внутри барабанной полости по отношению
к наружному. Слуховые косточки (стремя, наковальня, молоточек)
составляют цепь, передающую звуковые колебания и соединяющую
барабанную перепонку с закрытым вторичной барабанной перепон-
кой окном преддверия, ведущим в полость внутреннего уха. Рукоятка
молоточка сращена с барабанной перепонкой, а его головка сочленена
с телом наковальни. Длинный отросток наковальни сочленяется с го-
ловкой стремечка, основание которого входит в окно преддверия. Ко-
сточки покрыты слизистой оболочкой. Две мышцы (напрягающая ба-
рабанную перепонку и стременная) регулируют движение косточек.
Внутреннее ухо, расположенное в пирамиде височной кости, состо-
ит из перепончатого лабиринта, который залегает в костном лабирин-
те. Между обоими лабиринтами имеется пространство, заполненное
перилимфой. Три костных полукружных канала лежат в трех взаимно
перпендикулярных плоскостях: сагиттальной, горизонтальной, фрон-
тальной. Каждый канал имеет по две ножки, одна из которых (ампу-
лярная костная ножка) перед впадением в преддверие расширяется,
образуя ампулу. Соседние ножки переднего и заднего каналов соединя-
ются, образуя общую костную ножку, поэтому три канала открываются
в преддверие пятью отверстиями. Костная улитка образует 2,5 витка
вокруг горизонтально лежащего стержня-веретена.
Перепончатый лабиринт, повторяющий форму костного,
заполнен эндолимфой. Лабиринт состоит из двух частей: вестибуляр-
ного и улиткового. Вестибулярный лабиринт — периферический
отдел статокинетического анализатора (орган равновесия) — состоит
из двух мешочков: эллиптического (маточка) и сферического, которые
сообщаются между собой, а также трех полукружных протоков, зале-
гающих в одноименных костных каналах (рис. 191). Одна из ножек
каждого протока, расширяясь, образует перепончатые ампулы. Участ-
ки стенки мешочков, выстланные чувствительными рецепторными
клетками, называются пятнами, аналогичные участки ампул — гре-
бешками.
Эпителии пятен содержат воспринимающие (рецепторные) воло-
сковые клетки, на верхних поверхностях которых имеется по 60–
80 волосков (микроворсинок), обращенных в полость лабиринта.
414
ОРГАНЫ ЧУВСТВ
Кроме волосков, каждая клетка снабжена одной ресничкой. Поверх-
ность клеток покрыта студенистой мембраной, содержащей кристал-
лы углекислого кальция (статолиты). Мембрана поддерживается ста-
тическими волосками волосковых клеток.
Нервные окончания разветвляются, окружая наподобие чаш ре-
цепторные клетки, формируют синапсы с их телами. Рецепторные
клетки пятен воспринимают изменения силы тяжести, прямолиней-
Рис. 191. Орган равновесия и детали его строения:
А — полукружные протоки и эндолимфатические мешочки преддверного лабиринта
(костная стенка лабиринта частично удалена), Б — ориентация полукружных протоков
преддверного лабиринта внутреннего уха, В — расположение ампулярного гребешка и
пятна в стенке перепончатого лабиринта, Г — строение пятна (на продольном разрезе),
Д — строение ампулярного гребешка (на продольном разрезе).
Стрелками показаны места расположения структур в лабиринте: 1 — полукружные
каналы, 2 — преддверие, 3 — сферический мешочек, 4 — пятно сферического
мешочка, 5 — эндолимфатический проток, 6 — эллиптический мешочек, 7 — ампула,
8 — отолитовый аппарат, 9 — стаконии, 10 — мембрана стаконий,
11 — поддерживающие клетки, 12, 13 — волосковые сенсорные клетки,
14 — ампулярный гребешок, 15 — купол, 16 — латеральный полукружный канал,
17 — передний полукружный канал, 18 — задний полукружный канал
11
11
11
22
12
12
33
13
13
44
14
14
55
15
15
66
А
77
Б
88
В
99
Г
10
10
Д
16
16
17
17
18
18
415
ПРЕДДВЕРНО-УЛИТКОВЫЙ ОРГАН
ные движения и линейные ускорения. Ампулярные гребешки выстла-
ны аналогичными волосковыми клетками и покрыты желатинообраз-
ным куполом, в который проникают реснички. Они воспринимают
изменение углового ускорения.
При изменении силы тяжести, положения головы, тела, при уско-
рении движения мембрана скользит, а купол смещается. Это приво-
дит к напряжению волосков, что вызывает изменение активности раз-
личных ферментов волосковых клеток и возбуждение мембраны,
которое в конечном итоге передается ядрам мозжечка, спинному моз-
гу и коре теменной и височной долей больших полушарий, где нахо-
дится корковый центр анализатора равновесия.
Улитковый лабиринт — периферический конец слухового ана-
лизатора — залегает в костной улитке. Костный спиральный канал
разделяет проток на три части, занимая среднюю из них: верхняя —
так называемая лестница преддверия, нижняя — барабанная лестни-
ца. В них находится перилимфа. Улитковый проток заполнен эндо-
лимфой и представляет собой соединительнотканный мешок длиной
около 3,5 см. Улитковый проток на поперечном разрезе имеет треу-
гольную форму. На барабанной стенке и по всей длине улиткового ка-
нала располагается воспринимающий звуки спиральный орган (кор-
тиев). По всей его длине тянется в виде спирали покровная
мембрана — лентовидная пластинка желеобразной консистенции, ка-
саясь вершин его рецепторных волосковых клеток, лежащих на бази-
лярной соединительнотканной мембране. Мембрана образована при-
мерно 24 тысячами тонких радиальных коллагеновых волокон, длина
которых возрастает от основания улитки к ее вершине.
Рецепторные клетки несут на своей поверхности слуховые воло-
ски (микроворсинки), верхушки которых прикрепляются к описан-
ной покровной пластинке. К телам волосковых клеток подходят нерв-
ные окончания, образующие с ними синапсы. Тела афферентных
нейронов (первые нейроны) залегают в спиральном ганглии, располо-
женном в толще спиральной костной пластинки. Высокие звуки раз-
дражают только волосковые клетки, расположенные на нижних за-
витках улитки, а низкие звуки — волосковые клетки вершины улитки
и часть клеток на нижних завитках.
Звуковые волны через наружный слуховой проход достигают ба-
рабанной перепонки. Ее колебания передаются через цепь слуховых
косточек на окно преддверия, что вызывает передвижение перилим-
фы и воспринимается в улитковом протоке эндолимфой. Благодаря
этому происходит волнообразное движение основной мембраны, ко-
торая в зависимости от частоты и интенсивности звука колеблется по
всей своей длине. Эти колебания вызывают в волосковых клетках
определенные химические процессы, в результате которых генериру-
416
ОРГАНЫ ЧУВСТВ
ются нервные импульсы. В конечном итоге импульсы проводятся к
коре височной доли больших полушарий мозга, где расположен цент-
ральный (корковый) конец слухового анализатора.
Человек способен воспринимать звуковые колебания от 16 Гц
(16 колебаний в секунду) до 21 000 Гц. С возрастом эта величина сни-
жается в два-три раза — до 5000 Гц у старых людей. Некоторые жи-
вотные способны воспринимать колебания до 200—300 тыс. Гц, на-
пример летучие мыши — до 210 000 Гц, дельфины — до 280 000 Гц.
Сила звука измеряется в децибелах. Так, если принять абсолютную
тишину за 0, то шелест падающих листьев вызывает 10 децибел, ше-
пот — 20, обычная беседа — 60, движущийся автомобиль — от 60 до
90, интенсивное дорожное движение — 100—110, рок-музыка в испол-
нении оркестра — 110—120, а работающий двигатель реактивного са-
молета — 140. Шум вредно действует на орган слуха и на психику че-
ловека, вызывая психоэмоциональный стресс.
ОРГАН ОБОНЯНИЯ
Обонятельная область слизистой оболочки носа, которая у взро-
слого человека занимает 250—300 мм2 и расположена в слизистой
оболочке верхней носовой раковины и лежащей на этом уровне зоны
носовой перегородки, покрыта обонятельным эпителием. Обонятель-
ные клетки (их около 40 млн) имеют центральные и периферические
отростки. Короткий периферический отросток (дендрит) заканчива-
ется утолщением (обонятельная булава), на вершине которой распо-
лагается по 10—12 подвижных обонятельных ресничек. Центральные
отростки — аксоны — собираются в обонятельные нити (20—40), про-
ходящие через решетчатую пластинку одноименной кости и направ-
ляющиеся в мозг. Молекулы пахучих веществ взаимодействуют с ре-
цепторными белками ресничек, предварительно растворяясь в секрете
обонятельных желез, что генерирует нервный импульс. Он распро-
страняется по обонятельным нервам и в конечном итоге достигает
коркового центра обонятельного анализатора, который располагается
в крючке и парагиппокампальной извилине больших полушарий го-
ловного мозга. Несмотря на то, что существует всего девять четко раз-
личимых групп запахов, человек способен различить около трех ты-
сяч. Человек ощущает запах при концентрации около 500 млн молекул
в 1 м3 воздуха. У многих животных обоняние развито значительно
лучше, чем у человека. Так, например, собака различает до 100 000 за-
пахов при концентрации вещества около 200 000 молекул в 1 м3 воз-
духа, т. е. в 2500 раз меньше, чем человек. А самец бабочки «ночной
павлиний глаз» способен уловить запах неоплодотворенной самки на
расстоянии более 10 км!
417
ОРГАН ВКУСА
ОРГАН ВКУСА
Орган вкуса у человека образован примерно 2000 вкусовых почек,
расположенных в толще многослойного эпителия боковых поверхно-
стей желобоватых, листовидных и грибовидных сосочков языка, а так-
же в слизистой оболочке нёба, зева и надгортанника (рис. 192). На вер-
шине вкусовой почки имеется отверстие, которое ведет в маленькую
ямку, образованную верхушками вкусовых клеток. На верхней поверх-
ности каждой вкусовой клетки имеется около 40—50 микроворсинок.
Нервные волокна заходят во вкусовую почку и образуют множество
синапсов со вкусовыми клетками. Растворенное вещество, обладающее
вкусом, проникает во вкусовые почки через отверстие на его вершине,
реагирует с рецепторными белками цитолеммы микроворсинок, в ре-
зультате чего возникает нервный импульс. Для возникновения сладко-
го вкуса достаточно содержания в продукте 0,5% сахара, соленого —
0,25% соли, горького — 0,002% и кислого — 0,001% кислоты.
Импульс передается по ветвям VII, IX, X пар черепных нервов, че-
рез ряд подкорковых центров к корковому концу вкусового анализа-
тора, расположенному в коре парагиппокампальной извилины, крюч-
ке и аммоновом роге больших полушарий головного мозга.
7
4
3
2
5
6
1
5
6
почки (схема):
живающая клетка;
инки; 5 – эпители-
чание; 7 – нервное
Рис. 192. Строение вкусовой
почки (схема):
1 — вкусовая клетка,
2 — поддерживающая клетка,
3 — вкусовая пора,
4 — микроворсинки,
5 — эпителиальные клетки,
6 — нервное окончание,
7 — нервное волокно
418
ОРГАНЫ ЧУВСТВ
КОЖА
Кожа выполняет многообразные функции: защитную, терморегу-
ляционную, дыхательную, обменную, является депо крови и органом
осязания (анализатор общей чувствительности). Железы кожи выра-
батывают пот, кожное сало. С потом у человека в течение суток
в обычных условиях выделяется около 500 мл воды, солей, конечных
продуктов азотистого обмена. Кожа активно участвует в обмене вита-
минов, особенно важен синтез в коже витамина D под влиянием уль-
трафиолетовых лучей. Площадь кожного покрова взрослого человека
достигает 1,5—2 м2. Эта поверхность является рецепторным полем
осязания, болевой, температурной чувствительности и важнейшей
эрогенной зоной.
Кожа состоит из эпидермиса и дермы (рис. 193). Эпидермис — это
многослойный плоский ороговевающий эпителий, толщина которого
зависит от выполняемой функции. Так, на участках, подвергающихся
постоянному механическому давлению, его толщина достигает 0,5—
2,3 мм (например, ладони, подошвы); на груди, животе, бедре, плече,
предплечье, шее толщина не превышает 0,02—0,05 см. Пигментация
кожи зависит от количества меланоцитов — пигментных клеток. Пиг-
ментация у представителей разных рас различна.
Дерма, или собственно кожа, толщиной 1—2,5 мм образована сое-
динительной тканью. В ней различают сосочковый и сетчатый слои.
Рис. 193. Строение кожи:
1 — эпидермис,
2 — роговой слой,
3 — базальный слой,
4 — сосочковый слой,
5 — сальная железа,
6 — соединительнотканные
волокна (коллагеновые,
эластические, ретикулярные)
и клетки, 7 — пучки
миоцитов, 8 — волосяная
луковица, 9 — дольки
жирной ткани,
10 — сетчатый слой,
11 — корень волоска,
12 — потовая железа
11
11
11
22
12
12
33
44
55
66
77
88
99
10
10
419
КОЖА
Благодаря наличию сосочков на поверхности кожи видны гребешки,
разделенные бороздками. Сложное переплетение гребешков и бороз-
док образует рисунок, индивидуальный для каждого человека и не
меняющийся в течение всей его жизни. В сосочковом слое имеются
гладкие мышечные клетки. В сетчатом слое залегают корни волос, по-
товые и сальные железы. Подкожная клетчатка содержит жировую
ткань. Этот слой играет важную роль в терморегуляции и является
жировым депо организма.
Волосы — производное эпидермиса (рис. 194). Почти вся кожа
покрыта волосами. Всего на теле человека от 200 тыс. до 1 млн во-
лос. Исключение составляют ладони, подошвы, переходная часть
4
5
6
7
8
9
10
12
11
13
3
2
1
Рис. 194. Строение волоса
(схема):
1 — волосяная сумка,
2 — кора волоса,
3 — мозговое вещество волоса,
4 — кутикула, 5 — наружное
корневое влагалище,
6 — два слоя внутреннего
корневого влагалища,
7 — волосяная воронка,
8 — базальный (ростковый) слой
эпидермиса, 9 — роговой слой
эпидермиса, 10 — сальная
железа, 11 — мышца,
поднимающая волос,
12 — луковица волоса,
13 — сосочек волоса
(по В.Г. Елисееву и др.)
420
ОРГАНЫ ЧУВСТВ
губ, головка полового члена и малые половые губы. Наибольшее
число волос обычно на голове. Характер оволосения зависит от
пола, возраста и относится к вторичным половым признакам. Во-
лос имеет выступающий над поверхностью кожи стержень и ко-
рень, лежащий в толще кожи. Длина стержня колеблется от 1—2 мм
до 1,5 м, а толщина от 0,005 до 0,6 мм. Корень волоса расположен
в волосяном фолликуле. Волосы меняются в сроки от 2—3 месяцев
до 2—3 лет. Длина волос изменчива, связана с генетическими фак-
торами, полом, возрастом, волосы растут со скоростью около 0,2 мм
в сутки.
Подобно волосам ногти также являются производными эпидер-
миса. Ноготь представляет собой роговую пластинку, лежащую на
ногтевом ложе, ограниченную у основания и с боков ногтевыми вали-
ками. Ногти чувствительностью не обладают и кажутся поэтому, как
и волосы, «мертвыми», т. е. как бы живущими независимо от тела. По
форме и естественному цвету пытаются зачастую определить свойст-
ва человека, его судьбу. Ногти растут со скоростью около 0,15 мм
в сутки, на пальцах кистей сменяются каждые три, а на пальцах стоп —
каждые 4,5 мес. На протяжении жизни человек отстригает примерно
4 метра ногтей!
К железам кожи относятся потовые, сальные и молочные. По-
следние описаны ранее. Количество потовых желез около 2—2,5 млн,
они представляют собой простые трубчатые железы. Секрет потовых
желез — пот, который на 98% состоит из воды и 2% органических
и неорганических веществ (минеральные соли, мочевина, мочевая
кислота). С потом теряется большое количество воды, микроэлемен-
тов. При испарении пота теплоотдача усиливается, что является од-
ним из важных механизмов терморегуляции. Потовая система — это
не только наша система охлаждения, но и своеобразный половой при-
знак, несущий запаховую информацию. В течение суток при обычном
режиме человек выделяет около 0,7 л пота, однако в жарком климате
потоотделение может достигнуть 4 л в сутки, при интенсивной физи-
ческой нагрузке — до 10 л.
Сальные железы отсутствуют лишь на ладонях и подошвах, наи-
большее количество их на голове, спине. Вырабатываемое ими
кожное сало, будучи бактерицидным, не только смазывает волосы
и эпидермис, но в известной мере предохраняет его от микробов.
В период полового созревания у мальчиков функция сальных же-
лез активизируется, что связано с влиянием мужских половых гор-
монов.
421
КОЖА
Кожная чувствительность
Кожа иннервируется чувствительными нервами, отходящими от
спинномозговых и черепных нервов, а также волокнами вегетативных
нервов, подходящих к сосудам, гладким мышечным волокнам и желе-
зам. Кожа весьма обильно снабжена нервными окончаниями. В зави-
симости от характера воспринимаемого раздражения различают
терморецепторы, механорецепторы и ноцирецепторы. Первые вос-
принимают изменения температуры, вторые — прикосновения к коже,
ее сдавливание, третьи — болевые раздражения. Эти виды кожной
чувствительности в совокупности с проприорецепцией и болевой
чувствительностью всего тела могут быть объединены в соматовисце-
ральную чувствительность, которая отличается от описанных выше
признаков (зрение, слух, обоняние, вкус). Рецепторы соматовисце-
ральной чувствительности разбросаны по всему телу и не образуют
обособленных органов чувств. Афферентные нервные волокна, по ко-
торым распространяются импульсы от указанных рецепторов, явля-
ются дендритами чувствительных клеток, залегающих в спинномоз-
говых узлах и чувствительных узлах черепных нервов.
Осязание (механорецепция) включает восприятие ощущений
давления, прикосновения, вибрации, щекотки, которые воспринимают-
ся лишь в определенных осязательных точках кожи. В среднем на 1 см2
кожи приходится около 170 чувствительных нервных окончаний. Наи-
большая плотность осязательных точек в коже губ и подушечках паль-
цев, наименьшая — на спине, плечах, бедрах. В коже человека преобла-
дают рецепторы прикосновения. По-видимому, каждый отдельный
рецептор воспринимает определенное осязательное ощущение, но при
воздействии на кожу различных механических стимулов одновременно
реагирует несколько типов рецепторов. Терморецепторы воспринима-
ют соответствующие раздражения, причем различные точки кожи вос-
принимают тепло или холод, последние преобладают. Наиболее чувст-
вительна к температурным раздражителям кожа лица, на 1 см2 которой
приходится от 16 до 19 холодовых точек, в то время как вся кожа лица
воспринимает тепло. На 1 см2 кожи кисти приходится 1—5 холодовых
точек и лишь 0,4—0,5 тепловых.
Количество болевых точек кожи значительно больше, чем так-
тильных (примерно в 9 раз) и температурных (примерно в 10 раз).
Болевые ощущения воспринимаются ноцирецепторами. Время реаги-
рования кожи различно для разных ощущений: 0,9 сек. для боли,
0,12 сек. для осязания, 0,16 сек. для температурных. Особенно развита
чувствительность кисти и пальцев: так, кожа пальца способна воспри-
нять вибрацию с амплитудой 0,02 мкм.
422
РАСТЕНИЯ
РАСТЕНИЯ
К царству растений относят все фотосинтезирующие эукариоти-
ческие организмы. Кроме того, некоторые одноклеточные организмы,
такие как эвгленовые, способны к фотосинтезу на свету (как расте-
ния), а в темноте поглощают готовые органические вещества (как жи-
вотные клетки). Такой «смешанный» тип питания называется миксо-
трофным.
НИЗШИЕ РАСТЕНИЯ
Растения подразделяют на низшие и высшие. Тела низших не расчле-
нены на вегетативные органы. Характерной особенностью низших расте-
ний является крайняя простота организации органов размножения, как
правило, они имеют одноклеточное строение. Низшие растения по-
явились раньше высших, и в настоящее время они чрезвычайно широко
распространены. Низшие растения очень важны для нормального тече-
ния энергетических процессов в живой природе. Они обладают огром-
ной биомассой и высокой репродуктивной способностью. Они являются
основой трофической цепи большинства водных сообществ.
До недавнего времени к низшим растениям относили бактерии,
сине-зеленые водоросли, миксомицеты (слизевики), грибы, лишайни-
ки и некоторых других. Однако грибы являются гетеротрофами и во-
обще не могут относиться к растениям. Миксомицеты, бактерии, даже
фотосинтезирующие, и сине-зеленые водоросли (цианобактерии) —
прокариоты: они не имеют оформленного ядра и не могут считаться
растениями. Лишайники — это симбиоз гриба и водоросли. Они вы-
делены в отдельные крупные таксономические (систематические)
группы. Низшие растения — различные водоросли.
ВОДОРОСЛИ
Водоросли обитают в воде. Однако не все растения, живущие в во-
доемах, являются водорослями. Значительная часть их — это высшие
растения, которые имеют все присущие им вегетативные органы, при-
чем водный образ жизни ведут представители всех групп высших ра-
стений, кроме голосеменных. Однако все они происходят от наземных
форм и являются вторичноводными. Они попали в воду, осваивая но-
вые места обитания. В отличие от них водоросли всегда существовали
в воде.
Клетки водорослей практически не отличаются друг от друга. По-
этому большинство водорослей не имеют выраженных тканей (за
423
ВОДОРОСЛИ
исключением высокоорганизованных бурых водорослей, но и у них
ткани малочисленны и слабо дифференцированы). Тело водорослей
не расчленено на вегетативные органы и представляет собой единое
слоевище, которое может иметь самую разнообразную рассеченность.
Некоторые формы посредством ризоидов прочно прикрепляются
к субстрату, что защищает их от течения воды или волн. Но эти «кор-
неподобные» структуры не являются корнями.
Водоросли морфологически разнообразны. Встречаются чрезвы-
чайно простые одноклеточные, а также сложнорасчлененные. Послед-
ние очень похожи на высшие растения, их структуры внешне напоми-
нают стебли, листья и даже плоды. Варьируют размеры — от
микроскопической хлореллы до гигантского макроцистиса, достигаю-
щего в длину 60 м. Выделяют несколько основных групп водорослей,
различающихся сложной морфологической организацией.
1. Амебоидная структура — одноклеточные водоросли, которые
способны образовывать псевдоподии.
2. Монадная структура — одноклеточные водоросли, имеющие
жесткую оболочку, благодаря которой они имеют постоянную форму
тела. Эти водоросли перемещаются с помощью жгутиков.
3. Коккоидная структура напоминает монадную. В отличие от
нее, они лишены псевдоподии или жгутиков, поэтому клетки не пере-
мещаются самостоятельно, а пассивно переносятся током воды.
4. Пальмеллоидная структура образуется при постоянном или
временном объединении нескольких коккоидных клеток в общую
слизистую массу.
5. Нитчатая структура — одиночная или разветвленная нить, ко-
торая ведет свободный или прикрепленный образ жизни. Клетки де-
лятся лишь в одной плоскости, поэтому не образуется слой.
6. Пластинчатая структура — однослойная или многослойная
пластинка.
7. Сифональная структура — весь таллом — это одна гигантская
многоядерная клетка.
Бентосные (донные) водоросли прикрепляются своими ризоида-
ми ко дну, подводным камням, скалам или затонувшим предметам.
В прозрачных водах морей и океанов водоросли живут на глубинах,
превышающих 150 м. Глубже проникает небольшое количество света.
Его недостаточно для синтеза органических веществ, поэтому фото-
трофных организмов там нет. Бентосные водоросли живут лишь на
узкой прибрежной полосе и на возвышениях дна. Небольшие водоро-
сли, свободно плавающие в толще воды, образуют продуцентную
основу большинства водных биоценозов — фитопланктон.
Свет, проходя через толщу воды, постепенно рассеивается, поэто-
му в этих слоях живут лишь водоросли, имеющие вспомогательные
424
НИЗШИЕ РАСТЕНИЯ
пигменты. В результате этого вместо зеленой окраски водоросли
меняют цвет, при этом характер пигментов зависит от глубины оби-
тания организма. Различают золотистые, желто-зеленые, зе-
леные, красные (багрянки), бурые, эвгленовые, харовые, диа-
томовые и пирофитовые водоросли.
ВНИМАНИЕ!
Сине-зеленые водоросли (цианобактерии) не являются настоящими во-
дорослями, т. к . они относятся к прокариотам.
Размножение водорослей
Водоросли увеличивают свою численность путем вегетативного,
бесполого и полового размножения.
Вегетативное размножение происходит путем деления однокле-
точных водорослей или отделения частей от многоклеточного слоеви-
ща. Нитчатые формы расщепляются на две новые нити или распада-
ются на одно- или многоклеточные части. Слоевища истинно
многоклеточных водорослей могут разделяться благодаря механиче-
скому воздействию потоков воды, водных обитателей, деятельности
человека и т. д .
Многие водоросли образуют специальные структуры для вегета-
тивного размножения. У некоторых бурых водорослей на талломах
образуются своеобразные «почки», которые отламываются и дают на-
чало новым слоевищам. Харовые водоросли образуют одно- или мно-
гоклеточные клубеньки, которые после периода зимнего покоя прора-
стают в новые растения. Определенные клетки некоторых нитчатых
водорослей при наступлении неблагоприятных условий утолщают
оболочку и накапливают в протопласте большой запас питательных
веществ. Эти клетки способны долго находиться в состоянии покоя
и пережидать опасную ситуацию.
Бесполое размножение осуществляется спорами, которые развива-
ются или в обычных вегетативных клетках, или в спорангиях. Развитие
спор сопровождается делением ядра, которое может происходить не-
сколько раз в зависимости от количества образующихся спор (у различ-
ных водорослей ядро может делиться путем митоза или мейоза). Они
в дальнейшем окружаются более или менее плотной оболочкой и выхо-
дят через образующееся отверстие в оболочке материнской клетки.
Зооспоры снабжены жгутиками и могут активно передвигаться
(рис. 195). Апланоспоры имеют плотную оболочку и лишены жгути-
ков, они неподвижны. Гипноспоры имеют очень толстую оболочку.
Они способны длительно покоиться. Большинство водорослей обра-
зуют зооспоры. В других случаях при определении спор указывается
425
ВОДОРОСЛИ
их численность в материнской клетке или спорангии — моноспоры,
биспоры, тетраспоры и т. д.
Половое размножение всегда сопровождается слиянием двух га-
плоидных клеток. В результате этого образуется диплоидная зигота, из
которой развивается водоросль или же формируются зооспоры. Холо-
гамия — это слияние двух обычных вегетативных клеток (некоторые
колониальные жгутиковые). Конъюгация — это слияние вегетативных
клеток, не имеющих жгутиков (некоторые зеленые водоросли).
У остальных водорослей сливаются клетки — гаметы, образующиеся
в гаметангиях. Подвижные гаметы мужские, неподвижные — женские.
Выделяют три типа полового размножения — изогамию, анизога-
мию и оогамию.
Изогамия — слияние двух совершенно одинаковых гамет.
Анизогамия или гетерогамия — слияние гамет, различающихся
между собой по размерам или подвижности.
Оогамия — наиболее высоко организованный тип полового раз-
множения. В нем участвуют гаметы, отличающиеся по своим разме-
рам и по подвижности. Крупная неподвижная гамета — яйцеклетка,
которая содержит запас питательных веществ, необходимый для по-
следующего развития зиготы; значительно более мелкая гамета —
сперматозоид или антерозоид, снабженный жгутиком, способный ак-
тивно передвигаться в поисках яйцеклетки.
Гаметы образуются в особых структурах, являющихся, по сущест-
ву, половыми органами. Женские — оогонии (не путать с одноименны-
ми первичными женскими половыми клетками животных), муж-
ские — антеридии. За исключением харовых водорослей, которые
À
Á
Â
Ã
Ä
Рис. 195. Зооспоры:
А, Б — ночесветка — общий вид (А), зооспора (Б); В — макрозооспора улотрикса;
Г — микрозооспора того же вида; Д — дербезия
426
НИЗШИЕ РАСТЕНИЯ
имеют многоклеточные половые органы, у остальных водорослей ор-
ганы полового размножения состоят из одной клетки.
У некоторых водорослей яйцеклетки и сперматозоиды могут обра-
зовываться на разных талломах. Такие формы называют двудомными,
в отличие от однодомных, у которых на одном слоевище развиваются
гаметы обоих типов. Это относится к водорослям, гаметы которых от-
личаются одна от другой. У изогамных форм сливающиеся гаметы
могут образовываться на одном растении (гомоталличные) или на
разных — гетероталличные. Для различения морфологически одина-
ковых талломов, продуцирующих разные типы гамет, используют
простые обозначения — плюс (+) и минус (–) талломы. Аналогично
называют и гаметы, которые они образуют.
Чередование поколений. Некоторые водоросли способны на од-
ном талломе образовывать органы бесполого и полового размноже-
ния. Такие водоросли называют гаметоспорофитами. Если же органы
полового и бесполого размножения образуются на разных растениях,
происходит чередование поколений. Растения, на которых развива-
ются половые органы, называют гаметофитами, растения, образую-
щие органы бесполого размножения, — спорофитами.
Чередование поколений определяет и смену гаплоидной и дипло-
идной стадий жизни водорослей. В отличие от высших растений,
у которых соответствующие стадии строго закономерно сменяют друг
друга, у низших редукционное деление ядра, уменьшающее вдвое чи-
сло хромосом, случается на разных этапах развития, в связи с чем вы-
деляют несколько типов редукции.
Гаметическая редукция — уменьшение числа хромосом проис-
ходит непосредственно при образовании гамет.
Спорическая редукция — уменьшение числа хромосом проис-
ходит непосредственно при образовании спор.
Зиготическая редукция — первое деление мейоза.
Экологические формы водорослей
Выделяют следующие основные экологические формы водоро-
слей: 1) пресноводные водоросли; 2) водоросли морей и океанов;
3) бентосные (донные) водоросли; 4) фитопланктон; 5) водоросли
ледников; 6) водоросли термальных источников; 7) наземные водоро-
сли; 8) почвенные водоросли.
Значение водорослей
Водоросли появились в археозойскую эру. Первые примитивные
водоросли бурно развивались. В результате образовались огромные
427
ВОДОРОСЛИ
донные отложения, дошедшие до нашего времени в виде массивов
горных пород. И в настоящее время водоросли наряду с кораллами
активно участвуют в образовании рифов (красные водоросли лито-
тамнии). Помимо влияния на геологические процессы, водоросли на-
сытили кислородом воду и атмосферный воздух, что позволило суще-
ствовать аэробным организмам.
Водоросли очень важны для существования водных сообществ как
продуценты первичной органической массы. Однако условия для су-
ществования бентосных водорослей отсутствуют в большинстве рай-
онов. В первую очередь это связано с большой глубиной и отсутстви-
ем в связи с этим света, необходимого для фотосинтеза, поэтому
бо
льшая часть водорослей глубоководных зон представлена план-
ктонными формами. Суммарная сухая масса водорослей значительно
уступает массе наземных растений, относительная масса водорослей
незначительно меньше, чем относительная масса водных животных,
Однако огромная скорость воспроизводства делает их основой всех
трофических цепей. Микроскопические водоросли размножаются
столь быстро, что их просто не успевают съесть питающиеся ими жи-
вотные.
ВНИМАНИЕ!
На долю водорослей приходится 30—50% ежегодной массы органиче-
ского вещества, синтезируемого сообща всеми фототрофными организ-
мами.
Вместе с тем именно высокая скорость размножения микроскопи-
ческих водорослей часто является причиной серьезных экологиче-
ских нарушений. Это может привести к цветению воды, в результате
чего в ней резко снижается содержание кислорода, который использу-
ют аэробные бактерии-сапрофиты, утилизирующие огромные скопле-
ния мертвых клеток водорослей. Это часто приводит к массовой гибе-
ли водных организмов.
Роль водорослей в жизни человека разнообразна. Во многих стра-
нах некоторые виды употребляют в пищу. В частности, красный мор-
ской салат изготовляют из красной водоросли порфиры. Морской ка-
пустой называют бурую водоросль ламинарию, в которой ценят не
только вкусовые качества, но и способность выводить радионуклиды
из организма человека. Кроме того, ламинария содержит большое ко-
личество витаминов и минеральных элементов. Эти и многие другие
водоросли едят не только в сыром виде, но и активно используют для
приготовления различных блюд. Из слоевищ водорослей (особенно
красных) добывают агар, который широко используют в пищевой
428
ВЫСШИЕ РАСТЕНИЯ
промышленности для приготовления различного рода желе, мармела-
дов, пастилы и других продуктов. Кроме того, агар применяют при
производстве бумаги, на биотехнологических предприятиях — в каче-
стве отвердителя питательных сред, на которых культивируются
микроорганизмы, и в микробиологических лабораториях.
Из водорослей получают альгиновую кислоту. Ее широко исполь-
зуют в пищевой промышленности благодаря способности образовы-
вать гели, а также во многих технологических процессах из-за очень
высокой склеивающей способности. Водоросли очень богаты калием,
поэтому в прибрежных районах их используют в качестве удобрения.
Высокое содержание йода долгое время делало водоросли основным
промышленным источником получения этого ценного элемента.
Благодаря малым размерам и огромной скорости размножения
микроскопические водоросли активно используются в генной и кле-
точной инженерии. Значительный интерес для биотехнологии пред-
ставляет одноклеточная водоросль хлорелла. Она неприхотлива, быс-
тро размножается и способна в больших количествах накапливать
ценные продукты.
ВЫСШИЕ РАСТЕНИЯ
Высшие растения имеют четко дифференцированные ткани, кото-
рые выполняют специализированные функции. Тело высших расте-
ний расчленено на вегетативные органы, главные из которых — побег
и корень.
ВНИМАНИЕ!
Наличие большого числа тканей и расчленение тела на вегетативные
органы является важной отличительной особенностью высших расте-
ний.
Другая особенность высших растений — наличие многоклеточных
органов полового и бесполого размножения. Органы полового раз-
множения — гаметангии: мужские — антеридии, женские — архего-
нии защищены оболочкой, образованной бесплодными клетками, что
отличает половые органы высших растений от многоклеточных гаме-
тангиев — водорослей, в которых все клетки способны трансформиро-
ваться в гаметы. В антеридиях образуется большое количество мел-
ких мужских гамет (сперматозоидов), неподвижные мужские гаметы
большинства семенных называют спермиями. В архегониях развивает-
ся лишь одна крупная неподвижная яйцеклетка. В отличие от муж-
ских гамет, яйцеклетки снабжены значительным запасом питатель-
429
ВЫСШИЕ РАСТЕНИЯ
ных веществ. Степень развития гаметангиев высших растений
напрямую связана с уровнем организации гаметофита: чем он выше,
тем лучше развиты половые органы. У мхов они многочисленны, у
папоротникообразных их меньше, у голосеменных они подвергаются
значительной редукции, а у некоторых из них и у всех цветковых во-
все не образуются. Зигота, образовавшаяся в результате слияния
мужской и женской гамет, у высших растений дает начало зародышу.
Напомним, что у низших растений образовавшиеся в результате деле-
ния зиготы клетки сразу же используются для построения слоевища
и в последующем мало изменяются.
ВНИМАНИЕ!
Зародыши генетически детерминированы на специализацию в опреде-
ленном направлении.
У всех высших растений имеются две жизненные фазы, законо-
мерно сменяющие друг друга, — гаметофит и спорофит, которые со-
ставляют жизненный цикл высшего растения. Гаметофит, развиваю-
щийся из споры, представляет собой поколение, способное
размножаться половым путем, т. е. посредством слияния гаплоидных
клеток-гамет с образованием диплоидной зиготы. Гаметы высших ра-
стений всегда образуются в результате митоза, что принципиально
отличает их от половых клеток животных, поэтому тело гаметофита
построено из гаплоидных клеток. Фазу гаметофита еще называют га-
метофазой, или гаплофазой. На одном растении одновременно могут
развиваться мужские и женские половые органы. Такой гаметофит
называют однодомным. У двудомных растений гаметофиты формиру-
ют либо только мужские органы, либо только женские.
Спорофит, развивающийся из зиготы, размножается посредством
спор, тело образовано диплоидными клетками (соответственно, дру-
гое название этого поколения — диплофаза). Спорофит не образует
половые органы. Споры развиваются в особых многоклеточных —
спорангиях.
ВНИМАНИЕ!
Спорофит образован диплоидными клетками, но его гаплоидные споры
всегда образуются в процессе мейоза из материнских клеток-предшест-
венниц.
Количество образовавшихся при этом дефинитивных спор зави-
сит от того, какое это растение — равноспоровое или разноспоровое.
У равноспоровых в результате мейоза возникают четыре споры (тетра-
430
ВЫСШИЕ РАСТЕНИЯ
да), причем все они имеют одинаковое строение и размеры, и на раз-
вивающихся из них гаметофитах формируются как мужские, так и
женские половые органы. У разноспоровых образуются споры двух
типов — мегаспоры и микроспоры. Более крупные мегаспоры развива-
ются в мегаспорангиях, которые формируются на мегаспорофиллах.
При мейозе образуется лишь одна мегаспора, три меньшие клетки по-
гибают. Мегаспоры традиционно называют женскими спорами, хотя
употребление этого термина в прямом смысле некорректно, посколь-
ку спорофит представляет собой бесполое поколение, поэтому от-
дельные особи изначально нельзя делить на мужские и женские,
а развивающийся из них гаметофит формирует только женские поло-
вые органы — архегонии.
В процессе мейоза образуются четыре микроспоры, которые раз-
виваются в микроспорангиях на микроспорофиллах. Их традиционно
считают мужскими спорами, поскольку на развивающихся из них га-
метофитах формируются только мужские половые органы (антери-
дии). Гаметофит равноспоровых, как правило, хорошо развит, и его
фотосинтетическая активность вполне обеспечивает половые органы
органическими веществами. Напротив, у разноспоровых гаметофит
в значительной мере редуцирован. Часто он не выходит за пределы
споры, а потому не является фотосинтетическим. Необходимые для
развития половых органов органические вещества, которые образу-
ются в значительно меньших количествах, чем у разноспоровых, из-
начально запасаются в споре за счет спорофита.
В зависимости от преобладания гаплоидной или диплоидной фазы
высшие растения делят на две группы. Первая — растения, у которых
преобладает гаметофит, а спорофит развит слабо (мохообразные).
Вторая группа значительно больше: она включает растения, у кото-
рых превалирует спорофит, гаметофит при этом в большей или мень-
шей степени редуцируется (папоротникообразные, голосеменные
и покрытосеменные).
ТКАНИ РАСТЕНИЙ
Ткани формируются только у многоклеточных организмов. У ра-
стений ткани развиваются из меристем и состоят из клеток и синтези-
руемого ими межклеточного вещества. Ткани имеют сходное происхо-
ждение, строение и совместно выполняют общие функции. Выделяют
много различных типов растительных тканей, однако их количество
и степень дифференцированности напрямую зависит от систематиче-
ского положения растения. Так, у простых многоклеточных водоро-
слей клетки мало отличаются одна от другой. У более сложно органи-
зованных бурых водорослей уже имеются проводящие, механические,
431
ТКАНИ РАСТЕНИЙ
ассимиляционные и запасающие ткани. У высших растений насчиты-
вается несколько десятков (у покрытосеменных до 80) четко выра-
женных тканей.
В отличие от тканей животных, растительные ткани не образуют
внутренние органы. Ткани растений состоят из одного или несколь-
ких типов клеток. По этому признаку их подразделяют на две группы:
простые и сложные. Специализированные клетки могут быть разбро-
саны поодиночке или группами по всему телу растения в виде идио-
бластов (например, твердые склереиды в спелых плодах груши).
Главная функция, выполняемая тканями, со временем может изме-
няться, и даже не один раз. Например, по элементам ксилемы сначала
транспортируется вода с растворенными в ней веществами от корня
к листьям, а затем жесткие оболочки клеток выполняют только меха-
ническую функцию.
ВНИМАНИЕ!
Если ткани животных состоят только из живых клеток и межклеточного
вещества, то ткани растений могут содержать большее количество мерт-
вых клеток, чем живых. Более того, мертвые клетки (точнее, их оболочки)
зачастую выполняют основную функцию ткани (например, ксилема).
Образовательные ткани (меристемы). Меристемы состоят из жи-
вых недифференцированных клеток, способных постоянно делиться.
В онтогенезе этот тип растительных тканей возникает первым в ре-
зультате деления зиготы. На ранних этапах развития весь зародыш
состоит из меристем. Затем из них образуются все ткани растения.
Сами меристемы состоят из клеток двух типов. Клетки первого типа
называют инициальными. Они способны неограниченно долго раз-
множаться (аналогичные клетки животных называют стволовыми).
После деления одна дочерняя клетка сохраняет свойства инициаль-
ной, а другая, продолжая делиться, детерминируется в клетку опреде-
ленной ткани и приступает к начальным этапам дифференцировки.
Именно они представляют собой клетки второго типа. Количество
инициальных клеток обычно велико, иногда всего одна. Внешне ме-
ристематические клетки — это похожие друг на друга относительно
мелкие клетки с тонкими стенками, способными растягиваться. Рас-
положенное в центре ядро окружено небольшим количеством цито-
плазмы. Характерно сильное развитие гранулярного эндоплазматиче-
ского ретикулума, что свидетельствует об усиленном синтезе белков
в клетке, множество митохондрий и мелких вакуолей. Образователь-
ные ткани растений располагаются в разных местах. Их подразделяют
на несколько групп (рис. 196).
432
ВЫСШИЕ РАСТЕНИЯ
Верхушечные, или апикальные, ме-
ристемы расположены на верхушках
(апексах) осевых органов — стебля или
корня. С помощью этих меристем веге-
тативные органы растений растут в
длину.
Латеральные меристемы также ха-
рактерны для осевых органов, где они
располагаются концентрически в виде
муфты. Специализированные клетки
располагаются внутри, ближе к сердце-
вине, и снаружи — ближе к поверхности
тела. Первичные латеральные меристемы образуются из апикальных
и дают начало всем тканям растения. У древесных форм они сохраня-
ются в течение всей жизни в виде камбия, обеспечивая вторичное
утолщение. У травянистых форм латеральные меристемы быстро ис-
чезают, поэтому вторичное утолщение у них не происходит.
Интеркалярные (вставочные) меристемы представляют собой
группы клеток, уже вставших на путь дифференциации, но способных
размножаться. Интеркалярные меристемы имеются в основаниях мо-
лодых листьев. Они характерны для злаков, где располагаются в ниж-
них частях междоузлий, окруженных влагалищем листа. Эта особен-
ность позволяет подниматься полегшим после сильного ветра злакам:
изгиб побега образуется именно в указанных зонах.
Раневые меристемы восстанавливают повреждения тела.
Покровные ткани выполняют роль пограничного барьера, отделя-
ющего лежащие ниже ткани от окружающей среды. Основные функ-
ции покровных тканей: 1) защита от высыхания; 2) защита от попада-
ния вредных микроорганизмов; 3) защита от солнечных ожогов;
4) защита от механических повреждений; 5) регуляция обмена ве-
ществ между растением и окружающей средой; 6) восприятие раздра-
жения. Выделяют три типа покровных тканей: первичную, вторичную
и третичную.
Первичная покровная ткань (эпидерма, эпидермис), состоящая из
живых клеток, образуется из апикальных меристем. Эпидерма покры-
вает молодые растущие стебли и листья и защищает их от высыхания.
àÿ ìåðèñ-
î Òóòàþê)
1
2
3
2
Рис. 196. Схема расположения различных
меристем в растении:
1 — верхушечная меристема,
2 — интеркалярная меристема,
3 — боковая (латеральная) меристема
(по Тутаюк)
433
ТКАНИ РАСТЕНИЙ
Поэтому, кроме устьиц, все клетки эпидермы плотно соединены меж-
ду собой. Вся поверхность покрыта слоем кутина и растительных вос-
ков — кутикулой. Она отсутствует только на растущих корнях и под-
водных частях растений. В эпидерме имеются волоски (одноклеточные,
многоклеточные, железистые).
Устьица обнаружены только в эпидерме высших растений. Они
регулируют обмен воды и газов. Устьица представляют собой группу
клеток, в совокупности образующих устьичный аппарат (рис. 197).
Это две замыкающие клетки и примыкающие к ним побочные клетки.
Замыкающие клетки отличаются от окружающих их клеток формой,
наличием большого количества хлоропластов и неравномерно утол-
щенными стенками. Обращенные друг к другу клетки толще осталь-
ных. Обычно устьица располагаются на нижней поверхности листа.
У листьев, плавающих на поверхности воды, — на верхней. Устьица
1
23
5
6
7
8
9
11
10
7
7
7
6
2
3
Рис. 197. Строение устьичного аппарата:
1 — замыкающие клетки устьица, 2 — побочные клетки, 3 — основные клетки эпидермы,
4 — клетки губчатого мезофилла, 5 — устьичная щель, 6 — подустьичная полость,
7 — межклетники мезофилла, заполненные воздухом, 8 — хлоропласты, 9 — ядро,
10 — обращенные друг к другу толстые стенки замыкающих клеток, 11 — тонкие стенки
замыкающих клеток
4
434
ВЫСШИЕ РАСТЕНИЯ
листьев злаков часто расположены с обеих сторон равномерно, по-
скольку такие листья сравнительно равномерно освещаются.
Вторичная покровная ткань (перидерма) сменяет эпидерму, когда
зеленый цвет однолетних побегов становится коричневым. Перидер-
ма многослойная, состоит из центрального слоя камбиальных клеток
(феллогена), который откладывает наружу клетки феллемы, а внутрь —
феллодерму. Феллоген откладывает наружу клетки пробки (рис. 198).
Как только слой пробки оказывается достаточно мощным, зеленые
побеги меняют цвет на коричневый или бурый, у однолетних побегов
это обычно происходит в конце лета или осенью. Слой пробки непо-
1
2
3
4
5
6
7
Рис. 198. Развитие перидермы у зизифуса обыкновенного:
1 — кутикула, 2 — эпидермальный волосок, 3 — эпидермис, 4 — феллема,
5 — феллоген, 6 — феллодерма, 7 — первичная кора (по Александрову)
435
ТКАНИ РАСТЕНИЙ
стоянен. Периодически в нем происходят разрывы, которые сообща-
ются с расположенными рядом межклетниками. При этом на поверх-
ности образуются небольшие бугорки, называемые чечевичками, че-
рез них пространства межклетников сообщаются с атмосферным
воздухом.
Третичная покровная ткань (корка) так же, как и вторичная, ха-
рактерна только для древесных растений. Феллоген многократно за-
кладывается в более глубоких слоях коры, а ткани, которые оказыва-
ются снаружи от него, со временем отмирают, образуя более или
менее толстый слой — корку. Основной ее функцией является защита
от механических и термических повреждений.
Паренхима представлена группой тканей, которые заполняют
пространства внутри тела растения между проводящими и механиче-
скими тканями; для нее характерно наличие развитых межклетников
(рис. 199). Через устьица или чечевички они связаны с атмосферным
воздухом и обеспечивают оптимальный газовый состав внутри расте-
ния. Особенно хорошо развиты межклетники у растений, которые
произрастают на заболоченной почве, где газообмен затруднен. Такую
паренхиму часто называют аэренхимой.
1
2
3
4
5
Рис. 199.
Воздухоносная
паренхима в стебле
рдеста блестящего:
1 — кутикула,
2 — эпидерма,
3 — клетки
воздухоносной
паренхимы,
4 — воздухоносные
полости,
5 — эндодерма
(по Тутаюк,
с изменениями
и дополнениями)
436
ВЫСШИЕ РАСТЕНИЯ
В ассимиляционной паренхиме (хлоренхиме) хлоропласты располо-
жены не беспорядочно, а выстраиваются вдоль стенки, не затеняя
друг друга. Ассимиляционные ткани имеются только в освещенных
частях растения, причем они отделены от окружающей среды про-
зрачной эпидермой. Как только эпидерма сменяется непрозрачными
вторичными покровными тканями, хлоренхима исчезает.
Запасающая паренхима является вместилищем запасных органи-
ческих веществ (рис. 200). Они откладываются только в вегетацион-
ный период, а расходуются в период покоя и при подготовке к очеред-
ной вегетации. Вместилищем запасов могут быть как обычные органы
(побег, корень), так и специализированные — корневища, клубни, лу-
ковицы и др. Все семенные растения запасают энергетически ценные
вещества в семенах (в семядолях или эндосперме).
Механические ткани. Все растительные клетки обладают механи-
ческими свойствами. Это обеспечивается жесткой оболочкой клетки
и ее тургором. Но в растении есть ткани, для которых механические
свойства являются основными. Это колленхима и склеренхима. Обыч-
но они функционируют, взаимодействуя с другими тканями, образуя
внутри тела растения своеобразный каркас. В связи с этим их часто
называют арматурными.
Колленхима образована только живыми клетками, вытянутыми
вдоль оси органа. Они имеют неравномерные утолщения оболочки.
Клетки колленхимы функциональны только при наличии тургора.
При дефиците воды колленхима неэффективна, в результате растение
временно завядает (например, обвисающие листья огурцов в жаркий
1
2
Рис. 200А. Запасающая ткань
семядоли гороха. Крахмал и алейрон
(по Александрову)
Рис. 200Б. Запасающая ткань
в эндосперме пшеницы:
1 — алейроновый слой, 2 — ткань,
содержащая крахмал (по Александрову)
АБ
437
ТКАНИ РАСТЕНИЙ
день). После наполнения клеток водой функции колленхимы восста-
навливаются (рис. 201).
Склеренхима образована мертвыми клетками с очень толстыми стен-
ками, которые и выполняют механическую функцию. В зависимости от
формы различают два типа клеток склеренхимы — волокна и склереиды.
Волокна имеют сильно вытянутую форму с очень толстыми стенками
и небольшой полостью. Во флоэме их называют лубяными волокнами,
а в ксилеме — волокнами либриформа. Склереиды, или каменистые клет-
ки, округлые или ветвистые, покрыты мощными оболочками.
Выделительные ткани. У животных шлаки выводятся из организ-
ма, у растений ненужные вещества изолируются в живом протопла-
сте, в мертвых клетках или в межклеточных пространствах. Растения,
в отличие от животных, лишены целостной выделительной системы.
В зависимости от расположения выделительные ткани могут быть на-
ружными и внутренними.
Наружные выделительные ткани. Железистые волоски являются
производными эпидермы, к ним относятся и жгучие волоски крапи-
вы. Гидатоды осуществляют гуттацию — выделение избыточной
À
Á
Â
Ã
1
2
Рис. 201. Типы колленхимы:
А — пластинчатая колленхима в однолетнем побеге дуба черешчатого
под формирующейся пробковой тканью, Б — пластинчатая (1) и уголковая (2)
колленхимы в стебле баклажана, В — уголковая колленхима в черешке листа бегонии,
Г — рыхлая колленхима в черешке листа лопуха большого
(по Тутаюк)
438
ВЫСШИЕ РАСТЕНИЯ
воды в условия пониженной транспирации и высокой влажности.
Нектарники выделяют жидкость, содержащую большое количество
(от 7 до 87%) моно- и олигосахаридов. Пищеварительные железки
имеются у насекомоядных растений (росянка, непентис и др.). Соле-
вые железы есть у растений, произрастающих на засоленных почвах.
Внутренние выделительные ткани, как правило, не выводят вещества
за пределы организма, а накапливают их в себе.
Проводящие ткани. В отличие от предыдущих, эти ткани слож-
ные, т. к. состоят из различно дифференцированных клеток. Кроме
собственно проводящих элементов, здесь имеются механические, вы-
делительные и запасающие. Эти ткани объединяют все органы расте-
ния в единую систему. Выделяют два типа проводящих тканей — кси -
лему и флоэму (рис. 202).
Проводящие элементы ксиле-
мы образованы мертвыми клет-
ками. По ним осуществляется
дальний транспорт воды и раство-
ренных в ней веществ от корня к
листьям. Если в стенке проводя-
щей клетки нет сквозных перфо-
раций, ее называют трахеидой.
Трахеиды сообщаются между со-
бой посредством пор, которые
представляют собой углубления
во вторичной оболочке до первич-
ной. Если в стенке есть отвер-
стия — это сосуд. Вода передвига-
ется по трахеидам с меньшей
скоростью, чем по сосудам, потому
что у них нигде не прерывается
первичная оболочка.
Проводящие элементы флоэмы
сохраняют живой протопласт.
ÀÁ
ÂÃ
Ä
Å
3
2
1
3
2
1
1
1
2
Рис. 202. Схема строения разных типов
проводящих пучков: А — коллатеральный
открытый пучок, Б — коллатеральный
закрытый пучок, В — биколлатеральный
открытый пучок, Г — биколлатеральный
закрытый пучок, Д — концентрический
пучок с внутренней ксилемой,
Е — концентрический пучок с наружной
ксилемой; 1 — флоэма, 2 — ксилема,
3 — камбий
439
ОРГАНЫ РАСТЕНИЙ
Транспорт между клетками осуществляется через отверстия, располо-
женные на клеточных оболочках. Основными из них являются прово-
дящие, называемые ситовидными элементами. По ним происходит
дальний транспорт от фотосинтезирующих листьев к корню. У выс-
ших растений различных групп встречаются два типа ситовидных
элементов. У папоротникообразных и голосеменных они представле-
ны ситовидными клетками. Ситовидные элементы покрытосеменных
называют ситовидными трубками. В зрелых клетках отсутствуют
ядра. Зато рядом с ситовидной трубкой имеется клетка-спутница,
которая образуется вместе с ситовидной трубкой в результате мито-
тического деления общей материнской клетки и имеет ядро.
Обычно ксилема и флоэма располагаются в теле растения в опре-
деленном порядке, образуя слои или проводящие пучки. В зависимо-
сти от строения различают несколько типов проводящих пучков.
В коллатеральном открытом пучке между ксилемой и флоэмой нахо-
дится камбий, обеспечивающий вторичный рост. В биколлатеральном
открытом пучке флоэма располагается с двух сторон от ксилемы. За-
крытые пучки лишены камбия, поэтому они не способны к вторично-
му утолщению. Встречаются два типа концентрических пучков, в ко-
торых или флоэма окружает ксилему, или ксилема — флоэму.
ОРГАНЫ РАСТЕНИЙ
Дифференцированные органы присущи только высшим растени-
ям. У первых наземных растений почти все клетки содержали хлоро-
пласты. Но для фотосинтеза необходимы и углекислый газ и вода.
Низшие растения легко получают эти вещества из воды, в которой
они обитают, на суше это невозможно, т. к. бо
льшая часть углекислого
газа находится в воздухе, а воды — в почве. В результате возникла
полярность; часть растения тяготела к поверхности почвы, а затем по-
грузилась в нее полностью, поглощая воду из почвенных растворов.
Однако ее клетки лишились хлоропластов. Другая часть растения
приняла вертикальное положение, оставаясь на поверхности, в клет-
ках надземной части сохранились хлоропласты, которые обеспечива-
ли все растение органическими веществами. Так появились два ос-
новных вегетативных органа — корень и побег. Возникли и развились
высокоспециализированные ткани, которые сообща обеспечивали
нормальную жизнедеятельность всего растения.
Корень
Корень — один из двух основных вегетативных органов высшего
растения. Он обладает радиальной симметрией и способен к неогра-
440
ВЫСШИЕ РАСТЕНИЯ
ниченному росту в длину, т. к . имеет апикальную меристему (рис. 203).
Именно корень, специализированный для поглощения воды из почвы
и ее транспорта к фотосинтезирующей надземной части, резко изме-
нил общий уровень внутренней организации первых высших расте-
ний, что привело к их дальнейшему развитию на поверхности суши.
Строение корня зависит от его возраста. В молодом корне имеют-
ся несколько зон, которые отличаются одна от другой анатомически
и функционально. На верхушке корня располагается точка роста,
11
ÀÁ
I
II
III
IV
V
108
1
2
3
4
5
6
7
9
8
8
7
4
5
6
3
2
1
Рис. 203. Схема строения кончика корня (продольный разрез):
А — схематическое изображение: I — корневой чехлик, II — зона инициальных клеток,
III — зона меристемы, IV — зона растяжения, V — зона корневых волосков (зона
всасывания); 1 — перицикл, 2 — незрелые элементы флоэмы, 3 — зрелые элементы
флоэмы, 4 — эндодерма без поясков Каспари, 5 — незрелые элементы ксилемы,
6 — ризодерма, 7 — первичная кора, 8 — эндодерма с поясками Каспари, 9 — зрелые
элементы ксилемы, 10 — корневые волоски, 11 — центральный цилиндр; Б — клеточное
изображение: 1 — корневой чехлик, 2 — зона инициальных клеток, 3 — калиптроген
(меристема корневого чехлика), 4 — зона деления клеток, 5 — зона роста клеток,
6 — зона корневых волосков (зона всасывания), 7 — зона проведения,
8 — формирующиеся боковые корни (по Полувому и Тутаюк)
441
ОРГАНЫ РАСТЕНИЙ
образованная апикальной меристемой. Для защиты растущего корня
от механического повреждения твердыми частицами почвы служит
корневой чехлик, который имеется у большинства растений (отсутст-
вуя лишь у некоторых водных форм и паразитов). Чехлик образован
тонкостенными рыхло расположенными живыми клетками. Они пос-
тоянно слущиваются и заменяются новыми. Из отслоившихся клеток
образуется слизь, которая смазывает корень, облегчая его продвиже-
ние в почве. Корневой чехлик определяет и положительный геотро-
пизм корня. Напомним: геотропизм — это способность растущих ор-
ганов растений принимать определенное положение по отношению
к центру Земли. Положительный геотропизм — рост корней вниз, от-
рицательный — рост стебля вверх.
Непосредственно под чехликом расположена зона деления — пос-
тоянно делящиеся клетки, которые обеспечивают апикальный рост.
Дочерние клетки откладываются не только внутри, но и снаружи, по-
полняя популяцию постоянно погибающих клеток корневого чехли-
ка. Следующий участок корня — зона растяжения: здесь корень уве-
личивается в объеме и растягиваются. Начинает формироваться
ризодерма, и всасывается небольшое количество воды. Однако основ-
ное количество почвенных растворов поглощается в следующей зоне
всасывания, которая особенно важна для жизни растения, потому что
именно она снабжает все клетки водой и минеральными веществами.
Всасывание осуществляется специализированной эпидермальной
тканью — ризодермой, которая расположена на поверхности корня.
Ризодерма покрыта тонкими волосками, представляющими собой вы-
росты стенок клеток с цитоплазмой. Средняя длина волосков один-
два миллиметра, но у разных растений может колебаться от 0,05 до
10 мм. Волоски увеличивают поверхность всасывания примерно в
18 раз. Ризодерма активно влияет на процессы всасывания, избира-
тельно поглощая необходимые для растения вещества. Зона всасыва-
ния постоянно присутствует в молодом корне, она очень динамична.
Нежная ризодерма постоянно погибает, этот процесс идет в апикаль-
ном направлении, но со стороны зоны растяжения в нее постоянно
включаются новые клетки. Поскольку все это сопровождается ростом
корня, зона поглощения перемещается в почве, и корневые волоски
получают доступ к новым источникам воды и минеральных веществ.
Ниже ризодермы располагается первичная кора, состоящая из жи-
вых паренхиматозных клеток, покрытых тонкими оболочками. Пер-
вичная кора отделена от ризодермы слоем особых клеток — экзодер-
мой, а от осевого цилиндра — эндодермой. Экзодерма первоначально
регулирует транспорт, а после гибели ризодермы становится покров-
ной тканью корня. При этом функция всасывания вначале резко
ослабляется, а затем вовсе исчезает, поэтому данный участок корня
442
ВЫСШИЕ РАСТЕНИЯ
специализируется на транспорте веществ (зона проведения). Клетки
эндодермы имеют характерные утолщения, которые окружают клетку
в виде пояса — пояски Каспари. Благодаря суберину, имеющемуся
в волосках, они оказывают водоотталкивающее действие. Клетки эн-
додермы, смыкаясь в области поясков Каспари, окружают осевой ци-
линдр, создавая водонепроницаемое кольцо, которое вынуждает тран-
спортирующиеся по межклетникам растворы переходить в
проводящие структуры осевого цилиндра. Наружный слой осевого
цилиндра образован меристематической тканью — перициклом, под
ним располагается прокамбий, из клеток которого дифференцируют-
ся проводящие элементы растения.
Первичная структура характерна для молодых корней и корней,
в которых отсутствуют латеральные меристемы (папоротникообраз-
ные и однодольные покрытосеменные). Эти органы не способны
к вторичному утолщению. Однако в корнях голосеменных и двудоль-
ных покрытосеменных растений латеральные меристемы присутству-
ют в виде камбия, перицикла и феллогена. Клетки камбия не образу-
ют сплошной слой, а располагаются группами между ксилемой и
флоэмой. Откладывая наружу вторичную флоэму и внутрь вторич-
ную ксилему, камбий обеспечивает вторичное утолщение корня, кото-
рое со временем может стать значительным. Вторичные утолщения
корня и стебля схожи, однако годовые слои прироста в корне не выра-
жены. Феллоген образуется в перицикле, он откладывает снаружи
клетки, которые вскоре отмирают и образуют пробку. Оказавшиеся
снаружи от пробки живые клетки первичной коры также отмирают,
после чего корень становится буро-коричневым.
Корень формирует боковые ответвления, которые развиваются из
перицикла. Обычно они формируются в молодом корне на некотором
расстоянии от апекса. Сначала на поверхности осевого цилиндра воз-
никает бугорок из делящихся клеток перицикла. По мере его увеличе-
ния формируется апикальная меристема. Затем зачаточный корешок
прорывает слой эндодермы, а также первичную кору и становится бо-
ковым корнем с собственными апексом и чехликом. В нем развивают-
ся проводящие элементы осевого цилиндра. В отличие от боковых
корней, придаточные корни образуются на стеблях, листьях и на ста-
рых корнях.
Все корни одного растения образуют корневую систему (рис. 204).
В стержневой корневой системе хорошо развит главный корень, кото-
рый имеется только у семенных растений. Он развивается из зароды-
шевого корешка, который выделяется среди других корней. Такая си-
стема характерна для большинства двудольных растений. Мочковатая
корневая система образована придаточными корнями, главный ко-
рень неразличим. Мочковатую корневую систему имеют злаки.
443
ОРГАНЫ РАСТЕНИЙ
В зависимости от распределения корней в толще почвы выделяют
поверхностные, глубинные и универсальные корневые системы. Корне-
вые системы располагаются поверхностно у большинства однолетних
растений, а также у некоторых многолетних (например, злаков, какту-
сов и др.). Особенно это характерно для растений тропических лесов,
у которых бо
льшая часть корневой системы располагается на глубине
до 30 см, и растений, произрастающих в болотистой местности. Глу-
боко в почве располагаются корни многих растений, ксерофитов, жи-
вущих в условиях дефицита влаги. Их корни могут достигать уровня
грунтовых вод (рис. 205).
Функции корня разнообразны, главная — поглощение почвенных
растворов.
Для облегчения восприятия процесса всасывания почвенных рас-
творов напомним строение почвы. Она богата минеральными компо-
нентами: различные фракции песка, пыль, глина и гумус, имеющий
биогенное происхождение. Гумус образуется в результате переработ-
ки некоторыми живыми организмами детрита — мертвых корней, ли-
стьев, веток и других органических остатков. Гумус связывает мине-
Ðèñ. 68 . Êîðíåâàÿ ñèñòåìà:
À ñòåðæíåâàÿ: 1 ãëàâíûé êîðåíü; 2 áîêîâûå êîðíè; Á
ìî÷êîâàòàÿ;  ñìåøàííàÿ (àëëîðèçíàÿ): 1 ñåìÿäîëè;
2 íàäñåìÿäîëüíàÿ ÷àñòü ñòåáëÿ;3 ïîäñåìÿäîëüíàÿ ÷àñòü
ñòåáëÿ (ãèïîêîòèëü); 4 ïðèäàòî÷íûå êîðíè; 5 áîêîâûå êîð-
íè; 6 ãëàâíûé êîðåíü (À ïî Òóòàþê; Á, Â ïî Õðæàíîâñêîìó)
1
2
3
4
5
6
2
1
À
Á
Â
Рис. 204. Корневая система:
А — стержневая: 1 — главный корень, 2 — боковые корни;
Б — мочковатая, В — смешанная (аллоризная):
1 — семядоли, 2 — надсемядольная часть стебля,
3 — подсемядольная часть стебля (гипокотиль),
4 — придаточные корни, 5 — боковые корни,
6 — главный корень
(А — по Тутаюк; Б, В — по Хржановскому)
444
ВЫСШИЕ РАСТЕНИЯ
Ã
3
2
1
Â
I
I
I
I
I
I
I
V
V
I
V
Á
À
2
1
15ì
Р
и
с
.
2
0
5
.
Г
л
у
б
и
н
а
п
р
о
н
и
к
н
о
в
е
н
и
я
в
п
о
ч
в
у
к
о
р
н
е
й
н
е
к
о
т
о
р
ы
х
р
а
с
т
е
н
и
й
:
А
—
в
е
р
б
л
ю
ж
ь
я
к
о
л
ю
ч
к
а
в
п
у
с
т
ы
н
е
:
1
—
п
о
ч
в
а
;
2
—
г
р
у
н
т
о
в
ы
е
в
о
д
ы
,
Б
—
с
о
с
н
а
н
а
с
ф
а
г
н
о
в
о
м
б
о
л
о
т
е
;
В
—
н
е
к
о
т
о
р
ы
е
к
у
л
ь
т
у
р
н
ы
е
р
а
с
т
е
н
и
я
:
I
—
к
а
р
т
о
ф
е
л
ь
,
I
I
—
г
о
р
о
х
,
I
I
I
—
р
о
ж
ь
,
I
V
—
п
ш
е
н
и
ц
а
,
V
—
п
о
д
с
о
л
н
е
ч
н
и
к
,
V
I
—
с
а
х
а
р
н
а
я
с
в
е
к
л
а
;
Г
—
н
е
к
о
т
о
р
ы
е
р
а
с
т
е
н
и
я
с
т
е
п
е
й
:
1
—
ф
а
л
ь
к
а
р
и
я
,
2
—
ш
а
л
ф
е
й
,
3
—
в
е
р
о
н
и
к
а
с
и
з
а
я
(
А
,
Б
—
п
о
Р
о
д
и
о
н
о
в
о
й
и
с
о
а
в
т
.
;
В
—
п
о
К
о
р
ч
а
г
и
н
о
й
,
Г
—
п
о
Г
е
н
к
е
л
ю
)
445
ОРГАНЫ РАСТЕНИЙ
ральные частицы, в результате чего формируются плотные гранулы,
обильно пронизанные порами. Такая структура позволяет почве по-
глощать и удерживать значительное количество воды, обеспечивая
оптимальное соотношение воды и воздуха. Вода в почве может быть
доступной или недоступной для растения. Недоступная вода связана
почвенными коллоидами, минеральными компонентами почвы, а так-
же находится в мелких капиллярах. Чрезмерное содержание солей
в почвенных растворах не позволяет корням растений всасывать воду,
при этом вода может даже покидать растение.
В зависимости от условий, растения по-разному снабжаются во-
дой. По этому принципу их делят на ряд экологических групп.
Гидратофиты обитают в воде. Первичные гидратофиты — водоро-
сли, которые лишены корней и поглощают воду всей поверхностью
тела. Вторичные гидратофиты — высшие растения, предки которых
вторично заселили водную среду.
Гигрофиты — наземные растения, которые живут в условиях
обильного увлажнения. Дефицит воды для них губителен.
Ксерофиты обитают в жарком засушливом климате, поэтому они
постоянно испытывают недостаток влаги. Многие ксерофиты приспо-
собились запасать воду в своих тканях (например, алоэ, кактусы).
Мезофиты — большинство наземных растений растут при регу-
лярном увлажнении.
В корне синтезируются многие органические вещества, которые
необходимы растению, в частности аминокислоты (подробнее об этом
будет рассказано ниже). Корень заякоривает в земле и удерживает на
поверхности надземную часть растения, которая зачастую имеет гро-
мадную массу. Корень активно взаимодействует с различными по-
чвенными организмами, в частности с грибами. Взаимоотношения
между растением и грибом обоюдовыгодные (симбиоз). Ассоциация
корня растения с грибом называется микоризой. Она встречается
у громадного большинства семенных растений, отсутствуя лишь
у водных и паразитических растений, а также некоторых других (гре-
чишных, осоковых, крестоцветных).
Выделяют два типа микоризы — эктомикоризу и эндомикоризу.
Эктомикориза (эктотрофная микориза) широко распространена сре-
ди древесных растений средней полосы, например хвойные, береза,
бук, дуб, ива, некоторые кустарники и травы. Гифы гриба плотным
чехлом окутывают молодые корни, образуя гифовую мантию. Гифы
по межклетникам могут проникать в ткани корня, не входя в живые
клетки. Эндомикориза (эндотрофная микориза) встречается гораздо
чаще и развивается иначе: гифы не образуют чехол вокруг корня, но
проникают глубоко в кору корня, причем, в отличие от эктомикори-
зы, внедряются в живые клетки паренхимы. Там гифы образуют клуб-
446
ВЫСШИЕ РАСТЕНИЯ
ки, которые клетка впоследствии переваривает. Эндомикориза харак-
терна для большинства покрытосеменных.
Некоторые растения вступают в симбиоз с азотфиксирующими
бактериями, хотя это явление встречается гораздо реже, чем микори-
за. Все организмы нуждаются в регулярном поступлении азота извне.
Молекулярный азот недоступен для эукариот. Некоторые прокарио-
ты способны фиксировать атмосферный азот. Их делят на свободно-
живущие и симбиотические. К первой группе относят анаэробные бак-
терии — клостридии и аэробные — азотобактер, которые обитают
в почве. Отдельные фотосинтезирующие бактерии и сине-зеленые
водоросли сопрягают процесс азотфиксации с фотосинтезом.
Из симбиотических азотфиксирующих бактерий наиболее извест-
ны представители рода ризобиум, образующие клубеньки (рис. 206)
на корнях бобовых, — клубеньковые бактерии. Такое сожительство
чрезвычайно полезно для обоих симбионтов. Гетеротрофные бакте-
рии получают от растения необходимые им органические вещества,
а сами фиксируют атмосферный азот и передают его корню в виде
легкоусвояемых растением соединений. Корневые клубеньки форми-
руются эндогенно или экзогенно. При эндогенном способе клубеньки
образуются из перицикла, при экзогенном, который чаще встречает-
ся, — из паренхимы первичной коры молодого корня.
Азотфиксирующие бактерии не только снаб-
жают легкодоступными соединениями азота ра-
стение-хозяина, но в последующем значительно
обогащают ими почву. Так, например, люцерна
может за год накопить до 600 кг N/га, поэтому
бобовые часто используют с этой целью в сево-
обороте.
Часто из-за функциональной специализации
корни подвергаются весьма разнообразным
метаморфозам, в связи с чем их делят на не-
сколько типов: запасающие, втягивающиеся
(контрактильные), корни-присоски (гаусто-
рии), дыхательные (пневматофоры), ходульные,
досковидные корни, а также корневые отпрыски.
Запасающие корни содержат большое коли-
чество паренхимы, в живых клетках которой на-
капливаются необходимые вещества. Паренхи-
1
Рис. 206. Клубеньки на корнях люпина,
общий вид корневой системы:
1 — клубеньки (по Хржановскому и соавт.)
447
ОРГАНЫ РАСТЕНИЙ
ма может находиться в коре, ксилеме или в сердцевине. У многих
двулетних образуется сложная структура — корнеплод. В его форми-
ровании наряду с корнем принимает участие стебель, причем доля
участия этих органов у разных растений варьирует. Например, у репы
или свеклы корнеплод в основном образован стеблем, на корень при-
ходится лишь его самая нижняя часть. Корнеплод морковь, за исклю-
чением верхней части, состоит из корня. При утолщении боковых или
придаточных корней образуются корневые клубни, например у георги-
на, батата, чистяка и др. У орхидей корни снаружи покрыты веламе-
ном, который образован несколькими слоями крупных мертвых кле-
ток, имеющих сетчатые или спиральные утолщения оболочек, что
предотвращает их смятие. Пустые клетки веламена образуют капил-
ляры, по которым вода может всасываться и временно запасаться.
Втягивающиеся (контрактильные) корни имеются у многих лу-
ковичных, клубнелуковичных и корневищных растений (гладиолус,
крокус и др.). Такие корни находятся среди обычных, они способны,
сокращая свою длину, тянуть за собой луковицу или корневище и тем
самым погружают ее глубже в почву.
Корни-присоски (гаустории) имеют паразитические растения.
Они служат для проникновения в тело растения-хозяина, откуда вы-
сасывают необходимые для паразита вещества.
Дыхательные корни (пневматофоры) имеются у древесных тро-
пических растений, которые обитают на заболоченных морских побе-
режьях, в частности у мангровых. У них затруднена аэрация корней,
поэтому часть боковых подземных корней, проявляя отрицательный
геотропизм, растут вверх и поднимаются над почвой (вернее, над во-
дой). Для дыхательных корней характерны многочисленные чечевич-
ки и развитая система межклетников, которые обеспечивают газооб-
мен между растением и атмосферным воздухом.
Ходульные корни служат своеобразной опорой для тропических
деревьев, растущих на заболоченных почвах или в полосе прилива.
Эти корни по происхождению придаточные, они растут от стволов на
уровне воды, идут вниз и прочно укрепляют дерево в зыбком иле. Хо-
дульные корни также характерны для мангровых, но они особенно
развиты у баньяна, одно дерево которого, постепенно разрастаясь
вширь, может занять площадь в несколько сотен квадратных метров.
Досковидные корни характерны для крупных деревьев, растущих
в тропических лесах. Они представляют собой боковые корни, кото-
рые отходят от дерева у самой поверхности почвы.
Корневые отпрыски служат для вегетативного размножения. Они
развиваются в надземные побеги из образующихся на корнях прида-
точных почек. Корневые отпрыски встречаются, как правило, у мно-
голетних растений (сирень, слива, осина, вьюнок, осота и др.).
448
ВЫСШИЕ РАСТЕНИЯ
Побег
Побег относится к числу главных вегетативных органов растения.
Поскольку побег является осевым органом, у него присутствует апи-
кальная меристема, которая обеспечивает неограниченное нарастание
в длину. Такой рост сопровождается более или менее регулярным вет-
влением побега, что приводит к формированию системы побегов. Вет-
вление позволяет многократно увеличить фотосинтезирующую
поверхность и обеспечить растение органическими веществами.
У различных растений наблюдается несколько типов ветвления: ди-
хотомическое, моноподиальное, симподиальное (рис. 207).
Дихотомическое (вильчатое) ветвление имело место у давно
вымерших высших растений. В настоящее время такое ветвление
наблюдается у наиболее примитивных форм папоротниковидных и
плауновидных. Его принцип заключается в повторяющемся через
определенные интервалы разделении надвое верхушечной точки ро-
ста. При этом каждая из них дает начало новому побегу. Если каждый
из них развивается одинаково, такую дихотомию называют равной
(изотомия). Если преимущественно развивается одна из ветвей, ди-
хотомия называется неравной (анизотомией). При крайней выражен-
ности анизотомии разница между дочерними побегами настолько за-
метна, что прослеживается ось, которая находится не в выпрямленном,
а в зигзагообразном положении. Этот тип называется дихоподием, он
встречается у многих папоротникообразных.
Моноподиальное ветвление, или моноподий, характеризуется пол-
ным выпрямлением оси. При этом типе верхушечный рост происхо-
дит непрерывно, боковые же ветви закладываются под апексом. У них
также наблюдается непрерывный верхушечный рост и моноподиаль-
ное ветвление, при котором последовательно образуются боковые
ветви третьего, четвертого и последующих порядков, но скорость ро-
ста отходящих ветвей заметно уменьшается, чем рост основной оси.
Моноподиальное ветвление широко распространено среди хвойных
(ель, сосна, кипарис и др.).
Симподиальное ветвление (симподий) происходит от моноподиаль-
ного. Главная ветвь развивается относительно недолго, затем она или
погибает, или ее перевешивает образующаяся под верхушкой боковая
ветвь, которая после этого становится главной, но опять же лишь на
время, пока ее не «перевершинит» другая боковая ветвь. В итоге фор-
мируется ось, состоящая из последовательно развившихся боковых
ветвей. Такой тип ветвления характерен для цветковых растений
и особенно четко выражен у древесных форм.
Некоторые растения (например, пальмы) не ветвятся, но при этом
сохраняют верхушечный рост.
449
ОРГАНЫ РАСТЕНИЙ
В зависимости от выполняемой функции различают побеги двух
типов: вегетативные и генеративные. Вегетативные побеги осуществ-
ляют воздушное питание и обеспечивают синтез органических ве-
ществ из неорганических. В генеративных побегах фотосинтез чаще
всего не происходит, в них образуются спорангии, которые обеспечи-
I
I
I
1
2
3
4
À
Á
Â
Ã
Ä
Å
Р
и
с
.
2
0
7
.
Т
и
п
ы
в
е
т
в
л
е
н
и
я
с
п
о
р
о
ф
и
т
о
в
в
ы
с
ш
и
х
р
а
с
т
е
н
и
й
:
I
—
с
х
е
м
а
с
п
о
с
о
б
о
в
в
е
т
в
л
е
н
и
я
с
п
о
р
о
ф
и
т
о
в
в
ы
с
ш
и
х
р
а
с
т
е
н
и
й
:
А
—
р
а
в
н
а
я
д
и
х
о
т
о
м
и
я
(
и
з
о
т
о
м
и
я
)
,
Б
—
н
е
р
а
в
н
а
я
д
и
х
о
т
о
м
и
я
(
а
н
и
з
о
т
о
м
и
я
)
,
В
—
д
и
х
о
п
о
д
и
й
,
Г
—
м
о
н
о
п
о
д
и
й
,
Д
—
с
и
м
п
о
д
и
й
,
Е
—
л
о
ж
н
а
я
д
и
х
о
т
о
м
и
я
,
I
I
—
в
е
т
в
л
е
н
и
е
у
п
р
е
д
с
т
а
в
и
т
е
л
е
й
р
а
з
л
и
ч
н
ы
х
г
р
у
п
п
в
ы
с
ш
и
х
р
а
с
т
е
н
и
й
:
1
—
п
л
а
у
н
,
2
—
к
и
п
а
р
и
с
,
3
—
я
б
л
о
н
я
,
4
—
с
и
р
е
н
ь
(
п
о
Т
а
х
т
а
д
ж
я
н
у
,
с
и
з
м
е
н
е
н
и
я
м
и
и
д
о
п
о
л
н
е
н
и
я
м
и
)
450
ВЫСШИЕ РАСТЕНИЯ
вают размножение растения (к та-
ким побегам относят и цветок). Ве-
гетативный побег включает в себя
стебель, представляющий собой
ось побега; листья, которые явля-
ются уплощенными боковыми
органами побега; а также почки, яв-
ляющиеся зачатками молодых по-
бегов, они обеспечивают нараста-
ние побега и его ветвление. Для
побега характерна метамерность,
т. е . чередование одинаковых ча-
стей — метамеров (рис. 208).
Листья располагаются на сте-
бле в определенном порядке, в со-
ответствии с типом листорасполо-
жения; участок стебля на уровне
отхождения листа называется
узлом. Если основание листа пол-
ностью окружает стебель, узел на-
зывается закрытым, если нет —
открытым. Промежутки стебля
между соседними узлами называ-
ют междоузлиями. Метамер — уча-
сток побега, который включает
в себя узел с находящимся под ним
междоузлием, а также расположен-
ные на этом отрезке листья и по-
чки.
В зависимости от механических свойств стебля различают не-
сколько морфологических типов побегов: прямостоячие, припод-
нимающиеся, восходящие, лазящие, вьющиеся (лианы), ползучие и ле-
жачие.
Стебель
Стебель служит опорой для фотосинтезирующих органов (листь-
ев), а также обеспечивает транспортную связь между ними и корнем.
Кроме главных функций, стебель может выполнять и некоторые до-
полнительные, в частности клетки паренхимы молодых стеблей со-
держат большое количество хлоропластов и активно участвуют в фо-
тосинтезе. В стеблях многолетних растений часто откладываются
запасные питательные вещества, а у суккулентов — вода (например,
3
4
2
5
1
Рис. 208. Строение побега: 1 — листья;
2 — стебель; 3 — узлы; 4 — метамер
побега; 5 — междоузлие (по Тутаюк,
с изменениями и дополнениями)
451
ОРГАНЫ РАСТЕНИЙ
кактусы). Длина стебля напрямую зависит от длины междоузлий.
Узлы коротких стеблей настолько сближены друг с другом, что ме-
ждоузлия могут быть практически неразличимы.
Чаще всего стебель обладает радиальной симметрией. Форма его
обычно цилиндрическая, часто граненая, у некоторых растений сте-
бли совершенно сплющенные.
Стебель имеет апикальную меристему, рост его верхушечный. На
вегетативном апексе закладываются клеточные структуры, которые
затем войдут в состав стебля и остальных органов побега (листьев
и почек). Апекс побега существенно отличается от гладкого апекса
корня: у первого через определенные промежутки времени на поверх-
ности закладываются зачатки листьев — листовые примордии в виде
небольших валиков. Промежуток времени, который необходим для
закладки последующего листового зачатка, называется пластохрон,
его величина широко варьирует у различных растений и зависит от
периода вегетации.
Молодой стебель по строению сходен с молодым корнем. Снару-
жи он покрыт эпидермой, причем, в отличие от эпидермы, покрываю-
щей корень, имеются устьица. Под эпидермой располагается первич-
ная кора. Первичная кора стебля отделена менее четко, чем в корне.
Это вызвано тем, что в стебле эндодерма выражена гораздо слабее, в
ней часто откладываются крахмальные зерна. Поэтому пограничный
слой клеток, расположенных между первичной корой и осевым ци-
линдром, называется крахмалоносным влагалищем.
Осевой цилиндр, или стела, — совокупность проводящих структур
осевых органов растения в комплексе с ассоциированной паренхимой.
Стела имеется как в корне, так и в стебле. Однако строение стел раз-
личных осевых органов неодинаково, еще больше оно варьирует у ра-
стений из разных таксономических групп. Наиболее примитивна про-
тостела, которая была широко распространена среди вымерших
растений, в настоящее время встречается в стеблях многих папорот-
никообразных и в корнях почти всех высших растений. В исходном
виде протостела представляет собой сплошной тяж ксилемы, распо-
ложенный в центре, который по периферии окружен также сплош-
ным слоем флоэмы. Такой тип протостелы называют гаплостелой.
Сердцевина впервые появляется в сифоностеле, где по периферии
сердцевины расположены проводящие структуры ксилемы и флоэмы.
Трубчатое строение увеличивает механическую прочность конструк-
ции, что дало возможность растениям значительно увеличить свои
размеры и массу. Эта стела встречается у многих папоротников.
В эустеле (эвстеле) проводящие элементы образуют пучки, в ко-
торых снаружи обычно лежит флоэма, а внутри — ксилема. Если меж-
ду ксилемой и флоэмой имеется прослойка камбия, проводящий пу-
452
ВЫСШИЕ РАСТЕНИЯ
чок называется открытым, если нет — закрытым. Эустелой обладают
подавляющее большинство семенных растений. Большинство одно-
дольных покрытосеменных и некоторые травянистые двудольные
обладают атактостелой. В ней проводящие пучки без видимого по-
рядка разбросаны в коре и сердцевине осевого органа, поэтому на сре-
зе их можно обнаружить на любом участке.
Развивающийся стебель утолщается. В зависимости от участия
камбия утолщение может быть первичным или вторичным. Первичное
утолщение представляет собой увеличение объема стебля за счет рас-
тяжения клеток, составляющих первичные ткани. Это особенно важ-
но для травянистых форм, которые не обладают камбием и вторич-
ным ростом (почти все однодольные). Вторичное утолщение
осуществляется за счет деятельности камбия и приводит к существен-
ному увеличению диаметра стебля.
Проводящие элементы осевого цилиндра обеспечивают дальний
транспорт веществ. По ксилеме передвигается вода и растворенные
в ней вещества, в направлении от корня к листьям. Постоянный ток
воды обеспечивается транспирацией через раскрытые устьица. У раз-
ных растений значительно варьирует скорость движения воды. Она
может достигать нескольких десятков метров в час. По флоэме осу-
ществляется транспорт органических веществ, причем, в отличие от
ксилемы, он идет через живые клетки. Скорость движения веществ по
флоэме значительно ниже, чем по ксилеме — около одного метра
в час. По флоэме пластические вещества от органов, где они синтези-
руются, оттекают к корню, к цветкам и плодам (у покрытосеменных)
или спорангиям. По ксилеме также транспортируются растворенные
углеводы по направлению к развивающимся меристемам. Это проис-
ходит весной в период начала вегетации, когда фотосинтезирующие
зеленые органы еще отсутствуют, органические соединения по ксиле-
ме доставляются из мест, где они запасались к интенсивно распускаю-
щимся почкам.
Вторичное утолщение продолжается в течение всей жизни расте-
ния и за много лет может привести к формированию ствола, имеюще-
го колоссальные размеры: диаметр может превышать 10 м. У деревь-
ев, растущих в дождевых тропических лесах, интенсивность деления
камбиальных клеток очень мало изменяется в течение года, поэтому
у них почти не выражена слоистость древесины. У деревьев, произра-
стающих в условиях переменного климата, где сезонные колебания
температуры и влажности очень велики, активность камбия зависит
от времени года. Наиболее интенсивное деление инициальных клеток
здесь отмечается весной, когда из почек активно развиваются моло-
дые побеги, на которых имеются листья. В этот период в основном
откладываются проводящие воду элементы — трахеиды (бо
льшая
453
ОРГАНЫ РАСТЕНИЙ
часть древесины образована именно ими) и членики сосудов, имею-
щих большой внутренний объем и тонкие оболочки. Летом инициаль-
ные клетки камбия делятся реже и дают начало толстостенным трахе-
идам и волокнам. Осенью деятельность камбия ослабляется, а позднее
вовсе прекращается; зимой камбий не функционирует. В результате
образовавшаяся за год древесина неоднородна, и ее можно подразде-
лить на раннюю и позднюю, которые, чередуясь, совместно формиру-
ют ежегодный слой прироста в виде кольца.
Только молодые элементы ксилемы и флоэмы, расположенные не-
далеко от камбиального слоя, осуществляют транспорт веществ. Кси-
лема сохраняет способность проводить воду в течение нескольких
лет, затем проведение прекращается, ксилема выполняет только опор-
ную функцию. Флоэма функционирует обычно в течение одного сезо-
на вегетации. На смену утратившим способность проводить вещества
старым элементам камбий откладывает новые слои древесины и луба.
При этом старая флоэма участвует в формировании корки, а старая
ксилема оказывается в глубине ствола. Слои молодой древесины, ле-
жащие около камбия, называют заболонной древесиной (заболонью),
кнутри от нее лежит ядровая древесина. Заболонная древесина более
светлая, чем ядровая, хотя у многих деревьев это не наблюдается.
Почка
Почка — орган побега, который обеспечивает его верхушечный на-
рост и ветвление. В состоянии покоя почка является зачаточным побе-
гом, у него сильно укороченная ось — зачаток стебля, заканчивающий-
ся конусом нарастания (рис. 209). На оси вегетативной почки
располагаются зачаточные листья, причем в их пазухах уже находятся
зачатки пазушных почек следующего поколения. Зимующие почки по-
крыты специализированными чешуями, представляющими собой ви-
доизмененные наружные листья. Они предохраняют почку от высыха-
ния и отчасти от перепадов температуры. Такие почки называют
закрытыми. Почка, лишенная защитных чешуй, называется открытой.
По расположению почки могут быть верхушечными и пазушными.
Верхушечные почки обеспечивают апикальный рост побега. Самую
первую верхушечную почку можно обнаружить еще в зародыше. Впо-
следствии из нее образуются все органы побега. Пазушные почки на-
ходятся в пазухах листьев, положение их на стебле напрямую зависит
от листорасположения. Придаточные почки можно обнаружить на
стеблях, но, в отличие от пазушных, придаточные никак не связаны
с листьями и обычно находятся в междоузлиях. У некоторых расте-
ний придаточные почки образуются на листьях. Такими почками раз-
множается комнатное растение бриофиллюм (каланхоэ). У бегоний
454
ВЫСШИЕ РАСТЕНИЯ
придаточные почки развиваются из эпидермальных клеток, располо-
женных поблизости от перерезанных жилок листа. Такие почки на
листьях могут также образовывать многие папоротники.
Придаточные почки, кроме побега, образуются и на корнях. Раз-
вившиеся из них в дальнейшем побеги называют корневыми отпры-
сками, которые играют важную роль в вегетативном размножении
многих растений.
Многие многолетние растения образуют спящие почки. Они воз-
никают как пазушные или придаточные почки, но не развиваются
в побег, а впадают в состояние покоя и пребывают в нем в течение
длительного времени, сохраняя жизнеспособность. Сигналом для вы-
хода спящей почки из состояния покоя является повреждение или
снижение активности нормальных почек. Хорошо известно появле-
ние молодой поросли на пнях или на стволах после значительной об-
резки ветвей. У голосеменных спящие почки образуются редко, по-
этому на пнях ели или сосны не образуется молодая поросль. Кроме
вегетативных, у растений имеются еще вегетативно-генеративные
и генеративные почки.
Лист
Лист — это орган, специализированный для фотосинтеза
(рис. 210). Самыми первыми листьями семенных растений являются
семядоли. Остальные листья развиваются из зачатков — листовых
4
Á
I
À
2
1
II
À
Á
4
3
1
Рис. 209. Строение почек
бузины:
I — вегетативные почки,
II — генеративные почки;
А — внешний вид
и расположение на стебле,
Б — продольный разрез;
1 — почечные чешуи,
2 — конус нарастания,
3 — зачаточное соцветие,
4 — листья
(по Корчагиной, с изменениями
и дополнениями)
455
ОРГАНЫ РАСТЕНИЙ
примордиев, которые через определенные промежутки времени за-
кладываются в периферической зоне апикальной меристемы стебля
в виде бугорков. В зависимости от ряда факторов листья могут быть
более или менее крупными, но у них отсутствует бесконечный рост,
поэтому размер листа ограничен. Почти у всех растений лист живет
и функционирует определенное время, а затем он опадает со стебля.
Растения, у которых круглый год имеются листья, называют вечнозе-
леными, среди них встречаются деревья, кустарники и травы, все они
широко распространены во многих зонах. Несмотря на название, ли-
стья таких растений постепенно сменяются более молодыми. Вечно-
зеленые растения могут произрастать в географических зонах, где нет
зимы, или же в умеренных широтах. В последнем случае растения,
зимующие с листьями, имеют структуры, предохраняющие их от
чрезмерных потерь влаги. Например, игловидные листья хвойных
(хвоинки) имеют плотную консистенцию. Снаружи они покрыты
толстым слоем кутикулы, устьица глубоко погружены в углубления
эпидермы, и над устьичной щелью образуется наплыв кутикулы.
Многолетние растения, у которых в течение определенного сезона
года листовой покров отсутствует, называют листопадными. Времен-
ная потеря листьев — это защитная реакция, позволяющая растению
успешно пережить неблагоприятные условия окружающей среды. Ра-
стения, произрастающие в условиях сильной летней засухи, летом
сбрасывают листву, что приводит к уменьшению транспирации и эко-
номии воды.
Листья снабжают все растение органическими веществами. Усть-
ичная транспирация служит основой непрерывного транспорта ве-
ществ по проводящим структурам растения. Лист функционирует
очень активно, что довольно быстро приводит к его старению и отми-
ранию. В стареющем листе сначала уменьшается синтез углеводов,
затем происходит деградация хлоропластов и разрушается хлоро-
4
3
2
1
Рис. 210. Поперечный
разрез листа:
1 — кожица,
2 — столбчатая
паренхима,
3 — губчатая
паренхима,
4 — сосудисто-
волокнистый пучок
456
ВЫСШИЕ РАСТЕНИЯ
филл. Каротиноиды, которые до этого маскировались хлорофиллом,
становятся заметными. Они определяют желтую или красную окра-
ску старых листьев. В тканях листа накапливается большое количест-
во минеральных солей, попавших сюда, будучи растворенными в воде.
Одновременно из старого листа активно выводятся органические ве-
щества, которые могут быть полезными для других органов растения,
что еще больше ослабляет лист. У вечнозеленых растений оттекаю-
щие вещества идут на нужды развивающихся почек, поэтому опада-
ние листьев часто совпадает с распусканием почек. Старение листа
завершается его отпадением от стебля.
Лист снаружи покрыт прозрачной эпидермой. Под эпидермой рас-
полагается рабочая ткань листа — паренхима, или хлоренхима (мезо-
филл), которая занимает основной внутренний объем листовой пла-
стинки. Разделяют два типа паренхимы (мезофилла), которые
отличаются степенью развития межклетников и формой клеток — па-
лисадный мезофилл (столбчатая паренхима) и губчатый мезофилл
(губчатая паренхима). Палисадный мезофилл лежит под верхней эпи-
дермой. Его клетки содержат большое количество хлоропластов и до-
вольно плотно примыкают друг к другу, хотя небольшие межклетни-
ки, обеспечивающие газообмен, имеются. Оптимально освещаемые
в светлое время суток клетки палисадного мезофилла синтезируют
основную часть органических веществ, которые растение получает
в результате фотосинтеза.
Ниже палисадного мезофилла и ближе к нижней эпидерме лежит
губчатый мезофилл. Его округлые клетки располагаются более рыхло,
поэтому между ними имеются хорошо развитые межклетники, кото-
рые через устьица сообщаются с атмосферным воздухом. По меж-
клетникам к фотосинтезирующим клеткам доставляется углекислый
газ и удаляется кислород. Губчатый мезофилл содержит меньше хло-
ропластов, поэтому нижняя поверхность листа светлее верхней.
Разветвленная сеть проводящих пучков, которые располагаются
среди фотосинтезирующих клеток, обеспечивает нормальное функ-
ционирование мезофилла листа. Проводящие пучки, окруженные со-
путствующими клетками, называются жилками. Наиболее крупные
из них рельефно выдаются на нижней поверхности листа, а неболь-
шие находятся в толще мезофилла. Характер распределения жилок
в листе называется жилкованием, оно может быть сетчатым (у дву-
дольных), дуговидным и параллельным (у однодольных) и промежу-
точным.
Главная часть листа — листовая пластинка, чаще всего плоская.
Форма листовой пластинки удивительно разнообразная и причудли-
вая, при этом она служит очень важным систематическим показате-
лем. Самая простая пластинка цельная или рассеченная, рассечение
457
ОРГАНЫ РАСТЕНИЙ
может быть перистым или пальчатым, в результате чего образуются
перисто- и пальчато-рассеченные или перисто- и пальчато-лопастные
листовые пластинки. Лист одной листовой пластинки называется
простым. Его пластинка может быть рассеченной, но не до черешка.
Если на черешке располагаются две и более листовые пластинки, лист
сложный. Ось, на которой сидят пластинки, называется рахисом. Фор-
ма сложного листа весьма разнообразна. В зависимости от располо-
жения листовых пластинок на рахисе выделяют перистые, пальчатые,
тройчатые и другие типы сложных листьев.
Основание — часть листа, которая сочленяется со стеблем. В неко-
торых случаях оно выделяется слабо, но иногда сильно разрастается
и в виде влагалища со всех сторон охватывает стебель. Листовые вла-
галища защищают пазушные почки, у злаков также и вставочную ме-
ристему. Кроме того, влагалища поддерживают стебель. У многих ра-
стений в области основания имеются прилистники. Если прилистники
зеленые, они могут фотосинтезировать, причем в некоторых случаях
именно они обеспечивают растение пластическими веществами. Если
листовая пластинка непосредственно переходит в основание, лист на-
зывается сидячим (например, ирис или злаки). Если же между пла-
стинкой и основанием имеется черешок, лист черешковый (например,
береза).
Наиболее крупные листья — у растений, растущих в нижних яру-
сах дождевых тропических лесов, где сочетается оптимальная влаж-
ность и температура при умеренной освещенности. Плавающие ли-
стья водных растений также могут достигать огромных размеров.
Например, кувшинка Виктория обладает самыми большими среди
растений листьями, которые достигают двух метров в поперечнике
и способны удерживать на плаву груз до 40 кг. Напротив, у многих
ксерофитов мелкие листья развились в качестве приспособления к за-
сушливой среде обитания.
Последовательное положение листьев на стебле называется ли-
сторасположением. Выделяют несколько типов листорасположения:
спиральное, двурядное, мутовчатое и супротивное. Двурядное листо-
расположение часто у однодольных. Листья располагаются в одной
плоскости, охватывая своими основаниями стебель в области узла.
При мутовчатом листорасположении на одном узле располагаются
сразу несколько листьев. Супротивное листорасположение — вариант
мутовчатого, но на каждом узле имеются по два листа, пространствен-
но противопоставленных друг другу.
Метаморфозы. В зависимости от выполняемых функций листья
приобретают различные морфологические формы. В некоторых слу-
чаях листья частично или полностью подвергаются метаморфозу.
У кактусов и барбариса они становятся колючками, защищающими
458
ВЫСШИЕ РАСТЕНИЯ
растение. У тыквенных наряду с нормальными встречаются листья,
трансформирующиеся в усики. Усики листового происхождения име-
ются также у многих бобовых, при этом у гороха только верхушечная
часть листа превращается в усик. У насекомоядных растений лист ви-
доизменяется в аппарат, которым растение ловит насекомых и перева-
ривает их с помощью ферментов пищеварительных желез, компенси-
руя тем самым дефицит азотистых веществ. Часто листовой черешок
уплощается и принимает форму листовой пластинки, такой метамор-
фоз называется филлодием.
Не только листья, но и побеги в целом могут подвергаться мета-
морфозу. У дикой яблони, боярышника и многих других имеются ко-
лючки побегового происхождения. Растения, испытывающие недо-
статок влаги, используют в качестве фотосинтезирующего органа не
листья с их активной транспирацией воды, а уплощенные побеги. Та-
кие листоподобные побеги, имеющие зеленую окраску, называют
филлокладиями. Они, например, имеются у комнатного растения
иглицы. Часть побегов винограда видоизменяется в усики, с помощью
которых растение цепляется за опору и, не имея мощного стебля, под-
держивает свое тело высоко над землей. Многие растения, произра-
стающие в засушливых зонах, где постоянно не хватает влаги, при-
способились запасать воду в тканях собственных органов. Такие
растения называют суккулентами. У листовых суккулентов (алоэ)
вода запасается в листьях, у стеблевых суккулентов (кактус) —
в стебле.
Существует много типов подземных побегов, их происхождение
обычно связано с метаморфозом. Корневище представляет собой под-
земный побег. В нем могут запасаться питательные вещества, у многих
растений корневища служат для вегетативного размножения. На кор-
невище отсутствуют листья, но часто видны метамеры с узлами и ме-
ждоузлиями, что позволяет отличать их от корней. Из пазушных и
придаточных почек корневища развиваются надземные побеги, или с
помощью их корневище ветвится. Корневище нарастает по направле-
нию к надземной части побега, а с противоположного конца подзем-
ный побег постепенно отмирает. На корневище образуется множество
придаточных корней. Каудекс является видоизмененным подземным
побегом. Как и корневище, он запасает питательные вещества, но,
в отличие от корневища, каудекс не отмирает с нижнего конца, а по-
степенно переходит в главный корень. Такие подземные образования
имеются например, у василька, одуванчика, люцерны и др. Столоны
тоже представляют собой подземные побеги. Побеги могут быть над-
земными и подземными. Картофель имеет подземные столоны.
Хорошо развитые надземные столоны земляники называются уса-
ми (плетями). С помощью усов земляника вегетативно размножает-
459
ОРГАНЫ РАСТЕНИЙ
ся, так как из верхушечной почки развивается и молодое дочернее
растение, которое затем укореняется и теряет связь с материнским.
Луковица — это видоизмененный побег, имеющий очень короткую
продольную ось. Стебель луковицы сильно расширяется, образуя
донце, на котором располагаются толстые и очень сочные листья, ох-
ватывающие луковицу со всех сторон.
Особым типом видоизмененных побегов являются соцветия. Они
развиваются из вегетативно-генеративных почек и, в отличие от чи-
сто вегетативных побегов, чаще всего обладают ограниченным верху-
шечным ростом, потому что их апикальные меристемы образуют
цветки. Поэтому у многолетних растений соцветия отмирают после
того, как на них созреют плоды. Соцветия различаются между собой.
Если апикальная меристема полностью используется для формирова-
ния верхушечного цветка, соцветие называют закрытым, если же
длительное время сохраняется в деятельном состоянии — открытым.
В зависимости от наличия листьев выделяют облиственные (на-
пример, у фуксии), брактеозные (с мелкими чешуевидными листья-
ми-прицветниками, как у сирени и ландыша) и голые (пастушья сум-
ка) соцветия. Наиболее распространенным и самым примитивным
типом простых соцветий является кисть. Она представлена более
или менее длинной осью, на которой располагаются одиночные цвет-
ки с четко выраженными цветоножками. Различные варианты кистей
могут отличаться наличием или отсутствием апикальных меристем
(бывают открытыми и закрытыми), наличием или отсутствием листь-
ев (облиственные, брактеозные и голые), или количеством цветков
в соцветии (их может быть всего два или довольно много).
В тех случаях, когда на хорошо выраженной главной оси распола-
гаются цветки с цветоножками разной длины (у нижних они гораздо
длиннее, чем у цветков, расположенных выше), но при этом все цветки
находятся в одной плоскости, соцветие называется щитком. Если у си-
дящих на удлиненной главной оси цветков отсутствуют цветоножки,
соцветие называют колосом, или початком, если главная ось сильно
утолщена. Соцветия с сильно укороченной главной осью могут быть
зонтиком, если у цветков имеются цветоножки, или головкой, если
цветоножки не выражены. Наиболее сложным типом простых со-
цветий является корзинка сложноцветных, у которых цветки распола-
гаются на короткой и уплощенной главной оси. С боков и снизу все
соцветие окружено видоизмененными верховыми листьями, образую-
щими обертку. Часто цветки в корзинке разные: по периферии распо-
лагаются стерильные, но крупные и ярко окрашенные цветки, служа-
щие для привлечения насекомых-опылителей, а ближе к центру лежат
генеративные. Обычно первыми распускаются цветки, находящиеся
на периферии, а затем процесс идет центростремительно.
460
ВЫСШИЕ РАСТЕНИЯ
Сложные соцветия отличаются от простых тем, что у них на глав-
ной оси располагаются не отдельные цветки, а дополнительно ветвя-
щиеся оси второго, третьего и последующих порядков. Как и простые
соцветия, сложные подразделяются на несколько групп. Двойные ки-
сти отличаются от простых тем, что у них на длинной главной оси
располагаются не цветки, а простые кисти. Аналогично выделяют та-
кие соцветия, как сложный колос, сложный зонтик. Похожие на двой-
ные кисти метелки (или сложные кисти) ветвятся сильнее, нижние
оси у них развиты лучше верхних.
Соцветия имеются только у покрытосеменных, у которых они ши-
роко распространены. Большое количество цветков, расположенных
поблизости, имеют больше шансов быть опыленными, чем одиночные
цветки. Это привлекает насекомых-опылителей, которые опыляют не
один цветок, а сразу несколько, увеличивая тем самым выход семян.
ОТДЕЛ МОХООБРАЗНЫЕ
Мохообразные — единственный отдел высших растений, у кото-
рых гаметофит является преобладающим поколением. Растение, ко-
торое мы представляем себе как мох, и является гаметофитом. Мохо-
образные отличаются относительно простым строением. Их тело
расчленено на стебель и лист, у наиболее простых тело представляет
собой слоевище (таллом), уплощенный в дорсовентральном направ-
лении. Однако все вегетативные органы мохообразных не могут счи-
таться истинными, поскольку они принадлежат гаметофиту и негомо-
логичны настоящим органам спорофита. Поэтому их «стебли» и
«листья» именуют каулидиями и филлидиями, соответственно. Мохо-
образные не имеют корней и прикрепляются к субстрату нитевидны-
ми ризоидами, которые у различных форм могут быть одно- или мно-
гоклеточными. Поглощение воды и других необходимых веществ
растением осуществляется не только ризоидами, но и через всю по-
верхность тела, чему в значительной мере способствует полное (или
почти полное) отсутствие на поверхности защитной кутикулы. Это
вынуждает их обживать места с повышенной влажностью.
У большинства мохообразных отсутствует проводящая ткань.
Кроме того, у них отсутствуют или слабо развиты и большинство дру-
гих тканей, в частности механические, что ограничивает размеры ра-
стений несколькими сантиметрами. Водные мохообразные использу-
ют воду в качестве дополнительной опоры и нередко превышают
полметра в длину.
Развитие мохообразных представляет собой чередование гапло-
идной и диплоидной фаз (рис. 211). Преобладающим поколением яв-
461
ОТДЕЛ МОХООБРАЗНЫЕ
ляется гаметофит. Он развивается из споры, ядро которой имеет га-
плоидный набор хромосом, поэтому все клетки гаметофита тоже
гаплоидные. Первоначально из споры развивается протонема, имею-
щая вид ветвящейся нитчатой водоросли или пластинки. Спора не
содержит запаса питательных веществ, поэтому молодой гаметофит
должен их синтезировать самостоятельно посредством фотосинтеза.
Наличие необходимой для этого хлоренхимы определяет зеленую
окраску гаметофита.
У печеночных мхов многократное деление верхушечной клетки
дает начало пластинчатым структурам, на которых впоследствии ра-
зовьются половые органы. У листостебельных мхов гаметофит имеет
вид облиственных побегов, они развиваются из почек, которые фор-
мируются на протонеме. На гаметофорах образуются половые орга-
ны — женские архегонии и мужские антеридии. Чаще всего на одном
растении развиваются органы только одного пола — однодомные мхи,
но нередко встречаются и двудомные (на одной особи формируются
как женские половые органы, так и мужские). Антеридии представля-
ют собой овальные тельца, окруженные тонкой однослойной оболоч-
кой. Они наполнены сперматогенными клетками, которые при мито-
тическом делении дают начало двум подвижным сперматозоидам,
снабженным двумя жгутиками. Архегоний устроен сложнее и обычно
представляет собой структуру, по своей форме напоминающую бу-
тылку. В утолщенной части, называемой брюшком, находится круп-
ная яйцеклетка, которая также образуется в результате митоза. Вну-
три суженной шейки в один ряд располагаются шейковые клетки,
одна из которых — брюшная шейковая клетка — находится над яйце-
Ðèñ. 144. Ñõåìà æèçíåííîãî öèêëà ïå÷åíî÷íîãî ìõà
ïåëëèè (Pellia)
(ïî Ãðèíó è ñîàâò. , ñ èçìåíåíèÿìè)
Рис. 211. Схема жизненного цикла печеночного мха пеллии
(по Грину и соавт., с изменениями)
462
ВЫСШИЕ РАСТЕНИЯ
клеткой. Оплодотворение возможно только в капельно-жидкой воде.
Через шейку сперматозоиды проникают в брюшко архегония и опло-
дотворяют находящуюся там яйцеклетку. В результате образуется ди-
плоидная зигота, которая по прошествии определенного периода по-
коя дает начало диплоидному поколению — спорофиту.
Спорофит мохообразных называется спорогон, он устроен наибо-
лее просто среди всех высших растений. Спорогон представляет со-
бой коробочку (спорангий), которая посредством ножки переходит
в тело гаметофита. Спорогон мохообразных не содержит хлорофилла,
поэтому все необходимые для развития вещества получает от гамето-
фита посредством проникающей в его ткани нижней расширенной ча-
сти ножки — гаустории. Внутри коробочки многочисленные материн-
ские клетки спор делятся мейотически и дают начало гаплоидным
спорам, посредством которых осуществляется бесполое размножение.
В коробочке печеночных мхов среди спор находятся пружинки — эла-
теры. При повышении влажности элатеры раскручиваются, а при по-
нижении вновь скручиваются, при этом споры перемешиваются и
разрыхляются, что способствует выталкиванию их из коробочки по-
сле того, как она раскрывается. Споры, выпадающие из коробочки,
подхватываются ветром и переносятся на большие расстояния. По-
давляющее большинство погибают, попадая в неблагоприятную сре-
ду, но количество их столь велико, что часть обязательно попадает во
влажную почву, где из них прорастает протонема, дающая начало мо-
лодому гаметофиту. Жизненный цикл спорового растения представ-
лен на рис. 211 .
У мохообразных также широко распространено вегетативное раз-
множение. У печеночников образуются выводковые тельца и выводко-
вые корзиночки, а у листостебельных мхов из структур гаметофита
могут образовываться вторичные протонемы.
Значение мохообразных в природе в основном связано с участи-
ем в почвообразовании. Сами они не испытывают потребности в пи-
тательном субстрате, поскольку не обладают корнями и необходимые
им вещества способны всасывать всей поверхностью тела. Поэтому
мохообразные одними из первых (наряду с лишайниками) заселяют
места, доселе лишенные растительности, постепенно покрывая их
сплошным ковром. Мохообразные очень редко поражаются микроор-
ганизмами, крайне неохотно поедаются насекомыми, птицами и тра-
воядными животными, и, несмотря на медленный рост (несколько
миллиметров в год), в итоге они привели к образованию огромных
залежей торфа, который образуется из-за неполного гниения посте-
пенно отмирающих нижних частей растений. Мохообразные участву-
ют в регуляции водного баланса, так как они задерживают воду и
переводят ее в состояние грунтовой. Нередко это приводит к забола-
463
ОТДЕЛ ПАПОРОТНИКООБРАЗНЫЕ
чиванию почв, что ухудшает их продуктивность. Мохообразные нака-
пливают в своем теле соли тяжелых металлов и радионуклиды.
В хозяйственной деятельности человека широко используется
торф. Россия занимает первое место в мире по залежам торфа. Чаще
всего торф используют в сельском хозяйстве в качестве органическо-
го удобрения. Кроме того, он является хорошим топливом и широко
применяется в строительной индустрии благодаря высоким теплои-
золирующим свойствам. Сфагнум обладает бактерицидными свойст-
вами и хорошо впитывает, что позволяло врачам широко применять
его во время Второй мировой войны для лечения гнойных ран. Ко-
нечно, сегодня это достояние истории медицины.
ОТДЕЛ ПАПОРОТНИКООБРАЗНЫЕ
У папоротникообразных преобладающим поколением является
спорофит, его строение значительно сложнее, чем у мохообразных. Га-
метофит, напротив, в значительной мере упростился. Спорофиты па-
поротникообразных имеют прекрасно развитую проводящую систему.
ВНИМАНИЕ!
Папоротникообразные были первыми сосудистыми растениями на Земле.
Однако проводящие элементы ксилемы у них, как правило, не со-
суды, а трахеиды. Флоэма состоит из ситовидных клеток, у которых
ситовидные поля разбросаны по всей поверхности клеточной стенки.
В подавляющем большинстве случаев в пучках между флоэмой и кси-
лемой отсутствует камбий, поэтому у них не образуются вторичные
утолщения.
В спорангиях на листьях (спорофиллах) споры развиваются из по-
верхностных клеток. Такие листья у одних форм могут одновременно
фотосинтезировать, а у других они специализируются и морфологи-
чески отличаются от трофофиллов. В последнем случае спорофиллы
обычно собираются в колоски, или стробилы, занимающие на побеге
терминальное положение. Внутри спорангия из спорогенной ткани
развиваются споры. Сам спорангий имеет стенку, которая предохра-
няет спорогенную ткань от высыхания.
В зависимости от соотношения размеров формирующихся спор
папоротникообразные подразделяют на равноспоровые и разноспоро-
вые. У равноспоровых все споры одинаковые, прорастающий из них
гаметофит однодомный. Такие споры образуются в спорангии из ма-
теринских клеток спор, которые делятся мейотически, в результате
464
ВЫСШИЕ РАСТЕНИЯ
чего образуются тетрады (четыре совершенно одинаковые споры).
У разноспоровых формируются спорангии двух типов — микроспо-
рангии и мегаспорангии. В микроспорангиях из материнских клеток
спор образуется большее количество мелких спор — микроспоры, из
которых впоследствии прорастают мужские гаметофиты с антеридия-
ми. В мегаспорангиях, также мейотически из материнских клеток
спор, образуется одна очень крупная мегаспора. Сформировавшаяся
зрелая спора защищена двумя оболочками: внутренняя — эндоспорий,
наружная — экзоспорий.
Гаметофит значительно редуцирован. Чаще всего он выглядит
нежной зеленой пластинкой или нитью (заросток). У некоторых
форм гаметофит остается в споре, не фотосинтезирует, а живет за счет
питательных веществ, накопленных в процессе формирования споры.
На нижней поверхности заростка образуются нитевидные ризоиды
и развиваются малочисленные половые органы (часто единичные) —
архегонии и антеридии. У равноспоровых на одном заростке форми-
руются половые органы обоих типов (однодомный гаметофит), у раз-
носпоровых из микроспоры прорастает мужской гаметофит, на
котором развиваются антеридии, а из мегаспоры прорастает женский
гаметофит, дающий начало архегониям. Огромное количество дву-
или многожгутиковых сперматозоидов выходят из антеридия, про-
никают в архегонии, и один из них сливается с яйцеклеткой, в ре-
зультате чего образуется диплоидная зигота. Оплодотворение у
папоротникообразных возможно лишь при наличии капельно-жидкой
воды. Поэтому большинство папоротникообразных обитают в местах
с большой влажностью.
Зигота многократно делится, в результате чего образуется много-
клеточный зародыш, состоящий из зародышевых органов (корешка,
стебелька и листочка), а также ножки, проникающей в ткань гамето-
фита. Через ножку зародыш, не способный самостоятельно синтези-
ровать органические вещества, получает их из фотосинтезирующего
гаметофита. Гаметофит большинства папоротникообразных живет не-
долго, в течение нескольких недель, при появлении на развивающем-
ся спорофите функционирующих корней и листьев он постепенно от-
мирает (исключением являются плауны, у которых гаметофит живет
до нескольких лет).
Большинство современных папоротникообразных — это травяни-
стые растения, в тропических лесах произрастают и древовидные
формы. В предыдущих геологических эпохах древовидные папорот-
никообразные составляли основу лесов. В условиях теплого и очень
влажного климата они заселяли многочисленные болота, конкурен-
ция за наиболее освещенные места способствовала возникновению
массивных растений высотой до 30 м и нескольких метров в диаме-
465
ОТДЕЛ ПАПОРОТНИКООБРАЗНЫЕ
тре. Древовидные формы найдены в отложениях девонского периода
палеозойской эры. В следующем, каменноугольном, периоде древо-
видные папоротникообразные доминировали среди живших тогда ра-
стений. Именно их остатки сформировали залежи каменного угля.
В наше время живут различные представители трех основных
классов этого отдела: папоротниковидные, клинолистовидные (члени-
стые) и плауновидные. Кроме них, в настоящее время на Земле живут
еще четыре вида из класса псилотовых, имеющих очень простую орга-
низацию.
Класс папоротниковидные
Тело папоротниковидных расчленено на корень и побег. Зароды-
шевый корешок рано отмирает, и корни папоротниковидных отраста-
ют на стебле и листьях, т. е. они являются придаточными.
Стебли папоротниковидных, за исключением древовидных форм,
развиты относительно слабо и часто почти полностью располагаются
в почве. Они могут быть ползучими или вьющимися (их называют
корневищами), или прямостоячими (у древовидных форм). У разных
форм длина стебля варьирует от нескольких миллиметров до 25 м.
Укороченные стебли некоторых папоротников напоминают клубни.
Стебель многих папоротников ветвится.
Основная часть биомассы растения — это листья. Как правило,
они очень большие (у некоторых видов длина листьев может превы-
шать 30 м). Листья большинства папоротниковидных выполняют две
функции: фотосинтетическую и репродуктивную. Однако отдельные
формы обладают специальными функциями. Часть листьев служит
для развития спорангиев. Эти листья лишены хлорофилла и не осу-
ществляют фотосинтез. Такие листья называют фертильными, или
спорофиллами. На других листьях — трофофиллах — спорангии не
образуются. Они благодаря фотосинтезу снабжают органическими
соединениями все тело растения, в том числе фертильные листья со
спорангиями.
Бо
льшая часть папоротников — равноспоровые растения, имеется
много разноспоровых (например, сальвиниевые). Споры имеют ти-
пичное строение. Покровы образуют две оболочки: внутренняя — ин-
тина и наружная — экзина. Большое количество спор рассеиваются,
но лишь небольшая часть спор попадает в благоприятные условия,
и после определенного периода покоя они прорастают.
Из споры прорастает гаметофит. У равноспоровых папоротников
он представляет собой растение с хорошо развитой ассимилирующей
тканью и способен сам снабжать себя необходимыми органическими
соединениями. Однако гаплоидная фаза папоротников развивается
466
ВЫСШИЕ РАСТЕНИЯ
только в условиях достаточного увлажнения. Гаметофит имеет вид
уплощенной зеленой пластинки, часто сердцевидной формы. Обычно
гаметофит очень маленький — всего около 0,5 см в диаметре. На обра-
щенной к субстрату поверхности гаметофита образуются многочи-
сленные нитевидные ризоиды, которые связывают его с субстратом
и способствуют поглощению воды.
Как правило, на одном гаметофите имеются антеридии и архего-
нии, т. е . гаметофит однодомный. У большинства папоротников муж-
ские органы возникают несколько раньше женских. Количество поло-
вых органов, образующихся на одном гаметофите папоротниковидных,
относительно невелико.
Оплодотворение папоротниковидных осуществляется только в ка-
пельно-жидкой воде. Подвижные сперматозоиды, снабженные много-
численными жгутиками, выходят из антеридиев и, привлекаемые
веществами, которые выделяют архегонии, подплывают к ним, прони-
кают через шейку в брюшко и сливаются с находящейся там яйце-
клеткой. В результате образуется зигота с диплоидным набором хро-
мосом. Несмотря на количество оплодотворенных яйцеклеток, лишь
одна зигота развивается и дает начало многоклеточному зародышу.
Остальные погибают и не мешают развитию молодого спорофита, ко-
торый первоначально нуждается в значительном количестве органи-
ческого материала. Вскоре у растущего зародыша образуются корни и
фотосинтезирующие органы, после чего он способен самостоятельно
жить независимом от материнского гаметофита. Сам гаметофит вско-
ре после этого отмирает. Жизненный цикл равноспорового папорот-
ника представлен на рис. 212.
Рис. 212. Схема жизненного цикла равноспорового папоротника
(по Грину и соавт., с изменениями)
467
ОТДЕЛ ПАПОРОТНИКООБРАЗНЫЕ
Папоротниковидные — это наиболее процветающая группа папо-
ротникообразных. Класс насчитывает не менее 10 000 видов, заселяю-
щих разнообразные экологические ниши. Папоротниковидные в
основном заселяют влажные места, т. к. оплодотворение происходит
в воде. Однако актиниоптерис южный является выраженным ксеро-
фитом и заселяет засушливые области африканского и азиатского
континентов. Среди папоротников есть и вторично водные формы,
например сальвиния и др. Эпифитные формы живут на стволах дере-
вьев, используя их в качестве опоры. Среди папоротников немало
лиан. В тропических лесах произрастают ныне малочисленные древо-
видные папоротники.
Класс плауновидные
Тело плауновидных расчленено на побег и корень. Корневая сис-
тема образована придаточными корнями, отходящими от подземного
корневища. Надземная часть побега травянистая, хотя среди предко-
вых форм имелись древовидные. У плауновидных дихотомическое
(равное и неравное) ветвление. Осевые цилиндры могут быть различ-
ных типов, в процессе индивидуального развития происходит смена
одного другим. Листья на стебле располагаются по спирали, в отдель-
ных случаях наблюдается супротивное или мутовчатое листорасполо-
жение. Листья плауновидных небольшие — микрофилия. Листья пла-
уновидных отличаются от листьев всех других высших растений, это
выросты стебля. Листья могут быть вегетативными — трофофиллы
или спороносными — спорофиллы. Спорофиллы образуют скопления
в виде спороносных зон, собранных в средней части стебля, или стро-
бил на концевых участках веточек.
Среди плауновидных есть как равноспоровые, так и разноспоровые
формы. Плауновые являются равноспоровыми. Одиночные спорангии
развиваются в пазухах спорофиллов, которые собраны в стробилы.
Развитие спорангия и созревание продолжается от нескольких месяцев
до нескольких лет. Зрелый спорангий раскрывается в сухую погоду.
Спорангии разноспоровых плауновидных различные. Крупные ме-
гаспорангии развиваются на мегаспорофиллах, мелкие микроспоран-
гии — на микроспорофиллах. При этом оба типа спорофиллов разви-
ваются на одном и том же растении, будучи собранными в стробилы.
Размеры спор колеблются от 20 до 100 мкм — микроспоры, от 100 до
1500 мкм — мегаспоры. Количество микроспор всегда очень велико —
до миллиона в спорангии. Количество мегаспор гораздо меньшее де-
сятки или сотни, у некоторых видов в мегаспорангии развивается
одна мегаспора. У многих разноспоровых из спорангиев выходят не
споры, а женские и мужские гаметофиты.
468
ВЫСШИЕ РАСТЕНИЯ
У равно- и разноспоровых плауновидных дальнейшее развитие
споры происходит различно. У равноспоровых возможны два вариан-
та прорастания спор: у одних спора прорастает через несколько дней
после очень короткого периода покоя, у других спора долго находится
в состоянии покоя и прорастает лишь через несколько лет. Споры по-
степенно погружаются в субстрат на несколько сантиметров. Из-за
отсутствия под землей света гаметофит лишен возможности осу-
ществлять фотосинтез, и он является сапрофитом, вступая в симбиоз
с почвенными грибами. Гаметофит растет до 15 лет. На верхней сторо-
не гаметофита развиваются половые органы — однодомные заростки.
В результате оплодотворения в капельно-жидкой воде образуется ди-
плоидная зигота. Период покоя отсутствует, и зигота начинает де-
литься. Наземная часть молодого спорофита зеленеет, в ней постепен-
но накапливается большое количество фотосинтезирующей ткани,
достаточное для самостоятельного существования.
Иначе прорастают мега- и микроспоры всех папоротникообразных.
Гаметофит разноспоровых плауновидных наиболее упрощенный. Муж-
ской гаметофит остается в микроспоре. Женский гаметофит развива-
ется внутри мегаспоры благодаря наличию в ней большого количества
питательных веществ, за счет которых гаметофит растет и развивается.
Чаще всего мегаспоры выходят из мегаспорангия, но у многих видов
гаметофит может развиваться при нахождении мегаспоры в спорангии.
У разноспоровых плауновидных развитие зародыша значительно
варьирует. Полностью сформированный зародыш состоит из стебель-
ка с верхушечной точкой роста, двух листочков, корешка и ножки,
с помощью которой зародыш получает из гаметофита все необходи-
мые вещества. Со временем корешок и листочек прорывают ткань
гаметофита, выходят наружу и, достигнув достаточного уровня раз-
вития, снабжают спорофит водой с минеральными веществами и про-
дуктами фотосинтеза. После этого заросток постепенно отмирает.
Количество современных плауновидных значительно меньше, чем
папоротниковидных. В начале мезозойской эры абсолютное боль-
шинство плауновидных, в том числе все крупные формы, вымерли, не
выдержав конкуренции с более высокоорганизованными голосемен-
ными растениями. Среди ископаемых было много форм, способных
к вторичному утолщению.
Современные плауновидные имеют небольшие размеры — от не-
скольких миллиметров до нескольких десятков сантиметров. Побег
некоторых тропических видов прямостоячий, он достигает двух ме-
тров в высоту, длина побега лиан может превышать 20 м. Подавляю-
щее большинство плауновидных обитает во влажных местах, т. к. им
нужна вода для оплодотворения. В то же время плауновидным необ-
ходимо обычно очень мало света, поэтому они занимают самый ниж-
469
ОТДЕЛ ПАПОРОТНИКООБРАЗНЫЕ
ний ярус в лесу. Они успешно живут в разнообразных климатических
условиях — от влажных тропиков до умеренных широт, в тундре или
высоко в горах. Существуют эпифиты и ксерофиты, способные вы-
держивать значительный дефицит влаги и произрастать на сильно ос-
вещенных местах.
В настоящее время на Земле насчитывается около 400 видов пла-
уновых, 700 видов селягинелловых и 64 вида полушниковых. Значе-
ние плауновидных в природе невелико. Немногие служат пищей
травоядных животных. Плауны, содержащие сильнодействующее
вещество, которое останавливает передачу нервного импульса через
синапс подобно яду кураре, вообще не поедаются. Лишь некоторые
беспозвоночные изредка используют их без вреда для себя. Плауно-
видные почти не используются человеком. Споры плаунов ранее
применялись в медицине в качестве присыпки, ими пересыпали пи-
люли. Некоторые виды полушников украшают аквариумы, а многие
тропические селягинеллы, обладающие красивой окраской, выращи-
вают в оранжереях.
Класс клинолистовидные, или членистые
Самый малочисленный класс папоротникообразных — членистые.
В него входит только один порядок — хвощовые с единственным се-
мейством (хвощовые), к которому относится единственный род —
хвощ, насчитывающий 32 вида. Представители этого класса имеют
выраженное расчленение побега на узлы и междоузлия. От узла на-
земного побега отходит мутовка веточек, которые часто принимают за
листья. Но листья хвощовых сильно редуцируются, поэтому фото-
синтезирует освещенная надземная часть стебля. Травянистый побег
вторично не утолщается.
Все современные хвощовые являются равноспоровыми растениями.
Спорангии развиваются на особых, присущих только хвощовым,
структурах — спорангиофорах. Побеги, на которых они образуются,
у большинства видов наряду со спорогенной функцией обладают и
фотосинтетической. Такие вечнозеленые и многолетние побеги име-
ют очень высокую механическую прочность. У других видов (напри-
мер, хвощ лесной, хвощ полевой и др.) зеленые однолетние побеги двух
типов — чисто вегетативные и спорогенные. На последних вначале
образуются стробилы, но после созревания спор и опадания строби-
лов, эти побеги нельзя отличить от обычных вегетативных. И нако-
нец, у третьих (например, хвощ полевой) однолетние побеги изначаль-
но четко дифференцируются морфологически и функционально. При
этом спорогенные побеги, имеющие большую толщину, лишены хло-
рофилла и имеют розовато-бурую окраску. Они не ветвятся, появля-
470
ВЫСШИЕ РАСТЕНИЯ
ются ранней весной вместе с вегетативными побегами или несколько
раньше их и отмирают в тот же год, обычно сразу же после спороно-
шения.
Стробилы хвощей образуются на верхушке главного побега. Про-
растание споры начинается с деления ее ядра. Первоначально моло-
дой гаметофит напоминает нитчатую водоросль, но со временем он
становится сложно устроенным пластинчатым с многочисленными
уплощенными зелеными выростами, направленными вверх. На ниж-
ней поверхности заростка формируются бесцветные нитевидные ри-
зоиды, снабжающие растение водой и минеральными веществами.
В зависимости от условий развития размеры заростков варьируют от
миллиметра до нескольких сантиметров. У одних растений гаметофит
двудомный, у других — однодомный. Мужские гаметофиты меньше
женских. Оплодотворение происходит в капельно-жидкой воде. Зиго-
та дает начало зародышу, который состоит из корешка, стебелька и
двух-трех листочков. Вначале зародыш окружен тканью гаметофита,
по мере развития корешок прободает ее и попадает в почву, где начи-
нает активно выполнять свои функции. Зародышевый корешок живет
недолго. Из первичного побега сначала образуется корневище, от уз-
лов которого отходят придаточные корни, а затем надземные побеги.
Самые ранние ископаемые остатки предковых форм обнаружены в
отложениях начала девонского периода палеозойской эры. Хвощовые
достигли максимального развития и разнообразия в каменноугольный
период. Тогда были широко распространены очень похожие на совре-
менные хвощи древовидные каламиты, достигавшие двадцатиметро-
вой высоты, которые наряду с другими крупными папоротникообраз-
ными образовывали огромные леса. В триасовом периоде мезозойской
эры Землю заселили более высокоорганизованные семенные растения.
Большинство споровых, в том числе хвощовые, вымерли.
Несмотря на небольшое разнообразие видов, хвощовые широко
распространены во всех географических зонах, кроме Австралии
и Новой Зеландии (хотя в настоящее время искусственным путем
они занесены и туда).
Значение хвощей в природе и хозяйственной деятельности чело-
века невелико. Травоядные животные в естественных условиях не по-
едают большинство из них. Большое количество в составе кормов до-
машних животных может привести к гибели животных, т. к. хвощи
содержат токсичные вещества. Хвощи являются сорняками, и очень
трудно очищать от них поля. Молодые спороносные побеги и клу-
беньки, содержащие много крахмала, употребляют в пищу. Некоторые
биологически активные соединения используют при изготовлении
лекарств. Жесткие стебли хвощей используют для полировки различ-
ных материалов, а также для чистки металлической посуды.
471
СЕМЕННЫЕ РАСТЕНИЯ
СЕМЕННЫЕ РАСТЕНИЯ
Главная особенность семенных растений — это их размножение се-
менами. Семя — маленькое растение, имеющее все главные вегета-
тивные органы, снабженное запасом питательных веществ и надежно
защищенное собственной оболочкой. Количество семян значительно
меньшее, чем количество спор, но бо
льшая их часть продолжает раз-
виваться, потому что, будучи защищенными оболочками, они способ-
ны переживать неблагоприятные условия часто довольно долго, а при
наступлении необходимых перемен очень быстро прорастают и раз-
виваются в самостоятельное растение. Однако абсолютное большин-
ство слабо защищенных спор не попадает в благоприятные условия и,
не имея возможности пережидать в состоянии покоя, гибнет. Семя
развивается из семяпочки после оплодотворения находящейся там яй-
цеклетки. У семенных растений осуществляется внутреннее оплодот-
ворение, это позволяет им обойтись без воды. При внутреннем опло-
дотворении не нужны подвижные мужские гаметы, так как они
транспортируются к яйцеклетке пыльцевой трубкой. Семязачаток
представляет собой функционирующий мегаспорангий — нуцеллус,
окруженный стерильными мегаспорангиями, которые, сливаясь, обра-
зуют защитный покров — интегумент. Семязачаток связан с мате-
ринским спорофитом посредством халазы, через которую он получает
необходимые вещества. Противоположный полюс имеет отверстие
в интегументе — микропиле, через него в семязачаток проникают
оплодотворяющие неподвижные спермии.
Семенные растения размножаются семенами, но они все равно
образуют споры. Как и у других высших растений, у семенных после-
довательно сменяются половая (гаметофит) и бесполая (спорофит)
фазы жизненного цикла. Спорофит всегда преобладает, а гаметофит
(особенно мужской) редуцируется и развивается внутри споры.
ВНИМАНИЕ!
Все семенные растения являются разноспоровыми.
В процессе размножения у семенных растений появился новый
важный этап — опыление — перенос различными способами проросших
микроспор (пыльцевых зерен): либо непосредственно на семязачаток
у голосеменных, либо на рыльце пестика у покрытосеменных. После
этого в пыльцевом зерне образуется пыльцевая трубка, по которой не-
подвижные спермии доставляются непосредственно к микропиле се-
мязачатка. Интересно, что если мужской гаметофит в составе пыльце-
вого зерна транспортируется из места своего возникновения
472
ВЫСШИЕ РАСТЕНИЯ
к семязачатку, то образование мегаспоры, развитие из нее женского
гаметофита, оплодотворение находящейся в нем яйцеклетки и даль-
нейшее развитие зародыша и всего семени происходит стационарно
без нарушения связи с материнским спорофитом через халазу.
Большинство исследователей считают, что семенные растения име-
ют общее происхождение от одной из ветвей разноспоровых папорот-
никообразных. Самое древнее семя было найдено Д. Петтитом и Ч. Бе-
ком в отложениях верхнего девона. Вероятно, к этому времени и
появились первые семенные растения, которые были голосеменными.
ОТДЕЛ ГОЛОСЕМЕННЫЕ
В этот отдел входят только древесные наземные формы. Побег
большинства голосеменных прямостоячий (деревья и кустарники),
он может быть стелющимся или даже лиановидным (гнетум, эфедры).
Размеры растений широко варьируют от небольших кустарничков до
гигантских деревьев, достигающих 120 м в высоту и 12 м в диаметре
(секвойядендрон гигантский). Многие голосеменные долгожители,
например секвойядендрон гигантский, достигает за более чем 3000
лет колоссальных размеров.
Спорофит расчленен на вегетативные органы — побег и корень.
Главный корень почти всегда четко выражен в течение всей жизни ра-
стения (в отличие от папоротникообразных). От главного корня отхо-
дят боковые, а у некоторых примитивных форм голосеменных могут
образовываться и придаточные корни.
Стебель имеет сложное строение. Осевой цилиндр (эустель) содер-
жит коллатеральные проводящие пучки. Ситовидные клетки флоэмы
содержат ядра, а ситовидные поля рассеяны по боковым стенкам кле-
ток. Ксилема представлена трахеидами, лишь у оболочкосеменных
в древесине имеются настоящие сосуды. Проводящие элементы кси-
лемы и флоэмы образуют коллатеральные проводящие пучки. Нали-
чие камбия делает возможным вторичное утолщение стебля.
Анатомическое строение листьев сложнее, чем папоротникообраз-
ных. У голосеменных более совершенное строение проводящих пуч-
ков и хлоренхимы. У низших голосеменных, а также у произрастаю-
щих в зонах с постоянно высокой температурой листья крупные
рассеченные. У некоторых форм они имеют верхушечную или вста-
вочную меристему, за счет которой могут достигать значительной
длины (например, листья вельвичии удивительной растут в течение
всей очень продолжительной жизни, достигая нескольких метров
в длину). У высокоорганизованных голосеменных листья имеют
меньшие размеры и цельную листовую пластинку, часто в виде чешуи
473
ОТДЕЛ ГОЛОСЕМЕННЫЕ
или хвои. Большинство голосеменных являются вечнозелеными расте-
ниями, даже те из них, которые произрастают в условиях резко конти-
нентального климата с суровыми зимами. Для этого растения вырабо-
тали ряд адаптаций, повышающих их зимостойкость. Но есть и
листопадные формы (например, сибирская лиственница), сбрасываю-
щие к наступлению холодов листву. Лиственница является самым хо-
лодоустойчивым растением).
Голосеменные, как и все семенные растения, являются разноспоро-
выми. Мегаспорофиллы и микроспорофиллы у них собраны в груп-
пы — стробилы, или шишки. Стробилы образованы либо только
мегаспорофиллами — мегастробилы, либо только микроспорофилла-
ми — микростробилы. Их еще называют женскими и мужскими стро-
билами, соответственно. В зависимости от того, какие стробилы раз-
виваются на спорофите, последние могут быть однодомными, если
имеются мегастробилы и микростробилы, или двудомными, если
образуются стробилы только одного типа. Напомним, это никогда не
отмечается у спорофитов мохообразных или папоротникообразных.
Размеры стробил разных голосеменных имеют размеры от несколь-
ких сантиметров до метра, соответственно варьирует и их масса.
Стробилы саговников одиночные, у большинства других голосемен-
ных они собраны в группы.
В микроспорангиях голосеменных развивается большое количест-
во микроспор. Микроспора, окруженная экзиной и интиной, начинает
развиваться еще в микроспорангии. При этом мужской гаметофит го-
лосеменных растений, по сравнению с папоротникообразными, зна-
чительно редуцирован. Половые органы у первых не формируются
в полной мере, а вместо полноценного антеридия появляется антери-
диальная клетка, дающая начало сперматогенной клетке, из которой
при делении образуются два неподвижных спермия растений, у низ-
ших голосеменных — два подвижных сперматозоида.
Развиваясь внутри микроспоры, гаметофит увеличивает свой
объем. Пыльцевая трубка голосеменных служит для переноса непод-
вижных спермиев к яйцеклеткам, после опыления — для доставки
проросшей микроспоры (пыльцевого зерна) на семязачаток.
Мегаспорангий голосеменных развивается внутри семязачатка и
никогда его не покидает. Семязачатки лежат голо, они ничем не по-
крыты, что и дало название всему отделу. Они всегда связаны с мате-
ринским спорофитом, от которого получают все необходимые пита-
тельные вещества. Женский гаметофит просто устроен, хотя и меньше,
чем мужской. Женский гаметофит в течение всего своего развития
находится внутри мегаспорангия, где на нем формируются весьма
слаборазвитые архегонии (обычно по два). В каждом архегонии рас-
положена одна яйцеклетка. У некоторых голосеменных на женских
474
ВЫСШИЕ РАСТЕНИЯ
гаметофитах не образуются архегонии. В результате опыления спер-
мии из попавшего на семязачаток пыльцевого зерна с мужским гаме-
тофитом попадают на мегаспорангий с помощью пыльцевой трубки
через микропиле.
Голосеменные появились в девонский период палеозойской эры,
они были первыми семенными растениями. Полагают, что именно
они послужили причиной вымирания менее организованных папо-
ротникообразных, размножение которых всецело зависело от присут-
ствия воды. Голосеменные достигли наибольшего расцвета в мезозой-
скую эру, где они доминировали в растительных сообществах.
Появление и широкое распространение значительно более прогрес-
сивных цветковых растений привело к вымиранию большинства го-
лосеменных.
Современные голосеменные насчитывают около 800 видов. Одна-
ко их биомасса чрезвычайно велика во многих растительных сообще-
ствах. Особенно это относится к хвойным, которые широко распро-
странены на Земле, и особенно в областях с умеренным климатом, где
растения имеют ландшафтное значение. К голосеменным растениям
относят давно вымершие семенные папоротники и беннеттитовые,
а также современные саговниковые, гнетовые, гинкговые и хвойные.
Класс хвойные
Хвойные — наиболее распространенные среди всех голосеменных
растений. В природных сообществах по своему значению они прочно
занимают второе место после покрытосеменных. Хвойные чрезвычай-
но широко используются в хозяйственной деятельности человека.
Подавляющее большинство хвойных являются деревьями, но имеется
и небольшое количество кустарников. Один вид — паразитаксус опа-
ленный — является паразитом. Все хвойные имеют моноподиальный
тип ветвления с хорошо выраженным верхушечным побегом, у кото-
рого в течение всей жизни растения сохраняется апикальная меристе-
ма. Именно она обеспечивает неограниченный верхушечный рост.
При механическом повреждении верхушки главного побега и утрате
апикальной меристемы рост растения в высоту приостанавливается,
но затем начинает усиленно развиваться ближайшая к верхушке бо-
ковая ветвь, которая становится главным побегом. При этом может
нарушаться правильная форма кроны.
Многие хвойные — долгожители. Секвойядендроны гигантские,
произрастающие в Северной Америке, могут жить 3000 лет, а возраст
одного из экземпляров другого североамериканского долгожителя —
сосны долговечной — оценивается в 4900 лет! Секвойядендрон дости-
475
ОТДЕЛ ГОЛОСЕМЕННЫЕ
гает высоты более 100 м при толщине ствола 12 м, за что его называют
«мамонтовым деревом».
Древесина составляет бо
льшую часть объема стебля, кора и серд-
цевина развиты относительно слабо. Вся ксилема хвойных состоит из
трахеид, сосуды отсутствуют. Проводящая способность ксилемы
хвойных очень высока, этому способствует наличие в трахеидах окай-
мленных пор с торусами. Проводящие элементы флоэмы (ситовид-
ные клетки) лишены сопровождающих клеток, а ситовидные поля
располагаются на боковых стенках. Между ксилемой и флоэмой нахо-
дится камбий, обеспечивающий вторичное утолщение стебля. В стеб-
лях большинства хвойных образуются хорошо выраженные кольца
ежегодного прироста древесины. Многократное заложение перидер-
мы и деятельность феллогена приводят со временем к образованию
на периферии стебля толстого слоя корки. Характерной особенно-
стью класса является слабое развитие паренхимы в стебле и каналы
(смоляные ходы).
Листья у хвойных всегда цельные. У подавляющего большинства
хвойных продольные размеры листа многократно превышают попе-
речные. Эти сильно вытянутые игольчатые листья называют хвоей.
Реже у хвойных встречаются чешуевидные листья, у некоторых ви-
дов, обитающих в условиях теплого климата (араукариевые, подокар-
повые) — крупные листья, имеющие широкую листовую пластинку.
За редким исключением листья многолетние. Однако некоторые
виды, например лиственница, к зиме сбрасывают хвою, вообще исклю-
чают какую-либо потерю воды через устьица в течение зимы, у других
видов опадают ветви вместе с находящимися на них листьями.
Корневая система хвойных чаще всего стержневого типа, развита
хорошо.
Отдельные особи хвойных могут быть однодомными или двудом-
ными, но у них всегда однополые стробилы. Обычно они собраны
в группы, которые часто называют шишками.
Микростробил, устроенный очень просто, представляет собой уко-
роченную ось с расположенными на ней по спирали микроспорофил-
лами, на которых образуются микроспорангии. Из находящейся вну-
три спорангия спорогенной ткани возникают материнские клетки
спор. Каждая из них в ходе мейозы, дает начало четырем микроспо-
рам. Каждая спора окружена двумя оболочками — интиной и экзи-
ной. У микроспор многих хвойных в результате расхождения интины
и экзины образуются воздушные мешки. Развитие мужского гамето-
фита начинается, еще когда микроспоры находятся внутри микро-
спорангия. Мужской гаметофит сосны развивается так. После двух
первых делений ядра микроспоры образуются две небольшие протал-
лиальные клетки и крупная антеридиальная. Проталлиальные клетки
476
ВЫСШИЕ РАСТЕНИЯ
вскоре отмирают. На одном растении образуется громадное количест-
во микроспор. Хвойные являются ветроопыляемыми растениями. На-
личие воздушных мешков способствует переносу пыльцы.
Семязачатки развиваются на мегастробилах, которые, как и ми-
кростробилы, собраны в группы. На оси шишки находятся кроющие
чешуи, в пазухах которых расположены семенные чешуи. Они явля-
ются сильно упрощенными мегастробилами, названными так потому,
что на их верхних сторонах расположены семязачатки. У многих
хвойных обе чешуи (кроющая и семенная) свободны, однако часто
они срастаются между собой. Одна из них доминирует, другая реду-
цируется. Чаще всего преимущественно развита семенная чешуя (на-
пример, у сосновых).
На верхних сторонах семенных чешуи находятся семязачатки.
Чаще всего их два, реже один или более двух. Семязачаток состоит из
двух структур: нуцеллуса (мегаспорангия) и его покрова — интегу-
мента. В нуцеллусе большинства хвойных растений образуется един-
ственная материнская клетка споры (мегаспороцит). В результате
мейоза из нее образуются четыре мегаспоры, расположенные в виде
цепочки — одна над другой. Из тетрады развивается лишь одна мега-
спора — нижняя, расположенная дистальнее других от микропиле,
а остальные три погибают. Развитие женского гаметофита начинается
с последовательной серии митотических делений ядра мегаспоры. На
верхней части женского заростка (его называют первичным эндоспер-
мом) формируются чаще всего два сильно редуцированных архегония
с крупными яйцеклетками.
Опыление сосны происходит обычно в конце весны — начале лета,
когда семенные чешуи шишки немного расходятся, открывая при
этом семязачатки. В образовавшуюся щель проникает пыльца, кото-
рая летает в воздухе. Через микропиле семязачатка выделяется опы-
лительная жидкость, к которой прилетит пыльца. Затем в течение не-
скольких минут опылительная жидкость всасывается семязачатком
и вместе с ней пыльцевые зерна перемещаются по направлению к ну-
целлусу. После этого семенные чешуи плотно смыкаются и остаются
в таком состоянии вплоть до момента созревания семян, а мужской
гаметофит продолжает свое развитие.
Антеридиальная клетка делится, в результате чего образуется не-
большая генеративная клетка и более крупная клетка трубки. Затем
лопается наружный покров микроспоры — экзина, и из нее выходит
пыльцевая трубка, окруженная интиной. Пыльцевая трубка хвойных
служит только для доставки неподвижных спермиев яйцеклетке, на-
ходящейся в брюшке архегония. За некоторое время до этого генера-
тивная клетка делится, образуя стерильную клетку-ножку, которая
незадолго до оплодотворения отмирает, и сперматогенную клетку, по-
477
ОТДЕЛ ГОЛОСЕМЕННЫЕ
сле деления которой появляются два спермина. Следуя по ходу раз-
росшейся пыльцевой трубки через шейку архегония в его брюшко,
спермины выходят из трубки, один из них оплодотворяет яйцеклетку.
Сразу же после оплодотворения в зиготе ядро несколько раз по-
следовательно делится. Сформированный зародыш хвойных состоит
из корешка, подсемядольного лена (стебелька) и семядолей, количе-
ство которых значительно варьирует от двух (у тиссовых) до 18 у не-
которых сосновых (рис. 213).
Интегумент семязачатка трансформируется в покров семени.
Обычно покровы семян хвойных твердые, они надежно защищают за-
родыш от неблагоприятных воздействий факторов окружающей сре-
ды. Кроме того, из семенной чешуи, с которой семя все еще остается
связанным, образуется тонкое крылышко, благодаря которому семя
после выпадания из шишки подхватывается ветром и зачастую пере-
носится на большое расстояние от материнского растения. Прораста-
ние семян хвойных всегда начинается после периода покоя.
I
III
II
6
7
8
5
3
2
1
4
Ðèñ. 197. Ñåìÿ ñîñíû:
I îáðàçîâàíèå ñåìåíè; II ñåìÿ â ïðîäîëüíîì ðà
III çðåëîå ñåìÿ ñ êðûëûøêîì; 1 èíòåãóìåíò; 2
äîëè; 3 òî÷êà ðîñòà çàðîäûøà; 4 ïîäñåìÿäî
êîëåíî; 5 ýíäîñïåðì; 6 êîðåøîê; 7 íóöåëëóñ; 8
ñîê (ïî Êîìàðíèöêîìó è ñîàâò., ñ èçìåíåíèÿìè è
íåíèÿìè)
Рис. 213. Семя сосны:
I — образование семени, II — семя в продольном
разрезе, III — зрелое семя с крылишком;
1 — интегумент, 2 — семядоли, 3 — точка роста
зародыша, 4 — подсемядольное колено,
5 — эндосперм, 6 — корешок, 7 — нуцеллус;
8 — подвесок (по Комарницкому и соавт.,
с изменениями и дополнениями)
478
ВЫСШИЕ РАСТЕНИЯ
Для большинства хвойных нехарактерно вегетативное размножение.
Ископаемые остатки хвойных обнаружены в отложениях камен-
ноугольного периода палеозойской эры. Их расцвет приходится на
мезозой. В юрском и меловом периодах хвойные часто доминировали
в растительных сообществах и были представлены максимальным ко-
личеством видов. Появление цветковых приостановило гегемонию
хвойных, однако они прекрасно приспособились и к нынешним усло-
виям многих географических районов. При этом хвойные часто доми-
нируют в растительных сообществах (например, в таежных лесах).
Хвойные являются самыми распространенными среди голосемен-
ных растений. Они не только часто встречаются, их виды весьма раз-
нообразны. В настоящее время на Земле произрастают около 600 ви-
дов, которые объединяют в 55 родов и 7 семейств.
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
В этот отдел входят самые высокоорганизованные растения. У них
имеются совершенно новые структуры, осуществляющие как половое,
так и бесполое размножение, — цветок. Преобладающее поколение —
автотрофный спорофит. Гаметофит редуцирован, причем степень ре-
дукции максимальна во всем растительном мире. Кроме того, гамето-
фит никогда не бывает самостоятельным, он живет за счет спорофита,
т. е . является гетеротрофным. Выделяют три основные морфологиче-
ские группы покрытосеменных: деревья, кустарники и кустарнички.
Цветковые растения, будучи более совершенными по сравнению
с другими растениями по способу размножения, быстро распростра-
нились по Земле и приспособились к самым разнообразным экологи-
ческим факторам. Одной из наиболее важных форм адаптации стало
появление травянистых растений. В зависимости от продолжительно-
сти жизни травянистые растения подразделяют на однолетние, дву-
летние и многолетние. Однолетние отмирают сразу же после разви-
тия плодов. Двулетние обычно в первый год жизни образуют только
вегетативные органы, а во второй цветут и плодоносят. Многолетние
травы живут и плодоносят несколько лет, но при этом каждый год все
надземные побеги у них отмирают, а подземные зимуют и на следую-
щий год дают начало новому поколению надземных побегов. Некото-
рые растения прошли путь обратной трансформации травянистых
форм в древесные (например, пальмы).
У покрытосеменных тело расчленено на главные вегетативные ор-
ганы — побег и корень. В зависимости от условий обитания побег
имеет разнообразное строение. Он может быть прямым, лежачим,
вьющимся, лазящим, цепляющимся и т. д. Разнообразны и метамор-
479
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
фозы побегов. Их проводящие ткани — самые совершенные среди
всех растений. Ксилема имеет сосуды, которые обеспечивают более
быстрый транспорт воды и растворенных в ней веществ от корня по
стеблю и далее в листья. Проводящие элементы флоэмы представле-
ны безъядерными ситовидными трубками. У покрытосеменных ли-
стья, покрывающие стебли, разнообразны по размерам, степени рассе-
ченности, форме листовой пластинки и т. д .
Корневая система всегда начинается с зародышевого корешка се-
мени и возникающими от него боковыми корнями. Существует два
типа корневой системы цветковых: стержневая и мочковатая.
Цветок
Цветок — это укороченный видоизмененный побег, состоящий из
нескольких частей (рис. 214).
Цветок занимает апикальное положение, но при этом он может
располагаться на верхушке главного и бокового побега. Цветонож-
ка — относительно удлиненное междоузлие — связывает цветок с
остальным растением. У многих видов цветовых она отсутствует или
сильно укорочена, это сидячие цветки. Расширенная дистальная часть
цветоножки называется цветоложем, обычно оно уплощенное, но
иногда может быть вогнутым или, наоборот, выпуклым. Цветоложе
является сильно укороченной осью цветка, и в пазухах очень ее ко-
ротких междоузлий располагаются все органы цветка.
Околоцветник образован чашечкой и венчиком. У подавляющего
большинства растений они присутствуют в цветке одновременно, та-
кой околоцветник называют двойным, чаще всего имеется только ча-
шечка или только венчик — такие цветки простые. Цветок неболь-
4
5
6
3
2
1
Рис. 214. Схема строения
цветка:
1 — цветоножка,
2 — цветоложе,
3 — чашелистик,
4 — лепесток,
5 — тычинка,
6 — пестик
(по Хржановскому и соавт.)
480
ВЫСШИЕ РАСТЕНИЯ
шого числа видов лишен околоцветника и поэтому называется
беспокровным, или голым.
Чашечка образована различным количеством чашелистиков, кото-
рые происходят из обычных вегетативных листьев. Они часто зеле-
ные и фотосинтезируют. Основной функцией чашелистиков является
защита развивающихся частей цветка до его распускания. При отсут-
ствии венчика чашелистики бывают лепестковидными, они ярко
окрашены (например, у некоторых лютиковых). Чашелистики могут
быть обособлены друг от друга или срастаться между собой.
Венчик образован различным количеством лепестков. Их происхо-
ждение, как правило, связано с вегетативными листьями, но у боль-
шинства видов они представляют собой уплощенные и разросшиеся
стерильные тычинки. Вблизи основания лепестков иногда образуют-
ся дополнительные структуры, которые в совокупности называют
привенчиком. Лепестки венчика, как и чашелистики, могут срастаться
между собой краями (сростнолепестный венчик) или оставаться сво-
бодными (свободнолепестный венчик). Нередко сростнолистная ча-
шечка соседствует со свободнолепестным венчиком, у других свобод-
ные чашелистики сочетаются со сросшимися лепестками венчика.
Венчик развит у цветков, опыляемых насекомыми. У ветроопыля-
емых покрытосеменных венчик развит относительно слабо или реду-
цирован.
Тычинки (андроцей), являющиеся микроспорофиллами, состоят
из тычиночной нити и пыльника, образованного микроспорангиями.
Отдельные микроспорангии называются пыльцевыми гнездами. Сово-
купность тычинок одного цветка называют андроцеем. Количество
тычинок в одном цветке у разных покрытосеменных широко варьиру-
ет от одной до нескольких сотен. Часто тычинки, расположенные в
одном цветке, имеют различное строение по форме или длине тычи-
ночных нитей. Кроме того, все тычинки или несколько из них могут
срастаться между собой — это однобратственный андроцей (если сра-
стаются все тычинки); двубратственный, если одна из тычинок не
срослась с остальными; многобратственный, если тычинки срастают-
ся, образуя несколько групп. Разнообразие тычинок связано с особен-
ностями опыления цветков.
Плодолистики (гинецей) являются мегаспорофиллами. У покры-
тосеменных растений пыльцевые зерна попадают не на семязачаток,
как это происходит у голосеменных, а на рыльцевую область. При
этом семязачатки скрыты в полости плодолистика и надежно защи-
щены его стенками. В типичных случаях плодолистик состоит из
трех частей: завязи, столбика и рыльца. Если завязь не срастается
с окружающими стерильными частями цветка, а возвышается над
цветоложем, ее называют верхней (бобовые, пасленовые и др.). Если
481
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
завязь полностью обрастает элементами околоцветника или цветоло-
жем, она называется нижней (например, тыквенные). Промежуточ-
ные варианты называют полунижней завязью (например, земляника).
Нижняя завязь считается более прогрессивной, поскольку в ней семя-
зачатки, кроме стенок завязи, покрыты дополнительным защитным
покровом. Иногда плодолистики называют карпеллами, имея в виду,
что из них впоследствии разовьется плод. Другое название — пестик,
но его чаще применяют для характеристики всех плодолистиков од-
ного цветка. Однако правильнее называть всю совокупность плодоли-
стиков, находящихся в одном цветке, гинецеем. В зависимости от того,
насколько тесно связаны плодолистики между собой, различные типы
гинецеев подразделяют на две большие группы: апокарпный (все пло-
долистики свободны) и ценокарпный, если боковые стенки соседних
плодолистиков срастаются между собой.
Нектарники — особые железки, продуцирующие жидкие секреты,
богатые растворимыми сахарами, — нектар, привлекающий питаю-
щихся им насекомых опылителей, поэтому нектарники особенно хо-
рошо развиты у энтомофильных (опыляемых насекомыми) растений.
Нектарники могут быть вполне самостоятельными структурами, но
нередко они являются видоизменяющий частью цветка. Нектарники
лучше всего развиты у двудольных.
Большинство цветков имеют и тычинки, и плодолистики, поэтому
их называют обоеполыми. Но существует множество видов, цветки
которых содержат только тычинки или только плодолистики. Первые
принято называть мужскими, вторые — женскими. У некоторых по-
крытосеменных на одном растении одновременно или последователь-
но развиваются мужские и женские цветки. Такие формы называют
однодомными (например, огурец). У других одна особь имеет цветки
только одного пола, и в соответствии с этим отдельные экземпляры
подразделяют на мужские и женские, а вид в целом называют двудом-
ным. При этом мужские экземпляры значительно отличаются от жен-
ских не только по строению цветка, но и по ряду других признаков
(например, конопля, тополь, ива и другие).
Вокруг продольной оси большинства цветков можно провести не-
сколько плоскостей, каждая из которых разделит этот цветок на рав-
ные половины с одинаковым набором одинаковых органов. Такие
цветки называют актиноморфными, или правильными (подобный тип
симметрии в животном мире называется радиально-симметричным).
Однако у многих покрытосеменных относительно продольной оси
возможно провести лишь одну плоскость, делящую цветок на две рав-
ные половины. Такой цветок называют зигоморфным, или неправиль-
ным (у животных подобный тип именуется билатерально-симметрич-
482
ВЫСШИЕ РАСТЕНИЯ
ным). Наконец, можно встретить цветки, которые невозможно
разделить даже на две равные половины. Это асимметричные цветки.
Формула и диаграмма цветка. Формула цветка представляет со-
бой краткую запись, в которой в зашифрованной форме обозначены
все части цветка, а также указаны их численность и особенности. Вна-
чале указывают пол цветка. Если в нем имеются только тычинки, но
нет плодолистиков, то он обозначается как мужской — (стрела Мар-
са), или тычиночный, если, наоборот, имеются только плодолистики, —
женский — (зеркало Венеры), или пестичный. Обоеполые (гермафро-
дитные) цветки, имеющие как тычинки, так и плодолистики, либо не
указываются вообще, либо обозначаются совмещенным значком (вста-
вить символ совмещенного пола!). Затем обозначается тип симметрии
цветка. Если он имеет радиальную симметрию, то он обозначается зна-
ком S или знаком плюс, заключенным в кружочек — . Такой цветок
в ботанике называется актиноморфным, или правильным, как, напри-
мер, у розы или тюльпана. Билатерально-симметричный цветок обо-
значается вертикальной или косой стрелкой ␘ или вертикальной ли-
нией с точками по обеим сторонам —
∙
|.
. Билатерально-симметричный
цветок называется зигоморфным, например у гороха. Наконец, у неко-
торых растений цветки асимметричные, т. е. вокруг продольной оси
у них вообще нельзя провести ни одной плоскости, которая делила бы
цветок на две равные половины. Асимметричный цветок обозначается
зигзагообразной стрелкой —
.
Затем в формуле указываются отдельные части (члены) цветка.
Для обозначения каждого такого члена используются буквенные сим-
волы, соответствующие их латинскому названию. Количество одно-
именных членов выражается цифрами, если их не более 12, а если
количество превышает 12 или непостоянное, то знаком бесконеч-
ность — f. Если члены цветка срастаются между собой, то цифру, обо-
значающую их численность, помещают в скобки, например (5). Если
срастаются не все одноименные члены, то сросшиеся помещаются в
скобки, а свободные — вне: например (3) + 2 указывает, что три члена
сросшиеся, два несросшиеся. Встречаются цветки, у которых одно-
именные члены, например тычинки, имеют разную длину. В этом слу-
чае более длинные части выделяются размером обозначающих их
цифр: например, запись 2+3 говорит, что два члена имеют б
ольшую
длину, чем три другие.
Чашечка обозначается Са (Calyx), венчик — Со (Corolla), простой
околоцветник — Р (Perigonium), андроцей — A (Androceum), гине-
цей — G (Gynoeceum). Если одноименные члены цветка располагают-
ся несколькими кругами, то последовательно записывают количество
их в каждом круге, соединяя знаком +, например Р3+3
. Кроме того, ха-
рактеризуя завязь, необходимо показать, является ли она верхней или
483
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
нижней. Для этого над (если завязь нижняя) или под (если завязь
верхняя) цифрой, показывающей количество плодолистиков в гине-
цее, ставится черточка. В качестве примера приведем формулу цветка
яснотки белой — Ca(5), Co(2+3), А2+2
, G(2). Это говорит о том, что цветок
у яснотки зигоморфный, имеет сложный околоцветник, состоящий из
пяти сросшихся чашелистиков и такого же количества лепестков вен-
чика, расположенных в виде двух кругов, причем лепестки также
сросшиеся. Андроцей состоит из двух и более тычинок, причем две из
них выделяются большей длиной. Гинецей образован двумя сросши-
мися плодолистиками, завязь — верхняя. Несколько иной будет фор-
мула цветка лютика едкого: *Са5 Со5 А f G f, т. е . цветок обоеполый,
актиноморфный со сложным околоцветником, состоящим из пяти
свободных чашелистиков и такого же количества свободных лепест-
ков. Тычинок и плодолистиков много, завязь верхняя.
Для схематического обозначения цветка используют диаграмму,
которая представляет собой проекцию поперечного разреза цветка.
Диаграмма более информативна, чем формула, потому что она более
четко и зримо показывает взаимное расположение в цветке всех его
членов. Ось соцветия в диаграмме обозначают маленьким кружком
сверху, а кроющий лист — серповидной дугой с килем внизу. У верху-
шечных цветков ось не обозначают. Так же, как и кроющий лист, обо-
значают прицветники и чашелистики, а лепестки венчика — серпо-
видными дугами, но без киля. Символом тычинок в диаграмме
является фигура, напоминающая пыльник, а плодолистика — завязь,
кроме того, в завязи могут быть обозначены семязачатки. Сросшиеся
члены в диаграмме объединяются общими линиями (см. рис. 214).
Фотопериодизм — это реакция растений на длину светового дня,
т. е . фотопериода. В зависимости от внутренней реакции на продол-
жительность фотопериода организмы делят на длиннодневные и ко-
роткодневные. Длиннодневные растения (растения длинного дня) за-
цветают только при длинном световом дне (все растения, цветущие
летом, — рожь, пшеница, ячмень, укроп и др.) . Короткодневные ра-
стения (растения короткого дня) зацветают при коротком времени
суточного освещения. это все осенние цветы (хризантемы, рис, коно-
пля, соя и др.). Существуют также растения, для которых необходимо
чередование различных фотопериодов. Это длиннокороткодневные
и короткодлиннодневные растения. Наконец, есть растения, для кото-
рых продолжительность суточного освещения вообще не играет ника-
кой роли, они прекрасно цветут в любое время года (например, гра-
нат, на котором цветки образуются и весной, и летом, и осенью). Такие
растения являются нейтральными: по отношению к длине дня у них
переход к зацветанию не связан со световыми сигналами, они стиму-
лируются только внутренними факторами.
484
ВЫСШИЕ РАСТЕНИЯ
Фотопериодизм представляет собой адаптацию, которая позволя-
ет растению приспособиться к абиотическим факторам конкретной
географо-климатической области. Организм может переждать небла-
гоприятные условия, а при наступлении благоприятных — зацвести.
У разных видов цветение наступает в определенном возрасте и
в определенное время года. Однолетние зацветают в первый (и един-
ственный) год жизни, двулетние — на второй. У деревьев первое цве-
тение наступает позже. Оказывают влияние условия окружающей
среды: в южных широтах — раньше, чем в северных; произрастающие
на открытом месте — раньше, чем растущие в лесу, в теплом климате
цветение начинается раньше, чем в холодном. Продолжительность
цветения разных растений также неодинакова. Цветки некоторых ра-
стений распускаются и закрываются по нескольку раз, в зависимости
от погоды, времени суток и др. Способ опыления влияет на время рас-
пускания цветка. Например, растения, опыляемые ночными насеко-
мыми, распускаются с наступлением темноты. Многие растения, про-
израстающие в относительно стабильных условиях, закладывают
цветки и цветут в течение всего года. Такие растения — ремонтант-
ные (например, кофе, цитрусовые и др.) — одновременно закладыва-
ют и распускают цветки и развивают плоды.
Микроспорогенез и мужской гаметофит. В микроспорангии мате-
ринские клетки микроспор делятся мейозом, из каждой возникают четы-
ре гаплоидные микроспоры, покрытые двуслойной оболочкой (внутрен-
ней интиной и наружной экзиной). Гаметофиты цветковых максимально
редуцированы, особенно мужской гаметофит, который состоит из мини-
мального количества клеток, способных обеспечить половой процесс.
Цикл его формирования происходит в течение двух митозов; первый
всегда в микроспоре, когда она еще находится в пыльцевом гнезде ми-
кроспорангия. В результате митоза I образуются две клетки разных раз-
меров: маленькая генеративная клетка занимает пристенное положение;
более крупная — вегетативная. В дальнейшем генеративная клетка по-
степенно вдавливается в вегетативную и постепенно отшнуровывается
от ее поверхности, оказываясь внутри нее. Все это время оболочка ми-
кроспоры остается интактной, она называется не микроспорой, а пыль-
цевым зерном. Второй митоз осуществляет только генеративная клетка,
которая делится внутри вегетативной, из нее образуются два неподвиж-
ных спермия. Вскрываются микроспорангии в сухую погоду.
Семязачаток и мегаспорогенез. В отличие от голосеменных, се-
мязачаток цветковых растений окружен двумя интегументами. По-
кровы защищает находящийся под ними единственный фертильный
мегаспорангий (нуцеллус). Интегументы не полностью обрастают ну-
целлусом, неприкрытой остается его верхушка с отверстием — микро-
пиле (пыльцевход), через которое в последующем проникнет пыльце-
485
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
вая трубка, несущая неподвижные спермин. Hoжка семязачатка,
связывающая его с плодолистиком, называется фуникулюсом, а пере-
ход фуникулюса в семязачаток — халазой. Все указанные структуры
(один или два интегумента с микропиле, нуцеллус, халаза и фунику-
люс) составляют семязачаток. В мегаспорангии семязачатка форми-
руется единственная материнская клетка мегаспор, после ее мейоти-
ческого деления образуются четыре гаплоидные мегаспоры.
Развивается лишь одна из них, а остальные разрушаются.
Рис. 215. Мегаспорогенез и развитие женского гаметофита — зародышевого мешка:
А — развитие семязачатка, возникновение единственной археспориальной клетки
(материнской клетки мегаспор): 1 — археспориальная клетка в семязачатке,
Б — археспориальная клетка, В — редукционное деление археспориальной клетки
на две, Г — деление на четыре клетки (четыре мегаспоры), Д — дегенерация трех
мегаспор (они сплющиваются) и усиленное развитие одной мегаспоры, Е — начало
прорастания мегаспоры — ее ядро делится на два, Ж, З — двукратное деление двух
ядер с последующим образованием восьми ядер (на каждом полюсе по четыре ядра),
И — развившийся зародышевый мешок (женский заросток, или женский гаметофит),
на верхнем полюсе три обособленные клетки составляют лицевой аппарат: в центре
крупная яйцеклетка (3), рядом расположены две синергиды (2). Обособленные
у основания зародышевого мешка три клетки (5) являются антиподами, в центре
зародышевого мешка — два неслившихся полярных ядра (4)
(по Баранову, с изменениями и дополнениями)
486
ВЫСШИЕ РАСТЕНИЯ
Женский гаметофит (зародышевый мешок). Развитие женского
гаметофита цветковых растений начинается с усиленного роста объе-
ма оставшейся в нуцеллусе мегаспоры (рис. 215). Ее ядро делится
митотически, образуя сначала два ядра (первое деление), которые
расходятся к полюсам мегаспоры: одно ядро — к микропилярному по-
люсу, другое — к халазному). Затем эти ядра синхронно делятся (вто-
рое деление). Образовавшиеся в результате этого четыре ядра син-
хронно делятся еще раз (третье и последнее деление), образуя в
конечном итоге восемь гаплоидных ядер. В результате у каждого из
полюсов оказывается по четыре ядра.
В дальнейшем от каждой из двух полярных четверки ядер к центру
отходит по одному ядру, которые сливаются, образуя вторичное (цент-
ральное) ядро зародышевого мешка. Затем цитоплазма обособляется во-
круг ядер гаметофита, который из ядерной стадии развития переходит
в клеточную. Три ядра, оставшиеся вблизи халазного полюса, преобра-
зуются в три клетки (антиподы), которые питают гаметофит. Три ядра
вблизи микропиле отделяются клеточными перегородками, образуя от-
дельные клетки: крупную центральную яйцеклетку и две боковые клет-
ки синергиды. Вся цитоплазма, расположенная между антиподами с од-
ной стороны и клетками яйцевого комплекса с другой (яйцеклетка
и две синергиды), называется центральной клеткой. В ней находится
диплоидное вторичное (центральное) ядро.
На этом этапе женский гаметофит уже полностью сформирован
и состоит из шести гаплоидных клеток (одной яйцеклетки, двух си-
нергид и трех антипод) и одной диплоидной (центральной). Его стро-
ение внешне напоминает мешочек, поэтому женский гаметофит по-
крытосеменных называется зародышевым мешком.
Женский гаметофит цветковых имеет самое простое строение сре-
ди всех растений — для его полного формирования необходимо всего
восемь (!) клеток, которые образуются в результате трех делений ядра
мегаспоры. При этом половые органы (архегонии) у них отсутствуют.
По мере усложнения общей организации систематической группы про-
исходит уменьшение гаметофита и усиление спорофита.
Опыление — это перенос пыльцевых зерен на рыльце пестика,
у голосеменных пыльцевые зерна при опылении попадают непосред-
ственно на семязачаток. Пыльца переносится на рыльце пестика пу-
тем самоопыления или перекрестного опыления. При самоопылении
исключен обмен генетической информацией, поскольку пыльцевые
зерна попадают на рыльце пестика либо из одной из тычинок этого же
цветка, либо с другого цветка, расположенного на том же растении.
Это приводит к появлению чистых линий гомозиготных популяций
в пределах одного вида, неспособных обмениваться мутировавшими
генами, поэтому процессы видообразования в этих популяциях идут
487
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
самостоятельно. Постоянное самоопыление возникает в тех случаях,
когда невозможен перенос пыльцы с одного растения на другое. Слу-
чайное самоопыление происходит, когда цветки обмениваются между
собой пыльцой, но возможно попадание на рыльце пыльцевых зерен,
образованных в собственном цветке. У покрытосеменных в обычных
условиях рыльце опыляется чужой пыльцой, но если по каким-либо
причинам этого долго не происходит, в последний момент (когда пло-
долистик еще сохраняет способность воспринимать пыльцевые зер-
на) на рыльце попадает собственная пыльца. Тогда самоопыление яв-
ляется вынужденным событием, но лучше произвести самоопыление
и сформировать семена, чем не опылиться вовсе и остаться без семян,
т. е . без потомства. Опыление пыльцой другого цветка, расположен-
ного на том же растении также является самоопылением. Это способ
единственно возможный у форм с однополыми цветками, поскольку
у них тычинки и плодолистики находятся в разных цветках.
Перекрестное опыление — перенос пыльцы одного растения на
рыльце другого. Этот тип опыления встречается более часто, чем са-
моопыление между разными особями одного вида происходит обмен
аллелями, что приводит к увеличению доли гетерозиготных организ-
мов. Безусловно, перекрестное самоопыление имеет большие преиму-
щества по сравнению с самоопылением, поскольку возникшие мута-
ции свободно распространяются в пределах популяции. У двудомных
мужские и женские цветки находятся на разных растениях (мужских
и женских). К двудомным относятся тополь, ива, осина, конопля и др.
При самонесовместимости полноценное развитие пыльцевых зерен,
попавших на рыльце собственного цветка, становится невозможным.
Способы опыления у перекрестноопыляемых цветковых растений
весьма разнообразны. Их можно разделить на две группы. Первая: пе-
ренос пыльцы осуществляется главным образом насекомыми, а также
некоторыми позвоночными (птицы и летучие мыши). Соответствен-
но, различают энтомофилию, орнитофилию и зоофилию. Вторая груп-
па: пыльца переносится абиотическими факторами — ветром и реже
водой, в связи с чем различают анемофилию и гидрофилию. Растения,
опыляемые животными, обычно имеют ярко-окрашенные крупные
цветки. Мелкие цветки, как правило, собраны в соцветия, что зри-
тельно их увеличивает. Для привлечения опылителей служит нектар
или большое количество пыльцы, которую опылители охотно поеда-
ют.
Оплодотворение. После попадания пыльцы на рыльце начинается
прорастание пыльцевого зерна. Обычно это происходит, когда семяза-
чатки уже полностью сформированы и готовы к оплодотворению. Эк-
зина прорывается в области борозды или поры прорастания, и начи-
нает формироваться пыльцевая трубка, которая растет, проникает
488
ВЫСШИЕ РАСТЕНИЯ
в рыльце через столбик движется по направлению к завязи, где нахо-
дится семязачаток. Трубка растет на кончике. В растущую пыльцевую
трубку из пыльцевого зерна перемещается ядро клетки-трубки, а так-
же спермин (рис. 216).
Обычно развитие пыльцевой трубки происходит при уже сформи-
рованных семязачатках в завязи. Достигнув завязи, пыльцевая трубка
через микропиле проникает внутрь одного из находящихся там семя-
зачатков. Там пыльцевая трубка направляется к яйцевому аппарату,
проникает в одну из синергид и освобождает спермин. Один из спер-
миев сливается с яйцеклеткой, в результате чего образуется зигота.
Другой спермий сливается с полярными ядрами центральной клетки,
образуя триплоидное ядро. Зигота дает начало зародышу, а из трипло-
идной центральной клетки образуется ткань эндосперма. Механизм
оплодотворения цветковых, открытый отечественным ученым
С.Г. Навашиным (1898), называется двойным оплодотворением.
Развитие семени. Оплодотворенный семязачаток развивается в
семя, из зиготы возникает зародыш, из триплоидной центральной
клетки — эндосперм, из интегументов — семенная кожура, из стенок
завязи цветка — стенки плода.
4
5
6
7
8
9
10
11
12
13
14
3
2
1
Рис. 216. Схема двойного
оплодотворения:
1 — рыльце, 2 — столбик,
3 — завязь, 4 — семяпочка,
5 — зародышевый мешок,
6 — яйцевой аппарат,
7 — антисоды, 8 — два полярных
ядра, 9 — прорастающее
на рыльце пыльцевое зерно,
10 — пыльцевая трубка —
на кончике виднеются два
спермия, 11 — пыльник,
12 — тычиночная нить,
13 — венчик, 14 — чашечка
(по Тутаюк)
489
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
Развитие эндосперма. Эндосперм всех покрытосеменных образу-
ется из триплоидной центральной клетки. Эндосперм цветковых пол-
ностью отличается от первичного эндосперма голосеменных. У них
первичный эндосперм представляет собой гаплоидную вегетативную
ткань женского гаметофита, где накапливаются питательные вещест-
ва семени. Эндосперм у покрытосеменных происходит из триплоид-
ной центральной клетки, в образовании которой участвуют вторичное
ядро женского гаметофита и ядро спермия. У одних цветковых (зла-
ки, ситниковые, магнолиевые и др.) эндосперм сильно разрастается
и занимает большую часть семени, оттесняя на периферию маленький
зародыш (рис. 217). У других (тыквенные, бобовые, сложноцветные
и др.) весь эндосперм поглощается зародышем и используется его
семядолями, которые становятся самыми большими структурами
зрелого семени.
Семя
Зрелые семена по своим размерам, форме и окраске различных
цветковых растений, например семена кокосовой пальмы (кокосовые
орехи), очень велики, семена орхидей исчисляются микрометрами.
Семя снаружи покрыто прочной семенной кожурой, которая отграни-
чивает зародыш и запасные вещества от окружающей среды и защи-
щает его от внешних воздействий. В зрелом семени находится заро-
дыш со всеми зачатками вегетативных органов: зародышевый корешок,
семядоли, подсемядольное колено — гипокотиль (участок зародыша от
3
5
4
1
2
À
Á
Рис. 217. Схема строения семени с эндоспермом (у кукурузы)
и семени без эндосперма (у фасоли):
А — семя фасоли обыкновенной, Б — семя кукурузы; 1 — кожура семени,
2 — эндосперм с наружным алейроновым слоем, расположенным под кожурой,
3 — зародышевая почечка, 4 — зародышевый корень, 5 — семядоли
(у кукурузы щиток — одна семядоля) (по Тутаюк)
490
ВЫСШИЕ РАСТЕНИЯ
семядолей до корешка), зародышевый побег — зародышевая почечка.
Строение зрелого семени зависит от числа семядолей и наличия эндо-
сперма. Эндосперм хорошо развит в семенах злаков, у которых он за-
нимает значительную часть.
Зрелое сухое семя какое-то время находится в состоянии покоя,
в течение которого оно не прорастает. У разных семян сохраняется спо-
собность прорастать в течение различного времени. Рекордсмены — се-
мена люпина, которые сохранили жизнеспособность, будучи извлечен-
ными из ледника Аляски, где они пролежали около 10 000 лет!
Плод
Развитие плода. После оплодотворения развивается плод, кото-
рый имеется только у цветковых растений. После опыления сущест-
венно меняется направление транспортных потоков, и питательные
вещества следуют к развивающимся плодам. У травянистых растений
(особенно у однолетних) практически все синтезируемые органиче-
ские вещества используются развивающимися семенами и плодами,
что ведет к истощению других тканей растений. Созревание плода на-
чинается с прекращения его роста, разложения хлорофилла и дубиль-
ных веществ, накопления в вакуолях пигментов, определяющих ха-
рактерную для данного вида окраску плодов, разрушение пектинов,
содержавшихся в оболочках клеток, приводит к смягчению стенок
плода. В сочных плодах уменьшается содержание кислот и увеличи-
вается содержание сахаров. Поэтому спелые съедобные плоды имеют
более приятный вкус, чем незрелые. Созревший плод начинает отми-
рать: в плод обычно не поступают новые вещества, не делятся и не
растут его клетки, постепенно ткани плода разрушаются и сгнивают.
У большинства цветковых растений созревший плод опадает, он от-
мирает уже на грунте.
Строение плода. В образовании плода участвуют различные части
цветка, в первую очередь завязь. Стенка плода всегда состоит из трех
слоев: наружного — экзокарпия (эпикарпия), среднего — мезокарпия
и внутреннего — эндокарпия. Их совокупность называется перикарпи-
ем (околоплодником). У разных растений строение стенок плода имеет
свои отличительные особенности. Производительность семян и пло-
дов разных цветковых неодинакова. Относительное соотношение
массы и количества семян к массе родительского растения большее
у травянистых растений. Если семена мелкие, то на одном растении
их может образовываться очень много (например, у лебеды до 500 000
семян на одном растении). Еще больше семян продуцируют некото-
рые деревья (например, тополь и ива образуют свыше миллиона се-
мян на одном плодоносящем дереве), однако соотношение общей мас-
491
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
сы семян и дерева будет намного меньше. Масса семян и плодов
широко варьирует у разных растений. Например, зерновка пшеницы
весит меньше одного грамма, а масса одного плода тыквы может до-
стигать сотен килограммов.
Классификация плодов. Существует большое количество вариан-
тов классификации плодов, каждый из которых основывается на ка-
ком-то одном признаке (морфологическом, эволюционном и др.).
В зависимости от особенностей строения (морфологическая класси-
фикация) плоды подразделяют на простые, сборные и соплодия.
Простые плоды могут быть сухими и сочными, вскрывающимися
и невскрывающимися. К ним относятся листовка, боб, стручок, стру-
чочек и коробочка. Листовка вскрывается и становится похожей на
лист, отсюда и название. Если плод включает в себя не одну, а не-
сколько листовок, он называется многолистовкой (например, у маг-
нолии, калужницы и др.). Листовка может быть не только сухой, но
и сочной (например, многолистовка лимонника или однолистовка де-
генерии). Боб — сухой одно- или многосеменной плод, одновременно
вскрывается по брюшной и спинной сторонам, благодаря чему его
створки, раскрываясь, закручиваются, разбрасывая при этом семена
в разные стороны, тем самым способствуя их расселению. Такими
плодами обладают представители семейства бобовых. Стручок — су-
хой, многосеменной плод, его семена расположены двумя рядами по
обе стороны от тонкой ложной перегородки. Если длина стручка пре-
вышает его ширину не более чем в два раза, его называют стручочком.
Стручки и стручочки имеются у крестоцветных. Коробочка — сухой
плод, образованный в результате полного сращения нескольких пло-
диков.
К сухим невскрывающимся односеменным плодам относят орехо-
видные плоды: орех, орешек, желудь, семянку, крылатку и зерновку.
Орех — плод с жестким деревянистым околоплодником. Такие же
плоды имеет и лещина. Мелкие орехи называют орешками, они фор-
мируются, например, у гречихи. Плод, состоящий из многих орешков,
называется многоорешком, или сборным орешком (например, у люти-
ка, лапчатки, гравилата, липы и др.), такие плоды относят к сборным
плодам. Желудь отличается от ореха менее жестким околоплодником,
который у своего основания окружен чашевидной плюской, образо-
вавшейся из защитного покрова цветка. Семянка имеет кожистый
околоплодник, более мягкий, чем у ореха или желудя. Такие плоды
характерны для представителей семейства сложноцветных (напри-
мер, подсолнечника). Крылатка представляет собой своеобразный
тип плода, околоплодник которого по степени жесткости соответству-
ет семянке, но, в отличие от нее, у крылатки по краям околоплодника
имеется тонкий крыловидный кожистый или перепончатый вырост
492
ВЫСШИЕ РАСТЕНИЯ
(например, ильм; крылатка вяза становится дробной). Зерновка имеет
околоплодник, настолько тесно прилегающий к семенной кожуре, что
срастается с ней. Злаковые (пшеница, рожь, ячмень и др.) имеют зер-
новку.
Сочные плоды — это ягода, тыквина, яблоко, земляничина, ко-
стянка. Костянка и костянковидные плоды имеют околоплодник,
дифференцированный на тонкий экзокарпий, толстый мясистый
и сочный мезокарпий и многослойный деревянистый эндокарпий,
образующий косточку. Наиболее распространены однокосточковые
костянки (персик, абрикос, вишня, слива и др.), но существуют и мно-
гокосточковые костянки, которые называют сборными костянками
(например, ежевика, малина и др.), их относят к сборным плодам.
Встречаются костянки не только сочные, но и сухие (например, пло-
ды миндаля, грецкого ореха и кокосовой пальмы являются сухими
костянками).
Ягода и ягодообразные плоды, в отличие от костянки, не имеют
косточки. Индивидуальные особенности строения ягод различных ра-
стений (виноград, смородина, баклажан, крыжовник и др.) весьма
разнообразны. Тыквина — одна из разновидностей ягоды, она харак-
терна для тыквенных (огурец, тыква, дыня, арбуз и др.).
Яблоко — разновидность плода, в образовании которого, кроме за-
вязи, участвуют другие части цветка: разросшееся цветоложе, основа-
ния чашелистиков, лепестков, нижние части тычинок. Яблоко харак-
терно для яблони, груши, айвы, боярышника, рябины и др.
Земляничина — разросшееся сочное цветоложе, на котором располага-
ются простые плодики — орешки.
Сборные плоды. К сборным плодам относится сборная костянка
ежевики и малины, многоорешек или сборный орешек лютика, лап-
чатки, гравилата, липы и др., плод лотоса, который представляет со-
бой мясистое цветоложе, в которое погружены орешки; земляничина
с многочисленными орешками.
Соплодия. В отличие от плодов, соплодия образуются из несколь-
ких цветков (например, у свеклы) или соцветий (ананас, инжир и др.),
нередко соплодие включает в себя и ось соцветия.
Распространение семян и плодов. Для растения важно, чтобы со-
зревшие семена оказались на большем или меньшем расстоянии от
родительского растения и там проросли. Это важно по многим причи-
нам. В первую очередь, этот фактор ограничивает или исключает вну-
тривидовую конкуренцию, которая обычно более жесткая, чем меж-
видовая, поскольку особи одного вида нуждаются в одинаковых
условиях произрастания, на ограниченной территории они мешают
друг другу. Кроме того, это облегчает перекрестное опыление. Нако-
нец, именно перенос семян обеспечивает расселение растения, что
493
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
приводит к освоению ими новых территорий, а также обогащает видо-
вое разнообразие растительных сообществ.
Могут переноситься отдельные семена, плоды, соплодия, отдель-
ные части растения и даже целые растения (например, перекати-по-
ле). Переносимые структуры называют диаспорами. Относительно
немногие растения распространяют свои семена самостоятельно,
у большинства это осуществляют животные, человек или физические
факторы (ветер или вода). Из-за специализированного приспособле-
ния к конкретному типу переноса возникло такое разнообразие мор-
фологических особенностей различных плодов.
Роль человека — распространителя растений проявляется разноо-
бразно. Во-первых, человек может распространять семена и плоды
всеми способами, которые были описаны выше для зоохории (поедая
съедобные диаспоры, перенося их на своей одежде и т. д .) . Во-вторых,
человек распространяет растения во время путешествий, пересылая
почту и товары, производя передислокацию войск и т. д . В-третьих,
человек часто осознанно распространяет семена в соответствии со
своими хозяйственными нуждами или для внедрения новых растений
в уже имеющиеся фитоценозы.
Прорастание семян. Лишь у относительно небольшого числа по-
крытосеменных семена прорастают сразу после завершения созрева-
ния. Бóльшая часть таких растений произрастает в условиях постоян-
но теплого климата без резких температурных скачков. Однако
подавляющему большинству цветковых растений прорастанию пред-
шествует период покоя. Если условия неблагоприятны для прораста-
ния, семена находятся в состоянии вынужденного покоя, который
сопровождается максимальным замедлением физиологических про-
цессов в структурах семени. Вынужденный покой позволяет семени
дождаться сочетания благоприятных условий внешней среды и про-
расти. Однако внешние факторы не всегда стабильны, и благоприят-
ные условия нередко вскоре сменяются более суровыми. Поэтому се-
мена растений, произрастающих в географических зонах с сезонными
изменениями климата, должны прорастать только тогда, когда усло-
вия будут устойчиво благоприятными в течение более или менее дли-
тельного времени, чтобы проросток успел развиться в молодое расте-
ние и окрепнуть до наступления холодов или засухи. До этого семена
находятся в состоянии физиологического покоя. Данное состояние от-
личается от вынужденного покоя тем, что физиологические процессы
не заторможены, а, напротив, могут быть активными (дыхание, рост
тканей зародыша и др.), но прорастание при этом не происходит. Для
того чтобы вывести семена из состояния физиологического покоя, не-
обходимо воздействие низких температур, что происходит при насту-
плении холодов. Если ранее низкая температура повышается, семена
494
ВЫСШИЕ РАСТЕНИЯ
Á
Â
À
5
6
7
8
9
1
0
1
1
1
2
1
3
1
4
1
5
1
6
1
7
4
3
2
1
I
I
I
1
4
Р
и
с
.
2
1
8
.
Т
и
п
ы
п
р
о
р
а
с
т
а
н
и
я
с
е
м
е
н
и
:
А
—
с
х
е
м
а
с
т
р
о
е
н
и
я
с
е
м
е
н
и
,
Б
—
э
п
и
г
е
а
л
ь
н
о
е
(
н
а
д
з
е
м
н
о
е
)
п
р
о
р
а
с
т
а
н
и
е
,
н
а
п
р
и
м
е
р
,
у
п
о
д
с
о
л
н
е
ч
н
и
к
а
(
э
н
д
о
с
п
е
р
м
а
н
е
т
)
и
л
и
у
к
л
е
щ
е
в
и
н
ы
(
э
н
д
о
с
п
е
р
м
и
м
е
е
т
с
я
)
,
В
—
г
и
п
о
г
е
а
л
ь
н
о
е
(
п
о
д
з
е
м
н
о
е
)
п
р
о
р
а
с
т
а
н
и
е
,
н
а
п
р
и
м
е
р
,
у
о
г
о
р
о
д
н
ы
х
б
о
б
о
в
(
э
н
д
о
с
п
е
р
м
а
н
е
т
)
;
I
—
э
п
и
г
е
а
л
ь
н
о
е
п
р
о
р
а
с
т
а
н
и
е
(
у
д
л
и
н
я
е
т
с
я
г
и
п
о
к
о
т
и
л
ь
)
,
I
I
—
г
и
п
о
г
е
а
л
ь
н
о
е
п
р
о
р
а
с
т
а
н
и
е
(
у
д
л
и
н
я
е
т
с
я
э
п
и
к
о
т
и
л
ь
)
;
1
—
э
п
и
к
о
т
и
л
ь
,
2
—
с
е
м
я
д
о
л
я
,
3
—
к
о
р
е
ш
о
к
,
4
—
э
н
д
о
с
п
е
р
м
,
5
—
п
л
ю
м
у
л
а
,
6
—
с
е
м
е
н
н
а
я
к
о
ж
у
р
а
,
7
—
г
и
п
о
к
о
т
и
л
ь
,
8
—
и
с
к
р
и
в
л
е
н
н
ы
й
г
и
п
о
к
о
т
и
л
ь
,
9
—
д
в
е
с
е
м
я
д
о
л
и
,
1
0
—
с
е
м
е
н
н
а
я
к
о
ж
у
р
а
,
1
1
—
б
о
к
о
в
ы
е
к
о
р
н
и
,
1
2
—
п
л
ю
м
у
л
а
,
1
3
—
ф
о
т
о
с
и
н
т
е
з
и
р
у
ю
щ
и
е
с
е
м
я
д
о
л
и
,
1
4
—
в
ы
п
р
я
м
л
е
н
и
е
,
к
о
н
т
р
о
л
и
р
у
е
м
о
е
ф
и
т
о
х
р
о
м
о
м
,
1
5
—
п
е
р
в
ы
е
з
е
л
е
н
ы
е
л
и
с
т
ь
я
,
1
6
—
и
с
к
р
и
в
л
е
н
н
а
я
п
л
ю
м
у
л
а
,
1
7
—
б
о
к
о
в
ы
е
к
о
р
н
и
(
п
о
Г
р
и
н
у
и
с
о
а
в
т
.
,
с
и
з
м
е
н
е
н
и
я
м
и
)
495
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
перестают готовиться к прорастанию и впадают в состояние вторич-
ного покоя. Это предотвращает нежелательное преждевременное про-
растание семян при кратковременном повышении температуры во
время зимней оттепели.
Прорастание семян возможно только при благоприятном сочета-
нии температуры, влажности и аэрации. Для семян разных растений
интенсивность этих факторов может быть различной, и она зависит
от условий произрастания растений данного вида. Например, семена
северных растений прорастают при более низких температурах (пше-
ница при t от 0 до +7 °С), а южные — при более высоких (семена паль-
мы при t от +20 до +25 °С). Процесс прорастания семян осуществля-
ется в несколько этапов: набухание семян, проклевывание,
гетеротрофный рост проростка и его переход к автотрофному пита-
нию. Вначале из семени всегда появляется корешок. Вслед за корнем
начинает развиваться побег, причем оба эти органа обладают противо-
положной реакцией на силу тяжести: у корня — положительный гео-
тропизм, у побега — отрицательный. Это обеспечивает правильную
ориентацию вегетативных органов в пространстве. Все время, начи-
ная с проклевывания семени и до формирования фотосинтезирующих
вегетативных органов, развивающееся растение питается гетеротроф-
но за счет питательных веществ, запасенных в семени (гетеротроф-
ный этап). Когда побег достигает поверхности почвы, под действием
света в его тканях образуются хлоропласты, в результате чего моло-
дой побег зеленеет и постепенно переходит к автотрофному (фото-
трофному) питанию (рис. 218).
Вегетативное размножение цветковых
Вегетативное размножение является одним из типов бесполого
размножения, которое отличается от размножения спорами тем, что
его осуществляют не отдельные клетки (споры), а многоклеточные
структуры — части одного из вегетативных органов — корня или по-
бега. Подавляющее большинство цветковых растений может размно-
жаться вегетативным путем. Нередко этот способ увеличения числен-
ности популяции является единственным, т. е . некоторые растения не
образуют семян. Вегетативное размножение присуще, кроме покрыто-
семенных, и другим высшим растениям (мохообразным, папоротни-
кообразным), но крайне редко встречается среди голосеменных, для
которых является основным размножение семенами. Широкое рас-
пространение вегетативного размножения среди цветковых позволи-
ло им не только успешно освоить многие экологические ниши, но
и стать самой процветающей группой растений. Цветковые растения
выработали различные способы вегетативного размножения.
496
ВЫСШИЕ РАСТЕНИЯ
Размножение луковицами. Луковица — это видоизмененный по-
бег, у которого стебель сильно укорочен и похож на донце. От него
тесно отходят мясистые листья, содержащие большое количество за-
пасных питательных веществ, в пазухах листьев развиваются пазуш-
ные почки. На вершине донца находится верхушечная почка междо-
узлия, в луковице очень короткие почки. Снаружи луковица покрыта
сухими омертвевшими листьями. Луковица не впадает в состояние
полного покоя, поэтому в ней постоянно осуществляется рост, правда,
очень медленный. При активизации роста на донце образуются при-
даточные корни. Из верхушечной почки развивается надземный по-
бег. Луковичные растения широко распространены на высокогорных
альпийских лугах. Многие культурные растения размножаются луко-
вицами (лук, чеснок, тюльпаны, гиацинты, нарциссы и др.).
Размножение клубнелуковицами. Клубнелуковица — видоизме-
ненный подземный побег, внешне похожий на луковицу, но без мяси-
стых листьев. Запасные вещества накапливает в коротком вздутом
стебле. Снаружи клубнелуковица покрыта сухими пленчатыми ли-
стьями — остатками прошлогодних надземных вегетативных листьев.
Корни придаточные, способные укорачиваться, поэтому по оконча-
нии вегетационного периода они сокращаются, они втягивают клуб-
нелуковицу, углубляя ее в почве. Количество клубнелуковиц увели-
чивается путем образования нескольких дочерних (деток). Гладиолус,
шафран и другие растения образуют клубнелуковицы.
Размножение клубнями. Клубни образуются в результате утолще-
ния корня или подземной части побега, соответственно различают
корневые и стеблевые клубни. В течение вегетационного периода
в клубне накапливаются питательные вещества. Из перезимовавшего
клубня развивается надземный побег, а сам клубень атрофируется
и заменяется несколькими новыми. Это отличает клубни от клубне-
луковиц, которые образуются на основе старых. Корневые клубни
представляют собой вздувшиеся придаточные корни. Они имеются у
чистяка, батата, георгина и др. В области отхождения этих корней от
старого стебля находятся пазушные почки, из которых впоследствии
развиваются новые растения. Типичный пример стеблевых клубней —
клубни картофеля.
Размножение мясистыми корнями. Сильно утолщенные мяси-
стые корни образуются у различных представителей зонтичных, ор-
хидных и других семейств. Чаще всего одновременно утолщаются
и придаточные корни, которые могут образовывать разветвления;
мясистые главные корни встречаются реже.
Размножение корнеплодами. В составе корнеплода выделяют:
головку, которая образуется из побега и представлена короткой ро-
зеткой укороченных междоузлий; утолщенной шейкой, которая явля-
497
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
ется разросшимся подсемядольным коленом; мясистым корнем, кото-
рый всегда представляет собой стержневой разросшийся главный
корень. В процессе вегетационного периода в главный корень форми-
рующегося корнеплода поступают питательные вещества, однако ло-
кализация этих веществ может быть различной. В соответствии с
этим различают корнеплоды с запасающей паренхимой в ксилеме
(репа, редька, редис), корнеплоды с запасающей паренхимой во флоэ-
ме (петрушка, морковь), корнеплоды с запасающей паренхимой, воз-
никшей от деятельности нескольких добавочных слоев камбия (све-
кла). Доля запасных веществ в корнеплодах может быть очень
высокой, например до 20% в корнеплодах сахарной свеклы. Корне-
плоды характерны для двулетних растений. В первый год у них обра-
зуется мощная вегетативная масса, которая синтезирует много пита-
тельных веществ, откладываемых в корнеплоде. На следующий год
перезимовавший корнеплод использует запасные вещества для цвете-
ния и образования семян.
Размножение усами. Ус представляет собой надземный ползучий
побег с пленчатыми листьями и пазушными почками, способный
в определенных местах укореняться и образовывать укороченный ро-
зетковидный побег, от которого также отходят новые усы. Некоторое
время розеточные растения остаются связанными с родительским и
получают от него питательные вещества, но укоренившись, они уже
сами обеспечивают себя всем необходимым. К этому моменту связы-
вающий стебель обычно отгнивает. Размножение усами весьма эф-
фективно: один экземпляр земляники за два года дает начало пример-
но 200 новым растениям), а куст земляники за год заселяет 1,5 м2
окружающей территории.
Размножение корневищами. Корневище представляет собой ви-
доизмененный подземный побег, растущий горизонтально. В отличие
от корня, корневище несет чешуевидные листья и почки, из которых
развиваются надземные побеги или боковые ответвления. На узлах
образуются придаточные корни. Корневище заканчивается конусом
нарастания, обеспечивающим верхушечный рост: подобно типичному
побегу, там находится верхушечная почка, а не корневой чехлик, как
на корне. На поперечном срезе корневище подобно стеблю. Корневи-
ще может быть укороченным и вздутым (например, у канны, купены,
ириса) или удлиненным и тонким (например, у пырея ползучего,
мяты, астры, камыша, мятлика и др.). Укороченные корневища содер-
жат много запасных веществ. Корневища широко распространены
в природе, они имеются у многих многолетних травянистых, встреча-
ются среди кустарников и даже у древесных форм (бамбук). Кроме
цветковых растений, корневищами обладают все папоротникообраз-
ные. Корневища позволяют растению переждать под землей неблаго-
498
ВЫСШИЕ РАСТЕНИЯ
приятные условия среды, например зиму или засуху, при этом над-
земная часть побега отмирает. Для вегетативного размножения
достаточно небольшого участка корневища, содержащего почку, поэ-
тому корневищные растения очень быстро распространяются, захва-
тывая большие территории. Так, ежегодный прирост одного корневи-
ща пырея ползучего достигает 30—40 см. Корневища хорошо
укрепляют почву, поэтому такие растения используются для укрепле-
ния оврагов, оползней, сыпучих берегов и т. д .
Размножение корневыми отпрысками. У многих древесных и не-
которых травянистых цветковых на корнях образуются придаточные
почки, из которых развиваются надземные побеги, образующие кор-
невую поросль. Отделившись от родительского организма, они стано-
вятся самостоятельными растениями. Их называют корнеотпрыско-
выми (слива, вишня, тополь, липа, сирень, хрен, ежевика и др.) .
Образование корневых отпрысков активизируется после поврежде-
ния корня.
Размножение отводками. Отводок — это однолетний побег, при-
жатый к почве и в этом месте присыпанный землей. В присыпанной
части вскоре развиваются придаточные корни и формируется новое
растение. Отводки могут образовываться в естественных условиях,
особенно у лиан. Вегетативное размножение отводками широко ис-
пользуют в сельском хозяйстве, таким способом размножают виног-
рад, инжир, гранат, крыжовник и многие другие культурные растения.
От многолетнего побега пригибают к земле заранее выбранные здоро-
вые сильные однолетние побеги, специально пришпиливают их и за-
сыпают почвой.
Размножение делением кустов. Этот способ вегетативного раз-
множения используют для увеличения количества посадочного мате-
риала ценных сортов кустарников. Для этого куст делят на несколько
частей, каждая из которых должна иметь корневую систему и надзем-
ные побеги. Обычно это делают весной или ближе к осени.
Размножение черенками. Очень часто для искусственного вегета-
тивного размножения используют черенки — отрезки отделенные от
родительского растения. В зависимости от принадлежности к тому
или иному вегетативному органу различают корневые, стеблевые
и листовые черенки.
Размножение прививкой. Этот способ вегетативного размноже-
ния основан на искусственной пересадке части вегетативного органа
одного растения на другое, поэтому его называют трансплантацией.
Пересаженная (прививаемая) часть растения-донора называется
привоем, а растение-реципиент — подвоем (дичком). Основной за-
дачей прививки является тесное сближение камбия привоя и под-
воя, после чего живые клетки поверхностей среза дедифференциру-
499
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
ются и становятся меристемами. Они активно делятся и заполняют
пространство между раневыми поверхностями срезов паренхимной
тканью. Затем клетки формируют между собой многочисленные
плазмодесмы, которые их связывают и в итоге дифференцируются в
проводящие элементы, общие для привоя и подвоя. После этого па-
зушные почки привоя получают необходимые вещества и развива-
ются в боковые побеги. Прививки и другие способы вегетативного
размножения представляют собой удобный способ размножать цен-
ные сорта культурных растений, которые при этом сохраняют все
свои свойства.
Интересный и многообещающий способ вегетативного размноже-
ния — клонирование, суть которого состоит в получении идентичного
потомства из единичных клеток, взятых от донорского растения. Та-
ким образом можно производить неограниченное количество поса-
дочного материала с необходимыми качествами, при этом свободного
от вирусов и болезней.
Несмотря на распространенность, вегетативное размножение ме-
нее прогрессивно, чем половое, поскольку при этом не происходит не
только обмена генетического материала, как при половом размноже-
нии, но даже его рекомбинации как при размножении спорами. В ре-
зультате образуются точные копии материнского организма с иден-
тичными наборами аллелей.
Основные различия между классами двудольных и однодольных
представлены в табл. 27.
Таблица 27
Основные различия между представителями классов двудольных
и однодольных покрытосеменных растений
(по А.Л. Тахтаджяну, с дополнениями)
Класс двудольные, или магнолиопсиды
(Dicotyledones, aut Magnoliopsida)
Класс однодольные, или лилиопсиды
(Моnоcotyledones, aut Liliopsida)
12
Зародыш обычно с двумя семядолями,
которые, как правило, прорастают над-
земно. Иногда зародыш с одной семядо-
лей (например, у чистяка, некоторых
зонтичных), редко зародыш с тремя-че-
тырьмя семядолями (дегенерия и идио-
спермум). Семядоли обычно с тремя
главными проводящими пучками. Конус
нарастания располагается между семядо-
лями, часто он окружен зачаточными
листьями, вместе с которыми образует
верхушечную почечку
Зародыш обычно с одной семядолей,
которая занимает концевое положение,
продолжая ось зародыша. Семядоли
обычно с двумя главными проводящими
пучками. Конус нарастания и почечка
смещены и располагаются сбоку от про-
дольной оси зародыша. В большинстве
случаев подземное прорастание
500
ВЫСШИЕ РАСТЕНИЯ
12
Боковые вегетативные побеги имеют два
предлиста — профилла (самые нижние,
недоразвитые листья) и соответствую-
щий им прицветничек (брактеолей), рас-
положенные латерально, за исключением
некоторых примитивных групп (напри-
мер, кирказоновые, нимфейные и др.,
у которых они непарные и расположены
вентрально)
Предлистья и брактеоли непарные (оди-
ночные) и расположены на вентральной
стороне побега или реже парные и распо-
ложены латерально (многие представи-
тели семейств лилейных, осоковых, зла-
ковых и др.)
Листья обычно с перистым или реже
с пальчатым жилкованием, иногда жил-
кование дуговидное или параллельное;
обычно имеются свободные концы жи-
лок (жилкование незамкнутое). Черешок
обычно ясно выражен, и листья редко
имеют влагалищное основание. Листо-
вых следов обычно один — три, иногда
больше
Листья обычно с параллельным жилко-
ванием, однако главные жилки соединя-
ются между собой короткими боковыми
жилочками; реже жилкование дуговид-
ное и очень редко пальчатое или пери-
стое; свободных концов жилок, как пра-
вило, не бывает (жилкование обычно
замкнутое). Листья обычно не расчлене-
ны на черешок и пластинку, часто с вла-
галищным основанием. Число листовых
следов обычно большое
Проводящая система стебля обычно эу-
стелическая и состоит из одного кольца
проводящих пучков, реже сифоностели-
ческая. Однако в обоих случаях развит
камбий, обеспечивающий вторичное на-
растание стебля в толщину. Камбий от-
сутствует у некоторых семейств (напри-
мер, нимфейных). Во флоэме обычно
имеется паренхима (отсутствует у неко-
торых семейств, например у кирказоно-
вых и лютиковых). Кора и сердцевина
обычно хорошо дифференцированы
Проводящая система стебля обычно
атактостелическая, состоит из многих
отдельных пучков. Иногда проводящие
пучки образуют два и большее количест-
во колец. Проводящие пучки обычно ли-
шены камбия (иногда остаточный пучко-
вый камбий присутствует, например,
у красоднева, или гемерокаллиса).
Во флоэме нет паренхимы. Обычно нет
ясно дифференцированных коры и сер-
дцевины
Первичный (зародышевый) корешок
обычно развивается в главный корень, от
которого отходят более мелкие вторичные
(боковые) корни, в результате формиру-
ется стержневая корневая система (но
у многих травянистых форм корневая
система мочковатая). Чехлик и эпидерма
имеют в онтогенезе общее происхождение
(за исключением порядка нимфейных)
Первичный корешок рано отмирает, за-
меняясь системой адвентивных (прида-
точных) корней, обычно образующих
мочковатую корневую систему. Чехлик
и эпидерма имеют в онтогенезе разное
происхождение
Древесные или травянистые растения
(травянистые растения возникли из дре-
весных). Иногда вторичные древовидные
формы (например, саксаул)
Обычно травы, иногда вторичные древо-
видные формы (например, пальмы). Пер-
вично древесные растения отсутствуют
Цветки 5- или реже 4-членные и лишь
у некоторых, преимущественно прими-
тивных, групп бывают 3-членные (на-
пример, у анноновых и кирказоновых)
Цветки обычно 3-членные, иногда 4- или
2-членные, но никогда не бывают 5-член-
ными
Продолжение табл. 27
501
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ
12
Нектарники разных типов часто пред-
ставляют собой видоизмененные тычин-
ки, редко бывают септальными
Нектарники преимущественно септаль-
ные, т. е . расположены на перегородках
завязи
При делениях материнской клетки ми-
кроспор клеточные перегородки закла-
дываются преимущественно по симуль-
танному типу
При делениях материнской клетки ми-
кроспор клеточные перегородки закла-
дываются преимущественно по сукцес-
сивному типу
Оболочка пыльцевых зерен обычно трех-
бороздная или производных от нее типов
(однобороздная только у немногих при-
митивных групп)
Оболочка пыльцевых зерен обычно од-
нобороздная или производных от нее
типов (чаще всего однопоровая), но ни-
когда не бывает трехбороздной
Эндосперм обычно целлюлярный или
нуклеарный, редко гелобиальный
Эндосперм гелобиальный или нуклеар-
ный, очень редко целлюлярный
Периоды жизни растений. Жизнь растения можно разделить на
несколько периодов. Эмбриональный период начинается с момента об-
разования зиготы и завершается созреванием семени. Ювенильный
период длится от прорастания семени до перехода к автотрофному
питанию. В течение этих первых двух периодов питание гетеротроф-
ное, за счет родительского спорофита. Виргинальный период продол-
жается от начала автотрофного питания до первого цветения. Его
продолжительность у разных покрытосеменных широко варьирует —
от нескольких недель у однолетних трав до десятилетий у многих де-
ревьев. Время от первого до последнего цветения называется перио-
дом зрелости и размножения, или генеративным периодом. Именно
в этом периоде растение обильно плодоносит и оставляет потомст-
во — плоды с семенами. У многих древесных растений этот этап длит-
ся долго — от десятилетий до столетий, а у некоторых долгоживущих
деревьев даже тысячелетия! Однако рано или поздно любое растение
постепенно прекращает плодоносить, начинается период увядания,
который продолжается вплоть до естественной смерти организма.
Этот последний этап жизни растения называется периодом старости
и отмирания. Общая продолжительность жизни разных растений
очень широко варьирует, гораздо шире, чем у разных животных.
Классификация покрытосеменных
Цветковые — наиболее распространенная группа растений. Это
проявляется огромным разнообразием видов: так, на Земле произра-
стают не менее 250 000 видов, каждый год ученые обнаруживают
и описывают все новые и новые виды.
Окончание табл. 27
502
ВЫСШИЕ РАСТЕНИЯ
Отдел цветковые растения подразделяют на два класса: класс дву-
дольные, или магнолиопсиды, и класс однодольные, или лилиопсиды.
В класс двудольные входит около 180 000 видов, которые объединяются
в 10 000 родов и 325 семейств. Класс однодольные объединяет около
60 000 видов, 3000 родов и 65 семейств. Между этими классами имеет-
ся много отличий, однако ни одно из них не является всеобъемлющим.
Основные признаки основных семейств одно- и двудольных цвет-
ковых приведены в табл. 28.
Таблица 28
Основные признаки важнейших семейств одно- и двудольных цветковых
С
е
м
е
й
с
т
в
о
Формула цветка
Особенности цветка, соцветие
Плод
Представители
1234
С
л
о
ж
н
о
ц
в
е
т
н
ы
е
*Ca0Co(5)A(5)G1
Мелкие цветки собраны в со-
цветие корзинку, окруженную
оберткой, краевые цветки вы-
полняют функцию венчика.
Цветки трубчатые и язычковые
(асимметричные); их располо-
жение в корзинке разное: 1) все
трубчатые, 2) все язычковые,
3) центральные трубчатые, по
краям язычковые, 4) централь-
ные трубчатые, по краям ворон-
ковидные
Семянка,
орешек
Большинство представите-
лей — травы. Есть лекарствен-
ные и масличные. Одуванчик,
цикорий, василек
К
р
е
с
т
о
-
ц
в
е
т
н
ы
е
*Ca4Cо4A2+4G1
Четырехчленный околоцветник;
соцветия — кисть, реже в виде
щитка
Стручок,
стручочек
Одно-, дву- и многолетние
растения. Много культурных
форм. Репа, редис, турнепс,
редька, брюква, капуста и др.
Р
о
з
о
ц
в
е
т
-
н
ы
е
*Ca5Cо5AfGf
*Ca5Cо5AfG1
Одиночные цветки
Костянка
сборная,
сборный
орешек
Травы, кустарники, деревья.
Шиповник, малина, земляни-
ка, слива, яблоня, груша и др.
Б
о
б
о
-
в
ы
е
*Ca(5)Co5A(9)+1G1
Кисть, головка
Боб
Фасоль, горох, чечевица, ара-
хис, клевер, люцерна, люпин
П
а
с
л
е
-
н
о
в
ы
е
*Ca(5)Co(5)A5G1
Одиночные цветки или соцве-
тия — кисть, завиток
Ягода,
коробоч-
ка
Травы и деревья. Много куль-
турных и лекарственных
форм. Баклажаны, томаты,
перец, картофель, паслен
503
ГРИБЫ
1234
З
л
а
к
о
в
ы
е
*O(2)+2A3G1
Околоцветник состоит из двух
цветковых чешуй и двух цветко-
вых пленочек. Мелкие цветки
собраны в соцветия — сложный
колос, метелка, початок
Зерновка
Много важнейших зерновых
культур. Пшеница, рожь, яч-
мень, кукуруза, рис, овес и др.
Л
и
л
е
й
-
н
ы
е
*Ca6Co6A6G1
Цветки одиночные или собраны
в соцветие
Ягода,
коробоч-
ка
Большинство представителей —
многолетние травы. Спаржа,
алоэ, тюльпан, чеснок, лук
ГРИБЫ
Грибы составляют отдельное царство эукариот. Их долго относили
к растениям, но детальный анализ организации грибов говорит о том,
что это самостоятельная группа, сочетающая в себе признаки расте-
ний и животных. С растениями грибы сближает способность к не-
ограниченному росту и тенденция к формированию плодовых тел.
Кроме того, у грибов отсутствует фагоцитоз. Они поглощают необхо-
димые вещества через всю поверхность тела, которая значительно чем
у животных. Подобно животным у грибов отсутствуют пластиды. Тип
питания гетеротрофный. В качестве запасных веществ откладывается
гликоген, а не крахмал. Как и растения, грибы обладают клеточной
стенкой, образованной хитином, характерным для некоторых живот-
ных, например для кутикулы членистоногих. Все это и позволило вы-
делить отдельное царство грибов.
Протопласт грибов покрыт оболочкой. У некоторых она разделена
поперечными перегородками — это септированные, или членистые,
грибы, у других оболочка не разделена — асептированные, несептиро-
ванные, или нечленистые, грибы. Отдельные компартменты септиро-
ванных грибов, в свою очередь, разделены неполными перегородками,
пронизанными отверстиями, через которые цитоплазма соседних от-
секов свободно перетекает из одного в другой. Перегородки не связа-
ны с делением ядра, некоторые могут содержать одно или несколько
ядер, в других вовсе их нет.
Тонкая трубочка, имеющая многоядерный протопласт, называется
гифой гриба. Некоторые грибы не имеют гифовой организации, они
состоят из отдельных клеток (например, дрожжи). Цитоплазма содер-
жит все органеллы, свойственные эукариотической клетке. Отдельные
гифы увеличиваются благодаря верхушечному росту, они могут вет-
виться. Гифы способны объединяться в продольные группы, образуя
более крупные тяжи — ризоморфы до нескольких метров длиной и
Окончание табл. 28
504
ВЫСШИЕ РАСТЕНИЯ
нескольких миллиметров толщиной. Такие гифы развиты у опят. Бо-
лее плотные сплетения гиф формируют склероции, из которых фор-
мируются органы плодоношения. Совокупность гиф гриба составляет
его мицелий, или грибницу. Из мицелия также образуются плодовые
тела грибов, которые в обыденной речи собственно и называют гриба-
ми. При этом плотно переплетенные группы гиф образуют ложную
ткань, или плектенхиму.
Питание грибов. Все грибы являются гетеротрофами, они нужда-
ются в поступлении готовых органических веществ. Грибы способны
усваивать минеральные вещества из окружающей среды, но органиче-
ские они должны получать в готовом виде. В зависимости от потреб-
ности в тех или иных веществах определенный вид грибов заселяет
оптимальный для него субстрат. Грибы не могут усваивать крупные
частички пищи путем фагоцитоза, они всасывают только жидкости.
Для грибов характерно внешнее пищеварение: они сначала выделяют
в окружающую среду, содержащую пищу, ферменты, которые вне ор-
ганизма расщепляют полимеры до усваиваемых мономеров, затем мо-
номеры всасываются через всю поверхность гиф в цитоплазму.
Грибы могут быть симбионтами, паразитами или сапрофитами.
Симбионты вступают во взаимовыгодные отношения с растениями
в форме микоризы. При этом гриб получает от растения необходимые
ему органические соединения, в свою очередь снабжая это растение
минеральными веществами и водой. Паразиты используют другие
организмы в качестве источника нужных веществ односторонне, на-
нося ему вред. Чаще всего хозяевами паразитических грибов являют-
ся высшие растения, но ими также могут быть животные, человек,
а также другие виды грибов. Грибы, которые способны вести исклю-
чительно паразитический образ жизни (т. е . живут только на живом
хозяине), называют облигатными паразитами. Формы, способные пе-
риодически менять образ жизни с сапрофитного на паразитический
и обратно, называют факультативными паразитами. Облигатные па-
разиты не приводят к смерти хозяина, поскольку для них это будет
также губительно. Факультативные паразиты чаще всего убивают хо-
зяина, т. к. они в дальнейшем могут жить и на мертвых остатках, но
уже как сапрофиты. Сапрофиты питаются органическими фрагмен-
тами погибших организмов. Они (наряду с сапрофитными бактерия-
ми) составляют блок редуцентов биоценоза. Такими грибами являют-
ся пеницилл и мукор, известные в качестве плесени (соответственно,
голубого или белого цвета), покрывающей продукты питания, дрож-
жи и многие другие грибы.
Размножение грибов. Грибы размножаются половым и бесполым
способами. Бесполое размножение может быть вегетативным и собст-
венно бесполым. Вегетативное размножение осуществляется путем
505
ГРИБЫ
почкования гиф или отдельных клеток (например, дрожжи). Образую-
щиеся почки постепенно отделяются, растут и со временем начинают
почковаться. Гифы некоторых грибов могут распадаться на отдельные
клетки, каждая из которых впоследствии дает начало новому мицелию.
Собственно бесполое размножение осуществляется спорами, зооспора-
ми и конидиями, которые образуются на специальных гифах.
Половое размножение грибов осуществляется разными способами
(рис. 219). Слияние гаплоидных клеток путем изогамии, анизогамии
(гетерогамии) и оогамии с образованием зиготы, которая покрывает-
ся толстой оболочкой, некоторое время пребывает в состоянии покоя,
после чего прорастает. У многих грибов сливаются только клетки, на-
ходящиеся на разных типах мицелия, обозначаемых как «+» или «–»,
причем их внешнее строение одинаковое, но в пределах своих групп
половой процесс невозможен. Такие грибы называют гетероталлич-
ными, а те, которые имеют только один тип мицелия, — гомоталлич-
ными. Подчеркнем — разные типы мицелия гетероталличных грибов
не дает основания считать их мужскими или женскими. У высших
грибов половой процесс упрощен. У сумчатых грибов могут сливать-
ся не отдельные клетки, а половые органы — гаметангиогамия. Опло-
дотворение также может осуществляться посредством мелких непод-
вижных клеток спермаций. Такой процесс называется сперматизацией.
Половой процесс базидиальных грибов представляет собой слияние
участков вегетативных гиф. Такая форма полового размножения на-
зывается соматогамией. При этом вначале сливается цитоплазма
(плазмогамия), а затем ядра (кариогамия). Грибы, у которых половой
Рис. 219. Типы полового процесса у грибов
(по Горленко)
506
ВЫСШИЕ РАСТЕНИЯ
процесс отсутствует, относят к группе несовершенных грибов. Однако
эта группа во многом является искусственной, так как по мере обна-
ружения у того или иного вида несовершенных грибов полового про-
цесса этот вид относят к определенной систематической группе.
Экологические формы грибов. Грибы заселяют тот или иной суб-
страт в зависимости от потребности в конкретных веществах и спо-
собности синтезировать ферменты, расщепляющие эти вещества.
Большинство грибов тяготеют к влажным местам. Многие грибы за-
селяют различные виды почв — это почвенные грибы. Они участвуют
в почвообразовательных процессах. Часто почвенные грибы вступают
в симбиоз с корнями высших растений, образуя микоризу. Грибы,
предпочитающие жить на навозных кучах или богатой гумусом почве,
являются копрофилами. Грибы, использующие в качестве пищевого
субстрата древесину, называют ксилофитами. Некоторые из них пита-
ются мертвой древесиной (поваленные деревья, ветви, листья и т. д .),
поэтому являются сапрофитами. Другие используют древесину жи-
вых растений, т. е . являются паразитами. Большинство паразитиче-
ских грибов в качестве хозяев используют различные высшие расте-
ния. Многие поражают животных и человека. Отдельной
экологической группой являются грибы, которые вместе с водоросля-
ми образуют лишайники.
Систематика грибов. Различают следующие отделы: хитридиоми-
коты, оомикоты, зигомикоты, аскомикоты, базидиомикоты и несо-
вершенные грибы. Первые три отдела относятся к низшим грибам, ас-
комикоты и базидиомикоты — к высшим грибам, а несовершенные
грибы представляют собой сборную группу грибов, половой процесс
которых еще не установлен.
Отдел аскомикота. Все высшие грибы имеют членистые гифы.
Асковые грибы — одна из самых многочисленных групп грибов (30%
всех грибов), к ним относится около 30 000 видов. Их отличает огром-
ное морфологическое разнообразие: микроскопические одноклеточ-
ные почкующиеся формы (например, дрожжи) и формы, образующие
крупные плодовые тела (например, строчки). Размножение происхо-
дит половым и бесполым путями. Бесполое размножение осуществля-
ется конидиями. Половое размножение проходит в форме гаметангио-
гамии, без образования дифференцированных гамет. Сумчатые грибы,
широко распространенные в природе, способны жить на различных
субстратах и активно участвуют в почвообразовательных процессах.
Они могут вызывать порчу древесины. Часть видов, являясь парази-
тами растений (спорынья), грибов, лишайников, животных, а также
человека, приводят к развитию ряда заболеваний.
Различные виды трюфелей имеют великолепный вкус и аромат, за
что их высоко ценят гурманы. Соответствующим образом приготов-
507
ГРИБЫ
ленные сморчки и строчки также съедобны. Дрожжи незаменимы
в хлебопечении и бродильных производствах, другие виды использу-
ют для получения биологически активных веществ (антибиотики, ви-
тамины, ферменты, алкалоиды, гиббереллины и др.). Многие виды
широко используются для генетических исследований.
Отдел базидиомикота. Всего насчитывается свыше 30 000 видов
базидиальных грибов. Базидиальные, как и сумчатые, являются выс-
шими грибами. Их мицелии имеют членистое строение гиф. Бесполое
размножение осуществляется конидиями, но оно происходит редко.
Половое размножение осуществляется в виде соматогамии, при кото-
рой две вегетативные одноядерные клетки гаплоидного мицелия сли-
ваются. Большинство видов являются гетероталличными, соответст-
венно, у них соматогамия происходит только между гифами с
противоположными знаками «+» и «–» . У базидиальных грибов не
образуются половые органы. Органы полового спороношения — бази-
диоспоры — развиваются экзогенно в особых структурах — базидиях.
У большинства видов базидии образуются на плодовых телах в спо-
роносном слое. Попав на подходящий субстрат, базидиоспора прора-
стает и постепенно формируется мицелий, состоящий из гиф с гапло-
идными ядрами. Разрастаясь, гифы сталкиваются с гифами другого
мицелия, и, если они оказываются с противоположными знаками,
происходит соматогамия с образованием дикарионных гиф.
Базидиальные грибы распространены очень широко в природе.
Среди них имеются микроскопические формы и формы с очень круп-
ными плодовыми телами. Почти все шляпочные грибы являются ба-
зидиальными. По способу питания они могут быть сапрофитами и па-
разитами. Многие виды вступают в симбиоз с корнями растений,
образуя микоризу. Большое количество шляпочных грибов имеют
большое хозяйственное значение, так как они съедобны. Ядовитые
формы (бледная поганка, различные мухоморы и др.) содержат очень
сильные яды, поэтому они опасны для человека.
Значение грибов. Грибы играют важную роль в жизни природных
сообществ. Они участвуют в круговороте веществ в природе. Грибы-са -
профиты перерабатывают различные органические остатки, повышая
плодородие почв. Подстилочные сапрофиты разлагают лесную под-
стилку, которая образуется из опавших листьев, сучьев и т. д . Велика
роль грибов в качестве симбионтов высших растений (микориза). Не-
которые виды муравьев (листорезы) специально культивируют грибы
в особых отсеках своих муравейников. Однако многие грибы являются
паразитами и вызывают тяжелые заболевания растений и животных.
Ряд грибов продуцируют вещества, подавляющие рост высших расте-
ний. Среди грибов встречается даже сверхпаразитизм, когда на одном
паразите развивается другой, его собственный паразит.
508
ВЫСШИЕ РАСТЕНИЯ
Грибы имеют большое значение в хозяйственной деятельности че-
ловека. Многие шляпочные грибы съедобны, они употребляются
в пищу. Создаются специальные хозяйства по промышленному выра-
щиванию шампиньонов и вешенок. Пока для этого пригодны лишь
немногие виды. Различные микроскопические виды грибов широко
используются для получения биологически активных веществ.
Антибиотики. Некоторые грибы выделяют антибиотики. Еще
в 70-х годах XIX в. отечественные ученые В.А . Манассеин и А.Г. По-
лотебнов использовали плесень пенициллов для лечения сифилиса и
кожных заболеваний. Открытие в 1928 г. английским ученым А. Фле-
мингом пенициллина открыло новую эру в медицине. С тех пор анти-
биотики позволили спасти более двух миллиардов (!) людей от раз-
личных бактериальных инфекций. Активные штаммы пеницилла
открыты отечественным исследователем З.В. Ермольевой и ее со-
трудниками (1942).
Алкалоиды, полученные из склероциев спорыньи, применяют для
лечения сердечно-сосудистых и нервных заболеваний. В бродильном
производстве и хлебопечении невозможно обойтись без дрожжей.
Грибы являются продуцентами различных ферментов, что объясняет-
ся особенностями их питания. Плесневые грибы используют для при-
готовления некоторых ценных сортов сыра (рокфор, камамбер, ита-
льянская горгонцола и др.) . Использование в биотехнологическом
производстве грибов позволяет получить вещества, необходимые для
сельского хозяйства. Например, из дрожжей получают кормовой бе-
лок. Другие грибы продуцируют фитогормоны, например гибберел-
лин. Эти производства выгодны, поскольку в качестве пищевого суб-
страта используют отходы производства, в том числе и углеводороды
нефти. Вместе с сапрофитными бактериями грибы используют для
очистки сточных вод. Ряд грибов содержат весьма опасные токсичные
вещества. Некоторые являются паразитами человека, вызывающие
ряд заболеваний кожи (стригущий лишай, паршу, дерматиты и др.),
дыхательной и других систем.
Большой вред приносят возбудители заболеваний сельскохозяйст-
венных растений (фитофтора, спорынья, головневые, ржавчинные
и др.). Так, в 1845 г. в Ирландии широкое распространение фитофторы
привело к гибели урожая картофеля, что повлекло за собой голод и по-
следующую панику. В результате к 1851 г. численность населения стра-
ны уменьшилась на 2 млн человек. Милдью, или ложная мучнистая
роса, была завезена в Европу из Америки и быстро уничтожила почти
все виноградники Франции. Спорынья поражает во время цветения
хлебные злаки. В образующихся при этом рожках (склероциях) содер-
жится много сильнодействующих алкалоидов, которые воздействуют
на нервную систему. Употребление в пищу продуктов, приготовленных
509
ЛИШАЙНИКИ
из пораженного зерна, вызывает отравление. Паразитические грибы вы-
зывают заболевания домашних животных, например аспергиллез птиц,
развивающийся в легких, заболевания рыб и др. Многие сапрофитные
грибы портят меховые и кожаные изделия, ткани на основе природных
волокон (шелковые, шерстяные, льняные, хлопчатобумажные).
ЛИШАЙНИКИ
Лишайник — симбиоз гриба и водоросли. Это совершенно новые
морфологические формы, не похожие ни на грибы, ни на водоросли.
Лишайники синтезируют лишайниковые кислоты, которые другие ор-
ганизмы не вырабатывают. Лишайники обладают особыми способами
размножения.
Компоненты лишайника. Лишайник образован гетеротрофным
грибом, который составляет микобионт лишайника, и автотрофной
водорослью, которая составляет фикобионт. Основная часть объема
слоевища (90—95%) приходится на гифы гриба (рис. 220). Микоби-
онтами чаще всего являются асковые (сумчатые) грибы, реже базиди-
альные. Строение гиф лишайника имеет ряд особенностей. Гифы чле-
нистые, при этом отверстия, соединяющие соседние компартменты,
имеются не только в поперечных, но и в продольных перегородках.
1
2
3
4
Á
À
Рис. 220. Гифы лишайников:
А — строение гиф, Б — расширения гиф на месте расположения поперечной
перегородки; 1 — ядро, 2 — вакуоли, 3 — резервные вещества, 4 — плазмодесма
(по Голубковой)
510
ВЫСШИЕ РАСТЕНИЯ
Клеточные стенки гиф сильно утолщены, что предотвращает потерю
воды. В теле лишайника имеются особые двигающие гифы, которые
могут перемещать клетки водоросли в нужное место. Выделенные
и культивированные на питательных средах микобионты совершенно
не похожи на слоевище лишайника. Вне лишайника микобионт в при-
роде выжить не может.
В составе водорослевого компонента (фикобионта) лишайника
встречаются сине-зеленые водоросли и различные представители эука-
риотических водорослей: зеленые, желтые и бурые. Около половины
всех известных лишайников (примерно 10 000 видов) в качестве фико-
бионта имеет хлорококковую водоросль требуксию. Будучи в составе
лишайника, водоросли изменяются, резко замедляется их рост в связи
с тем, что значительная часть синтезированных органических веществ
поглощается грибом, в цитоплазме практически отсутствуют запасные
трофические включения, несмотря на то, что фотосинтез сохраняется,
как и у свободно живущих особей. В отличие от микобионта, водоросли
прекрасно выживают и в свободном состоянии. Окруженная гифами
гриба водоросль не может поглощать воду и неорганические вещества
извне, поэтому ей приходится добывать их из гиф.
Морфология лишайников. Тело лишайника не дифференцирова-
но на органы и представляет собой слоевище (таллом). Часто лишай-
ники окрашены в различные цвета пигментами, которые откладыва-
ются в гифах. В зависимости от внешнего вида таллома различают
накипные, листоватые и кустистые лишайники. Накипные лишайни-
ки имеют вид тонкого налета или корочки, прочно срастающейся
с поверхностью субстрата. Листоватые лишайники выглядят как
уплощенная округлая пластинка. Самые высокоорганизованные —
кустистые лишайники — напоминают разветвленный кустик, при-
крепленный к субстрату основанием своего таллома.
Размножение лишайников. Каждый из компонентов таллома спо-
собен размножаться самостоятельно. Большинство лишайников раз-
множаются преимущественно вегетативным путем. Высыхая, слое-
вище становится очень хрупким, от него легко отламываются кусочки,
которые затем подхватываются ветром и разносятся. Половое размно-
жение характерно для микобионта лишайников, оно сопровождается
формированием плодовых тел, которые часто напоминают миниатюр-
ные шляпочные грибы.
Экология лишайников. Лишайники заселяют практически любые
субстраты. Различают эпилитные лишайники, которые живут на по-
верхности камней, эпигейные — на почве, эпиксильные — на разлагаю-
щейся древесине, эпифитные — на коре деревьев, и многие другие.
Лишайники нуждаются в освещении. Водоросли в лишайнике сильно
затенены гифами гриба, поэтому значительная часть света ими не ис-
511
ЛИШАЙНИКИ
пользуется. Часто лишайники живут там, где растения жить не могут,
в то же время более комфортабельные места обитания остаются неос-
военными. Это происходит потому, что лишайники примерно в 15 раз
менее эффективно ассимилируют органические вещества, чем расте-
ния, поэтому они не могут конкурировать с последними и заселяют
недоступные для растений места. Лишайники способны долгое время
переносить недостаток влаги. Они поглощают не только жидкую воду,
но и пары воды из воздуха. Некоторые заселяют затопляемые места и
живут под водой. Лишайники выносливы к колебаниям температуры,
они могут переносить высокие (+50 ... +60 °С) и низкие (ниже –50 °С)
температуры. Причем фотосинтез может проходить при отрицатель-
ных температурах (даже при –25 °С), что абсолютно невозможно для
растений. Оптимальные температуры видоспецифичны, но у боль-
шинства видов фотосинтез происходит наиболее интенсивно при
+10 ... +25 °С. Лишайники активно реагируют на загрязненность воз-
духа, они поселяются лишь в экологически чистых местах. Быстрее
всех растут кустистые и листоватые формы, например кустистый ли-
шайник рамалина сетчатая при средней влажности за год вырастает
на 36,5 мм. Значительно медленнее растут накипные лишайники (за
год всего 0,25—0,5 мм). Бо
льшая часть слоевищ, несмотря на малые
размеры, имеют возраст около 50 лет.
Значение лишайников в природе велико, так как они растут даже
там, где не могут жить растения. Первыми заселяя безжизненные суб-
страты, лишайники участвуют в процессах почвообразования, что де-
лает возможным дальнейшее поселение растений. Лишайники погло-
щают минеральные вещества из каменистого субстрата и включают
их в биологический круговорот. Талломы лишайников создают усло-
вия для формирования сообществ различных организмов. Лишайни-
ки выделяют вещества, которые подавляют рост гриба. При этом де-
ревья, покрытые лишайниками, меньше страдают от разрушительной
деятельности грибов, питающихся древесиной.
Лишайниками питаются северные олени. Участие лишайников
в хозяйственной деятельности человека незначительно. Используют-
ся пигменты, содержащиеся в лишайниках, например лакмус, кото-
рым определяют рН среды. Исландский мох используется в парфю-
мерии и в медицине. Промышленное производство, сопровождаемое
вредными выбросами в атмосферу, приводит к сокращению числен-
ности лишайников, особенно в городах. Освоение нефтяных и газо-
вых месторождений приводит к загрязнению огромных территорий,
отчего страдают, в первую очередь, лишайники. Обычный след от гу-
сениц прошедшего по тундре вездехода наносит лишайниковому по-
крову раны, которые будут зарастать много десятилетий. Лишайники
нуждаются в срочной и долговременной защите.
512
ЖИВОТНЫЕ
ЖИВОТНЫЕ
Животные — гетеротрофные эукариоты. Для жизни им необходи-
мы органические вещества, которые они получают извне с пищей.
В биоценозах животные входят в блок консументов. Поэтому сущест-
вование животных возможно лишь в тесной связи с продуцентами
(растениями). Некоторые жгутиконосцы обладают хлоропластами и
могут на свету фотосинтезировать, а в темноте поглощать готовые ор-
ганические вещества. Чередование автотрофного и гетеротрофного
питания называется смешанным (миксотрофным) питанием. По ви-
довому разнообразию животные значительно превосходят другие
царства. Известно более двух миллионов видов, и каждый год откры-
вают все новые, в основном это беспозвоночные.
Царство животных разделяют на два подцарства: одноклеточные
и многоклеточные, каждое из которых включает в себя несколько
типов.
ПОДЦАРСТВО ОДНОКЛЕТОЧНЫЕ,
ИЛИ ПРОСТЕЙШИЕ
Главная особенность представителей подцарства одноклеточные —
это одноклеточное строение организма. Редко некоторые простей-
шие напоминают многоклеточные образования. Например, ряд про-
стейших образуют колонии. Однако они не являются истинно
многоклеточными, поскольку их клетки располагаются в один слой
и не дифференцированы на ткани.
Клетки простейших, будучи эукариотическими, содержит все ком-
поненты и органеллы, свойственные этим клеткам. Кроме того, они
обладают разнообразными специфическими структурами, свойствен-
ными только им. Это сократительные вакуоли, светочувствительное
пятно (глазок, или стигма), трихоцисты, мионемы, клеточный рот,
клеточная глотка, порошица и др. Цитоплазма простейших обычно
разделяется на два слоя: более плотный наружный — эктоплазму и
внутренний, содержащий большинство органелл и ядро, — эндоплаз-
му. Между двумя слоями у свободноживущих пресноводных форм
имеется одна или две сократительные вакуоли, необходимые для ре-
гуляции водно-солевого равновесия в клетке. Наличие органелл дви-
жения обеспечивает клетке не только подвижность, но и позволяет ей
захватывать пищу.
Многие простейшие имеют уплотненный слой цитоплазмы —
пелликулу, которая обеспечивает защиту клетки. Ряд простейших
образуют защитную капсулу из плотного органического вещества
513
ПОДЦАРСТВО ОДНОКЛЕТОЧНЫЕ, ИЛИ ПРОСТЕЙШИЕ
(например, псевдохитина у раковинных амеб) или неорганического
(соединений кальция, кремния, стронция и др. у лучевиков и морских
солнечников). Чаще всего одноклеточные имеют одно ядро (гаплоид-
ное — у споровиков и жгутиконосцев, диплоидное или полиплоид-
ное — макронуклеус инфузорий), но встречаются и многоядерные ор-
ганизмы (плазмодии, инфузории).
Размножение простейших осуществляется половым и бесполым
путями. У разных форм можно встретить все варианты полового про-
цесса: изогамию, анизогамию и оогамию. Только у самых примитив-
ных простейших (амеб) половое размножение неизвестно. Бесполое
размножение осуществляется путем митоза ядра и последующего раз-
деления цитоплазмы. У асимметричных простейших некоторые
структуры при этом достаются лишь одной из дочерних клеток, а дру-
гая впоследствии сама формирует их.
Важная особенность простейших — их сложный жизненный цикл
(не путать с клеточным циклом!), в котором закономерно чередуются
половой процесс и следующее за ним одно- или многократное бесполое
размножение. Жизненный цикл многих простейших довольно слож-
ный, его стадии широко варьируют у разных представителей.
Характерная особенность простейших — их способность образо-
вывать цисты (инцистироваться) при наступлении неблагоприятных
условий окружающей среды. В процессе инцистирования микроорга-
низм округляется, образует защитную оболочку и становится непод-
вижным. В состоянии покоя он может длительно сохранять свою
жизнеспособность. При этом его метаболизм сильно замедляется, что
весьма экономит питательные вещества. При наступлении благопри-
ятных условий из цисты выходит нормальный микроорганизм, кото-
рый называется трофозоитом, или вегетативной формой. У простей-
ших выделяют две основные жизненные формы: активный трофозоит
с органеллами движения и неподвижная циста.
Это позволяет простейшим благополучно выживать в чрезвычай-
но тяжелых условиях. Простейшие обитают практически везде, где
есть вода; вне водной среды длительное время ни одна клетка вообще
не может существовать. Простейшие обитают в пресных и соленых
водоемах, во влажной почве и других субстратах, в термальных источ-
никах и даже на снегу. Бóльшая часть простейших (около 3500 видов)
является паразитами растений и животных; описаны случаи сверхпа-
разитизма, когда в теле одноклеточного паразита находится другой,
его собственный паразит. В организме человека могут развиваться
25 видов простейших, многие из которых являются возбудителями
тяжелых заболеваний (малярия, сонная болезнь, дизентерия, ряд ве-
нерических заболеваний и др.) .
514
ПОДЦАРСТВО ОДНОКЛЕТОЧНЫЕ, ИЛИ ПРОСТЕЙШИЕ
Простейшие составляют подцарство одноклеточные, которое
включает следующие типы: 1) саркомастигофоры; 2) споровики;
3) книдоспоридии; 4) микроспоридии и 5) ресничные, или инфузо-
рии. Общая численность видов простейших по разным оценкам коле-
блется от 10 000 до 50 000 и более. Подавляющее большинство их
имеют размеры в пределах от 3 до 150 мкм, однако некоторые рако-
винные саркодовые достигают 2—3 см в диаметре, а некоторые много-
ядерные плазмодии могут быть еще больше
ТИП САРКОМАСТИГОФОРЫ
К этому типу относятся два класса: саркодовые и жгутиковые, ко-
торые перемещаются с помощью псевдоподий.
Класс саркодовые. Главная особенность саркодовых — движение
с помощью псевдоподий. Большинство представителей класса — оби-
татели моря (около 80%), остальные живут в пресных водоемах, во
влажной почве или являются паразитами. Выделяют три подкласса:
корненожки, лучевики и солнечники. Все они имеют относительно
простое строение. Подавляющее большинство ведет одиночный образ
жизни, однако известны и колониальные формы, например некото-
рые лучевики.
Корненожки являются самыми примитивными простейшими. Их
цитоплазма не дифференцирована, чаще всего одноядерные. Размеры
клеток колеблются от 10 мкм до 3 мм. Наиболее известный предста-
витель подкласса — обыкновенная амеба протей, которая обитает в
любом пресном водоеме. Ее размеры около 0,5 мм. Амеба перемещает-
ся с помощью псевдоподий, которые временно образуются благодаря
перетеканию цитоплазмы из одного участка клетки в другой. Ско-
рость передвижения — около 200 мкм/мин. Если амеба при этом на-
тыкается на мелких простейших, бактерий, одноклеточных водоро-
слей, мелкие органические фрагменты органики, она обтекает их и
фагоцитирует. Образовавшаяся пищевая вакуоль сливается с первич-
ными лизосомами, где органические частицы расщепляются и посту-
пают в цитозоль, а непереваренные остатки выбрасываются наружу
посредством экзоцитоза через любой участок плазматической мем-
браны. Подобно всем пресноводным микроорганизмам, амеба имеет
сократительную вакуоль, посредством которой удаляется излишняя
вода. Амеба размножается только бесполым путем.
Среди паразитов человека наиболее известна дизентерийная аме-
ба — возбудитель амебиаза (амебной дизентерии). Эти простейшие
попадают в организм человека в виде цист, которые он заглатывает, не
соблюдая правила гигиены (немытые руки, овощи, фрукты, сырая
вода). Исходно дизентерийная амеба живет в толстой кишке и пита-
515
ТИП САРКОМАСТИГОФОРЫ
ется обитающими там бактериями. Такую форму амебы называют
мелкой вегетативной, или формой минута, ее поперечные размеры
составляют 12—30 мкм. При ослаблении защитных сил организма
форма минута превращается в более крупную (до 50 мкм) патоген-
ную, или тканевую форму магна. Ее еще называют крупной вегета-
тивной формой. Форма магна разрушает кишечный эпителий и стен-
ки кровеносных сосудов, вызывая образование язв и кровотечение;
один из основных симптомов этого заболевания — кровавый понос.
При этом амебы питаются эритроцитами хозяина. Без лечения чело-
век может умереть, при эффективном лечении тканевая форма прев-
ращается в мелкую вегетативную, которая инцистируется. Огромное
количество цист (до 300 млн в сутки) выводится с фекалиями и попа-
дает в окружающую среду, заражая ее. При диагностике следует раз-
личать цисты дизентерийной и кишечной амеб: у амебы дизентерий-
ной в цисте четыре ядра, а у кишечной — восемь. Кишечная амеба, как
и ротовая амеба, безвредна.
Отряды раковинных амеб и фораминифер также являются корне-
ножками. Раковинные амебы образуют защитные раковины. В них
имеется отверстие — устье, через которое высовываются псевдоподии.
Эти амебы живут в пресных водоемах. Весьма многочисленные фора-
миниферы обитают в морях. Опадающие на дно пустые известковые
раковины со временем сформировали мощные залежи известняка, ко-
торые использует человек. Фораминиферы вызывают значительные
геологические преобразования.
Класс жгутиконосцы, или жгутиковые. Представители этого
класса имеют постоянную форму тела. У одних форм это обусловлено
жесткой оболочкой клетки, которая состоит из хитиноподобного ве-
щества или из целлюлозы; у других поверхностный слой эктоплазмы
уплотняется, образуя пелликулу. Она достаточно эластична, но благо-
даря ей не могут образовываться псевдоподии. Поэтому для пере-
движения и захвата пищи жгутиковые используют жгутики, их ко-
личество — от одного до нескольких тысяч. Жгутики обеспечивают
значительно более высокую подвижность, чем псевдоподии. Жгути-
ковые могут быть гетеротрофами или миксотрофами. Последние со-
держат хлоропласты и совмещают фотосинтез с потреблением гото-
вых органических веществ. У сапрофитов растворенные вещества
поступают в цитоплазму через всю поверхность тела микроорганизма.
Твердые частицы поступают через клеточный рот, или цитостом
(анимальное питание). Обычно цитостом располагается у основания
жгутика, винтообразное движение которого привлекает в эту область
взвешенные частицы. Клеточный рот ведет в узкий канал — клеточ-
ную глотку (цитофаринкс), заканчивающийся в эндоплазме, где от
него по мере поступления пищи отшнуровываются пищевые вакуоли.
516
ПОДЦАРСТВО ОДНОКЛЕТОЧНЫЕ, ИЛИ ПРОСТЕЙШИЕ
Если цитостома нет, то у основания жгутика на определенном участке,
где отсутствует пелликула, фагоцитируются твердые частицы. Сокра-
тительные вакуоли имеют строение более сложное, чем у саркодовых.
Размножение жгутиковых происходит половым и бесполым
образом. Бесполое размножение сопровождается митотическим деле-
нием ядра, после чего разделяется вся клетка. При этом жгутик мо-
жет отделяться, тогда обе дочерние особи формируют его заново или
он отходит к одной из клеток, а вторая отстраивает его заново. Поло-
вой процесс известен в основном у фотосинтезирующих колониаль-
ных форм (анизогамия или оогамия), реже — у одиночных (изога-
мия). У всех жгутиковых, размножающихся половым путем,
зиготическая редукция числа хромосом, т. е. два первых деления зи-
готы представляет собой мейоз. Диплоидный набор хромосом у этих
организмов имеет только зигота, а все остальное время набор хромо-
сом остается гаплоидным.
Жгутиконосцы бывают одиночными свободноживущими или па-
разитическими и колониальными. Колонии объединяют клетки оди-
накового строения, имеющие общее происхождение. Увеличение ко-
лонии происходит за счет митотического деления образующих ее
клеток. Чаще всего колония имеет форму шара, однако встречаются
древовидные колонии.
Жгутиконосцы подразделяют на растительные и животные.
Растительные жгутиконосцы включают в себя одиночные и коло-
ниальные формы, содержащие хлоропласты и поэтому способные
к фотосинтезу. Растительные жгутиконосцы обитают в соленой и
пресной воде, в крупных водоемах они входят в состав планктона.
Среди них отсутствуют паразитические формы. Особенностью пред-
ставителей этого подкласса (эвглены, динофлагелляты) является на-
личие светочувствительного глазка (стигмы). Наиболее известными
представителями являются эвглена и колониальный вольвокс.
Животные жгутиконосцы объединяют исключительно гетеро-
трофных жгутиковых. Различные представители подкласса ведут
одиночный или колониальный образ жизни. Многие из них паразиты,
в том числе и человека, вызывающие ряд тяжелых заболеваний.
Африканские трипаносомы являются возбудителями тяжелейшей
сонной болезни. Переносчиком паразита являются кровососущие мухи
цеце, а природным резервуаром — дикие копытные, в последнее вре-
мя — прежде всего человек, а также домашние животные. Для парази-
та характерен сложный цикл развития. Первая стадия проходит в пи-
щеварительной системе мухи цеце, а вторая — в крупном позвоночном
животном или человеке. Со слюной мухи-переносчика при укусе три-
паносома попадает в кровь и лимфу человека, после чего поражается
мозг. Первый симптом легкая лихорадка, затем появляется мышечная
517
ТИП СПОРОВИКИ
слабость, утомляемость, сонливость. Развивается глубокое истощение
организма, сонливость сменяется продолжительными периодами ко-
матозного состояния, в течение которых тело больного нередко охва-
тывают судороги. Западноафриканский вариант сонной болезни мо-
жет развиваться годами (до 10 лет). Без лечения обычно исход
летальный, хотя известны редкие случаи спонтанного выздоровления.
Восточноафриканский трипаносомоз развивается гораздо быстрее
и заканчивается смертью уже на шестом месяце заболевания. Болезнь
до сих пор распространена в странах тропической Африки, гамбий-
скими трипаносомами ежегодно заражаются около 10 000 людей,
а родезийскими более 1500. Вакцины малоэффективны, поскольку
паразит в ходе развития в организме человека после каждого размно-
жения изменяет свои антигенные свойства. Существуют относитель-
но эффективные медикаментозные средства, позволившие в значи-
тельной мере понизить смертность.
Лейшмании переносятся москитами. В зависимости от локализа-
ции, лейшманиозы человека бывают дерматотропными, они поража-
ют кожу и висцеротропными, поражающими внутренние органы.
Трихомонады и лямблии являются паразитами человека. Влага-
лищный трихомоноз вызывает влагалищная трихомонада — воспале-
ние женских и мужских половых органов. Известна кишечная трихо-
монада, которая обитает в толстой кишке и питается бактериями.
Лямблия является возбудителем лямблиоза. Паразит обычно заселяет
верхние отделы тонкой кишки, прежде всего двенадцатиперстную
кишку, и поглощает пищу из химуса. Интенсивное размножение пара-
зита приводит к тому, что значительная часть поверхности слизистой
оболочки оказывается покрытой слоем лямблий, что нарушает всасы-
вательную функцию тонкой кишки. Заболевание чаще всего развива-
ется у детей.
ТИП СПОРОВИКИ
В этот тип входят около 2000 видов простейших, все они являются
паразитами, что нашло отражение в упрощении строения клетки.
Споровики не имеют сократительных вакуолей, органелл движения и
клеточного рта. Обмен веществ осуществляется через всю поверх-
ность клеток. Споровики имеют сложный жизненный цикл, в кото-
ром последовательно чередуются половое и бесполое размножение.
Половому процессу предшествует бесполое размножение в виде ши-
зогонии — многократного митотического деления ядер в общей цито-
плазме, объем которой при этом увеличивается (такая многоядерная
структура называется шизонтом). Затем образуются гаметы, которые
сливаются (копулируют), зигота обычно формирует плотную оболоч-
518
ПОДЦАРСТВО ОДНОКЛЕТОЧНЫЕ, ИЛИ ПРОСТЕЙШИЕ
ку, поэтому она называется ооцистой. Ее первое деление всегда мейоз,
поэтому у споровиков происходит зиготическая редукция наборов
хромосом. После мейоза образовавшиеся гаплоидные клетки одно-
кратно или многократно митотически делятся. Такой тип бесполого
размножения получил название спорогонии, а образовавшиеся клетки
называются спорозоитами. Таким образом, в обобщенном виде жиз-
ненный цикл споровиков включает в себя шизогонию, половой про-
цесс и спорогонию. У разных представителей типа жизненный цикл
имеет свои особенности. Тип делится на два класса: грегарины и кок-
цидиеобразные.
Класс кокцидиеобразные включает внутриклеточных паразитов
позвоночных. Жизненный цикл сложен, он часто происходит со сме-
ной хозяев. При этом хозяин, в организме которого осуществляется
бесполое размножение, называется промежуточным хозяином, а тот,
в котором паразит размножается половым путем, называется оконча-
тельным, или дефинитивным, хозяином. Половой процесс осуществ-
ляется путем оогамии.
Очень опасен отряд кровяных споровиков. Наиболее известный из
них малярийный плазмодий — возбудитель малярии, которая широко
распространена в странах с тропическим и субтропическим клима-
том. Жизненный цикл кровяных споровиков проходит со сменой хо-
зяев. Окончательным хозяином являются кровососущие насекомые
(комары), в организме которых осуществляется половое размножение
и спорогония. Бесполое размножение шизогония и образование гамон-
тов происходит в организме промежуточных хозяев, которыми явля-
ются позвоночные животные (и человек).
Человек заражается при укусе самки малярийного комара, кото-
рый и является дефинитивным хозяином. Вместе со слюной комара в
кровь попадают очень мелкие (длиной 5—8 мкм) спорозоиты. С током
крови они переносятся в печень, проникают в гепатоциты, где прохо-
дят преэритроцитарную, или тканевую, стадию развития. Паразит
размножается путем шизогонии, и каждый шизонт дает начало огром-
ному (до 5000) количеству тканевых мерозоитов. Мерозоиты покида-
ют разрушенный гепатоцит и проникают в эритроциты, где проходит
эритроцитарная стадия развития. В эритроцитах паразит размножа-
ется шизогонией, в это время шизонт питается гемоглобином. После
разрушения эритроцита мерозоиты оказываются в плазме крови. Кро-
ме них, в плазму также попадают продукты их жизнедеятельности,
которые, как и сами мерозоиты, являются антигенами. Поэтому они
вызывают активную реакцию иммунной системы, которая начинает
борьбу с антигеном. Клинически это проявляется лихорадкой. Про-
должительность приступа обычно от 6 до 10 часов. Симптомы прекра-
щаются, когда мерозоиты проникают в другие эритроциты, где они
519
ТИП СПОРОВИКИ
вновь недоступны для иммунных молекул и клеток человека, поэтому
иммунные реакции не происходят. Развитие паразита внутри эритро-
цитарной стадии повторяется несколько раз. Каждый раз новые
мерозоиты выходят из эритроцитов в плазму синхронно, что сопрово-
ждается приступами лихорадки. Продолжительность внутриэритро-
цитарной стадии шизогонии для Р. malaria — 72 ч, а для других видов
48 ч. Человек является для малярийного плазмодия лишь проме-
жуточным хозяином, а для полного развития паразита необходимо
половое размножение, которое проходит в организме окончательного
хозяина — комара. Поэтому после многократных повторений эритро-
цитарной стадии развиваются не мерозоиты, а незрелые половые
клетки — гамонты (гаметоциты) — макрогамонты и микрогамонты.
На этом развитие малярийного плазмодия в организме человека за-
канчивается, для продолжения жизненного цикла плазмодии должны
попасть в организм комара (рис. 221).
Рис. 221. Цикл развития малярийного
плазмодия рода Plasmodium:
1 — спорозоиты, 2–4
—
шизогония
в печени (2 — спорозоит,
внедрившийся в печеночную клетку,
3 — растущий шизонт
с многочисленными ядрами,
4 — шизонт, распадающийся на
мерозоиты), 5–10 — эритроцитарная
шизогония (5 — молодой шизонт
в форме кольца, 6 — растущий шизонт
с псевдоподиями, 7–8
—
деление ядер
внутри развивающегося шизонта,
9–10 — распад шизонта на мерозоиты
и выход их из эритроцита, кроме
мерозоитов, видны зерна пигмента),
11 — молодой макрогамонт,
11а — молодой микрогамонт,
12 — зрелый макрогамонт,
12а — зрелый микрогамонт,
13 — макрогамета, 14 — микрогамонт,
15 — образование микрогамет
(флагелляция), 16 — копуляция,
17 — зигота, 18 — подвижная зигота
(оокинета), 19 — проникновение
оокинеты сквозь стенку кишечника
комара, 20 — превращение оокинеты
в ооцисту на наружной стенке
кишечника комара, 21, 22 — растущая
ооциста с делящимися ядрами,
23 — зрелая ооциста со спорозоитами
и остаточным телом,
24 — спорозоиты, покидающие
ооцисту, 25 — спорозоиты в слюнной
железе комара (по Хейсину)
520
ПОДЦАРСТВО ОДНОКЛЕТОЧНЫЕ, ИЛИ ПРОСТЕЙШИЕ
Гамонты попадают в желудок комара во время его кормления кро-
вью человека. В организме комара гамонты созревают и превращают-
ся в зрелые гаметы. В процессе созревания макрогамонты видоиз-
меняются в женские гаметы (макрогаметы), ядра микрогамонтов
несколько раз делятся митотически и дают начало пяти-шести микро-
гаметам. У споровиков зиготическая редукция (только зигота дипло-
идная, и первое деление ее ядра мейотическое), поэтому все клетки на
других стадиях развития, в том числе и гаметоциты, гаплоидные, их
ядра могут делиться только митотически. После этого происходит ко-
пуляция макро- и микрогамет. Образовавшаяся зигота (оокинета) ак-
тивно перемещается и проникает под эпителий стенки желудка кома-
ра, где фиксируется и превращается в ооцисту. Сначала ядро зиготы
делится мейотически, после чего образовавшиеся гаплоидные ядра
многократно делятся митотически. В результате спорогонии в ооци-
сте образуются до 10 000 спорозоитов. Стенка созревшей ооцисты ло-
пается, и наполняющие ее спорозоиты попадают в полости тела кома-
ра. Оттуда спорозоиты мигрируют в органы комара, главным образом
в его слюнные железы. Именно тогда комар способен при укусе вне-
сти паразита в кровь человека. С момента попадания гаметоцитов
с кровью в организм комара до заселения слюнных желез спорозоита-
ми обычно проходит от 7 до 45 дней.
ТИП ИНФУЗОРИИ, ИЛИ РЕСНИЧНЫЕ
В этот тип входят наиболее высокоорганизованные простейшие,
которые отличаются двумя главными особенностями: они движутся
посредством многочисленных ресничек — обладают одновременно
двумя ядрами: мелкого диплоидного микронуклеуса и крупного высо-
кополиплоидного макронуклеуса (гетероядерность). Микронуклеус
обеспечивает сохранение генетической информации и передачу ее
при размножении. Макронуклеус не участвует в размножении, но в
интерфазе именно в этом ядре происходит транскрипция РНК. Сле-
довательно, макронуклеус управляет жизнедеятельностью неделя-
щейся клетки. Тип включает в себя два класса: класс ресничные ин-
фузории и класс сосущие инфузории. Представители первого класса
остаются покрытыми ресничками в течение всей жизни (за исключе-
нием состояния цисты), а сосущие инфузории имеют реснички толь-
ко на определенных этапах онтогенеза (рис. 222).
Всего насчитывается около 6000 видов инфузорий.
Большинство ресничных инфузорий ведут свободный образ жиз-
ни (плавающие и прикрепленные к субстрату), однако встречаются
и паразиты (экто- и эндопаразиты). Наиболее изученным представи-
телем класса является инфузория туфелька, которая обитает в стоя-
521
ТИП ИНФУЗОРИИ, ИЛИ РЕСНИЧНЫЕ
чих пресных водоемах. Она покрыта снаружи
пелликулой и поэтому имеет постоянную форму
тела. У сложно организованных инфузорий
реснички неодинаковы по размеру, они могут
концентрироваться в определенных местах и
сливаться в комплексы (наиболее сложные ком-
плексы ресничек встречаются вблизи цитосто-
ма). От расположенного в эктоплазме базально-
го тельца каждой реснички по направлению
к переднему концу тела проходит кинетодес-
мальная фибрилла. Соседние фибриллы формируют тяж — кинето-
десму, вместе с базальными тельцами одного ряда ресничек они со-
ставляют кинетическую единицу. Все это обеспечивает согласованное
движение ресничек при движении и захвате пищи. В глубине экто-
плазмы находятся продольные тяжи сократительных микрофиламен-
тов — мионем. Их сокращение позволяет простейшему в определен-
ных пределах временно изменять форму тела.
Инфузории имеют углубление — перистом, или предротовую во-
ронку, ведущую в клеточный рот (цитостом). У наиболее прими-
тивных инфузорий цитостом располагается на переднем конце тела,
у более высокоорганизованных он находится на брюшной стороне.
Цитостом продолжается в узкий канал — клеточную глотку (цито-
фарингс), который заканчивается в эндоплазме, где от глотки от-
шнуровывается пищеварительная вакуоль. Затем она вовлекается в
ток цитоплазмы и совершает внутри клетки путь по определенной
траектории. Затем непереваренные частицы выводятся путем экзо-
цитоза наружу. Инфузории являются единственными простейшими,
имеющими постоянное место выброса неусвоенных продуктов — по -
рошицу.
У большинства инфузорий на границе между экто- и эндоплазмой
располагаются две сократительные вакуоли, которые устроены значи-
Рис. 222. Инфузория туфелька:
1 — реснички, 2 — пищеварительные вакуоли,
3 — микронуклеус, 4 — ротовое отверстие, 5 — глотка,
6 — порошица в момент выбрасывания непереваренных
остатков пищи, 7 — сократительная вакуоль
(центральный резервуар и радиально расположенные
приводящие каналы), 8 — макронуклеус,
9 — трихоцисты
(по Полянскому и Стрелкову)
522
ПОДЦАРСТВО ОДНОКЛЕТОЧНЫЕ, ИЛИ ПРОСТЕЙШИЕ
тельно сложнее, чем у других простейших. В состав каждой из них вхо-
дит центральный резервуар (собственно вакуоль), вокруг которой на-
ходятся радиально расположенные 5—7 приносящих и один выносящий
канальцы. Вакуоли попеременно сокращаются. Частота сокращений
вакуолей зависит от концентрации солей в окружающей воде.
Инфузории размножаются половым и бесполым способами. При
бесполом размножении сначала микронуклеус делится митотически,
а полиплоидный макронуклеус делится простой перетяжкой. Затем
клетка делится на две дочерние, которые обычно заново формируют
ротовые аппараты и другие недостающие структуры. Бесполое раз-
множение может повторяться многократно, однако для инфузорий
необходим и половой процесс. Инфузории не формируют гаметы, по-
этому у них нет и зиготы. Вместо этого между тесно сблизившимися
половыми партнерами образуется относительно узкий цитоплазмати-
ческий мостик, соединяющий обе клетки. Однако клетки не сливают-
ся полностью. Такой половой процесс называется конъюгацией. Наря-
ду с этим макронуклеус редуцируется и распадается, а микронуклеус
делится мейотически. Из образовавшихся в результате мейоза четы-
рех гаплоидных ядер три разрушаются, а оставшееся четвертое делит-
ся митозом, образуя два гаплоидных ядра, называемых пронуклеуса-
ми. Затем половые партнеры обмениваются пронуклеусами, т. е .
происходит обмен генетическим материалом. При этом один из про-
нуклеусов переходит через цитоплазматический мостик в другую
клетку и сливается с оставшимся там пронуклеусом, образуя дипло-
идное ядро. То же самое происходит и в другой клетке. После этого
цитоплазматический мостик разрывается и клетки расходятся, обла-
дая видоизмененным генетическим материалом. После расхождения
клеток ядро каждой из них неоднократно делится, и вновь реконстру-
ируются макронуклеусы.
Среди паразитических инфузорий только балантидий развивается
в организме человека; болезнь, которую он вызывает, называется ба-
лантидиозом. Этот крупный микроорганизм (ширина клетки до
70 мкм, длина до 200 мкм) имеет овальную форму, покрыт ресничками
и обладает всеми основными структурами, которые характерны для
инфузорий. Заражение происходит при проглатывания цист. Их
источником могут быть свиньи, поэтому работники свиноферм боле-
ют чаще. У человека паразит заселяет толстую кишку, где из цист вы-
ходят вегетативные формы, которые могут жить как комменсалы, пи-
таясь кишечной флорой, но иногда они разрушают слизистую
оболочку кишки и лежащие под ней сосуды. Это вызывает образова-
ние язв слизистой оболочки, в особо тяжелых случаях приводит даже
к перфорации стенки кишки и перитониту. Болезнь протекает в форме
тяжелого колита и сопровождается длительным кровавым поносом.
523
ТИП КИШЕЧНОПОЛОСТНЫЕ
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ТЕОРИИ ПРОИСХОЖДЕНИЯ
МНОГОКЛЕТОЧНЫХ ОРГАНИЗМОВ
Наиболее известны гипотезы Э. Геккеля, И.И . Мечникова и И. Хад-
жи. По мнению Э. Геккеля, многоклеточные произошли от высокоор-
ганизованных однослойных колониальных шаровидных простейших.
Появление двухслойности Геккель связывал с инвагинацией, по ана-
логии со способом образования у ланцетника двухслойной гаструлы
из однослойной бластулы. Клетки, оказавшиеся при этом внутри,
должны были видоизмениться и переваривать пищевые частицы, ко-
торые поступали в образовавшуюся первичную кишечную полость.
Клетки, расположенные снаружи, сохраняли реснички, с помощью
которых такое многоклеточное животное плавало. Геккель назвал
свой гипотетический организм гастреей.
И.И . Мечников, подобно Геккелю, считал, что многоклеточные
происходят от колониальных жгутиконосцев типа вольвокса. Однако
возникновение двухслойности он связывал не с инвагинацией стенки
внутрь, а с миграцией отдельных клеток из наружного слоя внутрь.
После этого, по мнению Мечникова, прорывался первичный рот, че-
рез который в первичную кишечную полость поступала пища и усва-
ивалась клетками внутреннего слоя. Эти же клетки осуществляли
и половой процесс. Клетки наружного слоя сохранили функцию дви-
жения, восприятие сигналов внешней среды и защиту. Ученый назвал
свой гипотетический организм фагоцителлой.
Иован Хаджи считал, что многоклеточность возникла сразу, путем
разделения на клетки некоторых специализированных участков цито-
плазмы. Из сократительных вакуолей могли возникнуть органы выде-
ления, из мионем — мускулатура и т. д .
ТИП КИШЕЧНОПОЛОСТНЫЕ
Кишечнополостные — водные двухслойные многоклеточные живот-
ные с радиальной симметрией тела. Клетки, которые происходят из двух
зародышевых листков — эктодермы и энтодермы, дифференцируются.
Симметрия тела называется радиальной потому, что вокруг продольной
оси животного можно провести неограниченное количество плоскостей,
и каждая из них делит тело на две симметричные половины. Остальные
животные имеют билатеральную симметрию тела, при которой вокруг
продольной оси можно провести только одну плоскость (сагиттальную),
которая разделит тело на две равные половины — правую и левую, тогда
как все остальные плоскости делят тело на несимметричные половины
(например, верхнюю и нижнюю или переднюю и заднюю).
524
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Кишечнополостные разделяются на два морфологических типа:
у полипов преобладают продольные размеры, у медуз — поперечные.
Медузы ведут одиночный образ жизни, полипы могут быть одиноч-
ными или колониальными. Колонии состоят из одинаковых (моно-
морфных) или из разных (полиморфных) особей. Развитие колонии
начинается с одной особи, которая в дальнейшем почкуется, образо-
вавшиеся новые особи не отделяются, а остаются связанными между
собой. Колонии разных кишечнополостных могут быть весьма много-
численными, они насчитывают тысячи особей. Медузы всю жизнь
проводят, свободно плавая в толще воды, водные потоки переносят их
на значительные расстояния, следовательно, они входят в состав ма-
кропланктона. Большинство из них живут в соленой воде, но есть и
пресноводные формы (например, краспедакуста). Одиночные полипы
ведут прикрепленный к определенному субстрату образ жизни, хотя
большинство из них способны медленно перемещаться. Колониаль-
ные формы полипов, как правило, фиксированы более жестко к суб-
страте и не могут перемещаться по дну. Хотя и здесь имеются исклю-
чения: колониальные сифонофоры ведут свободноплавающий образ
жизни.
Общий план организации тела одинаков у всех кишечнополост-
ных: они представляют собой двухслойный мешок с одним отверсти-
ем, которое сообщает гастральную полость с окружающей средой. Оба
слоя — наружный (эктодерма) и внутренний (энтодерма) — разделе-
ны бесструктурной мезоглеей, в которой имеются клеточные элемен-
ты. В зависимости от функциональной специализации, клетки экто-
дермы подразделяют на кожно-мышечные, стрекательные, нервные
и интерстициальные, или промежуточные — стволовые клетки, из ко-
торых образуются все другие клеточные типы. Энтодерма состоит из
клеток двух типов: жгутиковых и железистых. Дифференцировка
клеток позволяет говорить о появлении у кишечнополостных тканей.
Однако их клетки обычно не образуют значительных скоплений, как
в тканях более высокоорганизованных животных.
Кишечнополостные размножаются половым и бесполым способа-
ми. Бесполое размножение происходит путем почкования и стробиля-
ции. Кишечнополостные обладают выраженной способностью к реге-
нерации — восстановлению утраченных частей тела. При половом
размножении образуется двухслойная личинка — планула, покрытая
ресничками, благодаря этому она активно плавает.
В тип кишечнополостные входит примерно 9000 видов. Все они
водные животные, заселяющие водоемы с соленой и пресной водой.
В типе три класса: гидроидные, сцифоидные медузы и кораллы (ко-
ралловые полипы).
ТИП КИШЕЧНОПОЛОСТНЫЕ
525
Класс гидроидные полипы. Наибо-
лее изученный представитель класса —
гидра. Сегодня известно 29 видов гидр,
из них 8 встречаются в пресных водое-
мах стран СНГ. Гидра — это небольшой
полип цилиндрической формы (около
1 см длиной). Ее тело — двухслойный
мешок с ротовым отверстием, окру-
женным венчиком из 6—12 щупалец.
Подошва прикрепляет гидру к суб-
страту (рис. 223). Она способна мед-
ленно передвигаться, скользя на подо-
шве, или относительно быстро, как бы
«кувыркаясь».
Эктодерма состоит из эпителиаль-
но-мускульных, стрекательных, нерв-
ных и промежуточных клеток (рис.
224). Наиболее многочисленны эпите-
лиально-мускульные, в их базальной
части расположено мышечное воло-
конце, ориентированное по продоль-
ной оси тела. Тело гидры укорачивает-
ся при сокращении этих клеток.
Наличие стрекательных клеток —
характерная особенность всех кишеч-
нополостных. Каждая стрекательная
клетка имеет капсулу (книду), в кото-
рой находится спирально скрученная
полая нить, из клетки наружу выдает-
ся чувствительный волосок — книдо-
циль, окруженный микроворсинками
(рис. 225). Прикосновение к книдоци-
лю вызывает возбуждение клетки, в
результате чего нить выбрасывается из
капсулы.
%
$
#
"
!
Рис. 223. Продольный разрез
стебельчатой гидры:
1 — ротовой конус (гипостом),
2 — ротовое отверстие,
3 — мужские гонады,
4 — гастральная полость,
5 — яйцеклетка, 6 — подошва,
7 — стебелек, 8 — почка,
9 — желудочный отдел
(по Бриану)
Рис. 224. Участок среза гидры при большом увеличении:
1 — стрекательные клетки, 2 — интерстициальные
клетки, 3 — эктодермальные эпителиально-мускульные
клетки, 4 — нервные клетки, 5 — базальная мембрана,
6 — энтодермальные эпителиально-мускульные клетки,
7 — железистые клетки (по Кестнеру)
526
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Нервные клетки расположе-
ны в эктодерме диффузно, но
чаще всего они находятся во-
круг рта и на подошве. Отрост-
ки нервных клеток сообщаются
между собой. Такой диффузный
тип нервной системы — наибо-
лее примитивный из всех из-
вестных, так как все клетки на-
ходятся на поверхности тела
и не защищены. Кроме того,
у гидры отсутствуют нервные
центры. Однако даже столь
примитивная нервная система
позволяет гидре реагировать на
изменения внешней среды и вы-
рабатывать условные рефлексы.
Интерстициальные, или проме-
жуточные, клетки, окружен-
ные другими клетками, чаще
всего эпителиальными, являют-
ся стволовыми, поэтому они
всегда остаются недифференцированными и дают начало всем дру-
гим клеткам (эктодермы и энтодермы).
Энтодерма состоит из клеток двух основных типов: эпителиально-
мускульных пищеварительных и железистых. Первые имеют по
1–3 жгутика. Железистые клетки выделяют в пищеварительную по-
лость ферменты, расщепляющие органические молекулы. Эктодерма
и энтодерма разделены слоем мезоглеи.
Питание гидры. Гидра питается мелкими водными животными,
в основном рачками: дафниями, циклопами, а также простейшими.
Гидра парализует свою добычу стрекательными клетками, после чего
подтягивает щупальцами к ротовому отверстию и заглатывает. Пита-
тельные вещества усваиваются двумя способами. Пищевые частицы
могут непосредственно захватываться клетками, имеющими жгутики,
путем фагоцитоза. Это внутриклеточное пищеварение, свойственное
еще простейшим. У гидр и других кишечнополостных с возникнове-
нием полости тела появилась возможность и полостного пищеваре-
ния. Оказавшись в пищеварительной полости, проглоченное живот-
ное подвергается воздействию ферментов, которые выделяют
железистые клетки энтодермы. После этого переваренные вещества
всасываются клетками энтодермы и через мезоглею диффундируют
в эктодерму. Непереваренные остатки выводятся через рот.
Рис. 225. Различные типы стрекательных
капсул: А — пенетранта с невыбрашенной
стрекательной нитью, Б — пенетранта
с выброшенной стрекательной нитью;
1 — стрекательная нить, 2 — шипики,
3 — стилеты, 4 — шейка, 5 — крышечка;
В — пенетранта, стрекательная нить
которой внедрилась в покровы добычи,
Г — вольвента, Д — глютинанта
стрептолина, Е — глютинанта стреолина
(по Шульце)
ТИП КИШЕЧНОПОЛОСТНЫЕ
527
Размножение и развитие. Гидры размножаются бесполым и поло-
вым путями. Бесполое размножение осуществляется почкованием. При
этом на средней части тел (пояс почкования) периодически образу-
ются бугорки, которые развиваются в молодых гидр. Постепенно на
вершине бугорка прорывается рот и образуются щупальцы. Некото-
рое время молодая гидра сохраняет связь с родительской особью, за-
тем отделяется от нее и живет самостоятельно. Почкование у гидр
происходит летом, причем одновременно могут развиваться несколь-
ко почек. К половому размножению животные приступают осенью
с наступлением холодов. Гидры раздельнополые животные. Гаметы
у гидроидных полипов образуются из промежуточных клеток экто-
дермы. Мужские гонады располагаются в верхней, а женские — в ниж-
ней части тела гидры. Оплодотворение яйцеклеток происходит осе-
нью в теле матери, после чего они покрываются плотной оболочкой
и в таком состоянии зимуют, а родительский организм к зиме погиба-
ет. Весной из покоящихся оплодотворенных «яиц» развиваются мо-
лодые гидры.
У гидры немного врагов, самый опасный из них — прудовик.
Колониальные гидроидные полипы. Большинство гидроидных
полипов ведут колониальный образ жизни. Колония формируется из
одной особи в результате ее почкования. При этом дочерние полипы
не отрываются, а навсегда остаются связанными с родительской осо-
бью и в дальнейшем сами приступают к почкованию, в результате это-
го возникают древовидные колонии, состоящие из нескольких сотен
и даже тысяч особей — гидрантов. Преимуществом колониальной ор-
ганизации является высокая эффективность питания и защиты. Ги-
дранты колонии омываются значительно большим количеством воды,
а наличие многочисленных стрекательных клеток на щупальцах
исключают возможность освобождения добычи. Захваченная пища
становится достоянием всей колонии, так как питательные вещества
через каналы переносятся от одного гидранта к соседним. Кроме того,
сообща легче защищаться от врагов.
Размножение колониальных гидроидных происходит со сменой
полипоидного и медузоидного поколений. При этом полипы размно-
жаются только бесполым путем, а медузы — половым. Для медуз ха-
рактерно наличие мощного слоя мезоглеи между эктодермой и энто-
дермой. По периферии диска расположены многочисленные щупальца,
несущие у основания большое количество стрекательных клеток и ор-
ганы равновесия — статоцисты. Они являются замкнутыми пузырь-
ками, заполненными жидкостью и выстланными чувствительным
эпителием. Внутри каждого пузырька находятся кристаллы карбона-
та кальция. При изменении положения тела медузы в пространстве
они смещаются и воздействуют на волоски сенсорных клеток, кото-
528
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
рые вызывают мышечные сокращения, приводящие к восстановле-
нию нормального положения. Некоторые гидроидные медузы имеют
глазки. У медуз значительно более сложно, чем у полипов, устроена
нервная система. Кроме обычного сплетения, нервные клетки концен-
трируются по периферии зонтика.
Внизу выделяется длинный ротовой стебелек, или хоботок (ма-
нубриум), на конце которого имеется ротовое отверстие, окруженное
четырьмя лопастями — щупальцами. После ротового отверстия начи-
нается глотка, переходящая в желудок, от которого к периферии ку-
пола отходят радиальные каналы. По краю купола между наружным
и внутренним нервными кольцами проходит кольцевой канал, соеди-
няющий между собой все радиальные каналы.
Класс сцифоидные медузы. По своему строению сцифоидные ме-
дузы похожи на гидроидных медуз. Размеры первых обычно более
крупные — поперечник зонтика цианеи достигает двух метров, более
сложная нервная система: наряду с нервным кольцом одних имеются
ганглии — узловые скопления нервных клеток. Сцифоидные медузы
раздельнополые. Медузы широко распространены в Мировом океане.
Все медузы — хищники. Они питаются планктоном и мелкими рыба-
ми, которых парализуют стрекательными клетками. Некоторые меду-
зы могут парализовать и человека, после чего он утонет. Опасны для
человека: кубомедузы, обитающие у берегов Австралии и Индонезии;
цианеи, часто встречающиеся в Балтийском и Баренцевом морях;
дальневосточные медузы — крестовики, физалии.
Класс кораллы. Кораллы составляют наиболее крупный класс ки-
шечнополостных, в него входят около 6000 видов. Его представители
имеют только полипоидную организацию и не проходят стадию меду-
зы. Большинство из них ведут колониальный образ жизни, однако
имеются и одиночные формы (актинии). Размножение коралловых
полипов происходит половым и бесполым путями. Бесполое размно-
жение — почкование, половое — с помощью гамет, которые образуют-
ся в половых железах. Они раздельнополы и формируют гонады толь-
ко одного типа.
Полипы имеют наружный скелет, чаще он известковый, реже —
органический. У многих коралловых полипов (например, благородно-
го коралла) скелет очень красивый, и его широко используют как
украшение. Поэтому некоторые кораллы оказались на грани уничто-
жения и нуждаются в особой охране. Коралловые полипы — основ-
ные рифообразующие организмы. Кроме них, рифы образуют некото-
рые водоросли, но их роль в этом процессе скромнее. Результатом их
деятельности является Большой Барьерный риф, который тянется
вдоль восточного берега Австралии на протяжении более 1400 км.
Кроме того, коралловые рифы образуют атолловые острова, которые
ТИП ПЛОСКИЕ ЧЕРВИ
529
со временем заселяются растениями и животными. Коралловые поли-
пы обитают в теплых морях, особенно там, где температура не опуска-
ется ниже +20 °С. Они способны жить лишь на небольшой глубине
(до 50 м), при этом формируются характерные для коралловых рифов
экологические сообщества.
ТИП ПЛОСКИЕ ЧЕРВИ
В этот тип входят самые примитивные многоклеточные трехслой-
ные животные. В отличие от двухслойных кишечнополостных, у пло-
ских червей между эктодермой и энтодермой формируется третий заро-
дышевый листок — мезодерма. Форма тела плоских червей уплощенная,
у них выделяются передний и задний концы, но нет головы и хвоста.
Вокруг продольной оси можно провести лишь одну плоскость (сагит-
тальную), которая делит тело животного на две половины. Такой тип
симметрии называется двусторонней, или билатеральной; впервые он
появляется у плоских червей. В то же время у плоских червей отсутст-
вует сегментация тела. Гистологическая организация гораздо более
сложная, чем у кишечнополостных, прослеживается четкая сепарация
тканей, которые образуют внутренние органы.
Кожно-мускульный мешок образует стенку тела. Мешок состоит
из эпителиального покрова, расположенных под ним мышечных во-
локон и разделяющей их соединительной ткани. У свободноживущих
плоских червей эпителий ресничного типа, у паразитических форм
ресничек нет. У плоских червей эпителиальная и мышечная ткани
обособлены. Мышечные клетки образуют сплошные слои, покрываю-
щие тело червя наподобие мешка. Снаружи располагается слой коль-
цевых мышц, внутри — слой продольных. Имеются также мышечные
пучки, соединяющие спинную и брюшную части животного — дорсо-
вентральные (спинно-брюшные) мышцы. Хорошо организованная
мышечная система позволяет плоским червям совершать более слож-
ные движения по сравнению с кишечнополостными. У плоских чер-
вей полость тела, расположенная между стенкой тела и внутренними
органами, заполнена паренхимой, поэтому этих животных относят
к бесполостным. Через паренхиму осуществляется транспорт веществ
в пределах тела червя, в ней откладываются запасные вещества, кроме
того, паренхима обеспечивает опору внутренних органов.
Внутренние органы плоских червей образуют пищеварительную,
выделительную, нервную и половую системы. Кровеносная дыхатель-
ная системы и скелет отсутствуют. Газообмен осуществляется путем
двусторонней диффузии газов через покровы тела, при этом плоская
форма тела очень выгодная, поскольку обеспечивает большую пло-
щадь газообмена. Диффузно по тканевой жидкости паренхимы рас-
530
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
пространяются вещества внутри тела червя, а постоянство формы
тела обеспечивается кожно-мускульным мешком и паренхимой.
Пищеварительная система дифференцирована на эктодермаль-
ную переднюю кишку (глотку) и энтодермальную среднюю, которая
заканчивается слепо. У плоских червей отсутствует присущая более
высокоорганизованным животным задняя кишка, поэтому у них нет
и анального отверстия. Выведение непереваренных частиц осуществ-
ляется в обратном направлении через рот. Через переднюю кишку по-
глощается пища, в средней кишке она расщепляется, и необходимые
вещества всасываются в тканевую жидкость. У крупных форм для до-
ставки питательных веществ к удаленным от кишечника органам
средняя кишка образует многочисленные инвагинации, которые не-
посредственно подходят к органам. У ленточных червей пищевари-
тельная система редуцирована, и питательные вещества всасываются
через всю поверхность тела.
У плоских червей более совершенная нервная система. Нервная
система плоских червей — лестничного типа (ортогон). Наиболее
принципиальным является: 1) появление нервных ганглиев и ство-
лов; 2) погружение их вглубь тела, что значительно повышает защиту;
3) выраженная цефализация нервных центров, которая проявляется
в увеличении размеров мозгового ганглия, расположенного в перед-
ней части тела, и усилении его роли; 4) уменьшение количества нерв-
ных центров по мере совершенствования организации.
Органы чувств развиты в основном у свободноживущих форм
и практически отсутствуют у взрослых паразитов.
Выделительная система образована протонефридиями, которые
представляют собой систему ветвящихся канальцев, пронизывающих
все тело червя и заканчивающихся особыми мерцательными клетками
звездчатой формы. Внутри каждой такой клетки имеется каналец,
в просвет которого обращен пучок ресничек. Благодаря биению ресни-
чек тканевая жидкость с растворенными в ней продуктами распада по-
ступает в каналец мерцательной клетки и далее в многоклеточный ка-
налец. Канальцы объединяются в более крупные и открываются на
поверхности тела червя экскреторными (выделительными) отверстия-
ми, через которые выделяемая жидкость выбрасывается из организма
червя наружу. У многих представителей типа на конце тела имеется
расширение выделительной трубочки — мочевой пузырь, в котором вы-
деляемая жидкость может скапливаться. Протонефридии плоских чер-
вей обеспечивают не только удаление продуктов диссимиляции, но и
выводят из организма излишнюю воду, действуя наподобие сократи-
тельных вакуолей простейших.
Половая система. Большинство плоских червей гермафродиты, у них
одновременно имеются органы мужской и женской половых систем.
ТИП ПЛОСКИЕ ЧЕРВИ
531
Строение половой системы очень сложное, у разных представителей
типа оно может отличаться друг от друга. Чаще всего мужская половая
система состоит из различного количества семенников, семяпроводов,
которые объединяются в семяизвергательный канал, и совокупительно-
го органа (цирруса). Женская половая система более сложная. Она со-
стоит из яичников, желточников (недоразвитые яичники, поставляющие
питательные вещества для яиц); яйцеводов; влагалища, которое откры-
вается в половую клоаку. При оплодотворении совокупительный орган
вводит сперматозоиды в половую клоаку, где и происходит слияние га-
мет. Зигота окружается желточными клетками, снаружи их покрывает
скорлупа. Таким образом, яйца большинства плоских червей имеют
сложное многоклеточное строение, при этом питательные вещества ак-
кумулируются не в яйцеклетке, а за ее пределами в желточных клетках;
такие яйца называют экзолецитальными. Многие плоские черви, особен-
но паразиты, способны продуцировать огромное количество яиц.
Эмбриональное развитие плоских червей различное у разных пред-
ставителей типа. У паразитических форм (гельминты) оно происходит
со сменой одного или нескольких проме-
жуточных хозяев. Такие черви называют
биогельминтами.
Тип плоские черви подразделяется на
пять классов: ресничные черви, сосальщи-
ки, моногенеи, ленточные черви и цесто-
дообразные. Из них только ресничные
ведут свободный образ жизни, а осталь-
ные являются паразитами. Тип насчиты-
вает около 10 000 видов.
Класс ресничные черви, или турбел-
лярии (рис. 226). Это единственный
класс плоских червей, в который входят
Рис. 226. Схема строения трехветвистой
турбеллярии:
1 — щупальцевидные выросты, 2 — глаза,
3 — мозговой ганглий, 4 — передняя ветвь
кишечника, 5 — продольный нервный ствол,
6 — поперечные нервные перемычки,
7 — глотка, 8 — глоточный карман,
9 — семяпровод, 10 — ротовое отверстие,
11 — семенники, 12 — совокупительный
орган, 13 — задняя ветвь кишечника,
14 — половое отверстие, 15 — яйцевод,
16 — половая клоака, 17 — копулятивная
сумка, 18 — желточники, 19 — яичник .
Слева удалены семенники,
справа — желточники и яичник (по Граффу)
532
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
только свободноживущими формами. Большинство из них обитают
в соленых или пресных водоемах, немногие ведут наземный образ
жизни. Класс насчитывает около 3000 видов. Наиболее известные —
представители отряда планарий — это белая, черная, траурная, бурая
и др., всего около 100 видов. Их численность может быть очень боль-
шой, особенно при наличии обильной растительности. Размеры тур-
беллярии широко варьируют от нескольких миллиметров до 30 см.
Внешнее строение турбеллярии, обычное для плоских червей, описа-
но выше.
Кожно-мускульный мешок снаружи покрыт однослойным реснит-
чатым эпителием, в котором заложено много желез. Благодаря защит-
ной слизи, покрывающей тело червя, турбеллярии практически защи-
щены от поедания другими животными. Реснички эпителия и
сокращение мускулатуры обеспечивают перемещение турбеллярии,
они могут ползать по донному субстрату и активно плавать.
Пищеварительная система обычна для плоских червей. Рот распо-
ложен на брюшной стороне тела. Большинство турбеллярий питают-
ся различными мелкими беспозвоночными животными.
Выделительная система образована протонефридиями. Лучше
всего выделительная система выражена у пресноводных форм, кото-
рая обеспечивает им поддержание нормального водно-солевого рав-
новесия.
Нервная система имеет обычное для плоских червей строение.
Органы чувств у турбеллярии развиты значительно лучше, чем
у других классов плоских червей. Осязание осуществляется всей ко-
жей, кроме того, на переднем конце тела имеется пара чувствитель-
ных щупалец, которыми животное исследует поверхность субстрата.
По всей поверхности кожи рассеяны клетки, снабженные более длин-
ными и неподвижными ресничками (сенсиллами), которые восприни-
мают механические и химические воздействия извне. Два или более
глаза расположены вблизи мозгового ганглия или же по краям перед-
ней части тела. У некоторых турбеллярий имеются органы равнове-
сия — статоцисты.
Половая система турбеллярии гермафродитная. Мужские гаметы
образуются в семенниках. От каждого семенника отходит тонкий се-
мявыносящий канал, который впадает в семяпровод. Семяпроводы
соединяются и образуют семяизвергательный канал, расположенный
внутри совокупительного органа. Женская половая система состоит
из половых желез — яичников, желточников и половых протоков. Из
яичников яйцеклетки поступают в яйцеводы, туда же открываются
и протоки желточников, по которым поступают богатые питательны-
ми веществами желточные клетки. Яйцеводы объединяются и образу-
ют непарное влагалище, которое открывается в половую клоаку.
ТИП ПЛОСКИЕ ЧЕРВИ
533
У большинства турбеллярий происходит перекрестное оплодотворе-
ние, при котором половые партнеры передают друг другу мужские по-
ловые клетки. Сперма поступает через совокупительный орган, кото-
рый вводит ее в половую клоаку. У ресничных червей происходит
внутреннее оплодотворение. Оплодотворенные яйцеклетки окружают
желточные клетки, имеющие запас питательных веществ для яйца,
после чего формируется защитная скорлупа.
Развитие турбеллярии прямое, т. е . из яйца выходит организм, по-
хожий на взрослое животное. Определенное время личинка плавает
в составе планктона, после чего она дифференцируется в маленького
червя.
Турбеллярии могут размножаться и бесполым путем. При этом на
теле червя появляется поперечная перетяжка, постепенно разделяю-
щая животное на две части.
Ярко выражена способность турбеллярии к регенерации. Малень-
кие фрагменты размером с десятую или даже сотую часть тела плана-
рий образуют заново целый организм. При недостатке кислорода или
при резком повышении температуры воды планарии сами распадают-
ся на куски, которые регенерируют после наступления благоприят-
ных условий. Это явление называется аутотомией.
Класс сосальщики. Все сосальщики (их около 4000 видов) явля-
ются эндопаразитами беспозвоночных и позвоночных животных.
Около 40 видов живут в теле человека, поражая различные органы.
Наиболее распространен печеночный сосальщик. Внешнее строение
сосальщиков мало отличается от строения турбеллярии. Размеры
обычно невелики — несколько миллиметров, однако есть и более
крупные формы. Так, печеночный сосальщик может вырасти до 5 см,
а некоторые паразиты рыб достигают полутора метров в длину. Фор-
ма тела чаще всего листовидная. Червь имеет две присоски: ротовая
расположена на переднем конце тела, а более крупная брюшная — не-
сколько кзади на брюшной стороне. Присоски — это мелкие ямки в
теле, окруженные мощной мускулатурой, благодаря сокращению ко-
торой паразит присасывается к стенкам органа хозяина. Внутреннее
строение во многом сходно со строением ресничных червей (рис. 227).
На покровах нет ресничек, снаружи тело покрыто защитными выде-
лениями железистых клеток, которые предохраняют червя от разру-
шительного действия ферментов пищеварительных соков хозяина.
Эти черви ведут малоподвижный образ жизни. Форма тела изменяет-
ся путем его сокращения и вытягивания.
Пищеварительная система такая же, как и у турбеллярий, однако
ротовое отверстие находится на переднем конце и окружено ротовой
присоской.
534
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Органы чувств у взрослых червей пра-
ктически не развиты, что связано с их пара-
зитическим образом жизни. У свободно-
плавающих личинок имеются кожные
сенсиллы, у некоторых форм — глазки.
Выделительная система представлена
протонефридиями, при этом выделитель-
ные каналы объединяются в мочевой пузырь, который открывается
выделительным отверстием наружу.
Строение половой системы различается у разных сосальщиков.
Мужская половая система образована двумя семенниками, располо-
женными кзади от брюшной присоски. От них отходят два семяпро-
вода, которые объединяются в извитой семяизвергательный канал,
пронизывающий совокупительный орган. Женская половая система
образована одним яичником, который у печеночного сосальщика
сильно ветвится. От него отходит короткий яйцевод, который впада-
ет в мешкообразный оотип, куда открываются протоки желточни-
ков и скорлуповых желез. От оотипа отходит матка, которая откры-
вается вблизи от совокупительного органа в половую клоаку. При
оплодотворении совокупительный орган вводит семя в половую
клоаку, оттуда оно через матку попадает в оотип, где и происходит
слияние гамет. Яйца поступают в матку и постепенно продвигаются
по ней к выходу.
Для сосальщиков характерен сложный жизненный цикл, который
происходит со сменой одного или нескольких промежуточных хозяев
и с чередованием поколений. Взрослый гермафродитный червь — ма-
рита — паразитирует во внутренних органах позвоночных животных.
íèêè
íûå
êà; 9
1
3
4
6
7
8
9
10
11
12
13
14
15
2
5
16
17
18
Рис. 227. Схема строения сосальщика:
1 — ротовая присоска, 2 — глотка, 3 — пищевод,
4 — половое отверстие, 5 — брюшная присоска,
6 — желточники, 7 — лауреров канал, 8 — оотип,
9 — желточный проток, 10 — ветви кишечника,
11 — семенники, 12 — экскреторный пузырь,
13 — матка, 14 — семяпроводы, 15 — скорлуповые
железы, 16 — семяприемник,17 — яичник,
18 — совокупительный орган (по Смиту)
ТИП ПЛОСКИЕ ЧЕРВИ
535
Оплодотворенные яйца выходят с фекалиями из организма хозяина,
для правильного развития они должны попасть в воду, где из яйца
выходит личинка — мирацидий, покрытая ресничками и способная
активно плавать. Мирацидий активно ищет своего первого промежу-
точного хозяина за счет фото- и хемотаксиса. У всех сосальщиков та-
ким хозяином становится моллюск определенного вида для каждого
паразита. Обнаружив моллюска, мирацидий прикрепляется к нему и
проникает в его ткани. Внутри моллюска мирацидий теряет реснички
и превращается в бесформенную неподвижную личинку — спороци-
сту, содержащую зародышевые клетки, из которых образуются новые
личинки. Зародышевые клетки делятся, что приводит к увеличению
популяции. Это называется полиэмбрионией. Таким образом, споро-
циста размножается партеногенетически без оплодотворения и дает
начало большому количеству личинок следующего поколения — ре-
диям, которые выходят из нее. Редии, способные двигаться, имеют
пищеварительную систему и питаются, засасывая с помощью муску-
листой глотки ткани моллюска. Редии также размножаются партено-
генезом, давая начало другим редиям. В дальнейшем редии, размно-
жаясь, дают начало следующей личинке — церкарии, которая похожа
на взрослого червя (мариту). Важной особенностью церкария являет-
ся наличие у него подвижного хвоста. Церкарии выходят из моллю-
ска и активно плавают с помощью хвоста, но не питаются, используя
накопленные вещества. Церкарий прикрепляется к прибрежной траве
и инцистируется, превращаясь в адолескария, которого затем прогла-
тывает окончательный хозяин.
Многие сосальщики паразитируют в организме человека, вызывая
заболевания — трематодозы.
Печеночный сосальщик распространен повсеместно. Он вызыва-
ет заболевание фасциолез. Марита паразитирует в желчных протоках
и желчном пузыре человека, изредка — в протоке поджелудочной же-
лезы у крупных наземных позвоночных и человека, которые являют-
ся для паразита окончательным хозяином. Этот червь крупный, он
достигает 5 см в длину. Его промежуточный хозяин — малый прудо-
вик, в котором паразит развивается в течение 30—60 дней, после чего
выходит церкарий, который инцистируется на прибрежной траве,
становясь адолескарием. При поедании травоядными животными
водных растений происходит заражение. Человек может заразиться
при употреблении воды, в которой находится оторвавшийся адоле-
скарий.
Класс ленточные черви, или цестоды. Все цестоды — эндопарази-
ты, по строению и жизнедеятельности они более адаптированы
к паразитизму, поэтому их можно считать эталоном среди многокле-
точных паразитов. Личиночные стадии проходят в организме проме-
536
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
жуточных хозяев, которыми могут быть как позвоночные, так и бес-
позвоночные животные, а взрослые особи живут в просвете
кишечника позвоночных. Число видов ленточных червей превыша-
ет 3000.
Тело цестод похоже на тонкую ленту, оно подразделяется на голов-
ку, шейку и стробилу, состоящую из большего или меньшего количест-
ва нитчато соединенных между собой члеников (их еще называют цеп-
нями). Очень редко тело цельное. Окраска тела светлая и однотонная.
Червь прикрепляется к стенке кишки своей головкой, или сколексом,
на которой имеются обычно четыре присоски, а у некоторых помимо
присосок имеются и крючья. Червь, имеющий и присоски, и крючья,
называется вооруженным, если имеются только присоски — невоору-
женным. За головкой следует нерасчлененная шейка, от которой кзади
постоянно отшнуровываются членики — проглоттиды, их совокуп-
ность называется стробилой. Количество члеников определяет длину
цепня, у разных представителей оно широко варьирует от трех до
нескольких тысяч. Соответственно, неодинакова и длина червей: от
1 мм до 10 м, изредка до 20 м. Членики не зависят друг от друга, по-
скольку у каждого из них имеется полный набор необходимых для
жизни структур, тем более что крайнее упрощение организации этих
червей сводит к минимуму численность внутренних органов. Наи-
меньшие размеры имеют самые молодые членики. В дальнейшем они
растут, при этом изменяется их внутреннее строение, на конце строби-
лы оказываются самые старые членики. Когда цепень достигает мак-
симальных размеров, от его заднего конца начинают отрываться чле-
ники, которые затем выходят наружу.
Кожно-мускульный мешок цестод устроен по типу сосальщиков.
Отсутствие пищеварительной системы — характерная особенность
ленточных червей. Поглощение необходимых веществ осуществляет-
ся через всю поверхность тела. Цестоды обитают в тонкой кишке, где
пищевые вещества подвергаются ферментативному расщеплению пи-
щеварительными соками хозяина. Червь всасывает уже переработан-
ные продукты, поэтому внутреннее пищеварение им попросту не нуж-
на, так как эту функцию за них выполняет пищеварительная система
хозяина. Крупные размеры некоторых цестод приводят к истощению
хозяина, тем более что жизнь паразита может длиться десятилетиями.
Учитывая бескислородную среду обитания, цестоды имеют анаэроб-
ный тип дыхания, поэтому у них в паренхиме в качестве запасного
вещества накапливаются гранулы гликогена. Продукты неполного
расщепления субстрата цепни выделяют в полость кишечника, что от-
равляет организм хозяина.
Нервная система развита очень слабо. Органы чувств представле-
ны только чувствительными осязательными клетками, расположен-
ТИП ПЛОСКИЕ ЧЕРВИ
537
ными в эпителии. Выделительная система — протонефридиального
типа.
Половая система ленточных червей всегда гермафродитная, она
развита значительно лучше всех других внутренних органов
(рис. 228). Строение половой системы зависит от расположения чле-
ника в стробиле. У крупных цепней первые половые органы появля-
ются на расстоянии нескольких десятков или даже сотен члеников
(у бычьего цепня, например, у двухсотого) от сколекса. Поскольку
вначале развиваются мужские половые органы, такие членики назы-
вают мужскими. Мужская половая система состоит из многочислен-
ных мелких семенников, рассеянных в паренхиме, от которых отходят
тонкие семявыносящие протоки, сливающиеся между собой, образуя
семяпровод, который направляется к одной из боковых частей члени-
ка и там проходит через мускулистый совокупительный орган. При
сокращении мышц этот орган выдвигается и входит в половую клоа-
ку. После того как мужская половая система полностью сформирова-
лась, в проглоттиде начинают появляться женские половые органы, и
членик называется гермафродитным. Женская система состоит из
крупного яичника, от которого отходит яйцевод, впадающий в оотип.
Туда же впадает проток желточника и скорлуповых желез. Трубчатое
влагалище соединяет оотип с половой клоакой. Кроме того, от оотипа
отходит мешковидная матка. У разных цестод возможно перекрест-
ное оплодотворение и самооплодотворение. Яйца поступают в матку.
Ðèñ. 844 . Ñòðîåíèå ïîëîâîãî
àïïàðàòà øèðîêîãî ëåíòåöà
(Diphyllobofhrium laturn):
A ãåðìàôðîäèòíûé ÷ëåíèê (èç Øóëüöà è
Ãâîçäåâà); Á ñõåìà ñòðîåíèÿ æåíñêîé ïîëîâîé
ñèñòåìû (ïî Ñìèòó); 1 ñîâîêóïèòåëüíûé
îðãàí; 2 ñåìÿïðîâîä; 3 ñåìåííèêè; 4 ÿè÷íèê;
5 îîòèï; 6 æåëòî÷íèêè; 7 æåëòî÷íûé ïðî-
òîê; 8 ìàòêà; 9 âëàãàëèùå;10 îòâåðñòèå
ìàòêè. Ñëîæíûå ÿéöà (11), ñíàáæåííûå êðûøå÷-
êîé, âûõîäÿò â âîäó, ãäå â íèõ ðàçâèâàåòñÿ
ñâîáîäíàÿ ëè÷èíêà êîðàöèäèé (12)
Рис. 228. Строение полового аппарата
широкого лентеца (Diphullobofhrium latum):
А — гермафродитный членик (из Шульца
и Гвоздева), Б — схема строения женской по-
ловой системы (по Смиту);
1 — совокупительный орган, 2 — семяпровод,
3 — семенники, 4 — яичник, 5 — оотип,
6 — желточники, 7 — желточный проток,
8 — матка, 9 — влагалище, 10 — отверстие
матки. Сложные яйца (11), снабженные
крышечкой, выходят в воду, где в них
развивается свободная личинка —
корацидий (12)
538
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
По мере поступления яиц матка постепенно растягивается, при этом
от нее отходят боковые ответвления, количество и форма которых
специфичны для каждого вида, что очень важно при диагностике
гельминтоза. Основной объем проглоттиды занимает разросшаяся
матка, набитая яйцами, а яичник и все органы мужской половой сис-
темы дегенерируют и разрушаются. Такие членики, расположенные в
конце стробилы, называются зрелыми. Постепенно они отрываются и
выделяются из организма хозяина.
Развитие. У цестод сложный жизненный цикл со сменой промежу-
точных хозяев — они являются биогельминтами. Половозрелый червь
живет в просвете кишечника, прикрепившись к стенке кишки голов-
кой. Яйца попадают во внешнюю среду, где развивается личинка сфе-
рической формы, снабженная шестью крючьями, — онкосфера. Из обо-
лочки яйца онкосфера не выходит, пока не будет проглочена
промежуточным хозяином. Оказавшись в пищеварительном тракте
промежуточного хозяина, онкосфера освобождается от оболочки яйца.
С помощью крючьев она прободает стенку кишки, проникает в прос-
вет кровеносного сосуда и разносится с током крови по органам и тка-
ням промежуточного хозяина. Там онкосфера превращается в финну,
которая в дальнейшем пребывает в состоянии покоя, дожидаясь попа-
дания в организм окончательного хозяина. Обычно это происходит,
когда окончательный хозяин поедает промежуточного хозяина. Ока-
завшись в кишечнике окончательного хозяина, финна превращается в
молодого цепня. У разных видов в развитии могут быть отклонения от
описанной схемы. Промежуточное развитие может происходить в вод-
ной среде, со сменой нескольких промежуточных хозяев и т. д . У мно-
гих цестод окончательным или промежуточным хозяином является
человек. Вызываемые ими заболевания называются цестодозами.
Свиной солитер, или вооруженный цепень, распространен повсе-
местно. Во взрослом состоянии он является возбудителем тениоза,
заражение финнами вызывает заболевание цистицеркоз. Половозре-
лая особь обитает в тонкой кишке человека и поглощает питательные
вещества из химуса. Длина стробилы достигает 2—3 метров. Сколекс,
кроме четырех присосок, имеет двойной венчик из 22—32 кутикуляр-
ных крючьев. Признаком гермафродитных члеников является не-
большое количество ответвлений матки в зрелом членике — от семи
до двенадцати. Промежуточным хозяином является свинья, реже —
другие позвоночные: кошки, собаки и человек. Человек может про-
глотить яйца, например, при употреблении сырой воды. Онкосферы
оседают в мышцах, реже в легких, глазах, мозге и других органах.
Примерно через два месяца из онкосфер развиваются финны — ци-
стицерки. Заражение финнами происходит при употреблении сырого
или плохо обработанного мяса, зараженного финнами. В кишке окон-
ТИП ПЛОСКИЕ ЧЕРВИ
539
чательного хозяина головка финны выворачивается и прикрепляется
к стенке, после чего от шейки отшнуровываются молодые проглотти-
ды. Значительно более опасно заболевание цистицеркозом, поскольку
с током крови онкосферы заносятся в различные органы, развиваю-
щиеся финны вызывают мучительные боли, воспалительные процес-
сы и могут нарушать деятельность органа. Так, например развитие
финн в глазном яблоке приводит к слепоте, а в мозге — к смерти. Уда-
ление финн возможно только хирургическим путем.
Бычий солитер, или невооруженный цепень, вызывает заболева-
ние тениаринхоз, которое распространено повсеместно. Окончатель-
ным хозяином паразита является только человек, у которого он засе-
ляет тонкую кишку. Бычий солитер отличается от свиного
значительно большей длиной стробилы (до 10 м и более), наличием
только четырех присосок и отсутствием на сколексе крючьев. Наби-
тая яйцами матка зрелого членика имеет 17—35 боковых ответвлений.
В течение года один цепень образует и выделяет в окружающую среду
около 2500 члеников. Развитие типично, промежуточным хозяином
является крупный рогатый скот. В организме человека финны этого
цепня не развиваются.
Эхинококк является возбудителем эхинококкоза, которое распро-
странено повсеместно, преимущественно в районах с развитым жи-
вотноводством. Половозрелая особь заселяет тонкую кишку различ-
ных представителей семейства собак — шакалов, волков, лисиц,
домашних собак и др. Размеры взрослого червя около 5 мм. Помимо
присосок сколекс имеет двойной венчик крючьев, а стробила состоит
из трех-четырех члеников. Промежуточными хозяевами могут быть
различные млекопитающие, включая человека. С током крови онкос-
феры оседают в головном мозге, печени и др., где развиваются в фин-
ну. Финна эхинококка медленно, но постоянно растет: через месяц
она достигает 1 мм, а через 5 месяцев — 1 см в поперечнике. Через
несколько лет финна у человека достигает размеров детской головки,
а у крупных копытных — нескольких десятков килограммов. Извест-
ны случаи, когда размеры пузыря превышали 60 кг. При этом в перво-
начальном двуслойном пузыре финны образуются дочерние пузыри
с многочисленными головками. Эхинококк является одним из наибо-
лее опасных паразитов человека. Крупная, постоянно увеличивающа-
яся в размерах финна механически сдавливает окружающие ткани
печени, вызывая ее атрофию. Кроме того, ядовитые продукты жизне-
деятельности эхинококка отравляют организм. Возможно только хи-
рургическое лечение эхинококкоза.
Альвеококк — возбудитель альвеококкоза, многокамерного, или
альвеолярного, эхинококкоза, которое встречается значительно реже,
чем эхинококкоз. Область распространения паразита велика, в Рос-
540
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
сии — преимущественно на Урале и в Сибири. Половозрелая особь
обитает в тонкой кишке у тех же хозяев, что и эхинококк, однако раз-
меры взрослого альвеококка несколько меньшие, не более 3 мм. Ско-
лекс имеет крючья, расположенные на хоботке, стробила состоит из
3—4 члеников. Промежуточными хозяевами главным образом явля-
ются мышевидные грызуны, иногда человек. Финна чаще всего разви-
вается в печени, реже в легких, лимфатических узлах, мозге и др. ор-
ганах. В отличие от финны эхинококка, в которой дочерние пузыри
почкуются внутрь основного пузыря, у финны альвеококка почкова-
ние осуществляется только наружу. В результате образуется постоян-
но растущий узел, состоящий из мелких, лишенных жидкости пу-
зырьков, в каждом из них находится сколексы, которые постепенно
проникают в окружающие ткани и разрушают их. Альвеококкоз еще
более опасен, чем эхинококкоз. При хирургическом удалении финны
необходимо удалять большое количество тканей пораженного органа.
Широкий лентец вызывает дифиллоботриоз. Ареал распростране-
ния паразита очень широк, он живет в водоемах, где проходит проме-
жуточное развитие паразита. В России природные очаги находятся в
прибрежных населенных пунктах рек Северо-Запада и Сибири, Вол-
ги и Днестра. Половозрелая особь живет в тонкой кишке человека
и многих плотоядных млекопитающих. Широкий лентец является од-
ним из самых крупных гельминтов человека: длина взрослого червя
может превышать 10 м. На сплющенном с боков сколексе яйцевидной
формы длиной около 2,5 мм имеются две присасывательные борозды
(ботрии), нет крючьев. Заполненная матка образует многочисленные
петли, которые видны в зрелом членике в виде темноокрашенной ро-
зетки. Длина зрелого членика заметно меньше его ширины, поэтому в
десятиметровой стробиле может насчитываться до 4000 члеников,
тогда как стробила бычьего солитера почти такой же длины состоит
лишь из немногим более 1000 члеников. Через отверстие матки ог-
ромное количество яиц попадают в полость кишки и выделяются с
фекалиями из организма хозяина. Для дальнейшего развития яйца
должны попасть в воду, где в них начинают формироваться личин-
ки — корацидии, покрытые ресничками и способные медленно пла-
вать. Через некоторое время личинку заглатывает первый промежу-
точный хозяин — рачок-фильтратор циклоп. В нем корацидий
сбрасывает реснички, прободает стенку кишечника рачка, оказывает-
ся в полости его тела, где изменяется в процеркоид.
Дальнейшее развитие личинки возможно только в организме вто-
рого промежуточного хозяина, которым является рыба, проглотившая
зараженного рачка. Процеркоид мигрирует из кишечника рыбы в ее
мышцы и там превращается в плероцеркоида, имеющего вид уплощен-
ного белого червячка. Плероцеркоиды не погибают, если зараженную
ТИП КРУГЛЫЕ ЧЕРВИ
541
им рыбу съест другая рыба, — он осядет в ее мышцах и во внутренних
органах. Поэтому хищные рыбы в течение своей жизни могут нако-
пить огромное количество плероцеркоидов. Особенно часто заражен-
ными оказываются щуки, налимы и судаки, реже — окуни и ерши.
Окончательный хозяин заражения плероцеркоидами – тот, кто
употребляет в пищу сырую (непроваренную, непрожаренную, а также
непросоленную) рыбу или икру. Проглоченный плероцеркоид при-
крепляется к стенкам тонкой кишки с помощью ботрий и вырастает
в половозрелую особь.
ТИП КРУГЛЫЕ ЧЕРВИ
Круглые черви являются многоклеточными трехслойными живот-
ными с билатеральной симметрией тела. Тело имеет цилиндрическую
форму с круглым сечением на поперечном разрезе. Сегментация тела
отсутствует, но в большей степени, нежели у плоских червей, выраже-
на цефализация. Жизненно важные органы сосредоточены на перед-
нем конце тела. У круглых червей впервые появляется полость тела,
т. е. обособленное пространство между стенкой тела (кожно-мускуль-
ный мешок) и внутренними органами. У плоских червей это про-
странство заполнено рыхлой клетчаткой — паренхимой. Полость тела
у них является первичной (псевдоцель), поскольку эмбрионально про-
исходит из бластоцеля и представляет собой пространство между эк-
тодермой (покровом тела) и энтодермой кишки. Появление полости
выгодно для животного. Во-первых, она заполнена жидкостью, по ко-
торой значительно более эффективно диффундируют питательные
вещества от кишечника к тканям, продукты распада — от тканей к ор-
ганам выделительной системы, кислорода — от покровов тела к тка-
ням, углекислого газа — от тканей к поверхности тела и т. д . Во-вто-
рых, заполняющая полость тела жидкость давит изнутри на его стенку,
обеспечивая внутреннюю опору (внутренний гидроскелет).
Дыхательная и кровеносная системы отсутствуют у круглых чер-
вей, поэтому газообмен идет через покровы тела, а транспорт веществ
между тканями осуществляется посредством диффузии через полост-
ную жидкость. Черви имеют пищеварительную, выделительную,
нервную и половую системы.
В пищеварительной системе имеется задняя кишка и анальное от-
верстие. Благодаря этому пищеварительная система круглых червей
значительно более совершенная, чем у представителей описанных ра-
нее типов. Сквозная пищеварительная трубка дает возможность пи-
щевым массам проходить по ней в одностороннем направлении, а не
в двустороннем, как у плоских червей. Поэтому различные отделы
пищеварительной системы специализированы.
542
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Выделительная система, развитая слабо, образована протонефри-
диями или видоизмененными кожными железами.
Нервная система ортогонального типа состоит из окологлоточно-
го нервного кольца и отходящих от него нервных стволов, которые
связаны между собой многочисленными комиссурами. Органы чувств
развиты слабо.
Половая система организована более просто, чем у плоских чер-
вей. Большинство круглых червей раздельнополые, причем обычно у
них выражен половой диморфизм.
Тип включает в себя классы: собственно круглые черви,
брюхо ресничные, киноринхи, волосатики и ко-
ловратки. Ниже мы подробно рассмотрим организацию класса
собственно круглые черви.
Класс собственно круглые черви, или нематоды. К нематодам от-
носят несколько десятков тысяч видов. Эти организмы заселяют пра-
ктически все экологические ниши: любые водоемы, почву. Значитель-
ное количество видов являются паразитами растений, животных и
человека.
Форма, тело нематод веретенообразное, суживающееся к обоим
концам. На переднем конце располагается рот, анальное отверстие от-
крывается вблизи заднего конца тела на
брюшной поверхности. Позади рта на
брюшной поверхности имеется выдели-
тельное отверстие. У самок имеется еще
одно отверстие — половое, расположенное
на брюшной поверхности примерно в
средней части тела (рис. 229).
Рис. 229. Вскрытая аскарида (самка):
1 — губы, 2 — нервное кольцо, 3 — глотка,
4 — фагоцитарные клетки, 5 — «пищевод»
(передний отдел средней кишки), 6 — средняя
кишка, 7 — боковая линия, 8 — яйцевод, 9 — матка,
10 — яичник, 11 — влагалище, 12 — брюшной валик
гиподермы (по Стрелкову)
ТИП КРУГЛЫЕ ЧЕРВИ
543
Стенку тела образует кожно-мускульный мешок, покрытый снару-
жи сложно устроенной многослойной кутикулой, которая имеется
и у паразитических, и у свободноживущих видов. Кутикула выполня-
ет защитную роль — создает опору для лежащей под ней мускулатуры
кожно-мускульного мешка. Под кутикулой располагается гиподер-
ма — эпителий кожно-мускульного мешка, который имеет клеточное
строение или сливается в единую цитоплазматическую массу — син -
цитий. Гиподерма выделяет вещества, образующие кутикулу. Вдоль
тела гиподерма формирует четыре продольных валика: два по бокам
тела и два — по средней линии спинной и брюшной сторон. В отличие
от многослойных мышц плоских червей, мышечные волокна нематод
располагаются только продольно, будучи разделенными валиками ги-
подермы на четыре продольные ленты. Отсутствие кольцевой муску-
латуры ограничивает подвижность червя, который способен лишь из-
гибаться и медленно перемещаться на боку.
Пищеварительная система. На протяжении пищеварительной
трубки имеет место четкая функциональная специализация органов.
Ротовое отверстие, расположенное на переднем конце тела, обычно
окружено выростами — губами (у аскариды их три). Отверстие ведет
в ротовую полость (стому), которая затем продолжается в глотку (пи-
щевод). Все эти органы составляют переднюю кишку. В стенке пище-
вода расположены многочисленные железы, которые оказывают пер-
воначальное воздействие на пищу. Пищевод продолжается в длинную
среднюю кишку. Ее стенка образована лишь одним слоем эпителиаль-
ных клеток, лежащих на тонкой базальной мембране. В средней киш-
ке расщепляется пища и всасываются вещества, которые затем диф-
фундируют в полостную жидкость и распространяются по тканям.
Паразитические нематоды, обитающие в бескислородной среде поло-
сти кишечника, получают АТР путем гликолиза. За средней кишкой
следует короткая задняя, заканчивающаяся на брюшной стороне зад-
ней части тела анальным отверстием. У самцов в нее впадает семя-
провод, поэтому через анус у них выделяются не только экскременты,
но и половые клетки.
Выделительная система. У аскариды вся выделительная система
образована одной гигантской клеткой. У нематод отсутствуют прото-
нефридии. Это связано с отсутствием у них клеток, снабженных рес-
ничками или жгутиками. Напомним: в протонефридиях такие клетки
(пламенные) обязательно имеются.
Нервная система состоит из центрального и периферического
отделов. Центральный образован нервным кольцом, окружающим
глотку, и отходящими от него нервными стволами. Периферический
отдел составляют нервные ветви, отходящие от центров и сплетения
(рис. 230).
544
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
У нематод органы чувств выражены слабо, поскольку они ведут
либо паразитический, либо малоподвижный образ жизни. Нематоды
обладают чувствительными осязательными бугорками или щетинка-
ми, расположенными на переднем конце тела вокруг рта.
Половая система. Нематоды раздельнополые с выраженным по-
ловым диморфизмом. Они размножаются только половым путем.
У самца закладываются парные половые органы, однако в процессе
онтогенеза чаще всего они становятся непарными. У самок сохраня-
ются парные половые органы.
Опишем типичную половую систему аскариды. Мужская половая
система состоит из длинного, очень тонкого нитевидного семенника, в
котором образуются безжгутиковые сперматозоиды. Семенник перехо-
дит в семяпровод, который заканчивается коротким семенным пузырем,
в котором скапливаются сперматозоиды. Семенной пузырек переходит
в семяизвергательный канал, а тот, в свою очередь, впадает в заднюю
кишку неподалеку от анального отверстия. У нематод внутреннее опло-
дотворение. Женская половая система аскариды начинается двумя ни-
тевидными яичниками, в которых образуются и растут ооциты. Яични-
ки переходят в яйцеводы, по которым ооциты транспортируются в еще
более широкие трубчатые матки. В матке ооциты делятся мейотически,
в результате чего образуются яйцеклетки, которые окружаются толстой
оболочкой. В матке происходит и оплодотворение, причем одновремен-
но можно обнаружить ооциты и яйцеклетки на разных стадиях разви-
тия. Обе матки переходят в короткое непарное влагалище, открывающе-
еся половым отверстием на брюшной стороне тела.
Начальные этапы развития происходят в матке, дальнейшее раз-
витие — в окружающей среде. Паразитические черви (гельминты),
которые не нуждаются для личиночного развития в промежуточном
хозяине, называются геогельминтами. Среди нематод, заселяющих
различные экологические ниши, есть свободноживущие формы
Рис. 230. Схема нервной системы аскариды с брюшной стороны:
1 — ротовые сосочки с осязательными окончаниями и иннервирующими их нервами,
2 — окологлоточное нервное кольцо, 3 — боковые головные ганглии, 4 — брюшной
нервный ствол, 5 — боковые нервные стволы, 6 — кольцевые нервы, 7 — задний
ганглий, 8 — чувствительные сосочки с соответствующими нервами, 9 — анальное
отверстие, 10 — спинной нервный ствол (по Брауну)
ТИП КОЛЬЧАТЫЕ ЧЕРВИ
545
и много паразитических. Многие виды способны жить в организме
человека. Поэтому нематоды имеют большое медицинское значение.
Опишем наиболее важные для человека виды.
Аскарида человеческая — возбудитель аскаридоза. Половозрелые
особи живут в тонкой кишке человека, при этом самки достигают
40 см в длину, а самцы — 20—25 см при ширине 3—4 мм. У самок зад-
ний конец тела прямой, у самцов он загнут на брюшную сторону. За-
остренное на концах тело паразита упирается в стенки кишки, препят-
ствуя проходящим массам химуса механически вынести его наружу.
В течение суток самка аскариды откладывает в просвет кишечника до
240 000 овальных яиц, покрытых толстой крупнобугристой оболоч-
кой. Развитие эмбриона начинается еще в половых путях самки, но
дальнейшее формирование личинки требует кислорода. С фекалиями
яйца аскарид выносятся из кишечника хозяина и засоряют окружаю-
щую среду (почву, грунтовые воды и т. д.) . В присутствии кислорода
(первая кислородная стадия развития) и в условиях высокой влажно-
сти при температуре +20—25 °С в яйце развивается личинка. В про-
цессе развития личинка линяет, но не покидает оболочки яйца, пока не
будет проглочена хозяином. В таком состоянии яйца способны зара-
жать человека, т. е. они становятся инвазионными. Заражение происхо-
дит через грязные руки, при употреблении немытых овощей, ягод,
фруктов и сырой воды из открытых источников. Когда проглоченное
яйцо попадает в пищеварительный тракт человека, из яйца выходит
личинка, которая прободает стенку кишки, проникает в кровеносный
сосуд и с током венозной крови попадает в легкие, где в альвеолах про-
ходит вторая (кислородная) стадия развития личинки. Эта стадия со-
провождается двумя линьками. По мере развития личинка постепенно
мигрирует из альвеол в бронхиолы и более крупные бронхи и далее по
направлению к гортани. Личинка перемещается за счет движения рес-
ничек эпителия, дыхательных путей человека. Оказавшись в гортани,
личинка раздражает слизистую оболочку, вызывая кашель, в результа-
те чего она попадает в ротовую полость и проглатывается со слюной.
Так личинка возвращается в тонкую кишку, где развивается в половоз-
релую особь. Аскариды распространены повсеместно, особенно при
использовании в качестве удобрений фекалий.
ТИП КОЛЬЧАТЫЕ ЧЕРВИ
Кольчатые черви являются многоклеточными трехслойными жи-
вотными, имеющими билатеральную симметрию тела. Организация
кольчатых червей значительно более сложная, чем у представителей
рассмотренных ранее типов. Это первые сегментированные живот-
ные. Их тело подразделяется на три отдела: передний представлен
546
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
головной лопастью — простомиумом; средний отдел — основная
часть — сегментированное туловище, и задняя — пигидия, или аналь-
ная лопасть. Сегментация гомономная, т. е . все сегменты туловища
одинаковы (рис. 231). Выражена цефализация — обособление голов-
ного отдела на переднем конце тела путем слияния передних сегмен-
тов с одновременным увеличением относительной массы мозгового
ганглия. У кольчатых червей имеется вторичная полость тела (цело
м).
Напомним: первичная полость — это остаток бластоцеля, т. е. про-
странства, которое в ходе эмбрионального развития возникает между
эктодермой и энтодермой кишки. В отличие от нее, вторичная по-
лость развивается в мезодерме, из которой образуется большинство
тканей и органов. Она выстлана специальным однослойным плоским
эпителием — мезотелием. У плоских червей мезодерма покрывает из-
нутри стенку тела (мышечную часть кожно-мускульного мешка) и
снаружи — расположенные в целоме органы пищеварительной систе-
мы (все оболочки их стенок, кроме эпителия). Целом, в отличие от
первичной полости тела, разделяет не эктодерму и энтодерму, а раз-
личные структуры мезодермы — среднего зародышевого листка. Вто-
ричная полость кольчатых червей разделена на большое количество
парных целомических мешков, количество которых соответствует ко-
Рис. 231. Поперечный разрез из средней части тела дождевого червя:
1 — кутикула, 2 — эпидермис, 3 — слой кольцевой мускулатуры, 4 — слой
продольной мускулатуры, 5 — целомический эпителий, 6 — метанефридий,
7 — щетинка, 8 — мезентерий, 9 — брюшной сосуд, 10 — субневральный сосуд,
11 — брюшная нервная цепочка, 12 — хлорагогенные клетки, 13 — полость кишки,
14 — сосудистый плексус, 15 — тифлозоль, 16 — сосуд тифлозоля, 17 — спинной
сосуд (по Петрушевскому)
ТИП КОЛЬЧАТЫЕ ЧЕРВИ
547
личеству сегментов тела, так как в каждом сегменте имеется свой це-
ломический мешок. Целомические мешки разных сегментов отделены
друг от друга тонкими септами-перегородками. Пара мешков одного
сегмента соприкасается своими стенками над и под кишкой, образуя
здесь двухслойную перегородку. Эта перегородка не только разделяет
целомические мешки, но и поддерживает кишку, охватывая ее со всех
сторон, а также как бы подвешивает ее, образуя верхнюю и нижнюю
брыжейки (мезентерии). Между листками брыжеек к кишке подхо-
дят кровеносные сосуды и нервы. Целом заполнен жидкостью и вы-
полняет функцию гидроскелета. Целом осуществляет выделительную
функцию, там диффузно транспортируются вещества и созревают
гаметы.
Пищеварительная система состоит из передней, средней и задней
кишки. Пищеварительный тракт сквозной, он начинается ротовым
отверстием и заканчивается анальным. Каждый отдел имеет свою
функциональную специализацию. В отличие от круглых червей, стен-
ка кишки кольчецов образована не одним слоем клеток, а нескольки-
ми, причем все слои (кроме самого внутреннего — эпителия) сформи-
рованы из мезодермы.
У кольчатых червей отсутствует дыхательная система, газооб-
мен осуществляется через всю поверхность тела. У многощетинко-
вых имеются выросты тела — аналоги примитивных жабр. Транс-
порт веществ между тканями осуществляется циркулирующей
кровью, а не путем диффузии через тканевую жидкость. Кровеносная
система состоит из полых сосудов, которые пронизывают все тело
червя и образуют сплетения во всех его тканях. Кровеносные сосуды
возникают в связи с появлением вторичной полости тела, а остатки
первичной полости трансформируются в сосуды. Обе полости отде-
лены одна от другой, поэтому кровеносная система кольчатых чер-
вей замкнутая.
Кровеносная система состоит из двух больших сосудов (брюшного
и спинного), которые проходят под и над кишкой. Оба сосуда сообща-
ются между собой посредством кольцевых сосудов, которые имеются
в каждом сегменте тела. Кольчатые черви лишены сердца. Кровь дви-
жется по сосудам за счет периодической пульсации стенок спинного
сосуда, по которому кровь движется по направлению к переднему
концу тела. У малощетинковых, кроме спинного сосуда, сокращаются
и некоторые кольцевые сосуды. Кровь переносит по организму пита-
тельные вещества от органов пищеварения и кислород, поступающий
через покровы тела. Дыхательный пигмент, связывающий кислород
всех беспозвоночных, содержится не в клетках, он растворен в плазме
крови, дыхательный пигмент (гемоглобин) позвоночных находится
в эритроцитах.
548
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Выделительная система представлена метанефридиями, которые
расположены попарно в каждом сегменте тела. В отличие от прото-
нефридиев, которые образуют сеть соединенных между собой трубо-
чек, пронизывающих все тело животного, метанефридии представля-
ют собой короткие трубочки, не связанные между собой. Каждая из
них начинается во вторичной полости воронкообразным отверстием
и заканчивается отверстием на поверхности тела. Метанефридии —
это канал, соединяющий целом с окружающей средой, поэтому его от-
носят к целомодуктам. Продукты распада поступают в целомическую
жидкость, заполняющую вторичную полость тела, и постепенно нака-
пливаются. Через метанефридии жидкие продукты обмена удаляются
из тела червя в окружающую среду.
Нервная система состоит из пары слившихся узлов, образующих
«головной мозг»; двух нервных стволов, которые соединяют его с пер-
вой парой узлов брюшной нервной цепочки, огибая при этом с двух
сторон глотку (окологлоточное кольцо). Брюшная нервная цепочка
образована ганглиями, расположенными попарно в каждом сегменте
туловища червя. Оба ганглия связаны между собой и с ганглиями со-
седних сегментов.
Органы чувств представлены осязательными структурами, глаза-
ми (не у всех) и обонятельными ямками.
Половая система. Среди кольчецов имеются как раздельнополые,
так и гермафродитные формы. У разных представителей типа разви-
тие идет прямо или с метаморфозом, в результате которого образует-
ся плавающая личинка — трохофора, не похожая на взрослого червя.
Тип кольчатых червей подразделяется на два подтипа: беспоя-
сковые ипоисковые, которые насчитывают в общей сложности
около 10 000 видов. Согласно эволюционной теории, кольчатые черви
произошли от низших червей. Исходную группу составляли много-
щетинковые, от которых в процессе перехода к пресноводному образу
жизни образовались малощетинковые, а от них, в свою очередь, пияв-
ки, ведущие паразитический образ жизни. По мнению систематиков,
от кольчатых червей произошли членистоногие. Следует указать, что
эволюционная теория разделяется не всеми учеными.
Класс малощетинковые насчитывает около 3400 видов. Живут
они не только в водоемах, но и в почве, причем размеры почвенных
форм могут быть очень большими. Например, австралийский земля-
ной червь достигает трех метров в длину. Самые мелкие виды имеют
длину не более 0,5 мм. Наиболее известным видом является широко
распространенный дождевой червь, который имеет красноватую окра-
ску и достигает 20—30 см в длину.
Отсутствуют пальпы и параподии. Щетинки имеются, но их мало,
а размеры невелики. Сегменты передней части тела крупнее задних.
ТИП КОЛЬЧАТЫЕ ЧЕРВИ
549
Щетинки способствуют передвижению червя, так как увеличивают
сцепление его тела с субстратом. Утолщение тела (поясок) лучше ви-
ден со спинной стороны. Задняя часть тела дождевого червя уплоще-
на, и когда червь ночью частично выползает из норки в поисках пищи,
это утолщение держит его за край норки.
Эпителий кожно-мускульного мешка выделяет наружу тонкий
слой эластичной и прозрачной кутикулы. Слизь, выделяемая эпите-
лием, покрывает кожу, увлажняя ее, что необходимо для нормального
газообмена. Если покровы тела червя высыхают, он быстро погибает
от удушья. Расположенные под эпителием мышечные волокна обра-
зуют два слоя: наружный — кольцевой и более мощный внутренний —
продольный. Сокращение волокон позволяет червю совершать слож-
ные движения. При этом попеременно и согласованно работает не
только вся мускулатура, но и мышцы отдельных сегментов. В резуль-
тате определенные участки тела червя то последовательно сужаются
(при этом червь вытягивается вперед и раздвигает комочки почвы),
то расширяются, тело укорачивается и утолщается, уплотняя почву
вокруг. Таким образом, червь проникает в почву наподобие колышка,
что и определило название этих животных.
Пищеварительная система хорошо дифференцирована. Передняя
кишка подразделяется на глотку, пищевод (у некоторых имеется зоб)
и мускульный желудок. Ротовое отверстие переходит в ротовую по-
лость, которая ведет в объемистую глотку. В полость глотки открыва-
ются протоки многочисленных слюнных желез, секрет которых со-
держит ферменты и увлажняет пищу. Из глотки пища попадает в
пищевод. У дождевых червей в стенках пищевода имеются известко-
вые железы, которые удаляют излишки солей кальция из крови. Из
желез соли кальция поступают в полость пищевода и нейтрализуют
там гуминовые кислоты, большое количество которых имеется в по-
едаемых червями гниющих листьях. Передняя кишка заканчивается
широким желудком, в котором пища перетирается благодаря сокра-
щениям мощной мускулатуры его стенок. Перетиранию пищи способ-
ствуют песчинки, которые находятся в полости желудка.
На дорсальной стороне средней кишки имеется глубокое желобо-
видное впячивание, ориентированное вдоль органа, — тифлозоль,
который увеличивает переваривающую и всасывающую поверх-
ность. В стенке кишки имеется тонкая прослойка мускулатуры, со-
стоящая из наружного продольного и внутреннего кольцевого слоев.
Между базальной мембраной кишечного эпителия и мышечными
волокнами залегает густая сеть кровеносных капилляров. В средней
кишке биополимеров пищи расщепляются и всасываются образо-
вавшиеся продукты. Короткая задняя кишка заканчивается аналь-
ным отверстием.
550
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Дыхательная система отсутствует, газообмен осуществляется пу-
тем диффузии через всю поверхность кожи благодаря наличию в по-
верхностных слоях стенки тела густой сети кровеносных сосудов.
Кровеносная и нервная системы имеют типичное для кольчатых
червей строение.
Выделительная система представлена метанефридиями.
Органы чувств развиты плохо. Они образованы собранными в
группы чувствительными клетками — сенсиллами или отдельными
чувствительными клетками, которые рассеяны в коже. Глаза отсутст-
вуют, но дождевые черви способны реагировать на свет благодаря на-
личию в коже светочувствительных клеток.
Половая система малощетинковых кольчецов гермафродитна, по-
ловые железы расположены лишь в некоторых сегментах тела
(рис. 232). У дождевого червя в X и XI сегментах тела находятся две
пары семенников. Семяпроводы открываются на брюшной стороне
мужским половым отверстием. Они не связаны с метанефридиями и
представляют собой самостоятельные целомодукты. Женская поло-
вая система представлена парой яичников, локализованных в XIII сег-
менте. В них развиваются ооциты. Короткие яйцеводы открываются
женским половым отверстием в XIV сегменте.
У дождевых червей оплодотворение перекрестное. При копуляции
половые партнеры тесно соприкасаются друг с другом своими брюш-
ными частями, каждый половой партнер выделяет слизистый футляр,
который сливается с таким же футляром другого червя, образуя общую
слизистую муфту, охватывающую их обоих. Каждый партнер выделяет
1
34
6
7
8
9
10
11
12
13
14
15
2
5
16
17
18
19
20
IXXXIXIIXIIIXIVXV
XVI
Рис. 232. Схема половой системы
дождевого червя:
1 — передний семенной мешок,
2 — семенные капсулы,
3 — средний семенной мешок,
4 — задний семенной мешок,
5 — диссепимент XII/XIII,
6 — диссепимент XIII/XIV,
7 — часть диссепимента XIII/XIV,
8 — женская половая воронка,
9 — женский половой проток,
10 — брюшная нервная цепочка,
11 — брюшной ряд щетинок,
12 — боковой ряд щетинок,
13 — мужское половое
отверстие, 14 — семяпровод,
15 — яйцевой мешок,
16 — яичник,
17 — семявыносящий проток,
18 — мужские половые воронки, 19 — семенник, 20 — семяприемники.
IX—XVI
—
сегменты. На правой стороне семенные мешки вырезаны,
а семенные капсулы вскрыты
ТИП ЧЛЕНИСТОНОГИЕ
551
из своих мужских половых отверстий по капельке спермы. В результа-
те они попеременно выполняют роль и самца и самки, получая сперму
другой особи. Поэтому гермафродитизм не препятствует обмену гене-
тическим материалом. После копуляции железы пояска выделяют сли-
зистую муфту, в которую сначала откладываются одно — три яйца. По-
сле этого муфта застывает и превращается в кокон, защищающий яйца.
Развитие олигохет прямое. Из яйца выходит молодой червь, внеш-
не сходный со взрослой особью, который сразу же начинает самосто-
ятельную жизнь. Кроме полового размножения, у малощетинковых
кольчецов возможно и бесполое размножение путем архитомии. При
этом тело червя делится на две части, после чего в каждой из них до-
страиваются недостающие структуры. Все олигохеты обладают высо-
кой регенераторной способностью, поэтому при разделении особи из
разрезанных частей образуются полноценные новые черви.
Значение малощетинковых кольчатых червей в природе очень велико.
Земляные черви, являющиеся одними из самых многочисленных живот-
ных на планете, заселяют почти все географические зоны. Они являются
пищей для многих животных, увеличивают плодородие почвы, улучшая
ее структуру. Черви, обитающие на территории одного гектара, в течение
года заглатывают и выносят на поверхность до 30 тонн (!) переработан-
ной почвы. Пробивая себе дорогу в плотной почве, червь разрыхляет ее,
а затаскивая в норки опавшие листья — обогащает гумусом.
Ч. Дарвин в книге «Образование растительного слоя деятельнос-
тью дождевых червей и наблюдения над образом жизни последних»
(1881) приводит интересные данные. На одном гектаре почвы обитает
от 60 до 133 тыс., а в отдельных местах до 0,5—2 млн дождевых чер-
вей. За сутки каждый из них пропускает через себя количество земли,
равное собственной массе. На одном гектаре дождевые черви перера-
батывают около 250 кг земли, а там, где их особенно много, — до
2—3 тонн. Общее количество почвы, переработанной червями за год,
образует слой толщиной около 3 мм.
ТИП ЧЛЕНИСТОНОГИЕ
Этот тип наиболее разнообразный среди других систематических
групп. Он включает более полутора миллионов (!) видов, т. е. больше,
чем общее количество видов всех остальных организмов, вместе взя-
тых. Каждый год зоологи открывают сотни новых видов, и по прибли-
зительным оценкам общее количество видов превышает два миллио-
на членистоногих!
Членистоногие — многоклеточные животные с билатеральной
симметрией и вторичной полостью тела. В процессе онтогенеза сег-
ментированный целом смешивается с остатками первичной полости
552
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
тела, поэтому у взрослых особей полость тела смешанная — миксо-
цель. Выражена сегментация тела. Однако, в отличие от гомономной
сегментации кольчатых червей, у членистоногих она гетерономная,
т. е. строение сегментов разных частей тела различное. У некоторых
видов сегменты сливаются, образуя отделы тела: голову, грудь и
брюшко, которые также могут сливаться между собой; у некоторых
сливаются все отделы, образуя монолитное тело.
Голова состоит из головной лопасти — акрона и четырех сегмен-
тов. На акроне большинства членистоногих имеются придатки — уси-
ки, или антенуллы. Количество сегментов в груди и в брюшке широко
варьирует. Тело покрыто жесткой кутикулой, образующей экзоскелет
(наружный скелет). Кутикула покрывает тело снаружи и выстилает
начальные отделы передней и задней кишок, половых протоков и, др.
Кутикула состоит из наружного слоя — эпикутикулы и расположен-
ной под ней протокутикулы. Эпикутикула обеспечивает защиту от
потери влаги, что дает возможность членистоногим, в отличие от
кольчатых червей, длительное время жить без воды или в сухой по-
чве. Протокутикула придает прочность покровам. Ее основой являет-
ся хитин, обеспечивающий кутикуле гибкость. У ракообразных и
многоножек кутикула дополнительно пропитана солями и становится
благодаря этому более жесткой. Наличие легких и прочных покровов
позволяет животному быстро передвигаться. В результате членисто-
ногие освоили землю и воздушное пространство. Однако жесткая ку-
тикула нерастяжима и поэтому сдерживает рост животного. Поэтому
периодически происходит линька, в ходе которой старая кутикула от-
деляется и сбрасывается, а под ней к этому времени уже образуется
новая. Новая кутикула некоторое время мягкая и растяжимая, поэто-
му животное может увеличивать свои размеры. Благодаря смешанной
полости тела возможен ускоренный рост. Тело может заполняться
большим количеством жидкости, из-за чего его размер быстро увели-
чивается. После этого покровы тела затвердевают, и дальнейшее уве-
личение размера невозможно до следующей линьки.
Главной особенностью членистоногих является наличие у них под-
вижных конечностей. Конечности членистоногих функционируют эф-
фективно, так как состоят из нескольких звеньев, подвижно соеди-
ненных между собой суставами, и способны совершать движения
вокруг нескольких осей. Важно, что конечности, расположенные на
разных частях тела, специализированы.
Мышечная система значительно более совершенна, чем у других
беспозвоночных. Во-первых, членистоногие имеют поперечнополоса-
тую мышечную ткань. Во-вторых, у них нет кожно-мускульного меш-
ка, а соматическая мускулатура образована пучками мышечных
волокон, которые прикрепляются изнутри к наружному скелету
ТИП ЧЛЕНИСТОНОГИЕ
553
и обеспечивают движения отдельных частей тела. В зависимости от
выполняемой функции мышцы делят на различные группы (сгибате-
ли — разгибатели, приводящие — отводящие, сфинктеры — дилятато-
ры, синергисты — антагонисты).
В пищеварительной системе различают дифференцированные пе-
реднюю, среднюю и заднюю кишки. Средняя кишка энтодермальная,
передняя и задняя — эктодермальные и выстланы кутикулой. У мно-
гих членистоногих имеется развитый ротовой аппарат, в разные отде-
лы пищеварительного тракта открываются протоки многочисленных
мелких и крупных желез.
Хорошо развитые дыхательная и кровеносная системы обеспечи-
вают высокую подвижность членистоногих.
Дыхательная система. Можно выделить три основных типа дыха-
ния: жаберное, легочное и трахейное. Жабры, имеющиеся у первично-
водных членистоногих (ракообразных), представляют собой видоиз-
мененные конечности или их выросты. Легкие, которые также
образуются на основе конечностей, имеются у наземных форм. Тра-
хейное дыхание — наиболее совершенное, оно представлено системой
многочисленных ветвящихся трубочек, пронизывающих все тело жи-
вотного, по которым проходит воздух. Обычно у различных члени-
стоногих имеется только один тип органов дыхания.
Кровеносная система — незамкнутого типа. По сосудам и лакунам
течет гемолимфа, которая представляет собой кровь, смешанную с
тканевой жидкостью. Дыхательный пигмент растворен в гемолимфе.
Клеточные элементы — фагоциты. Движение гемолимфы осуществ-
ляется благодаря сокращениями сердца, расположенного на спинной
стороне тела. Гемолимфа из сердца направляется в сосуды, а из них
изливается в полостные лакуны, окружающие внутренние органы, и
заполняет их. Заполненная гемолимфой полость тела членистоногих
называется гемоцелем. У членистоногих с трахейным типом дыха-
тельной системы кровь обеспечивает только транспорт питательных
веществ, органы снабжаются кислородом непосредственно из трахей.
У остальных форм кровеносная система доставляет к органам и пита-
тельные вещества, и кислород.
Выделительная система образована коксальными железами, пе-
рикардиальными клетками, мальпигиевыми сосудами, жировым те-
лом и др.
Нервная система организована как у кольчатых червей в виде нерв-
ной брюшной цепочки. При этом увеличена роль надглоточных гангли-
ев, которые сообща образуют головной мозг, состоящий из трех отде-
лов: переднего — протоцеребрума, среднего — дейтоцеребрума и
заднего — тритоцеребрума. Отмечается тенденция к олигомеризации
ганглиев брюшной нервной цепочки, что выражается в уменьшении ко-
554
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
личества узлов за счет их слияния. Обычно очень хорошо развиты мно-
гочисленные органы чувств, обеспечивающие животному восприятие
основных сигналов внешней среды (свет, температуру, звук и др.) .
Половая система. Членистоногие раздельнополые, у них часто вы-
ражен половой диморфизм. Бесполое размножение отсутствует, спо-
собность к регенерации выражена слабее, чем у кольчатых червей.
Тип членистоногие подразделяется на четыре подтипа: три — л о -
битообразные, жабродышащие, трахейные и хе-
лицеровые.
Класс ракообразные. Ракообразные — единственный класс под-
типа жабродышащих. Они являются первичноводными животными,
поэтому в качестве органов дыхания у них имеются жабры, являющи-
еся выростами конечностей. Представители этого класса отличаются
от остальных членистоногих наличием двух пар усиков. Экологиче-
ские формы ракообразных весьма разнообразны, встречаются мор-
ские и пресноводные виды. Часть из них ведут донный образ жизни,
другие плавают в толще воды, входя в состав зоопланктона. Известны
сидячие ракообразные, неподвижно прикрепленные к донному суб-
страту. Некоторые виды являются паразитами. Лишь немногие пред-
ставители класса ведут наземный образ жизни. Всего известно около
25 000 видов.
Внешнее строение разнообразно. Тело расчленено на три отдела:
голову, грудь и брюшко. Строение головы у разных форм имеет ряд
особенностей. У десятиногих раков (речной рак, омары, лангусты,
крабы и др.) голова сливается с грудью, образуя головогрудь. На сег-
ментах головы располагаются различные парные и непарные придат-
ки. Непарные придатки: верхняя губа, которая представляет собой
складку кутикулы. Парные придатки — это антенны и видоизменен-
ные конечности. На акроне находится пара первых антенн — антенну-
лы, служащие для осязания и обоняния.
Конечности, расположенные в разных отделах тела, специализи-
руются для выполнения определенных функций, поэтому их строе-
ние сильно различается. Конечности первого сегмента видоизменя-
ются во вторую пару антенн, значительно более длинных, чем
антеннулы акрона. Антенны выполняют функцию осязания, у лангу-
стов очень длинные бичевидные антенны помогают животному обо-
роняться, а некоторым видам (например, водяным блохам) помогают
плавать. Остальные три сегмента головы также несут конечности, но
они обычно видоизменяются в ротовые части и осуществляют пер-
вичную обработку пищи. Конечности второго сегмента головы пре-
образуются в верхние челюсти, которые также называют жвалами
(мандибулами). Конечности III и IV головных сегментов образованы,
ТИП ЧЛЕНИСТОНОГИЕ
555
соответственно, первой и второй парами нижних челюстей — мак-
силл. Конечности грудных сегментов также специализируются.
У речного рака конечности первых трех пар грудных сегментов
преобразованы в ногочелюсти, которые помогают животному удер-
живать пищу. Остальные ножки служат для передвижения либо по
твердому субстрату (ходильные ноги), либо для плавания. От брюш-
ных сегментов отходят конечности только у высших раков, они спе-
циализируются для дыхания или размножения, у самцов и самок они
имеют различное строение. Последняя пара брюшных конечностей
видоизменилась в уплощенные пластинки, которые вместе с анальной
лопастью (тельсоном) образуют широкий плавник. Когда рак быст-
рыми движениями подгибает под себя брюшко, он способен относи-
тельно быстро плыть в обратном направлении (задом наперед).
Покровы тела образуют жесткий панцирь. У речного рака в обла-
сти головогруди слившиеся пластинки формируют головогрудной
щит — карапакс. В кутикуле ракообразных отсутствует водонепрони-
цаемый слой, а наружный слой кутикулы пропитан солями, что зна-
чительно повышает ее жесткость. Кутикула содержит множество раз-
нообразных пигментов, которые определяют окраску животного.
Внутренняя поверхность кутикулы образует углубления и гребни,
к которым прикрепляются скелетные мышцы. В течение некоторого
времени после линьки покровы остаются мягкими, поэтому в этот пе-
риод животное беззащитно, но оно может расти.
Постепенно покровы пропитываются солями и
затвердевают, благодаря этому рак защищен, но
его рост прекращается.
Пищеварительная система (рис. 233). Рото-
вое отверстие прикрыто верхней губой, сзади
нижней губой. В рот поступает пища, уже ча-
стично измельченная челюстями, затем она по-
ступает в толстостенный мускулистый пищевод,
а по нему в желудок. Желудок речного рака объ-
емистый, он подразделяется на два отдела — пе-
редний кардиальный и задний пилорический.
В кардиальном отделе имеются три кутикуляр-
'
&
%
$
#
"
!
Рис. 233. Пищеварительный канал речного рака:
1 — желудок, 2 — пилорическая часть желудка,
3 — слепой дорсальный вырост средней кишки,
4 — средняя кишка, 5 — валик, отделяющий среднюю
кишку от задней, 6 — задняя кишка, 7 — проток печени,
8 и 9 — гребни на стенке желудка
(по Гексли)
556
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ные пластинки, с помощью которых пища перетирается, там же она
подвергается химической обработке секретами печени. В пилориче-
ском отделе желудка происходит сортировка пищевых частиц.
Мелкие поступают в среднюю кишку и в печень, где окончательно пе-
рерабатываются. Более крупные из желудка направляются непосред-
ственно в заднюю кишку и без дальнейшей обработки выводятся че-
рез анальное отверстие.
Секрет печени содержит пищеварительные ферменты, а также ве-
щества, которые эмульгируют жиры пищи. Таким образом, печень
также выполняет роль поджелудочной железы. В клетках самой пече-
ни происходит внутриклеточное расщепление мелких частиц. Задняя
кишка в виде прямой трубки, которая тянется через все брюшко, за-
канчивается анальным отверстием.
Пища ракообразных разнообразна: речной рак всеядный, он поеда-
ет водную растительность, живую добычу, падаль и т. д . Плавающие
рачки, образующие зоопланктон, являются фильтраторами. Часть ра-
кообразных ведет паразитический образ жизни.
Дыхательная система. Жабры — специализированные структуры
газообмена речного рака — расположены в жаберных камерах голово-
груди. Вода постоянно поступает в жаберные камеры благодаря быст-
рым машущим движениям отростка максилл и омывает жабры.
Кровеносная система открытая (рис. 234). Движение гемолимфы
по сосудам обусловлено сокращениями сердца, расположенного на
спинной стороне тела в груди или в брюшке (это зависит от располо-
жения жабр). Сердце имеет вид трубки с отверстиями (остиями). Во
время систолы остии закрываются, гемолимфа продвигается в арте-
рии, которые отходят от сердца. Артерии разветвляются на более мел-
кие, из них гемолимфа изливается в полостные лакуны, расположен-
"
#
$
%
&
'
!
#
Рис. 234. Схема кровеносных сосудов и главнейших лакун у десятиногого рака:
1 — сердце, 2 — перикардий, 3 — передняя аорта, или глазная артерия, 4 — сяжковая
артерия, 5 — нижняя брюшная (поднервная) артерия, 6 — брюшной венозный синус,
7 — приносящие жаберные сосуды, 8 — нисходящая артерия, 9 — верхняя брюшная
артерия, 10 — жаберно-сердечные каналы. Печеночные артерии на схеме
не изображены, стрелками обозначено движение гемолимфы (из Иванова и соавт.)
ТИП ЧЛЕНИСТОНОГИЕ
557
ные между внутренними органами. Протекая по лакунам, гемолимфа
отдает кислород и питательные вещества тканям и насыщается про-
дуктами обмена. Затем гемолимфа поступает в жабры, где происходит
газообмен. Гемолимфа, насыщенная кислородом, оттекает от жабр в
полость перикардия и через остии попадает в сердце. Дыхательные
пигменты (гемоглобин или гемоцианин) растворены в гемолимфе.
Выделительная система представлена парными видоизмененны-
ми метанефридиями — целомодуктами. Их выводные каналы откры-
ваются либо у основания антенн (антеннальные железы), либо у осно-
вания второй пары максилл – максиллярные железы (за зеленоватый
цвет их также называют зелеными). У высших раков, в том числе
у речного рака, имеются антеннальные железы, у всех остальных —
максиллярные.
Нервная система образована окологлоточным нервным кольцом
и брюшной нервной цепочкой (рис. 235). Передний отдел представ-
лен сложно организованным надглоточным ганглием «головным моз-
гом», состоящим из парных ганглиев: переднего (протоцеребрума),
иннервирующего глаза, среднего (дейтоцеребрума), иннервирующего
антеннулы, и заднего (тритоцеребрума), который иннервирует вто-
рую пару антенн. Окологлоточные коннективы соединяют «головной
мозг» с подглоточным ганглием. Брюшные нервные стволы сближа-
ются, а соседние ганглии, находящиеся в одном сегменте, сливаются
между собой; кроме того, сливаются и ганглии, находящиеся в разных
í
1 íàäãëî
êîííåêòèâ
âè íåðâà
íûé ãàíãë
öåïî÷êè; 8
ïî÷êè, ÷åð
9 ïÿòûé
ëèé;111
11 ëîæí
&
'
!
"
#
$
%
!
"
#
$
%
Рис. 235. Общая топография нервной системы
речного рака:
1 — надглоточный ганглий (мозг),
2 — окологлоточные коннективы,
3–5
—
внутренняя, средняя и наружная ветви
нерва первой пары ходильных ног,
6 — подглоточный ганглий, 7 — первый грудной
ганглий брюшной нервной цепочки,
8 — продольная щель брюшной нервной
цепочки, через которую проходит нисходящая
артерия, 9 — пятый грудной ганглий,
10 — первый брюшной ганглий,
11–13 — нервы, отходящие от брюшных
ганглиев, 11 — ложный нерв ноги,
12 — дорсолатеральный нерв,
13 — вентральный нерв, 14 — пятый брюшной
ганглий, 15 — последний (шестой) брюшной
ганглий, 16 — нерв, идущий к антеннальной
железе, 17 — пищеводный ганглий
симпатической нервной системы
(из Кейма)
558
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
сегментах, из-за чего длина нервной цепочки и количество узлов
в ней уменьшаются. Наряду с соматической, имеется и вегетативная
(симпатическая) нервная система, которая состоит из головного отде-
ла и симпатического нерва с ганглиями. Вегетативная часть регулиру-
ет деятельность внутренних органов. Нервная ткань ракообразных
синтезирует нейрогормоны, обеспечивающие гуморальную регуля-
цию жизнедеятельности.
Органы чувств представлены глазами, органами осязания, органа-
ми химического чувства и органами равновесия. Глаза могут быть
сложными и более простыми. Сложные, или фасетированные, глаза
состоят из многочисленных мелких глазков — омматидиев, каждый
из которых функционирует отдельно и охватывает определенную
часть окружающего пространства. В результате рак воспринимает
картину в виде мозаичного изображения. Отдельный омматидий со-
стоит из светопреломляющих линз и чувствительных клеток, от кото-
рых отходят отростки, формирующие зрительный нерв. Глазки разде-
лены между собой тонкими прослойками пигмента. Парные сложные
глаза сидят на глазных стебельках. Всегда непарный простой глаз рас-
полагается на нижней поверхности головы между антеннулами. Ме-
ханические раздражения воспринимаются волосками, покрывающи-
ми тело и конечности, прежде всего антенны и антеннулы. Органами
химического чувства являются волоски, расположенные на антенну-
лах. Большинство ракообразных не имеют органов равновесия, одна-
ко у речного рака в основании антеннулы имеется статоцист.
Половая система раздельная, часто выражен половой диморфизм.
Речной рак откладывает яйца в конце осени — начале зимы. Перед
этим самка линяет. Ракообразные заботятся о потомстве. Некоторое
время после вылупления молодые рачата остаются под брюшком ма-
тери, которая их защищает. У речного рака развитие проходит без ме-
таморфоза, и из яйца выходит маленький рак.
Значение ракообразных в природе велико. Крупные ракообразные
являются санитарами водоемов, поедая погибших животных. Мелкие
создают основу зоопланктона, которым питаются большое количество
водных организмов. Многие ракообразные (речной рак, омары, лангу-
сты, многие креветки, крабы и др.) используются человеком в пищу.
Некоторые ракообразные являются промежуточными хозяевами мно-
гих опасных гельминтов. Например, веслоногий рачок циклоп служит
первым промежуточным хозяином личинки широкого лентеца.
В подтип трахейные входят наземные и вторично водные члени-
стоногие, у которых осуществляется трахейный газообмен. Подтип
объединяет два класса: насекомые и многоножки.
Класс насекомые — самый многочисленный не только среди чле-
нистоногих, но и среди всех животных вообще. Известно не менее по-
ТИП ЧЛЕНИСТОНОГИЕ
559
лутора миллионов видов насекомых. Великолепные адаптивные
возможности позволили насекомым освоить все оболочки биосферы:
гидросферу, верхнюю часть литосферы и даже нижние слои атмосфе-
ры. Насекомые являются единственными беспозвоночными, которые
имеют крылья и способны летать произвольно, что позволило им ос-
воить и воздушное пространство. Ни одна другая группа беспозво-
ночных не может соперничать с насекомыми по разнообразию адапта-
ции к различным условиям обитания.
Тело насекомых четко разделено на голову, грудь и брюшко, под-
вижно соединенные между собой. Голова состоит из акрона и слив-
шихся четырех сегментов головы. На голове располагается ротовое
отверстие, глаза и четыре пары придатков. Пара антенн, которые го-
мологичны антеннулам (первой паре антенн) ракообразных, являют-
ся придатками. Антенны насекомых называют усиками, они выполня-
ют функцию органов чувств, в первую очередь осязания и обоняния.
Многие насекомые обладают очень тонким обонянием.
Придатки сегментов головы — это видоизмененные конечности,
которые располагаются вокруг ротового отверстия и в совокупности
образуют ротовой аппарат, в состав которого входят мандибулы (верх-
ние челюсти). Мощные мандибулы называют жвалами; первая пара
нижних челюстей, или первые максиллы; вторая пара нижних челю-
стей, или вторые максиллы, которые вместе образуют непарную ниж-
нюю губу. К ротовому аппарату также относится и верхняя губа, пред-
ставляющая собой вырост кутикулы. Строение ротового аппарата
зависит от характера пищи: он может быть грызущим, лижущим, ла-
кающим, сосущим, колющим и т. д.
Грудь состоит из трех сегментов: переднегрудь, среднегрудь и зад-
негрудь. Все они несут по одной паре ходильных конечностей, а по-
следние два сегмента еще и по паре крыльев. Членики конечности
соединены между собой подвижными суставами, что позволяет жи-
вотному совершать разнообразные движения. Крылья насекомых не
связаны с конечностями, они представляют собой выросты покровов
тела, куда продолжаются нервные волокна и элементы трахейной си-
стемы, хорошо заметные на крыле в виде жилок. Чаще всего крылья
различные, хотя у стрекоз они почти одинаковы. У полужесткокры-
лых (клопов) передняя пара крыльев частично затвердевает, а у жест-
кокрылых (жуков) затвердевает полностью и превращается в жесткие
надкрылья, которые защищают заднюю пару настоящих крыльев.
У двукрылых (комары и мухи) полноценными крыльями становится
передняя пара, тогда как задняя видоизменяется в жужжальца.
Брюшко может содержать разное количество сегментов. Конечно-
сти на брюшке обычно отсутствуют, но у тараканов они сохраняются
в виде придатков.
560
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Тело насекомых покрыто трехслойной кутикулой, которая выде-
ляется поверхностным эпителием. Кутикула имеет обычное для на-
земных членистоногих строение, снаружи она покрыта водонепрони-
цаемой мембраной. В покровах имеются многочисленные волоски
и различные железы.
Скелетные мышцы у насекомых развиты очень хорошо. Общее ко-
личество мышечных пучков может достигать двух тысяч. Для сравне-
ния: у человека насчитывается всего около 600 скелетных мышц. Раз-
нообразие мышц позволяет насекомым совершать очень сложные
движения. Все части тела подвижны, особенно конечности и голова,
мускулатура брюшка организована проще. Особое значение для насе-
комого имеют крыловые мышцы, обеспечивающие машущие движе-
ния крыльев в полете. Особенностью мышц является то, что на каж-
дый нервный импульс волокна реагируют серией сокращений, что
позволяет животному совершать очень большое число взмахов кры-
льев, например некоторые комары совершают свыше тысячи взмахов
в секунду.
Пищеварительная система хорошо развита (рис. 236). Строение
каждого отдела зависит от характера пищи. Ротовая полость, распо-
ложенная на голове, окружена частями ротового аппарата. В нее впа-
дают протоки слюнных желез. Слюнные
железы гусениц выделяют жидкость, ко-
торая на воздухе превращается в прочную
шелковую нить. Глотка продолжается в
пищевод, который у многих видов (на-
пример, пчел) внизу образует расшире-
4
êà;
)è
9
êà;
ñî-
àð
àð-
'
!
"
&
%
$
#
"
!
#
$
Рис. 236. Пищеварительная система
черного таракана:
1 — глотка, 2 — слюнные железы,
3 — пищевод, 4 — зоб, 5 — пилорические
придатки, 6 — средняя кишка, 7 — тонкая
кишка, 8 — толстая кишка (тонкая (7)
и толстая (8) вместе составляют заднюю
кишку), 9 — слепой вырост толстой кишки,
10 — прямая кишка, 11 — клоакальное
расширение, 12 — мальпигиевы сосуды,
13 — жевательный желудок, 14 — резервуар
слюнной железы, 15 — выводной проток,
16 — непарный проток
(по Веберу)
ТИП ЧЛЕНИСТОНОГИЕ
561
ние — зоб, в котором пища задерживается, перемешивается и подвер-
гается воздействию пищеварительных ферментов. У хищных
насекомых, а также у питающихся твердой пищей (например, тарака-
нов, саранчи и др.) за зобом следует жевательный желудок, мускули-
стая стенка которого образует твердые выросты, перетирающие грубо
измельченную частями ротового аппарата пищу до состояния гомо-
генной кашицы. В средней кишке биополимеры расщепляются, и вса-
сываются питательные вещества. Для увеличения функциональной
поверхности в начале средней кишки имеются пилорические придат-
ки — несколько слепых выпячиваний. Кроме того, в стенке средней
кишки имеются многочисленные складки. На границе между средней
и задней кишкой у насекомых имеются многочисленные мальпигие-
вы сосуды. Задняя кишка заканчивается прямой кишкой, которая пе-
ред анальным отверстием образует клоакальное расширение, служа-
щее для накопления фекалий.
Насекомые питаются разнообразной пищей, но обычно у них
встречается определенная специализация. Различные насекомые по-
требляют растительную и животную, жидкую и грубую пищу, которая
может быть свежей или гниющей. В кишечнике термитов живут сим-
биотические микроорганизмы, которые расщепляют целлюлозу пое-
даемой древесины. Многие виды являются паразитами.
Дыхательная система насекомых трахейного типа. По бокам двух
задних грудных и брюшных сегментов имеются до десяти пар отвер-
стий — стигм, которые ведут в крупные трахейные стволы. Внутри
тела насекомого стволы многократно ветвятся, образуя тонкие тру-
бочки — трахеи, сообщающиеся между собой на брюшной стороне.
Самые мелкие трахеи — трахеолы — тонкостенные, что облегчает га-
зообмен. Трахеи пронизывают все тело насекомого, а их конечные
ветви даже проникают внутрь отдельных клеток. Для газообмена
с атмосферным воздухом животное совершает дыхательные движе-
ния, при этом брюшко пульсирует более или менее часто.
Кровеносная система насекомых незамкнутая. Сердце, располо-
женное на спинной стороне под слоем мышц в полости перикардиаль-
ного синуса, имеет вид длинной трубки со слепым задним концом.
Перегородки, расположенные по длинной оси, разделяют сердце на
несколько камер с отверстиями (остиями) в боковых стенках и клапа-
нами. Передний конец трубки продолжается в аорту, куда поступает
гемолимфа после сокращения сердца, обратный ток жидкости невоз-
можен из-за клапанов. Аорта направляется к голове, где открывается.
Из головы гемолимфа стекает по синусам в грудь и брюшко, омывает
внутренние органы и затем изливается в полость перикардиального
синуса, окружающего сердце. Гемолимфа поступает в полость сердца
562
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
благодаря сокращению парных крыловидных мышц, расположенных
по ходу сердца под ним.
Частота сердечных сокращений зависит от функциональной на-
грузки. Так, например, сердце бабочки бражника в покое сокращается
60 раз, а в полете — до 150 раз в минуту. Относительная простота
строения кровеносной системы насекомых (например, отсутствие
периферических сосудов) связана с тем, что тканевой газообмен вы-
полняет дыхательная трахейная система. Поскольку газообмен во
всем теле обеспечивают трахеи, кровеносная система служит только
для транспорта питательных и биологически активных веществ к ор-
ганам и тканям и удаления из них продуктов распада. Гемолимфа со-
держит мало дыхательного пигмента, поэтому чаще всего она бесцвет-
ная. Форменные элементы крови малочисленны, в основном это
фагоциты.
Выделительная система представлена мальпигиевыми сосудами,
перикардиальными клетками, нефроцитами и жировым телом. Маль-
пигиевы сосуды располагаются на границе между средней и задней
кишкой. У разных видов варьирует количество сосудов от 2 до 200
(у черного таракана от 80 до 100) у взрослых особей, у личинок —
меньше. Каждый сосуд — это тонкая трубочка, свободный конец кото-
рой заканчивается слепо, а другой впадает в полость кишечника. Из
гемолимфы, омывающей снаружи мальпигиевы сосуды, поглощаются
растворенные продукты распада и переводятся в нерастворимую фор-
му — кристаллы мочевой кислоты, которые поступают в полость со-
судов и постепенно перемещаются по направлению к полости кишки.
Через стенки сосудов освободившаяся вода всасывается обратно в ге-
молимфу, а обезвоженные продукты выделения выводятся наружу че-
рез анальное отверстие вместе с фекалиями. Такое строение выдели-
тельной системы очень выгодно для насекомых, так как выводит из
организма практически сухие вещества, что существенно снижает по-
терю воды и позволяет им обитать в очень жарком климате в услови-
ях дефицита воды.
Жировое тело обладает выделительной функцией. В нем запаса-
ются питательные вещества и накапливаются продукты обмена, при-
чем, попав в жировые клетки, эти вещества остаются в них навсегда
и вообще не выводятся из организма.
Нервная система образована головным мозгом и брюшной нерв-
ной цепочкой (рис. 237). Головной мозг — это сложно организован-
ный надглоточный ганглий. Его верхнюю часть составляет протоцере-
брум с большими зрительными долями, иннервирующими глаза.
К протоцеребруму относятся и грибовидные тела, управляющие наи-
более сложными движениями и рефлекторной деятельностью. Ниже
располагается дейтоцеребрум, его основную часть составляют обоня-
ТИП ЧЛЕНИСТОНОГИЕ
563
тельные доли, иннервирующие усики.
Тритоцеребрум образует самую ниж-
нюю часть головного мозга и бо
льшую
часть окологлоточных коннективов, ве-
дущих к подглоточному ганглию. Этот
отдел мозга развит значительно хуже
остальных, так как у насекомых нет вто-
рой пары антенн. Брюшная нервная це-
почка состоит из крупного подглоточ-
ного ганглия, иннервирующего части
ротового аппарата, трех менее крупных
грудных ганглиев, иннервирующих главным образом мышцы конеч-
ностей и крыловые мышцы и самых мелких брюшных ганглиев, кото-
рые иннервируют сегменты брюшка. У многих насекомых соседние
ганглии сливаются между собой, при этом возникают более крупные
скопления нервных клеток.
Кроме соматической нервной системы, у насекомых хорошо раз-
вита вегетативная симпатическая нервная система. Она начинается от
нервов, отходящих от головного мозга, и имеет собственные ганглии.
Симпатическая нервная система управляет деятельностью внутрен-
них органов. Во всех отделах нервной системы насекомых имеются
нейросекреторные клетки, обеспечивающие гуморальную регуляцию
деятельности организма.
Поведение многих насекомых очень сложное. Особенно это отно-
сится к общественным насекомым (муравьям, пчелам и др.), обладаю-
щим сложными механизмами коммуникации, у которых информация
передается от одной особи к другой.
íèÿ
êàíà:
!
Рис. 237. Схема строения нервной системы
насекомого:
1 — протоцеребрум, 2 — нейросекреторные
клетки, 3 — оптическая область мозга,
4 — дейтоцеребрум, 5 — антеннальный нерв,
6 — тритоцеребрум, 7 — кардиальные тела,
8 — прилежащие тела, 9 — окологлоточные
коннективы, 10 — подглоточный ганглий,
11 — нервы, идущие к ротовым конечностям,
12 — ганглии грудных сегментов, 13 — ганглии
брюшных сегментов, 14 — непарный нерв
симпатической системы
(из Вюрмбаха)
564
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Органы чувств насекомых обычно многообразные и очень хорошо
развитые. Насекомые воспринимают все основные типы раздраже-
ний — механические, вкусовые, обонятельные, звуковые, световые —
и способны определять положение тела в пространстве. Отсутствие
некоторых из этих функций обычно вызвано особенностями образа
жизни того или иного вида насекомых. Механические раздражения
воспринимают сенсиллы, расположенные, главным образом, на уси-
ках. Вкусовые сенсиллы, локализованные на органах ротового аппа-
рата и на дистальных члениках ходильных ног, являются органами
вкуса. Обонятельные сенсиллы располагаются в основном на антен-
нулах и на челюстных щупиках. У многих насекомых обоняние разви-
то великолепно, например самец шелкопряда обнаруживает неопло-
дотворенную самку по запаху на расстоянии нескольких километров.
Большинство насекомых способно воспринимать звуки, но органы
слуха имеются у тех насекомых, которые сами издают звуки, напри-
мер у кузнечиков имеются тимпанальные органы, расположенные на
ногах.
Глаза насекомых могут быть простыми или сложными. Простые
глаза имеются как у личинок, так и у взрослых особей, однако у по-
следних они закладываются заново, поскольку в процессе онтогенеза
личиночные глаза дегенерируют. Значительно более совершенными
являются сложные глаза, образованные большим количеством от-
дельных глазков — омматидиев. Каждый омматидий в составе слож-
ного фасеточного глаза функционирует как отдельный глаз, воспри-
нимая небольшую часть пространства. При этом насекомое получает
изображение окружающего мира в виде сложной мозаики из отдель-
ных крохотных картинок. Общее количество омматидиев в сложном
глазу у разных видов широко варьирует. Например, у муравья их 8—9,
у комнатной мухи — около 4000, а у стрекоз — до 28 000 в каждом
глазу. Этот показатель зависит от того, насколько важно зрение для
данного вида насекомых. Глаза насекомых не могут настраиваться на
разноудаленные предметы, поскольку у них отсутствует процесс ак-
комодации, поэтому зрение многих насекомых относительно нечет-
кое. Насекомые способны воспринимать коротковолновый свет (даже
ультрафиолетовый), но не воспринимают длинноволновой свет.
Половая система насекомых раздельнополая. У многих видов вы-
ражен половой диморфизм, проявляющийся в разных размерах тела,
крыльев или каких-либо придатков (например, длинные усики сам-
цов жука-дровосека или жвалы самца жука-оленя). Мужская половая
система образована парными семенниками. Семяпроводы соединяют-
ся, образуя непарный семяизвергательный канал. Перед тем как сое-
диниться, семяпроводы образуют расширения — семенные пузырьки,
в которые впадают придаточные железы. Семяизвергательный канал
ТИП ЧЛЕНИСТОНОГИЕ
565
проходит через совокупительный орган, который вместе с анальным
отверстием открывается в клоаку (рис. 238).
Женская половая система состоит из парных яичников и половых
путей: яйцевые трубки, парный яйцевод, непарная матка, влагалище
(рис. 239). Во влагалище также открывается мешковидный семяпри-
емник, в котором хранится сперма после спаривания, причем у неко-
торых насекомых сперма может сохраняться очень долго. Так, напри-
мер, матка пчел после спаривания всю жизнь (около пяти лет)
откладывает оплодотворенные яйца. У насекомых, откладывающих
яйца в древесину или в другие твердые структуры, имеется специаль-
ный орган — яйцеклад, который представляет собой видоизмененные
брюшные конечности. В течение жизни самка откладывает огромное
количество яиц, например пчелиная матка откладывает две-три тыся-
чи яиц в сутки, а в течение всей жизни — до полутора миллионов.
Еще более плодовитой является царица термитов, которая живет до
десяти лет, откладывая в сутки до 30 000 яиц! Личинка не всегда раз-
вивается из оплодотворенного яйца. Так, например, у общественных
насекомых (пчел, муравьев и др.) из оплодотворенных яиц развива-
À
Á
1
3
4
2
6
5
3
2
1
3
4
2
Рис. 238. Женская половая
система черного таракана:
1 — концевая нить, 2 — яйцевые
нити яичника, 3 — яйцевод,
4 — придаточные железы
(из Миала и Дени)
Рис. 239. Мужская половая система черного таракана.
Вид с брюшной (А) и спинной (Б) сторон:
1 — семенник, 2 — семявыносящий проток,
3 — разветвление семявыносящего протока,
4 — семяизвергательный канал, 5 — придаточные
железы, 6 — семенные пузырьки (из Миала и Дени)
566
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ются только самки, а мужские особи — из неоплодотворенных. У тлей
вообще нет самцов, вся популяция представлена самками, размножа-
ющимися партеногенезом.
Развитие насекомых может быть прямым или непрямым, т. е . оно
осуществляется либо с метаморфозом, либо без него. Развитие без ме-
таморфоза (развитие с неполным метаморфозом, или превращением,
или гемиметаболическое) проходит без стадии куколки. При этом из
яйца выходит личинка — нимфа, внешне похожая на имаго; она имеет
некоторые особенности. Например, обычно не развиты крылья, поло-
вые органы и др. Личинка питается, периодически линяет и быстро
растет, с каждой последующей линькой ее строение становится все
более похожим на взрослую особь. Этот тип развития характерен для
тараканов, полужесткокрылых, прямокрылых.
Развитие с полным метаморфозом (полным превращением, или го-
лометаболическое) — это непрямое развитие. Из яйца выходит личин-
ка, совершенно не похожая на взрослую особь. Личинка активно пита-
ется, значительно увеличивает свои размеры и массу, например после
выхода из яйца вес гусеницы тутового шелкопряда за 22—30 дней воз-
растает в 8—10 тысяч раз. Рост личинки всегда сопровождается линь-
ками. После личиночной стадии у насекомых с полным метаморфозом
следует стадия куколки. Эта стадия сопровождается почти полным по-
коем, при этом животное не питается и все жизненные процессы осу-
ществляются за счет накопленных ранее веществ. У куколки происхо-
дит полная перестройка строения, кроме нервной системы. Ткани
разрушаются, превращаясь в бесформенную полужидкую массу. Затем
все органы заново формируются, оболочка куколки лопается и из нее
выходит взрослое насекомое. Метаморфозом насекомых управляют
гормоны. Развитие с полным метаморфозом характерно для наиболее
высокоорганизованных насекомых (двукрылых, перепончатокрылых,
чешуекрылых, жесткокрылых). Такое развитие весьма полезно, так как
позволяет разделить экологические ниши между взрослыми особями
и личинками, что исключает конкуренцию между ними.
Онтогенез насекомого с полным метаморфозом представляет со-
бой чередование некоторых стадий: яйцо ␚ личинка ␚ куколка ␚
имаго. У насекомого без метаморфоза: яйцо ␚ личинка ␚ имаго.
Все насекомые подразделяются примерно на 40 отрядов.
Насекомые без метаморфоза
(с неполным превращением)
Отряд Вши. Мелкие, первично бескрылые насекомые, которые
имеют уплощенное тело и цепкие конечности. Строение ротовых ор-
ганов своеобразно, тип — колюще-сосущий. Вши являются эктопара-
ТИП ЧЛЕНИСТОНОГИЕ
567
зитами млекопитающих. На человеке паразитируют головная вошь
(вызывает педикулез, переносчик возбудителя возвратного тифа),
платяная вошь (переносчик возвратного и сыпного тифа), лобковая
вошь, или плошица (вызывает чесотку). Известно около 300 видов
вшей.
Отряд термиты — общественные насекомые, они совместно стро-
ят гнезда (термитники), состоящие из надземной (часто выше челове-
ка) и подземной частей. В составе общины несколько каст: половые
особи, рабочие и солдаты. Только половые особи имеют крылья, у ра-
бочих особей передние и задние крылья рано отпадают. Термиты пое-
дают древесину, которая расщепляется в кишечнике микро-
организмами-симбионтами; они причиняют вред, разрушая книги,
мебель, деревянные конструкции строений и т. п . Известно около
2500 видов термитов.
Отряд Клопы, или полужесткокрылые. Крылья этих насекомых
различные: вначале передние толстые и жесткие, их концы, а также
задние крылья — тонкие перепончатые. Ротовой аппарат колюще-
сосущий. Характерны пахучие железы. Имеются разные экологиче-
ские формы: наземные обитают в лесной подстилке, почве и на ра-
стениях, водные (гладыш, водяной скорпион и др.), есть паразиты
(постельный клоп и др.) и хищные. Среди клопов много вредителей
растений: вредная черепашка — вредитель злаков, крестоцветные
клопы повреждают огородные культуры и др. Известно около
4000 видов клопов.
Отряд Тараканы. Крылья тараканов неодинаковые: передние (над-
крылья) кожистые, задние — более тонкие перепончатые. Самки мо-
гут быть бескрылыми. Тело уплощено, ротовой аппарат — грызущего
типа. На конце брюшка имеются хорошо развитые брюшные конечно-
сти (церки). Тараканы — всеядные насекомые. Они обитают в почве,
под камнями, в подстилке, в человеческих жилищах (прусак, черный
таракан). Синантропные виды могут распространять возбудителей
некоторых заболеваний и яйца гельминтов, загрязняя пищу. Известно
около 3600 видов тараканов.
Отряд Равнокрылые. Обе пары крыльев одинаковые — тонкие,
с редкой сетью жилок. Ротовой аппарат колюще-сосущий. У многих
видов происходит чередование поколений со сменой растений-хозяев
или органа растения, на котором происходит питание. Ряд видов —
опасные вредители культурных растений (тли, в том числе виноград-
ная филлоксера, червецы, листоблошки, цикады и др.). Известно свы-
ше 30 000 видов равнокрылых.
Отряд Прямокрылые. Передние крылья — длинные кожистые, в
покое они выпрямлены; задние — более нежные, складываются вее-
ром. Иногда крылья недоразвиты. Ротовой аппарат — грызущего типа.
568
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Задние ноги обычно прыгательные. Представители: кузнечики, саран-
ча, сверчки, медведки. Есть опасные вредители растений (главным
образом — различные виды саранчи). Перелетная саранча и пустын-
ная саранча иногда образуют стаи колоссальных размеров, занимаю-
щих площадь в сотни гектаров. В 1962 г. в Туркмению из Ирана про-
никла стая, фронт распространения которой составил 160 км.
Известно около 20 000 видов прямокрылых.
Насекомые с метаморфозом
(с полным превращением)
Отряд Стрекозы. Передние и задние крылья одинаковые со слож-
ной сетью жилок. Ротовой аппарат — грызущего типа. Стрекозы актив-
ные хищники, их личинки ведут водный образ жизни, взрослые особи
имеют очень крупные сложные глаза и ловят добычу на лету. Предста-
вители: красотка, коромысло. Известно более 4500 видов стрекоз.
Отряд Жесткокрылые, или жуки. Крылья неодинаковые: передние
в виде твердых хитиновых пластинок (надкрылья), которые защища-
ют перепончатые задние крылья. Ротовой аппарат — грызущего типа.
Личинки имеют три пары грудных конечностей, иногда недоразви-
тых. Куколки свободные. Экологические формы разнообразны. Боль-
шинство видов ведут наземный образ жизни, некоторые живут в воде
(водолюбы, плавунцы и др.). Есть растительноядные и хищные фор-
мы. Многие являются опасными вредителями сельскохозяйственных
растений (майский и колорадский жуки, короеды, долгоносики и др.),
при этом и личинки, и взрослые особи. Однако некоторые виды по-
лезны (божьи коровки питаются тлями и червецами, жужелицы унич-
тожают насекомых-вредителей и т. д .). Жесткокрылые — самый круп-
ный отряд насекомых, известно более 250 000 видов.
Отряд Чешуекрылые, или бабочки. Насекомые имеют две пары
крупных крыльев, покрытых видоизмененными волосками — чешуй-
ками. Чешуйки могут быть оптическими, которые преломляют свет
(крыло приобретает перламутровый оттенок) или ярко окрашены,
при этом образуется характерный рисунок поверхности крыла. Рото-
вые органы преобразуются в длинный сосущий хоботок, у некоторых
видов они редуцируются. Их червеобразные личинки (гусеницы) име-
ют три пары грудных конечностей и пять пар брюшных ложных но-
жек. Ротовой аппарат — грызущего типа. В зависимости от формы
усиков отряд разделяют на два подотряда: булавоусые (дневные)
и разноусые (ночные) бабочки. Куколки открытого типа. Взрослые
особи питаются нектаром цветов, гусеницы — растительной или жи-
вотной пищей. Личинки многих видов наносят большой ущерб сель-
скохозяйственным растениям, например земляные совки, капустная
ТИП ЧЛЕНИСТОНОГИЕ
569
белянка, пяденица, коконопряды, шелкопряды и др. Тутовый шелко-
пряд одомашнен человеком (шелковичный червь). Уже тысячи лет
известно около 100 000 видов.
Отряд Блохи. Вторично бескрылые насекомые, тело которых сжа-
то с боков. Они не имеют сложных глаз. Ротовой аппарат — сосущего
типа. Блохи являются паразитами позвоночных животных, могут пе-
реходить на человека. Блохи переносят возбудителей опасных заболе-
ваний (чумы, эндемического сыпного тифа). Известно более 1000 ви-
дов блох.
Отряд Перепончатокрылые. Насекомые обычно имеют две пары
прозрачных перепончатых крыльев, задние крылья меньше передних,
сцеплены с ними в единую пластинку. Ротовой аппарат — грызущего
или лакающего типа. В зависимости от характера соединения груди
и брюшка, различают сидячебрюхих (брюшко широким основанием
соединено с грудными сегментами) и стебельчатых (грудь соединена
с брюшком тонким стебельком — сильно суженным вторым сегмен-
том брюшка). У самок высших форм яйцеклад преобразуется в жало
с ядовитым аппаратом. Личинки обычно не имеют конечностей, но
у пилильщиков есть три пары грудных и 6—8 пар брюшных ложных
ножек (ложногусеницы). Куколки свободные. Имеются растительно-
ядные и хищные виды. Высшие формы — общественные насекомые.
Среди них много опасных вредителей растений (пилильщики, рого-
хвостки, орехотворки и др.) . Ряд видов полезны для человека (медо-
носная пчела одомашнена несколько тысяч лет тому назад; наездники
используются для биологической борьбы с вредителями). Муравьи
уничтожают вредных насекомых и участвуют в почвообразовании,
шмели являются опылителями важных культур (например, клевера).
Известно более 150 000 видов перепончатокрылых.
Отряд Двукрылые. Самый высокоорганизованный отряд насеко-
мых. Имеют одну пару прозрачных или окрашенных перепончатых
крыльев (переднюю). Рудиментарные задние крылья превращаются
в жужжальца. Ротовой аппарат — лижущего или колющего типа, у не-
которых форм ротовые органы недоразвиты, поэтому взрослые особи
вообще не питаются (например, оводы). Среди взрослых особей
встречаются растительноядные или хищные. Личинки обитают в воде
(комары, мошки и др.), в гниющих органических остатках (например,
мухи) или паразитируют (оводы, вольфартова муха и др.). Многие
двукрылые являются важными опылителями цветковых растений.
Личинки мотылей служат кормом для рыб и водоплавающих птиц,
а взрослые особи — кормом для насекомоядных птиц и летучих мы-
шей. Однако многие виды двукрылых опасны, они переносят возбу-
дителей тяжелых заболеваний и являются паразитами. Мухи перено-
сят на лапках возбудителей инфекционных заболеваний и яйца
570
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
гельминтов. Мухи цеце являются кровососами и переносят возбуди-
телей сонной болезни, сибирской язвы и др. Малярийные комары пе-
реносят малярийного плазмодия, москиты — лейшманий, слепни —
некоторых трипаносом, возбудителей сибирской язвы, туляремию
и т. п . Известно около 80 000 видов двукрылых.
Насекомые активно участвуют в жизни природных сообществ.
Они обладают великолепными адаптационными способностями, по-
этому занимают разнообразные экологические ниши и участвуют во
многих биологических процессах. Многие виды очень быстро размно-
жаются и образуют огромную биомассу, что делает их незаменимым
источником пищи для рыб, амфибий, рептилий, птиц, млекопитаю-
щих. Особенно важно участие насекомых-опылителей в жизни расте-
ний. Многие виды растений на протяжении тысячелетий обитания
совместно с насекомыми выработали адаптации к определенным на-
секомым-опылителям. Эти растения не могут опыляться другими на-
секомыми, так, например, клевер опыляется в основном шмелями,
имеющими длинный хоботок.
Насекомые являются санитарами природы, так как они утилизи-
руют остатки растений и погибших животных. Многие виды насеко-
мых живут в почве и активно участвуют в ее образовании и улучша-
ют структуру почвы. Многие насекомые — хищники, которые
нападают не только на мелких, но и на относительно крупных жи-
вотных. Например, огромные массы южноамериканских кочующих
муравьев эситонов или южноафриканских дорилин перемещаются,
сметая на пути все живое. Растительноядные насекомые и их личин-
ки (например, личинки жуков или гусеницы бабочек) сильно повре-
ждают различные органы растений, что иногда приводит к их
гибели.
Велика роль насекомых в хозяйственной деятельности человека.
Некоторые виды насекомых человек одомашнил очень давно, напри-
мер медоносную пчелу или тутового шелкопряда. Во многих странах
особенно Юго-Восточной Азии, насекомых широко употребляют
в пищу, так как их мясо очень богато белками и многими полезными
для человека веществами. Многие насекомые являются вредителями
сельскохозяйственных растений, что приводит к огромным потерям.
Огромные стаи саранчи уничтожают на своем пути всю раститель-
ность. Борьба с ними обходится очень дорого, а использование ядови-
тых веществ сильнее губит окружающую среду, чем сами вредители.
Сегодня вырабатываются новые методы биологической борьбы, для
чего используют других насекомых, которые, в свою очередь, уничто-
жают вредителей. Например, наездники откладывают яйца в тело
вредителей, и личинки в последующем паразитируют на них. Многие
насекомые являются паразитами человека, например слепни, песча-
ТИП ЧЛЕНИСТОНОГИЕ
571
ные мухи, комары, блохи, вши и др. Часто насекомые переносят воз-
будителей заболеваний: домашняя муха и муха цеце (она к тому же
и кровосос), тараканы и многие другие.
Класс паукообразные. В этот класс входят более 36 000 видов.
Все они ведут наземный или вторично водный образ жизни, дышат
только атмосферным воздухом с помощью трахей или легких. Подав-
ляющее большинство паукообразных являются хищниками. Часть
видов паразиты, причем некоторые из них являются возбудителями
заболеваний (чесоточный зудень — возбудитель чесотки) или пере-
носчиками возбудителей опасных болезней (некоторые клещи явля-
ются переносчиками вируса энцефалита). Обычно паукообразные
ведут одиночный образ жизни, однако известны виды пауков, образу-
ющие крупные колонии до 10 000 особей. Многие виды паукообраз-
ных обладают сильнейшим ядом, опасным для человека. Большинст-
во современных форм ведут наземный образ жизни, однако предки
современных видов были водными животными.
Для паукообразных характерно слияние головных и грудных сег-
ментов, в результате чего образуется единая головогрудь, состоящая
из семи слившихся сегментов (рис. 240). Головогрудь несет шесть пар
конечностей, первая пара видоизменилась в хелицеры, которыми жи-
вотное прокалывает и разрывает добычу. Хелицеры — гомологи ан-
тенн. Вторая пара конечностей (являющаяся гомологами мандибул)
видоизменилась в педипальпы, которые удерживают пищу и манипу-
лируют ею, и несет на себе чувствительные структуры. Остальные че-
тыре пары конечностей являются ходильными ногами. У паукообраз-
ных отсутствуют придатки акрона — антеннулы (усики). Членистое
брюшко имеют сольпуги и скорпионы,
брюшко последних заканчивается тельсо-
ном, несущим ядовитую иглу. У клещей все
сегменты тела сливаются между собой, оно
вообще не расчленено. Брюшные конечно-
#
"
!
Рис. 240. Ротовые органы паука-крестовика:
1 — конечный когтевидный членик хелицеры,
2 — основной членик хелицеры, 3 — педипальпа,
4 — жевательный вырост основного членика
педипальпы, 5 — основной членик ходильной ноги
(по Иванову)
572
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
сти видоизменились в легкие и паутинные бородавки. На брюшке
имеются наружные отверстия органов дыхания — легочных мешков и
трахей, а также анальное и половое отверстия.
Густой секрет, выделяемый паутинными железами, быстро засты-
вает на воздухе и превращается в нити паутины. Каждая из пары бо-
родавок выделяет определенный тип паутины, затем мелкие нити со-
единяются в одну более или менее толстую нить, в зависимости от
числа функционирующих в тот момент бородавок. Нить может быть
гладкой и очень прочной, более тонкой, но липкой, или особенно тон-
кой, из которой образуется кокон. Механическая прочность паутины
в несколько раз больше, чем у шелковой нити. Пауки используют па-
утину не только для изготовления ловчих сетей, строение которых
разнообразно и строго организовано, но и для построения стенок ко-
конов, окружения пойманной жертвы, для передвижения. Кроме пау-
ков, клещи тоже имеют паутинные железы.
Тело паукообразных покрыто хитинизированной кутикулой обыч-
ного строения, наружный слой которой состоит из липопротеинов,
защищающих от потери воды. На примере паука-крестовика рассмо-
трим строение различных систем.
Пищеварительная система. В передней кишке выделяют глотку,
пищевод и сосательный желудок, расположенные в головогруди. Ро-
товое отверстие ведет в широкую мускулистую глотку, которая про-
должается в тонкий пищевод, переходящий позади головного мозга в
расширенный сосательный желудок. К его стенке прикрепляются хо-
рошо развитые мышцы; сокращаясь, они растягивают желудок, поэто-
му пища всасывается. В глотку открываются протоки слюнных желез,
слюна содержит активные ферменты. При укусе ферменты вводятся в
тело жертвы и растворяют ее ткани. Через некоторое время паук вы-
сасывает полупереваренную кашицу, оставляя лишь пустую оболочку
от жертвы. Таким образом, пищеварение у паука частично происходит
вне его организма.
Средняя кишка разделена на желудок (не путать с сосательным
желудком!), расположенный в головогруди, и тонкую кишку, которая
находится в брюшке. От желудка отходит тонкая кишка, лежащая в
брюшке, окруженная разветвлениями печени. Печень вырабатывает
пищеварительные ферменты, ее клетки фагоцитируют и расщепляют
мелкие частички пищи. Протоки печени впадают в тонкую кишку.
Кишка открывается в мешковидную клоаку, которая кзади переходит
в короткую заднюю кишку, заканчивающуюся анальным отверстием.
Дыхательная система образована легочными мешками (легкими)
и/или трахеями. Легочный мешок — это углубление в теле, через тон-
кие стенки которого происходит газообмен между гемолимфой и воз-
духом, поступающим в легочный мешок через отверстия дыхалец,
ТИП ЧЛЕНИСТОНОГИЕ
573
расположенных на брюшке. Легочное дыхание имеется у скорпионов
и низкоорганизованных пауков. У большинства пауков имеются и
легкие, и трахеи. Трахеи представляют собой тонкие ветвящиеся или
неветвящиеся трубочки. Трахеи лучше приспособлены к воздушному
газообмену, чем легкие.
Незамкнутая кровеносная система состоит из сердца и сосудов.
У организмов с легочным дыханием она более сложная, у трахей-
ных — проще, так как трахеи обеспечивают газообмен в тканях, а ге-
молимфа только распределяет питательные вещества и доставляет их
к тканям и клеткам. Наиболее простое кровообращение у клещей. Их
сердце, окруженное перикардием, расположено в передней части
брюшка над кишечником. Сердце имеет форму трубки с отверстиями
(остиями) в боковых стенках. От сердца отходят передняя, задняя
аорты и боковые артерии. Из артерий гемолимфа изливается в по-
лость тела, протекает по расположенным между внутренними органа-
ми лакунам, разносит необходимые вещества, после чего возвращает-
ся в полость перикардия и через остии поступает в сердце. У многих
паукообразных гемолимфа бесцветная, у некоторых видов может со-
держать пигменты, например гемоцианин.
Выделительная система образована мальпигиевыми сосудами,
коксальными железами и специализированными выделительными
клетками. Мальпигиевы сосуды, отходящие от средней кишки, чаще
всего ветвятся. Основное вещество, которое выделяется, — малораст-
воримый гуанин, что уменьшает потерю воды. Кроме того, гуанин вы-
деляется клетками клоаки. Выделяемые вещества поступают в по-
лость кишки и выводятся через анальное отверстие. Коксальные
железы (обычно одна или две пары желез) хорошо развиты только
у совсем молодых особей, позже они более или менее редуцируются.
Нервная система образована брюшной нервной цепочкой, имею-
щей разнообразное строение. В связи с редукцией антеннул в голов-
ном мозге отсутствует дейтоцеребрум, иннервирующий их. Имеются
протоцеребрум, который иннервирует глаза, и тритоцеребрум, иннер-
вирующий хелицеры. Ганглии брюшной нервной цепочки часто объе-
диняются, образуя более или менее выраженную ганглиозную массу.
Органы чувств развиты по-разному. Для пауков наиболее важно
осязание. Многочисленные осязательные волоски — трихоботрии —
рассеяны по всей поверхности тела, особенно их много на педипаль-
пах и ходильных ногах. Волоски воспринимают непосредственно ко-
лебания воздуха или паутины, чутко реагируя на них. Лировидные
органы, рассеянные по всему телу, являются органами химического
чувства. Органы зрения паукообразных — простые глаза, количество
которых варьирует у разных видов от 2 до 12. У пауков они располо-
жены в виде двух дуг на головогрудном щите, а у скорпионов одна
574
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
пара глаз спереди и еще несколько пар — по бокам. Несмотря на зна-
чительное количество глаз, зрение у паукообразных слабое.
Половая система. Все паукообразные раздельнополые, у них часто
выражен половой диморфизм. Так, у большинства пауков самка на-
много крупнее самца. Строение половой системы у разных видов раз-
личное. Половые протоки всегда парные.
Мужская половая система паука-крестовика парная. Два семенни-
ка расположены в нижней части передней трети брюшка. Спереди
они переходят в семяпроводы, которые открываются в непарный се-
менной мешок. Наружное половое отверстие, как и у всех паукообраз-
ных, находится на I сегменте брюшка. Педипальпы являются совоку-
пительным органом самца. Они на своих концах имеют копулятивный
придаток. Женская половая система паука-крестовика образована
двумя яичниками, лежащими в брюшке непосредственно под кишкой.
Впереди яичники переходят в парные яйцеводы, которые, в свою оче-
редь, сливаются, образуя непарную матку. Семяприемники, которые
открываются наружу самостоятельными отверстиями, сообщаются с
маткой. Спаривание паукообразных часто сопровождается ритуаль-
ным поведением, которое может быть достаточно сложным. Более
крупная самка может принять самца за свою добычу и съесть. (Инте-
ресно: один из пауков назван каракуртом — черной вдовой.)
Развитие большинства паукообразных прямое, без личиночных
стадий. Самки пауков плетут из паутины специальные коконы, куда
помещают яйца. Скорпионы живородящие, многие виды заботятся
о потомстве.
Паразитологическое значение паукообразных очень велико. Неко-
торые из них, особенно клещи, являются переносчиками ряда заболе-
ваний или сами являются возбудителями заболевания. Возбудитель
чесотки — повсеместно распространенный клещ чесоточный зудень.
Представители семейств иксодовых и аргасовых клещей, ведущие эк-
топаразитический образ жизни, — переносчики заболеваний. Из ик-
содовых наиболее опасен таежный клещ, который переносит вирус
таежного или весенне-летнего энцефалита. Клещ обитает в таежных
лесах восточнее Урала, Сибири и Дальнего Востока, он встречается и
в некоторых зонах европейской части России. Значение таежных кле-
щей в переносе вирусного энцефалита выяснил еще в 30-х годах XX в.
Е.Н. Павловский. Вирус энцефалита открыл академик Л.А . Зильбер.
Представители рода дермасентор переносят спирохет, вызывающих
клещевой сыпной тиф, возбудителей туляремии, а также энцефалита,
риккетсиозов и бруцеллеза. Эти клещи обитают в лесах и степях За-
падной Сибири и Забайкалья. Собачий клещ, живущий в лесных мас-
сивах европейской части России, переносит возбудителей весенне-
летнего энцефалита и туляремии.
ТИП МОЛЛЮСКИ
575
ТИП МОЛЛЮСКИ
Моллюски — это многоклеточные трехслойные билатерально-сим-
метричные животные, обладающие вторичной полостью тела. У части
моллюсков в течение онтогенеза смещаются органы, из-за чего тело
становится асимметричным. Вторичная полость тела в ходе онтогене-
за значительно редуцируется. Остатки целома окружают сердце, обра-
зуя перикардий, и половые железы. Промежутки между внутренними
органами заполнены соединительной тканью — паренхимой. Несег-
ментированное тело моллюсков подразделяется на голову, туловище
и ногу. У некоторых из них голова и нога отсутствуют. Вокруг основа-
ния туловища проходит обширная кожная складка — мантия. Про-
странство между мантией и туловищем образует мантийную полость,
в которой расположены жабры, открываются отверстия пищевари-
тельной, выделительной и половой систем. Вместе с сердцем эти орга-
ны составляют мантийный комплекс органов. Мантия выделяет нару-
жу защитную раковину. У некоторых видов она редуцируется.
Пищеварительная система состоит из хорошо дифференцирован-
ных отделов, у большинства видов в глотке расположена терка — ра-
дула, служащая для размельчения пищи, и печень.
Дыхательная система представлена жабрами или легкими.
Кровеносная система незамкнутая, поскольку объединены первич-
ная и вторичная полости тела. Кровь течет не только по кровеносным
сосудам, но и по полостным лакунам и синусам, омывая внутренние
органы и транспортируя питательные вещества и растворенные газы.
Пульсирующее сердце обеспечивает движение крови. Сердце состоит
из предсердий, количество которых соответствует количеству жабр и
желудочка. При сокращении сердца кровь выталкивается в аорту, из
него поступает в более мелкие артерии, а из них — в полость тела.
Пройдя по лакунам, кровь насыщается продуктами обмена и углекис-
лым газом, становится венозной и попадает в органы дыхания, где
очищается от СО2 и обогащается О2. После этого кровь поступает
обратно в сердце — сначала в предсердия, а из них — в желудочек.
Выделительная система представлена почками, которые своими
внутренними концами сообщаются с полостью перикардия.
Половая система может быть гермафродитной или раздельно-
полой.
Нервная система разбросанно-узлового типа.
Развитие примитивных моллюсков проходит со стадией личинки
трохофоры, у других образуется личинка — парусник.
Моллюски — очень древняя и самая многочисленная после члени-
стоногих группа животных. Они достигли наибольшего расцвета 50—
60 млн лет тому назад в палеогене. Тип включает около 130 000 видов.
576
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Класс брюхоногие, или улитки. Брюхоногие, или улитки, явля-
ются самым многочисленным классом подтипа раковинных моллю-
сков, их около 90 000 видов. Класс подразделяется на три подкласса:
переднежаберных, заднежаберных и легочных. Большинство из них
обитают в морях и океанах, но многие перешли в пресные водоемы
или ведут наземный образ жизни. Некоторые представители класса
являются паразитами. Размеры брюхоногих варьируют от нескольких
миллиметров до десятков сантиметров; так, хемифусус из подкласса
переднежаберных имеет раковину длиной 60 см. Тело расчленено на
голову, туловище и ногу, имеется цельная раковина. Характерная осо-
бенность улитки — выраженная асимметрия тела.
На голове располагаются одна-две пары щупалец и одна пара глаз,
которые могут находиться на концах щупалец или у их основания.
Там же находится и ротовое отверстие. Хорошо развитая нога образу-
ет широкую уплощенную ползательную поверхность — подошву, по-
средством которой животное медленно ползет по субстрату. Многочи-
сленные кожные железы подошвы выделяют густую клейкую слизь,
которая удерживает моллюска и предохраняет нежную ткань подо-
швы от механических повреждений в процессе движения. Туловище
располагается поверх ноги в виде спирально закрученного внутрен-
ностного мешка. Над туловищем образуется кожная складка — ман-
тия, которая направляется вниз и прикрывает мантийную полость.
Мантия выделяет кнаружи раковину, которая закручена в виде ко-
нической спирали. Многие виды имеют роговую крышечку, которой
моллюск прикрывает отверстие устья, после того как он втягивает
тело в раковину. Ее стенка снаружи покрыта тонким слоем органиче-
ского вещества, под ним лежит значительно более толстый фарфоро-
видный известковый слой, самый внутренний — перламутровый слой.
Органические вещества раковины синтезируются моллюском, а неор-
ганические поглощаются из пищи и из окружающей воды. У некото-
рых свободноплавающих и паразитических форм раковина редуциру-
ется.
Пищеварительная система (рис. 241). В передней кишке выделя-
ют ротовую полость, глотку и пищевод. Ротовая полость переходит
в глотку, имеющую толстые мускулистые стенки. На границе между
ротовой полостью и глоткой расположены роговые челюсти. В глотке
имеется мускулистый валик — язык, поверх которого лежит радула.
Ротовая полость и глотка выстланы кутикулой, а радула покрыта
твердыми зубцами, расположенными рядами. В процессе питания
моллюска язык вместе с радулой выдвигается из ротовой полости
и сдирает пищевые частицы. В глотку открываются выводные прото-
ки слюнных желез. Глотка продолжается в пищевод. Средняя кишка
начинается расширенной частью — желудком. Его окружает печень,
ТИП МОЛЛЮСКИ
577
которая выполняет функцию пищеварительной железы и фагоцити-
руют мелкие частицы пищи. От желудка отходит тонкая кишка, кото-
рая переходит в заднюю кишку, заканчивающуюся анальным отвер-
стием, открывающимся в мантийную полость.
Значительная часть улиток питается растительной массой, кото-
рую соскабливают при помощи радулы. Некоторые моллюски (на-
пример, виноградная улитка) наносят вред культурным растениям.
Имеются хищные и паразитические виды.
Дыхательная система большинства брюхоногих образована жа-
брами или непарным легким (подкласс легочные).
Незамкнутая кровеносная система состоит из сердца, которое рас-
полагается над задней кишкой в полости перикардия, и сосудов.
У примитивных переднежаберных сердце состоит их двух предсердий
и одного желудочка. У других правое предсердие редуцировано. При
сокращении желудочка артериальная кровь выталкивается в аорту,
которая вскоре разделяется на две ветви. У брюхоногих хорошо раз-
вита сосудистая система со сплетениями во всех частях тела. Из кон-
цевых сосудов кровь изливается в лакунарные пространства, где она
постепенно становится венозной. Затем кровь оттекает к органам ды-
хания, где насыщается кислородом. После этого артериальная кровь
поступает в предсердие, а оттуда — в желудочек.
Рис. 241. Вскрытая
виноградная улитка:
1 — губное щупальце,
2 — глазное щупальце,
3 — глотка,
4 — церебральный ганглий,
5 — легкое, 6 — легочная
вена, 7 — перерезанное
легочное отверстие,
8 — анальное отверстие,
9 — отверстие мочеточника,
10 — прямая кишка,
11 — мочеточник,
12 — предсердие,
13 — желудочек сердца,
14 — перикардий,
15 — почка, 16 — желудок,
17 — печень, 18 — гермафродитная железа,
19 — гермафродитный проток, 20 — белковая
железа, 21 — семяприемник, 22 — канал
семяприемника, 23 — яйцесемяпровод,
24 — семяпровод, 25 — мешок любовных
стрел, 26 — пальцевидные железы, 27 — бич,
28 — пенис, 29 — слюнные железы
(из Бриана)
578
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Выделительная система образована двумя почками, у взрослой
особи обычно остается только левая, а вторая редуцируется. Почка
состоит из ресничной воронки, обращенной в полость перикардия,
и канальца-мочеточника, который открывается в мантийную полость
рядом с анальным отверстием.
Нервная система. В типичном случае пять пар ганглиев в совокуп-
ности образуют нервную систему разбросанно-узлового типа.
Органы чувств воспринимают тактильные (щупальца, располо-
женные на голове), химические (осфрадии, которые находятся у осно-
вания жабр), световые (глаза на концах задней пары щупалец или
у их основания) раздражения и равновесие (статоцисты).
Половая система. Среди брюхоногих имеются раздельнополые
и гермафродитные формы. Половая система раздельнополых состо-
ит из половой железы (яичника или семенника) и половых протоков
(семяпровода или яичника). У мужской особи рядом с половым от-
верстием располагается совокупительный орган. У самки от яични-
ка отходит яйцевод, на котором образуется расширение — матка,
и боковое ответвление — семяприемник. Непарная половая железа
гермафродитов называется гермафродитной железой, в ней имеются
мужской и женский отделы. От гонады отходит общий гермафро-
дитный проток. У всех брюхоногих оплодотворение перекрестное,
внутреннее. У легочных развитие прямое, из яйца выходят крошеч-
ные улитки.
Значение брюхоногих в жизни человека невелико. В Западной
Европе употребляют в пищу виноградную улитку, для чего этих мол-
люсков даже специально разводят на фермах. Раковины улиток, со-
держащие перламутровый слой, используются для изготовления
украшений и пуговиц. Еще в Древнем Риме очень высоко ценились
ткани, окрашенные пурпуром, который добывали из пурпурной желе-
зы морских улиток мурексов. Растительноядные наземные брюхоно-
гие повреждают культурные посадки (виноградная улитка, слизни и
др.). Многие виды являются промежуточными хозяевами дигенетиче-
ских сосальщиков (например, в организме малого прудовика прохо-
дит личиночное развитие печеночный сосальщик).
Класс двустворчатые. Это второй по численности класс, насчиты-
вающий около 20 000 видов. Раковина состоит из двух створок, соеди-
ненных между собой. Двустворчатые ведут исключительно водный
образ жизни, заселяя соленые и пресные водоемы. Размеры раковины
варьируют от небольших до полутора метров. Масса тридакны ги-
гантской достигает 250 кг. Тело сплюснуто с боков и полностью скры-
то двумя створками раковины. Обычно сохраняется двусторонняя
симметрия. Характерна редукция головы, поэтому тело состоит толь-
ко из туловища и ноги. У некоторых видов нога имеет ползательную
ТИП МОЛЛЮСКИ
579
поверхность — подошву — и служит для передвижения моллюска.
Однако нога большинства двустворчатых сплюснута с боков и зао-
стрена на конце, она активно используется моллюском для закапыва-
ния в ил. Нога многих видов частично (мидии) или полностью (устри-
цы) редуцируется в связи с неподвижным образом жизни. Тело с
боков покрыто мантией, которая ограничивает мантийную полость
с находящимися в ней ногой и жабрами, у роющих форм свободные
края мантии срастаются, оставляя несколько отверстий. Через ввод-
ной сифон вода с пищевыми частицами поступает в мантийную по-
лость, омывая жабры; через другой — выводной — сифон отфильтро-
ванная вода выходит из мантийной полости. Мантия выделяет
компоненты раковины, состоящей из двух створок, гибко соединен-
ных между собой. Створки захлопываются благодаря сокращению
мощных замыкательных мышц. Песчинка, попадающая в промежуток
между мантией и раковиной, постепенно обволакивается слоями пер-
ламутра и превращается в жемчужину. У некоторых видов раковина
редуцирована.
Пищеварительная система имеет особенности, связанные с редук-
цией головы и отсутствием тех органов, которые в ней обычно долж-
ны находиться, — глотки с радулой и слюнными железами. Ротовое
отверстие расположено впереди ноги. В связи с отсутствием глотки,
ротовая полость продолжается в пищевод, который открывается в же-
лудок. По бокам от желудка располагается парная печень, протоки
которой впадают в полость желудка, от которой отходит средняя
кишка. В ней завершается расщепление пищи и всасывание мономе-
ров. После этого непереваренные остатки переходят в заднюю кишку.
Важная особенность двустворчатых: задняя кишка обычно пронизы-
вает насквозь желудочек сердца.
Дыхательная система образована жабрами.
Кровеносная система незамкнутая. Сердце, расположенное на
спинной стороне туловища, лежит в полости перикардия. Сердце со-
стоит из двух предсердий и одного желудочка. Из желудочка кровь
поступает в две аорты: переднюю и заднюю. Из артерий кровь посту-
пает в соединительнотканные лакуны. Омывая внутренние органы,
кровь из артериальной становится венозной и, оттекая от разных ча-
стей тела, собирается в венозный синус, лежащий под перикардием.
Из синуса кровь по приносящим жаберным сосудам направляется
к жабрам, где происходит газообмен. По выносящим жаберным со-
судам артериальная кровь поступает в предсердия, а из них — в же-
лудочек.
Выделительная система образована двумя видоизмененными по-
чками, расположенными в задней части тела ниже кишки. Перикар-
дий также выполняет выделительную функцию.
580
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Нервная система — разбросанно-узлового типа, некоторые узлы
сливаются между собой. В результате двустворчатые имеют три пары
ганглиев: цереброплевральные, педальные и висцеропариетальные.
Органы чувств развиты слабо, что связано с упрощенным строени-
ем и малоподвижным образом жизни. Отсутствуют головные щупаль-
ца и глаза. Однако глаза могут образовываться вдоль края мантии
(у гребешка) или в области сифонов (у сердцевидки). Обычно имеют-
ся органы химического чувства — осфрадии и два статоциста.
У подавляющего большинства видов половая система раздельно-
полая. Гонады, расположенные в передней части тела, частично захо-
дят в основание ноги. Яйцеводы или семенники, в зависимости от
пола, самостоятельно открываются в мантийную полость по бокам от
ноги. Оплодотворение наружное. Развитие происходит с метаморфо-
зом, т. е . проходит стадия личинки, которая оседает на донный суб-
страт и со временем превращается в молодого моллюска. Планктон-
ные личинки обеспечивают расселение этих моллюсков.
Большинство двустворчатых ведут малоподвижный образ жизни;
частично или полностью зарываясь в донный грунт, они выставляют
наружу сифоны и фильтруют воду в поисках пищи. Некоторые фор-
мы не способны перемещаться, поскольку прикрепляются ко дну с
помощью прочных нитей биссуса. Однако некоторые виды подвиж-
ные. Морской гребешок при опасности начинает часто хлопать створ-
ками раковины и уплывает.
Значение двустворчатых моллюсков в природе очень велико, так
как они являются фильтраторами. Пропуская через себя воду, они
очищают ее, поэтому в местах скоплений двустворчатых вода обычно
чистая. Одна устрица за один час фильтрует около десяти литров
воды, а мидии, заселяющие 1 м2 поверхности дна, в течение суток
фильтруют до 280 м3 воды.
Хозяйственное значение двустворчатых может быть как полез-
ным, так и вредным. Многие виды (устрицы, мидии, гребешок и др.)
человек употребляет в пищу из-за диетического мяса, богатого микро-
элементами. Поэтому их специально разводят. Хозяйства по выращи-
ванию морского гребешка имеются на российском побережье Дальне-
го Востока. Велико значение раковин двустворчатых в качестве
украшений (жемчуг, перламутровые раковины). Некоторые моллю-
ски приносят вред. Дрейссена, оказавшись в водопроводных трубах,
ограничивает движение воды. Другой двустворчатый моллюск — ко-
рабельный червь, или шашень, — проделывает длинные ходы в дере-
вянных предметах, например в деревянных сваях или деревянных ча-
стях кораблей, из-за чего может появиться течь.
Класс головоногие. Головоногие — наиболее высокоорганизован-
ные моллюски. Их ископаемые остатки найдены в отложениях кем-
ТИП МОЛЛЮСКИ
581
брийского периода палеозойской эры. Сегодня существует 700 видов
головоногих. Все они обитают в море, большинство плавают в толще
воды, заселяя различные глубины — от мелководий до нескольких ки-
лометров, некоторые виды ползают по дну. Размеры варьируют от не-
скольких сантиметров до 18 метров, последние весят не менее двух
тонн (гигантские кальмары архитеутис). У головоногих хорошо выра-
жена билатеральная симметрия. Тело подразделяется на голову, туло-
вище и ногу. Важная особенность строения — выросты ноги — щу-
пальца, которые окружают ротовое отверстие. Оставшаяся часть ноги
видоизменилась в воронку на брюшной стороне тела. У наиболее вы-
сокоорганизованных форм число щупалец 8—10. Внутренняя сторона
щупалец несет многочисленные присоски. С помощью щупалец жи-
вотные охотятся, захватывают и удерживают добычу, перемещаются
по дну, защищаются от врагов, контактируют с другими особями, ко-
пулируют и т. д. Мантия на спинной стороне срастается со стенкой
тела, а на брюшной стороне между мантией и телом образуется ман-
тийная полость, куда открываются анальное, половые и почечные от-
верстия, а также расположены жабры. Мантийная полость сообщает-
ся с окружающей средой через брюшное отверстие. При этом вода,
находящаяся в мантийной полости, с силой выбрасывается во внеш-
нюю среду через воронку. Таким образом, головоногие моллюски пе-
ремещаются в толще воды реактивным способом, при этом скорость
передвижения может быть очень высокой. Раковина имеется только
у немногих видов головоногих моллюсков (например, у наутилуса).
Кожа образована однослойным цилиндрическим эпителием, ле-
жащим на базальной мембране, под которым лежит соединительная
ткань (кутис). Под верхним волокнистым слоем располагается слой
пигментных клеток. Головоногие моллюски очень быстро меняют
свою окраску.
Мышечная система развита очень хорошо, она образована много-
численными пучками гладких миоцитов, однако мышечные волокна
мантии и щупалец имеют поперечную исчерченность.
Пищеварительная система имеет сложное строение. Ротовое от-
верстие, окруженное щупальцами, ведет в мускулистую глотку, где
имеются две мощные роговые челюсти (напоминающие клюв попу-
гая), которыми моллюск разрывает добычу. Протоки слюнных желез
открываются в глотку. Сзади от глотки отходит пищевод, который пе-
реходит в желудок, куда впадают и протоки печени. От печеночных
протоков отходят гроздевидные выросты, которые представляют со-
бой своеобразную поджелудочную железу. По печеночным протокам
в желудок попадают секреты печени и поджелудочной железы, содер-
жащие пищеварительные ферменты. От желудка отходит тонкая киш-
ка, которая переходит в прямую кишку, заканчивающаяся анальным
582
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
отверстием, открывающимся в мантийную полость на специальном
сосочке брюшной поверхности туловища. В самом конце прямой
кишки в нее впадает проток чернильного мешка. Эта железа имеется
только у головоногих моллюсков, она вырабатывает коричнево-чер-
ное вещество, которое при опасности выбрасывается животным и
сильно окрашивает воду. Сам моллюск при этом обычно меняет окра-
ску и уходит незамеченным.
Головоногие являются хищниками, охотящимися на любых жи-
вотных, с которыми могут справиться, демонстрируя при этом отмен-
ный аппетит. Свою добычу они схватывают щупальцами и затем
умерщвляют мощными челюстями и ядом слюнных желез.
Дыхательная система образована жабрами. Поверхность газооб-
мена у всех жаберных лепестков велика (у каракатицы около 1700 см2),
что приводит к поступлению большого количества кислорода в кровь.
Жабры омываются свежей водой, которая нагнетается в мантийную
полость за счет сокращений мускулатуры мантии.
Кровеносная система головоногих организована значительно
сложнее и совершеннее, чем у других классов моллюсков. Для нее ха-
рактерно сильное развитие кровеносных сосудов, причем кровь из ар-
терий в основном поступает в капилляры микроциркуляторного ру-
сла, после чего не изливается в лакунарные пространства полости
тела, как это всегда происходит при незамкнутой кровеносной систе-
ме моллюсков, а направляется непосредственно в вены. Таким обра-
зом, кровеносная система головоногих моллюсков почти замкнутая.
Сердце состоит из желудочка и расположенных по его бокам предсер-
дий. От переднего конца желудочка отходит головная (или передняя)
аорта, от заднего конца — брюшная аорта. В стенках органов артерии
распадаются на капилляры, образующие хорошо развитое микроцир-
куляторное русло. Из капилляров кровь направляется в вены. Кровь
собирается в приносящие жаберные сосуды, которые идут к жабрам.
Насыщенная кислородом артериальная кровь оттекает из жабр по вы-
носящим сосудам и поступает в предсердия сердца. При контакте
с кислородом кровь головоногих моллюсков приобретает синий цвет
благодаря наличию дыхательного пигмента гемоцианина.
Выделительная система представлена почками, количество кото-
рых соответствует числу жабр. Кроме того, выведение излишних со-
лей и продуктов обмена происходит в жабрах.
Нервная система организована очень сложно. В центральной
нервной системе имеются крупные ганглии — церебральные, педаль-
ные, плевральные и париетальные, расположенные вокруг пищевода,
при этом образуется общая окологлоточная масса нервной ткани
(«головной мозг»), защищенная головной хрящевой капсулой. Общая
относительная масса нервной ткани головоногих сопоставима с отно-
ТИП ХОРДОВЫЕ
583
сительной массой головного мозга рыб. Благодаря высокой организо-
ванной центральной нервной системе головоногие моллюски способ-
ны образовывать в течение жизни условные рефлексы и иметь
сложное поведение. Хорошо организованный вегетативный (симпа-
тический) отдел, состоящий из ганглиев и нервных сплетений, регу-
лирует деятельность внутренних органов.
Органы чувств разнообразны и хорошо развиты. Особенно совер-
шенны органы зрения, способные к аккомодация на предметы, нахо-
дящиеся на различном расстоянии. Очень крупного размера (до 40 см
в диаметре) глаза располагаются по бокам головы. Имеются и вне-
глазные светочувствительные клетки, расположенные внутри тела
моллюска. С их помощью животное может определять наиболее осве-
щенные места.
В головной капсуле расположена пара статоцистов, которые опре-
деляют положение тела в пространстве. Осфрадии или парные обоня-
тельные ямочки, расположенные под глазами, являются органами
обоняния. Вкус определяется чувствительными клетками, которые
находятся по краям ротового отверстия и на присосках.
Половая система. Головоногие раздельнополые животные, иногда
у них выражен половой диморфизм. Непарная половая железа распо-
ложена в половом отделе целома, где накапливаются половые клетки.
От половой железы отходят один или два половых протока. Эмбрио-
нальное развитие прямое. Многие головоногие заботятся о потомстве.
Например, самка осьминога сооружает гнездо и после откладки яиц
ничего не ест, постоянно нагнетая свежую воду в гнездо и охраняя
его. Вскоре после выхода детенышей из яиц самка погибает.
Многие головоногие являются объектом промысла из-за вкусного
мяса. Рудиментарный остаток раковины каракатиц используется в ка-
честве ингредиента для производства некоторых лекарств. Жидкость
чернильного мешка каракатиц используют в качестве чернил и для
изготовления китайской туши и краски сепии.
ТИП ХОРДОВЫЕ
Тип Хордовые объединяют главные черты их организации:
1) первичный осевой скелет — хорда. У низших хордовых она
функционирует всю жизнь, у более высокоорганизованных — только
на ранних стадиях онтогенеза, после чего хорда заменяется сегменти-
рованным позвоночным столбом;
2) полая нервная трубка, расположенная над хордой;
3) пищеварительная трубка, расположенная под хордой; перед-
ний отдел (глотка) сообщается с внешней средой посредством жабер-
ных щелей, которые пронизывают ее с боков. У первичноводных хор-
584
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
довых жаберные щели сохраняются в течение всей жизни, у наземных
форм — только на ранних этапах онтогенеза.
Ряд признаков также характерны для типа:
1) хордовые — многоклеточные трехслойные, вторичноротые, вто-
ричнополостные, животные с билатеральной симметрией тела; жи-
вотные;
2) для хордовых характерна сегментация.
Тип делится на три подтипа: личиночно-хордовые, бес-
черепные и черепные.
Подтип Бесчерепные. Все бесчерепные являются мелкими мор-
скими животными, обитающими на мелководьях с илистым или пес-
чаным дном. Они зарываются в донный грунт, оставляя на поверхно-
сти только передний конец тела. Для них характерна относительно
примитивная организация: отсутствие головы, наличие атриальной
(околожаберной) полости. Мышечная, выделительная и половая сис-
темы сегментированы. Сердце отсутствует, нервная трубка не диффе-
ренцированная, органы чувств развиты слабо. Известно около 30
видов, все они относятся к классу головохордовые. Рассмотрим орга-
низацию бесчерепных на примере ланцетника.
Ланцетник имеет уплощенное с боков полупрозрачное тело, сужа-
ющееся к обоим концам (рис. 242). Голова не выражена, вдоль спины
тянется невысокий тонкий спинной плавник, который кзади расши-
ряется, переходя в заостренный хвостовой плавник. На переднем кон-
це тела имеется предротовое отверстие, окруженное щупальцами.
Кзади от него вдоль боков снизу имеются складки, которые срастают-
ся, формируя атриальную полость, которая в задней трети тела сооб-
щается с внешней средой через атриопор. В области хвостового плав-
ника находится анальное отверстие.
Двуслойные покровы тела образованы однослойным эпидерми-
сом, покрытым кутикулой и нижележащим кориумом, или кутисом,
образованным студенистой тканью. В эпидермисе встречаются бока-
ловидные клетки (железистые).
Хорда в виде эластичного тяжа простирается вдоль всего тела до
его переднего конца дальше нервной трубки. Хорда окружена слоем
соединительной ткани, от которой кверху отходят выросты, защища-
ющие нервную трубку. Соединительнотканные волокнистые отростки
этой оболочки образуют миосепты, разделяющие мышцы, и создают
каркас в области жаберных щелей. Опорные структуры имеются в
плавниках, стенках предротовой воронки и щупальцах.
Мускулатура на всем протяжении тела сегментирована. Отдель-
ные мышечные группы (миомеры) разделены миосептами. Миомеры
представляют собой отдельные, вставленные один в другой конусы.
Соматическая мускулатура, образованная поперечнополосатой мы-
ТИП ХОРДОВЫЕ
585
шечной тканью, иннервируется периферическими нервами, отходя-
щими от нервной трубки. Простая организация мышечной системы
позволяет ланцетнику совершать лишь простые движения.
Пищеварительная и дыхательная системы ланцетника анатомиче-
ски связаны между собой. Ротовое отверстие располагается на перед-
нем конце тела в глубине предротовой воронки, которая окружена
щупальцами и кольцевой перепонкой (парус), которая нагнетает воду
в глотку. Пищеварительный тракт начинается длинной глоткой, боко-
вые стенки которой пронизаны многочисленными жаберными щеля-
ми (до 150 пар). Вода поступает через ротовое отверстие в глотку, вы-
ходит из нее через жаберные щели в околожаберную полость и
выводится во внешнюю среду через атриопор. Газообмен между про-
9
15
8
17
À
56
10
2
14
16
7
4
3
1
10
11
12
13
14
1
3
4
5
6
7
2
16
Á
Рис. 242. Строение ланцетника:
А — общая схема, Б — поперечный срез
в области глотки; 1 — плавниковая складка,
2 — метаплевральные складки,
3 — миомеры, 4 — миосепты, 5 — хорда,
6 — нервная трубка, 7 — глазки Гессе,
8 — предротовые щупальца, 9 — парус,
10 — глотка, 11 — атриальная полость,
12 — эндостиль, 13 — наджаберная
бороздка, 14 — печеночный вырост,
15 — анальное отверстие, 16 — гонады,
17 — атриопор
(по Левушкину и соавт.)
586
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ходящей водой и кровью, которая течет по тонким сосудам перегоро-
док, осуществляется, когда вода омывает межжаберные перегородки.
Кроме того, кислород поступает в организм через всю поверхность
тела (кожный газообмен). Взвешенные в воде пищевые частицы де-
трита и фитопланктон задерживаются в глотке и попадают в не диф-
ференцированную на отделы кишку, которая заканчивается анальным
отверстием вблизи основания подхвостового плавника. В начальном
отделе кишки от ее брюшной стороны отходит слепо заканчивающий-
ся печеночный вырост. Расщепление веществ и всасывание осуществ-
ляется в кишке и в печени. При этом происходит кишечное пищева-
рение в просвете кишки и в полой печени, а также внутриклеточное
переваривание.
Замкнутая кровеносная система устроена весьма примитивно,
сердце отсутствует (рис. 243). Венозная кровь собирается в венозный
синус, от которого вперед отходит брюшная аорта, стенки которой пе-
риодически сокращаются и гонят кровь в парные приносящие жабер-
ные артерии, отходят от брюшной аорты. В связи с тем, что основание
приносящих жаберных артерий расширено и пульсирует, эти сосуды
называют «жаберными сердцами». Жаберные артерии не распадаются
на капилляры. Кровь, протекающая по жаберным сосудам, обогащает-
ся кислородом и становится артериальной. Из перегородок выходят
выносящие жаберные артерии, впадающие в спинную аорту, которая
по своему ходу отдает сосуды ко всем органам, где происходит газооб-
мен, и артериальная кровь отдает кислород и становится венозной.
Оттекающая от передней части тела кровь собирается в передние кар-
динальные вены, от задней части тела — в задние кардинальные вены,
впадающие в венозный синус.
Венозная кровь, оттекающая от органов пищеварения, собирается
в подкишечную вену, которая направляется в печень, в ее стенках
(у ланцетника печень полая!) распадается на капилляры, образуя во-
Рис. 243. Схема кровеносной системы ланцетника:
1 — сонная артерия, 2 — корни спинной аорты, 3 — левый ковьеров проток,
4 — спинная аорта, 5 — левая задняя кардинальная вена, 7 — подкишечная вена,
8 — вена околожаберной полости, 9 — воротная система печени, 10 — брюшная
аорта, 11 — левая передняя кардинальная вена, 12 — жаберные артерии,
13 — расширенные основания жаберных артерий (по Нирштрассу)
ТИП ХОРДОВЫЕ
587
ротную систему печени. Напомним обычный путь крови: артерия ␚
капилляры ␚ вена; если при этом вена по своему ходу повторно обра-
зует микроциркуляторное русло и распадается на капилляры, то обра-
зуется воротная система, при этом кровь идет по иному пути: артерия
␚ капилляры ␚ вена ␚ капилляры ␚ вена. Проходя через печень,
кровь очищается от вредных веществ, а из моносахаридов синтезиру-
ется нерастворимый гликоген. Очищенная кровь оттекает от печени
по печеночной вене, впадающей затем в венозный синус, из которого,
как мы уже говорили, венозная кровь поступает в брюшную аорту.
Кровь ланцетника бесцветна, так как не содержит форменных эле-
ментов и дыхательных пигментов. Газы растворены непосредственно
в плазме и в процессе газообмена (жаберного и тканевого) диффунди-
руют в соответствии с градиентом концентрации.
Выделительная система образована метанефридиями, сходными
с органами выделения кольчатых червей. Каждый метанефридий
представляет собой короткую трубочку, обращенную одним концом
в целом, а другим — в атриальную полость. Мерцательный волосок
направляет поступившую из целома в трубочку метанефридия жид-
кость, которая выводится через отверстие в околожаберную полость,
а из нее во внешнюю среду. Метанефридии находятся над глоткой.
Нервная система подразделяется на центральную и перифериче-
скую. Центральная нервная система представлена расположенной
над хордой полой нервной трубкой, которая образуется в эмбрио-
нальном периоде в процессе нейруляции из дорсальной эктодермы.
На спинной стороне трубки хорошо заметен след срастания ее краев
(иногда срастание неполное). У ланцетника нервная трубка не раз-
делена на головной и спинной мозг, как это происходит у черепных
животных.
Периферическая нервная система образована нервами, отходящи-
ми от нервной трубки, и их волокнами.
Органы чувств развиты слабо, это связано с малоподвижным обра-
зом жизни. В эпидермисе рассеяны чувствительные клетки. Настоя-
щих глаз нет, однако имеются светочувствительные глазки Гессе.
К области нейропора прилегает орган обоняния — ямка Келликера.
Половая система образована многочисленными (около 25 пар) го-
надами, расположенными в полости тела вдоль атриальной полости
на уровне задней половины глотки и начального отдела кишки. Лан-
цетники — раздельнополые животные. Гаметы попадают в атриаль-
ную полость в результате разрыва стенки тела и половой железы.
Оплодотворение и развитие происходят во внешней среде. Личинки
вначале свободно плавают при помощи ресничек, которые покрывают
все их тело, и питаются планктоном. На поздних этапах развития ли-
чинка опускается на дно.
588
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ПОДТИП ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ
Класс Хрящевые рыбы
Класс объединяет рыб, у которых в течение всей жизни сохраняет-
ся хрящевой скелет. У них по 5—7 жаберных отверстий, кожа покрыта
плакоидной чешуей, отсутствует плавательный пузырь. Эти рыбы
обитают в морях и океанах, в пресные водоемы они заходят редко.
Размеры их от нескольких десятков сантиметров до 18 метров. Всего
насчитывается около 600 видов хрящевых рыб, которые подразделя-
ют на два подкласса: пластинчато-жаберные и химеро-
вые, или цельноголовые.
Подкласс Пластинчатожаберные. Большинство хрящевых рыб,
включая акул и скатов, относятся к этому подклассу.
У акул форма тела торпедообразная, туловище немного сплюсну-
то в дорсовентральном направлении. У скатов тело значительно боль-
ше уплощено дорсовентрально, что очень выгодно для донного образа
жизни. Подавляющее большинство скатов — обитатели дна, и лишь
немногие виды, например скат манта, ведут пелагический образ жиз-
ни. Тело нечетко разделено на голову, туловище и хвост. У большин-
ства видов на переднем конце головы имеется вырост — рострум
(рыло). По бокам головы располагаются глаза, несколько позади и
выше них находятся брызгальца. Щелевидное ротовое отверстие на-
ходится на нижней стороне головы, впереди рта видны парные ноз-
дри, а позади — пять пар вертикальных жаберных щелей. Туловище —
это часть тела от последней жаберной щели до отверстия клоаки,
которое находится на нижней поверхности тела вблизи хвоста. Туло-
вище заканчивается хвостом.
Конечности представлены парными или непарными плавниками.
Грудные и брюшные плавники парные, оба спинных плавника — перед-
ний и задний — непарные. У самцов акул на внутренних частях брюш-
ных плавников имеются пальцевидные выросты, которые способству-
ют копуляции. Лопасти хвостового плавника неодинаковые: верхняя
значительно больше, в нее заходит часть позвоночного столба. Гетеро-
цельный тип — основной орган движения (движитель) хвостового плав-
ника, боковые движения которого определяют поступательное движе-
ние тела. Такой способ перемещения более чем в 1,5 раза эффективнее
гребного винта судов. Парные плавники позволяют рулить, а непарные
обеспечивают равновесие тела. Хрящевые рыбы лишены плавательно-
го пузыря, поэтому они без движения погружаются в воду. У малопод-
вижных скатов хвостовой плавник редуцирован.
Тело покрыто двухслойной кожей. В эпидермисе много желези-
стых клеток, секрет которых выделяется на поверхность кожи. В дер-
ПОДТИП ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ
589
ме развиваются чешуи, покрывающие кожу и выполняющие защит-
ную функцию. Чешуя хрящевых рыб плакоидная, каждая чешуйка
представляет собой округлую пластинку, на которой возвышается зу-
бец, направленный назад. Зубец снаружи покрыт тонким слоем эма-
ли. Переходя на челюсти, чешуи становятся зубами.
Хрящевой скелет, который не окостеневает в течение всей жизни
рыбы, подразделяется на осевой и добавочный. Осевой включает
опорные структуры позвоночного столба и черепа, добавочный — хря-
щевые элементы поясов и свободных конечностей (рис. 244).
Осевой скелет. Позвоночный столб, образованный соединен-
ными между собой позвонками, в эмбриональном периоде заменяет
хорду, которая у взрослых особей сохраняется лишь между телами
позвонков, а также в самих телах. Каждый позвонок состоит из тела и
двух пар дуг: верхние ограничивают отверстие спинномозгового кана-
ла и нижние дуги. Тело позвонка имеет в центре отверстие, через ко-
торое проходит хорда. В позвоночном столбе выделяют два отдела:
туловищный и хвостовой. В туловищном отделе к коротким боковым
отросткам нижних дуг прикрепляются ребра.
Череп подразделяется на мозговой отдел, который защищает го-
ловной мозг, и висцеральный, образующий скелет переднего отдела
пищеварительной системы и опору для жаберного аппарата. Оба от-
дела имеют различное происхождение и объединяются в ходе эмбри-
онального развития.
Мозговой череп состоит из мозговой коробки, капсулы ор-
ганов чувств и скелета рострума. В мозговом черепе выделяют не-
сколько отделов: затылочный, слуховой, обонятельный, глазницы,
дно и крышу. Рострум образован хрящом ложковидной формы.
Висцеральный череп состоит из челюстного аппарата,
подъязычной дуги и жаберных дуг. Он образуется из висцеральных
дуг в стенках передней части кишечника, создает опору для него и
жаберного аппарата. Первые две пары висцеральных дуг редуцируют-
Рис. 244. Скелет акулы (схематично):
1 — позвонки, 2 — верхние дуги, 3 — нижние дуги, 4 — ребра, 5 — черепная коробка,
6 — обонятельная капсула, 7 — слуховая капсула, 8 — жаберная дуга,
9 — подъязычная дуга, 10 — нёбно-квадратный хрящ, 11 — меккелев хрящ,
12 — радиалии, 13 — плечевой пояс, 14 — тазовый пояс, 15 — базалии
(по Шмальгаузену)
590
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ся, от них остаются губные хрящи. Третья пара дуг преобразуется
в челюстную дугу, которая с каждой стороны формирует по две пары
хрящей: верхний парный небно-квадратный хрящ выполняет функ-
цию верхней челюсти, нижний парный меккелев хрящ является ниж-
ней челюстью. Половинки обоих хрящей, срастаясь между собой,
образуют цельные верхнюю и нижнюю челюсти. Подъязычная дуга,
возникающая из четвертой пары висцеральных дуг, состоит из двух
парных хрящей — гиомандибуляре (подвесок), и гиоида, а также одно-
го непарного — копулы, которая соединяет между собой гиоиды.
Подъязычная дуга подвижно связывает висцеральный череп с мозго-
вым. Позади подъязычной дуги находится остаток жаберной щели —
брызгальце и пять пар жаберных дуг (по числу жаберных щелей, у не-
которых акул их может быть 6—7). На каждой жаберной дуге (кроме
последней) находятся жабры.
Добавочный скелет — скелет плавников (парных и непар-
ных). Парные плавники пояса имеют дополнительную опору в теле,
которые создают поясы конечностей. Различают пояса передних и
задних конечностей.
ВНИМАНИЕ!
У всех рыб конечности представлены плавниками.
Мышечная система образована поперечнополосатыми скелетны-
ми мышцами, гладкие мышцы залегают в стенках внутренних органов
и сосудов. Скелетные мышцы подразделяются на мышцы туловища,
головы и конечностей (плавников). Лучше всего развита мускулатура
туловища, на долю которой приходится значительная часть массы
тела. Туловищные мышцы рыб сегментированы. Сегменты отделены
друг от друга тонкими соединительнотканными септами. Каждый сег-
мент — S-образно изогнутый миомер. Попеременно сокращаясь, мыш-
цы правой и левой сторон выполняют функцию локомоторного орга-
на, обеспечивающего поступательное движение вперед. Мышцы
головы приводят в движение челюстной аппарат, а мышцы конечно-
стей управляют движениями плавников. У некоторых видов специа-
лизированные мышечные пластинки преобразуются в электрические
органы; электрический скат способен генерировать разряд мощно-
стью около 70 В.
Пищеварительная система состоит из пищеварительной трубки и
пищеварительных желез (рис. 245). Пищеварительный тракт подраз-
деляется на ротовую полость, глотку, желудок, тонкую и толстую
кишки. Ротовое отверстие, расположенное на нижней стороне головы,
ограничено челюстями, покрытыми многочисленными зубами (обра-
ПОДТИП ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ
591
зуются из плакоидных чешуй). Они
располагаются в несколько рядов
(обычно 5—6, иногда до 15), имеют
коническую форму и обращены на-
зад, что позволяет рыбе эффективно
удерживать добычу. В течение жиз-
ни рыбы несколько раз сменяются
(за 10 лет у акулы может смениться
до 24 000 зубов).
Ротовая полость переходит в
глотку, которая сообщается с внеш-
ней средой через жаберные щели в ее
стенках. У гигантской и китовой акул, питающихся планктоном, на
жаберных дугах имеются густые тычинки, которые образуют цедиль-
ный аппарат. Однако большинство хрящевых рыб — хищники, питаю-
щиеся живой добычей или погибшими водными животными. У рыб
отсутствуют слюнные железы, поскольку вода постоянно поступает в
ротовую полость и глотку. Глотка переходит в короткий пищевод, ко-
торый, в свою очередь, открывается в V-образно изогнутый желудок.
В стенках желудка расположены многочисленные железы, выделяю-
щие компоненты желудочного сока. Частично переработанная пище-
вая кашица из желудка поступает в очень короткую тонкую кишку,
в которую впадают протоки двух крупных пищеварительных желез:
поджелудочной и печени.
Печень образует две или три лопасти и имеет значительно боль-
шие размеры, чем поджелудочная железа, у акул на нее приходится до
25% от всей массы тела. Печеночная желчь поступает в желчный пу-
зырь, где накапливается и некоторое время сохраняется. В печени
!
"
#
$
%
&
'
#
"
!
$
%
&
'
Рис. 245. Общее расположение внутренних
органов колючей акулы:
1 — венозный синус, 2 — предсердие,
3 — желудочек, 4 — артериальный конус,
5 — брюшная аорта, 6 — приносящие
жаберные артерии, 7 — кювьеров проток,
8 — боковая вена, 9 — желудок, 10 — тонкая
кишка, 11 — толстая кишка (просвечивает
спиральный клапан), 12 — прямая кишка,
13 — клоака, 14 — ректальная железа,
15 — печень, 16 — желчный пузырь,
17 — желчный проток, 18 — поджелудочная
железа, 19 — селезенка, 20 — правая почка
(левая не изображена)
592
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
откладывается много жира, что повышает плавучесть этих рыб, заме-
няя плавательный пузырь. Из тонкой кишки пищевая кашица перехо-
дит в широкую толстую кишку. В нем имеется спиральный клапан,
представляющий собой винтообразную складку стенки кишки, увели-
чивающую всасывающую поверхность. В толстой кишке заканчивает-
ся переработка пищевых веществ и происходит всасывание образо-
вавшихся при этом продуктов. Толстая кишка заканчивается прямой
кишкой, которая открывается в клоаку. Туда же впадают половые про-
токи и протоки мочевой системы.
Дыхательная система (рис. 246). Жабры расположены на межжа-
берных перегородках, которые отходят от жаберных дуг. По обеим
сторонам перегородки располагаются многочисленные выросты —
жаберные лепестки. Газообмен происходит при прохождении воды,
поступающей через ротовое отверстие в глотку, выходит через жабер-
ные щели во внешнюю среду, омывая при этом жаберные лепестки.
Ток воды всегда односторонний. Тонкие стенки жаберных лепестков
густо оплетены кровеносными сосудами, несущими венозную кровь,
после насыщения кислородом от жабр оттекает артериальная кровь.
Газообмен акул происходит пассивно в процессе движения. При
этом вода все время проникает через открытый рот в глотку и покида-
ет ее через жаберные отверстия. При снижении скорости движения,
тем более при остановке акулы, вода перестает поступать в рот, и про-
цесс насыщения крови кислородом замедляется. Поэтому акулы вы-
нуждены постоянно двигаться. Скаты, которые большую часть време-
Рис. 246. Схема, поясняющая, как происходит дыхание у акулы. Изменения давления
указаны относительно наружного давления: А — наполнение ротовой полости и глотки
водой, Б — изгнание воды через жаберные щели; 1 — вода, входящая через брызгальце,
2 — вода, входящая через рот, 3 — дно ротовой полости и глотки опускается,
4 — объем ротовой полости и глотки увеличивается, 5 — низкое давление,
6 — парабронхиальные полости, 7 — высокое давление, 8 — дно ротовой полости
и глотки поднимается, 9 — объем ротовой полости и глотки уменьшается
ПОДТИП ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ
593
ни проводят лежа на донном субстрате, активно засасывают воду за
счет периодического расширения полости глотки. Мускулатура рото-
вой полости и стенок глотки, сокращаясь, выдавливает воду, которая
омывает жаберные лепестки из глотки через жаберные щели, при
этом закрыто ротовое отверстие.
Кровеносная система образована сердцем и сосудами, составляю-
щими один круг кровообращения (рис. 247). Общая схема сосуди-
стой системы принципиально одинакова у всех хордовых. Имеются
два основных сосуда — брюшная и спинная аорты, по брюшной аорте
кровь движется по направлению к голове, а по спинной — к хвосту.
Сердце рыб состоит из двух камер — предсердия и желудочка. Отно-
сительные размеры сердца хрящевых рыб — самые маленькие среди
всех позвоночных. Сердце выполняет функцию насоса, обеспечиваю-
щего движение крови по артериям. Сердце рыб перекачивает только
венозную кровь, которая поступает в предсердие из прилегающего
к нему тонкостенного венозного синуса. Дистальный отдел желудоч-
ка хрящевых рыб (область перехода его в аорту) образует артериаль-
ный конус. У хрящевых рыб общий относительный объем крови по
отношению к массе тела ниже, чем у других позвоночных животных.
Кровь состоит из плазмы и форменных элементов (эритроцитов,
лейкоцитов и тромбоцитов). Основным органом образования фор-
менных элементов являются селезенка и почки. В селезенке, кроме
того, распадаются старые эритроциты. Эритроциты — ядерные клет-
ки. Кровь рыб содержит относительно мало эритроцитов и поэтому
имеет низкую кислородную емкость.
У рыб имеется примитивная лимфатическая система, образован-
ная лимфатическими сосудами; лимфатические узлы отсутствуют.
Лимфатическая система наряду с венозной обеспечивает дренаж тка-
Рис. 247. Кровеносная система акулы:
1 — левая сонная артерия, 2 — выносящая жаберная артерия, 3 — левый корень спинной
аорты, 4 — левая передняя кардинальная (яремная) вена, 5 — спинная аорта, 6 — левая
задняя кардинальная вена, 7 — хвостовая вена, 8 — воротная система почек, 9 — левая
боковая вена, 10 — воротная вена печени, 11 — воротная система печени,
12 — печеночная вена, 13 — сердце, 14 — левый кювьеров проток, 15 — брюшная аорта,
16 — приносящая жаберная артерия (по Шмальгаузену, с изменениями и дополнениями
594
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ней, удаляя скапливающуюся в них жидкость, которая поступает из
крови.
Выделительная система представлена парой удлиненных туло-
вищных почек (мезонефросов), которые располагаются в полости тела
вдоль позвоночного столба. В ходе эмбрионального развития сначала
закладываются головные почки (предпочки, или пронефросы) и лишь
затем они сменяются туловищными почками. Предпочка, внешне по-
хожая на метанефридии кольчатых червей или бесчерепных, образо-
вана большим количеством канальцев, каждый из которых своим
свободным концом открывается во вторичную полость тела, а проти-
воположным — в общий для всех канальцев выводной пронефриче-
ский канал. Свободный конец нефростома расширен в виде воронки,
куда из полости тела поступает жидкость. Предпочка удаляет вещест-
ва, растворенные в полостной жидкости. Пронефрос функционирует
только у зародышей и сменяется более совершенной туловищной по-
чкой, которая закладывается позади предпочки.
Канальцы туловищной почки начинаются слепыми расширения-
ми (боуменовы капсулы), окружающими сосудистый клубочек. Боу-
менова капсула вместе с сосудистым клубочком (мальпигиевое или
почечное тельце) и вместе с канальцем образует структурно-функци-
ональную единицу почки — нефрон. У хрящевых рыб в нефроне еще
сохраняется воронка (нефростом), но ее функциональная роль неве-
лика. Кровь, проходя под давлением через сосуды клубочка, теряет
часть плазмы, которая как бы выжимается в полость боуменовой кап-
сулы, являясь первичной мочой. Туловищная почка обеспечивает
фильтрацию крови, а не полостной жидкости, как это происходит
в головной почке. Это более эффективно, поскольку именно кровь
обеспечивает дальний транспорт веществ, в том числе токсичных.
Первичная моча из капсулы поступает в каналец нефрона, прохо-
дя по которому освобождается от нужных организму веществ (они
возвращаются обратно в кровь), но концентрация продуктов жизне-
деятельности становится выше. Объем вторичной мочи меньше объ-
ема первичной мочи за счет реабсорбции. У хрящевых рыб конечным
продуктом азотистого обмена является мочевина. Из нефрона вто-
ричная моча поступает в собирательные трубочки, которые впадают
в парные мочеточники. В туловищной почке мочеточником является
пронефрический канал пронефроса — мезонефрального канала (про-
тока), или вольфова канала (проток). Мочеточники тянутся вдоль
брюшной стороны почек, затем соединяются между собой и впадают
общим отверстием в клоаку. У самцов протоки мочевой и половой си-
стем связаны между собой.
Половая система представлена парными половыми железами и
половыми протоками. Мужская половая система анатомически связа-
ПОДТИП ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ
595
на с мочевой системой. Протоки парных семенников проходят через
вещество почки и впадают в вольфов проток, который у самцов вы-
полняет две функции: мочеточника и семяпровода. Канальцы перед-
него отдела почки не участвуют в образовании мочи и сообщаются
с семявыносящими канальцами, соответственно, вся передняя часть
почки, окружающая начальный сильно извитой отдел семяпровода,
называется придатком семенника. Оба вольфова протока соединяют-
ся и впадают в мочеполовой синус, который открывается на вдаю-
щемся в клоаку мочеполовом сосочке. У самцов почти редуцируется
мюллеров проток, но его остатки образуют парные мешки — семяпри-
емники, которые также впадают в мочеполовой сосочек.
Женская половая система полностью отделена от мочевой. Воль-
фов проток самок хрящевых рыб выполняет только функцию моче-
точника, почки не связаны с половой системой. Парные яичники,
в отличие от семенников, не сообщаются непосредственно с половы-
ми протоками, а выделяют яйцеклетки в полость тела, откуда они по-
падают в мюллеровы протоки, эмбрионально развивающиеся в ре-
зультате расщепления пронефрического протока на два: один из них
становится вольфовым протоком, а другой — мюллеровым. Подобное
образование мюллерова протока свойственно хрящевым рыбам, но
у других позвоночных они могут возникать в эмбриогенезе самостоя-
тельно, параллельно с вольфовыми протоками. У самок мюллеровы
протоки превращаются в яйцеводы. Передний конец, расположенный
вблизи яичника, расширяется, образуя воронку яйцевода, которая
соответствует нефростому головной почки. В начальном отделе яйце-
водов находятся скорлуповые железы, секрет которых формирует
оболочку яиц. Задние расширенные отделы яйцеводов иногда некор-
ректно называют маткой, они открываются в клоаку самостоятельны-
ми отверстиями, не сообщаясь с мочеточниками.
Развитие. У хрящевых рыб оплодотворение внутреннее, при этом
сперма выталкивается в клоаку и с помощью копулятивных органов
вводится в клоаку самки, а из нее в ее половые пути. Стенка яичника
разрывается, яйцеклетка сначала попадает в полость тела, а затем че-
рез воронку, которая к этому моменту увеличивается в размерах, про-
никает в просвет яйцевода. В передних отделах яйцевода за счет се-
крета скорлуповых желез образуется оболочка яйца, там же она
оплодотворяется, затем оформленное яйцо благодаря перистальтике
стенок мюллерова протока передвигается по направлению к «матке».
Яйца акул очень крупные, покрытые плотной оболочкой и часто
имеют сложную форму. Обычно яйца откладываются поодиночке в
укромных местах. Лишь немногие хрящевые рыбы (например, поляр-
ная акула) мечут многочисленную мелкую икру. Яйцо рыб относится
к телолецитальному типу, т. е . содержит очень большое количество
596
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
желтка. Развитие практически полностью проходит внутри яйца, так
что из него обычно выходит вполне сформированное животное. У мно-
гих хрящевых рыб оплодотворенное яйцо остается в половых путях,
где эмбрион вначале питается за счет запасов желтка, однако в даль-
нейшем стенки желточного мешка срастаются со стенками «матки», где
это яйцо находится. Между развивающимся эмбрионом и матерью
образуется общая сеть кровеносных сосудов, через которые происходит
обмен веществ. Эта структура напоминает плаценту млекопитающих.
По окончании эмбрионального развития молодое животное выходит из
половых путей и сразу же начинает самостоятельную жизнь.
Нервная система с точки зрения анатомии подразделяется на цен-
тральную и периферическую, а физиологически — на соматическую и
вегетативную (рис. 248). К центральной нервной системе (ЦНС) от-
носится спинной и головной мозг, все прочие нервные структуры,
расположенные за пределами головного и спинного мозга, образуют
периферическую нервную систему. Нервная трубка рыб подразделя-
ется на спинной мозг (задняя часть) и головной мозг (передняя).
Общее строение ЦНС рыб достаточно примитивна, их мозг очень
маленький, у хрящевых рыб всего 0,006—0,44% массы тела, у млеко-
питающих — 0,3—3,0%; слабо развит передний мозг и др. Спинной
Рис. 248. Головной мозг акулы сверху, сбоку и снизу:
1 — раковина мозжечка, 2 — обонятельная луковица, 3 — мозжечок, 4 — нижние
доли, 5 — промежуточный мозг, 6 — эпифиз, 7 — гипофиз, 8 — зрительные доли,
9 — продолговатый мозг, 10 — спинной мозг, 11 — средний мозг, 12 — подъязычный
нерв, 13 — спинной нерв, 14 — предобонятельный нерв, 15 — заднее сосудистое
сплетение, 16 — язычный мешок, 17 — передний мозг,
I, II, III, IV, V, VI, VII, VIII, IX, X, XI — головные нервы (по Маринелли)
ПОДТИП ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ
597
мозг представляет собой тонкий шнур, расположенный в позвоноч-
ном канале, образованном верхними дугами позвонков, которые за-
щищают спинной мозг от повреждений.
В эмбриогенезе головной мозг возникает в виде расширения пе-
реднего отдела нервной трубки. Вначале передний участок нервной
трубки образует вздутие — стадия одного мозгового пузыря, делят
этот пузырь затем две неполные поперечные перетяжки на три пузы-
ря (стадия трех мозговых пузырей), в дальнейшем образовавшиеся
пузыри дифференцируются на специализированные отделы головно-
го мозга. Из переднего пузыря образуются два отдела — больший ко-
нечный мозг и промежуточный мозг; средний отдел не делится, он
становится средним мозгом; задний пузырь преобразуется в ромбо-
видный мозг, который, в свою очередь, подразделяется на задний мозг
и продолговатый мозг, последний у большинства позвоночных, кроме
млекопитающих, без видимых границ переходит в спинной мозг.
У млекопитающих в состав заднего мозга входит мост и мозжечок,
у всех остальных позвоночных мост не является самостоятельным от-
делом головного мозга, так как между спинным мозгом и головным
мозгом связи малочисленные. Остатки невроцеля превращаются
в желудочки головного мозга. Среди хрящевых рыб головной мозг на-
иболее дифференцирован у акуловых в связи с высоким уровнем раз-
вития органов чувств.
Продолговатый мозг является продолжением спинного мозга и во
многом сходен с ним по строению. От продолговатого мозга отходят
шесть из десяти пар черепных нервов. Именно в продолговатом мозге
находятся центры, управляющие деятельностью внутренних органов,
опорно-двигательного аппарата и органов чувств (равновесия, слуха,
вкуса, боковой линии). Через продолговатый мозг проходят нервные
связи между спинным и головным мозгом, однако у рыб их немного.
У хрящевых рыб хорошо развит мозжечок, осуществляющий коор-
динацию и регуляцию движений, поэтому у быстро плавающих акул
он очень велик. Наиболее крупный и развитый отдел головного моз-
га — средний мозг. Две крупные зрительные доли образуют его основ-
ную часть. Средний мозг является интегрирующим центром нервной
системы рыб. В средний мозг поступают импульсы от органов чувств,
происходит анализ и синтез, эфферентные импульсы направляются
в другие отделы головного мозга и в спинной мозг. Головной мозг
хрящевых, костных рыб и амфибий, в котором высшим центром ин-
теграции нервной деятельности является средний мозг, относят к их-
тиопсидному типу именно в области среднего мозга образуется изгиб,
который имеется и у других позвоночных.
В состав промежуточного мозга входят зрительные бугры (тала-
мусы), к которым подходят зрительные тракты, образующиеся после
598
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
частичного перекреста зрительных нервов. В таламусы позвоночных
поступает информация от всех органов чувств, там находится центр
координации движений. В гипоталамусе — нижней части промежу-
точного мозга — залегают многочисленные ядра, в которых располо-
жены, помимо типичных нейронов, нейросекреторные клетки, синте-
зирующие нейрогормоны. В промежуточном мозге имеются два
придатка: верхний — эпифиз (эндокринная железа) и нижний, обра-
зующий заднюю долю, — гипофиз. Передняя доля гипофиза незави-
симо от задней формируется путем впячивания эктодермы ротовой
бухты (карман Ратке), у хрящевых рыб она сообщается с ротовым
эпителием.
Гипоталамус вместе с гипофизом образуют гипоталамо-гипофи-
зарную систему, которая через эндокринные железы регулирует дея-
тельность внутренних органов. Неполная перегородка разделяет боль-
шой (конечный) мозг на два относительно крупных полушария, при
этом нервные клетки имеются не только в их основании и в боковых
стенках, но и частично в крыше. К полушариям прилегают обоня-
тельные луковицы, являющиеся первичным центром обоняния.
Основная масса серого вещества сосредоточена в базальных ядрах —
полосатых телах, лежащих в вентральном отделе полушарий. В отли-
чие от высших позвоночных, серое вещество полушарий располагается
внутри белого, которое расположено снаружи. Основной функцией
конечного мозга рыб является обработка обонятельных импульсов.
Он также участвует в регуляции двигательной активности и поведе-
ния, поскольку связан с промежуточным и средним мозгом.
У всех рыб спинной мозг полностью заполняет спинномозговой
канал, у более высокоорганизованных позвоночных проявляется тен-
денция к сокращению длины спинного мозга. Спинной мозг сегмен-
тирован на всем протяжении.
Вегетативная нервная система у хрящевых рыб раз-
вита относительно слабо, анатомически она образована отдельными
ганглиями, которые разбросаны вдоль всего позвоночного столба.
Нейроны этих ганглиев своими отростками связаны с центрами, рас-
положенными в спинном мозге, и с внутренними органами.
Периферическая нервная система состоит из череп-
ных и спинномозговых нервов, их окончаниями и нервными узлами.
У рыб одиннадцать пар черепных нервов: I — обонятельные, II —
зрительный, III — глазодвигательный, IV — блоковый, V — тройнич-
ный, VI — отводящий, VII — лицевой, VIII — слуховой, IX — языко-
глоточный, X — блуждающий. Добавочный (XI) и подъязычный
(XII) зачаточные, их корешки отходят позади десятой пары. Все че-
репные нервы, начиная с тройничного, отходят от продолговатого
мозга.
ПОДТИП ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ
599
Спинномозговые нервы метамерно образуются в резуль-
тате слияния передних и задних (спинных и брюшных) корешков
спинного мозга (правых корешков с правыми, а левых — с левыми).
Каждая пара нервов иннервирует скелетные мышцы и покровы соот-
ветствующего сегмента тела. Наличие парных конечностей приводит
к тому, что в эмбриогенезе нарушается сегментация мускулатуры, ко-
торая обеспечивает движения конечностей, соответственно нарушает-
ся и сегментация спинномозговых нервов. При этом волокна несколь-
ких спинномозговых нервов смешиваются, образуются два сплетения:
плечевое и пояснично-крестцовое.
У хрящевых рыб очень быстро – на следующий день - затухают
условные рефлексы.
Органы чувств развиты хорошо. Органы зрения, располо-
женные по бокам головы, снабжены глазодвигательными мышцами,
обеспечивающими большую подвижность глаз, что особенно важно,
учитывая неподвижное соединение туловища с головой. Плоская ро-
говица глаз оптимально функционирует в водной среде, шарообраз-
ный хрусталик преломляет свет; у акул имеется мигательная пере-
понка, закрывающая глаз наподобие век.
Орган слуха образован только внутренним ухом, защищен-
ным хрящевой капсулой черепа. Внутри капсулы находится запол-
ненный эндолимфой перепончатый лабиринт, состоящий из трех
полукружных каналов, овального и круглого мешочков. Три полу-
кружных канала вместе с овальным мешочком образуют орган равно-
весия. В некоторых участках нижней части перепончатого лабиринта
расположены слуховые пятна и гребни, выстланные нейросенсорны-
ми клетками, воспринимающими звуки. У рыб отсутствует улитка,
имеется лишь ее зачаток в виде бокового расширения круглого ме-
шочка. Звуковые колебания воды сначала вызывают вибрацию голо-
вы рыбы, после чего колебания передаются эндолимфе и воспринима-
ются чувствительными волосковыми клетками.
Обонятельные мешки, расположенные на голове и открывающие-
ся на нижней поверхности рыла отверстиями-ноздрями, являются
органами обоняния. У хрящевых рыб очень тонкое обоняние:
акулы улавливают запах раненой добычи за несколько километров.
Органы боковой линии имеются только у рыб и водных
амфибий, поэтому их называют шестым чувством. Они расположены
на голове и по бокам вдоль тела. У рыб эти органы находятся на дне
ямок или в каналах, они снабжены рецепторами, воспринимающими
колебания воды, разницу в давлении. Это позволяет на значительном
расстоянии улавливать колебания, которые совершают другие живот-
ные. Таким образом, акула обнаруживает добычу на расстоянии около
600
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
300 м и способна обходить погруженные в воду предметы, что позво-
ляет ориентироваться в полной темноте, реагировать на изменение
глубины и т. д.
В подклассе пластинчатожаберные выделяют два отряда — акулы
и скаты. В более крупный отряд акулы входит около 250 видов и око-
ло 10 семейств. Все представители имеют веретенообразную форму
тела, жаберные щели, открывающиеся по бокам передней части туло-
вища над грудными плавниками; гетероцеркальный хвостовой плав-
ник. Большинство ведут пелагический образ жизни, т. е. постоянно
плавают в толще воды на разной глубине. Размеры акул широко варь-
ируют от 20 см до 18 м. Значительно более крупные виды давно вы-
мерли, но их остатки — зубы (хрящевой скелет не сохраняется) — за-
легают в донных отложениях древних морей. Акулы — хищники, при
этом сила, с которой акулы сжимают челюсти, не имеет аналогов во
всем животном мире. Наиболее крупные акулы (китовая и гигант-
ская) питаются планктоном. Ряд видов акул (большая белая, голубая,
мако, акула-молот и др.) нападают на людей. Мясо и плавники неко-
торых акул (например, сельдяной) употребляются в пищу, а печень
содержит много жира и богата витамином А, поэтому акулы являются
объектом промысла. За редким исключением акулы обитают в соле-
ных водоемах.
Скаты имеют тело уплощенное в дорсовентральном направлении
с хорошо развитыми грудными плавниками, под которыми открыва-
ются отверстия жаберных щелей. Большинство скатов — малопод-
вижные животные, они лежат на дне, сливаясь с грунтом. Многие
виды имеют на хвосте ядовитый шип, которым они могут нанести
укол при дотрагивании до него. Такой укол очень болезненный и
опасный. Донные скаты питаются моллюсками и ракообразными, по-
этому у них тупые зубы. Очень редко скаты ведут пелагический образ
жизни, например очень крупный скат манта.
Класс Костные (Костистые) рыбы
Большинство представителей надкласса рыб относятся к классу
костные рыбы. Известно около 25 000 современных рыб, однако
ихтиофауна больших глубин до сих пор еще изучена недостаточно хо-
рошо, поэтому многие виды неизвестны.
У костных рыб образован костный скелет, однако у осетровых, на-
ряду с костями имеются и хрящевые элементы скелета.
Подобно хрящевым, тело костных рыб подразделяется на голову,
туловище и хвост. Отсутствие шеи, хорошо обтекаемое гладкое тело
без выступов способствует активному движению в воде. Форма тела
может быть торпедовидной (тунец, сельдь), змеевидной (угорь), лен-
ПОДТИП ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ
601
товидной (рыба-сабля), стреловидной (щука), сплющенной (лещ, кам-
бала). Глаза располагаются на голове, впереди глаз расположены обо-
нятельные отверстия. Брызгальце имеется только у осетровых рыб.
Жаберные дуги находятся в жаберной полости и прикрыты костными
жаберными крышками. Парные плавники выполняют функцию орга-
нов движения и рулей, поддерживающих тело в нужном положении
и направляющих его при движениях. Строение и расположение плав-
ников зависит от образа жизни рыбы. Например, хорошо развитые
грудные плавники позволяют илистому прыгуну уверенно ползать по
суше, а летучим рыбам пролетать по воздуху значительные расстоя-
ния (до 800 м). Брюшные плавники смещаются далеко вперед (на-
пример, у тресковых) или видоизменены в присоску (например, у
бычков). Непарные плавники — спинной, анальный и хвостовой.
У некоторых рыб имеются дополнительные плавники. Основная
функция непарных плавников — стабилизация тела в пространстве.
Хвостовой плавник является главным органом движения рыбы. Раз-
личная форма хвостового плавника зависит от скорости плавания.
У разных видов в связи с определенным образом жизни некоторые
плавники редуцированы. Так, у угря отсутствуют брюшные плавники,
а у мурены — грудные.
Покровы тела представлены кожей, покрытой чешуей. Кожа со-
стоит из многослойного эпидермиса и лежащей под ней плотной дер-
мы. В эпидермисе находится много одноклеточных желез, выделяю-
щих бактерицидную слизь, защищающую рыбу от патогенных
микроорганизмов, участвует в выделении продуктов обмена и водно-
солевом обмене, ускоряют свертывание крови и уменьшают трение
рыбы о воду. Слизистые клетки синтезируют видоспецифические ве-
щества, позволяющие рыбам узнавать друг друга на расстоянии.
Слизь некоторых рыб токсична, иногда у них имеются ядовитые же-
лезы. В нижних слоях эпидермиса находятся пигментные клетки —
хроматофоры, благодаря этому многие рыбы пестрые. Некоторые
виды способны менять окраску тела, например лежащая на дне камба-
ла приобретает цвет донного субстрата. Окраска меняется в период
размножения (у лососевых и др.), при изменении температуры, соста-
ва воды, из-за эмоционального состояния рыбы (страх) и т. п . Степень
развития чешуи у разных видов различная, у некоторых рыб (напри-
мер, у сомов) чешуя вторично отсутствует. Костные чешуйки образу-
ются в кориуме. Чешуя улучшает гидродинамические свойства рыбы.
В коже располагаются различные рецепторы, в дерме образуются по-
кровные кости черепа и кости пояса передних конечностей.
Скелет костных рыб образован костями, но у осетровых в течение
жизни сохраняются хрящевые элементы (рис. 249). Костный скелет
более эффективен, он более прочный и легкий, чем хрящевой. Кост-
602
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ные элементы рыб возникают двояко. Кость развивается из хряща
(первичные, или замещающие, кости) и из очагов окостенения в дерме
(вторичные, или покровные, кости). Их также называют накладными,
поскольку они могут накладываться на хрящевые элементы скелета,
например в черепе осетровых).
Осевой скелет. Позвоночный столб состоит из двух отделов:
туловищного и хвостового. В отличие от хрящевых рыб, ребра которых
ограничивают полость тела лишь сверху, у костных ребра длинные, они
ограничивают полость тела еще и с боков. Череп устроен значительно
сложнее, чем у хрящевых рыб. Висцеральный череп образован челюст-
ной, подъязычной и жаберной дугами, а также костями жаберной крыш-
ки. Первичные челюсти костных рыб, развивающиеся из небно-ква-
дратного и меккелева хрящей, соединяясь, формируют челюстной
сустав, тогда как сами челюсти образованы вторичными костями. Нали-
чие вторичных челюстей позволяет значительно усложнить способы за-
хвата и удержания пищи, приспосабливаться к определенному типу
пищи. Это объясняет различное строение ротового аппарата и, соответ-
ственно, видовое разнообразие. У хрящевых рыб ротовой аппарат устро-
ен однотипно, поэтому они не могут потреблять столь же разнообразную
пищу, как различные костные рыбы. У них пять жаберных дуг, но по-
следняя развита хуже остальных. Подвижное соединение мозгового и
висцерального отделов черепа у костных рыб осуществляется подъя-
зычной дугой, при этом гиомандибуляре образует сустав со слуховым
отделом мозгового черепа. Жаберная крышка, имеющаяся только
у костных рыб, прикрывает с боков жаберные дуги. Она состоит из че-
тырех плоских накладных костей: крышки, предкрышки, подкрышки
и межкрышки. Жаберная крышка прикрепляется к гиомандибуляре.
Рис. 249. Скелет костистой рыбы (окуня):
1 — кости черепа, 2 — основные элементы спинного плавника, 3, 4 — лучи спинного
плавника, 5 — последние позвонки, удерживающие хвостовой плавник,
6 — хвостовые позвонки, 7 — основные элементы анального плавника,
8 — туловищные позвонки, 9 — ребра с придатками, 10 — кости и лучи брюшного
плавника, 11 — кости и лучи грудного плавника, 12 — жаберная крышка,
13 — верхняя и нижняя челюсти (по Баклашовой)
ПОДТИП ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ
603
Добавочный скелет устроен проще, чем у хрящевых рыб.
Он образован костями свободных конечностей и их поясов. Пояс пе-
редних конечностей (грудных плавников) образован небольшими ло-
паткой, коракоидом и цепочкой вторичных костей, самая крупная из
которых — клейтрум, имеющая серповидную форму (эту кость назы-
вают ключицей), которая соединяется с задним отделом мозгового че-
репа. Пояс задних конечностей (тазовый пояс) образован двумя срос-
шимися костями, свободно лежащими в толще мышц, к которым
прикрепляются костные лучи брюшных плавников. Непарные конеч-
ности не имеют поясов. Кроме этих костей, опору для соматической
мускулатуры формируют многочисленные тонкие межмышечные
(мускульные, или туловищные) косточки, которые расположены в
толще мышц. По своему происхождению они являются окостеневши-
ми сухожилиями.
Мышечная система у костных рыб развита лучше, чем у хрящевых,
что позволяет им первым совершать гораздо более сложные движения.
Она представлена мышцами туловища, головы и плавников. Основную
массу составляет сегментированная мускулатура туловища, обеспечива-
ющая движение рыбы. Мышцы головы (висцеральные) управляют дви-
жениями челюстей и жаберных крышек. Несегментированная мускула-
тура плавников изменяет положение парных плавников и стабилизирует
непарные. Соматическая мускулатура образована поперечнополосатыми
мышечными волокнами, которые дифференцированы на красные и бе-
лые (их больше). Скопления специализированных мышечных волокон
$
%
&
'
!
"
#
$
!
"
#
!
"#
$
%
&
'
Рис. 250. Общее расположение внутренних органов щуки:
1 — венозный синус, 2 — предсердие, 3 — желудочек сердца, 4 — брюшная аорта,
5 — луковица аорты, 6 — приносящие жаберные артерии, 7 — кювьеров проток,
8 — жабра, 9 — желудок, 10 — двенадцатиперстная кишка, 11 — тонкая кишка,
12 — прямая кишка, 13 — анальное отверстие, 14 — печень, 15 — желчный пузырь,
16 — желчный проток, 17 — поджелудочная железа, 18 — селезенка,
19 — плавательный пузырь, 20 — почка, 21 — мочеточник, 22 — мочевой пузырь,
23 — мочеполовой сосочек, 24 — мочевое отверстие, 25 — половая железа,
26 — половое отверстие
604
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
образуют электрические органы некоторых рыб, которые могут генери-
ровать разряды до 600 В (электрический угорь, электрический сом), спо-
собные оглушить жертву или врага и даже убить их.
Пищеварительная система костных рыб имеет те же отделы, что
и у хрящевых (рис. 250). Положение ротового отверстия зависит от
способа питания рыбы. У планктоноядных форм рот расположен
у хищников спереди, у бентосоядных (подбирающих пищу со дна) —
снизу; встречаются и различные переходные варианты. Ротовая по-
лость снабжена многочисленными зубами, которые имеются у многих
костей черепа. Все зубы однотипны, они наклонены по направлению
к глотке, поэтому рыбы могут только удерживать добычу или отры-
вать от нее куски (например, пираньи), но не могут ее жевать. Обычно
зубы прирастают к поверхности кости, иногда они соединяются с ней
подвижно. В течение жизни изношенные зубы заменяются новыми.
У рыб, питающихся растительной пищей или планктоном, зубы в ро-
товой полости отсутствуют. Как и у всех рыб, у костных отсутствует
настоящий язык. Имеющаяся вместо языка складка слизистой обо-
лочки лишена собственных мышц и не способна выдвигаться, у неко-
торых видов на ней могут находиться зубы. Пронизанная жаберными
щелями, глотка активно участвует в процессах питания.
Поступление пищи в ротовую полость осуществляется за счет
движения жаберных крышек, вода вместе с пищей засасывается в ро-
товую полость, из нее затекает в глотку и выталкивается из нее через
жаберные щели, а пища остается в глотке, поскольку ее не пропуска-
ют жаберные тычинки, расположенные на жаберных дугах. У некото-
рых видов (карповых и др.) на задней жаберной дуге расположены
глоточные зубы, которые перетирают пищу. Слюнные железы отсут-
ствуют. Короткий пищевод переходит в желудок, который имеется не
у всех видов, он отсутствует у карповых, бычков и др. Желудок пере-
ходит в тонкую кишку, в которую впадают протоки печени и подже-
лудочной железы. Из тонкой кишки расщепляется больше пищевых
веществ и осуществляется всасывание. Тонкая кишка костных рыб
значительно длиннее, чем у хрящевых, она образует множество пе-
тель (первая из них называется двенадцатиперстной кишкой), что
увеличивает поверхность всасывания.
Относительная масса печени костных рыб меньше, чем хрящевых
(1—8% от массы тела), но она выполняет те же функции. Имеется
желчный пузырь. Отсутствует анатомически оформленная поджелу-
дочная железа, ее структуры диссоциированы в ткани печени. Она
выполняет экзокринную и эндокринную функции. У представителей
надотряда костистых эндокринные клетки обособляются в виде
островков Лангерганса. Тонкая кишка переходит в толстую. Спираль-
ный клапан имеется только у осетровых и двоякодышащих рыб.
ПОДТИП ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ
605
У разных видов широко варьирует общая длина пищеварительно-
го тракта, что связано с характером питания: у растительноядных она
длиннее (у толстолобика в 6—15 раз превышает длину тела),
у всеядных рыб короче (кишечник карася длиннее тела в 2—3 раза),
а у хищных рыб он совсем короткий (у судака или щуки всего 0,6—
1,2 длины тела). Клоака имеется только у двоякодышащих рыб, у всех
остальных кишечник открывается анальным отверстием наружу.
Большинство костных рыб имеют плавательный пузырь, который
развивается как вырост дорсальной стенки передней части пищевари-
тельного тракта. В зависимости от того, сохраняется ли связь пузыря
с пищеводом, рыбы могут быть открытопузырными или закрытопу-
зырными. Плавательный пузырь заполнен воздухом, что уменьшает
удельный вес рыбы и позволяет ей зависать в воде. Хрящевые рыбы,
лишенные плавательного пузыря, этого делать не могут. Рыбы могут
произвольно изменять объем газа в пузыре, что ведет к уменьшению
или увеличению их плавучести.
Дыхательная система представлена жабрами. По сравнению с
хрящевыми рыбами у костных жаберный аппарат отличается рядом
особенностей. Частично (у осетровых) или полностью (у костистых)
редуцированы межжаберные перегородки, поэтому жаберные лепест-
ки расположенные непосредственно на жаберных дугах, омываются
водой со всех сторон. Только у костных рыб имеется жаберная крыш-
ка, которая активно участвует в акте дыхания, нагнетая воду в рото-
вую полость, и позволяет интенсифицировать газообмен даже при не-
подвижном положении рыбы. Газообмен рыб очень эффективный.
Некоторые виды высших костных рыб способны усваивать до 85%
растворенного в воде кислорода (хрящевые рыбы — 70—75%). Очень
важен кожный газообмен. Многие виды имеют дополнительные орга-
ны газообмена для усвоения кислорода воздуха (двоякодышащие, ла-
биринтовые и др.). При недостатке кислорода рыбы «глотают» воз-
дух, после этого кислород всасывается в капилляры, которые оплетают
стенки кишечника.
Кровеносная система костных рыб во многом сходна с системой
хрящевых рыб. Редуцирована передняя пара жаберных артерий. Зна-
чительные изменения в организации кровеносной системы имеются у
рыб, дышащих атмосферным воздухом, у двоякодышащих рыб имеет-
ся второй круг кровообращения.
У большинства видов гемоглобин находится в эритроцитах, при
этом относительное его содержание невелико (примерно 0,5—4 г/кг
массы тела, тогда как у млекопитающих 5—25 г/кг).
606
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Лимфатическая система развита значительно лучше, чем у хря-
щевых. Выражена система сосудов, по которым лимфа оттекает от
внутренних органов. Лимфатические сосуды впадают в вены.
Выделительная система образована лентовидными мезонефриче-
скими почками. Мочеточники объединяются в выводной канал, кото-
рый открывается не в клоаку, а на конце мочеполового сосочка. В не-
фронах осетровых сохраняется воронка (нефростом), однако у более
высокоорганизованных форм она отсутствует. Основной конечный
продукт азотистого обмена не мочевина (как у хрящевых рыб), а бо-
лее токсичный аммиак, поэтому костные рыбы выделяют большие
объемы мочи, так как аммиак требует гораздо большего разбавления.
Половая система костных рыб полностью отличается от системы
хрящевых рыб. Мужская половая система образована парными се-
менниками. В отличие от хрящевых, семявыносящие протоки семен-
ников костных рыб самостоятельные, они открываются общим поло-
вым отверстием позади анального отверстия. Таким образом, мужские
половые протоки костных рыб не имеют никакого отношения к воль-
фовым протокам, которые у них являются только мочеточниками.
Женская половая система образована парными полыми яичника-
ми, занимающими в теле примерно такое же положение, что и семен-
ники. Вытянутая часть оболочки яичника образует проток, через ко-
торый созревшие яйца (икринки) выходят либо непосредственно
наружу через самостоятельное непарное половое отверстие, лежащее
позади анального отверстия. У самок костных рыб мюллеровы прото-
ки редуцируются.
Развитие. У большинства костных рыб оплодотворение внешнее, от-
кладывание икры называется нерестом. У некоторых видов он сопрово-
ждается сложным половым поведением и изменением строения рыбы.
Многие костные рыбы проявляют заботу о потомстве (цихлиды, тиля-
пия, самцы лабиринтовых рыб, трехиглой колюшки и др.) . Проходные
рыбы для нереста мигрируют из моря в реки, где когда-то сами появи-
лись на свет. Количество икринок широко варьирует от единичных до
огромного (луна-рыба мечет до 30 млн икринок). Икра некоторых видов
свободно плавает в воде, других — прикрепляется к водным растениям,
поверхности донных камней и другим подводным предметам. Размеры
икринок маленькие, они богаты желтком (телолецитальные яйца). Раз-
витие костных рыб чаще всего осуществляется прохождением личиноч-
ной стадии. Среди костных рыб имеются и живородящие, например ак-
вариумные гуппи, меченосцы, моллиенезии и др. Много икринок
и молоди гибнет, поэтому до взрослой особи доживает лишь незначи-
тельная часть потомства.
Нервная система разделена, как и у хрящевых рыб, на несколько от-
делов, но головной мозг более примитивный (рис. 251). Конеч-
ПОДТИП ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ
607
ный мозг меньше, он не делится даже не-
полной перегородкой, поэтому его полость
едина и не образует боковых желудочков.
Нервные элементы имеются только в осно-
вании и боковых стенках конечного мозга
(крыша образована эпителием). Относи-
тельно крупный средний мозг является ин-
тегративным центром (ихтиопсидный тип
мозга). Мозжечок значительных размеров,
особенно у подвижных видов (активных
хищников и др.). У малоподвижных рыб
(камбал и др.) маленький мозжечок, но хо-
рошо развиты конечный и продолговатый
мозг, что важно для обоняния и осязания.
Спинной мозг не отличается от хрящевых рыб. Вегетативная
нервная система имеет более сложное строение, чем у хряще-
вых рыб. Расположенные вблизи позвоночного столба ганглии соеди-
няются в два симпатических ствола. Парасимпатический отдел обра-
зован вегетативными волокнами блуждающего нерва.
Органы чувств аналогичны таковым хрящевых рыб. Орган зре-
ния имеет обычное для рыб строение. Среди рецепторов различают
палочки, которые реагируют на слабый свет, но не могут создать чет-
кого изображения, и колбочки, нуждающиеся в большом количестве
света (цветовое зрение). У некоторых рыб зрение вторично редуци-
ровано.
Орган слуха — внутреннее ухо, сочетающее рецепторы, вос-
принимающие звуковые волны (слуховой анализатор) и изменение
положения тела. Внутреннее ухо имеет обычное для позвоночных
строение: внутри костного лабиринта слуховой капсулы (его вну-
тренние стенки не костные, а хрящевые!) залегает перепончатый ла-
биринт. Рецепторы, воспринимающие звук, расположены в круглом
мешочке. Рыбы воспринимают низкочастотные звуки боковой лини-
ей, а высокочастотные — внутренним ухом.
&
'
$
%
!
"
#
Рис. 251. Головной мозг окуня:
1 — обонятельная капсула, 2 — обонятельные доли,
3 — передний мозг, 4 — средний мозг, 5 — мозжечок,
6 — продолговатый мозг, 7 — спинной мозг,
8 — глазничная ветвь тройничного нерва,
9 — слуховой нерв, 10 — блуждающий нерв
(по Суворову)
608
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Органы обоняния представлены обонятельными мешками,
ноздри расположены между ртом и глазами. Многие рыбы имеют
очень тонкое обоняние, например лососи чувствуют воду родной реки
(в которой они появились на свет из икринки) на расстоянии 800 км
и безошибочно находят дорогу туда для нереста.
Орган вкуса — вкусовые почки разбросаны по всей поверхно-
сти кожи, особенно много их в слизистой оболочке ротовой полости,
глотки, на жаберных лепестках и усиках. Рыбы определяют вкусы
сладкий, соленый, кислый и горький. Химические раздражители так-
же воспринимают свободные окончания тройничного, блуждающего
и спинномозговых нервов.
Органы боковой линии имеют типичное строение, они
образованы погруженным в кожу каналом, который тянется вдоль
всего тела и через отверстия сообщается с окружающей водой.
Для многих видов рыб важны органы, воспринимающие электри-
ческое и магнитное поля. Эти органы расположены на всей поверхно-
сти тела, но больше всего их на голове и вблизи нее. Особенно важно
восприятие электромагнитных сигналов для рыб, обитающих в мут-
ной воде, где ориентация с помощью зрения неэффективна.
Поведение костных рыб сложнее, чем хрящевых, условные реф-
лексы у них вырабатываются легче, а затухают медленнее. Это позво-
лило костным рыбам приспосабливаться к разнообразным условиям
обитания и привело к разнообразию видов.
Класс костные рыбы подразделяется на четыре подкласса: хряще-
костные, лучеперые, кистеперые и двоякодышащие.
Хрящекостные являются наиболее древней и примитивной
группой костных рыб. По внешнему виду они напоминают акул,
тело покрывают костные «жучки». Хорда сохраняется в течение всей
жизни. Позвонки лишены тела. Первичный хрящевой череп снару-
жи покрыт накладными костями, однако жаберная крышка костная.
Плавательный пузырь связан с кишечником, в котором развит спи-
ральный клапан. Подкласс содержит всего один отряд — осетровые,
многие представители которых являются ценными промысловыми
рыбами.
Наиболее распространенный подкласс современных рыб — луче-
перые, на их долю приходится более 90% видов. Они распростране-
ны во всех водоемах (кроме термальных источников и некоторых
других) и занимают разнообразные экологические ниши. Скелет лу-
чеперых почти полностью костный. Тело покрыто костными чешуя-
ми, которые черепицеобразно накладываются друг на друга. В под-
класс лучеперые входят три надотряда: костные ганоиды, многоперые
и костистые (самые высокоорганизованные рыбы). Двоякодышащие
имеют одно или два ячеистых легких, которые сообщаются с брюш-
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
609
ной стороной пищевода. Легкие не гомологичны плавательному пу-
зырю костистых рыб (у двоякодышащих отсутствует). Кровеносная
система похожа на кровеносную систему наземных позвоночных.
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
К надклассу Наземные позвоночные относятся наземные, или вто-
ричноводные, позвоночные животные. Их пятипалые конечности
приспособлены для перемещения по твердой поверхности, у вторич-
новодных форм они видоизменяются в ласты или плавники, некото-
рые формы (червяги, змеи, некоторые ящерицы) вторично утратили
конечности. Органы дыхания (легкие) осуществляют газообмен меж-
ду атмосферным воздухом и кровью, в связи с этим у них имеется
малый, или легочный, круг кровообращения.
Класс Земноводные, или Амфибии
Амфибии — наиболее примитивные наземные животные. В яйце
отсутствует амниотическая полость, поэтому вместе с круглоротыми
и рыбами земноводные относятся к анамниям, они размножаются в
воде, где проходят начальные этапы их развития. Взрослые особи бо-
лее или менее связаны с водой, их система защиты организма от поте-
ри воды несовершенная. Это наиболее малочисленный класс позво-
ночных животных, насчитывающий примерно 2100—2600 видов.
Область их обитания ограничена прибрежными областями пресных
водоемов, морские виды отсутствуют. Класс включает три отряда:
хвостатые, безногие и бесхвостые, представители кото-
рых различаются между собой главным образом внешним строением.
Внешнее строение неодинаково у разных отрядов. У хвостатых от-
четливо различаются голова, туловище и хвост, соответственно, у бес-
хвостых отсутствует хвост, а у безногих — конечности.
Покровы тела земноводных голые, они лишены роговых или кост-
ных структур. Наряду с легкими, кожа является важным органом га-
зообмена, поэтому она всегда сохраняется влажной. Кислород снача-
ла растворяется в жидкости, покрывающей кожу, после чего
диффундирует из нее в кровь. То же самое происходит и с СО2, но в
обратном направлении. Это объясняет наличие в коже большого ко-
личества многоклеточных желез (у рыб кожные железы одноклеточ-
ные) эпидермального происхождения, погруженных в толщу дермы.
Секрет желез способствует газообмену и содержит бактерицидные
вещества, предохраняющие организм от внедрения инфекций. Неко-
торые железы выделяют ядовитый секрет, который защищает живот-
ное от нападения хищников. Верхний слой эпидермиса амфибий
610
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
(особенно жаб, длительно находящихся вне воды) ороговевает. Под
многослойным эпидермисом располагается соединительнотканная
дерма, в которой находятся пигментные клетки, определяющие окра-
ску кожи животного. Через кожу легко проникает вода, поэтому, на-
ходясь в воде, амфибии не пьют. Кожа прикреплена к телу не на всем
протяжении, а на определенных участках, пространства между кото-
рыми — лимфатические мешки — заполнены тканевой жидкостью.
Скелет, как и у всех позвоночных, подразделяется на осевой и до-
бавочный со всеми отделами, которые уже были перечислены при
описании скелета рыб. Строение скелета амфибий имеет особенности,
связанные с наземным образом жизни (рис. 252). Позвоночный
столб значительно больше дифференцирован на отделы, чем у рыб.
В нем выделяют отделы: шейный (один позвонок), туловищный (от 7
у бесхвостых до 100 и более у безногих), крестцовый (один позвонок)
и хвостовой (позвонки у хвостатых многочисленны, а у бесхвостых
Рис. 252. Скелеты: А — саламандры, Б — лягушки; 1 — череп, 2 — шейный позвонок,
3 — туловищные позвонки, 4 — ребра, 5 — крестцовый позвонок, 6 — хвостовые
позвонки (у лягушки — уростиль), 7 — плечевой пояс, 8 — плечо, 9 — локтевая кость,
10 — лучевая кость, 11 — запястье, 12 — пясть, 13 — фаланги пальцев,
14 — подвздошные кости таза, 15 — бедро, 16, 17 — большая и малая берцовые
кости, 18 — подплюсна, 19 — плюсна, 20 — фаланги пальцев
(по Левушкину и соавт.)
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
611
они срастаются в единую кость — уростиль). Наличие шейного по-
звонка очень важно, так как, в отличие от рыб, земноводные не могут
быстро разворачивать свое тело, а шейный позвонок увеличивает под-
вижность головы, которая у земноводных крайне мала: они не могут
ее поворачивать, но способны наклонять благодаря подвижному со-
членению шейного позвонка с черепом. К туловищным позвонкам
прикрепляются короткие ребра, которые у амфибий и не формируют
грудную клетку. Настоящие ребра есть только у безногих амфибий,
у хвостатых они имеют меньшие размеры, а у бесхвостых срастаются
с поперечными отростками позвонков. К единственному крестцовому
позвонку прикрепляются подвздошные кости. У позвонков хвостово-
го отдела есть верхние и нижние дуги, у бесхвостых они срастаются
в одну кость — уростиль. Благодаря наличию на телах позвонков со-
членовных поверхностей и сочленовных отростков позвонки прочно
соединены между собой, а весь позвоночный столб достаточно под-
вижный. Это особенно важно для передвижения хвостатых амфибий
в воде за счет боковых изгибов тела (наподобие рыб) без участия ко-
нечностей. Позвоночный столб может выполнять и вертикальные
движения.
Череп амфибий отличается большим количеством хрящей. Ко-
личество покровных (накладных) костей мало. Череп сочленяется с
шейным позвонком двумя мыщелками. Покровные кости образуют
дно, крышу и боковые стенки мозгового черепа. Висцеральный череп
также в основном хрящевой. В течение всей жизни сохраняется мек-
келев и небно-квадратный хрящи. Функционируют вторичные челю-
сти, образованные межчелюстными (предчелюстными) и верхнече-
люстными костями, которые расположены в виде костной дуги
кнаружи от небно-квадратного хряща. Череп амфибий плоский и ши-
рокий, что связано с особенностями нагнетания воздуха в легкие.
Редукция жаберного дыхания привела к радикальному преобразо-
ванию подъязычной дуги, при этом ее нижняя часть — гиоид — совмес-
тно с жаберными дугами видоизменяется в подъязычный аппарат,
к которому прикрепляются мышцы дна ротовой полости. Верхняя
часть подъязычной дуги — гиомандибуляре (подвесок) — перемещена
в полость среднего уха (она гомологична брызгальцу — жаберной
щели между челюстной и подъязычной дугами) и превращается
в слуховую косточку — стремечко, которое одним концом упирается
в центральную часть барабанной перепонки, а другим — в мембрану
овального окна внутреннего уха. Стремечко воспринимает колебания
барабанной перепонки и передает их на мембрану овального окна.
Жаберная крышка отсутствует.
Добавочный скелет состоит из костей поясов и свободных
частей конечностей. Пояс передней конечности (плечевой пояс) пред-
612
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ставляет собой состоящую из трех ветвей дугу, обращенную своей
вершиной к брюшной стороне. Спинная часть образована лопаткой,
брюшная — коракоидом («вороньей костью»), кпереди от него лежит
предкоракоидальный хрящ (предкоракоид), на который у бесхвостых
налегает тонкая ключица. Соединяясь между собой, все эти кости
образуют суставную впадину, куда входит головка плечевой кости.
Внутренние неокостеневшие концы коракоидов и прокоракоидов обе-
их сторон (правой и левой) соединяются между собой по средней ли-
нии. Впереди от этого соединения располагается предгрудина, а сза-
ди — грудина. Как и у рыб, кости плечевого пояса амфибий
расположены в толще мышц, которые связывают их с осевым скеле-
том, но сам пояс непосредственно не связан с осевым скелетом. Пояс
обеспечивает опору для свободной конечности.
Свободная конечность амфибий, как и всех наземных животных,
является пятипалой. Она служит опорой, позволяет приподнять тело
над поверхностью и обеспечивает перемещение в воздушной среде,
которая практически не поддерживает тело, поэтому всем наземным
животным постоянно приходится преодолевать силу тяжести (чего не
нужно делать рыбам). Наличие пяти пальцев не является главным
признаком пятипалой конечности. У многих четвероногих число
пальцев меньше пяти или даже они вообще вторично отсутствуют.
Передняя и задняя свободные конечности состоят из трех отделов:
проксимального (одна кость), промежуточного (две кости) и дисталь-
ного (относительно большое число костей). В типичном случае прок-
симальный отдел передней конечности образован плечевой костью,
задний — бедренной; промежуточный отдел передней конечности —
двумя костями (локтевой и лучевой), нижний — большеберцовой
и малоберцовой; дистальный отдел передней конечности состоит из
трех частей — запястья, пясти и фаланг пальцев, задний — из пред-
плюсны, плюсны и фаланг пальцев.
У представителей разных классов наземных позвоночных имеют-
ся особенности строения той или иной свободной конечности, но все
они носят вторичный характер. У всех амфибий проксимальный от-
дел свободной передней конечности — плечевая кость, промежуточ-
ный — локтевая и лучевая кости у хвостатых и единая кость предпле-
чья (она образуется в результате сращения локтевой и лучевой костей)
у бесхвостых.
Наиболее сложный отдел свободной передней конечности — ди-
стальный — образован запястьем, пястьем и фалангами пальцев. За-
пястье формируют два ряда мелких костей, часть из них срастается
между собой. К дистальному ряду костей запястья примыкают пять
пястных костей, дистальные концы которых сочленяются с фаланга-
ми пальцев. Пояс задних конечностей (тазовый пояс), в отличие от
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
613
плечевого, сочленяется непосредственно с осевым скелетом, а имен-
но — с его крестцовым отделом. У амфибий этот отдел состоит из од-
ного позвонка. Столь неравноценные способы прикрепления поясов
передней и задней конечностей связаны с тем, что для амфибий наи-
более важная — задняя конечность, которая обычно крупнее перед-
ней, особенно у бесхвостых.
Тазовый пояс всех наземных позвоночных состоит из трех парных
костей: подвздошных, седалищных и лобковых с крестцовым позвон-
ком (или с несколькими позвонками у более высокоорганизованных
позвоночных) сочленяются подвздошные кости. Соединяясь между
собой, все три кости тазового пояса образуют вертлужную впадину,
в которую входит головка бедренной кости, образуя тазобедренный
сустав. Особенностью тазового пояса амфибий является наличие
большего количества хрящевых элементов. Свободная задняя конеч-
ность имеет типичное строение: бедро, голень и стопа. Стопа подраз-
деляется на предплюсну, плюсну и фаланги пальцев. У хвостатых ам-
фибий голень состоит из большой и малой берцовых костей,
у бесхвостых они срастаются в одну кость голени.
Мышечная система сегментированная только у безногих, у хвоста-
тых и бесхвостых амфибий скелетная мускулатура утрачивает сег-
ментацию, что связано с развитием пятипалых конечностей. Если у
рыб движения плавников обеспечиваются главным образом мышца-
ми, расположенными на туловище, пятипалая конечность двигается
за счет мышц, расположенных в ней самой. Мускулатура конечностей
бесхвостых амфибий развита очень хорошо, особенно в задних конеч-
ностях, что связано со способом передвижения. Сегментированная
мускулатура имеется только в области позвоночного столба.
Пищеварительная система начинается обширной ротоглоточной
полостью, в которую ведет щелевидное ротовое отверстие. В общую
ротоглоточную полость также открываются отверстия внутренних
ноздрей (хоан), слуховых (евстахиевых) труб, гортанная щель, а так-
же протоки слюнных желез. Слюна земноводных лишена пищевари-
тельных ферментов и не оказывает на пищу химического воздейст-
вия, а только смачивает пищевой комок, облегчая его проглатывание.
На межчелюстных, верхнечелюстных костях, сошнике, а у некото-
рых и на нижней челюсти располагаются простые конические зубы,
которые своими основаниями прикрепляются к кости, а вершиной
обращены назад. Все зубы одинаковые, поэтому они только удержи-
вают добычу. У некоторых амфибий, например жаб, вообще отсутст-
вуют зубы. Амфибии имеют настоящий язык, который располагает-
ся на дне ротоглоточной полости. Язык покрыт клейким веществом
и полезен при ловле добычи, особенно функционален подвижный
язык, способный выдвигаться из ротоглоточной полости за счет
614
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
сокращения собственной мускулатуры и схватывать мелких жи-
вотных.
Проглатывание пищи сопровождается перемещением глазных
яблок вниз, при этом они давят на пищевой комок и проталкивают
его из ротоглоточной полости в короткий, растяжимый пищевод.
Пищевод только проводит пищу в желудок, где она задерживается,
размельчается благодаря сокращениям мускулатуры желудка, пропи-
тывается желудочным соком и подвергается воздействию ферментов.
Из желудка пищевая кашица (химус) поступает в кишку, которая
подразделяется на тонкую и толстую. В начальный отдел тонкой
кишки — двенадцатиперстную кишку — поступают секре-
ты печени и поджелудочной железы.
Поджелудочная железа в виде рыхлого тяжа располагается в бры-
жейке кишечника. Ее выводные протоки впадают в желчный проток
печени. Печень амфибий крупная, имеется желчный пузырь. Отдел
тонкой кишки, следующий за двенадцатиперстной кишкой, имеет
меньший диаметр, образует несколько петель и переходит в толстую
кишку, которая заканчивается прямой кишкой. В толстой кишке вса-
сывается бо
льшая часть воды, содержащаяся в химусе, и формируют-
ся каловые массы. Прямая кишка открывается в клоаку. Брыжейка
прикрепляется к спинной стороне стенки полости тела. В брыжейке
тонкой кишки расположена селезенка, которая является кроветвор-
ным органом.
Пищеварительная система амфибий отличается от аналогичной
системы рыб большей длиной пищеварительного тракта, кроме того,
конечный отдел толстой кишки открывается в клоаку. Земноводные
питаются мелкими беспозвоночными, однако крупные лягушки напа-
дают на мелких грызунов, птиц, змей и других амфибий, в том числе
и менее крупных особей своего вида.
Дыхательная система амфибий принципиально отличается от ды-
хательной системы рыб. Она обеспечивает газообмен в воздушной сре-
де. Жаберное дыхание имеется только у личинок, постоянно оби-
тающих в воде, но невозможно у взрослых животных, так как низкая
влажность воздуха исключает газообмен через наружные тонкостенные
жабры — они бы просто засохли. Дыхательная система амфибий наибо-
лее простая среди наземных позвоночных. Она состоит из ротоглоточ-
ной полости (верхние воздухоносные пути) и гортанно-трахеальной
полости (нижние пути), которая переходит в мешковидные легкие.
Гортань сообщается с глоткой через гортанную щель, которую с боков
ограничивают голосовые связкипокрытые складками слизистой обо-
лочки, натянутые между парными черпаловидными хрящами. Голосо-
вые связки вибрируют под действием воздуха, выходящего из легких,
тем самым участвуют в голосообразовании. Легкие — тонкостенные
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
615
мешки с ячеистой внутренней по-
верхностью за счет выростов стенок.
Стенки легких оплетены кровенос-
ными сосудами, которые приносят
венозную кровь и уносят артериаль-
ную, насыщенную кислородом.
У всех наземных позвоночных
органом воздушного дыхания явля-
ется легкое, поверхность газообме-
на которого располагается не сна-
ружи (как в жабрах), а внутри тела,
поэтому их функциональная по-
верхность не высыхает (рис. 253).
В процессе эмбрионального разви-
тия легкое формируется как слепой вырост переднего (глоточного)
отдела пищеварительной трубки, поэтому и у взрослых особей остает-
ся связанным с глоткой. Воздух доставляется в легкое по воздухонос-
ным (дыхательным) путям, которые связывают легкое, расположен-
ное внутри тела животного, с окружающим атмосферным воздухом.
Дыхательные пути выводят из легкого использованный воздух. Верх-
ние дыхательные пути, связывающие глотку с окружающей средой, —
это носовая полость, носоглотка и ротоглотка. Те, что проводят воздух
от глотки к легким, называют нижними дыхательными путями — это
гортань, трахея и бронхиальное дерево. Дыхательная система назем-
ных позвоночных анатомически и функционально подразделяется на
два отдела — воздухоносные пути и респираторный отдел. Воздухо-
носные пути осуществляют двусторонний транспорт воздуха, но не
участвуют в самом газообмене, респираторный отдел осуществляет
газообмен между внутренней средой организма (кровью) и атмосфер-
ным воздухом. Однако воздух и кровь — это разные агрегатные состо-
яния вещества — газ и жидкость, следовательно, непосредственный
обмен веществами между ними затруднен. Поэтому кислород воздуха
вначале растворяется в жидкости, которая тонкой пленкой выстилает
снаружи функциональную поверхность респираторного отдела, а по-
сле этого диффундирует в кровь. То же самое происходит и с углекис-
лым газом, но в обратном направлении: он из крови диффундирует в
поверхностную жидкость, после чего переходит в воздух. Таким обра-
зом, газообмен осуществляется пассивно в соответствии с градиентом
концентрации, т. е . из области с высокой концентрацией в область с
низкой концентрацией. В стенках воздухоносных путей имеются мно-
гочисленные слизистые железы, секрет которых не только увлажняет
воздух, но и задерживает механические частицы, содержащиеся в воз-
духе. Двустороннее движение воздуха по дыхательным путям способ-
Рис. 253. Легкие амфибий:
1 — легкие, 2 — их внутренняя
структура, 3 — гортань
(по Левушкину и соавт.)
616
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ствует его увлажнению. При дыхании воздух не заменяется полно-
стью, он лишь частично разбавляется свежим. Значительно более
высокое содержание О2 в атмосферном воздухе, по сравнению с во-
дой, делает газообмен у наземных позвоночных более эффективным.
Легочный тип дыхания требует совершенно иного способа венти-
ляции, чем у дышащих жабрами. Система жаберных крышек стано-
вится ненужной, поэтому жаберный аппарат наземных животных ви-
доизменяется, его скелетные элементы частично становятся хрящами
гортани. Вентиляция легких осуществляется за счет движений дыха-
тельных мышц, участвующих в акте дыхания, что обеспечивает пери-
одическое поступление и удаление воздуха из легких.
У амфибий в ходе дыхательного акта опускается дно ротовой по-
лости, атмосферный воздух засасывается и через отверстия ноздрей
попадает в полость ротоглотки, а затем через отверстие гортани
в легкие. Когда легкие заполнены, открываются отверстия ноздрей
и наружу выпускают излишний воздух. Для более эффективной
деятельности мышц дна ротовой полости большое значение имеет
характерный широкий и плоский череп амфибий. Общая функцио-
нальная поверхность респираторного отдела земноводных невелика:
соотносится с поверхностью тела примерно 2 : 3. Поэтому для амфи-
бий важен и кожный газообмен. Тонкая влажная кожа амфибий лег-
ко пропускает газы, например, у зеленой лягушки через кожу прони-
кает около 51% кислорода и удаляется около 86% углекислого газа,
тогда как через легкие — только 14%. Газообмен частично осуществ-
ляется через слизистую оболочку ротовой полости в процессе акта
дыхания. У жаб, имеющих относительно сухие покровы, большее
значение имеет легочный газообмен, тогда как у многих амфибий,
постоянно живущих в воде, кожный газообмен — основной, а иногда
даже единственный. У личинок амфибий газообмен осуществляется
с помощью наружных жабр, которые у большинства видов исчезают
в ходе метаморфоза, однако у сирен и протеев жабры сохраняются и
у взрослых особей.
Кровеносная система амфибий организована значительно слож-
нее, чем у рыб. Это связано с наземным образом жизни (рис. 254).
В связи с редукцией жаберного аппарата и переходом к легочному га-
зообмену у амфибий имеется второй (легочный) круг кровообраще-
ния. У всех наземных позвоночных сердце имеет два предсердия:
в одно поступает венозная кровь по венам большого (системного)
круга, а в другое — артериальная кровь по венам малого круга. Коли-
чество желудочков у разных классов неодинаково: один у амфибий
и большинства пресмыкающихся или два у крокодилов, всех птиц
и млекопитающих. Каждый круг кровообращения начинается в желу-
дочке, а заканчивается в предсердии.
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
617
!
"
#
!
"
#
$
%
&
'
$
%
&
'
!
"
#
$
%
&
'
!
!
!
!
!
!
"
!
#
Р
и
с
.
2
5
4
.
С
х
е
м
а
о
с
н
о
в
н
ы
х
к
р
о
в
е
н
о
с
н
ы
х
с
о
с
у
д
о
в
х
в
о
с
т
а
т
о
й
а
м
ф
и
б
и
и
(
в
и
д
с
б
о
к
у
)
:
1
—
в
и
с
о
ч
н
а
я
а
р
т
е
р
и
я
,
2
—
в
н
у
т
р
е
н
н
я
я
с
о
н
н
а
я
а
р
т
е
р
и
я
,
3
—
с
и
с
т
е
м
н
а
я
д
у
г
а
а
о
р
т
ы
,
4
—
а
р
т
е
р
и
а
л
ь
н
ы
й
п
р
о
т
о
к
,
5
—
л
е
г
о
ч
н
а
я
а
р
т
е
р
и
я
,
6
—
п
е
р
е
д
н
я
я
к
а
р
д
и
а
л
ь
н
а
я
,
и
л
и
в
н
у
т
р
е
н
н
я
я
,
я
р
е
м
н
а
я
в
е
н
а
,
7
—
з
а
д
н
я
я
к
а
р
д
и
а
л
ь
н
а
я
в
е
н
а
,
8
—
з
а
д
н
я
я
п
о
л
а
я
в
е
н
а
,
9
—
с
п
и
н
н
а
я
а
о
р
т
а
,
1
0
—
ж
е
л
у
д
о
к
,
1
1
—
с
е
л
е
з
е
н
к
а
,
1
2
—
ч
р
е
в
н
о
-
б
р
ы
ж
е
е
ч
н
а
я
а
р
т
е
р
и
я
,
1
3
—
б
р
ы
ж
е
е
ч
н
ы
е
а
р
т
е
р
и
и
(
м
н
о
ж
е
с
т
в
е
н
н
ы
е
)
,
1
4
—
п
о
д
в
з
д
о
ш
н
а
я
а
р
т
е
р
и
я
,
1
5
—
х
в
о
с
т
о
в
а
я
а
р
т
е
р
и
я
,
1
6
—
х
в
о
с
т
о
в
а
я
в
е
н
а
,
1
7
—
п
о
д
в
з
д
о
ш
н
а
я
в
е
н
а
,
1
8
—
п
о
п
е
р
е
ч
н
а
я
п
о
д
в
з
д
о
ш
н
а
я
в
е
н
а
,
1
9
—
в
о
р
о
т
н
а
я
в
е
н
а
п
о
ч
к
и
,
2
0
—
б
р
ю
ш
н
а
я
в
е
н
а
,
2
1
—
к
и
ш
к
а
,
2
2
—
п
о
д
ж
е
л
у
д
о
ч
н
а
я
ж
е
л
е
з
а
,
2
3
—
а
о
р
т
н
а
я
в
е
н
а
п
е
ч
е
н
и
,
2
4
—
п
е
ч
е
н
о
ч
н
а
я
в
е
н
а
,
2
5
—
п
о
д
к
л
ю
ч
и
ч
н
а
я
в
е
н
а
,
2
6
—
п
о
д
к
л
ю
ч
и
ч
н
а
я
а
р
т
е
р
и
я
,
2
7
—
л
е
г
о
ч
н
а
я
в
е
н
а
,
2
8
—
н
а
р
у
ж
н
а
я
я
р
е
м
н
а
я
в
е
н
а
,
2
9
—
л
е
г
о
ч
н
а
я
д
у
г
а
,
3
0
—
V
д
у
г
а
а
о
р
т
ы
,
3
1
—
с
о
н
н
а
я
а
р
т
е
р
и
я
,
3
2
—
б
о
к
о
в
а
я
в
е
н
а
г
о
л
о
в
ы
,
3
3
—
л
е
г
к
о
е
,
3
4
—
п
е
ч
е
н
ь
,
3
5
—
п
о
ч
к
а
618
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Малый круг начинается сосудами, отходящими от желудочка,
и заканчивается сосудами, впадающими в правое предсердие. Боль-
шой круг начинается сосудами, отходящими от желудочка, и заканчи-
вается сосудами, впадающими в левое предсердие. От желудочка
отходят артерии, которые в малом круге несут венозную кровь,
а в большом — артериальную, в предсердие впадают вены: в малом
круге — приносящие артериальную кровь, в большом — венозную.
От желудочка отходят артериальные жаберные дуги, число кото-
рых соответствует числу жаберных дуг висцерального черепа. Эмбри-
онально закладываются шесть пар артериальных дуг, первые две пары
жаберных дуг входят в состав черепа (первая полностью редуцирует-
ся, а вторая преобразуется в челюсти), поэтому аналогичные пары ар-
терий исчезают, и даже у рыб сохраняется всего четыре пары жабер-
ных артерий. У наземных позвоночных редукция жаберного аппарата
явилась причиной уменьшения числа жаберных дуг: третья пара пре-
образуется в сонные артерии, четвертая — в системные дуги аорты
(самые крупные сосуды большого круга), пятая пара редуцируется,
поскольку функционально она аналогична дугам аорты; шестая пара
превращается в легочные артерии.
Трехкамерное сердце амфибий состоит из двух предсердий, раз-
деленных перегородкой (у безногих и хвостатых перегородка непол-
ная, а у бесхвостых — полная), и одного желудочка (рис. 255). К серд-
цу относят и венозный синус (пазуха), сообщающийся с правым
предсердием, и артериальный конус, который соединяет желудочек
с сосудистой системой. Между предсердиями и желудочком распола-
гается общее отверстие, снабженное клапанами, которые препятству-
ют обратному затеканию крови в предсердия при сокращении (систо-
ле) желудочка. Сердце лежит в полости перикарда.
Артериальный конус разделен на два артериальных ствола, каж-
дый из которых, в свою очередь, разделен продольными перегородка-
Рис. 255. Схема вскрытого сердца лягушки:
1 — правое предсердие, 2 — левое
предсердие, 3 — желудочек, 4 — клапаны,
закрывающие общее отверстие, ведущее
из обоих предсердий в желудочек,
5 — артериальный конус, 6 — общий
артериальный ствол, 7 — кожно-легочная
артерия, 8 — дуга аорты, 9 — общая сонная
артерия, 10 — сонная «железа»,
11 — спиральный клапан артериального
конуса
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
619
ми на три сосуда: общую сонную артерию, кожно-легочную артерию
и системную дугу. Кровь поступает в желудочек из обоих предсердий
через общее отверстие. Единственный желудочек не разделен, поэто-
му в его полости смешивается венозная и артериальная кровь. Вдоль
всей поверхности артериального конуса располагается спиральный
клапан.
Сердечный цикл. Кровь из предсердий поступает в желудочек и ча-
стично смешивается в его полости, однако в правой части желудочка
сосредоточена преимущественно венозная кровь, поступившая из
правого предсердия, в левой — преимущественно артериальная, по-
ступившая из левого предсердия; в центре — смешанная кровь. В на-
чале систолы желудочка преобладающая венозная кровь из его пра-
вой части попадает в отверстия кожно-легочных артерий. В это время
спиральный клапан закрывает отверстия других сосудов. Дальнейшее
сокращение миокарда желудочка способствует открытию спирально-
го клапана отверстиям системных дуг, куда из центральной части же-
лудочка направляется смешанная кровь. Эта кровь не поступает в ле-
гочные артерии, так как они уже заполнены кровью. Далее спиральный
клапан сдвигается дальше, открывая отверстия сонных артерий, поэ-
тому в них попадает наиболее артериализованная кровь из левой ча-
сти желудочка, тогда как все другие сосуды уже заполнены кровью и
больше ее не принимают. Обратному затеканию крови из сосудов в
сердце препятствуют полулунные клапаны, которыми снабжено вход-
ное отверстие артериального конуса.
По выходе из сердца кожно-легочные артерии, несущие преиму-
щественно венозную кровь, вскоре разделяются на легочную артерию,
по которой кровь течет в легкие, и кожную артерию, которая ветвится
в коже спинной поверхности тела. Системные дуги аорты, несущие
смешанную кровь, направляются вверх и в стороны, отдавая затылоч-
но-позвоночную и подключичную ветви, кровоснабжающие передние
конечности; затем дуги, описав полукруг и направляются вниз и объ-
единяются в общий сосуд — спинную аорту, которая направляется на-
зад, отдавая ветви к внутренним органам. По артериям течет смешан-
ная в разных пропорциях артериальная и венозная кровь. Наличие
спирального клапана позволяет разделить кровь, а артерии распреде-
ляют ее по органам, при этом наиболее чистая артериальная кровь по-
падает в головной мозг и органы чувств; смешанная кровь направля-
ется во все остальные части тела, а наиболее венозная кровь течет
в сосудах малого круга кровообращения. Однако кровь в артериях не
полностью разделяет на артериальную и венозную, поэтому все орга-
ны получают смешанную кровь.
В тканях артерии распадаются на капилляры, образующие микро-
циркуляторное русло, где происходит тканевой газообмен. Кровь от-
620
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
текает от тканей по венам. Движение крови по большому кругу
можно так: желудочек ␚ системные дуги и сонные
артерии ␚ микроциркуляторное русло ␚ задняя
и передние полые вены ␚ венозный синус ␚ пра-
вое предсердие.
Малый круг кровообращения. Кровь из легочных артерий поступа-
ет в легкие, насыщается там и становится артериальной, после чего
она собирается в легочные вены, которые впадают непосредственно в
левое предсердие. Путь крови по малому кругу схематически можно
представить следующим образом: желудочек ␚ кожно-ле-
гочные артерии ␚ легочные артерии ␚ капилля-
ры легких ␚ легочные вены ␚ левое предсердие.
Относительно простая организация кровеносной системы, кото-
рая транспортирует по сосудам смешанную кровь, вместе со слабораз-
витой дыхательной системой препятствует интенсивному метаболиз-
му. Сердце амфибий сокращается редко, например, у травяной
лягушки всего 40—50 сокращений в минуту, что примерно в десять
раз меньше, чем у птицы такой же массы, поэтому у амфибий малая
скорость кровообращения и низкое давление крови. Из-за низкого
уровня обмена веществ амфибии менее подвижны по сравнению
с другими позвоночными.
Лимфатическая система развита относительно хорошо. Движение
лимфы по сосудам осуществляется за счет сокращения мышц и вну-
тренних органов, кроме того, имеются небольшие лимфатические сер-
дца, которые, периодически сокращаясь, перекачивают лимфу в кро-
веносные сосуды. Большое количество лимфы находится в подкожных
лимфатических мешках.
Выделительная система взрослых амфибий представлена парой
туловищных почек — мезонефросов, которые, в отличие от рыб, не
лентовидной формы, а овальные и весьма компактны. Вольфов про-
ток является мочеточником, который впадает в клоаку. В клоаку от-
крывается обширный мочевой пузырь, куда поступает и где временно
накапливается моча. При переполнении пузыря он опорожняется
в клоаку, оттуда моча выводится наружу.
Почки амфибий удаляют из крови продукты обмена и поддержи-
вают водно-солевое равновесие. Количество нефронов в почке зави-
сит от того, насколько тесно связано животное с водой. Водные амфи-
бии часть продуктов обмена выделяют через жабры и покровы тела
в окружающую воду. Окончательным продуктом азотистого обмена
у амфибий является мочевина. Через почки удаляется излишняя вода,
поступающая через кожу. У водных личинок амфибий основным про-
дуктом азотистого обмена является аммиак, в виде раствора которого
выводится через жабры и кожу.
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
621
Половая система. Мужская половая система образована
двумя округлыми семенниками. Над ними расположены жировые
тела, которые служат источником питательных веществ для развива-
ющихся сперматозоидов, поэтому осенью жировые тела значительно
более крупные, чем весной, когда образуется много гамет. По семявы-
носящим канальцам половые продукты проходят через переднюю
часть почки и попадают в вольфов проток, который у амфибий (так
же, как и у хрящевых рыб) совмещает функции мочеточника и семя-
провода. Вольфовы протоки впадают в клоаку, но незадолго до этого
каждый из них образует небольшое расширение — семенной пузырек,
в котором сперма временно накапливается. Подобно семенникам
и жировым телам, семенные пузырьки вне периода размножения
уменьшаются. В мужской половой системе амфибий нет собственных
половых протоков, также отсутствуют копулятивные органы.
Женская половая система образована двумя яичника-
ми, над которыми также лежат жировые тела. Размер яичников суще-
ственно меняется в зависимости от сезона, значительно увеличиваясь
к периоду размножения. Созревшие яйца выходят из яичника через
разрыв фолликулярной оболочки и оказываются в полости тела, от-
куда затем поступают в воронку яйцевода. Парный мюллеров проток
служит яйцеводом, одним концом (воронкой) он открывается в по-
лость тела, а другим — в клоаку. В период размножения яйцеводы зна-
чительно удлиняются, их стенки утолщаются.
Для многих амфибий характерно брачное поведение, часто сопро-
вождающееся голосовыми сигналами. Это необходимо для стимуля-
ции одновременного выхода половых продуктов у половых партне-
ров. Оплодотворение может быть внутренним или наружным.
Развитие подавляющего большинства амфибий проходит в воде,
некоторые виды вынашивают оплодотворенные яйца в своем теле.
Мезолецитальные яйца содержат относительно немного желтка, по-
этому происходит радиальное дробление. Для амфибий типично раз-
витие с метаморфозом, при этом из яйца выходит личинка, которая
по своему строению значительно ближе к рыбам, чем к взрослым ам-
фибиям. Она имеет характерную рыбообразную форму и перемещает-
ся благодаря продольным изгибам тела. Органами дыхания личинки
сначала служат наружные жабры (выросты кожи), позже прорывают-
ся жаберные щели, открывая внутренние жабры, после чего наруж-
ные жабры редуцируются. На ранних этапах отсутствуют конечности.
В ходе развития у личинки перестраиваются дыхательная, кровенос-
ная, выделительная и пищеварительная системы. Постепенно разви-
ваются конечности. Метаморфоз завершается формированием миниа-
тюрной копии взрослой особи, при этом у бесхвостых редуцируется
хвост. Для амбистом характерна неотения, т. е . у них размножаются
622
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
личинки, которые длительное время принимали за самостоятельный
вид, поэтому у них есть свое название — аксолотль. Такая личинка
более крупная, чем взрослая особь.
Нервная система. Центральная нервная система. Го-
ловной мозг ихтиопсидного типа, главным интегрирующим центром
является средний мозг, однако по сравнению с рыбами головной мозг
амфибий прогрессирует. Передний мозг значительно крупнее и более
четко дифференцирован. Относительно крупный конечный мозг раз-
делен полной перегородкой, имеются отдельные латеральные желу-
дочки, каждый из которых сообщается сзади с полостью третьего же-
лудочка. Крыша и боковые стенки переднего мозга целиком
образованы нервной тканью. У земноводных формируется первичный
свод мозга — архипаллиум (свод мозга имеют лишь двоякодышащие
рыбы). Расположенные впереди обонятельные доли слабо отделены
от полушарий мозга.
Промежуточный мозг маленький, сверху от него отходит эпифиз,
а снизу — воронка, связанная с гипофизом. Вместе с гипоталамусом
гипофиз образует единую гипоталамо-гипофизарную систему, регу-
лирующую деятельность эндокринных желез. Средний мозг амфибий
крупный, особенно его крыша. Средний мозг является интегрирую-
щим центром ЦНС амфибий. Мозжечок очень маленький, что связа-
но с простотой движений амфибий. Большое значение имеет ретику-
лярная формация — диффузное скопление серого вещества в переднем
отделе спинного мозга и стволе головного мозга. У низших позвоноч-
ных недостаточно развиты двигательные проводящие пути головного
мозга, управляющие деятельностью мышц, ретикулярная формация
выполняет эту функцию.
Спинной мозг имеет сегментированное строение: у видов, облада-
ющих конечностями (хвостатые и бесхвостые), в его передней и зад-
ней частях имеются расширения, соответствующие отхождению ко-
решков, которые участвуют в образовании нервных сплетений (см.
ниже). Спинной мозг амфибий короче позвоночника и занимает не
всю длину спинномозгового канала.
Периферическая нервная система образована череп-
ными и спинномозговыми нервами, их окончаниями — корешками и
узлами. От головного мозга амфибий отходят десять пар черепных
нервов, XI пара (добавочный нерв) у них отсутствует, а XII пара
(подъязычный нерв, который иннервирует мышцы языка) отходит от
спинного мозга. Передние ветви спинномозговых нервов образуют
два сплетения: плечевое и пояснично-крестцовое. Появление нервных
сплетений обусловлено развитием свободных пятипалых конечно-
стей: происходящее при этом нарушение сегментации зачатков ске-
летной мускулатуры влечет за собой такое же изменение сегментации
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
623
нервных пучков, которые эти мышцы иннервируют. Нервы, не участ-
вующие в образовании сплетений, сохраняют сегментарность. Задние
ветви сплетений не образуют.
Вегетативная нервная система состоит из симпатиче-
ского и парасимпатической частей. Центры симпатической части на-
ходятся в спинном мозге, периферический отдел образует симпатиче-
ские стволы, состоящие из цепи узлов, связанных между собой
волокнами, расположенных по сторонам от позвоночного столба. Па-
расимпатическая часть представлена волокнами, входящими в состав
черепных нервов.
Органы чувств. У подавляющего большинства земноводных име-
ются органы зрения. Зрение амфибий адаптировано к наземной
жизни: выпуклая роговица и двояковыпуклый хрусталик, что позво-
ляет различать предметы в воздушной среде на довольно большом
расстоянии. Роговица амфибий уплощается, когда животное попадает
в воду. Аккомодация глаза осуществляется путем перемещения хру-
сталика вперед за счет специальной ресничной мышцы. У земновод-
ных имеются подвижные веки, которые, смыкаясь, увлажняют рого-
вицу, предотвращая ее высыхание, а также очищают ее. Наряду с
верхним и нижним веками имеется третье веко, расположенное в пе-
реднем углу глаза. Земноводные реагируют только на движущиеся
предметы.
Орган слуха устроен гораздо сложнее, чем у рыб, поскольку
звук в воздушной среде распространяется значительно медленнее,
чем в воде. Поэтому орган слуха состоит не только из звуковосприни-
мающего аппарата — внутреннего уха, но и среднего уха, которое
улавливает звуки и передает их внутреннему уху. Среднее ухо пред-
ставляет собой небольшую барабанную полость, заполненную возду-
хом и выстланную слизистой оболочкой. Через отверстие слуховой
(евстахиевой) трубы барабанная полость сообщается с полостью глот-
ки, уравнивая давление воздуха в барабанной полости и атмосферно-
го воздуха. Снаружи барабанная полость прикрыта тонкой барабан-
ной перепонкой. Барабанная полость гомологична брызгальцу рыб.
Слуховая косточка — стремечко — осуществляет передачу звука.
Стремечко гомологично гиомандибуляре рыб. Одним концом оно
упирается в барабанную перепонку, а другим — в мембрану овального
окна внутреннего уха. Звуковые волны, достигая барабанной пере-
понки, заставляют ее вибрировать, что передается на стремечко, кото-
рое в свою очередь передает вибрацию на мембрану, разделяющую
барабанную полость и перепончатый лабиринт, это приводит к пере-
мещению эндолимфы в перепончатом лабиринте. Движения эндо-
лимфы возбуждают чувствительные клетки, вызывая генерацию
нервного импульса, который направляется по волокнам VIII пары че-
624
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
репных нервов в соответствующие центры головного мозга. По нерву
передаются импульсы от органа равновесия, рецепторный аппарат ко-
торого также расположен в полости внутреннего уха (вестибулярный
аппарат). Среднее ухо, адаптированное к восприятию звука в воздуш-
ной среде, имеется у всех наземных позвоночных животных.
Органы обоняния представлены обонятельными капсула-
ми, которые сообщаются с внешней средой через наружные ноздри,
с глоткой через внутренние ноздри (хоаны), поскольку поток воздуха
в процессе дыхания идет именно через них. Многочисленные обоня-
тельные нервы направляются в обонятельные доли переднего мозга.
Органы боковой линии имеются только у личинок, но у взрослых со-
храняются только у водных хвостатых и немногих постоянноводных
бесхвостых. Этот орган чувств у амфибий организован проще, чем
у рыб, поскольку чувствительные клетки попросту лежат в коже, а не
в специальном канале, как у рыб. Вся поверхность кожи осуществляет
осязание.
Поведение земноводных весьма примитивное, условные реф-
лексы медленно вырабатываются и быстро угасают. Специализация
рефлексов невелика, поэтому лягушка не может сформировать за-
щитный рефлекс отдергивания одной лапки, и при раздражении од-
ной конечности она дергает обеими лапками.
Нервная система и органы чувств земноводных сыграли важную
роль в разработке учения о физиологии нервной деятельности. На ля-
гушках экспериментировал И.М . Сеченов, изложивший результаты
работы в книге «Рефлексы головного мозга» (1863). В конце XVIII в.
лягушачьи лапки, подвешенные на проволоке, подсказали Л. Гальва-
ни идею «животного электричества», которая способствовала возник-
новению электрофизиологии.
Класс земноводных состоит из трех отрядов: хвостатые, бесхво-
стые и безногие.
Отряд хвостатые амфибии включает в себя около 300 видов, у ко-
торых имеются конечности примерно одинаковой длины и сжатый
с боков или округлый хвост. У некоторых видов (сирены) отсутству-
ют задние конечности. Значительная часть безногих бо
льшую часть
времени проводит в воде или даже вообще не выходит на сушу (около
200 постоянноводных видов). Тело наземных форм обычно укорочен-
ное, мощные конечности удлинены. Виды, обитающие преимущест-
венно в воде, более крупные. Так длина самого крупного современно-
го земноводного — гигантской саламандры — достигает 1,5 м. Но их
конечности короткие и слабые. Большинство хвостатых обитают в
зонах с умеренным климатом в западном и восточном полушариях, но
практически отсутствуют в южном полушарии.
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
625
Тело безногих амфибий червеобразное, конечности отсутствуют.
Хвост очень короткий, поэтому клоака открывается почти на самом
конце тела. Голая кожа богата железами, выделяющими на поверх-
ность тела обильную слизь. Глаза рудиментарны, барабанная пере-
понка отсутствует, слуховой нерв развит очень слабо. В толще кожи
имеются костные фрагменты, которые, возможно, являются остатка-
ми панциря древнейших земноводных (стегоцефалов). За исключени-
ем двух постоянноводных видов, все безногие ведут подземный образ
жизни, прокладывая себе норы во влажной почве и гниющей подстил-
ке. Пищей для них служат почвенные беспозвоночные, которых они
находят благодаря выраженному весьма тонкому обонянию. Некото-
рые виды живут в гнездах муравьев и питаются их личинками. У зем-
новодных выражена забота о потомстве. Известно одно семейство
безногих — червяги, насчитывающее около 60 видов, большинство ко-
торых обитают в тропических областях Центральной и Южной Аме-
рики, некоторые виды распространены в тропических зонах Африки
и Азии.
Наиболее высокоорганизованный отряд — бесхвостые амфибии,
куда входит большинство видов земноводных (около 1800). Тулови-
ще всех бесхвостых короткое и широкое, шея короткая, почти не вы-
раженная, конечности хорошо развитые, причем задние значительно
лучше передних, что обеспечивает характерное для них передвижение
по твердой поверхности скачками. Хвост отсутствует, хвостовые по-
звонки сливаются в одну косточку — уростиль.
Бесхвостые земноводные широко распространены на всех конти-
нентах, кроме Антарктиды. Большинство видов постоянно живут
вблизи водоемов, однако некоторые жабы приспособились жить
даже в пустыне, где размножаются в короткий период дождей, после
чего зарываются в грунт и впадают в спячку. Многие виды ведут
древесный образ жизни, для этого на кончиках пальцев у них име-
ются присоски, с помощью которых они удерживаются на листьях.
Они размножаются в дождевой воде, которая скапливается во влага-
лищах листьев, дуплах и других местах. Особенно многочисленны
древесные бесхвостые амфибии в дождевых тропических лесах.
Большинство бесхвостых земноводных питается насекомыми и дру-
гими беспозвоночными. Многие из них уничтожают большое коли-
чество вредителей сельскохозяйственных культур (например, жаба
поедает множество голых слизней), что приносит существенную
пользу. Мясо лягушек используется в пищу во многих странах (на-
пример, во Франции), что негативно сказывается на их численности
в естественных условиях. Реже в пищу употребляют яйца бесхво-
стых амфибий.
626
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Класс Пресмыкающиеся, или Рептилии
Рептилии, птицы и млекопитающие образуют группу высших по-
звоночных — амниот, которые способны размножаться без внешних
источников воды благодаря наличию амниотической полости, огра-
ниченной зародышевой амниотической оболочкой. Рыбы и земновод-
ные составляют группу низших позвоночных — анамний, у которых
размножение осуществляется в водоемах, т. к. оболочки яйца у них не
способны защитить развивающийся эмбрион от потери воды на суше.
Поэтому ареалы анамний ограничены водоемами или прибрежными
территориями. Наличие амниотической оболочки позволяет амнио-
там не зависеть от водоемов в период размножения, поэтому они рас-
селены гораздо более широко и занимают самые разнообразные
экологические ниши. Наиболее примитивны пресмыкающиеся —
единственные холоднокровные амниоты. Большинство современных
рептилий обитают на суше, в том числе в пустынях и в других местах
с очень сухим климатом. Некоторые виды вторично перешли к водно-
му образу жизни.
Внешнее строение рептилий разнообразно, в типичных случаях
тело подразделяется на голову, туловище и хвост. Тело черепах за-
ключено в костный панцирь, надежно защищающий тело животного.
Анатомия. Кожные покровы рептилий значительно более толстые
и сухие, чем у амфибий. Наружный слой многослойный ороговеваю-
щий эпителий (эпидермис), который у многих видов формирует утол-
щения — чешуи. Форма чешуи отличается не только у разных видов,
но и зависит от расположения на теле одного и того же животного.
Когти, которыми вооружены пальцы, являются производными эпи-
дермиса. Самый поверхностный роговой слой эпидермиса постоянно
слущивается. Клеточный состав пополняется за счет стволовых кле-
ток, которые расположены в прилегающем к дерме ростковом (маль-
пигиевом) слое. Змеи периодически линяют, в процессе линьки ста-
рый эпидермис сходит с тела наподобие чулка. В сухой коже рептилий
очень мало желез, лучше железы развиты у некоторых черепах. Под
эпидермисом располагается кутис, у некоторых ящериц в нем образу-
ются очаги окостенения, которые делают кожу более прочной. У чере-
пах такие окостенения формируют костный панцирь, состоящий из
спинного щита — карапакса и брюшного — пластрона.
Покрытая толстым ороговевающим эпидермисом, кожа рептилий
плохо пропускает воду. Однако через кожу вода все-таки испаряется,
например крокодилы, которым не нужно экономить воду, теряют та-
ким образом до 75% выделяемой воды. В отличие от кожи лягушек,
кожа рептилий плотно срастается с телом, не образуя лимфатических
мешков.
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
627
Скелет. Позвоночный столб подразделяется на четыре отдела:
шейный, пояснично-грудной, крестцовый и хвостовой кожи. В шей-
ном отделе всегда более одного позвонка, что придает шее (и голове!)
бóльшую подвижность по сравнению с амфибиями. Это важно для
животного, поскольку оно может не только наклонять или поднимать
голову (как лягушки), но и поворачивать ее во все стороны, обозревая
при этом окрестности. Не меньшее значение имеет обособление пер-
вых двух шейных позвонков в атланто-осевой комплекс, который по-
зволяет амниотам поворачивать голову вокруг продольной оси тела.
У рептилий и птиц череп соединяется с атлантом посредством лишь
одного затылочного мыщелка.
Грудные позвонки рептилий не отличаются от поясничных, поэто-
му эти два отдела объединяют в единый — пояснично-грудной. К ка-
ждому позвонку прикрепляется пара ребер. У ящерицы 22 пояснич-
но-грудных позвонка, все они несут ребра, но только первые пять пар
снизу присоединяются к грудине, образуя настоящую грудную клет-
ку. Остальные ребра не срастаются с грудиной. Своими свободными
концами они заканчиваются в мышцах стенки тела. У черепах — всего
восемь пар ребер, все они срастаются с расположенными выше пла-
стинами карапакса. У змей ребра не срастаются с грудиной, что связа-
но с ползающим образом жизни. Крестцовый отдел позвоночника
пресмыкающихся представлен двумя позвонками, к их поперечным
отросткам присоединяются подвздошные кости.
Позвонки хвостового отдела не имеют ребер. Их количество у раз-
ных видов широко варьирует и зависит от длины хвоста. У некоторых
ящериц посередине тела хвостового позвонка имеется хрящевая не-
окостеневающая прослойка, которая разделяет его на переднюю и
заднюю половины. В случае опасности мускулатура хвоста рефлек-
торно сокращается и переламывает тело позвонка по линии хрящевой
прослойки, при этом часть хвоста, расположенная сзади, отламывает-
ся и беспорядочно изгибается за счет сокращений мышц, что вводит
в замешательство врага и позволяет ящерице уйти. В дальнейшем
хвост регенерирует. Это называется аутотомией. У черепах все по-
звонки, кроме шейных и хвостовых, неподвижно срастаются с пласти-
нами карапакса.
Череп. Первичный череп почти полностью окостеневает. Кна-
ружи от него имеется большое количество покровных костей, входя-
щих в состав дна, крыши и боковых стенок. В висцеральном черепе
окостеневает небно-квадратный хрящ, который становится квадрат-
ной костью. Кости черепа змеи имеют многочисленные подвижные
соединения, благодаря чему они могут проглатывать добычу, значи-
тельно превосходящую по толщине саму змею (например, птичьи
628
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
яйца). Подъязычная дуга преобразуется в подъязычный аппарат, а ги-
омандибуляре функционирует как слуховая косточка — стремечко.
В плечевом поясе имеются ключица (отсутствует у кроко-
дилов) и надгрудинник, которые увеличивают прочность соединения
правой и левой сторон плечевого пояса. Соединение ребер с грудиной
и формирование грудной клетки обеспечивает непосредственное сое-
динение плечевого пояса с осевым скелетом. Плечевой пояс змей ре-
дуцирован, у черепах ключицы и надгрудинник срастаются с пластро-
ном. Т азовый пояс, образованный подвздошной, седалищной и
лобковой костями, не имеет существенных особенностей. Строение
свободных передней и задней конечностей типичное, в области ко-
ленного сустава расположена коленная чашечка. Проксимальный ряд
костей предплюсны срастается с берцовыми костями, а кости дисталь-
ного ряда соединяются с костями плюсны, поэтому вместо голено-
стопного сустава имеется межпредплюсневый сустав, соединяющий
проксимальный и дистальный ряды костей предплюсны.
Мышечная система сегментирована только в области позвоночно-
го столба, остальная часть мускулатуры представлена мышцами, рас-
положенными вокруг суставов. Мышцы тазового пояса прикрепляют-
ся главным образом не к костям таза, а к хвостовым позвонкам.
Скорость передвижения пресмыкающихся по твердому субстрату
значительно большая, чем у амфибий. Увеличение числа шейных по-
звонков сопровождается дифференцировкой мускулатуры шеи, по-
этому голова весьма подвижная. Поскольку имеется грудная клетка,
развиваются межреберные мышцы, необходимые для реберного типа
дыхания.
Пищеварительная система. Ротовая полость четко отделена от
глотки; ротовая полость крокодилов и черепах отделена от носовой
посредством костного нёба. Железы ротовой полости развиты лучше,
чем у амфибий. Подвижный язык, снабженный собственной мускула-
турой, участвует в процессе добывания пищи; змеи и многие ящери-
цы с помощью языка определяют запахи. Строение всех зубов одина-
ковое, поэтому рептилии не могут жевать и заглатывают пищу
целиком. У черепах зубы отсутствуют, вместо них имеются острые
роговые чехлы, покрывающие челюстные кости. Обычно зубы прира-
стают к кости, и только у крокодилов они расположены в альвеолах,
что делает их более устойчивыми.
Пищевод более длинный, чем у земноводных, что зависит от зна-
чительно более длинной шеи. Стенки желудка снабжены хорошо раз-
витой мускулатурой, что обеспечивает его перистальтику. Из желудка
химус поступает в тонкую кишку, начинающуюся двенадцатиперст-
ной кишкой, в которую впадают протоки печени и поджелудочной
железы. В области перехода тонкой кишки в толстую имеется неболь-
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
629
шой вырост — зачаточная слепая кишка. Слепая кишка у анамний от-
сутствует. Задний отдел толстой кишки образован прямой кишкой,
которая открывается в клоаку. Питание рептилий очень разнообраз-
ное, среди них имеются хищники и травоядные. У рептилий относи-
тельно медленный обмен веществ, поэтому они способны длительное
время голодать.
Дыхательная система более совершенная, чем у амфибий, что
проявляется более выраженной дифференцировкой воздухоносных
путей и значительным увеличением функциональной поверхности га-
зообмена в легких. Воздухоносные пути подразделяют на верхние —
носовую полость и нижние — гортань, трахею и бронхи. Стенки горта-
ни поддерживают парные черпаловидные и непарный перстневидный
хрящи. Трахея открыта благодаря кольцевидным хрящам в ее стенках.
На внутренней поверхности легких рептилий имеется сложная сеть
перегородок, которые формируют множество небольших внутренних
ячеек, совокупная поверхность которой гораздо бóльшая, чем в меш-
ковидном легком амфибий. Наиболее сложно устроены легкие чере-
пах и крокодилов.
Поступление и удаление воздуха у рептилий происходит не за счет
заглатывания, а благодаря сокращению межреберных мышц, изменя-
ющих объем грудной клетки. У рептилий отсутствует кожное дыха-
ние, тип дыхания называется реберным, поскольку покровы тела рез-
ко ограничивают свободную диффузию газов.
Эмбриональное развитие пресмыкающихся полностью происхо-
дит в яйце и не связано с естественными водоемами, поэтому жабер-
ное дыхание у них отсутствует, а газообмен в яйце осуществляется за
счет алантоиса и желточного мешка.
Кровеносная система. Сердце трехкамерное, но предсердия раз-
делены полной перегородкой, при этом предсердно-желудочковое от-
верстие не общее, как у амфибий, а индивидуальное в каждом пред-
сердии. Венозный синус входит в состав правого желудочка,
артериальный конус редуцирован, поэтому правая и левая артериаль-
ные дуги, а также легочная артерия отходят от желудочка самостоя-
тельно. В желудочке имеются внутренние перегородки, которые пре-
пятствуют смешиванию артериальной и венозной крови в его полости.
У рептилий отсутствуют кожные вены, поскольку нет кожного газо-
обмена. Более сложно устроено сердце крокодилов, у них имеется
полная перегородка, разделяющая полость желудочка на две изолиро-
ванные части. По сравнению с амфибиями у рептилий бóльшая часто-
та сердечных сокращений, более высокий сердечный индекс, выше
давление крови и быстрее кровоток. В крови рептилий содержится
в два раза больше эритроцитов и значительно больше кислородная
630
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
емкость крови. Все это делает интенсивность обмена веществ пример-
но в 5—10 раз более высокой, чем у земноводных.
Лимфатическая система. Лимфатические сердца
немногочислен-
ны. Лимфатические сосуды впадают главным образом в яремные вены.
У всех амниот выделительная система представлена гораздо более
совершенной тазовой почкой, или метанефросом. Туловищная почка
(мезонефрос), характерная для взрослых анамний, у амниот функци-
онирует только в течение небольшого периода эмбрионального разви-
тия, после чего она заменяется метанефросом, который у взрослой
особи функционирует в течение всей жизни. Таким образом, у эмбри-
онов амниот последовательно сменяют друг друга три поколения
почек: пронефрос, мезонефрос и метанефрос, образующиеся из обще-
го тяжа нефрогенной ткани, причем каждая из них (начиная с мезоне-
фроса) формируется позади предыдущей почки. Нефрон является
морфофункциональной единицей метанефроса, его строение было
описано в разделе, посвященном выделительной системе человека.
В отличие от нефронов мезонефроса, здесь всегда отсутствует ворон-
ка, а канальцы значительно более длинные, поэтому обратное всасы-
вание из первичной мочи веществ, необходимых организму, которые
проходят через фильтрационный барьер и в результате оказываются
в первичной моче, протекает гораздо более эффективно. Конечный
продукт азотистого обмена пресмыкающихся — нерастворимая моче-
вая кислота, что позволяет рептилиям существенно экономить воду.
Половая система. Мужская половая система представ-
лена семенниками и половыми протоками. Семяпроводы открывают-
ся в мочеточник на своей стороне тела незадолго до его впадения
в клоаку. Выросты стенки клоаки образуют совокупительный орган,
который у ящериц и змей является парным, у черепах и крокодилов
непарным, а у гаттерий вообще отсутствует. Во время полового акта
совокупительные органы выпячиваются наружу, один из них вводит-
ся в клоаку самки, куда затем вводится сперма.
Женская половая система образована парными яични-
ками и половыми путями. Созревшие яйцеклетки попадают в полость
тела и оттуда проникают в воронку одного из яйцеводов, которые од-
ним своим концом (воронкой) открываются в полость тела, а дру-
гим — в клоаку. Функцию яйцевода выполняют мюллеровы протоки
(у самок вольфовы протоки не сохраняются). Проходя по яйцеводу,
яйцеклетка окружается оболочками, образующимися из веществ, вы-
деляемых клетками эпителия: в верхнем отделе (маточной трубе) —
яичным белком, в среднем (матке) — кожистой скорлупой, которая
иногда пропитана известью. Оплодотворение яйцеклетки должно
произойти до образования скорлупы, поэтому у рептилий оплодотво-
рение внутреннее.
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
631
Развитие начинается в яйцеводах и полностью завершается в яйце.
Напомним: рептилии являются амниотами, у них зародыш в яйце
окружен оболочками, а скорлупа не дает яйцу высохнуть. Яйцо реп-
тилий содержит много желтка, который скапливается у вегетативного
полюса. Дробление дискоидальное. Развитие идет без метаморфоза,
поэтому из яйца выходит сформированное животное. Самка отклады-
вает яйца в подходящих для этого местах. Некоторые виды проявля-
ют заботу о потомстве. Вторичноводные пресмыкающиеся для раз-
множения выходят на сушу и там откладывают яйца (морские
черепахи) или становятся яйцеживородящими (морские змеи).
Строение нервной системы более сложное, чем у амфибий, это
связано с адаптацией к постоянному наземному обитанию. В наи-
большей степени усложнился головной мозг. Его самым круп-
ным отделом является передний мозг, прежде всего полушария
конечного мозга. Основная масса серого вещества полушарий сосре-
доточена в полосатых телах, расположенных в основании конечного
мозга, у рептилий они являются главными координаторами нервной
деятельности. Интегрирующим центром головного мозга являются
полосатые тела переднего (конечного) мозга (зауропсидный тип моз-
га). Кроме рептилий, мозг такого типа имеют птицы.
В полушариях переднего мозга рептилий дифференцированы обо-
нятельные и теменные доли. У рептилий на поверхности полушарий
имеются отдельные участки серого вещества — кора, это зачаток вто-
ричного свода, или неопаллиума. Кора рептилий примитивная, ее на-
зывают древней корой. Крупные полушария конечного мозга прикры-
вают промежуточный мозг. У рептилий (особенно у гаттерий) очень
хорошо развит теменной орган, строение которого напоминает глаз.
Средний мозг относительно невелик. Мозжечок развит хорошо, по-
этому рептилии могут совершать сложные движения. Продолговатый
мозг образует второй изгиб, который имеется у всех амниот, в том чи-
сле птиц и млекопитающих. Первый изгиб расположен в области
среднего мозга. Общая масса головного мозга рептилий примерно
равна массе спинного мозга. Спинной мозг имеет обычное строение,
он короче позвоночного столба, поэтому задняя часть спинномозгово-
го канала занята нервными волокнами и оболочками. Большое значе-
ние имеет ретикулярная формация спинного мозга и ствола головно-
го мозга, регулирующая функцию двигательных проводящих путей.
Периферическая нервная система. От головного
мозга отходят одиннадцать пар черепных нервов, XII пара — подъ-
язычный нерв — отходит от продолговатого мозга (у амфибий он от-
ходит не от ствола головного мозга, а от спинного мозга, поэтому его
нельзя считать черепным нервом). Однако XI пара (добавочный нерв)
соединена с X парой, поэтому общее количество черепных нервов
632
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
равно одиннадцати. Спинномозговые нервы образуют сплетения
(плечевое и пояснично-крестцовое, или тазовое), нервы которых ин-
нервируют кожу и мышцы конечностей.
Органы чувств. Орган зрения характеризуется наличием в
ресничной мышце поперечнополосатых мышечных волокон, эта мышца
не только перемещает хрусталик вперед и назад, но также изменяет его
кривизну. Глазодвигательные мышцы вращают глазные яблоки репти-
лий. Особенно эффектно это получается у хамелеонов, глаза которых
движутся независимо друг от друга. Глаза большинства рептилий при-
крыты веками (верхнее, нижнее и третье в виде мигательной перепон-
ки), у гекконов и змей подвижных век нет, у змей они сращены между
собой. Гекконы время от времени роговицу облизывают, а змеи периоди-
чески удаляют поверхностный слой сросшихся век во время линьки.
Орган обоняния развит очень хорошо. У ящериц и змей хорошо
развит якобсонов орган, полость которого сообщается с верхней стен-
кой полости рта. Чтобы этот орган начал функционировать, животное
периодически высовывает язык, который контактирует с летучими
веществами воздуха и частицами грунта. После этого животное втяги-
вает язык и прижимает его к каналу якобсонова органа, который ана-
лизирует химический состав веществ.
Орган слуха состоит из среднего и внутреннего уха, в пере-
пончатом лабиринте обособлена улитка. У некоторых змей между
ноздрями и глазами располагаются парные ямки, с помощью которых
животное различает инфракрасное излучение живой добычи, даже не
видя ее. Это позволяет улавливать разницу температур до тысячных
долей градуса. Змеи, которые обладают таким термолокатором, назы-
вают ямкоголовыми (гремучие змеи и др.).
Поведение пресмыкающихся намного сложнее, чем амфибий, что
связано с более высокой организацией нервной системы, прежде все-
го переднего мозга. Наряду с инстинктами, очень важна условно-реф-
лекторная деятельность, которая обеспечивает адаптацию к постоян-
но меняющимся условиям обитания.
В настоящее время известно около 6000 видов рептилий, которые
подразделяются на четыре отряда: клювоголовые, чешуйча-
тые, крокодилы и черепахи. Огромное количество видов
пресмыкающихся вымерло еще в меловом периоде мезозойской эры
по не вполне ясным причинам.
Отряд клювоголовые представлен единственным современным ви-
дом — гаттерией. Это животное одно из наиболее древних рептилий,
оно появилось задолго до динозавров — примерно 220 млн лет назад.
Гаттерия внешне похожа на крупную (до 75 см в длину) ящерицу, од-
нако отличается от нее рядом примитивных черт. У гаттерий лучше,
чем у других пресмыкающихся, развит теменной орган, в котором
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
633
имеются все структуры глаза: роговица, хрусталик и сетчатка. Темен-
ной глаз открывается на верхней стороне головы между теменными
костями. Доказано, что теменной глаз способен реагировать на свет.
Барабанная перепонка и барабанная полость отсутствуют. У молодых
особей зубы располагаются не только на челюстных и небных костях,
но и на сошнике, как у амфибий (кстати, это единственный случай
среди рептилий). Гаттерии обитают только на островах, лежащих
к югу и востоку от Новой Зеландии.
Самый крупный и процветающий (около 6000 видов) — отряд че-
шуйчатые. Он подразделяется на три подотряда: ящерицы, хамелео-
ны и змеи. Подотряд ящерицы объединяет рептилий, имеющих разное
строение. Тело большинства из них вытянутое, оно заканчивается
длинным хвостом. Чаще всего хорошо развиты конечности, но суще-
ствует немало безногих ящериц (веретеница, желтопузик и др.), по
внешнему виду похожих на змей. Многие ящерицы способны к ауто-
томии: в случае опасности они отбрасывают часть хвоста. Размеры
ящериц широко варьируют от нескольких сантиметров до 5 м в длину
и свыше 160 кг массой (вараны острова Комодо). Одна из ящериц
(ядозуб) выделяет яд, который вводится при укусе зубами, располо-
женными на нижней челюсти.
Подотряд хамелеоны внешне несколько похожи на ящериц. Строе-
ние тела максимально приспособлено к древесному образу жизни, ко-
торый ведут эти рептилии. Конечности заканчиваются пальцами, ко-
торые срастаются в клешнеобразно противопоставленные группы,
очень удобные для плотного обхватывания веток. Хамелеоны активно
используют при движении длинный цепкий хвост, на котором они
могут висеть, прицепившись к ветке. При этом глазодвигательные
мышцы каждого из глаз функционируют совершенно автономно, по-
этому движения обоих глаз не зависят друг от друга. Очень длинный
язык (длина превышает длину тела) с присоской на конце схватывает
добычу. Хамелеоны способны быстро перераспределять пигмент кожи
и изменять ее окраску, приближая к окружающему фону, благодаря
чему животное трудно заметить среди ветвей. Хамелеоны особенно
многочисленны на Мадагаскаре, обитают в лесах.
Подотряд змеи включает в себя специализированных рептилий, не
имеющих конечностей. От безногих ящериц змеи отличаются отсут-
ствием грудины, плечевого пояса, у многих отсутствует и тазовый
пояс. Немногие виды имеют рудиментарные остатки таза, а удавы —
зачатки задних конечностей. Кости черепа связаны между собой рас-
тяжимыми связками, которые в момент проглатывания добычи рас-
ходятся, пропуская пищу. Барабанная перепонка и полость среднего
уха отсутствуют, веки сросшиеся. Развито только одно — правое —
легкое. Отсутствует мочевой пузырь. Среди змей много ядовитых. Яд
634
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
поступает в ранку при укусе через канал, который проходит вдоль
особых ядовитых зубов, отличающихся от остальных большей дли-
ной. У неядовитых змей такие зубы отсутствуют. Змеиный яд входит
в состав многих лекарственных препаратов, поэтому змей отлавлива-
ют для получения яда. Большое количество змей истребляются, что
привело к сокращению их численности. Между тем змеи очень полез-
ны, так как они уничтожают грызунов, которые вредят посевам и
разносят возбудителей различных заболеваний. Все змеи питаются толь-
ко животной пищей. Змеи широко распространены на всех континентах,
кроме Антарктиды. Сегодня известно около 2700 видов змей. Длина тела
змей очень широко варьирует, слепозмейки достигают 10 см в длину,
длина анаконды или сетчатого питона может превышать 11 м.
Отряд крокодилы — наиболее высокоорганизованный среди репти-
лий. Крокодилы имеют обтекаемое тело, сплюснутое в области туло-
вища, но сжатое с боков в области хвоста. Зубы расположены в от-
дельных альвеолах (единственные среди пресмыкающихся!), развито
вторичное костное нёбо. Внутренние отверстия ноздрей открываются
не в ротовую, а в носовую полость (носоглоточный ход), а та, в свою
очередь, — в глотку. При этом пища, находящаяся во рту, не мешает
дыханию.
Полная перегородка имеется не только между предсердиями, но и
между желудочками, поэтому сердце крокодилов четырехкамерное.
Все крокодилы — хищники. Крупные виды (нильский, гребнистый) на-
падают и на человека. Крокодилы являются объектом промысла. Осо-
бенно высоко ценится их кожа, а мясо употребляют в пищу. Некон-
тролируемое уничтожение привело к резкому снижению численности
этих рептилий, поэтому крокодилов искусственно разводят на фермах.
Известно 25 видов современных крокодилов. Они обитают во влажных
тропических областях обоих полушарий, заселяя стоячие или медлен-
нотекущие пресные водоемы, реже крокодилы живут в морях. Самые
крупные особи (нильский крокодил) достигают более 10 м в длину.
Отряд черепахи — наиболее специализированная группа репти-
лий. Их туловище защищено костным панцирем, в который могут
втягиваться голова, конечности и хвост. Панцирь состоит из верхнего
щита — карапакса, который защищает тело сверху и с боков, и нижне-
го — пластрона. У кожистых черепах панцирь вместо роговых щитков
покрывает относительно мягкая кожа. У черепах хорошо развиты
мышцы конечностей, шеи и хвоста, однако мышцы туловища слабые,
что связано с наличием защитного панциря. Жесткий панцирь обезд-
виживает грудную клетку, поэтому черепахи не способны осуществ-
лять реберное дыхание. Они засасывают воздух, опуская дно ротовой
полости, наподобие того, как это делают амфибии. Среди черепах
имеются растительноядные и плотоядные. По образу жизни различа-
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
635
ют сухопутных и водных черепах. Большинство видов сухопутных че-
репах обитают в географических зонах с теплым климатом или в пу-
стынях. Известно около 330 видов современных черепах, образующих
4 подотряда: скрытошейные, бокошейные, морские и мягкокожистые.
Класс Птицы
Птицы — единственный класс высших позвоночных, освоивших
воздушную среду. Внешнее и внутреннее строение птиц во многом за-
висит от способности летать. Это, в первую очередь, морфологиче-
ские адаптации, облегчающие массу тела. Кроме того, птицы — тепло-
кровные животные, у них развитая нервная система и выражена
забота о потомстве. Класс насчитывает около 8000 видов, которые
входят в 35—40 отрядов.
Внешнее строение. Тело птиц обтекаемое, благодаря длинной шее го-
лова относительно маленькая, но очень подвижная. На челюстях имеет-
ся роговой клюв, зубы отсутствуют. Передние конечности превратились
в крылья, которые не участвуют в передвижении по твердому субстрату.
Опору для всего тела образуют только задние (нижние) конечности,
с помощью которых птицы перемещаются по твердому субстрату. Паль-
цы нижних конечностей снабжены когтями. Строение конечностей зави-
сит от образа жизни птицы. Хвост редуцирован, но это незаметно из-за
хорошо развитого хвостового оперения. Часть птиц (пингвины, страусы,
киви и др.) вторично утратили способность к полету, поэтому их тулови-
ще тяжеловесное, а крылья маленькие. Все тело птицы, кроме клюва
и части нижних конечностей, покрыто перьями (у белой совы, белой ку-
ропатки и др. нижние конечности тоже оперены).
Анатомическое строение птиц способствует полету. Кожные по-
кровы. Кожа птиц тонкая, почти лишена желез, исключением являет-
ся копчиковая железа, выделяющая жирный секрет, который смазы-
вает перья и делает их водонепроницаемыми (она лучше всего развита
у водоплавающих птиц). У некоторых видов (например, дроф) копчи-
ковая железа неразвитая, поэтому их перья намокают. Роговой клюв,
когти и перья, которые имеются только у птиц, являются производ-
ными эпидермиса кожи. Лишь у немногих видов (в основном нелета-
ющих) перья равномерно покрывают всю поверхность кожи, у других
перья располагаются только на определенных участках кожи — пте-
рилиях и отсутствуют на аптериях. Выделяют несколько типов перь-
ев, отличающихся по строению и функциям.
Контурные перья — наиболее сложные (рис. 256). Осевую
основу такого пера образует упругий стержень, который начинается
под кожей полым цилиндрическим очином, а над кожей переходит
в более длинную часть — ствол. Вершина очина, погруженная в кожу
636
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
открывается отверстием, через которое происходит кровоснабжение
пера; остатки этих сосудов сохраняются в полости очина в виде дуж-
ки. Вдоль ствола пера по обе его стороны располагаются опахала. Ка-
ждое опахало образовано многочисленными боковыми пластинками,
отходящими от ствола, — бородками I порядка. По обе стороны этих
бородок отходят более мелкие бородки II порядка. Они снабжены
крючочками, которыми сцепляются друг с другом, формируя тонкую,
но прочную пластинку опахала, которая в полете не пропускает воз-
дух. При механических ударах эластичные бородки сгибаются и сгла-
живают удар, но сцепление бородок при этом может нарушиться
и птица клювом восстанавливает целостность опахала. Контурные пе-
рья (маховые, кроющие, рулевые, надхвостье и др.) определяют обте-
каемость тела в полете, защищают тело от повреждений и смачивания
водой, участвуют в терморегуляции. Черепицеобразно налагаясь друг
на друга, они формируют сплошную плоскость. При опускании крыла
над его верхней поверхностью образуется разреженное воздушное
пространство, которое поднимает тело птицы вверх.
Пуховые перья лишены бородок II порядка и не образуют
опахала. Основная их функция — теплозащитная. Нитевидные
перья лишенные бородок, выполняют осязательную функцию.
Рис. 256. Строение контурного пера: А — общий вид, Б — схема строения опахала
(сильно увеличено), В — очин вскрыт, чтобы показать дужку пера; 1 — очин,
2 — стержень, 3 — бородки, 4 — бородочки, 5 — крючочки, 6 — наружное опахало,
7 — внутреннее опахало, 8 — пуховая часть опахала, 9 — стержень, 10 — ствол,
11 — отверстие очина, 12 — дужка пера (по Карташеву и соавт., с дополнениями)
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
637
У многих птиц (насекомоядных, ночных) в углах рта имеются перья,
видоизмененные в более жесткие щетинки. Перьевой покров регуляр-
но обновляется в ходе линек. Перья часто пигментированы, благодаря
чему они очень яркие. Основная цель окраски — участие в брачных
ритуалах, возможность слиться с окружающим фоном и др.
Скелет отличается прочностью и легкостью костей, так как они
содержат много воздуха, что очень важно для полета (рис. 257).Осе-
вой скелет характеризуется сращиванием костей в жесткие ком-
плексы. Позвоночный столб состоит из шейного, грудного, пояснич-
ного, крестцового и хвостового отделов. Шейный отдел позвоночника
птиц развит лучше, чем у других позвоночных. Это делает шею (и го-
Рис. 257. Скелет голубя:
1 — шейные позвонки, 2 — грудные позвонки,
3 — хвостовые позвонки, 4 — копчиковая кость,
5 — спинная часть ребра с крючковидным
отростком, 6 — брюшная часть ребра,
7 — грудина, 8 — киль грудины, 9 — лопатка,
10 — коракоид, 11 — ключица, 12 — плечо,
13 — лучевая кость, 14 — локтевая кость,
15 — пясть, 16 — II палец, 17 — III палец,
18 — IV палец, 19 — подвздошная кость,
20 — седалищная кость, 21 — лобная кость,
22 — бедро, 23 — голень, 24 — цевка,
25 — I палец, 26 — IV палец (по Наумову)
638
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
лову) очень подвижной. Птица может повернуть голову на 180°,
а совы могут совершить почти целый оборот, развернув голову на
270°! Для сравнения: человеку не стоит пытаться повернуть голову
даже на 90° во избежание серьезных травм. Первые два шейных по-
звонка отличаются от других, образуя атланто-осевой комплекс. По-
добно рептилиям, череп птиц сочленяется с атлантом посредством
одного затылочного мыщелка.
Грудные позвонки срастаются между собой, образуя спинную
кость. К каждому грудному позвонку присоединяется пара ребер, ко-
торые у птиц состоят из двух частей — спинной и брюшной, между
собой сочлененных подвижно. Снизу ребра соединяются с грудиной.
Такое строение грудной клетки позволяет приближать или удалять
грудину от позвоночного столба в процессе дыхания в полете. У киле-
вых птиц грудина несет широкую костную пластинку — киль, к кото-
рому прикрепляются мощные грудные мышцы, обеспечивающие опу-
скание крыла в полете. У нелетающих птиц (кроме пингвинов) киль
отсутствует. Все поясничные, крестцовые (их два) и часть ближайших
хвостовых позвонков сливаются, образуя сложный крестец, который
неподвижно соединяется с последним грудным позвонком. Хвосто-
вые позвонки сливаются между собой, образуя копчиковую кость,
к которой по бокам прикрепляются рулевые перья хвоста. Укороче-
ние хвостового отдела облегчает тело птицы и повышает его компакт-
ность.
Череп отличается большим объемом мозгового отдела, крупны-
ми глазницами. Его кости очень тонкие, они срастаются, из-за чего
череп очень легкий и прочный. Отсутствие зубов уменьшает вес чере-
па. Вторичное нёбо отсутствует.
Добавочный скелет имеет ряд особенностей, способствую-
щих полету. Плечевой пояс образован парными костями: лопаткой,
коракоидом и ключицей. Длинная саблевидно изогнутая лопатка ле-
жит поверх грудной клетки и способна подвижно перемещаться над
ребрами, не стесняя при этом движения крыльев. Хорошо развитый
коракоид нижним концом соединяется с грудиной, а верхним — с ло-
паткой, формируя совместно впадину для плечевого сустава. Обе
ключицы срастаются в непарную вилочку (дужку), концы которой со-
единяются с передними концами коракоидов, тем самым сглаживая
толчки при взмахах крыльев в полете.
Свободная передняя конечность птицы видоизменилась в крыло.
Ее скелет своеобразен, он состоит из типичных для пятипалой конеч-
ности отделов. Плечо образовано длинной плечевой костью, предпле-
чье — мощной локтевой и более тонкой лучевой, кисть сильно отли-
чается от кисти других наземных позвоночных. В проксимальном
ряду запястья имеются только две косточки. Дистальный ряд сраста-
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
639
ется с костями пясти, образуя две удлиненные кости. Редуцированы
пальцы, из которых сохраняются только три: II, III и IV, при этом
II палец имеет только одну проксимальную фалангу, к которой при-
крепляются перья, образующие крылышко, III — две фаланги, IV —
одну фалангу. Соединение звеньев крыла позволяет им двигаться
только в одной плоскости — плоскости крыла, поэтому птица может
только складывать и расправлять крыло.
Тазовый пояс образован парными подвздошными, седалищными и
лобковыми костями, которые с каждой стороны срастаются между со-
бой, образуя тазовые кости. Подвздошная кость каждой из них сра-
стается со сложным крестцом, что обеспечивает опору для задних ко-
нечностей, которые несут на себе всю массу тела птицы вне полета.
Все три кости образуют вертлужную впадину, с которой сочленяется
головка бедренной кости. Главная особенность тазового пояса птиц
состоит в том, что лобковые кости обеих тазовых костей не срастают-
ся между собой, оставляя впереди открытое пространство, что важно
для птиц, так как позволяет проходить крупным яйцам, покрытым
жесткой скорлупой. Такой тип строения таза называется открытым.
Исключением являются африканские страусы, у которых лобковые
кости срастаются между собой и таз закрытый.
Скелет свободной задней конечности состоит из бедра, голени
и стопы. В коленном суставе имеется сесамовидная кость — надко-
ленник (коленная чашечка). Голень имеет сложное строение — она
образована сросшимися большой и малой берцовыми костями (руди-
ментарная малоберцовая срастается с мощной большеберцовой),
к которым прирастает дистальный ряд костей предплюсны (две ко-
сти), при этом образуется костный комплекс — голено-предплюсна.
Дистальный ряд костей предплюсны сливается с костями плюсны,
образуя свойственную птицам длинную кость — цевку, или плюсно-
предплюсну. С дистальным концом цевки сочленяются проксималь-
ные фаланги пальцев, при этом количество пальцев у птиц варьирует,
например, большинство птиц имеют на нижних конечностях по четы-
ре пальца, казуары — три, а африканский страус — по два (единствен-
ный среди птиц).
Мышечная система птиц более дифференцирована, чем у репти-
лий. Особенно развиты мышцы конечностей, при этом в передней ко-
нечности — крыле — в основном располагаются сухожилия, а сами
мышцы вынесены из крыла и начинаются на костях туловища. Наи-
более развита мышца, опускающая крыло, — грудная мышца, которая
является самой крупной у килевых птиц (ее масса достигает 20% от
массы тела), что объяснимо, поскольку для того, чтобы с большой си-
лой опустить крыло в полете, требуется значительно больше усилий,
чем для того, чтобы это крыло поднять. Для прикрепления огромных
640
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
грудных мышц, обычных для позвоночных костных структур, тулови-
ща недостаточно, поэтому у летающих птиц и пингвинов имеется
крупный вырост грудины — киль. Значительно развита мускулатура
задних конечностей, особенно у бескилевых птиц (страусы и киви),
которые перемещаются исключительно с их помощью. У птиц хоро-
шо развита подкожная мускулатура, за счет которой птица может
поднимать перья (нахохливаться).
Пищеварительная система имеет типичное для высших позвоноч-
ных строение. Основные особенности связаны с приспособлением
к полету. У птиц отсутствуют зубы, что значительно уменьшает массу
тела. Однако у них имеется легкий и прочный роговой клюв, состоя-
щий из верхней части — надклювья и нижней — подклювья (рис. 258).
Клюв очень подвижен и способен совершать разнообразные и слож-
ные движения. Он служит не только для захвата, удержания и из-
!
"
!
"
#
$
%
&
'
Рис. 258. Клювы птиц: 1 — зеленоклювого тукана, 2 — колпицы,
3 — большеклювого попугая, 4 — тупика, 5 — водореза, 6 — большого пестрого
дятла, 7 — фламинго, 8 — большого кроншнепа, 9 — утки, 10 — шилоклювки,
11 — ибиса, 12 — клеста, 13 — марабу, 14 — козодоя-лягушкорота (по Наумову)
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
641
мельчения пищи, но и выполняет множество других функций. С по-
мощью клюва птица чистит, чинит поврежденные перья, укладывает
их, наносит секрет копчиковой железы. Клюв — грозное оружие в
драках с сородичами, при нападении и отражении агрессии врага.
С помощью клюва птицы сооружают гнезда, нередко очень сложной
конструкции (ремез, многие ткачиковые и др.), клюв необходим при
сооружении дупла. Некоторые птицы (например, определенные виды
вьюрков) клювом манипулируют «орудиями труда» (иголкой какту-
са), выковыривая насекомых из щелей. Форма клюва птиц весьма
разнообразна, она зависит от характера пищи и того, каким образом
птица ее добывает.
Подвижный язык имеется у всех птиц, но его форма широко варьи-
рует, например у гусиных он широкий и плоский, у хищных — корот-
кий и твердый, у голубя — ороговевший и заостренный на конце, у вер-
тишеек — длинный и тонкий и т. д. Слюнные железы у некоторых птиц
развиты хорошо, например у стрижей-саланганов, которые строят свои
гнезда из затвердевшей слюны (деликатес китайской кухни), нередко
они частично или полностью (у козодоев) редуцированы.
Пищевод очень длинный из-за длинной шеи, у многих птиц (голу-
би, куриные, попугаи и др.) в нижней части пищевода имеется расши-
рение — зоб, который отсутствует у других позвоночных. В зобе пища
временно накапливается и частично перерабатывается ферментами,
которые поступают из нижележащего железистого желудка. У неко-
торых видов (голуби, фламинго и др.) в период выкармливания клет-
ки слизистой оболочки зоба быстро делятся, слущиваются и образу-
ют жирную массу («птичье молочко»), которую птица скармливает
птенцам. У многих видов, например вороновых, зоба нет. Пищевод
переходит в желудок, который подразделяется на два отдела: перед-
ний — железистый и задний — мускульный. В железистом отделе
пища подвергается химическому воздействию желудочного сока.
Стенки мускульного желудка значительно более толстые за счет мощ-
ной мускулатуры. Изнутри он выстлан плотной кутикулярной вы-
стилкой, которую образует затвердевший секрет расположенных в
нем желез. В его полости находятся мелкие камешки, специально
проглатываемые птицей. Мощные сокращения мускулистых стенок
перемешивают пищу, которая при этом перетирается жесткими стен-
ками и камешками, выполняющими функцию жерновов. Таким обра-
зом птица компенсирует отсутствие зубов. Степень развития му-
скульного желудка зависит от типа пищи. Он развит наиболее
у куриных и других птиц, питающихся грубой пищей. Напротив,
у хищных и насекомоядных видов мускульный желудок развит слабо.
От мускульного желудка отходит двенадцатиперстная кишка, в
петле которой располагается поджелудочная железа. У большинства
642
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
птиц крупная печень снабжена желчным пузырем, но у некоторых
(например, у голубя) его нет, желчный проток открывается в двенад-
цатиперстную кишку, но отдельно от протоков поджелудочной желе-
зы. Длинная тонкая кишка переходит в толстую кишку, на границе
между ними имеется пара слепых выростов, каждый из которых явля-
ется слепой кишкой. У птиц отсутствует прямая кишка, и толстая
кишка непосредственно открывается в клоаку, на спинной стороне ко-
торой имеется слепой вырост — фабрициева сумка, являющаяся цен-
тральным лимфоидным органом, здесь образуются В-лимфоциты.
Отсутствие прямой кишки облегчает вес птицы, но при этом стано-
вится невозможным накапливать фекалии в кишечнике, поэтому пти-
цы очень часто испражняются. Для птиц характерна высокая скорость
пищеварения, например, сова за 3 часа полностью переваривает мышь,
а домовый воробей — зерно. Быстрее перевариваются насекомые:
жук — за 1 час, гусеница — за 15 минут.
Дыхательная система имеет очень сложное строение. Нижние
воздухоносные пути представлены гортанью, которая поддерживает-
ся черпаловидными и перстневидным хрящами, очень длинной тра-
хеей и системой бронхов. От бронхов III порядка — парабронхов —
отходит множество тонкостенных ячеистых бронхиол. Бронхиолы,
оплетенные кровеносными сосудами, образуют морфофункциональ-
ную структуру легкого, а в своей совокупности составляют респира-
торный отдел.
Характерной особенностью дыхательной системы птиц является
наличие тонкостенных воздушных мешков, которые представляют со-
бой выросты бронхов, расположенных между внутренними органами,
а их отростки проникают под кожу, между мышцами и в полости ко-
стей (рис. 259). Легочные мешки значительно превосходят по объему
относительно небольшие легкие, они играют очень важную роль в
осуществлении крайне своеобразного дыхания птиц. Птицы осу-
Ðèñ.
îðãà
1òðàõåÿ;2
ðåäíèå è
3
5
2
1
4
Рис. 259. Общая схема
органов дыхания птиц:
1 — трахея, 2 — легкие,
3 — мезобронх, 4, 5 — передние
и задние воздушные мешки
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
643
ществляют типичное для амниот реберное дыхание. При вдохе воздух
через ноздри засасывается в ротовую полость и через гортанную щель
попадает в полость гортани, затем проходит по трахее, бронхам, по-
ступает в задние легочные мешки и частично в легкие. Воздух, кото-
рый ранее находился в самих легких, выходит из них в передние меш-
ки. Выдох сопровождается уменьшением объема грудной клетки, при
этом мешки сжимаются и богатый кислородом атмосферный воздух
из задних мешков выдавливается в парабронхи легких, где происхо-
дит газообмен, а использованный воздух из передних мешков — в тра-
хею и через нее наружу.
В легких птиц воздух движется только в одном направлении, что
отличает их дыхательную систему от всех других наземных позвоноч-
ных. Односторонний постоянный поток свежего воздуха делает газо-
обмен в легких птиц непрерывным, тогда как у других наземных по-
звоночных диффузия газов прекращается, когда концентрации газов
в крови и легочном воздухе сравниваются. Поэтому у птиц содержа-
ние СО2 более низкое, а содержание О2 в артериальной крови выше,
чем в выдыхаемом воздухе. Более совершенный механизм газообмена
позволяет птицам сохранять активность даже при небольшом содер-
жании О2 в воздухе, что позволяет птицам летать на очень большой
высоте. Ранее считалось, что дыхательный акт у птиц в полете опреде-
ляется не реберным дыханием, а работой крыльев, однако сейчас до-
казано, что взмахи крыльев и дыхательные движения происходят не-
зависимо друг от друга и несинхронно.
Гортань птиц не участвует в голосообразовании, эту функцию
у них выполняет нижняя гортань, расположенная в области разделе-
ния трахеи на два бронха, соответственно, гортань, расположенная
перед трахеей, называется у птиц верхней гортанью.
Дыхательная система служит птицам не только для газообмена, но
и позволяет регулировать теплоотдачу. При повышении температуры
внешней среды резко возрастает частота дыхательных актов (полип-
ноэ) птиц, что влечет за собой испарение жидкости из воздухоносных
путей и, соответственно, охлаждение организма.
Кровеносная система. Артериальная и венозная кровь полностью
разделены, причем не только в сердце, но и в сосудистом русле, это
принципиально отличает птиц от амфибий и рептилий. Сердце птиц
состоит из двух половин, при этом в правой половине (правые пред-
сердие и желудочек) только венозная кровь, а в левой (левые предсер-
дие и желудочек) — только артериальная, ни в одной из камер кровь
не смешивается. В правое предсердие впадают полые вены, которыми
заканчивается большой круг кровообращения, от правого желудочка
отходит одна легочная артерия, с которой начинается малый круг.
В левое предсердие впадают легочные вены, от левого желудочка от-
644
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ходит единственная — правая — дуга аорты. Сердце лежит в полости
перикарда (околосердечной сумки).
Артериальная система большого круга крово-
обращения начинается единственной сохранившейся дугой аорты
(правой), отходящей от левого желудочка сердца.
ВНИМАНИЕ!
Все ткани птиц снабжаются чистой артериальной кровью.
Венозная система отличается частичной редукцией ворот-
ной системы почек. Венозная кровь от головы, шеи, крыла и грудных
мышц собирается в правую и левую передние полые вены, которые
впадают в переднюю часть правого предсердия. Венозная кровь от
задней части тела собирается в непарную заднюю полую вену, в кото-
рую также впадает печеночная вена. Для птиц характерен высокий
сердечный индекс, у мелких и более подвижных он больший, чем у
крупных. Работа сердца значительно интенсивнее, особенно у мелких
видов. Так, например, у синицы московки массой 8 г частота сердеч-
ных сокращений составляет примерно 1037 ударов в минуту. В полете
работа сердца значительно усиливается, что связано с резким увели-
чением потребности работающих мышц в О2. У птиц по сравнению
с рептилиями увеличен относительный объем крови, ее давление в со-
судистом русле выше. Кислородная емкость крови примерно в два
раза большая, чем у рептилий. Общее количество гемоглобина очень
велико, поскольку у птиц высокое содержание эритроцитов в крови.
Все эти показатели, а также полное разделение артериальной и веноз-
ной крови в сосудах позволяет птицам иметь очень высокий уровень
обменных процессов. У птиц температура тела постоянная, т. е . птицы
являются гомойотермными животными. У разных птиц температура
тела неодинакова (от 39 до 45 °С).
Постоянная температура тела делает возможным стабильную ра-
боту внутриклеточных ферментных систем, что способствует посто-
янной высокой активности организма. Это дает теплокровным живот-
ным преимущество перед холоднокровными, т. к. последним для
нормального течения биохимических реакций необходима определен-
ная температура окружающей среды, при которой могут оптимально
функционировать внутриклеточные ферменты.
В лимфатической системе отсутствуют лимфатические сердца.
Много лимфатических сосудов впадает в вены таза.
Выделительная система представлена парой очень крупных тазо-
вых почек. Более крупные, чем у рептилий и млекопитающих, разме-
ры почек объясняются весьма активным метаболизмом птиц. Общее
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
645
количество нефронов в почке достигает нескольких десятков тысяч,
что намного больше, чем у рептилий. Основным продуктом азотисто-
го обмена является мочевая кислота. Мочевой пузырь отсутствует.
Мужская половая система представлена двумя семен-
никами и половыми протоками; совокупительный орган имеется
лишь у немногих видов. Размеры семенников сильно увеличиваются
к периоду размножения (например, у скворца — в 1500 раз). Семя-
проводы впадают в клоаку. Функциональный совокупительный ор-
ган, похожий на орган крокодилов и черепах, имеется только у бески-
левых, гусиных и тинаму. У других видов сперма поступает в половые
пути самки при тесном соприкосновении клоак самца и самки. Опло-
дотворение внутреннее.
Женская половая система состоит из яичников и поло-
вых путей. Обычно сохраняется только одна половина органов — ле-
вый яичник и левый яйцевод. У немногих видов (куриных, попугаев,
сов, дневных хищников) правый яичник может иметься, но правый
яйцевод практически отсутствует. Богатые желтком яйцеклетки по-
падают из яичника в полость тела, а затем поступают в воронку яйце-
вода. В проксимальном отделе яйцевода (маточной трубе) вокруг зи-
готы сначала образуется толстый слой белковой оболочки (белок
яйца), который выделяют железистые клетки эпителия, а потом две
тонкие подскорлуповые оболочки. Следующий, более широкий отдел
(матка) содержит железы, которые секретируют жесткую наружную
известковую оболочку. Стенки дистального отдела яйцевода (влага-
лища) имеют хорошо развитую мускулатуру, с помощью которой
яйцо выдавливается в клоаку и выходит наружу.
Яйца птиц могут быть очень крупными, поэтому редукция поло-
вых органов правой стороны является для птиц полезной, поскольку
развивающиеся яйца следуют по яйцеводу строго поочередно. Если
бы оба яйцевода функционировали одинаково, очень крупные яйца
попросту не смогли бы разминуться. Таз птиц открытого типа, что по-
зволяет пропускать большие яйца. Птицы, которые гнездятся в хоро-
шо защищенных местах (например, в дуплах), откладывают белые
яйца, а те, кто строит открытые гнезда, — яйца, окрашенные в цвета
окружающего фона, что делает их менее заметными.
Развитие птиц проходит в яйце за счет очень большого количества
желтка. Дробление — дискоидального типа. В отличие от пресмыкаю-
щихся, птицы в течение всего периода эмбрионального развития со-
гревают яйца теплом своего тела (высиживают). Большинство птиц
откладывают яйца в специально построенные гнезда, некоторые пти-
цы (дятлы, многие попугаи и др.) выдалбливают дупла или занимают
чужие. Реже яйца откладываются без всякой подстилки. По заверше-
нии развития птенец пробивает изнутри скорлупу яйца и выходит на-
646
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ружу (вылупляется). Птицы заботятся о потомстве. Только что вылу-
пившиеся птенцы могут быть развитыми по-разному. У одних птиц
(куриных, гусиных, дроф, бескилевых и др.) из яйца выходят зрячие
птенцы, покрытые пухом и способные вскоре самостоятельно ходить
и клевать пищу, — это выводковые птенцы. У других (воробьиные, го-
луби, стрижи, дятлы и др.) вылупляются совершенно беспомощные
птенцы гнездового, или птенцового, типа. У них закрытые глаза, голая
кожа.
Нервная система птиц организована очень высоко. Крупный го-
ловной мозг превышает массу спинного. Увеличение размеров го-
ловного мозга связано со значительным укрупнением полушарий
конечного мозга, масса которых соответствует всем остальным его
отделам или даже превосходит их. У птиц плохое обоняние, поэто-
му их обонятельные доли очень малы. Свод мозга первичный, с не-
большими участками серого вещества. Его строение в основном
сходно с пресмыкающимися, основная масса серого вещества полу-
шарий сосредоточена в полосатых телах, которые являются основ-
ным интегрирующим центром нервной системы (зауропсидный
тип). Особенно хорошо развиты базальные ядра полосатого тела,
осуществляющие ассоциативные реакции. Промежуточный мозг
небольшой, эпифиз мал, но гипофиз развит хорошо. В среднем моз-
ге выделяются очень крупные зрительные доли, где происходит
высший анализ сигналов, полученных от органов чувств. У млеко-
питающих эту функцию выполняет кора полушарий конечного моз-
га. Мозжечок развит значительно лучше, чем у рептилий, поэтому
птицы могут совершать сложнейшие координированные движения,
что очень важно в полете.
Строение спинного мозга типичное сегментированное. В зо-
нах отхождения нервов, формирующих сплетения, имеются утолще-
ния спинного мозга. Длина спинного мозга взрослых особей короче
позвоночного столба.
Периферическая нервная система. От головного
мозга отходят двенадцать пар черепных нервов, при этом XI пара (до-
бавочный нерв) не полностью отделена от X пары (блуждающего
нерва). От спинного мозга посегментно отходят передние и задние ко-
решки, которые, соединяясь, образуют парные спинномозговые не-
рвы. Передние ветви некоторых нервов формируют плечевое и пояс-
нично-крестцовое сплетения, волокна которых иннервируют кожу
и мышцы конечностей.
Органы чувств. Орган зрения развит очень хорошо, посколь-
ку для большинства птиц зрение — основной способ ориентации. Гла-
за очень большие, особенно у ночных птиц. Птицы отличаются наибо-
лее совершенным среди всех позвоночных животных двойным
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
647
механизмом аккомодации глаза. При этом хрусталик не только меня-
ет свою кривизну, но также изменяется расстояние между хрустали-
ком и сетчаткой, что создает оптимальное фокусирование лучей на
сетчатке глаза.
Орган слуха состоит из среднего и внутреннего уха, улитка
развита лучше, чем у пресмыкающихся. В барабанной полости имеет-
ся одна слуховая косточка (стремечко), обе слуховые трубы объеди-
няются и открываются в глотку общим отверстием. Барабанная пере-
понка натянута ниже уровня кожи на дне воронки, представляющей
собой зачаток наружного слухового прохода. Это позволяет более
тонко улавливать звуковые волны, концентрировать их и направлять
к барабанной перепонке. Некоторые птицы (например, яванский са-
ланган) способны к эхолокации, поэтому могут летать в полной тем-
ноте в пещерах.
Обоняние почти не развито, хорошо различать запахи могут лишь
киви, которые разыскивают пищу в подстилке,и грифы-индейки, на-
ходящие падаль по запаху, и некоторые другие птицы.
Развитая нервная система птиц обусловливает очень сложное по-
ведение, в том числе социальное. Они легко приобретают условные
рефлексы. Птицы обладают наиболее сложной среди всех позвоноч-
ных инстинктивной деятельностью.
Современные птицы относятся к подклассу настоящих птиц, или
веерохвостых. У них имеется крупная грудина, отсутствуют брюшные
ребра и свободные пальцы на передней конечности, укороченный
хвост, в котором располагаются в виде веера рулевые перья. Настоя-
щие птицы делятся на три надотряда: бескилевые (бегающие), пин-
гвины (плавающие) и килевые (летающие).
К надотряду бескилевые, или страусовые, птицы относятся не ле-
тающие птицы, передвигающиеся путем шага или бега. На мощных
длинных задних конечностях уменьшено число пальцев. Только киви
четырехпалые, другие виды имеют по три пальца, а африканский
страус — два. Передние конечности развиты очень слабо, у киви они
почти полностью редуцированы. Недоразвит и пояс передних конеч-
ностей. Грудина лишена киля. Копчиковая кость очень мала. Кости
слабее пневматизированы, чем у других птиц. Перья не дифференци-
рованы на функциональные группы. Копчиковая железа отсутствует.
Самцы обладают функционирующим копулятивным органом. Бески-
левые птицы, кроме киви, имеют очень крупные размеры. Бескилевые
птицы обитают главным образом в южном полушарии (Южная Аме-
рика, Африка, Австралия и Новая Зеландия). Современные виды вхо-
дят в четыре отряда: африканские страусы, американские страусы
(нанду), австралийские страусы (казуары), бескрылые (киви). К на-
стоящему времени киви почти полностью истреблены. Страусы име-
648
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
ют большое промысловое значение, используются их прочная кожа,
мясо и яйца. Часто страусов разводят на фермах.
Надотряд пингвины включает в себя около 15 видов своеобразных
птиц, прекрасно адаптированных к долгому пребыванию в море.
У них не выражена пневматизация костей. Как и у бескилевых, отсут-
ствуют аптерии, поэтому перья равномерно и плотно покрывают кожу.
Несмотря на то что пингвины не могут летать, у них хорошо развит
киль грудины, что необходимо для прикрепления мощных грудных
мышц. Между пальцами задних конечностей имеется плавательная
перепонка. Большинство видов пингвинов гнездятся круглыми коло-
ниями. Пингвины распространены в южном полушарии, главным
образом в Антарктике. Кроме того, их можно встретить у южных бе-
регов Южной Америки, Африки и Австралии. Один вид гнездится на
Галапагосских островах вблизи экватора. Все современные виды пин-
гвинов входят в один отряд пингвины с одним семейством пингвино-
вых. Пингвины не имеют хозяйственного значения.
Надотряд килегрудые птицы объединяет большинство видов (око-
ло 8500) современных птиц. В основном это летающие птицы, но не-
которые виды вторично утратили способность к полету. Характерны
облегченные пневматичные кости, грудина с развитым килем. У вто-
рично нелетающих видов грудина плоская и без киля. Копчиковая
кость выражена. Дифференцированные на функциональные типы пе-
рья располагаются лишь по птерилиям. На контурных перьях имеют-
ся сомкнутые опахала. Многие виды килегрудых важны для хозяйст-
венной деятельности человека. Некоторые из них добываются
во время охоты, другие одомашнены (куры, индейки, утки, гуси,
цесарки и др.).
Класс Млекопитающие
Млекопитающие представляют собой наиболее высокоорганизо-
ванную группу высших позвоночных. У них лучше других развит го-
ловной мозг, особенно его лобные доли. Для млекопитающих харак-
терно сложное поведение, основанное не только на безусловных, но
и на условных рефлексах, поэтому кора больших полушарий весьма
у многих млекопитающих образует складки — извилины. Развитая
условно-рефлекторная деятельность позволяет млекопитающим быс-
тро приспосабливаться к изменяющимся условиям внешней среды,
что делает этот класс самым процветающим среди других позвоноч-
ных. Важным преимуществом является внутриутробное развитие
плода, живорождение и вскармливание потомства молоком. Млеко-
питающим присуща забота о детенышах, они охраняют и обучают их,
подготавливая к самостоятельной жизни. Многие виды являются
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
649
коллективными животными, что способствует передаче информации
от наиболее опытных особей молодым. Как и птицы, млекопитающие
теплокровные животные, что позволяет им заселять обширные терри-
тории и занимать разные экологические ниши, постоянно сохраняя
высокую активность, в том числе в холодное время года. Насчитыва-
ется более 4500 видов млекопитающих.
Внешнее строение разнообразно и зависит от образа жизни.
Наша задача во многом облегчается тем, что строение наиболее со-
вершенного вида млекопитающих — человека разумного — подробно
описано (см. раздел «Человек») выше. Поэтому здесь мы рассмотрим
лишь общие черты строения и функционирования млекопитающих.
Кожные покровы устроены сложно. Это связано с их активным
участием в терморегуляции. Ороговевающий эпидермис состоит из
нескольких слоев. Производными эпидермиса являются волосы, ког-
ти, ногти (отличаются от когтей тем, что они уплощены), копыта, рога
(за исключением костных рогов оленей), чешуи, иглы. Различные
кожные железы имеют эпидермальное происхождение. Волосы по-
крывают почти всю поверхность кожи, образуя характерный для мле-
копитающих волосяной покров. Отсутствие волос у ящеров, броне-
носцев, китов, дельфинов является вторичным. Различают несколько
типов волос. Длинные, толстые и жесткие волосы — остевые — защи-
щают кожу от повреждений, позволяют воде скатываться с тела, не
смачивая его. Пуховые волосы короче и мягче, они составляют основу
волосяного покрова. Густой подшерсток задерживает вблизи кожи
много воздуха, способствуя согреванию тела. Очень толстые и жест-
кие вибриссы, которые расположены на голове и выполняют функ-
цию осязания, являются специализированными волосами. В ходе се-
зонных линек волосяной покров периодически меняется. Зимний мех
более густой, отдельные волоски длиннее, чем летом, у многих млеко-
питающих выражена определенная сезонная окраска шерсти, обычно
сходная с окружающим фоном. На дистальных фалангах пальцев
большинства млекопитающих в зависимости от образа жизни разви-
ваются когти (лазающие и хищники), ногти (человекообразные обе-
зьяны) или копыта, которые развиваются не на всех, а на двух (у пар-
нокопытных) или одном (у непарнокопытных) пальце.
В коже млекопитающих имеется большое количество разнообраз-
ных желез эктодермального происхождения. Основные — потовые
и сальные железы. Потовые железы выделяют жидкий пот, который
предохраняет тело от перегревания. Их много у лошадей, очень мало
у собак и кошек и вовсе нет у китообразных (в водной среде испаре-
ние невозможно). Сальные железы открываются в волосяную сумку,
их секрет покрывает тонким слоем волосы, придавая им водоотталки-
вающие свойства; кроме того, слой жира покрывает снаружи эпидер-
650
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
мис. Потовые и сальные железы могут видоизменяться. Часто пот
имеет запах, причем характерный для определенного вида; кроме того,
запах самцов отличается от запаха самок, поэтому секрет потовых же-
лез очень важен для коммуникации между особями, а хищникам,
у которых хорошо развито обоняние, позволяет отыскать добычу.
Иногда потовые железы видоизменяются в пахучие, которые выделя-
ют сильно пахнущий секрет. Лучше всего пахучие железы развиты
у скунса, росомахи, ондатры, выхухоли.
Особенно важны млечные, или молочные, железы, которые пред-
ставляют собой видоизмененные потовые железы. Эти железы в пол-
ной мере развиты только у самок, они функционируют лишь опреде-
ленное время в период лактации после рождения детенышей.
Выделяемый млечными железами секрет — молоко — является основ-
ной пищей на ранних стадиях постнатального развития детенышей
млекопитающих. С этим связано и название всего класса. У большин-
ства млекопитающих выводные протоки млечных желез открываются
на сосках, количество которых у разных видов различное (у обезьян,
овец, слонов и др. по два соска, а у мелких грызунов, насекомоядных
и др.
—
до 24), но, как правило, это четное число. У однопроходных
соски отсутствуют, при этом молоко выделяется на поверхности тела,
и детеныши его слизывают.
Скелет млекопитающих имеет типичное строение, характерное
для высших позвоночных. Четко различают пять отделов позво-
ночного столба: шейный, грудной, поясничный, крестцовый
и хвостовой. У подавляющего большинства видов шейный отдел со-
стоит из семи позвонков, лишь ламантин (шесть шейных позвонков)
и различные ленивцы (от шести до десяти позвонков) являются
исключениями. Первые два позвонка образуют атланто-осевой ком-
плекс, который обеспечивает подвижность головы. Затылочная кость
черепа сочленяется с атлантом посредством двух затылочных мыщел-
ков. Длина шеи зависит не от количества позвонков, а от длины их
тел. Так, и длинное тело жирафа, и короткая шея мыши имеют по семь
позвонков. Грудной отдел состоит из большего числа позвонков — от
9 до 24. Грудные позвонки, ребра и грудина в совокупности образуют
выраженную грудную клетку, которая защищает органы дыхания,
сердце, крупные кровеносные сосуды и обеспечивает реберный тип
дыхания. Жесткая грудная клетка делает грудной отдел позвоночного
столба малоподвижным. Количество поясничных позвонков варьиру-
ет у разных видов млекопитающих от 2 до 9. Крестцовые позвонки
срастаются между собой, образуя единую кость — неподвижный кре-
стец. Количество крестцовых позвонков чаще всего 4, однако их мо-
жет быть 3 (у хищных), 2 (у утконоса) или более 4 (до 10), но истинно
крестцовыми являются только первые два позвонка, тогда как все
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
651
остальные — это хвостовые позвонки, сросшиеся с крестцом. Количе-
ство хвостовых позвонков у разных млекопитающих широко варьиру-
ет, у человекообразных обезьян все хвостовые позвонки срастаются
в одну кость — копчик. Значение хвостового отдела для млекопитаю-
щих велико. У некоторых обезьян хвост выполняет функцию пятой
конечности. У других хвост представляет собой балансир во время
бега или прыжка, кенгуру во время прыжка использует мощный хвост
в качестве опоры. В хвосте может откладываться жир (у сусликов).
У многих видов хвост позволяет выразить эмоции (собака, виляя хво-
стом, демонстрирует радость, а кошки — ярость).
У взрослых особей череп полностью окостеневает и не содержит
хрящевых элементов. Отдельные кости черепа срастаются в более
крупные костные образования, мозговой отдел срастается с висце-
ральным. В итоге череп млекопитающих, по существу, состоит из
сплошного костного монолита, отдельные элементы которого соеди-
нены между собой неподвижно, кроме нижней челюсти, которая под-
вижно соединяется с височной костью посредством височно-нижне-
челюстного сустава. Подъязычная кость связана с черепом посредством
мышц. Отдельную группу костей составляют слуховые косточки, рас-
положенные в полости среднего уха. Характерные для нижней челю-
сти других позвоночных квадратная и сочленовная кости у млекопи-
тающих находятся в полости среднего уха, где они выполняют
функции слуховых косточек: сочленовная кость видоизменилась
в молоточек, а квадратная — в наковальню. Таким образом, в среднем
ухе млекопитающих находится не одна слуховая косточка (стремеч-
ко), как у всех остальных четвероногих, а три (молоточек, наковальня
и стремечко), подвижно соединенные между собой суставами. Подъ-
язычный аппарат представлен подъязычной костью, которая распола-
гается между над- и подподъязычными мышцами и обеспечивает со-
гласованные движения глотки и гортани.
Добавочный скелет. Пояс передней конечности образован
лопаткой и ключицей. Самостоятельный коракоид имеется только
у однопроходных (утконоса, ехидны), у всех остальных он срастается
с лопаткой, образуя ее клювовидный отросток. Ключица имеется
только у млекопитающих, плечевой сустав которых многоосный, спо-
собный совершать движения вокруг трех осей (приматы, грызуны,
зайцеобразные, рукокрылые, насекомоядные, сумчатые). Животные,
у которых движения в плечевом суставе осуществляются преимуще-
ственно вокруг одной оси, не имеют ключицы (большинство хищных,
хоботных, копытных, китообразных). Пятипалая свободная верхняя
конечность имеет типичное строение: плечо (плечевая кость), пред-
плечье — локтевая и лучевая кости, кисть состоит из запястья, пясти,
652
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
фаланг пальцев. У летучих мышей все пальцы, кроме I, сильно удли-
нены, между ними натянута кожистая перепонка крыла.
Пояс задних конечностей состоит из парных тазовых костей. Ка-
ждая из них образуется в результате сращения подвздошной, седа-
лищной и лобковой костей. Лобковые кости, соединяясь между со-
бой, образуют симфиз, поэтому таз млекопитающих закрытый. Скелет
свободной задней конечности состоит из бедра (бедренная кость), го-
лени (большая и малая берцовые кости) и стопы (кости предплюсны,
плюсны и фаланг пальцев). У млекопитающих локтевой сустав на-
правлен назад, а коленный сустав — вперед (этот сустав укрепляет са-
мая крупная сесамовидная кость — надколенник).
Строение конечности зависит от образа жизни животного, напри-
мер, у роющих видов конечность мощная и короткая. Млекопитаю-
щие, которые бегают медленно, при ходьбе опираются на всю стопу
(медведи, обезьяны и др.). Быстроногие звери опираются на пальцы
(собака, волк, лисица, шакал, кошачьи). Самые быстрые бегуны опи-
раются на фаланги пальцев, при этом количество пальцев у них
уменьшается до двух — III и IV пальцы (у парнокопытных) или до
одного — III палец (у непарнокопытных). Конечности млекопитаю-
щих располагаются под туловищем, а не по его бокам, как у пресмы-
кающихся, что позволяет поддерживать тело над поверхностью земли
и способствует более эффективному перемещению.
Мышечная система млекопитающих очень сложная, она образова-
на большим количеством мышц. Наиболее мощные мышцы кисти и
стопы вынесены на более крупные кости предплечья или голени, тог-
да как к фалангам пальцев направляются лишь сухожилия. Мышцы
пояса передней конечности прикрепляются к костям плечевого пояса
и грудной клетки, мышцы пояса задней конечности прикрепляются к
костям таза, а не хвостовых позвонков, как у пресмыкающихся. Важ-
ной особенностью скелетной мускулатуры млекопитающих является
наличие у них куполообразной диафрагмы, которая разделяет груд-
ную и брюшную полости. Хорошо развита подкожная мускулатура,
иногда она образует под кожей сплошной слой (остатком такой му-
скулатуры являются мимические мышцы головы и подкожная мыш-
ца шеи человека). Подкожная мускулатура смещает участки кожи,
поднимает шерсть (ощетинивание), иглы (у ежей и дикобразов), осу-
ществляет движение вибрисс, образует основу губ и щек.
Пищеварительная система отличается особой сложностью строе-
ния и многообразием частных особенностей, которыми обладает тот
или иной вид. Спереди ротовая полость ограничена подвижными гу-
бами (они отсутствуют у однопроходных и китообразных), с боков —
щеками, сверху — нёбом, снизу — диафрагмой рта, задняя стенка от-
сутствует и через отверстие зева ротовая полость сообщается с
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
653
глоткой. Для млекопитающих характерно разделение ротовой поло-
сти на преддверие (пространство между губами и зубами) и собствен-
но ротовую полость (все остальное пространство), в которой находит-
ся язык. У грызунов, приматов и некоторых других млекопитающих
преддверие ротовой полости настолько обширно, что образует защеч-
ные мешки. Не у всех млекопитающих губы расположены впереди зу-
бов, у некоторых видов (например, слепыш, слепушонка) губы, нахо-
дящиеся позади зубов, предохраняют ротовую полость от попадания
в нее земли, когда животное прогрызает зубами свои норы в плотной
почве.
В альвеолах верхней и нижней челюстей располагаются зубы, при-
чем для млекопитающих характерна гетеродонтность, т. е . зубы раз-
делены на функциональные группы: резцы, имеющие долотообраз-
ный режущий верхний край коронки; клыки, с заостренной конической
коронкой; предкоренные (малые коренные, ложные коренные, премо-
ляры), с жевательной поверхностью и бугорками на ней; коренные
(моляры), жевательная поверхность которых обширнее и имеет боль-
ше бугорков. Для разных видов млекопитающих характерен опреде-
ленный набор зубов, графическая запись которого называется зубной
формулой. Смена зубов чаще всего происходит один раз в жизни: ран-
ние (молочные) зубы сменяются постоянными. Резцы грызунов рас-
тут в течение всей жизни, поэтому животное вынуждено даже перио-
дически грызть твердые предметы, чтобы сточить зубы до оптимальной
длины. Малодифференцированные зубы имеют насекомоядные (зем-
леройка, крот, выхухоль и др.) и неполнозубые (броненосцы, муравь-
еды, ленивцы), причем у последних зубы лишены эмали и корней или
могут даже вообще отсутствовать. Зубы отсутствуют у ящеров. У ки-
тообразных зубатых китов (кашалотов), а также различных дельфи-
нов они многочисленные, но одинаковые, а у усатых китов отсутству-
ют. Дифференцированные зубы позволяют не только отрывать куски
пищи, но и пережевывать их, что облегчает пищеварение.
У млекопитающих хорошо развит язык, посредством которого жи-
вотные схватывают пищу (бычьи, жирафы, муравьеды и др.), пьют
(лакают), перемешивают пищевые массы в ротовой полости, передви-
гают пищевой комок в глотку и т. д . Слюнные железы наиболее круп-
ные (корова в течение суток выделяет до 56 л слюны), тогда как у
китообразных они вообще не развиты. Слюна содержит ферменты,
расщепляющие углеводы (амилазу и мальтазу), поэтому в ротовой
полости происходит не только механическая, но и частичная химиче-
ская переработка пищи. У летучих мышей-вампиров в слюне раство-
рены антикоагулянты, препятствующие свертыванию крови в ранке,
которой они питаются; слюна некоторых землероек токсична. Глотка
дифференцирована на три отдела: носоглотка (проводит только воз-
654
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
дух), ротоглотка (проводит воздух и пищу) и гортанная часть глотки
(проводит только пищу). Длина пищевода соотносится с длиной шеи.
У большинства видов в зависимости от характера питания желудок
делится на несколько отделов. Наиболее сложно устроен желудок
жвачных, состоящий из четырех отделов: рубца, сетки, книжки и сы-
чуга. Пережеванная и пропитанная слюной пища из пищевода снача-
ла попадает в рубец, где под действием живущих там бактерий проис-
ходит брожение. Мышечные сокращения стенок желудка перемещают
пищевую массу из рубца в сетку. Из сетки пища отрыгивается обрат-
но в ротовую полость, где повторно пережевывается, орошается слю-
ной, после чего вновь проглатывается. Проглоченная полужидкая пи-
щевая масса из пищевода попадает в книжку, а из нее в сычуг. Такое
строение желудка позволяет жвачным перерабатывать очень грубые
корма; кроме того, сами бактерии рубца перевариваются вместе с ра-
стительной пищей, обеспечивая животное белком.
Кишечник подразделяется на тонкую и толстую кишку, в тонкой
кишке расщепляются полисахариды (кроме целлюлозы), белки и
жиры. В начальный отдел тонкой кишки — двенадцатиперстную киш-
ку — открываются протоки печени и поджелудочной железы (часто
она не обособлена анатомически и диффузно располагается в толще
брыжейки тонкой кишки), которые обычно предварительно объеди-
няются в общий проток. В толстой кишке всасывается вода, формиру-
ются каловые массы и накапливаются. Хорошо выражена слепая
кишка, причем ее длина зависит от характера пищи, то же самое отно-
сится и ко всей толстой кишке. Толстая кишка наиболее развита у ра-
стительноядных видов. Животные не способны самостоятельно усва-
ивать целлюлозу. Однако микроорганизмы, населяющие слепую
кишку, делают это. Толстая кишка заканчивается хорошо выраженной
прямой кишкой, открывающейся наружу анальным отверстием. У од-
нопроходных (утконоса, ехидны) кишечник открывается в клоаку.
Общая длина кишечника млекопитающих всегда больше длины тела,
например, у хищника ласки в 2,5 раза, а у травоядной овцы — в 29 раз.
Дыхательная система состоит из дыхательных путей и респира-
торного отдела. Дыхательные пути проводят воздух, очищают его от
пыли и микроорганизмов, согревают и увлажняют; в респираторном
отделе происходит газообмен между кровью и легочным воздухом.
Верхние дыхательные пути включают в себя носовую полость и часть
глотки (носоглотку и ротоглотку), нижние — гортань, трахею и систе-
му бронхов разного калибра. В носовой полости, отделенной от рото-
вой твердым нёбом, имеется система носовых ходов, благодаря кото-
рым поверхность полости значительно увеличивается. Здесь воздух
нагревается, очищается и увлажняется; кроме того, в носовой полости
имеется обонятельный отдел. Гортань имеет более сложное строение,
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
655
чем у других позвоночных, она выполняет две функции — проведение
воздуха и голосообразование. Стенки гортани укреплены не только
перстневидным и черпаловидными хрящами, но и характерным для
млекопитающих крупным щитовидным хрящом; кроме того, имеется
эластический хрящ — надгортанник, прикрывающий вход в гортань
во время глотания.
Морфофункциональной единицей легкого млекопитающих является
легочный ацинус. Газообмен в легких млекопитающих происходит
в альвеолярных ходах и альвеолах. Количество легочных альвеол
очень велико. Через легкие выделяются некоторые продукты обмена.
Дыхательные пути участвуют в терморегуляции, например собаки
при повышении температуры воздуха начинают более глубоко ды-
шать, при этом испаряется слюна, охлаждая животное. У млекопита-
ющих дыхательные движения осуществляются двумя способами: за
счет изменения объема грудной клетки (реберное дыхание) и за счет
деятельности диафрагмы (диафрагмальное дыхание).
Кровеносная система млекопитающих, как и птиц, характеризует-
ся полным разделением артериальной и венозной крови, поэтому ор-
ганы снабжаются чистой артериальной кровью. Четырехкамер-
ное сердце состоит из двух предсердий и двух желудочков.
Правая половина сердца, содержащая только венозную кровь, полно-
стью изолирована от левой половины, в которой находится артери-
альная кровь. Венозный синус и артериальный конус отсутствуют.
У млекопитающих два круга кровообращения: большой и малый.
От левого желудочка отходит единственная системная дуга аорты —
левая. У млекопитающих отсутствует воротная система почек, но
имеется непарная и полунепарная вены, которые являются рудимен-
тами задних кардинальных вен. Малый круг имеет ряд особенно-
стей.
Кровеносная система млекопитающих отличается высоким сер-
дечным индексом, большой частотой сердечных сокращений, быст-
рым кровотоком, большим объемом циркулирующей крови, высоким
давлением крови в сосудах. Важная особенность крови млекопитаю-
щих — отсутствие ядер в зрелых эритроцитах, что очень выгодно для
организма, так как позволяет заполнить всю клетку дополнительным
количеством гемоглобина. Все это наряду с весьма эффективной сис-
темой газообмена делает уровень обменных процессов очень высоким
(примерно в 20 раз выше, чем у рептилий). Млекопитающие наряду с
птицами являются гомойотермными животными.
Хорошо развита лимфатическая система, состоит из лимфатиче-
ских капилляров и сосудов, которые проходят через лимфатические
узлы. Основная масса лимфы вливается в кровеносное русло (в ярем-
ные вены).
656
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Выделительная система млекопитающих образована двумя ком-
пактными тазовыми почками, расположенными в брюшной полости.
Различают корковое и мозговое вещество почки, имеющие разную
окраску. На переднем конце почки располагается надпочечник (эндо-
кринная железа). Морфофункциональной единицей почки млекопита-
ющих является нефрон маммального типа. Между проксимальным и
дистальным отделами имеется петля Генле, которая участвует в кон-
центрировании мочи. В почках млекопитающих очень большое коли-
чество нефронов, например в обеих почках мыши их около 10 000,
а у кролика — 285 000. Продуктом азотистого обмена млекопитающих
в основном является мочевина; объем мочи относительно больший,
чем у рептилий или птиц. Вторичная моча собирается в почечной ло-
ханке, от которой начинается мочеточник, впадающий в мочевой пу-
зырь, в нем моча накапливается. Моча выводится через мочеиспуска-
тельный канал, который у самцов объединен с дистальным отделом
половых путей и проходит через губчатое тело полового члена. Моче-
испускательный канал самок — самостоятельная трубка, открываю-
щаяся наружным отверстием. У однопроходных мочеточники впада-
ют в мочеполовой синус, оттуда моча попадает в клоаку. Кроме почек,
выделительную функцию у млекопитающих выполняют легкие, кожа
(потовые железы) и толстая кишка.
Половая система. Мужская половая система состоит из
парных семенников, половых протоков, семенных пузырьков, пред-
стательной железы и полового члена. Семенники у однопроходных,
отдельных неполнозубых и насекомоядных, слонов и китообразных
остаются в брюшной полости в течение всей жизни. У самцов других
групп они выходят через паховый канал из брюшной полости, предва-
рительно все слои брюшной стенки выпячиваются, образуя мошонку,
которая имеется только у млекопитающих. В мошонке находятся се-
менники и их придатки, начальная часть семяпроводов, сосуды и
нервы. Семяпроводы, которые происходят из вольфовых протоков,
очень длинные, поскольку они возвращаются в брюшную полость
и в области таза впадают в мочеиспускательный канал, предваритель-
но образовав боковые ответвления — семенные пузырьки. Мочеиспу-
скательный канал самцов проходит через губчатое тело полового чле-
на, у основания которого располагается предстательная железа.
В ходе полового акта благодаря сокращению гладких мышечных
клеток сперма проталкивается и с силой выбрасывается из полового
члена во влагалище самки. Строение полового члена млекопитающих
значительно сложнее, чем копулятивного органа крокодилов и чере-
пах. Он образован двумя пещеристыми и губчатым телом, заканчива-
ющимся головкой. У многих млекопитающих (хищных, ластоногих
и др.) в соединительной ткани между пещеристыми телами распола-
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
657
гается длинная кость бакулюм, которая упрочняет половой член и
значительно увеличивает надежность эрекции.
Женская половая система образована парными яични-
ками и половыми путями. Яичники всегда находятся в брюшной по-
лости. Яйцеводы, возникшие из мюллеровых протоков, непосредст-
венно не связаны с яичниками (как и у всех других позвоночных),
они открываются в брюшную полость отверстиями воронок, располо-
женными вблизи яичников. Яйцеводы млекопитающих лучше других
позвоночных дифференцированы на отделы. Первый отдел, заканчи-
вающийся воронкой, называется маточной (фаллопиевой) трубой.
Затем следует более широкий отдел — матка, которая с каждой сторо-
ны тела впадает в непарное влагалище. В дистальный отдел также от-
крывается наружное отверстие женского мочеиспускательного кана-
ла. Там же находится гомолог пениса — клитор, который состоит из
двух пещеристых тел и головки, губчатого тела нет.
Развитие. Оплодотворение, как и у всех амниот, внутреннее.
Оплодотворение яйцеклетки и начальные этапы развития проходят в
маточных трубах. Дальнейшее развитие млекопитающих, принадле-
жащих к разным систематическим группам, происходит по-разному.
Яйца млекопитающих содержат мало желтка (изолецитальные яйца),
дробление асинхронное. Для раннего развития зародыша млекопита-
ющих характерна компактизация, в ходе которой бластомеры внезап-
но сближаются и контактируют друг с другом.
Наиболее примитивные — однопроходные — откладывают
яйца, покрытые скорлупой, однако, в отличие от рептилий и птиц,
яйцо содержит не зиготу, а зародыш, находящийся на средней стадии
развития. Первая половина развития зародыша происходит в поло-
вых путях матери. Достигнув определенного состояния, зародыш раз-
рушает изнутри скорлупу яйца и выходит из него, при этом у ехидн
он оказывается в сумке, куда самка помещает яйцо, а у утконосов —
в гнезде (у них нет сумки). Как и все млекопитающие, однопроходные
выкармливают детенышей молоком, которое у них стекает прямо по
шерсти, поскольку протоки млечных желез не образуют соска.
У сумчатых полное развитие в половых путях не происходит,
поэтому рождается очень маленький эмбрион. Питаясь молоком ма-
тери, детеныш быстро заканчивает свое развитие.
Развитие высших млекопитающих — плацентарных — цели-
ком проходит в женских половых путях. Оплодотворенная в маточ-
ной трубе яйцеклетка (зигота) там же приступает к дроблению, од-
новременно перемещаясь в полость матки. Там наружный слой
зародыша — трофобласт, на поверхности которого образуются вор-
синки хориона, — срастается со стенкой матки, слизистая оболочка
которой к тому моменту сильно утолщена. Со временем между разви-
658
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
вающимся зародышем и стенкой матки образуется общая связующая
структура — плацента, через которую происходит обмен веществ меж-
ду матерью и зародышем. По завершении внутриматочного развития
плацентарные млекопитающие рождают хорошо развитого детеныша
и некоторое время вскармливают его молоком. Детеныши могут ро-
ждаться зрячими, подвижными и покрытыми шерстью, например
жвачные, или слепыми, голыми и беззащитными, например хищные.
Отличия могут быть даже среди близких видов, например зайчата
рождаются зрячими и вскоре после рождения могут бегать, тогда как
крольчата слепые, голые и беспомощные. Млекопитающие проявля-
ют выраженную заботу о потомстве, при этом детеныши выкармлива-
ются сначала молоком, потом пищей взрослых. Родители охраняют
детенышей и вместе с другими взрослыми особями обучают их. Одна-
ко через определенное время молодые самцы чаще всего изгоняются,
чтобы предотвратить кровосмешение.
Нервная система. Центральная нервная система мле-
копитающих — наиболее сложная среди всех позвоночных. Головной
мозг очень крупный, по своей массе он значительно превосходит
спинной мозг. Так, например, у хищных головной мозг соотносится со
спинным в пропорции 4:1, у приматов — 8:1, у человекообразных
обезьян — 20—25:1, а у человека — 45:1. Особенно велик конечный
мозг, масса полушарий которого у ежей составляет 48% от массы все-
го головного мозга, у белок — 53%, у волков — 70%, у дельфинов —
75%. Увеличение достигается путем масштабного разрастания свода,
а не полосатых тел, как это происходит у птиц. Свод у млекопитаю-
щих вторичный (неопаллиум), вся его поверхность покрыта корой,
образованной серым веществом.
Кора полушарий конечного мозга (большого мозга) млекопитаю-
щих подразделяется на древнюю кору, которая имеет наиболее прими-
тивное однослойное строение, похожее на участки коры полушарий
рептилий и птиц; старую кору, которая также однослойная, но отде-
лена от нижележащей подкорки корковой пластинкой; и многослой-
ную новую кору, которая имеет наиболее сложное строение. Соотно-
шение перечисленных участков у разных видов млекопитающих
различно и зависит прежде всего от сложности поведения. Наиболее
развита новая кора у хищных, приматов и особенно у человека, у ко-
торого на долю новой коры приходится около 96% всей коры, при
этом новая кора отделена от старой и древней коры зонами средней
(промежуточной) коры (перипаллиокортикальной и периархикорти-
кальной, соответственно). В коре полушарий большого мозга распо-
лагаются центры высшей нервной деятельности, обеспечивающие на-
иболее сложные поведенческие реакции, которые не заложены
изначально, а развиваются в течение жизни, максимально эффектив-
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
659
но приспосабливая животное к быстро меняющимся условиям внеш-
ней среды. В коре располагаются центры анализаторов.
У млекопитающих кора конечного мозга является ведущим отде-
лом головного мозга, который интегрирует деятельность ЦНС. Мозг
такого типа называют маммальным. У малоподвижных млекопитаю-
щих (например, ленивцев) поверхность полушарий почти гладкая,
но у большинства видов разросшаяся кора образует более или менее
выраженные складки (извилины), разделенные бороздами. Базаль-
ные ядра (ядра полосатого тела и некоторые другие) у млекопитаю-
щих подчинены коре и выполняют роль подкорковых центров,
контролирующих движения и мышечный тонус. Обонятельные лу-
ковицы развиты хорошо, особенно у тех видов, для которых обоня-
ние является ведущим органом чувств. В крыше среднего мозга обо-
соблена характерная для млекопитающих пластинка четверохолмия,
состоящая из двух бугорков, из которых верхняя пара связана со
зрительным, а нижняя — со слуховыми анализаторами. У всех про-
чих классов позвоночных крыша среднего мозга разделена продоль-
ной бороздой на два бугра (двухолмие). Проводящие пути локализо-
ваны в ножках среднего мозга. Размеры этого отдела головного
мозга у млекопитающих невелики, что связано с его подчиненно-
стью высшим центрам, прежде всего коре. От среднего мозга отхо-
дят III и IV пары черепных нервов. Полость среднего мозга пред-
ставлена узким сильвиевым водопроводом, соединяющим между
собой третий и четвертый желудочки.
Ромбовидный мозг разделен на задний и продолговатый. Задний
мозг образован мозжечком и хорошо выраженным мостом, что связа-
но с более высокой организацией проводящих путей млекопитающих,
значительная часть которых проходит через мост. Очень хорошо раз-
витый мозжечок состоит из средней части — непарного червя и боко-
вых отделов — полушарий, также имеются клочок и узелок. Таким
образом, наряду с древними (клочок, узелок) и старыми (передняя
часть червя) структурами, у млекопитающих имеются более молодые
(полушария и задняя часть червя). При этом более старые структуры
получают информацию от органа равновесия и спинного мозга (вес-
тибуло- и спинно-мозжечковые пути), тогда как молодые — от коры
полушарий конечного мозга (корково-мостомозжечковые пути). На-
ряду с полушариями большого мозга, мозжечок снаружи покрыт ко-
рой (более простого строения, чем полушария большого мозга) серого
вещества, в ней локализуются наиболее молодые центры, управляю-
щие координацией сложных движений. На поверхности полушарий
мозжечка образуются складки — листики мозжечка, которые увели-
чивают поверхность коры. Более простыми функциями управляют
ядра, расположенные в мозжечке медиально.
660
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
Спинной мозг имеет характерное сегментарное строение,
в переднем и заднем отделах образует утолщения (шейное и пояснич-
но-крестцовое соответственно). У взрослых особей длина спинного
мозга меньше длины позвоночного столба.
Периферическая нервная система. От головного
мозга отходят 12 пар черепных нервов. У млекопитающих XI пара
(добавочный нерв) полностью дифференцирована. Спинномозговые
нервы (передние ветви) образуют нервные сплетения.
Вегетативная нервная система развита очень хорошо.
Центры парасимпатической части находятся в вегетативных ядрах со-
ответствующих черепных нервов, а также в крестцовом отделе спин-
ного мозга. Периферический отдел включает вегетативные волокна
черепных нервов вместе с промежуточными ганглиями. Центры сим-
патической части находятся в боковых рогах серого вещества некото-
рых сегментов спинного мозга. Периферический отдел — это симпа-
тические стволы, симпатические ганглии и сплетения.
Органы чувств у большинства млекопитающих развиты очень хо-
рошо, частичная или полная редукция того или иного анализатора
носит вторичный характер и связана с образом жизни.
Орган зрения имеется у большинства млекопитающих. Стро-
ение глаза типично для наземных позвоночных (выпуклая роговица
и двояковыпуклый хрусталик), аккомодация менее совершенная, чем
у птиц, осуществляется только путем изменения кривизны хрустали-
ка под воздействием ресничной мышцы. У мелких грызунов (напри-
мер, мыши) аккомодация вообще отсутствует. Не все млекопитающие
могут различать цвета, лесной хорек и многие другие животные
лишены цветового зрения, другие виды различают лишь отдельные
цвета, и только очень немногие млекопитающие (высшие приматы
восточного полушария) имеют полноценное цветовое зрение. Боль-
шинство видов способно различать только движущиеся предметы.
Это связано с тем, что у млекопитающих преимущественно развиты
другие органы чувств. Более острое зрение имеют обитатели обшир-
ных открытых территорий и ночные животные. Лесные млекопитаю-
щие видят хуже. У некоторых видов, преимущественно подземных
(слепыша, некоторых кротов), зрение атрофировалось, а редуциро-
ванные глаза полностью закрыты перепонкой.
Орган слуха развит очень хорошо, его строение намного
сложнее, чем у других позвоночных. Только у млекопитающих имеет-
ся наружное ухо — воронка из эластического хряща, покрытая кожей.
Хорошо развитые ушные мышцы позволяют млекопитающим повора-
чивать ушные раковины по направлению к источнику звука, опреде-
ляя его местонахождение. Это дает возможность ориентироваться без
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
661
участия зрения. В барабанной полости находятся три слуховые ко-
сточки — молоточек, наковальня и стремечко, подвижно связанные
между собой суставами, поэтому звукопередающий аппарат млекопи-
тающих совершеннее. Во внутреннем ухе имеется полностью сформи-
рованная улитка, в которой развит звуковоспринимающий кортиев
орган. Особенно хорошо орган слуха развит у ночных видов и хищ-
ных, часто слух является основным органом чувств (например, у ко-
шачьих).
Некоторые могут ориентироваться с помощью эхолокации (лету-
чие мыши, землеройки, ластоногие, дельфины и др.). Острота эхоло-
кации очень высока, например летучие мыши активно летают в тем-
ноте, уклоняясь от столкновений с предметами и разыскивая пищу,
а дельфины с высокой точностью определяют местонахождение кося-
ков рыб на расстоянии до 3 км.
Орган обоняния развит очень хорошо, для большинства
млекопитающих он является основным в процессе ориентации. Обо-
нятельная капсула крупная, область обонятельного эпителия обшир-
ная. У сумчатых, грызунов, копытных в обонятельной капсуле обо-
собляется отдел, открывающийся в небно-носовой канал (якобсонов
орган). Обоняние вторично редуцировано у китов, однако у тюленей
оно развито очень хорошо. Острое обоняние помогает млекопитаю-
щим ориентироваться по запахам, оставленным другими животными,
и находить предметы под землей.
Поведение млекопитающих наиболее сложное среди всех жи-
вотных, оно основывается на условных рефлексах. У многих видов
молодые особи обучаются, т. е . получают сведения, которые накапли-
вались в популяции в течение многих поколений. Например, слоны,
ведомые самой опытной слонихой, узнают место расположения всех
жизненно важных мест, которые могут быть далеко удалены друг от
друга, и в последующем находят дорогу самостоятельно. Высокое раз-
витие высшей нервной деятельности позволяет млекопитающим про-
являть великолепную экологическую пластичность. Поэтому они су-
мели занять самые разнообразные экологические ниши и в настоящее
время являются доминирующим классом среди позвоночных.
Современные млекопитающие подразделяются на три подкласса:
яйцекладущие, низшие звери и настоящие звери.
Подкласс яйцекладущие, или первозвери, включает в себя наибо-
лее примитивных млекопитающих, которые сочетают в себе призна-
ки разных классов позвоночных. Терморегуляция несовершенна,
поэтому температура тела непостоянна. Выкармливают своих дете-
нышей молоком, однако сосков у них нет и выводные отверстия мо-
лочных желез собраны на особых железистых полях. Выделяемое
662
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
молоко детеныши слизывают с шерсти. Млечные железы функцио-
нируют у обоих родителей. Ряд признаков сближает первозверей
с рептилиями и птицами. У них также имеется крупный самостоя-
тельный коракоид и прокоракоид. В тазовом поясе имеются сумча-
тые кости (у утконоса нет сумчатых костей). Кости черепа не обра-
зуют швов. У взрослых особей отсутствуют зубы. Челюсти покрыты
роговым клювом, который у утконоса широкий, наподобие утиного,
а у ехидн длинный и тонкий. Мягкие губы отсутствуют. Обладают
клоакой, в которую открываются выводные отверстия пищевари-
тельной системы и мочеполового аппарата. Головной мозг развит
хуже, чем у других млекопитающих, отсутствует мозолистое тело,
соединяющее полушария большого мозга. Первозвери размножают-
ся не путем живорождения, а откладыванием яиц, которые заключа-
ют в себе оформленный эмбрион, еще не завершивший свое разви-
тие. Ехидны размещают отложенное яйцо внутри сумки у самки.
Утконосы не имеют сумки, поэтому откладывают яйца в норе и, по-
видимому, насиживают их. Откладывание крупных яиц, богатых
желтком, приводит у самок к функциональной редукции правой по-
ловины половых путей, как у птиц. Первозвери входят в один от-
ряд — однопроходные, который подразделяется на семейство ехидн
(с двумя видами настоящих ехидн и тремя видами проехидн) и се-
мейство утконосов с единственным видом — утконосом. Эти крайне
своеобразные животные заселяют исключительно территории Ав-
стралии, Тасмании и Новой Гвинеи.
Подкласс низшие звери характеризуется более высокой организа-
цией, чем первозвери. Имеются зубы, однако в течение жизни сменя-
ется только один предкоренной зуб, тогда как остальные остаются по-
стоянными. Для большинства видов характерны мягкие губы.
Мочеполовой аппарат и пищеварительный тракт открываются нару-
жу самостоятельными выводными отверстиями, поэтому клоака от-
сутствует. В головном мозге нет мозолистого тела. Температура тела
выше, чем у первозверей, но ниже, чем у плацентарных, и непостоян-
ная. Размножение происходит путем живорождения, однако эмбрион
не завершает полного развития в половых путях матери (поскольку
плацента отсутствует или развита очень слабо). Поэтому беремен-
ность длится недолго (у гигантского кенгуру 30 суток), а детеныши
рождаются очень маленькими. Поскольку питание детеныша осу-
ществляется пассивно (молоко впрыскивается из соска в ротовую по-
лость), его гортань сильно приподнята и прижата к хоанам, что пред-
отвращает попадание молока в дыхательные пути в момент кормления.
Молочные железы у самцов нефункциональны. Кожистая сумка на-
ходится на брюхе (у вомбатов — на спине). Сумки нет у южноамери-
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
663
канских опоссумов. Вне зависимости от наличия сумки имеются две
сумчатые кости, которые прикрепляются к лобковому сочленению
тазовых костей, они укрепляют стенку брюшной полости, но не саму
сумку. Половые пути самки парные. У самцов многих видов также
раздвоен половой член. К низшим зверям относится лишь один от-
ряд — сумчатые, включающий в себя около 240 видов из 9 семейств,
которые занимают разнообразные экологические ниши. Подавляю-
щее большинство сумчатых обитает в Австралии и на прилегающих
к ней островах, некоторые виды заселяют Южную и Центральную
Америку и только один вид живет в Северной Америке.
Подкласс высшие звери, или плацентарные, наиболее обширен, он
объединяет наиболее высокоорганизованных млекопитающих. Они
живородящие, родившийся детеныш достаточно развит, чтобы само-
стоятельно сосать материнское молоко. Развитость детенышей зави-
сит от продолжительности беременности. Беспомощные детеныши
(например, у хищных) рождаются в хорошо защищенных убежищах
(обычно в норах), а подвижные детеныши (например, у парнокопыт-
ных и непарнокопытных) рождаются на открытой поверхности. Столь
значительное развитие эмбриона невозможно без образования надеж-
ной двусторонней связи между зародышем и матерью, которой у этой
группы млекопитающих является плацента. Отсутствует сумка и
сумчатые кости. Влагалище непарное. Молочные зубы сменяются по-
стоянными. Большие полушария головного мозга покрыты корой,
образованной серым веществом, которая у большинства видов обра-
зует складки — извилины (особенно у хищных и приматов). Полуша-
рия связаны между собой с помощью мозолистого тела. Протоки мо-
лочных желез открываются на поверхности соска. Обычно количество
сосков (их бывает от одной пары до шести пар и более) примерно со-
ответствует числу детенышей в одном помете. У видов с большим ко-
личеством детенышей в одном помете обычно имеется много сосков,
располагающихся в два длинных ряда на нижней стороне тела. Функ-
ционируют молочные железы лишь у самок и только в период кор-
мления потомства. Активизация функции молочных желез происхо-
дит непосредственно перед родами, этот процесс находится под
контролем эндокринного аппарата. Отсутствует клоака и роговой
клюв. У большинства видов (кроме китообразных и некоторых дру-
гих) имеются мягкие губы. Температура тела постоянная. Все эти
прогрессивные адаптации позволили плацентарным млекопитающим
заселить разнообразные экологические ниши, включая водную среду
и воздушное пространство. В настоящее время это наиболее процве-
тающая группа позвоночных, представители которой имеют самый
разный внешний вид.
664
ГЕНЕТИКА
ГЕНЕТИКА
Генетика изучает наследственность и изменчивость. Наследствен-
ность — это свойственная всем организмам способность передавать
свои признаки потомству, что приводит к биологическому сходству
между родителями и их потомством, а также обеспечивать возмож-
ность индивидуального развития сообразно с постоянно меняющими-
ся условиями среды. Изменчивостью, напротив, представляет собой
способность организмов приобретать различного рода изменения. Со-
гласно эволюционному учению, все имеющиеся признаки у любого
организма не присутствовали изначально, а появились путем мутаци-
онного преобразования других признаков, имевшихся ранее у предко-
вых форм. Универсальным материальным субстратом наследственно-
сти и изменчивости является нуклеиновая кислота. Для клеточных
организмов это всегда ДНК, для вирусов, наряду с ДНК, эту роль мо-
жет выполнять и РНК. Реализация информации осуществляется мо-
лекулами-посредниками, роль которых выполняет РНК. Каждая из
этих молекул представляет собой комплементарную копию опреде-
ленного участка молекулы ДНК, который принято называть геном.
Ген — это определенная последовательность ДНК или РНК, которая
отвечает за образование единичной полипептидной цепи. Термин
цистрон является функциональным эквивалентом гена. Признак пред-
ставляет собой любую дискретную характеристику организма — мор-
фологическую, физиологическую, биохимическую, этологическую и
т. д. Например, признаками являются цвет волос и глаз, группа крови
и резус-фактор, личностные качества: темперамент, характер, способ-
ности и др. Для возникновения любого из них необходима совместная
и строго согласованная деятельность не одного, а многих генов. При-
знак может иметь альтернативное (неодинаковое, противоположное)
проявление. В генетике каждый тип реализации одного и того же
признака называется аллелем, а гены, которые определяют развитие
признаков, имеющих альтернативное проявление, называют аллель-
ными или аллеломорфными. Такое разнообразие в проявлении одних
и тех же признаков возникло из-за мутаций генов. Каждый из алле-
лей (аллеломорфов), согласно хромосомной теории наследственно-
сти, располагается в хромосоме, занимая определенный ее участок —
локус.
Несмотря на то что у многих генов может быть большое количест-
во аллелей, их присутствие в генетическом аппарате всегда строго
определено. Каждый набор хромосом содержит один полный «ком-
плект» генов, отвечающих за развитие всех признаков организма. При
этом в гаплоидном состоянии всегда имеется лишь один аллель гена
каждого признака, в диплоидном — два, также и в полиплоидном со-
665
ГЕНЕТИКА
стоянии количество аллелей соответствует числу наборов хромосом.
Аллели в диплоидном или полиплоидном ядре могут быть одинако-
выми или разными. Если они одинаковые, то такое состояние называ-
ется гомозиготным, если разные — гетерозиготным. Диплоидные на-
боры хромосом свойственны соматическим клеткам большинства
эукариот. Появление диплоидности связано с половым размножени-
ем, смысл которого состоит в том, что новый организм получает оди-
наковое количество генетического материала от каждого из родите-
лей. При этом происходит комбинация генов; появляющиеся новые
сочетания признаков делают популяции значительно более гетероген-
ными, чем при бесполом размножении, что, в свою очередь, в значи-
тельной степени ускоряет эволюционный процесс.
В процессе полового размножения образуются гаметы (у живот-
ных в результате мейоза, а у растений в результате митоза), содержа-
щие по одному набору хромосом. В результате оплодотворения про-
исходит восстановление диплоидного набора. При этом хромосомы,
имеющие одинаковую форму и размеры, а также идентичную локали-
зацию в них генов, контролирующие одни и те же признаки, можно
условно разделить на пары, в каждой из которых одна хромосома по-
лучена от матери, а вторая от отца. Хромосомы каждой пары называ-
ют гомологичными. Число этих пар соответствует числу хромосом в
гаплоидном наборе. Хромосомы, гены которых определяют половые
признаки, являются половыми хромосомами, все остальные хромосо-
мы называются аутосомами. Кариотип — набор хромосом клетки,
который характеризуется их числом, размерами, формой и особенно-
стями строения. Представление набора хромосом на диаграмме также
называется кариотипом. Кариотип определяют путем микроскопиче-
ского изучения клетки во время ее митотического деления в метафа-
зе. Кариотипы разных организмов различаются между собой.
Геном — это совокупность всех генов, содержащихся в гаплоидном
наборе хромосом. Если же наборов хромосом два и больше, совокуп-
ность всех генов называют генотипом. Часто под генотипом подразу-
мевают аллельную пару рассматриваемого гена. При этом обычно
проявляется действие только одного аллеля. Тот аллель, который
проявляется в фенотипе как при гомозиготном, так и при гетерози-
готном генотипе, является доминантным. Напротив, рецессивный ал-
лель проявляется в фенотипе только в том случае, если генотип гомо-
зиготный. Совокупность всех проявленных признаков организма,
которые сформировались в процессе его индивидуального развития,
называется фенотипом. Элементарная единица фенотипа — феном.
Каждый из феномов контролируется аллелями одного гена. Фенотип
возникает в результате взаимодействия между генотипом и окружаю-
щей средой. В зависимости от условий окружающей среды возможна
666
ГЕНЕТИКА
некоторая вариабельность в проявлении одного и того же признака,
что связано с адаптацией к этим условиям. В зависимости от интен-
сивности действия фактора генотип будет обеспечивать строго опре-
деленное развитие фенотипа. Все возможное разнообразие сформиро-
вавшихся при этом фенотипов — это норма реакции, которая
индивидуальна для каждого организма и зависит от генотипа. Генети-
ческий контроль обусловливает границы нормы реакции. Норма ре-
акции — это потенциальная реактивность живой системы, которая
осуществляется при ее взаимоотношении с различными факторами
внешней и внутренней среды. Совокупность генов и их аллелей груп-
пы организмов называется генофондом. В зависимости от иерархиче-
ского положения группы выделяют генофонды популяции, вида, эко-
системы и биосферы.
Методы общей генетики. Гибридологический анализ, разра-
ботанный Г.Менделем, позволяет анализировать наследование от-
дельных признаков при половом размножении. Это основной метод
исследования наследственности и изменчивости. Основные положе-
ния метода состоят в следующем. 1 . Производится скрещивание го-
мозиготных родительских организмов, принадлежащих к одному
виду. 2. Скрещиваемые родительские особи должны различаться по
исследуемым признакам, наследование которых изучается отдельно
от других признаков родителей и их потомства. 3. Проведение реци-
прокного скрещивания, при котором различающиеся между собой
родительские организмы меняются местами по изучаемым аллелям.
4. Тщательное проведение статистического анализа всех классов
расщепления (т. е. проявления в потомстве гибридов особей с раз-
личными генотипами и фенотипами) у гибридов первого и последу-
ющих поколений. В настоящее время наряду с классическим гибри-
дологическим методом, предложенным Менделем, используются
также и другие. Например, методы отдаленной гибридизации, гибри-
дизации отдельных соматических клеток растений и животных, ги-
бридизации молекул ДНК и РНК, гибридизации клеток и вирусов
(трансдукция), гибридизации клеток и отдельных молекул ДНК
(трансформация).
Мутационный метод позволяет изучать наследование измене-
ний, вызванных мутациями. Используя различные виды мутагенов,
можно искусственно индуцировать мутации и затем изучить их с по-
мощью гибридологического метода. Вместе с гибридологическим
методом мутационный метод объединяют в метод генетического
анализа.
Цитологический метод используется для исследования кле-
точных структур, определяющих наследственность.
667
ГЕНЕТИКА
Математический метод позволяет рассчитать возможные ре-
зультаты эксперимента. Незаменим при исследовании наследования
количественных признаков и изменчивости.
Методы генетики человека. Применение самых распространен-
ных методов общей генетики, таких как гибридологический или экс-
периментально-мутационный, невозможно для изучения закономер-
ностей наследственности и изменчивости человека. Методические
трудности при изучении наследственности человека: 1) невозмож-
ность произвольного скрещивания в эксперименте; 2) позднее насту-
пление половой зрелости; 3) малое число потомков в каждой семье;
4) невозможность уравнивать условия жизни для потомства; 5) отсут-
ствие точной регистрации проявления наследственных признаков
в семьях и отсутствие гомозиготных линий; 6) большое число хромо-
сом. Наиболее значимыми методами являются следующие.
Генеалогический метод (метод родословных) заключается
в анализе наследования и проявления изучаемого признака в ряду по-
колений, при этом информативность метода возрастает по мере уве-
личения представленных поколений. Для этого составляют родослов-
ные, которые представляют собой последовательную, графически
изложенную совокупность сведений о предках того или иного челове-
ка, характеризующих его происхождение. Анализ полученной при со-
ставлении родословной информации позволяет определить тип на-
следования интересующего признака или болезни. Метод позволяет
выяснить, сколько генов вызывают проявление признака или болез-
ни, а также в каких хромосомах локализованы эти гены — в аутосомах
или половых хромосомах.
Близнецовый метод основан на изучении генетических законо-
мерностей у близнецов. Он позволяет сопоставить значимость гено-
типа и внешних факторов в проявлении признака.
Цитогенетический метод основан на изучении кариотипа в
норме и при патологии. При этом используют методы дифференци-
альной окраски хромосом, гибридизации соматических клеток, куль-
туры тканей in vitro (т. е. выращивание тканей вне организма на пита-
тельной среде). Метод позволяет диагностировать генетические
заболевания, связанные с нарушением строения и числа хромосом,
а также помогает определить локализацию генов в хромосомах.
Популяционный метод позволяет проследить распространен-
ность генов, отдельных аллелей и мутаций в различных более или
менее изолированных популяциях. Чаще всего при этом основыва-
ются на законе Харди — Вайнберга, согласно которому частоты ге-
нотипов в поколениях остаются неизменными, если на их сочетание
не воздействуют посторонние факторы (мутации, отбор, дрейф ге-
нов и др.).
668
НАСЛЕДСТВЕННОСТЬ
Онтогенетический метод позволяет определить присутствие
рецессивных аллелей у гетерозигот. Поскольку определенные генети-
ческие заболевания развиваются только в определенном возрасте,
возникает необходимость изучить фенотипические проявления в те-
чение индивидуального развития человека.
Метод моделирования. Общий смысл этого метода выражает
закон гомологических рядов наследственной изменчивости, сформу-
лированный великим отечественным генетиком Н.И . Вавиловым.
Согласно этому закону, виды и роды генетически близких организмов
в одинаковых условиях демонстрируют изменение признаков в сход-
ном направлении. Зная ряды измененных признаков у одного вида,
можно предвидеть такие же изменения у другого вида.
НАСЛЕДСТВЕННОСТЬ
ВЗАИМОДЕЙСТВИЕ АЛЛЕЛЬНЫХ ГЕНОВ
Различные состояния гена, при которых признак выражается по-
разному, называют аллелями. Общее количество возможных аллелей
не всегда ограничивается одной парой, часто имеет место множест-
венный аллелизм. В диплоидной клетке одновременно могут присут-
ствовать только два аллеля гена, один из которых получен от отца,
а другой от матери. Если аллели одинаковые, то генотип считается
гомозиготным, если разные — то гетерозиготным (эти понятия были
предложены В. Бэтсоном в 1902 г.) . Гаметы содержат гаплоидный на-
бор хромосом, поэтому они несут только по одному аллелю каждого
гена (правило чистоты гамет В. Бэтсона). В зависимости от количе-
ства признаков в гетерозиготном состоянии число различных типов
образующихся гамет выражается формулой 2n
, где n — число призна-
ков у гетерозиготной особи.
Взаимодействие аллелей одного гена осуществляется по принципу
доминирования (преобладания действия определенного аллеля в ал-
леломорфной паре), который может выражаться тремя способами:
полное доминирование, неполное доминирование и кодоминирова-
ние. При полном доминировании в фенотипе гетерозигот проявляется
только один аллель из аллеломорфной пары. Такой аллель называют
доминантным и обозначают заглавной буквой, например А. Другой
аллель может проявиться в фенотипе лишь при гомозиготном геноти-
пе. Этот аллель называется рецессивным и обозначается строчной
буквой, например а (понятия «доминантный» и «рецессивный» были
предложены Менделем), Гомозиготный генотип обозначается АА или
аа, а гетерозиготный — Аа. Например, желтая окраска семян гороха
669
ВЗАИМОДЕЙСТВИЕ АЛЛЕЛЬНЫХ ГЕНОВ
является доминантной, и при наличии такого аллеля семена оказыва-
ются желтыми как у гомозигот, так и у гетерозигот. Зеленая окраска
возможна лишь у гомозигот.
При неполном доминировании самостоятельное выражение аллеля
возможно только у гомозигот. У гетерозигот проявляются оба аллеля,
и признак приобретает промежуточное состояние между двумя алле-
лями. Например, если в гомозиготном состоянии один аллель опреде-
ляет красную окраску цветка, а другой белую, то в гетерозиготном
состоянии у гибрида проявится розовая окраска. Это пример проме-
жуточного наследования.
При кодоминировании возможно объединение у гетерозигот двух
аллелей, которые отдельно друг от друга являются доминантными.
Оказавшись вместе, оба аллеля совместно участвуют в выражении
признака, который при этом приобретает не промежуточное значение
(как в случае неполного доминирования), а совершенно самостоя-
тельное. Кодоминирование имеет место при наследовании группы
крови и выражается в следующем. В зависимости от наличия в мем-
бранах эритроцитов крови агглютиногенов и агглютининов в плазме
выделяют четыре группы крови системы АВО: О, А, В и АВ. Этот
признак определяют три аллеля одного гена: i, I
A
иIB
.
При этом ал-
лель i является рецессивным и проявляется в фенотипе только в со-
стоянии генотипа I (группа крови О). Аллели IА и IВ в генотипах IAi и
IBi являются доминантными по отношению к аллелю i (при этом гено-
типы IАIА и IАi определяют группу А; генотипы IBIB и IBi определяют
группу В), но в генотипе IАIB они кодоминантны и проявляются сов-
местно, образуя отдельную группу крови — АВ.
Наследование при полном доминировании. Моногибридное
скрещивание. Моногибридным называют скрещивание, при кото-
ром родительские организмы различаются по одной паре аллелей.
Первым эксперименты по моногибридному скрещиванию провел
Мендель. С этих опытов и началась генетика. Для своих эксперимен-
тов Мендель избрал горох. Основными мотивами для этого стало:
разнообразие сортов, каждый из которых отличался от других четко
различимыми признаками; самоопыление гороха, из-за чего можно
было не опасаться случайного перекрестного опыления, зато легко
можно проводить искусственное опыление согласно замыслу экспе-
римента; горох легко выращивать.
Мендель выбрал из многих чистых линий 22 с хорошо выражен-
ными контрастными различиями. На них он поэтапно изучал харак-
тер наследования семи признаков: окраска (желтая и зеленая) и фор-
ма (гладкая и морщинистая) семян, окраска плодов (зеленые и
желтые) и их форма (плоские и выпуклые с перетяжками), располо-
жение цветков (пазушные и верхушечные) и их окраска (красные
670
НАСЛЕДСТВЕННОСТЬ
и белые), высота побега (высокие и карликовые). В своей работе Мен-
дель использовал созданный им гибридологический метод и прово-
дил статистический анализ полученного цифрового материала. Уда-
чей Менделя явилось то, что у гороха в диплоидном наборе семь пар
хромосом и в каждой из них находятся гены, кодирующие один из
изучаемых им признаков. Но сам Мендель об этом не знал, так как
нахождение носителя наследственной информации тогда было неиз-
вестно. Для того чтобы исключить самоопыление, Мендель своевре-
менно удалял тычинки, после чего переносил пыльцу с другого расте-
ния. Так он подбирал родительские пары и искусственно их скрещивал.
В дальнейшем естественным образом происходило оплодотворение
семязачатков исследуемых растений гороха, после чего развивались
плоды с семенами полученных гибридов. Их исследование показало,
что у всех гибридов присутствовал признак только одного из родите-
лей. Позднее (1902) Бэтсон и Сондерс предложили обозначать ги-
бриды первого Поколения F1, родители обозначаются Р. На основа-
нии этих данных было сформулировано правило доминирования
и первый закон Менделя — закон единообразия гибридов первого поко-
ления. Согласно этому закону, гибриды первого поколения имеют
одинаковое проявление изучаемого признака, причем проявляется
вариант признака только одного родителя — доминантный.
Продолжая свои эксперименты, Мендель допустил самоопыление
у гибридов первого поколения и в результате получил гибриды второ-
го поколения — F
2
. Изучая их, Мендель обнаружил, что большая часть
из них несет тот же признак одного из родителей, который безальтер-
нативно присутствовал у гибридов первого поколения. Однако, наря-
ду с этим, у значительно меньшей части гибридов проявился «исчез-
нувший» признак другого родителя, который отсутствовал у всех
гибридов F1. Проведя статистический анализ гибридов второго поко-
ления, Мендель обнаружил, что варианты альтернативного проявле-
ния исследуемых признаков у них встречаются в соотношении 3:1.
Но, может быть, на выражение признака оказывает воздействие пол
родителей? Для большей достоверности Мендель также производил
реципрокные скрещивания. Их смысл состоит в том, что производятся
два скрещивания родителей, несущих изучаемый признак — допу-
стим, окраску цветка. В первом скрещивании пыльца с красных цвет-
ков искусственно переносилась на рыльца белых цветков. Во втором
производили рокировку родителей: пыльца с белых цветков перено-
силась на красные цветки. При этом результаты не изменились.
Мендель пришел к выводу, что каждое растение содержит по два
внутренних наследственных фактора, определяющих проявление
признака. У чистых линий эти факторы одинаковые, а у гибридов раз-
ные, причем от каждого из родителей растения получили по одному
671
ВЗАИМОДЕЙСТВИЕ АЛЛЕЛЬНЫХ ГЕНОВ
фактору через гаметы. Соответственно, гаметы содержат по одному
фактору признака. Оказавшись у гибридов вместе, наследственные
факторы не сливаются, а существуют отдельно, однако проявляется
только один фактор — доминантный, а другой — рецессивный — оста-
ется неактивным. Дальнейшее размножение гибридов приводит к по-
вторному появлению признака, отсутствующего у гибридов первого
поколения, в соотношении 3:1. Это происходит в результате того, что
в гаметы попадает только по одному внутреннему фактору, и дальней-
шее случайное слияние гамет, несущих эти факторы, приводит к обра-
зованию организмов либо с двумя одинаковыми факторами, либо с
двумя разными. При этом рецессивный признак проявляется только
у тех растений, у которых таких факторов два. Если в организме со-
держится по два доминантных фактора или один доминантный и один
рецессивный, в любом случае проявляется доминантный фактор.
В. Иогансен (1909) предложил называть наследственные факторы ге-
нами, а их альтернативные варианты — аллелями. Итогом этих выво-
дов стал второй закон Менделя — закон расщепления, по которому
в потомстве гибридов первого поколения происходит расщепление
в проявлении изучаемого признака: у части потомства проявляется
доминантный вариант признака, а у другой части — рецессивный.
Эти законы Менделя легко проиллюстрировать графически:
Р
♀АА
×
♂аа
␙
Гаметы Р: (А)
(а)
F1
Aa — 100%
Аа×Аа
Гаметы F1:
(А)и(а)(А)и(а)
␙
F2
АААаАааа
Как видно из приведенной схемы, в результате скрещивания гомо-
зигот (чистых линий, при размножении в себе не дающих расще-
пления) все 100% гибридов первого поколения (F1) имеют гетеро-
зиготный генотип и одинаковый фенотип. При моногибридном
скрещивании гетерозиготы дают два типа гамет: (А) и (а). Случайная
встреча гамет при оплодотворении дает три типа генотипа: АА (25%),
Аа (50%) и аа (25%), которые дают два типа фенотипа: генотипы АА
и Аа проявляют доминантный фенотип, а генотип аа — рецессивный.
Легко подсчитать, что доминантный фенотип будут иметь 75% гибри-
дов F2, а рецессивный только 25%, что в итоге и дает менделевское
расщепление 3:1.
672
НАСЛЕДСТВЕННОСТЬ
Для облегчения записи возможных генотипов английский генетик
Р.К. Пеннет предложил использовать таблицу, которая получила на-
звание решетка Пеннета. В ней по вертикали в крайнем левом стол-
бце в отдельных ячейках записывают аллели, содержащиеся во всех
возможных типах женских гамет, а в ячейках верхней строки — муж-
ских. В ячейки, расположенные на пересечении, вносят аллели соот-
ветствующего генотипа. Например, если скрещиваются особи с гено-
типами Аа × Аа, то каждая из них продуцирует гаметы двух типов:
А и а. Графически это будет выглядит так:
Гаметы ♂
Гаметы ♀
Аа
АА
А
А
а
аА
а
а
а
Генотип организма с рецессивным выражением признака всегда
гомозиготен. Доминантный же аллель проявляется и у гомозигот,
и у гетерозигот. Используя анализирующее скрещивание, можно точно
определить генотип интересующей особи с доминантным фенотипом.
Его сущность заключается в скрещивании такой особи с родитель-
ской особью, гомозиготной по рецессивному аллелю. Если генотип
был АА, то, скрещиваясь с аа, все потомство будет гетерозиготами Аа
и все будут иметь доминантный фенотип. Если исследуемая особь
была гетерозиготной и имела генотип Аа, то половина потомства бу-
дет иметь генотип Аа с доминантным фенотипом, а половина — гено-
тип аа с рецессивным фенотипом.
Дигибридное и полигибридное скрещивание. Проведя первую серию
опытов и исследовав характер наследования отдельных признаков,
Мендель стал изучать механизмы их совместного наследования. Для
этого он также использовал гибридологический метод. Скрещивание
родительских пар, различающихся между собой по многим призна-
кам, называется полигибридным скрещиванием. Вначале Мендель
изучил наследование одновременно двух альтернативных признаков,
такое скрещивание называется дигибридным. В одном из опытов Мен-
дель скрещивал гомозиготные растения, имеющие желтые и гладкие
семена (ААВВ), с также гомозиготными растениями, но с зелеными и
морщинистыми семенами (aabb). Желтый цвет и гладкая поверх-
ность семян доминируют над зеленой окраской и морщинистой по-
верхностью. Как и в случае с моногибридным скрещиванием, в F1
проявились лишь доминантные аллели. Однако в F2, наряду с имею-
щимися у родителей признаками (желтой окраской гладких семян
и зеленой морщинистых), у гибридов появились новые сочетания
673
ВЗАИМОДЕЙСТВИЕ АЛЛЕЛЬНЫХ ГЕНОВ
признаков: часть семян имела желтую окраску наряду с морщинистой
поверхностью, а еще одна часть — зеленую окраску в сочетании с
гладкой поверхностью.
Мендель отдельно просчитал частоту проявления каждого при-
знака: желтая окраска, зеленая окраска, гладкая поверхность, мор-
щинистая поверхность. Анализ показал, что независимо от формы
семян на каждые три семени желтой окраски в среднем приходится
по одному семени зеленой окраски. Точно так же и для другой пары
признаков: на каждые три семени с гладкой поверхностью независи-
мо от окраски семян приходится по одному морщинистому семени.
Он пришел к выводу, что каждый признак проявляется самостоя-
тельно и его наследование не зависит от другого. Таким образом,
Мендель сделал свое главное открытие — доказал дискретность
(независимость) наследственных факторов. Это выразилось в тре-
тьем законе Менделя: законе независимого наследования признаков
(независимого комбинирования генов), согласно которому каждый
признак наследуется независимо от наследования других признаков.
Графически расщепление при дигибридном скрещивании выглядит
так:
Р
♀ ААВВ
×
♂ ааbb
␙
Гаметы Р:
(АB)
(аb)
F1
AaBb (желтые и гладкие) — 100%
Аа
×
Аа
Гаметы F1:
(АB) (Ab) (aB) и (аb)
␙
F2
А-В (9 — желтые гладкие),
A-bb (3 — желтые морщинистые),
ааВ (3 — зеленые гладкие),
aabb (1 — зеленые морщинистые)
При скрещивании двойных гетерозигот (дигетерозигот) образу-
ется 16 возможных генотипов, дающих проявление в четырех типах
фенотипов. Математически расщепление в F2 можно выразить следу-
ющей формулой: х = (3+1)n
, где n — число пар признаков. При моно-
гибридном скрещивании будет х = (3+1)1 = 4 возможных генотипа и
два фенотипических типа в соотношении 3:1; при дигибридном: х =
(3+1)2
= 16 генотипов и четыре фенотипических типа — 9:3:3:1; при
тригибридном: х = (3+1)3 = 64 генотипа и восемь фенотипов —
27:9:9:9:3:3:3:1 и т. д . Для вычисления количества фенотипических ти-
пов в F2 употребляют формулу: х = 2
n
, где n — число пар признаков
674
НАСЛЕДСТВЕННОСТЬ
в гетерозиготном состоянии (это число равно количеству гамет, кото-
рые образует эта особь).
ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ
До сих пор мы рассматривали наследование признаков, которые
контролируются взаимодействием аллелей одного гена. При этом фе-
нотип гибрида зависит от наличия аллелей, их сочетания и типа до-
минирования. Наряду с этим имеет место взаимодействие нескольких
генов, расположенных не в гомологичных хромосомах, причем сов-
местно они определяют выражение только одного признака. Несмо-
тря на название, такой тип наследования также определяется аллеля-
ми. Только при аллельном взаимодействии учитываются аллели
одного гена, а при неаллельном взаимодействии — аллели нескольких
разных генов. Известно несколько типов взаимодействия разных ге-
нов: комплементарность, эпистаз и полимерия.
Комплементарность имеет место, когда доминантные аллели раз-
ных генов, объединившись в генотипе гибридов, дают новое феноти-
пическое проявление, отсутствующее у родительских форм. Напри-
мер, таким образом наследуется окраска цветков у душистого горошка.
У этого растения цветки, в зависимости от сочетания доминантных
аллелей двух комплементарных генов, могут иметь красную или бе-
лую окраску. Красные цветки имеют особи, обладающие одновремен-
но обоими доминантными аллелями — А и В (их взаимодействие об-
условливает образование пигмента). Если отсутствует хотя бы один
из них, то независимо от сочетания аллелей в генотипе пигмент не
образуется и цветки остаются белыми. Соответственно, если скрещи-
вать родителей с генотипами AAbb и ааВВ (они будут иметь белую
окраску цветков), то гибриды первого поколения будут дигетерозиго-
тами (АаВb) и унаследуют от каждого родителя по доминантному ал-
лелю каждого гена. Такое сочетание комплементарных генов приво-
дит к появлению новой формы признака, отсутствующей у обоих
родителей, — пигментации цветка. У гибридов второго поколения
произойдет расщепление, в результате чего 9 из 16 будут обладать
обоими доминантными аллелями генов — А и В — и, соответственно,
иметь окрашенные цветки. У оставшихся шести будет присутствовать
лишь один из доминантных аллелей, а у одной особи будут только
рецессивные аллели обоих генов. Следовательно, у всех семи пигмент
образоваться не сможет и цветки останутся неокрашенными.
Другим примером, иллюстрирующим взаимодействие комплемен-
тарных генов, является наследование формы гребня у кур, точнее
у петухов. Этот признак контролируется двумя генами, локусы кото-
675
ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ
рых находятся в разных хромосомах. Если в генотипе имеется один
или два доминантных аллеля одного гена (А-) и два рецессивных
другого (bb), то развивается гороховидный гребень; если присутству-
ет один или два доминантных аллеля гена В- и два рецессивных алле-
ля гена аа, сформируется розовидный гребень. Одновременное при-
сутствие в генотипе доминантных аллелей обоих генов (А-В-)
приводит к развитию у петуха ореховидного гребня. Если в генотипе
имеются только рецессивные аллели обоих генов (aabb), гребень
имеет листовидную форму.
Эпистаз. При таком взаимодействии аллель одного гена подавля-
ет действие аллеля другого гена. Различают доминантный эпистаз
(в случае, если А>В) и рецессивный (если аа>В или aa>bb). Рассмо-
трим проявление доминантного эпистаза, например окраски перьев
у кур. У большинства кур наличие пигмента перьев определяется до-
минантным аллелем гена С, тогда как его рецессивный аллель с детер-
минирует отсутствие пигмента и, как следствие, белое оперение. Вме-
сте с тем имеется эпистатический ген, доминантный аллель которого
I блокирует ген С и, соответственно, делает невозможным синтез пиг-
мента. Наличие аллеля I при любом сочетании других аллелей об-
условливает белое оперение. Рецессивный аллель ii не влияет на син-
тез пигмента.
Р
♀ CCII
×
♂ ccii
(белые перья) ␙ (белые перья)
Гаметы Р:
(CI)
(ci)
F1
CcIi — 100%
(белые перья)
Гаметы F1:
(CI) (Ci) (cI) и (ci)
␙
F2
13—C-I-
:
3—C-ii
(белые перья)
(окрашенные перья)
или
Мужские гаметы
Женские гаметы
CI
Ci
cI
ci
CI
ССII (бел.)
CCIi
(бел.)
CeII (бел.) CcIi (бел.)
Ci
CCIi (бел.) Ccii (окр.) CcIi (бел.) Ccii (окр.)
cI
CcII (бел.) CcIi (бел.) ccII (бел.)
ccIi (бел.)
ci
CcIi (бел.) Ccii (окр.) ccIi (бел.)
ccii (бел.)
676
НАСЛЕДСТВЕННОСТЬ
Полимерия. При полимерии доминантные аллели разных генов
одинаково действуют на проявление одного признака. Выделяют два
типа полимерии. При кумулятивной полимерии доминантные алле-
ли разных генов совместно усиливают степень проявления одного
признака. Такое взаимодействие генов имеет место главным образом
при детерминации количественных признаков (например, цвета
кожи или роста человека). В случае некумулятивной полимерии сте-
пень проявления признака аллелями не определяется, тогда как вы-
ражение самого признака зависит не от количества доминантных
аллелей, а от наличия доминантных аллелей вообще. Иными слова-
ми, если доминантные аллели имеются (пусть даже один), доми-
нантный фенотип проявляется в полной мере, а если генотип гомо-
зиготен по рецессивным аллелям всех взаимодействующих генов
(т. е. доминантные аллели отсутствуют), то проявляется рецессив-
ный фенотип.
Некумулятивная полимерия определяет наследование качествен-
ных признаков. Примером кумулятивной полимерии будет генетиче-
ская детерминация роста. Этот признак у человека определяется вза-
имодействием нескольких пар генов: Аа, Вb, Сс. Индивидуумы,
обладающие наибольшим количеством доминантных аллелей, дости-
гают наибольшего роста. При этом каждый доминантный аллель вза-
имодействующих генов действует равноценно и усиливает воздейст-
вие доминантных аллелей других генов. Если в генотипе преобладают
рецессивные аллели рост оказывается небольшим. Поэтому самыми
высокими будут лиц» с генотипом ААВВСС, а самыми низкорослы-
ми — с генотипом aabbcc. Чаще всего встречаются носители среднего
значения признака (т. е. люди среднего роста), тогда как крайние вы-
ражения (люди маленького или высокого роста) этого признака про-
являются значительно реже. Еще одним примером кумулятивной по-
лимерии является наследование цвета кожи. У человека степень
содержания пигмента (меланина) в коже определяется двумя парами
независимо расщепляющихся генов: ВВСС — темная кожа (высокое
содержание пигмента) и bbcc — белая кожа. Любые три аллеля тем-
ной кожи определяют темную кожу, любые два — смуглую, один —
светлую.
Некумулятивную полимерию легко проиллюстрировать на приме-
ре наследования формы плода у пастушьей сумки, которые могут
иметь треугольную или яйцевидную форму. Признак контролируется
аллелями двух пар полимерных генов: А-а и В-b, расположенных в
разных парах хромосом. При этом наличие в генотипе хотя бы одного
доминантного аллеля этих генов — А - или В-
—
приводит к образова-
нию треугольных плодов. И только у двойных гомозигот aabb плоды
яйцевидной формы.
677
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ
Выразим это в схеме скрещивания:
Р
♀ААbb × ♂ааBB
(треугольная форма)
(треугольная форма)
␙
Гаметы Р:
(Аb)
(аB)
F1
AaBa (треугольная форма) — 100%
Аа
×
Аа
Гаметы F1:
(АB) (Ab) (aB) и (аb)
␙
F2
15
:1
(треугольная форма) (яйцевидная форма — aabb)
9 A-B, 3 A-bb, 3 aab-
ХРОМОСОМНАЯ ТЕОРИЯ
НАСЛЕДСТВЕННОСТИ
Мендель обнаружил, что наследование отдельных признаков мо-
жет происходить независимо друг от друга. Великий ученый не был
понят и признан при жизни. В 1900 г. выявленные им закономерности
наследования открыли повторно три исследователя (Г. де Фриз,
К. Корренс и Э. Чермак). Вскоре после переоткрытия законов Мен-
деля (1902) сразу два исследователя В. Саттон и Т. Бовери предполо-
жили, что гены находятся в хромосомах. Превратить эту гипотезу в
теорию позволили опыты, произведены американским генетиком и
будущим лауреатом Нобелевской премии Т.Х . Морганом (1910) и его
сотрудниками К. Бриджесом, А.Г. Стертевантом и др. Морган для сво-
их исследований избрал плодовую мушку дрозофилу, поскольку муш-
ка легко и быстро разводится в лабораторных условиях, она обладает
множеством отчетливых альтернативных признаков, которые легко
изучать методом гибридологического анализа. Однако Морган обна-
ружил, что наследование некоторых признаков у дрозофил противо-
речит обнаруженным Менделем закономерностям, поскольку насле-
довались они не самостоятельно, а совместно. Таким образом было
открыто сцепленное наследование, а также явление кроссинговер.
Сцепленное наследование и кроссинговер. Морган скрещивал
мух, различавшихся по цвету тела и длине крыльев. Обычно дрозофи-
лы имеют серую окраску и нормальные крылья. Вместе с тем имеются
мутантные особи с черным телом и недоразвитыми крыльями. Оба
эти признака контролируются рецессивными аллелями. Как и ожида-
лось, скрещивание серых мух с нормальными крыльями и черных
с зачаточными дало единообразие гибридов первого поколения по до-
678
НАСЛЕДСТВЕННОСТЬ
минантным аллелям: все потомство имело серую окраску и нормаль-
ные крылья. Далее Морган предпринял анализирующие скрещива-
ния. Их результаты оказались совсем неожиданными. В первом
скрещивании он брал самцов из F1 и скрещивал их с гомозиготными
по обоим генам самками. В F
а
(потомстве от анализирующего скрещи-
вания) он получил не ожидаемые четыре фенотипических класса (что
следовало из открытых Менделем законов), а только два: половина
мух независимо от пола имела серое тело и нормальные крылья, а
другая — черное тело и зачаточные крылья. Морган сделал вывод, что
самец F1 продуцировал не четыре типа гамет, как при обычном диги-
бридном скрещивании, а только два. Причем каждая из таких гамет
содержала лишь изначальные родительские сочетания аллелей (раз-
умеется, имеются в виду генотипы родителей самца F1
, участвовавше-
го в этом анализирующем скрещивании). Во втором анализирующем
скрещивании Морган получил Fa от самок из F1 и гомозиготных по
исследуемым генам самцов. Результаты оказались совсем иными, чем
в предыдущем скрещивании. В этом случае в потомстве были все че-
тыре фенотипических класса: серые с нормальными крыльями, серые
с зачаточными крыльями, черные с нормальными крыльями и черные
с зачаточными крыльями. Таким образом, проявились возможные их
сочетания, как в случае с расщеплением при дигибридном скрещива-
нии. Однако частота встречаемости признаков здесь была иной. Ожи-
даемое менделевское расщепление для дигибридного скрещивания
должно быть 1:1:1:1 для каждого упомянутого фенотипического клас-
са, но Морган получил 41,5% серых мух с нормальными крыльями,
41,5% черных с зачаточными крыльями, 8,5% серых с зачаточными
крыльями и 8,5% черных с нормальными крыльями. Следовательно,
самцы и самки от F1 продуцируют неодинаковое количество различ-
ных типов гамет. Самцы производят только два типа гамет в одинако-
вых пропорциях, причем аллели исследуемых генов в них сцеплены и
всегда наследуются совместно. У самок образуется четыре типа гамет,
однако в разных соотношениях: в 83% (41,5% + 41,5% = 83%) случаев
аллели исследуемых генов наследуются также сцепленно, а в 17%
(8,5% +8,5% = 17%) — образуются их рекомбинантные сочетания.
ВНИМАНИЕ!
Морган доказал, что наследственные факторы — гены — располагаются
в хромосомах, причем гены, определяющие у дрозофил цвет тела и раз-
витие крыльев, находятся в одной хромосоме.
У гомозигот в гомологичных хромосомах присутствуют одинако-
вые аллели, а у гетерозигот — разные. Гены, расположенные в одной
679
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ
хромосоме, наследуются сцепленно. У самцов это сцепление абсолют-
ное, а у самок — частичное, поэтому в процессе мейоза у самок возмо-
жен частичный обмен одинаковыми участками у гомологичных (не-
сущих одинаковые гены) хромосом, что приводит к образованию
рекомбинантных сочетаний аллелей. Цитологическим основанием
для этого послужило наблюдение Янссенсом (1909) за образованием
перекреста (хиазм) в процессе профазы первого деления мейоза. Та-
кой обмен между гомологичными хромосомами получил название
кроссинговер (от англ. crossingover — перекрест). В дальнейшем Стер-
тевант предположил, что чем ближе гены расположены в хромосоме,
тем меньше вероятность кроссинговера между ними и больше вероят-
ность того, что они будут наследоваться сцепленно. С другой сторо-
ны, просчитав частоту кроссинговера, можно судить о расстоянии
между исследуемыми генами в хромосоме и даже определить их рас-
положение и удаленность друг от друга. В результате у генетиков по-
явилась возможность составления генетических карт.
Гены, расположенные в одной хромосоме, образуют группу сцепле-
ния. Если не произошел кроссинговер, они попадают в одну гамету и
наследуются вместе. Количество групп сцепления соответствует ко-
личеству хромосом в гаплоидном состоянии ядра. Из этого следует,
что открытые Менделем закономерности наследования раскрывают
индивидуальное поведение не отдельных генов, а отдельных хромо-
сом. Причиной тому послужило совпадение: гены, определяющие вы-
ражение изучаемых Менделем признаков, находились в разных хро-
мосомах и распределялись в гаметах независимо друг от друга.
В профазе первого деления мейоза у некоторых хромосом может
произойти перекрест — кроссинговер, в результате чего гомологичные
хромосомы обмениваются участками. Таким образом происходит ре-
комбинация аллелей генов. Гаметы, в которые затем попадают хромо-
сомы, претерпевшие кроссинговер, называют кроссоверными, гаметы
с неизмененными хромосомами называют некроссоверными.
Наследование, сцепленное с полом. Явление, при котором при-
знаки по-разному наследуются в зависимости от пола, также было от-
крыто и изучено Т. X . Морганом. Скрещивая мух, отличающихся по
цвету глаз, и проводя затем реципрокные скрещивания, он обнару-
жил, что наследование этих признаков по-разному проявляется у сам-
цов и самок. У дрозофил красная окраска глаз доминирует над белой.
Скрещивание красноглазой самки и белоглазого самца дало в F1 еди-
нообразие — все мухи были красноглазыми. Гибриды F2 также дали
привычное расщепление — 3:1. Однако в F2 все самки были красно-
глазыми, а среди самцов половина была красноглазых и половина бе-
логлазых. Реципрокное скрещивание дало иные результаты, также
противоречащие менделевскому расщеплению. При скрещивании го-
680
НАСЛЕДСТВЕННОСТЬ
мозиготных белоглазых самок и красноглазых самцов в F1 вообще не
было ожидаемого единообразия потомства: все самки оказались крас-
ноглазыми, а все самцы — белоглазыми. Иными словами, сыновья на-
следовали признак матери, а дочери — признак отца (такое наследова-
ние генетики называют крисс-кросс — крест-накрест). В F2 же
независимо от пола соотношение красноглазых и белоглазых мух
было одинаковым.
Морган объяснил это, сопоставив результаты скрещиваний с осо-
бенностями кариотипа дрозофил. В диплоидном ядре этих мух содер-
жится четыре пары хромосом, причем три пары у самцов и самок
между собой не отличаются по содержащимся в них генам, а четвер-
тая пара одинаковая только у самок — у нее имеются две Х-хромосо-
мы (генотип самок — XX), у самцов же Х-хромосома только одна,
а другая — не похожая на нее Y-хромосома (генотип самцов — XY,
генетики обозначают половые хромосомы такими значками: X — —,
Y — —). Хромосомы, отличающиеся у самцов и самок, получили на-
звание половые хромосомы. Морган предположил, что ген, определяю-
щий окраску глаз, у дрозофил расположен именно в Х-хромосоме.
Поскольку у самцов такая хромосома лишь одна, у них не происходит
взаимодействия аллелей, а реализуется любой аллель, имеющийся в
доставшейся единственной Х-хромосоме. У самок таких хромосом
две, соответственно, они получают не один (как самцы), а два аллеля,
поэтому реализация заложенных в них генов осуществляется соглас-
но принципу доминирования. Ген, контролирующий цвет глаз, нахо-
дится в Х-хромосоме, т. е. он сцеплен с полом, поэтому у гетерозигот-
ных самок оказывается красная окраска (поскольку она доминирует).
Самцы же всегда имеют только один аллель в своей единственной
Х-хромосоме. Поскольку другая половая хромосома — Y-хромосома —
не содержит такого гена, самцы всегда оказываются в «гомозиготном»
состоянии. Однако употреблять здесь понятие «гомозиготность»
нельзя, поскольку оно относится к диплоидной стадии и обозначает
парное состояние. Поэтому явление, при котором диплоидный орга-
низм обладает только одним аллелем какого-либо признака, получи-
ло обозначение гемизиготное состояние.
Наследование признаков, ограниченных полом и зависимых от
пола. Развитие многих признаков связано с половой принадлежно-
стью. Если локусы генов расположены в половых хромосомах, то их
наследование будет сцеплено с полом (окраска глаз у дрозофил, гены
дальтонизма и гемофилии у человека и др.). Кроме этого, существу,
ют признаки, гены которых находятся не в половых хромосомах, а в
аутосомах, развитие которых у представителей разного пола неодина-
ково. Если такие признаки проявляются только у представителей од-
ного пола, то они будут называться ограниченными полом. Например,
681
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ
быки имеют гены молочной продуктивности и передают их по на-
следству своим дочерям, но у самих быков эти гены не проявляются.
В других случаях признаки проявляются у обоих полов, однако у од-
ного из них они выражены интенсивнее. Например, и бараны и овцы
обладают рогами, но у баранов они развиты лучше. Признаки, кото-
рые по-разному проявляются у представителей мужского и женского
пола, называют зависимыми от пола.
Определение пола. Почти всем эукариотам свойственно половое
размножение. У гермафродитов одновременно присутствуют муж-
ские и женские гонады. Такие организмы не нуждаются в половых
партнерах и размножаются самостоятельно, например большинство
плоских червей. Однако при этом потомство получает генетический
материал лишь одного родителя, и такой половой процесс мало чем
отличается по значимости от бесполого размножения. Гораздо более
прогрессивным является полное разделение полов, при котором одни
организмы — самки — специализируются на производстве яйцекле-
ток, а другие — самцы — на производстве сперматозоидов. Половая
принадлежность организма может определяться на разных этапах его
развития, в связи с чем выделяют три типа: прогамное, сингамное
и эпигамное.
Прогамное определение пола осуществляется до оплодотворе-
ния в процессе оогенеза. Так происходит, в частности, у коловраток.
При этом образуются яйцеклетки разных размеров — крупные и мел-
кие. После оплодотворения из крупных яиц развиваются самки, а из
мелких — самцы.
Сингамное определение пола, происходящее при оплодотворе-
нии, определяется половыми хромосомами. Этот тип является наибо-
лее распространенным. Существует несколько вариантов хромосом-
ного определения пола. У многих организмов при мейозе мужские
особи продуцируют гаметы двух типов: половина из них будет содер-
жать Х-хромосому, другая — Y-хромосому, тогда как все гаметы жен-
ских особей будут одного типа — каждая из них содержит только
Х-хромосому. По этой причине у человека (и других организмов с по-
добным разделением половых хромосом) мужской пол является гете-
рогаметным, а женский — гомогаметным. Существуют примеры, ког-
да гетерогаметным оказывается не мужской, а женский пол. Так дело
обстоит у птиц и бабочек. Для того чтобы выделить женскую гетеро-
гаметность, половые хромосомы в этом случае обозначают другими
буквами — Z и W. Соответственно, самцы будут иметь генотип ZZ,
а самки — ZW.
Различие между половыми хромосомами далеко не всегда служит
причиной хромосомного определения пола. Половую принадлеж-
ность может детерминировать наличие или отсутствие одной хромо-
682
НАСЛЕДСТВЕННОСТЬ
сомы. Так наследуется пол у некоторых беспозвоночных (представи-
телей жесткокрылых, полужесткокрылых и прямокрылых насекомых,
пауков, многоножек и круглых червей). Впервые это было обнаруже-
но у клопа протенор. Соматические клетки самок этого насекомого
содержат четное количество хромосом — 14, из которых 12 являются
аутосомами, а 2 названы Х-хромосомами. У самцов также присутству-
ют 12 аутосом, но Х-хромосома всего одна, поэтому общее количество
хромосом становится нечетным — 13. Соответственно, во всех яйце-
клетках будет находиться по 7 хромосом (одна из которых Х-хромо-
сома), тогда как одна половина сперматозоидов также будет содер-
жать 7 хромосом (с Х-хромосомой), а другая — только 6 хромосом
(только аутосомы без Х-хромосомы). Детерминация женского пола
происходит при слиянии гамет, имеющих по 7 хромосом, две из кото-
рых Х-хромосомы, что соответствует генотипу самки. Мужские особи
развиваются при слиянии с яйцеклеткой «облегченного» сперматозо-
ида, не имеющего Х-хромосомы.
Пол также может определять плоидность (число хромосом в набо-
ре) не только половых хромосом, но и всех хромосом вообще. Такой
способ известен для перепончатокрылых насекомых. Например, мат-
ка медоносной пчелы по своему усмотрению (оценив размеры сото-
вой ячейки) откладывает оплодотворенные и неоплодотворенные
яйца. Из оплодотворенных яиц в дальнейшем развиваются матки или
рабочие пчелы (в зависимости от ухода и кормления личинок), имею-
щие диплоидный набор хромосом во всех клетках. Из неоплодотво-
ренных, а потому гаплоидных, яиц развиваются самцы — трутни.
Такой механизм делает ненужным присутствие в кариотипе специфи-
ческих половых хромосом. Развитие организма из оплодотворенной
яйцеклетки называется партеногенезом.
Эпигамное (метагамное) определение пола зависит не от
присутствия, соотношения или плоидности хромосом, а от интен-
сивности действия факторов окружающей среды, что может расце-
ниваться как модификационная изменчивость. Примером тому яв-
ляется детерминация пола у крокодилов. Из отложенных самкой
яиц в зависимости от температуры окружающей среды могут вылу-
пляться или юные самцы, или самки. Весьма оригинально определя-
ется пол у морского червя бонеллии (тип кольчатые черви). Если
его личинка развивается в одиночестве, то из нее вырастает подвиж-
ная самка, ведущая самостоятельный образ жизни. Однако если ли-
чинка оказывается поблизости от взрослой самки, то она оседает на
ее хоботок и под влиянием выделяемого самкой гормона развивает-
ся в микроскопического самца. Затем этот самец проникает в поло-
вые пути самки и живет там как паразит, в связи с чем его строение
значительно упрощено.
683
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ
У некоторых животных в течение жизни возможно изменение
пола. Так происходит у коралловых рыб лаброидов. Обычно они жи-
вут небольшими группами, состоящими из самца и гарема самок.
В случае гибели самца одна из самок начинает проявлять поведенче-
скую активность самца и через некоторое время трансформируется
в него морфологически.
У ряда видов половое размножение осуществляется путем парте-
ногенеза, в результате чего рождаются исключительно самки. Так раз-
множаются тли, а из позвоночных — скальная ящерица.
Определение пола у растений осуществляется под контролем по-
ловых хромосом или аутосом. Как правило, гетерогаметным является
мужской пол, имеющий генотип XY, тогда как генотип женских осо-
бей XX. Однако, например, у земляники и некоторых других цветко-
вых гетерогаметен женский пол. Кроме генетических детерминант, на
определение пола у многих растений активно влияют факторы внеш-
ней среды. Например, низкие температуры, высокое содержание в по-
чве доступных соединений азота, высокая влажность почвы и атмо-
сферного воздуха, освещение растений коротковолновым светом,
обработка растений этиленом или окисью углерода (СО), а также фи-
тогормоном ауксином усиливают женскую сексуализацию многих ра-
стений (прежде всего двудомных). Напротив, высокие температуры,
подкормка калийными удобрениями, обработка фитогормоном гиббе-
реллином, освещение длинноволновым светом стимулируют маску-
линизацию растений.
ИЗМЕНЧИВОСТЬ
На Земле нет двух совершенно одинаковых жизненных форм —
между ними всегда имеются определенные качественные или количест-
венные различия. Наибольшее сходство между собой проявляют особи
одного вида, однако и в пределах одного вида все организмы имеют вы-
раженные индивидуальные особенности (варианты нормы). Даже особи
с одинаковыми генотипами, появившиеся в результате вегетативного
размножения, различаются между собой. И клонирование не дает абсо-
лютного сходства, поскольку на образовавшиеся организмы по-разному
действуют факторы окружающей среды, вызывая формирование инди-
видуального фенотипа особи в соответствии с нормой реакции.
Изменчивость является обязательным и необходимым условием
индивидуального развития особи. Действительно, в течение онтогене-
за любого многоклеточного организма непрерывно происходит изме-
нение его внутреннего и внешнего строения. Особенно серьезные пре-
образования происходят в эмбриональном периоде. Однако и в пост-
684
ИЗМЕНЧИВОСТЬ
эмбриональном периоде организм постоянно изменяется, вплоть до
наступления смерти. Изменения происходят даже в течение неболь-
шого периода жизни. Например, обильная пища или голодание при-
водит к изменению массы тела. Различные биологические ритмы
вызывают неодинаковое состояние организма в зависимости от вре-
мени года или суток.
Материальные основы изменчивости целесообразно разделить на
несколько уровней. Ранее мы рассматривали и обосновывали законо-
мерности наследования признаков, при этом постоянно указывая на
гетерогенность самих признаков. Действительно, подавляющее боль-
шинство признаков имеет многочисленные альтернативные проявле-
ния, которые определяются количеством аллелей и способами взаи-
модействия между генами. Кроме того, в случае плейотропии один
ген может оказывать воздействие на проявление нескольких призна-
ков. В зависимости от проявления различают несколько типов измен-
чивости. Если признаки изменились у одной особи, изменчивость на-
зывают индивидуальной, если сразу у группы — групповой. Появление
изменений под воздействием действующих на определенной террито-
рии внешних факторов называется географической изменчивостью.
При этом изменение признака у популяций, расположенных поблизо-
сти, обычно менее резкое, нежели у популяций, значительно удален-
ных друг от друга географически. Такое постепенное изменение выра-
жения признака называется клинальной изменчивостью.
Радикальное преобразование какого-либо признака называется
качественной изменчивостью, а пропорциональное (признак изменя-
ется лишь в определенной степени) — количественной. Если имеет
место воздействие определенных специфических факторов внешней
среды на генетический материал, инициирующее изменение призна-
ков, то изменчивость называется направленной. Соответственно, спон-
танные изменения представляют ненаправленную изменчивость. Он-
тогенетическая изменчивость представляет собой преобразования
организма в процессе индивидуального развития. Однако, несмотря
на столь разнообразные формы проявления, не все измененные при-
знаки передаются по наследству. В соответствии с особенностями на-
следования изменений принято выделять два типа изменчивости: не-
наследственную и наследственную.
НЕНАСЛЕДСТВЕННАЯ (ФЕНОТИПИЧЕСКАЯ,
ИЛИ МОДИФИКАЦИОННАЯ) ИЗМЕНЧИВОСТЬ
В зависимости от условий обитания на организм действуют раз-
личные факторы. Кроме того, интенсивность действия одних и тех же
685
НЕНАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ
факторов может быть различной (температура, влажность, освещен-
ность, количество и качество пищи и др.). Каждый вид занимает опре-
деленную экологическую нишу в соответствии с особенностями своей
организации и по возможности не покидает ее, однако невозможно
создать абсолютно одинаковые условия даже для двух отдельных ор-
ганизмов. Имеющийся у любого организма генетический материал,
несмотря на свою уникальность, не возник внезапно — он длительно
формировался у предыдущих предковых форм в процессе постоянной
борьбы за существование и отбора (выживания) носителей наиболее
адаптивных сочетаний генов. Поэтому на уровне генома у каждого ор-
ганизма изначально заложена возможность изменяться в течение
жизни, приспосабливаясь к конкретным условиям обитания. Эти из-
менения не обязательно проявятся у потомства, если условия будут
несколько иными. Их называют ненаследственными, или модифика-
циями. Ч . Дарвин назвал такую изменчивость определенной ввиду
предсказуемости многих преобразований, или групповой, из-за сход-
ного направления изменения рассматриваемого признака у всей ис-
следуемой группы. Например, можно с уверенностью предположить
появление загара у впервые появившихся на пляже курортников, если
до этого их кожа долгое время не подвергалась воздействию прямых
лучей солнца. У них же могут возникнуть ожоги в результате слиш-
ком продолжительного воздействия этих лучей в первый день отдыха.
Наряду с групповым характером проявления, модификации также
могут быть индивидуальны. Загар появляется у всех, и все же он не-
одинаковый — это определяется внутренними свойствами организма.
Также тщетно отыскать два яблока абсолютно одинаковой формы,
размера и веса в магазине, несмотря на то, что все плоды будут иметь
характерные для данного сорта особенности.
А. Вейсман (1913) отреза
л хвосты у белых мышей, после чего
скрещивал бесхвостых животных. Если следовать логике наследова-
ния приобретенных признаков, то удаление хвоста у родителей до
размножения неминуемо должно отразиться на длине хвостов потом-
ства. Однако А. Вейсман, проведя тщательный статистический ана-
лиз, пришел к выводу, что длина хвоста у потомства вследствие при-
жизненного удаления его у родителей (причем до размножения) не
уменьшается. Многие жители экзотических стран имеют свое пред-
ставление о красоте, часто не понимаемое другими. Для этого они
всячески изменяют свою внешность. Например, девочкам обувают
ногу в деревянную колодку, дабы она оставалась маленькой, или с
детства надевают на шею кольца, чтобы постепенно удлинить ее, под-
пиливают зубы или отрезают клитор, — вариантов очень много! Не
остаются в стороне и мужчины: они могут удлинять губы, вставляя
в преддверие ротовой полости деревянную пластинку или протыкать
686
ИЗМЕНЧИВОСТЬ
носы. Во многих странах молодым людям наносят шрамы в виде ри-
туального узора. Однако дети у таких людей неизменно рождаются
обычными — с нормальными стопами, губами, носами и нетронутой
кожей. Следовательно, ни одно из таких приобретенных в ходе жизни
«украшений» невозможно передать по наследству. Каждому из
них для этого приходится прибегать к механическим вмешательствам
извне.
Норма реакции. Для нормального существования организм ну-
ждается в определенном спектре факторов окружающей среды, со-
вокупность которых составляет специфическую для каждого вида
экологическую нишу. Однако действующие на организм внешние
факторы постоянно изменяют свою интенсивность. Меняются как
абиотические (освещенность, температура, влажность и др.), так
и биотические (состав и количество пищи, различные типы взаимо-
отношений и др.) факторы. Если бы организм получал жестко очер-
ченную установку на неизменность всех своих признаков, он был бы
не способен приспосабливаться и, соответственно, выживать. Но,
к счастью, на самом деле наследуется способность пластично фор-
мировать признак сообразно действующим условиям. Например,
при изменении температуры окружающей среды изменяется актив-
ность биохимических реакций, листья поворачиваются вслед за дви-
жением солнца по небосводу, добиваясь тем самым оптимального
освещения, и т. д.
Масса тела человека может широко варьировать и составлять от
нескольких десятков до нескольких сотен килограммов, также чело-
век может жить при разной температуре воздуха. Однако способность
изменяться не абсолютна и для каждого фактора имеется максималь-
ная и минимальная интенсивность действия, за пределами которых
происходят необратимые изменения в организме, влекущие за собой
смерть от истощения или ожирения в первом случае и от переохла-
ждения или перегрева — во втором.
Диапазон возможных фенотипических изменений признака (мо-
дификаций) называется нормой реакции. Причем для разных призна-
ков ее размах может быть не только широким, но и узким, например,
цвет радужной оболочки глаза или определение пола у крокодилов
в зависимости от температуры: пол может быть или мужским, или
женским, но никаким другим. Для каждого организма границы нормы
реакции очерчены генотипом, преодолеть которые невозможно. Учи-
тывая это, легко понять, почему люди обладают разными возможно-
стями или почему спортсмены при одинаковых тренировках и сход-
ном питании демонстрируют непохожие результаты. Однако
непосредственное проявление признака зависит от среды. Так, спор-
тсмены физически не смогут постоянно показывать наивысшие ре-
687
НЕНАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ
зультаты. Пика формы они достигают только к соревнованиям, к ко-
торым они себя целенаправленно готовят, строго соблюдая режим,
тем самым добиваясь нужных изменений состояния организма.
Окраска шерсти у гималайского кролика зависит от температуры
воздуха. Так, при 30 °C крольчата вырастают полностью белыми,
а если температуру снизить до 20 °С, у кроликов появляется пятна
вокруг носа, а также чернеют лапы, уши и хвост. Добиться роста чер-
ной шерсти в нужном месте можно искусственно, если выбрить ее на
интересующем участке кожи и охлаждать эту область, например,
льдом. Также в зависимости от температуры изменяется окраска ле-
пестков на цветках примулы — с белой при температуре 30—35 °С на
красную при 15—20 °С. У водного растения стрелолиста в воде, на ее
поверхности и в воздухе образуются листья различной формы.
Механизм проявления признака можно рассматривать как взаимо-
действие генотипа с окружающей средой.
Интенсивность одного и того же признака у разных особей прояв-
ляется неодинаково интенсивно. Например, ультрафиолетовые лучи
вызывают потемнение кожи, однако степень пигментации может ши-
роко варьировать от едва заметной до почти черной. Поэтому для
оценки степени выраженности изучаемого признака генетики исполь-
зуют понятие экспрессивность. Этот показатель зависит от генотипа
(пигментация кожи интенсивнее проявляется у обладателей больше-
го числа доминантных аллелей, контролирующих этот признак). На-
ряду с генотипом на экспрессивность также влияет среда: загар на-
прямую зависит от полученного количества ультрафиолетовых лучей.
Поэтому фенотипическое проявление признака определяется взаимо-
действием генотипа со средой.
Обладание определенным геном не всегда означает, что он про-
явится в фенотипе. Так, у дрозофил мутировавший ген Lobe (L) вы-
зывает уменьшение размера глаз, причем с варьирующей экспрессив-
ностью (от почти нормальных глаз до полного их отсутствия). Но
этот ген проявляется не у всех носителей его, а только у 75%. Следо-
вательно, у 25% мух этот ген не проявится вовсе. Относительная доля
особей, у которых проявился исследуемый признак, называется пене-
трантностью. В разных случаях пенетрантность имеет неодинаковое
значение, например, у врожденного вывиха бедра она составляет лишь
20%, у сахарного диабета — 65%, а у аллелей гена, определяющего
группу крови, пенетрантность полная. Оба термина — «экспрессив-
ность» и «пенетрантность» — ввел в науку отечественный генетик
Н.В. Тимофеев-Ресовский в 1927 г.
Типы модификаций. Большинство из полученных в течение жиз-
ни преобразований являются адаптивными, потому что, изменяясь,
организм подстраивается под окружающую среду. В частности, жи-
688
ИЗМЕНЧИВОСТЬ
вотные, линяя, оптимально изменяют густоту шерсти и ее окраску.
Пигментация кожи защищает ее от ультрафиолетовых лучей. Оду-
ванчики, произрастающие в долине, обычно имеют высокий цвето-
нос, поскольку температура ночью уменьшается значительно. По
мере увеличения высоты над уровнем моря цветонос становится ко-
роче; у горных форм его длина минимальная. Это также является
адаптацией, поскольку в горах ночью холодно и плотно окруженный
листьями цветок значительно лучше защищен на коротком цвето-
носе.
Вместе с тем возможно также появление неадаптивных модифика-
ций. Обычно они возникают, если организм оказался в необычных
для его вида условиях. Так, у водяной гречихи развитие во влажном
воздухе приводит к появлению листьев, плавающих на воде. Неадап-
тивные модификации называют морфозами. Нередко они фенотипи-
чески похожи на известные для этого вида мутации и называются фе-
нокопиями таких мутаций. Известно немало фенокопий у дрозофил,
например воздействие на мух соединениями бора приводит к отсутст-
вию глаз, серебра — к появлению желтой окраски тела, а ртути —
к появлению тонких щетинок. Возможен и обратный эффект, когда
внешнее воздействие приводит к проявлению нормального фенотипа
у носителей мутировавшего гена. Например, увеличение длины кры-
льев у дрозофил с мутацией зачаточных крыльев при воздействии на
них высокой температурой. Все эти изменения проявляются только
при воздействии определенного фактора (физического, химического
или биологического). Если это воздействие прекращается, то фено-
тип возвращается к своему нормальному состоянию. Фенокопии со-
храняются в течение всей жизни только тогда, когда преобразующий
внешний фактор действует в период эмбрионального развития изме-
няемого органа. Но и тогда измененный признак не передается по на-
следству.
Иногда особенности, приобретенные в ходе онтогенеза под воздей-
ствием внешней среды, проявляются у потомства. Так, воздействие на
куколок самок колорадского жука высокой или, наоборот, низкой
температурой приводит к изменению окраски взрослых насекомых.
Этот признак проявляется у потомства в течение нескольких поколе-
ний, после чего возвращается к своему обычному фенотипу. Это при-
мер длительных модификаций.
Модификационная изменчивость имеет большое значение и для
хозяйственной деятельности человека. Зная особенности реагирова-
ния домашних животных и культурных растений на внешние факто-
ры, можно направленно изменять условия содержания или возделы-
вания, чтобы получить максимально высокий эффект.
689
КОМБИНАТИВНАЯ ИЗМЕНЧИВОСТЬ
НАСЛЕДСТВЕННАЯ (ГЕНОТИПИЧЕСКАЯ)
ИЗМЕНЧИВОСТЬ
Сюда относят те изменения признаков, которые передаются по на-
следству и впоследствии проявляются у потомства. Ч . Дарвин назвал
такой тип изменчивости неопределенной, или индивидуальной, т. к. не-
возможно определить, какие появятся изменения; кроме того, они
всегда индивидуальны. Различают два типа наследственной изменчи-
вости: комбинативную и мутационную.
КОМБИНАТИВНАЯ ИЗМЕНЧИВОСТЬ
Этот тип изменчивости возникает при сочетании имеющихся ге-
нов и их аллелей в процессе осуществления различных этапов поло-
вого размножения. При этом не происходит химических преобразо-
ваний молекул ДНК. Поэтому комбинативная изменчивость не
приводит к появлению новых генов или их аллелей — у потомков
проявляются признаки родителей и их предков, но в разных сочета-
ниях. Наименьшей единицей рекомбинаций генетического материала,
вызывающей появление новых сочетаний, является рекон, который
соответствует паре нуклеотидов двухцепочечной молекулы ДНК или
одному нуклеотиду в одноцепочечных молекулах нуклеиновых кис-
лот вирусов. Рекон нельзя разделить в процессе кроссинговера, он
всегда передается целиком. Комбинация наследственного материала
у эукариот достигается тремя способами: 1. Рекомбинацией генов в
процессе кроссинговера во время профазы первого деления мейоза,
когда гомологичные хромосомы обмениваются участками, в результа-
те появляются хромосомы с новыми сочетаниями аллелей (если в го-
мологичных хромосомах содержатся различные аллели гена — напри-
мер, зеленая или желтая окраска семян; если же аллели одинаковые,
то, несмотря на обмен участками, выражение признака не изменится).
2. Независимым расхождением хромосом в анафазе первого деления
мейоза, когда материнские и отцовские хромосомы расходятся в до-
черние клетки в случайном порядке, что приводит к разнообразным
комбинациям этих хромосом, и в результате все образовавшиеся при
мейозе гаметы имеют между собой генетические различия. 3 . Случай-
ным характером встреч гамет при оплодотворении.
У прокариот отсутствует половое размножение. Однако при опре-
деленных условиях и у них также имеет место рекомбинация наслед-
ственной информации, причем как ДНК нуклеоида, так и цитоплаз-
матической ДНК — плазмид.
Таким образом, комбинативная изменчивость приводит к тому,
что каждая зигота имеет уникальный набор наследственной инфор-
мации. Этим можно объяснить различия между потомками одних ро-
690
ИЗМЕНЧИВОСТЬ
дителей. Рекомбинация генетического материала имеет чрезвычайно
важное значение в эволюционном процессе, поскольку она создает
неисчерпаемое разнообразие генотипов, что делает популяцию гете-
рогенной. Появление неодинаковых, а следовательно, неравноценных
организмов одного вида открывает широкие возможности для естест-
венного отбора оставлять лишь наиболее удачные сочетания наслед-
ственных признаков. Поскольку новые организмы со временем также
включаются в половое размножение, процесс совершенствования ге-
нетического состава идет непрерывно.
МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ
В отличие от комбинативной изменчивости, обязательным услови-
ем мутационной изменчивости является качественное изменение на-
следственного субстрата. В результате происходит образование новых
аллелей или, напротив, утрата уже имеющихся. Это приводит к появ-
лению у потомков принципиально новых признаков, отсутствующих у
родителей. Основные положения теории мутаций изложил Г. де Фриз
(1901—1903). Он ввел термин мутация для обозначения скачкообраз-
ного, прерывного изменения наследственного признака. Основные по-
ложения его теории выглядят следующим образом: 1. Мутации возни-
кают внезапно, без промежуточных стадий, как скачкообразное
изменение признака. 2. Появившиеся новые формы проявляют устой-
чивость и передаются по наследству. 3 . Мутации отличаются от нена-
следственных изменений тем, что не образуют непрерывных рядов и не
группируются вокруг определенного «среднего типа»; мутации — это
качественные изменения. 4 . Мутации очень разнообразны, среди них
есть как полезные для организма и вида, так и вредные. 5 . Возможность
обнаружения мутаций зависит от числа проанализированных особей. 6 .
Одинаковые мутации могут возникать неоднократно.
Мутационной изменчивости подвержены все клеточные организ-
мы и вирусы. В настоящее время термином «мутация» принято обо-
значать любые изменения наследственного материала, передавае-
мые по наследству. Признак, который имелся до изменения,
называется диким, а измененный — мутантным.
Классификация мутаций. Единой классификации мутаций не
существует, и разделение наследственных изменений на группы
осуществляется по многим показателям. По характеру изменения
наследственного материала мутации подразделяют на генные, хромо-
сомные и геномные. В зависимости от направления мутации бывают
прямыми (из дикого типа возникает мутантный) и обратными (ревер-
сии), когда мутация приводит к ранее существовавшему дикому типу.
При этом важно помнить, что строго дикого типа в природе вообще не
691
МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ
бывает, поскольку любой ныне дикий аллель когда-то так же появил-
ся в результате мутации аллеля более раннего дикого типа. По прояв-
лению в гетерозиготном состоянии генотипа мутации могут быть
доминантными (если мутантный аллель всегда проявляется) и рецес-
сивными (если мутантный аллель проявляется только в гомозиготном
состоянии; большинство мутаций, сохранившихся в ходе естествен-
ного отбора, являются рецессивными). В зависимости от участия
внешних факторов различают спонтанные мутации, которые случай-
но возникают в природных условиях, и индуцированные мутации, воз-
никшие на фоне направленного воздействия каких-либо факторов на
генетический материал. По степени влияния на жизнедеятельность
организма мутации делят на полезные, нейтральные и вредные (край-
ним выражением такого рода мутаций являются летальные мутации).
Такие характеристики мутаций являются универсальными, т. е . они
применимы ко всем организмам.
Для отдельных форм также применимы более конкретные класси-
фикации. В зависимости от локализации в клетке мутации могут быть
ядерными и цитоплазматическими (изменения ДНК митохондрий и
пластид у эукариот, а у прокариот — плазмид). В зависимости от типа
клеток: мутации, происходящие в соматических клетках, называют
соматическими (этот тип мутаций не передается потомству при поло-
вом размножении), происходящие в половых клетках — генеративны-
ми. По фенотипическому проявлению мутации подразделяют на мор-
фологические, физиологические, биохимические, поведенческие и др.
Существует немало более частных классификаций мутаций, основан-
ных на каком-либо узком показателе.
Генные (точечные) мутации, или трансгенации, представ-
ляют собой неопределяемые цитологическими методами химические
изменения нуклеиновой кислоты в пределах отдельных генов. Эти из-
менения могут выражаться в нарушении пар нуклеотидов и сдвиге
рамки считывания. В результате при транскрипции появляется изме-
ненная тРНК и, соответственно, полипептид с иной последовательно-
стью аминокислот при трансляции и, следовательно, иной структурой
и свойствами. Генные мутации часто являются причиной наследст-
венных болезней, связанных с изменением обмена веществ. Наимень-
шая часть молекулы нуклеиновой кислоты, изменение которой при-
водит к появлению нового признака (или преобразованию уже
существующего), называется мутоном. Мутон соответствует паре
нуклеотидов в двухцепочечной молекуле ДНК или одному нуклеоти-
ду в одноцепочечной молекуле нуклеиновой кислоты у вирусов. Со-
ответственно этому генные мутации, охватывающие один сайт генно-
го локуса, называют односайтовыми, несколько — многосайтовыми.
В результате генных мутаций происходит изменение аллелей генов
692
ИЗМЕНЧИВОСТЬ
и их количество в генофонде популяции (и вида в целом) возрастает.
Это приводит к множественному аллелизму. Это понятие относится
исключительно к генофонду, поскольку отдельная особь не может
содержать более двух разных аллелей отдельного гена даже в гетеро-
зиготном состоянии. Примером тому служит серия аллелей, опреде-
ляющих окраску глаз у дрозофилы: красная (дикий тип), белая, виш-
невая, абрикосовая, эозиновая, цвета слоновой кости и т. д . (всего
более десяти). Генные мутации являются самыми распространенны-
ми — у растений и животных имеется до 10% половых клеток. Одна-
ко вероятность мутации отдельного гена очень мала — в среднем
10–5
— 10–7
.
Генные мутации выражаются двумя способами: при изменении
пар нуклеотидов и при сдвиге рамки считывания. Изменение нукле-
отидной последовательности может быть в виде транзиции или тран-
сверсии. В случае транзиции одно пуриновое основание в паре ну-
клеотидов заменяется на другое пуриновое, а пиримидиновое,
соответственно, на другое пиримидиновое. Например:
А (пурин) Т (пиримидин) ␚ G (пурин) С (пиримидин),
GC→AT,ТА→CGиCG→AT.
При трансверсии, напротив, пуриновое основание замещается пи-
римидиновым, а пиримидиновое, соответственно, пуриновым:
А (пурин) Т (пиримидин) ␚ С (пиримидин) G (пурин),
AT→ТА,GC →CG,CG →AT.
Мутации, сопровождаемые изменением или заменой оснований в
нуклеиновой кислоте, составляют примерно 20% общего числа ген-
ных мутаций. Значительно чаще происходят мутации, обусловленные
сдвигом рамки считывания. Их механизм состоит в выпадении имею-
щихся или во вставке лишних пар нуклеотидов.
Хромосомные мутации (перестройки, или аберрации)
представляют собой внутрихромосомные изменения или межхромо-
сомные обмены. Хромосомные перестройки встречаются только у эу-
кариот, поскольку лишь они обладают дискретными линейными
хромосомами. Они могут возникать как спонтанно, так и под воздей-
ствием мутагенов. В ходе любых хромосомных перестроек сначала
происходит разрыв хромосомы, а затем осуществляется соединение
фрагментов. Причем выделенные фрагменты либо удаляются и утра-
чиваются, либо встраиваются в ту же или в другую хромосому.
Внутрихромосомные перестройки представляют собой различно-
го типа структурные изменения в пределах одной хромосомы. По-
теря участка хромосомы называется делецией (или нехваткой) —
ABCDEFGH ␚ ABCEFGH (выделен утраченный ген). Впервые деле-
693
МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ
цию одной из хромосом у дрозофилы обнаружил К. Бриджес (1917),
это было первым открытием явления хромосомных мутаций. В ре-
зультате этой делеции у мух появляются вырезки края крыла. Не-
хватка концевого участка хромосомы называется дефишенси —
ABCDEFGH ␚ ABCDEFG (выделен утраченный ген). Нехватки
хромосом могут быть большими и малыми. Большие обычно деталь-
ны в гомозиготном (или гемизиготном) состоянии, если отсутствует
участок единственной Х-хромосомы у мужчин). Причина этого состо-
ит в том, что при делеции утрачивается слишком большое количество
генов, определяющих развитие органов и реализацию жизненно важ-
ных функций организма. Жизнеспособность гетерозигот объясняется
тем, что имеется возможность проявления генов, локализованных в
неповрежденной гомологичной хромосоме. У человека делеция в ко-
ротком плече пятой хромосомы в гетерозиготном состоянии служит
причиной болезни «кошачьего крика». Это заболевание сопровожда-
ется характерным «мяукающим» криком младенцев, а также малым
размером головы (микроцефалия) и умственной отсталостью. Боль-
ные дети очень редко доживают до 10—12 лет. Известны и другие тя-
желые заболевания, которые развиваются у человека при делеции
хромосом (также в гетерозиготном состоянии).
Возникновение делеций связано с разрывами в хромосоме. Если
это происходит не на конце, а в средней части хромосомы, то разо-
рванные участки соединяются и хромосома укорачивается. Выпаде-
ние одновременно обоих концевых участков приводит к образованию
кольцевой молекулы, поскольку утрачиваются теломеры, препятству-
ющие слипанию концов хромосомы, как это имеет место у прокариот.
Утраченные фрагменты хромосом разрушаются при делении клетки,
т. к . они не обладают центромерами.
Дупликации (или повторы) представляют собой повторение како-
го-либо участка хромосомы. Повторяющиеся участки образуют тан-
демы, которые могут быть прямыми, если последовательность генов
в них идет в одном направлении — ABCBCDEFGH, или обращенны-
ми (инвертированными), если последовательность генов в повторяю-
щихся участках противоположна — ABCCBDEFGH. Дупликация,
расположенная на конце хромосомы, называется концевой —
ABABCDEFGH. Многократные повторы (более двух раз) называют
мультипликациями (амплификациями) — ABCDCDCDEFGH (во
всех примерах выделены повторяющиеся участки). Примером фено-
типического проявления дупликации может служить уменьшение
размеров глаз у дрозофилы (в связи с уменьшением числа омматиди-
ев в сложных фасеточных глазах насекомого).
694
ИЗМЕНЧИВОСТЬ
Основной причиной появления дупликаций (и делеций тоже!) яв-
ляется разрыв хромосом. К этому могут привести разные факторы:
ионизирующая радиация, химические мутагены, вирусы и др. Однако
основной причиной разрывов и последующих мутаций генетики счи-
тают неравный кроссинговер. Он возникает при неправильной конъ-
югации гомологичных хромосом в фазе зиготены профазы первого
деления мейоза. Дупликации значительно менее опасны для жизнеде-
ятельности организма, нежели делеций, поскольку здесь не происхо-
дит утраты генетической информация. Повторы генов могут даже
быть полезны. Например, парные гены определяют синтез полипеп-
тидов гемоглобина человека.
Инверсии представляют собой внутрихромосомные перестройки,
в процессе которых происходит поворот участка хромосомы на
180° — ABCDEFGH ␚ ABCFEDGH (выделен инвертированный уча-
сток). В зависимости от того, захватывается ли область центромеры,
инверсии подразделяют на парацентрические (если центромера не
включается в оборачиваемый участок хромосомы) и церицентриче-
ские (если центромера также разворачивается). Такие перестройки,
как правило, происходят в средней части хромосом и не захватывают
область теломеры. Это связано с тем, что теломера препятствует сли-
панию хромосом, а срединные участки, разорвавшись, демонстрируют
высокую способность слипаться концами. Часто инверсии приводят
к летальному исходу в рецессивном состоянии. Кроме того, эти мута-
ции подавляют кроссинговер у гетерозигот (точнее, образовавшиеся
в результате кроссинговера гетерозигот рекомбинации делают орга-
низм нежизнеспособным), что может указывать на наличие инверсий.
У гомозигот инверсии не препятствуют кроссинговеру.
В хромосоме может быть не одна, а две инверсии, перекрывающи-
еся полностью, частично или не перекрывающиеся вовсе. В случае
одиночного кроссинговера при парацентрической инверсии образует-
ся одна хроматида с двумя центромерами и одна без центромеры. Обе
структуры утратятся в ходе анафазы, поэтому в результате мейоза
образуются не четыре гаметы, а две гаметы. При перицентрической
инверсии (также у гетерозигот) перенос центромер не происходит и
расхождение хромосом в анафазе не нарушается. Однако у части хро-
мосом происходит делеций, что также уменьшает количество гамет
с полным набором генов.
Межхромосомные перестройки представляют собой одно- или
двустороннее перемещение участков с одной хромосомы на другую,
а также взаимодействие негомологичных хромосом. Взаимный (или
реципрокный) обмен фрагментами между негомологичными хромо-
сомами называется транслокациями. Это явление было обнаружено
Дж. Беллингом (1915) в опытах на бархатных бобах. Позднее
695
МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ
К. Штерн (1926) выявил перенос фрагмента Y-хромосомы на Х-хро-
мосому у дрозофил. Схематично обмен генов при транслокации вы-
глядит следующим образом: если одна хромосома имеет последова-
тельность генов ABCDEFGH, а другая MNOPQR, то после
реципрокного обмена последовательность генов в хромосомах будет
MNOCDEFGH и ABPQR (выделены перемещенные гены). Приве-
денный случай демонстрирует обмен фрагментами одинаковой дли-
ны, однако так же может происходить перенос неравных по длине
участков. При некоторых транслокациях возможен перенос фрагмен-
та, содержащий центромеру; тогда появляется хромосома с двумя
центромерами, а также фрагменты без центромер, которые впоследст-
вии утратятся в процессе деления клетки.
Следствием транслокаций является изменение групп сцепления
генов, т. е. гены, ранее находившиеся в разных хромосомах и, соот-
ветственно в разных группах сцепления, при транслокациях оказы-
ваются сцепленными. Это приводит к тому, что гены негомологич-
ных хромосом наследуются сцепленно. При этом жизнеспособными
оказываются лишь те гаметы животных и споры растений, которые
несут родительские сочетания хромосом. Нарушение групп сцепле-
ния делает возможным обнаружение этих мутаций генетическими
методами. Участки гомологичных хромосом, несущие аллели одина-
ковых генов, в биваленте (в профазе первого деления мейоза) распо-
лагаются один напротив другого — именно они и притягиваются
друг к другу. Если произошла транслокация, то конъюгация хромо-
сом происходит иначе, поскольку в результате взаимного обмена
участками между негомологичными хромосомами у гетерозигот по
транслокации притягиваются не только две гомологичные хромосо-
мы, но также две другие, с одной из которых произошел обмен. Это
происходит из-за того, что гомологичные участки в изменившихся
группах сцепления оказываются во всех четырех хромосомах, и они,
как им и положено, взаимно притягиваются. Таким образом, по го-
мологичным участкам у гетерозигот по транслокациям конъюгиру-
ют не две, а четыре хромосомы. В результате этого образуются не
биваленты (состоящие из двух d-хромосом), а квадриленты (состоя-
щие из четырех d-хромосом). При этом конъюгирующие хромосомы
образуют характерную фигуру в виде креста. Транслокации подав-
ляют кроссинговер.
В отличие от кроссинговера, когда обмениваются гомологичными
участками гомологичные хромосомы, транслокации приводят к обра-
зованию хромосом с нарушенными последовательностями не алле-
лей, а отдельных генов. В результате расхождения таких хромосом из
шести возможных сочетаний четыре гаметы оказываются с нарушен-
ными хромосомами (делециями или дупликациями) и лишь две из
696
ИЗМЕНЧИВОСТЬ
них — с полноценными. Этим объясняется тот факт, что гетерозиготы
по транслокациям часто бывают стерильными — большинство проду-
цируемых ими гамет дефектны.
Транспозиции представляют собой перемещение участка в преде-
лах одной хромосомы, что приводит к изменению последовательно-
сти генов в группе сцепления, либо односторонний перенос этого
участка в другую хромосому. Такие переносы осуществляются с уча-
стием подвижных генетических элементов, которые имеются как у
прокариот, так и у эукариот. Открытие подвижных генетических эле-
ментов принадлежит Б. Мак-Клинтоку (1947), который изучал хро-
мосомные разрывы у кукурузы.
Робертсоновские перестройки представляют собой слияние двух
негомологичных хромосом, в результате чего образуется одна более
крупная хромосома. Эти преобразования названы по имени У.В. Ро-
бертсона, объяснившего уменьшение числа хромосом возможным их
слиянием. Противоположный процесс — разделение хромосомы на
две — встречается значительно реже.
Геномные мутации характеризуются изменением числа хромосом,
которые могут быть некратными или кратными. Некратное измене-
ние числа хромосом в диплоидном наборе называется гетероплоиди-
ей, или анэуплоидией. Это может сопровождаться отсутствием одной
из хромосом — моносомия по данной паре хромосом или всей пары
гомологичных хромосом — нуллисомия. Наличие одной или несколь-
ких лишних хромосом называется полисемией, которую, в свою оче-
редь, подразделяют на трисомию, если одна хромосома лишняя, те-
трасомию — при наличии двух лишних хромосом, и т. д. Название в
данном случае определяется количеством гомологичных хромосом,
например, если к двум имеющимся добавляется одна лишняя, то это
трисомия, если лишних две, то всего таких гомологичных хромосом
четыре и нарушение называется тетрасомией, и т. д. Все эти измене-
ния отражаются и на фенотипе, т. к. сопровождаются либо недостат-
ком, либо, соответственно, избытком генов. Причиной возникнове-
ния гетероплоидии является нарушение расхождения хромосом в
процессе мейоза. Если гомологичные хромосомы или хроматиды не
разошлись, то в одну из гамет попадут сразу две хромосомы, а в дру-
гую ни одной. Соответственно, при участии таких гамет в оплодотво-
рении образуется зигота с измененным числом хромосом. Явление
гетероплоидии было обнаружено К. Бриджесом в опытах по изуче-
нию наследования сцепленных с полом признаков у дрозофилы.
Гетероплоидия возможна как у аутосом, так и у половых хромосом.
Часто она сопровождается серьезными заболеваниями и может слу-
жить причиной летального исхода. В частности, моносомия у споро-
фитов растений обычно летальна. У дрозофил моносомия по четвер-
697
МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ
той хромосоме приводит к появлению более мелких и менее
фертильных мух, а моносомия по второй или третьей хромосомам
у тех же мух вызывает летальный исход, что указывает на неравно-
ценность расположенных в этих хромосомах генов. Воздействие по-
лисомии на споры растений неодинаково. Так, в микроспорах гамето-
фит не развивается, а в мегаспорах лишняя хромосома не оказывает
влияния на развитие женского гаметофита.
Неправильное расхождение хромосом возможно не только в про-
цессе мейоза, но также и митоза. Дальнейшее деление таких клеток
приводит к увеличению их числа. Нахождение в организме клеток од-
ного типа с различными свойствами называется мозаицизмом. Отно-
сительная доля измененных клеток зависит от того, на какой стадии
дробления произошло неправильное расхождение хромосом: чем
раньше это произошло, тем больше будет измененных клеток в разви-
вающемся организме. Тогда, как в случаях нарушения расхождения
хромосом при мейозе, образуются гаметы, последующее участие кото-
рых в оплодотворении приведет к образованию организма, все клетки
которого будут изменены.
Кратные изменения числа наборов хромосом выражаются полипло-
идией. При гаплоидии все хромосомы в кариотипе представлены в
единственном числе. Изменение числа наборов хромосом далеко не
всегда говорит о мутации. Многие исследователи вообще не считают
мутациями хромосомные изменения. Например, у высших растений в
обязательном порядке происходит закономерное чередование гапло-
идного поколения (гаметофит) и диплоидного (спорофит). Известны
гаплоидные формы у грибов и водорослей. У многих простейших
(жгутиконосцы, споровики) первое деление зиготы представляет со-
бой мейоз, в результате которого образуются гаплоидные клетки.
Даже среди многоклеточных организмов могут иметься особи с га-
плоидным набором хромосом (самцы перепончатокрылых — пчел,
муравьев и др.). Наконец, гаплоидные клетки (гаметы) образуются
у всех организмов, размножающихся половым путем. Индуцирован-
ное получение гаплоидных организмов имеет прикладное значение,
поскольку в гаплоидных клетках легче дифференцировать полезные
и вредные мутации. А дальнейший перевод гаплоидов в диплоиды по-
зволяет получать растения с нужными качествами. Для получения
индуцированных гаплоидных организмов используют различные ме-
тоды: искусственное опыление пыльцой с предварительно убитыми
ядрами или пыльцой другого вида; задержкой опыления (для стиму-
ляции деления неоплодотворенной яйцеклетки). Изредка в семенах
растений развиваются два зародыша (по аналогии с животными их
называют близнецами), один из которых является нормальным ди-
плоидным, а второй — гаплоидным.
698
ИЗМЕНЧИВОСТЬ
Полиплоидия приводит к кратному увеличению числа наборов
хромосом. Полиплоидными считают организмы, имеющие более двух
наборов хромосом. Различают сбалансированные полиплоиды, содер-
жащие четное количество наборов — 4n, 6n, 8n, 10n и т. д., и несбалан-
сированные с нечетным количеством наборов — 3n, 5n, 7n, 9n и т. д .
У несбалансированных полиплоидных организмов часто имеют место
дефекты в процессе мейоза из-за невозможности правильной конъ-
югации нечетного количества хромосом и нарушений при их расхо-
ждении.
Как и гаплоидия, полиплоидия значительно более распростране-
на у растений, особенно цветковых, где их доля составляет почти по-
ловину (большинство культурных растений полиплоидны). Для го-
лосеменных растений полиплоидия нехарактерна. У многоклеточных
животных полиплоидия всего организма встречается лишь у некото-
рых гермафродитов (например, у земляных червей) и у форм, раз-
множающихся партеногенетически (некоторые чешуекрылые, жест-
кокрылые, полужесткокрылые, ракообразные, рыбы и земноводные).
Кроме того, у многих животных имеет место полиплоидия клеток
отдельных органов (например, печени млекопитающих, слюнных
желез и мальпигиевых сосудов некоторых насекомых), что получило
название эндополиплоидии. Климат влияет на степень распростра-
ненности полиплоидных организмов — их становится больше по
мере приближения к арктическим и антарктическим широтам и в
условиях высокогорья.
Увеличение числа одинаковых геномов называется автополи-
плоидией. Примером тому является многократное увеличение набо-
ров хромосом в макронуклеусе инфузорий (более тысячи раз).
Автополиплоидия может возникать спонтанно и искусственно.
Спонтанная может происходить в случаях нарушений расхождения
хромосом в процессе мейоза при спорогенезе. Образовавшиеся в ре-
зультате этого не гаплоидные, а диплоидные споры впоследствии
продуцируют также диплоидные гаметы. Если плоидность увеличи-
вается в соматических клетках, то это приводит к появлению
мозаицизма, когда в организме одновременно присутствуют и ди-
плоидные, и полиплоидные клетки. Причиной этому может быть
репликация хромосом, которая не сопровождается последующим
разделением клеток.
Индуцированное увеличение плоидности вызывает интерес селек-
ционеров, поскольку позволяет получить высокопродуктивные расте-
ния. Полиплоидные формы высших растений имеют бо
льшие разме-
ры, чем гаплоидные. Однако увеличение плоидности приводит к
росту продуктивности лишь до определенных пределов, превышение
которых может привести к обратному результату — появлению более
699
МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ
слабых растений. Обычно для этого используют вещества, препятст-
вующие расхождению d-хромосом, например колхицин, который спо-
собен связываться с тубулином и блокировать тем самым полимери-
зацию необходимых для образования веретена деления микротрубочек.
Экспериментально можно добиться появления триплоидности у три-
тонов, если воздействовать на яйца аномально высокой или, наоборот,
низкой температурой.
Наличие нескольких наборов хромосом вносит определенные из-
менения в процесс мейоза. У полиплоидных организмов гомологич-
ных хромосом не две, а больше (пропорционально кратности генома).
В результате этого возникают не биваленты, а, например, квадрилен-
ты (если геном тетраплоиден). У несбалансированных полиплоидных
форм гораздо чаще проявляются нарушения мейоза, из-за чего они
почти всегда оказываются стерильными. Это обстоятельство, вредное
для растения, может оказаться весьма полезным для потребительских
нужд человека (например, триплоидные бананы с плодами без семян
или триплоидные арбузы без косточек).
В отличие от автополиплоидных организмов, у аллополиплоидных
умножается геном не одного вида, а разных. Они возникают в резуль-
тате гибридизации различных видов и родов — отдаленной гибриди-
зации. Полученные при этом гибриды с разными наборами хромосом
называют отдаленными гибридами, а их полиплоидные формы соот-
ветственно являются аллополиплоидами. Полиплоидные отдаленные
гибриды также могут быть бесплодными, но это случается реже, чем у
автополиплоидных. В начале 20-х годов XX в. отечественный цитоге-
нетик Г. Д. Карпеченко впервые получил плодовитый межродовой
гибрид, скрещивая редьку с капустой. Оба скрещиваемых растения
имеют в диплоидном наборе по 18 хромосом (в гаплоидном по 9).
Спорофитное поколение гибрида, названного рафанобрассикой, име-
ло в соматических клетках 36 хромосом, половина из которых редеч-
ные и половина капустные. Фенотип рафанобрассики совмещал при-
знаки обоих родителей, но в неприемлемом для сельского хозяйства
сочетании, поскольку побег был близок к редьке, а корень — к капу-
сте. Поэтому, несмотря на научный успех, этот гибрид не получил хо-
зяйственного распространения.
Хромосомные болезни человека. У человека известно большое
количество наследственных заболеваний. Многие из них вызваны из-
менением численности хромосом (как половых, так и аутосом), у но-
ворожденных они встречаются очень часто. В частности, из каждых
1000 живых новорожденных 3—4 имеют хромосомные нарушения.
В 35—40% случаях врожденных пороков развития «виноваты» имен-
но такие мутации. Далеко не все эмбрионы с хромосомными наруше-
ниями рождаются — в среднем из-за них происходит 40% спонтанных
700
ИЗМЕНЧИВОСТЬ
абортов и 6% всех мертворожденных. В последнее время доля детей
с хромосомными аномалиями неуклонно возрастает. Это связано с
воздействием неблагоприятных факторов (химических веществ, про-
никающей радиации, курением, алкоголизмом родителей и др.) .
Обычно проявившийся синдром называют по имени исследователя,
который первым его описал (табл. 30).
Мутагенез. В зависимости от природы происхождения все мута-
ции делят на спонтанные и индуцированные. Спонтанные мутации
возникают в естественных условиях обитания организма. На их появ-
ление не оказывается никакого воздействия извне, они всегда неожи-
данны и непредсказуемы, и действительные причины таких мутаций
остаются неизвестными. Характерной особенностью спонтанных му-
таций является то, что они крайне редки. У разных генов частота
спонтанных мутаций неодинакова. Кроме того, обнаружены мута-
торные гены, способные оказывать воздействие на появление мута-
ций в других генах.
Индуцированные мутации возникают под воздействием внешних
факторов — мутагенных, или мутагенов. Их делят на физические, хи-
мические и биологические.
Физические мутагены составляют высокоэнергетичные частицы
крайне малой величины, из-за чего они обладают высокой способно-
стью глубоко проникать в ткани и вызывать молекулярные наруше-
ния. Следствием этого является появление в тканях заряженных ча-
стиц — ионов, обладающих высокой реакционной активностью и
способных вызывать вторичные изменения генетического материала.
Особенно опасными являются свободные радикалы, образующиеся
из воды внутренней среды клетки. Наиболее известными физически-
ми мутагенами являются ионизирующая радиация (α-, β-, γ-лучи, Х-
лучи — рентгеновские лучи, а также потоки протонов и нейтронов)
и ультрафиолетовые лучи. Наиболее активно действует ионизирую-
щая радиация, обладающая гораздо более высокой проникающей
способностью. В результате возникают генные и различные типы
хромосомных мутаций. Похожее воздействие на генетический мате-
риал оказывает также облучение потоками нейтронов и протонов.
Ультрафиолетовые лучи обладают меньшей энергией, поэтому они
оказывают воздействие лишь на поверхностные ткани. При этом
образуются димеры тимидина, которые приводят к нарушениям
нуклеотидной последовательности в процессе репликации ДНК.
Первые индуцированные мутации были получены в 1925 г. отече-
ственными микробиологами Г. А . Надсоном и Г. С. Филипповым в
результате экспериментального облучения дрожжей ионизирующей
радиацией. Это привело к увеличению наследственных форм микро-
701
МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ
организма. У человека удваивается частота мутаций при получении
ионизирующей радиации в дозе 0,5—1,5 Гр (50—150 рад.). Техноген-
ные катастрофы, нарушения правил техники безопасности при ис-
пользовании радиоактивных веществ и ядерных отходов в значитель-
ной степени увеличили риск радиационного облучения. В связи с
этим вызывают интерес вещества, обладающие антимутагенной
активностью, которые способны снижать вредное воздействие иони-
зирующей радиации. К таким веществам относят радиопротекторы,
главным образом серосодержащие аминокислоты — метионин,
цистин, цистеин, а также ряд пуриновых и пиримидиновых производ-
ных (метилурацил, калия оротат, инозин, рибоксин). Физические му-
тагены используются селекционерами для индуцирования мутаций
при выведении новых сортов растений. В качестве источника γ-лучей
в лабораторных условиях обычно используют радиоактивный ко-
бальт (60Со).
Таблица 29
Хромосомные болезни человека
Название
синдрома
Причина
Частота
Проявление
Aнoмалии аутосом
Синдром
Дауна
Трисомия
по 21-й
хромосоме
1 : 500—700
новорожден-
ных; носите-
лей в четыре
раза больше,
но они гибнут
во внутри-
утробный
период
Умственная отсталость, изменение стро-
ения лица (монголоидность), у 40% име-
ют место различные пороки сердца;
больные редко живут более 20 лет и
крайне редко имеют детей
Синдром
Патау
Трисомия
по 13-й
хромосоме
1 : 5000—7000
новорожден-
ных
Расщепление губы (заячья губа), расще-
пление нёба (волчья пасть), пороки
развития головного мозга, глазных
яблок и внутренних органов (особенно
сердца, почек и половых органов), поли-
дактилия (многопалость); около 90%
детей погибают в течение первого года
жизни
Синдром
Эдвардса
Трисомия
по 18-й
хромосоме
1 : 7000—
10 000 новоро-
жденных
Нарушения в развитии всех систем орга-
нов; около 90% детей погибают в течение
первого года жизни
Синдром
«кошачий
крик»
Частичная
моносо-
мия
по 5-й хро-
мосоме
1:40000—
50 000 новоро-
жденных
Плач очень высокого тона
702
ИЗМЕНЧИВОСТЬ
Название
синдрома
Причина
Частота
Проявление
Аномалии половых хромосом
Синдром
Шерешев-
ского —
Тернера
Комплекс
половых
хромосом
ХО (моно-
сомия)
1 : 2500 ново-
рожденных;
носителей го-
раздо больше,
но они гибнут
во внутри-
утробный
период
Женский фенотип. Разнообразные нару-
шения физического и (иногда) умствен-
ного развития. Диагноз ставится, если
одновременно имеется гипогонадизм и
недоразвитие половых признаков (как
первичных, так и вторичных), врожден-
ные соматические пороки развития, низ-
кий рост
Синдром
Клайн-
фельтера
Комплекс
половых
хромосом
XXY
1 : 500 мужчин Мужской фенотип. Нарушения проявле-
ния первичных (недоразвитые умень-
шенные яички с дегенерированным
сперматогенным эпителием) и вторич-
ных половых признаков, умственная
отсталость (иногда), олигоспермия
(поэтому обычно бесплодны)
Трисомия
X
Комплекс
половых
хромосом
XXX
1 : 700 ново-
рожденных
девочек
Аномалии развития половых органов
и скелета, умственная отсталость, одна-
ко симптомы выражены не всегда, пони-
женная репродуктивная способность
Трисомия
XY
Комплекс
половых
хромосом
XXXY
Очень редко
То же, что и у XXY, но более выражено
Тетрасо-
мия X
Комплекс
половых
хромосом
ХХХХ
Очень редко
То же, что и у XXX, но более выражено
Тетрасо-
мия XY
Комплекс
половых
хромосом
XXXXY
Описано
более 100 слу-
чаев
То же, что и у XXXY, но более выражено
Пентасо-
мия XX
Комплекс
половых
хромосом
ХХХХХ
Единичные
случаи
То же, что и у ХХХХ, но более выражено
Пентасо-
мия XY
Комплекс
половых
хромосом
XXXXXY
Единичные
случаи
То же, что и у XXXXY, но более
выражено
Дисомия
XY
Комплекс
половых
хромосом
XYY
1 :1000
Более высокий рост, слабые мышцы и
импульсивное поведение (среди заклю-
ченных доля мужчин с таким генотипом
составляет около 2%); все фенотипиче-
ские отклонения обычно выражены
слабо
Продолжение табл. 29
703
МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ
Название
синдрома
Причина
Частота
Проявление
Трисомия
XY
Комплекс
половых
хромосом
XYYY
Очень редко
Выраженные половые и соматические
нарушения
Синдром
тестику-
лярной
феминиза-
ции
Наруше-
ние разви-
тия поло-
вых при-
знаков
Женский фенотип наружных половых
органов, но развиваются нормальные
яички; большая физическая сила, сопо-
ставимая с мужской (из-за высокого со-
держания тестостерона, обладающего
анаболическими свойствами); детей
иметь не могут
Андроге-
ниталь-
ный
синдром
То же
Мужской фенотип наружных половых
органов, но развиваются нормальные
яичники и другие внутренние женские
половые органы; детей иметь не могут
Химические мутагены должны обладать следующими свойствами:
1) — высокой проникающей способностью; 2) — свойством изменять
коллоидное состояние хромосом и 3) — определенным действием на
изменение гена или хромосомы. В зависимости от действия их подра-
зделяют на две группы: мутагены, действующие только на реплициру-
ющуюся ДНК (акридиновые красители и аналоги азотистых основа-
ний), и мутагены, действующие как на реплицирующуюся, так и на
покоящуюся ДНК (алкилирующие соединения — нитрозогуанидин,
метилметансульфонат и этилметансульфонат). Химические вещества,
индуцирующие мутации, были обнаружены в 30-х годах XX в. в экс-
периментах с дрозофилой. Ими оказались йод, аммиак, формалин,
азотистый иприт и др. В последующие годы было обнаружено боль-
шое количество других химических мутагенов, а также вещества
антимутагены, которые нейтрализуют или ослабляют воздействие
мутагенов. Антимутагены специфично действуют в отношении кон-
кретного мутагена. Из наиболее известных можно назвать ненасы-
щенные жирные кислоты (особенно полиненасыщенные), тониновую
кислоту, витамины, обладающие антиоксидантной активностью (ви-
тамины С, А, Е), катехин (содержащийся в чае, особенно зеленом,
и кофе).
Биологические мутагены — это, главным образом, вирусы, вызыва-
ющие наследственные изменения генетического материала у прока-
риот и эукариот. Кроме вирусов, мутации могут вызывать транспози-
руемые генетические элементы, а также микроорганизмы, выделяющие
токсины (прежде всего плесневые грибы). В последнем случае имеет
место не прямое воздействие биологического мутагена на генетиче-
Окончание табл. 29
704
СЕЛЕКЦИЯ
ский материал, а опосредованное — через выделяемые химические ве-
щества.
Установлено, что эффект индивидуального действия отдельных
мутагенов можно усилить, сочетая их с одновременным воздействием
других факторов. Так, К. В . Ватти и М.М . Тихомирова дополнитель-
но подвергали облученных рентгеновскими лучами дрозофил воздей-
ствию высокой температуры (+37 °С) и обнаружили более высокую
частоту мутаций по сравнению с вызываемыми только облучением.
При этом сама по себе высокая температура не индуцирует мутации.
Значение мутаций. Мутации так же, как и рекомбинации, дают
новые состояния генотипов. Однако, в отличие от последних, мута-
ции приводят к образованию новых аллелей и даже генов. Следова-
тельно, они являются причиной любого качественного изменения ге-
нофонда, что определяет микро- и макроэволюционные процессы.
Для хозяйственной деятельности человека мутации (особенно инду-
цированные) важны в качестве метода, позволяющего получить раз-
нообразие племенного материала с последующим отбором наиболее
ценных форм.
СЕЛЕКЦИЯ
Селекция — это процесс выведения новых и улучшения уже суще-
ствующих форм растений, животных или микроорганизмов. Селекция
не создает новых видов, она лишь изменяет уже существующие (ди-
кие) формы организмов в соответствии с хозяйственными или иными
потребностями человека.
История селекции. Растениеводство и животноводство имело ме-
сто уже в эпоху неолита. Вначале были одомашнены собаки, которые
помогали на охоте и охраняли жилище. Самыми первыми домашни-
ми сельскохозяйственными животными стали овцы, козы, коровы,
свиньи, ослы. Их предков человек одомашнил около 10 000 лет назад.
Позднее (за 2—3 тыс. лет до н. э .) одомашнили лошадь, буйвола, вер-
блюда, северного оленя, кур. Вначале животноводство было ограниче-
но лишь районами с теплым тропическим климатом, но со временем
распространилось повсеместно. В Европе домашние животные появи-
лись в III тысячелетии до н. э. В Южной Америке до прихода евро-
пейцев индейцы одомашнили лам, в Северной Америке была одомаш-
нена лишь индейка, а скотоводства не было вовсе. Еще на заре
сельскохозяйственной деятельности человек бессознательно прово-
дил селекцию одомашненных растений и животных. Она выражалась
в отборе самых продуктивных особей для размножения. 2000 лет на-
зад в древнем Риме были написаны трактаты поэта Вергилия, писате-
705
СЕЛЕКЦИЯ
ля и агронома Колумеллы, ученого Варрона, где имелись указания,
как следует вести отбор растений. Постепенно вырабатывались прие-
мы разведения, позволяющие увеличить производительность, кото-
рые передавались из поколения в поколение. В результате к началу
XIX в. удалось вывести множество сортов растений и пород живот-
ных. К настоящему времени одомашнены десятки видов растений,
около 40 видов животных, среди которых различные млекопитающие,
птицы (куры, индейки, цесарки, голуби и др.), рыбы (карп, аквариум-
ные рыбки), насекомые (пчелы, тутовый шелкопряд) и микроорга-
низмы. Продуктивность культурных форм многократно выше, чем у
их диких предков. Например, предки крупного рогатого скота — ди-
кие туры — кормили телят не более 3—4 месяцев, тогда как у коров
молочных пород лактация длится до 10 месяцев в году с регулярными
высокими удоями. Настриг шерсти от одного барана асканийской по-
роды составляет 30,6 кг за год!
Методы селекции. Для выведения новых форм и улучшения су-
ществующих селекционеры используют методы, основанные на гене-
тических закономерностях. Основными методами являются искусст-
венный отбор и гибридизация, искусственный мутагенез, полиплоидия,
гетерозис. В селекции микроорганизмов используют приемы генной
инженерии.
Искусственный отбор является наиболее старым методом. Дарвин
различал две формы искусственного отбора: бессознательный и есте-
ственный. В каждой группе есть лучшие и худшие по хозяйственному
признаку особи. Человек давно понял, что каждый организм способен
в той или иной мере передавать потомству и полезные и вредные при-
знаки. Суть бессознательного отбора состоит в сохранении для раз-
множения наиболее продуктивных особей и уничтожении (например,
для пищи) худших. В результате такого отбора формируются породы
и сорта с усредненными характеристиками (например, высокая удой-
ность и крупные размеры одновременно). Особенностью методиче-
ского отбора является то, что селекционер заранее представил цель
и систематически отбирает на племя особей с нужными ему качества-
ми (например, наиболее удойных коров для получения новой молоч-
ной породы или наиболее крупных особей для получения мясной по-
роды). При этом образуются высокоспециализированные породы и
сорта.
При массовом отборе сохраняют группу особей с желательными
признаками, после чего от них получают потомство. Потомство в та-
ком случае будет неоднородным, и периодически отбор придется по-
вторять, чтобы выбраковывать менее продуктивные экземпляры. Та-
кой отбор наиболее часто применяют в селекции растений. Массовый
отбор может быть эффективен лишь для признаков, которые кон-
706
СЕЛЕКЦИЯ
тролируются одним или немногими генами. При индивидуальном от-
боре селекционер ориентируется на генотип и выделяет отдельных
особей, чтобы получить от них потомство. Это позволяет получить
генетически однородную популяцию, которая состоит из гомозигот-
ных по нужному признаку особей — чистую линию.
Отбор не всегда дает нужные результаты. Часто имеет место не ге-
нетически закрепленное изменение признака, а модификация. В та-
ких случаях нужные признаки теряются у потомков, и необходимо
повторять отбор. Необходимо учитывать коэффициент наследуемо-
сти, который показывает долю генотипической изменчивости в по-
томстве. Например, отбор для племенного стада лучших несушек без
учета признаков их родителей малопродуктивен, поскольку коэффи-
циент наследуемости этого признака при массовом отборе составляет
всего около 0,25. На яйценоскость сильно влияет среда, поэтому куры,
полученные от лучших и худших по этому признаку матерей, несли
примерно одинаковое количество яиц. При этом коэффициент насле-
дуемости признака размера яиц гораздо выше (0,75), и большинство
дочерей кур, несших крупные яйца, также несли крупные яйца; доче-
ри матерей, несших мелкие яйца, проявляли тот же признак.
Гибридизация представляет собой скрещивание разнородных в
генетическом отношении родительских форм. С ее помощью можно
добиться внесения в геном ценных генов и получить нужную комби-
нацию признаков (например, если сорт обладает высокой продуктив-
ностью, но подвержен заболеваниям, то скрещивание с устойчивыми
формами делает потомство более выносливым). Потомство таких ро-
дителей называется гибридным, а особи — гибридами. Гибридизация
обязательно сочетается с последующим отбором наиболее удачных
фенотипов среди потомства для разведения. В зависимости от степе-
ни близости родителей различают внутривидовую и отдаленную ги-
бридизацию. Внутривидовая гибридизация — это направ-
ленное скрещивание между собой особей различных внутривидовых
форм (подвидов, сортов, пород, чистых линий), обладающих интерес-
ными для селекционера признаками. Отдаленная гибридиза-
ция — это скрещивание особей разных видов, позволяющее совме-
стить в гибриде ценные качества неродственных групп. Иногда
оказываются необходимы возвратные скрещивания с одной из роди-
тельской форм. Этот метод часто сопровождается трудностями, связан-
ными с генетической несовместимостью родителей. Полученные ги-
бриды могут быть стерильными (например, гибрид лошади и осла,
гибридные самцы от яка и крупного рогатого скота). Положительным
последствием гибридизации является гетерозис.
Гетерозис (гибридная мощность) представляет собой повышенную
продуктивность и жизнеспособность гибридов первого поколения. По
707
СЕЛЕКЦИЯ
этим показателям гибриды значительно превосходят родительские
формы. Гетерозис очень эффективен — межлинейные гибриды сорго
на 40—80% урожайнее исходных сортов и обычных межсортовых ги-
бридов, лука — на 30—45%, кукурузы — на 25—30%. Селекционеры
вывели линии двойных гибридов, обладающие цитоплазматической
мужской стерильностью. У растений таких линий пыльца не образует-
ся, но при опылении пыльцой нормальной родительской линии на них
обычным образом развиваются семена. Следовательно, на растениях
с цитоплазматической мужской стерильностью образуются только ги-
бридные семена. Явление гетерозиса свойственно не только растени-
ям, но и животным. Гибриды одно- и двугорбого верблюдов превосхо-
дят родителей размерами и силой; чрезвычайно сильны и выносливы
мулы — гибриды кобылы с ослом; продуктивность гибридов яка
и крупного рогатого скота значительно выше, чем у родителей. Гетеро-
зис проявляется не во всех признаках и зависит от направления скре-
щивания, условий выращивания гибридов. Лучшие результаты дают
скрещивание линий, проверенных на способность образовывать высо-
копродуктивные гибриды.
Классификация различных типов гетерозиса у растений: 1) р е -
продуктивный гетерозис выражается в лучшем развитии
генеративных органов, что приводит к повышению урожайности се-
мян и плодов; 2) соматический гетерозис обеспечивает
мощное развитие вегетативной массы; 3) приспособительный,
или адаптивный, гетерозис проявляется в повышении общей
жизнеспособности, что подтверждает адаптивную направленность
этого явления.
Во всех случаях гетерозис носит временный характер, поэтому у
гибридов последующих (после F1) поколений продуктивность возвра-
щается к состоянию исходных форм. Гетерозис широко используется
в селекции растений и животных, однако механизм гибридной мощ-
ности до сих пор не раскрыт.
Искусственный мутагенез стимулирует образование мутаций по-
средством воздействия мутагенными факторами. Это стимулирует ге-
нетическую гетерогенность имеющейся группы и тем самым предо-
ставляет селекционеру обширный материал для отбора. Этот метод
чаще используют для выведения новых сортов растений.
Полиплоидия. Не зная генетических механизмов этого явления, се-
лекционеры прошлого использовали его при выведении сортов пше-
ницы, овса, хлопчатника, картофеля и в цветоводстве. Сейчас это
можно делать направленно. Воздействие на клетки раствором колхи-
цина разрушает микротрубочки веретена деления и не позволяет хро-
мосомам разойтись. В результате появляются клетки с удвоенным (по
сравнению с обычным состоянием) набором хромосом. Если воздей-
708
СЕЛЕКЦИЯ
ствовать колхицином на определенные клетки семян, проростков ра-
стений, а также гаметы и эмбрионы животных, можно получить поло-
вые и соматические клетки с нужным набором хромосом. Полиплоидия
может выражаться нарушениями мейоза, слиянием соматических
клеток, удвоением хромосом в неделящихся клетках. Полиплоиды
могут образовываться в результате объединения генетического мате-
риала организмов разных видов, например, гибрид капусты и редьки,
полученный Г.Д. Карпеченко.
Разработаны методы трансформации растений и животных путем
использования клонированных генов. При этом можно получить
организм с генами других видов, например мышь с геном соматотроп-
ного гормона человека или крысы. Таким путем можно переносить
гены полезных белков растениям.
СЕЛЕКЦИЯ РАСТЕНИЙ
Селекционеры растений выводят новые сорта культурных форм и
улучшают уже имеющиеся сорта. Сорт — это группа растений одного
вида, устойчиво проявляющаяся в определенных условиях возделы-
вания некоторыми признаками, полезными для человека. Однако при
изменении благоприятных условий сорт теряет культурные свойства,
и признак возвращается к дикому типу. Основными методами селек-
ции растений являются разные способы искусственного отбора и ги-
бридизации. Широко используется явление гетерозиса, для его ини-
циации и управления составляют определенную схему скрещиваний.
Например, принудительное самоопыление растений, которые в обыч-
ных условиях опыляются перекрестно (кукуруза), приводит к осла-
блению потомства, но последующая гибридизация этих линий дает
высокоурожайное потомство. Высокоэффективным методом являет-
ся отдаленная гибридизация, ее широко использовал И. В . Мичурин,
сумевший вывести более 300 новых сортов. Для достижения скрещи-
ваемости он применял: предварительное вегетативное сближение
(привитый черенок выдерживался несколько лет на подвое, после
чего его цветы опыляли пыльцой нужного растения, т. е . проводили
собственно гибридизацию); использование посредника (скрещивал
сначала миндаль с полукультурным персиком, а полученный ги-
брид — с культурным персиком); опыление смесью пыльцевых зерен,
что облегчало восприятие плодолистиком пыльцы чужеродного вида.
Хорошие результаты дают экспериментальная полиплоидия,
искусственный мутагенез, методы культуры клеток и культуры тка-
ней, клеточная и генная инженерия. Многие селекционеры изучают
явление сомаклональной изменчивости — увеличение уровня наслед-
709
СЕЛЕКЦИЯ РАСТЕНИЙ
ственной изменчивости при размножении растений посредством ре-
генерации в культуре соматических клеток. Создан ряд генетически
модифицированных сортов культурных растений (соя, кукуруза, кар-
тофель и др.). Они содержат в геноме чужеродные гены, благодаря
которым значительно увеличивается продуктивность, устойчивость
к заболеваниям, паразитам и неблагоприятным условиям. Такие сорта
используются при изготовлении продуктов питания, но существуют
опасения в их безвредности (поэтому информация о содержании мо-
дифицированных ингредиентов должна присутствовать на упаковке
продукта).
Таблица 30
Центры происхождения культурных растений
(по Н.И . Вавилову)
Наименование
центра
Географическое
положение
центра
Примеры окультуренных растений
Южноазиатский
тропический
Тропическая Ин-
дия, Индокитай,
Южный Китай,
острова Юго-
Восточной Азии
Около 50% культурных растений, среди них
многие плодовые и овощные культуры, ци-
трусовые, баклажан, черный перец, огурец,
рис и др.
Восточно-
азиатский
Центральный
и Восточный
Китай, Япония,
Корея, Тайвань
Около 20% культурных растений, среди них
слива, вишня, соя, гречиха, просо и др.
Юго-Западно-
азиатский
Малая Азия,
Средняя Азия,
Иран, Афганис-
тан, Юго-Запад-
ная Индия
14% культурных растений, среди них некото-
рые формы зерновых (пшеницы, ржи), бобо-
вые, чеснок, виноград, лен, конопля, многие
плодовые (абрикос, груша) и др.
Средиземномор-
ский
Страны побере-
жья Средиземно-
го моря
11% культурных растений, среди них многие
овощные (капуста), маслины, сахарная
свекла, многие кормовые (клевер, чечевица)
и др.
Абиссинский
Абиссинское
нагорье Африки
(территория
Эфиопии)
Твердая пшеница, ячмень, зерновое сорго,
один вид банана и др.
Центральноаме-
риканский
Южная Мексика
Кукуруза, некоторые тыквенные, фасоль,
какао, табак, длинноволокнистый хлопчат-
ник и др.
Южноамерикан-
ский (Андий-
ский)
Часть района
Андского горного
хребта вдоль
западного побе-
режья Южной
Америки
Ананас, картофель, некоторые лекарствен-
ные (хинное дерево, кокаиновый куст) и др.
710
СЕЛЕКЦИЯ
Большое значение имеет генетическое разнообразие диких пред-
ковых форм окультуренного растения. Согласно учению Н.И. Вави-
лова о центрах происхождения культурных растений, исторической
родиной культурных форм является та географическая область, где
произрастает наибольшее количество предковых форм. Разные куль-
туры имеют неодинаковые области происхождения, при этом они
обычно совпадают с древними очагами зарождения земледелия. Из
них окультуренные растения широко распространились и стали ис-
ходным материалом для селекции новых сортов. Открытые Вавило-
вым центры перечислены в табл. 30. Учение Вавилова развивалось
и после внесения ряда уточнений. В настоящее время насчитывают
не 7, а 12 первичных центров происхождения культурных растений.
СЕЛЕКЦИЯ ЖИВОТНЫХ
Задача селекции животных — создание новых и улучшение суще-
ствующих пород домашних животных. Порода — это созданная раз-
личными методами селекции группа животных одного вида, которая
отличается генетически устойчивыми морфофизиологическими каче-
ствами, полезными человеку. Подобно культурным растениям, пород-
ные качества домашних животных проявляются только в определен-
ных условиях содержания, но при изменении этих условий порода их
теряет, и признак возвращается к дикому типу. Исключением являют-
ся собаки, которые, оставшись без внимания человека, не восстанав-
ливают фенотип предков, а превращаются в полиморфную группу
(«дворняги»). Особенностью селекции животных является то, что
для них свойственно только половое размножение. Поэтому многие
методы селекции растений для них неприменимы. Необходимо учи-
тывать и поведенческие реакции животных. Основными методами яв-
ляются различные формы отбора и гибридизации, но при подборе ро-
дительских пар важно учитывать хозяйственные признаки, которые
у самцов не проявляются в фенотипе, но влияют на продуктивность
породы (яйценоскость, жирность молока, продолжительность лакта-
ции и т. д .). При этом оценка производителей осуществляется по
внешним признакам (экстерьер), родословным и по качеству потом-
ства. Широкое применение метода искусственного осеменения позво-
ляет использовать половые клетки особо выдающихся по своим каче-
ствам самцов-производителей спустя много лет после их смерти
(глубокозамороженную сперму хранят в особых хранилищах). Для
получения максимального приплода от наиболее ценных самок разра-
ботаны методы стимуляции одновременного созревания нескольких
яйцеклеток путем введения определенных гормонов. После оплодо-
711
СЕЛЕКЦИЯ ЖИВОТНЫХ
творения эти яйцеклетки (ранние эмбрионы) пересаживают менее
продуктивным самкам, которые вынашивают генетически ценный
плод. Первым этапом селекции является приручение выбранного жи-
вотного. Это называют одомашниванием. Географические области, где
произошло одомашнивание того или иного вида, называют центрами
доместикации. Обычно они территориально совпадают с основными
центрами древних цивилизаций (там же находятся и центры проис-
хождения культурных растений). На культурные формы стабилизи-
рующая форма естественного отбора действует в значительно мень-
шей степени, поэтому признаки, которые в дикой природе являются
для вида вредными, не элиминируются, если они полезны человеку.
Типы скрещивания. Эффективность отбора усиливается правиль-
но проведенными скрещиваниями, которыми можно добиться сочета-
ния в потомстве нужных свойств или избавиться от ненужных. Скре-
щивания делят на два типа: близкородственное и неродственное.
Близкородственное скрещивание, или инбридинг, приводит к гомози-
готности популяции и позволяет при необходимости разложить попу-
ляцию на гомозиготные линии. Родственными называют родитель-
ские пары, находящиеся между собой в родстве: брат — сестра;
отец — дочь; мать — сын; двоюродные братья и сестры и т. д . Скорость
гомозиготизации тем выше, чем более близкими родственниками ока-
зываются подобранные родительские пары. В тех случаях, когда при-
знак контролируется несколькими генами, гомозиготизация идет
медленнее. В большинстве случаев рецессивные аллели, которые в
обычных условиях находятся в популяции в гетерозиготном состоя-
нии, отрицательно воздействуют на организм. Поэтому инбридинг
постепенно приводит к снижению устойчивости к факторам среды
и вырождению — инбредной депрессии. Начинают проявляться мно-
гие генные заболевания. Одновременно происходит выравнивание
линий по большинству признаков. Инбридинг не позволяет получить
абсолютно гомозиготные линии, потому что всегда будут гетерозиго-
ты по половым хромосомам (XX) и (XY); кроме того, всегда есть ве-
роятность возникновения мутации. Отбор на первых этапах инбри-
динга дает более значительный эффект в нужную сторону, отбор на
последующих близкородственных скрещиваниях менее эффективен,
но при этом возрастает гарантия закрепления нужных свойств в гено-
типе потомства. Инбредная депрессия часто используется для полу-
чения эффекта гетерозиса.
Неродственное скрещивание, или аутбридинг, приводит к повыше-
нию гетерозиготности группы. Чаще всего при этом скрещивают осо-
би разных популяций, не имеющих общих предков в 4—6 поколениях.
Такое скрещивание очень распространено, поскольку потомство пер-
вого поколения наиболее жизнеспособно и продуктивно — проявля-
712
БИОТЕХНОЛОГИЯ
ется гетерозис. Однако если в гибридном организме объединяют не-
сколько нужных наследственных свойств или стараются закрепить
определенное одно, то следует учитывать, что гибриды первого поко-
ления проявляют фенотипическое однообразие, а в последующих по-
колениях происходит расщепление. Поэтому необходимо тщательно
продумывать систему дальнейших скрещиваний и отбора. Разновид-
ностью аутбридинга является межпородное скрещивание, или кросс-
бридинг. В селекции животных широко применяют отдаленную ги-
бридизацию.
Выбор типа скрещивания зависит от поставленной задачи. Для
племенного (заводского) разведения сельскохозяйственных живот-
ных и растений и селекционной работы применяют как инбридинг,
так и аутбридинг. Если же ставится задача увеличения продуктивно-
сти имеющихся пород и сортов, используют их промышленное скре-
щивание. При этом не ведется работа по улучшению культурных
свойств этой группы, а лишь достигаются сиюминутные результаты.
БИОТЕХНОЛОГИЯ
Многие микробы человек научился использовать в различных
целях: производстве сырья для медикаментов (различные антибио-
тики, гормоны, интерферон, витамины В2, В12 и др., незаменимые
аминокислоты, например, лизин, ферменты и т. д .), пищевых (заме-
нители сахара — сорбит, ксилит) и кормовых добавок (аминокисло-
ты и кормовой белок — кормовые дрожжи содержат до 60% белка),
очистке промышленных отходов, в различных пищевых производст-
вах (хлебопечение, кисломолочные продукты, виноделие, пивоваре-
ние) и др. Микробы используются и в непищевой промышленности.
Они помогают выделять из бедных руд металлы, переводя их в рас-
творы. Полученная таким способом медь в 2—3 раза дешевле, чем
при использовании традиционных способов; кроме того, это эколо-
гично, поскольку снимается проблема выброса ядовитых газов, об-
разования отвалов пустой породы и т. п. Искусственное культивиро-
вание микроорганизмов для получения необходимых человеку
веществ называется биотехнологией. Преимуществами микробиоло-
гического синтеза являются: естественный способ получения про-
дукта — все вещества синтезируются клетками; дешевизна — микро-
бы культивируются на отходах производства (пищевого или
нефтехимического); высокая производительность; быстрое размно-
жение; высокая скорость синтеза; отсутствие прямого контакта ра-
ботников предприятия с микробами-продуцентами; относительная
простота селекции и др.
713
ЭВОЛЮЦИЯ
Селекция микроорганизмов. Наиболее продуктивные штаммы
микроорганизмов получают путем отбора. Поскольку многие из них
являются прокариотами, уровень мутабильности у них значительно
выше, чем у эукариот. Для того чтобы добиться еще большей генети-
ческой гетерогенности, широко применяют методы искусственного
мутагенеза, воздействуя мутагенами. После этого производят отбор
наиболее продуктивных клеток и клонируют их, выращивая на пита-
тельной среде. Высокоэффективные продуценты антибиотиков полу-
чены в СССР под руководством С. И. Алихиняна.
Генная инженерия. С помощью методов генной инженерии мож-
но манипулировать генами и переносить их из одного организма в
другой для получения нужных свойств. Получившая новый ген клет-
ка начинает продуцировать закодированный в этом гене белок, кото-
рый затем можно выделить и использовать. Таким способом удалось
перенести в геном бактерии кишечной палочки ген человеческого
инсулина. Полученный биотехнологический инсулин не отличается
от человеческого. Также в бактериальную клетку пересажен ген ин-
терферона — вещества, которое выделяется при вирусной инфекции.
Клеточная инженерия — это метод получения клеток с нужными
качествами путем гибридизации. Например, опухолевая клетка мо-
жет размножаться, а лимфоцит — выделять специфические иммуно-
глобулины — антитела. Путем гибридизации можно соединить в од-
ной клетке их способности. Полученная таким образом клетка будет
размножаться и синтезировать моноклональные антитела к тому или
иному антигену.
ЭВОЛЮЦИЯ
Эволюция — это историческое развитие живой природы с момента
возникновения жизни и до настоящего времени. Сюда включают все
адаптивные изменения, которые возникли в результате мутаций гене-
тического материала и явились причиной существующих различий
между организмами. Этот термин был предложен швейцарским нату-
ралистом Шарлем Боннэ (1762), но первоначально им обозначали
эмбриональное развитие организма.
Основными характеристиками эволюции являются необрати-
мость, адаптивность и преемственность. Необратимость проявляется
в том, что однажды исчезнувший признак не может повториться в
прежнем виде. Например, китообразные, предки которых вели назем-
ный образ жизни, вторично освоили водный образ жизни. В их строе-
нии выделяют рыбообразные признаки: обтекаемая форма тела, пе-
редние конечности в форме плавника и редукция задних конечностей,
714
ЭВОЛЮЦИЯ
гомодентность (зубы не дифференцированы на резцы, клыки и корен-
ные). Однако эти животные сохраняют принципиальные черты орга-
низации млекопитающих: теплокровность, внутриутробное развитие
и вскармливание детенышей молоком, высокоразвитая нервная сис-
тема. У них повторно не возникли жабры, и газообмен осуществляет-
ся в легких, нет воздушного пузыря и чешуи на коже. Аналогичный
пример можно привести и у растений, которые в процессе повторного
освоения водной среды сохранили расчленение тела на вегетативные
органы и дифференцированные ткани, но талломная организация
и ризоиды, присущие водорослям, не смогли появиться повторно.
Адаптивное направление понятно при рассмотрении борьбы за су-
ществование и естественного отбора. Преимущество организму дает
лишь полезная мутация. Поэтому накапливаются только полезные
для особи, а следовательно, и для всего вида в целом, признаки.
Преемственность эволюционных преобразований выражается в
том, что новые признаки не возникают из ничего, а образуются путем
преобразования уже имеющейся структуры. Например, разделение
позвоночного столба на отделы (шейный, туловищный, крестцовый,
хвостовой) у земноводных было бы невозможно, если бы они не име-
ли позвоночника. Усложнение отделов головного мозга у млекопита-
ющих не произошло бы, не имей их предки мозг.
Таким образом, эволюцию живых организмов можно представить
как исторический необратимый процесс адаптивных изменений, про-
исходящий на всех уровнях организации живой природы, который
характеризуется общей прогрессивной направленностью. В эволюци-
онном учении выделяют три основных момента: возникновение жиз-
ни на Земле, последовательные преобразования отдельных групп ор-
ганизмов и возникновение человека с его социальным укладом.
Додарвиновский период. Указания на естественное происхожде-
ние живых организмов и развитие материи имеются в работах древне-
индийских и древнекитайских философов. Китайский философ Кон-
фуций настаивал на возникновении жизни от единого источника,
после чего она постепенно развертывалась и разветвлялась. Наиболее
широко такие мысли были распространены в культуре древней Гре-
ции среди основателей диалектической натурфилософии. Так, пред-
ставители ионийской школы считали, что мир возник естественным
путем. В более поздний период были популярны взгляды механисти-
ческого материализма. В поисках возможных причин происхождения
жизни были заложены элементарные начала биологических наук (бо-
таники, зоологии) и медицины. Наибольший вклад внес древнегрече-
ский мыслитель Аристотель (384—322 гг. до н. э .), который первым
составил классификацию животных, провел сравнительный анализ
их анатомии и эмбриологии. Подобно многим его предшественникам,
715
ЭВОЛЮЦИЯ
Аристотель не возражал против самозарождения организмов, напри-
мер, клопов из соков тела животных, вшей из сырого мяса или дожде-
вых червей из ила прудов. После Аристотеля самозарождение жизни
подкреплялось «свидетельствами» многих знаменитых философов:
Цицерона, Вергилия, Овидия, Сенеки, Плиния, Апулея, Плутарха и
др. Идеи Аристотеля наложили отпечаток на мировоззрение многих
последующих поколений греческих и римских мыслителей и при
этом оказались на редкость живучими. Его авторитет был столь ве-
лик, что в течение последующих полутора тысяч лет (вплоть до
XVI в.) редко кто отваживался возражать его положениям. В средние
века эволюционные взгляды не были распространены. В Европе го-
сподствовали положения креационизма, основанные на Библии. Со-
гласно им, все живое признавалось созданным Богом и изменяться не
могло. И лишь с наступлением эпохи Возрождения возрос интерес
к идеям античных мыслителей, отличных от Аристотеля; с этим свя-
зывают начало формирования эволюционных идей в Европе.
Становление эволюционных идей. На раннем этапе (до конца
XVIII в.) преобладали умозрительные построения, основанные пре-
жде всего на данных ботаники и зоологии, которые в то время интен-
сивно развивались. Для упорядочения полученных сведений необхо-
дима была единая систематизация материала. Шведский натуралист
Карл Линней (1707—1778) первым увидел, что вид может быть уни-
версальной структурной единицей живой природы. Он ввел бинар-
ную номенклатуру, согласно которой каждый вид получал двойное
название — родовое и видовое, т. е . обозначался двумя словами, из
которых первым было имя прилагательное, обозначавшее род, а вто-
рым шло имя существительное, которым назывался этот вид. Линней
установил иерархию единиц систематики — таксонов. Например, по-
сле вида шел более крупный таксон — род, при этом ученый полагал,
что роды были созданы по отдельности, а виды представляют собой
лишь их варианты (отсюда и смысл бинарной номенклатуры). При
всем этом Линней не высказывал эволюционных идей. Он был уверен
в том, что видов насчитывается ровно столько, сколько их было созда-
но изначально. Его систематика во многом была искусственной, по-
скольку, объединяя виды, он часто руководствовался не родством,
а произвольными внешними признаками. Например, растения он раз-
делил на 24 класса, причем в первых 23 из них он распределил цвет-
ковые растения, руководствуясь при этом количеством тычинок
(в первом были однотычинковые, во втором — двутычинковые), их
размерами, сростностью и т. п . К последнему (тайнобрачные) он от-
нес все растения, у которых нет цветка, — водоросли, мхи, папоротни-
ки, хвощи, плауны и грибы с лишайниками.
716
ЭВОЛЮЦИЯ
По мере накопления биологических знаний умозрительные по-
строения сменялись более или менее убедительными теориями. Часть
ученых склонялась к выводу о возможности изменяемости, трансфор-
мации организмов. Поэтому их представления получили название
трансформизма. Одним из первых трансформистов был Жорж Луи
Бюффон (1707—1788), считавший, что разные животные возникли не
одновременно и имеют различное происхождение. Среди причин, вы-
зывающих изменения, он выделял воздействие внешней среды, пре-
жде всего климата и пищи, а также гибридизацию. Особое значение
Бюффон придавал воздействию человека, который одомашнил мно-
гие животные и растения. Он считал, что под воздействием климата
и пищи постепенно создаются разнообразные варианты изменений,
которые могут усовершенствоваться или же подвергаться дегенера-
ции. Таким образом, Бюффон предвосхитил учение о главных направ-
лениях эволюции — усложнении и упрощении организации. Идеи
трансформизма разделяли многие выдающиеся мыслители второй
половины XVII — начала XIX в. Среди них были Р. Гук, Д. Дидро, И.
В. Гете, Эразм Дарвин и др. Немало сторонников идей трансформиз-
ма было в России, наиболее активно эти принципы отстаивал
М. В. Ломоносов. Однако сторонники трансформизма не смогли пра-
вильно объяснить причины изменяемости видов.
Эволюционная теория Ламарка. Создателем первой целостной
эволюционной теории является французский ученый Жан Батист
Ламарк (1744—1829). Свои взгляды он изложил в книге «Философия
зоологии» (1809). Теория Ламарка основывалась на обширных бота-
нических и зоологических фактах. По мнению ученого, жизнь возни-
кла из неживой материи под стимулирующим воздействием света,
тепла и электрических разрядов. Но первопричиной материи Ламарк
считал Творца, который однажды запустил процесс движения всего
мироздания, дальнейшее развитие происходило по естественным при-
чинам. Иными словами, создание мира — это первичный закон разви-
тия, который находится в сфере религии, тогда как ход частных про-
цессов определяют вторичные законы развития, которые и составляют
предмет естествознания. При этом на первых порах существовали
только простые формы жизни, которые постепенно и очень медленно
изменялись во все более и более высокоорганизованные растения
и животные, дошедшие до наших дней.
Ламарк утверждал, что все организмы происходят от непохожих
на них более примитивных предковых форм. Ученый пришел к выво-
ду о плавной эволюции организмов, исключая само понятие вида:
есть только особи, которые постоянно и очень медленно изменяются,
поэтому каждое последующее поколение пусть незначительно, но от-
личается от родителей. Невозможность одномоментной фиксации
717
ЭВОЛЮЦИЯ
процесса Ламарк объяснял слишком малой скоростью процесса. Та-
ким образом, взгляды Ламарка на вид радикально отличались от лин-
неевских. Если последний постулировал реальный, но неизменный
вид, то Ламарк утверждал противоположное: настоящее состояние
организма — это всегда переходная стадия к более совершенному.
Подобно многим ученым своего времени, Ламарк был виталистом,
т. е. верил, что всем организмам присуще внутреннее стремление к со-
вершенству, которое позволяет преодолевать возникающие трудно-
сти. Ламарк разделил животный мир на 14 классов, различающихся
по своей сложности, поставив на вершину человека. Отрицая реаль-
ность существования вида, он считал, что каждый из классов нахо-
дится в постоянном движении в направлении усложнения, чтобы пе-
рейти на более высокую ступень организации. Таким образом, живая
природа никогда не бывает статичной, поскольку все организмы пос-
тоянно находятся на пути вверх, стремясь приблизиться к высшему
уровню — человеку. Причем этот процесс (процесс градации) был за-
пущен при возникновении жизни и никогда не прерывался. Ученый
объяснял причины постоянной изменяемости внутренним стремле-
нием живого организма к совершенствованию и дополнительными
моментами, которые изложил в своих трех законах.
Первый из них — закон прямого приспособления — имел отноше-
ние к растениям и низшим животным, которые не перемещаются и
зависят от внешних факторов — питания, врагов, температуры, влаги,
освещения и т. д . Попав в неблагоприятную обстановку, они не могут
перейти в лучшее место, а потому вынуждены приспосабливаться к
тому, что есть. Например, растение, которое растет на хорошей, доста-
точно увлажненной почве и хорошо освещено, отличается от такого
же растения, попавшего на пересохший каменистый грунт, да еще и в
тени. В каждом случае растение должно приобрести ту организацию,
которая позволит жить максимально хорошо.
Второй закон — закон упражнения и неупражнения органов — от-
носится к подвижным животным. При изменении условий таких жи-
вотные изменяют привычки, подстраиваясь под ситуацию. А это вле-
чет за собой более частое употребление того или иного органа и его
упражнение. Например, птицы, что бродят вдоль берега в поисках
пищи, должны вытягивать ноги из ила, чтобы не увязнуть в нем. В ре-
зультате у них вытягиваются ноги. Подобным же образом можно объ-
яснить и длинную шею жирафа: ему постоянно приходится тянуться
к листве деревьев, которой он питается, поэтому шея находится в по-
стоянном упражнении и удлиняется. Ученый считал, что по нервам
движутся тонкие жидкости (флюиды), направляясь к функциониру-
ющему, а потому упражняемому органу. Силой воли животное на-
правляет эту жидкость к нужному органу, который при этом усиленно
718
ЭВОЛЮЦИЯ
питается и совершенствуется. Поэтому длинные рога у самцов оленей
отрастают в результате дополнительного притока к голове нужных ве-
ществ. Соответственно, при неупражнении органа происходит отток
от него флюидов, отчего тот атрофируется и даже исчезает. Этим Ла-
марк объяснял отсутствие крыльев у части насекомых или конечно-
стей у змей.
Оба эти закона показывают изменения, которые происходят у от-
дельных особей, но ничего не говорят о потомстве. Для этого Ламарк
предложил свой третий закон — закон наследования приобретенных
признаков. По этому закону все полезные для организма качества,
возникшие в результате прямого приспособления или упражнения
(или же неупражнения) органа, в том же виде передаются по наслед-
ству. Соответственно, потомок, унаследовавший от родителя измене-
ния признака, тренируя орган, в еще большей степени развивает его.
Следовательно, потомство всегда отличается от родителей, а потому
стабильного вида просто не может существовать.
Теория Ламарка была по-настоящему эволюционистской, по-
скольку ступени его лестницы связаны между собой историческим
родством. Он обосновывал развитие организации от простой к слож-
ной, что приводит к постоянному совершенствованию и является
причиной образования множества разновидностей организмов. Дру-
гой его заслугой было то, что он выделял адаптивную направлен-
ность развития, обусловленную выработкой приспособлений к
внешней среде. Вместе с тем Ламарк не смог разобраться в причинах
возникающих изменений. Оправданием тому служит отсутствие в то
время знаний закономерностей наследования. Признаки, которые
приобретаются в течение жизни без изменения генетического мате-
риала, — то, что мы сейчас называем модификациями, не могут на-
следоваться. Например, дети великого певца не обязательно унасле-
дуют вокальные данные и мастерство отца. Однако, по Ламарку,
такое было бы возможно. Стремление к совершенствованию означа-
ло признание существования в организмах некой таинственной
силы, определявшей развитие органа в ответ на внешний фактор.
Это вносило идеалистическую составляющую в его внешне матери-
алистическую теорию.
Последним этапом додарвиновского периода была первая полови-
на XIX в., когда бурно развивалась биология, возникли сравнитель-
ная морфология и анатомия, эмбриология, палеонтология и была со-
здана клеточная теория. Одним из основателей сравнительной
морфологии является немецкий поэт и ученый И.В. Гете, разработав-
ший учение о морфологическом типе. Большой вклад в формиро-
вание сравнительной анатомии внес французский ученый Этьен
Жоффруа Сент-Илер. Ему принадлежит концепция единого типа
719
ЭВОЛЮЦИЯ
строения, в соответствии с которой разные животные имеют подоб-
ные органы.
Другой французский исследователь Жорж Кювье обосновал
принцип корреляций (соотносительности), по которому ни одна из
частей организма не может измениться без того, чтобы не изменились
другие. Например, если зубы травоядного животного приспособлены
к перетиранию грубой пищи, то они должны быть крупными и распо-
лагаться на тяжелой массивной челюсти. Соответственно этому
укрупнение головы влечет усиление мускулатуры шеи и т. д . Органы
не только согласованы между собой, но и зависят от образа жизни.
Кювье был основоположником палеонтологии. Он открыл и описал
около 40 видов вымерших животных. Палеонтологические находки
неопровержимо свидетельствовали об историческом изменении видо-
вого состава фауны Земли, что требовало объяснения. Кювье предло-
жил теорию катастроф, согласно которой на Земле периодически
происходят мощные геологические процессы, которые приводят к вы-
миранию некоторых видов организмов, после чего создаются новые
виды и заселяют освободившееся пространство. Причины возникно-
вения этих видов Кювье не объяснял.
Общность внутренней организации живых организмов доказыва-
ла клеточная теория Т. Шванна, появившаяся в середине XIX в. Со-
гласно этой теории, все существа независимо от систематического по-
ложения имеют клеточное строение, причем все клетки имеют
принципиально сходное строение и образуются путем деления (ос-
новные положения клеточной теории описаны в разделе «Клетка»).
Важной предпосылкой эволюционной теории явилось развитие эм-
бриологии. Изучение онтогенеза показало, что все организмы возни-
кают из зиготы, которая затем делится и дает начало многим популя-
циям дифференцированных клеток. Было обнаружено глубокое
сходство зародышей на ранних этапах развития, причем высшие жи-
вотные в эмбриогенезе последовательно повторяли строение зароды-
шей предковых форм. Это нашло отражение в теории параллелизма.
Значительный вклад в становление эмбриологии внес отечественный
ученый К. Бэр, заметивший, что эмбрион высокоорганизованного
животного похож не на взрослую форму более примитивного живот-
ного, а на его эмбрион. Другой отечественный исследователь X. Пан-
дер открыл зародышевые листки, из которых развиваются зачатки
органов эмбриона.
Эволюционная теория Ч. Дарвина. Английский ученый Чарлз
Роберт Дарвин (1809—1882) не только предложил концепцию эво-
люционного преобразования организмов, но и попытался обосновать
причины их возникновения. Эволюционная теория закрепилась во
время путешествия на корабле «Бигль». Во время путешествия Дар-
720
ЭВОЛЮЦИЯ
вин наблюдал и документировал особенности фауны и флоры по-
сещаемых земель. Собранные им фактические данные заставили
усомниться в неизменяемости видов. Палеонтологические находки
указывали на историческую связь между вымершими животными и
современными. Дарвин обнаружил большие различия в видовом со-
ставе Южной и Северной Америки. Например, в Северной Америке
нет страусов, броненосцев, тапиров, ленивцев, обитающих в Южной
Америке. У Дарвина возникло предположение, что причиной этого
могло бы быть географическое разделение ранее единого органиче-
ского мира непреодолимыми препятствиями, вроде Мексиканского
плоскогорья. Поэтому животные и растения по обе стороны преграды
стали развиваться по-разному, что и предопределило возникшие ви-
довые различия. На Галапагосских островах были обнаружены жи-
вотные, которые существенно отличаются от материковых форм. Дар-
вин усомнился в том, что для каждого острова было создана
специальная разновидность, скорее всего, они сами изменились в ту
или иную сторону под воздействием факторов, которые еще надлежа-
ло установить. Особенно были заметны различия между вьюрками,
которые на каждом острове имели особенности в строении клюва, но
при этом наблюдались переходные формы. По возвращении домой
Дарвин не сомневался в том, что виды изменяются.
Искусственный отбор, происхождение культурных форм расте-
ний и животных. Для того чтобы лучше обосновать эволюционное
учение, Дарвин предвосхитил публикации глобальных выводов уче-
нием о доместикации (происхождении культурных форм растений и
животных). Уже в то время мало кто сомневался, что домашние расте-
ния и животные произошли от диких предков, о чем свидетельствова-
ло их внешнее сходство. Стараясь объяснить причины этого, Дарвин
обратил внимание на то, что в каждой группе домашних растений или
животных имеется несколько, а иногда даже множество форм. Напри-
мер, известны сотни пород собак, которые отличаются друг от друга
гораздо сильнее, нежели от своего предка — волка. Если организация
диких форм всегда в максимальной степени соответствует тем усло-
виям, в которых живет организм, то домашние формы нередко обла-
дают вредными для самостоятельного выживания качествами. На-
пример, домашние свиньи имеют столь значительный запас жира, что
с трудом передвигаются, и в естественной среде они неминуемо поги-
бли бы. Поскольку вся история домашних животных связана с чело-
веком, то роль последнего в становлении возникших особенностей
выглядела несомненной. Но в чем она выражалась?
Существовавшие в то время гипотезы объясняли это гибридизаци-
ей и «внезапным порождением» (т. е . мутациями). Однако Дарвин
пришел к выводу, что это лишь факторы, но не причина. Гибридиза-
721
ЭВОЛЮЦИЯ
ция не может создать те породы, которые возникли от одного-единст-
венного предкового вида. Например, все домашние голуби берут на-
чало от дикого скалистого голубя, а куры — от дикой индийской
курицы. А так как дикий вид один, то и гибридизации не может
быть — кого с кем скрещивать?! Не убеждало Дарвина и «внезапное
порождение», которое периодически случалось, у культурных форм.
Внезапное возникновение нового признака является лишь предпо-
сылкой возникновения новой породы, но не ее причиной. В дикой
природе также возникают подобные изменения, но мы не увидим ди-
ких баранов с короткими ногами, волков с короткими и кривыми но-
гами, как у таксы. Такие животные просто бы погибли вскоре после
рождения, потому что новый признак существенно снижал мобиль-
ность. Между тем для человека такие признаки вызывают интерес,
поэтому измененные животные положили начало новым породам.
Дарвин считал предпосылками эволюции культурных растений
и животных наследственность, в которой он выделял несколько форм:
неопределенную (индивидуальную), определенную (групповую),
коррелятивную и комбинативную. Неопределенная наследственность
имеет место лишь у отдельных особей, у которых возникает внезапно.
Каждый организм в чем-то индивидуален — все отличаются по разме-
рам, окраске, форме и т. д . Нам это знакомо по мутационной изменчи-
вости. Определенная наследственность всегда групповая. Можно
предположить, что если отощавшее стадо перевести на хорошее паст-
бище, все особи скоро наберут вес. Точно так же при голодании они
все похудеют. В генетике такая наследственность называется моди-
фикационной. Коррелятивная, или соотносительная, изменчивость,
по Дарвину, представляет собой комплексное изменение признаков.
Например, домашняя утка летает меньше дикой, но больше ходит, по-
этому у нее кости ног увеличиваются, а кости крыла — уменьшаются.
Комбинативная изменчивость возникает при скрещивании разных
сортов растений или пород животных. При этом у потомства возника-
ет некоторая комбинация признаков, присущая родителям.
Каким бы ни было важным значение той или иной формы наслед-
ственности, ни одна из них не может самостоятельно привести к обра-
зованию новых видов, потому что при бессистемном размножении
возникший признак постепенно бы затух. По мнению Дарвина, основ-
ной причиной эволюции культурных форм был искусственный отбор,
который совершался человеком. При этом человек руководствовался
не полезностью закрепляемого признака для самого организма, а ис-
ключительно своими потребностями — хозяйственными или эстети-
ческими. Поэтому признаки, часто ненужные или даже вредные для
организма (коротконогость, излишняя масса тела, голая кожа, махро-
вость цветка и т. д .), оказываются востребованными человеком. Ос-
722
ЭВОЛЮЦИЯ
мысленно отбирая для скрещивания только тех особей, которые обла-
дали нужными свойствами, человек постепенно добивался стойкого
закрепления признака в потомстве, которое уже в дальнейшем разво-
дилось в чистоте. В результате появились многочисленные культур-
ные формы, которые в силу возникших изменений не смогли бы са-
мостоятельно существовать, но, находясь на попечении человека,
длительно сохраняются в измененном виде. Таким образом, искусст-
венный отбор — процесс создания новых культурных форм путем по-
следовательного и систематического размножения особей с наиболь-
шей выраженностью полезных для человека признаков и строгой
изоляции всех остальных особей.
Дарвин различал сознательный и бессознательный искусственный
отбор. Сознательный, или методический, отбор происходит, если че-
ловек, производящий отбор, — селекционер, четко представляет себе
цель и планомерно отбирает для размножения нужные особи. Тогда
новая форма появляется относительно быстро, например куры с пе-
рьями на голове в виде бороды были выведены за шесть лет. Основ-
ным условием сознательного отбора является большое количество
исходных особей, среди которых отбираются производители. Чем
меньше особей, тем дольше формируется новый сорт или порода. При
бессознательном отборе человек изначально не ставит особых целей,
но при этом постоянно выбраковываются худшие особи. Например,
отбирая животное для пищи, забивают в первую очередь менее цен-
ные. На семена обычно оставляют лучшие плоды от самых мощных
растений.
Основные положения эволюционного учения. Толчок к форму-
лировке теории эволюции Дарвину дала книга священника Мальтуса
«Трактат о народонаселении», которая вышла в свет в 1778 г. В ней
автор убедительно описал мрачные перспективы перенаселения чело-
веческого общества, которое может случиться, если рождаемость ни-
чем не сдерживать. Перенеся рассуждения Мальтуса на все остальные
организмы, Дарвин счел их универсальными. Для примера он взял
слонов, которые отличаются низкой репродукцией: одна слониха ро-
ждает за всю жизнь (несколько десятилетий) не более шести детены-
шей, часть из которых гибнет. Однако если все детеныши выживут
и такая же судьба постигнет всех их потомков, то уже через 750 лет
будут одновременно жить 19 млн особей! Что же говорить о наиболее
плодовитых организмах. Например, рыба-луна мечет за сезон около
300 млн икринок. Дождевик образует 7,0 × 1011 спор. Не говоря уже
о бактериях, которые в благоприятной обстановке размножаются в ге-
ометрической прогрессии. В реальной жизни такого не бывает, пото-
му что значительная часть потомства гибнет от негативных факторов
(недоедания, хищников, болезней, наводнениий, пожаров и др.) .
723
ЭВОЛЮЦИЯ
Тем не менее численность вида поддерживается. Например, буревест-
ник, который несет за сезон всего одно яйцо, остается многочислен-
ной птицей.
Обнаружив, что реально всегда рождается больше потомства, чем
оно может выжить, Дарвин понял, что после рождения любой орга-
низм вступает в борьбу за жизнь с неблагоприятными факторами
природы. Ученый назвал такие взаимоотношения борьбой за сущест-
вование. Наиболее активная борьба идет между особями одного вида,
поскольку они обладают сходными потребностями и близкими воз-
можностями, или между близкими видами, тогда более сильный вы-
тесняет слабого. Например, серая крыса вытесняет менее крупную и
неагрессивную черную крысу, азиатский таракан благодаря своей ак-
тивности вытесняет более крупного черного таракана. Дарвин пони-
мал борьбу за существование не в виде обычной драки, а метафорич-
но, подразумевая под этим активность, направленную на выживание.
Выживание наиболее сильных и активных особей в борьбе за сущест-
вование Дарвин назвал естественным отбором. После гибели менее
подготовленных сородичей сильным остаются все пищевые ресурсы,
но более важно то, что именно они участвуют в половом размножении
и оставляют потомству свои признаки. Выжившие особи несут наибо-
лее удачное сочетание признаков, что дает возможность виду изме-
няться при изменении окружающих факторов. Согласно Дарвину,
в этом состоит основное творческое значение естественного отбора.
Основная мысль эволюционной теории Дарвина выражается концеп-
цией выживания наиболее приспособленных особей, передающих
«удачные» изменения потомству. Особенности искусственного и ес-
тественного отбора приведены в табл. 31.
Таблица 31
Сравнение искусственного и естественного отбора
(по Георгиевскому, с изменениями и дополнениями)
Показатель
для сравнения
Эволюция культурных
форм (искусственный
отбор)
Эволюция природных видов
(естественный отбор)
Материал для отбора Индивидуальная наслед-
ственность (в том числе
резкие уклонения)
Индивидуальная наследствен-
ная изменчивость (в основном
незначительные уклонения)
Фактор отбора
Человек
Борьба за существование на
фоне факторов живой и нежи-
вой природы
Характер действия
отбора
Накопление изменений в последовательном ряде поколений
Скорость действия
отбора
Действует быстро (мето-
дический отбор)
Действует медленно, эволюция
идет постепенно
724
ЭВОЛЮЦИЯ
Показатель
для сравнения
Эволюция культурных
форм (искусственный
отбор)
Эволюция природных видов
(естественный отбор)
Формы отбора
Сознательный; бессозна-
тельный
Движущий; стабилизирующий
Результаты отбора
Создание форм, полезных
человеку; образование
сортов и пород
Образование приспособлений
к окружающей среде; образо-
вание видов и более крупных
таксонов
Свою эволюционную теорию Дарвин изложил в книге «Происхо-
ждение видов путем естественного отбора, или Сохранение благопри-
ятствуемых пород в борьбе за жизнь». Кроме этого, Дарвин опублико-
вал еще два капитальных труда: «Изменения домашних животных и
культурных растений» (1868), положения которого позже составили
теоретическую основу научной селекции, и «Происхождение челове-
ка и половой отбор» (1871), где автор изложил свои взгляды естест-
венного происхождения человека.
МИКРОЭВОЛЮЦИЯ
Ю.А . Филипченко предложил разделить дарвиновское эволюци-
онное учение на два процесса: микроэволюцию и макроэволюцию.
Микроэволюция представляет собой совокупность необратимых про-
цессов, которые происходят внутри вида в пределах популяций. Они
ведут к дивергенции внутри вида и возникновению нового вида. Уче-
ние о микроэволюции составляет основу современного дарвинизма.
Макроэволюция представляет собой процессы дивергенции на уровне
таксонов выше вида, которые ведут к образованию новых родов, се-
мейств, отрядов и т. д .
Объектом эволюции является особь. Именно на уровне отдельной
особи имеют место различные типы наследственной изменчивости.
Неравноценность особей является основным условием осуществле-
ния естественного отбора и эволюции. Элементарной единицей эволю-
ции является популяция — совокупность особей одного вида, которая:
обитает на территории, входящей в ареал вида; находится в относи-
тельной изоляции от других популяций своего вида; обладает выра-
женной устойчивостью; проявляет способность к эволюции, посколь-
ку состоит из особей с неодинаковой приспособленностью к среде
обитания.
Элементарным эволюционным материалом служит мутация. Воз-
никшие изменения только тогда имеют эволюционный смысл, если
Окончание табл. 31
725
МИКРОЭВОЛЮЦИЯ
они передаются по наследству. И .И . Шмальгаузен обнаружил, что
любые мутации ведут к нарушению сбалансированности функций ор-
ганизма. Поэтому в доминантном состоянии они оказываются вред-
ными и не поддерживаются сразу естественным отбором. Однако ре-
цессивные мутации сохраняются у гетерозигот и передаются по
наследству. При последующем размножении на протяжении ряда по-
колений генотип перестраивается, и включение в фенотип нового ал-
леля уже не вызовет нарушений. Все неудачные варианты фенотипа
будут немедленно удалены отбором.
ВНИМАНИЕ!
Ареной эволюционного процесса является биогеоценоз. Ни одна особь,
популяция или вид не могут самостоятельно существовать без многочи-
сленных связей с организмами других видов.
Предпосылки (элементарные факторы) эволюции
Дарвин считал основными предпосылками эволюции наследст-
венную изменчивость и безудержное стремление организмов к раз-
множению. Позднее к ним добавили еще несколько факторов: нена-
следственную (модификационную) наследственность, изоляцию,
миграцию и динамику численности популяций. Об изменчивости
рассказано в соответствующих разделах генетики, поэтому здесь мы
рассмотрим только последние три.
Изоляция. Для осуществления микроэволюции необходимо, что-
бы вид был гетерогенен, т. е . состоял из популяций, в которых могли
бы накапливаться мутировавшие аллели. Для этого популяция долж-
на быть отделена какими-нибудь барьерами, препятствующими скре-
щиванию с особями других популяций этого вида. Сама по себе изо-
ляция не может создать новых форм, но она обеспечивает условия
для дивергенции. Различают два основных способа изоляции — гео-
графический и биологический.
Географический способ изоляции обеспечивает территориальное
разделение популяций. Она обеспечивается различными преградами
природного ландшафта (непреодолимые водные преграды, горные
хребты, пустыни, лесные массивы и т. д .), которые делят ареал вида на
изолированные фрагменты. Изоляция может также возникнуть, если
на промежуточных территориях внутри ареала гибнут популяции, но
сохраняются по его периферии. Так появились, в частности, европей-
ский и дальневосточный виды ландыша.
Биологический способ изоляции создает условия, которые не допу-
скают скрещивания, а если оно происходит, исключают развитие нор-
726
ЭВОЛЮЦИЯ
мального потомства. Выделяют несколько типов биологической изо-
ляции: экологическую, этологическую, морфофункциональную и
генетическую. За исключением этологической, все остальное имеет
сходное значение у растений и у животных.
Экологическая изоляция обеспечивается различными условиями
или сроками размножения, поэтому ее делят на биотопическую и се-
зонную. Биотопическая форма достигается различным место-
обитанием близких видов. Например, одни лягушки размножаются
в ручьях, а другие — в прудах и болотах. Сезонная форма экологиче-
ской изоляции возникает при разделении по сезонам полового раз-
множения. При этом даже небольшой временной сдвиг исключает
встречу потенциальных половых партнеров и их скрещивание. Так,
прудовая и озерная лягушки, которые часто обитают вместе, размно-
жаются при разной температуре воды, поэтому не могут скрещивать-
ся. У севанской форели известно пять рас, каждая из которых мечет
икру в разные месяцы года.
Этологическая изоляция наблюдается при несоответствии полово-
го поведения у особей разных видов. Например, у журавлей неодина-
ковые брачные танцы, движения которых понятны лишь партнеру
своего вида. Этот вид изоляции имеется только у животных. Морфо-
физиологическая изоляция связана с особенностями организации и
функционирования органов репродуктивной системы, в связи с этим
выделяют морфологический и физиологический типы. Морфоло-
гическая изоляция происходит, если строение половых орга-
нов или размеры и форма тела делают невозможным скрещивание.
Физиологическая изоляция препятствует осуществлению
оплодотворения (если половой акт все же произошел) или нарушает
развитие зародыша. При этом спермины могут погибнуть в половых
путях самки другого вида или не смогут проникнуть в ее яйцеклетку.
У растений при попадании на рыльце плодолистика пыльцы других
видов чужеродная пыльца погибает сразу или вскоре после прораста-
ния пыльцевой трубки. Генетическая изоляция обеспечивается разли-
чиями генетического материала у самца и самки. Это несоответствие
проявляется на разных этапах развития и может выражаться в невоз-
можности оплодотворения, гибели эмбрионов, нежизнеспособности
потомства или его стерильности (из-за нарушения мейоза гаметоге-
нез гибридного потомства неосуществим). Механизмами генетиче-
ской изоляции являются хромосомные перестройки, несовместимость
экспрессии отдельных генов после их мутации, полиплоидия.
Все типы изоляции увеличивают вероятность межродственного
скрещивания, повышая этим степень инбридинга в популяции. Изо-
ляция стимулирует процессы специализации и ведет к освоению но-
вых экологических ниш.
727
МИКРОЭВОЛЮЦИЯ
Миграция представляет собой процесс, противоположный изоля-
ции. Его сущность состоит в перемещении организмов или генератив-
ного материала (например, спор, семян, плодов и т. д .) . Эволюцион-
ное значение миграции заключается в распространении новых аллелей
и обновлении генофонда популяций, а также расширении ареала и
создании новых популяций. Миграции способствуют поддержанию
устойчивости вида, потому что межпопуляционные скрещивания
обеспечивают выравнивание генофонда. Известны два способа мигра-
ции — поток генов и интрогрессия генов. Поток генов происходит при
скрещивании особей разных популяций. При этом мигрирующие осо-
би могут включаться в состав популяции или только спариваться, по-
сле чего продолжать одиночную жизнь. Поток генов обеспечивает
распространение аллелей и обогащает генофонд. Интрогрессия генов
представляет собой обмен генов в результате межвидового скрещива-
ния. У растений межвидовое скрещивание происходит чаще, чем
у животных.
Динамика численности популяций (популяционные волны). Ни
одна популяция не может поддерживать свою численность на неиз-
менном уровне. Время от времени число особей увеличивается или
снижается из-за действия различных факторов. Масштаб изменения
численности может быть разным — от нескольких особей до кратного
исчисления (в десятки и сотни раз от исходного). Такие колебания
С.С. Четвериков назвал волнами жизни. Основной причиной изме-
нения численности популяций является кормовая база. При доста-
точном количестве пищи численность популяции возрастает, при де-
фиците пищи — снижается за счет гибели менее сильных и больных
особей. Популяционные волны служат причиной изменения границ
ареала. При увеличении численности ареал расширяется, увеличива-
ется плотность заселения. Когда численность падает, границы ареала
могут сужаться или он становится дробным, что приводит к геогра-
фической изоляции территориально удаленных популяций. Эволю-
ционное значение популяционных волн состоит в том, что они спо-
собствуют гетерогенности популяций. При увеличении численности
популяции в ней пропорционально возрастает концентрация мутаций
(если особей стало больше в 10 раз, то и мутаций станет в 10 раз боль-
ше). Последующее снижение численности будет сопровождаться ги-
белью особей с вредными мутациями и выживание носителей полез-
ных. Повторный всплеск численности повлечет распространение
полезных аллелей, что благоприятно для вида.
Движущие силы эволюции. Причинами или движущими силами
эволюционного процесса являются открытые Дарвиным борьба за су-
ществование и естественный отбор. Они ведут к выживанию наибо-
лее приспособленных особей, которые участвуют в размножении
728
ЭВОЛЮЦИЯ
и передают свои признаки потомству, тем самым увеличивая количе-
ство носителей наиболее удачных генотипов.
Борьба за существование
Борьба за существование представляет собой совокупность про-
цессов, в которые вступает организм с целью выживания и оставления
потомства. По мнению Дарвина, борьбу за существование вызывают
противоречия между потребностью организмов к неограниченному
размножению при ограниченных средствах к существованию. Борьба
за существование начинается в момент оплодотворения и даже до
него. Она затрагивает гаметогенез, конкуренцию спермиев, условия
оплодотворения и т. д . Этот процесс происходит на всех этапах онто-
генеза. Развитие зародыша зависит от того, насколько правильно и
последовательно включаются механизмы эмбриогенеза — дробления,
гистогенеза, органогенеза. При многоплодной беременности эмбрио-
ны конкурируют за необходимые для развития вещества, и детеныши
рождаются неодинаковыми. Борьба за существование позволяет раз-
решить противоречие между количеством родившихся и количеством
выживших особей, что обеспечивает оптимальную внутреннюю
структуру популяции и вида в целом. Борьба может быть внутриви-
довой и межвидовой. Внутривидовая борьба носит наиболее острый
характер, потому что в нее вступают особи с одинаковыми потребно-
стями и близкими возможностями. Различают две основные формы
борьбы за существование: конкуренцию и прямую борьбу. Между
ними не всегда удается провести четкую грань, поэтому часто такое
деление условно.
Конкуренция — это любые антагонистические отношения, в кото-
рые вступают организмы для достижения цели, без нанесения друг
другу серьезных повреждений. Это наиболее распространенный спо-
соб борьбы за существование, поскольку в ней проявляются главные
противоречия между организмами. Конкуренцию обусловливают
сходные потребности и одинаковые возможности их реализации. По-
этому в нее вступают организмы, которым есть что делить. Например,
волк и лисица могут конкурировать за пищу. Но этого незачем делать
воробью и кабану. В зависимости от цели различают трофическую,
топическую и репродуктивную конкуренцию. Каждая из них может
быть внутривидовой и межвидовой, а также групповой и индивиду-
альной.
Трофическая конкуренция основана на потребности организмов
в одинаковой пище. Внутривидовая конкуренция у животных обычно
возникает при высокой плотности популяции, а также в условиях
сталкивания пищевых интересов у видов с территориальным укладом
729
МИКРОЭВОЛЮЦИЯ
жизни. Примером одиночной внутривидовой конкуренции будет кон-
куренция за пищу между тиграми, охотничьи участки которых пере-
крываются. Групповой вариант такой конкуренции будет между
семьями волков, живущих на соседних участках. Межвидовая трофи-
ческая конкуренция происходит, например, между тигром и леопар-
дом, которые могут охотиться на одинаковую добычу, но в разное вре-
мя суток. Групповой вариант — прайд львов и стая гиен, оказавшиеся
поблизости.
Трофическая конкуренция есть и у растений. Она выражается
в конкуренции за минеральные вещества, свет и влагу, которые необ-
ходимы для фотосинтеза и жизнедеятельности. В лесу конкуренция
выражается в стремлении деревьев быстрее вырасти, чтобы не ока-
заться в тени более высокорослого дерева. Вблизи наиболее крупных
деревьев тропического леса нельзя встретить другие особи подобного
размера, потому что они не могут получить нужное количество сол-
нечного света. Но если дерево окажется поваленным бурей, то за ос-
вободившееся пространство будут конкурировать несколько деревьев
меньшего размера. Эту борьбу выиграет тот экземпляр, который быс-
трее вырастет и затенит своих конкурентов. Получая достаточно све-
та, это дерево сумеет наилучшим образом подготовить генеративный
материал, что может оказаться не по силам менее развитым особям.
Конкуренция не только позволяет выявить возможности организмов,
но и обусловливает совершенствование морфофизиологической орга-
низации.
Топическая конкуренция происходит, когда совместно обитающие
организмы подвергаются воздействию одинаковых факторов нежи-
вой природы (температура, ландшафт, недостаточное или избыточное
увлажнение, засоленность и т. д .). При этом организмы конкурируют
не с этими факторами, а между собой. Например, чтобы замаскиро-
ваться зимой, нужно иметь светлую окраску меха или перьев. При
этом животное становится малозаметным. Но если не будет врага, жи-
вотное выживет на снегу независимо от окраски. Более того, темный
мех был бы предпочтительнее, потому что он способствует сохране-
нию тепла. Конкуренция на фоне абиотических факторов чрезвычай-
но важна. Ее не сможет выдержать неподготовленный организм. На-
пример, если животное вовремя не перелиняло, то оно станет
заметным на снегу и возрастет вероятность быть съеденным хищ-
ником.
Репродуктивная конкуренция охватывает отношения между орга-
низмами в борьбе за осуществление размножения. Ее формы могут
быть весьма разнообразными, у животных они основываются обычно
на брачном поведении. У многих видов распространена конкуренция
за полового партнера (чаще всего самку), выражающаяся в турнир-
730
ЭВОЛЮЦИЯ
ных боях (олени), токовании (глухари, тетерева) и др. Репродуктив-
ная конкуренция часто носит групповой характер. Самец и самка
обычно вместе защищают потомство. Нередко птицы сбиваются
в стаю и гнездятся поблизости, чтобы при опасности объединиться.
У растений репродуктивная конкуренция выражается в борьбе за
опылителя. Для этого у многих имеются крупные яркие цветки или
соцветия. Опылителя также можно привлечь пыльцой и нектаром.
Образование большого количества спор или семян увеличивает шан-
сы на попадание в благоприятные условия и способствует расселе-
нию. Конкуренция выражается в способах распространения генера-
тивного материала. Те растения, у которых эти признаки выражены
лучше, имеют больше шансов на выживание потомства. Репродуктив-
ная конкуренция стимулирует совершенствование тех адаптаций, ко-
торые связаны с размножением.
Прямая борьба подразумевает непосредственные отношения с
факторами живой и неживой природы. Прямая борьба может привес-
ти к физическому ущербу и даже гибели организма.
Прямая борьба с биотическими факторами выражается в борьбе
с другими организмами (как своего вида, так и другого) за пищу и
возможность размножаться. Прямая трофическая борьба происходит
при непосредственном контакте хищника и жертвы, между хищника-
ми за пищу, между паразитом и хозяином. Подобно конкуренции, она
может быть групповой и индивидуальной. Примером групповой борь-
бы будет столкновение за добычу между двумя стаями волков, инди-
видуальной — схватка между двумя леопардами за добычу. Крайней
формой является каннибализм. Прямая репродуктивная борьба про-
исходит при конфликте за полового партнера, место размножения,
пищу для потомства. Самцы морских котиков постоянно охраняют
свою территорию с самками, при этом постоянно вступая в схватку
с другими самцами. У микроорганизмов распространена борьба меж-
ду штаммами посредством выделяемых антибиотиков (аллелопатия).
Не всегда можно отличить конкуренцию от прямой борьбы. Нередко
участник драки гибнет не сразу, а спустя некоторое время от болезни
или истощения.
Прямая борьба с абиотическими факторами связана с защитой от
вредных воздействий факторов неживой природы. В отличие от топи-
ческой конкуренции, этот вид прямой борьбы связан с непосредст-
венным действием фактора. Так, если белая зимняя окраска обеспечи-
вает незаметность на снегу (топическая конкуренция), то зимний мех
должен быть прежде всего густым и теплым, чтобы защитить от холо-
да (прямая борьба). Лиственница, в отличие от других хвойных дере-
вьев, сбрасывает к зиме листву. Тем самым она в максимальной степе-
731
МИКРОЭВОЛЮЦИЯ
ни оказывается подготовленной к зимовке и способна выдерживать
сверхнизкую температуру воздуха.
Борьба за существование является необходимой предпосылкой
для осуществления естественного отбора. Она демонстрирует нерав-
ноценность особей, выявляя слабых и наиболее подготовленных,
а также определяет характер и направление действия отбора. В итоге
это обеспечивает сохранение и доминирование наиболее жизнеспо-
собных популяций в пределах вида в случае внутривидовой борьбы
или же более конкурентоспособного вида над менее жизнеспособным,
который занимает ту же экологическую нишу, если борьба носит меж-
видовой характер.
Естественный отбор
Все организмы различаются между собой по тем или иным анато-
мо-физиологическим критериям (физическим возможностям, актив-
ности, обучаемости и т. д .). Это очень важно для эволюционного про-
цесса, поскольку делает популяции гетерогенными. Посредством
различных способов борьбы за выживание проявляется реальный
уровень приспособленности организма. Задачей естественного отбора
является удаление слабых и выживание самых сильных особей. От-
бор осуществляется на всех этапах индивидуального развития и даже
до образования зиготы, потому что в размножении участвуют только
особи, доказавшие свою жизнеспособность. Отбор наиболее эффек-
тивно действует по отношению к доминантным аллелям. Естествен-
ный отбор, который направлен на сохранение определенных феноти-
пов, называется положительным. Отбор, удаляющий из популяции
фенотипы, называется отрицательным. Естественный отбор характе-
ризуется вероятностью, накапливающим и интегрирующим действи-
ем, адаптивной направленностью.
Вероятностный характер отбора определяет степень возмож-
ности его осуществления. Вероятностность выражается в статис-
тичности и стохастичности. Статистичность характеризует пред-
полагаемую встречаемость события, происходящего в группе
организмов — популяции. Выживать должны наиболее сильные и ак-
тивные особи, поэтому статистически можно ожидать, что зиму пере-
живут именно они. Стохастичность показывает, насколько реально
предполагаемое событие может осуществиться в условиях действия
случайного фактора. Например, сильное и здоровое животное, кото-
рое выгодно отличается своими физическими данными, неожиданно
погибает в лесном пожаре или становится жертвой инфекции. В та-
ком случае ожидаемого события (т. е. выживания сильного) не прои-
зойдет.
732
ЭВОЛЮЦИЯ
Накапливающее действие отбора состоит в постепенном сумми-
ровании тех или иных полезных признаков, что ведет к усилению
органа и его функции. Такая особенность отбора была замечена
В. О . Ковалевским, который отмечал постепенное увеличение тела и
изменение зубов у предковых форм лошадиных.
Интегрирующее действие отбора заключается в постепенном объ-
единении возникших изменений в целостную систему. Поскольку лю-
бое изменение оказывает воздействие на другие признаки, оно в той
или иной мере изменяет весь организм. Так, легочное дыхание влечет
за собой изменение строения сердца, появление второго круга крово-
обращения, редукцию жаберного аппарата и др.
Адаптивная направленность отбора определяется тем, что отби-
раются только особи, обладающие наилучшими адаптациями.
Способы осуществления естественного отбора. Сущность естест-
венного отбора состоит в недопущении к размножению менее приспо-
собленных, а значит, менее ценных особей. Это достигается либо ги-
белью более слабых, либо конкуренцией с более сильными. Отбор
через гибель (элиминацию) особей может осуществляться избира-
тельно и неизбирательно. При избирательной элиминации на первый
план выступает уровень приспособленности к внешним факторам,
когда выживают наиболее приспособленные особи. Это наиболее цен-
ная для эволюции форма отбора. Она может быть прямой и косвен-
ной. Прямая избирательная элиминация выражается в гибели
особи в результате непосредственного действия фактора. Например,
хищнику легче догнать зайца, который бегает медленнее, поэтому та-
кие особи элиминируются, а выживают более быстроногие зайцы.
Примером действия абиотического фактора является гибель медве-
дей, которые не смогли достаточно хорошо подготовиться к спячке и
не накопили достаточного количества жира. Особи в популяции раз-
личаются по способности находить пищу, выдерживать голодание,
жажду и т. д.
При косвенной избирательной элиминации гибель особи
происходит не от первоначально действующего фактора, а от его по-
следствий. Примером косвенной элиминации будет гибель зайца, ко-
торый сумел избежать гибели от хищника, но получил ранения. Впо-
следствии инфицированные раны могут послужить действительной
причиной смерти животного, но эта причина опосредована ранее на-
несенными повреждениями.
Неизбирательная элиминация приводит к гибели особей независи-
мо от уровня их адаптивности, потому что зависит от случайных при-
чин или форс-мажорных обстоятельств. Например, гибель в результа-
те стихийного бедствия, отстрела охотниками и т. п . Однако и в этом
733
МИКРОЭВОЛЮЦИЯ
случае более приспособленная особь перед случайной смертью имеет
больше шансов оставить потомство и передать ему свои признаки.
Естественный отбор осуществляется путем выживания и раз-
множения одних особей за счет гибели других.
Формы естественного отбора. Введенное Дарвиным понятие ес-
тественного отбора впоследствии было развито многими учеными,
в том числе отечественными. И. И. Шмальгаузен выделил две формы
естественного отбора — движущий и стабилизирующий.
Движущий (ведущий) отбор обусловливает сохранение тех осо-
бей, адаптации которых соответствуют изменившимся условиям. На-
пример, при общем похолодании выживают особи с более густым и
теплым шерстным покровом. Таким образом, движущий отбор сохра-
няет мутации, смещающие норму реакции генотипа в нужном (полез-
ном) направлении. Известны три способа проявления движущего от-
бора: направленный, дизруптивный и транзитивный.
Направленный отбор происходит в условиях однонаправленного
изменения одного или нескольких внешних факторов. Например, при
использовании антибиотиков гибнет большинство микроорганизмов,
но наиболее устойчивые к этим антибиотикам выживают и, размно-
жившись, восстанавливают численность популяции. Для борьбы
с ними приходится увеличивать концентрацию антибиотика, но это
также не уничтожит все микроорганизмы. Таким образом, происходит
отбор, направленный на выживание микроорганизмов с мутационны-
ми отклонениями в сторону устойчивости к данному антибиотику.
Так появились штаммы, устойчивые к пенициллину, тетрациклину
и многим другим антибиотикам. Часто результат направленного отбо-
ра становится заметным очень скоро. Например, популяция комнат-
ной мухи всего за два поколения повышает устойчивость к дусту ДДТ
в 4000 раз! Направленный отбор, осуществляемый человеком, состав-
ляет основу искусственного отбора, поэтому он также дает быстрые
результаты.
Дизруптивный, или разрывающий, отбор направлен на сохранение
носителей крайних адаптивных уклонений за счет гибели особей с
фенотипами средней нормы реакции. Такой отбор происходит при
длительном воздействии разнонаправленных внешних факторов.
Примером дизруптивного отбора является появление рас у человека,
каждая из которых отражает оптимальную приспособленность к опре-
деленным экологическим факторам.
Транзитивный, или переходный, отбор имеет место в случае ин-
тенсивного размножения ранее малочисленной фенотипической
группы особей, адаптации которых более соответствуют резко изме-
нившимся условиям среды. В основе транзитивного отбора лежит
«транзитивный полиморфизм» — существование внутри вида различ-
734
ЭВОЛЮЦИЯ
ных фенотипических форм. Например, во время промышленной рево-
люции в странах Западной Европы произошло резкое ухудшение эко-
логической обстановки в индустриальных городах, связанное с
обильным задымлением. Отдыхающие на стволах бабочки березовой
пяденицы в подавляющем большинстве (примерно 98%) имеют свет-
лую окраску и на светлых стволах березы малозаметны. Но когда кора
деревьев потемнела от заводских выбросов, светлые бабочки стали
выделяться, и почти все были съедены птицами. Однако вид не погиб,
поскольку выжили темноокрашенные особи, которые ранее были
немногочисленными, поскольку склевывались птицами. На темных
стволах их окраска оказалась адаптивной, и они численно стали пре-
обладать в популяции. Это явление получило название индустриаль-
ного меланизма. Первые темноокрашенные особи были обнаружены
в Манчестере в 1848 г., а к 1895 г. их в популяции было уже 98%.
Транзитивный отбор, в отличие от дизруптивного, не расчленяет
популяцию на отдельные формы. Он также не накапливает полезные
изменения, как это происходит при направленной форме движущего
отбора. Транзитивный отбор обратим: когда экологическая обстанов-
ка в Европе улучшилась, в популяциях березовой пяденицы вновь
возросла доля светлоокрашенных бабочек.
Стабилизирующий, или центростремительный, отбор обеспечива-
ет выживание особей со средней выраженностью признака за счет
элиминации особей с любыми отклонениями от этого. Этот вид отбо-
ра удерживает норму реакции в определенных рамках и не позволяет
ей расшириться или сместиться в ту или иную сторону. Выделяют три
формы стабилизирующего отбора: канализирующий, нормализую-
щий и балансированный.
Канализирующий отбор позволяет закрепить модификации, кото-
рые наиболее адаптивны в действующих условиях обитания. Он обес-
печивает выживание особей с наиболее устойчивым онтогенезом,
способных противостоять случайным изменениям среды. Это сопро-
вождается сужением нормы реакции. Примером такого отбора явля-
ется устойчивость растений к осенней оттепели. Если потепление
спровоцирует цветение, то последующие холода погубят генератив-
ный материал и растения не оставят потомство. Поэтому у них выра-
ботались адаптации, позволяющие сделать индивидуальное развитие
независимым от случайных колебаний внешних факторов. Это назы-
вается автономизацией онтогенеза.
Нормализующий (поддерживающий) отбор также ведет к сохране-
нию средних значений нормы, отсекая крайние варианты, но при этом
не происходит сужения нормы реакции. В качестве примера предста-
вим гнездование птиц, у которых выведено разное количество птен-
цов: 3, 5 и 7. Если пищи много, то во всех гнездах птенцы будут разви-
735
МИКРОЭВОЛЮЦИЯ
ваться хорошо и выживут. В этой ситуации преимущество
у «многодетных» родителей — они оставляют больше потомства. При
недостаточной кормовой базе большой выводок обеспечить пищей
сложно, и повышается вероятность гибели всех птенцов. В изменив-
шихся условиях получают преимущество уже те пары, в гнезде кото-
рых птенцов мало. Однако птицы не всегда могут предугадать количе-
ство пищи, поэтому они будут откладывать различное количество
яиц, в зависимости от генетических особенностей родительских пар.
Балансированный (уравновешивающий) отбор происходит, когда
имеются несколько внутривидовых форм, различающихся между со-
бой по приспособленности к тем или иным условиям. Так, у наземной
улитки цепи различные особи имеют разнообразную окраску ракови-
ны — от светло-желтой и розовой до темно-коричневой и красной.
В зависимости от сезона преимущество получают улитки с той окра-
ской раковины, которая максимально сливается с окружающим фо-
ном. Осенью и весной это будут бурые и розовые особи, незаметные
на опавшей листве, летом — зеленые. В каждом случае адаптивная
окраска позволяет выжить той или иной части популяции, тогда как
выпадающие из фона улитки уничтожаются хищниками (певчим
дроздом). В результате в любых условиях вид поддерживает свою
численность.
Несмотря на то что движущий и стабилизирующий отборы проти-
воположны по своему результату, они тесно взаимосвязаны. При рез-
ком изменении условий обитания осуществляется движущий отбор,
который позволяет выжить особям с адаптивными отклонениями. Но
когда условия станут относительно стабильными, в дело вступает ста-
билизирующий отбор, поддерживающий оптимальную для этой ситу-
ации норму реакции генотипа.
Существование естественного отбора сейчас мало кем подвергает-
ся сомнению. Имеется немало экспериментальных доказательств. На-
пример, в одной из бухт Англии в течение нескольких лет измеряли
ширину головогрудного щита (карапакса) у обитавших там крабов.
Это совпало с постройкой мола, которая уменьшила циркуляцию
воды и вызвала ее помутнение. В таких условиях преимущественно
выживали особи с узким щитом, а крабы с широким карапаксом поги-
бали, потому что у них жабры сильнее засорялись меловой взвесью.
Другим доказательством служит пример с березовой пяденицей, ко-
торый был рассмотрен выше.
Адаптации (приспособления) и адаптациогенез
Адаптацией считается любая особенность особи, популяции, вида
или сообщества организмов, которая способствует успеху в конкурен-
736
ЭВОЛЮЦИЯ
ции и обеспечивает устойчивость к абиотическим факторам. Это по-
зволяет организмам существовать в данных условиях среды и остав-
лять потомство. Критериями адаптации являются: жизнеспособность,
конкурентоспособность и фертильность. Жизнеспособность — это
способность организма жить и нормально развиваться в условиях
среды. Конкурентоспособность — это способность организма доби-
ваться успеха в борьбе за средства жизни. Фертильность — это спо-
собность организмов нормально размножаться.
Виды адаптации. Адаптации делят на аккомодации и эволюцион-
ные адаптации. Аккомодации представляют собой обратимый процесс.
Они возникают при резком изменении условий среды. Например, при
переселении особи попадают в новую для них обстановку, но посте-
пенно привыкают к ней. Эволюционная адаптация необратима, и воз-
никшие изменения генетически закрепляются. Сюда относят все при-
способления, на которые действует естественный отбор. Например,
покровительственная окраска или быстрый бег. Приспособления так-
же делят на организменные и видовые. Организменные адаптации
подразделяют на морфологические, физиологические, биохимические
и этологические.
Морфологические адаптации проявляются в покровительствен-
ной окраске, позволяющей быть незаметным среди окружающего
фона; предостерегающей окраске, которая предупреждает потенци-
ального врага о наличии защитных механизмов; мимикрии — внеш-
нем сходстве беззащитных видов с видом, обеспеченным защитой;
маскировке — особенности строения тела, позволяющей быть похо-
жим на окружающие предметы; преимуществах строения — опти-
мальные пропорции тела, расположение и густота волосяного или
перьевого покрова и т. п.
Физиологические адаптации обеспечивают функциональные преи-
мущества организма. Их условно разделяют на статические (посто-
янные физиологические параметры — температура тела, водно-соле-
вой баланс, концентрация сахара и т. п .) и динамические (адаптации
к колебаниям действия фактора — изменение температуры, влажно-
сти, освещенности, магнитного поля и т. п .).
Биохимические адаптации обеспечивают оптимальное течение
биохимических реакций в клетке (упорядочение ферментативного ка-
тализа, специфическое связывание газов дыхательными пигментами,
синтез нужных веществ в определенных условиях и т. п .).
Этологические адаптации представляют собой поведенческие ре-
акции, направленные на выживание отдельных особей и вида в целом.
Такими реакциями являются: поведение при поиске пищи и полового
партнера, спаривание, выкармливание потомства, избегание опасно-
сти и защита жизни в случае угрозы, агрессия и угрожающие позы
737
МИКРОЭВОЛЮЦИЯ
и многие другие. Некоторые поведенческие реакции наследуются
(инстинкты), другие приобретаются в течение жизни (условные
рефлексы). У различных организмов соотношение инстинктивного
и условно-рефлекторного поведения неодинаково. Например, у бес-
позвоночных и низших хордовых преобладает инстинктивное поведе-
ние, а у высших млекопитающих (приматов, хищных) — условно-
рефлекторное. Высший уровень поведенческой адаптивности,
основанный на механизмах высшей нервной деятельности, имеется
у человека.
Видовые адаптации обнаруживаются при анализе группы особей
одного вида. Основными из них являются различные конгруэнции,
уровень мутабильности, внутривидовой полиморфизм, уровень чи-
сленности и оптимальная плотность населения.
Конгруэнции представляют собой все морфофизиологические и
поведенческие особенности, которые способствуют существованию
вида как целостной системы. Репродуктивные конгруэнции обес-
печивают размножение. Некоторые из них непосредственно связаны
с репродукцией (соответствие половых органов, приспособления
к вскармливанию и др.), тогда как другие лишь опосредованно (раз-
личные сигнальные признаки: зрительные — брачный наряд, ритуаль-
ное поведение; звуковые — пение птиц, рев самца оленя во время гона
и др.; химические — различные аттрактанты, например, феромоны на-
секомых).
К конгруэнциям относят все формы внутривидовой кооперации —
конституциональной, трофической и репродуктивной. Конститу-
циональная кооперация выражается в согласованных дейст-
виях организмов в неблагоприятных условиях, которые повышают
шансы на выживание. Например, овцы в холодное время сбиваются
в тесную группу.
Трофическая кооперация состоит в объединении организ-
мов с целью более продуктивного добывания пищи. Например, стая
волков гораздо эффективнее охотится, нежели отдельная особь. При
этом у многих видов имеет место разделение обязанностей: одни осо-
би отделяют выбранную жертву от основного стада и гонят ее в заса-
ду, где затаились их сородичи и т. д .
Репродуктивная кооперация повышает успешность раз-
множения и способствует выживанию потомства. У многих птиц осо-
би собираются на токовищах, и в таких условиях облегчается поиск
потенциального партнера. То же самое происходит на нерестилищах,
лежбищах ластоногих и др. Вероятность опыления у растений повы-
шается, когда они растут группами и расстояние между отдельными
особями невелико.
738
ЭВОЛЮЦИЯ
Мутабильность представляет собой частоту возникновения мута-
ций в единицу времени (количество поколений) и на один ген. Для
каждого вида характерна своя частота, которая определяется уровнем
стабильности генетического материала и устойчивостью к мутагенам.
Мутации делают популяции гетероморфными и дают материал для
осуществления отбора. Для вида опасна как чрезмерно высокая, так и
недостаточная мутабильность. В первом случае возникает угроза це-
лостности вида, а во втором — невозможность осуществления отбора.
Внутривидовой полиморфизм обусловливает уникальность сочета-
ния аллелей у разных особей. Причиной полиморфизма являются
половое размножение, которое обеспечивает комбинативную измен-
чивость, и мутации, изменяющие субстрат наследственности. Поддер-
жание внутривидового полиморфизма обеспечивает устойчивость
вида и его существование в различных условиях.
Уровень численности определяет крайние значения количества
особей вида. Снижение численности ниже порогового уровня ведет
к гибели вида. Это связано с невозможностью встречи партнеров, на-
рушением внутривидовых адаптаций и др. Чрезмерное увеличение
численности также вредно, поскольку подрывает кормовую базу, спо-
собствует накоплению в популяции больных и ослабленных особей,
приводит к развитию стресса. Оптимальная плотность населения по-
казывает специфические для каждого вида особенности сосущество-
вания особей. Многие организмы предпочитают одиночный образ
жизни и встречаются лишь для спаривания (тигры, леопарды, самцы
слонов и др.) . У других сильно выражен инстинкт коллективности,
поэтому они нуждаются в высокой численности. Например, самые
многочисленные группы среди позвоночных образовывали американ-
ские странствующие голуби, стаи которых насчитывали миллиарды
(!) особей. После того как человеком была подорвана их численность,
странствующие голуби перестали размножаться — и вид исчез.
Адаптации не бывают идеальными или универсальными — ка-
ждая из них облегчает выполнение лишь определенной функции.
Например, длинные крылья стрижа, позволяющие ему быстро ле-
тать, затрудняют взлет с ровной поверхности. Есть птицы, которые
поедают ос и пчел, а также мух, мимикрирующих под них. Поэтому
любой адаптивный признак оказывается целесообразным лишь в
определенной среде. При резком изменении условий обитания чрез-
мерно развитые признаки могут оказаться нецелесообразными и
принести вред организму. Поэтому после глобальных экологических
катастроф преимущественно погибают высокоспециализированные
виды (например, динозавры в меловом периоде палеозойской эры).
То же самое происходит при отрицательном действии биотических
факторов, например гигантский торфяной олень был полностью
739
МИКРОЭВОЛЮЦИЯ
уничтожен хищниками из-за своих огромных рогов, сделавших его
малоподвижным.
Адаптациогенез. Происхождение адаптации связано с мутацион-
ной изменчивостью, которая ведет к появлению новых признаков
или изменению старых, и действием отбора. Однако признак должен
не только появиться, но и быть наследственно закреплен, а также
распространен в популяции. В процессе образования адаптации вы-
деляют три фазы: инадаптивную, преадаптивную и постадаптивную.
Инадаптивная фаза имеет место при резких изменениях условий
среды обитания. При этом может погибнуть значительная часть по-
пуляции, но часть особей, фенотип которых наиболее соответствует
сложившимся условиям, выживает и дает потомство, в генотипе ко-
торого будут закреплены предпосылки нужных качеств. Преадап-
тивная фаза наступает после элиминации особей с неподготовлен-
ным фенотипом. Она определяется генетическими факторами
(мутационной и комбинативной изменчивостью) совместно с дейст-
вием естественного отбора. В эту фазу предпосылки преобразуются
в реальную адаптацию, которая отвечает возникшим условиям сре-
ды. При этом отбор обязательно меняет свое направление (напри-
мер, стабилизирующий сменяется движущим). Постадаптивная
фаза представляет собой постепенное совершенствование появив-
шейся адаптации. При этом признаки, которые были раньше полез-
ны, могут оказаться ненужными. В таком случае происходит их по-
степенная редукция.
Критерии и признаки вида
Понятие вида является одним из главных в биологии. Однако не-
возможно дать определение вида лишь по одной его особенности. Для
этого необходимо рассмотреть основные его критерии, главными из
которых являются морфологический, физиолого-биохимический,
эколого-географический и генетико-репродуктивный.
Морфологический критерий характеризует вид по внешнему и вну-
треннему строению его особей. Этот критерий не абсолютен, потому
что существуют морфологически сходные виды-двойники (например,
шесть самостоятельных видов комаров из рода Anopheles). Физиоло-
го-биохимический критерий демонстрирует особенности физиологи-
ческих процессов, которые присущи разным видам. И этот критерий
не всегда точен, поскольку многие процессы в природе универсальны
(генетический код, биосинтез белка и др.). Эколого-географический
критерий характеризует вид в соответствии с территорией его обита-
ния (географическая зона) и занимаемой им экологической нишей.
Оба эти аспекта включаются в понятие адаптивной ниши (не путать
740
ЭВОЛЮЦИЯ
с экологической нишей!). Поскольку существуют виды-космополиты,
которые широко распространены на разных континентах (земляные
черви, комары, мыши; среди растений — береза, ель и др.), этот крите-
рий также не абсолютен. Генетико-репродуктивный критерий осно-
ван на механизмах генетической изоляции, препятствующей скрещи-
ванию между особями разных видов. Однако межвидовые гибриды
все-таки могут быть. Вдобавок этот критерий неприменим к организ-
мам с бесполым размножением.
Таким образом, ни один из перечисленных критериев не может
быть единственным и всеобъемлющим. Вид можно охарактеризо-
вать лишь по совокупности всех критериев. Основными признаками
вида, отражающими особенности этого уровня организации жизни,
являются дискретность, целостность, устойчивость, численность и
историчность. Дискретность показывает факт существования обо-
собленности рассматриваемого вида от других видов в соответствии
с перечисленными выше критериями. Целостность демонстрирует
сложную внутреннюю структуру вида, представляющую собой хоро-
шо организованную систему, в которой внутривидовые формы (по-
пуляции, расы, подвиды) объединены различного типа внутривидо-
выми адаптациями. Численность демонстрирует, что вид состоит не
из одной особи, а из множества. Устойчивость показывает способ-
ность вида существовать длительное время. Причиной этому служит
смена поколений, при этом продолжительность жизни особи огра-
ничена, но после нее остается потомство, которое также участвует
в размножении. Историчность показывает относительность устой-
чивости вида. На Земле существовало много видов растений, живот-
ных и других организмов, которые не сохранились до наших дней.
Так же непостоянны и ныне живущие виды. Именно устойчивость
и историчность вида вызывают наибольший интерес среди эволюци-
онистов.
Видообразование
Образование нового вида представляет собой конечный результат
микроэволюционного процесса. В зависимости от того, как возникает
вид, различают два основных способа видообразования: аллопатриче-
ский и симпатрический. Аллопатрическое (географическое) видообра-
зование происходит в случае глубокого преобразования внутривидо-
вых форм, изолированных территориально (реки, моря, леса, горы
и т. п .). Этот способ видообразования основан на дивергенции и при-
водит к адаптивной радиации (расхождению форм, приспосабливаю-
щихся к разным экологическим нишам). Известным примером дивер-
741
МАКРОЭВОЛЮЦИЯ
гентного видообразования являются галапагосские вьюрки, описанные
Дарвином.
Симпатрическое видообразование происходит на одном ареале с
материнской популяцией без территориальной изоляции. Причиной
возникновения нового вида в этом случае служит межвидовая гибри-
дизация, когда в потомстве объединяются два разных генотипа. Если
гибридная форма оказывается фертильной и конкурентоспособной,
действие естественного отбора приведет к появлению самостоятель-
ного вида. Этот способ видообразования называется синтезогенезом,
или сетчатой эволюцией. Таким образом возникла культурная слива:
при гибридизации терна (2n = 16) и алычи (2n = 8) при последующей
полинлоидизации (у сливы 2n = 48). Другим путем симпатрического
видообразования являются появление хромосомных перестроек и по-
липлоидия. Например, у хризантем все виды имеют кратное число
хромосом: 9, 18, 27, 36, 45...
Иногда рассматривают экологическое видообразование, которое
осуществляется через формирование в пределах популяции экотипов,
которые затем преобразуются в самостоятельные виды. Основными
факторами такого видообразования служат различные формы эколо-
гической изоляции.
Микроэволюция повышает уровень относительной приспосо-
бленности организмов к внешним условиям и создает многообразие
видов.
МАКРОЭВОЛЮЦИЯ
Макроэволюция представляет собой совокупность процессов, ко-
торые ведут к образованию надвидовых таксонов: рода, отряда, клас-
са, типа. Четкой границы между микро- и макроэволюцией не сущест-
вует, поскольку процесс дивергенции, стимулирующий появление
новых видов, продолжается без перерыва и в возникших формах. По-
этому макроэволюция не имеет собственных механизмов и осуществ-
ляется через процессы микроэволюции.
Формы макроэволюции. Различают несколько форм макроэво-
люции: филитическая, дивергентная, конвергентная, параллелизм и
сетчатая эволюция. Филитическая эволюция происходит в случае
последовательного преобразования единого ствола или отдельной
ветви филогенетического древа. Монофилия означает развитие так-
сона от одной предковой формы, а полифилия — от двух и более.
Большинство специалистов склоняются к монофилическому разви-
тию организмов от общей предковой формы, основываясь на уни-
версальных механизмах наследственности и изменчивости, клеточ-
742
ЭВОЛЮЦИЯ
ной организации, общности основных физиологических процессов.
Таксоны могут возникать и путем полифилии. Дарвин допускал та-
кую возможность только в отношении образования царств. Он счи-
тал, что растения и животные произошли от четырех-пяти родона-
чальных форм.
Дивергенция (основной путь эволюции) — это независимое обра-
зование различных признаков у родственных групп. Например, у всех
рыб газообмен осуществляется через жабры, позвоночный столб диф-
ференцирован лишь на туловищный и хвостовой отделы, соматиче-
ская мускулатура сегментирована и т. д . Частная адаптация к опреде-
ленным условиям приводит к появлению гомологичных органов,
которые имеют одинаковое происхождение, но выполняют различные
функции (например, передние конечности лошади, летучей мыши,
обезьяны и кита).
Конвергенция (схождение признаков) — это независимое форми-
рование сходных признаков у неродственных организмов, которые
обитают в одинаковой среде. Например, все животные, живущие
в лесу, имеют сходную защитную окраску, которая сезонно меняется.
При конвергентной эволюции у неродственных организмов образу-
ются аналогичные органы, которые имеют разное происхождение, но
выполняют одинаковую функцию. К таким органам относят колюч-
ки у растений, имеющие листовое (барбарис), прилистниковое (бе-
лая акация), побеговое (боярышник) происхождение или образуют-
ся из кончиков проводящих жилок (осот); у животных — плавники
рыб и китов, крылья у насекомых, птиц и летучих мышей и др.
Параллелизм (параллельное развитие) проявляется в сходном раз-
витии нескольких родственных организмов, которые первоначально
дивергировали, но обитают в одинаковых условиях. Примером явля-
ются сходные черты внешнего строения у разных семейств ластоно-
гих: тюленя, моржа и морского котика. Генетическое обоснование па-
раллельной эволюции дал Н. И. Вавилов в законе гомологических
рядов наследственной изменчивости. Согласно этому закону, «виды
и роды, генетически близкие, характеризуются сходными рядами на-
следственной изменчивости с такой правильностью, что, зная ряд
форм в пределах одного вида, можно предвидеть нахождение парал-
лельных форм у других видов и родов. Чем ближе генетически распо-
ложены в общей системе роды и линнеоны (совокупность морфоло-
гически сходных и близкородственных групп растений не обязательно
равноценных, но объединяемых в общий вид), тем полнее сходство
в рядах изменчивости». Сетчатая эволюция основана на механизмах
синтезогенеза, описанных выше при обсуждении симпатрического
видообразования. Макроэволюционные преобразования такого типа
могут осуществляться посредством гибридизации, симбиогенеза
743
НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
и трансдукции. Г ибридизация — это объединение генетических
признаков разных таксонов. Симбиогенез — это функциональное
объединение неродственных организмов на основе получения взаим-
ной пользы. Трансдукция — это перенос наследственной инфор-
мации вирусом из одного организма в другой.
НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
Основными направлениями эволюции являются ароморфоз,
идиоадаптация и катаморфоз (дегенерация).
Ароморфоз, или морфофизиологический прогресс, представляет со-
бой крупномасштабные морфофизиологические изменения, которые
ведут к общему подъему организации. Такие адаптации оказываются
полезными в самых разнообразных условиях среды. Примерами аро-
морфозов являются изменения, которые произошли у водных живот-
ных, перешедших к наземному образу жизни: разделение венозной
и артериальной крови, легочное дыхание, дифференциация позвоноч-
ного столба и появление конечностей пятипалого типа. У растений
ароморфозы привели к образованию тканей и вегетативных органов,
пыльцевой трубки, и семенному размножению и др. Особенностью
ароморфозов является то, что они охватывают все системы организ-
ма. Например, появление теплокровности обеспечило интенсифика-
цию деятельности всех клеток.
Идиоадаптация, или алломорфоз, представляет собой развитие
частных адаптаций, которые приспосабливают организм к конкрет-
ным условиям среды. Поэтому идиоадаптивные преобразования яв-
ляются более узкими, чем ароморфные. Идиоадаптации возникают на
основе исходной ароморфной организации и ведут к освоению новых
экологических ниш. Например, комплекс преобразований, возникших
у земноводных при выходе на сушу, являются ароморфозами, но в
дальнейшем земноводные приспособились жить в различных услови-
ях, вырабатывая для этого частные адаптации: преимущественно вод-
ные формы (тритоны, саламандры) сильно зависят от кожного дыха-
ния и часто имеют выросты на теле, увеличивающие поверхность
газообмена; древесные лягушки имеют на лапках присоски, позволя-
ющие им удерживаться на листьях и т. д . При идиоадаптации аро-
морфные преобразования не утрачивают своего значения: так, для
всех млекопитающих характерны основные признаки, выработанные
ароморфозами, — теплокровность, вскармливание детенышей моло-
ком, развитый головной мозг и др.
744
ЭВОЛЮЦИЯ
Катаморфоз (дегенерация), или морфофизиологический регресс,
охватывает все изменения, которые ведут к упрощению организации
и образа жизни организма. Нередко такие преобразования оказыва-
ются адаптивными, особенно для паразитических и малоактивных
форм. Например, утрата ленточными червями пищеварительной по-
лости является адаптацией, поскольку эти животные обитают в тон-
кой кишке, где ферменты хозяина расщепляют полимеры пищи. По-
этому вся поверхность тела червя действует на манер слизистой
оболочки кишки и всасывает нужные вещества. Пищеварительный
тракт в таких условиях паразиту попросту не нужен. У сидячих форм
редуцируются органы передвижения, у пещерных и роющих — орга-
ны зрения и т. д . Дегенерация одних структур при этом сопровожда-
ется выработкой частных приспособлений (идиоадаптация). Напри-
мер, у паразитических червей образуются прикрепительные структуры
(присоски, крючья), у сидячих полихет параподии передних сегмен-
тов разрастаются и участвуют в захвате пищи. Морфофизиологиче-
ский регресс всегда вторичен — он направлен на упрощение уже
существующей организации, если условия среды упрощаются (на-
пример, ленточные черви получают вещества, уже расщепленные до
мономеров). В природе такие изменения очень распространены.
В частности, паразитические виды имеются в 2500 родах покрытосе-
менных растений, во всех типах и классах простейших, в 13 из
24 классов червей, в 5 из 9 классов членистоногих! Поэтому морфо-
физиологический регресс — это адаптивный процесс, который повы-
шает уровень приспособленности организма в соответствующей нише.
БИОЛОГИЧЕСКИЙ ПРОГРЕСС
И БИОЛОГИЧЕСКИЙ РЕГРЕСС
Учение о биологическом прогрессе разработал отечественный уче-
ный А.Н . Северцов, который разделил понятия морфофизиологиче-
ского и биологического прогресса. Морфофизиологический про-
гресс — ароморфоз — заключается в усложнении организации, тогда
как биологический прогресс включает в себя все преобразования, ко-
торые отвечают трем критериям, выделенным Северцовым: нарастаю-
щее увеличение численности особей; расселение особей в новые места
обитания, ведущее к расширению ареала; появление внутривидовых
форм с последующей их эволюцией. Биологический прогресс охваты-
вает не только ароморфоз и идиоадаптацию, но и катаморфоз (все эти
термины предложены Северцовым), если возникшие изменения по-
лезны для вида и обеспечивают его процветание. Общее направление
прогрессивной эволюции живой природы называется орогенезом.
745
ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ
Биологический регресс представляет собой процессы, противопо-
ложные биологическому прогрессу. Он характеризуется уменьшени-
ем численности особей, сужением ареала, постепенным уменьшением
видового многообразия и внутривидовых форм. Как и биологический
прогресс, биологический регресс проявляется не только в упрощении
организации, но и во всеусложнении, если это снижает защищенность
группы. По мнению И. И. Шмальгаузена, возможность и вероятность
биологического регресса зависят от направления адаптивной специа-
лизации группы. В соответствии с этим он выделил два направле-
ния — теломорфоз и гиперморфоз.
Теломорфоз — это направление эволюции в сторону узкой специали-
зации к условиям обитания. Такие адаптации часто становятся тупико-
выми и в дальнейшем не прогрессируют. Они могут сопровождаться
частичной или полной редукцией органов и признаков или крайней
формой специализации (миноги, глубоководные рыбы, змеи, хамелео-
ны, кроты, ленивцы и др.) . Формирование крайних адаптаций называ-
ется телогенезом. Теломорфоз усиливается в случае ограничения среды
обитания (например, у роющих форм — кротов, слепышей и др.) . Узкая
специализация еще не означает скорой гибели вида или группы — этого
не произойдет, если не случится резкого изменения условий обитания.
Гиперморфоз — это чрезмерное увеличение размеров организма
или отдельных органов без корреляции с другими частями организма.
Множество фактов гиперморфоза собрано палеонтологами. Среди
них гигантские динозавры, саблезубые кошки с огромными клыками,
торфяной олень титанотерий с трехметровыми рогами и др. Совре-
менным примером гиперморфоза являются жирафы, длинная шея
которых позволяет легко доставать листья высоко расположенных
веток деревьев. Однако жирафы не в состоянии щипать траву
и исчезновение деревьев неминуемо приведет к их гибели.
ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ
Процесс эволюции отмечается рядом закономерностей. Во-пер-
вых, эволюция отличается прогрессивной направленностью (ароге-
нез). Усложнение организации происходит значительно чаще, чем
упрощение. Усложнение наиболее выражено у человека. Арогенез
проявляется в нарастании ускорения развития жизни. Если на появ-
ление аэробных организмов ушло более 2 млрд лет, то на развитие
человека пришлось лишь около 2 млн лет. Во-вторых, для историче-
ского развития организмов характерна смена направлений эволюции.
Ароморфоз, дающий начало новым, более высокоорганизованным
группам, закономерно сменяется выработкой частных адаптаций по-
746
ЭВОЛЮЦИЯ
средством идиоадаптации или адаптивной дегенерации. Причем ал-
ломорфоз происходит гораздо чаще, чем ароморфоз. Переход от спе-
циализации к ароморфозу возможен через смену и расширение
функций, а также через неотению. В-третьих, эволюцию отличает не-
обратимость и неравномерность. Дарвин считал положение о не-
обратимости применимой как для видов, так и для более крупных
групп. Впоследствии Л. Долло развил его, поэтому принцип необра-
тимости назван законом Долло. Принцип неравномерности эволю-
ции обосновывает различную скорость адаптивных преобразований
тех или иных групп. Такую неравномерность назвали «мозаичной
эволюцией». Дж. Симпсон выделил три формы эволюции в зависи-
мости от скорости преобразований: брадителическую, горотеличе-
скую и тахителическую. Брадителическая эволюция осуществляется
наиболее медленно. Горотелическая эволюция осуществляется сред-
ними темпами. Тахителическая эволюция отличается высокой скоро-
стью. Она часто ведет к массовому видообразованию, которое случа-
ется, если ароморфная группа попадает в новую среду обитания и
бурно осваивает различные экологические ниши. Высокую скорость,
большой масштаб и прогрессивность тахителической эволюции
Симпсон выразил в термине «квантовая эволюция». Скорость эво-
люционных преобразований зависит от внутренних факторов груп-
пы и действующих на нее внешних факторов. К внутренним фак-
т о рам относят интенсивность мутационного процесса (чем она
выше, тем эволюция идет быстрее), возможность гибридизации,
продолжительность смены поколений (чем быстрее сменяются по-
коления, тем быстрее эволюционный процесс). Внешними факто-
рами являются факторы внешней среды. Частое изменение усло-
вий обитания стимулирует движущий отбор и способствует
видообразованию. Результатом эволюции является возникновение
адаптаций и их совершенствование.
ДОКАЗАТЕЛЬСТВА ЭВОЛЮЦИИ
Эволюционные преобразования во всех случаях незаметны для
прямого наблюдения. Однако существование этого процесса доказы-
вается множеством общих признаков, которые имеются у организмов
разных систематических групп. Среди доказательств исторического
развития организмов и их филогенетической связи главными явля-
ются: палеонтологические, сравнительно-эмбриологические, сравни-
тельно-анатомические, клеточные, генетико-биохимические и биогео-
графические.
Палеонтологические доказательства получены в результате иссле-
дования ископаемых остатков вымерших организмов. Установлены
747
ДОКАЗАТЕЛЬСТВА ЭВОЛЮЦИИ
основные этапы исторического развития в ходе геологических перио-
дов, темпы и направления. Исследование остатков показало, что эво-
люция шла по пути постепенного усложнения организации. Палеон-
тологические находки выявили предковые формы современных
организмов, они показали последовательность их появления на Земле
и исчезновения.
Сравнительно-эмбриологические доказательства демонстриру-
ют общность гаметогенеза и ранних стадий онтогенеза у представи-
телей разных систематических групп, а также показывают соотно-
шение филогенеза и онтогенеза. Первым это сходство заметил
Аристотель. В 1828 г. Бэр показал, что эмбрионы разных системати-
ческих групп имеют между собой гораздо больше сходства, чем
взрослые формы тех же видов. При этом общая закономерность со-
стоит в последовательности появления признаков: сначала появля-
ются общие признаки типа, затем класса, отряда и последними по-
являются признаки вида. Таким образом, в индивидуальном
развитии особи можно наблюдать основные этапы исторического
развития всего вида. Наибольшее сходство эмбрионов на ранних
этапах и дивергенция на поздних стадиях показывают эволюцион-
ную дивергенцию вида. Выявленную Бэром взаимосвязь индивиду-
ального (онтогенеза) и исторического (филогенеза) развития Дар-
вин назвал законом зародышевого сходства. В 1864 г. Мюллер
предположил, что филогенетические изменения проявляются в он-
тогенезе двумя способами. Во-первых, эмбриогенез у различных
групп идет одинаково до тех пор, пока у более высокоорганизован-
ного представителя появится новый признак, который отсутствует
у предковой формы. Во-вторых, у более высокоорганизованных
форм развитие идет дольше, потому что прибавляются новые ста-
дии, отсутствующие у более примитивных форм. На основании вы-
водов Мюллера Геккель сформулировал биогенетический закон: он-
тогенез есть краткое и быстрое повторение филогенеза. Общие
признаки эмбрионов он назвал ценогенезами (образование желтка
в яйце, яйцевых оболочек, аллантоиса и др.), а признаки предковых
форм — палингенезами (формирование жаберных щелей и дуг, хор-
ды, однокамерного сердца и др.) .
Сравнительно-анатомические доказательства показывают общие
признаки строения разных групп. Такая общность проявляется в ор-
ганизации систем органов у различных позвоночных, в развитии го-
мологичных органов, рудиментарных структурах (например, у чело-
века насчитывается свыше 100 рудиментов) и атавизмах (проявление
признаков предковых форм).
Сравнительно-физиологические доказательства основаны на общ-
ности основных функций различных организмов: питании, дыхании,
748
ЭВОЛЮЦИЯ
кровообращении, рефлекторной деятельности и др. Клеточные дока-
зательства основываются на положениях клеточной теории.
Генетико-биохимические доказательства демонстрируют удиви-
тельное сходство молекулярной организации живых существ. Это от-
носится к материальному субстрату наследственности (нуклеиновые
кислоты), основным физиологическим процессам клетки и осуществ-
ляемым в ней биохимическим реакциям. Организмы используют
одни и те же мономеры (аминокислоты, нуклеотиды и др.) для по-
строения биомолекул. У всех одинаков генетический код. Геномы раз-
ных организмов имеют между собой много общего. Например, геном
человека примерно на 90% совпадает с геномом мыши и лишь на 1%
отличается от генома шимпанзе. Присутствием сохранившихся генов
далеко отставленных групп объясняется явление рекапитуляции —
проявление общих признаков на ранних стадиях эмбриогенеза, а так-
же рудиментов и атавизмов.
Биогеографические доказательства основаны на особенностях раз-
вития организмов, длительное время изолированных от других. При-
мером является эволюция сумчатых в Австралии, которые за дли-
тельное время освоили те же экологические ниши, что и плацентарные
млекопитающие на других континентах.
ГИПОТЕЗЫ ПРОИСХОЖДЕНИЯ ЖИЗНИ
НА ЗЕМЛЕ
Вопрос о происхождении жизни является наиболее дискуссион-
ным в биологии. Точного ответа на него получить нельзя, но сущест-
вует множество гипотез. Наиболее известными теориями являются:
креационизм, самопроизвольное зарождение из неживой материи, не-
прерывность жизни, гипотеза панспермии, гипотеза биохимической
эволюции. Многие из этих гипотез руководствуются одинаковыми
фактами, но с разной их интерпретацией.
Гипотеза божественного происхождения жизни (креационизм) осно-
вывается на образовании жизни и живых существ Создателем. В Би-
блии имеется прямое описание процесса и достаточно подробно указа-
на последовательность событий (Книга Бытия, 1.1 — 26). Практически
в каждом религиозном учении есть догма о сверхъестественном проис-
хождении жизни. Эта гипотеза принимает жизнь как данность и не
предпринимает попыток объяснения факта возникновения жизни за-
конами естественной природы.
Гипотеза самопроизвольного зарождения жизни возникла в антич-
ные времена в качестве альтернативы креационизму. Сторонниками
спонтанного зарождения жизни были Фалес, Анаксимандр, Анакри-
мен, Эмпедокл, Аристотель и др.
749
ГИПОТЕЗЫ ПРОИСХОЖДЕНИЯ ЖИЗНИ НА ЗЕМЛЕ
Гипотеза непрерывности жизни (гипотеза стационарного состоя-
ния) снимает сам вопрос о ее зарождении. Приверженцы этой гипоте-
зы уверены, что Земля существовала вечно, а вместе с ней существо-
вала жизнь. Они исключают развитие и преобразование организмов.
При этом виды существовали всегда, но могут в определенных усло-
виях исчезнуть.
Гипотеза панспермии также не пытается объяснить момент и усло-
вия возникновения жизни. Ее сторонники не считают Землю вечной,
а наличие на ней жизни объясняют попаданием из Вселенной, где она
была всегда. Доказательствами внеземного происхождения жизни
признаются «свидетельства» контактов с инопланетянами, НЛО, на-
скальные изображения объектов, похожих на космические корабли.
Современная наука не позволяет строго подтвердить или опроверг-
нуть факт существования внеземной жизни.
Гипотеза биохимической эволюции объясняет появление жизни
процессами, которые подчиняются законам химии и физики. Основ-
ные положения гипотезы биохимической эволюции состоят в после-
довательности следующих событий: 1) возникновение органических
соединений; 2) образование сложных биополимеров; 3) появление
первичных живых организмов. После того как температура на поверх-
ности Земли опустилась ниже 100 °С, вода из парообразного состоя-
ния перешла в жидкое, образовав Мировой океан. Отсутствие в ат-
мосфере молекулярного кислорода облегчало естественное появление
органических веществ, которые создали нужные предпосылки появ-
ления живой материи. Синтез органических веществ из неорганиче-
ских в условиях, которые предположительно соответствовали усло-
виям первобытной Земли, экспериментально доказан. В дальнейшем
из простых органических соединений должны были возникнуть более
сложные вещества, а те, в свою очередь, каким-то образом преобразо-
вались в живую систему.
По мнению А. И . Опарина (1924), решающее значение в этом пе-
реходном процессе имели белки, которые в силу своей амфотерности
притягивают воду и могут создать вокруг себя оболочку. Предполага-
лось, что такие частично обособленные комплексы в дальнейшем сли-
вались в более крупные — коацерватные капли. В результате происхо-
дило отделение первичных коллоидов от водного окружения. Опарин
считал, что коацерваты способны к примитивному обмену, в результа-
те которого в них избирательно накапливались определенные вещест-
ва, а на границе коацервата с окружающей водой выстраивались мо-
лекулы сложных углеводородов — липидов. Образовавшаяся при
этом пограничная мембрана еще в большей степени стабилизирует
750
ЭВОЛЮЦИЯ
систему, в которой могли бы осуществляться некоторые биохимиче-
ские реакции. Самовоспроизведение коацерватов Опарин объяснил
фрагментацией особо крупных капель на более мелкие идентичные
капли. Дальнейшее развитие процесса должно было привести к созда-
нию простейших форм гетеротрофных организмов. Г. Мёллер (1929)
считал более важным участие нуклеиновых кислот, а не белков.
Экспериментально доказана возможность репликации небольших
участков нуклеиновых кислот без участия белков-ферментов. Также
подтверждена ферментативная активность некоторых РНК. Это по-
зволяет предположить, что в ранних живых системах роль ферментов
выполняли нуклеиновые кислоты, а не белки.
Основная аргументация этой теории направлена на возможность
синтеза и устойчивости возникшей системы. Но она не объясняет по-
явления биохимических реакций, сложной взаимоподчиненной сис-
темы «нуклеиновые кислоты — белок», а также не может убедительно
объяснить самовоспроизводство образовавшихся систем. Исходя из
положений теорий Опарина и его сторонников, можно ожидать, что
кусок мяса (в котором находятся настоящие клетки, причем органи-
зованные куда совершеннее, чем коацерваты, и содержащие все необ-
ходимые для жизни вещества) способен инициировать образование
живых организмов (или отдельных клеток). А это уже напоминает
взгляды Ван Гельмонта, которому для создания мышей было доста-
точно грязной рубашки, темного шкафа и горсти пшеницы. Ученые
могут синтезировать основные органические соединения или выде-
лить их из организмов, однако еще ни одному человеку не удалось
вдохнуть жизнь в химические вещества. Возможность возникновения
такой сложнейшей организации как живая материя в результате слу-
чайных взаимодействий молекул, по образному высказыванию астро-
нома Фреда Хойла, «столь же нелепа и неправдоподобна, как утвер-
ждение, что ураган, пронесшийся над мусорной свалкой, может
привести к сборке Боинга-747». В соответствии с теорией вероятно-
стей подобные события столь же ожидаемы, как и возможность напи-
сания без единой ошибки Библии обезьяной, которой предоставили
пишущую машинку.
РАЗВИТИЕ ОРГАНИЧЕСКОГО МИРА
Возраст Земли ученые оценивают в 4,5—5 млрд лет. Примерно
3,5 млрд лет назад появились живые организмы. Предполагаемая по-
следовательность эволюционных преобразований изложена в табл. 32,
эволюции человека — в табл. 33.
751
РАЗВИТИЕ ОРГАНИЧЕСКОГО МИРА
Т
а
б
л
и
ц
а
3
2
О
с
н
о
в
н
ы
е
э
т
а
п
ы
э
в
о
л
ю
ц
и
и
ж
и
в
ы
х
о
р
г
а
н
и
з
м
о
в
(
п
о
З
а
х
а
р
о
в
у
и
с
о
а
в
т
.
,
с
и
з
м
е
н
е
н
и
я
м
и
и
д
о
п
о
л
н
е
н
и
я
м
и
)
Э
р
а
(
п
р
о
д
о
л
ж
и
-
т
е
л
ь
н
о
с
т
ь
,
м
л
н
л
е
т
)
П
е
р
и
о
д
(
п
р
о
-
д
о
л
ж
и
т
е
л
ь
-
н
о
с
т
ь
,
м
л
н
л
е
т
)
Н
а
ч
а
л
о
(
м
л
н
л
е
т
н
а
з
а
д
)
К
л
и
м
а
т
и
с
р
е
д
а
Р
а
з
в
и
т
и
е
о
р
г
а
н
и
ч
е
с
к
о
г
о
м
и
р
а
Ж
и
в
о
т
н
ы
е
Р
а
с
т
е
н
и
я
1
2
3
4
5
6
А
р
х
е
й
с
к
а
я
,
9
0
0
3
5
0
0
А
к
т
и
в
н
а
я
в
у
л
к
а
н
и
ч
е
-
с
к
а
я
д
е
я
т
е
л
ь
н
о
с
т
ь
.
А
н
а
-
э
р
о
б
н
ы
е
у
с
л
о
в
и
я
ж
и
з
н
и
в
м
е
л
к
о
в
о
д
н
о
м
д
р
е
в
н
е
м
м
о
р
е
.
Р
а
з
в
и
т
и
е
к
и
с
л
о
-
р
о
д
с
о
д
е
р
ж
а
щ
е
й
а
т
м
о
-
с
ф
е
р
ы
П
о
я
в
л
е
н
и
е
ж
и
з
н
и
н
а
З
е
м
л
е
.
С
о
х
р
а
н
и
в
ш
и
е
с
я
с
л
е
д
ы
ж
и
з
-
н
и
н
е
з
н
а
ч
и
т
е
л
ь
н
ы
.
О
б
н
а
р
у
ж
е
н
ы
о
с
т
а
т
к
и
б
а
к
т
е
р
и
й
,
а
н
а
э
-
р
о
б
н
ы
х
а
в
т
о
т
р
о
ф
н
ы
х
п
р
е
д
ш
е
с
т
в
е
н
н
и
к
о
в
с
и
н
е
-
з
е
л
е
н
ы
х
в
о
д
о
р
о
с
л
е
й
,
н
а
с
т
о
я
щ
и
х
в
о
д
о
р
о
с
л
е
й
.
П
е
р
в
ы
е
с
т
р
о
м
а
т
о
л
и
-
т
ы
.
О
т
д
е
л
ь
н
ы
е
н
а
х
о
д
к
и
п
р
о
к
а
р
и
о
т
и
ч
е
с
к
и
х
о
р
г
а
н
и
з
м
о
в
в
п
о
р
о
д
а
х
.
П
о
я
в
л
е
н
и
е
п
р
и
м
и
т
и
в
н
ы
х
э
у
к
а
р
и
о
т
(
п
р
о
с
т
е
й
-
ш
и
е
)
П
р
о
т
е
р
о
з
о
й
-
с
к
а
я
(
р
а
н
н
е
й
ж
и
з
н
и
)
,
2
0
0
0
2
6
0
0
±
1
0
0
П
о
в
е
р
х
н
о
с
т
ь
З
е
м
л
и
п
р
е
д
с
т
а
в
л
я
л
а
с
о
б
о
й
г
о
-
л
у
ю
п
у
с
т
ы
н
ю
.
К
л
и
м
а
т
х
о
л
о
д
н
ы
й
;
ч
а
с
т
ы
е
о
л
е
-
д
е
н
е
н
и
я
,
о
с
о
б
е
н
н
о
о
б
-
ш
и
р
н
о
е
в
с
е
р
е
д
и
н
е
п
р
о
-
т
е
р
о
з
о
я
.
В
к
о
н
ц
е
э
р
ы
с
о
д
е
р
ж
а
н
и
е
с
в
о
б
о
д
н
о
г
о
к
и
с
л
о
р
о
д
а
в
а
т
м
о
с
ф
е
-
р
е
—
д
о
1
%
.
А
к
т
и
в
н
о
е
о
б
р
а
з
о
в
а
н
и
е
о
с
а
д
о
ч
н
ы
х
п
о
р
о
д
Ш
и
р
о
к
о
е
р
а
с
п
р
о
с
т
р
а
н
е
н
и
е
п
р
о
-
с
т
е
й
ш
и
х
.
В
о
з
н
и
к
л
и
в
с
е
т
и
п
ы
б
е
с
-
п
о
з
в
о
н
о
ч
н
ы
х
ж
и
в
о
т
н
ы
х
.
Ш
и
р
о
к
о
р
а
с
п
р
о
с
т
р
а
н
е
н
ы
п
р
о
с
т
е
й
ш
и
е
,
г
у
б
-
к
и
,
к
и
ш
е
ч
н
о
п
о
л
о
с
т
н
ы
е
,
р
а
з
л
и
ч
-
н
ы
е
ч
е
р
в
и
,
п
р
е
д
к
и
т
р
и
л
о
б
и
т
о
в
и
и
г
л
о
к
о
ж
и
х
.
П
р
е
д
п
о
л
о
ж
и
т
е
л
ь
н
о
—
п
е
р
в
ы
е
п
р
е
д
с
т
а
в
и
т
е
л
и
х
о
р
д
о
-
в
ы
х
—
б
е
с
ч
е
р
е
п
н
ы
е
Р
а
с
п
р
о
с
т
р
а
н
е
н
ы
п
р
е
и
м
у
щ
е
с
т
в
е
н
н
о
з
е
л
е
н
ы
е
в
о
д
о
р
о
с
л
и
.
П
о
я
в
л
е
н
и
е
к
р
а
с
н
ы
х
в
о
д
о
р
о
с
л
е
й
752
ЭВОЛЮЦИЯ
1
2
3
4
5
6
П
а
л
е
о
з
о
й
с
к
а
я
(
д
р
е
в
н
е
й
ж
и
з
-
н
и
)
,
3
4
0
±
1
0
К
е
м
б
р
и
й
-
с
к
и
й
(
К
е
м
б
р
и
й
)
,
8
0
±
2
0
5
7
0
±
2
0
Р
а
н
н
е
к
е
м
б
р
и
й
с
к
о
е
о
л
е
-
д
е
н
е
н
и
е
с
м
е
н
я
е
т
с
я
в
н
а
-
ч
а
л
е
у
м
е
р
е
н
н
о
в
л
а
ж
-
н
ы
м
,
а
з
а
т
е
м
с
у
х
и
м
т
е
-
п
л
ы
м
к
л
и
м
а
т
о
м
.
А
к
-
т
и
в
н
о
е
н
а
с
т
у
п
л
е
н
и
е
м
о
р
я
,
с
м
е
н
и
в
ш
е
е
с
я
е
г
о
о
т
с
т
у
п
л
е
н
и
е
м
в
к
о
н
ц
е
п
е
р
и
о
д
а
Р
а
с
ц
в
е
т
м
о
р
с
к
и
х
б
е
с
п
о
з
в
о
н
о
ч
-
н
ы
х
,
и
з
к
о
т
о
р
ы
х
6
0
%
н
а
х
о
д
о
к
—
т
р
и
л
о
б
и
т
ы
.
П
о
я
в
л
е
н
и
е
о
р
г
а
н
и
з
-
м
о
в
с
м
и
н
е
р
а
л
и
з
о
в
а
н
н
ы
м
с
к
е
л
е
-
т
о
м
Д
и
в
е
р
г
е
н
т
н
а
я
э
в
о
-
л
ю
ц
и
я
в
о
д
о
р
о
с
л
е
й
;
в
о
з
н
и
к
н
о
в
е
н
и
е
м
н
о
-
г
о
к
л
е
т
о
ч
н
ы
х
ф
о
р
м
О
р
д
о
в
и
к
-
с
к
и
й
(
О
р
д
о
-
в
и
к
)
,
5
5
±
1
0
4
9
0
±
1
0
0
Р
а
в
н
о
м
е
р
н
о
у
м
е
р
е
н
н
ы
й
в
л
а
ж
н
ы
й
к
л
и
м
а
т
с
п
о
-
с
т
е
п
е
н
н
ы
м
п
о
в
ы
ш
е
н
и
-
е
м
с
р
е
д
н
е
й
т
е
м
п
е
р
а
т
у
-
р
ы
.
В
н
а
ч
а
л
е
п
е
р
и
о
д
а
б
о
л
ь
ш
а
я
ч
а
с
т
ь
с
у
ш
и
з
а
н
я
т
а
м
о
р
е
м
,
з
а
т
е
м
в
с
в
я
з
и
с
и
н
т
е
н
с
и
в
н
ы
м
г
о
р
о
о
б
р
а
з
о
в
а
н
и
е
м
о
с
в
о
-
б
о
ж
д
е
н
и
е
о
т
в
о
д
ы
з
н
а
-
ч
и
т
е
л
ь
н
ы
х
т
е
р
р
и
т
о
р
и
й
П
о
я
в
л
е
н
и
е
п
е
р
в
ы
х
п
о
з
в
о
н
о
ч
-
н
ы
х
—
б
е
с
ч
е
л
ю
с
т
н
ы
х
.
О
с
т
а
т
к
и
п
е
р
в
ы
х
к
о
р
а
л
л
о
в
ы
х
п
о
л
и
п
о
в
.
Г
о
-
с
п
о
д
с
т
в
о
т
р
и
л
о
б
и
т
о
в
,
и
г
л
о
к
о
ж
и
х
;
в
о
з
н
и
к
н
о
в
е
н
и
е
н
о
в
ы
х
к
л
а
с
с
о
в
и
в
ы
м
и
р
а
н
и
е
н
е
к
о
т
о
р
ы
х
г
р
у
п
п
б
е
с
п
о
з
в
о
н
о
ч
н
ы
х
И
с
к
л
ю
ч
и
т
е
л
ь
н
о
е
р
а
з
н
о
о
б
р
а
з
и
е
в
о
д
о
-
р
о
с
л
е
й
,
п
р
о
ц
в
е
т
а
н
и
е
в
с
е
х
о
т
д
е
л
о
в
С
и
л
у
р
и
й
-
с
к
и
й
(
С
и
л
у
р
)
,
3
5
±
1
0
4
3
5
±
1
0
В
н
а
ч
а
л
е
с
у
х
о
й
к
л
и
м
а
т
,
з
а
т
е
м
в
л
а
ж
н
ы
й
с
п
о
с
т
е
-
п
е
н
н
ы
м
п
о
т
е
п
л
е
н
и
е
м
.
И
н
т
е
н
с
и
в
н
о
е
г
о
р
о
о
б
р
а
-
з
о
в
а
н
и
е
(
С
к
а
н
д
и
н
а
в
-
с
к
и
е
г
о
р
ы
,
С
а
я
н
ы
)
,
в
о
з
-
н
и
к
н
о
в
е
н
и
е
п
е
р
в
ы
х
к
о
-
р
а
л
л
о
в
ы
х
р
и
ф
о
в
П
ы
ш
н
о
е
р
а
з
в
и
т
и
е
к
о
р
а
л
л
о
в
и
т
р
и
л
о
б
и
т
о
в
.
П
о
я
в
л
е
н
и
е
д
р
е
в
н
е
й
-
ш
и
х
ч
е
л
ю
с
т
н
о
р
о
т
ы
х
р
ы
б
(
б
о
л
ь
-
ш
е
р
о
т
ы
е
)
и
п
е
р
в
ы
х
д
ы
ш
а
щ
и
х
а
т
-
м
о
с
ф
е
р
н
ы
м
в
о
з
д
у
х
о
м
н
а
з
е
м
н
ы
х
ж
и
в
о
т
н
ы
х
—
с
к
о
р
п
и
о
н
о
в
.
В
ы
м
и
-
р
а
н
и
е
н
е
к
о
т
о
р
ы
х
г
р
у
п
п
к
о
р
а
л
л
о
в
В
к
о
н
ц
е
п
е
р
и
о
д
а
в
ы
-
х
о
д
р
а
с
т
е
н
и
й
н
а
с
у
ш
у
—
п
о
я
в
л
е
н
и
е
п
с
и
л
о
ф
и
т
о
в
.
П
е
р
в
ы
е
с
п
о
р
о
в
ы
е
с
о
с
у
д
и
-
с
т
ы
е
р
а
с
т
е
н
и
я
П
р
о
д
о
л
ж
е
н
и
е
т
а
б
л
.
3
2
753
РАЗВИТИЕ ОРГАНИЧЕСКОГО МИРА
Д
е
в
о
н
с
к
и
й
(
Д
е
в
о
н
)
,
5
5
±
1
0
4
0
0
±
1
0
К
л
и
м
а
т
х
а
р
а
к
т
е
р
и
з
у
е
т
-
с
я
с
м
е
н
о
й
с
у
х
и
х
и
д
о
-
ж
д
е
в
ы
х
с
е
з
о
н
о
в
.
О
л
е
д
е
-
н
е
н
и
е
н
а
т
е
р
р
и
т
о
р
и
и
с
о
в
р
е
м
е
н
н
ы
х
Ю
ж
н
о
й
А
м
е
р
и
к
и
и
Ю
ж
н
о
й
А
ф
-
р
и
к
и
.
П
о
л
н
о
е
о
с
в
о
б
о
-
ж
д
е
н
и
е
о
т
м
о
р
я
С
и
б
и
р
и
и
В
о
с
т
о
ч
н
о
й
Е
в
р
о
п
ы
П
о
я
в
л
е
н
и
е
р
ы
б
в
с
е
х
и
з
в
е
с
т
н
ы
х
с
и
с
т
е
м
а
т
и
ч
е
с
к
и
х
г
р
у
п
п
.
А
д
а
п
т
и
в
-
н
а
я
р
а
д
и
а
ц
и
я
р
ы
б
.
П
о
я
в
л
е
н
и
е
а
м
м
о
н
и
т
о
в
.
В
ы
м
и
р
а
н
и
е
з
н
а
ч
и
-
т
е
л
ь
н
о
г
о
к
о
л
и
ч
е
с
т
в
а
б
е
с
п
о
з
в
о
н
о
ч
-
н
ы
х
и
б
о
л
ь
ш
и
н
с
т
в
а
б
е
с
ч
е
л
ю
с
т
-
н
ы
х
.
О
с
в
о
е
н
и
е
ж
и
в
о
т
н
ы
м
и
с
у
ш
и
:
п
а
у
к
и
,
к
л
е
щ
и
и
д
р
у
г
и
е
ч
л
е
н
и
с
т
о
-
н
о
г
и
е
.
В
к
о
н
ц
е
п
е
р
и
о
д
а
п
е
р
в
ы
е
н
а
з
е
м
н
ы
е
п
о
з
в
о
н
о
ч
н
ы
е
—
с
т
е
г
о
ц
е
-
ф
а
л
ы
Р
а
з
в
и
т
и
е
,
а
з
а
т
е
м
в
ы
-
м
и
р
а
н
и
е
п
с
и
л
о
ф
и
т
о
в
.
В
о
з
н
и
к
н
о
в
е
н
и
е
о
с
-
н
о
в
н
ы
х
г
р
у
п
п
с
п
о
р
о
-
в
ы
х
р
а
с
т
е
н
и
й
:
м
о
х
о
-
о
б
р
а
з
н
ы
х
и
п
а
п
о
р
о
т
-
н
и
к
о
о
б
р
а
з
н
ы
х
(
п
а
п
о
-
р
о
т
н
и
к
о
в
и
д
н
ы
х
,
п
л
а
-
у
н
о
в
и
д
н
ы
х
,
х
в
о
щ
е
-
в
и
д
н
ы
х
)
,
а
т
а
к
ж
е
п
р
и
-
м
и
т
и
в
н
ы
х
г
о
л
о
с
е
м
е
н
-
н
ы
х
(
с
е
м
е
н
н
ы
е
п
а
п
о
-
р
о
т
н
и
к
и
)
.
В
о
з
н
и
к
н
о
-
в
е
н
и
е
г
р
и
б
о
в
К
а
м
е
н
н
о
-
у
г
о
л
ь
н
ы
й
(
К
а
р
б
о
н
)
,
6
5
±
1
0
3
4
5
±
1
0
В
с
е
м
и
р
н
о
е
р
а
с
п
р
о
с
т
р
а
-
н
е
н
и
е
л
е
с
н
ы
х
б
о
л
о
т
.
Р
а
в
н
о
м
е
р
н
о
т
е
п
л
ы
й
в
л
а
ж
н
ы
й
к
л
и
м
а
т
с
м
е
н
я
-
е
т
с
я
в
к
о
н
ц
е
п
е
р
и
о
д
а
х
о
л
о
д
н
ы
м
и
с
у
х
и
м
.
П
е
-
р
и
о
д
з
а
в
е
р
ш
а
е
т
с
я
о
б
-
ш
и
р
н
ы
м
о
л
е
д
е
н
е
н
и
е
м
Ш
и
р
о
к
о
е
р
а
с
п
р
о
с
т
р
а
н
е
н
и
е
ф
о
р
а
-
м
и
н
и
ф
е
р
,
к
о
р
а
л
л
о
в
,
м
о
л
л
ю
с
к
о
в
,
а
к
у
л
,
д
в
о
я
к
о
д
ы
ш
а
щ
и
х
р
ы
б
.
Р
а
с
-
ц
в
е
т
и
а
д
а
п
т
и
в
н
а
я
р
а
д
и
а
ц
и
я
з
е
м
-
н
о
в
о
д
н
ы
х
.
П
о
я
в
л
е
н
и
е
п
е
р
в
ы
х
р
е
п
т
и
л
и
й
—
к
о
т
и
л
о
з
а
в
р
о
в
,
л
е
т
а
ю
-
щ
и
х
н
а
с
е
к
о
м
ы
х
,
л
е
г
о
ч
н
ы
х
м
о
л
л
ю
-
с
к
о
в
.
С
о
к
р
а
щ
е
н
и
е
ч
и
с
л
е
н
н
о
с
т
и
т
р
и
л
о
б
и
т
о
в
и
с
т
е
г
о
ц
е
ф
а
л
о
в
Н
а
с
у
ш
е
«
к
а
м
е
н
н
о
-
у
г
о
л
ь
н
ы
е
л
е
с
а
»
с
п
р
е
о
б
л
а
д
а
н
и
е
м
с
п
о
-
р
о
в
ы
х
р
а
с
т
е
н
и
й
,
п
о
-
я
в
л
е
н
и
е
п
е
р
в
ы
х
х
в
о
й
н
ы
х
.
В
б
о
л
о
т
а
х
и
п
р
и
б
р
е
ж
н
ы
х
р
а
й
о
-
н
а
х
м
е
л
к
и
х
м
о
р
е
й
н
а
к
о
п
л
е
н
и
е
б
о
л
ь
ш
о
-
г
о
к
о
л
и
ч
е
с
т
в
а
р
а
с
т
и
-
т
е
л
ь
н
ы
х
о
с
т
а
т
к
о
в
П
е
р
м
с
к
и
й
(
П
е
р
м
ь
)
,
5
0
±
1
0
2
8
0
±
1
0
Р
е
з
к
а
я
з
о
н
а
л
ь
н
о
с
т
ь
к
л
и
м
а
т
а
.
З
а
в
е
р
ш
е
н
и
е
г
о
р
о
о
б
р
а
з
о
в
а
т
е
л
ь
н
ы
х
п
р
о
ц
е
с
с
о
в
к
а
р
б
о
н
а
.
О
т
-
с
т
у
п
л
е
н
и
е
м
о
р
е
й
и
ф
о
р
-
м
и
р
о
в
а
н
и
е
п
о
л
у
з
а
м
к
н
у
-
т
ы
х
в
о
д
о
е
м
о
в
.
Р
и
ф
о
-
о
б
р
а
з
о
в
а
н
и
е
В
м
о
р
я
х
д
о
м
и
н
и
р
у
ю
т
б
е
с
п
о
з
в
о
-
н
о
ч
н
ы
е
и
а
к
у
л
ы
.
Б
ы
с
т
р
о
е
р
а
з
в
и
-
т
и
е
и
а
д
а
п
т
и
в
н
а
я
р
а
д
и
а
ц
и
я
р
е
п
т
и
-
л
и
й
,
в
о
з
н
и
к
н
о
в
е
н
и
е
з
в
е
р
о
з
у
б
ы
х
и
т
р
а
в
о
я
д
н
ы
х
п
р
е
с
м
ы
к
а
ю
щ
и
х
с
я
.
П
о
я
в
л
е
н
и
е
ж
у
к
о
в
.
В
ы
м
и
р
а
н
и
е
т
р
и
л
о
б
и
т
о
в
,
с
т
е
г
о
ц
е
ф
а
л
о
в
и
с
о
-
к
р
а
щ
е
н
и
е
ч
и
с
л
а
о
т
р
я
д
о
в
д
р
у
г
и
х
б
е
с
п
о
з
в
о
н
о
ч
н
ы
х
и
р
я
д
а
п
о
з
в
о
н
о
ч
-
н
ы
х
И
с
ч
е
з
н
о
в
е
н
и
е
«
к
а
-
м
е
н
н
о
у
г
о
л
ь
н
ы
х
л
е
-
с
о
в
»
з
а
с
ч
е
т
в
ы
м
и
р
а
-
н
и
я
д
р
е
в
о
в
и
д
н
ы
х
п
а
п
о
р
о
т
н
и
к
о
о
б
р
а
з
-
н
ы
х
.
П
о
я
в
л
е
н
и
е
х
в
о
й
н
ы
х
и
р
а
с
п
р
о
-
с
т
р
а
н
е
н
и
е
и
х
в
С
е
-
в
е
р
н
о
м
п
о
л
у
ш
а
р
и
и
754
ЭВОЛЮЦИЯ
1
2
3
4
5
6
М
е
з
о
з
о
й
с
к
а
я
(
с
р
е
д
н
е
й
ж
и
з
-
н
и
)
,
1
6
5
Т
р
и
а
с
о
в
ы
й
(
Т
р
и
а
с
)
,
4
0
±
5
2
3
0
±
1
0
О
с
л
а
б
л
е
н
и
е
к
л
и
м
а
т
и
ч
е
-
с
к
о
й
з
о
н
а
л
ь
н
о
с
т
и
,
с
г
л
а
-
ж
и
в
а
н
и
е
т
е
м
п
е
р
а
т
у
р
-
н
ы
х
р
а
з
л
и
ч
и
й
.
Н
а
ч
а
л
о
д
в
и
ж
е
н
и
я
м
а
т
е
р
и
к
о
в
Н
а
ч
а
л
о
р
а
с
ц
в
е
т
а
р
е
п
т
и
л
и
й
—
н
а
-
ч
и
н
а
е
т
с
я
«
в
е
к
д
и
н
о
з
а
в
р
о
в
»
;
а
д
а
п
-
т
и
в
н
а
я
р
а
д
и
а
ц
и
я
р
е
п
т
и
л
и
й
,
п
о
я
в
-
л
я
ю
т
с
я
ч
е
р
е
п
а
х
и
,
к
р
о
к
о
д
и
л
ы
и
д
р
.
В
о
з
н
и
к
н
о
в
е
н
и
е
п
е
р
в
ы
х
м
л
е
к
о
п
и
-
т
а
ю
щ
и
х
(
я
й
ц
е
к
л
а
д
у
щ
и
е
и
с
у
м
ч
а
-
т
ы
е
)
,
н
а
с
т
о
я
щ
и
х
к
о
с
т
и
с
т
ы
х
р
ы
б
Р
а
с
п
р
о
с
т
р
а
н
е
н
ы
п
а
-
п
о
р
о
т
н
и
к
о
в
и
д
н
ы
е
,
х
в
о
щ
е
в
и
д
н
ы
е
и
п
л
а
-
у
н
о
в
и
д
н
ы
е
.
В
ы
м
и
р
а
-
ю
т
с
е
м
е
н
н
ы
е
п
а
п
о
-
р
о
т
н
и
к
и
.
О
б
и
л
и
е
с
а
г
о
в
н
и
к
о
в
ы
х
и
х
в
о
й
н
ы
х
Ю
р
с
к
и
й
(
Ю
р
а
)
,
6
0
1
9
0
—
1
9
5
±
5
К
л
и
м
а
т
,
в
н
а
ч
а
л
е
в
л
а
ж
-
н
ы
й
,
с
м
е
н
я
е
т
с
я
к
к
о
н
ц
у
п
е
р
и
о
д
а
з
а
с
у
ш
л
и
в
ы
м
в
о
б
л
а
с
т
и
э
к
в
а
т
о
р
а
.
Д
в
и
ж
е
н
и
е
к
о
н
т
и
н
е
н
т
о
в
,
ф
о
р
м
и
р
о
в
а
н
и
е
А
т
л
а
н
-
т
и
ч
е
с
к
о
г
о
о
к
е
а
н
а
В
о
к
е
а
н
е
п
о
я
в
л
е
н
и
е
н
о
в
ы
х
г
р
у
п
п
м
о
л
л
ю
с
к
о
в
,
в
т
о
м
ч
и
с
л
е
г
о
л
о
в
о
н
о
-
г
и
х
,
а
т
а
к
ж
е
и
г
л
о
к
о
ж
и
х
.
В
м
о
р
е
д
о
м
и
н
и
р
о
в
а
н
и
е
г
о
л
о
в
о
н
о
г
и
х
и
к
о
с
т
и
с
т
ы
х
р
ы
б
.
В
ы
м
и
р
а
н
и
е
д
р
е
в
-
н
и
х
х
р
я
щ
е
в
ы
х
р
ы
б
.
Г
о
с
п
о
д
с
т
в
о
п
р
е
с
м
ы
к
а
ю
щ
и
х
с
я
н
а
с
у
ш
е
,
в
о
к
е
а
-
н
е
и
в
о
з
д
у
х
е
.
О
б
и
л
и
е
н
а
с
е
к
о
м
ы
х
.
В
к
о
н
ц
е
п
е
р
и
о
д
а
п
о
я
в
л
е
н
и
е
п
е
р
-
в
о
п
т
и
ц
—
а
р
х
е
о
п
т
е
р
и
к
с
а
Ш
и
р
о
к
о
р
а
с
п
р
о
с
т
р
а
-
н
е
н
ы
п
а
п
о
р
о
т
н
и
к
и
и
г
о
л
о
с
е
м
е
н
н
ы
е
.
П
о
-
я
в
л
е
н
и
е
п
о
к
р
ы
т
о
с
е
-
м
е
н
н
ы
х
р
а
с
т
е
н
и
й
.
П
о
я
в
л
я
е
т
с
я
х
о
р
о
ш
о
в
ы
р
а
ж
е
н
н
а
я
б
о
т
а
н
и
-
к
о
-
г
е
о
г
р
а
ф
и
ч
е
с
к
а
я
з
о
н
а
л
ь
н
о
с
т
ь
М
е
л
о
в
о
й
(
М
е
л
)
,
1
3
6
±
5
1
3
6
±
5
В
о
м
н
о
г
и
х
р
а
й
о
н
а
х
З
е
м
л
и
п
о
х
о
л
о
д
а
н
и
е
к
л
и
м
а
т
а
.
В
ы
р
а
ж
е
н
н
о
е
о
т
с
т
у
п
л
е
н
и
е
м
о
р
е
й
,
с
м
е
н
и
в
ш
е
е
с
я
о
б
ш
и
р
-
н
ы
м
у
м
е
н
ь
ш
е
н
и
е
м
п
л
о
-
щ
а
д
и
М
и
р
о
в
о
г
о
о
к
е
а
н
а
и
н
о
в
ы
м
п
о
д
н
я
т
и
е
м
с
у
ш
и
.
И
н
т
е
н
с
и
в
н
ы
е
г
о
р
о
о
б
р
а
з
о
в
а
т
е
л
ь
н
ы
е
п
р
о
ц
е
с
с
ы
(
А
л
ь
п
ы
,
А
н
д
ы
,
Г
и
м
а
л
а
и
)
П
о
я
в
л
е
н
и
е
н
а
с
т
о
я
щ
и
х
п
т
и
ц
и
п
л
а
ц
е
н
т
а
р
н
ы
х
м
л
е
к
о
п
и
т
а
ю
щ
и
х
.
В
в
о
д
о
е
м
а
х
п
р
е
о
б
л
а
д
а
н
и
е
к
о
с
т
и
-
с
т
ы
х
р
ы
б
.
Р
а
с
ц
в
е
т
н
а
с
е
к
о
м
ы
х
.
В
ы
м
и
р
а
н
и
е
а
м
м
о
н
и
т
о
в
,
к
р
у
п
н
ы
х
р
е
п
т
и
л
и
й
и
п
р
и
м
и
т
и
в
н
ы
х
м
е
з
о
-
з
о
й
с
к
и
х
м
л
е
к
о
п
и
т
а
ю
щ
и
х
Р
е
з
к
о
с
о
к
р
а
щ
а
е
т
с
я
ч
и
с
л
е
н
н
о
с
т
ь
п
а
п
о
-
р
о
т
н
и
к
о
о
б
р
а
з
н
ы
х
и
г
о
л
о
с
е
м
е
н
н
ы
х
.
Д
о
м
и
н
и
р
о
в
а
н
и
е
п
о
к
р
ы
т
о
с
е
м
е
н
н
ы
х
р
а
с
т
е
н
и
й
П
р
о
д
о
л
ж
е
н
и
е
т
а
б
л
.
3
2
755
РАЗВИТИЕ ОРГАНИЧЕСКОГО МИРА
К
а
й
н
о
з
о
й
с
к
а
я
(
н
о
в
о
й
ж
и
з
н
и
)
,
6
6
±
3
П
а
л
е
о
г
е
н
о
-
в
ы
й
(
н
и
ж
н
е
-
т
р
е
т
и
ч
н
ы
й
)
,
4
1
±
2
6
6
±
3
У
с
т
а
н
а
в
л
и
в
а
е
т
с
я
т
е
-
п
л
ы
й
р
а
в
н
о
м
е
р
н
ы
й
к
л
и
м
а
т
.
И
н
т
е
н
с
и
в
н
о
е
г
о
р
о
о
б
р
а
з
о
в
а
н
и
е
(
К
а
в
-
к
а
з
,
К
р
ы
м
,
П
а
м
и
р
,
Г
и
-
м
а
л
а
и
,
А
н
д
ы
и
д
р
.
)
В
м
о
р
я
х
б
о
л
ь
ш
о
е
к
о
л
и
ч
е
с
т
в
о
к
о
-
р
а
л
л
о
в
и
м
о
л
л
ю
с
к
о
в
.
Ш
и
р
о
к
о
р
а
с
п
р
о
с
т
р
а
н
я
ю
т
с
я
к
о
с
т
и
с
т
ы
е
р
ы
б
ы
,
з
а
н
и
м
а
ю
щ
и
е
п
р
е
с
н
о
в
о
д
н
ы
е
в
о
д
о
е
м
ы
и
м
о
р
я
,
в
ы
м
и
р
а
ю
т
м
н
о
-
г
и
е
ф
о
р
м
ы
г
о
л
о
в
о
н
о
г
и
х
м
о
л
л
ю
-
с
к
о
в
.
Н
а
с
у
ш
е
с
р
е
д
и
п
о
з
в
о
н
о
ч
н
ы
х
:
х
в
о
с
т
а
т
ы
е
и
б
е
с
х
в
о
с
т
ы
е
а
м
ф
и
б
и
и
,
к
р
о
к
о
д
и
л
ы
,
я
щ
е
р
и
ц
ы
,
з
м
е
и
и
ч
е
-
р
е
п
а
х
и
.
П
о
я
в
л
я
ю
т
с
я
м
н
о
г
и
е
о
т
р
я
-
д
ы
м
л
е
к
о
п
и
т
а
ю
щ
и
х
,
в
т
о
м
ч
и
с
л
е
п
р
и
м
а
т
ы
.
В
о
з
н
и
к
а
е
т
о
б
о
с
о
б
л
е
н
-
н
ы
й
ц
е
н
т
р
р
а
з
в
и
т
и
я
р
а
с
т
и
т
е
л
ь
н
о
-
с
т
и
и
ж
и
в
о
т
н
ы
х
в
Ю
ж
н
о
й
А
м
е
р
и
-
к
е
.
Ш
и
р
о
к
а
я
д
и
в
е
р
г
е
н
ц
и
я
п
т
и
ц
.
Р
а
с
ц
в
е
т
н
а
с
е
к
о
м
ы
х
Г
о
с
п
о
д
с
т
в
о
п
о
к
р
ы
т
о
-
с
е
м
е
н
н
ы
х
р
а
с
т
е
н
и
й
;
с
о
х
р
а
н
я
е
т
с
я
з
н
а
ч
и
-
т
е
л
ь
н
о
е
к
о
л
и
ч
е
с
т
в
о
г
р
у
п
п
,
в
о
з
н
и
к
ш
и
х
в
м
е
л
о
в
о
м
п
е
р
и
о
д
е
.
С
о
с
т
а
в
ф
л
о
р
ы
б
л
и
-
з
о
к
к
с
о
в
р
е
м
е
н
н
о
м
у
;
в
к
о
н
ц
е
п
е
р
и
о
д
а
п
о
-
я
в
л
я
ю
т
с
я
т
а
й
г
а
и
т
у
н
д
р
а
Н
е
о
г
е
н
о
в
ы
й
(
в
е
р
х
н
е
т
р
е
-
т
и
ч
н
ы
й
)
,
2
3
2
5
±
2
Д
в
и
ж
е
н
и
е
к
о
н
т
и
н
е
н
т
о
в
;
о
б
о
с
о
б
л
я
ю
т
с
я
К
а
с
п
и
й
-
с
к
о
е
,
С
р
е
д
и
з
е
м
н
о
е
,
Ч
е
р
н
о
е
и
А
р
а
л
ь
с
к
о
е
м
о
р
я
А
н
т
р
о
п
о
-
г
е
н
о
в
ы
й
,
1
,
5
—
2
,
0
1
,
5
–
2
Х
а
р
а
к
т
е
р
н
ы
н
е
о
д
н
о
-
к
р
а
т
н
ы
е
с
м
е
н
ы
к
л
и
м
а
-
т
а
.
К
р
у
п
н
ы
е
о
л
е
д
е
н
е
-
н
и
я
С
е
в
е
р
н
о
г
о
п
о
л
у
ш
а
-
р
и
я
П
о
я
в
л
е
н
и
е
и
р
а
з
в
и
т
и
е
ч
е
л
о
в
е
к
а
.
Ж
и
в
о
т
н
ы
й
м
и
р
п
р
и
о
б
р
е
т
а
е
т
с
о
в
-
р
е
м
е
н
н
ы
е
ч
е
р
т
ы
Р
а
с
т
и
т
е
л
ь
н
ы
й
м
и
р
п
р
и
о
б
р
е
т
а
е
т
с
о
в
р
е
-
м
е
н
н
ы
й
о
б
л
и
к
,
ф
о
р
-
м
и
р
у
ю
т
с
я
н
ы
н
е
ж
и
-
в
у
щ
и
е
с
о
о
б
щ
е
с
т
в
а
756
ЭВОЛЮЦИЯ
Т
а
б
л
и
ц
а
3
3
О
с
н
о
в
н
ы
е
э
т
а
п
ы
э
в
о
л
ю
ц
и
и
ч
е
л
о
в
е
к
а
В
р
е
м
я
Х
а
р
а
к
т
е
р
н
ы
е
п
р
и
з
н
а
к
и
С
т
а
д
и
я
П
р
е
д
с
т
а
в
и
т
е
л
и
О
б
ъ
е
м
м
о
з
г
а
М
е
с
т
о
о
б
и
т
а
н
и
я
2
5
м
л
н
л
е
т
Д
р
е
в
е
с
н
ы
й
о
б
р
а
з
ж
и
з
н
и
.
С
т
а
д
н
о
с
т
ь
.
С
п
о
с
о
б
-
н
о
с
т
ь
м
а
н
и
п
у
л
и
р
о
в
а
н
и
я
п
р
е
д
м
е
т
а
м
и
О
б
щ
и
е
п
р
е
д
к
и
а
н
-
т
р
о
п
о
м
о
р
ф
н
ы
х
о
б
е
з
ь
я
н
и
г
о
м
и
н
и
д
Д
р
и
о
п
и
т
е
к
и
О
к
о
л
о
5
0
0
с
м
3
А
ф
р
и
к
а
,
А
з
и
я
9
м
л
н
л
е
т
Д
а
л
ь
н
е
й
ш
е
е
р
а
з
в
и
т
и
е
с
т
а
д
н
о
с
т
и
.
И
с
п
о
л
ь
з
о
-
в
а
н
и
е
п
р
е
д
м
е
т
о
в
в
к
а
ч
е
с
т
в
е
о
р
у
д
и
й
д
л
я
з
а
-
щ
и
т
ы
и
д
о
б
ы
в
а
н
и
я
п
и
щ
и
П
р
о
т
о
а
н
т
р
о
п
ы
Д
р
е
в
н
е
й
ш
и
е
а
в
с
т
р
а
л
о
п
и
т
е
-
к
и
О
к
о
л
о
5
0
0
с
м
3
А
ф
р
и
к
а
,
А
з
и
я
4
,
5
—
5
И
с
п
о
л
ь
з
о
в
а
н
и
е
р
а
з
л
и
ч
н
ы
х
п
р
е
д
м
е
т
о
в
П
р
е
г
о
м
и
н
и
д
ы
А
в
с
т
р
а
л
о
п
и
т
е
-
к
и
О
к
о
л
о
5
0
0
с
м
3
А
ф
р
и
к
а
,
А
з
и
я
2
—
2
,
5
м
л
н
л
е
т
И
з
г
о
т
о
в
л
е
н
и
е
п
е
р
в
ы
х
о
р
у
д
и
й
(
г
а
л
е
ч
н
а
я
к
у
л
ь
т
у
р
а
)
.
П
е
р
е
х
о
д
н
а
я
с
т
а
д
и
я
к
ф
о
р
м
и
р
о
в
а
-
н
и
ю
с
о
в
р
е
м
е
н
н
о
г
о
т
и
п
а
ч
е
л
о
в
е
к
а
Ч
е
л
о
в
е
к
у
м
е
-
л
ы
й
О
к
о
л
о
6
0
0
с
м
3
А
ф
р
и
к
а
1
—
2
м
л
н
л
е
т
П
о
д
д
е
р
ж
а
н
и
е
о
г
н
я
.
П
р
о
с
т
ы
е
ф
о
р
м
ы
к
о
л
л
е
к
-
т
и
в
н
о
й
д
е
я
т
е
л
ь
н
о
с
т
и
,
ф
о
р
м
и
р
о
в
а
н
и
е
р
е
ч
и
в
в
и
д
е
о
т
д
е
л
ь
н
ы
х
в
ы
к
р
и
к
о
в
Д
р
е
в
н
е
й
ш
и
е
л
ю
д
и
(
а
р
х
а
н
т
р
о
п
ы
,
и
л
и
п
и
т
е
к
а
н
т
р
о
п
ы
)
Ч
е
л
о
в
е
к
п
р
я
-
м
о
х
о
д
я
щ
и
й
8
0
0
—
1
1
0
0
с
м
3
А
ф
р
и
к
а
,
Ц
е
н
т
-
р
а
л
ь
н
а
я
и
З
а
-
п
а
д
н
а
я
Е
в
р
о
п
а
,
В
о
с
т
о
ч
н
а
я
А
з
и
я
,
И
н
д
о
н
е
з
и
я
4
0
–
5
0
0
т
ы
с
.
л
е
т
С
о
ч
е
т
а
н
и
е
п
р
и
з
н
а
к
о
в
а
н
т
р
о
п
о
и
д
о
в
и
с
о
в
р
е
-
м
е
н
н
о
г
о
ч
е
л
о
в
е
к
а
.
В
ы
с
о
к
а
я
к
у
л
ь
т
у
р
а
и
з
г
о
-
т
о
в
л
е
н
и
я
о
р
у
д
и
й
.
Д
о
б
ы
в
а
н
и
е
о
г
н
я
.
С
о
в
е
р
-
ш
е
н
с
т
в
о
в
а
н
и
е
п
л
е
м
е
н
н
ы
х
о
т
н
о
ш
е
н
и
й
—
з
а
-
б
о
т
а
о
б
л
и
ж
н
и
х
Д
р
е
в
н
и
е
л
ю
д
и
(
п
а
-
л
е
о
а
н
т
р
о
п
ы
)
Н
е
а
н
д
е
р
т
а
л
ь
-
ц
ы
Д
о
1
5
0
0
с
м
3
А
ф
р
и
к
а
,
Е
в
р
о
п
а
,
А
з
и
я
М
е
н
е
е
4
0
т
ы
с
.
л
е
т
Ф
о
р
м
и
р
о
в
а
н
и
е
ч
е
л
о
в
е
к
а
с
о
в
р
е
м
е
н
н
о
г
о
т
и
п
а
.
Р
а
з
в
и
т
ы
е
с
о
ц
и
а
л
ь
н
ы
е
о
т
н
о
ш
е
н
и
я
.
О
д
о
м
а
ш
-
н
и
в
а
н
и
е
р
а
с
т
е
н
и
й
и
ж
и
в
о
т
н
ы
х
.
Н
а
с
т
о
я
щ
а
я
р
е
ч
ь
.
П
о
я
в
л
е
н
и
е
и
с
к
у
с
с
т
в
а
С
о
в
р
е
м
е
н
н
ы
е
л
ю
д
и
К
р
о
м
а
н
ь
о
н
ц
ы
О
к
о
л
о
1
4
0
0
—
1
5
0
0
с
м
3
А
ф
р
и
к
а
,
Е
в
р
о
п
а
,
А
з
и
я
,
А
м
е
р
и
к
а
,
А
в
с
т
р
а
л
и
я
757
ПОЛОЖЕНИЕ ЧЕЛОВЕКА В ПРИРОДЕ
ПОЛОЖЕНИЕ ЧЕЛОВЕКА В ПРИРОДЕ
Человек относится к типу хордовых, подтипу позвоночных, клас-
су млекопитающих, подклассу плацентарных одноутробных, отряду
приматов, подотряду человекоподобных приматов, секции узконо-
сых обезьян Нового Света, надсемейству человекообразных прима-
тов, семейству людей, виду «человек разумный», подвиду «совре-
менный человек». В состав надсемейства человекообразных приматов
входит семейство гоминид, куда относятся современный человек раз-
умный и ископаемые люди. Человек разумный имеет общие черты
с другими позвоночными. Основные из них: 1. Наличие внутреннего
скелета, основу которого составляет осевой скелет, состоящий из мета-
мерных позвонков. 2 . Разделение черепа на два отдела: мозговой, явля-
ющийся вместилищем головного мозга, и лицевой, служащий скеле-
том для органов чувств и начальных отделов пищеварительного и
дыхательного путей. 3 . Гомологичные пары пятипалых конечностей,
скелет которых прикрепляется посредством поясов к осевому скелету.
4. Центральная нервная система состоит из спинного и головного моз-
га. Последний представлен пятью отделами, содержащими полости —
желудочки. 5 . Периферическая нервная система состоит из 12 пар че-
репных нервов (I, II и VIII являются нервами органов чувств, III, IV,
VI и XII — двигательными, а V, VII, IX, X, XI — смешанными), а также
смешанных спинномозговых нервов. Последние отходят сегментарно
от спинного мозга двумя корешками. 6 . Дифференцированные органы
чувств. 7. Выстланная серозной оболочкой полость тела, разделенная
на 3 отдела: перикард, плевру и брюшину. 8. Наружные (кожные) по-
кровы состоят из эпидермиса и дермы. 9. Органы пищеварения начи-
наются расположенной на головном конце тела полостью рта, снаб-
женной зубами, которая переходит в глотку, затем идут пищевод,
расширенный желудок и кишечник, снабженный двумя крупными же-
лезами — печенью и поджелудочной. 10. Органы дыхания состоят из
воздухоносных путей и легких, в которых происходит газообмен.
11. Замкнутая кровеносная система состоит из артерий, микроцирку-
лярного русла и вен. Наличие центрального органа кровообращения —
сердца, разделенного (как у всех наземных позвоночных) на артери-
альную и венозную половины. 12. Органы выделения, представленные
почками и мочевыводящими путями (мочеточники, мочевой пузырь,
мочеиспускательный канал). 13. Парные половые железы, в них обра-
зуются гаметы (половые клетки) и синтезируются половые гормоны.
14. Железы внутренней секреции.
Человек относится к млекопитающим, которые обладают рядом
признаков, отличающих их от других позвоночных. Это, в первую
очередь, наличие молочных желез, волосяного покрова, кожных
758
ЭВОЛЮЦИЯ
желез, четырехкамерного сердца с одной левой дугой аорты и по-
стоянной температуры тела. Наряду с описанными человек облада-
ет рядом морфологических признаков, общих с другими млекопита-
ющими: 1. Череп, сочленяющийся с I шейным позвонком двумя
мыщелками. 2 . Семь шейных позвонков, I и II позвонки отличаются
от остальных. 3 . Гетеродонтные зубы, сочленяющиеся с челюстями с
помощью неподвижных зубоальвеолярных сочленений. Смена молоч-
ных зубов постоянными. Наличие резцов, клыков и коренных зубов.
4. Наличие трех слуховых косточек в полости среднего уха и развито-
го наружного уха. 5 . Наличие оформленных губ, в толще которых рас-
положены мышцы, и щек. 6 . Наличие слюны, содержащей ферменты,
расщепляющие крахмал. 7 . Усложненная пищеварительная трубка с
расширенным желудком. 8 . Наличие диафрагмы, разделяющей груд-
ную и брюшную полости. 9 . Легкие, построенные из ацинусов, состо-
ящих из альвеол. 10. Отсутствие ядер в зрелых эритроцитах. 11 . На-
личие новой коры (неокортекса) головного мозга. 12 . Развитие
гортани с голосовыми связками. 13. Окаймленные ресницами веки.
Человек принадлежит к отряду приматов. С большинством при-
матов человека сближает ряд общих морфофункциональных осо-
бенностей: 1. Относительно крупный головной мозг с развитым ко-
нечным мозгом и корой полушарий большого мозга, увеличение
лобной, височной и затылочной долей конечного мозга, ассоциатив-
ных зон лобных и теменных долей. 2 . Пятипалая хватательная кисть с
противопоставляющимся большим пальцем. Очень подвижные верх-
ние конечности с развитыми ключицами и способностью к пронации
и супинации. 3 . Плоские ногти на пальцах кистей и стоп. 4 . Наличие
папиллярных линий и узоров на ладонях и подошвах. 5 . Увеличенный
мозговой отдел черепа. 6. Относительно большая продолжительность
жизни. 7 . Опорная стопа. 8 . Слабое развитие органов обоняния, хоро-
шее — органов слуха и зрения. 9 . Высокая степень гомологии ДНК
человека и приматов. Так, у человека и шимпанзе около 90% сходных
генов. 10. Низкая плодовитость, компенсируемая сильно развитой за-
ботой о потомстве. 11 . Строение внутренних органов. 12 . Строение
мимической мускулатуры.
В то же время человек по своему строению существенно отличает-
ся от приматов и, в том числе, от человекообразных обезьян. Основ-
ные отличительные признаки: 1. Очень крупный (абсолютно и отно-
сительно) мозг, превосходящий в 3—4 раза мозг человекообразных
обезьян, развитие и дифференцировка областей, связанных с члено-
раздельной речью и мышлением (лобная и височная доли, нижняя те-
менная область). 2 . Изменение пропорций конечностей — удлинение
ног по сравнению с руками. 3 . S -образная форма позвоночника с чет-
ко выраженными шейным и поясничным лордозами. 4 . Особое разви-
759
ПОЛОЖЕНИЕ ЧЕЛОВЕКА В ПРИРОДЕ
тие и расположение некоторых мышц (например, малоберцовых)
и связок нижних конечностей в связи с прямохождением. 5 . Форма
таза. 6 . Уплощенная в переднезаднем направлении грудная клетка.
7. Сводчатая стопа с массивным большим пальцем и некоторой ре-
дукцией остальных. 8 . Полное противопоставление большого пальца
кисти остальным. 9 . Особенности строения гортани в связи с членораз-
дельной речью (например, голосовая мышца). 10. Относительное уве-
личение мозгового отдела черепа и уменьшение лицевого в связи с
редукцией жевательных мышц и челюстей. 11 . Уменьшение волосяно-
го покрова. 12 . Сильное развитие папиллярных узоров на коже под-
ушечек пальцев кисти. 13. Увеличение периода детства. У человека
соотношение длительности детства и общей продолжительности жиз-
ни1:5,уприматов1:6—1:13.
Все современные люди относятся к одному виду, который широко
расселен по Земле. Представители различных рас и популяций дают
при смешении плодовитое потомство.
ЭКОЛОГИЯ
Экология — это биологическая дисциплина, изучающая взаимоот-
ношения организма со средой обитания. Этот термин в 1866 г. ввел
в науку Э. Геккель. На любой организм воздействуют биотические
и абиотические факторы. Совокупность всех биотических факторов
(факторов живой природы), действующих на определенной террито-
рии, составляют биоценоз, или сообщество. Если же учитываются еще
абиотические факторы (неживая природа, или биотоп), то это будет
биогеоценоз, или экосистема (термин «биогеоценоз» в 1940 г. ввел
В.Н. Сукачев). Биоценозы имеют видовую, пространственную и тро-
фическую структуры. Видовую структуру образуют виды, входящие
в него со своей численностью и биомассой. Для каждого биоценоза
характерна особая видовая структура. В зависимости от видового раз-
нообразия биоценозы могут быть богатыми или бедными видами. Бо-
гатые видами сообщества обычно образуются под действием опти-
мальных для жизни условий (например, влажные тропические леса).
Молодые сообщества беднее видами, чем сложившиеся. Виды с наи-
большей численностью популяций называют доминантными, а те из
них, участие которых наиболее важно для сообщества, называют
эдификаторами (дубы в дубравах, ель и сосна в хвойных лесах
и т. д.). Виды, популяции которых насчитывают небольшое количест-
во особей, называют малочисленными.
Пространственная структура биоценоза формируется фитоцено-
зом, расчлененным на вертикальные составляющие — ц е ноэле-
760
ЭКОЛОГИЯ
менты (ярусы) и горизонтальные — микрогруппировки. Вертикаль-
ное распределение растительности позволяет наиболее эффективно
использовать солнечный свет. Каждый из ярусов создает условия для
жизни определенных животных, адаптации которых соответствуют
данной среде. Горизонтальная расчлененность фитоценоза (мозаич-
ность) выражается в существовании микрогруппировок со своим ви-
довым составом, отличным от группировок, расположенных побли-
зости.
Трофическую структуру создают сложные пищевые связи.
АБИОТИЧЕСКИЕ ФАКТОРЫ
Многочисленные факторы неживой природы называют абиотиче-
скими. В зависимости от природы они подразделяются на климатиче-
ские, эдафические (почвенные), топографические и другие факторы
физической природы (волны, морские течения, огонь и др.).
В отдельных зонах обитания климат может иметь свои особенно-
сти. Это называют микроклиматом.
Климатические факторы включают в себя свет, температуру,
влажность, ветер, давление воздуха и его состав. Свет является источ-
ником энергии для фотосинтеза, который лежит в основе всех энерге-
тических процессов подавляющего большинства сообществ (за
исключением тех, где продуцентами являются хемотрофы). Главным
источником света является Солнце. Движение Земли вокруг своей
оси обеспечивает периодическую смену светлого и темного времени
суток, что лежит в основе образования суточных ритмов. Лишь на эк-
ваторе длина дня и ночи постоянна (около 12 часов), тогда как в бо-
лее низких или высоких широтах существуют сезонные изменения
продолжительности дня и организмы синхронизируют свою актив-
ность с текущим временем года. Весеннее увеличение светлого време-
ни суток стимулирует деятельность половых желез. Цветение расте-
ний наступает лишь при определенной продолжительности дня,
в связи с чем они делятся на короткодневные и длиннодневные. Реак-
ция организмов на продолжительность светового дня (фотопериод)
называется фотопериодизмом. Растения могут существовать лишь
при наличии освещения, поэтому большая часть дна Мирового океана
не заселена растениями (на глубину свыше 200 метров достаточное
для фотосинтеза количество света не проникает). Однако животные,
будучи гетеротрофами, менее зависимы от света, поэтому среди них
встречаются как дневные, так и ночные животные. Некоторые виды
живут при отсутствии света, например пещерные, роющие и глубоко-
водные формы.
761
АБИОТИЧЕСКИЕ ФАКТОРЫ
Температура дает возможность осуществляться биохимиче-
ским реакциям в клетке, поэтому она влияет на все жизненные про-
цессы. Температура организма не может упасть ниже нуля, поскольку
тогда кристаллики льда повредят клетки. Губительна и чрезмерно вы-
сокая температура, вызывающая денатурацию структуры белка. Глав-
ным источником тепла является солнечная радиация, теплокровные
организмы (птицы и млекопитающие) используют энергию биохими-
ческих реакций. В зависимости от способности поддерживать темпе-
ратуру тела различают пойкилотермных и гомойотермных животных.
Жизнедеятельность первых (все беспозвоночные, круглоротые, рыбы,
амфибии, рептилии) зависит от внешнего тепла, при понижении тем-
пературы внешней среды у них пропорционально снижается актив-
ность. У гомойотермных (теплокровных) животных температура тела
постоянна за счет тепла, выделяемого при метаболизме, который у
них очень высок и относительно постоянен. Способность выдержи-
вать крайние значения температурного фактора у разных организмов
неодинакова. Многие организмы выдерживают замораживание (неко-
торые земноводные зимой промерзают), а для других (кишечная па-
лочка, тихоходки и др.) даже температура абсолютного нуля (–273 °С)
не приводит к гибели. При снижении температуры тела основные
процессы в организме снижаются до такой степени, что видимые про-
явления жизни исчезают (анабиоз).
Влажность воздуха обеспечивается насыщением его водяным па-
ром. Наибольшая влажность имеется вблизи водных источников,
а также во влажных тропических лесах, где растения выделяют много
водяного пара в процессе транспирации. Степень насыщенности опре-
деляет количество выпадаемых осадков, что определяет характер фи-
тоценоза. Если количество осадков за год не превышает 250 мм, неза-
висимо от других факторов развивается пустынный ландшафт, на
котором могут произрастать немногие ксерофиты. Такие зоны назы-
вают аридными. Если осадков около 2000 мм в год, то это влажные,
или гумидные, зоны. В умеренных зонах годовое количество осадков
составляет 250—750 мм в год. Существуют сезонные колебания влаж-
ности, причем зимой при сильных морозах количество пара в воздухе
снижается. Многие организмы выработали адаптации, способствую-
щие запасанию воды или снижающие ее потерю. Среди них — запаса-
ние жира (верблюд, пустынная крыса), самой воды (суккуленты —
кактус, алоэ), снижение транспирации, сбрасывание листьев, спячка
и т. д . Многие пустынные животные вообще не пьют, довольствуясь
водой, поступающей с пищей. Круговорот воды в процессе выпадания
осадков и последующего испарения называется гидрологическим
циклом.
762
ЭКОЛОГИЯ
Ветер обеспечивает циркуляцию воздушных масс, а вместе с
ними — перераспределение водяного пара и осадков. Таким же обра-
зом транспортируются кислород, выделяемый растениями, загрязня-
ющие вещества и др. Ветер играет значительную роль в распростране-
нии генеративного материала (спор, семян) и микроорганизмов.
Давление воздуха и его состав оказывают серьезное
влияние на видовой состав сообщества. Атмосферное давление сни-
жается по мере увеличения высоты, и уже на высоте 5450 м давление
вдвое меньше, чем на уровне моря. Одновременно происходит сниже-
ние количества О2. В таких условиях организмы испытывают гипок-
сию, что приводит к недостаточному снабжению тканей О2, быстрому
утомлению и недомоганию. У растений увеличивается транспирация,
и они теряют много воды. Газовый состав воздуха постоянен (N2
—
78,09%, С2 — 20,95%, СО2 — 0,03—0,04%), некоторое снижение относи-
тельного содержания кислорода происходит в полярных районах.
Содержание вредных примесей увеличивается за счет выбросов пред-
приятий. Наряду с выхлопными газами автомобилей это создает
плотный туман, смешанный с дымом, — смог.
Эдафические (почвенные) факторы — это значение почвы (слой,
расположенный над горными породами земной коры) на организмы.
В.В . Докучаев показал, что в почве постоянно идут разнообразные
физические, химические и биологические процессы. Он выделил пять
почвообразующих факторов — климат, геологическая основа, образу-
емая материнской породой, рельеф местности, живые организмы, вре-
мя. В состав почвы входят четыре главных компонента: минеральная
основа — 50—60%, органические вещества (гумус) — до 10%, вода —
25—35%, воздух — 15—25%.
Топографические факторы определяются рельефом местности. Ос-
новным фактором является высота над уровнем моря, с увеличением
которой снижается средняя температура (и увеличивается ее суточ-
ный перепад), атмосферное давление и концентрации газов, но увели-
чивается скорость ветра, количество осадков и солнечная радиация.
Это приводит к появлению вертикальной зональности. При этом в
характеристику склона входит его крутизна (уклон более 35° делает
невозможным образование почвы — она смывается — и жизнь расте-
ний), экспозиция (обращенность на ту или иную сторону света).
Лишь некоторые факторы оказываются жизненно важными — это
лимитирующие факторы. У каждого организма есть крайние границы
реакции на действие этих факторов, которые определяются нормой
реакции генотипа. Наиболее комфортные условия действия фактора
называют оптимумом. Зоны выражения фактора, отклоняющиеся от
оптимума, но не нарушающие жизнедеятельности организма, называ-
ют зонами нормы. Дальнейшее смещение в сторону недостаточного
763
БИОТИЧЕСКИЕ ФАКТОРЫ
или избыточного действия фактора приводит к снижению адаптив-
ных возможностей организма и нарушает его жизнедеятельность (на-
рушение размножения, замедление роста, физиологическую актив-
ность и др.). Такие зоны называют зонами пессимума. Например, для
развития яйца аскариды оптимум является t +25 °C, крайними значе-
ниями +12° и +40°. Величины крайних значений действия фактора
у разных организмов неодинаковы. Те из них, кто способен без ощу-
тимого вреда переносить значительные колебания действия фактора,
имеют приставку эври- и называются эвритопными. Организмы, су-
ществующие в узких пределах колебания интенсивности фактора,
имеют приставку стено- и называются стенотопными. Для большин-
ства организмов оптимальной является температура +15 ... +30 °С, но
у некоторых животных оцепенение наступает уже при +15 °С. Есть
микроорганизмы, способные жить в термальных источниках, и похо-
лодание для них губительно. Соответственно, эвритермными являют-
ся те организмы, которые устойчивы к колебаниям температуры (на-
пример, многие насекомые, сезонно переживающие значительные
перепады температуры). Ксенобионтные организмы, напротив, не мо-
гут переносить такие перепады, например, веслоногий рачок копилия
удивительная не выдерживает изменения температуры за пределами
+23 ... +29 °С. Эвриоксибионтные и стеноксибионтные организмы
аналогично реагируют на содержание кислорода, эвригалинные и сте-
ногалинные — на колебания соленности воды. Если организм устой-
чив к изменениям целого комплекса факторов внешней среды, его на-
зывают эврибионтным, а если неустойчив — стенобионтным.
БИОТИЧЕСКИЕ ФАКТОРЫ
Живые организмы не только подвергаются влиянию абиотических
факторов, но и сами оказывают взаимное воздействие друг на друга.
При этом большое значение имеют видовое разнообразие биоценоза,
численность популяций в нем и биомасса (общее количество массы,
принадлежащей всей совокупности особей). Все разнообразие взаи-
моотношений между организмами (коакции) можно разделить на
прямые и косвенные. Прямые отношения проявляются в непосредст-
венном воздействии организмов друг на друга, например отношения
«хищник — жертва». Косвенные отношения влияют на организмы
опосредованно, например фитоценоз влияет на абиотические факто-
ры (температура, влажность, освещение и др.), поэтому абиотические
условия леса и степи разные. Наиболее прочными связями, объединя-
ющими совместно живущие организмы, являются трофические, или
пищевые. Пищевой, или трофический, фактор может различаться по
764
ЭКОЛОГИЯ
количеству, качеству (составу) и доступности. Каждый организм пи-
тается за счет других организмов, но при этом и сам является пищей
для других. Например, траву едят травоядные животные, тех, в свою
очередь, поедают плотоядные. Но и хищники также включены в тро-
фические связи, поскольку любой из них или сам станет жертвой бо-
лее сильного хищника, либо после естественной смерти будет разла-
гаться под действием сапрофитных организмов (бактерий и грибов),
которые обогащают почву соединениями, необходимыми для рас-
тений.
Любой биоценоз включает в себя три блока: продуценты, консу-
менты и редуценты. Продуценты — это зеленые растения, синтезиру-
ющие органические вещества из неорганических, используя для этого
энергию света. Они создают первичную органическую массу, за счет
которой живут они сами и все остальные участники биоценоза.
В определенных условиях роль продуцентов выполняют бактерии-хе-
мотрофы, которые используют для синтеза органических веществ
энергию, выделяющуюся в ходе химических реакций неорганических
соединений. Животные, которые питаются растениями или поедают
других животных, составляют блок консументов, подразделяющихся
на консументы первого (травоядные), второго (хищники, питающие-
ся травоядными), третьего и последующего порядков (крупные хищ-
ники, которые могут справиться с менее крупными хищниками). Кон-
сументы первого порядка регулируют численность продуцентов, но
при этом они потребляют в пищу не более 5—10% растительной мас-
сы, тогда как все остальное, а также остатки самих консументов под-
вергаются разложению сапрофитами, которые составляют в биоцено-
зе блок редуцентов. Они используют для своей жизнедеятельности
органические соединения, содержащиеся в остатках, и переводят про-
дукты деструкции в абиотическую среду, обогащая ее веществами, ко-
торые необходимы продуцентам.
Любой организм проявляет выраженную избирательность в отно-
шении состава пищи, поскольку каждому из них необходимы опреде-
ленные вещества. В зависимости от разнообразия рациона различают
виды, способные питаться одним видом растения или животного (мо-
нофаги, или стенофаги), и виды, поглощающие широкий ассортимент
пищи (полифаги, или эврифаги). Промежуточное положение занима-
ют олигофаги, кормовая база которых более или менее ограничена.
Трофические связи позволяют наиболее полно использовать свето-
вую энергию, которая переводится растениями в химические связи
органических соединений. При переходе веществ от одного блока к
другому значительная часть энергии теряется в виде энтропии, что
соответствует второму закону термодинамики. Поэтому суммарная
биомасса вышестоящего блока всегда меньше нижестоящего. Напри-
765
БИОТИЧЕСКИЕ ФАКТОРЫ
мер, биомасса растений в биоценозе значительно превышает биомас-
су травоядных животных, но биомасса последних гораздо больше, чем
у плотоядных хищников. Совсем иная ситуация в биоценозах морей.
Там биомасса растений значительно уступает биомассе животных.
Однако противоречия здесь нет, просто основная часть морских про-
дуцентов приходится на мелкие водоросли, образующие фитоплан-
ктон. Количество крупных талломных водорослей невелико, потому
что водоросли (как и все остальные растения) могут жить только
в условиях достаточной освещенности. Между тем основная часть
акватории Мирового океана глубоководная и поэтому не заселена во-
дорослями, которым приходится заселять узкие прибрежные зоны
мелководья. Но и фитопланктон имеет относительно небольшую био-
массу, потому что его постоянно поедают животные-фильтраторы,
образующие зоопланктон. Однако фитопланктон размножается на-
столько быстро, что его просто не успевают полностью съесть.
Организмы в сообществе, объединенные трофическими связями,
образуют цепи питания. Первым звеном в такой цепи являются про-
дуценты, следующими — консументы того или иного порядка. В каче-
стве примера можно привести такую цепь: растения ␚ насекомые ␚
птицы ␚ хищные птицы; или фитопланктон ␚ зоопланктон ␚ мел-
кие рыбы ␚ более крупные рыбы ␚ рыбоядные ластоногие. Обычно
цепь питания включает в себя не более четырех-пяти звеньев, что свя-
зано с потерей энергии на каждом уровне. Все организмы, образующие
сообщество, являются взаимополезными, поскольку изъятие из цепи
или сети одного вида неизбежно приведет к нарушению цепи пита-
ния. Так, в Китае пытались уничтожить воробьев, результатом чего
стало взрывное размножение насекомых-вредителей, которые ранее
уничтожались этими птицами. Закономерное снижение биомассы
каждого последующего блока цепи питания получило название пра-
вила экологической пирамиды. При этом различают пирамиду массы
(собственно изменение биомассы организмов) и пирамиду энергии
(изменение количества энергии). При фотосинтезе растения исполь-
зуют 0,1—0,5% световой энергии Солнца, падающей на землю. Каж-
дый последующий блок цепи питания использует всего 5—15% энер-
гии, полученной от предыдущего звена. Если расположить друг над
другом энергетическую продуктивность питания по ходу пищевой
цепи, то как раз получится экологическая пирамида энергии. В сред-
нем один гектар леса в течение года воспринимает примерно 2,1 ×
× 109 кДж солнечной энергии. Но если сжечь образовавшуюся за это
время растительную биомассу, освободится всего 1,1 × 106 кДж — 0,5%
от первоначальных цифр, что и будет отражать реальную продуктив-
ность фототрофов. За год растения Земли синтезируют около
100 млрд т органических веществ, содержащих 1,8 × 1018 кДж энергии.
766
ЭКОЛОГИЯ
Типы взаимоотношений между организмами. В зависимости от
реакции выделяют гомотипические и гетеротипические взаимоотно-
шения. Гомотипические коакции представляют собой взаимоотноше-
ния между особями одного вида. Они выражаются, например, в сов-
местной защите детенышей и кормового участка. Соответственно,
гетеротипическими коакциями являются взаимоотношения между
представителями разных видов. Эти отношения могут проявляться
различными способами, основными из которых являются антибиоз
(включает конкуренцию, хищничество и паразитизм) и симбиоз
(включает кооперацию, мутуализм и комменсализм). Различные фор-
мы конкуренции и кооперации были рассмотрены выше в разделах,
посвященных борьбе за существование, адаптациям.
Хищничество представляет собой наиболее распространенную
форму взаимоотношений между животными, при которых особи од-
них видов преследуют, убивают и затем пожирают особей других ви-
дов. Хищники и жертвы вырабатывают адаптации, которые первым
облегчают поиск и ловлю добычи, а вторым позволяют остаться в жи-
вых. При этом некоторые хищники могут долго бежать (волки), дру-
гие способны развивать большую скорость и на короткой дистанции
догоняют жертву (гепарды) или же, затаившись, совершают резкий
рывок и убивают жертву (леопарды). Адаптации жертв могут быть
морфологическими (покровительственная окраска, мимикрия, проч-
ные покровы, шипы и др.), физиологическими (выделение ядовитых
или отпугивающих веществ) и поведенческими (активная оборона,
затаивание, убегание). Развитие адаптации у хищников и их жертв не
приводит к безусловному преимуществу ни у одного из них, что важ-
но для обоих. Если бы хищники, получив возможность всегда доби-
ваться успеха на охоте, уничтожили бы все свои кормовые объекты,
то, оставшись без пищи, они погибли бы от голода. Соответственно и
потенциальные жертвы, избежав нападения хищников, сильно раз-
множатся и подорвут свою кормовую базу, в результате чего погибнут
не только хищники, но и они сами. Поэтому итогом сосуществования
является такое состояние системы «хищник — жертва», при котором
поддерживается более или менее постоянный уровень поедания хищ-
никами своих жертв, дающий возможность длительного сосущество-
вания этих популяций в составе биоценоза.
Паразитизм — это такая форма межвидовых взаимоотношений,
при которой один организм (паразит) живет за счет другого (хозяи-
на) и при этом наносит ему ущерб, но обычно не приводит к гибели.
В зависимости от места пребывания на хозяине различают эктопара-
зитов (на поверхности тела, например, блохи) и эндопаразитов (вну-
три тела хозяина, например, гельминты). По времени нахождения па-
разита на хозяине паразитизм бывает истинным, или постоянным
767
БИОТИЧЕСКИЕ ФАКТОРЫ
(вши, живущие на хозяине постоянно), и периодическим (личинки
оводов — паразиты, а взрослые особи живут свободно). Паразитизм
широко распространен среди растений и животных, а вирусы выража-
ют крайнюю форму паразитизма, поскольку вообще не способны осу-
ществлять свои физиологические функции вне клетки-хозяина. Дли-
тельное сосуществование системы «паразит — хозяин» приводит к
выработке адаптации (главным образом у паразита). К их числу отно-
сится сложный цикл развития паразита со сменой промежуточного
хозяина. Это обеспечивает расселение паразита. Паразиты очень ред-
ко приводят к гибели хозяина, поскольку не заинтересованы в этом:
гибель хозяина лишает пищи и паразита. Организмы, которые не
всегда являются паразитами, называют факультативными. Облигат-
ные паразиты всегда ведут паразитический образ жизни.
Симбиоз представляет собой разные формы тесного сожительства
организмов разных видов, не приносящие вреда никому из них. В бо-
лее узком понимании используют термин мутуализм, которым обо-
значают сожительство, из которого оба вида извлекают пользу, но
лишь один из них может развиваться только в присутствии другого,
тогда как другой может жить самостоятельно. Примерами таких взаи-
моотношений является сожительство актинии и некоторых рыбок,
обитающих среди ее щупалец, рака-отшельника и той же актинии.
Актиния получает остатки пищи, но защищает своими стрекательны-
ми клетками своего сожителя. При этом актиния может жить и само-
стоятельно, а сожитель — нет. Взаимную пользу получают бобовые
растения и азотфиксирующие бактерии, образующие на их корнях
клубеньки. Это пример взаимовыгодного симбиоза, при котором оба
сожителя не могут нормально развиваться друг без друга.
Комменсализм — питание одного вида остатками пищи другого
вида (например, гиены и другие падальщики), не принося ему вреда;
нейтрализм — взаимную независимость совместно обитающих видов;
форезия — использование одними видами других в качестве транс-
портного средства; синойкия — использование чужих гнезд и нор или
другого организма в качестве жилища; протокооперация — совмест-
ное проживание с целью повышения эффективности адаптации,
например коллективное гнездование нескольких видов птиц, способ-
ствующее защите всего поселения от хищников; аменсализм — поло-
жение, при котором один вид не может нормально питаться и размно-
жаться в присутствии другого; интерференция — непреднамеренное
подавление жизнедеятельности одним видом другого, например пти-
цы, поедающие семена шишек, разрушают их, чем вызывают гибель
обитающих в шишках насекомых.
Антропогенный фактор. В настоящее время природа испытывает
чрезвычайно большое воздействие человека. Оно может быть нега-
768
ЭКОЛОГИЯ
тивным и положительным. Человеческий фактор чаще всего является
причиной отрицательных изменений в сообществах. Это выражается
в усилении давления на окружающую среду в процессе различного
рода производств (сточные воды, выбросы токсичных газов и задым-
ление), роста городов и увеличении их числа и др. Особое значение
имеет постоянно увеличивающееся население Земли, уже превышаю-
щее 7 млрд человек. Ежегодный прирост составляет примерно 2% в
год (75—85 млн человек). Все увеличивающиеся потребности людей
влекут за собой освоение все новых территорий для хозяйственных
нужд, прокладку средств коммуникации (подземные и подводные ка-
бели, провода и др.). При этом происходит уменьшение площади аре-
алов организмов. Все это приводит к вымиранию видов — каждый
день с лица Земли исчезает несколько видов растений и животных.
Из-за вырубки тропических лесов изменяется климат, при этом
длительные засухи сменяются проливными дождями, вызывающими
губительные наводнения. Развивающаяся в результате неправильного
землепользования эрозия почвы снижает ее плодородие, а образовав-
шиеся овраги изменяют ландшафт и также приводят к гибели сооб-
ществ. Постоянно расширяется площадь пустынь и образуются
новые. Большой вред приносит непосредственное истребление жи-
вотных. Это явилось причиной исчезновения многих видов (стелле-
рова корова, дронт, тур, квагга, странствующий голубь и др.). Виды
растений и животных, которым грозит вымирание, занесены в Крас-
ную книгу и подлежат охране. В местах их естественного обитания
создаются заповедники (запрещена любая хозяйственная деятель-
ность), заказники (влияние человека сильно ограничено) и нацио-
нальные парки (обширные территории, где охраняемые зоны переме-
жаются с хозяйственными объектами).
Некоторые животные сумели выработать адаптации, позволяю-
щие им обитать рядом с человеком и находить там пищу. Численность
популяций серых крыс многократно превышает число самих горожан.
То же самое относится и к тараканам.
РАЗВИТИЕ ЭКОСИСТЕМ
Экосистема — это устойчивая система живых организмов и окру-
жающей среды. В пределах этой системы осуществляется поток энер-
гии и круговорот веществ. Следует помнить, что экосистема — это
любое сообщество организмов в абиотическом окружении, тогда как
биогеоценоз всегда связан с участком Земли. То есть такие экосисте-
мы, как пробирка с питательной средой, заселенная бактериями, под-
водная лодка в плавании, аквариум, — биогеоценозом не являются.
769
РАЗВИТИЕ ЭКОСИСТЕМ
Для него необходима четко ограниченная часть земной поверхно-
сти — открытой или под водой. Однако любой биогеоценоз является
экосистемой.
Экосистема является открытой системой, динамично изменяю-
щейся под воздействием различных внутренних и внешних факторов.
Формирование биогеоценоза начинается с заселения первыми орга-
низмами ранее безжизненных территорий. Такими организмами не
могут быть животные (им там нечего есть, поскольку они гетеротро-
фы) и высшие растения (для их произрастания нужна почва с гуму-
сом). Обычно новые участки осваиваются лишайниками, которые
представляют самодостаточную систему, состоящую из продуцента
(водоросли) и консумента (гриба). Эти организмы создают пионерные
сообщества и постепенно из их остатков и материнской породы нака-
пливается слой почвы, достаточный для поселения высших растений.
Чаще всего первыми поселяются споровые (мхи и папоротникообраз-
ные), а после появляются различные травы, кустарники и деревья.
Постепенная смена одних видов другими в процессе формирования
экосистемы называется экологической сукцессией. В результате сук-
цессии формируется устойчивое самовозобновляющееся сообщество,
находящееся в равновесии с окружающей средой. Такое сообщество
называется климаксным. Различают первичную и вторичную сукцес-
сию. Первичная сукцессия представляет собой все вышеописанные
этапы формирования биогеоценоза на ранее безжизненном участке.
Вторичная сукцессия — это восстановление экосистемы, поврежден-
ной до этого каким-либо фактором (вырубка леса человеком, пожар,
буря, наводнение, снежная лавина и т. п .). Вторичная сукцессия идет
гораздо быстрее, поскольку после повреждения экосистемы может со-
храниться генеративный материал (споры, семена, почвенные живот-
ные и др.). Но она часто приводит к смене сообщества в экосистеме,
поскольку изменяются условия обитания. При этом также проходят
ряд стадий. Например, после пожара или вырубки леса на оголенной
территории быстро заселяются травянистые растения. Затем появля-
ются всходы деревьев, чьи семена разносятся ветром (береза, осина,
сосна), и постепенно восстанавливается лес. Его видовой состав мо-
жет быть иным, чем ранее, и часто меняется. Например, если в берез-
няке появляются всходы ели, то теневыносливая ель постепенно вы-
растет настолько, что затенит светолюбивые березы и они погибнут.
Теория сукцессии была разработана Клементсом (1916). Постепен-
ные изменения в процессе сукцессии ведут к стабилизации биогеоце-
ноза. Формирующаяся экосистема характеризуется высокой про-
дуктивностью, которая снижается с образованием климаксного
сообщества. Поэтому продуктивность старых лесов ниже, чем моло-
дых. Это объясняется тем, что в формирующейся системе еще не все
770
ЭКОЛОГИЯ
экологические ниши заняты. Процесс их освоения сопровождается
увеличением видового разнообразия и усложнением пищевых цепей.
Увеличивается и биомасса сообщества. Освоение экологических ниш
усложняет структуру сообщества. Это может выражаться в ярусно-
сти, характерной для старого леса, где различные организмы могут
найти много специфических мест обитания. Постоянно возрастающая
конкуренция ведет к сужению специализации по экологическим ни-
шам. В молодых экосистемах много видов с быстрой скоростью раз-
множения, в климаксных больше видов, которые размножаются мед-
ленно.
Причиной устойчивости климаксного сообщества является са-
морегуляция. Она выражается в том, что организмы, сосуществую-
щие на одной территории, не уничтожают полностью друг друга,
а лишь ограничивают численность на оптимальном в данных усло-
виях уровне.
Смена биогеоценоза. Эволюция биогеоценоза со временем ведет
к его смене. Причины для этого могут быть внешними и внутренни-
ми. Примерами внешних факторов, изменяющих экосистему, могут
быть различные природные бедствия или воздействие человека. Вну-
тренние факторы также разнообразны. Например, постройка бобрами
плотины ведет к затоплению более или менее обширных территорий.
При этом гибнут деревья и изменяется фитоценоз — энергетическая
основа сообщества. Другим примером может служить зарастание озе-
ра, что приводит к нарушению круговорота веществ. Из-за недостатка
кислорода органические остатки не окисляются до конца и скаплива-
ются на дне в виде ила. Это ведет к обмелению водоема и постепенно
он превращается в болото с другим видовым составом биоценоза.
Дальнейшая эволюция может привести к образованию на месте воз-
никшего болота осокового луга, а затем леса.
Агроценозы. Активное воздействие человека на живую природу
приводит к возникновению искусственных биогеоценозов. Среди них
наиболее распространены агроценозы — сельскохозяйственные эко-
системы. Это поля, пастбища, сады, пруды и др. В отличие от естест-
венных сообществ, в агроценозах биотические компоненты подби-
раются в соответствии с хозяйственными или эстетическими
потребностями человека. Часто практикуются монокультуры, т. е. за-
селение искусственно обработанной территории каким-то одним ви-
дом растения или животного. Ранее существовавшее на этом месте
сообщество обычно уничтожается. Главная задача агроценоза — дать
наибольшую продукцию, а это характерно для ранней стадии сукцес-
сии. Для освобождения места для агроценоза используются различ-
ные способы: выжигание, корчевание, осушение, затопление и др. Ча-
сто при этом происходит эрозия почвы (разрушение плодородного
771
КРУГОВОРОТ ВЕЩЕСТВ С УЧАСТИЕМ ЖИВЫХ ОРГАНИЗМОВ
слоя и последующее удаление его водными потоками и ветром), сни-
жение ее плодородности.
КРУГОВОРОТ ВЕЩЕСТВ С УЧАСТИЕМ
ЖИВЫХ ОРГАНИЗМОВ
Биогенная миграция атомов
В природе постоянно происходит перемещение различных ве-
ществ, создающее циркуляцию химических элементов и неорганиче-
ских соединений. Различают два основных круговорота веществ:
большой (геологический) и малый (биотический). Большой кругово-
рот, который длится очень долго (миллионы лет), заключается в по-
степенном разрушении горных пород, после чего растворимые веще-
ства сносятся в Мировой океан, где образуют пласты морских
отложений. Когда в ходе геотектонических изменений перемещаются
и сталкиваются материки, происходит подъем морского дна, осадоч-
ные напластования вновь оказываются на поверхности суши, и про-
цесс повторяется. Если в транспорте веществ значительную роль иг-
рают живые организмы, имеет место биогеохимический круговорот.
Он является частью большого круговорота и происходит на уровне
биогеоценоза.
Суть его в том, что растения аккумулируют в себе минеральные
вещества из почвы и воздуха и используют полученные элементы для
синтеза органических веществ. Посредством цепей питания элементы
мигрируют в организмы консументов и после распада, осуществляе-
мого редуцентами, вновь возвращаются в неорганическую среду (по-
чву, воду, атмосферу). Полный оборот химических веществ из абиоти-
ческой среды через растительные и животные организмы обратно в
неорганическую с использованием световой энергии называется био-
геохимическим циклом. В соответствии с направлениями миграции ве-
ществ различают два типа круговорота: газовые с обширными резер-
вуарами неорганических веществ в атмосфере и океанах (круговорот
азота, кислорода, углекислого газа и воды) и осадочные, которые
образуют менее крупные и обычно локальные резервуары в земной
коре (круговороты кальция, фосфора, железа).
Живые организмы (живое вещество) в круговоротах выполняют
газовую, концентрационную, биохимическую, окислительно-восста-
новительную, энергетическую, деструкционную функции.
Газовая функция проявляется в выделении организмами некото-
рых газов и их преобразовании. Растения в процессе фотолиза разла-
гают воду и выделяют кислород, причем в таком большом количестве,
что примерно пятая часть атмосферы состоит из биогенного О2.
772
ЭКОЛОГИЯ
В процессе дыхания организмы выделяют СО2
.
Многие прокариоты
могут восстанавливать газы (например, N2, H2S).
Концентрационная функция проявляется в избирательном погло-
щении и накоплении организмами определенных химических элемен-
тов (С, Н, О, N, P, S и др.). При отмирании организмов эти вещества
освобождаются, но остаются на месте, в результате чего постепенно
образуются их скопления.
Биохимическая функция выражается в том, что в ходе метаболизма
в организмах происходят разнообразные биохимические реакции, ко-
торые глубоко изменяют строение реагирующих веществ.
Окислительно-восстановительная функция осуществляется в про-
цессе окисления организмами одних веществ с образованием различ-
ных соединений (оксидов, солей и др.), а также восстановления дру-
гих (азот, серное железо и др.).
Энергетическая функция состоит в усвоении автотрофными орга-
низмами внешней энергии и последующей передаче ее по пищевой
цепи. В основе этого лежит деятельность фототрофов и хемотрофов.
Деструкционная функция проявляется в разложении организмов
после их смерти. При этом органические вещества, входившие в со-
став тела организма, разрушаются до неорганических соединений, ко-
торые через фототрофов возвращаются в биологический круговорот.
Биогенная миграция атомов химических элементов может осу-
ществляться двумя способами — микроорганизмами и многоклеточны-
ми организмами. При этом миграция с участием микроорганизмов бо-
лее значительна. Особое место занимает участие человека, которое
часто рассматривают в качестве третьего способа биогенной миграции.
Круговорот воды. Испаряясь с поверхности суши и моря, вода
входит в состав атмосферы и переносится воздушными массами на
значительные расстояния. Когда пар конденсируется, образуется
жидкая вода, которая выпадает в виде осадков. Круговорот воды спо-
собствует осуществлению большого круговорота. Кроме того, в круго-
ворот воды включаются ювенильные воды, которые поступают на по-
верхность земной коры при извержении вулканов и гейзеров.
Выделяют воду и живые организмы (транспирация растений, с потом,
мочой, фекалиями и т. п .).
Круговорот углерода. Углерод создает основу любого органиче-
ского соединения благодаря своим химическим свойствам. Как и у
любого другого элемента, круговорот углерода осуществляется по
большому и малому циклам. Малый (биотический) круговорот угле-
рода тесно связан с живыми организмами. Основным резервуаром
этого элемента является СО2 атмосферы. В таком виде его утилизиру-
ют фотосинтезирующие организмы, используя атом углерода для по-
строения органических веществ. Фиксированный в органических сое-
773
КРУГОВОРОТ ВЕЩЕСТВ С УЧАСТИЕМ ЖИВЫХ ОРГАНИЗМОВ
динениях углерод впоследствии потребляется консументами разных
трофических уровней, поэтому общая масса углерода в живых орга-
низмах очень высока. В процессе дыхания и деятельности деструкто-
ров (редуцентов) углерод вновь переходит в СО2 и возвращается
в атмосферу. Ежегодно в экосистемы включается около 12% СО2
атмосферы, поэтому на весь малый (биотический) цикл миграции
этого элемента приходится 8 лет. Некоторая часть углерода направля-
ется по другому пути и накапливается в органических остатках в виде
залежей ископаемых (каменный уголь, торф).
Круговорот азота. Азот является самым распространенным га-
зом в составе атмосферного воздуха (около 80%). Значительная
часть его находится в форме неорганических (нитриты, нитраты, ам-
монийные соли) и органических (аминокислоты, белки, азотистые
основания нуклеиновых кислот. Сущность биотического (малого)
круговороты азота состоит в следующем. Молекулярный азот не мо-
жет усваиваться эукариотами. Однако это под силу некоторым про-
кариотам, имеющим особый ген (nif-ген). Они могут связывать N2
и включать его в состав нитритов и нитратов, за что получили назва-
ние нитрифицирующих бактерий. Они могут находиться в свобод-
ном состоянии или быть в симбиозе с корнями растений (клубень-
ковые бактерии). Связанный таким образом азот доступен для
растений, он используется ими для построения нужных органиче-
ских веществ и передается по цепям питания. После отмирания ра-
стений органические остатки, содержащие азот, перерабатываются
редуцентами (гнилостные бактерии) до аммиака. Затем некоторые
хемосинтезирующие бактерии переводят его сначала в состав азоти-
стой, а затем азотной кислот. Наряду с процессом нитрификации
(связывания азота) происходит и обратный процесс — денитрифи-
кация, который представляет собой последовательное восстановле-
ние азота из химических соединений в молекулярное состояние, по-
сле чего он вновь попадает в атмосферу. Часть азота выводится из
биогенной миграции, оседая в составе отложений. Однако это ком-
пенсируется азотом, поступающим в атмосферу в процессе вулкани-
ческой деятельности. Общее время прохождения малого (биотиче-
ского) цикла миграции азота составляет 110 лет.
Круговорот фосфора. Содержание фосфора в органических соеди-
нениях гораздо меньшее, чем углерода и азота, но значение его высоко.
В составе ортофосфорной кислоты он входит в состав фосфолипидов
и нуклеиновых кислот. Остатки неорганического фосфата связаны
между собой макроэргическими связями, которые при разрыве выде-
ляют энергию, используемую для осуществления биохимических реак-
ций в клетке. Поэтому молекулы нуклеозидтрифосфата (АТР и др.)
составляют энергетический потенциал клетки. Отложения фосфора
774
ЭКОЛОГИЯ
всецело связаны с жизнедеятельностью живых организмов. Ископае-
мые остатки неравномерно скапливаются в земной коре. Постепенно
вымываясь, соединения фосфора попадают в экосистемы и включают-
ся в трофические цепи. Недостаточное содержание этого элемента в
культивируемых почвах компенсируется внесением фосфорных удо-
брений.
Круговорот серы. Сера входит в состав немногих органических
соединений (например, серосодержащие аминокислоты), но ее при-
сутствие необходимо для жизнедеятельности организма. Растения ис-
пользуют серу в виде сульфатов, которые образуются из отложений
сульфидов в результате деятельности микроорганизмов. В абиотиче-
скую среду сера возвращается после деструкции органики редуцента-
ми. Избыток соединений серы угнетает фотосинтез и вызывает гибель
растений в промышленных зонах.
Содержание
КЛЕТОЧНАЯТЕОРИЯ..........................................................3
ПРОКАРИОТИЧЕСКИЕИЭУКАРИОТИЧЕСКИЕКЛЕТКИ...................5
ХИМИЧЕСКАЯОРГАНИЗАЦИЯКЛЕТКИ.....................................6
СТРОЕНИЕКЛЕТКИ..........................................................11
Биологическиемембраны........................................................12
Поверхностныйкомплекс....................................................16
Межклеточныесоединения..................................................22
Микроворсинки.............................................................25
Ядро.........................................................................26
Хромосомыиядрышки................................................29
Кариотип...........................................................33
Цитоплазма......................................................................34
Гиалоплазма.................................................................34
Органеллы..................................................................34
Органеллыобщегозначения............................................35
Немембранныеорганеллы..............................................35
Цитоскелет.........................................................35
Клеточныйцентр....................................................38
Рибосомы............................................................40
Мембранныеорганеллы................................................41
Митохондрии........................................................41
Эндоплазматическаясеть.............................................44
КомплексГольджи....................................................46
Лизосомы............................................................52
Пероксисомы.........................................................55
Специальныеорганеллы.....................................................56
Ресничкиижгутики..................................................56
Включения...........................................................59
Целостныереакцииклетки.......................................................59
Эндоцитоз...................................................................60
Пиноцитоз...................................................................60
Фагоцитоз...................................................................62
Внутриклеточныебиохимическиереакции.......................................63
Синтезбелка................................................................63
Основныереакциитканевогообмена.........................................67
Экзоцитоз...................................................................71
Путивосприятияипередачиинформацииклеткой............................71
Жизненныйпутьклеток.........................................................73
Клеточныйцикл.................................................................74
Интерфаза...................................................................76
Митоз...........................................................................79
Мейоз...........................................................................83
776
СОДЕРЖАНИЕ
ОСОБЕННОСТИСТРОЕНИЯРАСТИТЕЛЬНОЙКЛЕТКИ....................90
Делениерастительнойклетки(цитокинез).......................................95
Фотосинтез......................................................................97
Световаястадияфотосинтеза...............................................100
Темноваястадияфотосинтеза...............................................103
СТРОЕНИЕ И ФУНКЦИОНИРОВАНИЕ ПРОКАРИОТИЧЕСКОЙ
КЛЕТКИ.......................................................................105
ВИРУСЫ......................................................................111
ТКАНИ........................................................................115
Эпителиальныеткани...........................................................115
Соединительныеткани..........................................................117
Рыхлаяволокнистаясоединительнаяткань..................................124
Плотнаяволокнистаясоединительнаяткань.................................127
Тканисоспециальнымисвойствами.............................................128
Хрящеваяткань.............................................................128
Костнаяткань..............................................................129
Мышечныеткани...............................................................133
Нервнаяткань..................................................................138
ОРГАНЫ,СИСТЕМЫИАППАРАТЫОРГАНОВ.............................146
ОСОБЕННОСТИ СТРОЕНИЯ, РОСТА И РАЗВИТИЯ ЧЕЛОВЕКА. . . . . . . . . . . 147
БИОЛОГИЧЕСКАЯ И ПСИХОСОЦИАЛЬНАЯ СУЩНОСТЬ ЧЕЛОВЕКА . . . 155
ОПОРНО-ДВИГАТЕЛЬНЫЙАППАРАТ......................................157
Пассивнаячастьопорно-двигательногоаппарата.................................157
Скелетиегосоединения........................................................162
Скелеттуловища...........................................................167
Череп.......................................................................175
Скелетконечностей.........................................................185
Костиверхнейконечности..................................................186
Костинижнейконечности...................................................189
Активнаячастьопорно-двигательногоаппарата..................................195
Скелетныемышцы..........................................................195
Элементыбиомеханики.....................................................201
Мышцыразличныхобластей................................................203
Работоспособность,работа,утомлениеиотдых..................................216
Физическаяактивность.....................................................222
ВНУТРЕННИЕОРГАНЫ......................................................223
ПИЩЕВАРИТЕЛЬНАЯСИСТЕМА...........................................225
Полостьрта.....................................................................228
Глоткаипищевод...............................................................235
Желудок.......................................................................237
777
СОДЕРЖАНИЕ
Тонкаякишка...................................................................239
Печеньижелчныйпузырь......................................................241
Поджелудочнаяжелеза.........................................................246
ФУНКЦИИПИЩЕВАРИТЕЛЬНОЙСИСТЕМЫ.............................252
Пищаихарактерпитания.......................................................253
Белки......................................................................255
Жиры(липиды)............................................................257
Углеводы...................................................................259
Балластныевещества.......................................................262
Свободныерадикалы...........................................................263
Витамины......................................................................263
Жирорастворимыевитамины...............................................264
Водорастворимыевитамины................................................265
ВитаминыгруппыВ........................................................266
Витаминоподобныевещества................................................268
Потребностьввитаминах.......................................................269
Минеральныевещества.........................................................270
Вода............................................................................272
ПИЩЕВАРЕНИЕ..............................................................273
Пищеварениевполостирта.....................................................274
Пищеварениевжелудке.........................................................275
Пищеварениевтонкойкишке...................................................276
Пищеварениевтолстойкишке..................................................277
ДЫХАТЕЛЬНАЯСИСТЕМА..................................................278
Гортань.........................................................................280
Легкие..........................................................................286
Плевра.....................................................................289
Средостение....................................................................289
Функциядыхательнойсистемы.................................................289
МОЧЕПОЛОВОЙАППАРАТ..................................................294
Мочевыеорганы................................................................294
Функцияпочек.............................................................301
Физическиеихимическиесвойствамочи....................................301
ПОЛОВЫЕСИСТЕМЫ........................................................303
Биологическаяисоциальнаясущностьполачеловека............................303
МУЖСКИЕПОЛОВЫЕОРГАНЫ.............................................305
Внутренниемужскиеполовыеорганы...........................................305
Простата...................................................................308
Наружныемужскиеполовыеорганы............................................310
Механизмэрекции..............................................................312
778
ЖЕНСКИЕПОЛОВЫЕОРГАНЫ.............................................313
Внутренниеженскиеполовыеорганы............................................313
Матка......................................................................316
Влагалище..................................................................317
Наружныеженскиеполовыеорганы.............................................317
Овариально-менструальныйцикл...............................................319
Молочнаяжелеза...........................................................321
Половоесозревание.............................................................322
Гаметогенез.....................................................................323
Половойциклчеловека.........................................................324
ПОЛОСТЬЖИВОТА.БРЮШИНА...........................................327
СЕРДЕЧНО-СОСУДИСТАЯСИСТЕМА.......................................331
КРОВЕНОСНАЯСИСТЕМА..................................................331
Сердце.....................................................................334
Функциисердца............................................................337
Кровоснабжениетелачеловека..............................................341
Функциясердечно-сосудистойсистемы.....................................346
Регуляцияфункцийсердечно-сосудистойсистемы...........................349
Лимфатическаясистема.........................................................350
ЛИМФОИДНАЯ СИСТЕМА (органы кроветворения
ииммуннойсистемы)..........................................................351
Костныймозг...................................................................355
Тимус..........................................................................356
Лимфоидная ткань стенок органов
пищеварительнойидыхательнойсистем.........................................356
Лимфатическиеузлы...........................................................358
Селезенка......................................................................359
Неспецифическаясопротивляемостьорганизма.................................360
ЭНДОКРИННЫЕЖЕЛЕЗЫ..................................................361
Гипофиз........................................................................365
Щитовиднаяжелеза............................................................365
Надпочечник...................................................................366
Паращитовидныежелезы.......................................................366
Панкреатическиеостровки......................................................367
НЕРВНАЯСИСТЕМА.........................................................367
ЦЕНТРАЛЬНАЯНЕРВНАЯСИСТЕМА(ЦНС)...............................369
Спинноймозг...................................................................369
Головноймозг..................................................................370
Оболочкиспинногоиголовногомозга.......................................381
СОДЕРЖАНИЕ
779
Периферическаянервнаясистема...............................................383
Вегетативная(автономная)нервнаясистема(ВНС).............................386
Высшаянервнаядеятельность...................................................392
Особенностивысшейнервнойдеятельностичеловека........................393
Типынервнойдеятельности.................................................394
Интегративнаяфункциянервнойсистемы.......................................395
Ритмымозга................................................................395
Сонибодрствование........................................................396
Сознаниеимышление......................................................396
Членораздельнаяречь.......................................................396
Научениеипамять..........................................................397
Поведение..................................................................399
Мотивация.................................................................400
Интеллект..................................................................402
ОРГАНЫЧУВСТВ............................................................403
Органзрения...................................................................405
Преддверно-улитковыйорган(органслухаиравновесия)........................411
Органобоняния................................................................416
Органвкуса....................................................................417
Кожа...........................................................................418
Кожнаячувствительность...................................................421
РАСТЕНИЯ....................................................................422
НИЗШИЕРАСТЕНИЯ........................................................422
Водоросли......................................................................422
Размножениеводорослей...................................................424
Экологическиеформыводорослей...........................................426
Значениеводорослей.......................................................426
ВЫСШИЕРАСТЕНИЯ........................................................428
Тканирастений.................................................................430
Органырастений...............................................................439
Корень.....................................................................439
Побег.......................................................................448
Стебель.....................................................................450
Почка......................................................................453
Лист........................................................................454
Отделмохообразные............................................................460
Отделпапоротникообразные....................................................463
Класспапоротниковидные..................................................465
Классплауновидные........................................................467
Классклинолистовидные,иличленистые....................................469
Семенныерастения.............................................................471
ОтделГолосеменные............................................................472
Классхвойные..............................................................474
СОДЕРЖАНИЕ
780
СОДЕРЖАНИЕ
Отделпокрытосеменные,илицветковые.........................................478
Цветок.....................................................................479
Семя.......................................................................489
Плод.......................................................................490
Вегетативноеразмножениецветковых.......................................495
Классификацияпокрытосеменных..........................................501
Грибы..........................................................................503
Лишайники.....................................................................509
ЖИВОТНЫЕ..................................................................512
ПОДЦАРСТВООДНОКЛЕТОЧНЫЕ,ИЛИПРОСТЕЙШИЕ.................512
ТипСаркомастигофоры.........................................................514
ТипСпоровики.................................................................517
ТипИнфузории,илиРесничные................................................520
ПОДЦАРСТВОМНОГОКЛЕТОЧНЫЕ........................................523
Теориипроисхождениямногоклеточныхорганизмов.............................523
ТипКишечнополостные.........................................................523
ТипПлоскиечерви..............................................................529
ТипКруглыечерви.............................................................541
ТипКольчатыечерви...........................................................545
ТипЧленистоногие.............................................................551
Насекомыебезметаморфоза(снеполнымпревращением)....................566
Насекомыесметаморфозом(сполнымпревращением)......................568
ТипМоллюски.................................................................575
Типхордовые...................................................................583
ПодтипПозвоночные,илиЧерепные............................................588
КлассХрящевыерыбы......................................................588
КлассКостные(Костистые)рыбы...........................................600
НадклассНаземныепозвоночные...............................................609
КлассЗемноводные,илиАмфибии..........................................609
КлассПресмыкающиеся,илиРептилии.....................................626
КлассПтицы...............................................................635
КлассМлекопитающие.....................................................648
ГЕНЕТИКА....................................................................664
НАСЛЕДСТВЕННОСТЬ.......................................................668
Взаимодействиеаллельныхгенов...............................................668
Взаимодействиенеаллельныхгенов.............................................674
Хромосомнаятеориянаследственности..........................................677
ИЗМЕНЧИВОСТЬ.............................................................683
Ненаследственная (фенотипическая, или модификационная)
изменчивость...................................................................684
Наследственная(генотипическая)изменчивость.................................689
781
СОДЕРЖАНИЕ
Комбинативнаяизменчивость...................................................689
Мутационнаяизменчивость.....................................................690
СЕЛЕКЦИЯ....................................................................704
Селекциярастений.............................................................708
Селекцияживотных............................................................710
БИОТЕХНОЛОГИЯ...........................................................712
ЭВОЛЮЦИЯ..................................................................713
Микроэволюция................................................................724
Предпосылки(элементарныефакторы)эволюции...........................725
Борьбазасуществование....................................................728
Естественныйотбор.........................................................731
Адаптации(приспособления)иадаптациогенез..............................735
Критерииипризнакивида..................................................739
Видообразование...........................................................740
Макроэволюция................................................................741
Направленияэволюции.........................................................743
Биологическийпрогрессибиологическийрегресс...............................744
Основныезакономерностиэволюции............................................745
Доказательстваэволюции.......................................................746
ГипотезыпроисхожденияжизнинаЗемле.......................................748
Развитиеорганическогомира...................................................750
Положениечеловекавприроде..................................................757
ЭКОЛОГИЯ...................................................................759
Абиотическиефакторы..........................................................760
Биотическиефакторы...........................................................763
Развитиеэкосистем.............................................................768
Круговоротвеществсучастиемживыхорганизмов...............................771
Биогеннаямиграцияатомов.................................................771
Все права защищены. Книга или любая ее часть не может быть скопирована, воспроизведена в
электронной или механической форме, в виде фотокопии, записи в память ЭВМ, репродукции или каким-
либо иным способом, а также использована в любой информационной системе без получения разрешения
от издателя. Копирование, воспроизведение и иное использование книги или ее части без согласия
издателя является незаконным и влечет уголовную, административную и гражданскую ответственность.
Научно-популярное издание
Билич Габриэль Лазаревич
Зигалова Елена Юрьевна
БИОЛОГИЯ ДЛЯ ПОСТУПАЮЩИХ В ВУЗЫ
Директор редакции Е. Капьёв
Руководитель медицинского направления О. Шестова
Ответственный редактор О. Ключникова
Художественный редактор Е. Анисина
Подписано в печать 19.03 .2018. Формат 70x1001/16 .
Печать офсетная. Усл. печ. л. 63,52.
Тираж
экз. Заказ
ООО «Издательство «Э»
123308, Москва, ул. Зорге, д. 1. Тел.: 8 (495) 411-68-86.
;ндіруші: «Э» А?Б Баспасы, 123308, МAскеу, Ресей, Зорге кBшесі, 1 Cй.
Тел.: 8 (495) 411-68-86.
Тауар белгісі: «Э»
?азаEстан Республикасында дистрибьютор жAне Bнім бойынша арыз-талаптарды EабылдаушыныG
Bкілі «РДЦ-Алматы» ЖШС, Алматы E., Домбровский кBш., 3«а», литер Б, офис 1.
Тел.: 8 (727) 251-59-90/91/92. Интернет-магазин: www.book24.kz
;німніG жарамдылыE мерзімі шектелмеген.
Сертификация туралы аEпарат сайтта ;ндіруші «Э»
Сведения о подтверждении соответствия издания согласно законодательству РФ
о техническом регулировании можно получить на сайте Издательства «Э»
;ндірген мемлекет: Ресей
Сертификация EарастырылмаMан
Оптовая торговля книгами Издательства «Э»:
142700, Московская обл., Ленинский р-н, г. Видное,
Белокаменное ш., д. 1, многоканальный тел.: 411-50 -74 .
По вопросам приобретения книг Издательства «Э» зарубежными оптовыми
покупателями обращаться в отдел зарубежных продаж
International Sales: International wholesale customers should contact
Foreign Sales Department for their orders.
По вопросам заказа книг корпоративным клиентам,
в том числе в специальном оформлении, обращаться по тел.:
+7 (495) 411-68 -59 , доб. 2261.
Оптовая торговля бумажно-беловыми
и канцелярскими товарами для школы и офиса:
142702, Московская обл., Ленинский р-н, г. Видное-2,
Белокаменное ш., д. 1, а/я 5. Тел./факс: +7 (495) 745-28-87 (многоканальный).
Полный ассортимент книг издательства для оптовых покупателей:
Москва. Адрес: 142701, Московская область, Ленинский р-н,
г. Видное, Белокаменное шоссе, д. 1 . Телефон: +7 (495) 411-50 -74 .
Нижний Новгород. Филиал в Нижнем Новгороде. Адрес: 603094,
г. Нижний Новгород, ул. Карпинского, д. 29, бизнес-парк «Грин Плаза».
Телефон: +7 (831) 216-15-91 (92, 93, 94).
Санкт-Петербург. ООО «СЗКО». Адрес: 192029, г. Санкт-Петербург, пр. Обуховской Обороны,
д. 84, лит. «Е». Телефон: +7 (812) 365-46-03 / 04. E-mail: server@szko.ru
Екатеринбург. Филиал в г. Екатеринбурге. Адрес: 620024,
г. Екатеринбург, ул. Новинская, д . 2щ. Телефон: +7 (343) 272-72-01 (02/03/04/05/06/08).
Самара. Филиал в г. Самаре. Адрес: 443052, г. Самара, пр-т Кирова, д. 75/1, лит. «Е».
Телефон: +7(846)207-55-50 . E-mail: RDC-samara@mail.ru
Ростов-на-Дону. Филиал в г. Ростове-на-Дону. Адрес: 344023,
г. Ростов-на -Дону, ул. Страны Советов, д. 44 А. Телефон: +7(863) 303-62-10.
Центр оптово-розничных продаж Cash&Carry в г. Ростове-на -Дону. Адрес: 344023,
г. Ростов-на -Дону, ул. Страны Советов, д. 44 В. Телефон: (863) 303-62-10. Режим работы: с 9-00 до 19-00 .
Новосибирск. Филиал в г. Новосибирске. Адрес: 630015,
г. Новосибирск, Комбинатский пер., д. 3 . Телефон: +7(383) 289-91-42 .
Хабаровск. Филиал РДЦ Новосибирск в Хабаровске. Адрес: 680000, г. Хабаровск,
пер. Дзержинского, д. 24, литера Б, офис 1. Телефон: +7(4212) 910-120.
Тюмень. Филиал в г. Тюмени. Центр оптово-розничных продаж Cash&Carry в г. Тюмени.
Адрес: 625022, г. Тюмень, ул. Алебашевская, д. 9А (ТЦ Перестройка+).
Телефон: +7 (3452) 21-53-96/ 97/ 98.
Краснодар. Обособленное подразделение в г. Краснодаре
Центр оптово-розничных продаж Cash&Carry в г. Краснодаре
Адрес: 350018, г. Краснодар, ул. Сормовская, д. 7, лит. «Г». Телефон: (861) 234-43-01(02).
Республика Беларусь. Центр оптово-розничных продаж Cash&Carry в г. Минске. Адрес: 220014,
Республика Беларусь, г. Минск, пр-т Жукова, д. 44, пом. 1 -17, ТЦ «Outleto».
Телефон: +375 17 251-40-23; +375 44 581-81-92. Режим работы: с 10-00 до 22-00 .
Казахстан. РДЦ Алматы. Адрес: 050039, г. Алматы, ул. Домбровского, д. 3 «А».
Телефон: +7 (727) 251-59-90 (91,92). Интернет-магазин: www.book24.kz
Украина. ООО «Форс Украина». Адрес: 04073 г. Киев, ул. Вербовая, д. 17а.
Телефон: +38 (044) 290-99-44 . E-mail: sales@forsukraine.com
Полный ассортимент продукции Издательства «Э» можно приобрести в книжных
магазинах «Читай-город» и заказать в интернет-магазине www.chitai-gorod.ru.
Телефон единой справочной службы 8 (800) 444 8 444. Звонок по России бесплатный.
В Санкт-Петербурге: в магазине «Парк Культуры и Чтения БУКВОЕД», Невский пр-т, д. 46.
Тел.: +7(812)601-0-601, www.bookvoed.ru
Розничная продажа книг с доставкой по всему миру. Тел.: +7 (495) 745-89-14.
РЕДАКЦИЯ РЕКОМЕНДУЕТ
НАШИ НОВИНКИ
https://eksmo.ru/b2b/