






























































































































Author: Ижболдина Л.А.
Tags: биология клетки и субклеточных частиц цитология ботаника экология атлас монография флора фауна очерки определитель водорослей байкал
ISBN: 5-9554-0015-X
Year: 2007




СПРАВОЧНИКИ И ОПРЕДЕЛИТЕЛИ
ПО ФАУНЕ И ФЛОРЕ ОЗЕРА БАЙКАЛ
Серия основана в 1995 г.
GUIDES AND KEYS ТО IDENTIFICATION
OF FAUNA AND FLORA OF LAKE BAIKAL
The series is initiated in 1995
RUSSIAN ACADEMY OF SCIENCES
SIBERIAN DIVISION
Limnological Institute
MINISTRY OF EDUCATION
OF RUSSIAN FEDERATION
IRKUTSK STATE UNIVERSITY
Institute of Biology
L.A. IZHBOLDINA
GUIDE AND KEY TO BENTHONIC
AND PERIPHYTON ALGAE
OF LAKE BAIKAL
(MEIO- AND MACROPHYTES)
WITH SHORT NOTES ON THEIR ECOLOGY
Editors
OLEG A. TIMOSHKIN (Doctor Sc., Biology)
SERGEI I. GENKAL (Doctor Sc., Biology)
NOVOSIBIRSK
NAUKA-CENTER’
2007
РОССИЙСКАЯ АКАДЕМИЯ НАУК
СИБИРСКОЕ ОТДЕЛЕНИЕ
Лимнологический институт
МИНИСТЕРСТВО ОБРАЗОВАНИЯ
РОССИЙСКОЙ ФЕДЕРАЦИИ
ИРКУТСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Научно-исследовательский институт биологии
Л.А. ИЖБОЛДИНА
АТЛАС И ОПРЕДЕЛИТЕЛЬ
ВОДОРОСЛЕЙ БЕНТОСА
И ПЕРИФИТОНА ОЗЕРА БАЙКАЛ
(МЕЙО- И МАКРОФИТЫ)
С КРАТКИМИ ОЧЕРКАМИ ПО ИХ ЭКОЛОГИИ
Ответственные редакторы
доктор биологических наук О.А. ТИМОШКИН
доктор биологических наук С. И. ГЕНКАЛ
НОВОСИБИРСК
“НАУКА-ЦЕНТР”
2007
УДК 576.8
ББК 28.591
И31
Ижболдина Л.А. Атлас и определитель водорослей бентоса и перифитона
озера Байкал (мейо- и макрофиты) с краткими очерками по их эколо-
гии / Л.А. Ижболдина. — Новосибирск: Наука-Центр, 2007. — 248 с., [0.5] л.
вкл. — (Справочники и определители по фауне и флоре озера Байкал).
ISBN 5-9554-0015-Х.
Монография представляет собой одну из книг серии “Справочники и определители по фауне
и флоре озера Байкал”, в которой приведены сведения по систематике, синонимике, морфологии
и экологии бентосных водорослей и перифитона оз. Байкал. Описано 137 видов мейо- и макро-
фитов, обитающих в озере, среди которых наиболее разнообразны зеленые и синезеленые водо-
росли. Даны иллюстрированные диагнозы всех таксонов и ключи для их определения. Особое
внимание уделено эндемичным родам и видам. Рассмотрены особенности вертикального и гори-
зонтального распределения водорослей. Описан характер сезонных изменений в составе и коли-
чественном развитии донной флоры.
Книга рассчитана на ботаников, альгологов, гидробиологов, преподавателей и студентов био-
логических факультетов вузов.
Табл. 6. Ил. 109. Библиогр.: 160 назв.
Перевод на английский Е.М. Тимошкиной
Izhboldina L.A. Guide and key to benthonic and periphyton algae of Lake Baikal
(meio- and macrophytes) with short notes on their ecology / L.A. Izhboldina. — No-
vosibirsk: Nauka-Center, 2007. — 248 p. — (Guides and keys to identification of fauna
and flora of Lake Baikal).
This monograph provides knowledge on the systematics, synonymy, morphology and ecology of bent-
honic algae and periphyton of Lake Baikal. It has been found that the lake is inhabited by 137 meio- and
macrophyte species among which most common are green and blue-green algae. This work includes illustra-
ted diagnoses of all taxa and keys to their identification. Special attention is focused on endemic genera and
species. Peculiarities of vertical and horizontal distribution of these algae, and seasonal variations in their
composition and quantitative distribution are considered.
The book is intended for botanists, algologists, hydrobiologists, teachers and students of biological
faculties.
Tabl. 6. Ill. 109. Ref. 160.
English version by E.M. Timoshkina
Редакционная коллегия
академик РАН М.А. Грачев (главный редактор),
доктора биологических наук О.А. Тимошкин (зам. главного редактора),
Г. Ф. Мазепова, В.В. Дрюккер, Т.Я. Ситникова, Е.В. Лихошвай,
кандидаты биологический наук Т.И. Земская, Н.Г. Мельник, НА. Рожкова,
Р.М. Камалтынов
Рецензенты
доктор биологических наук С.В. Пыжьянов
кандидаты биологических наук Г.В. Помазкина, НА. Бондаренко
Утверждено в печать Ученым советом Лимнологического института СО РАН
Издание осуществлено при финансовой поддержке Сибирского отоеления РАН,
Байкальского музея ВСФ СО РАН и частных спонсоров
© Л.А. Ижболдина, 2007
© Российская академия наук, 2007
© Перевод. Е.М. Тимошкина, 2007
© Гравюра. Б.И. Лебединский, 2007
© Составление и редактирование тома.
О.А. Тимошкин, 2007
© Разработка серии “Справочники и опре-
делители по фауне и флоре озера Бай-
кал». О.А. Тимошкин, 1995
ISBN 5—9554—0015—X © Оформление. “Наука-Центр», 2007
Посвящается светлой памяти
моего Учителя
Заслуженного деятеля науки РСФСР,
известного байкаловеда
профессора Михаила Михайловича Кожова
CONTENTS
PREFACE ...................................................................... 8
Chapter 1. HISTORY OF PHYTOBENTHOS AND PERIPHYTON RESEARCH ON
LAKE BAIKAL......................................................... 10
Chapter 2. COLLECTION AND TREATMENT PROCEDURES............................... 25
Chapter 3. SYSTEMATIC PART................................................... 27
Division Cyanophyta.................................................. —
Class Chroococceae ................................................. —
Order Chroococcales.............................................. 28
Class Hormogoneae ................................................. 31
Order Stigonematales.............................................. —
Order Nostocales ................................................ 32
Order Oscillatoriales ........................................... 54
Division Chrysophyta ............................................... 69
Class Chrysocapsineae .............................................. —
Order Chrysocapsales ............................................. —
Order Hydrurales ................................................ 73
Class Chrysothrichineae ............................................ —
Order Chrysothrichales............................................ —
Division Bacillariophyta ........................................... 74
Class Pennatophyceae................................................ —
Order Raphales................................................... 75
Division Rhodophyta ................................................ 78
Class Florideophyceae............................................... —
Order Nemaliales.................................................. —
Division Chlorophyta ............................................... 81
Class Tetrasporinophyceae.......................................... 82
Order Tetrasporales............................................... —
Class Ulotrichophyceae............................................. 86
Order Ulotrichales................................................ —
Order Oedogoniales.............................................. 168
Class Siphonocladophyceae......................................... 171
Order Cladophorales............................................. 172
Order Sphaeropleales............................................ 205
Class Zygnematophyceae (Conjugatophyceae)........................... —
Order Zygnematales.............................................. 206
-Division Charophyta................................................ 215
Class Charophyceae................................................ 216
Order Charales.................................................... —
INFORMATION ON THE AUTHOR................................................... 224
REFERENCES ................................................................. 236
GLOSSARY ................................................................... 243
ALPHABETIC INDEX OF LATIN NAMES............................................. 246
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ ............................................................... 8
Глава 1. ИСТОРИЯ ИССЛЕДОВАНИЯ ФИТОБЕНТОСА ОЗЕРА БАЙКАЛ ... 10
Глава 2. МЕТОДЫ СБОРА И ОБРАБОТКИ ПОЛЕВОГО МАТЕРИАЛА.................... 25
Глава 3. СИСТЕМАТИКА ВОДОРОСЛЕЙ БЕНТОСА И ПЕРИФИТОНА ОЗЕРА
БАЙКАЛ............................................................ 27
Отдел Cyanophyta................................................... —
Класс Chroococceae............................................... —
Порядок Chroococcales......................................... 28
Класс Hormogoneae............................................... 31
Порядок Stigonematales......................................... —
Порядок Nostocales............................................ 32
Порядок Oscillatoriales...................................... 54
Отдел Chrysophyta................................................. 69
Класс Chrysocapsineae............................................ —
Порядок Chrysocapsales ........................................ —
Порядок Hydrurales............................................ 73
Класс Chrysothrichineae.......................................... —
Порядок Chrysothrichales....................................... —
Отдел Bacillariophyta............................................. 74
Класс Pennatophyceae............................................. —
Порядок Raphales.............................................. 75
Отдел Rhodophyta.................................................. 78
Класс Florideophyceae............................................ —
Порядок Nemalialea............................................. —
Отдел Chlorophyta................................................. 81
Класс Tetrasporinophyceae ...................................... 82
Порядок Tetrasporales.......................................... —
Класс Ulotrichophyceae ......................................... 86
Порядок Ulotrichales........................................... —
Порядок Oedogoniales......................................... 168
Класс Siphonocladophyceae....................................... 171
Порядок Cladophorales........................................ 172
Порядок Sphaeropleales....................................... 205
Класс Zygnematophyceae (Conjugatophyceae)........................ —
Порядок Zygnematales ........................................ 206
Отдел Charophyta.................................................. 215
Класс Charophyceae............................................. 216
Порядок Charales............................................... —
СВЕДЕНИЯ ОБ АВТОРЕ....................................................... 224
БИБЛИОГРАФИЧЕСКИЙ СПИСОК.................................................. 236
СЛОВАРЬ ТЕРМИНОВ, ВСТРЕЧАЮЩИХСЯ В ТЕКСТЕ.................................. 243
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ................................... 246
ПРЕДИСЛОВИЕ
Исследования, проведенные в первой половине прошлого столетия сотруд-
никами Российской академии наук, Иркутского государственного университе-
та (ИГУ), Биолого-географического научно-исследовательского института при
ИГУ (БГНИИ), с 1975 г. переименованного в Научно-исследовательский ин-
ститут биологии (НИИБ), и другими ботаниками, показали, что в донных фи-
тоценозах прибрежной зоны Байкала преобладают водоросли. Были описаны
видовой состав и основные особенности распределения фитобентоса в летний
период [Мейер, 1930; и др.]. Изучение этой группы растений началось в
БГНИИ под руководством профессора М.М. Кожова. С 1961 по 1985 г. был об-
следован фитобентос (водоросли мейофиты — высота талломов 0.5-2 мм, и
макрофиты — высота талломов более 2 мм), вдоль всего прибрежья озера про-
ведены круглогодичные наблюдения за развитием водорослей, свидетельствую-
щие о наличии сезонной динамики видового состава в зоне литорали (глубина
0-20 м). Сделаны первые шаги в определении продукции массовых видов, изу-
чении их химического состава. С 1961 по 1996 г. организованы регулярные на-
блюдения за состоянием фитобентоса у юго-восточного побережья озера — в
районе сброса сточных вод Байкальского целлюлозно-бумажного комбината
(БЦБК), позволившие выявить характер влияния последних на донные фито-
ценозы. В 1997 г. исследования фитобентоса начаты в Лимнологическом инсти-
туте СО РАН (ЛИН) и с 2000 г. проводятся режимные наблюдения у северо-за-
падного побережья Юж. Байкала.
В настоящей работе изложены результаты изучения водорослей Байкала —
мейо- и макрофитов. Представлены краткая морфологическая характеристика
и систематика водорослей Байкала, приведены краткие диагнозы отделов,
классов, семейств, родов и видов, даны ключи для определения таксонов, отме-
чены отклонения от известных диагнозов, описаны экология, особенности рас-
пространения видов в оз. Байкал. Все виды и вариации иллюстрированы ри-
сунками, кроме того, в оформлении книги использованы работы художника
Ю.В. Ижболдина. При составлении систематического списка водорослей и
ключей для определения таксонов за основу взяты “Определитель пресновод-
ных водорослей СССР” (т. 2-4, 10, 13, 14); “Определитель зигнемовых водо-
рослей России” [Рундина, 1988], подготовленные сотрудниками Ботанического
института им. В.Л. Комарова. При описании водорослей порядка Oedogoniales
мы пользовались работой Н.Н. Воронихина [1951], а видов порядка Tetraspora-
Предисловие
9
les — работами О.А. Коршикова [1953], Л.И. Курсанова и М.М. Забелиной
[1953]. Кроме того, при описании ряда диагнозов и ареалов водорослей исполь-
зованы монографии и статьи других российских и зарубежных авторов [Матвн
энко, 1965; Комаренко, Васильева, 1975, 1978; Мошкова, 1979; Топачевский,
Масюк, 1984; Мошкова, Голлербах, 1986; Музафаров, Эргашов, Халилов, 1988;
Водоросли..., 1989; Pascher, 1914; Hoek, 1963; К. Starmach, 1972].
При проведении исследований фитобентоса оз. Байкал большую помощь
автору оказали дирекция НИИ биологии в лице профессора О.М. Кожовой,
сотрудники института В.М. Максимова, А.А. Гуменюк, Н.А. Вологдина, аква-
лангисты Н.С. Резников, А.М. Мурахвери, Б.А. Дмитриев, В.И. Максимов,
Ю.Г. Наумов, В.А. Гомбрайх, М.С. Развозжаев, А.Ю. Верномудров, В.И. Се-
мейкин, студенты биологического факультета ИГУ и команды экспедицион-
ных катеров; научные сотрудники Ботанического института РАН, Заслужен-
ный деятель науки, профессор М.М. Голлербах и кандидат биологических наук
Н.В. Сдобникова консультировали по вопросам систематики синезеленых и зе-
леных водорослей; сотрудник Лимнологического института СО РАН доктор
биологических наук О.А. Тимошкин оказывал постоянное содействие в подго-
товке рукописи к публикации.
Всем перечисленным лицам автор приносит самую искреннюю благодар-
ность.
Научное редактирование рукописи и подготовка ее к печати частично
профинансированы программой президиума РАН “Биоразнообразие и дина-
мика генофондов... ”, проект № 11.14 “Анализ биоразнообразия беспозво-
ночных на примере озера Байкал и водоемов Прибайкалья ” (руководитель
О.А. Тимошкин).
ГЛАВА
ИСТОРИЯ ИССЛЕДОВАНИЯ ФИТОБЕНТОСА
ОЗЕРА БАЙКАЛ
Фитобентос Байкала исследуется с начала прошлого века. Первые данные
опубликованы в работе В.Ч. Дорогостайского [1906], исследовавшего участок
западного побережья озера от истока р. Ангары до о. Ольхон и на восточном бе-
регу — район Туркинских минеральных вод. В ней отмечена оригинальность
донной флоры и указано на наличие зональности в ее вертикальном распреде-
лении. Автор выделил зону Ulothrix zonata Kg. и Draparnaldia на глубине 0.5-2
м. Во второй зоне (глубины от 2 до 50 м) живут в основном диатомеи, а в тре-
тьей (глубины более 50 м) — только диатомеи. Впервые приведен список
встреченных видов донных водорослей. По инициативе В.Ч. Дорогостайского в
1916 г. была организована гидробиологическая экспедиция, взятая под покро-
вительство Российской академии наук. В работе экспедиции участвовали
В.А. Яшнов, К.И. Мейер, Г.Ю. Верещагин, А.Г. Генкель, Л.А. Зенкевич,
Л.Л. Россолимо, И.И. Месяцев. Г.Ю. Верещагин [1918], обследовавший ряд
районов Байкала летом 1916 г., отметил на глубинах от 0.5 до 2 м на камнях зону
улотрикса, а глубже 2м — зону, занимаемую драпарнальдиями. Он же подчер-
кивает ряд особенностей в характере распределения фитобентоса в Чивыркуй-
ском заливе, у Мысовой и в бух. Бабушка. Позднее И.И. Месяцев, Л.А. Зенке-
вич и Л.Л. Россолимо [1922] отметили своеобразие в распределении донной
флоры Чивыркуйского залива. Ими впервые обнаружено скопление Cladophora
на черном органическом иле у входа в бух. Фертик и Онгоконскую.
В.Ч. Дорогостайский [1923], изучив характер вертикального распределения
бентоса в Байкале, выделяет 4 зоны: литораль (глубины от 0 до 5 м), сублито-
раль (5-50 м), переходную зону (50-300 м), абиссаль (более 300 м). Характе-
ризуя эти зоны, он отмечает, что в литорали сильное освещение вызывает оби-
льное развитие водорослей (в основном зеленых), а в более спокойных мес-
тах — высшей водной растительности. В сублиторали, где влияние берега менее
заметно, более постоянны температуры, а света проникает достаточно, живут
харовые водоросли и донные диатомеи, а зеленые водоросли встречаются реже.
Из высших растений в этой зоне растут виды Potamogeton, Ceratophyllum и др. В
переходной зоне, куда свет проникает в незначительном количестве, наблюда-
ются почти одни диатомеи. Характеризуя горизонтальное распределение бен-
тоса в оз. Байкал, В.Ч. Дорогостайский выделяет семь провинций (централь-
ную, южную, северную, селенгинскую, маломорскую, островную и ангар-
скую), различающихся по характеру донной фауны и флоры. Специфичность
Глава 1. История исследования фитобентоса озера Байкал
11
донной флоры впервые установил К.И. Мейер [1922а, б, 1925а б, 1927, 1930;
Мейер, Рейнгард, 1925; Meyer, 1930]. Он описал целый ряд новых, эндемичных
для озера видов зеленых водорослей семейства Chaetophoraceae (Harv.) De Toni
et Levi и Cladophoraceae (Hass.) Wittr. Em. Обследовав участок западного побе-
режья от истока Ангары до о. Ольхон, К.И. Мейер [1922а] делит его по характе-
ру донной флоры на три части. Участок от с. Лиственичного до р. Голоустная
является типичным, открытым для всех ветров побережьем озера. Здесь слабо
развиты высшие растения и наиболее разнообразно представлена флора диато-
мей. В вертикальном распределении макрофитов К.И. Мейер выделяет три
растительных зоны: улотрикса (на глубине от 0 до 1.5 м), тетраспоры (1.5-2.5
м) и драпарнальдий (2.5—12—20 м). Отмечено своеобразие донной флоры в
бух. Анга, во внутренней части которой обильно развита высшая водная расти-
тельность, и в прол. Ольхонские Ворота, отличающемся по характеру донной
флоры от вышерассмотренных районов.
Исследования, проведенные на Сев. Байкале в 1926 г., позволили изучить
фитобентос на участке от мыса Котельниковского на западном берегу до
губы Хакусы — на восточном. Было отмечено отсутствие эндемичных видов
рода Draparnaldia Вогу севернее губы Богучанская, что К.И. Мейер связал с
влиянием многочисленных рек, впадающих здесь в озеро. В этот же период
К.И. Мейером и Л.В. Рейнгардтом [1925] были опубликованы результаты обра-
ботки сборов В.Н. Сукачева, проведенных им на Байкале в 1914-1915 гг. Дан
список видов, обнаруженных в районе Ольхонских Ворот, у Ушканьих остро-
вов, у Котельниковского маяка, пос. Култук и вблизи устья р. Снежная. Описа-
но два новых эндемичных вида рода Draparnaldia. Наиболее полно результаты
исследований К.И. Мейера представлены в монографии “Введение во флору
водорослей озера Байкала” [1930]. Подчеркнув оригинальность донной флоры,
К.И. Мейер первым описал особенности ее распределения вдоль всего байка-
льского побережья, выделив ряд следующих областей, отличающихся по харак-
теру фитобентоса.
Открытый коренной Байкал. К открытым районам Байкала К.И. Мейер от-
носит и прол. Мал. Море и Баргузинский залив. По вертикали в открытом Бай-
кале фитобентос располагается поясами, сменяющими друг друга по мере уве-
личения глубин. Первый растительный пояс на глубинах от 0 до 0.5-1.5м обра-
зует Ulothrix zonata. На каменистом грунте этот пояс выражен вдоль всего
побережья. На нитях U. zonata и на камнях живут многочисленные диатомовые
водоросли. На глубинах от 1.5 до 2-2.5 м, образуя второй растительный пояс,
развиваются Tetraspora cylindrica (Wahl.) Ag., var. bullosa C. Meyer и Didymosphe-
nia geminata (Lingb.) M. Schmidt, с многочисленными разновидностями. Этот
пояс развит вдоль побережья менее равномерно, а местами Tetraspora отсутст-
вует. Далее, на глубине до 10-12 м, расположен третий растительный пояс, об-
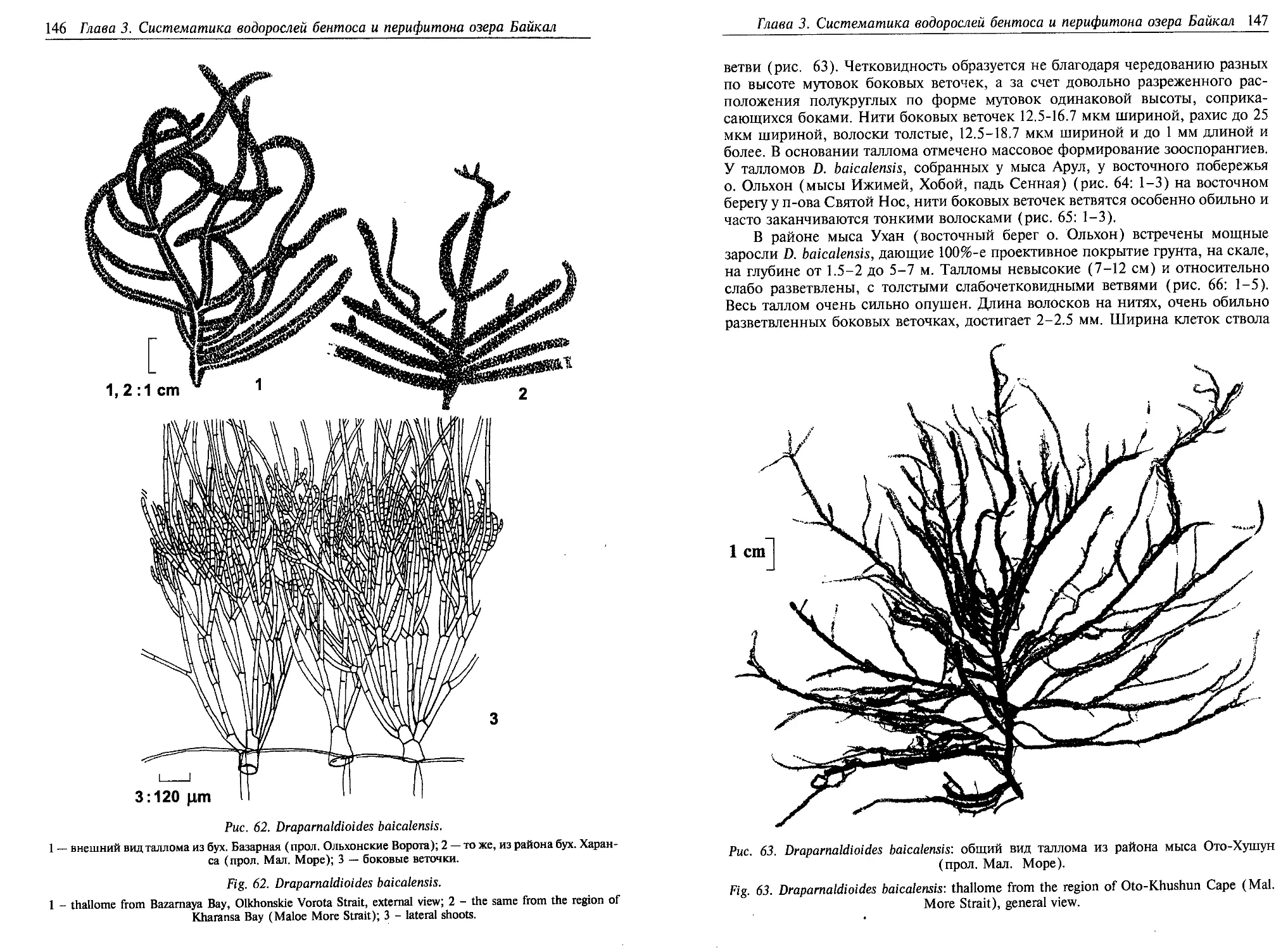
разованный эндемичными видами рода Draparnaldia, выделенными впоследст-
вии в эндемичный байкальский род [Мейер, Скабичевский, 1969; Скабичев-
ский, 1976а]. К.И. Мейер [1930] подчеркивает, что виды этого рода встречаются
исключительно в открытом Байкале и исчезают перед устьями рек всюду, где
сказывается их влияние. На Сев. Байкале эти виды не были встречены севернее
мыса Курлы. Не заходят Draparnaldia и в губы Ая, Богучанская, не были найде-
ны виды этого рода на мелководье около о. Лиственичный. Вместе с Draparnal-
dia растут и многие эндемичные виды семейства Cladophoraceae, синезеленые
водоросли родов Tolypothrix Kutz., Calothrix (Ag.) V. Poljansk., Stratonostoc Elenk.
и богатейшая флора диатомей.
12
Глава 1. История исследования фитобентоса озера Байкал
Пролив Ольхонские Ворота. Здесь на всех глубинах преобладают мягкие
грунты и лишь на мысах, где берег образован скалами, развиты три раститель-
ных пояса, характерные для литорали открытых побережий озера. На мягком
грунте на глубине от 1.5 до 16 м развит мощный пояс Chaetomorpha baicalensis
Meyer, вместе с которой растут Chara sp., Stratonostoc verrucosum (Vauch.)
Elenk., Sphaeronostoc pruniforme (Ag.) Elenk. и большое количество диатомей
[Яснитский, 1928; Мейер, 1930]. Третий пояс на глубинах от 16 до 36 м образуют
одни диатомовые водоросли.
Чивыркуйский залив. Донная растительность его резко отличается от Бай-
кальской и носит общесибирский или сибирско-байкальский характер [Вере-
щагин, 1935, 1940; Кожов, 1936,1960,1973] и лишь в створе залива, где ощутимо
влияние Байкала, донная флора носит байкальский характер.
Район впадения р. Селенги, Ангаро-Кичерское мелководье, бухта Анга. В этих
районах масса осадочного материала, отлагающегося при впадении рек в озеро,
постоянный прибой, достигающий большой силы, создают неблагоприятные
условия для поселения здесь донной растительности. Лишь в тихих местах жи-
вет богатая флора диатомей.
С 1923 г. на Байкале начали вести исследования биологи Иркутского уни-
верситета. Ботаник В.Н. Яснитский [1928] обследовал фитобентос прол. Оль-
хонские Ворота, отметил его своеобразие и составил первую карту-схему рас-
пределения фитобентоса. Им же [Яснитский, 1934] изучен половой процесс у 4
видов рода Draparnaldia, отмечено наличие морфологической и физиологиче-
ской дифференцировки полов. Образование зооспор у исследованных видов
Draparnaldia не встречено. Целый ряд водорослей семейства Cladophoraceae,
найденных ранее [Мейер, 1930] в литоральной зоне озера, М.М. Кожов [Буров,
Кожов, Талызин, 1933] обнаружил в районе о. Богучан, в бухтах Аяя, Фролиха и
Лаканда в сублиторали на глубинах до 35 м. Данные о распределении в Байкале
макрофитов имеются и в других работах М.М. Кожова [1931, 1934, 1936, 1962].
А.П. Скабичевский [1931, 1934, 1936], проводивший исследования донной
флоры в Южном (район пос. Бол. Коты) и Северном (бухты Аяя, Лаканда и
Фролиха) Байкале, описал новый род Gemmiphora Skabitsch., целый ряд новых
видов и разновидностей зеленых водорослей из семейства Cladophoraceae. В гу-
бах Сев. Байкала он выделил и описал два глубинных растительных пояса [Ска-
бичевский, 1934]. Четвертый, занимающий глубины от 11-16 до 20-35 м, обра-
зуют Cladophora meyeri Skabitsch., Chaetocladiella microscopica C. Meyer et Ska-
bitsch. Кроме того, на этих глубинах могут встречаться Cladophora floccosa
С. Meyer, С. kursanovii Skabitsch., С. pulvinata С. Meyer, Chaetomorpha monilifor-
mis Skabitsch., Ch. curta Skabitsch., Chaetocladiella pumila (C. Meyer) Meyer et
Skabitsch., эпифитные диатомовые водоросли. Пятый пояс располагается на
глубинах от 20-35 до 60-78-116 м на сильно заиленном песке, где обнаружены
Chaetomorpha curta и масса диатомей. Было отмечено [Скабичевский, 1966,
1968], что в разных районах озера два последних пояса развиты неодинаково.
Лучше они выражены вдоль восточного берега, для которого характерен поло-
гий уклон дна и преобладание мягких грунтов. Этот же автор [Скабичевский,
1937] изучал половой процесс у Tetraspora cylindrica var. bullosa. Было показано,
что данный вид является строго раздельнополым организмом с физиологиче-
ской дифференцировкой полов. Морфологическая дифференцировка между
гаметами противоположного пола отсутствует.
Таким образом, в результате исследований, проведенных в начале прошло-
го столетия, был изучен видовой состав фитобентоса оз. Байкал. Установлено,
Глава 1. История исследования фитобентоса озера Байкал
13
что макрофиты живут на глубине до 60-116 м и в их вертикальном распределе-
нии ярко выражена зональность. Выделено пять растительных поясов, каждый
из которых характеризуется определенными доминирующими видами и глуби-
нами их обитания. Отмечен ряд особенностей в горизонтальном распределении
макрофитов. Выявлены виды, характерные для Сев. и Юж. Байкала. Получены
данные по биологии пяти эндемичных видов.
Целый ряд статей был опубликован в 50-70-х годах прошлого столетия.
В.Н. Яснитский [1952], рассматривая причины гигантизма байкальских водо-
рослей, ведущую роль отводит “низкой температуре воды, обусловленной гран-
диозными размерами нашего озера”.
В статье А.П. Скабичевского [1966], написанной им по материалам 1927-
1928 гг. и ряда последующих лет, рассмотрены особенности распределения дон-
ной растительности в районе Бол. Котов, им же опубликован целый ряд других
работ [Скабичевский, 1965, 1968, 1969а,б, 1970, 1975, 19766, 1978, 1984], уточня-
ющих систематическое положение отдельных видов и родов водорослей или
особенности их морфологии и распределения в Байкале.
Все исследования фитобентоса Байкала до 1961 г. носили качественный
характер и в основном были выполнены в летний период. С 1961 г. на Байкаль-
ской биостанции БГНИИ под руководством профессора М.М. Кожова были
начаты регулярные круглогодичные наблюдения за развитием донных биоце-
нозов, обследован фитобентос вдоль всего побережья озера. С 1997 г. регуляр-
ные исследования фитобентоса проводятся Лимнологическим институтом
СО РАН. В результате проведенных исследований в Байкале обнаружено 137
видов и разновидностей водорослей, относящихся к шести отделам низших
растений (табл. 1).
Наиболее разнообразно представлены в Байкале зеленые и синезеленые
водоросли.
М.М. Кожов [1962, 1973] среди населения Байкала выделяет по происхож-
дению две главные экологические группы: лимнофилов и лимнореофилов. В
современном Байкале к первой группе он относит виды и формы, которые
встречаются в прибрежно-соровой зоне озера и в окружающих Байкал мелко-
водных озерах и других водоемах эвтрофного типа. Эти виды со времен вселе-
ния их в оз. Байкал не изменились. Г.Ю. Верещагин [1930,1935,1939] называет
комплекс таких видов сибирским. Среди донных водорослей мейо- и макрофи-
Таблица 1
Распределение водорослей по таксономическим группам
Отдел Класс Порядок Подпоря- док Семейство Род Вид Разновид- ность
Cyanophyta 2 4 3 8 15 43 (1) 7
Chrysophyta 2 3 — 3 3 4(1) —
Bacillariophyta 1 1 1 1 2 5(2) —
Rhodophyta 1 1 — 1 1 2 —
Chlorophyta 4 6 3 И 21(5) 55 (24) 16 (12)
Charophyta 1 1 — 2 2 5 —
Всего... 11 16 7 26 44(5) 114 (28) 23 (12)
Примеч а н и e. В скобках указано число эндемичных родов, видов и разновидностей.
14
Глава 1. История исследования фитобентоса озера Байкал
тов к этому комплексу могут быть отнесены обычные для Палеарктики виды
родов Oscillatoria, Cladophora (С. aegagropila, C.fracta'), Stigeoclonium, Bulbochae-
te, Oedogonium, Spirogyra, Zygnema, Mougeotia. К лимнореофилам (по Г.Ю. Ве-
рещагину [1930, 1939] — байкало-сибирский биогеографический комплекс)
принадлежат палеарктические виды, широко распространенные в крупных
проточных озерах и быстрых реках Сибири. Такие виды живут в литорали и
сублиторали открытых прибрежий Байкала. К этой группе могут быть отнесены
виды родов Stratonostoc, Calothrix, Tolypothrix, Schizothrix, Ulothrix, Stigeoclonium,
Cladophora (C. glomerata).
По мнению многих исследователей [Верещагин, 1935, 1939; Кожов, 1957,
1962, 1973; Старобогатов, 1970; Порфирьева, 1977; и др.], исходными для фор-
мирования эндемичной фауны и флоры Байкала служили представители си-
бирско-евразиатской фауны и флоры из окружающих Байкал и имевших с ним
связь водоемов с высоким содержанием кислорода и сравнительно постоянны-
ми температурами. Г.Ю. Верещагин [1930] выделяет эндемиков Байкал, не име-
ющих родственной связи с сибирскими видами, в третий, байкальский, комп-
лекс. В настоящее время в Байкале обнаружено 40 эндемичных видов и разно-
видностей донных мейо- и макрофитов. Особенно ярко эндемизм выражен у
зеленых водорослей (52.2 % всех встреченных видов и разновидностей), среди
них 5 эндемичных родов (Ireksokonia, Myxonemopsis Meyer, Draparnaldioides
Meyer et Skabitsch., Gemmiphora Skabitsch., Chaetocladiella Meyer et Skabitsch.) и
36 видов и разновидностей. Наибольшее число эндемиков — среди представи-
телей семейств Chaetophoraceae и Cladophoraceae. Семейство Chaetophoraceae
представлено в Байкале 27 видами и разновидностями, среди которых 20 энде-
миков.
Особый интерес представляют 9 массовых видов эндемичного рода Drapar-
naldioides, имеющих короткий период вегетации и доминирующих в зоне лито-
рали в весенне-осенний период. К.И. Мейер [1930, 1969] и А.П. Скабичевский
[1969, 1976а] считают, что эволюция представителей этого рода шла в Байкале
по линии Stigeoclonium Kutz. — Ireksokonia — Draparnaldioides, а виды широко
распространенного рода Draparnaldia составляют параллельную байкальской
линию развития. Отличительными чертами представителей эндемичного рода
Draparnaldioides является большой размер талломов (до 25-30 см), наличие
мощного ризоидного или слизистого чехла, защищающего водоросли от силь-
ного волнового воздействия, и сетчато продырявленный хлоропласт, занимаю-
щий всю высоту клетки [Мейер, Скабичевский, 1969]. У большинства видов
рассматриваемого рода есть целый ряд морфологических форм, что, возможно,
является результатом межвидового скрещивания близкородственных видов,
одновременно обитающих на одной территории.
Нельзя исключать и влияния разных экологических условий произраста-
ния. Очевидно, автохтонный процесс видообразования эндемичной донной
флоры Байкала продолжается и в наши дни. Водоросли семейства Cladophora-
ceae представлены в Байкале относительно мелкими (высота 0.2-1 см) теневы-
носливыми (многие из них живут на глубинах до 50-70 м) видами, вегетирую-
щими и размножающимися в Байкале круглый год. Обнаружено 23 вида и раз-
новидности этого семейства, среди которых 15 эндемиков. Последние не имеют
сходства с видами, широко распространенными в пресных водоемах Палеарк-
тики. А.П. Скабичевский [1978] подчеркивает, что по одностороннему гребен-
чатому ветвлению некоторые эндемичные виды Cladophora напоминают аль-
пийские и морские виды, однако говорить о связи пресноводных видов этого
рода с морскими можно лишь в генетическом смысле. Предками байкальских
Глава 1. История исследования фитобентоса озера Байкал
15
Cladophora А.П. Скабичевский [1978] считает обитателей горных водоемов, а
эволюция Cladophoraceae в Байкале, по его мнению, шла по двум направлени-
ям: по пути образования подушечковидных или шарообразных талломов и по
пути редукции ветвления и образования хетоморфообразных Chaetocladiella.
М.М. Кожов [1973] подчеркивает, что значительную роль в процессе обра-
зования эндемичных видов играли выходцы из древних крупных бассейнов
Центральной Азии и из водоемов палеогеновой Ангариды. В настоящее время в
оз. Хубсугул (Монголия), имеющем через речную систему связь с оз. Байкал,
обнаружен ряд эндемичных и близкородственных байкальским видов семейст-
ва Cladophoraceae [Загоренко, Ижболдина, 1976, 1977, 1983].
Вдоль открытых прибрежий Байкала донные фитоценозы образованы эн-
демичными видами и водорослями байкало-сибирского комплекса (лимноре-
филами). В зоне литорали эндемики составляют в разных растительных поясах
от 28.2 до 35.1 % от общего числа встреченных видов. В фитоценозах сублитора-
ли, где видовое разнообразие водорослей снижается, доля эндемичных видов
увеличивается до 46.6-69.2 % (табл. 2).
Все встреченные в мейо- и макрофитобентосе и перифитоне Байкала водо-
росли являются типичными обитателями пресных холодных вод, но некоторые
из сибирских и байкало-сибирских видов могут обитать и в водоемах с разной
степенью засоления, минерализации и в горячих источниках (табл. 3). Извест-
но [Голлербах, Полянский, 1951], что лишь очень немногие водоросли могут
жить и в морской, и в пресной воде. В Байкале 18 % от встреченных видов спо-
собны расти в солоноватой и соленой воде, а 6 видов встречаются и в минера-
льных источниках. Среди эвригалинных водорослей преобладают зеленые во-
доросли, а среди эвритермных — синезеленые водоросли родов Oscillatoria и
Phormidium.
По продолжительности периода вегетации выделены две группы водорос-
лей [Ижболдина, 1975]. Это развивающиеся в течение круглого года виды родов
Stratonostoc, Calothrix, Tolypothrix, Oscillatoria, Schizothrix, Gemmiphora, Chaetop-
Таблица 2
Число эндемичных (1) и сибирских и байкало-сибирских (2) видов и разновидностей мейо- и
макрофитов бентоса и перифитона оз. Байкал
Отдел Всего видов и разновидно- стей в оз. Байкал Растительный пояс
I II III IV V
1 2 1 2 1 2 I 2 1 2 1 2
Cyanophyta 1 51 — 9 — 33 1 39 1 10 1 2
Chrysophyta 1 3 1 3 1 2 1 2 — — — —
Bacillariophyta 2 3 1 2 2 3 — — — — — —
Rhodophyta — 2 — 2
Chlorophyta 36 33 9 12 27 26 31 16 13 5 8 2
Charophyta — 5 — — — 1 — 4 — 1 — —
Всего видов и разновидностей 40 97 11 28.2 28 71.8 30 32J 65 68.4 33 35.1 61 64.9 14 46.6 16 53.4 9 69.2 4 ЗОЛ
Примечание. В числителе — число видов и разновидностей, в знаменателе — доля (в %)
этих видов от общего числа видов в поясе.
16
Глава 1. История исследования фитобентоса озера Байкал
Таблица 3
Мейо- и макрофиты бентоса и перифитона оз. Байкал, обитающие не только в пресных водах
Отдел, вид Пресноводные виды, встречающиеся и в
солоноватых водах соленых водах минеральных источниках горячих источ- никах
Cyanophyta
Microcystis muscicola — — — +
Calothrix parietina + — — —
Gloeothrichia pisum + — — —
Oscillatoria sancta + — — —
0. limosa 4- — — —
0. lacustris •— + —
0. amoena — —. — 4-
0. formosa — — + 4-
Phormidium purpurascens — — +
Ph. tenue — — + +
Ph. ambiguum + — — +
Ph. uncinatum — — — +
Ph. papiraceum — + — +
Lyngbya major — — — +
L. aestuari + + — +
Microcoleus subtorulosus — — — +
Chlorophyta
Ulothrix zonata — 4- — —
U. tenuissima — 4- — —
Chaetophora elegans 4- — — —
Cladophora aegagropila 4- — — —
C. fracta f. fracta 4- — — —
C. globulina 4- — 4- —
C. glomerata var. glomerata 4- 4- — —
C. glomerata var. crassior + 4- — —
Sphaeroplea annulina 4- — — —
Spirogyra decimina f. jurgensii 4- — — —
S. decimina f. longata — — 4- —
S. weberi f. weberi — 4- — —
S. hassallii — — 4-
S. calospora f. calospora 4- — — —-
Mougeotia genuflexa f. genuflexa — — 4- —
M. laetewirens 4- — — —
Charophyta
Nitella opaca 4- — — —
N. flexilis 4- — — —
Chara tomentosa 4- — — —
Всего... 19 8 6 11
Глава 1. История исследования фитобентоса озера Байкал
17
hora, Cladophora, Chaetocladiella и
виды с коротким периодом вегета-
ции (представители родов Ulothrix,
Tetraspora, Tetrasporopsis, Stigeocloni-
um, Ireksokonia, Draparnaldioides,
Batrachospermum и др.). Массовое
развитие сезонных видов наблюда-
ется в зоне литорали, где встречены
представители всех известных для
Байкала отделов низших растений.
В сублиторали золотистые, диато-
мовые (образующие крупные коло-
нии) и красные водоросли отсутст-
вуют. Представители синезеленых и
зеленых водорослей обнаружены на
глубинах от 0.1 до 70-100 м. Макси-
мальным диапазоном глубин обита-
ния отличаются зеленые водоросли
семейства Cladophoraceae, вегетиру-
ющие в озере круглый год и перено-
сящие пониженные температуры и
освещенность. Это Cladophora floc-
cosa (0.5-50 м), С. floccosa var. irre-
gularis (4.5-40 м), C. kursanovii
(1-60 м), С. comp acta (1.5-70 м),
С. meyeri (3-67 м), С. aegagropila
(3-65 м), Chaetomorpha curta
(10-116 м).
В зоне литорали наименее раз-
нообразно мейо- и макрофиты
представлены в I растительном поя-
се, испытывающем на себе активное
волновое воздействие в период от-
крытой воды и влияние понижен-
ных температур в подледный пери-
од. В разных районах озера в этом
поясе обнаружено всего 11 видов во-
дорослей, вегетирующих круглый
год (табл. 4). Среди них синезеле-
ные и зеленые водоросли, из кото-
рых всего 5 эндемичных видов се-
мейства Cladophoraceae. Более раз-
нообразно представлены сезонные
виды, но и среди них число эндеми-
ков (6 видов) невелико. Значитель-
но богаче видовой состав водорос-
лей II и III растительных поясов,
имеющих высокий (0.71) коэффи-
циент видового сходства и в этих
поясах по числу видов преобладают
18
Глава 1. История исследования фитобентоса озера Байкал
водоросли с коротким периодом вегетации (сезонные виды). Наиболее богат
видовой состав сезонных видов во II поясе, где, очевидно, проявляется явление
краевого эффекта. В рассматриваемых поясах по числу видов, так же, как и в
I поясе, преобладают синезеленые и зеленые водоросли. Среди последних зна-
чительно возрастает доля эндемичных видов, формирование которых, очевид-
но, наиболее активно шло в зоне литорали, откуда некоторые теневыносливые,
медленно растущие многолетние виды (Cladophora floccosa var. floccosa, C. floc-
cosa var. irregularis, C. kursanovii) проникли и в зону сублиторали, где нашли
благоприятный для своего поселения грунт.
На мягком грунте вдоль открытых прибрежий Байкала в глубинной зоне
трех первых поясов водоросли обычно отсутствуют. В двух поясах сублитораль-
ной зоны (IV и V) по числу видов преобладают вегетирующие круглый год во-
доросли, хорошо переносящие пониженную освещенность и в большинстве
своем многолетние растения, способные к размножению в течение всего года.
Богаче видовой состав водорослей IV пояса, что происходит благодаря разви-
тию здесь в летний период сезонных видов. В обоих поясах наиболее разнооб-
разно представлены зеленые водоросли, на 70-80 % являющиеся эндемиками
Байкала.
Круглогодичные наблюдения за развитием фитобентоса, выполненные на
участке прибрежья, типичном для северо-западного побережья Юж. Байкала
(район пос. Бол. Коты), позволили выделить в развитии макрофитов литорали
четыре биологических сезона, различающихся по структуре донных фитоцено-
зов и их фитомассе [Ижболдина, 1970, 1974а, 1975, 1985, 1989, 1990а, б].
Зима (январь—март) — период, когда в Байкале представлены вегетирую-
щие в течение круглого года виды родов Stratonostoc, Calothrix, Tolypothrix, Oscil-
latoria, Schizothrix, Cladophora, Chaetomorpha, Chaetocladiella, Gemmiphora и др.
Весна (апрель — первая половина июня) — начало вегетации весенне-лет-
них видов из родов Ulothrix1, Tetraspora, Tetrasporopsis, Ireksokonia, Draparnaldio-
ides (D. simplex C. Meyer et Skabitsch., D. dorogoshankinii C. Meyer et Skabitsch. и
их формы) и др.
Лето (вторая половина июня — сентябрь) — период массовой вегетации
весенне-летних видов, а также появление летних представителей рода Drapar-
naldioides (D. baicalensis C. Meyer et Skabitsch., D. pilosa C. Meyer et Skabitsch., D.
vilosa C. Meyer et Skabitsch.), а также Cladophora glomerata L., Didymosphenia ge-
minata (Lyngb.) M. Schmidt и летне-осенних видов — Ulothrix zonata Kg., Dra-
parnaldioides arenaria C. Meyer et Skabitsch., D. arnoldi C. Meyer et Skabitsch.,
D. lubrica C. Meyer et Skabitsch. и их формы.
Осень (октябрь — декабрь) — период, когда вместе с ветегирующими круг-
лый год видами встречаются немногочисленные представители летне-осенних
видов — Ulothrix zonata, Draparnaldioides arenaria, D. arnoldi, D. lubrica, D. pumila.
В весенне-осенний период макрофиты трех первых поясов продуцируют
основную массу водорослей, включающихся ежегодно в круговорот веществ.
При этом в фитоценозах обычно доминируют эндемики, а в зимний период
они играют в сообществах подчиненную роль и лишь в районах массового раз-
вития Chaetocladiella pumila С. Meyer et Skabitsch., Cladophora compacta и C. floc-
cosa доминируют. Наиболее разнообразно представлены и обильно развиты
макрофиты в летний период. У массовых сезонных видов отмечен один пик
lUlothrix zonata обильно развит в весенне-летний период, а в небольшом количестве встреча-
ется в зоне прибоя до ноября.
Глава 1. История исследования фитобентоса озера Байкал
19
фитомассы, сроки достижения которого у всех летне-осенних видов в разные
годы неодинаковы и могут быть сдвинуты в ту или иную сторону на 10-20 дней
и более.
Круглогодичные наблюдения, проведенные за развитием фитобентоса в
районе пос. Бол. Коты (Юж. Байкал), показали, что в I растительном поясе на-
блюдается один пик фитомассы (в конце июня — начале июля) в период мас-
сового развития Ulothrix zonata, максимальная фитомасса которого в разных ча-
стях побережья колеблется от 100 до 400 г/м2 . В это время прибойная зона при-
влекает к себе массу беспозвоночных животных, находящих себе здесь пищу
(скопление бактерий, эпифитные, в основном диатомовые, водоросли) и убе-
жище [Кожов, 1960; Каплина, 1974; Окунева, 1974, 1976, 1989]. Численность
гаммарид, например, в период развития U. zonata увеличивается в 88 раз в зоне
глубин от 0.3 до 1 м [Каплина, 1974]. Во II поясе прослежены два разных по ве-
личине пика фитомассы. Первый пик наблюдали в период массового развития
Tetraspora cylindrica var. bullosa в июле, второй обусловлен массовым развитием
Cladophora glomerata и отмечен в середине сентября. В разные годы максималь-
ная фитомасса Tetraspora колеблется от 200 до 600 г/м2, а С. glomerata — от 80
до 100 г/м2. В III поясе смена доминантных видов за весенне-осенний период
происходит трижды. В конце июня — первой половине июля здесь доминируют
Draparnaldioides simplex и D. goroschankinii, максимальная фитомасса которых
колеблется в отдельные годы от 50 до 180 г/м2. В середине сентября отмечена
максимальная фитомасса (700-1200 г/м2) у D. baicalensis. У D. arenaria и D. аг-
noldii она обычно не превышает в это время 250 г/м2. У D. pumila и D. lubrica
фитомасса достигает максимальной величины в конце сентября — декабре и
колеблется в разные годы от 60 до 280 г/м2.
В зимний период водоросли в зоне литорали представлены менее разнооб-
разно; фитомасса их обычно ниже, чем у весенне-осенних видов и не имеет за-
метных сезонных и межгодовых колебаний, но может существенно отличаться
в разных районах озера [Ижболдина, 1974а, 1976а, 1979,1987]. Фитомасса сине-
зеленых водорослей обычно не превышает 0.5-20, реже 70 г/м2, и лишь на уча-
стках с обильным развитием Stratonostoc может достигать 80-130 г/м2, реже
(район пади Мал. Сенная, мыс Тонкий у губы Богучанская) 160-180 г/м2. У зе-
леных водорослей этот показатель колеблется в среднем от 5-10 до 25-35 г/м2,
достигая в некоторых районах с обильным развитием Chaetocladiella pumila
С. Meyer et Skabitsch., Cladophora compacta, C. floccosa, C. kusnetzowii, Chaetomor-
pha baicalensis 50-270 г/м2 [Ижболдина, 1990а].
В IV и V растительных поясах, где преобладают вегетирующие круглый год
виды, фитомасса водорослей обычно ниже, чем в литорали, и не имеет замет-
ных сезонных и межгодовых колебаний. На мягком грунте в IV поясе она ко-
леблется в разных районах озера от 0.1 до 3.8 г/м2 и изредка, в местах обильного
развития С. floccosa, может достигать 49 г/м2. В V растительном поясе на мяг-
ком грунте средняя фитомасса водорослей меньше — 0.03-0.46 г/м2 — и лишь в
немногих районах в основном восточного побережья Сев. Байкала, где обильно
развита Chaetomorpha curta, она достигает 23.5 г/м2. На твердом грунте (галька,
камни, щебень), изредка встречающемся в этих поясах, водоросли развиты бо-
лее обильно и средняя фитомасса их колеблется в разных участках побережья
от 14 до 28 г/м2. Вдоль западного берега, где значительные площади дна зани-
мают скалы, под крутым утлом уходящие на глубину, количественный учет
бентоса с помощью обычных гидробиологических орудий лова провести невоз-
можно. Отдельные пробы, которые удалось взять аквалангистам с отвесных
20
Глава 1. История исследования фитобентоса озера Байкал
скал на глубинах до 40-60 м, свидетельствуют об обильном обрастании послед-
них Cladophora Kursanovii, которая местами сплошной коркой покрывает скалы.
Фитомасса ее в сублиторали достигает 50-70 г/м2.
Дальнейшее исследование фитоценозов сублиторали с применением под-
водной фотосъемки позволит дать более полную качественную характеристику
и количественную оценку фитобентоса скальных подводных склонов западно-
го побережья оз. Байкал.
Изучение особенностей горизонтального распределения фитобентоса по-
казало, что по структуре донных фитоценозов и количественному развитию
макрофитов восточное и западное побережья различаются более заметно, чем
разные части озера. Это вполне естественно, поскольку побережья существен-
но различаются по геолого-геоморфологическому строению дна [Лут, 1964;
Бекман, Деньгина, 1969; Карабанов, 1990] и другим физико-географическим
условиям [Вотинцев, 1961; Верболовдр., 1965; Шимараев, 1977; Фиалков, 1983].
Вдоль большей части западного побережья, где развит абразионный склон, об-
разованный кристаллическими породами, донные фитоценозы имеют боль-
шую фитомассу и видовое многообразие, чем на абразионно-аккумулятивном
шельфе, характерном для восточного берега [Ижболдина, 19906,1992; Кравцо-
ва, Ижболдина и др., 1991]. Коэффициент видового сходства (S) донной флоры
вдоль открытых прибрежий Байкала в разных его частях и в прол. Мал. Море
колеблется от 0.63 до 0.75. Наиболее велико видовое сходство водорослей
(S = 0.81) западного и восточного прибрежий Сев. Байкала. В прол. Мал. Море
обнаружено максимальное по сравнению с другими районами озера число (6)
разновидностей эндемичных зеленых водорослей, что, видимо, обусловлено
наличием здесь многочисленных бухт, отличающихся по своим экологическим
условиям [Верболов, 1959; Патрикеева, 1959; Бекман, 1965; Россолимо, 1959;
Толмачев, 1959; Вотинцев, 1961; Кожов, 1962; Кожова, Ижболдина, 19956].
Ушканьи острова, в донной флоре которых преобладают эндемичные виды,
имеют высокий коэффициент видового сходства (S - 0.62) с донной флорой
прол. Мал. Море и фитоценозами открытых прибрежий западного (S - 0.59-
0.64) и восточного (S = 0.64-0.77) берегов. У западного берега эндемичные во-
доросли выступают, как правило, в роли доминантных и субдоминантных видов.
Вдоль восточного субдоминантными, а иногда и доминантными видами являют-
ся широко распространенные в Палеарктике виды водорослей. Вероятно, лито-
ральная зона прибрежий с абразионным шельфом была более благоприятным
местом для формирования специфичной донной флоры оз. Байкал. Представи-
тели последней, как и донные беспозвоночные [Верещагин, 1940; Кожов, 1962,
1973], в процессе эволюции заселили и сублиторальную зону. Интересен харак-
тер распространения в Байкале эндемичных зеленых водорослей Chaetomorpha
baicalensis, Chaetocladiella pumila, Draparnaldioides baicalensis, Cladophora compacta
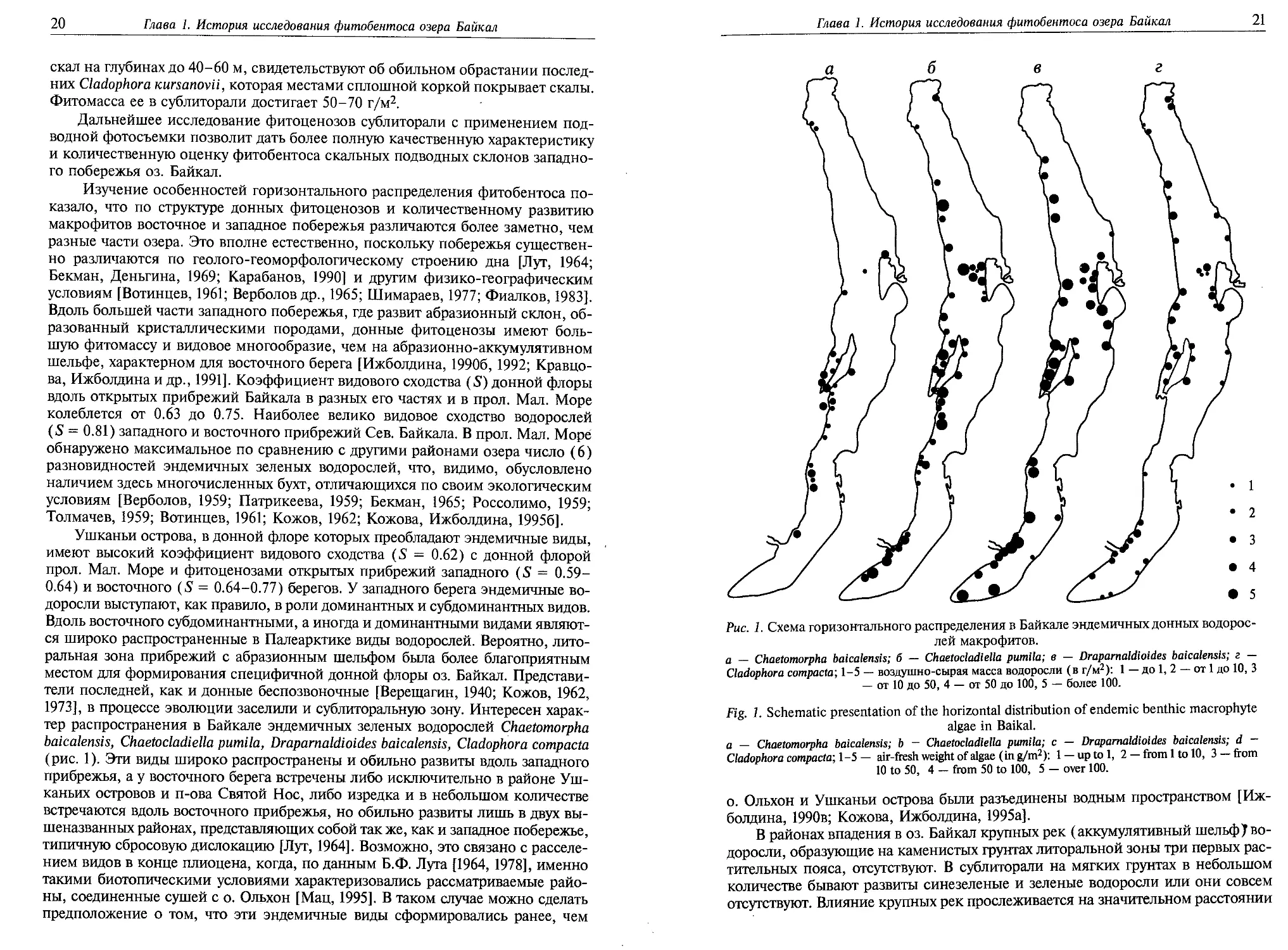
(рис. 1). Эти виды широко распространены и обильно развиты вдоль западного
прибрежья, а у восточного берега встречены либо исключительно в районе Уш-
каньих островов и п-ова Святой Нос, либо изредка и в небольшом количестве
встречаются вдоль восточного прибрежья, но обильно развиты лишь в двух вы-
шеназванных районах, представляющих собой так же, как и западное побережье,
типичную сбросовую дислокацию [Лут, 1964]. Возможно, это связано с расселе-
нием видов в конце плиоцена, когда, по данным Б.Ф. Луга [1964, 1978], именно
такими биотопическими условиями характеризовались рассматриваемые райо-
ны, соединенные сушей с о. Ольхон [Мац, 1995]. В таком случае можно сделать
предположение о том, что эти эндемичные виды сформировались ранее, чем
Глава 1. История исследования фитобентоса озера Байкал
21
Рис. 1. Схема горизонтального распределения в Байкале эндемичных донных водорос-
лей макрофитов.
а — Chaetomorpha baicalensis; б — Chaetocladiella pumila; в — Draparnaldioides baicalensis; г —
Cladophora compacta; 1-5 — воздушно-сырая масса водоросли (в г/м2): 1 — до 1, 2 — от 1 до 10, 3
— от 10 до 50, 4 — от 50 до 100, 5 — более 100.
Fig. 1. Schematic presentation of the horizontal distribution of endemic benthic macrophyte
algae in Baikal.
a — Chaetomorpha baicalensis; b — Chaetocladiella pumila; c — Draparnaldioides baicalensis; d —
Cladophora compacta; 1-5 — air-ftesh weight of algae (in g/m2): 1—uptol, 2 —from Ito 10, 3 —from
10 to 50, 4 — from 50 to 100, 5 — over 100.
о. Ольхон и Ушканьи острова были разъединены водным пространством [Иж-
болдина, 1990в; Кожова, Ижболдина, 1995а].
В районах впадения в оз. Байкал крупных рек (аккумулятивный шельф) во-
доросли, образующие на каменистых грунтах литоральной зоны три первых рас-
тительных пояса, отсутствуют. В сублиторали на мягких грунтах в небольшом
количестве бывают развиты синезеленые и зеленые водоросли или они совсем
отсутствуют. Влияние крупных рек прослеживается на значительном расстоянии
22
Глава 1. История исследования фитобентоса озера Байкал
от их устья и выражается в снижении фитомассы и обеднении видового состава
донных водорослей мейо- и макрофитов и уменьшении среди них числа энде-
мичных видов [Ижболдина, 19816,19826,1984]. Минимальное (10 видов) видовое
разнообразие фитоценозов отмечено на Селенгинском мелководье (табл. 5). В
районах впадения рек Кичеры и Верх. Ангары видовой состав водорослей более
разнообразен (35 видов и разновидностей). Однако здесь так же, как и в районе
Селенгинского мелководья, по числу видов преобладают синезеленые водорос-
ли. Индекс видового сходства водорослей в этих районах равен 0.26.
Особенности в распределении фитобентоса в мелководных заливах, бухтах,
сорах отмечены многими исследователями [Верещагин, 1918; Месяцев, Россо-
лимо, Зенкевич, 1922; Яснитский, 1928; Мейер, 1930; Кожов, 1947, 1962; Гага-
рин, 1977; Вотякова, 1981; Азовский, Паутова, Ижболдина, 1990в, и др.]. Схемы
распределения донных фитоценозов в зал. Чивыркуйский, в южной части Мал.
Моря и прол. Ольхонские Ворота и в Посольском соре приводит Л.А. Ижбол-
дина [1990в]. Ею же дана количественная оценка фитобентоса этих районов.
Отмечена малочисленность видового состава водорослей мейо- и макрофитов в
зал. Провал и в Посольском соре. Донные фитоценозы зал. Провал имеют ми-
нимальный коэффициент видового сходства (S = 0.04-0.24) с фитоценозами
других районов озера, что скорее всего обусловлено отрицательным влиянием
поступающих в залив вод р. Селенги. Донная флора Посольского сора также
существенно отличается от таковой вдоль открытых прибрежий озера
Таблица 5
Число видов водорослей мейо- и макрофитов в разных районах оз. Байкал
Район Всего ви- дов и раз- новид- ностей Суапо- phyta Chryso- phyta Bacillario- phyta Rhodo- phyta Chloro - phyta Charo- phyta
Ангаро-Кичерское мелководье 35(3) 20(1) — 1 — 13(2) 1
Сев. Байкал 71(29) Западн 23(1) ый берег 3(1) 5(2) — 38(25) 2
Мал. Море 63(30) 19(1) 3(1) 2(2) — 35(26) 4
Ольхонские Ворота 43(17) 10 2(1) 1 — 26(16) 4
Сред. Байкал 65(33) 17(1) 3(1) 5(2) — 38(27) 2
Юж. Байкал 71(30) 21(1) 4(1) 5(2) — 39(27) 2
Сев. Байкал 58(27) Восточн 17(1) ый берег 2(1) 5(2) — 32(23) 2
Чивыркуйский залив 42(14) 16(1) 2(1) 1 — 21(12) 2
Ушканьи острова 37(20) 9 2(1) 3(2) — 23(19) —
Сред. Байкал 52(25) 11(1) 3(1) 3 — 36(23) —
ЗаЛ. Провал 16(2) Ю(1) — — 2 4(1) —
Селенгинское мелководье Ю(4) 5(1) 1(1) — — 4(2) —
Посольский сор 15(3) 10(1) — — 1 4(2) —
Юж. Байкал 59(22) 18(1) 3(1) 5(1) 1 31(19) 2
Примечание. В скобках указано число эндемичных видов.
Глава 1. История исследования фитобентоса озера Байкал
23
(S = 0.19-0.28). Несколько выше видовое сходство фитоценозов Посольского
сора и Чивыркуйского залива (S — 0.42) и мелководных бухт Анта и Богучан-
ская (S = 0.65). Значительно различается состав донной флоры Селенгинского
и Ангаро-Кичерского мелководий (S = 0.26).
Фитоценозы Чивыркуйского залива, где встречено 42 вида водорослей,
среди которых 14 эндемиков, наибольшее видовое сходство имеют с флорой
восточного прибрежья Сев. Байкала (S = 0.67) и проливов Мал. Море (S - 0.62)
и Ольхонские Ворота (S = 0.65).
В глубоко вдающихся в берег бухтах, расположенных вдоль открытых при-
брежий озера (Анга, Богучанская, Фролиха и др.), и под прикрытием мысов на
мягких грунтах литорали развиты обычно высшие водные растения (Potamoge-
ton, Ranunculus, Myriophyllum и др.), вместе с которыми растут и широко рас-
пространенные в Палеарктике летние (Cladophora fracta, С. globulina, Phizoclo-
nium sp., Sphaeroplea annulina) и вегетирующие круглый год (Chara sp., Nitella
sp., Cladophora aegagropila и др.) виды водорослей, здесь же встречаются много-
численные эндемики Байкала (Draparnaldioides simplex, Ireksokonia formosa,
Chaetomorpha baicalensis') [Кожов, 1931, 1962; Паутова, 1973, 1975; Ижболдина,
1990в]. Несмотря на то, что эти бухты расположены на значительном расстоя-
нии друг от друга и даже на разных берегах озера, коэффициент видового сход-
ства фитоценозов довольно высок — 0.68, что обусловлено, очевидно, сходст-
вом экологических условий в рассматриваемых бухтах (преобладающий тип
грунта, уклон дна, защищенность от волнового воздействия, влияние речного
стока и др.). В сублиторали таких бухт развит обычный для Байкала комплекс
из 11-14 видов водорослей. В более открытых и более подверженных волновому
воздействию бухтах (Бабушка, Песчаная, Сеногда, Лаканда, Хакусы и др.), где
в литорали преобладает мягкий грунт, водоросли отсутствуют или представле-
ны менее разнообразно (15-19), чем в бухтах, глубоко вдающихся в берег
(26-29 видов). В сублиторали многих открытых бухт преобладают крупнозер-
нистые слабозаиленные пески, иногда с галькой, и донная флора представлена
8-11 видами водорослей мейо- и макрофитов. В таких районах фитоценозы, от-
личающиеся иногда по составу доминантных и субдоминантных видов, имеют
высокий коэффициент видового сходства (S = 0.73). Флора открытых и глубо-
ко вдающихся в берег бухт имеет более низкий коэффициент видового сходства
(0.3-0.5).
Самые первые шаги сделаны с целью определения продуктивности ряда
массовых видов донных водорослей [Бочкарев, Вотинцев, Яснитский, 1950; Во-
тинцев, 1963; Ижболдина и др., 1978а, 1979,19816,19826,1984а]. Ежегодно в ли-
торали основную фитомассу продуцируют водоросли с коротким периодом
развития. Отмечено, что в общей продукции сезонных водорослей литорали
значительная доля (76.7 %) приходится на продукцию Ulothrix zonata, ниже она
у видов Draparnaldioides (23.1 %). Доля продукции Tetraspora cylindrica var. bul-
losa, у которой сухое вещество составляет всего 0.7 % от сырой массы водорос-
ли, ничтожна — 0.2 %. В настоящее время расчет годовой продукции проведен
для массовых видов на участке, типичном для северо-западного прибрежья
Юж. Байкала. Для расчета продукции массовых видов для всего озера этих дан-
ных недостаточно, поскольку в разных частях озера неодинаковы как площадь
литорали и пригодность ее для поселения макрофитов, так и сами виды, отли-
чающиеся друг от друга по продуктивности. Для определения общей продук-
ции макрофитов необходимо провести аналогичные выполненным исследова-
ния в разных частях байкальского прибрежья.
24
Глава 1. История исследования фитобентоса озера Байкал
Положено начало изучению химического состава донных водорослей [Гуме-
нюк, 1986, 1987; Ижболдина, Корнакова, Гуменюк, 1988; Элементный состав...,
1994]. Отмеченная для макрофитов Байкала избирательная способность к акку-
муляции металлов свидетельствует о возможности использования байкальских
водорослей в качестве индикаторов среды их обитания.
Режимные наблюдения за развитием донной флоры проведены в 1961—
1996 гг. НИИ биологии на одном из типичных участков юго-восточного побе-
режья Байкала в районе БЦБК. Изучена изменчивость донных фитоценозов,
свидетельствующая о наличии значительных естественных межгодовых колеба-
ний их фитомассы и относительной стабильности структуры сообществ [Ко-
жов, Ижболдина и др., 1965, 1969; Ижболдина, 1970, 1974; Кожова, Окунева,
Ижболдина и др., 1974; Ижболдина, 1982а]. В районе поступления в озеро про-
мышленных очищенных сточных вод (ПОСВ) БЦБК, сбрасываемых в озеро с
1966 г., фитоценозы дна находятся в неустойчивом состоянии. Наиболее замет-
но воздействие ПОСВ сказалось на развитии эндемиков Tetraspora cylindrica
var. bullosa и Draparnaldioides pilosa. Эти водоросли, ранее сплошным ковром
покрывавшие в районе сброса ПОСВ каменистый грунт на глубине от 1 до
3-3.5 м, с 1981 г. образуют здесь разреженные неравномерные заросли или со-
всем отсутствуют [Ижболдина, 1985, 1992; Кожова, Ижболдина и др., 19956].
В 1982 г. в этом районе отмечено обильное развитие Oscillatoria, свидетельству-
ющее о возможном вторичном загрязнении этого района озера. Отмеченные
нарушения в структуре донных фитоценозов связаны, очевидно, и с качеством
стоков и с поступлением последних в поверхностные слои воды (1978-1982 гг.),
что привело к длительному повышению их концентрации в зоне развития мак-
рофитов. Исследования, проведенные А.Б. Ошаровым с соавт. [1983], показали,
что неразведенные сточные воды подавляют важные физиологические процес-
сы водорослей. В неразведенных стоках через неделю содержание азота в тал-
ломах снижается до 40—60 %. При 50-кратном разбавлении ПОСВ подавляется
фосфорный обмен. Отмечено, что в разные годы воздействие стоков на процес-
сы ассимиляции неодинаково. Наиболее устойчивым к воздействию ПОСВ
оказался доминирующий в зоне прибоя Ulothrix zonata, который в годы депрес-
сии Tetraspora и Draparnaldioides занимал участки дна, свободные от их зарос-
лей. В районе сброса условно чистых вод (УЧВ) БЦБК совсем исчезли эндеми-
ки Tetraspora cylindrica var. bullosa (с 1976 г.) и D. pilosa (с 1979 г.), снизилась
фитомасса Didymosphenia geminata [Кожова, Ижболдина, Бокова, 1998]. В небо-
льшом количестве здесь появились виды, не характерные для открытых при-
брежий озера — Oedogonium sp., Spirogyra sp., Batrachospermum moniliforme.
Очевидно, при организации мониторинга донных фитоценозов необходи-
мо обращать внимание на количественное развитие эндемичных видов родов
Tetraspora и Draparnaldioides. При этом следует иметь в виду, что для оценки со-
стояния донных фитоценозов всего озера режимные наблюдения за их состоя-
нием необходимо провести в разных частях побережья, различающихся по
условиям обитания донной растительности, по видовому составу донной фло-
ры и структуре растительных сообществ.
Необходимы углубленные исследования биологии эндемичных байкаль-
ских видов, что поможет решить проблему их эволюции. Кроме того, чрезвы-
чайно важным представляется изучение роли донных водорослей мейо- и мак-
рофитов в жизни донных беспозвоночных и некоторых рыб.
ГЛАВА
МЕТОДЫ СБОРА
И ОБРАБОТКИ ПОЛЕВОГО МАТЕРИАЛА
В настоящей работе речь идет о водорослях мейо- (размер от 0.1 до 2 мм) и
макрофитах (более 2 мм) Байкала. Представители подавляющего большинства
видов прочно прикреплены к субстрату с помощью ризоидов и имеют относите-
льно небольшие размеры (до 10-25 см), поэтому сбор их с помощью водных
граблей, скребков, якорьков, используемых ботаниками на водоемах, где преоб-
ладают высшие водные растения [Катанская, 1981; Руководство..., 1983; и др.), на
Байкале не эффективен. Лишь на глубине от 0 до 1.5-3 м качественный сбор
макрофитов можно проводить с помощью щипцов Рубцова. При количествен-
ном учете фитобентоса в литорали Байкала мы пользовались преимущественно
методом пробных площадок, широко применяемом при учете бентоса как в кон-
тинентальных, так и в морских водоемах [Браун, 1957; Барашков, 1965; Блинова,
Возжинская, 1971; Катанская, 1981; Кокин, 1982; Распопов, 1985].
Пробы отбирали на разрезах, равномерно расположенных перпендикуляр-
но к берегу. Число разрезов зависит от того, насколько разнообразны экологи-
ческие условия на обследуемом участке озера (крутизна и протяженность мел-
ководной террасы, преобладающий на ней тип грунта, близость рек и степень
подверженности разных частей побережья волновому воздействию и т.д.). Под
водой направление разреза указывал уложенный по дну капроновый шнур, раз-
меченный через каждые Юм. Проплывая над шнуром, аквалангист определяет
ширину пояса, образованного изучаемым видом или видами, и глубинные гра-
ницы пояса. Затем в центре пояса и вблизи его глубинных границ (глубоковод-
ной и мелководной) берут по одной пробе, стараясь разместить рамку на зарос-
лях, наиболее типичных для данной глубины. Если ширина пояса более 6-10 м,
число проб увеличивают. При отборе пробы аквалангист с участка дна, ограни-
ченного учетной рамкой площадью 0.1 м2, переносит весь грунт с обрастанием
в мешок из плотной ткани, завязывает его и пробу с помощью лебедки подни-
мают на палубу экспедиционного судна.
Поскольку водоросли весьма неравномерно распределены в литорали Бай-
кала, одновременно с учетом фитомассы проводят учет проективного покры-
тия грунта водорослями. Мы делали это с помощью железной рамы площадью
0.25 м2, разделенной на 16 квадратов [Ижболдина, Верномудров, Максимов,
1978, 1984; Ижболдина, Максимов, Мурахвели и др., 1978]. Заросли, занимаю-
щие один квадрат, составляют 6.2 % проективного покрытия дна в раме. В зоне
зарослей изучаемого вида аквалангист через каждые 10 м 4 раза перекладывает
26
Глава 2. Методы сбора и обработки полевого материала
раму в обе стороны от шнура, указывающего направление разреза, и считает
число квадратов в раме, занятых зарослями водорослей. Все данные о грунте,
ширине растительного пояса, его глубинных границах, проективном покрытии
дна водорослями, передают по переговорному устройству на поверхность, где
их фиксируют в полевом дневнике, или данные аквалангист под водой заносит
на специальную пластину и с нее позднее переносят их в полевой журнал. На
мягком грунте сбор фитобентоса можно проводить методом пробных площа-
док, с помощью всасывающего эрлифтного пробоотборника (ВЭП) [Развозжа-
ев, 1988] и с помощью дночерпателя Петерсена. Сравнительная оценка разных
способов отбора количественных бентосных проб [Ижболдина, Максимова,
Гуменюк и др., 1990] показала следующее. Репрезентативные количественные
данные могут быть получены на иле и заиленном мелком песке с помощью
дночерпателя Петерсена, на крупнозернистом слабозаиленном песке с помо-
щью ВЭП, на каменистом грунте путем отбора проб аквалангистами из учет-
ной рамки в мешки.
На борту экспедиционного судна камни из мешков перекладывают в тазы с
водой, где с камней с помощью острого ножа тщательно отделяют все водорос-
ли. Пробу промывают через капроновое сито, переносят в емкость для хране-
ния и фиксируют 4%-м раствором формалина. Для того, чтобы талломы Dra-
parnaldioides, многие из которых покрыты слоем слизи, сохранили свою форму
и массу, необходимо, чтобы объем фиксирующего раствора был раза в 2 боль-
ше объема водорослей. Для сохранения водорослями зеленой окраски в пробу
можно добавить раствор медного купороса до появления голубой окраски фик-
сирующей жидкости [Малый практикум..., 1976].
В лаборатории пробу промывали от формалина, растения разбирали по ви-
дам, определяли фенологические фазы развития водорослей, отмечали степень
обрастания эпифитами1. Перед взвешиванием поверхностную воду с водорослей
убирали обсушиванием их на фильтровальной бумаге. Водоросли, талломы ко-
торых окружены слизистым чехлом, обсушивали на плотном (№ 73) капроно-
вом сите, помещенном на фильтровальную бумагу. Крупные формы взвешивали
на аналитических, мелкие — на торзионных весах2. Для определения общей фи-
томассы водорослей в том или ином растительном поясе средневзвешенную фи-
томассу растений (в г/м2), определенную по методу пробных площадок, умно-
жали на общую площадь пояса, учитывая при этом проективное покрытие грун-
та водорослями. Для оценки степени видового сходства фитоценозов в разных
растительных поясах и районах озера использовали коэффициенты видового
сходства Серенсена S = 2 с/а + Ь, где а и b — числа видов, обнаруженных в каж-
дом из сравниваемых районов, с - число общих для них видов. Коэффициент
встречаемости (V) видов определяли по формуле V =---, где а — число проб,
п
в которых данный вид зарегистрирован, п — число всех проб [Константинов,
1986]. Масштаб дан для эндемичных байкальских водорослей.
1 При определении систематического положения ряда Cyanophyta для окрашивания влагалищ
использовали хлор-цинк-иод (20 г хлорида цинка, 6.5 г иодида калия, 1.3 г кристаллического иода в
10.5 мг воды), окрашивающий целлюлозу в синий цвет.
2 В настоящей работе приведена воздушно-сырая масса водорослей.
ГЛАВА
СИСТЕМАТИКА ВОДОРОСЛЕЙ БЕНТОСА
И ПЕРИФИТОНА ОЗЕРА БАЙКАЛ
ОТДЕЛ CYANOPHYTA
Водоросли одноклеточные, колониальные, многоклеточные, окрашенные
в сине-зеленый цвет различных оттенков, реже в оливково-зеленый, жел-
то-зеленый, розовый или фиолетовый, но никогда не бывают ярко-зеленые
или коричневые. При наличии массы газовых вакуолей клетки под микроско-
пом кажутся почти черными. Протопласт разделен на окрашенный перифери-
ческий слой — хроматоплазму, содержащий пигменты, и бесцветную внутрен-
нюю часть — центроплазму1. Слизистые оболочки (влагалища) часто толстые,
бесцветные или желтые, до коричневых; реже красные, синеватые или фиоле-
товые. Подвижные жгутиковые стадии в цикле развития отсутствуют. Полово-
го процесса нет, размножение вегетативное и бесполое. Встречаются стадии
покоя — споры и гормоспоры (гормоцисты). Для многоклеточных нитчатых
форм характерно образование особых пограничных клеток — гетероцист.
В Байкале найдены представители двух классов.
I. Водоросли одноклеточные и колониальные; расположение клеток в коло-
ниях беспорядочное или более или менее правильное, очень редко ните-
видное, но ни талломов, ни настоящих нитей не образуется. Эндоспоры,
экзоспоры и гетероцисты отсутствуют..................Chroococceae
II. Водоросли образуют нити; клетки в нитях обычно тесно соединены друг с
другом и связаны плазмодесмами, образуя трихом (строение многоклеточ-
ное); трихомы часто окружены влагалищем; гормогонии в громадном боль-
шинстве случаев образуются; гетероцисты имеются или отсутствуют. Эндос-
поры и экзоспоры отсутствуют, у многих образуются споры и гормоспоры ..
..................................................... Hormogoneae
Класс CHROOCOCCEAE
Водоросли одноклеточные и колониальные свободно плавающие; реже при-
крепленные. Колонии слизистые, реже — из плотно сомкнутых клеток. Располо-
жение клеток в колонии беспорядочное или более-менее правильное, очень ред-
ко — нитевидное. Плазматическая связь между клетками отсутствует. Клетки
’Морфологического оформленного ядра у синезеленых водорослей нет, в связи с чем их отно-
сят к подцарству прокариотических водорослей [Водоросли..., 1989].
28 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
обычно не дифференцированы на основание и вершину. Размножение исклю-
чительно вегетативное.
В Байкале известны представители одного порядка.
ПОРЯДОК CHROOCOCCALES
Клетки одиночные или соединены в свободно плавающие или свободно
лежащие на дне слизистые колонии, разнообразного, но не трубчато-студени-
стого вида.
ПОДПОРЯДОК STEREOMETREAE
Колонии объемные, слизистые, разнообразной формы и размера. Клетки в
колониях расположены во всей толще слизи или только одним перифериче-
ским слоем, окруженным слизистой оболочкой вокруг.
СЕМЕЙСТВО MICROCYSTIDACEAE
Колонии, преимущественно неправильных очертаний, состоящие из гомо-
генной слизи с клетками, обычно без собственных слизистых оболочек.
В Байкале найдены представители 2 родов.
I. Клетки шаровидные, делящиеся во всех направлениях пространства.
1. Колонии разнообразной формы, сплошные или неправильно продыряв-
ленные. Клетки в слизи колонии разбросаны без порядка.... Microcystis
II. Клетки эллипсоидные или палочковидно-цилиндрические и очень редко
шаровидные, но в таком случае делятся всегда только в одном направлении
пространства.
1. Колонии разнообразной формы и размера. Клетки удлиненные, всегда
разбросаны в слизи колонии без порядка............... Aphanothece
Род Microcystis (Kutz., 1833) Elenk. (incl. Aphanocapsa Nag.)
Колонии слизистые, микроскопические и макроскопические, преимуще-
ственно бесформенные, реже более или менее шаровидные или удлиненные.
Колониальная слизь мягкая или довольно плотная. Клетки шаровидные, иног-
да слегка эллипсоидные, расположены в слизи без порядка. Деление клеток по
всем направлениям пространства.
В Байкале найдено 2 вида.
I. Водоросли, свободно живущие в воде или на суше, иногда плотно прилегают
к субстрату, но не внедряются в него.
1. Клетки с газовыми вакуолями не более 3 мкм в диаметре.
А. Колонии сплошные, с несколькими дочерними колониями в общей
оболочке, клетки 2-3 мкм в диаметре.............. М. ichthyoblabe
2. Клетки без газовых вакуолей.
А. Колонии сливаются в бесформенные массы, клетки 2-4 мкм в диа-
метре.............................................М. muscicola
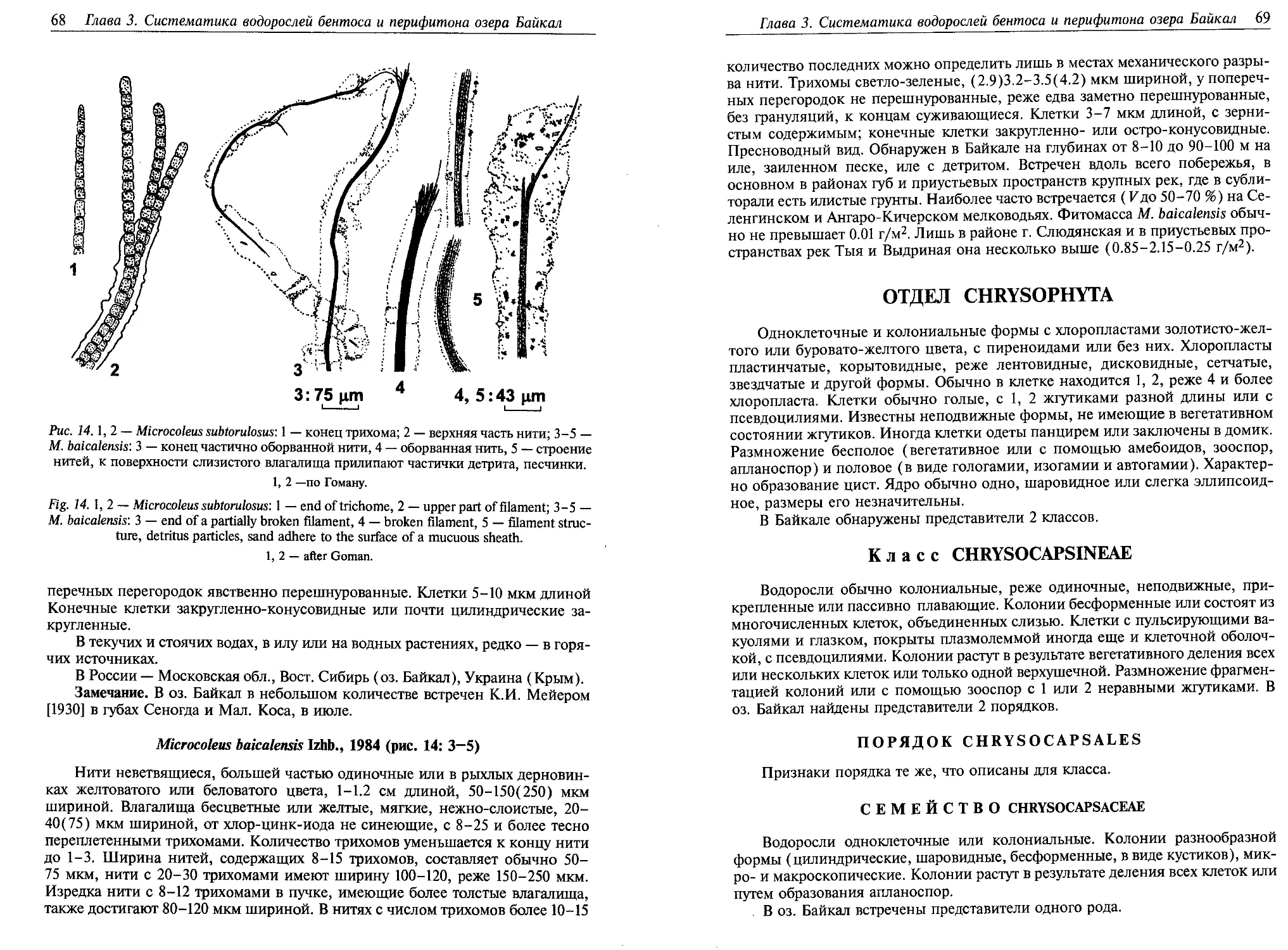
Microcystis ichthyoblabe Kutz. (рис. 2: 1, 2)
Колонии микроскопические, более или менее округлые, сплошные, с об-
щей наружной студенистой оболочкой и дочерними колониями внутри. Клет-
ки с газовыми вакуолями.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 29
Рис. 2. 1, 2 — Microcystis ichthyoblabe; 3, 4 — М. muscicola; 5 — Aphanothece stagnina; 6,7 —
Sphaeronostoc kihlmani: 6 — колонии в натуральную величину, 7 — молодая колония
(большое увеличение); 8-10 — Fischerella major: 8 — участок нити со спорой, 9, 10 —
ветвящиеся нити.
По: 1 — Кютцингу, 2 — Тейлингу, 3 — Вилле, 4 — Комаренко, 5 — Смису, 6, 7 — Косинской, 8-10 —
Гоману.
Fig. 2. 1, 2 — Microcystis ichthyoblabe; 3, 4 — М. muscicola; 5 — Aphanothece stagnina; 6,7 —
Sphaeronostoc kihlmani: 6 — colonies, full size, 7 — juvenile colony (high magnification);
8-10 — Fischerella major: 8 — segment of filament with a spore, 9,10 — branching filaments.
After: 1 — Kiitzing, 2 — Teiling, 3 — Ville, 4 — Komarenko, 5 — Smise, 6, 7 — Kossinskaya, 8-10 —
Goman.
В планктоне стоячих и текущих вод, вызывает иногда цветение воды.
Местами в Европейской части России и в Сибири. В Байкале отмечена
В.Ч. Дорогостайским [1906] без указания места нахождения.
Замечания. Нами ввд встречен в небольшом количестве в районе Анга-
ро-Кичерского мелководья на глубине 18-25 м на иле с детритом. Размер слож-
ных колоний неправильных очертаний достигает 0.5 мм.
30 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Microcystis muscicola (?) (Menegh.) Elenk. (=Aphanocapsa muscicola (Menegh.)
Wille и некоторые другие) (рис. 2: 3, 4)
Колонии, сливающиеся в слизистую или студенистую бесформенную, ши-
роко распростертую массу сине-зеленого цвета. Клетки шаровидные, 2-4 мкм
в диаметре, тесно скученные, сине-зеленые или других оттенков.
На влажных скалах, стенах, дереве, а также в теплицах и в воде холодных и
горячих источников. В Определителе пресноводных водорослей СССР [Гол-
лербах и др., 1953] вид отмечен для Байкала без указания его местонахождения
в озере и условий обитания.
Замечания. Нами найдены колонии Microcystis неправильных очертаний,
некоторые из них достигают 2.5 мм. Клетки диаметром 3.8 мкм. Вид обнаружен
вдоль всего прибрежья на глубинах от 1.6 до 102 м на заиленном песке и иле
[Ижболдина, 1990в]. Встречаемость М. muscicola вдоль большей части при-
брежья менее 10 %. Несколько выше она вдоль западного берега Северного
(V = 23-29 %) и Среднего Байкала (V = 15-16 %), а на Ангаро-Кичерском
мелководье достигает 40-60 %. Фитомасса этой водоросли в разных районах
озера колеблется от 0.001 до 0.1 г/м2 Фитомасса более 0.1 г/м2 была отмечена на
глубинах от 3 до 35 м в районе мыса Орео (0.41-0.76 г/м2) и на глубине 3.5-15 м
в районе мыса Харгантуй (5.6 г/м2).
Род Aphanothece (Nag., 1849) Elenk.
Колонии слизистые, микроскопические, в основном бесформенные. Ко-
лониальная слизь гомогенная, различной консистенции, бесцветная или окра-
шенная. Клетки эллипсовидные, до палочковидно-цилиндрических, очень
редко шаровидные. Деление клеток только перпендикулярно длинной оси. В
слизи клетки расположены беспорядочно.
Виды Aphanothece встречаются в самых разнообразных водных и наземных
местообитаниях. В Байкале найден один вид.
Aphanothece stagnina (Spreng) В.-Peters, et Geith emend.
(рис. 2: 5)
Колонии студенистые, довольно крепкие, шаровидные, полушаровидные,
эллипсоидные или цилиндрические, до 4 см в поперечнике. Клетки цилиндри-
ческие, 3-5 мкм шириной, 5 мкм длиной, более-менее сине-зеленые, преиму-
щественно рыхло, реже густо расположенные.
В сточных водах на камнях, иле, растениях или в свободно плавающем со-
стоянии. Повсюду.
Замечания. В Байкале найдена К.И. Мейером [1930] в зал. Провал. М.Ю. Бек-
ман [1959] залежи A. stagnina обнаружены в оз. Загли-Нур, расположенном в
вершине бух. Загли и сообщающимся с ней протоком. Нами вид встречен в гу-
бах Слюдянской, Онокочанской, Сеногда, вблизи устьев р. Тыя и на Анга-
ро-Кичерском мелководье. У восточного берега озера найдена в губе Дагарской
и в бух. Змеиная (Чивыркуйский залив), в Посольском соре и в зал. Провал на
глубине от 2 до 4 м, на заиленном песке и иле. Колонии до 2.1 мм в поперечни-
ке, клетки цилиндрические, сине-зеленые. Встречаемость A. stagnina обычно
не превышает 5 %, лишь на Ангаро-Кичерском мелководье она достигает
15-17 %, а в Посольском соре — 52 %.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 31
Класс HORMOGONEAE
Водоросли нитевидные. Нити свободные, очень редко срастающиеся с бо-
ков. Клетки в нити тесно соединены друг с другом посредством плазмодесм,
образуют “трихомы”, характерные только для этого класса. Гетероцисты име-
ются или отсутствуют. В Байкале найдены представители 3 порядков.
I. Гетероцисты имеются, хотя иногда встречаются очень редко.
1. Нити с настоящим ветвлением, иногда сопровождаемые и ложным
...............................................Stigonematales
2. Ветвление только ложное или нити совсем не разветвленные.
А. Нити на всем протяжении одинакового строения и даже в том случае,
если базальные их части прилегают к субстрату, разница между восходя-
щими и прилежащими их частями незначительна или отсутствует ....
.......................................................Nostocales
II. Гетероцисты всегда отсутствуют................Oscillatoriales
ПОРЯДОК STIGONEMATALES
Ветвление настоящее — неправильно-боковое (типичное), правильно-ди-
хотомическое или ложнодихотомическое. Иногда настоящее ветвление сопро-
вождается ложным. Нити с однорядными или многорядными трихомами обра-
зуют дерновники неопределенного облика или шаровидные колонии. Нити
свободные или срастаются боками, образуя пластинку или прямостоячие пуч-
ки. Основная часть таллома может отличаться от боковых ветвей, которые мо-
гут расширяться или суживаться к вершинам, редко имеют волосковидные
окончания. Характерно почти постоянное наличие гетероцист и гормогониев.
В Байкале встречен представитель одного семейства.
СЕМЕЙСТВО STIGONEMATACEAE
Ветвление боковое, более или менее неправильное (настоящее), иногда
сопровождаемое ложным. Боковые ветви однородные, никогда не заканчиваю-
щиеся терминальной гетероцистой и не образующие волосков.
В Байкале встречен представитель одного рода — Fischerella.
Род Fischerella (Born, et Flah.) Gom., 1895
Нити стелющиеся, образующие тонкие или толстые войлочные, подушко-
образные или корковидные дерновинки. Ветвление настоящее, обычно одно-
стороннее, боковые ветви уже основной нити. Редко наблюдаются случаи лож-
ного ветвления. Влагалища гомогенные или слоистые. Трихомы основных ни-
тей чаще многорядные, состоящие из более или менее округлых клеток.
Трихомы ветвей обычно однородные. Гетероцисты боковые или интеркаляр-
ные, из удлиненных клеток. Имеются гормогонии и споры.
В Байкале найден один вид.
Hscherella major Gom. (рис. 2: 8—10)
Дерновинки толстые, серовато-коричневой или черной окраски. Основ-
ные нити — стелющиеся, 8-16 мкм шириной, с широкими желто-коричневы-
ми слоистыми влагалищами. Клетки основных нитей почти шаровидные или
32 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
цилиндрические, 6-8 мкм шириной. Ветви прямостоящие, тесно переплетают-
ся, 6-12 мкм шириной. Клетки ветвей квадратные, до короткодисковидных,
слегка расширяющиеся на концах трихомов, 4-10 мкм шириной. Гетероцисты
цилиндрические или почти квадратные. Гормогонии длинные, слегка палице-
образные. Споры 10-14 мкм длиной и 7-10 мкм шириной.
На влажных стенах, старом дереве, а также на стеблях водяных растений.
Эстония, Украина, Байкал, Якутия.
Замечание. В Байкале в небольшом количестве встречена К.И. Мейером
[1930] в июле в зал. Провал, около Костылей, а также в протоках и старых рус-
лах р. Кичера.
ПОРЯДОК NOSTOCALES
Водоросли этого порядка имеют всегда однорядные трихомы, не ветвящи-
еся или ложно разветвленные, с гетероцистами и нередко со спорами. Трихомы
обычно по одному в каждом влагалище, редко по нескольку. Трихомы одина-
ковой ширины на всем протяжении или они симметрично расширяются или
суживаются к обоим концам, или они асимметричные, суживающиеся от осно-
вания к вершине.
В Байкале найдены представители 2 подпорядков.
ПОДПОРЯДОК SYMMETREAE
Нити и трихомы симметричные, одинаковой ширины на всем протяжении
или симметрично утолщающиеся или утончающиеся к обоим концам.
В Байкале обнаружены представители 3 семейств.
I. Трихомы симметричные, реже асимметричные, по одному в каждом влага-
лище.
1. Трихомы не ветвящиеся, одинаковой ширины на всем протяжении, реже
суживаются к обоим концам.
А. Влагалища большей частью расплывающиеся. Трихомы всегда собра-
ны в макроскопические, реже микроскопические, колонии различного
облика........................................... Nostocaceae
Б. Влагалища хорошо оформленные, очень редко расплывающиеся
.............................................Nodulariaceae
2. Трихомы с ложными ветвлениями, иногда оба конца нитей расширенные
или суженные. Рост чаще апикальный.............Scitonemataceae
СЕМЕЙСТВО NOSTOCACEAE
Представители семейства имеют хорошо оформленные колонии макро-
или реже микроскопические, шаровидные, распростертые, нитевидные, сли-
зистые или студенистые, мягкие и твердые, с мягким или крепким перидер-
мом, плотные или полые внутри. Трихомы на периферии колоний имеют
ясно заметные, иногда окрашенные и слоистые влагалища. Внутри колоний
последние обычно расплывающиеся и бесцветные. Трихомы извилистые, ко-
роткие или длинные, густо или рыхло переплетающиеся, иногда радиально
расположенные. Вегетативные клетки разнообразной формы, гетероцисты
интеркалярные, редко терминальные, шаровидные или эллипсоидные, раз-
множение посредством гормогониев и спор. Последние шаровидные или эл-
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 33
липсоидные, гладкие или с шагреневой оболочкой. У некоторых видов споры
неизвестны.
Для Байкала известны представители 2 родов.
I. Колонии макроскопические, очень редко микроскопические, с крепким пе-
ридермом.
1. Колонии шаровидные, реже эллипсоидные от 1 мм до 1 см и более в по-
перечнике ........................................Sphaeronostoc
2. Колонии в молодых стадиях развития шаровидные, затем неправильно
распростертые, твердые или мягкие....................Stratonostoc
Род Sphaeronostoc Elenk.
Колонии на всех стадиях развития более или менее шаровидные или, реже,
эллипсоидные, от 1 мм до 5 см в поперечнике, внутри плотные или полые, мяг-
кие или твердые, студенистые или хрящеватые, с крепким перидермом. Трихо-
мы рыхло или тесно лежащие, с влагалищами, хорошо заметными на перифе-
рии колоний. Вегетативные клетки бочонкообразные, реже шаровидные или
эллипсоидные, гетероцисты почти шаровидные, иногда по нескольку рядом.
Споры образуются редко.
В Байкале найдено 4 вида этого рода.
I. Колонии макроскопические, от 1 мм до 2.5-5 см в поперечнике.
1. Клетки с газовыми вакуолями...........................S. kihlmani
2. Клетки без газовых вакуолей.
А. Колонии шаровидные или эллипсоидные, гладкие.
а. Колонии до 1 см в поперечнике, с тесно лежащими трихомами.
а. Трихомы на концах не суживающиеся...............£ coeruleum
б. Колонии от 1 до 5 см в поперечнике, с рыхло лежащими трихомами. .
.................................................S.prumiforme
Б. Колонии более или менее шаровидные, бугорчатые.
а. Трихомы длинные, радиально расположенные от центра к периферии,
с расплывающимися внутри колоний влагалищами .... S. zetterstedtii
Sphaeronostoc kihlmani (Lemm.) Elenk.
(=Nostoc kihlmani L., Nostoc planctonicum Poretzky et Tschemow)
(рис. 2: 6-7)
Колонии шаровидные, реже удлиненные, с крепким перидермом, бирюзо-
вой окраски, 100-600 мкм в диаметре, реже 2-4.5 мм. Влагалища незаметные,
трихомы тесно переплетающиеся — 4-7.5 мкм шириной. Бочонкообразные
или почти шаровидные клетки заполнены газовыми вакуолями. Гетероцисты
шаровидные, реже эллипсоидные, 6-10 мкм в диаметре, иногда по нескольку
рядом, споры неизвестны.
В озерах и сфагновых болотах, свободно плавают в воде. Указывается для
Кавказа, Зап. и Вост. Сибири и Дальнего Востока.
Замечание. В Байкале вид найден К.И. Мейером [1930] в июле в зал. Про-
вал среди зарослей водно-болотной растительности. Нами [Ижболдина, 1990]
единичные колонии встречены в районе Ангаро-Кичерского мелководья, на
глубине 3 м, на заиленном песке.
34 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Sphaeronostoc coeruleum (Lyngb.) Elenk. {—Nostoc coeruleum Lyngb.)
(рис. 3: 1, 2)
Колонии шаровидные, гладкие, с крепким перидермом, ярко-сине-зеленые,
реже коричневатые, 5-6 мкм, реже до 1 см, в диаметре. Влагалища обычно за-
метные, трихомы тесно переплетающиеся, 5-7 мкм шириной. Клетки и гетеро-
цисты почти шаровидные. Гетероцисты до 10 мкм в диаметре. Споры неизвестны.
В стоячих и медленно текущих водах, на водяных растениях, свободно пла-
вает в воде или лежит в илу на дне водоемов. Во многих местах Европейской
части России, на Кавказе, в Вост. Сибири, в Сред. Азии. В Байкале найден
К.И. Мейером [1930] в Посольском соре — в июле.
Рис. 3. 1,2 — Sphaeronostoc coeruleum: 1 — колонии; 2 — трихомы с гетероцистами; 3-5 —
Stratonostoc linckia f. linckia: 3 — разрез колонии, 4 — отдельные мелкие колонии, 5 —
проростки гормогониев; 6 — Stratonostoc linckia f. piscinale; 7 — Stratonostoc linckia f. cameum.
По: 1-3 — Косинской, 4 — Кондратьевой, 5 — Горбуновой, 6, 7 — Фреми.
Fig. 3.1, 2 — Sphaeronostoc coeruleum: 1 — colonies, full size, 2 — trichomes with heterocysts;
3-5 — Stratonostoc linckia f. linckia: 3 — colony section, 4 — separate small colonies, 5 —
hormogone sprouts; 6 — Stratonostoc linckia f. piscinale; 7 — Stratonostoc linckia f. carneum.
After: 1-3 — Kossinskaya, 4 — Kondrat’eva, 5 — Gorbunova, 6, 7 — Fremi.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 35
Sphaeronostoc pruniforme (Ag.) Elenk. (=Nostoc pruniforme Ag.) (рис. 4: 1—4)
Колонии шаровидные или эллипсоидные, гладкие, 1-1.5 см в диаметре,
реже до 4 см ширины и 5 см длины, мягкие внутри, иногда с центральной по-
лостью, с крепким перидермом, ярко-сине-зеленые, оливковые, до черно-ко-
ричневых. Влагалища бесцветные или желтоватые, хорошо заметны. Трихомы
расположены радиально от центра к периферии, 4-6 мкм ширины, рыхло пе-
реплетающиеся. Клетки коротко-бочонкообразные или несколько удлинен-
Рис. 4. Sphaeronostoc pruniforme.
1,2 — колонии в натуральную величину; 3 — поперечный разрез колонии; 4 — трихомы с гетеро-
цистами и спорами. По: 1, 4 — Косинской, 3 — Фреми.
Fig. 4. Sphaeronostoc pruniforme.
1,2 — colonies, full size; 3 — cross-section of the colony; 4 — trichomes with heterocysts and spores. After:
1, 4 — Kossinskaya, 3 — Fremi.
36 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
ные. Гетероцисты почти шаровидные, 6-7 мкм в диаметре. Споры около
10 мкм в диаметре.
В прудах и озерах, свободно плавает в воде или лежит на илистом дне водо-
ема. В Европейской части России, в Вост, и Зап. Сибири, в Сред. Азии и на Да-
льнем Востоке.
Замечания. В Байкале массовое развитие S. pruniforme в губе Крутой (Чи-
выркуйский залив) наблюдал Г.Ю. Верещагин [1918]. В бухтах прол. Ольхонские
Ворота обильное развитие вида отмечено В.Н. Яснитским [1928]. К.И. Мейер
[1930] подчеркивает, что S. pruniforme встречается в сорах (Посольский, Исток-
ский), заливах (Баргузинский, Чивыркуйский, Мухор) и в мелководных, глу-
боко вдающихся в берег бухтах (Харгантуй, Курминская). Кроме того, в боль-
шом количестве встречен им около ст. Мысовой. Нами [Ижболдина, 1990а]
кроме вышеперечисленных районов вид обнаружен в бух. Хужир-Нуго, в губе
Мужинайской, у мыса Коврижка и в бух. Песчаная. Единичные талломы най-
дены на Селенгинском мелководье. Колонии обычно шаровидные или слегка
приплюснутые, 3-3.5 см в диаметре. В районе бух. Базарной (прол. Ольхон-
ские Ворота) размер многих колоний достигает 6-8 см в диаметре. В бухтах
прол. Мал. Море и Чивыркуйского залива встречаемость S. pruniforme не пре-
вышает 23-35 %, а фитомасса колеблется от 0.5 до 40 г/м2. В бухтах Ольхонских
Ворот встречаемость вида больше (48-60 %) и фитомасса в разных бухтах со-
ставляет от 5 до 211 г/м2.
Sphaeronostoc zetterstedtii (Aresch.) Elenk. (=Nostoc zetterstedtii Aresch.)
(рис. 5: 1—4)
Колонии мелкобуторчатые и лопастные, с крепким перидермом, более или
менее шаровидные, реже с почти гладкой поверхностью, 0.5-1.5 (2.5)1 см в по-
перечнике, темно-зеленой или оливково-бурой окраски. Влагалища желтые
или бесцветные. Хорошо заметны лишь на периферии колонии. Трихомы ра-
диально расположенные, 3.5-5 мкм шириной, на поверхности переплетающи-
еся. Клетки бочонкообразные, почти шаровидные или несколько удлиненные.
Гетероцисты 6-10 (15) мкм шириной, иногда по нескольку рядом. Споры ша-
ровидные, 5-7 мкм в диаметре.
В литоральной, реже элиторальной зонах озер и в болотах. Нередко снача-
ла прикрепляется, затем свободно лежит на дне водоема.
В Европейской части России, на Кавказе (оз. Севан), в Вост. Сибири, в
Сред. Азии и на Дальнем Востоке.
Замечание. В Байкале вид встречен нами [Ижболдина, 1990в] в районе
прол. Мал. Море у мыса Харгантуй, в губе у мыса Курминский, у мысов Мужи-
най, Арул и в зал. Мухор. Растет на глубине от 3.5 до 24 м, как на твердом, так и
на мягком грунте.
Род Stratonostoc Elenk.
Колонии в молодости шаровидные, затем более или менее распростертые,
с тенденцией к плоскостному росту. Перидерм мягкий или крепкий. Влагали-
ща бесцветные или окрашенные, заметные только у периферии колоний. Три-
хомы рыхло или тесно переплетающиеся, иногда расположены радиально. Ве-
гетативные клетки чаще всего бочонкообразные, споры шаровидные или эл-
липсоидные, с гладкими или покрытыми мелкими шипиками оболочками.
1 Здесь и далее: приведен размах колебаний размеров; в скобках - размеры особей, встре-
чающихся единично.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 31
Рис. 5. Sphaeronostoc zetterstedtii.
1,2 — колонии в натуральную величину; 3 — детальное изображение трихомов; 4 — разрез при не-
большом увеличении через два соседних бугорка с характерным для этого вида радиальным распо-
ложением. По: 1, 3 — Косинской, 4 — Голлербаху.
Fig. 5. Sphaeronostoc zetterstedtii.
1,2 — colonies, full size; 3 — detailed presentation of trichomes; 4 — section (through two neighbouring
tubercles with typical for this species radial location of trichomes, low magnification) After: 1,3 —
Kossinskaya, 4 — Gollerbach.
В Байкале найдено 3 вида.
I. Колонии с мягким, иногда почти незаметным перидермом.
1. Колонии сначала шаровидные, потом распростертые, разнообразного об-
лика.
А. Споры с гладкой оболочкой, шаровидные или эллипсоидные. Клетки
очень разнообразны по форме....................................S. linckia
II. Колонии с крепким перидермом.
1. Колонии более 1 см в поперечнике.
А. Колонии распростертые, волнисто-гладкие или бугорчатые.
а. Колонии свободные, плотные, кожистые, волнисто-пластинчатые
или складчато-бугорчатые....................................S. commune
б. Колонии прикрепленные к субстрату, распростерто-вздутые, толстые
или мягкие внутри, бугорчатые или складчато-волнистые, часто по-
лые .......................................................S. verrucosum
Stratonostoc linckia (Roth) Elenk. (=Nostoc linckia (Roth) Born, et Flah.)
f. linckia (рис. 3: 3—5)
Колонии сначала шаровидные, прикрепленные, далее неправильно рас-
простертые, слизистые, свободно плавающие, слабо-лиловатой, синевато-зе-
леной, грязно-зеленой или коричневатой окраски. Влагалища бесцветные,
ясно заметные на периферии колоний. Трихомы извиты, густо переплетающи-
еся. Клетки бочонкообразные, длина их равна или немного больше или мень-
ше ширины. Гетероцисты и споры почти шаровидные. Споры с гладкой корич-
невой, реже бесцветной оболочкой.
38 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Stratonostoc linckia forma linckia (рис. 3: 3-5)
Трихомы 3.5-4.0 (4.7) мкм шириной, клетки бочонкообразные, эллипсо-
идные или шаровидные, 2-5 мкм шириной, 5-6 мкм длиной Гетероцисты
5-6 мкм в диаметре, споры 6-7 мкм шириной и 7-8 мкм длиной, с гладкой ко-
ричневой или редко бесцветной оболочкой.
В стоячих, реже в текучих водах, сначала прикрепление на подводных
предметах, затем плавает свободно.
В России почти повсеместно.
Замечание. В оз. Байкал вид найден К.И. Мейером [1930] в зал. Провал в
лагуне у пос. Оймур в июле.
Stratonostoc linckia forma piscinale (Kutz.) Elenk.
(=Nostoc piscinale Kutz.) (рис. 3: 6)
Отличается от типа бугорчатой поверхностью колоний, рыхлым располо-
жением трихомов и бесцветными оболочками спор. Трихомы 3.7-4 мкм шири-
ной, гетероцисты 4.5-6 мкм в диаметре, споры 6-8 мкм шириной.
В ряде мест Европейской части России, на Кавказе, в окрестностях оз.
Байкал и на Камчатке.
Замечание. Для оз. Байкал приводится М.М. Голлербахом [Голлербах и др.,
1953] без указания места нахождения. Нами не встречен.
Stratonostoc linckia forma rivulare (Kutz.) Elenk.
(=Nostoc rivulare Kutz.)
Колонии сначала шаровидные, потом неправильно разорванные. Влагали-
ща желтые, заметны только на периферии колонии. Трихомы рыхло переплета-
ются (3) — 4-4.2 (4.5) мкм шириной, гетероцисты обычно удлиненные или
бочонкообразные, 6-8 мкм шириной и 7-10 мкм длиной, с гладкой бесцветной
или коричневой оболочкой.
В стоячих или текучих водах, сначала прикрепленно, затем плавают сво-
бодно.
Бассейн р. Сятка, Кавказ, Камчатка, оз. Байкал.
Замечание. Для Байкала приводится М.М. Голлербахом [Голлербах и др.,
1953] без указания места нахождения. Нами не встречен.
Stratonostoc linckia forma carneum (Ag.) Elenk.
(—Nostoc carneum Ag.) (рис. 3: 7)
Характеризуется разнообразной окраской колоний с преобладанием оттен-
ка мяса и красно-коричневого оттенка, рыхлым расположением трихомов и эл-
липсоидной формой спор.
Колонии в старости распростертые, кожистые, иногда пузырчато-бугорча-
тые. Влагалища бесцветные, трихомы рыхло переплетаются, 3-4 мкм шири-
ной. Клетки почти шаровидные, бочонкообразные, удлиненные. Длина их мо-
жет превышать ширину до 3 раз. Гетероцисты удлиненные, 6 мкм шириной,
споры 6 мкм шириной, 8-10 мкм длиной.
В стоячих водах, сначала прикреплены, затем свободно плавают. В Евро-
пейской части России, в Сибири, в горах Сред. Азии и на Камчатке.
Замечание. В оз. Байкал найдена К. И. Мейером [1930] в небольшом коли-
честве в зал. Провал, в лагуне около пос. Оймур и в Истокском соре, в июле.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 39
Stratonostoc commune (Vauch.) Elenk.
Колонии долго остаются шаровидными, достигая размера вишни, потом
распростертые, неправильно лопастные и складчато-бугорчатые, до 6-7 см в
поперечнике, сине-зеленые, желтые, фиолетово-коричневые, до черных. Вла-
галища толстые, темно-коричневые, хорошо заметные ближе к периферии.
Трихомы тесно переплетающиеся, бледно-оливково-зеленые, 4-6 мкм шири-
ной. Клетки коротко-бочонкообразные или почти шаровидные, около 5 мкм
длиной. Гетероцисты по нескольку рядом, обычно шаровидные, около 7 мкм
в диаметре. Споры такой же величины, как вегетативные клетки. Встречаются
редко.
Stratonostoc commune forma commune
В сухих, преимущественно степных и полупустынных местах, на сухих лугах.
В России распространен широко, особенно в южных районах. В Байкале
не найден.
Stratonostoc commune forma sphaericum (Vauch.) Elenk.
(=Nostoc sphaericum Vauch.) (рис. 6: 3—5)
Колонии до 6-7 см в поперечнике. Влагалища обычно незаметные. Трихо-
мы 4-5 мкм шириной, гетероцисты около 6 мкм в диаметре. Споры эллипсоид-
ные, около 5 мкм шириной и 7 мкм длиной, с гладкой коричневой оболочкой.
В стоячих водах на подводных растениях, на влажных мхах, на омываемых
водой скалах, иногда в поверхностном слое почв.
В России повсеместно.
Замечания. Приводится для Байкала К.И. Мейером [1930]. Найдена им в
губе Анга и около Ушканьих островов. Нами [Сравнительная оценка..., 1990]
встречена в бухтах Базарной и Хужир-Нуго, где свободно лежит на мягком
грунте на глубине 3-5 м. Колонии до 5 см в поперечнике, крупнобугорчатые,
большей частью полые.
Stratonostoc verrucosum (Vauch.) Elenk. (=Nostoc verrucosum Vauch.) (рис. 6: 1, 2)
Колонии сначала шаровидные или полушаровидные, крупнобуторчатые,
потом распростертые, складчато-волнистые в старости, внутри мягкие, 1-3,
реже до 10 см в поперечнике, черно-зеленые, оливковые, до темно-коричневых,
с крепким перидермом. Влагалища желто-коричневые, заметны на краях коло-
ний. Трихомы 3-5 (6) мкм шириной, у периферии колоний густо переплетают-
ся. Клетки трихомов коротко бочонкообразные, гетероцисты почти шаровид-
ные, 6-8 мкм в диаметре, споры эллипсоидные, около 5 мкм шириной и 7 мкм
длиной, с гладкой темной оболочкой. В стоячих и текучих водах, сначала плотно
прикрепляется к грунту и подводным предметам, затем может плавать свободно.
Широко распространен в России, на Кавказе, в Крыму, в Сред. Азии.
Замечания. По наблюдению К.И. Мейера [1930], байкальский S. verruco-
sum отличается от типовой формы большим размером трихомов (5-6 мкм
шириной) и гетероцист (7.2-8.2 мкм шириной). Обычный для Байкала вид,
растет на глубине 2-5 м на камнях, где иногда образует обширные заросли.
Растет вместе с Chaetocladiella pumila С. Meyer et Skabitsch. и Tetraspora cylind-
rica (Wahlenb.) Ag. var. bullosa C. Meyer. На мягком грунте вид в массе найден
К.И. Мейером [1930] в бух. Харин-Ирги, где развит вместе со Sphaeronostoc
prumiforme на глубине 10-20 м, сплошным слоем покрывая дно бухты. Кроме
того, был встречен в прол. Мал. Море, в Чивыркуйском и Баргузинском зали-
40 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 6. 1,2 — Stratonostoc verrucosum: 1 — детальное изображение трихомов, 2 — общий
вид колоний в натуральную величину; 3-5 — Stratonostoc commune f. sphaericum: 3 — сре-
занные большие колонии, 4 — общий вид колонии, 5 — вид колоний из субстрата.
По: 1, 2 — Фреми, 3-5 — Тюре.
Fig. 6. 1, 2 — Stratonostoc verrucosum: 1 — detailed presentation of trichomes, 2 — full-sized
colonies, general view; 3-5 — Stratonostoc commune f. sphaericum: 3 — cut-away large
colonies, 4 — general view of the colony, 5 — colonies viewed from the substratum.
After; 1, 2 — Fremi, 3-5 — Ture.
вах и в бух. Аяя. Обильное развитие вида в бухтах Харин-Ирги и Загли отме-
чено также В.Н. Яснитским [1928]. А.П. Скабичевский [1934] обнаружил
S. verrucosum в бух. Аяя на глубине 12-14 м. При наших исследованиях [Иж-
болдина, 1990в] вид обнаружен вдоль всего побережья, исключая Селенгин-
ское и Ангаро-Кичерское мелководья, на глубинах от 0.8 до 35 м. Наиболее
часто S. verrucosum встречается (V = 30-50 %) на твердом грунте на глубинах
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 41
от 1.5 до 10-12 м, где в качестве субстрата может также использовать талломы
Cladophora compacta и С. floccosa. Средняя фитомасса S. verrucosum вдоль за-
падного прибрежья больше (4.95 г/м2), чем вдоль восточного (0.66 г/м2). У
восточного берега лишь в районе губы Максимиха и у о. Лиственичный фито-
масса данного вида достигает соответственно 25.4 и 32.2 г/м2. На мягком
грунте фитомасса водорослей вдоль всего прибрежья невелика (0.9-2.8 г/м2)
и лишь в бухтах южной части Мал. Моря она достигает 83 г/м2 в губе Курма и
63 г/м2 в бух. Шибетейская. В последней S. verrucosum образует мощные за-
росли вместе с Chaetomorpha baicalensis.
В ряде районов Байкала (бухты Дагарская, Аяя, Фролиха, мыс Амнунда-
кан, губа Богучанская, бух. Бабушка, падь Улан, падь Орео, падь Широкая)
S. verrucosum вместе е другими водорослями образует IV растительный пояс.
СЕМЕЙСТВО NODULARIACEAE
Нити прямые или слабо изогнутые, одиночные или соединенные в дерно-
винки, прикрепленные или свободно плавают. Влагалища мягкие или крепкие,
тонкие или толстые, гомогенные или слоистые, очень редко расплывающиеся,
содержат всегда по одному трихому. Трихомы симметричные или асимметрич-
ные, с интеркалярным или верхушечным ростом. Гетероцисты интеркалярные
и базальные. Споры лежат вне связи с гетероцистами.
В Байкале найден представитель одного рода.
Род Aulosira Kirchn., 1878
Нити симметричные, прямые или изогнутые, одиночные или, реже, соеди-
нены в дерновинки. Влагалища крепкие. Трихомы с интеркалярным или верху-
шечным ростом, гетероцисты интеркалярные. Споры удлиненно-цилиндриче-
ские, вне связи с гетероцистами.
В Байкале встречен один вид.
Aulosira implexa Bom. et Flah. (рис. 7: 6)
Нити прямые или изогнутые, обычно собраны в пучки, 7-17 мкм (чаще
12 мкм) шириной и 5-10 мм длиной. Влагалища тонкие, бесцветные. Трихомы
8-9 мкм шириной, сине-зеленые, у поперечных перегородок слегка перешну-
рованные. Клетки цилиндрические, длина их равна или меньше ширины. Гете-
роцисты квадратные или цилиндрические. Споры цилиндрические, 8-9 мкм
шириной и 16-34 мкм длиной, располагаются рядами от 2 до 32 подряд.
В стоячих водах. Латвия, Вост. Сибирь.
Замечание. В Байкале в небольшом количестве встречена К. И. Мейером
[1930] в бух. Анга среди зарослей водной растительности, кроме того, в сборах
В.Н. Яснитского встречена в Онгоконской бухте.
СЕМЕЙСТВО SCYTONEMATACEAE
Строение нитей и боковых ветвей обычно симметричное. Ветвление лож-
ное. Гетероцисты интеркалярные и базальные. Влагалища гомогенные или
слоистые, крепкие или ослизняющиеся. Иногда наблюдается образование гор-
могониев, реже спор.
В Байкале встречаются представители одного рода.
42 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 7. 1, 2 — Tolypothrix distorta f. distorta — ветвящиеся нити с гетероцистами; 3 —
Т. distorta f. penicillata; 4 — Т. helicophyla; 5 — Т. limbata; 6 — Aulosira implexa.
По: 1, 5, 6 — Фреми, 2 — Кондратьевой, 3 — Косинской, 4 — Леммерману.
Fig. 7. 1, 2 — Tolypothrix distorta f. distorta — branching filaments with heterocysts; 3 —
T. distorta f. penicillata; 4 — T. helicophyla; 5 — T. limbata; 6 — Aulosira implexa.
After: 1, 5, 6 — Fremi, 2 — Kondrat’eva, 3 — Kossinskaya, 4 — Lemmerman.
Род Tolypothrix Kutz., 1843 ex Bomet et Flax., 1887
Различно изогнутые ветвящиеся нити, растущие в виде кустиков. Ветвле-
ние ложное. Ложные ветви преимущественно двойные. Влагалища крепкие,
гомогенные или слоистые с параллельными или косо расходящимися слоями.
Трихомы к концам часто расширяющиеся, с дифференцированными клетка-
ми. Гетероцисты интеркалярные, чаще базальные, нередко по нескольку ря-
дом. Гормогонии конечные. Споры встречаются редко, одиночные или це-
почками.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 43
В Байкале встречено 4 вида, ведущих подводный образ жизни.
А. Влагалища с неровными волнистыми краями, нити 7-11 мкм шириной
....................................................Т. helicophila
Б. Влагалища с ровными неволнистыми краями, нити 4-18 мкм шириной
а. Нити до 3 мм длиной
а. Нити 10-15 мкм шириной.....................Т. limbata
б. Нити до 2-3 см длиной
а. Нити 4-10 мкм шириной ......................T.tenue
р. Нити 8-15 мкм шириной (у F. penicillata — до 18 мкм шириной),
влагалища в средних частях нити окрашены в желто-коричневый
цвет...........................................Т. distorta
Tolypothrix helicophila Lemm. (рис. 7: 4)
Дерновинки маленькие, плотно прикреплены к субстрату. Нити 7-11 мкм
шириной, обильно ветвистые. Влагалища слизистые, узкие, бесцветные, сна-
ружи волнистые. Трихомы 4-5 мкм шириной, у поперечных перегородок не
перешнурованные. Клетки квадратные или длина их меньше или больше ши-
рины. Гетероцисты одиночные, обычно цилиндрические, реже почти квадрат-
ные, бесцветные.
В стоячих водах на подводных растениях и створках раковин.
Вост. Сибирь (Байкал).
Замечание. В Байкале найден К.И. Мейером [1930] в бух. Половинной
(западное побережье Юж. Байкала) на глубине 1.25 м. Ширина трихомов
7.5 мкм.
Tolypothrix limbata Thur. (рис. 7: 5)
Дерновинки хлопьевидно-кустистые, сине-зеленые, 10-15 мкм шириной,
2-3 мм длиной, обильно ветвящиеся. Влагалища бесцветные или желто-корич-
невые, толстые, слоистые, с ровными краями. Трихомы 5.5-9 мкм шириной,
клетки бочонковидной формы, длина их равна или немного более ширины. Ге-
тероцисты одиночные или по 2 ряда.
В стоячих и текучих водах среди других водорослей.
Латвия, Эстония, Россия (Кольский полуостров, Вост. Сибирь), горы
Сред. Азии.
Замечания. В Байкале встречен К.И. Мейером [1930], где, иногда в значите-
льном количестве развит.на камнях на глубине 2-5 м. Найден им в падях Сен-
ная, Фролиха, у ст. Мысовая, у мыса Ирексокон, а также в Нижнеангарском
соре и в зал. Мухор. В бух. Аяя Т. limbata в качестве субстрата использует тал-
ломы Chara sp. и Nitella sp., развитые у северного крутого берега бухты. Нами в
небольшом количестве встречен в районе пос. Бол. Коты.
Tolypothrix tenuis Kutz.
Дерновинки хлопьевидно-кустистые, реже кистевидные, сине-зеленые,
оливковые или темно-бурые. Ветвление обильное, одиночное, редко двойное.
Ветви идут в том же направлении, что и основная нить, под некоторым углом
к ней. Влагалища узкие, бесцветные, редко слегка темноватые. Гетероцисты
квадратные, закругленные или цилиндрические, обычно по 4-7, реже по
3 подряд.
44 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Tolypothrix tenuis forma tenuis
В Байкале не найден.
В стоячих и медленно текущих водах.
В России почти повсеместно.
Tolypothrix tenuis forma polymorpha (Lemm.) Kossinsk.
Нити 15-17 мкм шириной, трихомы 12-13 мкм шириной, гетероцисты квад-
ратные, закругленные или цилиндрические, большей частью по 4-7 подряд.
Река Волга (близ Саратова), Вост. Сибирь (оз. Байкал, оз. Сартлан).
Замечание. В Байкале найдена К.И. Мейером [1930] в небольшом количе-
стве в губе Куркутской (прол. Ольхонские Ворота).
Tolypothrix distorta (Fl. Dan.) Kutz.
Дерновинки кустистые или кустисто-кисточкообразные, от ярко-сине-зе-
леной до темно-коричневой окраски. Ветви идут в том же направлении, что и
основная нить, отклоняясь от нее под углом 45° или прилегая к ней. Влагалища
узкие, в средней части таллома окрашенные в желто-коричневый цвет. Трихо-
мы ярко-сине-зеленые или оливковые. Клетки трихомов квадратные или дли-
на их менее ширины. Гетероцисты шаровидные или продолговатые, одиноч-
ные или по 2-3, реже по 4 рядом.
Tolypothrix distorta forma distorta (рис. 7: 1, 2)
Дерновинки кустистые, до 3 см длины. Нити 8.5-15 мкм шириной, трихо-
мы 6-12 мкм шириной, ярко-сине-зеленые или оливковые. Клетки короткие,
посередине нити иногда квадратные, 3.6-5 мкм длиной, с зернистым содержи-
мым. Гетероцисты шаровидные до 8 мкм в диаметре, или продолговатые до
15 мкм, одиночные, реже по 2-3 рядом.
В стоячих и медленно текущих водах, а также в выжимках из мхов.
В России встречается почти повсеместно.
Замечания. По К.И. Мейеру [1930], вид распространен по всему Байкалу
на глубине 2-5 м, на камнях, часто поселяется внутри колоний Cladophora
compacta, образуя с ней одну подушечку. В зал. Провал найден на высших вод-
ных растениях. А.П. Скабичевский [1934,1966,1978] отмечает Т. distorta f. dis-
torta как вид, обычный для III растительного пояса. При наших исследовани-
ях [Ижболдина, 1990в] вид был встречен вдоль всего прибрежья, исключая
районы впадения крупных рек, на глубине от 1.5 до 25 м, на твердом грунте.
Наиболее обильно вид развит на глубинах от 1.5 до 6 м. Встречен нами среди
ветвей Cladophora compacta, С. floccosa, С. meyeri, Chaetocladiella pumila. Чаще
(7 = 25-40 %) и в большем количестве (0.25 г/м2) вид встречается вдоль за-
падного прибрежья. У восточного прибрежья эти показатели ниже и состав-
ляют соответственно 10-20 % и 0.06 г/м2. Лишь в нескольких пунктах запад-
ного (мысы Бол. Коса, Харгантуй, Ото-Хушун, о. Угунгой) и восточного
(бух. Сосновка, у р. Солзан) прибрежий фитомасса Т. distorta f. distorta колеб-
лется от 13 до 47 г/м2, а у мыса Харгантуй во II поясе достигает 87 г/м2. Круг-
логодичные наблюдения за развитием водорослей показали, что в нояб-
ре-марте, когда в Байкале отсутствуют виды водорослей, образующих в ве-
сенне-летний период верхний растительный ярус, размер талломов Т. distorta
f. distorta больше (5-6 мм в высоту), чем летом (2-3 мм выс.). В июне-сен-
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 45
тябре часто встречаются талломы, верхняя часть которых состоит из пустых
влагалищ, на последних в массе поселяются сувойки рода Vorticella. В августе
в районе пади Ниж. Хомуты и у мыса Онгурен встречены талломы Т. distorta f.
distorta с массой гормогониев.
Tolypothrix distorta forma penicillata (Ag.) Kossinsk. (рис. 7: 3)
Дерновинки кустисто-кисточкообразные, ложные нити более или менее
прилегают к основной нити. Нити 8-20 мкм шириной, трихомы 4-13 мкм
шириной. Конечная клетка почти шаровидная. Гетероцисты одиночные или по
4 подряд.
Преимущественно в быстротекущей воде, реже по берегам озер и прудов.
Россия (острова Арктики, европейская часть, оз. Байкал, Дальний Вос-
ток), Сред. Азия.
Замечания. Впервые для Байкала отмечена К.И. Мейером [1930]. Найдена
им около пос. Берла и в бухтах Аяя и Лаканда. Нами эта форма встречена во
многих районах озера (губа Слюдянская, мыс Онокочанский, о. Лиственич-
ный, о. Угунгой, в прол. Мал. Море, губа Томпа), на глубине от 1.2 до 7м на
камнях. Целые заросли этой водоросли обнаружены у о. Богучан на глубине
1.2-1.5 м, на камнях. Кроме того, найдена в сборах О.А. Тимошкина и
Т.Я. Ситниковой в прол. Ольхонские Ворота (бухты Тутайская, Базарная) и в
Чивыркуйском заливе (бух. Змеиная). В июле-сентябре часто встречаются
талломы с гормогониями и пустыми влагалищами в верхней части дерновинок.
Фитомасса редко превышает 1 г/м2.
ПОДПОРЯДОК ASIMMETREAE
Нити и трихомы в типе асимметричные: утончающиеся или утолщающие-
ся от основания к вершине.
СЕМЕЙСТВО RIVULARIACEAE
Трихомы утончающиеся от основания к вершине и заканчивающиеся
обычно волоском, гетероцисты базальные, реже интеркалярные. Ложные ветви
обычно с базальными гетероцистами, симподиального типа, по одному или
пучками по нескольку вместе. У некоторых видов в основании нитей над гете-
роцистой образуются споры.
В оз. Байкал встречены представители 3 родов.
I. Нити одиночные или помногу вместе в форме налетов, ковриков, дернови-
нок, хлопьев и т.п., никогда не образуют студенистых сферических колоний
...........................................................Calothrix
II. Нити соединены в студенистые шаровидные или полушаровидные колонии,
внутри которых располагаются радиально.
1. Споры отсутствуют постоянно..........................Rivularia
2. Споры образуются.................................Gloeothrichia
Род Calothrix (Ag., 1824) V. Poljansk., 1937, sensu lat. (incl. Dichothrix
Zanard., Sacconema Borzi, Rivulariopsis (Kirchn.) Woronich.)
Трихомы утончаются от основания к вершине постепенно или резко, пере-
ходя или не переходя в волосок. Нити неветвящиеся или ветвящиеся, одиноч-
ные или помногу вместе, извитые или параллельно расположенные. Колонии в
46 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
виде налетов, ковриков, дерновинок, хлопьев. Ветви могут оставаться в слизи
материнской нити, образуя разветвленные кустики, или обособляются от нее.
Некоторые виды образуют споры.
В оз. Байкал найдено 5 видов.
I. Нити неветвящиеся или ветвящиеся, обычно не образующие разветвленных
кустиков. Обычно одно влагалище заключает один трихом, реже — неско-
лько.
1. Влагалища обычно без широкой остуденевшей периферической части. Нити
более 100 мкм длиной
А. Нити и трихомы утончаются от самого основания более или менее посте-
пенно, иногда у основания луковицеобразно расширенные.
а. Окончания трихомов разнообразны: они то незначительно утончены, то
несут волосок, переход к которому постепенен или часто внезапен
.....................................................С. kossinskajae
б. Окончания трихомов никогда не бывают столь разнообразными. При на-
личии волоска переход к нему чаще постепенен.
а. Трихомы обычно заканчиваются волосками, которые сбрасываются
при образовании гормогониев.
+ Нити у основания в большинстве случаев луковицеобразно расшире-
ны.
х Нити у основания большей частью внезапно и сильно вздутые (до
3 раз) ........................................С. kawraiskyi
хх Нити у основания не столь резко и сильно вздутые.
о Ширина нитей над расширенным основанием 3.5-12 мкм.
Л Клетки средней части нитей не бывают сильно удлиненными.
□ Основания нитей и трихомов сильно и постоянно луковице-
образно расширены. Нити одиночные или помногу, часто
расположены звездообразно ......................С. stellaris
О □ Луковицеобразные расширения основания нитей не столь
сильны и непостоянны. Нити одиночные или помногу, но
не образуют звездообразных групп.
§ Нити приблизительно до 0.5 мм длиной.
- Нити обычно до 0.5 мм длиной, большей частью помногу,
тесно сближенные, перпендикулярные к субстрату. Влага-
лища тонкие, неслоистые......................С. braunii
++ Нити у основания в большинстве случаев без луковицеобразного
расширения.
х Влагалища окрашенные, иногда местами бесцветные.
о Влагалища всегда узкие, неслоистые, не бывающие темно-ко-
ричневыми. Нити обычно расположены параллельно, неветвя-
щиеся ........................................... С. braunii
оо Влагалища большей частью широкие, слоистые, иногда тем-
но-коричневые, часто с воротничками. Нити нередко ветвя-
щиеся. В местах ветвления влагалища одевают 2 трихома
.............................................С. parietina
Calothrix kossinskajae N. Poljansk. (рис. 8: 4—16)
Нити в группах, реже одиночные, до 200 мкм длиной, у основания иногда
несколько расширенные, 4-6 (7.5 мкм), в средней части 3-5 (5.7 мкм и на
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 8. 1 — Calothrix braunii; 2 — С. parietina; 3 — С. stellaris; 4-16 — С. kossinskajae,
вершины нитей с разнообразно заканчивающимися верхушками трихома; 17 — Rivularia
borealis, колония в вертикальном разрезе.
По; 1, 3 — фреми, 2, 4-16 — Полянскому, 17 — Куку.
Fig. 8. 1 — Calothrix braunii; 2 — С. parietina; 3 — С. stellaris; 4-16 — С. kossinskajae, fila-
ment apices with a variety of trichome tips; 17 — Rivularia borealis colony, vertical section.
After: 1, 3 — Fremi, 2, 4-16 — Poljansky, 17 — Kukk.
конце 1.5-3.8 мкм шириной Влагалища бесцветные, тонкие, неслоистые, про-
должающиеся дальше окончания трихомов.
Трихомы сине-зеленые или бледно-зеленые, у клеточных перегородок
перешнурованные или ровные, иногда слабо расширенные у основания,
(3)3.8-5.3 (6) мкм, посередине 2.3-4.6 мкм и на конце (не считая волосков)
0.8-3.4 мкм шириной, заканчивающиеся округленной или конусовидной
клеткой или постоянно или внезапно переходящие в волосок. Иногда воло-
сок оторван от трихома и сидит на конце влагалища, гетероцисты базальные,
обычно одиночные, полушаровидные, реже слегка вытянутые, 2.3-5.3 мкм
длиной, 3.8- 5.3 (6) мкм шириной.
48 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
В стоячих и текучих водах, эпифитно на водяных растениях, раковинах
моллюсков, на твердом субстрате.
Европейская часть России и территории СНГ, с севера до юга, не часто.
Замечания. Вид найден во многих районах Сев. Байкала [Ижболдина,
1990в], где растет на глубинах от 0.8 до 7 м, прикрепляясь к камням. Образует
плотные дерновины диаметром 0.5-1.5 мм. Характерно разнообразие оконча-
ний трихомов. Наиболее часто (V = 58-77 %) вид встречается вдоль западного
прибрежья Сев. Байкала на участке от Ангаро-Кичерского мелководья до губы
Слюдянская. Средняя фитомасса С. kossinskajae в районах обнаружения вида
колеблется от 0.001 до 7-10 г/м2, а в отдельных пробах достигает 200 г/м2
(губа Сеногда) и 982 г/м2 (губа Слюдянская).
Calothrix kawraiskyi Schmidle
Нити соединены в плотные дерновники 0.5-0.75 мм высоты, не ветвящие-
ся, прямые, реже согнутые на конце, у основания обычно сильно и внезапно
вздутые (до 12 мкм шириной). Имея над этим расширением около 4 мкм
ширину, к концу постепенно утончаются и заканчиваются очень длинным тон-
ким волоском. Влагалища нерасщепленные, тонкие, бесцветные. Вегетативные
клетки квадратные или длина их больше или меньше ширины. Гетероцисты ба-
зальные, полушаровидные или удлиненно округлые, бесцветные.
В озерах, на подводных предметах. Латвия, оз. Байкал.
Замечание. В Байкале найден К.И. Мейером [1930] в зал. Провал, в лагуне
около пос. Оймур, в июле.
Calothrix stellaris Born, et Hah. (рис. 8: 3)
Нити одиночные или в группах, часто расположены звездообразно, изо-
гнутые, у основания луковицеобразно расширенные, 13-21 мкм, выше 6.5-
12 мкм шириной. Влагалища тонкие, бесцветные, обычно неслоистые. Трихо-
мы в местах клеточных перегородок слабо перешнурованные, у основания рас-
ширенные, над этим расширением 6-7 мкм шириной, заканчивающиеся воло-
ском. Вегетативные клетки в нижней части трихома короткие, выше почти
квадратные. Гетероцисты базальные, почти полушаровидные, одиночные или
по 2-3 подряд, обычно шире вегетативных клеток.
В стоячих и текучих водах, на высших водных растениях и водорослях,
твердом субстрате.
Эстония, Грузия, Россия (оз. Байкал).
Замечание. В оз. Байкал найдена К.И. Мейером [1930] в июле в зал. Провал
на высших водных растениях.
Calothrix braunii Bom. et Flail, (рис. 8: 1)
Нити не ветвящиеся, приблизительно до 500 мкм длиной, образующие си-
не-зеленые или коричневые дерновники или встречающиеся одиночно. Нити у
основания луковицеобразно расширенные, (7)9-11.2(13) мкм шириной, в
средней части 4.5-8.5 мкм шириной, обычно прямые, помногу вместе. Влага-
лища обычно бесцветные (лишь в нижней части нитей желтоватые или корич-
невые), тонкие, неслоистые, продолжаются далее трихомов. Трихомы в местах
клеточных перегородок слабо перешнурованы, у основания (4.5) 5-7 мкм
шириной, в средней части 2.5-5 мкм шириной, переходящие в волосок или за-
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 49
канчивающиеся закругленно конусовидной или закругленной клеткой. Вегета-
тивные клетки короткие или длина их равна ширине. Гетероцисты базальные,
одиночные, шаровидные или полушаровидные.
В стоячих и текучих водах, на камнях, на раковинах, в перифитоне.
В России и по территории стран СНГ распространен широко — с севера до
юга.
Замечания. Для Байкала вид приводится К.И. Мейером [1930]. Найден им в
районе падей Жилище и Смородовая. Нами отмечен как вид, характерный для
открытых прибрежий юго-западной части озера. В течение года растет на кам-
нях, на глубине от 2 до 10-15 м. Диаметр колоний может достигать 2-2.5 мм
[Ижболдина, 19906].
Calothrix parietina (Nag.) Thur. (рис. 8: 2)
Нити приблизительно до 1 мм длиной, помногу вместе, тесно сближенные,
образующие коричнево-бурые, более или менее широко распростертые налеты
или корочки или встречающиеся одиночно. Нити в основании (7) 9-
18.5 (23) мкм шириной, в средней части (5.5) 7-14 (16) мкм шириной, нередко
ветвящиеся. Влагалища обычно желто-коричневые, в местах ветвления неред-
ко на некотором протяжении одевающие 2 трихома, обычно слоистые. Пери-
ферические слои часто несколько расходятся, образуя воротнички. Трихомы у
основания перешнурованные в местах клеточных перегородок, выше не пере-
шнурованы. В нижней части трихомы (4)4.5-9(13) мкм шириной, посередине
3.5-7(10.5) мкм шириной, на конце’переходящие в волосок. Вегетативные
клетки короткие или удлиненные. Гетероцисты базальные, одиночные, реже
по 2-3 рядом, обычно полушаровидные, реже интеркалярные.
В стоячих и текучих, пресных и солоноватых водах, на камнях, часто в зоне
прибоя.
В России и странах СНГ распространен широко — с севера до юга.
Замечания. Для Байкала вид впервые отмечен К.И. Мейером [1930]. Най-
ден им в большом количестве в районе пос. Бол. Коты в июле. По нашим на-
блюдениям, вид характерен для всего озера, живет в течение всего года на кам-
нях, на глубине 2-10 м. Кроме того, обнаружен в притоках Юж. Байкала (реки
Бол. Коты, Жилище, Черная, Крестовка) [Кожова, Ижболдина, 1994].
Кроме 5 вышеперечисленных видов рода Calothrix К.И. Мейер [1930] при-
водил для Байкала еще 2 вида: С. epiphytica W. et G.S. West, C. fusca (Kutz.)
Bom. et Flax., имеющих колонии микроскопических (до 200-350 мкм длиной)
размеров. Первый из них найден в зал. Провал, в лагуне около пос. Оймур на
корнях Phragmites communis Trin., второй, развивающийся обычно в слизи дру-
гих водорослей, встречен в июле около о. Лиственичный.
При наших исследованиях [Ижболдина, 1990в] вдоль открытых прибрежий
озера на глубинах от 1.5-2 до 5-8 м на твердом грунте обнаружен Calothrix sp.,
определить видовую принадлежность которого в настоящее время не представ-
ляется возможным. Встречаемость вида не превышает обычно 20 %, реже дости-
гает 30 % (у западного побережья Юж. Байкала) — 39 % (у восточного побе-
режья Сев. Байкала). Вдоль западного прибрежья озера и у Ушканьих островов
фитомасса Calothrix sp. составляет в среднем 0.7 г/м2, а вдоль восточного при-
брежья в прол. Мал. Море она колеблется от 0.1 до 0.3 г/м2. Отмечены три райо-
на, где Calothrix sp. развит особенно обильно. Это о. Угунгой (5 г/м2), мыс Бол.
Коса (9.6 г/м2) и мыс Валукан, где средняя фитомасса во II поясе составила
22.8 г/м2, а в III — 9.8 г/м2. В районах владения крупных рек вид не найден.
50 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Род Rivularia (Roth) Ag., 1824, emend Thur.
Трихомы к вершине постепенно утончаются и переходят в волосок. Нити
соединены в студенистые колонии более или менее правильной шаровидной
или полушаровидной формы, прикрепленные или, реже, свободно плавающие.
Нити внутри колонии располагаются радиально. Споры всегда отсутствуют.
В Байкале найдены 2 вида.
I. Колонии обычно не инкрустированы известью.
1. Колонии обычно более 0.5 мм в диаметре. Газовые вакуоли отсутствуют.
А. Колонии мягкие. Нити при надавливании легко отделимы друг от друга,
а. Колонии до 3 мм в диаметре. Влагалища бесцветные, неслоистые.
а. Колонии до 0.5 мм в диаметре. Клетки короткие или одинаковой
длины и ширины. Гетероцисты около 4 мкм в диаметре ....
..................................................R. borealis
р. Колонии до 3 мм в диаметре. Клетки удлиненные. Гетероцисты
10-12 мкм диаметром................................R.aquatica
Rivularia borealis Р. Richt. (рис. 8: 17)
Колонии мягкие, шаровидные или полушаровидные, до 0.5 мм в диаметре,
иногда сливающиеся и широко распростертые. Нити до 200 мкм длиной, рас-
положены рыхло и при надавливании легко отделяются друг от друга. Влагали-
ща бесцветные, неслоистые, в старости расплываются. Трихомы 4 мкм шири-
ной, переходящие в волосок. В основании нитей ширина клеток вдвое превос-
ходит длину, выше клетки квадратные. Гетероцисты базальные, одиночные,
шаровидные. Гормогонии помногу один за одним.
В стоячих водах на водяных растениях.
Эстония, Нов. Земля, оз. Байкал.
Замечание. Найдена в Байкале К.И. Мейером [1930] в бух. Анга в конце
июня.
Rivularia aquatica (de Wild.) Geitl. sensu lato (=Rivularia giobiceps G.S. West.)
(рис. 9: 14—17)
Колонии полушаровидные, до шаровидных, до 3 мм в диаметре, мягкие,
коричневые, реже оливково-зеленые, не инкрустированы известью. Трихомы в
интеркалярной меристемальной зоне 7-9 мкм шириной, в основании до 9 мкм
шириной, на старых нитях тесно, не более 4-6 мкм шириной, заканчивающие-
ся длинным волоском. Вегетативные клетки в нижней части трихома удлинен-
ные, в меристемальной зоне короткие. Гетероцисты базальные, обычно оди-
ночные, реже по 2, почти шаровидные, 10-12 мкм диаметром.
В стоячих водах, на водных растениях.
В разных районах России, не часто.
Замечание. В оз. Байкал найдена К.И. Мейером [1927] в Ангаро-Кичерском
соре.
Род Gloeothrichia J. Agardh., 1824
Постепенно утончающиеся к вершине и обычно переходящие в волосок
трихомы, соединены в студенистые колонии шаровидной или полушаровид-
ной формы, сплошные или полые, прикрепленные или свободно плавающие.
Нити внутри колонии располагаются радиально.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 51
Рис. 9. 1-13 — Gloeotrichia pisum: 1, 13 — простые колонии, 2 — сложные колонии, 3 —
облик колоний, эпифитирующих на Hydrocharis morsus ranae, 4 — группа спороносных
нитей в колонии, 5, 8 — нити с молодой спорой, 6 — нить со зрелой спорой, 7 — молодая
нить без споры, 9-11 — основания спороносных нитей, в которых трихом разрушен, 12 —
колонии в натуральную величину, 13 — фрагмент колонии в разрезе с основанием
спороносных нитей; 14-17 — Rivularia aquatica: 14 — макроскопическая форма колоний,
эпифитирующих на стебле водного растения, 15 — отдельный трихом из колонии (верхний
конец не изображен), 16, 17 — основания 7 трихомов (большее увеличение).
По: 1-6 — Косинской, 14, 15 — Уэсту, 16, 17 — Гейтлеру и Рутнеру.
Fig. 9. 1-13 — Gloeotrichia pisum: 1, 13 — simple colonies, 2 — complex colonies, 3 — external
view of colonies-epiphytes of Hydrocharis morsus ranae, 4 — a group of sporiferous filaments in
the colony, 5, 8 — filaments with a juvenile spore, 6 — filament with a mature spore, 7 — juvenile
filament without a spore, 9-11 — bases of sporiferous filaments with a destroyed trichome, 12 —
colonies, full size, 13 — section of a colony fragment with a base of sporiferous filaments; 14-17 —
Rivularia aquatica: 14 — macroscopic colonies — epiphytes on an aquatic plant stem, 15 — a sep-
arate trichome from the colony (upper tip not shown), 16,17 — bases of 7 trichomes (high mag-
nification).
After: 1-6 — Kossinskaya, 14, 15 — West, 16, 17 — Gaitler & Rutner.
52 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Единственно достаточно общим и постоянным отличием от Rivularia явля-
ется способность к спорообразованию.
В оз. Байкал найдены 2 вида.
I. Колонии обычно твердые. Нити при надавливании с трудом отделимы друг
от друга......................................................G. pisum
II. Колонии более мягкие. Нити при надавливании легче отделимы друг от друга.
1. Волоски не выступают из студня колоний. Трихомы состоят из большого
числа клеток и переходят в волосок.
А. Оболочка спор гладкая. Влагалища вокруг спор мешковидно расши-
ренные, часто с перетяжками............................. G. natans
Gloeothrichia pisum (Ag.) Thur. {=Rivularia pisum Ag.) (рис. 9: 1—13)
Колонии шаровидные, реже чечевицеобразные, 1-2 мм, реже до 1 см в
диаметре, обычно твердые, коричневые, темно-зеленые или сине-зеленые, ино-
гда несколько инкрустированные известью. Нити расположены тесно и при на-
давливании с трудом отделимы друг от друга. Влагалища узкие, обычно бесцвет-
ные, реже темноватые или коричневые. Трихомы у основания 4-11.2 мкм
шириной, с интеркалярной меристемальной зоной, заканчиваются длинным во-
лоском. Вегетативные клетки квадратные или длина их больше или меньше ши-
рины. Гетероцисты базальные, обычно одиночные, шаровидные, эллипсоидные,
полушаровидные, редко конусовидные. Споры гладкие, цилиндрические, пря-
мые (реже изогнутые), 60-400 мкм длиной, (7) 8-15 мкм шириной. Наружная
оболочка споры плотно склеена с внутренним слоем влагалища.
В стоячих, реже текучих, водах, обычно прикрепляется к подводным расте-
ниям, реже плавает свободно.
Вид широко распространен в России и на территории СНГ.
Замечания. Найдена в оз. Байкал К.И. Мейером [1930] — в Чивыркуйском
заливе, около о. Бакланий. При наших исследованиях [Ижболдина, 1998в]
встречена во многих районах озера на мягком грунте (заиленный песок, ил) на
глубине от 2-5 до 70-100 м. Наиболее часто встречается на Селенгинском
(V = 63 %) и Ангаро-Кичерском (V = 38 %) мелководьях, в заливах Чивыркуй-
ский (7 = 52-92 %) и Мухор (V до 62 %) и в Посольском соре (Удо 63 %). Фи-
томасса водоросли и ее численность наиболее высоки в сорах и мелководных
заливах. В зал. Мухор численность G. pisum на глубине 2 м составила
65 280 экз/м2, а фитомасса 11.6 г/м2. В Чивыркуйском заливе в IV растительном
поясе численность колоний достигает 500 тыс. экз/м2, а фитомасса 34.6 г/м2. В
Посольском соре средняя фитомасса рассматриваемого вида составляет 5.8 г/м2,
а в отдельных участках достигает 19 г/м2. Вдоль открытых прибрежий озера, в
том числе и на вышеупомянутых мелководьях, фитомасса G. pisum не превыша-
ет обычно 0.1-0.5 г/м2. Максимального размера колонии G. pisum достигали в
зал. Мухор (3.1 мм) и в бух. Харанса (2.25 мм в диаметре). В других районах
озера диаметр колоний обычно не превышает 0.5-0.8 мм. В летних сборах
встречены в основном колонии со спорами.
Gloeothrichia natans (Hedw.) Rabenh. {Rivularia natans (Hedw.) Welw.)
(рис. 10: 1—7)
Колонии более или менее правильно шаровидные, до 2 см в диаметре, час-
то сливающиеся в бесформенную слизистую массу, мягкие, сине-зеленые,
грязно-оливково-зеленые, буро-коричневые, иногда слабо инкрустированы
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 53
Рис. 10. Gloeotrichia natans.
1 — нить в стадии Pseudorivularia с интеркалярной меристемальной зоной и утонченным ниже нее
трихомом; 2 — нить в стадии Eugloeotrichia (со зрелой спорой); 3 — основание и интеркалярная
меристемальная зона одной нити (в клетках последней — газовые вакуоли); 4 — основание нити в
стадии Pseudorivularia; 5-7 — начало спорообразования. По Полянскому.
Fig. 10. Gloeotrichia natans.
1 — filament at a Pseudorivularia stage with an intercalary meristemal zone and thinned trichome below it;
2 — filament at a Eugloeotrichia stage (with a mature spore); 3 — base and intercalary meristemal zone of
one filament (the cells of the latter contain gas vacuoles); 4 — filament base at a Pseudorivularia stage;
5-7 — beginning of spore formation. After Poljansky.
54 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
известью. Нити расположены рыхло и легко отделимы друг от друга. Влагали-
ща вокруг споры расширенные, обычно с извилистыми контурами, желтые,
коричневые, иногда с фиолетовым оттенком, реже бесцветные. Трихомы у
основания 5-13.5 мкм шириной, заканчивающиеся длинным тонким и ломким
волоском. Гетероцисты базальные, одиночные, шаровидные, полушаровид-
ные, эллипсоидные. Споры цилиндрические, 40-250 мкм длиной, 6-18 мкм
шириной, с влагалищами до 39.6 мкм шириной
В стоячих водах, сначала прикреплена к водным растениям, затем свобод-
но плавает.
В России и странах СНГ широко распространена.
Замечания. По наблюдению К.И. Мейера [1930], G. natans в Байкале явля-
ется наиболее распространенным эпифитом. В большом количестве встречает-
ся в сорах и реках, где поселяется на водных и водно-болотных растениях (Ra-
nunculus, Polygonum, Potamogeton, Myriophyllum, Phragmites communis). Найдена в
зал. Провал, в Посольском, Истокском и Нижнеангарском сорах и в протоках
и старых руслах р. Кичера. Встречена также в зал. Мухор и в бух. Анга.
При наших исследованиях [Ижболдина, 1976, 1990в], вид, кроме того,
встречен в проливах Мал. Море (бухты Курма, Хужир-Нуго) и Ольхонские Во-
рота (бухты Базарная, Куркутская, у мысов Имыш-Таме и Тыгтэ) на Potamoge-
ton perfoliatus L. и Polygonum amphibium L. Найден в Чивыркуйском заливе
(бух. Онгоконская, о. Мал. Калтыгей, по линии между мысами Катунь и Ирка-
на) и в Посольском соре на Potamogeton perfoliatus. На Селенгинском мелково-
дье свободно лежит на дне на глубинах от 5 до 35-40 м.
ПОРЯДОК OSCILLATORIALES
Трихомы всегда без гетероцист и обычно без спор, однорядные, с влагали-
щами или без них, не ветвящиеся или образующие ложные, реже — настоящие
ветвления. Трихомы по одному или помногу в каждом влагалище, одинаковой
ширины на всем протяжении или могут утончаться к концам или утолщаться,
или асимметричные, суживающиеся от основания к вершине и заканчивающи-
еся волоском.
Порядок объединяет всех безгетероцистных представителей класса Ноппо-
goneae.
Для Байкала известно 2 семейства.
I. Трихомы по одному в каждом влагалище или без него.
1. Трихомы цилиндрические.
А. Трихомы прямые или разнообразно изогнутые, не образуют ложных
ветвлений. Конечные клетки трихомов разнообразны по форме и ино-
гда образуют на вершинах трихома особые колпачки (калиптры)
..............................................Oscillatoriaceae
II. Трихомы по нескольку или помногу в одном влагалище. Ложное ветвление
трихомов имеется.....................................Schizothrichacea
СЕМЕЙСТВО OSCILLATORIACEAE
Прямые или разнообразно изогнутые нити (иногда спиралевидные), с вла-
галищами или без них. Влагалище содержит один трихом. Трихомы одной ши-
рины на всем протяжении, лишь некоторые суживаются или расширяются к
концам. Конечные клетки обычно отличаются от других клеток трихома по
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 55
своей форме, образуют иногда колпачки (калиптры). Ложное ветвление, гете-
роцисты и споры отсутствуют. В Байкале встречены представители 3 родов.
I. Трихомы обычно без влагалищ.
1. Трихомы длинные, прямые или разнообразно изогнутые, образующие
иногда спираль.......................................Oscillatoria
II. Трихомы с влагалищами.
1. Влагалища мягкие, слизистые, нередко расплывающиеся . . . Phormidium
2. Влагалища крепкие.
А. Нити соединены в разнообразные, но не пучковатые дерновинки или
свободные..................................................Lyngbya
Род Oscillatoria Vauch., 1803 ex Gomont., 1892
Трихомы сине-зеленые, реже — желто-зеленые, красные или фиолетовые,
прямые или изогнутые, иногда к концам слабоспиралевидные или крючкооб-
разно согнутые. Трихомы одиночные или соединены в дерновинки в виде ко-
жистых пленок, реже — пучков. Влагалища отсутствуют. Трихомы имеют оди-
наковую ширину на всем протяжении или утончаются к концам. Клетки обыч-
но цилиндрические, иногда перешнурованы у клеточных перегородок, редко
бочонкообразные. Клетки равной длины и ширины или длина может быть зна-
чительно больше или меньше ширины. Содержимое клеток однородное или
зернистое, зернышки могут быть равномерно расселены по клетке или собра-
ны рядами у поперечных перегородок. Встречаются газовые вакуоли. Конеч-
ные клетки могут быть закругленные, головчатые, заостренные, нередко с ка-
липтрой (колпачковидно утолщенной оболочкой наружных стенок). Нити
способны к движению, напоминающему качание маятника.
В Байкале найдено 9 видов, из которых 7 идентифицированы.
I. Ширина клеток в 3-11 раз превосходит длину, трихомы к концам несу-
живающиеся или суживающиеся.
1. Трихомы у поперечных перегородок перешнурованные.
А. Конечные клетки чаще слабоголовчатые, с более или менее утолщен-
ной наружной оболочкой. Трихомы (7) 10-20 мкм шириной...........
......................................................О. sancta
2. Трихомы у поперечных перегородок не перешнурованные, прямые, реже
изогнутые, но на концах не согнутые и не спиралевидные. Клетки без ва-
куолей, придающих им ячеистый вид.
А. Ширина трихомов 11-23 мкм, клетки очень короткие, длина их
2-5 мкм..................................................О. limosa
II. Длина клеток от 2 до 3 раз меньше или значительно больше ширины, реже
длина их равна ширине. Трихомы к концам не суживающиеся.
1. Длина клеток в 1-3 раза меньше ширины.
А. Клетки явственно бочонкообразные, с газовыми вакуолями.......
................................................... О. lacustris
Б. Клетки без газовых вакуолей, с явственной зернистостью у поперечных
перегородок.
а. Дерновинки сине-зеленые, более или менее слизистые. Трихомы. . .
6—10 мкм шириной, прямые или изогнутые, но не спиралевидные
.....................................................О. tenuis
III. Длина клеток равна ширине, превосходит ее или меньше не более чем в
3 раза, но трихомы явственно суживающиеся к обоим концам.
56 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
1. Трихомы неспиралевидно извитые, не желто-зеленые, без газовых ваку-
олей.
А. Конечные клетки головчатые.
а. Длина клеток в 2-4 раза больше ширины, конечные клетки без ка-
липтры.................................................О. splendida
б. Клетки почти квадратные. Конечные клетки с калиптрой.........
.....................................................О. атоепа
Б. Конечные клетки более или менее конусовидные.
а. Трихомы свыше 4 мкм шириной В содержимом клеток зерна отсут-
ствуют.
+ Клетки квадратные или длина их до 2 раз меньше ширины — трихо-
мы 4-6 мкм шириной....................................О. formosa
Oscillatoria sancta (Kutz.) Gom. (рис. 11: 14, 15)
Дерновинки черновато-стального оттенка, блестящие, тонкие, слизисто-
студенистые. Трихомы 10-20 мкм шириной, у поперечных перегородок пере-
шнурованные, на концах слегка суженные, прямые, с грануляциями у попереч-
ных перегородок. Клетки 2.5-6 мкм длиной. Конечные клетки плосковато-по-
лушаровидные, слегка головчатые, с утолщенной оболочкой.
В стоячих пресных и солоноватых водах, на подводных предметах.
Широко распространена в России и странах СНГ.
Замечание. В оз. Байкал в небольшом количестве найдена К.И. Мейером
[1930] в прол. Мал. Море у мыса Кобылья Голова в августе.
Oscillatoria limosa Ag. (рис. И: 18-21)
Дерновинки черновато-сине-зеленые, до коричневых. Трихомы более или
менее прямые, (11) 13-16 (22) мкм шириной, у поперечных перегородок не пе-
решнурованные, с грануляциями. Длина клеток 2-5 мкм. Трихомы прямые или
слабо утончающиеся к концу. Конечные клетки плоско закругленные, с утол-
щенной оболочкой.
В стоячих и медленно текущих водах, в илу озер.
Широко распространена по всей территории России и стран СНГ.
Замечание. В Байкале в небольшом количестве найдена К.И. Мейером
[1930] в Нижнеангарском соре, около среднего устья.
Oscillatoria lacustris (Kleb.) Geitl. (Trichodesmium lacustre Kleb.)
(рис. 11: 6-13)
Трихомы соединены в свободно плавающие чешуйкообразные связки или
пучки из параллельно расположенных нитей, неравномерно выступающих на
обоих концах. Трихомы прямые, 5-7(8) мкм шириной, клетки бочонкообраз-
ные, 4-7 мкм длиной, с газовыми вакуолями. Конечные клетки закругленные,
иногда удлиненные и слегка утонченные на конце до 12 мкм длиной.
В планктоне пресных и соленых водоемов, эпифитно на водных растениях,
реже в бентосе болот, прудов и озер.
В разных районах России, нередко.
Замечание. В оз. Байкал найдена К.И. Мейером [1930] в бух. Анга, где в зна-
чительном количестве эпифитирует на высших водных растениях Potamogeton
perfoliatus, Р. pectinatus L., Myriophyllum spicatum L., Ranunculus divaricatus Schrank.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 57
Рис. 11.1-5 — Oscillatoria атоепа; 6-13 — О. lacustris: 6 — пучок нитей (небольшоеуве-
личение), 7, 8,13 — концы нитей, 9-12 — короткие нити, 10 — верхние клетки изобра-
жены без содержимого; 14, 15 — О. sancta; 16 — О. tenuis; 17 — О. formosa; 18-21 —
О. limosa; 22-24 — О. splendida.
По: 1, 7, 8, И, 12,16-18, 23 — Косинской, 2,14, 22 — Гоману, 3 — Фреми, 4,15, 21 — Комаренко,
5, 24 — Кондратьевой, 6, 9, 10 — Клебану, 19 — Шкорбатову, 20 — Анисимовой.
Fig. 11. 1-5 — Oscillatoria атоепа; 6-13 — О. lacustris". 6 — tuft of filaments (low
magnification), 7, 8, 13 — filament tips, 9-12 — short filaments, 10 — upper cells without
showing their inside; 14-15 — 0. sancta; 16—0. tenuis; 17 — O. formosa; 18-21 — O. limosa;
22-24 — O. splendida.
After: 1, 7, 8,11,12,16-18, 23 — Kossinskaya, 2,14, 22 — Goman, 3 — Fremi, 4,15, 21 — Komarenko.
5, 24 — Kondrat’eva, 6, 9, 10 — Kleban, 19 — Shkorbatov, 20 — Anisimova.
Oscillatoria tenuis Ag. (рис. 11: 16)
Дерновинки ярко-, реже желто-сине-зеленые, плоские, слизистые. Трихо-
мы прямые, 6-10 мкм шириной, клетки 2.6-5 мкм длины. Трихомы ярко-си-
не-зеленые, не сильно перешнурованы у поперечных перегородок, большей
частью с грануляциями, конечные клетки более или менее полушаровидные,
иногда с немного утолщенной оболочкой.
58 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Oscillatoria tenuis forma tenuis
Трихомы 6-10 мкм шириной, клетки у поперечных перегородок не сильно
перешнурованные, 2.6-5 мкм длиной.
В водоемах со стоячей чистой и загрязненной водой, прикрепляется к под-
водным предметам или плавает свободно.
Один из самых распространенных в России и странах СНГ видов этого рода.
Замечания. Для Байкала приводится К. И. Мейером [1930]. Найдена им в
прибрежной зоне Кичеро-Ангарского сора. По сообщению К. И. Мейера,
В.Н. Яснитский наблюдал ее в бух. Онгоконская. Нами [Ижболдина, 1990]
встречена в августе в районе впадения р. Кичера.
Oscillatoria tenuis forma tergestina (Kutz.) Elenk.
Трихомы 4-6 мкм шириной.
В разных районах России.
Замечание. В Байкале найдена К. И. Мейером [1930] в восточной части Ки-
черо-Ангарского сора у берегов, где скапливается ил.
Oscillatoria splendida Grev.
Дерновинки ярко-сине-зеленые или оливково-зеленые. Трихомы прямые
или изогнутые, 2-3 мкм шириной, у поперечных перегородок не перешнуро-
ванные, обычно с хорошо выраженной грануляцией. К концам трихомы посте-
пенно суживающиеся, согнутые, спиралевидно извилистые или крючкообраз-
но изогнутые. Конечные клетки головчатые, почти шаровидные, без калиптры.
Oscillatoria splendida forma splendida (рис. 11: 22—24)
Трихомы бледно-сине-зеленые, прямые или изогнутые, 2-3 мкм шириной.
Длина клеток 3-9 мкм.
В стоячих водах, в илу, на водных растениях, а также в загрязненной воде.
В России и странах СНГ распространена широко.
Замечания. В Байкале найдена К.И. Мейером [1930] в районе пос. Култук, в
июле. Кроме того, приводится В.Н. Яснитским (для Онгоконской бухты). При
наших исследованиях [Ижболдина, 1982а, 1985] вид встречен в районе мысов
Утулик, Немчинова, Кочериковский, Бол. Солонцовый, Улан-Нур, в губах Да-
гарская и Фролиха. Обильное развитие О. splendida наблюдали в июле 1976 и
1982 гг. в районе сброса промышленных очищенных сточных вод БЦБК. Яр-
ко-сине-зеленые дерновинки водорослей покрывали грунт на глубинах от 2 до
3-4 м, а фитомасса их в отдельных участках достигала 0.7-3.2 г/м2.
Oscillatoria атоепа (Kutz.) Gom. (рис. 11: 1—5)
Дерновинки сине-зеленые. Трихомы прямые или слегка согнутые, 2.5-
5 мкм шириной, у поперечных перегородок едва заметно перешнурованные
или неперешнурованные, обычно с грануляциями, к концам суживающиеся.
Конечные клетки головчатые, ширококонусовидные, с калиптрой. Длина кле-
ток 2.5-4.2 мкм, равна или чуть меньше ширины.
В стоячих горячих и холодных водах, а также в илу, на подводных растени-
ях и в болоте, реже — в горячих источниках.
В России и на территории СНГ, с севера до юга, довольно часто.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 59
Замечания. В Байкале найдена К.И. Мейером [1930] в устье Селенги, около
ст. Танхой и мыса Рытый. В районе мыса Заворотный обнаружена на Drapar-
naldioides pumila С. Meyer et Skabitsch. Нами [Ижболдина, 1990в] встречена во
многих районах озера (в бухтах прол. Мал. Море, на восточном берегу о. Оль-
хон, против падей Варначка, Бол. Коты, Орео, Слюдянская, у мысов Немчино-
ва и Ухан). Растет обычно на твердом грунте в зарослях других водорослей на
глубине от 0.5 до 10 м.
Oscillatoria formosa Вогу (рис. 11: 17)
Дерновинки темно-сине-зеленые. Трихомы ярко-сине-зеленые, прямые,
4-6 мкм шириной, у поперечных перегородок слегка перешнурованные, часто
с легкой зернистостью, к концам суженные и загнутые. Длина клеток 2.5-
5.7 мкм. Конечные клетки тупо- или, реже, более или менее закругленно-кону-
совидные, без калиптры.
В стоячих водах, на твердом грунте, в илу озер, а также в горячих и холод-
ных, минеральных и в том числе серных источников.
Широко распространен по всей России и странам СНГ, за исключением
Крайнего Севера.
Замечания. В Байкале найдена К.И. Мейером [1930] в августе около
ст. Мысовая. Кроме того, встречена К.И. Мейером в сборах В.Н. Сукачева из
района о. Бол. Ушканий и ст. Култук.
Oscillatoria sp. 1
Трихомы сине-зеленые, соединены в спирально завернутые пучки. Шири-
на трихомов 5-5.3 мкм. Трихомы не перешнурованы у клеточных перегородок,
с грануляциями, к концам постепенно суженные. Длина клеток равна ширине,
несколько меньше или больше ее. Толщина трихомов на зауженных концах
3-3.5 мкм, конечные клетки закругленно-конусовидные, иногда с калиптрой.
Вид близок к О. brevis (Kutz.) Gom. и О. deflexoides Elenk. et Kossinsk. От пер-
вой отличается способностью трихомов соединяться в спирально закрученные
пучки, большей длиной клеток и присутствием калиптры; от последней — разно-
образной длиной клеток, закругленно-конусовидными конечными клетками.
В Байкале вид встречен в районе падей Бол. Коты, Орео, у мыса Немчино-
ва, вблизи устья р. Бол. Осиновка. Растет на глубине от 0.2 до 10 м, наиболее
часто встречается на глубине от 0.2 до 1.5 м. В июне, июле встречается в зарос-
лях Ulothrix zonata Kg.
Кроме вышеописанных видов в наших сборах обнаружен ряд водорослей
этого рода, определение видовой принадлежности которых в настоящий мо-
мент не представляется возможным. Условно эти виды объединены под назва-
нием Oscillatoria sp. 2. Встречены они (V = 2-10 %) в основном вдоль открытых
прибрежий Байкала во II и III растительном поясе, на твердом грунте, чаще
(V = 14-78 %) — вдоль западного побережья Сред. Байкала. Высока вероят-
ность того, что при дальнейшем исследовании список видов этого рода будет
расширен.
Род Phormidium Kutz., 1843 ex Gomont, 1892
Трихомы в мягких, слизистых, часто расплывающихся влагалищах, облада-
ющих способностью склеиваться в слизистые или пленчато-кожистые дерно-
винки, прикрепляющиеся нижней стороной к субстрату. Наблюдается движе-
ние нитей. В Байкале найдено 8 видов.
60 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
I. Трихомы у поперечных перегородок не перешнурованы или лишь слабо пе-
решнурованы, концы их часто крючкообразно согнутые или головчатые.
Нити более 50 мкм длиной
1. Трихомы не свыше 3 мкм шириной
А. Дерновинки более или менее красного или фиолетового оттенка.
а. Трихомы у поперечных перегородок не перешнурованные, большей
частью с 2 зернышками.............................Ph. purpurascens
Б. Дерновинки других оттенков, чаще более или менее сине-зеленые.
а. Трихомы у поперечных перегородок слабо перешнурованы, 1-2 мкм
шириной. Конечные клетки без калиптры...................Ph. tenue
2. Трихомы свыше 3 мкм шириной Конечные клетки без яйцевидного или
шаровидного колпачка, закругленные.
А. Трихомы к концам не суженные, 4-7.5(9.5) мкм шириной В воде.
а. Трихомы у поперечных перегородок слегка перешнурованные, иногда
с грануляциями. Влагалища от хлор-цинк-иода чаще синеют........
.................................................Ph. ambiguum
Б. Трихомы к концам более или менее суженные, слабо спиралевидно
изогнутые.
а. Конечные клетки часто головчатые.
+ Длина клеток обычно в 2-3 раза менее ширины, реже они почти
квадратные. Влагалища крепкие или ослизняющиеся..............
...............................................Ph. uncinatum
б. Конечные клетки никогда не головчатые.
а. Трихомы с грануляциями у поперечных перегородок.
+ Трихомы 3-5 мкм шириной, у поперечных перегородок не пере-
шнурованные. Клетки почти квадратные или несколько удли-
ненные ........................................Ph. inundatum
р. Трихомы без грануляций у поперечных перегородок.
+ Длина клеток обычно больше ширины ..............Ph. сопит
++ Длина клеток обычно меньше ширины.
— Конечные клетки прямые, тупо-конусовидные. Длина кле-
ток не более чем в 2 раза меньше ширины.
А Ширина трихомов 3-5 мкм.................Ph. papiraceum
ДА Ширина трихомов 5.5-10.5 мкм.............Ph. crouanii
Phormidium purpurascens (Kutz.) Gom. (рис. 12: 2)
Дерновинки пурпуровые, до коричнево-фиолетовых, кожистые. Трихомы
грязно-фиолетовые, искривленные, 1.5-2.5 мкм шириной, у поперечных пере-
городок не перешнурованные, обычно с 2 зернышками, к концам не суженные.
Влагалища расплывающиеся, от хлор-цинк-иода не синеют. Клетки 2-4.5 мкм
длиной, конечные клетки закругленные, без калиптры.
Phormidium purpurascens forma purpurascens (Kutz.) Gom. (рис. 12: 2)
Трихомы сильно искривленные, 1.5-2.5 мкм шириной.
В стоячих водах (на скалах и в горячих источниках).
Вост. Сибирь (оз. Байкал), Камчатка, горы Сред. Азии.
Замечание. В Байкале найдена К.И. Мейером [1930] около пади Барун-Бу-
дэ, на восточном берегу о. Ольхон, в июле.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 61
Рис. 12.1 — Phormidium papiraceum; 2 — Ph. purpurascens; 3-7 — Ph. ambiguum:
3 — трихом без влагалища, 4, 7 — трихом с тонким влагалищем, 5 — трихом
с толстым влагалищем, 6 — трихом с толстым и частично остуденевшим
влагалищем; 8 — Ph. crouanii; 9 — Ph. inundatum; 10 — Ph. uncinatum; 11 — Ph.
corium; 12, 13 — Ph. tenue.
По: 1, 2, 8, 9, И — Гоману, 3-6 — Фреми, 7 — Комаренко, 10 — Полянскому, 12,13 —
Косинской.
Fig. 12. 1 — Phormidium papiraceum; 2 — Ph. purpurascens; 3-7 — Ph. ambiguum:
3 — trichome without sheath, 4, 7 — trichome with a thin sheath, 5 — trichome
with a thick sheath, 6 — trichome with a thick and partially jellified sheath; 8 — Ph.
crouanii; 9 — Ph. inundatum; 10 — Ph. uncinatum; 11 — Ph. corium; 12, 13 — Ph.
tenue.
After: 1, 2, 8, 9, 11 — Goman, 3-6 — Fremi, 7 — Komarenko, 10 — Poljansky, 12, 13 —
Kossinskaya.
62 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Phormidium tenue (Menegh.) Gom. (рис. 12)
Дерновинки ярко-сине-зеленые, кожистые, распростертые. Трихомы ярко
(реже бледно)-сине-зеленые, прямые или слабо искривленные, 1-2 мкм ши-
риной, у поперечных перегородок слабо перешнурованные, к концам сужива-
ющиеся. Длина клеток (1.5) 2.5-5 мкм. Грануляции у поперечных перегородок
отсутствуют. Конечные клетки удлиненно-конусовидные, без калиптры. Вла-
галища тонкие, расплывающиеся, от хлор-цинк-иода не синеющие.
В стоящих водах, в горячих и минеральных источниках.
Широко распространен по территории России и стран СНГ.
Замечания. Вид встречен нами [Ижболдина, Галкин, 2000] в сборах
В.В. Тахтеева в горячих источниках Прибайкалья. В Чивыркуйском заливе в
бух. Змеиная, где растет при температуре воды 46 °C, в бух. Хакусы — при
48 °C, в источнике на мысе Горячинский — при 55 °C. Кроме того, нити Ph. te-
nue встречены в основании талломов Draparnaldioides goroschankinii С. Meyer et
Skabitsch., собранных у западного побережья Юж. Байкала.
Phormidium ambiguum Gom. (рис. 12: 3-7)
Дерновинки ярко-сине-зеленые, иногда с черноватым или желтоватым
оттенком, обычно распростертые. Трихомы искривленные, сине-зеленые,
4-6 мкм шириной, у поперечных перегородок слегка перешнурованные, к
концам не суженные. Содержимое их иногда с газовыми вакуолями или с гра-
нуляциями у поперечных перегородок. Влагалища тонкие, крепкие или ос-
лизняющиеся, иногда толстые и более-менее слоистые, от хлор-цинк-иода
обычно синеющие.
Phormidium ambiguum forma ambiguum
Трихомы 4-6 мкм шириной, клетки 1.5-1.9 мкм длиной. Содержимое кле-
ток с газовыми вакуолями. Конечные клетки закругленные, без калиптры.
В стоячих пресных и солоноватых и в текучих водах, в горячих источниках.
В разных районах России и стран СНГ, не редко.
Замечания. В Байкале в небольшом количестве встречен К.И. Мейером
[1930] в зал. Мухор. Нами встречен в губе Анга, где был обильно развит (фито-
масса до 13 г/м2) среди талломов Draparnaldioides pumila С. Meyer et Skabitsch.,
на камнях, на глубине 1.5 м, в сентябре.
Phormidium uncinatum (Ag.) Gom. (рис. 12: 10)
Дерновинки черно-зеленые, до коричнево-черных, или ярко-сине-зеленые,
распростертые или плавающие и кустисто-раздельные. Трихомы сине-зеленые
или грязновато-зеленые, прямые или слабо согнутые, (4) 5.5-9 (9.5) мкм
шириной, у поперечных перегородок не перешнурованные, часто с грануляция-
ми. Концы трихомов сужены, крючковидно загнуты или слабо спиралевидно
изогнуты. Длина клеток обычно в 2-3 раза меньше ширины. Конечные клетки
головчатые, обычно с калиптрой.
В текучих и стоячих водах, реже на влажной почве и в горячих источниках.
В разных районах России и стран СНГ, часто.
Замечания. В Байкале найден К.И. Мейером [1930] в районе ст. Мысовая.
Нами [Ижболдина, 1990в] встречен в районе губы Таланка, на глубине 3.5 м, на
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 63
камнях, в губе Улан-Нур, где растет вместе с Lyngbya major Menegh. на заилен-
ном песке на глубине 23 м, около мысов Кочериковский (глубина 4 м, камни),
Бол. Солонцовый (глубина 20 м, галька) и Бол. Коса, где растет на глубине 2 м
среди зарослей Tolypothrix distorta. В последнем районе наблюдали на талломах
Ph. uncinatum массовое формирование гормогониев.
Phormidium inundatum Kutz. (рис. 12: 9)
Дерновинки сине-зеленые, с черноватым оттенком, кожистые. Трихомы
сине-зеленые, 3-5 мкм шириной, у поперечных перегородок не перешнуро-
ванные, с грануляциями, к концам суженные, не согнутые. Влагалища тонкие,
ослизняющиеся, от хлор-цинк-иода синеют. Конечные клетки тупо-конусо-
видные, без калиптры.
Phormidium inundatum forma inundatum
Дерновинки сине-зеленые, с черноватым оттенком. Трихомы 3-5 мкм
шириной, клетки 4-8 мкм длиной. Влагалища от хлор-цинк-иода синеют.
В текучих и стоячих водах и на влажных скалах.
В разных районах России, редко.
Замечания. В Байкале найден К.И. Мейером [1930] у мыса Ижимей. Нами
вид встречен в районе мысов Дыроватый и Бол. Солонцовый, в губе Бол. Коса
и у пади Ниж. Хомуты, на глубине от 2 до 18 м. Во всех районах на талломах
Ph. unindatum отмечено формирование гормогониев [Ижболдина, 1990в].
Phormidium corium (Ag.) Gom. (рис. 12: И)
Дерновинки черно- (до коричнево-зеленых) или сине-зеленые, кожистые,
распростерты. Трихомы 3-4.5 мкм шириной, у поперечных перегородок не пе-
решнурованы, к концу прямые, коротко сужены. Длина клеток 3.4-8 мкм. Гра-
нуляций у поперечных перегородок нет, конечные клетки тупо-конусовидные,
реже закругленные, без калиптры. Влагалища тонкие, от хлор-цинк-иода
обычно синеют. Преимущественно в текучих, реже в стоячих водах, на влаж-
ных стенах, соломенных крышах, у основания древесных стволов и в почвах.
В разных районах России и странах СНГ, нередко.
Замечания. Впервые отмечен в Байкале К.И. Мейером [1930]. Найден им
около мысов Рытого и Заворотного, где растет в сообществе с Cladophora meyeri
и Tolypothrix distorta. Нами [Ижболдина, 1990в] встречен в губе Богучанской, на
глубине 2.5 м, среди харовых водорослей.
Phormidium papiraceum (Ag.) Gom. (рис. 12: 1)
Дерновинки черно-зеленые, блестящие, распростертые, тонкие, кожистые.
Трихомы сине-зеленые, тесно сплетающиеся, 3-5 мкм шириной, у поперечных
перегородок перешнурованы, к концам сужены. Длина клеток 2-4 мкм, грану-
ляций у клеточных перегородок нет. Конечные клетки тупо-конусовидные, без
калиптры. Влагалища тонкие, редко ослизняющиеся, от хлор-цинк-иода обыч-
но синеют.
В текучих и стоячих водах, в соленых водоемах, в горячих источниках и на
влажной земле. В России и странах СНГ, редко.
Замечание. В Байкале найден К.И. Мейером [1930] около пади Жилище, в
июле.
64 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Phormidium crouanii Gom. (рис. 12: 8)
Дерновинки сине-зеленые, с черноватым оттенком, тонкие, кожистые.
Трихомы 7.5-10.5 мкм шириной, слабо искривленные, располагающиеся поч-
ти параллельно друг к другу. Длина клеток 4-8 мкм. Трихомы у поперечных пе-
регородок не перешнурованы, грануляции отсутствуют. Конечные клетки ту-
по-конусовидные, без калиптры. Влагалища тонкие, расплывающиеся, от
хлор-цинк-иода не синеющие.
В стоячих и текучих водах.
В разных районах России и странах СНГ, редко.
Замечания. В Байкале был найден К.И. Мейером [1930] в районе мыса
Ирексокон, в июле. Нами [Ижболдина, 1990в] встречен в районе губы Слюдян-
ской на глубине 3.5 м, среди зарослей Draparnaldioides pumila и Tolypothrix dis-
torter и у мыса Кочериковский, где на глубине 4 м вместе с Ph. crouanii встрече-
ны нити Ph. uncinatum.
Род Lyngbya Ag.
Трихомы с крепкими не склеивающимися влагалищами. Нити одиночные
или образуют дерновинки разной формы, обычно симметричные, редко обна-
руживают асимметрию. Влагалища бесцветные или желто-коричневые, тонкие
или с возрастом толстые, иногда слоистые. У лингбий с толстыми влагалищами
отмечали ложное ветвление и присутствие 2 трихомов в одном влагалище.
В оз. Байкал найдено 2 вида.
I. Нити обычно образуют дерновинки.
1. Дерновинки кустистые.
А. Длина клеток в 3-8 раз меньше ширины.
а. Трихомы 11-17 мкм шириной, у поперечных перегородок не пере-
шнурованные (редко слабо перешнурованные), к концам утончен-
ные. Конечные клетки слегка головчатые, со слабо утолщенной обо-
лочкой ................................................L. major
2. Дерновинки другой формы или нити одиночные.
А. Длина клеток меньше ширины.
а. Трихомы сине-зеленые или оливковые, (6.6)10-16(24) мкм шири-
ной. Длина клеток в 3-6 раз меньше ширины. Конечные клетки за-
кругленные, плосковатые или плосковато-конусовидные...........
..................................................L. aestuarii
Lyngbya major Menegh. (рис. 13: 5)
Нити длинные, прямые, соединены в черно-зеленые кустистые дерновин-
ки. Трихомы темно-сине-зеленые, 11-17 мкм шириной, у поперечных перего-
родок не перешнурованные или слабо перешнурованные, с грануляциями, к
концам слабо утолщенные, слегка головчатые. Влагалища бесцветные, широ-
кие, слоистые, от хлор-цинк-иода не синеющие.
Lyngbya major forma major
Трихомы 11-17 мкм шириной, образуют кустистые дерновинки. Длина
клеток 2-5 мкм.
В стоячих водах, в горячих источниках.
В разных районах России и странах СНГ, нечасто.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 65
Рис. 13. 1-4 — Lyngbya aestuarii: 1 — нить с расширяющимся кверху трихомом, 2 —
нити с фрагментами трихома, 3,4 — освобождение фрагментов нитей; 5 — L. major;
6-8 — Schizothrix sp.: 6 — вид дерновинок в натуральную величину, 7 — характер
ветвления дерновинок (х 10), 8 — строение трихомов.
По: 1 — Косинской, 2 — Комаренко, 3, 4 — Кондратьевой, 5 — Фреми.
Fig. 13. 1-4 — Lyngbya aestuarii: 1 — filament with a trichome expanding upwards, 2 —
filaments with trichome fragments, 3, 4 — release of filament fragments; 5 — L. major; 6-8 —
Schizothrix sp.: 6 — fiill-sized view of swards, 7 — branching pattern of swards (x 10), 8 —
structure of trichomes.
After: 1 — Kossinskaya, 2 — Komarenko, 3-4 — Kondrat’eva, 5 — Fremi.
66 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Замечания. В Байкале в большом количестве обнаружена К.И. Мейером в
районе пади Нугды, где вместе с Chaetomorpha baicalensis С. Meyer, Spirogyra sp.
и Oedogonium sp. устилает все дно бухты. Нами [Ижболдина, 1990в] встречена в
районе падей Улан и Улан-Нур, на глубинах от 10 до 20 м, на заиленном песке в
зарослях харовых водорослей. В прол. Мал. Море, у мыса Ото-Хушун, найдена
на глубине 1.9 м, на мелких камнях, занесенных песком. Длина встреченных
нитей достигала 2-2.5 см, ширина 20-21 мкм. Во многих нитях наблюдали гор-
могонии и большие (до 200-300 мкм) участки влагалищ без трихомов, сохра-
няющиеся, очевидно, какое-то время после выхода гормогониев.
Lyngbya aestuarii (Mert.) Liebm. (рис. 13: 1-4)
Нити одиночные или в дерновинках от коричневого до темно-сине-зеле-
ного оттенка, почти прямые. Трихомы (8)10-16(24) мкм шириной, у попереч-
ных перегородок не перешнурованные. Часто с грануляциями. Влагалища сна-
чала тонкие, бесцветные, потом широкие, темно-коричневые, слоистые, не-
ровные с поверхности, от хлор-цинк-иода не синеющие. Конечные клетки
плосковатые или плосковато-закругленные, с утолщенной оболочкой, обычно
слабо суженные или совсем не суженные.
Lyngbya aestuarii forma aestuarii
Нити одиночные или соединены в дерновинки, почти прямые или искрив-
ленные. Трихомы (8)10-16(24) мкм шириной, клетки 2.1-2.6-5.6 мкм длиной,
с грануляциями у поперечных перегородок, конечные клетки плосковатые или
плосковато-закругленные.
В стоячих пресных, особенно же в солоноватых и соленых водах и в горя-
чих источниках.
В России и странах СНГ широко распространена.
Замечания. Ранее найдена в оз. Байкал К.И. Мейером [1930] в марте подо
льдом в районе Маритуя. Нами [Ижболдина, 1990в] встречена в августе в райо-
не пади Улан-Нур и в сентябре в районе пади Бол. Солонцовая на глубинах
15-25 м. Во многих нитях отмечены гормогонии.
СЕМЕЙСТВО SCHIZOTHRICHACEAE
Нити большей частью ложно ветвящиеся, реже неветвящиеся. Влагалища
преимущественно широкие, бесцветные или окрашенные, крепкие или мягкие,
иногда ослизняющиеся и расплывающиеся, содержащие по нескольку или по-
многу трихомов, реже по 1-2 трихома.
В оз. Байкал найдены представители 2 родов.
I. Влагалища крепкие.
1. Влагалища содержат обычно много или несколько трихомов, реже 1-2.
Нити более или менее сильно ветвящиеся ...............Schizothrix
II. Влагалища мягкие, иногда расплывающиеся.
1. Влагалища содержат очень много тесно сближенных трихомов.........
......................................................Microcoleus
Род Schizothrix (Kutz., 1843) Gomont., 1892
Нити тесно переплетающиеся, образуют кожистые дерновинки или соеди-
нены в подушки, кустики, прямостоящие пучки с более или менее обильным
ветвлением. Влагалища обычно крепкие, бесцветные или окрашенные в жел-
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 67
тый, коричневый, красный, реже фиолетовый или синеватый цвет, узкие или
широкие, в большинстве случаев на вершинах закрытые, к концам суживаю-
щиеся, содержащие помногу, реже по 1-2, трихома.
Включает виды, развивающиеся в воде (но не в планктоне) — преимуще-
ственно в ручьях и в зоне прибоя озер.
В оз. Байкал найден 1 вид.
Schizothrix sp. (рис. 13: 6—8)
Нити соединены в обильно ветвящиеся, не инкрустированные известью
сине-зеленые кустистые дерновинки, высота которых колеблется в разных
районах Байкала от 2-2.5 до 8-10 мм. Влагалища бесцветные или желтоватые,
крепкие, неслоистые, тонкие (2.7-5 мкм шириной), содержат 1-3, иногда до
6-10 переплетающихся между собой трихомов. Ширина нити с одним трихо-
мом 11-13 мкм. Трихомы 6.6-8.3 мкм шириной, у поперечных перегородок
слабо перешнурованы, без грануляций, иногда в клетках видна зернистость.
Клетки обычно квадратные, но иногда их длина может быть несколько меньше
или больше ширины (6-8.3, реже 13.5 мкм длиной). Конечные клетки закруг-
ленные или округло-конусовидные.
Вид обнаружен вдоль большей части байкальского побережья на глубинах от
1.5 до 15, реже до 40 м, на твердом грунте, реже на песке [Ижболдина, 1976а,
1990в]. Schizothrix sp. часто растет среди талломов Tolypothrix distorta, Cladophora
compacta, C. floccosa, C. floccosa var. irregularis. Вдоль западного прибрежья Сред,
и Юж. Байкала, вдоль восточного прибрежья Ср. Байкала, у Ушканьих островов
и п-ова Святой Нос встречаемость Schizothrix sp. несколько выше (20-40 %),
чем в других частях прибрежья (V = 2-10 %). На Селенгинском и Ангаро-Ки-
черском мелководьях, в Посольском соре и зал. Провал этот вид не найден.
Обычно фитомасса Schizothrix sp. во II и III растительных поясах колеблется от
0.01 до 4 г/м2. Вдоль западного прибрежья Юж. Байкала средняя фитомасса Schi-
zothrix sp. выше (2.2 г/м2), а на отдельных участках (у пади Емельяниха, у
мыса Бакланий) она достигает 24.2 и 34.9 г/м2 соответственно.
Род Microcoleus Desmaz., 1823
Нити часто склеивающиеся или одиночные, неветвящиеся или скудно вет-
вящиеся. Влагалища обычно бесцветные, мягкие, слизистые, нередко расплы-
вающиеся, содержат очень много тесно расположенных, часто канатообразно
перекручивающихся и к концам большей частью суженных трихомов, без ка-
липтры на конечных клетках.
В оз. Байкал найдено 2 вида.
I. Обитают преимущественно в воде.
1. Трихомы 2-10 мкм шириной.
А. Длина клеток равна или несколько меньше ширины.
а. Трихомы 6-10 мкм шириной, помногу во влагалище . . М. subtorulosus
Б. Длина клеток больше ширины.
а. Трихомы (2.0)3.2-3.5(4.2) мкм шириной, помногу во влагалище . . .
..................................................М. baicalensis
Microcoleus subtorulosus (Breb.) Gom. (рис. 14: 1, 2)
Дерновинки сине-стального оттенка. Влагалища сильно слизистые, от
хлор-цинк-иода синеющие, содержат помногу прямых, к концам иногда посте-
пенно суживающихся трихомов. Трихомы сине-зеленые, 6-10 мкм шир., у по-
68 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 14.1, 2 — Microcoleus subtorulosus: 1 — конец трихома; 2 — верхняя часть нити; 3-5 —
М. baicalensis: 3 — конец частично оборванной нити, 4 — оборванная нить, 5 — строение
нитей, к поверхности слизистого влагалища прилипают частички детрита, песчинки.
1, 2 —по Гоману.
Fig. 14. 1,2 — Microcoleus subtorulosus: 1 — end of trichome, 2 — upper part of filament; 3-5 —
M. baicalensis: 3 — end of a partially broken filament, 4 — broken filament, 5 — filament struc-
ture, detritus particles, sand adhere to the surface of a mucuous sheath.
1, 2 — after Goman.
перечных перегородок явственно перешнурованные. Клетки 5-10 мкм длиной
Конечные клетки закругленно-конусовидные или почти цилиндрические за-
кругленные.
В текучих и стоячих водах, в илу или на водных растениях, редко — в горя-
чих источниках.
В России — Московская обл., Вост. Сибирь (оз. Байкал), Украина (Крым).
Замечание. В оз. Байкал в небольшом количестве встречен К.И. Мейером
[1930] в губах Сеногда и Мал. Коса, в июле.
Microcoleus baicalensis Izhb., 1984 (рис. 14: 3-5)
Нити неветвящиеся, большей частью одиночные или в рыхлых дерновин-
ках желтоватого или беловатого цвета, 1-1.2 см длиной, 50—150(250) мкм
шириной. Влагалища бесцветные или желтые, мягкие, нежно-слоистые, 20-
40(75) мкм шириной, от хлор-цинк-иода не синеющие, с 8-25 и более тесно
переплетенными трихомами. Количество трихомов уменьшается к концу нити
до 1-3. Ширина нитей, содержащих 8-15 трихомов, составляет обычно 50—
75 мкм, нити с 20-30 трихомами имеют ширину 100-120, реже 150-250 мкм.
Изредка нити с 8-12 трихомами в пучке, имеющие более толстые влагалища,
также достигают 80-120 мкм шириной. В нитях с числом трихомов более 10-15
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 69
количество последних можно определить лишь в местах механического разры-
ва нити. Трихомы светло-зеленые, (2.9)3.2-3.5(4.2) мкм шириной, у попереч-
ных перегородок не перешнурованные, реже едва заметно перешнурованные,
без грануляций, к концам суживающиеся. Клетки 3-7 мкм длиной, с зерни-
стым содержимым; конечные клетки закругленно- или остро-конусовидные.
Пресноводный вид. Обнаружен в Байкале на глубинах от 8-10 до 90-100 м на
иле, заиленном песке, иле с детритом. Встречен вдоль всего побережья, в
основном в районах губ и приустьевых пространств крупных рек, где в субли-
торали есть илистые грунты. Наиболее часто встречается (Удо 50-70 %) на Се-
ленгинском и Ангаро-Кичерском мелководьях. Фитомасса М. baicalensis обыч-
но не превышает 0.01 г/м2. Лишь в районе г. Слюдянская и в приустьевых про-
странствах рек Тыя и Выдриная она несколько выше (0.85-2.15-0.25 г/м2).
ОТДЕЛ CHRYSOPHYTA
Одноклеточные и колониальные формы с хлоропластами золотисто-жел-
того или буровато-желтого цвета, с пиреноидами или без них. Хлоропласты
пластинчатые, корытовидные, реже лентовидные, дисковидные, сетчатые,
звездчатые и другой формы. Обычно в клетке находится 1, 2, реже 4 и более
хлоропласта. Клетки обычно голые, с 1, 2 жгутиками разной длины или с
псевдоцилиями. Известны неподвижные формы, не имеющие в вегетативном
состоянии жгутиков. Иногда клетки одеты панцирем или заключены в домик.
Размножение бесполое (вегетативное или с помощью амебоидов, зооспор,
апланоспор) и половое (в виде гологамии, изогамии и автогамии). Характер-
но образование цист. Ядро обычно одно, шаровидное или слегка эллипсоид-
ное, размеры его незначительны.
В Байкале обнаружены представители 2 классов.
Класс CHRYSOCAPSINEAE
Водоросли обычно колониальные, реже одиночные, неподвижные, при-
крепленные или пассивно плавающие. Колонии бесформенные или состоят из
многочисленных клеток, объединенных слизью. Клетки с пульсирующими ва-
куолями и глазком, покрыты плазмолеммой иногда еще и клеточной оболоч-
кой, с псевдоцилиями. Колонии растут в результате вегетативного деления всех
или нескольких клеток или только одной верхушечной. Размножение фрагмен-
тацией колоний или с помощью зооспор с 1 или 2 неравными жгутиками. В
оз. Байкал найдены представители 2 порядков.
ПОРЯДОК CHRYSOCAPSALES
Признаки порядка те же, что описаны для класса.
СЕМЕЙСТВО CHRYSOCAPSACEAE
Водоросли одноклеточные или колониальные. Колонии разнообразной
формы (цилиндрические, шаровидные, бесформенные, в виде кустиков), мик-
ро- и макроскопические. Колонии растут в результате деления всех клеток или
путем образования апланоспор.
. В оз. Байкал встречены представители одного рода.
70 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Род Tetrasporopsis Lemm.
Колонии макроскопические, бесформенные, в виде трубок, продырявлен-
ных мешков или пластинок, сначала прикрепленных к грунту, позднее свобод-
но плавающих. Клетки овальные, размещены поодиночке или попарно. Хлоро-
пластов 1, 2, корытовидных или в виде подковы, без пиреноида, с глазком и
2 пульсирующими вакуолями. Рост колонии благодаря делению всех клеток
колонии, зооспоры одножгутиковые.
Tetrasporopsis reticulata С. Meyer, 1927 (рис. 15: 1-6)
Молодые, прикрепленные к грунту талломы, имеют вид маленькой (2-
5 мм) трубочки, в стенке которой уже образовалось несколько отверстий. Раз-
растаясь, трубочка превращается в пузырь со стенками сетчато-продырявлен-
ными. Позднее таллом на вершине разрывается и Tetrasporopsis принимает вид
очень тонкой, скользкой и слизистой на ощупь пленки с бахромчатыми краями
до 5-15 см в длину и ширину. Таллом темно- или светло-бурый, иногда олив-
ково-бурый. В пленке плотной прозрачной слизи расположены в один слой
клетки овальной формы, лежащие поодиночке или по 2. Клетки без оболочки,
с одним желтым или желто-бурым хлоропластом подкововидной формы, в
клетках по 2 сократительные вакуоли. Размножаются клетки продольным деле-
нием. Клетки Т. reticulata могут превращаться в зооспоры, имеющие один
длинный жгутик, красный нитевидный глазок, прилегающий к хлоропласту.
Клетки 6-8 мкм шириной и около 10(20) мкм длиной, зооспоры 8 мкм
шириной и 12-13 мкм длиной.
От Т. fuscesceus отличается крупными размерами, сетчато-продырявлен-
ным талломом и прикрепленным образом жизни.
Эндемик Байкала, растет на глубине 2-10 м всюду, где на этой глубине
имеются камни.
Замечания. При наших исследованиях [Ижболдина, 1990в] вид встречен во
многих районах озера в июне-августе, преимущественно на глубине 1-3.5 м,
реже до 5-15 м. Встречаемость вида обычно не превышает 10-15 %. На мягком
грунте в качестве субстрата может использовать талломы харовых водорослей,
ветви лютика и урути. В Юж. Байкале Т. reticulata вегетирует в июне-июле, в
августе встречаются лишь обрывки талломов.
Мы наблюдали выход зооспор. Движение их беспорядочное, но спокойное,
плавное, вращательное вокруг оси. Ярко выражен положительный фототаксис.
Зооспоры в течение минуты собираются на более освещенной стороне капли.
Кроме Т. reticulata нами встречен в Байкале вид, по ряду признаков отли-
чающийся как от рассмотренного выше вида, так и от Т. fuscesceus.
Tetrasporopsis sp. (рис. 16: 13, 14)
Колонии имеют вид полых тонкостенных мешков с продырявленными
стенками. Общий размер колоний не превышает обычно 1 см, чаще меньше. В
пленке слизи в один слой расположены овальные клетки, лежащие обычно по
2 рядом. В клетке один желто-бурый чашевидный хлоропласт. Длина клеток
6.2-6.6 мкм, ширина - 4.4-4.9 мкм. Встречаются одиночные клетки, имеющие
длину 8-8.3 мкм и ширину 5.8 мкм. Tetrasporopsis sp. — довольно распростра-
ненный в Байкале эпифит. Развит в июне-октябре. Встречен нами [Ижболди-
на, 1974а,б, 1976] на талломах Cladophora glomerata, С. compacta, Chaetocladiella
pumila, Didymosphenia geminata. Встречаемость вида 5-10 %, а фитомасса редко
достигает 1 г/м2.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 71
Рис. 15. 1-6 — Tetrasporopsis reticulata: 1 — молодые колонии, 2 — колонии, разорванные
в верхней части, 3 — таллом колонии при слабом увеличении, 4 — неподвижные клетки
внутри колонии, 5, 6 — зооспоры; 7-9 — Hydrurus foetidus: 7 — внешний вид колонии,
8 — кончик ветви, 9 — зооспора.
По: 3-5 — Мейеру, 8 — Бертольду, 9 — Клебсу.
Fig. 15. 1-6 — Tetrasporopsis reticulata: 1 — juvenile colonies, 2 — colonies broken in their
upper part, 3 — colony thallome (low magnification), 4 — non-motile cells inside colony, 5,
6 — zoospores; 7-9 — Hydrurus foetidus: 7 — external view of the colony, 8 — tip of branch,
9 — zoospore.
After: 3-5 — Meyer, 8 — Bertold, 9 — Klebs.
72 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 16. 1-12 — Chrysothallus baicalensis: 1-7 — стадии развития таллома, 8-10 — внеш-
ний вид таллома, 11 — неподвижные споры, 12 — часть таллома после выхода зооспор;
13, 14 — Tetrasporopsis sp.: 13 — общий вид колонии на ветвях Cladophora, 14 — непод-
вижные клетки внутри колонии.
1-12 — по Мейеру.
Fig. 16. 1-12 — Chrysothallus baicalensis: 1-7 — stages of thallome development, 8-10 —
external view of thallome, 11 — non-motile spores, 12 — part of thallome after zoospore release;
13, 14 — Tetrasporopsis sp.: 13 — general view of the colony on Cladophora branches, 14 —
non-motile cells inside of the colony.
1-12 - after Meyer.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 73
ПОРЯДОК HYDRURALES
Колонии макроскопические, прикрепленные, имеют вид обильно разветв-
ленных слизистых кустиков дб 20-30 см длины. В клетках один крупный бурый
хлоропласт. Рост таллома верхушечный, делятся клетки, находящиеся на конце
верхушечных тяжей. Клетки, находящиеся в боковых ответвлениях таллома,
способны превращаться в зооспоры тетраэдрической формы.
СЕМЕЙСТВО HYDRURACEAE
Включает лишь один род.
Род Hydrurus Ag.
Колонии имеют вид перистых тяжей. Клетки округлые, яйцевидные, позд-
нее округло-эллипсоидные или веретено'видные, размещены наиболее густо по
периферии колонии. В каждой клетке один крупный, корытовидный хлоро-
пласт с голым пиреноидом в средней части. Ядро одно в центре клетки.
Лишь один вид.
Hydrurus foetidus (Vill.) Kirchn. (рис. 15: 7-9)
Колонии от 1 до 30 см длиной и больше, клетки 6-10 мкм в диаметре.
В быстротекучих, реже стоячих холодных водах, в высокогорном поясе, в
арктических районах.
Замечания. По данным К.И. Мейера [1930], в большом количестве растет в
истоке Ангары, где образует на камнях целые заросли, кроме того, найден им в
речке в пади Сенной. Нами в большом количестве Н. foetidus встречен в прито-
ках Байкала — речках Бол. Котинка, Крестовка, Безымянная, Слюдянка, в
июле — первой декаде сентября. В конце сентября встречаются отдельные по-
луразрушенные талломы.
Класс CHRYSOTHRICHINEAE
Многоклеточные прикрепленные или свободноплавающие талломы в виде
многорядных простых или ветвящихся нитей, однослойных, реже многослой-
ных пластинок, распростертых по субстрату, не заключенных в слизистый че-
хол. Клетки разрастаются в одном, двух и трех направлениях. Размножение од-
ножгутиковыми зооспорами.
ПОРЯДОК CHRYSOTHRICHALES
Пресноводные, реже солоновато-водные и морские формы. Наиболее вы-
сокоорганизованные представители золотистых водорослей. Признаки те же,
что описаны для класса.
СЕМЕЙСТВО THALLOCHRYSIDACEAE
Нитчатые многорядные или паренхиматозные, прикрепленные, реже сво-
бодноплавающие талломы в виде пластинок, растянутых по субстрату. Размно-
жение одножгутиковыми зооспорами.
74 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Род Chrysothallus С. Меуег, 1930
Таллом в виде многоклеточных округлых или округло-лопастных пласти-
нок желто-бурого цвета, разной величины и формы. Пластинка однослойна и
образована сжатыми с боков призматическими клетками, содержащими под-
ковообразной формы хлоропласт желто-бурого цвета, лежащий у наружной
выпуклой стенки. В клетке видны 2 маленькие сократительные вакуоли и мно-
гочисленные зерна лейкозина. Размножение при помощи зооспор.
В оз. Байкал описан один вид. Эпифит на Didymosphenia geminata.
Chrysothallus baicalensis C. Meyer, 1930 (рис. 16: 1—12)
Размер пластинок колеблется от 40-50 до 150 мкм и более. Ширина клеток
около 10 мкм, высота около 2 мкм.
Найден К.И. Мейером [1930] в оз. Байкал, в районе мыса Котельников-
ский, где эпифитирует на Didymosphenia geminata. Для Украины приводится
А.М. Матвиенко [1965]. Найден в болотах, где эпифитирует на разных водорос-
лях (Oedogonium, Cladophora и др.) и высших водных растениях.
Замечание. Нами [Ижболдина, 19766] встречен во многих районах прол.
Мал. Море, где эпифитирует на D. geminata, Chaetocladiella pumila С. Меуег,
Cladophora meyeri Skabitsch. и C. glomerata L., на глубине 1.5-5, реже 15 м.
ОТДЕЛ BACILLARIOPHYTA
Одноклеточные микроскопические водоросли, часто собранные в различ-
ного рода колонии. Наружная оболочка кремниевая, состоящая из двух поло-
винок. Стенки панциря структурные. Протопласт клетки состоит из прото-
плазмы, ядра, хлоропластов бурого цвета. Продукты ассимиляции — масло и
волютин. Основной способ размножения — вегетативное деление клеток на-
двое. Половой процесс изогамный, гетерогамный и оогамный. Вегетативные
клетки диплоидные.
В Байкале представлены колониальные водоросли класса Pennatophyceae.
Класс PENNATOPHYCEAE
Клетки одиночные или собраны в колонии. Панцири имеют обычно сим-
метричное строение, с пояска линейные, таблитчатые или клиновидные, пря-
мые, иногда S-образно изогнутые, часто со вставочными ободками и септами.
Створки овальные, ланцетные, линейные, булавовидные и др. Структурные
элементы на створках расположены обычно симметрично по отношению к
продольной оси и состоят из точек, образующих поперечные ряды. Иногда на-
блюдаются поперечные ребра, слизевые поры, многочисленные мелкие шипи-
ки. По продольной оси створки обычно проходит осевое поле, часто расширя-
ющееся в центре створки в среднее поле. У многих видов вдоль середины осе-
вого поля расположен щелевидный шов. Каналовидный шов обычно сдвинут к
краю створки, находится в киле или в крыловидном выросте створки. Размно-
жение вегетативным делением клетки надвое, известно образование ауксоспор,
сопровождающееся половым процессом. Вегетативные клетки диплоидные.
В Байкале встречены представители одного порядка, образующие крупные,
прикрепленные к грунту колонии.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 75
ПОРЯДОК RAPHALES
Панцирь продольно или поперечно симметричный. Створки от линейных
до широкоэллиптических. Шов всегда хорошо развит, может быть щелевид-
ным или каналовидным. Иногда щелевидный шов имеется только у полюсов
створки, короткий. Имеются поперечные штрихи, ребра, иногда — ареолы.
Вставочные ободки и септы встречаются редко.
ПОДПОРЯДОК DIRAPHINEAE
Шов щелевидный, имеется на обеих створках, развит хорошо, имеются
центральный и конечный узелки. В Байкале найдены представители одного се-
мейства.
СЕМЕЙСТВО NAVICULACEAE
Разнообразные по форме клетки одиночные или образуют разнообразной
формы колонии. Створки и концы створок разнообразной формы. Обе створки
имеют хорошо развитый щелевидный шов, состоящий из наружной и внутрен-
ней щелей с центральным и конечными узелками. Обычно шов находится на
середине створки, реже смещен к краю или располагается в киле, иногда его
трудно обнаружить. Структура обеих створок одинаковая, представлена штри-
хами и ребрами или ареолами, расположенными поперечными рядами. Осевое
поле линейное или ланцетное, часто наблюдается центральное поле. Хлоро-
пласты обычно в виде 1-2 крупных пластинок, реже в виде длинных лент или
мелкозернистые. В Байкале встречены представители 2 родов.
I. Панцирь по одной оси гетеропольный.
1. Панцирь асимметричный по поперечной плоскости, с пояска клиновид-
ный.
А. Поперечные штрихи, ребра грубопунктирные......Didymosphenia
Б. Поперечные штрихи нежно пунктирные...............Gomphonema
Род Didymosphenia М. Schmidt
Клетки на длинных разветвленных слизистых ножках, образуют кустико-
видные колонии коричневого или беловатого цвета до 1-2 см высоты. Пан-
цирь с пояска клиновидный. Вставочные ободки и септы отсутствуют. Створ-
ки клиновидные, резко асимметричные по отношению к поперечной плоско-
сти и едва заметно асимметричные по отношению к продольной плоскости.
Края створок посередине выпуклые, концы головчато отшнурованные, го-
ловной конец шире базального. Поперечные штрихи грубые, из крупных то-
чек, образующих продольные ряды. Осевое поле линейное, среднее поле уд-
линенно-овальное, асимметричное, с одной или несколькими точками на
брюшной стороне. Два пластинчатых хлоропласта прилежат к поясковым сто-
ронам.
В Байкале известно 2 вида.
I. Края створок гладкие.......................................D. geminata
II. Края створок с зубцами .....................................D. dentata
76 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Didymosphenia geminata (Lyngb.) М. Schmidt. (Gomphonema geminata Lyngb.)
(рис. 17: 4, 5)
Створки клиновидные, едва асимметричные по продольной оси, со слабо
выраженными спинной и брюшной сторонами, посередине сильно расширен-
ные, перед концами суженные, длиной 48-135 мкм, шириной 24-43 мкм. Кон-
цы широко закругленные, головчатые. Штрихи радиальные, грубопунктирные,
10 в 10 мкм, срединные неодинаковой длины. Шов расположен по продольной
оси, слегка изогнутый, довольно широкий, полярные узелки отодвинуты от
концов створки, полярные щели длинные, изогнуты к спинной стороне. Осе-
вое поле линейное, среднее поле немного расширено. Брюшная сторона сред-
него поля с 2-4 изолированными точками.
Вид сильно изменчив по размерам панциря и форме створок.
Североальпийский вид, часто в быстротекучих водотоках. Повсеместно.
Иногда образует крупные макроскопические колонии.
Замечания. К.И. Мейер [1930] приводит D. geminata как характерный для
коренного Байкала вид. Встречена им от Култука до Нижнеангарска, везде,
где в береговой зоне имеются камни. Вместе с Tetraspora cylindrica в летний
период образует на глубине 1.5-2.5 м II растительный пояс. Вид охотно селит-
ся на высших водных растениях. Нами [Ижболдина, 1990в; Кожова, Ижбол-
дина, Бокова, 1998] D. geminata встречена на глубинах от 0.3 до 5, реже 7-11 м.
Наиболее часто она растет на глубине 2-2.5 м. Развитие D. geminata с мая до
октября, наиболее обильно — в августе-сентябре. Вид входит в состав разных
фитоценозов, обнаруженных на каменистой литорали в указанной зоне глу-
Рис. 17. 1 — Gomphonema quadripunctatum; 2 — G. firmum; 3 — Didymosphenia dentata;
4, 5 — D. geminata, 5 — колонии на ветвях Ranunculus sp.
По: 1, 2 - Скабичевский, 3 - Дорогостайский, 4 - Густедт.
Fig. 17. 1 — Gomphonema quadripunctatum; 2 — G. firmum; 3 — Didymosphenia dentata;
4, 5 — D. geminata, 5 — colonies on Ranunculus sp. branches.
After: 1, 2 - Skabitschewski; 3 - Dorogostayski; 4 - Gustedt.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал Т1
бин. В силу малой удельной массы D. geminata редко входит в состав субдоми-
нантных видов и лишь на участке западного побережья Сев. Байкала от
р. Томпа до губы Онокочанской, где сильно влияние рек Верх. Ангары, Киче-
ры и Тыи, и Tetraspora cylindrica var. bullosa развита слабо, Didymosphenia до-
минирует в фитоценозах II пояса.
Didymosphenia dentata (Dorogostaisky) Skv. (Gomphonema dentata Dorogostaisky;
Didymosphenia dentata f. elongata Skv.; D. dentata f. subcapitata Skv.; D. dentata
var. subcapitata f. curta Skv.) (рис. 17: 3)
Створки клиновидные, слегка асимметричные по продольной оси, со сла-
бо выраженными спинной и брюшной сторонами; край створки усажен корот-
кими толстыми заостренными зубцами, 3.5-5 в 10 мкм. Длина створки 50—
78.5 мкм (чаще 70-80 мкм), ширина 28-54 мкм. Концы широко закругленные,
головной конец обычно головчатый, иногда широко клювовидно оттянут.
Штрихи радиальные, грубопунктирные, 6.5-10 в 10 мкм. На середине длинные
штрихи чередуются с короткими.
Вид, близкий к D. geminata, отличается присутствием зубцов на краю
створки. Эндемик Байкала.
Замечания. По данным К.И. Мейера [1930], широко распространена по
всему озеру, растет вместе с D. geminata, но в северной половине Байкала она
встречается чаще. Как характерный для открытых прибрежий Байкала вид упо-
минает А.П. Скабичевский [1966].
Род Gomphonema Ag.
Клетки одиночные, прикрепляются к субстрату студенистыми ножками,
реже образуют ветвистые колонии. Панцирь асимметричный по отношению к
поперечной плоскости, с пояска клиновидный, со створки булавовидный, лан-
цетный, реже почти линейный. Концы закругленные, головчатые, иногда за-
остренные; верхний и базальный обычно неодинаковые. Поперечные штрихи
нежные, пунктирные, линеолированные, очень редко грубые, в виде ребер.
Осевое поле обычно узкое, среднее поле небольшое, округлое, большей частью
с одной или несколькими изолированными точками. Вставочные ободки и
септы имеются только у 3 видов. Пластинчатые хлоропласты прилегают к пояс-
ковым сторонам и частично заходят под створки. В Байкале встречено 3 вида
[Скабичевский, 1984].
Gomphonema quadripunctatum (Qstr.) Wisl. (=G. quadripunctatum var. genuina
Skv. et Meyer; G. quadripunctatum var. genuina f. tumida Skv. et Meyer;
G. olivaceum var. quadripunctatum (Qstr.) (рис. 17: 1)
Створки ланцетные, длиной 31-100 мкм, шириной 7.5-20 мкм. Концы за-
кругленные, головной конец более широкий и плоско закругленный, с септой.
Штрихи линеолированные, на середине створки радиальные, 12-16 в 10 мкм, к
концам створки параллельные. Осевое поле линейное, среднее поле большое,
округлое, с 4-10 изолированными точками.
Замечания. Особо обильное развитие G. quadripunctatum отмечает К. И. Мей-
ер в г. Тань, где данный вид растет в значительно большем количестве, чем D. ge-
minata, развивающаяся на этих же глубинах.
78 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Gomphonema flrmum Skv. (рис. 17: 2)
Образует ветвистые колонии, характеризующиеся тонкими ветвями. Длина
створок 70—133 мкм, ширина 19-24 мкм, штрихов 11-12 в 10 мкм. Растет в
литорали Байкала среди других водорослей. К.И. Мейером [1930] обнаружена у
западного и восточного берега. А.П. Скабичевским [1934,1965] встречена в губах
Аяя и Якшакан и в других местах восточного прибрежья. Эндемик Байкала.
Gomphonema ventricosum Greg. (=G. ventricosum forma curtum Skv.)
Размеры клеток широко варьируют. По замечанию А.П. Скабичесвского
[1983], вид нуждается в дальнейших исследованиях. Длина створки 20-56 мкм,
ширина 8,5-13 мкм. Растет в литорали Байкала на ножках и даже на клетках
Didymosphenia. Вид встречен К.И. Мейером [1930] по всему Байкалу от Култука
до Нижнеангарска.
ОТДЕЛ RHODOPHYTA
Окраска в типе красная (разных оттенков), может быть голубоватой, олив-
ково-зеленой, желтоватой, сине-зеленой. У пресноводных форм красный цвет
наблюдается редко, преобладают указанные тона. Водоросли колониальные (в
виде соединенных слизью клеток) или многоклеточные (в виде нитей, кусти-
ков, шнуров и т.п.). Клетки одноядерные или многоядерные, с одним или мно-
гими хлоропластами различной формы. Вегетативное размножение у однокле-
точных форм простым делением на две, у многоклеточных — частями слоеви-
ща. Половой процесс — оогамия. У большинства видов с половым размножени-
ем имеется изоморфная и гетероморфная смена половой и бесполой форм раз-
вития. В жизненном цикле Rhodophyta отсутствуют подвижные стадии.
В оз. Байкал встречены представители одного класса.
Класс FLORIDEOPHYCEAE
Талломы многоклеточные, однорядно-нитчатые или более сложного стро-
ения. В основе строения всех форм лежит клеточная разветвленная нить. Рост
апикальный, клетки одно- или многоядерные. Хлоропласты различной формы,
пристенные, единичные или многочисленные. Пиреноиды у низкоорганизо-
ванных форм. Вегетативное размножение редкое, бесполое — тетраспорами,
биспорами, полисифами, реже моноспорами. Половое размножение оогамное.
Имеется чередование гаметофита и спорофита.
ПОРЯДОК NEMALIALES
Таллом нитчатый, одно- или многорядно-нитчатый, простой или разветв-
ленный. Нити гонимобласта развиваются от оплодотворенного карпогона.
Ауксиллярные клетки отсутствуют. Бесполое размножение преимущественно
моноспорами.
В оз. Байкал встречены представители одного семейства.
СЕМЕЙСТВО BATRACHOSPERMACEAE
Талломы обильно разветвленные. Кора из ризоидоподобных клеточных
нитей плотно прилегает к клеткам центральной оси. Мутовки веточек ограни-
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 79
ченного роста, рыхлые, заключенные в слизь или срастающиеся. Карпоспоры
конечные. Водоросли исключительно пресноводные.
В Байкале найдены представители одного рода.
Род Batrachospermum Roth, 1800
Талломы до 40 см длины, слизистые, рыхлые, цилиндрической формы, с
мутовчато расположенными боковыми веточками предельного роста, часто
придающими таллому четковидный характер. Центральная ось из одного ряда
цилиндрических и бочонкообразных клеток, попеременно разветвленная. Му-
товки боковых веточек образуют многократно и в основном дихотомически
разветвленные нити, верхушечные клетки которых часто заканчиваются воло-
ском. Вокруг центральной оси коровой слой из ризоидоподобных нитей. Поло-
вые растения одно-, дву- и многодомные. Сперматангии на верхушечных клет-
ках мутовочных веточек. Карпогенные ветви разной длины, из 3-13 клеток,
трихогина крупная. Гонимобласты четко отграниченные, клубочковидные, со-
стоящие из коротких нитей с конечными карпоспорангиями.
В Байкале растут 2 вида Batrachospermum.
I. Мутовки хорошо развитые, более 200 мкм в диаметре, из многократно раз-
ветвленных нитей. Гонимобласты среди мутовочных веточек, по 1, 2 или по
нескольку в мутовке. Трихогина различной формы.
1. Гонимобласты по 1-2 в центре каждой мутовки, обычно крупные.
А. Трихогина лишена отчетливой ножки, обратно-конусовидная, до почти
цилиндрической. Гонимобласты от 140 мкм в диаметре и более.
а. Моноспорангии имеются. Гонимобласты обычно недоразвитые. Клет-
ки центральной оси 120-360 мкм длиной .................В. vagum
2. Гонимобластов помногу в каждой мутовке, разбросанных, обычно мелких.
А. Трихогина булавовидная или урнообразная. Боковые веточки карпо-
гонной ветви длинные, покрывающие карпогон и гонимобласт.
а. Моноспорангии отсутствуют. Гонимобласты нормально развиты.
б. Ветви почти одинаковой ширины. Мутовочные веточки прямые, со-
стоят из клеток почти одинаковой формы. Волоски разной длины
...............................................В. moniliforme
Batrachospermum vagum (Roth) Ag. (-B. vagum var. Keratophyllum (Bory) Sirod.)
(рис. 18: 4, 5)
Талломы 2-25 см длиной, сильно слизистые, ярко-зеленые, с голубоватым
оттенком, обильно разветвленные, однодомные. Мутовки эллипсоидные и ди-
сковидные, 300-700(1000) мкм в диаметре, мутовочные веточки из 6-12 кле-
ток, утончающиеся к верхушке, нижние клетки 20-60 мкм длиной, 7-15 мкм
шириной. Волоски редкие или многочисленные. Клетки центральной оси
120-360 мкм длиной. Моноспорангии 9-14 мкм длиной, 7.5-10 мкм шириной,
по 1-2 на концах веточек. Карпогон 30-60 мкм длиной, 5-8 мкм шириной
внизу. Карпоспорангии до 16 мкм длиной, до 13 мкм шириной Гонимобласты
часто недоразвитые.
В проточных и стоячих водоемах со свежей водой.
По всей территории России и странах СНГ.
Замечание. Вид встречен К.И. Мейером [1930] в Байкале в зал. Провал у
дер. Дубинине.
80 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 18. 1-3 — Batrachospermum moniliforme: 1 — часть мутовочной веточки с карпогоном
и сперматангиями, 2 — мутовочная веточка с гонимобластом, 3 — общий вид таллома;
4, 5 — В. vagum: 4 — мутовочная и 2 вторичные боковые веточки, 5 — карпогон с
трихогиной и 2 спермациями.
По: 1, 2 — Кюлину, 4, 5 — Стармаху.
Fig. 18. 1-3 — Batrachospermum moniliforme: 1 — fragment of a whorled shoot with a
carpogonium and spermatangia, 2 — whorled shoot with gonimoblast, 3 — general view of
thallome; 4, 5 — B. vagum: 4 — whorled and 2 secondary lateral shoots, 5 — carpogonium with
a trichogyne and 2 spermatia
After: 1, 2 — Culin, 4, 5 — Starmach.
Batrachospermum moniliforme Roth (=B. corbula Sirod.; B. moniliforme
var. isoeticola; B. pyramidale Sirod.; B. densum Sirod.)) (рис. 18: 1—3)
Талломы 2-6 см длиной, сильно слизистые, желто- или оливково-серые или
буроватые, обильно разветвленные, с главными осями, однодомные. Мутовки
шаровидные, эллипсоидные, бочонкообразные 500-800 (1000) мкм в диаметре.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 81
Мутовочные веточки из (6) 8-16(20) клеток, последние в нижней части вето-
чек 20-60 мкм длиной, 5-10(12) мкм шириной. Волоски редкие или обильные,
разной длины. Клетки центральной оси (100)300-800 мкм длиной. Карпогон-
ные ветви 6-7-клеточные, карпогон 20-40 мкм длиной, 4-6 мкм шириной
внизу. Трихогина булавовидная или урнообразная. Гонимобласты многочис-
ленные, шаровидные, 70-120 мкм в диаметре. Карпоспорангии яйцевидные
или грушевидные, 10-13 мкм длиной, 6-10 мкм шириной.
В быстро текущих ручьях и родниках и в озерах.
Европейская и Азиатская части России, Крым, Кавказ.
Замечания. Вид встречен в оз. Байкал К.И. Мейером [1930] в зал. Провал,
около дер. Дубинине. Нами [Ижболдина, 1990в] обнаружен в притоке Байкала
р. Бол. Котинка (северо-западное побережье Юж. Байкала) и в районе ее
устья, в Посольском соре — в районе устья р. Абрамиха. В 1981 г. В. moniliforme
обнаружен у юго-восточного побережья озера в районе сброса УЧ В БЦБК, в
июле [Ижболдина, 1985]. В августе на мутовочных веточках развиты карпого-
ны, сперматангии, гонимобласты.
ОТДЕЛ CHLOROPHYTA
Водоросли имеют хлоропласт чисто-зеленого цвета, разнообразную форму
талломов (одноклеточные, колониальные, многоклеточные, неклеточные).
Клетки одеты оболочкой, в составе которой обычно преобладает целлюлоза.
Хлоропласты от одного до нескольких, разнообразной формы, часто с пирено-
идами. Клетки одно- или многоядерные. Размножение бесполое (с помощью
зооспор, апланоспор, акинет) и половое (изо-, гетеро- и оогамное). Некото-
рые водоросли имеют правильную смену половой и бесполой форм развития.
В оз. Байкал найдены представители 5 следующих классов.
Класс TETRASPORINOPHYCEAE
Водоросли одноклеточные или колониальные. Колонии разнообразных
очертаний, микроскопические и крупные, иногда слизистые, но никогда не
бывают нитевидными. Хлоропласты зеленые, обычно чашевидные. Половой
процесс (когда он имеется) - копуляция гамет.
Класс ULOTRICHOPHYCEAE
Водоросли в виде простых или разветвленных нитей, у некоторых форм с
резким различием между основным стволом и боковыми ветвями, иногда с во-
лосками, часто окруженные слизью. Хлоропласт обычно один (реже их много)
пластинчатый (постенный), иногда продырявленный, но не спирально извитой.
Класс SIPHONOCLADOPHYCEAE
Нити простые или разветвленные, без резко выраженного основного ство-
ла, у некоторых форм соединены в виде шаров, в которых располагаются ради-
ально. Хлоропласт один, сетчато-продырявленный или в виде многочисленных
пластинок. У нитчатых форм хлоропласты кольчатые, с зазубренными краями.
Половой процесс — чаще изогамия, а также гетеро- и оогамия.
82 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Класс OEDOGONIOPHYCEAE
Нити не разветвленные или разветвленные, с одноклеточными щетинка-
ми, луковицеобразно вздутыми у основания. Для нитчатых форм характерно
деление вегетативных клеток с образованием колпачков. Хлоропласт стенко-
положный, сетчатый или в виде анастомозирующих между собой лент. Поло-
вой процесс - оогамия.
Класс ZYGNEMATOPHYCEAE
Водоросли одноклеточные, колониальные или многоклеточные (нитча-
тые, очень редко ветвящиеся). Хлоропласты осевые или пристенные, по одно-
му или по нескольку в клетке. Подвижные жгутиковые стадии в процессе раз-
множения отсутствуют. Вегетативное размножение - делением клеток и фраг-
ментацией талломов у нитчатых форм. Половое размножение - конъюгация, в
процессе которой происходит слияние хлоропластов 2 клеток, одиночных или
находящихся в одной или разных нитях.
Класс TETRASPORINOPHYCEAE
Этот класс представлен в Байкале одним порядком.
ПОРЯДОК TETRASPORALES
Талломы одноклеточные или колониальные. Клетки шаровидные, оваль-
ные, веретеновидные, располагающиеся в колониях по периферии слизи или
неравномерно внутри нее. Слизистые чехлы клеток гомогенные или слоистые,
внутренняя оболочка протопластов целлюлозная, наружная — пектиновая.
Хлоропласт чашевидный, мешкообразный, звездообразный, с пиреноидами
или без них, ядро одно. Размножение изо- или анизогамное.
Порядок представлен в Байкале одним семейством и одним родом.
СЕМЕЙСТВО TETRASPORACEAE
Клетки с 2 или 4 псевдоцилиями, в слизистых колониях разнообразной ве-
личины и формы (шаровидные, цилиндрические, грушевидные или бесфор-
менные), прикрепленные или свободноплавающие. Хлоропласт чашевидный,
с пиреноидом или без него, с 2 сократительными вакуолями. Бесполое размно-
жение — зооспорами, половое — изо- или анизогамное.
Для оз. Байкал известны представители одного рода.
Род Tetraspora Link., 1809
Колонии от микроскопических до макроскопических, прикрепленные или
свободно плавающие, разнообразной формы. Клетки шарообразные, по 1, 2
или 4. Передняя часть клеток с псевдоцилиями обращена к внешней стороне
колоний. Хлоропласт чашевидный, с одним пиреноидом, ядро одно, пульсиру-
ющих вакуолей 2 или они отсутствуют. Бесполое размножение — с помощью
двухжгутиковых зооспор, половое — двухжгутиковыми изогаметами. Предста-
вители этого рода предпочитают тихие воды.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 83
В оз. Байкал обнаружены 2 вида рода Tetraspora.
I. Колонии прикрепленные, достигают макроскопических размеров.
1. Колонии в виде длинных тонких слизистых тяжей, прикрепленных к грун-
ту тонкой ножкой........................................Т. cylindrica
2. Колонии иной формы без ножки.
А. Колонии в виде слизистых, простых или неправильно-бугристых меш-
ков, иногда сетчато-продырявленных........................Т. lubrica
Tetraspora cylindrica (Wahl.) Ag.
Колонии в виде слизистых цилиндрических трубок, прикрепленных к суб-
страту тонкой ножкой. Клетки шаровидные, 4 мкм в диаметре, с псевдоцилия-
ми, не выступающими из слизи колоний. Известны акинеты.
В озерах, лужах. В России и странах СНГ повсеместно.
В оз. Байкал встречены две вариации этого вида.
Tetraspora cylindrica var. bullosa C. Meyer, 1927 (рис. 19: 1-4)
Водоросль имеет форму длинной светло-зеленой трубки, прикрепленной к
камням с помощью тонкой ножки, расширяющейся в небольшую подошву. С
возрастом трубка принимает вид мешка с волнистыми гофрированными стен-
ками. Мешок на вершине разрывается и край его становится клочковатым, что
отличает данную вариацию от типа. Длина таллома колеблется от 2.5 (молодые
талломы) до 20 см. Ширина колоний от 0.3-0.5 до 3 см. Клетки имеют харак-
терное для Tetraspora строение и диаметр 10-15 мкм.
Широко распространена по всему озеру, где на камнях, на глубине от
(1)1.5 до 2-2.5 м образует (вместе с Didymosphenia geminata) II растительный
пояс, впервые описанный для Байкала К.И. Мейером [1927].
Замечания. Половой процесс у Т. cylindrica var. bullosa изучен А.П. Скабиче-
ским [1937], отметившим, что в районе пос. Бол. Коты (северо-западное побере-
жье Юж. Байкала) массовое развитие этого вида наблюдается в конце июня —
первой половине июля. Во второй половине июля — начале августа Tetraspora
отмирает. Установлено, что Т. cylindrica var. bullosa — строго раздельнополый ор-
ганизм с физиологической дифференцировкой полов, показано отсутствие мор-
фологических различий между гаметами противоположного пола. В.Н. Яснит-
ским [1952] в этом же районе Байкала были встречены талломы, имеющие раз-
мер, значительно больший, чем указывает для вариации К.И. Мейер. Талломы
достигали 1.0-1.9 м в длину и 33-37-см в ширину. По нашим наблюдениям [Иж-
болдина, 1990а], более крупные, чем указывает К.И. Мейер, талломы встречают-
ся вдоль западного побережья Юж. и Сред. Байкала и в прол. Мал. Море. В оста-
льных районах озера размер талломов не превышает таковые, приведенные
К.И. Мейером [1930]. Водоросль найдена нами на глубинах от 1 до 10 м, а в райо-
не пади Жилище она обнаружена на уступах скалы, занесенной песком и илом
на глубине 30-35 м [Ижболдина, 1975]. Массовое развитие ее наблюдали на тех
же глубинах (1-2.5 м), что приводит К.И. Мейер, на камнях.
Известно [Мейер, 1930; Скабичевский, 1966], что Tetraspora не образует
сплошных зарослей вдоль прибрежья, а растет на камнях отдельными пятнами,
находящимися иногда на значительном расстоянии друг от друга. Вдоль откры-
тых прибрежий Байкала, где в литорали преобладают мягкие грунты, Tetraspora
появляется лишь там, где на глубине 1.5-3.0 м растут Nitella, Myriophyllum spica-
turn L., Ranunculus sp., ветви которых могут служить для нее субстратом [Иж-
84 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 19. Tetraspora cylindrica var. bullosa.
1 — внешний вид таллома; 2 — талломы в качестве субстрата используют Nitella sp.; 3 — деление
клеток; 4 — гаметы; 5 — зиготы.
Fig. 19. Tetraspora cylindrica var. bullosa.
1 — external view of thallome; 2 — thallomes used Nitella sp. as substratum; 3 — cell division;
4 — gametes, 5 — zygotes.
болдина, Верномудров, Максимов, 1984]. В районах мелководий против устьев
крупных рек — Селенга, Кичера, Верх. Ангара, а также во внутренних частях
Чивыркуйского и Баргузинского (зал. Култук) заливов и бухт Анга и Богучан-
ская тетраспора не встречена. Круглогодичные наблюдения за развитием Tet-
raspora показали, что в районах выхода теплых ключей она появляется подо
льдом в апреле [Ижболдина, 1970]. Массовое развитие вида вдоль всего при-
брежья начинается после таяния льда в мае — июне и в разных районах озера
происходит не одновременно. Однако массовое размножение ее во всех частях
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 85
озера заканчивается к 15-20 августа, и позднее встречаются лишь отдельные
полуразрушенные талломы. Т. cylindrica var. bullosa дает ежегодно один пик фи-
томассы, обычно в конце июля — начале августа — в период созревания гамет
[Ижболдина и др., 1984]. В это время вдоль западного побережья Юж. Байкала
масса водорослей составляет в среднем 270-360 г/м2, а в отдельных случаях мо-
жет достигать 2-3 кг/м2.
К.И. Мейер [1930] отметил, что на внутренней стороне трубчатого таллома
Tetraspora, особенно в верхней разрушающейся части, развивается эпифитная
водоросль Chaetonema irregulare Novak, из сем. Aphanochaetaceae. Талломы,
пронизанные плетями Ch. irregulare, обнаружены им во многих районах озера.
Tetraspora cylindrica var. sp. (рис. 20, 1, 2)
Вместе с Т. cylindrica var. bullosa вдоль западного берега Юж. и Сред. Бай-
кала, в прол. Мал. Море, у восточного берега о. Ольхон и у Ушканьих островов
встречаются талломы Tetraspora в виде цилиндрических сильно сетчато проды-
Рис. 20. Tetraspora cylindrica var. sp.
1 — общий вид таллома; 2 — расположение клеток в колонии; 3, 4 - Tetraspora lubrica: 3 — общий
вид колонии, 4 — расположение клеток в колонии. 3, 4 — по Смису.
Fig. 20. Tetraspora cylindrica var. sp.
1 — general view of thallome; 2 — location of cells in the colony; 3,4 — Tetraspora lubrica: 3 - general view
of the colony, 4 - location of cells in the colony. 3, 4 — after Smise.
86 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
рявленных мешков, прикрепляющихся к субстрату с помощью тонкой ножки
(рис. 20: 1, 2). Клетки шарообразные, с характерным для Tetraspora строением,
лежат по 4, 10-11 мкм в диаметре. Псевдоцилии не выступают из слизи коло-
ний. От Т. cylindrica var. bullosa отличается сетчато-продырявленным талломом
и чуть меньшим размером клеток, от Т. lubrica присутствием тонкой ножки и
более плотным расположением клеток.
Tetraspora lubrica (Oth.) Ag. (рис. 20: 3, 4)
Колонии желто-зеленые, мешковидные, сетчато-продырявленные, до 20
см длиной. Клетки шаровидные, имеют псевдоцилии, не выступающие из сли-
зи колоний.
В России и странах СНГ повсеместно в стоячих водоемах.
Замечание. В Байкале в небольшом количестве вид обнаружен К.И. Мейе-
ром [1930] на глубине 1, 2 м у мысов Красный, Хадартуй, Онгурен, Ижимей и
Барун-Будэ.
Класс ULOTRICHOPHYCEAE
ПОРЯДОК ULOTRICHALES - УЛОТРИКСОВЫЕ
Водоросли многоклеточные, нитчатые, разнонитчатые, пластинчатые или
мешковидные, реже одноклеточные, преимущественно прикрепленные. Клет-
ки цилиндрические. На поверхности многих клеток образуются волоски
(хеты) или щетинки. Хлоропласт пристенный, пластинчатый или дисковид-
ный, поясковидный или кольцевидный. Цельный или продырявленный, с од-
ним или несколькими пиреноидами или без них. Центральный звездчатый хло-
ропласт встречается только у празиоловых. Ядро одно. Вегетативное размноже-
ние — путем разрыва таллома на фрагменты или акинетами, бесполое —
2-4-жгутиковыми зооспорами и апланоспорами. Половой процесс изогамный,
гетерогамный, оогамный. Для некоторых видов характерна правильная смена
половой и бесполой форм развития.
В Байкале встречены представители 4 семейств.
1. Талломы нитчатые, без волосков и щетинок........Ulotrichaceae
2. Слоевище в виде макроскопических разветвленных кустиков. Конечные клет-
ки веточек заостренные или заканчиваются одно- или многоклеточными во-
лосками .........................................Chaetophoraceae
3. Слоевище в виде однослойных дисков или разнообразно ветвящихся по суб-
страту нитей. Клетки с волосками................Aphanochaetaceae
4. Талломы макроскопические, одно- или двуслойные, волоски или щетинки
отсутствуют. Хлоропласт центральный, звездчатый.....Prasiolaceae
СЕМЕЙСТВО ULOTRICHACEAE — УЛОТРИКСОВЫЕ
Талломы многоклеточные, в виде простых, обычно однорядных нитей,
реже одноклеточные, клетки одноядерные, хлоропласт пристенный, кольце-
видный, поясковидный с одним или несколькими треноидами. Все клетки, за
исключением базальной, способны к делению надвое. Бесполое размноже-
ние — 2- и 4-жгутиковыми зооспорами, апланоспорами, гипноспорами; поло-
вое — изогамное, гаметы 2-жгутиковые. В оз. Байкал встречены представители
одного рода.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 87
Род Ulothrix Kutz., 1833 — Улотрикс
Нити однорядные, прикрепленные к субстрату базальной клеткой или сво-
бодноживущие. Клетки цилиндрические. Оболочка клеток разной толщины,
сплошная, недвустворчатая. Хлоропласт поясковидный или кольцевидный, с 1
или несколькими пиреноидами.
В Байкале встречены 2 вида.
I. Нити от 10 мкм и более.
А. Оболочки клеток тонкие, неслоистые.
а. Клетки широкого диапазона размерности, (6.5) — 10-22(34) мкм
шириной................................................U. tenuissima
Б. Оболочки клеток толстые, слоистые (только в молодом состоянии иногда
тонкие).
а. Клетки (11)30-45(83)мкмшириной ....................... U.zonata
Ulothrix tenuissima Kutz. (=U. tenuis Kiltz.; Hormiscia tenuis De-toni)
(рис. 21: 10)
Нити длинные, прямые, темно-зеленые. Клетки цилиндрические, 10-
20 мкм шириной. Оболочки клеток тонкие, хлоропласт широкий, с (1)2-4
крупными пиреноидами. Вид изучен недостаточно.
В пресных минерализованных стоячих и особенно проточных водах.
Россия, Кавказ, Казахстан, Узбекистан, нередко.
Замечание. В Байкале встречен [Ижболдина и др., 19906] на глубине от 0.1
до 1.5 м, на камнях, где растет вместе с Ulothrix zonata. Развит в мае-июле.
Ulothrix zonata (Web. et Mohr.) Kutz. {Conferva zonata Web. et Mohr.;
Hormiscia zonata (Web. et Mohr.) Aresch.) (рис. 21: 1-9)
Нити прикрепленные, зеленые или желто-зеленые. Клетки нитей цилинд-
рические или слегка бочонкообразные, (11)30-45(83) мкм шириной. Молодые
нити 10-11 мкм шириной. Длина клеток равна ширине, меньше или больше ее.
У молодых нитей оболочки тонкие, у старых — толстые, слоистые. В молодых
клетках хлоропласт поясковидный, позднее кольцевидный, с несколькими
(до 20) крупными пиреноидами.
Предпочитает хорошо аэрируемые чистые пресные воды.
Ulothrix zonata var. zonata (рис. 21: 1—9)
Нити на всем протяжении одинаковой толщины. Клетки 20-30(38) мкм
шириной, по длине равные ширине или до 2 раз меньше ее.
В проточных, реже стоячих водах. Повсеместно.
Замечания. В Байкале встречен многими исследователями [Дорогостай-
ский, 1906; Яснитский, 1928; Верещагин, 1930; Кожов, 1931, 1947; Скабичев-
ский, 1934; и др.]. К.И. Мейер [1927,1930] подчеркивал, что U. zonata встречает-
ся вдоль всего побережья, где в прибойной зоне есть твердый грунт. Первые
нити U. zonata появляются в апреле на свободных ото льда участках дна у уреза
воды. Массовое развитие начинается в мае, после освобождения озера ото
льда, в мае — августе на глубинах от 0 до 1.5 м U. zonata образует растительный
пояс, названный К.И. Мейером [1927, 1930] поясом U. zonata. В этот период
встречаемость U. zonata на глубине от 0 до 1,5 м составляет 50-70 %. Вдоль се-
веро-западного прибрежья Юж. Байкала максимальная фитомасса вида отме-
88 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 21. 1-9 — Ulothrix zonata: 1 — верхушка молодой нити, 2 — базальная часть нити,
3 — фрагмент взрослой нити, 4 — кольцевидный хлоропласт с пиреноидом, 5 — зооспо-
ра, 6 — стадии прорастания зооспор, 7 — гаметы, 8 — слияние гамет, 9 — зигота; 10 —
U. tenuissima, фрагменты нитей; 11-14 — Chaetophora elegans: 11 —общий вид таллома на
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 89
чена в конце июля [Ижболдина, Верномудров, Максимов, 1978]. В это время
средняя фитомасса водорослей колеблется в разные годы от 100 до 210 г/м2. К
началу сентября массовое развитие U. zonata заканчивается и средняя масса его
не превышает 16-21 г/м2. На крупных валунах и скалах у уреза воды нити U. zo-
nata можно встретить до ноября. Молодые нити обнаружены и в подледный пе-
риод в январе под прорубью на глубине 0.7 м.
На нитях U. zonata в громадном количестве поселяются диатомеи, список
которых, по данным К.И. Мейера [1930], может включать до 22 видов, относя-
щихся к 10 родам. Появление зарослей U. zonata привлекает в прибрежную зону
массу макро- и мезобентических животных, находящих себе здесь благоприят-
ные условия для жизни [Каплина, 1974; Кожов, 1962; Окунева, 1974,1976]. Чис-
ленность гаммарид, например, в период развития U. zonata увеличивается в
зоне глубин 0.3-1 м в 88 раз по сравнению с зимой, а биомасса их возрастает
при этом в 11 раз [Каплина, 1974].
ПОДПОРЯДОК CHAETOPHORINEAE
СЕМЕЙСТВО CHAETOPHORACEAE
Таллом обычно состоит из 2 частей: горизонтальной и вертикальной, обра-
зованных ветвящимися или простыми нитями. Горизонтальная часть состоит
из нитей, тесно расположенных в одной плоскости и срастающихся боковыми
спорами в однослойную паренхимную пластину. Вертикальная часть представ-
лена простыми или обильно ветвящимися нитями, многие из которых закан-
чиваются одно- или многоклеточными волосками. При недоразвитии горизон-
тальной части от нижних клеток прямостоячих нитей отходят ризоидные выро-
сты, с помощью которых они и прикрепляются к субстрату. Хлоропласт
преимущественно кольцевидный или в виде полуцилиндра, цельный или сет-
чато-продырявленный. Бесполое размножение 2- или 4-жгутиковыми зооспо-
рами, половой процесс — изо- или гетерогамный. Гаметы 2- или 4-жгутиковые.
ПОДСЕМЕЙСТВО CHAETOPHOROIDEAE
Нити ветвящиеся, реже простые, с хорошо выраженной ветвящейся глав-
ной нитью. Реже нити распростертые по субстрату или частично приподнима-
ющиеся над ним. Конечные клетки нитей заостренные, реже закругленные или
вытягивающиеся в одно- или многоклеточный волосок.
В Байкале встречены представители 6 родов.
субстрате (при малом увеличении), 12 — нижняя часть таллома, 13 — верхушечная часть
таллома, 14 — образование акинет.
По: 1,5-9 — Локхорсту и Вроману, 2, 3,10 — Газену, 4 — Рич, И, 14 — Мошковой, 12,13 — Тиффани.
Fig. 21.1-9 — Ulothrix zonata: 1 — apex of a juvenile filament, 2 — basal part of filament, 3 —
fragment of a mature filament, 4 — ring-shaped chloroplast with a pyrenoid, 5 — zoospore, 6 —
stages of zoospore germination, 7 — gametes, 8 — gamete fusion, 9 — zygote; 10 —
U. tenuissima, filament fragments; 11-14 — Chaetophora elegans: 11 — general view of thallome
on substratum (low magnification), 12 — lower part of thallome, 13 — apical part of thallome,
14 — akinete formation.
After: 1, 5-9 — Lokhorst & Wroman, 2, 3,10 — Gazen, 4 — Rich, 11,14 — Moshkova, 12,13 — Tiffany.
90 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
I. Боковые ветви не отличаются от главных нитей. Водоросли зеленые с
хлоропластами.
1. Прямостоячие нити заключены в обильную слизь, формирующую таллом
от шаровидной до неправильной формы................... Chaetophora
2. Прямостоячие разветвленные нити не заключены в слизь.
А. Талломы в виде разветвленных кустиков, объединенных в дерновинки
или обособленные. Хлоропласт цельный или продырявленный, с не-
ровными краями, с 1 или несколькими пиреноидами .... Stigeoclonium
Б. Талломы в виде обособленного разветвленного кустика. Хлоропласт
сетевидный, занимающий всю высоту клетки, с многими крупными
пиреноидами; эндем Байкала..........................Ireksokonia
II. Боковые ветви отличаются от главной нити.
I. Талломы обильно разветвлены и ясно дифференцированы на ствол и си-
дящие на нем сильно разветвленные ветви. Все они покрыты короткими
боковыми веточками, которые могут быть одиночными, супротивными
или по 3-4 в мутовках. Конечные клетки веточек часто оканчиваются
многоклеточным волоском.
А. Клетки главных нитей с тонкими оболочками.
а. Хлоропласт в виде тонкой сеточки, занимает всю полость клетки.
Вокруг главного ствола и крупных ветвей развит чехол из обильно
разветвленных ризоидов, развивающихся из базальных клеток бо-
ковых веточек; эндем Байкала...............Draparnaldioides
б. Хлоропласт в виде узкого или широкого продырявленного кольца.
Ризоидный покров отсутствует...................Draparnaldia
Б. Клетки главных нитей с толстыми слоистыми оболочками; эндем Бай-
кала ..............................................Myxonemopsis
Род Chaetophora Schrank., 1789
Таллом слизистый, шаровидный, полушаровидный или бородавчатый. В
основании таллома расположены шаровидные клетки, от которых отходят ра-
диально расположенные главные нити. В средней части таллома ветвление ни-
тей менее обильно, чем в верхней. Конечные клетки заостренные или перехо-
дят в длинный многоклеточный волосок. Клетки нитей цилиндрические или
несколько вздутые. Бесполое размножение 4-жгутиковыми зооспорами, апла-
носпорами, гипноспорами, акинетами. Половое размножение 2-жгутиковыми
гаметами.
В Байкале обнаружен один вид.
Chaetophora elegans (Roth) Ag. (=Rivularia elegans Roth)
(рис. 21: 11-14)
Таллом в виде шаровидных и полушаровидных слизистых образований.
Нити поднимающиеся от подошвы вверх, расходятся радиально. Конечные
разветвления нитей никогда не собраны в виде плотных кисточек. Клетки по-
дошвы шаровидные, клетки главных нитей цилиндрические или слегка бочон-
кообразные, 9.6-12 мкм шириной, по длине в 3-10 раз больше, у конечных вет-
вей цилиндрические, 7.2-10.8 мкм шириной, по длине в 1.5-2 раза больше. Ко-
нечные клетки конусовидно заостренные, редко вытянутые в волосок.
В стоячих и проточных водах, на водных цветковых растениях, на твердом
субстрате. Повсеместно.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 91
Замечания. В Байкале обнаружена К.И. Мейером [1930] в Посольском Соре
на Phragmites communis Trin. и Ranunculus divarucatus Schrank., в зал. Провал -
в лагуне около Оймура и в зал. Мухор. Нами, кроме того, вид встречен в Исток-
ском соре, в бухтах Чивыркуйского залива, а также в районе губы Слюдянская
на Cladophora kursanovii Skabitsch., в губе Онокочанской на Nitella sp. и в районе
пади Орео на стебельках Didymosphaenia geminata.
Род Stigeoclonium Kutz., 1843
Талломы состоят из стелющихся по субстрату и восходящих разветвленных
нитей, оканчивающихся многоклеточными или одноклеточными волосками.
Иногда конечные клетки заостренные или закругленные. Хлоропласт пристен-
ный, цельный или сетчато продырявленный, с одним или многими пиреноида-
ми. В верхней части таллома хлоропласт выстилает стенки клеток почти полно-
стью, в средней части и в основании таллома он занимает обычно только сред-
нюю часть клеток в виде узкого кольца. Бесполое размножение - 2- и
4-жгутиковыми зооспорами, половое — изогамное.
В Байкале найдено 4 вида.
I. Таллом с хорошо развитыми прямостоячими нитями.
1. Клетки главных нитей не отличаются по форме и величине от клеток вет-
вей. Ветвление супротивное или очередное. Конечные клетки веточек за-
остренные, реже вытянутые в 1-3-клеточный волосок ......... S.tenue
2. Клетки главных нитей по форме и величине отличаются от клеток главных
ветвей. Последние в нижней части таллома только цилиндрические.
А. Ветви 2-го порядка очередные или супротивные, сближенные по неско-
льку, оканчивающиеся шиловидно заостренной клеткой или длинным
многоклеточным волоском...............................£ flagelliferum
Б. Ветви 1-го порядка обычно супротивные. Ветви 2-го порядка преимуще-
ственно очередные, заканчивающиеся короткой заостренной клеткой.
Волоски обычно отсутствуют..............................S. атоепит
II. Таллом с редуцированными прямостоячими нитями.
1. Прямостоячие нити из 3-5 и более клеток, простые, реже скудно ветвящи-
еся, волоски в основании не вздутые.........................£ farctum
1. Stigeoclonium tenue (Ag.) Kutz. (=Draparnaldia tenuis Ag.;
Myxonema tenue (Ag.) Rabenh.; Stigeoclonium irregulare Kutz.;
£ tenue var. irregulare (Kutz.) Rabenh.; £ (tenue) klebsii Heering;
£ (tenue) pascheri Heering; £ (tenue) westii Heering;
£ stellare Kutz.; £ gracile Skvortz.) (рис. 22: 1—12)
Таллом кустиковидный, до 5 см высотой. Ветвление очередное или супро-
тивное. Клетки главных нитей цилиндрические или слабо вздутые в средней
части нитей, 6-15 мкм шириной, по длине в 3-5(6) раз больше. У ветвей изо-
диаметрические, 5-6 мкм шириной.
Stigeoclonium tenue var. tenue (=£ tenue var. genuinum Kirchn.)
Клетки главных нитей с тонкими бесцветными оболочками. Конечные
клетки шиловидно заостренные или заканчиваются 1-3-клеточными волосками.
В стоячих и проточных водах. Одна из наиболее обычных, повсеместно
распространенных водорослей.
92 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 22. 1-12 — Stigeoclonium tenue: 1 — ветвь с формирующимися макрозооспорами,
2 — формирующиеся микрозооспоры, 3 — прорастающая зооспора в воде, 4 — макро-
зооспора, 5 — микрозооспора, 6 — покоящаяся клетка, образовавшаяся из одной
микрозооспоры, 7 — прорастающая клетка, 8 — вышедшая макрозооспора, 9 —
гаметы, 10 — копуляция и зигота, 11 — прорастающие макрозооспоры, 12 — характер
ветвления таллома, 13 — Stigeoclonium flagelliferum; 14-16 — Stigeoclonium farctum var.
farctum: 14 — общий вид молодого слоевища сверху, 15 — фрагмент слоевища, 16 —
стелющаяся нить с прямостоячими веточками.
По: 1-7 — Klebs, 8-11 — G.S.West, 12 — Газену, 13,15 — Мошковой [1979], 14,16 — Бертхольду.
Fig. 22. 1-12 — Stigeoclonium tenue: 1 — branch with forming macrozoospores, 2 — forming
microzoospores, 3 — germinating zoospore in water, 4 — macrozoospore, 5 — microzoospore,
6 — resting cell formed from one microzoospore, 7 — germinating cell, 8 — released
macrozoospore, 9 — gametes, 10 — copulation and zygote, 11 — germinating macrozoospores,
12 — thallome branching pattern; 13 — Stigeoclonium flagelliferum; 14-16 — Stigeoclonium
farctum var. farctum: 14 — general top view of a juvenile thallome, 15 — fragment of thallome,
16 — trailing filament with erect shoots.
After: 1-7 - Klebs, 8-11 - G.S. West, 12 - Gazen, 13,15 - Moshkova [1979], 14,16 - Berthold.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 93
Замечания. В Байкале встречена нами вдоль северо-западного побережья
южной части озера в августе — сентябре, на глубине 1-2.5 м, на камнях. В те же
сроки развита в заметном количестве в притоке Байкала р. Бол. Коты [Кожова,
Ижболдина, 1994].
Stigeoclonium атоепит Kiitz. (=5. атоепит f. biforme Collins;
Мухопета атоепит (Kiitz.) Hazen) (рис. 23)
Кустики светло-зеленые, пушистые, 0.5-1.5(15 см) высотой. Главные нити
обильно разветвленные. Ветви 1-го порядка преимущественно супротивные,
сближенные по 2-3 пары, ветви 2-го порядка обычно короткие, супротивные
Рис. 23. Stigeoclonium атоепит var. атоепит.
По Газе ну.
Fig. 23. Stigeoclonium атоепит var. атоепит.
After Gazen.
94 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
или очередные. Клетки главных нитей в нижней части таллома 11.4-18(38) мкм
шириной, по длине в 1.5-8 раз больше. Ветви первого порядка в основании
6-11.4 мкм шириной, ветви 2-го порядка 5-7 мкм шириной. На концах нитей
волоски обычно отсутствуют.
В проточных и стоячих водах, довольно часто.
Замечание. В Байкале в небольшом количестве встречена К.И. Мейером
[1930] в Ангаро-Кичерском соре, на глубине 1.6 м, среди зарослей Equisetum li-
mosum L. и в районе г. Сеногда в августе на глубине 1.6 м.
Stigeoclonium flagelliferum Kutz. (рис. 22: 13)
Ярко-зеленые кустики, скользкие на ощупь, до 2-5(15) см высоты. Ветви
1-го порядка обильно разветвленные, супротивные, часто сближенные, по
2-5 пар. Клетки главных нитей в средней части таллома 14-23(25) мкм
шириной, по длине в 3-8 (12) раз больше. Ветви 2-го порядка 9-12(15.2) мкм
шириной, по длине в 4-6 (10) раз больше.
В реках, ручьях, реже в прудах на камнях и различных растительных субст-
ратах.
В России довольно часто.
Замечания. В Байкале вид встречен нами в бух. Анга на стебельках Didymos-
phenia geminata в августе, у о. Лиственичный - на нитях Cladophora floccosa
С. Meyer в конце июня, в прол. Мал. Море у мыса Курминский на талломах
Chaetocladiella pumila С. Meyer et Skabitsch., в сентябре [Ижболдина, 1976а].
Кроме того, вид встречен у юго-восточного побережья Байкала, в районах
сброса УЧВ и ПОСВ БЦБК [Ижболдина, 1985].
Stigeoclonium farctum Berth. (=5. huberi Heering; S', najadeanum Skvortz.)
(рис. 22: 14-16)
Таллом в виде подушковидных дерновинок, до 1,5 мм высотой. Ветвящи-
еся в плоскости нити срастаются боками в плотно прижатую к субстрату па-
ренхимную пластину и отходящих от нее коротких прямостоячих, простых
или слабо ветвящихся нитей, состоящих иногда из 1-5 клеток. Между прямо-
стоячими нитями находятся длинные многоклеточные волоски, вырастаю-
щие из клеток подошвы. Слегка вздутые клетки подошвы 6-12 мкм шириной,
8-12 мкм длиной. Цилиндрические клетки прямостоячих нитей 4-8,6 мкм
шириной, 6-16 мкм длиной.
В стоячих и проточных водах, широко распространенный вид.
Замечания. В Байкале найден К.И. Мейером [1930] в зал. Мухор. При на-
ших исследованиях встречен в районе пади Орео и мыса Крест на стебельках
Didymosphenia geminata и у о. Угунгой (прол. Мал. Море) на глубине 2.5 м на
поверхности колоний Stratonostoc verrucosum и на ветвях Chaetocladiella pumi -
la. Кроме того, в 1981, 1988 гг. встречен у юго-восточного побережья озера в
районе поступления в него условно-чистых вод БЦБК [Ижболдина, 19766,
1985а].
Род Ireksokonia Meyer, 1927
Тонкие ветвящиеся нити высотой 10-12 см, прикрепляющиеся к субстрату
с помощью мощной системы ризоидов, развивающихся у нижних клеток тал-
лома. Ветвление нитей моноподиальное, ветви очередные или супротивные.
Клетки нитей обычно тонкостенные, цилиндрические, в средней части слегка
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 95
перетянутые у поперечных перегородок. Длина клеток у главных и крупных
ветвей в 1.5-3 раза больше ширины. Клетки тонких боковых ветвей квадратные
или длина их в 1.5-2 раза более ширины. Конечные клетки закругленные или
переходят в длинный тонкий волосок. Хлоропласт в виде тонкой нежной се-
точки, с многочисленными пиреноидами, занимает всю высоту клетки. В клет-
ках, граничащих с волоском, хлоропласт в виде пластинки с мелкими отверсти-
ями и несколькими пиреноидами. Ядро одно, довольно крупное. Бесполое раз-
множение - зооспорами, половое - изогамное.
Этот род близок к роду Stigeoclonium, от которого отличается размерами
таллома, строением хлоропласта, мощной ризоидной системой. Большое сход-
ство с родом Draparnaldioides С. Meyer et Skabitsch., а именно D. simplex, от ко-
торого отличается менее выраженной дифференциацией таллома на ствол и
боковые ветви. Эндем Байкала. Встречен один вид.
Ireksokonia formosa С. Meyer (1927)
(рис. 24: 1-6)
Образует длинные, до 10-12 см высотой, тонкие нити, растущие дерновин-
ками на камнях на глубине 2-5 м. Нити сильно ветвятся. Ветвление моноподи-
альное, ветви располагаются большей частью одиночно, изредка по 2 супро-
тивно. Закладываются они в акропетальном порядке. Клетки главных и круп-
ных боковых ветвей изодиаметрические или длина их превосходит ширину в
1.5-3 раза. Они слегка вздуты, на поперечных перегородках слегка перетянуты.
Оболочки клеток тонкие, прозрачные, середина клетки занята крупной вакуо-
лей клеточного сока; в тонком стенкоположном слое плазмы лежит хлоро-
пласт. Он имеет вид нежной и тонкой сеточки с широкими отверстиями, зани-
мающей всю клетку (рис. 24: 2). В нем заключены многочисленные крупные
пиреноиды. По мере приближения к концам ветвей хлоропласт принимает вид
пластинки, продырявленной мелкими отверстиями и содержащей несколько
пиреноидов. Ядро одно, довольно крупное.
Рост ветвей происходит главным образом на концах ветвей, где под щетин-
кой находится зона роста, построенная из энергично делящихся клеток. Эти же
клетки дают начало щетинкам. Клетки, превращающиеся в щетинку, сильно
вытягиваются, хлоропласт их принимает вид очень редкой, бледной сеточки с
мелкими пиреноидами. Концы щетинок почти всегда обломаны. Талломы
Ireksokonia formosa прикрепляются к субстрату при помощи мощной системы
ризоидов, развивающейся из нижних клеток (рис. 24: 3). Ризоиды возникают в
большом количестве, по 1-2 из каждой клетки. Они состоят из длинных тонко-
стенных клеток, цилиндрических или слегка вздутых. Размеры клеток Ireksoko-
nia formosa: толщина их в нижних частях нитей 120-135 мкм, в середине
50-80 мкм, на концах нитей 14-27 мкм, ширина щетинок 9-15 мкм. Толщина
ризоидов 18-45 мкм. Гаметангии развиваются на концах ветвей (рис. 24: 4).
Клетки ветвей делятся поперечными и продольными перегородками и вздува-
ются, вследствие чего нити принимают четковидную форму. По внешнему
виду и строению многогнездные гаметангии Ireksokonia походят на гаметангии
видов рода Draparnaldioides.’Толщина гаметангиев 22.5-35 мкм.
По К.И. Мейеру [1930], распространена в северной части Байкала, на глу-
бине 2-5 м, где встречается довольно редко. К.И. Мейер рассматривает I. for-
mosa как форму, связывающую род Stigeoclonium с эндемичным байкальским
родом Draparnaldioides и, возможно, исходную для них.
96 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Замечания. Вид ранее встречен [Скабичевский, 1966; Поповская, Скаби-
чевский, 1968] в районе Ушканьих островов и пос. Бол. Коты. При наших ис-
следованиях он обнаружен вдоль всего западного прибрежья Байкала и в сред-
ней и северной частях восточного, на камнях, на глубине 2-8(15) м. Встречае-
мость вида не превышает обычно 10 %, а фитомасса — 16 г/м2. В мелководных
бухтах и заливах, где обычно преобладает мягкий грунт, I. formosa в качестве
субстрата использует высшие водные растения родов Potamogeton L. и Myriop-
hyllum L., талломы крупных водорослей родов Nitella Ag. и Chara L. В июле
Рис. 24. Ireksokonia formosa.
1 — ветвление нитей в верхней части таллома; 2 — клетка ствола; 3 — ризоиды в основании тал-
лома; 4 — гаметангии; 5 — зооспорангии разной формы; 6 — зооспорангий, образовавшийся из
клетки крупной ветви. 2-3 — по К.И. Мейеру [1927].
Fig. 24. Ireksokonia formosa.
1 — filament branching in the upper part of thallome; 2 — spine cell; 3 — rhizoids at the thallome base;
4 — gametangia; 5 — zoosporangia of various shapes; 6 — zoosporangium formed of a large branch cell.
2-3 — after Meyer [1927].
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 97
встречаются стерильные талломы, в конце августа—начале сентября наблюда-
ли массовое формирование гаметангиев и выход гамет. Длина отдельных гаме-
тангиев может достигать 1.2 мм. Одновременно с гаметангиями и на одних с
ним веточках наблюдается формирование зооспорангиев. Последние развива-
ются из клеток как мелких, так и крупных ветвей (рис. 24: 5-6). Длина зооспо-
рангиев 90-180 мкм, ширина - 50-75 мкм. Выход зооспор пронаблюдать не
удалось.
Род Myxonemopsis Meyer, 1930
Таллом кустиковидный, 2 см высотой, обильно разветвлен. Главный ствол
и крупные ветви состоят из цилиндрических или бочонкообразно перешнуро-
ванных клеток с толстыми слоистыми оболочками. Ствол ветвится моноподиа-
льно. Расположение крупных ветвей очередное или супротивное. Ствол и
крупные ветви, в свою очередь, покрыты длинными, обильно разветвленными
боковыми веточками, с очередным или супротивным расположением нитей.
Клетки этих нитей боковых веточек короткие, вздутые, иногда почти шарооб-
разные, а к концам нитей становятся цилиндрическими и заканчиваются за-
остренными клетками или длинным волоском. Таллом прикрепляется к грунту
с помощью мощной ризоидальной системы, напоминающей таковую у Irekso-
konia formosa. Хлоропласт сетчато продырявленный, с 2-4 пиреноидами. Раз-
множение не прослежено. Рассматриваемый род близок к роду Ireksokonia, но
отличается длинными обильно разветвленными боковыми веточками, напоми-
нающими таковые у рода Draparnaldioides, но от последнего отличается отсут-
ствием мутовок боковых веточек. По мнению К.И. Мейера [1930], Myxonemop-
sis надо рассматривать как боковое ответвление Ireksokonia formosa. Эндем
Байкала. Встречен один вид.
Myxonemopsis crassimembranacea Meyer, 1930
(рис. 25: 1-4)
Невысокие (около 2 см) пушистые кустики. Клетки главного ствола и
крупных боковых ветвей 90-150 мкм шириной, ширина боковых веточек в
нижней части 70 мкм, в средней — 30-40 мкм, на концах ветви — 15 мкм; клет-
ки волосков — 7.5-11 мкм. Оболочки клеток главного ствола бесцветные, сло-
истые, 5.5-7.5 (10) мкм толщиной. На концах ветвей, где клетки становятся
цилиндрическими, толщина клеточных оболочек не более 1.5 мкм.
Поданным К.И. Мейера [1930] и других исследователей [Поповская, Ска-
бичевский, 1968], редкий для Байкала вид. К.И. Мейером обнаружен у мыса
Рытый на глубине 2 м, на камнях в августе.
Род Draparnaldioides Meyer et Skabitsch., 1976
Кустиковидные талломы высотой до 10-40 см. От главного ствола отходят
крупные ветви 2-го и 3-го порядка. Все они несут боковые веточки, распола-
гающиеся мутовками, реже одиночные или супротивные. Боковые веточки
образованы ветвящимися нитями, которые обычно заканчиваются длинными
прозрачными волосками. У одних видов в боковых веточках четко выражена
средняя ось — рахис, выдающийся за пределы боковой веточки; у других ви-
дов рахиса нет или он быстро становится незаметным, переходя в разветвле-
ние. Ствол и крупные ветви таллома состоят из одного ряда цилиндрических
98 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 25. Myxonemopsis crassimembranacea.
1 — одна клетка главного ствола; 2, 3 — части боковых ветвей; 4 — одна боковая ветвь. По
К.И. Мейеру [1930].
Fig. 25. Myxonemopsis crassimembranacea.
1 — one cell of the main spine; 2, 3 — fragments of lateral shoots; 4 — one lateral branch. After Meyer
[1930].
или бочонковидно перешнурованных круглых клеток. Длина их равна шири-
не, больше или меньше. Клетки боковых веточек мелкие, цилиндрические.
Хлоропласт клеток ствола и крупных ветвей в виде тонкой сеточки с много-
численными пиреноидами, прилегающей к клеточным стенкам и занимаю-
щей всю полость клетки. В боковых веточках хлоропласт в виде сплошной
или слабо продырявленной пластинки с несколькими пиреноидами. Крупные
клетки в основании рахиса имеют обычно сетчато-продырявленный хлоро-
пласт.
Вокруг клеток главного ствола и крупных ветвей развивается ризоцдный
покров. Бесцветные, лишенные хлоропласта ризоидные нити, развиваются из
базальных клеток боковых веточек. Разрастаясь, они вытягиваются вниз
по стволу, ветвятся, переплетаются с ризоидами других боковых веточек. В ре-
зультате образуется рыхлый ризоцдный покров, наиболее толстый в нижних
частях таллома. Половое размножение, изученное В.Н. Яснитским [1934], анизо-
гамное, со строго выраженной гетероталличностью. Многогнездные гаме-
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 99
тангии развиваются из клеток нитей боковых веточек, в которых происходит
вначале поперечное, а затем продольные деления. В каждой клеточке много-
гнездного гаметангия формируется одна 4-жгутиковая гамета со стигмой и се-
тевидным хлоропластом. Бесполое размножение мало изучено.
Род Draparnaldioides близок к роду Draparnaldia, но, по мнению К.И. Мей-
ера и А.П. Скабичевского, филогенетически с ним не связан. В Байкале эволю-
ция шла по линии Stigeoclonium — Ireksokonia — Draparnaldioides, a Draparnaldia
составляет параллельную байкальской линию развития. Род Draparnaldioides
отличается от рода Draparnaldia сетевидным хлоропластом, занимающим всю
высоту клетки, наличием ризоидного покрова. Важными диагностическими
признаками для разграничения видов служат форма боковых веточек и нали-
чие или отсутствие в них рахиса. Для определения видов следует отбирать рас-
тения, вступившие в стадию формирования гаметангиев, а в них — боковые
ветви из средней части таллома.
Эндемик Байкала. Всего встречено 9 видов.
I. Рахис в боковых веточках выражен ясно.
1. Боковые веточки слабо разветвлены.
А. Боковые веточки простые, в общих очертаниях ланцетные, ветвление
их двустороннее. Рахис заканчивается тонким волоском .... D. simplex
Б. Боковые веточки узкие, сжатые с ветвями, расположенными лишь у их
основания. Рахис резко выражен, заканчивается толстым длинным
волоском.................................................D. arenaria
2. Ветвление боковых веточек более обильное.
А. Боковые веточки в очертании округлые, веерообразно разветвлены. Ра-
хис длинный, заканчивающийся толстым волоском или короткий (по-
сле разрушения гаметангия), с округлой клеткой на конце D. arnoldii
Б. Боковые веточки узкоромбические, ветвление их неправильно двояко-
перистое. Рахис длинный, заканчивается тонким волоском...........
.................................................D. goroschankinii
В. Боковые веточки широкояйцевидные или полукруглые. Рахис заканчи-
вается толстым волоском. От рахиса в акропетальном порядке отходят
ветви, загибающиеся кверху................................D. pumila
П. Рахис в боковых веточках короткий, незаметный, быстро переходящий в ветви.
1. Боковые веточки компактные, в очертании округлые, образованы сильно
ветвящимися нитями. Волоски образуются в небольшом относительно
количестве ..................................................D. vilosa
2. Боковые веточки округлые, обильно разветвленные, концы нитей закан-
чиваются длинными толстыми волосками. Нити боковых веточек плотно
прилегают друг к другу, изогнуты дугообразно, выпуклой стороной обра-
щены наружу .................................................D. pilosa
3. Боковые веточки округло-треугольной формы, имеют довольно толстый
рахис, состоящий обычно из 1-2 клеток, от которых сразу начинаются
ветви более или менее одинакового размера, особенно обильно ветвящие-
ся в верхней части .......................................D. baicalensis
4. Боковые веточки в очертании округло-яйцевидные, очень густые благода-
ря чрезвычайно обильному ветвлению составляющих ее нитей. Рахис ко-
роткий, состоит из 7-8, иногда из 2-3 клеток и заканчивается пучком ве-
точек с длинными волосками...................................D. lubrica
100 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Draparnaldioides simplex (Meyer) Meyer et Skabitsch. (=Draparnaldia simplex
C. Meyer, 1922; Draparnaldiella simplex Meyer et Skabitsch., 1969)
(рис. 26: 1-3; 27: 1-11)
Густые, сильно и тонко разветвленные кустики высотой 15-20 см, сли-
зистые на ощупь. Таллом прикреплен к субстрату с помощью неясно вы-
раженной подошвы. Сразу над местом прикрепления начинается ветвление
главного ствола. Внизу оно более редкое и ветви толстые и длинные. Выше
ветвление становится густым и ветви приобретают кистевидный характер.
Ветвление ствола мутовчатое. Мутовку образуют 3-5 ветвей, чаще их 4.
Боковые веточки расположены поодиночке или по 2 супротивно. Они слабо
разветвлены и нити их заканчиваются длинными волосками. Между мутов-
ками крупных ветвей располагаются добавочные ветви с довольно густым
ветвлением нитей. В нижних частях ствола клетки имеют толстую оболочку и
длина их меньше (в 1.5- 2 раза) ширины. В средней и верхней частях таллома
длина клеток равна ширине или превосходит ее в 2-4 раза. Размеры клеток
главного ствола и крупных ветвей в нижних частях таллома: 200-370 мкм
шириной, 100-180 мкм длиной; в средних частях таллома — 80-180 мкм
шириной, на концах ветвей — 25-45 мкм шириной, 55-90 мкм длиной;
боковых веточек в основании рахиса — 10-35 мкм толщиной, в верхней части
нитей — 10-12.5 мкм, волоски 7.5-18 мкм. Ризоидный покров у D. simplex
выражен слабо и развит лишь в нижних частях таллома, где невозможно
рассмотреть строение боковых веточек без удаления ризоидного чехла. Все
нити боковых веточек, кроме основания рахиса, превращаются в гаметангии,
ширина которых колеблется от 23 до 32 мкм. Выход гамет совершается ночью
или в предрассветные часы. Гаметы, образующиеся в огромном количестве,
выходят в течение 2-3 ночей. Пустые гаметангии разрушаются и на стволе и
крупных ветвях остаются лишь короткие основания рахисов. Из-за этого
таллом сильно изменяется, делается прозрачным (рис. 26: 1-3). Гаметы имеют
широкогрушевидную форму и снабжены 4 жгутиками. Длина гамет 8-10.6 мкм.
Гаметы имеют короткий бесцветный носик, мелкий палочковидный глазок и
сетчато-продырявленный хлоропласт с 1-3 пиреноидами. Ядро расположено
ближе к заднему концу клетки.
Эндемик Байкала, D. simplex широко распространена по всему озеру,
являясь наиболее частым представителем рода.
Draparnaldioides simplex forma simplex
Ствол обычно образован мутовками из 4 ветвей, расположенных на неко-
тором расстоянии друг от друга.
Широко распространена по всему Байкалу на глубине 2.5-10 м.
К.И. Мейер [1930] выделял 2 формы D. simplex, подчеркивая, что, возможно,
это формы роста.
Draparnaldioides simplex forma trijurca Meyer, 1930
Ветви крупные, с трехчленными, относительно редко расположенными
мутовками, с небольшим числом добавочных ветвей.
Оз. Байкал, Чивыркуйский залив, мыс Халтыгей.
Draparnaldioides simplex forma pentafiirca Meyer, 1930
Пятичленные тесно расположенные мутовки с густым ветвлением. Кустики
лохматые.
Оз. Байкал, падь Сенная на о. Ольхон.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 101
Рис. 26. Draparnaldioides simplex.
1 — на некоторых ветвях в основании таллома начался выход гамет; 2 — в основании таллома выход
гамет закончен; 3 — внешний вид таллома после массового выхода гамет.
Fig. 26. Draparnaldioides simplex.
1 — some branches at the thallome base started to release gametes; 2 — gamete release at the thallome base
finished; 3 — external view of thallome after mass gamete release.
102 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 27. Draparnaldioides simplex.
1 — ветви в верхней части таллома; 2 — боковые веточки; 3 — добавочная веточка с гаметангиями;
4 — ветвление главного ствола; 5 — боковые веточки в основании крупной ветви (ризоидный че-
хол убран); 6 — фрагмент таллома после выхода гамет и разрушение гаметангиев; 7, 8 — гаметы;
9 — слияние гамет; 10 — зигота; 11 — зооспорангии. 2-4, 6, 7 — по К.И. Мейру [1930].
Fig. 27. Draparnaldioides simplex.
1 — branches in the upper part of thallome; 2 — lateral shoots; 3 — extra shoot with gametangia; 4 —
branching of the main spine; 5 — lateral shoots at the base of a large branch (rhizoid cover is removed);
6 — thallome fragment after gamete release and disintegration of gametangia; 7, 8 — gametes; 9 — gamete
fusion; 10 — zygote; 11 — zoosporangia. 2-4, 6, 7 — after Meyer [1930].
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 103
Замечания. В.Н. Яснитский [1934], изучавший размножение байкальских
Draparnaldioides, отмечает, что D. simplex начинает свое развитие подо льдом, а
в конце мая в Байкале встречаются вполне развитые, но стерильные талломы.
К середине августа развитие вида заканчивается и растения быстро отмирают.
Зиготы, которые удалось наблюдать В.Н. Яснитскому, всегда были двуядерны.
Наши исследования показали, что D. simplex растет на глубинах от 2.5 до 15 м,
на камнях. Встречается она и на мягком грунте, где в качестве субстрата ис-
пользует талломы харовых водорослей и высшие водные растения. Появление
D. simplex в Байкале отмечено нами [Ижболдина, 1970, 1976в] подо льдом в ап-
реле, а в районах выхода теплых ключей — в марте. Разные части озера могут
различаться по срокам появления и массового развития стерильных талломов,
но созревание гаметангиев и выход гамет во всем озере происходит в первой
половине июля, позднее встречаются лишь полуразрушенные талломы
D. simplex. Длина гаметангиев достигает 400-500 мкм. Бесполое размножение
изучено слабо. Часто на одних талломах с гаметангиями развиты зооспорангии
с микрозооспорами. Это клетки округлой или овальной формы, достигающие
50-55 мкм в диаметре. Выход зооспор наблюдать не удалось.
Draparnaldioides goroschankinii Meyer et Skabitsch. (=Draparnaldia goroschankinii
Meyer, 1922; Draparnaldiella goroschankinii Meyer et Skabitsch., 1969)
(рис. 28: 1-5; 29: 1-6)
Светло-зеленые слизистые сильно разветвленные кустики высотой 10-
15 см. Прикрепляются к субстрату при помощи неясно выраженной подошвы.
Главный ствол обильно разветвлен. Ветвление ствола моноподиальное, реже
мутовчатое. Мутовка состоит из 3-5 крупных ветвей. Боковые веточки оди-
ночные, супротивные или по 3-4 в мутовке, узкоромбической формы, с ясно
выраженным рахисом, всегда выдающимся за пределы веточки и заканчи-
вающимся обычно длинным тонким волоском. Ветвление боковой веточки
неправильное, двоякоперистое. Клетки главного ствола цилиндрические или
слегка бочонковидные, с тонкой оболочкой, в основании таллома 300-350 мкм
шириной, в средней части 200-280 мкм шириной, концы ветвей 30-40 мкм
шириной. Клетки боковых веточек 12-17.5 мкм шириной, волосков 5-8 мкм
шириной. Гаметангии 16-30 мкм шириной. Ризоидный покров выражен слабо
и развит лишь в нижних частях таллома.
Эндемик Байкала, широко распространен преимущественно в южной час-
ти озера, на глубине 2-3 м.
Draparnaldioides goroschankinii forma goroschankinii
Узкоромбическая форма боковых веточек.
Широко распространена преимущественно в Юж. Байкале.
Замечания. По нашим данным, в 1 см от конца ветвей ширина клеток
ствола 60-200 мкм, длина их равна ширине или чуть больше или меньше ее.
Конечные клетки главного ствола и крупных ветвей 15-25 мкм шириной.
Клетки в основании рахиса 12-37 мкм шириной, а в нижней части таллома их
ширина достигает 60-90 мкм. Нити в верхней части боковых веточек
(6)8-12(17) мкм шириной, волоски на конце нитей 3.6-8(10) мкм шириной и
до 1 мм длиной. Толщина ризоидного покрова в нижних частях таллома
достигает 150-250 мкм. Исследованиями В.Н. Яснитского [1934] установлено,
что D. goroschankinii появляется в Байкале подо льдом, половой процесс ее
104 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 28. Draparnaldioides goroschankinii.
1 — строение боковых веточек в верхней части таллома; 2 — боковые веточки в средней части таллома;
3 — боковые веточки в основании ствола молодого таллома (ризоцдный чехол убран); 4 — клетка ствола;
5 — основание молодого таллома. 2, 4 — по К..И. Мейру [1930], 5 - по Л.А. Ижболдиной [1976].
Fig. 28. Draparnaldioides goroschankinii.
1 — structure of lateral shoots in the upper thallome part; 2 — lateral shoots in the middle part of thallome; 3 — lat-
eral shoots at the spine base of a juvenile thallome (rhizoid cover is removed); 4 — spine cell; 5 — base of
a juvenile thallome. 2, 4 — after Meyer [1930], 5 - after Izhboldina [1976].
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 105
I____I
6:30 цт
Рис. 29. Draparnaldioides goroschankinii.
1 — фрагмент ветви с гаметангиями; 2 — гаметангий; 3 — гаметы; 4 — копулярция гамет; 5 —
зиготы через 3-5 мин после слияния клеток; 6 — зооспорангий. 2 — по К.И. Мейру [1930], 1, 6 - по
Л.А. Ижболдиной [1976].
Fig. 29. Draparnaldioides goroschankinii.
1 — fragment of a branch with gametangia; 2 — gametangium; 3 — gametes; 4 — gamete copulation;
5 — zygotes 3-5 minutes after cell fusion; 6 — zoosporangium. 2 — after Meyer [1930], 1, 6 - after
Izhboldina [1976].
106 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
приходится на первую половину июля и к середине июля она отмирает.
Вдоль северо-западного побережья Юж. Байкала отмечены [Яснитский, 1934;
Ижболдина, 1978] значительные межгодовые колебания в количественном
развитии D. goroschankinii. При наших исследованиях [Ижболдина, 1990в]
отмечено, что в северной части озера формирование гаметангиев начинается
на 10-15 дней позднее. Гаметангии 10-30 мкм шириной и 62-350 мкм длиной.
Гаметы 6-10.5 мкм длиной и 7.8-8.8 мкм шириной. Иногда из клеток крупных
ветвей формируются образования, напоминающие зооспорангии у рода Dra-
parnaldia Вогу [Ижболдина, 19766]. Зооспорангии грушевидной формы, 36.6-
40 мкм длиной, 12.5-18.3 мкм шириной. Выход зооспор пронаблюдать не
удалось. Зооспорангии развиты на тех же талломах, что и гаметангии.
Вид широко распространен в Байкале и при наших исследованиях не был
обнаружен лишь на участке восточного побережья между ст. Танхой на юге и
о. Лиственичный на севере. Растет на твердом грунте на глубине (1)2.5-5(13)
м. На мягком грунте в качестве субстрата может использовать харовые водо-
росли и высшие водные растения.
Draparnaldioides goroschankinii {огтз. plumosa Meyer (1930) (рис. 30; 31: 1—4)
Широкоромбическая форма боковых веточек, которые очень обильно раз-
ветвлены, благодаря чему таллом принимает более пушистый характер.
В Байкале найдена К.И. Мейером [1930] в июне 1925 г. в губе Половинной
на глубине 3 м.
Рис. 30. Draparnaldioides goroschankinii f. plumosa, общий вид таллома.
Fig. 30. Draparnaldioides goroschankinii f. plumosa, general view of thallome.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 107
1-4:125 цт
Рис. 31. Draparnaldioides goroschankinii f. plumosa.
1 — фрагмент верхней части таллома с формирующимися гаметангиями; 2 — боковые веточки в
основании таллома (ризоидный покров убран); 3 — зооспорангии и гаметангии на боковых ве-
точках из основания таллома; 4 — клетки главного ствола в самом основании таллома, ризоиды.
Fig. 31. Draparnaldioides goroschankinii f. plumosa.
1 — fragment of upper thallome part with growing gametangia; 2 — lateral shoots at the thallome base
(rhizoid cover is removed); 3 — zoosporangia and gametangia on lateral shoots at thallome base; 4 —
cells of the main spine at the very base of thallome, rhizoids.
108 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Замечания. Найдена нами [Ижболдина, 1990в] в районе пади Бол. Коты, в
губе Половинная, у о. Угунгой (в прол. Мал. Море) и на Сев. Байкале в губе
Том па и у мыса Мужинай. Растет в мае-июле на глубине 3-5(15) м, на твердом
грунте или использует в качестве субстрата харовые водоросли и высшие
водные растения.
Клетки главного ствола в самом основании таллома 100-120 мкм шириной,
но уже на 1 см выше их ширина увеличивается до 250-280 мкм. В средней части
таллома ширина клеток ствола и крупных ветвей 190-230 мкм, а в 1 см от
верхнего конца ветви - 68-112(137) мкм. Конечные клетки крупных ветвей
20-37 мкм шириной. В верхней и средней частях таллома клетки боковых
веточек в основании рахиса 20-30 мкм шириной, а в нижней части таллома -
50-60 мкм. Размножение происходит одновременно с типовой формой в
первой половине июля. Гаметангии 12-25 мкм шириной, 190-380 мкм длиной.
Зооспорангии округлой или овальной формы, 50-75 мкм шириной, 86-112 мкм
длиной.
Draparnaldioides arenaria Meyer et Skabitsch. (=Draparnaldia arenaria Meyer,
1922; Draparnaldiella arenaria Meyer et Skabitsch., 1969) (рис. 32, 33)
Обильно разветвленные темно-зеленые кустики высотой 10-20 см. Ветвле-
ние главного ствола моноподиальное, реже мутовчатое. Боковые веточки в
верхней и средней частях таллома узкие, сжатые, с ветвями, расположенными
лишь у их основания. Рахис резко выражен, заканчивается длинным толстым
волоском. В основании ствола и крупных ветвей боковые веточки сильно раз-
растаются и определить по ним вид водоросли трудно. Боковые веточки на
клетках ствола и ветвей расположены обычно мутовками, по 2-3, реже одиноч-
ные. Главный ствол и крупные ветви состоят из бочонковидных клеток с тон-
кой оболочкой, у основания таллома 300-340 мкм шириной, в средней части
главного ствола 180-200 мкм шириной, конечные клетки ветвей 50-70 мкм
Рис. 32. Draparnaldioides arenaria, общий вид.
Fig. 32. Draparnaldioides arenaria, general view.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 109
Рис. 33. Draparnaldioides arenaria f. arenaria.
1 — строение таллома на конце ветви; 2 — то же, в средней части ветви; 3 — боковые веточки в основа-
нии таллома (ризоидный чехол убран); 4 — боковые веточки; 5 — гаметангий на рахисе боковой веточ-
ки; 6 — после разрушения гаметангия рахис становится коротким; 7 — гаметангий и зооспорангий на
рахисе боковой веточки. 4 — по К.И. Мейру [1930].
Fig. 33. Draparnaldioides arenaria f. arenaria.
1 — thallome structure at the end of the branch; 2 — the same in the middle part of the branch; 3 — lateral
shoots at the base of thallome (rhizoid cover removed); 4 — lateral shoots; 5 — gametangium on the lateral
shoot of the rachis; 6 — after gametangium destruction of the rachis becomes short; 7 — gametangium and
zoosporangium on the lateral shoot of the rachis. 4 — after Meyer [1930].
ПО Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
шириной. Нити боковых веточек 22-45 мкм шириной, волоски на концах ни-
тей 11-25 мкм шириной. Гаметангии 45-60 мкм шириной. Ризоидный покров
развит приблизительно в нижней трети таллома.
Эндемик Байкала. По данным К.И. Мейера [1930], широко распростране-
на в Байкале, преимущественно в северной его части.
Draparnaldioides arenaria forma arenaria
Узкие, сжатые и скудно ветвящиеся в нижней части боковые веточки,
прижатые к клеткам ствола и ветвей.
Широко распространена преимущественно в Сев. Байкале, на глубине
3-15 м.
Замечания. В.Н. Яснитский [1934], изучавший размножение D. arenaria,
отмечает, что в районе пос. Бол. Коты развитие вида начинается с июля и
продолжается до начала зимы. Максимум развития в сентябре-октябре, а
отдельные талломы встречены им до конца января. Половой процесс наблю-
дали в конце августа-начале ноября, т.е. во время максимального развития
D. arenaria. Гаметы широкогрушевидной формы, с 4 жгутиками, 5-10 мкм
длиной, 4-7 мкм шириной, длина жгутиков 10-11 мкм. Зиготы имеют удли-
ненно-грушевидную форму и 8 жгутиков. Длина зиготы 12-14 мкм, ши-
рина 7-8 мкм. Зиготы сохраняют подвижность в течение 10-15 мин.
Проведенные нами исследования [Ижболдина, 1990а] позволили расширить
границы ареала D. arenaria. Отсутствует она вдоль восточного побережья южной
и средней частей озера, испытывающих на себе влияние крупных рек Выдрина,
Снежная, Мишиха, Селенга, Баргузин. Появление молодых талломов D. arenaria
во всех частях озера наблюдали во второй половине июня. С середины июля,
когда высота талломов достигает 6-8 см, на рахисах — самых крупных нитях бо-
ковых веточек, формируется гаметангий, опоражнивающийся обычно к концу
августа. В это время начинается массовое формирование гаметангиев на осталь-
ных нитях боковых веточек, а рахис становится коротким после разрушения зоо-
спорангия. Часто на рядом расположенных веточках рахис может быть разной
длины (рис. 33: 5-6). В середине сентября отмечали массовый выход гамет. От-
дельные плодущие экземпляры встречены до октября. Длина гаметангиев может
достигать 400-700 мкм, ширина 60-75 мкм. На одних нитях с гаметангиями мо-
гут быть развиты зооспорангии (рис. 33: 7). Последние встречены на талломах
из района пади Широкая и губы Харанса в прол. Мал. Море.
Draparnaldioides arenaria forma plumosa Meyer, 1930 (рис. 34: 1—4)
Сильное перистое ветвление таллома вследствие того, что оттопыренные
мутовки боковых веточек располагаются близко друг к другу и в каждой мутов-
ке одна или 2 веточки развивают свой рахис в длинную ветвь.
Эндемик Байкала. В небольшом количестве встречена К.И. Мейером
[1930] в Курме (прол. Мал. Море).
D. arenaria f. plumosa нами встречена [Ижболдина, 1990а] также в районе
бух. Курма, на глубине 4-7 м, и в районе пади Бугульдейка, на глубине 2-5 м.
Draparnaldioides arenaria forma elegans Izhboldina, forma nov. (рис. 35, 36)
Мутовки боковых веточек в верхней и средней частях таллома удалены на
значительное (370-600 мкм) расстояние и не соприкасаются друг с другом
боками.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 111
Рис. 34. Draparnaldioides arenaria f. plumosa.
1 — общий вид таллома; 2 — боковые веточки на конце ветви; 3 — фрагмент боковой веточки из ос-
нования крупной ветви; 4 — у 2 боковых веточек рахис продолжает расти, образуя новую ветвь.
Fig. 34. Draparnaldioides arenaria f. plumosa.
1 — general view of thallome; 2 — lateral shoots at the branch tip; 3 — fragment of the lateral shoot at the
base of a large branch; 4 — rachis on two lateral shoots continues to grow forming a new branch.
112 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 35. Draparnaldioides arenaria f. elegans,
внешний вид таллома в натуральную вели-
чину.
Fig. 35. Draparnaldioides arenaria f. elegans,
a full-size external view of thallome.
Оз. Байкал, против пади Бол. Коты, на глубине 7-15 м, на песке. Растет
прикрепляясь к талломам Nitella flexilis (L.) Ag.
Draparnaldioides arenaria forma pilosissima Izhboldina, forma nov.
(рис. 37: 1-3; 38: 1-2)
Невысокие (2-2,5 см) и сильно опушенные талломы. Боковые веточки
располагаются близко друг к другу и заканчиваются толстыми и длинными (до
2 мм) волосками.
Оз. Байкал, к югу от устья р. Бугульдейка. Растет прикрепляясь к ветвям
Myriophyllum spicatum, на глубине 2-4 м.
Draparnaldioides arnoldii Meyer et Skabitsch. (=Draparnaldia arnoldii Meyer,
1925; Draparnaldiella arnoldii Meyer et Skabitsch., 1969) (рис. 39; 40)
Сильно разветвленные, густые и слизистые кустики высотой 10-15 см.
Расположение крупных ветвей на стволе супротивное или очередное. Таллом
прикреплен к камням круглой подошвой. Боковые веточки располагаются на
стволе и ветвях мутовками - по 2-3 в мутовке, и сидят на каждой клетке ствола
или через 1-2.
Боковая веточка округлая, с хорошо выраженным рахисом. Иногда корот-
ким, иногда длинным, но всегда заканчивающимся на верхушке закругленной
клеточкой без щетинки. Веточка имеет сложное двоякоперистое ветвление,
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 113
Рис. 36. Draparnaldioides arenaria f. elegans.
1 — расположение боковых веточек на конце маленькой ветви; 2 — боковые веточки в основании
той же ветви; 3 — боковые веточки в средней части главного ствола, ризоидный чехол убран; 4 —
клетки боковых веточек; 5 — клетки главного ствола в основании таллома, ризоиды.
Fig. 36. Draparnaldioides arenaria f. elegans.
1 — position of lateral shoots at the tip of a small branch; 2 — lateral shoots at the base of the same branch;
3 — lateral shoots in the middle part of main spine, rhizoid cover removed; 4 — cells of the lateral shoots;
5 — cells of main spine at thallome base, rhizoids.
114 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 37. Draparnaldioides arenaria f. pilosissima.
1 — боковые веточки на конце ветви; 2 — боковые веточки ниже на той же ветви; 3 — общий вид
таллома.
Fig. 37. Draparnaldioides arenaria f. pilosissima.
1 — lateral shoots at the branch tip; 2 — lateral shoots below on the same branch; 3 — general view of
thallome.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 115
Рис. 38. Draparnaldioides arenaria f. pilosissima.
1 — боковые веточки очень тесно расположены на клетках ствола; 2 — боковые веточки на круп-
ной ветви.
Fig. 38. Draparnaldioides arenaria f. pilosissima.
1 — lateral shoots tightly located on the spine cells; 2 — lateral shoots on a large branch.
116 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
1
2:1 ст
i
j
Рис. 39. Draparnaldioides
arnoldii f. arnoldii. Общий вид
таллома.
1 — слабое увеличение (фото
О.А. Тимошкина), 2 — таллом в
натуральную величину.
Fig. 39. Draparnaldioides
arnoldii f. arnoldii, general view
of thallome.
1 — low magnification (photo by
O.A. Timoshkin), 2 — a full-size
thallome.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 117
Рис. 40. Draparnaldioides arnoldii.
1-3 — строение таллома в разных частях крупной ветви; 4 — на рахисе боковой веточки сформиро-
ван гаметангий; 5-6 — после разрушения гаметангия рахис в боковых веточках становится
коротким, а — гаметангий; б — зооспорангий; в — ризоиды. 5 — по К.И. Мейеру [1930].
Fig. 40. Draparnaldioides arnoldii.
1-3 — structure of thallome in various parts of a large branch; 4 — gametangium formed on the rachis of a
lateral shoot; 5-6 — after gametangium desintegration rachis on lateral shoots becomes short, a —
gametangium; 6 — zoosporangium; в — rhizoids. 5 — after Meyer [1930].
118 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
придающее ей веерообразный характер. Клетки главного ствола и крупных вет-
вей цилиндрические, слегка перетянутые у поперечных перегородок. Их шири-
на 170- 180 мкм у основания таллома, 90-100 мкм - в средней части таллома,
нити боковых веточек 25-45 мкм шириной. Гаметангии 40-60 мкм шириной.
Ризоидный покров развит хорошо, окутывает 2 нижние трети таллома.
К.И. Мейер [1930] отмечает близость к D. arenaria, от которой D. arnoldii
отличается двоякоперистым ветвлением боковых веточек, ризоидным покро-
вом, одевающим 2 нижние трети таллома, коротким рахисом, заканчивающим-
ся закругленной клеткой.
Эндемик Байкала. Найден К.И. Мейером [1930] во всех частях озера, но
встречается редко и в небольшом количестве.
Draparnaldioides arnoldii forma arnoldii Meyer et Skabitsch.
Боковые веточки округлые, веерообразно разветвлены, имеют хорошо вы-
раженный рахис, заканчивающийся округлой клеткой.
В небольшом количестве встречен во всех частях озера на глубине 2.5-15 м.
Замечания. По нашим данным, D. arnoldii развита в Байкале в те же сроки
(конец июня — октябрь), что и D. arenaria. Найдена вдоль большей части по-
бережья на глубине от 1.5-2 до 5-10 м, на камнях. Сезонные наблюдения за
развитием данного вида, как и других видов Draparnaldioides, свидетельствуют
об усложнении строения боковых веточек от вершины к основанию таллома.
В молодых (верхних) частях талломов ризоидный покров и у D. arenaria и
D. arnoldii отсутствует и боковые веточки D. arnoldii похожи по своему строе-
нию на таковые у D. arenaria, т.е. они слабо разветвлены и в них обычно хоро-
шо выражен рахис (рис. 39: Г, 40:1). В июле-августе у обоих видов рахис в бо-
ковых веточках длинный и заканчивается толстой щетинкой. Формирование
гаметангиев начинается на самых крупных нитях боковых веточек, каковыми
у рассматриваемых видов являются рахисы. На остальных, более тонких,
ветвях гаметангии развиваются позднее. После выхода гамет во второй поло-
вине сентября оболочки гаметангиев разрушаются и рахис с закругленной
клеткой на конце становится значительно короче остальных ветвей боковой
веточки, т.е. становится типичным, по описанию К.И. Мейера, для D. агпоГ
dii. Таким образом, к числу признаков, отличающих рассматриваемые виды,
может быть отнесена разная степень ветвления боковых веточек и ризоидный
покров, одевающий клетку ствола и крупных ветвей на одну нижнюю треть
таллома у D. arenaria и на 2 нижние трети у D. arnoldii. У встреченных нами
талломов D. arnoldii ширина клеток ствола в нижней части 170-280 мкм, в
средней — 90-100(207) мкм, в верхней — 40-100 (118) мкм, конечных клеток
ветвей — 35-45 мкм, в верхней части нитей боковых веточек — 12.5-31 мкм,
волосков 12.5-31 мкм. Гаметангии 35-60(70-100) мкм шириной и 370-
750 мкм длиной. Одновременно с гаметангиями и на одних с ними талломах
могут быть развиты зооспорангии округлой или овальной формы 32-50 мкм
шириной (рис. 40: 4, 6).
В Мал. Море — у о. Хынок, у пади Сенной на восточном берегу о. Оль-
хон — встречены талломы D. arnoldii, у которых нити боковых веточек заканчи-
ваются очень длинными (до 2.5 мм) волосками, отчего талломы кажутся опу-
шенными. Толщина ризоидного покрова в средней части таллома 200-
250 мкм, а в основании талломов из районов Мал. Моря — 500 мкм, на талло-
мах с мыса Амундакан — 1000 мкм.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 119
Кроме вышеописанной типичной формы К.И. Мейером встречены еще
2 формы, самостоятельное значение которых, по его мнению, требует допол-
нительных исследований.
Draparnaldioides arnoldii forma compacta Meyer, 1930
Имеет более низкие, чем тип, талломы — 6-7 см, более густое и обильное
ветвление, ветви короткие. Мутовки боковых веточек сидят на стволе через
2-3 клетки.
Местонахождение вида не указано.
Замечания. D. arnoldii f. compacta встречена нами в районе пади Сенной
(восточный берег о. Ольхон) и севернее мыса Туркинский у пос. Гремячинск.
Боковые веточки сидят на стволе часто и смыкаются боками (рис. 41: 1-4; 42:
1-4). Толщина ризоидного чехла в основании талломов достигает 300-600 мкм.
На талломах поселяется масса сувоек рода Vorticella и диатомей родов Navicula,
Synedra и Cymbella.
Draparnaldioides arnoldii forma gracilis Meyer, 1930
Имеет более длинные и тонкие ветви, широко раздвинутые и менее густые
мутовки боковых веточек.
Замечания. D. arnoldii f. gracilis (рис. 43:1; 44:1-3) встречена нами в августе
в Мал. Море — в губе Курминской, на глубине 5 м, в прол. Ольхонские Воро-
та — в бухте Базарной, у уреза воды, в губе Амундакан, на глубине 2.5 м, на
камнях. Длина волосков на нитях боковых веточек достигает 1.5-2.5 мм. Гаме-
тангии 30-44 мкм шириной и до 500 мкм длиной.
Draparnaldioides pumila Meyer et Skabitsch. (=Draparnaldia pumila Meyer, 1926;
Draparnaldiella pumila Meyer et Skabitsch., 1969) (рис. 45: 1—6; 46: 1—7)
По описанию К. И. Мейера [1930], представляет слизистые кустики высотой
7-10 см, прикрепляющиеся к субстрату маленькой круглой подошвой. Главный
ствол хорошо прослеживается на протяжении всего кустика и утончается от
основания к вершине от 2-3 до 0.5 мм. От ствола отходят ветви, ширина
которых 0.5 мм. Иногда в основании главного ствола образуется несколько
крупных ветвей такого же диаметра, как ствол. Они в свою очередь ветвятся, что
делает ветвление таллома более обильным. Боковые веточки широкояйце-
видные или полукруглые, с четко выраженным рахисом, заканчивающимся
длинным волоском, ширина которого может быть больше, чем у рахиса. Раз-
растаясь, рахисы могут давать начало новым ветвям. Боковые веточки рас-
полагаются обычно мутовками по 4, которые, соприкасаясь краями, образуют
сплошной покров вокруг ствола. Клетки ствола и крупных ветвей цилин-
дрические или слегка бочонкообразные, в основании таллома 250 мкм шири-
ной, в средней части его 135-175 мкм, в верхушечной 45-50 мкм. Длина
клеток равна ширине, до 1.5 раза больше или меньше ее. Клетки боковых
веточек цилиндрические, 10-13.5 мкм шириной. Конечные клетки переходят
в волосок, диаметр которого может превышать ширину самой нити. Ризоид-
ный покров развит хорошо и появляется уже на расстоянии 1.5-2 см от конца
ветви.
Эндемик Байкала. К.И. Мейером [1930] отмечен как вид, характерный для
северной части озера. Там же обнаружен А.П. Скабичевским [1934].
120 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 41. Draparnaldioides arnoldii f. compacta (севернее мыса Туркинский).
1 — внешний вид таллома; 2-4 расположение боковых веточек в разных частях крупной ветви.
Fig. 41. Draparnaldioides arnoldii f. compacta (north of Turkinsky Cape).
1 — external view of thallome; 2-4 — position of lateral shoots in various parts of a large branch.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 121
2
Начало рис. 42.
Beginning of Fig. 42.
122 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 42. Draparnaldioides arnoldii f. compacta (падь Сенная на о. Ольхон).
1 — боковые веточки на конце ветви; 2 — та же ветвь ниже; 3 — рахис боковой веточки прорастает в
новую ветвь; 4 — боковые веточки с главного ствола. От клеток в основании веточек отходят ризоиды.
Fig. 42. Draparnaldioides arnoldii f. compacta (Sennaya Pad on Olkhon Island).
1 — lateral shoots at the tip of the branch; 2 — the same branch below; 3 — rachis of the lateral shoots grows
into a new branch; 4 — lateral shoots on the main spine. Rhizoids off the cells at the base of the shoots.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 123
Замечания. По нашим данным [Ижболдина, 1990в], вид широко распро-
странен вдоль западного прибрежья Байкала. Вдоль восточного берега встре-
чен на севере между мысами Ирексокон и Каткова и на юге — от Муринской
банки до мыса Шаманский. Вид появляется в конце июля, массовое развитие в
сентябре — начале октября на глубине (1)2.5-5(11) м, на камнях. В конце сен-
тября — октябре наблюдали формирование гаметангиев. Отдельные талломы
встречаются в ноябре — декабре. В августе на талломах D. pumila поселяется
масса сувоек рода Vorticella. Ширина клеток ствола в основании таллома может
достигать 290 мкм, в средней части — 300 мкм, в верхней — 87-168 мкм, конеч-
ные клетки 20-50 мкм; в основании рахиса боковой веточки — 18.5-37.5 мкм,
волосков на нитях веточки — 12-16.8 мкм. Толщина ризоидного покрова в ос-
новании таллома 200-400 мкм, в средней части — 125-300 мкм, в 1-2 см от вер-
шины клетки ствола оплетают единичные ризоиды.
На Сев. Байкале — в районе губ Амундакан, Богучанской, Слюдянской,
Онокочанской и в Чивыркуйском заливе — встречены талломы D. pumila высо-
той 4-5.7 мм. Главный шнуровидный ствол достигает в диаметре 4-5 мм, а
единичные крупные ветви 2-3 мм. Они не утончаются к вершине (рис. 47:1-3)
и усажены массой тонких (0.5-1 мм) и коротких 3-5(10) мм ветвей. Толщина
нитей боковых веточек 13-20 мкм и высота последних в 1.5 раза больше, чем у
типа. Размер и форма клеток ствола и строение боковых веточек типично для
D. pumila [Ижболдина, 197бв].
Draparnaldioides pilosa Meyer et Skabitsch. (=I)raparnaldia pilosa Meyer, 1926;
Draparnaldiella pilosa Meyer et Skabitsch., 1969) (рис. 48: 1—3)
По К.И. Мейеру [1930], слизистые сильно разветвленные кустики высотой
15-20 см. Многочисленные длинные шнурованные ветви, постепенно утон-
чающиеся, к концам становятся нитевидными. На стволе и крупных ветвях
развита масса коротких ветвей, от 1-2 см до 11.4 см длиной. Присутствие мел-
ких веточек придает кустику лохматый вид. Боковая веточка округлая; ветвле-
ние нитей обильное, рахис незаметен, так как быстро переходит в ветви, кото-
рые по своему размеру не уступают рахису. Лишь на очень молодых ветвях ра-
хис можно проследить. Нити ветвей боковой веточки плотно прилегают друг к
другу, изогнуты дугообразно и выпуклой стороной обращены наружу. Концы
нитей заканчиваются длинными толстыми щетинками, придающими таллому
волосистый вид. Боковые веточки собраны в мутовки по 3 и сидят довольно
рыхло, но соприкасаются краями. Клетки главного ствола цилиндрические,
слегка перетянутые у поперечных перегородок, 160-180 мкм шириной в ниж-
них частях таллома, 100-150 мкм шириной в средних и в окончаниях —
40-55 мкм шириной. Толщина нитей и щетинок в боковых веточках
10.7-12.9 мкм. Формирующиеся гаметангии 17-21 мкм шириной. Ризоидный
покров развит очень сильно и особенно в нижней части таллома, где его тол-
щина может достигать 1-1.5 мм.
По К.И. Мейеру [1930], распространена по всему Байкалу, встречена им в
июле — августе.
Замечания. По нашим данным [Ижболдина, 1990в], D. pilosa широко рас-
пространена вдоль открытых прибрежий озера, на камнях, на глубине
(0.6)1.5-3.5(5) м в июне—сентябре. В конце августа — начале сентября в рай-
оне мыса Разбор (Сев. Байкал) и мыса Немчинова (Юж. Байкал) встречены
талломы D. pilosa с формирующимися гаметангиями 20-25 мкм шириной
(рис. 49). В районе пади Бол. Коты в августе наблюдали на талломах D. pilosa
124 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 43. Draparnaldioides arnoldii f. gracilis, общий вид таллома.
Fig. 43. Draparnaldioides arnoldii f. gracilis, general view of thallome.
массовое развитие зооспорангиев весьма разнообразной формы, заполненных
яйцевидными клетками (рис. 50). Проследить выход зооспор не удалось.
К.И. Мейер [1930] отмечал большое сходство D. pilosa и D. vilosa, высказы-
вая предположение о том, что, возможно, правильнее смотреть на D. vilosa как
на форму D. pilosa, от которой первая отличается характером ветвления и раз-
мерами таллома, наличием немногочисленных волосков. Наши данные также
подтверждают сходство этих видов. Более того, нами встречен целый ряд
Draparnaldioides, обладающих морфологическими признаками нескольких од-
новременно развивающихся видов со слабо выраженным рахисом. Возможно,
это результат межвидового скрещивания близкородственных видов. Установ-
ление видовой принадлежности таких водорослей требует более глубоких ис-
следований на молекулярном уровне. В настоящей работе мы приведем лишь
краткое морфологическое описание встреченных нами форм.
Талломы D. pilosa, развитые вдоль юго-восточного побережья озера, не
превышают 10-15 см в высоту, чаще они короче (рис. 51). На концах ветвей
боковые веточки очень слабо разветвлены и в них хорошо виден рахис
(рис. 52: 1). В средней части таллома строение боковых веточек аналогично
описанному для данного вида К.И. Мейером [1930]. Однако в некоторых бо-
ковых веточках хорошо выражен рахис, что придает сходство с D. vilosa
(рис. 52: 2). В нижней части таллома, где рахис представлен 1-2 клетками, бо-
ковые веточки по строению походят на D. baicalensis (рис. 52: 4), но более
обильно ветвятся от самого основания веточки и несут массу длинных, до
1-1.5 мм, волосков.
В районе бух. Хакусы (Сев. Байкал) встречены талломы D. pilosa высотой
18-22 см. Кустики не очень обильно разветвлены, но строение боковых вето-
чек типично для вида, а размер клеток ствола и нитей боковых веточек не-
сколько больше, чем указывает для этого вида К.И. Мейер [1930] (рис. 53:1-4).
Вблизи устья р. Большая встречены обильно разветвленные талломы D. pilosa
высотой 6-10 см, с массой коротких (2-5 см) и тонких, почти нитевидных,
ветвей (рис. 54: 1-4). Строение боковых веточек имеет сходство с таковыми у
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 125
Рис. 44. Draparnaldioides arnoldii f. gracilis.
1 — расположение боковых веточек на концах коротких (длиной 1, 2 см) ветвей; 2 — боковые ве-
точки в основании той же ветви; 3 — боковые веточки на крупной ветви, гифовый чехол убран.
Fig. 44. Draparnaldioides arnoldii f. gracilis.
1 — location of lateral shoots at the tips of short (1, 2 cm long) branches; 2 — lateral shoots at the base of
the same branch; 3 — lateral shoots on a large branch, hypha is withdrawn.
126 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
1-6:1 ст
Рис. 45. Draparnaldioides pumila, внешний вид талломов из разных районов оз. Байкал.
1 — ст. Танхой; 2 — бух. Анга; 3 — падь Бол. Коты; 4 — о. Угунгой в прол. Мал. Море; 5 — мыс Ва-
лукан; 6 — бух. Фролиха.
Fig. 45. Draparnaldioides pumila, external view of thallomes from different regions of Lake
Baikal.
1 — Tankhoy Station, 2 — Anga Bay, 3 — Bolshye Koty Settlement, 4 — Ugungoy Island in Maloe More
Strait, 5 — Valukan Cape, 6 — Frolikha Bay.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 127
Рис. 46. Draparnaldioides pumila.
1 — строение боковых веточек на маленькой (0.5-1 см длиной) ветви; 2-4 — боковые веточки в
основании той же ветви; 5 — боковые веточки в средней части крупной ветви; 6 — рахис боковой
веточки прорастает в новую ветвь; 7 — боковые веточки из основания таллома. 2, 3 — по
К.И. Мейеру [1930].
Fig. 46. Draparnaldioides pumila.
1 — structure of lateral shoots on a small (0,5-1 cm long) branch; 2-4 — lateral shoots at the base of the
same branch; 5 — lateral shoots in the middle part of a large branch; 6 — rachis on a lateral shoot grows
into a new branch; 7 — lateral shoots on the thallome base. 2, 3 — after Meyer [1930].
128 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Данные о размерах Draparnaldioides pilosa в разных районах оз. Байкал
SO
Л
ZJ
S
=?
ю
Л
н
Ширина нитей боковых веточек, мкм щетинки £ 2 22-7 00^22 ое Г** I 1 1 1 1 I т—I о । 1Л чп “7 I Н о чп 22 । “ Н Н rid s s d d s
в верхней части О\ Г*» f*» f*» о гч’ 2 щ 2 od ш об in о оо сп «—। in I I -и 1— •— гч »—> w 1 ГЧ *П Itf-jlll 1|11 гч !• in о in in о гч гч о S 2 гч гч гч об ”
рахис в основа- нии ш ш о f** f** (Q г-ч ,а о СП СП Т «н IO и 7 II 1 ГЧ in г* 1 00 00 гч S 01 Я*
Ширина клеток ствола и крупных ветвей, мкм конечные клет- ки ЧП ЧП Н ЧП о ° 5 § о ЧП Г. Г~ ЧП ПП ill 1 чг 1 1 1 1 чп чп о ' _| 00 еч — Г-Г; £ < СП СП СП £** СП
в 1-1.5 см от | вершины 90-157 93-118 62-112 87-120 157-188 93-157 94-145 87-112 62-94 87-100 87-112
в средней части Q О 1П О О СП о so Г*» 00 Г*. in г*» ГЧ 00 о о <п о so 00 ГЧ »— гч гч сп сп сп гч гч сп гч го 1 1 1 1 1 III I’ll ©о©©«п о о © о гч о О СП о г*» г*» г** 00 00 00 о 00 OS ^ч^СПГЧГЧ гч — Я— ГЧ ч- гч
в основании таллома 160-180 180-280 300-460 244-343 250-337 275-343 240-343 100-280 207-281 340-500 130-280 318-330
Высота талло- ма, см 8й2 2 Я ° оо Я । । । । । iii <П 40 Г** о\ С^чоОО
Район сброса По К.И. Мейеру [1930] Мыс Немчинова Падь Култук Падь Клюевка Нижнее Изголовье п-ова Святой Нос Мыс Ото-Хушун Мыс Амундакан У р. Большая Мыс Якшакан Падь Варначка Бух. Хакусы Бух. Базарная
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 129
Рис. 47. Draparnaldioides pumila.
1 — из района г. Богучанской; 2 — из района мыса Амундакан; 3 — из Чивыркуйского залива.
Fig. 47. Draparnaldioides pumila.
1 — from the region of Boguchanskaya Bay; 2 — from the region of Amundakan Cape; 3 — from
Chivyrkuy Bay.
D. vilosa, но отличается от них наличием массы длинных волосков. Размер кле-
ток ствола и крупных ветвей ближе к D. vilosa. Большие, чем приводит для
D. pilosa К.И. Мейер [1930], размеры клеток ствола, крупных ветвей и нитей бо-
ковых веточек, мы отмечали у талломов D. pilosa из других районов озера
(табл. 6).
Draparnaldioides vilosa Meyer et Skabitsch. (=Draparnaldia vilosa Meyer, 1926;
Draparnaldiella vilosa Meyer et Skabitsch., 1969) (рис. 48: 4—7)
По К.И. Мейеру [1930], сильно ветвистые кустики высотой 6-10 см. Ветв-
ление очень густое. Главный ствол и крупные ветви в свою очередь усажены
многочисленными короткими ветвями длиной 1-2 мм и шириной 0.5 мм. Бо-
ковые веточки компактные, в очертании округлые, образованы сильно ветвя-
щимися нитями, некоторые из них заканчиваются длинными волосками такой
же ширины, что и нити. Волосков образуется немного. У молодых боковых ве-
130 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 48. 1-3 — Draparnaldioides pilosa:
1 — общий вид таллома, 2 — клетки
ствола, 3 — боковая веточка; 4-7 —
D. vilosa: 4 — общий вид таллома,
5 — молодая боковая веточка, 6,7 —
боковые веточки.
1-7 — по К..И. Мейеру [1930].
Fig. 48. 1-3 — Draparnaldioides pilosa: 1 — general view of thallome, 2 — spine cells, 3 — lat-
eral shoot; 4-7 — D. vilosa: 4 — general view of thallome, 5 — lateral juvenile shoot, 6, 7 — lat-
eral shoots.
1-7 - after Meyer [1930].
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 131
3:125 |im
Рис. 49. Draparnaldioides pilosa, район мыса Разбор (Сев. Байкал).
1 — общий вид талллома; 2 — боковые веточки в верхней части таллома, гаметангии; 3 — боковая
веточка в основании крупной ветви.
Fig. 49. Draparnaldioides pilosa, region of Razbor Cape (Northern Baikal).
1 — general view of thallome; 2 — lateral shoots in the upper part of thallome, gametangia; 3 — lateral
shoot at the base of a large branch.
132 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 50. Draparnaldioidespilosa, район пади Бол. Коты (Юж. Байкал).
1,2 — общий вид таллома, 2 — после выхода гамет нижняя часть таллома полуразрушена; 3 — боко-
вые веточки из верхней части таллома с зооспорангиями; 4 — боковая веточка в основании крупной
ветви, зооспорангии.
Fig. 50. Draparnaldioides pilosa: from the region of Bolshye Koty Pad (Southern Baikal).
1,2 — general view of thallome, 2 — after gamete release the lower part of thallome is partially desintegrated;
3 — lateral shoots from the upper part of thallome with zoosporangia; 4 — lateral shoots at the base of a large
branch, zoosporangia.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 133
Рис. 51. Draparnaldioides pilosa, район мыса Немчинова (юго-восточное побережье
оз. Байкал), общий вид таллома.
Fig. 51. Draparnaldioides pilosa from the region of Nemchinova Cape (south-eastern coast of
Lake Baikal), general view of thallome.
точек рахис хорошо выражен, у сформировавшихся веточек он короткий и не-
заметный.
Мутовки боковых веточек соприкасаются краями, образуя вокруг клеток
ствола сплошной покров. Клетки главного ствола и крупных ветвей цилиндри-
ческие, у поперечных перегородок слегка перетянутые, в основании таллома
300 мкм шириной, в 2 см от конца — 200 мкм шириной, на коротких ветвях —
47 мкм шириной. Рахис в основании боковой веточки 26 мкм шириной,
нити — 10-15 мкм шириной. Гаметангии 26 мкм шириной. Ризоидный покров
появляется уже на расстоянии 0.5 см от концов ветвей.
Редкий для Байкала вид, встречен К.И. Мейером [1930] в районе мысов
Толстый, Немнянка, Мужинай, Тонкий, у ст. Мысовая.
134 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 52. Draparnaldioides pilosa, юго-восточное побережье оз. Байкал.
1 — боковые веточки на конце короткой ветви; 2 — в основании той же ветви; 3 — строение боковой
веточки; 4 — боковые веточки на крупной ветви, ризоидный чехол убран. 3 — по К..И. Мейру [1930].
Fig. 52. Draparnaldioides pilosa (south-eastern coast of Lake Baikal).
1 — lateral shoots at the end of a short branch; 2 — at the base of the same branch; 3 — structure of a lateral
shoot; 4 — lateral shoots on a large branch, rhizoid cover withdrawn. 3 — after Meyer [1930].
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 135
Рис. 53. Draparnaldioides pilosa.
1 — фрагмент таллома из района бух. Хакусы; 2 — боковые веточки на короткой маленькой ветви,
некоторые веточки, разрастаясь, дают начало новой ветви; 3-4 — боковые веточки из нижней час-
ти таллома.
Fig. 53. Draparnaldioides pilosa.
1 — fragment of thallome from the region of Khakusy Bay; 2 — lateral shoots on a small short branch, when
growing some shoots give rise to a new branch; 3, 4 — lateral shoots from the lower part of thallome.
136 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 137
Рис. 54. Draparnaldioides pilosa.
1 — общий вид таллома из района вблизи устья р. Большая (Сев. Байкал); 2 — боковые веточки с
мелких ветвей; 3 — боковая веточка в средней части таллома; 4 — боковая веточка в основании
главного ствола.
Fig. 54. Draparnaldioides pilosa.
1 — general view of thallome from the region near mouth of Bolshaya River (Northern Baikal); 2 — lateral
shoots from fine branches; 3 — lateral shoot in the middle part of thallome; 4 — lateral shoot at the base of
main spine.
Замечания. Нами D. vilosa найдена в губе Таланка — у мыса Тонкий (конец
июля) и у мыса Повалишина и о. Лиственичный (середина сентября). Кроме
того, в районе мыса Рытый встречены талломы Draparnaldioides, по ряду
признаков близкие к D. vilosa, но не идентичные ей. Это сильноветвистые
кустики высотой 6-11 см, усажены короткими ветвями 3-6 мм длиной (рис. 55:
1-4). По размеру клеток и строению боковых веточек в разных частях таллома
вид ближе всего к D. vilosa, но отличается от нее наличием массы длинных
щетинок и довольно четко выраженным рахисом в боковых веточках в средней
части таллома. Отмечено массовое формирование гаметангиев.
В районе пади Жилище встречены обильно разветвленные талломы Drapar-
naldioides высотой 10-13.5 см (рис. 56: 1-3). Мутовки боковых веточек сидят
через 2-4 клетки, но, благодаря чрезвычайно обильному ветвлению веточек,
138 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 55. Draparnaldioides vilosa, мыс Рытый, Сев. Байкал.
1 — общий вид; 2 — боковые веточки на конце ветви длиной 2 мм; 3 — боковые веточки в средней
части ветви длиной 0.5-1 см; 4 — строение боковой веточки на крупной ветви.
Fig. 55. Draparnaldioides vilosa, Rytyi Cape, Northern Baikal.
1 — general view; 2 — lateral shoots at the branch tip 2 mm long; 3 — lateral shoots in the middle part of a
0.5-1 cm long branch; 4 — structure of a lateral shoot on a large branch.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 139
Рис. 56. Draparnaldioides vilosa, район пади Жилище, Юж. Байкал.
1 — общий вид таллома; 2 — боковые веточки на коротких ветвях; 3 — боковая веточка в основании
крупной ветви.
Fig. 56. Draparnaldioides vilosa, region of Zhilishche Pad, Southern Baikal.
1 — general view of thallome; 2 — lateral shoots on short branches; 3 — lateral shoot at the base of a large
branch.
140 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
соприкасаются боками. По внешнему виду таллома вид похож на D. vilosa, но в
отличие от нее у данной водоросли рахис в боковых веточках просматривается
во всех частях таллома. У очень многих веточек рахис разрастается и дает начало
новым коротким ветвям (рис. 56: 3), благодаря чему весь таллом так сильно
разветвлен. Кроме того, на концах ветвей многие нити боковых веточек закан-
чиваются волосками, как у D. pilosa. Ширина клеток ствола описываемого
вида - 400-413 мкм в основании таллома, 318-340 мкм — в средней его части,
120-176 мкм в 1-2 см от вершины ветвей и конечных клеток ветвей —
30-40 мкм. Ширина нитей боковых веточек 10-12(18) мкм, рахиса в основании
боковых веточек в разных частях таллома от 20-25 мкм на концах ветвей до
37-50 мкм в основании таллома. Размеры клеток встреченной Draparnaldioides,
а также наиболее обильное ветвление нитей в верхней части боковых веточек
придают виду сходство с D. baicalensis.
Draparnaldioides baicalensis Meyer et Skabitsch. (=Draparnaldia baicalensis Meyer,
1922; Drapamaldiella baicalensis Meyer et Skabitsch., 1969) (рис. 57: 1—12; 58—66)
Сильно разветвленные кустики темно-зеленого цвета, 15-25(40) см вы-
сотой. Весь таллом одет толстым слоем прозрачной слизи. Главный ствол,
прикрепляющийся к субстрату при помощи круглой подошвы, в нижних час-
тях имеет 3-4 мм ширины, в верхней части таллома ствол и крупные ветви
1.5-2 мм ширины и на концах — до 0.5-2 мм ширины. Боковые веточки округ-
ло-треугольной формы, обильно разветвлены. Рахис состоит обычно из 1-2
клеток, от которых начинается масса ветвей почти одинакового размера, очень
обильно ветвящихся, особенно в своей верхней части. Обычно мутовки образу-
ют 2-3 веточки. Мутовки неодинаковой величины и расположены на некото-
ром расстоянии друг от друга, вследствие чего ветви таллома D. baicalensis име-
ют узловатый вид. Клетки ствола и ветвей цилиндрические, слабо перетянутые
у клеточных перегородок, ширина их 400-450 мкм у основания таллома,
240-360 мкм — в средней части, ближе к концам ветвей — 110-150 мкм, конеч-
ной клетки — около 75 мкм, клетки боковых веточек — 12-16.5 мкм, волос-
ков — 9.5-12.5 мкм, гаметангии — 16-22.5 мкм. Ризоидный покров сильно раз-
вит, утолщается от верхней части ветвей к основанию таллома, где может дос-
тигать 3 мм ширины.
По К.И. Мейеру [1930], вид характерен для западного прибрежья Байкала,
встречен им на участке от ст. Маритуй до мыса Котельниковский. На восточ-
ном берегу D. baicalensis найдена лишь на Муринской банке, у Ушканьих ост-
ровов и в Баргузинском заливе - в губе Бученкова.
Замечания. По наблюдению В.Н. Яснитского [1934], у западного берега
Юж. Байкала D. baicalensis вегетирует в июле-октябре, а плодущие экземпляры
встречаются с конца августа. Гаметы 5-10 мкм длиной, 4-6 мкм шириной, зи-
готы, сохраняющие подвижность в течение 5-10 мин, 12-14 мкм длиной,
7-8 мкм шириной (рис. 57: 11-13). По нашим данным [Ижболдина, 1970,
1990в], вид начинает вегетировать в июне, сначала на глубине 1.5-2 м появля-
ются единичные молодые обильно ветвящиеся талломы с ровными нечетко-
видными ветвями, утончающимися к вершине. Строение боковых веточек в
разных частях таллома типично для вида (рис. 58:1-5). В августе талломы име-
ют четковидные ветви (рис. 59) и образуют на глубине 3-6 м и более, на кам-
нях, целые заросли. Массовый выход гамет происходит обычно в середине сен-
тября. В это же время на талломах D. baicalensis в районе пади Сенной (на
о. Ольхон), в губе Заворотной, у Ушканьих островов и в губе Харанса (прол.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 141
10-12 :15 цт
Рис. 57. 1—12. Draparnaldioides baicalensis: 1 — клетка рахиса, 2 — образование волоска,
3-6 — образование ризоидов, 7 — часть боковой веточки с гаметангиями, 8 — клетка
ствола, 9 — боковая веточка, 10 — гаметы, 11 — спаривающиеся гаметы, 12 — зигота;
13 — Draparnaldioides lubrica, боковая веточка.
1-9,13 - по К.И. Мейру [1930].
Fig. 57. 1-12 — Draparnaldioides baicalensis: 1 — rachis cell, 2 — formation of a hair, 3-6 —
formation of rhizoids, 7 — part of a lateral shoot with gametangia, 8 — spine cell, 9 — lateral shoot,
10 — gametes, 11 — pairing gametes, 12 — zygote; 13 — Draparnaldioides lubrica, lateral shoot.
1-9,13 — after Meyer [1930].
Мал. Море) обнаружены образования, похожие на зооспорангии у рода
Draparnaldia Вогу (рис. 60; 1-4). Это бочонкообразные, круглые, яйцевидные
или грушевидные клетки нитей боковых веточек, заполненные зернистым со-
держимым и достигающие 33-80 мкм длиной и 22-55 мкм шириной (в самой
широкой части). Выход зооспор проследить не удалось. Обычно зооспорангии
развиты на талломах, где гаметангиев нет или они только начинают формиро-
ваться [Ижболдина, 19766].
142 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 58. Draparnaldioides baicalensis.
1 — общий вид молодого таллома; 2 — боковые веточки на конце ветви; 3 — боковые веточки в
верхней части таллома; 4 — боковые веточки на крупной ветви в средней части таллома; 5 — боко-
вые веточки в нижней части ствола, ризоцдный покров убран.
Fig. 58. Draparnaldioides baicalensis.
1 — general view of juvenile thallome; 2 — lateral shoots at the branch end; 3 — lateral shoots in the upper
part of thallome; 4 — lateral shoots on a large branch in the middle part of thallome; 5 — lateral shoots in
the lower part of the spine, rhizoid cover withdrawn.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 143
Рис. 59. Draparnaldioides baicalensis, общий вид таллома в августе.
Fig. 59. Draparnaldioides baicalensis, thallome in August, general view.
В районе пос. Бол. Коты встречены талломы D. baicalensis, у которых
толщина слабочетковидных ветвей во всех частях таллома может достигать
3-4 мм (рис. 61:1-4). В таких талломах длина клеток ствола и крупных ветвей
обычно меньше ширины, а высота боковых веточек на крупных ветвях может
достигать 0.8-1.0 мм. В районе проливов Ольхонские Ворота (бух. Базарная) и
Мал. Море (бух. Харанса, о. Угунгой), в бух. Заворотная встречены талломы
D. baicalensis, имеющие толстые, очень слизистые на ощупь шнуровидные
ветви 3-4 мм в диаметре. Очень редко характерная для вида четковидность
прослеживается на концах толстых ветвей (рис. 62: 1, 2). Строение боковых
144 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 60. Draparnaldioides baicalensis (Ушканьи острова).
1, 2 — боковые веточки с зооспорангиями в верхней (1) и средней (2) частях таллома; 3 — боковая
веточка с зооспорангиями и гаметангиями из нижней части таллома; 4 — зооспорангии.
Fig. 60. Draparnaldioides baicalensis (Ushkany Islands).
1, 2 — lateral shoots with zoosporangia in the upper (1) and middle (2) parts of thallome; 3 — lateral shoot
with zoosporangia and gametangia from the lower part of thallome; 4 — zoosporangia.
веточек типично для D. baicalensis, но их высота (до 1.5 мм) значительно
больше, чем в других районах Байкала. Больше (до 25 мкм) ширина нитей
боковых веточек и рахиса (до 70 мкм). Толстый (до 0.7-0.8 мм) ризоидный
чехол окружает клетки ствола и крупных ветвей, делает их прочными и хорошо
видными. В материалах О.А. Тимошкина (сентябрь 2000 г.), собранных в
районе мыса Ото-Хушун (прол. Мал. Море) встречен таллом D. baicalensis
высотой 18 см, имеющий как толстые шнуровидные, так и слабочетковидные
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 145
Рис. 61. Draparnaldioides baicalensis (район пади Бол. Коты, Юж. Байкал).
1 — общий вид таллома; 2 — боковые веточки на конце маленькой тонкой ветви; 3 — боковые ве-
точки на ветви 2 мм шириной; 4 — боковые веточки на ветви 3 мм шириной.
Fig. 61. Draparnaldioides baicalensis (region of Bolshye Koty, Southern Baikal).
1 - thallome, general view; 2 - lateral shoots at the tip of a small thin branch; 3 - lateral shoots on the
branch 2 mm wide; 4 - lateral shoots on a 3 mm wide branch.
146 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 62. Draparnaldioides baicalensis.
1 — внешний вид таллома из бух. Базарная (прол. Ольхонские Ворота); 2 — то же, из района бух. Харан-
са (прол. Мал. Море); 3 — боковые веточки.
Fig. 62. Draparnaldioides baicalensis.
1 - thallome from Bazamaya Bay, Olkhonskie Vorota Strait, external view; 2 - the same from the region of
Kharansa Bay (Maloe More Strait); 3 - lateral shoots.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 147
ветви (рис. 63). Четковидность образуется не благодаря чередованию разных
по высоте мутовок боковых веточек, а за счет довольно разреженного рас-
положения полукруглых по форме мутовок одинаковой высоты, соприка-
сающихся боками. Нити боковых веточек 12.5-16.7 мкм шириной, рахис до 25
мкм шириной, волоски толстые, 12.5-18.7 мкм шириной и до 1 мм длиной и
более. В основании таллома отмечено массовое формирование зооспорангиев.
У талломов D. baicalensis, собранных у мыса Арул, у восточного побережья
о. Ольхон (мысы Ижимей, Хобой, падь Сенная) (рис. 64: 1-3) на восточном
берегу у п-ова Святой Нос, нити боковых веточек ветвятся особенно обильно и
часто заканчиваются тонкими волосками (рис. 65: 1-3).
В районе мыса Ухан (восточный берег о. Ольхон) встречены мощные
заросли D. baicalensis, дающие 100%-е проективное покрытие грунта, на скале,
на глубине от 1.5-2 до 5-7 м. Талломы невысокие (7-12 см) и относительно
слабо разветвлены, с толстыми слабочетковидными ветвями (рис. 66: 1-5).
Весь таллом очень сильно опушен. Длина волосков на нитях, очень обильно
разветвленных боковых веточках, достигает 2-2.5 мм. Ширина клеток ствола
Рис. 63. Draparnaldioides baicalensis: общий вид таллома из района мыса Ото-Хушун
(прол. Мал. Море).
Fig. 63. Draparnaldioides baicalensis: thallome from the region of Oto-Khushun Cape (Mai.
More Strait), general view.
148 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 64. Draparnaldioides baicalensis из района пади Сенной на о. Ольхон и у мыса Арул.
1 — общий вид таллома; 2 — боковая веточка на крупной ветви таллома; 3 — то же, мыс Арул.
Fig. 64. Draparnaldioides baicalensis from Sennaya Pad on Olkhon Island and opposite of Aral
Cape.
1 — thallome, general view; 2 — lateral shoot on a large thallome branch; 3 — the same, Anil Cape.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 149
2:100 цш
3
Рис. 65. Draparnaldioides baicalensis из района Ушканьих островов.
1 — общий вид таллома; 2 — боковая веточка в верхней части таллома; 3 — боковые веточки на
крупной ветви, формирующиеся гаметангии.
Рис. 65. Draparnaldioides baicalensis from the region of Ushkany Islands.
1 — general view of thallome; 2 — lateral shoots in the upper part of thallome; 3 — lateral shoots on a large
branch, growing gametangia.
150 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
2:100 |im 3:23 цт
Рис. 66. Draparnaldioides baicalensis (район мыса Ухан, у восточного берега о. Ольхон).
1 — внешний вид таллома; 2 — боковые веточки в верхней части таллома; 3 — зооспорангии (а) и
гаметангии (б); 4, 5 — боковые веточки в нижней части таллома.
Fig. 66. Draparnaldioides baicalensis (region of Ukhan Cape near eastern coast of Olkhon
Island).
1 — thallome, general view; 2 — lateral shoots in the upper part of thallome; 3 — zoosporangia (a) and
gametangia (6); 4, 5 — lateral shoots in the lower part of thallome.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 151
Окончание рис. 66.
End of Fig. 66.
152 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
составляет в основании 300-360 мкм, в средней части 210-280 мкм, на конце
ветви 90-160 мкм. В основании таллома обнаружены зооспорангии (рис. 66).
Наиболее обильно D. baicalensis развита вдоль западного берега Юж. и
Сред. Байкала и у восточного берега в районе Ушканьих островов и п-ова Свя-
той Нос. На участке северо-западного побережья Юж. Байкала, расположен-
ном между падями Жилище и Сенная, фитомасса D. baicalensis в середине сен-
тября составляет 900-1600 г/м2 [Ижболдина, Максимов, Мурахвери, 1978а].
Draparnaldioides lubrica Meyer et Skabitsch. (=Draparnaldia lubrica Meyer, 1925;
Draparnaldiella lubrica Meyer et Skabitsch., 1969) (рис. 57: 13; 67-75)
Сильно разветвленные темно-зеленые кустики 15-20 см высотой, со шну-
ровидными слизистыми и упругими ветвями 2-3 мм в толщину. Главный ствол
прикрепляется к грунту круглой подошвой. И ствол, и крупные ветви дают в
свою очередь начало новым ветвям, на которых развиты короткие тонкие ветви
до 1 см длиной и 0.5-1 мм шириной. Боковые веточки округло-яйцевидные,
очень обильно разветвлены (рис. 57:13). Рахис короткий, иногда из 2-3 клеток
и заканчивается пучком веточек. Мутовки боковых веточек смыкаются краями,
образуя вокруг клеток ствола сплошной покров. Клетки ствола и ветвей ци-
линдрические, слабо перешнурованные у клеточных перегородок, с толстыми
оболочками 200-240 мкм ширины, в средней части таллома 120-150 мкм
ширины, короткие (1см длины) ветви 60-70 мкм ширины. Толщина нитей бо-
ковых веточек 9-10 мкм, гаметангиев 17.5-21 мкм. Ризоидный покров появля-
ется почти на самой верхушке ветвей и утолщается к основанию таллома.
К.И. Мейер [1930] отмечает сходство в строении боковых веточек D. lubrica и
D. baicalensis, от которой рассматриваемый вид отличается наличием сплош-
ных шнуровидных упругих и слизистых ветвей, несущих массу коротких и тон-
ких ветвей.
В Байкале встречена К.И. Мейером [1930] около Мысовой и у Карги, отде-
ляющей Посольский сор от Байкала, у мыса Тонкий. А.П. Скабичевским [1966]
единичные экземпляры D. lubrica обнаружены в районе пади Бол. Коты. Автор
отмечает, что вид встречается крайне редко и не каждый год.
Draparnaldioides lubrica forma ramulifera Meyer (1930)
Отличается присутствием многочисленных, коротких (0.5 см) ветвей, по-
крытых еще совсем неразвитыми мутовками боковых веточек.
В Байкале встречена К.И. Мейером
[1930] у Мысовой и у Карги, отделяю-
щей Посольский сор от Байкала.
Замечания. При наших исследо-
ваниях D. lubrica обнаружена вдоль за-
падного берега озера в районе пади
Бол. Коты, где она растет одновремен-
но с D. baicalensis и на одних с ней
Рис. 67. Draparnaldioides lubrica, общий
вид таллома.
Fig. 67. Draparnaldioides lubrica, general
view of thallome.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 153
глубинах. Талломы D. lubrica имеют массу коротких ветвей и характерное для
вида строение боковых веточек, очень сильно разветвленных, смыкающихся
краями и образующих сплошной покров вокруг клеток ствола (рис. 67; 68:
1-4). Однако в основании таллома клетки ствола шире (280-320 мкм), чем
указывает для вида К.И. Мейер [1930]. Больше (12.0-14.6 мкм) ширина
Рис. 68. Draparnaldioides lubrica.
Боковые веточки: 1 — на короткой и тонкой ветви, 2 — в верхней части крупной ветви, 3 — в
средней части таллома, 4 — в основании таллома.
Fig. 68. Draparnaldioides lubrica.
Lateral shoots: 1 — on a short and thin branch, 2 — in the upper part of a large branch, 3 — in the middle
part, 4 — at the base of thallome.
154 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
волосков на нитях боковых веточек, а на рахисе крупных нитей боковой
веточки она может достигать 19-24 мкм. Кроме того, в районе пади Бол. Коты
встречены крупные талломы, на которых есть шнуровидные и слабочетко-
видные ветви (рис. 69), усаженные короткими ветвями. На коротких тонких
ветвях строение боковых веточек имеет большое сходство с таковыми у
D. baicalensis, но нити веточек более обильно разветвлены и на них отмечено
формирование зооспорангиев (рис. 70: 1, 2). В средней и нижней частях
таллома боковые веточки типичны для D. lubrica (рис. 70: 3, 4), но размер
клеток ствола во всех частях таллома как у D. baicalensis. В нижней части
таллома ширина клеток в основании хорошо выраженного рахиса, состоящего
из 8-9 клеток, может достигать 80-90 мкм.
Одновременно с вышеописанными в Байкале в массе развиты обильно
разветвленные талломы Draparnaldioides с массой длинных шнуровидных вет-
вей (рис. 71), с боковыми веточками, по строению характерными для D. lubrica
(рис. 57: 13). Однако размер клеток в любой части таллома больше, чем у
D. lubrica, и соответствует таковому у D. baicalensis.
Рис. 69. Draparnaldioides lubrica, внешний вид таллома из района пади Бол. Коты.
Fig. 69. Draparnaldioides lubrica, thallome from the region of Bolshye Koty Settlement, gen-
eral view.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 155
Рис, 70. Draparnaldioides lubrica.
1 — строение боковой веточки на короткой и тонкой ветви; 2 — зооспорангии; 3, 4 — боковая ве-
точка в средней части крупной ветви (3) и в самом основании таллома (4); rh — ризоиды в осно-
вании веточки; g — формирующиеся гаметангии, z - зооспорангии.
Fig. 70. Draparnaldioides lubrica.
1 — structure of a lateral shoot on a short and thin branch; 2 — zoosporangia; 3,4 — lateral shoot in the
middle part of a large branch (3) and lateral branch immediately at the base of thallome (4); rh — rhizoids
at the base of the shoot; g — growing gametangia, z - zoosporangia.
156 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 71. Draparnaldioides lubrica, общий вид таллома.
Fig. 71. Draparnaldioides lubrica, thallome, general view.
В районе пади Бугульдейка на ветвях Myriophyllum spicatum найдены невы-
сокие (5-6 см) обильно ветвящиеся талломы Draparnaldioides со слизистыми
на ощупь шнуровидными ветвями (рис. 72). Строение боковых веточек имеет
сходство как с D. baicalensis, только веточки более обильно разветвлены
(рис. 72: 2, 3), так и с D. lubrica (рис. 72: 4). Размер клеток ствола как у
D. baicalensis и в основании таллома может достигать 500 мкм. Ширина много-
численных длинных (до 1.5 мм) волосков может достигать 18.5 мкм, что боль-
ше, чем у D. lubrica и D. baicalensis.
В прол. Ольхонские Ворота на глубине 40 м О.А. Тимошкиным встречен
таллом Draparnaldioides высотой 18 см (рис. 73), имеющий значительное мор-
фологическое сходство с D. lubrica (шнуровидные слизистые ветви, наличие
многочисленных коротких ветвей, строение боковых веточек), но отличаю-
щийся от этого вида более толстыми клетками ствола в основании таллома и
толщиной нитей боковых веточек (до 15-18.7 мкм) и волосков (18.7-25 мкм).
Оригинальный таллом Draparnaldioides высотой 10 см встречен в бух.
Базарной в прол. Ольхонские Ворота (рис. 74: 1-5). Наличие шнуровидных,
скользких на ощупь и упругих ветвей и строение боковых веточек в основании
таллома, мощный ризоидный покров придают ему сходство с D. lubrica.
Строение боковых веточек в верхней части таллома и размер клеток ствола и
крупных ветвей делают его похожим на D. baicalensis. Наличие массы длинных
волосков и относительно рыхло расположенные мутовки боковых веточек
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 157
1:1 ст
Рис. 72. Draparnaldioides lubrica.
1 — общий вид таллома; 2-4 — боковые веточки в верхней (2), средней (3) и нижней (4) частях
таллома.
Fig. 72. Draparnaldioides lubrica.
1 — thallome, general view; 2-4 — lateral shoots in the upper (2), middle (3) and lower (4) parts of
thallome.
свидетельствуют о сходстве вида с D. pilosa. Похожий таллом был встречен в
районе губы Солонцовой, но главный ствол и крупные ветви, одетые мощным
(до 300 мкм) ризоидным чехлом, значительно толще (3-4 см), чем у таллома
из губы Базарной (рис. 74: 6). Соответственно выше и очень обильно раз-
ветвлены боковые веточки в средней и нижней частях таллома (рис. 74: 8, 9).
158 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 73. Draparnaldioides lubrica, общий вид таллома (из района прол. Ольхонские
Ворота).
Fig. 73. Draparnaldioides lubrica, thallome, general view (from the region of Olkhonskie
Vorota Strait).
Строение боковых веточек на концах тонких ветвей типично для D. baicalensis
(рис. 74: 7). Размер клеток ствола и круглых ветвей также ближе к таковому у
D. baicalensis. Однако от последней он отличается прочным талломом, не
раздавливающимся при нажиме на покровное стекло (как D. baicalensis) и
чрезвычайно обильным ветвлением боковых веточек, начинающемся в самом
основании их.
D. lubrica f. ramulifera найдена нами в Юж. Байкале у ст. Танхой, Култук и
в районе пади Бол. Коты в августе - сентябре на глубине 2.5-3 м на камнях
(рис. 75: 1, 2).
Заканчивая раздел, посвященный роду Draparnaldioides, необходимо отме-
тить следующее. Учитывая значительное морфологическое сходство, можно
выделить несколько групп видов. К.И. Мейер делит все виды по строению
боковых веточек на 2 группы. Виды первой группы (D. simplex, D. arenaria,
D. arnoldii, D. goroschankinii, D. pumila) имеют хорошо выраженный рахис —
главный ствол боковой веточки. Виды второй группы (D. pilosa, D. vilosa,
D. baicalensis, D. lubrica) лишены рахиса. Учитывая возрастающую сложность
талломов, К.И. Мейер [1930] располагает виды рассматриваемого рода в еле-
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 159
дующем порядке: D. simplex, D. arenaria, D. arnoldii, D. goroschankinii, D. pumila,
D. pilosa, D. vilosa, D. baicalensis, D. lubrica.
Позднее H.S. Forest [1957] отмечал, что у байкальских драпарнальдий
можно ожидать широкие морфологические вариации с постепенными пе-
реходами. Учитывая это, он считал, что все байкальские драпарнальдии при-
надлежат к единой популяции. Однако он признает все же 3 следующих вида:
D. arenaria, к которой он относит также D. arenaria f. plumosa, D. simplex,
D. goroschankinii, D. goroschankinii f. plumosa; D. arnoldii, куда включает 2 формы
этого вида и D. vilosa; D. baicalensis, к которой относит D. lubrica и ее форму,
D. pilosa и D. pumila.
Нам представляется логичным выделение трех групп видов, обладающих
значительным морфологическим сходством, наличием переходных форм, по-
являющихся, вероятно, благодаря близкородственному скрещиванию, име-
ющих общий ареал обитания видов.
К двум первым группам мы относим виды, у которых боковые веточки
расположены на клетках ствола довольно редко и не образуют вокруг него
сплошного покрова.
К первой группе принадлежат D. simplex f. simplex, D. simplex f. trifurca,
D. simplex f. pentafurca, D. goroschankinii f. goroschankinii, D. goroschankinii f.
plumosa. Виды и формы расположены в ряд по степени усложнения строения у
них боковых веточек. Виды этой группы появляются в Байкале подо льдом и
заканчивают вегетацию после массового выхода гамет к середине июля. Из них
наиболее широко распространена D. simplex.
Ко второй группе видов мы относим D. arenaria f. arenaria, D. arenaria f.
plumosa, D. arenaria f. elegans, D. arenaria f. pilosissima, D. arnoldii f. arnoldii,
D. arnoldii f. gracilis, D. arnoldii f. compacta. Развитие видов этой группы в
Байкале начинается в конце июня — начале июля и продолжается до ноября.
Но массовое развитие видов заканчивается к началу октября. Оба вида широко
распространены в Байкале, но наиболее часто встречаются у западного берега и
у восточного в северной части озера.
К третьей группе мы относим водоросли с погруженными в слизь разной
плотности и очень обильно разветвленными боковыми веточками, сопри-
касающимися друг с другом своими краями и сплошь покрывающими клетки
ствола и ветвей. Виды этой группы обычно лишены резко выраженного рахиса.
Он состоит из 1-2 или 12 клеток и не возвышается над боковой веточкой,
теряясь в ее ответвлениях. Хорошо заметен он лишь в молодых боковых
веточках, расположенных в верхней части таллома. Исходным видом в этой
группе может быть D. pumila, у которой рахис хорошо выражен в молодых
боковых веточках и достаточно ясно различим даже в боковых веточках в
основании таллома. Далее по степени усложнения строения боковых веточек и
укорачивания в них рахиса виды могут быть расположены следующим образом:
D. vilosa, D. lubrica, D. lubrica ramunlifera, D. pilosa, D. baicalensis. Виды этой
группы развиты в Байкале с июля до глубокой осени.
Значительное сходство видов D. arenaria — D. arnoldii, D. vilosa — D. pi-
losa — D. pumila, D. baicalensis — D. lubrica первым отмечал К.И. Мейер [1930].
Предложенное Форестом [Forest, 1957] объединение видов Draparnaldioides
представляется нам необоснованным. Несомненно, вопрос о путях эволюции
видов рассматриваемого рода еще ждет своего решения.
160 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 74. Draparnaldioides lubrica (?).
1 — общий вид таллома из района бух. Базарной (Ольхонские Ворота); 2 — боковая веточка на
конце ветви; 3 — боковая веточка в средней части таллома, 4, 5 — боковые веточки на крупных
ветвях в основании таллома.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 161
Fig. 74. Draparnaldioides lubrica (?).
1 - general view of thallome from the region of Bazamaya Bay (Olkhonskie Vorota); 2 - lateral shoot at the
tip of the branch; 3 - lateral shoot in the middle of thallome; 4, 5 - lateral shoots on large branches at the
base of thallome.
162 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
7, 8:100 цт
Продолжение рис. 74.
Continuation of Fig. 74.
6 - общий вид таллома из губы Со-
лонцовой; 7 - боковые веточки на
конце ветви; 8,9 — боковые веточки в
основании таллома.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 163
Окончание рис. 74.
End ofFig. 7 4.
6 — general view of thallome from Solontsovaya Bay; 7 - lateral shoots at the end of a large branch; 8,9-
the same at the base of thallome.
164 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 75. Draparnaldioides lubrica f. ramulifera.
Внешний вид таллома: 1 — из района ст. Танхой, 2 — из района пади Бол. Коты.
Fig. 75. Draparnaldioides lubrica f. ramulifera.
General view of a thallome from: 1 — the region of Tankhoy station; 2 — the region of Bolshye Koty
Settlement.
Фото 1. Sphaeronostoc pruniforme (Ag.) Elenk.
1-3 — общий вид колоний (уменьшено); 4 — трихомы в центре колоний.
Фото 1-3 — О.А. Тимошкина; 4 — А.Н. Галкина.
Photo 1. Sphaeronostoc pruniforme (Ag.) Elenk.
1-3 — general view of the colonies (reduced); 4 — trichomes in the center of the colony.
Photos by 1-3 — O.A. Timoshkin; 4 — A.N. Galkin.
Фото 2. Schizothrix sp. среди ветвей Cladophora glomerata (L.) Kutz.
Фото А.Н. Галкина.
Photo 2. Schizothrix sp. among the branches of Cladophora glomerata (L.) Kutz.
Photo by A.N. Galkin.
Фото 3. Batrachospermum moniliforme Roth. Фото O.A. Тимошкина.
Photo 3. Batrachospermum moniliforme Roth. Photo by O.A. Timoshkin.
Фото 4. Tetraspora cylindrica (Wahl.) Ag. var. sp. Фото O.A. Тимошкина.
Photo 4. Tetraspora cylindrica (Wahl.) Ag. var. sp. Photo by O.A. Timoshkin.
Фото 5. Ulotrix zonata Kiitz.
Видны пустые зооспорангии. Фото А.Н. Галкина.
Photo 5. Ulotrix zonata Kiitz.
Empty zoosporangia are clearly seen. Photo by A.N. Galkin.
Фото 6. Draparnaldioides
simplex Meyer et Skabitsch.
forma simplex.
В нижней части таллома
начался выход гамет.
Фото О.А. Тимошкина
с гербария Л.А. Ижболдиной.
Photo 6. Draparnaldioides
simplex Meyer et Skabitsch.
forma simplex.
Discharge of gametes begins
in the lower part of the thallome.
Photo by O.A. Timoshkin,
from herbarium of L.A. Izhboldina.
Фото 7. Draparnaldioides
arenaria Meyer et Skabitsch.
forma arenaria.
Фото O.A. Тимошкина
с гербария Л.А. Ижболдиной.
Photo 7. Draparnaldioides
arenaria Meyer et Skabitsch.
forma arenaria.
Photo by O.A. Timoshkin,
from herbarium of L.A. Izhboldina.
Фото 8. Draparnaldioides
arenaria Meyer et Skabitsch.
forma elegans Izhboldina.
Фото O.A. Тимошкина
с гербария Л.А. Ижболдиной.
Photo 8. Draparnaldioides
arenaria Meyer et Skabitsch.
forma elegans Izhboldina.
Photo by O.A. Timoshkin,
from herbarium of L.A. Izhboldina.
Фото 9. Draparnaldioides arnoldii
Meyer et Skabitsch.
forma arnoldii.
Фото O.A. Тимошкина
с гербария Л.А. Ижболдиной.
Photo 9. Draparnaldioides arnoldii
Meyer et Skabitsch.
forma arnoldii.
Photo by O.A. Timoshkin,
from herbarium of L.A. Izhboldina.
Фото 10. Draparnaldioides arnoldii
Meyer et Skabitsch.
forma arnoldii - фрагменты ветвей.
Фото O.A. Тимошкина
с гербария Л.А. Ижболдиной.
Photo 10. Draparnaldioides arnoldii
Meyer et Skabitsch.
forma arnoldii - branch fragments.
Photo by O.A. Timoshkin,
from herbarium of L.A. Izhboldina.
Фото 11. Draparnaldioides pumila
Meyer et Skabitsch. из разных
районов оз. Байкал.
Фото O.A. Тимошкина с гербария,
Л.А. Ижболдиной.
Photo 11. Draparnaldioides pumila
Meyer et Skabitsch. from different
regions of Lake Baikal.
Photo by O.A. Timoshkin,
from herbarium of L.A. Izhboldina.
Фото 12. Draparnaldioides pumila
Meyer et Skabitsch.
Фото O.A. Тимошкина
с гербария Л.А. Ижболдиной.
Photo 12. Draparnaldioides pumila
Meyer et Skabitsch.
Photo by O.A. Timoshkin,
from herbarium of L.A. Izhboldina.
Фото 13. Draparnaldioides pumila
Meyer et Skabitsch. из района острова
Богучан (Сев. Байкал).
Фото О.А. Тимошкина
с гербария Л.А. Ижболдиной.
Photo 13. Draparnaldioides pumila
Meyer et Skabitsch. from the region
of Boguchan Island (Northern Baikal).
Photo by O.A. Timoshkin,
from herbarium of L.A. Izhboldina.
Фото 14. Draparnaldioides pumila
Meyer et Skabitsch. из района острова
Богучан (Сев. Байкал).
Фото О.А. Тимошкина
с гербария Л.А. Ижболдиной.
Photo 14. Draparnaldioides pumila
Meyer et Skabitsch. from the region
of Boguchan Island (Northern Baikal).
Photo by O.A. Timoshkin,
from herbarium of L.A. Izhboldina.
Фото 15. Draparnaldioides pumila
Meyer et Skabitsch. из района мыса
Амундакан (Сев. Байкал).
Фото О.А. Тимошкина
с гербария Л.А. Ижболдиной.
Photo 15. Draparnaldioides pumila
Meyer et Skabitsch. from the region
of Amundakan Cape (Northern Baikal).
Photo by O.A. Timoshkin,
from herbarium of L.A. Izhboldina.
Фото 16. Draparnaldioides baikalensis
Meyer et Skabitsch.
Фото O.A. Тимошкина
с гербария Л.А. Ижболдиной.
Photo 16. Draparnaldioides baikalensis
Meyer et Skabitsch.
Photo by O.A. Timoshkin,
from herbarium of L.A. Izhboldina.
Фото 17. Draparnaldioides baikalensis Meyer et Skabitsch.
Собран 16 августа 1983 г., западное побережье Южного Байкала.
Фото О.А. Тимошкина с гербария Л.А. Ижболдиной.
Photo 17. Draparnaldioides baikalensis Meyer et Skabitsch.
Collected on August 16, 1983; Southern Baikal, westen coast.
Photo by O.A. Timoshkin, from herbarium of L.A. Izhboldina.
Фото 18. Draparnaldioides pilosa (Meyer) Meyer et Skabitsch.
Собран 14 сентября 1976 г., район пади Широкая, глубина 4 м, обломки скал.
Фото О.А. Тимошкина с гербария Л.А. Ижболдиной.
Photo 18. Draparnaldioides pilosa (Meyer) Meyer et Skabitsch.
Collected on September 14, 1976; depth 4 m; rocks; region of pad' Shirokaya.
Photo by O.A. Timoshkin, from herbarium of L.A. Izhboldina.
Фото 19. Draparnaldioides baikalensis Meyer et Skabitsch. в естественной обстановке.
Фото O.A. Тимошкина, Д.В. Антипенко, сделано на южном побережье (в заливе) у мыса Ото-Хушун,
Малое Море, глубина около 2 м.
Photo 19. Draparnaldioides baikalensis Meyer et Skabitsch. in the natural environment.
Photo by O.A. Timoshkin, D.V. Antipenko, made on the southern coast (in a small bay) of Oto-Khushun Cape,
Maloe More strait, depth about 2 m.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 165
Род Draparnaldia Вогу, 1808
Таллом кустиковидный, слизистый на ощупь, состоит из главной оси и
крупных боковых ветвей, покрытых, в свою очередь, маленькими боковыми
веточками. Прикрепляется к субстрату с помощью базальной клетки и много-
численных ризоидов. Главный ствол и крупные ветви состоят из цилиндриче-
ских или слегка бочонковидных клеток. Хлоропласт проходит по середине кле-
ток, поясковидный или сетчато-продырявленный, занимающий всю клетку и
содержащий много пиреноидов. Боковые веточки сидят на стволе и крупных
ветвях мутовками и образованы сильноветвящимися нитями, которые часто за-
канчиваются длинным прозрачным волоском. Главная ось боковых веточек —
рахис - у одних видов четко выражена и проходит через всю веточку, у других
она довольно быстро становится незаметной, переходя в разветвление. Беспо-
лый процесс - с помощью 4-жгутиковых зооспор, половой — изогамный или
гетерогамный. Известно образование акинет и апланоспор.
Преимущественно в холодных проточных, реже стоячих водах, в обраста-
ниях камней, затонувшей древесины, высших водных растений.
В Байкале встречено 2 вида:
1. Боковые веточки имеют ясно выраженную ось ..............D. plumosa
2. Боковые веточки не имеют ясно выраженной оси.......... D. glomerata
Draparnaldia plumosa (Vauch.) Ag. (=Batrachospermum plumosum Vauch.;
Conferva mutabilis Roth; Draparnaldia mutabilis (Roth) Cedercr.)
(рис. 76: 3).
Кустики длиной 7-10 см. Боковые веточки широколанцетные, имеют хо-
рошо выраженный рахис 6-10 мкм шириной, заканчивающийся волоском
3.8-5 мкм шириной. Клетки главного ствола цилиндрические или слегка пере-
тянутые, 38—107 (114) мкм шириной. Клетки боковых веточек 5-10 мкм
шириной, хлоропласт кольцевидный, занимает среднюю часть клетки.
Draparnaldia plumosa var. plumosa (Vauch.) Ag.
Боковые ветви плотные, в очертании широколанцетные.
В реках, прудах, озерах, водохранилищах. Повсеместно. Вегетирует с вес-
ны до осени.
Замечание. В небольшом количестве найдена К.И. Мейером [1930] в
Нижнеангарском соре на севере оз. Байкал и в губе Крохалиной (Чивыр-
куйский залив).
Draparnaldia glomerata (Vauch.) Ag. (рис. 76: 1, 2).
Кустики 8-10 см высотой. Боковые веточки широкояйцевидные, округ-
лые, полуокруглые. Четко выраженного рахиса нет, он быстро теряется в раз-
ветвлениях. Клетки главного ствола слабо перетянутые, реже цилиндрические
(20)32—(99)125 мкм шириной. Клетки боковых веточек 6-10 мкм шириной.
Хлоропласт кольцевидный, занимает середину клетки.
Draparnaldia glomerata var. glomerata (=var. genuina Kirchn.)
Боковые веточки обильно разветвленные, рыхлые, расположенные близко
друг к друту.
В реках, ручьях, озерах. Повсеместно.
166 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 76. 1, 2 — Draparnaldia glomerata var. glomerata: 1 — средняя часть главного ствола;
2 — нижняя часть с ризоидными выступами; 3 — D. plumosa var. plumosa', 4-7 —
Coleochaete pulvinata var. pulvinata: 4 — фрагмент слоевища, 5 — зрелый оогоний,
покрытый корой, 6 — стадии прорастания ооспоры, 7 — карпоспоры.
По: 1-3 — Мошковой; 4-7 — Принсгейм.
Fig. 76. 1, 2 — Draparnaldia glomerata var. glomerata: 1 — middle part of main spine; 2 —
lower part with rhizoid protrusions: 3 — D. plumosa var. plumosa. 4-7 — Coleochaete pulvinata
var. pulvinata: 4 - fragment of thallome, 5 - mature oogonium, covered by bark, 6 - stages of
oospore hutching, 7 - carpospores.
After: 1-3 — Moshkova; 4-7 — Princeheim.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 167
Замечание. Стерильные талломы D. glomerata высотой 4-6 см встречены в
июле-августе в притоке оз. Байкал р. Бол. Котинка и в районе ее впадения.
СЕМЕЙСТВО COLEOCHAETACEAE
Таллом в виде прикрепленных к субстрату нитей, иногда срастающихся
боками. Иногда образуются прямостоячие нити и тогда таллом подушковидный,
до нескольких миллиметров длиной. Клетки различной формы, преимуществен-
но цилиндрические. Некоторые клетки несут щетинки, окруженные у основания
воротничком. Хлоропласт пластинчатый, с 1, 2 пиреноидами, ядро одно. Бесполое
размножение двужгутиковыми зооспорами. Половой процесс оогамный.
Эпи- и эндофиты, поселяются на высших водных растениях и водорослях,
в пресных водоемах.
Род Coleochaete Breb., 1844
Признаки рода те же, что описаны для семейства.
В Байкале встречен один вид.
Coleochaete pulvinata А. Вг. (рис. 76: 4—7)
Таллом полушаровидный — до 2 мм высоты. Нити прямостоячие, ветвя-
щиеся, радиально расходящиеся от клеток подошвы, образованной ползучими
по субстрату нитями. Клетки цилиндрические, реже многоугольные, 10- 43 мкм
шириной, щетинки встречаются часто. Зооспоры образуются в верхушечных
клетках прямостоячих нитей. Оогонии на ранних стадиях развития бутылко-
видные, позднее шаровидные, до 150 мкм в диам. Ооспоры до ПО мкм шири-
ной. Антеридии бутылковидные, 7-17 мкм шириной, 17.5-42.4 мкм длиной.
В стоячих водах — озерах, болотах, реже в проточных водах, на водных
цветковых растениях, камнях, раковинах моллюсков.
Coleochaete pulvinata var. pulvinata
Таллом полушаровидный, до 2 мм высоты, с хорошо развитыми прямо-
стоячими нитями. Клетки до 20-43 мкм шириной, оогонии до 150 мкм в
диаметре, ооспоры 80-110 мкм в диаметре. Антеридий 17 мкм шириной, по
длине до 2.5 раза больше.
В озерах, реках, болотах.
Замечания. В притоке оз. Байкал — р. Кичера — вид встречен К.И. Мейе-
ром [1930] в июле 1926 г. Им же обнаружены в Байкале три микроскопические
водоросли этого рода: С. orbicularis Prinahsh. в Посольском соре, С. soluta
Breb. и С. scutata Breb. в бух. Анга в августе 1916 г.
ПОДПОРЯДОК PRASIOLINEAE
Таллом однорядно и многорядно нитевидный или однослойный, ленто-
видный и пластинчатый, с ризоидами. Таллом образуют округленно-угловатые
клетки, сближенные друг с другом группами, а весь таллом кажется разбитым
на участки из 4 клеток. Хлоропласт один, центральный, звездчатый, с одним
пиреноидом. Вегетативное размножение — фрагментацией таллома, беспо-
лое — акинетами. Половой процесс — гетерогамия, гаметы двужгутиковые.
Встречается во всех частях света, в том числе и в арктических и антарктических
районах. Подпорядок включает одно семейство.
168 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
СЕМЕЙСТВО PRASIOLACEAE (Rabenh.) Borli
В оз. Байкал встречен представитель одного рода.
Род Prasiola С.А. Ag., 1822
Таллом однорядный, лентовидный или пластинчатый, прикрепленный.
Клетки округленно-угловатые, располагаются довольно правильными квадрат-
ными участками, отграниченными друг от друга утолщенными частями оболо-
чек. Ризоиды короткие, отходят чаще от клеток в основании таллома.
В оз. Байкал найден один вид.
Prasiola sp.
Таллом пластинчатый, курчаво-складчатый, до 0.6-1 см длиной. Клетки 8-
12 мкм шириной и 15-17 мкм длиной. Хлоропласт с одним пиреноидом.
Встречена на глубине от 1 до 12-15 м на урути. В июне большая часть тал-
лома образована молодыми клетками, размер которых не превышает 4.8-
5.2 мкм. Водоросль вегетирует до декабря. В феврале-апреле изредка встреча-
ются полуразрушенные пластинки Prasiola sp. с пустыми клетками, обильно
покрытыми эпифитами.
В небольшом количестве встречен вдоль западного и вдоль восточного по-
бережья Сев. и Сред. Байкала.
Вид более всего напоминает Prasiola crispa (Lightf.) Menegh., от которой
отличается местообитанием.
ПОРЯДОК OEDOGONIALES
Таллом нитчатый, простой или ветвистый. Нити прикреплены к субстрату
с помощью базальной клетки, оторвавшиеся от субстрата свободно плавают.
Нарастание нити происходит в результате своеобразного деления клеток, при
котором образуется две дочерние клетки. Одна из них полностью покрыта обо-
лочкой материнской клетки, другая — новообразовавшейся оболочкой, при-
крытой сверху останками старой оболочки в виде слегка выступающего кол-
пачка. В результате повторных делений таких колпачков становится несколько.
По наличию колпачков виды этого порядка легко отличаются от других нитча-
тых водорослей. Клетки одноядерные, хлоропласт пристенный, сетчатый, с не-
сколькими пиреноидами. Размножение фрагментацией нитей, зооспорами, с
венчиком жгутиков на переднем конце. Зооспоры образуются по одной в клет-
ке. Половой процесс оогамный. Виды одно- и двудомные. Отмечен половой
диморфизм.
СЕМЕЙСТВО OEDOGONIACEAE
Признаки те же, что описаны для порядка. В Байкале встречены предста-
вители 2 родов.
1. Нити не разветвленные................................Oedogonium
1. Нити разветвленные, с одноклетными щетинками, луковицеобразно вздуты-
ми у основания ......................................Bulbochaete
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 169
Род Oedogonium Link., 1820
Нити неветвящиеся. Клетки цилиндрические, иногда по краям волнистые.
Базальные клетки с ризоидальными выростами, иногда полушаровидные. Вер-
хушечные клетки закругленные, заостренные, реже вытянутые в щетинку. Од-
но- или двудомные.
В Байкале обнаружено 3 вида.
I. Вегетативные клетки цилиндрические, с волнистой или узловатой оболочкой
1. Вид двудомный, с карликовыми мужскими растеньицами.
А. Оогонии почти шаровидные, одиночные или по 2, вскрываются кры-
шечкой. Ооспоры шаровидные или почти шаровидные, гладкие.........
...................................................О. undulatum
II. Вегетативные клетки цилиндрические с прямой гладкой оболочкой
1. Виды двудомные, с карликовыми мужскими растеньицами.
А. Оогонии вскрываются небольшой щелью (порой). Отверстие оогония
на экваториальной линии или несколько выше ее. Ооспоры почти ша-
ровидные, гладкие..................................O.flavescens
2. Виды без карликовых мужских растеньиц.
А. Оогонии вскрываются небольшой щелью, отверстие оогония лежит на
экваториальной линии. Ооспоры почти шаровидные, гладкие........
......................................................О. sociale
Oedogonium undulatum (Breb.) A. Br. (рис. 77: 2, 3)
Двудомный. Вегетативные клетки 15-22 мкм шириной, оболочка клеток
волнистая. Оогонии почти шаровидные или эллипсоидально-шаровидные,
48-56 мкм шириной, 50-75 мкм длиной, одиночные или по 3, открываются
крышечкой. Ооспоры шаровидные или почти шаровидные, 42-50 мкм шири-
ной, 42-52 (60) мкм длиной. Карликовые мужские растеньица удлиненно
обратно-конусовидные, 8-10 мкм шириной, 18-65(70) мкм высотой.
Озера (на макрофитах), пруды, болота, реки, канавы.
Замечание. В Байкале встречен К.И. Мейером [1930] среди зарослей вод-
но-болотной растительности в Ангаро-Кичерском соре в июле.
Oedogonium flavescens (Hass.) Wittr. (рис. 77: 1)
Двудомный. Ширина вегетативных клеток 18-23 мкм. Оболочка клеток глад-
кая. Оогонии эллипсоидально-шаровидные или почти шаровидные, 49-52 мкм
шириной, 51-60 мкм длиной. Ооспоры шаровидные, не всегда вполне заполняю-
щие оогонии, 45-49 мкм шириной, 45-49 мкм длиной, оболочка гладкая. Карли-
ковые мужские растеньица слегка согнутые, высота их 36-45 мкм, ширина ножки
11-12 мкм.
Растет в болотах, в озерах.
Замечания. В Байкале найден К. И. Мейером [1930] в бух. Анга в начале ав-
густа, среди зарослей высших водных растений.
Oedogonium sociale Wittr. (рис. 77: 4)
Двудомный. Ширина вегетативных клеток мужских и женских нитей оди-
наковая, 9-16 мкм. Оогонии почти шаровидные, 30-38 мкм шириной, 32-
42 мкм длиной, одиночные. Ооспоры шаровидные, почти заполняющие по-
170 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 77.1 — Oedogonium flavescens; 2-3 — Oedogonium undulatum; 4 — Oedogonium sociale;
5 — Bulbochaete radinospora.
По: 1 — Витгроку; 2 — Роллу, 3-5 — Хирну.
Fig. 77. 1 — Oedogonium flavescens-, 2-3 — Oedogonium undulatum-, 4 — Oedogonium sociale;
5 — Bulbochaete radinospora.
After: 1 - Wittrock, 2 - Roll, 3-5 - Him.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 171
полость оогония, (26)28-35 мкм шириной, 28-35 мкм длиной, оболочка
гладкая. Антеридии 1-5-клеточные, 12-14 мкм шириной, 8-11 мкм длиной.
Растет в прудах.
Замечания. Вид встречен в Байкале К.И. Мейером [1930] в бух. Анга среди
зарослей водной растительности. При наших исследованиях встречены обрывки
стерильных нитей Oedogonium, идентифицировать которые в настоящее время
невозможно. Условно мы объединили такие виды под названием Oedogonium sp.
Водоросли найдены в бух. Анга, в Баргузинском заливе — у с. Макарово. У
открытого юго-восточного побережья обрывки Oedogonium sp. обнаружены в
районах поступления в озеро условно чистых и промышленных очищенных
сточных вод БЦБК (вблизи рек Бол. Осиновка и Солзан) [Ижболдина, 1985в].
Кроме того, Oedogonium sp. найден в сборах В.В. Тахтеева из районов горячих
источников Прибайкалья, у мыса Котельниковский, в пади Горячинской, в
бух. Хакусы. Водоросль встречена на некотором удалении от места выхода
термальных вод, где температура последних не превышает 20-23 °C [Ижбол-
дина, Галкин, 2000].
Род Bulbochaete С. Ag., 1817
Нити ветвящиеся, плотно прикрепляющиеся к субстрату с помощью ри-
зоидов. Щетинки, развивающиеся по клеткам, длинные, волосовидные, у
основания луковицеобразно вздутые.
В Байкале встречен один вид.
Bulbochaete rhadinospora Wittr. sec. Hirn,
(рис. 77: 5)
Гинандроспоровые. Вегетативные клетки 15-22 мкм шириной, длина их в
1.5-2 раза больше. Оогонии 29-37 мкм шириной, 60-80 мкм высотой, антеридии
7-9 мкм шириной, 6-8 мкм длиной. Клетки андроспорангиев 13-15 мкм шири-
ной, 16-23 мкм длиной.
Замечания. В Байкале обнаружен К.И. Мейером [1930] в июле в Ангаро-
Кичерском соре, где в небольшом количестве развит среди зарослей водно-
болотной растительности. Нами [Ижболдина, 2001] виды этого рода встречены
в водоемах Байкальского заповедника.
Класс SIPHONOCLADOPHYCEA
Водоросли прикрепленные или свободноплавающие, многоклеточные, в
виде нитей или разветвлены. Оболочка клеток трехслойная, состоящая из хи-
тинового, пектинового и целлюлозного слоев (последний иногда отсутствует).
Хлоропласт постенный, в виде полого цилиндра, сетчато-продырявленный, в
виде поперечных кольцевых полос или мелких дисков. Пиреноиды и ядра
обычно многочисленны. Размножение вегетативное, бесполое, 2-4-жгутико-
выми зооспорами. Половой процесс чаще изогамия, гетеро- и оогамия. Зоо-
споры и гаметы развиваются в любых клетках таллома.
В Байкале встречены представители 2 порядков: Cladophorales и Sphaero-
pleales.
172 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
ПОРЯДОК CLADOPHORALES
Нити простые или ветвящиеся, прикрепленные или свободноплавающие,
часто образующие дерновинки. Ветвление обильное, ветви многоклеточные,
реже короткие, иногда из 1-3 клеток. Хлоропласт в виде сетчато-продырявлен-
ного цилиндра, с равномерно расположенными пиреноидами. Ядра рассеяны
по всему протопласту или расположены по спиральной линии на одинаковом
расстоянии друг от друга. Размножение вегетативное, бесполое, половое.
СЕМЕЙСТВО CLADOPHORACEAE
Признаки семейства те же, что описаны для порядка. Водоросли этого се-
мейства обладают большой морфологической изменчивостью. В Байкале встре-
чены представители 5 родов.
I. Талломы изоморфные, из однотипных нитей.
1. Нити очень короткие, из нескольких клеток, тесно сомкнутые в плот-
ные дерновинки.......................................Gemmiphora
2. Нити более длинные, образующие дерновинки разнообразного строе-
ния или свободноживущие.
А. Нити всегда довольно обильно ветвящиеся на всем протяжении. Бо-
ковые ветви длинные.
а. Пиреноиды имеются.
а. Ветви отходят не под прямым углом. Геликоиды не образуются
или образуются редко. Зооспоры и гаметы имеются..........
....................................Cladophora (частично)
Б. Нити неветвящиеся или скудно ветвящиеся на всем протяжении. Бо-
ковые ветви очень короткие, нередко в виде простых бугорков.
а. Нити прикрепленные или свободноплавающие. Ветвление всегда
отсутствует. Зооспоры, как правило, четырехжгутиковые......
.............................................Chaetomorpha
б. Нити обычно свободноплавающие. Ветвление скудное или отсут-
ствует. Зооспоры двужгутиковые.
а. Нити одинаковой ширины на всем протяжении. При ветвлении
эвекция отсутствует..........................Rhizoclonium
р. Нити к основанию утончаются. При ветвлении эвекция отчетли-
вая .................................Cladophora (частично)
II. Талломы гетероморфные, из нитей двух типов, стелющихся по субстрату и от-
ходящих вверх.
1. Нити ветвящиеся только у самого основания, далее на всем протяжении
без ветвей. Неветвящиеся прямостоячие нити довольно длинные
...............................................Chaetocladiella
2. Прямостоячие нити, как правило, ветвящиеся. В воде............
............................................Cladophora (частично)
Род Gemmiphora Skabitsch., 1931
Водоросль образует плотные темно-зеленые дерновинки, не более 1.5 мм
высотой. Таллом нитевидный, изоморфный, нити плотно сомкнутые так, что
ризоиды их срастаются краями. Узкая внизу и расширяющаяся кверху нить
состоит не более чем из 8 клеток. Ветвление нечастое. Оно начинается с
врастания одной из клеток нити в оболочку нижележащей клетки, которая
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 173
разрастается вверх и вбок, вследствие чего получается нить с раздвоенным
верхом, а при многократном повторении — нить с кистью ветвей на верхушке
(рис. 78: 6). Все клетки нити могут образовывать направленные вниз ризоиды.
От одной нити может отходить несколько вторичных ризоидов. Оболочка клеток
толстая и явственно слоистая. Хлоропласт в виде овальных пластинок, зани-
мающих всю клетку. Пиреноиды отсутствуют. Запасное питательное вещество —
масло в виде капелек разного размера — скапливается в верхнем и нижнем
концах клетки. Ядра (от 2 до 10) расположены посредине клетки в один
более-менее прямой ряд. В апикальной клетке ядра разбросаны по всей клетке.
Зооспоры и половой процесс не обнаружены. Вегетативное размножение обо-
лочковыми спорами, которые образуются в верхней части нитей путем отшну-
ровывания протопласта с 1-2 ядрами и внедрения их в стенку клетки. Споры
разрастаются и округло выпячиваются наружу. Некоторые из этих спор могут
прорастать, не отделяясь от материнской нити. Акинеты продолговатой, реже
шаровидной, формы, образуются из верхних клеток нитей. Акинета имеет более
темную окраску, чем обычные клетки, и содержит до 10 ядер, разбросанных по
всей клетке.
В Байкале найден один вид.
Gemmiphora compacta Skabitsch., 1931 (рис. 78: 1-9).
Признаки вида те же, что описаны для рода. Клетки в нижней части нити
11.2-39.6 мкм шириной, в средней 30.8-64 мкм, в верхушечной 58.8-134.4 мкм.
Длина клеток в средней части нити 57-207 мкм, ризоидная клетка 115—
320 мкм длиной.
В Байкале найдена А.П. Скабичевским [1931] в районе пади Бол. Коты, где
на глубине 1.5-3.5 м заполняет трещинки и углубления камней. Позднее [По-
повская, Скабичевский, 1968] G. compacta отмечена им как обычный вид
III растительного пояса (глубины от 3.5 до 10-12 м) вдоль восточного берега в
районах губы Бирея, мыса Понгонье, губ Якшакан, Язовка, Давша и на участке
побережья от ст. Танхой до мыса Половинка и у западного берега у мысов Ко-
тельниковский, Заворотный, Узур, в губе Песчаная, у мысов Сытый, Пещерка
и скалы Два брата (южнее пади Жилище).
Замечания. Нами [Ижболдина, 1976] вид встречен также в губе Дагарской, в
бухтах Богучанская и Бабушка, у мысов Тонкий, Бол. Коса, Кочериковский, во
многих районах прол. Мал. Море и вдоль юго-восточного побережья озера на
участке между устьями рек Утулик и Хара-Мурин, где на камнях может давать
до 20-25 % покрытия грунта. В перечисленных районах вид растет на глубине
от 1.5-2 до 7-8 м. Обнаружен в Байкале в течение круглого года. В разных час-
тях нитей G. compacta хлоропласт может иметь форму сетчато-продырявленной
пластинки [Ижболдина, 1965, 1981].
Род Chaetomorpha Kutz., 1845
Нити неразветвленные, прикрепленные к грунту в течение всей жизни или
на начальной стадии развития. Нити растут одиночно или образуют дерновины
разной плотности. Вегетативные клетки цилиндрические или бочонкообраз-
ные, различной длины. Базальная клетка обычно более узкая, чем нить и за-
канчивается дисковидно расширенным или коралловидно разветвленным ри-
зоидом. Оболочка клеток крепкая, с хорошо заметной слоистостью. Хлоро-
пласт сетчато-продырявленный, цилиндрический, с многочисленными ядрами
174 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
7, 8 :100 цт
Рис. 78. Gemmiphora compacta.
1 — часть дерновинки; 2 — нижняя часть нити, образование вторичного ризоида; 3 — нить с двумя
вторичными ризоидами; 4 — врастание протопласта верхней клетки в оболочку нижележащей; 5
— дальнейшая стадия; 6 — нить с разветвлением наверху; 7 — споры (почки); 8 — прорастающие
в нити споры; 9 — дальнейшая стадия прорастания. 1-9 — по Скабичевскому.
Fig. 78. Gemmiphora compacta.
1 — part of thallus; 2 — lower part of filament, formation of secondary rhizoid; 3 — filament with two
secondary rhizoids; 4 — protoplast of the upper shoot grows into the sheath of the lower one; 5 — the
following stage; 6 — filament branching at the top; 7 — spores (buds); 8 — spores growing into filaments;
9 — subsequent stage of in-growth. 1-9 — after Skabitschevsky.
и пиреноидами. Размножение фрагментацией нитей, акинетами, 4-жутиковы-
ми зооспорами. Половой процесс — копуляция двужгутиковых изогамет.
Преимущественно в морях, а также в солоноватых и пресных водах.
В Байкале найдено 4 вида.
I. Клетки менее 100 мкм шириной. Нити короткие (0.57-1.74 мкм длиной)
.................................................................Ch. solitaria
II. Клетки от 100 мкм шириной и более.
1. Нити короткие (не более 3.5 см) всегда прикрепленные.
А. Клетки цилиндрические, без перетяжек у поперечных перегородок, пре-
имущественно 250-300 мкм шириной ..............................Ch. curta
Б. Клетки явственно бочонкообразные, преимущественно 450-600 мкм
шириной..................................................Ch. moniliformis
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 175
2. Нити длинные, прикрепленные или свободно плавающие.
А. Нити свободно плавающие, клетки до 350 мкм шириной.
а. Клетки 250-350 мкм шириной, длина их равна ширине или до 2 раз
меньше ее (только у старых клеток длина может быть больше шири-
ны) ................................................Ch. baicalensis
Chaetomorpha solitaria Skabitsch., 1931 (=Cladochaete microscopica (Meyer)
Meyer et Skabitsch., 1965; Chaetocladiella microscopica (C. Meyer)
Meyer et Skabitsch., Chaetomorpha solitaria, Skabitsch.
[Загоренко, Ижболдина, 1983] (рис. 79: 1-5)
Нити одиночно растущие, прямые, очень короткие, 0.57-1.74 мм длиной
и 38.8-77.6 мкм шириной, состоящие из 8-16, чаще 14 клеток. Нить может от
основания к середине расширяться, а выше опять суживается. Нити из
цилиндрических, слабо перетянутых клеток. Длина клеток увеличивается
вверх и всегда больше ширины. Иногда в нижней части нити длина может
быть равна ширине или меньше ее. Верхняя клетка округлая. Ризоид диско-
видный, с изрезанными краями. Клеточная оболочка толстая (до И мкм
шириной), слоистая, хлоропласт сетчатый, с большим числом пиреноидов.
Зооспорангии образуются из клеток в верхней части нити, 44.2-76.6 мкм
шириной, раскрываются посередине, сбоку, округлым окошечком. Выхода
зооспор наблюдать не удалось.
В Байкале найдена против пади Бол. Коты, на глубине 40-55 м, на камнях
и мелких камешках.
Замечания. Из Байкала вид описан А.П. Скабичевским [1931]. Позднее
[Мейер, Скабичевский, 1965; Скабичевский, 1966] Ch. solitaria была отнесена
к роду Cladochaete С. Meyer et Skabitsch., переименованному впоследствии
из-за номенклатурных соображений в род Chaetocladiella [Скабичевский,
1968], и отождествлена с видом Chaetocladiella microscopica (С. Meyer) Meyer et
Skabitsch. Последний, по мнению авторов [Мейер, Скабичевский, 1965],
наряду с типичными экземплярами, разветвленными у основания, может
иметь вид и простой неразветвленной нити, соответствующей облику ти-
пичных представителей рода Chaetomorpha. А.П. Скабичевский [1965, 1966]
считает, что именно эти одиночные неразветвленные нити Chaetocladiella
microscopica и были первоначально описаны под названием Chaetomorpha
solitaria. Мы считаем [Загоренко, Ижболдина, 1983], что описание А.П. Ска-
бичевского, данное им для Chaetomorpha solitaria, не соответствует таковому
для Chaetocladiella microscopica. Дело в том, что неразветвленные экземпляры
последней по своему облику и по размерам не отличаются от нитей раз-
ветвленных экземпляров этого вида. Одиночные нити, как и разветвленные,
часто расширяются в средней части, никогда не зауживаются, но часто
загибаются на вершине. Клетки у нее цилиндрической формы, равной длины
и широкие, а в верхней части длина их меньше ширины. В местах клеточных
перегородок нити перетянуты, форма конечной клетки всегда округлая.
Приведенное описание полностью совпадает с описанием, данным для этого
вида К.И. Мейером [1927, 1930], но отличается от приводимого для Chae-
tomorpha solitaria А.П. Скабичевским, где отмечено сужение нити в верхней
части и превышение длины над шириной. Поэтому мы посчитали сом-
нительным, что при первоописании Ch. solitaria автор имел дело с оди-
ночными неразветвленными нитями Ch. microscopica. Большой объем ма-
териала, просмотренный нами к настоящему времени, позволяет нам ут-
176 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 79. 1-5 — Chaetomorpha solitaria: 1 — нити в натуральную величину, 2, 3 — нити на
разной стадии роста, 4 — на конце клетки пустые зооспорангии, 5 — рост нити после
разрушения зооспорангия возобновляется; 6-11 — Chaetomorpha moniliformis: 6 — нити
в натуральную величину, 7 — молодые нити, 8 — нижняя часть нити с ризоидом, 9 —
участок клеток средней части нити, 10 — то же, соседние перетяжки отдельной группы
клеток, 11 — верхняя часть нити. 3 — по А.П. Скабичевскому [1936].
Fig. 79. 1-5 — Chaetomorpha solitaria: 1 — filaments, natural size, 2, 3 — filaments at
different growth stages, 4 — empty zoosporangia at the cell end, 5 — filament growth after
disintegration of zoosporangium is resumed; 6-11 — Chaetomorpha moniliformis: 6 —
filaments, full size, 7 — juvenile filaments, 8 — lower part of filament with rhizoid, 9 —
fragment of cells in the middle part of filament, 10 — the same. Neighbouring cell division of
some cells, 11 — upper part of filament. 3 — after Skabitschevsky [1936].
верждать, что в Байкале наряду с одиночными неразветвленными нитями
Chaetocladiella microscopica развиваются одиночные неразветвленные нити, не
имеющие отношения к этому виду. А соответствие таких нитей диагнозу,
данному А.П. Скабичевским для Ch. solitaria, свидетельствует о том, что они
являются ничем иным, как Ch. solitaria.
Напомним, что в одной из последних работ А.П. Скабичевский [1976а]
ограничивается предположением о том, что Ch. solitaria, возможно, — лишь
одна из модификаций Ch. microscopica.
Учитывая вышеизложенное, мы считаем Chaetomorpha solitaria самостоя-
тельным видом. Chaetomorpha solitaria имеет сходство только с Ch. curta Skabitsch.
[Скабичевский, 1969а], от которой хорошо отличается значительно меньшими
размерами клеток и их формой, в том числе и формой базальной клетки.
Сходство этих видов ранее было подчеркнуто и А.П. Скабичевским [1936].
Вид широко распространен в сублиторальной зоне Байкала, встречен нами
в марте, июне-октябре. Кроме Байкала найден в оз. Хубсутул (Монголия), в
северной и южной частях озера.
На песчаных грунтах с глубинами 8-30 м.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 177
Chaetomorpha curta Skabitsch., 1969 (=Сй. baicalensis С. Meyer,
var. curta Skabitsch., 1936) (рис. 80: 1—17)
Нити темно-зеленые, неразветвленные, короткие (до 3.5 см длины),
единичные, редко по 2-3 вместе, прямые или слегка изогнутые, нередко к
середине расширяющиеся, а у основания суженные. Клетки цилиндрической
формы, длина их чаще равна ширине, реже несколько меньше или больше ее.
Редко клетки слабобочонкообразные. Верхушечная клетка куполообразно за-
кругленная. Базальная клетка уже основных, с широкой дисковидной по-
дошвой, поэтому основание нити имеет вид вазы. Иногда суженными могут
быть 2 клетки. Оболочка клеток слоистая, до 32 мкм толщиной. Клетки нити
128-470 мкм шириной, чаще 250-300 мкм шириной. Молодые нити 20-
100 мкм шириной. Зооспорангии образуются в верхней части нити, слегка
бочонкообразные, открывающиеся круглым отверстием в середине клетки.
Зооспоры лежат плотно, заполняя всю полость клетки. У талломов с глубины
60 м, лежащих на иле, нижняя часть нитей разрушена, ризоид отсутствует.
Если по каким-либо причинам конец нити обламывается, на нем может
образоваться апикальная куполообразная клетка. Тогда получается свободно
лежащая нить, оба конца которой соответствуют вершине. Продолжительность
жизни предположительно более 1 года.
Обитает Ch. curta в сублиторали (глубины 20-70 м) оз. Байкал, образуя
V растительный пояс [Скабичевский, 1936]. Позднее [Поповская, Скаби-
чевский, 1968] она была обнаружена на глубине 78 м у мыса Понгонье, 116 м в
губе Давша и на 86 м у мыса Котельниковский.
А.П. Скабичевский [1936, 1969] указывает на близость данного вида с
Ch. solitaria, от которой отличается значительно большими размерами. От
Ch. baicalensis отличается прикрепленным образом жизни, меньшей длиной
нитей и в среднем меньшей их толщиной. Ch. curta является как бы про-
межуточной формой между Ch. baicalensis и Ch. solitaria.
Замечания. А.П. Скабичевский [1969а] отмечает, что вид встречается в
Байкале в двух состояниях нити: прикрепленные к грунту с помощью ризоида,
и свободно лежащие на дне. Первые встречаются на глубинах от 15 до 30 м на
крупнозернистом песке, гальке, гравии, мелких камешках, вторые — на или-
сто-песчаном грунте на глубинах от 30 до 116 м.
Нами [Ижболдина, 1990в] Ch. curta встречена в сублиторальной зоне
многих районов озера. Средняя ширина прикрепленных к песчинкам нитей
колеблется в разных районах Байкала от 140-150 до 200-250, реже 300 мкм.
Причем в Юж. Байкале нити Ch. curta обычно тоньше, чем в северной части
озера. Длина нитей, как правило, не превышает 1-2 см.
В проливах Ольхонские Ворота (бух. Харин-Ирги) и Мал. Море (бух-
ты Харанса, Тутырхейская, острова Угунгой, Зумугой, мыс Ото-Хушун), а
также в бух. Анга, у о. Бакланий (вблизи бух. Песчаной) у мысов Арул,
Онгурен и против падей Широкая, Улан-Нур и Бугульдейка, на заиленном
песке обнаружены свободно лежащие нити длиной 0.5-1.2, реже 3.5 см
(в бух. Харин-Ирги). Эти нити очень хрупкие, прямые или надломленные. На
обоих концах остро- или тупозакругленные клетки (рис. 80: 10-16). Един-
ственный раз была встречена такая нить с ризоидом, прикрепленным к
песчинке на домике ручейника. Ширина нитей в вышеперечисленных районах
колеблется от 375 до 425 мкм, достигая в отдельных случаях 450-590 мкм.
Наблюдения за продолжительностью жизни прикрепленных и свободно
лежащих нитей показали следующее. Прикрепленные к песчинкам нити в
178 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 80. 1-17 — Chaetomorpha curta: 1 — нижняя часть нити с ризоидом, 2 — участок
клетки в средней части нити, 3 — верхняя часть нити, 4 — верхняя часть нити с
опорожнившимся зооспорангием, 5 — молодые талломы, 6 — общий вид нити перед
образованием зооспорангиев, 7 — талломы с опорожнившимися зооспорангиями, 8 —
зооспорангий разрушен и рост нити продолжается, видна делящаяся апикальная клетка,
9 — оба конца нити обломлены, 10 — на концах свободно лежащей на дне нити
образовались апикальные клетки, 11 — нити из района бух. Лаканда в натуральную
величину, 12 — фрагмент свободно лежащей нити из прол. Ольхонские Ворота, 13 —
короткая нить с двумя апикальными клетками, 14-16 — концы нитей, 17 — нити Ch.
curta из прол. Ольхонские Ворота (бух. Харин-Ирги) в натуральную величину. 18-23 —
Ch. baicalensis: 18, 19 — зооспорангии, 20 — часть нити, 21 — концы нитей с разной
формой апикальной клетки, 22 — фрагмент нити с апикальной закругленной клеткой,
23 — нити в натуральную величину.
По: 1-4 — Скабичевскому, 18-20 — Мейеру.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 179
аквариумах с регулярной сменой байкальской воды живут более 3 лет. За это
время дважды было отмечено в октябре формирование зооспорангиев. Пустые
зооспорангии сохраняются до 6 мес. В марте-апреле рост нити возобновляется.
В озере талломы Ch. curta с зооспорангиями встречены в июле-октябре. На
одной крупной песчинке может расти до 15-30 нитей Ch. curta, среди которых
совсем молодые, с формирующимися и пустыми зооспорангиями. Очевидно,
процесс размножения в разных талломах протекает не одновременно. Под-
тверждением тому служат молодые стерильные талломы Ch. curta, в течение
года обнаруживаемые в Байкале и свободно лежащие на грунте талломы
Ch. curta, собранные в районе о. Бакланий Камень в июне 1982 г., вегетировали
в условиях аквариума до октября 1987 г. За это время увеличилось число нитей
и уменьшилась их длина. Зооспорангии не встречены.
К.И. Мейер [1930] в небольшом количестве обнаружил в бух. Аяя короткие
(до 5 мм) нити Chaetomorpha, которые были определены им как Ch. аёгеа
(Dillw.) Kutz. Позднее найденные здесь нити описаны А.П. Скабичевским
[1936, 1969] как Ch. curta и отличаются от Ch. аёгеа по цвету хлоропласта, длине
нитей и базальной клетки. Очевидно, определение этой водоросли как
Ch. аёгеа было ошибочным, на возможность чего указывают К.М. Виноградо-
ва, М.М. Голлербах и др. [1980]. Нами Ch. аёгеа в оз. Байкал не встречена.
Chaetomorpha moniliformis Skabitsch., 1936
Нити короткие (до 2 см), толстые ((236)450-600(667) мкм), расширяющие-
ся снизу вверх и достигающие постоянной ширины у 5-7-й клетки. Благодаря пе-
ретянутости нитей у клеточных перегородок, они принимают четковидную фор-
му. Четковидность лучше выражена в верхней и средней частях нити. Длина кле-
ток сильно варьирует, но обычно она меньше ширины. Верхушечная клетка тупо
закругляется или слегка заостренная. Базальная клетка с расширенным в виде ле-
пешки ризоидом в основании имеет форму обратного конуса. Хлоропласт сетча-
то-продырявленный, интенсивно-зеленого цвета. Зооспорангии, образующиеся
из верхних клеток нити, 480-690 мкм шириной, 225-372 мкм длиной.
Эндемик Байкала, растет на глубине 6-35 (38) м, на песке. Встречен
А.П. Скабичевским [1936, 1968] в бухтах Аяя, Лаканда, Фролиха и у Ушканьих
островов, где вместе с другими водорослями образует IV растительный пояс.
Наши исследования [Ижболдина, 1990в] позволили значительно расши-
рить границы распространения вида в Байкале. В литоральной зоне на камени-
стых грунтах Ch. moniliformis растет обычно среди талломов Chaetocladiella
pumila Меуег et Skabitsch.
Fig. 80.1 -17 — Chaetomorpha curta: 1 — lower part of filament with rhizoid, 2 — cell fragment
in the middle part of filament, 3 — upper part of filament, 4 — upper part of filament with an
empty zoosporangium, 5 — juvenile thallomes, 6 — filament before formation of zoosporangia,
general view, 7 — thallomes with empty zoosporangia, 8 — zoosporangium disintegrated and
filament continues to grow, dividing apical cell is seen, 9 — both filament ends broken, 10 —
apical cells formed at the ends of filament lying free on the bottom, 11 — filaments from the
region of Lakanda Bay, full size, 12 — fragment of a free-lying filament from Olkhonskie Vorota
Strait, 13 — a short filament with 2 apical cells, 14-16 — filament tips, 17 — Ch. curta filaments
from Olkhonskie Vorota Strait (Kharin-Irgi Bay), full size; 18-23 — Ch. baicalensis: 18, 19 —
zoosporangia, 20 — fragment of filament, 21 — filament ends with different forms of apical cell,
22 — filament fragment with a rounded apical cell, 23 — filaments, full size.
After: 1-4 — Skabitschevsky, 18-20 — Meyer.
180 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Вдоль западного прибрежья Ch. moniliformis обнаружена от бух. Песчаной
до мыса Онгурен. Наиболее обильное развитие вида наблюдали в бух. Хара-Ун-
дурская (о. Ольхон) в III (фитомасса 20 г/м2) и IV (фитомасса 92 г/м2) расти-
тельных поясах, на твердом грунте. В других районах фитомасса обычно менее
0.5 г/м2. У восточного берега Ch. moniliformis обнаружена на участке от мыса
Тонкий до губы Фролиха. Наиболее обильно развита в районе мысов Амунда-
кан и Разбор, где ее фитомасса в III поясе составляет 2.8 и 2.4 г/м2 соответст-
венно. Наиболее часто Ch. moniliformis встречается в районе Ушканьих остро-
вов во II (7= 22 %) и III (V = 26 %) растительных поясах. В других районах
озера встречаемость вида обычно менее 5 %. Пустые и полуразрушенные зоо-
спорангии наблюдали в августе-сентябре. В аквариуме с байкальской водой
талломы Ch. moniliformis живут более 1 года.
Chaetomorpha baicalensis С. Meyer (Meyer, 1922)
(рис. 80: 18-23)
Нити светло-зеленые, длинные (до 3-4 см), ломкие, разнообразно изогну-
тые. Переплетаясь между собой, образуют на глубине 1-2 м обширные дерно-
вины. Клетки нити цилиндрические, отношение длины к ширине сильно варь-
ирует. Клеточная оболочка толстая, слоистая. Хлоропласт в виде сетчато-про-
дырявленного цилиндра с многочисленными мелкими пиреноидами. Ширина
нитей 250-350 мкм. Все клетки нити могут превращаться в зооспорангии. При
этом они слегка вздуваются и участки нитей с зооспорангиями принимают чет-
кообразную форму.
Эндемик Байкала. По данным К.И. Мейера [1930], образует обширные за-
росли на мягком грунте на глубине 1-2 м. Найдена им у Ушканьих островов, в
пади Нугды, в прол. Ольхонские Ворота и в Мал. Море — против мыса Хадар-
туй и у мыса Ото-Хушун.
Замечания. В.Н. Яснитский [1928] обнаружил Ch. baicalensis в бухтах Ха-
рин-Ирги и Загли (прол. Ольхонские Ворота), где на глубине 1.5-16 м она об-
разует растительный пояс. Им же прослежена в южной части прол. Мал. Море
до о. Хубын. При наших исследованиях [Ижболдина, 1990в] Ch. baicalensis об-
наружена во многих пунктах вдоль западного побережья Байкала (бух. Богу-
чанская, мыс. Онгурен, против падей Орео, Улан, Улан-Нур, Песчаная, Сен-
ная, Бол. Коты), на глубине от 3 до 23 м. Против пади Улан она найдена на глу-
бине 68 м. В прол. Мал. Море Ch. baicalensis прослежена нами от мыса Кобылья
Голова до губы Харанса и мыса Ото-Хушун, в районе которого длина нитей
достигала 10-16 см. В бухтах Тутырхейской, Харгойской, Семисосенной и Ша-
ра-Шулун Ch. baicalensis толстым (до 0.5 м) слоем покрывает песчаный грунт
на глубине от 1.5 до 10 м, а фитомасса ее колеблется от 4 до 39 г/м2, в то время
как в других районах озера она редко превышает 1, 2 г/м2. В бух. Шибетейской
Ch. baicalensis, как и вдоль открытых прибрежий озера, растет среди зарослей
харовых водорослей. У восточного берега вид встречен у мыса Орлова и в губе
Сорожья (п-ов Святой Нос), и в районе губы Максимиха (Баргузинский
зал.) — Ch. baicalensis развита в Байкале в течение всего года. В аквариуме с
байкальской водой отдельные талломы жили более 3 лет.
Род Rhizoclonium Kutz., 1843
Водоросль образует свободноплавающие пучки из тесно переплетенных ни-
тей, реже растет прикрепленно. Нити грубые, без ветвей или скудно ветвящиеся.
Ветви обычно короткие, из 1-2 клеток, редко длинные. Вторичного ветвления
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 181
не бывает. Характерно развитие коротких ризоидных ветвей. Клетки нити ци-
линдрические, длинные, реже короткие. Оболочка клеток у разных видов разной
толщины. Хлоропласт сетчатый, иногда из множества неправильных овальных
хлоропластов, с одним пиреноидом в каждом. Ядер от одного до нескольких. Ве-
гетативное размножение — за счет разрыва нитей, с помощью акинет, представ-
ляющих собой клетки неправильной формы с сильно утолщенными стенками.
В морских, соленых, солоноватых и пресных водах.
В Байкале обнаружен один вид.
Rhizoclonium sp. (рис. 81)
Представляет собой тонкие крепкие нити длиной от 1-1.5 до 2.5-3, реже
6-10 см. Нити неприкрепленные, спутанные, построены из цилиндрических
клеток шириной 50-55 мкм, длина их колеблется от 55 до 150, чаще 70-93 мкм.
В одной нити размер клеток может сильно изменяться. Наблюдаются утолще-
ния одного или нескольких участков нити, захватывающее обычно 3-6, реже
10-12 клеток. Ширина нити в утолщенных местах достигает 70-90 мкм.
Вид близок к Rh. hieroglychicum, отличается большей шириной нитей, от-
сутствием ризоидов и вегетативных ветвей.
Rhizoclonium sp. найден нами в районе губ Богучанская (глубина 10 м), Да-
гарская (12 м), Томпа (от 10 до 35 м), Улан (глубина 38 м). В губах Бабушка и
Песчаная он встречен на глубинах от 1.5 до 30 м, в Чивыркуйском заливе — на
глубине 20 м, где растет вместе с Cladophora floccosa var. irregularis, на Анга-
ро-Кичерском мелководье — вблизи устья р. Верх. Ангара, где рос переплета-
ясь с талломами Cladophora aegagropila (L.) Rabenh.
Замечания. Rhizoclonium sp. растет обычно среди зарослей харовых водорос-
лей и Ch. baicalensis, иногда среди ветвей Cladophora glomerata L. и так перепле-
тается с талломами последних, что его трудно отделить, не повредив нити. Во
многих нитях в августе — сентябре наблюдали образование, напоминающие
споры почки, описанные ранее А.П. Скабичевским [1931] для Gemmiphora
compacta. Однако ни уплотнения хлоропласта в отпочковавшихся клетках, ни
прорастания последних в новую нить не замечено. Возможно, это акинеты
Rhizoclonium sp.
Род Cladophora Kutz., 1843
Рыхлые кустистые дерновинки разнообразного облика (плотные подушко-
образные, полушаровидные, шаровидные колонии или бесформленные скоп-
ления ветвящихся нитей). Таллом изоморфный или гетероморфный. Органы
прикрепления отсутствуют или в виде простого дисковидного ризоида, или ко-
ралловидных разрастаний. Ветвление боковое, одностороннее, поочередное,
супротивное. Инсерция ветвей апикальная или латеральная.
Часто наблюдается эвекция и, как следствие, ее псевдодихотомия. Клетки
обычно вытянутые, длина их превышает ширину до 20 раз. Хлоропласт сетча-
то-продырявленный или в виде многочисленных пластинок с многими пире-
ноидами. Вегетативное размножение фрагментацией талломов и акинетами,
бесполое — с помощью 2- или 4-жгутиковых зооспор, половое — изогамное.
У некоторых видов отмечена гетероталличность. Известна изоморфная смена
половой и бесполой форм развития.
Представители рода распространены почти повсеместно. В Байкале найде-
но 11 видов и вариаций.
182
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 81. Rhizoclonium sp.
Fig. 81. Rhizoclonium sp.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 183
I. Талломы изоморфные, кустистые, с органами прикрепления или без них.
1. Органы прикрепления всегда отсутствуют.
А. Клетки основных нитей обычно свыше 60 мкм, но не менее 50 мкм
шириной.
а. Дерновинки неправильно обратно-конусовидные, 2-4 мм длиной.
.. Клетки в нижней части кустиков 80-90 мкм шириной..........
.................................................С. kusnetzowii
2. Органы прикрепления хорошо выражены, но у некоторых видов развиты
только на начальных стадиях развития, затем талломы отрываются и рас-
тут в свободноплавающем состоянии.
А. Органы прикрепления в виде простых дисковидных ризоидов. Нити
короткие (до 7 см длиной), постоянно прикрепленные к субстрату.
а. Нити 0.5-2.5 см длиной, обильно ветвятся, собранные в шары или
дерновинки. Ветвление в нижней части талломов всестороннее, в
верхней — одностороннее, гребенчатое.
а. Дерновинки хлопьевидные, реже шаровидные, 0.5-1.5 см в диа-
метре, клетки в нижней и средней частях нитей 35-140 мкм
шириной, конечные 15-65 мкм шириной.................С. floccosa
р. Дерновинки округлой, полушаровидной или шаровидной формы,
1—2.5 мм в диаметре. Клетки в нижних частях нитей 72-117 мкм
шириной, конечные около 50 мкм шириной .............С. meyeri
т. Дерновинки шарообразной формы, до 0.5 см в диаметре. Клетки в
нижней части таллома 27-32 мкм шириной, конечные 18-27 мкм
шириной; ширина зооспорангиев 30-32 мкм ............С globus
Б. Органы прикрепления в форме коралловидных ризоидов. Нити длин-
ные, прикрепленные к субстрату постоянно или на начальных стадиях
роста. Дерновины крупные, кустистые, рыхлые, реже шаровидные или
в виде бесформенных скоплений.
а. Инсерция ветвей латеральная, псевдодихотомия встречается редко.
а. Дерновинки преимущественно шаровидные, реже хлопьевидные и
бесформенные. Нити обильно ветвящиеся. Базальные ризоиды
многочисленные. Клетки часто палицеобразные, 125-200 мкм, на
конце нитей 30-70 мкм шириной....................С. aegagropila
р. Дерновинки в виде спутанных пучков или тяжей с более или ме-
нее параллельным расположением скудно ветвящихся нитей. Ба-
зальные ризоиды редкие. Клетки преимущественно цилиндриче-
ские.
+ Клетки 16-27(38) мкм, на вершине нитей 13-21 мкм шириной
...........................................C.globulina
б. Инсерция ветвей апикальная, поперечные перегородки в местах от-
хождения ветвей косые и часто переходят в горизонтальное положе-
ние, вызывая псевдодихотомию.
а. Клетки основных осей 45-85 мкм шириной, апикальные 16-32 мкм
шириной. Ветвление обильное. Только в пресной воде . . С. flacra
р. Клетки основных нитей 65-275 мкм шириной, апикальные 19-
91 мкм шириной. Ветвление скудное, почти отсутствует или оби-
льное. В пресной, солоноватой (до 15 %о) и морской воде....
...............................................С. glomerata
II. Талломы гетероморфные, из ризоидных нитей, стелющихся по субстрату
и отходящих от них вверх простых или ветвящихся нитей.
184 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
1. Восходящие нити обильно ветвятся, не более 2.5 см высотой.
А. Дерновинки не более 10(15) мм в высоту. Эндемики Байкала.
а. Дерновинки до 1 мм высотой. Клетки восходящих нитей 100—
130 мкм шириной.................................. С. kursanovii
б. Дерновинки 2-5 мм высотой. Клетки восходящих нитей менее
100 мкм шириной.
а. Клетки восходящих нитей в нижних частях 32-40 мкм шириной.
Ветвление особенно обильное в верхних частях нитей, кистевид-
ное. Нити плотно сжатые, образуют очень плотные полушаро-
видные подушечки, темно-зеленого цвета..........С. compacta
р. Клетки восходящих нитей в нижних частях 63-80 мкм шириной.
Ветвление равномерно обильное, одностороннее. Талломы в
виде округлых рыхлых подушечек сероватого цвета С pulvinata
Cladophora kusnetzowii С. Meyer, 1930 (рис. 82: 1—6 )
Темно-зеленые дерновинки неправильно-треугольной формы, 2-4 мм высо-
той. Строение таллома изоморфное, ризоиды отсутствуют. От верхнего рас-
ширенного конца нижней клетки, служащей основанием таллома, отходят 3-5
ветвей, которые в свою очередь образуют в верхней части мутовки по 3-5 клеток.
В периферической части таллома ветвление одностороннее или супротивное.
Клетки сильно вытянутые, ширина их в нижней части кустика 80-90 мкм, в
средней — 45-65 мкм, на концах — 30-37 мкм. Вегетативное размножение с
помощью плетей, вырастающих из нижних клеток таллома. Плети в виде тонких
нитей первое время не ветвятся, затем начинают ветвиться и отрываются от
материнского таллома. Кроме того, талломы по мере разрастания разламываются.
Эндемик Байкала. Найден К.И. Мейером [1930] в зал. Мухор (прол. Мал.
Море), где сплошным слоем покрывает дно на глубине 3-4 м. Растет вместе с
Lemna и Nostoc. На поверхности С. kusnetzowii часто встречается эпифит^
Lyngbya kuetzingii (Kutz.) Schmidle.
Замечания. Нами [Ижболдина, 1990а] С. kusnetzowii прослежена в Мал.
Море — от зал. Мухор до губы Харанса. В прол. Ольхонские Ворота в неболь-
шом количестве обнаружена у мысов Харин-Ирги, Тыгтэ, Харганте, Хален и в
бух. Куркутской на глубине от 1 до 15(36) м. Отмечена сезонность в развитии
С. kusnetzowii [Ижболдина, 1976а]. В июне в зал. Мухор найдена масса мелких
(2-2.8 мм высотой) дерновинок, в конце сентября длина хлопьевидных дерно-
винок достигает 5-7 мм. Фитомасса этой водоросли в зал. Мухор на глубине
2-3 м составляет в среднем 40 г/м2. В условиях аквариума с байкальской водой
талломы С. kusnetzowii вегетируют на протяжении 2-3 лет. Верхние концы мно-
гих конечных клеток могут утолщаться. Иногда несколько таких утолщений
наблюдается в одной клетке, которая принимает четковидную форму [Ижбол-
дина, 1988]. Возможно, мы наблюдаем образование акинет, прорастание кото-
рых проследить не удалось.
Cladophora floccosa С. Меуег, 1927 (рис. 83: 1—7 )
Образует небольшие кустики шарообразной или неправильной формы до
1 см высотой и 1-1.5 см диаметром, прикрепленных к камням или песчинкам
простым лепешковидным ризоидом. Строение таллома изоморфное. От ризо-
идной клетки отходит нить, ветвящаяся всесторонне, давая начало простым
длинным нитям, которые, достигнув значительной длины, образуют густое
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 185
Рис. 82. 1—6 — Cladophora kusnetzowii: 1 — ветвление таллома, 2-3 — внешний вид
таллома в натуральную величину в июне (2) и в сентябре (3), 4, 5 — конец ветвей, 6 —
образование акинеты (?) (большое увеличение); 7-13 — Cladophora aegagropila: 7 —
участок таллома свободноплавающего растения, 8, 9 — участки талломов из шаро-
видных дерновинок, 10 — нить с ризоидами, образованными из апикальных клеток,
11 — участок таллома подушкообразной дерновинки с большим числом ризоидов и
инверсиями полярности, 12 — коралловидный ризоид, 13 — внешний вид колонии в
натуральную величину.
По: 1, 4 — Мейеру, 7-12 — Хуку, 13 — Кютцингу.
Fig. 82. 1-6 — Cladophora kusnetzowii: 1 — thallome branching, 2, 3 — full-sized thallome in
July (2) and September (3), general view, 4, 5 — end of branches, 6 — akinete formation (?)
(large magnification), 7-13 — Cladophora aegagropila: 7 — thallome fragment of a free-
swimming plant, 8, 9 — thallome fragments from globose swards, 10 — filament with rhizoids
composed of apical cells, 11 — fragment of thallome of a pillow-shaped sward with a large num-
ber of rhizoids and polarity inversions, 12 — coral-shaped rhizoid, 13 — external view of the
colony, full size.
After: 1, 4 — Meyer, 7-12 — Hoek, 13 — Kiitzing.
186 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
одностороннее ветвление гребенчатого типа. Новые ветви в свою очередь вы-
растают в длинные нити, дающие вновь гребенчатое ветвление. В результате
образуется хлопьевидный кустик, состоящий из нитей с несколькими яруса-
ми ветвления. Бывает и разностороннее ветвление в верхних частях нитей.
Иногда талломы состоят лишь из одного яруса гребенчато разветвленных ни-
тей. Клетки цилиндрические, ширина их в нижних частях нитей 35-120 мкм,
в средних — 35-140 мкм, апикальных клеток 15-64 мкм. Оболочка тонкая или
толстая, слоистая. Клетки в нижней части таллома могут превращаться в аки-
неты. Зооспорангии бочонкообразные, 80-140 мкм шириной, располагаю-
щиеся цепочками в верхних частях кустика. Иногда нить с зооспорангиями
спирально закручивается.
Cladophora floccosa var. floccosa C. Meyer, 1927 (рис. 83: 1—7 )
Ветвление в верхней части кустика только одностороннее, гребенчатое.
Клетки в нижних и средних частях нитей 35-70 мкм шириной, конечные клет-
ки 15-25 мкм шириной. Оболочка утолщенная, слоистая только у нижних и
акинетообразных клеток.
В Байкале — на камнях и крупнозернистом песке на глубине 0.5-5 м.
А.П. Скабичевским [1968, 1976] встречена во многих бухтах озера на глубине
30-35(50) м.
Рис. 83. Cladophora floccosa var. floccosa.
1, 3-5 — характер ветвления таллома; 2 — молодые талломы; 6 — зооспорангии; 7 — внешний вид
таллома в натуральную величину. По: 1, 2 — Скабичевскому, 4 — Мейеру.
Fig. 83. Cladophora floccosa var. floccosa.
1, 3-5 — thallome branching pattern; 2 — juvenile thallome; 6 — zoosporangia; 7 — external view of
thallome, full size. After: 1, 2 — Skabitschevsky, 4 — Meyer.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 187
Замечания. При наших исследованиях [Ижболдина, 1988, 1990в] вид встре-
чен вдоль большей части прибрежья. Отсутствует он на Ангаро-Кичерском и
Селенгинском мелководьях, в зал. Култук (Баргузинский залив), изредка (V=
до 10 %) встречается в Посольском соре, проливах Мал. Море и Ольхонские
Ворота. Вдоль остальной части прибрежья С. floccosa является одним из наи-
более распространенных видов, обнаружена во всех растительных поясах, но
наиболее часто встречается (И = 40-60 %) во II и III. Обычно фитомасса ко-
леблется от 0.01 до 10-16 г/м2 и составляет в среднем 0.74 г/м2 во II и 2.91 г/м2 в
III поясах. К.И. Мейер [1930] подчеркивает, что С. floccosa встречается главным
образом в наиболее мелких и хорошо прогреваемых местах. Наши наблюдения
показывают, что водоросль здесь развита наиболее обильно. Значительные за-
росли ее вдоль западного берега обнаружены в районе мыса Курлы (153 г/м2,
III пояс), губы Сеногда (100 г/м2, III пояс), у мыса Котельниковский (42 г/м2,
III пояс) и в районе пос. Сенная на о. Ольхон (23 г/м2, III пояс). Вдоль восточ-
ного берега — это мыс Разбор (33 г/м2, III пояс), о. Бол. Ушканий (78 г/м2, а в
отдельных случаях — до 600 г/м2, II пояс), у о. Лиственичный (30 г/м2, II пояс)
и у ст. Танхой (22 г/м2, III пояс).
Вид развит в Байкале в течение круглого года. Экспериментальные исследо-
вания показали, что С. floccosa — многолетнее растение. Темп роста молодых,
слабо разветвленных талломов составляет в среднем 140 мкм в месяц. Во все се-
зоны года в Байкале присутствуют как стерильные талломы С. floccosa, так и
талломы с зооспорангиями в разной стадии формирования или пустые. В зоо-
спорангии превращаются от 1-2 до 10-12 клеток на конце нити. Отмеченные
А.П. Скабичевским [1966] в районе Бол. Котов талломы С. floccosa с одним яру-
сом ветвей встречены нами вдоль всего побережья и особенно часто на глубинах
более 10-15 м. Кроме того, в разных районах озера (ст. Маритуй, мыс. Телеграф-
ный, падь Сенная на о. Ольхон, мыс Каткова, губа Иринда, р. Бол. Осиновка,
мыс Кочериковский) в разные годы мы встретили талломы С. floccosa, у которых
ветви 2-го и 3-го порядка не ветвились и достигали длины 2-3 см, а в отдельных
случаях даже 7 см. Как правило, такие талломы стерильны и чрезвычайно обиль-
но покрыты эпифитами. Причина такого роста нитей не ясна.
Кроме Байкала вид встречен [Загоренко, Ижболдина, 1976] в оз. Хубсугул.
Cladophora floccosa var. irregularis Skabitsch., 1930 (рис. 84: 1-7)
Ветвление в верхней части таллома обычно всестороннее. Ширина клеток
в нижних частях нитей 60-120 мкм, в средней — 80-140 мкм, апикальных кле-
ток 40-64 мкм. Клеточные оболочки толстые, слоистые.
Оз. Байкал, на глубине 4.5-40 м.
Замечания. Растет обычно в тех же районах, что и типовая форма. Наиболее
обильное развитие отмечено в районе о. Бол. Ушканьего, во II и III поясах, где
встречаемость ее составляет 75-87 %, а фитомасса 83-57 г/м2. В течение года в
Байкале присутствуют как стерильные талломы, так и растения с зооспорангия-
ми на разной стадии развития. В сборах О.А. Тимошкина из бух. Загли-Нур
(прол. Ольхонские Ворота) ризоиды у талломов С. floccosa var. irregularis, сво-
бодно лежащих на мягком грунте на глубине 7-10 м, обычно разрушены
(рис. 85). Ветви очень длинные, скрученные, маловетвящиеся, утончающиеся к
концу. Ветвление в разных частях таллома редко разностороннее. В основании и
в средней части нитей ширина их составляет 75-90 мкм. Апикальные клетки
25-40 мкм шириной.
188 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Cladophora meyeri Skabitsch., 1976 (= С. humilis С. Meyer, non C. humilis Kiitz.)
(рис. 86: 1-8)
Очень маленькие, до 1-1.5 мм высотой, талломы в виде шаровидных или
полушаровидных кустиков, прикрепленные к камням и песчинкам при помо-
щи простого ризоида, расширенного в виде небольшой лепешки. Таллом обыч-
но ветвится. Ветвление в нижней части таллома всестороннее, в верхней обыч-
но одностороннее. Клетки более или менее цилиндрические, в нижней части
таллома (50)72-117 мкм шириной, преимущественно короткие, в верхних час-
тях 32-90 км шириной, по длине в 1.5-2(5) раз больше ширины. Апикальные
клетки 27-50 мкм шириной, ризоидная клетка 125 мкм шириной. Толстая
слоистая оболочка у клеток в нижней части таллома и у зооспорангиев. Зоо-
Рис. 84. Cladophora floccosa var. irregularis.
1 — неразветвленная веточка с участком опо-
рожнившихся зооспорангиев, загнутых в спи-
раль; 2 — дихотомическое ветвление; 3 —
разветвленная веточка в верхней части талло-
ма; 4-6 — ветвление таллома; z — зооспоран-
гии после выхода зооспор. 1-3 — по
Скабичевскому.
Fig. 84. Cladophora floccosa var. irregularis.
1 — non-branched shoot with a fragment of empty
zoosporangia, coiled; 2 — dichotomous branching;
3 — branched shoot in the upper part of thallome;
4-6 — thallome branching; z — zoosporangia after
zoospore release. 1-3 — after Skabitschevsky.
Z
4-6:250 pirn
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 189
Рис. 85. Cladophora floccosa var. irregularis.
Характер ветвления талломов из района бухты Загли-Нур (прол. Ольхонские Ворота), где талломы,
свободно лежащие на песке, обычно лишены ризоидов.
Fig. 85. Cladophora floccosa var. irregularis.
Thallome branching pattern from the region of Zagli-Nur Bay (Olkhonskie Vorota Strait), thallomes lying
free on the sand are generally devoid of rhizoids.
спорангии бочонкообразные, 72-94 мкм шириной, располагаются в верхней
части нитей цепочками.
Эндемик Байкала, растет на мелких камнях и песчинках. Найден К.И. Мей-
ером [1930] на глубине 3-10 м во многих районах Сев. Байкала.
190 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Cladophora meyeri var. meyeri (рис. 86: 1—8)
Кустики 1-1.5 мм высотой. Клетки в нижних частях нитей 72-117 мкм
шириной, в верхних — 54-90 мкм шириной, по длине в 1.5-2(5) раза больше.
Апикальные клетки около 50 мкм шириной.
Оз. Байкал, на глубине 3-40 м.
Замечания. А.П. Скабичевский [1934, 1966, 1968] отметил, что С. meyeri
обычно входит в состав водорослей, образующих в Байкале IV растительный
пояс, занимающий глубины от 10-12 до 35 м. При наших исследованиях [Иж-
Рис. 86. 1-8 — Cladophora meyeri var. meyeri: 1 — молодое растение, 2 — участок нити с
зооспорангиями, 3-5, 7 — молодые талломы, 6 — кустики в натуральную величину, 8 —
характер ветвления в разных частях таллома; 9-12 — С. meyeri var. gracilior: 9 — общий
вид таллома в натуральную величину, 10 — сильно ветвящийся кустик, 11 — отдельная
нить, 12 — верхняя часть нити с зооспорангиями.
1-3,10-12 — по Мейеру.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 191
болдина, 1981в, 1990b] вид обнаружен на глубинах от 3 до 67 м, но наиболее час-
то встречается (V = 50-100 %) в зоне глубин 15-35 м. С. meyeri растет вдоль
большей части побережья, где на глубинах ее обитания есть крупнозернистый
заиленный песок. Не встречена она лишь в Истокском и Посольском сорах, в
зал. Провал, на Селенгинском мелководье и на север от него до мыса Толстый,
на Ангаро-Кичерском мелководье.
Fig. 86. 1-8 — Cladophora meyeri var. meyeri: 1 — young alga, 2 — fragment of filament with
zoosporangia, 3-5, 7 — juvenile thallomes; 6 — bushes, full size, 8 — branching pattern in
various parts of thallome; 9-12 — C. meyeri var. gracilior: 9 — general view of thallome, full
size, 10 — highly branched bush, 11 — separate filament, 12 — upper part of filament with
zoosporangia.
1-3, 10-12 — after Meyer.
192 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Ранее [Верещагин, 1908; Месяцев, Зенкевич, Россолимо, 1922; Кожов,
1962] в глубине бухт Онгоконской и Фертик (Чивыркуйский залив) было
отмечено наличие черного рыхлого и сильно маркого ила, “обязанного своим
происхождением Cladophora, которая здесь исключительно многочисленна”.
При наших [Ижболдина, 1990в] исследованиях в этих губах также обна-
ружены целые залежи Cladophora meyeri и масса ее полуразрушившихся
черных талломов. Фитомасса С. meyeri, не превышающая обычно вдоль
открытых прибрежий озера 1-3, редко 10-15 г/м2, здесь достигает 140 г/м2.
Кроме Чивыркуйского залива скопления С. meyeri встречены в районе пади
Широкая и у р. Бугульдейка. Это — медленно растущие многолетние расте-
ния. Максимальный прирост отдельных нитей за 7 мес наблюдения составил
500 мкм [Ижболдина, 1988]. Зооспорангии в разной стадии формирования и
опустошенные встречаются на талломах в течение всего года. Молодые слабо
разветвленные талломы (рис. 86: 1, 4, 5) найдены в разных районах озера в
июне-сентябре.
Кустики С. meyeri большей частью - шарообразной формы и внешне напо-
минают Cladophora kozhowii Zagor. et Izhbol. [Загоренко, Ижболдина, 1977], в
массе развивающуюся в оз. Хубсугул (Монголия), но отличаются от последней
присутствием ризоида, характером ветвления и упругими, но не хрупкими тал-
ломами.
Cladophora meyeri var. gracilior (C. Meyer) Hollerb.
(=C humilisNAr. gracilior C. Meyer, 1927) (рис. 86: 9-12)
Кустики до 2,5 см высоты. Клетки в нижних частях таллома 50-80 мкм
шириной, в верхних — 32-40 мкм шириной, по длине в 2-5 раз и более превос-
ходят ширину. Апикальные клетки до 27 мкм шириной.
В оз. Байкал, на глубине 2-5 м, встречена К.И. Мейером [1930] в губах
Фролиха, Мал. Коса, Онгоконская.
Замечания. Вид найден А.П. Скабичевским [1934,1968] в губах Аяя, Фроли-
ха, Лаканда. Нами [Ижболдина, 1990в] данная вариация встречена у западного
берега в бух. Бихин, в губе Харанса (Мал. Море) и у восточного берега в районе
губ Таланка, Дагарская, Амундакан на Сев. Байкале, кроме того, в Чивыркуй-
ском заливе, у Ушканьих островов. В Юж. Байкале — у ст. Боярск, Переемная
и Танхой. Растет на глубинах от 6 до 24 м, на крупнозернистом песке, гальке,
заиленном песке.
Cladophora globulus С. Meyer (рис. 87: 11—13)
Дерновинки в виде шариков, до 0.5 см в диаметре, состоящие из тесно пе-
реплетенных нитей. Во внутренней части колонии ветвление всестороннее, в
верхней части нитей — одностороннее, кистевидное. Клетки в основном ци-
линдрические, отношение длины к ширине варьирует. Толщина клеток в ниж-
них частях нитей 27-32 мкм, на концах ветвей клетки 18-27 мкм шириной.
Зооспорангии формируются из конечных клеток 30-32 мкм шириной, с тол-
стой слоистой оболочкой. После выхода зооспор клетка, лежащая ниже зоо-
спорангия, прорастает и рост нити возобновляется.
В Байкале — в губах Аяя и Фролиха, на глубине 10 м. Вид недостаточно
изучен.
Замечания. А.П. Скабичевским [1934] вид найден на глубине 10 м в губе
Аяя. Кроме того, нами [Ижболдина, 1990в] встречен вдоль восточного берега в
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 193
районе губ Дагарская, Фролиха, Лаканда, Максимиха, у мыса Орловский и у
Нижнего Изголовья полуострова Святой Нос. У западного берега С. globulus
обнаружена в HI, IV и V поясах в районе мысов Южный Кедровый, Бол. Со-
лонцовый, Анютха, Рытый, Кочериковский, Онгурен, Ухан и Ижимей. Фито-
масса редко превышает 1 г/м2.
Рис. 87. 1-10 — Cladophora kursanovii: 1-4 — молодые талломы, 5 — стелющиеся нити,
6 — восходящая нить, 7 — участок таллома со стелющейся нитью, от которой отходят
восходящие нити и вторичные ризоиды; 8-10 — верхние участки ветвящихся нитей;
11-13 — С. globulus: 11 — общий вид таллома в натуральную величину, 12 — отдельная
нить, 13 — зооспорангий.
По: 2, 3, 7-10 — Скабичевскому, 5, 6, 12,13 — Мейеру.
Fig. 87. 1-10 — Cladophora kursanovii: 1-4 — juvenile thallomes, 5 — trailing filaments, 6 —
ascending filament, 7 — fragment of thallome with a trailing filament with ascending filaments
and secondary rhizoids running from it, 8-10 — upper parts of branched filaments; 11-13 —
C.globulus: 11 — general view of thallome, full size, 12 — separate filament, 13-zoosporangium.
After: 2, 3, 7-10 — Skabitschevsky, 5, 6,12, 13 — Meyer.
194 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Cladophora aegagropila (L.) Rabenh. (= Conferva aegagropila L.; Aegagropila sauteri
(Nees) Kiitz.; Ae. holsatica Kiitz.; Ae. armeniaca (Wittr.) Heering; Cladophora
aegagropila var. sauteri (Nees) Rabenh.; C. aegagropila var. bulnheimii Rabenh.;
C. sauteri (Nees) Kiitz.; C. profunda Brand) (рис. 81: 7-13)
Дерновинки свободноплавающие или прикрепленные. Плавающие дерно-
винки обычно шаровидные или удлиненно-овальные, до 21 см в диаметре, плот-
ные снаружи и рыхлые внутри, состоящие из многочисленных радиально распо-
ложенных талломов. Реже они хлопьевидные или бесформенные. Строение тал-
лома изоморфное. Ярко выражена инверсия полярности. Наблюдается
отрастание ветвей и ризоидов на обоих полюсах клетки и образование коралло-
видных ризоидов по вершине ветвей. Каждая клетка может дать до 4 ветвей. Ин-
серция ветвей латеральная, эвекция наблюдается редко. Клетки цилиндрические
или палицеобразные. У основной оси 125-200 мкм шириной, по длине в 1.5-15
раз больше. Конечные ветви 30-100 мкм шириной, апикальные, 30-70 мкм
шириной, по длине в 5-24 раза больше. Оболочка клеток в основании таллома до
20 мкм шириной, у конечных ветвей 2.5-8 мкм шириной. Размножение преиму-
щественно вегетативное, путем деления таллома, возможно апланоспорами и
зооспорами.
В пресных или, очень редко, солоноватых озерах.
В Байкале найдена К.И. Мейером [1930] в Нижнеангарском соре и в Чи-
выркуйском заливе.
Замечания. Нами [Ижболдина, 1990в] С. aegagropila найдена на Ангаро-Ки-
черском мелководье, на глубине 3 м, в губе Дагарской, на глубинах от 14 до
65 м, и в Чивыркуйском заливе, где ее заросли занимают значительные площа-
ди на глубинах 5-15 м. В вершине залива на глубине 5-15 м фитомасса водо-
рослей достигает 5-19 г/м2. Дерновинки С. aegagropila хлопьевидные, высотой
от 1.5 до 12.5 мм, чаще 3.6-4.1 мм. Ширина верхнего расширенного конца ни-
тей клетки может достигать 80-123 мкм, конечные клетки 33-66 мкм.
Cladophora globulina (Kiitz.) Kiitz. (=Conferna globulina Kiitz.; Cladophora
lacustris Kiitz.; C. fracta var. lacustris (Kiitz.) Brand) (рис. 88: 1-7)
Дерновинки свободноплавающие, в виде бесформенных пучков из перепу-
танных между собой нитей. Строение таллома изоморфное. Нити длинные,
обычно неветвящиеся или скудно ветвящиеся, при массовом образовании аки-
нет ветвление может быть обильным. Инсерция ветвей латеральная, у старых
растений отмечено псевдодихотомическое ветвление. Поперечная перегород-
ка, отделяющая ветвь от основной оси, часто смещается в базальную клетку
ветви. Короткие ветви заканчиваются клеткой в виде шипика. Рост интерка-
лярный, базальные ризоиды коралловидные, редкие. Клетки цилиндрические у
основания оси, 16-27(38) мкм шириной, по длине в 3-11 раз больше, апикаль-
ные 13-21.5 мкм шириной, по длине в 3-8 раз больше, у конечных ветвей
13.5-16 мкм шириной, по длине в 2-7 раз больше.
В пресных, минерализованных и солоноватых водах. Широко распростра-
ненный в Палеарктике вид.
Замечания. В Байкале найден К.И. Мейером [1930] в Баргузинском заливе
около мыса Холодянка и в Чивыркуйском заливе около бух. Крохалиная.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 195
Рис. 88. Cladophora globulina.
1-4 — участок нитей разной степени разветвленности; 5, 6 — участки нитей со вздувшимися клет-
ками; 7 — характер ветвления нитей. Разное увеличение. По Хуку.
Fig. 88. Cladophora globulina.
1-4 — fragment of filament with a different degree of branching; 5-6 — filament parts with swollen cells;
7 — branching pattern of filaments. Various magnification, after Hoek.
Cladophora fracta (Miill. ex Vahl.) Kiitz. (рис. 89: 1—3)
Дерновинки прикрепленные или свободноплавающие, рыхлые. Строение
таллома изоморфное. Нити обильно и равномерно ветвящиеся. Инсерция вет-
вей апикальная, косая до горизонтальной. Базальные ризоиды коралловидные.
Клетки цилиндрические, у основных осей 45-85 мкм шириной, по длине в
1.5-14 раз больше, у конечных ветвей 17.5-38 мкм шириной, по длине в 3-17
раз больше, апикальные 16-32 мкм шириной, по длине в 3.5-25 раз больше.
Размножение фрагментацией таллома и зооспорами.
Вид чрезвычайно полиморфный. Близок к С. glomerata, от которого от-
личается характером ветвления, размерами клеток и экологией. Описание
дано с учетом последней монографии рода Cladophora [Hoek, 1963], в кото-
рой автор различает 2 разновидности этого вида, одна из них встречена в
оз. Байкал.
Cladophora fracta var. fracta (= Conferva fracta Mull ex. Vahl.; C. patens Ag.;
Cladophora viadrina Kiitz.; Aegagropila agardhii Kiitz.)
Дерновинки свободноживущие. Базальные ризоиды редкие. Клетки основ-
ных ветвей 45-85 мкм шириной, конечных ветвей 17.5-38 мкм шириной, апи-
кальные, 16-27 мкм шириной. Размножение с помощью акинет.
В пресных, более или менее эвтрофированных водоемах; выносит лишь
слабую степень засоления.
196 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 89. 1-3 — Cladophora fracta var. fracta: 1,2 — характер ветвления вегетативных нитей
(разное увеличение), 3 — прорастание акинет; 4-7 — С. glomerata var. glomerata: 4 — тал-
лом акропетального строения, 5 — общий вид таллома из оз. Байкал, 6, 7 — части талломов
акропетального строения; 8-11 — С. glomerata var. crassior: 8 — прикрепленный старый
таллом ветвится только в основании, 9 — базальная часть таллома, 10,11 — части молодых
растений слабо выраженного акропетального строения.
1-4, 6-11 — по Хуку.
Fig. 89. 1-3 — Cladophora fracta var. fracta: 1,2 — branching pattern of vegetative filaments
(various magnification), 3 — akinete germination; 4-7 — C. glomerata var. glomerata: 4 —
thallome of acropetal structure, 5 — general view of thallome from Lake Baikal, 6, 7 — thallome
parts of acropetal structure; 8-11 — C. glomerata var. crassior: 8 — sessile old thallome branching
only at the base, 9 — basal part of thallome, 10, 11 — parts of juvenile algae with poorly defined
acropetal structure.
1-4, 6-11 - after Hoek.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 197
Замечания. В Байкале найдена К.И. Мейером [1930] в сорах, заливах и
реках около берега. В р. Селенга, около мыса Облом, в зал. Провал, в лагуне
около сора Оймур, в Истокском соре, в зал. Мухор. В Истокском соре на
талломах в большом количестве развит эпифит Lyngbya kuetzingii (Kutz.)
Schmidle. Нами [Ижболдина, 1990в] этот вид также встречен во многих
районах Байкала (в губах Богучанской, Анге, Улан, Берхин, Максимихе,
Орео, в Чивыркуйском заливе и ряде других районов), где он растет в
основном среди зарослей Nitella sp. и Chaetomorpha baicalensis. Обнаружена
С. fracta var. fracta на глубинах от 2.5-3 до 15-30 м. Фитомасса ее в зарослях
харовых водорослей может достигать 23 г/м2.
Cladophora glomerata (L.) Kutz,
(рис. 89: 4-11)
Кустики рыхлые, прикрепленные или свободноплавающие и лежащие
на дне. Строение таллома изоморфное. Рост апикальный и интеркалярный.
Ветвление разнообразное — обильное или скудное. Инсерция ветвей апи-
кальная, косая, до горизонтальной, изредка у плавающих нитей латеральная.
Базальные ризоиды коралловидные. Клетки цилиндрические, у основных осей
65-275 мкм шириной, по длине в 1.5-12 раз больше, у конечных ветвей
19-100 мкм шириной, по длине в 1.5-15 раз больше, апикальные часто слегка
заостренные, 19-91 мкм шириной, по длине в 1.5-21 раз больше. Размножение
с помощью акинет и зооспор.
В пресноводных и солоновато-водных (до 15 %о) водоемах и морской воде.
Описание составлено по последней монографии рода Cladophora [Hoek,
1963], в которой автор в пределах вида различает только 2 разновидности.
Cladophora glomerata var. glomerata (= Conferva glomerata L.;
C. fontinalis Dill.; C. rigida Hudson; C. vaginata Ducluzeau;
Cladophora fasciculata Kutz.; C. fluitans Kutz.; C. declinata Kutz.;
C. brachystelecha Rabenh.) (рис. 89: 4—7)
Дерновинки разнообразной формы. Рост в основном апикальный. Клетки
основных осей 100-275 мкм шириной, конечных ветвей — 22-100 мкм шири-
ной, апикальные — 21-91 мкм шириной.
Широко распространен в Байкале [Ижболдина, 1970, 1974а, 19766, 1990в],
растет в зоне глубин от 1-1.5 до 3, реже 5 м, на камнях.
Замечания. Появляясь в середине июля, кустиковидные талломы дости-
гают максимальной высоты 5-7 см в августе — начале сентября. Отмечено, что
С. glomerata var. glomerata наиболее обильно развита в тех районах озера, где
отсутствует Chaetocladiella pumila С. Meyer et Skabitsch. Таким районом явля-
ется восточное побережье Юж. и Ср. Байкала (исключая Селенгинское мелко-
водье и район впадения р. Баргузин), где встречаемость вида во II и III поясах
колеблется от 21-26 до 30-39 %, а средняя фитомасса составляет 3.5 г/м2.
Вдоль остальной части побережья и встречаемость вида (1.5-8 %) на рас-
сматриваемых глубинах и его фитомасса (0.21 г/м2) ниже. Характерно массовое
развитие на талломах С. glomerata var. glomerata эпифитов. В районе сброса
сточных вод БЦБК на водоросли в большом количестве поселялись сувойки
рода Vorticella [Ижболдина, 1982].
198 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Cladophora glomerata var. crassior (Ag.) Hoek
C= Conferva crispata var. crassior Ag.; C. bullosa L.; C. capillaris L.;
C. crispata Roth; Cladophora crispata (Roth) Kutz.; C. callicoma Kutz.;
C. longissima Kiitz.; C. canalicularis Rabenh.;
C. fracta var. patens f. gossypina (Draparn. ex Kiitz.) Rabenh.;
C. insignis var. abbreviata Kiitz.; C. insignis f. crispata Grun. ex Rabenh.
(рис. 89: 8-11)
Характерны тенденция к интеркалярному росту, подавленность процессов
ветвления и спорообразования. Ветвление преимущественно разбросанное.
Клетки основных нитей 65-165 мкм шириной, конечных ветвей 16-70 мкм
шириной, апикальные 19-55 мкм шириной.
Отличить эту разновидность от типовой можно только в условиях культуры
[Hoek, 1963].
Замечания. В Байкале найдена К.И. Мейером [1930] в районе станций Бо-
ярск и Маритуй, в последнем случае - в гавани подо льдом.
Cladophora kursanovii Skabitsch., 1976
(=Aegagropila pigtnaea C. Meyer, 1927) (рис. 87: 1-10)
Дерновинки в виде маленьких, 1-2 мм диаметром и около 1 мм высотой,
подушечек (на мелких камнях и крупных песчинках) и корковидных наростов
в несколько квадратных сантиметров на камнях и скале. Строение талломов
гетероморфное. Стелющиеся нити, прикрепленные к грунту с помощью ризои-
дов, дают восходящие вверх короткие, неправильно и обильно ветвящиеся
нити, которые, переплетаясь между собой, образуют на камнях корковидные
наросты.
Клетки стелющихся нитей бочонкообразные или почти шаровидные, 125-
399 мкм шириной. Длина их меньше ширины или до 2 раз больше ее.
Восходящие нити цилиндрические или более-менее вздутые, 100-130(250) мкм
шириной, по длине равные ширине или в 2-4 раза больше. Апикальные клетки
более или менее конической формы, 63-90 мкм шириной. Хлоропласт в виде
сетчато-продырявленного цилиндра, с многочисленными пиреноидами. Зоо-
спорангии развиваются из любой клетки.
Эндемик Байкала, на глубине 5-10 м.
Замечания. А.П. Скабичевским [1965, 1966, 1968] С. kursanovii найдена во
многих районах озера. Проведенные нами [Ижболдина, 1967, 1976а, 1990в]
исследования свидетельствуют о том, что эта водоросль является одним из
наиболее широко распространенных в Байкале эндемичных видов. Она отсут-
ствует лишь на Ангаро-Кичерском и Селенгинском мелководьях, в зал. Провал
и единично встречается в прол. Ольхонские Ворота и в Посольском соре. На
твердом грунте фитомасса водоросли колеблется в разных районах озера от 0.01
до 10 г/м2, а на отдельных участках может достигать 38-43 г/м2 (Ушканьи
острова, мыс Бакланий, губа Хара-Ундурская) и даже 160 г/м2 (падь Идэбэ на
о. Ольхон) и 300 г/м2 (губа Сеногда, мыс Тыя). На мягком грунте фитомасса
С. kursanovii значительно ниже (0.001-0.2 г/м2), лишь в Баргузинском заливе у
с. Марково она достигает в IV поясе 5.6 г/м2 и севернее устья р. Выдриная
3.8-30 г/м2.
С. kursanovii найдена на глубинах от 1.5 до 40-60 м, но наиболее часто
встречается (V =20-50 %) в III и IV поясах на глубинах от 3.5 до 35 м. Площадь
корковидных наростов С. kursanovii достигает 5-6, реже 60-70 см2. В литораль-
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 199
ной зоне водоросль растет обычно на боковых и часто на нижней стороне кам-
ней, неплотно прилегающих ко дну. Способна вегетировать, оказавшись под
корковой формой губки Lubomirskia baicalensis Dyb. Вдоль северо-западного
прибрежья Юж. Байкала крупные корковидные наросты С. kursanovii с тем-
но-зеленым хлоропластом обнаружены Б.А. Дмитриевым на отвесных скалах
на глубине 60-62 м. Способность вида развиваться в условиях слабого освеще-
ния позволяет предположить, что эта водоросль распространена и на больших
глубинах, не доступных сегодня для обследования. С. kursanovii - растение
многолетнее, хорошо развивается в аквариумах с байкальской водой, при по-
стоянной смене последней.
Cladophora compacta (С. Meyer) С. Meyer, 1976
{=Aegagropila compacta C. Meyer, 1926) (рис. 90: 1—5)
Дерновинки в виде плотных подушечек, 2-10(15) мм диаметром, 2-5 мм
высотой. Строение таллома гетероморфное. Стелющиеся по субстрату ветвя-
щиеся нити дают побеги, радиально расходящиеся вверх. Во внутренних частях
подушечки нити построены из длинных клеток и ветвятся слабо, ближе к
периферии наблюдается обильное одностороннее гребенчатое ветвление. Из
конечных ответвлений нитей образуются зооспорангии. После выхода зооспор
зооспорангии разрушаются и рост нити возобновляется, “надстраивается но-
вый этаж” гребенчато разветвленных нитей. Клетки восходящих нитей цилин-
дрические, 32-40 мкм шириной, к периферии укорачивающиеся. Апикальные
клетки 25-32 мкм шириной. Хлоропласт в виде продырявленного цилиндра.
Зооспорангии по 5-10 на концах ветвей, 45-60 мкм шириной. Зооспоры 15
мкм длиной и 8-10 мкм шириной.
Эндемик Байкала. Растет на твердом субстрате, образуя иногда целые за-
росли, на глубине 2-20, местами до 38 м.
Замечания. Вдоль западного берега и у Ушканьих островов, где каменистые
грунты литорали слабо заилены и занимают значительную часть литорали,
подушечки С. compacta обычно крупнее и развиты в большем количестве
(фитомасса от 3 до 37 г/м2), чем вдоль восточного побережья (фитомасса
0.05-3 г/м2), для которого характерны пологий уклон дна и значительное
заиливание каменистого грунта. Размерный состав подушечек С. compacta
довольно разнообразен [Ижболдина и др., 1985]. В любой период года встре-
чаются подушечки высотой от 0.5 до 10 мм с площадью основания от 1 до
200 мм2. Зооспорангии в разной стадии формирования прослеживаются также
во все сезоны года. Мы наблюдали формирующиеся зооспорангии на молодых
(1-2 мм высотой) и еще слабо разветвленных кустиках. В ноябре — мае
зооспорангии встречаются чаще, чем в июне — октябре. На вертикальном
разрезе каждой колонии хорошо видна смена зон, образованных слабо и
сильно разветвленными нитями. Последние обычно сильно забиты илом и
песком. С. compacta — многолетние растения. В районе о. Бол. Ушканьего
подушечки С. compacta обнаружены на глубине 50-70 м [Ижболдина, 1974в].
Cladophora pulvinata (С. Meyer) С. Meyer, 1976
(=Aegagropila pulvinata C. Meyer, 1926) (рис. 90: 6—9)
Талломы в виде рыхлых округлых серовато-зеленых подушечек, 10 мм в
диаметре, 2-5 мм высотой. Строение таллома гетероморфное. От стелющихся
по субстрату нитей отходят вертикальные побеги с обильным односторонним
ветвлением. Клетки стелющихся нитей короткие, толстостенные, восходящие,
200 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 90.1-5 — Cladophora compacta: 1 — внешний вид таллома в натуральную величину, 2
— характер ветвления таллома, 3,4 — кистеобразно ветвящиеся нити с зооспорангиями,
5 — отдельная клетка; 6-9 — С. pulvinata: 6 — общий вид таллома в натуральную величи-
ну, 7 — ветвление таллома, 8 — ветви с зооспорангиями, 9 — зооспорангии.
3-5, 8, 9 — по Мейеру.
Fig. 90. 1-5 — Cladophora compacta: 1 — external view of thallome, full size, 2 — thallome
branching pattern, 3,4 — branching filaments with zoosporangia, bundle-shaped, 5-separate
cell; 6-9 — C. pulvinata: 6 — general view of thallome, full size, 7 — thallome branching, 8 —
branches with zoosporangia, 9 — zoosporangia.
3-5, 8, 9 — after Meyer.
63-80 мкм шириной в нижних частях, 35-50 мкм шириной в верхних частях
нитей, апикальные клетки 27-36 мкм шириной, длинные, с закругленной вер-
хушкой. Хлоропласт цилиндрический, сетчато-продырявленный, с крупными
пиреноидами. Зооспорангии в большом числе образуются на концах нитей,
54-72 мкм шириной и 90-126 мкм длиной, с толстой слоистой оболочкой, от-
крывающиеся одним отверстием.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 201
Эндемик Байкала, растет на камнях на глубине 1.5-5 м и более. Вид встре-
чен К.И. Мейером в районах Ср. и Сев. Байкала.
Замечания. У восточного берега Сев. Байкала найдена А.П. Скабичевским
[1934,1968], где входит в состав IV растительного пояса, занимающего глубины
от 11—13 до 20-35 м, реже 40 м. Нами [Ижболдина, 1974в, 1990в] вид встречен во
многих районах озера, где на глубине 15-35 м есть круглозернистый песок,
галька, мелкие камешки. Вдоль западного берега С. pulvinata найдена в районе
мысов Кочерикова, Онгурен, Арул, в прол. Мал. Море (мыс. Харгантуй,
губа. Хара-Ундурская, у о. Угунгой) — у мысов Идебе и Ухан на Ольхоне, про-
тив падей Улан, Анга, у мысов Бугульдейка, Арка и в бухтах Бабушка и Песча-
ная. Вдоль восточного побережья она встречена в северной части озера в рай-
оне губ Якшакан, Лаканда, Сосновка, а также у Ушканьих островов и п-ова
Святой Нос. Фитомасса С. pulvinata обычно не превышает 2 г/м2 и лишь в рай-
оне губы Хара-Ундурская она составила 9.8 и у мыса Онгурен на твердом грун-
те в IV поясе достигала 31.8 г/м2. Одновременно со стерильными талломами
постоянно встречаются кустики с зооспорангиями в разной стадии формиро-
вания или пустыми. В условиях аквариума с регулярной сменой байкальской
воды талломы С. pulvinata живут по нескольку лет.
Род Chaetocladiella С. Meyer et Skabitsch., 1968
(= Chaetomorphora Kiitz., pr. p.; Cladochaete C. Meyer et Skabitsch., 1965,
non Cladochaete Saccardo)
Водоросли нитевидные, изоморфного или гетероморфного строения с про-
стым ризоидом. При гетероморфном строении стелющаяся часть нитей разно-
образно ветвится и образует вторичные ризоиды. Вертикально восходящие
нити не ветвящиеся. Клетки цилиндрические, длина их равна или в 1.5 раза
меньше или до 4 раз больше ширины. Хлоропласт в виде сетчато-продырявлен-
ного цилиндра, с многочисленными пиреноидами и ядрами, зооспоры (гаме-
ты?) 2-жгутиковые.
В Байкале найдено 3 вида.
I. Таллом изоморфного строения.
1. Нити неветвящиеся, отходящие от ризоидной клетки в числе 1-5....
.................................................Ch. microscopica
2. Нити ветвящиеся только при основании таллома на протяжении первых
2-6 клеток....................................................Ch. litoralis
II. Таллом гетероморфного строения................................. Ch.pumila
Chaetocladiella microscopica (C. Meyer) C. Meyer et Skabitsch., 1968
(= Chaetomorpha microscopica C. Meyer, 1927; Cladochaete microscopica
(C. Meyer) C. Meyer et Skabitsch., 1965) (рис. 91: 1-13)
Одна-пять, чаще 4 неветвящихся нити отходят от одной ризоидной клет-
ки. По направлению к вершине нити постепенно утолщаются до 140-160 мкм,
могут загибаться и даже спирально закручиваться. Апикальная клетка округ-
лая. Хлоропласт сетчато-продырявленный, с многочисленными пиреноидами,
зооспорангии (гаметангии?) образуются в клетках на концах нитей. Ширина
зооспорангиев колеблется от 108 до 150 мкм.
Эндемик Байкала, растет на глубине 7-10 м на камнях.
Замечания. А.П. Скабичевский [1934, 1966, 1968] относит эту водоросль к
числу видов IV и V растительных поясов.
202 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 91. Chaetocladiella microscopica.
1 — зооспорангий; 2 — ризоид; 3 и 13 — талломы с зооспорангиями; 4, 5 — наиболее часто встре-
чающаяся форма таллома из 4 нитей; 6-12 — талломы с разным числом и направленностью нитей;
8 — одна из нитей развита значительно сильнее других и закручена в клубок; И — на вертикально
расположенной нити после разрушения зооспорангиев рост нити возобновился. По: 1-4 — Мейеру,
5-8 — Скабичевскому.
Fig. 91. Chaetocladiella microscopica.
1 — zoosporangium; 2 — rhizoid; 3 and 13 — thallomes with zoosporangia; 4, 5 — most common form of
thallome with 4 filaments; 6-12 — thallomes with various number of filaments; 8 — one of the filaments
more developed and curled like a ball; 11 — filament continues to grow on a vertically running stalk after
destroying of zoosporangia. After: 1-4 — Meyer, 5-8 — Skabitschevsky.
При наших исследованиях C. microscopica обнаружена вдоль большей части
открытых прибрежий озера, где в зоне сублиторали есть мягкий грунт. Не
найдена она лишь в районе Селенгинского мелководья и к северу от него до
мыса Тонкий, встречаемость вида в разных участках прибрежья колеблется от
14-16 до 45-55 %. Лишь вдоль восточного берега Сев. Байкала этот показатель
достигает 83 %, а в проливах Мал. Море (V = 5.5 %) и Ольхонские Ворота
(V = 2.3 %) вид встречается редко. Фитомасса водоросли невелика (0.001-
0.08 г/м2) и только в двух районах у мыса Бол.Солонцовый и на Мурине кой
Банке она достигала 0.51 и 0.36 г/м2. Наблюдения за развитием С. microscopica
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 203
показали, что это многолетние медленно растущие растения. На каждом
талломе образование зооспор происходит неоднократно. Пустые зооспорангии
сохраняются на протяжении 5-6 мес, затем разрушаются и рост нитей возоб-
новляется.
Chaetocladiella litoralis (Skabitsch.) С. Meyer et Skabitsch., 1968
(—Chaetomorpha litoralis Skabitsch., 1931; Cladochaete litoralis (Skabitsch.)
C. Meyer et Skabitsch., 1965) (рис. 92: 1—3)
Светло-желтые дерновинки до 2 см длиной. От 2-6 клеток, идущих после
ризоидальной, отходят вертикальные нити, разнообразно изогнутые, но не вет-
вящиеся. В основании вертикальных нитей образуются вторичные ризоиды. В
нижней части нитей клетки цилиндрические, 28-76.5 мкм шириной, в средней
и верхней частях нити бочонкообразно вздутые, 44.8-75.6 мкм шириной, сужи-
вающиеся к вершине, до 36.4 мкм шириной. Хлоропласт сетчатый, с многочис-
ленными ядрами. Зооспорангии 61.65-72.8 мкм шириной, 42-106.4 мкм дли-
ной Зооспоры продолговато-яйцевидной формы, 8.5-10.2 мкм шириной, 17-
18.6 мкм длиной, с 4 короткими (11.9-13.6 мкм длиной) жгутами.
6, 7,10: 800 цт
Рис. 92. 1-3 — Chaetocladiella litoralis'. 1 — нижняя разветвленная часть таллома, 2 —
верхняя часть нити, 3 — зооспоры; 4-11 — С. pumila'. 4, 5 — стелющиеся нити, 6 —
ризоид, 7 — заложение восходящих нитей, 8 — хлоропласт, 9 — зооспора, 10, 11 —
общий вид таллома (разное увеличение).
По: 1-3 — Скабичевскому, 4-9, 11 — Мейеру.
Fig. 92. 1-3 — Chaetocladiella litoralis: 1 — lower branched fragment of thallome, 2 — upper
part of filament, 3 — zoospores; 4-11 — C. pumila: 4,5 — trailing filaments, 6 — rhizoid, 7 —
formation of ascending filaments, 8 — chloroplast, 9 — zoospore, 10, 11 — general view of
thallome (various magnification).
After 1-3 — Skabitschevsky, 4-9, 11 — Meyer.
204 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Эндемик Байкала, обнаружена А.П. Скабичевским [1931] в районе пади
Бол. Коты, на глубине 0.5-0.75 м, во второй половине лета, когда на камнях в
прибойной зоне заканчивается развитие Ulothrix zonata.
Chaetocladiella pumila (C. Meyer) C. Meyer et Skabitsch., 1968
(= Chaetomorpha pumila C. Meyer, 1926; Cladochaete pumila (C. Meyer)
C. Meyer et Skabitsch., 1965) (рис. 92: 4—11)
Дерновинки до 2-4 см высотой. Таллом гетероморфный. Ветвящиеся нити
стелются по субстрату. Восходящие нити неветвящиеся, 286-360 мкм шири-
ной. Длина клеток равна ширине, чуть больше ее или в 1.5-2 раза меньше.
Оболочка толстая, слизистая, иногда слоистая. Хлоропласт светло-зеленый,
сетчато-продырявленный, со многими пиреноидами или в виде округло-
многоугольных дисков, соединенных тонкими нитями. В зооспорангий могут
превратиться все клетки неветвящихся нитей.
Эндемик Байкала, растет на глубине 2-4 м на камнях, которые обычно по-
крывает сплошным ковром. Распространен преимущественно по западному
берегу и чаще на юге, чем на севере.
Замечания. А.П. Скабичевский [1970] отметил, что стелющиеся нити зна-
чительно тоньше (115-130 мкм шириной), чем восходящие (260-430 мкм ши-
риной). Мелкие веточки в основной части кустика 50-60 мкм шириной. Длина
восходящих нитей 3-4 см, иногда до 7 см. Вторичные ризоиды, описанные
К.И. Мейером [1930], А.П. Скабичевским не встречены. Наиболее пышные
заросли Ch. pumila отмечены им в районе Ушканьих островов.
Наши исследования [Ижболдина, 1974в, 19766, 1990в] подтверждают сде-
ланные ранее [Мейер, 1930; Скабичевский, 1966] наблюдения о том, что
Ch. pumila имеет разорванный ареал обитания и помогают определить грани-
цы ареала. Вид встречен нами на глубинах от 1 до 23 м. Наиболее часто встре-
чается во II (V = 30-55 %) и III (V = 15-43 %) растительных поясах. Вдоль за-
падного берега вид обнаружен нами от мыса Шары-Жалгай до пади Емелья-
ниха. Севернее ее, до пади Ниж. Хомуты, Ch. pumila в наших сборах не
встречена. Севернее пади Ниж. Хомуты заросли рассматриваемого вида про-
слежены до мыса Бол. Коса, в том числе вдоль всего побережья о. Ольхон. Се-
вернее единичные талломы Ch. pumila обнаружены лишь у мыса Тонкий (у
губы Богучанской) и у мыса Слюдянский. Вдоль восточного берега в южной и
средней частях озера вид не найден. Появляется он в районе п-ова Святой
Нос и очень обильно развит в районе Ушканьих островов. Далее на север он
встречен лишь в губе Фролиха, где в небольшом количестве (фитомасса
1,5 г/м2) развит в III растительном поясе. Средняя фитомасса Ch. pumila в раз-
ных районах западного побережья колеблется от 13 до 25 г/м2 во II и 9-15 г/м2
в III поясе, а в районе мысов Крест, Хадартуй и Ото-Хушун она достигает со-
ответственно 99.5; 251; 57.5 г/м2. У восточного побережья, в районе Ушканьих
островов, средняя фитомасса водорослей составляет во II поясе 42 г/м2 при
встречаемости 83 % и в третьем поясе — 26.5 г/м2 при встречаемости 63 %.
В районе мыса Ото-Хушун, далеко выдающемся в море и защищающем
расположенное южнее побережье от северных ветров, на талломах Ch. pumila
длина неветвящихся нитей достигает 7-13 см. По наблюдению К.И. Мейера
[1930] и по нашим данным [Ижболдина, 1981а], вид вегетирует в озере круглый
год. Осенью (в конце сентября-октябре) на талломах — масса пустых зооспо-
рангиев. На зиму остаются нижняя, сильно разветвленная, часть таллома и ко-
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 205
роткие (0.5-1 см длиной), обломленные на вершине прямостоячие нити. Вес-
ной (в марте—апреле) рост этих неветвящихся нитей возобновляется.
ПОРЯДОК SPHAEROPLEALES
Порядок включает одно семейство, структура хлоропластов которого схожа
с Ulotrichales, но отличается многоядерным талломом.
СЕМЕЙСТВО SPHAEROPLEACEAE
Нити неветвящиеся, цилиндрические, образующие рыхлые бесформенные
скопления тины. Таллом изоморфный. Клетки нитей цилиндрические, длина
их превышает ширину в 20-25, реже в 90 раз. Оболочка на боковых стенках
тонкая, поперечные перегородки часто утолщенные. Хлоропласт в виде проды-
рявленных колец, опоясывающих клетку, лежащих на некотором расстоянии
друг от друга и соединенных между собой перемычками с пиреноидами. Ядер
несколько в каждом кольце. Вегетативное размножение распадением нитей на
фрагменты. Бесполое размножение известно для одного вида (S. wilmanii).
Половое размножение оогамное. Известен один род.
Род Sphaeroplea Ag., 1824
Характеристика та же, что для семейства. В Байкале найден один вид.
Sphaeroplea annulina (Roth) Ag. (= Conferva annulina Roth;
Sphaeroplea treviranii Kiitz.; S. braunii Kiitz.; S. leibleinii Kiitz.;
S. crassisepta Klebahn; S. annulina var. crassisepta Heinr.) (рис. 93: 1-10)
Нити 24-72 (170) мкм шириной. Длина клеток в 8-20( 35) раз больше ши-
рины. Поперечные перегородки тонкие, редко утолщенные, с неправильными
разрастаниями. Хлоропласты многочисленные, по 20-32 в клетке, с 2-10 пире-
ноидами. Ооспоры шаровидные, располагающиеся в 1 или несколько рядов,
11-20(38) мкм диаметром, с толстой оболочкой, несущей полые тупокониче-
ские выросты, основания которых соединены правильной полигональной се-
тью тонких складок.
В неглубоких пресных и солоноватых водоемах, искусственных бассейнах,
оранжереях, на рисовых полях. Космополит.
В Байкале найдена К.И. Мейером [1930] в небольшом количестве в бух. За-
воротной, в начале августа.
Класс ZYGNEMATOPHYCEAE (CONJUGATOPHYCEAE)
Одноклеточные, колониальные или многоклеточные (нитчатые) водорос-
ли. Нитчатые формы иногда образуют короткие ответвления. Водоросли при-
креплены к субстрату ризоидом или свободноплавающие. Клеточная оболочка
цельная или состоит из 2 и более сегментов, однослойная или дифференциро-
вана на внутренний и внешний слой с гладкой или скульптурной оболочкой.
Хлоропласты зеленые, крупные, осевые или пристенные, по одному или по не-
скольку в клетке, с пиреноидами. Ядро одно. Вегетативное размножение путем
деления клетки на 2. Половое размножение — конъюгация (изо-, гетеро- и
оогомия), при которой происходит слияние протопластов двух клеток, находя-
206 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 93. Sphaeroplea annulina.
1-5 — строение хлоропластов; 6 — форма поперечных перегородок; 7 — участок нити в момент оп-
лодотворения яйцеклеток; 8 — ооспора; 9,10 — прорастающие зооспоры. По: 1-4, 8 — Фричу, 5 —
Клебану, 6 — Хейнрихеру, 7 — Ольтмансу, 9, 10 — Кону.
Fig. 93. Sphaeroplea annulina.
1-5 — structure of chloroplasts; 6 — shape of transverse walls; 7 — part of filament at the moment of
ovigerm fertilization; 8 — oospore; 9,10 — germinating zoospores. After: 1-4, 8 — Fritsch, 5 — Klebahn,
6 — Heinricher, 7 — Oltmanns, 9, 10 — Kone.
щихся в одной или разных нитях. В последнем случае нити предварительно
связываются между собой копуляционным каналом. Зиготы одеваются пло-
ской оболочкой, прорастая после периода покоя 1, 2 или 5 проростками.
В оз. Байкал встречены представители одного порядка.
ПОРЯДОК ZYGNEMATALES
Клетки соединены в длинные неразветвленные нити, редко с короткими
ответвлениями из одной-немногих клеток, свободноплавающие или прикреп-
ленные к субстрату ризоидом. Клетки по форме всегда цилиндрические, кле-
точная оболочка цельная, одно- или двуслойная, гладкая, без пор. Хлоропла-
сты зеленые, осевые и пристенные, звездчатые, дисковидно-пластинчатые,
мешковидные, спирально изогнутые или параллельные продольной оси клет-
ки, от 1 до 16. Ядро одно, пиреноиды многочисленные, беспорядочно располо-
женные по всему хлоропласту. Размножение вегетативное, бесполое и половое.
Конъюгация лестничная и боковая.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 207
В оз. Байкал встречены представители 3 семейств.
I. Хлоропласт осевой, звездчатый, с радиальными выростами, реже дисковид-
ный, без выростов. Ядро в центре клетки ...............Zygnemataceae
II. Хлоропласт осевой, реже пристенный, широкопластинчатый. Ядро примы-
кает сбоку к хлоропласту, реже в центре клетки ...........Mougeotiaceae
III. Хлоропласт пристенный, спирально извитой, слегка извитой или почти па-
раллельный продольной оси клетки. Ядро в центре клетки . . . Spirogyraceae
СЕМЕЙСТВО ZYGNEMATACEAE
Нити неразветвленные, редко с короткими ответвлениями, прикреплен-
ные к субстрату с помощью ризоида или свободно плавающие. Поперечные
стенки клеток гладкие. Хлоропласты осевые, звездчатые, многолучевые, по 1-6
в клетке, чаще по 2. В каждом хлоропласте 1 пиреноид. Ядро в центре клетки, в
протоплазматическом мостике между хлоропластами. Размножение в основ-
ном зигоспорами, реже апланоспорами или акинетами. Зигоспора формирует-
ся обычно в полости одного из гаметангиев, реже в конъюгационном канале.
В оз. Байкал встречен представитель одного рода.
Род Zygnema Ag., 1824 (=Zygogonium Kiitz., pr.p.;
Zygnemopsis (Skuja) Trans., pr. p.; Neozygnema Yamagishi)
Нити преимущественно неразветвленные, свободно плавающие или при-
крепленные к субстрату. Вегетативные клетки цилиндрические, до 60 мкм
шириной, короткие или длина их до 4 раз превышает ширину. Звездчатые,
многолучевые хлоропласты обычно по 2 в клетке, с пиреноидом в центре. Ядро
одно в центре клетки между хлоропластами. Конъюгация лестничная, реже бо-
ковая. Зигоспоры формируются обычно в одном из гаметангиев, иногда в
конъюгационном канале. Бесполое размножение акинетами и апланоспорами.
В оз. Байкал встречен один вид.
Zygnema cruciatum (Vauch.) Ag. (=Z. inconspicuum Czurda;
Z. inconspicuum Czurda emend Skv.; Z. mucosum Skv.) (рис. 94: 1-2)
Вегетативные клетки 30-36(39) мкм шириной, по длине равные ширине
или до 2(4) раз больше. Конъюгация лестничная. Зигоспоры формируются в
одном из гаметангиев, широкоэллиптические, 29-40 мкм шириной, 30-
60 мкм длиной. Партеноспоры цилиндрически-яйцевидные, 30-35 мкм ши-
риной, 30-60 мкм длиной.
В стоячей воде, лужах, прудах, канавах, болотах, реже в водоемах со слабо
проточной водой, зарастающих участках рек и озер.
Повсеместно.
Замечания. В оз. Байкал Z. cruciatum встречена в июне в Истокском соре
[Мейер, Рейнгардт, 1925]. Нами обрывки стерильных нитей найдены в Байкале
в бухтах прол. Ольхонские Ворота и в материале О.Т. Русинек из водоемов
Прибайкалья (р. Осиновка, озера Очки, Кривое, Метковские).
СЕМЕЙСТВО SPIROGYRACEAE
Нити неразветвленные, реже прикрепленные. Хлоропласты пристенные,
от 1 до 16 в клетке, лентовидные, обычно спирально извитые или параллельные
продольной оси клетки. Пиреноиды многочисленные, расположены беспоря-
208 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 94.1, 2 — Zygnema cruciatum: 1 — лестничная конъюгация, 2 — акинета и зигоспоры в
двух положениях; 3-10 — Mougeotia laetevirens, различные положения зигоспоры, лест-
ничная конъюгация; 11-16 — М. genuflexa f. genuflexa: 11 — ложная конъюгация, 12 —
ранняя стадия лестничной конъюгации, 13,14 — лестничная конъюгация, 15,16 — боко-
вая конъюгация.
По: 1 — Скворцову, 2 — Transeau, 3 — Волковой, Даниловой, 4 — Transeau, 5, 6 — Skuja, 7-10 —
Gauthier Lievre, 11, 12 — Рундиной, 13, 14 — Krieger, 15, 16 — Fott.
Fig. 94.1,2 — Zygnema cruciatum: 1 — staircase conjugation, 2 — akinete and zygospores in two
positions; 3-10 — Mougeotia laetevirens, different positions of zygospore, staircase conjugation:
11-16 — M. genuflexa f. genuflexa: 11 — false conjugation, 12 — early stage of staircase
conjugation, 13, 14 — staircase conjugation, 15, 16 — lateral conjugation.
After: 1 — Skvortsov, 2, 4 — Transeau, 3 — Volkova, Danilova, 5-6 — Skuja, 7-10 — Gauthier Lievre,
11-12 — Rundina, 13, 14 — Krieger in: Kolkwitz, Krieger, 15, 16 — Fott.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 209
дочно. Ядро крупное, в центре клетки на плазматических тяжах, соединенных с
постенным слоем протоплазмы. Конъюгация лестничная и боковая. Конъюги-
рующие нити обычно спирально извитые. Размножение зигоспорами, реже ап-
ланоспорами и акинетами.
В Байкале найдены представители одного рода.
Род Spirogyra Link., 1820 (=Coufera Mull., pr. p.; Mougeotia Ag. pr. p.;
Zygnema Ag., pr. p.; Rhynchonema Kutz.; Degagnya Conard, Temnogyra Lewis)
Нити длинные, неветвистые, свободно плавающие или прикрепленные с
помощью ризоида к субстрату. Вегетативные клетки до 200 мкм шириной, по
длине менее ширины, равные ей или до 25 раз больше, хлоропласты обычно в
виде спирально извитых лент, с многочисленными пиреноидами, расположен-
ными по оси ленты. Ядро одно. Конъюгация — лестничная или боковая или
обе одновременно. Зигоспоры эллипсоидные, эллипсоидно-цилиндрические,
шаровидные, линзовидные. Мезоспорий структурированный.
В оз. Байкал найдено 5 видов.
По строению поперечных перегородок Л.А. Рундина [1988] условно делит
род Spirogyra на 3 секции, не имеющие таксономического значения, но облег-
чающие пользование ключом при определении видов.
В оз. Байкал встречены представители двух секций.
СЕКЦИЯ CONJUGATA
Поперечные стенки клеток гладкие.
I. Размножение преимущественно зигоспорами, редко апланоспорами.
1. Зигоспоры правильно-эллипсоидные (двухосно-эллипсоидные), в попе-
речном сечении (вид сверху) круглые.
А. Экзоспорий и мезоспорий гладкие.
а. Хлоропласт постоянно один.
* Клетки без конъюгационной пары, обычно невздутые.
+ Воспринимающие клетки невздутые.
х Конъюгация лестничная.
о Зигоспоры эллипсоидные.
А Вегетативные клетки (17)20-38(42) мкм шириной.......
......................S. deciminaf. juergensii (частично)
хх Конъюгация лестничная и боковая.
о Конъюгация преимущественно боковая, вегетативные клетки
20-55(62) мкм шириной
А Вегетативные клетки (24)26-38(42) мкм шириной....
......................S. decimina f. longata (частично)
++ Воспринимающие клетки вздутые
х Воспринимающие клетки вздутые со стороны конъюгационно-
го канала.
о Конъюгация лестничная
А Конъюгационный канал образован равномерно обеими
клетками...............S. decimina f. Juergensis (частично)
* * Клетки без конъюгационной пары, вздутые
х Конъюгация боковая и лестничная
+ Воспринимающие клетки невздутые ....................
..........................S', decimina f. longata (частично)
210 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
СЕКЦИЯ SALMACIS
Поперечные стенки клеток складчатые.
I. Оболочка зигоспор гладкая.
1. Хлоропластов 1-2.
А. Хлоропласт один, редко их 2.
а. Воспринимающие клетки невздутые или слабо вздутые со всех сторон,
х Конъюгация лестничная или преимущественно лестничная.
А Вегетативные клетки (19)23-32(36) мкм шириной ..........
....................................... S. weberi f. weberi
2. Хлоропластов (1) 2-4.
А. Воспринимающие клетки веретеновидно вздутые.
а. Конъюгация лестничная и боковая.
А Вегетативные клетки (26) 30-35 мкм шириной......S. hassallii
И. Оболочка зигоспор скульптурированная.
1. Экзоспорий скульптурированный, мезоспорий гладкий.
А. Воспринимающие клетки невздутые или слабо вздутые.
а. Концы стерильных клеток невздутые......S. calospora f. calospora
Spirogyra decimina (Mull.) Kiitz.
Вегетативные клетки (17)20-38(42) мкм шириной, по длине в 1.5-12 раз
больше. Ризоиды латеральные, разветвленные, многочисленные. Поперечные
стенки гладкие, хлоропласт 1 (2). Конъюгация лестничная и боковая. Конъю-
гационный канал при лестничной конъюгации образован обеими клетками.
Воспринимающие клетки невздутые или слабо вздутые, до 43.5 мкм шириной.
Клетки без конъюгационной пары невздутые или вздутые, до 55-78 (95) мкм
шириной. Зигоспоры эллипсоидные, нередко с более или менее заостренны-
ми вершинами, реже цилиндрически-эллипсоидные, почти шаровидные или
неправильной формы (16.5) 18-44 мкм шириной (26)35-83(113) мкм длиной.
Экзоспорий тонкий, бесцветный, гладкий, мезоспорий толстый, от темного
до желто-коричневого. Партеноспоры 29-38 мкм шириной, 36-64 мкм дли-
ной.
Широко распространенный, эврибионтный, полиморфный вид.
В оз. Байкал найдены 2 формы этого вида.
Spirogyra decimina forma juergensii (Kiitz.) V. Poljansk.
(=SI. juergensii Kiitz., non sensu Skuja, 1924; S. quinina (Ag.)
Kiitz. f. juergensii (Kiitz.) Kirchn.) (рис. 95: 12—14)
Вегетативные клетки 24-32 (36) мкм шириной, по длине в 2-8 раз больше.
Конъюгация лестничная. Зигоспоры от эллипсоидных до эллипсоидно-цилинд-
рических, иногда почти шаровидные, 22-35(38) мкм шириной, (35)40- 81 мкм
длиной. Партеноспоры (32)35-38 мкм шириной, (35)43.5-46 мкм длиной.
В стоячих и текучих водах, нередко в лужах и канавах, иногда на рисовых
полях и в солоноватых водах. Повсюду.
Замечание. В оз. Байкал найдена К.И. Мейером и Б.В. Рейнгардтом [1925] в
Истокском соре в июне.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 211
Рис. 95. 1, 2 — Spirogyra decimina f. longata: 1 — партеноспоры, 2 — боковая конъюгация;
3-7 — S. weberi var. weberi: 3-6 — разная степень вздутия воспринимающих клеток,
лестничная конъюгация, 7 — боковая конъюгация; 8-11 — 5. hassallii: 8 — боковая
конъюгация, 9 — вегетативная нить, 10, 11 — лестничная и боковая конъюгации;
12-14 — S. decimina f. juergensii: 12, 13 — лестничная конъюгация, 14 — партеноспоры;
15 — S. calospora f. calospora.
По: 1-7,13-15 - Л.А. Рундиной [1998], 8, 9 - Пти; 12 - Jao [19356], 10,11 - Gauthier-Lievre.
Fig. 95. 1, 2 — Spirogyra decimina f. longata: 1 — parthenospores, 2 — lateral conjugation;
3-7 — S. weberi var. weberi: 3-6 — various degree of swelling in perceiving cells, staircase
conjugation, 7 — lateral conjugation; 8-11 — S. hassallii: 8 — lateral conjugation, 9 —
vegetative filament, 10, 11 — staircase and lateral conjugation; 12-14 — S. decimina f.
Juergensii: 12,13 — staircase conjugation, 14 — parthenospores; 15 — S. calospora f. calospora.
After. 1-7,13-15 — Rundina, 8, 9 — Ptie; 12 — Jao, 10, 11 — Gauthier-Lievre.
212 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Spirogyra decimina forma longata (Vauch.) V. Poljansk. (=SI. longata (Vauch.)
Kiitz.; S. longata var. reversa (Hass.) Kirchn. (?); S. indica (Rao) Krieger)
(рис. 95: 1-2)
Вегетативные клетки (24)26-38 мкм шириной, по длине в 2-5 (10) раз бо-
льше. Конъюгация боковая, редко лестничная, иногда совмещенные* в одной
паре нитей. Конъюгационный канал при боковой конъюгации выдается на
1/4-1/2 ширины клетки. Воспринимающие клетки невздутые. Зигоспоры эл-
липсоидные, нередко с более или менее заостренными вершинами (18)25—
40(43) мкм шириной, (26)41-79(100) мкм длиной.
В стоячих и текучих водах, в болотах, в минеральных и серных источниках,
на рисовых полях. Повсюду.
Замечания. В оз. Байкал найдена К.И. Мейером [1930] в губе Анга, среди
зарослей высших водных растений, в августе.
Spirogyra weberi Kiitz.
Вегетативные клетки (19)23-35(39) мкм шириной, по длине в (2)3-16 раз
больше. Поперечные стенки складчатые, хлоропласт 1(2). Конъюгация лест-
ничная, реже боковая, иногда совмещенные в одной паре нитей. Воспринима-
ющие клетки при лестничной конъюгации невздутые, слабо или сильно взду-
тые. Клетки без конъюгационной пары невздутые или слегка вздутые. Зигоспо-
ры эллипсоидные, реже цилиндрически-эллипсоидные, с более или менее
закругленными вершинами, 24-49(54) мкм шириной, (20)40-105(122.5) мкм
длиной. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, от
желтого до коричневого, двухслойный, гладкий. Партеноспоры элипсоидные
или почти шаровидные, 25-35 мкм шириной, 32-67 мкм длиной.
Широко распространенный, полиморфный вид.
В оз. Байкал встречена типовая форма.
Spirogyra weberi forma weberi (=S. weberi var. elongata (Spree) Rabenh.;
.S’. weberi var. genuina (Kiitz.) Kirchn.; S. laxa Kiitz.; S. semigrnata Jao;
S. tolosana Comere (?)) (рис. 95: 3-7)
Вегетативные клетки (19)23-32(36) мкм шириной. Конъюгация лестнич-
ная, редко боковая, иногда совмещенная в одной паре нитей. Конъюгацион-
ный канал при лестничной конъюгации образован обеими клетками равномер-
но. Воспринимающие клетки невздутые или слабо вздутые. Зигоспоры 21-
40.5(43.5) мкм шириной, (26)40-93.5(120) мкм длиной.
В стоячих и текучих водах, в родниках, в заводях и старицах рек, в соленых
озерах и на рисовых полях. Развивается в кислых нейтральных и щелочных во-
дах и при малом содержании азота. Повсюду.
Замечания. В оз. Байкал найдена К.И. Мейером [1930] в бух. Анга, в авгу-
сте, среди зарослей высших водных растений.
Spirogyra hassallii (Jenner) Petit (рис. 95: 8—11)
Вегетативные клетки (26)30-35 мкм шириной, по длине в 4-13 раз больше.
Поперечные стенки складчатые, хлоропластов 2-4. Конъюгация боковая, редко
лестничная. Конъюгационный канал при лестничной конъюгации образован
обеими клетками. Воспринимающие клетки веретновидно вздуты со всех сто-
рон, до 49-50 мкм шириной. Клетки без конъюгационной пары, невздутые. Зи-
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 213
госпоры (30)35-54 мкм шириной, 52-169(223) мкм длиной. Экзоспорий тон-
кий, бесцветный, гладкий. Мезоспорий толстый, от желтого до желто-коричне-
вого, двухслойный, гладкий. Партеноспоры 38-40 мкм шириной, 47-72 мкм
длиной, встречаются редко.
В стоячих, реже текучих водах, прудах, минеральных источниках, в том
числе радоновых.
Россия: европейская часть — почти повсюду, Вост. Сибирь (Республика
Саха, оз. Байкал), Дальний Восток, Украина (окрестности Киева), Узбеки-
стан, Казахстан.
Замечания. В оз. Байкал найдена К.И. Мейером [1930] в бух. Анга, в авгу-
сте, среди зарослей высших водных растений.
Spirogyra calospora Cleve (рис. 95: 15)
Вегетативные клетки 20-56 мкм шириной, по длине в 4-14 раз больше.
Поперечные клетки складчатые. Хлоропластов 2-3, конъюгация лестничная,
конъюгационный канал образован обеими клетками. Воспринимающие клет-
ки невздутые или слабо вздутые. Зигоспоры от эллипсоидных до цилиндриче-
ски-эллипсоидных, 26-60 мкм шириной, 52-130 мкм длиной. Экзоспорий
толстый, бесцветный, двухслойный. Мезоспорий тонкий, от желтого до ко-
ричневого, двухслойный, гладкий.
Spirogyra calospora forma calospora (=S. protecta Wood; S. punctata Petit,
non sensu Cleve; S. cleveana Trans.) (рис. 95: 15)
Вегетативные клетки (26)29-42 мкм шириной. Зигоспоры 30-51 мкм
шириной, 52-125 мкм длиной.
В стоячих и текучих водах, водохранилищах, прудах, оросительных кана-
лах, иногда при солености 5.7 %о.
Россия: Кольский п-ов, окрестности Москвы, устье Волги, западное побе-
режье Каспийского моря, Вост. Сибирь (Сев.Байкал, Республика Саха), Чу-
котка, Колыма, Эстония, Латвия, Узбекистан, Туркмения.
Замечания. Кроме 5 вышеописанных видов Spirogyra Л.А. Рундина [1998]
приводит для окрестностей Байкала еще 2 вида: 5. majuscula Kiitz. — для Тур-
кинских минеральных вод и S. setiformis (Roth) Kiitz.— без указания места на-
хождения.
Стерильные нити Spirogyra отмечены для Байкала многими авторами [Яс-
нитский, 1928; Мейер, 1930; Кожов, 1947,1962; Вотякова, 1981; и др.]. К.И.Мей-
ером [1930] они встречены в мелководных бухтах западного (материкового) бе-
рега прол. Ольхонские Ворота, в Посольском соре и на Ангаро-Кичерском
мелководье и в соре. В.Н. Яснитский [1928] обнаружил Spirogyra sp. в бухтах
прол. Ольхонские Ворота, а Н.Е. Вотякова [1981] — в дельте р. Селенги. При
наших [Ижболдина, 19766,1990в] исследованиях Spirogyra sp. найдена в мелко-
водных бухтах Чивыркуйского залива и прол. Мал. Море, в бухтах Дагарской и
Сеногда, у о. Лиственичный и в приустьевых пространствах рек Тыя, Курлы,
Тошка. Очевидно, что представители Spirogyra встречаются обычно в закрытых
мелководных хорошо прогреваемых бухтах. Однако в 1976 и 1982 гг. Spirogyra
sp. была обнаружена на участке открытого юго-восточного побережья Байкала,
расположенном в районе сброса в озеро более теплых, чем озерные, услов-
но-чистых вод Байкальского целлюлозно-бумажного комбината [Ижболдина,
1985а].
214 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Кроме того, стерильные обрывки нитей Spirogyra sp. были обнаружены в
сборах О.Т. Русинек из водоемов Байкальского заповедника (реки Осиновка,
Мишиха, Первая Шестипалиха, Кривое) и в материалах В.В. Тахтеева, собран-
ных в районе горячих источников Прибайкалья (падь Давше, мыс Котельни-
ковский). Spirogyra sp. встречена в водотоках с температурой воды не более
23 °C [Ижболдина, Галкин, 2000; Ижболдина, 2001].
СЕМЕЙСТВО MOUGEOTIACEAE
Нити обычно неразветвленные, свободно плавающие, прикрепленные к
субстрату. Поперечные стенки гладкие, хлоропласт 1 (редко 2), осевой, плас-
тинчатый, изменяющий положение при разной интенсивности освещения.
Пиреноиды от одного до многих, расположены или беспорядочно или в ряд по
центру хлоропласта, иногда пиреноиды отсутствуют. Ядро одно. Размножение
зигоспорами, апланоспорами, акинетами, иногда с развитием одной из этих
форм. Формирование зигоспоры в конъюгационном канале, иногда с внедре-
нием в полости обоих гаметангиев.
В оз. Байкал найдены представители одного рода.
Род Mougeotia Ag., 1824 (=Serpentinaria Gray; Mesocarpus Hass.;
Staurospermum Kiitz.; Craterospermum A. Br.; Pleurocarpus A. Br.; Debarya Wittr.,
pr. p.; Gonatonema Wittr.; Zygnema kg. pr. p.; Mougeotiella Yamagishi)
Вегетативные клетки в нитях до 63 мкм шириной, по длине до 20 раз боль-
ше ширины. Хлоропласт один (редко 2) в клетке, осевой, Широкопластинча-
тый, изменяющий положение при разной интенсивности освещения. Конъю-
гация лестничная, реже боковая, зигоспоры разнообразной формы. Бесполое
размножение с помощью акинет, апланоспор, партеноспор.
В оз. Байкал найдено 2 вида.
I. Зигоспоры имеются.
А. Мезоспорий гладкий.
а. Формирование зигоспоры только в конъюгационном канале.
* Конъюгация боковая, редко лестничная...........М. genuflexa
* * Конъюгация только лестничная.
+ Зигоспоры короткоцилиндрические (катушковидные).
х Вегетативные клетки (22)28-42 мкм шириной . ... М. laetevirens
Mougeotia genuflexa (Dillw.) Ag. (рис. 94: 11-16)
Вегетативные клетки 15-40 мкм шириной, по длине в 2-9( 12) раз больше,
часто коленчато изогнутые. Хлоропласт с (2)4-8 беспорядочно расположен-
ными пиреноидами. Конъюгация редкая, обычно боковая, реже лестничная.
При лестничной конъюгации зигоспора формируется в конъюгационном ка-
нале.
Зигоспоры угловато-округлые, квадратно-округлые, трапециевидно-округ-
лые (обычно при боковой конъюгации), эллипсоидные или шаровидные,
20-40 мкм шириной и 30-44 мкм длиной.
Вид недостаточно изучен из-за редкой встречаемости зигоспор.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 215
Mougeotia genuflexa forma genuflexa (= Zygnema genuflexum Ag.;
Pleurocarpus mirabilis A. Br.; Mesocarpus genuflexus Ag.;
M. pleurocarpus De Вагу) (рис. 94: 11—16)
Вегетативные клетки (24)29-40 мкм шириной, по длине в 2-5,5(12) раза
больше. Хлоропласт с 4-8 пиреноидами. Зигоспоры 29-40 мкм шириной,
36-44 мкм длиной.
В стоячих и текучих водах, в горных речках и минеральных (радоновых)
источниках.
Россия: европейская часть, Зап. Сибирь, Прибайкалье; Украина, Казах-
стан, Ср. Азия.
Замечания. В оз. Байкал в большом количестве найдена К.И. Мейером
[1930] в бух. Анга, среди зарослей высших водных растений, в зал. Провал око-
ло с. Оймур и в Нижнеангарском соре.
Mougeotia laetevirens (A. Br.) Wittr. (= Craterospermum laetevirens A. Br.;
Mougeotia laetevirens var. varians Wittr. f. minor Kissel.) (рис. 94: 3-10)
Вегетативные клетки (22)28-42 мкм шириной, по длине в 3-12 раз боль-
ше. Хлоропласты с многочисленными пиреноидами. Конъюгация лестничная.
Зигоспоры в конъюгационном канале короткоцилиндрические, обычно с во-
гнутыми боковыми сторонами, сверху и снизу круглые, 38-60 мкм шириной,
(29)36-60(75) мкм длиной. Конъюгирующие клетки коленообразно изогнуты.
Мезоспорий гладкий, желто-коричневый, с 2 линиями разрыва.
В стоячих, заболоченных, реже текучих водах.
Россия, повсюду.
Замечания. В оз. Байкал встречена К.И. Мейером [1930] в большом коли-
честве в губе Анга, в августе, среди высших водных растений.
Стерильные нити Mougeotia sp. найдены К.И. Мейером [1930] на Анга-
ро-Кичерском мелководье и в соре. Н.Е. Вотякова [1981] приводит Mougeotia sp.
для дельты р. Селенги. Нами найдена в сборах О.Т. Русинек из водоемов Бай-
кальского заповедника (р. Переемная, оз. Кривое) и в материале, собранном
В.В. Тахтеевым в районе горячих источников Прибайкалья, в пади Давше и у
мыса Котельниковский [Ижболдина, Галкин, 2000]. Mougeotia sp. обнаружена
в водотоках с температурой воды, не превышающей 23 °C.
ОТДЕЛ CHAROPHYTA
Крупные многоклеточные зеленого цвета водоросли членисто-мутовчатого
строения. Рост побегов осуществляет верхушечная клетка. Клетки таллома од-
ноядерные и многоядерные. Хлоропласты в виде многочисленных зерен без
пиреноидов. Вегетативное размножение клубеньками — специализированны-
ми участками таллома. Бесполое размножение отсутствует. Половой процесс
оогамный. Оогонии образованы рядом вытянутых клеток, спирально обвола-
кивающих яйцеклетку. Шаровидные антеридии состоят из щиткообразных
клеток, снаружи соединенных между собой зазубренными краями и сложно
устроенных внутри. Сперматозоиды спирально извитые, с 2 жгутами. Зигота
прорастает после периода покоя с редукцией числа хромосом. Чередования
форм развития в онтогенезе нет.
216 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Класс CHAROPHYCEAE
Оогонии однотипного строения, с 5 обволакивающими клетками, спира-
льно завитыми снизу вверх, справа налево.
В классе один порядок.
ПОРЯДОК CHARALES
Талломы высотой до 0.5 м, часто инкрустированы известью. Основные по-
беги (стебли) ветвятся. От основания стебля отходят ризоиды, проникающие в
грунт и служащие для прикрепления. Стебли состоят из чередующихся узлов и
междоузлий. Узлы образуют короткие одноядерные клетки, междоузлия образу-
ют длинные многоядерные клетки, покрывающиеся у некоторых видов одно-
слойной корой из продольно вытянутых клеток, усеянных одноклеточными ши-
пиками. В стеблевых узлах располагаются мутовки коротких побегов (листья),
также имеющих членистое строение. В пазухах листьев возникают боковые вет-
ви стебля. Размножение вегетативное и половое. Виды однодомные и двудом-
ные. Оогонии с одной яйцеклеткой имеют покров из 5 спиральных завитых кле-
ток, завернутых, начиная снизу, справа налево. Верхние концы этих клеток об-
разуют ясно выраженную шейку, и каждый несет по 1-2 клетки, слагающихся в
коронку из 5 клеток в один ярус, из 10 клеток в 2 яруса. Антеридии шаровидные.
Ооспоры более или менее эллипсоидные, с коричневыми, до почти черных,
стенками, на поверхности которых видны спирально извитые ребра.
В Байкале найдены представители 2 семейств.
I. Коронка оогония из 10 клеток в 2 яруса по 5. Стебли и листья без коры. Ли-
стья сложные, членисто-вильчатые, иногда неправильные, а в том случае с
более или менее обособленной основной осью члеников и многочисленны-
ми листочками в ее узлах, реже простые..........Семейство Nitellaceae
1. Листья правильно однократно или многократно вильчатые, реже простые.
Антеридии на вершине члеников в развилках листа, оогонии сбоку, у од-
нодомных видов под антеридиями. Ооспоры в поперечном сечении эл-
липтические .........................................Род Nitella
II. Коронка оогония из 5 клеток в один ярус. Стебли и листья с корой или без
коры. Листья сложные, членисто-линейные, с основной осью члеников и
кучками одноклеточных листочков в ее узлах.
1. Кора имеется или отсутствует. Прилистники всегда имеются, иногда руди-
ментарные. Листья из 4 и более члеников. Листочки разной длины, по 4 и
более в узле..................................Семейство Characeae
А. Растения однодомные и двудомные, у первых оогонии располагаются
выше антеридиев. Кора хорошо развитая, иногда только на стебле или
совсем отсутствует...................................Род Chara
Род Nitella Ag., 1824 emend. A. Br., Leonh. (=Nitella Ag. pr. p.)
Обычно 2 ветви в пристеблевых узлах. Листья сложные, правильно одно-
кратно или многократно вильчатые, состоящие из равновеликих одноклеточ-
ных члеников. Плодоносящие листья обычно собраны в крупные рыхлые, мел-
кие плотные головки. Одиночные антеридии развиваются на вершине члени-
ков в развилках листа. Оогонии располагаются тут же сбоку, у однодомных
видов — непосредственно под антеридием. Ооспоры эллипсоидные или шаро-
видные, в поперечном сечении эллиптические.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 217
В оз. Байкал найдены 2 вида.
I. Конечные листовые членики только одноклеточные. Листья однократно ви-
льчатые (реже простые). Коронка оогония быстро опадающая.
1. Гаметангии без слизи. Растения однодомные и двудомные.
А. Мутовки одинарные, из одного круга однотипных листьев.
а. Растение двудомное.................................N. ораса
б. Растения однодомные. Мутовки из 6-7(8) листьев ...N. flexilis
Nitella ораса (Bruz.) Ag. (—Chara flexilis Smith) (рис. 96: 9-16)
Растение двудомное, 20-30 см высотой, грязно-зеленое, иногда почти чер-
ное, не инкрустировано известью. Стебли тонкие, междоузлия сильно вытяну-
ты, до 9 см длиной. Мутовки из 6-7 длинных листьев, плодоносящие мутовки
из укороченных листьев. Листья однократно вильчатые, с (1)2-3 члениками
второго порядка. Вторые членики на конце обычно переходят в короткий за-
остренный тупой кончик. Гаметангии в развилке листьев. Оогонии по (1)2(3),
эллипсоидные, 430-700 мкм длиной (без коронки), 375-610 мкм шириной.
Спиральные клетки образуют 7-8 извивов. Коронка быстро опадающая, рыхло
сложенная, 50 мкм высотой, 75 мкм длиной. Ооспоры эллиптические, темной
каштаново-коричневой окраски, 345-470(500-520) мкм длиной, 315-450 мкм
шириной. Антеридии 670-775 мкм в диаметре.
В пресных, преимущественно стоячих водоемах, повсеместно. Иногда в
солоноватых водах.
Замечания. Для Байкала приводится К.И. Мейером [1930]. Найдена
Л.В. Рейнгардтом в районе Култука.
Nitella flexilis (L.) Ag. (—Chara flexilis L.(ex. p.); C. commutata Rupr.)
(рис. 96: 1—8)
Растение однодомное, до 20-40 см высотой, чисто-зеленого цвета, не инк-
рустированное известью. Стебли сильно ветвящиеся, крепкие и гибкие. Меж-
доузлия обычно в 1.5-2 раза длиннее листьев, мутовки из 6-7(8) основных ли-
стьев, к которым иногда добавляются 1-3 дополнительных (акцессорных) лис-
та, реже плодоносящие листья, собранные в головки. Листья однократно
вильчатые или двучленно-простые, с( 1)2(3) члениками 2-го порядка. Первые
членики длинные, вторые одноклеточные, закругленные или конусовидные на
конце. Гаметангии объединенные на основных листьях. Оогонии одиночные
или по 2-3, эллипсоидные, 550-583 мкм длиной (без коронки), 500-610 мкм
шириной, спиральные клетки образуют (6)8-9 извивов, коронка рыхлая, бы-
стро опадает, 50 мкм высотой и 85 мкм шириной. Ооспоры эллиптические,
480-575 мкм длиной, 400-500 мкм шириной, темно-красно-коричневые. Анте-
ридии 500-625(800) мкм в диаметре.
В пресных преимущественно мелких водоемах, реже в текучих и солонова-
тых водах.
Nitella flexilis var. flexilis (рис. 96: 1-8)
Плодоносящие мутовки преимущественно распростертые. Конечные лис-
товые членики тупые или постепенно суженные. Гаметангии развиваются поч-
ти одновременно. Спиральные клетки оогониев образуют 8-9 извивов, ооспо-
ры с 6-8 ребрами.
218 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 96. 1-8 — Nitella flexilis: 1 — общий вид растения, 2, 3 — плодоносящие листья, 4,
5 — концы листочков, 6 — коронка, 7 — ооспора, 8 — оогоний; 9-16 — N. ораса: 9 — об-
щий вид части мужского растения, 10 — общий вид части женского растения, 11 — пло-
доносящий лист с антеридием, 12 — плодоносящий лист с оогониями, 13, 14 — концы
листочков, 15 — коронка, 16 — ооспора.
По: Голлербаху, Красавиной.
Fig. 96. 1-8 — Nitella flexilis: 1 — general view, 2, 3 — bearing leaves, 4, 5 — tips of leaflets,
6 — corona, 7 — oospore, 8 — oogonium; 9-16 — Nitella opaca: 9 — general view of a part of
male alga, 10 — general view of a part of female alga, 11 — bearing leaf with antheridium, 12 —
bearing leaf with oogonia, 13, 14 — tips of leaflets, 15 — corona, 16 — oospore.
After: Gollerbach, Krasavina.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 219
В пресных, преимущественно мелких водоемах, а также в озерах, реже в те-
кучих и солоноватых водах. Повсеместно.
Замечания. В оз. Байкал обнаружена нами [Ижболдина, 1984,1990в) вдоль
юго-западного побережья в районе падей Бол. Коты и Варначка, на глубине от
3 до 18-20 м. Массовое появление плодоносящих мутовок наблюдали в конце
июля — начале августа.
Кроме 2 вышеназванных видов в Байкале найдены стерильные талломы
Nitella, что не позволило определить их вид. К.И. Мейером [1930] в большом
количестве Nitella sp. обнаружена в районе пади Нугды и в губе Аяя. При наших
исследованиях стерильные талломы Nitella sp. встречены преимущественно у
западного побережья Сред. Байкала, а также в губах Сев. Байкала — Слюдян-
ской, Богучанской, Дагарской, Аяя. В районе мысов Улан-Хапсагай и Имыш-
Таме (прол. Ольхонские Ворота) высота талломов Nitella sp., образующих на
глубине 9-18 м мощные заросли, достигает 50 см. Фитомасса Nitella sp. дости-
гает здесь максимальной (523 г/м2) известной для Байкала величины [Ижбол-
дина, 1976а, 1990в].
СЕМЕЙСТВО CHARACEAE AG. EMEND HOLLERB.
Растения часто инкрустированные известью, мутно-зеленые, сероватые,
реже прозрачно-зеленые. Стебли и листья с корой или без коры. Прилистники
имеются. Листья членисто-линейного строения. Коронка оогония из 5 клеток в
один ярус.
В оз. Байкал найдены представители одного рода.
Род Chara (Vaillant, 1719) L.
Листья в большинстве из 2-7 члеников. Венчик прилистников одно- или
двурядный. Листочки по 3-7 в каждом листовом узле. Однодомные и двудом-
ные растения. У однодомных гаметангии объединенные, реже раздельные.
У двудомных оогонии располагаются над антеридиями. У подавляющего боль-
шинства видов стебли и листья в той или иной степени покрыты корой.
В Байкале встречены 3 вида.
I. Венчик прилистников двурядный.
1. Стеблевая кора двухполосная, число коровых трубок вдвое больше числа
листьев в вышестоящей мутовке (реже неправильная, местами трех-
полосная).
А. Первичные (основные) коровые трубки более крупные и выступающие,
чем вторичные (промежуточные), вследствие чего коровые шипы или
узловые клетки находятся на выступающих частях коры.
а. Листья нормально развитые.
а. Растение двудомное ............................С. tomentosa
р. Растение однодомное.
* Стеблевая и листовая кора хорошо развиты.
+ Ооспоры черные (или темно-коричневые).
х Коровые шипы отсутствуют или совсем слабо развитые и тог-
да всегда одиночные.
о Ооспоры свыше 500 мкм длиной............С. contraria
2. Стеблевая кора трехполосная, число коровых трубок втрое больше числа
листьев в вышестоящей мутовке.
220 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
А. Растения двудомные.
а. Стеблевые шипы только в форме округлых или заостренно-бородав-
чатых клеток.
* На погруженных в ил корневых частях имеются сложные крупные
клубеньки; стенки клеток очень тонкие, так что при высушивании
растение делается плоским и нежным, как бумага......С. fragifera
Chara contraria A. Br. (рис. 97: 12-20)
Растение однодомное, обычно 5-20 см, редко до 100 см высотой, умерен-
но-кустистое, зеленовато-серое, сильно инкрустировано. Стебли 400-1100 мкм
в диаметре. Междоузлие в 2-4 раза длиннее листьев. Кора правильно двухпо-
лосная. Шипы одиночные, тупые, короткие. Прилистников по 2 пары на каж-
дый лист, хорошо развиты. Мутовки из 6-8(10) листьев. Листья 5-12 (30) мм
длиной, обычно из 3-5 члеников с корой и голого 2-3-клеточного концевого
членика. Верхушечная клетка короткая, конусовидная, заостренная. Листочки
обычно по 5-7 вокруг листовых узлов, заостренные или тупые, чаще рудимен-
тарные в форме округлых или заостренно-бородавчатых клеток. Гаметангии
всегда объединенные, одиночные, реже парные, на 2-4 нижних листовых уз-
лах. Оогонии 650-975 мкм длиной (без коронки), 375-575 мкм шириной; спи-
ральные клетки образуют 13-16 извивов, коронка плотная, 110-199 мкм высо-
той, 180-360 мкм шириной. Ооспоры эллипсоидные, до цилиндрически-эл-
липсоидных, 525-725 мкм длиной, 285-425 мкм шириной, черные, с 11-14
тонкими ребрами. Антеридии 255-450 мкм в диаметре.
В пресноводных, преимущественно непроточных водоемах.
В России повсеместно.
Замечание. В Байкале найдена К.И. Мейером [1930] в Истокском соре, в
июле.
Chara fragifera Durieu (рис. 97: 1-11)
Двудомное растение, (7)10-15(50) см высотой, обильно кустистое, неин-
крустированное, зеленое. Стебли не более 400 мкм в диаметре. Междоузлия
обычно равны по длине листьям. Кора правильно трехполостная, с почти рав-
новеликими коровыми трубками. Шипы полностью рудиментарные, в виде
шаровидных или заостренно-бородавчатых клеток. Прилистники в двуряд-
ном венчике, по 2 пары на каждый лист. Мутовки из 6-9 листьев. Последние
обычно 1-2 см длиной, у вытянутых форм до 3.5 см длиной, из 7-11, чаще
8-10 члеников с корой. Концевой членик из 1-2 клеток, верхушечная клетка
конусовидная, заостренная. Листочки сильно редуцированы, у женских рас-
тений в числе 8, у мужских растений их 5. Гаметангии одиночные, реже пар-
ные, на 3-5(7) нижних листовых узлах. Оогонии удлиненно-эллипсоидные,
(675)875-1000(1200) мкм длиной (без коронки), (450)525-675(800) мкм
шириной. Спиральные клетки образуют 12-14 извивов, коронка 105-175 мкм
высотой, 175-200 мкм шириной. Ооспоры 550-700(800) мкм длиной, (300)
345-450 мкм шириной, каштаново-коричневые, до почти черных, с 9-12 яв-
ственными ребрами. Антеридии 450-575(640) мкм в диаметре.
В пресных водах, преимущественно в прудах, реже в озерах.
Россия (оз. Байкал), Прибалтика, Белоруссия, Киргизия, Узбекистан,
Туркмения.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 221
Рис. 97. 1-11 — Chara fragifera: 1 — общий вид части растения, 2 — основание мутовки,
3 — часть стебля с коровыми шипами, 4 — поперечный разрез стебля, 5, 6 — окончания
листьев, 7 — клубеньки, 8 — коронка, 9 — плодоносящий женский листовой узел, 10 —
ооспора, 11 — плодоносящий мужской листовой узел; 12-20 — Ch. contraria: 12 — общий
вид части растения, 13,16 — части стебля и основания мутовки с различно выраженными
прилистниками и коровыми шипами, 14,15 — конечные членики листа, 17 — плодонося-
щий листовой узел, 18 — поперечный разрез стебля, 19 — оогоний, 20 — ооспора.
По: Голлербаху, Красавиной.
Fig. 97. 1-11 — Chara fragifera: 1 — part of alga, general view, 2 — base of whorl, 3 — fragment
of the stalk with cortical spinules, 4 — stalk, cross-section, 5,6 — ends of leaves, 7 — tubercles,
8 — corona, 9 — bearing female leaf knot, 10 — oospore, 11 — bearing male leaf knot; 12-20 —
Ch. contraria: 12 — fragment of alga, general view, 13, 16 — parts of stalk and whorl base with
cortical spinules, 14, 15 — terminal segments of leaf, 17 — bearing leaf knot, 18 — stalk,
cross-section, 19 — oogonium, 20 — oospore.
After: Gollerbach, Krasavina.
222 Глава 3. Систематика водорослей бентоса и перифитона озера Байкал
Рис. 98. Chara tomentosa.
1 — общий вид части растения; 2 — основание мутовки; 3 — часть стебля; 4 — поперечный разрез
стебля; 5, 6 — листовые узлы с плодоношением; 7 — ооспора; 8 — коронка; 9 — оогоний. По: Гол-
лербаху, Красавиной.
Fig. 98. Chara tomentosa.
1 — fragment of alga, general view; 2 — whorl base; 3 — fragment of stalk; 4 — stalk, cross section; 5,6 —
bearing leaf knots; 7 — oospore; 8 — corona; 9 — oogonium. After; Gollerbach, Krasavina.
Глава 3. Систематика водорослей бентоса и перифитона озера Байкал 223
Замечание. В Байкале найдена К.И. Мейером [1930] в бух. Харин-Ирги
(прол. Ольхонские Ворота) в начале августа.
Chara tomentosa L. (=С. ceratophylla Wallr.; C. latifolia Willd.) (рис. 98: 1—9)
Растение двудомное, до 60 см высотой. Стебли до 2 мм в диаметре, ветвят-
ся умеренно. Междоузлия в 1.5-3 раза длиннее листьев. Кора правильно двух-
полосная. Шипы одиночные или парные, обычно широко яйцевидно заост-
ренные. Прилистники в двурядном венчике, по 2 пары на лист. Мутовки из
6-8 листьев, 2-3 см длиной, обычно из 2-3 члеников с корой и голого двукле-
точного концевого членика. Гаметангии обычно одиночные, иногда парные,
на 2-3 нижних листовых узлах. Оогонии 750-1000 мкм длиной (без коронки),
675-800 мкм шириной Спиральные клетки образуют 14-15 извивов. Коронка
200-300 мкм высотой, 400-500 мкм шириной. Ооспоры эллипсоидные, 765-
900 мкм длиной, 525-645 мкм шириной, темно-коричневые с 13-14 ребрами.
Антеридии 900-1425 мкм в диаметре.
В пресных и солоноватых водах, преимущественно в непроточных водо-
емах. Повсеместно.
Замечание. В Байкале заросли Ch. tomentosa обнаружены К.И. Мейером
[1922] в оз. Нур, расположенном в вершине бух. Загли, сообщающимся с ней
протоком.
Кроме 3 вышеописанных видов во многих районах Байкала обнаружены
стерильные талломы Chara sp. Г.Ю. Верещагин [1918] наблюдал заросли Chara
sp. в бух. Крохалиной (Чивыркуйский зал.). К.И. Мейером [1930] она обнару-
жена в бух. Харин-Ирги (прол. Ольхонские Ворота) и в бухтах Чивыркуйского
залива, В.Н. Яснитским [1928] — в прол. Ольхонские Ворота, где на глубине от
1,5 до 16 м образовывала вместе с Chaetomorpha целые заросли. На развитие
Chara sp. в губах и защищенных от господствующих ветров участках байкаль-
ского побережья указывает и М.М. Кожов [1931, 1947, 1962]. Нами стерильные
талломы Chara sp. встречены кроме вышеуказанных районов в губе Богучан-
ской, у пади Орео, у о. Угунгой, у мыса Ото-Хушун. В зал. Мухор (прол. Мал.
Море) в августе встречены талломы Chara sp. с оогониями [Ижболдина, 1990].
Фитомасса водоросли в разных участках прол. Ольхонские Ворота колеблется
от 5-10 до 230 г/м2, в Чивыркуйском заливе — от 1 до 230 г/м2, а южнее о. Бак-
ланий и в бух. Змеиная может достигать 1500 г/м2.
СВЕДЕНИЯ ОБ АВТОРЕ
ИЖБОЛДИНА ЛЮДМИЛА АЛЕКСАНДРОВНА — кандидат биологиче-
ских наук, старший научный сотрудник. Много лет работала в Научно-исследо-
вательском институте биологии при Иркутском государственном университете, с
1997 г. — в Лимнологическом институте РАН. Область научных интересов —
экология, биология, систематика, продуктивность донных водорослей мейо- и
макрофитов Байкала и окружающих его водоемов. Имеет 86 научных публика-
ций, в том числе несколько монографий.
225
Рис. 1. Байкальский берег, северо-западное побережье.
Рис. 2. Скала Кобылья Голова в районе пади Черной. Южный Байкал.
Рис. 3. Мысы западного побережья озера.
Рис. 4. Хвойный лес на обрывистом скалистом берегу Байкала.
Рис. 5. Северо-западное побережье озера. Вид из района пади Черная.
40
230
Рис. 6. Волны разрушают байкальские берега. Сосна, нависшая над водой, обречена.
231
Рис. 7. Галечный материал при волнении озера разрушает скальные берега.
Рис. 9. Одно из главных зданий (вид сбоку) Байкальской биостанции ИГУ в пос. Большие Коты (Южный Байкал).
Й
234
Рис. 10. Территория биостанции ИГУ. Пришла весна.
235
Рис. 11. Одно из общежитий научных сотрудников на территории биостанции ИГУ.
библиографический список
Азовский М.Г., Паутова В.Н., Ижболдина Л.А. К флоре гидрофитов озера Байкал // Бо-
тан. жури. — 1983. — № 10. — С. 1392-1397.
Барашков Г.К. О методике количественного учета литоральных водорослей // Распреде-
ление и состав промысловых водорослей Баренцева моря. — М.; Л., 1965. — С. 8-12.
Бекман М.Ю. Некоторые закономерности распределения и продуцирования массовых
видов зообентоса в Малом Море // Тр. Байкал, лимнол. ст. АН СССР. — 1965. —
Т. 17. - С. 342-381.
Бекман М.Ю., Деньгина Р.С. Население бентали и кормовые ресурсы Байкала // Биоло-
гическая продуктивность водоемов Сибири. — М., 1969. — С. 42-47.
Блинова Е.И., Возжинская В.Б. Морские макрофиты и растительные ресурсы океана //
Основы биологической продуктивности океана и ее использование. — М., 1971. —
С. 137-171.
Бочкарев П.Ф., Вотинцев К.К., Яснитский В.Н. Об энергии фотосинтеза некоторых мак-
рофитов озера Байкал // Докл. АН СССР. — 1950. — Т. 70, № 3. — С. 519-522.
Браун Д. Методы исследования и учета растительности. — М., 1957. — 316 с.
Буров В.С., Кожов М.М., Талызин Ф.Ф. и др. Материалы к распределению грунтов и фа-
уны прибрежной полосы Северного Байкала // Изв. Биол.-геогр. НИИ при Иркут,
ун-те. — 1933. — Т. 6, вып. 1. — С. 154-175.
Верболов В.И. Течения в Малом Море // Исследование Малого Моря. — М.; Л., 1959. —
С. 34-53. — (Тр. Байкал, лимнол. ст. АН СССР; Т. 17).
Верболов В.И., Сокольников В.М., Шимараев М.Н. Гидрометеорологический режим и
тепловой баланс озера Байкал. — М.; Л., 1965. — 362 с.
Верещагин Г.Ю. Отчет о работах, произведенных на Байкале во время командировки от
Императорской Академии Наук летом 1916 года // Труды комиссии по изучению
озера Байкала. — 1918. — Т. 1, вып. 1. — С. 1-53.
Верещагин Г.Ю. Маршруты Байкальской экспедиции Академии наук СССР 1925-
1929 гг. //Докл. АН СССР. - 1930. - № 3. - С. 227-266.
Верещагин Г.Ю. Два типа биологических комплексов Байкала // Тр. Байкал, лимнол. ст.
АН СССР. - 1935. - Т. 6. - С. 199-212.
Верещагин Г.Ю. Происхождение и история фауны и флоры Байкала в связи с вопросом
о происхождении их в континентальных водах вообще // Изв. Гос. геогр. об-ва. —
М.; Л., 1939. - Т. 71, вып. 9. - С. 1277-1293.
Верещагин Г.Ю. Происхождение и история Байкала, его фауны и флоры // Тр. Байкал,
лимнол. ст. АН СССР. - 1940. - Т. 10. - С. 72-239.
Виноградова КЛ., Голлербах М.М., Зауер Л.М., Сдобникова Н.В. Определитель пресно-
водных водорослей СССР. Зеленые водоросли — Chlorophyta. Классы сифонокладо-
вые, сифоновые. Красные водоросли — Rhodophyta. Бурые водоросли — Phaeophy-
ta. — Л., 1980. — Вып. 13. — 248 с.
Библиографический список
237
Водоросли: Справочник / С.П. Вассер, Н.В. Кондратьева, Н.П. Масюк и др. — Киев,
1989. - 606 с.
Воронихин Н.Н. Эдогониевые водоросли Европейского Севера СССР. Oedogoniales //
Тр. Ботан. ин-та им. В.Л. Комарова. — М.; Л., 1951. — Сер. И, вып. 7. — С. 415-480.
Вотинцев К.К. Гидрохимия озера Байкал // Тр. Байкал, лимнол. ст. АН СССР. — 1961. —
Т. 20. - С. 311.
Вотинцев К.К. Метод склянок и истинная продукция водорослей // Тр. Лимнол. ин-та
СО АН СССР. - 1963. - Т. 3(23). - С. 113-119.
Вотякова Н.Е. Систематическая характеристика водорослей обрастаний // Экология
растительности дельты реки Селенги. — Новосибирск, 1981. — С. 61—73.
Гагарин П.К. Водная растительность прибрежных участков // Лимнология прибреж-
но-соровой зоны Байкала. — Новосибирск, 1974. — С. 142-148.
Голлербах М.М., Полянский В.И. Определитель пресноводных водорослей СССР. — М.,
1951. — Вып.1: Общая часть. — 199 с.
Голлербах М.М., Косинская Е.К., Полянский В.И. Определитель пресноводных водорос-
лей СССР. — М., 1953. — Вып.2: Синезеленые водоросли. — 652 с.
Голлербах М.М., Красавина Л.К. Определитель пресноводных водорослей СССР. — Л.,
1983. — Вып. 14: Харовые водоросли — Charophyta. — 190 с.
Гуменюк А.А. К вопросу о химическом составе макрофитов озера Байкал // Тезисы 4-й
конференции молодых ученых. — Иркутск, 1986. — С. 48.
Гуменюк А.А. Зольный состав массовых видов макрофитов озера Байкал // Тезисы 5-й
конференции молодых ученых. — Иркутск, 1987. — С. 28.
Дорогостайский В.Ч. Материалы для альгологии оз.Байкал и его бассейна // Изв.
Вост.-Сиб. отд. Император. Рус. геогр. о-ва. — 1906. — Т. 35, № 3. — С. 1-44.
Дорогостайский В.Ч. Вертикальное и горизонтальное распределение фауны Байкала //
Сиб. тр. / Иркут, ун-т. — 1923. — Вып. 4. — С. 103-131.
Загоренко Г.Ф., Ижболдина Л.А. Нахождение байкальской эндемичной кладофоры в
озере Хубсугул (МНР) // Новые материалы по фауне и флоре Байкала. — Иркутск,
1976. - С. 49-58.
Загоренко Г.Ф., Ижболдина Л.А. Новый вид рода Cladophora Kutz. // Новости системати-
ки низших растений. — Л., 1977. — С. 20-24.
Загоренко Г.Ф., Ижболдина Л.А. О систематическом положении Chaetomorpha solitaria
Skabitsch. (Chlorophyta) // Новости систематики низших растений. — Л., 1983. —
С. 18-22.
Забелина М.М., Киселев И.А., Прошкина-Лавренко А.И., Шешукова В.С. Определитель
пресноводных водорослей СССР. Диатомовые водоросли. — М., 1951. — Т. 4. — 619 с.
Ижболдина Л.А. О бентосной макрофлоре открытого Байкала // Изв. Биол.-геогр. НИИ
при Иркут, ун-те. — Иркутск, 1965. — Т. 18, вып. 1-2. — С. 35-49.
Ижболдина Л.А. Заметки по биологии некоторых видов макрофитов открытого Байка-
ла // Изв. Биол.-геогр. НИИ при Иркут, ун-те. — Иркутск, 1967. — С. 9-11.
Ижболдина Л.А. Бентосные макрофиты открытых вод Южного Байкала // Бентос и
планктон Южного Байкала. — Иркутск, 1970. — С. 13-41. — (Изв. Биол.-геогр.
НИИ при Иркут, ун-те; Т. 23, вып. 1).
Ижболдина Л.А. Макрофиты Южного Байкала и продукция некоторых видов, домини-
рующих в литорали района Больших Котов // Продуктивность Байкала и антропо-
генные изменения его природы. — Иркутск, 1974а. — С. 111-116.
Ижболдина Л.А. Состояние макрофитов в районе Утулик — Мурино // Продуктивность
Байкала и антропогенные изменения его природы. — Иркутск, 19746. — С. 181-191.
Ижболдина Л.А. Макрофиты островов Ушканьего архипелага // Продуктивность Байка-
ла и антропогенные изменения его природы. — Иркутск, 1974в. — С. 117-125.
Ижболдина Л.А. Сезонная динамика биомассы макрофитов Южного Байкала // Круго-
ворот вещества и энергии в озерных водоемах. — Новосибирск, 1975. — С. 107-111.
Ижболдина Л.А. Новые данные о бентосных водорослях озера Байкал // Новые материа-
лы по фауне и флоре Байкала. — Иркутск, 1976а. — С. 4-13.
Ижболдина Л.А. Макрофиты пролива Малое Море // Новые материалы по фауне и фло-
ре Байкала. — Иркутск, 19766. — С. 14-34.
238
Библиографический список
Ижболдина Л.А. Новые данные о представителях эндемичного байкальского рода Dra-
parnaldiella С. Meyer et Skabitsch. 11 Новые материалы по фауне и флоре Байкала. —
Иркутск, 1976в. — С. 35-49.
Ижболдина Л.А. К характеристике представителей семейства кладофоровых в озере Бай-
кал. — Иркутск, 1981а. — Рукопись деп. в ВИНИТИ, № 3085.81. — 34 с.
Ижболдина Л.А. Особенности количественного распределения макрофитобентоса вдоль
открытых побережий Байкала // Круговорот вещества и энергии в водоемах: Пятое
Всесоюз. Лимнол. совещ. Тез. докл. — Лиственичное-на-Байкале, сентябрь, 1981. —
Иркутск, 19816. — Вып. 1. — С. 65-67.
Ижболдина Л.А. Биомасса и продукция макрофитобентоса открытых побережий озера
Байкал // Тезисы докладов 4-го съезда ВГБО. — Киев, 1981в. — Ч. 1. — С. 117-118.
Ижболдина Л.А. Макрофитобентос // Состояние сообществ Южного Байкала. — Ир-
кутск, 1982а. — С. 58-69.
Ижболдина Л.А. Макрофитобентос Селенгинского мелководья оз. Байкал // Проблемы
экологии Прибайкалья: Тез. докл. к Всес. научн. конф. — Иркутск, 19826. — Ч. 3. —
С. 68-69.
Ижболдина Л.А. Новый вид рода Microcoleus Desmaz. из озера Байкал // Новости систе-
матики низших растений. — М.; Л., 1984. — С. 20-22.
Ижболдина Л.А. Количественное распределение эндемичных видов макрофитов вдоль
открытых побережий озера Байкал // Круговорот вещества и энергии в водоемах:
Структура и продуктивность растительных сообществ (фитопланктон, фитобентос,
высшая водная растительность). — Иркутск, 1985а. — Вып. 11. — С. 31-32.
Ижболдина Л.А. Макрофитобентос как один из показателей импактного мониторинга
озера Байкал // Совершенствование регионального мониторинга состояния озера
Байкал. - Л., 19856. - С. 215-223.
Ижболдина Л.А. Особенности распределения в озере Байкал макрофитов, вегетирующих
круглый год // Актуальные проблемы современной альгологии: Тез. докл. Всесоюз.
конф. — Чебоксары, 1987. — С. 107-108.
Ижболдина Л.А. Новые данные по биологии ряда видов семейства Cladophoraceae
(Hass.) Cohn, из озера Байкал // Новое в изучении флоры и фауны Байкала и водо-
емов его бассейна. — Иркутск, 1988. — С. 53-62.
Ижболдина Л.А. Характер распределения эндемичных видов донных водорослей в озере
Байкал // Тезисы докладов по споровым растениям Средней Азии и Казахстана. —
Ташкент, 1989. — С. 21-22.
Ижболдина Л.А. Макрофитобентос открытых побережий озера // Подводные ландшаф-
ты Байкала. — Новосибирск, 1990а. — С. 142-166.
Ижболдина Л.А. К вопросу о распределении мейо- и макрофитобентоса в озере Байкал
(водоросли) // Биология внутренних вод. — Л., 19906. — С. 33-37. — (Информ, бюл.;
№ 88).
Ижболдина Л.А. Мейо- и макрофитобентос озера Байкал (водоросли). — Иркутск,
1990в. - 175 с.
Ижболдина Л.А. Результаты исследований бентоса Байкала // Экологические исследова-
ния Байкала и Байкальского региона. — Иркутск, 1992. — Ч. 1. — С. 137-151.
Ижболдина Л.А. Мейо- и макрофиты // Флора и фауна заповедников. Флора и фауна
водоемов и водотоков Байкальского заповедника. — М., 2001. — Вып. 92. — С. 15-18.
Ижболдина Л.А., Верномудров А.Ю., Максимов В.И. Продукция Ulothrix zonata Kg. в озе-
ре Байкал // Новые материалы по фауне и флоре Байкала. — Иркутск, 1978. —
С. 56-61.
Ижболдина Л.А., Верномудров А.Ю., Максимов В.И. Биомасса и продукция Tetraspora
cylindrica (Wahlenb.) Ag. var. bullosa C.Meyer в озере Байкал // Экологические иссле-
дования озера Байкал и Прибайкалья. — Иркутск, 1984. — С. 10-17.
Ижболдина Л.А., Галкин А.Н. Мейо- и макрофиты (водоросли) горячих источников
Прибайкалья // Сохранение биологического разнообразия геотермальных рефугиев
Байкальской Сибири. — Иркутск, 2000. — С. 7-8.
Ижболдина Л.А., Коневецкая М.С., Максимова В.М. Макрофитобентос юго-восточного
побережья озера Байкал на участке между устьями рек Снежная и Селенга // Эколо-
гические исследования озера Байкал и Прибайкалья. — Иркутск, 1984. — С. 17-24.
Библиографический список
239
Ижболдина Л.А., Корнакова Э.Ф., Гуменюк А.А. Характеристика элементного состава
макрофитов северо-западного побережья Южного Байкала // Первая Всесоюз.
конф, по рыбохозяйственной токсикологии (Юрмала). — Рига, 1988. — Ч. 1. —
С. 163-164.
Ижболдина Л.А., Максимова В.М. Биомасса и продукция ряда макрофитов, вегетирую-
щих в оз. Байкал в течение года // Гидробиологические и ихтиологические исследо-
вания в Восточной Сибири: Чтения памяти проф. М.М.Кожова. — Иркутск, 1979. —
Вып. 3. - С. 6-13.
Ижболдина Л.А., Максимов В.И. Продукция видов рода Draparnaldiella С. Meyer et Ska-
bitsch. вдоль западного побережья Среднего Байкала // Проблемы экологии При-
байкалья: Тез. докл. к Всесоюз. науч. конф. — Иркутск, 1982. — С. 96-97.
Ижболдина Л.А., Максимова В.М., Гуменюк А.А., Кусовникова Т.Г. Сравнительная оцен-
ка разных способов отбора количественных бентосных проб в озере Байкал. — Ир-
кутск, 1990. - Рук. деп. ВИНИТИ 21.03.90, № 1513 - В90.
Ижболдина Л.А., Максимов В.И., Мурахвери А.М. и др. Биомасса и продукция видов
рода Draparnaldiella С. Meyer et Skabitsch в Байкале// Гидробиологические и ихтио-
логические исследования в Восточной Сибири: Чтения памяти проф. М.М. Кожо-
ва. — Иркутск, 1978. — Вып. 2. — С. 47-56.
Ижболдина Л.А., Семейкин В.И., Максимова В.М. Новые сведения о Cladophora compacta
(С. Meyer) С. Meyer из озера Байкал. — Иркутск, 1985. — Деп. в ВИНИТИ 16.12.86,
№ 8548-В.
Каплина Г.С. Макрозообентос каменистых грунтов литорали оз. Байкал и его сезонная
динамика (данные 1963-1968 гг.) (район Больших Котов) // Продуктивность Байка-
ла и антропогенные изменения его природы. — Иркутск, 1974. — С. 126-137.
Карабанов Е.Б. Структура подводных ландшафтов // Подводные ландшафты Байкала. —
Новосибирск, 1990. — С. 3-66.
Катанская В.М. Высшая водная растительность континентальных водоемов СССР: Ме-
тоды изучения. — Л., 1981. — 187 с.
Кожов М.М. К познанию фауны Байкала, ее распределения и условий обитания // Изв.
Биол.-геогр. НИИ при Иркут, ун-те. — 1931. — Т. 5, вып. 1. — 170 с.
Кожов М.М. Гидрологические и гидробиологические исследования в Баргузинском за-
ливе на Байкале в 1932 г. // Изв. Биол.-геогр. НИИ при Иркут, ун-те. — 1934. — Т. 6,
вып. 1. — С. 9-83.
Кожов М.М. Моллюски озера Байкал // Тр. Байкал, лимнол. ст. АН СССР. — М.; Л.,
1936. - Т. 8. - 352 с.
Кожов М.М. О распространении современной байкальской фауны вне Байкала // Тр.
Карел, фил. АН СССР. - 1956. - Вып. 5. - С. 39-46.
Кожов М.М. О генезисе основных экологических комплексов в современной байкаль-
ской фауне // Изв. Биол.-геогр. НИИ при Иркут, ун-те. — Иркутск, 1957. — Т. 27,
вып. 1-4. — С. 68-83.
Кожов М.М. О видообразовании в озере Байкал // Бюл. МОИП. Отд. биологии. — М.,
1960. - Т. 65. - С. 39-47.
Кожов М.М. Биология озера Байкал. — М., 1962. — 315 с.
Кожов М.М. Становление и пути эволюции фауны озера Байкал // Проблемы эволю-
ции. — Новосибирск, 1973. — Т. 3. — С. 5-30.
Кожов М.М., Ижболдина Л.А., Каплина Г.С. и др. Бентос литорали и сублиторали оз.
Байкал вдоль юго-восточных берегов // Гидробиол. журн. — 1965. — Т. 1, № 4. —
С. 3-11.
Кожов М.М., Ижболдина Л.А., Каплина Г.С. и др. Бентос юго-восточного побережья
оз. Байкал // Биологическая продуктивность водоемов Сибири. — М., 1969. —
С. 29-36.
Кожова О.М., Ижболдина Л.А. Фитоценозы реки Большие Коты (приток Байкала, Рос-
сия) //Альгология. - 1994. — Т. 4, № 3. — С. 84-87.
Кожова О.М., Ижболдина Л.А., Бокова И.К. Экология Didymosphenia geminata (Lyngb.)
М. Schmidt в озере Байкал // Альгология. — 1998. — Т. 8, № 2. — С. 132-139.
Кожова О.М., Ижболдина Л.А.; Гуменюк А.А., Максимова В.М. Пространственная
структура и динамика донных фитоценозов литорали вдоль открытых побережий
240
Библиографический список
озера Байкал // Природные ресурсы, экология и социальная среда Прибайкалья.
Состояние донных сообществ и рыб Байкала. — Иркутск, 1995а. — С. 58-61.
Кожова О.М., Ижболдина Л.А., Гуменюк А.А., Максимова В.М. Макрофиты Байкала: со-
став, распространение, мониторинг // Проблемы экологии Прибайкалья: Чтения
памяти проф. М.М.Кожова. — Новосибирск, 19956. — С. 61-66.
Кожова О.М., Окунева Г.Л., Ижболдина Л.А., Каплина Г.С. Оценка распределения бен-
тоса в Южном Байкале (район Утулик — Мурино) по критерию Фишера // Продук-
тивность Байкала и антропогенные изменения его природы. — Иркутск, 1974. —
С. 181-191.
Кокин К.А. Экология высших водных растений. — М., 1982. — 157 с.
Комаренко Л.Е., Васильева И.И. Пресноводные диатомовые и синезеленые водоросли
водоемов Якутии. — М., 1975. — 423 с.
Комаренко Л.Е., Васильева И.И. Пресноводные зеленые водоросли водоемов Якутии. —
М., 1978. - 283 с.
Константинов А.С. Общая гидробиология. — М., 1986. — 472 с.
Коршиков О.А. Визначник прюноводних водоростей Украшсько! РСР. Подкласс Прото-
коков. — Кшв, 1953. — Т. 5. — 439 с.
Кравцова Л.С., Ижболдина Л.А., Акиншина Т.В. и др. Количественная характеристика
донных биоценозов литорали оз.Байкал с использованием ландшафтного райониро-
вания // Тез. докл. съезда ВГБО. — Мурманск, 1991. — С. 179-180.
Курсанов Л.И., Забелина М.М. Определитель низших растений. — М., 1953. — Т. 1.
Лут Б.Ф. Геоморфология Байкала // Геоморфология дна Байкала и его берегов. — М.,
1964. - С. 5-123.
Лут Б.Ф. Геоморфология Прибайкалья и впадины озера Байкал. — Новосибирск, 1978.
- 213 с.
Малый практикум по низшим растениям / Н.П. Горбунова, Е.С. Клюшникова, Н.А. Ко-
марницкий и др. — М., 1976. — 216 с.
Матвиенко А.М. Определитель пресноводных водорослей СССР. Золотистые водорос-
ли. — М., 1954. — Вып.З. — 188 с.
Матвшэнко О.М. Визначник пр!сноводних водоростей Украшсько! РСР. Золотист! во-
дорост! Chrysophyta. — Кшв, 1965. — Ч. 1. — 367 с.
Мац В.Д. Развитие Байкальской впадины: хронология. Трансформации зоогеографиче-
ских барьеров // Вторая Верещагинская Байкальская конференция: Тез. докл. и
стенд, сообщений. Иркутск, 1995. — С. 130-131.
Мейер К.И. Новые виды зеленых водорослей оз. Байкал // Ботан. материалы / Институт
споровых растений главного ботан. сада РСФСР. — 1922а. — Т. 1, вып. 1. — С. 13-15.
Мейер К.И. Материалы по флоре водорослей оз. Байкал // Журн. Моск, отд-ния Русск.
Ботан. о-ва. — М., 19226. — № 4. — С. 1-27.
Мейер К.И. О водорослях Северного Байкала // Русск. арх. протистологии. — М.,
1927. - Т. 4, вып. 1-4. - С. 93-118.
Мейер К.И., Рейнгард Л.В. К флоре водорослей оз.Байкала и Забайкалья // Бюлл.
МОИП, 1925. - Т. 33. - Вып.3-4. - С. 201-243.
Мейер К.И. Введение во флору водорослей озера Байкала // Бюл. МОИП. Отд. биоло-
гии. — М., 1930. — Т. 39, вып. 3-4. — С. 179-396.
Мейер К.И., Скабичевский А.П. Новый род из Cladophoraceae // Новости систематики
низших растений. — Л., 1965. — С. 41-44.
Мейер К.И., Скабичевский А.П. О систематическом положении видов рода Draparnaldia
Вогу из оз. Байкал // Новости систематики низших растений. — Л., 1969. —
Вып. 6. - С. 38-47.
Месяцев И., Зенкевич Л., Россолимо Л. Предварительный отчет о работах Байкальской
экспедиции Зоологического Музея Московского университета летом 1917 г. // Тр.
Комис, по изучению оз. Байкал. — 1922. — Т. 1, вып. 2. — С. 162-179.
Мошкова Н.О. Визначник прюноводних водоростей Украшсько! РСР. Улотриксов! во-
дорост! — Ulotrichales. Кладофоров! водорост! — Cladophorales. — Кшв, 1979. —
Т. 6. - 497 с.
Мошкова Н.А., Голлербах М.М. Определитель пресноводных водорослей СССР. Зеле-
ные водоросли. Класс Улотриксовые. — Л., 1986. — Вып. 10(1). — 360 с.
Библиографический список
241
Музафаров А.М., Эргашев А.Э., Халилов С. Определитель синезеленых водорослей
Средней Азии. — Ташкент, 1988. — Т. 2. — С. 405-892.
Окунева Г.Л. Сезонные изменения мезобентоса на каменистой литорали (район
пос. Большие Коты) // Продуктивность Байкала и антропогенные изменения его
природы. — Иркутск, 1974. — С. 137-152.
Окунева Г.Л. Мезо- и микробентос в районе Больших Котов (Южный Байкал) // Новые
материалы по фауне и флоре Байкала. — Иркутск, 1976. — С. 116-142.
Окунева Г.Л. Гарпактициды озера Байкал. — Иркутск, 1989. — 150 с.
Ошаров А.Б., Назарова Г.Д., Мыльникова Э.В., Алексеева Н.Р. Влияние промстоков
БЦБК на физиологические показатели бентосных водорослей // Экология Южного
Байкала. — Иркутск, 1983. — С. 268-275.
Патрикеева Г.И. Донные отложения Малого Моря // Тр. Байкал, лимнол. ст. АН
СССР. - М.; Л., 1959. - Т. 17. - С. 205-254.
Паутова В.Н. Высшая водная растительность на мелководьях озера Байкал // Кругово-
рот вещества и энергии в озерах и водохранилищах. Краткое содержание докла-
дов. — Иркутск, 1973. — Вып. 1. — С. 142-143.
Паутова В.Н. Высшая водная растительность на мелководьях озера Байкал // Кругово-
рот вещества и энергии в водоемах. — Новосибирск, 1975. — С. 142-144.
Поповская Г.И., Скабичевский А.П. О распределении донной растительности Байкала //
Докл. АН СССР. - 1968. - Т. 181, № 3. - С. 723-726.
Порфирьева Н.А. Планарии озера Байкал. — Новосибирск, 1977. — 206 с.
Распопов И.М. Высшая водная растительность больших озер Северо-Запада СССР. —
Л., 1985. - 199 с.
Развозжаев М.С. Всасывающий эрлифтный пробоотборник // Открытия, изобрете-
ния. - 1988. - № 36. - С. 152. - 1427210.
Россолимо Л.Л. Некоторые черты температурного режима Малого Моря // Тр. Байкал,
лимнол. ст. АН СССР. - М.; Л., 1959. - Т. 7. - С. 7-33.
Рундина Л.А. Зигнемовые водоросли России (Chlorophyta: Zygnematophyceae, Zygnema-
tales). - СПб., 1998 - 451 с.
Руководство по методам гидробиологического анализа поверхностных вод и донных от-
ложений / Под ред. В.А. Абакумова. — Л., 1983. — 239 с.
Скабичевский А.П. О новых зеленых водорослях из оз.Байкал // Изв. Биол.-геогр. ин-та
при Иркут, ун-те. — 1931. — Т. 5, вып. 4. — С. 69-76.
Скабичевский А.П. О распределении донной растительности в губах Аяя, Фролиха и Ла-
канда на Байкале // Изв. Биол.-геогр. НИИ при Иркут, ун-те. — 1934. — Т. 4,
вып. 1. — С. 103-115.
Скабичевский А.П. Заметки по альгологии Байкала // Изв. Биол.-геогр. НИИ при Ир-
кут. ун-те. — 1936. — Т. 7, вып. 1-2. — С. 32-41.
Скабичевский А.П. К вопросу о половом процессе у Tetraspora cylindrica Ag. var. bullosa
C. Meyer // Ботан. журн. — 1937. — T. 22, № 6. — С. 546-553.
Скабичевский А.П. О некоторых водорослях семейства Cladophoraceae из Байкала // Но-
вости систематики низших растений. — М.; Л., 1965. — С. 44-50.
Скабичевский А.П. О распределении донной растительности Байкала в окрестностях Бо-
льших Котов // Бюл. МОИП. Отд. биол. — 1966. — Т. 71, № 6. — С. 108-119.
Скабичевский А.П. Изменение названия рода Cladochaete // Новости систематики низ-
ших растений. — М.; Л., 1968. — С. 73-74.
Скабичевский А.П. Основные черты распределения донных водорослей Байкала // Вто-
рое совещ. по вопросам круговорота вещества и энергии в озерных водоемах. — Ли-
ственичное-на-Байкале. — 1969а. — Ч. 2. — С. 17-18.
Скабичевский А.П. Новый вид рода Chaetomorpha Kiitz. // Новости систематики низших
растений. — М.; Л., 19696. — С. 47-52.
Скабичевский А.П. К морфологии Chaetocladiella pumila (С. Meyer) С. Meyer et Ska-
bitsch. // Новости систематики низших растений. — М.; Л., 1970. — С. 27-32.
Скабичевский А.П. Основные черты распределения донной (водорослевой) раститель-
ности Байкала и методы исследования в продуктивности // Круговорот вещества и
энергии в озерных водоемах. — Новосибирск, 1975. — С. 102-107.
242
Библиографический список
Скабичевский А.П. Изменение названия рода Draparnaldiella С. Meyer et Skabitsch. и во-
прос о систематическом положении его видов // Бюл. МОИП. Отд. биол. — 1976а. —
№ 5. - С. 114-115.
Скабичевский А.П. Семейство кладофоровых во флоре озера Байкал // Бюл. МОИП.
Отд. биол. - 19766. - Т. 81, № 2. - С. 139-143.
Скабичевский А.П. Особенности состава и систематических взаимоотношений водорос-
лей Байкала // Круговорот вещества и энергии в водоемах. Элементы биотического
круговорота. Некоторые вопросы систематики. — Лиственичное-на-Байкале,
1977. - С. 325-328.
Скабичевский А.П. Флора и растительность // Проблемы Байкала. — Новосибирск,
1978. - С. 146-157.
Скабичевский А.П. Виды рода Gomphonema Ag. (Bacillariophyta) озера Байкал // Новости
систематики низших растений. — М.; Л., 1984. — С. 51-62.
Скабичевский А.П., Вотинцев К.К., Флоренсов Н.А. К истории изучения фитопланктона
и донных водорослей Байкала // Бюл. МОИП. Отд. биол. — 1979. — Т. 84, вып. 1.
Состояние макрофитобентоса в районе Больших Котов (Южный Байкал) по данным
1982-1983 гг. / Л.А. Ижболдина, В.М. Максимова, В.И. Семейкин и др. — Иркутск,
1984. - 20 с. - Рук. деп. в ВИНИТИ 13.3.85, № 1804-85.
Сгаробогатов Я.И. Фауна моллюсков и зоогеографическое районирование континенталь-
ных водоемов земного шара. — Л., 1970. — 370 с.
Топачевский А.В., Масюк Н.П. Пресноводные водоросли Украинской ССР. — Киев,
1984. - 333 с.
Толмачев В.А. О гидрохимии Малого Моря // Тр. Байкал, лимнол. ст. АН СССР. — М.;
Л., 1959. -Т. 17. - С. 137-191.
Фиалков В.А. Течения прибрежной зоны озера Байкал. — Новосибирск, 1983. — 191 с.
Шимараев М.Н. Элементы теплового режима оз.Байкал. — Новосибирск, 1977. — 147 с.
Элементный состав массовых видов макрофитов озера Байкал / О.М. Кожова, Л.А. Иж-
болдина, В.И. Резчиков и др. — М., 1994. — 22 с. — Деп. ВИНИТИ 3.08.94,
№ 2108-В.94.
Яснитский В.Н. Некоторые результаты гидробиологических исследований на Байкале
летом 1925 г. // Докл. АН СССР. - 1928. - № 18-19. - С. 353-358.
Яснитский В.Н. К вопросу о морфологической и физиологической дифференцировке
полов у некоторых видов Draparnaldia Вогу // Бюл. МОИП. Отд. биол. — 1934. —
Т. 43, № 2. - С. 171-186.
Яснитский В.Н. Явление гигантизма во флоре озера Байкал // Изв. Биол.-геогр. НИИ
при Иркут, ун-те. — 1952. — Т. 13, вып. 2. — С. 3-8.
Forest M.S. The remarkable Draparnaldia species of lake Baical, Siberia. — “Castanea” // J.
southern Appalachian Botan. Club. Norman. — 1957. — Vol. 22, N 4. — P. 126-132.
Hoek C. Van den. Revision of the European species of Cladophora. — Leiden, 1963. — 248 p.
Meyer K.I. Alge nonnulae novae baicalensis 2 // Not. Sist. Inst. Crypt. Hort. Bot. Petropol. —
1924. - N 3. - P. 107-108.
Meyer K.I. Sur 1’endemisme de la flore algologique du lac Baical. Rev. Algologique. — 1925a. —
Bd 5, N 2. - S. 110-117.
Meyer K.I. Untersuchungen uber die Algenflora des Baicalsees // Der Deutsch. Bot. Gesell-
schaft Ber. - 1926. - Bd 44, N 7. - S. 410-419.
Meyer K.I. Einige neue Algenformen aus dem Baicalsee // Arch. Protistenkunde. — 1930a. —
Bd 72, N 2. - S. 44-51.
Pascher A. Die Siisswasser-flora Deutschlands, Osterreichs und der Schweiz. Chrysophyceae
III. Ulotrichales, Microsporales, Oedogoniales. — Jena, 1914. — H. 6. — 226 S.
Starmach K. Chlorophyta. Ill: Zielenice nitkowate; Ulothrichales, Ulvales, Prasiolales, Sphae-
ropleales, Cladophorales, Chaetophorales, Trentepohliales, Siphonales, Dichotomosip-
honales (With Keys for the identification of filamentous green algae mentioned in the
Volume). - Warszawa; Krakow, 1972. - 750 S.
СЛОВАРЬ ТЕРМИНОВ
Акинеты — клетки с толстыми оболочками и большим количеством запас-
ных веществ, служащие для бесполого размножения и переживания небла-
гоприятных условий.
Антеридий (гаметангий) орган, в котором развиваются антерозоиды —
мужские гаметы, снабженные жгутиками.
Апланоспоры — клетки без жгутов, выполняющие роль зооспор.
Ауксилярные клетки — клетки, богатые питательными веществами, из кото-
рых развивается гонимобласт (после слияния карпогона с А.к. у красных водо-
рослей).
Ауксоспора — спора роста, спора полового размножения, образующаяся в
результате копуляции вышедших из створок панцирей двух протопластов диа-
томовых водорослей, а затем прорастающая в клетки, резко отличающиеся по
размерам от исходных.
Ареолы — регулярно повторяющиеся мельчайшие отверстия в стенках пан-
циря диатомовых водорослей.
Боковые веточки — невысокие (до 1 мм, реже до 2 мм высотой) разветвлен-
ные веточки, располагающиеся обычно мутовками вокруг клеток ствола и
крупных ветвей у зеленых водорослей родов Draparnaldia и Draparnaldioides.
Боковая конъюгация — способ полового процесса у зеленых водорослей
класса Zygnematophyceae, характеризующийся тем, что содержимое мужской
клетки переливается в соседнюю женскую клетку той же нити через образую-
щийся конъюгационный канал, где оба протопласта сливаются, образуя зиготу.
Влагалище — слизистое или студенистое образование у многих синезеле-
ных водорослей, заключающее в форме чехла или футляра один или несколько
трихомов.
Газовые вакуоли — полости в протоплазме клеток синезеленых водорослей,
наполненные азотом и придающие клетке в проходящем свете микроскопа чер-
но-бурый или почти черный цвет.
Гаметы — половые клетки со жгутиками или без них, сливающиеся при
оплодотворении.
Гаметангии — половые органы растений, в которых развиваются гаметы.
Гетероцисты — характерные для нитчатых синезеленых водорослей своеоб-
разные клетки, образующиеся из вегетативных клеток в разных местах трихома,
отличающиеся хорошо выраженной двухслойной оболочкой и содержимым,
244
Словарь терминов
всегда лишенным ассимиляционных пигментов, газовых вакуолей и зерен за-
пасных веществ.
Гипноспора — спора водорослей, покрывающаяся особо утолщенной обо-
лочкой внутри материнской клетки и способная длительный срок находиться в
состоянии покоя. В отличие от акинет оболочка материнской клетки не прини-
мает участия в образовании оболочки гипноспоры.
Гонимобласты — особые нитевидные выросты, развивающиеся из оплодо-
творенного карпогона красных водорослей и образующие на своих ответвле-
ниях карпоспорангии.
Гормоспоры (гормоцисты) — у синезеленых водорослей — однорядно- мно-
гоклеточные ’тельца, одетые в отличие от гормогониев толстой оболочкой,
представляющей собой разросшееся влагалище.
Гормогонии — фрагменты трихомов синезеленых водорослей, способные к
произвольному активному движению и прорастанию в новые особи; служат
для размножения.
Зигота — клетка, образующаяся в результате слияния двух клеток при по-
ловом процессе у различных водорослей.
Зигоспора — зигота с оформившимися слоями оболочки, приобретшая
определенную форму и цвет, заполнившаяся запасными питательными веще-
ствами, в частности большим количеством масла.
Карпогон — одноклеточный женский половой орган красных водорослей
(оогоний) в виде бутылочки с вытянутой верхней (трихогиной) и раздутой
нижней (брюшной) частью, в которой помещается одно женское ядро (диф-
ференцированной яйцеклетки нет).
Карпоспоры — споры у красных водорослей, образующиеся из зиготы и да-
ющие начало новому растению. К. представляют собой голые, лишенные жгу-
тиков клетки, обычно неподвижные.
Конъюгация — форма полового процесса, заключающаяся в слиянии про-
топластов двух равноценных клеток, не имеющих морфологической диффе-
ренцировки на мужские и женские половые элементы.
Коронка оогония — образование на верхушке оогония харовых водорослей,
представляющее собой сросшиеся верхушки коровых клеток, спиралевидно
обрастающих яйцеклетку.
Мезоспорий — средняя оболочка зигоспор у зигнемовых водорослей. В ме-
зоспории содержатся хитин, спорополленин, хромовая и плавиковая кислоты,
он устойчив к ферментам и ацетолизу.
Оогамия — тип полового процесса, при котором крупная и неподвижная
женская половая клетка (яйцеклетка) оплодотворяется небольшой и обычно
подвижной мужской половой клеткой.
Оогоний — женский половой орган многих водорослей, обычно однокле-
точный и лишь у харовых водорослей многоклеточный.
Ооспора — зигота с многослойной оболочкой, возникшая в результате
оогамного полового процесса.
Партеноспора — спора, развившаяся из диплоидной неоплодотворенной
яйцеклетки без редукционного деления ядра.
Перидерм — уплотненный поверхностный слой слизи, окружающий коло-
нии некоторых синезеленых водорослей из рода Nostoc.
Пиреноид — клеточная органелла многих водорослей, представляют собой
бесцветные плотные образования белковой природы, расположены внутри
хлоропласта или на нем.
Словарь терминов
245
Полисифонный — многорядный, сложенный из члеников, которые состоят
из длинных клеток (сифонов).
Псевдоцилии — длинные, очень тонкие выросты клеток, в отличие от жгу-
тиков неподвижны; встречаются у представителей рода Tetraspora и др.
Рахис — главная ось боковой веточки у зеленых водорослей родов Drapar-
naldia и Draparnaldioides.
Ризоиды — волосковидные образования из одной или нескольких клеток,
расположенных в один ряд; служат для прикрепления таллома к субстрату.
Септы — неполные кремниевые перегородки у диатомовых водорослей,
образующиеся в результате разрастания вставочного ободка в полость клетки.
Спора — специальная клетка, служащая для бесполого размножения и пе-
реживания неблагоприятных условий.
Сперматангии — клетки, функционирующие как мужские органы размно-
жения у красных водорослей, в которых образуются спермации.
Спермации — безжгутиковые пассивно-подвижные мужские половые клет-
ки (встречаются у некоторых водорослей).
Таллом (слоевище) — особая форма тела у низших растений, характеризую-
щаяся отсутствием разделения на типичные органы высших растений (стебель,
корень и лист).
Трихомы — соединение клеток в однорядные или реже многорядные ните-
видные образования у синезеленых водорослей из класса Hormogoniophyceae.
Трихогина — узкая верхняя часть женских половых органов красных водо-
рослей (карпогона), обычно удлиненная, трубчатая, способствует удержанию
прилипающих к ней спермациев.
Хроматоплазма — окрашенная периферическая часть протопласта клеток
синезеленых водорослей, где сосредоточены пигменты.
Хлоропласты — внутриклеточные органеллы растительной клетки — зеле-
ные пластиды, в которых осуществляется процесс фотосинтеза.
Центроплазма — центральная часть клеток синезеленых водорослей, в про-
топласте которых диффузно распределены хроматиновые и запасные вещества.
Центроплазма синезеленых водорослей является аналогом ядра.
Эвекция — ускоренный рост боковой ветви, принимающей положение и
облик главной нити, тогда как последняя смещается в сторону в форме боково-
го ветвления.
Экзоспоры — споры, развивающиеся на поверхности образующего их орга-
на и по мере созревания отчленяющиеся от него. Встречаются только у синезе-
леных водорослей класса Chamaesiphoneae.
Экзоспорий — наружная оболочка зигоспор, покрывающая эписпорий. Со-
стоит из пектина и целлюлозы.
Эндоспора — спора, формирующаяся эндогенно, часто внутри спорангиев.
Эндоспорий — тонкая внутренняя оболочка зигоспоры.
алфавитный указатель
ЛАТИНСКИХ НАЗВАНИЙ
Aphanothece (Nag.) Elenk. 30
— stagnina (Spreng.) В.-Peters, et Geitl.
emend. 30
Aulosira Kirchn. 41
— implexa Bom. et Flah. 41
Batrachospermum Roth 79
— moniliforme Roth 80
— vagum (Roth.) Ag. 79
Bulbochaete C. Ag. 171
— radinospora Wittr. sec. Him. 171
Calothrix (Ag.) V. Poijansk. sensu lat. 45
— braunii Bom. et Flah. 48
— kawraiskyi Schmidle. 48
— kossinskajae V. Poijansk. 46
— parietina (Nag.) Thur. 49
— stellaris Bom. et Flah. 48
Chara L. 219
— contraria A. Br. 220
— fragifera Durieu. 220
— tomentosa L. 223
Chaetocladiella Meyer et Skabitsch. 201
— litoralis (Skabitsch.) C. Meyer et Ska-
bitsch. 203
— microscopica (C. Meyer) C. Meyer et
Skabitsch. 201
— pumila (C. Meyer) C. Meyer et Ska-
bitsch. 204
Chaetomorpha Kiitz. 173
— baicalensis C. Meyer 180
— curta Skabitsch. 177
— moniliformis Skabitsch. 179
— solitaria Skabitsch. 175
Chaetophora Schrank. 90
— elegans (Roth) Ag. 90
Chrisothallus C. Meyer 74
— baicalensis C. Meyer 74
Cladophora Kiitz. 181
— aegagropila (L.) Rabenh. 194
— compacta (C. Meyer) C. Meyer 199
— floccosa C. Meyer 184
---var. floccosa 186
---var. irregularis Skabitsch. 187
— fracta (Miill. ex Vahl.) Kiitz. 195
---f. fracta 195
— globulina (Kiitz.) Kiitz. C. Meyer 194
— globulus C. Meyer 192
— glomerata (L.) Kiitz. 197
---var. glomerata 197
---var. crassiori (Ag.) Hoek. 198
— kursanovii Skabitsch. 198
— kusnetzowii C. Meyer 184
— meyeri Skabitsch. 188
---var. meyeri 190
---var. gracilior (C. Meyer) Hellerb.
192
— pulvinata (C. Meyer) C. Meyer 199
Coleochaete Breb. 167
— pulvinata 167
Didymosphenia M. Schmidt. 75
— geminata (Lyngb.) M. Schmidt. 76
— dentata Dorogostaisky 77
Draparnaldia Bory 165
— glomerata (Vauch.) Ag. 165
— plumosa (Vauch.) Ag. 165
Draparnaldioides C. Meyer et Skabitsch. 97
— arenaria C. Meyer et Skabitsch. 108
---f. arenaria 109, 110
---f. elegans Irhb. f. nov. 110
Алфавитный указатель латинских названий
247
---f. pilosissima Irhb. f. nov. 112, 114,
115
---f. plumosa C. Meyer 110
— arnoldii C. Meyer et Skabitsch. 112
------ f. arnoldii 116-118
---f. compacta C. Meyer 119-122
---f. gracilis C. Meyer 119
— baicalensis C. Meyer et Skabitsch. 140
— goroschankinii C. Meyer et Skabitsch.
103
---f. goroschankinii 103
---f. plumosa C. Meyer 106
— lubrica C. Meyer et Skabitsch. 152
---f. ramulifera 152
— pilosa C. Meyer et Skabitsch. 123
— pumila C. Meyer et Skabitsch. 119
— simplex C. Meyer et Skabitsch. 100,102
---f. simplex 100
---f. pentafurca C. Meyer 100
---f. trifurca C. Meyer 100
— vilosa C. Meyer et Skabitsch. 129
Fischerella (Bom. et Flah.) Gom. 31
— major Gom. 31
Gemmiphora Skabitsch. 172
— compacta Skabitsch. 173
Gloeothrichia Agardh. 50
— natans (Hedw.) Rabenh. 52
— pisum (Ag.) Thur. 52
Gomphonema Ag. 77
— quadripunctatum (Oestr.) Wislouch.
77
— firmum Skabitsch. 78
— ventricosum Greg. 78
Hydrurus Ag. 73
— foetidus (Vill.) Kirchn. 73
Ireksokonia C. Meyer 95
— formosa C. Meyer 95
Lyngbya Ag. 64
— aestuarii (Mert.) Liebm. 66
— major Menegh. 64
Microcoleus Desmaz. 67
— baicalensis Izhb. 68
— subtorulosus (Breb.) Gom. 67
Microcystis (Kiitz.) Elenk. 28
— ichthyoblabe Kiitz. 28
— muscicola (Menegh.) Elenk. 30
Myxonemopsis C. Meyer 97
— crassimembranacea C. Meyer 97
Mougeotia Ag. 214
— genuflexa (Dillw.) Ag. 214
---f. genuflexa 215
— laetevirens (A. Br.) Wittr. 215
Nitella Ag. emend A. Br., Leonh. 216
— flexilis (L.) Ag. 217
— opaca (Bruz.) Ag. 217
Oedogonium Link. 169
— flavescens (Hass.) Wittz. 169
— sociale Wittr. 169
— undulatum (Breb.) A. Br. 169
Oscillatoria Vauch. 55
— атоепа (Kiitz.) Gom. 58
— formosa Bory 59
— lacustris (Kleb.) Geitl. 56
— limosa Ag. 56
— sancta (Kiitz.) Gom. 56
— splendida Grev. 58
— tenuis Ag. 57
---f. tenuis 58
---f. tergestiana (Kiitz.) Elenk. 58
Oscillatoria sp. 1. 59
Phormidium 59
— ambiguum Gom. 62
— corium (Ag.) Gom. 63
— crouanii Gom. 64
— inundatum Kiitz. 63
— papiraceum (Ag.) Gom. 63
— purpurasceus (Kiitz.) Gom. 60
— tenue (Menegh.) Gom. 62
— uncinatum (Ag.) Gom. 62
Prasiola sp. 168
Rhizoclonium Kiitz. 180
— sp. 181
Rivularia (Roth.) Ag. emend Thur. 50
— aguatica (de Wild.) Geitl. sensu lat. 50
— borealis P. Richt. 50
Schizothrix (Kiitz.) Gom. 66
— sp. 67
Sphaeronostoc Elenk. 33
— coeruleum (Lyngb.) Elenk. 34
— kihlmani (Lemm.) Elenk. 33
— pruniforme (Ag.) Elenk. 35
— zetterstedtii (Aresch.) Elenk. 36
Sphaeroplea Ag. 205
— annulina (Roth) Ag. 205
Spirogyra Link. 209
— calospora Cleve 213
— decimina (Miill.) Kiitz. 210
---f. jurgensis (Kiitz.) V. Poijansk. 210
---f. longata (Vauch.) V. Poijansk. 212
— hassallii (Jenner) Petit 212
248
Алфавитный указатель латинских названий
— weberi Kiitz. 212
----f. weberi 212
Stigeoclonium Kiitz. 91
— amoenum Kiitz 94
— farctum Berth. 94
— flagelliferum Kiitz 94
— tenue (Ag.) Kiitz 91
Stratonostoc Elenk. 36
— commune (Vauch.) Elenk. 39
----f. sphaericum (Vauch.) Elenk. 39
— linckia (Roth.) Elenk. 37
----f. carneum (Ag.) Elenk. 38
----f. linckia 38
----f. piscinale (Kiitz ) Elenk. 38
----f. rivulare (Kiitz ) Elenk. 38
— verrucosum (Vauch.) Elenk. 39
Tetraspora Link. 82
— cylindrica (Wahl.) Ag. 83
----var. bullosa C. Meyer 83
----var. sp. 85
— lubrica (Roth.) Ag. 86
Tetrasporopsis Lemm. 70
— reticulata C. Meyer 70
— sp. 70
Tolypothrix Kiitz. 42
— distorta (Fl. Dan.) Kiitz. 44
----f. distorta 44
----f. penicillata (Ag.) Kossinsk. 45
— helicophyla Lemm. 43
— limbata Thur. 43
— tenuis Kiitz. 43
----f. polymorpha (Lemm.) Kossinsk. 44
Ulothrix Kiitz. 87
— tenuissima Kiitz. 87
— zonata (Web. et Mohr.) Kiitz. 87
----f. zonata 87
Zygnema Ag. 207
— cruciatum Vauch. Ag. 207
Научное издание
Ижболдина Людмила Александровна
АТЛАС И ОПРЕДЕЛИТЕЛЬ ВОДОРОСЛЕЙ БЕНТОСА
И ПЕРИФИТОНА ОЗЕРА БАЙКАЛ (МЕЙО- И МАКРОФИТЫ)
С КРАТКИМИ ОЧЕРКАМИ ПО ИХ ЭКОЛОГИИ
Редактор Н.А. Воронина
Художник И. Чевыкалов
Корректор Е.В. Литвиненко
Оператор электронной верстки В.Е. Постникова
Сдано в набор 10.02.05. Подписано в печать 28.11.06.. Бумага офсетная. Гарнитура Times ЕТ.
Формат 70 х 108Vi6. Уч.-изд. л. 20,7. Усл. печ. л. 21,7 + 0,5 л. вкл. на мел. бум.
Зак. 4, тираж 600 экз.
ООО Издательство “Наука-Центр”. 630090, Новосибирск, пр. акад. М.А. Лаврентьева, 6.
Тип. РПЦ «Вета», 630033, г. Новосибирск, ул. Мира, 62.