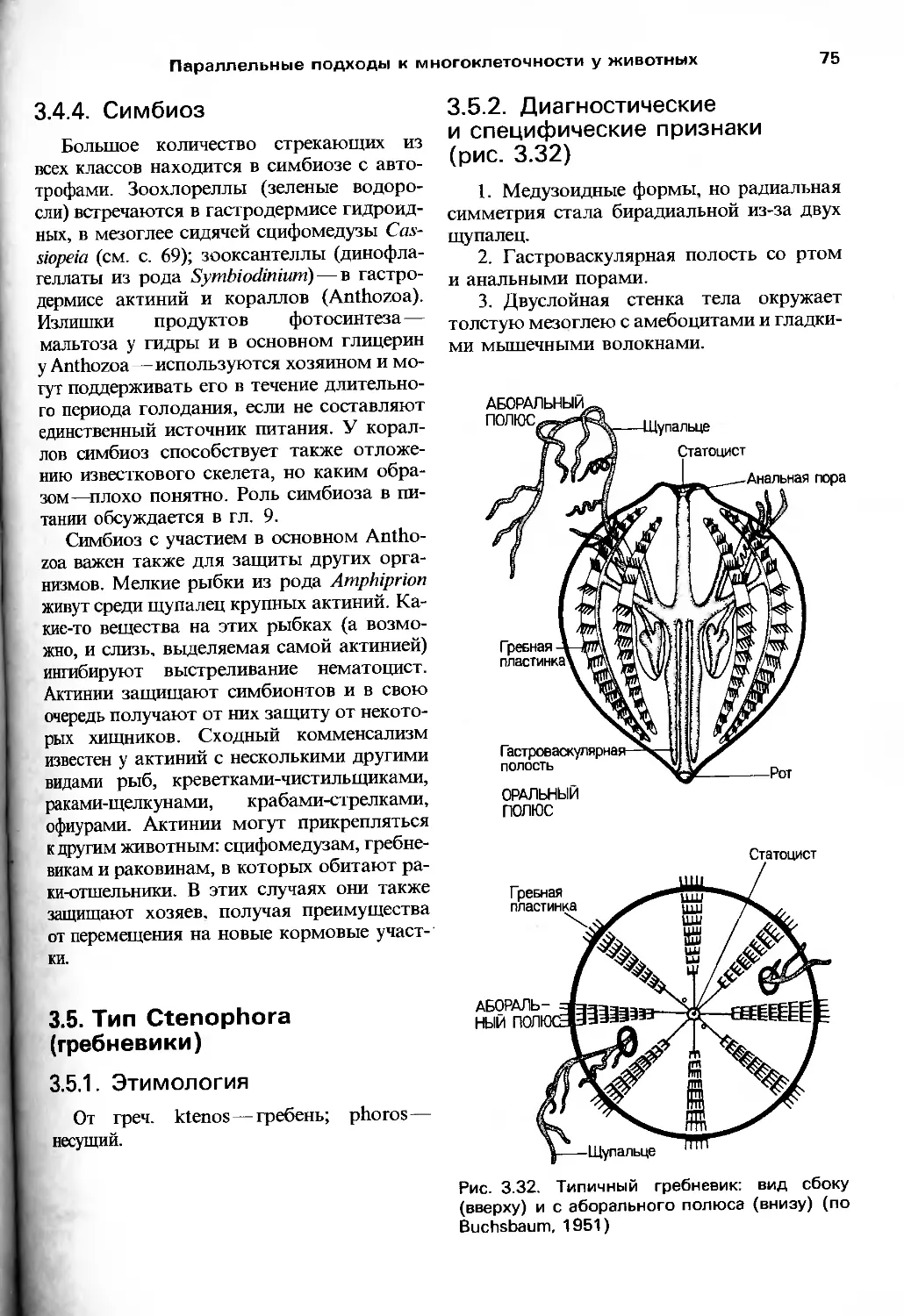
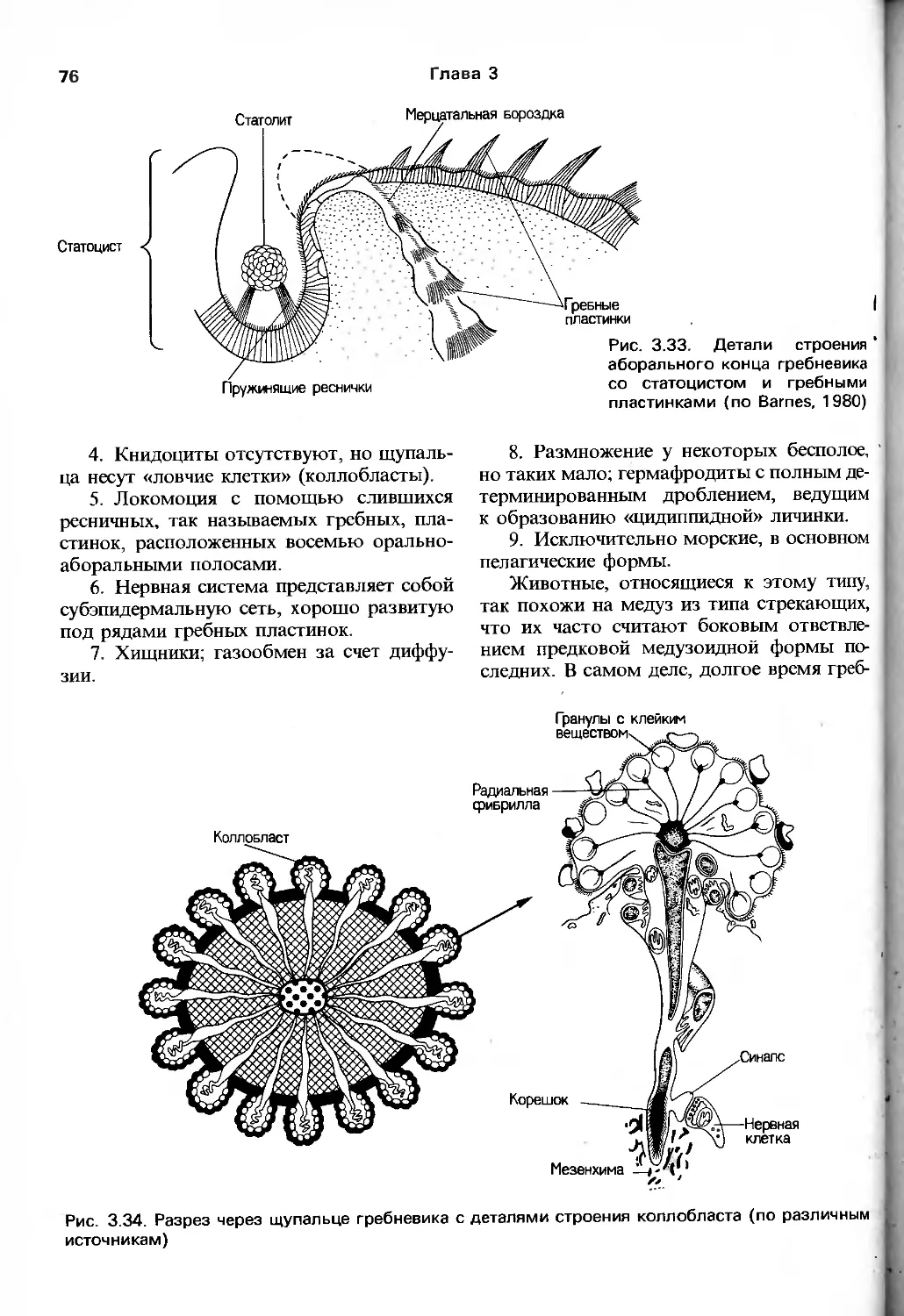
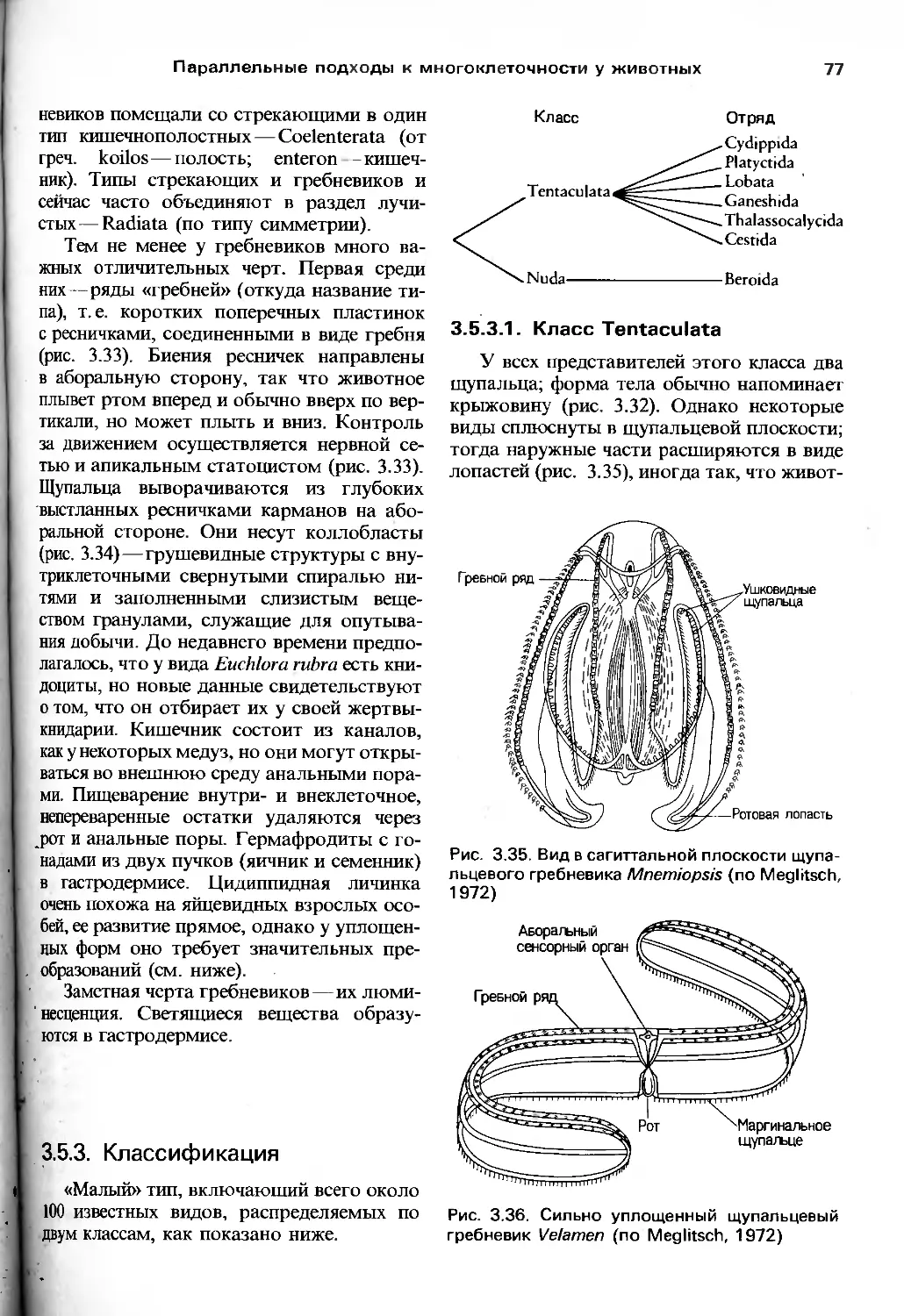
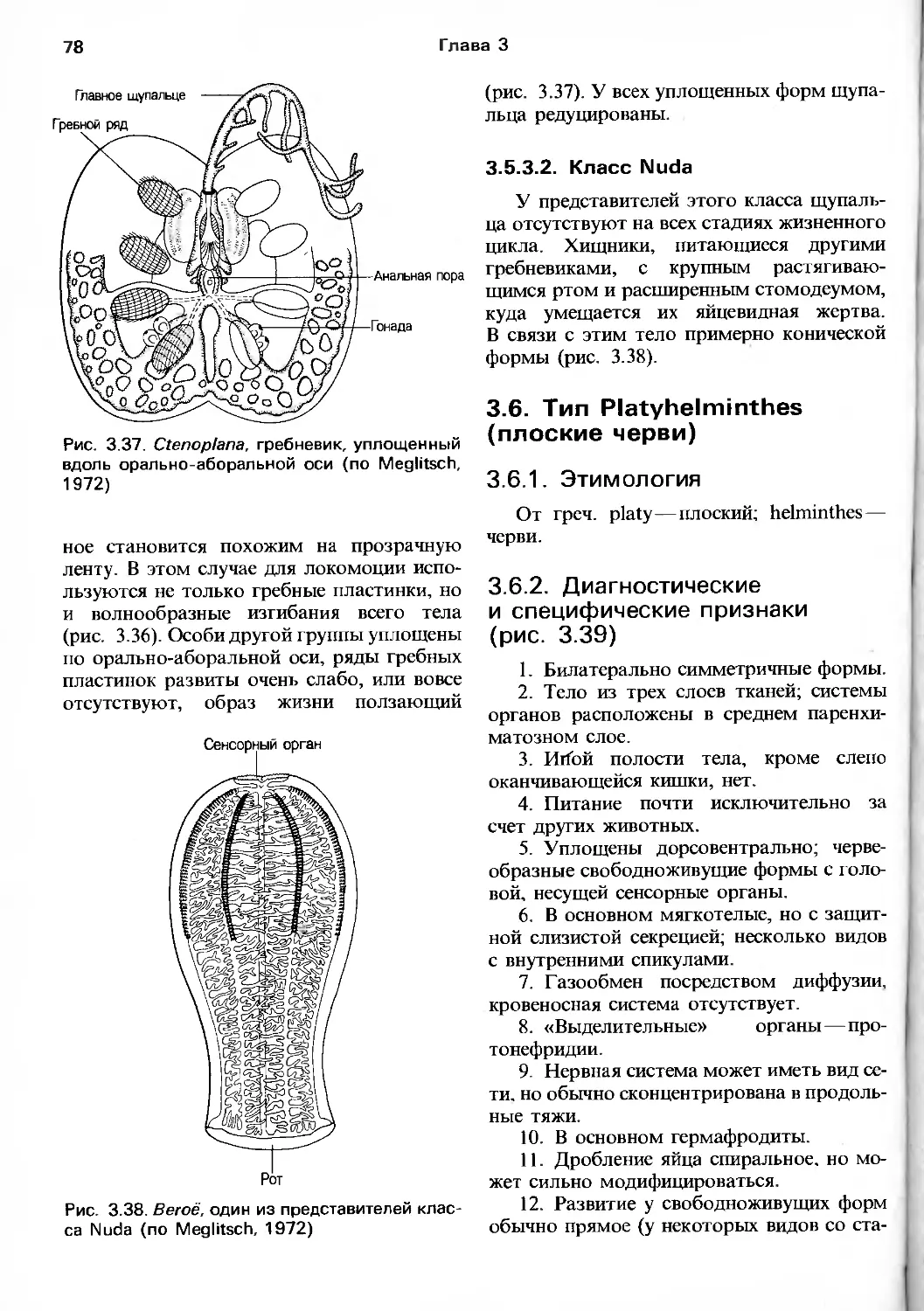
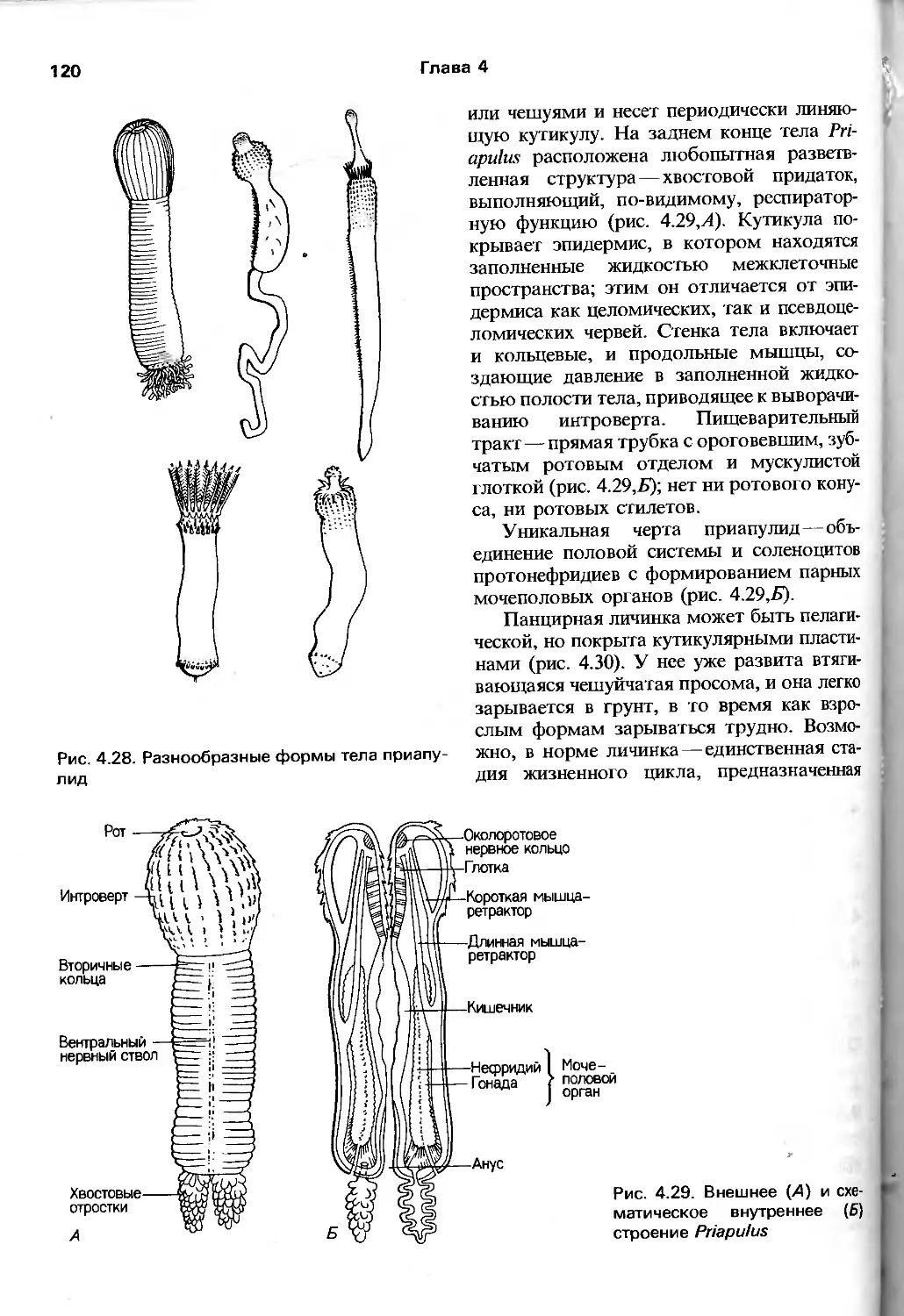
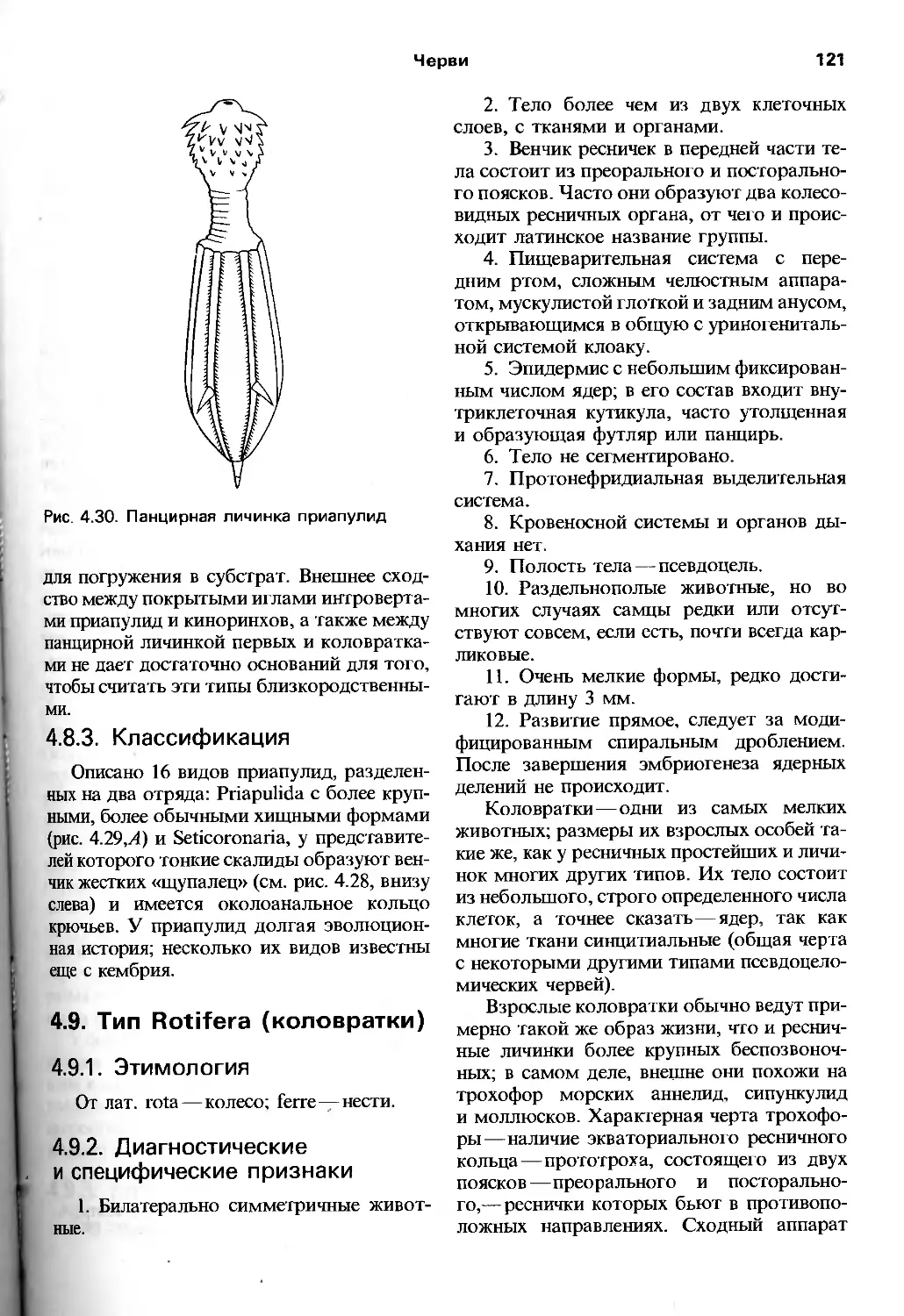
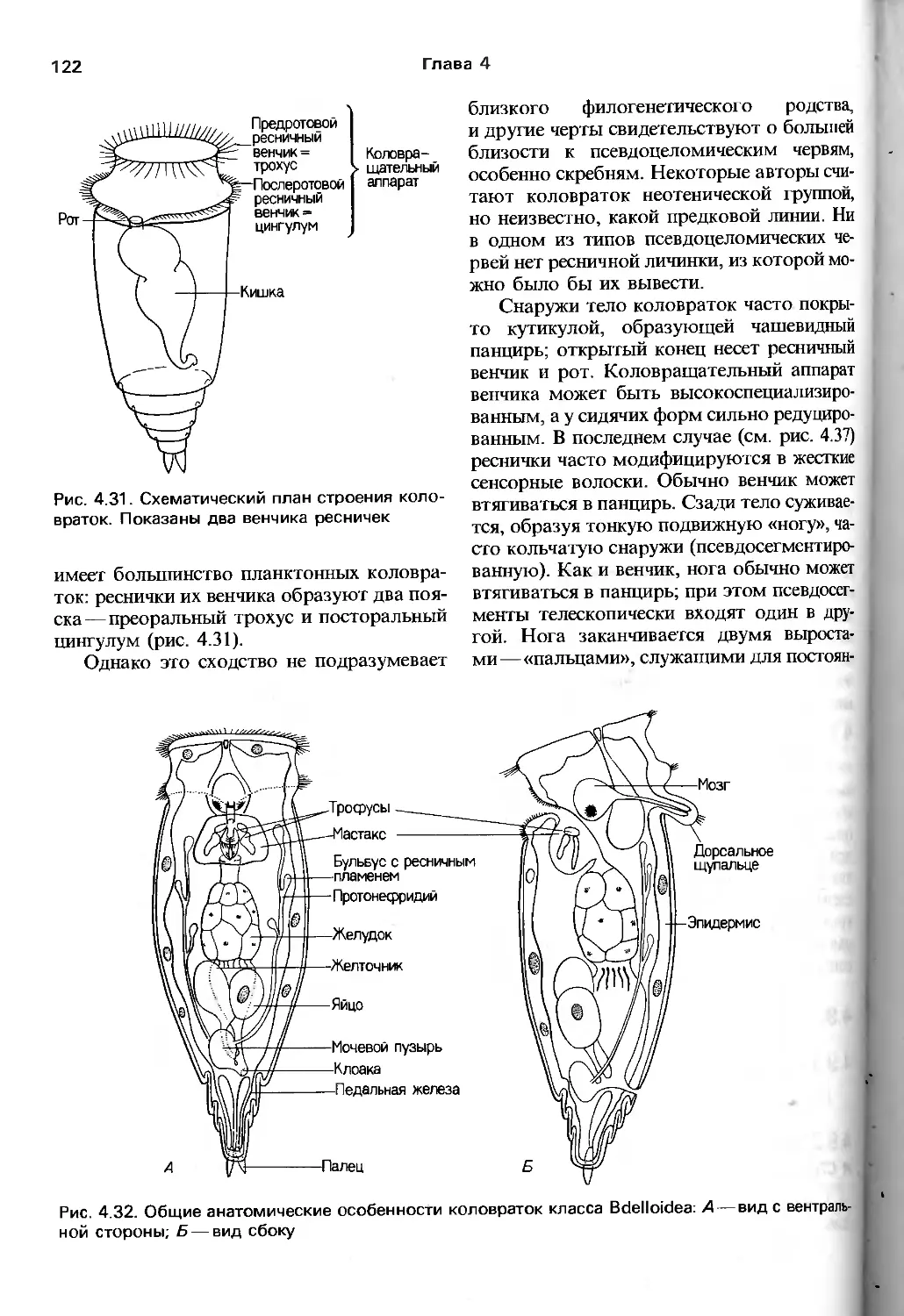
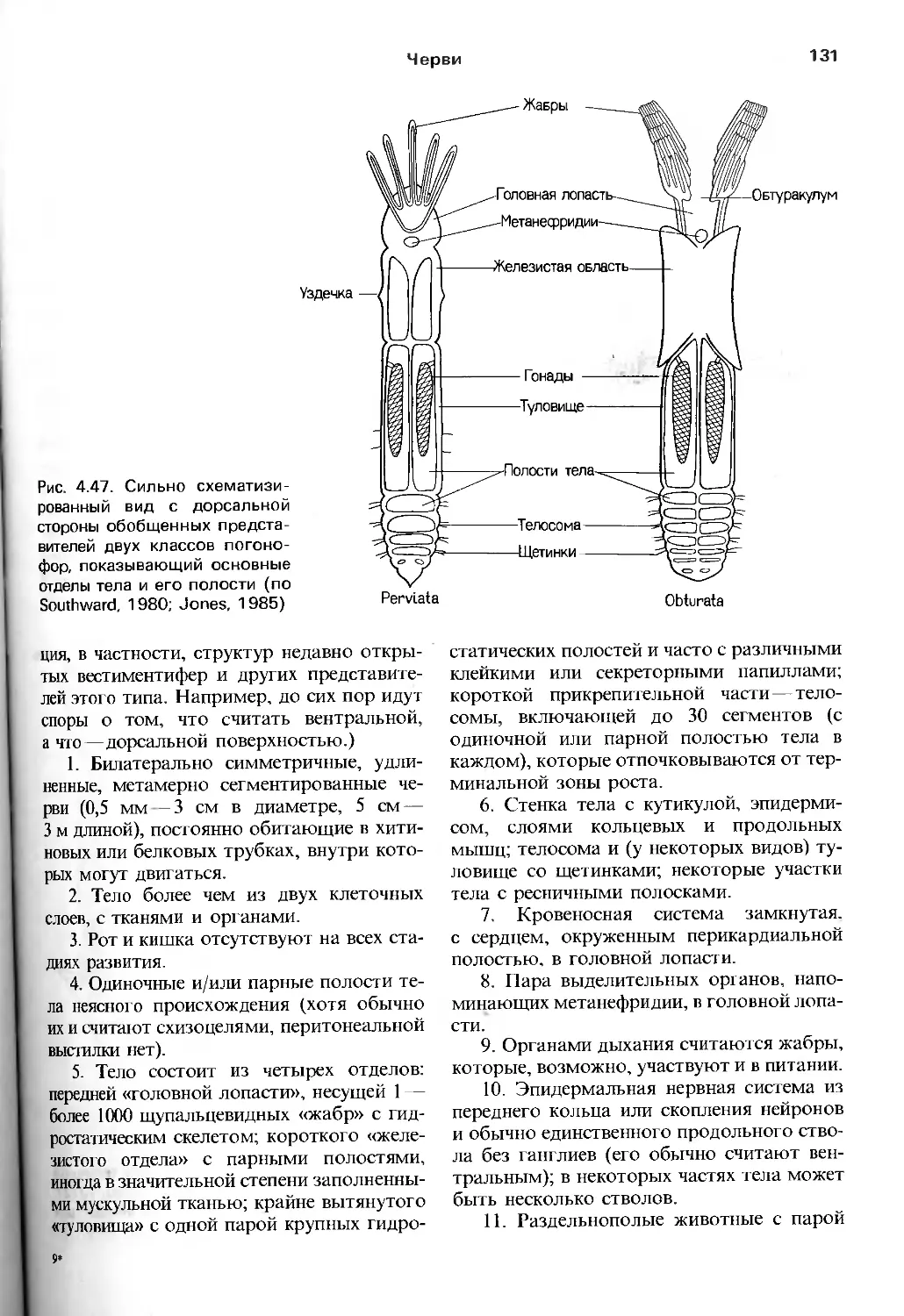
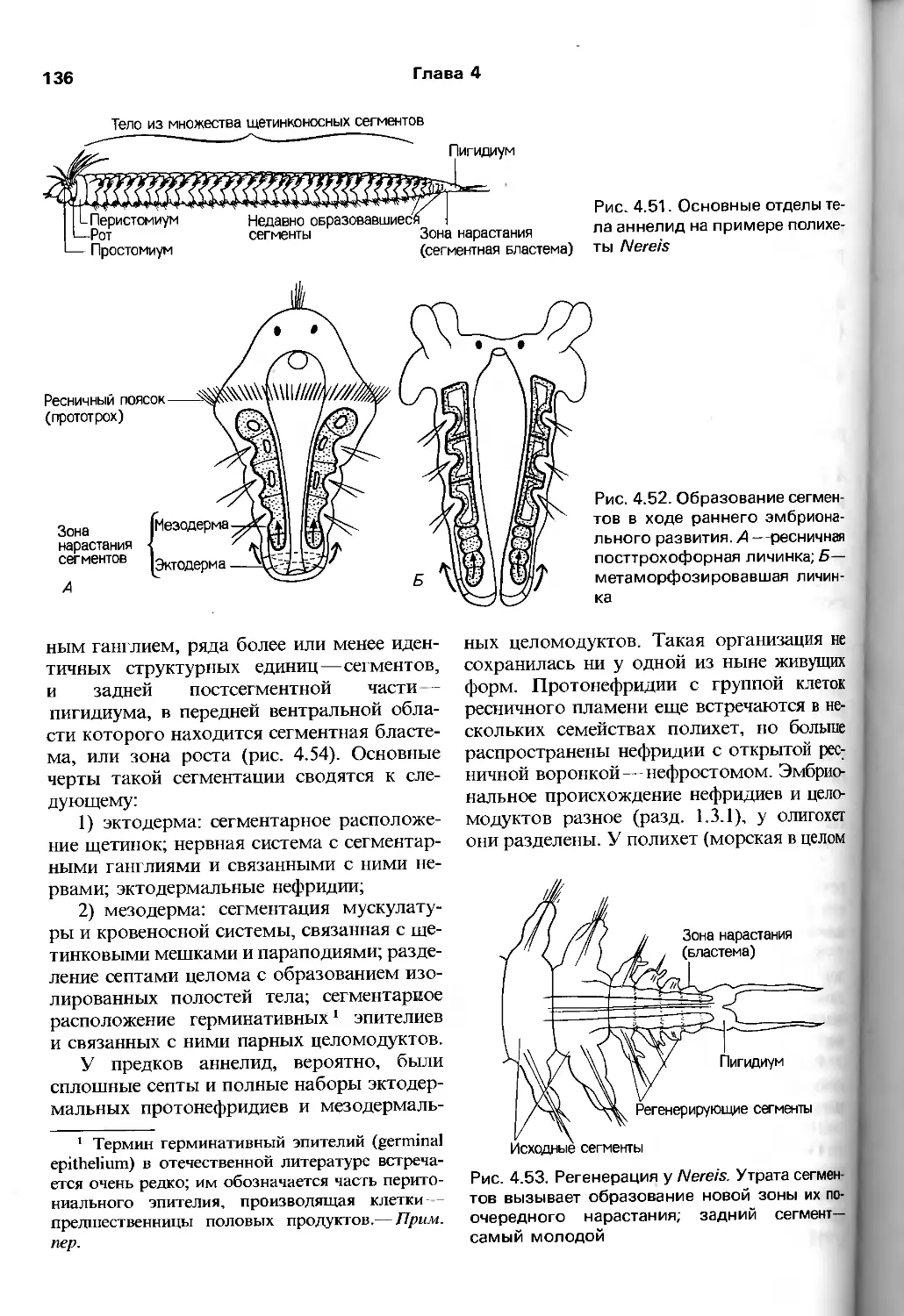
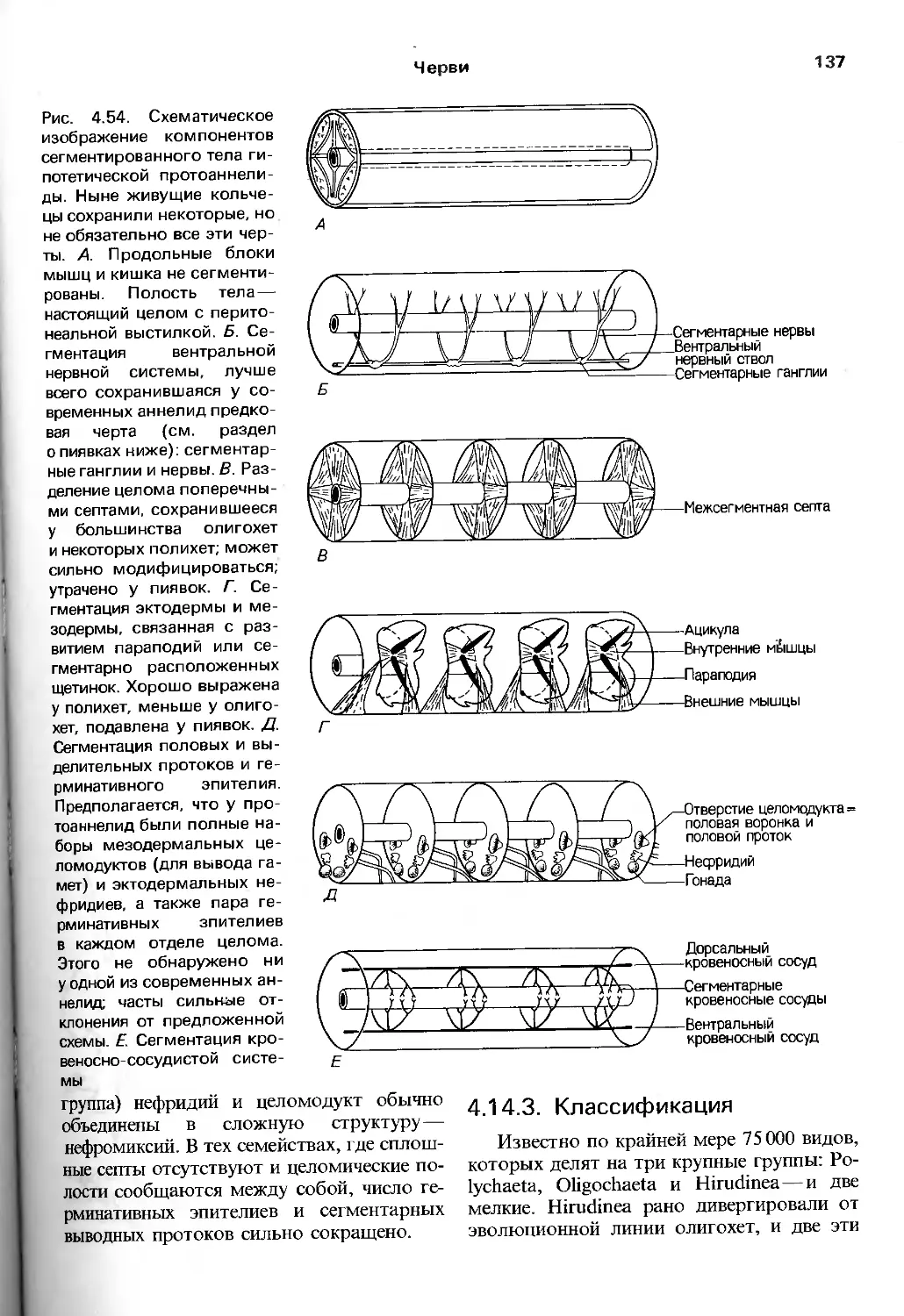
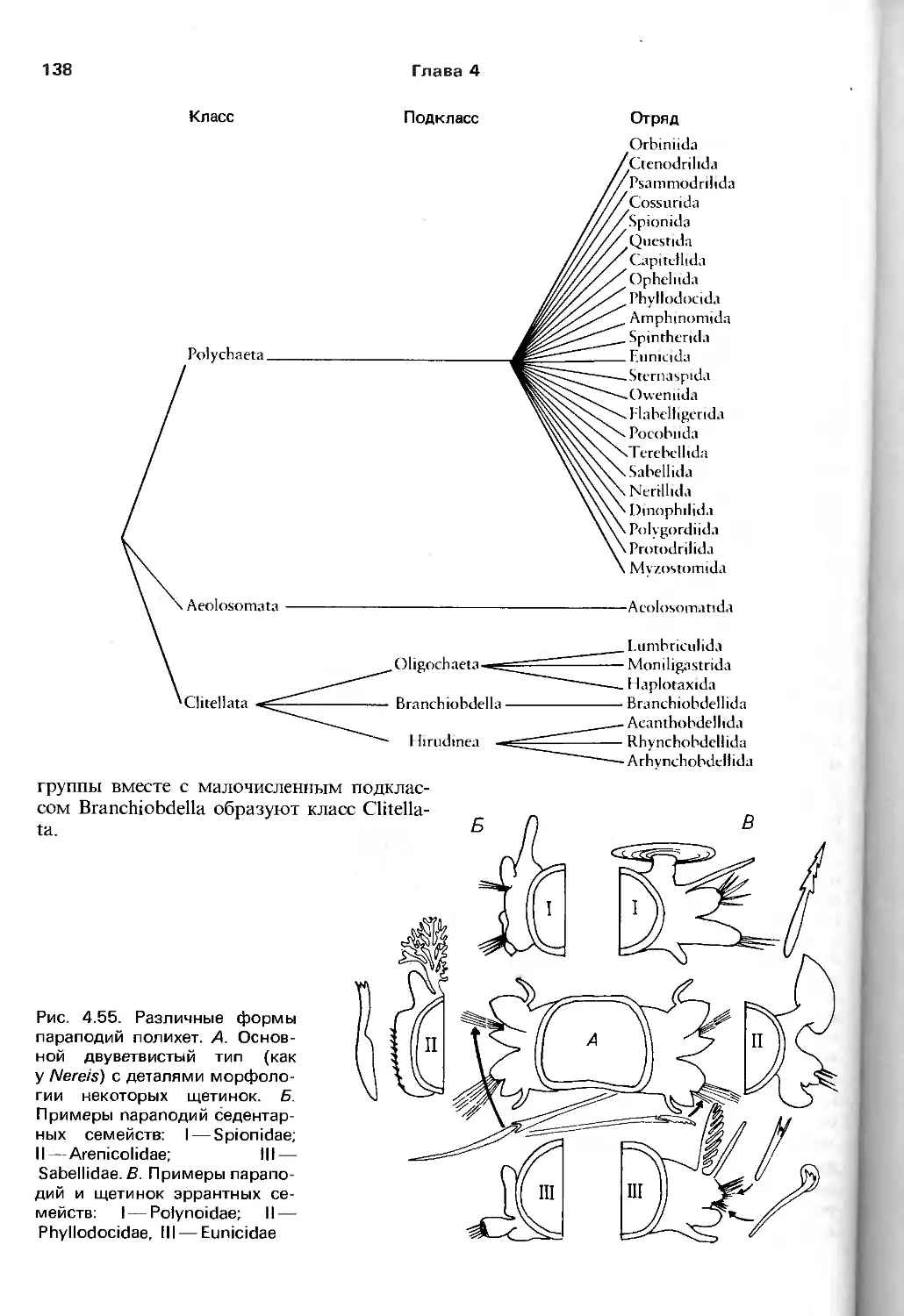
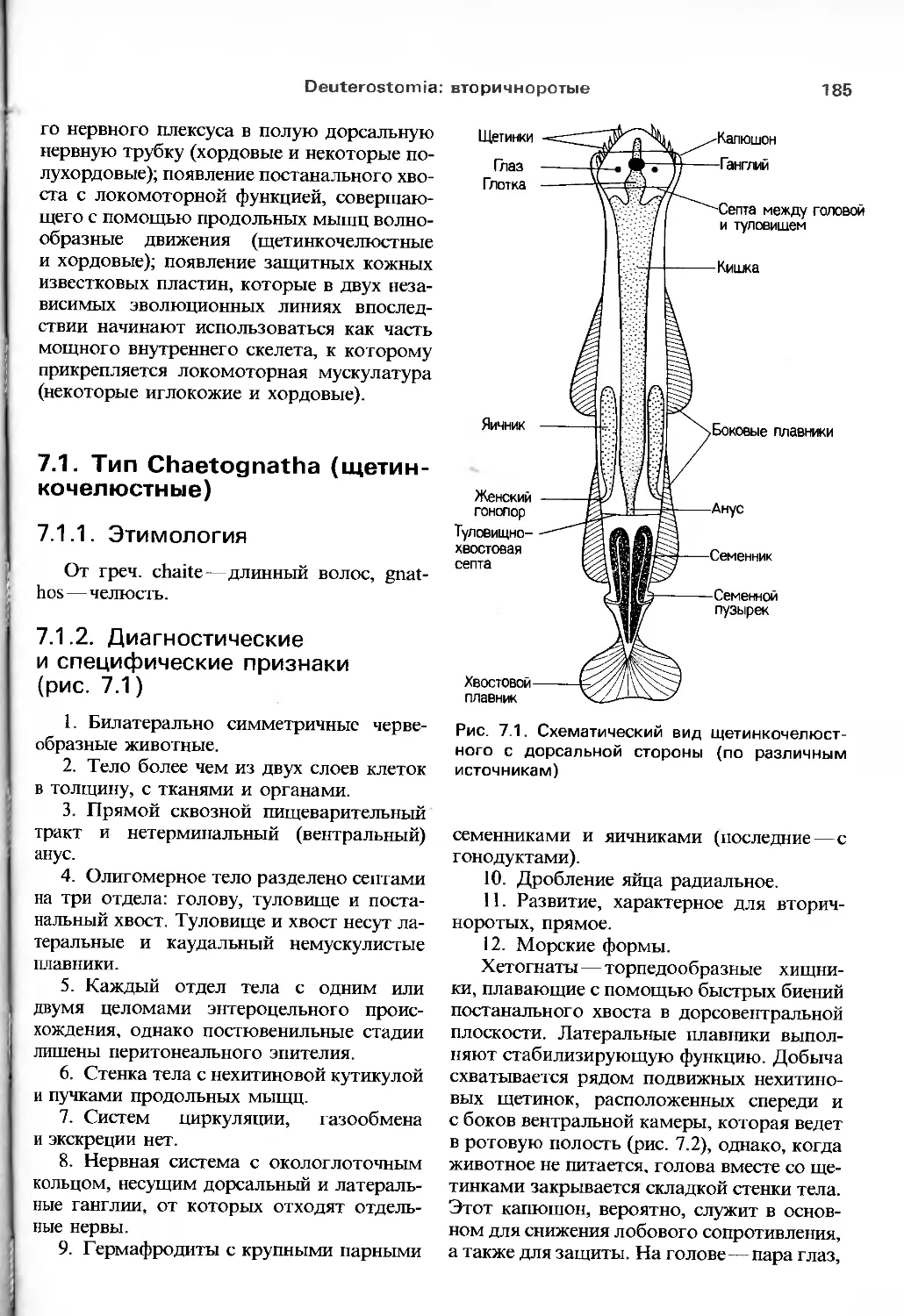
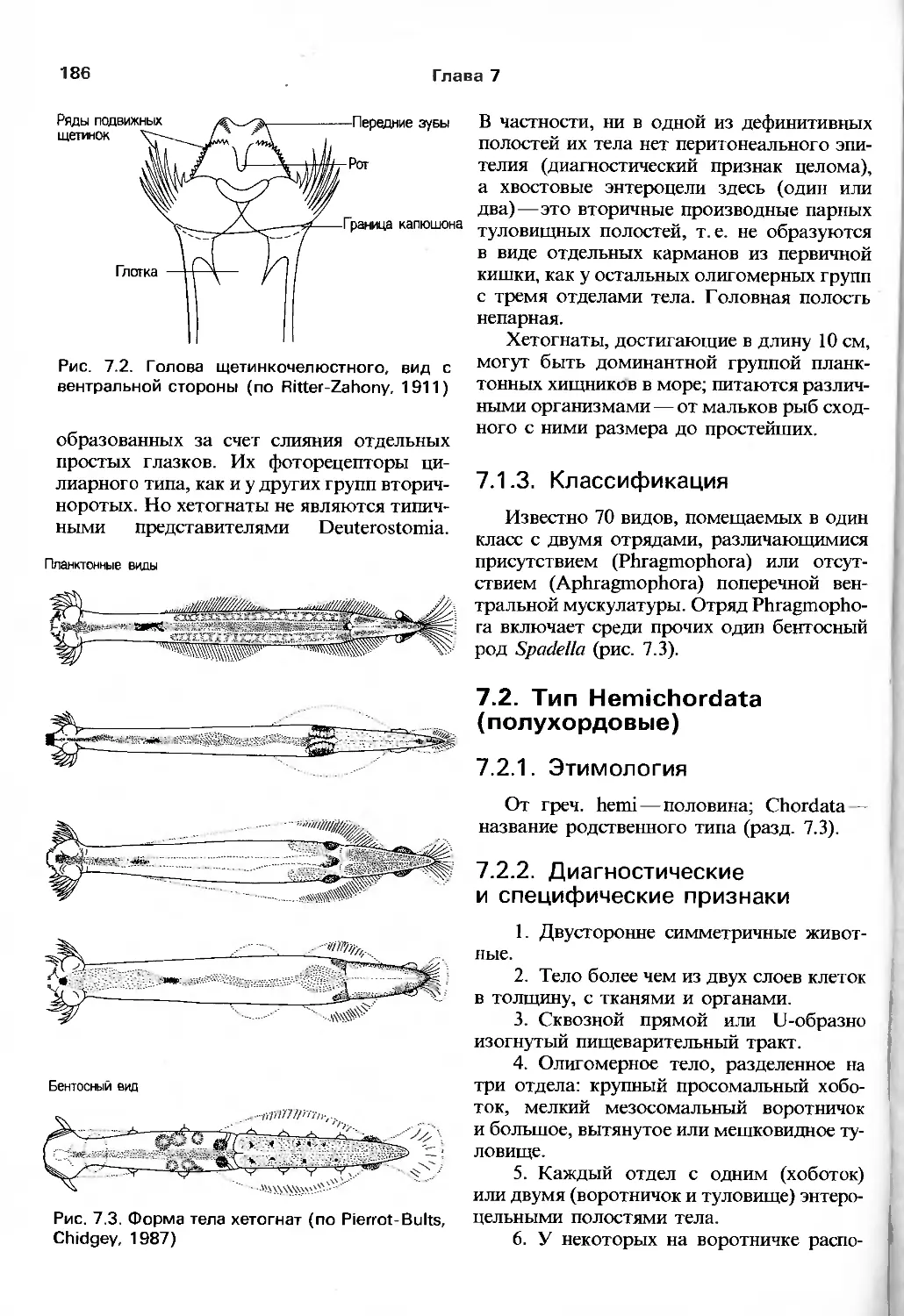
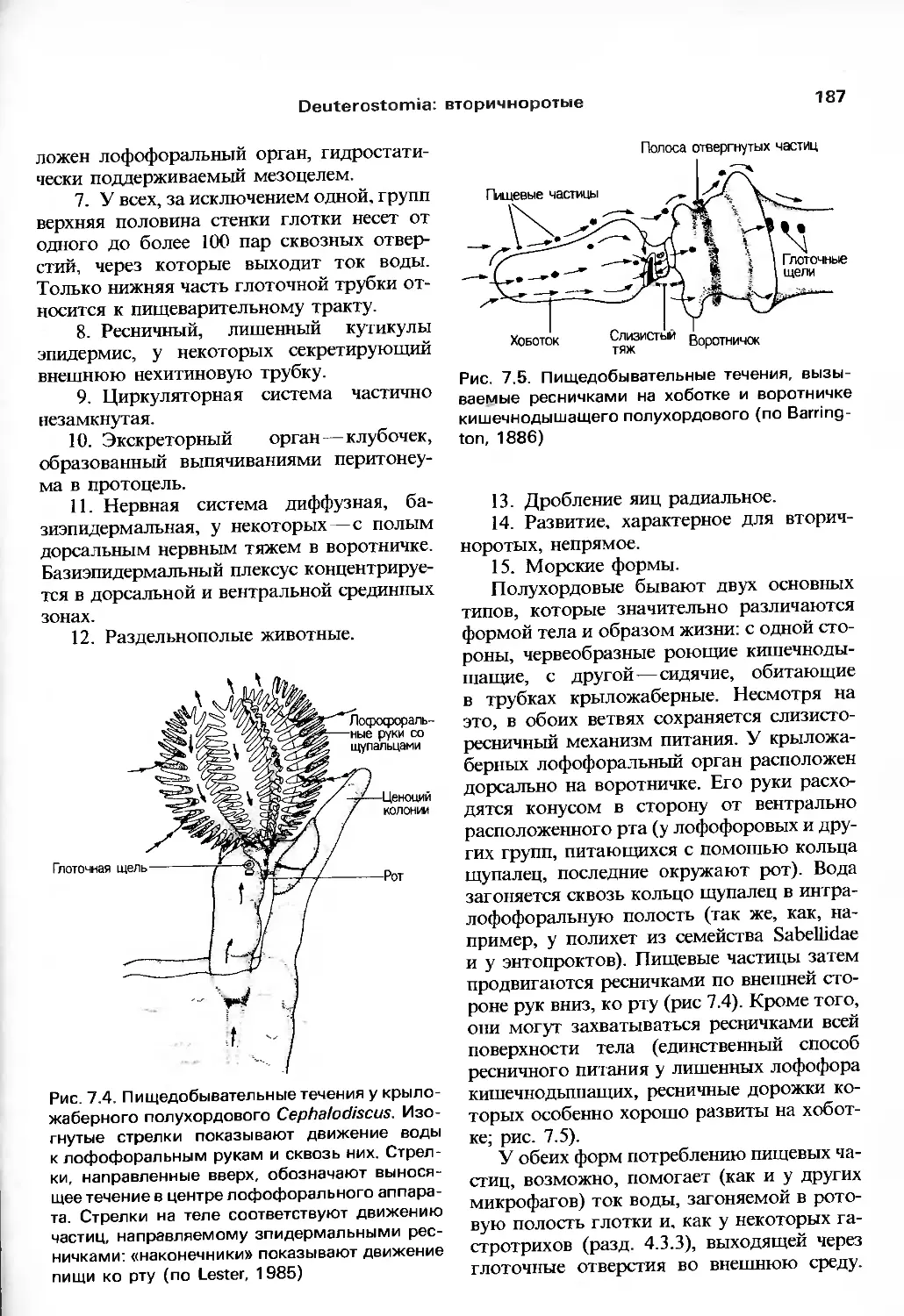
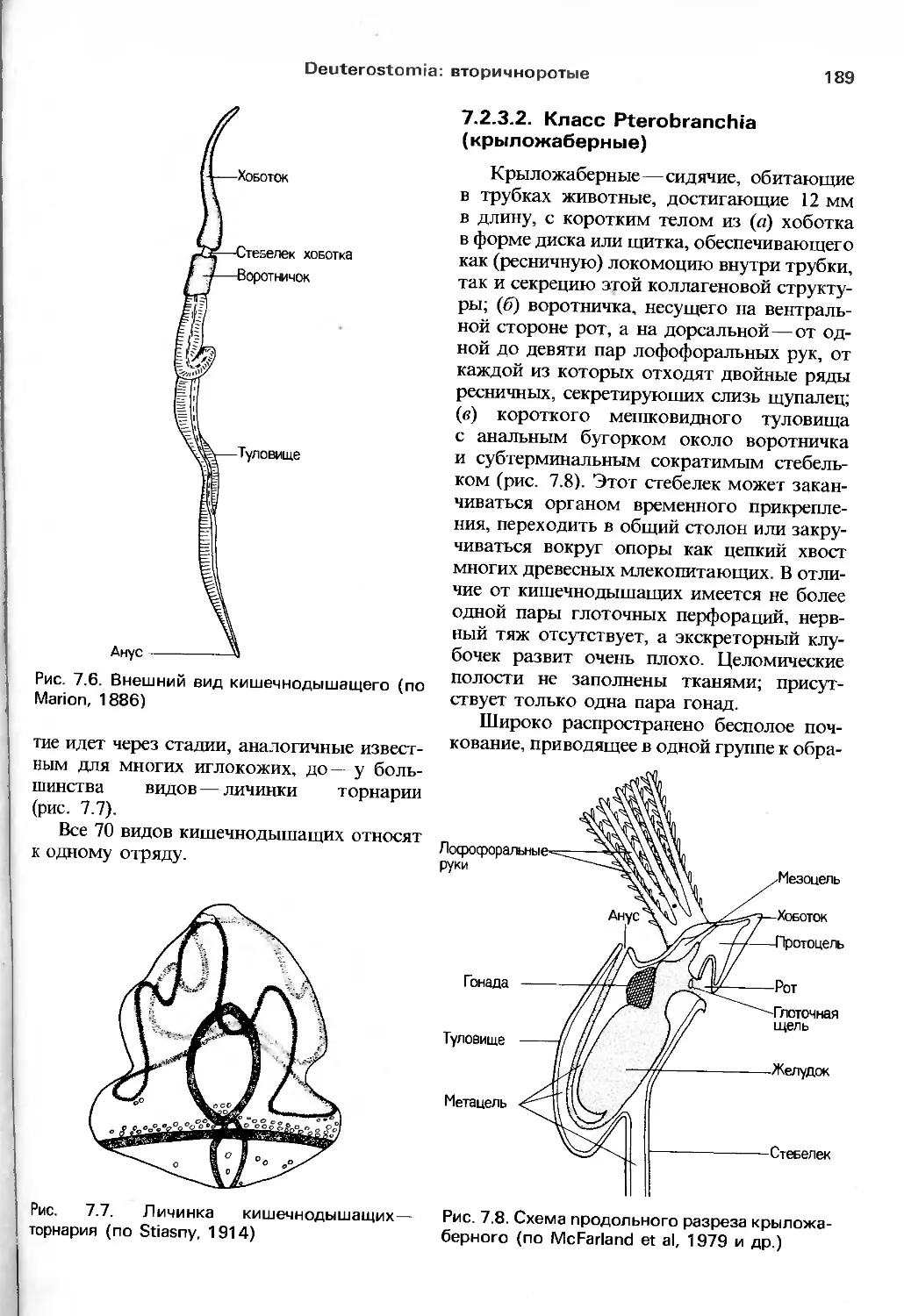
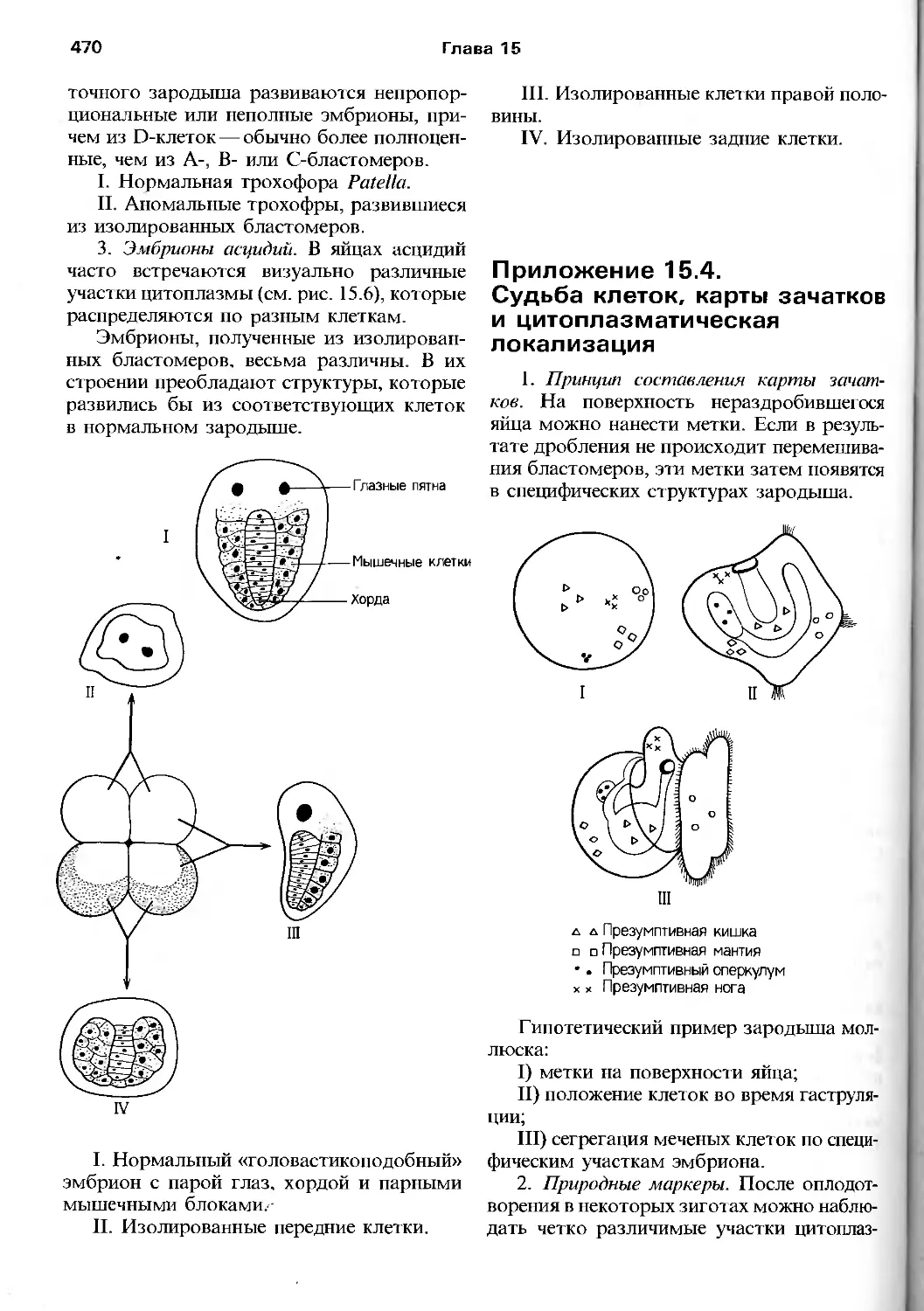
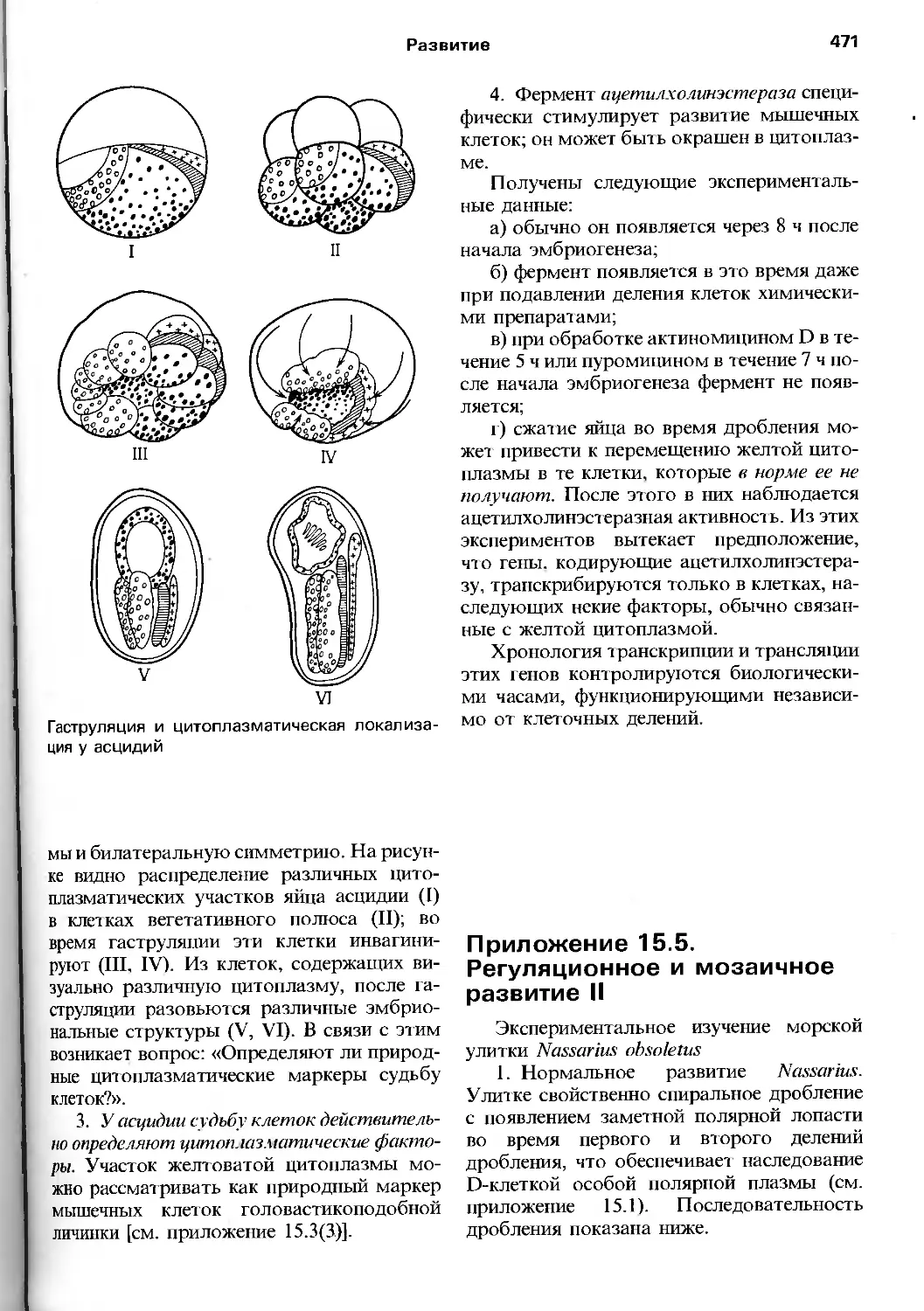
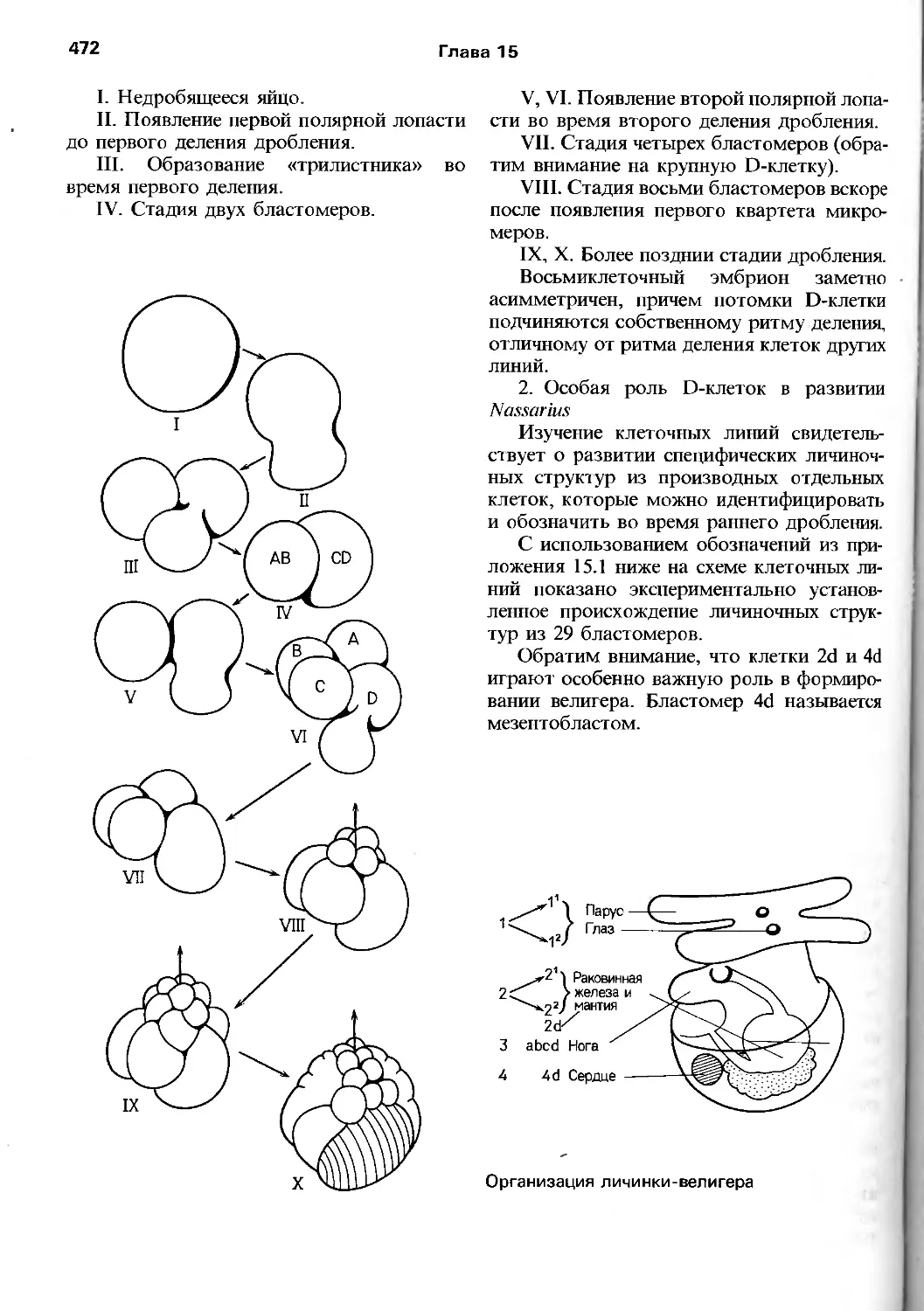
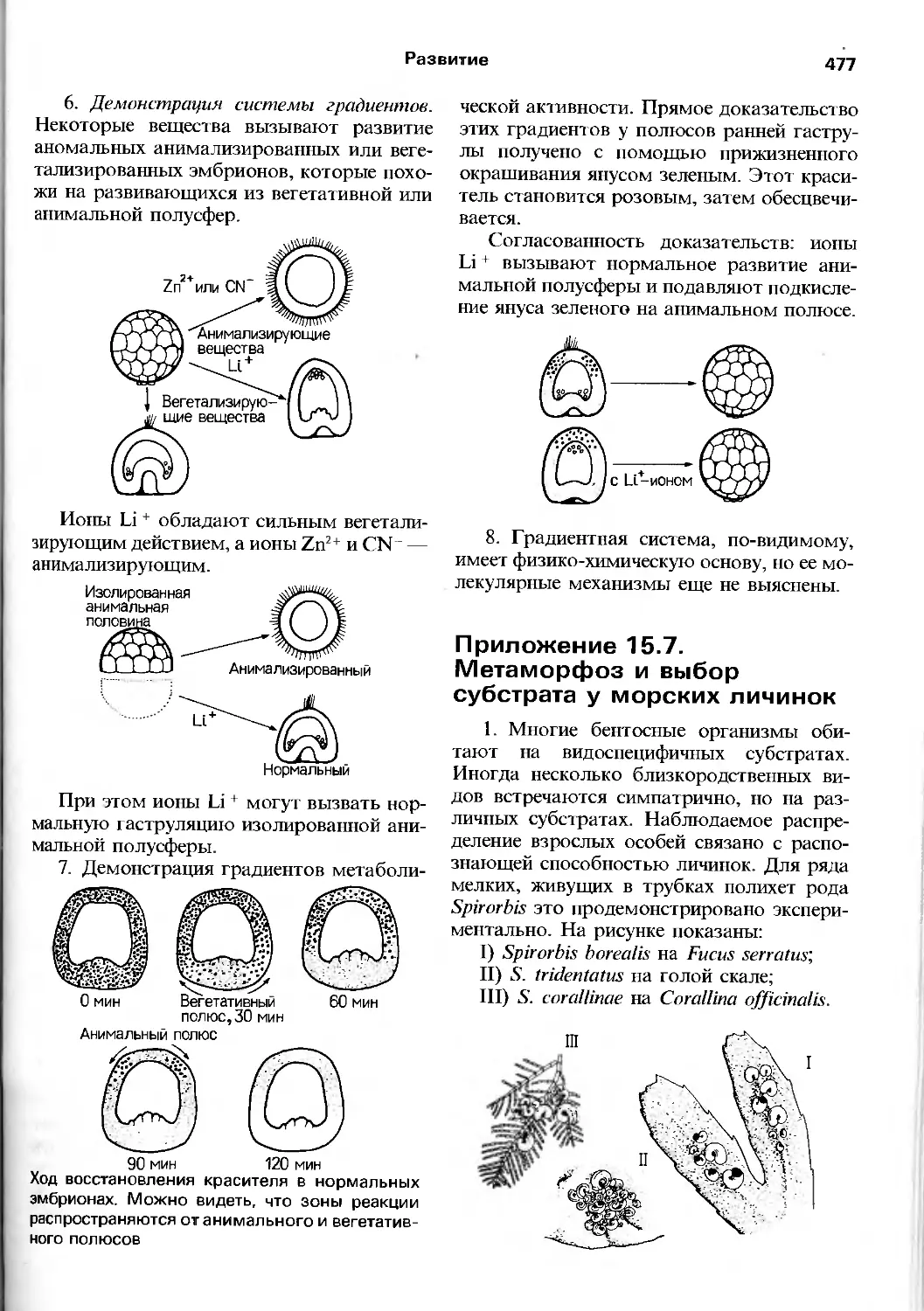
/




































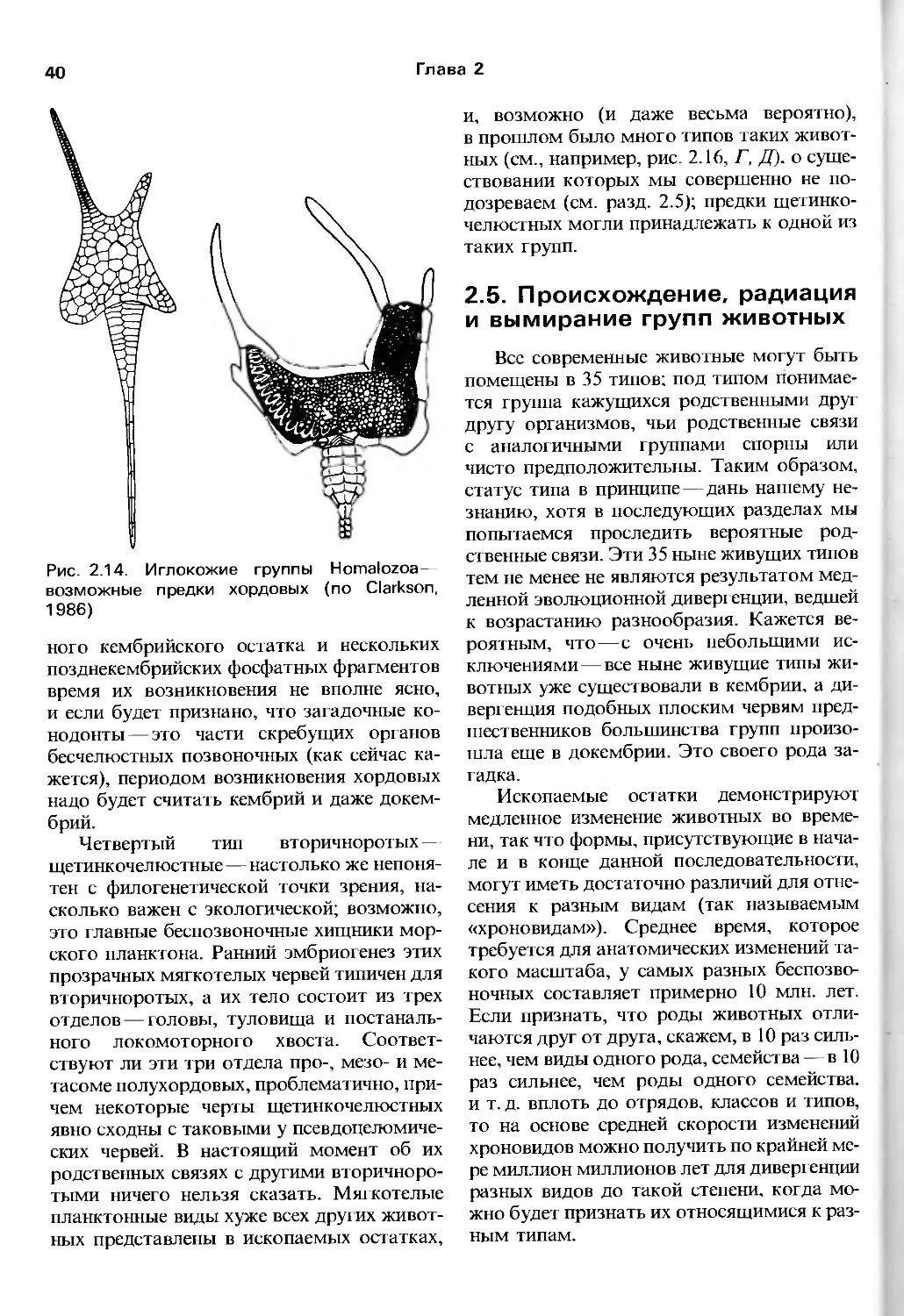

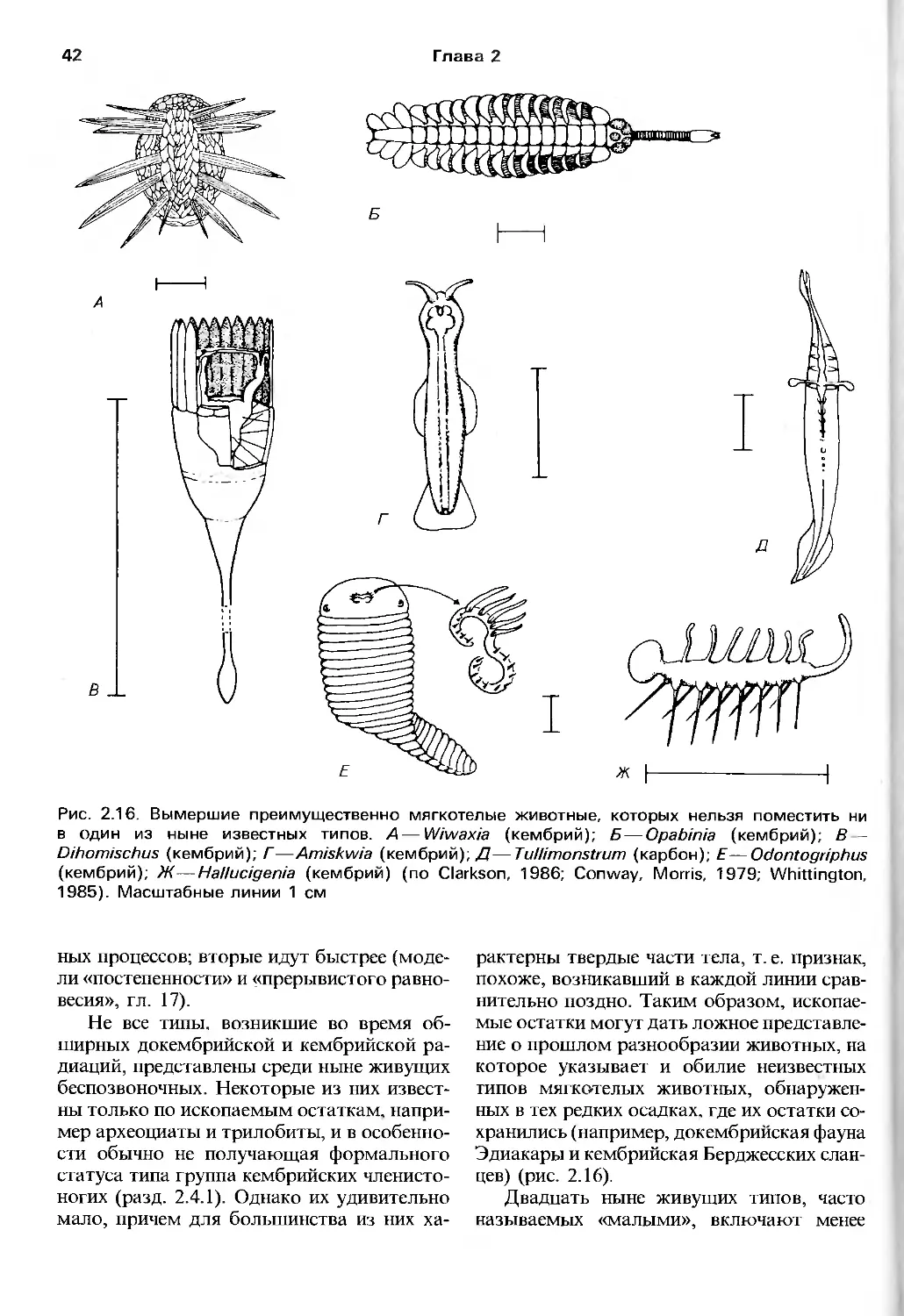
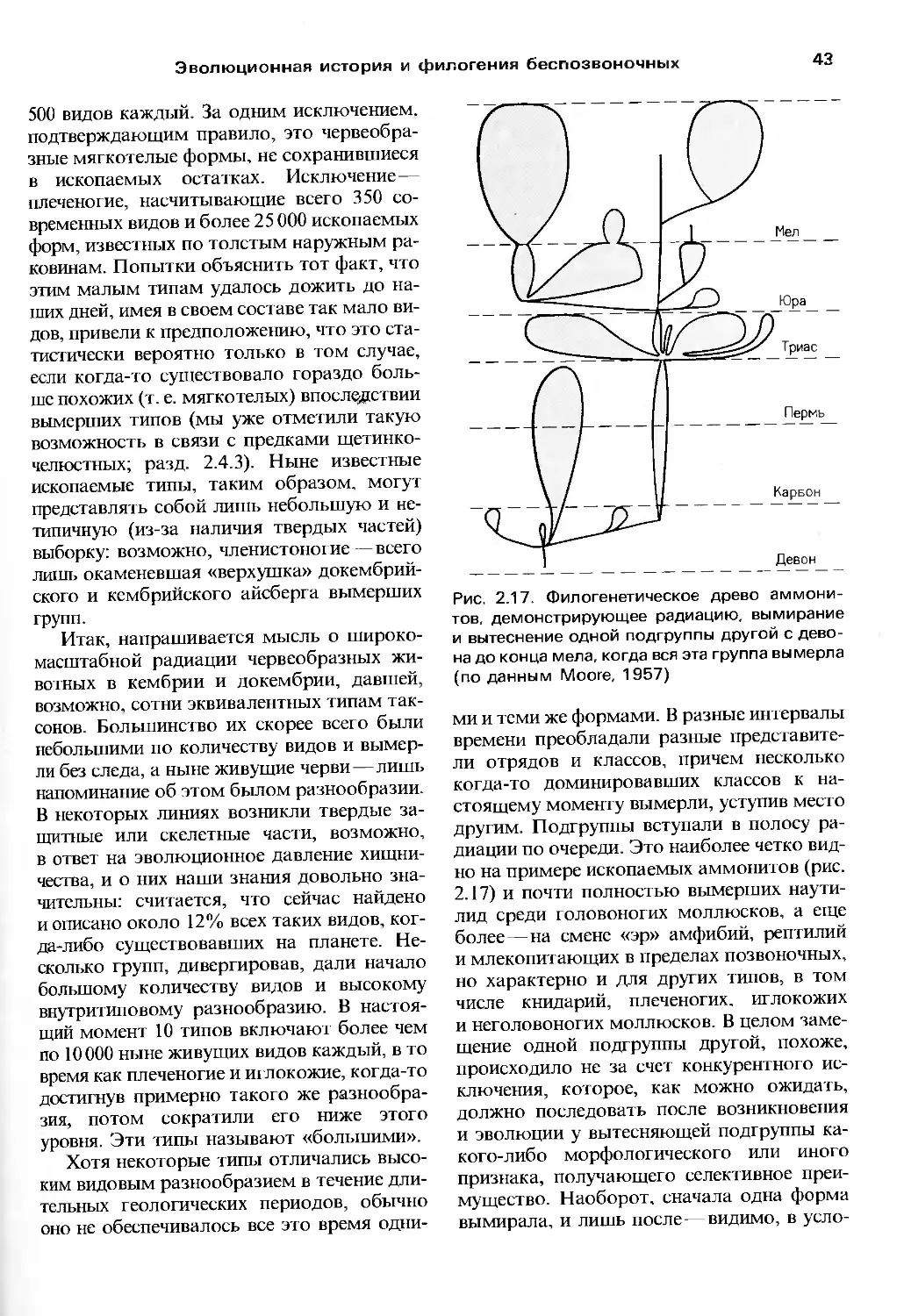
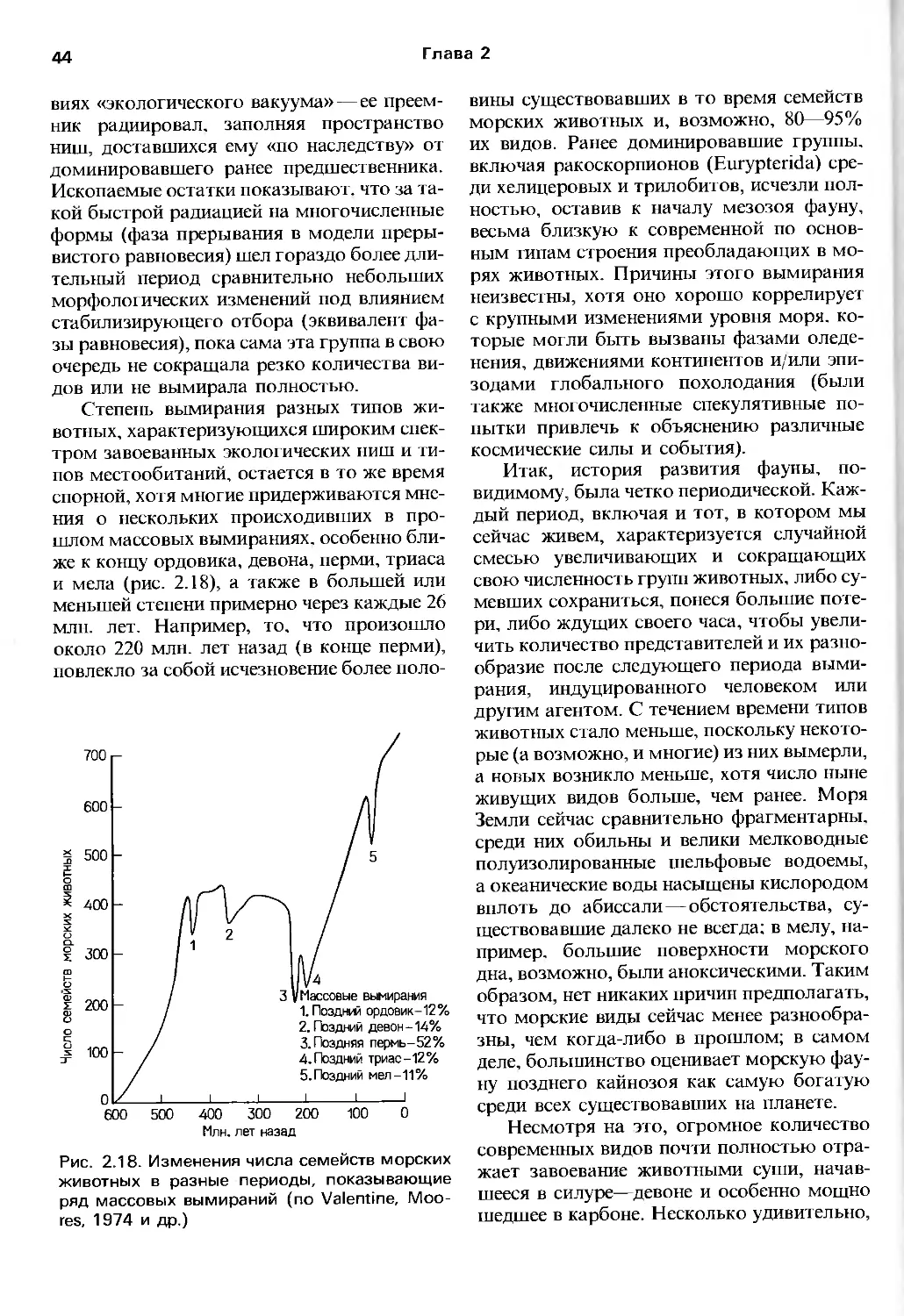









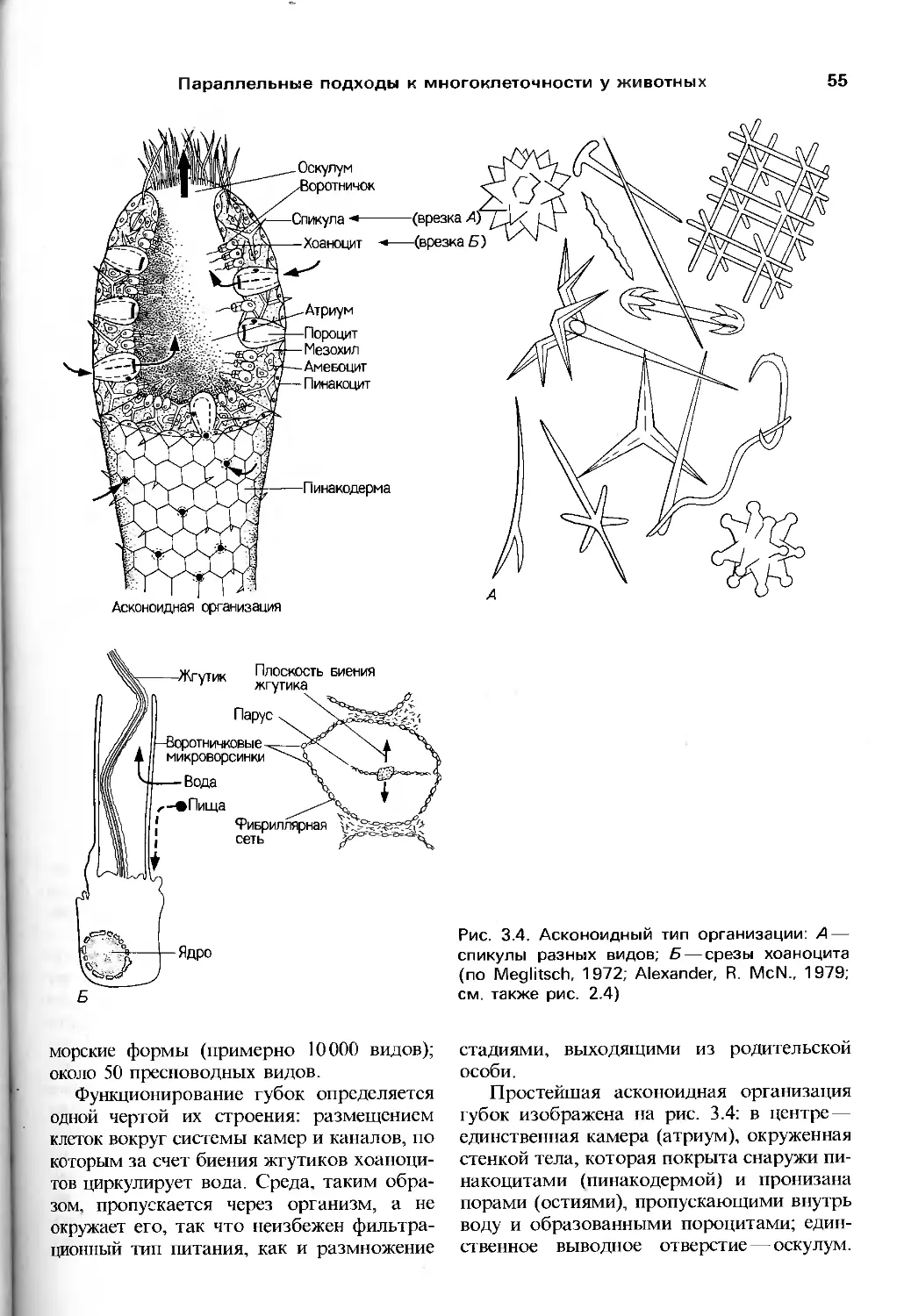
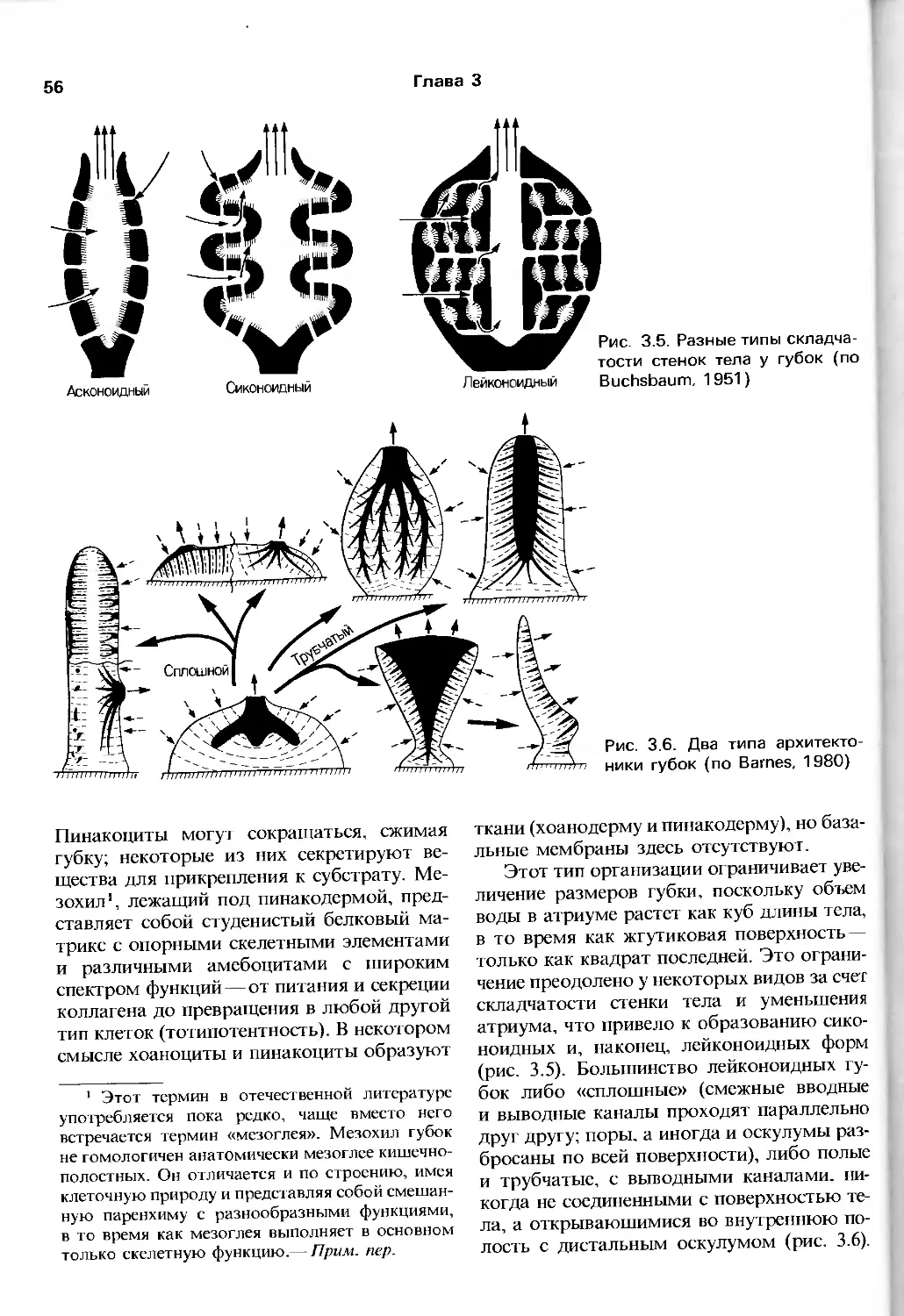






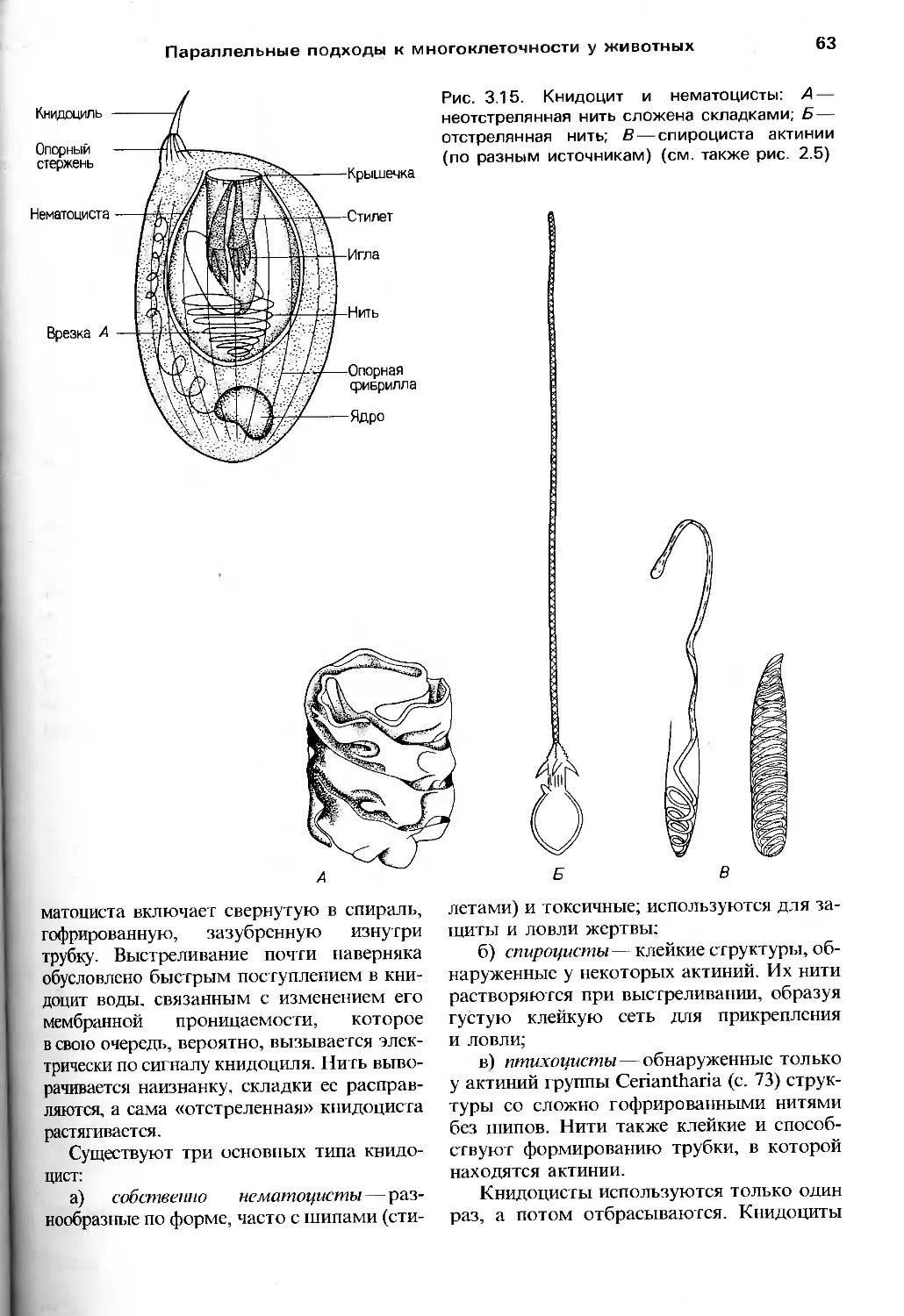








































































































































































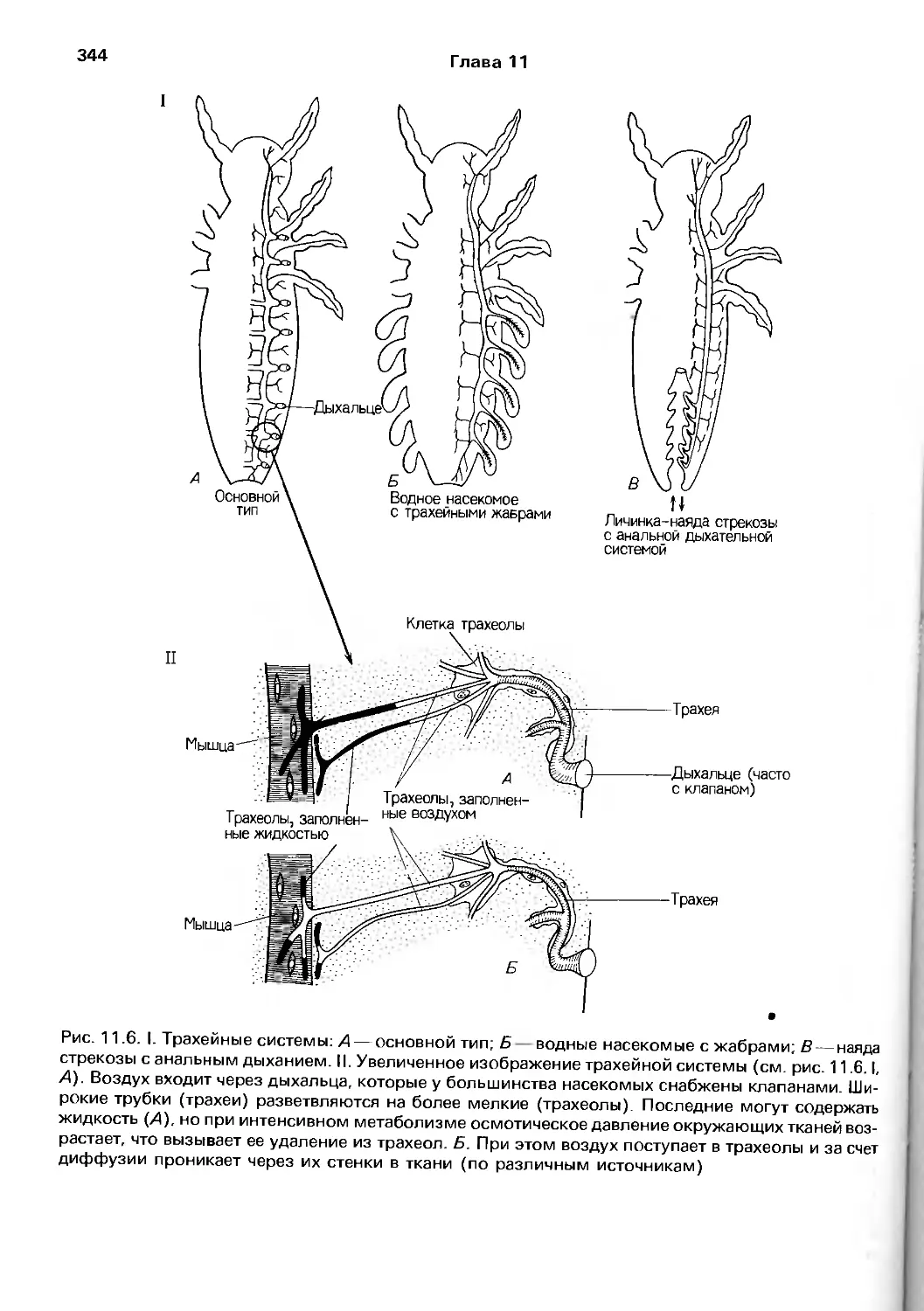


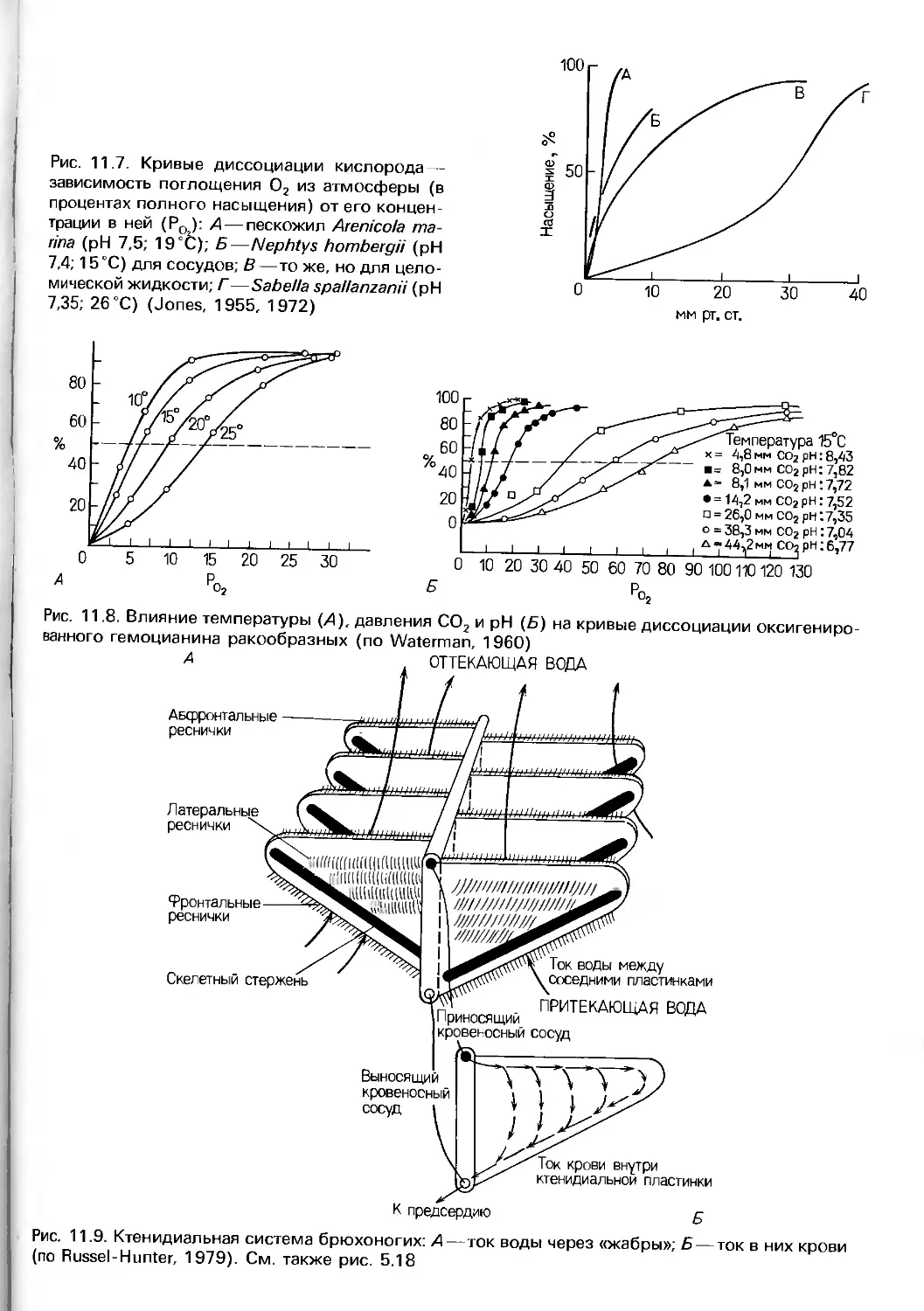

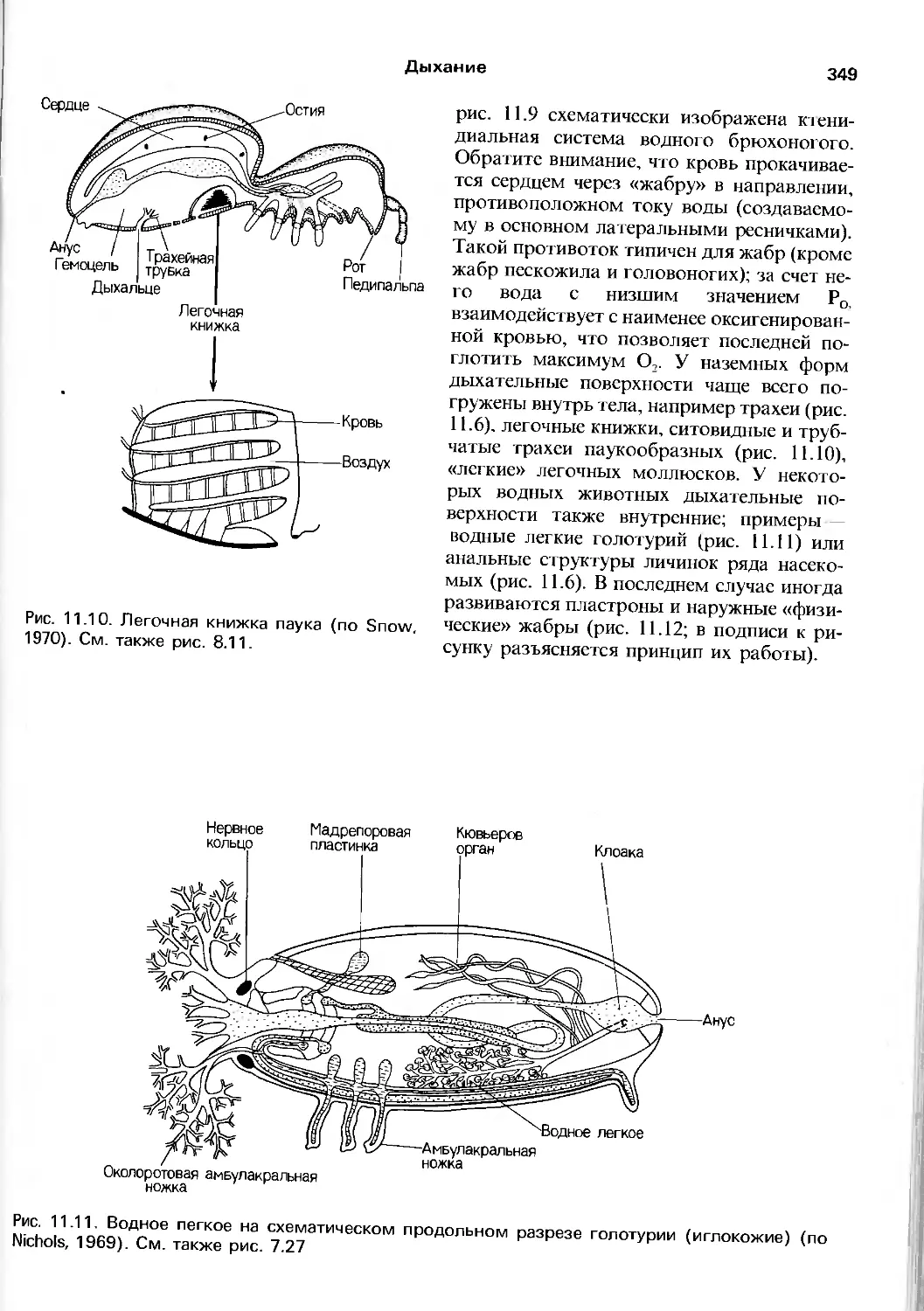








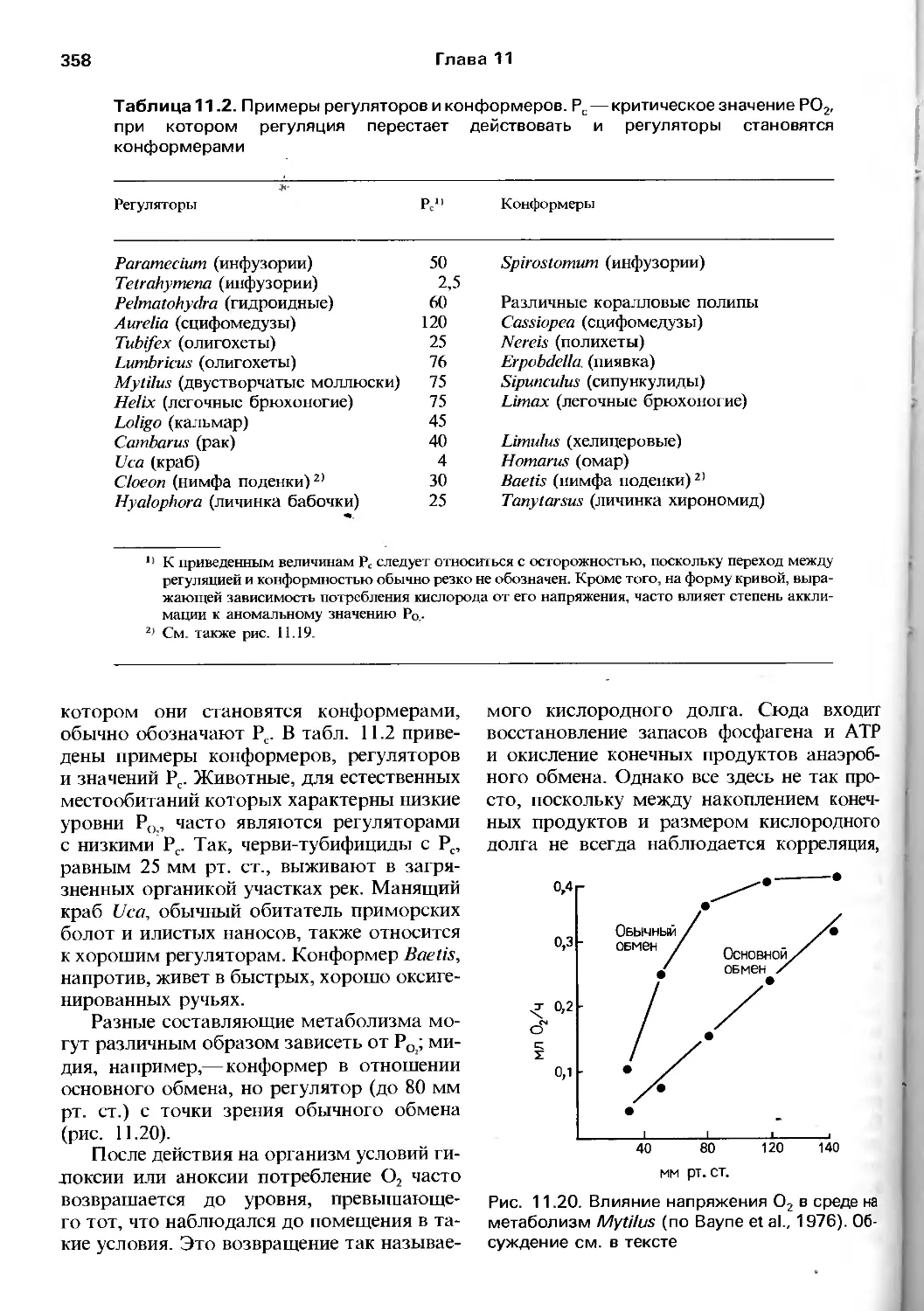




































































































































































Author: Барнс Р. Кейлоу П. Олив П. Голдинг Д.
Tags: invertebrata беспозвоночные зоология биология издательство мир
ISBN: 5-03-001496-9
Year: 2009




Text
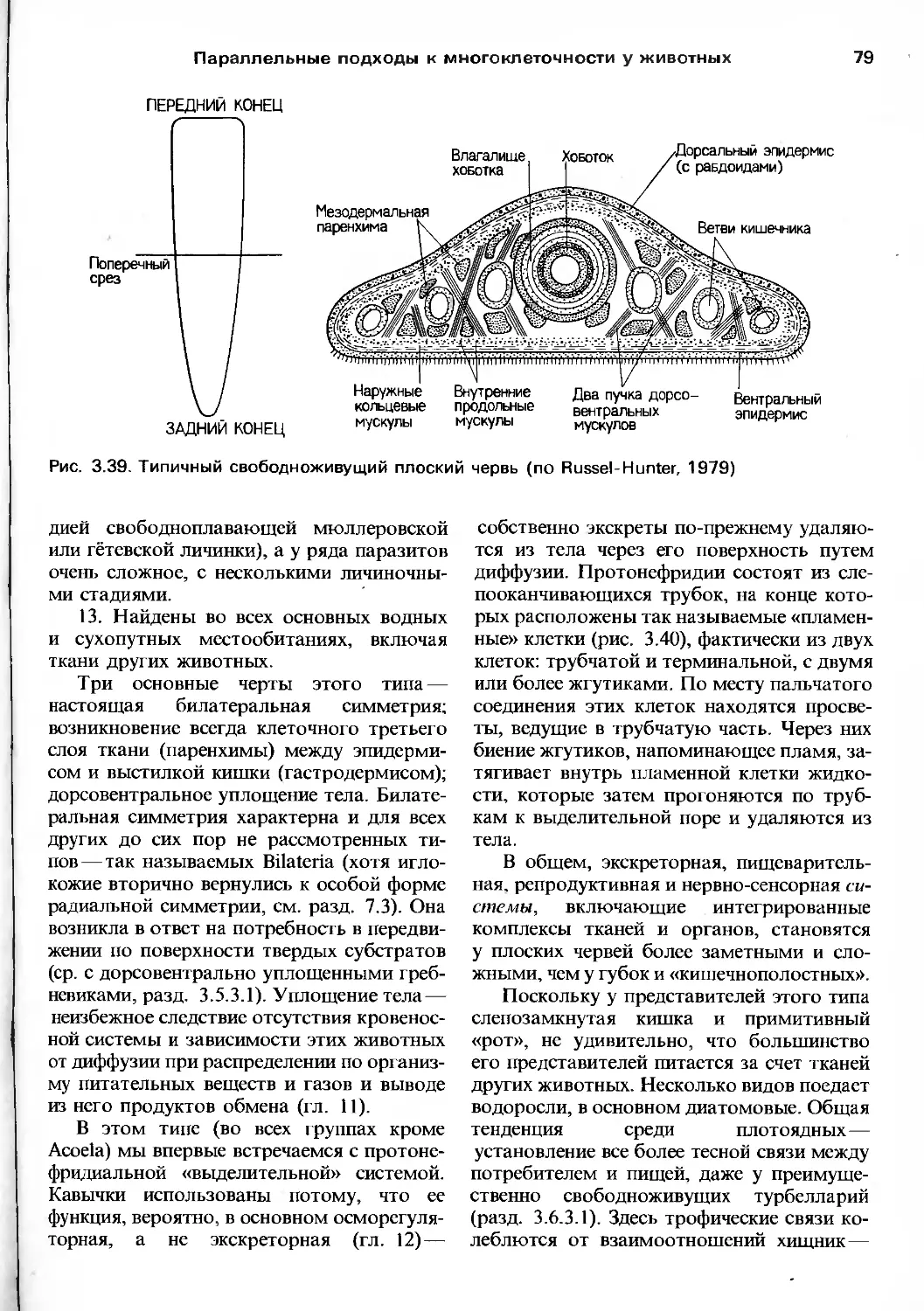
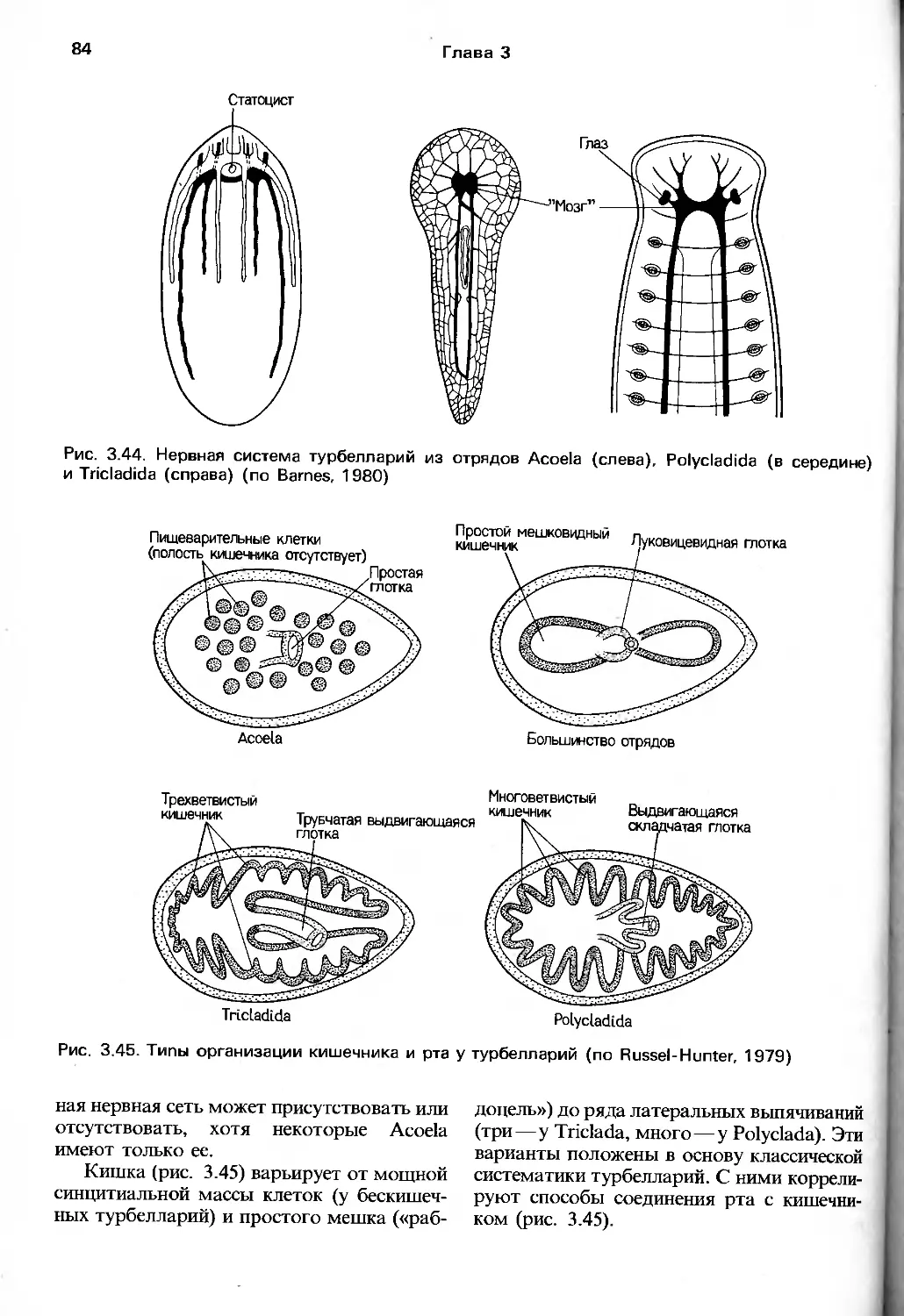
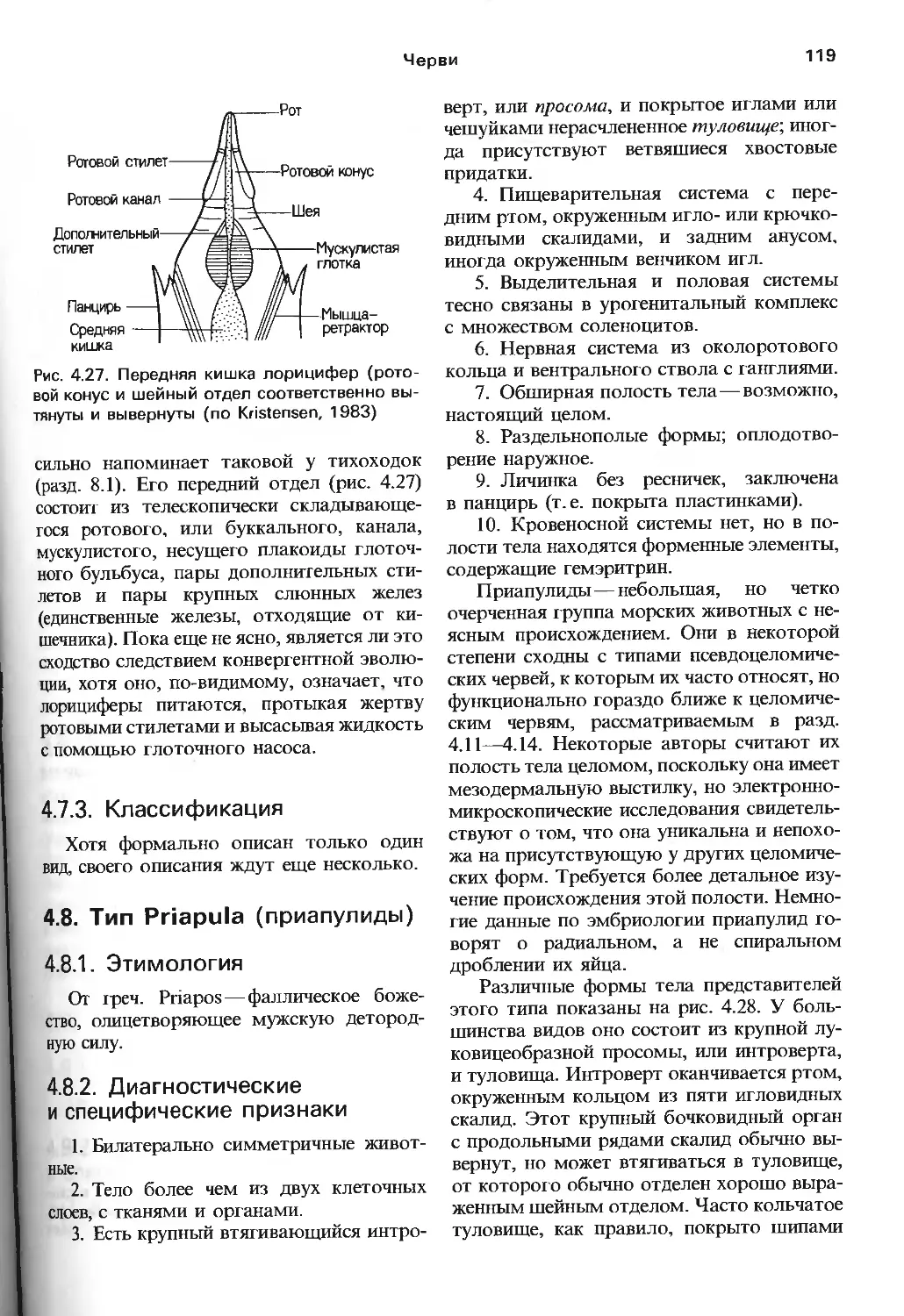
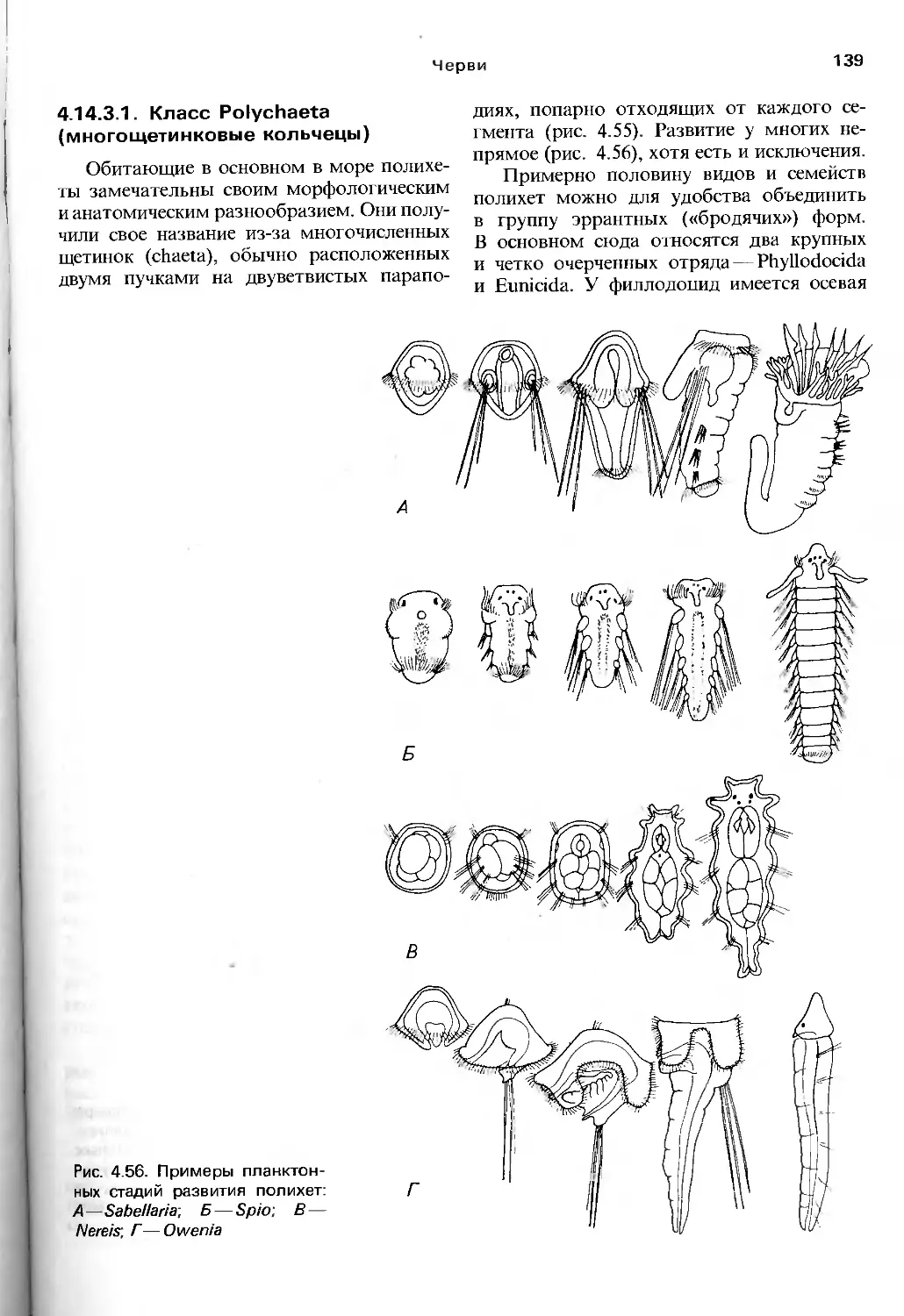
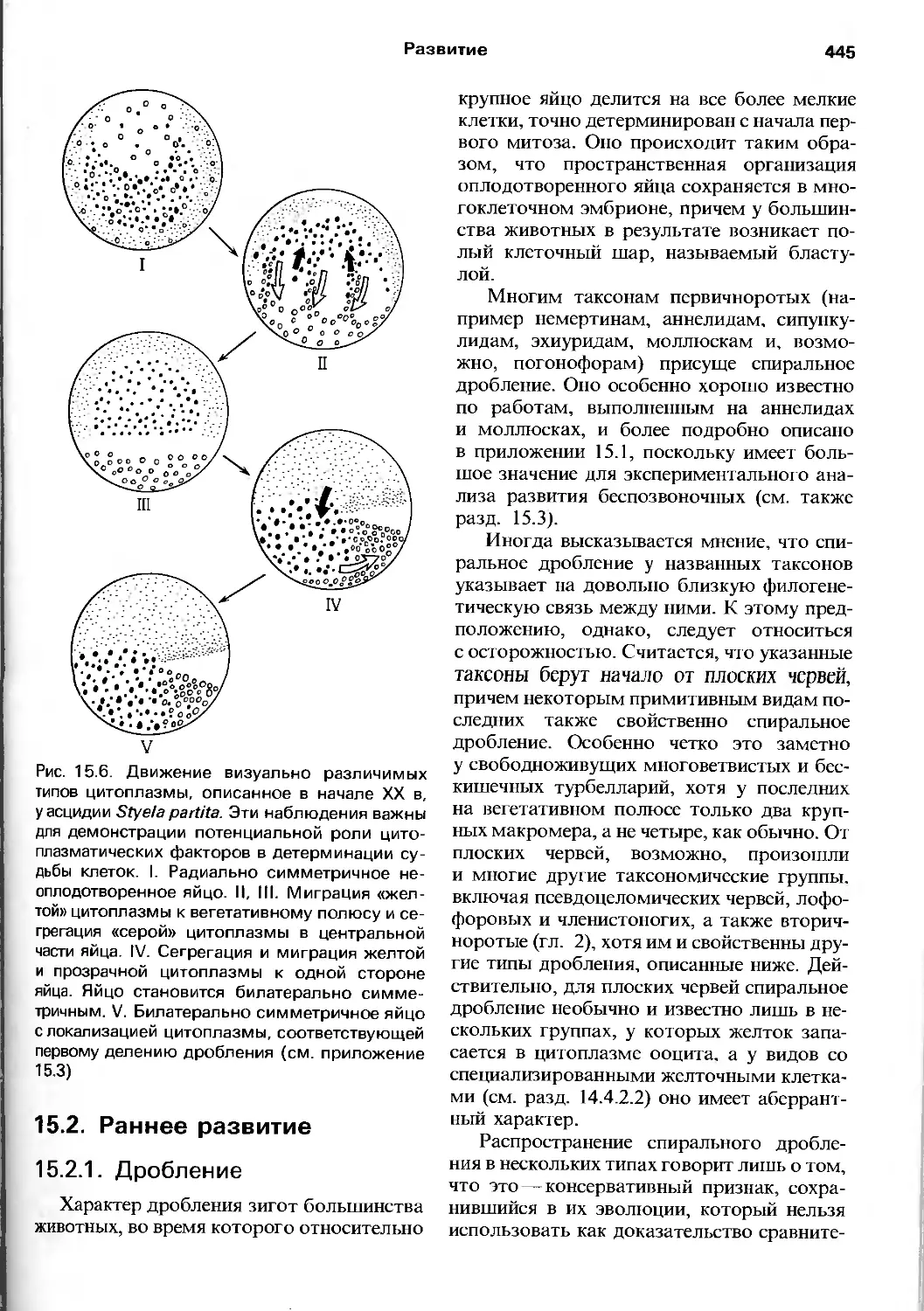
сннъ.
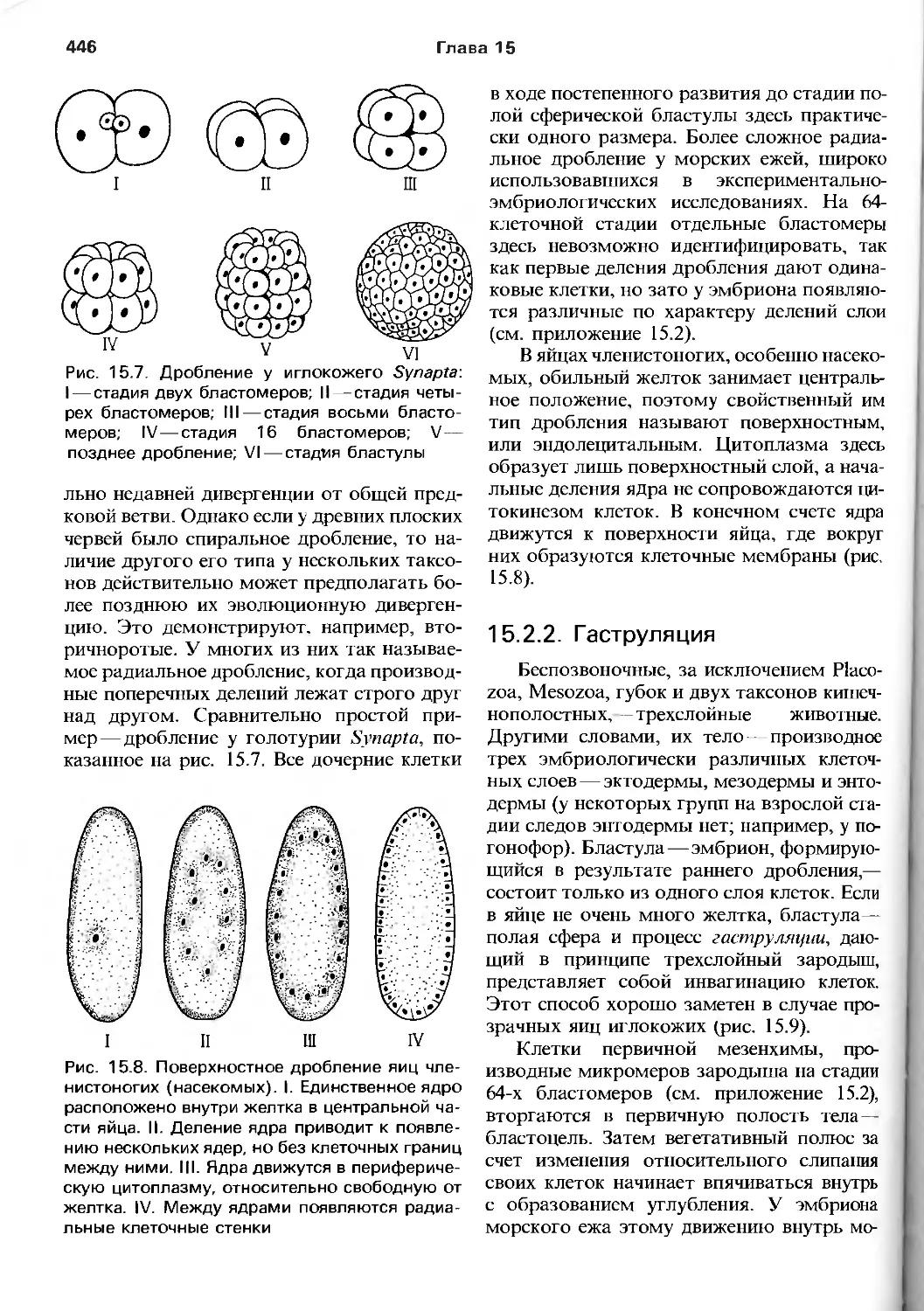
" , - 1
О SO О
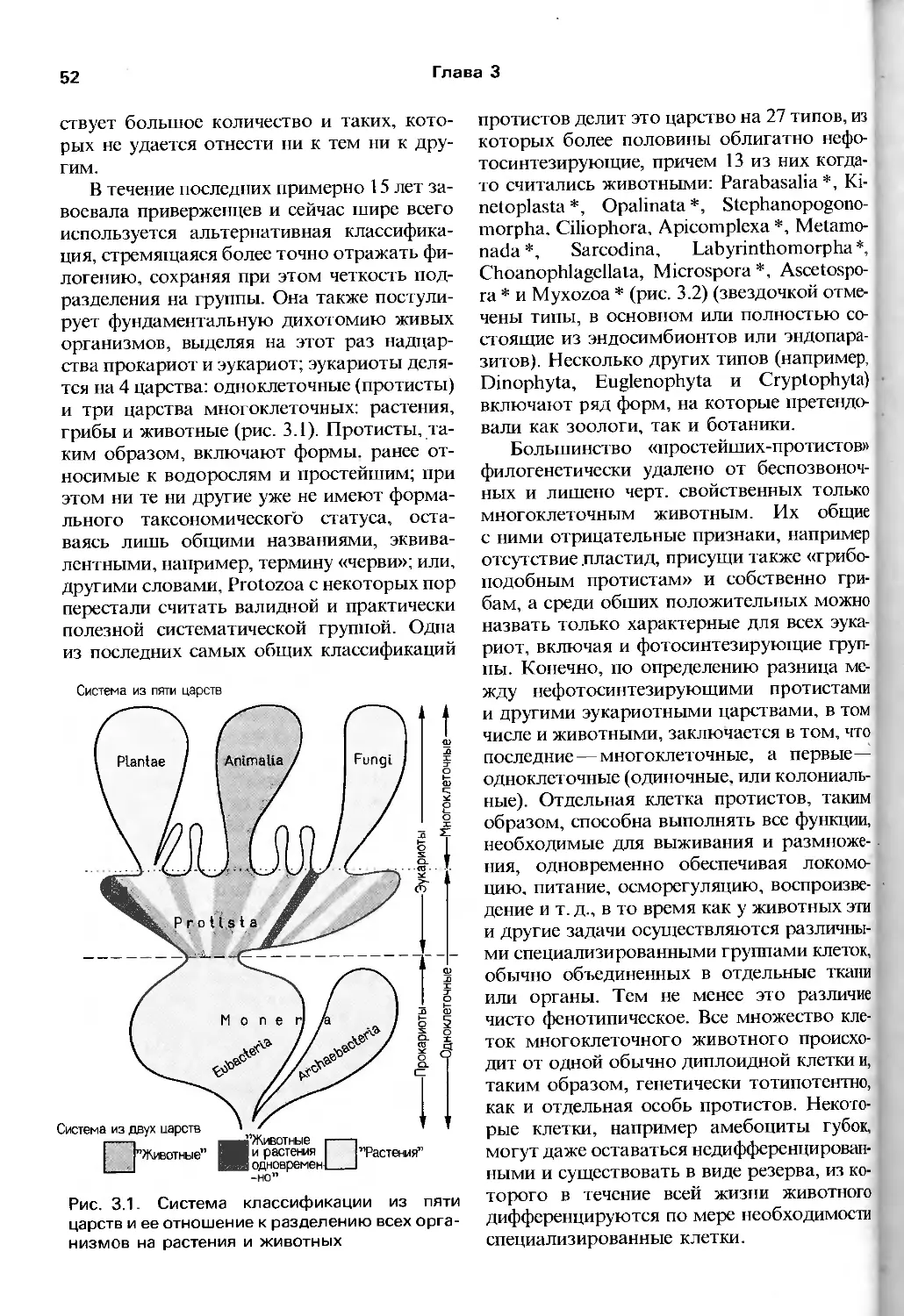
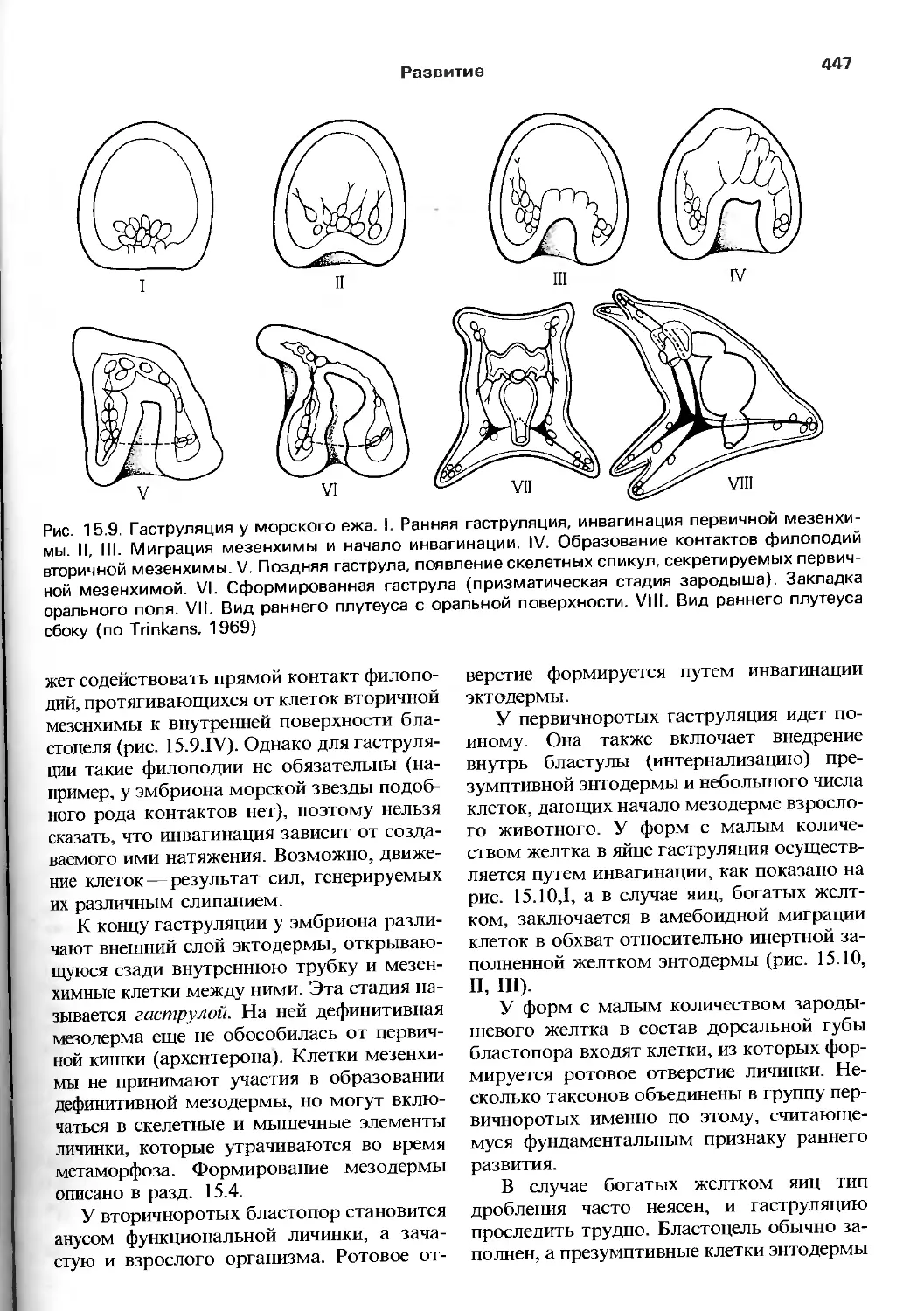
БЕСПОЗВОНОЧНЫЕ
THE INVERTEBRATES
A NEW SYNTHESIS
R.S.K. BARNES
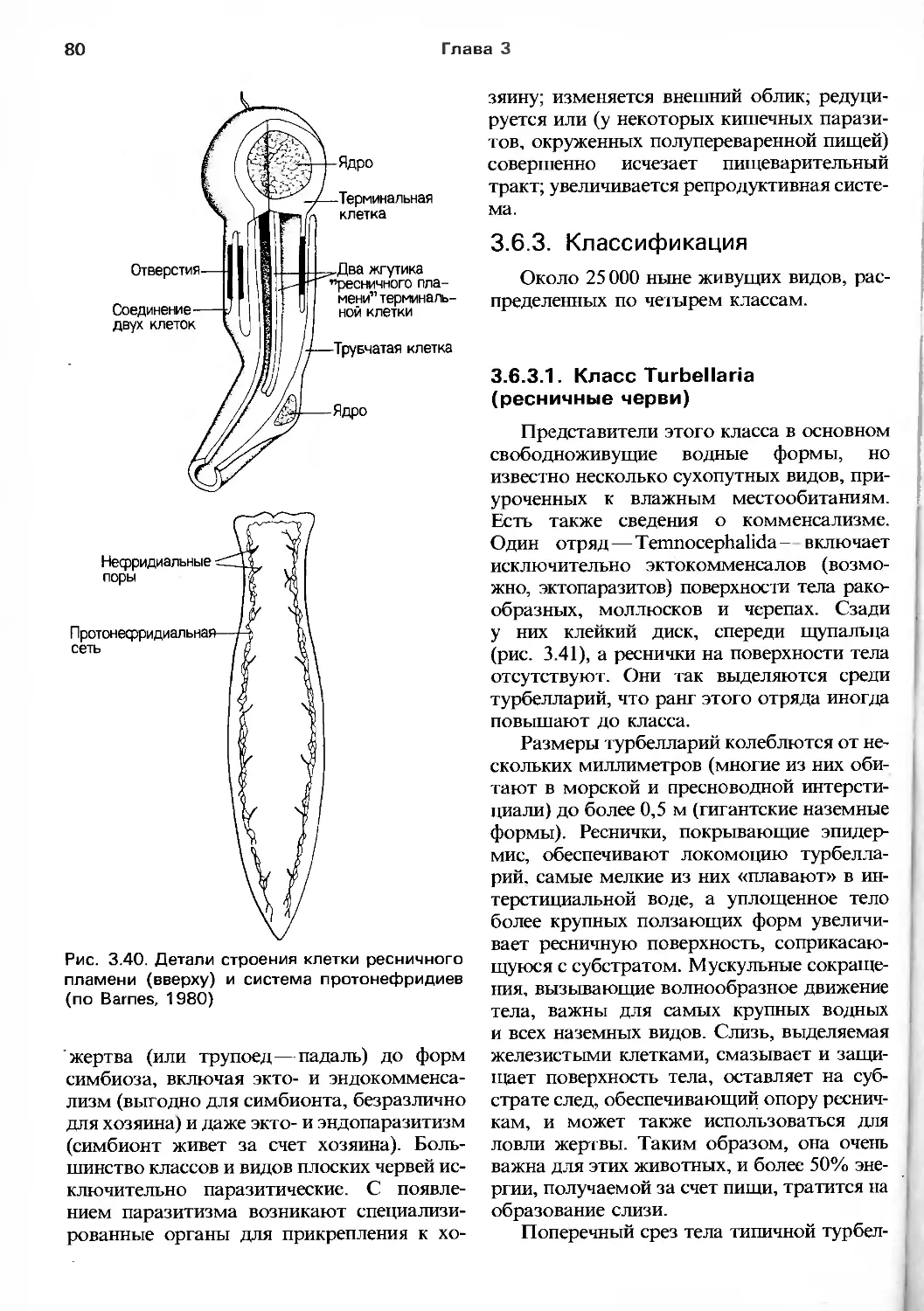
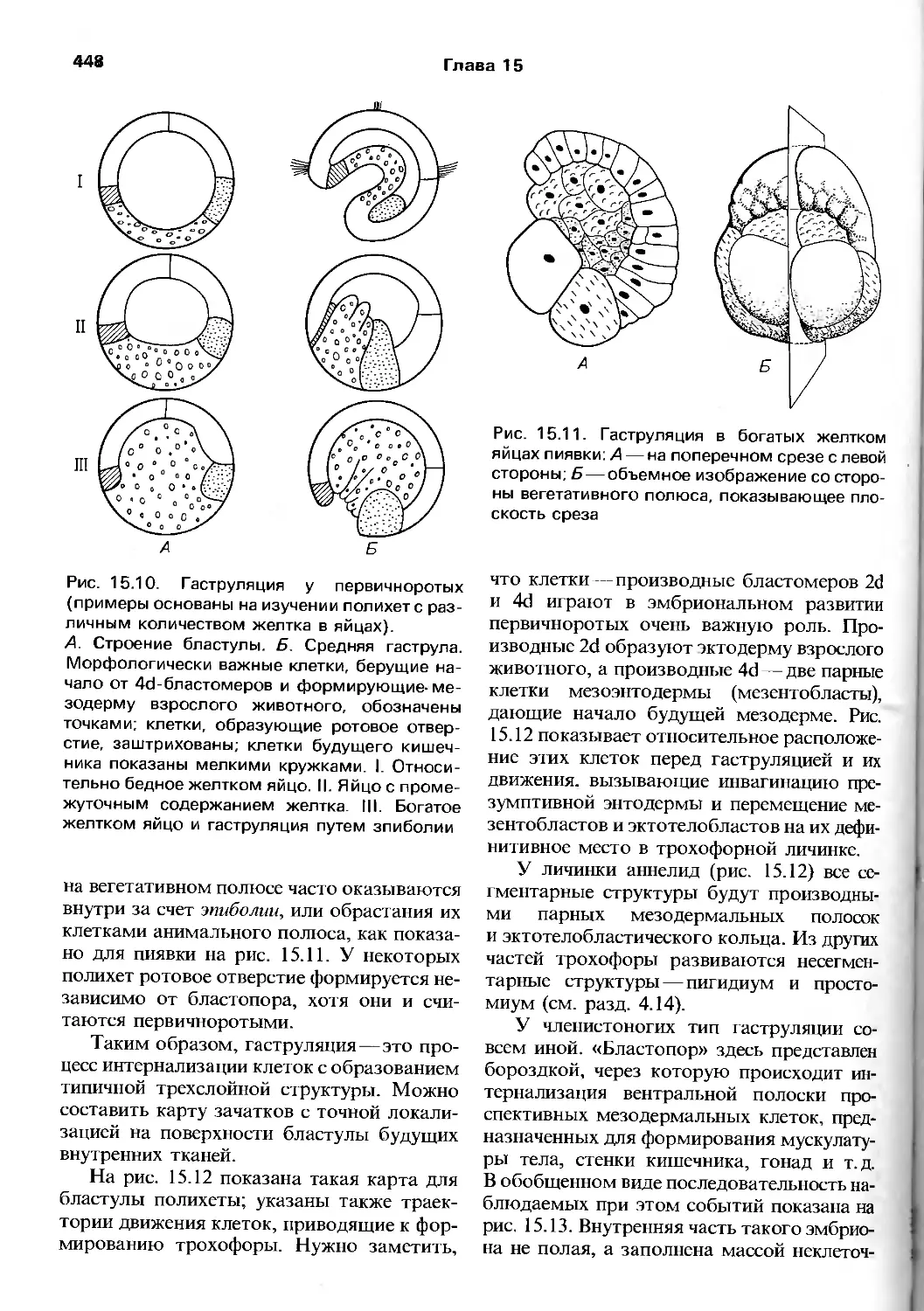
Department of Zoology and St Catharine's Colledge,
University of Cambridge, UK
P. CALOW
Department of Zoology, University of Sheffield, UK
P.J.W. OLIVE
Department of Zoology,
University of Newcastle upon Tyne, UK
WITH A CHAPTER CONTRIBUTED BY
D.W. GOLDING
Department of Zoology,
University of Newcastle upon Tyne, UK
BLACKWELL SCIENTIFIC PUBLICATIONS
Oxford London Edinburgh
Boston Palo Alto Melbourne
Р. БАРНС, П. КЕИЛОУ, П. ОЛИВ, Д. ГОЛДИНГ
НОВЫЙ ОБОБЩЕННЫЙ
подход
Перевод с английского
М.А. Сафроновой,
канд. биол. наук И. В. Успенского,
канд. биол. наук А. Б. Цетлина
под редакцией
канд. биол. наук Б. Я. Виленкина
МОСКВА «МИР» 1992
ББК 28.6
Б53
УДК 592
Авторы: Барнс Р., Кейлоу П., Олив П., Голдинг Д.
Беспозвоночные: Новый обобщенный подход: Пер.
с англ.— М.: Мир, 1992.— 583 с, ил.
ISBN 5-03-001496-9
В руководстве по зоологии беспозвоночных, написанном известными
английскими учеными, впервые обобщены сведения по эволюции, систематике, морфологии
и физиологии беспозвоночных. Книгу характеризует полнота охвата предмета и
наглядность изложения. Рассмотрены эволюция и систематика беспозвоночных, а также
их функциональная биология: питание, дыхание, движение, выделение и осморегуля-
ция, репродукция, онтогенез, нервная и эндокринная системы. Приведен словарь
терминов.
Для студентов-биологов, специалистов, интересующихся биологией
беспозвоночных, преподавателей биологии в школе.
Б 1907000000-^0101_92 ББК 28.6
041(01)—91
Редакция литературы по биологии
ISBN 5-03-001496-9 (русск.) © 1988 by Blackwell Scientific Publications
ISBN 0-632-01638-8 (англ.) /^Р 1л i. * л ^перевод на русский язык, Сафронова М. А
Успенский И. В., Цетлин А. Б., 1992
С ччь№
ПРЕДИСЛОВИЕ К РУССКОМУ ПЕРЕВОДУ
Задачи, построение и многие другие
особенности предлагаемой читателю книги
вполне объяснены в предисловии ее
авторов. Остается сказать лишь несколько слов
о том, какое, по нашему мнению, место
может занять эта книга в отечественной
зоологической литературе и чем она отличается
от большинства наших руководств по
зоологии и сравнительной физиологии
беспозвоночных.
Характерная ее особенность состоит
в том, что равное внимание уделено всем
группам беспозвоночных, независимо от
количества входящих в них видов, т. е. пла-
козои, гнатостомулиды и форониды
описаны так же подробно, как и моллюски или
членистоногие. При этом вопросы
филогении и происхождения тех или иных систем
органов животных рассматриваются как
второстепенные для данной книги, хотя
в ряде случаев они изложены достаточно
полно. Часто, говоря о происхождении той
или иной группы, авторы приводят
различные гипотезы, обращая внимание читателя
на то, что эти проблемы еще далеки от
окончательного разрешения.
Там, где это возможно, строение
животных описывается с функциональной точки
зрения, что, безусловно, помогает читателю
разобраться в многообразии
беспозвоночных. Большая удача книги — рисунки и
схемы, которые не дублируют, а дополняют
текст.
И, пожалуй, основное отличие от
распространенных у нас руководств в том, что
описывается строение именно живых
животных. Образно говоря, эта книга—
путеводитель скорее по зоопарку или
аквариуму, чем по анатомической коллекции.
Некоторые представления авторов
могут показаться читателю непривычными.
Например, ими принимается так
называемая архицеломатная теория происхождения
вторичноротых, описывается система
брюхоногих моллюсков, считающаяся многими
архаичной, и т. п. Однако это не затрудняет
чтения книги и делает ее, пожалуй, особо
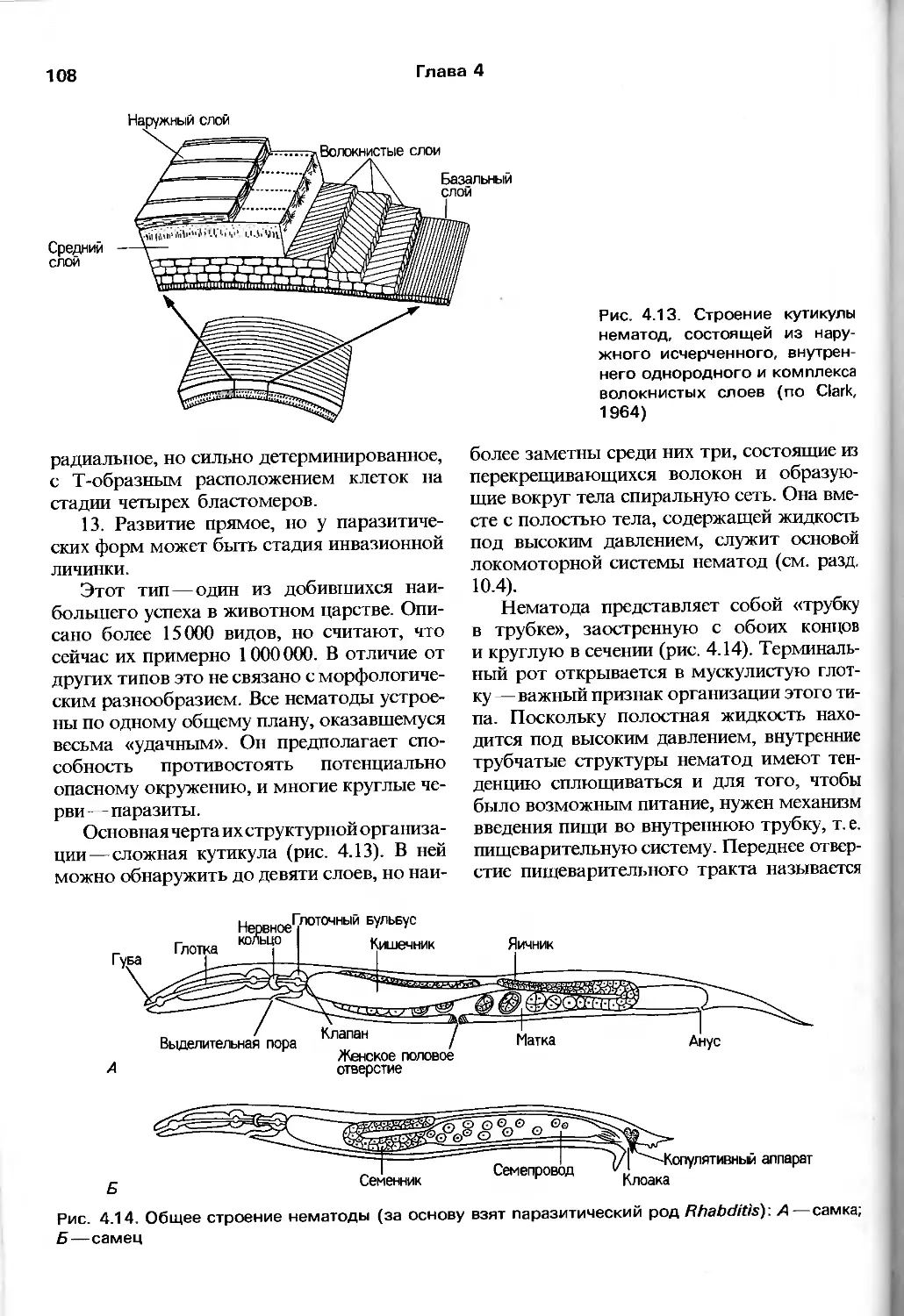
интересной в качестве источника
«альтернативных» точек зрения на важные
зоологические проблемы.
Как уже говорилось, в предлагаемой
книге идет речь о живых животных, и в этом
отношении она уникальна среди
распространенных у нас учебников и руководств
по зоологии. Сведения об организации
процессов жизнедеятельности у
беспозвоночных не столь обширны и не так равномерно
охватывают разные типы и классы, как
данные об их анатомии и морфологии. В этом
отношении сравнительные эмбриология,
физиология, биохимия еще не достигли
такой цельности, как сравнительная
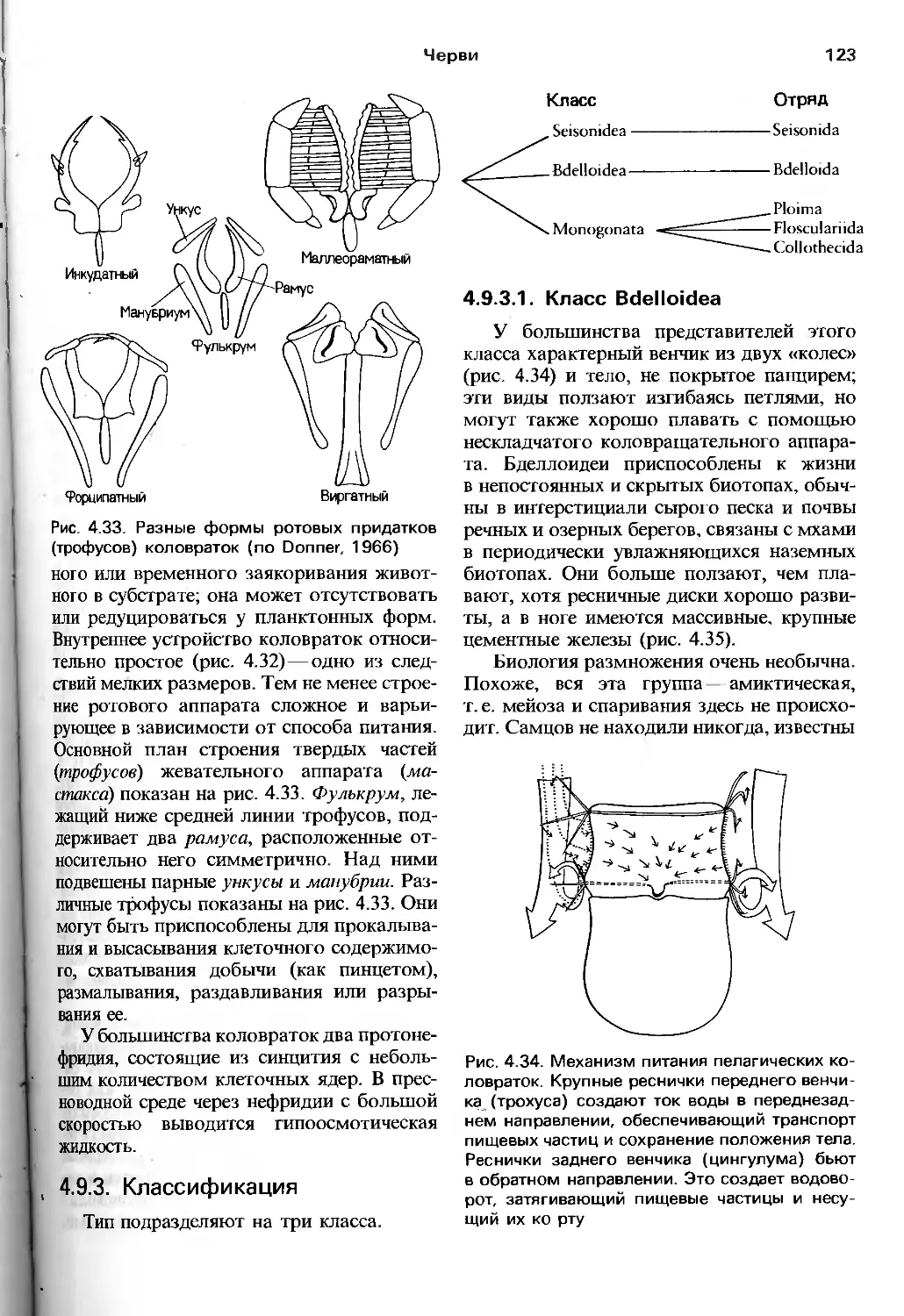
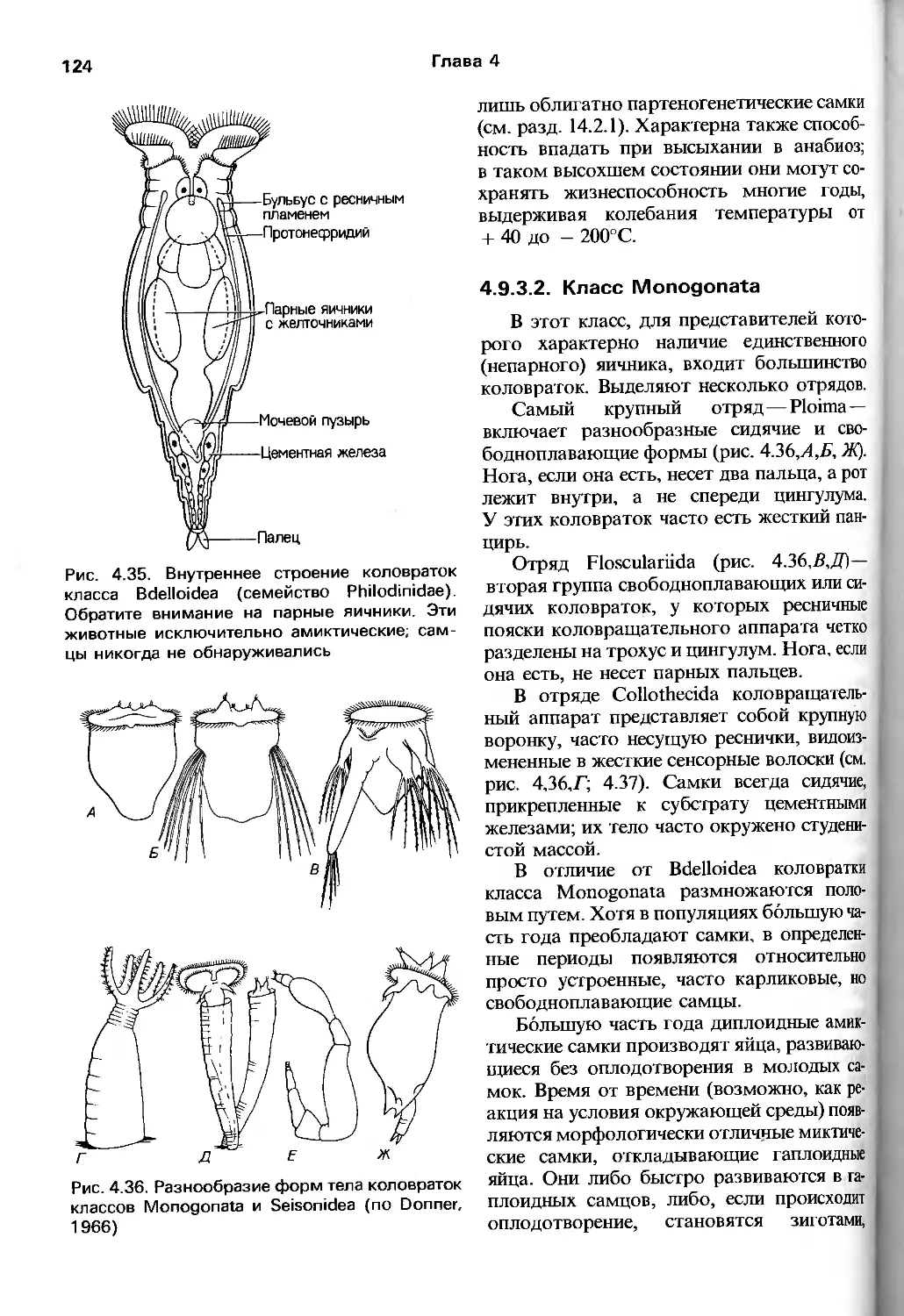
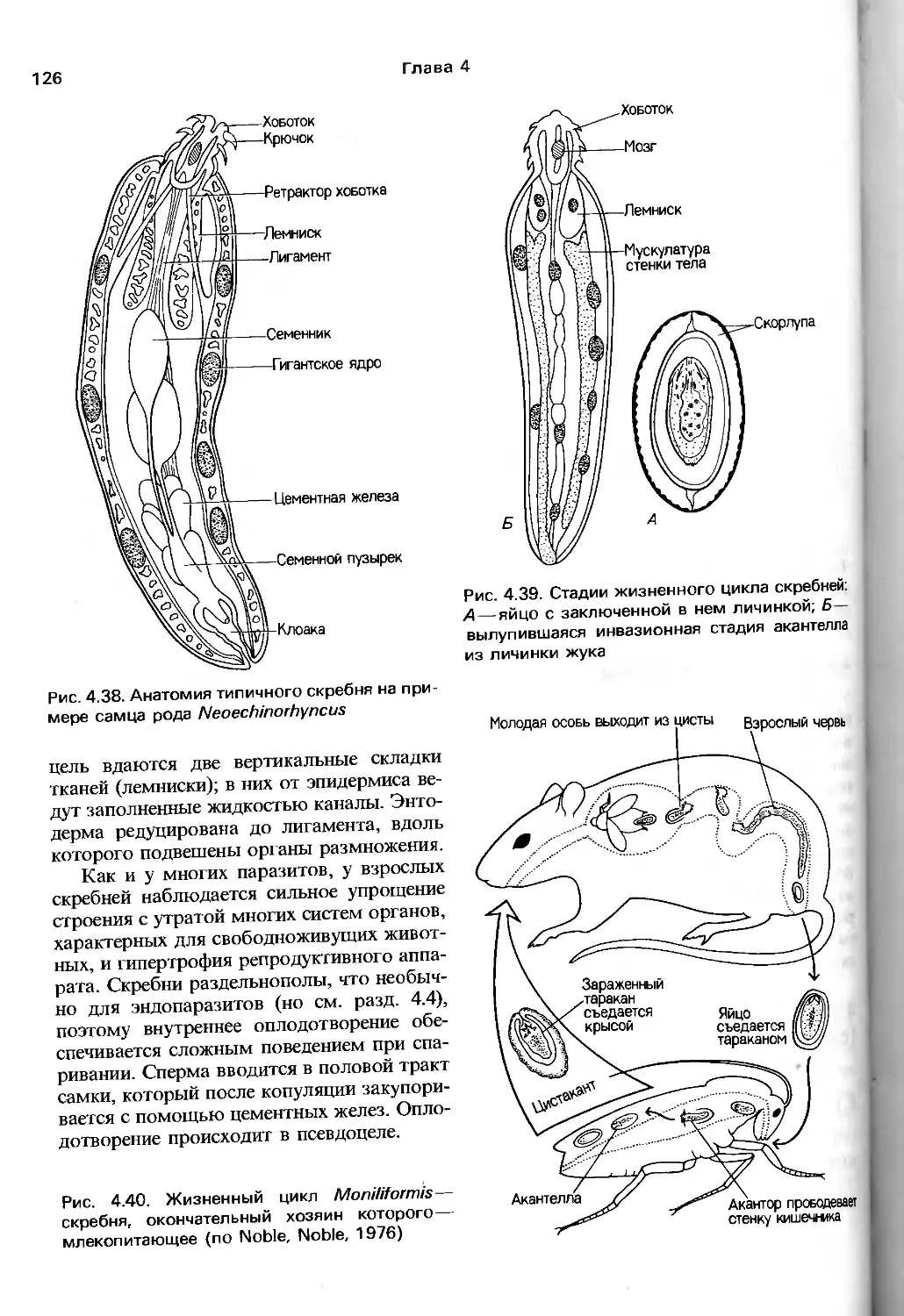
анатомия, что отмечают и авторы руководства.
Соответствующие разделы (гл. 9—17)
заслуживают внимания по двум причинам.
Во-первых, здесь довольно полно собраны
современные сведения о механизме
процессов жизнедеятельности у представителей
тех групп беспозвоночных, где
исследования такого рода достаточно продвинулись
вперед. Во-вторых, что более важно для
зоолога, авторы сумели показать, что
далекие от традиционных зоологических
вопросов работы по физиологии, биохимии,
молекулярной генетике, электрофизиологии
и т.п. имеют непосредственное отношение
к пониманию поведения, питания,
адаптации видов к внешним условиям, их
эволюции и прочим предметам, непосредственно
интересующим зоологов. Можно
надеяться, что предложенная манера подачи и
обсуждения нетрадиционного материала бу-
6
Предисловие к русскому переводу
дет способствовать преодолению той
неприязни к новейшим биологическим
подходам, которую часто проявляют зоологи
классического направления.
Эволюционный метод авторов четко
прослеживается в том. что они наряду
с устройством и возможностями
эволюционирующего объекта демонстрируют
и внешние силы, создающие давление
отбора. Другими словами, значительное
внимание уделено экологическим вопросам, т.е.
рассматривается большое количество
экологических классификаций, стратегий и
других до некоторой степени умозрительных
построений. Нельзя, вероятно, утверждать,
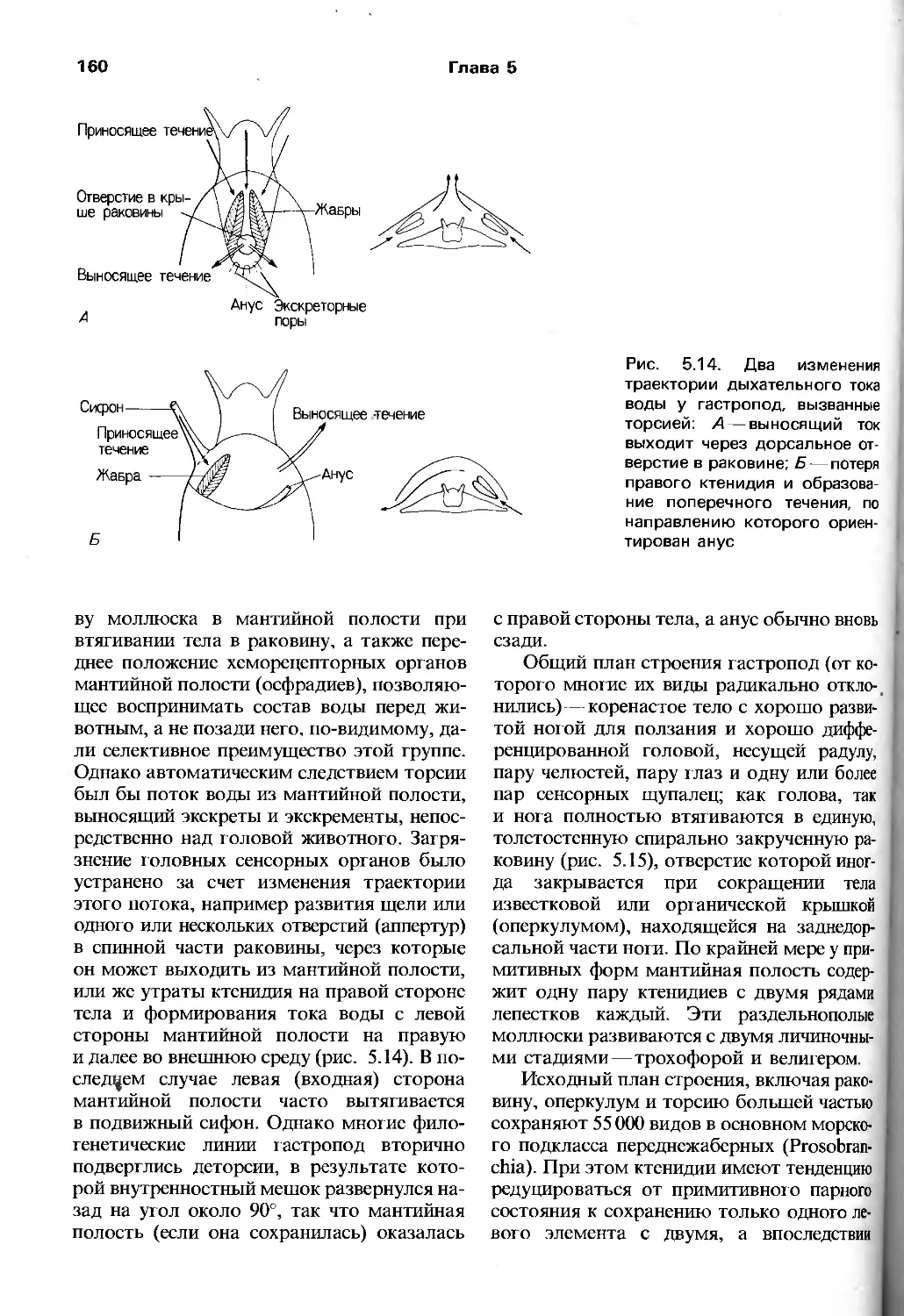
что экология в ее современном состоянии
вносит весомый вклад в целостное
понимание эволюционного процесса. Тем не менее
соответствующие разделы книги
способствуют развитию представления о зоологии
как о науке о живых организмах,
взаимодействующих с разнообразными
биотическими и абиотическими факторами
окружающей их среды.
А. Б. Цетлин
Б. Я. Виленкин
ПРЕДИСЛОВИЕ
Существует несколько вполне
доступных учебников, посвященных
беспозвоночным, поэтому появление еще одного
требует некоторых пояснений. Уже
опубликованные книги попадают в одну из двух
категорий: либо это систематические обзоры,
последовательно рассматривающие
различные типы животных (например, R. D.
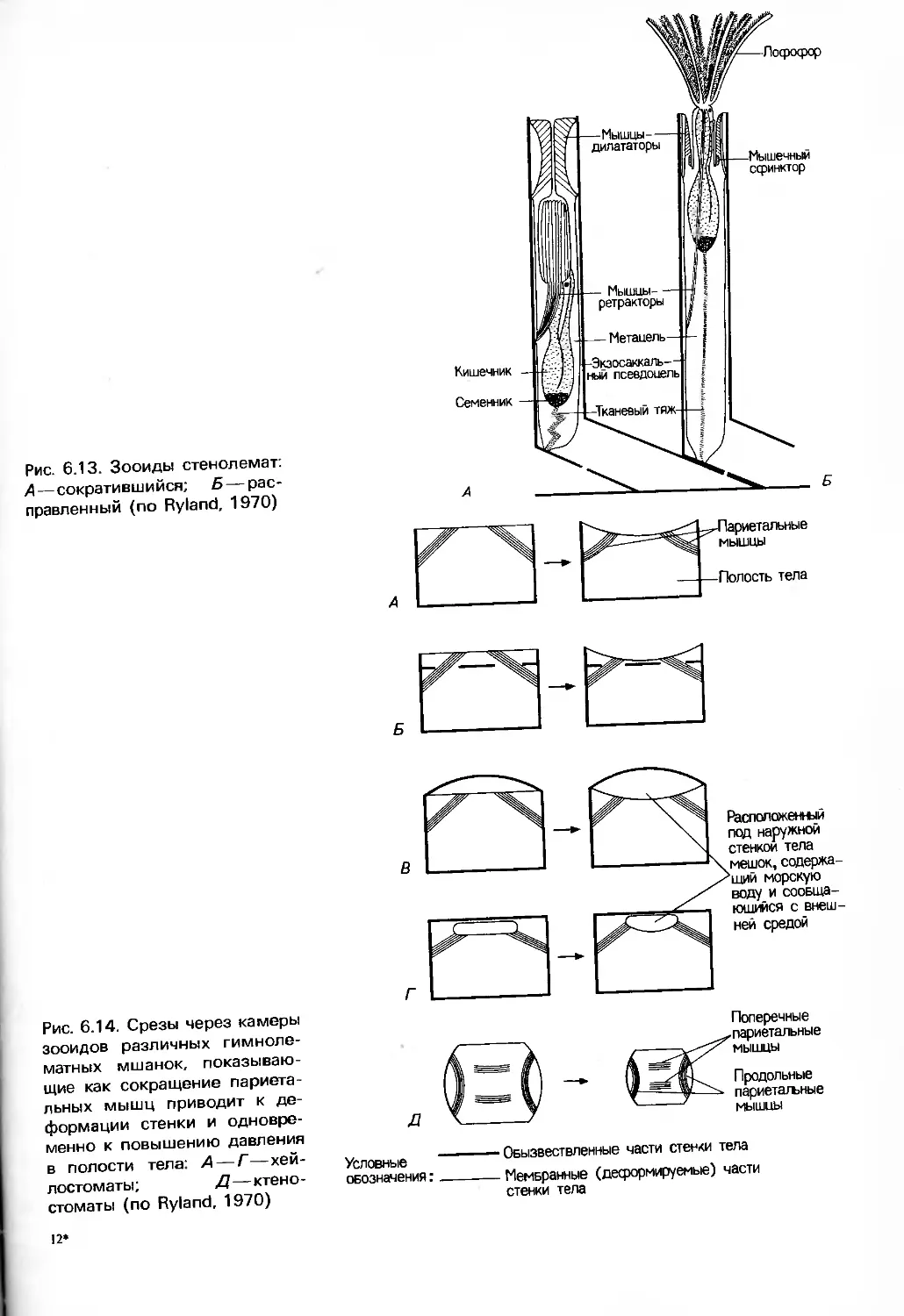
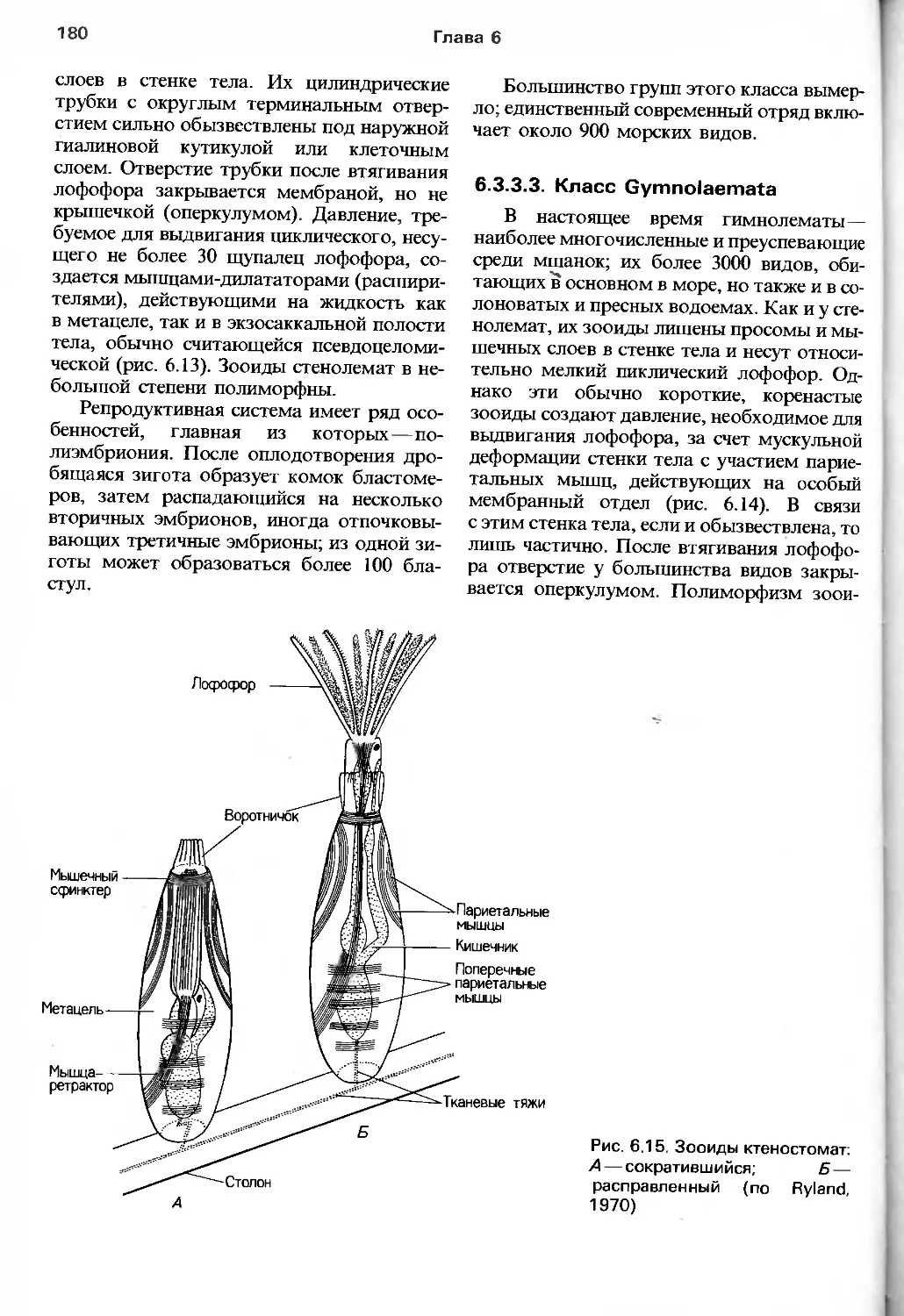
Barnes, "Invertebrate Zoology", Saunders, 1987),
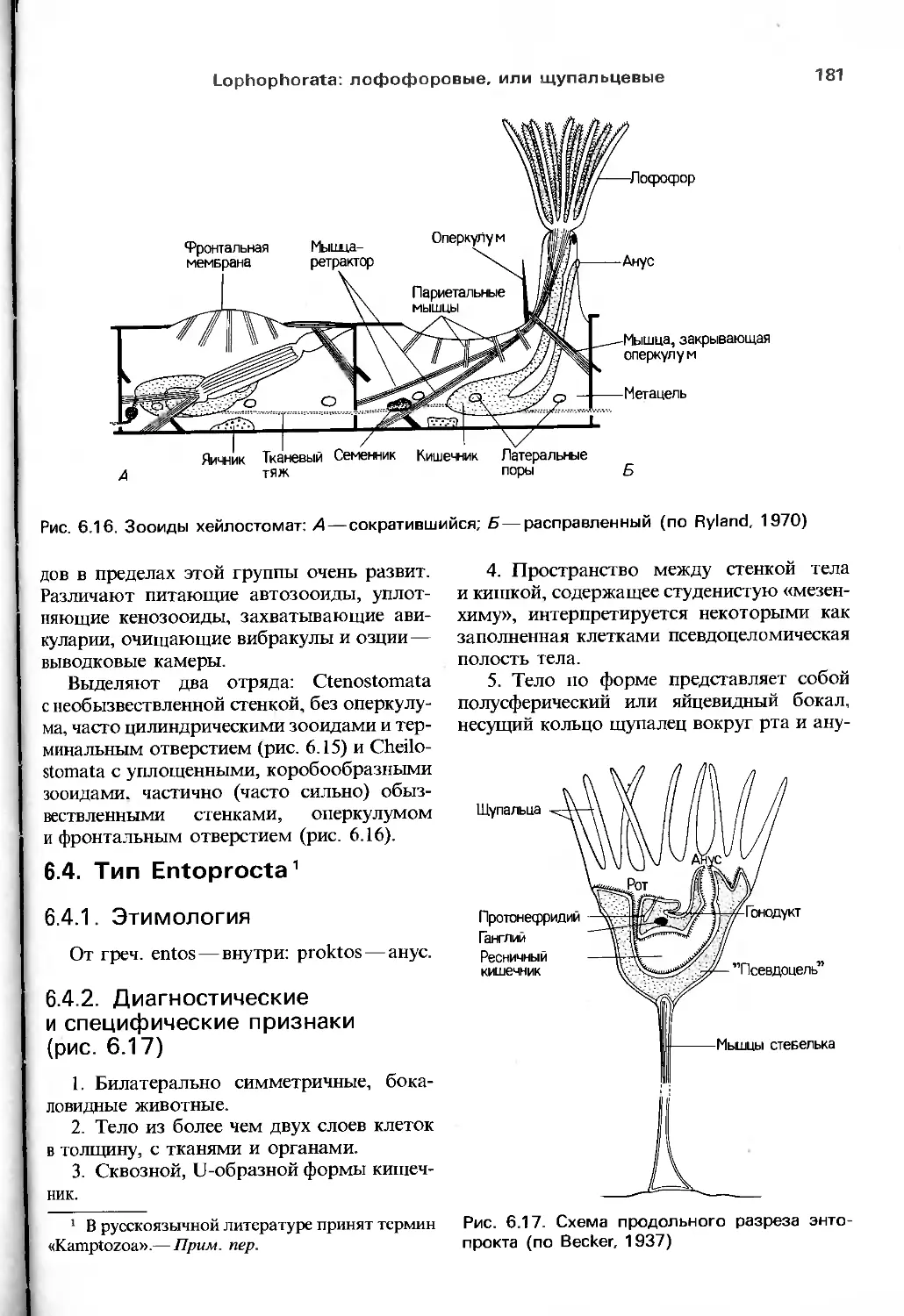
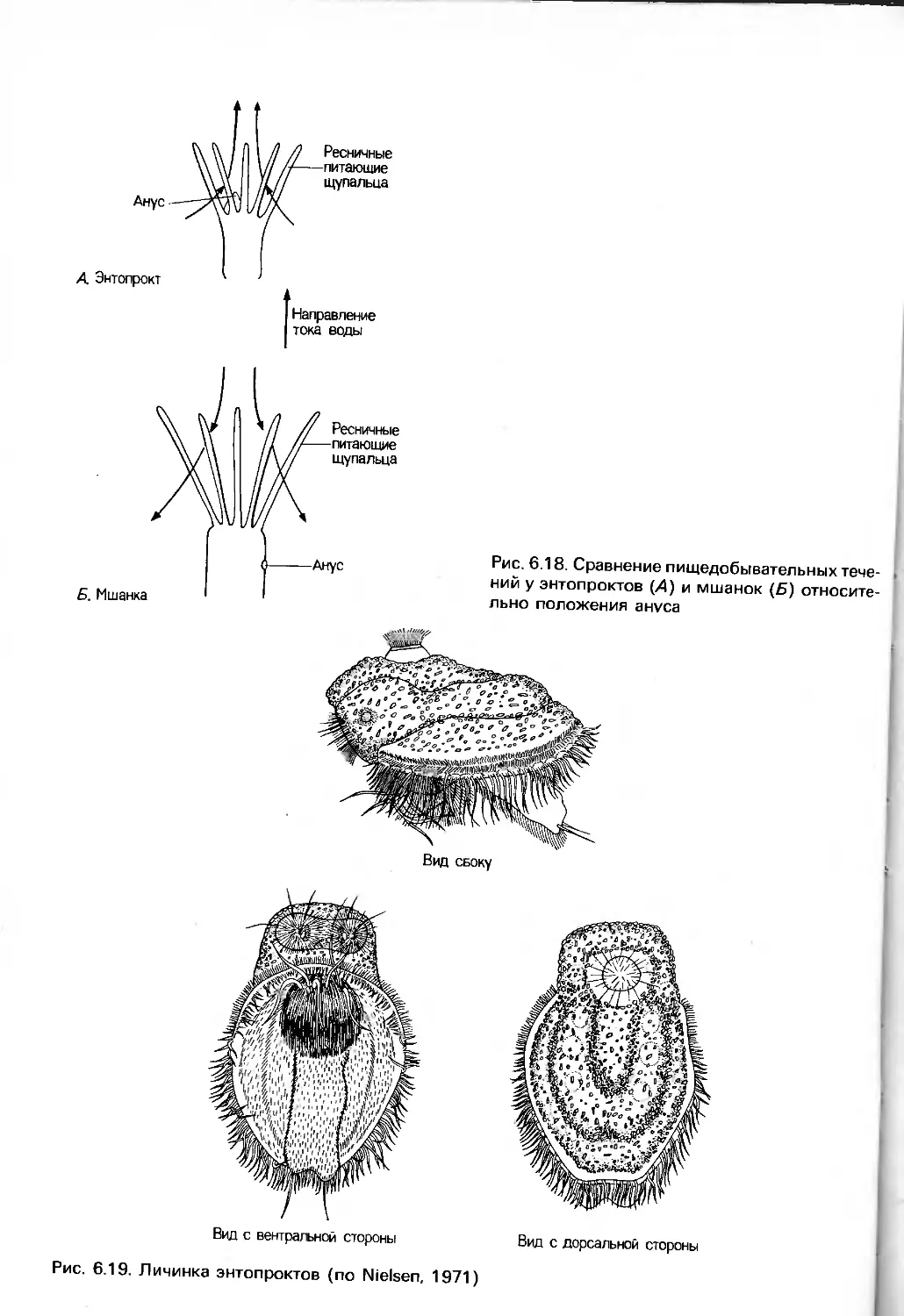
либо функциональные исследования тех или
иных анатомических и физиологических
«систем» (дыхательной, двигательной,
координационной и т. д.) в основном
хорошо изученных групп (например, Е. J.W.
Barrington, "Invertebrate Structure and
Function", Nelson, 1979). Курсы беспозвоночных,
таким образом, требуют по крайней мере
по одной из книг обеих категорий.
Однако в общем за последние 25 лет
произошло сокращение учебного времени,
отведенного специально отдельным
группам, частично для того, чтобы высвободить
время для новых предметов, а также в связи
с потерей популярности систематических
обзоров разнообразия организмов по
сравнению с временами классической зоологии.
В конечном итоге и каждая пара
существующих учебников, и даже многие
отдельные работы содержат гораздо больше
информации, чем это требуется для краткого
курса. Утопая в деталях, студенты не в
состоянии за деревьями увидеть леса.
Поэтому мы и предприняли попытку
объединить под одной обложкой основную
информацию о разнообразии
беспозвоночных и различных их функциональных
системах, информацию, которая, как нам
кажется, действительно требуется для
университетского курса. Нашей главной
проблемой стал отбор того, что можно выкинуть,
а не того, что оставить, и здесь мы
постарались критически оценить существенные
черты каждой группы или системы, обратив
внимание читателя именно на них. Далее,
мы твердо уверены, что эволюционный
подход—стержень понимания всех
аспектов биологии, но, к сожалению, слишком
мало авторов смотрит на животных не как
на застывшие механические формы.
Поэтому везде, где это возможно, мы вели речь об
эволюции, ставя перед собой цель
охарактеризовать разнообразие и функции
беспозвоночных на фоне давления отбора и
селективных преимуществ групп в прошлом
и теперь. Это также повлияло на подбор
и обработку материала. Эта книга, таким
образом, не обобщает уже опубликованные
сведения, а, как мы надеемся, является
новым критическим взглядом на основные
вопросы биологии беспозвоночных.
Поскольку, как отмечалось выше, с
каждым десятилетием зоологические курсы все
меньше и меньше останавливаются на
отдельных типах животных, мы считаем здесь
уместным выступить в защиту широкого
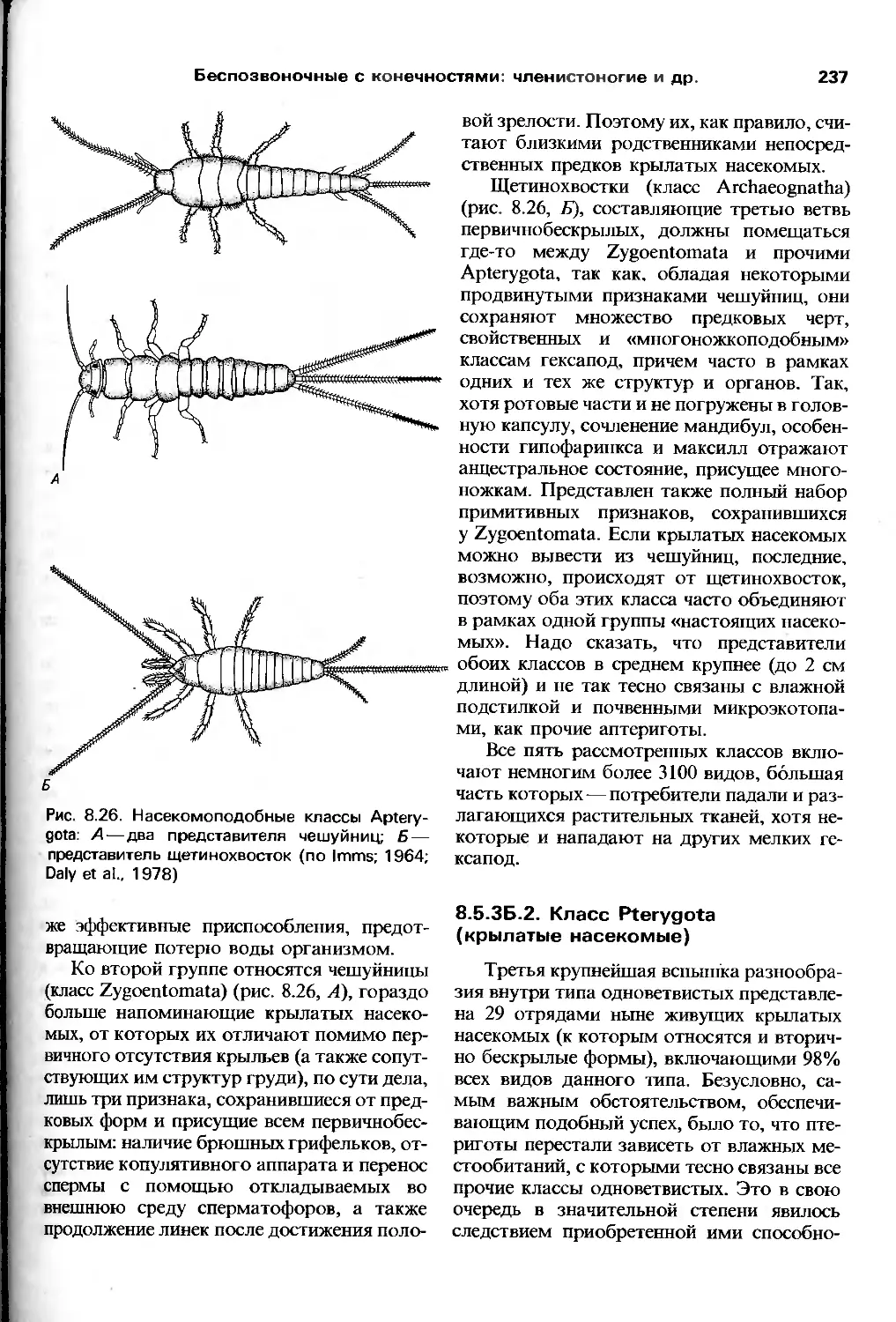
изучения беспозвоночных в высшей школе.
Множество наших современных
представлений о биологических процессах в целом
возникло в ходе исследований именно этих
животных; достаточно вспомнить
плодовую мушку в связи с генетикой и кальмара
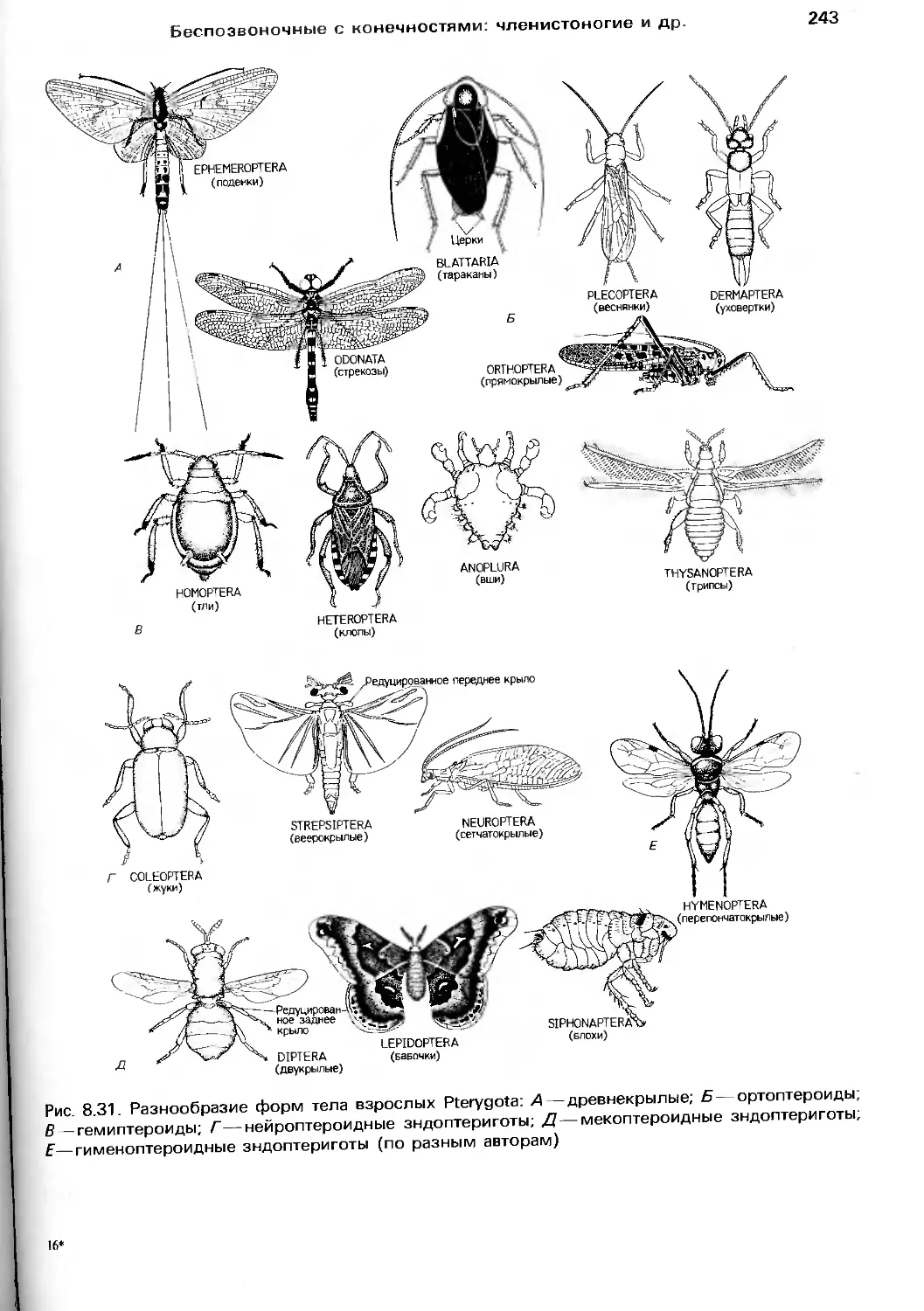
в связи с нейрофизиологией, чтобы все
стало ясно. Однако до сих пор число детально
изученных типов животных, не говоря уже
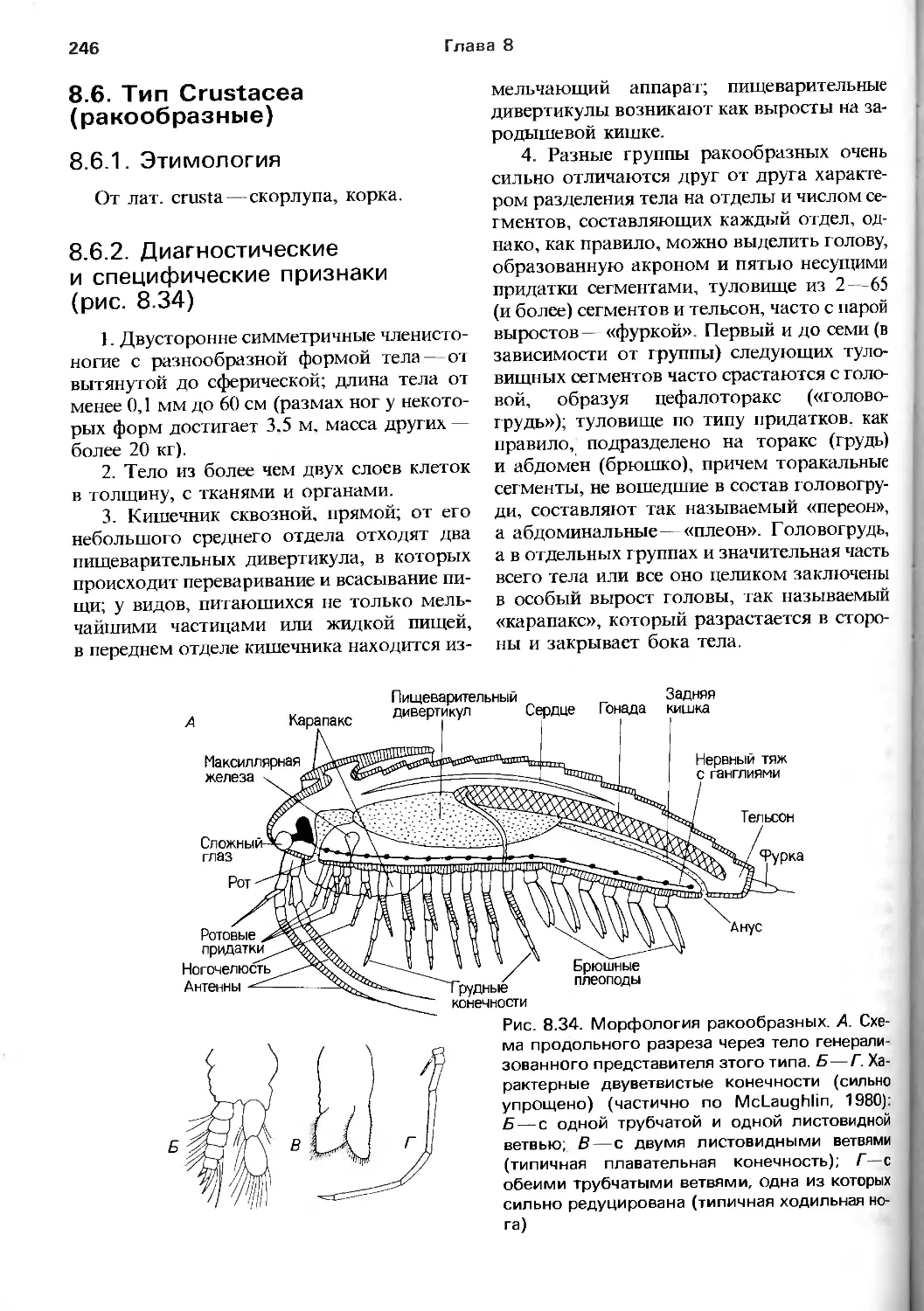
о видах, крайне мало и, конечно, не
отражает всего разнообразия структур и
функций беспозвоночных. Мы верим, что в
будущем множество обобщений возникнет как
раз при изучении групп, не привлекающих
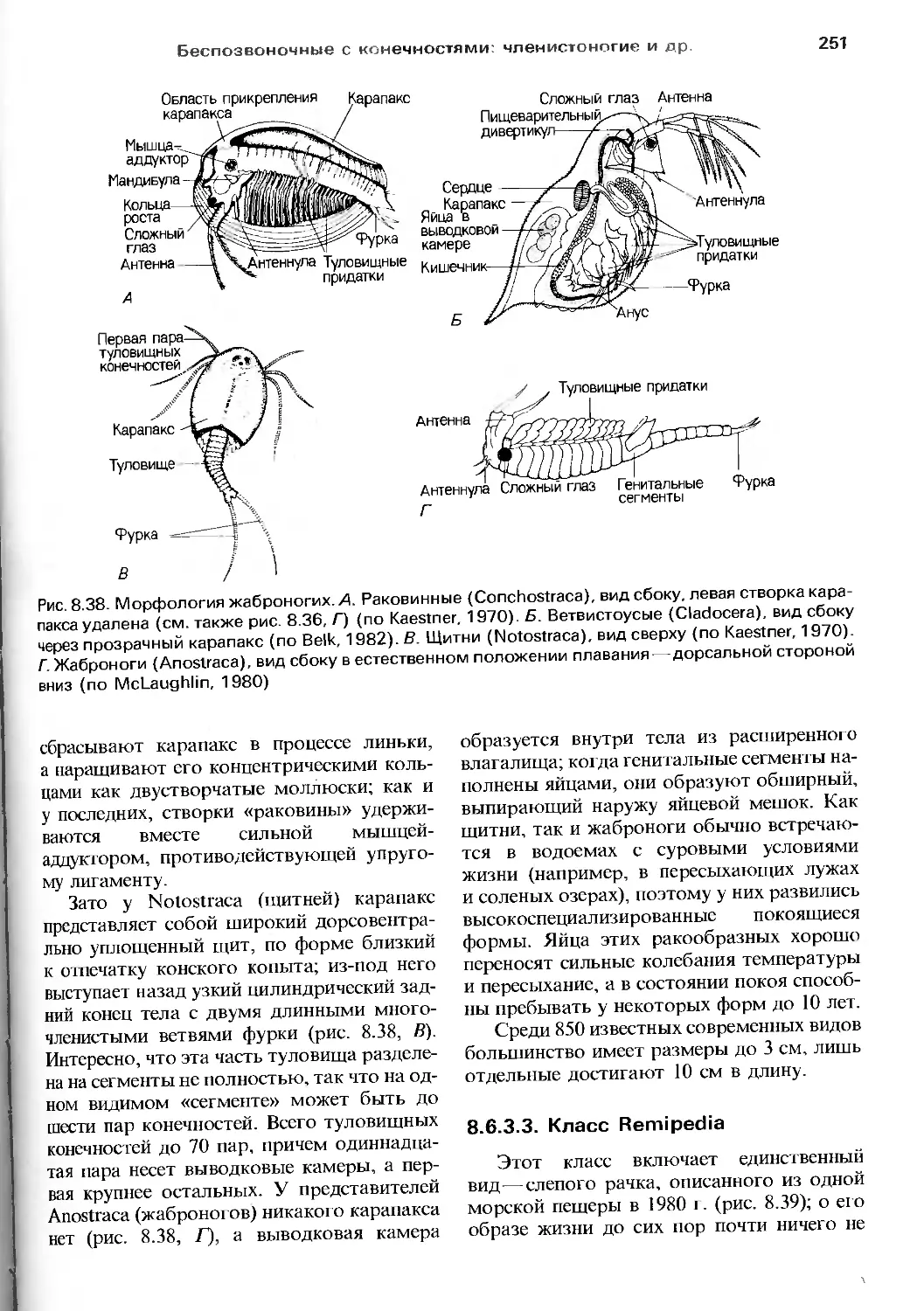
пока всеобщего внимания, и что без оценки
разнообразия, так же, как и единства жизни,
невозможно получить адекватного пред-
8
Предисловие
ставления как о биологии в целом, так и
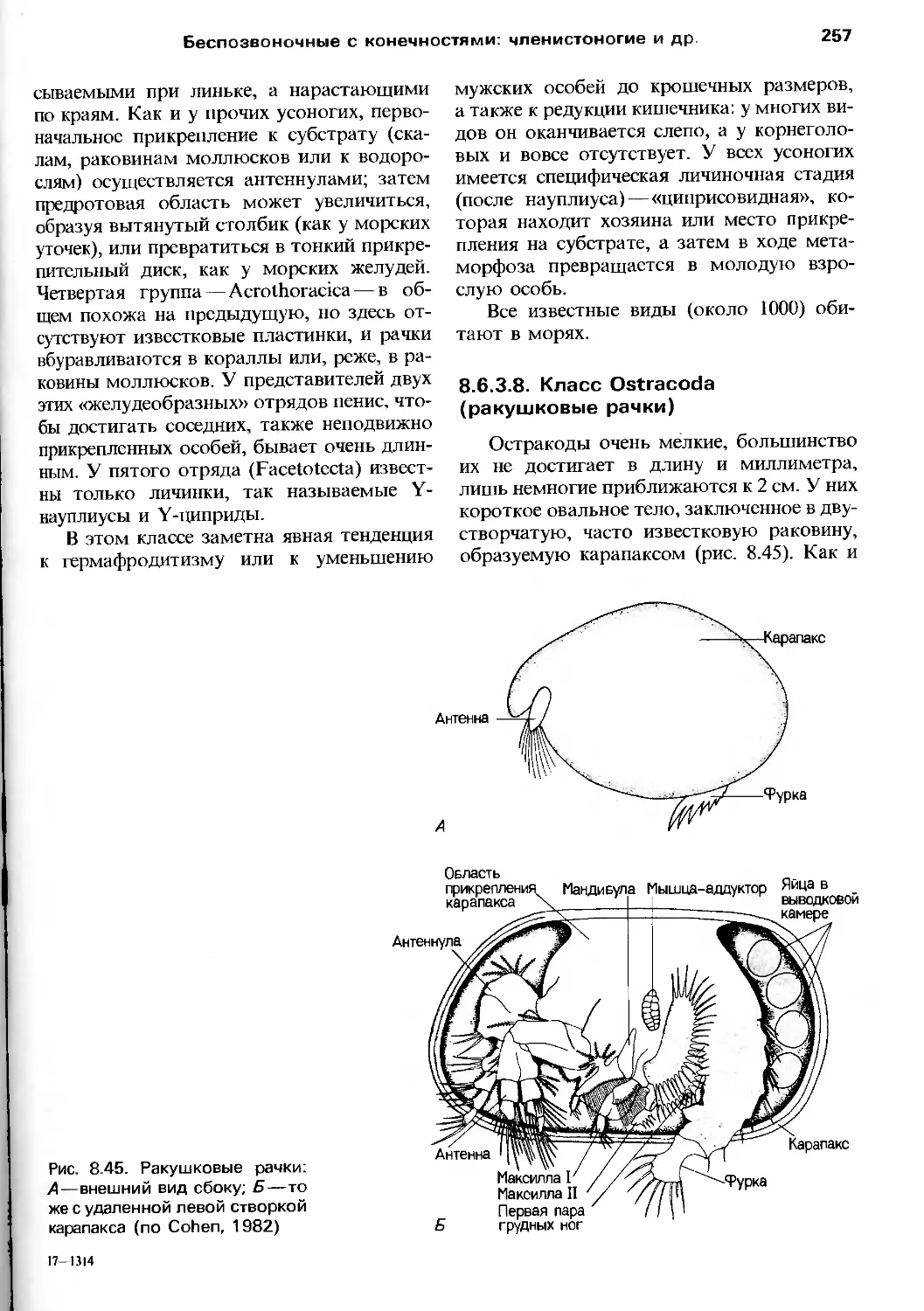
о том, в какой степени наши современные
знания опираются на крошечную
необъективную выборку.
За исключением гл. 16, эта книга—плод
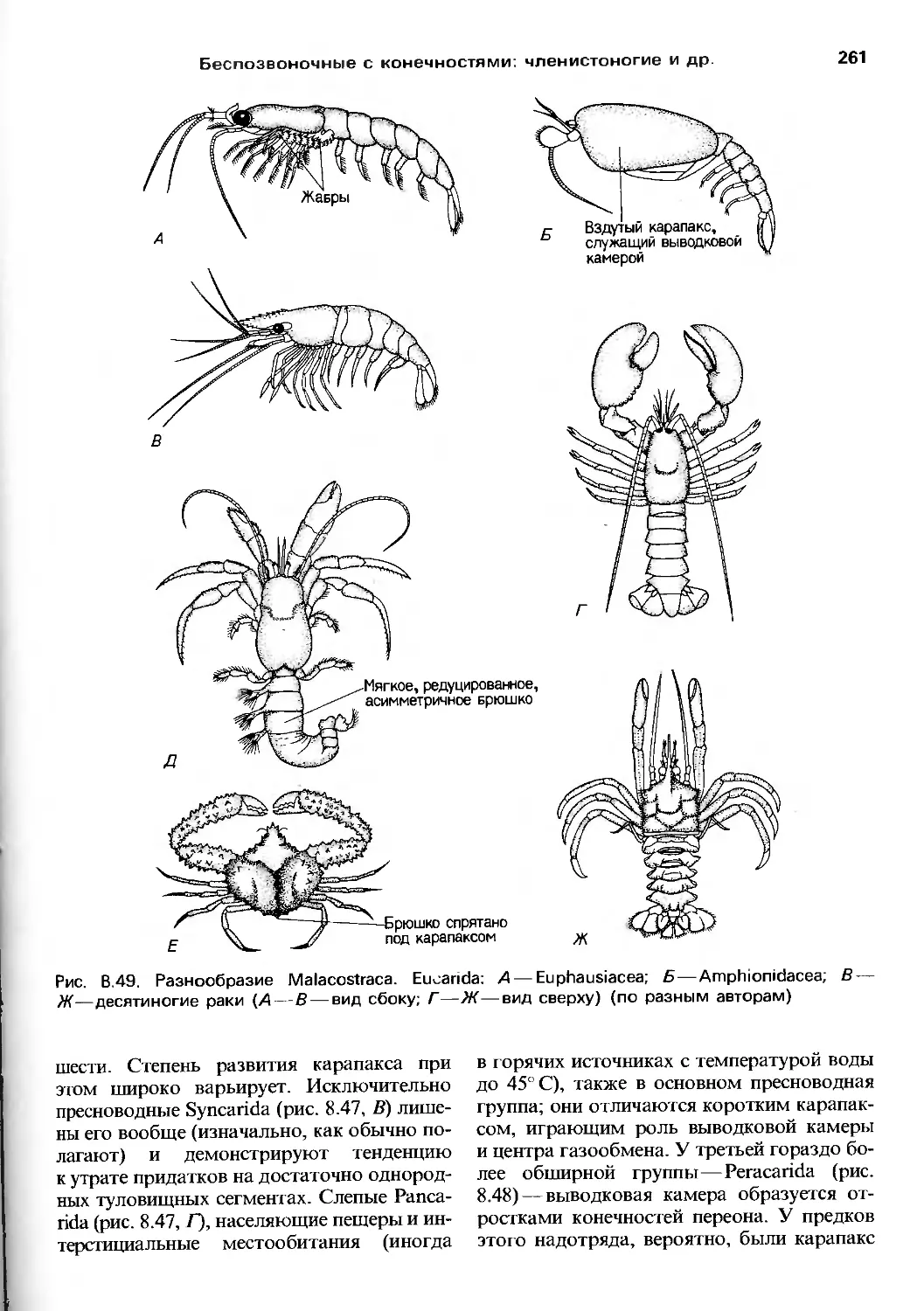
совместных усилий трех авторов. Хотя
практически первые наброски разных глав
или их частей были написаны каждым из
них поодиночке, весь текст затем
перерабатывался в ходе совместного критического
обсуждения: мы все втроем принимаем на
себя ответственность за гл. 1—15, а также за
гл. 17. Однако нет книги, которая была бы
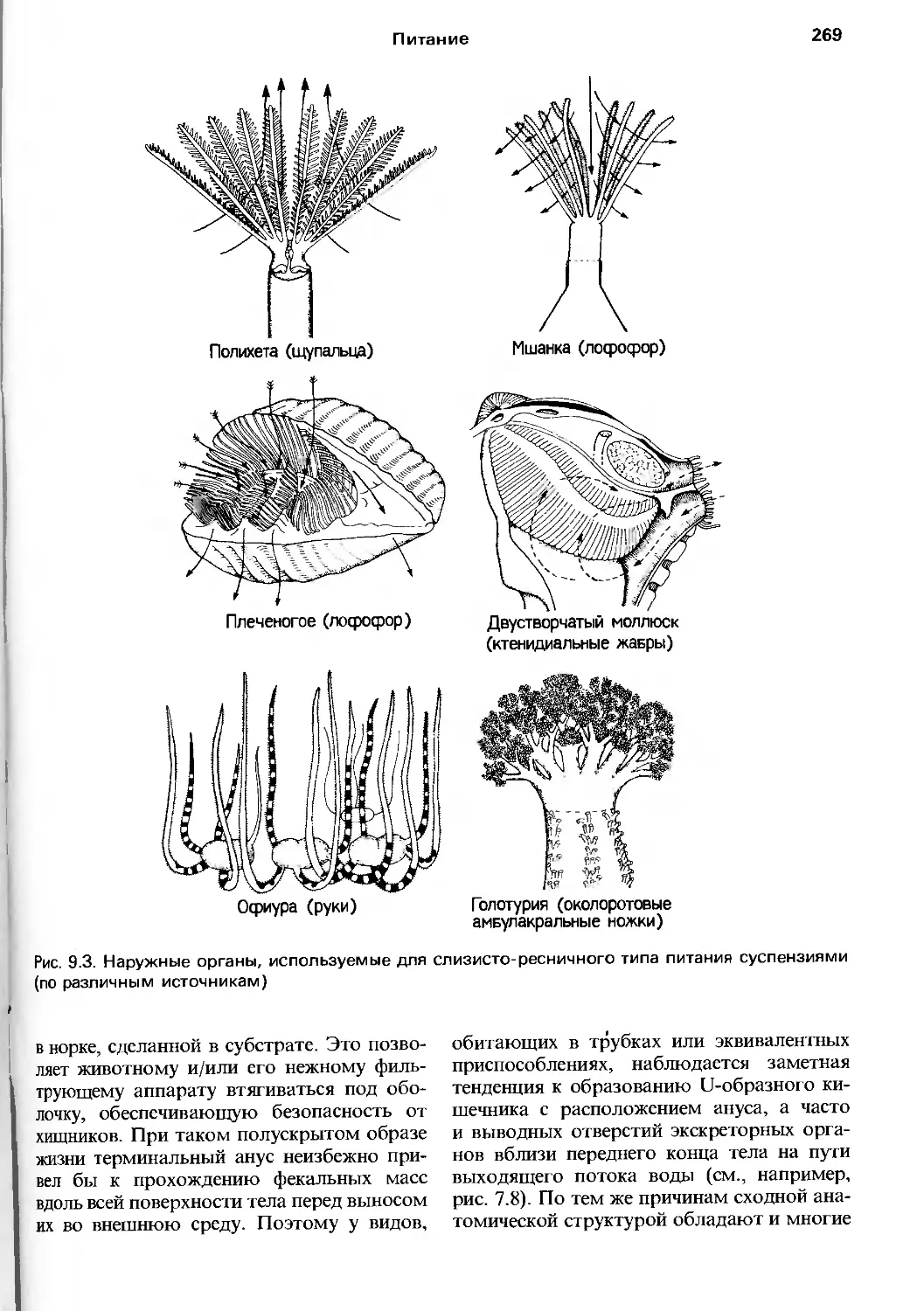
произведением только ее авторов, и мы
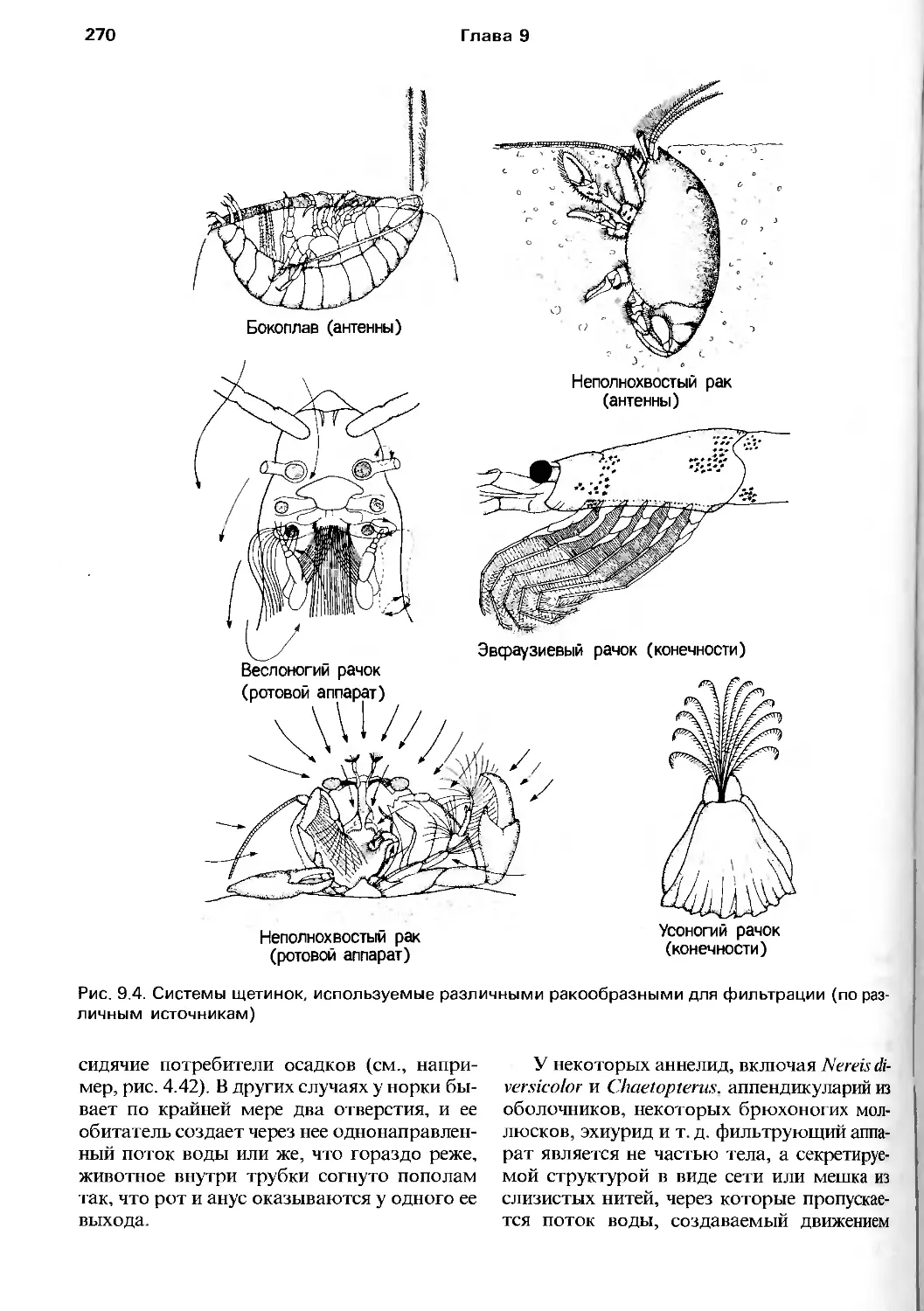
очень признательны множеству людей,
которые помогали нам во время ее
подготовки. В частности, мы хотели бы выразить
нашу благодарность Д. Голдингу, взявшему
на себя гл. 16, и высоко оценить работу
Хелен Крейтон, которая вместе с Б. Фостер-
Смитом и П. Кингстоном снабдили текст
рисунками. Несколько наших коллег были
так любезны, что ознакомились с
набросками материала книги: Г. Беннет-Кларк
прочел всю работу, а Б. Бейн, Дж. Коэн, С. Ко-
нуэй-Моррис, П. Кроган, М. Джамгоз,
Д. Джордж, П. Гиббс, Р. Хьюс, П. Миллер,
Т. Ньюберри, Д. Николе, Дж. Райленд,
Р. Сид, С. Тайлор, и П. Уиллмер — разные
ее части. Многие другие предоставили нам
ту или иную информацию и поделились
своими мнениями. Их усилия спасли нас от
фактических ошибок и текстовых
погрешностей. Мы не рассчитываем на то, что
в книге не осталось ни одной неточности
и ни одного отступления от общепринятых
положений, не в последнюю очередь—
из-за нашей неуступчивости перед лицом
справедливой критики. Р. Кемпбеллу
и С. Роллисону из издательства "Blackwell
Scientific Publications" мы весьма
признательны за всестороннюю помощь, советы,
участие и административное содействие.
Наш долг перед собственными семьями
может оценить только тот, кто когда-либо
посвяшал большую часть своего
«свободного времени» такого рода работе.
Большинство иллюстраций в книге
основано на уже опубликованных в научной
литературе, хотя все они так или иначе
переработаны. Ссылки на оригинальные
источники даны в соответствующих подписях
к рисункам и в списке, прилагаемом в конце
книги.
Р.С.К.Б.
П.К.
П.Дж.У.О.
ЧАСТЬ I
ЭВОЛЮЦИОННЫЕ ВОПРОСЫ
Основная идея, пронизывающая наш
обзор разнообразия беспозвоночных (часть II)
и их функциональной биологии (часть
III),—давление отбора и преимущества,
отразившиеся на этих животных в прошлом
и продолжающие формировать их
биологию в настоящее время. В рамках вводной
части мы кратко описываем эти
могущественные эволюционные факторы.
Слово «эволюция» означает просто
«изменение», а изменения можно
анализировать с двух различных точек зрения,
которые в общем относятся друг к другу, как
причина к следствию или как механизм
действия к его проявлению: а) наличие
процессов, в конечном итоге обусловливающих
наблюдаемые изменения; б) наличие общей
схемы или последовательности
происшедших с течением времени изменений.
Фактически хотя обычно считается, что Чарлз
Дарвин продемонстрировал факт эволюции,
он лишь предложил ее правдоподобный
механизм—естественный отбор,— который
мог бы объяснить эволюционные
изменения, признававшиеся и до него. Как уже
говорилось, эволюционное (или
«филогенетическое») древо типов беспозвоночных и
процесс естественного отбора связаны друг
с другом, но на практике можно видеть
несопоставимость результатов и большое
количество разногласий между, с одной
стороны, популяционными генетиками,
изучающими процессы отбора на живых
организмах, и, с другой—таксономистами,
строящими филогенетические схемы и
пытающимися понять причины возникновения
новых таксонов надвидового уровня.
Здесь мы рассмотрим оба этих подхода
в основном раздельно; в этом смысле гл.
1 помимо того, что является введением
в книгу в целом, описывает отбор как
механизм изменений (этот аспект иногда
называют «специальной теорией эволюции»),
в то время как в гл. 2 обсуждаются
филогенетические связи групп беспозвоночных
(«общая теория эволюции») и
возникновение разнообразия во времени. Тем не менее
в пределах каждой из этих глав мы сочли
уместным представить часть материала
другой главы, например комментируя
такой противоречивый вопрос, как способ
возникновения классов и типов
беспозвоночных.
ГЛАВА I
ВВЕДЕНИЕ: ОСНОВНОЙ ПОДХОД И ПРИНЦИПЫ
1.1. Почему беспозвоночные?
Эта книга о беспозвоночных, т. е.
животных без позвоночника. Такое определение,
основанное на отсутствии, а не наличии
характерного признака, необычно и
подразумевает отклонение от стандартного типа,
у которого этот признак имеется. Если бы
стандарта или нормы не было, подобного
рода определение вряд ли имело бы смысл.
Когда Аристотель разделил животных
на полнокровных и бескровных,
подразумевалось, что присутствие крови—норма. Он
верил в эволюцию живого к некой
совершенной форме, предполагающей наличие
крови, и обобщил эту мысль в
иерархической системе живых существ, называемой
«Лестница природы» (Scala naturae),
отражающей развитие от бескровного
состояния к полнокровному, являющемуся целью
(табл. 1.1).
Аналогичным образом, когда Ламарк
(известный своим учением о
благоприобретенных признаках) впервые отделил
беспозвоночных от позвоночных (в своей «Syste-
me des Animaux sans Vertebres», Paris, 1801),
подразумевалось, что последние—норма.
Вероятно, это опять же вытекало из
своеобразной эволюционной теории Ламарка,
которая предполагала, что приобретенные
признаки наследуются в соответствии
с принципом не только выживаемости, но
и прогрессивного развития к некой высшей
форме, к которой ближе всего подошли
позвоночные и человек.
Современная зоология отказалась от
концепций направленной эволюции
(телеологии), однако деление на позвоночных
и беспозвоночных сохранилось и оказало
влияние на многие поколения студентов.
Это удивительно, потому что проводимая
граница едва ли естественна и даже не
вполне определенна; она отделяет группу,
включающую много типов (беспозвоночные), от
части одного типа (у некоторых
представителей типа Chordata нет настоящего
позвоночника!)
Тем не менее есть две другие важные
причины для разделения учебного курса на
зоологию позвоночных и беспозвоночных.
Первая—историческая; Ламарк создал
прецедент, от которого, с тех пор как он
утвердился как метод подхода к предмету,
оказалось трудно отказаться. Вторая, и,
вероятно, более существенная—
сохранившееся до сих пор ощущение, что,
поскольку у нас самих есть позвоночник,
животные с этим признаком достойны
большего внимания, чем следует просто из их
таксономического статуса.
Обращаясь здесь к биологии
беспозвоночных, мы тем самым сохраняем это
разделение, но не из-за какой-то философской
приверженности теории направленной
эволюции или представлению о
фундаментальных биологических различиях между
позвоночными и беспозвоночными. Наша
позиция скорее прагматическая. Мы хотим
показать, что:
1) у всех живых организмов есть общие
основные черты строения и
функционирования;
2) черты варьируют, и группы таксонов,
объединенные общими их «вариантами»,
называют типами;
3) варианты эволюционировали и,
следовательно, должны быть связаны общим
происхождением;
4) в пределах каждой морфо-
физиологической системы животные
приспособились к окружающим экологическим
условиям в ходе естественного отбора (в ка-
Введение: основной подход и принципы
Таблица 1.1. Лестница жизни, или Лестница природы (Scala naturae), no
Аристотелю
Живородящие
Яйцеродящие
Черверодяшие
Возникают из зародышевой
слизи, путем почкования
или самозарождения из
неживого
Самозарождаются из
неживого
С
совершенными
яйцами
С
несовершенными
яйцами
Полнокровные
1. Человек
2. Волосатые четвероногие
(наземные млекопитающие)
3. Cetacea (морские
млекопитающие)
4. Птицы
5. Чешуйчатые четвероногие
и безногие (рептилии и
амфибии)
6. Рыбы
Бескровные
7. Malaria (головоногие)
8. Malacostraca (ракообразные)
9. Насекомые
10. Ostracoderma (моллюски
кроме головоногих)
11. Зоофиты
кой степени эти микроэволюционные
процессы могут быть связаны с макроэволю-
ционными изменениями, отмеченными
в пп. 2 и 3, вопрос спорный, и мы вернемся
к нему позже).
При рассмотрении этих проблем было
бы целесообразно каким-либо образом
ограничить материал, и мы сделали это на
основе исторического прецедента. Более
того, колоссальное разнообразие
беспозвоночных позволяет полнее
проанализировать пп. 2—4. Однако прежде всего
необходимо остановиться на основных чертах,
общих для всех живых организмов (п. 1), их
отличиях от неживой природы, их
возникновении. Этому посвящено несколько
следующих разделов.
1.2. Свойства живых существ
1.2.1. Введение
В химическом составе живых
организмов обнаружено менее трети всех
встречающихся в природе элементов (табл. 1.2).
Около 75% веса всех животных составляет
вода, 50% сухой массы—углерод с
небольшим количеством кремния или без него.
Земная кора, напротив, состоит более чем
на 20% из кремния и менее чем на 0,1% из
углерода.
Несмотря на ограниченное количество
химических элементов, обнаруженных в
живых организмах, составляющие их
молекулы очень разнообразны по структуре
и функциям. Это объясняется тем, что
атомы углерода в отличие почти от всех прочих
элементов (только кремний несколько
напоминает его в этом отношении) способны
соединяться друг с другом в разнообразные
цепочки и кольца. Такими имеющими
углеродную основу молекулярными блоками
организмов являются: сахара,
аминокислоты, жирные кислоты и нуклеотиды; они
в свою очередь объединяются в
макромолекулы, образуя полисахариды, белки, липи-
ды и нуклеиновые кислоты. Свободные
органические вещества подобного уровня
сложности образуются в неживых системах
крайне редко (с. 14).
12
Глава I
Таблица 1.2. Элементы, обнаруженные в
живых организмах
Элемент Символ Приблизи- Атомный
тельная номер
атомная
масса, Д
Водород "
Углерод1}
Азот1}
Кислород"
Фтор
Натрий 2)
Магний 2)
Кремний
Фосфор 2)
Сера 2)
Хлор2)
Калий г)
Кальций
Ванадий
Хром
Марганец
Железо 2)
Кобальт
Медь
Цинк
Селен
Олово
Иод
Н
С
N
О
F
Na
Mg
Si
Р
S
CI
к
Ca
V
Cr
Mn
Fe
Co
Cu
Zn
Se
Sn
[
1
12
14
16
19
23
24
28
31
32
35
39
40
51
52
55
56
59
64
65
79
119
127
1
6
7
8
9
11
12
14
15
16
17
19
20
23
24
25
26
27
29
30
34
50
53
!) Четыре первых по частоте встречаемости в
живых организмах элемента.
2) Следующие по частоте встречаемости в живых
организмах элементы.
Еще более глубокое различие между
живым и неживым заключается в способе
организации органического вещества. В
живых системах макромолекулы образуют
мембраны, ограничивающие другие
неслучайные скопления макромолекул,
реагирующих друг с другом в процессе
упорядоченного метаболизма. Эти «упаковки»—
клетки. Множество клеток образует
многоклеточный организм, в котором они
объединены в хорошо управляемое и
организованное структурно-функциональное целое.
Само существование, устойчивость такой
упорядоченности и сложности долгое время
считались специфической и даже загадочной
чертой организмов, созданных и
поддерживаемых мистической жизненной силой,
поскольку неживая природа подчиняется
физическому Второму закону термодинамики,
гласящему, что порядок и организация—
нестабильны. Энтропия, т. е.
неупорядоченность, должна постепенно возрастать во
всех реакциях и процессах.
Тем не менее сейчас признается, что
устойчивая организация биологических
систем основана на двух отнюдь не
таинственных чертах, присущих всему живому. Они
являются решающими для понимания
основных принципов биологии, и мы постоянно
будем к ним обращаться.
1. Организмы запрограммированы.
2. Этим программам подчинены
рабочие системы и подсистемы, которые
открыты для поступления вещества и энергии.
1.2.2. Программа
Если взять самый фундаментальный
уровень, генетическая программа
контролирует свойства белков, определяя типы
и последовательность аминокислот, из
которых они образованы. Для животных
характерны только 20 аминокислот, но
цепочка, включающая всего 100 из них (это
немного для белка), может иметь в принципе
20 10° различных конфигураций! Это
объясняет большое разнообразие белков.
Некоторые из них относятся к ферментам,
контролирующим все процессы обмена
веществ, другие используются как
строительный материал клеток организма в целом.
Сама программа закодирована в ну-
клеотидной последовательности ДНК.
Четыре типа нуклеотидов (аденин, тимин,
гуанин и цитозин) определяют 20 различных
аминокислот, так что однозначного
соответствия первых последним быть не может.
Только комбинация как минимум трех
нуклеотидов дает достаточное число
альтернативных сочетаний (43 = 64); такой три-
плетный код универсален для живого,
причем лишние альтернативы (около 40)
обусловливают его избыточность (одна
аминокислота кодируется несколькими
триплетами) и наличие особой «пунктуации». Тем не
менее прямой передачи информации от
ДНК синтезирующимся белкам не
происходит. Сначала она «переписывается» в виде
Введение: основной подход и принципы
13
ДНК
РНК
Белок
Ядерная
мемврана
Рис. 1.1. Схема синтеза белка на
молекулярном уровне (по Callow, 1976): гяРНК —
гетерогенная ядерная РНК, впоследствии
преобразующаяся в мРНК
молекул РНК (сходны с ДНК, но тимин
заменен урацилом), которые переносят ее
в цитоплазму, где на содержащих особый
тип РНК частипах — рибосомах—служат
матрицами для упорядоченной сборки
определенных аминокислот.
Аминокислоты переносятся и «вставляются» на место
другим типом РНК—транспортной. Вся
эта сложная система схематически
изображена на рис. 1.1.
1.2.3. Открытость
Упорядоченные системы организмов
вплоть до их макромолекул постоянно
подвергаются «атакам энтропии», но их
разрушающиеся компоненты могут замещаться
в соответствии с информацией,
заключенной в генетических программах. Это
возможно только в том случае, если существуют
постоянный приток упорядоченного
«сырья» (в своей знаменитой книге "What is
Life?", вышедшей в 1944 г., Эрвин Шредингер
назвал это поглощением негэнтропииХ) и
отток утратившего организацию материала
(экскретов) и энергии (в основном тепла).
Таким образом, организмы и
составляющие их клетки должны быть открытыми
системами, а это подразумевает, что даже
у нерастущих животных идет постоянная
замена клеток и/или молекул—то, что Ше-
нхеймер (Schoenheimer) в своей книге "The
Dynamic State of Body Constituents" (1946)
определил как динамическое устойчивое
состояние тела.
1.2.4. Эволюция путем
естественного отбора —
неизбежное следствие
возобновления систем
за счет репликации
Целые организмы также могут
заменяться себе подобными, т.е.
воспроизводиться путем репликации генетической
программы. Это предполагает отделение
многоклеточных или одноклеточных структур
(пропагул), несущих полную или неполную
генетическую программу (геном)
родителей. Многоклеточные пропагулы всегда
имеют более или менее полные копии
родительского генома; репродуктивные
процессы с их участием называются бесполыми
или вегетативными. Одноклеточные
пропагулы (гаметы), как правило, содержат лишь
часть родительской наследственной
программы и должны слиться с другими
гаметами, чтобы она стала полной (процесс
оплодотворения); только после этого
начинается развитие. Это называют половым
размножением.
Таким образом, речь идет о системах,
реплицирующих геном для воспроизведения
организма. Однако геном не всегда
копируется точно. Даже при бесполом
размножении мутации вызывают его изменения,
а при половом процессе дополнительная
«перетасовка» генов, связанная с мейозом,
и их «перемешивание» при оплодотворении
обусловливают значительные различия
между родителями и потомством.
Изменчивость генетической программы ведет к
разнообразию формы и функций фенотипов,
что в свою очередь влияет на способ их
взаимодействия с окружающей средой,
а следовательно, определяет шансы особей
на выживание и скорость их
воспроизведения. Из этого следует, что генетические
программы, лучше всего обеспечивающие
выживание и плодовитость в данной
окружающей среде, т.е. наиболее адаптированные
к ней, станут чаще встречаться. Более того,
поскольку мир конечен и ресурсы, необхо-
14
Глава I
димые для жизненных процессов,
ограничены, эти программы будут иметь тенденцию
замещать другие, менее выигрышные. Так
схематически выглядит процесс эволюции
путем естественного отбора, который
впервые четко описан Чарлзом Дарвином в его
книге «Происхождение видов» (1859). Для
характеристики этого процесса он
использовал заимствованную у Герберта Спенсера
формулировку «выживание наиболее
приспособленных». Однако из сказанного выше
ясно, что приспособленность представляет
собой свойство одного генетически
детерминированного признака распространяться
в популяции за счет других, а ото требует
как выживания, так и плодовитости.
1.3. Происхождение жизни
Самая фундаментальная черта живых
систем — их способность сохранять
упорядоченное и организованное состояние за
счет процессов запрограммированной
репликации и воспроизведения. Эволюция
путем естественного отбора является
автоматическим следствием этого. Но каким
образом возникла такая система? Открытие
того, как возникли органические молекулы,
составляющие живое (с. 11), дает только
частичный ответ на этот вопрос. Мы должны
представить себе, как они организовались
в самореплицирующиеся системы.
Углеродсодержащие молекулы, из
которых состоят организмы, когда-то считали
настолько специфическими и уникальными,
что возможность их синтеза связывалась
только с живыми существами. Таким
образом проводилось разграничение между
органическими (= из живого) и
неорганическими веществами. Первая брешь в
разделяющей их стене была пробита Велером,
синтезировавшим в 1832 г. очень простую
органическую молекулу (мочевины) из циа-
ната аммония (неорганическая молекула)
в результате простого ее нагревания. Это
положило начало рациональному и
научному подходу к химии живого, а тем самым —
современной биохимии и молекулярной
биологии. Хотя, конечно, контролируемый
синтез мочевины—это еще очень далеко от
спонтанного образования полисахаридов,
липидов, белков и нуклеиновых кислот,
необходимого для возникновения живых
систем.
1.3.1. Добиологический синтез
органических полимеров
О первоначальной земной атмосфере
известно очень мало, но, возможно, она
возникла в ходе «дегазации» планеты и
была очень похожа на газовую смесь,
выделяемую вулканами. Если это так, то в ней
почти наверняка не было кислорода (см. гл. 11).
Эксперименты показали, что при этих
условиях любое энергетическое воздействие —
освещение, ударные волны,
ультрафиолетовая радиация (поскольку отсутствовал
кислород, не было и озонового экрана,
отфильтровывающего волны этой длины из
солнечного спектра) или тепло
вулканического пепла привело бы к добиологическо-
му синтезу разнообразных «органических»
мономеров: Сахаров, аминокислот и даже
нуклеотидов. При благоприятных
обстоятельствах, например при высоких
концентрациях неорганических полифосфатов, они
могут объединяться в длинные цепочки,
образуя, в частности, полинуклеотиды и
полипептиды. Все эти вещества, вероятно,
накапливались в древнем океане, образуя так
называемый «первичный бульон».
В этом добиолегическом мире шел
своего рода отбор, поскольку молекулы,
способные полимеризоваться быстрее других
и/или оказавшиеся более стабильными,
получали большее распространение. Но
темпы таких изменений были медленными
и вряд ли могли привести к возникновению
принципиально новых структур, поскольку
образование каждого полимера
происходило независимо и не закреплялось в
генетической памяти. Следующий за этим шаг не
представлял особой трудности, поскольку
некоторые полимеры, однажды
образовавшись, могут влиять на образование других
полимеров. Полинуклеотиды, в частности,
способны определять
последовательность нуклеотидов, действуя как матрица
для полимеризации. Если один полинуклео-
тид служит матрицей для
комплементарного ему другого полинуклеотида, который
в свою очередь является матрицей для
первого, можно говорить о множестве моле-
Введение: основной подход и принципы
15
кул, связанных чем-то вроде генетической
памяти. Полинуклеотиды,
воспроизводящиеся наиболее эффективно,
увеличиваются в количестве по сравнению с прочими,
т.е. имеют селективное преимущество.
Матричным системам свойственны
ошибки репликации, и новые
полинуклеотиды, образующиеся за счет «мутаций»,
вероятно, конкурировали с другими за
ограниченное количество мономерных
структурных блоков. Поскольку дезоксирибону-
клеозиды (предшественники нуклеотидов)
синтезируются сложнее рибонуклеозидов
и поскольку РНК играет центральную роль
в современном белковом синтезе, широко
(но не всеми) признана гипотеза, согласно
которой первыми самореплицирующимися
полимерами были короткие цепочки РНК.
1.2.3. Происхождение
и эволюция клеток
Следующий шаг по направлению к
системам, иллюстрируемым рис. 1.1,
представить труднее всего. При нормальной
температуре спонтанная репликация, описанная
выше, протекала бы медленно и с высоким
процентом ошибок. Включение в этот
процесс репликазы—белка, способного
катализировать репликацию, существенно бы
его ускорило. Каким образом это
произошло— не ясно, но, возникнув, такая
система получила преимущества. Более того,
должны быть определенные преимущества
в окружении матрицы и репликазы
оболочкой, чтобы выгоды от их взаимодействия не
могли использоваться другими, немного
отличными матрицами-конкурентами. Так
появилась клетка и разница между
генотипом и фенотипом. На эти примитивные
клетки отбор должен был действовать
таким образом, что те из них, у которых
взаимодействие между генотипом и фенотипом
стимулировало скорость репликации и ее
точность, распространялись быстрее
других. Хотя трудно точно представить себе
ход событий, но именно в ходе
взаимодействия и отбора возникли и
совершенствовались сложные системы, включающие ДНК
и различные формы РНК.
Первые клетки были мелкими и просто
устроенными. Отчасти они напоминали
современных бактерий, так называемых
прокариот. В некоторых из них шло
дальнейшее формирование мембран для отделения
генетической информации от остального
объема, что, вероятно, давало
преимущества, поскольку обеспечивало лучшую
защиту генетического материала от
повреждений. Такие клетки, так называемые про-
тоэукариоты, возможно, позднее
приобрели цитоплазматические органеллы, из
которых наибольшее значение имели
митохондрии. Последние очень сходны со свободно-
живущими прокариотами, напоминая их
размерами, формой, наличием собственной
ДНК и размножением путем деления
надвое. Поэтому сейчас считается, что они
возникли за счет симбиоза между мелкими
прокариотами, похожими на ныне
живущую бактерию Рагасоссш, и более
крупными, содержащими ядра протоэукариотами.
Разрушая эукариотические клетки, можно
показать, что весь механизм аэробного
метаболизма связан именно с
митохондриями, так что этот предполагаемый симбиоз
развивался параллельно накоплению в
земной атмосфере кислорода вследствие
фотосинтетической активности древних циано-
бактерий.
1.3.3. Почему самозарождение
жизни не происходит все время?
Если крупные биомолекулы и даже
клетки возникли однажды, правомерен вопрос,
почему этого не происходит постоянно.
Вероятно, ответ заключается в том, что живые
существа сами создали условия, в которых
самозарождение невозможно. Например,
кислород, продукт их обмена веществ,
образовавшись, должен разрушать
органические молекулы, из которых построено
живое. В его присутствии органические
полимеры окисляются до простых
неорганических составляющих. Следовательно, когда
свободного кислорода накопилось
достаточно много, существование «первичного
бульона» стало невозможным. Кроме того,
сложные органические молекулы в его
составе, вероятно, были превосходным
источником питания для первых организмов
и поедались или разрушались скорее, чем
синтезировались вновь.
16
Глава I
1.4. Уровни организации
организмов
Вряд ли вся совокупность
физиологических процессов, когда они «втиснуты» в
одну клетку (как у протистов; разд. 3.1), может
работать столь же эффективно, как в случае
их разделения между различными клетками
многоклеточного организма. Многоклеточ-
ность обеспечивает большее пространство
для реакций, а разделение функций между
клетками означает сведение к минимуму
физиологических противоречий по крайней
мере в пределах каждой из них. Таким
образом, в сторону многоклеточности, т. е.
происхождения царства Animalia, должно было
действовать значительное давление отбора.
Следующая глава обобщает основные
черты беспозвоночных и соображения об их
родственных связях и эволюции. Она
иллюстрирует различные возможные тенденции
развития, постепенно раскрывающие
физиологический потенциал многоклеточных
животных, в частности:
1) эволюцию клеточной дифференпи-
ровки;
2) пространственное распределение
клеток одного типа по тканям, а затем
организацию их в органы (совокупности клеток,
обслуживающих общую функцию);
3) эволюцию сквозного кишечника,
обеспечивающего большую специализацию
разных отделов тела;
4) развитие заполненной жидкостью
полости тела, которая позволяет кишке и
другим органам (например, сердцу) работать
независимо от мускулатуры стенки тела,
способствует диффузионному
распределению питательных веществ, и, действуя как
гидростатический скелет (разд. 10.5),—
более эффективной локомоции;
5) эволюцию специфических систем
переноса питательных элементов и
дыхательных газов между тканями,
позволяющую избежать размерных ограничений,
связанных с диффузионным
распределением этих веществ (см. гл. 11);
6) эволюцию конечностей, значительно
расширяющую возможности локомоции,
особенно на суше и в воздухе.
Легко показать, как естественный отбор
приводит к совершенствованию функций
в пределах одного уровня организации. Но
обеспечивает ли он переход с одного уровня
на другой? Происходят ли эти переходы
постепенно за счет непрерывных мелких
изменений, улучшающих функционирование
и приспособленность организмов, или
путем скачков между уровнями, больше
объясняющихся случайностью, чем
естественным отбором? Эти альтернативы
рассмотрены соответственно как гипотезы
постепенности и прерывистого равновесия. Без
сомнения, эволюция беспозвоночных
представляет собой отчасти прерывистый
процесс (гл. 2). Но поскольку такие скачки
обычно происходят в геологических
масштабах времени, т.е. в течение нескольких
миллионов лет, они могли произойти и под
действием естественного отбора. Похоже,
что его давление время от времени сильно
менялось, что вызывало значительные
колебания темпов эволюции. Так что
прерывистый характер эволюции не исключает
действия дарвиновских механизмов. Этот
вопрос горячо дебатировался в
эволюционной экологии (Calow, 1983) и мы позже
вернемся к нему.
1.5. План книги
В этой главе кратко рассмотрено то, что
именно мы считаем основными признаками
живых систем.
Это организованные системы,
поддержание которых зависит от программ,
репликации и открытости.
Отсюда почти логически вытекают
черты животных.
Они потребляют пищевые ресурсы из
окружающей среды и используют их так,
что это обеспечивает выживание и
плодовитость.
Различные уровни организации
животных эволюционировали, и в пределах типов
действовал естественный отбор,
повышающий адаптацию процессов получения и
утилизации ресурсов. После предварительного
обзора разных уровней организации в
следующей главе мы более детально опишем
их в части II. Это даст основу для более
глубокого рассмотрения поведения и
физиологии беспозвоночных в части III, где мы
сосредоточим внимание на отдельных аспек-
Введение: основной подход и принципы
17
тах их функционирования. Таким образом,
в части II последовательно обсуждаются
разные типы, а в части III принят
межтиповой, функциональный подход к
беспозвоночным. Читатель, таким образом, может
выбирать, заняться ли ему в основном
систематикой (часть II), используя часть III
как источник дополнительной информации,
или, наоборот, обратиться к
функциональной биологии беспозвоночных (часть III),
пользуясь частью II как «указателем»
упоминающихся таксонов. В любом случае обе
эти части составляют одно целое и
призваны дать полное и всестороннее
представление об организмах беспозвоночных.
Центральна. иа.ун4*шц
б!бл1отека ХДУ 3
1.6. Дополнительная
литература
Calow Р. 1976. Biological Machines. Edward
Arnold, London.
Calow P. 1983. Evolutionary Principles. Blackie,
Glasgow.
Maynard Smith J. 1986. The Problems of Biology.
Oxford University Press, Oxford.
Smith DC, Douglas A.E. 1987. The Biology of
Symbiosis. Edward Arnold, London.
ГЛАВА С-
ЭВОЛЮЦИОННАЯ ИСТОРИЯ
И ФИЛОГЕНИЯ БЕСПОЗВОНОЧНЫХ
Современные животные —результат
эволюционной истории; невозможно
полностью понять современную биологию без
оценки ее прошлого и ограничений,
наложенных им на возможности и образ жизни
организмов. В этой главе мы опишем
основные черты эволюционной истории
животного царства, в том числе его
происхождения.
Слишком часто филогенетические
реконструкции представляют собой довольно
бесплодное занятие—остроумные, но
малоперспективные попытки объединять
планы строения в разные комбинации с целью
получения удовлетворительной, т. е. более
или менее логичной картины — что-то
вроде составления картинки из кусочков,
большинство которых утеряно. Для заполнения
пробелов в таких схемах предлагались
гипотетические формы, часто без учета того,
что такие постулируемые животные
должны были выживать, добывать пищу,
избегать хищников и размножаться. Причиной
бесплодности нескольких из этих попыток
было отчасти использование
исключительно анатомического подхода, нескольких
«ключевых признаков» (часто
эмбриологических по своей природе), исследование
которых на более широком спектре животных
выявило их меньшую определенность
и диагностическую ценность, чем
предполагалось изначально.
Здесь мы постарались дать современную
филогенетическую оценку накопленных
данных, исходя из возможного действия
отбора в разные периоды в прошлом и
вероятных реакций на его давление живших
тогда организмов. Мы также
сосредоточили внимание на том, что говорят
ископаемые находки о характере увеличения
разнообразия и ходе вымирания,
продемонстрировав, как наши знания о прошлом влияют
на понимание эволюционного процесса.
Именно в этой области у палеонтологов
возобновились эволюционные споры,
связанные с широко известными гипотезами
«макроэволюции» и «прерывистого
равновесия».
Читатели должны обратить внимание
на две вещи. Во-первых, в этой главе
неизбежны ссылки на некоторые анатомические
черты, которые описаны в части II. Мы
считаем более логичным дать общий обзор
перед детальным описанием отдельных
групп, даже если это подразумевает, что
некоторые особенности строения и концепции
будут описаны позднее. Пока же
характеристики различных типов сведутся к
необходимому минимуму. Во-вторых, как
подчеркивается в этой главе, филогенетические
связи животных остаются спорными
вопросами, а в нашей книге не ставится задача
обсуждения всех противоположных точек
зрения. Мы просто приводим те
представления, которые кажутся нам наиболее
правдоподобными, не вдаваясь в детальное
обоснование всех принимаемых точек зрения.
Главы части II также содержат только
конкретную информацию (например,
относительно фундаментальных различий между
типами членистоногих), и те, кто интересуется
принципами таксономического разделения,
должны обратиться к спискам
«дополнительной литературы», рекомендованной
в конце этой и других глав.
2.1. Введение
Очевидно, что многоклеточные
животные, как и представители двух других
царств многоклеточных—грибы и
растения,—произошли от одноклеточных (или
Эволюционная история и филогения беспозвоночных. 19
Charnia Sprtggina Cyctomedusa Rangea Praecambridium
Рис. 2.1. Докембрийские (из отложений Эдиакары) животные (по Glaessner, Wade, 1966)
неклеточных) эукариотных протистов. Но
на этом ясность кончается. Большинство
типов животных, представленных в
ископаемых остатках, впервые появляются
«полностью сформированными» уже в
кембрии, около 550 млн. лет назад. Это
касается и таких продвинутых, анатомически
сложных типов, как трилобиты, иглокожие,
плеченогие и моллюски. Докембрийские
остатки животных немногочисленны, но
обычно считается, что книдарии и
сегментированные черви тогда уже существовали
(рис. 2.1). Ископаемые находки, таким
образом, ничего не говорят о
возникновении и ранней дивергенции разных типов, за
исключением того, что эти события должны
датироваться докембрием, вероятно,
периодом 1200—900 млн. лет назад. Сама
жизнь, как считается, возникла 3500 млн.
лет назад.
Множество споров вызывает вопрос,
какую группу животных считать наиболее
примитивной и от какой группы протистов
ее можно вывести. Если не учитывать
слишком фантастических гипотез, три типа
считались и считаются основными
претендентами на роль предковой группы
животных: губки (Porifera), кишечнополостные (в
особенности Cnidaria) и плоские черви (Р1а-
tyhelminthes). Ранние сторонники
примитивности той или иной из этих групп
впоследствии стремились вывести из нее две
остальные, а в конечном итоге и все прочие
известные типы. Сравнительно рано губок
начали считать довольно специфической
изолированной группой, возможно
неродственной другим животным, и поместили
в особое подцарство Parazoa. отдельно от
всех остальных типов, кроме вымерших
Archaeocyatha. Вопрос тем самым был
сведен к тому, произошли ли
кишечнополостные от плоских червей или наоборот?
Первые животные были, по-видимому,
мелкими, состояли из относительно
небольшого числа клеток и не имели твердых
частей. Поскольку в подавляющем
большинстве случаев сохраняются ископаемые
остатки организмов именно с твердыми
структурами—раковинами, пластинками
или скелетом, было бы нереальным
ожидать, что палеонтологические находки
когда-нибудь помогут определить предка
животных; в высшей степени невероятно, что
такая форма сохранилась. Таким образом,
зоологи вынуждены опираться лишь на
сравнение структуры и функций живых
представителей разных типов животных
и протистов и строить гипотезы о том,
какие из существующих признаков живых
организмов следует считать примитивными
2*
20
Глава 2
чертами, дошедшими до наших дней. Здесь
надо помнить, что современные
представители всех групп организмов отделены
сроком по крайней мере 600 млн. лет, а
возможно, и более, от момента возникновения
каждой из них, а значит, и всевозможными
изменениями в биохимии, физиологии,
эмбриологии, анатомии и т. д., которые
могли произойти за этот период.
Большинство ранних споров
характеризуется приверженностью к ныне непризна-
ваемым «законам», например к формуле:
«онтогенез повторяет филогенез» (гл. 17),
выдумыванием чисто гипотетических
промежуточных стадий, отвечающих скорее
требованиям, продиктованным выдвинутой
автором гипотезой, чем потребностям
успешного выживания, а также серией
остроумных кабинетных спекуляций,
которые, учитывая природу вопроса, в принципе
нельзя было ни проверить, ни
опровергнуть. В качестве реакции на этот
первоначальный поток спекуляций и контрспекуляций
некоторые современные биологи считают
любую попытку реконструировать
филогенез пустой и ненаучной: если мы никогда не
узнаем, правилен ли данный ответ,
бесполезно ставить и сам вопрос! Хотя этот взгляд
в общем можно считать корректным,
маловероятно, что пытливый человеческий
разум позволит сковать себя такими рамками.
Если строго придерживаться определения
биологии как науки, отвергающей любую
область, в которой гипотезы нельзя
проверить экспериментально, многие
современные биологические исследования, в том
числе все, связанные с событиями
прошлого, потеряли бы право на существование.
Свидетельства, полученные в результате
сравнительного изучения живых
организмов и ископаемых находок, можно
использовать в качестве аргументов в пользу того
или иного пути филогенеза, и именно на
таких данных основан наш последующий
обзор. Тем не менее читатель должен
помнить, что альтернативные интерпретации не
только возможны, но и фигурируют в
литературе.
Одна из основных черт сравнительно
нового подхода к разгадке тайн
филогенетической истории—падение доверия к
нескольким считавшимся решающими «ключевым
признакам», таким, как тип личинки или
сегментация. В этой связи некоторые,
возможно, удивятся, прочтя на следующих
страницах, что, например, глоточные щели—
структура для удаления попадающей в
пищеварительный тракт воды, свойственная
не только хордовым и полухордовым, но
и неродственным им гастротрихам; что
«специфически нематодный» способ локо-
моции с использованием продольно
несжимаемых скелетных элементов, продольной
мускулатуры и последовательных
волнообразных изгибов присущ и головохордо-
вым (и у тех и у других мышцы с
отростками, идущими к нервному стволу, а не
наоборот); что киноринхи сегментированы так
же, как многие членистоногие. Даже
структуры, подобные нематоцистам, характерны
не только для книдарии, но присутствуют
и у некоторых протистов.
Животные, сталкиваясь с одинаковым
давлением отбора, находили сходные пути
решения возникающих проблем. Поэтому
все шире распространяется мнение, что
различные уровни структурной организации
достигались в результате конвергенции
параллельно различными филогенетическими
линиями («кладонами»). Пример —
сегментация. Во многих кладонах
наблюдается сериальное повторение мускульной
и связанных с ней систем органов вдоль
вытянутого в длину тела, т. е. разная степень
метамерной сегментации, обычно
связанной с локомоторными структурами. Когда
это сочеталось с развитием покровов тела,
используемых в качестве наружного
скелета, эволюционные ограничения приводили
к появлению внешне сходных животных:
наружный скелет должен быть
сегментированным, а конечности, если имеются,
членистыми. Таким образом, логической или
эволюционной необходимости родственных
связей между всеми сегментированными
животными нет. Это справедливо и для
животных с членистыми конечностями. Для
установления их возможного родства
нужно использовать другие критерии.
2.2. Первые животные
Теоретически многоклеточные
организмы могли возникнуть из протистов тремя
Эволюционная история и филогения беспозвоночных.
21
А Симвиотический
fl Разделение на клетки
Рис. 2.2. Возможные пути эволюции
многоклеточное™ животных начиная от протистов
способами. Во-первых, разные типы
последних могли образовать сложный
организм симбиотически по аналогии с
предполагаемым путем возникновения эукариот-
ной клетки из разных прокариот и
лишайников из грибов и водорослей (рис. 2.2,А).
Второй возможный способ заключается
в том, что продукты деления одного
протиста остаются вместе и многоклеточность
возникает через стадию колониальности
(рис. 2.2,Б). При этом каждый отдельный
протист был бы эквивалентен одной клетке
многоклеточного организма и сами проти-
сты считались бы истинно
одноклеточными. Третий способ—образование у
многоядерного протиста внутренних мембран
вокруг каждого ядра, ограничивающих сферу
его влияния определенным участком тела,
т.е. формирование самостоятельных
отделов (компартментов) последнего (рис.
2.2,В). В этом случае многоклеточный
организм было бы лучше считать «клеточным»,
а предкового
протиста—неклеточным, а не одноклеточным.
Первый из трех рассмотренных
механизмов сталкивается с серьезными
генетическими проблемами. Каким образом
генетически различные протесты интегрируются
в единый репродуцирующийся организм?
Даже два или три разных симбионта в
составе лишайников вынуждены
размножаться по отдельности, а затем вновь
объединяться. Что касается третьего
возможного механизма, никаких указаний на
внутреннюю компартментацию ныне живущих
протистов, а значит, и сравнительных
данных, позволяющих предположить, что она
могла иметь место в прошлом, нет. Тем не
менее следует сказать, что если бы какой-то
современный многоядерный протист и был
разделен внутри на отделы, биологи,
вероятно, отнесли бы его к многоклеточным
организмам, а не к протистам. Вместе с тем
известно, что многие протесты, бактерии
и цианобактерии (синезеленые водоросли)
образуют колонии (рис. 2.3), в некоторых из
которых наблюдается дифференцировка на
разные клеточные типы. Неудивительно,
что в пользу этого второго способа
высказывается большинство биологов и
существует множество указаний на возможные
пути эволюции многоклеточное™ через
колониальную стадию.
В самом деле, трудно найти различия
между колонией протистов и
многоклеточным организмом. Не у всех организмов,
которые классически считаются
многоклеточными, хорошо развита координация
деятельности клеток, а клеточная
дифференцировка, как уже говорилось, встречается не
только у них. Зачастую дело в традиции
и удобстве. Из 27 типов протистов,
признаваемых одной недавней классификацией, 16
включают колониальные виды, а в трех
уровень организации нескольких или всех
видов признается истинно
многоклеточным. Не считая животных, от протистов
произошли более шести ныне живущих
групп многоклеточных, и нет никаких
оснований утверждать, что многоклеточность
животных могла возникнуть лишь
однажды, даже если все они имеют общие
цитологические и биохимические черты (как нет
никакой необходимости в предпринимав-
22
Глава 2
Choanoflagellata [у,
Baclllarlophyta
Xanthophyta
Chlorophyta
Sarcodlna
Chrysophyla
Рис. 2.З. Колониальные
представители нескольких типов
протистов (по разным
источникам)
шихся попытках, напротив, вывести все их
типы из протистов независимо). *
2.2.1. Губки
Среди всех животных губок удобнее
всего рассматривать как колонии протистов,
а не как многоклеточных. Если бы на
протяжении многих лет не было традицией
включать их в царство Animalia, можно было бы
отнести их к Protista параллельно с
сохранением в пределах этого царства
многоклеточных водорослей. Функции отдельных
клеток губок мало скоординированы
(нервной координации нет); отсутствует какая-
либо система симметрии этих особей-
колоний; разные типы клеток не образуют
совместно четко определенных тканей или
систем органов, базальные мембраны
отсутствуют. Губок называют животными
с клеточным уровнем организации в
противоположность тканевому уровню
кишечнополостных и уровню систем органов
плоских червей. То, что единица организации
здесь—отдельная клетка, подтверждается
способностью дезагрегированных губок
к самовосстановлению и к регенерации из
очень мелких фрагментов.
Один тип протистов — воротничковые
жгутиконосцы (Choanoflagellata) включает
одиночных или колониальных
представителей, практически идентичных наиболее
характерному типу клеток губок — хо-
аноцитам (рис. 2.4). Некоторые хоанофла-
геллаты живут, прикрепившись к субстрату,
многие секретируют мембранную или
кремнеземную оболочку или панцирь. Они
питаются, насколько это известно, тем же
способом, что и хоаноциты. Поэтому
большинство зоологов полагает, что губки
произошли от колоний хоанофлагеллат. Тот
факт, что хоаноцитоподобные клетки
обнаружены у других животных, возможно,
говорит о родстве с этими жгутиконосцами не
только Porifera.
Строение губок настолько своеобразно,
что вывести из его плана какой-либо из
ныне живущих типов животных невозможно.
Основываясь на этом, некоторые считают
губок ранней неудачной «попыткой»
перехода к многоклеточности. Ничего
подобного! Губки—в высшей степени процветаю-
Эволюционная история и филогения беспозвоночных
23
Жгутик
Воротничок
Хоаноцит
Воротничковый
жгутиконосец
Рис. 2.4. Морфологическое сходство между
хоаноцитами губок и воротничковыми
жгутиконосцами
щая морская группа с большим числом
современных видов, чем у иглокожих, и почти
с таким же, как у морских аннелид: они
составляют существенную часть морской
фауны, начиная с кембрия. Нельзя
рассматривать их кажущуюся просто гу как результат
неспособности к развитию систем органов,
свойственных другим группам животных,
поскольку она может быть прямым
результатом «неживотного» образа жизни.
Губки— прикрепленные, сидячие и
совершенно неподвижные фильтраторы. В
самом деле, функции их скелетного аппарата
прямо противоположны таковым у всех
других типов животных: он служит для
предотвращения движения и дает телу жесткую
опору. Окружающая вода прогоняется
через каналы и камеры губок за счет
неупорядоченного биения жгутиков хоаноцитов;
если бы эти протоки не были жесткими и
неподвижными, локальные падения давления
воды превращали бы всю их систему в
препятствие, а не в приспособление для
пропускания максимального количества
жидкости. Если тело неспособно передвигаться,
нервная система, например, оказывается
«безработной». Защита от хищников
обеспечивается не их обнаружением и
избеганием, а неприятными на вкус веществами
и волокнистой или игольчатой структурой
скелета. С этой же точки зрения трудно
представить, каким образом какая-либо из
других систем органов, присущих более
организованным животным, могла бы
увеличить приспособленность и выживаемость
губок. Также совсем необязательно считать
губок сравнительно рано возникшей
многоклеточной формой. Древнейшие
ископаемые остатки их спикул датируются более
поздним периодом, чем, например, остатки
сложных сегментированных видов, и
вполне возможно, что губки возникли позже
некоторых других групп.
Таким образом, это один из
альтернативных способов организации животных,
а не эволюционный «брак».
2.2.2. Кишечнополостные
Так же, как и губок, кишечнополостных
(т. е. Cnidaria и Ctenophora) обычно считали
крайне своеобразной тупиковой ветвью
эволюции. Предположение, согласно которому
они не дали начала какому-либо другому
типу (тоже, как и в случае с губками),- -
просто иной способ признания их общего
плана строения настолько удачным, что
никакие коренные изменения в нем, похоже, не
могли привести к большему успеху.
Особенности его радиальная симметрия,
тканевый уровень организации с координацией
функций клеток, обеспечиваемой
диффузной сетью лишенных оболочки отростков
нейронов, и наличие только двух клеточных
слоев со студенистой и часто неклеточной
мезоглеей между ними. Характерны
внутриклеточные органеллы—нематоцисты (у
Cnidaria) или коллобласты (у Ctenophora),
отвечающие за защиту и нападение.
Структуры такого типа известны только у
некоторых протистов (например, у Myxozoa, Mi-
crospora и Dinophyta) (рис. 2.5).
Форма тела кишечнополостных в
основном напоминает чашу, иногда уплощенную
или удлиненную, с простой полостью
внутри, сообщающейся с внешней средой
единственным отверстием (рот-анус). Это. а
также двуслойное клеточное строение дало
ранним исследователям филогенеза повод
отмечать, что такое устройство аналогично
стадии гаструлы в эмбриогенезе животных.
Глава 2
Книдоциль
Крышечка
(оперкулум)
Трувка
Нематоциста
Ядро
•Нить (трувка)
Крышечка
Рис. 2.5. Подобные нематоцистам органеллы стрекающих кишечнополостных и протистов из
групп Myxozoa и Microspora. Нематоцисты книдарий: А — книдоцит с неотстреленной нематоци-
стой; Б—отстреленная нематоциста. В. Спора представителя Myxozoa с неотстреленной органел-
лой. Г,Д. Споры представителей Microspora с неотстреленными и (£) с отстреленными органел-
лами (по Нутап, 1940; Calkins, 1926; Wenyon, 1926 и др.)
Поскольку в дальнейшем двуслойная га-
струла становится трехслойной за счет
образования третьего зародышего листка
(мезодермы), кишечнополостных считали
реликтовыми предковыми формами, от
которых произошли все остальные,
«трехслойные», типы животных. Предполагались
три стадии эволюции последних: а) полый
клеточный шар (= бластула), форма,
известная для некоторых колониальных
жгутиконосцев; б) шар, по каким-то
причинам ставший двуслойной чашей (= гастру-
ла = кишечнополостные); в) появление
мезодермы и с" ней—«трехслойных» форм.
Эта эмбриологическая аналогия, возможно,
остроумна, но нет никаких свидетельств
в пользу того, что зародышевые листки
в той или иной мере эквивалентны клеткам
внешне сходного взрослого организма.
Эмбриологические понятия двух и трех
листков не следует применять к морфологии
взрослых животных. Как уже говорилось
выше, кишечнополостные вряд ли были
предками какой-либо другой группы, хотя
предпринимались попытки вывести плоских
червей из планулы (расселительной стадии
некоторых
книдарий)—свободноплавающей, непитающеися, лишенной полости
бластулы, которая, хотя и радиально
симметрична, вытянута вдоль продольной оси.
Кстати, основной план строения
кишечнополостных таков, что и они вряд ли
могли произойти от какой-либо другой
известной группы животных. Предка их, таким
образом, вероятнее всего, следует искать
среди протистов, хотя невозможно решить,
Эволюционная история и филогения беспозвоночных
25
какой именно (если вообще такие есть) из
выживших типов последних стоит к нему
ближе всего. Поскольку клетки книдарий
типично жгутиковые, а не ресничные, как
у большинства животных, причем пищевые
частицы поглощаются путем амебоидного
фагоцитоза, предполагалось, что протесты,
о которых идет речь, скорее всего были
гетеротрофными жгутиковыми. Хотя
интенсивно обсуждались вопросы о том, какая
форма тела колониальных протистов или
«протокишечнополостных» могла быть
исходной для группы и какая фаза—полип
или медуза—более примитивна, дело не
пошло дальше предположений, и нет
особого смысла спорить о достоинствах каждой
конкретной схемы.
У большинства книдарий полип может
размножаться бесполым почкованием, и
у многих видов сохраняется тканевый
контакт между отпочковывающимися
дочерними и родительской особью. Все такие
полипы генетически идентичны и связаны между
собой, вследствие чего возможен их
полиморфизм: некоторые могут
специализироваться на питании или защите, другие—на
половом размножении и т. д. В
определенном смысле специализация отдельных
полипов и сохранение медуз в составе
колоний—процесс, в целом эквивалентный
развитию систем органов у большинства
других животных. Колонии полипов в
результате почкования могут достигать крупных
размеров (например, у многих кораллов),
так же как и отдельные полипы и медузы,
потому что каждый из двух их клеточных
слоев контактирует с морской водой
(наружный слой—с находящейся во внешней
среде, внутренний—с водой в гастроваску-
лярной полости), а значит, диффузия
дыхательных газов, продуктов выделения и т. д.
не зависит от массы тела. То же
справедливо и для губок, но никак не для плоских
червей (см. ниже).
И еще одна заслуживающая упоминания
особенность кишечнополостных.
Известные их виды, за небольшим исключением,—
хищники, ни один из них не может питаться
водорослями. Они поглощают в основном
планктонных животных, случайно
касающихся их щупалец с нематоцистами или
коллобластами. Предковая форма,
вероятно, питалась бактериями и протистами (см.
гл. 9); несколько ныне живущих видов
имеют сходный рацион, но расцвет
кишечнополостных в позднедокембрийских
морях, вероятно, был следствием
переключения на более крупную добычу в условиях
завоевания толщи воды различными
группами животных (в основном
членистоногими), образовавшими зоопланктон. Таким
образом, еще одно сходство с губками —
вероятность возникновения и особенно
адаптивной радиации после появления
других групп животных.
Стоит отметить, что, хотя мы и следуем
здесь традиционному представлению о
родстве Ctenophora и Cnidaria, все более
распространяется мнение, что их сходство —
результат конвергенции. «Мезоглея» ктено-
фор, возможно, ближе к мезенхиме плоских
червей, чем к мезоглее книдарий; клетки
ктенофор также многоресничные, а не од-
ножгутиковые.
2.2.3. Плоские черви
Современные плоские черви (Platyhel-
minthes) — обширная и разнообразная
группа, в которой сейчас преобладают
паразитические виды. Свободноживущие турбел-
ларии, от которых обычно принято
выводить паразитических сосальщиков и ленте-
цов,— билатерально симметричные
организмы с плоским, лишенным полости
телом, приводимым в движение ресничками.
Имеются системы органов, в том числе
и покрытые оболочкой нервные волокна,
организованные у всех видов, кроме
некоторых Acoela, в отчетливые продольные
стволы. Среди ныне живущих турбелларий план
строения этих мелких Acoela (рис. 2.6) и
нескольких родственных им групп ближе
всего к предковому, хотя сами они не
обязательно дали начало всем прочим плоским
червям. (Одно из затруднений, упомянутых
в разд. 2.1, возникающее при
эволюционных построениях, основанных на
современных формах, связано с тем, что все
выжившие группы могли значительно дивергиро-
вать от вымерших предковых форм, даже
сохранив на первый взгляд множество
примитивных черт. Таким образом, лучше
считать эти ныне живущие организмы анало-
26
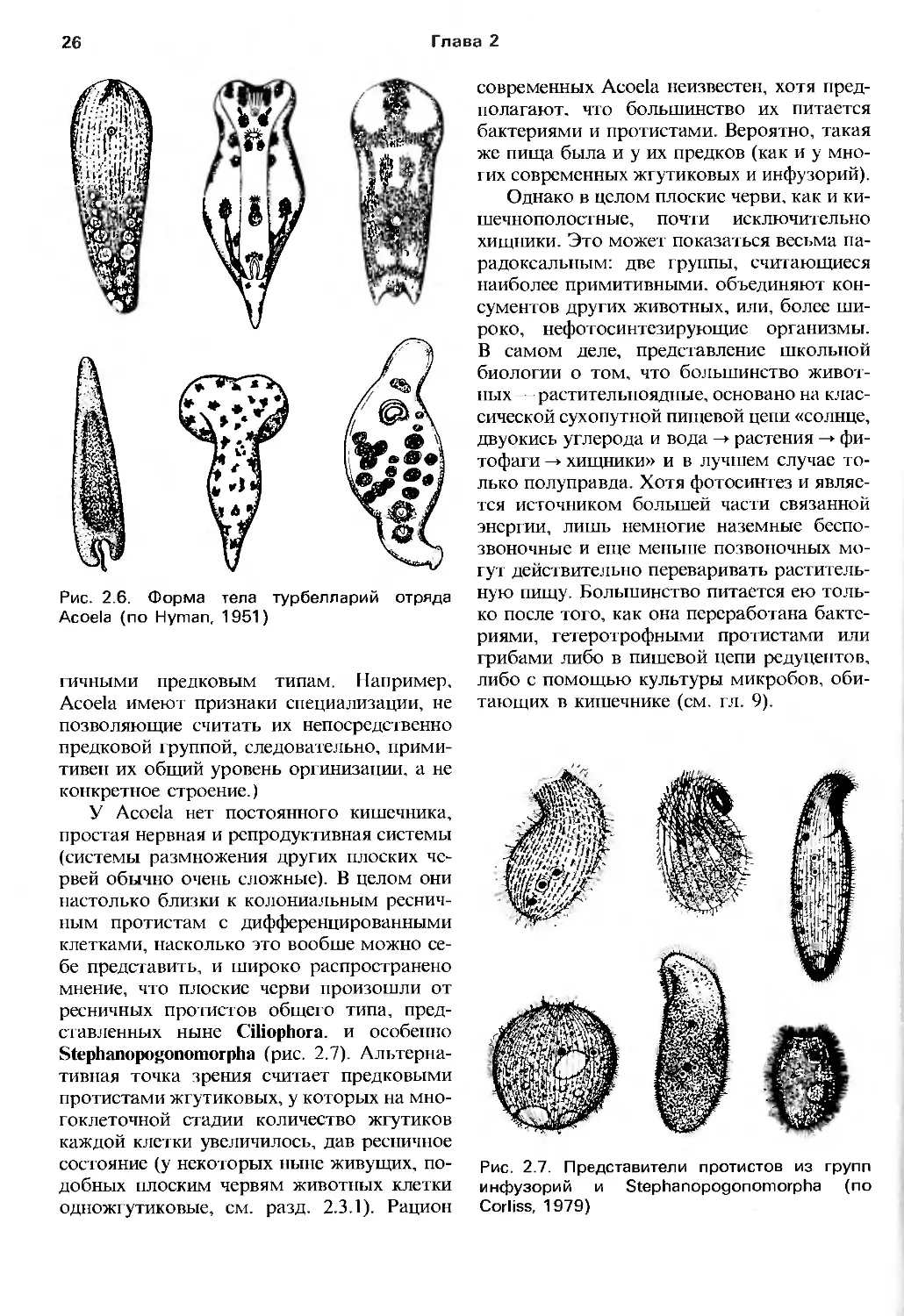
Глава 2
Рис. 2.6. Форма тела турбелларий отряда
Acoela (по Hyman, 1951)
гичными предковым типам. Например,
Acoela имеют признаки специализации, не
позволяющие считать их непосредственно
предковой группой, следовательно,
примитивен их общий уровень оргинизации, а не
конкретное строение.)
У Acoela нет постоянного кишечника,
простая нервная и репродуктивная системы
(системы размножения других плоских
червей обычно очень сложные). В целом они
настолько близки к колониальным
ресничным протистам с дифференцированными
клетками, насколько это вообше можно
себе представить, и широко распространено
мнение, что плоские черви произошли от
ресничных протистов общего типа,
представленных ныне Ciliophora. и особенно
Stephanopogonomorpha (рис. 2.7).
Альтернативная точка зрения считает предковыми
протистами жгутиковых, у которых на
многоклеточной стадии количество жгутиков
каждой клетки увеличилось, дав ресничное
состояние (у некоторых ныне живущих,
подобных плоским червям животных клетки
одножгутиковые, см. разд. 2.3.1). Рацион
современных Acoela неизвестен, хотя
предполагают, что большинство их питается
бактериями и протистами. Вероятно, такая
же пища была и у их предков (как и у
многих современных жгутиковых и инфузорий).
Однако в целом плоские черви, как и
кишечнополостные, почти исключительно
хищники. Это может показаться весьма
парадоксальным: две группы, считающиеся
наиболее примитивными, объединяют кон-
сументов других животных, или, более
широко, нефотосинтезирующие организмы.
В самом деле, представление школьной
биологии о том, что большинство
животных— растительноядные, основано на
классической сухопутной пищевой цепи «солнце,
двуокись углерода и вода -» растения -*
фитофаги -> хищники» и в лучшем случае
только полуправда. Хотя фотосинтез и
является источником большей части связанной
энергии, лишь немногие наземные
беспозвоночные и еще меньше позвоночных
могут действительно переваривать
растительную пищу. Большинство питается ею
только после того, как она переработана
бактериями, гетеротрофными протистами или
грибами либо в пишевой цепи редуцентов,
либо с помощью культуры микробов,
обитающих в кишечнике (см. гл. 9).
Рис. 2.7. Представители протистов из групп
инфузорий и Stephanopogonomorpha (по
Corliss, 1979)
Эволюционная история и филогения беспозвоночных
27
И напротив, морские фотосинтезирую-
щие организмы—почти исключительно
одноклеточные протисты, сравнительно легко
перевариваемые животными. Трудность
питания ими вытекает из пространственных
ограничений. Морской фотосинтез
приурочен к сравнительно тонкому
поверхностному слою воды, освещенному солнцем, а
большинство групп животных (возможное
исключение— кишечнополостные, если
медуза была их предковой формой) скорее всего
эволюционировало в связи с донными
осадками и осталось бентосными формами. Они
зависели от поступления сверху мертвых
водорослевых тканей, уже
колонизированных редуцентами (в основном бактериями)
и питающимися бактериями протистами.
Поэтому предковый рацион должен был
включать в первую очередь нефотосинтези-
рующие организмы из пищевых цепей
редуцентов. По мере эволюции животных для
потребления стали доступны более крупные
и энергетически более ценные организмы,
но. за исключением мелководных и
физически неблагоприятных морских окраин, это,
по-видимому, произошло только тогда,
когда они сами смогли освоить толщу воды
и начать питаться фотосинтезирующими
протистами. Следовательно, прямое
потребление живых фотосинтезирующих
организмов возникло, вероятно, в филогении
животных относительно поздно как
специализированный тип питания.
2.2.4. Placozoa и Mesozoa
Серьезными кандидатами на
независимую эволюцию от протистов являются
только две другие группы животных—
Placozoa и Mesozoa. Можно считать, что
обе они ближе к колониальным протистам,
чем к многоклеточным животным,
поскольку лишены нервных, мускульных,
пищеварительных и скелетных клеток, хотя
детальных сведений о каждой из этих групп мало.
Placozoa выглядят и ведут себя как крупные,
плоские, покрытые жгутиками клеточные
амебы. Их тела лишены какой-либо
симметрии и состоят из нескольких тысяч клеток,
расположенных в два слоя,— «дорсальный»
и «вентральный» с клетками-волокнами
в матриксе между ними. Уровень их
организации эквивалентен наблюдаемому у губок,
и некоторые считали их неотеническими
личинками последних. Тем не менее
равновероятно, что это — группа жгутиковых
протистов, перешедшая к многоклеточности
(возможно, сравнительно недавно).
Известно, что они питаются протистами путем
наружного пищеварения с последующей
абсорбцией его продуктов.
Mesozoa также лишены каких-либо
тканей и органов, однако имеют определенную
форму тела и организацию. Две достаточно
различные и, возможно, неродственные
между собой группы этого типа объединяют
крошечных билатерально симметричных
ресничных эндопаразитов морских
беспозвоночных. Их тело может состоять только
из 20 30 ресничных клеток, окружающих
одну или более репродуктивных, однако
у некоторых видов развитие внутри хозяина
приводит к потере наружных ресничных
клеток и образованию плазмодия. Их
билатеральная симметрия и общая
червеобразная форма тела позволили высказать
предположение, что одна или обе группы
Mesozoa произошли от плоских червей и что
простота их строения—следствие
паразитической дегенерации, но против этого говорят
особенности расположения их клеток и
жизненного цикла, а также крайнее
своеобразие репродуктивных процессов. Возможно,
предков этого мелкого загадочного типа
следует искать среди ресничных протистов,
однако никакой информацией на этот счет
зоологи пока не располагают.
2.3. Производные
плоских червей
Возможно, многоклеточные животные
возникали от протистов от 3 до 5 6 раз,
если принимать во внимание только
группы, дожившие до наших дней, но почти
наверняка только от одной из этих линий —
билатерально симметричных плоских
червей— произошли все остальные типы
животных (см., например, Sleigh, 1979). У
многих этих потомков развились новые
системы органов, такие, как кровеносная система
и гидростатический или твердый скелет,
а системы, унаследованные от предков,
стали более специализированными; все «усо-
28
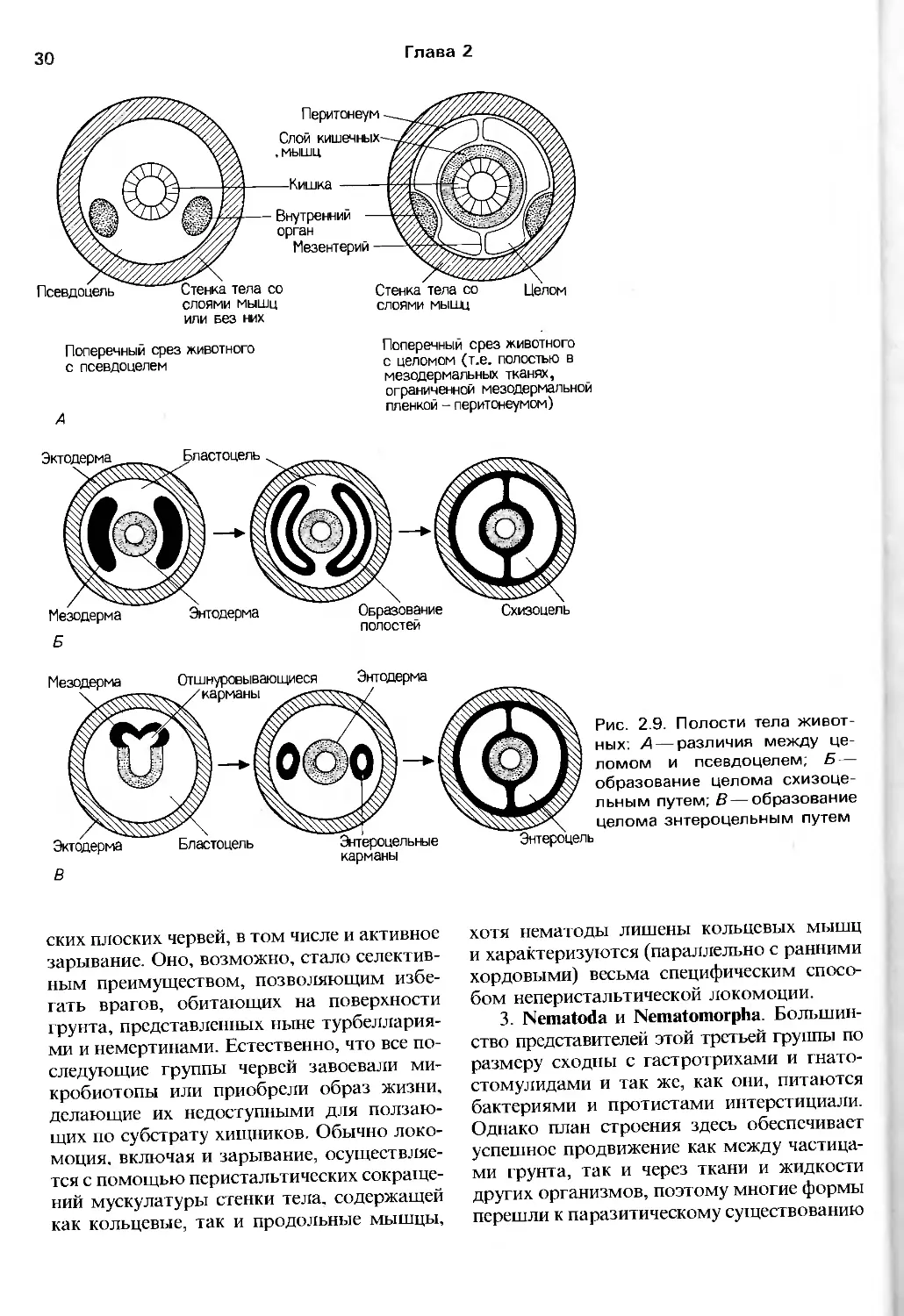
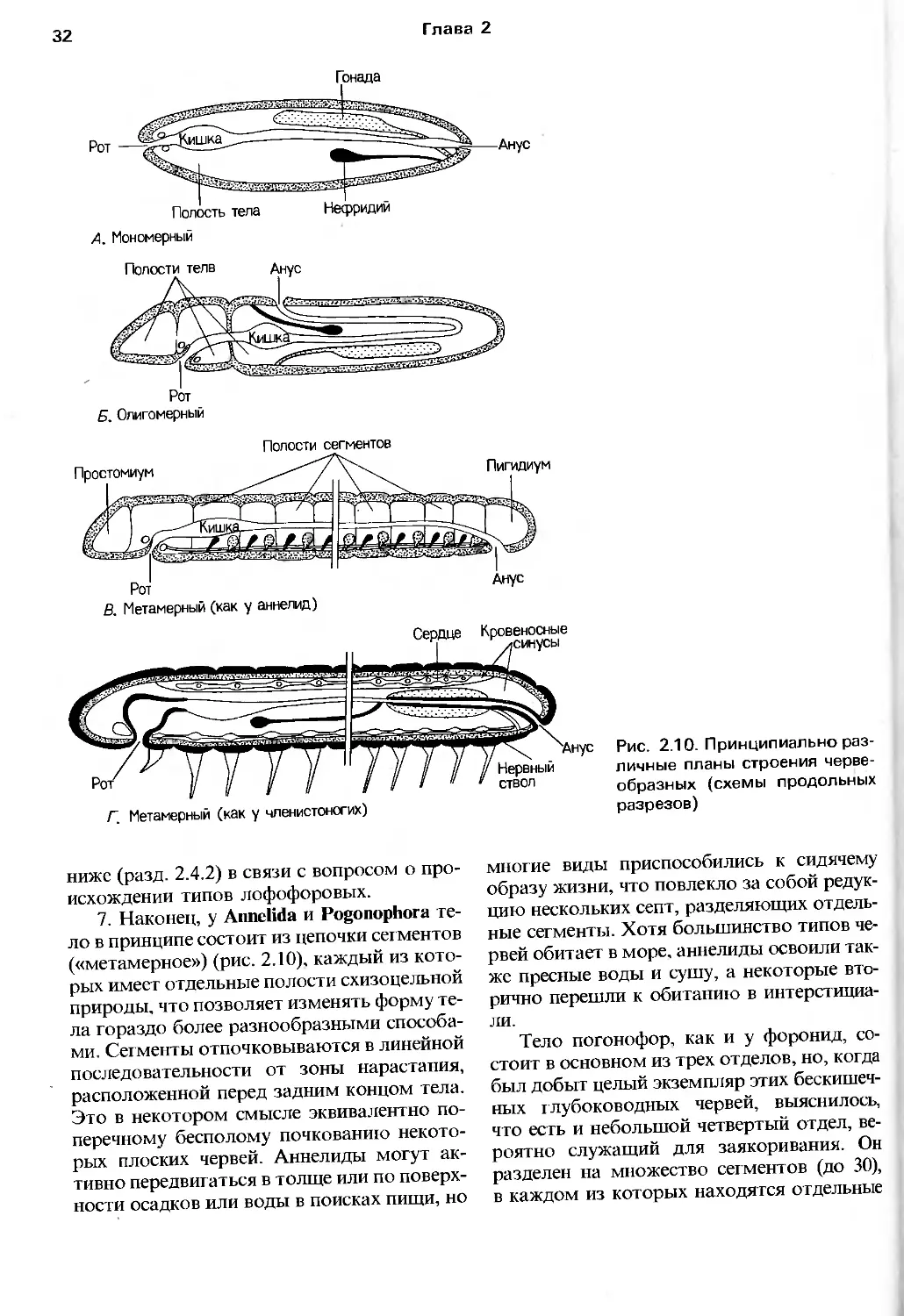
Глава 2
03
СО
СО
g га <я
ы in l о. К
о tS га о о
JS Ь с -22 S
о. о. оо 2:
р га оо » га 9 - <_,
- i-| 8 f £ ° с о I ^1-s
■:3 J
411
-c -g 5- ra
Q.b't- ra-c 0> fc-'c l."__ — —
J*r:a-£:oa-Oz>omLULUcocOQ_
I
b_L
Радиация
артропод
CO -•-*
5 О) га
tie
Ф О -C
X Ш О
X
Радиация ЛОфО
форовых
Радиация
вторично-
ротых
I
Радиация предковых, подобных плоским червям животных
т
Рис. 2.8. Радиации, давшие начало современным типам животных (см. также рис. 2.20)
вершенствования» в общем вытекают из
неспособности диффузии справляться с
транспортом веществ через крупное сплошное
тело и из ограничений локомоторной системы
плоских червей. Несмотря на это, основной
план строения последних остается
очевидным. Непосредственно от подобного
плоским червям организма, вероятно,
произошли моллюски и группа других
«червеобразных» типов, в то время как членистоногие,
лофофоровые и вторичноротые стали
производными этих вторичных групп червей
(рис. 2.8).
2.3.1. Группы червеобразных
Червя можно определить как
билатерально симметричное, безногое животное
с мягким телом, длина которого в 2 —3 раза
превышает ширину. Исходя из этого,
примерно 16 типов животных включают
только червей (не считая Platyhelminthes), а еще
в нескольких группах известны отдельные
червеобразные виды. Хотя свидетельства
присутствия червей, такие, как норы,
трубки, следы ползания в осадочных породах,
в том числе и докембрийских,
многочисленны, остатки самих мягкотелых животных
сохраняются редко. Известно, что приапу-
лиды и сегментированные черви были
обильны в кембрии, однако отнесение другого
ископаемого материала к конкретным ныне
живущим типам вызывает разногласия.
Возможно, несколько других современных
типов червей также имеют кембрийский или
докембрийский возраст. Ископаемые
остатки в целом малопригодны для установления
родственных связей между ними и во
взглядах на филогению этих животных
много расхождений. Без сомнения, черви не
Эволюционная история и филогения беспозвоночных
29
образуют естественной группы; общее у них
только то, что все они тем или иным путем
произошли от свободноживущей формы,
сходной с Platyhelminthes. Однако можно
выделить семь крупных группировок,
объединяемых преимущественно образом
жизни и экологией, а часто также и некоторой
степенью потенциального
филогенетического родства (не включая щетинкочелюст-
ных, которые будут рассмотрены в одном
из следующих разделов вместе с другими
вторичноротыми, и пятиусток, вторично
утративших конечности и, вероятно,
родственных членистоногим).
1. Более крупные свободноживущие тур-
белларии — потомки ранних плоских червей
и родственная им группа немертин (Nemer-
tea). Это плоские, имеющие сплошное тело
(т.е. «ацеломические») хищники, медленно
ползающие по субстрату с помощью
вентральных щетинок или (более крупные
формы) за счет волнообразных мускульных
сокращений, проходящих по вентральной
поверхности. Поскольку ограничения,
связанные с диффузией дыхательных газов и
других веществ, требуют, чтобы хотя бы одно
из измерений тела оставалось небольшим,
увеличение размеров возможно только за
счет удлинения или разрастания в ширину.
Вследствие этого крупные формы
лентовидны или листовидны; некоторые достигают
большой длины —одна, лишь наполовину
раскрученная немертина—до 30 м. Хотя
эта группа животных в основном морская,
в ней много пресноводных и несколько
сухопутных форм; образ жизни
представителей также разнообразен: среди них есть
свободноплавающие виды, комменсалы и
паразиты. Эти турбелларии и немертины —
самые крупные и самые активные
животные из тех, у кого сохранился предковый
план строения без полости тела. Они не
могут быстро зарываться в субстрат, но,
несмотря на это, успешно питаются весьма
разнообразными животными.
2. Gnathostomula и Gastrotricha также
сохранили общий план строения предковых
плоских червей, хотя каждая их эпидерма-
льная клетка несет только одну ресничку.
Мелкие гнатостомулиды и гастротрихи
(длина тела обычно меньше 2 мм)
специализировались на обитании в интерстициали
песчаных осадков, где перемещаются с
помощью эпидермальных ресничек, не
сдвигая при этом песчинок. Некоторые
населяют относительно глубокие,
бескислородные слои, и несколько видов, — вероятно,
облигатные анаэробы. Представители
обоих типов питаются бактериями и
протестами, либо свободноплавающими в интер-
стициальной воде, либо связанными с
частицами органического детрита.
3—7. Все остальные группы червей
имеют одну общую черту, отсутствующую
у Platyhelminthes и у двух производных от
них групп, рассмотренных выше. Речь идет
о заполненной жидкостью полости тела,
играющей роль гидростатического скелета.
В разных эволюционных линиях у нее
различная форма и эмбриональное
происхождение, и, вероятно, она возникала
несколько раз независимо. Тем не менее по способу
развития можно выделить три основных
типа этой полости (рис. 2.9): «псевдоцель»,
часто образующийся из сохранившегося бла-
стоцеля, т.е. полости внутри бластулы,
обычно исчезающей во время гаструляции;
«схизоцель», возникающий путем
расхождения массы мезодермальных клеток;
«энтеронель», который формируется за счет
отшнуровывания карманов от анхентерона,
или первичной кишки :. Четвертый тип
полости тела, неизвестный у ныне живущих
червей (вероятно, был у некоторых их
вымерших групп—см. разд. 2.4.1), но
присутствующий у ряда нечервеобразных типов,—
«гемоцель», т. е. система кровеносных
синусов.
В большинстве случаев этот
гидростатический скелет делает возможными формы
локомоции, недоступные для ацеломиче-
1 В отечественной литературе принято
деление полости тела по способу ее происхождения на
первичную и вторичную, или целом. Первичная
полость тела (протоцель) образуется за счет
расхождения экто- и энтодермы и лишена
собственной выстилки. В отличие от нее целом имеет ме-
зодермальную выстилку, которая может
возникать двумя способами: а) энтероцельным, т. е. от-
шнуровыванием карманов от первичной кишки,
и б) телобластическим, т. е. из мезодермы,
образованной делением клеток-телобластов.
Различают также смешанный тип полости тела —
схизоцель.— Прим. пер.
30
Глава 2
Псевдоцель
Перитонеум
Слой кишечных
мышц
•Кишка
Внутренний
орган
Мезентерий
Стенка тела со
слоями мышц
или вез них
Поперечный срез животного
с псевдоцелем
Стенка тела со
слоями мышц
Целом
Поперечный срез животного
с целомом (т.е. полостью в
мезодермальных тканях,
ограниченной мезодермальной
пленкой - перитонеумом)
Эктодерма
Бластоцель
Энтодерма
Образование
полостей
Схизоцель
Мезодерма
Отшнуровывающиеся Энтодерма
'карманы
Бластоцель
Энтероцельные
карманы
Энтероцель
Рис. 2.9. Полости тела
животных: А — различия между
целомом и псевдоцелем; Б —
образование целома схизоце-
льным путем; В — образование
целома знтероцельным путем
ских плоских червей, в том числе и активное
зарывание. Оно, возможно, стало
селективным преимуществом, позволяющим
избегать врагов, обитающих на поверхности
грунта, представленных ныне турбеллария-
ми и немертинами. Естественно, что все
последующие группы червей завоевали
микробиотопы или приобрели образ жизни,
делающие их недоступными для
ползающих по субстрату хищников. Обычно локо-
моция, включая и зарывание,
осуществляется с помощью перистальтических
сокращений мускулатуры стенки тела, содержащей
как кольцевые, так и продольные мышцы,
хотя нематоды лишены кольцевых мышц
и характеризуются (параллельно с ранними
хордовыми) весьма специфическим
способом неперистальтической локомоции.
3. Nematode и Nematomorpha.
Большинство представителей этой третьей группы по
размеру сходны с гастротрихами и гнато-
стомулидами и так же, как они, питаются
бактериями и протистами интерстициали.
Однако план строения здесь обеспечивает
успешное продвижение как между
частицами грунта, так и через ткани и жидкости
других организмов, поэтому многие формы
перешли к паразитическому существованию
Эволюционная история и филогения беспозвоночных 31
на протяжении всего или части жизненного
цикла. Паразитические виды могут быть
намного крупнее свободноживующих—
некоторые нематоды достигают в длину
8 м. Особенности их локомоторной
системы в наличии обширного псевдоцеля,
заполненного жидкостью под высоким
давлением и окруженного покрытой коллагено-
вой кутикулой стенкой тела, содержащей
только продольные мышцы. (Недавно у
одной нематоды обнаружены кольцевые
мышцы, и можно допустить, что предковая
для этой группы форма была устроена
сходным образом.) Коллагеновые волокна
образуют своеобразную решетку,
препятствующую изменению длины тела.
Сокращения продольной мускулатуры,
противодействующие сопротивлению
гидростатического псевдоцеля, ограниченного этой
решеткой, приводят к синусоидальным
изгибаниям тела в дорсовентральной
плоскости, за счет чего черви могут довольно
быстро передвигаться в узких заполненных
жидкостью каналах.
4. У четвертой группы (Priapula, Kino-
rhyncha и. возможно. Loricifera) также
обширный псевдоцеломический '
гидростатический скелет и тело, покрытое кутикулой,
которая в данном случае линяющая и
хитиновая, а в стенке тела присутствуют
кольцевые мышцы. У мелких киноринхов
кутикула сегментированная и наблюдается также
некоторая сегментация внутренних
органов, например мускульной и нервной
систем. Приапулиды и киноринхи активно
закапываются в грунт в поисках добычи,
раздвигая перед собой частицы осадка с
помощью выворачивающегося за счет
увеличения давления в полости тела его переднего
конца, дифференцированного в сократимый
роющий орган. Его шипы цепляются за
окружающий субстрат, а сокращения
продольных мышц затем подтягивают
остальное тело, натягивая его на «головной»
отдел. Еще мало известно о типе Loricifera,
открытом позже других (описан только
в 1983 г.), хотя, похоже, исходя из общего
строения, он должен принадлежать к
обсуждаемой группе. У его представителей
1 Точная природа полости тела приапулид
спорна; она у них весьма специфического типа.
сходный выворачивающийся покрытый
шипами передний отдел, но взрослые лорици-
феры, вероятно, являются комменсалами
или эктопаразитами других интерстициаль-
ных животных.
Возможно, от подобного плоским
червям предка этой группы произошли
бескишечные и вторично упрощенные
паразитические скребни (Acanthocephala) и свободно-
живущие коловратки (Rotifera). Во всяком
случае, два этих типа многие считают
родственными между собой. Коловратки — в
основном пресноводные формы,
передвигающиеся И; или питающиеся мелкими
частицами с помощью ресничного коловра-
щательного аппарата у переднего конца
тела. Как и у некоторых других животных
с псевдоцелем, у них синцитиальные органы
с постоянным числом ядер («эвтелия»).
Весьма вероятно, учитывая в основном
специализированные формы размножения
коловраток, что они, как и все псевдоцеломи-
ческие группы с мелкими размерами тела,
произошли от гораздо более крупных
червей путем педоморфоза.
5. Sipuncula и Echiura приспособились
к сидячему образу жизни и питанию
осадком. Их крупное мешковидное тело
находится в постоянной норке, трещине или
подобного рода убежище. Как у приапулид
и других представителей предыдущей
группы, здесь обширная полость тела, но не
псевдоцель, а схизоцель. Сипункулиды
и эхиуриды питаются, используя
специализированный передний отдел тела: у первых
он удлиненный выворачивающийся и
втягивающийся, несет терминальные лопасти
или щупальца для сбора детрита; у вторых
это растяжимый и сократимый хоботок
с вентральным желобом и расширенным,
покрытым ресничками, собирающим пищу
концом. До некоторой степени
представители обоих этих типов напоминают
гигантских односегментных аннелид.
6. У групп 3—5, описанных выше,
единственная обширная полость тела (т.е. они
«мономерны») независимо от способа ее
образования. У шестой группы —
Phorona — тело состоит из трех отделов,
с собственной полостью каждый («олиго-
мерное» строение) (рис. 2.10). Эти сидячие
фильтраторы будут детально рассмотрены
32
Глава 2
Гонада
Полость тела Нефридий
А. Мономерный
Полости телв Анус
Рот
Б. Олигомерный
Простомиум
Полости сегментов
В. Метамерный (как у аннелид)
Анус
Пигидиум
Сердце Кровеносные
-синусы
Г. Метамерный (как у членистоногих)
"Анус Рис. 2.10. Принципиально раз-
Нервный личные планы строения черве-
ствол образных (схемы продольных
разрезов)
ниже (разд. 2.4.2) в связи с вопросом о
происхождении типов лофофоровых.
7. Наконец, у Annelida и Pogonophora
тело в принципе состоит из цепочки сегментов
(«метамерное») (рис. 2.10), каждый из
которых имеет отдельные полости схизоцельной
природы, что позволяет изменять форму
тела гораздо более разнообразными
способами. Сегменты отпочковываются в линейной
последовательности от зоны нарастания,
расположенной перед задним концом тела.
Это в некотором смысле эквивалентно
поперечному бесполому почкованию
некоторых плоских червей. Аннелиды могут
активно передвигаться в толще или по
поверхности осадков или воды в поисках пищи, но
многие виды приспособились к сидячему
образу жизни, что повлекло за собой
редукцию нескольких септ, разделяющих
отдельные сегменты. Хотя большинство типов
червей обитает в море, аннелиды освоили
также пресные воды и сушу, а некоторые
вторично перешли к обитанию в интерстициа-
ли.
Тело погонофор, как и у форонид,
состоит в основном из трех отделов, но, когда
был добыт целый экземпляр этих
бескишечных глубоководных червей, выяснилось,
что есть и небольшой четвертый отдел,
вероятно служащий для заякоривания. Он
разделен на множество сегментов (до 30),
в каждом из которых находятся отдельные
Эволюционная история и филогения беспозвоночных 33
полости тела, и после достаточно бурных
споров среди систематиков погонофоры
сейчас признаются родственными аннели-
дам.
2.3.2. Моллюски
У моллюсков нет ни гидростатического
целомического скелета, ни признаков мета-
мерной сегментации, однако из-за такого
же типа личинки — трохофоры, что и у
сегментированных целомических аннелид
(типу личинки в свое время придавалось
первостепенное значение при установлении
филогенетического родства), в течение многих
лет считалось, что у них когда-то были и це-
ломическая полость тела, и сегменты, но
и то и другое утрачено в процессе
эволюции. Правда, у ряда моллюсков имеется
несколько пар некоторых органов (у монопла-
кофор, например, восемь пар мыщц-
ретракторов ноги, шесть пар
выделительных органов и пять-шесть пар жабр), но эти
пары не приурочены к определенным
участкам тела, которые можно было бы считать
остатками сегментов. И у других несегмен-
тированных животных бывают
расположенные в линейной последовательности
множественные пары органов, но при этом
никогда не предполагалось, что эти формы
произошли от метамерно
сегментированных предков (их иногда называют «псевдо-
метамерными»). У моллюсков целомиче-
ская полость окружает сердце, но более
вероятно, что она возникла в пределах данной
группы как пространство, в котором оно
может биться, и не является остатком
когда-то более обширной полости, служившей
гидростатическим скелетом.
Таким образом, нет никаких
убедительных доказательств изначального
присутствия и последующей редукции или утраты
этого признака, и моллюсков, как правило,
уже не считают близкими родственниками
аннелид (см., например. Trueman, Clarke,
1985). В самом деле, первые моллюски,
похоже, мало чем отличались от плоских
червей с дорсальным защитным покровом
(белковым с рядом известковых чешуек или
пластин). Как и ныне живущие Platyhelmin-
thes, они ацеломические и несегментирован-
ные. Как и у крупных плоских червей и не-
мертин (за исключением
специализированных видов), их движение осуществляется за
счет ресничек или волн мускульных
сокращений, проходящих вдоль плоской
вентральной поверхности.
Помимо спинной защитной раковины
другой основной чертой, отличающей пред-
кового моллюска от плоских червей, было,
по-видимому, наличие в ротовой полости
зубчатого языковидного органа — радулы.
Он позволял соскребать пленки и колонии
водорослей с твердых поверхностей, и
широко признается, что первые моллюски
питались именно водорослями-протистами
в мелководных местообитаниях. Из-за
развития твердого покрова на дорсальной
стороне (единственной поверхности, открытой
воздействию токов воды, а также хищников
и волн) газообмен через поверхность тела,
как и в случае плоских червей, сильно
ограничивается. Таким образом, параллельно
эволюции раковины должно было идти
развитие специфических органов дыхания
(жабр) и циркуляторной системы для
транспорта газов по телу. Парные жабры
моллюсков, своеобразные и специфические для
этого типа органы, называемые ктенидия-
ми, изначально располагались сзади в
мантийной полости, скрытой раковиной. Две
другие вымершие группы— Hyolitha и Wi-
waxiida—похоже, реагировали на давление
отбора, благоприятствующее появлению
защитного покрова тела, сходным с
моллюсками способом.
От такого покрытого раковиной,
напоминающего плоского червя животного в
ходе адаптивной радиации возникли весьма
разнообразные формы, описанные в гл. 5,
включая и наиболее сложно устроенные не
только среди потомков плоских червей, но
и среди всех беспозвоночных. Тем не менее
некоторые виды сохранили древний способ
питания за счет соскабливания водорослей.
Этот исходный образ жизни может быть
сравнительно легко адаптирован к
наземному существованию, и моллюски
составляют одну из очень немногих групп
животных, обладающих ферментами,
способными разрушать стойкие структурные
углеводы морских и наземных растений.
3—1314
34
Глава 2
2.4. Более отдаленные
потомки плоских червей
2.4.1. Членистоногие
Основные анатомические отличия
червей с сериально повторяющимися по
длине тела органами от членистоногих—
наличие у последних конечностей и
твердого (членистого) суставчатого наружного
скелета. Эти отличия не столь существенны,
как может показаться на первый взгляд.
Сначала, вероятно, возникли конечности.
Некоторые современные животные,
например онихофоры и тихоходки,— мягкотелые
червеобразные организмы с вентральными
или почти вентральными нечленистыми
конечностями и гидростатическим скелетом.
Онихофоры покрыты гибкой хитиновой
кутикулой, тихоходки — наружными кутику-
лярными пластинами. Таким образом,
сравнительно легко продемонстрировать,
как эволюционировали кутикулярные
защитные структуры, пока не покрыли тело
целиком, образовав суставные сочленения
между различными наборами сериально
повторяющихся элементов (так же, как,
например, у неродственных членистоногим
киноринхов) и члениками конечностей и
в итоге взяв на себя функции скелета,
заменившего исходный гидростатический. В
одной из групп членистоногих—у хелицеро-
вых—выпрямление конечностей может
осуществляться гидравлически, и только их
сгибание происходит с помощью экзоске-
летно-мускульной системы.
Такая «артроподизация» предкового
червя шла, похоже, еще в докембрии во время
основной радиации этой группы (рис. 2.11),
и многочисленные весьма разнообразные
и временами крайне причудливые варианты
сегментированных артроподоподобных
животных известны уже с кембрия (рис. 2.12).
Если бы они дожили до наших дней, типов
членистоногих было бы не меньше, если не
больше, чем среди червеобразных. Вероят-
Рис. 2.11.
1979)
Радиация «артроподоподобных» животных в докембрии/кембрии (по Whittington,
Эволюционная история и филогения беспозвоночных.
35
£ С вентральной стороны
Ж. Вентролатеральныи вид
Рис. 2.12. Животные, напоминающие членистоногих, не относящиеся к филогенетическим
линиям трилобитов, хелицеровых, ракообразных и одноветвистых: А — Burgessia (кембрий); Б —
Mimetaster (девон); В — Marrella (кембрий); Г—Branchiocaris (кембрий); Л—Johoia (кембрий);
Е—Anomalocaris (кембрий); Ж—Sarotrocercus (кембрий) (по Manton, Anderson, 1979; Whitting-
ton, 1985). Масштабные линии 1 см
нее всего, предковая группа червей, доживи
до наших дней, не была бы отнесена к анне-
лидам—основному современному типу
сегментированных червеобразных. Очень
может быть, например, что гидростатическая
полость тела предка членистоногих
представляла собой гемоцель, а не сегментарно
разделенный схизоцель, характерный для
аннелид, и тело его было не
сегментированным, а мономерным с множественными
линейно расположенными парами
локомоторных органов. Этих предков можно
представить себе в виде удлиненных, внешне
похожих на киноринхов червей с кровеносной
GUI'
36
Глава 2
стемой (возникшей по той же причине, что
и у моллюсков, см. разд. 2.3.2), сильно
расширившейся для выполнения
гидростатических функций. Существуют также
фундаментальные эмбриологические различия
между аннелидами и разными группами
членистоногих, которые также развиваются
по разным схемам (см., например, Manton,
Anderson, 1979).
Лишь немногие из большого числа
групп членистоногих пережили кембрий,
а из них только четыре —девон. Так,
трилобиты, существовавшие по крайней мере 300
млн. лет, вымерли в конце Перми. До наших
дней сохранились только три группы —
хелицеровые (Chelicerata), ракообразные
(Crustacea) и одноветвистые (Uniramia),
отличающиеся в основном строением и
расположением конечностей и других придатков,
а также эмбриологией и внутренним
строением, причем до такой степени, что можно
говорить об их независимом
происхождении от червеобразного предка. Похоже
даже, что по крайней мере ракообразные и
одноветвистые — потомки совершенно
различных групп червей (Abele, 1982). (Иногда
Pycnogona считают четвертой группой
такого же статуса; в противоположном случае
их включают в Chelicerata.)
«Атроподизированными» предками всех
этих современных линий были, возможно,
удлиненные передвигающиеся по субстрату
животные с большим числом сегментов,
несущих по паре придатков, морские — в
случае хелицеровых и ракообразных, и,
вероятно, наземные — в случае одноветвистых.
Последняя группа до силура неизвестна.
Многие представители этих трех линий
сохранили удлиненную предковую форму
тела, в разной степени дифференцированного
на отделы (вместе с придатками),
соответствующие голове и туловищу, просоме
и опистосоме или голове, груди и брюшку
и т. д. Однако у других число сегментов
редуцировано за счет утраты или слияния
и еще более сократилось число пар ног.
Возникновение многих линий, например
морских и пресноводных планктонных
ракообразных и насекомых среди
одноветвистых, предположительно связано с педо-
морфозом. Веслоногие рачки, например,
могли произойти от планктонной личинки
крупного, удлиненного, бентосного предка.
Как уже говорилось, обитание в толще
воды дает селективное преимущество — тут
относительно много корма (из-за близости
фотосинтезирующего слоя) по сравнению
с донными осадками, питательность
которых зависит от оседающего
неиспользованного органического материала.
Три ныне живущие группы считаются
родственными членистоногим и, возможно,
представителями первой стадии «артропо-
дизации». Две из них—онихофоры (Опу-
chophora) и тихоходки (Tardigrada)
упоминались выше, третья — пятиустки (Pentasto-
ma). Все три отличаются от членистоногих
отсутствием их основных признаков—
наружного скелета и членистых
конечностей. Тело не сегментировано, хотя пары
мясистых, заканчивающихся коготками
ножек расположены вдоль всего тела или его
части, причем каждой паре соответствует
отдельный ганглий нервного ствола. У
тихоходок четыре пары таких ножек, у
онихофор—14—43, у пятиусток можно говорить
о двух сильно редуцированных парах (три
пары у ранней личинки). У онихофор другие
органы также сериально повторяются — по
паре на каждый «сегмент» с ножками.
Пятиустки — червеобразные
эндопаразиты, и их предполагаемое вторичное
упрощение затрудняет филогенетическую
интерпретацию— некоторые современные
исследования наводят на мысль об их близости
к ракообразным. Хотя гидростатический
скелет всех трех групп гемоцель,
характерный, вероятно, для предковых червей,
и у пятиусток, и у тихоходок наблюдается
кратковременное присутствие энтероцель-
ных карманов (три пары у первого типа,
две — у второго), т. е. признака,
характерного для лофофоровых и вторичноротых (см.
ниже). Эти «протоартроподы» являются
в сущности червями с мягким телом и
закладывающимися чертами «членистоного-
сти». Сейчас они обитают в основном на
суше (несколько видов тихоходок—в море),
хотя известны кембрийские морские
формы, напоминающие онихофор. Многие
исследователи считают онихофор близкими
по происхождению к одноветвистым.
Природа сегментации членистоногих,
протоартропод и киноринхов (и позвоноч-
Эволюционная история и филогения беспозвоночных
37
ных), таким образом, совершенно отлична
от таковой у аннелид. У первых она
вытекает из мономерного строения с
множеством пар конечностей (или у
позвоночных—мышечных блоков, обеспечивающих
плавание) и последующей сериализацией
связанных с движением скелетных,
мышечных, нервных и сосудистых элементов, а
у вторых основана на линейной цепочке
частично независимых единиц, каждая из
которых имеет собственные полости тела
и наборы органов, включая у примитивных
форм и гонады (рис. 2.10). Метамерные
группы членистоногих, вероятно, возникли
независимо от предковой группы или групп
неметамерных червей. Тем не менее
конечный результат эволюции членистоногих
и аннелид внешне сходен: тело состоит из
пресегментного переднего отдела («акрон»
у первых, «простомиум» у вторых), ряда ме-
тамерных сегментов, добавляющихся сзади
по мере роста и/или развития, и конечного
постсегментного отдела («тельсона» у
членистоногих, «пигидиума» у аннелид). У ки-
норинхов новые сегменты по мере развития
добавляются сходным образом.
2.4.2. Лофофоровые
Все потомки плоских червей,
рассматривавшиеся выше (за одним исключением),
имеют общие черты развития': а)
дробление яйцеклеток спиральное (иногда —с
модификациями); б) из бластопора возникает
рот, т.е. это—первичноротые животные; в)
ход дальнейших преобразований бластоме-
ров определяется на очень ранней
эмбриональной стадии (16-клеточной бластулы),
т.е. развитие детерминированное; г) если
гидростатический скелет или полости
внутри или вокруг органов — целомические
(ограничены мезодермальной выстилкой),
то они образуются путем расхождения
массы мезодермальных клеток, т. е. схизоцель-
ным путем (разд. 2.3.1, см. также разд.
15.2—15.4). Исключение составляет группа
червей, кратко упомянутая в разд. 2.3.1,
1 Из них первая — спиральное дробление -
наименее характерна; она присутствует только
в семи типах и не выражена, например, у брюхо-
ресничных, нематод и приапулид.
с телом, разделенным на три отдела, со
своей собственной полостью каждый,—
Phorona.
Хотя они и первичноротые. дробление
яиц у них радиальное и
недетерминированное, и по крайней мере некоторые из
полостей и мезодермальных тканей образуются
так, что это можно считать модификацией
энтероцельного способа (разд. 2.3.1; см.
разд. 15.2 - 15.4). Ныне живущие форони-
ды — мелкие сидячие черви, обитающие
в хитиновых трубках. Первый отдел их тела
(просома) очень мелкий, нависает над ртом.
Второй (мезосома)—также небольшой, но
несет крупный лофофор и содержит полость
(мезоцель), функционирующую как
гидравлическая система последнего. Лофофор
включает в себя полые, покрытые
ресничками щупальцевидные выросты стенки тела
вокруг рта, которые в первую очередь
используются для питания взвесями. Самый
крупный отдел тела — несегментированная,
лишенная придатков метасома с
полостью—метацелем, служащей основным
гидростатическим скелетом. Все три
полости тела связаны между собой порами.
Похоже, что от червей, подобных этим,
произошли два других типа современных лофо-
форовых и, возможно, вторичноротые. И
у лофофоровых, и у вторичноротых
радиальное недетерминированное дробление,
тройная возникающая энтероцельным
путем полость тела, причем некоторых
лофофоровых можно считать вторичноротыми
(рот образуется не из бластопора, а путем
вторичного прорывания кишки). Другие
общие черты включают личинок с системой
восходящих выстланных одноресничными
клетками полосок для сбора пищи вокруг
рта, трансформирующихся при
метаморфозе в руки лофофора с такими же клетками,
мозг в мезосоме и мезодерму,
возникающую из энтодермы или клеток
вегетативного полюса бластулы, что отличает ее от
мезодермы других билатерально
симметричных животных, развивающейся из губ
бластопора.
Плеченогие (Brachiopoda), появившиеся
в кембрии, утратили мелкий передний
отдел тела (просому); их метасома по
сравнению с метасомой форонид сильно
редуцирована, однако мезосома очень крупная
38
Ордовик
Л
у.
X
^
J^
Глава 2
Кемврий
Х.Г
Рис. 2.13. Возможное полифилетическое происхождение плеченогих как волна «брахиоподиза-
ции» различных форонидоподобных червей в докембрии/кембрии (по Wright, 1979)
и поддерживает хорошо развитый лофо-
фор. Это сидячие формы с телом,
покрытым вместо трубки известковой или
фосфатной раковиной, похожей на раковину
двустворчатых моллюсков, только с
дорсальной и вентральной, а не боковыми
створками. У некоторых плеченогих ее выросты
укрепляют мощный спирально
закрученный лофофор. Вероятно, разные группы
этого типа — потомки форонидоподобного
предка, а разные форониды прошли в
докембрии стадию «брахиоподизации»,
подобную радиации, давшей предка
членистоногих (рис. 2.13).
Ныне живущие форониды могут
размножаться бесполым путем, а один из их
видов — почкованием, что является нормой
для третьей группы лофофоровых —
мшанок (Bryozoa). У них колонии из тысяч
связанных между собой особей (зооидов)
образуются в ходе неоднократного
бесполого почкования особи-основательницы
и ее потомков точно так же, как у
некоторых книдарий (разд. 2.2.2). Для некоторых
мшанок характерен и полиморфизм
зооидов. По сравнению с предковым форонидо-
подобным червем отдельные зооиды очень
мелкие (увеличение размеров — свойство
целой колонии), часто лишены просомы
и секретируют раковину, домик или
студенистый матрикс, покрывающий их так, что
остается только отверстие, через которое
лофофор высовывается для сбора пищи
и втягивается для безопасности. Почти все
мшанки сидячие и могут рассматриваться
как крошечные колониальные варианты фо-
ронид. Они появились в ископаемых
остатках сравнительно поздно: не ранее
ордовика.
Родственные отношения мшанок с
другими группами лофофоровых остаются
спорными, отчасти потому, что способ
образования их полостей тела аберрантен:
у нескольких видов они формируются зано-
Эволюционная история и филогения беспозвоночных
39
во во время метаморфоза. Некоторые
предполагают, что мшанки близки к неясной
группе Entoprocta, с которыми их
объединяет высокоспециализированный тип
фоторецепторов. Эти спирально дробящиеся
крошечные сидячие животные несут
расположенные по кругу щупальца, как и лофо-
форовые, однако строение их не
соответствует структуре настоящего лофофора,
а полость тела здесь нецеломическая,
заполнена мезенхимой. Другие предполагают
родство с Entoprocta коловраток, но
свидетельств близости этого типа с какой-либо
конкретной группой слишком мало.
2.4.3. Вторичноротые
Парадоксально, но хотя четыре типа
вторичноротых — полухордовые (Hemichor-
data), иглокожие (Echinodermata), хордовые
(Chordata) и щетинкочелюстные (Chaetog-
natha) — образуют группу с большим
количеством общих черт, об их связях между
собой почти ничего не известно. В целом
можно считать, что различия между форони-
доподобными лофофоровыми и
полухордовыми крыложаберными выражены нечетко.
Тело последних разделено на три отдела,
каждый из которых содержит
образовавшиеся энтероцельным путем полости
(единственная в просоме, и парные в мезо- и ме-
тасоме), а мезосома несет крупный
гидравлически управляемый лофофоральный
аппарат. Единственные существенные
различия заключаются в том. что просома кры-
ложаберных хорошо выражена и
используется при движении, ее полость (протоцель)
образует гидростатический скелет, и
развитие животного происходит с образованием
вторичного рта. Из-за этого сходства
широко распространено мнение о близости
общего плана строения крыложаберных
и предковой формы вторичноротых; при
этом другая группа полу
хордовых—червеобразные кишечнодышащие, - вероятно,
утратила лофофор в связи с возвращением
к свободноживущему роющему образу
жизни.
Хотя иглокожие выглядят совершенно
иначе из-за раннего развития у них
сферической формы тела с почти радиальной
симметрией (в связи с сидячим существованием
и питанием за счет фильтрации), строение
их полостей тела ясно указывает на
исходную тройственную их организацию,
сходную с наблюдаемой у лофофоровых и
полухордовых. Один протоцель образует осевой
синус, левые мезоцель и протоцель
вместе — амбулакральную систему, правый
мезоцель сильно редуцирован у всех
иглокожих, за исключением аберрантных
организмов, а два метацеля дают основные отделы
полости тела. Иглокожие — одна из многих
групп, появившихся впервые с уже
характерным для типа обликом, начиная с
кембрия, хотя и не все ранние формы имеют
пятилучевую симметрию, свойственную
современным классам. О предковом плане
строения можно только догадываться.
Одна из наиболее правдоподобных гипотез
выводит иглокожих из сидячих, близких
к крыложаберным животных, у которых
лофофор с его гидравлической системой мезо-
целя развился в систему радиальных
каналов амбулакральной системы; у некоторых
современных крыложаберных у лофофора
пять основных стволов с каждой стороны.
Помимо черт, характерных для всех
вторичноротых, общий признак иглокожих
и хордовых—система твердых защитных,
содержащих кальций пластинок мезодерма-
льного происхождения в коже; у некоторых
вымерших иглокожих, похоже, были
образования, эквивалентные глоточным щелям,
присутствующим также у ныне живущих
полухордовых (и ряда гастротрихов).
Неудивительно поэтому, что большинство
попыток вывести хордовых из других групп
животных помещает иглокожих неподалеку
от точки их происхождения, но, как и в
случае перехода полухордовые/иглокожие, все
предложенные филогенетические схемы
страдают недостатком потенциальных
промежуточных форм; им нет подходящих
аналогов среди ныне живущих и ископаемых
видов, если не считать своеобразных Нота-
lozoa кембрия -девона (рис. 2.14),
лишенных какой бы то ни было симметрии,
которых некоторые считают предками
хордовых. Другие исследователи выводят
последних путем педоморфоза из личинок
полухордовых/иглокожих. Древнейший
ископаемый материал, точно относящийся к
хордовым, датируется ордовиком, хотя из-за од-
40
Глава 2
Рис. 2.14. Иглокожие группы Homalozoa—
возможные предки хордовых (по Clarkson,
1986)
ного кембрийского остатка и нескольких
позднекембрийских фосфатных фрагментов
время их возникновения не вполне ясно,
и если будет признано, что загадочные ко-
нодонты — это части скребущих органов
бесчелюстных позвоночных (как сейчас
кажется), периодом возникновения хордовых
надо будет считать кембрий и даже
докембрий.
Четвертый тип вторичноротых —
щетинкочелюстные—настолько же
непонятен с филогенетической точки зрения,
насколько важен с экологической; возможно,
это главные беспозвоночные хищники
морского планктона. Ранний эмбриогенез этих
прозрачных мягкотелых червей типичен для
вторичноротых, а их тело состоит из трех
отделов — головы, туловища и
постанального локомоторного хвоста.
Соответствуют ли эти три отдела про-, мезо- и ме-
тасоме полухордовых, проблематично,
причем некоторые черты щетинкочелюстных
явно сходны с таковыми у псевдоцеломиче-
ских червей. В настоящий момент об их
родственных связях с другими вторичноро-
тыми ничего нельзя сказать. Мягкотелые
планктонные виды хуже всех других
животных представлены в ископаемых остатках,
и, возможно (и даже весьма вероятно),
в прошлом было много типов таких
животных (см., например, рис. 2.16, Г, Д). о
существовании которых мы совершенно не
подозреваем (см. разд. 2.5); предки
щетинкочелюстных могли принадлежать к одной из
таких групп.
2.5. Происхождение, радиация
и вымирание групп животных
Все современные животные могут быть
помещены в 35 типов; под типом
понимается группа кажущихся родственными друг
другу организмов, чьи родственные связи
с аналогичными группами спорны или
чисто предположительны. Таким образом,
статус типа в принципе—дань нашему
незнанию, хотя в последующих разделах мы
попытаемся проследить вероятные
родственные связи. Эти 35 ныне живущих типов
тем не менее не являются результатом
медленной эволюционной дивергенции, ведшей
к возрастанию разнообразия. Кажется
вероятным, что—с очень небольшими
исключениями— все ныне живущие типы
животных уже существовали в кембрии, а
дивергенция подобных плоским червям
предшественников большинства групп
произошла еще в докембрии. Это своего рода
загадка.
Ископаемые остатки демонстрируют
медленное изменение животных во
времени, так что формы, присутствующие в
начале и в конце данной последовательности,
могут иметь достаточно различий для
отнесения к разным видам (так называемым
«хроновидам»). Среднее время, которое
требуется для анатомических изменений
такого масштаба, у самых разных
беспозвоночных составляет примерно 10 млн. лет.
Если признать, что роды животных
отличаются друг от друга, скажем, в 10 раз
сильнее, чем виды одного рода, семейства — в 10
раз сильнее, чем роды одного семейства,
и т. д. вплоть до отрядов, классов и типов,
то на основе средней скорости изменений
хроновидов можно получить по крайней
мере миллион миллионов лет для дивергенции
разных видов до такой степени, когда
можно будет признать их относящимися к
разным типам.
Эволюционная история и филогения беспозвоночных
41
Вероятно, первые животные произошли
от предков-протистов около 1000 млн. лет
назад, т. е. произведенный выше расчет
должен был бы привести нас к выводу, что их
потомки дивергируют с образованием
нескольких различных типов еще примерно
через 1,1 х 10 12 лет. К настоящему
моменту их потомки должны были бы отличаться
друг от друга, как, например, разные
семейства жуков. Даже если допустить средний
временной интервал между
последовательными хроновидами равным всего 1 млн. лет
и сократить коэффициент различия между
видами и родами, родами и семействами
и т. д. с 10 до 5, потребовалось бы более
3000 млн. лет для того, чтобы предковые
плоские черви дивергировали в несколько
типов. Однако типы, известные нам
сегодня, уже существовали и отличались друг
от друга не меньше, чем сейчас, всего через
400 млн. лет после вероятного
возникновения многоклеточных животных, и, по-
видимому, дивергировали за гораздо более
короткий промежуток времени.
Неудивительны слова одного современного автора
(Day, «Genesis on Planet Earth», 1984):
«Внезапное появление кембрийской фауны
представляет собой одну из крупнейших
биологических загадок... Типы, представленные
тогда, возникли быстро, происхождение их
неясно».
Очевидно, что-то не так в наших
эволюционных взглядах. Оказывается, не все
филогенетические изменения протекают так
медленно, как отражено в ископаемых
остатках близких видов. Временами
происходило взрывоподобное преобразование
морфологии, такое быстрое, что
промежуточные его стадии не представлены в
отложениях. Возможно также, что оно шло в
относительно небольших популяциях
(скорость потенциальных эволюционных
изменений обратно пропорциональна размеру
популяции), а это также уменьшает
возможность сохранения в ископаемом материале.
Педоморфоз (см. выше), например, в ходе
смены одного поколения другим может за
несколько лет привести к такого масштаба
изменениям взрослой формы, что в
результате потомство и родителей будут считать
принадлежащими к разным отрядам или
классам. (Таксономические категории, та-
КлассА
Класс В
КлассС
Класс D
Возрастающая
морфологическая дивергенция
Класс А
Класс В
Рис. 2.15. Одно и то же гипотетическое
филогенетическое древо, представленное в двух
разных формах:/!—классификация на основе
традиционных пиний, выделяемых по
морфологическому сходству и различию; Б—
классификация в соответствии с принципами
кладистики
ким образом, часто указывают на степень
морфологических различий и совсем
необязательно — на удаленность от общего
предка; такое положение хотели бы исправить
филогенетики «кладистской школы»;
удайся им это, типов и классов в том
объеме, в каком мы их понимаем сейчас, если
бы и осталось, то очень мало— рис. 2.15.)
Вспышки филогенетических изменений
часто были связаны с сильной дивергенцией
на уровне видов, родов, семейств и даже
отрядов от одной формы. Такая «адаптивная
радиация» происходит очень быстро в
геологическом масштабе времени, и этот
способ эволюции за короткий период
нескольких новых форм от одной предковой привел
к: а) представлениям о том, что процессы,
контролирующие изменения внутри одной
популяции (микроэволюцию), качественно
отличаются от приводящих к образованию
новых таксонов высшего ранга
(макроэволюции); б) теориям о количественно
различных скоростях микро- и макроэволюцион-
42
Глава 2
В _1_
д
ж
Л
Рис. 2.16. Вымершие преимущественно мягкотелые животные, которых нельзя поместить ни
в один из ныне известных типов. А— Wiwaxia (кембрий); Б — Opabinia (кембрий); В -
Dihomischus (кембрий); Г—Amiskwia (кембрий); Д—Tullimonstrum (карбон); Е—Odontogriphus
(кембрий); Ж—Hallucigenia (кембрий) (по Clarkson, 1986; Conway, Morris, 1979; Whittington,
1985). Масштабные линии 1 см
ных процессов; вторые идут быстрее
(модели «постепенности» и «прерывистого
равновесия», гл. 17).
Не все типы, возникшие во время
обширных докембрийской и кембрийской
радиации, представлены среди ныне живущих
беспозвоночных. Некоторые из них
известны только по ископаемым остаткам,
например археоциаты и трилобиты, и в
особенности обычно не получающая формального
статуса типа группа кембрийских
членистоногих (разд. 2.4.1). Однако их удивительно
мало, причем для большинства из них
характерны твердые части тела, т. е. признак,
похоже, возникавший в каждой линии
сравнительно поздно. Таким образом,
ископаемые остатки могут дать ложное
представление о прошлом разнообразии животных, на
которое указывает и обилие неизвестных
типов мягкотелых животных,
обнаруженных в тех редких осадках, где их остатки
сохранились (например, докембрийская фауна
Эдиакары и кембрийская Берджесских
сланцев) (рис. 2.16).
Двадцать ныне живущих типов, часто
называемых «малыми», включают менее
Эволюционная история и филогения беспозвоночных
43
500 видов каждый. За одним исключением,
подтверждающим правило, это
червеобразные мягкотелые формы, не сохранившиеся
в ископаемых остатках. Исключение—
плеченогие, насчитывающие всего 350
современных видов и более 25 000 ископаемых
форм, известных по толстым наружным
раковинам. Попытки объяснить тот факт, что
этим малым типам удалось дожить до
наших дней, имея в своем составе так мало
видов, привели к предположению, что это
статистически вероятно только в том случае,
если когда-то существовало гораздо
больше похожих (т. е. мягкотелых) впоследствии
вымерших типов (мы уже отметили такую
возможность в связи с предками щетинко-
челюстных; разд. 2.4.3). Ныне известные
ископаемые типы, таким образом, могут
представлять собой лишь небольшую и
нетипичную (из-за наличия твердых частей)
выборку: возможно, членистоногие—всего
лишь окаменевшая «верхушка» докембрий-
ского и кембрийского айсберга вымерших
групп.
Итак, напрашивается мысль о
широкомасштабной радиации червеобразных
животных в кембрии и докембрии, давшей,
возможно, сотни эквивалентных типам
таксонов. Большинство их скорее всего были
небольшими по количеству видов и
вымерли без следа, а ныне живущие черви—лишь
напоминание об этом былом разнообразии.
В некоторых линиях возникли твердые
защитные или скелетные части, возможно,
в ответ на эволюционное давление
хищничества, и о них наши знания довольно
значительны: считается, что сейчас найдено
и описано около 12% всех таких видов,
когда-либо существовавших на планете.
Несколько групп, дивергировав, дали начало
большому количеству видов и высокому
внутритиповому разнообразию. В
настоящий момент 10 типов включают более чем
по 10000 ныне живущих видов каждый, в то
время как плеченогие и иглокожие, когда-то
достигнув примерно такого же
разнообразия, потом сократили его ниже этого
уровня. Эти типы называют «большими».
Хотя некоторые типы отличались
высоким видовым разнообразием в течение
длительных геологических периодов, обычно
оно не обеспечивалось все это время одни-
Мел
tL_Z_J _ _ _Юра_
Триас
Пермь
Карвон
Девон
Рис. 2.17. Филогенетическое древо
аммонитов, демонстрирующее радиацию, вымирание
и вытеснение одной подгруппы другой с
девона до конца мела, когда вся эта группа вымерла
(по данным Moore, 1957)
ми и теми же формами. В разные интервалы
времени преобладали разные
представители отрядов и классов, причем несколько
когда-то доминировавших классов к
настоящему моменту вымерли, уступив место
другим. Подгруппы вступали в полосу
радиации по очереди. Это наиболее четко
видно на примере ископаемых аммонитов (рис.
2.17) и почти полностью вымерших наути-
лид среди головоногих моллюсков, а еще
более—на смене «эр» амфибий, рептилий
и млекопитающих в пределах позвоночных,
но характерно и для других типов, в том
числе книдарий, плеченогих, иглокожих
и неголовоногих моллюсков. В целом
замещение одной подгруппы другой, похоже,
происходило не за счет конкурентного
исключения, которое, как можно ожидать,
должно последовать после возникновения
и эволюции у вытесняющей подгруппы
какого-либо морфологического или иного
признака, получающего селективное
преимущество. Наоборот, сначала одна форма
вымирала, и лишь после—видимо, в уело-
44
Глава 2
виях «экологического вакуума»—ее
преемник радиировал, заполняя пространство
ниш, доставшихся ему «по наследству» от
доминировавшего ранее предшественника.
Ископаемые остатки показывают, что за
такой быстрой радиацией на многочисленные
формы (фаза прерывания в модели
прерывистого равновесия) шел гораздо более
длительный период сравнительно небольших
морфологических изменений под влиянием
стабилизирующего отбора (эквивалент
фазы равновесия), пока сама эта группа в свою
очередь не сокращала резко количества
видов или не вымирала полностью.
Степень вымирания разных типов
животных, характеризующихся широким
спектром завоеванных экологических ниш и
типов местообитаний, остается в то же время
спорной, хотя многие придерживаются
мнения о нескольких происходивших в
прошлом массовых вымираниях, особенно
ближе к концу ордовика, девона, перми, триаса
и мела (рис. 2.18), а также в большей или
меньшей степени примерно через каждые 26
млн. лет. Например, то, что произошло
около 220 млн. лет назад (в конце перми),
повлекло за собой исчезновение более поло-
' Массовые вымирания
1.Поздний ордовик-12%
2. Поздний девон-14%
3. Поздняя пермь-52%
4.ГЪздний триас-12%
5.Поздний мел-11%
400 300 200
Млн. лет назад
Рис. 2.18. Изменения числа семейств морских
животных в разные периоды, показывающие
ряд массовых вымираний (по Valentine, Moo-
res, 1974 и др.)
вины существовавших в то время семейств
морских животных и, возможно, 80—95%
их видов. Ранее доминировавшие группы,
включая ракоскорпионов (Eurypterida)
среди хелицеровых и трилобитов, исчезли
полностью, оставив к началу мезозоя фауну,
весьма близкую к современной по
основным типам строения преобладающих в
морях животных. Причины этого вымирания
неизвестны, хотя оно хорошо коррелирует
с крупными изменениями уровня моря,
которые могли быть вызваны фазами
оледенения, движениями континентов и/или
эпизодами глобального похолодания (были
также многочисленные спекулятивные
попытки привлечь к объяснению различные
космические силы и события).
Итак, история развития фауны, по-
видимому, была четко периодической.
Каждый период, включая и тот, в котором мы
сейчас живем, характеризуется случайной
смесью увеличивающих и сокращающих
свою численность групп животных, либо
сумевших сохраниться, понеся большие
потери, либо ждущих своего часа, чтобы
увеличить количество представителей и их
разнообразие после следующего периода
вымирания, индуцированного человеком или
другим агентом. С течением времени типов
животных стало меньше, поскольку
некоторые (а возможно, и многие) из них вымерли,
а новых возникло меньше, хотя число ныне
живущих видов больше, чем ранее. Моря
Земли сейчас сравнительно фрагментарны,
среди них обильны и велики мелководные
полуизолированные шельфовые водоемы,
а океанические воды насыщены кислородом
вплоть до абиссали—обстоятельства,
существовавшие далеко не всегда; в мелу,
например, большие поверхности морского
дна, возможно, были аноксическими. Таким
образом, нет никаких причин предполагать,
что морские виды сейчас менее
разнообразны, чем когда-либо в прошлом; в самом
деле, большинство оценивает морскую
фауну позднего кайнозоя как самую богатую
среди всех существовавших на планете.
Несмотря на это, огромное количество
современных видов почти полностью
отражает завоевание животными суши,
начавшееся в силуре—девоне и особенно мощно
шедшее в карбоне. Несколько удивительно,
Эволюционная история и филогения беспозвоночных
45
ЦАРСТВА Мопега
I Protista
nr4-!-&jn9'L
*S* /n\Plantae
Anlmalia
ЖИВОТНЫЕ
Кишечнополостные Плоские черви
Губки / Яервеовразные
/^ \ I /V\/МОЛЛЮСКИ
/ ». I / / /\^,ЛофОсроровые
/ / \ / / // V-Вторичноротые
/ / j*. I / /s ^^Лг Ракообразные
/ / / l^iV""""""' -4-Хелицеровые
I \"Членистоногие" / / /
\ Одноветвистые / /
Рис. 2.19. Число ныне живущих видов разных
царств и типов животных (по данным Barnes,
1984)
что эта колонизация, похоже,
осуществлялась в основном из литоральной зоны
непосредственно на сушу, а не по воде—из
моря в эстуарий, дальше в реку (озеро)
и т. д.; при этом большинство наземных
животных содержат много жидкости и
устойчивы к высыханию, а не наоборот.
(Основными группами, использовавшими менее
«популярный» водный путь, были
позвоночные и некоторые моллюски.) Пресные
воды заселялись главным образом
формами, уже адаптированными к наземным
условиям. Мелкие интерстициальные про-
тисты, инфузории, нематоды, тихоходки
и т.д. никогда не покидали водных пленок,
окружавших частицы осадка или почвы
и могут считаться наземными только в том
смысле, что их микроместообитание
находится на суше. Какой из двух путей ее
заселения они использовали, неизвестно и,
возможно, непознаваемо. На суше гораздо
больше, чем в воде, препятствий к
распространению, а это привело к массовому
видообразованию в пределах сравнительно
немногих успешно освоивших сушу типов
(по той же причине гораздо больше бентос-
ных морских организмов, чем
пелагических). Многие наземные виды
представляют собой географически сменяющие друг
друга варианты одного основного плана
строения. Сухопутные хелицеровые и одно-
ветвистые, особенно насекомые, сейчас
доминируют среди беспозвоночных планеты
по числу видов (рис. 2.19), так что за
последние 350 млн. лет центр разнообразия
организмов на видовом и родовом уровнях
переместился из моря, где и произошло все
филогенетическое развитие, рассмотренное
в этой главе, на сушу.
2.6. Резюме
и эволюционный обзор
Около 1 млрд. лет назад несколько
линий гетеротрофных протистов, обитавших
на морском дне, вероятно, пересекли крайне
плохо выраженную границу между коло-
ниальностью и многоклеточностью. дав
начало «животным». В течение следующих
нескольких сотен миллионов лет потомки
одной из этих линий дали ряд
крупномасштабных радиации, вызванных, по-
видимому, в основном наличием
неиспользованных источников пищи, давлением
хищничества, а также экологическими
и эволюционными преимуществами,
связанными с увеличением размеров тела (см.
разд. 9.1). Эти радиации привели к
широкому разнообразию бентосных черве- и
моллюскообразных животных, от некоторых
червей эволюционировали членистоногие,
лофофоровые и вторичноротые (рис. 2.20),
так что 600 млн. лет назад, вероятно, уже
существовали все известные типы и
множество неизвестных. С того времени разные
типы и входящие в них классы и отряды
46
Глава 2
Platyhelminthes
Gastrotricha и
Gnathostomula
Псевдоцелом и чес кие
группы
Arthropoda Nemertea Mollusca
(См. ниже)
1. Эвтелия I
2. Внутреннее
оплодотворение
З.Интерстициальные
1. Внутреннее
оплодотворение
формы
1.Внутреннее
оплодотворение
7. Метамерия
2. Псевдоцель
становится
циркуляторной
системой
3. Наружный
скелет
Внутреннее
оплодотворение
Конечности
Annelida,
Echiura и
Pogonophora
Sipuncula Lophophorata Deuterostomia
1. Метамерия
2. Циркуляторная
система
3. Прямой кишечник
с терминальным
анусом
4. Щетинки
1-Ц-образный
кишечник
1. Линяющая
кутикула
ч.Схизоцель
2. Гермафродитизм 2.Гермафродитизм 2.Псевдоцель
3. Утрата первичной З.Утрата первичной
личинки
личинки
4.Моноцилиарный
эпидермис
5-Интерстициальные
формы
1.Циркуляторная 1-Циркуляторная
система система
„ t- - 2.Кишка
2.Специализирован- 2.Сквознои
кишечный хоботок ниК с терминаль-
^ _ - ным анусом
Затрата первичной ^£»™к
личинки стерминаль-
4.Сквозной кишечник ным анусом 4.Радула
с терминальным
I анусом
3. Ктенидии
5. Моди'Фи ци рован ное
дроолениь
б.Дорсапьная
раковина
_L
Предковые, подобные плоским червям животные
Мономерип Тело без полости
Раздельнополость Нет кровеносной системы
Трохофороподобная Спиральное дробление
личинка
Слепо заканчивающая- Первичноротость
ся кишка
Наружное
оплодотворение
Д етерм и н и рован ное
развитие
(См. ниже)
1. Вторичноротые
1. Модифицированная
личинка
2. Лофофор
З.Энтероцель
4. Циркуляторная
система
5. Недетерминированное
развитие
6.Радиальное дробление
7. Олигомерия (три отдела)
8. Сквозной U-образный
кишечник
Э. Гермафродитизм, широко
„распространено бесполое
почкование
Псевдоцеломические и членистоногие
Группа лофофоровых/вторичноротых
III
I 1
Рис. 2.20. Гипотетические схемы родственных отношений между потомками близких к плоским
червям форм с указанием предковых признаков (сохранившиеся у потомков, кроме особо
оговоренных случаев) и основных вторичных черт специализации (по крайней мере исходных) в разных
группах или кладонах. Подробно связи в пределах линий псевдоцеломических/членистоногих
и лофофоровых/вторичноротых показаны отдельно
приобретали и теряли экологическое
значение. Палеоэкологи, например, на основе
форм с твердыми легко переходящими
в ископаемое состояние остатками
выделяют три сменявшие друг друга крупные
фауны, доминировавшие в морях: кембрий-
ско-ордовикскую (трилобиты и другие
ранние членистоногие, беззамковые плеченогие
и моноплакофоровые моллюски), позднепа-
леозойскую, или ордовикско-пермскую
(раковинные головоногие, морские лилии,
замковые плеченогие, мшанки класса Stenolae-
mata), и современную (мезозойско-кай-
нозойскую), характеризующуюся ведущей
Эволюционная история и филогения беспозвоночных
47
ролью брюхоногих и двустворчатых
моллюсков, ракообразных класса Malacostraca,
морских ежей, мшанок класса Gymnolaema-
ta и позвоночных. Каждая из этих фаун, по-
видимому, формировалась в мелководной
среде обитания, а позже перемещалась (?)
в более глубоководные зоны внешнего
шельфа и континентального склона.
Одной из важнейших стадий в радиации
типов строения животных было завоевание
толщи воды, позволяющее использовать на
месте фитопланктонную продукцию
поверхностных слоев моря. Основным
фактором, позволившим животным оторваться
от морского дна, был педоморфоз (см. гл.
17), который дал врожденные
репродуктивные преимущества, заключающиеся в
сокращении времени генерации. Изменения
в относительной хронологии процессов
развития (т. е. мутации «регуляторных генов»),
сопровождающие педоморфоз, являются
мощным средством быстрых и крупных
морфологических преобразований: за
несколько поколений анатомия половозрелой
особи может измениться до такой степени,
что предковую форму и ее потомка в
системах классификации, основанных на
морфологических различиях, будут относить
к разным классам или даже типам.
Известно несколько современных морских
личинок, у которых, по-видимому, в связи с
длительностью их периода оседания на дно
развитие гонад начинается еще до
метаморфоза; недавно наблюдали, как личинки
одного из видов гребневиков выметывали
вполне жизнеспособные гаметы (как
некоторые двустворчатые моллюски—икру);
это, возможно, указывает на способ
возникновения педоморфических групп и на
механизм ряда крупных «макроэволюционных
событий».
Последний этап в формировании
разнообразия животных имел место около 300—
400 млн. лет назад, когда животные с
конечностями начали колонизацию суши. По
сравнению с морем, где обитали их предки,
суша представляла собой в высшей степени
расчлененную с географической точки
зрения среду с множеством препятствий для
расселения организмов. Таким образом,
видообразование у наземных форм стало идти
в неизвестных прежде масштабах: хотя к
новым окружающим условиям смогло
приспособиться очень мало крупных групп,
подавляющее большинство видов
современных животных —наземные.
Довольно любопытная черта истории
больших типов животных —то, что. хотя
составляющие их подгруппы и имели
тенденцию сменять друг друга во времени,
похоже, это очень редко происходило за счет
конкурентного исключения. Напротив,
предшествовавшая группа сначала
вымирала или резко сокращала численность,
и лишь затем шла радиация последующей
для заполнения, очевидно, вакантных ниш.
Возможно, с этим перекликается то
обстоятельство, что, согласно современным
экологическим данным, известно очень мало
фактов вытеснения анатомически сложно
устроенными животными морфологически
более простых из общих местообитаний:
членистоногие и аннелиды, например, и по
сей день сосуществуют в осадках с
«примитивными» приапулидами и сипункулидами
без каких-либо признаков конкурентного
исключения.
Таким образом, следует избегать
соблазна считать, скажем, плоских червей и
кишечнополостных в каком-либо смысле
«ниже» ракообразных или хордовых,
отличающихся гораздо более сложным строением
и поведением. Разнообразные типы
животных, возникшие в результате крупных до-
кембрийской и кембрийской радиации,
соответствуют более или менее одновозраст-
ным альтернативным планам строения, не
демонстрируя постепенного возрастания
приспособленности и «совершенства».
Другими словами, вся жизнь
эволюционировала от ранних докембрийских прокариот;
некоторые из их потомков сейчас сложно
устроенные дубы, слоны и люди, но они ни
в коей мере не вытеснили прокариот, и
бактерии до сих пор успешно доминируют
в большинстве типов местообитаний!
Вымирание тех или иных типов и их
подгрупп, о котором упоминалось выше, по-
видимому, определялось в значительной
степени случайными причинами, как и то,
какие конкретно формы придут им на
смену. Как уже говорилось, группы не
вымирали из-за того, что их план строения в каком-
то смысле был хуже, чем у других; целые на-
48
Глава 2
боры неродственных между собой и хорошо
приспособленных животных, похоже,
становились жертвами радикальных
изменений окружающей среды, природа которых
хотя и неизвестна, но хорошо коррелирует
с крупными изменениями уровня моря
и глобальной температуры. Возможно,
многим типам с небольшим количеством
входящих в их состав видов не- удалось
пережить эти критические периоды, а более
крупные группы, сохранившиеся, хотя и со
значительными потерями в разнообразии,
исключительно благодаря большей
численности и более широкому географическому
и экологическому распространению своих
представителей, получили в результате
возможность новой радиации. Итак, хотя и не
возникает вопроса о том. является ли
экологической реальностью межвидовая
конкуренция, она, как правило, тем
интенсивнее, чем больше сходства между
конкурирующими видами. Следовательно, ничего
удивительного, если выживание в ходе
геологической истории таксонов высокого
ранга, таких, как отряды, классы и типы,
было по большей части делом случая.
Радиация | |_
Л
пт
S -■■•«- •
Радиация
III
Л
Фаза
равновесия или
"стазис"
|Прерыв
Рис. 2.21. Схематическое изображение двух
радиации потомков одной филогенетической
линии, демонстрирующее фазы прерывания
непрерывности и равновесия, а также
уменьшение разнообразия группы, вызванное
массовой смертностью из-за экологических
причин (Е) и восстановления высокого давления
отбора при насыщении местообитаний (S); см.
текст
Представители групп, переживших
основные периоды вымирания, сразу после
этого должны были освоить сравнительно
свободные местообитания, в которых
давление отбора, похоже, на время сильно
снижалось. В таких условиях последующая
радиация могла идти без заметных
ограничений, приводя к широкому
«экспериментальному» разнообразию, сокращавшемуся
только позже в ходе вымирания,
начинавшегося после нового насыщения
местообитаний с установлением относительно
высокого давления отбора. Отбор в этом случае
можно рассматривать как ограничитель
разнообразия, формирующегося во время
каждой радиации после стадии
непосредственного расселения (рис. 2.21, см. также
рис. 2.11). Именно пережившие отбор линии
в пределах каждой группы или подгруппы,
доминировавшей в период между
экологически обусловленными фазами массового
вымирания, в большей или меншей степени
давали начало новому эволюционному
циклу.
Таким образом, случай решал, какая
пьеса будет поставлена, определял
потенциальный состав исполнителей и открывал
занавес; отбор прослушивал кандидатов на
вакантные роли и следил за непрерывными
исправлениями, вносимыми в сценарий
и характеры персонажей.
2.7. Дополнительная
литература
Для иллюстрации филогенетической
точки зрения, противоположной
высказанной здесь, особенно в отношении
родственных связей гребневиков, мшанок и щетин-
кочелюстных, а также возможного места
возникновения основных групп животных
см. Nie/хеп С. 1985. Animal phylogeny in the
light of the trochaea theory. Biol. J. Linn. Soc.
Lond., 25, 243—299. (Этот автор
предполагает, что предковые формы большинства
эволюционных линий животных были ра-
диально симметричными планктонными
организмами, а билатеральная симметрия
сформировалась независимо у лофофоро-
вых/вторичноротых и у первичноротых.)
Эволюционная история и филогения беспозвоночных 49
Abele L.G. (Ed.) 1982. The Biology of Crustacea
(Vol. 2). Embryology, Morphology and Genetics.
Academic Press, New York.
Barnes R.S.K. (Ed.) 1984. A Synoptic Classification
of Living Organisms. Blackwell Scientific
Publications, Oxford.
Bryce D. 1986. Evolution and the New Phylogeny.
Llanerch, Dyfed.
Clarkson E.N.K. 1986. Invertebrate Palaeontology
and Evolution, 2nd edn. Allen & Unwin, London.
Cohen J., Massey B. D. 1983. Larvae and the origins
of major phyla. Biol. J. Linn. Soc, Lond., 19,
321 -8.
Conway Morris S., George J.D.. Gibson R., Piatt
H.M. (Eds) 1985. The Origins and Relationships
of Lower Invertebrates. Clarendon Press,
Oxford.
Glaessner M.F. 1984 The Dawn of Animal Life.
Cambridge University Press, Cambridge.
Goldsmith D. 1985. Nemesis. The Death Star and
Other Theories of Mass Extinction. Walker, New
York.
Henxon E.D. 1977. The Origin and Early Evolution
of Animals. Wesley University Press, Middleton,
Connecticut.
House M. R. (Ed.) 1979. The Origin of Major
Invertebrate Groups. Academic Press, London.
Manton S.M., Anderson D.T. 1979. Polyphyly and
the evolution of arth ropods. In: M. R. House
(Ed.) The Origin of Major Invertebrate Groups,
pp. 269 321. Academic Press, London.
McKerrow W.S. (Ed.) 1978. The Ecology of Fossils.
Duckworth, London. Scientific American 1982.
The Fossil Record and Evolution, Freeman, San
Francisco.
Sleigh M.A. 1979. Radiation of the eukaryote
Protista. In: M. R. House (Ed.) The Origin of Major
Invertebrate Groups, pp. 23—53. Academic Press,
London.
Trueman E. R.. Clarke M. R. (Ed.) 1985. The Mollus-
ca (vol. 10). Evolution. Academic Press, Orlando.
Valentine J. W. (Ed.) 1985. Phanerozoic Diversity
Patterns. Princeton University Press, Princeton.
New Jersey.
IVhittington H.B. 1985. The Burgess Shale. Yale
University Press, New Haven.
ЧАСТЬ
ТИПЫ БЕСПОЗВОНОЧНЫХ
В главах этой части мы
проиллюстрируем и кратко охарактеризуем все
известные современные типы беспозвоночных,
а также входящие в их состав классы.
Этот систематический обзор гораздо
менее детален, чем в других учебниках по
зоологии беспозвоночных, так как вместо
описания всех анатомических черт разных
типов животных мы сосредоточили внимание
только на тех жизненно важных признаках
каждой группы, которые должен знать
каждый студент, т. е. на определяющих
индивидуальность каждого типа или класса, их
эволюционные преимущества или
экологическую роль. Приводятся также списки
диагностических черт, облегчающие
сравнение отдельных типов.
В этой части мы постарались
подчеркнуть разнообразие планов строения
беспозвоночных, отраженное основным
таксономическим делением животного царства.
При этом всем классам уделено одинаковое
внимание независимо от числа входящих
в них видов. Этот «уклон» исправлен тем,
что в части III преобладает материал о
более крупных, хорошо известных и, без
сомнения, более важных группах.
Деление на типы, классы и т.д.,
принятое в этой части, основано на работе Барнса
(Barnes, 1984, «A Synoptic Classification of
Living Organisms», Blackwell Scientific
Publication, Oxford). Как известно, до сих пор
продолжается открытие новых таксонов
животных—с 1980 г. описаны новый тип
Loricifera и два новых класса,
предположительно из хорошо известных типов (Remipe-
dia из ракообразных и Concentricycloidea из
иглокожих). Таким образом, хотя в книге
полностью охвачены все известные в 1987 г.
классы беспозвоночных, нет оснований
считать, что человеку уже знакомы все главные
группы животных, живущих ныне на
планете. Похоже, что более детальное
исследование таких малоизученных биотопов, как
глубоководная и интерстициальная зоны,
позволит обнаружить новые, совершенно не
похожие на известные типы существ.
Последнее слово о разнообразии животных
еще долго не будет сказано!
ГЛАВА О
ПАРАЛЛЕЛЬНЫЕ ПОДХОДЫ
К МНОГОКЛЕТОЧНОСТИ У ЖИВОТНЫХ
Protozoa
Porifera
Placozoa
Coelenterata: Cnidaria и Ctenophora
Platyhelminthes
Mesozoa
Группы животных, рассматриваемые
в этой главе, в основном относятся к таким,
которые, согласно предположению,
высказанному в гл. 2, могли возникнуть
непосредственно в рамках царства Protista,
поскольку они обладают принципиально
различными чертами организации. Таким
образом, их можно рассматривать как
параллельные эволюционные линии,
достигшие многоклеточности независимо друг от
друга.
3.1. Protozoa (простейшие)
До начала 1970-х гг. было принято
помещать все организмы в одно из двух царств:
фотосинтезирующие или растущие в земле
формы относили к растениям, а нефотосин-
тезирующие и способные к свободному
перемещению—к животным. Является
организм про- или эукариотным, одно- или
многоклеточным, считалось при этом
фундаментальном подходе к делению всего
живого неважным. В рамках подобной
классификации многоклеточные растения потом
объединили в группу Metaphyta, в то время
как одноклеточные фотосинтезирующие
формы (включая также различных
прокариот) были названы Protophyta или Algae.
Соответственно многоклеточные животные
составили группу Metazoa, а
одноклеточные — Protozoa.
Такая система из двух царств явно
страдала многими недостатками, два из
которых— важнейшие: во-первых, она не
отражала реальную филогенетическую
дихотомию: нет никаких данных,
подтверждающих, что цианобактерии, диатомеи,
багрянки, мухоморы и хвойные, например,
составляют одну естественную группу, в то время
как амебы, кишечные жгутиконосцы, губки
и иглокожие—другую. Впрочем, гакая
искусственность не была бы серьезной
проблемой, если считать цели классификации
чисто прагматическими: два царства
должны представлять собой лишь четко
очерченные, удобные, хотя и произвольные
взаимоисключающие подразделения
организмов. Но второй недостаток как раз и
заключается в том, что категории растений
и животных реально не различаются. В
пределах группы явно близкородственных
организмов (эвгленовые жгутиконосцы,
например) часть видов одного рода можно
отнести к растениям, часть — к животным,
а еще несколько форм приходится считать
и теми и другими одновременно.
Некоторые динофлагеллаты удовлетворяют свои
пищевые потребности на 5% за счет
фотосинтеза и на 95% — за счет гетеротрофного
питания. Более того, известны
жгутиконосцы, способные менять царство, к которому
они были отнесены первоначально, на
противоположное, просто оставаясь сутки
в темноте! Таким образом, система из двух
царств не только не отражает
филогенетического родства, но и не является
работающей схемой, поскольку, хотя множество
организмов и можно с уверенностью назвать
«животными» или «растениями», суще-
4»
52
Глава 3
ствует большое количество и таких,
которых не удается отнести ни к тем ни к
другим.
В течение последних примерно 15 лет
завоевала приверженцев и сейчас шире всего
используется альтернативная
классификация, стремящаяся более точно отражать
филогению, сохраняя при этом четкость
подразделения на группы. Она также
постулирует фундаментальную дихотомию живых
организмов, выделяя на этот раз надцар-
ства прокариот и эукариот; эукариоты
делятся на 4 царства: одноклеточные (протисты)
и три царства многоклеточных: растения,
грибы и животные (рис. 3.1). Протисты,
таким образом, включают формы, ранее
относимые к водорослям и простейшим; при
этом ни те ни другие уже не имеют
формального таксономического статуса,
оставаясь лишь общими названиями,
эквивалентными, например, термину «черви»; или,
другими словами. Protozoa с некоторых пор
перестали считать валидной и практически
полезной систематической группой. Одна
из последних самых общих классификаций
Система из пяти царств
Рис. 3.1. Система классификации из пяти
царств и ее отношение к разделению всех
организмов на растения и животных
протистов делит это царство на 27 типов, из
которых более половины облигатно нефо-
тосинтезирующие, причем 13 из них когда-
то считались животными: Parabasalia*, Ki-
netoplasta *, Opalinata*, Stephanopogono-
morpha. Ciliophora, Apicomplexa *, Metamo-
nada*, Sarcodina, Labyrinthomorpha *,
Choanophlagellata, Microspore*, Ascetospo-
ra * и Myxozoa * (рис. 3.2) (звездочкой
отмечены типы, в основном или полностью
состоящие из эндосимбионтов или
эндопаразитов). Несколько других типов (например,
Dinophyta, Euglenophyta и Cryptophyta)
включают ряд форм, на которые
претендовали как зоологи, так и ботаники.
Большинство «простейших-протистов»
филогенетически удалено от
беспозвоночных и лишено черт, свойственных только
многоклеточным животным. Их общие
с ними отрицательные признаки, например
отсутствие .пластид, присущи также «грибо-
подобным протестам» и собственно
грибам, а среди обших положительных можно
назвать только характерные для всех
эукариот, включая и фотосинтезирующие
группы. Конечно, по определению разница
между нефотосинтезирующими протестами
и другими эукариотными царствами, в том
числе и животными, заключается в том, что
последние—многоклеточные, а первые—
одноклеточные (одиночные, или
колониальные). Отдельная клетка протистов, таким
образом, способна выполнять все функции,
необходимые для выживания и размноже-.
ния, одновременно обеспечивая локомо-
цию, питание, осморегуляцию,
воспроизведение и т. д., в то время как у животных эти
и другие задачи осуществляются
различными специализированными группами клеток,
обычно объединенных в отдельные ткани
или органы. Тем не менее это различие
чисто фенотипическое. Все множество
клеток многоклеточного животного
происходит от одной обычно диплоидной клетки и,
таким образом, генетически тотипотентно,
как и отдельная особь протистов.
Некоторые клетки, например амебоциты губок,
могут даже оставаться
недифференцированными и существовать в виде резерва, из
которого в течение всей жизни животного
дифференцируются по мере необходимости
специализированные клетки.
Рис. 3.2. Разнообразие форм тела протистов (по Sleigh et al., 1984)
54
Глава 3
1(Г
10"'
й Ю"
10"'
Более интенсивный
метаволизм
Беспозвоночных при тех
же размерах
10"
10"а ТО"6 10"
Масса тела,г
10
Рис. 3.3. Отношение между скоростью
метаболизма в покое и размерами тела для
протестов и беспозвоночных, демонстрирующее
более высокие метаболические потребности
многоклеточных по сравнению с одноклеточными
тех же размеров (по Schmidt-Nielsen, 1984)
(1 ккал = 4,2 кДж)
Неудивительно, что тело протистов
обычно намного мельче, чем у животных,
хотя это и не всегда так. Размеры
значительного числа сложно устроенных
животных, состоящих из тысяч клеток, например
среди лофофоровых, членистоногих или
хордовых, менее 1 мм вдоль самой длинной
оси, а у большинства представителей псев-
доцеломических групп—менее 2 мм. С
другой стороны, клетки некоторых ресничных
протистов (тип Ciliophora) достигают 5 см
в длину, а псевдоподии некоторых Хепо-
phyophora (тип Sarcodina) могут
охватывать поверхность диаметром 25 см.
Единственный способ, которым неколониальные
протисты отвечают на давление отбора,
благоприятствующее увеличению размеров
(см., например, разд. 9.1),— разрастание
самой клетки, часто сопровождающееся
полиплоидией или появлением нескольких ядер;
при этом токи цитоплазмы формируют
впоследствии внутриклеточную систему
циркуляции. В известном смысле крупных
размеров протистам достичь сравнительно
легче, поскольку интенсивность
метаболизма у одноклеточных ниже, чем у
многоклеточных (рис. 3.3).
Таким образом, организмы, ранее
относимые к типу или полцарству Protozoa,
представляют собой неоднородный набор
протистов, отчасти эквивалентных одной из
клеток тела животных, отчасти — целом)
животному. Слово «эквивалентных» в
предыдущем предложении очень важно,
поскольку у ныне живущих протистов более
длинная самостоятельная эволюционная
история, чем у различных многоклеточных
организмов, и только тип Choanoflagellata
сохранил некоторые признаки,
позволяющие говорить о его прямом родстве с
животными (гл. 2).
3.2. Тип Porifera (губки)
3.2.1. Этимология
От лат. porus — пора; ferre—нести.
3.2.2. Диагностические
и специфические признаки
(рис. 3.4)
1. Общей характерной симметрии нет
даже у видов, форма которых может иногда
зависеть от локальной экологии.
2. Многоклеточные, но с небольшим
числом типов клеток; тканевая организация
развита очень слабо; органы отсутствуют
и координация функций клеток слабая;
нервной системы нет.
3. Характерны хоаноциты,
обеспечивающие циркуляцию воды и питание.
4. Иногда имеется сложный скелет либо
из известковых или кремнеземных спикул,
либо из белковых (коллагеновых = спонги-
новых) волокон, либо из элементов обоих
типов.
5. Клетки располагаются вокруг
водяных камер или каналов разной степени
сложности; настоящая полость тела или кишка
отсутствуют.
6. Питание фильтрационное; газообмен
путем диффузии.
7. Только сидячие (прикрепленные)
формы.
8. Размножение половое или бесполое;
известна раздельнополость, но обычен
гермафродитизм; дробление полное; личинки
планктонные.
9. Исключительно водные, в основном
Параллельные подходы к многоклеточное™ у животных
55
Оскулум
Юротничок
■Спикула ■*—
Хоаноцит
Атриум
Пороцит
Мезохил
Амеюцит
Пинакоцит
Пинакодерма
-(врезка А,
-(врезка Б)
Асконоидная организация
-Жгутик
ютничковые
микроворсинки
Плоскость Биения
жгутика
Парус
Фибриллярная \_i>=R
сеть ~^'
Рис. 3.4. Асконоидный тип организации: А —
спикулы разных видов; Б— срезы хоаноцита
(по Meglitsch, 1972; Alexander, R. McN., 1979;
см. также рис. 2.4)
морские формы (примерно 10000 видов);
около 50 пресноводных видов.
Функционирование губок определяется
одной чертой их строения: размещением
клеток вокруг системы камер и каналов, по
которым за счет биения жгутиков хоаноци-
тов циркулирует вода. Среда, таким
образом, пропускается через организм, а не
окружает его, так что неизбежен
фильтрационный тип питания, как и размножение
стадиями, выходящими из родительской
особи.
Простейшая асконоидная организация
губок изображена на рис. 3.4: в центре —
единственная камера (атриум), окруженная
стенкой тела, которая покрыта снаружи пи-
накоцитами (пинакодермой) и пронизана
порами (остиями), пропускающими внутрь
воду и образованными пороцитами;
единственное выводное отверстие — оскулум.
56 Глава 3
Асконоидныи
Сиконоидныи
Рис. 3.5. Разные типы
складчатости стенок тела у губок (по
Лейконоидный Buchsbaum. 1951)
1Д У$£¥&Л
Рис. 3.6. Два типа
архитектоники губок (по Barnes, 1980)
Пинакоциты могут сокращаться, сжимая
губку; некоторые из них секретируют
вещества для прикрепления к субстрату. Ме-
зохил1, лежащий под пинакодермой,
представляет собой студенистый белковый ма-
трикс с опорными скелетными элементами
и различными амебоцитами с широким
спектром функций — от питания и секреции
коллагена до превращения в любой другой
тип клеток (тотипотентность). В некотором
смысле хоаноциты и пинакоциты образуют
1 Этот термин в отечественной литературе
употребляется пока редко, чаще вместо него
встречается термин «мезоглея». Мезохил губок
не гомологичен анатомически мезоглее
кишечнополостных. Он отличается и по строению, имея
клеточную природу и представляя собой
смешанную паренхиму с разнообразными функциями,
в то время как мезоглея выполняет в основном
только скелетную функцию.— Прим. пер.
ткани (хоанодерму и пинакодерму), но база-
льные мембраны здесь отсутствуют.
Этот тип организации ограничивает
увеличение размеров губки, поскольку объем
воды в атриуме растет как куб длины тела,
в то время как жгутиковая поверхность —
только как квадрат последней. Это
ограничение преодолено у некоторых видов за счет
складчатости стенки тела и уменьшения
атриума, что привело к образованию сико-
ноидных и, наконец, лейконоидных форм
(рис. 3.5). Большинство лейконоидных
губок либо «сплошные» (смежные вводные
и выводные каналы проходят параллельно
друг другу; поры, а иногда и оскулумы
разбросаны по всей поверхности), либо полые
и трубчатые, с выводными каналами,
никогда не соединенными с поверхностью
тела, а открывающимися во внутреннюю
полость с дистальным оскулумом (рис. 3.6).
Параллельные подходы к многоклеточности у животных
57
Это разделение по форме тела не отражает
систематики губок (см. ниже). Наиболее
распространена лейконоидная организация,
что свидетельствует о ее эффективности.
Она возникала в ходе эволюции губок
неоднократно, иногда, по-видимому, через
промежуточное сиконоидное состояние. Лейко-
ноидные формы в своем развитии часто
сначала проходят асконоидную и сиконоид-
ную стадии.
Хоаноциты крайне важны для
метаболизма губок. Они прогоняют через них воду
за счет биения своих жгутиков, а также
улавливают мелкие пищевые частицы,
заносимые внутрь тела токами воды, когда вода
проходит через их «воротнички»,
представляющие собой венчики из микроворсинок
(рис. 3.4). Более крупные пищевые частицы
могут поглощаться амебоцитами. Как
хоаноциты, так и амебоциты передают
захваченные частицы пищи другим клеткам, и,
похоже, основными пищеварительными
структурами служат амебоциты, а не
хоаноциты.
Губки размножаются бесполым путем,
высвобождая небольшие фрагменты тела
или скопления необходимых для развития
клеток, в основном амебоцитов. Например,
геммулы (рис. 3.7, А), производимые
пресноводными губками, состоят из археоцитов
(крупных специализированных
амебоцитов), покрытых прочной твердой оболочкой
из спонгина и спикул, позволяющей им
зимовать. Половое размножение, хотя
обычно гермафродитное (черта, кореллирующая
с сидячим образом жизни), часто включает
перекрестное оплодотворение. При этом
спермий выходит через оскулум одной
особи, проникает в пору другой, улавливается
ее хоаноцитом и передается яйцеклетке.
Развитие до личиночной стадии чаше всего
протекает в материнском организме.
Личинка—обычно паренхимула со сплошным
телом, покрытым жгутиками, кроме одного
из полюсов (рис. 3.7, £), но иногда — полая
амфибластула, одна из полусфер которой
состоит из мелких жгутиковых клеток,
а другая — из макромеров без жгутиков
(рис. 3.7, В)—покидает родительское тело
и короткий период времени перед
оседанием на дно ведет планктонный образ
жизни.
Микропиле
Внутренняя
мембрана
Археоциты
Бластоцель
Макромер
Рис. 3.7. А. Геммула пресноводной губки. Б.
Паренхимула. В. Амфибластула (no Meglitsch,
1972)
58
Глава 3
Неудивительна высокоразвитая
способность губок к регенерации (даже из одной
клетки), обусловленная автономностью их
клеточных комплексов.
3.2.3. Классификация
Примерно 10000 описанных на
сегодняшний день видов распределены по
четырем классам.
3.2.3.1. Класс Hexactinellida
(шестилучевые губки)
Представителей этого класса часто
называют стеклянными губками. Их скелет
образован кремнеземными,
преимущественно шестилучевыми спикулами (рис.
3.8), откуда их название; тело часто похоже
на вазу в среднем 30—40 см высотой (рис.
3.9). Преобладают сиконоидные формы.
Пинакоциты, присутствующие во всех
других классах, здесь отсутствуют, и наружные
покровы представляют собой сетевидный
синцитий, образованный соединенными
псевдоподиями амебоцитов.
Исключительно морские виды, встречающиеся
преимущественно на больших глубинах. Следует
отметить, что некоторые авторы выделяют
их в отдельный подтип или даже тип (Sym-
plasma).
3.2.3.2. Класс Calcarea (известковые
губки)
Представители этого класса отличаются
присутствием спикул из карбоната каль-
Рис. 3.8. Кремнеземные спикулы губок класса
Hexactinellida (no Alexander, 1979)
Рис. 3.9. Стеклянная губка корзинка Венеры
(по Barnes, 1980)
ция—кальцита или арагонита.
Встречаются все три типа организации ас-
коноидная, сиконоидная и леиконоидная.
Высота большинства видов менее 10 см.
Только морские формы.
3.2.3.3. Класс Demospongiae
(обыкновенные губки)
Самый крупный класс, объединяющий
90% всех известных видов. Скелет состоит
из кремнеземных спикул и/или спонгиновых
волокон. Одно семейство (Spongiidae)
включает обычные туалетные губки со
скелетом только из спонгина. Организация
только леиконоидная, некоторые формы
достигают значительных размеров (например,
тропические Spheciospongia—более 1 м
в диаметре и в высоту). Окраска часто
блестящая.
Среди представителей этого класса так
называемые сверлящие губки способны
делать отверстия в известняковых
образованиях—кораллах или раковинах
моллюсков. Есть и пресноводные виды, в
основном из семейства Spongillidae.
Параллельные подходы к многоклеточности у животных
5В
3.2.3.4. Класс Sclerospongiae
(коралловые губки)
Класс содержит очень небольшое число
специализированных морских видов,
обитающих в гротах и туннелях, связанных
с коралловыми рифами в разных частях
света. Его представители напоминают
обыкновенных губок, но отличаются сложным
скелетом из кремнеземных спикул и спонгино-
вых волокон, внутри тонкого
поверхностного слоя живой ткани, лежащего на
массивном основании из карбоната кальция (рис.
ЗЛО). Все виды лейконоидные.
Класс
Hcxactinellida
Calcarca
\Demospongue
\ Sclerospongiae
Отряд
Amphidiscosida
Hexactinosida
Lychniscosida
Lyssacmosida
Clathnnida
Leucettida
Leucosolennda
Sycettida
Inozoida
Sphinctozoida
Homosclerophonda
Chonstida
Spirophonda
Lithisnda
Hadromenda
Axmellida
Agelasida
Halichondrida
Poecilosclenda
Petrosiida
Haplosclenda
Verongnda
Dictyoceratida
Dendroceratida
.Ceratoporellida
_ Tabulospongida
3.3. Тип Placozoa
(пластинчатые)
3.3.1. Этимология
От греч. plakos—плоский; zoon —
животное.
3.3.2. Диагностические
и специфические признаки
(рис. 3.11)
1. Осей симметрии нет; форма может
меняться, как у амеб.
2. Нет отдельных тканей или органов.
3. Нет полости тела или
пищеварительной полости.
4. Отсутствует система нервной
координации.
5. Тело в форме толстой пластины,
которая может двигаться в любом
направлении в своей плоскости.
6. Единственный наружный слой
жгутиковых клеток окружает заполненный
жидкостью мезохил, содержащий сеть
звездчатых клеток-волокон.
7. Морские формы.
Верхняя поверхность плоского диско-
видного тела образована тонким слоем
чешуевидных клеток, многие из которых
несут жгутик. Более толстый нижний слой
состоит из двух типов клеток—столбчатых
жгутиковых, перемежающихся
железистыми безжгутиковыми (рис. 3.12). Последние
секретируют на добычу (разнообразных
протистов) ферменты, частично
переваривающие ее вне тела. Продукты
переваривания поглощаются теми же железистыми
клетками. Нижние жгутиковые клетки
обеспечивают скользящее движение животных;
изменения формы тела, делающие их
похожими на крупных амеб, обеспечиваются
координированными сокращениями и
расслаблениями клеток-волокон в мезохиле.
Размножение обычно бесполое—как
делением, так и почкованием. Известно и
половое размножение, но оно и последующее
развитие зародыша очень плохо изучены.
Яйцеклетки, вероятно, образующиеся из
клеток нижнего слоя, по-видимому,
находятся в мезохиле; в воде, где находились
животные, обнаруживались спермин, но
процессы сперматогенеза и оплодотворения
никогда не наблюдались.
Тело пластинчатых состоит из
нескольких тысяч клеток и достигает диаметра
3 мм, хотя в нем представлены всего 4
клеточных типа, и эти животные относятся
к простейшим из известных представителей
многоклеточных. Клетки содержат меньше
60
Глава 3
Известковый
скелет
Рис. 3.10. Губка класса Sclerospongiae с деталями строения верхней поверхности (по Bergquist,
1978)
Дорсальный (чешуйчатый)
клеточный слои
Жгутики
Мезохил
Полость, заполненная
жидкостью
Вентральный
клеточный слой
Рис. 3.11. Общий вид Placozoa (no Barnes, Рис. 3.12. Схематический срез Placozoa (по
1980) Margulis, Schwartz, 1982)
Параллельные подходы к многоклеточное™ у животных
61
ДНК, чем у всех прочих животных
(количества того же порядка, что у бактерий),
хромосомы очень мелкие, меньше 1 мкм в
длину. Почти сто лет эти формы считали пла-
нулоподобными личинками губок или
стрекающих, но в конце 1960-х гг. было
обнаружено, что они могут достигать половой
зрелости, ив 1971 г, для них был описан новый
тип.
Пластинчатых чаще всего обнаруживали
в морских аквариумах, а в естественных
условиях находили на литорали. Вероятно,
они широко распространены, но их трудно
заметить.
3.3.3. Классификация
Описан только один вид (возможно, их
два). Этот тип часто помещают в отдельное
подцарство фагоцителлообразных (Phago-
cytellozoa), название которого неудачно,
поскольку их способ питания обычно не
фагоцитоз.
3.4. Тип Cnidaria (стрекающие:
гидроидные, сцифомедузы,
актинии, кораллы)
3.4.1. Этимология
От греч. cnide — крапива.
3.4.2. Диагностические
и специфические признаки
(рис. 3.13)
1. Радиально симметричные животные
со щупальцами, окружающими рот, на
одном из концов тела.
2. Есть внутренняя пищеварительная
(«гастроваскулярная») полость,
открывающаяся во внешнюю среду только ртом.
3. Две основные формы тела: медуза,
приспособленная к пелагическому образу
жизни, и бентосный полип. Полипы бывают
колониальными, а в пределах колонии
полиморфными.
4. Стенка тела образована двумя
тканями— эпидермисом и гастродермисом1,
1 В советской литературе эти ткани принято
называть эктодермой и энтодермой
соответственно.— Прим. ред.
состоящими из разного числа типов клеток.
Между этими двумя тканями находится
бедная клетками мезоглея. Органы развиты
слабо.
5. Во всех классах типа присутствуют
так называемые книдоциты со
стрекательными органеллами (в основном — не-
матоцистами), не встречающиеся больше
ни в одной из групп многоклеточных.
6. Могут быть скелетные элементы из
хитина и/или карбоната кальция.
7. Нервная система в виде одной или
двух сетей.
8. Большинство -хищники;
газообмен за счет диффузии.
9. Размножение бесполое и половое;
последнее часто включает формирование
почти билатерально симметричной
покрытой ресничками личинки планулы.
10. Только водные формы,
большинство- морские.
В жизненных циклах стрекающих могут
участвовать две разные формы
организации— относительно подвижная медуза,
несущая гонады, и сидячий полип, который
исходно их, по-видимому, не имел (рис.
3.13). Полип трубчатый, на одном конце
у него рот, на другом - базальный диск,
с помощью которого животное приклепляе-
тся к субстрату. У медуз рот расположен на
конце хоботка—манубриума,
прикрепленного к нижней поверхности зонтика
наподобие ручки. Полипы отпочковывают медуз
бесполым путем, а медузы образуют
полипы в результате полового размножения,
часто с промежуточной стадией планулы -
сплошной (без полости внутри) покрытой
ресничками личинки (см. гл. 14, приложение
14.5).
Это — очень общее описание, и в
пределах типа много исключений: не все медузы
подвижны, не все полипы сидячие,
некоторые полипы несут гонады, некоторые
медузы отпочковывают медуз, в ряде жизненных
циклов отсутствует стадия медузы, в
других—стадия полипа, иногда развитие
протекает без планулы.
Клеточная организация полипа и
медузы в основном одинакова, различия в ней
касаются частностей: у некоторых полипов
мезоглея—тонкий, в основном
неклеточный слой, а у медуз она толще, часто более
62
Глава 3
ОРАЛЬНЫЙ ПОЛЮС
Щупальца
Эпидермис
Гастродермис
Гастроваскулярная
полость
Мезоглея —
АБОРАЛЬНЫЙ ПОЛЮС
УМБРЕЛЛА
Гастроваскулярная полость
ПОЛИП
1
1
Рот
Стебелек
,
Парус
Эпидермис
Мезоглея
Гастродермис
-Щупальца
СУБУМБРЕЛЛА
МЕДУЗА
Рис. 3.13. Два типа формы тела стрекающих (по Fingerman, 1976)
волокнистая, иногда в ней присутствуют
бродячие клетки (амебоциты).
Обобщенная схема строения стенки
тела, типичной для гидроидных полипов,
приведена на рис. 3.14.
Эпидермис состоит из пяти основных
типов клеток:
1. Эпителиально-мышечные:
столбчатые «покровные» клетки (иногда
чешуйчатые) с базальным расширением,
содержащим сократимые миофибриллы,
ориентированные параллельно орально-абораль-
ной оси тела, т.е. продольные мускулы.
2. Интерстициальные: содержат
крупные ядра и мало цитоплазмы; способны
образовывать другие типы клеток.
3. Стрекательные клетки, или книдо-
циты: содержат способные выворачиваться
структуры—нематоцисты (рис. 3.15, см.
также рис. 2.5), чаще называемые книдоци-
стами. У гидр и медуз на одном конце этих
клеток расположена жесткая щетинка —
книдоциль, устроенная так же, как и
жгутик. У актиний ее нет, но иногда
присутствует конический комплекс ресничек,
вероятно, выполняющий те же функции. Не-
Мезоглея
Пищеварительная
вакуоль
Клетки, секрети
рующие слизь
Псевдоподии
Ядро эпителиально-
мускульной клетки
Сенсорная клетка
Книдоцит
Книдоциль
Пищеварительные
мускульные клетки
Клетки, секре-
тирующие ферменты ^"^ Нервная клетка
ГАСТРОДЕРМИС ЭПИДЕРМИС
Клетки, секретирующие
слизь
Интерстициальная клетка
Рис. 3.14. Продольный срез стенки тела в классе Hydrozoa (no Barnes, 1980)
Параллельные подходы к многоклеточное™ у животных
63
Книдоциль
Опорный
стержень
Нематоциста
Крышечка
Врезка А
Рис. 3.15. Книдоцит и нематоцисты: А —
неотстрелянная нить сложена складками; Б—
отстрелянная нить; В—спироциста актинии
(по разным источникам) (см. также рис. 2.5)
Опорная
фибрилла
матоциста включает свернутую в спираль,
гофрированную, зазубренную изнутри
трубку. Выстреливание почти наверняка
обусловлено быстрым поступлением в
книдоцит воды, связанным с изменением его
мембранной проницаемости, которое
в свою очередь, вероятно, вызывается
электрически по сигналу книдоциля. Нить
выворачивается наизнанку, складки ее
расправляются, а сама «отстреленная» книдоциста
растягивается.
Существуют три основных типа книдо-
цист:
а) собственно
нематоцисты—разнообразные по форме, часто с шипами
(стилетами) и токсичные; используются для
защиты и ловли жертвы;
б) спироцисты—клейкие структуры,
обнаруженные у некоторых актиний. Их нити
растворяются при выстреливании, образуя
густую клейкую сеть для прикрепления
и ловли;
в) птихоцисты—обнаруженные только
у актиний группы Ceriantharia (с. 73)
структуры со сложно гофрированными нитями
без шипов. Нити также клейкие и
способствуют формированию трубки, в которой
находятся актинии.
Книдоцисты используются только один
раз, а потом отбрасываются. Книдоциты
64
Глава 3
могут регенерировать из интерстициальных
клеток; развивающийся книдоцит
называется книдобластом.
4. Слизисто-железистые клетки:
вырабатывают слизь для защиты, ловли добычи
и приклеивания.
5. Сенсорно-нервные клетки:
удлиненной формы, расположенные под прямым
углом к поверхности эпидермы. Более или
менее тождественны мультиполярным
нейронам (гл. 16). Соединяясь с другими
нейронами, образуют неправильную сеть, или
плексус (см. разд. 16.4.1). У некоторых
стрекающих (не гидр) две нервные сети в
эпидерме, у других — одна в эпидермисе,
другая— в гастродермисе (см. ниже).
Гастродермис состоит из: а) питательных
мышечных клеток, поглощающих пищевые
частицы для последующего
внутриклеточного пищеварения. Они, как и
эпителиально-мышечные, способны к сокращению, но
их сократимые части ориентированы под
прямым углом к орально-аборальной оси,
образуя слой кольцевой мускулатуры. С
помощью жгутиков перемешивают пищу в га-
строваскулярной полости; б) железистых
клеток, вырабатывающих ферменты,
необходимые для пищеварения в гастроваску-
лярной полости; в) клеток, секретирующих
слизь; г) у некоторых стрекающих
(например, актиний) — книдоцитов. У медуз и
актиний гастродерма сложно-складчатая.
Локомоция происходит с помощью
мышечных сокращений, преодолевающих
давление жидкости в гастроваскулярной
полости, которая выполняет функции
гидростатического скелета (см. гл. 10), как, вероятно,
и мезоглея. Движения у полипов обычно
ограничены изгибанием и скольжением;
у медуз они гораздо разнообразнее. Но
даже последние могут активно перемещаться
только в орально-аборальной плоскости
благодаря хорошему развитию кольцевой
мускулатуры по нижнему краю зонтика,
позволяющей животным двигаться в
основном вертикально; тогда как горизонтальное
перемещение происходит пассивно за счет
океанических течений. Тем не менее
некоторые медузы (Cubomedusae, см. с. 69)
способны к быстрому активному движению
в горизонтальной плоскости аборальной
поверхностью вперед.
Следует отметить способность
стрекающих к регенерации. Она использована
эмбриологами для изучения общих
процессов клеточного деления и дифференциров-
ки, в частности у гидры, которую легко
разводить в лаборатории (см. разд. 15.5). У нее
орально-аборальная полярность
восстанавливалась, даже если животное разрезали на
несколько кусков вдоль этой оси. Щупальца
всегда возникали на ближайшем ко рту
целого животного конце такого фрагмента.
Похоже, что необходимая для
этого информация связана с орально-
аборальным градиентом какого-то
вещества, различаемого клетками и
вызывающего у них соответствующую реакцию
(деление и дифференцировку) в зависимости от
его концентрации, а следовательно, от
уровня вдоль градиента, на котором
расположены клетки.
Митозы происходят вдоль всей оси тела,
но существует градиент их скорости—от
высокой у орального полюса, сразу под ги-
постомом, т. е. бугорком, над которым
располагается рот (см. рис. 3.16), до
низкой —у аборального. Таким образом,
поддерживается постоянный «ток» клеток от
орального к аборальному полюсу и к
концам щупалец. В результате со своих мест
ОРАЛЬНЫЙ ПОЛЮС
'4 дня
Пип остом
2дня
Формирующаяся-
почка
Движение
\ метки незна-
J чительно или
отсутствует
20 дней
дней
■Сформировавшаяся
почка
АБОРАЛЬНЫЙ ПОЛЮС
Рис. 3.16. Перемещения клеток гидры (по
Campbell. 1967)
Параллельные подходы к многоклеточное™ у животных
65
вытесняются клетки, которые отмирают
(происходит «линька») и используются для
формирования новых почек. Таким
образом идет непрерывное обновление клеток —
в течение нескольких недель все они у
одной особи замещаются новыми, что
позволяет животным быть как бы бессмертными
(разд. 13.2.4).
3.4.3. Классификация
Описано около 10000 ныне живущих
видов. Их распределение по таксонам
высшего ранга зависит от того, какая стадия—
полип или медуза доминирует в жизненном
цикле. В классе Hydrozoa (гидроидные)
стадия полипа обычно преобладает, в классе
Scyphozoa (сцифомедузы) — все наоборот.
В третьем классе Anthozoa все формы—
полипоиды, а медуз, вероятно, нет вовсе.
Однако термин «полипоид» использован
в данном случае сознательно, поскольку эти
животные, хотя и сидячие, имеют много
общих черт со сцифомедузами: толстую
клеточную мезоглею, ввернутый внутрь ману-
бриум (так называемый стомодеум), га-
стродермальные гонады. Известны и
сидячие медузоидные формы (см. ниже). Так что
Anthozoa вполне можно считать сидячими
медузами без настоящей полипной стадии!
Тем не менее одна из существующих
классификаций рассматривает Anthozoa как
(коралловые) полипы, а отсутствие у них
медузной стадии—как настолько
серьезный признак, что их помещают в
отдельный от гидроидных и сцифомедуз подтип
(табл. 3.1). Те же аргументы можно
использовать, если считать Anthozoa медузами,
лишенными полипной стадии. Такой
подход имеет свои преимущества: он позволяет
выделить у Anthozoa несколько классов,
которые, как мы увидим, сильно различаются.
Однако отсутствие в жизненном цикле
медузы или полипа не такая уже редкость и
у представителей других классов, следова-
Класс
Подкласс
Hydrozoa -
„Scyphozoa -
Отряд
Athecata
Milleporina
Stylasterina
Chondrophora
Thecata
Limnomedusae
Trachylina
Siphonophora
Actinulida
Stauromedusae
Coronatae
Semaeostomeae
Rhizostomeae
- [Cubozoa] •
'Anthozoa
Cenantipatharia
Alcyonaria
Zoantharia'
- Cubomedusae
Antipathana
Ceriantharia
Protoalcyonaria
Stolonifera
Telestacea
Gastraxonacea
Gorgonacea
Coenothecalia
Alcyonacea
Pennatulacea
Actiniana
Zoanthinaria
Scleractinia
Corallimorpharia
Ptychodactiaria
5-1314
66
Глава 3
Таблица 3.1. Классификация стрекающих
Классическая
Альтернативная
Класс Hydrozoa
Класс Scyphozoa
Класс Anthozoa
Подкласс Alcyonaria
[Подкласс Zoantha-
ria1»!
Подтип Medusozoa
Класс Hydrozoa
Класс Scyphozoa
Класс Cubozoa
Подтип Anthozoa
[Класс Ceriantipatha-
ria1»]
Класс Alcyonaria
Класс Zoantharia
"Включая Ceriantharia.
[ ] Некоторые считают таксон искусственным
(Schmidl, 1972).
тельно, ему можно было бы не придавать
слишком большого значения и у Anthozoa.
Поэтому здесь мы используем более
классическую схему, делящую стрекающих на
три крупных класса — Hydrozoa, Scyphozoa
и Anthozoa, признавая ее, впрочем, не
вполне удовлетворительной.
3.4.3.1. Класс Hydrozoa
(гидроидные)
Hydra—хорошо всем известная гидра
(рис. 3.16) не является типичным
представителем этого класса, так как она: а)
одиночная; б) лишена медузной стадии. Гораздо
более характерна Obelia (рис. 3.17). Три
момента характеризуют полипную стадию
этих животных: а) они состоят из
нескольких полипов (модулей), связанных со
столонами (стелющаяся гидрориза и восходящий
гидрокаулус), так что представляют собой
колониальные, или модульные, организмы;
б) не все полипы одинаковы: некоторые, со
щупальцами, выполняют
пищеварительную функцию (гастрозоид, или трофозоид);
другие (гонозоиды)—репродуктивную, т.е.
отпочковывают медуз, третьи,
отсутствующие у Obelia, но обычные у других
гидроидных,—булавовидные, несущие батареи кни-
доцитов и служащие для защиты
(дактилозоиды, с. 70); в) присутствует опорный
скелет из хитина (иногда обызвествленного),
называемый перисарком; ткань, которую
он окружает (ценосарк), охватывает общую
для всей колонии полость тела. У разных
таксонов могут быть значительные как
количественные, так и качественные различия
в этих чертах.
У медузы Obelia много характерных для
гидромедуз черт. Она мелкая (0,5 см), ее
гонады эпидермального происхождения,
яичники и семенники находятся на разных
особях. Однако на внутреннем краю зонтика
присутствует только рудиментарный парус
(велум), гораздо лучше развитый у других
видов гидромедуз (ср. рис. 3.18). Гастродер-
мис без мускулов. Вместо них
специализированные мускулы по краю внутренней
поверхности зонтика образуют кольцевые
слои, создающие движущую силу. Они
сокращаются, выталкивая воду вниз, а
медузу— вверх. Серии сокращений напоминают
пульсацию. Иногда парус образует узкое
отверстие, создающее струю воды.
Мускулы работают, преодолевая сопротивление
мезоглеи, которая, хотя и неклеточная,
однако содержит белковые волокна,
придающие ей упругость. Движение вниз пассивно.
Нервная система также
специализирована—плексус образует два нервных кольца
у нижнего края зонтика, иногда даже с
ганглиями. Последние связаны с сенсорными
органами, расположенными также по краю
зонтика: глазками (реагируют на свет)
и статоцистами (реагируют на ускорение)
(рис. 3.19).
У некоторых видов медуза не плавает
свободно, а остается прикрепленной к
полипу, высвобождая только гаметы. Гонозоид
в этом случае называется гонофором. В
случае Tubularia от гастрозоида не отделяется
не только медуза (гонофор), но и планула.
Вместо них высвобождается личинка-
актинула—округлая пелагическая поли-
поидная форма (рис. 3.20). У некоторых
видов прикрепленная медуза редуцируется
и остаются только гонады (например, у
гидры). Прикрепленность и редукция медуз,
вероятно, возникали у гидроидных несколько
раз независимо. С другой стороны, в ряде
случаев из жизненного цикла исчезает
полип, а не медуза (рис. 3.21).
Колониальность и полиморфизм
достигли своего предельного выражения у
некоторых своеобразных пелагических видов:
знаменитого португальского кораблика
(Physalia) с заполненным газом поплавком
Параллельные подходы к многоклеточное™ у животных
67
Рот
Гипостом
Стебелек
Щупальце
Гидротека
Гидрокаулус
Гастрозоид
Гонада
Гидрориза столона -
Рис. 3.17. Жизненный цикл Obelia (по Barnes, 1980)
Молодая колония
Зонтик
Стеьелек —
Гастроваскулярная
полость
Радиальный канал
Кольцевой канал-
Рис. 3.18. Гидромедуза рода
Bougainvillea (no Bullough,
1958)
Ротовое щупальце
Рот
Глазок
Парус
Батареи
нематоцист
(видоизмененная медуза) и свисающими
вниз щупальцами; отходящими от корми-
диев (пучков модифицированных полипов,
рис. 3.22,А), а также парусника (Velelld) с его
блюдцеобразным поплавком,
(видоизмененный гастрозоид), несущим гонозоиды
и дактилозоиды (рис. Ъ.22,Б).
Наконец, некоторые колониальные
гидроидные секретируют внутренний эпидер-
мальный известковый скелет. Это так назы-
5«
68
Глава 3
Сувумврелла
Сувумвреллярное
нервное
кольцо
Парус
Конкреция
Кольцевой канал
Экзумвреллярное
нервное
кольцо
Сенсорный
эпителий
Сенсорная
ресничка
Рис. 3.19. Статоцист
гидромедузы (no Barnes, 1980)
Выходящая актинула
Перисарк
Колония гидроидов Прикрепленная актинула
Рис. 3.20. Жизненный цикл Tubularia (no Barnes, 1980)
Параллельные подходы к многоклеточное™ у животных
69
Рис. 3.21. Жизненный цикл
гидроида Aglaura, не
имеющего полипной стадии (по Barnes,
1980)
ваемые гидрокораллы (отряды Milleporina
и Stylasterina) (рис. 3.23). Это прямостоячие
и обрастающие формы, достигающие
иногда значительных размеров. Их
многочисленные дактилозоиды могут причинять
сильную боль, поэтому некоторые
представители данной группы называются
огненными или жгучими кораллами.
3.4.3.2. Класс Scyphozoa
(сцифомедузы, сцифоидные)
Типичный для сцифоидных жизненный
цикл—у рода Amelia (рис. 3.24,Б).
Доминирует медуза (рис. Ъ.2Л,А) с диаметром
зонтика в среднем 10 см, у некоторых видов он
достигает 30 см. Если не принимать во
внимание размер, сцифомедузы очень похожи
на гидромедуз, описанных выше (с. 66).
Различия следующие: а) манубриум несет
щупальца (короткие у Amelia, но очень
длинные у некоторых других видов); б) нет
паруса (кроме Cubomedusae; см. ниже); в)
мезоглея толстая и волокнистая, с
блуждающими амебоцитами; г) нервные
кольца редки, контроль за пульсацией
осуществляется краевыми скоплениями нейронов,
расположенных в мелких булавовидных
структурах—ропалиях, несущих также ста-
тоцисты и иногда глазки (рис. 3.25); д)
гонады гастродермального происхождения.
Большинство сцифомедуз—хищники, но
некоторые, в том числе и Amelia,—
фильтраторы. У этого вида молодь
фильтрует воду краями щупалец, а взрослые
особи улавливают планктон с помощью
слизистых тяжей, секретируемых нижней
поверхностью зонтика. Вся она и лопасти ману-
бриума покрыты жгутиками, которые гонят
пищевые частицы ко рту.
Медузы обычно раздельнополы.
Оплодотворенные яйца развиваются в планулу,
которая образует полипоидную сцифисто-
му; та почкуется поперечным
расщеплением, давая ювенильные медузы — зфиры
(рис. 3.24). Как сцифистома, так и эфира—
питающиеся стадии.
Существуют аберрантные сцифоидные:
некоторые не образуют сцифистому, у
других сцифистома быстро формирует гонады,
но отсутствует медуза. Ставромедузы
живут, прикрепившись стебельком к морским
водорослям (рис. 3.26). Упомянутые выше
кубомедузы—быстрые пловцы, наносящие
очень сильные ожоги. У них есть парус и га-
стродермальные гонады, так что их часто
считают промежуточными формами между
гидромедузами и сцифоидными, помещая
в отдельный класс Cubozoa. Часто
встречающаяся в Карибском море Cassiopeia
Хитиновые септы
Газовая
/камера
Уровень воды
Дактилозоид
(со щупальцами)
Щупальце
(вытягивается
до 9 м)
"Парус"
Газовые трувки + выросты
энтодермы
Пневматофор, разделенный на камеры
Три ряда
дактилозоидов
Рис. 3.22. A. Physalia и детали строения ее кормидия. Б. Разрез через тело Velella (по разным
источникам)
Гастрозоид
Дактилозоид
Скелет
?/:ма&слШь&'-ъЖ .v«Lv^a6«№;
о О ®
© о
о
Рис. 3.23. Участок колонии
гидрокоралла Millepora:
слева— вертикальный разрез,
справа—вид сверху (по Rus-
sel- Hunter, 1979)
Ропалии
Свободноплавающие самец и самка медузы
(половое размножение)
яСцифистома
Рис. 3.24. Медуза (А) и жизненный цикл (Б) Aurelia (no Fingerman, 1976)
Пигментное пятно
Рис. 3.25. Ропалий Aurelia
в разрезе (по Hyman, 1940)
-Статоцист
72
Глава 3
Отверстие
сувумврел-
лярной ямки
Положение
гонад внутри
Вторичные
щупальца
Ротовой
стебелек
Стебелек
Прикрепительный диск
Рис. 3.26. Haliclystus—представитель отряда
Stauromedusae (no Bullough, 1958)
стродермиса с мезоглеей внутри, разделяют
гастроваскулярную полость. Некоторые из
них, так называемые полные,
прикрепленные к стомодеуму, но имеют в районе рта
отверстия в стенках для облегчения
циркуляции гастроваскулярной жидкости; другие,
неполные, к стомодеуму не прикреплены
(рис. 3.27).
Края мезентериев представляют собой
трехлопастные тяжи (рис. 3.28). Наружные
лопасти покрыты ресничками, а
центральная несет книдоциты и клетки,
вырабатывающие ферменты. Для Anthozoa
характерно, что нематоцисты лишены крышечки
(нить часто вырывается из нематоцисты
просто через трехклапанное отверстие).
Книдоциль также отсутствует (см. с 62).
Септы могут нести и свободные нити—
аконции, которые закручиваются вокруг
добычи и участвуют в ее умерщвлении с
помощью многочисленных книдоцитов.
Мезоглея толстая, волокнистая, клеточная.
Мускулатура в основном гастродермального
прикрепляется куполом вниз на тихом
мелководье мангровых зарослей. У нее
множество вторичных ртов, открывающихся
непосредственно в гастроваскулярную
полость. Мелкие животные, уловленные
поверхностью оборчатых «рук», перемещаются
к этим ртам по слизистым тяжам.
3.4.3.3. Класс Anthozoa
Представители этого
класса—полипоиды, но (см. также выше, с. 059) крупнее
и сложнее по строению, чем полипы в
других классах (рис. 3.27). Их уплощенный
оральный диск несет щупальца и щелевидный
рот, ведущий в латерально уплощенный
стомодеум. В одном или в обоих «углах»
рта может присутствовать ресничный
желобок, сифоноглиф, ведущий в стомодеум.
Сифоноглифы, вероятно, направляют воду
в гастроваскулярную полость, поддерживая
в ней гидростатическое давление. Вместе
с щелевидной формой рта наличие сифоно-
глифов придает симметрии Anthozoa черты
бирадиальности, т.е. уже невозможно
разрезать их тело на две эквивалентные
половины вдоль любого диаметра. Мезентерии
(или септы), т. е. выступающие складки га-
'от
Щупальце
1ерсрорация
в мезентерии
Стомодеум
Мезентериаль-
ная нить
Мускул-ре -
трактор
Аконция
Неполные
вторичные
мезентерии
Полные
первичные
мезентерии
Гонада
Подошвенная
пластинка
СифОНОГЛИфЫ
■Неполный
третичный
мезентерий
Рис. 3.27. Схема строения типичной актинии.
Внизу—поперечный разрез на уровне стомо-
деума (по Alexander, 1979)
Парвплельные подходы к многоклеточное™ у животных
73
Отверстие
в септе
Продольный
мускул
Столбик
Радиальный
мускул
Подошва
■Аконция
Рис. 3.28. Строение мезентерия (по Fretter, Graham, 1976)
происхождения; продольные мускулы-
ретракторы расположены в мезентериях,
а кольцевые—в гастродермисе внутренней
трубки. Размножение как бесполое—
почкованием (иногда образуются
колонии—см. ниже), поперечным
расщеплением и разрывом (см. гл. 14), так и
половое—с гонадами гастродермального
происхождения в некоторых или во всех
мезентериях. Известны раздельнополые и герма-
фродитные формы. Оплодотворение
наружное или внутреннее; в последнем случае
часть развития может протекать внутри
материнского организма (примитивное
живорождение). Обычно образуется планула,
планктотрофная или лецитотрофная (см.
гл. 14), время существования которой
варьирует. Рот и мезентерии могут начать
формироваться внутри личинки, но
щупальца обычно не образуются до оседания на
Дно.
Актинии—самые обычные одиночные
Anthozoa; они распространены повсеместно
в прибрежных водах. Обычно они
прикреплены к твердым субстратам в литоральной
зоне. У некоторых, например, Metridium,
хорошо развита мускулатура вокруг
орального диска, образующая сфинктер таким
образом, что щупальца втягиваются
внутрь, когда актиния сжимается во время
отлива (рис. 3.29) для защиты и экономии
энергии на дыхании. Обычный
североамериканский и европейский вид Stomphia cocci-
пеа может при угрозе отрываться и плавать,
изгибая тело. Несколько других видов
плавают за счет ударов щупалец.
Представители семейства Minyadidae пелагические; они
плавают вниз щупальцами с помощью
газового пузыря. Ceriantharia также
представители одиночных Anthozoa,
приспособленные к существованию на илистом
дне. Они живут в слизистых трубках,
образованных птихоцистами (см. с. 63).
Известны также различные
колониальные кораллообразующие Anthozoa.
Каменистые склерактиниевые кораллы состоят
из полипов, соединенных латерально и
«сидящих» на известковом «наружном скелете»
(рис. 3.30). Это обеспечивает опору и
защиту, так как когда полипы втягиваются, они
74
Глава 3
Рис. 3.29. Metridium senile на разных стадиях
сокращения тела (по Robbins, Snick, 1980, с
изменениями)
Наружный
скелет
W
Рис. 3.30. Продольный разрез через тело
особи коралла из отряда Scleractinia (no Barnes,
1980)
лишь слегка выступают над скелетной
пластиной. Октокораллы (или восьмилуче-
вые—у них всего восемь щупалец) также
образуют разветвленные колонии полипов,
латерально соединенных цененхимой,
состоящей из толстой мезоглеи и связующих
гастродермальных трубок (рис. 3.31).
Мезоглея содержит амебоциты, секретирующие
скелет из СаСОэ. Таким образом, здесь
скелет внутренний. У некоторых в цененхиме
находится центральный опорный стержень
из белка или известковых спикул. В
последнем случае его выделяют из так
называемого благородного коралла (Corallium)
и обрабатывают, получая всем известные
коралловые ювелирные изделия.
И наконец, группа Alcyonaria—это
мягкие кораллы, образующие мягкие
кожистые колонии, достигающие размеров более
1 м. Их цененхима также содержит
известковые спикулы, но их здесь меньше, чем
у других восьмилучевых.
Форма всех этих кораллов значительно
варьирует и зависит от типа модульного
роста.
Полое перистое
щупальце
Эпидермис на поверх-
ности колонии \
Стомсдеум
Мезоглея
Спикулы внутреннего
.скелета
Гастродермальные трувки
между полипами
V1
Молодой полип
Рис. 3.31. Продольный разрез
через полип восьми лучевого
коралла (по Russel- Hunter,
1979)
Параллельные подходы к многоклеточности у животных
75
3.4.4. Симбиоз
Большое количество стрекающих из
всех классов находится в симбиозе с авто-
трофами. Зоохлореллы (зеленые
водоросли) встречаются в гастродермисе
гидроидных, в мезоглее сидячей сцифомедузы
Cassiopeia (см. с. 69); зооксантеллы (динофла-
геллаты из рода Symbiodiniurri) — в
гастродермисе актиний и кораллов (Anthozoa).
Излишки продуктов фотосинтеза—
мальтоза у гидры и в основном глицерин
у Anthozoa—используются хозяином и
могут поддерживать его в течение
длительного периода голодания, если не составляют
единственный источник питания. У
кораллов симбиоз способствует также
отложению известкового скелета, но каким
образом—плохо понятно. Роль симбиоза в
питании обсуждается в гл. 9.
Симбиоз с участием в основном
Anthozoa важен также для защиты других
организмов. Мелкие рыбки из рода Amphiprion
живут среди щупалец крупных актиний.
Какие-то вещества на этих рыбках (а
возможно, и слизь, выделяемая самой актинией)
ингибируют выстреливание нематоцист.
Актинии защищают симбионтов и в свою
очередь получают от них защиту от
некоторых хищников. Сходный комменсализм
известен у актиний с несколькими другими
видами рыб, креветками-чистильщиками,
раками-щелкунами, крабами-стрелками,
офиурами. Актинии могут прикрепляться
к другим животным: сцифомедузам,
гребневикам и раковинам, в которых обитают
раки-отшельники. В этих случаях они также
защищают хозяев, получая преимущества
от перемещения на новые кормовые
участки.
3.5. Тип Ctenophora
(гребневики)
3.5.1. Этимология
От греч. ktenos—гребень; phoros—
несущий.
3.5.2. Диагностические
и специфические признаки
(рис. 3.32)
1. Медузоидные формы, но радиальная
симметрия стала бирадиальной из-за двух
щупалец.
2. Гастроваскулярная полость со ртом
и анальными порами.
3. Двуслойная стенка тела окружает
толстую мезоглею с амебоцитами и
гладкими мышечными волокнами.
Рис. 3.32. Типичный гребневик: вид сбоку
(вверху) и с аборального полюса (внизу) (по
Buchsbaum, 1951)
76
Глава 3
Статолит
Мерцательная Бороздка
Статоцист <
Гревные
пластинки
Пружинящие реснички
Рис. 3.33. Детали строения
аборального конца гребневика
со статоцистом и гребными
пластинками (по Barnes, 1980)
4. Книдоциты отсутствуют, но
щупальца несут «ловчие клетки» (коллобласты).
5. Локомоция с помощью слившихся
ресничных, так называемых гребных,
пластинок, расположенных восемью орально-
аборальными полосами.
6. Нервная система представляет собой
субэпидермальную сеть, хорошо развитую
под рядами гребных пластинок.
7. Хищники; газообмен за счет
диффузии.
8. Размножение у некоторых бесполое,
но таких мало; гермафродиты с полным
детерминированным дроблением, ведущим
к образованию «цидиппидной» личинки.
9. Исключительно морские, в основном
пелагические формы.
Животные, относящиеся к этому типу,
так похожи на медуз из типа стрекающих,
что их часто считают боковым
ответвлением предковой медузоидной формы
последних. В самом деле, долгое время греб-
Гранулы с клейким
веществом-,,
Колловласт
Нервная
клетка
Рис. 3.34. Разрез через щупальце гребневика с деталями строения коллобласта (по различным
источникам)
Параллельные подходы к многоклеточности у животных
77
невиков помещали со стрекающими в один
тип кишечнополостных—Coelenterata (от
греч. koilos—полость; enteron
—кишечник). Типы стрекающих и гребневиков и
сейчас часто объединяют в раздел
лучистых— Radiata (по типу симметрии).
Тем не менее у гребневиков много
важных отличительных черт. Первая среди
них—ряды «гребней» (откуда название
типа), т.е. коротких поперечных пластинок
с ресничками, соединенными в виде гребня
(рис. 3.33). Биения ресничек направлены
в аборальную сторону, так что животное
плывет ртом вперед и обычно вверх по
вертикали, но может плыть и вниз. Контроль
за движением осуществляется нервной
сетью и апикальным статоцистом (рис. 3.33).
Щупальца выворачиваются из глубоких
выстланных ресничками карманов на або-
ральной стороне. Они несут коллобласты
(рис. 3.34)—грушевидные структуры с
внутриклеточными свернутыми спиралью
нитями и заполненными слизистым
веществом гранулами, служащие для
опутывания добычи. До недавнего времени
предполагалось, что у вида Euchlora rubra есть кни-
доциты, но новые данные свидетельствуют
о том, что он отбирает их у своей жертвы-
книдарии. Кишечник состоит из каналов,
как у некоторых медуз, но они могут
открываться во внешнюю среду анальными
порами. Пищеварение внутри- и внеклеточное,
непереваренные остатки удаляются через
рот и анальные поры. Гермафродиты с
гонадами из двух пучков (яичник и семенник)
в гастродермисе. Цидиппидная личинка
очень похожа на яйцевидных взрослых
особей, ее развитие прямое, однако у
уплощенных форм оно требует значительных пре-
. образований (см. ниже).
Заметная черта гребневиков—их
люминесценция. Светящиеся вещества
образуются в гастродермисе.
3.5.3. Классификация
«Малый» тип, включающий всего около
100 известных видов, распределяемых по
двум классам, как показано ниже.
Класс
Tcntaculata
Nuda-
Отряд
Cydippida
Platyctida
Lobata
Ganeshida
Thalassocalycida
Cestida
- Beroida
3.5.3.1. Класс Tentaculata
У всех представителей этого класса два
щупальца; форма тела обычно напоминает
крыжовину (рис. 3.32). Однако некоторые
виды сплюснуты в щупальцевой плоскости;
тогда наружные части расширяются в виде
лопастей (рис. 3.35), иногда так, что живот-
Гревной ряд
Ротовая лопасть
Рис. 3.35. Вид в сагиттальной плоскости
щупальцевого гребневика Mnemiopsis (no Meglitsch,
1972)
Аворальный
сенсорный орган
Гревной pi
Маргинальное
щупальце
Рис. 3.36. Сильно уплощенный щупальцевый
гребневик Velamen (no Meglitsch, 1972)
78
Глава 3
Главное щупальце
Гревной
Анальная пора
Гонада
Рис. 3.37. Ctenoplana, гребневик, уплощенный
вдоль орально-аборальной оси (по Meglitsch,
1972)
ное становится похожим на прозрачную
ленту. В этом случае для локомоции
используются не только гребные пластинки, но
и волнообразные изгибания всего тела
(рис. 3.36). Особи другой группы уплощены
по орально-аборальной оси, ряды гребных
пластинок развиты очень слабо, или вовсе
отсутствуют, образ жизни ползающий
Сенсорный орган
Рис. 3.38. Вегоё, один из представителей
класса Nuda (no Meglitsch, 1972)
(рис. 3.37). У всех уплощенных форм
щупальца редуцированы.
3.5.3.2. Класс Nuda
У представителей этого класса
щупальца отсутствуют на всех стадиях жизненного
цикла. Хищники, питающиеся другими
гребневиками, с крупным
растягивающимся ртом и расширенным стомодеумом,
куда умещается их яйцевидная жертва.
В связи с этим тело примерно конической
формы (рис. 3.38).
3.6. Тип Platyhelminthes
(плоские черви)
3.6.1. Этимология
От греч. platy—плоский; helminthes —
черви.
3.6.2. Диагностические
и специфические признаки
(рис. 3.39)
1. Билатерально симметричные формы.
2. Тело из трех слоев тканей; системы
органов расположены в среднем
паренхиматозном слое.
3. Иной полости тела, кроме слепо
оканчивающейся кишки, нет.
4. Питание почти исключительно за
счет других животных.
5. Уплощены дорсовентрально;
червеобразные свободноживущие формы с
головой, несущей сенсорные органы.
6. В основном мягкотелые, но с
защитной слизистой секрецией; несколько видов
с внутренними спикулами.
7. Газообмен посредством диффузии,
кровеносная система отсутствует.
8. «Выделительные» органы—про-
тонефридии.
9. Нервная система может иметь вид
сети, но обычно сконцентрирована в
продольные тяжи.
10. В основном гермафродиты.
11. Дробление яйца спиральное, но
может сильно модифицироваться.
12. Развитие у свободноживущих форм
обычно прямое (у некоторых видов со ста-
Параллельные подходы к многоклеточное™ у животных
79
ПЕРЕДНИЙ КОНЕЦ
(
Поперечный
срез
ЗАДНИЙ КОНЕЦ
Мезодермальная
паренхима
Дорсальный эпидермис
(с равдоидами)
Ветви кишечника
Наружные
кольцевые
мускулы
Внутренние
продольные
мускулы
Два пучка дорсо-
вентральных
мускулов
Вентральный
эпидермис
Рис. 3.39. Типичный свободноживущий плоский червь (по Russel-Hunter, 1979)
дней свободноплавающей мюллеровской
или гётевской личинки), а у ряда паразитов
очень сложное, с несколькими
личиночными стадиями.
13. Найдены во всех основных водных
и сухопутных местообитаниях, включая
ткани других животных.
Три основные черты этого типа —
настоящая билатеральная симметрия;
возникновение всегда клеточного третьего
слоя ткани (паренхимы) между
эпидермисом и выстилкой кишки (гастродермисом);
дорсовентральное уплощение тела.
Билатеральная симметрия характерна и для всех
других до сих пор не рассмотренных
типов— так называемых Bilateria (хотя
иглокожие вторично вернулись к особой форме
радиальной симметрии, см. разд. 7.3). Она
возникла в ответ на потребность в
передвижении по поверхности твердых субстратов
(ср. с дорсовентрально уплощенными
гребневиками, разд. 3.5.3.1). Уплощение тела —
неизбежное следствие отсутствия
кровеносной системы и зависимости этих животных
от диффузии при распределении по
организму питательных веществ и газов и выводе
из него продуктов обмена (гл. 11).
В этом типе (во всех группах кроме
Acoela) мы впервые встречаемся с протоне-
фридиальной «выделительной» системой.
Кавычки использованы потому, что ее
функция, вероятно, в основном осморегуля-
торная, а не экскреторная (гл. 12) —
собственно экскреты по-прежнему
удаляются из тела через его поверхность путем
диффузии. Протонефридии состоят из сле-
пооканчивающихся трубок, на конце
которых расположены так называемые
«пламенные» клетки (рис. 3.40), фактически из двух
клеток: трубчатой и терминальной, с двумя
или более жгутиками. По месту пальчатого
соединения этих клеток находятся
просветы, ведущие в трубчатую часть. Через них
биение жгутиков, напоминающее пламя,
затягивает внутрь пламенной клетки
жидкости, которые затем прогоняются по
трубкам к выделительной поре и удаляются из
тела.
В общем, экскреторная,
пищеварительная, репродуктивная и нервно-сенсорная
системы, включающие интегрированные
комплексы тканей и органов, становятся
у плоских червей более заметными и
сложными, чем у губок и «кишечнополостных».
Поскольку у представителей этого типа
слепозамкнутая кишка и примитивный
«рот», не удивительно, что большинство
его представителей питается за счет тканей
других животных. Несколько видов поедает
водоросли, в основном диатомовые. Общая
тенденция среди плотоядных—
установление все более тесной связи между
потребителем и пищей, даже у
преимущественно свободноживущих турбелларий
(разд. 3.6.3.1). Здесь трофические связи
колеблются от взаимоотношений хищник—
80
Глава 3
Отверстия-
Соединение -J
двух клеток V I
-Ядро
-Терминальная
клетка
,Два жгутика
"ресничного
пламени"
терминальной клетки
-Трувчатая клетка
Ядро
Нефридиальные
поры
Протонефридиальная-
сеть
Рис. 3.40. Детали строения клетки ресничного
пламени (вверху) и система протонефридиев
(по Barnes, 1980)
жертва (или трупоед—падаль) до форм
симбиоза, включая экто- и эндокомменса-
лизм (выгодно для симбионта, безразлично
для хозяина) и даже экто- и эндопаразитизм
(симбионт живет за счет хозяина).
Большинство классов и видов плоских червей
исключительно паразитические. С
появлением паразитизма возникают
специализированные органы для прикрепления к
хозяину; изменяется внешний облик;
редуцируется или (у некоторых кишечных
паразитов, окруженных полупереваренной пищей)
совершенно исчезает пищеварительный
тракт; увеличивается репродуктивная
система.
3.6.3. Классификация
Около 25 000 ныне живущих видов,
распределенных по четырем классам.
3.6.3.1. Класс Turbellaria
(ресничные черви)
Представители этого класса в основном
свободноживущие водные формы, но
известно несколько сухопутных видов,
приуроченных к влажным местообитаниям.
Есть также сведения о комменсализме.
Один отряд—Temnocephalida- включает
исключительно эктокомменсалов
(возможно, эктопаразитов) поверхности тела
ракообразных, моллюсков и черепах. Сзади
у них клейкий диск, спереди щупальца
(рис. 3.41), а реснички на поверхности тела
отсутствуют. Они так выделяются среди
турбелларий, что ранг этого отряда иногда
повышают до класса.
Размеры турбелларий колеблются от
нескольких миллиметров (многие из них
обитают в морской и пресноводной интерсти-
циали) до более 0,5 м (гигантские наземные
формы). Реснички, покрывающие
эпидермис, обеспечивают локомоцию
турбелларий. самые мелкие из них «плавают» в ин-
терстициальной воде, а уплощенное тело
более крупных ползающих форм
увеличивает ресничную поверхность,
соприкасающуюся с субстратом. Мускульные
сокращения, вызывающие волнообразное движение
тела, важны для самых крупных водных
и всех наземных видов. Слизь, выделяемая
железистыми клетками, смазывает и
защищает поверхность тела, оставляет на
субстрате след, обеспечивающий опору
ресничкам, и может также использоваться для
ловли жертвы. Таким образом, она очень
важна для этих животных, и более 50%
энергии, получаемой за счет пищи, тратится на
образование слизи.
Поперечный срез тела типичной турбел-
Параллельные подходы к многоклеточности у животных
81
Класс
Подкласс
Turbellaria •
. Monogenea -
•чTrematoda ■
. Aspidogastrea
.Digenea
Cestodai
^ Cestoda-<
^Eucestoda
ларии приведен на рис. 3.42 (см. также
рис. 3.39): видны реснички (у более крупных
форм они могут отсутствовать на спинной
поверхности) и несколько мускульных
слоев—из кольцевых, продольных, косых
и поперечных волокон. Эти волокна не
исчерченные. Если учесть все сказанное о
слизи, неудивительно, что железистые клетки
такие крупные. Они часто расположены
в паренхиме и имеют выводной проток,
открывающийся на внешнюю поверхность
тела. Заметны также различные
палочковидные структуры—рабдоиды, одни из них во-
Рис. 3.41. Представитель отряда Temnocephali-
da
Отряд
Nemertodermatida
Acoela
Catenulida
Macrostomida
Proplicastomata
Lecithoepitheliata
Prolecithophora
Neorhabdocoela
Temnocephalida
Proseriata
Tricladida
Polycladida
Monopisthocotylea
Polyopisthocotylea
Aspidobothria
Strigeidida
Azygiida
Echinostomida
Plagiorchiida
Opisthorchiida
Amphftinida
Gyrocotylida
Caryophylhda
Spathebothriida
Trepanorhynchida
Pseudophyllida
Lecanicephalida
Aporida"
TetraphylHda
Diphyllida
Litobothrida
Nippotaenida
Proteocephalida
Cyclophyllida
Щупальце
Глотка
Кишка
Репродуктивная
система
Прикрепительный
диск
82
Глава 3
§ШШ Щ— Равдоиды
——Дорсальный эпидермис
■■-.-■•■■-■■•.-■. :.--з;—Базальная мембрана
-Кольцевые мышцы
»-v. <t«»4 •—■ —Пигмент
О
оооооооооо о
о оо о ооооооо
i/PffffOOP,
-Диагональные мышцы
-Продольные мышцы
Железистая клетка
Эпидермис
Мышечный слой
Паренхима
Гастрсдермис
■Дорсовентральная мышца
•Тело мышечной клетки
Равдоиды
о о о о ооооооо
оооооооооооо
■Вентральный
эпидермис
Реснички
Рис. 3.42. Поперечный разрез через турбелларию с деталями строения рабдоидов, дорсальной
и вентральной поверхностей и выстилки кишечника (по различным источникам)
зникают из эпидермальных, другие (так
называемые рабдиты)—из железистых
клеток в паренхиме. Их функции окончательно
не ясны, но они выбрасываются, когда
животное потревожено, и служат для защиты
(см. разд. 13.2.1.2) и/или разрушаются,
образуя вокруг него толстый слой слизи.
Они есть почти у всех турбелларий.
Пространство между внутренними органами
заполняет паренхима—скопление
неправильной формы клеток. Среди более крупных
клеток встречаются помельче, с крупным
Параллельные подходы к многоклеточное™ у животных
83
Наземная форма
Tri.cladi.da
Пресноводная форма
TricLadida
Голова Catenulida
-Ресничная ямка
—"Мозг"
ядром и тонким слоем цитоплазмы,
похожие по структуре и функциям на интерсти-
циальные клетки стрекающих. Они
называются необластами и играют важную роль
в замене износившихся тканей, а также в
регенерации после бесполого размножения
или повреждений (см. разд. 15.5.2).
Подвижность турбелларий привела
к определенной цефализации, т.е.
обособлению головного
отдела—модифицированной передней части тела,
первой попадающей в новые условия при
Клетка Пигментные
сетчатки клетки
Рис. 3.43. Цефализация (специализация
головного конца) у турбелларий и детали
строения «глазных пятен» (по различным
источникам)
движении (рис. 3.43). Обычно здесь
появляются глаза (два или много), устроенные
по типу пигментного бокала (разд. 16.7.3).
На голове также заметны ресничные
полоски, погруженные в желобки. Вероятно,
это—хеморецепторы, помогающие
находить пищу. Для ихпобслуживания развилось
переднее скопление нервной ткани—
«головной мозг». От него отходят пары
продольных нервных стволов, количество
которых сокращается у некоторых групп до
двух вентральных (рис. 3.44). Эпидермаль-
6*
84
Глава 3
Статоцист
Глаз
'Мозг'
Рис. 3.44. Нервная система турбелларий из отрядов Acoela (слева), Polycladida (в середине)
и Tricladida (справа) (по Barnes, 1980)
Пищеварительные клетки
(полость кишечника отсутствует)
Простая
к&икМеШК0ВадНЫЙ Луковицевидная глотка
Acoela
Большинство отрядов
Трехветвистый
кишечник Трувчатая выдвигающаяся
глотка
Многоветвистыи
кишечник Выдвигающаяся
цчатая глотка
TricLadida Polycladida
Рис. 3.45. Типы организации кишечника и рта у турбелларий (по Russel-Hunter, 1979)
ная нервная сеть может присутствовать или
отсутствовать, хотя некоторые Acoela
имеют только ее.
Кишка (рис. 3.45) варьирует от мощной
синцитиальной массы клеток (у
бескишечных турбелларий) и простого мешка («раб-
доцель») до ряда латеральных выпячиваний
(три—у Triclada, много—у Polyclada). Эти
варианты положены в основу классической
систематики турбелларий. С ними
коррелируют способы соединения рта с
кишечником (рис. 3.45).
Параллельные подходы к многоклеточности у животных
Яичник
Семенники
Яичник
1Т0ЧНИК
Сперма Яйцеклетка
Рис. 3.46. Системы размножения турбелларий
Сперматогонии
Гонады хорошо выражены (за
исключением Acoela) и связаны с внешней средой
выводными трубками, на которых может
быть несколько расширений с различными
функциями (рис. 3.46). Половая система
самца всегда состоит из многих
семенников, сперматозоиды обычно двужгутико-
вые. У самки может быть либо много
яичников, где образуются эндолецитальные
яйца, либо единственный яичник и
множество отдельных желточников,
производящих желточные клетки; в этом случае яйца
эктолецитальные (разд. 14.4).
Турбелларий без желточников называют
архоофорными, с желточниками—нео-
офорными.
Другая заметная черта турбелларий —
их способность к бесполому размножению,
в основном путем поперечного деления,
коррелирующая с ярко выраженной
способностью к регенерации. При этом, как и
у стрекающих, наблюдается полярность
(здесь—переднезадняя), возможно,
основанная на осевом физиологически-
эмбриональном градиенте. И наконец, при
голодании Triclada способны сильно умень-
86
Глава 3
шаться, иногда от размеров взрослой особи
до размеров только что вылупившегося
животного, а начав питаться, вырастают
заново. Предполагалось, что этот процесс
омолаживает ткани Triclada, делая их
практически бессмертными.
3.6.3.2. Сосальщики
У следующих двух паразитических
классов настолько много общих черт, что
раньше они считались одним классом (и
рассматриваются так до сих пор в некоторых
учебниках). Отличие в том, что представители
класса Monogenea в основном
эктопаразиты с простым жизненным циклом и одним
хозяином (обычно—рыбой), в то время как
в классе Trematoda почти все виды—
эндопаразиты. Более того, у
представителей основного подкласса трематод—
Digenea сложный жизненный цикл со
сменой двух или более хозяев. Внешне по
форме тела сосальщики напоминают турбелла-
рий, но есть и важные отличия:
характерный для последних ресничный клеточный
эпидермис заменен безресничным
синцитием (тегументом) (рис. 3.47); появилась
мускулистая глотка, с помощью которой
пища активно вытягивается из тканей
хозяина; она открывается в двуветвистый
кишечник (рис. 3.48); у взрослых особей
отсутствуют специальные сенсорные органы.
Класс Monogenea
(моногенетические сосальщики)
Представители этой группы — как
правило, эктопаразиты рыб, хотя из этого
правила много исключений: некоторые из них
обитают в полостях тела, открывающихся
наружу—во рту, на жабрах, в мочеполовой
системе, а несколько видов обнаружены
даже в целоме; есть паразиты амфибий,
рептилий и головоногих моллюсков.
Как и следует ожидать в случае
эктопаразитов, которым необходимо
прикрепляться к наружной поверхности тела часто
подвижного хозяина, как на переднем, так
и на заднем конце тела моногеней заметны
органы прикрепления, включая присоски,
крючья и зажимы (рис. 3.49). Сзади они
особенно крупные, и их строение положено
в основу разделения этого класса на два
отряда: MoHophisthocotylea—с цельным
задним фиксаторным аппаратом и парными
прикрепительными органами спереди, Ро-
/yophisthocotylea—с расчлененным задним
фиксаторным аппаратом, но единственной
ротовой присоской или парой
выворачивающихся буккальных присосок.
Гемидесмосома
Гликокаликс
Ммкротрувочка Гликокаликс
Микроворсинка
Интерстициальное
вещество
Митохондрия
Секреторные
тельца Т2
Секреторные
тельца Т1
Базальная
плазматическая
мембрана
Интерстициальное
вещество
Клетка паренхимы
Тело клетки
погруженного эпителия
с тельцами Та
Щелевой контакт
Рис. 3.47. Схема строения покровов моногенетического сосальщика (А) и трематоды (Б): ШЭР-
шероховатый эндоплазматический ретикулум
Ротовая присоска
Глотка
Пищевод
Семенной пузырек
Сумка цирруса
Семепровод
Семенник
Матка
Оотип
Скорлуповая железа
Желточные железы
Мочевой пузырь
Рис. 3.48. Схема строения генерализованной двуустки
Передний прикрепительный
орган
Прикрепительные железы
Рот
Глотка
Скорлуповая
железа
-Двуветвистый кишечник
Половое отверстие
Циррус
Предстательные железы
Присоска
Семевыносящий
канал
Выделительный
канал
Лауреров канал
Семеприемник
Яичник
ОРАЛЬНАЯ
СТОРОНА
■Желточный проток
Яйцевод
Желточный
резервуар
Присоска
Кишка
■Эмбрион
Яичник
-Семенники
Краевые
крючья
Рот
Нефридиальная пора
.Копуляционное
отверстие
Семепровод
Копуляционное
отверстие
Яичник
.елточный проток
Генитально-кишечный
канал
Кишка
Протонеорридии
Присоска
Прикрепительный диск "Якорь" Прикрепительный диск
"Якорь"
А Б
Рис. 3.49. Моногенетические сосальщики: А— Gyrodactylus из отряда Monopisthocotylea
Polystoma из отряда Polyopisthocotylea (no Meglitsch, 1972)
88
Глава 3
Прикрепительная
поверхность
Рот
Железистые клетки глотки
Выделительная пора
Внутренний
резервуар
Внешний
резервуар
Семеприемник
Яичник
Выносящий канал
сперматосрора
Внешний мускул
Семеизвергательный
канал
■Отверстие влагалища
Влагалище
Краевой крючочек
- ^—Кишечник
I
-Реснички
Прикрепительный
диск
Рис. 3.50. Типичный жизненный цикл моногенетического сосальщика на примере паразита рыб
Entobdella soleae (по различным источникам)
Параллельные подходы к многоклеточное™ у животных
89
Жизненный цикл моногеней
относительно прост. Яйца защищены скорлупой, при
вьшуплении высвобождается
свободноплавающая, покрытая ресничками личинка—
онкомирацидий. Она служит для
расселения на новых хозяев (рис. 3.50).
Класс Trematoda
Представители этого класса —
эндопаразиты, многие из которых
вызывают заболевания человека и домашних
животных. Как и у моногеней, есть органы
прикрепления, но у представителей
крупного подкласса Digenea (дигенетические
сосальщики) они расположены вокруг рта
(ротовая присоска) и посередине брюшной
поверхности (брюшная присоска). В
физиологии этих животных тегумент играет важную
роль. Он обеспечивает защиту, особенно от
ферментов—у видов, обитающих в
кишечнике, но также и от антител хозяина (см.
разд. 13.2.2.5). Через него непосредственно
от хозяина могут быть получены некоторые
питательные вещества, в частности
аминокислоты.
Жизненный цикл дигенетических
сосальщиков сложен: хозяин взрослой
(половозрелой) особи называется окончательным; это
позвоночное. Хозяева, в которых
развиваются другие стадии цикла, способные к
бесполому размножению, называются
промежуточными; их может быть от одного до трех
(один—обычно брюхоногий моллюск).
Типичный жизненный цикл выглядит
так (рис. 3.51):
1. Яйцо, покрытое скорлупой с
крышечкой, попадает в кишечник хозяина и
выводится наружу с фекалиями (а у некоторых
дигенетических сосальщиков — через
мочевой пузырь с мочой).
2. Из него вылупляется свободноживу-
ВЗРОСЛАЯ ОСОБЬ
Яичник РУмка цирруса
Желточники
Температура
Осмотическое давление
Температура
Св<?трСОг
,р02
Циста
метацеркари!
ЦЕРКАРИЯ
Поведение
Фототаксис
Геотаксис
Обнаружение улитки:
хемотаксис?
Паразитическая кастрация,
Общий метаболизм,
Иммунитет Беспозвоночного
Рис. 3.51. Типичный жизненный цикл трематоды (по Smyth, Halton, 1983)
90
Глава 3
щая, покрытая ресничками личинка—
мирацидий.
3. Ее съедает моллюск, или она сама
внедряется в него через эпидермис. Затем,
потеряв реснички, паразит проникает в
пищеварительную железу хозяина.
4. Там он превращается в спороцисту.
5. Спороциста содержит зародышевую
массу, из которой митотически возникает
несколько дочерних спороцист и/или редия.
6. Последняя содержит зародышевые
клетки, которые аналогичным образом
развиваются в личинки—церкарии.
7. Они покидают хозяина, прободая его
ткани, и становятся свободноживущими.
8. Церкарии проникают в
окончательного хозяина или съедаются им (жизненный
цикл завершен), или же находят
промежуточного хозяина из позвоночных и там ин-
цистируются, образуя метацеркарию.
9. Если промежуточный хозяин,
содержащий метацеркарию, будет съеден
окончательным хозяином, цикл завершен.
Половозрелые паразитические особи
производят гигантское количество яиц
(например, печеночная двуустка—около
20000 в день). Более того, каждая
выходящая из яйца личинка размножается в ходе
дальнейших бесполых стадий. Такая
высокая репродуктивная способность
(возможно, связанная с тем, что эндопаразиты по
определению окружены сверхобильной
пищей—тканями хозяина) компенсирует
массовую гибель расселительных стадий при
случайной их передаче от одного хозяина
к другому.
Schistosomatidae (шистосомовые) —
семейство дигеней, выделяющееся по двум
причинам. Во-первых, оно включает виды,
вызывающие у человека опасное (иногда—
смертельное) заболевание шистосомоз.
В эндемичных областях им может страдать
большая часть населения; это—один из
самых распространенных гельминтозов
человека. Во-вторых, шистосомовые
раздельнополы, чем резко отличаются от других
плоских червей. Почему раздельнополость
возникла у паразитов, где шансы на встречу
особей разных полов проблематичны,
остается загадкой. Тем не менее, когда
встреча происходит, между ними возникает
тесная взаимосвязь: у самца по всей длине
Вентральная присоска самки
Вентральная присоска
самца
Ротовая присоска
самца
Вентральный (гинекосрор
ный) желов
Рис. 3.52. Schistosoma mansoni (no Meglitsch,
1972)
тела проходит вентральный желоб, в
которой помещается более тонкая самка.
Взрослых животных часто обнаруживают
именно в такой ассоциации (рис. 3.52).
Представители подкласса Aspidogastrea
в чем-то сходны как с дигенетическими, так
и с моногенетическими соеалыциками
(рис. 3.53). У них нет заднего прикрепите-
Присоска
Ячейка
Рис. 3.53. Представитель подкласса
Aspidogastrea (по Meglitsch, 1972)
Параллельные подходы к многоклеточное™ у животных
91
льного аппарата, но присутствует либо
крупная присоска, покрывающая всю
вентральную поверхность тела, либо ряд
вентральных присосок. Это эндопаразиты,
обнаруженные в кишечнике рыб и рептилий,
а также в перикардиальной и почечной
полостях моллюсков. Оцнако их жизненный
цикл прост; он включает двух, а иногда
всего одного хозяина.
3.6.3.3. Класс Cestoda
Представители этого класса, обычно
известные под названием ленточных червей,
приспособились к паразитированию в
кишечнике позвоночных, и, таким образом,
стали самой специализированной группой
Platyhelminthes. Окруженные массой
полупереваренной пищи хозяина, они утратили
и рот, и кишечник. Вместо этого, как и у
сосальщиков (и даже в большей степени), те-
гумент (рис. 3.54) приобрел способность
к непосредственной абсорбции пищи. Он
покрыт микроскопическими выростами
(микротрихами), увеличивающими его
поверхность, а в своей толще содержит
большое количество митохондрий, что
свидетельствует об интенсивности происходящих
здесь процессов активного транспорта. На
поверхности тегумента также может
происходить переваривание пищи с помощью
ферментов хозяина—так называемое
мембранное пищеварение.
Сложно устроенный орган
прикрепления—сколекс с присосками и крючьями
(рис. 3.55) — удерживает червя в потоке
пищи, создаваемом кишечной
перистальтикой. Жизнь в длинном трубчатом
пространстве с движущейся пищевой средой
благоприятствовала эволюции особой формы
тела—стробилы, состоящей из одинаковых
члеников (проглоттид),
отпочковывающихся вблизи сколекса (стробилизация),
созревающих и постепенно отделяющихся на
противоположном конце тела в виде
мешочков, набитых яйцами. В средней части
этого цикла в них развивается половая
система (рис. 3.55), очень похожая на
половую систему дигеней. Если возможно,
происходит перекрестное оплодотворение, но
известно самооплодотворение между двумя
проглоттидами в одной стробиле или даже
ЙШШ
ю*
Микротрихи
•Наружная
плазматическая
мемврана
1иноцитозный
пузырек
Внутренняя
плазматическая
мемврана
Базальная мемврана
Соединительный
Митохондрия
Аппарат Гольджи
Эндоплазматический
ретикулум
Ядро
Рис. З.Б4. Схематизированный разрез через
покровы цестоды (по Meglitsch, 1972)
в одной проглоттиде. Характерно, что
образование и развитие сегментов у
кольчатых червей и членистоногих происходит
в противоположном по сравнению с цесто-
дами направлении, т. е. с заднего конца тела
вперед, а не спереди назад (см. с. 37).
Оплодотворенные яйца заполняют матку,
и в зрелых проглоттидах мало что остается,
кроме этой растянутой набитой яйцами
матки. Такая «конвейерная» система
размножения обеспечивает поразительную
плодовитость. Например, взрослый бьмий
цепень (Taenia saginata) может состоять из
700—1000 проглоттид, теряя от 3 до 10 из
них в день; по 100000 яиц в каждой.
Яйца из зрелой проглоттиды
вылупляются либо во внешней среде, либо после
проглатывания этой проглоттиды
хозяином. Из них выходит личинка онкосфера;
иногда она окружается ресничной
мембраной (эмбриофором) и в этом случае назы-
92 Глава 3
Матка
Семенники SZZ-:
Матка
вается корацидием. Эта стадия в
промежуточном хозяине может развиваться в
разные типы личинок, основные из которых:
1. Процеркоид—мелкая веретеновид-
ная форма с массивным телом и крючьями
на заднем конце.
2. Плероцеркоид—массивная форма
с дефинитивным сколексом, но без
зародышевых крючьев процеркоида, из которого
она развивается.
3. Цистицеркоид, состоящий из
переднего пузыря с ввернутым внутрь сколек-
Рис. З.ББ. Форма тела
ленточных червей с деталями
строения различных отделов и тремя
типами сколексов (по
различным источникам)
сом и хвостовидного заднего отдела с
личиночными крючьями.
4. Цистицерк—пузырь с единственным
ввернутым внутрь сколексом.
Другие личиночные типы представляют
собой варианты этих четырех. Иногда
происходит бесполое размножение сколексов
пузыревидной стадии в промежуточном
хозяине. Когда его съедает окончательный
хозяин, личинка попадает в кишечник и
превращается во взрослого червя. На рис. 3.56,
А—В показано несколько типичных жизнен-
Параллельные подходы к многоклеточное™ у животных 93
Жизнеспособны
яйца проходят
сквозь кишечник
At #о
Онкосфера •:-"•■■•■• Цистицерк
Рис. 3.56. Жизненные циклы цестод: А— Taenia saginata: Б—Diphyllobothrium dendriticum
(с. 090); B—Hymenolepsis папа (с. 091) (no Smyth, 1962)
ных циклов, каждый из которых включает
некоторые из этих стадий.
Не следует думать, что,раз ленточные
черви прочно прикреплены сколексом, они
не могут передвигаться внутри кишечника
хозяина. Их мускулатура под тегументом
состоит из кольцевых и продольных, а в
паренхиме—из продольных, поперечных
I и дорсовентральных волокон. Более того,
в сколексе расположено переднее скопление
нервной ткани, от которого отходят два
латеральных ствола, тянущиеся назад вдоль
всей стробилы. Могут присутствовать
дорсальные, вентральные и дополнительные
латеральные стволы, а также связывающие
продольные стволы кольцевые комиссуры.
Карликовый цепень (Hymenolepis) способен
мигрировать на крупные расстояния в
кишечнике хозяина, вероятно, благодаря этой
сложной нервно-мускульной системе.
Обычно класс ленточных червей делят
на два подкласса — Eucestoda и Cestodaria.
В первый (собственно ленточные черви)
входят 12 отрядов, сильно отличающихся
друг от друга формой тела и хозяевами.
К последним относится широкий спектр
теплокровных и холоднокровных
позвоночных. Специального упоминания
заслуживают представители отряда Caryophyllida
(рис. 3.57), паразитирующих на пресновод-
94
Глава 3
Д=-0,56°С
рН-6,7
Эмбриональное
развитие
8 дней при 25°С
+ 02
Корацидий
Рис. З.Б6 (продолжение)
ных рыбах (окончательный хозяин) и
ракообразных (промежуточный хозяин). У
взрослых форм сколекс слабо развит, а в
нечленистой «стробиле» находится единственный
набор мужских и женских половых органов
(один яичник, много семенников, но один
копулятивныи орган). Эти черви обитают
в полости тела, а не в кишке хозяина. Их
считают педоморфной личинкой (см.
гл. 17) неизвестной группы ленточных
червей. У представителей Cestodaria есть
общие черты как с эвцестодами, так и с
сосальщиками (рис. 3.58). У них отсутствуют
как рот и кишечник, так и стробилизация
и сколекс, зато присоски
«моногенетического» типа. Несмотря на это, их личинка напо-
в
Влагалище
Рис. 3.57. Schistocephalus
solidus—представитель отряда Caryophyllida, встречающийся
в полости тела колюшек (по Smyth, 1962) Семелриемник
Рис. 3.58. Gyrocotyle—представитель
класса Cestodaria (no Barnes, 1980)
96
Глава 3
минает онкосферу, но с 10 крючьями вместо
6. Они паразитируют в целомической и
кишечной полостях пластиножаберных и
примитивных костистых рыб.
3.7. Тип Mesozoa
3.7.1. Этимология
От греч. mesos — средний; zoon—
животное (такое название предложено
потому, что этих животных считали
промежуточными формами между
многоклеточными животными, т. е. Metazoa, и
одноклеточными— Protozoa).
3.7.2. Диагностические
и специфические признаки
1. Билатерально симметричные формы.
2. Органов нет.
3. Полости тела, включая
пищеварительную, отсутствуют.
4. Очень небольшое количество клеток
расположено всего в два «слоя»—
наружный ресничный слой и внутренняя
клетка или клетки с репродуктивной
функцией.
5. Некоторые клетки развиваются
внутри других.
6. Эндопаразиты беспозвоночных на
некоторых стадиях своего жизненного цикла.
7. Сложный жизненный цикл,
включающий половую и бесполую репродуктивные
фазы.
8. Морские животные.
3.7.3. Классификация
В этот тип входит 50 видов из двух
довольно различных групп—Rhombozoa и Ог-
thonecta, между которыми, вполне
возможно, нет родства, поэтому ниже они
рассматриваются порознь.
Класс Отряд
__-Dicyemida
• РгттЬптл Т 1т Hirjfiniifi
. Orthonecta Orthonectida
3.7.3.1. Класс Rhombozoa
Это паразиты почек головоногих мол- (
люсков. Их червеобразное тело длиной
0,5—7,5 мм состоит из 20—30 ресничных
соматических клеток (у каждого вида их
строго постоянное число), расположенных
по спирали вокруг одной удлиненной
осевой (аксиальной) клетки (рис. 3.59). Внутри
последней находятся от одной до многих
(более 100) клеток-аксобластов, дающих
начало половому и бесполому
поколениям.
Жизненный цикл сложный и не изучен
до конца, но, по-видимому, описанное
выше строение свойственно двум совершенно
различным стадиям жизненного цикла,
размножающимся бесполым (нематоген) и
половым (ромбоген) путями. Они
различаются дальнейшей судьбой аксобластов.
Ромбоген—стадия, характерная для взрослого
головоногого моллюска; аксобласты его
аксиальной клетки образуют «инфузориге-
ны», эквиваленты гонад, высвобождающие
яйца и/или спермин. Оплодотворение,
происходящее внутри осевой клетки, приводит
к инфузориевидной личинке, которая вскоре
уже содержит полный набор клеток, и
дальнейшее развитие личинки происходит уже
за счет их роста и дифференцировки, без
делений. Внутриклеточно на этой стадии уже
заложены будущие герминативные клетки.
Инфузориевидная личинка покидает
ромбоген и выводится из моллюска с его мочой.
Обладая отрицательной плавучестью (из-за
высокой концентрации инозитолгексафо-
сфата магния в двух специальных клетках),
она опускается на морское дно. Затем
неизвестным способом (возможно, на более
поздних стадиях собственного развития или
превратившись в другую личиночную
стадию) она инфицирует молодого
головоногого моллюска и, проникнув в его почку,
развивается в нематоген. Внутри аксиаль-
Ресничная соматическая клетка
Рис. 3.59. План строения тела представителей
класса Rhombozoa (no Lapan, Morowitz, 1972)
Параллельные подходы к многоклеточное™ у животных
97
Незрелая
червеовразная
личинка
Аксовласт
Рис. 3.60. Жизненный цикл
представителей класса Rhom-
bozoa (no Margulis. Schwartz,
1982)
Герминативные
клетки
Инсрузориевидная
личинка
Ресничная
соматическая
клетка
Аксиальная
клетка
Незрелая
инсрузориевидная
личинка
ной клетки последнего аксобласты через
стадию червеобразной личинки формируют
новые нематогены; таким путем может
накопиться много бесполых поколений. В
конце концов, когда плотность нематогенов
в почках становится высокой и/или хозяин
достигает половой зрелости, они либо
трансформируются в ромбогены, либо
производят их новое поколение (рис. 3.60).
Питание всех эндопаразитических стадий
происходит, вероятно, за счет абсорбции
соматическими клетками питательных веществ,
растворенных в моче хозяина, однако
никаких подробностей этого неизвестно.
Описано только пять родов, входящих
в 2 отряда.
3.7.3.2. Класс Orthonecta
Ортонектиды—эндопаразиты
широкого круга морских беспозвоночных: плоских
Червей, немертин, аннелид, двустворчатых
моллюсков и иглокожих. Для половой
стадии характерен половой диморфизм; это
свободноживущие мелкие формы (размер
самцов около 0,1 мм, самок чуть больше;
рис. 3.61). И самцы и самки по строению
несколько напоминают взрослых Rhombo-
zoa—кольцевой наружный слой ресничных
соматических клеток окружает
многочисленные яйца или спермин. Осевая клетка
отсутствует. Самцы выделяют сперму в
море, она проникает в самок и оплодотворяет
содержащиеся в них яйца. Затем внутри
тела самки развивается инвазионная личинка.
Яйцеклетки
Сперма
Рис. 3.61. Представители класса Orthonecta:
самка (А) и самец (б) (no Atkins, 1933)
7—1314
98
Глава 3
Вегетативные ядра
Рис. 3.62. Плазмодий ортонектид (по Caullery,
Mesnil. 1901)
состоящая из слоя ресничных клеток,
окружающих несколько герминативных клеток.
После высвобождения личинок и
инвазии тканей хозяина наружные ресничные
клетки утрачиваются, а герминативные
размножаются, образуя многоядерный
плазмодий (рис. 3.62). От него отделяются
фрагменты, образующие бесполым путем
новые плазмодии внутри хозяина.
Некоторые клетки плазмодия затем дают особей
одного или обеих полов, которые покидают
хозяина и завершают жизненный цикл.
Известны только 3 рода, образующие
один отряд.
3.7.4. Родственные связи
У некоторых эндопаразитов
отсутствуют различные системы органов,
характерные для свободноживущих форм, от
которых они предположительно произошли,
и простота строения Mesozoa раньше
рассматривалась как результат такой
связанной с паразитизмом дегенерации. Однако
если это так, Mesozoa представляют собой
крайнюю стадию данного процесса—они
не только лишены вообще каких-либо
органов, но и могут состоять менее чем из 30
клеток. Кроме того, некоторые их
особенности, в частности внутриклеточное
развитие половых и бесполых личиночных форм
Rhombozoa, не имеют аналогов в животном
царстве. Нет никаких свидетельств родства
Rhombozoa с какой-либо известной группой
животных, т. е. они вполне могли
независимо произойти от протистов (см. гл. 2),
причем гораздо позже, чем другие группы
многоклеточных, поскольку их хозяева
относятся к сравнительно продвинутым
группам.
Orthonecta—несколько менее
аберрантные формы, и их происхождение от плоских
червей вполне возможно, хотя и далеко не
очевидно.
3.8. Дополнительная
литература
Bergquisi P.R. 1978. Sponges. Hutchinson, London
[Porifera].
Fry W. G. (Ed.) 1970. The Biology of Porifera.
Academic Press, New York [Porifera].
Grell K. G. 1982. Placozoa. In: Parker S. P. (Ed.)
Synopsis and Classification of Living Organisms,
(Vol. 1), p. 639. McGraw-Hill, New York
[Placozoa].
Hyman L. H. 1940. The Invertebrates (Vol. 1).
Protozoa through Ctenophora. McGraw-Hill, New
York [Porifera, Cnidaria, Ctenophora, Mesozoa].
Hyman L. H. 1951. the Invertebrates (Vol. 2). Platy-
helminthes and Rhynchocoela. McGraw-Hill,
New York [Platyhelminthes].
Kaestner A. 1967. Invertebrate Zoology (Vol. 1).
Wiley, New York [Porifera, Chidaria,
Ctenophora, Platyhelminthes, Mesozoa].
Lapan E.A., Morowitz H. 1972. The Mesozoa.
Scient. Am., 227, 94—101 [Mesozoa].
Mackie G. O. (Ed.) 1976. Nutritional Ecology and
Behavior. Plenum, New York [Cnidaria].
Miller R.L. 1971. Trichoplax adhaerens Schulze
1883: Return of an Enigma. Biol. Bull mar. Biol.
Lab., Woods Hole, 141, 374 [Placozoa].
Morris S. C, Geordge J. D., Gibson R., Piatt EM.
1985. The Origins and Relationships of Lower
Invertebrates. The Systematics Association, Special
Volume No. 28. Clarendon Press, Oxford.
Reeve M.R.. Walker M. A. 1978. Nutritional
ecology of ctenophores—a review of past research.
Adv. mar. Biol. 15, 246—287 [Ctenophora].
Schmidt H. 1972. Die Nesselkapseln der Anthozoen
und ihre Bedeutung fur die phylogenetische
Systematic Helgolander Wiss. Meeresunters., 23,422—
458 [Platyhelminthes].
Schockaert E.R., Ball I.R. 1981. The Biology of the
Turbellaria. Junk, The Hague [Platyhelminthes].
Smyth T.D. 1977. Introduction to Animal
Parasitology, 2nd edn. Wiley, New York [Platyhelminthes].
ГЛАВА *Т
ЧЕРВИ
Nemertea
Gnathostomula
Gastrotricha
Nematoda
Nematomorpha
Kinorhyncha
Loricifera
Priapula
Rotifera
Acanthocephala
Sipuncula
Echiura
Pogonophora
Annelida
Все включенные в эту главу 14 типов —
первичноротые производные плоских
червей, хотя общих специфических черт
лишены и не составляют естественную группу
родственных животных (см. гл. 2). Общим
для ее представителей, помимо
червеобразной формы тела, является скорее
отсутствие некоторых черт, характерных для
других групп или типов. Можно сказать, что
эти черви не имеют ног, не покрыты
раковиной, не вторичноротые, не несут лофофора.
У червеобразных форонид есть лофофор
и они рассмотрены в гл. 6; кишечнодыша-
шие и
щетинкочелюстные—вторичноротые, поэтому речь о них пойдет в гл. 7;
пятиустки обсуждаются со своими
предполагаемыми сородичами-членистоногими
в гл. 8. Таким образом, практически 14
описываемых ниже групп—простые животные,
сохранившие более или менее неизменной
червеобразную предковую форму тела
(длина в 2—3 —1500 раз превышает
ширину), круглые или уплощенные в сечении.
Хотя для этих «червей» не разработано
общепринятой системы родства, широко
признается ее деление на четыре группы
типов (см., например, рис. 2.20). У первой
и второй групп (Nemertea и Gnathostomu-
la/Gastrotricha) идентичный с предками—
плоскими червями—план строения и
относительно небольшое таксономическое
разнообразие в пределах каждой группы; для
обеих характерна высокая специализация,
которая делает невозможным их
объединение с Platyhelminthes.
Гастротрихи, возможно, могут служить
связующим звеном с гораздо более крупной
и разнообразной третьей группой типов,
известной как Aschelminthes, или псевдоце-
ломические черви. В нее входит семь типов,
распадающихся на две подгруппы.
Kinorhyncha, Loricifera и Nematomorpha на
некоторых стадиях жизненного цикла имеют
выворачивающийся интроверт ', несущий
окруженный стилетами и кругами «скалид»
ротовой конус; у Priapula есть сходный
интроверт со скалидами, но без ротовых
стилетов. Nematoda, похоже, также связаны
с этой подгруппой через недавно описанный
из Бразилии вид с аналогичным
интровертом и игловидными скалидами; кроме того,
у Nematoda и Nematomorpha одинаковая
высокоспециализированная мускульно-
локомоторная система. И напротив,
Rotifera и Acanthocephala лишены интроверта,
1 В отечественной литературе для интроверта
и хоботка обычно используется один термин —
хоботок. Поскольку между интровертом и
хоботком существуют выраженные функциональные
и морфологические различия, здесь и далее под
интровертом понимается передняя часть тела,
которая может выворачиваться и втягиваться,
а под хоботком — специальный, иногда сложно
устроенный орган.— Прим. пер.
V
100
Глава 4
имеют внутриклеточную кутикулу,
локомоторный и/или клейкий «хоботок» и лемни-
ски. У коловраток последние две
структуры— только в отряде Bdelloidea. Если не
брать в расчет интроверт и панцирную
личинку, Priapula—довольно нетипичные
Aschelminthes; возможно, по форме тела
и репродуктивному типу они близки
к предкам. Другие типы псевдоцеломиче-
ских червей можно объединить по
устройству стенки тела, природе глотки и полости
тела, феномену эвтелии (постоянство
клеточного состава), внутреннему
оплодотворению с копуляцией, отсутствию
способности к регенерации частей тела и бесполому
размножению. Для многих также
характерны протонефридиальная система, линька
кутикулы, ее утолщение с образованием
защитного панциря, мелкие размеры
непаразитических форм, сегментация кутикулы (и
связанных с ней элементов стенки тела),
флоскулы и дуогландулярные
прикрепительные железы.
Четвертая группа также включает
разнообразных по признакам червей. Однако
Annelida, Pogonophora и Echiura
объединяют такие черты, как схизоцельная
полость тела', хитиновые щетинки, замкнутая
кровеносная система, органы выделения
типа метанефридиев; представители первых
двух типов метамерные, в то время как
эхиуриды явно мономерны. Также моно-
мерны Sipuncula, у которых «метанефри-
дии» и схизоцель часто считаются
признаками, сближающими их с четвертой
группой типов, хотя щетинок и замкнутой
кровеносной системы нет. Только у Sipuncula
есть интроверт, но в отличие от короткого,
боченковидного, покрытого иглами
интроверта Aschelminthes, у них он представляет
собой длинную тонкую трубку,
оканчивающуюся лопастями или щупальцами для
собирания детрита.
Положение Sipuncula в группе схизоцело-
мических2 червей, как и Priapula среди
1 Под схизоцелем здесь понимается
вторичная полость тела, образованная расхождением
мезодермальных клеток, см. рис. 2.9,Б.— Прим.
пер.
2 Схизоцеломические черви — вторичнопо-
лостные черви, у которых целом образован схи-
зоцельным способом.— Прим. пер.
Aschelminthes, аберрантно (в отношении
одних черт типичное, в отношении других—
резко отличное от остальных типов).
В обеих этих группах одни анатомические
признаки могут быть истолкованы в пользу
параллельного приобретения одинакового
уровня организации, а другие—для
доказательства филогенетического родства. То,
что один автор считает железой и ее
протоком, другой рассматривает как глотку и
ротовой канал (этот пример—две точки
зрения на строение личинок нематоморф).
В некоторых типах, в частности, Nemato-
morpha среди псевдоцеломических червей
и Pogonophora среди схизоцеломических
выделяют группы совершенно различного
происхождения. Одну группу нематоморф
можно вывести из мерметидных нематод,
в то время как другую связывают с кино-
ринхами и лорициферами; не исключено,
что один класс (или подтип) погонофор
близкородствен аннелидам, в то время как
другой возник независимо, и схизоцель
в нем развился параллельно!
Ясно, что преждевременно объединять
какие-либо из этих групп червей в меньшее
число более крупных типов. Следует
провести еще много исследований, прежде чем
можно будет сделать определенные
филогенетические выводы, поэтому в данной главе
мы будем рассматривать все 14 групп как
независимые таксоны одного
систематического ранга.
4.1. Тип Nemertea (немертины)
4.1.1. Этимология
От греч. Nemertes—средиземноморская
нимфа, дочь Нерея и Дориды.
4.1.2. Диагностические
и специфические признаки
(рис. 4.1)
1. Билатерально симметричные,
червеобразные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Сквозной кишечник с терминальным
анусом.
Черви
101
Отверстие для
выворачивания
ховотка
Латеральный
нерв
Гонады
Мускул-ретрактор
ховотка
Ринхоцель
Хоботок
Кишечные
дивертикулы
Анус
Рис. 4.1. Схематическое изображение
внутреннего строения немертины с дорсальной
стороны (по нескольким источникам, главным
образом по Pen nak, 1978)
4. Полости тела нет, функции скелета
выполняет паренхима.
5. Наружный ресничный эпидермис
лишен кутикулы.
6. Тело дорсовентрально уплощено,
часто с сериально повторяющимися органами
(например, кишечными карманами, прото-
нефридиями, гонадами).
7. Есть кровеносная система, но
дыхательная система отсутствует.
8. Нервная система с головным мозгом
и обычно тремя продольными стволами.
9. Имеется хоботок эктодермального
происхождения, способный выворачиваться
и втягиваться в продольную дорсальную
полость (ринхоцель), которая открывается
наружу вблизи рта.
10. Дробление яиц спиральное.
Немертины, длина тела которых
колеблется от менее 0,5 мм до более 30 м, могут
быть названы «сверхплоскими червями»,
так как основные черты их строения в
принципе те же, что и у крупных турбелларий
(см. разд. 3.6.3.1). Тем не менее они
отличаются от этой группы тремя основными
признаками; во-первых, более эффективно
работающим сквозным кишечником; во-
вторых, замкнутой кровеносной системой,
ток крови (правда, нерегулярный) в которой
обеспечивается сокращениями стенок
кровеносных сосудов и движениями тела;
в-третьих, характерным хоботком,
совершенно изолированным от кишечника и не
являющимся его частью (рис. 4.2).
Этот хоботок помещается в трубчатой
полости — ринхоцеле, проходящей почти
Отверстие для-
выворачивания
ховотка
-Анус
Рис. 4.2. Схема продольного разреза немертины, демонстрирующая ринхоцель и хоботок (А -
втянутый; Б—вывернутый) (по Gibson, 1982)
102
Глава 4
Рис. 4.3. Личинка немертин—пилидий (по
Kershaw, 1983)
вдоль всего тела и имеющей такое же
строение, что и стенки последнего. Давление,
возникающее в ринхоцеле при сокращении
мышц его стенок, выворачивает хоботок.
Втягивается он с помощью продольных
мускулов, тянущихся от его заднего конца
к заднему краю ринхоцеля. Хоботок
используется в основном для ловли добычи, а
у наземных видов также для быстрого
перемещения: в вывернутом состоянии он
прикрепляется к земле впереди животного,
которое затем как бы подтягивается на нем
вперед.
Локомоция у немертин осуществляется
в целом так же, как у турбелларий: они
скользят за счет биения ресничек по
выделяемой слизи, а крупные формы
перемещаются благодаря волнообразным
мышечным сокращениям вентральной
поверхности тела. Некоторые виды могут плавать,
волнообразно извиваясь в дорсовентраль-
ной плоскости.
Размножение происходит как бесполым
путем (фрагментацией), так и половым,
причем гонады являются временными
структурами, формирующимися из
скоплений дедифференцированных мезенхимных
клеток, которые окружаются мембранами,
а в зрелом состоянии связываются с
внешней средой временными протоками.
Большинство видов раздельнополые, хотя ряд
наземных и пресноводных форм —
гермафродиты, способные к
самооплодотворению. Развитие обычно прямое, хотя
у представителей одного отряда известна
личиночная стадия (рис. 4.3);
оплодотворение, как правило, наружное.
Почти все немертины — хищники,
питающиеся широким набором организмов,
от простейших до моллюсков,
членистоногих и рыб. К этому типу относится
длиннейшее из всех известных животных—Lineus
longissimus, обычно достигающее 30 м, а
в некоторых случаях и вдвое больше в
полностью вытянутом состоянии.
4.1.3. Классификация
Около 900 известных видов
распределены по двум классам.
Класс Отряд
Palaeonemertea
Heteronemertea
Hoplonemertea
Bdellonemertea
4.1.3.1. Класс Anopla
У аноплид центральная нервная система
расположена в стенке тела, которая часто
включает три мускульных слоя: либо два
слоя кольцевых мышц и
один—продольных (между ними), как в отряде
Palaeonemertea, либо два слоя продольных и
между ними один—кольцевых, как в отряде
Heteronemertea. Хоботок аноплид устроен
относительно просто, не имеет регионарной
дифференцировки или стилетов, хотя несет
большое количество эпителиальных раб-
доидов того же типа, что и у турбелларий;
ринхоцель открывается совершенно
изолированным от рта отверстием.
Большинство представителей этого
класса (рис. 4.4)—морские бентосные
животные, но три вида—пресноводные, а еще
несколько обитают в солоноватых водах.
Heteronemertea—единственная группа
немертин, у которой есть личиночная стадия.
4.1.3.2. Класс Enopla
У эноплид центральная нервная система
расположена изнутри относительно
мускулатуры стенки тела. Мускулатура
двуслойная; хоботок дифференцирован на отделы;
центральный из них представляет собой ко-
Черви
103
Рис. 4.4. Форма тела представителей класса
Anopla (no Gibson, 1982)
Рис. 4.5. Форма тела представителей класса
Enopla (no Gibson, 1982)
роткую мышечную луковицу, несущую
один или более стилетов (за исключением
аберрантного комменсального рода Mala-
cobdella, представители которого питаются
путем фильтрации и выделены в отдельный
отряд Bdellonemertea). Кишечник эноплид
сложный и открывается наружу общим
с ринхоцелем отверстием. У большинства
представителей класса (отряд Hoplonemer-
tea) кишка снабжена многочисленными
парами боковых ответвлений (дивертикулов),
в то время как у Bdellonemertea крупная
глотка служит фильтрующим органом и
содержит ресничные сосочки, улавливающие
пищевые частицы без помощи слизи.
Хотя большинство эноплид—морские
бентосные формы, есть несколько
пелагических видов, плавающих в толще воды и на
поверхности (рис. 4.5), а также ряд
пресноводных и наземных представителей.
Известны комменсалы (например, из ат-
риальной полости оболочников) и
паразиты.
4.2. Тип Gnathostomula
(гнатостомулиды)
4.2.1. Этимология
От греч. gnathos—челюсть; stoma—рот.
4.2.2. Диагностические
и специфические признаки
(рис. 4.6 и 4.7)
1. Билатерально симметричные,
червеобразные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Кишечник слепо замкнут.
104
Глава 4
-Рострум
Кишечник
Семенники
Мужское
половое
отверстие
Рис. 4.6. Анатомия гнатостомулиды из отряда
Filospermoidea (no Sterrer, 1982)
4. Полость тела отсутствует, функции
скелета выполняет слабо развитая
паренхима.
5. Кровеносной и дыхательной систем
нет.
6. Выделительные органы простые дву-
клеточные протонефридии, напоминающие
эпидермальные клетки тем, что
терминальная клетка несет только одну ресничку.
7. Эпидермис без кутикулы, из однорес-
ничных клеток.
8. Диффузная эпидермальная нервная
система.
9. Сложный ротовой аппарат
образован парными челюстями и единственной ба-
зальной пластинкой.
10. Синхронные гермафродиты
(мужские и женские гонады созревают
одновременно).
11. Дробление яиц спиральное.
12. Развитие прямое.
13. Морские интерстициальные формы
часто бескислородных песков.
У этих крошечных (менее 1 мм в длину)
прозрачных червей в принципе такое же
строение тела, что и свободноживущих тур-
белларий (разд. 3.6.3.1) и гастротрихов
(разд. 4.3). От обеих этих групп они
отличаются высокоспециализированной
мускулистой глоткой со сложной системой
челюстей, которая служит для сбора бактерий,
протестов и грибов с поверхности песчинок
(рис. 4.8). В поисках этой добычи
гнатостомулиды движутся в интерстициальных
порах, плавая или скользя с помощью своего
ресничного эпидермиса (реснички бьют
вперед или назад) или за счет резких
сокращений трех или четырех пар продольных
мышц, образующих мускулатуру стенки
тела. Единственный яичник образует за цикл
размножения одно крупное яйцо, вероятно,
всегда оплодотворяемое внутри тела. По-
видимому, в течение жизни некоторые гна-
Ресничная ямка
Базальная пластинка-
Челюсти -
Глотка -
Яичник
Бурса
Влагалище
Семенник
Мужское половое
отверстие
Рис. 4.7. Анатомия гнатостомулиды из отряда
Bursovaginoidea (no Sterrer, 1982)
Черви
105
Базальная пластинка
^-Челюсти
Рис. 4.8. Базальная пластинка и парные
челюсти гнатостомулид (по Sterrer, 1982)
тостомулиды проходят превращение от
бесполой питающейся стадии до половой непи-
тающейся.
Этот тип был описан только в 1956 г.
вместе с первым его представителем. Такое
позднее открытие объясняется не тем, что
гнатостомулиды редки—их может быть до
600000 особей в 1 м3. Причина—слабая
изученность бескислородных слоев морских
осадков и то, что гнатостомулиды сильно
деформируются в результате фиксации.
4.2.3. Классификация
Около 100 описанных видов помещены
в два отряда в пределах одного класса:
сильно вытянутые Filospermoidea (рис. 4.6)
с длинным рострумом, по без передних
парных сенсорных органов, пениса или семе-
приемника (бурсы), и короткие Bursovagi-
noidea (рис. 4.7) без удлиненного рострума,
но с парными сенсорными щетинками,
ресничками на переднем конце тела, с пенисом
и бурсой. Несомненно, еще многие виды
ждут своего описания.
4.3. Тип Gastrotricha
(брюхоресничные черви)
4.3.1. Этимология
Отгреч. gaster—желудок; thrix—волос.
4.3.2. Диагностические
и специфические признаки
(рис. 4.9)
1. Билатерально симметричные,
червеобразные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Сквозной кишечник с
субтерминальным анусом.
4. Полость тела отсутствует (несмотря
на существовавший ранее
противоположный взгляд).
5. Кровеносной и дыхательной систем
нет.
6. Наружная нехитинизированная
кутикула, обычно нересничный дорсальный
эпидермис и обильно покрытый ресничками
вентральный. Слои кутикулы поднимаются
над каждой ресничкой как футляр.
7. Нервная система из пары связанных
дорсальной комиссурой ганглиев по бокам
глотки и пары продольных стволов.
8. Тело покрыто кутикулярными
чешуйками, иглами или крючками и несет до
250 прикрепительных трубочек.
Комиссура
Мускулистая
глотка
Ганглий
Глоточная пора
Семенник
Яичник
■Прямая кишка
■Прикрепительная
труБОчка
Рис. 4.9. Схематическое изображение
внутреннего строения брюхоресничного с
дорсальной стороны (по нескольким источникам)
106
Глава 4
и " I Jlllllllllllllllll
" "V / \
« || \ /II iiimIiii\
II и \ / \
II И \ / \
ц || \ /lOIIHlllllllllllHllllllV
■ ч \ / \
II " \ / \
„ И \ /i'iiiiiiiiiiiiiiiiiiiiiiiiiiiiR
" " \ I '
и !{ | In iiiiiiiiiHiiiiiiiiiiii
" II / \ /
II n / ViiiMlliliinilliiiil^
Рис. 4.10. Схематические изображения двух
брюхоресничных с вентральной стороны,
демонстрирующие разные типы расположения
ресничек (по Нутап, 1951)
9. Гермафродиты или партеногенети-
ческие формы.
10. Яйца высвобождаются через
разрывы в стенке тела, дробятся билатерально-
радиально, но развитие
детерминированное.
11. Развитие прямое.
12. Водные формы, обитают в интерсти-
циали, на поверхности водоемов, изредка —
в планктоне.
У гастротрихов более или менее
прозрачное, дорсовентрально уплощенное тело
до 4 мм в длину (обычно менее 1 мм).
Спереди расположены сенсорные органы того
же типа, что и у гнатостомулид отряда Bur-
savaginoidea (парные щетинки, реснички
и сенсорные ямки), у некоторых—глазные
пятна; сзади тело может заканчиваться
мощной вилочкой (фуркой) или тонким
хвостом. Выделяющие клейкое вещество
трубочки обеспечивают временное
прикрепление к поверхностям. Хотя стенка
тела содержит кольпевые и продольные
мускулы, гастротрихи движутся скользя с
помощью ресничек, которые на их
вентральной стороне часто расположены
неравномерно, образуя, например, поперечные или
продольные полосы характерным для
разных родов способом (рис. 4.10).
Питаются гастротрихи бактериями,
протистами и детритом, загоняя пищу в рот
биением ротовых ресничек или с помощью
работающей как насос глотки. Как и у
нематод, глотка в сечении трехлучевая и
выстлана единственным слоем мышечно-эпи-
телиальных клеток. Как у коловраток и ки-
норинхов, у пресноводных видов пара про-
тонефридиев с соленоцитами.
4.3.3. Классификация
Известно более 400 видов,
распределяемых между двумя отрядами одного класса;
у ремневидных морских Macrodasyida (рис.
4. 11) глотка перфорирована двумя порами,
служащими, вероятно, для вывода воды,
поступившей внутрь в процессе питания,
как у крыложаберных (разд. 7.2.3.2, см.
также разд. 9.2.5); пресноводные и морские
Chaetonotida (рис. 4.12) обычно
веретенообразны, лишены глоточных пор; если
присутствуют прикрепительные трубочки, то
только сзади; могут размножаться партено-
генетически.
Рис. 4.11. Формы тела брюхоресничных
отряда Macrodasyida (no Grasse, 1965; Hummon,
1982)
Черви
107
Рис. 4.12. Формы тела брюхо-
ресничных отряда Chaetonotida
(по Grasse, 1965; Hummon,
1982)
4.4. Тип Nematoda
(круглые черви)
4.4.1. Этимология
От греч. пета—нить; eidos — форма.
4.4.2. Диагностические
и специфические признаки
1. Билатерально симметричные,
червеобразные формы с тенденцией к
радиальной симметрии вокруг продольной оси.
2. Тело из двух клеточных слоев, с тка-
, нями и органами.
3. Сложная кутикула.
4. Кольцевые мышцы в стенке тела,
отсутствуют.
5. Полость тела—псевдоцель, обычно
развивается из бластоцеля.
6. Мускулистая кишка проходит от рта
на переднем конце через мускулистую
глотку к субтерминальному анусу.
7. Продольная мускулатура разделена
на четыре зоны вентральным, двумя
латеральными и дорсальным эпидермальными
валиками.
8. Нервная система из продольных
тяжей, расположенных в вентральном
и дорсальном эпидермальных валиках и
непосредственно контактирующих с
мышечными клетками.
9. Поперечный срез представляет
собой круг; полостная жидкость всегда
находится под большим давлением.
10. Кровеносной системы нет.
11. Выделительная система без клеток
с ресничным пламенем или нефридиев.
Экскреторные трубочки в одной или
нескольких клетках-ренеттах.
12. Дробление яйца не спиральное и не
108
Глава 4
Наружный слой
Волокнистые слои
Средний
слой
радиальное, но сильно детерминированное,
с Т-образным расположением клеток на
стадии четырех бластомеров.
13. Развитие прямое, но у
паразитических форм может быть стадия инвазионной
личинки.
Этот тип — один из добившихся
наибольшего успеха в животном царстве.
Описано более 15000 видов, но считают, что
сейчас их примерно 1000 000. В отличие от
других типов это не связано с
морфологическим разнообразием. Все нематоды
устроены по одному общему плану, оказавшемуся
весьма «удачным». Он предполагает
способность противостоять потенциально
опасному окружению, и многие круглые
черви —паразиты.
Основная черта их структурной
организации—сложная кутикула (рис. 4.13). В ней
можно обнаружить до девяти слоев, но наи-
Рис. 4.13. Строение кутикулы
нематод, состоящей из
наружного исчерченного,
внутреннего однородного и комплекса
волокнистых слоев (по Clark,
1964)
более заметны среди них три, состоящие из
перекрещивающихся волокон и
образующие вокруг тела спиральную сеть. Она
вместе с полостью тела, содержащей жидкость
под высоким давлением, служит основой
локомоторной системы нематод (см. разд.
10.4).
Нематода представляет собой «трубку
в трубке», заостренную с обоих концов
и круглую в сечении (рис. 4.14).
Терминальный рот открывается в мускулистую
глотку—важный признак организации этого
типа. Поскольку полостная жидкость
находится под высоким давлением, внутренние
трубчатые структуры нематод имеют
тенденцию сплющиваться и для того, чтобы
было возможным питание, нужен механизм
введения пищи во внутреннюю трубку, т. е.
пищеварительную систему. Переднее
отверстие пищеварительного тракта называется
НервноеГлоточный БУЛЬБУС
Глотка
кольцо
Кишечник
Яичник
Семенник
■Копулятивный аппарат
Клоака
Рис. 4.14. Общее строение нематоды (за основу взят паразитический род Rhabditis): A—самка;
Б—самец
Черви
109
Передняя часть глотки Задняя часть глотки
Стома
Рис. 4.15. Механизм работы глоточного
насоса нематод. Диаметр переднего отдела глотки
намного меньше, чем у заднего отдела. Клапан
сзади закрывается при расширении глотки,
и жидкость втекает в ее переднюю часть. Когда
глотка сокращается, задний клапан
открывается, и жидкость течет назад из-за большего
диаметра заднего отдела (по Croll, Matthews,
1977)
стома. Оно ведет в мускулистую глотку, на
другом конце которой находится клапан.
Глотка закачивает пищу против градиента
давления, причем однонаправленный поток
в сторону кишечника обеспечивается
соотношением объемов передней и задней
частей глотки (рис. 4.15).
Значительная часть полости тела занята
парными, сериально организованными
репродуктивными органами, семенниками
или яичниками, которые часто спирально
закручены. Расположение их выводных
отверстий показано на рис. 4.14.
Нематоды обычо раздельнополы,
оплодотворение внутреннее. «Яйца»,
откладываемые самкой, фактически представляют
собой оплодотворенные зиготы или ранние
эмбриональные стадии. Ооциты второго
порядка движутся по яйцеводам к семе-
приемнику, где происходит
оплодотворение. Затем они покрываются слоем
белковой скорлупы, вырабатываемой маткой,
а сама зигота секретирует две внутренние
яйцевые оболочки. «Яйца» бывают крайне
устойчивы к внешним воздействиям и
образуются у некоторых паразитических видов
в огромных количествах.
Строение стенки тела лучше всего видно
на поперечном срезе. Оно выделяет этот
тип среди всех прочих (рис. 4.16). Под слоем
сложной кутикулы лежит эктодермальный
эпидермис, включающий клетки нервной
и выделительной систем, основные
элементы которых расположены в четырех
продольных валиках. Дорсальный и
вентральный валики содержат крупные дорсальный
и вентральный нервы. В промежутках
между эпидермальными валиками лежат
клетки продольной мускулатуры; их небольшое
количество строго фиксировано.. После са-
Латеральный
Мышечные клетки Глотка нервный ствол
Матка
Выделительный
канал
Рис. 4.16. Строение нематоды на поперечном срезе:/)—объемное изображение головы,
демонстрирующее треугольную организацию рта, губ и связанных с ними сенсорных органов; Б —
поперечный срез в области глотки; В — поперечный срез в средней .части тела
110
Глава 4
Тело мышечной клетки
Иннервационный otdoctoj
мышечной клетки
Сократимая часть
мышечной клетки
Нервные волокна
Кутикула
Эпидермис Гиподермальный валик
Рис. 4.17. Объемное изображение
нервно-мускульного комплекса
нематод. Показаны сократимые
части мышечных клеток, их тела
и иннервационные отростки,
устанавливающие прямые синаптиче-
ские связи с нервными волокнами
в латеральных, дорсальном и
вентральном стволах
мых ранних стадий эмбрионального
развития рост тела происходит за счет
увеличения размеров клеток, а не их числа.
Мышечные клетки правильнее было бы
назвать нейромоторными комплексами
(рис. 4.17). Сократимые волокна каждой
клетки расположены дистально и
прилегают к эпидермису. Удлиненный
иннервационный отросток непосредственно
контактирует с нервными тяжами дорсального
или вентрального валиков в зависимости от
положения мышечной клетки. Каждая
такая клетка дает несколько отростков, концы
которых образуют синапсы с другими
мышечными и нервными волокнами. Таким
образом, вероятно, формируется система,
обеспечивающая одновременное
сокращение всех мышечных клеток. Связь их
отростков с нервами—необычная черта,
поскольку у большинства животных отростки
отходят наоборот—от нервных к
моторным клеткам (см. разд. 16.4.2).
У нематод известны различные
сенсорные приспособления. К ним относятся
головные папиллы (сосочки), амфиды (хемо-
чувствительные ямки, часто связанные
с передними амфидными железами),
головные щетинки, соматические щетинки,
хвостовые папиллы и спикулы, глазки и фазми-
ды (рис. 4.18). Фазмиды представляют
собой расположенные в хвостовом отделе
сенсорные ямки, похожие на амфиды,
причем иногда те и другие связаны с железами.
Нематод делят на два класса в зависимости
от отсутствия или наличия фазмид.
Класс
Головная папилла
Головная щетинка
Соматическая
щетинка
Различные мужские
сенсорные органы
Сосочки
Хвостовые
папиллы
Амфида
Глазок
Гемизонид
Внутренние
рецепторы
Соматическая
щетинка
-Спикулы
Фазмиды
Рис. 4.18. Обобщенная схема, показывающая
разные типы сенсорных органов,
обнаруженных у нематод. Ни у одного вида нет их всех (по
Croll, Matthews, 1977)
Adenophorea
Secernentea
Отряд
Enoplida
Isolaimida
Mononchida
Dorylaimida
Trichocephalida
Mermithida
Muspiceida
Araeolaimida
Chromadorida
Desmoscolecida
Desmodorida
Monhysterida
Rhabditida
Strongylida
Ascaridida
Spirurida
Camallanida
Diplogasterida
Tylenchida
Aphelenchida
Черви
111
Экскреторная система круглых червей—
пример исключительной экономии числа
клеток, характерной для типа. Исходно в ее
состав входят одна или две
специализированные клетки-ренетты. В более
продвинутом случае формируется система трубочек
внутри этих железистых клеток-ренетт,
которая у некоторых форм утрачена (см. разд.
12.3.1).
Однотипный план строения нематод,
отражающийся на их внешнем облике, может
быть приспособлен к огромному
разнообразию внешних условий. Лучше всего это
иллюстрирует разнообразие их механизмов
питания или жизненных циклов
паразитических форм. Многие свободноживущие
нематоды питаются бактериями, которых они
поглощают в виде взвеси, или детритом.
Есть и хищники, поедающие в основном
других нематод или микроорганизмы.
У них возникли разнообразные зубы или че-
люстеподобные пластинки на передней
поверхности стомы. Эти «челюсти»
протыкают жертву, содержимое которой затем
высасывается с помощью работающей как
насос глотки.
Ряд свободноживущих
видов—фитофаги. Некоторые заглатывают грибы
и дрожжевые клетки целиком, но
большинство протыкает клетки растений и
высасывает содержимое. Для того чтобы питаться
таким образом, многие
нематоды-фитофаги располагают в ротовой полости
специальным приспособлением: копьевидным
образованием для вскрытия клеток или
полым стилетом, который в совокупности
с глоточным насосом используется как
всасывающий шприц.
Эти механизмы питания
свободноживущих видов могут быть использованы и
паразитическими формами. Например,
полости кишечника позвоночных наполнены
богатой смесью бактерий и разрушенных
клеток, т.е. питательной средой для нематод,
заглатывающих бактерий. Челюстные
пластинки могут быть использованы для
разрыва выстилки кишки и высвобождения
форменных элементов крови, которыми
питаются нематоды-паразиты.
Ослабление позвоночных, вызываемое
инвазией их пищеварительной системы
нематодами, частично зависит от биологии
питания последних. Детская острица (Ente-
robius vermicularis) относительно безвредна,
пока питается бактериями в кишечнике,
и становится патогенной, когда ее
популяция возрастает настолько, что вызывает
закупоривание пищеварительного тракта.
Свайник двенадцатиперстной кишки (Anky-
lostoma duodenale) более опасен, так как
вызывает кишечные кровотечения, питаясь
клетками крови. Человек, в котором живет
100 таких червей, может терять до 50 мл
крови в день.
Стома всех нематод-фитопаразитов
вооружена стилетом или «копьем». Многие
из них образуют постоянные цисты, в
которых неподвижные самки питаются
гигантскими клетками. Черви сами вызывают
образование таких клеток растением-
хозяином, вероятно, воздействуя на
продуцирование им веществ, стимулирующих
рост, или их ингибиторов. Наиболее
важные роды фитонематод—Heteroda и Meloi-
dogyne.
Паразитизм возникал в филогенезе
нематод неоднократно. Этому
способствовали некоторые основные черты организации
типа, а именно:
1) сложная непроницаемая и прочная
кутикула;
2) внутреннее оплодотворение и
способность производить высокоустойчивые к
внешним воздействиям яйца;
3) микрофагия, т.е. питание
мелкодисперсным материалом;
4) мелкие размеры тела;
5) химическое разнообразие,
включающее механизмы, необходимые для
избегания защитных систем хозяина.
Паразитизм у многих беспозвоночных
ведет к сильному видоизменению взрослых
форм и развитию специализированных
жизненных циклов со стадиями,
способными к бесполому размножению, как,
например, у многих паразитических плоских
червей (разд. 3.6.3.2 и 3.6.3.3). Однако у
нематод этого не произошло. Взрослые особи
«преадаптированы» к паразитическому
образу жизни, и способ их развития остался
неизменным. Исходный жизненный цикл
изображен на рис. 4.19, А. Он включает
в себя спаривание самцов и самок и
высвобождение хорошо защищенных «яиц». Раз-
112
Глава 4
Б. Strongyloldes
\\ Взрослая самка
||в кишечнике
Яйца, Образующиеся
ерез ^^рартеногенетически
LAIffT 'трахею в ^ -
*\\ ю кро- кишечник „ т
. носнои Паразитизм
n -JE^IZ^_ ''_ Вылупление
Прониййт; через ко*у ' в кишечнике
или загпатыва.гсй
* * Почва йгтголии
Д Ancylostoma
По кровеносной
системе к легким
Ж| «ченнчйцикл
СВ Ь0ДИСШЩ«(ИХ 1
.осовей т
Взрослые особи
" - кишечнике
ОСОБИ
)Яйца
Iх кишечник Паразитизм
1 гВыделение с
Проникает через"кад^.йлй^ "*", "фекалиями
заглатыва*,^.-
черезели- Ч Почва и фекалии f
ЗИСТуЮ BF4^ J-*"^****'
%
Г. AscarLs л
Через трахею / /Взрослые особи
в кишечник! Lb кишечнике
Оплодотворенные яйца
Д. Enteroblus Q(l„*
Яйца
^
inec
фекалиями
г
в кишечник] 1~|
По Кровей
Ч\Яосной \
'к л^Тим Паразитизм
«Паразитизм в____тВ—делеН|
кишечнике^Почва иТФекалия
^брекагми] г
-^ JP<1в яйч3
^Поглощение
\с пищей
Яйца
■ Выделение с фекалиями
Проникает через/Г
слизистую /<Почва'и
.1Лу1
ление
в кишечнике/.
Поглощение £, в яйце
с пищей
Рис. 4.19. Жизненные циклы нематод: А — основной цикл с четырьмя личиночными стадиями;
Б—Д—модифицированные циклы видов, приспособившихся к зндопаразитизму (по СгоН
и Matthews, 1977). L, — L4—различные личиночные стадии
витие включает четыре личиночные стадии.
Из этого жизненного цикла можно
вывести ряд более приспособленных к
паразитизму, которые тем не менее никоим
образом не следует считать последовательными
эволюционными стадиями.
Наименее модифицированный
жизненный цикл включает личинку, живущую
свободно в почве. Станут взрослые формы
паразитами или нет, зависит от случая.
Например, особи Strongyloldes на протяжении
всего цикла могут быть свободноживущи-
ми, но некоторые личинки третьей стадии
проникают в млекопитающее-хозяина через
кожу и мигрируют через трахею к
кишечнику, где живут размножающиеся партеноге-
нетически самки (см. гл. 14). Яйца либо
начинают жизненный цикл во внешней среде,
Черви
113
A. Oxyspirura
Выходит в зову,
мигрирует в
глазницу i
Съедается с
насекомым
L.
Инцистируется
в полости тела
зрослые особи в глазнице
Яйца проходят через
слезные/носовые
протоки в кишечник
латывание
полости тела
Б. Thelazia
3frS---
^/111 \1°нктиваг
I Млекопитающее
Выходит 1Ълвзную~\
жидкость \
Взрослые особи в конъ-
" «ктивальных мешках
Яйца
Движется к
ротовым частям
мухи
В. РагаШагча
Взрослые особи в подкожной
ткани, мигрируют на поверхность кожи
•\Яйца в кровоточащих
франках
Г. Stephanofilaria
Заглатывается
со слезной
Чжидкостью,
проходит из
кишечника в яйцевые
фолликулы
Взрослые особи в
подкожной ткани
Лошадь
Выходит в~р^Й^кстда" "Л
насекомое питается \
" Кровососущее
BN. насекомое.
.Микрофиларии
■ув изъязвлениях
Д. Onchocerca
<Л) ([Взрослые особи
т подкожной ткани
Е. Diroftlaria
Микрофиларии
] в дерме
Млекопитающее
Выходит фйЪтёдующем
у Мошка
'уЛ'Взрослые особи в сердце
Выходит при следующем
кровососании
Комар
Микрофиларии
крови
кровососании|
Рис. 4.20. Жизненные циклы зндопаразитических нематод, включающие смену хозяев: А —
Oxyspirura; Б—Thelazia; В—Parafilaria; Г—Stephanofilaria; Д—Onchocerca; Е—Dirofilaria (no
Croll, Matthews, 1977)
либо дают начало новым инвазионным
личинкам третьей стадии (рис. 4.19,£).
У свайника Ankylostoma сходный
жизненный цикл, но без свободноживущих
взрослых форм. Как самцы, так и самки
обитают в кишечнике (рис. 4Л9,В). Первая
и вторая личиночные стадии — типичные
рабдитовидные личинки-микрофаги,
питающиеся в почве. Третья, инвазионная,
стадия инцистируется и не питается.
У необычно крупной человеческой
кишечной нематоды—аскариды (Ascaris) сво-
бодноживущая личинка исчезает. Яйца
выделяются с фекалиями наружу, но личинки
первой и второй стадий остаются инцисти-
рованными внутри яиц, которые
заглатываются хозяином с загрязненной пищей.
Личинки третьей стадии вылупляются
8-1314
114
Глава 4
в просвете кишки, но не остаются там.
Через кровеносную систему они мигрируют
к легким, а оттуда обратно в кишечник (рис.
4.19,7"). Такой сложный жизненный цикл
может отражать его эволюционную
историю, в которой был промежуточный
хозяин, утраченный впоследствии.
В сходных жизненных циклах паразитов
кишечника человека Trichurns и Enterobins
(рис. 4.19,Д) отсутствуют признаки двойной
миграции перед поселением в кишечнике.
Самки Enterobius могут вызывать очень
сильный зуд вокруг анального отверстия. Он
приводит к расчесыванию и в результате—
к прямому повторному заражению и
переносу паразитов через пальцы к другим
потенциальным хозяевам.
Некоторые круглые черви,
паразитирующие в позвоночных, имеют
промежуточных хозяев и используют
взаимоотношения животных в пищевой цепи (рис. 4.20).
Другой пример—жизненные циклы нема-
тод-мермитид: личинка паразитирует в
насекомых, а взрослые особи свободноживу-
щие (ср. тип Nematomorpha. разд. 4.5).
4.5. Тип Nematomorpha
(волосатики)
4.5.1. Этимология
От греч. nematos—нить; morpha—
форма.
4.5.2. Диагностические
и специфические признаки
(рис. 4.21)
1. Билатерально симметричные,
тонкие удлиненные черви (диаметр менее 3 мм,
длина от 10 см до 1 м).
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Кишечник сквозной прямой, часто
дегенерирует, вероятно, никогда не
функционирует.
4. Тело мономерное с псевдоцеломиче-
ской полостью, часто заполненной
мезенхимой.
5. Стенка тела состоит из гибкой кол-
лагеновой кутикулы, эпидермиса и слоя
Рис. 4.21. Форма тела взрослых волосатиков:
А — Gordioidea, Б—Nectonematida (no Margu-
lis, Schwartz, 1982; Fewkes, 1883)
продольных мышц; слой кольцевых мышц
отсутствует.
6. Кровеносная, выделительная и
дыхательная системы отсутствуют.
7. Нервная система расположена в
эпидермисе, состоит из переднего кольца
и двух продольных стволов без ганглиев.
8. Раздельнополые формы, с
единственной удлиненной гонадой или парой
гонад; оплодотворение внутреннее с
помощью сперматофоров.
9. Взрослые особи свободноживущие,
живут недолго.
Черви
115
Крючковидная
скалида
Кутикулярное
кольцо
11—Ротовой конус
со стилетами
Ротовой канал
Кишечник
Стилеты
Псевдоцел!
Кутикулярн!
КОЛЬЦОСепта
Кишечник >.
Рис. 4.22. Личинка Gordioidea: A — с
вывернутым интровертом, Б—с втянутым интровертом
(по Нутап, 1951; Реппак, 1978)
10. Личиночные стадии заражают
членистоногих (или, редко, пиявок) и
развиваются в их гемоцеле; у ювенильных особей
три ротовых стилета и три круга крючко-
видных скалид (рис. 4.22).
11. Обитатели пресных вод или влажной
почвы. Один род—морской.
Строение тела взрослых волосатиков
и нематод (разд. 4.4) в принципе сходно.
Как и последние, они передвигаются за счет
волн, проходящих вдоль тела в дорсовен-
тральной плоскости. Морские виды и
самцы пресноводных и наземных групп
наиболее подвижны. Более крупные
пресноводные самки обычно ведут относительно
оседлый образ жизни, они лежат, свернувшись
в спираль или в кольцо, на субстрате.
Наиболее очевидное отличие волосатиков от
нематод— дегенерация у первых кишечника
и/или его отверстий, что связано с
редукцией взрослой стадии до короткоживущей
репродуктивной и расселительной фазы
жизненного цикла.
Питающаяся фаза — паразитические
ювенильные особи, которые, хотя и имеют
кишечник, скорее всего получают большую
часть, а то и всю пищу за счет абсорбции
через поверхность тела из гемоцеля хозяина.
Личинки и ювенильные особи совершенно
непохожи на нематод; они напоминают
взрослых киноринхов (разд. 4.6) и лорици-
фер (разд. 4.7) наличием ротовых стилетов,
скалид и строением передней части
кишечника (рис. 4.22). Когда личиночный
интроверт вывернут (рис. 4.22,А) с помощью
поперечной септы, изолирующей от его
полости переднюю часть псевдоцеля, эти
стилеты и скалиды, по-видимому, способствуют
проникновению через ткани хозяина в гемо-
цель. Происходит ли это через покровы или
через кишечник, пока не ясно.
4.5.3. Классификация
В настоящее время известно 250 видов,
помещаемых в два отряда одного класса.
Наиболее многочисленная группа
пресноводные Gordioidea (рис. 4.21,А),
паразиты одноветвистых. Для них характерен
единственный (вентральный) нервный
ствол, парные гонады и полость тела, почти
целиком заполненная мезенхимными
клетками, по крайней мере в пререпродуктив-
ный период. Небольшое число известных
морских Nectonematida (рис. 4.21,Б) —
паразиты десятиногих раков — отличается
вторым (дорсальным) нервным стволом,
единственной оформленной (у самцов) или
диффузной (у самок) гонадой и псевдоце-
лем, незаполненным мезенхимой. Они несут
также двойной ряд латеральных щетинок,
проходящий вдоль всего тела, что
увеличивает ундулирующую поверхность во время
плавания. Некоторые зоологи считают
родство между этими двумя отрядами очень
отдаленным, выводят нектонематид из
нематод и сближают гордиоидей с киноринха-
ми и лорициферами.
116
Глава 4
4.6. Тип Kinorhyncha
4.6.1. Этимология
От греч. kinema—движение; rynchos—
рыло.
4.6.2. Диагностические
и специфические признаки
(рис. 4.23)
1. Билатерально симметричные,
червеобразные, но довольно короткие формы.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Пищеварительная система
трубчатая, с задним анусом, мускулистой глоткой
и выступающим ротовым конусом,
содержащим ротовую полость.
4. Тело снаружи разделено на
определенное количество (13 или 14) сегментов,
или «зонитов».
5. Эпидермис с игольчатой кутикулой,
Ротовые стилеты
Ротовой конус
Скалиды-
Шейная глас-
тинка(плацида)-
Сенсорное пятно
Рис. 4.23. Наружное строение киноринхов: А —
Cyclorhagida: (I) вид с вентральной стороны;
(II) вид спереди с втянутым интровертом; Б—
Homalorhagida: (I) виде вентральной стороны;
(II) вид спереди с втянутым интровертом,
защищенным пластинками третьего зонита (по
Higgins. 1983)
включающей единственную дорсальную
и парные вентральные пластинки на
каждом зоните.
6. Полость тела—псевдоцель,
происходящий из сохраняющегося бластоцеля'.
7. Стенка тела с кольцевыми и
диагональными мышцами.
8. Выделительная система из протоне-
фридиев в 11-м зоните; соленоциты с
парными ресничками и многочисленными
ядрами.
9. Нервная система из переднего около-
кишечного кольца и вентрального ствола,
на котором группы ганглионарных клеток
расположены в соответствии с
сегментацией эпидермиса и мускулатуры.
10. Мелкие животные; почти всегда
входят в состав морской мейофауны.
11. Наружных ресничек нет.
Киноринхи—одна из наименее
изученных групп животных. Они явно близки
к другим типам псевдоцеломических
червей, особенно—к Loricifera и Nematomor-
pha.
Наиболее характерная черта типа—
покрытая иглами кутикула, отчетливо
подразделяющаяся на определенное число
поперечных зонитов. Сегментация кутикулы
влияет на эпидермис, мускулатуру и
нервную ткань, придавая им метамерную
структуру, которая тем не менее не гомологична
наблюдаемой у целомических червей-
аннелид (разд. 4.14).
Все киноринхи—крошечные формы
мейофауны морских илистых осадков.
Описано около 100 видов, основное строение
которых сходное.
Ротовой конус со стилетами образует
выворачивающийся интроверт, который
может втягиваться во второй, «шейный»,
сегмент. Интроверт вооружен
расположенными по кругу загнутыми скалидами.
Туловищный отдел состоит из 10
одинаковых туловищных зонитов, каждый из
которых несет единственную дорсальную
пластинку—тергит, и пару вентральных—
стерншпы. Мускулатура стенки тела
уплощает туловише так, что спинная и брюшные
1 Эмбриональное развитие этих животных
пока до конца не изучено, поэтому трудно
сказать, как образуется их полость тела.—Прим:
пер.
Черви
117
Мозг
Слюнная железа
Ротовой конус
Скалида
Трихоскалида
Глотка
Пищеварительная
железа
■Средняя кишка
-Половые органы
Нефридий
Рис. 4.24. Внутреннее строение обобщенного
киноринха с вытянутым интровертом
пластинки сближаются, и давление в псев-
доцеле выворачивает интроверт. Животные
передвигаются, выталкивая его вперед, зая-
кориваясь скалидами, а затем подтягивая
все тело с помощью мышц-ретракторов.
Некоторые черты внутреннего строения
киноринхов (рис. 4.24) также общие с
другими типами псевдоцеломических червей.
Эпидермис частично синцитиальной
структуры образует продольные среднедорсаль-
ный и латеральные валики (как у нематод)
и заходит в самые крупные иглы кутикулы.
Его утолщения—«подушечки» вдаются
в псевдоцель. Хорошо развита мускулистая
глотка, треугольная в поперечном сечении,
что характерно для многих
псевдоцеломических червей.
4.6.3. Классификация
В основе деления киноринхов на два
отряда лежит способ втягивания интроверта.
У Cyclorhagida втягивается только первый
зонит, который защищают крупные
пластины второго зонита (см. рис. 4.23,Л). В
отряде Homalorhagida втягиваются оба
первых зонита, обычно защищаемые
вентральными пластинами третьего (рис. 4.23,Б).
4.7. Тип Loricifera
4.7.1. Этимология
От лат. lorica-
ferre—нести.
-грудной щит или латы;
4.7.2. Диагностические
и специфические признаки
(рис. 4.25)
1. Билатерально симметричные
животные.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Прямой сквозной кишечник и
терминальный анус.
4. Есть полость тела.
5. Тело делится на три отдела:
передняя выворачивающаяся голова или ин^
Скалиды
Панцирь
Рис. 4.25. Вид с дорсальной
стороны лорициферы (по Kri-
stensen, 1983 с упрощениями)
Ротовой конус
Рот
Ротовой стилет
Вытянутый интроверт Шейнь1й отдел
118
Глава 4
Ротовой конус
Скалиды
Сегментированный
шейный отдел
Панцирь
Хвостовые ножки
Рис. 4.26. А. Вид с дорсальной
стороны личинки последней
стадии лорициферы. Б. Вид
с вентральной стороны ее
локомоторных игл (по Kristensen,
1983 с упрощениями)
троверт с девятью кругами направленных
назад игло- или зубовидных скалид и
вытягивающимся ротовым конусом,
окруженным 8—9 твердыми ротовыми стилетами;
короткий наружно сегментированный
шейный отдел с рядами пластинок; туловище,
покрытое кутикулярным панцирем из
шести продольных пластин, несущим
направленные вперед полые иглы. У взрослых
особей два передних отдела могут втягиваться
в туловищный панцирь; пластинки шейного
отдела, по-видимому, служат защитным
покровом после такого втягивания.
6. Пищеварительный тракт с крупным
мускулистым глоточным бульбусом
(луковицей) и телескопически раздвигающимся
ротовым каналом; весь тракт, вероятно,
выстлан кутикулой.
7. Одна пара гонад; раздельнополые
животные.
8. Хорошо развитый мозг и
вентральный нервный ствол с ганглиями.
9. Личиночная стадия похожа на
уменьшенную взрослую особь (рис. 4.26),
если не считать отсутствия ротовых
стилетов и наличия двух типов локомоторных
органов: а) 2—3 пар игл на передневентра-
льной части панциря, с помощью которых
животное может ползать; б) пары
напоминающих ножки подвижных хвостовых
придатков, используемых для плавания.
Несколько возрастных стадий личинки разделено
линьками.
10. Интерстициальные формы морских
ракушечно-гравийных осадков.
Строение этих крошечных (менее 0,3 мм)
животных известно пока лишь в общих
чертах; нет никакой информации об их
эмбриологии и образе жизни. Впервые они были
описаны в 1983 г. Одна из причин столь
позднего открытия, вероятно, то, что
лорициферы прочно прикрепляются к частицам
осадка или другим организмам и не
обнаруживаются с помощью обычных для сбора
мейобентоса методик; другая причина, по-
видимому, сходство животных во втянутом
состоянии с личинками коловраток или
приапулид.
Наиболее характерная черта строения—
деление тела на передние интроверт и
шейный отдел и заднее туловище, покрытое
панцирем. У взрослого животного вся
передняя часть может быть втянута в
туловищный панцирь, в то время как личинка
способна втянуть только интроверт в шейный
отдел. Эта особенность очень близка к
тому, что наблюдается у приапулид, личинок
волосатиков, нематод рода Kinochidus
и особенно у киноринхов (разд. 4.6). Как
у последних, так и у лорицифер «голова»
выворачивается за счет давления в полости
тела и втягивается специфической мышцей-
ретрактором, в то время как ротовой конус
с его стилетами втягивается и вытягивается
независимо с помощью мускулов. Функции
выдвижения и втягивания переднего конца
тела у лорицифер неизвестны; в других
группах это обеспечивает продвижение
в осадках или тканях животных-хозяев,
ловлю и поглощение добычи. Возможно,
лорициферы — эктопаразиты.
Пищеварительный тракт по строению
Черви
119
Роговой стилет
Ротовой канал
Дополнительный
стилет
Панцирь
Средняя
кишка
Мускулистая
глотка
Мышца-
ретрактор
Рис. 4.27. Передняя кишка лорицифер
(ротовой конус и шейный отдел соответственно
вытянуты и вывернуты (по Kristensen, 1983)
сильно напоминает таковой у тихоходок
(разд. 8.1). Его передний отдел (рис. 4.27)
состоит из телескопически
складывающегося ротового, или буккального, канала,
мускулистого, несущего плакоиды
глоточного бульбуса, пары дополнительных
стилетов и пары крупных слюнных желез
(единственные железы, отходящие от
кишечника). Пока еще не ясно, является ли это
сходство следствием конвергентной
эволюции, хотя оно, по-видимому, означает, что
лорициферы питаются, протыкая жертву
ротовыми стилетами и высасывая жидкость
с помощью глоточного насоса.
4.7.3. Классификация
Хотя формально описан только один
вид, своего описания ждут еще несколько.
4.8. Тип Priapula (приапулиды)
4.8.1. Этимология
От греч. Priapos—фаллическое
божество, олицетворяющее мужскую
детородную силу.
4.8.2. Диагностические
и специфические признаки
1. Билатерально симметричные
животные.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Есть крупный втягивающийся
интроверт, или просома, и покрытое иглами или
чешуйками нерасчлененное туловище;
иногда присутствуют ветвящиеся хвостовые
придатки.
4. Пищеварительная система с
передним ртом, окруженным игло- или крючко-
видными скалидами, и задним анусом,
иногда окруженным венчиком игл.
5. Выделительная и половая системы
тесно связаны в урогенитальный комплекс
с множеством соленоцитов.
6. Нервная система из околоротового
кольца и вентрального ствола с ганглиями.
7. Обширная полость тела—возможно,
настоящий целом.
8. Раздельнополые формы;
оплодотворение наружное.
9. Личинка без ресничек, заключена
в панцирь (т.е. покрыта пластинками).
10. Кровеносной системы нет, но в
полости тела находятся форменные элементы,
содержащие гемэритрин.
Приапулиды—небольшая, но четко
очерченная группа морских животных с
неясным происхождением. Они в некоторой
степени сходны с типами псевдоцеломиче-
ских червей, к которым их часто относят, но
функционально гораздо ближе к целомиче-
ским червям, рассматриваемым в разд.
4.11—4.14. Некоторые авторы считают их
полость тела целомом, поскольку она имеет
мезодермальную выстилку, но электронно-
микроскопические исследования
свидетельствуют о том, что она уникальна и
непохожа на присутствующую у других целомиче-
ских форм. Требуется более детальное
изучение происхождения этой полости.
Немногие данные по эмбриологии приапулид
говорят о радиальном, а не спиральном
дроблении их яйца.
Различные формы тела представителей
этого типа показаны на рис. 4.28. У
большинства видов оно состоит из крупной
луковицеобразной просомы, или интроверта,
и туловища. Интроверт оканчивается ртом,
окруженным кольцом из пяти игловидных
скалид. Этот крупный бочковидный орган
с продольными рядами скалид обычно
вывернут, но может втягиваться в туловище,
от которого обычно отделен хорошо
выраженным шейным отделом. Часто кольчатое
туловище, как правило, покрыто шипами
120
Глава 4
%
Рис. 4.28. Разнообразные формы тела приапу-
лид
или чешуями и несет периодически
линяющую кутикулу. На заднем конце тела Pri-
apulus расположена любопытная
разветвленная структура—хвостовой придаток,
выполняющий, по-видимому,
респираторную функцию (рис. 4.29,А). Кутикула
покрывает эпидермис, в котором находятся
заполненные жидкостью межклеточные
пространства; этим он отличается от
эпидермиса как целомических, так и псевдоце-
ломических червей. Стенка тела включает
и кольцевые, и продольные мышцы,
создающие давление в заполненной
жидкостью полости тела, приводящее к
выворачиванию интроверта. Пищеварительный
тракт—прямая трубка с ороговевшим,
зубчатым ротовым отделом и мускулистой
глоткой (рис. 4.29,5); нет ни ротового
конуса, ни ротовых стилетов.
Уникальная черта при any ли
д—объединение половой системы и соленоцитов
протонефридиев с формированием парных
мочеполовых органов (рис. 4.29,5).
Панцирная личинка может быть
пелагической, но покрыта кутикулярными
пластинами (рис. 4.30). У нее уже развита
втягивающаяся чешуйчатая просома, и она легко
зарывается в грунт, в то время как
взрослым формам зарываться трудно.
Возможно, в норме личинка—единственная
стадия жизненного цикла, предназначенная
-Околоротовое
нервное кольцо
■Глотка
-Короткая мышца-
ретрактор
-Длинная мышца-
ретрактор
-Кишечник
-Нефридий I Моче-
- Гонада ( половой
I орган
Анус *
Рис. 4.29. Внешнее {А) и
схематическое внутреннее (Б)
строение Priapulus
Черви
121
Рис. 4.30. Панцирная личинка приапулид
для погружения в субстрат. Внешнее
сходство между покрытыми иглами
интровертами приапулид и киноринхов, а также между
панцирной личинкой первых и
коловратками не дает достаточно оснований для того,
чтобы считать эти типы
близкородственными.
4.8.3. Классификация
Описано 16 видов приапулид,
разделенных на два отряда: Priapulida с более
крупными, более обычными хищными формами
(рис. 4.29,А) и Seticoronaria, у
представителей которого тонкие скалиды образуют
венчик жестких «щупалец» (см. рис. 4.28, внизу
слева) и имеется околоанальное кольцо
крючьев. У приапулид долгая
эволюционная история; несколько их видов известны
еще с кембрия.
4.9. Тип Rotifera (коловратки)
4.9.1. Этимология
От лат. rota — колесо; ferre—нести.
4.9.2. Диагностические
и специфические признаки
1. Билатерально симметричные
животные.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Венчик ресничек в передней части
тела состоит из преорального и
посторального поясков. Часто они образуют два
колесовидных ресничных органа, от чего и
происходит латинское название группы.
4. Пищеварительная система с
передним ртом, сложным челюстным
аппаратом, мускулистой глоткой и задним анусом,
открывающимся в общую с уриногениталь-
ной системой клоаку.
5. Эпидермис с небольшим
фиксированным числом ядер; в его состав входит
внутриклеточная кутикула, часто утолщенная
и образующая футляр или панцирь.
6. Тело не сегментировано.
7. Протонефридиальная выделительная
система.
8. Кровеносной системы и органов
дыхания нет.
9. Полость тела — псевдоцель.
10. Раздельнополые животные, но во
многих случаях самцы редки или
отсутствуют совсем, если есть, почти всегда
карликовые.
11. Очень мелкие формы, редко
достигают в длину 3 мм.
12. Развитие прямое, следует за
модифицированным спиральным дроблением.
После завершения эмбриогенеза ядерных
делений не происходит.
Коловратки—одни из самых мелких
животных; размеры их взрослых особей
такие же, как у ресничных простейших и
личинок многих других типов. Их тело состоит
из небольшого, строго определенного числа
клеток, а точнее сказать—ядер, так как
многие ткани синцитиальные (общая черта
с некоторыми другими типами псевдоцело-
мических червей).
Взрослые коловратки обычно ведут
примерно такой же образ жизни, что и
ресничные личинки более крупных
беспозвоночных; в самом деле, внешне они похожи на
трохофор морских аннелид, сипункулид
и моллюсков. Характерная черта трохофо-
ры—наличие экваториального ресничного
кольца—прототроха, состоящего из двух
поясков—преорального и
посторального,— реснички которых бьют в
противоположных направлениях. Сходный аппарат
122
Глава 4
Предротовой
ресничный
венчик =
трохус
Послеротовой
ресничный
венчик =
цингулум
Кишка
Коловра-
> тательный
аппарат
Рис. 4.31. Схематический план строения
коловраток. Показаны два венчика ресничек
имеет большинство планктонных
коловраток: реснички их венчика образуют два
пояска— преоральныи трохус и посторальный
цингулум (рис. 4.31).
Однако это сходство не подразумевает
близкого филогенетического родства,
и другие черты свидетельствуют о большей
близости к псевдоцеломическим червям,
особенно скребням. Некоторые авторы
считают коловраток неотенической группой,
но неизвестно, какой предковой линии. Ни
в одном из типов псевдоцеломических
червей нет ресничной личинки, из которой
можно было бы их вывести.
Снаружи тело коловраток часто
покрыто кутикулой, образующей чашевидный
панцирь; открытый конец несет ресничный
венчик и рот. Коловращательный аппарат
венчика может быть
высокоспециализированным, а у сидячих форм сильно
редуцированным. В последнем случае (см. рис. 4.37)
реснички часто модифицируются в жесткие
сенсорные волоски. Обычно венчик может
втягиваться в панцирь. Сзади тело
суживается, образуя тонкую подвижную «ногу»,
часто кольчатую снаружи (псевдосегментиро-
ванную). Как и венчик, нога обычно может
втягиваться в панцирь; при этом
псевдосегменты телескопически входят один в
другой. Нога заканчивается двумя
выростами—«пальцами», служащими для постоян-
Трофусы
1астакс
Булызус с ресничным
пламенем
Протонефридий
Желудок
•Желточник
Яйцо
Мочевой пузырь
Клоака
-Педальная железа
Мозг
Дорсальное
щупальце
—Эпидермис
Палец
Рис. 4.32. Общие анатомические особенности коловраток класса Bdelloidea: A—виде
вентральной стороны; Б — вид сбоку
Черви
123
Инкудатный С\\
МануБриум\л
Фулькрум
Форципатный Виргатный
Рис. 4.33. Разные формы ротовых придатков
(трофусов) коловраток (по Donner, 1966)
ного или временного заякоривания
животного в субстрате; она может отсутствовать
или редуцироваться у планктонных форм.
Внутреннее устройство коловраток
относительно простое (рис. 4.32)—одно из
следствий мелких размеров. Тем не менее
строение ротового аппарата сложное и
варьирующее в зависимости от способа питания.
Основной план строения твердых частей
(трофусов) жевательного аппарата (ма-
стакса) показан на рис. 4.33. Фулькрум,
лежащий ниже средней линии трофусов,
поддерживает два рамуса, расположенные
относительно него симметрично. Над ними
подвешены парные ункусы и манубрии.
Различные трофусы показаны на рис. 4.33. Они
могут быть приспособлены для
прокалывания и высасывания клеточного
содержимого, схватывания добычи (как пинцетом),
размалывания, раздавливания или
разрывания ее.
У большинства коловраток два протоне-
фридия, состоящие из синцития с
небольшим количеством клеточных ядер. В
пресноводной среде через нефридии с большой
скоростью выводится гипоосмотическая
жидкость.
, 4.9.3. Классификация
Тип подразделяют на три класса.
Класс
Seisonidea -
Bdelloidea -
Отряд
-Seisonida
-Bdelloida
.Ploima
-Flosculariida
-Collothecida
Monogonata
4.9.3.1. Класс Bdelloidea
У большинства представителей этого
класса характерный венчик из двух «колес»
(рис. 4.34) и тело, не покрытое панцирем;
эти виды ползают изгибаясь петлями, но
могут также хорошо плавать с помощью
нескладчатого коловращательного
аппарата. Бделлоидеи приспособлены к жизни
в непостоянных и скрытых биотопах,
обычны в интерстициали сырого песка и почвы
речных и озерных берегов, связаны с мхами
в периодически увлажняющихся наземных
биотопах. Они больше ползают, чем
плавают, хотя ресничные диски хорошо
развиты, а в ноге имеются массивные, крупные
цементные железы (рис. 4.35).
Биология размножения очень необычна.
Похоже, вся эта группа амиктическая,
т. е. мейоза и спаривания здесь не
происходит. Самцов не находили никогда, известны
Рис. 4.34. Механизм питания пелагических
коловраток. Крупные реснички переднего
венчика (трохуса) создают ток воды в переднезад-
нем направлении, обеспечивающий транспорт
пищевых частиц и сохранение положения тела.
Реснички заднего венчика (цингулума) бьют
в обратном направлении. Это создает
водоворот, затягивающий пищевые частицы и
несущий их ко рту
124
Глава 4
Бульвус с ресничным
пламенем
Протонеорридий
Парные яичники
с желточниками
Мочевой пузырь
Цементная железа
Палец
Рис. 4.35. Внутреннее строение коловраток
класса Bdelloidea (семейство Philodinidae).
Обратите внимание на парные яичники. Эти
животные исключительно амиктические;
самцы никогда не обнаруживались
Рис. 4.36. Разнообразие форм тела коловраток
классов Monogonata и Seisonidea (no Donner,
1966)
лишь облигатно партеногенетические самки
(см. разд. 14.2.1). Характерна также
способность впадать при высыхании в анабиоз;
в таком высохшем состоянии они могут
сохранять жизнеспособность многие годы,
выдерживая колебания температуры от
+ 40 до - 200°С.
4.9.3.2. Класс Monogonata
В этот класс, для представителей
которого характерно наличие единственного
(непарного) яичника, входит большинство
коловраток. Выделяют несколько отрядов.
Самый крупный отряд—Ploima—
включает разнообразные сидячие и
свободноплавающие формы (рис. 436,А,Б, Ж).
Нога, если она есть, несет два пальца, а рот
лежит внутри, а не спереди цингулума.
У этих коловраток часто есть жесткий
панцирь.
Отряд Flosculariida (рис. 4.36,В,Д)—
вторая группа свободноплавающих или
сидячих коловраток, у которых ресничные
пояски коловращательного аппарата четко
разделены на трохус и цингулум. Нога, если
она есть, не несет парных пальцев.
В отряде Collothecida коловращатель-
ный аппарат представляет собой крупную
воронку, часто несущую реснички,
видоизмененные в жесткие сенсорные волоски (см.
рис. 4,36,7"; 4.37). Самки всегда сидячие,
прикрепленные к субстрату цементными
железами; их тело часто окружено
студенистой массой.
В отличие от Bdelloidea коловратки
класса Monogonata размножаются
половым путем. Хотя в популяциях большую
часть года преобладают самки, в
определенные периоды появляются относительно
просто устроенные, часто карликовые, но
свободноплавающие самцы.
Большую часть года диплоидные
амиктические самки производят яйца,
развивающиеся без оплодотворения в молодых
самок. Время от времени (возможно, как
реакция на условия окружающей среды)
появляются морфологически отличные
мистические самки, откладывающие гаплоидные
яйца. Они либо быстро развиваются в
гаплоидных самцов, либо, если происходит
оплодотворение, становятся зиготами,
Черви 125
Рис. 4.37. Сидячая коловратка отряда Collothe-
cida. Обратите внимание на
модифицированные реснички коловращательного аппарата
дающими начало амиктическим самкам.
Таким образом, миктические самки
являются факультативно партеногенетическими
(см. разд. 14.2.1).
4.9.3.3. Класс Seisonidea
Этот мелкий класс морских коловраток
обитает на жабрах ракообразных—Nebalia
и некоторых равноногих. Сюда входит
только один род Seison (рис. 4.36,5). Его
представители относительно крупны (несколько
миллиметров), у них сильно
редуцированный коловращательный аппарат и
выступающий мастаке. Яичники, как и у видов
Bdelloidea, парные, но (что не типично для
коловраток) происходит обычное половое
размножение с участием нормально
развитых самок и самцов, встречающихся в
популяциях с одинаковой частотой.
4.10. Тип Acanthocephala
(скребни)
4.10.1. Этимология
От греч. akantha—шип; kephale—
голова.
4.10.2. Диагностические
и специфические признаки
1. Билатерально симметричные,
червеобразные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Пищеварительной системы нет.
4. Сегментация тела отсутствует, но
иногда выражена внешняя поперечная коль-
чатость.
5. Выступающий хоботок вооружен
крючьями.
6. Полость тела—псевдоцель.
7. Синцитиальный эпидермис с
небольшим фиксированным числом относительно
крупных ядер.
8. Нервная система с единственным
передневентральным ганглием и парными
или одиночными нервами, отходящими
к органам.
9. Иногда присутствуют нефридии.
10. Кровеносной и дыхательной систем
нет.
11. Раздельнополые животные с
внутренним оплодотворением и
живорождением.
12. Инвазионная личинка проникает
в промежуточного хозяина—насекомое.
13. Взрослые особи паразитируют в
пищеварительном тракте позвоночных.
Общее строение тела скребней показано
на рис. 4.38. Большинство видов мелкие (от
1 мм до нескольких сантиметров длиной),
но некоторые достигают длины 1 м.
Наиболее характерная черта — хоботок с
изогнутыми шипами. Он вместе с шейным
отделом может вворачиваться, но обычно
образует постоянный аппарат
прикрепления к тканям хозяина. Как и у других псев-
доцеломических червей, некоторые ткани
состоят из четко фиксированного
небольшого количества клеток.
Стенка тела включает тонкую кутикулу,
покрывающую синцитиальный эпидермис,
содержащий внутриклеточную кутикулу
и небольшое количество четко
расположенных и довольно крупных ядер. Положение
этих гигантских ядер—хороший
диагностический признак. Под эпидермисом
находится тонкий слой кольцевых и продольных
мышц, окружающих псевдоцель. В псевдо-
126
Глава 4
Хоботок
Крючок
Ретрактор ховотка
Лемниск
Лигамент
Семенник
-Гигантское ядро
Цементная железа
Семенной пузырек
Клоака
Рис. 4.38. Анатомия типичного скребня на
примере самца рода Neoechinorhyncus
цель вдаются две вертикальные складки
тканей (лемниски); в них от эпидермиса
ведут заполненные жидкостью каналы.
Энтодерма редуцирована до лигамента, вдоль
которого подвешены органы размножения.
Как и у многих паразитов, у взрослых
скребней наблюдается сильное упрощение
строения с утратой многих систем органов,
характерных для свободноживущих
животных, и гипертрофия репродуктивного
аппарата. Скребни раздельнополы, что
необычно для эндопаразитов (но см. разд. 4.4),
поэтому внутреннее оплодотворение
обеспечивается сложным поведением при
спаривании. Сперма вводится в половой тракт
самки, который после копуляции
закупоривается с помощью цементных желез.
Оплодотворение происходит в псевдоцеле.
Рис. 4.40. Жизненный цикл Moniliformis—
скребня, окончательный хозяин которого—
млекопитающее (по Noble, Noble, 1976)
Скорлупа
Рис. 4.39. Стадии жизненного цикла скребней:
А—яйцо с заключенной в нем личинкой; Б—
вылупившаяся инвазионная стадия акантелла
из личинки жука
Молодая особь выходит из цисты взрослый червь
Акантор прОБодевает
стенку кишечника
Черви
127
У многих специализированных
паразитов строение личинки позволяет судить
о филогенетическом родстве взрослых
особей, однако у скребней это не так. Их
личинка также высокоспециализирована и сходна
с взрослыми животными. После
внутреннего оплодотворения ранние личиночные
стадии протекают в покрытом прочной
скорлупой «яйце» (рис. 4.39,А). Оно выводится
из хозяина с фекальными массами и, чтобы
происходило дальнейшее развитие,
должно быть съедено
насекомым—промежуточным хозяином. В нем личинка
вылупляется и проникает в гемоцель, где
превращается в следующую личиночную
стадию — акантеллу (рис. 4.39,Б). Такая
личинка может инцистироваться в насекомом,
и после того как его проглотит
позвоночное, развивается в этом окончательном
хозяине во взрослого скребня. Рис. 4.40
иллюстрирует жизненный цикл Moniliformis—
паразита мышей, крыс, кошек и собак.
4.10.3. Классификация
Известно 1000 видов, распределяемых
по трем отрядам единственного класса. Два
отряда составляют паразиты пресноводных
рыб, а третий—наземных четвероногих.
4.11. Тип Sipuncula
(сипункулиды)
4.11.1. Этимология
От лат. sipunculus—маленькая дудка.
4.11.2. Диагностические
и специфические признаки
(рис. 4.41 и 4.42)
1. Билатерально симметричные,
червеобразные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Мускулистая U-образная кишка со
ртом и анусом, последний расположен дор-
сально на передней части тела.
4. Тело подразделяется на переднюю
часть—интроверт, заканчивающийся ртом,—
и более толстое туловище сзади.
5. Тело не сегментировано.
Рот—&>
Интроверт—V \
Рис. 4.41. Внешний вид типичной сипункулиды
Мозг
Нервное кольцо
Вентральный
нервный ствол
Ретракторы
-Задняя кишка и анус
Кишечник
Щупальца
Щупальцевый
мешок
Пищевод
Рис. 4.42. А. Внутренее строение сипункулиды;
левая сторона тела удалена. Б. Детали
строения гидравлической системы щупалец и
интроверта
6. Полость тела схизоцель, но без
внутренних септ.
7. Наружный эпителий покрыт
кутикулой, без волосков и щетинок.
128
Глава 4
8. Кровеносной или
дифференцированной дыхательной систем нет.
9. Нервная система с передним мозгом,
околопищеводным нервным кольцом и
вентральным нервным стволом без ганглиев.
10. Единственный нефридий или пара не-
фридиев.
11. Развитие со спиральным дроблением
и обычно трохофорной личинкой, которая
у некоторых видов сменяется океанической
«пелагосферой»—личинкой, характерной
только для данного типа.
Тело четко разделено на два отдела:
передний, интроверт, втягивается в
находящееся сзади туловище мощными мышцами-
ретракторами (см. рис. 4.42) и
выворачивается за счет гидравлического давления,
создаваемого туловищной мускулатурой.
Локальное расслабление кольцевых мышц
может вызывать расширение тела,
закрепляющее животное в субстрате при
выворачивании интроверта.
Сипункулиды в
целом—неизбирательные детритофаги, захватывающие
пищу с помощью щупалец, окружающих рот
на вершине интроверта. Щупальца
управляются гидравлической системой,
изолированной от общей полости тела. Из каждого
из них пара протоков ведет в кольцевой
канал, от которого отходят один или два
мешочка, расположенных параллельно оси
интроверта. Эти дорсальный и вентральный
компенсационные мешочки могут
сжиматься и служат гидравлическими
резервуарами для системы щупальцевых каналов
(рис. 4.42, Б).
Многие виды живут во временных
норках, прорытых в илистых субстратах, но
некоторые заселяют пустые раковины
моллюсков или трубки червей. Сверлящие формы
проделывают ходы в известковых
коралловых отложениях. Разнообразие форм
иллюстрирует рис. 4.43.
Нервная система сипункулид напоминает
известную для аннелид и эхиурид: передний
ганглий (мозг) в интроверте,
околопищеводное нервное кольцо и вентральный
нервный ствол (однако без признаков
сегментарных ганглиев).
Обширная гидравлическая полость тела
служит также для накопления гаметоцитов,
выпадающих в нее из просто устроенных
Рис. 4.43. Разнообразные формы тела
сипункулид
гонад на ранних стадиях дифференцировки
и хранящихся там до тех пор, пока
количество зрелых гаметоцитов не станет
достаточно большим. Затем гаметоциты через
нефридии выводятся наружу. Эта схема ви-
I
Рис. 4.44. Способы эмбрионального развитая
сипункулид: (I) через трохофорную личинку;
(II) через непитающуюся трохофору и
характерную для данного типа пелагосферу; (III)
через крупную планктотрофную пелагосферу,
характерную для открытого океана; (IV) через
непитающуюся, богатую желтком личинку
с подавлением пелагических личиночных
стадий (по Rice, 1985 с изменениями)
Черви
129
доизменена у крошечной формы Golfingia
minuta—протандрического гермафродита,
производящего крупные яйца,
развивающиеся без оплодотворения. Это—
следствие мелких размеров животного, а
в целом способ размножения сипункулид
характерен для всех крупных морских
беспозвоночных.
Из яйца у большинства сипункулид
выходит пелагическая личинка, близкая к
личинке некоторых аннелид. У некоторых
видов эта трохофора прямо развивается в
молодую особь, но чаще претерпевает
метаморфоз до следующей личиночной
стадии— пелагосферы, которая может быть
и бентосной, хотя обычно пелагическая.
Четыре способа развития сипункулид
проиллюстрированы рис. 4.44.
4.11.3. Классификация
Все 250 известных видов можно
объединить в один класс и отряд.
4.12. Тип Echiura (эхиуриды)
4.12.1. Этимология
От греч. echis — змея; ига — хвост.
4.12.2. Диагностические
и специфические признаки
1. Билатерально симметричные,
червеобразные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Мускулистая кишка с передним и
задним отверстиями.
4. Тело с крупным вытягивающимся
и сократимым передним выростом, или
«хоботком», с латеральными складками,
ведущими ко рту.
5. Единственная неразделенная полость
тела—схизоцель—расположена между
мышцами стенки тела и кишкой.
6. Тело несегментировано.
7. Единственная пара щетинок
расположена спереди вентрально, иногда в другом
месте.
8. Замкнутая кровеносная система с
дорсальным и вентральным сосудами; в одной
группе сосудистая система незамкнутая.
9. Нервная система с околопищеводным
кольцом и вентральным нервным стволом
без выраженных ганглиев.
10. Выделительная система включает до
400 неметамерно расположенных нефри-
диев.
11. Развитие со спиральным дроблением
и трохофороподобной личинкой.
Эхиуриды—несегментированные цело-
мические черви, родственные кольчецам
(разд. 4.14), к которым их прежде и
относили. Наиболее их характерная черта—
хоботок, где находится передняя часть
нервной системы; возможно, он гомологичен
простомиуму аннелид (рис. 4.45). Парные
вентральные щетинки или крючья на
переднем отделе тела — также черта,
напоминающая аннелид, как и нефридии с
ресничной воронкой, расположение мускульных
слоев в стенке тела и строение
пищеварительной системы. Тем не менее эхиуриды
отличаются от кольчецов полным
отсутствием сегментации или метамерии. Можно
считать их близкими к аннелидной линии
филогенеза. В самом деле,
онтогенетические данные говорят о возможной
метамерии на ранних стадиях эволюции этой
группы.
Все эхиуриды—морские животные,
ведущие сидячий образ жизни в мягких
осадках. Большинство питается детритом и
обитает в постоянных норах; при этом хоботок
служит инструментом неизбирательного
сбора пищи; вдоль его желобка она
направляется ко рту (см. рис. 9.6). Разнообразие
форм тела иллюстрирует рис. 4.46. Для
рода Urechis характерен другой способ
питания. Эти эхиуриды живут в глубоких U-
образных норках, через которые они
прогоняют воду за счет волновых
перистальтических сокращений мышц.
Биология размножения эхиурид—
характерная для крупных целомических
червей, обитающих в морской среде.
Животные раздельнополы, и целом служит
полостью, в которой гаметоциты развиваются
перед массовым выходом через нефридии
во время «нереста».
Один из родов эхиурид отличается
специализированным типом полового
размножения, который обсуждается в разд.
14.2.3.3. Самцы здесь карликовые и живут
9—1314
130
Глава 4
Желовок
Щетинка
Туловище
Вентральный
нервный
ствол
«Хоботок
Кишечный сосуд
Передняя кишка
Нефридий
Вентральный
нервный ствол
Вентральный
кровеносный сосуд
Средняя кишка
Анальный мешок
Гонада
Прямая кишка
Рис. 4.45. Внешнее и
внутреннее строение эхиурид:
А — вид с вентральной
стороны Echiurus; Б — вид
в разрезе с дорсальной
стороны
на хоботке самок. Наличие карликовых
самцов ранее считалось чертой,
характерной только для отряда Bonellida, но
недавно этот феномен был обнаружен и у других
эхиурид; это, возможно, свидетельствует
Рис. 4.46. Разнообразные формы тела эхиурид
о том, что он возникал в пределах данного
типа неоднократно.
4.12.3. Классификация
Описано 150 видов, распределяемых по
трем отрядам в зависимости от
расположения мускульных слоев в стенке тела,
количества нефридиев, замкнутого или
незамкнутого характера кровеносной системы.
У представителей одного из отрядов, Хе-
nopneusta, задняя часть кишки
функционирует как дыхательный орган.
4.13. Тип Pogonophora
4.13.1. Этимология
От греч. pogon — борода; phoros—
носитель.
4.13.2. Диагностические
и специфические признаки
(рис. 4.47)
(Следует отметить, что интерпретация
различных анатомических черт погонофор
все еще не устоялась, как и их гомологиза-
Черви
131
Уздечка
Рис. 4.47. Сильно
схематизированный вид с дорсальной
стороны обобщенных
представителей двух классов
погонофор, показывающий основные
отделы тела и его полости (по
Southward. 1980; Jones, 1985)
Жавры
Головная лопасп
'етанефридии
■Железистая область
Гонады
-Туловище
Полости тела-
■Телосома
Щетинки
Овтуракулум
Pervlata
Obturata
ция, в частности, структур недавно
открытых вестиментифер и других
представителей этого типа. Например, до сих пор идут
споры о том, что считать вентральной,
а что—дорсальной поверхностью.)
1. Билатерально симметричные,
удлиненные, метамерно сегментированные
черви (0,5 мм —3 см в диаметре, 5 см —
3 м длиной), постоянно обитающие в
хитиновых или белковых трубках, внутри
которых могут двигаться.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Рот и кишка отсутствуют на всех
стадиях развития.
4. Одиночные и/или парные полости
тела неясного происхождения (хотя обычно
их и считают схизоцелями, перитонеальной
выстилки нет).
5. Тело состоит из четырех отделов:
передней «головной лопасти», несущей 1 —
более 1000 щупальневидных «жабр» с
гидростатическим скелетом; короткого
«железистого отдела» с парными полостями,
иногда в значительной степени
заполненными мускульной тканью; крайне вытянутого
«туловища» с одной парой крупных
гидростатических полостей и часто с различными
клейкими или секреторными папиллами;
короткой прикрепительной части—тело-
сомы, включающей до 30 сегментов (с
одиночной или парной полостью тела в
каждом), которые отпочковываются от
терминальной зоны роста.
6. Стенка тела с кутикулой,
эпидермисом, слоями кольцевых и продольных
мышц; телосома и (у некоторых видов)
туловище со щетинками; некоторые участки
тела с ресничными полосками.
7. Кровеносная система замкнутая,
с сердцем, окруженным перикардиальной
полостью, в головной лопасти.
8. Пара выделительных органов,
напоминающих метанефридии, в головной
лопасти.
9. Органами дыхания считаются жабры,
которые, возможно, участвуют и в питании.
10. Эпидермальная нервная система из
переднего кольца или скопления нейронов
и обычно единственного продольного
ствола без ганглиев (его обычно считают
вентральным); в некоторых частях тела может
быть несколько стволов.
11. Раздельнополые животные с парой
9*
132
Глава 4
Кутикула
Кровеносны!
сосуды жавры
Кровеносные
сосуды пиннулы
Полость тела
Эпидермис
Реснички
Пиннула
Рис. 4.48. Поперечный срез жабры Perviata,
демонстрирующий две пиннулы и расположение
ресничек (по Ivanov, 1963)
удлиненных гонад в туловище; мужские
гонады производят сперматофоры.
12. Оплодотворение считается
наружным, развитие непрямое, яйца и личинки не
покидают материнской трубки.
13. Морские, обычно глубоководные
(100—4000 м) формы.
Хотя впервые эти обитатели морских
глубин обнаружены в 1900 г., известно о них
мало. Например, только в 1964 г. получен
целый экземпляр погонофоры (ранее мета-
мерная телосома была неизвестна), а до
этого времени их обычно считали
родственными лофофоровым и вторичноротым из-
за как будто олигомерного строения тела,
состоящего из трех отделов. Обнаружение
сегментированной, несущей щетинки
терминальной части и первых вестиментифер
в 1969 г. привело к изменению прежних
представлений и сближению их с
кольчецами. В самом деле, некоторые авторы
склоняются к тому, чтобы поместить один или
оба класса погонофор в тип аннелид, но, как
-Пиннулы
Жавры
Головная лопасть
Уздечка
Железистая
Область
Реснички
Непарные
папиллы
Туловище
А
Постаннулярные
Телосома^
Рис. 4.49. Погонофоры класса Perviata: A—схематическое изображение внешнего вида типичной
первиаты (сильно укорочено), показывающее ее основные черты (по George, Southward, 1973);
у живой погонофоры тело ориентировано прямо и вертикально; Б—Г—передние концы трех
первиат с разным количеством и расположением жабр (по Ivanov, 1963)
Черви
133
отмечено выше, этот и многие другие
вопросы еще ждут своего разрешения.
Погоно'форы обитают в прямых, плотно
их облегающих трубках, образуемых
железистым отделом и утолшаемых секретами
туловища. Внутри них они удерживаются
с помощью: а) туловищных и/или телосо-
мальных щетинок; б) некоторыми
туловищными папиллами; в) валиками или
крыльями железистого отдела. Жабры выставлены
из трубки в окружающую воду и сильно
варьируют по количеству и расположению.
Часто их основания слиты вместе или
отходят от языковидной или гребенчатой
структуры. У представителей самого обширного
класса они расположены так, вероятно, для
того, чтобы образовать центральную
трубчатую полость; у видов с единственным
толстым «щупальцем» тот же эффект
достигается за счет спирального скручивания
последнего (рис. 4.49,В). В эту полость
вдаются парные ряды «пиннул» —
удлиненных выростов эпидермальных
клеток, сбоку от которых находятся длинные
эпидермальные реснички (рис. 4.48).
Функцией этих ресничек, вероятнее всего,
является создание нисходящего тока воды в
полости, образованной жабрами, однако роль
пцннул неясна.
Отсутствие кишечника породило
множество спекуляций относительно способа
питания погонофор. Наружное переваривание
пищи маловероятно, поскольку не
обнаружено соответствующих секретирующих
ферменты клеток. Всасывание
растворенных органических веществ возможно, но
наличие в среде достаточных их концентраций
проблематично. Не исключено, что мелкие
частицы органики, приносимые током воды
в межжаберную полость, могут
улавливаться пиннулами и поглощаться
посредством пиноцитоза; известно, что эпидермис
по крайней мере нескольких видов несет
микроворсинки, сходные с присутствующими
на эпителии средней кишки других
животных. К настоящему времени доподлинно
Рис. 4.49 (продолжение)
134 Глава 4
известно только то, что виды, живущие
вблизи подводных гидротермальных фума-
рол или участков просачивания холодных
вод, удовлетворяют свои потребности в
основном (если не целиком) за счет
внутриклеточных хемоавтотрофных бактерий, ме-
таболизирующих восстановленные
соединения серы и метан, поступающие из жерл,
причем аналогичные бактерии
присутствуют у изученных видов погонофор
класса Perviata.
4.13.3. Классификация
Сто видов погонофор делят на два клас-
Класс
Perviata
Obturata ■
Отряд
.Athecanephna
.Thccancphria
-Vestimentifera
4.13.3.1. Класс Perviata
Эти типичные погонофоры
сравнительно мелкие (диаметр менее 3 мм, длина
менее 85 см); они живут в трубках,
заякоренных в мягких осадках. У них немного (1—
250) удлиненных жабр и выражен косой
валик— «уздечка»; окружающий железистый
отдел (рис. 4.49). Возможно, он удерживает
переднюю часть тела в устье трубки.
Описано 95 видов, разделяемых на два отряда.
4.13.3.2. Класс Obturata
Единственный отряд Vestimentifera (рис.
4.50), составляющий этот класс, включает
червей гораздо больших размеров (1—3 см
в диаметре, длина некоторых особей более
2 м) с более короткими и более
многочисленными жабрами (свыше 1000),
расположенными на «обтуракулуме» — части тела,
прикрывающей отверстие трубки после
того, как животное в нее втягивается. В
отличие от первиат у вестиментифер нет
щетинок на туловище и уздечки, а последняя
заменена двумя крупными крыльями,
которые сходятся вдоль средней дорсальной
линии и тянутся вперед над основанием обту-
ракулума.
По мере исследования новых зон проса-
Жавры
Реснички
Овтуракулум
-Железный
отдел
-Туловище
-Телосома
Рис. 4.50. Схематическое изображение
внешнего вида типичной вестиментиферы (по
Southward, 1982)
чивания и гидротермальных жерл число
открываемых видов вестиментифер
возрастает почти экспоненциально. О находке
первого вида было объявлено в 1969 г.,
второго—в 1975 г., третьего — в 1981 г., еще
шесть видов описаны в 1986, вероятно,
около дюжины новых видов описываются в
настоящее время. Хотя в одной из последних
публикаций таксономический ранг обтурат
поднят до уровня самостоятельного типа
с двумя классами и тремя отрядами ввиду
Черви
135
того, что сведения и суждения об этой
группе постоянно меняются и будут меняться
в дальнейшем по мере получения нового
материала, мы предпочитаем сохранить
здесь традиционную систему из одного
класса и отряда, пока классификация обту-
рат как-то стабилизируется.
4.14. Тип Annelida
(кольчатые черви, кольчецы)
4.14.1. Этимология
От лат. annellus или annelus —
уменьшительное от anulus—кольцо.
4.14.2. Диагностические
и специфические признаки
1. Билатерально симметричные,
червеобразные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и органами.
3. Мускулистый пищеварительный
тракт со ртом и анусом.
4. Тело сегментировано (сегментация
может быть внешне незаметна, но всегда
видна по нервной системе).
5. Имеются пресегментный простомиум
с нервным ганглием и постсегментный пи-
гидиум.
6. Полость тела представляет собой ряд
схизоцелей, нечетко выражена у форм
с передней и задней присосками.
7. Полость тела часто разделена
поперечными септами, но они часто неразвиты
или плохо различимы в нескольких или во
всех сегментах.
8. Наружный эпителий покрыт
кутикулой и несет эпидермальные щетинки
(единичные или в пучках), за исключением
экземпляров с передней и задней
присосками.
9. Стенка тела мускулистая, часто со
сплошными слоями кольцевых мышц и
четырьмя полосами продольных мышц.
10. Кровеносная система замкнутая.
11. Нервная система с пресегментным
надпищеводным ганглием,
околопищеводным кольцом и вентральным нервным
стволом с сегментарными ганглиями.
12. Сегментарные выводные протоки
мезодермального и эктодермального
происхождения могут быть объединены,
ограничены одним или несколькими сегментами
и/или частично редуцированы.
13. В разной степени выражена цефали-
зация.
14. Развитие со спиральным
дроблением, но возможны его модификации и га-
струляция путем эпиболии у форм с
богатыми желтком яйцами.
15. У морских форм планктонное
развитие со свободноживущей трохофорной
личинкой, но эта стадия иногда
инкапсулируется; у пресноводных и наземных форм -
внутри яиц, покрытых прочными
оболочками.
В структуре аннелид сочетаются
гидростатическая и функциональная особенности
целомического уровня организации с
сегментированным строением тела. В самом
примитивном случае сегментация
отражается на эктодерме и мезодерме и включает
посегментное разделение целома септами.
У многих аннелид тем не менее такой план
строения исчез или модифицирован, а септы
или целом вторично утрачены.
Сегментация (рис. 4.51, см. также
рис. 4.54) возникает в онтогенезе из двух
парных мезодермальных зон роста и
соответствующей эктодермальной полоски
перед пигидиумом, т. е. задним отделом,
через который энтодерма сообщается с
внешней средой. Новые сегменты образуются
спереди от пигидиума так, что последний
сформировавшийся сегмент всегда
оказывается задним (рис. 4.52, А). У некоторых
полихет первые три сегмента образуются
очень рано и одновременно и могут
специализироваться для жизни в планктоне, но
последующие возникают постепенно один за
другим (рис. 4,52,2?). Их число может
увеличиваться в течение всей жизни, хотя
обычно достигает определенного предела.
У взрослых полихет и олигохет
образование новых сегментов можно индуцировать
поперечным перерезанием, причем утрата
последних сегментов приводит к
формированию нового пигидиума и препигидиаль-
ной зоны роста (рис. 4.53) (см. разд. 15.5).
В наиболее общих чертах тело аннелид
можно представить себе состоящим из пре-
сегментного простомиума с надпищевод-
136
Глава 4
Тело из множества щетинконосных сегментов
Пигидиум
-Перистомиум
-Рот
Простомиум
Недавно образовавшиеся
сегменты Зона нарастания
(сегментная Бластема)
Ресничный поясок
(прототрох)
Зона Мезодерма
нарастания <
сегментов |эктодерма
А
Рис. 4.51. Основные отделы
тела аннелид на примере полихе-
ты Nereis
Рис. 4.52. Образование
сегментов в ходе раннего
эмбрионального развития. А —ресничная
посттрохофорная личинка; Б—
метаморфозировавшая
личинка
ным ганглием, ряда более или менее
идентичных структурных единиц—сегментов,
и задней постсегментной части —
пигидиума, в передней вентральной
области которого находится сегментная
бластема, или зона роста (рис. 4.54). Основные
черты такой сегментации сводятся к
следующему:
1) эктодерма: сегментарное
расположение щетинок; нервная система с
сегментарными ганглиями и связанными с ними
нервами; эктодермальные нефридии;
2) мезодерма: сегментация
мускулатуры и кровеносной системы, связанная с ще-
тинковыми мешками и параподиями;
разделение септами целома с образованием
изолированных полостей тела; сегментарное
расположение герминативных • эпителиев
и связанных с ними парных целомодуктов.
У предков аннелид, вероятно, были
сплошные септы и полные наборы эктодер-
мальных протонефридиев и мезодермаль-
1 Термин герминативный эпителий (germinal
epithelium) в отечественной литературе
встречается очень редко; им обозначается часть перито-
ниального эпителия, производящая клетки -
предшественницы половых продуктов.— Прим.
пер.
ных целомодуктов. Такая организация не
сохранилась ни у одной из ныне живущих
форм. Протонефридии с группой клеток
ресничного пламени еще встречаются в
нескольких семействах полихет, но больше
распространены нефридии с открытой рес:
ничной воронкой—нефростомом.
Эмбриональное происхождение нефридиев и
целомодуктов разное (разд. 1.3.1), у олигохет
они разделены. У полихет (морская в целом
Зона нарастания
(властема)
Пигидиум
Регенерирующие сегменты
Исходные сегменты
Рис. 4.53. Регенерация у Nereis. Утрата
сегментов вызывает образование новой зоны их
поочередного нарастания; задний сегмент—
самый молодой
Черви
137
Рис. 4.54. Схематическое
изображение компонентов
сегментированного тела
гипотетической протоаннели-
ды. Ныне живущие
кольчецы сохранили некоторые, но
не обязательно все эти
черты. А. Продольные блоки
мышц и кишка не
сегментированы. Полость тела—
настоящий целом с перито-
неальной выстилкой. Б.
Сегментация вентральной
нервной системы, лучше
всего сохранившаяся у
современных аннелид предко-
вая черта (см. раздел
о пиявках ниже):
сегментарные ганглии и нервы. В.
Разделение целома
поперечными септами, сохранившееся
у большинства олигохет
и некоторых полихет; может
сильно модифицироваться;
утрачено у пиявок. Г.
Сегментация эктодермы и
мезодермы, связанная с
развитием параподий или се-
гментарно расположенных
щетинок. Хорошо выражена
у полихет, меньше у
олигохет, подавлена у пиявок. Д.
Сегментация половых и
выделительных протоков и
герминативного эпителия.
Предполагается, что у про-
тоаннелид были полные
наборы мезодермальных це-
ломодуктов (для вывода
гамет) и эктодермальных не-
фридиев, а также пара
герминативных зпителиев
в каждом отделе целома.
Этого не обнаружено ни
у одной из современных
аннелид; часты сильные
отклонения от предложенной
схемы. £ Сегментация
кровеносно-сосудистой
системы
группа) нефридий и целомодукт обычно
объединены в сложную структуру—
нефромиксий. В тех семействах, где
сплошные септы отсутствуют и целомические
полости сообщаются между собой, число
герминативных эпителиев и сегментарных
выводных протоков сильно сокращено.
^?V^r-AV
^
Ф^Р^)^¥
Сегментарные нервы
Вентральный
нервный ствол
Сегментарные ганглии
Межсегментная септа
Ацикула
Внутренние мышцы
Параподия
Внешние мышцы
тверстие целомодукта =
половая воронка и
половой проток
Нефридий
Гонада
Дорсальный
кровеносный сосуд
■Сегментарные
кровеносные сосуды
Вентральный
кровеносный сосуд
4.14.3. Классификация
Известно по крайней мере 75000 видов,
которых делят на три крупные группы: Ро-
lychaeta, Oligochaeta и Hirudinea—и две
мелкие. Hirudinea рано дивергировали от
эволюционной линии олигохет, и две эти
138
Глава 4
Класс
Подкласс
Polychaeta.
Отряд
Orbiniida
Ctenodrilida
Psammodnlida
Cossurida
Spionida
Questida
Capitcllida
Ophcluda
Phyllodocida
Amphinomida
Spinthcnda
F.umcida
Stcrnaspida
Owemida
Flabelligenda
Pocobnda
Terebellida
Sabellida
Nertllida
Dinophilida
Polygordiida
Protodrilida
Mvzostomida
vAeolosomata
-Acolosomatida
'Clitellata
„Oligochaeta-
Branchiobdella
Hirudinea
группы вместе с малочисленным
подклассом Branchiobdella образуют класс Clitella-
ta.
Рис. 4.55. Различные формы
параподий полихет. А
Основной двуветвистыи тип (как
у Nereis) с деталями
морфологии некоторых щетинок. Б.
Примеры параподий седентар-
ных семейств: I — Spionidae;
II—Arenicolidae; III —
Sabellidae.fi. Примеры
параподий и щетинок эррантных
семейств: I — Polynoidae; II —
Phyllodocidae, ill — Eunicidae
. Lumbriculida
-Moniligastrida
. Haplotaxida
• Branchiobdellida
- Acanthobdellida
• Rhynchobdcllida
- Arhvnchobdcllida
Черви
139
4.14.3.1. Класс Polychaeta
(многощетинковые кольчецы)
Обитающие в основном в море полихе-
ты замечательны своим морфологическим
и анатомическим разнообразием. Они
получили свое название из-за многочисленных
щетинок (chaeta), обычно расположенных
двумя пучками на двуветвистых парапо-
диях, попарно отходящих от каждого
сегмента (рис. 4.55). Развитие у многих
непрямое (рис. 4.56), хотя есть и исключения.
Примерно половину видов и семейств
полихет можно для удобства объединить
в группу эррантных («бродячих») форм.
В основном сюда относятся два крупных
и четко очерченных отряда — Phyllodocida
и Eunicida. У филлодоцид имеется осевая
Рис. 4.56. Примеры
планктонных стадий развития полихет:
ASabellaria; Б—Spio; В—
Nereis; Г—Owenia
140
Глава 4
Челюсти
Парагнаты
Простомиук
Глотка
(втянута)
Рис. 4.57. Хоботок у отряда Phyllodocida — мускулистая глотка, которая выворачивается за счет
давления в целоме и втягивается при сокращении мышц-ретракторов (А—Г). Она часто, какуЛ/е-
reis, вооружена белковыми челюстями
Передний конец
Передняя -
поверхность
Рукоятка
Передний конец
(клещи)
Мандивулы
Максиллы(М)
Рис. 4.58. Челюстной аппарат
у отряда Eunicida. А.
Изолированный челюстной комплекс.
Б. I Челюсти втянуты в
мускулистую вентральную глотку. II.
Вывернутые челюсти (по Olive,
1980)
Черви
141
Рис. 4.59. Разнообразие форм
тела эррантных полихет; I и II —
вид животного с дорсальной
и вентральной сторон
соответственно
глотка-хоботок (передняя часть
пищеварительного канала), которая может
гидравлически выворачиваться и втягиваться мыш-
цами-ретракторами. Часто она вооружена
небольшим числом челюстей из
отвердевшего белка с высоким содержанием ионов
тяжелых металлов, но никогда не обы-
звествленных (рис. 4.57).
Эвнициды напоминают филлодоцид
внешне, но у них не осевая глотка-хоботок,
а выворачиваемая буккальная масса',
вооруженная сложным набором обызвествлен-
ных челюстей, парой мандибул,
напоминающих по форме долото, и несколькими
парами максилл (рис. 4.58). Разнообразие
форм эррантных полихет
проиллюстрировано рис. 4.59. У большинства хорошо
развита голова с разнообразными сенсорными
органами и сложным мозгом. Многие—
хищники, но есть и детритофаги, фильтра-
торы и всеядные формы. Хорошо развиты
параподии с вентральной и дорсальной
ветвями, укрепленными жестким белковым
стержнем—ацикулой.
Для некоторых эррантных полихет
характерна эпитокия, т.е. своего рода
«роение», связанное с половым размножением.
Это часто сочетается со сложным метамор-
1 Вместо термина «буккальная масса» (buccal
mass) в отечественной литературе принято
употреблять термин «вентральная мускулистая
глотка».— Прим. пер.
фозом взрослых животных. Эпитокия
может быть двух типов (рис. 4.60 и 4.61). При
эпигамии весь червь трансформируется
в «роевую» (эпитокную) форму, в то время
как при схизогамии (столонизации) задние
сегменты половозрелого животного
отделяются, становясь мигрирующими
несущими гаметы столонами;
Остальных полихет часто искусственно
и по разным принципам объединяют в
группы седентарных («сидячих») форм и «ар-
хианнелид»; внутри них родственные связи
не прослеживаются. Большинство биологов
предпочитает оперировать четко
очерченными семействами, а не отрядами.
У седентарных полихет обычно
наблюдаются регионарные различия в строении
сегментов и четко выраженные
торакальный («грудной») и абдоминальный
(«брюшной») отделы (рис. 4.62). Все они
микрофаги, питающиеся осадком или взвесью и не
способные передвигаться за счет извивов
тела или с помощью параподии, что
характерно для эррантных видов (см. разд. 10.5).
Их разнообразие обусловлено
значительными различиями в образе жизни.
Некоторые живут в норах, другие—в трубках из
секретируемого пергаментоподобного
материала или карбоната кальция, иногда—
из песчинок. В нескольких семействах
наблюдается функциональная конвергенция
с олигохетами. Такие черви роются
в мягком субстрате, либо заглатывая его,
142
Глава 4
Рис. 4.60. Эпитокия: эпигамия. Половозрелые особи претерпевают метаморфоз, роятся и
погибают: А — немодифицированная атокная форма Nereis; Б— метаморфозировавшая эпитокная
форма Nereis (Heterohereis); В—схема жизненного цикла
Рис. 4.61. Эпитокия: схизогамия: А — образование единственного столона за счет модификации
задних сегментов; Б—образование множества столонов путем терминального почкования;
терминальный столон—самый старший; В—схема жизненного цикла
Черви
143
Рис. 4.62. Разнообразие форм
тела седентарных полихет.
Мелкие рисунки показывают
характерный образ жизни (см.
также рис. 9.6)
Рис. 4.63. Разнообразие форм
и размеров тела семейств
полихет, ранее объединявшихся
в группу архианнелид. Сейчас
считается, что они вторично
упрощены из-за мелких
размеров
144
Глава 4
либо «облизывая» песчинки. У них
небольшой простомиум без выступающих
сенсорных органов, простые щетинки на
редуцированных параподиях, хорошо развитая
кольцевая мускулатура и сплошные септы.
Представители нескольких семейств
собирают мелкие частицы или органический
осадок и доставляют их ко рту мягкими
хватательными щупальцами (см. рис. 9.6),
которые часто отходят от простомиума.
Живущие в трубках черви некоторых
относительно продвинутых семейств имеют
венчик жестких простомиальных щупалец,
функционирующий как настоящий
фильтрационный аппарат (рис. 9.3). Другие
типы пищедобывательной специализации
обсуждаются в гл. 9 (см., например, рис. 9.5).
Архианнелиды (рис. 4.63) представляют
собой группу неродственных друг другу
крошечных полихет, которые независимо
приспособились к обитанию в интерстициа-
ли морских песков.
4.14.3.2. Класс Clitellata
(поясковые кольчецы)
Подкласс Oligochaeta
(малощетинковые кольчецы)
Олигохеты- в основном наземные
и пресноводные аннелиды; некоторые виды
вторично перешли к обитанию в морской
среде, особенно в эстуариях и интерсти-
циали.
При локомоции олигохеты используют
гидростатические свойства полностью се-
птированного целома и метамерной
организации. Таким образом, у них сохранился
план строения, близкий к приписываемому
протоаннелидам (см. рис. 4.53), однако сам
подкласс не является примитивным. Для
олигохет характерен синхронный
гермафродитизм, вынашивание небольшого
количества крупных, богатых желтком яиц
в защитной и питающей структуре—
коконе, секретируемом пояском, и сильно
редуцированное число гонад. Расположение
гениталий используют для разделения
олигохет на три отряда.
Функционально и экологически
существуют в принципе два типа олигохет: в
основном водные микродрилы и сухопутные
мегадрилы, или земляные черви.
Функциональная анатомия земляных
червей на удивление постоянна и может быть
проиллюстрирована на примере Lumbricus
terrestris (рис. 4.64,А).
Поперечный разрез (рис. 4.64,£)
демонстрирует следующие детали строения:
водонепроницаемая кутикула, смазываемая
выделениями бокаловидных секреторных
клеток эпидермиса; эпидермис; нервный слой;
сплошной слой кольцевой мускулатуры;
полосы продольных мышц; одиночные или
расположенные небольшими группами (как
у Lumbricus) щетинки; целом, разделенный
сплошными септами и выстланный перито-
неальным эпителием; мускулатура
кишечной стенки; энтодермальная выстилка
пищеварительного тракта.
План строения червя метамерный
(сегментарный), но передний отдел сильно
специализирован, особенно — строение
пищеварительной и половой систем
(рис. 4.64,А,Б; рис. 4.65). У земляных
червей сложное псевдокопуляционное
поведение, при котором спаривающиеся животные
покрываются общей слизью, а сперма
переносится наружно в семеприемники или
семенные мешки в области пояска (рис. 4.66).
Оплодотворение происходит внутри
кокона, причем после одного спаривания червь
может произвести много яйцевых коконов.
Олигохеты-микродрилы мельче
земляных червей. К ним относят несколько
семейств, обитающих в основном в пресных
водах, хотя Enchytraeidae—в основном
сухопутная группа. Некоторые живут даже
в морских глубинах. Форма тела здесь
более разнообразна, чем у земляных червей,
у некоторых имеются выступающие
волосовидные щетинки, напоминающие
щетинки полихет (рис. 4.67). Биология
размножения часто высокоспециализирована.
Распространен бесполый процесс в форме
спонтанного разделения (схизогенез); у
некоторых видов настоящее половое
воспроизведение неизвестно совсем.
Подкласс Hirudinea (пиявки)
Специфические черты пиявок
определяются развитием присосок, позволяющих
им плотно прикрепляться обоими своищ
концами. После этого тело функционирует
как единая гидростатическая система. Соот-
Черви
145
Простомиум Отверстие
семеприемника
Поясок Щетинки
Половые щетинки
Эпидермис + кутикула
Кольцевые мышцы
Продольные мышцы
Щетинки
Кишечник
Нефридий
Целом
Вентральный
нервный ствол
Рис. 4.64. А Внешнее строение земляного червя Lumbricus terrestris, вид с вентральной стороны.
Б. Схема поперечного среза этого же вида
Надпищеводный Кровеносная Семенники
Подпищеводный Нефридий Семенные Яичники
ганглий пузырьки
Рис. 4.65. Внутреннее строение передних сегментов Lumbricus terrestris
10—1314
146
Глава 4
А. Копуляция
Б. Откладка яиц
В. Оплодотворение
3
Г. Образование кокона
Рис. 4.66. Поведение Lumbricus
terrestris при спаривании. А.
Животные покидают свои
норки и образуют покрытую
слизью пару. Во время копуляции
происходит взаимный обмен
сперматозоидами, сперма
переходит из семенных
пузырьков в семеприемники, как
показано стрелками. Б. Зрелое
яйцо выходит из женского
полового протока и переносится
назад к пояску. В. Секретируе-
мый пояском кокон вокруг
небольшого количества яиц
движется вперед; яйца
оплодотворяются, когда он проходит
мимо отверстия семеприемника.
Г. Кокон с развивающимися
эмбрионами откладывается
в почву
Простомиум
Рот
Поясок
Задняя
присоска
Рис. 4.67. Разнообразие форм тела олигохет-
микродрил
Рис. 4.68. Внешнее строение пиявки Hurudo.
Вторичная кольчатость показана слева, а
настоящая сегментация — справа
Черви
147
Дорсальный кровеносный сосуд
Продольная мышц.
Кольцевая мышца
Косая мышца
Нефридий
Кутикула
Эпидермис
Семепровод
Семенник
Ботриоидальная ткань
Кишечник
Вентральный нервный ствол
Рис. 4.69. Поперечный срез Hi-
rudo. Целомическая полость
заполнена паренхиматозной
тканью. Пространство между
ее клетками может быть
организовано в хорошо
выраженные синусы.
ветственно сегментация целома, эктодермы
и мезодермы по большей части утрачена.
Функционально пиявки напоминают
плоских червей, но используют преимущества
целомических синусов (для создания
нагнетающего усилия), нервной системы с
сегментарными ганглиями (для координации
сложной локомоторной и поведенческой
активности) и замкнутой кровеносной
системы с дыхательными пигментами.
У всех пиявок ровно 33 сегментарных
ганглия. Передняя присоска образована
сегментами I—IV, а задняя —
XXV—XXXIII (рис. 4.68). Щетинки и
наружная сегментация не выражены, хотя тело
разделено вторичными кольцами, не
совпадающими с границами сегментов.
Поперечный разрез показывает, что пространство
между мезодермой и мускулистой стенкой
кишки заполнено рыхлой «паренхимопо-
Глотка
Мышца-расширитель
глотки
Пенис
Яичник
Влагалище
Семенник
Рис. 4.70. Внутренее строение
Hirudo, вид с дорсальной
стороны. Кишка показана
отдельно
Семепровод
Вентральный
нервный ствол
Сегментарный
ганглий
Слюнная железа
■Дорсальный сосуд
Слепой вырост кишки
(цекум)
ю*
148
Глава 4
Рис. 4.71. Разнообразие форм тела пиявок;
правый рисунок иллюстрирует характерную
способность этих червей сильно изменять
форму своего тела
добной» ботриоидальной тканью'
(рис. 4.69). Некоторые внутренние органы,
особенно герминативные эпителии и
нефридии, все же отражают предковую
метамерию (рис. 4.70).
Пиявки—гермафродиты с несколькими
парами семенников, парой яичников и
единственным половым отверстием.
Оплодотворение внутреннее, и при размножении
происходит перенос сперматозоидов между
партнерами, часто сопровождающийся
сложным копуляционным поведением. У
многих видов сложно устроенные
сперматозоиды вводятся «подкожно» и мигрируют
к яичникам между клетками животного-
реципиента. После спаривания взрослые
пиявки откладывают многочисленные
коконы, с одним или несколькими яйцами
каждый. Некоторые крупные виды живут по
нескольку лет, но большинство — только
один год, зимуя в эмбриональном
состоянии внутри кокона.
Разнообразие формы тела пиявок
иллюстрирует рис. 4.71. Все они плотоядные, но
не все наружные паразиты или
кровососущие формы. Классификация основана на
строении ротовых частей и способе
питания.
Acanthobdellida—отряд своеобразных
примитивных эктопаразитов лососевых
рыб. Передней присоски здесь нет,
несколько сегментов несут щетинки, целом
сегментарный. Таким образом, они представляют
собой связующее звено между уровнями
организации олигохет и пиявок.
= Подкласс Branchiobdella
К этому подклассу относятся
крошечные эктопаразиты пресноводных
ракообразных. Тело состоит из 15 или 16 сегментов,
из которых четыре первых слиты и
образуют цилиндрическую голову с присоской.
Эти малоизученные поясковые кольчецы,
похоже, представляют независимую
эволюционную линию, идущую от олигохетопо-
добного предка и во многих отношениях
параллельную линии пиявок (рис. 4.72).
4.14.3.3. Класс Aeolosomata
Этих очень мелких аннелид ранее
считали примитивными олигохетами, ,но сейчас
их эволюцию признают независимой от
линии поясковых. Некоторые крошечные
формы обитают в интерстициали пресных и
солоноватых водоемов. Гермафродиты;
яичник всего в одном сегменте, а семенники—
1 Термином «ботриоидальная ткань» здесь
обозначена паренхима, происходящая из целоми-
ческой выстилки.— Прим. пер.
Рис. 4.72. Branchiobdella.
Внешнее строение
типичного вида
Рис. 4.73.
Aeolosomata. Типичный
вид
Черви
149
в сегментах, расположенных спереди и
сзади от него. «Поясок» образован
вентральными железами и не гомологичен
дорсальному пояску Clitellata.
Типичный представитель класса
изображен на рис. 4.73. У него покрытый
ресничками простомиум, образующий основной
локомоторный орган, и довольно длинные
волосовидные щетинки. У Aeolosomata, как
и в семействах архианнелид, наблюдается
структурное упрощение, связанное с
мелкими размерами, а родственные связи с
другими аннелидами неизвестны. В класс входит
всего около 25 видов.
4.15. Дополнительная
литература
Bird A.F. 1971. The Structure of Nematodes.
Academic Press, New York [Nematoda].
Boaden P.J.S. 1985. Why is a gastrotrich? In:
Conway Morris S. el al. (Ed.) The Origins and
Relationships of Lower Invertebrates, pp. 248—260.
Clarendon Press, Oxford [Gastrotricha].
Croll N.A. 1976. The Organisation of Nematodes.
Academic Press, London [Nematoda].
Croll N. A., Mathews B.G. 1977. Biology of
Nematodes. Blackie, London [Nematoda].
Dales R.P. 1963. Annelids. Hutchinson, London
[Annelida].
D'Hondt J.-L. 1971. Gastrotricha. Oceanogr. mar.
Biol., Ann. Rev., 9, 141 192 [Gastrotricha].
DonnerJ. 1966. Rotifers (transl. Wright H.G.S.).
Warne, London [Rotifera].
Edwards С A., Lofty J. R. 1972. The Biology of
Earthworms. Chapman & Hall, London
[Annelida].
Gibson R. 1972. Nemerteans, Hutchinson, London
[Nemertea].
Hyman L.H. 1951. The Invertebrates (Vol. 2). Pla-
tyhelminthes and Rhynchocoela. McGraw-Hill,
New York [Nemertea].
Hyman L.H. 1951. The Invertebrates (Vol.3).
Acanthocephala, Aschelminthes and Entoprocta.
McGraw-Hill. New York [Gastrotricha.
Nematoda, Nematomorpha, Kinorhyncha, Priapula,
Rotifera. Acanthocephala].
Ivanov A. V. 1963. Pogonophora. Academic Press,
London.
Kaestner A. 1967. Invertebrate Zoology (Vol. 1).
Wiley, New York [Nemertea, Gastrotricha,
Nematoda, Nematomorpha, Kinorhyncha, Priapula,
Rotifera, Acanthocephala, Sipuncula, Echiura,
Annelida].
Kristensen R.M. 1983. Loricifera, a new phylum
with aschelminthes characters from the meiobent-
hos. Z. zool. Syst. Evolutionsforsch., 21, 163 -180
[Loricifera].
Mill P. 1978. Physiology of the Annelids. Academic
Press, London [Annelida].
Norrevang A. (Ed.) 1975. The Phylogeiiy and
Systematic Position of Pogonophora. Parey, Hamburg
[Pogonophora].
Rice M. E., Todorovic M. 1975. Proceedings of the
International Symposium on the Biology of
Sipuncula and Echiura. Smithsonian Inst., Washington
[Sipuncula, Echiura].
Sterrer W. \912. Systematics and evolution within
the Gnathostomulida. Syst. Zool., 21, 151—173
[Gnathostomula].
Sterrer W., Mainitz M., Reiger R.M. 1985.
Gnathostomulida: enigmatic as ever. In: Conway Morris
S. et al. (Ed.). The Origins and Relationships of
Lower Invertebrates, pp. 181 —199. Clarendon
Press, Oxford [Gnathostomula].
ГЛАВА %J
МОЛЛЮСКИ
Моллюски, или мягкотелые,— хорошо
очерченный и своеобразный тип, не
имеющий ни близкого сходства, ни
филогенетического родства с какой-либо из ныне
живущих групп, если не считать сохранения рада
признаков, указывающих на то, что их
предками были плоские черви. В гл. 2 они
и были описаны как плотные плоские черви
со спинными защитными щитками
(панцирем). Тем не менее большинство видов
этого типа очень далеко дивергировало от
формы и образа жизни плоских червей.
Действительно, представители в высшей
степени преуспевающего типа моллюсков
достигли такого же морфологического
разнообразия, что и у целого набора типов,
описанных в других главах, поэтому
заслуживают отдельной главы.
5.1. Тип Mollusca
5.1.1. Этимология
От лат. molluscus — мягкий (орех или
гриб).
5.1.2. Диагностические
и специфические признаки
(рис. 5.1)
1. Билатерально симметричные
животные.
2. Тело из более чем двух слоев клеток
в толщину, с тканями и органами.
3. Сквозной кишечник.
4. Полости тела нет, за исключением
кровеносных синусов.
5. Тело несегментировано и весьма
изменчиво по форме, но в основном
закрученное и часто коническое, обычно вытянутое
в дорсовентральном направлении, так что
образуется «внутренностный мешок». Как
правило, спереди располагается голова,
несущая глаза и сенсорные щупальца. В
большинстве случаев с вентральной стороны
имеется массивная с уплощенной подошвой
нога, а сзади — мантийная полость, но все
это бывает сильно модифицировано.
6. Защитная наружная дорсальная
раковина из белка (конхиолина), упрочненного
известковыми спикулами, или из 1—8
известковых пластин, секретируемых
эпидермисом спинной и боковых сторон тела
(мантией). Иногда раковина вторично
редуцирована, покрыта мягкими тканями,
утрачена вовсе или увеличена настолько, что
скрывает все тело.
7. Несущая зубы, хитиновая, языковид-
ная терка (радула), способная выдвигаться
из ротовой полости наружу; соскобленные
ею частицы попадают на слизистый тяж,
который втягивается в желудок с помощью г
стебелька, помещающегося в специальном I
влагалище (см. рис. 9.17 и разд. 9.2.5). |
8. Газообмен осуществляется одной или .
несколькими парами ктенидиальных жабр,
расположенных в мантийной полости
(иногда они редуцированы).
9. Кровеносная система незамкнута;
сердце окружено перикардиальной
полостью мезодермального происхождения,
сквозь которую проходит кишка.
10. Пара мешковидных «почек»,
открывающихся проксимально в перикардиаль-
ную полость и впадающих в мантийную
полость.
11. Нервная система с
околопищеводным нервным кольцом и двумя парами ган-
глионизированных продольных стволов.
Иногда ганглии сильно сближены.
12. Типична одна пара гонад с
протоками, впадающими в мантийную полость.
Моллюски
151
Рис. 5.1. Схематичный
продольный разрез
обобщенного моллюска
Раковина
Гонада—
Пищеварительный дивертикул
Стевелек
Сенсорное
щупальце
Радула
Продольные Нога
нервные стволы
Сердце в перикардиальной
/ полости
Почка
Анус
Мантийная
4 »^ Ктенидий
Дыхательное
течение воды
У примитивных форм половые продукты
выводятся через перикардиальную полость
и почки.
13. Дробление яиц спиральное.
14. Развитие непрямое, с личиночными
стадиями трохофоры и велигера (рис. 5.2).
Наблюдаются случаи вторичного прямого
развития.
Моллюски, насчитывающие почти
100 000 видов, вероятно, второй после
нематод (тысячи видов которых еще ожидают
своего открытия и описания) тип животных
по численности. Их преуспевание связано не
столько с какими-либо особыми
анатомическими или экологическими свойствами,
сколько с исключительной пластичностью
и адаптационными возможностями
основного плана строения. Перечисленные выше
признаки типа модифицируются и очень
широко варьируют в разных его классах.
Эту пластичность можно
проиллюстрировать, с одной стороны, изменчивостью
функций какой-либо одной структуры тела
моллюсков (например, раковина помимо
защиты может обеспечивать плавучесть,
быть роющим органом или пластинкой
внутреннего скелета), а с другой стороны,
множеством структур, способных
адаптироваться к выполнению какой-либо одной
функции (захватывать пищу могут несущие
реснички щупальца, пальпы, сильно
увеличенные жабры, руки, вооруженные
присосками, радулярные зубы и т.д.).
По форме тела моллюски варьируют от
цилиндрических, сильно вытянутых вдоль
переднезадней оси, обитающих в вырытых
ими ходах «червей» без ноги и раковины до
почти сферических, лишенных головы
животных, заключенных внутри крупной
двустворчатой раковины. При этом длина
видов составляет от 2 мм у планктонных и ин-
терстициальных форм до более 20 м у
гигантских кальмаров (включая руки). В
последнем случае это результат удлинения
дорсовентральной плоскости предков.
Моллюски занимают все основные типы
местообитаний и включают представителей
всех известных типов питания. К ним
относятся как наиболее, так и наименее
подвижные формы свободноживуших беспозво-
Рис. 5.2. Личинки
моллюсков: А—трохофора; Б —
велигер (по Нуглап, 1967)
152
Глава 5
Одонтофор
Радулярная
терка
Мышцы:
а - мышца-протрактор
радулы,
б- мышца-протрактор
одонтофора,
в - мышца-ретрактор
одонтофора,
г - мышца-ретрактор
радулы
Сокращение мышц "а" и
«Б" выдвигает одонтофор
и радулярную терку изо
рта (II)
Продолжение сокращения
мышцы "Б", расслабление
па" и сокращение "г"
обеспечивают движение терки
поверх одонтофора (III), a
сокращение «в» возвращает
одонтофор и терку назад
в ротовую полость (как I)
Рис. 5.3. Соскабливание
пищи радулой (по Russel—
Hunter, 1979 и др.
источникам)
ночных. В этот тип, вместе с самыми
«умными» беспозвоночными помещены
животные с почти не развитыми органами чувств
и мозгом. («Если бы Создатель
действительно щедро наделил свое лучшее создание
собственным обликом, креационист,
несомненно, сделал бы вывод, что Бог похож на
кальмара».— Diamond, 1985.)
Как говорилось в гл. 2, план строения
моллюсков в общем такой же, как у плоских
червей (разд. 3.6), но с двумя
существенными дополнениями (и несколькими другими,
вытекающими из них): радулой и
дорсальной раковиной. Радула (рис. 5.3)
помещается в заднем вентральном кармане ротовой
полости и включает хрящевой скелетный
элемент — одонтофор, на котором
закреплена подвижная радулярная терка с
поперечными рядами обращенных назад острых
зубов, одновременно образующих и
продольные ряды. Мышцы-протракторы могут
вызывать как выдвигание одонтофора
наружу из ротовой полости, так и некоторое
движение терки относительно ее базальной
опоры, а также поднимание отдельных
зубов. В результате зубы скребут по
поверхности, отделяя от нее пищевые частицы,
которые в процессе втягивания мышцами-
ретракторами одонтофора и радулы внутрь
попадают в ротовую полость. Хотя эти
хитиновые зубы могут минерализоваться SiO,
и Fe304, они сильно изнашиваются,
поэтому вся радула медленно смешается вперед
по одонтофору, вынося еще не
использовавшиеся зубы в «рабочую зону», в то время
как на проксимальном конце нарастает
новая терка. Следовательно, радула-
главным образом орган собирания,
соскабливания и сверления, но у ряда хищных
видов она модифицирована в аппарат для
ловли добычи за счет увеличения размера
отдельных зубов (с соответственным
уменьшением их числа в рядах) и иногда
соединения с зубами ядовитых желез.
Спинное защитное покрытие дорсовен-
трально уплощенных предковых
моллюсков было, возможно, белковой или
хитиновой кутикулой, укрепленной известковыми
«косточками» или чешуйками, секретируе-
мыми эпидермисом. У большинства их по-
Моллюски
153
томков содержание карбоната кальция
в этой структуре сильно увеличено; она
представлена одной или несколькими
крупными известковыми пластинами,
покрытыми тонким слоем конхиолинового перио-
стракума. В типичном случае кальцит или
арагонит откладывается различными
слоями (каждый — в органическом матриксе).
Различают наружный призматический слой
и внутренние перламутровые. Появившись,
раковина эволюционировала, причем
главное направление роста моллюсков
перестало быть переднезадней осью, как обычно
у червеобразных животных, и сменилось
дорсовентральным, так что раковина и
животное, заключенное у большинства
современных форм внутри нее, приняли более
или менее коническую форму, иногда
вытянутую, и при этом закрученную в плоскую
или винтовую спираль. Мантия, секрети-
рующая раковину, обычно
распространяется в виде складки поверх незащищенных
частей тела, поэтому раковина с помощью
мышц-ретракторов может надвигаться на
голову и вентральную локомоторную ногу,
обеспечивая безопасность животного.
Одновременно нога может прикрепляться
к субстрату, действуя как присоска.
Наличие спинного защитного покрова
имеет прямые последствия для газообмена
через поверхность тела. Развитию
раковины должно сопутствовать преобразование
специализированной непокрытой ею
области в жабру одновременно с эволюцией
системы циркуляции дыхательных газов по
телу (если таковая не сформировалась
ранее). Охватывание раковиной и мантией
тела моллюска особенно заметно сбоку и
(или) сзади, где мантийная полость
замкнута (рис. 5.4). В ней расположены одна или
более пар ктенидиальных жабр, каждая из
которых состоит из уплощенной
продольной оси, по обеим сторонам которой
находятся плоские треугольные лепестки (фила-
менты). Каждый из них поддерживается
мелким хитиновым тяжем, идущим вдоль
фронтальной поверхности. Реснички на
поверхности жабр гонят воду между
отдельными филаментами, через которые в
противотоке движется кровь (см. рис. 11.9).
Обычно вода входит в мантийную полость
по бокам тела и выбрасывается наружу дор-
Прямая кишка
Опорный
_к/ • ,,, ,, „ ХИТИНОВЫЙ
Приносящий Ц&- M[/\J VM1* стержень
веносный сосуд —'
Выносящий
кровеносный
сосуд
Рис. 5.4. Поперечный разрез мантийной
полости обобщенного моллюска; показаны ктени-
диальные жабры и направления входящего
водного тока, образующегося в результате
действия их ресничек
сально вдоль средней линии. Анус и почки
открываются в эту полость в зоне
выносящего течения.
Однако, как подчеркивалось выше,
природа и развитие радулы, раковины и ктени-
диев, как и большинство других
анатомических признаков моллюсков, значительно
варьируют в различных классах этого типа,
поэтому дальнейшая их характеристика
будет дана ниже по отдельности. Поскольку
именно моллюски среди всех
беспозвоночных составляют излюбленный объект ней-
робиологических исследований, сведения
о них приведены также в гл. 16.
5.1.3. Классификация
Тип моллюсков состоит из 8 классов,
два из которых —гастроподы (брюхоногие)
и бивальвии (двустворчатые) включают
более 98% их современных известных видов.
5.1.3.1. Класс Chaetodermomorpha
Хетодермоморфы— своеобразная
группа лишенных раковины червеобразных
моллюсков, сильно вытянутых вдоль передне-
задней оси. Они роют и заселяют
вертикальные ходы в мягких морских осадках. Их
цилиндрическое тело длиной от 2 до 14 см
располагается головным концом вниз,
в толщу осадка, который поглощается
терминальным ртом. Мантийная полость,
несущая одну пару ктенидиев (с двумя рядами
филаментов каждый), расположена на
заднем конце тела (рис. 5.5), т. е. в устье хода.
Нога отсутствует, мантия покрывает все
тело; поэтому движения животных нетипич-
Класс
Подкласс
Надотряд Отряд
Chaetodermomorpha -
Neomeniomorpha
Monoplacophora
Polyplacophora -
Prosobranchia,
Gastropoda "4
Heterobranchia
\Bivalvia
Protobranchia
Lamellibranchia
iScaphopoda „
(Cephalopoda
Nautiloidea ■
Coleordea
* В настоящее время классификация
брюхоногих не устоялась, поскольку старая,
разработанная до 1930 г. система, основанная на разных
уровнях организации, постепенно уступает место
Pulmonata-
- Caudofoveata
-Aplotegmentaria
. Pachytegmentaria
- Tryblidiida
. Lepidopleurida
Ischnochitonida
• Acanthochitonida
Docoglossida
Pleurotomanida
Amsobranchida
Lepetellida
Nentida
Architaenioglossa
Ectobranchida
Neotaenioglossa
Heteroglossa
Stenoglossa
Archaeopulmonata
Basommatophora
Stylommatophora
Onchidiida
Gymnomorpha'^— Sqleolifera
Opisthobranchia<
Allogastropoda-
Ctenidiobranchia -
Palaeobranchia —
Pteriomorpha
Rhodopida
Cephalaspida
Anaspida
Saccoglossa
Nudibranchia
Pleurobranchomorpha
Umbraculomorpha
Pyramidellomorpha
Nucuhda
Solemyida
Arcida
Mytilida
Ptenida
Palaeoheterodonta<;Trl6cnllda
Heterodonta -
Anomalodesmata «
Unroniida
_Venenda
-Myida
, Pholadomyida
- Poromyida
-Dentalida
. Siphonodentalida
Nautilida
Sepiida
Teuthida
Octopoda
Vampyromorpha
более филогенетически обоснованным подходам.
Приводимая нами схема — скорее один из
примеров современных классификаций, ни одна из
которых пока не получила общего признания.
Моллюски
155
Ганглий
Гонада Перикардиальная полость
Рот
Железисто
кутикулярная
пластинка
Радулярный Дивертикул
мешок кишечника
Анус
Сенсорный
орган
Мантийная
ПОЛОСТЬ о |- с /^
Рис. 5.5. Схема
продольного разреза через передний
Ктенидий (А) и задний (Б) концы хе-
тодермоморфного
моллюска (по Boss, 1982)
ное и происходит за счет
перистальтического сокращения хорошо развитой
мускулатуры стенки тела. Вместо раковины
эпидермис секретирует хитиновую кутикулу, в
которой закреплены налегающие друг на
друга обращенные назад, заостренные
чешуевидные пластинки, покрывающие все тело.
Некоторые системы органов, характерные
для моллюсков, отсутствуют; очень плохо
различимая голова не несет глаз и
сенсорных щупалец; нет экскреторных органов
и половых протоков (гонады выводят свои
продукты через перикардиальную полость);
некоторые виды лишены радулы. Однако на
голове находится терминальная хитиновая
пластинка, ассоциированная с различными
железами (рис. 5.6), функция которой пока
неясна.
Семьдесят видов этих раздельнополых
питающихся осадками моллюсков
образуют один отряд.
Задний коней
Ктенидий
Передний конец
Кутикулярная пластинка
Задний конец
Рис. 5.6. Форма тела хето-
дермоморф: А — внешний
вид; Б— образ жизни
в морском осадке (по Ну-
man, 1967; Jones, Baxter,
1987)
Задний
конец
Кутикулярная
пластинка
Передний
конец
Ктенидий
156 Глава 5
5.1.3.2. Класс Neomeniomorpha
Представители этого класса внешне
сходны с хетодермоморфами отсутствием
раковины, а практически и головы,
червеобразно вытянутым в переднезаднем
направлении телом, отсутствием
экскреторных органов, половых протоков, иногда
радулы (рис. 5.7). Однако они отличаются во
многих других отношениях и обычно
считаются стоящими ближе к раковинным
моллюскам.
Их тело длиной от 1 мм до 30 см сжато
с боков и несет продольный вентральный
желобок, в котором расположены один или
более мелких гребней, рассматриваемых
как сильно редуцированная нога. Мантия
покрывает все тело, кроме этого желобка,
и образует один или более слоев отдельных
известковых пластинок или спикул под
кутикулой. У переднего конца желобка
располагаются ресничная ямка и сдвинутый
к вентральной стороне рот, а у
противоположного конца находится задневентральная
мантийная полость без ктенидиев. Часто
развиты вторичные жабры в виде складок
или сосочков (рис. 5.8).
Неомениоморфы—хищники,
питающиеся книдариями, на которых их обычно
находят. Способ их питания, однако, не
совсем ясен ввиду частой редукции или
отсутствия радулы. У некоторых видов передняя
часть пищеварительного тракта может
выворачиваться и захватывать пищу. Хотя
мускулатура стенки тела хорошо развита,
движение происходит не за счет сокращения
мышц, а в результате скольжения с
помощью ресничек на выдвигающихся гребнях
вентрального желобка. Все виды
гермафродиты; копуляция происходит с помощью
стилетов; у некоторых видов сперма
партнера накапливается в семеприемниках.
Из 180 видов все морские и разделяются
на два отряда в зависимости от числа слоев
известковых образований в мантии и
присутствия или отсутствия эпидермальных
сосочков.
5.1.3.3. Класс Monoplacophora
Считалось, что моноплакофоры
вымерли в девоне, однако в 1952 г. живые их
экземпляры были обнаружены в одном из
океанических желобов Тихого океана.
Изучение материала из других районов привело
к описанию в целом 8 видов из одного
отряда.
Передний
конец
Ресничная
ямка
Рис. 5.7. Форма тела неомениоморф: А— внешний вид; Б— образ жизни, ползание по колонии
гидроида (по Jones. Baxter, 1987)
Моллюски
157
Ганглий
Рис. 5.8. Схема продольного
разреза неомениоморфного
моллюска (по Boss, 1982)
Все это мелкие (3 мм — 3 см) морские
раздельнополые потребители донных
осадков с телом, покрытым со спины единой
конической или чашеобразной раковиной, под
которой расположена округлая
слабомускулистая нога, окруженная сзади и с боков
широкой мантийной полостью (рис. 5.9).
Внутри нее по бокам ноги расположено
пять или шесть пар ктенидиев с одним
рядом филаментов каждый. Других
органов также по нескольку пар: шесть пар
дольчатых почек, открывающихся независимо
друг от друга по бокам мантийной полости,
две пары гонад и восемь пар мышц-
ретракторов ноги (рис. 5.10). Голова разли-
Рис. 5.9. Моноплакофорный моллюск с (А)
вентральной и (Б) боковой сторон (по Lemche,
Wingstrand, 1959)
Кишка Гонада Перикардиальная полость
сие чима, но плохо развита, не несет сенсорных
ад- щупалец (за исключением расположенных
ко- около рта) и глаз. Радула развита хорошо.
юд Анус открывается в заднем конце мантий-
ку- ной полости.
;ов
9)- 5.1.3.4. Класс Polyplacophora
но (хитоны, или панцирные)
им
га- Овальные или более вытянутые, дорсо-
jo- вентрально уплощенные хитоны отлича-
мо ются восемью расположенными в линию
ги, и последовательно налегающими друг на
щ- друга спинными пластинами панциря. Ман-
т- тия вокруг них развивается в широкий
«пояс», с кутикулой, часто несущей иглы,
чешуйки или щетинки. У некоторых видов
он частично или полностью покрывает
спинные пластины (рис. 5.11). Разделение
дорсального защитного покрытия на
отдельные элементы позволяет хитонам
сворачиваться в шар, как некоторым
многоножкам и мокрицам.
Голова плохо развита и спрятана под
передней частью мантийного пояса; глаза
и сенсорные щупальца отсутствуют, но
радула крупная и несет в каждом поперечном
ряду помногу зубов. Большая часть
вентральной поверхности тела занята мощной
вытянутой мускулистой ногой, с помощью
которой хитоны медленно ползают по
твердому субстрату. Нога и пояс могут
использоваться как сильная присоска, плотно притя-
i гивающая животных к субстрату. Узкая
мантийная полость окружает ногу со всех
сторон, кроме непосредственно передней,
и содержит 6—88 пар ктенидиев с двумя
д\ рядами филаментов каждый: сзади в нее вы-
he, ходит анальное отверстие и протоки одной
пары почек. Последние крупные и, как
Педальный Мантийная
желсеок полость
Педальный нервный ствол
Околопищеводное
нервное кольцо
Почки
Латеральный
нервный ствол
Мышцы-ретракторы
ноги
Ктенидии
2йЭ/ Сердце
Мантийная
полость
Анус
Рис. 5.10. Схема
анатомического строения моноплакофор;
вид с вентральной стороны
с удаленными ногой и стенкой
тела. Видны сериально
повторяющиеся внутренние органы
и ктенидии (по Lemche, Wing-
stand, 1959)
Мелкие
непокрытые участки
дорсальных
пластин
Рис. 5.11. Вид с дорсальной
стороны четырех моллюсков-
полиплакофор. показывающий
различную степень покрытия
пластинок панциря мантией
(по источникам в Нуглап, 1967)
Моллюски
159
Гонодукт
Экскреторная пора
Анус
Рис. 5.12. Схема анатомического строения обобщенного панцирного моллюска; вид с
вентральной стороны с удаленными ногой, стенкой тела и кишкой (по Hescheler, 1900)
Голова
Ктенидии
Мантийная полость
Гонада
Гонопор
Сердце и перикардиальная
полость
и единственная (сросшаяся) гонада (с
парными половыми протоками и половыми
порами), часто занимают всю длину тела (рис.
5.12). Нервная система устроена
относительно просто; ее стволы, к примеру, не ган-
глионизированы.
Хитонов 550 видов, все они морские
и раздельнополые; большинство питается
соскабливая водоросли. Они могут быть от
3 мм до 40 см длиной. Имеются 3 отряда,
которые различаются по расположению
жабр в мантийной полости, присутствию
или отсутствию прикрепительных зубов на
пластинах панциря и степени покрытия
пластин панциря мантийным поясом.
5.1.3.5. Класс Gastropoda
(брюхоногие)
Гастроподы—обширная и
разнообразная группа, общий признак которой—
поворот (торсия) внутренностного мешка
в процессе развития примерно на 180°
против часовой стрелки, так что мантийная
полость занимает переднее положение, и
вперед соответственно открываются протоки
почек и анальное отверстие: брюхоногие —
как бы «скрученные» животные (рис. 5.13).
Это заметно по асимметричному развитию
двух мышц-ретракторов ноги и/или
неравномерному росту (см. разд. 15.4.1).
Возможность размешать хорошо развитую голо-
Нервная
система
Кишка
Ктенидии
Приносящее
течение
Выносящее течение"
А Б
Рис. 5.13. Процесс торсии у гастропод: А—до
торсии; Б — после торсии
160 Глава 5
Приносящее течент
Отверстие в
крыше раковины
Выносящее течение
А
Сифон
Приносящее
течение
Жавра
Жавры
Анус Экскреторные
горы
Рис. 5.14. Два изменения
траектории дыхательного тока
воды у гастропод, вызванные
торсией: А — выносящий ток
выходит через дорсальное
отверстие в раковине; Б — потеря
правого ктенидия и
образование поперечного течения, по
направлению которого
ориентирован анус
ву моллюска в мантийной полости при
втягивании тела в раковину, а также
переднее положение хеморецепторных органов
мантийной полости (осфрадиев),
позволяющее воспринимать состав воды перед
животным, а не позади него, по-видимому,
дали селективное преимущество этой группе.
Однако автоматическим следствием торсии
был бы поток воды из мантийной полости,
выносящий экскреты и экскременты,
непосредственно над головой животного.
Загрязнение головных сенсорных органов было
устранено за счет изменения траектории
этого потока, например развития щели или
одного или нескольких отверстий (аппертур)
в спинной части раковины, через которые
он может выходить из мантийной полости,
или же утраты ктенидия на правой стороне
тела и формирования тока воды с левой
стороны мантийной полости на правую
и далее во внешнюю среду (рис. 5.14). В
последнем случае левая (входная) сторона
мантийной полости часто вытягивается
в подвижный сифон. Однако многие
филогенетические линии гастропод вторично
подверглись деторсии, в результате
которой внутренностный мешок развернулся
назад на угол около 90°, так что мантийная
полость (если она сохранилась) оказалась
с правой стороны тела, а анус обычно вновь
сзади.
Общий план строения гастропод (от
которого многие их виды радикально откло-,
нились)—коренастое тело с хорошо
развитой ногой для ползания и хорошо
дифференцированной головой, несущей радулу,
пару челюстей, пару глаз и одну или более
пар сенсорных щупалец; как голова, так
и нога полностью втягиваются в единую,
толстостенную спирально закрученную
раковину (рис. 5.15), отверстие которой
иногда закрывается при сокращении тела
известковой или органической крышкой
(оперкулумом), находящейся на заднедор-
сальной части ноги. По крайней мере у
примитивных форм мантийная полость
содержит одну пару ктенидиев с двумя рядами
лепестков каждый. Эти раздельнополые
моллюски развиваются с двумя
личиночными стадиями—трохофорой и велигером.
Исходный план строения, включая
раковину, оперкулум и торсию большей частью
сохраняют 55 000 видов в основном
морского подкласса переднежаберных (Prosobran-
chia). При этом ктенидии имеют тенденцию
редуцироваться от примитивного парного
состояния к сохранению только одного
левого элемента с двумя, а впоследствии
Моллюски
161
Рис. 5.15. Спиральные раковины гастропод (по
источникам в Нутап, 1967)
лишь с одним рядом лепестков. С
исчезновением правого ктенидия коррелирует
потеря правой почки, правого предсердия
сердца и правого осфрадия.' Большинство
переднежаберных раздельнополые, хотя
некоторые—гермафродиты с чередованием
мужской и женской фаз. В основном это —
бентосные «улитки», поедающие
водоросли (блюдечки, литторины, каллиостомы,
маргаритесы), неподвижные колониальные
животные (каури) или илоядные формы, но
одна группа (отряд Stenoglossa) включает
много хищников с радулой, несущей лишь
несколько крупных клыковидных зубов
в каждом ряду. Несколько видов-
фильтраторов питаются, используя
наружные слизистые сети (рис. 9.5) или
увеличенные ктенидии (параллелизм с бивальвия-
ми). Среди переднежаберных и вообще
среди всех гастропод блюдечки (отряд Doco-
glossida) образуют весьма изолированную
группу с особым строением раковины,
радулы и пищеварительного тракта.
Во втором подклассе гастропод, Hete-
robranchia, происшедшем от
переднежаберных, наблюдаются несколько тенденций
развития: а) редукция, погружение под
мягкие ткани или утрата раковины; б)
утрата оперкулума; в) деторсия; г) утрата пред-
ковых ктенидиев и замещение их
вторичными органами газообмена; д) развитие
синхронного гермофродитизма. Два основных
надотряда гетеробранхий—это морские
заднежаберные (Opisthobranchia: около
1000 видов) и в основном наземные и
пресноводные легочные (20000 видов). У первых
(голожаберники, морские зайцы,
крылоногие и др.) ктенидии обычно замещается
вторичными жабрами или развивается
газообмен через поверхность тела, часто
покрытую сосочками. Большинство их видов, как
планктонных, так и донных—хищники,
включая эндо- и эктопаразитов, но
некоторые потребляют водоросли (часто—сосут
их сок), а одна группа планктонных филь-
траторов кормится, используя секретируе-
мую слизистую сеть и реснички на широкой
крыловидной ноге (рис. 9.5). Бывает до
четырех пар головных щупалец.
Легочные (Pulmonata) характеризуются
в основном преобразованием мантийной
полости в воздушное легкое с сократимым
отверстием—пневмостомом. Большинство
их питается наземными растениями; у всех
исчезли личиночные стадии, так что
эмбриональное развитие идет прямо до
молодых улиток или слизней. Многие
легочные сохраняют спиральную гастроподную
раковину (обычно тонкостенную), но
некоторые представители наземного отряда Sty-
lommatophora ее утратили (наземные
слизни).
Третий надотряд гетеробранхий—
Gymnomorpha—также состоит из слизней,
но три эти, возможно неродственные, ветви
морских или наземных гастропод (в общей
сложности около 200 видов)
характеризуются различным смешением признаков
легочных и заднежаберных. То же можно
сказать о надотряде Allogastropoda (500
видов). Очевидно, что Heterobranchia—это
гастроподы, для которых типично
параллельное развитие многих «прогрессивных»
признаков, и распутать их взаимосвязи
затруднительно.
В целом брюхоногие включают около
77000 улитко- и слизнеподобных видов
(рис. 5.16), достигающих 60 см высоты. Их
можно разделить на 23 отряда.
11—1314
162 Глава 5
Рис. 5.16. Форма тела различных гастропод (в основном по источникам в Нутап, 1967)
5.1.3.6. Класс Bivalvia
(двустворчатые)
В сущности, бивальвии—это сжатые
с боков моллюски, полностью
помещающиеся между парой створок раковины.
Будучи защищены таким образом, они
малоподвижны или вообще неподвижны, иногда
сцементированы с субстратом или
прикреплены к нему другим способом. Как и у
других животных, «спрятанных» внутри
толстого защитного покрова, голова этих
моллюсков сильно редуцирована (рис. 5.17).
Радулы, глаз и сенсорных щупалец нет, хотя
отсутствие головных органов чувств может
быть эффективно восполнено щупальцами,
а иногда и глазами, располагающимися по
краю мантии. Две створки раковины
являются боковыми, сочленены вдоль
дорсальной средней линии и открываются
пассивно за счет действия упругого
дорсального лигамента. Сохранение их в закрытом
состоянии требует сокращения двух (иногда
их число редуцировано до одного)
мускулов-замыкателей (аддукторов), которые
способны к так называемому запирательно-
му тонусу, т.е. могут длительное время
оставаться в сокращенном состоянии, не
требуя непрерывного расхода энергии и че-
Моллюски
163
Зувы раковины
Пищеварительный дм-
Рот вертикул -
Гонада Лигамент Влагалище
-^ \ I стебелька Почка
-Сердце
-Мышца-аддуктор
-Анус
Выводной
хисрон
Вводной сисрон
Место
расположения жавры
Ктенидиальная
жавра
Ротовая лопасть
Рис. 5.17. Схема продольного
разреза двустворчатого
моллюска: А—с удаленной
жаброй (для ясности); Б—с
жаброй в естественном
положении и указанием траектории
пищевых токов воды (по
нескольким источникам) Б
редующихся сокращений и расслаблений
отдельных волокон.
Собственно тело, у самых крупных
видов достигающее 1 м в длину,
располагается внутри раковины, в дорсальной ее
части, а крупная мантийная полость занимает
оставшееся вентральное и латеральное
пространство. Когда мускулы-замыкатели
расслаблены, створки несколько
приоткрываются, позволяя току воды проходить
через полость и (у форм, закапывающихся
в мягкий осадок) выдвигаться наружу
сжатой с боков ноге. Часто вентральные и
боковые края мантии срастаются вместе,
оставляя только отверстие для ноги и
мелкие отверстия для циркуляции воды. В
районе последних мантия часто вытянута,
образуя вводной и выводной сифоны,
иногда способные втягиваться внутрь раковины.
На основе происхождения и функций
ктенидиев различают три крупные
категории бивальвий. У примитивных форм
(подкласс Protobranchia) парные ктенидии
выполняют главным образом исходную
функцию газообмена, а питание, если оно не
зависит полностью от симбиотических хе-
моавтотрофов, осуществляется при
помощи лопастей с обеих сторон рта. У одного
отряда (Nuculida) они образуют длинные
щупальца, которые, двигаясь по
поверхности окружающего осадка, подгоняют
пищевые частицы к ротовым лопастям, которые
сортируют их перед поеданием (см.
рис. 9.6). Эти виды питаются частицами
осадка с помощью слизисто-ресничного
механизма. Однако у преобладающего
большинства видов (надотряды Pteriomorpha,
Palaeoheterodonta и Heterodonta) парные
ктенидии очень сильно увеличены и
сложены в поперечном сечении W-образно.
образуя органы фильтрационного питания
(рис. 5.18). Пищевой материал в виде
суспензии засасывается в мантийную полость
вместе с приносящим током воды, его
частицы отфильтровываются, когда этот ток
проходит сквозь жабры, а затем
передаются ко рту с участием ротовых лопастей,
т. е. (слизисто-)ресничного механизма.
И наконец, у «перегородчатожаберных»
видов третьей группы (отряд Poromyida надо-
тряда Anomalodesmata) жабры сильно
редуцированы и формируются мышечные
«насосные» перегородки, с помощью которых
и увеличенного вводного сифона мелкие
животные засасываются внутрь мантийной
полости, где захватываются мускулистыми
ротовыми лопастями и поедаются.
Двустворчатые, насчитывающие 20000
видов,— это обычные бентосные животные
морей и пресных водоемов. Большинство
из них раздельнополые и характеризуются
непрямым развитием, хотя личинки
некоторых пресноводных бивальвий аберрантные,
поскольку приспособлены к паразитирова-
нию на рыбах. Различают 11 отрядов.
164
Глава 5
Ктенидиальные
филаменты
Мантийная
полость
Камера
выносящего течения (КВТ)
Камера приносящего
течения (КПТ)
Ктенидиальная ось
Ктенидиальный филамен'
КПТ.
Отсрильтрованная
пища гонится
фронтальными ресничками
Приносящий кровеносный
сосуд
мй кровеносный
сосуд
Течение воды,
вызванное ла-
, теральными
Гресничками
| латеро-фронтальные
Реснички .J фронтальные —
I латеральные —
Рис. 5.18. Поперечный разрез
двустворчатого моллюска,
иллюстрирующий
фильтрационное питание с помощью ктени-
диальных жабр (по Russell-
Hunter, 1979)
5.1.3.7. Класс Scaphopoda
(лопатоногие)
Скафоподы—удлиненные,
цилиндрические моллюски (длина 2—150 мм), почти
полностью окруженные мантией, секрети-
рующей единую трубчатую известковую
раковину, открытую с обоих концов. Они
закапываются в мягкие морские осадки,
оставляя более узкий конец трубки-
раковины несколько выступать над
поверхностью дна (рис. 5.19). Из более широкого
вентрального отверстия (апертуры)
раковины выдвигаются коническая или
цилиндрическая роющая нога и мелкая хоботообра-
зная голова, лишенная глаз и сенсорных
Респираторный ток воды /*
Рис. 5.19. Образ жизни моллюсков-скафопод
Моллюски
165
Отверстие в раковине
Гонада
Кишечник
Голова
Мантийная полость
Мантия
Каптакулы
Рис. 5.20. Схема продольного разреза лопато-
ногого (по различным источникам)
щупалец. Однако имеются радула,
единственная срединная челюсть и собранные
в пару пучков узкие булавовидные
сократимые филаменты — ловчие щупальца
(каптакулы). Мелкие пищевые частицы осадка
передаются ко рту вдоль каптакул с
помощью ресничек, а более крупные
приклеиваются к липким расширениям на их концах
и переносятся прямо в рот (рис. 9.6).
Трудно определить направление, в
котором вытянуто тело. Если считать, что анус
и мантийная полость находятся сзади, как
у гастропод и цефалопод, то скафоподы
сильно удлинены дорсовентрально (рис.
5.20). Однако мантийную полость и анус
часто рассматривают как вентральные, а
в таком случае скафоподы вытянуты вдоль
переднезадней оси тела. Мантийная
полость простирается вдоль задней высоты (или
вентральной длины) раковины и лишена
ктенидиев. Вода попадает внутрь ее сквозь
дорсальное (или заднее) отверстие за счет
биения ресничек мантии и периодически
выбрасывается наружу через то же отверстие
при помощи мышечного сокращения.
Газообмен идет через стенки мантийной
полости. Все виды раздельнополые с одной
непарной гонадой, продукты которой
выводятся через правую почку.
Лопатоногих 350 видов; их делят на два
отряда в зависимости от числа и формы
каптакул и формы ноги.
5.1.3.8. Класс Cephalopoda
(головоногие)
Целиком морские головоногие—
анатомически и поведенчески наиболее
совершенные представители моллюсков и,
возможно, вообще всех безпозвоночных.
Они также включают самых крупных
современных беспозвоночных, гигантских
кальмаров, достигающих общей длины 20 м.
Диагностическими признаками цефалопод
являются структуры, образованные тем,
что у других моллюсков было бы ногой,
которая здесь отсутствует. Передний отдел
эмбриональной ноги развивается в
несколько хватательных щупалец или рук вокруг
рта, а задний формирует мускулистую
воронку вокруг части отверстия мантийной
полости. Вода может входить в последнюю
по краям головы и с силой выталкиваться
через эту подвижную воронку за счет
мышечного сокращения. Цефалоподы — в
основном пелагические животные,
плавающие, используя реакцию выталкиваемой
струи, за подвижной добычей, которую
ловят щупальцами (руками). Как и гастропо-
ды, головоногие удлинены в дорсовентра-
льном направлении, но в связи с их
плавающим образом жизни исходно вентральный
отдел функционально становится передним,
а дорсальный внутренностный мешок
фактически образует задний конец тела
животного (рис. 5.21). Мантийная полость, таким
образом, открывается спереди, поэтому при
быстром движении цефалоподы движутся
задом наперед. При более медленной локо-
моции ориентация воронки может
обеспечивать движение в различных
направлениях. Добыча, пойманная руками,
измельчается парой клювовидных дорсовентраль-
ных челюстей, а затем радулой. Некоторые
руки у самцов модифицированы для
копуляции (переноса сперматофоров). Большая
часть видов раздельнополые, с одной
гонадой и прямым развитием. Удивительно,
учитывая их крупные размеры, что все
головоногие, за исключением наутилоидей,
относительно короткоживущие, размножаю-
166
Глава 5
Голова
Передняя (дорсальная) сторона
Вентральная
(передняя)
сторона
Дорсальная
(задняя)
сторона
Отверстие мантийной
Задняя (вентральная) сторона
Рис. 5.21. Схематичный вид сбоку головоногого моллюска, показывающий изменение
ориентации тела (в скобках) по сравнению с анцестральными формами (без скобок)
тся один раз и затем умирают
(моноцикличность— см. разд. 14.3).
Другие признаки этой группы заметно
различаются у представителей двух ныне
живущих подклассов. У шести современных
видов Nautiloidea единая наружная
плоскоспиральная раковина с множеством
расположенных в ряд друг за другом камер, из
которых последняя обитаема, хотя тонкий
тяж живой ткани—сифон—тянется сквозь
другие камеры (рис. 5.22). По мере роста
животного откладывается новый материал
раковины, добавляющий примерно
цилиндрическую секцию к старой раковине. Затем
животное продвигается вперед, секретируя
сзади новую перегородку. Таким образом
добавляется новая камера к уже
существующим. Наутилоидеи—плохие пловцы. Вода
выталкивается из мантийной полости
только за счет сокращения мышц воронки, и,
чтобы оставаться взвешенными в толще
воды, они используют плавучесть раковины,
которая не зависит от давления и
обеспечивается необитаемыми камерами. Сифон
поглощает воду, исходно содержащуюся в
камере, и вместо нее секретирует газ, что дает
нейтральную плавучесть (см. разд. 12.2.2).
Рот окружает большое количество (80—90)
не несущих присосок ловчих щупалец. Из
них четыре модифицированы для передачи
Капюшон Глаз
Щупальца
Воронка
Раковина
Щупальца Рад ла
'\Капюшон/
Воронка
Ганглии
Мантийная полость
Ктенидии
Б
Камеры
раковины
Сифон
Гонада
Сердце
Рис. 5.22. Наутилоидеи: А —
внешний вид; Б—
схематический продольный
разрез (по Boss, 1982 и др.)
Моллюски
167
сперматофоров. Имеется по две пары кте-
нидиев и почек. Нервная система и глаза
устроены относительно просто.
У Coleoidea (внутрираковинных)
заметна тенденция к редукции раковины,
коррелирующая с более активным плаванием.
У этой группы вода выталкивается из
мантийной полости с большой силой за счет
одновременного сокращения мощных
кольцевых мышц стенки мантийной полости,
координируемого крупными звездчатыми
ганглиями, от которых отходят гигантские
нервные волокна тем большего диаметра,
чем больше расстояние между иннервируе-
мой мышцей и ганглием. Каракатицы
(отряд Sepiida) сохраняют известковую
раковину как орган плавучести (разд. 12.2.2),
но она уменьшена в размере и помещается
внутри тела животного. У кальмаров
(отряды Teuthida и Vampyromorpha) раковина
редуцирована в еще большей степени—до
тонкого внутреннего хрящевого элемента,
не связанного с плавучестью, а у
большинства осьминогов (отряд Octopoda) она
полностью утрачена. Многие пелагические ко-
леоидеи имеют обтекаемую или торпедо-
видную форму (рис. 5.23) и должны
непрерывно плавать, чтобы оставаться в толще
воды, за счет волнообразного движения
боковых плавников, обеспечивающих
медленную, но эффективную локомоцию.
Некоторые виды, однако, выработали альтерна-
Рис. 5.23. Форма тела голово-
ногих-колеоидей (по
различным источникам)
168
Глава 5
Клюв Радула Внутренний Пищеварительные
остаток дивертикулы
раковины
Рис. 5.24. Схема
продольного разреза колеоидей (по щуГ1альца
Boss, 1982 и др.)
Воронка
Мантийная Ктенидий
полость
Анус
тивный способ поддержания плавучести:
путем замещения тяжелых двухвалентных
катионов на ионы аммония в расширенной
перикардиальной полости одновременно
с развитием студенистых тканей с низкой
плотностью. Осьминоги в основном
адаптированы к донному существованию, хотя
у некоторых их форм кожистая перепонка
между руками может использоваться как
парус для дрейфа в придонных течениях.
У таких видов редуцированные или
рудиментарные ктенидий. В отличие от наути-
лоидеи у колеоидей одна пара массивных
складчатых ктенидиев, одна пара почек
и восемь коротких рук, вооруженных
присосками (две из них у самцов обычно
модифицированы для переноса сперматофоров).
У кальмаров и каракатиц, кроме того, еще
две удлиненные щупальцевидные ловчие
руки, несущие на концах подушки присосок
(рис. 5.24).
Однако, возможно, наиболее
отличительные признаки 650 видов колеоидей—
замкнутая кровеносная и в высшей степени
Нервы к
рукам
Нерв к —
щупальцу
Верхний
вуккальный
ганглий
иеревральный
ганглий
Зрительный
нерв
исцеральныи
нерв
Брахиальный ганглий J \ Задний нерв воронки
Педальный ганглий Передний нерв воронки
Ресничная
мышца
Хрусталик
Роговица
Радужка
Рис. 5.25. Мозг (А) и глаза
(б) колеоидей (по Wells,
1982; Kaestner, 1967) Б
Сетчатка
.Зрительные нервы
I
Моллюски
169
концентрированная нервная системы,
последняя—с хорошо развитыми глазами.
Ктенидии лишены ресничек (вода движется
через мантийную полость за счет
мускульного сокрашения, обеспечивающего
реактивную локомоцию), но содержат
капилляры, по которым кровь без системы
противотока прокачивается жаберными сердцами.
Околопищеводное нервное кольцо
расширено в мозг, устроенный сложнее, чем у
каких-либо иных беспозвоночных. Общее
строение парных глаз, иннервирующихся от
мозга, такое же, как у позвоночных: с
роговицей, радужковой диафрагмой,
хрусталиком и сетчаткой (рис. 5.25). Однако в
отличие от позвоночных фоторецепторы такого
глаза обращены в сторону падающего
света. Нервная система контролирует также
многочисленные поверхностные хромато-
форы, которые (что необычно) управляются
мелкими мышцами и поэтому могут
реагировать очень быстро, приводя к почти
мгновенному изменению окраски.
5.2. Дополнительная
литература
Fretter V., Реаке J. (Eds) 1975 1978. Pulmonates (2
Vols). Academic Press, London.
Hughes R. N. 1986. A Functional Biology of Marine
Gastropods. Croom Helm, Beckenham.
Hyman L. H. 1967. The Invertebrates (Vol. 6):
Mollusca 1. McGraw-Hill, New York.
Kaestner A. 1967. Invertebrate Zoology (Vol. 1).
Wiley. New York.
Morton J. E. 1979. Molluscs, 5th edn. Hutchinson,
London.
Purchon R.D. 1968. The Biology of the Mollusca.
Pergamon Press, Oxford.
Runham N. W., Hunter P.J. 1970. Terrestrial Slugs.
Hutchinson, London.
Solem A. 1974. The Shell Makers. Wiley, New York.
Wells M.J. 1962. Brain and Behaviour in Cephalo-
pods. Heinemann, London.
Wells M.J. 1978. Octopus. Chapman & Hall,
London.
Wilbur K.M. (Ed.) 1983—1987. The Mollusca (12
Vols.). Academic Press, New York.
ГЛАВА \J
LOPHOPHORATA: ЛОФОФОРОВЫЕ,
ИЛИ ЩУПАЛЬЦЕВЫЕ
Phorona
Brachiopoda
Bryozoa
Entoprocta
Обычно к лофофоровым относят три
типа: Phorona, Brachiopoda и Bryozoa. Общим
признаком этих групп, как указывает их
название, является лофофор, т.е.
циклический, подковообразный, в виде сложного
завитка или складчатого кольца орган с
покрытыми ресничками полыми щупальцами,
функционирующий как аппарат для
фильтрации пищевых частиц из толщи воды.
Этот лофофор, окружающий рот (но не
анус), гидростатически поддерживается
собственной полостью тела.
В своей основе тело лофофоровых
состоит из трех отделов с очень мелкой прео-
ральной частью (просомой), небольшим
вторым отделом (мезосомой), несущим рот
и лофофор, и обширным третьим (метасо-
мой), включающим остальные системы
органов, а также анус у основания лофофора.
Далее, общий отличительный признак
лофофоровых— их «гонады», которые
представляют собой не оформленные органы,
а просто рыхлые перитонеальные
скопления половых клеток. У большинства
лофофоровых просома утрачена и, даже если
присутствует, крайне мала. Ее значение для
понимания родственных взаимоотношений
между типами этих животных гораздо
важнее, чем в их жизни.
Хотя лофофоровые первичноротые
в том смысле, что их рот формируется из
бластопора, они имеют и ряд характерных
черт вторичноротых (гл. 7), включая план
строения с тремя отделами (у некоторых
вторичноротых просома хорошо развита),
каждый из которых с собственной
полостью тела, в некоторых случаях
образующейся энтероцельно (разд. 15.2—15.4).
Общие признаки включают также радиальное
и недетерминированное дробление и ба-
зиэпидермальную нервную систему. У
некоторых вторичноротых известны даже
временные перитонеальные «гонады» и мезо-
сомальный лофофор. Поэтому
лофофоровых рассматривают как группу,
занимающую промежуточное положение между
первично- и вторичноротыми (см. разд. 2.4.3).
Лофофоровые—прикрепленные или
сидячие животные, и их тело защищено секре-
тируемым наружным покровом. Он бывает
в виде хитиновых трубок, внутри которых
животные свободно двигаются, или же
студенистых, хитиновых или известковых
раковин или домиков, постоянно соединенных
с эпидермисом.
Таким образом, лофофоровые
представляются хорошо очерченной группой, и
некоторые считают, что всех их следует
включать в один тип Lophophorata. Эта точка
зрения не стала общепризнанной, с одной
стороны, из-за продолжающихся споров по
поводу взаимоотношений мшанок с
остальными группами, а с другой—из-за
особенностей Entoprocta, которые остаются плохо
понятными. Служащие для питания
щупальца энтопроктов не подходят под
определение лофофора, но при этом сами
животные отчасти сходны с мшанками, хотя у них
спиральное, детерминированное дробление,
явно не свойственное лофофоровым.
Поэтому включение энтопроктов в эту
главу во многом условно.
Lophophorata: лофофоровые. или щупальцевые 171
6.1. Тип Phorona (форониды)
6.1.1. Этимология
От родового названия Phoronis, одного
из эпитетов египетского божества Изиды.
6.1.2. Диагностические
и специфические признаки
(рис. 6.1)
1. Билатерально симметричные,
червеобразные животные.
2. Тело из более чем двух слоев клеток
в толщину, с тканями и органами.
3. Сквозной, U-образной формы
кишечник.
4. Олигомерное тело из трех отделов,
каждый со своим единым целомом.
5. Просома очень мелкая, мезосома
тоже, но с крупным лофофором,
поддерживаемым мезоцелем; метасома крупная и
удлиненная.
6. Стенка тела без кутикулы, с
мышечными слоями.
7. Замкнутая кровеносная система с ге-
Подковоовразно
.расположенные
лофОфоральные
щупальца
Эпистом
•Анус
Мезоцель
Протоцель
Нефридий
Метацель
Секретируемая
труБка
Желудок
Тонада"-
Рис. 6.1. Схема продольного разреза
форониды (по Emig, 1979)
Рис. 6.2. Различные формы лофофора у форо-
нид (по Emig, 1979)
моглобином, заключенным в форменные
элементы.
8. Одна пара напоминающих метане-
фридии органов (у личинок—прото-
нефридии).
9. Нервная система диффузная, базиэпи-
дермальная.
10. Дробление яиц радиального типа.
11. Развитие непрямое, обычно с
личиночной стадией — актинотрохой.
12. Морские формы.
Форониды—сидячие черви, постоянно
обитающие в секретируемых ими
хитиновых трубках, частично погруженных
в мягкий осадок или, реже, прикрепленных
к твердому субстрату. Достигают в длину
50 см. но обычно — менее 20 см, причем
большую часть длины тела занимает
метасома, или туловище. Кроме этого,
единственная заметная внешне часть тела —
крупный и топографически терминальный
лофофор—аппарат для фильтрационного
питания, который может нести до 1500
образованных мезосомой щупалец,
расположенных в совокупности подковой (рис.
6.2); у видов с особенно большим числом
щупалец свободные концы «подковы»
закручиваются в спирали.
172
Глава 6
Рис. 6.3. Личинка форонид—актинотроха (по
Emig. 1979)
Большинство
форонид—гермафродиты, некоторые—раздельнополые.
Половые клетки образуются в перитонеуме, а
затем попадают в метацель, откуда
выводятся во внешнюю среду через нефридии.
Оплодотворение наружное, хотя яйца и
личинки (рис. 6.3) могут «инкубироваться»
в полости, образованной лофофором, или
внутри трубки в течение 40 — 75% периода
развития. Один вид способен размножаться
бесполым способом путем поперечного
деления и почкования, другой — только
поперечным делением.
У форонид целый ряд признаков, общих
как с остальными типами лофофоровых
(разд. 6.2 и 6.3), так и с вторичноротыми
(гл. 7), и их часто считают близкими по
происхождению с обеими этими группами (гл.
2). Некоторые даже выступают за отнесение
их к собственно вторичноротым, несмотря
на развитие рта из б ласт опора.
6.1.3. Классификация
Известно только десять видов,
помещаемых в два рода одного семейства.
6.2. Тип Brachiopoda
(плеченогие)
6.2.1. Этимология
От греч. brachion—плечо; pous—нога.
6.2.2. Диагностические
и специфические признаки
(рис. 6.4)
1. Билатерально симметричные
животные.
2. Тело из более чем двух слоев клеток
в толщину, с тканями и органами.
3. Кишечник либо сквозной, U-
образной формы, либо вторично
замкнутый на конце.
4. Олигомерное, состоящее из двух
отделов тело, каждый отдел с единым, в
основном энтероцельным по происхождению
цел омом.
5. Просома отсутствует, мезосома
мелкая, но с крупным лофофоральным
комплексом, который поддерживается мезоце-
лем (см. рис. 9.3), а у некоторых видов—
известковыми выростами раковины; мета-
сома мелкая.
6. Тело целиком, за исключением
стебелька, заключено внутри двустворчатой
раковины, которая может быть
сцементирована с субстратом, прикреплена к камню,
заякорена в мягком осадке с помощью
стебелька или вообще не прикреплена. Мелкое
тело животного располагается в задней
части раковины, большая часть внутреннего
пространства которой занята лофофором.
7. Незамкнутая кровеносная система
с сердцем или сердцами.
8. Одна или две пары метанефридиепо-
добных органов.
9. Нервная система из
околопищеводного нервного кольца с ганглиями, от
которого отходят отдельные нервы.
10. Бесполого размножения нет;
большей частью раздельнополые, без
оформленных гонад, но с четырьмя скоплениями
половых клеток, связанными с перитонеу-
мом. Гаметы поступают в метацель и
выводятся во внешнюю среду через нефридии.
11. Дробление яиц радиальное.
12. Морские формы.
Lophophorata: лофофоровые, или щупальцевые 173
Пищевой желоБок
Боковая рука ло-
фофора
Гонада
Нервное кольцо
Желудок
Полость тела
Раковина
Мантия
Центральная рука
лофофора
Рот
Гонада
Нефридий
Мускулы раковины
Стебелек
Рис. 6.4. Схема продольного
разреза обобщенного плечено-
гого
Современные брахиоподы, являющиеся
остатком некогда очень крупного типа (335
ныне живущих видов — 26000 вымерших),
по своему внешнему виду и образу жизни
удивительно схожи с двустворчатыми
моллюсками (разд. 5.1.3.6). Обе группы—
прикрепленные или в основном
неподвижные животные-фильтраторы,
помещающиеся внутри двустворчатой раковины, се-
кретируемой эпидермальной мантией и
покрытой органическим периостракумом.
Однако фильтрационный аппарат плечено-
гих—циклический, петлевидный или
спиральный лофофор (а не ктенидиальные
жабры), а две створки их раковины —
дорсальная и вентральная (а не
латеральные, как у бивальвий). Следующая точка
конвергенции—редукция имевшегося
у предков головного отдела (у брахиопод—
утрата просомы) как следствие полного
заключения тела внутрь толстой наружной
раковины.
Все ныне живущие брахиоподы —
относительно мелкие (раковина менее 10 см
в длину и ширину) бентосные животные.
В эволюционном смысле они, вероятно,
эквивалентны раковинным форонидам (гл.
2). Действительно, их план строения мог
возникнуть от таких червеообразных
организмов полифилетически (рис. 2.13).
6.2.3. Классификация
Различают два класса, дивергировавших
друг от друга по многим признакам.
Класс
Jnarticulata-
Articulata -
Отряд
. Lingulida
Acrotretida
. Rhynchonellida
• Terebratulida
-Thecideidida
6.2.3.1. Класс Inarticulate
(беззамковые)
Беззамковые брахиоподы, как
свидетельствует их название, характеризуются
створками раковины, состоящими из
фосфата кальция и хитина и соединенными
друг с другом только с помощью мышц.
В задней части тела между створками
выходит стебелек, формирующийся как часть
тела животного из личиночной мантии. Он
содержит вырост метацеля и собственные
мышцы; в некоторых группах, где стебелек
отсутствует, вентральная створка
сцементирована с субстратом. В отличие от других
брахиопод анус у беззамковых есть (рис.
6.5), лофофор относительно простой,
поддерживающих его скелетных элементов
(выростов раковины) нет.
174
Глава 6
Щетинки
Раковина
Кишечник
Мышцы раковины-
Лофосрораль-
ная рука
Рот
Гонада
Анус
Нефридий
Целомичес-
кая полость
j-Кутикула
tf-Мышца
Рис. 6.5. Схема продольного разреза
беззамкового плеченогого
Рис. 6.6. Различная форма тела беззамковых
плеченогих (по источникам в Нутап, 1959)
Стевелек
Мышцы раковины
Рис. 6.7. Схема продольного
разреза замкового
плеченогого (по Moore, 1965)
Слепо замкнутый
I кишечник
ЛофОфор
Раковина
Нефридий Гонада
Внутренний вид
дорсальной створки,
показывающий
опорные известко
вые структуры
лофофора
Lophophorata: лофофоровые, или щупальцевые
175
Личинка—уменьшенное
свободноплавающее подобие взрослых животных с
раковиной и лофофором, реснички которого
обеспечивают движение в толще воды.
Метаморфоза при оседании на дно не
происходит. В ходе развития мезодерма
формируется из архентерона энтероцельным
способом, а полости тела — схизоцельно внутри
образовавшихся энтероцельно мезодер-
мальных клеточных масс.
К беззамковым относят 47 видов,
разделяемых на два отряда: роющие Lingulida
и сидячие, блюдечковидные Acrotretida
(рис. 6.6).
6.2.3.2. Класс Articulate (замковые)
У замковых раковины из карбоната
кальция, створки которых сочленяются
с помощью зубов на вентральной створке,
входящих в углубление на дорсальной.
В вентральной створке находится также
вырез или отверстие, через которое выходит
стебелек (если он есть). Дорсальная створка
часто несет направленные внутрь выросты,
поддерживающие крупный, уложенный
сложными петлями или спиральный лофофор
(рис. 6.7). Ни собственных мышц, ни
выроста метацеля в стебельке нет. Он образуется
не из мантии, а за счет одного из трех
отделов тела личинки. Эта личиночная стадия
(рис. 6.8) сильно отличается от взрослых
животных и развивается с метаморфозом.
В отличие от беззамковых кишечник на
конце замкнут; как мезодерма, так и полости
тела образуются энтероцельно.
Замковые формы появились в процессе
эволюции брахиопод относительно поздно
(в ордовике); сохранилось почти 300 их
видов, помещаемых в три отряда на основе
структуры лофофора и поддерживающих
его известковых образований.
6.3. Тип Bryozoa (мшанки)
6.3.1. Этимология
От греч. bryon мох; zoon—животное.
6.3.2. Диагностические
и специфические признаки
(рис. 6.9)
1. Модульные, колониальные
животные; каждая колония состоит из отдельных
особей (зооидов), число которых может
доходить до миллиона. Колонии
формируются за счет бесполого размножения
(почкования) особи-основательницы. Ткани
каждого зооида контактируют с тканями его
непосредственных соседей.
■Лосрофораль-
ные щупальца
Глотка
Мышца-
ретрактор
Яичник
Защитное
покрытие
Пилорическая
часть
Полость тела
Семенник
■Тканевый тяж
Рис. 6.8. Личинка замкового плеченогого (по Рис. 6.9. Схема продольного разреза обобщен-
Lacaze-Duthier, 1861) ной мшанки
176
Глава 6
2. Зооиды часто полиморфны;
различают их питающие, защитные,
репродуктивные и др. типы.
3. Колонии в целом гермафродитные,
хотя зооиды могут быть гермафродитными
или раздельнополыми.
Приводимые ниже особенности
относятся к отдельным зооидам.
4. Билатерально симметричные формы.
5. Тело более чем из двух слоев клеток
в толщину, с тканями и органами.
6. Сквозной, U-образной формы
кишечник.
7. Тело олигомерное, из двух или трех
отделов, каждый—со своей полостью тела,
формирующейся de novo в ходе
метаморфоза; некоторые с дополнительной метасо-
мальной полостью тела.
8. Просома присутствует только у
одной группы, мелкая; мезосома тоже мелкая,
с относительно мелким циклическим или
подковообразным лофофором, который
поддерживается гидростатически мезоце-
лем. Метасома крупная, мешковидная.
9. Тело замкнуто или погружено в
хитиновую, студенистую или известковую
трубку, в домик или общий матрикс; сквозь
отверстие в этой оболочке может
выдвигаться (за счет гидростатического давления)
или втягиваться (с помощью мышц-
ретракторов) лофофор.
10. Циркуляторной и экскреторной
систем нет.
11. Нервная система в виде ганглия
между ртом и анусом и отходящих от него
околопищеводного нервного кольца и
отдельных нервов.
12. Гонады «перитонеальные», гаметы
попадают в метапель и выходят во
внешнюю среду через «целомопоры», связанные
с лофофором.
13. Дробление яиц радиальное, обычно
они развиваются под защитой
родительского организма.
14. Развитие обычно непрямое.
Мшанки похожи на крохотных (до 0,5
мм) форонид (разд. 6.1), формирующих за
счет бесполого размножения (почкования)
(рис. 6.10) обширные столонообразующие,
ковро-, древо- или листовидные колонии.
Сходство с другими типами лофофоровых
(разд. 6.1 и 6.2) в наличии лофофора для
фильтрационного питания, олигомерного
плана строения тела, радиального
дробления, внешне аналогичной гонадной системы
с половыми клетками, развивающимися из
клеток эпителиальной выстилки полости
тела. Вопрос о происхождении последней до
сих пор дебатируется, что связано с
отсутствием как ее эмбриологических
предшественников, так и преемственности между
зародышевыми листками и тканями
взрослого животного. В ходе метаморфоза
ткани личинок подвергаются гистолизу, а
затем полностью реорганизуются в тело
взрослой особи. Мезодерма и полость тела
остальных лофофоровых в основном энте-
роцельные и образуются из зародышевой
кишки; однако у мшанок, если она и
имеется, то исчезает в процессе метаморфоза.
Поэтому прямых свидетельств мезодерма-
льного происхождения полостей взрослых
животных нет и целомическии статус,
который им обычно приписывают, базируется
в основном на аналогии с форонидами
и брахиоподами. Единства взглядов здесь
нет: альтернативная школа предлагает
рассматривать полости тела мшанок как
эквивалент предположительно псевдоцело-
мической полости энтопроктов (разд. 6.4).
Как группа, мшанки—весьма
преуспевающая линия фильтраторов, причем на
сегодняшний день более, чем какие-либо иные
лофофоровые. За несколькими
исключениями, колонии неподвижные, покрывающие
относительно твердый субстрат или
прикрепленные к нему. Исключения—
пресноводная форма Cristatella, способная
медленно ползать (около 10 см в день) с
помощью мускулистой «ноги», и морская
форма Selenaria maculata, передвигающаяся
значительно быстрее (до 1 м/ч) на длинных
щетинках своих авикуларных зооидов (см.
разд. 6.3.3.3). S. maculata обитает на
мелководьях коралловых рифов; показано, что
она движется к освещенным солнцем
участкам. Колония зеленого цвета, поэтому
возможно, что такое поведение связано с
присутствием в ее тканях симбиотических зоок-
сантелл, как и у многих других рифовых
беспозвоночных (разд. 9.1).
Lophophorata: лофофоровые. или щупальцевые
177
Рис. 6.10. Форма колоний мшанок (по различным источникам)
6.3.3. Классификация
Большинство из 4000 описанных
видов—морские формы, но один класс и
отдельные виды в двух других классах—
пресноводные.
Класс
Phylactolacmata -
Stcnolacmata
Gymnolacmata-
Отряд
-Plumatellida
- Cyclostomata
- Ctenostomata
-Cheilostomata
12—1314
178
Глава 6
6.3.3.1. Класс Phylactolaemata
Филактолематы — относительно
неспециализированные мшанки. Тело зооидов
сохраняет предположительно анцестраль-
ное разделение на три отдела. Просомаль-
ная тканевая складка, эпистом, с
гидростатическим протоцелем нависает над
ротовым отверстием, как у форонид. В стенке
тела хорошо развиты слои продольной
и кольцевой мускулатуры. Сокращение
кольцевых мышц создает давление,
приводящее к выдвиганию крупного лофофора,
несущего до 120 щупалец, расположенных
в форме подковообразного гребня (рис.
6.11). Зооиды мономорфные,
цилиндрические, одни из наиболее крупных у мшанок.
Полости тела соседних особей часто
связаны между собой.
Все виды пресноводные, эпидермис се-
кретирует необызвествленный хитиновый
или студенистый чехол (солей кальция
в пресной воде гораздо меньше, чем в
морской). В процессе эволюции мшанки этого
класса выработали механизм образования
покоящихся стадий для распространения
и/или зимовки. Такие стадии весьма
своеобразны: на тяже ткани, идущем от кишки
к стенке тела, скопления эпидермальных
Рис. 6.11. Расправленный зооид
филактолемат, изображенный прозрачным (по Реппак,
1978)
Рис. 6.12. Статобласты филактолемат (по Ну-
man, 1959)
и «перитонеальных» клеток развиваются
в «статобласты» (рис. 6.12), т.е. богатые
желтком клетки, обычно заключенные в
хитиновые дископодобные створки. Они
могут оставаться прикрепленными к стенке
тела и обеспечивать восстановление колонии
in situ после ее отмирания или же
высвобождаться при жизни или после гибели зооида
в окружающую среду. В последнем случае
они способны плавать и могут переноситься
на большие расстояния. Статобласты
крайне устойчивы к замерзанию и высыханию.
В умеренных широтах после зимовки они
«прорастают» в зооидов-основателей.
Один отряд этого класса включает
около 50 видов.
6.3.3.2. Класс Stenolaemata
Зооиды стенолемат также
цилиндрические, но лишены просомы и мышечных
Лофосрор
Рис. 6.13. Зооиды стенолемат:
А—сократившийся; б —
расправленный (по Ryland, 1970)
Полость тела
Рис. 6.14. Срезы через камеры
зооидов различных гимноле-
матных мшанок,
показывающие как сокращение
париетальных мышц приводит к
деформации стенки и
одновременно к повышению давленин
в полости тела: А — Г—хей-
лостоматы; Д—ктено-
стоматы (по Ryland, 1970)
Расположенный
под наружной
стенкой тела
мешок,
содержащий морскую
воду и
сообщающийся с
внешней средой
Поперечные
париетальные
мышцы
Продольные
париетальные
мышцы
Условные
обозначения:
■ Овызвествленные части стенли тела
- Мемвранные (десрормируемые) части
стенки тела
12*
180
Глава 6
слоев в стенке тела. Их цилиндрические
трубки с округлым терминальным
отверстием сильно обызвествлены под наружной
гиалиновой кутикулой или клеточным
слоем. Отверстие трубки после втягивания
лофофора закрывается мембраной, но не
крышечкой (оперкулумом). Давление,
требуемое для вьщвигания циклического,
несущего не более 30 щупалец лофофора,
создается мышцами-дилататорами
(расширителями), действующими на жидкость как
в метацеле, так и в экзосаккальной полости
тела, обычно считающейся псевдоцеломи-
ческой (рис. 6.13). Зооиды стенолемат в
небольшой степени полиморфны.
Репродуктивная система имеет ряд
особенностей, главная из которых—по-
лиэмбриония. После оплодотворения
дробящаяся зигота образует комок бластоме-
ров, затем распадающийся на несколько
вторичных эмбрионов, иногда отпочковы-
вающих третичные эмбрионы; из одной
зиготы может образоваться более 100
бластул.
Большинство групп этого класса
вымерло; единственный современный отряд
включает около 900 морских видов.
6.3.3.3. Класс Gymnolaemata
В настоящее время гимнолематы—
наиболее многочисленные и преуспевающие
среди мшанок; их более 3000 видов,
обитающих в основном в море, но также и в
солоноватых и пресных водоемах. Как и у
стенолемат, их зооиды лишены просомы и
мышечных слоев в стенке тела и несут
относительно мелкий циклический лофофор.
Однако эти обычно короткие, коренастые
зооиды создают давление, необходимое для
вьщвигания лофофора, за счет мускульной
деформации стенки тела с участием
париетальных мышц, действующих на особый
мембранный отдел (рис. 6.14). В связи
с этим стенка тела, если и обызвествлена, то
лишь частично. После втягивания
лофофора отверстие у большинства видов
закрывается оперкулумом. Полиморфизм зоои-
ЛофОфор
Мышечный
Сфинктер
Метацель
Мышца-
ретрактор
Париетальные
мышцы
Кишечник
Поперечные
париетальные
мышцы
Тканевые тяжи
-Столон
Рис. 6.15. Зооиды ктеностомат:
А — сократившийся; Б—
расправленный (по Ryland,
1970)
Lophophorata: лофофоровые. или щупальцевые
181
Фронтальная Мышца-
мемврана ретрактор
-Лосрофор
Анус
Мышца, закрывающая
оперкулум
Метацель
Яичник Тканевый Семенник Кишечник Латеральные
тяж поры Б
Рис. 6.16. Зооиды хейлостомат: А—сократившийся; Б—расправленный (по Ryland, 1970)
дов в пределах этой группы очень развит.
Различают питающие автозооиды,
уплотняющие кенозооиды, захватывающие ави-
куларии, очищающие вибракулы и озции—
выводковые камеры.
Выделяют два отряда: Ctenostomata
с необызвествленной стенкой, без оперкулу-
ма, часто цилиндрическими зооидами и
терминальным отверстием (рис. 6.15) и Cheilo-
stomata с уплощенными, коробообразными
зооидами. частично (часто сильно) обыз-
вествленными стенками, оперкулумом
и фронтальным отверстием (рис. 6.16).
6.4. Тип Entoprocta1
6.4.1. Этимология
От греч. entos — внутри: proktos — анус.
6.4.2. Диагностические
и специфические признаки
(рис. 6.17)
1. Билатерально симметричные,
бокаловидные животные.
2. Тело из более чем двух слоев клеток
в толщину, с тканями и органами.
3. Сквозной, U-образной формы
кишечник.
4. Пространство между стенкой тела
и кишкой, содержащее студенистую
«мезенхиму», интерпретируется некоторыми как
заполненная клетками псевдоцеломическая
полость тела.
5. Тело по форме представляет собой
полусферический или яйцевидный бокал,
несущий кольцо щупалец вокруг рта и ану-
Щупальца
Протонефридий
Ганглий
Ресничный
кишечник
Гонодукт
'Псевдоцель"
Мышцы стебелька
1 В русскоязычной литературе принят термин
«Kamptozoa».— Прим. пер.
Рис. 6.17. Схема продольного разреза энто-
прокта (по Becker, 1937)
Анус
Л Энтопрокт
Б. Мшанка
/I /) Ресничные
у/ /f- питающие
111/ щупальца
Направление
тока воды
Ресничные
питающие
щупальца
Анус
Рис. 6.18. Сравнение пищедобывательных
течений у энтопроктов (А) и мшанок (£)
относительно положения ануса
Вид своку
Вид с вентральной стороны Вид с дорсальной стороны
Рис. 6.19. Личинка энтопроктов (по Nielsen, 1971)
Lophophorata: лофофоровые. или щупальцевые
183
са; прикрепляется к субстрату сократимым
стебельком.
6. Стенка тела с кутикулой, но без
мышечных слоев.
7. Циркуляторной и газообменной
систем нет.
8. Одна пара или множество (у
пресноводных видов) протонефридиев.
9. Нервная система в виде ганглия
между ртом и анусом и отдельных отходящих
от него нервов.
10. Щупальца невтягивающиеся, но
могут сокращаться и загибаться внутрь,
заполняя межщупальцевую полость.
11. Дробление яиц спиральное.
12. Развитие непрямое.
Энтопрокты (внутрипорошицевые)—
мелкие (0,5—5,0 мм высоты), одиночные
или колониальные животные, временно или
чаще постоянно прикрепленные к
субстрату, в том числе и к другим организмам. Все
они фильтраторы. От 6 до 36 выростов
стенки тела образуют щупальца с
ресничками, улавливающими частицы пищи из воды
и передающими их в слизи ко рту. У форм,
создающих собственные токи воды для
питания, она идет снаружи между
щупальцами в окруженное ими пространство, а затем
из него вверх (рис. 6.18): это—направление,
обратное наблюдаемому у остальных лофо-
форовых. Анус на вершине конического
возвышения выводит остатки пищи в зону
центрального выносящего течения.
Широко распространенное бесполое
размножение почкованием может приводить
к возникновению модульных колоний.
Вероятно, все энтопрокты—гермафродиты.
Внешне однополые особи соответствуют
различным половым стадиям гермафродит-
ного организма. Сперма выбрасывается
в воду, но оплодотворение, видимо, всегда
внутреннее. Личинка—планктотрофная
или лецитотрофная трохофора (рис. 6.19),
в большинстве случаев метаморфозирую-
щая во взрослое животное, но у некоторых
видов последнее развивается из образуемой
личинкой почки.
Взаимоотношения этого мелкого типа
с другими спорны. Некоторые
интерпретируют обширную «мезенхимную» ткань как
заполненный гемолимфой псевдоцель
и объединяют энтопроктов с другими псев-
доцеломическими типами (гл. 4); другие
считают их близкими мшанкам на основе
сходства процессов развития, а также
неясного происхождения полости тела (разд.
6.3.2).
6.4.3. Классификация
Описано 150 видов, помещенных в один
класс и один отряд. Все, за исключением
одного пресноводного рода,—морские
формы.
6.5. Дополнительная
литература
Emig С. С. 1979. British and Other Phoronids.
Academic Press, London [Phorona].
Hyman L.H. 1951. The Invertebrates (Vol. 3).
Acanthocephala, Aschelminthes and Entoprocta.
McGraw-Hill, New York [Entoprocta].
Hyman L. H. 1959. The Invertebrates (Vol. 5).
Smaller Coelomate Groups. McGraw-Hill, New York
[Phorona, Brachiopoda, Bryozoa].
Nielsen C. 1971. Entoproct life-cycles and the ento-
proct/ectoproct relationship. Ophelia 9, 209 341
[Entoprocta].
Rudwick M.J.S. 1970. Living and Fossil Brachio-
pods. Hutchinson, London [Brachiopoda].
Ryland J. S. 1970. Bryozoans. Hutchinson, London
[Bryozoa].
Wright A.D. 1979. Brachiopod radiation. In: House
M. R. (Ed.) The Origin of Major Invertebrate
Groups, pp. 235—252. Academic Press, London
[Brachiopoda].
ГЛАВА /
DEUTEROSTOMIA: ВТОРИЧНОРОТЫЕ
Chaetognatha
Hemichordata
Echinodermata
Chordata
О том, что вторичноротые—
естественная группа родственных друг
другу типов, говорит наличие у них ряда общих
эмбриологических, структурных и
биохимических признаков, а также эволюционных
тенденций, неизвестных в других группах.
Например, в ходе раннего эмбрионального
развития большинства вторичноротых: а)
бластопор не образует рта, являющегося
в результате вторичным отверстием кишки
(откуда термин «вторичноротые»); б)
дробление идет по радиальному типу; в)
судьба каждой клетки в процессе развития
не определена окончательно до
относительно поздних стадий морфогенеза
(недетерминированное развитие); г) полость тела
формируется путем отделения от
эмбриональной кишки серии «энтероцельных»
карманов (см., например, рис. 2.9). Все это в
совокупности типично для вторичноротых, хотя
отдельные из названных признаков
(особенно б и г) известны и в других типах. Далее,
к характерным чертам вторичноротых
относятся фоторецепторы ресничного (ци-
лиарного) типа (в отличие от рабдомерных
фоторецепторов первичноротых),
преобладание одноресничных клеток в эпидермисе,
креатинфосфат как депо фосфатов (у
первичноротых более обычен в этой роли арги-
нинфосфат) и практически полное
отсутствие хитина.
Предок вторичноротых был, вероятно,
похож на современных крыложаберных
полухордовых (см. разд. 7.2.3.2), которые
сами напоминают форонид (см. разд. 6.1).
По-видимому, он представлял собой
короткое сидячее червеобразное животное с оли-
гомерным телом, состоявшим из трех
отделов, второй из которых (мезосомальный)
нес лофофор, а третий (метасомальный)
заключал временные скопления перитонеаль-
ных клеток, вероятно образующие гонады.
Их нервная система, по-видимому,
состояла в основном из диффузного субэпидер-
мального плексуса, с одним или более
продольными тяжевидными утолщениями.
В отличие от всех известных лофофорат
(однако по аналогии с некоторыми гастро-
трихами, см. разд. 4.3.3) ток воды,
транспортирующий захваченные лофофором
пищевые частицы в передний отдел
кишечника, выходит наружу через несколько
боковых отверстий, ведущих из глотки
непосредственно на поверхность тела. Таким
образом кишка предков вторичноротых
сообщалась с внешней средой не только ртом
и анусом.
В большинстве эволюционных линий
вторичноротых лофофор утрачен или
сильно модифицирован. В связи с развитием
в некоторых группах прикрепленного
образа жизни (иглокожие, некоторые
полухордовые) или личиночного
свободноплавающего существования (большинство
хордовых) исчезло исходное разделение тела на
три отдела. У двух групп, наконец,
глоточные перфорации модифицированы для
обеспечения фильтрационного питания
(ранние хордовые) или газообмена (более
поздние хордовые и кишечнодышащие
полухордовые). К другим эволюционным
тенденциям, неизвестным у первичноротых,
относятся: сворачивание субэпидермально-
Deuterostomia: вторичноротые 185
го нервного плексуса в полую дорсальную
нервную трубку (хордовые и некоторые
полухордовые); появление постанального
хвоста с локомоторной функцией,
совершающего с помощью продольных мышц
волнообразные движения (щетинкочелюстные
и хордовые); появление защитных кожных
известковых пластин, которые в двух
независимых эволюционных линиях
впоследствии начинают использоваться как часть
мощного внутреннего скелета, к которому
прикрепляется локомоторная мускулатура
(некоторые иглокожие и хордовые).
7.1. Тип Chaetognatha
(щетинкочелюстные)
Щетинки
Глаз
Глотка
7.1.1. Этимология
От греч. chaite-
hos—челюсть.
-длинный волос, gnat-
7.1.2. Диагностические
и специфические признаки
(рис. 7.1)
1. Билатерально симметричные
червеобразные животные.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и органами.
3. Прямой сквозной пищеварительный
тракт и нетерминальный (вентральный)
анус.
4. Олигомерное тело разделено септами
на три отдела: голову, туловище и
постанальный хвост. Туловище и хвост несут
латеральные и каудальный немускулистые
плавники.
5. Каждый отдел тела с одним или
двумя целомами энтероцельного
происхождения, однако постювенильные стадии
лишены перитонеального эпителия.
6. Стенка тела с нехитиновой кутикулой
и пучками продольных мыщц.
7. Систем циркуляции, газообмена
и экскреции нет.
8. Нервная система с окологлоточным
кольцом, несущим дорсальный и
латеральные ганглии, от которых отходят
отдельные нервы.
9. Гермафродиты с крупными парными
Яичник
Женский
гонопор
Туловищно-
хвостовая
септа
Капюшон
Ганглий
Септа между головой
и туловищем
Кишка
Боковые плавники
Семенной
пузырек
Хвостовой
плавник
Рис. 7.1. Схематический вид щетинкочелюст-
ного с дорсальной стороны (по различным
источникам)
семенниками и яичниками (последние—с
гонодуктами).
10. Дробление яйца радиальное.
11. Развитие, характерное для вторич-
норотых, прямое.
12. Морские формы.
Хетогнаты—торпедообразные
хищники, плавающие с помощью быстрых биений
постанального хвоста в дорсовентральной
плоскости. Латеральные плавники
выполняют стабилизирующую функцию. Добыча
схватывается рядом подвижных
нехитиновых щетинок, расположенных спереди и
с боков вентральной камеры, которая ведет
в ротовую полость (рис. 7.2), однако, когда
животное не питается, голова вместе со
щетинками закрывается складкой стенки тела.
Этот капюшон, вероятно, служит в
основном для снижения лобового сопротивления,
а также для защиты. На голове—пара глаз,
186
Глава 7
Ряды подвижных дч Зг Передние зувы
щетинок V-^_^^ А^ \\ .
^R^____y_V__ й<-—Граница капюшона
Глотка —I \— / /
Рис. 7.2. Голова щетинкочелюстного, вид с
вентральной стороны (по Ritter-Zahony, 1911)
образованных за счет слияния отдельных
простых глазков. Их фоторецепторы ци-
лиарного типа, как и у других групп вторич-
норотых. Но хетогнаты не являются
типичными представителями Deuterostomia.
Планктонные виды
Бентосный вид
Рис. 7.3. Форма тела хетогнат (по Pierrot-Bults,
Chidgey, 1987)
В частности, ни в одной из дефинитивных
полостей их тела нет перитонеального
эпителия (диагностический признак целома),
а хвостовые энтероцели здесь (один или
два)—это вторичные производные парных
туловищных полостей, т.е. не образуются
в виде отдельных карманов из первичной
кишки, как у остальных олигомерных групп
с тремя отделами тела. Головная полость
непарная.
Хетогнаты, достигающие в длину 10 см,
могут быть доминантной группой
планктонных хищников в море; питаются
различными организмами — от мальков рыб
сходного с ними размера до простейших.
7.1.3. Классификация
Известно 70 видов, помещаемых в один
класс с двумя отрядами, различающимися
присутствием (Phragmophora) или
отсутствием (Aphragmophora) поперечной
вентральной мускулатуры. Отряд
Phragmophora включает среди прочих один бентосный
род Spadella (рис. 7.3).
7.2. Тип Hemichordata
(полухордовые)
7.2.1. Этимология
От греч. hemi—половина; Chordata -
название родственного типа (разд. 7.3).
7.2.2. Диагностические
и специфические признаки
1. Двусторонне симметричные
животные.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и органами.
3. Сквозной прямой или U-образно
изогнутый пищеварительный тракт.
4. Олигомерное тело, разделенное на
три отдела: крупный просомальный
хоботок, мелкий мезосомальный воротничок
и большое, вытянутое или мешковидное
туловище.
5. Каждый отдел с одним (хоботок)
или двумя (воротничок и туловище) энтеро-
цельными полостями тела.
6. У некоторых на воротничке распо-
Deuterostomia: вторичноротые
187
ложен лофофоральный орган,
гидростатически поддерживаемый мезоцелем.
7. У всех, за исключением одной, групп
верхняя половина стенки глотки несет от
одного до более 100 пар сквозных
отверстий, через которые выходит ток воды.
Только нижняя часть глоточной трубки
относится к пищеварительному тракту.
8. Ресничный, лишенный кутикулы
эпидермис, у некоторых секретирующий
внешнюю нехитиновую трубку.
9. Циркуляторная система частично
незамкнутая.
10. Экскреторный орган—клубочек,
образованный выпячиваниями перитонеу-
ма в протоцель.
11. Нервная система диффузная, ба-
зиэпидермальная, у некоторых—с полым
дорсальным нервным тяжем в воротничке.
Базиэпидермальный плексус
концентрируется в дорсальной и вентральной срединных
зонах.
12. Раздельнополые животные.
ЛофОфораль-
-ные руки со
щупальцами
-?В5? \зЬ, JwirSv / колонии
Глоточная щель f-—g\Ii*~—У /
"' ""v4 и
if. -1
Рис. 7.4. Пищедобывательные течения у крыло-
жаберного полухордового Cephalodiscus.
Изогнутые стрелки показывают движение воды
к лофофоральным рукам и сквозь них.
Стрелки, направленные вверх, обозначают
выносящее течение в центре лофофорального
аппарата. Стрелки на теле соответствуют движению
частиц, направляемому зпидермальными
ресничками: «наконечники» показывают движение
пищи ко рту (по Lester, 1985)
Полоса отвергнутых частиц
Рис. 7.5. Пищедобывательные течения,
вызываемые ресничками на хоботке и воротничке
кишечнодышащего полухордового (по Barring-
ton, 1886)
13. Дробление яиц радиальное.
14. Развитие, характерное для вторич-
норотых, непрямое.
15. Морские формы.
Полухордовые бывают двух основных
типов, которые значительно различаются
формой тела и образом жизни: с одной
стороны, червеобразные роющие кишечноды-
шащие, с другой—сидячие, обитающие
в трубках крыложаберные. Несмотря на
это, в обоих ветвях сохраняется слизисто-
ресничный механизм питания. У крыложа-
берных лофофоральный орган расположен
дорсально на воротничке. Его руки
расходятся конусом в сторону от вентрально
расположенного рта (у лофофоровых и
других групп, питающихся с помощью кольца
щупалец, последние окружают рот). Вода
загоняется сквозь кольцо щупалец в интра-
лофофоральную полость (так же, как,
например, у полихет из семейства Sabellidae
и у энтопроктов). Пищевые частицы затем
продвигаются ресничками по внешней
стороне рук вниз, ко рту (рис 7.4). Кроме того,
они могут захватываться ресничками всей
поверхности тела (единственный способ
ресничного питания у лишенных лофофора
кишечнодышащих, ресничные дорожки
которых особенно хорошо развиты на
хоботке; рис. 7.5).
У обеих форм потреблению пищевых
частиц, возможно, помогает (как и у других
микрофагов) ток воды, загоняемой в
ротовую полость глотки и, как у некоторых га-
стротрихов (разд. 4.3.3), выходящей через
глоточные отверстия во внешнюю среду.
188
Глава 7
Такой однонаправленный поток—более
эффективная система, чем у лофофоровых,
которые, например, должны периодически
выбрасывать через рот избыток воды,
захваченной вместе с пищевыми частицами.
Хотя обеспечение питания было, вероятно,
исходной функцией глоточных перфораций
(см. разд. 9.2.5), сохраняющейся у
крыложаберных, у кишечнодышащих
(потребляющих также частицы осадка) ток воды
стал обслуживать газообмен и количество
глоточных пор сильно увеличено.
Как следует из названия типа,
полухордовые имеют ряд черт, общих с хордовыми.
В целом последние характеризуются
четырьмя признаками: присутствием хорды,
глоткой, перфорированной щелями или
порами, постанальным хвостом и дорсальной
нервной трубкой (разд. 7.4). Из них у
кишечнодышащих есть по крайней мере два—
перфорированная глотка и дорсальная
нервная трубка, а возможно, и третий: у юве-
нильных особей некоторых видов описан
постанальный хвост. Однако если у щетин-
кочелюстных и хордовых хвост в принципе
локомоторная структура, о постанальном
отделе гемихордат этого сказать нельзя.
(Действительно, такой отдел свойствен
многим беспозвоночным, особенно с
кишкой U-образной формы, и ювенильный
«хвост» кишечнодышащих, вероятно,
гомологичен стебельку крыложаберных.) Хорда
у гемихордат определенно отсутствует,
хотя за нее в течение многих лет ошибочно
принимали направленное вперед
выпячивание кишки, отходящее от ротовой полости.
Таким образом, некоторым полухордовым
все же свойственна половина специфических
признаков хордовых, что оправдывает
данное им название.
Как указывалось выше, ряд признаков
объединяет крыложаберных с лофофоровы-
ми, особенно с форонидами (разд. 6.1),—
мезосомальный лофофор, поддерживаемый
гидростатическим давлением в мезоцеле,
трехраздельная система энтероцельных
полостей тела, диффузный базиэпидермаль-
ный нервный плексус и т.д. Единственные
важные отличия крыложаберных состоят
в развитии вторичного рта и парной
природе мезо- и метацелей. Таким образом, две
главные группы полухордовых в
анатомическом отношении представляют собой
звено, связующее первичноротых
лофофоровых и другие группы вторичноротых.
Предполагается, что именно крыложаберные,
а не форониды являются предковыми
формами для обеих филогенетических линий.
7.2.3. Классификация
Известно 100 видов полухордовых,
разделяемых на три класса.
Класс Отряд
Enteropneusta Helminthomorpha
/ Rhabdopleurida
С- Pterobranchia «^rrZTCZ Cephalodiscida
\ Planctosphaeroidea Planctosphaerida
7.2.3.1. Класс Enteropneusta
(кишечнодышащие)
Одиночные, подвижные, удлиненные
черви длиной до 2,5 м; роют ходы в мягких
субстратах или обитают под камнями, где
питаются частицами осадка или
взвешенной в воде пищей. Тело состоит из (а)
длинного хоботка, реснички которого создают
основную движущую силу при локомоции
животного и образуют транспортную
систему для пищевых частиц; (б) короткого,
лишенного лофофора воротничка; (в) очень
длинного туловища с терминальным
анусом и многочисленными глоточными
порами (рис. 7.6), количество которых
увеличивается всю жизнь. В глоточной стенке эти
перфорации имеют U-образную форму
и поддерживаются системой скелетных
перегородок, но на поверхность тела
выходят в виде мелких, округлых, дорсальных
отверстий. Нервный тяж и экскреторный
клубочек хорошо развиты, но полость тела
в основном заполнена мышечными
волокнами и соединительной тканью —
производными перитонеума,
заменяющими большую часть мускулатуры стенки
тела.
Имеется как половое, так и бесполое
размножение (фрагментация); много пар
гонад, расположенных в передней части
туловища, открываются, каждая своим
протоком. Оплодотворение наружное. Разви-
Deuterostomia: вторичноротые 189
-Хоботок
■/гевелек ховотка
-Воротничок
-Туловище
Анус
Рис. 7.6. Внешний вид кишечнодышащего (по
Marion, 1886)
тие идет через стадии, аналогичные
известным для многих иглокожих, до — у
большинства видов—личинки торнарии
(рис. 7.7).
Все 70 видов кишечнодышащих относят
к одному отряду.
7.2.3.2. Класс Pterobranchia
(крыложаберные)
Крыложаберные—сидячие, обитающие
в трубках животные, достигающие 12 мм
в длину, с коротким телом из (а) хоботка
в форме диска или щитка, обеспечивающего
как (ресничную) локомоцию внутри трубки,
так и секрецию этой коллагеновой
структуры; (б) воротничка, несущего на
вентральной стороне рот, а на дорсальной—от
одной до девяти пар лофофоральных рук, от
каждой из которых отходят двойные ряды
ресничных, секретируюших слизь щупалец;
(в) короткого мешковидного туловища
с анальным бугорком около воротничка
и субтерминальным сократимым
стебельком (рис. 7.8). Этот стебелек может
заканчиваться органом временного
прикрепления, переходить в общий столон или
закручиваться вокруг опоры как цепкий хвост
многих древесных млекопитающих. В
отличие от кишечнодышащих имеется не более
одной пары глоточных перфораций,
нервный тяж отсутствует, а экскреторный
клубочек развит очень плохо. Целомические
полости не заполнены тканями;
присутствует только одна пара гонад.
Широко распространено бесполое
почкование, приводящее в одной группе к обра-
ЛофОфоральные-
руки
Мезоцель
Хоботок
Лротоцель
Стебелек
Рис. 7.7. Личинка кишечнодышащих— Рис. 7.8. Схема продольного разреза крыложа-
торнария (по Stiasny, 1914) верного (по McFarland et al, 1979 и др.)
Рис. 7.9. Одиночные крыложаберные рода Се- Рис. 7.10. Трохофороподобная личинка крыло-
phalodiscus питаются, цепляясь за выступы об- жаберных (по Schepotieff, 1909)
щего ценоция (по Lester, 1985)
Рис. 7.11. Часть колонии рабдоплеврид (А) и увеличенный отдельный зооид (Б) (по Grasse, 1948
и др.)
Deuterostomia: вторичноротые
191
зованию колоний с общим столоном из
стебельков, а в другой—к клонам отдельных
особей, живущих в общей оболочке—
«ценоции» (рис. 7.9). О половом
размножении известно мало, но личиночная стадия не
такая, как у кишечнодышащих (рис. 7.10).
Десять видов крыложаберных
распределены между двумя отрядами. В отряд Се-
phalodiscida входят неколониальные
формы, либо ползающие по поверхности
колоний гидроидов, либо живущие внутри
общего «ценоция» и поднимающиеся к его
отверстиям для добывания пищи (рис. 7.9). У них
одна пара глоточных перфораций, от
четырех до девяти пар лофофоральных рук и
одна пара гонад. Отряд Rhabdopleurida
объединяет колониальные формы (рис. 7.11)
с непарной гонадой, одной парой
лофофоральных рук и без глоточных щелей.
7.2.3.3. Класс Planctosphaeroidea
Этот класс выделен, чтобы поместить
в него (хочется надеяться, только временно)
гигантские личинки (до 2,2 см в диаметре),
напоминающие своей общей формой
личинки кишечнодышащих (рис. 7.12). Их
находят в образцах планктона из
Атлантического океана с начала 1930-х гг. Поскольку
Рис. 7.12. Planctosphaera. вид с вентральной
стороны (по Spengel, 1932)
взрослая форма пока еще не известна, их
статус и особенности остаются
неопределенными.
7.3. Тип Echinodermata
(иглокожие)
7.3.1. Этимология
От греч. echinos—еж; derma—кожа.
7.3.2. Диагностические
и специфические признаки
(рис. 7.13)
1. Взрослые—с пятилучевой
симметрией, в большинстве случаев
действительно радиальные, некоторые билатеральные.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и органами.
3. Кишечник сквозной (у некоторых
вторично слепозамкнутый, у одной группы
отсутствует).
4. Форма тела в высшей степени
разнообразна. Анцестральные и современные
сидячие формы со сферическим или
чашеобразным телом, прикрепленные абораль-
но к субстрату стебельком с обращенным
вверх ртом, окруженным пятью руками.
Производные свободноживущие формы без
стебелька, с телом, ориентированным так,
что рот обращен вниз, к субстрату или
(реже) вперед; с различимыми руками или без
них, иногда билатерально симметричные.
5. Какой-либо дифференцированной
головы нет.
6. Исходно с олигомерной, состоящей
из трех парных целомов полостью тела.
Метацели («соматоцели») образуют ее
основную часть. У немногих форм с прямым
развитием полость тела образуется схизо-
цельно.
7. Водно-сосудистая (амбулакральная)
система, формирующаяся в основном из
левого мезоцеля (левого «гидроцеля») и
частично из левого протоцеля (левого «аксо-
целя»), содержит не связанную прямо
с внешней средой морскую воду и
гидравлически управляет движениями трубкообра-
зньгх локомоторных (амбулакральных)
ножек и/или захватывающих пищу щупалец
(«подий»). Часть этой системы образует ко-
192
Глава 7
Околоротова!
кольцевая
система
Кишечник
•Ампулы
Амбулакральные
ножки
Мышцы
Иглокожее на стебельке
Мадрепоровая Анус
пластинка
Вторично
СВОБОДНОПОДВИЖНОе
иглокожее
Гонада
Условные обозначения:
• Нервы
9 Амвулакра л ьная
система
• Гёмальная система
fS^d Известковые структуры
Рис. 7.13. Схемы разрезов двух обобщенных иглокожих (по Nichols, 1962)
Околоротовая рот
кольцевая
система
льцевои канал вокруг пищевода, от
которого начинаются радиальные каналы, идущие
вдоль каждой руки.
8. Имеется мезодермальная субэпидер-
мальная система известковых «косточек»
или пластинок, часто с выступающими на
поверхность бугорками или шипами.
9. Экскреторных органов нет.
10. Циркуляторная «гёмальная»
система выражена слабо; ее функции в
значительной мере выполняются целомическими
жидкостями.
11. Нервная система субэпидермальная,
в форме околопищеводного нервного
кольца, от которого начинаются диффузные
нервы, идущие вдоль каждого амбулакра.
12. Обычно раздельнополые.
13. Дробление яиц радиальное.
14. Развитие типичное для вторичноро-
тых, как правило, непрямое, с ресничной,
билатерально симметричной личинкой
(рис. 7.14).
15. Морские формы.
Иглокожие — в высшей степени
своеобразная группа с тремя весьма
характерными чертами: особой симметрией,
известковым мезодермальным скелетом, водно-
сосудистой (амбулакральной) и другими
целомическими системами. Все эти три
особенности могут быть прямо связаны с их
происхождением от сидячих фильтраторов,
которые, от каких бы предков ни
происходили, имеют тенденцию к развитию
радиальной симметрии ловчего аппарата (разд.
9.1). У иглокожих этот процесс привел
к формированию круга из пяти часто
дихотомически ветвящихся рук. У предковых
форм это были эквиваленты и, возможно,
производные лофофоральных рук
животных, подобных крыложаберным
полухордовым (разд. 7.2.3.2). Как и у последних,
гидравлическая система рук и ресничных
щупалец на них поддерживается в основном
давлением в мезоцеле. Некоторые
эволюционные линии иглокожих вторично стали
свободноживущими; большинство этих
форм как бы перевернулось «вверх ногами»,
так что исходно обращенная вверх несущая
рот и руки поверхность стала
контактировать с субстратом, а пищедобывающие
щупальца превратились в локомоторные.
Относительно небольшие анатомические
изменения сформировали из них
амбулакральные ножки (общая адаптация при этом
Deuterostomia: вторичноротые
193
А. Виттелария морских лилий
Рот
В. Осриоплутеус осриур
Я Брахиолария морских звезд
Скелетные спикулы
Анус
Г. Эхиноплутеус морских ежей
Кишечник
Д. Аурикулария голотурий
Рис. 7.14. Личинки иглокожих (no Barnes, 1980)
изменении функций - появление при
каждой ножке гидравлического резервуара,
ампулы, позволяющей ей индивидуально
сокращаться и вытягиваться). При этом руки
часто становились крупнее и шире (как
у морских звезд) или объединялись с телом,
заворачиваясь вокруг него (как у морских
ежей и голотурий). В любом случае пять рук
все еще определяют план строения тела, так
что у большинства современных свободно-
живущих форм радиальная симметрия
сохраняется, и, следовательно, они могут
двигаться в любом направлении (пятилучевая
Анус
Е. Долиолария голотурий
симметрия была не у всех ранних групп
иглокожих; известны спирально
симметричные и даже асимметричные формы—
см. рис. 2.14).
Сидячие животные нуждаются в
механической опоре тела и защите его от хищников
и действия волн. У иглокожих это
обеспечивается отложением в дермисе системы
«косточек», каждая из которых представляет
собой пористую решетку из карбоната
кальция, ведущую себя как единый кристалл
кальцита. Отдельные «косточки» часто
увеличиваются в размерах, принимая форму
13—1314
194
I лава 7
пластинок, иногда сцепляюшихся с другими
с образованием жесткого, почти наружного
панциря. Как и у других панцирных или
раковинных животных (например,
двустворчатых моллюсков, плеченогих) имевшаяся
у предков голова утрачена. Покровный, как
правило ресничный, эпидермис иногда
исчезает, и части панциря оказываются
внешней границей тела. Известковые структуры
могут вторично выполнять локомоторную
функцию, замещая в этом отношении
амбулакральные ножки. Мышцы, идущие от
одних «косточек» к другим, обеспечивают
движение по субстрату с помощью рук или
игл (см. гл. 10).
Хотя тело иглокожих мономерное, у них
развиваются три пары энтероцельных
карманов (иногда в модифицированной
форме), как и у олигомерных животных с тремя
отделами тела, т.е. форм, которым, как
принято считать, родственны иглокожие
(см. разд. 15.4.1). Как и у лофофоровых,
и полухордовых, метацели («соматоцели»)
формируют основную полость тела, а мезо-
цели (у иглокожих — один из них), или
«гидроцели»,—гидравлическую систему рук,
похожих на лофофоральные; правый мезо-
цель обычно редуцируется до мелкого
сократимого мешочка. Правый протоцель
(«аксоцель») образует аксиальный (осевой)
синус, т. е. пространство вокруг части гема-
льной системы, функции которой неясны,
в то время как левый объединяется с мезо-
целем в амбулакральную систему. Прото-
цели и мезоцели этих олигомерных
энтероцельных животных связаны с внешней
средой порой. Согласно классическим
взглядам, эта связь сильно развита у
иглокожих как средство изменения объема
жидкости в амбулакральной системе.
Считается, что через общий аксогидропор в виде
пористой (мадрепоровой) пластинки
происходит водообмен с внешней средой
(фактически в амбулакральной системе находится
не целомическая жидкость, а морская вода
с небольшим количеством целомоцитов).
От этой пластинки так называемый
«каменистый канал» ведет к околопищеводному
кольцу. В действительности прохождение
воды через мадрепоровую пластинку ни
разу не наблюдалось, однако она,
возможно, обеспечивает выравнивание
гидростатического давления внутри и вне
животного.
Таким образом, своеобразие иглокожих
не столько в самом их анатомическом
строении, сколько в том, что это
единственная процветающая группа свободноживу-
щих животных, происшедшая от
прикрепленных сидячих предков, которые, по-
видимому, были способны двигать лишь
несущими щупальца руками и изгибать
стебелек.
7.3.3. Классификация
Хотя известно около 6250 современных
видов иглокожих, нынешнее их
разнообразие значительно меньше, чем было в
палеозое: сейчас осталось только 6 из 24 классов
этого типа.
Класс Отряд
Isocnnida
Comatulida
Millericrinida
Bourgueticrinida
Cyrtocrinida
Platyasterida
Paxillosida
Valvatida
Spmulosida
Forcipulatida
Euclasterida
Oegophiurida
Phrynophiunda
Ophiurida
Crinoidea
Asteroidea
Ophiuroidea
^Concentricydoidea
4Echinoidea
\Holothuroidea«
- Peripodida
Cidaroida
Echinothuroida
Diadematoida
Pedinoida
Salenoida
Phymosomatoida
Arbacioida
Temnopleuroida
Echmoida
Holectypoida
Clypeasteroida
Cassiduloida
Spatangoida
Neolampadoida
Holasteroida
, Dendrochirotida
. Dactylochirotida
. Aspidochirotida
. Elasipoda
. Apodida
«Molpadiida
Deuterostomia: вторичноротые
195
7.3.3.1. Класс Crinoidea
(морские лилии)
Криноидеи — это единственные
современные иглокожие, сохранившие анцестра-
льную ориентацию тела со ртом,
обращенным вверх и окруженным слизисто-
ресничными, несущими мелкие щупальца
(ножки, или подии) руками, фильтрующими
пищу из толщи воды. Хотя в принципе рук
пять, они могут неоднократно разделяться,
давая от 10 до 200 ложных рук с
многочисленными боковыми ответвлениями (пин-
нулами) на каждой. По их обращенной
вверх поверхности проходит
амбулакральный желобок (рис. 7.15). Частицы пищи
захватываются ресничными, секретирующи-
ми слизь ножками и продвигаются вдоль
ресничных амбулакральных желобков ко
рту и далее в U-образный кишечник (рис.
7.16). Амбулакральная и другие целоми-
ческие системы простые, без ампул,
управляющих ножками, и мадрепоровой
пластинки. Каменистые каналы (часто
многочисленные) открываются в соматоцели.
Ножки вытягиваются за счет сокращения
снабженных сфинктерами каналов
амбулакральной системы.
Шаровидное тело постоянно или
временно прикреплено к субстрату с помощью
несократимого аборального стебелька
длиной до 1 м, часто несущего мутовки так
называемых цирр, с терминальным
прикрепительным диском или системой цирр,
способных удерживаться за морское дно или
заякориваться в нем. У многих
современных видов после короткого периода
прикрепленного существования животные
становятся подвижными и плавают с помощью
СвоБоднопоцвижная
Рис. 7.15. Форма тела
криноидеи (по Danielsson, 1892;
Carpenter, 1866)
Стебельковая
196
Глава 7
Амбулакральная
'система
Осевой орган
и синус
Кишка
Цирры
Рис. 7.16. Схематический
разрез тела и части руки свобод-
ноживущей морской лилии (по
Nichols. 1962)
перьевидных рук, лишь временно
прикрепляясь к субстрату аборальными циррами.
Однако анцестральное положение
обращенного вверх рта при этом сохраняется. Руки,
тело и стебелек несут сильно обызвествлен-
ные пластинки, занимающие большую
часть их объема, так что пространство,
приходящееся на живые ткани, мало.
Ткань гонад распределена диффузно.
Гаметы образуются в области перитонеума
внутри рук, развиваются в мелких
проксимальных «гонадах» и высвобождаются
через разрывы в стенках пиннул. Характерная
личинка виттелария после метаморфоза
превращается в миниатюрное стебельковое
подобие взрослых животных.
В классе 625 видов. Различают пять
отрядов, в основном по деталям строения
стебелька и его прикрепительной системы.
7.3.3.2. Класс Asteroidea
(морские звезды)
Астероидеи в целом характеризуются
уплощенным телом, постепенно
переходящим в пять (иногда до 40) рук. Они могут
быть короткими и широкими, придавая
животным пятиугольные очертания, или же
тонкими и длинными (рис. 7.17). Животные
свободноподвижные, рот и
амбулакральные борозды расположены на нижней
поверхности тела, а мадрепоровая пластинка
и анус (если он есть)—аборально.
Амбулакральные борозды хорошо развиты и
защищены с каждой стороны двойным рядом не
слившихся друг с другом пластинок, через
вырезы в которых направляются протоки,
соединяющие ампулы с локомоторными
амбулакральными ножками, часто с
присосками на концах. В целом известковый
скелет развит относительно слабо, хотя несет
внешние бугорки и шипы, иногда
расположенные в определенном порядке.
Некоторые из шипов модифицированы в педицел-
ларии (рис. 7.18)—группы, обычно из трех
мелких «косточек», действующих как
ножницы или щипцы и за счет этого
очищающих животное от организмов, пытающихся
осесть на его поверхности.
Полости тела, включая амбулакральную
систему, обширны; соматоцели образуют
мелкие выросты (папулы), проходящие на
аборальной стороне сквозь стенку тела
и служащие для газообмена (рис. 7.19).
Некоторые астероидеи—гермафродиты;
известно несколько форм размножения
и онтогенеза, включая бесполое
почкование, своеобразную заботу о потомстве,
прямое (особенно в высоких широтах) и
непрямое развитие с личиночными стадиями
бипиннария и брахиолария (рис. 7.14,£).
Большинство видов трупоеды и хищники,
питаюшиеся сидячими животными, но
известны и детритофаги, и фильтраторы.
При макрофагии добыча заглатывается
целиком или желудок, который очень
крупный, выворачивается через рот и
охватывает ткани жертвы. От желудка отходит па-
Deuterostomia: вторичноротые
197
■M0
Рис. 7.17. Форма тела астероидей (по источникам в Нутап, 1955)
ра слепых выростов в каждую руку. в 1962 г. род Platasterias сочли сохранив-
Шесть отрядов, включающих 1500 ви- шимся представителем давно вымерших
дов, различают по типам педицелларий, ра- иглокожих из класса Somasteroidea. Однако
сположению амбулакральных пластин сейчас его принято считать специализиро-
и форме амбулакральных ножек. Открытый ванной астероидеей из отряда Platyasterida.
198
Глава 7
Характерные типы
Структура окончании стебельковых педицелларии
Рис. 7.18. Педицелларии (по Mortensen, 1928 -
1951; Hyman, 1955)
Рот Нерв Гемальная ральная
система система
Рис. 7.19. Схематический разрез тела и части
руки морской звезды (по Nichols, 1962)
7.3.3.3. Класс Ophiuroidea
(змеехвостки, или офиуры)
Как и морские звезды, которым они
родственны, офиуры плоские, свободноживу-
щие иглокожие со ртом на нижней стороне
тела. Однако у этой группы тело мелкое,
дискообразное и резко отграниченное от
длинных узких рук (рис. 7.20). Известковый
скелет занимает большую часть их объема
и также хорошо развит в центральном
диске. Пластинки рук слиты между собой,
образуя продольный ряд сочлененных друг
с другом «позвонков», способных двигаться
за счет «межпозвонковых» мышц. С
помощью такого механизма офиуры могут
ходить на своих руках; центральный диск при
этом часто приподнят над субстратом.
Соответственно амбулакральные ножки
лишены локомоторных функций и только
помогают рукам опираться на субстрат; в
основном они используются для захвата пищи.
Амбулакральная борозда погружена
внутрь скелетных пластин. В отличие от
астероидей мадрепоровая пластинка
у офиур расположена на оральной стороне
тела.
Многие офиуры—слизисто-ресничлые
фильтраторы, у некоторых из них пять
основных рук дихотомически ветвятся.
Однако среди змеехвосток обычны и детритофа-
ги, и трупоеды. У всех имеется крупный
желудок, но кишечник и анус отсутствуют.
Между желудком и оральной поверхностью
находятся 10 впячиваний нижней
поверхности тела—бурс (рис. 7.21). Это органы
газообмена. Вода попадает в них через
расположенные вокруг рта щели за счет биения
ресничек или мышечного накачивания.
В бурсы открываются гонады и здесь же
часто развиваются яйца. У некоторых видов
эмбрионы прикреплены к стенке бурс, так
что происходит своего рода
«живорождение». Развитие прямое или с личиночной
стадией—офиоплутеусом (рис. 7.14,5).
Около 2000 видов разделяют на три
отряда.
7.3.3.4. Класс Concentricycloidea
В 1986 г. по девяти экземплярам одного
нового вида, найденным на затопленной
древесине на глубине около 1000 м у
побережья Новой Зеландии, был описан новый
класс необычных медузоподобных
иглокожих. Тело Xyloplax medusiformis диаметром
8 мм имеет форму плоского диска,
окруженного шипами (рис. 7.22). Рук и каких-
либо признаков пищеварительного тракта
нет. Верхняя поверхность диска покрыта
чешуевидными пластинками, а нижняя несет
два концентрических кольца амбулакраль-
Deuterostomia: «торичноротые
199
Рис. 7.20. Форма тела различных ocbnvp (no источникам в Hvman. 1955}
Диск
Полость тела
Желудок
Гонада
Бурса
Каменистый канал
и осевой орган
Позвонки
Мадрепоровая Рот Нерв Гемальная Амвулакральная
пластинка система система
Рис. 7.21. Схематический разрез лиска и части руки осЬиуры(по Nichols. 1962)
200
Глава 7
Рис. 7.22. Вид с дорсальной (А) и вентральной
(б) сторон представителя концентрициклоидей
Xyloplax (no Baker et al, 1986)
ных каналов, от которых отходит
единственное краевое кольцо амбулакральных
ножек. Пять пар гонад; прямое развитие
в молодь, сходную по строению со
взрослыми особями, идет, по-видимому, в
бурсах.
7.3.3.5. Класс Echinoidea
(морские ежи)
Эхиноидеи—сферические или вторично
уплощенные свободноживущие иглокожие
без рук (рис. 7.23) (руки в определенном
смысле включены в состав тела). Для них
характерен жесткий панцирь из
соединенных друг с другом скелетных пластинок.
Каждая амбулакральная и
интерамбулакральная зоны несут в принципе по две
вертикальные дуговидные полосы таких
пластинок, идущих от аборального полюса ко
рту. На панцире находятся педицелларии
(см. разд. 7.3.3.2) и подвижные иглы, с
помощью которых некоторые виды могут
передвигаться по поверхности субстрата
или зарываться в него. Помимо этого локс-
моция происходит за счет амбулакральных
ножек, выходящих сквозь поры в
амбулакральных пластинках.
А Вид сьоку
1
■\
77^"
В Вид своку
Г Вид с оральной стороны Л Вид с аворальной стороны
Вид своку
Рис. 7.23. Форма тела морских ежей: А, Б—«правильные»; В—Д—«неправильные» (по
нескольким источникам, в основном в Hyman, 1955)
Deuterostomia: вторичноротые
201
Проксимальные
концы зувов
Мышцы
Пирамиды, удерживающие
зуьы
Дистальные концы зувов
Рис. 7.24. Аристотелев фонарь (по Нутап,
1955)
Расположенный на нижней стороне тела
рот снабжен характерным грызущим
аппаратом из пяти крупных и нескольких мелких
пластинок— «аристотелевым фонарем»
(рис. 7.24), с помощью которого
пережевываются водоросли и сидячие животные. Для
этого «фонарь» выдвигается изо рта.
У «правильных» видов эпифауны рот
находится в центре нижней поверхности тела,
а анус—посередине аборальной.
Некоторые виды эхиноидей роются в рыхлых
осадках и вторично стали билатерально
симметричными («неправильные» морские ежи).
У таких форм анус заметно смещен к
«заднему концу» тела, а рот иногда отчасти
сдвинут к «переднему». Они детритофаги,
в связи с чем аристотелев фонарь
редуцируется или отсутствует, а питание
осуществляется с помощью специализированных
слизисто-ресничных амбулакральных
ножек.
Обширная полость тела (рис. 7.25)
образует выросты в виде сильно
модифицированных амбулакральных ножек (у
«неправильных» ежей) или в виде
околоротовых жабр (у «правильных» видов),
служащих для газообмена. Четыре (у
«неправильных») и пять (у «правильных» форм)
крупных гонад, вдающихся в полость тела,
открываются наружу аборальными гонопора-
ми. У некоторых наблюдается забота о
потомстве, однако у большинства развитие
непрямое с личинкой офиоплутеусом
(рис. 7.14,Г)-
Классификация по отрядам базируется
в основном на вымерших формах; в 15
современных отрядах около 950 ныне
живущих видов.
7.3.3.6. Класс Holothuroidea
(голотурии, или морские огурцы)
Голотурии лишены рук (как и у морски>
ежей они включены в тело), а их
билатерально симметричное тело, сильно вытянутое
вдоль орально-аборальной оси, лежит как
бы на боку. При этом одна его сторона
с тремя рядами ножек, амбулакрами,
(тривиум) является «вентральной», а
противоположная, с двумя амбулакрами (биви-
ум)—«дорсальной» (рис. 7.26). Стенка тела
также необычная, кожистая, с хорошо
развитыми кольцевыми и пятью продольными
Гонада
Кишка
Аристотелей
сронарь
Рис. 7.25. Схематический
разрез морского ежа (по Nichols,
1962)
Мадрепоровая
пластинка
Анус
Каменистый канал
и осевой орган
Амбулакральная
система
202
t Лг|ва ,'
Рис. 7.26. Форма тела голотурий (по нескольким источникам, в основном в Hyman, 1955)
мышечными пучками. Известковый скелет булакральная система поддерживает от 8 до
редуцирован до изолированных микроско- 30 пальцевидных, разветвленных или щит-
пических «косточек». ковидных ротовых щупалец, использую-
Вокруг находящегося «спереди» рта ам- щихся для питания детритом или, реже, для
Deuterostomia: вторичноротые
203
Мадрепоровая Гонада
пластинка
Полость тела
Клоака
-Анус
Водное легкое
Рис. 7.27. Схематический разрез голотурии (по Nichols, 1962)
фильтрования (рис. 9.3 и 9.6). Сквозной,
часто очень длинный пищеварительный тракт
заканчивается «сзади» прямой кишкой или
клоакой и затем анусом. В клоаку у многих
видов открывается пара обширных впячи-
ваний, лежащих в полости тела; это
разветвленные водные легкие. Вода заполняет
их и с силой выталкивается обратно,
обеспечивая газообмен (рис. 7.27). Некоторые
голотурии при раздражении выбрасывают
заднюю кишку включая водные легкие
через анус (эвисцерация). У нескольких видов
при раздражении выбрасываются
специальные расположенные около легких клейкие
или содержащие токсин кювьеровы
трубочки. Мадрепоровая пластинка лежит
свободно в полости тела.
Локомоция очень медленная, у
большинства видов с помощью
амбулакральных ножек, которые часто разбросаны по
поверхности тела, а не ограничены
определенными зонами (амбулакрами).
Некоторые группы полностью лишены ножек
и движутся за счет перистальтических
сокращений мышц, отталкиваясь от осадка
заякоривающимися в нем, выступающими
из тела мелкими известковыми
«косточками». Некоторые голотурии—сидячие
животные, использующие амбулакральные
ножки больше для прикрепления, чем для
локомоции. У ряда глубоководных форм
сильно увеличенные и удлиненные
амбулакральные ножки функционируют как
ходули (см. рис. 7.28). Большинство видов—
представители эпифауны, хотя некоторые
роются в рыхлых осадках, а одна группа
включает планктонные формы.
Обычно непарная гонада открывается
гонопором около ротовых щупалец.
Распространена забота об эмбрионах, которые
у некоторых форм развиваются в полости
тела, а у одного вида—в яичнике.
Некоторые—протандрические гермафродиты.
Если отсутствует забота о потомстве,
развитие непрямое с личиночными стадиями ви-
телларии (или аурикуларии) и затем долио-
ларии (рис. 7.14,Д).
Амвулакральные ножки
Рис. 7.28. Голотурии, идущие по
океаническому дну с помощью крупных удлиненных
амбулакральных ножек
204
Глава 7
По форме ротовых щупалец, характеру
амбулакральных ножек и форме тела
выделяют шесть отрядов, объединяющих около
1150 ныне живущих видов.
7.4. Тип Chordata (хордовые)
7.4.1. Этимология
От лат. chorda—струна (из кошачьей
кишки).
7.4.2. Диагностические
и специфические признаки
Тип хордовых включает три подтипа, из
которых два относятся к беспозвоночным,
поэтому обсуждаются в настоящей книге.
Эти два подтипа достаточно сильно
отличаются друг от друга и будут рассмотрены
по отдельности. Тем не менее у них
следующие общие признаки.
1. Билатерально симметричные
животные.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и органами.
3. Кишечник сквозной, анус не
терминальный.
4. Тело в принципе мономерное, без
различимой головы, конечностей или
челюстей.
5. Обширная глотка со стенками,
перфорированными насквозь глоточными
щелями, число которых варьирует от нескольких
до очень многих.
6. Слизисто-ресничные фильтраторы
(см. разд. 9.2.5); ток воды, приносящий
пищевые частицы, покидает тело через
глоточные щели.
7. Обычно глоточная область окружена
вторичной стенкой тела, образующей
атриальную полость, связанную с
окружающей средой единственной порой (см.
рис. 9.14).
8. Эпидермис не секретирует наружной
кутикулы и не несет ресничек.
9. На некоторых или на всех стадиях
жизненного цикла присутствует внутренний
дорсальный скелетный тяж—хорда.
10. На некоторых или на всех стадиях
жизненного цикла дорсальнее хорды
проходит нервная трубка.
11. На некоторых или на всех стадиях
жизненного цикла органом активного
плавания служит мускулистый постанальный
хвост.
12. Циркуляторная система частично не
замкнута.
Глоточные
пер
А Глотка Желудок Нервная трубка
Сердце
Эндостиль □ Атриальное отверстие
Пищевой желовок
Просвет глотки
Эндостиль
Атриум
Срез на уровне А - В
Глоточная и первичная
стенки тела
Вторичная стенка тела
Рис. 7.29. План строения
обобщенного беспозвоночного
хордового
Deuterostomia: вторичноротые
205
13. Дробление яйца радиальное.
14. Развитие, характерное для вторич-
норотых, обычно недетерминированное
и непрямое.
15. Морские формы.
7.4А. Подтип Urochordata
(личиночнохордовые,
или оболочники)
7.4А.1. Диагностические
и специфические признаки
1. Хорда, нервная трубка и
постанальный хвост (если есть) только на личиночной
стадии и у одной неотенической группы.
2. Целомической полости нет.
3. Экскреторных органов нет.
4. Мышечные и другие структуры не
сегментированы.
5. Тело целиком заключено внутри се-
кретируемой туники, панциря или
«домика», обычно состоящих из целлюлозы и
белка и содержащих мигрировавшие из тела
клетки, а у некоторых форм—
экстра-корпоральные кровеносные сосуды.
6. Пищеварительный тракт U-образной
формы с обширной глоткой, обычно
занимающей большую часть объема тела.
7. Нервная система в виде ганглия,
расположенного между ртом и атриальным
отверстием, с отходящими от него
отдельными нервами.
8. Гермафродиты, обычно с непарными
семенником и яичником.
Оболочники—сидячие или свободножи-
вущие. одиночные или колониальные филь-
траторы с признаками хордовых только на
стадии свободноплавающей личинки.
7.4А.2. Классификация
Около 1400 видов оболочников
распределены между тремя классами.
Класс Отряд
„ Aplousobranchia
. Phlebobranchia
. Stohdobranchia
.Aspiraculata
Ascidiacea
Thaliacea
Larvacea -
Pyrosomida
Doliolida
Salpida
-Copelata
7.4A.2.1 Класс Ascidiacea
(асцидии)
Этот класс включает около 1250 сидячих
бентосных видов. Обычно имеется свобод-
ноживущая планктонная, похожая на
головастика личинка (рис. 7.30), которая в
конце концов оседает на дно и прикрепляется
головным концом к субстрату. Затем
неравномерный рост в течение метаморфоза
фактически приводит к повороту передне-
задней оси тела на 180°. так что рот и
окружающий его жаберный сифон туники или
панциря оказываются на противоположном
точке прикрепления конце тела (рис. 7.31).
Этот жаберный сифон ведет через
окруженное щупальцами отверстие в обширную
глотку, обычно перфорированную
большим количеством мелких щелей (за
исключением глубоководных видов из отряда
Aspiraculata, у которых глотка
редуцирована, щели отсутствуют, а жаберный сифон
модифицирован в ряд ловчих выростов).
Щупальца вокруг отверстия, ведущего
в глотку, используются для того, чтобы
Туника Нервная трувка Атриальный Глотка Жаберный сиерон
СИфОН
Рис. 7.30. Анатомия головасти-
коподобной личинки асцидий
(по McFarland et al., 1979 и др.)
Прикрепительные
органы
Хорда Кишечник Сердце Эндостиль
206
Глава 7
Рис. 7.31. Метаморфоз голова-
стикоподобной личинки (по
Barnes, 1980)
предотвращать попадание в нее крупных
частиц. Атриальное отверстие также
оформлено в виде сифона, находящегося
рядом с жаберным сифоном (рис. 7.32).
Сердце расположено базально и, что
необычно, способно выталкивать кровь в двух
периодически сменяемых направлениях.
Кровь интересна тем, что в некоторых ее
клетках высока концентрация связанных
с сульфат-ионами тяжелых металлов,
особенно ванадия (может быть в 105—106 раз
выше, чем в морской воде), железа, ниобия.
Распространено бесполое размножение
почкованием. Оно происходит несколькими
способами от различных частей тела асци-
дий. Как и у неродственных асцидиям кни-
дарий и мшанок, отпочковавшиеся особи
могут оставаться связанными друг с другом
в колонии, однако полиморфизма зооидов
не наблюдается. Некоторые колониальные
формы окружены общим панцирем, у
многих общее атриальное отверстие, другие
связаны базальными столонами; часто у
колоний характерная форма. У некоторых
видов почкование начинается еще на
личиночной стадии. Однако большинство асцидий
одиночные; эти формы часто гораздо
крупнее зооидов колониальных видов, достигая
в высоту более 15 см (рис. 7.33).
Кроме аберрантных Aspiraculata
выделяют еще три отряда асцидий, в основном
по размещению гонад и структуре
глоточной стенки. У исключительно
колониальных Aplousobranchia гонады окружены
петлей кишечника, стенка глотки простая. У,
как правило, одиночных Stolidobranchia они
погружены в стенку тела и лежат вдоль
глотки с продольными складками и
внутренними перегородками. У также большей
частью одиночных Phlebobranchia гонады
расположены, как у Aplousobranchia, глотка
без складок, но с характерными
продольными перегородками внутри, образованными
раздвоенными сосочками.
Приносящее
течение
Жаверныи сисра
Ротовые
щупальца
Ганглии
Выносящее
if течение
-Атриальный
сифон
-Анус
-Панцирь или туниа
-Гонады
777777777777777777777777/^^
Форма атриальной
полости
Срез на уровне А - В
Просвет глотки
Рис. 7.32. Схематический разрез асцидий (по
нескольким источникам)
Deuterostomis: вторичноротие
207
Колониальные формы
Одиночные формы
Рис. 7.33. Форма тела и колоний асцидий (по Millar, 1970 и др.)
7.4А.2.2. Класс Thaliacea
(пелагические оболочники)
Талиацей 70 видов, все они планктонные
и используют ток воды, несущей пищевые
частицы, для реактивного движения.
Жаберное и атриальное отверстия
расположены на противоположных концах их верете-
новидного или боченкообразного тела.
У всех форм наблюдается бесполое
почкование от вентрального столона, отходящего
фазу за глоточным эндостилем.
Наблюдаются два различных образа
жизни. Отряд Pyrosomida (огнетелки)
включает колониальных талиацей, которые
помещаются в общем цилиндрическом
домике с центральным просветом, открытым во
внешнюю среду только с одного конца.
Каждый зооид расположен так, что
жаберное отверстие обращено вовне общего
цилиндра, а атриальное — в его внутреннюю
полость, куда, таким образом, попадают
все пищевые токи воды, и вся трубчатая
колония длиной до нескольких метров движе-
Овщая полость
колонии
Отдельные зооиды
Общий
панцирь
Ганглий
Жаверный
СИфОН
Перфориро-
Анус
Атриалыная
-. -Желудок.
^—Яичник
Семенник
Общая полость
Атриальный
СИфОН
колонии
Атриальный
СИфОН
Выносящее
течение
Глоточная Анастомозирующие и неполные
перегородка мышечные кольца
Ганглий
Рис. 7.34. Схематический
разрез колонии (А) и отдельного
зооида (б) пиросомы (по Gras-
se, 1948; Fraser, 1982)
серный сифон
Семенник
Желудок Сердце Атриум
Панцирь-чехол
Эндостиль
Одиночная Боковая
глоточная перфорация
Атриальньп
сифон
Жаберный
сифон
Столон
Мутовки развивающихся почек
Рис. 7.35. Схематичная
анатомия двух стадий жизненного
цикла сальп: А—
размножающееся половым
путем агрегированное
«поколение»; Б — размножающееся
бесполым путем одиночное
«поколение» (по Berill, 1950;
Fraser, 1982 и др.)
Deuterostomia: вторичноротые
209
Ганглий Глоточные Мышечные ободья
перфорации
Глотка
Жаберный—
СИфОН
Атриум
Атриальный
сифон
Анус
Эндостиль CeMeHJK Сердце Желудок Яичник
Развивающиеся почки
Рис. 7.36. Схематичная
анатомия двух стадий жизненного жаБернЬ|и
цикла боченочников: А—сифон
размножающееся половым
путем одиночное «поколение»;
Б—размножающееся
бесполым путем агрегированное
«поколение» (по Berrill. 1950;
Fraser, 1982 и др.) Б
Столон
Атриальный
сифон
Почки,
мигрирующие от столона
к кадофору
тся как единое целое, напоминая крупную
морскую змею (рис. 7.34). У отдельных
зооидов множество глоточных перфораций,
внешне сходных с имеющимися у асцидий.
Отряды Salpida (сальпы) и Doliolida (бо-
ченочники) — в основном одиночные
животные, хотя у них происходит чередование
одиночных форм и агрегаций различных
половых типов. У сальп (рис. 7.35) одиночная
стадия размножается бесполым путем,
давая цепочки особей, которые в конечном
итоге отделяются в виде колонии полового
«поколения». У долиолид половым путем
размножается, напротив, одиночная стадия,
образующая бесполое агрегированное
«поколение» (рис. 7.36). В обоих отрядах
наблюдается полиморфизм зооидов (на
различных стадиях жизненного цикла),
имеются мощные ленты или обручи мышц,
идущие вокруг тела и служащие для
выталкивания воды (вместо ресничного механизма ее
продвижения), более или менее прозрачный
слизистый чехол и небольшое число
крупных глоточных перфораций. У сальп всего
две огромные глоточные щели (рис.
9.15).
Личиночные стадии есть только у
боченочников; от них, как обычно, считается,
произошли путем педоморфоза
представители следующего класса.
7.4А.2.3. Класс Larvacea
(аппендикуларии)
Строение этих планктонных форм
характерно для личинок оболочников (рис.
7.37): мелкое тело с крупным хвостом, по
всей длине которого проходят хорда и
нервная трубка. Общая длина животных около
5 мм. Эпидермальные железы секретируют
14—1314
210
Глава 7
Эндостиль'
Глоточные щели
Яичник
Семенник
Кишечник
■Хорда
Рис. 7.37. Анатомия аппендикуларии (по Alld-
redge, 1976)
Задний
конец
тонкий студенистый (нецеллюлозный)
«домик», образующий внешний
фильтрационный аппарат (рис. 7.38). Вода поступает
в домик за счет биения хвоста через особые
сита, и пищевые частицы, в основном нано-
планктонные водоросли,
отфильтровываются тонкой сетью внутри студенистой
оболочки (см. рис. 9.5), а затем заглатывается
с помощью тока воды, создаваемого
глоточными ресничками. Вода выходит из
пищеварительного тракта через пару
глоточных щелей, открывающихся независимыми
отверстиями на поверхности тела. Роль
атриальной полости в результате играет
пространство внутри «домика». Когда его
сита и фильтры необратимо засоряются,
аппендикуларии могут покидать его через
«дверь» (в норме закрытую) и «надувают»
новый уже заложенный к этому моменту
домик.
Фильтрационная
система
Сито
Приносящее
течение
Животное внутри
домика
Вид своку
Передний
конец
Отверстие для выхода
из домика
Передний конец
Приносящее течение
Рис. 7.38. «Домик»
аппендикуларии и индуцируемые
питающие течения сквозь него (по
Hardy, 1956 и др.)
Задний конец
Приносящее течение
Выносящее
течение
Вид с дорсальной стороны
Deuterostomia: вторичноротые
211
Все аппендикуларии—одиночные
формы, размножение только половое. Один
вид, что необычно для урохордат,
раздельнополый. Всего один отряд, включающий
70 видов.
7.4Б. Подтип Cephalochordata -
(головохордовые,
или ланцетники)
7.4Б.1. Диагностические
и специфические признаки
(рис. 7.39)
1. Тело, сжатое с боков, рыбообразное.
2. Хорда проходит по всей его длине.
3. Полый дорсальный нервный ствол
тянется почти вдоль всего тела, не
расширяется спереди в мозг.
4. Постоянный постанальный хвост.
5. Тело с сериально повторяющимися
мышечными блоками, нервами,
выделительными органами и гонадами.
6. Полость тела формируется энтероце-
льно из множества сериально
повторяющихся карманов. Их вентральные части
Миомеры
Хорда
сливаются, а дорсальные вытесняются
мышцами.
7. Экскреторная система очень сходна
с протонефридиальной, но формируется
перитонеальными клетками.
8. Глоточный отдел покрыт вторичной
стенкой тела, образованной парой метапле-
вральных складок, растущих в вентральном
направлении и срастающихся по средней
вентральной линии тела.
9. Глотка обширная, занимающая
половину длины тела.
10. Раздельнополые животные с
многочисленными парными гонадами.
Головохордовые — мелкие (до 10 см
длиной) свободноживущие животные,
ведущие бентосный и малоподвижный образ
жизни, но способные плавать, меняя места
питания или спасаясь от хищников. При
этом хорда используется как несжимаемая,
но гибкая опора. Сокращение продольно
ориентированных мышц приводит к
волнообразному изгибанию тела, а не к его
укорочению (эквивалентно системе движения,
наблюдаемой у хетогнат и нематод).
Пищедобывательная система голово-
хордовых в сущности такая же, как у обо-
Мерцательный орган
Анус
Атриопор Гонады Эндостиль Глотка
Ротовые цирры
Миомеры
Просвет
глотки
Атриум
Вторичная
стенка тела
П1 Камера "плавникового луча"
Нервная трувка
Хорда
Глоточные
щели
Эндостиль
Миомеры
Рис. 7.39. Головохордовые: А— схематический продольный разрез; Б—прижизненный внешний
облик (по Young, 1962)
14*
212
Глава 7
Камеры
^плавниковых
лучей1'
Миомеры
Ж.
Мерцательный орган
Парус
Ротовые цирры
Эндостиль
Глотка
Рис. 7.40. Передний конец го-
ловохордового (по Young,
1962)
лочников. Ротовой отдел окружен рядом
щупалец (ротовыми циррами и веларными
щупальцами —рис. 7.40), которые
предотвращают попадание внутрь нежелательных
частиц. Глотка удлиненная, а не
бочкообразная и атриопор расположен на средней
вентральной линии тела сразу перед анусом.
Домика, окружающего тело и вытянутого
в сифоны, нет.
В некоторой степени головохордовые
являются связующим звеном между
беспозвоночными и третьим подтипом
хордовых— позвоночными (Vertebrata), особенно
с бесчелюстными, также лишенными
конечностей и челюстей. У личиночной стадии
миног, как и у ланцетника, слизисто-
ресничная фильтрующая глотка, хотя в
данном случае вода выталкивается из нее с
помощью мышечного сокращения. Однако
более очевидны отличия головохордовых
от позвоночных. Они состоят в отсутствии
дифференцированной головы с парными
сенсорными органами, мозгом и
защищающим его черепом, в особенностях строения
нервной трубки, к которой мышечные
клетки посылают свои отростки (формируя
ложные вентральные корешки), и
периферических нервных волокон, лишенных миелино-
вой оболочки, в очень тонком однослойном
эпидермисе. Тем не менее общие черты
головохордовых, вероятно, соответствуют
организации «протопозвоночных».
Очевидно также сходство ланцетников с
увеличенным неотеническим вариантом личинки
урохордат, во всем прочем похожих на
беспозвоночных.
7.4Б.2. Классификация
Известно 25 видов, помещаемых в один
класс и один отряд.
7.5. Дополнительная
литература
Aharino А. 1965. Chaetognaths. Oceanogr. mar.
Biol., Ann. Rev., 3, 115—194 [Chactognatha].
Baker A.N., Rowe F.W.E., Clark H.E.S. 1986.
A new class of Echinodermata from New Zealand.
Nature, Lond., 321, 862 864.
Barrington E.J. W. 1965. The Biology of Hemichor-
data and Protochordata. Oliver & Boyd,
Edinburgh [Hemichordata, Chordata].
Berrill N.J. 1950. The Tunicata. Ray Society,
London [Urochordata].
Hyman L. H. 1955. The Invertebrates (Vol. 4)
Echinodermata. McGraw-Hill, New York
[Echinodermata].
Hyman L.H. 1959. The Invertebrates (Vol. 5)
Smaller Coelomate Groups. McGraw-Hill, New York
[Hemichordata, Chaetognatha].
Nichols D. 1969. Echinoderms, 4th edn. Hutchinson,
London [Echinodermata].
Young J.Z. 1981. The Life of Vertebrates, 3rd edn.
Clarendon Press, Oxford [Chordata].
ГЛАВА О
БЕСПОЗВОНОЧНЫЕ С КОНЕЧНОСТЯМИ:
ЧЛЕНИСТОНОГИЕ И СХОДНЫЕ ГРУППЫ
Tardigrada
Pentastoma
Onychophora
Chelicerata
Uniramia
Crustacea
За исключением первичноротости, для
представителей шести рассматриваемых
в данной главе типов фактически общими
являются только два признака — парные
конечности по всей длине тела или какой-то
ее части (каждая пара иннервируется
обычно парными ганглиями, расположенными
на продольных нервных тяжах) и
псевдоцелом, часто заполненный гемолимфой и
потому называемый гемоцелем.
У представителей трех типов
(Tardigrada, Pentastoma и Onychophora) жесткого
наружного (экзо-) скелета нет; полость их тела
играет роль гидростатического скелета;
в сущности эти животные—черви с
мягкими, мясистыми, нечленистыми
конечностями, вооруженными коготками, т.е. с
пальцевидными выростами тела, приводимыми
в движение внешними по отношению к ним
мышцами. В гл. 2 мы допустили, что таких
«червей с конечностями» можно было бы
считать «протоартроподами» и реликтами
исходной волны «артроподизации»
(приобретения членистых конечностей) в
докембрии или в самом начале кембрия. Однако
если онихофоры и кажутся родственными
некоторым истинным членистоногим (од-
новетвистым), особенности тихоходок так
же сильно, если не сильнее, сближают их
с (андестральными?) псевдоцеломическими
животными, как и с другими
протоартроподами, а истинные родственные связи
пятиусток по меньшей мере туманны, в
значительной степени из-за их
специализированного паразитического образа жизни.
У «истинных» членистоногих (Crustacea,
Chelicerata и Uniramia) жесткий, членистый,
склеротизированный кутикулярный экзо-
скелет из хитина и белка, иногда
пропитанный карбонатом кальция (рис. 8.1). Он
покрывает все тело, включая членистые
конечности (слово артроподы, переводимое как
«членистоногие», происходит от греч. arth-
ron—сустав, podos—ноги) (рис. 8.2). Как
и у других животных, наличие кутикулы
затрудняет рост, который требует ряда линек.
Особенно остро эта проблема встает
именно у артропод, где кутикула выполняет
также роль скелета. Во время их линьки
старый скелет частично резорбируется, затем
сбрасывается, а новый мягкий скелет,
образующийся под ним, растягивается
(благодаря поступлению в тело воздуха или воды)
и отвердевает. В этот период животные
особенно уязвимы, поэтому линька часто
осуществляется в убежищах. Изначально экзо-
скелет, вероятно, представлял собой ряды
защитных кутикулярных пластин или
обручей, как, например, у киноринхов (разд. 4.6)
и некоторых тихоходок (разд. 8.1). Позднее
он адаптировался к выполнению скелетной
функции, частично заменив анцестральный
гидростатический псевдоцель. В некоторых
группах эта замена так и осталась
частичной, поскольку ряд членистоногих все еще
использует гидростатическое давление для
выпрямления конечностей, а систему экзо-
скелет-мышцы - только для сгибания их.
Исходно тело членистоногого мономер-
но, хотя с каждой парой ног связана
заметная метамерия стенки тела, экзоскелета
214
Глава 8
Эпидермис
Тонофивриллы,
пронизывающие
клетки эпидермиса
Эпикутикула
Отвердевшая и иногда
ОБЫзвествленная экзокутикула
Неовызвеотвленная и
гибкая эндокутикула
Эпидермис
Базальная мембрана
Железистая Клетка, Образующая
клетка щетинку
Рис. 8.1. Разрез кутикулярного зкзоскелета членистоногого, демонстрирующий различные слои
и внутренний вырост (аподему), выполняющий функцию внутреннего скелета (по Hackman, 1971
и др.)
и некоторых внутренних структур (см..
например, рис. 2.10). В сущности артропод-
ный план строения следующий: небольшой
передний отдел (акрон) и эквивалентная ему
задняя часть (тельсон), не несущие
конечностей, а между ними ряд промежуточных
отрезков (сегментов), каждый из которых
с парой конечностей и сериально
повторяющимся набором обслуживающих их
структур (мышечных, нервных, скелетных и т. д.).
При этом не связанные с конечностями
органы (например, выделительная и
репродуктивная системы) не метамеризуются. Во
многих линиях членистоногих
прослеживается значительное слияние несущих
конечности сегментов, утрата придатков и/или диф-
ференцировка различных отделов тела;
особенно широко распространено
преобразование передних конечностей в пищезахваты-
вающий аппарат. Однако кроме частичной
Плейрит
Членистая конечность
Рис. 8.2. Поперечный разрез
тела членистоногого,
демонстрирующий различные экзо-
скелетные пластинки и
членистую конечность. Число
отдельных члеников в конечности
в разных группах неодинаково;
в каждом типе они носят
особые названия
Беспозвоночные с конечностями: членистоногие и др.
215
или полной модификации кутикулы в экзо-
скелет (и вытекающих отсюда
морфологических особенностей, например, членистых
конечностей), отсутствия ресничек и
тенденции к развитию сложных глаз, у артропод
мало общих особенностей, поэтому весьма
вероятно, что членистоног ость—скорее
уровень организации, а не отличительный
признак единой филогенетической линии.
К артроподам относится подавляющее
большинство видов животных, главным
образом в результате успешного заселения
этой группой суши и той легкости, с
которой может идти видообразование у мелких
наземных организмов. Успех
членистоногих как обитателей суши, резко
отличающий их от большинства групп
беспозвоночных, вероятно, в значительной степени
обусловлен эволюцией влагосберегающих
органов выделения и газообмена, а также
развитием препятствующей испарению
непроницаемой эпикутикулы.
8.1. Тип Tardigrade
(тихоходки)
8.1.1. Этимология
От лат. tardus—медленный, gradu—
шаг.
8.1.2. Диагностические
и специфические признаки
(рис. 8.3)
1. Мелкие, коренастые, билатерально
симметричные животные.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и органами.
3. Кишечник сквозной, прямой.
4. Тело мономерное, хотя с четырьмя
парами коротких нечленистых конечностей
с коготками, служащих для ползания
(рис.8.3,^4); конечности обслуживаются
внешними по отношению к ним мышцами
и иннервируются сериально
повторяющимися парами нервных ганглиев.
5. Скелет гидростатический, образован
хорошо развитым псевдоцелем.
6. Тело покрыто кутикулой, но
мышечных слоев под эпителием нет; в целом нехи-
тинизированная кутикула сбрасывается при
линьке; ее поверхность часто скульптуриро-
вана шипиками и/или чешуйками; тело
пронизано сетью отдельных гладких
мышечных волокон.
7. Число клеток тела строго
фиксировано (эвтелия).
8. Мускулистая сосущая глотка
выстлана хитиновыми чешуйками (плакоидами);
Рис. 8.3. Строение тихоходок:
А. Внешний облик животных,
ползающих по поверхностиСтенка тела-
нитчатой водоросли (по
Marcus, 1929). Б. Схема
продольного разреза
генерализованной тихоходки (по Cuenot,
1949)
"Мальпигиевы
сосуды"
Стилет
216
Глава 8
для поражения добычи из ротовой полости
может выдвигаться пара стилетов.
9. Органов кровообращения и
газообмена нет.
10. У ряда видов выделительная
система состоит, вероятно, из трех «мальпигие-
вых сосудов».
11. Нервная система с головным
мозгом и вентральной парой стволов с
ганглиями, иннервирующими конечности.
12. Раздельнополые животные, иногда
наблюдается партеногенез; гонада
непарная.
13. Развитие прямое.
14. Свободноживущие обитатели интер-
стициали пресных и соленых вод, а также
водной пленки на наземных растениях;
ведут скрытный образ жизни.
Для тихоходок характерна
специфическая смесь признаков: как и у других про-
тоартропод, у них парные конечности с
Kofi ид СБОКУ
«боченка» у тихоходок (по
Рис. 8.4. Стадия
Morgan, 1982)
готками; как у лофофоровых и вторичноро
тых, в их развитии, хотя и кратковременно,
прослеживается стадия парных (пять пар)
энтероцельных карманов; с псевдоцеломи-
ческими формами их сближает общий
уровень организации и образ жизни.
Хотя ряд видов обитает в относительно
стабильных биотопах типа морского песка,
Вид о дорсальной
стороны
Вид с вентральной
стороны
Рис. 8.5. Разнообразие форм тела, скульптуры кутикулы, числа и формы коготков у тихоходок (по
Morgan, King, 1976 и др.)
Беспозвоночные с конечностями: членистоногие и др.
217
. для многих характерна жизнь во временных
водных пленках и текучей воде. У
последних для переживания неблагоприятных усло-
^ вий развились различные устойчивые ста-
|дии. Например, когда водная пленка на
листьях мха высыхает, через проницаемую
кутикулу тихоходок также происходит
испарение влаги. Так они могут терять большую
часть жидкости тела, превращаясь в мелкие
1 сухие «бочоночки» (рис. 8.4), которые
\ остаются жизнеспособными до 10 лет (воз-
' можно, и дольше). В этом состоянии их
потребление кислорода в 600 раз меньше
исходного, они способны выдерживать более
8 ч температуру — 272° С и
кратковременное нагревание до 150" С. Менее суровые
условия переносятся благодаря: (а) инци-
стированию (животное втягивает
конечности, сбрасывает кутикулу, как при линьке,
и сворачивается в шарик внутри кутикуляр-
ной капсулы); (б) образованию толстостен-
* ных покоящихся яиц. Возможно, что с
учетом таких анабиотических фаз продолжите-
' льность жизни тихоходок может превы-
I шать 60 лет.
Все тихоходки —сосущие организмы,
хотя их кормовой спектр широк: от клеток
водорослей или наземных растений до
таких интерстициальных или ведущих
скрытный образ жизни животных, как
коловратки, нематоды или даже другие тихоходки.
Несколько видов—паразиты, один из
них—в желудке брюхоногих моллюсков.
Животное прикладывается ротовым отвер-
\ стием к пищевому объекту и прокалывает
его двумя выдвигающимися при помощи
мышц стилетами, после чего благодаря
насасывающему действию глотки в желудок
перекачиваются жидкости и клеточные ор-
ганеллы. Глоточные плакоиды, вероятно,
служат для измельчения заглатываемых
твердых частиц.
1
i 8.1.3. Классификация
* 400 современных видов (длина тела
0,05—1,2 мм) слабо различаются
анатомически, если не считать орнаментации
кутикулы, формы и числа коготков (рис. 8.5); все
виды относятся к одному классу.
8.2. Тип Pentastoma
(пятиустки)
8.2.1. Этимология
От греч. pente—пять; stoma — рот.
8.2.2. Диагностические
и специфические признаки
(рис. 8.6)
1. Билатерально симметричные,
уплощенные, червеобразные животные.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и органами.
3. Кишечник сквозной, прямой.
4. Тело мономерное, хотя внешне
кольчатое; на переднем конце две пары «ног»
с коготками или просто две пары коготков
(рис. 8.7).
5. Псевдоцель в роли
гидростатического скелета.
6. Тело покрыто хитинизированной
кутикулой, пронизанной порами и
сбрасываемой при линьке; под эпителием залегают
слои кольцевой и продольной
поперечнополосатой мускулатуры.
7. Органов выделения, кровообращения
и газообмена нет.
8. Нервная система с головным мозгом,
ганглиями, иннервирующими каждый одну
«ногу» (иногда все пять ганглиев слиты
воедино), и вентральным нервным тяжем '.
9. Раздельнополые животные, одна или
две гонады.
10. Оплодотворение внутреннее путем
копуляции.
11. Развитие с тремя «личиночными
стадиями»; у первой из них три пары
лопастных, ногоподобных, нечленистых
придатков.
12. Паразиты дыхательных путей и
легких позвоночных; питаются кровью хозяев.
Как и у многих эндопаразитов,
основную часть тела пятиусток занимает половая
система и образуется огромное количество
мелких яиц. Оплодотворение внутреннее;
оплодотворенные яйца (одновременно—до
полумиллиона) до момента откладки
остаются в организме самки. Матка, где
1 Нервная цепочка лестничного типа с 4—5
парами ганглиев. Прим. пер.
218
Глава 8
"Педальные железы"
Репродуктивная
система
Кишка
J—Анус
"Нога"
Брюшной
нервный тяж
Стенка тела
Рис. 8.6. Внутреннее строение
самки пятиустки на
схематическом продольном разрезе
(преимущественно по Cuenot,
1949)
идет их накопление, фактически занимает
всю полость тела самки, увеличивающегося
при этом более чем в 100 раз. Развитие
включает последовательно три
«личиночные» или ювенильные стадии, первая из
которых завершается еще до выхода из яйца.
Последнее выходит из тела хозяина через
его пищеварительный тракт (все хозяева —
наземные или пресноводные хищные
позвоночные; 90% видов пятиусток—паразиты
пресмыкающихся). Если личинка, еще
находящаяся внутри яйца, проглатывается
промежуточным хозяином (всеядным или
растительноядным насекомым, рыбой или
четвероногим), она выходит из яйца и
активно движется через ткани позвоночного,
используя для этого три хитиновых стилета
Рис. 8.7. Разнообразная морфология
переднего конца тела пятиусток; видна степень
развития или редукции «ног» (вид с вентральной
стороны) (по Kaestner, 196B)
и короткие толстые конечности; достигнув
специфической области тела (печени и т. д.),
инцистируется и развивается во вторую
личиночную стадию. Если затем зараженный
промежуточный хозяин становится
добычей окончательного (рептилии, птицы или
млекопитающего), появляется третья
личинка, похожая на мелкий вариант взрослой
пятиустки, мигрирующая в легкие или
верхние дыхательные пути. У ряда видов во
время одной из последних личиночных
линек происходит утрата конечностей, на
месте которых остаются лишь терминальные
крючковидные коготки. У личинок многих
видов на каждой ноге изначально пара
коготков; на взрослой стадии их по одному.
8.2.3. Классификация
Описано примерно 100 видов этих
мелких (2—16 см длиной) «червей»; всех их
можно отнести к одному классу.
8.3. Тип Onychophora
8.3.1. Этимология
От греч. onychos—коготки; phoros—
носитель.
8.3.2. Диагностические
и специфические признаки
(рис. 8.8)
1. Билатерально симметричные
животные с удлиненно-цилиндрической,
червеобразной формой тела.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и органами.
3. Кишечник сквозной, прямой. Перед
ним пара ротовых придатков, каждый из
которых несет две когтевидные мандибулы
(образуют внутреннюю и внешнюю че-
Беспозвоночные с конечностями: членистоногие и др.
219
Рис. 8.8. Строение онихофор:
А—схема продольного
разреза; Б—внешний облик (по Se-
dawick, 1888; Cuenot. 1949)
Клеикая
-Антенна железа Сердце Гонада Гемоцель
Глаз
Ротовая
папилла
Выделительный
орган
А
Мозг Кишка Нога
Анус
люстные пластинки). Передняя и задняя
кишка с кутикулярной выстилкой;
выпячивания (дивертикулы) пищеварительного
тракта отсутствуют.
4. Тело с 14—43 парами коротких,
нечленистых, мясистых ног по всей своей
длине. Каждая нога — полое выпячивание
стенки тела с терминальной подушечкой, парой
коготков и внутренней мускулатурой (хотя
движение ноги вызывается сокращением
внешних, т.е. лежащих вне ее мышц);
каждой паре ног соответствует пара остий
сердца и пара выделительных органов.
5. В роли гидростатического скелета
хорошо развитый гемоцель; сердце трубчатое,
других циркуляторных сосудов нет.
6. Тело покрыто очень тонкой, гибкой,
хитинизированной кутикулой. Под
эпителием расположены слои кольцевой, косой
и продольной гладкой мускулатуры.
7. Органы выделения—сериально
повторяющиеся пары мешковидных желез;
передняя пара образует слюнные железы,
задняя—половые протоки.
8. Органы газообмена—просто
устроенные трубчатые трахеи, подходящие
пучками к многочисленным мелким
дыхальцам.
9. Нервная система с головным
мозгом и парой очень широко расставленных
вентральных нервных тяжей, соединенных
9—10 комиссурами в каждом из несущих
конечности «сегментов», но без отчетливых
ганглиев. Органы чувств—пара кольчатых
антенн с небольшим простым глазом в
основании каждой.
10. Раздельнополые животные, гонады
парные, оплодотворение внутреннее с
помощью сперматофоров.
11. Развитие прямое.
12. Свободноживущие наземные
формы.
Длительное время онихофоры вызывали
интерес исследователей главным образом
как живой пример промежуточной стадии
между уровнями организации червей и
членистоногих. Как и у червей, у онихофор
мягкое тело, гидростатический скелет,
ресничные выделительные каналы и слой
гладкой мускулатуры под эпителием. С
членистоногими их сближает наличие
конечностей, трахей, сердца с остиями, продольно
разделенных кровеносных синусов и
челюстей—производных придатков, в данном
случае коготков, которыми оканчиваются
ходильные ноги. Однако тонкая структура
их общих для артропод особенностей явно
указывает на независимую эволюцию
онихофор и всех известных групп
членистоногих. Если они и напоминают внешне,
например, древних одноветвистых, то предками
их считаться никак не могут. Трахеи
онихофор, в частности,— это просто устроенные
в основном неразветвленные трубочки,
открывающиеся пучками через
многочисленные (до 75) дыхальца (рис. 8.9),
разбросанные по поверхности каждого «сегмента»
с конечностями. Челюсти онихофор,
движущиеся в переднезадней плоскости,
действуют независимо друг от друга как
разрывающие пищу приспособления
(благодаря своим заостренным концам), а не в ка-
220
Глава 8
Трахейные
7труБочки
Дыхальце
Рис. 8.9. Дыхальце и пучок трахейных трубочек
(по Clarke, 1973)
честве жующих придатков. К другим
особенностям относится строение вентральных
нервных тяжей с их многочисленными ко-
миссурами, но без «сегментарных»
ганглиев.
Онихофоры—наземные животные,
связанные в основном с влажными
микроместообитаниями. Толщина их
проницаемой для воды кутикулы всего 1 мкм
(меньше, чем у тонкой эпикутикулы
членистоногих, см. рис. 8.1); дыхальца без
замыкающего механизма. Однако потерянная влага
может быть восполнена благодаря
выпячиванию через щели в кутикуле тонкостенных
пузырьков, соприкасающихся с влажными
поверхностями. Онихофоры — в основном
ночные хищники, обнаруживающие добычу
с помощью антенн и охотящиеся даже на
достаточно подвижных животных типа
кузнечиков, разбрызгивая на расстояние до
0,5 м слизеподобное вещество из желез,
открывающихся на парных ротовых сосочках
по бокам рта. Эта слизь почти мгновенно
застывает на воздухе, образуя очень
клейкую ловчую сеть, в которой запутывается
потенциальная добыча. Этот же прием
используется с целью защиты.
8.3.3. Классификация
Известно 70 видов онихофор длиной до
15 см, относящихся к одному классу и отря-
ДУ-
8.4. Тип Chelicerata
8.4.1. Этимология
От греч. chele—клешня; cerata — рога.
8.4.2. Диагностические
и специфические признаки
(рис. 8.10)1
1. Билатерально симметричные артро-
поды длиной от менее 1 мм до 60 см; форма
тела от удлиненной до почти сферической.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и органами.
3. Кишечник сквозной, прямой; от
среднего его отдела отходят от двух до
множества пар слепых пищеварительных
выростов (дивертикулов), секретирующих
ферменты и внутриклеточно
переваривающих и абсорбирующих пищу (они
возникают не как выпячивания эмбриональной
кишки, а в результате разделения желтка
зародыша до дифференцировки
пищеварительного тракта); рот на переднем конце тела
с вентральной стороны.
4. Тело разделено на два отдела:
передний («просома») образован акроном и
шестью сегментами, несущими придатки,
полностью или частично прикрыт карапаксом;
задний отдел («опистосома») без придатков
или же они сильно модифицированы.
5. Придатки одноветвистые; на просо-
ме это пара «хелицер» (клешневидных,
в форме ложных клешней или стилетов),
пара ного-, щупальце- или клешневидных «пе-
дипальп» и четыре пары ходильных ног (все
крепятся вблизи вентральной срединной
линии и у некоторых видов выпрямляются за
счет гидростатического давления в гемоце-
ле); антенн и челюстей нет.
6. Ротовой аппарат образован только
одной парой придатков (хелицерами), хотя
в измельчении пищи и направлении ее в рот
могут также участвовать медиальные
отростки базального членика других
конечностей («коксальные эндиты»).
7. Обычно (если не происходит
вторичной утраты) на просоме находятся простые
1 Морские пауки сильно отличаются от
других хелицеровых, и к ним не относятся многие из
перечисленных ниже признаков.
Беспозвоночные с конечностями: членистоногие и др.
221
пульпы Й1ЙГнные v Пищеварительные
пед а Та К^Ра™.КС„ желе3ь,Р Мальпигиев Сердце
-I - -I
Рис. 8.10 Схема продольного разреза тела обобщенного хелицерового
фединные и боковые глазки; в одной из
групп за счет скопления боковых глазков
формируются сложные глаза.
8. Опистосома иногда внешне
разделена на сегменты (до 12), а у некоторых
видов—на широкую переднюю «мезосому»
и узкую заднюю «метасому»; иногда несет
постанальный шип, жало или жгутик.
9. Выделительная система из слепо
оканчивающихся коксальных желез в про-
соме и/или разветвленных эндодермальных
мальпигиевых сосудов опистосомы,
отходящих от среднего отдела кишечника и
выводящих главным образом гуанин.
10. Экзоскелет необызвествленный;
иногда в просоме еще и пластинчатый мезо-
дермальный эндоскелет.
И. Органы дыхания связаны с
придатками опистосомы или их эмбриональными
зачатками; у морских форм это наружные
жаберные «книжки», у наземных—
внутренние легочные «книжки» и
производные от них ситовидные или трубчатые
трахеи.
12. В газообмене участвует циркулятор-
ная система; в гемолимфе обычно
содержится гемоцианин.
13. Нервная система с отдельными
ганглиями по всей длине тела или, чаще,
слившимися в один просомальный узел.
14. Раздельнополые животные. В
исключительно морских классах
оплодотворение наружное (хотя партнеры при
спаривании объединяются друг с другом,
совершая псевдокопуляцию), в главным образом
наземных группах— внутреннее (копуляция
или с помощью сперматофоров); половые
поры расположены на втором сегменте
опистосомы.
15. Ювенильные особи внешне
напоминают взрослых, обычно выходят из яйца
с полным набором конечностей.
16. Исходно морские бентосные
организмы; один класс очень успешно заселил
сушу и пресные воды.
Хелицеровые отличаются от двух
других типов членистоногих по ряду
важнейших признаков, хотя эволюция
мальпигиевых сосудов и трахей шла у них и у одновет-
вистых параллельно. Наиболее явная
отличительная черта — их придатки. Ни у
одного из хелицеровых нет челюстей или других
конечностей, способных перемещаться друг
относительно друга, пережевывая пищу.
Добыча схватывается и разрывается с
помощью клешней или когтей (на педипаль-
пах и/или хелицерах), но заглатывается
только в сильно размельченном или, как
правило, жидком виде. Чтобы в кишечник не
проникли крупные частицы, ротовое
отверстие обычно прикрыто щетинками.
Несмотря на это, хелицеровые — почти
исключительно хищники. Процесс их питания
состоит в удержании жертвы вблизи
ротового отверстия, впрыскивания в нее
пищеварительных ферментов и последующего
всасывания продуктов этого внешнего
переваривания. Если ферменты не вводятся в тело
222
Глава 8
жертвы, то переваривание или
механическое дробление пищи осуществляется
в предротовой области, огороженной кокса-
льными эндитами нескольких или всех
конечностей просомы, часто почти радиально
расходящихся вокруг ротового отверстия.
В связи с таким способом питания передняя
кишка преобразована в своего рода насос
или насосы, а средняя, особенно множество
ее дивертикулов (они могут занимать
большую часть объема тела),— в место
окончательного переваривания и всасывания.
Далее, у хелицеровых нет антенн, хотя педипа-
льпы, первые или вторые пары ходильных
ног могут модифицироваться для
выполнения соответствующей функции.
Среди хелицеровых прослеживаются
морфологические ряды, демонстрирующие
возможный путь эволюции трахейной
системы из наружных жабр. У меростомо-
вых—морской и, почти несомненно, анце-
стральной линии хелицеровых — органы
дыхания (ка# и у большинства других
морских животных)—наружные жабры. Они
развиваются из заднего края клапановид-
ных придатков опистосомы; биение послед-
ных создает ток воды над жаберными
листочками. У наземных паукообразных
воздушное дыхание, но в противоположность
одноветвистым (см. разд. 8.5) типичные для
них органы газообмена эмбриологически
связаны с задними краями зачатков описто-
сомальных конечностей (они не
развиваются в ходильные ноги, хотя паутинные
бородавки пауков и гребневидные органы
скорпионов представляют собой сильно
видоизмененные придатки опистосомы). Так,
например, образуются легочные книжки (рис.
8.11), гомологичные рядам жаберных
лепестков в карманах, погруженных внутрь
тела. Каждая такая книжка—это впячива-
ние со свисающими в него рядами
параллельных лепестков, слипнутся которым не
дают особые распорки; воздух между ними
перемещается благодаря диффузии, а
внутрь них заходит синус с гемолимфой.
У некоторых видов лепестки легочных
книжек удлиненные и напоминают скорее
трубочки, чем пластинки; в зависимости от
числа таких трубочек они называются
ситовидными (их много близко расположенных)
или трубчатыми (их мало) трахеями. Сле-
воздуха
Дыхальце
Рис. 8.11. Объемная схема внутреннего
строения легочной книжки паукообразных,
демонстрирующая циркуляцию крови и воздуха (по
Barnes, 1980)
довательно, эти трахейные системы в
принципе не что иное, как удлиненные,
погруженные внутрь тела и связанные с
конечностями жабры. У некоторых пауков
дополнительно развиты вторичные трахеи из
пустотелых внутренних выростов экзоскелета
(«аподем»—см. рис. 8.1).
8.4.3. Классификация
Описано 63 000 видов хелицеровых,
отнесенных к трем далеко отстоящим друг от
друга классам, из которых один некоторые
считают неродственным двум другим.
Класс Отряд
Merostomata Xiphosura
/ Scorpiones
/ /,Ur°pygi
/ y^/Schizomida
/ //s' Amblypygi
/ ^gJ^^^___Araneae
/ Arachnida ^CS^~—Ricinulei
L—' ^ Pseudoscorpiones
\ NNN^\^~""~--Solpugida
\ ^^ч\~^ Opiliones
\ \Ns\Notostigmata
\ \/\ Parasitiformes
\ \Acariformes
^Pycnogona Pycnogonida on
Беспозвоночные с конечностями: членистоногие и др.
223
8.4.3.1. Класс Merostomata
Хотя до пермского периода это была
доминирующая группа беспозвоночных, в
настоящее время меростомовые
представлены только четырьмя видами отряда Xipho-
sura (мечехвосты). Это крупные морские хе-
лицеровые с толстым карапаксом,
напоминающим очертаниями лошадиное копыто,
покрывающим крупную просому и
нависающим над ней спереди (ротовое
отверстие оказывается в центре вентральной
стороны) и с боков, прикрывая сверху все
конечности. Небольшая подвижно причлен-
ная опистосома представляет собой
плоскую пластину, частично входящую в вырез
карапакса и окаймленную с боков
мощными шипами; терминально она несет
длинный постанальный хвостовой шип (рис.
8.12). Хелицеры, педипальпы и все
ходильные ноги, кроме последней пары,
вооружены клешнями; последняя пара, несущая ряд
терминальных шипов или пластин,
используется для рытья в грунте (рис. 8.13).
В отличие от прочих современных хели-
церовых у мечехвостов почти полный набор
•Просома
Опистосома
-Хвостовой шип
Рис. 8.12. Вид мечехвоста с дорсальной
стороны (по Kaestner, 1968)
опистосомальных придатков: передняя их
пара небольшая и трубчатая, остальные
шесть пар — плоские пластины, при
помощи которых животное плавает,
перевернувшись вверх ногами. Первая из этих пар
пластин служит также половой крышечкой,
Рис. 8.13. Полусхематический
вид мечехвоста с вентральной
стороны; показаны различные
придатки (по Savory, 1935)
Карапакс
Опистосома
Рот Хелицера
Педипальпа
Придатки
опистосомы
Хвостовой шип-
224
Глава 8
а последние пять несут наружные жабры.
Другие особенности, присущие морским
формам, но нетипичные для хелицеровых
в целом,— отсутствие мальпигиевых
сосудов и внешнее оплодотворение. Среди всех
современных хелицеровых только у
мечехвостов имеются крупные, диффузные
сложные глаза достаточно своеобразного типа
по бокам карапакса. В отличие от
паукообразных гонады и пищеварительные
дивертикулы расположены в просоме.
Мечехвосты—ночные, бентосные
животные прибрежного мелководья; они
роются в рыхлых осадках и ползают по дну,
питаясь крупными моллюсками и полихета-
ми, раскусывая их коксальными эндитами
(«гнатобазами») и размельчая в
специализированном желудке1.
8.4.3.2. Класс Arachnida
(паукообразные)
Почти целиком наземные формы,
составляющие 98% современных видов
хелицеровых, с хорошо выраженными
приспособлениями к существованию на суше
(например, выделительная система с мальпи-
гиевыми сосудами; органы дыхания,
погруженные внутрь тела; кутикула,
непроницаемая для воды благодаря слою воска;
внутреннее оплодотворение). Внешний облик
паукообразных весьма разнообразен: от
удлиненных, хорошо вооруженных артропод
с крупными хватательными педипальпами,
отчетливой наружной сегментацией и опи-
стосомой, расчлененной на мезо- и метасо-
му (скорпионы, телифоны и т. д.), до почти
сферических форм с тонким экзоскелетом,
незаметной наружной сегментацией и менее
развитыми педипальпами (большинство
пауков, клещей и т.д.). Это разнообразие
находит отражение в делении класса на 13
отрядов (рис. 8.14). Скорпионовидная
форма тела, почти несомненно,— примитивное
состояние, поскольку она слабо отличается
от облика некоторых (вымерших)
ракоскорпионов из меростомовых, которые вели
пресноводный, а возможно, и земноводный
1 Жевательный желудок лежит в передней
части просомы и имеет кутикулярную выстилку
с грубыми зубцами.— Прим. пер.
образ жизни. Некоторые современные
паукообразные вновь вернулись в пресную
воду, а ряд клещей — в море.
В отличие от мечехвостов у
паукообразных нет сложных глаз и локомоторных
придатков на опистосоме; у них длинные
ходильные ноги без клешней', а половые
железы и пищеварительные дивертикулы
расположены в опистосоме. В нескольких
группах паукообразных общая тенденция
к облегчению экзоскелета привела к
отделению двух последних сегментов просомы от
карапакса (хотя на них и могут
присутствовать самостоятельные дорсальные
пластинки) и к развитию мягкой, гибкой опистосо-
мы. У большинства клещей эти два послед-
ных просомальных сегмента слиты с опи-
стосомой, и тело состоит из двух отделов,
отличных от наблюдающихся у прочих
хелицеровых. Однако в общем случае
сохраняется типичное деление на просому и опи-
стосому, разграниченные внутренней
диафрагмой или узким стебельком. Такое
разделение позволяет повышать давление
в просоме (для выпрямления конечностей),
не изменяя его в опистосоме.
Паукообразные—один из наиболее
преуспевающих классов животных. Их
образ жизни связан главным образом с
охотой на представителей другой не менее
преуспевающей группы—насекомых. К
замечательным приспособлениям для ловли
этих очень подвижных организмов
относятся ядовитые железы, секрет которых
может впрыскиваться в жертву, а также
образование паутинной нити- явление,
достигающее своей кульминации в постройке
ловчей сети пауками. Размеры
паукообразных колеблются от менее 0,1 мм до 18 см;
в классе свыше 62 000 видов. Известны
клещи, питающиеся растительными тканями
и детритом; некоторые клещи —паразиты.
8.4.3.3. Класс Pycnogona
(морские пауки)
Этих мелких бентосных морских
беспозвоночных с длиной тела менее 6 см ряд
зоологов считает неродственными хелице-
ровым, а возможно, и всем прочим
членистоногим. Первая пара конечностей этих
1 На просоме.— Прим. пер.
Беспозвоночные с конечностями: членистоногие и др.
225
Scorplones
(скорпионы)
Pseudoscorplones
(ложноскорпионы)
Uropygl (телифоны.или
скорпионопауки)
Amblypygl
(жгутоноше)
RlcinuLel
Агапеае (пауки)
Notostigmata
Рис. 8.14. Форма тела представителей различных отрядов паукообразных (по Savory, 1935; Huges,
1959 и др.)
15—1314
226
Глава 8
Рис. 8.15. Форма тела морских пауков (преимущественно по Hedgpeth, 1982)
животных, хелифоры, оканчивается, как и
у хелицеровых, клешнями; вторая, пальпы,
возможно, гомологична педипальпам; у
большинства видов присутствуют еще четыре
пары ходильных ног. Однако у некоторых
видов ходильных ног пять или шесть пар,
а между пальпами и первой парой
ходильных ног располагаются дополнительные
придатки—яйценосы. В отличие от прочих,
латеральных, придатков (ходильные ноги
причленены к мощным боковым выростам
тела) они крепятся вентрально (рис. 8.15).
У ряда видов хелифоры, пальпы и яйценосы
рудиментарные или отсутствуют, но
ходильные ноги всегда хорошо развиты и
бывают очень длинными (до 75 см).
Цефалоторакс, или головогрудь (просо-
ма?), не покрытая карапаксом, несет
центральный выступ с двумя парами простых
глаз и (спереди или вентрально) крупный
трубчатый невтягивающийся хоботок с
терминальным ротовым отверстием. Абдомен,
или брюшко (опистосома?),— крошечный
несегментированныи сосочек, лишенный
придатков. Органов выделения и дыхания
нет. Необычно то, что гонады, как и
пищеварительные дивертикулы, заходят внутрь
ходильных ног, где и созревают яйца. В
ходе псевдокопуляции гаметы выводятся из
множества половых пор (гонопоров),
расположенных у основания каждой ходиль-
Рис. 8.16 Протонимфон морских пауков (по
Kaestner, 1968)
Беспозвоночные с конечностями: членистоногие и др.
227
Глазной о I- л
Бугорок Сердце Гонада Абдомен
Сито из щетинок
Хоботок
Рис. 8.17. Схема продольного
разреза тела и хоботка
морского паука (по King, 1973)
ной ноги или их последних двух пар. После
оплодотворения самец собирает яйца и
носит в виде шаров, прикрепленных к яйцено-
сам. В итоге вылупляется своеобразная
личинка с тремя парами придатков- -
протонимфон (рис. 8.16), чаще всего
ведущая полупаразитический образ жизни на
поверхности или внутри стрекающих или
моллюсков.
Взрослые морские пауки питаются в
основном губками, книдариями, мшанками.
Часть тела добычи захватывается хелифо-
рами (если они имеются), а заглатывание
тканей жертвы осуществляется за счет
мощного насасывающего действия глотки.
Поеданию пищи способствует грызущее
действие трех зубов, расположенных на конпе
хоботка. В глотке пища измельчается
другими зубами или мощными щетинками;
у основания глотки воротничок из щетинок
обеспечивает попадание в пищевод только
мельчайших пищевых частиц (рис. 8.17).
Описано более 100 видов, относимых
к одному отряду.
8.5. Тип Uniramia
(одноветвистые)
8.5.1. Этимология
От лат. unus один и ramus—ветвь.
8.5.2. Диагностические
и специфические признаки
(рис. 8.18)
1. Двусторонне симметричные
членистоногие с разнообразной формой тела (от
■Анус
Срезы
дивертикулов гонад и
кишечника в
конечностях
рюшной нервный
ствол с ганглиями
очень вытянутой до почти шаровидной)
длиной от менее 1 м до 35 см.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и органами.
3. Кишечник сквозной, прямой, без
пищеварительных дивертикулов.
4. Тело разделено на два отдела—
«голову», образованную акроном и тремя
или четырьмя сегментами с придатками,
и «туловище» с парами ходильных ног.
В одном из подтипов в состав туловища
входит до 350 относительно однородных
сегментов, подавляющее большинство из
которых—с ходильными ногами; в другом
подтипе оно дифференцировано на «грудь»
(торакс) с тремя парами ног и «брюшко»
(абдомен), включающее до одиннадцати
сегментов с крайне видоизмененными
придатками, если таковые вообще имеются.
5. Придатки одноветвистые; на голове
находится по паре «антенн» (усиков),
«мандибул» (жвал) и «максилл» (челюстей), а
у ряда групп—и вторая пара максилл;
туловище несет только функциональные или
модифицированные ходильные ноги; хели-
цер или клешненосных конечностей нет.
6. Ротовых придатков (мандибулы
и максиллы) две или три пары, причем
элементы каждой пары работают в фазе или
в противофазе с образующими другую
пару; базальные членики максилл или вторая
пара максилл (в тех группах, где их две
пары) срастаются в пластинку (нижнюю губу,
или лабиум), ограничивающую предрото-
вую полость снизу.
7. По бокам головы находятся глазки,
часто образующие сложные глаза; иногда
простые глазки присутствуют и на средней
линии головы.
15*
228
Глава 8
МНОГОНОЖКА
Антенна
Боковые Максиллярная
k глазки железа Кишечник
Мозг
Ротовые
придатки
Гонада Сердце
Анус
Слюнная Дыхальца Мальпигие- Нервный Ноги Гонопор
железа вы сосуды тяж
НАСЕКОМОЕ
Антенна
Крыло
^/ Прямая
Гонада Сердце кишка
Церка -
Анус
Пенис
Класпер
Мальпигиевы
сосуды
Нервный ^Дыхальца
тяж
Ноги
Рис. 8.18. Схематические продольные разрезы генерализованных представителей двух подтипов
одноветвистых
8. Туловище наружно сегментировано,
голова — нет.
9. У большинства представителей
одного из подтипов на груди одна или две
пары крыльев.
10. В гемоцеле находится жировое тело,
часто тесно связанное с кишечником.
11. Выделительная система
представлена 0—2 парами максиллярных желез в
голове и 1—75 парами неветвящихся эктодерма-
льных мальпигиевых сосудов, отходящих
от задней кишки вблизи места ее
соединения со средней и выводящих главным
образом аммиак и/или мочевую кислоту.
12. Наружный скелет известковый или,
чаще, неизвестковый.
13. Органы газообмена парные в виде
ветвистых трубчатых трахей, через которые
диффундирует воздух; исходно они
открываются наружу парой дыхалец на каждом
сегменте с ходильными ногами, а
оканчиваются в теле трахеолами;
эмбриологически трахейная система не связана с
придатками.
14. Гемолимфа не переносит
дыхательных газов, и дыхательные пигменты в ней
отсутствуют (исключение—некоторые
личиночные стадии).
15. Раздельнополые формы;
оплодотворение внутреннее с помощью сперматофо-
ров или путем копуляции; исходно гонопо-
ры находятся на последнем туловищном
сегменте, в ряде случаев это состояние
претерпело модификации.
16. В ряде групп выделяется четкая
личиночная стадия, сильно отличающаяся от
взрослых форм; иногда особь выходит из
яйца с неполным набором сегментов (и ног)
и они добавляются при линьках по одному,
причем в некоторых случаях и после
достижения половой зрелости.
17. В основном наземные формы, неко
Беспозвоночные с конечностями: членистоногие и др.
229
торые пресноводные, очень мало
морских.
Возникшие, по-видимому, на суше одно-
ветвистые наряду с паукообразными,
которых они превосходят и по видовому
разнообразию, и по общей численности, сейчас
доминирующая группа наземных
беспозвоночных. Причины подобного успеха на
суше отчасти те же, что и у паукообразных.
В обеих группах независимо появились
системы газообмена, основанные на
диффузии газов из окружающей среды в ткани
и обратно, что гораздо эффективнее
предотвращает потери влаги, чем, к примеру,
респираторная система четвероногих
позвоночных, выдыхающих большое количество
водяных паров во время принудительной
вентиляции легких. Характерная для арах-
нид и одноветвистых выделительная
система с мальпигиевыми сосудами благодаря ее
связи с кишечником также, по крайней мере
потенциально, способствует экономии воды
за счет реабсорбции ее из продуктов
азотного обмена после их выведения в заднюю
кишку. Наружный кутикулярный скелет в
большей степени препятствует потере влаги,
нежели мягкие влажные покровы кольчецов
и моллюсков. И все же в большинстве
классов одноветвистых эти адаптации решают
проблему потери воды лишь частично, и их
представители связаны в основном с
влажными местообитаниями—в почве или
поблизости от нее. В частности, их
дыхальца не способны закрываться, некоторое
количество воды уходит с продуктами
азотного обмена (представленных главным
образом аммиаком), а кутикула относительно
проницаема. Лишь в одном классе потери
влаги удалось сократить еще сильнее—и
намного — за счет появления механизмов
закрывания дыхалец, водонепроницаемой
воскоподобной эпикутикулы и системы
извлечения воды из экскрементов в прямой
кишке. Именно этот класс, точнее крылатые
насекомые, стал самой крупной и
разнообразной группой одноветвистых.
Хотя перечисленные выше механизмы
экономии воды позволяют понять, почему
одноветвистые достигли по крайней мере не
меньшего процветания, чем паукообразные,
все же для объяснения гораздо больших
масштабов их успеха этого мало. Не
исключено, что значительную долю его можно
отнести на счет появления челюстей,
способных кусать и жевать. Унирамии могут
захватывать в пищеварительный тракт
твердую пищу и, следовательно, не ограничены
в своем рационе жидким кормом, а значит,
и добычей, пригодной для наружного
пищеварения. На деле это означает, что им
доступны растительные ткани, а суша как раз
и характеризуется прежде всего изобилием
этого относительно грубого материала,
который приходится откусывать и разжевы-
Мышца для
отведения назад
Мышца для
выдвижения
вперед
Передний
аддуктор
Мандивула
Верхняя гува
(лаврум)
Мандивула
Сросшиеся
максиллы
(лавиум)
Верхняя гува
(лаврум)
Мандивула
МаксиллаI
Максилла II
(лавиум)
Рис. 8.19. Ротовой аппарат одноветвистых. А Схема поперечного разреза через голову,
показывающая примитивное состояние, при котором мандибулы расположены вертикально, направлены
вниз и приводятся в движение примерно теми же мышцами, что и ходильные ноги. Б. Вид спереди
и снизу на предротовую полость двупарноногой многоножки: видна верхняя губа (лабрум),
образованная склеритами головной капсулы, и нижняя губа (лабиум), образованная сросшимися мак-
силлами (заметно горизонтальное положение мандибул). В. Вид сбоку головы насекомого:
видны три пары ротовых придатков и причленение мандибулы к головной капсуле в двух точках (1
и 2) (по Kaestner. 1968)
230
Глава 8
вать, прежде чем проглотить. В отличие от
почти исключительно плотоядных
паукообразных здесь лишь одна крупная
группа —губоногие—полностью состоит из
хищников, хотя большинство
представителей типа потребляет все же мертвое и
разлагающееся растительное вещество, а не
собственно живые растения (причины этого
рассмотрены в гл. 9).
Первоначально мандибула одноветви-
стых была ходильной конечностью первой
пары, у которой развивалась лишь базаль-
ная часть. Ее режущий край —широкий,
направленный внутрь вырост, неподвижно
соединенный с остальной частью жвалы у всех
форм, кроме двупарноногих и симфил.
Исходно мандибула причленяется к телу так
же, как и прочие ходильные ноги, т.е.
ориентирована перпендикулярно головной
капсуле (рис. 8.19,^4). Однако у
большинства одноветвистых ее положение
изменилось таким образом, что она располагается
параллельно вентральной поверхности
головы, т.е. развернута вперед. Поэтому ее
первоначально дорсальное соединение с
телом стало задним, и у некоторых форм
спереди возникла еще одна точка причленения
(см. рис. 8.19,5). В итоге движения жвал
ограничиваются в основном раздвиганием
и сдвиганием. Нижняя стенка или задний
край предротового пространства, в
котором работают ротовые
придатки,—результат слияния базальных частей максилл
или второй пары максилл (в группах, где
таковые имеются), также развившихся из
ходильных ног (рис. 8Л9,Б). Это совсем не
похоже на то, что мы видим в двух остальных
типах членистоногих, нижняя губа которых
представляет собой часть вентрального
экзоскелетного стернита. У многоножек
строение ротовых частей в целом довольно
однообразно, но среди крылатых
насекомых морфология мандибул и максилл
крайне варьирует, коррелируя с адаптивной
радиацией этих форм в широчайшем
диапазоне пищевых ниш (см. разд. 8.5.3Б.2).
8.5.3. Классификация
Представителей одноветвистых,
которых известно свыше миллиона видов,
можно разделить на два подтипа, один из них
с четырьмя классами, а другой—с шестью.
Однако общий план строения здесь
относительно постоянен, так что различия между
большинством классов выражены гораздо
слабее, чем во многих других крупных
типах, например среди моллюсков, хелицеро-
вых и ракообразных.
8.5.3А. Подтип Myriapoda
(многоножки)
Классы многоножек, вероятно,
отражают первую крупную адаптивную
радиацию внутри данного типа (или среди его
современных представителей); один из этих
классов дал затем второе важнейшее
ответвление, положившее начало классам
второго подтипа.
Главная отличительная особенность
многоножек в том, что их туловище
образовано рядом более или менее одинаковых
сегментов, каждый из которых, не считая од-
ного-двух последних и иногда первого,
несет пару ходильных ног; грудь и брюшко не
выражены. Можно назвать и другие общие
признаки подтипа, хотя они присущи не
только ему, но в некоторых случаях и первич-
нобескрылым насекомым (см. разд.
8.5.3Б.1). Так, имеется лишь одна пара ма-
льпигиевых сосудов (как правило,
вытянутых); на голове нет срединных глазков, но
присутствуют «органы Темешвари»,
воспринимающие, по-видимому, переносимые
воздухом химические соединения, а
возможно, и влажность; отдельные членики
антенн обслуживаются собственной
мускулатурой; нервная система не
сконцентрирована, и ганглии расположены во всех
туловищных сегментах. Кроме того, как уже
говорилось выше, механизмы экономии воды
развиты относительно слабо. Классы
различаются между собой в основном числом
пар максилл, расположением гонопоров,
числом тергитов и тем, какие сегменты
лишены ног.
Хотя в целом это сугубо наземная
группа, в каждом классе по нескольку видов
населяет морскую литораль.
Беспозвоночные с конечностями: членистоногие и др.
231
Класс
^ Pauropoda
Отряд
Scutigerida
Lithobiida
Scolopendrida
Gcophilida
Scolopendrelhda
Polyxenida
Glomendesmida
Oniscomorpha
Polyzdniida
Stemmiulida
Spirobolida
luliformida
Typhlogena
Chordeumatida
Polydesmida
Pauropodida
8.5.3A.1. Класс Chilopoda
(губоногие)
Губоногие —сильно или очень сильно
вытянутые, дорсовентрально уплощенные
многоножки с двумя парами максилл на
голове и туловищем из 15—181 и более
сегментов (их число всегда непарное) с
ногами и двух хвостовых безногих сегментов—
прегенитального и генитального (рис. 8.20).
Все ноги имеют сходное строение, хотя
иногда длина их возрастает спереди назад;
только первая пара (это диагностический
признак класса) видоизменена в
хватательные органы — крупные когтевидные крючья
с ядовитыми железами внутри (рис. 8.21).
Хотя у большинства видов количество
боковых глазков достаточно велико, а в
одной группе (отряд
Scutigerida—единственный случай среди многоножек)
имеются даже крупные сложные глаза, эти
хищные артроподы ведут ночной образ жизни
или обитают под поверхностью почвы,
обнаруживая добычу (олигохет и
членистоногих) с помощью антенн или, реже, ног.
Некоторые группы глаз лишены.
Как и у большинства многоножек,
перенос спермы происходит с помощью
наружного сперматофора, выдавливаемого из
хвостового гонопора лишь после сложного
ухаживания. Во всех группах, кроме одной,
у самца из того же отверстия выдвигается
прядильный аппарат, выделяющий неско-
Рис. 8.20. Форма тела губоногих (по Lewis,
1981)
лько паутинных нитей для подвешивания
сперматофора и направления к нему самки.
Некоторые виды охраняют яйца и даже
молодь после вылупления.
Необычно для представителей этого
подтипа то, что у многих видов потомство
выходит из яиц с полным набором
сегментов и ног (в том числе, как ни странно,
у форм с максимальным их числом), однако
в более типичном случае у свежевылупив-
шихся особей сегментов меньше, чем
у взрослых, иногда всего четыре.
Известно 3000 видов с длиной тела
до 27 см. Их разделяют на четыре отряда.
232
Глава 8
Антенна
МаксиллаI
Максилла II
Видоизмененная
первая пара
туловищных ног
Рис. 8.21. Сильно видоизмененная первая пара
туловищных конечностей губоногого (по Borror
et al., 1976)
8.5.3А.2. Класс Symphyla
Симфилы — мелкие (менее 8 мм длиной)
многоножки; общий план их строения такой
же, как у губоногих, т.е. имеется голова
с двумя парами максилл и туловище из
двенадцати (в данном случае) сегментов с
ногами и двух терминальных безногих,
последний из которых слит с тельсоном (рис. 8.22).
Прядильные органы также находятся на
последнем сегменте тела. Однако в отличие от
губоногих у симфил тергитов больше, чем
сегментов (до 24), что делает тело более
гибким. Гонопоры вторично открываются
в передней части тела (на третьем сегменте),
а первая пара ног не образует крючьев, хотя
Тергиты
Прядильный
аппарат
Рис. 8.22. Внешний вид симфил (по Kaestner,
1968). Обратите внимание, что тергитов
больше, чем пар ног
эти конечности заметно мельче
последующих—иногда вдвое. Еще одна необычная
для многоножек черта—единственная пара
коротких трахей, открывающихся
дыхальцами на голове.
Однако во многих других отношениях
этот класс напоминает представителей
подтипа Hexapoda, особенно первичнобескры-
лых насекомых. В принципе сходны,
например, их рртовые придатки, а рядом с
ногами на большинстве сегментов развиты
мелкие грифельки и коксальные мешочки,
которые способны выворачиваться наружу за
счет давления жидкости в гемоцеле и
используются для поглощения влаги из
окружающей среды. Поскольку, как и
большинство многоножек, симфилы выходят из яиц
с неполным составом сегментов, не
исключено, что Hexapoda произошли от симфило-
подобных предков в результате педоморфо-
за (некоторые двупарноногие и пауроподы
также вылупляются лишь с тремя парами
ног).
Среди 160 известных видов этих мелких,
слепых, мягкотелых и бледноокрашенных
многоножек большинство
растительноядные, питающиеся в основном мелкими
живыми корешками и обитающие в почве,
листовой подстилке, гниющей древесине, под
камнями и т. п. Все виды относят к одному
отряду.
8.5.3А.З. Класс Diplopoda
(двупарноногие)
Тело от короткого до очень сильно
вытянутого (рис. 8.23). Наиболее характерный
и бросающийся в глаза признак—наличие
«сдвоенных» сегментов. Туловище двупар-
ноногих состоит из безногого первого
сегмента, следующих за ним трех сегментов
с типичной для многоножек единственной
парой ног на каждом, а затем—ряда
«колец» (от пяти до 85 и более) с двумя парами
ног, ганглиев, остий сердца и т. д. в каждом,
после чего идет один или несколько
безногих сегментов, включая хвостовую зону их
прироста. Каждое сдвоенное сегментное
кольцо, из которых в основном и образовано
почти цилиндрическое или, реже,
уплощенное туловище, возникает вследствие
частичного или полного попарного слияния туло-
Беспозвоночные с конечностями: членистоногие и др.
233
Рис. 8.23. Разнообразие двупарноногих (по
разным авторам, главным образом по Blower,
1985)
вищных сегментов; у двух сегментов самое
меньшее может быть общий тергит. У
самцов многих видов одна—две пары ног
(конечности VII сдвоенного сегмента, или
задние конечности VII и передние VIII) в
различной степени видоизменены и образуют
копулятивные «гоноподии» (половые
ножки), забирающие сперму из гонопоров на III
сегменте и переносящие ее в
соответствующие отверстия самки. Другие виды для
переноса спермы используют мандибулы;
иногда самки и самцы тесно сближают свои
гонопоры, или же (классический для
многоножек способ) самец выделяет пакеты
спермы и паутинные направляющие нити. У
большинства диплопод в момент вылупления
всего семь туловищных «сегментов», а
новые добавляются всю жизнь—даже спустя
долгое время после наступления половой
зрелости.
К другим особенностям двупарноногих
относится лишь одна пара максилл,
большое число боковых простых глазков,
собранных в группы, которые на первый
взгляд можно принять за сложные глаза
(впрочем, у ряда видов глазки вовсе
отсутствуют), а также обызвествленный
наружный скелет (исключая представителей
одного отряда). Из всех одноветвистых
последний признак встречается лишь у
диплопод.
Класс выключает 10000 видов и делится
на 10 отрядов. Это медленно
передвигающиеся потребители растительных тканей
(обычно уже начавших разлагаться) в тех же
местообитаниях, что и симфилы, т. е.
гниющих стволах, листовой подстилке и т. п.
Самые крупные представители достигают
в длину 28 см.
8.5.3А.4. Класс Pauropoda
Эти крошечные существа (длиной менее
2 мм) обитают там же, где симфилы и ди-
плоподы; они надкусывают гифы грибов
и высасывают их содержимое с помощью
передней кишки, работающей наподобие
насоса. Их трудно заметить, хотя
численность их часто весьма высока. В смысле
плана строения они примерно так же
относятся к двупарноногим, как симфилы—к
губоногим. Так, у пауропод одна пара
максилл, первый и два последних сегмента
туловища лишены ног, гонопор находится
спереди (на третьем сегменте), у
туловищных сегментов наблюдаются начальные
этапы попарного слияния: общий тергит
полностью или отчасти покрывает
сегменты I и II, III и IV, V и VI и т. д. (рис. 8.24).
Сегментов всего 11—12, последний
сливается с тельсоном. У вылупившихся из яйца
особей сегментов мало и в большинстве
случаев три пары ног.
Однако пауропод нельзя считать просто
примитивными диплоподами, поскольку
у них есть и отличительные признаки.
Например, отсутствует сердце, а у
большинства и трахеи (если трахеи все же имеются,
234
Глава 8
ВИД СВЕРХУ
Рис. 8.24. Внешний вид пауропод (по Kaestner,
1968 и Borror et al, 1976). Обратите внимание,
что тергитов меньше, чем пар ног
они очень слабо развиты). Кроме того,
антенны ветвистые.
Все 500 видов класса слепые, мягкотелые
и бесцветные. Их относят к единственному
отряду.
8.5.3Б. Подтип Hexapoda
(насекомые)1
От многоножек представителей
данного подтипа отличает лишь один признак:
туловище разделено на грудь из трех
несущих ноги сегментов и брюшко из одиннад-
1 Вмировойи отечественной литературе
«насекомыми» в настоящее время чаще всего
называют группу, включающую отряды Thysanura
и Microcoryphia (объединяемые под названием
Apterygota, или первичнобескрылые насекомые),
а также всех Pterygota. В таком объеме этот
таксон принято именовать классом Insecta (иногда
используют и название Hexapoda). Отряды Diplu-
ra, Collembola и Protura либо противопоставляют
насекомым, объединяя их в класс Entognatha,
либо (как и здесь) считают самостоятельными
классами.— Прим. пер.
цати сегментов без ходильных ног, но
иногда с придатками различной формы, причем
абдоминальные сегменты могут исчезать
или сливаться, так что их число бывает
меньше одиннадцати. Кроме того, у всех ге-
ксапод две пары максилл, а гонопоры
занимают терминальное или субтерминальное
положение; в большинстве случаев имеются
срединные простые глазки, а также простые
глазки или сложные глаза по бокам головы.
8.5.3Б.1. Классы первично-бескрылых
насекомых (Apterygota)
Выделяют пять групп первичнобескры-
лых насекомых. По всей видимости, каждая
из них отделилась от прочих на очень
ранних стадиях эволюции насекомых (если они
вообще не произошли независимо от каких-
то предковых форм многоножек), поэтому
вполне можно говорить о самостоятельных
классах. Лишь у одного из них ряд черт
указывает на явное родство с крылатыми
насекомыми, шестым классом гексапод. Все
классы первичнобескрылых очень невелики
по объему и включают всего по одному
отряду, поэтому удобнее рассматривать их
вместе.
Эти пять классов можно разбить на три
группы, первая из которых
(двухвостки = Diplurata; ногохвостки = Oligoentoma-
ta и бессяжковые = Myrientomata)
характеризуется частично погруженными внутрь
головной капсулы ротовыми частями и еще
некоторыми особенностями, общими
скорее с многоножками, чем с прочими
насекомыми. По образу жизни, в том числе по
брачному поведению и местообитаниям,
эти формы также очень напоминают
многоножек, и неудивительно, что многие
энтомологи выступают за включение
двухвосток, ногохвосток и бессяжковых в подтип
Myriapoda. Среди сохранившихся у них
предковых черт—сочленение мандибул
с головой в одной точке, трехлопастный ги-
пофаринкс (непарный, похожий на язычок
орган, расположенный около слюнных
желез), выпячивающиеся коксальные
мешочки, брюшные грифельки, собственная
мускулатура в члениках антенн, органы Те-
мешвари (или очень близкие к ним
образования), некоторые типы абдоминальных
I
Беспозвоночные с конечностями: членистоногие и др.
235
Класс
Diplurata-
Надотряд
Oligoentomata —
Myrientomata —
Zygoentomata —
Archaeognathata
i Pterygota
Palaeoptera
Orthopteroidea
Hemipteroidea
Endopterygota
Отряд
- Diplura
-Collembola
- Protura
- Thysanura
- Microcoryphia
Ephemeroptera
Odonata
Blattaria
Mantodea
Isoptera
Zoraptera
Grylloblattaria
Dermaptera
Orthoptera
Phasmida
Embioptera
Plecoptera
Psocoptera
Mallophaga
Anoplura
Thysanoptera
Homoptera
Heteroptera
Coleoptera
Strepsiptera
Hymenoptera
Raphidioida
Neuroptera
Megaloptera
Mecoptera
Diptera
Siphonaptera
Trichoptera
Lepidoptera
придатков. Все они малы по размерам
(длина тела, как правило, не превышает 7 мм) *;
вероятно, по этой причине мальпигиевых
сосудов нет, а продукты азотного обмена
удаляются через эпителий средней кишки;
глаза и простые глазки имеются лишь
у представителей одного класса из трех.
Двухвостки (рис. 8.25, А) лишены черт
специализации, которые столь ярко
проявляются у остальных двух классов. У бессяж-
ковых (рис. 8.25, Б) антенны рудиментарны
(их функцию берут на себя грудные ноги
1 Известны представители двухвосток
длиной до 30 мм.— Прим. пер.
первой пары), а мандибулы превратились
в колющие стилеты. Как и у большинства
многоножек, число туловищных сегментов
после вылупления постепенно растет, пока
не достигнет полного взрослого набора (11
брюшных). Зато ногохвостки (рис. 8.25, В)
сохранили лишь пять брюшных сегментов
(и тельсон), а две пары брюшных придатков
у них преобразованы в особое устройство,
за которое этот класс и получил свое
название. Конечности четвертого
абдоминального сегмента образуют отросток, способный
с силой распрямляться под действием
давления полостной жидкости. В покое этот
отросток удерживается в согнутом положении
236
Глава 8
Церки
Антенно-
овразные
передние
ноги
Тельсон
Вид сверху
Придаток
IV сегмента
Вид своку
Рис. 8.25. Многоножкоподобные классы Apterygota: А—два представителя двухвосток; Б—
бессяжковое; В—различные ногохвостки; показаны видоизмененные брюшные придатки (по
Imms, 1964; Wallace, Mackerras. 1970)
зацепкой, образованной придатком
третьего сегмента. Кроме того, конечности
первого сегмента брюшка образуют крупную
вентральную трубку с выпячивающимися,
подобно коксальным мешочкам,
пузырьками; назначение этой структуры
окончательно не выяснено. Возможно, она
используется для пополнения запасов влаги из
окружающей среды. Ни в одном из трех классов
гонопоры не расположены на VIII (у самок)
или X (у самцов) сегментах брюшка, как
у всех прочих насекомых; отсутствуют так-
Беспозвоночные с конечностями: членистоногие и др.
237
Рис. 8.26. Насекомоподобные классы Aptery-
gota: A—два представителя чешуйниц; £ —
представитель щетинохвосток (по Imms; 1964;
Daly et al., 1978)
же эффективные приспособления,
предотвращающие потерю воды организмом.
Ко второй группе относятся чешуйницы
(класс Zygoentomata) (рис. 8.26, А), гораздо
больше напоминающие крылатых
насекомых, от которых их отличают помимо
первичного отсутствия крыльев (а также
сопутствующих им структур груди), по сути дела,
лишь три признака, сохранившиеся от пред-
ковых форм и присущие всем первичнобес-
крылым: наличие брюшных грифельков,
отсутствие копулятивного аппарата и перенос
спермы с помощью откладываемых во
внешнюю среду сперматофоров, а также
продолжение линек после достижения
половой зрелости. Поэтому их, как правило,
считают близкими родственниками
непосредственных предков крылатых насекомых.
Щетинохвостки (класс Archaeognatha)
(рис. 8.26, Б), составляющие третью ветвь
первичнобескрылых, должны помещаться
где-то между Zygoentomata и прочими
Apterygota, так как, обладая некоторыми
продвинутыми признаками чешуйниц, они
сохраняют множество предковых черт,
свойственных и «многоножкоподобным»
классам гексапод, причем часто в рамках
одних и тех же структур и органов. Так,
хотя ротовые части и не погружены в
головную капсулу, сочленение мандибул,
особенности гипофаринкса и максилл отражают
анцестральное состояние, присущее
многоножкам. Представлен также полный набор
примитивных признаков, сохранившихся
у Zygoentomata. Если крылатых насекомых
можно вывести из чешуйниц, последние,
возможно, происходят от щетинохвосток,
поэтому оба этих класса часто объединяют
в рамках одной группы «настоящих
насекомых». Надо сказать, что представители
обоих классов в среднем крупнее (до 2 см
длиной) и не так тесно связаны с влажной
подстилкой и почвенными микроэкотопа-
ми, как прочие аптериготы.
Все пять рассмотренных классов
включают немногим более 3100 видов, большая
часть которых—потребители падали и
разлагающихся растительных тканей, хотя
некоторые и нападают на других мелких
гексапод.
8.5.3Б.2. Класс Pterygota
(крылатые насекомые)
Третья крупнейшая вспышка
разнообразия внутри типа одноветвистых
представлена 29 отрядами ныне живущих крылатых
насекомых (к которым относятся и
вторично бескрылые формы), включающими 98%
всех видов данного типа. Безусловно,
самым важным обстоятельством,
обеспечивающим подобный успех, было то, что пте-
риготы перестали зависеть от влажных
местообитаний, с которыми тесно связаны все
прочие классы одноветвистых. Это в свою
очередь в значительной степени явилось
следствием приобретенной ими способно-
238
Глава 8
сти эффективно препятствовать потере
воды телом. Эволюция их современного
состояния, вероятно, включала целый ряд
этапов, начиная с выработки особых свойств
кутикулы и трахей.
Возможно, восковой слой эпикутикулы
первоначально играл адаптивную роль
водоотталкивающего гидрофобного покрова:
в местообитаниях, где существует риск
переувлажнения, мелкие животные могут
захватываться водной пленкой, и
поступление в тело воды представляет для них куда
более серьезную опасность, чем ее потеря;
максиллярные железы аптеригот, по-
видимому, должны откачивать ее
значительный избыточный объем. У некоторых но-
гохвосток возникли ряды покрытых воском
бугорков с водоотталкивающей функцией,
и, надо полагать, предок крылатых
насекомых имел аналогичные приспособления.
У него также вполне могла отсутствовать
трахейная система, как и у большинства
многоножкоподобных аптеригот, что
можно приписать, по-видимому, прежде всего
мелким размерам любого организма,
возникшего в результате педоморфоза—
особенно если его предок и сам по себе был
не особенно велик. Как только (вновь)
развились трахеи, гидрофобный восковой слой
смог распространиться по всей поверхности
тела, никак не препятствуя газообмену.
В то же время наземные членистоногие,
по-видимому, теряют через трахейную
систему больше воды, чем через кутикулу,
поэтому сам по себе водонепроницаемый
покров не дал бы нужного эффекта.
Обычно у птеригот имеются мускулы,
закрывающие дыхальца (бывают и
другие— открывающие их), например,
насекомые сухих местообитаний могут открывать
их лишь на очень короткое время. Иногда
потери воды через дыхальца сокращаются
и другими путями. Один из них—
накопление в течение длительного времени
образовавшейся двуокиси углерода в
негазообразной форме. Поскольку кислород из
пространства трахеи потребляется, не
замещаясь эквивалентным объемом
углекислого газа, возникает частичное разрежение,
обеспечивающее поступление новых порций
воздуха через дыхальца в виде
однонаправленного потока, что противодействует
диффузии водяных паров наружу.
Последнее усовершенствование системы
сохранения воды связано с задней кишкой
и заключается в механизме обратного
всасывания воды из экскрементов и продуктов
выделения по мере их прохождения через
этот отдел пищеварительного тракта. Реаб-
сорбция может осуществляться разными
путями; среди них широко распространен
способ активной секреции неорганических
и/или органических растворимых веществ
в систему межклеточных пространств
внутри бугорков или сосочков стенки прямой
кишки. В результате вода пассивно
диффундирует из просвета кишечника в эти
пространства по осмотическому градиенту;
ПРОСВЕТ ПРЯМОЙ КИШКИ
Кутикула
Межклеточное
пространство
Воронка
Клапаны
Абсорбция растворенных веществ
жидкости в воронке
\ Поступление воды по
^.осмотическому градиенту
-Секреция растворенных
веществ в межклеточное
пространство
Поступление воды после абсорбции
растворенных веществ в воронке
Рис. 8.27. Схематический разрез сосочка прямой кишки падальной мухи, показывающий систему
межклеточных полостей, в которые и из которых могут перемещаться растворенные вещества
и куда из просвета кишки вынуждена поступать вода, переходящая затем в гемоцель (с сильными
упрощениями по Gupta, Berridge, 1966)
Беспозвоночные с конечностями: членистоногие и др.
239
когда накапливающаяся жидкость
переносится в гемоцель, растворимые вещества
абсорбируются из нее и вновь
возвращаются в круговорот (рис. 8.27).
Освоение местообитаний с недостатком
влаги, ставшее возможным благодаря этим
усовершенствованным механизмам
экономии воды, должно было сильно сказаться
на биологии ранних птеригот, что и
обусловило формирование многих характерных
черт этой группы. Откладывание капелек
спермы или сперматофоров, например,
в относительно сухих условиях было бы
ненадежной стратегией, и у крылатых
насекомых сперма переносится от самца к самке
непосредственно путем копуляции, хотя
сперматозоиды по-прежнему чаще всего
заключены в сперматофор. У примитивных
форм самки сохраняют унаследованные от
предков парные гонопоры, и самцы имеют
парные совокупительные органы, однако
в большинстве отрядов половое отверстие
самок непарное, а парные гениталии самца
соответственно срастаются в единую
структуру. У самца развита также пара
кластеров, т.е. захватов, которые удерживают
самку, в то время как выворачивающийся
дистальный отдел его полового протока
(или обособленная гоноподия—половая
ножка) вводится через половое отверстие
самки, чтобы отложить один или несколько
сперматофоров или пакетов спермы в ее
совокупительную сумку или другой отдел
полового тракта. При высвобождении из спер-
матофора сперма, как правило, хранится—
порой на протяжении длительного
периода—в семеприемнике (сперматеке), покуда
не оплодотворит все яйца по мере их
откладки, причем сперматозоиды попадают
в яйцо через крошечные поры в его
защитной оболочке, характерной для птеригот.
У ряда насекомых самки спариваются всего
один раз в жизни, но полученная при этом
сперма может оплодотворить несколько
кладок яиц; ни один из видов не линяет
после достижения половой зрелости. У
насекомых часто есть яйцеклад, что дает
возможность откладывать яйца в особые, как
правило, укромные места; другие способны
с этой же целью вытягивать конец брюшка
в виде трубочки, тогда как у некоторых
перепончатокрылых (пчел, ос и др.) яйце-
Сегмент, несущий гонопорСы)
?
Яйцеклад
Непарный пенис
Класпер
Рис. 8.28. Модификации придатков брюшных
сегментов предковых форм одноветвистых,
способствующие откладке яиц и спариванию
крылатых насекомых (вид сбоку) (по Snod-
grass, 1935). Яйцеклад самки представляет
собой базальные части придатков VIII и IX
сегментов брюшка; гонопоры самца находятся на
X сегменте, однако у большинства насекомых
они смещаются вперед и кажутся структурой IX
сегмента. Следует отметить, что у многих Pte-
rygota сформировались вторичные
совокупительные органы и ложные яйцеклады
клад видоизменился в жало, утратив тем
самым связь с размножением. Кластеры и
гоноподия самца, как и яйцеклад самки
(рис. 8.28), произошли из анцестральных
придатков брюшных генитальных
сегментов; кроме часто встречающихся парных
церков (придатки XI сегмента), эти органы
спаривания и откладки яиц представляют
собой единственные функционирующие
у взрослых птеригот остатки целой серии
сегментарных придатков брюшка.
Однако больше, чем любой другой
чертой, крылатые насекомые выделяются
среди всех членистоногих наличием крыльев—
еще одного приобретения, ставшего
возможным лишь после преодоления
зависимости от влажных местообитаний. У взрослых
особей (имаго) второй и третий грудные
сегменты несут кроме ног по паре крыльев;
впрочем, одна или обе пары могут быть
редуцированы или совсем утрачены.
Происхождение этих крыльев, как и селективные
преимущества обладателей предшествую-
240
Глава 8
щих им структур пока не известны, однако
скорее всего крылья развились из лопасте-
видных боковых выростов грудных терги-
тов с богатой сетью трахей,
встречающихся, например, у некоторых чешуйниц
и многочисленных ископаемых гексапод
(см. разд. 10.6.3). Установлено, что у ныне
живущих птеригот крылья формируются из
выпячиваний стенки тела путем дорсолате-
рального разрастания эпидермальных
зачатков. Верхняя и нижняя поверхности
последних прирастают друг к другу на всем
протяжении, кроме участков вдоль системы
каналов, в которых циркулирует
гемолимфа и расположены трахеи и сенсорные
нервы. В конечном итоге клетки эпидермиса
секретируют тонкую кутикулярную
оболочку; когда происходит последняя линька (на
имаго) мелкие, несколько мясистые
крыловые зачатки расправляются под давлением
полостной жидкости, выстилка каналов
укрепляется кутикулой (они становятся
дефинитивными «жилками крыла»),
эпидермис дегенерирует и крыло превращается
в тонкую двуслойную кутикулярную
структуру. Естественно поэтому, что его
движениями управляют мышцы, размещенные
в других местах; при этом они не всегда
крепятся к основанию крыла. Кроме того,
последнее должно быть сочленено с грудью
так, чтобы описывать необходимую для
полета траекторию в виде эллипса или вось-
Рис. 8.29. Разнообразное
жилкование крыльев насекомых
(по разным авторам).
Примитивные крылья с
многочисленными жилками показаны
в верхней части рисунка, а
внизу представлен ряд,
демонстрирующий постепенное
сокращение количества жилок
Беспозвоночные с конечностями: членистоногие и др.
241
мерки. У мелких форм крылья могут
работать с частотой до тысячи взмахов в
секунду. Весьма необычно для животных то, что
летательные мышцы насекомых отвечают
на каждый полученный нервный
импульс несколькими сокращениями (см.
разд. 16.10.5). Крылья примитивных птери-
гот, как правило, относительно велики
и снабжены многочисленными жилками, но
во многих эволюционных линиях их
размеры и количество жилок сильно
сокращаются (рис. 8.29); это соответствует эволюции
от длиннотелых животных с устойчивым,
но неманевренным полетом к короткоте-
лым формам, у которых он неустойчив,
зато высокоманеврен. В тех же
эволюционных линиях возникла и способность
полностью складывать крыловую пластинку,
когда она не используется—тенденция,
достигшая своего наиболее полного развития
в видоизменении всех частей переднего
крыла в защитные покрышки для сложенных
задних крыльев. Полет насекомых
рассматривается в разд. 10.6.3.
Фактором, обеспечившим
окончательный эволюционный успех крылатых
насекомых, оказалось, по всей видимости,
разнообразие доступных им источников пищи.
Это нашло свое отражение в крайне
разнообразном строении ротового аппарата,
обусловленном модификацией различных его
придатков в разных отрядах, хотя в отличие
от большинства аптеригот (если не считать
чешуйниц) птериготы и представляют
собой единую группу, поскольку у всех них
эти придатки не погружены в головную
капсулу, мандибулы причленяются в двух
точках, а гипофаринкс однолопастный. Пред-
ковой формой этих придатков был, без
сомнения, стандартный ротовой аппарат
многоножек, приспособленный для кусания
и пережевывания, причем такое его
строение сохранилось и у некоторых
современных крылатых насекомых (рис. 8.30, А). Он
дал начало целому ряду направлений
эволюции колющих и сосущих структур,
рассмотрение которых мы ограничим здесь
лишь немногими примерами.
Так, клопы питаются в зависимости от
группы соками растений или животных
с помощью хоботка, образованного нижней
губой (вторая пара максилл), внутри
которой размещаются две пары развившихся из
мандибул и максилл стилетов (рис. 8.30, Б).
Мандибулярные стилеты прокалывают
ткани жертвы и позволяют максиллярным
проникнуть в ранку. На внутренней
поверхности последних находятся два канала—через
один из них выпускается слюна, через
второй высасывается жидкость. У комаров
(рис. 8.30, В) есть два дополнительных
стилета, образованные верхней губой (окру-
жаюшей пищевой канал) и гипофаринксом
(в ней находится проток для слюны).
Однако в обоих случаях нижняя губа играет
исключительно защитную роль и
складывается—телескопически или отгибаясь локте-
образно назад—по мере проникновения
остальных ротовых придатков в ткани.
Другие насекомые сосут более
легкодоступные жидкости—такие, как нектар или
выделения из ран и трещин покровов.
Бабочки, например, пьют нектар через
длинный хоботок, способный расправляться за
счет нагнетания гемолимфы (когда
давление ее падает, он сам сворачивается в
спираль благодаря своим упругим свойствам).
Хоботок образован частями максилл, все
же прочие ротовые придатки редуцированы
или бесследно исчезли (рис. 8.30, Г). У ряда
ночных бабочек его кончик заострен, несет
кисточку из длинных волосков и может
использоваться для прокалывания. У
комнатной мухи для захвата пищи служит
нижняя губа. Мандибулы здесь также
отсутствуют, но верхняя губа и гипофаринкс
погружены в углубление разросшейся нижней
губы, несущей на конце большой, мягкий
двулопастный «лабеллум», действующий
подобно губке (рис. 8.30, Д).
Есть насекомые с гигантскими
мандибулами, которые, однако, не принимают
никакого участия в питании, а служат для
защиты или для сражений между самцами (рис.
8.30,£); кроме того, по меньшей мере в
восьми отрядах отмечены виды с
рудиментарным ротовым аппаратом, вообще не
питающиеся на имагинальной стадии. У
некоторых птеригот, особенно с полным
метаморфозом или с водными ювенильными
стадиями, большая часть жизненного цикла
и вся активность, связанная с питанием,
завершается до наступления половой
зрелости, а взрослая особь представляет собой
16-1314
242
Глава 8
Голова спереди
Голова своку, При питании стилеты Разрез в
стилеты спрятаны выходят наружу плоскости X-Y
-Li
Голова своку Разрез в плоскости
X-Y
Lr mrr Li
Голова своку
Разрез в плоскости X-Y
mi-
Хоботок
Общий вид тела сверку
Голова своку
Разрез в плоскости
X-Y
Рис. 8.30. Примеры адаптивной радиации ротового аппарата крылатых насекомых (см. текст):
А— грызущие мандибулы жука (Coleoptera); Б— колюще-сосущий хоботок клопа (Heteroptera);
В — ротовой аппарат комара (Diptera); Г—хоботок нектароядной бабочки (Lepidoptera); Д—
лижущий ротовой аппарат комнатной мухи (Diptera); E—сильно разросшиеся мандибулы самца
жука-рогача (Coleoptera), используемые в схватках самцов, а не для питания (по ряду авторов,
главным образом по Borror et al., 1976). Lr—верхняя губа, md — мандибула, mx — максилла, Li-
нижняя губа, h — гипофаринкс
недолговечный «автомат» для расселения
и размножения.
Миллион видов крылатых насекомых
можно разделить на четыре основные
группы отрядов (рис. 8.31). Paleoptera, или древ-
некрылые (два отряда), отличаются
главным образом одним (примитивным)
признаком—неспособностью укладывать
сзади на брюшке крылья, которые у них
крупные и несут густую сеть продольных и
поперечных жилок. Неполовозрелые стадии
(наяды, нимфы), живущие долго и
многократно линяющие, развиваются только
в воде, и, следовательно, их адаптации
отличаются от имагинальных. У многих,
например, имеются наружные парные трахей-
Беспозвоночные с конечностями: членистоногие и др.
243
-РедуцировамД\-.
ное заднее ол
' крыло
LEPIDOPTERA
DIPTERA (бзбочки)
(двукрылые)
Рис. 8.31. Разнообразие форм тела взрослых Pterygota: А— древнекрылые; Б— ортоптероиды;
В — гемиптероиды; Г—нейроптероидные зндоптериготы; Д—мекоптероидные зндоптериготы;
Е—гименоптероидные зндоптериготы (по разным авторам)
16*
244
Глава 8
ные жабры—листовидные пластинки на
брюшке, напоминающие настоящие жабры,
за исключением того, что вместо кровос-
набжающих структур (циркуляторная
система одноветвистых не участвует в
переносе дыхательных газов) в них находится
густая сеть замкнутых трахей. Возможно, эти
образования развились из предковых
придатков абдоминальных сегментов. Все
прочие птериготы относятся к «новокрылым»,
т.е. в состоянии покоя способны плоско
укладывать крылья поверх брюшка.
Вторая группа (из 10 отрядов)—
ортоптероиды. Для этих стоящих у основа-
0D0NATA
DERMAPTERA
EPHEMEROPTERA
Наружный зачаток крыла
PLECOPTERA
0RTH0PTERA
HETEROPTERA
THYSANOPTERA
Г STREPSIPTERA
NEUROPTERA
C0LE0PTERA
В AN0PLURA
H0M0PTERA
^ЩЩЙ
DIPTERA
Ложноножки <
Д LEPIDOPTERA
SIPHONAPTERA
HYMENOPTERA
Рис. 8.32. Неполовозрелые формы различных групп Pterygota, изображенных на рис. 8.31: Л —
наяды древнекрылых; Б. В — ортоптероидные и гемиптероидные нимфы; Г—Е—личинки эндоп-
теригот (по разным авторам)
Беспозвоночные с конечностями: членистоногие и др. 245
ния линии новокрылых (Neoptera) форм
типичны грызущий ротовой аппарат, нескон-
центрированная нервная система, неветвя-
щиеся, длинные многочлениковые антенны,
пара церков на конце брюшка,
многочисленные мальпигиевы сосуды,
выпячивающиеся совокупительные органы. Нимфы
несколько раз линяют, постепенно меняясь
и приближаясь по облику к взрослому
насекомому (имаго). Все органы взрослой
особи, включая крылья, сложные глаза и т. п.,
развиваются по мере прохождения нимфа-
льных возрастов. Поэтому образ жизни
ювенильных особей примерно такой же, как
и у взрослых.
В шести отрядах следующей—
гемиптероидной—группы происходит
такое же постепенное изменение нимф до
взрослых форм, хотя в большинстве
случаев наиболее серьезная перестройка
организма приурочена к последней
неполовозрелой стадии (или стадиям). В
подавляющем большинстве эти насекомые питаются
жидкой пищей, и определенные части мак-
силл превратились в склеротизированные
стилеты. Для них характерно небольшое
число (четыре и менее) мальпигиевых
сосудов, отсутствие церков, сильно
редуцированное жилкование крыла, короткие
антенны с небольшим числом члеников и хорошо
выраженная гоноподия. У нимф никогда
нет простых глазков.
Последняя группа—эндоптериготы'
(11 отрядов) в чем-то сходна с предыдущей,
хотя у ее представителей имеются короткие
церки и может быть множество
мальпигиевых сосудов. Однако, если всем прочим но-
вокрылым свойственно постепенное
превращение первой нимфальной стадии в
имаго с развитием крыловых зачатков снаружи
тела, у эндоптеригот есть личиночная
стадия, резко отличающаяся от взрослой
особи по строению, питанию, а часто и по
местообитанию—уникальный случай среди
наземных членистоногих. У многих мух
и перепончатокрылых, к примеру, личинки
безногие, червеобразные, в то время как
у бабочек и других перепончатокрылых на
1 В отечественной литературе ее обычно
называют Holometabola, т. е. насекомые с полным
превращением.— Прим. пер.
Рис. 8.33. Вид куколок эндоптеригот с
вентральной стороны; куколка двукрылого (в
середине) заключена в «пупарий»—оболочку из
шкурки, сброшенной личинкой последнего
возраста (по Jeannel, 1960; Wallace, Mackerras,
1970)
их брюшке находятся дополнительные
вторичные ходильные ноги («ложные ножки»)
(рис. 8.32). При этом зачатки крыльев
развиваются под покровами и, следовательно,
снаружи неразличимы. Поскольку
личиночные формы так существенно отличаются от
взрослых, переход между ними включает
так называемое полное превращение (см.
разд. 15.4.2). Этот процесс разворачивается
на особой неактивной стадии—в «куколке»
(рис. 8.33); почти все ткани личинки
разрушаются путем гистолиза, взрослый
организм формируется заново, и
развивающиеся крылья впервые появляются на
поверхности тела. Некоторые куколки
сохраняют сброшенный последней личинкой
покров в качестве защитной оболочки (рис.
8.33, Б). В эволюционном отношении такая
стратегия экологического разделения
личинок и имаго представляется весьма
выигрышной: 85% всех видов крылатых
насекомых относятся к эндоптериготам. Личинки
ряда видов могут встречаться даже в таких
местообитаниях, которые для
преимущественно сухопутных животных в высшей
степени неблагоприятны, например в
донных осадках приливной зоны и временных
водоемах; личинка одного африканского
комарика способна после дегидратации
к криптобиозу, соперничая в этом
отношении с тихоходками (см. с. 0222).
246
Глава 8
8.6. Тип Crustacea
(ракообразные)
8.6.1. Этимология
От лат. crusta—скорлупа, корка.
8.6.2. Диагностические
и специфические признаки
(рис. 8.34)
1. Двусторонне симметричные
членистоногие с разнообразной формой тела- от
вытянутой до сферической; длина тела от
менее 0,1 мм до 60 см (размах ног у
некоторых форм достигает 3.5 м, масса других —
более 20 кг).
2. Тело из более чем двух слоев клеток
в толщину, с тканями и органами.
3. Кишечник сквозной, прямой; от его
небольшого среднего отдела отходят два
пищеварительных дивертикула, в которых
происходит переваривание и всасывание
пищи; у видов, питающихся не только
мельчайшими частицами или жидкой пищей,
в переднем отделе кишечника находится
измельчающий аппарат; пищеварительные
дивертикулы возникают как выросты на
зародышевой кишке.
4. Разные группы ракообразных очень
сильно отличаются друг от друга
характером разделения тела на отделы и числом
сегментов, составляющих каждый отдел,
однако, как правило, можно выделить голову,
образованную акроном и пятью несущими
придатки сегментами, туловище из 2- -65
(и более) сегментов и тельсон, часто с парой
выростов — «фуркой». Первый и до семи (в
зависимости от группы) следующих
туловищных сегментов часто срастаются с
головой, образуя цефалоторакс
(«головогрудь»); туловище по типу придатков, как
правило, подразделено на торакс (грудь)
и абдомен (брюшко), причем торакальные
сегменты, не вошедшие в состав
головогруди, составляют так называемый «переон»,
а абдоминальные—«плеон». Головогрудь,
а в отдельных группах и значительная часть
всего тела или все оно целиком заключены
в особый вырост головы, так называемый
«карапакс», который разрастается в
стороны и закрывает бока тела.
Пищеварительный Задняя
дивертикул Сердце Гонада кишка
Нервный тяж
с ганглиями
Ротовые
придатки
Ногочелюсть
Антенны
Брюшные
плеоподы
Рис. 8.34. Морфология ракообразных. А.
Схема продольного разреза через тело
генерализованного представителя этого типа. Б—Г.
Характерные двуветвистые конечности (сильно
упрощено) (частично по McLaughlin, 1980):
Б — с одной трубчатой и одной листовидной
ветвью; В — с двумя листовидными ветвями
(типичная плавательная конечность); Г—с
обеими трубчатыми ветвями, одна из которых
сильно редуцирована (типичная ходильная но-
га)
Беспозвоночные с конечностями: членистоногие и др.
247
5. Цилиндрические или листовидные
придатки в основе своей двуветвистые,
причем ветви, как правило, имеют разные
размеры и форму, а часто несут и вторичные
ответвления; к придаткам головы относятся
две пары антенн (первую называют «антен-
нулы»), одна пара мандибул и две пары
максилл; туловищные придатки широко
варьируют в числе, строении и регионарной
дифференцировке; исходно каждый сегмент
несет пару конечностей, хотя на брюшке
они нередко отсутствуют; хелицер нет, но
отдельные конечности могут оканчиваться
клешнями.
6. Первичных ротовых придатков три
пары (мандибулы и две пары максилл), но
во всех группах бывает от одной до трех
дополнительных пар — «ногочелюстей»,
отходящих от грудных сегментов в составе
головогруди; элементы каждой пары могут
двигаться в фазе или в противофазе с другими
парами.
7. На голове находятся срединные
простые глазки или сложные глаза по бокам
(часто на подвижных стебельках).
8. Обычно наружная сегментация
выражена на туловище (часто скрыта под ка-
рапаксом, иногда исчезает) и отсутствует на
голове.
9. Выделительная система
представлена слепо оканчивающимися антеннальными
и/или максиллярными железами.
10. Наружный скелет часто обызвеств-
лен.
11. Газообмен идет через внутреннюю
стенку карапакса, через всю поверхность
тела или с помощью жабр, развивающихся из
определенных частей грудных или
брюшных конечностей.
12. Для циркуляторной системы
характерно наличие гемоцианина. реже—других
пигментов.
13. Нервная система с парными
ганглиями в каждом сегменте; вентральные
тяжи и ганглии исходно раздельные, но
чаще слиты; иногда все грудные ганглии
сливаются в единую массу.
14. Раздельнополые животные, редко
гермафродиты; оплодотворение внутреннее
путем копуляции с помощью гоноподий
или пенисов; расположение гонопора
варьирует, часто он расположен на груди.
15. Яйца самка обычно носит на себе
или держит в специальных карманах; у
одних форм из яйца выходит особь с полным
набором сегментов, у других — личинка
«науплиус» лишь с тремя сегментами (рис.
8.35) (ряд высокоспециализированных
паразитических форм удалось отнести к
ракообразным только благодаря наличию у них
науплиуса).
16. По большей части морские формы,
хотя встречаются и пресноводные (13%),
реже — наземные (3%).
Ракообразные, по сути дела,
представляют членистоногих в морских
экосистемах: лишь 3% видов артропод мирового
океана не относится к этому типу. Они
преобладают в планктоне (и в пресноводном
также) и входят в число трех-четырех
важнейших бентосных групп, включая интер-
стициальный комплекс и макрофауну.
Кроме того, ряд видов ведет паразитический
образ жизни.
Процветание ракообразных можно
в определенной мере объяснить таким
общим для всех артропод признаком, как
наличие членистых конечностей. Их ноги
и/или плавательные придатки позволяют
быстро передвигаться и без труда осваивать
различные участки местообитания.
Несомненно, успеху способствует и другая важная
черта артропод—наружный скелет:
крупные, заключенные в крепкий панцирь
и мощно вооруженные формы гораздо
меньше подвержены риску поедания
хищниками, чем мягкотелые животные. Однако,
может быть, основная причина практически
повсеместного распространения водных
ракообразных— разнообразие формы тела,
хотя внутренняя анатомия их достаточно
сходна.
Глаз \\| \1/ АнтеннУЛЬ1
Рис. 8.35. Личинка-науплиус (по Green, 1961)
248
Глава 8
Если строение хелицеровых (разд. 8.4)
и насекомых (разд. 8.5.3Б) соответствует
некой общей схеме, а многоножки (разд.
8.5.ЗА), несмотря на все различия в числе
сегментов, неизменно сохраняют общую
форму тела, у ракообразных дело обстоит
совершенно иначе; такого понятия, как
типичный план строения тела ракообразных,
попросту не существует. У одних есть голова
и туловище, у других—голова, грудь
и брюшко, у некоторых—головогрудь,
переон и брюшко, а в одной крупной группе
лишь головогрудь и брюшко; в немногих
случаях брюшко отсутствует, а бывает, что,
по сути дела, нет и дифференцированной
головы или груди. Кроме того, число
сегментов в каждом из этих отделов также
варьирует и зависит от группы—иногда внутри
одного класса. Аналогичным образом,
конечности бывают самой разнообразной
формы—от ходильных ног, практически
таких же, как у хелицеровых и одноветви-
стых, до листовидных плавательных
придатков. Антенны могут играть роль
сенсорных органов, служить для локомоции или
сбора корма, действовать как средство
прикрепления к хозяину, выполнять роль
кластеров, удерживающих партнера при
спаривании, и т. д. У морского желудя,
паразитического веслоногого, раковинного жабро-
ногого краба и равноногого из семейства
антурид (рис. 8.36) едва ли есть какие-либо
внешние черты, свидетельствующие об их
принадлежности к одной и той же группе
животных.
Такая пластичность строения позволяет
ракообразным плавать, закапываться
в грунт, ползать, делать ходы в древесине,
прилепляться к скалам, охотиться,
объедать растения, питаться взвешенной пищей
и донными осадками (см. рис. 9.4),
паразитировать на животных различных типов (и
на других ракообразных)—так что,
вероятно, не будет преувеличением сказать, что
они занимают все возможные типы
экологических ниш в морях. Ракообразные—
прежде всего водные обитатели. Хотя
представители ряда их групп живут и на суше,
это все же не наземные животные в полном
смысле слова. В частности, у них
сохраняется та же система газообмена, что и у
водных сородичей, а это ограничивает распро-
Рис. 8.36. Разнообразие формы тела
ракообразных: А — морской желудь (Cirripedia). Б—
паразитическое веслоногое (Copepoda); В—
равноногое (Malacostraca); Г—раковинное
жаброногое (Branchiopoda); Д—краб
(Malacostraca) (по разным авторам)
странение лишь влажными
местообитаниями. Более того, некоторые виды, например
наземные крабы, вынуждены возвращаться
в воду для размножения, поскольку их цикл
развития по-прежнему включает фазу
водной личинки. Только среди мокриц
встречаются формы с сугубо сухопутными
приспособлениями, и среди наземных
ракообразных эта группа распространена
наиболее широко. В дополнение к жабрам на
плеоподах—характерному признаку этого
отряда (Isopoda) — у некоторых мокриц эти
придатки брюшка снабжены трахеевидны-
ми втачиваниями покровов, заходящими
в гемоцель конечностей. У других видов
выработались лишь механизмы, позволяющие
постоянно поддерживать жабры в
увлажненном состоянии, например направляя
на плеоподы капельки воды с поверхности
тела.
Беспозвоночные с конечностями: членистоногие и др.
249
8.6.3. Классификация
Почти 40000 видов, включаемых в этот
тип, распределены между девятью класса-
Класс
ми. Все шире распространяется традиция
объединять эти девять таксонов в четыре
более крупные группы (их можно считать,
например, подтипами); при этом относите-
Надотряд
Отряд
Cephalocarida -
Branchiopoda -
Remipedia
' Mystacocarida -
Branchiura
Copepoda-
4Cirripedia -
*Osiracoda-
' Malacostraca
Phyllocarida-
Hoplocarida -
Syncarida-
Pancarida -
Peracarida
Eucarida
- Brachypoda
Anostraca
Notostraca
Conchostraca
Cladocera
- Nectiopoda
- Derocheilocarida
-Arguloida
Calanoida
Cydopoida
Misophrioida
Mormonilloida
Harpacticoida
Monstrilloida
Siphonostomatoida
Poecilostomatoida
PTantulocaridida
Rhizocephala
Ascothoracica
Thoraeica
Acrothoracica
Facetotecta
Myodocopida
Cladocopida
Podocopida
Platycopida
Palaeocopida
Leptostraca
Stomatopoda
Anaspidacea
Stygocaridacea
Bathynellacea
Thermosbaenacea
Mysidacea
Cumacea
Spelaeogriphacea
Tanaidacea
Mictacea
Isopoda
Amphipoda
Euphausiacea
Amphiorridacea
Decapoda
250
Глава 8
льно неспециализированные классы Remi-
pedia и Malacostraca помещают в отдельные
монотипические подтипы, а остальные два
подтипа (возможно, обязанные своим
возникновением педоморфозу) включают
соответственно классы Cephalocari-
da + Branchiopoda (подтип Phyllopoda)
и Mystacocarida + Branchiura + Copepo-
da + Cirripedia + Ostracoda (подтип Maxil-
lopoda). Хотя Malacostraca часто называют
«высшими ракообразными», на самом деле
это довольно примитивная группа,
находящаяся где-то у основания ствола
ракообразных; наиболее продвинутые черты
организации заметны у Phyllopoda и Maxillopoda.
8.6.3.1. Класс Cephalocarida
Эти мелкие слепые рачки (длина их не
достигает и 4 мм) обитают на морском дне
и питаются детритом. О них ничего не было
известно до 1955 г., несмотря на то что
местами их численность может быть высокой.
Тело (рис. 8.37) делится на голову, грудь
и брюшко без каких-либо намеков на
образование головогруди или карапакса; все
восемь пар грудных ног похожи друг на друга
и вполне идентичны II паре максилл, также
состоящих из листовидной и трубчатой
части. Брюшко включает 11 сегментов и
лишено придатков, если не считать
сохранившихся на первом сегменте редуцированных
конечностей, к которым прикрепляются
Антенна
Грудь / Голова
Брюшко-£у I / /
UJ Грудаые Антен^7
Y-I придатки
Фурка \1
<1\
Рис. 8.37. Цефалокарида (вид сбоку) (по
Sanders, 1957)
яйцевые мешки. Цефалокариды —
гермафродиты (что для ракообразных
нетипично); у парных яичников и семенников
общий проток; развитие протекает постепенно
и сопровождается многочисленными
личиночными линьками.
Всего известно десять видов; тело у всех
вытянутое, цилиндрическое,
заканчивающееся тельсоном с длинной фуркой, каждая
ветвь которой несет щетинку. Все известные
формы относят к одному отряду.
8.6.3.2. Класс Branchiopoda
(жаброногие)
Жаброногие представляют собой
довольно разнообразную группу (рис. 8.38),
включающую главным образом
пресноводных рачков с мелкими или
рудиментарными придатками головы (за исключением,
как правило, антенн), не приросшими к
голове туловищными сегментами и с
туловищем, несущим ряд примерно одинаковых
конечностей, которые обычно
уменьшаются в размерах спереди назад (несколько
последних сегментов вообще их лишены).
Конечности эти несут жабры, и в типичном
случае представляют собой листовидные
плавательные и/или фильтрующие органы,
форма которых поддерживается в основном
давлением полостной жидкости, а не
кутикулой. Многие виды размножаются парте-
ногенетически и «вынашивают» на себе
яйца до выхода личинок. Отдельные
представители способны образовывать
покоящиеся стадии.
Жаброногих делят на четыре отряда,
сильно различающиеся по внешнему облику.
У раковинных (Conchostraca) и ветвисто-
усых, или водяных блох (Cladocera), тело
короткое, иногда почти округлое, антенны
выполняют локомоторные функции, фурка
в виде коготков, а выводковая камера
помещается дорсально под сдавленным с боков
карапаксом. Однако если у раковинных до
более 30 туловищных сегментов, а карапакс
покрывает все тело, включая голову, у вет-
вистоусых последний, хотя часто велик,
никогда не закрывает головы, а иногда
редуцирован до небольшой выводковой камеры;
туловищных конечностей всегда не более
шести пар (рис. 8.38, А, Б). Раковинные не
Беспозвоночные с конечностями: членистоногие и др.
251
Область прикрепления
карапакса
Карапакс
Мышцам
аддуктор
Мандивула
Кольца
роста
Сложный
глаз
Антенна
Первая пара
туловищных
конечностей
Сложный глаз
Пищеварительный^
дивертикул-
Антенна
Сердце -
Карапакс
Яйца в
выводковой
камере
.Антеннула Туловищные Кишечник^
^^Туловищные
" придатки
Туловищные придатки
Антенна
Антеннула Сложный глаз
Г
Генитальные
сегменты
Фурка
Рис. 8.38. Морфология жаброногих. А. Раковинные (Conchostraca), вид сбоку, левая створка
карапакса удалена (см. также рис. 8.36, Г) (по Kaestner, 1970). Б. Ветвистоусые (Cladocera), вид сбоку
через прозрачный карапакс (по Belk, 1982). В. Щитни (Notostraca). вид сверху (по Kaestner, 1970).
Г. Жаброноги (Anoslraca), вид сбоку в естественном положении плавания—дорсальной стороной
вниз (по McLaughlin, 1980)
сбрасывают карапакс в процессе линьки,
а наращивают его концентрическими
кольцами как двустворчатые моллюски; как и
у последних, створки «раковины»
удерживаются вместе сильной мышцей-
аддуктором, противодействующей
упругому лигаменту.
Зато у Notostraca (идитней) карапакс
представляет собой широкий дорсовентра-
льно уплощенный щит, по форме близкий
к отпечатку конского копыта; из-под него
выступает назад узкий цилиндрический
задний конец тела с двумя длинными
многочленистыми ветвями фурки (рис. 8.38, В).
Интересно, что эта часть туловища
разделена на сегменты не полностью, так что на
одном видимом «сегменте» может быть до
шести пар конечностей. Всего туловищных
конечностей до 70 пар, причем
одиннадцатая пара несет выводковые камеры, а
первая крупнее остальных. У представителей
Anostraca (жаброногов) никакого карапакса
нет (рис. 8.38, Г), а выводковая камера
образуется внутри тела из расширенного
влагалища; когда генитальные сегменты
наполнены яйцами, они образуют обширный,
выпирающий наружу яйцевой мешок. Как
щитни, так и жаброноги обычно
встречаются в водоемах с суровыми условиями
жизни (например, в пересыхающих лужах
и соленых озерах), поэтому у них развились
высокоспециализированные покоящиеся
формы. Яйца этих ракообразных хорошо
переносят сильные колебания температуры
и пересыхание, а в состоянии покоя
способны пребывать у некоторых форм до 10 лет.
Среди 850 известных современных видов
большинство имеет размеры до 3 см, лишь
отдельные достигают 10 см в длину.
8.6.3.3. Класс Remipedia
Этот класс включает единственный
вид—слепого рачка, описанного из одной
морской пещеры в 1980 г. (рис. 8.39); о его
образе жизни до сих пор почти ничего не
252
Глава 8
Антенна Головогрудь Боковые плавательные конечности
Фурка
Рис. 8.39. Ремипедия, вид
с вентральной стороны (по
Yager, Schram, 1986)
известно. Мелкое (меньше 3 см в длину)
вытянутое прозрачное тело состоит из
короткой, лишенной карапакса головогруди,
включающей голову и первый туловищный
сегмент, и длинного туловища из более чем
30 похожих друг на друга сегментов,
каждый из которых несет пару боковых
листовидных конечностей, служащих для
плавания дорсальной стороной вниз. Среди
придатков головы необычны палочковидные
выросты спереди от антеннул и
хватательные ротовые органы, в том числе
ногочелюсти. Сейчас специалисты собираются
описать еще восемь видов; все они обнаружены
в морских пещерных местообитаниях в
Атлантике.
8.6.3.4. Класс Mystacocarida
Мистакокариды (рис. 8.40) — крохотные
(менее 1 мм длиной) вытянутые, лишенные
пигментации интерстициальные морские
рачки. Их характерные признаки—прежде
всего разделение головы на небольшую
переднюю и обширную заднюю части,
а также туловище из 10 сегментов, первый
из которых несет ногочелюсти, хотя и не
срастается с головой. Головные придатки
крупные (они используются для локомо-
ции), а придатки брюшка или
редуцированы до мелких одночленистых структур
(например, на I] —V сегментах), или вовсе
отсутствуют; на тельсоне мощная
клещевидная фурка. К примитивным признакам
группы относятся парные туловищные
ганглии, которые лишь соприкасаются между
собой, но не слиты, отсутствие сложных
глаз, пищеварительных дивертикулов
(вероятно, по причине малых размеров тела).
Еще одна их любопытная черта уникальна
среди ракообразных: на каждом
туловищном сегменте находится пара
расположенных по бокам поперечных бороздок с
зубчатыми краями: назначение их не выяснено.
Все десять известных видов относят
к одному отряду.
Антеннула
Антенна
Ногочелюсть
Зуечатая
Бороздка
Тельсон
Рис. 8.40. Мистакокарида, вид сверху (по
Kaestner, 1970)
8.6.3.5. Класс Branchiura
(карпоеды, или карповые вши)
Представители этого
класса—небольшие (меньше 3 см длиной) формы,
часть жизненного цикла проводящие в
качестве эктопаразитов на морских и
пресноводных рыбах; они прокалывают покровы
хозяина мелкими мандибулами и питаются
жидкой пищей—чаще всего кровью, как
комары. Их сильно уплощенное дорсовентра-
льно тело состоит из головогруди (голова
и приросший к ней первый грудной
сегмент), трехсегментного переона и двуло-
пастного нерасчлененного брюшка;
головогрудь, а иногда и большая часть переона
покрыты обширным плоским карапаксом
в виде круглого, двудольчатого или напо-
Беспозвоночные с конечностями: членистоногие и др.
253
Антенны
Сложный глаз
Карапакс
Максиллярная присоска
.Дыхательные участки
■Максилла II
1лавательные конечности
Фурка
-Брюшко
ВИД С ВЕНТРАЛЬНОЙ СТОРОНЫ
Максиллярная
присоска
Максилла II
Карапакс
Брюшко
Б ВИД С ВЕНТРАЛЬНОЙ СТОРОНЫ fi ВИД С ДОРСАЛЬНОЙ СТОРОНЫ
Рис. 8.41. Морфология и разнообразие карпоедов (по Kaestner, 1970)
минающего наконечник стрелы щита,
простирающегося в стороны или в стороны
и назад (рис. 8.41) и несущего пару сложных
глаз.
Придатки головы могут быть либо
сильно уменьшены, либо превращены в органы
прикрепления к телу хозяина; они
заканчиваются крючьями, или же, как в случае мак-
силл I, крупными стебельчатыми
присосками. Все четыре пары грудных придатков,
включая конечности сегмента, полностью
или частично слитого с головой,
представляют собой плавательные ноги; на брюшке
придатки полностью отсутствуют.
Несколько необычно то, что самка не носит на
себе яйца до вылупления молоди, а
прикрепляет их к субстрату или к донной
растительности.
Единственный отряд насчитывает
порядка 150 видов.
8.6.3.6. Класс Copepoda (веслоногие)
Веслоногие доминируют в морском и
в несколько меньшей степени в
пресноводном планктоне. К ним относятся и
многочисленные интерстициальные формы, а
примерно четверть всех известных видов—
паразиты самых разных животных — от
губок до китов. Хотя в большинстве случаев
это мелкие рачки (меньше 2 мм длиной),
254
Глава 8
Просома
Головогрудь
Антеннула
Мандивула-
Антенна
Максилла I
Максилла II
Ногочелюсть Грудные%
.Сочленение
Уросома
плавательные
конечности
длина одного свободноживущего вида 2 см,
а одного эктопаразита— 0,3 м.
В типе тело веслоногих Состоит из
головы с хорошо развитыми ротовыми
придатками и антеннами, шестисегментной груди
с плавательными конечностями и брюшка
из пяти сегментов без придатков. Однако
сегменты могут срастаться между собой
самыми различными способами (по меньшей
мере один грудной сегмент всегда слит с
головой, а у многих форм в состав
головогруди входит и второй), так что исходное
разделение на головогрудь, переон и брюшко
не обязательно отражает фактическое
расчленение тела, если в нем вообще
сохраняются обособленные отделы. Так, у
паразитических видов можно видеть различные
стадии морфологической дегенерации, причем
в крайних случаях утрачивается всякая
внешняя сегментация и исчезают придатки.
У свободноживущих и ряда паразитических
форм наблюдается функциональное
разделение тела на заднюю область и
сочлененную с ней переднюю. За исключением ин-
терстициальных цилиндрических форм,
передний отдел («просома») овальный,
иногда удлиненно-овальный, в то время как
Рис. 8.42. Морфология
веслоногих. А. Внешний вид свобод-
ноживущей формы сбоку;
показаны придатки. Б. Место
сочленения просомы с уросомой
(показано стрелкой) у разных
веслоногих; грудные сегменты
пронумерованы, номера
сегментов, слитых с другими
отделами, взяты в скобки (по
Kaestner, 1970)
задний («уросома») тонко-трубчатый (рис.
8.42). Хотя в одной из групп граница между
ними и в самом деле совпадает с делением
на головогрудь и переон + брюшко, во
многих случаях она проходит между 111 и IV
сегментами переона (соответствующими
V и VI туловищным сегментам), так что
последний сегмент переона (и груди) входит
в состав уросомы. Карапакс и сложные
глаза у веслоногих всегда отсутствуют.
Паразитические формы могут иметь
самый разнообразный облик (рис. 8.43),
причем некоторые червеобразные или
мешковидные виды можно отнести к
ракообразным лишь по признакам ранних стадий их
жизненного цикла. Они часто носят на себе
яйца в виде длинных тяжей в отличие от
свободноживущих веслоногих, у которых
один-два овальных яйцевых мешка.
Около 8400 видов мировой фауны
группируют в девять отрядов, из которых один
помещен в данный класс лишь условно. Эти
слабоизученные рачки-тантулокариды—
крошечные эктопаразиты морских
ракообразных, живущие постоянно на теле
хозяина, прикрепившись к нему ротовым
диском; головные придатки у них отсут-
Беспозвоночные с конечностями: членистоногие и др.
255
Рис. 8.43. Разнообразие формы тела и его дегенерация у паразитических веслоногих (по разным
авторам)
ствуют, а из яйца выходит продвинутая ли-
чинка-танталус, осуществляющая
заражение. Самка, по сути дела, представляет
собой мешок с яйцами, удерживаемый на теле
хозяина личиночной головой, а свободно-
живущий самец развивается из массы де-
дифференцированной ткани внутри
личинки. Высказывалось предложение выделить
эти виды в самостоятельный класс Tantulo-
carida, возможно родственный Copepoda
или Cirripedia.
8.6.3.7. Класс Cirripedia (усоногие)
Усоногие —наиболее видоизмененная
группа ракообразных, представленная либо
сидячими формами, либо паразитами. Они
по сути лишены головы, у большинства
отсутствует и брюшко, а от наружной
сегментации в лучшем случае сохраняются лишь
следы. Крайняя специализация
наблюдается в отряде корнеголовых (Rhizocephala),
представители которого больше всего
напоминают гриб-трутовик с сетью тонких
трубочек, распространяющейся по всему телу
хозяина (почти всегда — десятиногого
рака), и наружным мешком, где
располагаются гонады (рис. 8.44, А).
Слабее всего структурная специализация
в отряде Ascothoracica (рис. 8.44, Б), куда
относятся паразиты книдарий и иглокожих.
У немногих видов здесь сохраняются даже
рудименты головы, несущей антеннулы
с клешнями (что совершенно нетипично для
ракообразных), и грудь с шестью парами
плавательных конечностей, покрытыми
двустворчатым карапаксом, из-под
которого наружу выступает пятисегментное
брюшко. Наиболее известные представители
усоногих—морские желуди и морские
уточки (отряд Thoracica)- прикрепленные
организмы; шесть пар их грудных ног
(«усиков») образуют фильтрующий аппарат для
сбора пищи, а бочковидный карапакс
усилен известковыми пластинками (от
нескольких до множества) (рис. 8.44, В, Г), не сбра-
256
Глава 8
Внутренние ветвящиеся отростки
стебелька (показаны лишь на
одной стороне нрава)
Тело паразита
Отогнутое Брюшко крава
Карапакс
Клешненосные
антеннулы
г Ротовой
конус
Мышца-аддуктор
Фурка
Грудные
конечности
Брюшко
Ротовой конус
Яйцевод Тело Антеннула
Основание
Г
Рис. 8.44. Усоногие. А. Вид с вентральной стороны краба, пораженного представителем корнего-
ловых (покровы краба показаны прозрачными). Б. Схематический вид сбоку представителя Asco-
thoracica (левая створка карапакса удалена). В. Морской желудь, вид сбоку (часть карапакса и его
пластинок удалены; см. также рис. 8.36). Г. Внешний вид двух морских уточек, прикрепившихся
к деревянному бруску (А и Б по Kaestner, 1970; б и Г по Zullo, 1982)
Беспозвоночные с конечностями: членистоногие и др.
257
сываемыми при линьке, а нарастающими
по краям. Как и у прочих усоногих,
первоначальное прикрепление к субстрату
(скалам, раковинам моллюсков или к
водорослям) осуществляется антеннулами; затем
предротовая область может увеличиться,
образуя вытянутый столбик (как у морских
уточек), или превратиться в тонкий
прикрепительный диск, как у морских желудей.
Четвертая группа — Acrothoracica — в
общем похожа на предыдущую, но здесь
отсутствуют известковые пластинки, и рачки
вбуравливаются в кораллы или, реже, в
раковины моллюсков. У представителей двух
этих «желудеобразных» отрядов пенис,
чтобы достигать соседних, также неподвижно
прикрепленных особей, бывает очень
длинным. У пятого отряда (Facetotecta)
известны только личинки, так называемые Y-
науплиусы и Y-циприды.
В этом классе заметна явная тенденция
к гермафродитизму или к уменьшению
мужских особей до крошечных размеров,
а также к редукции кишечника: у многих
видов он оканчивается слепо, а у корнеголо-
вых и вовсе отсутствует. У всех усоногих
имеется специфическая личиночная стадия
(после науплиуса) — «циприсовидная»,
которая находит хозяина или место
прикрепления на субстрате, а затем в ходе
метаморфоза превращается в молодую
взрослую особь.
Все известные виды (около 1000)
обитают в морях.
8.6.3.8. Класс Ostracoda
(ракушковые рачки)
Остракоды очень мелкие, большинство
их не достигает в длину и миллиметра,
лишь немногие приближаются к 2 см. У них
короткое овальное тело, заключенное в
двустворчатую, часто известковую раковину,
образуемую карапаксом (рис. 8.45). Как и
Антенна
Карапакс
Фурка
Область
прикрепления Мандибула Мышца-адцуктор Яйца в
карапакса \ I I выводи
Антеннула
Рис. 8.45. Ракушковые рачки:
А — внешний вид сбоку; Б—то
же с удаленной левой створкой
карапакса (по Cohen, 1982)
выводковой
камере
Карапакс
Максилла I
Максилла II
Первая пара
грудных ног
17—1314
258
Глава 8
у раковинных рачков (Conchostraca) и
двустворчатых моллюсков, створки
удерживаются в сомкнутом состоянии поперечной
мышцей-аддуктором, противодействующей
упругому лигаменту. Иногда присутствуют
зубы замка. Однако в отличие от
упомянутых групп остракоды сбрасывают раковину
и образуют ее заново при каждой линьке.
На их мешковидном теле не заметно
сегментации, но, если судить по придаткам,
половину его объема занимает то, что
раньше было головой. Сохраняется всего от
пяти до семи пар придатков, причем первые
четыре из них несомненно принадлежат
«голове» (антеннулы, антенны, мандибулы
и первые максиллы), а две задние (одна или
обе могут отсутствовать)—«груди»; что же
касается лежащей между ними пятой пары,
взгляды специалистов на ее природу
расходятся. Большинство считает эти придатки
второй парой максилл, некоторые —
грудными конечностями, т. е. ногочелюстя-
ми или первыми торакальными ногами.
Если это, в самом деле, вторые максиллы,
ракушковые рачки оказываются
единственной группой ракообразных, в которой
могут сохраняться только головные придатки.
Функции этих конечностей у разных форм
неодинаковы, но антенны обычно являются
основными плавательными органами,
а первые грудные конечности, если
присутствуют, часто бывают ходильными. Яйца,
как правило, вынашиваются до вылупления
под карапаксом, хотя ряд видов просто
прикрепляет их к субстрату, в том числе к
подводным растениям.
Остракод делят на пять отрядов (из них
один описан лишь по пустым раковинам).
Всего известно 5700 видов, по большей
части морских. Есть и несколько наземных
форм, перешедших из пресных водоемов
к жизни в слое лесного гумуса или
подстилки.
8.6.3.9. Класс Malacostraca
По объему этот класс намного
превышает все прочие и насчитывает примерно
23000 видов. По-видимому, такого
разнообразия форм тела нет больше ни в одном
классе животного мира; лишь один из 16
входящих сюда отрядов — Decapoda—
включает столь несхожие друг с другом
организмы, как крабы, речные раки, креветки
и раки-отшельники. Главный признак,
объединяющий представителей этого класса—
тело из головы, восьмисегментной груди
и брюшка из шести (редко семи) сегментов,
причем все перечисленные отделы
снабжены полным набором придатков.
Наблюдаемое разнообразие обусловлено тем, что
разное количество грудных сегментов (от
нуля до восьми) может объединяться с
головой в головогрудь, 0—3 пары грудных
придатков бывают преобразованы в ного-
челюсти. карапакс имеется или отсутствует
(как первично, так и вторично), а если он
есть, то покрывает от первых двух до всех
восьми грудных сегментов и до нескольких
брюшных.
К числу типичных черт специализации
Malacostraca относятся, во-первых,
развитие в передней кишке «желудка», где пища
тонко перемалывается, а все оставшиеся
крупные частицы отфильтровываются от
получающейся массы, которая затем
поступает в пищеварительные дивертикулы (рис.
8.46), а во-вторых,—превращение послед-
Мышцы, приводящие
пластинки в движение
Кардиальный
отдел желудка
Зубчатая
пластинка
Клапан
Пилорический
отдел желудка
Задняя кишка
Устье
пищеварительного
дивертикула
Пищевод
Рис. 8.46. Схематический
продольный разрез через желудок
десятиногого рака; видны
жевательные пластинки,
измельчающие проглоченную пищу,
а также фильтрующая система
пилорического отдела,
прикрывающая отверстие одного
из пищеварительных
дивертикулов (по Warner, 1977)
Беспозвоночные с конечностями: членистоногие и др.
259
Мышца-
аддуктор
Карапакс
VII ерюшной
сегмент
Тельсон
Хватательные ложно-
клешневидные
ноги
Грудные
конечности
Голова
Грудные
сегменты
Брюшные
сегменты
Хвостовой веер
Выводковая камера
с яйцами ~~
Рис. 8.47. Разнообразие Malacostraca. A. Phyllocarida, вид сбоку (карапакс как бы прозрачен). Б.
Hoplocarida, вид сверху. В. Syncarida. вид сбоку. Г. Pancarida, вид сбоку; карапакс изображенной
самки, используемый в качестве выводковой камеры, сильно увеличен и вздут (по разным
авторам)
ней пары брюшных придатков («уропод»)
и тельсона в хвостовой веер, которым
и оканчивается тело вместо более обычной
для ракообразных фурки. Характерно
также наличие пары хорошо развитых
сложных глаз, иногда вторично утрачиваемых.
Многие виды крупные, с сильно обызвеств-
ленными покровами, у некоторых
наблюдается значительная концентрация нервной
системы и сложное поведение.
Malacostraca—существенный компонент морского
нектона и бентоса; многие виды встречаются
в пресных ручьях, реках и озерах, другие
ведут наземный образ жизни (сюда относятся
все ракообразные, способные выживать не
только в постоянно увлажненных
местообитаниях).
Можно выделить шесть групп отрядов.
В двух из них сохранился исходный план
строения тела (т. е. голова, грудь и брюшко)
без всякой тенденции к образованию
головогруди или добавочных ротовых частей.
В одной группе (Phyllocarida, рис. 8.47. А)
наблюдается нетипичный для этого класса
признак: VII брюшной сегмент и фурка,
а уроподы отсутствуют. Наиболее
характерный внешний признак филлокарид—
крупный, сжатый с боков двустворчатый
карапакс, покрывающий грудь и восемь пар
ее листовидных конечностей, щетинки кото-
17*
260
Глава 8
Карапакс
Стевельчатые Карапакс
сложные глаза
Карапакс
Карапакс
Рис. 8.48. Разнообразие Malacostraca. Peracarida: А—Tanaidacea; Б — Mysidacea; В—
Spelaeogriphacea; Г—Cumacea; Д—Isopoda (равноногие раки; см. также рис. 8.36. б); £—
Amphipoda (бокоплавы) (А и Д—вид сверху, прочие—сбоку) (по разным авторам). Обратите
внимание на неодинаковую степень развития карапакса и разную форму придатков
рых окружают выводковую камеру. Во
второй группе—Hoplocarida (раки-богомолы)
(рис. 8.47, Б)—карапакс гораздо мельче
и закрывает лишь половину дорсовентраль-
но сплюснутой груди. Особенно
характерно, что первые пять пар грудных
конечностей здесь несут ложные клешни и не
связаны с локомоцией; вторая пара очень
крупная, хватательная, а последние три пары
у самок удерживают яйцевую массу. У ряда
видов конечности второй пары превращены
в «кулаки», способные выбрасываться со
скоростью 1 см/мс, нанося удар такой же
силы, что и пуля 22-го калибра.
В отличие от двух этих групп следующие
три надотряда характеризуются слиянием
по меньшей мере одного (как правило, а
самое большее—трех) грудного сегмента
с головой в головогрудь, соответственно
переоном из семи сегментов и брюшком из
Беспозвоночные с конечностями: членистоногие и др.
261
Вздутый карапакс,
служащий выводковой
камерой
Мягкое, редуцированное,
асимметричное Брюшко
Брюшко спрятано
под карапаксом
Рис. В.49. Разнообразие Malacostraca. Eucarida: А— Euphausiacea; Б—Amphionidacea; В-
Ж—десятиногие раки (А — В — вид сбоку; Г—Ж—вид сверху) (по разным авторам)
шести. Степень развития карапакса при
этом широко варьирует. Исключительно
пресноводные Syncarida (рис. 8.47, В)
лишены его вообще (изначально, как обычно
полагают) и демонстрируют тенденцию
к утрате придатков на достаточно
однородных туловищных сегментах. Слепые Рапса-
rida (рис. 8.47, Г), населяющие пещеры и ин-
терстициальные местообитания (иногда
в горячих источниках с температурой воды
до 45° С), также в основном пресноводная
группа; они отличаются коротким
карапаксом, играющим роль выводковой камеры
и центра газообмена. У третьей гораздо
более обширной группы—Peracarida (рис.
8.48) — выводковая камера образуется
отростками конечностей переона. У предков
этого надотряда, вероятно, были карапакс
262
Глава 8
и сложные глаза на стебельках, однако и то
и другое претерпело сильную редукцию
в разных эволюционных линиях, так что
сложные глаза стали по большей части
сидячими, а в двух крупных отрядах —
бокоплавов (Amphipoda) и равноногих (Iso-
poda) —исчез и карапакс. Перакариды
очень разнообразны по форме и местам
обитания: одни плавают, другие роют
норки, многие передвигаются, используя
придатки переона в качестве ходильных ног,
есть и паразитические формы. Тело может
быть вытянутым или коротким,
уплощенным дорсовентрально, сжатым с боков или
«креветкоподобным». Мокрицы (из отряда
равноногих) успешно заселили сушу (см.
разд. 8.6.2).
Последняя группа—Eucarida (рис.
8.49)—характеризуется включением всех
восьми сросшихся между собой грудных
сегментов в головогрудь, так что тело
состоит лишь из нее и брюшка, причем
головогрудь целиком заключена в карапакс,
простирающийся обычно по бокам вниз
с образованием вокруг грудных жабр
защитной камеры. Брюшные плеоподы, как
правило, используются для плавания, а
у самок десятиногих раков—для
удержания яйцекладок, в то время как грудные
ноги служат для питания, ловли и
удерживания добычи и для локомоции. В наиболее
обширном отряде—Decapoda (десятиногие
раки) — у особей три пары ногочелюстей
и прослеживается тенденция к редукции
брюшка, зашедшей дальше всего у крабов,
которые подгибают его под крупный, часто
очень широкий карапакс, так что при
взгляде сверху оно совершенно незаметно.
Среди ракообразных только у крабов
ширина тела подчас намного превышает его
длину.
8.7. Дополнительная
литература
Borror D.J., De Long D.M., Triplehorn С. А. 1976.
An Introduction to the Study of Insects, 4 th edn.
Holt, Reinhart & Winston, New York.
Boudreaux H.B. 1979. Arthropod Phylogeny with
Special Reference to Insects. Wiley, New York.
Chapman R. F. 1969. The Insects: Structure and
Function. English Universities Press, London.
Clarke K. U. 1973. The Biology of the Arthropoda.
Edward Arnold, London.
Cloudsley-Thompson J.L. 1968. Spiders, Scorpions,
Centipedes and Mites. Pergamon Press, Oxford
[Chelicerata, Myriapoda].
Daly H. V., Doyen J. Т., EhrlichP.R. 1978.
Introduction to Insect Biology and Diversity. McGraw-
Hill. New York.
Gupta A. P. (Ed.). 1979. Arthropod Phylogeny. Van
Nostrand Reinhold, New York.
Kaestner A. 1968. Invertebrate Zoology, (Vol. 2).
Wiley, New York [Onychophora, Tardigrada, Penta-
stoma, Chelicerata, Myriapoda].
Kaestner A. 1970. Invertebrate Zoology, (Vol. 3).
Wiley, New York [Crustacea].
King P. E. 1973. Pycnogonids. Hutchinson, London.
LewisJ.G.E. 1981. The Biology of Centipedes.
Cambridge University Press, Cambridge.
Little C. 1983. The Colonisation of Land.
Cambridge University Press, Cambridge [все наземные ар-
троподы].
Manton S.M. 1977. The Arthropoda. Oxford
University Press, Oxford.
McLaughlin P. A. 1980. Comparative Morphology
of Recent Crustacea, Freeman, San Francisco.
Ramazzotti G. 1972. II Phylum Tardigrada, 2nd edn.
Istituto Italiano di Idrobiologia, Pallanza.
Savory Т. Н. 1977. Arachnida. Academic Press, New
York.
SchramF.R. 1986. Crustacea. Oxford University
Press, New York.
Sedgwick A. 1888. A Monograph of the
Development of Peripatus capensis, and of the Species and
Distribution of the genus Peripatus. Clay, London.
ЧАСТЬ III
ФУНКЦИОНАЛЬНАЯ БИОЛОГИЯ
БЕСПОЗВОНОЧНЫХ
Если в предыдущей части было показано
морфологическое и биологическое
многообразие беспозвоночных, то в этой
основное внимание уделяется общим чертам их
функциональной анатомии, физиологии
и поведения. Совершенно ясно, что
независимо от строения, эволюции или экологии
все животные имеют некие общие
потребности, обеспечивающие, по крайней мере
потенциально, выживание отдельных
особей и сохранение их генов в следующих
поколениях. В связи с этим животные
располагают эквивалентными наборами
функциональных систем, позволяющих как
получать необходимые ресурсы и информацию,
так и обрабатывать полученное, особым
образом упорядочивая ею. Возникшие
в процессе отбора селективные
преимущества тех или иных планов строения, образов
жизни и местообитаний обусловливают
неодинаковые способы решения этих общих
для всех проблем, поэтому, как и в
предыдущей части, в главах этой функциональная
биология беспозвоночных рассматривается
с учетом различных факторов отбора и
оптимальных вариантов взаимодействия с
ними.
Некоторые потребности служат нуждам
непосредственного выживания, и в этих
случаях отбор часто очень сильно действует на
особей. Например, животным необходимо
находить, потреблять и ассимилировать
пищу, содержащую необходимое вещество
и энергию, нередко в условиях жесткой
конкуренции за эти ресурсы (гл. 9) и в то же
время избегать опасности стать пищей для
других животных (гл. 13). Другие задачи,
обеспечивающие любому организму саму
возможность существования, можно
считать возникшими и разрешенными в
эволюции большинства групп относительно рано,
поэтому сейчас они связаны в основном
с действием стабилизирующего отбора.
Так, большинство животных нуждается
в локомоторных системах, всем
необходимы подвижные части тела, чтобы
захватывать пищу, спасаться от хищников, избегать
неблагоприятные условия среды и т. д. (гл.
10). Нужны также газообмен с окружающей
средой и метаболические процессы, в
результате которых высвобождается энергия
(гл. 11). Поскольку химический состав тела
не такой, как у окружающей среды,
требуется регулировать состав и/или
концентрации химических веществ в организме, а
значит, и удалять «отходы» (гл. 12). Чтобы
многоклеточный организм поведенчески
и физиологически проявлял себя как единое
целое (гл. 16), необходимо получать и
оценивать информацию о внутренней и
внешней средах и, если нужно, реагировать на
нее, координируя во времени и
пространстве развитие и работу различных
функциональных систем тела. Наконец (гл. 14 и 15),
животные должны обладать стратегией
воспроизводства и жизненным циклом,
позволяющими передать максимум генов
последующим поколениям, а их зиготы или иные
пропагулы должны в свою очередь
развиваться в организмы, которые сами способны
к воспроизводству, к потреблению и
переработке добываемых ресурсов; иногда это
развитие включает своеобразные
личиночные стадии, приспособленные для распро-
264
Часть III
странения, отыскания специфических хозяев
или питания до метаморфоза во взрослую
размножающуюся форму.
Поскольку исследованиями по всем
этим направлениям затронуто очень
ограниченное число беспозвоночных, животные,
обсуждаемые ниже, составляют лишь
небольшую часть от тех, что были описаны
в ч. II. По вполне понятным причинам
экспериментальный материал относится
в основном к наиболее многочисленным
и относительно крупным среди
беспозвоночных (членистоногие, моллюски и др.).
Это, однако, послужит оправданием
нашему кажущемуся невниманию к так
называемым «большим типам», обусловленному
подходом, принятым в ч. II.
ГЛАВА d
ПИТАНИЕ
Питание животных представляет собой
до некоторой степени неразработанную
область исследования. Верно, что накоплен
огромный материал, например по механике
фильтрационного питания или о том, как
хищники охотятся за своими жертвами,
много известно о строении
пищеварительной системы и физиологии пищеварения, но
ответ на вопрос, почему животные
питаются той или иной пищей, предполагался
очевидным—поедается то, что можно добыть
и усвоить. Но почему большинство
примитивных типов животных почти
исключительно плотоядные? Почему так много
растительных тканей не потребляется? Почему
некоторые виды более «всеядны», чем
другие? И почему некоторые болотные птицы
предпочитают мидий червям?
В этой главе мы рассматриваем
филогенетические ограничения, связанные с
питанием животных (эволюционное прошлое),
различные механизмы питания (наследие
прошлого), а также факторы,
определяющие выбор той или иной пищи в рамках
широкого кормового спектра, и возможности
влияния жертвы на выбор хищника
(экологическое настоящее).
9.1. Введение: эволюция
способов питания животных
Все типы животных (может быть, за
исключением Uniramia) возникли в море,
очень стабильной и однородной среде. Если
не считать пограничные приливно-
отливные и непосредственно прибрежные
зоны, такие физические факторы, как
температура, соленость, ионный состав,
насыщение кислородом и т. д., не представляют
никакой опасности для выживания морских
организмов и не создают физиологических
ограничений для их биологической
активности. Поэтому жизнь обитателей моря
лимитируется всего двумя, может быть, тремя
факторами, из которых большинство, если
не все, связаны с питанием. Животные
нуждаются в достаточном количестве пиши,
должны избегать опасности стать пищей
для других и иметь определенное
пространство для удовлетворения первых двух
потребностей. Успешное выживание особей
определяется только этим. Конечно, если их
генам предписано существовать в течение
более длительного периода, чем жизнь
одной особи, необходимо также произвести
максимально возможное число потомков,
способных в свою очередь выживать и
производить себе подобных (см. гл. 14). Все
анатомические, физиологические,
биохимические или онтогенетические особенности
лишь механистически способствуют
повышению шансов на решение этих главных
жизненных задач.
Что же могло служить пищей для
первых животных, населявших поверхность до-
кембрийского морского дна (гл. 2)? Только
колониальные или одноклеточные бактерии
и протесты, которые во всех водоемах,
кроме самых мелководных, также были в
основном гетеротрофами. Достаточно света
для фотосинтеза проникает в толщу воды
примерно на 100 м в открытом море, а на
мелководье обычно на меньшую глубину
(20—30 м в береговой зоне, а иногда лишь
несколько сантиметров в очень мутных
водах с илистыми взвесями). Большая часть
континентального шельфа и все дно океана
вообще не знают солнечного света.
Первичная продукция органического вещества
фотосинтетическими морскими протестами
266
Глава 9
образуется только в поверхностных слоях
воды, сильно удаленных от основных бен-
тосных местообитаний. В наше время
большинство донных животных почти
полностью зависят в конечном счете от оседания
сверху мертвых и уже частично
разложившихся тканей и от организмов,
обусловливающих их разложение как во время
прохождения через водную толщу, так и после их
попадания на морское дно.
Только в прибрежных мелководных
зонах фотосинтезирующие организмы были
непосредственно доступны животным, либо
отфильтровывающим их из взвешенного
в воде материала, либо заглатывающим
прикрепленные формы. Если не считать
филогенетически изолированных губок,
питание путем фильтрации было в эволюции
животных гораздо более поздней
специализацией, чем обгладывание или ощипывание
(гл. 2). Вместе с тем хемосинтетическая
деятельность бактерий морского дна была, а
в некоторых местах все еше остается
важным источником первичной продукции.
Бактериальный фотосинтез и хемосинтез
характерны для анаэробных или
микроаэробных местообитаний, а также
восстановленных субстратов.
Поэтому неудивительно, что наиболее
просто устроенные из доживших до наших
дней плоских червей и. по-видимому, их
предковые формы были и остаются
потребителями в основном бактерий и
протестов, связанных тем или иным образом
с донными осадками и камнями. Пожалуй,
более удивительно то, что явно и скрыто
этот примитивный рацион доминировал
у всех форм, которым дали начало плоские
черви, даже у перешедших к наземному
существованию. Численность бактерий и
протестов в морских осадках огромна, но они
очень малы и часто относительно рассеяны
в пространстве, редко встречаясь плотными
скоплениями, хотя бывают и исключения,
которые мы рассмотрим позже. Эти
особенности имели два очень важных
последствия для образа жизни животных.
Во-первых, они должны обладать
определенной подвижностью, чтобы отыскивать
новые источники пищи, когда прежние
оказываются исчерпанными. Таким образом,
подвижность стала одним из условий
животного образа жизни. Во-вторых,
потребитель мелких, рассеянных в пространстве
организмов и сам должен быть относительно
мелким. Чем крупнее животное, тем выше
его метаболические потребности и тем
больше ему требуется пищи в единицу
времени. Это можно видеть на графике
зависимости между размером тела и расходом
энергии, необходимой для его существования
(рис. 9.1). Крупное животное не сможет
прокормиться бактериями или мелкими проти-
стами, поскольку не сумеет потребить
достаточное их количество в единицу времени.
Однако само увеличение размеров тела,
по-видимому, дает селективные
преимущества по трем причинам:
1) более крупные животные обычно
способны произвести больше потомства (т.е.
повышается репродуктивный потенциал);
2) больший размер может повысить
шансы не быть съеденным другими
организмами (т. е. повышается выживаемость);
3) более крупные животные, как
правило, могут оттеснить более мелких от
ограниченных источников общих пищевых
ресурсов (т. е. опять же повышается
выживаемость).
Кроме того, при одинаковой усваивае-
мости энергетически всегда выгоднее
поглощать крупные частицы данной пищи, чем
мелкие, а для этого тоже нужно обладать
большим размером.
Итак, с учетом селективных
преимуществ увеличения размеров неудивительно,
что большинство доживших до наших дней
1 мкг 1 мг 1 г
Размер тела
Рис. 9.1. Связь между размером тела и
интенсивностью метаболизма (по Philipson, 1981)
(см. также рис. 11.16)
Питание
267
плоских червей крупнее видов, которые
питаются бактериями, и пищей им служат
более крупные объекты, т. е. животные. В
наше время две наиболее значительные
исходные группы животных — билатеральные
плоские черви и радиальные
кишечнополостные — представлены практически лишь
плотоядными формами. Тем не менее
размер тела плоских червей ограничен
скоростью диффузии (разд. 11.4.1), поэтому
преимущества, связанные с дальнейшим
увеличением тела, делают выгодными любые
морфологические изменения, которые бы
сняли такое ограничение. Это
обстоятельство и стремление избежать хищничества со
стороны плоских червей, возможно, и
послужили двумя главными толчками,
приведшими к широкому разнообразию
червеобразных форм, появившихся на ранних
этапах эволюции животных.
Однако в самых мелких морских водах
в изобилии присутствуют протесты в виде
нитчатых или слоевищных, колониальных
или многоклеточных водорослей. Хотя
более крупные морские водоросли создают
ряд особых затруднений (см. ниже),
нитчатые формы и ювенильные стадии
макроскопических типов можно относительно легко
отделить от субстрата, к которому они
прикреплены, с помощью специального
соскабливающего органа (рис. 5.3), что делает
этот источник обильной и
концентрированной фотосинтезирующей ткани доступным
для потребления. Моллюски, одни из
ранних эволюционных производных плоских
червей, использовали именно этот
специализированный способ питания,
обезопасившись как от быстрых течений, характерных
для мелких вод, так и от хищников
дорсальной раковиной. Соскабливание с помощью
радулы является также эффективным
средством добывания в качестве пищи сидячих
или прикрепленных животных,
защищенных наружными раковинами, оболочками
или домиками, а также наземных растений,
и в течение своей последующей эволюции
моллюски распространили свой исходный
способ питания на большинство доступных
типов пищи.
Другие рано дивергировавшие от
плоских червей животные либо сохранили пред-
ковый кормовой рацион, либо развили
механизмы для концентрации взвесей мелких
частиц, проплывающих мимо или
осаждающихся из верхних слоев воды. Во многих
морских средах скорость осаждения и
количество органического материала,
взвешенного в воде, практически постоянны.
Значит, если консумент расположится так, что
обеспечит себе его перехват и
концентрирование, он уже не будет нуждаться в большой
подвижности или даже в подвижности
вообще, если не считать одной системы органов,
и, став сидячим или прикрепленным,
сможет сократить энергетические потребности
и, следовательно, необходимое количество
пищи на единицу веса тела. В результате
относительно крупные животные способны
существовать на рационе, состоящем в
основном из бактерий, протестов и мертвого
органического вещества.
Некоторые из этих ранних производных
плоских червей извлекали пищевые частицы
из водной взвеси, пропуская ее через некое
фильтрационное приспособление
(потребители суспензий); другие собирали эти
частицы и связанные с ними скопления
микроорганизмов уже после их оседания на
поверхность субстрата (потребители осадков);
третьи перехватывали частицы во время их
пассивного оседания еще до того, как они
достигали дна (потребители оседающих
частиц).
К потребителям суспензий относятся
губки, лофофоровые, беспозвоночные
хордовые, ряд представителей моллюсков,
кольчатых червей и членистоногих, в том
числе некоторые формы, которые позже
в ходе филогенеза перенесли с собой этот
способ питания в толщу воды вплоть до
зоны фотосинтеза. Пищеварительные
адаптации потребителей суспензий описаны в этой
главе ниже (разд. 9.2.5), а здесь мы
остановимся лишь на некоторых общих чертах их
питания. Всем им свойственны фильтр
определенного типа и либо особое
положение участков захвата пиши, позволяющее
перехватывать естественный (и
постоянный) водный поток, либо приспособления,
создающие собственный ток воды через
фильтр.
У губок одни и те же клетки—хоа-
ноциты—создают ток воды за счет биения
своих жгутиков и улавливают из нее пише-
268
Глава 9
Поток воды
Задержанная
пищевая частица
Пищеварительная
вакуоль
Амебоидная клетка
мезохила
Рис. 9.2. Хоаноциты губок (по
Brill, 1973)
вые частицы, задерживающиеся на
воротничке из микроворсинок вокруг жгутика
(рис. 9.2). Мелкий размер отверстий
образуемого микроворсинками фильтра
позволяет задерживать частицы, размер которых
меньше, чем у бактерий. Вполне очевидно,
что всем потребителям суспензий выгодно
фильтровать один и тот же объем воды не
более одного раза, и у губок это
обеспечивается ее пропусканием через множество
мелких пор, распределенных по всей
поверхности тела, и выведением через одно
или несколько крупных отверстий (см. разд.
3.2). Образующийся более сильный поток
относит профильтрованную воду от тела
губки (у колониальных и планктонных огне-
телок из оболочников с аналогичной
системой общий для всех зооидов выводимый
поток создает, кроме того, реактивную
силу, способствующую движению колонии
сквозь толщу воды; рис. 7.34).
В отличие от этого у разнообразных
потребляющих суспензии потомков плоских
червей для фильтрационного питания
адаптировались или образовались особые
органы—щупальца на голове или вблизи от нее
у аннелид, мезосомальный лофофор у фо-
ронид и их производных, сильно
увеличенные ктенидиальные жабры у двустворчатых
молюсков и т. д. (рис. 9.3). Однако принцип
действия здесь один и тот же. Биение
ресничек в определенных участках
фильтрующего органа создает поток воды, в то время
как другие ресничные поверхности
осуществляют захват и транспорт пищевых
частиц ко рту с помощью слизи. Хотя
захватываются лишь частицы определенного
размера, имеются дополнительные
ресничные зоны либо на самом фильтре, либо на
каком-нибудь вспомогательном органе,
в которых частицы сортируются на
потребляемые и отсеиваемые, часто в виде
псевдофекальной массы. Эквивалентная
система есть и у беспозвоночных хордовых
животных (разд. 9.2.5).
Однако у членистоногих фильтраторов
реснички отсутствуют в связи с наличием
экзоскелета, и вместо них функцию фильтра
выполняют щетинки на различных
конечностях (рис. 9.4). Поток воды, несущий
пищевые частицы, создается как бы
плавательными движениями фильтрующих или
других придатков. Фильтры из щетинок
обычно не такие мелкие, как ресничные, поэтому
захватывают только относительно крупные
частицы.
Все животные, использующие щупальце-
подобные структуры для питания
суспензиями (как и многие другие фильтраторы),
являются прикрепленными или сидячими,
а их ловчие щупальца расположены в
принципе радиально-симметрично, обычно
вокруг рта. Такая же функциональная система
наблюдается у первично радиальных книда-
рий (см. разд. 3.4). Тело обычно защищено
раковиной или трубкой или же помещается
Питание
Полихета (щупальца)
Мшанка (лофОфор)
Плеченогое (лофОфор)
Двустворчатый моллюск
(ктенидиальные жавры)
Офиура (руки)
Голотурия (околоротовые
амбулакральные ножки)
Рис. 9.3. Наружные органы, используемые для слизисто-ресничного типа питания суспензиями
(по различным источникам)
в норке, сделанной в субстрате. Это
позволяет животному и/или его нежному
фильтрующему аппарату втягиваться под
оболочку, обеспечивающую безопасность от
хищников. При таком полускрытом образе
жизни терминальный анус неизбежно
привел бы к прохождению фекальных масс
вдоль всей поверхности тела перед выносом
их во внешнюю среду. Поэтому у видов,
обитающих в трубках или эквивалентных
приспособлениях, наблюдается заметная
тенденция к образованию U-образного
кишечника с расположением ануса, а часто
и выводных отверстий экскреторных
органов вблизи переднего конца тела на пути
выходящего потока воды (см., например,
рис. 7.8). По тем же причинам сходной
анатомической структурой обладают и многие
270
Глава 9
Веслоногий рачок
(ротовой аппарат)
Эвсраузиевый рачок (конечности)
Неполнохвостыи рак
(ротовой аппарат)
Усоногий рачок
(конечности)
Рис. 9.4. Системы щетинок, используемые различными ракообразными для фильтрации (по
различным источникам)
сидячие потребители осадков (см.,
например, рис. 4.42). В других случаях у норки
бывает по крайней мере два отверстия, и ее
обитатель создает через нее
однонаправленный поток воды или же, что гораздо реже,
животное внутри трубки согнуто пополам
так, что рот и анус оказываются у одного ее
выхода.
У некоторых аннелид, включая Nereis di-
versicolor и Chaetopterus. аппендикуларий из
оболочников, некоторых брюхоногих
моллюсков, эхиурид и т. д. фильтрующий
аппарат является не частью тела, а секретируе-
мой структурой в виде сети или мешка из
слизистых нитей, через которые
пропускается поток воды, создаваемый движением
Голова
Крыловидные
параподии
Веерообразные
выросты
ВИД СБОКУ
А. Обитающая в трувках полихета Chaetopterus
Щупальца ноги
Поток
воды
Рот
Ресничная
дорсальная
Бороздка
Глотка
Секретируемый
слизистый мешок
ВИД СВЕРХУ
ПЕРЕДНЕЙ ЧАСТИ ТЕЛА
Слизистые нити
Крышечка -
Рэковинэ
" ^д^шЯШШЬ
Б. Червеовразный врюхоногий моллюск семейства Vermetldae
«я$£ЩЦ\\
Слизистый фильтр
Рот
В. Аппендикулария
Хвост
Тело
■з
s
н
0)
I
S
ID
-Секретируемая
слизистая сфера
или воронка
Г. Полихета Nereis dlversicolor
Раковина
-Крыловидный вырост
ноги и мантии
Д. Крылоногий врюхоногий моллюск
"морская вавочка"
Рис. 9.5. Фильтрационное питание с использованием секретируемых слизистых сетей (по различным источникам)
ISO
272
Глава 9
Желовок Хоботок
Сифоны
Хоботок (эхиуриды)
Каптакула
Оворчатые
гувы
Голова
Вводной сифон (и жавры)
(двустворчатый моллюск)
Пища, скопившаяся
в продольной Бороздке
Каптакулы (лопатоногий моллюск)
[упальца
Щупальца (полихеты)
Околоротовые щупальца
(модифицированные подии)
Голотурия (иглокожие)
rinipobepi ^uvii (ункулиды;
Ротовые
лопасти
Ротовые лопасти (двустворчатый моллюск
подкласса Protobranchia)
Рис. 9.6. Органы обитающих в грунте
беспозвоночных, используемые для слизисто-рес-
ничного питания осадками (по различным
источникам)
специальных образований, служащих у
других животных локомоторными органами.
Через определенные периоды времени эта
сеть вместе с собранной на ней пищей
съедается, и животное секретирует новый
фильтр (рис. 9.5). У некоторых кораллов,
других брюхоногих моллюсков и личинок
насекомых независимо образовались
системы захвата пищи, сходные с этой, но
использующие в основном естественные, а не
специально создаваемые потоки воды.
Сбор пищи с поверхности субстрата
потребителями осадков не требует таких
сложных приспособлений, как при питании
суспензиями, и, действительно, некоторые
черви просто заглатывают поверхностный
слой субстрата с помощью совершенно
неспециализированной ротовой области. Тем
не менее в ряде случаев, особенно у сипунку-
лид, голотурий и эхиурид, а также у
некоторых аннелид и полухордовых около
ротового отверстия развиты особые лопасти,
щупальца или растягивающийся хоботок,
покрытые ресничками и способные двигаться
по субстрату или в нем, что позволяет более
эффективно собирать пищевые частицы
вблизи норок, где живут такие формы (рис.
9.6). У потребителей осадков с лопастями
или щупальцами вокруг рта есть черты,
общие с потребителями суспензий (как отме-
Питание
273
чалось выше в связи с U-образным
кишечником), и некоторые из них могут
использовать оба этих способа питания. Однако две
проблемы касаются только потребителей
осадков: а) органический материал часто
составляет лишь малую часть последних;
б) основная часть самого органического
материала часто состоит из твердых и непере-
вариваемых остатков, накапливающихся
именно из-за использования их не
животными, а только бактериями (см. разд. 9.2.6).
В связи с этим, за исключением
мелководий, где на поверхности субстрата могут
обитать бентосные фотосинтезирующие
протесты, потребители осадков, вероятно,
зависят от бактерий, способных
превращать недоступную иначе органику в
съедобный материал, и от тех протистов и интер-
стициальных животных, которые сами
зависят от этих бактерий. Так или иначе, они
вынуждены либо поглощать большие
количества отложений (поэтому переоткладывают
бентосные осадки и являются мощными
агентами биотурбации), либо проводить
большую работу по их сортировке и отбору
пищи. Щупальца многих потребляющих
осадки червей способны, например,
вытягиваться на большие расстояния от норки, где
обитает животное. За исключением особо
«плодородных» участков, потребители
осадков не могут достигать плотности
населения, характерной для потребителей
суспензий, у которых лимитирующим
фактором часто является пространство, а не
пищевые ресурсы. Поскольку во многих
случаях в конечном счете приходится
полагаться на продуктивность бактерий, а не на
скорость оседания детритных частиц, а
продуктивность бактерий в свою очередь
может ограничиваться не только
поступлением углерода (например, дефицитом
питательных веществ в интерстициальных
водах), скорость роста потребителей
осадков часто ниже, чем у потребителей
суспензий.
Последняя группа животных,
питающихся веществом, поступающим из
поверхностных вод,— потребители оседающих
частиц, например некоторые стебельчатые
иглокожие с телом, прикрепленным к
морскому дну, но значительно
поднимающимся над ним, чтобы перехватить частицы
Система
прикрепле-
^^СЧСуБСтрат
тттптттЬтттттгттптт,
-rjnnmmiTiiniin",
Рис. 9.7. Стебельковая морская лилия
(иглокожие), питающаяся оседающими частицами (по
Clark, 1915)
детрита до того, как они «разбавятся»
донными отложениями (рис. 9.7). «Перехват»
производят несколько радиально
расположенных рук, несущих гидравлически
управляемые и покрытые слизью сосочки; затем
по ресничным бороздкам пища
направляется к ротовому отверстию,
расположенному в центре образованного основаниями
рук круга.
Судя по выжившим представителям
древних групп, на очень ранних этапах
истории многоклеточных животных уже,
вероятно, существовал еще один совершенно
особый способ питания. У некоторых
морских обитателей мелководья в
поверхностных тканях тела находятся симбиотические
оксифотобактерии (прохлорофиты и циано-
бактерии) или фотосинтезирующие проти-
сты (одноклеточные динофлагеллаты или
хлорофиты). Питание некоторых плоских
червей и книдарий, по-видимому,
полностью зависит от этих симбионтов, хотя
в большинстве случаев они не способны их
переваривать. Такие бескишечные турбел-
ларии, мягкие кораллы и медузы постоянно
(сидячие формы) или временно (подвижные
формы) находятся на полном солнечном
свету. Их симбионты фотосинтезируют
18-1314
274
Глава 9
&.
$
.Энтодерма
■ Г %
xv*
ф
4 $-*&'*1'' J**** ^v> ь
*Л Л £S
> !
-г
-у»
■ У
**" <">;
1 * &\#~
I
♦ S"
<"■"«
,.^"»"V'
Рис. 9.8. Электронная микрофотография водорослей-симбионтов в тканях книдарии (Muscatine
et al.. 1975)
обычным способом, но как источник
минерального питания частично используют
неорганические азот и фосфор, образующиеся
из органических соединений в ходе
метаболизма хозяина, а в качестве источника
углерода — углекислый газ, выделяющийся
в процессе его дыхания. По-видимому,
животное-хозяин каким-то образом делает
клеточные стенки симбионта более тонкими
и/или более проницаемыми, чем в норме,
и поэтому часть веществ, образующихся за
счет фотосинтеза (сахара, жиры и
аминокислоты), диффундирует в ткани хозяина, где
ассимилируется: продукты их распада
затем снова путем диффузии попадают
в клетки симбионта. Таким образом
животное «доит» этих автотрофов, плотность
которых может достигать 30 000 в 1 мм-1 ткани
хозяина (рис. 9.8). Другие виды морских
животных хотя и частично, но также зависят
от фотосинтезирующих симбионтов.
Аналогичным образом некоторые
глубоководные погонофоры целиком или
отчасти зависят от симбиотических хемоавто-
трофных бактерий, использующих
восстановленные соединения серы и метан,
выделяющиеся из жерл и фумарол морского дна.
Недавно было обнаружено, что различные
олигохеты и двустворчатые моллюски,
связанные с другими, также богатыми
сульфидами, морскими местообитаниями,
получают около 50% фиксированного углерода
от бактерий, окисляющих серу. Тот факт,
что симбионты участвуют в питании
представителей большинства типов животных,
причем особенно широко в группах,
считающихся наиболее близкими к предковым
формам, дает основание предполагать, что,
хотя симбиотическое происхождение
многоклеточное™ и спорно (гл. 2), своему
возникновению и успеху древние группы
животных могли быть в значительной степени
обязаны симбиозу и, вероятно, особенно
тогда, когда пищевые ресурсы были
ограничены (и в самом деле, хлоропласты зеленых
водорослей, по-видимому, произошли от
других, когда-то эндосимбиотических, эука-
риотных водорослей, а если это так,
симбиотическое объединение отдельных эука-
риотных организмов могло дать начало
некоторым другим эукариотам).
Способы питания и типы пищи,
описанные выше, были, по всей вероятности, в
течение сотен миллионов лет единственными
доступными свободно живущим животным,
и в наше время обитатели морей кормятся
Питание
275
в основном живыми и мертвыми
бактериями, протестами и друг другом. В конце
концов возникшие в море формы заселили
сушу, и, хотя наземные бактерии, протисты
и другие животные могли обеспечить им
практически такой же рацион (с поправкой
на неэффективность потребления суспензий
в наземных условиях), в новых
местообитаниях они столкнулись с новым, обильным,
однако совершенно особым источником
пищи — наземными бриофитами и
сосудистыми растениями. Серьезная проблема,
вставшая перед их потенциальными
потребителями, заключалась в том, что большая
часть биомассы растительного
организма—грубые неперевариваемые клеточные
стенки из целлюлозы и почти инертных
химически структурных полимеров типа
лигнина. Предки большинства животных
никогда не нуждались в целлюлазах или
сходных ферментах для усвоения пищи, и
первые наземные формы, во-видимому,
оказались абсолютно неприспособленными для
использования этого обильного, но
химически стойкого источника пищи. Только в
меловом периоде растительноядность широко
распространилась среди наземных
животных. Однако небольшое число морских
видов уже до некоторой степени решило очень
близкую проблему. Крупные водоросли
морских мелководий также содержат
сложные уллеводы, обеспечивающие
устойчивость к сильным движениям воды. Эти
макроскопические формы так же
потенциально пригодны в пищу, как и большинство
тканей наземных растений, но потреблять
этот в целом непривлекательный корм,
пока он живой, способны только две группы
морских животных.
Первая из них—это некоторые морские
ежи. Большинство потребителей морских
водорослей нуждаются в обязательном
предварительном расщеплении их прочных
полисахаридов бактериями до усвояемой
формы. К таким животным относятся
классические потребители осадков. Бактерии,
особенно анаэробы, способны разлагать
самые разнообразные органические
молекулы, в том числе из нефти и многих
пластмасс, однако, вместо того чтобы
использовать результаты этих процессов,
происходящих во внешней среде, некоторые морские
ежи перенесли их внутрь своего тела.
Специальный участок их кишечника содержит
культуру анаэробных бактерий. Заглатывая
грубые ткани морских водорослей, морские
ежи обеспечивают бактерий пищей; те
разлагают эти ткани, и животные-хозяева
переваривают как продукты метаболизма
бактерий, так и их самих. Следующая
проблема, связанная с питанием морскими
водорослями,— низкое содержание в них азота
на единицу веса, но в анаэробных условиях
некоторые бактерии способны фиксировать
атмосферный азот, растворенный в
морской воде, т. е., живя в кишечнике
иглокожих, могут повышать количество азота в их
преимущественно углеводной диете. Хотя
морские ежи как бы преадаптированы к
наземной растительной пище, ни один из них
не перешел к жизни на c>iui\ но многие из
наиболее успешных наземных фитофагов
решили эту проблему сходным образом.
Другая группа потребителей морских
водорослей—это брюхоногие моллюски,
и неудивительно, что при их исходном
образе жизни на мелководье, включающем
соскабливание пищи с субстрата, они
оказались среди немногих групп животных,
обладающих ферментами, необходимыми для
разложения большого числа полимерных
углеводов, в том числе и целлюлазами.
В отличие от иглокожих брюхоногие стали
процветающей группой наземных
фитофагов, что может засвидетельствовать любой
садовод.
Хотя у некоторых сухопутных
членистоногих есть целлюлазы, питание, основанное
на деятельности кишечных симбиотических
микроорганизмов, характерно для многих
успешных наземных фитофагов, включая
тараканов, термитов, некоторых жуков
и млекопитающих. Все они для
переваривания растительной пищи используют сим-
биотические бактерии и протистов,
обеспечивающих распад углеводных цепей и/или
поступление дополнительных количеств
органического азота. Эти так называемые
«травоядные» животные столь же зависимы
от бактерий и простейших, как и бескишеч-
ные плоские черви, сипункулиды и
питающиеся осадками аннелиды.
Однако не все части растения одинаково
химически устойчивы; например, молодые
18*
276
Глава 9
фотосинтезирующие листья для животных
доступнее. И все же лишь немногие из
фитофагов могут разрушать стенки
растительных клеток, чтобы добраться до их
содержимого, используя свой собственный
ферментный арсенал. Наземные
беспозвоночные, не способные разлагать целлюлозу
с помощью симбионтов или собственных
ферментов, выработали два особых метода
извлечения содержимого растительных
клеток или растительных соков. Гусеницы,
кузнечики и другие насекомые, питающиеся
листьями, откусывают их мелкие кусочки
или тонкие полоски и потребляют
содержимое клеток, разрушенных при разрывании
и последующем пережевывании тканей
растения. Целые клетки при этом в пищу
непригодны, поэтому лишь относительно
небольшая доля (в среднем одна треть)
откусанных частей усваивается и для
возмещения низкой эффективности питания
требуется переработать большое количество
корма. Подобная система может существовать
только благодаря обильной биомассе
растений. Второй способ основан на
проникновении в систему транспортировки
жидкостей внутри растения с помощью
колюще-сосущего ротового аппарата (рис. 9.9;
см. также рис. 8.30). Так питаются все
равнокрылые, например тли. Они избегают
устойчивых химически структурных
углеводов, однако иногда нуждаются в симбиоти-
ческих кишечных бактериях для увеличения
Стилеты
Рис. 9.9. Голова клопа, питающегося соками
растений; стилет введен в ткани растения (по
Barnes, 1980)
с их помощью количества азота в
разбавленной и бедной этим элементом кормовой
жидкости.
Растениям, используемым в пищу,
очевидно, были выгодны приспособления,
препятствующие потреблению фотосинтези-
рующих тканей, и подобно другим
прикрепленным организмам (морским
колониальным животным, например), мало
приспособленным к другим способам обороны, они
развили сложную систему механической
и химической защиты (разд. 9.2.4). Как
обычно бывает в таких случаях, это
положило начало «гонке» наступательных и
оборонительных вооружений—образованию
токсинов у кормовых объектов и
механизмов детоксикации или устойчивости у их
потребителей.
Только одна категория фитофагов
эффективно использует вещества
растительного происхождения без помощи симбио-
тических бактерий или протистов. Это те
виды, которые питаются веществами или
структурами, развившимися специально
для привлечения животных (нектаром,
плодами, семенами и т. д.), необходимых
растениям для опыления и/или распространения.
Хотя плоды и семена в основном
предназначены для позвоночных, членистоногие
и моллюски также иногда «приобщаются»
к этим ресурсам до того, как они съедены
птицами или млекопитающими.
Грибы с их хитиновыми клеточными
стенками и отсутствием прочных опорных
тканей не столь трудны для переваривания
животными и поэтому широко
используются ими. Некоторые насекомые в этом
плане заходят столь далеко, что собирают
растительные ткани, пережевывают их до
мягкой массы и затем используют ее как
субстрат для выращивания грибов, идущих
в конце концов в пищу. В какой-то степени
грибы играют ту же роль, что и водоросли
в питании предков наземных животных.
Несмотря на широко распространенное
представление о преобладании на суше
пищевых цепей типа растение->раститель-
ноядное животное->плотоядное животное,
сложившееся в связи с большим
вниманием, уделяемым в научных исследованиях
травоядным (особенно пастбищным
млекопитающим), основу большинства наземных
Питание
277
пищевых сетей составляют не живые
растительные ткани и не обгрызание их, а
разлагающиеся органические остатки. Основные
потоки энергии идут через животных,
использующих в пищу детрит и лесную
подстилку с участием бактерий, протистов и грибов.
Таким образом, наземные животные
недалеко ушли от своих морских предков в
смысле способности «перерабатывать»
различные типы корма, и даже сейчас фитофаги
потребляют менее 3% лесной продукции
в виде живого растительного материала.
9.2. Типы питания животных:
способы захвата
и переработки пищи
9.2.1. Классификация типов
питания
Влияние давних представлений о
важнейшей экологической роли цепей
питания, включающих обгрызание растений,
отражено даже в названиях классических
трофических уровней. Многие годы выделение
типов питания животных основывалось на
таких пищевых цепях и на систематической
принадлежности поедаемых организмов;
отсюда, например, «растительноядные»,
«плотоядные» и (промежуточные)
«всеядные» формы. Подобная классификация
порождает много неудобств. Во-первых,
исключительно растительноядных или
плотоядных видов относительно мало,
особенно если учитывать все стадии их
жизненного цикла, т. е. большинство животных
попадают в неопределенную категорию
«всеядных». Во-вторых, будучи основана на
старом представлении о разделении живых
организмов на два царства—растений и
животных, она не решает вопроса о том, куда
отнести виды, питающиеся бактериями
и протистами и составляющие довольно
обширную группу. В-третьих,
филогенетические связи видов добычи необязательно
важны для экологии питания ее потребителей.
Позже стало принято различать
категории консументов по способам питания.
Можно говорить о хищниках, паразитах,
обгрызающих и ощипывающих животных,
о потребителях суспензий и осадков, о
питающихся с помощью симбионтов. Эта
классификация не учитывает
систематического положения организмов,
используемых в пищу,— например, «обгрызатели»,
потребители суспензий и осадков могут
поедать бактерий, протистов, грибы,
растения и животных. Если способы питания
животных не делают различий между
царствами поедаемых организмов, не следует
делать это и нам при анализе биологии
питания. Поэтому в следующем разделе мы от
них абстрагируемся и более детально
рассмотрим особенности поглощения и
переваривания пищи.
9.2.2. Общие черты
У большинства беспозвоночных
«сквозной» кишечник с ротовым отверстием,
расположенным более или менее спереди
и служащим для захвата пищи, и анальным
отверстием, сдвинутым более или менее
назад и служащим для вывода из кишечника
во внешнюю среду непереваримых остатков
и экскретов. У некоторых животных
кишечник заканчивается слепо, и единственное
отверстие служит как для поглощения пищи,
так и для освобождения от «отходов».
Сюда относятся в основном книдарии и
плоские черви, но также и ряд других
животных, у большинства из которых такое
строение носит вторичный характер. У
других форм (некоторые паразиты, но также
и свободноживущие виды)
пищеварительный канал вообще отсутствует, и пища
поступает либо от внутренних симбионтов,
либо через наружную поверхность тела.
Много и отклонений от этих вариантов.
Например, у форм, ведущих прикрепленный
образ жизни, рот часто расположен в
центре верхней стороны тела, а не в его
передней части; у некоторых свободноживущих
форм, предки которых были
прикрепленными, например у свободно передвигающихся
иглокожих, рот находится в центре нижней
поверхности тела, а у пищеварительного
канала хордовых и ряда других животных
наружных отверстий множество (разд. 9.2.5).
С точки зрения развития
пищеварительный канал животных включает три отдела:
эктодермальную переднюю кишку (обра-
278
Глава 9
vПередняя
г кишка
.Средняя
лкишка
ныи дивертикул
Кишечник
Прямая
кишка
Задняя
'кишка
Анус
Рис. 9.10. Отделы пищеварительного тракта
обобщенного беспозвоночного (схема)
зуется при втягивании части наружной
поверхности тела), также эктодермальную
заднюю кишку и энтодермальную среднюю
кишку, где происходят переваривание и
всасывание, часто снабженную слепыми
дивертикулами. Каждый из этих основных
отделов разделен на функционально различные
части (рис. 9.10). Передняя кишка может
состоять из а) ротовой полости, в которую
ведет ротовое отверстие и открываются
протоки «слюнных желез» (иногда
специализированных на образовании клейких секретов,
антикоагулянтов или токсинов); б)
мускулистой глотки, иногда участвующей в
поглощении пищи, либо действуя как насос, либо
образуя выворачиваемый орган для
схватывания добычи, либо (как у хордовых) служа
фильтром при питании суспензией; в)
короткого пищевода и/или г)
накапливающего органа—зоба, особенно хорошо
развитого у животных, поглощающих через
длительные интервалы времени большие
количества пищи, постепенно поступающей
в среднюю кишку.
Средняя кишка обычно подразделяется
на а) мускулистый желудок, б) различные
слепые секреторные и/или всасывающие
отростки и в) кишечник. В желудке
происходит механическое измельчение и
сортировка проглоченной пищи; иногда в нем
имеется особый отдел для растирания пищевых
частиц. Иногда здесь же начинается
переваривание, но чаще переваривание и/или
всасывание протекают в различных
дивертикулах— слепых выростах, отходящих от
кишки сразу за желудком. У моллюсков и
ракообразных эти выросты превращаются
в крупные сложные органы - гепа-
топанкреасы.
У примитивных форм переваривание
идет в основном внутриклеточно, когда
частицы пищи фагоцитируются клетками
выстилки абсорбирующего отдела
пищеварительной системы и расщепляются в их
вакуолях. Такой способ питания преобладает
у губок, кишечнополостных, плоских червей
и ряда других животных, у которых пища
поступает в среднюю кишку в
тонкодисперсном состоянии. У более сложно
организованных животных, особенно у
поглощающих пищу отдельными крупными
массами, переваривание происходит в
основном с помощью ферментов, секретируемых
в просвет кишки, а его продукты затем
абсорбируются клетками ее стенки. У таких
видов дивертикулы имеют, как правило,
чисто секреторную функцию, и всасывание
происходит в кишечнике. Внеклеточное
переваривание способствует ферментной
специализации как отдельных клеток, так
и различных участков кишки, однако
приводит к необходимости производить большее
общее количество ферментов, поскольку
просвет кишки— крупная открытая
система, в которой нелегко поддерживать их
оптимальную для пищеварительных
процессов концентрацию.
Наконец, задний отдел
пищеварительного тракта (если он есть) представлен прямой
кишкой (ректумом), где может происходить
всасывание воды (у некоторых наземных
животных; см. рис. 8.27) и образование
фекальных масс перед их выведением наружу
через анальное отверстие.
Все животные нуждаются в получении из
пищи энергосодержащих веществ, исполь-
Питание
279
зуемых немедленно или несколько позже,
аминокислот, необходимых для синтеза
структурных и метаболических белков,
и ряда других химических элементов и
соединений, например витаминов,
участвующих в катализе биохимических реакций.
Другими словами, животные должны
поедать углеводы, жиры, белки и витамины и,
если они не могут всасываться сразу,
переваривать их до усвояемых форм. Виды,
пища которых бедна белком и/или
витаминами, обычно нуждаются в симбиотических
кишечных бактериях, синтезирующих эти
вещества, а потребители сложных
полимерных углеводов — в симбиотических
бактериях и протистах, разлагающих эти
вещества до более простых органических
молекул. Значительная часть фекалий состоит
из кишечных бактерий.
9.2.3. Охотники и паразиты
Охотники — это подвижные животные,
которые атакуют, убивают и потребляют
одну особь-жертву (почти всегда - также
подвижное животное) в данный момент
времени. Среди охотников различают три
крупных типа: преследователи (например,
кальмар) догоняют, хватают и
обездвиживают других быстро передвигающихся
животных; поисковики (многие гастроподы,
членистоногие и т. д.) отыскивают менее
подвижную, чем они сами, добычу; засадчи-
ки (такие, как пауки и богомолы) большую
часть времени относительно неподвижны
и хватают жертву, совершая редкие рывки
или прыжки.
Преследователи и засадчики (в меньшей
степени поисковики) имеют специальное
вооружение для схватывания и
обездвиживания жертвы. Как правило, это либо
органы, расположенные вокруг рта, например
клешневидные и ложноклешневидные
придатки у членистоногих, руки головоногих
с присосками или крючками, шипы у ще-
тинкочелюстных (рис. 9.11), либо сильные
челюсти, связанные с передней кишкой, как
у аннелид, где они находятся на
выворачиваемой глотке, которая с силой
«выстреливается» для захвата добычи. Некоторые
засадчики привлекают жертв с помощью
мимикрии. Например, у некоторых сифонофор
щупальца несут образования,
напоминающие веслоногих рачков, и их добычей
становятся в основном животные, питающиеся
копеподами (обычно другие ракообразные).
Пойманное животное либо заглатывается
целиком, либо разрывается с помощью
различных придатков и заглатывается по
частям, либо из его тела высасывается
жидкость. Если жертва заглатывается целиком,
передняя часть пищеварительного тракта
способна растягиваться в соответствии
с размерами пищи.
С другой стороны, многие поисковики
питаются относительно малоподвижными
животными, которые защищаются от
нападения с помощью наружных покровов из
карбоната кальция, целлюлозы, хитина
и т.д. (разд. 9.33 и 13.2.1). В связи с этим
у поисковиков иногда развиваются
специализированные способы хишничества: про-
буравливание защитных покровов
(например, с помощью радулы у моллюсков);
отламывание их кусочков и выворачивание
в образовавшееся отверстие своего
желудка, секреция на незащищенные ткани
ферментов и абсорбция продуктов наружного
пищеварения; заглатывание жертвы
целиком и раздавливание ее раковины в
желудке; высасывание отдельных полипов или
зооидов в колониях с помощью хоботков
или стилетов и глоточного насоса
(например, некоторые заднежаберные моллюски,
морские пауки и др.).
У некоторых охотников сосущий
механизм поглощения пищи. В дополнение к уже
перечисленным группам ряд хищных
насекомых и прежде всего пауки высасывают
жидкость из своих жертв либо
непосредственно, либо сначала впрыскивая в
пойманную добычу слюну, содержащую про-
теолитические ферменты (и парализующие
токсины), разжижающие ее ткани до такого
состояния, что их можно всосать.
Небольшой переход отделяет описанный тип
питания от эктопаразитизма, когда животное
питается жидкостями хозяина, не убивая
его самого. Категории охотников и
паразитов различаются нечетко, и их разделение
в значительной мере опирается на
относительные размеры потребителя и объекта
потребления. Пиявка, сосущая кровь крупного
млекопитающего, вряд ли может его убить.
280
Глава 9
НОГИ С ЛОЖНЫМИ
ПЕДИПАЛЬПЫ С КЛЕШНЯМИ ПЕДИПАЛЬПЫ С ЛОЖНЫМИ КЛЕШНЯМИ КЛЕШНЯМИ
^Клешни
Скорпион (хелицеровые)
Ложные
клешни - „ „
Жгутоногое (хелицеровые) рак-БОгомол (ракообразные)
Ложные клешн
Ложноскорпион (хелицеровые) Скорпионопаук (хелицеровые) Богомол (одноветвистые)
ВЫВОРАЧИВАЮЩАЯСЯ ГЛОТКА
НОГИ С КЛЕШНЯМИ
А
Челюсть Выворачивающаяся глотка
Ядовитая железа
Проток ядовитой железы
Полихета (аннелиды)
РУКИ С ПРИСОСКАМИ
~Клешни
Крав (ракообразные)
ГОЛОВНЫЕ ШИПЫ
Задние зувы Передние
Ловчие шипы
Головоногий моллюск Щетинкочелюстное
Рис. 9.11. Органы схватывания добычи (по различным источникам)
так как потеря крови будет для него
незначительна. Однако другой вид пиявок,
питающийся кровью мелкой прудовой
улитки, губит последнюю, поэтому должен
считаться охотником. Даже в случае животных
одинакового размера отнесение к той или
иной категории представляет трудности.
Некоторые планктонные полихеты
нападают на щетинкочелюстных и отъедают
у них передний конец тела. Жертвы от этого
не погибают и способны регенерировать
недостающую часть. Как квалифицировать
полихету—как охотника или как паразита?
Столь же проблематична ситуация и со
многими эндопаразитами. Например,
некоторые перепончатокрылые насекомые
проводят часть жизненного цикла в теле других
животных, что приводит к гибели
последних. Эти так называемые «паразитоиды»
съедают свою жертву изнутри наружу, а не
наоборот, и не так быстро, как классические
охотники. Такие насекомые откладывают
Питание
281
яйцо или яйца в особь-жертву (обычно
другое насекомое), и развивающиеся личинки
питаются тканями хозяина до своего
окукливания и метаморфоза. Имаго здесь —
типичные охотники, за исключением того,
что они только нападают на добычу, но не
поедают ее, а оставляют потомкам. И
в этом случае встает вопрос об
относительных размерах животных. Личинка
перепончатокрылого или двукрылого насекомого
велика по отношению к размеру жертвы,
в которой она паразитирует, поэтому
хозяин может быть крупным лишь настолько,
чтобы обеспечить возможность развития до
окукливания одной или нескольким
личинкам. В то же время взрослые формы,
скажем трематоды или нематоды, хотя и
питаются тканями хозяина по существу таким
же образом, очень малы по сравнению,
например, с млекопитающим-хозяином и не
вызывают его гибели. Поскольку оба типа
питания, не считая их крайних форм,
неразличимы, неудивительно, что во многих
группах мелких хищных беспозвоночных
имеются и паразитические виды.
Более четкую границу можно провести,
по крайней мере в отношении биологии
питания, между охотниками и паразитами,
питающимися жидкостями тела и/или
тканями своих жертв, с одной стороны, и
бескишечными эндопаразитами, обитающими
в пищеварительном тракте хозяина,—с
другой, хотя опять-таки эволюционный
переход между эндопаразитическими
самостоятельно переваривающими животными
и формами, всасывающими уже
переваренные хозяином продукты, не слишком велик.
Некоторые паразитические черви, особенно
цестоды и скребни, лишены кишечника, но
их наружная кутикула снабжена
микроворсинками (так называемыми микротриха-
ми), эквивалентными тем, которыми
покрыты поглощающие клетки кишечника
других животных. Внутри хозяина эти черви
абсорбируют продукты его пищеварения,
а их кутикулы, кроме того, защищают их от
влияния его расщепляющих ферментов.
В то время как другие типы «паразитов»,
рассмотренные выше, фактически являются
микрохищниками, цестоды и скребни—
истинные паразиты в обыденном
понимании этого слова, поскольку существуют за
счет ресурсов, добытых другим видом. То
же самое справедливо и в отношении
других кишечных паразитов (включая и
некоторых нематод с функционирующей
пищеварительной системой), потребляющих
только содержимое кишечника своего хозяина.
Однако другие живущие в кишечнике
животные питаются тканями его стенки и
кровью хозяина, поэтому относятся к
хищникам.
Виды, питающиеся тканями и
жидкостями тела- животных, потребляют легко
перевариваемый, богатый белком материал,
поэтому, как правило, секретируют протеа-
зы и имеют относительно короткий и
просто устроенный кишечник,
специализированный (если специализация вообще имеет
место) только в передней части с
образованием жевательного желудка (если жертва
заглатывается целиком), зоба для
накапливания запасов или насасывающей глотки.
Высокое содержание белка в животной
пище делает ее привлекательной и для ряда
неплотоядных животных на некоторых
критических стадиях их жизненного цикла.
Иногда самки двукрылых насекомых,
например комаров, нуждаются в крови, чтобы
получить белок, необходимый для развития
яиц, хотя самцы и личинки могут
обходиться без животной пищи.
9.2.4. Обгрызатели
и соскабливатели
Обгрызатели и соскабливатели —это
подвижные потребители неподвижной
добычи, использующие в пищу доступные для
них ткани, но обычно не убивающие при
этом свои одиночные или колониальные
жертвы. На суше источниками питания для
них являются растения и грибы, а в море—
колонии животных (таких, как книдарии,
мшанки или оболочники), бактерий и
многоклеточные водоросли. Добывание пищи
требует наличия твердых грызущих или
скоблящих ротовых частей типа радуляр-
ной ленты моллюсков (рис. 5.3),
аристотелева фонаря морских ежей (рис. 7.24), скле-
ротизованных челюстей насекомых (рис.
8.30) и т.д., а поиск корма здесь не
составляет главной проблемы (сильное отличие их
от большинства типов охотников), посколь-
282
Глава 9
Средняя
кишка
Анус
Б Trichomonas
В trichonympna
Рис. 9.12. Пищеварительный тракт термита, питающегося древесиной (А), и симбиотические
жгутиконосцы (Б, В) из его задней кишки (А по Morton, 1979; Б, В по Mackinnon, Hawes, 1961)
ку съедобный материал, как правило,
находится в изобилии. Трудности этой
категории животных в следующем: а) наличие
систем химической защиты у неподвижных
жертв; б) небольшая доля пригодной для
усвоения пищи на единицу веса всего
поглощенного материала из-за большого
количества в нем устойчивых структурных или
защитных элементов и часто низкое
содержание белка в усвоенной органике.
Проблема высокого содержания непере-
вариваемых углеводных полимеров в
растениях и водорослях (агар, альгин,
ламинарии, целлюлоза и др.) частично решается,
как уже говорилось, благодаря
специфическим ферментам и одновременному
удлинению пищеварительного тракта (особенно
средней кишки), увеличивающему размер
органов, переваривающих устойчивые
материалы, а частично — за счет симбиотиче-
ских бактерий и протестов, обитающих
в особых частях кишки (часто в задней
кишке, но иногда в зобе или желудке).
Нередко присутствует крупный зоб для
накопления запасов.
Кишечные симбионты разлагают
полисахариды анаэробным путем с
образованием жирных кислот и простых углеводов,
пригодных для всасывания. Такой способ
достигает своего наивысшего развития
у термитов, использующих в пищу
наиболее устойчивый из природных органических
материалов — древесину. У этих насекомых
задняя кишка длиннее всего остального
пищеварительного тракта (рис. 9.12) и
содержит плотную популяцию жгутиковых Ну-
permastigina. Эти протисты фагоцитируют
частицы древесины и сами содержат
симбиотические бактерии, которые, по-
видимому, в основном и отвечают за
расщепление целлюлозы. Другой компонент
дерева, лигнин, вероятно, не
переваривается. Равноногий рачок Limnoria также
питается древесиной, однако, по всей
вероятности, лишен кишечных симбионтов и
переваривает целлюлозу и гемицеллюлозы с
помощью собственных ферментов.
Структурные углеводы, если могут усваиваться,
служат богатым источником энергии, но не
дают почти ничего, кроме нее. Limnoria
получает необходимый белок из грибов,
живущих в потребляемой этим ракообразным
древесине, которая без таких
колонизировавших ее организмов-редуцентов не может
удовлетворить потребности животного.
Фактор размера также важен при
потреблении растительного материала с
относительно низкой питательной ценностью.
У крупных животных, например
позвоночных размером с кролика, низка удельная
Питание
283
скорость метаболизма, что снижает
потребность в энергии на единицу веса тела.
Благодаря большому объему они способны
накапливать в переваривающем отделе кишки
много растительного материала.
Млекопитающие, будучи гомойотермными
животными, поддерживают температуру,
необходимую для эффективной работы
пищеварительных ферментов. Поэтому они могут
существовать за счет грубых трав и подобных
им материалов, поглощаемых в больших
количествах. Мелкие животные, например
растительноядные беспозвоночные,
вынуждены питаться кормом более высокого
качества и обеспечивать себе доступ к
содержимому растительных клеток, протыкая их,
соскабливая целлюлозные стенки с
помощью радулы или прокусывая их своими
ротовыми частями. Поглощение больших
количеств пищи здесь, как правило,
невозможно просто из-за мелких размеров тела.
Даже мелкие птицы и млекопитающие не
могут существовать за счет малопитательного
корма: если это растительноядные
животные, их рацион ограничен богатыми
энергией семенами или сходными
образованиями.
У растений много разных средств
химической защиты, включая алкалоиды
(никотин, кокаин, хинин, морфин и кофеин), глю-
козиноляты, цианогенные гликозиды и тан-
нины. Эти вещества, вероятно, специально
образуются в их тканях, чтобы
воспрепятствовать потреблению последних, и иногда
сильно токсичны, являясь естественными
инсектицидами — например, у дерриса и
пиретрума. Таннины, высвобождаясь,
связывают белки, делая их неусвояемыми и
таким образом уменьшая белковую ценность
поглощенной пищи. Другие соединения
имитируют собственные гормоны и
феромоны фитофага, отрицательно влияя на его
рост, развитие, размножение или вызывая
неадекватные поведенческие реакции. У
некоторых растений механизм химической
защиты включается только во время
выедания, исключая ненужную трату ресурсов,
необходимых для роста и размножения:
недавно показано, что (по крайней мере в
одном случае) растения, даже не будучи
объектом нападения, могут реагировать
мобилизацией своей химической защиты на пора-
Овразования, имитирующие яйца
Рис. 9.13. На листьях и стеблях страстоцвета
находятся образования, имитирующие яйца
бабочек Heliconius. Это предотвращает откладку
яиц на растение (самки избегают листьев, на
которые уже отложены яйца, возможно,
в связи с тем, что только что вылупившиеся
личинки способны поедать друг друга) (no
Gilbert, 1982)
жение фитофагами соседних экземпляров
того же вида.
Другие защитные механизмы растений
бывают структурными, например
волосовидные выступы, выделяющие при
прикосновении липкие секреты или отпугивающие
вещества, или анатомическое
имитационное сходство здоровых листьев с листьями,
пораженными фитофагами (рис. 9.13), а
также симбиотическими, особенно заметными
в случае ассоциаций различных
покрытосеменных и других сосудистых растений с
муравьями. Муравьи изгоняют насекомых,
обгрызающих растение, «в обмен» на его
нектар и место для устройства гнезд.
Сходные структурные, химические и другие
системы защиты имеются и у морских
животных, страдающих от грызущих консумен-
тов, но они пока слабо изучены.
Тем не менее консументы коэволюцио-
нировали со своими неподвижными
жертвами в течение многих тысячелетий, и
некоторые соскабливатели и обгрызатели
приобрели способность обезвреживать,
избегать, связывать или экскретировать
специфические защитные метаболиты, обходить
284
Глава 9
структурные барьеры и ускользать от
внимания охраняющих растения муравьев
путем химической мимикрии под этих
насекомых. И все же в большинстве наземных
местообитаний, благоприятных для развития
растений, накоплено огромное количество
биомассы живых растений так же, как
в океане—водорослей. Это говорит о том,
что большая часть тканей макрофитов на
самом деле остается недоступной для
грызущих и скоблящих животных и/или не
используется ими и становится источником
пищи только после отмирания и
разложения.
Поедание макрофитов осуществляется
не только путем обгрызания и
соскабливания. Некоторые растительноядные
животные как в море (пример—заднежаберные
моллюски), так и на суше (пример—многие
клопы) обходят трудности, создаваемые
структурными углеводами, высасывая
содержимое отдельных клеток или внедряясь
своими трубчатыми ротовыми частями
в ксилему и флоэму сосудистых растений
(рис. 9.9 и 8.30). В последнем случае
гидравлическое давление в проводящих элементах
растения бывает достаточным для
закачивания жидкости непосредственно в
кишечник, например, тли, которая паразитирует
на растении-хозяине примерно так же. как
клещи или самки комаров—на животных.
Как отмечалось выше в отношении
плотоядных охотников, лишь небольшая
ступень отделяет эктопаразитизм на крупном
хозяине от эндопаразитизма, и некоторые
нематоды паразитируют в растениях так
же, как другие нематоды в животных, а
личинки ряда насекомых выгрызают ткани
растений, обитая внутри последних.
Однако даже при наличии целлюлаз
и эквивалентных им ферментов или симбио-
тических макроорганизмов в кишечнике (а
тем более при отсутствии тех и других)
переваривание и ассимиляция тканей
макрофитов, как правило, идут очень
неэффективно, а в случае питания жидкостями
поступление углеводов намного превышает
потребность в них. Поэтому фекальные
массы таких консументов обильны и
содержат много неассимилированного
органического вещества. В связи с этим они
являются важным экологическим путем доступа
к углероду, фотосинтетически
фиксированному живыми макрофитами, для других
категорий животных консументов, особенно
для потребителей осадков.
9.2.5. Питание взвесями
Основные механизмы, с помощью
которых животные, питающиеся суспензиями,
отфильтровывают пищевые частицы от
воды, были рассмотрены в разд. 9.1. У групп,
использующих для этого наружные секреты
неанатомического характера (рис. 9.5), по-
видимому, отсутствуют другие
специальные приспособления для улавливания
взвесей. Эти животные, по крайней мере по
строению своего кишечника, вероятно,
наиболее близки к охотникам-засадчикам,
улавливающим свои небольшого размера
жертвы сетями. В самом деле, эти два
способа питания не имеют четкой границы:
паук, плетущий тенета, может
рассматриваться и как потребитель «воздушной
суспензии», и как охотник-засадчик. Такое же
двойственное положение у морских книда-
рий; вокруг их центрального ротового
отверстия расположены радиально
симметричные кольца щупалец, а в фазе полипов
эти животные ведут прикрепленный образ
жизни; оба этих признака характерны для
потребителей суспензий, и, действительно,
жертвами здесь являются в основном зоо-
планктонные виды, взвешенные в воде.
Однако их особи улавливаются не совсем
пассивно, а индивидуально поражаются нема-
тоцистами, пусть даже сама жертва
способствует их выстреливанию, случайно
коснувшись щупальца, и книдария не нападает «по
собственной инициативе».
В отличие от этого у групп со слизисто-
ресничной системой фильтрационного
питания четко прослеживается специализация
пищеварительной системы. У большинства
таких видов с фильтрационным аппаратом,
либо свободно омываемым водой, либо
защищенным снаружи раковиной (например,
у плеченогих и двустворчатых моллюсков)
(рис. 9.3), передняя кишка представляет
собой короткую трубку, соединяющую
наружный фильтр с желудком. Однако у
хордовых передняя кишка сама является
местом, где происходит фильтрация, и поэто-
Питание
285
му сильно специализирована. Глоточная
стенка у этих животных пронизана по бокам
многочисленными мелкими отверстиями
(«стигмами»), которые, прободая стенку
тела, открываются на его поверхности. Вода
поступает в рот, оттуда в глотку, затем
проходит через стигмы и выводится обратно
в окружающую среду, т.е. ее поток
однонаправленный. Хотя сам по себе процесс
фильтрации глоткой и необычен, аналогичная
система известна и в других группах, что,
возможно, указывает на ее эволюционное
происхождение. Животные, питающиеся
мелкими частицами, часто используют для
втягивания их в рот поток воды, которая
затем должна как-то выводиться из
организма. У лофофоровых вода просто
периодически «выплевывается», и поглощение частиц
в эти моменты прекращается. У брюхорес-
ничных макродазиид (рис. 4. 9) и
полухордовых цефалодисцид (разд. 7.2.3.2) глотка
несет пару перфораций, ведущих на
поверхность тела, через которые поглощенная
вода выводится наружу без перерывов в
питании. Древнейшие хордовые, по-видимому,
специально приспособили такой поток
воды для отфильтровывания пищевых частиц,
что избавило их от необходимости
втягивать лофофороподобный орган при угрозе
со стороны хищников (что также прерывает
питание). Кишечнодышащие полухордовые
используют этот же поток для газообмена,
что свойственно и более поздним водным
хордовым.
Глотка питающихся суспензиями
хордовых очень велика и занимает большую
часть объема тела. Пронизанная тысячами
отверстий стенка тела в глоточной области
у многих видов почти исчезает и поэтому
заменяется вторичной стенкой,
образованной у головохордовых складками ткани, а
у туникат—секретируемой целлюлозной
оболочкой. В связи с этим собственно тело
целиком или частично окружено
морфологически внешней полостью, или атриумом,
между истинной и ложной стенками тела.
Сюда поступает вода, проходящая через
стигмы, и отсюда она выводится в
окружающую среду через единственное
отверстие, т. е. атриопор или атриальный сифон
(рис. 9.14). В сущности такая же система
наблюдается у позвоночных. Однако у рыб
жаберных щелей (= стигм), ведущих из
кишечника к поверхности тела, значительно
меньше, а сама стенка тела толще, поэтому
нет необходимости в дополнительной
ложной стенке, а поток воды используется
прежде всего для газообмена, а не для
поглощения пищевых частиц.
Специальная железа, эндостиль, распо-
Гипервранхиальный желовок (в котором
собирается слизистый тяж с пищевыми частицами)
Отверстия
АТРИУМ
Ток слизи
Латеральные
реснички
У^^%
О . ПРОСВЕТ
fti глотки JJ
\ %
/лЧ __^--:* фронтальные
Ток воды
АТРИУМ
реснички
Эндостиль (продуцирует слизь)
Рис. 9.14. Схема поперечного среза глотки беспозвоночного хордового, показывающая действие
системы фильтрационного питания (по различным источникам)
286
Глава 9
Ток
воды
Жаберный
СИфОН
Ганглий
Атриальная
полость
Атриальный
СИфОН
Задний
конец
Вид сверху
Атриальная полость
Ток воды
Задний конец
Глоточная
перегородка
Эндостиль
Слизистый мешок
Глоточная перегородка
(единственный остаток
дорсальной стенки глотки)
Оволочка
Передний
конец
Слизистый мешок
Вид своку
Рис. 9.15. У пелагических
оболочников (например, у сальп)
слизистый мешок перекрывает
оба крупных глоточных
отверстия (по Berrill, 1950, и др.)
ложенная вдоль вентральной средней линии
глотки, вырабатывает слизь, которая
тонкими слоями гонится вверх по
перфорированным боковым стенкам. Когда вода
вытекает наружу через стигмы, эти слои
перехватывают потенциальные пищевые
частицы в некоторых случаях размером до
0,5 мкм. В конце концов потоки слизи с
пищевыми частицами сходятся дорсально, где
образуют продольный тяж в гипербранхиа-
льном желобе. Движение в этой системе как
воды, так и слизи обеспечивается
полосками ресничек внутри глотки (исключение —
пелагические сальпы).
У пелагических оболочников поток воды
с пищевой взвесью обычно обеспечивает
и движение животного вперед, поскольку
ротовое и атриальное отверстия
расположены на противоположных концах тела.
В связи с этим реактивным типом движения
число стигм уменьшается до двух (у сальп),
слизь образует одну внутреннюю сеть
конической формы, подвешенную в полости
глотки (рис. 9.15), а проталкивание воды
обеспечивается мышечными кольцами,
охватывающими тело (рис. 7.35 и 7.36).
Не только у этих беспозвоночных
хордовых, но и у лофофоровых и моллюсков-
фильтраторов пища поступает из
фильтрующего органа (независимо от его типа)
в желудок в виде слизистого жгута,
несущего съедобные частицы. У всех животных со
слизисто-ресничной системой питания в
желудке кислая реакция. Это снижает вязкость
слизистого жгута, обеспечивая выход из
него пищевых частиц. Переваривание и
абсорбция питательных веществ происходят
в желудке и/или в дивертикулах средней
кишки, отходящих поблизости от него.
Непереваренные остатки попадают в обычно
короткий кишечник, где щелочная реакция
вновь увеличивает вязкость слизи и
способствует образованию фекальных комочков
или нитей.
Движущая сила, обеспечивающая
доставку слизистого жгута в желудок, также
обеспечивается ресничками, у хордовых
действующих на этот жгут непосредствен-
Питание
287
Рис. 9.16. Желудок мшанок со
слизисто-фекальным
стержнем, используемым для
подтягивания пищи в
пищеварительный тракт (no Gordon, 1975)
Желудок
Слизистый
тяж с пищей
Пилорус Вращающийся стержень
Реснички
Прямая кишка
Глотка
но, а у лофофоровых и моллюсков —
косвенно. У двух последних групп в
желудке находится вращающийся стержень,
наматывающий на себя слизистый жгут. У
лофофоровых он состоит из слизи и
фекальных масс, расположен в пилорическои части
желудка и вращается расположенными там
ресничками (рис. 9.16). Время от времени
использованный стержень поступает в
кишечник, превращается в фекальный комок,
выбрасывается наружу и заменяется новым.
У моллюсков этот стержень, известный
под названием кристаллического стебелька,
гораздо крупнее, сохраняется значительно
дольше и состоит из гиалинового мукопро-
теина. Он также вдвинут в желудок, но
выходит из специального дивертикула, так
называемого стебелькового мешка, и его
вращают выстилающие этот мешок реснички.
Кончик стебелька не только работает как
лебедка, накручивая на себя слизистый жгут
с пищевыми частицами (рис. 9.17), но и
постепенно растворяется в желудке,
высвобождая фермент амилазу. Параллельно
стебелек медленно выдвигается вперед в желудок
(опять-таки благодаря работе ресничек),
а секретируемыи проксимально материал
поддерживает его длину постоянной.
Поэтому кончик стебелька все время
упирается в стенку желудка, что создает эффект
вращающегося жернова, и служит для
отправки пищевых частиц из ставшего менее
вязким слизистого жгута в сортирующую
часть желудка, где реснички продолжают их
разделение.
9.2.6. Питание осадками
Вероятно, большинство современных
животных питаются детритом
(органическим материалом, состоящим из мелких
частиц) или подстилкой (из частиц большего
Пищевод
Слизистый
тяж с пищей
Зона
сортировки
Вход в пищеварительный
дивертикул Кишечник
Щиток
Кристаллический
стебелек
Стебельковый
мешок
Рис. 9.17. Желудок фильтрующего моллюска с кристаллическим стебельком, используемым для
подтягивания пищи в пищеварительный тракт (по Morton, 1979)
288
Глава 9
размера), но, несмотря на это, четкого
представления о природе этого рациона пока
нет. Лист, съедаемый, скажем, земляным
червем,— это не просто мертвый лист, а
целая экосистема. На его поверхности и в его
тканях обитают бактерии и грибы,
обеспечивающие разложение органики, различные
протисты, потребляющие деструкторов
(например, амебы, ресничные, гетеротрофные
жгутиковые), а также лишь немногим более
крупные нематоды и клещи, питающиеся
этими мелкими организмами. На
поверхности листа иногда оказываются фотосинте-
зирующие водоросли и цианобактерии,
мертвые и разлагающиеся остатки других
организмов (не в последнюю очередь — из
фекалий животных). Все эти живые и
мертвые компоненты могут быть поглощены
животным вместе с листом, и задача
состоит в том, чтобы определить, какие из них
перевариваются и ассимилируются и какие
в наибольшей степени удовлетворяют
метаболические запросы потребителя.
Это не обязательно означает, что
потребитель просто заглатывает основной
материал и переваривает все, что может. Теперь
известно, что некоторые потребители
осадков способны к значительно более
избирательному поглощению, чем представлялось
ранее (впрочем, лишь немногие животные
были настолько детально изучены с этой
точки зрения).
Избирательное поглощение, безусловно,
должно давать преимущества, поскольку
пищевая ценность различных компонентов
детрита весьма неодинакова. Сравнение
органики в осадке, заглатываемом
некоторыми морскими животными, не способными
к такому отбору, и их фекалий, а также
обработка известными ферментами из их
кишечника проб этого осадка показали, что
основная часть того небольшого
количества органического вещества, что
содержится в донных отложениях, остается таким
видам недоступной. Они усваивают менее
5—10% его детритного «пула». (На чисто
песчаных пляжах вся органика иногда
составляет менее 1% заглатываемого
материала. Слой лесной подстилки ею богаче,
но все равно часто лишь небольшая ее доля
пригодна в пищу.)
К тому времени, когда любой кусочек
органических остатков становится частью
поверхностного слоя подстилки или
детрита, он уже теряет большую часть своей
питательной ценности. Если это лист, то перед
сбрасыванием все растворимые вещества
переходят из него в остающиеся на
родительском растении ткани, а выщелачивание
приводит к потере других органических
веществ уже в первые часы после падения на
землю. Органические частицы попадают на
субстрат только пройдя через кишечник
консумента, где из них обычно извлекается
большинство питательных соединений,
а фекальные массы — один из основных
источников органического вещества в
осадках и почве. Поэтому органика в них—это
чаще всего устойчивые структурные,
скелетные или защитные соединения, проходящие
без изменений через пищеварительные
тракты большинства животных (у детрито-
фагов, как правило, отсутствуют целлюла-
зы).
Однако в случае свежих остатков других
организмов потребитель осадка имеет шанс
усвоить некоторые содержащиеся в них
органические вещества, хотя и тогда белка
скорее всего будет очень мало или он
окажется недоступен из-за присутствия танни-
нов (см. разд. 9.2.4). Вообще наиболее
вероятно, что потребители мертвой органики
(за исключением питающихся падалью)
существуют за счет живых микроорганизмов,
связанных с этими остатками, служащими
лишь удобным транспортным средством
для доставки пищи в кишечник. На суше и
в пресных водах грибы, разлагающие
подстилку, возможно, являются основной
пищей для потребителей осадков, а на
прибрежном морском мелководье наибольшую
питательную ценность, вероятно, имеют
обрастающие частицы грунта фотосинтези-
рующие одноклеточные протисты.
Показано, что у животных, способных выборочно
питаться только диатомовыми
водорослями, эффективность ассимиляции достигает
70%, в то время как у неизбирательных
потребителей органического материала она
менее 4%. Так или иначе, на основной
площади морского дна, где фотосинтез
невозможен и частицы, оседающие из верхних
слоев воды, редки, к тому времени, когда
они достигают субстрата, в них остаются
Питание
289
только устойчивые к разложению вещества,
и для консументов источником как энергии,
так и белка служат бактерии.
Пищеварительная система потребителей
осадков, как правило, не специализирована,
но кишечник часто удлинен (см. рис. 4.42
и 4.45), что обусловлено теми же
причинами, что и в случае обгрызающих и
соскабливающих животных. Как и у последних,
обычно образуется очень много фекальных
масс, питательная ценность которых для
других потребителей намного ниже, чем
у проглоченного материала.
9.2.7. Питание с участием
симбионтов
Хотя питание относительно немногих
видов животных полностью зависит от
фотосинтеза их эндосимбионтов, как это
показано на рис. 9.18 (см. также разд. 9.1),
некоторые представители таких
филогенетически различных групп, как губки, моллюски,
книдарии, хордовые и т. д., в той или иной
степени используют в пищу продукты
жизнедеятельности своих фотосинтезирующих
партнеров. Ими могут быть цианобактерии
(например, у некоторых губок и эхиурид),
прохлорофиты (у оболочников), а чаще ди-
нофлагеллаты (в виде зооксантелл во
многих морских беспозвоночных) или зеленые
водоросли, представленные зоохлореллами
(обычно у пресноводных видов). Некоторые
мадрепоровые кораллы удовлетворяют 2/3
своих метаболических потребностей за счет
внутриклеточных зооксантелл, и только
1/3 — за счет внешних источников питания,
включая типичное для кишечнополостных
улавливание зоопланктона. У некоторых
актиний зооксантеллы и приспособления
для ловли добычи расположены в
различных частях тела: «органы» с симбионтами
расправляются в дневное время, а
щупальца с нематоцистами — ночью (рис. 9.19).
Симбиоз может дополнять растительнояд-
ность. Мешкоязычные моллюски
высасывают клетки макроскопических зеленых (и
других) водорослей; некоторые их виды
могут сохранять в дивертикулах средней
кишки целые функционирующие
хлоропласт своей добычи (рис. 9.20), которые
захватываются пищеварительными клетками-
фагоцитами, но продолжают фотосинтези-
ровать в некоторых случаях еще более двух
месяцев. До половины фиксируемого
углерода передается моллюску, чего вполне
достаточно для удовлетворения его
энергетических потребностей. (Возможно,
хлоропласт некоторых эвглен имеют сходное
эволюционное происхождение.)
Несколько иные симбиотические
отношения существуют между растениями,
выделяющими нектар, и насекомыми,
которые посещают их для поедания этой бога-
Рис. 9.1 В. Схема потоков
углерода (С) и питательных
веществ (N) при симбиотических
взаимодействиях (по Barnes,
Hughes. 1982)
Гетеротрофное животное
Фотосинтезируюший
СИМБИОНТ
Органические
~ C + N
Органические
— C+N
i_
Неорганические
-C + N
Внешняя среда
\Органические
4 C + N
290
Глава 9
ДЕНЬ
LebrunLa danae
НОЧЬ
Lebrunia coralligens
Bunodeopsis antilliensls
Рис. 9.19. Использование различных частей тела тремя видами рифообразующих коралловых
полипов в разное время суток: органов, содержащих зооксантеллы, при дневном освещении (слева)
и щупалец, несущих нематоцисты, ночью (справа) (по Sebens, De Riemer, 1977)
той сахарами жидкости и в результате
обеспечивают опыление. У некоторых видов
имаго потребляют только нектар (чтобы
обеспечить себя энергией для полета), а
питающаяся фаза их жизненного цикла
представлена личинками, накапливающими
достаточно ресурсов для формирования
взрослого организма, который нуждается
Питание
291
г е..
4?.:
**
<>&■
At, 4Щ
9U I * '.
*i
Ш* *>
4#- J' «t>
• V
.4 '"'
., ~J>;*
Рис. 9.20. Электронная микрофотография функциональных хлоропластов в пищеварительном
дивертикуле мешкоязычного моллюска (Trench, 1975)
в течение своего недолгого существования сутствует на всех стадиях жизненного цикла
лишь в источнике энергии. (некоторые двустворчатые моллюски, оли-
Наконец, у ряда относительно крупных гохеты, полихеты и все погонофоры). Их
свободноживущих животных кишечник от- способ питания до сих пор остается загад-
292
Глава 9
кой, хотя в тканях большинства таких
видов найдены симбиотические хемоавто-
трофные бактерии. Известно, однако, что
по крайней мере у некоторых погонофор на
поверхности тела имеются микроворсинки,
эквивалентные покрывающим поверхность
тела у бескишечных паразитов, обитающих
в пищеварительном тракте других
животных. Следовательно, возможно, что
некоторые из этих свободноживущих червей
способны извлекать органические соединения
из окружающей воды. Относительно других
мягкотелых свободноживущих животных
предполагается, что их пища частично
состоит из растворенной органики, хотя это
признается не всеми отчасти из-за
необходимости при таком способе питания
достаточно высокой, нечасто встречающейся
концентрации в окружающей среде
органических веществ. Другая возможность—
наружное пищеварение с последующим
всасыванием переваренных продуктов
стенкой тела, однако соответствующих
железистых клеток пока не обнаружено.
9.3. Затраты и выигрыш
при различных способах
питания:
оптимальное фуражирование
9.3.1. Введение
Мы видим, что типы питания животных
строго ограничены их эволюционным
прошлым: представителю линии потребителей
суспензий нелегко стать
охотником-преследователем, и наоборот. Тем не менее
многие виды обладают некоторой
пластичностью, и некоторые «генералисты» могут
даже переключаться с одного рациона на
совершенно иной и менять способы
питания. Например, обитающая в эстуариях по-
лихета Nereis diversicolor способна питаться
взвесью (рис. 9.5,Г), осадком, быть
охотником, поедать падаль и соскабливать
фрагменты макроскопических водорослей (табл.
9.1).
Во время своей дневной активности
животные сталкиваются с различными
потенциально съедобными объектами, т.е. с
альтернативными пищевыми ресурсами.
Для большинства специализированных кон-
сументов это может означать выбор лишь
между различными особями одного и того
же вида жертвы, но обычно диапазон
возможных типов пищи шире. Таким образом,
в рамках ограничений, наложенных их
эволюционным прошлым, животные в
масштабах своего экологического времени
(каждую минуту, каждый час, каждый день)
вынуждены выбирать, использовать тот
или иной объект в качестве пищи или
отказаться от него в данных условиях в пользу
другого. Такие «решения» имеют важные
последствия, поскольку выигрыш от
потребления в пищу различных объектов
неодинаков, т. е. одни из них будут «питательнее»
других. Однако наиболее питательная пища
предпочитается и большим числом консу-
ментов, что ведет к более интенсивному ее
потреблению и в результате к истощению
этого ресурса. У других объектов
питательная ценность ниже, поэтому они
пользуются меньшим вниманием консументов,
а следовательно, более обильны и более
доступны в любом местообитании.
Питательная ценность (на единицу
поглощенной массы) — только один из
факторов, влияющих на выбор пищи, поскольку
кроме очевидной выгоды добывание ее
влечет за собой и определенные расходы.
Независимо от способа питания на поиск,
захват, переработку и переваривание добычи
должны быть затрачены время и энергия,
которые можно употребить на что-нибудь
другое. Расходы на процесс питания
неодинаковы при различных его способах, однако
во всех случаях отбор благоприятствует
стратегии, приносящей максимум «чистой
прибыли» (выигрыш минус расходы).
Пища— одно из первых условий
существования, поэтому любая особь, у которой эта
«чистая прибыль» наибольшая, будет иметь
селективное преимущество с точки зрения
и выживания, и воспроизведения перед
другими, поскольку:
а) сможет тратить меньше суммарного
времени на поиск пищи (и таким образом
уменьшит для себя риск оказаться жертвой
хищников);
б) будет быстрее расти или дорастать
до больших размеров (с соответствующими
репродуктивными преимуществами);
Питание 293
Таблица 9.1. Пищевые предпочтения Nereis diversicolor в порядке убывания
количества усвояемой энергии на пищевой объект
Тип питания и характер нищи Порядок пред- Порядковый номер
почтения в ряду убывания
усвояемой энергии
Поедание мертвых двустворчатых моллю- 1 1
сков Масота
Охота на живых олигохет Tubifex 2 2
Охота на живых рачков-бокоплавов Сого- 3 4
phium
Охота на живых личинок двукрылого насе- 4 3
комого Erioptera
Питание поверхностными частицами дон- 5 5
ных осадков
Питание суспензиями 6 6
Обгрызание живых зеленых водорослей Еп- 7 7
teromorpha
Обгрызание живых зеленых водорослей Ulva 8 8
Охота на живых брюхоногих моллюсков 9 9
Hydrobia
в) будет «здоровее», а значит,
устойчивее к паразитам и лучше защищенной от
хищников и т.п.;
г) извлечет большую пользу из
меньшего количества ресурсов при дефиците пищи.
Каким же образом могут животные
максимизировать «чистую прибыль» в единицу
времени? Эта проблема изучалась на
простых моделях стратегии оптимального
фуражирования животных с последующей
проверкой прогнозов, полученных в этих
моделях, с помощью экспериментов и
наблюдений. Обычно при постановке
экспериментов выделяют какую-нибудь одну
переменную, в данном случае—выбор жертвы.
Следует помнить, однако, что в
естественных условиях поведение животного в
каждый отрезок времени представляет собой
компромиссный ответ на множество
разнонаправленных воздействий. Максимизация
«чистой прибыли» от питания сама по себе
уже дает преимущества, но не менее
выгодны избегание хищников, уход за
потомством и т. д. Эти другие потребности часто
накладывают ограничения на процесс
питания, в результате чего потребление добычи
будет идти на уровне ниже оптимального,
т.е. животное может иметь очень
небольшой выбор и поглощать то, что удастся
найти за тот короткий период, который
останется ему для питания.
Кроме того, живая добыча отнюдь не
ждет, пока ее съест консумент! Фактически
стремление жертвы.не оказаться
потребленной значительно больше, чем стремление
хищника поглотить ее; это называют
принципом «жизнь/обед». Например, при
любой встрече между каракатицей и
креветкой для первой это лишь проблематичная
возможность поесть, для второй - вопрос
жизни и смерти, а значит, и давление
отбора для этих партнеров различно.
Следующие разделы будут
посвящены обсуждению способов максимизации
«чистой прибыли» консументами и ее
минимизации их жертвами, т. е. уменьшения ими
для себя опасности быть съеденными.
Подвижные хищники, имеюшие возможность
схватить или отвергнуть ту или иную особь-
жертву, представляют собой простейший
случай, и мы особенно подробно разберем
именно его. Потребители суспензий и
осадков сталкиваются с иными проблемами
и будут рассмотрены отдельно.
9.3.2. Теория оптимального
фуражирования
Любой съедобный объект имеет для
потребителя свою особую «питательную
ценность». Потребляемые вещества служат для
294
Глава 9
животного источником энергии и
материалом для роста соматических и
репродуктивных клеток и тканей, значит, питательная
ценность должна учитывать обе эти
потребности. К сожалению, они могут
проявляться в разное время года или на различных
стадиях жизненного цикла, т.е. в какой-то
момент главной может быть потребность
в энергии, в другой—в органическом азоте,
а у некоторых видов (например, у наземных
брюхоногих) бывают периоды, когда
больше всего прочего они нуждаются в
неорганических элементах или соединениях
(например, кальция).
В большинстве исследований в качестве
условной меры питательной ценности пищи
используется ее энергетическое содержание,
или калорийность; с помощью этого
упрощенного термина питательная ценность
может быть выражена как энергия, полученная
при поедании данного объекта (Ef). минус
все энергозатраты, связанные с его
поимкой, умерщвлением, обработкой и
перевариванием («затраты на переработку», Е,,).
Помимо этого различные потенциально
съедобные объекты встречаются в данном
местообитании с различной частотой, и,
если потребитель предпочитает
определенный тип объекта, он может активно искать
именно его, т. е. идти на «поисковые
затраты» (EJ. Чистая энергетическая прибыль
поэтому будет равна
Eg - Е„ - Es.
В свою очередь время, которое легче
измерить, можно подставить в это выражение
вместо энергии, и тогда питательная
ценность будет измеряться энергией,
полученной в единицу времени. Она составит E/Th,
если встреча с жертвой уже произошла, или
Е/(Т,, + TJ, если требуется поиск жертвы
(Th — «время перераб отки», Ts—
«поисковое время», а Е—доступное
количество энергии = Eg в выражении выше).
Возьмем сначала более простой случай,
когда отсутствуют затраты времени на
поиски. Потребитель случайно наталкивается
на различные потенциально съедобные
объекты в месте своего обитания, и возникает
вопрос: какой из них он должен съесть,
чтобы получить максимальное количество
энергии в единицу времени? По логике
следует ожидать, что он предпочтет добычу
с наибольшей питательной ценностью,
выраженной отношением Е/Т,, (или, если
в данный момент он нуждается в
определенном элементе N, с максимальным
отношением N/Th). Другими словами, выбирается
жертва, из которой можно извлечь
наибольшую пользу.
Эта простая модель приводит к ряду
уже проверенных прогнозов.
1. Потребитель будет поедать только
жертв с наибольшей питательной
ценностью, если частота встреч с ними настолько
велика, что включение в рацион других
жертв (с более низкой питательной
ценностью) уменьшит среднюю скорость
поглощения энергии.
2. Если, однако, частота встречи с
жертвами с наивысшей питательной ценностью
ниже, потребитель должен расширить
диапазон типов жертв, включив в их число
следующие по питательной ценности объекты
и т.д.
3. Если на распознавание различных
типов жертв требуется значительное время
(например, когда это возможно только при
соприкосновении с ними), следует
использовать в пищу жертвы с более низкой
питательной ценностью, но встречающиеся
чаще других, даже если более ценных жертв
остается много. Когда же идентификация
жертвы происходит мгновенно (например,
с помощью зрения), животное должно
отвергать жертвы с меньшей питательной
ценностью, если более ценная пища вполне
доступна. (Установление пищевой ценности
жертвы —просто дополнительный
компонент времени переработки.)
Все перечисленные прогнозы полностью
подтвердились, например, для зеленого
краба (Carcinus), питающегося мидиями
(Mytilus) различного индивидуального
размера (рис. 9.21—9.23; приложение 9.1).
Опыты по изучению взаимодействия
хищника и жертв, относящихся к разным
видам, проводились реже, однако полихета
Nereis diversicolor как потребитель очень
широкого диапазона пищи также служит
примером иерархического подхода к выбору
жертвы, соответствующего количеству
ассимилируемой энергии, заключенной в
каждом типе добычи (табл. 9.1).
Питание
295
12 3 4
Длина мидии, см
Рис. 9.21. Связь между питательной ценностью
(Б/Т,,) и выбором жертвы у краба Carcinus
maenas (шириной 6,0—6,5 см), питающегося
мидиями Mytilus edulis; см. текст (по Elner,
Hughes, 1 978)
20
10
X
Первые 5 дней
/ .Вторые 5 дней
\
\ ..-■
Л
\
\
'■•.Третьи 5 дней
2 4 6
Длина мидии, см
Рис. 9.22. Выбор жертвы крабами Carcinus
maenas, когда число мидий (Mytilus edulis)
предпочитаемого размера уменьшается в
результате их выедания; см. текст (по данным
Elner, Hughes, 1978)
Теперь можно усложнить базовую
модель, включив в нее разное время,
необходимое для поиска различных типов пищи.
Предположим, что Т,, постоянно для
данного типа и данного размера жертв, a Ts
зависит от частоты различных
потенциальных особей жертв в данном местообитании.
Пусть будут два типа жертв х и у, при этом
питательная ценностью д: больше, чем у г.
Тогда
Е(х)^ Е(у)
Th(x) Th(y)
Очевидно, что, если потребитель
обнаруживает жертву типа х, он всегда ее съедает, по-
20-
10-
30-
20-
' 10-
1
1
ои-
40-
^ зо-
го-
10-
:
60-
50-
40-
30-
20-
10-
I
В наличии
В рационе
Рис. 9.23. Влияние доли менее питательных
мидий (Mytilus edulis) на выбор добычи
крабами Carcinus maenas; см. текст (по данным Elner,
Hughes, 1978)
скольку не получает никакого
преимущества при отказе от нее в пользу жертвы у
(при прочих равных условиях). А если
сначала он встречается с жертвой у, — съест он
ее или откажется, чтобы продолжать
поиски более выгодной для него жертвы х?
С точки зрения максимизации чистой
энергетической прибыли решение зависит
только от частоты встречаемости х, т. е. от
величины Т„(х). Особь типа у, встреченная
животным-потребителем, должна быть
съедена, если
ЕЫ_>_ Е(х)
и отвергнута, если
Е(у) „
Т„Ы' Т„(дс) + Т.(дс)'
если
ЕМ
Т„(>) Т„(дс) + Т,(х)"
Прозрачный
пластик
Выдвижная
крышка
Выдвижная
крышка
Конвейерная лента
"Поисковое время"
для крупных
червей, с
А 40
0вщее число
пищевых
Объектов в минуту
Процент
предъявляемых крупных и
мелких червей
Крупные Мелкие
50,***, г^-,50
Рис. 9.24. Аппарат,
используемый для предъявления с
меняющейся частотой различных
пищевых объектов (мучных
червей) неподвижному
потребителю (большой синице); см.
текст (по Krebs et al., 1977)
Процент съеденных
крупных и мелких
червей
Крупные
53 ™ Мелкие
Мия ,47
50
Б 20
т50
49*та ГЩ
ттл 51
75
87
12
25
J3
88
50
г-п50
Г
18
тч-12
Fxl67
27
33
гл22
Рис. 9.25. Специализация на более крупных мучных червях при небольшом связанном с ними
поисковом времени, как в опытах В, Г и Д:
Е(мелк)
Т,,(мелк) "^ Ти(крупн) + Т5(крупн)'
Е(крупн)
.(по Krebs et al., 1977)
Питание
297
Иными словами, данная встреченная особь-
жертва должна съедаться, если за время,
затраченное на ее переработку, найти и съесть
другую, более благоприятную с точки
зрения полученной энергии жертву
невозможно (рис. 9.24 и 9.25, приложение 9.2).
Дальнейшее усложнение состоит в том,
что многие виды потенциальной пищи
распределены в виде компактных скоплений,
т.е. отдельными пятнами, например
организмы, обрастающие морские скалы,
муравьи в муравейниках, стаи мизид, заросли
крапивы и т. д. В этом случае консумент
сталкивается с проблемой другого рода:
любое подобное скопление, участок или
область локальной концентрации содержит
определенное количество пищи, поедание
которой может привести к уменьшению ее
обилия и снижению чистой прибыли для
потребителя. Как же долго следует
эксплуатировать данное пятно, прежде чем перейти
к следующему? Стоит ли сначала исчерпать
все его ресурсы или выгоднее прекратить
использование раньше, а если да, то в какой
момент?
Хотя внешне иные, но по сути такие же
проблемы встают и перед другими
животными-потребителями. Например, в высоких
широтах количество доступной пищи
зависит от сезонных климатических условий,
и животные должны решать, когда им
покидать географическую область с
уменьшающимися пищевыми ресурсами. Когда дол-
ЖЕ1Ы улетать ласточки? Ряд особых
взаимоотношений хищник—жертва также
относится к этой категории, особенно если
хищникам требуется значительное время,
чтобы потребить все ткани пойманной
жертвы, в том числе высосать из нее все
соки. Пчелы, питающиеся нектаром из цветка,
водяные клопы, сосущие жидкость тела
комариных личинок, и даже львы,
пожирающие антилопу,— во всех этих случаях
сначала скорость поглощения пищи велика, но по
мере того, как основная часть более
доступных и более питательных ее компонентов
усваивается, процесс потребления
замедляется. В какой момент животному следует
оставить старую жертву и переключиться
на новую? После того как извлечен весь
съедобный материал или не дожидаясь этого?
Когда потребитель начинает эксплуати-
ш
Т
(_
О-
<D
X
СП
'Х
о
х _^_^_^__—
X ^* '
Т уг
>- s
§ /
1= /
аз /
Z /
Z /
>- /
о "
т
Рис. 9.26. Изменение со временем скорости
получения энергии из ограниченного
источника; см. текст
ровать еще нетронутое до него пятно, будь
то географическая область или пойманная
особь жертвы, количество энергии,
извлекаемое в единицу времени, можно выразить
соотношением, представленным на рис.
9.26. Рано или поздно животное должно
покинуть этот источник пищи, перейдя на
другой участок или поймав другую жертву,
причем этот переход может стоить ему
много времени и энергии. Даже такое медленно
передвигающееся животное, как береговая
улитка Littorina, тратит в единицу времени
в 12 раз больше энергии на ползание, чем на
соскабливание водорослей на одном месте,
и, конечно, миграции на большие расстоя-
X
<D
т
>- ^~
с; /^
о /
>-х /
-« »--вб -—*-
Время Время, проведенное
миграции на пятне
Рис. 9.27. Потребление энергии при
эксплуатации пятна ресурса в зависимости от времени,
проведенного в его пределах и затраченного на
его достижение; см. текст
298
Глава 9
ш
ЕС
1_ X
CLtr-
QJ СП
Hi с
mz
Sg
х cl
го
is
?ГГС
О СП
SI
51
S* 1
^
,1
Среднее
время
миграции
Время, проведенное
на пятне
Оптимальный момент
покидания пятна
Рис. 9.28. Время нахождения
в пределах пятна,
обеспечивающее максимальную
суммарную прибыль на единицу
суммарного времени
фуражировки, можно найти, проведя
касательную от начала
координат к кривой получения
энергии; см. текст
ния обходятся ей очень дорого. Значит,
необходимо (с поправкой на различный
расход энергии) учитывать и время, требуемое
на передвижение как между участками, так
и внутри них.
Теперь, если мы соотнесем среднюю
чистую энергию, полученную со всех пятен
в данном местообитании, со временем и
учтем среднее время, Егеобходимое для
перемещения от одного пятна к другому (рис.
9.27), можно задать вопрос, какой период
времени, проведенный на пятне, даст
максимальный выигрыш в энергии за единицу
суммарного времени (т. е. включая
перемещение от одного пятна к другому и
пребывание в пределах пятна). Ответ дает
касательная, проведенная от начала координат
к кривой, отражающей общее количество
поглощенной энергии (рис. 9.28), т. е. линия,
идущая от начала координат к этой кривой
под максимальным углом. Таким образом,
потребитель, покидающий участок с
пищевыми ресурсами или прекращающий
поедать данную жертву, когда количество
поглощенной энергии в единицу времени
падает до этого среднего уровня,
максимизирует энергию, полученную в единицу
суммарного времени фуражирования.
Потребители должны прекратить использование
сокращающихся источников пищи в
оптимальный для этого момент!
Если это так, то следуют выводы:
а) когда пятна внутри местообитания
различаются по качеству, потребители
должны дольше оставаться на лучших пятнах
(рис. 9.29);
б) когда пятна качественно
приблизительно одинаковы, но расстояния между ними
различны, для потребителя выгоднее
дольше оставаться на данном пятне, если он
затратил больше времени, чтобы достичь его
(рис. 9.30) (т. е., если затраты на передвиже-
Средняя скорость
для всех пятен
Покинуть
раньше
Покинуть
позже
Время, проведенное
на пятне
Рис. 9.29. Влияние качества
пятна на оптимальное время,
которое следует провести на
различных пятнах в пределах
данной области; см. текст
Питание
299
Короткий Относительно
период недолго
миграции
между пятнами
Долгий период
миграции между
пятнами
Относительно
долго
Рис. 9.30. Влияние среднего периода миграции
между пятнами на оптимальное время,
которое следует провести на пятнах одинакового
качества; см. текст
ние выше, выигрыш от него будет меньше,
и потребитель должен остаться на каждом
пятне дольше).
9.3.3. Подвижные потребители
и их жертвы
Каким образом общие соображения,
изложенные выше, влияют на биологию
питания подвижных животных, пищей которым
служат отдельные объекты, и как могут их
жертвы минимизировать шансы быть
съеденными?
Очевидно, одно и то же количество
пищи бывает представлено либо большим
числом мелких объектов, либо несколькими
крупными, и эти два экстремальных случая
требуют различных затрат и дают
неодинаковую выгоду. Выгода от одной крупной
особи велика, однако такая жертва скорее
всего будет очень подвижна, поэтому,
чтобы поймать ее, потребуются большие
затраты времени и энергии. С другой
стороны, питание многими мелкими особями
может не требовать много времени на поимку
каждой из них, однако велика вероятность
долгих поисков добычи. Эти особенности
будут отражаться как на потребителе, так
и на его жертве.
Преследующие охотники обычно
большую часть своего суммарного времени
питания проводят в погоне за крупными
особями-жертвами, причем не всегда успешно.
На каждую потенциальную жертву
тратится много энергии, поэтому и прибыль
должна быть велика. Охотнику выгодно
минимизировать затраты на преследование,
а жертве—максимизировать их. Это ведет
к своего рода «гонке вооружений» между
хищником и жертвой. Первый ограничивает
себя таким кругом своих потенциальных
жертв, которые наиболее уязвимы для его
специфических форм преследования и
захвата, получая при этом максимальную
энергетическую компенсацию и все больше
приспосабливаясь к особенностям
ускользания своих жертв от преследования. В
результате максимализации чистой прибыли
преследующие охотники по своему рациону
становятся «специалистами».
Для жертв помимо очевидного давления
отбора в сторону увеличения скорости
и ловкости наиболее распространенным
способом уменьшения шансов быть
пойманными хищниками (не только
преследующими) является групповое
существование (стаями, стадами, косяками и т. п.). Это
дает целый ряд- преимуществ.
1. Одно из наиболее известных
наблюдений заключается в том, что большая группа
может обнаружить приближение хищника
и отреагировать на него быстрее, чем
мелкая группа или отдельная особь. Например,
мелкий бескрылый морской клоп Halobates
скользит по поверхностной пленке моря так
же, как водомерка по поверхности пресных
водоемов. Когда эти клопы не питаются,
они собираются во «флотилии» которые
очень четко реагируют на приближение
хищников (птиц, рыб и т. п.). Расстояние,
с которого «флотилия» реагирует избега-
300
Глава 9
i 4. 90-
Я *
8-1
ei бо -
га х
ш
о. о;
i g зо -
100 200
Число осовей во "флотилии"
Рис. 9.31. Зависимость расстояния, на котором
экспериментальная модель хищника вызывает
поведенческую реакцию у морского клопа На-
lobates, от числа его особей во «флотилии» (по
Treherne, Foster, 1980)
нием на хищника, зависит от ее размеров,
как показано на рис. 9.31: оно больше в
случае более крупных флотилий, хотя, конечно,
существует критический размер, начиная
с которого дистанция обнаружения уже не
возрастает (т. е. достигнут максимум).
2. Второе преимущество существования
в группе—это введение хищника в
замешательство. Преследующие охотники
одномоментно нападают на одну потенциальную
жертву, и мы уже видели, что им выгоднее
схватить особь с высокой питательной
ценностью. Группа может реагировать на
приближение хищника рассеянием особей в
разных направлениях. Это затрудняет
«нацеливание» на индивидуальную жертву и
приводит к переключению с цели на цель, когда
разные особи мелькают в поле его зрения.
3. Хищников может отпугивать
групповое защитное поведение животных, в то
время как подобное затруднение
отсутствует при встрече с одиночным
представителем того же вида. Классические примеры
групповой защиты относятся к
позвоночным, однако есть они и среди
беспозвоночных, причем особенно яркие—среди
общественных перепончатокрылых. Кроме того,
личинки некоторых пилильщиков при
нападении (например, хищных насекомых)
выделяют своими ротовыми частями липкий
секрет. Подобная реакция не мешает клопам-
щитникам атаковать отдельных личинок,
однако, нападая на их скопление, клопы
быстро покрываются липкими выделениями
и после этого едва могут двигаться.
4. Все три положительных эффекта
группового существования, описанные выше,
ведут к увеличению времени, которое
требуется хищнику для удачной поимки жертвы.
Это может привести к тому, что для него
окажется более выгодным переключение на
другую жертву, не относящуюся к
общественным животным. Четвертый эффект
группового существования относится к
вероятности для каждой данной особи в
группе оказаться жертвой удачного нападения:
чем крупнее группа, тем эта вероятность
ниже. Это было показано для клопа Halobcttes,
о котором упоминалось выше в связи с
дистанцией обнаружения хищников, когда
этим насекомым угрожают рыбы (рис.
9.32). Один клоп из 10 в группе подвергается
1/10 общего числа нападений,
направленных на одиночную особь, а если в группе
100 клопов, вероятность оказаться жертвой
составляет 1/100.
Этот эффект необязательно чисто
статистический. Часто вероятность быть
схваченным выше всего на периферии группы и
минимальна в ее центре. В ряде случаев
наблюдали непрерывное движение особей от
периферии к центру; при этом по краям
остаются животные, менее других
способные занять относительно безопасное
положение внутри «эгоистического стада». Так
происходит активная минимизация
индивидуальной вероятности быть съеденным.
В противоположность этому охотники
из категории поисковиков и засадчиков
о
* 0,011 1 1
1 10 100
Число осовей в группе
Рис. 9.32. Число нападений рыб на отдельных
особей Halobates в зависимости от числа
клопов в группе (по Treherne, Foster, 1980)
Питание
301
предпочитают добычу, которую легко
поймать из-за небольшого (по сравнению
с хищником) размера. Любая с легкостью
схваченная жертва тут же съедается, что
дает в сумме достаточно еды, и
поисковикам легче всего максимизировать свою
чистую прибыль, становясь по рациону ге-
нералистами. Затем, поскольку поисковое
время обратно пропорционально общему
обилию подходящей жертвы, в
местообитаниях, где мало добычи или в периоды
снижения ее количества, поисковики, по-
видимому, наименее специализированы
в смысле питания. Зато в периоды пика
обилия определенного вида-жертвы они могут
тратить непропорционально много
времени, потребляя именно его (становясь
временно «специалистами»), и переключаться
на другие виды, только когда их
численность также достаточно возрастет.
Особям видов, выедаемых
поисковиками, выгодно максимизировать поисковое
время охотника. Этого можно добиться
различными путями, увеличивающими
трудности обнаружения жертвы или
распознания в ней потенциальной пищи. Способы
следующие (рис. 9.33):
1) заселение таких микроместообитаний,
которые нелегко обследовать большинству
потенциальных хищников, т. е. жизнь в
щелях, под камнями, в норах и т.д.;
2) незаметность, т. е. особые
(покровительственные) форма тела, поза, окраска (или
запах), сливающиеся с фоном и таким
образом увеличивающие время распознавания
(некоторые виды способны изменять цвет
и рисунок тела так, чтобы сливаться с
несколькими типами фона; другие изменяют
свое окружение, чтобы оно более
эффективно скрывало их от хищника);
3) мимикрия, как и
покровительственный облик, увеличивает время
распознавания хищников за счет имитации
малосъедобного или вовсе несъедобного объекта
(мертвого листа, сучка, кусочка фекалий
и т. д., вида с плохим вкусом или
способного к агрессивной самозащите).
Увеличение необходимого поискового
времени не единственный способ
уменьшения пищевой ценности отдельной особи для
хищника; к другим относятся удлинение
периода переработки жертвы и уменьшение
энергии, содержащейся в единице веса
жертвы.
Увеличение времени переработки часто
достигается в ходе эволюции следующих
признаков (рис. 9.34):
1) защитного вооружения (жал,
челюстей и т.д.);
2) защитных покровов (раковин,
известковых или хитиновых пластин и т.д.);
3) специальных образований,
затрудняющих захват жертвы, манипулирование ею
и/или заглатывание (например, игл). Иглы
и подобные им выросты тела особенно
эффективны, поскольку увеличивают не
только время переработки, но и видимый
размер тела жертвы (при небольшом приросте
тканей) и таким образом исключают
животное из числа возможных объектов поедания
хотя бы некоторых хищников.
Содержание энергии и/или питательных
веществ на единицу веса или объема тела
может быть уменьшено путем
«разбавления» живых тканей инертными, мало
питательными или несъедобными веществами.
Вкусовая пригодность может быть снижена
присутствием особых веществ в некоторых
органах или во всем теле. В этом случае
хищнику необходим определенный уровень
научения методом «проб и ошибок», и
часто жертвы делают такое научение как
можно более быстрым и эффективным,
подкрепляя неприятный вкус яркой
предупреждающей окраской.
Увеличение времени на переработку,
снижение содержания энергии на единицу
веса и уменьшение вкусовой пригодности
(или явная токсичность) используются
также в качестве способов защиты пищевыми
объектами обгрызающих и
соскабливающих животных, функционально
эквивалентных охотникам-поисковикам, но
потребляющих прикрепленные организмы, часто
способные к фотосинтезу. Поисковики, как
правило, крупнее и живут дольше, чем их
жертвы-животные, а у соскабливателей
и обгрызателей обычно как раз наоборот —
их добыча крупнее их самих и долговечнее.
Таким образом, в то время как жертвам
охотников-поисковиков трудно
использовать какую-либо иную стратегию, кроме
научения своих долгоживущих
потребителей с помощью отпугивающих веществ из-
302
Глава 9
ОБИТАНИЕ В НОРАХ
Полихета
ПОКРОВИТЕЛЬСТВЕННЫЙ ОБЛИК
?!>, Клоп, сливающийся
с корой
Палочник
(насекомое) ,
МИМИКРИЯ
.... ,:--» - ^
J4
St .-•
5^, _ ^&э а*^ ^ Гусеницы бэбочки,
s -~А '%. . », ^^ имитирующие
3 >ж '' ^" V- птичий помет
Л- ■'■■ _* v
Рис. 9.33. Механизмы,
увеличивающие поисковое время
для потенциальных хищников
(из различных источников)
бегать их, долгоживущим растениям легче
и выгоднее просто убивать токсинами ко-
роткоживущих насекомых-фитофагов,
осуществляя, таким образом, отбор особей,
игнорирующих данный вид в качестве
пищевого ресурса.
Все эти защитные системы (см. также
разд. 13.2.1) создают для консумента-
генералиста такую ситуацию, когда ему
выгоднее искать другие типы жертв — более
заметные, менее защищенные, более
привлекательные на вкус. Однако успешное
избегание нападения со стороны генералистов
приводит к высокой численности
полагающихся в основном на пассивную
механическую или химическую защиту
прикрепленных или сидячих организмов, а это делает
их доступными для нападения «специали-
Питание
303
ИГЛЫ
ПОКРОВЫ
У
/"■/ |'П
Морской еж
■ "7 /I i 1 \ ч
Личинка ракообразного
ЖАЛА
Жало
Рис. 9.34. Структуры,
увеличивающие время переработки для
потенциальных хищников (из
различных источников)
._ Карапакс
Раковина
Ракообразное
Дорсальная створка
Вентральная створка
Плеченогое
Раковина
Скорпион (хелицеровые)
Переднежаберный моллюск
стов», эволюционно приспособленных для
преодоления таких барьеров. Обгрызатели
и соскабливатели, а также специалисты-
потребители форм, использующих
покровительственный облик, мимикрию и
отпугивающие вещества, принимали участие
в такой же эволюционной «гонке
вооружений» с поедаемыми видами, что и охотники-
преследователи с их чрезвычайно
подвижными жертвами. В самом деле,
специалисты-потребители, выработав механизмы,
которые делают неэффективными
химические методы защиты их жертв, могут даже
накапливать токсичные вещества в
собственном теле, используя их для защиты
против своих врагов.
9.3.4. Потребители суспензий
и осадков
Животные, улавливающие пищу путем
процеживания потока воды, очевидно,
тратят энергию не на преследование или
поиски добычи, а на фильтрацию и отбраковку
ненужных частиц.
Размеры отверстий в их фильтрах
обычно могут меняться лишь в масштабах
эволюционного времени, поэтому трудно
предполагать возможность отбора из воды
только частиц с наибольшей питательной
ценностью: фильтр специализирован на
размер, а не на съедобность пищи.
Прикрепленные фильтраторы могут максимизиро-
304
Глава 9
а
Содержание усвояемой энергии
во взвешенном материале, кДж/л
Рис. 9.35. Связь между концентрацией
съедобных частиц в воде и скоростью фильтрации
у полихеты Nereis diversicolor, питающейся
суспензией (по Pashley, 1985) (см. рис. 9.5, Г
и табл. 9.1)
вать чистую скорость потребления энергии,
изменяя только следующие параметры:
1) скорость фильтрации в зависимости
от относительного обилия в воде частиц
разного типа;
2) скорость отбраковки в зависимости
от относительной питательной ценности
частиц и затрат на отбраковку.
Скорость фильтрации должна
увеличиваться при повышении концентрации
частиц с высокой питательной ценностью
(рис. 9.35), однако здесь может быть
верхний предел, определяемый возможностями
пищеварительного тракта хищника
перерабатывать поступающий материал. При сли-
зисто-ресничном механизме это зависит от
движения слизистого тяжа внутри кишки,
обусловленного деятельностью ресничек.
У членистоногих, просеивающих пищу с
помощью щетинок, может произойти полное
заполнение полости кишки заглоченными
частицами. Как только это происходит,
скорость поглощения энергии ограничивается
временем прохождения частиц через
пищеварительный тракт и интенсивностью их
переваривания. Следовательно, скорость
фильтрации падает до величины,
способствующей поддержанию кишечника в
заполненном состоянии. Как и в ситуации
с хищниками, поедающими только
наиболее питательные части добычи, чтобы после
этого перейти к другой жертве (разд. 9.3.2),
потребителям суспензий, зависящим от
качества и обилия пищевых частиц, бывает
выгоднее быстро пропускать пищу через
кишечник, усваивая лишь часть ее, чем
медленно переваривать большую долю
заглоченного.
Затраты на отбраковку бывают высоки,
поэтому по мере роста затрат на
отбраковку частиц определенного типа можно
ожидать увеличения скорости их заглатывания.
Когда эти затраты очень велики, бывает
дешевле избавляться от даже полностью
несъедобных частиц через кишечник, а не с
помощью сортирующих органов. В этих
условиях потребитель суспензий может
столкнуться с дефицитом энергии, и ему иногда
выгоднее полностью прекратить питание до
изменения состава взвешенного в воде
вещества.
При подобных системах сбора пищи
возможность стать «специалистом» очень
мала (если вообще существует), разве
только в эволюционном масштабе времени и
в отношении определенного размерного
диапазона частиц. Например, у
пелагических оболочников очень тонкоячеистые
фильтры, способные задерживать бактерии:
с другой стороны, киты отцеживают
крупных планктонных эвфаузиид. Эти различия
связаны просто с величиной отверстий их
фильтров и относительными размерами
самих фильтраторов. Киты не могли бы
использовать фильтр, способный собирать
бактерий и протистов: им не удалось бы
одновременно продвигаться вперед и
фильтровать по той же самой причине, по которой
невозможно быстро протащить в воде
мелкую планктонную сеть и что-нибудь в нее
поймать. Вода просто проталкивается
вперед, почти не проходя через ячеи. Такая
избирательность у потребителей суспензий,
естественно, не позволяет регулировать
питательную ценность частиц, а проявляется
в основном у свободноживущих личинок,
имеющихся у многих видов, когда они
расселяются в места, способные обеспечить
прикрепленные взрослые стадии
необходимыми взвешенными пищевыми частицами.
Потребители осадков представляют со-
Питание
305
бой нечто среднее между потребителями
суспензий и объединенной категорией
охотников и обгрызателей-соскабливателей.
Питаясь неживым органическим веществом,
они потребляют пищу низкой питательной
ценности, которая соответственно очень
обильна и не может защищаться от
поглощения. Некоторые консументы этой
категории просто пропускают через кишечник
неорганический осадок, содержащий
органический компонент, поэтому сталкиваются
с такими же проблемами переполнения
кишечника, как и потребители суспензий.
Когда содержание усвояемого вещества в
осадке ниже определенного предела, может
быть энергетически выгоднее не поглощать
такой материал, а перейти в другое место.
Однако многие потребители осадков
способны к избирательному поглощению
(разд. 9.2.6) и могут вести себя как
охотники-поисковики с той разницей, что «поиск»
совершает не все животное, а очень
подвижные щупальца или подобные им
образования.
В целом живые существа в рационе
потребителей суспензий и осадков очень малы по
сравнению с размером самого потребителя,
и, хотя некоторые из поглощаемых видов
и располагают химическими средствами
защиты, большинство, по-видимому,
полностью лишены препятствующих выеданию
систем. Вместо этого они наделены
способностью быстро размножаться половым
и/или бесполым путем, которой более чем
достаточно для компенсации потерь от
хищничества. В этом они отчасти сходны
с добычей обгрызателей и соскабливателей,
обычно достаточно интенсивно растущей
или бесполо делящейся, чтобы возмещать
утраченные структуры типа листьев,
полипов или зооидов, так что только при
исключительно высокой скорости выедания
наблюдается гибель отдельных особей или
колоний.
9.4. Выводы
По всей видимости, первые животные
были мелкими потребителями бактерий,
протистов и друг друга, причем эти
исходные рационы не только сохранились
относительно неизменными в течение
миллионов лет, но и всегда оказывали влияние на
питание животных. Селективные
преимущества крупного размера способствовали
развитию механизмов, увеличивающих
заглатываемое количество этих мелких и
рассеянных в среде объектов, но не привели
к появлению принципиально новых
рационов. Потребление макрофитов, будь го
водоросли или растения, возникло
относительно поздно и без участия кишечных
микроорганизмов остается малоэффективным
не только из-за низкой питательности
тканей, богатых структурными углеводами, но
и в связи с защитными приспособлениями
этих во всех других отношениях легко
доступных объектов нападения.
Подразделение животных на
«растительноядных», «плотоядных» и «всеядных» не
способствует пониманию их биологии
питания: с этой точки зрения большинство
видов относятся к всеядным. Более
рациональна классификация по типам питания—
охотники/паразиты, соскабливатели/об-
грызатели, потребители осадков,
потребители суспензий, потребители продуктов
жизнедеятельности эндосимбионтов и т. д.
Такая система не считается с систематическим
положением поедаемых видов и
ориентируется скорее на потребителя, чем на
потребляемое.
Хотя и ограниченные в разной степени
своим эволюционным прошлым, все
животные тем не менее оказываются в масштабах
экологического времени перед выбором:
какую из особей-жертв предпочесть в данных
пространственно-временных условиях;
продолжать питаться на этом участке или
перейти на другой и т. д. По-видимому,
потребители не принимают случайных
«решений», и теория оптимизации дает модель,
довольно успешно предсказывающую
поведение животных в ходе фуражирования.
В рамках ограничений, создаваемых
факторами, не относящимися к качеству пищи
и обеспеченности ею (например,
хищничеством, биологией разможения), некоторые
животные, действительно, питаются таким
образом, чтобы максимизировать
получаемую прибыль (выгода минус затраты).
С другой стороны, отбор среди видов-
жертв ведет к максимизации относительных
расходов, связанных с использованием их
20—1314
306
Глава 9
в пищу, за счет особенностей поведения,
морфологии или биохимии. Как и
следовало ожидать, различные типы питания
подразумевают неодинаковые потенциальные
решения проблемы максимизации
прибыли, дающие селективное преимущество
либо генералистам. либо специалистам.
Пищевое поведение, очевидно,
ежедневно находится под давлением не менее
интенсивного отбора, чем другие, более
изученные аспекты биологии животных.
9.5. Дополнительная
литература
Barnard С. J. (Ed.). 1985. Producers and Scroungers.
Croom Helm, London.
Calow P. 1981. Invertebrate Biology: A Functional
Approach. Croom Helm, London.
Crawley M.J. 1983. Herbivory. Blackwell Scientific
Publications, Oxford.
Hodkinson L D., Hughes M. K. 1982. Insect
Herbivory. Chapman and Hall, New York.
Jennings D.H., Lee D.L. (Ed.) 1975. Symbiosis.
Cambridge University Press, Cambridge.
Jennings J. W. 1972. Feeding, Digestion and
Assimilation in Animals, 2nd edn. Macmillan, London.
Jorgenson C.B. 1966. Biology of Suspension
Feeding. Pergamon Press, Oxford.
Mason C.F. 1977. Decomposition. Edward Arnold,
London.
Morton J. 1979. Guts, 2nd edn. Edward Arnold,
London.
Owen J. 1980. Feeding Strategy. Oxford University
Press, Oxford.
Smith D.C., Douglas Л.Е. 1987. The Biology of
Symbiosis. Edward Arnold, London.
Taylor R.J. 1984. Predation. Chapman and Hall,
New York.
Townsend C.R.. Calow P. (Ed.) 1981. Physiological
Ecology. Blackwell Scientific Publications,
Oxford.
Vermeij G. 1987. Evolution and Escalation. An
Ecological History of Life. Princeton University Press,
Princeton.
Приложение 9.1.
Выбор мидий крабами
В результате наблюдений не
представляет большого труда определить время,
которое требуется крабу, чтобы разломать
раковину мидии и съесть ее содержимое (Th).
Количество энергии, полученное при
поедании моллюска (Е), определяется с помощью
калориметрической бомбы. Краб ломает
раковину с помощью своих клешней, на что
уходит значительное «время переработки»,
которое более или менее экспоненциально
увеличивается с размером мидии. Таким
образом, для открывания крупных раковин
требуется очень много времени, но им
соответствует большое значение Е, а мелкие
мидии можно открыть быстро, но они дают
и меньше энергии. Для краба данного
размера отношение E,Th изменяется с
изменением размера мидий, как показано на рис.
9.21. На этом же графике указан размерный
диапазон мидий, поедаемых крабами
рассматриваемого размера. Очевидно,
соотношение между теми мидиями, которые
съедены, и теми, что обладают наибольшей
предсказанной питательной ценностью, т. е.
прогноз 1 (стр. 294), оказывается верным.
В этом опыте каждая съеденная мидия
тут же заменялась другой такого же
размера, и таким образом частота мидий с
наибольшей питательной ценностью
поддерживалась на уровне достаточно высоком для
того, чтобы включение в рацион менее ценных
жертв привело к снижению средней
скорости потребления энергии. Во втором
эксперименте съеденные мидии не возмещались,
и частота наиболее питательных
моллюсков уменьшалась. Реакция на это крабов
показана на рис. 9.22: в соответствии с
прогнозом 2 (с. 294) они расширили диапазон
своих жертв, включив в него следующие по
ценности размеры жертв.
Крабы распознают размеры мидий,
прикасаясь к ним, и в третьем опыте изменяли
соотношение моллюсков разного размера.
Из рис. 9.23 следует, что в подтверждение
прогноза 3 (с. 294) в случае высокого
обилия менее ценных мидий они все же
употреблялись в пищу, хотя особи оптимального
размера были также многочисленны.
Таким образом, когда Carcinus
сталкивается с особями разного размера одного
типа жертвы, он способен выбрать тот
размер, который даст ему максимальное
количество энергии за единицу времени. В
какой-то мере он, вероятно, оценивает
питательную ценность мидий перед тем, как
питаться ими.
Питание
307
Приложение 9.2. Поисковое
время и выбор жертвы:
синица и мучные черви
Одна из трудностей, связанных с
проверкой вывода о том, что жертва будет
съедена, если искать другую менее выгодно,
заключается в проблеме правильной оценки
поискового времени: когда потребитель
передвигается в пределах своего
местообитания, как установить, ищет ли он
определенный тип жертвы или преследует иные
цели? Эта трудность была изящно обойдена
учеными из Оксфорда. Вместо того чтобы
дать потребителю возможность
перемещаться среди различных жертв, его
удерживали на месте и предлагали ему несколько
типов пищевых объектов с неодинаковой
частотой. Эффект получался такой же.
поскольку частота обнаружения пищи
(эквивалент поискового времени) регулировалась
скоростью ее движения мимо животного.
В этом опыте потребителем была птица
(большая синица, Parus), а жертвами —
кусочки мучных червей (личинок жуков-
хрущаков) двух размеров, причем больший
по размеру кусочек был вдвое богаче
энергией, чем меньший.
Мучные черви продвигались мимо
сидящей в клетке птицы на ленте конвейера
(рис. 9.24), и синица могла либо съесть,
либо проигнорировать любой потенциальный
пищевой объект. Однако, склевывая одного
из червяков, она упускала возможность
схватить тех, которые могли оказаться
доступны в течение следующих 5 - 15 с (время
переработки). Был проведен ряд опытов
с различными эквивалентами поискового
времени для более ценных крупных червей
и различными частотами предъявления
более и менее выгодных пищевых объектов
(рис. 9.25).
Согласно прогнозам модели
максимизации энергии в вариантах В, Г и Д (но не А
и Б), выбор падает на крупных червей.
В опытах это подтвердилось. Даже если
мелких жертв было в два раза больше, чем
крупных (вариант Д), когда поисковое
время для более выгодной пищи снижалось,
потребитель игнорировал менее выгодных
для него мелких жертв и склевывал в
основном крупных, причем переключение на
избирательность происходило при близком
к предсказанному соотношении числовых
параметров.
20*
ГЛАВА I О
БИОМЕХАНИКА И ДВИЖЕНИЕ
(ЛОКОМОЦИЯ)
В этой главе мы познакомимся с тем,
каким образом беспозвоночные могут
механически воздействовать на окружающую их
среду. В основном речь пойдет о типах ло-
комоции, однако те же механические
принципы применимы и к прикрепленным
животным, которые тратят энергию,
перемещая именно среду, а не самих себя.
Раздел, открывающий главу, посвящен
основным принципам механики и
терминологии. Затем обсуждаются биологические
аспекты генерирования механических сил
клетками животных. Общая их черта —
использование получаемой в ходе дыхания
энергии (см. гл. 11) для образования
перекрестных связей между белковыми
фибриллами.
Следующая тема—сравнительное
изучение локомоции животных. Обсуждается
влияние их относительного размера на
локомоторные системы и дается обзор
потенциальных возможностей в этом смысле
различных уровней структурной организации
беспозвоночных. Отдельно описываются
локомоторные системы мягкотелых
животных с так называемым гидростатическим
скелетом и форм с твердыми скелетными
элементами, в частности ракообразных
и насекомых.
10.1. Принципы локомоции
У беспозвоночных можно видеть
широкое структурное разнообразие. Многие из
них мягкотелые, у других развиты твердые
ткани или покровы. Фактически все они
способны совершать механическую работу,
в результате которой их тело перемещается
относительно окружающей его среды или
среда относительно фиксированного тела.
Энергия, затраченная на эту работу, должна
принести выгоду. Последняя может
выражаться в повышении интенсивности
питания или использования другого ресурса,
в снижении опасности со стороны
хищников, в избегании неблагоприятных
абиотических условий, в активизации расселения
и/или повышении вероятности контакта
с потенциальными партнерами для
полового размножения.
Когда животные передвигаются, они
подчиняются тем же основным принципам,
что и все подвижные объекты, т.е.
ньютоновским законам движения. Они гласят
следующее:
1) если тело находится в состоянии
покоя относительно своего окружения, оно
может быть приведено в движение только
за счет приложения внешней силы;
2) приложение неуравновешенной силы
к движущейся массе приводит либо к
ускорению, либо к замедлению движения
данной массы в зависимости от направления
действия силы;
3) каждое действие вызывает
противодействие, равное по силе и
противоположное по направлению;
4) энергия внутри замкнутой системы
остается постоянной, хотя может
переходить из одной формы в другую.
Эти принципы универсальны, и мы
проиллюстрируем их в общем виде до
рассмотрения различных типов движения
беспозвоночных, В приложении 10.1 приведены
основные законы, управляющие локомо-
цией животных, и определены некоторые
единицы измерения.
Когда животное движется, оно должно
преодолевать силы, стремящиеся
препятствовать этому движению; их называют
силами трения или лобового сопротивления
(поскольку противодействие среды, через
которую движется объект, стремится
замедлить его движение, речь также идет о си-
Биомеханика и движение (локомоция)
309
ле). Их принято выражать в единицах,
приводимых в приложении 10.1. В любой
системе имеется определенный запас энергии,
которая может быть превращена в
кинетическую, связанную с движением массы, но
в конечном счете рассеиваемую в виде
тепла. Это показано в приложении 10.2.
Механическая работа, совершаемая при
локомоции, обеспечивается энергией,
высвобождающейся при химических реакциях
в клетках тела животного (см. гл. 11),
однако химическая работа необязательно равна
по величине механической.
Человек может очень устать, стараясь
поднять слишком тяжелый для него
предмет, однако по определению он не
совершает при этом механической работы.
Разумеется, в его теле происходит
метаболическая работа: мышцы используют АТР, т. е.
энергию, запасенную в ходе обмена
веществ, и их усилия приводят к выделению
тепла. Поэтому при оценке эффективности
локомоции мы должны учитывать то, что
затраченная энергия не всегда вызывает
движение тела. Механическая работа
определяется величиной массы и расстоянием,
на которое она переместилась. Это можно
назвать полезной работой. Общие затраты
энергии при движении животного измерить
труднее, но их можно оценить по
интенсивности поглощения кислорода. Тогда
эффективность проделанной животным работы
(как и к.п.д. любой машины) выразится
отношением полезная работа
затраченная энергия
Мы увидим далее, что одни
локомоторные системы могут быть эффективнее, чем
другие, но в то же время иметь
ограниченную мощность, т. е. скорость выполнения
работы. Как и повсюду в биологии, здесь
наблюдается целый ряд компромиссов,
среди которых один из важнейших — между
эффективностью (к.п.д.) и абсолютной
мощностью.
Как показано в приложении 10.2,
существует инерция движения; кроме того, на
движущиеся организмы влияют силы,
действующие в направлении,
противоположном направлению перемещения,
обусловленные сопротивлением среды. Их роль
очень сильно зависит от вязкости (или
трения) последней, относительного размера
организма и его абсолютной скорости. Эта
взаимозависимость выражается числом Рей-
нольдса (Re):
v
где 1—длина движущейся системы; Vmax —
ее максимальная скорость; v — вязкость
среды (для воды v = 1 х 10~6 м2/с).
Число Рейнольдса — это мера
относительного влияния инерции и трения. Для
очень мелких движущихся систем типа
сперматозоидов, жгутиковых, инфузорий,
покрытых ресничками личинок и
крохотных беспозвоночных вроде коловраток
(разд. 4.9) число Рейнольдса очень мало,
где-то между Ю-1 и 10 '. В этих случаях
движение почти целиком обусловлено
силами трения. Некоторые более крупные
беспозвоночные достигают скоростей, при
которых число Рейнольдса значительно
больше единицы; их движение уже нельзя
объяснить лишь действием сил трения. Это
относится прежде всего к насекомым и
головоногим моллюскам.
В приложениях 10.1 и 10.2 генерируемые
силы считаются действующими в
направлении движения, однако они могут
прикладываться и в других направлениях. Такая
ситуация описана в приложении 10.3.
10.2. Генерирование
механических сил
клетками животных
Локомоция у всех животных
обусловлена изменением формы клеток,
противодействующим сопротивлению окружающей
среды, за счет использования энергии,
высвобождающейся в ходе метаболизма.
Локомоция обеспечивается тремя типами
клеток жгутиковыми, ресничными и
мышечными (рис. 10.1). Несмотря на
морфологические различия, механизмы, приводящие
к изменению их формы, сходны; все они
включают зависимое от АТР образование
поперечных мостиков между белковыми
фибриллами.
Жгутик—это трубчатый вырост
клеточной поверхности, окруженный мембраной,
310
Глава 10
Рис. 10.1. Различные типы двигательных клеток. А. Жгутиковые клетки: I — свободноживущее
жгутиковое простейшее; II — хоаноциты губок. Б. Ресничные клетки: I — свободноживущая
инфузория (простейшее); II — ресничные клетки животного. В. Мышечные клетки
Рис. 10.2. Схематическое
изображение реснички:
А—поперечный срез; Б —
продольный срез; В — сгибание
Жгутик
Ресничка
Рис. 10.3. Направление силы,
создаваемой биением жгутика
(,4) и реснички (Б)
Биомеханика и движение (локомоция)
311
являющейся продолжением мембраны,
покрывающей и всю клетку. Он содержит
пучок белковых фибрилл, так называемую ак-
сонему, которая, как показывает
электронная микроскопия, состоит из двух
центральных микротрубочек, окруженных кольцом
из девяти пар микротрубочек (рис. 10.2).
Изменение формы жгутика вызывается
скольжением наружных дублетов дру1 по другу
вследствие образования поперечных
мостиков между соседними микротрубочками.
Для образования каждого такого мостика
требуется энергия фосфатной связи,
поставляемая молекулой АТР (см. разд. 11). Тот
же самый механизм действует в ресничках.
Основное различие между этими
структурами касается их абсолютной длины,
характера размещения и биения. Длина волны,
пробегающей по жгутику, обычно меньше его
общей длины, поэтому он может
генерировать более или менее постоянную силу,
действующую параллельно его продольной
оси (рис. 10,3,^4).
В отличие от жгутика ресничка
относительно короткая, и цикл ее движений часто
включает жесткий сильный удар и гибкое
возвращение назад (рис. 10.3.£).
Постоянная движущая сила создается строго
последовательным и координированным
движением большого числа ресничек,
расположенных на обширных участках поверхности
в виде поясков или полос из многих тысяч
этих органелл.
Генерирование сил мышечными
волокнами обусловлено их способностью
становиться короче, преодолевая сопротивление.
Они состоят из трубчатых сократимых
элементов, миофибрилл, каждая из которых
диаметром примерно I мкм. в свою
очередь образованных белковыми филамента-
ми. расположенными в строго
определенном порядке. Во многих мышцах можно
выделить функциональную единицу, так
называемый саркомер, из совокупности
тонких и толстых филаментов, образованных
двумя разными белками —миозином и
актином (рис. 10.4).
Между миозиновыми и актиновыми фи-
ламентами могут возникать поперечные
мостики; их образование и изменения
формы «головки» миозиновои молекулы
приводят к скольжению тонких филаментов
Рис. 10.4. Строение мышцы при различном
увеличении:/! — целая иннервируемая мышца;
Б—отдельное мышечное волокно с нервным
окончанием (синапсом); В— функциональная
единица мышцы—саркомер; Г—поперечные
срезы саркомера под электронным
микроскопом
А
ШИПИ 1ГТП IIIIMMIII ГТТ
нищ" "-ни—чинпниж-
•W+IHMtHHH 1 t Н-
Dlllll 1П llllllllllllllll
Б
mf|Htf III tHII ИНН НИМ I I I III III t
it шипим'*' шин ч и
i ii
Рис. 10.5. Структура мышцы и генерирование
силы мышечными клетками: А — схема
строения поперечнополосатой мышцы; Б—схема
строения гладкой мышцы; В — генерирование
силы: I—строение мышцы при различной
степени сокращения; II—зависимость
развиваемой силы от относительной длины
312
Глава 10
вдоль толстых, пока этому движению не
воспрепятствует уравновешивающая сила,
действующая в противоположном
направлении. Значительное сокращение
мышечного волокна требует повторяющихся циклов
образования и разрыва поперечных
мостиков, что обеспечивается энергией
фосфатных связей АТР. Свойства мышечной
системы определяются скоростью укорачивания
саркомеров и их числом в одном ряду; от
этого зависит скорость сокращения всей
мышцы. Сила, действующая на единицу
поперечного сечения, зависит от числа
параллельно расположенных филаментов. По
мере увеличения степени их
взаимоперекрывания число свободных мест для образования
поперечных связей снижается и мощность
мышцы падает до нуля (рис. 10.5).
10.3. Движение
с помощью ресничек
Беспозвоночные (за исключением очень
просто устроенных Placozoa; разд. 3.3) не
используют для локомоции жгутиков, хотя
движение с их помощью сохранилось почти
у всех сперматозоидов животных. Однако
у губок жгутиковые клетки —
хоаноциты—прогоняют воду через камеры
и каналы тела.
Обычным локомоторным
приспособлением являются реснички, либо
покрывающие всю поверхность тела, либо
расположенные в виде поясов или полос, но такой
способ движения характерен только для
животных длиной менее 1 мм. Это связано
с тем, что движение отдельной реснички
строго фиксированно, и единственный
способ увеличения суммарной движущей силы
состоит в увеличении числа ресничек. Оно
ограничено площадью поверхности тела,
в то время как масса организма
увеличивается пропорционально его объему. Более
того, гидродинамический к. п. д. ресничных
организмов уменьшается с увеличением их
размеров, следовательно, более крупным
организмам, использующим этот тип
движения, требуется гораздо больше энергии
для достижения тех же относительных
скоростей.
У личинок многих беспозвоночных
реснички расположены в виде полос или поя-
Рис. 10.6. Личинки с полосами ресничек: А —
велигер брюхоногого моллюска; Б—плутеус
иглокожего (офиуры)
сов, длина или число которых возрастают
по мере роста животного (рис. 10.6).
Реснички гребневиков образуют сложные
гребные пластинки, обеспечивающие скорость
до 15 мм/с.
Работа ресничек организована таким
образом, что все они обычно имеют
одинаковые характер и частоту биения, но
движутся в разных фазах. Координированное
действие создает волну, пробегающую от
одной реснички к другой. Эта волна может
идти в направлении движущей силы или в
противоположном ему, так что каждая
ресничка совершает удар чуть позже той, которая
находится непосредственно перед ней. Во
время ударов отдельные реснички не гасят
усилие друг друга. Такой тип координации
их активности называется метахроналъной
ритмичностью (рис. 10.7).
Переход от ресничного к мышечному
движению произошел, по-видимому,
у близких к плоским червям предков
билатеральных форм, причем у некоторых сво-
бодноживущих плоских червей и немертин
встречаются оба типа локомоции.
Эпидермис свободноживущих турбелла-
рий и немертин обильно покрыт
ресничками. Самые мелкие их формы длиной около
Биомеханика и движение (локомоция)
313
Внешний
погранич-
Рис. 10.7. Метахрональное биение ресничек. А
Отдельные реснички в различных фазах цикла
биения. Б. Удар (I) и возвратное движение (II)
создают результирующую движущую силу во
внешнем пограничном слое
1 мм находятся на верхнем пределе
размерного диапазона животных, эффективно
использующих ресничную локомоцию.
Более крупные плоские черви (Triclada и Polyc-
lada), сохранившие реснички в качестве
основного средства передвижения (ползания),
способны к нему только благодаря плоской
форме тела — площадь их поверхности
увеличивается быстрее, чем квадрат линейных
размеров.
Самые крупные животные, ползающие
с помощью ресничек,— это, вероятно, не-
мертины, например вид Lineus longissimus,
длиной до нескольких метров при ширине
не более 1—2 мм, что обеспечивает ему
требуемую площадь поверхности тела. Однако
такой выход из положения открыт только
для водных организмов. На суше быстрая
потеря воды, неизбежная при такой
большой относительной поверхности, была бы
губительна. Мышечные механизмы
передвижения плоских червей и немертин
разнообразны: сюда относятся локомоторные
волны, пробегающие по нижней стороне
тела, перистальтические движения и
«шагания» с подтягиванием заднего конца к
переднему. Эти способы значительно сильнее
развиты в других типах животных и
описаны в следующих разделах.
10.4. Работа мышц
и скелетные системы
Химическая энергия может
использоваться клетками мышц таким образом, что
мышечные волокна укорачиваются, однако
эти изменения формы клеток—лишь часть
локомоторной системы у организмов со
скелетными структурами, передающими
генерируемые силы.
Выделяют два основных типа скелетов
животных: жидкие, свойственные
широкому кругу мягкотелых беспозвоночных,
и жесткие, используемые членистоногими
и некоторыми иглокожими. Различия
между формами с жестким и жидким скелетом
далеко не абсолютны. У многих типов,
кроме членистоногих, наблюдаются жесткие
скелетные элементы; примерами могут
служить иглы морских ежей, «позвонки» офиур
и ацикулы в параподиях полихет.
Аналогичным образом у членистоногих
для передачи сил часто используется
гидростатический скелет, например при движении
кишечника, а у паукообразных—для
распрямления членистых конечностей.
Жесткие скелеты артропод являются наружными
(экзоскелеты), т.е. мягкие ткани окружены
твердыми скелетными элементами. У
хордовых в качестве примитивной формы
внутреннего скелета развился упругий
стержень—хорда. На ее основе
сформировались хрящевые и костные скелеты высших
позвоночных.
Во всех локомоторных системах,
включающих мышцы, активность последних
ограничивается сокращением; для
восстановления их прежней длины требуется
особая сила. Чаще всего она обеспечивается
антагонизмом между различными группами
мышечных волокон, однако ее источником
может быть и работа ресничек (например,
у некоторых стрекающих) или энергия,
накопленная упругими тканями. Жидкие
скелеты мягкотелых беспозвоночных могут
использоваться по-разному, но в основе их
функционирования лежат одни и те же
принципы, рассмотренные в приложении 10.4.
Многие мягкотелые животные способны
значительно изменять свою форму. Этим
особенно отличаются немертины, однако
деформируемость имеет свои физические
ограничения, поскольку объем тканей
и обычно полостей тела остается
постоянным. У немертин нерастягивающиеся
волокна стенки тела расположены под углом
к его продольной оси. Если их ориентиро-
314
Глава 10
1 -
Максимальное
^сокращение
Максимальное/
вытягивание .
Овьем
10 20 30 40 50 60 70 80 90
10 20 30 40 50 60 70 80 90
10 20 30 40 50 60 70 80 90
Угол между волокном и продольной осью
вать параллельно этой оси, объем тела
станет нулевым, то же самое произойдет, если
расположить их к ней под углом 90'. Между
этими двумя нереальными крайностями
существует положение, при котором объем
тела максимален—угол волокон около 55'
относительно продольной оси. Отношение
между объемом, ограничиваемым этой
системой волокон, и их углом с продольной
осью тела показано на рис. 10.8. По
рис. Ю.8.А видно, что объем червя типа не-
мертины обычно меньше максимально
возможного в теории. При максимальном
сокращении или максимальном растяжении
червь будет в поперечном сечении круглым,
в прочих случаях — эллиптическим.
Изменения длины нескольких видов немертин
в рамках этих теоретических границ
представлены на рис. 10.8,£". Механика
движения нематод также определяется
свойствами волокон стенки тела. В отличие от
немертин эти черви относительно слабо
подвержены деформации; организованные
в сложную систему их волокна (см.
рис. 4.13) расположены под углом более 55
к продольной оси, а полостная жидкость
находится под высоким давлением.
При сокращении продольных мышц
угол прохождения волокон становится
больше, и ограниченный ими объем
уменьшается (рис. 10.8,В). Это в свою очередь
повышает внутреннее давление,
противодействующее усилию продольных мышц
и возвращающее их к прежней длине.
Поэтому у нематод в отличие от модели,
представленной в приложении 10.4. нет
кольцевых мышц.
Мягкотелые животные с косыми
мышечными волокнами, например пиявки,
способны устанавливать структурные волокна
Рис. 10.8. Физические свойства спиральной
системы неупругих волокон и изменение формы
червеобразного животного. А. Связь между
внутренним объемом и наклоном неупругих
волокон относительно оси цилиндра, вокруг
которого они закручены. Б. Пределы
деформируемости некоторых немертин, объем тела
которых меньше, чем теоретически допустимый
их системой волокон. В. Связь между углом
наклона волокон и объемом тела у нематод.
Поскольку у этих животных высокое
внутреннее давление, их тело в поперечном сечении
круглое. При сокращении продольных мышц
объем тела уменьшается, и вызываемое этим
увеличение давления противодействует
мышечному сокращению. Эти животные обычно
перемещаются синусоидальными движениями,
при которых контралатеральные (т. е. лежащие
на противоположных сторонах тела) мышцы
сокращаются поочередно (из Clark, 1964)
Биомеханика и движение (локомоция)
315
стенки тела под углом 55 к продольной
оси, становясь фактически твердыми.
Мышцы, действующие на жесткие или членистые
скелеты, передают силы от места их
генерации к точке их приложения к внешней среде.
Жесткий скелетный элемент работает как
рычаг, принципы которого рассмотрены
в приложении 10.5. Следует отметить,
однако, что в примере 7 приложения 10.4 речь
также идет о своего рода рычаге, но
основанном не на относительной длине, а на
относительной площади.
10.5. Локомоция мягкотелых
животных
10.5.1. Передвижение
по твердой поверхности
Многие мягкотелые беспозвоночные
способны передвигаться по твердому
субстрату, т. е. по такому, поверхность
которого, соприкасающаяся с телом животного, не
сдвигается по отношению к частям
субстрата, находящимся на расстоянии от этого
тела, когда животное прикладывает к
субстрату силу. Чтобы обеспечить локомоцию,
эта сила должна передаваться субстрату
через фиксированную точку, часто
называемую точкой опоры.
Во многих локомоторных системах
вдоль продольной оси тела параллельно
направлению движения распространяются
волны мышечного сокращения и
расслабления. Перемещение происходит либо в
направлении распространения этих волн, либо
в противоположном ему. Это показано
в приложении 10.6. Плоские черви,
некоторые книдарии и прежде всего брюхоногие
моллюски движутся благодаря волнам
мышечных сокращений поверхности тела,
прилегающей к субстрату (педальным
локомоторным волнам).
Педальные локомоторные волны легко
наблюдать на нижней поверхности тела
планарии или улитки, когда они ползут по
стеклянной пластинке, наклоненной под
небольшим углом. У наземной улитки Helix
можно видеть одновременно несколько
таких волн, проходящих через всю ногу в
направлении движения животного, но с
большей, чем перемещение самой улитки,
скоростью. У других брюхоногих, например
у морского блюдечка (Patella),
относительно небольшое число волн проходит в
направлении, противоположном движению
животного. Более того, здесь, как правило,
наблюдается разница в половину длины
волны между правой и левой сторонами
ноги. В результате различают педальные
локомоторные волны нескольких типов:
прямые и обратные, монотаксические и ди-
таксические.
Прямые монотаксические волны Helix,
длина которых намного меньше длины
ноги,— это как бы двигатель, работающий на
первой передаче: он обеспечивает
относительно низкую скорость движения и слабую
маневренность, однако способен
перемещать довольно большую массу или
преодолевать значительные силы сопротивления.
Дитаксические системы с волнами такой же
длины, что и длина ноги, или даже
большей, позволяют двигаться относительно
быстрее и более маневренно.
Нога моллюска — весьма сложно
устроенный орган. Основной его скелетный
элемент представлен сложной системой
кровеносных синусов, т.е., хотя сами жи-
316
Глава 10
вотные ацеломические, внутри их ноги
находится гидростатическая скелетная
система. Главные мышечные волокна здесь не
продольные, а передние и задние косые
с противоположным действием (рис. 10.9).
Принято было считать, что при
прохождении локомоторной волны каждый отрезок
ноги поочередно отделяется от поверхности
(как показано в приложении 10.6,^4 и Б), но
это маловероятно в связи с огромным
усилием, необходимым для разделения двух
липких влажных поверхностей.
Образование точек опоры и зон движения зависит от
свойств слизистого слоя под ногой. Слизь
ведет себя как упругое твердое тело в
условиях слабого сдвигового напряжения, а при
увеличении последнего — как вязкая
жидкость. В точках опоры она находится как раз
в первом состоянии, а под фронтом волны,
где сдвиговые силы больше,— во втором.
Некоторые улитки, включая Helix,
могут передвигаться и другим способом —
«галопом», при котором генерируются
обратные волны мышечных сокращений
с длиной волны, примерно равной длине
ноги. Дальнейшая модификация этого типа
локомоции состоит в поочередном
прикреплении к субстрату переднего и заднего
концов тела; примером может служить
передвижение с петлеобразным выгибанием
тела, характерное для пиявок (см. ниже).
У многих крупных плоских червей и
большинства немертин чередующиеся волны
сокращений кольцевых и продольных
мышц создают обратные
перистальтические волны, расширяющие локомоторные
возможности, создаваемые работой
поверхностных ресничек. Такая система лучше все-
, го развита у червей с септированным цело-
мом, особенно у земляных.
На рис. 10.10,Л показан механизм
движения земляного червя. Обратим внимание,
что, когда кольцевые мышцы расслаблены,
а продольные сокращены, сегменты тела не
перемещаются (см. приложение 10.6).
Сегменты, находящиеся перед самым
коротким сегментом, растягиваются,
отталкиваясь от субстрата этой точкой опоры,
тогда как задние сокращаются, обеспечивая
с ее помощью подтягивание (рис. 10.10,£).
Проскальзывание исключается за счет рас-
ставления прочных щетинок.
(11111 II Mill lllll IlllllllfX)
«r-r-T-flllllllllllllllHIIIID
"им ашммиппиипр
(гт I I I I zdiiiiiiiiiiTTTD
(ШШ^Д=гдД| 111111 ДПР
dlllllllltx I i I 1 СГПТШ&
c=i=r=tiiii I I МП i i i i 1-СПТП)
ст-г-т-тХШШШ!).! i i i д=Ш>
СШД i i i ПТПТПТТП iin
Б д
V
Рис. 10.10. А. Последовательные стадии
движения земляного червя. У каждого сегмента
постоянный объем, как на рис. А. II.
приложения 10.6. Когда продольные мышцы
сокращены, а кольцевые расслаблены, сегменты
неподвижны относительно земли; объем сегмента
фиксирован, и локомоторная волна
распространяется назад. Б. Силы, действующие на
неподвижный сегмент, заякоренный щетинками
Волны изменений давления,
возникающие при сокращении кольцевых и
продольных мышц, независимы друг от друга, так
как септы эффективно изолируют сегменты
(рис. 10.11). Наивысшее давление
отмечается в момент сокращения кольцевых мышц
и проталкивания сегмента сквозь субстрат.
В фазе заякоривания давление ниже. Такая
изоляция давлений невозможна у животных
с неразделенной полостью тела. Полагают,
что это одно из основных преимуществ
сегментации, способствовавших эволюции
метамерии у аннелид. Каждый сегмент
представляет собой отдельный
гидростатический элемент с постоянным объемом (см.
приложение 10.5). Хотя здесь рассмотрено
движение земляных червей по плоской
поверхности, те же принципы лежат в основе
их перемещения в толще почвы, к которому
они главным образом и адаптированы.
Совершенно по-другому обеспечивается
Биомеханика и движение (локомоция)
317
локомоция пиявок. Их передние и задние
присоски являются весьма эффективными
точками опоры. Все тело работает как
единственная гидростатическая система, причем
кишка — единственная заполненная
жидкостью полость. Прикрепление к субстрату
осуществляется поочередно передней и
задней присосками (рис. 10. \2,А).
Сходный способ передвижения
встречается у личинок некоторых насекомых.
Например, у гусениц бабочек петлеообразные
выгибания тела эквивалентны сокращению
продольных мышц (рис. \0Л2,Б). Полость
тела (целом) у пиявок фактически вытеснена
деформируемой ботриоидальной тканью,
так что с точки зрения архитектоники эти
Рис. 10.12. Шагающие движения, при которых
точками опоры служат попеременно передний
и задний концы животного: А— пиявка; Б—
гусеница (личинка бабочки)
-О
=о
BIIIHI МШИ IIIIIIIIIIIH ТПТШ
А В
~>
« »
Рис. 10.11. Изучение локомоции земляного
червя. А Система, позволяющая регистрировать
давление внутри сегментов А и В с помощью
вставленных в них пластиковых трубок,
непосредственно связанных с датчиком давления. Б.
Последовательные положения сегментов А и
В при движении червя. В. Волнообразное
изменение давления (по Seymour, 1969)
животные несегментированные и ацеломи-
ческие, хотя, безусловно, относятся к анне-
лидам, и их происхождение от
сегментированных целомических предков выдают
некоторые анатомические детали прежде
всего в строении нервной системы.
Эррантные полихеты используют способ
локомоции, отличающий их как от олиго-
хет, так и от пиявок. Она обеспечивается
здесь многочисленными конечностями,
концы которых движутся назад относительно
тела, перемещая при этом последнее вперед
по субстрату (см. приложение 10.5).
Когда полихета, например Nereis,
ползет медленно (рис. 10.13,Л), по ее парапо-
диям от хвоста к голове проходит метахро-
нальная волна активности; при этом фазы
Рис. 10.13. Схема локомоции полихеты Nereis:
А— медленное ползание; Б — быстрое
ползание. Для простоты движение щетинок не
показано. Они могут выдвигаться и втягиваться,
увеличивая длину «шага»
318
Глава 10
движения параподий правой и левой сторон
тела различаются ровно на половину длины
волны. Поскольку волна направлена
вперед, каждая конечность совершает сильное
отталкивание без риска интерференции
с толчком ближайшей сзади параподий, так
как последняя оттолкнулась чуть раньше.
При более быстром ползании или при
плавании Nereis к этой дитаксической прямой
локомоторной волне добавляется
двигательная сила, генерируемая сокращением
продольной мускулатуры стенки тела
(рис. 10.12,Ь). Активность продольных
мышц правой и левой сторон
распространяется так, что каждая параподия
совершает мошное отталкивание, находясь на
гребне синусоидальной волны,
пробегающей вперед по стенке тела синхронно с
волной движения параподий.
В этом случае силы, прикладываемые
к субстрату, генерируются не только
мускулатурой параподий, но и контралатераль-
ными продольными мышцами. При таком
типе движения в соответствии с общим
правилом прямые локомоторные волны
должны наблюдаться тогда, когда точки опоры
возникают в областях максимальной длины
продольных мышц.
У многих эррантных полихет
продольные мышцы играют в процессе ползания
незначительную роль, и движущая сила в
основном создается мускулатурой параподий,
пересекающей границы сегментов. Это
особенно характерно для червей, покрытых
чешуйками, и близких к ним форм, у которых
межсегментные септы сильно
редуцированы или утрачены. В локомоции здесь
участвуют как собственные мышцы параподий,
так и мышцы, входящие в них извне.
Щетинки выталкиваются наружу и
втягиваются обратно собственными мышцами,
а движение параподий вверх-вниз и вперед-
назад обеспечивается совместно
собственными и входящими мышцами, что
приводит к своего рода шаганию, и движение
некоторых полихет напоминает сложную
походку многоножек.
Уникальное локомоторное устройство
представляет собой амбулакральная
система иглокожих. Для морской звезды,
например, характерно наличие пяти лучей, в
каждом из которых проходит амбулакральный
канал, начинающийся от околоротового
кольца. Вдоль каждого радиального канала
находятся ряд резервуаров-ампул и
амбулакральные ножки (рис. 10.14,£). Мышцы
ампул и амбулакральных ножек действуют
антагонистически по отношению друг к другу:
сокращение мышц, сжимающих ампулу,
выталкивает воду в амбулакральную нож-
Ампула
''WW/'
•ЩЩ"'
Радиальный канал
■ Амвулакральная
ножка
Рис. 10.14. Амбулакральные
ножки и ампулы
амбулакральной системы иглокожих. А.
Общая организация. Б. Схема
поперечного среза луча морской
звезды, показывающая
амбулакральную систему. В. Цикл
шагающего движения
отдельной амбулакральной ножки (ее
мышцы-ретракторы не
показаны)
Биомеханика и движение (локомоция)
319
аш=х
с
СИ
жг
з=шш;
Ж)
ку, а сокращение ножки вызывает отток
воды обратно в ампулы. При этом отдельная
ампула не служит резервуаром только
одной «своей» ножки, поскольку наблюдается
перемещение жидкости между частями всей
системы, и при полном растяжении ножки
она может достигать большего объема, чем
соответствующая ей ампула в полностью
растянутом состоянии. Однако, как
правило, мышцы, сокращающие ампулу,
действуют как антагонисты тех, что
укорачивают ножку.
Амбулакральные ножки, вытягиваясь
под действием гидравлического давления,
могут совершать простые шагающие
движения; на их концах часто находятся липкие
присоски (рис. 10.14,5).
Особенность движения морских звезд —
отсутствие какого-либо заметного метах-
ронального ритма в движении
амбулакральных ножек.
10.5.2. Движение в толще
субстрата и в трубках
Роющим червям свойственно наличие
крупных полостей тела и по крайней мере
частичное отсутствие септированности.
Животные с таким строением способны
использовать различные способы локомоции
внутри нор (рис. 10.15). Перистальтическое
движение бывает четырех типов, поскольку
любая часть тела может одновременно
увеличиваться и в длину, и в обхвате. Эти
движения используются для прокачивания
воды через норку, для перемещения в ней или
для того и другого вместе.
Несегментированные и
сегментированные, но лишенные септ животные с
крупными полостями тела могут существенным
образом менять форму последнего, что
используется для внедрения в субстрат.
Примером служат сипункулиды, способные
делать это чрезвычайно быстро;
соответствующие движения показаны на рис. 10.16
вместе с графиком ^изменений внутреннего
давления. Эти животнЫе-^югут развивать ему ряд преимуществ. Сегментарное распо-
высокую мощность, но обьгчТ1о~в_течешге~~'ложение ганглиев улучшает нервную коор-
коротких периодов времени. При поддер- динацию; параподии со щетинками со-
жании высокого давления в целоме все здают точки опоры; имеется хорошо разви-
мышцы тела должны расходовать метабо- тая сосудистая система с сегментарными
лическую энергию для сохранения своей по- жабрами. С механической точки зрения ту-
кххшдш оттспш^
Рис. 10.15. Четыре типа перистальтических
движений, используемые обитающими в
трубках червями. В случаях А, Б и Г объем
сегментов не фиксирован. В случае В (Sabella)
сегменты разделены септами и имеют
постоянный объем. Перемещение тела по трубке при
перестальтических движениях
предотвращается за счет образования точек опоры
сегментами промежуточной длины
стоянной длины. Механическая работа
совершается за счет контролируемого
расслабления определенных мышц, что позволяет
растягиваться соответствующим участкам
тела (см. приложение 10.4). Мощность при
этом может быть очень высокой, но к.п.д.
будет низок, так как требуется
непрерывный мышечный тонус для поддержания
высокого давления в целоме. Полагают, что
эволюционное возникновение поперечных
септ у аннелид связано, по крайней мере
частично, с необходимостью предотвращать
растяжение стенок тела без использования
мышечной работы, что увеличивает
механический к.п.д. системы (ср. разд. 10.1).
Полихета Arenicola marina (пескожил) —
это сегментированный червь, по-видимому,
вторично приспособленный к роющему
образу жизни. Метамерное строение дает
320
Глава 10
(
)
(
>
)„
<=t
)«
-—^—
>
<И
>
С
^)vn
Рис. 10.16. А. Схема последовательных
движений сипункулиды при зарывании и
перемещении сквозь субстрат. Б. Изменение внутреннего
давления в различных фазах локомоторного
цикла. Обратите внимание, что максимальное
давление отмечено в момент проникновения
в субстрат (как и у земляных червей — рис
10.11), но при этом оно во всех частях тела
одинаково (по Trueman, Foster Smith, 1976)
ловищный целом пескожила неразделен,
что позволяет ему совершать самые
разнообразные движения и развивать высокую
мощность. Это дает ему возможность
внедряться в субстрат путем попеременного
терминального и пенетрационного заякорива-
ний (объяснение см. на рис. 10.17). Такой
же тип движения встречается у роющих кни-
дарий и на поздних стадиях закапывания
у двустворчатых моллюсков (рис. 10.18).
Пескожилы обычно обитают в
открытых J-образных трубках, являющихся
частью U-образной норовой системы
(рис. 9.32). Животное извлекает и поедает
песок в «забое» с помощью
соскабливающих и заглатывающих движений хоботка.
Скорость этой работы невысока, и во время
ее выполнения передний целом изолируется
от остальной полости тела глоточной се-
птой, клапаны которой позволяют затем
объединять его с туловищным целомом.
Высокое давление в последнем приводит
к вытягиванию хоботка при более
интенсивном рытье.
Обитающие в трубках черви,
сохранившие межсегментные септы (например, Sa-
bella), сталкиваются со специфической
проблемой. Им необходимо прокачивать воду
через трубку с помощью перистальтических
движений. Имея постоянный объем
сегментов, они могут использовать только
перистальтическую волну типа изображенной
на рис. 10.15,В; следовательно, их будет
сносить в направлении, противоположном
движению волны. Чтобы этого не
произошло, Sabella образует точки опоры сегмен-
Рис. 10.17. Стадии движения пескожила в субстрате. А. Терминальное заякоривание: передняя
часть тела расширена, а задняя подтягивается вперед. Б. Пенетрационное заякоривание: сегменты
передней части тела образуют заякоривающие выступы, хоботок раскапывает песок, и «голова»
продвигается вперед (по Trueman, 1975)
Биомеханика и движение (локомоция)
321
I II III
Рис. 10.18. Закапывание двустворчатого
моллюска. I. Створки раковины раздвигаются,
создавая опору для проталкивания ноги. II, III.
Конец ноги расширяется, и раковина
втягивается в песок (по Trueman, 1975)
тами промежуточной длины, раздвигая на
них шетинки. Остальные сегменты скользят
по очень гладкой выстилке трубки.
10.5.3. Плавание
Самые мелкие мягкотелые
беспозвоночные плавают за счет движущей силы
ресничек, как описано в разд. 10.3. Более
крупные животные используют работу мышц;
исключение составляют гребневики,
несущие ряды ресничных гребных пластинок.
Для плавания необходимо, чтобы сила,
генерируемая мышцами, воздействовала на
среду (обычно это вода), сквозь которую
животное движется. Для относительно
крупных животных число Рейнольдса (см.
разд. 10.1) больше единицы, так что
следует учитывать инерцию тела. Когда
плывет крупное животное с Re > 1, масса
окружающей воды обычно приводится в
вихревое движение. Такой тип плавания особенно
характерен для позвоночных, но многие
мягкотелые беспозвоночные, не
обладающие в воде нейтральной плавучестью, также
способны генерировать подъемную и
движущую силу за счет мышечной активности.
Для этого используются три основных
механизма: а) распространение назад волн по
телу животных с гладкой поверхностью; 6)
гребущие движения; в) реактивное
движение.
Относительно мелкие беспозвоночные
с Re < 1 не могут использовать вихреобра-
зования, тем не менее также способны
плавать. Их движение основано на различиях
в сопротивлении воды при разных
скоростях.
Основные работы по механике
волнообразного движения выполнены на угрях,
но аналогичные локомоторные волны
генерируются и гладкотелыми червеобразными
беспозвоночными, например пиявкой Hiru-
do. При плавании ее тело сплющено дорсо-
вентрально и по нему пробегают
направленные назад волны. Волна движется назад
со скоростью U (относительно земли),
которая выше скорости V перемещения тела
вперед. Принципы такого типа плавания
изложены в приложении 10.7.
Механизм плавания полихет, у которых
тело шероховатое, иной. Волна движется
в том же направлении, что и тело. Частично
это связано со сложной динамикой
жидкости, омывающей колеблющееся тело с
выступами, а частично с действием параподий.
совершающих мах назад в тот момент,
когда их достигает гребень проходящей по
телу волны (см. рис. 10.13,Ь).
Многие крупные позвоночные движутся
толчками, в перерыве между которыми
скользят по инерции; при этом энергия толчка
сохраняется в виде кинетической энергии
тела. Головоногие моллюски используют тот
же принцип в своем реактивном движении.
Вода с силой выталкивается из мантийной
полости при сокращении выстилающих ее
мышц. Поскольку вода имеет массу и
приобретает ускорение, направленное назад,
должна возникнуть сила противодействия,
разгоняющая животное вперед. Этот
принцип более полно объясняется в
приложении 10.8.
10.6. Локомоция
беспозвоночных
с членистыми конечностями
Ракообразные, хелицеровые и одновет-
вистые—три разнообразные и
процветающие группы беспозвоночных. Все они
характеризуются наличием твердого экзоскелета,
позволяющего иметь членистые
конечности. Успех этих трех групп может быть
частично связан с локомоторными
преимуществами, которые дает такое строение
последних. Кроме того, насекомые -это
единственные беспозвоночные, имеющие
21—1314
322
Глава 10
Рис. 10.19. А. Конечность типичного членистоногого. Обратите внимание на мощные мышцы в ее
базальной части, сухожилия и суставы. Б Характерное положение конечности членистоногого.
При движении вперед происходит вращение проксимального сустава в горизонтальной
плоскости и разгибание более дистальных суставов
крылья и способные к полету, чем отчасти
обусловлено их доминирование в наземных
местообитаниях. Несмотря на свое полифи-
летическое происхождение, ходильные
конечности наиболее высокоорганизованных
ракообразных, хелицеровых и одноветви-
стых устроены весьма сходно в результате
конвергентной эволюции. Они состоят из
ряда подвижно сочлененных элементов,
масса которых снижается по направлению
к дистальному концу (рис. 10.19,А).
Каждый сустав допускает движение только в
одной плоскости; это обусловливает сгибание
и разгибание конечности, а также вращение
ее плоскости в суставе, где происходит при-
членение к туловищу; последнее обычно
и определяет перемещение тела вперед.
Чаще всего оно как бы подвешено между
расставленными в стороны ногами
(рис. 10.19,£), так что при ходьбе центр
тяжести движется горизонтально.
За счет вращения конечности у
основания и сгибания и разгибания в остальных
суставах животное может перемещать
кончик ноги по прямой линии, параллельной
направлению движения. Конечность
работает как механический рычаг (см.
приложение 10.5), и у длинноногих артропод его
устройство таково, что большая сила
перемещает относительно малый груз на
большое расстояние, обеспечивая тем самым
быструю ходьбу.
Наиболее массивные двигательные
мышцы расположены не в конечностях,
а внутри туловища, и, поскольку животное
не подскакивает вверх-вниз при
перемещении вперед, основные изменения момента
затрагивают только конечность. Они
сводятся к минимуму за счет общих для артропод
особенностей строения: масса конечности
убывает от основания к дистальному концу.
Боковые волнообразные колебания тела,
как у полихет (см. рис. 10.13) и некоторых
многоножек (см. ниже), конечно, приводят
к изменениям момента, и их подавление,
по-видимому, оказало существенное
влияние на эволюцию членистоногих.
Хотя ходильные конечности насекомых
и продвинутых ракообразных, например
крабов, сходны по строению, их
эволюционная история существенно различается.
Конечности ракообразных происходят от
предковой структуры, выполнявшей
помимо локомоторной различные функции,
включая захват пищи и дыхание.
Биомеханика и движение (локомоция)
323
10.6.1. Локомоция ракообразных
Конечности ракообразных выводятся из
общего плана строения, включающего
несколько составных частей: два непарных ба-
зальных сегмента, листовидный эпиподит
и пару дистальных частей—эндоподит
и экзоподит (см. рис. 8.34,i>— Г). У
современных листоногих раков Cephalocarida
и Branchiopoda (см. рис. 8.37—8.39)
подобные конечности имеются на каждом нецефа-
лизованном торакальном сегменте и
участвуют не только в передвижении, но и
в питании.
Конечность ракообразных может
действовать как весло, поверхность которого
максимальна во время гребка, а при
возвратном движении существенно
уменьшается за счет складывания и изменения
положения щетинок (рис. 10.20), что снижает
сопротивление воды. Однако примитивные
ракообразные используют для плавания
также изменения объема пространства,
окружаемого конечностями. Вода
затягивается внутрь него по средней линии
(рис. 10.21) и выталкивается назад.
Однонаправленное ее движение создается за счет
того, что экзоподит действует как клапан.
Пищевые частицы задерживаются
щетинками эндоподита и передаются вперед со
щетинки на щетинку вдоль срединной
вентральной борозды по направлению ко рту.
Таким образом, метахрональный ритм
движения конечностей обеспечивает локомоцию,
питание, а возможно, и дыхание.
Отличительная черта локомоции
ракообразных—относительная легкость пере-
Рис. 10.20. Роль щетинок при создании
неодинакового сопротивления во время плавания
ракообразных, для которых значения числа Re
относительно низки: А— положение при
возвратном движении; Б—положение во время
локомоторного гребка (по Hesseid, Fowtner,
1981)
21*
Рис. 10.21. Конечности листоногих
ракообразных создают потоки воды, служащие для
плавания и питания. А. Вид снаружи правой
стороны тела. Б. Вид изнутри левой стороны тела.
Показаны щетинки, фильтрующие пищевые
частицы, которые затем передаются вперед ко
рту. Вода затягивается в срединную
вентральную полость, а затем проталкивается назад,
проходя через фильтры эндоподитов, т. е.
внутренних ветвей конечностей
хода от плавания к хождению. У многих
креветкоподобных Malacostraca
торакальные конечности — ходильные, а
абдоминальные (плеоподы)—плавательные
(рис. 10.22). Во многих эволюционных
линиях либо утрачивается способность к
хождению (см. рис. 8.49,А), либо снижается
роль плавательных плеоподов, как у
омаров и крабов (см. рис. 8.49, Г—£). У омаров
хорошо развита реакция избегания с
участием уроподов — последней пары
абдоминальных придатков. Мощные брюшные
мышцы сокращаются, вызывая резкое
сгибание туловища, и в результате загребания
воды уроподами животное отбрасывается
назад (см. разд. 16.10.4).
У крабов (Brachyura) брюшко
редуцировано, и большинство из них передвигаются
УУ~~^^[МЪ Тш) * (для реакции
СУ ггек п*^ извегания)
й I УЦ- Плаватель-
\ \^_ v- ные ноги
Ходильные (плеоподы)
ноги
Рис. 10.22. Креветкоподобный представитель
Malacostraca с ходильными и плавательными
конечностями
324
Глава 10
Рис. 10.23. Плавающий краб Callinectes. Его
задние конечности при плавании действуют как
гребные лопасти
на пяти парах грудных конечностей.
Несмотря на это, и здесь возможен переход к
плаванию за счет уплощения и модификации
последних грудных ног, как у Callinectes из
семейства Portunidae (рис. 10.23). Краб
плавает за счет вращения этих конечностей,
которые действуют как гребные лопасти.
Переходы между плаванием и
хождением в эволюции ракообразных—следствие
обитания в водной среде, в основном
ограничивающей распространение этих
животных. В морской воде безпозвоночные могут
иметь почти нейтральную плавучесть, а
высокая (по сравнению с воздухом) вязкость
среды не позволяет развивать большую
скорость. Строение ракообразных
позволяет некоторым представителям этого типа
передвигаться на суше, но другие
физические особенности исключают широкую
радиацию в наземных местообитаниях. К ней
оказались способны другие
членистоногие— хелицеровые и одноветвистые.
10.6.2. Наземная локомоция:
хождение
У животных, обитающих на суше,
плотность значительно выше, чем у
окружающего их воздуха, поэтому им необходимы
опорные структуры, и формы, которым
приходится часто или быстро
передвигаться, используют для опоры о землю
твердые скелетные элементы. Последние
должны противостоять существенным
деформирующим силам: сгибающим, т.е.
действующим перпендикулярно оси
конечности, и скручивающим ее вокруг собственной
оси.
Структурой, устойчивой в таких
условиях и имеющей минимальную массу,
является полый цилиндр. Быстрое
передвижение требует также гибких сочленений
(суставов) между жесткими элементами,
сухожилий для передачи сил, генерируемых
мышцами, и устройств для запасания
энергии. Кутикула членистоногих идеально
удовлетворяет всем этим требованиям.
Она состоит в основном из углеводно-
белкового комплекса —хитина. Это
прочное и гибкое вещество, но само по себе не
пригодное для формирования жестких
скелетных элементов. Жесткость может быть
повышена за счет образования связей
между составляющими его белками. В
результате такого «дубления» возникает белок,
называемый склеротином. Он может
сообщать кутикуле различную жесткость,
иногда сравнимую с жесткостью кости, в то
время как гибкий хитин образует суставы
конечностей. Как правило, движение в
каждом суставе ограничено только одной
плоскостью, что обеспечивается развитием
внутренних выступов.
Обычно сгибание требует сокращения
мышц, а разгибание часто происходит за
счет упругости мембраны сустава или с
участием гидравлического механизма, когда
сила, распрямляющая конечность,
передается через заполняющую ее гемолимфу.
Шагание наземных членистоногих
включает вращение направленной вбок оси ноги.
У двупарноногих очень много коротких
конечностей (рис. 8.23), движение которых
следует направленному вперед метахро-
нальному ритму, так что каждая из них
совершает толчок чуть позже, чем
находящаяся непосредственно за ней. Это как бы
движение «на малых оборотах»,
позволяющее развивать относительно большое
усилие при низкой максимальной скорости.
Двупарноногие — фитофаги, обитающие,
как правило, в гниющей древесине и
аналогичных субстратах, так что такой способ
передвижения вполне подходит для их
образа жизни.
Многие губоногие многоножки,
напротив, активные хищники, способные
передвигаться относительно быстро. На рис. 10.24
показано их движение с различными
скоростями. Характер локомоции здесь сходен
Биомеханика и движение (локомоция)
325
Рис. 10.24. Движение губоногих многоножек
с различной скоростью. Черными точками
обозначены концы конечностей, контактирующие
с землей (по Manton, 1965)
с описанием для Nereis (рис. 10.12). С
ростом скорости число конечностей,
касающихся земли, уменьшается, а изгибания
тела усиливаются.
Боковые волнообразные изгибы тела
служат для увеличения длины шага у
членистоногих с короткими конечностями, но это
связано с существенными изменениями
момента. Их можно избежать путем
удлинения каждой конечности при снижении их
числа, и в различных эволюционных линиях
артропод прослеживается именно такая
тенденция. У десятиногих раков их пять пар
(но для хождения часто используются
только три или четыре пары), у
паукообразных - четыре, а у насекомых — три.
Хождение требует поднятия ноги, ее движения
вперед, опускания и движения назад
относительно тела. Во время последней фазы
конец ноги упирается в землю, а центр
тяжести тела перемещается относительно него
вперед. Движения в различных суставах
позволяют при этом удерживать тело на
постоянной высоте над землей.
Рис. 10.25. Вид сбоку и траектории движения
конечностей скорпиона. Заметно, что
траектории отдельных конечностей не пересекаются
между собой (по Hesseid, Fowtner, 1981)
А Б В Г
Рис. 10.26. Траектории движения конечностей
различных членистоногих: И — краб; Б — омар;
В — паук; Г—насекомое (по Manton, 1952)
На рис 10.25 показан вид сбоку и
траектории движения конечностей скорпиона при
его движении вперед. Ноги не мешают друг
другу благодаря расположению их концов
на неодинаковых расстояниях от тела.
Траектории конечностей различны, что
требует сложной координации движений в
каждом суставе, причем они обычно не
пересекаются друг с другом, как показано на рис.
10.26. Большинство артропод движутся
вперед за счет вращения базального членика
конечности относительно тела (рис. 10.26,,Z>-
Г), однако крабы перемещаются вбок,
сгибая и разгибая ходильные ноги в дисталь-
ных суставах.
Если при малом числе конечностей часть
из них переставляется одновременно,
возникает проблема устойчивости. У
насекомых с тремя парами ног они должны
переставляться в такой
последовательности, чтобы в любой момент времени по
крайней мере три ноги опирались о землю,
а центр тяжести находился в пределах
треугольника, образуемого точками опоры.
Наиболее распространенные способы
хождения насекомых удовлетворяют этим
условиям. Две группы конечностей
переставляются поочередно, и каждая,
становясь на землю, образует устойчивую опору.
Метахрональная последовательность их
движений направлена вперед, и ноги с двух
сторон одного сегмента действуют в проти-
вофазе. То же самое наблюдается и у
животных с большим числом конечностей,
например у полихет-нереид (рис. 10.13) и
губоногих многоножек (рис. 10.24).
326
Глава 10
Такое «чередование треугольников»
может обеспечивать различные скорости локо-
моции: чем больше отношение периодов
движения конечности вперед и назад, тем
быстрее перемещается животное.
У некоторых насекомых подобного
«чередования треугольников» не наблюдается,
особенно если одна пара конечностей
специализирована для какой-либо особой
функции, как, например, у богомолов, у
которых первая их пара редко используется
для опоры.
10.6.3. Полет
Физические свойства усиленной склеро-
тином кутикулы позволили крылатым
насекомым выработать способность к полету.
Кутикула обладает почти идеальными
свойствами для построения
аэродинамической структуры. Она может образовывать
тонкую, легкую, гибкую мембрану,
усиленную склеротизованными жилками и
ребрами. Такая конструкция фактически является
гофрированной пластиной, но по своим
свойствам аналогично значительно более
тяжелой сплошной конструкции
аналогичных габаритов. Кроме того, упругие белки
способны запасать поставляемую
мышцами энергию, чтобы использовать ее для
генерации быстрых движений крыла.
Первые летающие насекомые появились
около 200 млн. лет назад, и с тех пор
механизм полета модифицировался и
совершенствовался. Поэтому у современных
насекомых структурные и аэродинамические
приспособления к полету значительно
разнообразнее, чем. скажем, у птиц и летучих
мышей. На рис. 10.27 показаны лишь
некоторые формы крыльев насекомых; они
изображены одинакового размера, но на самом
деле и по этому признаку существенно
различаются (см. также рис. 8.29).
Структурные приспособления,
сделавшие возможной эволюцию полета у
насекомых, изначально, по-видимому, служили
для терморегуляции. Многие современные
насекомые до сих пор используют крылья
для этой цели: бабочки повышают
температуру тела, поглощая ими солнечную
энергию, а перепончатокрылые (например,
пчелы и осы) «разогреваются» перед поле-
Рис. 10.27. Различные формы крыльев
насекомых. Изображения примерно одного размера,
но в действительности размеры этих видов
сильно различаются (см. также рис. 8.29)
том, быстро ими взмахивая. Возможно,
изменение относительных размеров привело
к тому, что крылья, первоначально
приспособленные для терморегуляции, приобрели
аэродинамические свойства.
В частности, для точной
терморегуляции необходимо движение крыловидных ,
придатков, так что их развитие должно
было бы сопровождаться появлением
соответствующей мускулатуры. Примитивные
летающие насекомые, по-видимому, были
способны лишь к планирующему полету,
Биомеханика и движение (локомоция)
327
II II
Нервные импульсы Нервные импульсы
VJvhrhnvhrvrh rrrrrrmrnvrrrmrr
Движение торакса Движение торакса
Ш III
Рис. 10.28. Структурные и нервно-мышечные
приспособления к полету у насекомых. А.
Прямые летательные мышцы, характерные,
например, для стрекоз: I — стрекоза; II —
поперечные срезы ее торакса с прямыми
летательными мышцами, прикрепленными
непосредственно к основаниям крыльев; III — запись
синхронных нервных импульсов и движений
торакса у таких насекомых. Б. Непрямая
летательная мускулатура, например, двукрылых:
I — муха; II — поперечные срезы торакса с
основными антагонистическими летательными
мышцами; III — запись асинхронных нервных
импульсов и движений торакса у насекомых
такого типа (по Pringle, 1975)
при котором требуются относительно
медленные движения крыльев для
регулирования стационарных аэродинамических сил.
Однако для понимания аэродинамики
многих ныне живущих насекомых необходимо
учитывать нестационарные и неустойчивые
условия.
Современные виды используют два типа
структурных приспособлений для
приведения крыльев в движение. Стрекозы
считаются одними из самых примитивных
ныне живущих крылатых насекомых. У них
крыло приводится в движение так
называемыми прямыми летательными мышцами,
прикрепляющимися к его основанию, как
показано на рис. 10.28.Л, причем
наблюдается однозначное соответствие между
каждым нервным импульсом, стимулирующим
эти мышцы, и соответствующим
движением крыльев и торакса (рис. 10.28,А. III).
Такую иннервацию летательных мышц
называют синхронной (см. разд. 16.10.5).
Однако у большинства насекомых
движение крыльев вызывается «непрямыми»
летательными мышцами, изменяющими
форму торакса, который посредством
особого сочленения с основаниями крыльев
вызывает их поднимание и опускание (рис.
10.28,Б). Нервные импульсы,
стимулирующие такие мышцы, не синхронизированы
с наблюдаемой частотой движений торакса
и соответственно крыльев (см. разд.
16.10.5). Это называется асинхронной
иннервацией летательных мышц. Нервные
импульсы, показанные на рис. 10.28,51, III,
поддерживают эти мышцы в состоянии
возбуждения, характеризующемся высоким
внутриклеточным содержанием ионов Са 2+. В
таком состоянии сокращение вызывается
стимуляцией расположенных в мышцах
рецепторов растяжения. При этом частота
биения крыльев зависит от физических
свойств торакальных сегментов.
Сокращение одной группы летательных мышц
приводит к растяжению мышц-антагонистов,
что в свою очередь вызывает их
сокращение, индуцируя каскад поочередных
сокращений мышц обеих групп, который не
прекратится, пока они находятся в
возбужденном состоянии.
Ранее считалось, что под действием
непрямых летательных мышц торакс
двукрылых насекомых как бы «перескакивает» из
одного стабильного состояния в другое—с
поднятыми или опущенными крыльями.
Однако оказалось, что кинематика
движения крыла не согласуется с этой моделью,
предсказывающей замедление движения
крыла по мере приближения к среднему
положению и роста напряжения в тораксе
с быстрым ускорением при переходе к
стабильным точкам (рис. 10.29,у4).
Схема, иллюстрирующая
альтернативную модель, изображена на рис. 10.29,5.
Эта модель предполагает, что движение
328
Глава 10
20 Номер кадра
5 Время, мс
Рис. 10.29. А. I — Изменение положения кончика крыла мухи, зафиксированное с помощью
сверхбыстрой киносъемки (4000 кадров в секунду); II — кривая, предсказанная на основе
простого механизма «перескакивания». Б. Полусхематическая модель причленения крыла у двукрылого
насекомого. Опускание крыла из положения А в положение Б поднимает дорсальную пластинку
торакса (скутум) и тем самым активирует дорсовентральные мышцы (ДВМ II), которые
сокращаются и поднимают крыло. Аксиллярные мышцы (Акс 3) могут увеличивать амплитуду
движения крыла (до положения В), что ведет к возрастанию энергии, запасенной в упругих белках
торакса при деформации. Это способствует быстрому подъему крыла, как показано на рис. 10.29,
А (по Miyan, Ewing, 1986)
мышц, опускающих крыло, вызывает
поднятие сочлененной с его основанием
дорсальной стенки торакса. Это приводит к
растяжению дорсовентральных мышц
(стимулирующему их сокращение) и накоплению
энергии в упругих компонентах системы,
высвобождающих ее при движении крыла
вверх.
Чтобы насекомое могло лететь, на него
должна действовать сила противодействия,
одна из составляющих которой равна весу
насекомого, а другая не меньше, чем сила
сопротивления воздуха при движении
сквозь него. Обе эти составляющие—
подъемная и тяговая силы—создаются
движением аэродинамической плоскости
в воздухе.
Результаты тщательных наблюдений за
движениями крыльев саранчи,
подтвержденные затем на других насекомых,
свидетельствуют о том, что крыло создает и
подъемную, и тяговую силы при движении как
вниз, так и вверх. Кончик крыла следует
почти по эллиптической траектории относи-
Биомеханика и движение (локомоция)
329
Рис. 10.30. Clap/fling-механизм генерирования вихрей. I. В верхнем положении крылья прижаты
друг к другу. II. Крылья начинают раздвигаться, причем их более жесткие передние края
расходятся первыми. Это вызывает циркуляцию воздуха вокруг крыла. III. Движение крыла вниз сообщает
воздуху ускорение; совершается работа и возникает сила противодействия с направленными
вперед и вверх составляющими (по Weis-Fogh, 1975)
тельно тела насекомого, что означает
зубчатую траекторию относительно
неподвижной точки отсчета при движении
животного вперед.
Однако подъемый коэффициент у
некоторых насекомых оказывается выше, чем
можно было бы ожидать от обычной
аэродинамической плоскости, исходя из
измеренных значений числа Рейнольдса. Такое
несоответствие особенно велико для мух-
журчалок и других насекомых, способных
парить неподвижно. Для понимания этих
явлений необходимо принять во внимание
нестационарные, неустойчивые силы,
создаваемые движением крыла. За летящим
насекомым образуются вихревые потоки
воздуха, и полет обусловлен силой
противодействия, возникающей при генерировании
вихревого движения вокруг поверхности
крыла. Более подробно этот принцип
разбирается в приложении 10.9.
Когда крылья насекомого смыкаются
и затем расходятся, передние, более жесткие
их края разъединяются первыми и воздух
устремляется в область низкого давления,
возникающую между крыльями (рис. 10.30).
Работа затрачивается на ускорение массы
воздуха, закручивающегося вокруг крыла,
а сила противодействия включает как
подъемную, так и тяговую составляющие.
Как только движение воздуха достигает
максимальной скорости, крылья перестают
совершать полезную работу до тех пор,
пока вихрь не будет «сброшен», что
произойдет при смене крылом направления
движения в нижней ei о точке. В приложении 10.9
аналогичным образом разбирается взлет
бабочки.
Насекомые настолько разнообразны,
что ко всем представителям этой группы
вряд ли может быть применена единая
теория полета. Для самых мелких форм типа
«мошкары» значение числа Рейнольдса
настолько низко, что они могут разгребать
воздух, как будто это вязкая жидкость.
У таких насекомых крылья покрыты
волосками и утрачивают свойства
аэродинамической поверхности (см. рис. 10.27). И
напротив, у крупных жуков Re достигает
23 000, и их жесткие надкрылья могут вести
себя как плоскости самолета.
10.6.4. Наземная локомоция:
прыгающие насекомые
Некоторые насекомые способны
прыгать. Для большинства из них это важная
реакция избегания; тот, кто хоть раз
пытался поймать блоху, знает, насколько
эффективно это непредсказуемое поведение.
330
Глава 10
Способность к прыжкам наиболее развита
у блох, кузнечиков и цикадок.
Чтобы подпрыгнуть, насекомое должно
подействовать на землю с такой силой,
чтобы сила противодействия сообщила
стартовую скорость его массе. Высота прыжка
определяется уравнением:
Кинетическая энергия (1/2 mV2) =
Потенциальная энергия в высшей точке прыжка
(mgh),
где m—масса насекомого;
V—стартовая скорость;
g—ускорение свободного падения;
h—высота прыжка.
V2
Следовательно, h = — и
2g
Кинетическая энергия
h = .
mg
Сила, с которой конец ноги прыгающего
насекомого воздействует на землю, имеет
вертикальную и горизонтальную
составляющие, как показано на рис. 10.31 для
кузнечика. Вертикальная составляющая равна
F-sin6.
Ускорение продолжается только до тех
пор, пока ноги насекомого касаются земли,
и стартовая скорость определяется силой
толчка и временем, в течение которого она
действует (см. также приложение 10.2). Это
время зависит от длины ног, и у всех
прыгающих форм они относительно длинные.
Рис. 10.31. Кузнечик до и во время прыжка.
При разгибании задней ноги на землю
действует сила. Чем длиннее ноги, тем дольше ее
действие, следовательно, больше ускорение
А Б В
Рис. 10.32. Прыжок блохи. А. Бедро
поднимается и в деформированных упругих белках
кутикулы запасается энергия. Б. Бедро «снимается
с предохранителя» при расслаблении мышцы.
Сила воздействия голени на землю сообщает
блохе начальную скорость, от которой зависит
высота прыжка
В разд. 10.2 было показано, что скорость
сокращения мышечных клеток ограничена их
физиологическими особенностями: у
летающих насекомых это ограничение
преодолевается с помощью непрямых
летательных мышц и способности запасать энергию
в упругих компонентах стенки тела при
деформации (см. разд. 10.6.3). Аналогичные
приспособления есть и у большинства
прыгающих насекомых. Элементы ноги
обобщенного насекомого показаны на рис.
10.19,/4, и мы будем ссылаться на этот
рисунок при описании механизма прыжка. Нога
блохи характеризуется удлиненными
тазиком и бедром. При подготовке к прыжку
бедро приподнимается в положение,
показанное на рис. 10.32,/4. Затем в участке
очень упругой кутикулы стенки торакса
запасается энергия. Хотя блохи неспособны
к полету, считается, что этот участок
произошел от гомологичного ему места у
летающих насекомых, причем для
деформации торакса используются прежние
летательные мышцы. Торакальные сегменты
удерживаются в таком напряженном
положении с помощью своего рода
«предохранителя» из выступов и пазов в покровах,
и блоха находится как бы во «взведенном»
состоянии, готовая к резкому
высвобождению запасенной энергии во время прыжка.
При расслаблении мышц, поднимающих
бедро, «предохранитель» снимается и
голень с большой силой ударяет о землю при
возвращении сегментов торакса в
первоначальное положение.
Биомеханика и движение (локомоция)
331
10.7. Выводы
В этой главе дано общее введение в
проблему локомоции у беспозвоночных.
Читатель должен уяснить себе значение
размеров: например, почему крупные животные
используют для движения мышечные
клетки, тогда как более мелким достаточно
ресничек или жгутиков.
Следует понимать, что двигательные
системы всех животных подчиняются одним
и тем же законам механики и что в передаче
силы всегда участвуют скелетные элементы.
Эта передача может осуществляться либо
посредством жидкости, заключенной в
замкнутом объеме, либо с помощью системы
жестких рычагов.
Некоторые беспозвоночные
медлительны и малоподвижны, другие перемещаются
очень быстро и маневренно; такие
животные, как головоногие моллюски и
насекомые, располагают весьма совершенными
локомоторными системами, одна из
которых основана на реактивном движении,
а другая—на использовании маховых
крыльев. Здесь изложены лишь элементарные
принципы; читателей, желающих более
подробно ознакомиться с проблемой, мы
отсылаем к приведенному ниже списку
дополнительной литературы.
Понимание механизмов локомоции
животных крайне важно для понимания
эволюционного происхождения
таксономических групп, поскольку любая
предполагаемая предковая форма должна быть
структурно оправданной. Она должна
«работать», подчиняясь основным законам
физики точно так же. как и любой ныне живущий
организм.
10.8. Дополнительная
литература
Alexander R. MeN. 1982. Locomotion of Animals.
Tertiary Level Biology. Blackie, Glasgow.
Clark R.B. 1964. Dynamics in Metazoan Evolution.
Clarendon Press. Oxford.
ElderH. Y., Trueman E. R. 1980. Aspects of Animal
Movement. Society for Experimental Biology
Seminar Series. Cambridge University Press,
Cambridge.
Hesseid C. F.,Fowtner C. R. (Eds) 1981. Locomotion
and Energetics in Arthropods. Plenum Press, New
York.
Kingsolver J.G. 1985. Butterfly engineering. Scient.
Am., 233 (2), 90 97.
Rainey R.C. (Ed.) 1984. Insect Flight. Blackwell
Scientific Publications, Oxford.
Trueman E. R. 1975. The Locomotion of Soft-Bodied
Animals. Edward Arnold, London.
Weis-Fogh T. 1975. Unusual mechanisms for the
generation of lift in flying animals. Scient. Am., 233
(5), 81- 87.
Приложение 10.1.
Термины и определения
механики
1. Сила проявляется в ее действии на
массу.
Сила стремится изменить направление
или скорость движения массы. Сила есть
масса х ускорение: F = М • а.
В единицах системы СИ сила величиной
1 ньютон (Н), приложенная к массе 1 кг,
сообщает ей ускорение 1 м/с.
2. Когда одно тело прикладывает силу
к другому, это равнозначно тому, что
второе тело прикладывает к первому
противоположно направленную силу равной
величины. Последняя называется силой
противодействия.
3. Когда сила двигает массу,
совершается механическая работа. Работа есть
сила х расстояние:
W = F-d,
W = (M-a)d.
В системе СИ единица измерения
работы—джоуль (Дж). Скорость совершения
работы—мощность, т.е. мощность—это
сила х скорость. Единица СИ для
измерения мощности — ватт (Вт); 1 Вт =
= 1 Дж/с.
Приложение 10.2.
Кинетическая энергия и трение
1. В приложении 10.1 сила F придает
нашей модели (осьминог в лодке) такое
ускорение, что
F = М • а.
Совершаемая работа будет зависеть от
расстояния, пройденного массой за время
действия данной силы: W = F • d.
332
Глава 10
Иллюстрация принципа равота = сила х расстояние
Иллюстрация концепции сил действия
и противодействия
2. Если бы лодка плыла без трения,
осьминог мог бы двигаться вечно за счет
кинетической энергии:
KE=fm-V2.
где V - скорость, м/с.
Лодка останавливается, поскольку
существует сила трения, вызывающая
замедление. Эту силу называют также
сопротивлением.
3. Кинетическая энергия рассеивается
в виде тепловой энергии. Чтобы продолжать
двигаться с постоянной скоростью,
осьминогу нужно прикладывать силу, равную
силе трения.
О
Лововое conpo-^ID
вление,или трение
Приложение 10.3.
Разложение сил на
составляющие
1. Любая сила действует в
определенном направлении, она является вектором.
2. Две силы, действующие на одну и ту
же массу, но в разных направлениях, можно
Силы, действующие в различных
направлениях, дают результирующую силу,
действующую в каком-то ином направлении
Биомеханика и движение (локомоция)
333
суммировать, получив результирующую
силу, действующую в своем собственном
направлении.
3. а) Направления действия сил можно
изображать линиями со стрелками,
начинающимися от массы.
б) Длина линии пропорциональна
величине силы.
в) Направление и относительную
величину результирующей силы можно
определить геометрически.
Геометрическое разложение сил. Sin 6 =
cos 6= F2/F
F./F,
4. Одна сила может быть разложена на
две составляющие, направленные
перпендикулярно друг другу. За направление одной
из составляющих обычно принимают
направление движения вперед. Другая
составляющая, действующая перпендикулярно
этому направлению, не имеет отношения
к движению, но тем не менее требует
метаболических затрат.
Выделение составляющей, действующей в
направлении движения, если дана сила,
действующая в ином направлении. В направлении
движения действует сила, равная F cos 6
Приложение 10.4.
Гидростатический скелет
I. Сокращение мышц, окружающих
замкнутую полость, заполненную
жидкостью, увеличивает давление этой жидкости:
Сила
Давление
Площадь
В системе СИ давление измеряется
в ньютонах на квадратный метр: Н/м2.
2. Таким образом, сила, действующая
на любую поверхность, составит F =
давление х площадь.
3. Давление действует одинаково на все
точки поверхности и направлено под
прямым углом к ней (см. рисунок).
I U Г Г7
Давление в жидкости действует во всех
направлениях
4. Внутреннему давлению в
гидравлической системе противостоит сила
противодействия ограничивающей поверхности.
Если сопротивление этой поверхности
в каком-то участке меньше, чем то. которое
необходимо для сдерживания давления,
движение жидкости приведет к изменению
формы поверхности. Это лежит в основе ло-
комоции всех животных с
гидростатическим скелетом.
На приведенном ниже рисунке мышцы
в правой и левой частях цилиндра
действуют как взаимные антагонисты.
ОТТй
t t , , , ,t_f
I ,
I I
Антагонизм между соединенными между
собой правым (R) и левым (L) компартментами
системы
334
Глава 10
5. Большая изменчивость системы
обеспечивается тем, что мышцы стенки тела
включают продольные и кольцевые
волокна.
Антагонистическая организация
(снаружи) и продольных мышц
кольцевых
.спив .(ШЛИ)
ш
IV
V
GMQ
VI
I-IV. Участки тела переменного объема: I —
кольцевые мышцы сокращаются, жидкость
выталкивается; II — кольцевые мышцы
расслабляются, жидкость входит; III — кольцевые
и продольные мышцы сокращаются, жидкость
выталкивается; IV—продольные и кольцевые
мышцы расслабляются, жидкость входит. V, VI.
Участки тела фиксированного объема: V—
продольные мыщцы расслабляются, объем не
меняется; VI — кольцевые мышцы
расслабляются, объем не меняется
6. Любой отрезок животного с
разделенным на изолированные отсеки телом
цилиндрической формы может в результате
претерпевать разнообразные деформации.
7. При изменении формы может
совершаться механическая работа. Она равна
произведению действующей силы (=
давление х площадь) на пройденное расстояние.
В приведенном ниже примере хоботок
выталкивается наружу при сокращении
большей по объему части тела.
Общая сила стенки тела
= Давление =
Сокращающаяся площадь at
Сила, действующая на конец хоботка
Площадь хоботка а2
- = Р = -.
а1 а2
Это так называемый гидростатический
рычаг (см. также приложение 10.5).
Совершенная работа равна J = Р х а, х d,.
а,
Приложение 10.5. Жесткие
скелеты — принцип рычага
1. Сила F может приводить в движение
груз М, вращая его вокруг точки опоры.
Если F—приложенная сила, a FR — сила
Биомеханика и движение (локомоция)
335
противодействия, они связаны равенством
F х d, = FR x d,,
где d, — путь, пройденный точкой
приложения силы F, a d, — путь, пройденный точкой
приложения силы FR.
Обратим внимание, что в такой системе
работа совершается силой,
противоположной по направлению приложенной.
2. Небольшая приложенная сила может
перемещать крупный груз, но количество
энергии должно остаться постоянным,
поэтому
—5 = Выигрыш в силе =
4i
'd,
Проигрыш в расстоянии.
3. Приложив большое усилие, можно
передвинуть маленький груз на
значительное расстояние.
Приложение 10.6
На рисунке изображено тело животного,
разделенное на пять отрезков (они могут
соответствовать или не соответствовать
настоящим сегментам), прикрепленных к
поверхности, а затем растягивающихся (А)
или сокращающихся (£). Волны мышечного
сокращения могут проходить по
мускулистой ноге, как показано на рис. /4(1) и £"(1),
или по цилиндрическому червеобразному
телу, как в случаях /4(H) и £(11).
А (I). Опора на короткие сегменты -
волна идет назад
Б (I). Опора на длинные сегменты -
волна идет вперед
с
J С
JEHU
3 С
J С
о=с
с
х=
С
о
о оз
Э
А (II). Опора на короткие
сегменты — волна идет
назад (как, например,
у дождевых червей)
С
Б (II). Опора на длинные сегменты —
волна идет вперад. Для червя
цилиндрической формы такой
тип движения трабует
использования щетинок
(однако см. также описание
движения по принципу
нереид)
336
Глава 10
Результаты одинаковы. Если точки
опоры возникают в областях с максимальной
длиной отрезков, животное передвигается
в одном направлении с волной мышечного
сокращения, т.е. волна прямая. Если,
напротив, точки опоры соответствуют
участкам сокращения продольных мышц, т. е. на
месте остаются самые короткие участки
тела, животное перемещается в направлении,
противоположном движению волны
мышечного сокращения, которая в таком
случае называется обратной.
Приложение 10.7.
Плавание мягкотелых червей
1. Плавание мягкотелых животных
можно анализировать, рассматривая
движение отдельного продольного отрезка их
тела.
2. Такой отрезок описывает
«восьмерку» с углом 9 между его продольной осью
и линией, перпендикулярной оси движения
всего тела. Мы рассмотрим только силы,
действующие на среднюю линию отрезка,
хотя это, конечно, упрощение.
. Ось движения
Поперечная
ось
Для понимания действуюших сил
необходимо знание правил разложения сил (см.
приложение 10.3).
3. Если отрезок тела находился в покое,
а затем начал двигаться под углом к оси
перемещения всего тела, он приложит силу
к массе окружающей воды и приведет ее
в движение (см. приложение 10.2).
л у Поперечная
Продольная ось
Одна из составляющих силы F, F",
вызовет движение воды вдоль поверхности тела,
а составляющая F' будет действовать под
прямым углом к этой поверхности,
сообщая воде ускорение.
Сила противодействия воды RF', равная
по величине и противоположная по
направлению силе F', может быть разложена на
составляющие, одна из которых параллельна,
а другая перпендикулярна оси движения
всего животного.
За время прохождения полного цикла
волнообразного движения составляющие,
перпендикулярные оси движения,
взаимоуничтожаются, а составляющие,
действующие вдоль этой оси, складываются.
По мере того как животное набирает
скорость, угол между направлениями
движения отдельных отрезков его тела и тела
целиком уменьшается. Этот угол называют
углом атаки (а).
Действительное направление
движения отрезка тела
с учетом скорости пере
мещения всего тела вперед
RF'
Биомеханика и движение (локомоция)
337
Это снижает силу противодействия,
направленную вдоль оси движения. Скорость
будет увеличиваться до тех пор, пока эта
сила не уравновесится силой лобового
сопротивления.
Приложение 10.8.
Реактивное движение
1. При реактивном движении сила,
действующая на воду, и сила противодействия
направлены вдоль линии движения
животного.
2. У головоногих моллюсков
сокращение мышц тела в стенке мантийной полости
повышает в последней давление. Таким
образом, в любой точке полости действует
одинаковая сила, направленная
перпендикулярно ее стенке.
Сила = Давление х Площадь.
3. При открывании сифона эта сила
обеспечивает ускорение массы воды,
выходящей из мантийной полости.
Произведенная работа =-~ m V2
(= кинетическая энергия).
Поскольку масса выталкиваемой воды
меньше, чем масса тела животного,
скорость этой воды выше, чем скорость
движения животного в противоположном
направлении. У реактивных двигателей с такими
свойствами низкий к. п. д.
4. Часть работы, производимой
мышцами, сжимающими мантийную полость,
расходуется на деформацию упругих стенок
тела; имеются также мышцы-антагонисты,
расширяющие полость до ее
первоначального объема.
Приложение 10.9. Вихревое
движение воздуха и полет
1. Полет насекомых основывается на
нестационарных воздушных потоках,
генерируемых движениями крыльев; важный
элемент этого процесса — образование
и сбрасывание вихрей.
В составе вихря масса текучего вещества
вращается в виде кольца или эллипса
вокруг центральной линии, как показано на
приведенном ниже рисунке. Вихревое
кольцо обладает моментом и энергией.
Индуцированный
поток
Вихревое .^
вращение /Оь.
Направленный
вниз момент
вихря
Схематическое изображение вихревого
кольца. Чтобы генерировать вихрь в текучей среде,
необходимо затратить работу
2. При полете движение крыльев
вызывает вращение воздуха вокруг их
аэродинамической поверхности; этот воздух
«сбрасывается» с крыла в виде вихревого кольца,
когда крыло изменяет направление
движения, наклон или форму.
Q.
Летящее насекомое поддерживается
в воздухе и движется вперед за счет силы
противодействия Q, возникающей в
результате «сбрасывания» вниз вихревых колец.
3. Вращение воздуха часто возникает
при разведении в стороны сомкнутых
крыльев (см. также рис. 10.30). Это иногда
называют механизмом "clap/fling".
22 1314
Глава 10
Почти вертикальный взлет бабочки (см.
рисунок) возможен благодаря
«сбрасыванию» вниз воздушного вихря (по Kingsol-
ver, 1985).
I. Крылья соединены наверху.
II, III. По мере разведения крыльев
возникает циркуляция воздуха вокруг их
передних краев (у большинства насекомых—
более жестких — см. характер жилкования
на рис. 10.27).
IV. Ускорение массы вращающегося
воздуха генерирует подъемную силу, и
бабочка отрывается от земли.
V, VI. Когда крылья достигают нижнего
положения, воздух «сбрасывается» с их
поверхности в виде вихревого кольца, момент
которого направлен вниз.
ГЛАВА 11
ДЫХАНИЕ
Потребность в кислороде считается
фундаментальным свойством всего живого. Он
поглощается через дыхательные
(респираторные) поверхности, такие, как жабры или
легкие, и после поступления в организм
используется для окисления органических
субстратов (в основном углеводов, хотя
и не только их) с выделением энергии,
необходимой для обеспечения процессов
метаболизма. Однако жизнь зародилась без
кислорода, поэтому аэробное дыхание
свойственно не всем организмам, и некоторые
из них до сих пор функционируют по
анаэробному типу. В этой главе
обсуждаются процессы аэробного и анаэробного
дыхания у беспозвоночных. Мы начнем с
разбора биохимических основ дыхания, а
затем, обратившись к его аэробному типу,
рассмотрим, как кислород извлекается из
окружающей среды и переносится в ткани
и как на его поглощение влияют факторы
окружающей среды.
11.1. Центральная роль АТР
в дыхании
Фосфорилированные нуклеотиды, и
особенно аденозинтрифосфат (АТР), играют
важную роль посредников в переносе
энергии от «топлива» (питательных веществ)
к энергопотребляющим процессам
метаболизма. Потенциальная энергия пищи
переносится на так называемые макроэргиче-
ские (высокоэнергетические) фосфатные
связи (обозначаемые ~ Р) следующим
образом:
Энергия из потребленной пищи +
А-Р~Р + Р,->А-Р~Р~Р,
где А — аденозин, а Р,.—неорганический
фосфат. Эта запасенная энергия может
затем быть использована в ходе метаболизма
с образованием из АТР аденозиндифосфата
(ADP) и Р,. Следует, однако, отметить, что
термин «макроэргическая фосфатная связь»
не вполне точен. Энергия не запасается в ко-
валентной связи между фосфатом и
остальной частью молекулы. Правильнее
говорить о том, что энергия этой связи отражает
разницу между энергетическим
содержанием молекул АТР и ADP.
11.2. Основной путь
катаболизма
Гликолиз и цикл трикарбоновых кислот
(ТКК) являются «становым хребтом»
катаболизма у всех типов беспозвоночных. Они
подробно изложены в других учебниках,
поэтому ниже дается лишь краткое их
описание.
На рис. 11.1 приведена очень упрощенная
схема этих двух путей. «Топливом» служит
Глюкоза -\
Лактат -<////ЛУ/>/дПируват
Л
V2ATP
>36АТР
Рис. 11.1. Очень упрощенная схема
метаболических путей, связанных с генерированием АТР
22*
340
Глава 11
глюкоза, получаемая непосредственно из
пищи или в ходе ферментативных
превращений других молекул пищи или резервов
тела.
Гликолиз происходит в цитоплазме и не
требует присутствия кислорода. Он дает
АТР в ходе прямых превращений субстрата,
как бы продвигающегося по этому
метаболическому пути «вниз» — в сторону
упрощения молекул. Это фосфорилирование на
уровне субстрата.
Цикл ТКК протекает в митохондриях
при участии кислорода. Он дает
восстановленный никотинамидадениндинук. геотид
(NADr), который в свою очередь
используется для образования АТР, отдавая
электроны (т. е. окисляясь с образованием N ADC)
в цитохромную транспортную систему, где
в роли их конечного акцептора выступает
кислород. Это—фосфорилирование,
сопряженное с транспортом электронов. На рис.
11.1 отражено, что участие кислорода
позволяет получить больше молекул АТР на
одну молекулу глюкозы, чем
бескислородная часть общего процесса.
11.3. Получение АТР
без кислорода
До появления в ходе эволюции фотоав-
тотрофов организмам приходилось
обходиться без кислорода. Кроме того,
экологическая аноксия (отсутствие О,) может
встречаться и в современных условиях; с ней
сталкиваются литоральные животные,
оказавшись на воздухе во время отлива, формы,
роющие ходы в восстанавливающих
субстратах, а также многие паразиты.
Определенные ткани могут находиться в аноксиче-
ских условиях, даже когда организм в
целом достаточно насыщен кислородом.
Например, в мышцах, участвующих в
реакциях избегания у двустворчатых
моллюсков, обычно протекает анаэробный
метаболизм.
Таким образом, «первые» организмы
были анаэробами, а некоторые остаются
ими и по сей день. Теоретически возможно
большое число анаэробных метаболических
путей, но лишь четыре из них были
использованы беспозвоночными. Различные пути
возникли в ходе эволюции для
удовлетворения определенных энергетических
потребностей.
Путь молочной кислоты (или лактатный
путь) изучен лучше других. Он показан на
рис. 11.1; NADr, образующийся при
гликолизе, вновь окисляется при восстановлении
конечного продукта этого метаболического
пути — пирувата—до лактата под
действием фермента лактатдегидрогеназы. Как
уже отмечалось, это неэффективный способ
получения АТР, но он позволяет
генерировать его с высокой скоростью в отсутствие
кислорода и обычно используется для
обеспечения интенсивной мышечной работы
у позвоночных, когда мышца может
временно оказаться в анаэробных условиях.
Лактатный путь встречается не во всех
тканях беспозвоночных. Он, по-видимому,
присутствует у насекомых в мышцах
конечностей, но отсутствует в летательных
мышцах, которые так обильно снабжены
трахеями и трахеолами (рис. 11.6,£), что редко
оказываются в условиях аноксии.
Опиновый путь близок к лактатному
и приспособлен для «взрывной» работы,
требующей быстрого, но не обязательно
эффективного синтеза АТР. При этом
углеводы катаболизируются в ходе гликолиза,
но вместо восстановления пирувата
происходит его восстановительная конденсация
с аминокислотой, приводящая к
образованию опина—производного аминокислоты:
Глюкоза + 2 аминокислоты + 3 ADP +
ЗР,->
2Н,0 + 2 опина + 3 АТР.
Можно выделить несколько опиновых
метаболических путей в зависимости от
участвующих аминокислот и ферментов.
Например, при резком сокращении мышц
мантии у головоногих образуется вещество,
называемое октопином. Другой опин—
стромбин — обнаружен у некоторых
двустворчатых моллюсков.
Путь янтарной кислоты (сукцинатный
путь). У некоторых организмов типа
двустворчатых моллюсков, обитающих в анок-
сических илах, или паразитов, населяющих
анаэробные среды своих хозяев, например
кишечник позвоночных, в ходе эволюции
сформировались анаэробные
метаболические пути, позволяющие получить на моле-
Дыхание
341
кулу глюкозы больше АТР, чем при опино-
вом и лактатном путях, но медленнее. При
этом энергия NAD,, полученного на первых
этапах гликолиза, используется для фосфо-
рилирования, сопряженного с транспортом
электронов, при котором конечным их
акцептором вместо кислорода служит фума-
рат. Общая схема этого процесса приведена
на рис. 11.2. Фосфоенолпируват (ФЕП) —
предшественник пирувата при гликолизе —
превращается в оксалоацетат за счет
присоединения СО, (карбоксилирования),
катализируемого ФЕП-карбоксикиназой, а
затем в фумарат. Фумарат окисляется до сук-
цината с участием электронотранспортной
системы, а сукцинат может метаболизиро-
ваться дальше до пропионата и других
летучих жирных кислот. Оксалоацетат,
фумарат и сукцинат являются промежуточными
соединениями цикла ТКК, однако там они
образуются в последовательности,
противоположной только что описанной.
Поэтому путь янтарной кислоты является как бы
обращенным циклом ТКК. Часть пирувата
в такой сукцинатной системе может
превращаться в лактат, ацетат, аланин
и этанол, т.е. широкий спектр конечных
продуктов. В общей сложности этот
процесс позволяет синтезировать от 4 до 6
молекул АТР на каждую молекулу глюкозы.
У двустворчатых моллюсков описана
сходная последовательность реакций,
дающая сукцинат в ходе восстановления окса-
лоацетата; последний, однако, образуется
не из ФЕП, а из аминокислоты (аспартата)
в реакции трансаминирования.
Фосфагены. Эти запасные соединения
получают группу ~ Р от АТР в периоды покоя
и отдают ее назад при интенсивной
нагрузке или аноксии:
Фосфоаргинин + ADP = Аргинин + АТР.
Такая система типична для
беспозвоночных, тогда как у позвоночных в
качестве донора используется фосфокреатин.
Исключениями являются иглокожие,
которые могут использовать оба фосфагена,
а также аннелиды, содержащие помимо фо-
сфоаргинина четыре других фосфагена.
Рис. 11.3 демонстрирует
филогенетическое распределение основных
метаболических путей (фосфагены не указаны). Все они
распространены очень широко. Так, либо
путь янтарной кислоты мог
эволюционировать из системы гликолиз-цикл ТКК, либо
наоборот, т. е. один путь мог возникнуть из
другого путем обращения направления
реакций (см. выше). Принято считать, что
в ранней биотической среде были широко
представлены аминокислоты. поэтому
Гликоген
-*NAD„
NAD„
3.
^ADP+
ADP+Pt
► АТР
- ADP+R
i АТР
Пиоуват Различные конечные
Оксалоацетат ""продукты
!
ADP+Pt
I Фумарат
ТКК
Система транспорта
электронов
♦ I
АТР
Сукцинат
Рис. 11.2. Вариант анаэробного
метаболического пути, используемый эндопаразитами
и двустворчатыми моллюсками,
переживающими длительные периоды гипоксии (по Са-
low, Townsend, 1981).
ФЕП—фосфоенолпируват
342
ARTHROPODA
Глава 11
ECHINODERMATA
CHORDATA
Мол (Янт)
NEMATODA
Мол Янт
Мол
Мол
ANNELIDA
SIPUNCULA
MOLLUSCA
Мол Янт
Опиновые?
PLATYHELMINTHES
Мол Янт
Окт Янт
Янт Окт (Мол)
Опиновые?
LOPHOPHORATA
Мол Опиновые
CNIDARIA
PORIFERA
Аминокислотные
Опиновые? (Янт)
Мол Опиновые'2
ПРЕДКОВЫЕ ПРОТИСТЫ
Рис. 11.3. Филетическое распространение
основных метаболических путей синтеза АТР (по
Livingstone, 1983, с изменениями). Мол — путь
молочной кислоты; янт — путь янтарной
кислоты; окт—октопиновый путь. Метаболические
пути, указанные в скобках, используются при
особых обстоятельствах
предполагается, что первые системы
синтеза АТР использовали их в качестве как
доноров, так и акцепторов электронов.
Исходя из этого, опиновые пути следует
считать примитивными. Ясно, что на ранних
этапах эволюции беспозвоночных
присутствовали все пути анаэробного
метаболизма, а их нынешнее распределение, по-
видимому, отражает различные
направления отбора, т.е. специфические адаптапии
к экологическим условиям, в которых
приходится функционировать организму.
Некоторые системы эволюционировали как
приспособления к «взрывной» работе (они
дают большие количества АТР, но не
обязательно высоко эффективны, поскольку
работают непродолжительно), другие,
наоборот, как способ более экономичного, хотя
и не такого быстрого получения АТР
в условиях длительной аноксии.
11.4. Поглощение кислорода
11.4.1. Главное—это диффузия
Аэробный метаболизм возможен только
при поступлении в ткани кислорода. В
основе этого процесса лежит диффузия, т. е.
движение молекул О, из области высокого
в область низкого парциального давления
(Ра)' в соответствии с законом Фика.
Скорость диффузии через ткани зависит от
градиента Р0, и от свойств самой ткани—
последние учитываются коэффициентом
диффузии. Исходя из оценок этих
коэффициентов и потребностей тканей в кислороде
можно рассчитать, что расстояние между
1 Парциальное давление газа, растворенного
в жидкости, обычно называют его
напряжением.— Прим. ред.
Дыхание
343
1 мм
о
А
Рис. 11.4. Поперечные срезы различных тур-
белларий (в одинаковом масштабе): А—
прямокишечная; Б— трехветвистая; В —
многоветвистая (по Alexander, 1971)
потребляющей кислород тканью и
дыхательной поверхностью не может превышать
1 мм. Это одна из причин плоской формы
тела крупных бесполостных турбелларий,
тогда как для более мелких бескишечных
и прямокишечных форм она не обязательна
(рис. 11.4). Однако многие сцифоидные
медузы и коралловые полипы достигают
очень крупных размеров, несмотря на
массивное строение. Это связано с тем. что
здесь наружный и внутренний слои ткани
тонкие и непосредственно контактируют
с окружающей водой или с гастроваскуляр-
ной полостью. Находящийся между ними
толстый слой мезоглеи в основном лишен
клеток, и его метаболические потребности
невелики. В мезоглее коралловых полипов
клеток больше, но она обычно тоньше, чем
у сцифомедуз, и благодаря сложной
складчатости гастродермиса (рис. 3.27) всегда
находится не далее 1 мм от воды,
циркулирующей в гастроваскулярной полости.
11.4.2. Циркуляторные системы
Возникновение в ходе эволюции систем
циркуляции жидкостей позволяет снять
ограничения, накладываемые законами
диффузии. Такие системы повышают
возможности транспорта 02, позволяя перейти
«одномиллиметровый предел», а за счет
быстрого удаления кислорода с
дыхательных поверхностей поддерживают крутой
градиент Р0,, тем самым увеличивая
скорость поглощения О, организмом.
Известны два основных типа циркуляторных
систем (рис. 11.5): незамкнутые с обширным
гемоцелем (бластоцелем или разросшимся
кровеносным сосудом), типичные для ар-
тропод и моллюсков, и замкнутые,
состоящие из артерий и вен, хорошо развитые
у аннелид. Для обоих типов систем
необходимы мышечные насосы, например
сократительные трубки у артропод. снабженные
Связка, поддерживающая
сердце
Перикардиальная
полость
Перикардиальная
мемврана
Остия
Крыловидная
мышца
Кишка
Рис. 11.5. Два типа
циркуляторных систем: А—
незамкнутая у членистоногого;
Б—замкнутая у полихеты
(Nereis)
Дорсальный параподиальный
сосуд
Кишечный
сосуд
Вентральный параподиальный
сосуд
-^^Дорсальныи сосуд
Дорсальный
кишечный сосуд
Кишечная сосудистая
сеть
Вентральный
кишечный сосуд
344
Глава 11
Рис. 11.6. I. Трахейные системы: А— основной тип; Б водные насекомые с жабрами; В — наяда
стрекозы с анальным дыханием. II. Увеличенное изображение трахейной системы (см. рис. 11.6.1,
А). Воздух входит через дыхальца, которые у большинства насекомых снабжены клапанами.
Широкие трубки (трахеи) разветвляются на более мелкие (трахеолы). Последние могут содержать
жидкость (А), но при интенсивном метаболизме осмотическое давление окружающих тканей
возрастает, что вызывает ее удаление из трахеол. Б. При этом воздух поступает в трахеолы и за счет
диффузии проникает через их стенки в ткани (по различным источникам)
Дыхание
345
парами отверстий (остий) с клапанами (рис.
11.5, /4), или пульсирующие кровеносные
сосуды (латеральные «сердца») у аннелид.
У ниркуляторных систем иглокожих и
полухордовых строение промежуточное; они
включают мелкие сосуды, впадающие в
более крупные синусы. Однако у голотурий
(иглокожие) хорошо развитая замкнутая
система. У мокриц в связи с выходом на сушу
сформировались более крупные и более
мускулистые, чем у других ракообразных,
сердца и сосудоподобные лакуны,
образующие почти замкнутую циркуляторную
систему. Незамкнутая система насекомых
практически не используется для
транспорта кислорода. Вместо этого у них
сформировалась обширная и сложная система
стволов, трахей и трахеол (рис. 11.6),
гарантирующая почти прямую доставку
газообразно! о кислорода ко всем активно мета-
болизирующим тканям (как она работает,
объясняется в подписи к рис. 11.6).
11.4.3. Кровь
Изначально кровь, по-видимому, была
бесцветной и близкой по составу к морской
воде, как у многих современных
брюхоногих и двустворчатых моллюсков. Такая
солевая среда может переносить
сравнительно небольшие количества кислорода,
поэтому кровь многих беспозвоночных содержит
его переносчики — специализированные
белки, называемые дыхательными
пигментами, способные обратимо связывать О,.
Например, в крови аннелид концентрация
растворенного кислорода составляет менее
1% об. (мл О,/100 мл крови), тогда как с
дыхательными пигментами связано 1 -10%
об. Структура этих пигментов сходна: все
они состоят из белковой части, соединенной
с простетической группой, содержащей
металл. Близкие по строению простетические
группы гемоглобина и хлорокруорина
называют гемом. Это порфирин, связанный
с одним атомом железа. У гемоцианина
простетическая группа образована не пор-
фирином, а полипептидом, связанным с
медью и серой. У гемэритрина это также не
гем, хотя и включает железо. Некоторые
свойства и распределение в животном
царстве дыхательных пигментов приведены
в табл. 11.1.
Количество переносимого кровью О, (ее
кислородная емкость) зависит от
содержания пигментов и от их способности
связывать и отдавать О, (сродства к кислороду).
Важный показатель — отношение количеств
кислорода, способного связаться с
пигментом и доступного в окружающей среде
(внешнее Р0). Это отношение
выражается так называемой кривой диссоциации
кислорода1, которую получают, насыщая
кровь кислородом при различных
значениях Р0 и измеряя связывание О, по
изменению оптических свойств пигмента.
Такие кривые часто, хотя и не всегда (см.
ниже), имеют S-образную форму (рис. 11.7).
Полезной характеристикой пигмента
является величина Ра, при которой он
насыщается кислородом наполовину (Р50).
Значения Р50 варьируют в зависимости от
температуры (ее снижение сдвигает кривую
диссоциации влево; рис. 11.8. А) и рН.
Уменьшение рН чаще всего понижает сродство
пигмента к кислороду, т. е. сдвигает кривую
вправо (рис. 11.8, Б). Это явление называют
эффектом Бора (нормальным, или
отрицательным). Рост концентрации СО,
вызывает снижение рН, приводя к эффекту
Бора, а поскольку содержание СО, выше
всего в тканях с высокой интенсивностью
метаболизма, высвобождение кислорода
стимулируется там, где он более всего
необходим. У некоторых беспозвоночных,
например у хелицерового Limulus
(мечехвоста), обнаружен обратный (аномальный,
или положительный) эффект Бора.
Мечехвост обитает на поверхности ила, где
периодическая нехватка О, и повышенный
уровень СО,, возможно, способствовали
выработке такой реакции на рН, которая
усиливает поглощение кислорода из среды, хотя
при этом затрудняется его высвобождение
в тканях. Не всем случаям обратного
эффекта Бора можно дать такое рациональное
объяснение, поэтому их и называют
«аномальными».
1 Такое название неудачно, поскольку
диссоциирует не сам кислород, а его комплекс с
пигментом, так что правильнее говорить «кривая
диссоциации оксигенированного пигмента».—
Прим. ред.
346 Глава 11
Таблица 11.1. Дыхательные пигменты (по Schmidt-Nielsen, 1979)
Пигмеи г
Описание
Молекулярная масса
Распространение в животном мире
Гемоцианнн Медьсодержащий белок, перено- 300 000
симый в растворенном состоя- 9000000
нии; синего цвета в оксигени-
рованной форме, бесцветный
в дезоксигенированной
Гемэритрин
Хлорокруорин
Железосодержащий белок;
находится внутри клеток; не
содержит порфирина; фиолетового
цвета
Белок, содержащий железопор- 2 750000
фириновый комплекс;
переносится в растворенном
состоянии; красный в
концентрированных растворах, зеленый
в разбавленных
Гемоглобин Белок, содержащий железопор- 17000
фириновый комплекс; перено- 300000
сится в растворенном
состоянии или внутри клеток;
наиболее часто встречающийся
у животных пигмент; красного
цвета
Моллюски: панцирные,
головоногие, переднежаберные и
легочные брюхоногие; отсутствует
у двустворчатых.
Ракообразные: крабы, омары.
Хелицеровые: Limulus, Euscorphis
Сипункулиды: все изученные
виды.
Полихеты: Magelona.
Приапулиды: Halicrvptus. Priapu-
lus.
Плеченогие: Lingula
Распространение ограничено
четырьмя семействами полихет:
Sabellidac, Serpulidae, Chlorhae-
midae, Ampharetidae.
Простетическая группа отдельно
от белка обнаружена у морских
звезд (Luidia, Astropecten)
Позвоночные.
Иглокожие: ю.кмурии.
Моллюски: Planorbix, Tivela.
Насекомые: Chironoimts, Gastrophi-
lus.
Ракообразные: Duphnia. Anemia.
Аннелиды: Lumbricus. Tubifex, Are-
nicolu. Spirorbis (у некоторых
видов гемоглобин, у чругих
хлорокруорин, остальные
лишены дыхательных пигментов), Se-
rpula (гемоглобин и
хлорокруорин).
Нематоды: Ascaris.
Плоские черви: паразитические
трематоды.
Инфузории: Paramecium. Tctrahy-
тепа
Различия по кривым диссоциации и по
значениям Р50 наблюдаются как внутри
отдельных классов пигментов, так и между
ними, что, наверняка, отражает адаптацию
пигментов к внутренней и внешней средам,
в которых им приходится функционировать
(см., например, рис. 11.7). Можно отметить
следующие корреляции.
1. Р5„ выше у животных, обитающих при
высоких внешних Р0 и имеющих
дыхательные поверхности, не представляющие
существенных препятствий для диффузии, так
что Р0> в их крови не очень отличается от
Р0 в среде. Такая ситуация характерна
для полихеты Sabella (рис. 11.7) с
дыхательным пигментом хлорокруорином (Р5(| * 30
мм рт.ст.). Это сублиторальное животное
обитает в трубке из ила и слизи, но
способно выставлять венчик из щупалец в
окружающую хорошо оксигенированную воду.
Кроме того, ею постоянно омывается
поверхность тела за счет перистальтических
движений мускулатуры его стенки. Другой
пигмент с низким сродством к кислороду—
Рис. 11.7. Кривые диссоциации кислорода —
зависимость поглощения 02 из атмосферы (в
процентах полного насыщения) от его
концентрации в ней (Р0г): А — пескожил Arenicola
marina (рН 7,5; 19°С); Б—Nephtys hombetgii (pH
7,4; 15°С) для сосудов; В—то же, но для цело-
мической жидкости; Г—Sabella spallanzanii (pH
7,35; 26 °С) (Jones, 1955, 1972)
10 20
мм рг. ст.
80
60
40
20
-
-
// /2у25°
i i i i i i i i i i i i
/о
10 15 20 25 30
Температура 15 С
х = 4,8 мм со2 рН: 8,43
■= 8,0 мм С02рН:7,82
а- 8,1 мм со2Рн:7,72
• = 14,2 мм С02рН: 7,52
d - 26,0 мм со2 рН :7,35
о = 38,3 мм СО2рН:7,04
u-44j2mmСО, рН: 6,77
0 10 20 30 40 50 60 70 80 90 100110 120 130
Рис. 11.8. Влияние температуры (А), давления С02 и рН (Б) на кривые диссоциации оксигениро-
ванного гемоцианина ракообразных (по Waterman, 1960)
А . ОТТЕКАЮЩАЯ ВОДА
f
АБфронтальные
реснички
Латеральные
реснички
Фронтальные
реснички
Скелетный стержень
Ток воды между
соседними пластинками
ПРИТЕКАЮЩАЯ ВОДА
Приносящий
кровеносный сосуд
Выносящий
кровеносный
сосуд
Ток крови внутри
ктенидиальнои пластинки
К предсердию
Рис. 11.9. Ктенидиальная система брюхоногих: А—ток воды через «жабры»; Б—ток в них крови
(по Russel-Hunter, 1979). См. также рис. 5.18
348
Глава 11
гемоцианин кальмара Loligo (Р5(1 % 50 мм
рт.ст.;. К этой же категории животных
относится эррантная роющая полихета Neph-
tys с гемоглобином в крови. Как видно из
рис. 11.7, у этого пигмента довольно низкое
значение Р50(«5—7 мм рт. ст.), но следует
отметить, что кривая диссоциации здесь
гиперболическая, поэтому по сравнению с S-
образной кривой со сходным Р50 пигмент
менее насыщен кислородом при более
высоких Р0. Иными словами, пигмент с S-
образной кривой полностью насыщается
кислородом при более низких Р0, чем
пигмент с таким же Р50, но с
гиперболической кривой диссоциации. Эта полихета
обитает в песке литоральной зоны; во время
прилива вода прокачивается через
сделанный ею ход с помощью мерцательного
движения ресничек. В таких условиях пигмент
должен быть способен как связывать, так
и отдавать О, при высоких значениях Р().
Когда наступает отлив, ход спадается
и происходит резкое снижение Р0 —по-
видимому, до такого уровня, что
поглощение кислорода становится вообще
невозможным, и животное скорее всего переходит
в этот период (около 6 ч) на анаэробный
метаболизм. Следует также отметить, что
у гемоглобина сосудов Nephtys сродство
к кислороду выше, чем у целомического
гемоглобина. Возможно, это связано с
поступлением О, через всю поверхность тела,
сначала в целом, а уже оттуда в сосудистую
систему. Ход кривых диссоциации
сосудистого и целомического гемэритринов сипун-
кулиды Dendrostomwn zostericolum как раз
противоположный: у целомического
пигмента сродство больше, чем у сосудистого.
У этого животного дыхательным органом
служат щупальца. Кислород диффундирует
через них в сосудистую систему, а уже
оттуда- -в целомическую жидкость.
2. Pj,, относительно низкое у животных
с затрудненной диффузией через
дыхательные поверхности, обитающих в средах с
высоким Р0, т.е. когда значение Р0 в
крови значительно ниже внешнего. К этой
категории относятся многие десятиногие раки
с Рх в пределах от 1 до 20.
3. Р5(, может быть очень низким (<1)
у животных, населяющих среду с низким
значением Р0. К этой категории
относятся, например, олигохеты-тубифициды
и личинки комаров-хирономид (и те и
другие с гемоглобином в качестве
дыхательного пигмента), встречающиеся в местах,
богатых (или загрязненных) органическими
веществами. Хирономиды живут в норках,
через которые активно прокачивают воду.
Однако они делают это не постоянно, так
что даже при насыщенности окружающей
воды кислородом внутри норки может
наступить гипоксия; в этих условиях пигмент
с высоким сродством к кислороду важен
для его поглощения и, возможно, для
запасания на короткие периоды аноксии. Низкое
значение Р50 характерно также для морского
литорального кольчеца пескожила, часто
роющего ходы в неаэрируемых илах (рис.
11.7). Как и хирономиды, он постоянно
прогоняет воду через норку, и пигмент с
высоким сродством к кислороду, возможно,
важен для кратковременного запасания О,.
Тем не менее маловероятно, что
запасенный таким образом кислород обеспечивает
пескожила в течение всего отливного
периода, так что в это время он, по-видимому,
способен частично переходить на
воздушное дыхание. Наконец, следует отметить,
что кривая диссоциации у пескожила
сравнительно мало чувствительна к низким
значениям рН, что также может быть
адаптацией к жизни в аноксических илах, часто
бывающих закисленными.
11.4.4. Органы дыхания
Еще одним фактором, ограничивающим
поглощение кислорода, является площадь
дыхательной поверхности. Для дыхания
плоских червей, длинных, тонких немертин,
нематод и некоторых аннелид достаточно
неспециализированной поверхности тела.
Однако дальнейшее увеличение размеров
или активности, а также эволюция
относительно малопроницаемых защитных
наружных покровов, в том числе раковины,
потребовали возникновения васкуляризован-
ных дыхательных поверхностей. У водных
форм широко распространены выросты
стенки тела, например «жабры» некоторых
полихет и членистоногих (рис. 11.6), ктени-
дии моллюсков, жаберные книжки у
мечехвоста, ножки некоторых иглокожих. На
Дыхание
349
Сердце
Остия
Анус
Гемоцель
Рот
Педипал'ьпа
Рис. 11.10. Легочная книжка паука (по Snow.
1970). См. также рис. 8.11.
рис. 11.9 схематически изображена юени-
диальная система водного брюхоногого.
Обратите внимание, что кровь
прокачивается сердцем через «жабру» в направлении,
противоположном току воды
(создаваемому в основном латеральными ресничками).
Такой противоток типичен для жабр (кроме
жабр пескожила и головоногих); за счет
него вода с низшим значением Р0,
взаимодействует с наименее оксигенирован-
ной кровью, что позволяет последней
поглотить максимум О,. У наземных форм
дыхательные поверхности чаще всего
погружены внутрь тела, например трахеи (рис.
11.6), легочные книжки, ситовидные и
трубчатые трахеи паукообразных (рис. 11.10),
«легкие» легочных моллюсков. У
некоторых водных животных дыхательные
поверхности также внутренние; примеры —
водные легкие голотурий (рис. 11.11) или
анальные структуры личинок ряда
насекомых (рис. 11.6). В последнем случае иногда
развиваются пластроны и наружные
«физические» жабры (рис. 11.12; в подписи к
рисунку разъясняется принцип их работы).
Нервное
кольцо
Мадрепоровая
пластинка
Кювьеров
орган
Клоака
Анус
Водное легкое
Околоротовая амбулакральная
ножка
■Амбулакральная
ножка
Рис. 11.11. Водное пегкое на схематическом продольном разрезе голотурии (иглокожие) (по
Nichols, 1969). См. также рис. 7.27
350
Глава 11
Водоотталкивающие
волоски (10 /мм )
Воздух
Слой воздуха-
Дыхальце
Кутикула
-Ткань
mtmimmimhff^utmmtmmmmmm
Трахея
Рис. 11.12. Пластрон. Часть поверхности тела покрыта гидрофобными волосками,
обеспечивающими ее несмачиваемость и постоянное присутствие слоя воздуха. В результате она может
действовать как несжимаемая жабра, в которую из воды диффундирует кислород. Густой слой
волосков препятствует поступлению в воздушный слой воды по мере использования кислорода, а, так
как объем слоя остается постоянным, Р0 в нем падает, вызывая диффузию 02 из окружающей
воды. У водного насекомого Aphelocheirus пластрон может выдерживать давление в несколько
атмосфер (по Ramsay, 1962; Eckert, Randall, 1983)
11.4.5. Вентиляция
Еще одним механизмом,
способствующим поглощению кислорода, является
вентиляция дыхательных поверхностей. Она
обеспечивает постоянное поступление к ним
кислорода, т.е. поддержание на их уровне
как можно более крутого градиента
концентраций последнего. Вентиляция
свойственна животным как со специализированными
дыхательными поверхностями, так и без
них, но отсутствует в трахейной системе
насекомых. Структура приспособлений,
осуществляющих вентиляцию, самая разная:
от ресничек на жаберных пластинках
двустворчатых моллюсков до мышечных
насосов, обслуживающих водные легкие
голотурий или анальные структуры у стрекоз.
Живущие в трубках полихеты используют
реснички и/или перистальтические движения
(см. выше). У ракообразных вентиляцию
обычно создают колебательные движения
придатков. При снижении Р0> во внешней
среде вентиляция обычно усиливается. На
рис. 11.13 показан аппарат, позволяющий
одновременно измерять характер
активности (путем регистрации изменений
электрического импеданса на респирометре) и
поглощение кислорода (с помощью
электродов) у водных пиявок. При снижении
Р0г в среде усиление двигательной
активности увеличивает поглощение О, по
сравнению с пиявками в состоянии покоя. При
этом упорядоченные движения могут так же
эффективно улучшать снабжение
кислородом, как и регулярная вентиляция.
11.5. Измерение метаболизма
Основная часть энергии, получаемой за
счет дыхания, в конечном счете
превращается в тепло (рис. 11.14). Поэтому, если
измерить количество выделяемого животным
тепла, можно довольно точно оценить
интенсивность метаболизма. Известны
устройства с очень чувствительными термоэле-
Дыхание
351
Дыхательная камера
Насыщение Покой Неупорядочен- Вентиляция
воздухом,% ное движение
К кислородочувст
вительному электроду
L
Пиявка в воде дышит через
поверхность тела
Резервуар с регуляцией
температуры и напряжения
кислорода
Nephelopsis obscura
100 0,21+0,02 0,54+0,09 0,52+0,10
50 0,04±0,01 0,36±0,05 0,32±0,04
25 0,03+0,01 0,18 + 0,01 0,10+0,07
0 0,02+0,01 0,0310,01 0,03+0,02
Erpobdetta punctata
100 0,27+0,02 0,59 + 0,11 0,54+0,09
50 0,05+0,01 0,35+0,07 0,55 + 0,05
25 0,03 + 0,01 0,13±0,04 0,20 + 0,03
0 0,02±0,01 0,06+0,01 0,04±0,01
Преобразователь
импеданса
I
Усилитель
I
Осциллоскоп-
самописец
Вентиляция
Неупорядоченное движение
Покой
Время, мин
Рис. 11.13. Схема аппарата, описанного в тексте (А), и некоторые полученные с его помощью
результаты (б. В). Б. Тилы движений, зарегистрированные путем измерения импеданса. В.
Интенсивность потребления 02 (мкл/мг сухой массы в час ± среднеквадратичное отклонение) у двух
видов пиявок при различных концентрациях 02 в среде и разных типах активности. Можно видеть,
что движение связано с повышенным потреблением кислорода при любых его концентрациях,
кроме самой низкой, причем существенной разницы в этом смысле между вентиляцией и
неупорядоченным движением нет (по Wrona, Davies, 1984)
Поступление
энергии
Дыхание
С02
ADP
.ыхание v. ,
/v
Синтез
CL
Механическая
работа
!<
Химическая
равота
-Тепло
Тепло
Тепло
-Тепло
Рис. 11.14. Схема использования энергии, получаемой из пищи. Процессы дыхания генерируют
АТР неэффективно, с потерей энергии в виде тепла; большая часть запасенной в АТР энергии
также рассеивается в виде тепла после совершения работы
ментами, способные зарегистрировать
малые количества тепла, выделяемые
беспозвоночными (их называют
микрокалориметрами), но используют их редко. На рис.
11.15 приведены результаты
микрокалориметрического эксперимента с водной олиго-
хетой Lumbriculus variegatus. На графике
можно различить: (I) аэробное состояние; (II)
аноксическое состояние; (III) состояние
отравленного животного. Заметно, что
интенсивность аэробного метаболизма
приблизительно вчетверо выше, чем аноксического.
Глава 11
Теплоизоляция
Экспериментальное
животное
i
23
Аноксия (8%C02+92%N2)
1
28
Время эксперимента
IV Температура термостата
"""'V—>'N"YV4>tV,'v,'',*w~>'v^^
Рис. 11.15. Прямой микрокалориметр. А. Устройство аппарата: очень чувствительные датчики (S,
и S2) измеряют температуру воды, втекающей в теплоизолированную камеру, в которой
находится экспериментальное животное, и вытекающей из нее. Б. Некоторые результаты, более
подробно описанные в тексте (по Gnaiger, 1983)
Поглощение кислорода легче измерить,
чем выделение тепла. Для этого имеются
различные методы, от химического
титрования О, в водном растворе до физического
измерения объема или давления газа.
Существуют также весьма чувствительные
электроды нескольких типов. Содержание
кислорода может быть определено либо в
закрытой системе, где находится животное,
либо по поступлению О, в открытую
систему и оттоку из нее. По определению эти
методы позволяют следить только за
аэробными процессами и поэтому чреваты
недооценкой дыхательного метаболизма, так как
иногда беспозвоночные используют
одновременно анаэробные и аэробные процессы
даже при достаточном поступлении
кислорода. Когда, как и при каких именно
условиях снижение Ра вызывает
переключение с аэробного метаболизма на
анаэробный, не вполне понятно; для выяснения этих
вопросов требуется больше данных,
полученных методом прямой калориметрии.
Тем не менее в большинстве работ
интенсивность метаболизма оценивается
именно по поглощению кислорода. На
основе этой информации ниже показано, как
различные факторы влияют на
дыхательный метаболизм. Однако интерпретация
проводимых результатов вызывает сложно-
Дыхание
353
сти, так как метаболизм беспозвоночных
настолько зависим от состояния животного
и от условий, в которых оно находится, что
данные разных опытов могут быть
несопоставимы друг с другом. Дыхательный
метаболизм подразделяют на основной обмен,
характеризующий организм в состоянии
покоя, обычный обмен, измеряемый при
обычном для животного уровне
активности, пищевой обмен у только что
питавшегося животного и активный обмен при
повышенной активности. Экспериментаторы
обычно стремятся определить основной
обмен, поскольку это наиболее
воспроизводимая оценка метаболизма.
11.6. Факторы, влияющие
на дыхание
Ниже перечисляются наиболее
изученные факторы, влияющие на дыхание у
беспозвоночных. Они расположены в порядке
снижения их связи с особенностями самого
организма, т. е. начиная с неотъемлемых от
него свойств и кончая внешними
влияниями.
11.6.1. Размер тела
Естественно ожидать, что крупные
беспозвоночные дышат более интенсивно, чем
мелкие, но, поскольку поглощение
кислорода ограничивается поверхностью (см.
выше), а поверхность тела пропорциональна
квадрату линейных размеров, тогда как его
масса—их кубу, следует также ожидать,
что зависимость между массой тела и
потреблением кислорода нелинейна.
Сравнение животных различного размера в
пределах данного вида или относящихся к
разным видам свидетельствует о том, что
интенсивность дыхания в покое возрастает
с увеличением массы, но с убывающей
скоростью. Это выражено следующим
уравнением:
Интенсивность дыхания = а(масса)ь,
(11.1)
где а и b—константы, причем b обычно
меньше 1. Прологарифмировав, получаем
log интенсивности дыхания =
К + b(log массы),
(11.2)
где К = log а. Поэтому, если построить
график зависимости логарифма поглощения
кислорода от логарифма массы тела,
получится линейное отношение с наклоном,
соответствующим b (рис. 11.16).
Разделив-уравнения 11.1 и 11.2 на массу
тела, получим:
Интенсивность дыхания на единицу
массы = а(масса)Ь|,
log интенсивности дыхания на единицу
массы = К + (b— I) (log массы).
Поскольку b меньше 1, величина Ь— 1 будет
отрицательной, так что график зависимости
логарифма интенсивности дыхания на
единицу массы от логарифма массы будет
прямой линией с отрицательным наклоном
(рис. 11.17). Следовательно, как и
ожидалось, интенсивность дыхания на единицу
массы тела снижается с возрастанием
последней. Если бы это отношение
определялось только увеличением поверхности тела
с возрастанием его массы без учета других
факторов, b всегда было бы 0,67, а Ь— 1
составляло бы —0,33. Объясняется это
просто. Дыхание, будучи зависимым от
площади поверхности (см. выше), должно (для
геометрически сходных тел) быть
пропорционально квадрату длины тела (I2). Масса
пропорциональна объему и, следовательно
(опять же при геометрическом подобии),
пропорциональна F. Поэтому дыхание
было бы пропорционально кубическому кор-
V*-
ню из массы {yj\3 = 1) в квадрате (= I2), т. е.
з
^/М* = м0,67. Однако реальные значения
b редко точно соответствуют 0,67 и обычно
составляют от 0,67 до 1 (рис. 11.16).
Считается,- что, хотя геометрические факторы
и вносят свой вклад в зависимость дыхания
от размера, они не являются
единственными или хотя бы главными факторами,
определяющими это соотношение. Остальные
предстоит выяснить.
11.6.2. Активность
При движении моллюск-блюдечко
потребляет приблизительно в 1,4 раза больше
кислорода, чем в покое; эта цифра
составляет около 2 для бокоплава Gammarus, от
3 до 4 для обитающих в эстуариях креветок
23—1314
354
Глава 11
1л г
1мл •
1мкл -
1нл
1пл
1фЛ
•
-
■\S
-
гУ
г/
>
23
лгулъ
9 п'тз1^ Наклон = 0,
7/по
i i _ i i
1пг
1нг
1 мкг 1 мг
Масса тела
1г
1кг
Номер Группа
на рис.А
Одноклеточные эктотермы
1 Бактерии
2 Гривы
3 Жгутиковые
4 Инфузории
5 Корненожки
Многоклеточные эктотермы
6 Нематоды
7 Микроракоовразные
8 Клещи
9 Ногохвостки
10 Термиты (личинки)
11 Энхитреиды
12 Жуки (личинки)
13 Термиты (личинки)
14 Муравьи (рабочие)
15 Дождевые черви (коконы)
16 Сенокосцы
п
1}
4
5
5
24
12
71
29
4
61
17
21
23
3
30
b
0,68
1,33
0,28
0,93
0,82
0,91
0,61
0,74
0,75
0,В7
0,67
0,49
1,14
1,00
0,69
Номер Группа
на рис. А
17 Двупарноногие многоножки
1В Пауки
19 Равноногие раки
20 Моллюски
21 Жуки (имаго)
22 Дождевые черви
(взрослые особи)
23 Макроракоовразные
п
77
6
40
6
14
1В
3
b
0,79
0,В1
0,69
0,76
0,81
0,76
0,В1
Рис. 11.16. А. Графики зависимости потребления 02 от массы тела для ряда таксонов (в двойных
логарифмических координатах). Б. Список таксонов, использованных при построении графиков
(А), с указанием наклона b прямой для каждого таксона (по Phillipson, 1981); п — число
изученных представителей
Дыхание
355
Palaemonetes, и может превышать 100 для
летящей саранчи. В этом нет ничего
неожиданного, так как движение требует биения
ресничек или жгутиков или же сокращения
мышц, что повышает интенсивность
метаболизма по сравнению с основным
обменом.
11.6.3. Питание
Потребление кислорода обычно
возрастает сразу после приема пищи, но вскоре
опять снижается. Например, у
ракообразного Macrobrachium rosenbergii эти колебания
составляют 10—40%. Такую дыхательную
реакцию на питание называют
специфическим динамическим действием (СДД),
специфическим динамическим эффектом (СДЭ)
или калоригенным эффектом.
В основе СДЭ лежат следующие
факторы:
1) энергетические затраты на
переработку пищи в кишке;
2) энергетические затраты на
расщепление и выделение лишних белков,
поступивших с пищей;
3) энергетические затраты на
использование веществ, полученных из пищи, для
синтеза компонентов новых тканей.
В исследовании, проведенном на мидии
Mytilus edulis (Bayne, Scullard, 1977),
зарегистрировано возрастание на 25%
потребления кислорода после приема пищи; с
энергозатратами на фильтрование пищи связано
более 80% этого прироста, на выделение
аммиака — менее 20%.
Длительное голодание обычно
сопровождается снижением потребления
кислорода. Это связано не только с отсутствием
СДЭ, но и с пониженной двигательной
активностью и даже с переходом на более
экономичный «поддерживающий»
метаболизм. Например, для разных видов морских
желудей, обитающих на различных уровнях
литорали и получающих неодинаковые
количества пищи, характерны разные
интенсивности потребления кислорода
(рис. 11.17). У Balanus crenatus и В. rostratus,
которым пища доступна непрерывно в
течение большей части приливно-отливного
цикла, интенсивность метаболизма самая
высокая. У видов нижне- и среднелиторальной
о*
2,5
2,0
3 1,5
Balanus glandula (сред)
Balanus crenatus (сув)
Balanus rostratus (сув)
Jtelanus cariosa
Chthamalus
nssus Chthamalus
(верх) data
(верх)
Polliclpes
polymerus
(сред)
0 0,5 1,0 1,5
Log массы (мг)
2,0
Рис. 11.17. Связь между логарифмом
потребления кислорода на единицу массы тела и
логарифмом этой массы для некоторых видов
морских желудей (по Newell, Branch, 1979);
верх, сред, ниж—верхняя, средняя, нижняя
литораль; суб—сублитораль
зон В. glandula и В. cariosa уровень
метаболизма промежуточный, тогда как виды
рода Chthamalus, населяющие верхнюю
литораль и питающиеся лишь короткое время на
протяжении каждого приливно-отливного
цикла, а в периоды квадратурных приливов
вообще несколько дней не
имеющие.доступа к пище, отличаются очень низким
уровнем потребления кислорода.
11.6.4. Температура
Изменения температуры среды могут
влиять на метаболизм, только если они
влияют на температуру тела, что обычно
свойственно беспозвоночным, но не
млекопитающим или птицам, поэтому первых
называют пойкилотермными (от греч. poiki-
1о—изменчивый), а вторых—
гомойотермными (от греч. homoio—
остающийся таким же). Однако многие
беспозвоночные, например глубоководные или
населяющие тропические районы океанов,
живут при постоянной температуре и,
строго говоря, не являются пойкилотермными.
Более общие термины, учитывающие
источник тепла, определяют
беспозвоночных как эктотермных, а млекопитающих
и птиц— как эндотермных животных. Но
и эта классификация не универсальна, так
как в летательных мышцах некоторых
насекомых образуется так много тепла (эндо-
термия), что оно может использоваться для
356
Глава 11
поддержания постоянной температуры
торакса (гомойотермность). Например,
шмели не могут летать, если температура их
летательных мышц ниже 30°С или выше 44°С.
По крайней мере 90% энергии,
затрачиваемой летящей пчелой, высвобождается в
тораксе в виде тепла, так что при интенсивном
полете температура там может подняться
на несколько градусов в считанные секунды.
Более того, есть данные, что во время
свободного полета шмели способны
поддерживать постоянную внутреннюю температуру
при различных внешних. Крупные пчелы (в
основном матки) могут летать даже при
0°С, поддерживая при этом в тораксе
температуру 30"С и выше. Такая регуляция
обеспечивается отчасти пассивно— с
помощью толстого слоя теплоизолирующих
волосков на тораксе, но главным образом
активно за счет работы, совершаемой при
полете.
Таким образом, ни один из
перечисленных терминов не может строго описать все
температурные характеристики
беспозвоночных, однако большинство их все же
относится к пойкилотермным эктотермам.
В пределах температурного интервала,
приемлемого для пойкилотермного
животного, интенсивность дыхания возрастает
с увеличением температуры окружающей
среды. Однако, как характерно для
химических реакций, такая зависимость нелинейна:
увеличение температуры на несколько
градусов вызывает повышение интенсивности
дыхания в несколько раз. Широко
используется индекс Q10, отражающий влияние
температуры на метаболизм. Он
определяется следующим образом:
Qio =
R'0/t2-'.
R,
(logJV-logRJlO
logQio = ,
i2 ti
где R, и R2— интенсивности метаболизма
(оцененные, например, по поглощению
кислорода) при температурах t, и t2
соответственно. Отсюда следует, что при
t2 — t, = 10°С величина Q,0 равна
отношению R2 и R,, так что Q1(l означает, во
сколько раз возрастает R при повышении
температуры на каждые 10°С. При резких сменах
температур Q,0 часто достигает 2 и более,
& 0,6
Q
5-
0,4
0,2
0,1
14
Сутки
21
28
Рис. 11.18. Акклимация у Mytilus. • —
постоянные условия 10°С; *—переход от 10
к 15°С; О — переход от 10 к 5°С. Пояснения
в тексте (по Widdows, Ваупе, 1971)
причем эта величина может меняться в
зависимости от температурного интервала.
Кроме того, непосредственная реакция
на изменение температуры бывает
непродолжительной. На рис. 11.18 показано, что
происходит при перенесении мидий Mytilus
edulis из среды с температурой 10°С в среду
с 5 или 15°С. Потребление кислорода при
15° резко возрастает, а затем постепенно
снижается до некоторого постоянного
уровня; при 5° оно также сначала резко
падает, а затем возрастает, выходя на плато.
Этот процесс называется акклимацией;
характерно, что значение Q10 после акклима-
ции ниже, чем в случае первоначальной
реакции, а при идеальной акклимации
стремится к 1. Такая реакция считается
адаптивной, так как при высоких температурах
позволяет экономить энергию, а при низких
поддерживать синтез АТР на достаточном
уровне. Акклимация такого рода была
обнаружена у различных плоских червей,
моллюсков, аннелид и иглокожих.
Акклимация может действовать и в
противоположном направлении, т. е. еще более
снижать потребление кислорода при низких
Дыхание
357
температурах и повышать его при высоких,
т.е. значения Ql0 после акклимации будут
больше, чем для непосредственной реакции.
Это характерно для некоторых блюдечек,
включая как пресноводные {Ancylus fluviati-
lis), так и морские {Patella aspera) виды. При
низких температурах такой тип акклимации
может быть адаптивным, обеспечивая
переход организма в диапаузу и позволяя
сохранять энергию в условиях зимнего
недостатка пищи и кислорода, связанного,
например, с замерзанием водоема. Стимуляцию
усиленного метаболизма при высоких
температурах объяснить труднее; возможно,
она связана с тонкими компенсаторными
механизмами.
Реакция, проиллюстрированная
рис. 11.18, называется положительной ак-
климацией, а описанная выше для
блюдечек— отрицательной, или обратной, аккли-
мацией. Масштабы происходящих
изменений неодинаковы у разных видов и могут
зависеть от состояния животного. Так, у
некоторых литоральных беспозвоночных
уровень основного обмена быстро и почти
полностью положительно акклимируется при
изменении температуры (Q1(l к, 1), тогда как
значение Ql0 для обычного обмена всегда
остается больше 1. Возможно, такая
реакция адаптивна, поскольку на этих животных
наиболее существенный перепад
температур действует в период между приливами,
когда они неподвижны и не питаются.
Низкие значения Q10 для потребления
кислорода покоящимися организмами
демонстрируют определенную экологическую
закономерность: для сублиторальных видов,
например Strongylocentrotus franciscanus и
Ammonia natalensis, а также роющих форм
нижней литорали типа Diopatria cuprea и
некоторых двустворчатых моллюсков, не
испытывающих значительных и быстрых
изменений температуры, характерны величины
Q10 > 1, а для обитателей литорали Littorina
littorea, Strongylocentrotus purpuratus, Maco-
ma balthica, Actinia equina и Bullia digitalis
Ql0 к 1. Но есть и много исключений;
например, у литорального моллюска Patella
vulgata значение Q,0 достаточно высокое, а
у сублиторальной полихеты Hyalinoecia—
низкое.
11.6.5. Концентрация
кислорода (Р0)
Водным беспозвоночным часто
приходится переносить низкие значения Ра,
например при загрязнении пресноводных
водоемов органическими веществами, при
выходе из-под воды во время отливов, в
озерах ниже термоклина или при обитании
в субстратах с восстанавливающими
свойствами. Некоторые животные реагируют на
это такой перестройкой вентиляции и
кровообращения, что потребление кислорода
остается на прежнем уровне (см., например,
рис. 11.19). Такие формы называют
регуляторами. У других видов потребление
кислорода изменяется параллельно Ра во
внешней среде. Их называют конформерами
(рис. 11.19). Регуляторы могут
поддерживать потребление кислорода на постоянном
уровне лишь в определенных пределах Р0;
критическое значение этого параметра, при
г
Baetis (конформер)
Leptophlebia
(регулятор^
Рс
\/\Рс
Ephemera
(конформер)
О 50 100 150 200 250 300
Напряжение 02, мм рт. ст.
Диапазон
конформной
1. реакции
Диапазон регуляции
Критическое напряжение
Напряжение 02 в среде
Рис. 11.19. Потребление кислорода в
зависимости от напряжения 02 в окружающей среде.
Вертикальной пунктирной линией показано
содержание 02 в воде, уравновешенное его
содержанием в атмосфере (по Fox et al., 1937)
358 Глава 11
Таблица 11.2. Примеры регуляторов и конформеров. Рс—критическое значение Р02,
при котором регуляция перестает действовать и регуляторы становятся
конформерами
Регуляторы Рс" Конформеры
Paramecium (инфузории)
Tetrahymena (инфузории)
Pelmatohydra (гидроидные)
Amelia (сцифомедузы)
Tubifex (олигохеты)
Lumbricus (олигохеты)
Mytilus (двустворчатые моллюски)
Helix (легочные брюхоногие)
Loligo (кальмар)
Cambarus (рак)
Uca (краб)
Cloeon (нимфа поденки)2'
Hyalophora (личинка бабочки)
50
2,5
60
120
25
76
75
75
45
40
4
30
25
Spirostomum (инфузории)
Различные коралловые полипы
Cassiopea (сцифомедузы)
Nereis (полихеты)
Erpvbdella. (пиявка)
Sipunculus (сипункулиды)
Limax (легочные брюхоногие)
Limulus (хелицеровые)
Homarus (омар)
Baetis (нимфа поденки)21
Tanytarsus (личинка хирономид)
" К приведенным величинам Рс следует относиться с осторожностью, поскольку переход между
регуляцией и конформностью обычно резко не обозначен. Кроме того, на форму кривой,
выражающей зависимость потребления кислорода от его напряжения, часто влияет степень аккли-
мации к аномальному значению Ра.
21 См. также рис. 11.19.
котором они становятся конформерами,
обычно обозначают Рс. В табл. 11.2
приведены примеры конформеров, регуляторов
и значений Рс. Животные, для естественных
местообитаний которых характерны низкие
уровни Р0,, часто являются регуляторами
с низкими Рс. Так, черви-тубифициды с Рс,
равным 25 мм рт. ст., выживают в
загрязненных органикой участках рек. Манящий
краб Uca, обычный обитатель приморских
болот и илистых наносов, также относится
к хорошим регуляторам. Конформер Baetis,
напротив, живет в быстрых, хорошо оксиге-
нированных ручьях.
Разные составляющие метаболизма
могут различным образом зависеть от Ра;
мидия, например,—конформер в отношении
основного обмена, но регулятор (до 80 мм
рт. ст.) с точки зрения обычного обмена
(рис. 11.20).
После действия на организм условий ги-
локсии или аноксии потребление 02 часто
возврашается до уровня,
превышающего тот, что наблюдался до помещения в
такие условия. Это возвращение так
называемого кислородного долга. Сюда входит
восстановление запасов фосфагена и АТР
и окисление конечных продуктов
анаэробного обмена. Однако все здесь не так
просто, поскольку между накоплением
конечных продуктов и размером кислородного
долга не всегда наблюдается корреляция,
0,4
Обычный
°.3г овмен
0,1
40
80
мм рт. ст.
120
140
Рис. 11.20. Влияние напряжения 02 в среде на
метаболизм Mytilus (no Bayne etal., 1976).
Обсуждение см. в тексте
Дыхание
359
а уровни АТР и фосфагена обычно
возвращаются к норме очень быстро, уже на
начальной стадии процесса. Поэтому следует
учитывать и другие перестройки, например
восстановление доаноксического
внутриклеточного рН, пониженного в результате
накопления кислых конечных продуктов,
или использование дополнительного
кислорода для насыщения жидкостей тела и
дыхательных пигментов.
В заключение следует отметить, что
избыток кислорода может быть так же
опасен, как и его недостаток. Радикалы
свободного кислорода (супероксид-радикалы)
способны денатурировать макромолекулы,
особенно белки. Это становится
существенным при очень высоких концентрациях О,,
например в тканях беспозвоночных,
содержащих симбиотические водоросли (у
многих обитателей коралловых рифов), где
кислород является конечным продуктом
фотосинтеза. Такие организмы выработали
клеточные механизмы защиты от
кислородного отравления, в частности ферменты су-
пероксиддисмутазу (СОД),
обезвреживающую опасный супероксид-радикал, и ката-
лазу, удаляющую Н,02, образуемую СОД.
На рис. 11.21 показано, что у некоторых
групп беспозвоночных активность СОД
повышается с увеличением концентрации
хлорофилла в тканях, т. е. с возрастанием сим-
биотической способности к высвобождению
кислорода. Однако эта зависимость
неодинакова для разных групп, что связано с раз-
Хлорофилл, мкг/мг велка
Рис. 11.21. Связь между активностью СОД
и концентрацией хлорофилла у основных
групп беспозвоночных коралловых рифов (по
Shick, Dykens, 1985)
личной локализацией симбионтов:
активность СОД наиболее высока у стрекающих
с исключительно или преимущественно
внутриклеточными зооксантелами
(цитоплазма непосредственно испытывает
воздействие кислорода), тогда как у оболочника
Didemnum симбиотические прохлорофиты
находятся не только вне клеток, но и вне
организма (в выстилке общей клоакальной
камеры), и избыточный кислород быстро
уносится током жидкости, не контактируя с
цитоплазмой клеток. Самая низкая
активность СОД среди стрекающих наблюдается
у видов, обитающих в затененных местах —
в пещерах или под скалами.
11.6.6. Соленость
Поскольку изменения солености воды
могут вызвать у морских беспозвоночных
активные поведенческие и физиологические
реакции, можно ожидать, что они будут
сопровождаться изменениями потребления
кислорода. Креветка Metapenaeus monocer-
cus встречается в условиях различной
солености. Потребление кислорода особями из
солоноватоводных популяций минимально
в 50%-ной морской воде и повышается при
изменении этого уровня как в гипо-, так и
в гиперосмотическую сторону. И напротив,
потребление кислорода особями из
морской популяции минимально в 100%-ной
морской воде и возрастает до максимума
в 25%-ной.
Затраты энергии на осморегуляцию и
активный транспорт ионов, по-видимому,
невелики, так что большая часть
наблюдаемых изменений, вероятно, связана с
реакциями всего организма, например с
переходом к другому типу активности. Это более
подробно обсуждается в работе Ньюэлла
(Newell, 1979).
11.7. Выводы
Термином «дыхание» обозначают
процессы высвобождения биологически
полезной энергии из питательных веществ,
главным образом углеводов. Эти реакции
по большей части, хотя и не
исключительно, аэробные, поэтому «дыханием»
называют также процесс потребления О, и из-
360
Гпава 11
бавления от ненужного С02. Его
биохимические аспекты иногда называют
«внутренним», или «тканевым дыханием»; а
физиологические— «внешним дыханием». В
настоящей главе рассмотрены механизмы,
действующие на обоих этих уровнях,—
системы, обеспечивающие получение О, из
окружающей среды и его доставку к
тканям, а также внешние и внутренние
факторы, влияющие на интенсивность
метаболизма.
11.8. Дополнительная
литература
Alexander R.McN. 1971. Size and Shape. Edwards
Arnold, London.
Bayne B.L., Scullard C. 1977. An apparent specific
dynamic action in Mytilus edulis. J. mar. biol.
Assoc, UK, 57, 371—378.
Calow P., Townsend C.R. 1981. Resource utilization
in growth. In: C. R. Townsend and P. Calow (Eds)
Physiological Ecology: an Evolutionary Approach
to Resource Utilization. Blackwell Scientific
Publications, Oxford.
Eckert R., Randall D. 1983. Animal Physiology, 2nd
edn. W. H. Freeman. New York.
Heinrich B. 1979. Bumble-bee Economics. Harvard
University Press, Cambridge, Massachusetts.
Jones J.D. 1972. Comparative Physiology of
Respiratory Mechanisms. University of Pennsylvania
Press, Philadelphia.
Livingstone D. R. 1983. Invertebrate and vertebrate
pathways of anaerobic metabolism: evolutionary
considerations. J. Geol. Soc. 140, 27 38.
Newell R. С 1979. Biology of Intertidal Animals, 3rd
edn. Marine Ecological Surveys Ltd., Kent.
Newell R. C, Branch C. 1979. The influence of
temperature on the maintenance of energy balance in
marine invertebrates. Adv. mar. Biol., 17, 329 -
346.
Phillipson J. 1981. Bioenergetic options and phyloge-
ny. In: C. R. Townsend. P. Calow (Eds)
Physiological Ecology: an Evolutionary Approach to
Resource Utilization. Blackwell Scientific
Publications, Oxford.
Ramsay J. A. 1962. A Physiological Approach to the
Lower Animals. Cambridge University Press,
London.
Schmidt-Nielsen K. 1979. Animal Physiology:
Adaptation and Environment. Cambridge University
Press, Cambridge.
Имеется перевод: Шмидт-Ниельсен К.
Физиология животных: приспособление и среда: в 2 т.—
М.: Мир, 1982.
ГЛАВА I £-
ЭКСКРЕЦИЯ, ИОННАЯ И ОСМОТИЧЕСКАЯ
РЕГУЛЯЦИЯ, ПЛАВУЧЕСТЬ
Вещества, количество которых
превышает метаболические потребности, должны
быть удалены из организма животного.
К ним относятся непереваримые
компоненты пищи, выводимые с фекалиями (гл. 9),
и СО,, удаляемый в процессе дыхания
(гл. 11). Однако существуют и другие
избыточные вещества — вода, различные ионы
и продукты распада лишних белков и
аминокислот. Именно последние
азотсодержащие соединения в физиологии обычно
называются продуктами выделения, или
экскреции (экскретами). Вместе с тем процессы
удаления лишних ионов, воды и
азотсодержащих веществ у беспозвоночных часто
настолько тесно связаны, что логично
рассматривать их вместе. Поэтому в этой главе мы
сначала опишем истинную экскрецию,
затем разберем проблемы, связанные с
регуляцией водно-ионного баланса, и, наконец,
рассмотрим структуру и функции
выделительных систем, связанных с ионной и
осмотической регуляцией, но не обязательно
с удалением азота.
12.1. Экскреция
Ненужные аминокислоты могут
появиться в организме при поступлении слишком
большого их количества с пищей или в
результате катаболизма белков. Обычно они
подвергаются дальнейшей окислительной
деградации с образованием кетокислот
и аммиака:
R
I
NH2—CHCOOH + 1/2 02 =
Аминокислота
R
I
= 0=ССООН + NH3 .
Кетокислота Аммиак
Кетокислоты могут использоваться в
других метаболических превращениях, а
аммиак, будучи крайне токсичным, должен
быть либо быстро удален из организма,
либо переведен в безвредное состояние (см.
ниже). Он легко растворим и легко
выводится в случае водных животных за счет
диффузии через покровы тела или
дыхательные поверхности. Если среди экскретов
преобладает аммиак, такой тип выделения
называют аммониотелией; он широко
распространен у водных беспозвоночных,
в том числе у водных личинок насекомых (в
отличие от наземных имаго; см. ниже).
У некоторых животных аммиак
превращается в менее токсичную мочевину:
NH2
I
с=о
I
NH2
Экскреция азота в виде мочевины известна
как уреотелия; такой тип выделения
типичен для млекопитающих. У беспозвоночных
он распространен слабо. Возможно, это
связано с тем, что, будучи еще токсичной,
мочевина требует для своего выведения
потери больших количеств воды. Тем не менее
уреотелия встречается у некоторых плоских
червей, аннелид и моллюсков.
Основным экскретом наземных
беспозвоночных (а также наземных позвоночных,
за исключением млекопитающих) является
мочевая кислота, а основанный на ней тип
выделения называется урикотелией. По
химическому строению она относится к
классу пуринов. Другие вещества этого класса
также используются как экскреты
некоторыми беспозвоночными (пуринотелия).
Появление в эволюции этих продуктов
выделения связано с их малотоксичностью
и низкой растворимостью, которая позво-
362
Глава 12
ляет удалять их в кристаллическом
состоянии и тем самым сокращать потери воды.
Урикотелия характерна для онихофор, од-
новетвистых и в меньшей степени для
ракообразных и моллюсков. У пауков основным
экскретом является другой пурин—гуанин.
^
НС
"Ч
1ST
Пурин
./"
HN
^
Мочевая кислота
HN'
сн
H,N
^
Гуанин
Исключение из указанной выше
эволюционной тенденции составляют
ракообразные. Даже у наземных равноногих раков
преобладает аммониотелия. Кутикулярные
покровы этих животных легко проницаемы,
поскольку почти лишены эпикутикулярного
воска, характерного для насекомых (разд.
12.2.5), и газообразный аммиак может
свободно удаляться через них путем диффузии.
Ткани ракообразных также, по-видимому,
менее чувствительны к действию аммиака,
чем у других животных. Кроме того, его
выделение через покровы способствует
формированию экзоскелета: щелочная среда,
возникающая на поверхности тела,
необходима для образования карбонат-иона из
растворенного СО, и осаждения СаСО,.
Моллюски также могут использовать- аммиак
для упрочнения раковины. Из других
наземных беспозвоночных непосредственное
выделение аммиака свойственно некоторым
олигохетам и многоножкам, но они, как
и равноногие раки, обитают во
влажной среде, где опасность
обезвоживания очень мала.
Еще один фактор, сыгравший роль
в эволюции экскреторных систем,—
экономия метаболических ресурсов. Так,
хотя пурины и менее токсичны, чем аммиак,
в их состав входит углерод—
потенциальный источник энергии.
Мочевина в этом смысле занимает промежуточное
положение. В табл. 12.1 сопоставляются
токсичность и потери воды и углерода
(энергии), связанные с каждым из трех
основных продуктов выделения. Однако,
насколько важны эти потери для «бюджета»
всего организма, неясно; возможно,
большее значение имеют энергозатраты
(выделение тепла) в метаболических процессах,
ведущих к образованию экскретов.
Важно понимать, что «продукты
выделения», особенно малотоксичные
(например, пурины), вовсе не обязательно должны
удаляться из организма. Так, и у наземных,
и у водных легочных моллюсков мочевая
Таблица 12.1. «Экономичность» экскретов. Число звездочек соответствует
приблизительной количественной оценке (Pilgrim, 1954)
C/N
Теплота сгорания Токсич- Потребность
ность в воде
кДж/моль кДж/моль N
Аммиак
Мочевина
Мочевая кислота
0
0,5
1,25
378
638
1932
378
319
483
***
**
*
***
**
*
Экскреция, ионная и осмотическая регуляция 363
кислота накапливается в организме по мере
его роста и старения. Обнаружено также
накопление мочевины в тканях тропического
легочного моллюска Bulimulus в период
летней диапаузы. Значение этих процессов
неизвестно, но помимо экскреторной функции
высокая концентрация мочевины в тканях
и жидкостях тела улитки может повышать
осмотическое давление, тем самым
препятствуя потере воды путем испарения. Не
исключено, что это способствует
переживанию длительных засушливых периодов.
Мочевая кислота накапливается также в
ходе эмбрионального развития клейдоических
яиц. Хорошим примером может служить
опять-таки легочный моллюск: содержание
мочевой кислоты в яйцах Lymnaea
возрастает с 0,5% их свежего веса при дроблении
до 4,5% при выходе личинки. Есть мнение,
что именно возникновение клейдоических
яиц в ходе эволюции привело к появлению
пуринотелии. Как бы то ни было, оба этих
признака сыграли важную роль в
завоевании суши.
12.2. Осмотическая и ионная
регуляция
Жидкости тела животных представляют
собой разбавленные солевые растворы,
основным электролитом которых является
хлорид натрия; иными словами, они
напоминают по составу морскую воду. Обычно
считается, что это следствие зарождения
жизни в море. Тем не менее между морской
водой и жидкостями тела имеются
существенные различия (табл. 12.2), и Макал-
лум (Macallum) в 1920-х гг. предположил,
что это отражает постепенное изменение
состава океана с момента возникновения
жизни до наших дней. Однако результаты па-
леохимических исследований говорят об от-
Таблица 12.2. Концентрации ионов в плазме крови или полостной жидкости, выраженные в
процентах от концентраций тех же жидкостей, диализованных против морской воды (по Barrington,
1979)
Na
Са Mg CI
so4
Книдарии
Amelia aurita
Иглокожие
Marthasterias glacialis
Оболочники
Salpa maxima
Аннелиды
Arenicola marina
Сипункулиды
Phascolosoma vulgare
Ракообразные
Maja squinado
Dromia vulgaris
Carcinus maenas
Pachygrapsus marmoratus "
Nephrops norvegicus
Моллюски
Pecten maximus
Neptunea antiqua
Sepia officinalis
99
100
100
100
104
100
97
110
94
113
100
101
93
106
111
113
104
110
125
120
118
95
77
130
114
205
96
101
96
100
104
122
84
108
92
124
103
102
91
97
98
95
100
69
81
99
34
24
17
97
101
98
104
101
102
100
99
102
103
104
87
99
100
101
105
47
100
65
92
91
66
53
61
46
69
97
98
22
" Этот грапсоидный краб—единственное представленное в таблице животное с гипоосмотической гемолимфой
(концентрация ионов — 86% их концентрации в морской воде).
364
Глава 12
сутствии существенных различии в составе
воды древних и современных океанов.
Таким образом, из табл. 12.2 следует,
что даже морским животным приходится
регулировать состав жидкостей своего тела.
У обитателей эстуариев и пресных вод эта
проблема стоит еще более остро, так как
они должны поддерживать в жидкостях
тела более высокие концентрации веществ,
чем во внешней среде. Обратная ситуация
наблюдается у животных, а) которые во
время жизни в пресной воде выработали
в своих жидкостях более низкую
концентрацию веществ, чем в морской воде, а затем
вернулись в море; б) обитающих в очень
концентрированной среде, например в
соленых озерах. Для наземных форм,
естественно, основная проблема - сохранение воды.
Теперь давайте более детально
рассмотрим вопросы, связанные с ионной и
осмотической регуляцией в четырех
перечисленных экологических ситуациях. Основные
термины и понятия, важные для понимания
этого материала, приведены в приложении
12.1.
12.2.1. Морская среда
У большинства морских
беспозвоночных жидкости тела изоосмотичны морской
воде, а сами животные являются осмокон-
формерами. Тем не менее, как уже
говорилось, ионный состав жидкостей их тела
довольно существенно отличается от состава
морской воды, так что необходима ионная
регуляция. Это иллюстрируется данными
табл. 12.2. Стоит отметить следующее, а)
Aurelia регулирует в основном
концентрацию ионов сульфата, поддерживая ее на
более низком уровне, чем в море. Возможно,
это связано с поддержанием плавучести:
ионы сульфата тяжелые, и их замена хлорид-
ионами снижает плотность медузы. То же,
по-видимому, справедливо для каракатицы
Sepia (см. также разд. 12.2.2). б)
Единственный ион, содержание которого способны
регулировать иглокожие,— это калий.
Возможно, это одна из причин, ограничивших
их распространение морской средой, в)
Уровень магния очень низок в жидкостях
тела членистоногих. Это может быть
связано с тем, что большинство членистоногих
очень активны, а магнии имеет свойства
анестетика. Однако у весьма подвижной
каракатицы Sepia концентрация магния не
понижена.
12.2.2. Отступление по поводу
плавучести
Водные животные с плотностью выше,
чем у воды, должны опускаться на дно,
поэтому плавающим формам выгодно
поддерживать свою плотность равной или
меньшей, чем у воды, иначе им придется
затрачивать дополнительную энергию, чтобы
не погружаться в глубину. Уже говорилось,
что этого можно достичь, по крайней мере
отчасти, регулируя свой ионный состав.
Ниже мы проиллюстрируем это новыми
примерами, а также обратим внимание на
другие, обеспечивающие плавучесть
приспособления.
У глубоководного кальмара Heliocran-
chia очень большая перикардиальная
полость, заполненная жидкостью. Она легче
морской воды за счет гораздо более
низкого содержания ионов натрия и очень
высокого—ионов аммония. Последние, являясь
конечным продуктом катаболизма белков,
диффундируют в перекардиальную
жидкость и удерживаются в ней за счет кислой
реакции. Кроме того, как и в случае Sepia
и Aurelia, анионы в этой жидкости
представлены почти исключительно хлоридом,
заменяющим тяжелые ионы сульфата.
Существуют и другие способы
повышения плавучести: а) удаление ионов без их
замещения, что, однако, приводит к гипоос-
мотическому состоянию жидкостей тела,
поддержание которого требует
соответствующих затрат; б) устранение тяжелых
компонентов; например, у некоторых
пелагических брюхоногих моллюсков (в
частности, птеропод и гетеропод) раковина
редуцирована или вовсе отсутствует; в)
увеличение содержания жиров (они легче воды)
распространено как у морских, так и у
пресноводных планктонных ракообразных; г)
использование заполненных газом
поплавков с мягкими стенками, как у книдарии
Physalia, или в виде твердостенных камер
раковины головоногих наутилоидей и ее
рудимента у каракатицы (рис. 12.1).
Экскреция, ионная и осмотическая регуляция
365
Заполненное газом
пространство
—Газовая
камера
Отверстие
трувки сифона
Жилая
камера
Рис. 12.1. А. Перегородчатая раковина моллюска Nautilus. По мере роста животного к ней
прибавляются одна за другой новые заполненные газом камеры. Сначала они заполнены жидкостью,
в которой растворен в основном NaCI. Затем Na+ удаляется посредством активного транспорта,
жидкость становится гипоосмотической по отношению к морской воде и удаляется, оставляя
заполненное газом пространство. Газ поступает внутрь камеры путем диффузии, давление l\l2
достигает 0,8 атм, как в окружающей воде и тканях животного, а у 02 оно в камере ниже, чем
вокруг нее. Поскольку у камеры жесткие стенки, давление газа остается около 0,9 атм, независимо
от глубины погружения животного. Б. Рудимент раковины у каракатицы имеет слоистую
структуру и заполнен газом приблизительно такого же состава и давления, что и у наутилуса. Однако
в наиболее старых, задних камерах (показанных черным) находится раствор NaCI. Количество
газа можно изменять вместе с объемом жидкости, что позволяет каракатице регулировать свою
плотность, а тем самым плавучесть и глубину погружения. Это обеспечивается за счет изменения
концентрации раствора NaCI в раковине: на небольшой глубине он близок по осмотическим
свойствам к морской воде, по мере погружения концентрация соли снижается, жидкость становится
гипоосмотичной морской воде и крови животного, а возникающее при этом осмотическое
давление уравновешивает гидростатическое давление, стремящееся заполнить «раковину» водой (по
Schmidt-Nielsen, 1979)
12.2.3. Пресноводная среда
Если морских животных поместить
в разбавленную морскую воду, то либо
осмотическое давление жидкостей их тела
уравняется с окружающим (осмоконформе-
рами), либо включатся механизмы,
препятствующие разбавлению их внутренней
среды (у осморегуляторов). Примеры
приведены на рис. 12.2. Обитатели эстуариев и
отчасти литорали в естественных условиях
периодически оказываются в разбавленной
воде, например во время отлива или после
сильного дождя. Среди таких животных
встречаются как осмоконформеры, так и ос-
морегуляторы.
Не следует, однако, считать осмокон-
формеров обязательно более стеногалин-
ными, чем осморегуляторы. Так,
осмотическое давление крови мидии Mytilus edulis
соответствует окружающему как в Северном
море (нормальная соленость), так и в
Балтийском (менее половины нормальной
солености). Одна из причин такой
толерантности в том, что эффективно регулируется
не внеклеточная, а внутриклеточная среда.
По-видимому, важную роль в этом
процессе играют аминокислоты: их концентрация
при разбавлении окружающей среды
возрастает, тем самым повышая осмотическое
давление внутри клеток. Выделение
аммиака мидиями усиливается при снижении
внешней солености. Это, вероятно,
стимулирует катаболизм белков и образование
свободных аминокислот под действием
осмотического стресса. Подтверждением
служит тот факт, что у мидий обнаружена
генетическая изменчивость по лейцинаминопеп-
тидазе — важному ферменту белкового
катаболизма, причем более активные его ва-
366
Глава 12
£
£
Эквивалентность между
■внутренней
и внешней
средой
Морская
вода
100 200 300 400 500 600
Концентрация во внешней среде, ммоль/л
Рис. 12.2. Связь концентраций солей в
жидкостях тела и среде обитания некоторых солоно-
водных животных (по Schmidt-Nielsen, 1979)
Q.J
So
0,7-
0,6 -
ge o,5
0,4
0,3
0,2
0,1
о
"f к- ей
с с: о
op.
"О
о
Si £ S£.
to
О
J3_
I
о
CD
s
и га
§
О)
о
с
«3
1
$
^
Рис. 12.3. Проницаемость экзоскелета
ракообразных из различных местообитаний (по
Ноаг, 1966)
рианты чаще встречаются в эстуариях, т. е.
в условиях пониженной солености.
Осморегуляторы могут контролировать
приток воды и отток ионов двумя
способами— с помощью малопроницаемых
покровов и/или активного выкачивания воды
и регуляции поступления ионов в жидкости
тела и их выведения. На рис. 12.3 показано,
что проницаемость экзоскелета
ракообразных ниже у видов, обитающих на литорали
или в слабосоленых водах.
Пресноводные животные находятся в
такой же ситуации, что и осморегуляторы при
пониженной солености, но им
приходится регулировать состав внутренней
среды постоянно. На рис. 12.4 показано, как
жидкости тела таких животных реагируют
на повышение солености. Обратите
внимание на значительные различия в
концентрации этих жидкостей в пресной воде—у
двустворчатого моллюска Anodonta и
ракообразного Daphnia она ниже, чем у бокопла-
ва Gammarus или клопа-гребляка Sigara, а
у этих животных она в свою очередь ниже,
чем у родственных им морских форм.
Возможно, это снижает затраты на
поддержание большой разницы осмотических
давлений внешней и внутренней сред.
Большинство пресноводных животных
100 200 300 400 500 600
Концентрация во внешней среде, ммоль/л
Рис. 12.4. То же, что на рис. 12.2, но для
пресноводных животных (по Schmidt-Nielsen, 1979)
Экскреция, ионная и осмотическая регуляция
367
регулируют состав внутренней среды за
счет низкой проницаемости эпидермиса
(рис. 12.3) и выделения большого
количества мочи. Если его выразить в процентах
к весу тела в сутки, эта величина будет
гораздо меньше 10% у большинства морских
беспозвоночных и намного больше у
пресноводных. Например, для Astacus и
Gammarus она составляет около 40%, для Daph-
nia—более 200%, а для пресноводного
двустворчатого моллюска Anodonta — более
400%. Моча пресноводных беспозвоночных
обычно гипоосмотична жидкостям тела,
так как в соответствующих отделах
«выделительных» органов (с. 375) из нее
избирательно извлекаются полезные ионы.
Несмотря на это, а также на низкую
проницаемость эпидермиса, некоторые ионы все же
теряются, и их запас должен восполняться.
Некоторые из них поступают с пищей, но
многие пресноводные беспозвоночные
способны и к прямому активному захвату
ионов из окружающей среды. В этом можно
убедиться, предварительно выдержав
животное в дистиллированной воде, чтобы
снизить концентрацию ионов в жидкостях
тела, а затем возвратив его в обычную
пресноводную воду. Очень скоро состав
жидкостей тела возвращается к норме, хотя в
течение всего эксперимента концентрация
ионов в среде ниже, чем в организме.
Следовательно, они должны активно поглощаться.
Эти процессы наиболее подробно изучены
у пресноводных ракообразных: речного
рака Astacus, водного ослика Asellus и боко-
плава Gammarus. Возможно, в активном
транспорте ионов какую-то роль играет вся
поверхность тела, но имеются
убедительные данные, что он идет в основном через
жабры.
Если выращивать личинок комара Aedes
aegypti в дистиллированной воде, уровень
натрия в их гемолимфе станет ниже 30%
нормы. При перенесении их в обычную
пресную воду концентрация натрия
восстанавливается за несколько часов.
Эксперименты позволили установить, что на 90%
поглощение этого иона происходит через
анальные сосочки, а остальное—в
основном через кишечник. У личинок стрекоз
аналогичную роль играют ректальные
жабры.
12.2.4. Гиперосмотическая среда
Некоторые морские беспозвоночные,
например креветка Palaemonetes, гипоосмо-
тичны морской воде (рис. 12.2). Обычно
считается, что такие животные происходят
от пресноводных форм, вернувшихся в
море. Жидкости тела некоторых грапсоидных
крабов также гипоосмотичны среде (см.
примечание к табл. 12.2), но, поскольку
возможность обитания их предков в
пресных водах исключена, происхождение ги-
поосмотичности в этом случае неясно.
Основная проблема для организма в
гиперосмотических условиях состоит в том.
чтобы не пропускать внутрь соли и не
выпускать воду. Ее приходится решать и
беспозвоночным, обитающим в более соленых
водах, чем морская. Ярким примером
может служить рачок Artemia, живущий в
соленых озерах. Он поддерживает состав
своей внутренней среды путем активной
регуляции: «выпивает» большое количество
воды и выделяет избыток ионов через
жабры во взрослом состоянии и через
специализированный шейный орган на
личиночных стадиях развития.
12.2.5. Наземная среда
В принципе главная физиологическая
проблема для наземных животных—
обезвоживание. Поверхность некоторых из
них, например земляных червей, слизней
и улиток, влажная, а их распространение
ограничено влажными местообитаниями
в почве и листовой подстилке, так что
назвать их истинно сухопутными нельзя.
После дождя среда в почве может стать даже
гипоосмотической, и некоторые нематоды
в таких условиях активно удаляют воду из
тканей (возможно, через кишечник).
Другую крайность представляют насекомые со
своим непроницаемым экзоскелетом,
способные обитать в очень сухих условиях.
Непроницаемость их покровов обусловлена
эпикутикулярным восковым слоем (рис.
12,5,у4) (см. также разд. 8.5.3Б.2). Если этот
слой удалить, потери воды за счет
испарения многократно возрастут. Значение эпи-
кутикулярного слоя для сохранения воды
показано также путем измерения скорости
368
Глава 12
writer
' (Кутикул!
Восковый
канал
Экзокутикула
Поровый
канал
«утикула
Эпидермис
CZ 10
f
?
с^Г^
24 26 28 30 32 34 36 38 40 42 44 46
Б Температура, °С
Рис. 12.5. А. Срез кутикулы насекомого с восковым эпикутикулярным слоем (no'Edney, 1974). Б.
Потери воды через кутикулу таракана в зависимости от температуры воздуха (по Beament, 1958)
испарения при разных температурах. Она
резко увеличивается при достижении
определенной температуры, соответствующей
точке плавления эпикутикулярного воска
(рис. 12.5.5).
Как уже отмечалось (с. 362), у
наземных ракообразных воскового
эпикутикулярного слоя нет, и они обитают в
основном во влажных условиях. Хотя мокрицы
обычно встречаются в сырых местах,
иногда им приходится переносить и воздействие
сухого воздуха. Неизбежные потери воды
при этом частично восполняются за счет
влажной пищи, которую потребляют эти
животные,—разлагающихся растительных
тканей; кроме того, некоторые их виды
способны пить воду и поглощать ее через анус.
Пустынному равноногому рачку Hemilepi-
stus избегать высыхания позволяют
поведенческие приспособления: в жаркое
дневное время он прячется в норках. Их глубина
достигает 30 см, в них гораздо прохладнее,
чем на поверхности, а относительная
влажность может составлять 95% (подробнее
см. Edney, 1957).
Насекомые также способны пить воду,
получать ее из пищи или в результате
метаболического окисления органических
молекул, например:
СН„0. + 60,->6СО, + 6Н,0.
Более того, некоторые наземные насекомые
и пауки поглощают водяные пары
непосредственно из атмосферы. Каким образом,
точно неизвестно; в различных группах
в этом процессе участвуют эпителии
ротовой полости, прямой кишки и, возможно,
трахейная система.
12.2.6. «Завоевание» суши
и пресных вод
Никто не сомневался в том, что
зарождение жизни и адаптивная радиация
организмов произошли в морской среде (гл. 2).
Завоевание других местообитаний шло как
прямо (из моря на сушу, из моря в пресные
воды), так и через промежуточные стадии
(из моря на сушу через пресные воды, из
моря в пресные воды через сушу). Кроме
того, выход из моря на сушу мог
происходить через литораль или интерстициаль.
Эти варианты схематически показаны на
рис. 12.6. Мы уже говорили о
несостоятельности теории Макаллума (с. 363), но все же
разумно предполагать, что предковые
образ жизни и местообитание отразились
на составе внутренней среды современных
Экскреция, ионная и осмотическая регуляция
369
Суша
Литораль
ЙИнтер-
встициаль
Море
Рис. 12.6. Пути «завоевания» суши (—) и
пресноводных местообитаний ( )
животных. Так, вполне вероятно, что
осмотическое давление жидкостей тела у
наземных животных, происшедших
непосредственно от морских предков, выше, чем
у потомков пресноводных форм (с. 365).
Данные, приведенные в табл. 12.3, в
основном подтверждают это предположение, но,
Таблица 12.3. Осмотическое давление крови некоторых наземных животных (Little, 1983, с
изменениями) 1)
как и следовало ожидать, не всегда.
Например, считается, что у сухопутного
десятиногого рака Holthuisana transversa
пресноводные предки, тем не менее осмотическое
давление его гемолимфы довольно высокое.
Хотя у мелких пресноводных ракообразных
осмотическое давление жидкостей тела
низко (< 300 мосмоль), пресноводные
десятиногие раки сохранили более высокие его
значения (> 500 мосмоль), возможно,
в связи с более крупными размерами.
Таким образом, высокое осмотическое
давление у наземных представителей этой группы
не противоречит их происхождению от
пресноводных форм.
Животные, вышедшие на сушу через ин-
терстициаль, например некоторые немерти-
ны, нематоды и аннелиды, вероятно,
пережили в своей эволюционной истории
условия, промежуточные между пресноводными
и наземными. Донные отложения забуфери-
вают соленость окружающей воды, однако
по мере продвижения из моря на сушу
соленость интерстициальной воды снижается,
так что почвенные животные, следовавшие
этим путем, вероятно, должны были
выработать эффективные механизмы ионной
регуляции. Поэтому вполне возможно, что
Вид
Немертины
Agronemertes dendyi
Аннелиды
Lumbricus terrestris
Моллюски
Eutrochatella tankervillei
Poteria lineata
Pseudocyclotus laetus
Helix pomatia
Pomatias elegans
Agriolimax reticulatus
Ракообразные
Talitrus sp.
Holthuisana transversa
Porcellio scaber
Cardisoma armatum
Coenobita brevimanus
" Приведены средние вели
дой.
Таксономическое
положение
Hoplonemertea
Oligochaeta
Prosobranchia
—"—
—■•—
Pulmonata
Prosobranchia
Pulmonata
Amphipoda
Decapoda
Isopoda
Decapoda
Осмотическое
давление,
мосмоль
145
165
67
74
ЮЗ
183
254
345
400
517
700
744
—"— 800
niiiu для активных животных во влажных услов
Возможный путь заселения
суши
Морская литораль
Пресные воды
—"—
—"—
Солоноватые воды
Соленые болота
Морская литораль
Соленые болота
Морская литораль
Пресные воды
Морская литораль
—"—
—"—
лях или в равновесии с пресной во-
24-1314
I
370
Глава 12
концентрация жидкостей их тела низка, как
и у форм с пресноводными предками.
Примером служит сухопутная немертина Argo-
nemertes dendyi, которая, как считается,
проникла на сушу именно через интерстициаль
и характеризуется низким осмотическим
давлением внутренней среды (табл. 12.3).
Даже из весьма ограниченного набора
данных, приведенных в табл. 12.3, можно
сделать вывод, что большинство
беспозвоночных вышли на сушу прямо из моря.
Хотя две важнейшие наземные группы —
насекомые и паукообразные — в этой
таблице не отражены, они, судя по всему,
также происходят непосредственно от морских
предков. В отличие от беспозвоночных
происхождение наземных позвоночных
исключительно пресноводное. Их покровы, как
и у наземных членистоногих, особенно
насекомых (разд. 8.5.3Б.2), эффективно
противостоят испарению. Мягкотелые
беспозвоночные избегают высыхания, выбирая для
обитания влажные условия, однако иногда
им, по-видимому, приходится переносить
частичные потери воды и повышение
внутреннего осмотического давления.
Животные с разбавленными жидкостями тела
(происходящие от пресноводных предков)
обычно не способны выдержать такое
повышение; возможно, белки,
эволюционировавшие в разбавленных растворах, легче
денатурируются в концентрированных.
Поэтому мягкотелые беспозвоночные
с пресноводным происхождением хуже
приспособлены к жизни на суше, чем
непосредственные потомки морских форм.
Завоевав сушу, некоторые
беспозвоночные (ряд немертин, многоножек,
насекомых, брюхоногих моллюсков и т. д.)
вторично переселились в пресную воду.
Насекомые в этом особенно преуспели: из
примерно 1 млн. их описанных современных
видов 25 000—35 000 проводят в пресной
воде хотя бы одну стадию жизненного цикла.
Так как кутикула взрослых особей
непроницаема, осмотические проблемы стоят для
них не так остро, как для личинок, покровы
которых гидрофильны и слабо хитинизиро-
ваны. Поэтому у них возникли сложные
механизмы регуляции, в том числе ионные
насосы, локализованные в специальных
структурах (см. ниже). Эти личинки, как и
мягкотелые беспозвоночные, вторично
приспособившиеся к пресноводным условиям,
испытывают, по-видимому, такое же давление
осмотических условий, что и у форм,
перешедших в пресные водоемы
непосредственно из моря. В обоих случаях можно
ожидать снижения концентраций внутренней
среды организма.
12.3. Выделительные системы
Специализированные «выделительные
системы» встречаются во всех типах
беспозвоночных, за исключением стрекающих
и иглокожих. У губок их тоже нет, но у
пресноводных видов имеются сократительные
вакуоли. «Выделительные системы»
присутствуют и у аммониотелических животных,
т. е. не всегда участвуют в выведении азота.
Их основная функция почти всегда
заключается в ионной и осмотической регуляции
и лишь иногда в истинной экскреции; то же
можно сказать и о сократительных
вакуолях губок, поэтому сам термин
«выделительные системы» взят нами в кавычки.
В этом разделе мы сначала рассмотрим их
строение, а затем функцию.
12.3.1. Строение
Исходя из особенностей развития,
можно выделить две основные категории
выделительных систем: нефридии - грубочки
эктодермального происхождения,
врастающие внутрь тела, и целомодукты—
мезодермалытые протоки, развивающиеся
от внутренних органов к поверхности.
Нефридии бывают двух основных типов.
(I) Протонефридии -это замкнутые
системы трубочек, заканчивающиеся клетками
с мерцательным пламенем у плоских червей
(разд. 3.6.2) и немертин, луковицами (буль-
бусами) с мерцательным пламенем у
коловраток и соленоцитами у Priapulus,
некоторых гастротрихов, полихет и архианнелид
(рис. 12.7). (II) Метанефридии,
оканчивающиеся ресничными воронками, характерны
для олигохет (рис. 12.8) и, возможно,
некоторых других групп.
Целомодукты — это трубчатые, иногда
ресничные выделительные структуры,
которые до того, как было изучено их эмбриона-
2
Экскреция, ионная и осмотическая регуляция
371
Жгутик
Рис. 12.7. А. Луковица с мерцательным
пламенем, замыкающая протонефридии
коловраток: I — продольный срез; II — поперечный
срез. Б, В. Соленоциты приапулид и брюхорес-
ничных соответственно (по Barrington, 1979)
льное развитие, часто не отличали от не-
фридиев. Они встречаются у онихофор,
членистоногих и моллюсков. У Peripatus по два
целомодукта (называемых коксальными
железами) почти в каждом сегменте (рис.
12.9), но у животных других групп их
обычно меньше. У ракообразных они
открываются в основании антенн (антеннальные же-
Рис. 12.8. Схема метанефридия аннелиды (см.
также рис. 12.15)
лезы; рис. 12.9) и/или вторых максилл (мак-
силлярные железы; рис. 12.9); у некоторых
одноветвистых они также связаны с максил-
лами, а у пауков протоки пары целомодук-
тов ведут на поверхность шестого сегмента
(коксальные железы, рис. 12.9). У
моллюсков «выделительные» целомодукты
(«почки») произошли из общего зачатка с
сердцем и гонадами, но в ходе эволюции четко
обособились от этих органов, как показано
на рис. 12.10. (Следует отметить, что, хотя
по определению эти целомодукты являются
«целомическими», т. е. их полость окружена
мембраной мезодермального
происхождения, они не связаны с целомом. На самом
деле животные с выделительными целомо-
дуктами лишены целомической полости, а
у целомических беспозвоночных
«целомодукты», связывающие целом с внешней
средой, не используются для экскреции!)
В состав некоторых выделительных
систем, называемых миксонефридиями или
нефромиксиями (рис. 12.11), входят как
эктодермальные, так и мезодермальные
компоненты. Они особенно распростране-
мж ГП
Рис. 12.9. Выделительные и половые целомодукты онихофор и некоторых членистоногих (по
Goodrich, 1945): аж—антеннальная железа; кж—коксальная железа; гп — гонопор; мж—
максиллярная железа
24*
372
Глава 12
Кишка (К)
Гонада
Перикардиальная
полость (П)
Сердце (С)
Почечно-перикардиаль-
ный канал
(Пч)
ПРИМИТИВНАЯ
ФОРМА
Мочеполовое \(например, неоменио-
отверстие уюроры)
Гонопор (г) Мочевы-
водящее м г
отверстие (м) ДВУСТВОРЧАТЫЕ ГОЛОВОНОГИЕ
ЭДНОПРЕДСЕРДНЫЕ
БРЮХОНОГИЕ
Рис. 12.10. Репродуктивные
и выделительные органы
моллюсков (по Goodrich, 1945)
ны у полихет, но «нефридии» форонид, си-
пункулид, эхиурид и плеченогих, по-
видимому, также относятся к этому типу.
«Выделительные системы» нематод
и крылатых насекомых совершенно иные.
У нематод это всегда одноклеточная
вентральная железа, расположенная в псевдо-
целе, с терминальной ампулой,
открывающейся порой на вентральной поверхности
тела. Она может быть связана с каналами,
образующими так называемую
Н-систему (рис. 12.12). Ее боковые каналы
внутриклеточные, а вся система
одноядерная. У насекомых органами выделения
служат мальпигиевы сосуды, слепые трубочки
(от одной до многих), открывающиеся в
пищеварительный тракт на границе средней
и задней кишки (рис. 12.13). Сходные
трубочки, возможно, также с выделительными
функциями присутствуют у многоножек,
некоторых паукообразных и тихоходок, но
возникли они, по крайней мере у пауков
и тихоходок, независимо, так что
представляют собой результат конвергентной
эволюции. Наконец, еще одной интересной
выделительной структурой является так
называемый клубочек кишечнодышащих полу-
Рис. 12.11. Схема строения нефромиксия
(= миксонефридия): М — мезодермальный
(= целомодуктовый) компонент; Э—экто-
дермальный (= нефридиальный) компонент;
Г—гаметы
Рис. 12.1 2. Выделительные системы нематод./! и Б— системы Н-типа с двумя вентральными
железистыми клетками; В—асимметричная система, состоящая из одной ветви Н-системы; Г—
Н-система без железистых клеток; Д и Е—укороченные Н-системы; Ж—укороченная и
редуцированная Н-система; 3—система из единственной вентральной железистой клетки (по Lee,
Atkinson. 1976)
Рис. 12.13. Мальпигиевы сосуды (М) и ректальная железа (РЖ) насекомых;-«движение
пищи; > движение воды и некоторых ионов (мальпигиевы сосуды и ректальная железа
расположены так, что вода и ионы могут циркулировать по кругу); ...> мочевая кислота (по Potts, Parry,
1964). Показана также ультраструктура клетки мальпигиевого сосуда (по Oschman, Berridge,
1971). Заметно большое количество митохондрий, что предполагает участие этой клетки в
активном транспорте
374
Глава 12
хордовых. Он образован выпячиванием
перитонеума в целом хоботка.
12.3.2. Функция
Итак, существует множество типов
различных по эмбриональному
происхождению «выделительных систем». Несмотря на
такое разнообразие, принцип их действия
сходен, поскольку в образовании
выделяемой жидкости участвуют лишь два
процесса.
1. Ультрафильтрация: жидкость под
давлением проходит через
полупроницаемую мембрану, задерживающую белки
и другие макромолекулы, но
пропускающую воду и низкомолекулярные
растворенные вещества.
2. Активный транспорт: движение
веществ против градиента их концентрации
с затратой энергии. Если оно направлено
в выделяемую жидкость, процесс называют
активной секрецией, если в
противоположную сторону,— активной реабсорбцией.
12.3.2.1. Системы, участвующие
в ультрафильтрации
Считается, что давление, необходимое
для ультрафильтрации, создается биением
КМ Л К М
Рис. 12.14. Антеннальная (зеленая) железа
рака и изменение содержания хлоридов в моче
по мере ее прохождения по этому органу:
КМ — концевой мешочек; Л—лабиринт; К—
почечный каналец; М — мочевой пузырь (по
Potts, Parry, 1964)
жгутиков или ресничек (мерцательного
пламени) в протонефридиях и «давлением
крови» в целомодуктах, например на уровне
концевого мешочка антеннальной (зеленой)
железы ракообразных (рис. 12.14) или
в сердце моллюсков. Из этих структур
ультрафильтрат поступает в канальцы, а
у моллюсков - через перикардиальную
полость в почки. Все низкомолекулярные
компоненты жидкостей тела переходят в
ультрафильтрат в концентрациях,
пропорциональных их содержанию в этих жидкостях.
Физиологически важные молекулы, в
частности глюкоза, а у пресноводных
беспозвоночных также ионы Na+, K+, CI и Са2+
в канальцах реабсорбируются (см.,
например, рис. 12.14), а токсичные и ненужные
вещества остаются в выделяемой жидкости.
Как уже отмечалось, этот избирательный
процесс связан с активным транспортом.
12.3.2.2. Системы, не участвующие
в ультрафильтрации
Фильтрация с помощью ресничек
Движение целомической жидкости
в «выделительные» каналы метанефридиев
и нефромиксиев обеспечивается биением.,
ресничек. По мере прохождения через эти
каналы состав жидкости изменяется за счет
активного транспорта (рис. 12.15).
Этим путем могут выводиться и
экскреты. Например, у земляных червей в
выделительных процессах участвует хлорагогенная
ткань эпителиального происхождения,
лежащая в целоме на поверхности средней
кишки. Хлорагогенные клетки запасают жиры
и углеводы, но, кроме того, в них
происходит дезаминирование, и образующиеся
аммиак и мочевина поступают в целом, а
оттуда в воронки нефридиев. Остатки,
образующиеся при распаде хлорагогенных
клеток, также могут удаляться через нефридиа-
льную систему. У пиявок функцию хлораго-
генной ткани выполняет ботриоидальная
ткань. У некоторых групп, например в роде
Theromyzon, экскреты из нее удаляются
через открытую систему нефридиев, однако
в других случаях, в частности у Glossiphonia
и Hirudo, воронки метанефридиев отделены
от выводных протоков перегородкой.
В этих случаях биение ресничек направлено
Экскреция, ионная и осмотическая регуляция
375
Узкий Широкий Мочевой
каналец каналец пузырь
200
1 150
СО
f «о
zr
I 50
0
Рис. 12.15. Осмотическое давление в
различных отделах метанефридия земляного червя
(по Potts, Parry, 1964)
из воронок внутрь целома и служит для
расселения образующихся в них амебоцитов.
Протоки открываются наружу и. возможно,
служат для ионной и осмотической
регуляции.
Мальпигиевы сосуды насекомых
Прямое поступление О, в ткани
насекомых через трахеи снизило роль
находящейся под давлением циркуляторной
системы. Поэтому давление жидкости внутри
и снаружи мальпигисвых сосудов примерно
одинаково, что исключает возможность
ультрафильтрации. Вместо этого ионы
калия и. возможно, также другие
растворенные вещества, активно секретируюгся
в просвет сосуда, а вода поступает туда за
счет осмоса. Секретируемые ураты (соли
мочевой кислоты) поддерживаются в
растворенном состоянии благодаря высокому
значению рН в дистальной части сосудов.
Из них жидкость поступает в заднюю
кишку, где происходит реабсорбция воды
(особенно интенсивно ректальными
железами; см. рис. 8.27), значение рН снижается,
а ураты осаждаются в виде мочевой
кислоты, которая выводится с фекалиями (см.
рис. 12.13).
12.3.2.3. Выделительная система
нематод
О принципах действия этой системы
известно мало, однако функции каналов
и железистых клеток почти наверняка
различны. Первые, по-видимому, участвуют
в ионной и осмотической регуляции, хотя
и лишены ресничек для создания тока
жидкости (обычно считается, что в процессе
участвуют ультрафильтрация и активный
транспорт). Вероятно, эта система не играет
существенной роли в выведении азота. У
некоторых нематод ее секреты, по-видимому,
обладают ферментативной активностью; их
источником скорее всего является
железистая клетка.
12.4. Выводы
Теперь читателю должно быть понятно,
что у беспозвоночных не только трудно, но
и бесполезно рассматривать экскрецию
(удаление азотистых отходов) отдельно от
ионной и осмотической регуляции и даже от
плавучести. Образование, транспорт и
удаление азотистых отходов на каком-то этапе
обязательно сопряжены с током и
фильтрацией жидкостей тела, даже если экскреты
в конечном итоге оказываются твердыми,
как, например, мочевая кислота. Поэтому
выделение тесно связано с движением
жидкостей и ионов, а значит, с процессами
осмоса, диффузии, активного транспорта
и ультрафильтрации. «Выделительные
системы» всегда участвуют в ионной и
осмотической регуляции, но не всегда —
собственно в экскреции. Несмотря на
разнообразие строения, их функционирование
основано на сходных процессах, и именно
на них сделан акцент в этой главе. И
наконец, замена ионов на ионы другой массы
или жидкости на газ влияет на плотность
ткани и, как теперь должно быть понятно,
является важным фактором,
контролирующим плавучесть водных беспозвоночных.
12.5. Дополнительная
литература
Burlington E.J. W. 1979. Invertebrate Structure and
Function, 2nd edn. Nelson. London.
Beament J. W. L. 1958. The effect of temperature on
the waterproofing mechanism of an insect. J. exp.
Biol., 35, 494 519.
Denton E.J., Gilpin-Brown J.B. 1961. The
distribution of gas and liquid within the cuttlebone. J. mar.
biol. Assoc, U.K., 41, 365 381.
Denton E.J., Gilpin-Brown J.B. 1966. On the
376
Глава 12
buoyancy of the pearly Nautilus, J. mar. biol.
Assoc, 46, 723—759.
Edney E. B. 1957. The Water Relations of Terrestrial
Arthropods. Cambridge University Press.
Cambridge.
Edney E. B. 1974. Desert artropods. In: G. W. Brown
(Ed.) Desert Biology, (Vol. 2). Academic Press,
New York.
Hoar W.S. 1966. General and Comparative
Physiology. Prentice Hall, New Jersey.
Home F. R. 1971. Accumulation of urea by a
pulmonale snail during aestivation. Сотр. Biochem.
Physiol., 38A, 565 570.
Koehn R.K. 1983. Biochemical genetics and
adaptations in molluscs. In: P. W. Hochachka (Ed.) The
Mollusca (Vol. 2), pp. 305—330. Academic Press,
New York.
Krogh A. 1939. Osmotic Regulation in Aquatic
Animals. Cambridge Univesity Press, Cambridge.
Lee D. L., Atkinson H. J. 1976. Physiology of
Nematodes, 2nd edn. Macmillan, London.
Little C. 1983. The Colonisation of Land. Cambridge
University Press, Cambridge.
Needham J. 1931. Chemical Embryology (3 vols).
Cambridge University Press, Cambridge.
Pilgrim R. L. C. 1954. Waste of carbon and energy in
nitrogen excretion. Nature (London), 173, 491—
492.
Potts W.F. W., Parry G. 1964. Osmotic and Ionic
Regulation in Animals. Oxford University Press,
London.
Schmidt-Nielsen K. 1972. How Animals Work.
Cambridge University Press, Cambridge.
Schmidt-Nielsen K. 1979. Animal Physiology:
Adaptation and Environment. Cambridge University
Press, Cambridge.
Имеется перевод: Шмидт-Ниельсен К.
Физиология животных: приспособление и среда: в
2 т.— М.: Мир, 1982.
Приложение 12.1.
Некоторые определения
Если растворы разделены барьером
(мембраной), проницаемой как для
растворителя, так и для растворенного вещества,
то происходит следующее.
1. Растворенное вещество (ионы)
переходит из более в менее концентрированный
раствор путем диффузии (движение ионов
подчиняется более сложным
закономерностям, если между разделенными мембраной
растворами существует разность
электрических потенциалов; это более подробно
обсуждается в гл. 16).
2. Растворитель переходит из менее
в более концентрированный раствор за счет
осмоса; силу, обеспечивающую этот
переход, называют осмотическим давлением.
Оно зависит от числа молекул
растворенного вещества, но не от их природы.
Процессы 1 и 2 продолжаются до тех
пор, пока градиент концентраций не
исчезнет.
Мембраны живых организмов обычно
лишь частично проницаемы для воды и
растворенных веществ, как показано на
рисунке, где А—ион; * результирующее
движение ионов; -» движение воды.
I
А А
А
-*■
А А
А А
А
■■»•
А А
II
А А
—
А
А А
•»
А
Организм (0) Внешняя
среда(ВС)
ВС
А А
А
А А
AAA
AAA
AAA
о
ВС
В случае I организм изоосмотичен
окружающей среде;
в случае II он гиие/тосмотичен ей, а
в случае III—гияоосмотичен по
отношению к среде.
Термины изотонический и изоосмотиче-
ский (и по аналогии остальные) часто
используют как синонимы, но это не вполне точ- ,
но, так как тоничность описывает реакцию (
объема клетки (организма); если организм
изоосмотичен внешней среде, он обычно,
хотя и не всегда, ей изотоничен. Так,
например, яйца морского ежа изотоничны изоос-
мотическому раствору NaCl, но не изоосмо-
тическому раствору СаС12 (более детальное
объяснение см. Potts, Parry, 1964).
Экскреция, ионная и осмотическая регуляция
377
О смоконфор мерами называют
животных, у которых осмотическая концентрация
жидкостей тела изменяется в зависимости
от среды, т. е. остается ей изоосмотич-
ной.
Осморегуляторами называют
животных, поддерживающих осмотическую
концентрацию внутренней среды отличной от
осмотической концентрации внешней
среды.
Ионная регуляция—это поддержание
концентрации ионов в жидкостях тела на
уровне, существенно отличающемся от их
концентрации во внешней среде.
Эвригалинными называют животных,
жизнеспособных в широком диапазоне
концентрации солей в окружающей воде.
Стеногалинные животные могут
переносить лишь ограниченные колебания
солености воды.
Концентрации. Легко понять, что такое
вес вещества, но число молей—это более
сложное понятие. Моль (грамм-
моль)— это молекулярная масса,
выраженная в граммах, или масса вещества, в
котором содержится число Авогадро
(6,023 х 1023) его молекул. Молярные
концентрации удобны, поскольку отражают
число молекул растворенного вещества,
а именно от него зависит осмотическое
давление.
Молярность—число молей
растворенного вещества в литре раствора.
Моляльность — число молей
растворенного вещества на килограмм чистого
растворителя.
ГЛАВА I О
ЗАЩИТА
В этой главе сначала дается
классификация опасностей, которым подвергаются
беспозвоночные, а затем рассматриваются
способы защиты от них. Разнообразие
возможных опасностей определяет широкий
круг затрагиваемых вопросов: от защиты
против патогенов до защиты от хищников
и даже от процессов старения, если,
конечно, такая защита возможна.
13.1. Классификация
опасностей
13.1.1. Две основные группы
На рис. 13.1 представлены примеры
кривых выживаемости, т. е. численности (или
доли) особей, родившихся приблизительно
в одно и то же время (относящихся к одной
когорте) и доживших до того или иного
возраста. Можно видеть, что в природных
популяциях смертность либо наиболее высока
у молоди, либо распределена довольно
равномерно по всем возрастным группам.
Гибель здесь, вероятно, вызывается главным
образом экологическими, т. е. внешними по
отношению к организму, факторами типа
хищничества, несчастных случаев или
болезней. Молодь обычно более уязвима
к ним, чем взрослые особи, но иногда
чувствительность всех возрастных групп
сходна. В лабораторных культурах можно
исключить большинство внешних факторов
смертности, и она наиболее высока у особей
самых старших возрастных групп. Это
свидетельствует об увеличении уязвимости
организма с возрастом, вызванным, вероятно,
внутренними факторами, т. е. процессами
старения.
13.1.2. Экологические (внешние)
факторы смертности
Хотя существует множество
разнообразных экологических причин смертности,
они могут быть объединены в четыре
основные группы: несчастные случаи, болезни,
хищничество и стрессы, вызываемые
окружающей средой. Первые три очевидны и не
нуждаются в специальном пояснении,
а стрессы возникают либо при отсутствии
какого-нибудь необходимого для
организма фактора, либо при наличии стрессора—
природного токсина или антропогенного
загрязнителя.
13.1.3. Старение
Старение на первый взгляд идет при
исключении внешних факторов смертности
и может поэтому рассматриваться как
процесс внутренней дегенерации
физиологических систем, клеток и молекул, который,
вероятно, прослеживается вплоть до
денатурации биологически важных веществ
(нуклеиновых кислот и белков) под влиянием
теплового шума, ковалентной сшивки
боковых цепей макромолекул, самоокисления
и т. д. Не следует, однако, полагать, что на
эти внутренние процессы не могут влиять
внешние факторы. На рис. 13.2 показано,
что облучение всего тела снижает
продолжительность жизни дрозофилы, причем
эффект зависит от дозы радиации. Однако
форма кривых выживаемости при этом не
меняется, и некоторые геронтологи
связывают укорочение жизни в данном случае
с ускорением старения, возможно
объясняющимся в свою очередь повреждением
Защита
379
И.Полевые популяции
О
Рис. 13.1. А Кривые
выживаемости полевых популяций
беспозвоночных; х'—отклонение
(%) от средней
продолжительности жизни (по lto, 1980). Б.
Кривая выживаемости и
распределение по возрастам
абсолютного числа погибших
особей и их доли от общей
численности для лабораторной
популяции Drosophila melanogaster
(по Lamb. 1977); кривая
выживаемости лабораторной
популяции перепончатокрылого
насекомого Nasonia vitripennis
(по Davies, 1983)
Б. Лабораторные популяции
Drosophua
melanogaster
100
0 50 100 150 200
10 20
Возраст, дни
макромолекул, особенно ДНК,
высокоэнергетическим облучением. С другой стороны,
добавки некоторых веществ, например
витамина Е, в культуральную среду нематод
или в пищу дрозофил удлиняют
продолжительность их жизни. Эти вещества,
вероятно, защищают макромолекулы от
повреждений (так, витамин Е, по-видимому,
относится к антиоксидантам).
13.1.4. Классификация
Как показано выше, смертность
организмов всегда в той или иной степени
зависит от условий окружающей среды, однако
некоторые ее причины более тесно связаны
с самим организмом, чем другие. В табл.
13.1 ее факторы классифицируются в
соответствии со степенью такой связи: от
нападения хищников, с одной стороны, до
процессов старения, с другой. Простота
исключения того или иного фактора в условиях
эксперимента уменьшается в обратном
порядке.
30 40 50
Возраст, дни
Рис. 13.2. Влияние тотального
гамма-облучения на самцов Drosophila melanogaster (no
Lamb, 1977)
380 Глава 13
Таблица 13.1. Классификация факторов смертности
Фактор
Несчастный Хищниче-
случай ство
Болезнь
Внешний Внутренний
стрессор (на- стрессор (напри-
пример, загря- мер, системные
знитель) нарушения)
Зависимость от
погибшего животного
Легкость исключения
данного фактора
в эксперименте
Реакция организма
'' X — низкая; XXX — вьк
X»
XXX »
;окая.
X
XXX
Защита
XX
XX
Иммунная
защита
XX
XX
Устойчивость,
репарация
XXX
X
Репарация
13.2. Защита
13.2.1. Против хищников
Все животные могут быть съедены
другими животными (см. гл. 9) и, чтобы
избежать этого, защищают себя разными
способами, объединяемыми в три основные
группы: избегание потенциальных хищников,
пассивная защита и активный отпор.
13 2 1.1. Избегание
Этот способ включает либо избегание
встреч с хищниками, либо незаметность для
них. Примером использования обеих
стратегий служит массовая вертикальная
миграция некоторых морских и пресноводных
планктонных животных. Хотя ее характер
обычно сложен, он часто включает уход
вниз от солнечного света в дневное время
и всплывание ночью. Тем самым животные
снижают вероятность быть замеченными
хищниками, руководствующимися при
охоте зрением (рис. 13.3, А). Такое поведение
особенно выражено у наиболее уязвимых
для хищничества видов и возрастных
классов. Более того, в результате уникального
исследования одного из видов веслоногих
в нескольких изолированных горных озерах
показано (Gliwicz, 1986), что вертикальные
миграции этого рачка происходят только
там, где обитают планктоноядные рыбы.
Удалось также пронаблюдать эту
миграцию в одном из озер на разных стадиях его
заселения такими рыбами. Через 12 лет
после первого их появления вертикальная
миграция была выражена слабо, но еще через
23 года четко прослеживалось погружение
рачков в дневное время. Однако
хищничество не служит единственным и
исчерпывающим объяснением эволюционного
возникновения вертикальных миграций, ибо
некоторые беспозвоночные опускаются в их
процессе на глубину, намного
превышающую необходимую для того, чтобы
скрыться от света. Кроме того, некоторые
мигрирующие зоопланктеры люминесцируют
по ночам, увеличивая этим свою
видимость. Другие возможные объяснения
вертикальных миграций включают
оптимизацию использования неравномерно
распределенной пищи (гл. 9), экономию энергии
и связь с горизонтальной миграцией.
Сходные вертикальные миграции, но
в меньшем масштабе, наблюдаются у
пресноводных беспозвоночных, обитающих на
погруженных камнях. Днем животные
находятся под ними в неактивном состоянии,
а ночью обычно перемещаются на
поверхность валунов, где ведут активный образ
жизни. Большое количество этих, как
правило, бентосных организмов можно
обнаружить пассивно плывущими в потоке
воды. Этот так называемый дрейф
беспозвоночных особенно выражен в сумерках (рис.
13.3, Б), вероятно, потому, что именно в это
время они выползают на омываемую
течением верхнюю часть камней, откуда легко
могут быть смыты.
Особые «реакции избегания»
представляют собой крайнюю форму локомоторно-
Защита 381
го ускользания от опасности. При них
используются либо обычные двигательные
стереотипы, либо специализированные
поведенческие ответы. Так, у каракатицы
Sepia чернильный мешок содержит жидкость
с гранулами меланина. Будучи атакован,
моллюск выталкивает ее наружу, создавая
«чернильную завесу», тут же изменяет свой
цвет на очень бледный и уплывает под
прямым углом к предшествовавшему
направлению движения. Двустворчатые
моллюски, например обычная прикрепленная
форма Cerastoderma, могут избегать
хищника за счет внезапных и быстрых сокращений
мышц ноги и аддукторов раковины.
Исключительно мощную реакцию избегания
моллюсков этого рода вызывает прикосновение
амбулакральных ножек морской звезды;
вероятно, она индуцируется какими-то
веществами, выпускаемыми иглокожим в воду.
Криптическая (маскирующая, или
покровительственная) окраска—еще один широ-
0
50
100
ко распространенный метод избегания
хищников у беспозвоночных (см. рис. 9.33).
Примеры ее можно, по-видимому,
обнаружить во всех типах животных, но наиболее
детально этот феномен изучен на
некоторых улитках и насекомых.
1. Исчерченность раковины у Сераеа пе-
moralis. У этого вида наземных улиток
широко варьируют цвет и узор раковины за
счет изменения ее фонового оттенка, а
также числа, ширины и яркости окраски полос.
Все эти варианты находятся под
генетическим контролем. Дрозды питаются
особями Сераеа, находя раковины с помощью
зрения и разбивая их о камни, так
называемые дроздовые наковальни. В середине
1950-х гг. Кейн и Шеппард (A. J. Cain,
P. M. Sheppard) показали, что окраска
раковин, разбитых дроздами, в среднем лучше
заметна, чем у живых улиток, обитающих
в том же районе. Различные узоры по-
разному заметны в зависимости от место-
1900 0100 0700 1300
1600 2200 0400 1000 1600 ч
Рис. 13.3. Вертикальные
миграции (А) веслоногого рачка
Calanus и (Б) более
глубоководной креветки-акантвфири-
ды (по Barnes, Hughes, 1982).
Влияние света и темноты на
миграции некоторых
пресноводных беспозвоночных (В)
в открытом водоеме и (Г)
в управляемой
экспериментальной системе (по Holt, Waters,
1967)
Г
о
о
S
§
я
о
о
я
о
о
я
о
о
я
о
о
CNJ
о
о
CD
о
о
а
382
Глава 13
Полосатая Светлая
светлая
Темная
Ж
Т*Ж;
уЛ
Ж
ж ж
ж т
'ж
ж
Ж'
ж
тт
'*ж
л
,тт
'Ж
ж
4
ЖЖЛ пЖТЛ
„л
л
о
т ЛЛл л' п
т л лл л л
л л л
-л6^
80
Рис. 13.4.
20 40
Процент весполосых
Частоты встречаемости различных
морф Сераеа в разных местообитаниях.
Светлые раковины с полосами малозаметны в
низкой растительности (Т—трава, Ж—живые
изгороди), тогда как темные однотонные
раковины труднее различимы в тенистых лесах (Л)
(по Calow. 1983; оригинальные данные из
работы Cain, Sheppard, 1954)
обитания и времени года. Например,
полосатые светлые раковины трудно разглядеть
в буйной растительности (луга, живые
изгороди), где взаимодействие проникающего
света и узких теней создает резко
контрастирующие темные и светлые участки. В
тенистых лесах труднее всего различить
темные раковины без полос (рис. 13.4).
2. Меланизм у березовой пяденины
(Bistort hetularia). У многих бабочек и других
насекомых сформировались на крыльях
и теле рисунки, делающие их незаметными
на деревьях, обросших лишайниками.
Промышленные загрязнения губят лишайники
и покрывают стволы копотью. В таких
районах ранее адаптированные к фону
светлые бабочки типичной формы замещены
темными, ме.шнистическими формами. Это
явление известно как индустриальный
меланизм (рис. 13.5). В 1950-е гг. Кеттлуэлл
(Н. В. D. Kettlewell) показал его связь у
березовой пяденицы с селективным выеданием
бабочек птицами. При прямом наблюдении
было установлено, что в загрязненных
местах типичные формы более уязвимы для
насекомоядных птиц, чем меланистические.
Рис. 13.5. Частоты трех морф березовой пяденицы (insularia и carbonaria — меланистические).
Несмотря на то что окраска insularia кажется промежуточной между типичной и carbonaria, она
контролируется другим генным локусом (по Sheppard, 1958)
Защита
383
и наоборот. Кеттлуэлл выпускал пядениц
в бочку, раскрашенную изнутри черными
и белыми вертикальными полосами.
Оказалось, что 65% бабочек предпочитали
совпадающий с ними по цвету фон (типичные —
белый, меланистические — черный), т.е.
соответствующий поведенческий механизм
позволяет им выбирать для отдыха такой
фон, на котором они наименее заметны.
(Как и следовало ожидать, эволюция по-
лосатости у улиток и меланизма у бабочек
протекала сложнее, чем представлено в
данном здесь кратком описании. За более
подробными сведениями можно обратиться
к генетическим работам, например Berry.
1977.)
И наконец, маскировка не всегда делает
животное незаметным, но и придает ему
иногда сходство с несъедобными
объектами. Многие насекомые напоминают внешне
сучки или листья растений. Личинки ранних
возрастов некоторых видов бабочек-
парусников хорошо заметны, но избегают
нападения, поскольку своей черной
окраской с белым «седлом» на спине похожи на
птичий помет (см. рис. 9.33)! Их узор резко
меняется, когда они становятся чересчур
крупными для такого способа маскировки.
Эриксен и др. (Erichsen et al., 1980)
провели оригинальные опыты по выяснению
влияния покровительственной окраски на
выбор жертвы насекомоядными птицами.
Большой синице (Parus major) предлагалось
выбрать между крупными и мелкими
личинками мучного хрущака (Tenebrio moli-
tor), помещенными в соломинки, причем
крупные особи находились в
непрозрачных соломинках, напоминающих веточки,
а мелкие—в прозрачных, что делало их
легко заметными. Энергетическая ценность
крупных личинок примерно вдвое выше,
поэтому они были более выгодной пищей,
но их обнаружение требовало больше
времени, потраченного птицей на
рассматривание и перебирание «веточек». В
экспериментах синицы выбирали мелких личинок, если
«веточек» было много, и переключались на
крупных, если мало.
Другие животные маскируются,
прикрепляя к себе материал из окружающей
среды. Например, крабы-пауки покрывают
кусочками водорослей и другими предметами
острые выросты своего экзоскелета.
Домики личинок ручейников, вероятно, также
используются для маскировки. Так, для
вида Potamophyiax cingulatus, у которого они
построены из листьев, вероятность стать
жертвой форели меньше, когда фоном
служит дно, покрытое листьями, а не песком.
По мере роста личинки листовой домик
сменяется песчаным, но в этом случае
вероятность поедания уже не зависит от
фонового субстрата, что, возможно, объясняется
низкими вкусовыми качествами нового
домика (см. ниже). (Более подробно см. Han-
sell, 1984.)
13.2.1.2. Пассивная защита
Животные могут физическими или
химическими методами «разубеждать»
хищников нападать на них.
Широко используются для этого
известковые защитные образования, например
спикулы губок, скелеты кораллов, трубки
аннелид, раковины моллюсков и плечено-
гих, домики некоторых лофофоровых,
панцири иглокожих. Хитинизированные
покровы членистоногих также могут
использоваться для защиты и в ряде случаев
укреплены соединениями кальция. Это особенно
заметно у сидячих усоногих рачков, покрытых
толстыми известковыми пластинками (см.
рис. 8.36, А).
Иногда неорганические включения
настолько снижают питательную ценность
тканей животного, что оно теряет свою
привлекательность как источник пищи.
Возможно, такова функция, по крайней мере
частично, известковых или кремнеземных
скелетов губок, известковых скелетов
кораллов, трубочек из гравия или песчинок,
окружающих тело некоторых полихет и личинок
ряда пресноводных насекомых (см. выше).
Удивительно, но лишь у небольшого числа
морских турбелларий в стенке тела
обнаруживаются известковые пластинки или шипы
(гл. 3). Их функция, по-видимому, опорная,
но есть основания полагать, что они
одновременно снижают питательную ценность
организма. Более того, кажется вполне
правдоподобным, что раковина моллюсков
произошла именно от таких образований.
Раковина — наиболее очевидное сред-
384
Глава 13
I
Рис. 13.6. Защитная скульптурированность
раковин моллюсков
ство физической защиты (см. рис. 9.34). Ее
толстая стенка, сильная
скульптурированность поверхности, закрывающееся устье
и спиральная форма эффективно
предохраняют многих морских бентосных гастропод
от рыб, крабов, омаров и других хищников,
способных разламывать раковины других
форм (рис. 13.6). Крышка, прикрывающая
устье раковины наземных улиток,
препятствует проникновению внутрь нее хищных
жуков. Сложная скульптура, включающая
разнообразные выросты, ребра, шишки
и шипы, служит не только для укрепления,
но и повышает эффективный размер, тем
самым снижая доступность животного для
хищников или увеличивая время
переработки жертвы. То, что у пресноводных
моллюсков нет толстостенных раковин со сложной
архитектурой, возможно, частично
объясняется отсутствием в их местообитаниях
специализированных моллюскофагов.
Кроме того, это может быть связано с меньшей
доступностью кальция в пресной воде.
Необычные защитные покровы
встречаются у асцидий. Их наружный
однослойный эпителий дополнительно окружен
туникой (оболочники по латыни—Tunicata;
см. разд. 7.4А), обычно достаточно
толстой, но сильно варьирующей по
консистенции— от мягкой и непрочной до грубой
и хрящеподобной. Она состоит из
волокнистого материала, основным компонентом
которого у многих, хотя и не у всех, асцидий
служит особая форма целлюлозы,
называемая туницином. Помимо этого в состав
туники входят белки и неорганические
включения, например кальций. Она бывает ва-
скуляризована и иногда содержит
амебоидные клетки, т. е. ее нельзя считать мертвым
покровом.
Защитные структуры не всегда секрети-
руются самим обороняющимся
организмом. Хороший пример обратной
ситуации— домики личинок ручейников. Они
построены из материалов, полученных из
окружающей среды. Сходным примером
может служить так называемый
«фекальный щит» личинок жука Cassida rubiginosa,
хотя в этом случае защита обеспечивается
отходами жизнедеятельности
обороняющегося. Речь идет о спрессованной массе
сброшенных покровов и фекалий, которую
личинка носит на прикрывающем ее спину
особом вилкообразном органе. Она может
маневрировать щитом, используя его для
защиты от нападения других насекомых,
в частности муравьев. Раки-отшельники
забираются в пустые раковины гастропод,
избавляясь таким образом от необходимости
создавать собственный экзоскелет. Их уро-
поды модифицированы, причем
увеличенная левая приспособлена для удерживания
за столбик раковины (см. рис. 8.49, Д).
Химические методы пассивной защиты
также широко распространены у
беспозвоночных. У многих видов ткани тела
содержат токсины. Так, до 0,3% веса одной из не-
мертин составляют нейротоксины.
Различные токсины, в том числе серную кислоту,
используют многочисленные
безраковинные брюхоногие моллюски, например зад-
нежаберники и легочные слизни. Некоторые
губки выделяют раздражающие вещества;
было показано, что экстракт из
пресноводных губок убивает мышей. Ряд животных
пользуется чужими токсинами. Так,
некоторые крабы «украшают» себя губками,
возможно, для маскировки, а возможно, с
целью использования образуемых ими токси-
Защита
385
нов. Сходным образом определенные виды
раков-отшельников обеспечивают себе
защиту, занимая раковины моллюсков с
прикрепленными к ним губками и
актиниями.
Турбелларии, судя по всему, не
выделяют токсинов в ткани своего тела, но зато
образуют рабдоиды—стержневидные
структуры, расположенные в эпидермисе
перпендикулярно поверхности тела и секре-
тируемые эпидермальными железистыми
клетками (разд. 3.6.3.1). При раздражении
червя они выбрасываются наружу.
Защитную роль рабдоидов легко
продемонстрировать экспериментально. Колюшки
охотно питаются олигохетой Tubifex tubifex
и могут выхватывать ее прямо из пинцета.
Однако, если предварительно покрыть
червя слизью, выделяемой турбелларией при
раздражении, рыба отказывается от такой
пищи. Если же Tubifex покрыт слизью,
оставляемой на субстрате спокойно
ползущим плоским червем, приманка съедается.
Слизь раздраженных турбелларии
содержит много рабдоидов, тогда как в слизи
непотревоженных животных их нет или очень
мало. Рабдоиды, вероятно, выполняют
и другие функции, например участвуют
в быстром образовании самой слизи или
служат антимикробными агентами.
Токсины обычны у насекомых, которые
в свою очередь могут «заимствовать» их от
растений, на которых питаются. Например,
кузнечик Poekilocerus bufonius поедает
молочаи, содержащие сложные токсины (так
называемые карденолиды), способные
нарушать сердечную деятельность.
Кузнечик извлекает их из своей пищи,
запасает в ядовитой железе, а при нападении
хищника выпускает содержащую эти
вещества жидкость. Если выдержать
кузнечиков на рационе без молочаев, содержание
карденолидов в выпускаемой жидкости
снижается в 10 раз. Бабочки-данаиды также
питаются молочаями, насыщая свои ткани
карденолидами, что делает их
несъедобными для птиц. Если исключить эти растения
из рациона их личинок, данаиды
оказываются безвредными для хищников. Токсичные
вещества типа карденолидов, не
вырабатываемые в организме насекомых, а
получаемые ими из растений, иногда называют кай-
ромонами, противопоставляя их алломонам,
т.е. токсинам, используемым самими
производящими их организмами (в данном
случае— растениями) (более детальное
описание см. Nordlund, Lewis, 1976).
Присутствие токсинов часто сочетается
с предостерегающей окраской. Такую
корреляцию можно встретить во многих типах
беспозвоночных, от немертин и слизней до
насекомых. Окраска обычно связана с
простым узором, часто включающим красный,
желтый, черный и белый цвета. Каждый
хорошо знаком с черными и желтыми
полосами пчел и ос.
Возникновение токсинов и
предостерегающей окраски в ходе эволюции—
непростой процесс. Их преимущество
проявляется только тогда, когда они
защищают от хищников, и в то же время
единственный способ для хищника научиться
распознавать несъедобное животное —
попробовать его на вкус. Одним из
механизмов может быть отбор сородичей, при
котором гибель одной особи с генами
признаков, предостерегающих о наличии токсина,
защищает особей, несущих такие же гены.
Гены могут распространяться аналогичным
способом, если их носитель с трудом
уязвим и способен выжить после обнаружения
его хищником или если хищник
отпугивается каким-либо неприятным стимулом еще
до нападения. У большинства насекомых
с предостерегающей окраской прочные
покровы, а ярко окрашенные слизни часто
испускают сильные запахи. Немертины
способны регенерировать ткани, уничтоженные
хищниками.
Нетоксичные животные могут
имитировать предостерегающие окраски; это
свойство называют бейтсовской мимикрией по
имени того, кто первым его
проанализировал. Поскольку предостерегающая окраска
у имитатора — обман, а у животного-
модели— правда, необходимо, чтобы а)
модель была несъедобной и ярко
окрашенной; б) модель встречалась чаще
имитатора, ибо, если она встречается редко, хищник
не усвоит взаимосвязи между внешним
обликом и средствами защиты; в)
имитаторы обитали рядом с моделями и были с
трудом от них отличимы. Еще одна форма
мимикрии называется мюллеровской; при ней
25—1314
386
Глава 13
/.
Ч"
ч
л
Б Г Е
Рис. 13.7. Бейтсовская мимикрия между бабочками Alcidis agarthyrsus (А) и имитирующей ееРа-
pilio lag (Б) и между североамериканским видом Limenitis archippus (В) и ее моделью—данаидой
Danaus plexippus (Г). Мюллеровская мимикрия между Podotricha telesiphe (Д) и Heliconius
telesiphe (£)
несъедобные виды конвергируют
морфологически, получая от этого взаимную
выгоду. Критерий 6 здесь не существенен,
а сходство критерий в не обязательно
должно быть столь точным. У различных
видов ос и пчел одинаковый полосатый
рисунок: это одно из проявлений мюллеров-
ской мимикрии. Многие двукрылые, в
частности мухи-журчалки и некоторые бабочки,
внешне походят на ос и пчел — это
бейтсовская мимикрия. Примеры для чешуекрылых
можно видеть на рис. 13.7.
Ход эволюции мюллеровской мимикрии
понять нетрудно. Существенной проблемой
при бейтсовской мимикрии остается
условие е, т. е. достаточно близкое сходство
между имитатором и моделью, чтобы оно
обеспечило надежную защиту. Как в таком
случае могли выжить формы,
промежуточные между немимикрирующим предком
и современным имитатором? Одним из
возможных путей может быть двухфазный
эволюционный процесс: установление
приблизительного, но достаточно адекватного
сходства в результате одной крупной
мутации с дальнейшей постепенной «подгонкой»
деталей путем отбора мелких генетических
изменений.
13.2.1.3. Активный отпор
Органы, использующиеся для захвата
и умертвления добычи, часто могут
применяться и для активной защиты от
хищников. Примером служат книдобласты
стрекающих. Как это ни парадоксально,
книдобласты видов, которыми питаются
голожаберные моллюски, могут заимствоваться
последними для собственной защиты.
Неразряженные нематоцисты съеденной кни-
дарии транспортируются ресничками
желудка не перевариваясь в выросты на его
дорсальной поверхности («рожки», или це-
расы^, дистальные концы которых, книдоса-
ки, открываются наружу (рис. 13.8).
Отстреливание нематоцист, по-видимому,
вызывается сокращением кольцевых мыщц кни-
досака. Запас нематоцист может быть
восстановлен приблизительно за 10 дней,
причем большинство голожаберников
используют лишь определенные их типы,
присутствующие у жертв. Некоторые турбелларии
аналогичным путем пользуются нематоци-
стами гидроидов, а гребневик Euchlora ги-
Ьга—нематоцистами медуз.
Хитинизированные челюсти и жала
членистоногих также служат примером средств
нападения, применяемых для защиты.
С другой стороны, жало иногда играет
только защитную роль, например у медонос-
Защита
387
Пищевод
Проток к пищеварительной
железе и "рожкам"
Нематоцисты
Ядро клетки
книдосака
Слой кольцевых-
мышц
Кишечник
Рис. 13.8. Желудок голожаберного моллюска со вскрытой дорсальной стенкой (.4) и книдосаки
(Б) (по Barnes. 1980)
ной пчелы. Оно представляет собой
модифицированный яйцеклад, не используемый
для откладки яиц, состоит из парных
зазубренных ланцетов и непарного стилета (рис.
13.9) и в состоянии покоя находится в
футляре внутри седьмого абдоминального
сегмента. Механизм действия жала разобран
в подписи к рисунку. Ядовитая жидкость се-
кретируется парой продолговатых желез
в брюшке и у медоносной пчелы содержит
помимо собственно яда ферменты,
вызывающие образование гистамина в тканях
жертвы.
Жуки рода Brachimts, известные под
названием жуков-бомбардиров,
разбрызгивают особую жидкость для отражения
атаки хищников — пауков, богомолов и даже
лягушек. Потревоженный жук выпускает ее
из пары желез, расположенных на кончике
брюшка. Последнее способно вращаться,
поэтому жидкость может быть направлена
точно в цель, фактически в любом
направлении. Ее действующие составляющие —
бензохиноны, синтезируемые взрывооб-
разно в момент разбрызгивания путем
окисления фенолов. Это сопровождается
хорошо слышимой детонацией, причем
жидкость извергается при температуре
100 С!
Большинство двупарноногих
многоножек передвигаются относительно медленно;
для своей защиты помимо толстого
известкового экзоскелета они используют также
набор отпугивающих желез (рис. 13.10). Их
выводные отверстия расположены по краям
тергитов, обычно по паре на сегмент, хотя
на некоторых сегментах полностью
отсутствуют. Состав секрета у разных видов
неодинаков; в него могут входить альдегиды,
хиноны, фенолы и синильная кислота.
Последняя синтезируется непосредственно
перед использованием при смешивании ее
предшественника и фермента,
образующихся в разных долях железы. Эта
жидкость токсична или репеллентна для других
мелких животных, а у крупных тропических
видов действует разъедающе на кожу
человека. Обычно она выделяется медленно, но
у некоторых многоножек выпускается под
большим давлением струей или в виде
брызг на расстоянии 10—30 см. Такое ее
извержение вызывается, по-видимому,
сокращением туловищных мышц,
прилежащих к секреторному мешку. Плотоядные
388
Глава 13
Ядовитый мешок
Квадратная
пластинка
Б Разрез
Щелочная
железа
А
Рис. 13.9. Жало медоносной пчелы (Apis mellifera). Стержень, состоящий из стилета и
зазубренных ланцетов, опускается при сокращении мышцы м,, а затем сокращение мощных мышц м2,
идущих от квадратной пластинки к передней части продолговатой пластинки, приводит к повороту
треугольной пластинки и к выталкиванию ланцета наружу. Втягивание ланцета происходит при
сокращении мышц м3. Мышцы по обе стороны жала работают поочередно, так что за счет
последовательных выдвиганий и втягиваний ланцетов они все глубже проталкиваются в тело жертвы.
Яд секретируется парой нитевидных кислых желез в брюшке. Их секрет накапливается в ядовитом
мешке, проток которого открывается у основания жала в ядовитый канал. Функция так
называемой щелочной железы точно неизвестна (по Imms. 1964)
губоногие многоножки передвигаются
быстрее, поэтому в меньшей степени
оснащены отпугивающими механизмами,
полагаясь в основном на спасение бегством и на
использование ядовитых коготков,
применяемых также для захвата добычи (рис.
8.21). Тем не менее некоторые их виды
обладают отпугивающими железами. Так,
некоторые литобиоморфы несут большое число
одноклеточных желез на последних четырех
парах ног, которыми они могут
отшвыривать врага, одновременно разбрасывая
Часть
железистого мешка
■Мышца,
окружающая выводной
проток
Рис. 13.10. Отпугивающие
железы двупарноногих
многоножек (по Cloudsley-Thompson,
1958)
Защита
389
клейкие капли секрета.
Педицелларии морских звезд и морских
ежей также относятся к органам,
сформировавшимся специально для защиты. Они
представляют собой клешневидные
придатки, используемые в основном для борьбы
с личинками, которые могут поселяться на
поверхности тела (рис. 7.18). Выделяют три
основных типа таких структур:
стебельчатые, сидячие и альвеолярные
(погруженные). Один из нескольких типов
педицелларии морских ежей содержит железы, секре-
тирующие яд, способный быстро
парализовать мелких животных и отпугнуть крупных
хищников. Аналогичные функции у зоои-
дов-авикуларий некоторых мшанок (разд.
6.3.3.3).
Интересная форма защиты —
«обескураживание» потенциальных хищников.
Узор на крыльях некоторых бабочек и
других насекомых напоминает глаза
позвоночного. В состоянии покоя «глаз» не видно, но
они мгновенно «открываются», стоит
только животное потревожить. Их присутствие,
возможно, связано и с отвлечением
внимания хищника на менее уязвимые части тела
или даже на органы защиты; например,
у некоторых ос белые абдоминальные пятна
находятся как раз около жала.
Эксперименты с отловленными птицами
продемонстрировали обе функции «глазных пятен».
Резко меняющаяся окраска жертвы также
может обескураживать хищника; быстрое
изменение цвета потревоженной
каракатицы—яркий пример такого рода.
13,2.2. Против внутренних врагов
Некоторые патогены могут проникать
через наружные защитные берьеры, но
любой организм располагает внутренней
линией обороны, способной противостоять
такому вторжению. У позвоночных это
иммунная система с антителами,
нейтрализующими специфические чужеродные
антигены. У беспозвоночных такой системы нет.
но и они, конечно же, располагают
внутренними средствами защиты, в основном —
фагоцитарными клетками, способными
распознавать и уничтожать чужеродный
материал.
13.2.2.1. Главное—отличать
«свое» от «чужого»
Классическим примером распознавания
«своего» служит реагрегация клеток у
губок. Продавливая кусочки губок через
ткань или помещая их в раствор этилендиа-
минтетрауксусной кислоты (ЭДТА), можно
разделить весь организм на отдельные
клетки. Смесь клеток разных видов или
даже клонов одного и того же вида даст видо-
специфичные и клоноспецифичные реагре-
гаты.
Опыты по пересадке тканей дают
сходные результаты. Например, горгонарии
(колониальные кораллы) отторгают ткани
других видов (ксенотрансплантаты) и даже
генетически отличных особей своего вида
(аллотрансплантаты), но ткани других
частей той же колонии
(автотрансплантаты) всегда приживаются.
13.2.2.2. Роль фагоцитов,
или амебоидных клеток.
в защите беспозвоночных
Все описанные выше реакции
характерны для колониальных и обрастающих
организмов, часто оказывающихся в условиях,
когда их жизненное пространство
ограничено и является объектом жесткой
конкуренции. Это, по-видимому,
благоприятствовало эволюции способности распознавания
«своего» как средства поддержания
собственной целостности. Следовательно,
такие типы систем самораспознавания могут
быть результатом особого давления
отбора, а вовсе не общей основой эволюции
иммунных систем в царстве животных.
Иммунологические исследования были
начаты на заре XX в., когда И. И. Мечников
ввел шипы розы под эпидермис личинки-
бипиннарии морской звезды и вскоре
обнаружил, что они «атакованы» амебоидными
клетками. Сходные результаты он получил
и при инъекции бацилл сибирской язвы
личинкам жука-носорога (Oryctes nasicornis).
На основе этих наблюдений Мечников
высказал предположение, что амебоидные
клетки, которые у многих примитивных
беспозвоночных участвуют в процессах
внутриклеточного пищеварения (разд. 9.2.2),
390
Глава 13
Таблица 13.2. Типы беспозвоночных, в которых зарегистрировано удаление чужеродных
материалов амебоидными клетками
Таксон животных
Губки
Аннелиды
Сипункулиды
Моллюски
Ракообразные
Одноветвистые
Иглокожие
Оболочники
Инъецированные частицы или вещество
Индийская тушь, кармин
Эритроциты
Редии и церкарии трематод
Индийская тушь, кармин
Частицы железа
Эритроциты
Чужеродные сперматозоиды
Частицы латекса
Бактерии
Кармин, индийская тушь
Эритроциты, дрожжи, бактерии
Диоксид тория
Кармин
Бактерии
Бактерии
Частицы латекса
Железо, сахариды
Аралдитовые имплантаты
Bacillus thuringiensis
Индийская тушь, кармин
Эритроциты, бактерии
Бычий сывороточный альбумин, бычий
гамма-глобулин
Клетки морского ежа (в морской звезде)
Кармин
Частицы стекла
Трипановый синий
Диоксид тория
Фагоцитоз
+
+
+
+
+
+
+
-f
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
Реакция
Инкапсулирование
+
+
+
+
сохранились у более высокоразвитых
животных в качестве внутренней системы
защиты. Действительно, фагоциты широко
распространены у беспозвоночных, и
эксперименты по введению живым организмам
чужеродного материала
продемонстрировали их способность удалять различные
чуждые организму частицы (табл. 13.2).
13.2.2.3. Как же фагоциты
отличают «свое» от «чужого»?
Фагоциты должны одновременно
«игнорировать» нормальные ткани своего
организма и поглощать чужеродные частицы.
По-видимому, они участвуют и в удалении
«своей» ткани, поврежденной в результате
присутствия ксенобиотика, например
загрязнителя.
О механизме этого процесса известно
сравнительно мало. Априорно можно
предположить, что распознавание происходит
при контакте фагоцита с его мишенью.
Поскольку маловероятно, что все чужеродные
частицы подают специфический сигнал
«убей», разумно предположить подачу
своими клетками особого сигнала «не
убивай». Нет данных о существовании у
беспозвоночных такой же развитой, как у
позвоночных, системы антител, служащих
промежуточным звеном при разпознавании
антигенов, но есть указания на опсониновую
активность жидкостей их тела (опсонины—
молекулы, связывающиеся с чужеродными
частицами, с одной стороны, и с
фагоцитами, с другой, что облегчает процесс
фагоцитоза). Так, амебоциты мечехвоста (хелице-
ровые) не обладают существенным бакте-
Защита
391
Обозначения:
i—1 Бивалентный опсонин
• Опсониновый рецептор
А Н-молекула ("своя")
V Анти-Н-рецептор
—«-Сигнал "не увивай"
Рис. 13.11. Две модели фагоцитарных
иммунных систем беспозвоночных; см. объяснение
в тексте (по Coombe et el., 1984)
рицидным действием без его сыворотки, но
в ее присутствии убивают кишечную
палочку. Сходным образом фагоцитоз
эритроцитов человека гемоцитами (амебоцитами
крови) осьминога Eledone cirrosa
происходит только после того, как эритроциты
обработаны его сывороткой. Экстракты из
многих беспозвоночных действуют как
агглютинины, связывая друг с другом
различные клетки и бактерии in vitro (табл. 13.2).
Возможно, они обладают и опсониновыми
свойствами, связывая чужеродные частицы
с поверхностью фагоцитов. (Более
подробное обсуждение этих механизмов см.
Coombe et al., 1984.)
Предполагаемые механизмы
распознавания клеток фагоцитами представлены на
рис. 13.11. Прямое узнавание (рис. 13.11,Л),
вероятно, осуществляется фагоцитами,
«патрулирующими» ткани беспозвоночных
с массивным телом, тогда как у животных
с заполненными жидкостями полостями
в качестве промежуточных факторов
фагоцитоза могут выступать опсонины (рис.
13.11,£).
13.2.2.4. Размножение связано
с определенными сложностями
Когда происходит внутреннее
оплодотворение (гл. 14), в организм животного
попадает сперма с чужеродным генотипом.
Оплодотворенные яйцеклетки и эмбрионы,
находящиеся в тканях матери (гл. 15), также
являются генетически наполовину
«чужими». Однако в норме они не должны
разрушаться ее иммунной системой—напротив,
материнский организм должен служить им
надежным хозяином. Неизвестно, как
конкретно половые клетки избегают
разрушения иммунной системой, но некоторое
представление об этом могут дать
результаты описанных ниже опытов.
Сперматозоиды аллогенных доноров не
фагоцитируются при введении в целомическую
полость земляных червей, однако
сперматозоиды млекопитающих или других видов
дождевых червей фагоцитируются. Самцы си-
пункулид не могут инкапсулировать яйца
того же вида, введенные в их целом; это
позволяет предположить, что половые клетки
обладают каким-то универсальным
механизмом, позволяющим им избегать атаки
иммунной системы
организма-хозяина— ведь в природе яйца в организме
самцов не встречаются! Те же яйца,
поврежденные химической, тепловой или
ультразвуковой обработкой, быстро инкапсулируются,
в то время как предварительно
замороженные яйца, хотя и нежизнеспособны,
инкапсуляции не подвергаются. Интересующихся
мы вновь отсылаем к работе Кумба и др.
(Coombe et al., 1984).
13.2.2.5. Защита паразитов
от иммунного ответа хозяина
Успех паразита зависит от его
способности защитить себя от иммунного ответа
хозяина. Этот вопрос был подробно
исследован на трематодах, у которых открыты
разнообразные способы защиты. Так, шисто-
сомы окружают свое тело антигенами,
близкими или идентичными антигенам
хозяина, которые либо синтезируются самим
паразитом (молекулярная мимикрия), либо
берутся у хозяина и связываются с
поверхностью паразита. Независимо от происхо-
392
Глава 13
ждения они маскируют собственные
антигены паразита, так что те более не вызывают
ответной реакции. Фасциолиды используют
иной механизм: они выделяют вещества,
токсичные для лимфоцитов и других клеток
иммунной системы хозяина. Кроме того,
гликокаликс покровов паразита с высокой
скоростью обновляется, что позволяет
трематодам «сбрасывать» антитела хозяина.
fТоксин
ОШСС
Энергия
-Токсин-ОН
У
Метаволизм
и экскреция
13.2.3. Реакция на загрязнители
Стрессоры окружающей среды и
вызываемые ими реакции столь разнообразны,
что невозможно обсудить все их достаточно
исчерпывающе. Некоторые способы
«физиологической защиты» от стресса,
вызываемого кислородом, соленостью и т.п.,
уже обсуждались в предыдущих главах
(например, разд. 11.6.5 и 12.2). Ниже мы
рассмотрим некоторые общие реакции
беспозвоночных на два типа загрязнителей:
ксенобиотики (органические токсины) и
тяжелые металлы.
13.2.3.1. Оксигеназа с широкой
субстратной специфичностью
(ОШСС) и ксенобиотики
Органические загрязнители типа
углеводородов, попадающих в среду при утечках
нефти, могут проникать в ткани морских
беспозвоночных. Они липофильны,
поэтому их метаболизация затруднена, и они
могут накапливаться в липидных отложениях
и липидных компонентах клеточных
мембран до такой концентрации, что это
нарушает биохимические процессы. Однако
у ряда морских беспозвоночных, в
частности у некоторых полихет, моллюсков и
ракообразных, имеется ферментная система,
способная окислять такие токсины путем
присоединения атомов кислорода, что
делает эти вещества более гидрофильными и,
следовательно, легче метаболизируемыми.
Эта система довольно неспецифична в
отношении субстратов, на которые
воздействует, и состоит из нескольких ферментов
и цитохромов, связанных в основном с
мембранами микросомной фракции эндоплаз-
матического ретикулума некоторых тканей
(например, пищеварительной железы или
Рис. 13.12. Упрощенная схема действия
системы ОШСС (по Calow, 1985)
гепатопанкреаса). Процесс окисления
требует больших затрат энергии; в самом
общем виде он представлен на рис. 13.12. Он
включает эпоксидирование, гидроксилиро-
вание и дезалкилирование. Работа системы
требует индукции, т. е. входящие в нее
ферменты образуются только при наличии
ксенобиотика. Например, содержание
соответствующего цитохрома в тканях мидий
возрастает после суточного воздействия
дизельного топлива и возвращается к норме
через 8 дней после снятия стресса.
Оксигеназы с широкой субстратной
специфичностью присутствуют и у
растительноядных насекомых, где они, вероятно,
участвуют в обезвреживании природных
органических токсинов, вырабатываемых
растениями для защиты от фитофагов.
Например, у насекомых-полифагов активность
ОШСС обычно выше, чем у стенофагов
(более специализированных в смысле питания),
no-видимому, в связи с воздействием более
широкого спектра токсичных соединений,
включая фенолы, хиноны, терпеноиды и
алкалоиды.
13.2.3.2. Металлотионеины
Тяжелые металлы типа ртути, кадмия,
меди, серебра и олова могут быть
исключительно токсичными для водных
беспозвоночных. Например, они способны
денатурировать ферменты, так как, взаимодействуя
с ними, изменяют их третичную структуру.
Защита
393
Однако некоторые морские
беспозвоночные, в том числе крабы, двустворчатые
моллюски, блюдечки и некоторые аннели-
ды, обезвреживают тяжелые металлы,
связывая их с особыми белками,
называемыми металлотионеинами. Это соединения
низкого молекулярного веса, богатые SH-
группами из-за высокого содержания
аминокислоты цистеина. SH-группа может
связывать или хелатировать металлы, что
делает их менее токсичными.
13.2.4. Репарация—защита
против старения?
В разд. 13.3.1 вскользь упоминалось,
что старение целого организма вызывается
накоплением повреждений на суборганиз-
менном уровне. Об этом свидетельствуют
следующие данные.
Разрушение тканей. Старение у
двукрылых насекомых сопровождается снижением
способности к полету, и оно в свою очередь
коррелирует с дегенеративными
изменениями в структуре летательных мышц.
Показано, что среднее число клеток в мозге
рабочих пчел падает с 522 при вылете из куколки
до 350 в возрасте 10 недель (рис. 13.13).
Липофусцин (пигмент старения).
Вероятно, это продукт пероксидирования липидов,
550 т
500
450-
400-
350-
10 20 30 40 50 60 70
Возраст, дни
Рис. 13.13. Снижение с возрастом числа клеток
мозга у пчел (по Rockstein, 1950)
ш
0,6
0,5
0,4
0,3
0,2
0,1
|-
4
^ i
i
i
}
i
• Самец
о Самка
i
10 20
Возраст,дни
30
40
Рис. 13.14. Накопление липофусцина
(измеренное по его флуоресценции) у Drosophila
melanogaster (no Biscardi. Webster. 1977)
образующийся при разрушении клеточных
мембран. Показано, что липофусцин
накапливается с возрастом в тканях нематод
и насекомых (рис. 13.14).
Работа ферментов. Данные,
полученные в основном при изучении нематод,
говорят о нарушении с возрастом структуры
и функции ферментов: они становятся более
чувствительными к тепловой денатурации
(что указывает на изменения в
молекулярной организации), проявляют аномальные
иммунологические свойства и снижают
каталитическую активность. Однако это не
относится к некоторым ферментам
нематод, а также ко многим изученным
ферментам других животных.
Основная причина всех этих нарушений
связана, вероятно, с молекулярными
процессами типа теплового шума,
вызывающими ошибки синтеза белков и других
метаболических превращений. В то же время
поврежденные белковые молекулы могут
в принципе в соответствии с генетической
программой заменяться новыми так же, как
394
Глава 13
3-г
2--
I i i_
j
^ >.о сто
;S £Jm За
bog O-O
Я cdcclc
о о
SI
m bo_ 5a E
— н° e[o ^
-o 3-m ош
e 0.0 o-o
Q> ё~е e^
0_фО OO
о о-с С1-c
§s
§r?
Dendrocoelum
tacteum
(короткоживущий
моноцикличный)
Polycelis tenius
(долгоживущий
полицикличный)
Рис. 13.1 Б. Скорость обновления клеток на
различных стадиях жизненного цикла двух
видов трехветвистых турбелларий—долго-
живущего и короткоживущего. Последний
вкладывает гораздо больше энергии в
размножение, чем первый. В обоих случаях число
клеточных делений снижается с возрастом, но
и целые клетки (в данном случае—путем
митоза) (гл. 1). Действительно, результаты
опытов на нематодах позволяют говорить,
что белки с аномальной аминокислотной
последовательностью обновляются
быстрее нормальных, но этот процесс
замедляется по мере старения червей. Клеточные
деления у взрослых особей характерны для
книдарий, плоских червей, аннелид и
моллюсков, но весьма ограничены у нематод
и насекомых. У пресноводных
трехветвистых турбелларий обновление клеток с
возрастом замедляется (рис. 13.15), причем
особенно заметно — с переходом у
моноцикличных форм к размножению (см. разд.
14.4). Последнее связано с ускорением
самого процесса старения. Таким образом,
биологический возраст, или жизнеспособность,
организма зависит от баланса между
возникновением дефектов и их заменой или
исправлением (репарацией). Неудивительно
поэтому, что процессы старения наименее
выражены у таких организмов, как
книдарий, у которых обновление тканей
происходит на протяжении всей жизни, и наиболее
заметны у нематод и насекомых, где оно
весьма ограничено (табл. 13.3). А среди
насекомых митотическое деление клеток
захватывает больше тканей и продолжается
дольше у видов с более продолжительной
жизнью, например у жуков.
у короткоживущеи формы митотическая
активность в репродуктивном возрасте падает
сильнее. Считается, что замедление обновления
тканей вызывается большим вкладом энергии
в размножение и сокращает жизнь, ускоряя
процессы старения (по Calow, Read, 1986)
Таблица 13.3. Старение у беспозвоночных
Группа
Подозревается
отсутствие старения
у некоторых видов
Подозревается
существование
старения у некоторых
видов
Доказано
старение некоторых
видов
Книдарий
Плоские черви
Моллюски
Нематоды
Аннелиды
Коловратки
Членистоногие
+
+
+
+
+
+
+
+
+
+
+
Защита
395
13.3. Выводы
Опасности, которым подвергаются
организмы, весьма разнообразны, и
неудивительно, что они вызывают широкий спектр
ответных реакций. Однако можно
предложить единую классификацию таких
опасностей в зависимости от их более или менее
тесной связи с организмом, находящимся
под угрозой. Сходным образом за внешним
разнообразием реакций можно различить
некоторые общие черты. Например, все
защитные механизмы независимо от того,
направлены они против хищников, паразитов,
микроорганизмов или изнашивания самого
организма, требуют больших затрат
вещества и энергии. Именно эти общие
принципы читатель должен постараться
уяснить из этой и других глав данной книги.
13.4. Дополнительная
литература
Berry R.J. 1977. Inheritance And Natural Selection.
New Naturalist No. 61. William Collins and Co.,
Glasgow.
Blest A.D. 1957. The function of eyespot patterns in
the Lepidoptera. Behaviour, 11, 209—256.
Calow P. 1981. Invertebrate Biology. Croom Helm,
London.
Calow P. 1983. Evolutionary Principles. Blackie,
Glasgow.
Coombe D. R., Ey P. L., Jerikin С R. 1984. Self/non-
self recognition. Q. Rev. Biol., 59, 231 255.
Danes I. 1983. Ageing. Edward Arnold. London.
Eisner Т., Van Tassell E., Carrel J. E. 1967.
Defensive use of a "faecal shield" by a beetle larva.
Science, N.Y., 158, 1471 - 1473.
Erichsen J. Т.. Krehs J.R., Houston A.I. 1980.
Optimal foraging and cryptic prey. J. anim. Ecol., 49,
271—276.
Gliwicz M.Z. 1986. Predation and the evolution of
vertical migration in zooplankton. Nature
(London), 320, 746—748.
Hansell M. H. 1984. Animal Architecture and
Building Behaviour. Longman, London.
Livingstone D.R., Moore M.N., Lowe D.M., Nas-
ci C, Farrar S. V. 1985. Responses of the
cytochrome P-450 monoxygenase system to diesel oil in
the common mussel, Mytilus edulis L., and the
periwinkle, Littorina littorea L. Aquatic Toxicology.
7, 79—81.
Nordlung D.A., Lewis W.J. 1976. Terminology of
chemical releasing stimuli in intraspecific and
interspecific interactions. J. chem. Ecol., 2, 211
220.
Rockstein M. 1950. The relation of cholinesterase
activity to change in cell number with age in the
brain of the adult worker bee. J. cell. сотр.
Physiol., 35. 11—23.
TheodorJ.L. 1976. Histo-incompatibility in a
natural population of gorgonians. Zool. J. Linn. Soc,
58, 173—176.
Turner J. R.G. 1984. Darwin's coffin and Dr. Pan-
gloss— do adaptationist models explain mimicry?
In: B. Shorrocks (Ed.) Evolutionary Ecology, pp.
313—361. Blackwell Scientific Publications,
Oxford.
ГЛАВА ■ 4
РАЗМНОЖЕНИЕ И ЖИЗНЕННЫЕ ЦИКЛЫ
В принципе многоклеточные эукарио-
ты—это диплоидные организмы,
размножающиеся половым путем, образуя
гаплоидные гаметы, пары которых при
слиянии дают диплоидную зиготу. Генотип
половых клеток (гамет) обычно отличается от
генотипа родительского организма
вследствие перераспределения генов в ходе мейо-
за. Более того, новый диплоидный
организм обычно (хотя и не всегда) образуется
при слиянии гамет различных генотипов.
Таким образом у многоклеточных
организмов в результате полового размножения
и оплодотворения возникает клетка с
новым, уникальным набором генов.
В этой главе рассмотрены процессы,
связанные с половым размножением, а
дальнейшее развитие и дифференцировка
зиготы и продуктов ее деления описаны в гл.
15. Хотя жизненные циклы животных
следуют одному основному плану,
особенности полового и бесполого размножения
(также широко распространенного среди
животных) сильно варьируют. Мы
рассмотрим это разнообразие, обусловливающие
его факторы и адаптивное значение
различных вариантов, включая селективные
преимущества полового размножения перед
бесполым. По ходу изложения станет ясно, что
различные особенности жизненных циклов
и размножения характерны для
определенных филогенетических линий, уровней
организации животных и экологических
ситуаций. Это позволит обсудить стратегии
размножения беспозвоночных в свете
современных теорий эволюции жизненных
циклов.
14.1. Введение
В гл. 1 высказывалась мысль о том, что
эволюция—это естественное следствие
самовоспроизведения систем путем
полуконсервативной репликации, причем у всех
организмов один и тот же носитель
генетической информации—ДНК, а наследственная
изменчивость возникает при неправильном
копировании последовательности
азотистых оснований в процессе репликации этой
молекулы. Однако эволюция
многоклеточных существ требует полуконсервативного
воспроизведения не просто клеток, а
организма в целом, т. е. процесса, называемого
размножением. У подавляющего
большинства форм он сопровождается генетической
рекомбинацией; в этом случае говорят о
половом размножении. Существуют
различные его системы, но их общей чертой
остается обмен генетическим материалом между
особями с разными генотипами.
В приложении 14.1 приведены схемы
трех различных половых процессов.
Генетический обмен у вирусов возможен при
смешанном заражении одной клетки-хозяина
вирусами разных штаммов. У бактерий
часть молекулы ДНК одного из партнеров
передается другому в ходе конъюгации,
после чего между гомологичными нитями
ДНК может происходить обмен. Все
многоклеточные эукариоты используют третью
из представленных систем полового
процесса.
У животных обычно образуются два
сильно различающихся типа гамет —
сперматозоид и яйцеклетка (яйцо) (разд.
14.4.2). Часто они происходят от
специализированных клеток, обособляющихся на
Размножение и жизненные циклы
Таблица 14.1. Репродуктивные признаки морских беспозвоночных
397
Признак
Развитие
Размер яиц
Плодовитость
Частота размножения
Число размножений за
жизнь
Продолжительность
жизни
(Время генерации)
Размер тела
Сперматозоиды
Оплодотворение
Репродуктивное
усилие 1}
Пелагическое
Мелкий:
~50 мкм
Высокая:
106
Низкая:
1 раз в год
Одно
Многолетняя
Много лет
Крупный:
длина > 1000 мм
Простые
Наружное без
1 планкотрофное
■< лецито*грофное
( смешанное
Однолетняя
хранения спермы
Большое; совершается
позже
Непелагическое
Крупный:
>1000 мкм
Низкая:
1
Высокая:
много раз в год, почти непрерывно
Множество
Меньшая
Несколько дней или недель
Мелкий:
< 1 мм
Сложной формы
Внутреннее или
включающее процессы
переноса и хранения спермы
Небольшое;
совершается раньше
" Репродуктивное усилие можно определить по формулам
ДЕГ.
где EG- энергия, вло-
Es "г EG AES + EG
женная в половые структуры, Es—энергия, вложенная в соматические ткани, АЕС—ежегодный прирост половых
тканей, AES- ежегодный прирост соматических тканей.
ранних стадиях эмбриогенеза, но в
некоторых группах могут возникать в результате
дедифференцировки соматических клеток.
Жизненные циклы многоклеточных
организмов разнообразны; например, у
многих растений чередуются гаплоидные и
диплоидные поколения (приложение 14.2)
с одинаковой или различной морфологией.
У животных такой смены поколений не
встречается, но известно четкое
чередование диплоидных фаз, размножающихся
половым и бесполым путями (разд. 14.2).
Несмотря на общие черты полового
процесса у всех животных, условия его
осуществления и детали жизненных циклов
различаются очень сильно. Отличительные
признаки последних в каждой группе
беспозвоночных часто в совокупности называют
несколько антропоморфным термином
«репродуктивная стратегия» (табл. 14.1).
14.2. Значение полового
и бесполого размножения
14.2.1. Бесполое размножение
в жизненных циклах
беспозвоночных
Половое размножение распространено
очень широко, но многие организмы могут
размножаться бесполым путем, т. е. без
генетической рекомбинации. Генотип
потомства при этом практически идентичен
родительскому. Бесполое размножение может
идти как путем разделения тела животного
398
Глава 14
0 0
А
Рис. 14.1. Схема двух типов бесполого
(амейотического) размножения: А —
фрагментация; Б—партеногенез
на многоклеточные части {почкование
и фрагментация), так и с образованием не-
оплодотворенных яиц {партеногенез)'. На
рис. 14.1 показаны оба этих механизма,
свойственные многим беспозвоночным.
Почкование и фрагментация особенно
распространены у мягкотелых форм —
губок, книдарий, плоских червей, немертин,
аннелид и некоторых иглокожих. Эти
способы размножения редко встречаются у
животных с твердыми покровами и не
обнаружены у моллюсков и членистоногих.
Фрагментация может означать либо простое
разделение организма на две части, каждая из
которых регенерирует недостающую (рис.
\4Л,А), либо распад на множество пропа-
гул. развивающихся впоследствии в целые
организмы (рис. 14.2). Она обычно сочетае-
1 Партеногенез часто относят к половому
(однополому) размножению.— Прим. ред.
тся с половым размножением в сложных
жизненных циклах с чередованием половых
и бесполых «поколений» (рис. 14.4,Л).
Затруднения, вносимые почкованием и
фрагментацией в формулировку понятия
«особь», особенно выражены у тех
организмов, где эти процессы идут не до конца, так
что возникают колонии из большого числа
структурных единиц. Колониальность
особенно распространена у книдарий, мшанок
и оболочников. Некоторые из таких
беспозвоночных показаны на рис. 14.3. По-
видимому, этих животных правильнее
считать состоящими не из особей, а из
модульных единиц.
Партеногенез у беспозвоночных
встречается также часто (нами под этим
термином понимается бесполое размножение
посредством неоплодотворенных яиц, но
некоторые авторы включают в это понятие
и способы, описанные в предыдущем
абзаце). Он включает либо модифицированный
мейоз с образованием диплоидного яйца,
для развития которого не требуется
слияния с мужской гаметой, либо развитие из
неоплодотворенных гаплоидных яиц
самцов, а из оплодотворенных диплоидных—
самок (такой способ называется арреното-
кия). Облигатный партеногенез, т. е. полное
отсутствие полового размножения,
встречается редко. Он предполагается, например,
у бделлоидных коловраток (см. разд.
4.9.3.1), самцы которых не обнаружены.
Чаще партеногенез циклически чередуется
с половым размножением. После одного
или нескольких бесполо размножающихся
поколений образуются особи разного пола,
причем слияние их гамет обычно дает
покоящиеся, устойчивые к внешним
воздействиям яйца (рис. \4А,А). Жизненные
циклы такого типа характерны для моного-
натных коловраток, многих мелких
пресноводных ракообразных и тлей. Диплоидная
особь, развивающаяся из
оплодотворенного покоящегося яйца, дает начало клону
генетически идентичных потомков. Скорость
роста популяпии при этом максимальна,
и такой жизненный цикл можно
рассматривать как приспособление к максимальному
использованию непостоянных пищевых
ресурсов в условиях, когда другие
биологические факторы ограничивают размеры тела.
Размножение и жизненные циклы
399
Л o<Xqu
Яйцо (п) ■
Сперматозоид (п)-
В
Взрослый
организм
Партеногенез или
фрагментация
L Бесполое 1
размножение
Половое размножение
Рис. 14.2. Пример множественной фрагментации: полихета Dodecaceria. A. I. Взрослая особь до
разделения на сегменты. II. Стадия регенерации: каждый сегмент формирует голову и хвост путем
прироста новых сегментов. Б. Образование первичных, а затем вторичных особей из одного
сегмента. В конечном счете этот сегмент оказывается в составе особи, прекратившей
фрагментацию. В. Схема жизненного цикла, включающего бесполое и половое размножение: бесполое
сменяется половым, и все бесполые особи в конце концов погибают (по Dehorne, 1933; Gibson, Clark,
1976)
На определенной стадии цикла появляются
особи, дающие потомков двух типов —
самцов и самок, способных к половому
размножению. Переход к нему бывает
эндогенно детерминирован, но чаще является
реакцией на изменение окружающих условий,
например на перенаселение, изменение
качества пищи или сокращение
продолжительности дня (см. также разд. 14.4.4). Переход
к половому размножению может
сопровождаться морфологическими изменениями
(рис. 14.4,2>,Б), а у тлей эти изменения
происходят и на бесполой стадии (рис. 14.4,7").
14.2.2. Типы полового
размножения
Более 99% беспозвоночных
размножаются половым путем на той или иной стадии
жизненного цикла, причем для
подавляющего большинства это единственный
способ воспроизведения. При нем всегда
происходит слияние относительно крупной и
неподвижной женской гаметы -
яйцеклетки— с более мелкой и подвижной
мужской— сперматозоидом. Такое различие
гамет называют анизогамией. Оно позволяет
различать мужские и женские функции,
которые не обязательно выполняются
разными особями. Виды, представленные самца-
400
Глава 14
Рис. 14.3. Примеры колониальных, или
«модульных», беспозвоночных./l. Гидроидный полип
Obelia: I — ветвящаяся колония; II—отдельный
полип. Б. Мшанка Membranipora: I — часть
колонии; II — отдельный зооид. В — оболочник
Sydnium: I—три колонии с отдельными
входными и общим выводным сифонами; II —
отдельный зооид
ми и самками, называют раздельнополыми:
если одна и та же особь одновременно или
последовательно выполняет мужские и
женские функции, животных называют
гермафродитами. Характеристики обоих типов
размножения представлены в табл. 14.2,
а их распространение в основных группах
беспозвоночных—в табл. 14.3. При
последовательном гермафродитизме смена пола
может наступать по достижении
определенного, видоспецифичного размера или
возраста, так что самцов и самок одновременно
не встречается; в других случаях этот
момент различен у разных особей в
популяции, что допускает наличие двух «чистых»
полов. Обе эти ситуации показаны на рис.
14.5.
14.2.3. Механизмы определения
пола
Эти механизмы у беспозвоночных
можно разделить на три основных типа:
а) материнский, б) генетический и в)
экологический.
14.2.3.1. Материнское
определение пола
В этом случае пол потомства
детерминируется самкой, образующей яйца двух
типов. Пример приведен в приложении
14.3(1). При такой системе размножения
неизбежен инбридинг, и все особи зрелой
популяции являются самками. Так, у пчел
матка способна контролировать
оплодотворение своих яиц; если оно произошло,
диплоидные яйца развиваются в стерильных
самок (рабочих особей), но при
определенных условиях способны дать и плодовитых
маток; неоплодотворенные яйца партеноге-
нетически развиваются в гаплоидных
самцов.
14.2.3.2. Генетическое
определение пола
У некоторых животных хромосомные
наборы самцов и самок заметно
различаются: у самок две половые хромосомы
идентичны (так называемые Х-хромосомы),
у самцов — различны (Х- и Y-хромосомы).
Пол зиготы определяется при
оплодотворении половой хромосомой, оказавшейся
в сперматозоиде. Это явление, называемое
гетерогамией, распространено у
беспозвоночных не так широко, как часто считают.
Несколько его разновидностей встречаются
О Покоящееся яйцо-
результат полового размножения
(диплоидное, 2 п)
Самка, не
образующая Эфиппий
Вылупление партеногенетической
/^■диплоидной самки
( I
V Ди
Вылупление
рачка из
эфиппия
Диплоидные яйца
х Несколько поколений
Изменение условии:
"температура,
- освещенность,
. пищевые ресурсы,
стресс
Диплоидные самки дают
размножающееся половым путем
потомство
\ \
Самцы Самки
Гаплоидные
сперматозоиды
I J
\>
Гаплоидные
яйцеклетки
®
Оплодотворение
Поколения,
размножающиеся вез полового процесса
Откладка
эфиппия
Самка,
образующая
Эфиппий
Мейоз/\
РинГамия
у^ Мейоз .
Одно половое
поколение
Появление
самцов и самок
Самец
2п) Покоящееся яйцо
ЛЕТО
Крылатые Бесполые
расселительные формы
Бескрылые Бесполые
формы,
обеспечивающие рост популяции
Крылатые самки,
дающие в
потомстве яйцекладущих
самок
ОСЕНЬ
ВЕСНА
-о-
ЗИМА
С^Зигота — "покоящаяся"
стадия
Неоплодотворенные
гаплоидные яйца
Рис. 14.4. Чередование партеногенеза и полового размножения в жизненных циклах
беспозвоночных. А. Элементы обобщенного жизненного цикла. Б. Жизненный цикл пресноводного ветви-
стоусого рачка Daphnia. В. Жизненный цикл коловратки Branchionus: за бесполой фазой следует
половая, когда неоплодотворенные самки откладывают мелкие яйца, из которых развиваются
самцы, а оплодотворенные самки откладывают более крупные яйца, представляющие собой
зимующую стадию жизненного цикла. Г. Жизненный цикл вишневой тли. Для партеногенетических
самок характерно живорождение, и только осенние особи откладывают яйца. Хорошо выражен
полиморфизм, особенно в образовании крылатых и бескрылых форм (£ — по Bell, 1982, Г—по
Dixon, 1973)
26-1314
402 Глава 14
Таблица 14.2. Системы полового размножения животных
Раздельнополость Каждая особь в популяции образует либо мужские, либо женские половые
клетки и имеет соответствующие вспомогательные железы и органы
Гермафродитизм Некоторые особи в популяции образуют как мужские, так и женские
половые клетки и имеют вспомогательные железы и органы, связанные как
с мужской, так и с женской воспроизводительными функциями в тот или
иной период жизни
Синхронный гермафродитизм
Взрослые животные одновременно образуют оба типа половых клеток. При
этом они могут либо обмениваться мужскими половыми клетками с
другим ор1анизмом (аутбридинг), либо размножаться
самооплодотворением
Последовательный гермафродитизм
Каждое животное выполняет функции сначала одного, затем другого пола.
1. Протандрия. Мужские функции развиваются и выполняются раньше,
чем женские. При активации женских функций мужские либо
утрачиваются, либо сохраняются (синхронный гермафродитизм)
2. Протогиния. Женские функции развиваются и выполняются раньше
мужских. В конечном итоге также может наблюдаться синхронный
гермафродитизм
3. Чередование. Протандрическое или протогиническое, но с последующим
обращением пола
Таблица 14.3. Типы полового размножения у беспозвоночных
Тип Класс Характер полового размножения
Губки
Mesozoa
Стрекающие
Гребневики
Плоские черви Турбелларии
Моногенеи
Трематоды
Цестоды
Гнатостомулиды
Все способны к половому размножению, но
многие также размножаются бесполым путем за
счет отделения фрагментов-геммул
Самооплодотворяющиеся гермафродиты с
чередованием полового и бесполого поколений
Раздельнополые, но часто размножаются
почкованием или фрагментацией. Иногда имеют
сложные жизненные циклы с пелагическими
медузами, образующими гаметы, и бентосны-
ми гидроидами, размножающимися бесполым
путем
Синхронные гермафродиты, по-видимому,
перекрестно-оплодотворяющиеся
Синхронные гермафродиты, по-видимому,
перекрестно-оплодотворяющиеся. Распространено
бесполое размножение фрагментацией, у
некоторых форм половой процесс не обнаружен
Синхронные гермафродиты; при множественных
заражениях, возможно, распространено
перекрестное оплодотворение. Иногда
самооплодотворяющиеся. На личиночных стадиях часто
размножаются множественной фрагментацией
Синхронные гермафродиты, обычно
самооплодотворяющиеся. Один род раздельнополый
Синхронные гермафродиты
Размножение и жизненные циклы
403
Продолжение табл. 14.3
Тип
Класс
Характер полового размножения
Немертины
Гастротрихи
Моллюски
Коловратки
Киноринхи
Скребни
Loricifera
Волосатики
Нематоды
Приапулиды
Сипункулиды
Эхиуриды
Аннелиды
Погонофоры
Форониды \
Мшанки /
Плеченогие
Внутрипорошицевые
Полухордовые
Иглокожие
Chaetodcrmomorpha
Monoplacophora
Панцирные
Лопатоногие
Брюхоногие
Neomeniomorpha
Двустворчатые
Головоногие
Bdelloidea
Monogonata
Seisonidea
Полихеты
Поясковые
Хордовые
Морские ежи
Голотурии
Морские лилии
Морские звезды
Офиуры
Оболочники
Почти все раздельнополые; некоторые
пресноводные формы гермафродиты
Синхронные гермафродиты, но распространен
партеногенез
Раздельнополые
Весьма различный характер размножения:
переднежаберные — в основном раздельнополые,
но иногда последовательные (протандриче-
ские) гермафродиты; заднежаберные- в
основном синхронные гермафродиты;
легочные— синхронные гермафродиты с
перекрестным оплодотворением
Гермафродиты
Раздельнополые
Облигатный партеногенез; самцы неизвестны
Циклический партеногенез; в половой фазе
раздельнополы (самиы карликовые)
Раздельнополые
Раздельнополые; иногда самцы карликовые
Обычно раздельнополые; иногда
последовательные протандрические гермафродиты, изредка
синхронные гермафродиты. Некоторые виды
размножаются фрагментацией
Всегда синхронные гермафродиты с
перекрестным оплодотворением
Раздельнополые
Синхронные гермафродиты; отдельные вилы
раздельнополые
В основном раздельнополые, изредка
гермафродиты
Синхронные или последовательные
гермафродиты
Раздельнополые
\ В основном раздельнополые, иногда синхронные
/ гермафродиты. Некоторые виды
размножаются фрагментацией
Синхронные гермафродиты
404
Глава 14
Продолжение табл. 14.3
Тип
Класс
Характер полового размножения
Ракообразные
Хелицеровые
Онихофоры
Тихоходки
Пятиустки
Одноветвистые
Жаброногие
Ракушковые
Веслоногие
Усоногие
Malacostraca
Раздельнополые, но распространен партеногенез;
у некоторых видов самцы не обнаружены
Раздельнополые
Торациковые обычно гермафродиты, хотя
некоторые виды раздельнополые с карликовыми
самцами; остальные раздельнополые с
карликовыми самцами
Обычно раздельнополые, но иногда
последовательные (протандрические) гермафродиты
Почти всегда раздельнополые
Почти всегда раздельнополые, но иногда
встречается партеногенез или арренотокия (самцы
обычно неизвестны)
у насекомых; оно подробно узучено на
примере плодовой мушки Drosophila tnelanoga-
ster. Как показано в приложении 14.3, у
самок здесь две Х-хромосомы, а у самцов—X
и Y. В ходе мейоза все яйцеклетки получают
по одной Х-хромосоме, а ровно половина
сперматозоидов—Y-хромосому. При
случайном слиянии гамет устанавливается
отношение полов 1:1. У многих
беспозвоночных гетерогамия не обнаружена, тем не
менее наблюдается фиксированное отношение
полов.
100
90
80
70
60
50
40
30
20
10
А
Б
i i i
Л
\
\ >v
i i I i"1 1 1 \-
Размер тела
(длина, число сегментов и т.д.)
Рис. 14.5. Типы обращения пола при
последовательном гермафродитизме:/!—ложная
раздельнополость; Б—несбалансированный
гермафродитизм
14.2.3.3. Экологическое
определение пола
Пол особи не всегда определяется до
или в момент оплодотворения; иногда он
зависит от окружающих условий во время
эмбрионального или личиночного
развития. Одним из наиболее известных
примеров служит эхиурида Bonellia viridis;
карликовые самцы этого червя паразитируют на
самках (разд. 4.12). В начале нынешнего
столетия было показано, что пол
свободноплавающих планктонных личинок не
фиксирован— оседая на дно, они становятся
самками, а попадая на крупный хоботок самки
или оказываясь вблизи от него,
превращаются в самцов (приложение 14.3).
Эксперименты показали, что самки
выделяют вещество, оказывающее сильное
маскулинизирующее действие на личинки:
в его отсутствие почти все они развиваются
в самок. У большинства других эхиурид
определение пола генетическое, но недавно
в семействах, не связанных близким
родством с Bonellia, были обнаружены
карликовые самцы, что может отражать
независимую эволюцию сходного механизма в
различных филетических линиях этой группы.
Эхиуриды раздельнополы, но у многих
последовательных гермафродитов смена пола
также сильно зависит от внешних условий.
Считается, что у таких животных в процес-
Размножение и жизненные циклы
405
I II III
Рис. 14.6. Определение пола у блюдечка
Crepidula fornicata: I — ранняя стадия — образование
пары; II, III — более поздние стадии —
образование сложных комплексов,
включающих самцов, самок и интерсексов
сах определения пола участвуют феромоны.
Рис. 14.6 показывает это на примере
блюдечка Crepidula fornicata. Животные
образуют плотные стопки, причем самые
нижние особи всегда превращаются в самок,
а верхние — в самцов; находящиеся между
ними животные обычно становятся
гермафродитами.
14.2.4. Адаптивное значение
различных способов
полового размножения
14.2.4.1. Зачем нужно
половое размножение?
Одна из задач эволюционной биологии
состоит в том, чтобы понять, почему
существуют различные типы размножения.
Почему, например, половое размножение
настолько преобладает над другими типами?
Ответ не так самоочевиден, как может
показаться. Половое размножение приводит
к разнообразию потомства и скрывает
проявление потенциально вредных мутаций, но
эти преимущества необходимо сравнить
с недостатками и «стоимостью» процесса.
Во-первых, снижается эффективность
передачи генов следующему поколению,
поскольку от каждого родителя потомку
достается лишь половина генома. Поскольку
приспособленность определяется именно
способностью к сохранению генов в ряду
поколений, речь идет об основной «статье
расходов» на половое размножение. Во-вторых,
необходимо тратить ресурсы на развитие
половых признаков и на брачное поведение;
наконец, риск для жизни во время поиска
партнера часто повышается.
Многие теории считают, что половое
размножение выгодно виду или группе
в эволюционном масштабе времени, а
бесполое дает кратковременное преимущество
на уровне отдельных особей (концепция
группового отбора). В последнее время
разработаны более приемлемые теории,
признающие селективное преимущество
полового размножения для особи, а не для
группы или вида.
Предполагалось, что у особей,
размножающихся половым путем, разнообразное
генетически потомство будет в среднем
лучше приспособлено к изменившимся
условиям среды в будущем, чем одинаковые
потомки бесполого организма. Согласно этой
теории, преимущество полового
размножения максимально в нестабильной среде и
теряется в относительно устойчивых
условиях. Однако анализ распространения
бесполого размножения среди
беспозвоночных, обитающих в различных биотопах, не
подтверждает такого прогноза. Оно
особенно часто встречается у организмов,
зависящих от изменчивых ресурсов, тогда как
половое размножение свойственно почти всем
животным самых стабильных
местообитаний. Так, бесполое размножение более
обычно для пресноводных форм, чем для
морских представителей тех же
таксономических групп.
Альтернативная теория предполагает,
что преимущество размножающихся
половым путем организмов обусловлено
конкурентоспособностью их разнообразных
потомков в сложно организованной среде
с интенсивной конкуренцией: однообразное
потомство бесполого животного не сможет
вытеснить их из всех возможных
местообитаний. Ее создатель Белл (Bell, 1982)
считает, что половое размножение должно
преобладать в стабильных, но сложных
биотопах, а бесполое —там, где численность вида
ниже максимальной, которую может
поддерживать данная среда, и возможен
экологический оппортунизм. В этих условиях
преимущество получают животные с
максимальной скоростью роста или увеличения
численности потомства.
Существуют и другие теории,
пытающиеся объяснить преимущество полового
размножения (рис. 14.7). Некоторые из них,
406
Глава 14
Половое размножение важно для сохранения видов
Нет
Половое размножение — историческое наследие,
не связанное в Настоящее время с отбором
Прогнозируется корреляция полового размножения с
таксономическим положением, но не с экологическими
параметрами.
НЕ СООТВЕТСТВУЕТ НАБЛЮДЕНИЯМ -отвергается
Да
Отбор, действующий в настоящее время,
поддерживает половое размножение в связи с его
преимуществами
Признается,но каков механизм?
2. Половое размножение поддерживается кратковременным
отбором на адекватную изменчивость
3. Половое размножение
преадаптирует потомство
к Новым условиям среды
в будущем
Прогнозируется полоеое
размножение главным образом
е нестабильных средах,
а бесполое — в стабильных
НЕ СООТВЕТСТВУЕТ
НАБЛЮДЕНИЯМ -
отвергается как общая
теория
Половое размножение —
следствие пространственной
и биотической
неоднородности среды. В таких
условиях общая приспособленность
разнообразного потомства
особей, размножающихся
половым путем, будет выше,
чем у однородного потомства
от бесполого резмножения,
т. е. половое размножение
дает селективное
преимущество отдельным особям.
Прогнозируется
преобладание полового размножения
в стабильных сложных
средах, а партеногенеза —
в простых и нарушенных
СООТВЕТСТВУЕТ
НАБЛЮДЕНИЯМ — принимается как
основа для общей теории
Половое размножение поддерживается
долговременной эволюцией при неадекватной современной
изменчивости |
Половое размножение поддерживается за счет
вымирания популяций, размножающихся бесполым
путем I
Вымирание обусловлено
неспособностью
адаптироваться к изменениям
занятых ниш
Преимущество только
на уровне популяции
Прогнозируется быстрое
вымирание и
преобладание полового
размножения в нестабильных
средах
НЕ СООТВЕТСТВУЕТ
НАБЛЮДЕНИЯМ -
отвергается
Вымирание обусловлено
накоплением вредных
алпелей в стабильных
средах
Прогнозируется большая
стабильность популяций
с половым резмноже-
нием в простых, быстро
меняющихся средах
НЕ СООТВЕТСТВУЕТ
НАБЛЮДЕНИЯМ -
отвергается
Рис. 14.7. Алгоритм выбора между альтернативными теориями селективного преимущества
полового размножения (по Bell, 1982)
например предположение, что половое
размножение является механизмом
предотвращения заражения всего потомства
паразитами, совпадают в своих прогнозах с более
общими теориями, в частности с теорией
Белла.
14.2.4.2. Гермафродитизм
и раздельнополость
Задаваясь вопросом, каковы
селективные преимущества полового размножения
по сравнению с бесполым, логично
спросить, чем определяется распространение
раздельнополости и гермафродитизма. При
этом следует учитывать как
таксономические, так и экологические аспекты.
Например, из табл. 14.3 видно, что в некоторых
таксономических группах преобладают
гермафродиты, а другие состоят почти
исключительно из раздельнополых форм.
Кроме того, гермафродитизм чаще
встречается в определенных условиях среды:
например, пресноводные и наземные аннели-
ды и моллюски гермафродиты, а их
морские сородичи в основном раздельнополы.
Аналогичным образом среди
глубоководных ракообразных гермафродитизм более
распространен, чем среди близких
мелководных форм. Разнообразие типов
полового процесса у беспозвоночных позволяет
предположить, что тот или иной из них
возник под влиянием естественного отбора,
поэтому можно попытаться найти
функциональное объяснение ситуаций, в которых
встречается гермафродитизм.
Размножение и жизненные циклы
407
Предложено несколько моделей
эволюции, три из которых вкратце рассмотрены
ниже.
Модель низкой плотности: если
плотность популяции низка, или организмы ведут
сидячий образ жизни, синхронный
гермафродитизм повышает вероятность
оплодотворения при неизбежно редких встречах
между особями, а если встреч не
происходит, делает возможным
самооплодотворение.
Модель преимущества размеров: если
успех спаривания у одного пола связан
с размерами тела, а у другого — нет,
возникает последовательный гермафродитизм.
Популяционно-генетическая модель:
при низкой численности популяции
повышается вероятность инбридинга и дрейфа
генов; в этих условиях гермафродитизм
повышает эффективную численность
популяции.
Была также предложена более общая
теория, включающая в себя все три
описанные модели. В ее основе лежит простое, но
глубокое наблюдение: при половом
размножении у любой особи только одна мать
и один отец. При этом ровно половина
генома зиготы вносится каждым из
родителей, поэтому и мужская, и женская
репродуктивные функции обеспечивают
одинаковый успех размножения.
Объем ресурсов, которые может
использовать организм, ограничен, и их следует
распределять на поддержание
жизнедеятельности и воспроизведение таким образом,
чтобы суммарная приспособленность особи
была максимальной. Определенная часть
затрат на половое размножение связана
с развитием вспомогательных структур —
протоков и желез, облегчающих движение
гамет, а также с повышенным риском
гибели при поисках партнера. Гермафродитам
приходится «тратиться» и на мужскую, и на
женскую репродуктивные функции,
поэтому у них общие расходы любого конечного
ресурса на размножение, по-видимому,
выше. Следовательно, можно ожидать, что
давление отбора благоприятствует
раздельнополости.
Однако ситуация меняется, если
«отдача» от затрат на какую-то из
репродуктивных функций падает: можно полагать, что
в таких условиях отбор будет
благоприятствовать гермафродитизму. Например,
пониженная «отдача» от энергозатрат на
женскую функцию, вероятно, свойственна
животным, выращивающим потомство в
выводковой камере с ограниченным внутрен-
ным пространством.
14.3. Разнообразие полового
размножения:
жизненные циклы
14.3.1. Введение
Существует много типов полового
размножения; наиболее важные признаки,
в сумме составляющие репродуктивную
стратегию, следующие:
1) максимальная потенциальная
продолжительность жизни;
2) число эпизодов размножения в
течение жизни;
3) характер расходования гамет;
накопление и массовое использование или
постепенное расходование по мере
образования;
4) степень синхронизации половых
циклов особей в популяции;
5) избирательность спаривания и
степень аутбридинга в популяции;
6) относительный размер и «стоимость»
гамет;
7) тип развития и степень
подверженности ювенильных и взрослых особей
давлению отбора;
8) относительная доля всех доступных
ресурсов, вкладываемых в размножение.
Жизненные циклы животных часто
классифицируют в зависимости от числа
эпизодов размножения в течение жизни и от
продолжительности половозрелой стадии.
Используется несколько
невзаимоисключающих систем, некоторые из которых
сравниваются в табл. 14.4. Различные типы
размножения также схематически
представлены в приложении 14.4.
14.3.2. Морские беспозвоночные
Глубокое влияние на тип размножения
животных оказывает среда их обитания.
Морские беспозвоночные могут высвобо-
408
Глава 14
Таблица 14.4. Системы классификации типов размножения
Размножение происходит одни раз в жизни
Моноцикличность " (монотелия): практически синонимы; второй термин
используется в основном исследователями аннелид
В эту категорию попадают все насекомые; их жизненные циклы далее классифицируются следующим
образом:
унивольтинный (однолетний):
мультивольтинный:
бивольтинный:
семивольтинный
Размножение происходит несколько раз в жнзни
Полицикличность (полителия)
Ежегодная полицикличность (полителия):
Непрерывная полицикличность (полителия):
одно поколение в год
много поколений в год
два поколения в год
одно поколение каждые два года
размножение в определенные периоды,
разделенные обычно однолетними перерывами
размножение идет относительно непрерывно. Если
продолжительность жизни составляет не более
года, такой жизненный цикл бывает неотличим от
унивольтинного
" В англоязычной литературе термину «моноцикличность» соответствует «semelparity», а
«полицикличность»—«iteroparity», но производные от них типа «семельпарность» и «итеропарность» в советской литературе не
привились — Прим. ред.
ждать гаметы прямо в окружающую среду,
где и происходит оплодотворение, но
пресноводные и наземные формы такой
возможности лишены. После наружного
оплодотворения из яиц морских беспозвоночных
часто развиваются подвижные планктонные
личинки (см. разделы по систематике и гл.
15). Это оказывает большое влияние на
стратегию размножения.
Почти во всех типах животных с
морскими представителями большинство или хотя
бы некоторые виды характеризуются
наружным оплодотворением и наличием
пелагических личинок. Такой жизненный цикл,
называемый пелаго-бентосным,
проиллюстрирован в приложении 14.5. Однако во
всех этих типах с пелагической личиночной
стадией встречаются также виды с
непелагическими, бентосными личинками, что
обычно считается прогрессивным
признаком, связанным с утратой или
модификацией личиночной стадии. В большинстве
типов животных имеются также виды,
пелагические личинки которых не питаются, а
существуют за счет запаса желтка,
полученного от родительского организма.
Перечисленные типы личинок называют планкто-
трофными пелагическими и лецитотрофны-
ми пелагическими, а развитие без
пелагической личиночной стадии — прямым. Это
разделение не абсолютно, и развитие
многих личинок смешанное. Наличие
пелагической личинки обычно считают
примитивным признаком. У более 70% всех морских
беспозвоночных умеренных зон планкто-
трофные пелагические личинки, что
подразумевает явные преимущества такого
способа развития в большинстве ситуаций,
заключающиеся, по-видимому, в следующем:
1) использование временного источника
пищи во время «цветений» фитопланктона;
2) заселение новых местообитаний;
3) расширение географического ареала;
4) избегание катастрофического
вымирания при исчезновении локальных
местообитаний;
5) избегание локальной конкуренции
и конкуренции между сибсами;
6) предоставление разнообразному
потомству максимального разнообразия
местообитаний.
Пелагическое развитие часто (но не
всегда) связано с наружным оплодотворением,
которое в свою очередь сочетается с еще
одним предположительно примитивным
признаком— простой формой сперматозоида
с круглой головкой вроде приведенного на
рис. 14.10. Гаметы такого типа встречаются
во всех группах животных, и две эти черты
характеризуют особый морской тип раз-
Размножение и жизненные циклы
409
Таблица 14.5. Примитивные и производные признаки морских беспозвоночных11
Исходные признаки
Производные признаки
Яйца высвобождаются непосредственно в воду,
пелагическое развитие
Наружное оплодотворение; форма
сперматозоидов примитивная
Небольшое количество желтка в яйце
Полное равномерное дробление
Бластула с бластоцелем
Гаструляция посредством инвагинации
Планктотрофная личинка
Автосинтетический оогенез
Размножение один раз в год в строго
определенный период, с четкой синхронизацией гаметоге-
неза у одной и у различных особей
Длительное хранение накопленных половых
клеток
Крупный размер тела, часто обширная полость
тела или широкие межклеточные пространства
Яйца не высвобождаются; часто вынашиваются
Внутреннее оплодотворение, часто
сопровождающееся хранением спермы; форма
сперматозоидов сложная
Большое количество желтка в яйце
Неравномерное дробление
Бластула более или менее заполнена
Гаструляция другого типа
Лецитотрофная личинка
Оогенез другого типа
Многократное, часто относительно непрерывное
размножение; сезонность и синхронизация га-
метогенеза не выражены
Частое использование небольших количеств
половых клеток без их накопления
Тело небольшое, с частично заполненной
полостью. Накопление гамет в межклеточных
пространствах невозможно
Некоторые данные из Jagersten, 1972.
множения. С ними связаны и некоторые
другие, а все вместе они составляют
комплекс коррелирующих между собой
особенностей жизненных циклов морских
беспозвоночных (табл. 14.5, «Исходные
признаки»). Производные, или прогрессивные,
признаки, развившиеся на основе этих
предположительно примитивных, также
перечислены в табл. 14.5. Они особенно
характерны для очень мелких морских
беспозвоночных, обитающих, например, в интерстициа-
ли песчаного дна.
При эволюции относительно крупных
размеров тела оказалось возможным
сохранение типа размножения, считающегося
примитивным. В типах животных без крупных
форм, например у киноринхов, а также
у мелких моллюсков, аннелид и иглокожих
(с крупными сородичами) наблюдаются
в основном продвинутые признаки. Они
встречаются также у морских
беспозвоночных, обитающих в экстремальных или
необычных условиях, например на большой
глубине или в полярных районах, а также
в эстуариях.
Важным свойством, коррелирующим
с «примитивным» типом размножения.
является массовое высвобождение гамет во
время ежегодных нерестов. Как правило,
морские беспозвоночные накапливают их
в полостях тела (часто в целоме), а затем
полностью расходуют в течение
единственного нерестового сезона. Одной из причин
эволюции продвинутого типа размножения
у мелких организмов, возможно, была их
неспособность запасать достаточное
количество гамет. Два различных типа
жизненных циклов с массовым и четко сезонным
нерестом рассмотрены в приложении
14.4(3). Более распространен ежегодный
нерест при продолжительности жизни два
года или более. Однако некоторые виды,
например полихеты семейства Nereidae и все
головоногие моллюски (за исключением
примитивных наутилоидей) нерестятся
всего раз в жизни. Их массовый нерест
сопровождается генетически
запрограммированной гибелью.
Для мягкотелых пресноводных и
наземных беспозвоночных также характерны
продвинутые репродуктивные признаки,
приведенные в табл. 14.5. Они описаны в
следующем разделе.
410
Глава 14
14.3.3. Пресноводные
и наземные беспозвоночные
У животных, обитающих в пресных
водах или на суше, нет выбора между планк-
тотрофным и лепитотрофным
пелагическим или прямым развитием. В этих средах
осмотические и другие ограничения не
допускают выделения в среду незащищенных
сперматозоидов и яйцеклеток, так что
оплодотворение должно быть внутренним.
Развивающиеся эмбрионы также требуют
защиты от потерь воды и осмотического
стресса, например с помощью
водонепроницаемых покровов или внутри кокона.
Следствием этого явилась утрата
примитивной пелагической личинки, и
размножение крупных мягкотелых неморских
беспозвоночных характеризуется следующими
признаками:
1) живорождение или откладка яиц,
защищенных непроницаемыми мембранами
или коконами;
2) внутреннее оплодотворение,
требующее непосредственного спаривания
партнеров;
3) структурно усложненные, часто
нитевидные сперматозоиды;
4) относительно большой вклад
материнских ресурсов в каждое яйцо, что
снижает плодовитость;
5) забота о потомстве или обеспечение
его пищей;
6) многократная или эпизодическая
откладка яиц с использованием внутреннего
запаса спермы для их оплодотворения;
7) гермафродитизм (особенно
характерен для мягкотелых беспозвоночных, но не
свойствен членистоногим).
Признаки некоторых пресноводных
животных выдают их недавнее происхождение
от морских предков. Так. пресноводные
двустворчатые моллюски, например
беззубка (Anodonta), очень сходны со своими
морскими сородичами. Они, в частности,
образуют довольно многочисленные глохи-
дии (модифицированные личинки-
велигеры), которые, однако, не являются
свободноживущими, а распространяются,
прикрепляясь к жабрам или коже рыб.
Жизненные циклы пресноводных
плоских червей, поясковых аннелид и легочных
моллюсков очень сходны. Все они -
синхронные гермафродиты (кроме
утративших способность к половому
размножению) и характеризуются сложным половым
поведением, высокоспециализированными
придаточными железами и способностью
защищать развивающиеся эмбрионы. В их
жизненные циклы входят продолжительные
периоды откладки яиц (см. приложение
14.4). Основной компромисс, на который
они вынуждены идти, между
плодовитостью и цродолжительностью жизни;
некоторые виды живут всего год или меньше,
другие, обычно более крупные,— несколько
лет.
Так как в пресноводной или наземной
средах колебания физических параметров
значительно сильнее, чем в море,
большинство обитающих в этих биотопах
беспозвоночных способны переходить в состояние
физиологического покоя, называемое диа-
паузой, при котором организм переносит
без вреда для себя потери воды и
экстремальные окружающие условия. Его
метаболические потребности снижены, он может
обходиться без внешних источников энергии
и за счет этого переживать периоды
отсутствия корма, характерные для умеренных
и полярных областей. У долгоживущих
видов, например некоторых легочных
моллюсков и поясковых кольчецов, диапауза
наступает во взрослом состоянии, а у мелких
форм она часто соответствует стадии яйца.
Так. многие популяции пиявок в течение
зимних месяцев представлены только
эмбрионами в коконах. Перезимовывание
в виде покоящихся яиц особенно
характерно для таких крошечных форм, как
коловратки и некоторые группы ракообразных,
в частности для водяных блох (Daphnia).
У этих организмов жизненные циклы
включают чередование половых и бесполых
поколений (рис. 14.4): из диапаузирующего
яйца, образующегося в результате
полового размножения, весной вылупляются
бесполые особи, которые дают начало летним
клонам, использующим обильные в это
время ресурсы. Крайняя форма диапаузы
наблюдается у бделлоидных коловраток: их
инцистированные яйца могут находиться
практически в состоянии анабиоза многие
годы, пока не попадут в подходящие для ро-
Размножение и жизненные циклы
411
ста условия. Это свойство, возможно,
связано с отсутствием полового
размножения: избегая неблагоприятных условий, они
живут только в оптимальной для себя
среде.
Насекомые ипаукообразные
приспособились к наземной жизни особенно успешно
(см. разд. 8.4.3.2 и 8.5.3Б.2); кроме того,
некоторые их личиночные, нимфальные или
половозрелые стадии (особенно у
насекомых) вторично заселили пресные воды.
Репродуктивная биология этих групп
отличается от свойственной мягкотелым
беспозвоночным одним очень важным признаком:
среди них очень редко встречаются
гермафродиты; в подавляющем большинстве
случаев возможно только половое
размножение, причем всегда с раздельнополостью.
Эволюционный успех здесь в значительной
мере связан с появлением
водонепроницаемых покровов, а также способностью
к образованию водонепроницаемых яиц.
Они должны быть оплодотворены до
откладки, поэтому у всех одноветвистых
внутреннее оплодотворение, часто связанное со
сложным брачным поведением.
Преобладание раздельнополости в этих группах,
вероятно, объясняется высокой
подвижностью животных.
Жизненные циклы несекомых
традиционно классифицируют по типу
развития— с неполным (экзоптериготы) или
полным (эндоптериготы) превращением (см.
гл. 15). Здесь, однако, будет использован
иной, функциональный подход. Жизненный
цикл насекомого можно рассматривать как
смену различных типов активности в юве-
нильном и имагинальном состояниях.
Поскольку развитие включает несколько
дискретных стадий, разделенных линьками,
каждая стадия может выполнять
специфические функции, основные из которых
перечислены ниже:
1) развитие и дифференцировка;
2) накопление пищи и других ресурсов;
3) расселение и поиск ресурсов;
4) спаривание и выбор партнера;
5) обеспечение ресурсами потомства;
6) выбор места для развития потомства;
7) откладка яиц.
Жизненные циклы некоторых
насекомых проанализированы с этой точки зрения
в приложении 14.6. Обратим внимание на
то, что накопление ресурсов и расселение
могут происходить на различных стадиях
развития, а также на отличие по этим
параметрам большинства насекомых от
морских беспозвоночных, у которых расселение
обычно происходит на ювенильной стадии,
а накопление ресурсов - во взрослом
состоянии.
Самый простой тип развития такой, при
котором переход от личинки к имаго
происходит постепенно, причем все стадии
используют одни и те же пищевые ресурсы.
Накопление их также идет на всех стадиях, но
взрослые особи, кроме того, выполняют
функции расселения, поиска ресурсов,
спаривания и откладки яиц. Такой тип
развития характерен для прямокрылых
(например, саранчи) и проиллюстрирован в
приложении 14.6(1).
Однако довольно часто личинки и имаго
различаются по характеру питания, и,
следовательно, давление отбора для них
неодинаково. Например, личинки падальной
мухи (двукрылое) используют обильный, но
временный источник пищи — мясо мертвых
животных. Найти его - задача
родительской особи, которая часто питается иначе
[см. приложение 14.6(2)]. Личинки
некоторых насекомых обитают в воде, так что
отличие их экологической ниши от имагина-
льной еще более выражено.
На таком несходстве, вероятно,
основана эволюция жизненных циклов, в которых
функции расселения и поиска ресурсов
полностью отделены от функции их
накопления, так что имаго почти или совсем не
питаются. Это разделение в филогенезе
насекомых возникало неоднократно и
независимо. Крайними его примерами могут
служить поденки, веснянки и ручейники. Их
личинки— водные хищники, а взрослые особи
не питаются и живут всего несколько часов,
в течение которых должны спариться и
отложить яйца в места, подходящие для их
развития [см. приложение 14.6(3)].
У чешуекрылых жизненный цикл
сходный, хотя взрослые бабочки все же
получают некоторое количество энергии,
потребляя нектар. Основная функция их личинок
(гусениц) —накопление как можно
большего количества доступных ресурсов с макси-
412
Глава 14
мальной скоростью. Переход ко взрослому
состоянию, в задачи которого входит
спаривание и поиск ресурсов, осуществляется
через непитающуюся стадию куколки
[приложение 14.6(3)]. Пищевые ресурсы бывают
доступны лишь в определенные сезоны, их
поиск может идти как в пространстве, так
и во времени, включая диапаузу на стадии
яйца, куколки или имаго, а также
миграцию.
Некоторые взрослые насекомые кроме
расселения, спаривания и поиска ресурсов
занимаются также накоплением последних.
При этом они только находят их, чтобы
передать потомству. Такой жизненный цикл
известен у некоторых прямокрылых,
жесткокрылых и двукрылых, но наиболее
характерен для перепончатокрылых [см.
приложение 14.6(4)]. Считается, что он сыграл
главную роль в эволюции эвсоциального
поведения пчел и ос [см. приложение
14.6(5)].
14.4. Контроль процессов
размножения
14.4.1. Первичные
и непосредственные
контролирующие факторы
Жизненные циклы, обсуждавшиеся
в предшествующих разделах, включают
сложные переключения клеточных
процессов, которые должны быть
скоординированы во времени и в пространстве как друг
с другом, так с внешними факторами, в
некоторых случаях обеспечивая необходимый
уровень синхронизации жизненных
отправлений особей в популяции. Принято
считать, что четкие циклы репродуктивной
активности в популяциях животных связаны
с циклическими изменениями внешних
условий, в определенные периоды более
благоприятных для размножения, чем
в другие. Это создает давление отбора, на-
1. Меняющиеся параметры внешней среды и
сигналы времени
Долгота Осовые химические
Температура дня Пища вещества
\ \ / /
2. Г Восприятие сигналов I
Центральная
нервная система
©Длинные или короткие дни1
Температура
Приливы и фазы луны
Взаимоотношение фаз
[Суммирование и интеграция}
Нейроэндокринная система:
ПРЕОБРАЗОВАНИЕ
1. Изменение условий
среды
2. Восприятие этих
изменений
3. Преобразование
сигнала
Прямое и косвенное
влияние на
лимитирующие процессы
Внешний цикл гаме-
тогенеза и
размножения
Эндокринная реакция
Пролиферация гамет *
^ Развитие гамет
Высвобождение
гамет
5. Навлюдаеиая цикличность Образования и высвобождения гамет
Рис. 14.8. Схематическое изображение цепочки контролирующих элементов, участвующих в
регуляции репродуктивного цикла, синхронизированного с внешними событиями (по Olive, 1985)
Размножение и жизненные циклы
413
правленного на синхронизацию
репродуктивной активности с такими оптимальными
периодами. Это давление обусловлено
первичными факторами, контролирующими
размножение. Они не обязательно
непосредственно регулируют репродуктивный
цикл. Гаметогенез (особенно оогенез)
иногда продолжается несколько месяцев, и для
правильного протекания этих клеточных
процессов, завершающихся размножением,
организм должен ориентироваться по
совершенно иным факторам внешней среды.
События, используемые для такой
регуляции, называют непосредственными
факторами. Воспринятая информация об
изменениях среды должна быть обработана в
центральной нервной системе и передана
к эффекторам в виде нервных, нейроэндо-
кринных или эндокринных сигналов (разд.
16.12). Последовательность таких команд
определяет высокоупорядоченный видоспе-
цифичный репродуктивный цикл (рис. 14.8).
Под их контролем находится дифференци-
ровка половых клеток и распределение
энергии между поддержанием жизни,
ростом и размножением.
14.4.2. Гаметогенез
Мужские половые клетки образуются
в результате сперматогенеза, а женские —
ЭМБРИОНАЛЬНОЕ
РАЗВИТИЕ
Сперматогониальная
стволовая клетка
Сперматогонии-
(митотическое
размножение)
Сперматоцит
I порядка
МЕЙОЗ I
Сперматоциты
II порядка
х2
Сперматиды
СПЕРМАТОГЕНЕЗ
Сперматозоиды
«4
ГАМЕТОГЕНЕЗ У САМКИ
Оогониальная стволовая
клетка
Оогонии (митотическое
размножение)
Ооцит I порядка
ВИТЕЛЛОГЕНЕЗ
МЕЙОЗ I
Ооцит II порядка +
полярное тельце
МЕИОЗ II
@ Яйцо + 3 полярных тельца
ГАМЕТОГЕНЕЗ У САМЦА
Рис. 14.9. Схематическое изображение основных клеточных процессов гаметогенеза
414
Глава 14
А Б В Г
Рис. 14.10. А. Примитивный сперматозоид
с круглой головкой. Б—Г. Более продвинутые
нитевидные типы сперматозоидов, которые
могут иметь различное внутреннее строение
оогенеза; схемы этих весьма различных
процессов показаны на рис. 14.9 и более
детально они рассмотрены ниже.
14.4.2.1. Сперматогенез
У большинства беспозвоночных
сперматогенез протекает быстро; мейозу
предшествуют многочисленные митотические
деления, что приводит к образованию очень
большого числа половых клеток. Каждый
сперматогоний превращается в спермато-
цит первого порядка, из которого
образуются четыре сперматиды. Сперматозоиды
формируются в результате дифференциров-
ки последних (рис. 14.9, А). У многих
морских беспозвоночных сперматозоиды
сходные, с круглой головкой (рис. 14.10, А), но
для некоторых характерно более сложное
строение (рис. 14.10, Б—Г); это обычно
наблюдается в жизненных циклах,
включающих хранение спермы, внутреннее
оплодотворение, а также тогда, когда у яиц прочные
защитные покровы (см. выше).
14.4.2.2. Оогенез
Оогенез — это продолжительный
процесс, в ходе которого в развивающемся
яйце откладываются запасы питательных
веществ (обычно в течение длительной
профазы мейоза), а мейоз завершается либо
непосредственно перед оплодотворением,
либо уже после него.
У некоторых морских беспозвоночных,
например сипункулид, эхиурид и некоторых
полихет, развивающиеся ооциты по
отдельности свободно плавают в целомической
жидкости—так называемый оогенез со.ш-
В II
Рис. 14.11. Солитарный, фолликулярный и ну-
триментарный типы оогенеза. А. Солитарный
тип: ооциты не связаны с вспомогательными
клетками, а свободно взвешены в жидкостях
тела. Характерен для эхиурид, сипункулид и
некоторых (по-видимому, немногих) полихет. Б.
Фолликулярный тип: ооциты сильно зависят
от покрывающих их соматических клеток. В.
Нутриментарный тип: ооциты связаны с
сестринскими клетками, происходящими из того
же зачатка, что и сам ооцит. В результате
неполного цитокинеза часто возникает синцитиа-
льный комплекс, в котором ооциты и
питающие клетки соединены цитоплазматическими
мостиками или канальцами. I. Пример ооцита
без покрывающих фолликулярных клеток (по-
лихета Diopatra). II. Пример ооцита,
окруженного фолликулярными клетками, что
характерно для двукрылых насекомых, например для
мухи Drosophila (см. также рис. 14.13)
Рис. 14.12. Синтез желтка в ооцитах.
Автосинтетический тип, характерный для солитарного
оогенеза. Низкомолекулярные
предшественники поглощаются микроворсиночной
поверхностью ооцита, а синтез высокомолекулярных
запасных веществ происходит в его
цитоплазме с участием информационных молекул,
образующихся в ядре. ЭР—эндоплазма-
тический ретикулум
Низкомолекулярные предшествен
ники желтка
Рис. 14.1 3. Клеточные процессы при развитии яйца двукрылых насекомых (например, у Drosophi-
la). I. При делении стволовой клетки образуется стволовая клетка следующего поколения и
половая клетка первого порядка. II—V. Половая клетка первого порядка делится четыре раза, но за
счет неполного цитокинеза образуется комплекс из 16 клеток, соединенных цитоплазматически-
ми мостиками. Обратим внимание, что у двух заштрихованных клеток на любом этапе деления
больше связей с соседними, чем у любых других. Одна из этих двух клеток превратится в ооцит,
остальные станут питающими. VI. Питающие клетки действуют как система генной
амплификации: образующиеся в их ядрах информационные молекулы (мРНК) поступают в цитоплазму
ооцита. Весь комплекс окружается фолликулярным эпителием из соматических клеток. VII. Ооцит
растет за счет питающих клеток. VIII. Зрелое яйцо полностью заполняет фолликул. Оно готово
к эмбриогенезу после оплодотворения и откладки
416
Глава 14
Высокомолекулярные
Низкомолекулярные составляющие
предшественники желтка
Эндо'пиноцитоз
Фолликулярные
клетки
Рис. 14.14. Гетеросинтетический тип синтеза запасных веществ, обычно (но не обязательно)
связанный с фолликулярным развитием ооцита. Сложные высокомолекулярные
предшественники желтка приносятся к ооциту жидкостями тела (кровью, гемолимфой, целомической
жидкостью и т.д.). Эти вещества синтезируются в неполовых клетках с использованием их
информационных молекул. Приведенная схема типична для некоторых насекомых
тарного типа (рис. 14.11, А). При этом
будущие запасные вещества поглощаются оо-
цитом в виде низкомолекулярных
предшественников (аминокислот, простых Сахаров
и моноглицеридов), из которых органеллы
его цитоплазмы синтезируют
высокомолекулярные вещества, в совокупности
называемые «желтком» (как показано на рис.
14.12). Такой характер накопления
последнего называют автосинтетическим; он
обычно (но не всегда) связан с солитарным
типом оогенеза (см. ниже).
Однако для большинства
беспозвоночных характерны фолликулярный или нутри-
ментарный типы оогенеза (рис. 14.11, Б, В),
при которых развивающийся ооцит тесно
связан с эпителиальным слоем
соматических клеток (называемых фолликулярными)
и/или со своими сестринскими клетками,
образующимися в результате неполного
цитокинеза при митотических делениях
материнской клетки (оогония) до начала
профазы мейоза. Развитие сложного яичника
такого типа детально рассматривается на рис.
14.13 на примере двукрылого насекомого
Drosophila. Накапливается все больше
данных о том, что при более сложных типах
оогенеза большинство
высокомолекулярных веществ, накапливающихся в
цитоплазме ооцита, синтезируется не им самим, а
соматическими клетками в других частях
организма и доставляется к ооциту
жидкостями тела или кровью по сосудам (рис. 14.14).
Такой тип оогенеза называют гетеросинте-
тическим. Впервые он был обнаружен у
насекомых, но затем оказался широко
распространенным, характерным, например, для
ракообразных и моллюсков.
Рис. 14.15. Схема строения турбелларии, показывающая обширную систему желез, участвующих
в образовании сложного «яйца», состоящего из оплодотворенной яйцеклетки, желточных клеток
и белковой оболочки. Стрелками показан путь движения яйца из яичника, в ходе которого оно
оплодотворяется и окружается совместно с питающими клетками белком перед выведением
наружу (см. также рис. 3.46)
Размножение и жизненные циклы
417
Ранее считалось, что для полихет
типичен автосинтетический солитарный оогенез,
но, как выяснилось, эта группа по
разнообразию и сложности вариантов оогенеза
и участвующих в нем структур не уступает
насекомым. У некоторых видов
обнаружены его фолликулярный и нутриментарный
типы, а у нереид (семейство Nereidae) он,
хотя и солитарный, но, вероятно, гетеросин-
тетический.
«Яйцо» турбелларий обычно
представляет собой сложное образование, состоящее
из почти не содержащего желтка ооцита
и питающих клеток с цитоплазмой,
заполненной желтком, синтезированным в
особом органе — желточнике (рис. 14.15); все
это упаковано в плотную белковую
оболочку. Такие яйца называют эктолецитальны-
ми; считается, что для плоских червей этот
признак — продвинутый, а примитивное
состояние— эндолецитальныи тип с желтком,
запасенным в цитоплазме ооцита; он
встречается у некоторых свободноживущих
турбелларий. У пресноводных и наземных ан-
нелид и моллюсков адаптации сходные:
откладываемые яйца окружены коконом,
содержащим питательный белок,
дополняющий вещества,
яйцеклетки.
запасенные в цитоплазме
14.4.3. Синхронное размножение
морских беспозвоночных
Наиболее распространенный тип
размножения морских беспозвоночных (разд.
14.3.2) требует очень точной синхронизации
репродуктивных процессов как в одном
организме, так и между особями одной
популяции. Эта точность бывает весьма
впечатляющей. В табл. 14.6 приведены даты
нереста тихоокеанского червя палоло (Polychae-
ta, Eunicida) за последние 100 лет; его
период жестко связан с третьей четвертью луны,
наступающей после определенной даты
в начале октября. Сроки соблюдаются
с точностью до одного дня, причем нерест
каждый раз начинается в одно и то же время
суток. Так же строго соблюдаются сроки
размножения у японской морской лилии
Comanthns japonicus; на рис. 14.16 показано,
что весь годовой цикл гаметогенеза у этих
животных строго синхронизирован.
Приведенные примеры представляют собой, по-
видимому, крайние случаи, но у большин-
Таблица 14.6. Подборка данных, иллюстрирующая синхронность всплывания для нереста
половозрелых червей палоло (Eunice viridis) у островов Самоа
Год
Третья четверть луны
Даты всплывания
Октябрь
Ноябрь
Октябрь
Ноябрь
П843
19 лет 11862
19 лет<^
19 лет*
1874
1893
1926
1927
1928
1929
1930
1943
1944
1945
16
15
31
31
27
17
5
24
14
20
8
27
14
14
30
29
26
15
4
23
13 \
19
7
26
("15/16
115/16
<Р'
131
[28
17
25
1445
20
28
14/15
7/9
Эти данные свидетельствуют о том, что а) нерест никогда не происходит ранее 8 октября; 6) он происходит
в октябре, если третья четверть луны наступает позднее 18 октября; в) каждые 19 лет нерест приходится на одно и то
же число. Кроме i oi о, время суток, когда взрослые особи всплывают на поверхность моря, также точно
фиксировано.
27—1314
418
Глава 14
250
1974
Рис. 14.16. Синхронное развитие яиц у морской лилии Comanthus japonicus. Цикл гаметогенеза
занимает 12 месяцев и заканчивается синхронным спариванием всех особей популяции. Оно
продолжается двое суток и строго коррелирует с фазами Луны. Горизонтальной полосой обозначено
время размножения гаметоцитов (по Holland et al., 1975)
ства морских беспозвоночных, относящихся
к любому типу, проявляется та или иная
степень синхронизации репродуктивных
процессов. Возникает вопрос—чем же
регулируются эти циклы? Ранее считалось, что
основными сигналами служат изменения
температуры окружающей среды, но для
обеспечения такой точности, как в двух
приведенных выше примерах, этого, очевидно,
недостаточно, и вообще у температурных
циклов слишком велика случайная
изменчивость (т.е. у сигнала слишком высок
уровень шума), чтобы они могли служить
единственным регулирующим фактором.
Море—сложная среда с множеством
ритмических изменений (табл. 14.7), и
репродуктивные циклы морских
беспозвоночных могут быть согласованы по фазе с
каждым из средовых циклов. Было показано,
что такие разные животные, как полихеты,
иглокожие и ракообразные, способны четко
реагировать на длину светового дня,
причем иногда не менее сложно, чем в случае
описанных ниже фотопериодических
реакций наземных насекомых. Кроме того,
у них известны непосредственные реакции
на смены фаз луны, а также эндогенные
ритмы с примерно лунной или приливной пе-
Таблица 14.7. Геофизические циклы в морской среде
Название
Периодичность
Метонов
Годичный
Лунный
Полулунный
Приливный
Суточный
Полуприливный
Повторение одинаковых сочетаний фаз Солнца и Луны 19 лет
Оборот Земли вокруг Солнца 1 год
Оборот Луны вокруг Земли 29,5 сут
Повторение одинаковых сочетаний фаз приливного 15 сут
и солнечного циклов (цикл
квадратурного/сизигийного приливов)
1 лунные сутки 24,8 ч
1 солнечные сутки 24 ч
Время между полной или малой водой и средней водой 12,4 ч
Результаты наблюдений говорят о том, что репродуктивная активность по крайней мере каких-то морских
беспозвоночных коррелирует с каждым из этих циклов. Это, однако, не позволяет выявить причинный фактор
(обсуждение в тексте).
Размножение и жизненные циклы
419
Часы
10 20 30 АО 50 60 70
Г
Месяцы
III
Циркадианные
компоненты
П
Лунные
компоненты
Годовые
компоненты
Модуляция факторами
внешней среды
Рис. 14.17. Схематическая модель высокосинхронизованного спаривания животных типа
морской лилии Comanthus (рис. 14.16) или полихеты Eunice (табл. 14.6). Предполагается, что
размножение запускается взаимодействием эндогенных ритмов, каждый из которых, возможно,
настраивается внешним времязадателем с годовой, лунной или суточной периодичностью. В
результате размножение происходит в течение одного или двух месяцев в год в дни, жестко связанные
с лунным циклом, и в строго определенное время суток (по Olive, 1985)
риодичностью, которые настраиваются
(«захватываются») цикличностью
соответствующих внешних процессов («времязада-
телей»), причем эти внутренние «часы»
очень точно «показывают» лунное или
приливное время. Накапливаются сведения,
говорящие о том, что в основе годичных
репродуктивных циклов могут также лежать
эвдогенные ритмы с периодом, близким
году (рис. 14.17).
Репродуктивные циклы морских
беспозвоночных включают чередование двух
основных процессов — образования и
высвобождения гамет. Эти циклы могут
регулироваться циклическими изменениями
образования гормонов. По характеру действия
можно выделить два основных типа
последних: гонадотропные и индуцирующие
нерест или созревание и/или активацию гаме-
тоцитов, как показано на рис. 14.18. Такой
тип эндокринного контроля размножения
возникал неоднократно и независимо в ходе
эволюции беспозвоночных, поэтому
вещества, его осуществляющие, могут иметь
у разных животных совершенно различную
молекулярную структуру. У иглокожих го-
надотропная функция выполняется двумя
гормонами, типичными также для
позвоночных,— прогестероном и эстроном, а
нерест вызывается каскадом сигналов,
начинающимся высвобождением нейросекре-
торного пептида их радиальных нервов, что
в свою очередь индуцирует образование
в яичнике простого регуляторного вещества
1-метиладенина. Последний стимулирует
сокращение мышц яичника, т. е. начало
нереста, а помимо этого — образование
третьего посредника на внутренней
поверхности мембраны ооцита, что в конечном счете
ведет к исчезновению его ядерной оболочки
и делает возможным оплодотворение.
Сходные функциональные
взаимоотношения обнаружены у полихет и моллюсков,
хотя участвующие в них вещества имеют
совершенно иную молекулярную природу.
По-видимому, эти гормональные
механизмы эволюционировали независимо.
Некоторые морские беспозвоночные
27«
Глава 14
- Активность гонадотропного гормона
■ Активность гормона нереста
Время
Соматический рост
Накопление
запасов
Мобилизация запасов
Накопление репродуктивных
клеток
Рассасывание яиц
Соматический рост
Накопление запасов
Рис. 14.18. Контроль годового репродуктивного цикла гонадотропным и индуцирующим нерест
гормонами: А обобщенная схема; Б— клеточные события, связанные с различными фазами
гормонального цикла
размножаются не циклически, а лишь раз
в жизни (см. выше). Участие эндокринной
системы в контроле репродуктивных
процессов у этих животных, по-видимому,
существенно отличается от описанного
выше—здесь происходит обычно
необратимое переключение с соматического роста на
размножение. Такой механизм
наблюдается у нереид. Созревание и часто
сопровождающий его соматический метаморфоз
зависят от постепенного снижения уровня
единственного гормона (см. гл. 16).
необходимого для регенерации и образования
сегментов, т. е. по мере созревания животного
его способность расходовать ресурсы на
данные процессы утрачивается. Это —
адаптивный признак, так как после
размножения животное неизбежно умирает, новые
сегменты все равно не могли бы повысить
его будущую плодовитость и, таким
образом, оказываются бесполезными.
У полителических червей (см. табл. 14.4)
ситуация иная, поэтому у них способность
к регенерационному росту не связана с
размножением так жестко. Характер
эндокринного контроля репродуктивных процессов
и общая репродуктивная стратегия
организма, по-видимому, тесно коррелируют, но
остается неясным, какой из этих двух
факторов является причиной, а какой —
следствием.
Сходная ситуация наблюдается у
моллюсков. Для большинства их видов
характерна полицикличность, и считается, что их
эндокринные системы соответствуют
общей схеме, приведенной на рис. 14.18.
Однако у большинства головоногих при
половом размножении происходит быстрое
массовое перераспределение резервов:
запасные метаболиты переходят в
развивающиеся половые клетки, и животные после
спаривания гибнут. Такое переключение
вызывается секретом глазных желез,
выполняющих таким образом гонадотропную
функцию; активность зрительного нерва
подавляет их секрецию. Изоляция желез или
перерезание зрительного нерва вызывает
преждевременное половое созревание и
в конечном итоге смерть, а переключение,
о котором идет речь, обычно необратимо.
Размножение и жизненные циклы
421
14.4.4. Циклы размножения
и диапауза в наземной
и пресноводной средах
Репродуктивная биология наземных
и пресноводных беспозвоночных в
значительной степени определяется их
способностью к внутреннему оплодотворению и
хорошей защищенностью откладываемых
яиц. С этим также связана способность
самок сохранять сперматозоиды и
оплодотворять яйца последовательно.
Большинство беспозвоночных, обитающих в
умеренных, бореальных и полярных широтах,
характеризуются сезонным размножением
с продолжительными периодами откладки
яиц, перемежающимися репродуктивным
покоем. Такой цикл характеризуется не
столько синхронностью размножения, сколько
закономерными переходами между
репродуктивными активным и неактивным
состояниями. Последнее может быть покоем,
т.е. непосредственной временной реакцией
на неблагоприятные окружающие условия,
прекращающейся при их улучшении:
факультативной диапаузой—непосредственной
реакцией на неблагоприятные условия
среды, которая не прекращается, пока не
пройдет определенный период времени;
облигатной диапаузой — прекращением
активности в определенное время года вне
зависимости от неблагоприятных условий,
для избежания которых оно бывает
приспособлено.
Состояния покоя и факультативной диа-
паузы наступают на взрослой стадии
легочных моллюсков, дождевых червей и
некоторых насекомых; облигатная диапауза,
напротив, чаще связана с более ранними
стадиями развития—эмбриональной,
личиночной или куколочной. Так, многие
популяции наземных и пресноводных
беспозвоночных в умеренных и бореальных областях
переживают зиму исключительно на стадии
яйца. Это явление особенно подробно
изучено на насекомых, причем установлены
некоторые факторы, его вызывающие.
Насекомых, которые активны летом и впадают
в диапаузу осенью, называют «длинноднев-
ными», а тех, что активны зимой, как,
например, тутовый шелкопряд (Bombyx то-
ri),—«короткодневными». Хотя эти
термины носят чисто описательный характер, они
вполне отражают суть дела, так как именно
продолжительность светового дня
регулирует переход из одного физиологического
состояния в другое.
Циклические изменения долготы
светового дня могут служить очень точным
источником информации о смене времен
года во всех областях, кроме тропических.
Ход этих изменений в течение года на
различных широтах показан на рис. 14.19;
обратим внимание, что два раза в год 21
марта и 21 сентября продолжительность
и дня. и ночи составляет ровно 12 ч в любой
точке земной поверхности. Часто животные
реагируют на изменение долготы дня
и используют эту информацию для
контроля процессов, имеющих годовую
периодичность. Это явление называется
фотопериодизмом. Реакция многих животных на
относительную долготу дня (или ночи)
обычно нелинейна, т. е. световой период,
меньший определенной критической
величины, воспринимается просто как короткий,
а превышающий некую величину — как
длинный. В этом можно убедиться,
поместив колонии насекомых в различные
фотопериодические условия и регистрируя
частоту наступления у них диапаузы (рис.
14.20). Точность определения животными
времени находится при этом в пределах 30
мин, что привлекло значительное внимание
физиологов, интересующихся природой
биологических часов.
Изучение различных насекомых
показало, что фотопериодизм, вероятно,
определяется различными процессами. У
некоторых видов для оценки эффективной
долготы дня, по-видимому, используется
внутренний циркадианный' осциллятор.
Наиболее отчетливо это демонстрируют
опыты, когда насекомых содержат в условиях
с фиксированным световым днем и ночью
различной продолжительности (от 12 и
менее до 72 ч и более). При этом некоторые
«дни» животные воспринимают как
длинные, а другие—как короткие, причем таким
1 В советской литературе более употребим
термин «циркадный», однако грамматически он
менее верен (от латинского "dies" = «день»
остается только одна буква).— Прим. ред.
24
22
20
18
16
14 -
a
-
—
-
~0°
-20°-<^Я
~щ//ll
'W П
"65/ /
_ 70°
ll i I i
70° J
I i I i I
65°
60° \
50° ^
40°\
""30°^
- Ю" ■
i I > I
70°
1 i 1 i
0
учХ4--!00
\\ \JL0°
\ \ёГ
\70°
1 , 11,1,
I II III IV V VI VII VIII IX X XI XII
Рис 14.19. Сезонная изменчивость долготы дня в зависимости от широты. Обратим внимание,
что в любой точке Земли дважды в год день и ночь продолжаются по 12 ч. Период изменений
долготы дня везде одинаков; от широты зависит только его амплитуда (по Saunders, 1977)
Естественная долгота дня
100 г-
60
20-
-
•
I
•<4
Б __—
6
Фоте
период
Л
\
\
\
1
1
12
<*-*"
\
\
v_
i i
18
(Св
зтлое время суток, ч)
Критическая долгота дня
Рис. 14.20. Схема фотопериодической реакции у (А) длиннодневных и (6) короткодневных
насекомых. Выраженность реакции оценивается по проценту особей, входящих в диапаузу при данном
фотопериоде
Размножение и жизненные циклы
423
L Т
I 8 24 СГ8:16
[I 8 36 СТ8:?8
III 8 Л8 СТ8:Л0
IV 8 60
111 1
тт
I^S
WWW^^^^^T
Т^^ S ^^^ \\\\\\4\\\4s ^ \\\Ч\\ЧУ
ж??
k\4\\4sS\44444\vR\4\\44\444\k^^
I
I II'
[ I. II..
Ill I ' I
I Г ',' '.I
Г.ч
Рис. 14.21. Данные, свидетельствующие об
участии свободнотекущего циркадианного
ритма в фотопериодических явлениях. А. Схема
резонансного опыта. Предполагается
существование некого эндогенного осциллятора
(показан в виде синусоиды) с
фоточувствительной фазой ф1. Различные световые режимы
изображены чередованием заштрихованных
и светлых полос. В примерах I — IV световой
период (С) фиксирован (8 ч), а темновой (Т)
варьирует; это дает различную
продолжительность циклов т. Обратим внимание, что в
примерах I и III ф1 всегда приходится на темновой
период, а в примерах II и IV—иногда на
световой (.). Это может восприниматься как
условия длинного дня. Б. Экспериментальная
демонстрация «резонанса», свидетельствующая
о существовании внутреннего осциллятора. На
графиках показан процент серых мясных мух
Sarcophaga, входящих в куколочную диапаузу
при выращивании в различных
фотопериодических режимах с длиной светового дня 4—
20 ч и эффективной продолжительностью
суток 20—72 ч. Явно заметен резонанс (по
Saunders, 1982)
образом, как будто есть осциллятор со
«свободнотекущим» циркадианным (т. е.
примерно 24-часовым) ритмом, имеющим
светочувствительную фазу, определенным
образом сдвинутую по отношению к началу
цикла, устанавливаемого либо «рассветом»,
либо «закатом». На рис. 14.21 показано, как
в таких «резонансных» опытах можно
добиться того, что эта фаза всегда будет
приходиться на светлое время «суток» даже при
424
Глава 14
х i-
IUU
60
20
-
/ч^
1 1 1 1 1 1
постоянной продолжительности светового
периода. Это возможно только при условии
«свободного течения» внутреннего нирка-
дианного ритма в течение нескольких
циклов. Однако у некоторых животных таких
резонансных эффектов обнаружить не
удается.
Например, у тли Megoura viciae в опыте,
аналогичном проиллюстрированному на
рис. 14.21, признаков резонанса не заметно.
Как следует из рис. 14.22, эти насекомые
используют для измерения времени
механизм типа «песочных часов». Переход от
партеногенетического бесполого
размножения к половому, характерный для
жизненного цикла тлей, обычно индуцируется
укорочением светового дня, что имеет явное
адаптивное значение, так как обильные
весенние и летние пишевые ресурсы к этому
времени почти иссякают. Фактически этот
переход вызывается увеличением
относительной продолжительности ночи,
измеряемой внутренними «песочными часами» (т. е.
определяющими истекший период
времени). Как только длина темнового периода
превысит 9,75 ч, возникает реакция на
«короткий день» и появляются половые
формы, даюшие начало зимующим в состоянии
диапаузы яйцам.
14.5. Размножение
и распределение ресурсов
Образ жизни и циклы размножения
беспозвоночных весьма разнообразны, но при
этом видовые репродуктивные особенности
обычно жестко фиксированы. Одна из
проблем современной биологии
беспозвоночных заключается в создании эволюционной
Рис. 14.22. Данные, свидетельствующие о том,
что тли Медоша viciae измеряют
продолжительность темнового времени суток с помощью
механизма типа «песочных часов». А. Процент
бескрылых (летних) партеногенетических
самок в колониях при различных фоторежимах
с фиксированной продолжительностью
светового времени (8 ч). Заметно отсутствие
резонанса. Б. Процент бескрылых
партеногенетических самок, появляющихся, когда после
наступления темноты возникает полуторачасовой
световой перерыв, а затем следует темновой
период различной длительности. В. Эффект
одночасового светового перерыва в различное
время очень длинной «ночи». Такой режим
воспринимается как «длинный день» только тогда,
когда свет подается через 8 ч после «заката».
Даже кратковременное прерывание темноты
в это время воспринимается как рассвет,
наступивший до истечения критического периода,
что оказывает глубокое влияние и физиологию
насекомого. Более ранний световой импульс
интерпретируется как новый «закат»; его
воздействие перекрывается следующей длинной
ночью (по Saunders, 1982)
Размножение и жизненные циклы
425
теории, объясняющей наблюдаемое в
природе разнообразие жизненных циклов.
Пока такой теории не существует, но уже
можно сформулировать некоторые ее общие
положения.
Наиболее признана в настоящее время
так называемая «демографическая теория
жизненного цикла». Она базируется на
следующих положениях:
1) естественный отбор действует на
отдельные признаки жизненного цикла;
2) естественный отбор направлен на
оптимизацию адаптивных стратегий (т.е. на
повышение приспособленности особей, где
под приспособленностью понимается
эффективность передачи генетического
материала последующим поколениям);
3) отдельные признаки жизненного
цикла могут эволюционировать независимо.
Более того, поскольку доступные
организму ресурсы ограничены, возникает
потенциальный конфликт между их
использованием для выживания и плодовитости,
который разрешается тем или иным
компромиссом.
Демографическая теория полагает, что:
1) увеличение репродуктивного усилия,
т.е. вклада ресурсов в размножение, ведет
к росту эффективности последнего, но в то
же время уменьшает затраты на
поддержание жизнеспособности самого организма;
2) увеличение репродуктивного усилия
ведет к повышению собственной
плодовитости и/или выживаемости потомства;
3) снижение вклада ресурсов в
соматическое развитие ведет либо к снижению
выживаемости особи, либо к замедлению
роста, а следовательно, к снижению
эффективности размножения в будущем;
4) возможен компромисс между
сиюминутной репродуктивной эффективностью
и остаточной репродуктивной ценностью,
определяемой как произведение
выживаемости и плодовитости в оставшийся период
жизни особи.
Там, где возможны компромиссы,
различные решения будут оптимальными
в разных условиях, однако адаптивная
изменчивость организма ограничена его
эволюционной историей. Например,
высказывалось предположение, что утрата стадии
личинки-велигера брюхоногими
моллюсками связана с очень глубокими
генетическими изменениями, сделавшими этот шаг эво-
люционно необратимым. Если это так,
следует ожидать резкого таксономического
расхождения групп гастропод,
сохранивших и утративших такую личиночную
стадию; это и подтверждается наблюдениями.
Важный для беспозвоночных
компромисс—между численностью и
выживаемостью потомства. Это иллюстрируется,
например, выбором между планктотрофным
и лецитотрофным типами развития или
между стратегиями заботы о потомстве или
отсутствия таковой. Одни и те же русурсы
можно вложить или в относительно
большое число «дешевых» потомков, или в
немногих, но «дорогих», каждый из которых
будет иметь более высокие шансы на
выживание. Этот выбор совершается обычно не
на индивидуальном уровне, а в ходе
эволюционной истории вида, так что вклад
ресурсов в каждого потомка — видоспецифичный
признак. Нередко даже близкородственные
виды по нему различаются.
Вторая важная
переменная—относительное количество доступных ресурсов,
вкладываемых в каждый акт размножения.
Следует предполагать, что любой вклад
в него снижает соматические возможности,
причем этот конфликт разрешается на
индивидуальном уровне. Репродуктивные
стратегии большинства беспозвоночных
отнюдь не случайны, и между накоплением
ресурсов и их вкладом в размножение
обычно проходит какое-то время. У этой
задержки своя «стоимость», связанная с
определенной вероятностью гибели особи до
начала размножения, а также с затратами на
сохранение запасов. Затраты могут быть
уравновешены следующими
преимуществами:
1) условия для выживания потомства
бывают непостоянными. Это часто
наблюдается, если молодь использует временный
источник пищи или менее устойчива к
определенным условиям среды;
2) у видов с наружным
оплодотворением его вероятность выше, если гаметы
выделяются всеми особями популяции
одновременно;
3) размножение бывает сопряжено с
риском. Если, например, в этот период живот-
426
Глава 14
ное подвергается повышенной опасности
нападения хищника, может быть выгоднее
сначала накопить большую часть или все
доступные ресурсы, а затем вложить их
в размножение;
4) если накопление жизненного опыта
родительской особью повышает шансы
потомства на выживание, этого может
оказаться достаточным для того, чтобы
перевесить стоимость позднего размножения.
Биология популяций и взаимодействия
между членами биоценоза в значительной
мере определяются особенностями
репродуктивной биологии составляющих их
особей. Жизненные циклы выражают характер
распределения ресурсов между
соматическими и репродуктивными функциями. От
него зависят плодовитость и
выживаемость, т. е. в конечном счете возрастная
структура популяции. К сожалению, в
настоящее время не понятны ни
функциональное значение всего разнообразия
жизненных циклов и типов размножения
беспозвоночных, ни давления отбора, ответственные
за их формирование; однако в этой области
уже появляются новые идеи.
Обычно считают, что признаки,
характеризующие жизненный цикл, подвержены
давлению отбора. С другой стороны,
необходимо признать, что бывают
филогенетические и функциональные ограничения,
препятствующие оптимальному ответу на это
давление. Трудности возникают в связи
с возможностью не одного, а нескольких
решений. Например, иногда
близкородственные организмы занимают сходные
экологические ниши, но существенно
различаются по характеру распределения своих
ресурсов. Так, у морских беспозвоночных
часто встречается ситуация, когда из двух
близких видов, обитающих в практически
идентичных условиях, один образует
небольшое число крупных яиц с большим
запасом желтка, а другой — множество мелких,
почти не содержащих желтка. Значит,
можно говорить о компромиссе между числом
и размером яиц, который не обязательно
зависит от общего вклада ресурсов в
размножение, причем не исключено, что
существует несколько компромиссных решений.
Вот примеры еще нескольких
компромиссов:
— вклад в одного потомка - число
потомков;
— вклад в одного потомка—
выживание потомства;
— число потомков данного размера—
выживание родительской особи;
— число потомков данного размера—
численность будущего потомства.
Один из подходов к пониманию
разнообразия типов размножения—выделение
совместно изменяющихся признаков
и поиск теорий, которые бы предсказывали
их коэволюцию. В качестве иллюстрации
можно рассмотреть виды, обычно
встречающиеся во временно
«недоиспользуемых» экологических нишах. Условия
существования в них краткосрочны и
непредсказуемы, и у видов, к ним приспособленных,
следовало бы ожидать появления
признаков, позволяющих производить
максимально возможное количество потомства. Им
могут быть противопоставлены виды кли--
максовых сообществ, где шансы на
выживание отдельной молодой особи ограничены,
а конкуренция за ресурсы сильна. Можно
предположить, что в этих двух
противоположных ситуациях естественный отбор
будет способствовать возникновению разных
наборов репродуктивных признаков,
причем конкретные признаки вида станут
отражать относительную интенсивность
различных, действовавших на него давлений
отбора. Эта идея оказала очень большое
влияние на развитие теории размножения, и,
хотя в настоящее время понятна ее
недостаточность для полного объяснения
эволюции репродуктивных признаков, она
заслуживает более детального обсуждения.
Для описания ситуаций, в которых
отбор идет преимущественно на образование
максимально возможного числа потомков,
используется термин «r-отбор», где г—
константа, определяющая скорость роста
популяции в уравнениях
dN
— = rN или N, = Nne .
dt
Отбор на максимальное значение г
благоприятствует раннему размножению,
высокой плодовитости и короткой
продолжительности жизни. Можно предполагать, что
«г-стратеги» характеризуются набором
Размножение и жизненные циклы
427
признаков, способствующих производству
максимального числа потенциальных
потомков, и что численность популяций таких
видов будет нестабильной, склонной к
сильным колебаниям. В условиях интенсивной
конкуренции, напротив, возможности для
роста численности невелики, и можно
ожидать корреляции максимальной
приспособленности с поздним размножением,
низкой плодовитостью и обеспечением
каждого из немногих потомков большим запасом
питательных веществ. Отбор такого типа
называется /f-отбором по константе К
в «логистическом» уравнении
Это уравнение описывает рост популяции,
плотность которой асимптотически
стремится к предельной для данной среды. Одна из
трудностей такого анализа связана с тем,
что величины г и К, строго говоря,
несравнимы. Мальтузианский параметр г можно
измерить или оценить при исследовании
реальной популяции, тогда как ее предельная
плотность для среды — абстрактная
безразмерная величина, которую определить на
практике нельзя.
Параметры роста популяции часто
выводятся из исследований выживаемости
и плодовитости особей. Один из подходов
здесь — использование демографических
таблиц, отражающих среднюю
выживаемость и среднюю плодовитость самок
каждой возрастной группы. Эти таблицы
можно составить либо при наблюдении за
популяцией в состоянии динамического
равновесия, где поддается определению
возраст любой особи, либо изучая
выживаемость и плодовитость данного возрастного
класса популяции. Получение таких данных
о беспозвоночных сопряжено с особыми
трудностями, поскольку возраст обычно
трудно определить, однако некоторые
методы все же дают нужную информацию;
например, прикрепленные организмы типа
морских желудей на каменистых берегах
можно метить и наблюдать в течение
нескольких лет. Пример демографической
таблицы для одного из североамериканских
видов этих ракообразных приведен
в табл. 14.8, где указаны средняя
выживаемость 1(х) и средняя плодовитость т|х) в
зависимости от возраста (х).
Сумма произведений 1( -т,,,, 21(х)тх,
представляет собой среднюю плодовитость
самки в течение всей ее жизни. Ее часто
обозначают R0. Время генерации — это средний
возраст особей в момент смерти; его можно
вычислить из данных таблицы по формуле
Z1wm<x) "
Мальтузианский параметр часто
приближенно получают из уравнения
lnR0
г^^р—, где R0 = Il(x,m,x).
Если значение г положительно (как в
табл. 14.8), популяция растет, если
отрицательно — она сокращается, а если равно
нулю—ее численность постоянна. Заметим,
что если г выводится из данных таблицы, то
величина К— нет.
Условия среды всегда изменчивы, но
в некоторых местообитаниях они
стабильнее, чем в других. В последнем случае
мелкие и слабые молодые особи будут более
подвержены гибели, чем крупные и хорошо
приспособленные взрослые. Поэтому
можно ожидать, что в стабильных условиях
отбор благоприятствует большой
продолжительности жизни с растянутым во
времени производством хорошо обеспеченного
запасом питательных веществ потомства.
И напротив, во флуктуирующих условиях
среды все возраста будут примерно
одинаково подвержены смертности. Эти доводы,
как и концепция г/Л^-отбора, позволяют
предполагать коэволюцию в стабильных
условиях таких признаков, как длительная
продолжительность жизни, низкая
плодовитость и поздний переход к размножению,
а в неустойчивых условиях — таких, как
короткая продолжительность жизни, высокая
плодовитость и раннее половое созревание.
Иными словами, можно ожидать, что
стабильность среды обитания будет
предсказуемым образом влиять на компромиссы
жизненного цикла, обсуждавшиеся выше.
Это не вполне подтверждается
наблюдениями. В рассмотренных в этой главе циклах
размножения морских беспозвоночных про-
428 Глава 14
Таблица 14.8. Демографическая таблица для морского желудя Balanus glandula (Hines, 1979)
Возраст (х), Выживаемость к дан- Среднее число яиц, 1,
мес ному возрасту, 1„ выметанное к
возрасту х, тх
х|к> 'И,,
Оценочная смертность О
до прикрепления
, 0
3
12
24
36
48
60
72
84
96
1,000
1,17х 10 4
2,04x10 5
3,84 х 10 6
2.05x10 6
1,28x10 6
7,33 х 10-"
4,18 х Ю-7
2,44х Ю-7
1,42 х Ю-7
0
0
20 504
66 814
113 125
140 742
159 435
170 892
176 922
180 540
0
0
0,418
0.256
0,231
0,180
0,117
0,075
0,043
0,026
= 1.346
0
0
5,016
6,153
8,335
8.641
7,008
5,151
3,629
2,469
Время генерации Т = Хх 1 (х)т ,х) = 46.402 мес
Скорость прироста популяции в месяц г
I'lRo
Т
= 0,006,
т. е., если популяция сохранится, ее численность будет возрастать медленно.
Выживаемое 1ь в данном случае оценивали по числу прикрепившихся особей на каждые 1000 выметанных яиц (на
более поздних стадиях- путем прямого наблюдения). Эти данные составляют столбец |„.
Плодовитость оценивали путем измерения базально! о диаметра (по отношению базального диаметра желудя
данного возраста и числа выметываемых яиц за один раз). Регистрировали также число нерестов за год.
Произведение этих величин дает оценку средней плодовитости в каждом возрасте (столбец шх).
слеживается явный компромисс между
размером яиц и числом потомков. Однако в
целом организмы с планктотрофным
пелагическим развитием, очень высокой
плодовитостью и массовой смертностью молоди
образуют во взрослом состоянии довольно
стабильные популяции, живут
относительно долго и отличаются крупными
размерами тела. И напротив, морские виды,
адаптированные к использованию временных
экологических ниш, часто отличаются
низкой плодовитостью, а также лецитотроф-
ным и прямым развитием, минующим
пелагическую стадию.
Чтобы понять, почему каждый вид
животных размножается так, а не иначе,
необходимо учесть ограничения,
накладываемые средой обитания, функциональной
морфологией организма на различных
стадиях развития и, самое главное, его
эволюционной историей, которая может
сужать круг доступных ему путей решения
этой проблемы.
14.6. Выводы
В последнее время интенсивно
разрабатывается теория жизненного цикла,
стремящаяся функционально объяснить
разнообразие типов размножения животных,
в частности беспозвоночных.
В этой главе мы кратко рассмотрели
некоторые данные, на которых базируются
эти теоретические построения. Очевидно,
что основной способ размножения всего
живого — половой, хотя он может
перемежаться периодами бесполого
воспроизведения в сложных жизненных циклах многих
книдарий, плоских червей, аннелид, мелких
ракообразных, насекомых и т.д.
Типы полового размножения и
распределения доступных ресурсов между ним
и соматическими нуждами поразительно
разнообразны. Это разнообразие
рассмотрено с таксономической, экологической
и функциональной точек зрения. Ясно, что
далеко не все возможные комбинации ре-
Размножение и жизненные циклы
429
продуктивных признаков встречаются
в каждой среде обитания или
таксономической группе. Так, развитие морских
беспозвоночных часто идет через стадию свобод-
ноживущей пелагической личинки, и с этой
особенностью связаны многие другие:
наружное оплодотворение, массовые
синхронные нересты, простое строение
сперматозоидов, яйца, бедные питательными
веществами.
У всех пресноводных и наземных видов,
а также в некоторых морских средах
пелагическое личиночное развитие отсутствует.
При этом обычно наблюдаются внутреннее
оплодотворение, хранение спермы и
наличие большого запаса питательных веществ
в яйцах.
Любой тип размножения не случаен
и обусловлен контролируемым
распределением ресурсов между репродуктивными
процессами. Если размножение животных
в популяции синхронизировано, должны
действовать механизмы восприятия и ней-
роэндокринной передачи информации из
внешней среды. В данной главе мы лишь
кратко коснулись этой проблемы; более
подробно она рассмотрена в гл. 16.
Приведенные здесь описания и анализ
репродуктивных процессов у
беспозвоночных послужат основой для более
детального разбора вопросов экологии и биологии
сообществ, а также индивидуального
развития (гл. 15).
14.7. Дополнительная
литература
Детальные сведения по размножению
беспозвоночных можно получить из двух
многотомных трудов и следующей серии обзорных томов:
Adiyodi К. С, Adiyodi R. G. (Eds) 1983.
Reproductive Biology of Invertebrates. Wiley, New York.
Vol. 1. Oogenesis, oviposition and oosorption.
Vol. 2. Spermatogenesis and sperm function.
Следующие тома ждут выхода в свет
Giese A.G., PearseJ.S. (Eds) 1974 1977.
Reproduction in Marine Invertebrates. Academic Press,
New York.
Vols I -IV и следующие.
Advances in Invertebrate Reproduction. Elsevier
Science, Amsterdam.
Vol. 1. Clark W., Adams T. S. (Eds) 1981.
Vol. 2. Engels W. (Ed.) 1984.
Vol. 3. Porchet M. (Ed.) 1986.
В следующих монографиях рассмотрены
различные аспекты размножения
беспозвоночных:
Begon M., Harper J.L., Townsend С. R. 1986.
Ecology: Individuals, Populations and Communities.
Blackwell Scientific Publications Oxford.
Имеется перевод: Бигон М.. Харпер Дж.,
Таунсенд К. Экология. Особи, популяции и
сообщества: в 2 т.— М.: Мир. 1989.
Bell G. 1982. The Masterpiece of Nature: The
Evolution and Genetics of Sexuality. University of
California Press, Berkeley.
Brady J. 1979. Biological Clocks (Studies in Biology,
104). Edward Arnold, London.
С alow P. 1978. Life Cycles. Chapman and Hall.
London.
Charnov E. 1982. The Theory of Sex Allocation. Pri-
nseton University Press, Princeton, New Jersey.
Cohen J. 1977. Reproduction. Butterworth, London.
Greenwood P. J., Adams J. 1987. The Ecology of Sex.
Edward Arnold, London.
Maynard-Smith J. 1978. The Evolution of Sex.
Cambridge University Press, Cambridge.
Saunders D.S. 1977. An Introduction to Biological
Rhythms. Blackie, Glasgow.
Приложение 14.1.
Системы полового
размножения (миксиса)
Половое размножение—это система
воспроизведения, обеспечивающая обмен
генетической информацией (носителем
которой является молекула ДНК) и появление
особи, чья генетическая информация будет
результатом рекомбинации родительских
генов.
1. Вирусы. Бактериофаги заражают
бактериальные клетки, вводя в них свою ДНК.
При этом белковая оболочка вируса
остается вне клетки. Вирусная ДНК внутри
бактерии содержит информацию для синтеза
новых белковых оболочек фага и лизиса
бактерии-хозяина.
A. Одновременное заражение разными
мутантами по одному и тому же
структурному гену не обеспечивает воспроизведение
фага, а одновременное заражение
мутантами по разным структурным генам дает
активные фаги.
Б. Мутантные фаги способны
размножаться в бактерии-хозяине.
B. При смешанном заражении фагами
430
Глава 14
Белки, кодируемые
генами А и В-
МУТАНТНЫЕ формы
не способны
синтезировать полный навор
генных продуктов
Белковая оволочка
Бактериофага
Мутант I, ген А
1асть ДНК
с генами А и В
Мутант II, ген А Мутант III, ген В
Б Заражение некомпле- Заражение
комплементарными мутантами ментарными
мутантами (инфекция
"дикого типа")
Т
Заражение
некомплементарными мутантами
Кроссинговер
не дает форм
дикого типа
Кроссинговер дает
вирусную частицу
дикого типа
с комплементарными мутациями
кроссинговер может привести к появлению фагов,
способных заражать бактерии «в
одиночку», т.е. к половому размножению.
2. Бактерии.
I. Разные штаммы бактерий способны
к конъюгации. При этом образуется
прикрепительный мостик (пилус), по которому
молекула ДНК (бактериальная хромосома)
передается от донора («самца») реципиенту.
II, III. Длина переданного отрезка ДНК
зависит от времени конъюгации, которую
можно прервать путем интенсивного
встряхивания культуры.
а
g_s)
е.
^ I
ш
Размножение и жизненные циклы
431
IV. Между дуплицированными
участками бактериальной хромосомы может
произойти обмен генами. При этом возникают
бактерии, совмещающие признаки обеих
родительских особей.
3. Половое размножение у высших
организмов. Диплоидные клетки имеют два
набора хромосом, в каждом из которых
содержится генетическая информация одного
из родителей. В ходе особого типа
клеточного деления, вызываемого мейозом, из
диплоидных клеток образуются гаплоидные
гаметы. Их генетическая информация
может быть идентична родительской или
отличаться от нее. если произошел кроссинго-
вер.
Гаплоидные половые клетки от двух
родителей (или иногда от одного родителя)
сливаются (это называют
оплодотворением) с образованием диплоидной зиготы.
Геном зиготы отличается от геномов
диплоидных клеток, давших начало гаметам.
В нем сочетаются генетические признаки
обоих родителей, каждый из которых
представлен одним набором хромосом.
Диплоидная животная клетка.
Гены последовательностей
а-е и А-Е находятся на
хромосомах, унаследованных
от обоих родителей.
Эта клетка гетерозиготна
а- - -
Ь- - -
е-#- -•
В ходе МЕИОЗА хромосомы
спариваются, и между их
хроматидами может
происходить генетический
овмен (кроссинговер)
МЕЙОЗ
В гаметах может оказаться родительская или рекомвинантная
последовательность генов
ГАМЕТЫ
Родительская Рекомвинантная Рекомвинантная Родительская
Частота кроссинговера зависит от длины ДНК, разделяющей
гены на хромосоме
432
Глава 14
Приложение 14.2.
Жизненные циклы и половое
размножение
1. Половое размножение у большинства
протистов. Пример—инфузория Рагате-
литься, формируя гаплоидный гаметофит.
На приведенном ниже рисунке показано
протекание этого процесса у папоротников.
У высших растений гаметофит обычно
редуцирован до очень небольшого числа
клеток.
Гаметофит
Конъюгация
2. Жизненный цикл всех
многоклеточных животных (и некоторых водорослей).
Взрослая особь состоит из нескольких
или многих (обычно различных)
диплоидных клеток. В многоклеточном теле
образуются гаплоидные гаметы. Их слияние
дает диплоидную зиготу. В результате ряда
митотических делений развивается новая
взрослая диплоидная особь.
Мейоз
Оплодотворение
2п Зигота
3. Для жизненных циклов многих
водорослей и всех высших растений характерно
чередование многоклеточных гаплоидной
и диплоидной фаз. Первая называется гаме-
тофитом; она образует (путем митоза)
спермин и/или яйцеклетки. Слияние гамет дает
зиготу. Последняя путем митозов
развивается в многоклеточный спорофит. В процессе
мейоза образуются гаплоидные споры. Не
сливаясь, они продолжают митотически де-
Оплодотворение
2п
У животных такого чередования
гаплоидного и диплоидного поколений
никогда не происходит. Чередование поколений,
размножающихся бесполым и половым
путями, ему не эквивалентно (см. рис. 14.4).
Приложение 14.3.
Системы определения пола
I. Пример материнского определения
пола: мелкая полихета Dinophilus gyrocilia-
tus.
Взрослая
особь 5
Кокон с круп
ными и мелки
ми яйцами
Осеменение
неполовозрелых
самок рано
созревшими
карликовыми самцами
Размножение и жизненные циклы
433
В яичниках образуются два типа яиц: из
крупных развиваются самки, из мелких —
рано созревающие карликовые самцы.
Осеменение самок происходит внутри кокона
на эмбриональной стадии. Все взрослые
особи—это осемененные самки.
2. Пример генетического определения
пола: плодовая мушка Drosophila.
Ж-хромосома
•хромосома
3. Экологическое определение пола.
У некоторых организмов пол особи
определяется после оплодотворения.
Примером может служить эхиурида ВопеШа vi-
ridis.
Все ресничные планктонные личинки
этого червя способны развиться как в самку
(все крупные взрослые особи—самки), так
и в карликового самца. Оседая на
незаселенные участки морского дна, они дают
самок, а оседая вблизи хоботка самки, под
влиянием выделяемых им секретов
превращаются в карликовых самцов. В
экспериментах выявлено также определенное
генетическое влияние на пол, но влияние среды
его обычно пересиливает.
АЛ. У самок диплоидный генетический
набор с двумя одинаковыми половыми
хромосомами —XX. Все яйцеклетки
идентичны по способности развиться в особь того
или иного пола.
II. У самцов диплоидный генетический
набор с неодинаковыми половыми
хромосомами—X и Y. Образуются
сперматозоиды двух типов: из яиц, оплодотворенных
сперматозоидами, несущими Y-хромосому,
разовьются самцы, а из оплодотворенных
сперматозоидами с Х-хромосомой —
самки.
Б. Пол потомка определяется тем,
какой из двух типов сперматозоидов
оплодотворил данную яйцеклетку. При такой
системе определения пола нормальное
соотношение полов будет 1:1.
Приложение 14.4.
Жизненные циклы
беспозвоночных
На каждом рисунке показан типичный
обладатель того или иного цикла. Ширина
полосы отражает количество ресурсов: ее
28—1314
434
Глава 14
L
1 год
1 год
1. МУЛЬТИВОЛЬТИННЫЙ ВИД
III. Неоднократные последовательные \ Ч~\ \
кладки \ Ч Ч Ч
возрастание означает их накопление,
уменьшение—их расходование.
1. Мультивольтинный вид.
В течение года сменяется несколько
поколений; в большинстве случаев в
пресноводных и наземных местообитаниях
умеренной зоны у животных имеется
репродуктивная диапауза. На рисунке показана диа-
пауза на взрослой стадии, но она часто
бывает также на стадиях яйца, личинки или
куколки.
2. Однолетний вид.
Многие виды умеренных областей
живут ровно год, т. е. за это время у них
сменяется только одно поколение.
Размножение может либо быть представлено, как
в примере II, однократным массовым
нерестом, либо продолжаться в течение
нескольких месяцев (III).
3. Многолетний вид.
Эти животные способны жить в течение
нескольких лет, весь этот период
накапливая ресурсы. Возможны две различные
стратегии вкладывания их в размножение.
Размножение и жизненные циклы
1-й год | 2 - й год | 3-й год
435
3. МНОГОЛЕТНИЕ ВИДЫ
IV. Полицикличный вид, размножающийся один раз в год
V. Полицикличный вид, размножающийся небольшими кладками несколько раз в год
ХЗ:
=Ос
VI. Моноцикличный вид, размножающийся один раз в конце жизни, длительность которой
определяется условиями среды: 2, 3 года или волее
Чаще всего животные (например, морские
звезды; IV) размножаются один раз каждый
год в определенный период. Размножение
может быть синхронным и массовым или
многократным несинхронным (например,
у земляного червя, V). Некоторые
многолетние животные моноцикличны, т.е.
размножаются один раз в жизни в возрасте не
менее одного года. Наиболее известные их
примеры—головоногие моллюски и поли-
хеты семейства Nereidae (VI).
Приложение 14.5.
Жизненные циклы морских
беспозвоночных
Яйца и сперматозоиды морских
беспозвоночных могут высвобождаться в
морскую воду, где и происходит
оплодотворение. Это оказывает глубокое влияние на
всю репродуктивную биологию этих
животных.
436
CNIDARIA
Планула
PLATYHELMINTHES
NEMERTEA
ANNELIDA
Полихета
Мюллер Ob екая
личинка
Пилидий
Трохофора
Глава 14
ECHINODERMATA
Морские ежи
Морские звезды
Бипиннария
Плутеус
Брахиолария
Нектохета
MOLLUSCA
Двустворчатые
Офиуры
Офоплутеус
Голотурии
Полиолария
Примеры взрослых и характерных личиночных форм основных групп морских беспозвоночных.
Взрослые животные изображены в значительно меньшем масштабе, чем личинки
Приложение 14.6.
Функциональный анализ
жизненных циклов насекомых
1. Ювенильные и взрослые стадии
используют сходные пищевые ресурсы.
Функции ювенильных стадий—
развитие и дифференцировка, накопление
ресурсов.
Функции имаго — накопление ресурсов,
расселение и поиск ресурсов, спаривание
и откладка яиц.
Пример: прямокрылые (саранча и
кузнечики).
Размножение и жизненные циклы
437
CRUSTACEA
LOPHOPHORATA
Форониды
Актинотроха
Мшанки
Цифонаутес
CHORDATA
Malacostraca Зоеа
HEMICHORDATA
Мегалопа
"Головастик"
Торнария
I Л Ш
Характерные жизненные циклы. I. Пелагобентосныи — с пелагической личиночной стадией.
II. Голобентосный — с утратой примитивного признака, т.е. пелагической личиночной стадии. III.
Голопелагический — весь жизненный цикл проходит в пелагиали. Во многих группах можно
встретить все три этих типа. Приведенные здесь примеры относятся к брюхоногим моллюскам
2. Ювенильные и взрослые стадии
используют различные пищевые ресурсы.
Функции ювенильных стадий—
развитие и накопление ресурсов.
Функции куколок—дифференцировка
и развитие.
Функции имаго — накопление ресурсов,
их поиск, спаривание и выбор мест для
развития потомства.
Пример: падальная муха и другие
двукрылые.
3. Взрослая стадия непродолжительна
и неспособна к накоплению ресурсов.
Функции ювенильных стадий—развитие и
накопление ресурсов; функции имаго—
спаривание, поиск ресурсов и расселение.
438
Глава 14
1. Водные личинки, например у поденок.
П. Наземные личинки, например у
чешуекрылых. Здесь взрослая стадия
продолжительная, и насекомые своим ротовым
аппаратом собирают нектар, используемый
как горючее для полета.
4. Имаго обеспечивают ресурсами
потомство.
Функции личинок: накопление ресурсов.
Функции имаго: расселение, поиск и
накопление ресурсов, спаривание, поиск мест
для развития потомства и откладка яип.
Пример: одиночные осы
(перепончатокрылые).
Все доступные потомству ресурсы
поставляются родителями.
5. Накопление ресурсов, развитие,
спаривание и откладка яиц разделены между
кастами.
Ювенильные стадии—использование
ресурсов и рост.
Стерильные рабочие особи—
накопление ресурсов.
Матки (фертильные самки)—
спаривание и выбор партнера, откладка
яиц, расселение и поиск ресурсов.
Самцы—спаривание; в накоплении
ресурсов участия не принимают.
Пример: медоносные пчелы.
ГЛАВА I О
РАЗВИТИЕ
Большинство многоклеточных
животных начинают жизнь с оплодотворенного
яйца, возникшего при слиянии двух очень
разных гаплоидных гамет. Слияние дает
диплоидную зиготу, и именно эта довольно
специфическая клетка развивается во
взрослый многоклеточный организм.
Последовательность ее ранних, точно
организованных делений называется дроблением; во
время него роста дочерних клеток не
происходит. Зигота особым способом
расщепляется на большое количество мелких клеток,
взаиморасположение которых в
пространстве соответствует организации
оплодотворенного яйца. Эта организация может
определить судьбу дочерней клетки на ранних
стадиях эмбриогенеза. У беспозвоночных
обнаружено несколько различных типов
дробления яйца, среди которых наиболее
известны спиральное у большинства пер-
вичноротых и радиальное у вторичноро-
тых.
Дальнейшее развитие эмбриона
включает его пространственную реорганизацию
в соответствии с планом строения
взрослого организма, а на более поздних стадиях—
дифференцировку клеток в зависимости от
их функций у личинки и взрослого
организма. Последовательность развития, таким
образом, включает следующие этапы:
1) оплодотворение;
2) активация метаболизма зиготы
и трансляция материнской
информационной РНК (мРНК);
3) дробление;
4) активация ядра зиготы и
транскрипция новых специфичных для нее мРНК;
5) органогенез;
6) дифференцировка.
Развитие часто включает метаморфоз,
когда адаптированная к определенным
условиям окружающей среды личинка
внезапно превращается в резко отличающееся
от нее взрослое животное, выполняющее
иногда совсем другие функции и
адаптированное к совершенно иным условиям (см.
гл. 14).
Многие беспозвоночные оказались
весьма ценными моделями для анализа
биохимических механизмов, участвующих в
клеточной дифференцировке и органогенезе.
Изучение развития важно и потому, что оно
проливает свет на филогенетические
взаимосвязи между животными.
В этой главе развитие беспозвоночных
будет рассматриваться в основном на
основе экспериментальных данных по
механизмам клеточной детерминации. Наш обзор
включает анализ событий, происходящих
в момент оплодотворения яйца, и его
реакцию на это. Описание типов дробления дает
основу для экспериментального изучения
клеточной детерминации, т.е.
формирования проспективного значения клеток в ходе
онтогенеза. Большое число описательного
материала по личиночным формам и
метаморфозу закладывает фундамент
экспериментальных исследований, раздвигающих
границы нашего понимания развития
и дифференцировки. В этой главе
обсуждается также регенерация у беспозвоночных,
т. е. процесс восстановления целой
структуры из ее фрагмента. Участвующие в этом
механизмы сходны с обеспечивающими
нормальное развитие.
15.1. Оплодотворение
и инициация развития
Оплодотворение—сложный процесс;
его основные компоненты таковы:
1) физический контакт гамет;
440
Глава 15
2) взаимодействие мембранной
поверхности, ведущее к слиянию двух клеток;
3) физиологическая реакция оболочки
яйца, препятствующая дальнейшему
проникновению спермы и обычно называемая
блокадой полиспермии;
4) активация метаболизма ооцита;
5) слияние пронуклеусов с
формированием нового диплоидного генома зиготы;
6) инициация раннего дробления.
В понимании общих принципов этого
процесса особую роль сыграли морские
беспозвоночные с наружным
оплодотворением.
Точная последовательность событий
зависит от степени зрелости «яйца» в момент
слияния гамет. У некоторых
беспозвоночных они сливаются до окончания мейоза
и отделения полярных телец. В других
случаях мейоз заканчивается до
оплодотворения. Например, у морского ежа он к этому
моменту завершен, а яйцеклетка морской
звезды вступает в метафазу I мейоза
непосредственно перед выметыванием (см.
гл. 16) и, если не будет оплодотворена, он
дальше не пойдет, т. е. полярные тельца
появляются уже после взаимодействия
между яйцом и сперматозоидом.
Большинство морских беспозвоночных
выметывают яйца и сперму в воду, где
и происходит оплодотворение. Можно
было бы ожидать, что между мужской и
женской гаметами существует какое-то
притяжение, но, по сути дела, это бывает
довольно редко. В морской воде сперматозоиды
становятся активными и совершают
хаотические движения, которые в случае
достаточно высокой концентрации яиц и
сперматозоидов приводят к физическому контакту
между гаметами. Лишь в некоторых
случаях, например у оболочников, ряда
моллюсков и колониальных гидроидов
Campanularia наблюдается хемокинез. У
упомянутых гидроидов сперма должна проникнуть
в яйцо через сравнительно узкое отверстие
в гонангий—колбообразной структуре,
окружаюшей яйца. Киносъемка показала,
что движения отдельных сперматозоидов
не хаотичны, а направлены к устью гонан-
гия (рис. 15.1). Вещество, экстрагированное
из этого устья, привлекает их, будучи
растворенным в агаре.
Рис. 15.1. Гонангий гидроида Campanularia
и траектории движения сперматозоидов,
зарегистрированные на кинопленке. Это один из
первых случаев демонстрации хемотаксиса
между яйцеклетками и сперматозоидами.
Данный вид, конечно, не исключение; другие
примеры можно найти среди оболочников и
моллюсков, но насколько широко распространено
подобное явление, неизвестно (по Muller,
1966)
Механизм самого обычного
оплодотворения яиц хаотически движущимися
сперматозоидами полностью еще не ясен. В
начале века было обнаружено, что
сперматозоиды морского ежа, помещенные в воду,
где находились яйца, временно лишаются
подвижности, как бы агглютинируясь. Это
вызвано диффузией в воде
высокомолекулярного вещества (из студенистой оболочки
яиц). Его назвали ферти.шзином, а
соответствующий белок-детектор на
сперматозоиде— антифертилизином. На поверхности
многих яиц находится видоспецифичный
коагулянт спермы, но только в некоторых
случаях он диффундирует в окружающую
среду, как у морского ежа.
В световой микроскоп можно увидеть
физические изменения, происходящие со
сперматозоидом некоторых
беспозвоночных при контакте с яйцом (рис. 15.2,Л).
Сначала его иглообразный отросток
соприкасается с цитоплазмой яйца. Этот
отросток— акросомная нить—развивается из
самой передней части сперматозоида—
акросомы. У других беспозвоночных он
отсутствует (рис. 15.2,£).
Развитие
441
I
II
III
IV
Рис. 15.2. Изменения в ходе оплодотворения по данным световой микроскопии. А. Морская
звезда Asterias. I. Приближение сперматозоида к яйцу, окруженному студенистой оболочкой. II.
Акросомная реакция. Кажется, что тонкая нить выбрасывается из мелкой, напоминающей пузырек
акросомы спереди сперматозоида. III. Акросомная нить соприкасается с мембраной яйцеклетки
и начинается кортикальная реакция, распространяющаяся от точки контакта. IV. Сперматозоид
поглощается коническим выступом цитоплазмы ооцита. Б. Морской еж Arbacia. I. Приближение
сперматозоида к студенистой оболочке. II. Прохождение сперматозоида через студенистую
оболочку. III. Контакт головки сперматозоида с желточной оболочкой; иногда, как показано на
рисунке, заметна очень мелкая акросомная нить. IV. Кортикальная реакция и внедрение в ооцит ядра
сперматозоида (по Austin, 1965)
С помощью электронной микроскопии
установлено, что некоторые аспекты
взаимодействия сперматозоид/яйцо сходны
у всех видов, поскольку все они включают
слияние мембран. На рис. 15.3 показаны
изменения, наблюдаемые в момент
контакта сперматозоида с яйцом у морского ежа
Arbacia и у полихеты Hydroides. В обоих
случаях сначала происходит слияние
мембраны акросомного пузырька и
плазматической мембраны сперматозоида, в
результате чего высвобождается ферментное
содержимое пузырька. Последующий лизис
обеспечивает расправление акросомных
трубочек или акросомной нити, их
проникновение через оболочки яйца и
установление контакта с его плазматической
мембраной. Дальнейшее слияние мембран
приводит к образованию единой оболочки вокруг
яйца и сперматозоида.
Начальные реакции, вызванные
контактом, иногда называют «блокадой
полиспермии». Существуют два компонента этого
явления. Первый—невидимый, хотя у
некоторых видов связан с изменением
электрического потенциала у поверхности яйца.
Второй—видимая кортикальная реакция,
которая через несколько секунд
распространяется радиально от места контакта
сперматозоида с яйцом. Она представляет
собой слияние мембраны кортикальных
гранул, формирующихся в позднем оогенезе,
с мембраной яйца и выход их содержимого
в перивителлиновое (околожелточное)
пространство (рис. 15.4).
Реакция такого типа почти
универсальна; ее результат- -выделение студенистой
оболочки оплодотворения, которая,
начиная с этой стадии, препятствует слиянию
яйца с другими сперматозоидами.
Теперь в яйце происходят глубокие
метаболические изменения. До
оплодотворения оно как бы пребывает в анабиозе.
Связывание сперматозоида с рецепторным
г- г ' ч$&ъ
YII VIII IX X
Рис. 15.3. Оплодотворение по данным электронной микроскопии. А. Акросомная реакция у
морского ежа Arbacia. Акросомная нить этого вида настолько мала, что с трудом различима в
световой микроскоп. I. Передняя часть головки сперматозоида под электронным микроскопом.
II. Слияние мембран акросомы и сперматозоида для высвобождения содержимого акросомы.
Б. Nereis ш
Рис. 15.4. Кортикальная реакция: отслаивание оболочки оплодотворения. А. Морской еж Arbacia.
I. Сперматозоид контактирует с плазматической мембраной яйца; кортикальные гранулы интакт-
ные. II. Кортикальные гранулы разрушаются, и отслаивается оболочка оплодотворения; реакция
постепенно распространяется от места контакта. III. Оплодотворенное яйцо. Обратите внимание,
что студенистая оболочка присутствует еще до оплодотворения IV. Схематическое представление
цепи событий по данным электронной микроскопии. Б. Полихета Nereis. I. Сперматозоид
приближается к яйцу; на этой стадии студенистого слоя нет. II. Постепенное распространение
кортикальной реакции от точки контакта. Наружная часть яйца состоит из крупных кортикальных ячеек,
которые разрушаются во время этого процесса. III. Оплодотворенное яйцо. У большинства видов
содержимое кортикальных ячеек выделяется через оболочку оплодотворения (внежелточную
оболочку), формируя студенистую оболочку (по Austin, 1965)
III. Удлинение акросомной нити. Б. Детали процесса оплодотворения у полихеты Hydroides. I.
Приближение сперматозоида к внешнему пограничному слою (ВПС) желточной оболочки. Этот
слой образуется из кончиков микроворсинок на поверхности ооцита. II, III. Слияние мембраны
акросомного пузырька с ВПС, вызывающее высвобождение содержимого акросомы. Начало
прободения желточной оболочки. IV. Продолжающееся проникновение головки сперматозоида и
удлинение многочисленных акросомных трубочек. V, VI. Контакт и окончательное слияние акросом-
ных трубочек с плазматической мембраной ооцита. VII, VIII. Проникновение сперматозоида в
ооцит по мере того, как цитоплазма последнего смещается к месту слияния мембран ооцита
и сперматозоида. IX. Дальнейшее проникновение. X. Ядро и митохондрии сперматозоида в
цитоплазме яйца. Обратите внимание на отверстие, оставшееся в желточной оболочке (с электронных
микрофотографий в кн. Colwin, Colwin, 1961)
444
Глава 15
Связывание сперматозоида с рецептором
40
60 -
__ внутриклеточных депо
Кортикальная реакция
_ Начало выделения кислоты и
основного поступления Г\1а+
- Превращение NAD в NADP
- Рост потребления кислорода
Формирование оболочки
Незначительное поступление Na
"Изменение мембранного потенциала
Высвобождение Са из
_ оплодотворения завершено
80 -
100
Увеличение внутриклеточного рН
(пониженная кислотность)
- Усиление синтеза белка
- Активация транспортных систем
200 -
300
400
500
600
800
1000 — Слияние ядер яйцеклетки
— и сперматозоида
— Инициация синтеза ДНК
2000 -
3000
4000
5000
6000
-Первое клеточное деление
0123456789 10
Время после осеменения, мин
1200
600
200
Оплодотворенные у
яйца
8 16 24 32 40 48
Врамя после добевления спермы, мин
Рис. 15.5. Последовательность событий после контакта мембран сперматозоида и яйца во время
оплодотворения у морского ежа (по Epel,-1977). На вставках приведены примеры
экспериментальных данных, подтверждающих эту схему: А — потребление кислорода (по Ohnishi, Sugiyama,
1963); Б—синтез белка (по Epel, 1967)
участком запускает последовательность
событий (рис. 15.5), которые выводят яйцо из
этого состояния. Уровень потребления
кислорода значительно увеличивается,
достигая пика в первую минуту после контакта
(когда образуется оболочка
оплодотворения), но по сравнению с неоплодотворенны-
ми яйцами остается гораздо более высоким
и в дальнейшем. Затем становятся
доступными «замаскированные» молекулы
информационной (матричной) РНК, накопленные
в цитоплазме яйца во время оогенеза, в
результате чего усиливается синтез белка.
Первое деление дробления происходит
через строго определенный период времени
после оплодотворения, однако еще до него
можно наблюдать важные изменения в
распределении цитоплазматических
компонентов яйцеклетки. У билатерально
симметричных животных эти движения
цитоплазмы формируют главные оси будущего
эмбриона, определяя передний, задний,
дорсальный и вентральный квадранты
зиготы. Впервые такое перераспределение было
обнаружено у асцидий; оно изображено на
рис. 15.6. Однако, как будет показано
в разд. 15.3, пространственному
распределению различных составных частей
цитоплазмы вряд ли стоит придавать слишком
важное значение.
Развитие
445
Рис. 15.6. Движение визуально различимых
типов цитоплазмы, описанное в начале XX в,
уасцидии Styela partita. Эти наблюдения важны
для демонстрации потенциальной роли цито-
плазматических факторов в детерминации
судьбы клеток. I. Радиально симметричное не-
оплодотворенное яйцо. II, III. Миграция
«желтой» цитоплазмы к вегетативному полюсу и
сегрегация «серой» цитоплазмы в центральной
части яйца. IV. Сегрегация и миграция желтой
и прозрачной цитоплазмы к одной стороне
яйца. Яйцо становится билатерально
симметричным. V. Билатерально симметричное яйцо
с локализацией цитоплазмы, соответствующей
первому делению дробления (см. приложение
15.3)
15.2. Раннее развитие
15.2.1. Дробление
Характер дробления зигот большинства
животных, во время которого относительно
крупное яйцо делится на все более мелкие
клетки, точно детерминирован с начала
первого митоза. Оно происходит таким
образом, что пространственная организация
оплодотворенного яйца сохраняется в
многоклеточном эмбрионе, причем у
большинства животных в результате возникает
полый клеточный шар, называемый
бластулой.
Многим таксонам первичноротых
(например немертинам, аннелидам, сипунку-
лидам, эхиуридам, моллюскам и,
возможно, погонофорам) присуще спиральное
дробление. Оно особенно хорошо известно
по работам, выполненным на аннелидах
и моллюсках, и более подробно описано
в приложении 15.1, поскольку имеет
большое значение для экспериментального
анализа развития беспозвоночных (см. также
разд. 15.3).
Иногда высказывается мнение, что
спиральное дробление у названных таксонов
указывает на довольно близкую
филогенетическую связь между ними. К этому
предположению, однако, следует относиться
с осторожностью. Считается, что указанные
таксоны берут начало от плоских червей,
причем некоторым примитивным видам
последних также свойственно спиральное
дробление. Особенно четко это заметно
у свободноживущих многоветвистых и
бескишечных турбелларий, хотя у последних
на вегетативном полюсе только два
крупных макромера, а не четыре, как обычно. От
плоских червей, возможно, произошли
и многие другие таксономические группы,
включая псевдоцеломических червей, лофо-
форовых и членистоногих, а также вторич-
норотые (гл. 2), хотя им и свойственны
другие типы дробления, описанные ниже.
Действительно, для плоских червей спиральное
дробление необычно и известно лишь в
нескольких группах, у которых желток
запасается в цитоплазме ооцита, а у видов со
специализированными желточными
клетками (см. разд. 14.4.2.2) оно имеет
аберрантный характер.
Распространение спирального
дробления в нескольких типах говорит лишь о том,
что это — консервативный признак,
сохранившийся в их эволюции, который нельзя
использовать как доказательство сравните-
446
Глава 15
Рис. 15.7. Дробление у иглокожего Synapta:
I — стадия двух бластомеров; II — стадия
четырех бластомеров; III — стадия восьми
бластомеров; IV — стадия 16 бластомеров; V —
позднее дробление; VI — стадия бластулы
льно недавней дивергенции от общей пред-
ковой ветви. Однако если у древних плоских
червей было спиральное дробление, то
наличие другого его типа у нескольких
таксонов действительно может предполагать
более позднюю их эволюционную
дивергенцию. Это демонстрируют, например, вто-
ричноротые. У многих из них так
называемое радиальное дробление, когда
производные поперечных делений лежат строго друг
над другом. Сравнительно простой
пример—дробление у голотурии Synapta,
показанное на рис. 15.7. Все дочерние клетки
I II III IV
Рис. 15.8. Поверхностное дробление яиц
членистоногих (насекомых). I. Единственное ядро
расположено внутри желтка в центральной
части яйца. II. Деление ядра приводит к
появлению нескольких ядер, но без клеточных границ
между ними. III. Ядра движутся в
периферическую цитоплазму, относительно свободную от
желтка. IV. Между ядрами появляются
радиальные клеточные стенки
в ходе постепенного развития до стадии
полой сферической бластулы здесь
практически одного размера. Более сложное
радиальное дробление у морских ежей, широко
использовавшихся в экспериментально-
эмбриологических исследованиях. На 64-
клеточной стадии отдельные бластомеры
здесь невозможно идентифицировать, так
как первые деления дробления дают
одинаковые клетки, но зато у эмбриона
появляются различные по характеру делений слои
(см. приложение 15.2).
В яйцах членистоногих, особенно
насекомых, обильный желток занимает
центральное положение, поэтому свойственный им
тип дробления называют поверхностным,
или эндолецитальным. Цитоплазма здесь
образует лишь поверхностный слой, а
начальные деления ядра не сопровождаются
цитокинезом клеток. В конечном счете ядра
движутся к поверхности яйца, где вокруг
них образуются клеточные мембраны (рис.
15.8).
15.2.2. Гаструляция
Беспозвоночные, за исключением Placo-
zoa, Mesozoa, губок и двух таксонов
кишечнополостных, — трехслойные животные.
Другими словами, их тело — производное
трех эмбриологически различных
клеточных слоев — эктодермы, мезодермы и
энтодермы (у некоторых групп на взрослой
стадии следов энтодермы нет; например, у
погонофор). Бластула—эмбрион,
формирующийся в результате раннего дробления,—
состоит только из одного слоя клеток. Если
в яйце не очень много желтка, бластула—
полая сфера и процесс гаструляции,
дающий в принципе трехслойный зародыш,
представляет собой инвагинацию клеток.
Этот способ хорошо заметен в случае
прозрачных яиц иглокожих (рис. 15.9).
Клетки первичной мезенхимы,
производные микромеров зародыша на стадии
64-х бластомеров (см. приложение 15.2),
вторгаются в первичную полость тела—
бластоцель. Затем вегетативный полюс за
счет изменения относительного слипания
своих клеток начинает впячиваться внутрь
с образованием углубления. У эмбриона
морского ежа этому движению внутрь мо-
Развитие
447
Рис. 15.9. Гаструляция у морского ежа. I. Ранняя гаструляция, инвагинация первичной
мезенхимы. II, III. Миграция мезенхимы и начало инвагинации. IV. Образование контактов филоподий
вторичной мезенхимы. V. Поздняя гаструла, появление скелетных спикул, секретируемых
первичной мезенхимой. VI. Сформированная гаструла (призматическая стадия зародыша). Закладка
орального поля. VII. Вид раннего плутеуса с оральной поверхности. VIII. Вид раннего плутеуса
сбоку (по Trinkans, 1969)
жет содействовать прямой контакт
филоподий, протягивающихся от клеток вторичной
мезенхимы к внутренней поверхности бла-
стоцеля (рис. 15.9.IV). Однако для гаструля-
ции такие филоподий не обязательны
(например, у эмбриона морской звезды
подобного рода контактов нет), поэтому нельзя
сказать, что инвагинация зависит от
создаваемого ими натяжения. Возможно,
движение клеток — результат сил, генерируемых
их различным слипанием.
К концу гаструляции у эмбриона
различают внешний слой эктодермы,
открывающуюся сзади внутреннюю трубку и мезен-
химные клетки между ними. Эта стадия
называется гаструлой. На ней дефинитивная
мезодерма еще не обособилась от
первичной кишки (архентерона). Клетки
мезенхимы не принимают участия в образовании
дефинитивной мезодермы, но могут
включаться в скелетные и мышечные элементы
личинки, которые утрачиваются во время
метаморфоза. Формирование мезодермы
описано в разд. 15.4.
У вторичноротых бластопор становится
анусом функциональной личинки, а
зачастую и взрослого организма. Ротовое
отверстие формируется путем инвагинации
эктодермы.
У первичноротых гаструляция идет по-
иному. Она также включает внедрение
внутрь бластулы (интернализацию) пре-
зумптивной энтодермы и небольшого числа
клеток, дающих начало мезодерме
взрослого животного. У форм с малым
количеством желтка в яйце гаструляция
осуществляется путем инвагинации, как показано на
рис. 15.10,1, а в случае яиц, богатых
желтком, заключается в амебоидной миграции
клеток в обхват относительно инертной
заполненной желтком энтодермы (рис. 15.10,
II, III).
У форм с малым количеством
зародышевого желтка в состав дорсальной губы
бластопора входят клетки, из которых
формируется ротовое отверстие личинки.
Несколько таксонов объединены в группу
первичноротых именно по этому,
считающемуся фундаментальным признаку раннего
развития.
В случае богатых желтком яиц тип
дробления часто неясен, и гаструляцию
проследить трудно. Бластоцель обычно
заполнен, а презумптивные клетки энтодермы
448
Глава 15
Рис. 15.10. Гаструляция у первичноротых
(примеры основаны на изучении полижете
различным количеством желтка в яйцах).
А. Строение бластулы. Б. Средняя гаструла.
Морфологически важные клетки, берущие
начало от 4d-6nacTOMepoB и
формирующие-мезодерму взрослого животного, обозначены
точками; клетки, образующие ротовое
отверстие, заштрихованы; клетки будущего
кишечника показаны мелкими кружками. I.
Относительно бедное желтком яйцо. II. Яйцо с
промежуточным содержанием желтка. III. Богатое
желтком яйцо и гаструляция путем зпиболии
на вегетативном полюсе часто оказываются
внутри за счет эпиболии, или обрастания их
клетками анимального полюса, как
показано для пиявки на рис. 15.11. У некоторых
полихет ротовое отверстие формируется
независимо от бластопора, хотя они и
считаются первичноротыми.
Таким образом, гаструляция—это
процесс интернализации клеток с образованием
типичной трехслойной структуры. Можно
составить карту зачатков с точной
локализацией на поверхности бластулы будущих
внутренних тканей.
На рис. 15.12 показана такая карта для
бластулы полихеты; указаны также
траектории движения клеток, приводящие к
формированию трохофоры. Нужно заметить,
Рис. 15.11. Гаструляция в богатых желтком
яйцах пиявки; А — на поперечном срезе с левой
стороны; Б—объемное изображение со
стороны вегетативного полюса, показывающее
плоскость среза
что клетки—производные бластомеров 2d
и 4d играют в эмбриональном развитии
первичноротых очень важную роль.
Производные 2d образуют эктодерму взрослого
животного, а производные 4d — две парные
клетки мезоэнтодермы (мезентобласты),
дающие начало будущей мезодерме. Рис.
15.12 показывает относительное
расположение этих клеток перед гаструляцией и их
движения, вызывающие инвагинацию пре-
зумптивной энтодермы и перемещение ме-
зентобластов и эктотелобластов на их
дефинитивное место в трохофорной личинке.
У личинки аннелид (рис. 15.12) все
сегментарные структуры будут
производными парных мезодермальных полосок
и эктотелобластического кольца. Из других
частей трохофоры развиваются
несегментарные структуры — пигидиум и просто-
миум (см. разд. 4.14).
У членистоногих тип гаструляции
совсем иной. «Бластопор» здесь представлен
бороздкой, через которую происходит ин-
тернализация вентральной полоски
проспективных мезодермальных клеток,
предназначенных для формирования
мускулатуры тела, стенки кишечника, гонад и т.д.
В обобщенном виде последовательность
наблюдаемых при этом событий показана на
рис. 15.13. Внутренняя часть такого
эмбриона не полая, а заполнена массой неклеточ-
Развитие
449
Бластопор
Прототрох
Мезодермальные
полоски
Эктотеловластическое
кольцо
Рис. 15.12. Гаструляция у первичноротых на примере полихет: А — вид с вентральной
поверхности^— виде левой стороны. I. Схематическое изображение простой карты зачатков. II. Гаструля-
ционные движения: сплошные стрелки показывают движение клеток к бластопору, пунктирные—
миграцию инвагинировавших клеток. Обратите внимание, что стомодеум, или рот, образован
инвагинацией клеток через переднюю часть бластопора. Мезодермальные полоски берут начало от
парных мезентобластов, мигрирующих через задний край бластопора. III. Сформированная тро-
хофора с парными мезодермальными полосками и эктотелобластическим кольцом. Эти два
элемента образуют сегментную бластему, которая сохраняется на передней стороне пигидиума на
протяжении всей взрослой жизни. Простомиум, перистомиум и пигидиум являются
несегментными структурами (частично по Anderson, 1964)
ного желтка. Одновременно инвагинируют
ганглионарные клетки сегментарной
нервной системы.
Эмбрион насекомого окружен «внезаро-
дышевыми оболочками», позволяющими
29—1314
провести интересные аналогии между
развитием «высших» и «низших»
беспозвоночных, с одной стороны, и «высших» и
«низших» позвоночных с другой.
450
Глава 15
Рис. 15.13. Гаструляция у насекомых. А.
Поперечный разрез. Б. Вид с вентральной стороны.
I. Раннее деление ядра. II. Поверхностное
дробление. Ill, IV. Инвагинация мезодермы. V,
VI. Сегментация нейрозктодермы, образование
внезародышевых оболочек. VII.
Сформированная гаструла (по Slack, 1983)
15.3. Экспериментальная
эмбриология беспозвоночных:
детерминация судьбы клетки
15.3.1. Введение
Сложное многоклеточное животное
происходит от одноклеточной,
недифференцированной тотипотентной структуры —
оплодотворенного яйца. В процессе
развития оно дает множество клеток, причем
отдельные их группы различаются по
химическому составу, морфологии и функциям.
Экспериментальная эмбриология—это
наука о том, как в развивающемся
эмбрионе возникают популяции
дифференцированных клеток со специфическими функциями.
Эмбрионы и личинки беспозвоночных
представляют собой наиболее удобные
модельные системы для анализа участвующих
в этом процессов.
В ходе раннего онтогенеза
жизнедеятельность развивающегося эмбриона
регулируют материнские гены. Однако затем
в действие вступает геном собственно
зиготы, а, поскольку со временем появляется
много различных типов клеток,
заключенная в ядре информация должна в каждом
случае особым образом контролироваться.
Потенциальных точек контроля много; они
включают один или несколько этапов такой
последовательности:
1) генетическая информация в ядре
зиготы;
2) распределение информации по
дочерним клеткам;
3) транскрипция генетической
информации;
4) популяция молекул мРНК;
5) перенос мРНК из ядра в
цитоплазму;
6) популяция мРНК в цитоплазме
дочерних клеток;
7) трансляция информации на
молекулах мРНК;
8) образование специфичных для клеток
белков;
9) функция специфических веществ.
В настоящее время общепринято, что
у большинства организмов ядра
дифференцированных клеток содержат ту же
генетическую информацию, что и ядро оплодот-
Развитие
451
Элиминируемый
хроматин
римордиальные
репродуктивные
клетки
П1 IV
Рис. 15.14. Пример диминуции хромосом во время раннего эмбрионального развития (Ascaris).
I. Первое деление дробления: хромосомы не утрачиваются. II. Элиминация хромосомного
материала во время деления передней клетки. III. Элиминация хромосомного материала и
перегруппировка клеток на стадии четырех бластомеров. Только одна клетка сохраняет полный набор
хромосом и тотипотентность. IV. На стадии гаструлы полный набор хромосом остается в двух при-
мордиальных половых клетках
воренного яйца. Доказательства этого
основаны на результатах
1) экспериментальной трансплантации
ядер из дифференцированных клеток в
цитоплазму энуклеированных яиц;
2) изучения цитологии хромосом во
время данного дробления;
3) сравнения характера исчерченности
гигантских хромосом некоторых
двукрылых и других насекомых;
4) изучения дифференцировки и редиф-
ференцировки во время регенерации (см.
также разд. 15.5);
5) анализа «отпечатков пальцев» ДНК
из разных тканей.
Однако из этого общего правила есть
исключения. Например, у нематоды Ascaris
только примордиальные половые клетки
эмбриона получают полный набор
хромосом, а в других клеточных линиях часть их
утрачивается во время раннего дробления
(рис. 15.14).
Если все ядра развивающегося
зародыша имеют одинаковый генотип, дифферен-
иировка должна выражать взаимодействие
цитоплазматической информации клетки
и ее ядра.
15.3.2. Мозаичное
и регуляционное развитие
В начале этого столетия эмбриологи-
экспериментаторы изучили возможности
нормального развития изолированных
клеток зародышей беспозвоночных. В ряде
случаев, например у морского ежа, каждый из
бластомеров на стадии четырех клеток
давал целую и совершенно нормальную ли-
чинку-плутеус (см. приложение 15.3,1).
Такое развитие назвали регуляционным.
В других случаях эмбрионы,
развивающиеся из каждого бластомера на четырех-
клеточной стадии, были весьма
неполноценными (см. приложения 15.3,2 и 15.3,3).
Такое развитие назвали мозаичным. Был
сделан вывод, ^то отдельные клетки
наследуют особый тип цитоплазмы,
определяющий их дальнейшую судьбу. Более
детальный анализ этих данных требует знания
судьбы различных блестомеров при
нормальном развитии, которую можно отразить
картой зачатков на бластуле или нераздро-
бившейся цитоплазме яйца. Такую карту
составляют, маркируя специфические зоны
29*
452
Глава 15
цитоплазмы и прослеживая судьбу меченых
клеток, а в случае асцидии Styela partita она
выявляется в ходе тщательных наблюдений
за яйцом после оплодотворения (см. рис.
15.6). Эти наблюдения говорят, что
благодаря движениям цитоплазмы яйцо
становится явно билатерально симметричным,
а среди его зон выделяется серповидный
участок цитоплазмы желтоватого цвета,
так называемый желтый серп. Его можно
наблюдать от ранних стадий дробления до
формирования мышечных блоков у личин-
ки-«головастика». Все презумптивные
мышечные клетки наследуют некоторое
количество желтой цитоплазмы, причем ни одна
из клеток, которая не станет мышечной, ее
не получает. Факторы, определяющие
судьбу клетки у беспозвоночных, более
детально обсуждаются в приложении 15.4.
Результаты экспериментов с асцидией
явились убедительным доказательством
роли локализации цитоплазматических
компонентов в детерминации судьбы клетки.
Опыты с яйцами насекомых также
показывают, что в процессе развития деятельность
ядер может определяться цитоплазмой,
в которую они попали. На рис. 15.8 показан
ход дробления. На заднем полюсе яйца
часто находится хорошо различимая
цитоплазма, так называемая полярная плазма.
Способностью формировать половые
клетки (т.е. оставаться тотипотентными)
обладают только ядра, оказавшиеся в клетках
с такой цитоплазмой. Эмбрионы с
удаленной полярной плазмой стерильны.
Полярная плазма определенной генетической
линии мухи Drosophila при введении в
переднюю часть яйца мухи другой линии
вызывает формирование из клеток этой области
гамет.
Однако развитие нельзя рассматривать
исключительно как результат особенностей
распределения цитоплазматических
детерминантов в яйце. Оно также включает
взаимодействие между клетками.
Эксперименты с улиткой Nassarius obsoletus (часто
употребляют также родовое название Ilyanassa)
продемонстрировали взаимодействие
цитоплазматических детерминантов и
межклеточных влияний, определяющее судьбу
клетки в ходе эмбриогенеза (более детально
см. приложение 15.5).
Nassarius obsoletus свойственно
спиральное дробление (см. приложение 15.1) с
появлением при первом и втором клеточных
делениях выступающей полярной лопасти.
Специфическая полярная плазма передается
только в D-бластомер. Эмбрионы с
удаленной полярной лопастью становятся
симметричными, определенная хронология
дробления в последовательности поколений
D-клеток нарушается, и развитие приводит
к весьма неполноценной «безлопастной»
личинке (см. приложение 15.5). У таких
личинок нет глаз, статоцистов, ноги, паруса,
раковины, сердца и оформленной кишки.
Однако некоторые продукты клеточной
дифференцировки можно распознать.
Схема клеточных поколений в приложении 15.1
подтверждает эти наблюдения. Многие из
зависящих от полярной лопасти структур не
являются производными клеток,
наследующих ее материал. Ясно, что судьба клеток,
происходящих от 1а и 1с, дающих глаза
или от 2d, из которых формируется
раковина, определяется не только их цитоплазма-
тической наследственностью, но и
взаимодействием с клетками D-линии во время
критического периода, длящегося от
появления 2d до формирования 4d.
Аналогичным образом клетки, компетентные дать
начало раковинной железе, являются
производными единственного бластомера 2d,
однако они будут выполнять свои функции,
только если клетка 2d контактно
взаимосвязана с другими клетками D-линии.
У эмбриона морского ежа судьба
различных клеточных слоев еще в большей
степени определяется их
взаиморасположением и не закреплена цитоплазматической
наследственностью. Основные элементы
личинки здесь можно проследить вплоть до
бластомеров последних стадий дробления
в соответствии со следующей схемой:
An 1—апикальный ресничный султан;
An 2 — оральное поле и стомодеум;
Veg 1—эктодерма;
Veg 2 — архентерон и целомические
мешки;
Микромеры—первичная и вторичная
мезенхима.
Взаимосвязи слоев рассмотрены в
приложении 15.6. Поперечное разделение
бластулы четко выявляет ее части с разным по-
Развитие
453
тенциалом развития, однако говорить
о простой мозаике этих частей нельзя. По-
видимому, каждый клеточный слой
развивается в соответствии с его положением
относительно двух градиентов,
направленных один от анимального полюса к
вегетативному, а другой—наоборот.
Некоторые эксперименты, приведшие к такой
концепции, обсуждаются в приложении 15.6.
Различия между регуляционным и
мозаичным эмбрионами, видимо,
закладываются в момент первого разделения
клеток со специфическими цитоплазматически-
ми детерминантами.
15.4. Личиночное развитие
и метаморфоз
Жизненный цикл многих животных
характеризуется дивергенцией экологических
ниш, занимаемых ювенильной и взрослой
стадиями. Особенно ярко это проявляется
у свободноживущих морских
беспозвоночных (см. гл. 14), паразитических видов и
некоторых насекомых. Такая особенность
развития требует более или менее резкого
перехода между образом жизни, к которому
была адаптирована личинка, к новому,
характерному для взрослого организма.
Часто это сопровождается существенным
изменением строения, так называемым
метаморфозом.
15.4.1. Метаморфоз морских
личиночных форм
Ресничные личинки морских
беспозвоночных адаптированы к пелагическому
образу жизни. Полосы и пояса ресничек,
обеспечивающие их движение, не подходят
взрослым формам (см. гл. 10). Для таких
животных характерен метаморфоз с
утратой ресничных поясов и переходом к
локомоции с помощью мышечных клеток.
У гастропод метаморфоз личинки-
велигера сопровождается постепенной
редукцией паруса, который в качестве
главного локомоторного органа в конечном итоге
заменяется ногой (см. рис. 15.15), поскольку
уже не в состоянии поддерживать
развивающуюся улитку. Метаморфоз двуствор-
Рис. 15.15. Закручивание (торсия) во время
развития личинки моллюска на примере вели-
гера брюхоногих. К началу торсии мантийная
полость находится сзади, а после нее—
спереди, что соответствует топологии
взрослой улитки. I. Ранний велигер с
асимметричной правой мышцей-ретрактором. II. Первая
фаза торсии — быстрый поворот на 90° за счет
сокращения мышцы-ретрактора. Это вызывает
частичное закручивание внутренностного
мешка относительно ноги. III. Вторая фаза
торсии—закручивание внутренностного мешка
еще на 90° относительно ноги за счет
неравномерного роста
чатых моллюсков часто происходит
быстрее, с резкой утратой паруса.
У полихет также существует
противоречие между характером локомоции личинки
454
Глава 15
и потребностями развития во взрослое
животное. Сегменты тела добавляются на
эмбриональной или личиночной стадии
постепенно (см. разд. 15.2.2) и берут начало от
парных вентральных тяжей в задней части
трохофоры. Мезодермальные клетки тяжей
являются производными парных мезен-
тобластов. образующихся из бластомера 4d
в ходе типичного спирального дробления
(см. разд. 15.2.1 и приложение 15.1). У этих
зачатков сегментов имеются
мезодермальные и эктодермальные элементы. Два тяжа
клеток, дающие мезодерму, разрастаются
в ряд тканевых масс, в которых
формируется целом. В отличие от описанных ниже
иглокожих у этих животных мезодерма
образуется не из полости архентерона. Такой
гип целома назван схизоцелем. Сегменты
формируются путем координированного
органогенеза мезодермальных и эктодерма-
льных тканей, в котором главную
индукционную и организационную роль играет
вентральный нервный ствол. Каждый
новый сегмент развивается спереди от пиги-
диума, увеличивая массу личинки. Часто
каждый сегмент во время пелагической
стадии жизни снабжен локомоторными или
плавательными приспособлениями.
Развитие иглокожих характеризуется
одной из наиболее впечатляющих форм
метаморфоза в животном царстве. Полностью
сформировавшаяся их личинка
билатерально симметрична, в то время как у взрослого
животного доминирует пятилучевая
симметрия, хотя билатеральная иногда
накладывается на нее вторично (см. разд. 7.3.2). Це-
ломические мешки иглокожих развиваются
из боковых выпячиваний вершины
архентерона через некоторое время после
завершения гаструляции (см. рис. 15.9). Такой
способ их формирования называется энтероце-
льным. Изучение ныне живущих иглокожих
говорит о том, что изначально было три
парных целомических мешка: аксоцель,
гидроцель и соматоцель. Их развитие и
последующий метаморфоз показаны на рис.
15.16. .У многих современных иглокожих
правые аксоцель и гидроцель редуцированы
или полностью исчезли. Левый гидроцель
разделяется на гидроцель и вырост,
образующий каменистый канал и гидропор (рис.
15.16, I- IV).
Эти примитивные целомические зачатки
показаны у девятидневной личинки-
плутеуса на рис. 15.16,V. Правый и левый
соматоцели распространяются вокруг
желудка и формируют полости тела взрослого
организма. Левый гидроцель преобразуется
в пятилучевой заполненный жидкостью
зачаток, из которого развивается
амбулакральная система взрослой особи.
Дефинитивное ротовое отверстие, определяющее
оральное поле морского ежа, будет
формироваться в центре кольца гидроцеля. Таким
образом, орально-аборальная ось
взрослого организма проходит примерно вдоль
лево-правосторонней оси плутеуса (рис.
15.16,VI). Морской еж или морская звезда
развивается из имагинальньтх дисков,
разрастающихся во время метаморфоза
параллельно сморщиванию и сбрасыванию экто-
дермального эпителия личинки (рис.
15.16, VII, VIII).
Животные с экзоскелетом не могут
расти и развиваться постепенно. Они должны
пройти через ряд линек, при каждой из
которых сбрасывается старый покров и
организм увеличивается в размерах вплоть до
затвердения нового наружного скелета. Это
хорошо заметно у ракообразных. Их
развитие часто включает ряд морфологически
различных личиночных стадий, которые
в конечном итоге линяют, давая взрослую
форму (см. приложение 14.5 и гл. 8). При
этом рост с линьками нередко
продолжается и после того, как сформировалась
морфология взрослого организма.
У морских беспозвоночных личинка
обеспечивает поселение организма в
пригодных для дальнейшего развития условиях.
..Это очень важная стадия в жизненном
цикле животного, особенно для видов,
ведущих во взрослом состоянии сидячий или
прикрепленный образ жизни, для которых
выбор подходящего субстрата бывает
решающим для выживания фактором.
Гидробиологи иногда наблюдают тучи
только что прошедших метаморфоз
личинок, как будто случайно оседающих на
непригодные для жизни субстраты. Однако
этот процесс не всегда случаен. Многие
работы говорят о точности, с которой
пелагическая личинка способна «выбрать»
благоприятный субстрат. При этом действуют
Развитие
455
Рис. 15.16. Развитие целомической системы морского ежа и метаморфоз его личинки во
взрослую форму. I. Формирование целомических мешков из выпячиваний у вершины архентерона
после окончания гаструляции. II. Парные цепомические мешки. Ill, IV. Разделение мешков.
V. Дифференцировка второго левого целома в гидроцель и комплекс каменистого канала-
гидропора, рост третьих (левого и правого) целомических мешков. У Psammechinus эта стадия
достигается примерно на девятый день планктонного развития. VI. Формирование пятилучевого
амбулакрального кольца, определяющего оральную поверхность развивающегося иглокожего,
и становление орально-аборальной оси. VII. Постепенное развитие орального и аборального
имагинальных дисков, сопровождающееся ростом ресничных локомоторных поясов плутеуса.
VIII. Эмбрион Psammechinus незадолго до завершения метаморфоза. Личиночные ткани будут
разрушены, зачатки взрослой особи объединяются, сформировав уменьшенную копию морского
ежа
следующие механизмы: 3) различение и выбор предпочтитель-
1) последовательность поведенческих ного субстрата;
актов, приводящая к контакту личинки 4) агрегационное поведение и хемосен-
с благоприятным субстратом; сорное распознавание субстратов, уже засе-
2) замедленный метаморфоз в отсут- ленных взрослыми и личиночными форма-
ствие такого субстрата; ми своего вида.
456
Глава 15
<TD
Относительный размер
Эмбриональное
развитие
ЯЙЦО
J
*f<=
^Mf^yjj.
Питание
1-й
личиночной
стадии
1
X
<fr
^-^
S
^~
/
1
•zJ
Krxj=r3=^
1
Питание
2-й личи
ночной
стадии
Питание
3-й
личиночной
стадии
Питание
4-й личи
ночной
стадии
Питание
5—й
личиночной
стадии
Неполовозрелые личинки
1-е
имаги-
нальное
питание
2-е
имаги-
нальное
питание
3-е
имаги-
нальное
питание
Размножение
ИПА1 U
Рис. 15.17. Схематическое представление роста и развития насекомых. А. Аметаболические
формы: метаморфоза нет. В этом примере рост продолжается и после достижения половой зрелости.
Б. Гем и метаболические формы: неполный или частичный метаморфоз. В этом примере крылья
появляются у личинки второго возраста в виде наружных зачатков. Полностью они формируются
после пятой (последней) линьки. Увеличение размера происходит в течение короткого периода
после каждой линьки. В. Голометаболические формы: полный метаморфоз. В этом примере
четыре личиночные, куколочная и имагинальная стадии. Рост происходит после каждой личиночной
линьки
I
Развитие
457
Г
Эмбриональное
развитие
Питание
1-й личин:
ночной
стадии
Питание
2- й личи
ночной
стадии
Питание
3-й
личиночной
стадии
Питание
4-й личи
ночной
стадии
Непитающаяся
5-я личиночная
стадия или
куколка
Имаго
Размножение и расселение
Иногда питание
ЯЙЦО
Зачатков [ Внутренние зачатки крыльев
крыльев нет ' г
ЛИЧИНКА
ЛИЧИНКА
Наружные
зачатки крыльев
КУКОЛКА
Крылья
ИМАГО
В
Рис. 15.17 (продолжение)
Например, сложная последовательность
поведенческих актов в процессе развития
наблюдается у личинки мидии Mytilus edu-
lis.
Установлено, что личинки многих видов
высокоизбирательно относятся к
заселяемым субстратам. Это иллюстрируют
описанные в приложении 15.7 эксперименты
с выбором поверхностей заселения
личинками близкородственных видов живущих
в трубках полихет Spirorbis, взрослые особи
которых обычно встречаются на различных
субстратах.
Циприсовидные личинки морских
желудей также демонстрируют перед оседанием
на субстрат ярко выраженное и сложное
поведение. Это позволяет им выбрать
удобные для метаморфоза места. Личинки
реагируют не только на текстуру поверхности
(предпочитают грубую и пористую), но и на
присутствие других морских желудей, их
личинок, а также остатков взрослых особей
собственного вида.
15.4.2. Развитие и метаморфоз
насекомых
Тип Uniramia включает большую группу
животных, обычно называемых
насекомыми (подтип Hexapoda). У них три
различных способа развития. Представители
близких к многоножкам бескрылых групп,
например ногохвостки и щетинохвостки,
развиваются постепенно через ряд линек. Их
строение при этом не претерпевает каких-
либо сильных изменений, поэтому говорить
о метаморфозе нельзя (рис. 15.17,^4). Такие
формы называют аметаболическими.
Полное дробление яиц у многих видов этих
животных— примитивная черта, однако у
некоторых оно, хотя сначала и полное, в
конечном счете приближается к типу,
характерному для крылатых насекомых (рис.
15.18).
У других насекомых во время развития
наблюдается более или менее выраженный
метаморфоз и фиксированное число
личиночных стадий, предшествующих взрослой
форме. В нескольких отрядах метаморфоз
не слишком резкий, поскольку преимагина-
льные формы, называемые «нимфами» или
«наядами» (если они водные), имеют
наружные зачатки крыльев. Развитие этих
насекомых (к ним относятся стрекозы и
кузнечики) называют гемиметаболическим, а их
самих— экзоптериготами. У кузнечиков и
саранчи, как показано на рис. 15. П.Б, нимфы
и имаго занимают одну и ту же нишу,
поэтому при метаморфозе последней
личиночной стадии в крылатое насекомое значи-
458
Глава 15
Рис. 15.18. Раннее развитие бескрылых
насекомых. Можно видеть, что дробление сначала
полное (ср. рис. 15.8)
тельной перестройки строения не требуется.
Если же ювенильные стадии обитают в
воде, изменение морфологии может быть
значительным, поскольку оно связано с
дивергенцией неполовозрелых особей и имаго по
нишам. Однако и такой тип метаморфоза
не столь впечатляющ, как у насекомых
с внутренними крыловыми зачатками
(эндоптеригот). Их развитие называют го-
лометаболическим. Между последней
личиночной и имагинальной стадиями здесь есть
промежуточная стадия куколки (рис. 15.17
и 8.33). Личинки голоматаболических
насекомых относятся к различным типам (рис.
15.19); им часто дают обобщенные
названия —гусеницы, черви и т.д. Как показано
в гл. 14 и на рис. 15.17,5, это
специализированные питающиеся стадии, в то время как
I II
V VI
Рис. 15.19. Примеры личинок голометаболи-
ческих насекомых: I—жужелицы (жуки); II —
жука-щелкуна («проволочник»); III —
долгоносика (жуки); IV—медоносной пчелы
(перепончатокрылые); V—бабочки (гусеница);
VI — падальной мухи (двукрылые). В каждом
случае морфология личинки и имаго
совершенно различна
Рис. 15.20. Система имагинальных дисков
личинки двукрылого (Drosophi/a). Обозначены
местоположение дисков в личинке и
развивающиеся из них имагинальные структуры. Можно
видеть, что пищеварительная система при
метаморфозе не заменяется
имаго выполняют функцию расселения
и размножения. На стадии куколки
движение и питание прекращаются, поскольку
происходит коренная структурная
перестройка организма.
У голометаболических насекомых во
время раннего эмбриогенеза
закладываются группы клеток, которые
дифференцируются только во время метаморфоза. Эти
имагинальные клетки, собранные довольно
рыхлыми гнездами или чаще оформленные
в виде компактных имагинальных дисков,
дают начало всему внешнему покрову
имаго с его сложной регионарной структурой,
слюнным железам и другим внутренним
органам. Схема расположения имагинальных
дисков у личинки двукрылого и их
соответствие имагинальным структурам показаны
на рис. 15.20. В современной биологии
особый интерес приобрела система
имагинальных дисков плодовой мушки Drosophila me-
lanogaster. В основном это обусловлено
богатством генетической информации,
накопленной по этому организму, однако
связано также и с преимуществами
использования имагинальных дисков в качестве
модели для анализа судьбы клеток, процессов
детерминации, дифференцировки и
органогенеза, которые обсуждались выше в разд.
15.3.
Развитие
459
15.4.3. Экспериментальный
анализ развития имагинальных
дисков Drosophila melanogaster
Имагинальные диски голометаболиче-
ских насекомых закладываются в раннем
эмбриогенезе, т. е. клетки должны рано
принять «решение», будут ли они
функционировать в теле личинки или останутся
недифференцированными в составе имагинальных
дисков. В последнем случае их дифференци-
ровка начнется под влиянием изменений
гормональной среды во время окукливания.
Несмотря на отсутствие дифференци-
ровки, имагинальные диски являются высо-
кодетерминированными структурами, по
свойствам больше напоминающими
мозаичных, чем регуляционных эмбрионов.
Однако, как показано ниже, имеются
данные о том, что в основе этой детерминации
лежит система градиентов. Путем
выборочного удаления частей диска с последующей
его имплантацией в личинку последней
стадии можно составить карту его зачатков. Во
время метаморфоза в соответствии с
удаленными частями будут развиваться
неполные имагинальные структуры. Карта
зачатков дисков конечностей, например, имеет
концентрическое строение (рис. 15.21,Л).
Из внешних зон развиваются
проксимальные элементы ноги, а из внутренних —
дистальные (рис. 15.21,/>,Z?). Диск
конечности можно разрезать и имплантировать его
фрагмент в молодую личинку. При этом
происходит компенсационный рост, и
недостающие части восстанавливаются. Однако
на морфологию получившейся конечности
влияет положение внутри диска исходных
фрагментов.
Если, например, диск конечности
разделен на секторы, как показано на рис.
\5.22.A, то верхняя медиальная четверть
(А), регенерируя, дает нормальный полный
диск, а нижняя медиальная (С) — зеркально
удвоенную нижнюю половину. Эти
результаты сопоставимы с градиентной теорией
организации. Можно предположить
существование некой организационной
иерархии, причем фрагменты, находящиеся на
высшей ее ступени, могут регенерировать
стоящие ниже их по рангу, а в обратном
направлении регенерация невозможна. Эта
Тазик
Вертлуг
Бедро
Голень
Лапка
В
Голень
Лапка
Рис. 15.21. Развитие конечности насекомого
из имагинального диска. А. Карта зачатков
имагинального диска, полученная путем
селективного удаления его частей. Б. Постепенное
развитие конечности путем разрастания
имагинального диска. В. Элементы
сформированной ноги
модель представлена схематически на рис.
15.22,£". Подобные модели ценны, но
следует заметить, что, воспроизводя
наблюдаемое, они не дают никаких ключей к
разгадке биологической природы
действующих механизмов. В то же время именно
система имагинальных дисков позволяет бы-
460
Глава 15
Верхняя
4 5 6 7 6 5 Л
Рис. 15.22. Доказательство градиентной, или иерархической, системы регионарной организации
имагинального диска конечности дрозофилы. А. I. Определение четвертей интактного диска.
II. Регенерация верхней медиальной четверти дает полный диск. III. Регенерация нижней
медиальной четверти дает зеркальную копию нижней половины диска. Б. Диск с контурными линиями
предполагаемой способности к развитию. В. Модель регионарной организации: участки высшего
ранга могут восстанавливать все недостающие части, а участки назшего ранга—только части не
превышающего их ранга
стро продвигаться вперед в понимании
молекулярных и генетических основ
детерминации судьбы клеток и у высших
организмов.
Когда эксперименты показывают, что
характер будущей дифференцировки клетки
предопределен и не подвержен влиянию ее
клеточного окружения, такую клетку часто
называют детерминированной в своем
развитии. Клетки имагинального диска
насекомых обладают именно этим свойством. То,
что они детерминированы еще до начала
биохимической дифференцировки, явилось
важным открытием биологии развития.
Диск можно извлечь из эмбриона-
донора и трансплантировать в другую
часть эмбриона-реципиента (рис. 15.23).
Обычно он продолжает развиваться в
соответствии со своим исходным положением
в теле мухи. Другими словами, если это
диск будущей конечности, его клетки будут
развиваться в конечность. Различают два
типа детерминации:
1) регионарная—всего диска;
Развитие
461
I II
Рис. 15.23. Регионарная организация имагина-
льного диска фиксирована (см. также
приложение 15.8). Диск конечности личинки
двукрылого (I) удален хирургически и (II)
имплантирован в брюшко эмбриона реципиента. После
завершения метаморфоза (III) у вышедшей из
куколки мухи на брюшке находится
дополнительная конечность
2) локальная, т.е. отдельных клеток
внутри диска, обеспечивающая дифферен-
цировку организованной интегрированной
структуры.
Ниже будет показано, что эти два типа
детерминации могут быть разграничены.
Клетки имагинального диска способны
к пролиферации (митотическому делению),
а его рост является естественной
составляющей развития. Диск способен также
к компенсационному росту, т.е. может
быть восстановлен из своего фрагмента (см.
рис. 15.22).
Гормональная среда взрослого
насекомого не препятствует пролиферации клеток
имагинального диска, но подавляет или не
стимулирует их дифференцировку.
Последняя происходит только в том случае, если
диск поместить в гормональную среду
личинки последней стадии или куколки.
Эти факты легли в основу многих
интересных экспериментов, результаты которых
часто оказывались совершенно
неожиданными.
Диск может оставаться
недифференцированным гораздо дольше, чем в норме,
если разводить его серийную культуру
внутри взрослой мухи. Низкий уровень линоч-
ного гормона в ее гемолимфе не тормозит
пролиферацию клеток диска, однако не
позволяет ему претерпевать метаморфоз.
Фрагменты диска можно
последовательно трансплантировать в ряду многих
поколений, в любой момент проверяя состояние
их детерминации путем пересадки тестовых
участков в личинку последней стадии.
Метод проиллюстрирован на рис. 15.24.
Тестируемые таким путем клетки имаги-
нальных дисков обычно сохраняют свою
исходную регионарную детерминацию,
превращаясь во время метаморфоза
в структуру (глаз, антенну, крыло и т.д.),
определяемую первоначальным
местоположением диска. Однако это правило знает
исключения: иногда фрагмент диска дает
начало структуре, соответствующей другой
части зародыша (приложение 15.8), т.е.
происходит транс детерминация. Особенно
интересно, что различные клетки диска
продолжают при этом вести себя
координирование, т.е. как составные части
нормальной, но иной карты зачатков.
Большое число наблюдений
свидетельствует, что некоторые трансдетерминации
встречаются часто, другие редко или
вообще никогда (см. приложение 15.8). Когда все
клетки диска неожиданно изменяют свое
состояние детерминации, создается
впечатление, что при этом действует хорошо
отлаженная система контроля. Ее генетическую
основу можно исследовать на нескольких
мутантах Drosophila (см. приложение 15.9).
Схема координированного развития диска,
соответствующая каждой специфической
Дальнейшее
культивирование
Рис. 15.24. Метод серийной трансплантации
имагинальных дисков. Диск извлекают из
личинки-донора, разрезают и помещают в имаго,
где он регенерирует. Регенерировавший диск
можно извлечь из хозяина, снова разрезать
и одну из его частей протестировать на
характер развития в личинке последнего возраста.
Другой фрагмент переносят во взрослого
хозяина для дальнейшего культивирования (см.
также приложение 15.8)
462
Глава 15
области тела, контролируется группой
генов, называемых гомеозисными.
Например, генетическая мутация Anten-
napedia изменяет регионарную
организацию таким образом, что на месте антенны
развивается неполная конечность. Чтобы
предотвратить развитие антеннального
диска в конечность, необходим нормальный
(«дикий») аллель гена Antennapedia (см.
приложение 15.9).
Гомеозисные гены контролируют также
дифференцировку других структур,
отражающих регионарную организацию.
Некоторые из таких генов и свойственные им
эффекты приведены в приложении 15.9. Все
гомеозисные гены способны превращать
один сегмент в другой или одну часть
сегмента в другую. Для исследования
регионарной и сегментарной организаций
животных эти гены являются крайне интересным
материалом. Так, известно, что некоторые
последовательности оснований у гомеозис-
ных генов Drosophila такие же, что и у
других генов животных самых различных
систематических групп. Эти
последовательности называют «гомеобоксами»; их
существование свидетельствует о раннем
эволюционном происхождении таких генов
регионарной организации.
Однако некоторые фундаментальные
вопросы до сих пор не получили
объяснения. Регионарная организация раннего
эмбриона несекомых запрограммирована
в яйцеклетке, т. е. это результат процесса га-
метогенеза. В дальнейшем различные
группы генов, по-видимому, транскрибируются
в имагинальных дисках в соответствии с их
местоположением в раннем эмбрионе.
Установлено, что в этом участвуют
гомеозисные гены, однако молекулярные основы
такой регионарной дифференцировки и
точная роль этих генов до сих пор не выяснены.
15.5. Регенерация
15.5.1. Введение
Регенерацию можно определить как
способность к восстановлению утраченных
случайно или в результате автотомии
частей тела путем компенсационного роста
и дифференцировки. Способность к
регенерации недостающих частей тела —
замечательная особенность многих
мягкотелых беспозвоночных, например губок,
стрекающих, плоских червей, немертин, анне-
лид и некоторых иглокожих. Для них
характерно бесполое размножение путем
фрагментации (см. гл. 14), очевидно тесно
связанной с регенерацией. Беспозвоночные
с твердыми или
высокоспециализированными покровами (например, членистоногие,
псевдоцеломические черви, моллюски)
обычно слабо регенерируют и часто не
способны к размножению путем
фрагментации. У членистоногих регенерация
происходит во время линьки и ограничена, как
правило, конечностями.
Процессы, происходящие во время
регенерации, сходны с происходящими при
нормальном развитии. К ним относятся
1) пролиферация
недифференцированных клеток (как у бластулы) и
формирование зачатка (бластемы);
2) морфогенез и организация клеток
в пространственную иерархию;
3) дифференцировка и экспрессия
специфических генов.
В связи с этим явление регенерации
представляет собой удобную модель для
изучения процессов развития. У некоторых
животных во время нее происходит дедиф-
ференцировка, не свойственная
нормальному развитию.
Регенерация означает реакцию
организма на утрату части тела, причем ответ
должен включать как пролиферацию бластемы,
так и развитие соответствующей структуры
из пролиферировавших клеток.
15.5.2. Природа регенерационной
бластемы
Новые клетки при регенерационном
росте могут происходить либо из резервной
популяции недифференцированных тотипо-
тентных клеток, либо из ранее
дифференцированных клеток путем их дедифференци-
ровки. Какой из этих альтернативных
вариантов осуществляется на самом деле, не
всегда ясно. Особенно хорошо выражена
способность к регенерации у стрекающих.
Они располагают запасом интерстициаль-
ных (1-)клеток, которые в норме образуют
Развитие
463
Интерстициальные (7)-клетки
Рис. 15.25. Поперечный разрез книдарий.
показывающий интерстициальные клетки и их
пролиферацию перед регенерацией
и постоянно замещают различные классы
Дифференцированных клеток, например
книдобласты (рис. 15.25).
У гидры эти I-клетки обычно
скапливаются в эктодерме перед бесполым
размножением почкованием. Такую же их
реакцию вызывает повреждение тела, причем
рана становится местом их конкурентного
привлечения и на ее месте они формируют
регенерационную бластему. У гидры в
отличие от некоторых других книдарий I-
клетки пролиферируют в течение всей
жизни, благодаря чему поддерживается
постоянный регенерационный потенциал.
Однако гипотеза, рассматривающая их как
самовозобновляющуюся популяцию,
обязательную для восстановительных процессов,
слишком упрощена. Если эти клетки
разрушить химическим путем, организм не
утрачивает способности к регенерации.
Регенерировать могут и фрагменты гидры,
обычно не содержащие I-клеток. Более того,
в благоприятной среде
дифференцированные клетки тканевых эксплантатов гидры
дедифференцируются в интерстициальные
клетки, размножаются и редифференци-
руются в клетки других типов. Это значит,
что обычно наблюдаемый путь к дифферен-
цировке через популяцию резервных клеток
не единственный.
Удивительная регенерационная
способность свободноживущих плоских червей
также связана с недифференцированными
резервными клетками, так называемыми
необластами. Этих животных используют
для изучения феномена регенерации уже
в течение более 100 лет. Из фрагментов,
полученных в результате поперечного
перерезания планарии, восстанавливаются две
целые особи (рис. 15.26,^4). Точно так же
целые черви регенерируют из мелких
фрагментов, включая и сагиттально отсеченные
(рис. 15.26,5).
Первая стадия
регенерации—формирование раневой бластемы и ее
последующее заселение необластами. Роль этих
резервных клеток была
продемонстрирована экспериментами по облучению и
трансплантации. Облучение рентгеновскими
лучами в дозе 3000 рад организм не убивает,
но препятствует пролиферации необластов.
После облучения животное утрачивает
способность к регенерации, однако она может
произойти, если в тело такого червя
имплантировать фрагмент необлученного
донора. Более того, если клетки имплантата
помечены, например по-другому окрашены,
можно видеть, что цвет регенерировавшего
фрагмента такой же, что у
имплантированной ткани (рис. 15.27).
Тотипотентные резервные клетки не
всегда участвуют в регенерации, и их
ведущая роль в этом процессе, доказанная для
планарии, может быть скорее исключением.
У аннелид регенерационная бластема
<5<\
7—\
Рис. 15.26. Регенерация у плоских червей. А.
Поперечное разрезание и последующая
регенерация передним или задним фрагментом
утраченной задней или передней части. Б.
Даже мелкие кусочки могут восстановить целого
червя, причем исходные ткани сохраняют свою
регионарную специфику
464
Глава 15
IV
Рис. 15.27. Доказательство особой роли кле-
ток-«необластов» при регенерации у планарии.
А. I. Особь облучена жесткими
рентгеновскими лучами. II. Передняя часть удалена.
III. Задний фрагмент не способен к
регенерации. Б. I. II. См. A. III. В среднюю часть тела
имплантирован небольшой фрагмент генетически
отличной особи. IV. Теперь голова может быть
регенерирована; генетически она идентична
имплантированному фрагменту
формируется по-иному: путем дедифферен-
цировки и перегруппировки
дифференцированных клеток эктодермы и мезодермы.
Например, заживление раны при утрате
каудальных сегментов идет за счет
миграции целомоцитов к поврежденной
поверхности и восстановления зон роста, которые
появляются в виде тяжей характерных
клеток со сферическими ядрами и четко
заметными ядрышками. У нереид дифференци-
ровка этой области нарастания сегментов
требует присутствия ростового гормона
мозга, а формирование сегментарных экто-
дермальных структур (сумок щетинок и па-
раподий) идет под индукционным влиянием
вентральной нервной цепочки (см. ниже,
а также приложение 15.10).
15.5.3. Регенерация
и регионарная организация
Многие мягкотелые беспозвоночные
способны целиком восстанавливать
регионарно организованную структуру из очень
мелкого фрагмента тела. Это
иллюстрируют рис. 15.26, а также рис. 14.2, на
котором изображено бесполое размножение по-
лихеты с помощью спонтанно автотомиро-
ванных фрагментов.
Общее для этих примеров —
восстановление целой структуры из ее части.
В каждом случае фрагмент сохраняет свою
исходную полярность, и компенсационный
рост возвращает его на прежнее положение
в общем плане строения организма. На рис.
15.26 фрагмент головы планарии
восстанавливает утраченную «хвостовую» область,
а «хвостовой» фрагмент—утраченную
голову. В составе целого каждой части
присуща регионарная специфика, которая
сохраняется при регенерации. У
сегментированных животных эта специфика выражена еще
четче и закреплена в линейной иерархии
сегментов. У эмбриона насекомых число
сегментов фиксировано, и на каждом
развиваются свойственные только ему
структуры. Специфика этих сегментов
контролируется или регулируется особыми
генетическими локусами (см. приложение 15.9).
Следует, однако, отметить, что гены bithorax
свидетельствуют о существовании
морфологических границ, дополняющих легче
распознаваемые сегментарные границы.
Членистоногие не могут
восстанавливать утраченные сегменты, а многие анне-
лиды, строение которых также
сегментарное, обладают такой способностью.
Каждый сегмент аннелид уникален и является
частью единого интегрированного целого;
это выражается, в частности, в том, что
многим аннелидам свойственно
фиксированное их число. Позиционная информация,
по-видимому, поступает от вентральной
нервной цепочки (см. приложение 15.10).
У полихет семейства Nereidae для
регенерации «хвостовой» области необходим
гормон мозга, но это представляется
вторичной адаптацией, связанной с облигатно
моноцикличным размножением данных
червей (см. разд. 14.3 и 14.4); тем не менее
скорость нарастания новых сегментов остается
зависимой от позиционного контроля
(приложение 15.10).
Развитие
465
15.5.4. Регенерация, рост
и размножение
Для регенерации требуются ресурсы
организма, и это, по-видимому, должно
приводить к их перераспределению в пользу
соматических функций за счет уменьшения
вклада в половое размножение.
Следовательно, существует потенциальный
конфликт между регенерацией и половым
размножением, и можно предполагать наличие
регуляторных механизмов, эффективно
контролирующих антагонизм между этими
процессами. Особенно это касается
моноцикличных организмов. Ресурсы,
необходимые для образования репродуктивных
тканей, должны расходоваться у них на регене-
рационный рост только в том случае, если
он обеспечивает компенсационное
увеличение выживаемости или жизнеспособности
потомства.
Подобный механизм следует
предполагать у моноцикличных нереид, которые, как
показано выше, способны к регенерацион-
ному росту после утраты задних сегментов.
Когда эти полихеты достигают
половозрелое™, все ресурсы полностью расходуются
на размножение, и после него черви
погибают. Значит, регенерация сегментов в
конце репродуктивного периода теряет смысл.
На этой стадии жизненного цикла регенера-
ционный рост подавляется эндокринным
механизмом. Во время полового
созревания постепенно снижается секреция
гормона мозга, и уменьшение его концентрации
в циркуляторной системе способствует
завершению гаметогенеза (см. разд. 14.4),
инициируя одновременно соматические
изменения, связанные с размножением (см.
рис. 16.35). Этот же гормон необходим для
каудальной регенерации, поэтому
половозрелые нереиды не способны
восстанавливать утраченные задние сегменты. У поли-
хет, размножающихся несколько лет
подряд, подобный эндокринный механизм не
встречается. Регенерационный рост здесь
идет и у половозрелых особей, поскольку
восстановленные сегменты в последующие
годы могут участвовать в размножении и их
репродуктивная ценность остается
высокой.
30-1314
15.6. Выводы: развитие
беспозвоночных и
генетическая программа
Экспериментальное изучение развития
беспозвоночных началось в конце XIX в.
Оно развивалось параллельно с
генетическими исследованиями, открытием
молекулярных основ клеточной наследственности
и быстрым расширением знаний в области
молекулярной биологии. На повестке дня
стоит выдающаяся по сложности задача
объединения этих дисциплин.
Беспозвоночные являются кладезем удобных
модельных систем для ее решения. На этом пути
уже достигнут значительный прогресс, и
большая часть материала, представленного
в настоящей главе, может быть
переосмыслена с точки зрения молекулярной
биологии.
Информационные молекулы животной
клетки—это последовательности мРНК,
синтезированные на матрице ДНК в ядре.
В первых разделах этой главы говорилось,
что почти у всех животных ядерная ДНК
зиготы полностью передается каждой
дочерней клетке во время раннего дробления.
Эти молекулы мРНК можно изучать
методами современной биохимии. Во время
оогенеза (см. гл. 14) молекулы материнской
мРНК накапливаются в цитоплазме ооци-
та. При оплодотворении они активируются
и обеспечивают ранний белковый синтез.
В конечном счете ядерный геном зиготы
становится источником генетической
информации всего организма. Однако в
процессах дифференцировки и морфогенеза
решающую роль сохраняет цитоплазма
ооцита. Мы видели, что у широкого круга
беспозвоночных функции клеточного ядра
и информация, которая в конечном счете
транскрибируется или транслируется,
детерминированы их предысторией в
эмбрионе.
Иногда синтез тех или иных ферментов,
по-видимому, определяется
специфическими цитоплазматическими факторами (см.,
например, развитие оболочника Styela).
В других случаях функциональные реакции,
вероятно, вызываются межклеточными
контактами. Роль таких взаимодействий
показана в экспериментах с эмбрионами
466
Глава 15
моллюсков и морских ежей. Ряд наиболее
удивительных открытий сделан при
изучении эмбрионов насекомых, особенно
плодовой мушки Drosophila melanogaster. в случае
которой обилие имеющихся генетических
сведений сочетается с удобством
постановки опытов. Вполне вероятно, что эмбрионы
беспозвоночных еще долго будут
оставаться в числе лучших модельных систем
для исследования сложных проблем
развития животных.
15.7. Дополнительная
литература
Berril N.J. 1971. Developmental Biology. McGraw
Hill, New York.
Brookbank J. W. 1978. Developmental Biology:
Embryos, Plants and Regeneration. Нафег and Row,
New York.
Browder L. W. 1984. Developmental Biology, 2nd
edn. Saunders, New York.
Epel D. 1977. The program of fertilisation. Scient.
Am. 237, 129 138.
Gehring W.J. 1985. The molecular basis of
development. Scientific American, 253, 137—146.
Oppenheimer S.B. 1980. Introduction to Embryonic
Development. Allyn and Bacon, New York.
Reverberi G. 1971. Experimental Embryology of
Marine and Freshwater Invertebrates.
Amsterdam.
Slack J. M. W. 1983. From Egg to Embryo.
Cambridge University Press, Cambridge.
Stearns L. W. 1974. Sea Urchin Development:
Cellular and Molecular Aspects. Dowden, Hutchinson
and Ross. Pennsylvania.
Приложение 15.1.
Спиральное дробление
Этот тип дробления характерен для
таких таксонов первичноротых, как немерти-
ны, аннелиды, сипункулиды, эхиуриды
и моллюски. Наиболее четко его можно
наблюдать у эмбрионов с относительно
небольшим количеством желтка, например
у морских полихет и моллюсков.
1. Неоплодотворенное яйцо радиально
симметрично относительно оси,
соединяющей анимальный (бедный желтком) и
вегетативный полюса (I). При этом все
сегменты, отсекаемые анимально-вегетативной
плоскостью, эквивалентны.
После оплодотворения яйцо
билатерально симметрично, и на нем можно
проследить основные плоскости эмбриона (II—
IV):
II) поперечная—разделяет переднюю
и заднюю части;
III) сагиттальная—разделяет левую
и правую половины;
IV) фронтальная — разделяет
дорсальную и вентральную стороны.
Развитие
467
2. Плоскости двух первых делений
являются продольными и делят пополам
углы между фронтальной и сагиттальной
плоскостями.
В результате образуются первые два
бластомера, АВ и CD (II), а затем первые
четыре бластомера А, В, С, D (III).
II III
D-бластомер обычно крупнее трех
остальных. Все последующие продукты
дробления могут быть идентифицированы
относительно этой клетки.
3. Плоскость третьего деления
поперечная; она проходит над экватором и
отделяет четыре меньшие клетки на анималь-
ном полюсе от четырех более крупных на
вегетативном (I).
30*
Четыре мелкие клетки называют
первым квартетом микромеров. Они
расположены над границами между макромерами,
поскольку плоскость дробления наклонена
относительно продольной оси эмбриона.
Если смотреть с анимального полюса,
кажется, что микромеры сдвинуты по часовой
стрелке (II).
Восемь бластомеров эмбриона
обозначают так:
la lb lc Id
1А IB 1С ID
Все последующие деления являются
поперечными и заключаются в разделении
существующих микромеров и отделении
новых квартетов микромеров в результате
неравного дробления макромеров. На рис. III
показано появление 16-клеточного
эмбриона после четвертого деления дробления.
Обозначения бластомеров на этой стадии
следующие:
деления первого jai j^i ici j^i
квартета: * »- * *
la2, lb2, lc2, Id2
второй квартет: 2a, 2b, 2c, 2d
till
макромеры: 2A, 2B, 2C, 2D
Плоскость каждого деления дробления
наклонена относительно продольной оси
таким образом, что направления сдвигов
микромеров по часовой и против часовой
стрелки чередуются.
468
Глава 15
4. Разработана условная система
обозначений, позволяющая идентифицировать
каждую клетку вплоть до стадии 64 бласто-
меров. Схема дробления (завершенная
только по наиболее важной линии D-клеток)
выглядит так:
и т.д.
Яйцо
и т.д.
Число клеток 12 4 8 16 32 64
Деление 1-е 2-е 3-е 4-е 5-е 6-е
дробления
Почему особенно важна линия D-
бластомера, объясняется далее в тексте
и приложениях 15.2 и 15.3.
Приложение 15.2.
Радиальное дробление
Этот тип дробления, отличный от
наблюдаемого у аннелид и моллюсков,
присущ, например, иглокожим и в измененном
виде хордовым. У морских ежей он
выражен очень четко.
1. Схема дробления. Оплодотворенное
яйцо билатерально симметрично (I). Пло-
IV
■Мезомеры (8)
1акромеры (4)
■Микромеры (4)
An 1 (16 клеток)
,п 2 (16 клеток)
'eg 1 (8 клеток)
'eg 2 (8 клеток)
Развитие
469
скости двух первых делений дробления
проходят вдоль анимально-вегетативной оси
и делят яйцо сначала на два. а затем на
четыре равных бластомера (II, III).
Третье деление дробления поперечное
и неравное, плоскость его проходит
непосредственно над экватором (IV) и отделяет
четыре «мезомера» от несколько более
крупных «макромеров».
Четвертое деление дробления в нижнем
и верхнем клеточных слоях идет по-
разному. В верхней половине оно
продольное и дает кольцо из восьми клеток, а в
нижнем ярусе поперечное и неравное, в
результате чего образуются четыре макромера
и четыре микромера. В этом примере
микромеры находятся на вегетативном
полюсе (V).
Два следующих деления в верхней
половине поперечные, а в нижней—продольное
и поперечное. В результате образуется
эмбриональная стадия из 64 бластомеров
(VI).
2. Идентификация бластомеров.
Поскольку первые четыре бластомера структурно
идентичны, идентифицировать отдельные
клетки невозможно. Однако могут быть
идентифицированы клеточные слои. На
стадии 64 бластомеров их условно обозначают
следующим образом:
первый анимальный слой = Anl = 16
верхних бластомеров;
второй анимальный слой = Ап2 =16
нижних бластомеров анимального
полушария;
первый вегетативный слой = Vegl =
кольцо из восьми макромеров под экватором;
второй вегетативный слой = Veg2 =
нижнее кольцо из восьми макромеров
вегетативного полушария;
микромеры = Mic = группа самых
мелких клеток на вегетативном полюсе.
3. Бластула. В результате дробления
формируется бластула (VII)—полый шар
из ресничных клеток.
Гаструляция начинается инвагинацией
микромеров (VIII). Она описана в разд.
15.2.2 и проиллюстрирована на рис. 15.9.
Приложение 15.3.
Регуляционное и мозаичное
развитие I
В конце XIX—начале XX вв. на
эмбрионах морских беспозвоночных стало
возможным изучать характер развития
отдельных клеток, изолированных на ранних
стадиях дробления (двух или четырех
бластомеров). Результаты оказались на
удивление различными.
1. Эмбрионы иглокожих. Из каждой
изолированной на стадии четырех
бластомеров клетки может развиться нормальный
(но мелкий) плутеус. Эти мелкие эмбрионы
были названы регуляционными.
Мелкий, но
/V/ нормальный
плутеус
1. Нормальный плутеус.
II. Результаты разделения бластомеров.
Из бластомера, изолированного на че-
тырехклеточной стадии, в результате
деления образуются два мезомера, макромер
и микромер, т.е. наблюдается схема
дробления нормального эмбриона (см.
приложение 15.2).
2. Эмбрионы моллюсков. Из
изолированных бластомеров двух- и четырехкле-
470
Глава 15
точного зародыша развиваются
непропорциональные или неполные эмбрионы,
причем из D-клеток — обычно более
полноценные, чем из А-, В- или С-бластомеров.
I. Нормальная трохофора Patella.
П. Аномальные трохофры, развившиеся
из изолированных бластомеров.
3. Эмбрионы асцидий. В яйцах асцидий
часто встречаются визуально различные
участки цитоплазмы (см. рис. 15.6), которые
распределяются по разным клеткам.
Эмбрионы, полученные из
изолированных бластомеров, весьма различны. В их
строении преобладают структуры, которые
развились бы из соответствующих клеток
в нормальном зародыше.
Глазные пятна
Мышечные клетки
Хорда
III. Изолированные клетки правой
половины.
IV. Изолированные задние клетки.
Приложение 15.4.
Судьба клеток, карты зачатков
и цитоплазматическая
локализация
1. Принцип составления карты
зачатков. На поверхность нераздробившегося
яйца можно нанести метки. Если в
результате дробления не происходит
перемешивания бластомеров, эти метки затем появятся
в специфических структурах зародыша.
I. Нормальный «головастикоподобный»
эмбрион с парой глаз, хордой и парными
мышечными блоками.--
II. Изолированные передние клетки.
л л Презумптивная кишка
n d Презумптивная мантия
•. Презумптивный оперкулум
х * Презумптивная нога
Гипотетический пример зародыша
моллюска:
I) метки на поверхности яйца;
II) положение клеток во время гаструля-
ции;
III) сегрегация меченых клеток по
специфическим участкам эмбриона.
2. Природные маркеры. После
оплодотворения в некоторых зиготах можно
наблюдать четко различимые участки цитоплаз-
Развитие
471
I II
Гаструляция и цитоплазматическая
локализация у асцидий
мы и билатеральную симметрию. На
рисунке видно распределение различных цито-
плазматических участков яйца асцидий (I)
в клетках вегетативного полюса (II); во
время гаструляиии эти клетки инвагини-
руют (III, IV). Из клеток, содержащих
визуально различную цитоплазму, после га-
струляции разовьются различные
эмбриональные структуры (V, VI). В связи с этим
возникает вопрос: «Определяют ли
природные цитоплазматические маркеры судьбу
клеток?».
3. У асцидий судьбу клеток
действительно определяют цитоплазматические
факторы. Участок желтоватой цитоплазмы
можно рассматривать как природный маркер
мышечных клеток головастикоподобной
личинки [см. приложение 15.3(3)].
4. Фермент ацетилхолинэстераза
специфически стимулирует развитие мышечных
клеток; он может быть окрашен в
цитоплазме.
Получены следующие
экспериментальные данные:
а) обычно он появляется через 8 ч после
начала эмбриогенеза;
б) фермент появляется в это время даже
при подавлении деления клеток
химическими препаратами;
в) при обработке актиномицином D в
течение 5 ч или пуромицином в течение 7 ч
после начала эмбриогенеза фермент не
появляется;
г) сжатие яйца во время дробления
может привести к перемещению желтой
цитоплазмы в те клетки, которые в норме ее не
получают. После этого в них наблюдается
ацетилхолинэстеразная активность. Из этих
экспериментов вытекает предположение,
что гены, кодирующие ацетилхолинэстера-
зу, транскрибируются только в клетках,
наследующих некие факторы, обычно
связанные с желтой цитоплазмой.
Хронология транскрипции и трансляции
этих генов контролируются
биологическими часами, функционирующими
независимо от клеточных делений.
Приложение 15.5.
Регуляционное и мозаичное
развитие II
Экспериментальное изучение морской
улитки Nassarius obsoletus
1. Нормальное развитие Nassarius.
Улитке свойственно спиральное дробление
с появлением заметной полярной лопасти
во время первого и второго делений
дробления, что обеспечивает наследование
D-клеткой особой полярной плазмы (см.
приложение 15.1). П осле довательность
дробления показана ниже.
472 Гла
I. Недробящееся яйцо.
II. Появление первой полярной лопасти
до первого деления дробления.
III. Образование «трилистника» во
время первого деления.
IV. Стадия двух бластомеров.
1 15
V, VI. Появление второй полярной
лопасти во время второго деления дробления.
VII. Стадия четырех бластомеров
(обратим внимание на крупную D-клетку).
VIII. Стадия восьми бластомеров вскоре
после появления первого квартета
микромеров.
IX. X. Более поздний стадии дробления.
Восьмиклеточный эмбрион заметно •
асимметричен, причем потомки D-клетки
подчиняются собственному ритму деления,
отличному от ритма деления клеток других
линий.
2. Особая роль D-клеток в развитии
Nassarius
Изучение клеточных линий
свидетельствует о развитии специфических
личиночных структур из производных отдельных
клеток, которые можно идентифицировать
и обозначить во время раннего дробления.
С использованием обозначений из
приложения 15.1 ниже на схеме клеточных
линий показано экспериментально
установленное происхождение личиночных
структур из 29 бластомеров.
Обратим внимание, что клетки 2d и 4d
играют особенно важную роль в
формировании велигера. Бластомер 4d называется
мезентобластом.
Организация личинки-велигера
Развитие
473
ЗА Кишка
За
2а1
2а2 __ Стомодеум
1а1 ——1а > Левый глаз
~~-----1а12 )
1а2 п
Парус
OD Кишка
ЗЬ
2Ь1
2b ^^-1Ь11 1 Пространство
1Ь <!Г 1h« > между глазами
1Ь2 J Парус
ЗС Кишка
Зс ^- Правая сторона ноги
2с1 L Область мантии
2с2 1с" 1
,1с1^^Г" 2 I Правый глаз
1C2^^^D^ к
Зр ' —== Кишка
, . " 4d Кишечник, сердце
, Левая сторона ноги
2d1 \ п
2 > Раковинная железа и
2d 1rin J соматическая
. .1^---" „ пластинка
1d2
Схема клеточных линий (Р — полярные тельца)
Бластула Гаструляция Трохофора
Перемещение клеток 2d и 4d в специфическое для них положение во время гаструляции
474
Глава 15
3. Организующая роль полярной
плазмы. Материал полярной лопасти во время
раннего дробления можно удалить
микрохирургическим путем. Результаты этого
впечатляющие.
Образующаяся безлопастная личинка
утрачивает всю структурную организацию,
сохраняя некоторые признаки клеточной
дифференцировки. Ниже изображены два
типичных примера таких личинок.
В классической серии экспериментов А.
Клемент изучал результаты
последовательности изъятия D-клетки во время
эмбриогенеза. Результаты сведены в приводимую
ниже таблицу.
Обобщение результатов последовательного удаления клеток D-линии у Nassarius (по А. С.
Clement, 1962)
Удаленная клетка Эмбрион Обнаруженные дефекты
I. D (см. приложение 15.1)АВС Как и у безлопастного эмбриона, утрачены:
кишка, сердце, раковина, нога, статоцисты, глаза
II. ID ABC 4- Id — »—
III. 2D ABC + Id + 2d —»—
IV. 3D ABC + Id + 2d + 3d Раковина развита в различной степени, утрачены
кишка и сердце
V. 4D ABC + Id + 2d + 3d + 4d Отсутствуют
4. Интерпретация: доказательство
индукционных взаимодействий. Исходя из
схемы клеточных линий и таблицы, можно
отметить следующее: глаза образуются из
производных клеток 1а и 1с, а раковинная
железа — из производных клетки 2d. Клетки
1а и 1с есть у всех экспериментальных
эмбрионов, но глаза формируются только
в опытах IV и V. Почему? Ответ: «Глаза
формируются только в присутствии клеток
D-линии, по крайней мере до образования
бластомера 3d».
Нормальная раковинная железа
развивается только в том случае, если клетка 2d
присутствует по крайней мере до
образования бластомера 3d.
Доказательство мозаичного развития:
глаза формируются из клеток 1а и 1с.
Разрушение их препятствует развитию глаз.
Вывод: только производные 1а и 1с
компетентны формировать глаза, но для
детерминации этого направления развития им
требуется индуктивное влияние клеток D-
линии.
Сердце обычно развивается из клетки
4d: только она компетентна обеспечить его
дифференцировку.
Развитие
475
Приложение 15.6.
Экспериментальный анализ
развития морского ежа
1. Клеточные слои. Дробление яйца
морского ежа описано в приложении 15.2.
Основные слои 64-клеточного зародыша
следующие:
Anl — 16 клеток,
Ап2—16 клеток,
Veg 1 — 8 клеток,
Veg 2 — 8 клеток,
микромеры.
Бластула Гаструла
Развивающийся плутеус
(призматическая стадия)
Взаимное соответствие клеточных слоев
бластулы, гаструлы и раннего плутеуса
2. Развитие изолированной половины
эмбриона. Изолированная анимальная
половина эмбриона превращается в
неразвивающуюся далее бластулу с аномально
крупным апикальным султаном ресничек.
Изолированная вегетативная половина дает
начало более или менее нормальному
эмбриону с чрезмерно развитым
кишечником. Такой эмбрион называется вегетализи-
рованным.
Эти наблюдения можно было бы
объяснить самодифференцировкои первичных
клеточных слоев («мозаичная» теория
развития), но дальнейшие эксперименты
показывают, что это не так.
Неразвивающаяся Бластула
(анимализированный эмврион)
Вегетализированный
эмбрион
3. Доказательство градиентной теории
развития.
Если изолированную анимальную
половину эмбриона объединить с четырьмя
микромерами, она развивается в нормальный
плутеус. На рисунке показано:
I) разделение эмбриона;
II) объединение анимальной полусферы
с четырьмя микромерами;
III) гаструляция;
IV) нормальный плутеус.
IV III
476
Глава 15
В эксперименте, описанном в п. 3, о
микромерах говорят, что они «индуцируют»
архентеронформирующую способность
клеток слоя Ап-2. Клетки Ап-2 становятся
компетентными дифференцироваться в ар-
хентерон, хотя в норме они таковыми не
являются.
4. Много экспериментов было
проведено по развитию изолированных слоев
эмбриона и сочетаний различных слоев.
Результаты одного из них описаны ниже.
Нормальные эмбрионы получаются при
«равновесии» анимальнои и вегетативной
тенденций развития:
Anl +- 4 микромера = нормальный плу-
теус;
An 2 + 2 микромера = нормальный плу-
теус;
Veg 1+1 микромер = вегетализирован-
ный эмбрион.
+4
микромера
+2,
микромера
+1
микромер
+1
■микромер
Нормальный
Нормалы
Славо
'вегета-
лизированный
]Вегета-
'лизированный
Развитие изолированных Сочетание с
клеточных слоев микромерами
Градиент от анимального полюса
5. Описанные выше результаты можно
интерпретировать как наличие двухгра-
диентной системы. Предполагается, что
в нормальном эмбрионе каждый клеточный
слой занимает определенное положение
относительно двух градиентов.
Градиентная теория развития.
Нормальное развитие требует
"равновесия" между двумя
градиентами
Градиент от вегетативного полюса
Развитие
477
6. Демонстрация системы градиентов.
Некоторые вещества вызывают развитие
аномальных анимализированных или веге-
тализированных эмбрионов, которые
похожи на развивающихся из вегетативной или
анимальной полусфер.
Ионы Li + обладают сильным вегетали-
зирующим действием, а ионы Zn2+ и CN~ —
анимализирующим.
Изолированная
анимальная
половина
Анимализированный
Нормальный
При этом ионы Li + могут вызвать
нормальную гаструляцию изолированной
анимальной полусферы.
7. Демонстрация градиентов метаболи-
О мин Вегетативный
полюс, 30 мин
Анимальный полюс
60 мин
90 мин 120 мин
Ход восстановления красителя в нормальных
эмбрионах. Можно видеть, что зоны реакции
распространяются отанимального и
вегетативного полюсов
ческой активности. Прямое доказательство
этих градиентов у полюсов ранней гастру-
лы получено с помощью прижизненного
окрашивания янусом зеленым. Этот
краситель становится розовым, затем
обесцвечивается.
Согласованность доказательств: ионы
Li+ вызывают нормальное развитие
анимальной полусферы и подавляют подкисле-
ние януса зеленого на анимальном полюсе.
с LI-ионом
8. Градиентная система, по-видимому,
имеет физико-химическую основу, но ее
молекулярные механизмы еще не выяснены.
Приложение 15.7.
Метаморфоз и выбор
субстрата у морских личинок
1. Многие бентосные организмы
обитают на видоспецифичных субстратах.
Иногда несколько близкородственных
видов встречаются симпатрично, но на
различных субстратах. Наблюдаемое
распределение взрослых особей связано с
распознающей способностью личинок. Для ряда
мелких, живущих в трубках полихет рода
Spirorbis это продемонстрировано
экспериментально. На рисунке показаны:
I) Spirorbis borealis на Fucus serratus;
II) S. tridentatus на голой скале;
III) S. corallinae на Corallina officinalis.
Глава 15
Эксперименты по выбору субстрата для
поселения личинками различных видов полихет
рода Spirorbis (no De Silva, 1962-)
Вид
Spirorbis borealis
S. tridentatus
S. corallinae
Субстрат
Fucus serratus
Corallina officinalis
Fucus serratus
Камень
F. serratus
Камень
С. officinalis
Камень
С. officinalis
F. serratus
Общее число
личинок
1297
18
457
295
0
52
0
55
63
2
Когда в эксперименте личинкам
предлагали «на выбор» два разных субстрата, у
видов обнаружились четкие различия в
предпочтениях (см. таблицу ниже).
2. Циприсовидная личинка морского
желудя (см. ниже) очень разборчива в
отношении места поселения.
Мускульная сумка
Масляные
гловулы
Рот
Цементная железа
Прикрепительный Антеннула
и сенсорный диски
Перед тем как окончательно выбрать
место для прикрепления, она передвигается
по субстрату, демонстрируя
исследовательское поведение, причем иногда уплывет,
чтобы выбирать в другом месте.
3. Многие факторы могут увеличить
вероятность поселения личинки морского
желудя на специфическом субстрате.
Так, поселение и метаморфоз
стимулируют:
1) шероховатая поверхность;
2) ямчатая или бороздчатая
поверхность;
3) остатки панцирей морских желудей;
4) присутствие поселившихся ниприсо-
видных личинок. Наиболее важны среди
этих факторов те, что связаны с
присутствием других морских желудей.
Непривлекательная поверхность может
стать привлекательной, если ее пропитать
экстрактом из тканей морского желудя. Ат-
трактантом служит белок, распознаваемый
циприсовидной личинкой уже на уровне
мономолекулярного слоя.
Развитие
479
Приложение 15.8.
Серийная трансплантация
имагинальных дисков
и открытие трансдетерминации
1. С помощью метода серийной
трансплантации можно многократно
тестировать характер детерминации фрагмента до-
норного имагинального диска.
о Тест 3
| и т.д.
Метод серийной трансплантации
Если характер детерминации донорного
диска сохраняется, каждый тестируемый
имплантат будет проявлять его свойства.
Это обычно и наблюдается.
2. Однако в случае многократных
тестирований имплантантов иногда
наблюдаются спонтанные изменения характера
детерминации. При этом возможны не все
переходы. На рисунке проиллюстрированы
некоторые из них, встречающиеся у
дрозофилы. Стрелки показывают направление
спонтанных изменений характера
детерминации.
Например, наблюдалось развитие из
дисков конечностей крыльев, антенн и
ротовых придатков, но из них никогда не
образуются глаза, жужжальца или гениталии.
Следует отметить, что дальнейшая
судьба спонтанно изменяется сразу у всех
клеток диска, однако они должны сохранять
локальную специфику, поскольку из них
формируются регионарно организованные
структуры. Такое спонтанное изменение
называется трансдетерминацией.
Приложение 15.9.
Генетический контроль
и регионарная организация:
гомеозисные гены
1. В культурах Drosophila иногда
встречаются аномальные или уродливые мухи,
у которых из имагинального диска
развиваются структуры несоответствующего или
неправильного типа. Например, из
имагинального диска антенны может
сформироваться нога.
480
Глава 15
Генетический анализ показывает, что
причиной таких аномалий является
мутация по одному генному локусу. Иначе
говоря, существуют гены, каким-то образом
поддерживающие или упорядочивающие
характер детерминации отдельных имаги-
нальных дисков. Эти контролирующие
гены названы гомеозисными.
Название
мутанта
Antennapedia
Ophthalmoptera
Hexaptera
Символ
Abtp
Opt.
Hx
Генный Эффект
локус
3^8
2—68
2
Приводит к
развитию ноги
вместо антенны (I)
Приводит к
развитию крыла из
глазного диска
(П)
Приводит к
развитию вместо
дорсального про-
торакса крыла,
иногда ноги (III)
Эти мутации доминантные, известны
и другие—рецессивные. В какой-то степени
эффекты этих мутаций сходны со
спонтанными изменениями при
трансдетерминации.
2. Гомеозисные гены, по-видимому,
участвуют в процессах, контролирующих
регионарную специфику. Это особенно
наглядно проявляется в случае генов
комплекса bithorax.
Комплекс bithorax—это ряд тесно
сцепленных генов третьей хромосомы
дрозофилы, влияющих не некоторые особенности
регионарной организации торакса или
передней части абдомена. Фактически его
мутанты демонстрируют ранее не различимые
морфологические границы.
Ниже на рисунке представлено
схематическое изображение мухи, рассеченной на
основные структурные элементы.
Гомеозисные мутанты по комплексу bithorax
свидетельствуют о том, что существует
регионарная граница между передней и задней
частями мезоторакса (второго
торакального сегмента), метаторакса (третьего
торакального сегмента) и первого
абдоминального сегмента.
Дорсальная сторона
Вентральная сторона
Проторакс Я Передний/задний
♦ метаторакс
Передний/задний
мезоторакс
Схема нормальной конфигурации торакальных
структур дрозофилы
Мутант bithorax (bx)—это рецессивный
ген, приводящий к тому, что на переднем
метатораксе (где в норме образуются
жужжальца) развивается передняя часть крыла,
как на переднем мезотораксе (I).
Развитие
481
Ген
Символ
Что меняется
Во что превращается
I. bithorax
II. postbithorax
III. Ultrabithorax
V. bithoraxoid
bx
pbx
Ubx
bxd
Передний метаторакс
(передняя часть жужжальца)
Задний метаторакс (задняя
часть жужжальца)
Метаторакс (жужжальце)
Первый абдоминальный
сегмент
Передний мезоторакс
(передняя часть крыла)
Задний мезоторакс (задняя
часть крыла)
Мезоторакс (крыло)
Метаторакс (жужжальце)
Обратим внимание, что двойное рецес-
bx pbx
сивное состояние (IV) сход-
bx pbx
но в отношении регионарной организации
с доминантным (III).
Ubx+
Приложение 15.10.
Позиционная информация
и каудальная регенерация
у аннелид
I. Число сегментов. Тело аннелид
состоит из следующих отделов (см. также
рис. 4.51):
1) простомиум (головная лопасть);
2) специфическое число сегментов;
3) пигидиум (постсегментная лопасть).
У некоторых полихет, как и у пиявок,
число сегментов невелико; например, у Cly-
140
120
100-
80
Дефинитивное число
сегментов
Дефинитив-,
ное число
сегментов •„
•/• °-
Nephtys
Longosetosa
1 2 3 4 5 6 7 8
Длина тела, см
Взаимосвязь между числом сегментов и
длиной тела у двух близкородственных видов
Nephtys
menella torquata—22, у Ophryotrocha pueri-
lis—25. Иногда сегментов значительно
больше, но тем не менее их число может
служить видовым признаком.
У молодых животных скорость
образования новых сегментов высокая, однако
у более старых особей рост в большей
степени идет за счет увеличения размеров
существующих сегментов. При ампутации
«хвостовых» сегментов восстанавливается
высокая скорость их формирования,
характерная для молодых червей. После
ампутации и заживления раны образуется новая
зона нарастания сегментов. У Nereis его
скорость прямо пропорциональна числу
удаленных сегментов.
j5 10 20 30
Число ампутированных сегментов
2. Специфика сегментов. Каждый
сегмент аннелиды ведет себя как часть
интегрированного целого, имеет особое
строение и специфику. Это наиболее ясно
выражено у живущего в трубке червя Chaetopte-
rus variopedatus, изображенного на рис. 9.5.
Однако один-единственный сегмент этой
внешне сложно организованной полихеты
способен регенерировать части тела
спереди и сзади от него, т. е. восстанавливать
целого червя!
31—1314
482
Глава 15
Стадия регенерации целого червя из
изолированного веероподобного сегмента Chaetopte-
rus variopedatus
Во время регенерации фрагментов червя
Clymenella torquata, во взрослом состоянии
состоящего из 22 сегментов, также можно
наблюдать регионарную специфику
каждого сегмента.
/4~fct~~~~13 сегментов _ _ - 15
II
£ШЗ~1е] "13^ментов|—^-^8^1107
Ш
STE£KEU I 9^-—13 сегментов"—" ^21 [jj
IV
На рисунке изображены:
I) интактный червь;
II—IV) компенсационная регенерация
13-сегментных фрагментов из разных
участков:
II) сегменты 3—15;
III) сегменты 6—18;
IV) сегменты 9—21.
Обратим внимание, что в каждом случае
регенерирующие сегменты сохраняют свое
изначальное положение в порядковой
иерархии.
3. Морфаллаксис. У некоторых полихет
утрата головных сегментов вызывает
морфологическую перестройку оставшихся.
Это явление иногда называют морфаллак-
сисом. Сегменты как бы изменяют свое
положение в иерархической организации тела.
Это наблюдается, например, у червей
рода Sabella. По-видимому, для них
характерно восстановление венца щупалец, время
от времени обкусываемого рыбами.
Тело у Sabella состоит из простомиума
со сложным венцом щупалец, перистомиу-
ма, фиксированного числа торакальных
сегментов с характерным расположением па-
раподий и многих идентичных
абдоминальных сегментов (I). Регенерироваться может
не более трех передних сегментов. Если по-
лихета теряет их больше, задние сегменты
утрачивают щетинки и превращаются в
торакальные (II IV).
На рисунке изображены:
I) интактная Sabella;
II) утрата простомиума, перистомиума
и всех торакальных сегментов;
III) ранние стадии регенерации и мор-
фалл аксиса;
IV) регенерация венца щупалец,
перистомиума и торакальных сегментов, в
которые превращается соответствующее
число абдоминальных.
Такой путь регенерации ловчих
щупалец—самый быстрый. Очевидно также, что
каждый сегмент имеет характерное
строение благодаря определенному положению
в переднезаднем градиенте тела
Развитие
483
4. Модель регенерации. Многие
наблюдения можно описать следующей простой
моделью:
а) позиционная ценность простомиума
равна нулю;
б) позиционная ценность пигидиума
равна 1 + видоспецифичное число
сегментов;
в) зона нарастания (бластема)
сегментов расположена у передней стороны
пигидиума;
г) скорость образования сегментов
пропорциональна разности между
позиционными ценностями пигидиума и последнего
сегмента и равна нулю, когда эта разность
составляет единицу;
д) образование сегментов продолжается
до тех пор, пока разность между
позиционными ценностями пигидиума и самого
молодого сегмента не станет равна единипе;
е) утрата каудальных сегментов
приводит к восстановлению пигидиума с
присущей ему высокой позиционной ценностью.
Эта модель проиллюстрирована на
примере полихеты Ophryotrocha. Она
применима к нормальному эмбриональному (А)
и регенерационному (Б) росту.
Эксперимент (В) наводит на мысль, что
нервный ствол несет позиционную
информацию. Отклонение его в сторону,
например в 9-м сегменте, вызывает
формирование дополнительного пигидиума. Этот
лишний «хвост» будет теперь образовывать
сегменты по описанным выше правилам.
Подобные двухвостые полихеты изредка
встречаются в природе и могут быть также
получены путем соединения фрагментов
двух особей. В любом случае скорость
образования сегментов у каждого
пигидиума подчиняется законам нормального
роста и пропорциональна разности между
позиционными ценностями бластемы и
последнего сегмента.
В
А. Нормальный рост Ophryotrocha. Ценность простомиума—0, пигидиума—25.
Последовательность сегментов во время роста устанавливается путем интеркаляции. Б. Регенерационный рост
Ophryotrocha. После удаления каудальной части между последним сегментом и восстановленным
пигидиумом существует позиционный разрыв. В. Индукция дополнительного пигидиума путем
хирургического нарушения вентрального нервного тяжа
31*
ГЛАВА I О
СИСТЕМЫ КОНТРОЛЯ
Первые главы этой части книги
посвящены отдельным функциональным
системам— питания, движения, дыхания и
т.д.,— однако наш главный тезис
заключается в том, что отбор действует не на
частные признаки, а на организм в целом. Все
гены особи в совокупности обеспечивают ей
успех или гибель.
Уже подчеркивалось, что эволюционное
преимущество имеет две составляющие:
выживание самого индивида и более
высокую вероятность оставления им потомства.
Обе они требуют усвоения информации из
внутренней и внешней среды и успешного
осуществления необходимых реакций на
нее. Уже говорилось, что на животных
постоянно действует противоречивое
давление среды. Каждый вид активности влияет
на другие и может, например, повышать
вероятность гибели особи, которая
вынуждена искать компромиссы и подстраховывать
свои действия. Поэтому для регуляции
развития от зиготы до размножающейся
взрослой формы (см. гл. 15), для получения
и обработки требуемых данных, а затем для
инициации, контроля и видоизменения
координированных реакций (если нужно
реагировать, чтобы вероятность
непосредственного выживания была максимально
возможной) организму необходимы особые
системы. Они должны действовать на всех
стадиях жизненного цикла. В данной главе
как раз и рассматриваются такие нейробио-
логические и эндокринные системы
контроля.
16.1. Введение
Изучать системы контроля у
беспозвоночных практически начали по трем
основным причинам. Во-первых, из чисто
научного любопытства, во-вторых, с целью
использования полученных данных в
экономике и медицине и, наконец, в связи с тем,
что эти системы представляют собой
удобные «модели» для исследования научения,
обратной связи и т. д.
Ряд видов («лабораторные крысы» мира
беспозвоночных), главным образом
насекомые и моллюски, широко использовались
для изучения фундаментальных проблем,
которое привело ко многим из наиболее
значительных достижений в области нейро-
биологии. Выдающуюся роль при этом
играли гигантские аксоны, или нервные
волокна, кальмара Loligo (приложение 16.1),
однако крупные нейроны с диаметром тела
до 1 мм обычны для моллюсков, и в нейро-
биологии стало почти рутинным делом
вводить до четырех электродов в одну такую
клетку, регистрируя ее импульсы,
воздействуя на нее током и т. д.
Нервные системы многих
беспозвоночных отличаются простотой, которая
позволила исчерпывающе описать их строение
и функциональные взаимоотношения
между их частями. В отношении позвоночных
это пока лишь отдаленная мечта.
Примерами могут служить вся нервная система
нематоды Caenorhabditis elegans, а также
периферические кардиальный и стоматогаст-
ральный ганглии ракообразных. Более
того, в разных особях можно
идентифицировать одни и те же индивидуальные нейроны,
что позволяет исследовать их строение,
физиологию и значение. Важна также
несвойственная позвоночным устойчивость
многих беспозвоночных к экспериментальным
воздействиям.
Изучение плодовой мушки дрозофилы
внесло исключительный вклад в понимание
генетики нейробиологии и поведения. Ха-
Системы контроля
485
Тонкая стеклянная
нить
■Капля раствора
электролита
Электроннолучевой
осциллоскоп
Кончик
микропипетки
1 мкм
а
, Электрометр
(входной сигнал
I от микроэлектрода)
-i- Заземление
Аксон кальмара
Рис. 16.1. Микроэлектрод и установка для регистрации внутриклеточных электрических
потенциалов (по Shepherd, 1983)
рактерно, что многие полученные мутанты
были прежде всего идентифицированы по
поведенческим отклонениям, что дало
материал для исследования морфологических,
биохимических и генетических основ
поведения. Для этих целей была также широко
использована нематода Caenorhabditis.
У нее шесть хромосом (у дрозофилы—
четыре) и только 3000—5000 генов.
Мутанты способны жить в питательной среде
даже при наличии крупных дефектов
(например, будучи парализованными); возможно
и их размножение, поскольку один из полов
является самооплодотворяющимся
гермафродитом. К 1984 г. было известно 228
мутаций по 14 генам, в том числе делающие
животных нечувствительными к
прикосновению.
Для изучения нервных систем сейчас
используется множество методов. Самые
передовые из них применяют
внутриклеточные электроды и осциллоскопы,
регистрирующие электрические потенциалы (рис.
16.1). Недавно разработана методика
«patch-clamp» (локальной фиксации), с
помощью которой к отверстию
микроэлектрода могут быть присоединены
мельчайшие фрагменты клеточной мембраны для
изучения функций индивидуальных
мембранных каналов. Широко используются
также световая и электронная микроскопия
в сочетании с методами введения в
отдельные нервные клетки красителей (например,
проционового желтого) или электроноплот-
ных материалов, что позволяет проследить
распространение отростков нейронов и их
контакты с другими клетками. В последние
годы наблюдается настоящий
информационный взрыв, связанный с применением
методов современной биохимии и
молекулярной биологии. Нейромедиаторы,
гормоны, рецепторы и т.д. были
идентифицированы химически, установлены кодирующие
их последовательности, к их молекулам
получены антитела.
Нейробиология особым образом
связана с поведением животных. Наблюдение за
поведением целого организма можно
сравнить с изучением «черного ящика» типа
калькулятора или компьютера. О многих
свойствах аппарата можно узнать,
наблюдая его в работе, сравнивая информацию на
его входе и выходе и т. д. Однако для
изучения его внутренней структуры, свойств
различных составных частей и механизмов их
функционирования необходимо «вскрытие
ящика», чем и занимается нейробиология.
Как будет показано, вместе с
эндокринологией она обеспечивает наиболее
перспективный подход к всесторонней оценке
поведения, физиологии и общей биологии
организмов.
486
Глава 16
16.2. Потенциалы
16.2.1. Мембраны
Согласно жидкостно-мозаичной
модели, плазматическая мембрана клетки
состоит главным образом из двойного слоя
(бислоя) фосфолипидных молекул,
гидрофильные концы которых направлены
наружу. Она обычно отделена от мембраны,
ограничивающей соседнюю клетку, элек-
тронопрозрачным межклеточным
пространством шириной около 20 нм. Бислой
образует главный барьер для диффузии
ионов и т. д. В него погружены молекулы
белков и гликопротеинов, многие из которых
пронизывают его полностью, поэтому
могут функционировать в качестве «каналов»
и «насосов» для перемещения ионов через
мембрану. Многие компоненты мембраны
свободно плавают внутри нее, другие
прочно заякорены на специфических участках
клеточной поверхности,
специализированных для восприятия стимулов, нервной
передачи и т.д.
Концентрация ионов в цитоплазме
и межклеточной жидкости зависит от
пассивного движения (диффузии) и активного
транспорта. В мембранах нервных волокон
находится «натриевый насос», отвечающий
за выведение из клетки ионов Na *" и
поступление в нее ионов К+. Это движение
происходит против электрохимического
градиента (см. ниже) и зависит от
метаболической энергии. Насос состоит из фермента,
Na+/K+-ATPa3bi, названного так из-за
способности катализировать распад АТР
и использовать высвобождающуюся
энергию.
16.2.2. Потенциалы
Наличие у плазматической мембраны
разности электрических потенциалов—
общая особенность живых клеток,
основанная на неодинаковом распределении ионов
и различной проницаемости мембраны.
Обычно наиболее важны при этом ионы
натрия и калия. Если мембрана проницаема
в основном для ионов К+,
сконцентрированных в клетке, их диффузия из нее ведет
к появлению отрицательного заряда внутри
(анионы оказываются в избытке) и к поло-
К+*-
-К+
Na+*-
. Na+
Рис. 16.2. Электрические потенциалы
клеточной мембраны, вызванные различной
проницаемостью для ионов, особенно для К+ (слева)
и Na+ (справа)
жительному заряду снаружи (рис. 16.2).
Поскольку противоположные заряды
притягиваются, а одноименные отталкиваются,
возникающий мембранный потенциал
задерживает дальнейший отток К+, пока сила,
обусловленная разностью концентраций,
уравновешивается силой, создаваемой
разностью потенциалов. Это состояние
системы называется электрохимическим
равновесием. И напротив, если ион Na+,
присутствующий главным образом вне клетки,
способен проникать в нее, то в момент
равновесия изнутри от мембраны установится
положительный заряд. Такие простые
изменения проницаемости являются основой
большинства сигнальных процессов в
нервной системе.
Потенциал, обычно свойственный
неактивной клетке, называется потенциалом
покоя. В основном он определяется
относительными концентрациями К+, поскольку
мембрана хорошо проницаема для этого
иона. Это легко продемонстрировать,
изменяя концентрацию К+ в окружающей среде.
Оказалось также возможным выдавить из
аксона кальмара цитоплазму и заменить ее
экспериментальным солевым раствором.
В результате естественная разность
концентрации ионов К+ меняет знак, что в
соответствии с ожиданиями изменяет на
противоположную полярность потенциала покоя.
Изменения мебранного потенциала
могут происходить спонтанно в связи с
работой особых каналов, через которые в клетку
проникают ионы Са+, а в сенсорных
клетках— из-за влияния стимулов окружающей
Системы контроля
487
среды. Они происходят также в местах
специфического контакта между нейронами,
называемых синапсами, где обычно
вызываются химическими веществами (нейроме-
диаторами), которые высвобождаются из
одной клетки и возбуждают или тормозят
активность другой.
16.2.3. Проведение градуальных
потенциалов
Главная функция нервных элементов —
передача информации из одной точки тела
в другую. Это обеспечивается проведением
изменений мембранного потенциала. Так
как нервное волокно обладает свойствами
электрического кабеля, вдоль него
распространяются локальные изменения
потенциала (рис. 16.3). Такое проведение
называется пассивным, или электротоническим,
а происходящие изменения потенциалов —
градуальными и декрементными
(затухающими). Их величина зависит от уровня
начального потенциала и постепенно
уменьшается по мере распространения тока по
мембране. В глазу мухи за время
проведения по аксону фоторецепторной клетки
такой потенциал уменьшается примерно
наполовину. Аналогичным был бы эффект от
броска камня в озеро. Высота
расходящихся волн зависит от размера камня и
расстояния от источника. Многие нервные
-Л0 r
-50 - \ \«Нейрон большого диаметра
m \ ^
z \ \.
-60 - \ ^^-^^
\>1елкий нейрон '
-701 1 1 1—
0 12 3
Расстояние от места деполяризации, мм
Рис. 16.3. Пассивное и декрементное
распространение деполяризации по крупным и
мелким аксонам. «Утечка» потенциала зависит
также от эффективности глиальной изоляции (по
Darnell et al., 1986)
клетки, «неспайковые нейроны», по-
видимому, функционируют только таким
образом, а все нейроны имеют аналогично
действующие участки.
16.2.4. Потенциал действия
Частичная деполяризация многих
аксонов вызывает быстрое изменение их
состояния, приводящее к исчезновению и перемене
знака разности потенциалов (приложение
16.2). Возникающий потенциал действия,
или «спайк», связан с пороговым эффектом,
т.е. наблюдается только тогда, когда
стимул достигает определенного
минимального уровня. Это ответ по типу «все или
ничего», поскольку дальнейшее увеличение
интенсивности стимуляции его не усиливает.
В организме потенциалы действия
вызываются спонтанными, рецепторными или
синаптическими потенциалами, которые
пассивно распространяются от мест
возникновения к способной активно реагировать
области мембраны, откуда вдоль нервного
волокна пойдет ток. Он деполяризует
прилежащие области мембраны, стимулируя их
активность, т.е. обеспечение дальнейшей
деполяризации и т.д. Таким образом,
потенциал действия активно
распространяется вдоль мембраны. Он не градуальный,
а соответствует принципу «все или ничего»,
проводится с постоянной величиной, а не
декрементно. Аналогией ему может
служить поджигание пороха запальным
фитилем.
Скорость проведения сильно зависит от
диаметра волокна, поэтому передача
импульсов в реакциях избегания и подобных
им у беспозвоночных обеспечивается
гигантскими волокнами. Морская аннелида
Myxicola держит по их размеру первенство:
у ее аксонов диаметром более 1 мм
скорость проведения 20 м/с. В некоторых случаях
эта скорость сильно увеличивается за счет
изоляции участков волокна глиальной
оболочкой (разд. 16.3.2). В результате
проведение происходит сальтаторно
(скачкообразно), т.е. от одного неизолированного
участка мембраны к следующему при
восстановлении каждый раз потенциала
действия. У гигантских волокон земляного
червя по две такие «горячие точки» на дорса-
488
Глава 16
льной поверхности в каждом сегменте; в
аксонах креветки—«золотых медалистах»
животного мира по скорости проведения
(200 м/с при 20° С) — эти точки
располагаются в местах разветвления аксонов. Миели-
новые оболочки, изолирующие нервное
волокно у позвоночных, у беспозвоночных не
обнаружены.
16.2.5. Спайковые
и неспайковые нейроны
Неспайковые нейроны описаны в
основном у членистоногих и обычно
соответствуют рецепторам (например,
фоторецепторам глаза насекомых) и интернейронам.
Неспайковые мотонейроны известны
только в системе иннервации мышц стенки тела
у нематоды Ascaris. Мы просто не знаем,
почему по некоторым нейронам
распространяются градуальные потенциалы, в то
время как по другим—потенциалы
действия. Это нельзя объяснить только длиной
волокон. Если они иннервируют окончания
ног у крупных беспозвоночных, потенциалы
действия им, несомненно, необходимы.
Однако в фоторецепторах усоногого рака по
волокнам длиной до 1 см
распространяются градуальные сигналы, тогда как во
многих нейронах с гораздо более короткими
волокнами, включая некоторые амакриновые
клетки (разд. 16.3.1),— потенциалы
действия, как и в крошечных ооцитах.
железистых клетках и т.д.
Во многих клетках, генерирующих
градуальные потенциалы, происходит почти
непрерывное высвобождение нейромедиа-
тора, и незначительные изменения
мембранного потенциала (для фоторецептора
мухи — всего 0,3 мВ) могут его усилить или
ослабить. Даже присутствуя в небольшом
числе, что типично для систем
беспозвоночных, такие нейроны способны обеспечить
плавный градуальный контроль, который,
по-видимому, требует гораздо большего
количества спайковых клеток с
суммированным выходом. Информация,
передаваемая с помощью потенциалов действия,
возможно, «ненамного сложнее чередования
точек и тире азбуки Морзе» (Adrian, 1932),
однако комбинирование частоты,
последовательности импульсов и
продолжительности активности обеспечивает очень тонкую
систему кодирования. Обычны
промежуточные варианты между двумя этими
крайностями: небольшой участок клеточной
мембраны может генерировать спайк,
который затем декрементно распространяется
к окончаниям нейрона. Расположенные
вдоль аксона и зависящие от мембранного
потенциала каналы для Са2+ могут
поддерживать этот потенциал, остающийся тем не
менее градуальным, сходные каналы в
мембране нервного окончания способны
усиливать поступающий градуальный сигнал.
16.3. Нейроны и их связи
16.3.1. Нейроны
Нервные клетки (нейроны)
характеризуются наличием удлиненных отростков
и способностью проводить электрические
потенциалы. Они традиционно
подразделяются на сенсорные (афферентные),
передающие информацию в центральную
нервную систему, мотонейроны (эфферентные,
или двигательные), передающие команды
из центра к эффекторам (мускулатуре,
железам и т. д.), и интернейроны, которые
связывают предыдущие два типа клеток
(локальные интернейроны часто отличают от
«релейных» клеток с более длинными
отростками). Можно добавить еще нейросекретор-
ные клетки, выделяющие в кровь гормоны.
Сейчас уже обнаружены клетки с
признаками как сенсорных, так и двигательных
нейронов, а также с другими сочетаниями
свойств.
Нейроны часто описывают, используя
термины функциональной морфологии
(рис. 16.4). Входная (или дендритная)
зона— это чувствительный рецепторный
участок сенсорных и постсинаптическая
область других нейронов. Здесь возникают
и отсюда распространяются по мембране
градуальные рецепторные или синаптиче-
ские потенциалы. У спайковых нейронов
есть зона генерации импульсов, где
достигающие порогового значения градуальные
потенциалы запускают потенциалы
действия. Аксон проводит потенциалы к
нервным окончаниям, где осуществляется синап-
тическая передача (т. е. связь с другой клет-
Системы контроля
489
Дендритная зона
Аксон
Передающие
окончания
Рецептор Контак'
Хеморецептор растяжения рецептор
насекомого ракообразного пиявки
Мотонейрон Нейросекреторная
насекомого клетка моллюска
Рис. 16.4. Нейроны с различными морфологией и функцией обладают сходной функциональной
организацией с характерной динамической полярностью. Некоторые нейроны монополярные,
т.е. от их тела отходит единственный отросток (пример — мотонейрон насекомых), другие би-или
мультиполярные
Тело клетки
Аксон
Рис. 16.5. А. Дендриты мотонейронов стома-
тогастрального ганглия ракообразного (и
многих других) одновременно получают и
передают информацию. Их выходные окончания
могут одновремено быть постсинаптическими
по отношению к другим волокнам. Однако
естественное проведение импульса по аксону
кой). Заметим, что расположение тела
нейрона не важно для этих аспектов его
физиологии.
Поскольку многие дендриты, а также
пресинаптические окончания могут как
получать, так и передавать информацию (рис.
16.5), такая концепция динамической
полярности нейронов требует уточнения. Более
того, аксоны некоторых нервных клеток,
вероятно, проводят импульсы в обоих
направлениях, а у других (амакриновых)
клеток отростка, соответствующего аксону,
нет, причем их разветвления, вероятно,
способны к независимой активности.
16.3.2. Глиальные клетки
Вторую категорию клеток,
присутствующих в нервной системе, называют ней-
роглией. Их роль механическая (укрепле-
носит односторонний характер. Б. Изополяр-
ный нейрон стрекающих. В. Нейрон
грибовидного тела полихеты. В таких нейронах
динамическая полярность, по-видимому, отсутствует.
Г. У амакриновых клеток, например, из
зрительной доли насекомого, отсутствует главный
отросток, соответствующий аксону
490
Глава 16
ние, разделение и защита нервных
элементов), электрическая (изоляция волокон
и увеличение скорости нервного
проведения) и метаболическая (контроль ионной
среды внутри нервной системы и
разрушение нейромедиаторов после их выделения
из нейронов).
16.3.3. Химические синапсы
Многие нейроны передают
информацию, секретируя химические вещества,
называемые нейромедиаторами и нейромоду-
ляторами. В отличие от секретов обычных
железистых клеток, распространяющихся
на большие расстояния, эти продукты
высвобождаются строго локализованно и
селективно в специальные места контакта
с другими клетками, называемые
синапсами.
Чаще всего химические синапсы
образуются между окончаниями нервных
волокон. Они могут включать
луковицеобразные концевые расширения или вздутия (ва-
рикозы) по всей длине волокна. У многих
синапсов настолько характерная структура
(рис. 16.6), что существование
функционального контакта обычно предполагается уже
на основе наблюдений в электронном
микроскопе, хотя подобная корреляция и
вызывает споры. Заслуживает внимания и тот
факт, что обнаруженный
электрофизиологическими методами «синаптический
контакт» между двумя клетками в
действительности может соответствовать тысячам
соединений (25000 в стоматогастральном
ганглии омара), различимых при помощи
электронной микроскопии.
Обычные нейромедиаторы, например
ацетилхолин, запасаются в синаптических
пузырьках, и принято считать, что их
выброс происходит за счет экзоцитоза, т. е.
слияния мембраны пузырька с мембраной
окончания и последующего выхода
содержимого в синаптическую щель. Слабое
выделение медиаторов, по-видимому,
происходит из окончаний даже в состоянии
покоя, но потенциал действия (или
деполяризация не более чем на 0,3 мВ в неспайковом
нейроне, например фоторецептора мухи)
вызывает одновременный экзоцитоз
множества пузырьков. Секреторные гранулы,
-**--#'
/*
Секреторная гранула
Синаптические пузырьки
Пресинаптическая
"Т-пластина"
Синаптическая шель
Постсинаптическое
уплотнение
-V*
9-4 л
_ » 4
Рис. 16.6. А. Пресинаптическое окончание
химического синапса у членистоногих. Диаметр
синаптических пузырьков, группирующихся
у пресинаптического утолщения, составляет
обычно 30—50 нм; секреторные гранулы, как
правило, находятся позади них и, возможно,
преобладают в объеме окончания. Многим
синапсам не свойственны некоторые из
указанных признаков. Например, у иглокожих ни
специализированных щелей, ни утолщений, ни
выростов не наблюдается. Б. Электронная
микрофотография синапса в церебральном ганглии
земляного червя Lumbricus. Заметно
диффузное пресинаптическое уплотнение. Стрелкой
показана синаптическая щель (* 40000,
масштаб 100 нм)
Системы контроля
491
являясь местом запасания пептидных ней-
ромедиаторов и модуляторов, также
высвобождают их путем экзоцитоза (см.
приложение 16.11). Фундаментальные механизмы
функционирования синапсов были изучены
на гигантских волокнах кальмара, пре- и
постсинаптические окончания которых
достаточно велики для введения
микроэлектродов. Льинас (Llinas) с соавторами
показал, что потенциал действия открывает
в пресинаптической мембране зависящие от
мембранного потенциала каналы для Са2+,
а выброс медиатора активируется
повышением уровня Са2+ внутри окончаний, а не
самой деполяризацией.
После высвобождения нейромедиатор
диффундирует через синаптическую щель
и связывается с белковыми рецепторными
молекулами постсинаптической мембраны.
После такой активации молекула-рецептор
может функционировать как селективный
ионный канал, причем крайне короткое
время—не более нескольких миллисекунд.
Ответ рецептора может заключаться и в
активации внутри клетки синтеза второго
посредника, например сАМР (циклический аде-
нозинмонофосфат), который обладает
более продолжительным метаболическим
воздействием на ионные каналы.
У отдельного нейрона обычно много си-
наптических входов с различными
медиаторами, вызывающими генерирование
возбуждающих (деполяризующих) и тормозных
(обычно гиперполяризующих) постсинапти-
ческих потенциалов. На нейроны действуют
также нейромодуляторы— вещества,
называемые так из-за своих менее четких
эффектов (см., например, разд. 17.10.5). Действие
медиаторов быстро прекращается
благодаря их ферментативной деградации (аце-
тилхолин, пептиды) или обратному
поглощению пресинаптическими окончаниями
(амины, аминокислоты).
Преимущество химических синапсов
заключается в их способности усиливать
сигнал. Например, нейромедиатор (возможно,
гистамин), выделяемый окончаниями фото-
рецепторных нейронов мухи, в ответ на
определенную деполяризацию вызывает
в 7—14 раз большее изменение
мембранного потенциала (гиперполяризацию в
данном случае) постсинаптических нейронов.
Другая важнейшая для функционирования
нервной системы черта химических
синапсов— высокая пластичность их действия;
действительно, есть данные, что на этом
основаны различные типы научения
(приложение 16.3).
16.3.4. Электрические синапсы
Один из наиболее спорных вопросов
в истории нейробиологии касается
механизма синаптической передачи. По иронии
судьбы, у рака как раз к тому времени,
когда вопрос наконец решался в пользу
химической гипотезы—главным образом на
основе работ по периферическим нервным
контактам у позвоночных (см. приложение
16.9), - в 1959 г. Фершпаном (Furshpan)
и Поттером (Potter) были открыты
электрические синапсы. Они позволяют клеткам
непосредственно стимулировать друг друга
без участия медиаторов и их рецепторов.
Передача зависит от щелевых контактов1,
в которых белковые молекулы образуют
пересекающие межклеточное пространство
полые цилиндры, через которые может течь
ток, что обеспечивает распространение
деполяризации непосредственно от одной
клетке к другой. Передача происходит
быстро, исключая синаптическую задержку,
составляющую для химических стимулов
около 0,4 мс, и во многих случаях может
происходить в любом направлении. Однако
иногда электрические синапсы действуют
как выпрямители, обеспечивая проведение
с низким сопротивлением только в одном
направлении.
Иногда такие синапсы способствуют
интеграции в единую функциональную единицу
нескольких клеток. Это может
синхронизировать действие всей их группы (рис. 16.7),
например в случае рецепторных клеток от-
1 Щелевые контакты не следует путать
с плотными контактами, в которых происходит
слияние внешних листков мембран. Плотвые
контакты препятствуют диффузии через
межклеточное пространство и являются структурной
основой гемато-энцефалического барьера. С
помощью последнего у насекомых, головоногих и
позвоночных регулируется химический состав
среды, окружающей нервную систему.
492
Глава 16
Рис. 16.7. Электрические синапсы придают
клеточной популяции свойства единого
функционального элемента. ХС—химический
синапс; ЭС—электрические синапсы
дельных элементов (омматидиев) сложного
глаза саранчи и нейросекреторных клеток,
синтезирующих нерестовый гормон у
моллюсков (как и в случае многих железистых
и мышечных клеток). Электрические
синапсы обычны для нейронов с гигантскими
волокнами; они обеспечивают высокую
скорость функционирования их системы,
работающей в результате как одна огромная
клетка (см. рис. 16.25).
Эмбриональные нейроны связаны
множественными щелевыми контактами,
многие из которых утрачиваются по мере
развития ионных каналов и химических
синапсов.
16.4. Организация
нервных систем
16.4.1. «Нейроидная» система
губок
У губок не может сформироваться
настоящей нервной системы, поскольку у них
вообще нет фиксированных «тканей»,
однако эти организмы способны сокращать
и отдельно оскулумы, и все тело. Таким
способом некоторые виды создают
обратный ток воды, прочищающий каналы,
а другие используют мощные, кажущиеся
координированными сокращения для
выбрасывания в окружающую среду личинок.
Многие реакции на стимуляцию, вероятно,
вызваны исключительно распространением
механических воздействий: миоциты,
сокращаясь, растягивают соседние клетки и т. д.
Однако у Rhabdocalyptus не исключено
электрическое проведение возбуждения,
поскольку электрический стимул генерирует
ответ, распространяющийся через весь
организм со скоростью 0,25 см/с и
вызывающий закрытие всех оскулумов.
Эпителиальное проведение (см. ниже) представляет
собой другой тип нейроидной функции.
16.4.2. Нервные сети
Нервная сеть—это диффузное
двумерное сплетение би- или мультиполярных
нейронов (рис. 16.8); такая организация
характерна для нервных систем книдарий.
Элементы сети расположены между
основаниями эпителиальных клеток поверх
мезоглеи. Они встречаются как в одном, так и
в обоих клеточных слоях и способны
проводить сигналы со скоростью от 10 до 100
см/с.
Характерная черта нервной сети—
распространение активности в любом
направлении из любой точки стимуляции.
Первые экспериментаторы вырезали из
стенки тела книдарий извилистые фигуры
и показывали, что через них могут распро-
шшкшпш
Эпидермис
Нейрон
Мезоглея
Гастродермис
Рис. 16.8. А. Вид с поверхности нервной сети
книдарий, состоящей главным образом из изо-
полярных и биполярных нейронов (детали
синапсов не показаны). Б. Поперечный разрез,
демонстрирующий нервную сеть в
эпидермисе. Когда такие сети находятся в обоих
эпителиальных слоях, они могут контактировать
через мезоглею
Системы контроля
493
страняться импульсы, объясняя это
симметричностью (т.е. двусторонним
проведением) синапсов. Однако были обнаружены
и обычные поляризованные контакты, и
пары взаимно противоположных
поляризованных синапсов. Возможно, в стебельке
колониальной книдарии Namonia клетки
соединены электрическими синапсами. В
некоторых сетях проведение активное, тогда
как в других—декрементное, т. е. сигнал
затухает с увеличением расстояния от точки
его возникновения (разд. 16.9.3).
Нервная сеть может быть регионарно
дифференцирована. Например, в
мезентериях актиний встречаются крупные
ориентированные по вертикали биполярные
нейроны, обеспечивающие быстрое
активное проведение. Более того, в одном и том
же эпителии иногда присутствуют две
различные нервные сети. Для сцифомедуз
(например, Aurelid) характерен ряд ганглиев,
расположенных по краю колокола. Их
сенсорная стимуляция обеспечивается
«диффузной нервной сетью», а двигательные
сигналы от них идут к плавательным
мышцам через быстро проводящую «нервную
сеть из гигантских волокон». Это первая
известная нервная сеть; ее описал в 1879 г.
Шафер (Schafer). Две сети покрывают
примерно одну и ту же область на нижней
поверхности колокола, но прямо между собой
не связаны.
Поведение книдарии, например актиний,
включает питание, плавание,
вскарабкивание на раковины комменсалов—раков-
отшельников, «хождение» на щупальцах,
движение базального диска и закапывание.
Сложность и интеграция этих действий,
если учесть крайне низкий уровень
организации нервной системы, поразительны.
У плоских червей присутствует ряд
нервных сетей с теми же функциональными
характеристиками, что и у книдарии. Две из
них, сочетающие сенсорные и двигательные
функции, иногда расположены внутри
вентрального эпидермиса и непосредственно
под ним, а третья, исключительно
двигательная,—под лежащим глубже мышечным
слоем. Описаны также хорошо различимые
нервные сети, связанные с дорсальной
поверхностью, глоткой и другими частями
пищеварительного тракта. В глубине тела они
соединяются с нервными стволами.
Нервные сети, вероятно, играют важную роль
и в других группах животных: у иглокожих,
в ноге моллюсков, в кишечнике (даже у
позвоночных).
16.4.3. Центральная
и периферическая
нервные системы
Между центральной нервной системой,
состоящей из нервных стволов и ганглиев,
и периферической нервной системой,
включающей нервы и рецепторы, существует
важное отличие. Первая является «пунктом
назначения» сенсорных сигналов, центром
синаптическои интеграции и источником
двигательного контроля, тогда как нервы
служат главным образом проводящими
путями (нервные сети сочетают обе эти роли).
Функция миниатюрных периферических
ганглиев промежуточная в том смысле, что они
всегда связаны с центральной нервной
системой и находятся под ее влиянием,
однако в значительной мере способны и к
независимой активности. Примером могут
служить ганглии мелких клешнеобразных
органов защиты (педицелларий) некоторых
иглокожих (рис. 7.18), кардиальные ганглии
ракообразных и параподиальные ганглии
полихет.
Считалось, что у беспозвоночных
клеточные тела практически всех сенсорных
нейронов расположены на периферии,
однако сейчас у многих групп они обнаружены
и в центральной нервной системе.
Любопытная черта нематод, некоторых плоских
червей и иглокожих (рис. 16.9, Г)
заключается в наличии вместо нервных волокон,
идущих от мотонейронов к мускульным
блокам, длинных аксоноподобных отростков
мышечных клеток, образующих постсинап-
тические окончания на поверхности
нервного ствола.
Эпителиальное проведение со скоростью
3—35 см/с играет важную роль у
гидромедуз и оболочников. В некоторых случаях
эпителий функционирует как единый рецеп-
торный орган с огромной поверхностью.
Так, у Sarsia механическая стимуляция
любой точки внешней поверхности колокола
возбуждает потенциал действия, который
494
Глава 16
Надпишеводный
ганглий
Подпищеводный
ганглий
Церебральный-
ганглий
Нервный
ствол
Комиссуры
_. N ^ Вентральный
ь Сегментарные^нервный
нервы ствол
Церевральный
ганглий
Плевральный
ганглий
Висцеральные
ганглии
Церебральный
ганглий
Околопищеводные
коннективы
Торакальный
ганглий
Аникальная
Гипоневральная
Эктоневральная
Д
Рис. 16.9. Разнообразие нервных систем беспозвоночных. А. Ортогональная организация парных
нервных стволов и сериально расположенных комиссур у плоских червей. Б. Сегментированный
вентральный нервный ствол пиявок не медуллярный, как у других аннелид, а четко
дифференцирован на ганглии, содержащие клеточные тела, и коннективы, образованные нервными
волокнами. Подпищеводный ганглий формируется несколькими сегментарными ганглиями. В. Для
моллюска Patella показаны основные ганглии и перекручивание висцеральных коннектив.
Организация нервной системы самых примитивных моллюсков ненамного выше, чем у плоских червей,
в то время как у некоторых головоногих моллюсков мозг и поведение такие же сложные, как и
у рыб. Г. Высококонцентрированная нервная система десятиногих раков. Д. Поперечный разрез
луча морской звезды. Нервные волокна эктоневральной системы слепо оканчиваются на
поверхности гипоневральной системы. Выделяемое вещество, вероятно, действует как «локальный
гормон» с диффузной активностью
распространяется через щелевые
межклеточные контакты по всему эпителию.
Обычно эпителиальное проведение дает входные
сигналы нервной системе (например, у
оболочника Oikopleura—через электрические
синапсы с эпидермальными сенсорными
нейронами). Оно также участвует в
двигательном контроле, передавая возбуждение,
возникающее в нервной системе, слою
мышечных волокон или ресничкам (например,
в глотке оболочников).
У гидромедуз вдоль края колокола
проходят два кольцевых нервных тяжа.
Внутренний, медуллярный, включает
равномерно рассеянные тела нейронов (главным
образом биполярных) с вытянутыми
параллельно друг другу отростками.
Медуллярные стволы у плоских червей могут быть
Системы контроля
495
расположены по ортогональному плану
(рис. 16,9, А), который считается у первично-
ротых основным. Передние участки их
иногда лишь слегка утолщены, но уже
у высших плоских червей и немертин
образуется четко дифференцированный
церебральный ганглий, или «мозг».
У большинства аннелид вдоль всей
длины тела проходит единый вентральный
нервный ствол, образованный путем попарного
объединения в каждом сегменте коннекти-
вами эктодермальных ганглионарных
рудиментов (рис. 16.9, Б). К периферии
сегментов от них отходят пары нервов. Спереди
развивается надпищеводный ганглий, а
у многих полихет хорошо развиты
сенсорные щупальца, глаза и ольфакторные
органы, отсылающие сигналы различным
частям этого «мозга» (рис. 4.59).
Членистоногим также свойственны сегментированные
нервные стволы, но во многих группах еще
сильнее проявляется тенденция
объединения сегментарных рудиментов в сложные
ганглии (рис. 16,9, Г). Обычно формируется
хорошо развитый мозг, состоящий из про-
то-, дейто- и тритоцеребрума. Нервная
система моллюсков состоит из ряда связанных
комиссурами и коннективами ганглиев, от
которых к периферии отходят нервы. У
некоторых гастропод наблюдается
интересное расположение коннектив в виде
восьмерки (рис. 16.9, В)—результат торсии. Во
многих группах ганглии имеют тенденцию
концентрироваться, образуя
околопищеводное кольцо.
У нематод нервное кольцо вокруг
пищевода соединяет несколько продольных
медуллярных нервных стволов (например,
у Ascaris—два главных и четыре более
мелких). У иглокожих, что хорошо заметно на
морских звездах, эктоневральная система
сохраняет примитивное эпидермальное
положение (рис. 16.9, Д) и совмещает
сенсорную и моторную функции. Радиальные
нервы простираются вдоль нижней
поверхности каждого луча, отходя от околоротового
нервного кольца. Для морских лилий
характерна глубже залегающая исключительно
моторная гипоневральная система, но
наиболее важна для них апикальная система на
аборальной стороне тела. У асцидий
нервные волокна идут к периферии от
единственного церебрального ганглия,
расположенного между сифонами, а у их «голова-
стикоподобной» личинки от этого ганглия
вдоль хвоста тянется полая нервная трубка,
по-видимому, гомологичная таковой у
позвоночных.
16.4.4. Дифференцировка
нейропиля
Различают по крайней мере три типа
гистологической организации нервных
стволов и ганглиев (рис. 16.10). Наиболее
обычны участки дифференцированного нейропиля.
Тела нейронов, часто окруженные глиаль-
ными клетками, оказываются
изолированными в коре, или периферической зоне.
Сложное переплетение их разветвляющихся
отростков формирует сердцевину, или ней-
ропиль, являющийся основной зоной синап-
тических контактов, хотя часть из них
локализована на телах клеток (например, у
моллюсков).
В нервных стволах нематод и
внутреннем нервном кольце медуз можно видеть
параллельное расположение нейронов. Их
тела гистологически не изолированы, а си-
наптические контакты образуются между
соседними волокнами. У нейронов нематод
обычно по одному отростку (неразветвлен-
ному у Caenorhabditis, с двумя точками
ветвления у большинства Ascaris),
возможные синаптические контакты которого
ограничены прилежащими волокнами.
Наконец, наиболее прогрессивный
островковый тип организации обнаружен
в мозге осьминога {Octopus). Тела клеток
расположены не только в коре, но и
рассеяны внутри нейропиля. Это напоминает
строение серого вещества у позвоночных.
16.4.5. Сегментарное строение
Передвижение многих удлиненных
животных происходит благодаря волнам
сокращений, распространяющимся в том или
ином направлении вдоль тела (например,
у аннелид; см. гл. 10). При этом
мускулатура представляет собой ряд блоков или
сомитов, способных к последовательному
возбуждению. Каждый сегмент снабжен
«стандартным набором» нервных структур,
496
Глава 16
Нейропиль
•i\,■ '.'■«'•i '♦* *
или коннектива
Соединительнотканная
оволочка
Кора с клеточными
телами
Ядро
Синапс
Рис. 16.10. Характер дифференцировки
неиропиля. А. Дифференцированный нейропиль.
Б. Параллельное расположение нейронов. В.
Нейропиль с островками Островковая организация. Г. Поперечный раз-
рез церебрального ганглия полихеты Nereis.
Т—клеточные тела нейронов: О—
соединительнотканная оболочка мозга; Н —
нейропиль; П — пучки нейросекреторных
аксонов, идущих к нейрогемальным зонам в
основании мозга (ср. разд. 16.11.3). * 125; масштаб
100 мкм (Golding, Whittle. 1974)
тел клеток
Кора с клеточными
телами
сериально повторяющихся вдоль всей
длины тела. У некоторых беспозвоночных
выявлены соответствующие структурные
единицы и их связи. «Принципиальная
схема» системы, контролирующей плавание
пиявки, приведена на рис. 16.11 (см. также
приложение 16.4).
Обычно эта система включает
мотонейроны с аксонами, идущими к мышцам,
локальные интернейроны для генерирования
соответствующего типа активности (разд.
16.10) и релейные интернейроны,
координирующие работу соседних сегментов (у
пиявки с помощью тормозных волокон). На рис.
16.11 не показаны сенсорные клетки, одни
из которых обеспечивают проприорецеп-
торную обратную связь с мышцами, а
другие, активируясь, инициируют плавание.
Сериальное повторение нервных
структур встречается не только у
сегментированных животных. Аналогично устроены
радиальные нервы иглокожих и осевые
стволы в щупальцах головоногих. Они
включают группы тел нейронов, зоны неиропиля
и билатеральные нервы, отходящие к
периферии. У нематоды Ascaris пять групп
мотонейронов (из одиннадцати клеток
каждая) сериально расположены вдоль тела.
Системы контроля
497
Рис. 16.11. Сегментарно повторяющийся комплекс нейронов (обозначены одинаковыми
цифрами), контролирующих плавание пиявки. В передний ганглий (слева) входят четыре интернейрона
с циклической активностью, обеспечивающие поочередную активацию возбуждающих
мотонейронов 3 и 4, индуцирующих сокращение соответственно дорсальных и вентральных мышц. Справа
показаны синаптические связи внутри и между ганглиями. Светлые утолщения — возбуждающие,
темные—тормозные химические синапсы; между клетками 33 и 28 находится также
выпрямляющий электрический синапс (по Friesen et al., 1976)
16.4.6. «Горячие линии»
Когда земляной червь, выставив из
норки передний конец тела, сталкивается с
вошедшей в поговорку «ранней пташкой»,
описанного выше межсегментного
распространения сигналов со скоростью около
3 см/с для спасения недостаточно —
требуется сквозной проводящий путь.
Такие пути часто состоят из «гигантских»
аксонов (разд. 16.2.4), образованных, в
частности у земляных червей, рядом клеток (по
одной на ганглий), некоторые из которых
продольно сливаются в многоклеточный
синцитий, а другие соединяются
электрическими синапсами. У гидромедуз пучок
связанных электрически гигантских
волокон обеспечивает очень быстрое проведение
возбуждения ко всем частям внутреннего
нервного кольца, а тем самым—
симметричные сокращения колокола,
необходимые для плавания. У других животных
с помощью таких путей осуществляется
общая стимуляция или торможение
сегментарных систем, конкретная работа которых
контролируется межсегментными связями
(рис. 16.28). Многие волокна внутри
нервных стволов проходят через несколько
сегментов и являются промежуточными по
функции между двумя описанными выше
крайностями.
16.4.7. Роль мозга
Передние ганглии, особенно у эволю-
ционно продвинутых беспозвоночных,
развиты лучше других. Такая тенденция—
часть более общего процесса цефализации,
т.е. дифференцировки головы.
Во-первых, это явление, несомненно, во
многом связано с неизбежной
концентрацией сенсорных органов на переднем конце
32—1314
498
Глава 16
тела подвижных билатерально
симметричных животных. Роль мозга в обработке,
интеграции сенсорных сигналов и
реагировании на них хорошо изучена у насекомых.
Протоцеребрум с крупными зрительными
долями получает информацию от глаз, дей-
тоцеребрум—от антенн с хорошо
развитыми хеморецепторами, а тритоцеребрум—от
передней части пищеварительного тракта.
Выдающаяся черта протоцеребрума многих
членистоногих, как и церебрального
ганглия высших полихет и даже плоских
червей,— наличие «грибовидных тел». Они
состоят из большого количества крошечных
нейронов (у насекомых диаметром менее
3 мкм), пучки аксонов которых образуют
«ножки». Эти клетки, по-видимому,
участвуют в интеграции информации,
поступающей от различных сенсорных органов,
расположенных на голове и других частях
тела.
Вторая функция передней части тела—
нервный контроль пищедобывающего
поведения. У аннелид, например, оно подавляется
при удалении над- и/или подпищеводного
ганглия. Физиология питания падальной
мухи была подробно описана Детье (De-
thier, 1976) в его книге «The Hungry Fly»
(«Голодная муха»). Распознавание пищи
при помощи хеморецепторов на тарсаль-
ных (дистальных) сегментах ног и на лабел-
луме (см. рис. 8.30, Д) вызывает
прохождение нервных импульсов к подпищеводному
ганглию и активацию его интернейронов,
а. затем мотонейронов, что индуцирует
выворачивание хоботка и всасывающие
движения глоточного насоса.
В-третьих, передние ганглии
функционируют как высшие центры главного контроля
рефлексов и спонтанной активности
«низших» уровней. У плоского червя Notoplana
во время локомоции пища попадает прямо
в рот, тогда как неподвижное животное
сначала направляет ее к переднему концу тела
для идентификации. Децеребрированная
особь не способна к последней форме
поведения и продолжает питаться, даже если
кишечник заполнен до конца. Эти эсперимен-
ты показывают, что мозг получает
различную сенсорную информацию, в данном
случае— о состоянии кишечника и движении.
Он принимает «стратегические решения»
и отдает «команды», как бы выключая или
включая рефлекторные реакции низших
центров, обладающих собственным
нервным аппаратом для организации
конкретной деятельности. Децеребрированные
особи у многих видов беспозвоночных обычно
характеризуются гиперактивностью.
В-четвертых, высшие центры
контролируют общее «возбуждение», или
«настроение», которое разносторонне влияет на всю
нервную систему животного. Например,
чувствительность к стимулам у нейронов
глаза саранчи значительно увеличивается
при поступлении возбуждающих сигналов
от мозга; одновременно стимулируются
моторные центры.
Наконец, мозг играет ключевую роль
в организации высших нервных функций
и высокоразвитых форм поведения,
например у осьминога (рис. 16.12). Работы Янга
(Young) продемонстрировали, что в
зависимости от типа сенсорной информации
в процессе научения участвуют различные
части мозга. Зрительные пути ведут от
зрительных к верхнефронтальным, а затем
к вертикальным долям, в то время как
тактильное научение связано с
нижнефронтальными, субфронтальными и вертикальными
долями. Внутри каждого из этих путей
информация переключается на ряд центров;
например, внутри вертикальных долей
различают четыре центра поступления
зрительной информации. Существуют также
волокна, идущие в обратном направлении от
вертикальных к зрительным долям.
В любой нервной системе «следы
памяти» присутствуют более чем на одном
уровне ее организации. Так, удаление
вертикальных долей существенно ослабляет
зрительную память, хотя некоторые ее следы
сохраняются в других долях. Более того,
информация, усвоенная одной стороной мозга
при одностороннем предъявлении
стимулов, через несколько часов закрепляется
другой его стороной.
Несмотря на выводы, которые можно
сделать из анализа механизма научения
в приложении 16.3, высшие нервные
функции, по-видимому, требуют огромного
количества нейронов. Например, такие
«наивысшие» центры, как вертикальные доли,
содержат 25 млн. крошечных «микронейро-
Системы контроля
499
Вертикальная
Суввертикальная
ДорсальнО'
вазальная
Медиально-
вазальная
Зрительная
комиссура
Рис. 16.12. Надпищеводная
область мозга осьминога на
продольном срезе (плоскость
сечения показана внизу) с
некоторыми из главных долей
(по Young, 1963)
Медиально-верхне -
фронтальная
Сувсрронтальная
Медиально-
нижнефронтальная
Буккальная
нов», многие из которых не имеют аксонов.
Эти доли облегчают процесс научения,
возможно формируя кодированное
отображение ситуации, что лежит в основе
замечательной способности осьминогов к
обобщению.
16.5. Спонтанная активность
16.5.1. Нервная инициация
Важнейший вклад в нейробиологию,
внесенный изучением рефлекторных дуг,
имел прискорбный побочный эффект.
Появилась тенденция считать, что нервная
система зависит от внешнего
стимулирования, как компьютер от источника тока.
Ничто не может быть дальше от истины— в
этом убеждают любые, даже самые
поверхностные наблюдения за поведением
организмов— от амебы до осьминога.
Способность нервной системы к спонтанной
активности, ее роль в инициации событий
является такой же решающей, как и ее свойство
реагировать на изменение окружающих
условий.
Часто активность с эндогенной
инициацией является ритмической, и, возможно, ее
проявление в течение коротких временных
интервалов обусловлено спонтанно
генерируемыми потенциалами действия (разд.
16.2.2). Участвующие в этом клетки
называются водителями ритма. Подобная
активность может также возникать в
результате взаимного и чередующегося
торможения двух или более нейронов. Синхронные
ритмические вспышки активности
(«мозговые волны»), охватывающие большое число
нервных клеток, хорошо известны у
позвоночных, включая и человека. У
беспозвоночных они также широко распространены
и обычно отличаются высокой частотой
(около 50 Гц), хотя у осьминога она ниже
(менее 25 Гц) и удивительно близка к
отмечаемой у позвоночных. Периоды других
ритмов значительно больше; как будет
показано, у них другая клеточная основа.
500
. Глава 16
16.5.2. Движение
Сжимание гидры в мелкий комок
происходит благодаря последовательным
спонтанным сокращениям эпидермиса. Они
вызываются серией регистрируемых на
поверхности тела электрических «импульсов
сокращения», инициируемых в
расположенной под гипЬстомом области эпидермаль-
ной нервной сети. После двукратной
обработки колхицином, избирательно
токсичным для нервных клеток, животное
становится совершенно неподвижным.
Возникнув, импульсы распространяются во всех
направлениях, вероятно, как по нервной
сети, так и по эпидермису.
Спонтанная активность ряда краевых
ганглиев сцифомедузы генерирует
ритмические сокращения колокола, необходимые
для плавания. Такую активность
проявляют одновременно все центры, но в
течение некоторого времени «задает темп»
сначала один ганглий, затем другой и т. д.
«Лидер» генерирует потенциалы действия с
интервалом, скажем, 2 с у животного
диаметром 2 см и 20 с у 20-сантиметрового.
Потенциалы распространяются по «нервной
сети гигантских волокон» нижней
поверхности колокола. Они стимулируют
плавательные мышцы и регулируют активность
других ганглиев, чтобы она не препятствовала
установившемуся ритму.
У гидромедуз (например, Polyorchis)
спонтанная активность, задающая ритм
плавания, генерируется гигантскими
нейронами внутреннего нервного кольца; по-
видимому, здесь присутствует
единственный водитель ритма. Сокращение щупалец
происходит благодаря спонтанной
активности ганглиев внешнего нервного кольца.
Добавляющиеся по мере роста новые
щупальца и ганглии ведут себя независимо до тех
пор, пока не сформируются их нервные
связи. В дальнейшем их активность
синхронизируется с активностью остальной
системы.
16.5.3. Автономные функции
Аннелида Arenicola живет в песчаной
норке (рис. 9.32), которую она вентилирует,
обеспечивая дыхание (разд. 11.4.3).
Периодические насасывающие движения с
интервалом около 40 мин прогоняют через
жабры червя воду или пузырьки воздуха. Это
поведение включает сложную
последовательность движений: сначала сдвигание к
хвостовому концу норки; затем перемещение
вперед в сочетании с волнообразными
перистальтическими сокращениями стенки
тела; и наконец - кратковременную смену
направления вентиляции на
противоположное. Эта активность не является
рефлекторным ответом на недостаток кислорода,
а генерируется находящимися в
вентральном нервном стволе многочисленными
скоординированными водителями ритма.
Точно так же спонтанные пищедобываю-
щие движения с более коротким
интервалом (около 7 мин) генерируются нервными
элементами в пищеводе.
Наиболее очевидная спонтанная
ритмическая активность животных наряду с дыха-
Перикардиальный орган
ОтЦНС
Эндокринные*-;
влияния
Сенсорная
Обратная
связь
Возбуждающее и
тормозящее
влияния из ЦНС
Рис. 16.13. Кардиальный ганглий омара
проявляет спонтанную активность и генерирует
сокращения сердца.
1—9—идентифицированные нейроны (по Shepherd, 1983)
Системы контроля
501
тельными движениями—биение сердца.
У многих форм ее первичный источник
локализован в мышечных клетках, и она
только модулируется нервной системой. Однако
у ракообразных биение сердца неврогенное
и контролируется кардиальным ганглием
(рис. 16.13). Все его клетки проявляют
спонтанную активность, хотя координируются
одной клеткой, действующей как водитель
ритма. Ганглий находится под влиянием
высших центров (через синаптические связи
с центральной нервной системой),
сенсорных нейронов сердца (по принципу обратной
связи) и эндокринной системы.
16.5.4. Биологические часы
Некоторые периодические явления дают
основание думать, что существует
механизм узнавания времени суток или
слежения за ходом времени. Он называется
биологическими часами (приложение 16.5). Такие
часы необходимы для животных,
ориентирующихся по солнцу, например для пчел
и муравьев. Медоносная пчела не только
может сообщать другим рабочим особям
направление на источник пищи и
расстояние до него, исполняя виляющий танец
перед входом в улей [как показал фон Фриш
(von Frisch) в 1950 г.], но и способна по
прошествии времени корректировать этот
задаваемый по солнцу азимут с учетом
движения светила по небосводу.
Многие животные реагируют на
изменение долготы дня, меняя, например,
характер развития или размножения с приходом
коротких осенних дней (см., например,
разд. 14.3.4). Это подразумевает
способность «измерять» длину дня и/или ночи,
являющуюся температурно-компенсатор-
ной (т. е. независимой от температуры), что
необычно для эктотермных
беспозвоночных. В противном случае в измерении
времени могли бы быть большие ошибки.
16.6. Рецепторы
16.6.1. Основная характеристика
Чувствительность к воздействиям
окружающей среды — общая характеристика
живых клеток, наблюдаемая даже в
отсутствие явной структурной дифференцировки.
Например, протист амеба обладает
фоточувствительностью, а у насекомых
восприятие продолжительности светового дня, по-
видимому, происходит в мозгу и не зависит
от глаз. Однако у большинства животных
сформировался ряд специализированных
рецепторных клеток, часто входящих в
состав многоклеточных сенсорных органов
(см. приложение 16.6). У подавляющего
большинства рецепторов чувствительная
часть клетки структурно дифференцирована
и несет реснички или микроворсинки (а в
некоторых случаях и те и другие) (см. рис.
16.16). Часто строение таких ресничек
видоизменено, хотя это, по-видимому, слабо
связано с их сенсорной модальностью.
Значение их, вероятно, заключается в
увеличении площади сенсорной клетки. В
некоторых случаях (например, в глазу рака) на
электронно-микроскопическом уровне
можно видеть внутримембранные частицы,
предположительно состоящие из
зрительного пигмента, которые концентрируются
в мембранах микроворсинок, являющихся,
по-видимому, чувствительным участком.
У других видов (например, у дрозофилы)
чувствительность скорее всего более
диффузная, так как распределение частиц внутри
микроворсинок и в окружающей их
плазматической мембране сходно.
16.6.2. Классификация
рецепторов
Аристотель различал так называемые
«пять чувств», которые затем были
дополнены другими, иногда довольно
экзотическими (например, чувствительностью к
магнитным полям). Типы чувствительности
называются сенсорными модальностями. Их
современная классификация основана на
физических свойствах соответствующих
стимулов (например, свет, механические,
химические воздействия и т.д.). Кроме
того, принято различать экстерорецепторы,
чувствительные к внешним раздражениям,
интерорецепторы, реагирующие на
внутренние факторы, и проприорецепторы,
сигнализирующие о движении или положении
мышц, суставов и т. д. Выделяют также
физические рецепторы, отвечающие на измен-
502
Глава 16
Статолит
Сувумвреллярное
нервное кольцо
Сенсорная
ресничка
Сенсорный
нейрон
Рис. 16.14. Статоцист книдарии (по Singla,
1975) (см. также рис. 3.19 и 3.25)
чивость внешней среды, и тонические
рецепторы, активность которых зависит от
абсолютного уровня стимуляции. Многие
рецепторы являются сочетанием двух этих
типов.
Изучая какую-либо сенсорную
модальность, можно получить информацию о
другой. Так, статоцисты, чувствительные к
силе тяжести, являются
специализированными механорецепторами. Они широко
распространены у беспозвоночных, например
обнаружены у осьминога, а у медуз
расположены по краю колокола. Этот орган
состоит из пузырька, содержащего одно или
несколько твердых образований, статоли-
тов (рис. 16.14). Когда статолит
перемещается под действием силы тяжести, он
касается сенсорных ресничек или изгибает их,
активируя тем самым рецепторные клетки.
У оболочника Oikopleura статолит заменен
меланиновой каплей, которая, будучи легче
воды, всплывает, указывая таким образом
направление силы тяжести.
Роль статоцистов была
продемонстрирована Крайдлем (Kreidl) в 1893 г. на
примере креветки. Эти животные во время
линьки заменяют песчинки, образующие их
статолиты, на новые. Крайдль подбросил
им вместо песчинок железные шарики, а
затем с помощью магнита заставлял креветок
плавать вверх ногами, на боку и т. д.,
поскольку магнитное поле имитировало
гравитационное. Хотя статоцисты чувствительны
к ускорению, у некоторых беспозвоночных
(например, у осьминога) имеется также
система каналов, аналогичных полукружным
каналам позвоночных, в которых
рецепторы активируются движением жидкости.
16.6.3. Специалисты
и генералисты
Рецепторы данной сенсорной
модальности обычно обладают повышенной
чувствительностью к определенному виду (или
диапазону) поступающей энергии, что
выражается в более низком пороге
раздражения и более интенсивном ответе на
стимуляцию. Однако одни из них являются
специалистами, а другие генералистами. У оль-
факторного специалиста спектр
воспринимаемых запахов крайне узкий; иногда
четкую реакцию вызывает лишь одно
соединение феромон (разд. 16.10.6), секретируе-
мый как химический сигнал другой особью
того же вида, или только запахи, связанные
с пищей. Ольфакторные генералисты
реагируют на широкий диапазон стимулов одной
модальности, но, поскольку у каждого из
них свой собственный характер
чувствительности, животное способно распознавать
вещества за счет специфического сочетания
активированных рецепторов.
16.6.4. Кодирование
интенсивности
Информация, поступающая от
рецепторов, обычно означает не только «да» или
«нет», но также и «сколько». Это
закодировано в частоте импульсов, генерируемых
в нервных волокнах, которая
пропорциональна логарифму интенсивности стимула
или какой-либо другой аналогичной
функции от него. Диапазон интенсивности
стимула, к которому чувствителен организм,
расширяется путем контроля
чувствительности рецептора центробежными
волокнами (проводящими импульсы к периферии),
иннервирующими, например, рецепторы
растяжения у ракообразных (см. рис. 16.24).
Это свойство особенно хорошо развито
у беспозвоночных. Наконец, различные
клетки могут работать в разных частях
широкого диапазона. В классической работе
Системы контроля
503
Николлса (Nicholls) с соавторами показано,
что механорецепторы в стенке тела пиявки,
клеточные тела которых обнаружены в
каждом ганглии вентрального нервного ствола,
относятся к нескольким типам. Клетки
«прикосновения» (Т) чувствительны даже
к току воды. Рецепторы «давления» (Р)
начинают реагировать, когда сила,
прикладываемая щупом, достигает 7 г. Наконец,
клетки «ноцицептора» (N) активируются
только в ответ на сильные повреждающие
стимулы.
16.6.5. Сенсорные клетки
и сенсорные нейроны
Многие сенсорные органы позвоночных
состоят из сенсорных клеток без волокон.
Через химические синапсы они передают
информацию окончаниям сенсорных
нейронов, аксоны которых идут в центральную
нервную систему. Несмотря на свою
промежуточную функцию, такие нейроны (по
недоразумению) раньше назывались
первичными, а периферические клетки—
вторичными. У большинства
беспозвоночных сам рецептор является нейроном.
Исключение составляют рецептор Лангерган-
са у оболочника Oikopleura и
фоторецепторы у книдарии Татоуа (рис. 16.15).
А Б В Г
Рис. 16.15. Сенсорные нейроны хеморецеп-
торных нухальных органов полихет (.Л) Nephtys
и (Б) Nereis. Сенсорные клетки Oikopleura (В)
и Tamoya (Г) образуют с сенсорными
нейронами электрический и химический синапсы
соответственно
16.7. Зрение
Зрение—это сенсорная модальность,
дающая максимум информации о мире.
Это справедливо и для многих наиболее
сложно устроенных беспозвоночных, о чем
свидетельствует строение их глаз.
16.7.1. Зрительные пигменты
Чувствительность к свету, т.е. узкому
участку спектра электромагнитного
излучения, обусловлена зрительным пигментом —
химическим веществом, поглощающим
лучистую и выделяющим в ответ на эту
свободную энергию. Родопсин—это
соединение, состоящее из каротиноида ретиналя
(производного витамина А) и белка опсина.
Оно распространено почти у всех групп
организмов и обнаружено, например, как у
водоросли Volvox, так и у человека.
Воздействие света превращает ретиналь из \\-цие
в 11-транс конфигурацию, а затем
диссоциация ретиналя и опсина приводит
к обесцвечиванию пигмента. Эти изменения
влияют на ионные каналы фоторецептор-
ной мембраны, стимулируя синтез вещества
типа сАМР (см. разд. 16.3.3), вероятно
cGMP (циклический гуанозинмонофосфат).
В ходе ферментативного синтеза пигмент
регенерируется.
Родопсин существует в различных
формах, определяемых типами ретиналя и
опсина. Светляки, активные в сумерках,
излучают как сигнал для других особей своего
вида желтый свет, в то время как их виды,
проявляющие активность в темноте,
светятся зеленым. Таким образом,
максимальная чувствительность ретиналевых
пигментов у различных видов точно
отрегулирована.
Цветовое зрение важно для многих
ракообразных, насекомых и паукообразных
(удивительно, что у осьминога его нет) и
основано на присутствии нескольких типов
рецепторов, пигменты которых
чувствительны к различным частям спектра.
Например, различные рецепторы пчел
воспринимают в желто-зеленой (540 нм), синей (440
нм) и ультрафиолетовой (340 нм) областях.
Бабочки чувствительны также к красному
свету. Большинство фоторецепторов мух
504
Глава 16
благодаря наличию родопсина и другого
пигмента обладают любопытной двойной
чувствительностью (в сине-зеленой и
ультрафиолетовой областях). Диапазон
спектральной чувствительности других
рецепторов зависит от экранирующих пигментов,
которые позволяют реагировать только на
определенные длины волн, «отсекая»
остальные.
Некоторые животные способны
различать плоскость поляризации света. У мух
эту функцию выполняют краевые элементы
сложного глаза. Характер поляризации
рассеянного света неба позволяет пчелам
определять положение солнца, даже если оно
скрыто облаками, и ориентироваться по
нему. Осьминог благодаря этой способности
«видит сквозь» маскирующую серебристую
чешую рыбы.
16.7.2. Ресничные
и рабдомерные глаза
Как показал Икин (Eakin), подавляющее
большинство фоторецепторов принадлежат
к этим двум типам (рис. 16.16). У одного из
них фоторецепторные органеллы
представлены ресничками или их
модифицированными мембранами (например, у
позвоночных). Набор микротрубочек в ресничке
может быть обычным или видоизмененным.
У второго типа фоторецепторные
органеллы—микроворсинки, образующие
правильные ряды, называемые рабдомами. Следует
обратить внимание, что дендриты, несущие
микроворсинки, часто оканчиваются
ресничкой или ее рудиментом, т.е. можно
предполагать, что ресничный комплекс
играет роль индуктора при развитии рабдо-
мов. Более того, у полихет семейства Serpu-
lidae на разных уровнях одной и той же
клетки обнаружены дефинитивные
ресничные и рабдомерные рецепторы. У книдарии
Polyorchis микроворсинки, образованные
клеточной и ресничной мембранами,
расположены вперемешку; некоторые из них, по-
видимому, имеют смешанную природу.
Многие фоторецепторные мембраны
обоих типов ежесуточно в массе
разрушаются и обновляются. У ночных пауков-
динопид рабдом занимает днем около 15%
объема рецепторной клетки, а через час
после захода солнца—до 90%. Этот избыток
разрушается в течение двух часов после
восхода. У плоских червей микроворсинки
в темноте дегенерируют и
восстанавливаются между пятой и десятой минутами
освещения (по-видимому, из мембранного
материала, накопленного в виде готовых
к использованию внутриклеточных
пузырьков).
Большинство ресничных элементов
является рецепторами «выключения».
Темнота приводит к открытию №+-каналов
(обычно), деполяризапии и выделению
медиаторов из окончаний. Рабдомерные
рецепторы, напротив, обычно реагируют на
«включение» (перечисленные эффекты
вызываются освещением). Эти различия легко
наблюдать у гребешка Pecten: верхний слой
рецепторов его глаза—ресничный, а
нижний —рабдомерный.
Ресничные рецепторы, например у поли-
хет-сабеллид и гребешка, по-видимому,
хорошо адаптированы к участию в «теневых
рефлексах»—защитных реакциях при
неожиданном снижении освещенности.
Однако у усоногих рачков при этом
используются рабдомерные рецепторы «включения».
Уменьшение освещенности, вызывая их
гиперполяризацию, блокирует выделение
тормозного медиатора и тем самым
активирует нейроны мозга. Однако из правила
всегда есть исключения. Например, у брю-
хоногого моллюска Onchidium
микроворсинковые рецепторы, по-видимому, реагируют
на «выключение». У сальп (оболочники)
такая реакция свойственна фоторецепторам
обоих типов. Принцип работы рецепторов
смешанного типа пока не выяснен.
Долгое время считалось, что тип
фоторецепторов имеет филогенетическое
значение: ресничный характерен для книдарии
и вторичноротых, а микроворсинковый—
для первичноротых. Однако сейчас
известны многочисленные исключения из этой
схемы. Например, у морской звезды
рецепторы микроворсинковые; в глазах гребешка
представлены оба типа: правый
личиночный глазок плоского червя Pseudoceros
состоит из нескольких микроворсинковых
рецепторов, а левый—из трех
микроворсинковых и одного ресничного.
У многих жгутиковых протистов часть
Системы контроля
505
Ресничные фоторецепторы
Равдомерные (микроворсинковые)
сроторецепторы
Кольчецы
Членистоногие
Щетинкочелюстные
Кольчецы /плоские черви
Стрекающие
Моллюски
Протисты
Ресничка
1икроворсинки
Рис. 16.16. Разнообразие ресничных и рабдомерных фоторецепторов (в основном по Eakin.
1968)
жгутика специализирована для выполнения
рецепторных функций, причем стигма
(пигментное пятно) обеспечивает дирекциона-
льную чувствительность (т. е. определение
направления на источник стимуляции).
В высшей степени удивительно наличие
у некоторых динофлагеллат оцеллоидов, т. е.
частей клетки, напоминающих простои глаз
с роговицей, хрусталиком и
кристаллическим сетчаткоподобным слоем,
подстилаемым пигментным бокалом, которые, как
предполагается, способны формировать
изображения.
506
Глава 16
16.7.3. Глазки и глаза
Уровень организации фоторецепторных
органов беспозвоночных весьма
неодинаков (рис. 16.17). Простейшие из них,
называемые глазками, дают информацию
только об интенсивности и направлении света.
При оценке эволюционного значения
различных типов структур следует соблюдать
осторожность, поскольку все данные
соответствуют ныне живущим видам. К тому же
последовательное усложнение может
наблюдаться внутри многих крупных
таксонов— выделяют до 40—65 «филетических
линий сложности» (Salvini-Plawen, Mayr,
1977). Например, среди фоторецепторных
органов книдарий встречаются и участки
пигментированного эпителия, и глаза с
хрусталиком (у Татоуа); У моллюсков это
и простые открытые глазные бокалы (у
блюдечек), и пятиклеточные глазки (у
некоторых голожаберных), и глаза головоногих
с 20 млн. рецепторов, выдерживающие
сравнение с нашими собственными.
Глаза, формирующие хорошо
сфокусированные изображения, называются
камерными. Во-первых, они характеризуются
обширным слоем рецепторов, составляющих
сетчатку (ретину), которая может быть
прямой или инвертированной в зависимости
от направления рецепторных полюсов
клеток к источнику света или от него
соответственно (рис. 16.18). Во-вторых, такие глаза
снабжены устройствами для фокусирования
света, хотя, если сетчатка толстая, а
хрусталик прилегает к ее поверхности,
изображение будет нечетким.
Хорошо развитое зрение имеет
решающее значение для хищников, например для
пауков-скакунов, которым необходимо
оценивать скорость движения жертвы (у пауков
все глаза состоят из пигментного бокала
с хрусталиком). Камерные глаза достигают
наивысшего развития у осьминога и
близких к нему форм (рис. 5.25,F).
Совершенство их выражается помимо прочего в
появлении век, подвижного зрачка,
изменяющего свою форму хрусталика, и внешних
мышц, которые при участии сигналов от
статоциста фиксируют зрение на интере-
суюшем предмете независимо от движений
тела. Головоногие моллюски—
рекордсмены по размеру глаз. У
гигантского кальмара Architeuthis их диаметр
достигает 40 см, а сетчатка, по всей вероятности,
содержит до Ю10 рецепторов (у человека —
108). Глаз наутилуса представляет собой
загадку, поскольку лишен и хрусталика, и
роговицы. Он функционирует как камера
с крошечным входным отверстием; при
этом чувствительность и разрешение низки.
16.7.4. Линзы и зеркала
У наземных видов (например, пауков) за
преломление "света отвечает главным
образом выпуклая роговица. У водных
животных эту функцию выполняют линзы
(хрусталики), дающие, вопреки ожиданиям,
лишь незначительную сферическую аберра-
Рис. 16.17. Уровни
организации глазков с пигментным
бокалом и глаз книдарий (А —
фоторецепторный эпителий
Laukartiara; В—Bougainvillia;
Г— Tamoya) и
переднежаберных моллюсков (Б—Patella;
B — Nerita; Г—Valvata). ПК-
пигментная клетка, СД—
сенсорный дендрит, СН —
сенсорный нейрон. Дендриты
несут реснички или
микроворсинки.
Системы контроля
507
Рис. 16.18. Сетчатка аннелид.
А. Прямая — глазной бокал по-
лихеты Vanadis (no Hermans,
Eakin, 1974). Б.
Инвертированная— глазные бокалы в мозгу
гголихеты Armandia с
единственным сенсорным
нейроном и одноклеточным
пигментным бокалом (по
Hermans, Cloney, 1966). В. Глазной
бокал пиявки Hirudo с
сенсорными нейронами,
содержащими вакуолеподобную «фаосо-
му», нельзя отнести к какому-
либо из этих типов. Фаосомы
могут быть образованы и
ресничными, и
микроворсинковыми фоторецепторами (по Hess,
1897)
цию. Маттиссен (Matthiessen) в 1886 г.
предположил, что у таких линз
коэффициент преломления изменяется от высоких
значений в центре до низких на периферии.
Это было подтверждено для головоногих
и ряда брюхоногих моллюсков, а также
некоторых веслоногих и полихет-альциопид
(и рыб). Очевидно, «существует только
один путь создания хорошей линзы с
использованием биологического материала»
(Land, 1984).
Зеркала важны для фокусирования света
в большинстве глаз и выполняют эту
функцию либо самостоятельно, либо в
сочетании с линзами. Отражая свет, они могут
направлять его сквозь слой рецепторов
в обратном направлении, фактически
удваивая эффективную освещенность (у
некоторых пауков); иногда на одном слое
рецепторов свет фокусируется линзой, а на
втором перефокусируется зеркалом (у
некоторых ракообразных).
16.7.5. Сложные глаза
Сложные глаза—это органы,
состоящие из ряда практически идентичных
элементов, омматидиев, расположенных в
геометрическом порядке. Их иногда
называют мозаичными глазами, поскольку
изображение формируется с участием всех
омматидиев, каждый из которых дает лишь
небольшую его часть. По-видимому, такие
глаза хорошо адаптированы к регистрации
движения, но их разрешающая способность
(за исключением мух) составляет в лучшем
случае Г, в то время как у позвоночных
и осьминога— Г. Если сетчатка
пигментированного глазного бокала вогнутая и дает
перевернутое изображение, сложные глаза
и их сетчатки всегда выпуклые, а
получаемое изображение прямое (рис. 16.19). Такие
глаза обнаружены у многих членистоногих,
за исключением пауков, диплопод и т.д.
Они присутствуют также у двустворки Area,
расположены на щупальцах полихет-
сабеллид, а у морских звезд образуют
«зрительные подушечки» на концах лучей.
Острота зрения сложных глаз зависит от
углового расхождения омматидиев.
Например, в глазу дафнии 22 омматидия,
направление каждого из которых отличается от
направления соседнего на 38°, а у краба Lepto-
grapsus их несколько тысяч с разницей
между соседними 1,5°. Разрешающая
способность в последнем случае выше, а
мозаичное изображение из мелких кусочков четче,
чем из крупных, но это коррелирует с
потерей чувствительности. В глазах некоторых
крабов наблюдаются мелкозернистые
полоски, образованные омматидиями с
малым угловым расхождением; они
аналогичны желтому пятну в глазах позвоночных.
У членистоногих каждый омматидий
представляет собой удлиненный элемент,
обычно состоящий из роговичной линзы,
кристаллического конуса, ряда рецептор-
ных (ретинулярных) клеток (у дафнии—4,
508
Глава 16
Рис. 16.19. А. Разница в организации глазного
бокала (слева) и сложного глаза (справа). Б.
Голова ктыря (двукрылые). Диаметр крупных
передних фасеток сложного глаза около 60
мкм (с любезного разрешения профессора М.
Land, Университет Суссекса)
у крабов и мух—8, у мечехвоста—от 10 до
15) и примыкающих к ним пигментных
клеток (рис. 16.20). Ретинулярные клетки
расположены как дольки в апельсине.
Упорядоченные наборы микроворсинок вдоль их
внутренних поверхностей образуют рабдо-
меры, а рабдомеры одного омматидия
составляют его рабдом.
Омматидии некоторых членистоногих
зависят от дифракции света. У эвфаузиевых
рачков и бабочек длинные, по форме
напоминающие пулю, кристаллические конусы,
согласно Экснеру (Ехпег, 1881), образуют
«линзовые цилиндры» с высоким значением
коэффициента преломления вдоль оси,
постепенно снижающимся по направлению
к периферии. Однако сейчас известно, что
в других глазах (например, у креветки
и омара) используется отражающая
оптика; при этом пирамидальные конусы
«посеребрены» снаружи слоями кристаллов.
Поверхность таких глаз образована
квадратными, а не шестиугольными фасетками.
В аппозиционных глазах поля зрения
соседних омматидиев обычно слегка
перекрываются, хотя у ракообразного Phronima
каждый омматидии срединных глаз
воспринимает свет от поля с углом 4°, тогда как
угол расхождения омматидиев составляет
только 0,5°. Возможно, наиболее
замечательные глаза у раков-богомолов. В то время
как у нас формируются два изображения
одного объекта, у этих животных—шесть,
поскольку в каждом их глазу находятся три
полосы омматидиев, ориентированных
Рис. 16.20. А. Продольный разрез омматидия
аппозиционного сложного глаза (по Wiggles-
worth, 1965). Б. Пути световых лучей в оптико-
суперпозиционных глазах при световой
(нижняя половина) и темновой (верхняя
половина) адаптации
Системы контроля
509
в одном направлении! Два последних
примера свидетельствуют о том, что никакой
анализ возможностей сложных глаз
неправомерен без учета «компьютерной»
способности мозга обрабатывать и
интерпретировать информацию, получаемую от
рецепторов.
Типы сложных глаз у членистоногих
различаются по степени и природе
функциональной изоляции омматидиев.
Аппозиционные глаза (рис. 16.20,Л),
обнаруженные, например, у крабов и пчел,
адаптированы к высокой освещенности. Пигментные
клетки в них экранируют рабдом от любого
света, не проходящего через роговичную
линзу данного омматидия.
Оптико-суперпозиционные глаза
(рис. 16.20,-6), например, у креветок,
омаров и бабочек, имеют широкую
«прозрачную зону» между кристаллическим конусом
и рабдомом, причем последний короче
первого. При сильном освещении они
функционируют как аппозиционные, обеспечивая
максимальную разрешающую способность.
Однако при слабом освещении рабдомы
получают свет, проходящий через несколько
соседних омматидиев, благодаря
изменению формы пигментных клеток или
миграции в них пигмента, если сами они остаются
неподвижными. Лучи от данного
источника, падая на разные омматидии, изменяют
направление, тем самым увеличивая
количество света, возбуждающего один рабдом,
что максимизирует чувствительность глаза
при тусклом освещении. Однако, вероятно,
в связи с рассеиванием света во время его
прохождения через мембраны
разрешающая способность глаза снижается.
В описанных выше типах глаз
невозможно разделить информацию, поступающую
от отдельных ретинулярных клеток одного
омматидия, поскольку рабдом здесь
«замкнутый», т. е. различные рабдомеры как бы
вставлены друг в друга микроворсинками.
Более того, каждая рецепторная клетка
образует синапсы на всех нейронах,
объединенных в мелкую группу («патрон»),
расположенную непосредственно под оммати-
дием. Совсем другая картина наблюдается
в нейросуперпозиционных глазах мух,
представляющих собой вершину развития
зрения у ченистоногих (приложение 16.7).
16.8. Обработка сенсорной
информации
16.8.1. Смысловое восприятие
мира
Информация о внешней и внутренней
средах должна быть не только собрана и
закодирована в изменениях мембранного
потенциала рецепторов, но и обработана, т. е.
видоизменена и преобразована так, чтобы
обеспечить адаптацию рассматриваемого
животного. Например, у саранчи простые
глазки на вершине головы дают слабо
сфокусированное изображение и
демонстрируют массовую конвергенцию—
информация от приблизительно 1000
рецепторов каждого глазка передается
сравнительно небольшим числом (25) нейронов
второго порядка. Во время полета это
обеспечивает быструю общую оценку положения
горизонта, тогда как сложные боковые
глаза, как мы увидим ниже, могут различать
детали окружающего.
Совершенство сенсорного и нервного
аппарата членистоногих проявляется в
распознавании ими признака или сочетания
признаков среди множества простых
альтернативных вариантов. Пчела научается
различать важные для нее участки, «как будто
делает их моментальные панорамные
снимки» (Collett, 1983). Некоторые
южноамериканские пчелы, опыляющие орхидеи,
ежедневно посещают в определенной
последовательности одни и те же цветы, положение
каждого из которых на постоянном
маршруте протяженностью около 20 км они
должны запомнить.
Главным аспектом сенсорной обработки
является выделение признаков —
распознавание или выбор в качестве
объекта внимания особенно важной для
животного информации. Так, у членистоногих
некоторые виды поведения стереотипны и ярко
выражены, например движение клешнями
при сексуальных демонстрациях у
манящего краба. Они играют роль «сигнальных
раздражителей», «высвобождающих»
специфические поведенческие реакции у особей
своего вида, ничего не знача для других
видов. По всей видимости, сенсорная и
нервная системы организованы так, чтобы раз-
510
Глава 16
личать, проводить и усиливать одни
сигналы, не реагируя на другие.
Интересно рассмотреть поведение
самцов некоторых журчалок. Когда в поле
зрения появляется потенциальный партнер,
самец летит ему на перехват. Чтобы
проложить правильный курс, нужно знать
расстояние до объекта, его скорость и
направление движения. Эти параметры, вероятно,
нельзя определить путем наблюдения,
поскольку быстро передвигающийся крупный
объект на большом расстоянии будет
выглядеть почти так же, как медленно
движущийся мелкий объект, находящийся рядом
с самцом. По-видимому, муха «допускает»,
что объект примерно такого же размера,
как она сама, и что он перемещается со
скоростью, близкой к ее собственной.
Расстояние, с которого он может быть замечен,
зависит от его размера, следовательно,
можно сделать вывод о расстоянии. Вместе
с направлением и скоростью движения
объекта в поле зрения оно позволяет (как в
компьютере с соответствующей программой)
проложить перехватывающий курс.
Конечно, если объект представляет собой
летящий вдалеке реактивный авиалайнер,
свадьба не состоится, однако в запасе еще
столько попыток!
16.8.2. Сенсорные системы
Информация, поступающая в
центральную нервную систему по сенсорным путям,
проходя через центры синаптического
взаимодействия, видоизменяется (рис. 16.21).
На данном уровне передачи представлен
ряд модулей — практически идентичных си-
наптических центров. Они часто связаны
локальными интернейронами и
посредством их способны влиять друг на друга.
Среди такого рода взаимодействий
наиболее изучено латеральное торможение,
впервые описанное Хартлайном (Hartline)
в 1949 г. в его классической работе по
мечехвосту (Limulus). Реакция омматидия на
данный уровень освещенности сильнее, если
он освещен один, поскольку одновременное
стимулирование нескольких соседних ом-
матидиев вызывает их взаимное
торможение. Организации сенсорных систем
свойственна также иерархия — информация
постепенно передается от низших центров
к высшим.
Зрительный путь насекомых связан
с тремя участками мозга—ламинарной,
медуллярной и лобулярной областями
зрительной доли, не считая протоцеребрума
(приложение 16.8). Каждому омматидию
соответствует модуль, или патрон, нейронов
Гривовидные тела
Релейные нейроны
Местные
От антенны нейроны
Гломерулы
Рис. 16.21. Ольфакторный
путь у таракана. Узелки
(гломерулы) являются синаптически-
ми центрами, состоящими из
триад, включающих входные
окончания, местные нейроны
и релейные нейроны,
передающие обработанную
информацию к высшим центрам (по
Boeckh et al., 1975)
Системы контроля
511
второго порядка в ламине и пучок клеток
третьего порядка в медулле. Этот принцип
ретинотопии означает, что характер
освещения глаза неоднократно отражается
в морфологически упорядоченных формах
активности на четырех различных уровнях
организации зрительной системы.
16.8.3. Конвергенция
и дивергенция
Конвергенция, свойственная глазкам
насекомых, типична для многих сенсорных
систем. С ней связана концепция сенсорного
поля—набора рецепторов, передающих
сенсорную информацию одной клетке или
одному центру нервного пути. Например,
у гигантского срединного волокна
земляного червя сенсорное поле переднее, т. е.
главная информация поступает к нему от
рецепторов, расположенных в передней части
тела, тогда как у пары боковых гигантских
волокон— главным образом задние
сенсорные поля. Поля обоих типов волокон
перекрываются в средней части тела.
Для сенсорных путей характерна также
дивергенция, поскольку информация от
одного рецептора или их группы передается
в центральную нервную систему по
множественным, или параллельным, путям,
которые могут быть использованы для
выделения и изоляции информации различного
типа. Дивергенция также характерна для
сенсорных систем, каждая из которых хотя
обычно и соответствует только одной
сенсорной модальности, имеет выход на
несколько моторных центров, влияя таким
образом на различные типы поведения.
16.8.4. Меченые линии
В отличие от эндокринных сигналов,
качественно отличающихся друг от друга
в связи с химическими особенностями
гормонов, в нервной системе передача
различных элементов информации происходит
с помощью внешне идентичных
последовательностей электрических изменений и
одного и того же химического медиатора.
Значение информации определяется
участвующими в передаче нервными элементами,
каждый из которых как бы по-своему
«меченый».
Этот принцип можно наблюдать в
случае реакции избегания у тараканов и
сверчков. Стремительная атака хищника—
жабы - создает ток воздуха, который
воспринимается сенсорными волосками на
анальных горках насекомого (см.,
например, рис. 8.31,£), удаление которых
снижает шансы на спасение. Волоски собраны
в несколько пучков, чувствительных к
ветру, дующему в разных направлениях.
Каждый пучок образует отдельный комплекс
соединений с гигантскими интернейронами,
доходящими до торакса, а различные
гигантские интернейроны косвенно
стимулируют специфические мотонейроны.
Например, интернейрон № 5 активируется только
струями воздуха от заднего квадранта этой
же стороны тела и возбуждает медленные
депрессорные мотонейроны (и мышцы)
этой стороны, что приводит к повороту от
источника ветра. Если церки в эксперименте
поменять местами, животное «глупеет»
и реагирует так, как будто поток воздуха
идет с другой стороны.
16.8.5. Выделение признаков
Одного фотона достаточно для того,
чтобы вызвать заметный скачок
мембранного потенциала фоторецептора. В
некоторых случаях единственная молекула
способна возбудить хеморецептор. Поскольку
у одного организма могут быть миллионы
рецепторов, понятно, что животное должно
отфильтровывать из поступающей
сенсорной информации значимые сигналы, считая
остальные лишними.
Как показано выше, в зрительную долю
саранчи заходят все ретинотопные
отростки. В конечном счете они образуют контакт
с нейроном, называемым гигантским лобу-
лярным детектором движения (LGMD),
имеющим (рис. 16.22) множество
веерообразно расходящихся дендритов,
позволяющих ему получать стимулы от
элементов, соответствующих любой части поля
зрения. LGMD возбуждается только в том
случае, если объект в поле зрения, во-
первых, достаточно мал, так что саранча не
реагирует, скажем, на движения раститель-
512
Глава 16
*/*. *
Рис. 16.22. Нейрон LGMD в зрительной доле саранчи, заполненный раствором хлорида кобальта.
Д — веерообразное расположение дендритов в зрительной доле; Т—тело клетки; А—аксон,
идущий в мозг (слева), х 80; масштаб 100 мкм (с любезного разрешения доктора Claire Rind,
Университет Ньюкасл-апон-Тайна)
ности. Информация о крупных объектах
частично отфильтровывается благодаря
латеральному торможению (разд. 16.8.2), а
частично— за счет активации параллельного
пути, оказывающего на LGMD тормозное
воздействие. Во-вторых, объект должен
двигаться. Пресинаптические по отношению
к LGMD элементы являются фазически-
ми—каждый из них, выпустив залп
импульсов, прекращает работу. Следовательно,
только движущийся объект, возбуждающий
целый ряд таких элементов, обеспечивает
последовательное поступление стимулов
к LGMD и его активацию. Наконец,
движение должно варьировать—неоднократная
стимуляция одних и тех же синаптических
входов LGMD вызывает нервную
депрессию и становится неэффективной.
16.8.6. Центробежный контроль
Информация передается по сенсорным
путям не в одном направлении; с любого
уровня нервные волокна могут
направляться назад, к более периферическим
центрам (т.е. центробежно), обеспечивая,
например, контроль по принципу
отрицательной обратной связи. Нейроны-детекторы
движения у саранчи находятся под
влиянием как тормозных, так и стимулирующих
центробежных волокон, идущих от мозга.
Когда саранча шевелит головой, нейроны
LGMD тормозятся, поскольку иначе любой
мелкий стационарный объект двигался бы
в поле зрения, активируя детекторную
систему. И напротив, когда общее
возбуждение организма центробежно повышается,
чувствительность этой системы к
визуальной стимуляции может возрасти.
16.9. Нервные основы
поведения
16.9.1. Независимые эффекторы
Некоторые действия являются
адаптивной реакцией клеток на самостоятельно
получаемые стимулы, т.е. клетки ведут себя
как независимые эффекторы. В эту
категорию входят некоторые железистые и
мышечные клетки; хроматофоры иглокожих,
непосредственно реагирующие на свет; кни-
доциты (стрекательные клетки книдарий),
Системы контроля
513
разряжающиеся при контакте с
чужеродным организмом (хотя на них действует
и то, насколько животное голодно).
Аналогичным образом координированное биение
ресничек обеспечивается механическими
взаимодействиями между смежными орга-
неллами,
Протисты являются независимыми
эффекторами по определению, однако
демонстрируют контрольные механизмы,
идентичные некоторым из действующих
в нервной системе. При встрече с
препятствием при движении вперед в Paramecium
возникает мембранная деполяризация, и
в клетку поступает Са24, что изменяет
направление биения ресничек на
противоположное. Стимуляция сзади увеличивает
проницаемость мембраны для К+, повышает
потенциал покоя и ускоряет биение.
16.9.2. Элементы поведения
Большинство эффекторов не относятся
к независимым. Обычно поведение
представляет собой результат спонтанной
активности, генерируемой в нервной системе,
или ответ на стимуляцию. Его элементы
принято относить либо к рефлексам, либо
к устойчивым комплексам действий.
Рефлекс— это стереотипный, относительно
простой двигательный ответ на
специфический стимул. Его сила и/или масштабы
зависят от интенсивности стимуляции.
Концепция устойчивых комплексов
действий возникла в ходе развития этологии,
сравнительного изучения видоспецифично-
го поведения, начатого Лоренцом и
другими. Такие комплексы представляют собой
стереотипные действия, часто весьма
сложные и не изменяющиеся в зависимости от
силы стимуляции. Они могут быть и
спонтанными, а если вызываются стимуляцией,
последняя действует только как пусковой
механизм. Один и тот же комплекс могут
вызывать совершенно различные стимулы;
например, рак принимает характерную позу
защиты в ответ на широкий диапазон
сигналов угрозы. Комплексы генетически
детерминированы и точное выполнение
определенной «программы» обеспечивается
организацией нервной системы. Обратные
связи, как таковые, их не контролируют.
Итак, две эти концепции различаются
главным образом ролью, которая
отводится стимулам. По отношению к
рефлексам она примерно такая же, как у человека,
играющего на пианино, а в случае
устойчивых комплексов действий ближе всего будет
аналогия с включением магнитофонной
записи.
16.9.3. Рефлекторные действия
У многих книдарий в ответ на слабые
прикосновения можно наблюдать
медленные локальные сокращения,
распространение которых зависит от интенсивности,
количества и частоты стимулов. Мы будем
называть такой тип реакции «облегченным-»
(употребляют также термины «декремент-
ный» и «инкрементный»), поскольку
одиночный стимул, даже если он не в состоянии
вызвать данный ответ, тем не менее
облегчает эту задачу следующему и т.д. (рис.
16.23). Классическое объяснение,
предложенное Пантином (Pantin), основано на
явлении пресинаптического облегчения
в нервной сети, за счет которого второй
и следующие за ним импульсы, достигая
синапса, запускают постсинаптический ответ.
Рис. 16.23. Облегченное сокращение полипов
у коралла Pontes в ответ на пять
последовательных идентичных электрических стимулов (по
Shelton, 1975)
33 I314
514
Глава 16
не возникающий при поступлении первого
импульса.
Примечательно, что облегченные
рефлексы, встречающиеся у гидроида Cordy-
lophora, опосредованы электрическими
импульсами в эпителии (разд. 16.4.3). Это
свидетельствует о возможности имитации
многих свойств химических синапсов щелевыми
контактами. Наконец, у некоторых
кораллов (рис. 16.23) такие рефлексы
обусловлены потенциалами в нервной сети со
сквозной проводимостью, т.е.
распространяющимися по всей колонии в ответ на каждый
стимул! Этот парадокс можно объяснить
постепенным снижением скорости
проведения в случает серии импульсов. Любой
следующий импульс будет неизбежно все
больше и больше отставать от предыдущего,
временной интервал между поступлением
двух импульсов в данную точку—
постепенно увеличиваться, а облегчение в
ее нервно-мышечных
синапсах—уменьшаться.
Рефлекс с участием рецептора
растяжения рака, как и коленный рефлекс человека,
является моносинаптическим, т.е. между
рецепторным и двигательным нейронами
находится только один химический синапс.
Этот рефлекс отвечает за сохранение позы,
Командное волокно
Рис. 16.24. Нейронные цепи рака, связанные
с рецептором растяжения. Если внешние
факторы препятствуют сокращению «рабочей»
и рецепторной мышц, вызванному активацией
командного волокна и мотонейрона, рецептор
активируется, что ведет к дальнейшей
стимуляции мышцы. Рецептор участвует также в моно-
синаптическом рефлексе на растяжение
(тормозная иннервация рецепторной клетки
центробежными волокнами не показана) (по
Kennedy, 1975)
Рис. 16.25. Нейронные цепи, участвующие
в реакции испуга у пиявки. Передача от
сенсорных Т-клеток через соеиннерней-
роны и ряд S-клеток к мотонейронам
осуществляется исключительно через
электрические синапсы. Только для нервно-мышечных
синапсов характерна химическая передача.
Показана одна сторона двух смежных ганглиев
поскольку, если соответствующая мышца
растягивается, активация рецептора и затем
связанного с ним мотонейрона вызывает
противодействующее мышечное
сокращение. Такая нервная цепь также
контролирует выполнение мышцами команд,
поступающих из центральной нервной системы
(рис. 16.24).
Во многих реакциях избегания или
испуга участвуют гигантские аксоны (разд.
16.2.4). У пиявки их нет, однако по ее
нервному стволу проходит путь с быстрой
сквозной проводимостью, обеспечивающий
резкое сокращение всего тела. Он состоит
из ряда «S-клеток», по одной на сегмент,
продольно направленные аксоны которых
связаны с S-клетками соседних сегментов
электрическими синапсами. Эта система
иллюстрирует важность электрических (т.е.
«быстрых») синапсов для реакции
избегания (рис. 16.25). Разные типы механорецеп-
торов стенки тела также образуют моноси-
наптические рефлекторные дуги с
мотонейронами «своего» сегмента.
Пожалуй, наиболее любопытный из всех
Системы контроля
515
Мозг
Сенсорное
волокно
Место стимуляции
Рис. 16.26. Рефлекторные дуги внутри зооидов Salpa. Каждая дуга состоит из сенсорного пути,
идущего от эпидермальной пластинки в мозг, двигательной иннервации эпидермиса и
эпителиального проведения к выходной пластинке (по Anderson, Bone, 1980)
известных рефлексов наблюдается у агрега-
ционной стадии некоторых сальп
(оболочники), которые образуют цепочки,
включающие до 20 «зооидов». Каждый зооид
связан с соседним двумя эпидермальными
пластинками, проводящими в
противоположных направлениях (рис. 16.26). Клетки
на «выходе» каждой пластинки
напоминают по ультраструктуре пресинаптические
окончания (но это эпителиальные клетки,
а не нейроны!). На «постсинаптической»
стороне, отделенной тонким слоем
внеклеточного вещества, находятся 6—12
ресничных рецепторных нейронов. Их стимуляция
вызывает прохождение импульсов по
аксонам в мозг и возбуждение там
мотонейронов, индуцирующих потенциал действия
в клетках эпидермиса, куда тянутся их
отростки. За счет эпителиального проведения
возбуждение распространяется к
«выходной» пластинке зооида и т.д.
16.9.4. Устойчивые комплексы
действий
Врожденная способность центральной
нервной системы генерировать и
координировать упорядоченные комплексы
двигательных действий продемонстрирована
у многих групп животных. «Втягивание
мантии и смыкание раковины у
двустворчатого моллюска My а,... копуляционные
движения ..., ритм полета, ходьбы ...,
дыхательные движения ..., биение сердца ...,
плавательные сокращения у медузы—показано,
что все это более или менее жестко (sic)
контролируется центром» (Bullock, 1977).
Заднежаберный моллюск Tritonia
реагирует на прикосновение хищной морской
звезды 30-секундным «припадком»
плавания. Это требует чередующихся
сокращений дорсальных и вентральных мышц с
участием либо сигналов, генерируемых
центральной нервной системой, либо
рефлекторной цепи—начальный стимул вызывает
сокращение дорсальных мышц, это по
принципу сенсорной обратной связи через
рефлекторную дугу стимулирует вентральные
мышцы и т. д. Исследования показали, что
типичная картина наблюдаемой активности
мотонейронов может быть воспроизведена
в мозгу in vitro при стимуляции
перерезанных сенсорных нервов. Это ясно
показывает, что в данном случае для
генерирования поведенческого ответа сенсорной
обратной связи, обеспечиваемой мышцами,
недостаточно.
16.9.5. Вариации на две темы
Многие элементы поведения заключают
в себе признаки как рефлексов, так и
устойчивых комплексов действий (приложение
16.9). У саранчи единственная клетка
рецептора растяжения, связанная с местами
сочленения крыльев, активируется в конце
каждого взмаха. Здесь по крайней мере
можно предположить систему,
функционирующую посредством рефлекторных дуг,
однако это не так, поскольку в
изолированных торакальных ганглиях и без сенсорной
стимуляции наблюдается активность,
характерная для полета. В то же время
импульсы, генерируемые рецепторами за
каждый цикл, оказывают тоническое
воздействие на скорость и силу взмаха крыла на
протяжении нескольких циклов.
Нейроны LGMD саранчи (разд. 16.8.5)
образуют синаптический выход на пару ней-
33»
516
Глава 16
Рис. 16.27. Нейронные цепи, контролирующие
прыжок саранчи, DCMD — нейрон-детектор
движения; С — С-нейрон; М — М-нейрон; Е.
F—мотонейроны соответственно разгибателя
и сгибателя; R,, R2—сенсорные нейроны,
обеспечивающие обратную связь при
одновременном сокращении. Жирные линии —
нервные пути, тормозящие сгибатели (по
Pearson, 1983)
ронов, называемых «контралатеральными
детекторами движения» (DCMD), отростки
которых спускаются в третий торакальный
ганглий, где каждый из них иннервирует два
нейрона — «С» и «М» (рис. 16.27). Синапсы
каждого С-нейрона на мотонейронах двух
антагонистических мышц—разгибателя
и сгибателя голени—находятся в бедрах
третьей пары ног.
Сначала С-клетка вызывает как
«взведение» ноги с подгибанием голени под тело
насекомого, так и совместное сокращение
двух названных мышц-антагонистов,
искривляющее эластичную кутикулу бедра.
Затем по принципу положительной
обратной связи кутикулярные рецепторы
напряжения действуют на мотонейроны,
усиливая сокращение мышцы-разгибателя.
Наконец, обратная связь, образуемая другой
группой рецепторов, в ответ на
одновременное сокращение стимулирует М-клетки.
Если это совпадает с продолжающимся
поступлением сигналов от DCMD и т.д., М-
клетки активируются, тормозя
мотонейроны сгибателя и тем самым индуцируют
прыжок с высвобождением запасенной
в искривленной кутикуле энергии. Ясно, что
сенсорная обратная связь, являясь
интегральной частью системы, контролирующей
прыжок, гарантирует неэффективность
внешней инициации последнего, пока его
механизм полностью не подготовлен (см.
разд. 10.6.4).
В заключение следует отметить
«непочтительность» природы к применяемым
нами категориям. Нервная система,
безусловно, обладает врожденной способностью
генерировать интегрированные двигательные
программы. Однако некоторые устойчивые
комплексы действий оказываются
«устойчивее» других. Например, одни быстрые
реакции сокращения тела у аннелид,
опосредованные гигантскими волокнами,
являются градуальными, в то же время как другие
осуществляются по принципу «все или
ничего». Исключение при их анализе эффектов
обратной связи — дурная традиция. Они
так же заложены в генетической программе,
как и организация центральной нервной
системы, являясь одним из главных средств,
с помощью которых устойчивые формы
поведения достигают поставленных природой
целей.
16.10. Организация
двигательного «выхода»
16.10.1. Аспекты организации
Системы нейронов, контролирующие
двигательный эффект (например,
сенсорные), обычно организованы иерархически.
Это хорошо заметно в цепях, управляющих
брюшными конечностями (плеоподами,
или плавательными ножками) рака (рис.
16.28). Различают нейроны трех уровней:
командные, интернейроны—генераторы
программ и мотонейроны.
16.10.2. Командные нейроны
У рака к каждому из сегментарных
ганглиев подходят как стимулирующие, так
и тормозные нейроны. Они впервые были
открыты у ракообразных Вирсмой (Wiers-
ma), который обнаружил, что активация
особых клеток запускает
координированную поведенческую программу.
Командный нейрон отвечает за принятие решения.
В то время как сенсорные пути диверги-
руют, обеспечивая стимуляцию множества
двигательных центров (разд. 16.8.3),
командные нейроны внутри двигательных
систем являются местом конвергенции и
интеграции входных сигналов от различных
сенсорных органов. Их активация или
отсутствие таковой определяют, начнется или
Системы контроля
Командный нейрон
517
|®>) Генератор программы
Мотонейрон
©УС/ 1СИМ1
нейрон
Межсегментный
релейный vr /ф\
Jtt
Рис. 16.28. Основные компоненты нейронных цепей, контролирующих движение плавательных
ножек рака (показаны клетки в двух смежных сегментарных ганглиях). Обычно наблюдается
более одного нейрона каждого типа (no Stein, 1971)
нет двигательное действие. Следовательно,
для его генерирования работа таких клеток
необходима и достаточна.
Гигантские волокна, управляющие
ударами брюшка у рака (приложение 16.9),
считались моделью командных структур.
Однако более детальное исследование
показало, что ответ на их прямую
экспериментальную стимуляцию не совсем такой, как
в природе: естественные удары обычно
более вертикальные и продолжительные.
Таким образом, гигантские волокна
определяют общий характер действия, но для его
точного выполнения необходимы
параллельные негигантские нервные пути.
Командные клетки обычно не
определяют «деталей» двигательной активности:
не устанавливают хронологии или порядка
прохождения импульсов в контролируемых
ими интер- и мотонейронах. Исключение
составляют гигантские волокна,
расположенные по краю колокола медуз, а также
известные у аннелид и других удлиненных
животных и отвечающие за практически
одновременные сокращения отдаленных друг
от друга частей тела, характерные для
реакции избегания.
Во многих двигательных системах ни
одна клетка или группа клеток, по-
видимому, не соответствует командному
элементу (см., например, рис. 16.27). Более
того, хотя маутнеровские клетки у
костистых рыб и водных амфибий являются
командными нейронами, вся эта концепция
плохо приложима к позвоночным.
16.10.3. Генерирование
программы
Генерирование программы
двигательного ответа часто осуществляется не
командными и не двигательными нейронами,
а расположенными между ними
интернейронами. Одна из наиболее обычных
двигательных программ связана с двумя
мышцами, работающими как антагонисты.
Нервная система должна гарантировать
единовременную стимуляцию только одной из
них. Одним из примеров может служить
замкнутая петля из интернейронов,
контролирующая плавание пиявки (рис. 16.11).
В каждом сегменте рака одному неспай-
ковому интернейрону свойственны
регулярные спонтанные колебания мембранного
потенциала, происходящие в фазе с биением
плавательных ножек. Деполяризация и
выход медиатора стимулируют мотонейроны
мышц, отвечающие за гребущий удар,
и тормозят мотонейроны, обеспечивающие
возвратное движение конечности. Хотя от
командных клеток зависит, будет активен
этот интернейрон или нет, за генерирование
программы действий отвечает именно он.
Это можно продемонстрировать, ускоряя
или замедляя колебания его потенциала
экспериментальным путем. Ритм оказыва-
518
Глава 16
MN2V - -«- - -d^MNV
UNlY -<-- ^(dMnV - - - 5>
_. Мышца-депрессор
i> сокращается
^_^ Мышца-леватор
■ i> расславляется
Мышца-деп рессор
расславляется
*^ Мышца-леватор
"t-^ сокращается
Рис. 16.29. Нейронная цепь, контролирующая
вентиляцию жабр у краба. Два неспайковых
интернейрона (IN,, IN2), поочередно
возбуждаясь, активируют мотонейроны мышцы-
депрессора (DMN) и мышцы-леватора (LMN)
соответственно. Прерывистые линии —
неактивные волокна; светлые утолщения —
возбуждающие синапсы, темные—тормозные
синапсы
ется перенастроенным — он не
соответствует временным характеристикам
предшествующего. Другие интернейроны
участвуют в межсегментной координации,
обеспечивающей метахрональный ритм биения
плавательных ножек.
В торакальном ганглии краба,
контролирующем вентиляцию жабр, обнаружена
сходная система: один неспайковый
интернейрон функционирует по описанной выше
схеме, а колебания потенциала второго не
совпадают с первым по фазе (рис. 16.29).
Работа обоих обеспечивает нормальные
респираторные движения.
Благодаря наличию нескольких
интернейронов, способных генерировать
различные программы, индивидуальные
мотонейроны могут быть использованы различным
образом, что делает ненужным
дублирование волокон, осуществляющих нервно-
мышечную иннервацию. Так, Берроус
(Burrows) обнаружил, что у саранчи на
единственный мотонейрон, идущий к мышце
ноги, влияют по крайней мере двенадцать
интернейронов.
16.10.4. Мотонейроны
Хотя за генерирование двигательной
программы часто ответственны
интернейроны, многие мотонейроны в результате
своих взаимодействий также включены
в этот процесс. У нематоды Ascaris вдоль
всего тела проходят пять крупных
интернейронов, обеспечивающие тоническую
стимуляцию плавания. Мотонейроны,
возбуждающие дорсальную мышцу в каждом
Рис. 16.30. Нейронная цепь, контролирующая плавание у Ascaris, обеспечивает поочередные
сокращения дорсальной (слева) и вентральной (справа) мышц. IN — интернейроны; DE, VE, DI, VI —
дорсальные и вентральные возбуждающие и тормозные мотонейроны соответственно. Светлые
утолщения — возбуждающие синапсы; темные утолщения—тормозные синапсы (по Stretton et al.,
1984)
Системы контроля
519
«сегменте» тела, одновременно
стимулируют тормозные нейроны вентральной
мышцы и через них тормозят вентральные
возбуждающие нейроны (рис. 16.30).
Активация последних оказывает аналогичное
влияние на дорсальные нейроны.
Чередующаяся активность дорсальных и
вентральных мотонейронов, зависящая по частоте
и интенсивности от степени их стимуляции
интернейронами, вызывает попеременные
сокращения соответствующих мышц.
Другие клетки образуют связи между
соседними участками тела и координируют их
активность.
То. что генерирование программы
двигательными элементами потребовало бы
дублирования последних, по-видимому,
вытекает из приведенных наблюдений,
поскольку плавание вперед и назад у аскариды
контролируют различные типы
мотонейронов. Возможно, в данном случае это
неважно, так как набор совершаемых червем
движений ограничен. Подобный вывод
справедлив в отношении многих
двигательных систем беспозвоночных.
Нервные элементы, участвующие в
реакциях избегания, в большинстве случаев не
подтверждают правила, согласно которому
один и тот же набор мотонейронов
используется в зависимости от входных сигналов
в различных целях. У кальмара гигантские
волокна участвуют в реактивном отплыва-
нии при испуге, тогда как более мелкие
управляют сокращениями мышц,
обеспечивающими дыхательные движения. У
медузы Aglantha реакция избегания выражается
в одном—трех мощных сокращениях
колокола, контролируемых восемью
гигантскими двигательными волокнами под его
нижней поверхностью и боковыми
мотонейронами меньшего диаметра. Обычные
плавательные сокращения управляются только
последними элементами. Аналогичным
образом у рака боковые гигантские волокна
возбуждают не только (через пару
центральных гигантских волокон) от пяти до
девяти быстрых «сгибающих» мотонейронов,
участвующих в пластичных реакциях
избегания, но и пару гигантских сегментарных
мотонейронов, ответственных
исключительно за удары брюшка (приложение 16.9).
16.10.5. Нервно-мышечная
иннервация
Контроль мышечной активности
мотонейронами связан с выделением химических
нейромедиаторов в нервно-мышечных
синапсах между нервными окончаниями и
мышечными волокнами (разд. 16.3.3 и
приложение 16.10). Очень важно также наличие
или отсутствие распространяющихся
потенциалов действия внутри отдельных
мышечных волокон и/или их передача через
щелевые контакты между ними. Способы
иннервации мышц и желез чрезвычайно
разнообразны. Так, сердце асцидий (оболочники)
и некоторых насекомых ее лишено, и его
сокращения миогенные, возможно, с
эндокринной модуляцией. И напротив, биение
сердца у ракообразных неврогенное и
контролируется только возбуждающими
мотонейронами. Соматическая мышца аскариды
находится под двойным (возбуждающим
и тормозным) контролем.
У саранчи разгибатель голени
контролируется четырьмя нервными волокнами.
Возбуждающие и тормозные волокна
выделяют медиаторы глутамат и гамма-
аминомасляную кислоту соответственно.
Кроме того, окончания нейрона DUM
(дорсальный непарно-срединный) секретируют
нейромодулятор октопамин (табл. 16.1).
Его собственный эффект, по-видимому,
слаб, а основная роль заключается в
придании мышце способности отвечать на
двигательное возбуждение более интенсивным
сокращением с быстрым расслаблением,
что важно для локомоции. В отсутствие ок-
топамина сокращение становится более
длительным и способствует сохранению
позы.
У двустворчатых моллюсков мышцы-
аддукторы очень выносливые — они могут
держать створки раковины сомкнутыми
в течение нескольких часов или даже дней,
обеспечивая защиту от хищников.
Сокращение происходит в результате активации
возбуждающих мотонейронов с выделением
ацетилхолина. Когда напряжение уже
развилось, мышца входит в состояние «замка»,
характеризующееся низким потреблением
кислорода и поддерживаемое независимо
от дальнейшей стимуляции. Расслабление
520 Глава 16
Таблица 16.1. Химические медиаторы беспозвоночных
Ней ро медиаторы
Ацетил холин
О
Н3С—С—О—(СН2)2—N*-(СН, )3
Глутамат (аминокислота) H3N*—СН—СНг—СН2—С—О
с=о
I
О
Октопамин (амин)
HOV^/
Гормоны
-СН—СНг—NHj*
I
ОН
Головной активатор гидры (пептид)
pGlu—Pro—Pro—Gly—Gly— Ser—I.ys—Val— He—Leu—Phe
1-метиладенин
Экдиэон (стероид)
Ювенильный гормон I
СООСН,
Феромон
Бомби коп
НННННННННННННННН
I I I I I I I I I I I I I I I I
н-с-с-с-с=с-с=с-с-с-с-с-с-с-с-с-с-он
I ! I I I I I I I I I I
ннн ннннннннн
индуцируется активацией тормозных
волокон, медиатором которых является, по-
видимому, 5-гидрокситриптамин.
У насекомых с «прямыми»
летательными мышцами импульсы в нервных волокнах
«отбивают ритм» сокращений. В
противоположность этому специализированные
«непрямые» летательные мышцы,
например у мух, сокращаются с частотой более
1000 раз в секунду, тогда как число
потенциалов действия, проходящих по нервным
и мышечным волокнам, составляет всего
5—10. Нервы обеспечивают лишь
тонический эффект, переводя мышцу в «активное
состояние». Сокращение и расслабление
являются внутренними «рефлексами»
мышечных волокон, которые индуцируются
воздействиями (растяжением и т. д.),
сопровождающими биение крыльев (см. разд.
10.6.3).
Системы контроля
521
16.11. Химическая
коммуникация
16.11.1. Спектр химической
коммуникации
Нейромедиаторы — это вещества, секре-
тируемые нейронами и обычно выделяемые
в непосредственной близости от других
клеток, на чью активность они влияют
(табл. 16.1). В результате нервное
воздействие высокоспецифично в пространстве
и времени. В противоположность этому
гормоны—это химические посредники,
вырабатываемые клетками, которые обычно
объединены в железы, и влияющие на
другие, отдаленные клетки, переносясь к ним
кровотоком или другими внутренними
жидкостями. Железы, секретирующие гормоны,
называются эндокринными, поскольку
выделяют свои продукты внутрь тела.
Секреты экзокринных желез, например слизь или
ферменты, выводятся наружу, обычно через
протоки.
Если по функции обычный нейрон
можно сравнить с телефоном, эндокринный
сигнал напоминает команду, поданную
через мегафон. Гормон часто действует сразу
по всему организму, хотя это не означает
отсутствия у него высокоспецифичной
активности. Эффект гормонального сигнала
зависит от рецепторов на клетках-мишенях.
Например, разные типы хроматофоров,
даже с одним и тем же пигментом, могут быть
чувствительными к разным гормонам.
Другие секретируемые вещества,
называемые феромонами, выделяясь в
окружающую среду, имеют биологическое значение
для других особей. У протестов, например,
они обеспечивают агрегацию амебоидных
клеток слизевиков. Крайне важна роль
феромонов у общественных насекомых,
которых можно сравнить с «подвижными
батареями экзокринных желез» (Е. О. Wilson,
1975).
16.11.2. Критерии эндокринной
активности
Существует ряд критериев,
позволяющих судить об эндокринной активности
органа. В частности, его хирургическое удале-
Рис. 16.31. По две личинки пятого и четвертого
возрастов Rhodnius в состоянии парабиоза.
Они соединены восковыми перемычками
с имаго и «подключены» к его
кровообращению. Личинки секретируют линочный и ювени-
льный гармоны, вызывая дополнительные
линьки имаго, приводящие к его ювенилизации
(по Wigglesworth, 1940)
ние должно прекращать действие
предположительно вырабатываемого им гормона,
а реимплантация данного органа или
инъекция экстрактов из него должны это действие
восстанавливать. Можно использовать
парабиоз или трансплантационный метод
(рис. 16.31). Альтернативный способ
включает сохранение клеток или органов in vitro.
Эффект внесения в культуральную среду
желез или их экстрактов можно
использовать для оценки предполагаемой
активности.
16.11.3. Нейросекреция
В 1928 г. Эрнст Шаррер (Е. Scharrer)
постулировал, что некоторые клетки,
названные нейросекреторными. проявляют
свойства как нейронов, так и клеток
эндокринных желез. Эта идея получила развитие
в исследованиях Берты Шаррер и других
авторов на беспозвоночных. Было показано,
что такие клетки у них широко
распространены. Особенно убедительные
подтверждения теории дали работы с насекомыми.
Тела нейросекреторных клеток обычно
собраны в центральной нервной системе
в ганглионарные ядра (рис. 16.32). Их
аксоны образуют вздутые окончания, тесно
связанные с циркуляторной системой. Эти
окончания могут быть объединены в офор-
522
Глава 16
f>^ .^МйУ -""V* y?i<t: v-_-:*.
At' <£*
4'CM»
Фг- .Г , ч>
* # ? «и
/=*. ,; • • ■ * й
>« -; :
Рис. 16.32. А. Нейросекреторные клетки (на врезке) в церебральном ганглии полихеты Harmot-
hoe. Цитоплазма заполнена секреторным материалом, окрашенным «по Гомори» в темный цвет
паральдегид-фуксином; ядра неокрашены; масштаб 100 мкм (Golding, Whittle, 1977). Б.
Электронная микрофотография нейросекреторной клетки земляного червя Lumbricus. Секрет состоит
из большого числа мелких гранул диаметром около 200 нм. Я — ядро с ядрышком; ШЭР—
шероховатый эндоплазматический ретикулум; * 6500, масштаб 1 мкм
мленное тело (нейрогемальный орган) или
рассеяны по нейрогемальной зоне на
поверхности ганглия или нерва (рис. 16.33).
Нейросекрет вырабатывается в телах
клеток, переносится по аксонам течением аксо-
плазмы и накапливается в окончаниях. Под
электронным микроскопом видно, что он
состоит из мельчайших секреторных гранул
диаметром обычно 100—200 нм, которые
отпочковываются аппаратом Гольджи.
Активность нейросекреторных клеток
возникает либо спонтанно, либо в
результате синаптического возбуждения. Она
вызывает прохождение потенциалов действия по
аксонам, поступление ионов Са2+ в
окончания и выделение гормона в кровяное русло
за счет экзоцитоза (приложение 16.11).
16.11.4. Размытая граница
Недавние открытия сгладили различие
между нейросекреторными клетками
Системы контроля
523
Гувной нерв-
Рис. 16.33. Церебральный
ганглий моллюска Lymnaea.
Показаны нервные тракты от двух
типов нейросекреторных
клеток к нейрогемальным зонам
поверхности комиссуры и
губным нервам. Дорсальные
тела— эндокринные органы
ненервной природы (по Joosse,
Geraerts, 1983)
и «обычными» нейронами. Нейроны,
напоминающие неиросекреторные клетки, были
описаны у книдарий и плоских червей,
лишенных сосудистых систем. У гидры
обнаружен головной активатор (табл. 16.1),
очищенный в 1981 г. Шаллер (Schaller) с
соавторами. Это вещество в концентрации
1 «гидра-эквивалент»/мл, т.е. менее
[0 ю моль/л, вызывает in vitro регенерацию
щупалец фрагментами тела. Интересно, что
с помощью антител к этому пептиду в
мозгу млекопитающих была обнаружена
система содержащих его нейронов.
Более того, некоторые описанные на
сегодняшний день нейроны функционируют
как классические неиросекреторные клетки
(т. е. секретируют гормоны), но лишены
характерной для них цитологии. Другие
нейроны, напротив, имея цитологию
типичных нейросекреторных клеток, не
выполняют их функций: вместо выделения секрета
в циркуляторную систему они
непосредственно контактируют с клетками-
мишенями.
16.11.5. Нейропептиды
Принципиальное сходство
нейросекреторных и обычных нейронов доказывается
секрецией многими, а возможно, и всеми
нейронами (включая и те, что образуют
обычные медиаторы) особых пептидов, хотя
первоначально это считали специфическим
свойством нейросекреторных клеток. Более
того, типичные неиросекреторные клетки
могут оказывать координированное
воздействие на физиологию и поведение
животного. В «сумковых клетках» моллюска Aplysia
нейросекрет синтезируется в виде
высокомолекулярного белка, или прогормона. В
секреторных гранулах он расщепляется на
более короткие фрагменты под действием
ферментов, так что вьщеляется не
единственный гормон, а «коктейль» пептидов.
Один из них — «гормон откладки яиц» —
через кровоток достигает гонад и
индуцирует икрометание. Другой стимулирует
нервные окончания и нейрогемальном
комплексе, обеспечивая их максимальную
активность. Ряд гормонов, включая гормон
откладки яиц, диффундирует по всему
абдоминальному ганглию и стимулирует
особые нейроны, запускающие
высокоспецифический тип поведения, сопровождающий
икрометание.
16.11.6. Нейроэндокринные
системы
Нервная, эндокринная и нейросекретор-
ная системы функционально не
изолированы друг от друга, а образуют
интегрированные (у эволюционно продвинутых
беспозвоночных—сложные) нейроэндокринные
системы. В рефлекс, включающий сенсорное
восприятие, обработку и интеграцию
интернейронами и контроль эффекторов, входит
эндокринный этап, опосредующий послед-
524
Глава 16
нюю его часть. Таким образом,
эндокринная система обычно контролируется
нервной, и наоборот, ее секреты часто
оказывают глубокое влияние на нервную систему
(приложение 16.12).
16.11.7. Феромоны
Первым идентифицированным
феромоном был бомбикол (см. табл. 16.1) —
вещество, секретируемое самками тутового
шелкопряда {Bombyx mori). В 1959 г.
кульминацией его исследований стала работа
Бутенандта (Butenandt) с сотрудниками,
связанная с экстрагированием продуктов
полумиллиона абдоминальных желез.
Потрясает то, что единственная молекула бом-
бикола, по-видимому, оказывает заметное
воздействие на ольфакторный рецептор
антенны самца, а 200 молекул, вызывающих
200 импульсов, достаточно для запуска
поведенческого ответа. Самец начинает
характерный зигзагообразный полет против
ветра.
Более позднее исследование
продемонстрировало важность смесей компонентов.
Сочетание химических веществ, возможно,
необходимо для возбуждения начального
эффекта, например свойственного
половому аттрактанту. Не исключено также, что
один из компонентов оказывает первичное
длиннодистанционное воздействие
(например, аттрактивное), а другой — вторичное,
на близком расстоянии (например,
инициирует копуляцию). Даже бомбикол секрети-
руется в сочетании с другим, сходным
соединением — бомбикалем, тормозяще
влияющим на полет. Вблизи друг от друга
самцы многих бабочек стимулируют друг
у друга секрецию полового феромона,
вызывающего'реакцию у самки. Изучение
подобных веществ, очевидно, имеет огромное
потенциальное значение для борьбы с
насекомыми-вредителями.
Феромоны очень важны для
регулирования кастовой структуры общественных
насекомых. Наиболее известный пример —
секреция маточного молочка мандибуляр-
ными железами матки медоносной пчелы.
Два его компонента—9-оксодекановая и 9-
гидроксидекановая кислоты—действуют
как синергисты, тормозя развитие яичников
у рабочих особей и постройку ими
специальных ячеек, в которых развиваются матки.
Когда матку удаляют из улья, это
торможение снимается, и вместо нее развиваются
новые активные в половом отношении
самки. Это же «молочко» важно как половой
аттрактант, а также во время роения и
основания новой семьи.
Вещества, секретируемые особями
одного вида, могут заметно влиять на другой и
в этом случае называются аллолюнами. На
них основаны взаимоотношения многих
симбиотических организмов. У некоторых
насекомых вещество, действующее как
феромон на особей своего вида, влияет как ал-
ломон на представителей другого,
например привлекая паразитов. У аннелиды
Nereis снижение концентрации гормона мозга
вызывает нерест (после которого животное
погибает), а паразиты-споровики (проти-
сты), «настраиваясь» на этот сигнал,
синхронизируют свое собственное размножение
с размножением хозяина.
У бабочки дубового шелкопряда Antherea
polyphemus, личинки которой питаются
листьями красного дуба, к размножению
приводит интересная последовательность
химических сигналов. Алломон, образуемый
листьями дуба, стимулирует у самки
выделение нейросекреторного гормона,
вызывающего выдвигание гениталий и секрецию
феромона. Он привлекает самцов, и
размножение происходит в месте, богатом пищей
для развития следующего поколения.
16.12. Роль эндокринной
системы
У беспозвоночных гормоны участвуют
в регуляции широкого спектра процессов.
Ниже будут описаны лишь некоторые из
этих механизмов.
16.12.1. Эндокринный пусковой
механизм
Некоторые гормоны выделяются в
ответ на специфический стимул и действуют
как «пусковой механизм», вызывая
быстрый ответ по принципу «все или ничего».
У морской звезды выметывание половых
продуктов у особей каждого пола может
быть вызвано инъекцией экстрактов радиа-
Системы контроля
525
льных нервов в полость тела взрослого
животного. Странно, что участвующий в этом
пептидный гормон («гонадостимулирую-
гдее вещество», или GSS),
распространенный по всем радиальным нервам и в
нервном кольце, по-видимому, секретируется
слоем субкутикулярных опорных клеток
(т. е. ненастоящими нейронами). Как он
транспортируется по нервным тяжам и
участвует ли в его переносе к тканям целомиче-
ская жидкость, вопрос спорный. С этой
необычной системой связано еще много
нерешенных проблем.
Гонадостимулирующее вещество
вызывает созревание гамет в яичниках или в их
фрагментах in vitro, что, казалось бы,
свидетельствует о его прямом действии. Однако
на самом деле GSS стимулирует синтез
и выделение второго вещества
фолликулярными клетками гонад. Это «индуцирующее
мейоз вещество» (MIS), как показал Кана-
тани (Kanatani) с сотрудниками в 1969 г.,
оказалось 1-метиладенином (табл. 16.1).
MIS любопытным образом сочетает
эндокринную и параэндокринную функции.
Во-первых, оно влияет на фолликулярные
клетки, которые его синтезируют, что
приводит к их отделению от гамет и
выделению последних через половые протоки. Во-
вторых, MIS диффундирует через
межклеточное пространство, индуцируя мейоз
ооцитов. Наконец, оно диффундирует в це-
ломическую жидкость и стимулирует
сокращение мышц, участвующих в выметывании
гамет, а у некоторых видов вызывает своего
рода заботу о потомстве.
16.12.2. Эндокринный баланс
Некоторые функции контролируются не
единственным гормоном, а балансом
антагонистических факторов. Углеводный,
солевой и водный обмен часто регулируется
именно таким путем, обеспечивающим
быструю обратимость эндокринного эффекта.
У ракообразных изменение окраски
носит физиологический характер, т.е. не
связано с количеством пигмента, а
обусловлено движением (или «миграцией») его
гранул в клетках, называемых хроматофорами
(рис. 16.34). Сами клетки при этом не
изменяют формы и положения.
Рис. 16.34. Хроматофоры с различной
степенью дисперсии пигмента
Гормоны, контролирующие
хроматофоры. обычно выделяются в ответ на внешние
стимулы (освещение фона и т. д.), но в
некоторых случаях их секреция подчиняется
циркадианному или приливно-отливному
ритму. Обычно наблюдаются два
антагонистических гормона, один из которых
вызывает рассеивание пигмента, а другой — его
концентрирование. Однако в некоторых
случаях на хроматофоры одного цвета
влияет множество факторов, что приводит
к различным окраскам тела. Гормоны
воздействуют также на миграцию ретинальных
пигментов, связанную с изменением
физиологии зрения сложного глаза (рис. 16.20,£).
16.12.3. Эндокринный контроль
развития у Nereis
Характер эндокринной регуляции у по-
лихет-нереид сравнительно прост и похож
526
Глава 16
на известный для немертин и некоторых
других беспозвоночных (но не для
большинства других аннелид). Как впервые
показал Деркон (Durchon) в 1948 г., децере-
брация этих червей приводит к
преждевременному созреванию. На последней стадии
жизненного цикла большинство видов
превращаются в гетеронереидную форму
(рис. 4.60), адаптированную к «роению»
и нересту в поверхностных морских водах.
Этот процесс также подавляется гормоном
мозга.
Децеребрация на ранних стадиях
жизненного цикла приводит к аномалиям
развития ооцитов или их дегенерации, а также
к неполному метаморфозу. Это позволяет
предполагать, что у гормона сложное,
двойственное действие, включающее
трофический эффект, поддерживающий раннее
развитие гамет и тела, и тормозное влияние
на процессы, связанные с последними
стадиями развития. У самок трофическое
влияние на гаметы может быть косвенным,
поскольку синтез специфических белков
желтка (вителлогенинов), необходимых для
развития ооцитов, происходит в целомоцитах.
Гормон необходим также для образования
новых сегментов тела (см. разд. 14.4.3).
На ранних стадиях жизненного цикла
уровень гормона высок, но впоследствии
интенсивность его секреции снижается. Это,
вероятно, зависит от окружающих условий.
Однако вещество, продуцируемое
созревающими гаметами, подавляет
эндокринную активность мозга, что в свою очередь
ускоряет развитие гамет, приводя к
выделению еще больших количеств их ингиби-
рующего вещества и т.д. (эффект
положительной обратной связи). Вопрос о том,
какая здесь связь прямая, а какая обратная,
спорный.
Считалось, что множество процессов,
участвующих в образовании сегментов, га-
метогенезе и метаморфозе, контролируется
простыми изменениями секреции
«мозгового гормона». Однако электронная
микроскопия позволяет предполагать, что мозг
нереид является источником множества
гормонов. Более того, недавно у этих и
Других нечленистоногих беспозвоночных
обнаружены экдистероиды (разд. 16.12.4), а
у пиявок они, вероятно, контролируют
линьку. Несомненно, эндокринные механизмы
у нереид намного сложнее, чем
представлялось ранее.
16.12.4. Эндокринный контроль
развития насекомых
В течение последних 50 лет насекомые
привлекают большое внимание
эндокринологов из-за скачкообразного характера их
развития, включающего ряд линек, и в
некоторых группах—метаморфоз от личинки
до куколки и имаго. Сложность этих
процессов у членистоногих отражается на
контролирующей их нейроэндокринной
системе (приложение 16.12).
Впервые значение мозга было
продемонстрировано Копецом (Корее) в
классических экспериментах 1917—1923 гг. (рис.
16.35). Гусеница непарного шелкопряда
будет окукливаться, только если испытывает
влияние мозга в течение некоторого
минимального (критического) периода.
Участвующий в этом проторакатропный
гормон (РТТН) необходим для линьки на
любой стадии жизненного цикла. У бабочки
Manduca] источником этого гормона, по-
видимому, является единственная нейросе-
креторная клетка с каждой стороны мозга,
а местами его хранения и выделения служат
нервные окончания внутри кардиального
и прилежащего тел (главным образом в
последнем) (приложение 16.12). Стимулы,
индуцирующие выделение гормона у разных
насекомых, неодинаковы. У клопа Rhodnius
пусковым сигналом служит растяжение
брюшка в результате питания кровью
(животное может быть «обмануто»
заполнением брюшка воздухом), а у Manduca—
достижение критической массы тела.
Целью недавней работы Исидзаки (Ishi-
1 В настоящее время установлено, что у
Manduca источником РТТН являются четыре
латеральные клетки (по две с каждой стороны мозга).
Их аксоны проходят через кардиальное тело, не
ветвясь, а затем по аллатальному нерву
достигают прилежащего тела, где дают
многочисленные разветвления, характерные для нейрогемаль-
ного органа. Местом выделения и запасания
гормона является только прилежащее тело (М.А
O'Brien et al., J. Neural Sci., 1988, v. 8 (9), pp.
3247—3257).— Прим. пер.
Системы контроля
527
Рис. 16.35. Эксперименты, демонстрирующие роль мозга в регуляции окукливания у Lymantra.
После критического периода его гормон уже выделился в достаточном количестве, поэтому деце-
ребрация или пигатура не могут блокировать развития
zaki) и Гилберта (Gilbert) с соавторами
была очистка РТТН. Из мозга бабочки Вот-
Ьух выделены два пептида с
молекулярными массами около 5000 и 22 000, способные
стимулировать секрецию экдизона прото-
ракальными железами. Меньший из них
состоит из двух аминокислотных цепей,
связанных дисульфидными мостиками. Это
послужило основанием для гипотез
относительно филогенетического значения
сходства этого соединения с инсулином и инсу-
линоподобными факторами роста
позвоночных. Установление физиологической
роли двух этих пептидов еще впереди.
Роль проторакальных желез
продемонстрирована в экспериментах на куколках
бабочки цекропиевого шелкопряда (Saturnia
cecropia). Если в изолированное брюшко
имплантировать активный мозг, оно не
в состоянии линять. Если же для этой пели
использовать одновременно активный мозг
и неактивные проторакальные железы,
развитие происходит. Это свидетельствует
о том, что гормон мозга стимулирует в
проторакальных железах или их гомологах се-
крепию второго фактора, стероида экдизона
(табл. 16.1). Карлсон (Karlson) с
соавторами использовал 1000 кг куколок
шелкопряда для получения 200 мг чистого вещества,
ставшего в 1963 г. первым химически
идентифицированным гормоном
беспозвоночных. В тканях экдизон превращается в 20-
гидроксиэкдизон, который и является
собственно гормоном линьки.
Дляподобных соединений, известныхпод
общим названием экдистероиды,
разработан ряд чувствительных количественных
анализов. Каллифоровый тест,
использованный Карлсоном, основан на
способности экдизона вызывать окукливание у
перевязанных личинок мухи. Сравнительно
недавно стал широко применяться
радиоиммунологический анализ. Антитела к экдизон-
белковому комплексу связывают
радиоактивный экдизон. Масштабы этого процесса
зависят от концентрации немеченого
экдизона, конкурирующего за места
связывания, и, таким образом, позволяют судить
о его количестве. У клопа Rhodnius
единственный пик экдизона наблюдается перед
каждой линькой (рис. 16.36).
РТТН—типичный пептидный гормон
в том смысле, что его воздействие на
рецепторы плазматических мембран
проторакальных желез индуцирует внутриклеточный
синтез сАМР. Стероиды, напротив,
проникают внутрь клетки и (предположитель-
но),связываясь с цитоплазматическими
белками, стимулируют генетический аппарат
ядра. Для исследования этой проблемы
ценным материалом являются мухи, так как их
слюнные железы содержат «гигантские»
528
Глава 16
Rhodnius
Линька
3 6 9 12 О
4-я стадия
3 6 9 12 15 18
5-я стадия
Дни 0 3 6 9
4-я стадия
3 6 9 12 15 18
5-я стадия
Рис. 16.36. Уровни в гемолимфе 20-гидроксиэкдизона (А) и ювенильного гормона (Б) в течение
последних личиночных стадий Rhodnius (no Baehr et al., 1978)
хромосомы, которые в 10 раз длиннее и
в 100 раз шире обычных. Во время развития
различные «диски» таких хромосом сильно
утолщаются, образуя «пуффы», где
повышенная активность генов выражается в
интенсивном синтезе РНК.
Экспериментальным путем была показана индукция экди-
стероидами появления специфической
последовательности ранних и поздних
хромосомных пуффов, идентичной той, что
обычно наблюдается перед линькой.
Третий гормон—ювенильный (JH)
(табл. 16.1)—секретируется прилежащими
телами (приложение 16.12). Три основные
формы JH — близкие по структуре
молекулы; каждое насекомое секретирует одну из
них или более. Роль JH наиболее четко
продемонстрирована в экспериментах с
удалением прилежащих тел и их трансплантацией
в особей, находящихся на разных стадиях
развития (рис. 16.37). На ранних этапах
жизненного цикла наблюдается высокий
уровень JH, который обеспечивает появление
после линьки (запускаемой экдизоном)
следующей личиночной стадии. У Rhodnius во
время пятой (последней) личиночной
стадии уровень JH сильно снижается (рис.
16.36), и в результате индуцируемой
Рис. 16.37. Эксперименты, демонстрирующие роль прилежащих тел у Bombyx. А. Нормальное
развитие: личинка пятого возраста, куколка и имаго. Б. Ускоренное развитие личинки третьего
возраста после удаления прилежащих тел. В. Гигантская личинка шестого возраста, полученная
путем имплантации активного прилежащего тела в личинку пятого возраста, развивается в
гигантскую куколку и имаго (по Turner, 1966)
Системы контроля
529
экдизоном линьки появляется имаго (или
сначала куколка, а затем имаго, как,
например, у бабочек).
На ранних стадиях развития ювениль-
ный гормон защищен от распада
специфическим белком гемолимфы, с которым он
связывается. Низкий уровень JH перед
последней линькой обеспечивается снижением
скорости его синтеза и образованием
специфического фермента, разрушающего
гормон, несмотря на его связь с белком.
У насекомых ювенильный гормон
участвует также в контроле размножения.
Вместе с родственными химическими
соединениями он используется в борьбе с
вредителями, обладая такими явными
преимуществами, как нетоксичность и способность
к биодеградации. Кроме того, насекомые
с трудом вырабатывают устойчивость
к веществу, от которого зависит их
нормальное развитие.
Гормон выхода секретируется мозгом
и запускает сложную последовательность
событий, происходящих при сбрасывании
насекомым старой кутикулы (через
некоторое время после образования эпидермисом
новой кутикулы под влиянием экдизона).
У сверчка Teleogryllus различают четыре
основные стадии этого процесса. Во-первых,
кутикула отслаивается и прикрепляется
к субстрату; затем насекомое выбирается из
старой шкурки; имагинальные крылья, ноги
и т. д. расправляются и приобретают
окончательную форму; наконец, старая
кутикула съедается. Эти стадии включают
стереотипную и координированную
последовательность по крайней мере 48 специфических
движений, повторяющихся не менее 4 ч.
Понятно, что о нейросекреторных клетках,
вырабатывающих гормон выхода, можно
говорить как о «командных нейронах»
(разд. 16.10.2).
Новая кутикула вскоре темнеет и
отвердевает. Ее «дубление» стимулируется ней-
росекреторным гормоном бурсиконом,
который секретируется после выхода из
старой шкурки мозгом или абдоминальными
ганглиями.
Интересно сравнить эндокринный
контроль развития у насекомых и других
членистоногих, особенно ракообразных. У них
такой же стероидный гормон линьки—
20-гидроксиэкдизон, секретируемый
эндокринными железами ненейронной
природы— Y-органами. Секреция стероида
контролируется нейросекреторной системой
глазного стебелька. Однако, если у
насекомых РТТН оказывает стимулирующее
действие, у ракообразных соответствующий
гормон угнетает работу Y-органов,
поэтому удаление глазного стебелька ускоряет
линьку или даже вызывает серию быстрых
линек. И наконец, недавно показано, что
у ракообразных есть JH. Он секретируется
антеннальными железами и его роль сходна
с таковой у насекомых.
Главное отличие заключается в
присутствии у ракообразных половых гормонов.
Как показал Шарнио-Коттон (Charniaux-
Cotton) в 1954 г., мужской гормон
секретируется андрогенными железами,
связанными с семепроводами, а один или несколько
женских гормонов — яичником. Эти
вещества стимулируют гаметогенез и диффе-
ренцировку вторичных половых признаков.
У большинства насекомых генотип, по-
видимому, влияет на половую дифференци-
ровку непосредственно, без участия
эндокринной системы.
16.13. Выводы
Мы завершим наш обзор регуляторных
систем у беспозвоночных двумя
примерами, свидетельствующими о прогрессе,
достигнутом за последние годы в объяснении
поведения и физиологии с точки зрения их
нервных механизмов.
Почвенная нематода Caenorhabditis ele-
gans была детально изучена Бреннером
(Brenner) и его коллегами в Кембридже.
Этот червь почти прозрачен, длина его
всего 1 мм, а жизненный цикл в лабораторной
культуре при 20°С длится 3,5 дня. Строение
его нервной системы было полностью
реконструировано по электронным
микрофотографиям серийных срезов.
У этого гермафродита ровно 302
нейрона, каждый из которых характеризуется
определенной формой и местоположением.
Более того, сейчас известны синаптические
контакты каждой клетки, т.е. построена
«принципиальная схема» нервной системы.
Обнаружено около 600 щелевых контактов
34—1314
530
Глава 16
и примерно 7000 химических синапсов, хотя
их число и положение могут варьировать.
Прослежены последовательность
клеточных поколений, предшествующих
в эмбриогенезе каждому нейрону, стадии
развития, на которых происходит их
специфическая дифференцировка, а также
изменения их синаптических связей.
Изучение морского заднежаберного
моллюска Pleurobranchia californica
Дейвисом (Davis) с соавторами в Калифорнии
оказалось наиболее успешным с точки
зрения выяснения нервных механизмов,
лежащих в основе поведения почти у любого
животного.
Различные формы поведения
организованы иерархически. Так, реакция избегания
важнее пищевой, ее стимуляция активирует
тормозные синапсы единственного
«командного» нейрона, контролирующего
питание. Мотивация питания зависит от
предшествующего потребления пищи, а
активируемые им сенсорные пути тормозят этот
командный нейрон. Конкурирующие
стимулы вызывают различные ответы,
зависящие от относительной силы стимуляции,
и животное должно обладать способностью
к принятию решения, поскольку в каждый
момент времени оно обычно занято только
одним делом. Основой этого является
интеграция сенсорной информации в нервной
системе. Например, если пищевой стимул
сопровождается стимулом для втягивания
ротовой лопасти, предпочтение будет
отдано первому, поскольку активируются два
нейрона, блокирующие двигательный
выход, ответственный за втягивание. Если же
пищевой стимул слабый или животное
насытилось, активируются другие цепочки
и доминирующим оказывается сигнал на
втягивание ротовой лопасти. В этих
процессах участвует и эндокринная система:
питание тормозится до тех пор, пока на нейроны
буккального ганглия влияет гормон
откладки яиц.
Поведение упорядоченно и адаптивно,
и этим оно четко отличается от
разнообразных конкурирующих влияний, с которыми
сталкивается животное. Изучение нервной
системы беспозвоночных открыло
некоторые свойства нервной организации,
лежащие в основе их поведения. Тем не менее,
несмотря на составление полных
«принципиальных схем» нейронных соединений, мы
до сих пор не знаем, что эти животные
«чувствуют» и чувствуют ли они вообще. Это
должно сделать нас осторожными в
прогнозах при будущем исследовании более
развитых нервных систем.
16.14. Дополнительная
литература
Bullock Т.Н., Horridge G.А. 1965. Structure and
Function in the Nervous System of Invertebrates.
Freeman, San Francisco.
Eaton R. C. (Ed.) 1984. Neural Mechanisms of
Startle Behaviour. Plenum Press, New York.
Halliday T. R., Slater P. J. B. (Eds) 1983. Animal
Behaviour. Vol. 1, Causes and Effects. Blackwell
Scientific Publications, Oxford.
Mill P. J. 1982. Comparative Neurobiology. Ed.
Arnold, London.
Nighnam K.L., Hill L. 1977. The Comparative
Endocrinology of the Invertebrates, 2nd edn. Ed.
Arnold, London.
Roberts A., BushB.M.H. (Eds) 1981. Neurones
Without Impulses: Their Significance for
Vertebrate and Invertebrate Systems. Cambridge
University Press, Cambridge.
SheltonG.A.B. (Ed) 1982. Electrical Conduction
and Behaviour in "Simple" Invertebrates.
Clarendon Press, Oxford.
Приложение 16.1.
Система гигантских волокон
кальмара Loligo
А. Аксоны двух гигантских нейронов
первого порядка сливаются по средней
линии тела и иннервируют несколько клеток
второго порядка в задней части мозга.
Большинство последних являются
мотонейронами, непосредственно управляющими
мышцами головы и воронки, однако
аксоны двух клеток идут к звездчатому
ганглию, где образуют синапсы с гигантскими
волокнами третьего порядка (показана
только одна пара) (по Joung, 1939).
Системы контроля
531
Гигантское
волокно 2-го
порядка
Проксимальный
синапс
Дистальныи
:инапс
Гигантский аксон кальмара"
Нервы от звездчатого
ганглия
Б. Звездчатый ганглий. Гигантские
волокна второго порядка образуют «дисталь-
ные» синапсы с волокнами третьего
порядка, а другие гигантские волокна образуют
с ними «проксимальные» контакты. Каждое
волокно третьего порядка возникает путем
слияния отростков нескольких клеток, и все
отходящие от звездчатого ганглия нервы
включают по одному такому волокну
(показано только пять нервов). Самый задний
и самый крупный нерв является излюблен-
Гигантское
волокно 3-го
порядка
Гигантское
волокно 2-го
порядка
1-й
порядок
2-Й
порядок
Воронка
Звездчатый
ганглий
3-й
порядок
ным объектом исследования нейробиоло-
гов.
В. Синаптические контакты между
гигантскими волокнами второго и третьего
порядков. Физиологическое соединение
двух этих элементов опосредуется
множеством контактов, в каждом из которых по-
стсинаптический отросток как бы зубцами
вставлен в пресинаптический участок.
Стрелки указывают направление потока
импульсов.
Приложение 16.2.
Потенциал действия
Открытие механизма возникновения
потенциала действия Ходжкином (Hodgkin)
и Хаксли (Huxley) из Плимута и Кембриджа
и другими привело к присуждению им
Нобелевской премии 1963 г. С помощью так
называемого метода «фиксации
потенциала» и путем замещения одного из ионов
веществом, для которого мембрана
непроницаема, были измерены токи
определенных ионов через мембрану при различных
потенциалах. Другой метод заключался
в избирательной блокировке ионных
каналов ядами: натриевых—тетродотоксином,
выделенным из иглобрюха, калиевых—
тетраэтиламмонием.
Возникновение потенциалов действия
обусловлено наличием в мембране ионсе-
лективных каналов с воротным
потенциалом. Натриевые каналы, по-видимому,
меняют свою форму при деполяризации, от-
34*
532
Глава 16
крывая «ворота», через которые ионы Na+
входят в аксон, приводя к еще большей
деполяризации мембраны. Это открывание—
явление кратковременное, вслед за ним
быстро наступает «натриевая инактивация».
Существуют также калиевые каналы
с воротным потенциалом. Они
открываются после натриевых, и диффузия ионов К +
через них (достаточно одного из каждых 107
ионов К+ на импульс) восстанавливает
потенциал покоя. За этим следует короткий
период «отдыха», в течение которого
мембрана рефрактерна, т. е. не способна
активироваться. В клетках многих моллюсков
основной вклад в возникновение потенциала
действия вносят Са2+-каналы. Известен
мутант дрозофилы shaker (трясущийся),
названный так из-за паралитических
вздрагиваний конечностей во время эфирной
анестезии. Калиевые каналы с воротным
потенциалом у него отсутствуют или
дефектны (затронутые этой мутацией гены
идентифицированы). В результате реполяризация
запаздывает, и потенциалы действия
растягиваются во времени.
501=^
I о |—\
£ - |д—t-Каналы для №+
§ -50 - I / \ \ \^Каналы для К+
i i i i
2 4
Время, мс
На графике показаны изменения
мембранного потенциала Р (ось ординат) в
любой точке аксона в зависимости от времени
(ось абсцисс) и обусловливающие их
последовательные изменения числа открытых
К+- и Na+-каналов (по Hodgkin, Huxly,
1952).
Приложение 16.3.
Клеточные механизмы
научения
Одни и те же типы научения обнаружены
во всем животном царстве, но только
недавно Канделом (Kandel) и другими были
исследованы их клеточные механизмы.
Мантийная полость моллюска Aplysia
имеет сифон и защитную пластинку. Легкое
прикосновение активирует сенсорный путь
от каждой из этих структур, нейроны
которого образуют синаптические контакты
непосредственно с мотонейронами,
управляющими защитной реакцией втягивания
ктенидия (моносинаптическая
рефлекторная дуга—см. разд. 16.9.3). Сенсорные
нейроны, иннервирующие хвостовую зону,
стимулируют интернейроны, образующие
синапсы с сенсорными окончаниями.
В пресинаптических окончаниях
сенсорных нейронов происходят изменения,
определяющие модификацию поведения, т.е.
научение. Частая стимуляция дыхательных
структур, вызывая неоднократное
проведение потенциалов действия в эти окончания,
приводит к градуальной инактивации Са2+-
каналов (разд. 16.3.3), уменьшению
количества выделяемого в ответ на каждый
стимул медиатора и ослаблению
поведенческой реакции. Эта простейшая форма
научения называется привыканием.
Если на «хвостовую» зону действует
мощный, угрожающий животному
раздражитель, ответ на широкий круг других
стимулов, включая и легкое прикосновение
к дыхательным структурам, усиливается.
Такая форма неассоциативного научения,
называемая сенсибилизацией, опосредуется
медиатором 5-гидрокситриптамином (5-
НТ), выделяемым интернейронами и
запускающим целый каскад метаболических
процессов в сенсорных окончаниях:
5-НТ -> Синтез сАМР -> Активация
фермента протеинкиназы -»Фосфорилирова-
ние специфических белков К+-каналов
мембраны -> Инактивация К+-
каналов -»Увеличение длительности
потенциалов действия -> Увеличение притока Са2+
внутрь клетки -> Усиление выброса
медиатора.
Системы контроля
533
Выработка условного рефлекса—это
одна из форм ассоциативного научения
(помните собак Павлова?). Так, если за серией
слабых стимуляций жаберной пластинки
каждый раз немедленно следует
угрожающее раздражение «хвоста», животное
начнет реагировать гораздо сильнее на
стимуляцию пластинки, чем, например, сифона.
Описанное сочетание стимулов приводит
к выделению из интернейронов 5-НТ и
синтезу сАМР во всех сенсорных окончаниях
как раз тогда, когда с приходом
потенциалов действия повышается уровень ионов
Са2+ в окончаниях только стимулированных
сенсорных нейронов пластинки. Считается,
что Са2+ активирует сАМР, а тот в свою
очередь усиливает выделение медиатора,
которого в результате высвобождается
значительно больше, чем во время
сенсибилизации.
Описанные выше формы научения
являются кратковременными—они
сохраняются в течение нескольких минут или,
в лучшем случае, часов. Однако применение
особых программ приводит к формирова-
От От От
£ сифона хвоста пластинки
А. Нервная система и дыхательная камера
Aplysia (вид сверху). 5. Синаптические связи
нейронов в абдоминальном ганглии,
использованные для изучения клеточных механизмов
процессов научения: С—сенсорные нейроны;
И — интернейрон; М — мотонейрон
нию долговременной памяти (у Aplysia—
более трех недель), обусловленной не
неустойчивыми метаболическими сдвигами,
а структурными изменениями. Число и
размер синаптических участков, а также число
синаптических пузырьков существенно
возрастают при долговременной
сенсибилизации и снижаются в ходе привыкания.
Приложение 16.4.
Становление дефинитивных
нейронных связей
Хотя окончательное строение нервной
системы невероятно сложное, она
развивается из простого эпителия. На конце
отростков эмбриональных клеток находятся
конусы амебоидного роста,
оканчивающиеся псевдоподиями. Формирование
«правильных» связей нейронов имеет решающее
значение для дальнейшей работы нервной
системы (разд. 16.8.4). Механизмы этого
процесса изучены у насекомых, в частности,
у кузнечиков, поскольку их эмбриональные
ганглии достаточно прозрачные, а число
клеток в них невелико. В результате
отдельные нейроны и даже их растущие аксоны
могут быть надежно идентифицированы.
В то время как одни клетки
развиваются, другие становятся избыточными
и элиминируются. Хотя различные ганглии
в нервном стволе эмбриона насекомых
содержат одинаковое число клеток,
некоторые клетки абдоминальных ганглиев
утрачиваются, а их аналоги в тораксе
развиваются для управления конечностями (они
сохраняются, даже если конечности удалить).
Аналогичным образом у гермафродитной
формы нематоды Caenorhabditis 302
нейрона, а у самцов в связи с репродуктивными
функциями сохраняются еще 79 нейронов.
Такой процесс запрограммированной гибели
клеток детерминирован генетически.
«Программа» развития дегенерирующих клеток
такая же, что и у их выживающих «сестер»,
однако у них включается «программа
самоубийства». Один из наиболее интересных
мутантов у Caenorhabditis (по гену ced-3)
в этом отношении дефектен. Клетки,
которые в норме должны погибать, выживают
и формируют функциональные связи; по-
видимому, на животном это не отражается!
534
Глава 16
vMP2 (€S>
А. Становление нейронных путей в
сегментарном ганглии кузнечика на седьмой день
двадцатидневного развития. Б. После удаления
пионерных нейронов U, и U2 нейрон аСС
дезориентируется. В. При удалении пионерных
нейронов MP, и dMP2 из этого же и соседнего
ганглия дезориентируется нейрон рСС (по Ва-
stiani et al., 1985)
vMP2
vMP2
dMP2 (0)=--==.-я<
Mp1vOW_-_- ';
Выросты различных клеток в ганглиях
эмбриона кузнечика появляются в четкой
последовательности; отростки,
заканчивающие развитие первыми, называются
пионерными волокнами. Прорастая от
ганглия к ганглию или от эпидермальных
сенсорных клеток в центральную нервную
систему, они следуют стереотипными путями.
Ориентация конусов роста, по-видимому,
определяется различными факторами их
окружения, но главным образом, вероятно,
положением базальных мембран и глиаль-
ных клеток.
Волокна, вырастающие позднее, часто
следуют за пионерными и в их отсутствие
(при экспериментальном удалении
отдельных эмбриональных клеток методами
лазерной микрохирургии) дезориентируются.
Если в эксперименте задержать развитие
клетки, ее аксон «отыщет» правильный
путь, несмотря на появление множества
дополнительных факторов.
Системы контроля БЗБ
Использование моноклональных
антител к центральной нервной системе
дрозофилы показало, что различные волокна
в составе нервного пучка имеют
одинаковую специфическую молекулу. По-
видимому, она является «фактором
узнавания», а нейрон «запрограммирован»
следовать сначала одному химическому
указателю, затем другому и т. д. Более того, хотя
конкретный, окончательно
сформированный нейрон может значительно различаться
своими дендритами у разных особей, он
в любом случае образует практически
одинаковые синаптические связи.
Приложение 16.5.
Клеточные механизмы
биологических часов
Среди наиболее известных ритмов
биологической активности некоторые связаны
с регулярным чередованием дня и ночи.
Циркадианный (circa dies—около суток)
ритм у моллюска Aplysia захватывается
(настраивается) изменениями в суточном
цикле освещенности, однако, если животное
содержать в неизменных условиях, он
сохраняется в течение нескольких дней, лишь
немного изменяя длину каждого цикла. Это
свойство циркадианного ритма дает
организму возможность реагировать на
факторы окружающей среды еще до их
наступления.
Ритмическая активность широко
распространена среди нейронов центральной
нервной системы и сохраняется в течение
нескольких недель даже при извлечении
ганглиев из тела и содержании их в культураль-
ной среде. Различные клетки имеют свои
собственные ритмы, однако в природе они
синхронизируются «главными часами»,
локализованными в «D-клетках» глаза (это не
фоторецепторы, а клетки, непосредственно
получающие от них информацию через
синапсы). Циркадианный ритм может быть
зарегистрирован в D-клетках
изолированного глаза, которые оказывают свое
влияние не за счет секреции гормонов, как когда-
то считалось, а через аксоны, проходящие
почти по всей нервной системе.
Были исследованы фундаментальные
механизмы работы биологических часов.
5 р До удаления глаза
х
С- L
5 г После удаления глаза
I
Время
Локомоторная активность Aplysia в течение дня
и ночи до удаления обоих глаз и через три дня
после него (по Strumwasser, 1974)
Обработка анизомизином, который
блокирует синтез белка в клетке, связываясь с
одной из субъединиц рибосом, уничтожает
ритм, а короткое воздействие этого
вещества «останавливает часы» на время, т. е.
заставляет их «отставать». Джаклет (Jac-
klet, 1981) следал вывод, что «суточный
синтез белка—основное условие хода цирка-
дианных часов». Соответствующие белки
могут быть ферментами протеинкиназами,
которые, как известно, влияют на ионные
каналы и, следовательно, на импульсную
работу других систем.
Приложение 16.6.
Сенсиллы насекомых
У насекомых некоторые рецепторные
нейроны связаны с кутикулярными
волосками, шипиками, пластинками, ямками и т. д.
Такие сенсорные органы называются
сенсиллами; тело буквально изобилует этими
536
Глава 16
Материнская клетка1
Кутикула
Дистальные сегменты
(сенсорные реснички)
Тормогенная
клетка
Трихогенная
клетка
Механо
рецептор
Сахарный
Рецептор
Водный
рецептор
-Катионный рецепт!
-Анионный рецепта]
Овкладочная
клетка
Развитие и строение сенсиллы насекомого. Тормогенная клетка секретирует материал для сочле-
новочной впадины, а трихогенная клетка—для волоскового стержня (обе они участвуют в
образовании лимфы); обкладочная клетка формирует чехол вокруг ресничек (по Dethier, 1976;
Hansen, 1978)
крошечными «антеннами». Все клетки,
входящие в состав сенсиллы, образуются путем
деления одной материнской клетки. У
нематод хеморецепторные органы, называемые
амфидами (рис. 4.18), развиваются сходным
образом.
Вкусовые или обонятельные хеморецеп-
торы у разных животных чрезвычайно
схожи. Дендрит (или «внутренний сегмент»)
каждого биполярного рецепторного
нейрона направлен к основанию этого органа, где
образует одну или более ресничек
(«наружный сегмент»). Проходя через множество
пор (до 15000 на волосок), химические
вещества достигают этих ресничек по особой
омывающей их жидкости («лимфе»).
Хорошо развитые механорецепторы
насекомых, как и у подавляющего
большинства других животных, ресничные.
Движение щетинки органа осязания деформирует
ресничку и тем самым стимулирует рецеп-
торную клетку. Кампаниформные сенсиллы,
состоящие из единственного нейрона,
связанного с жестким кутикулярным
колпачком, воспринимают напряжение в
кутикуле. Хордотональные сенсиллы дают
информацию о положении или движениях
суставов у насекомых и ракообразных. Они
состоят из рецепторов, каждых из которых
покрыт специализированной сколопоидной
клеткой, связанной с тяжами
соединительной ткани. Такие рецепторы служат, в
частности, сенсорными компонентами
тимпанальных органов слуха, встречающихся
в разных частях тела насекомых. Эти
органы состоят из модифицированных частей
трахейной системы и имеют воздушные
мешки с тимпанальными мембранами,
вибрация которых в ответ на звук стимулирует
связанные с ними рецепторы.
У насекомых, по-видимому, нет особых
структур, чувствительных к силе тяжести
и ускорению, и соответствующую
информацию они получают от многочисленных
сенсорных органов, связанных с суставами,
и т. д. Однако у мух задние крылья
превращены в булавовидные жужжальца,
которые быстро вибрируют во время полета
и по принципу гироскопа, используемого
в навигации, дают информацию о
поворотах тела.
Системы контроля
537
Приложение 16.7.
Нейросуперпозиционная
сетчатка мухи
Разрешающая сила нейросуперпози-
ционного глаза может быть в 100 раз выше,
чем у сложных глаз других типов. Рабдоме-
ры омматидия здесь остаются
разделенными, т. е. рабдом «открытый». Каждый раб-
домер получает свет, падающий под
определенным углом, а его аксон идет к
патрону, обшему не для всех рабдомеров данного
омматидия, а для ориентированных вдоль
той же оптической оси рабдомеров
соседних омматидиев. В результате лучи света от
одного источника падают на шесть
ретинулярных клеток смежных омматидиев. Их
нервные волокна, спирально закручиваясь,
образуют синапсы на одном патроне в
оптическом ганглии.
На рисунке показаны пути световых
лучей от двух точек в поле зрения (светлые
и темные линии), их проекции на ретинуляр-
ные клетки соседних омматидиев и
схождение (конвергенция) волокон активируемых
ими рецепторов на оптических патронах ла-
мины. Справа—световые лучи от одной
точки попадают на различные ретинуляр-
ные клетки смежных омматидиев, но
аксоны последних сходятся на одном патроне.
Из восьми ретинулярных клеток каждого
омматидия в нервной суперпозиции обычно
участвуют только шесть (по Strausfeld, Nas-
sel, 1981).
Ретинулярные
клетки сетчатки
Патрон ламины
Окончание аксона
ретинулярнои клетки
Постсинаптическая
монополярная клетка 2-го порядка
Ламина
538
Глава 16
Приложение 16.8.
Оптическая доля комнатной
мухи (Musca domestica)
«Сложность сетчатки насекомых
ошеломляет, обескураживает и не имеет аналогов
у других животных. Размышляя о
запутанной системе сложных, или фасеточных,
глаз, проникая в лабиринт нейронов и
соединяющихся волокон трех крупных
сегментов сетчатки,... исследователь
совершенно теряет голову» (Ramon, Cajal, 1937).
Все типы клеток ламины и большинство
образуемых ими связей сейчас
идентифицированы, и многие из них изучены электро-
физиологически. Достигнут также большой
прогресс в исследовании медуллы и лобулы
(рисунок из Strausfeld, Nassel, 1981).
Приложение 16.9.
Удары брюшка у рака
Удары брюшка у рака считают
типичными устойчивыми комплексами действий.
А. Упрощенная схема нервных цепей из
гигантских нейронов и «негигантских» проводящих
путей, присутствующих в каждом
абдоминальном сегменте и контролирующих удар брюшка
у рака. Эти системы активируются внешними
стимулами (стрелки). ЦГ—центральный
гигантский нейрон; БС—быстрый мотонейрон
мышцы—сгибателя; Г—гигантский
командный нейрон; ТМ—тормозный мотонейрон;
ТВ—тормозный вход рецептора растяжения;
ГМ — гигантский мотонейрон; MP—
мышечный рецептор растяжения; НП —
негигантский нервный путь; С—сенсорные
нейроны; СМ — стимулирующий мотонейрон.
Гигантское волокно и гигантские мотонейроны
связаны первыми известными науке
электрическими синапсами (см. разд. 16.3.4). 5. Удар
брюшка у рака (по Wine, Krasne, 1972)
Системы контроля
539
Неожиданный угрожающий стимул,
достаточно интенсивный для возбуждения
единственного импульса в гигантских волокнах,
инициирует быструю высокостереотипную
реакцию избегания—«удар брюшка» за
счет синхронного сокращения мышц-
сгибателей абдоминальных сегментов.
Действие совершается настолько быстро, что не
может быть модифицирано обратной
связью.
Активность в гигантских волокнах
тормозит как мотонейроны разгибателей
(антагонистов сгибателей), так и рецепторы
растяжения разгибателей (поскольку иначе
они помешали бы удару), пока сгибатели не
разовьют максимальную силу. Таким
образом, гигантские волокна действуют как
командные нейроны (разд. 16.10.2),
запускающие различные координированные
элементы двигательного акта.
Однако после подгибания брюшка
происходит быстрое распрямление тела за счет
сокращения мышц-разгибателей. Этот
«возвратный удар», будучи неизменной
частью естественной последовательности, не
генерируется центральной нервной
системой, а является рефлекторным. Как только
прекращается торможение мышечных
рецепторов, они стимулируются растяжением
и активируют разгибатели через описанную
ранее рефлекторную дугу (разд. 16.9.3).
Интересно, что торможение сгибателей,
сопутствующее обусловленному обратной
связью сокращению разгибателей,
стимулируется (хотя и с задержкой) гигантскими
волокнами.
Приложение 16.10.
Контроль мышечных
сокращений у ракообразных
У краба отдельные мышцы состоят из
волокон с различной структурой и
функциональными характеристиками. Некоторые из
них быстро генерируют потенциалы
действия и развивают напряжение, в то время
Быстрый
аксон
Медленный!
аксон 1
1
Л h Ь ,
О))))))))»)))
h b hri-^
■1ШШШ
1 "Ъ Ь Ъ*-^
(Ш1Ш1)
Ь \ ье^
:итш
Тормозный
аксон
Иннервация мышц у краба (по Atwood, 1973)
как другим свойственны в основном
градуальные потенциалы и более медленное
сокращение. Мышцы имеют сложную
полинейронную иннервацию и в отличие от
соматических мышц позвоночных «принятие
решения» о характере их работы часто
осуществляется периферией, где, как и у
многих других беспозвоночных, внутри инне-
рвируемой структуры сталкиваются
противоположные влияния.
Отростки единственного быстропрово-
дящего аксона контактируют главным
образом с быстрыми мышечными
волокнами; эти синапсы в ответ на повторную
стимуляцию вскоре «устают». Окончания
нервных волокон малого диаметра подходят
в основном к медленным мышечным
волокнам; в ответ на повторную стимуляцию
здесь наблюдается облегчение.
Следовательно, степень сокращения контролируется
не подключением дополнительных
двигательных единиц, как у позвоночных, а
градуальным управлением мышечными
волокнами. Кроме того, тормозные волокна
выделяют гамма-аминомасляную кислоту, как
в синапсах, образуемых с возбуждающими
окончаниями (и/>есинаптическое
торможение), блокируя выброс возбуждающего
медиатора глутамата, так и в синапсах на
мышечных волокнах (иостесинаптическое
торможение), что тормозит любое их
возбуждение.
540
Глава 16
Приложение 16.11.
Нейросекреция путем экзоци-
тоза
Секрет из нейросекреторных нейронов
выделяется, как и из большинства обычных
железистых клеток, путем экзоцитоза. Во
время этого процесса мембрана гранулы
сливается с плазматической мембраной
окончания аксона, что приводит к
диффузии ее содержимого в кровь. Впервые это
было установлено при изучении
беспозвоночных, в частности в работах Норманна
(Normann, 1965) по падальной мухе СаШ-
phora.
Позже исследования на аннелидах, а
затем на широком круге других
беспозвоночных показали, что секреторные гранулы из
«обычных» нервных окончаний (т.е.
образующих синаптические контакты с другими
клетками) также выделяются путем
экзоцитоза. Это означает, что синаптическим
окончаниям свойственны два различных
секреторных механизма.
Синаптические пузырьки, по-видимому,
формируются локально внутри окончаний
и захватывают обычные медиаторы,
например ацетилхолин, из клеточного сока, где
эти вещества синтезируются. Пузырьки
обычно выделяют свое содержимое в
местах синаптических утолщений (рис. 16.6).
Нейропептиды, напротив, синтезируются
и упаковываются в гранулы в теле клетки.
После переноса в нервное окончание
гранулы высвобождают свое содержимое путем
экзоцитоза. по-видимому, через
неспециализированные участки мембраны.
После экзоцитоза мембраны пузырьков
и гранул отделяются от плазматической
мембраны, переходя внутрь окончания
(эндоцитоз). Однако в то время как
пузырьки осуществляют круговорот (т. е. вновь
заполняются и используются), мембраны
гранул, вероятно, разрушаются.
V *
Г
6 -. *
•
г 'СГ
Ъ
А
ш.
А. Электронная микрофотография,
демонстрирующая экзоцитоз секреторных гранул из
синаптического окончания в церебральном
ганглии земляного червя Lumbricus. Стрелкой
показано выделяемое содержимое гранулы.
СГ—секреторные гранулы (* 60000, масштаб
100 нм). Б. В синаптических окончаниях
действуют два различных секреторных механизма
с участием пузырьков и гранул соответственно.
Ультраструктура нейросекреторных окончаний
идентична (Golding, Pow, 1987). ГЭР—
гладкий эндоплазматический ретикулум
I
Аппарат/
Гольджи
Секреторные
(Ж) гранулы
^ ®
Синаптические пузырьки^!
о о . о ^-
ОО о°
°ооо°о
Экзоцитоз *
Перикарион
^Деградация
Лизосома О
i Рециклиздция\ Q_) s
о°о0 W.
-жОж О
Эндоцитоз
Системы контроля
541
Приложение 16.12.
Нейроэндокринные системы
саранчи
Разнообразие и сложность нейроэндо-
кринных систем некоторых беспозвоночных
хорошо иллюстрирует пример саранчи.
Система 1. Диуретический гормон
синтезируется нейросекреторными клетками
мозга и транспортируется по аксонам в
запасающую долю кардиального тела (нейро-
гемальный орган), из которой выделяется
в кровь. Гормон стимулирует образование
мочевины малытигиевыми сосудами. Это
эндокринная система первого порядка—
только с одним гормональным этапом.
Система 2. Нервные волокна, идущие
к железистой доле кардиального тела,
образуют там типичные синаптические
контакты с железистыми клетками, работой
которых они управляют, выделяя нейромедиа-
тор октопамин. При стимуляции клетки
выделяют адипокинетический гормон
пептидной природы, идентифицированный Мор-
дью (Mordue) с соавторами в 1976 г. Он мо-
билизирует липиды жирового тела для
использования в качестве источника
энергии при полете.
Система 3. Другой гормон, секретируе-
мый нейросекреторными клетками
комплекса мозг/кардиальное тело, называется
проторакотропным (РТТН). Он
стимулирует секрецию проторакальными железами
стероидного гормона экдизона, который
после модификации в тканях запускает
линьку. Это эндокринная система второго по-
Мозг
Диуретический
гормон *'
.ардиальное тело
{запасающая доля)
Кардиальное тело
этезистая доля)
Прилежащее
"" тело
Адипокинетический
гормон
t-яПроторакот-
'"лропный
^гормон •'
Экдизон
Ювен ильный
гормон
Проторакаль-
ная железа
Мозг и эндокринные органы саранчи
(относительные размеры кардиального и
прилежащего тел сильно увеличены)
рядка—с двумя эндокринными этапами.
Система 4. Секреция ювенильного
гормона прилежащим телом, по-видимому,
контролируется как прямой иннервацией,
так и гуморальными факторами.
Обратная связь. Эндокринные системы
обычно действуют по принципу обратной
связи. Например, высокий уровень
циркулирующего 20-гидроксиэкдизона, влияя на
неиросекреторные клетки мозга, тормозит
синтез экдизона проторакальными
железами.
ГЛАВА 1 7
ЕЩЕ РАЗ ОБ ОСНОВНЫХ ПРИНЦИПАХ
17.1. Основные
физиологические черты фенотипов
Для построения новых соматическиих
и репродуктивных тканей, замены
отживших соматических тканей и получения
энергии для этих процессов животным
требуются ресурсы. В отличие от автотрофных
организмов, которые могут синтезировать
органические соединения из
неорганических, используя в качестве источника
энергии солнечный свет, животные должны
питаться другими организмами (см. гл. 9).
Эта их гетеротрофностъ требует развития
способности к передвижению для поиска
и добычи ресурсов, а также как средства
спасения от других животных. Появление
локомоции (см. гл. 10) обусловило коэво-
люционный отбор хищников и их жертв на
эффективность передвижения. Локомоция
также требует энергии (гл. 11). Кроме того,
ресурсы необходимо вкладывать в
различные процессы и структуры,
обеспечивающие защиту от других организмов (см. гл.
13).
Поступившие с пищей макромолекулы
для их усвоения тканями расщепляются на
субъединицы в ходе ферментативного
процесса, известного как пищеварение. Затем
эти субъединицы либо используются для
ресинтеза макромолекул (анаболизм), либо
распадаются с выделением необходимой
для жизнедеятельности энергии
(катаболизм). Излишек ресурсов, особенно
аминокислот, и «отработанные» тканевые белки
также катаболизируются и экскретируются
(см. гл. 12).
Главным энергетическим ресурсом для
метаболизма (анаболизма и катаболизма)
являются углеводы, обычно моносахариды,
которые могут запасаться в виде
полисахаридов (часто гликогена) и / или жира.
Энергия высвобождается в ходе их катаболиче-
ского окисления. В период между
возникновением жизни и появлением фотосинтеза,
когда атмосфера Земли была лишена
кислорода (гл. 1 и 2), оно шло путем переноса
электронов к органическим соединениям,
которые в восстановленной форме
накапливались как конечные продукты обмена.
С появлением кислородной атмосферы
стало возможным более полное и более
эффективное окисление, потребовавшее
поступления из внешней среды 02 и удаления из
организма С02 в качестве одного из
«отходов» жизнедеятельности. Аэробное дыхание
сейчас распространено в животном царстве
наиболее широко, хотя у некоторых видов
основу катаболизма все еще составляет
анаэробное дыхание. Универсальной
формой кратковременного запасания и
переноса энергии, выделяемой в катаболических
процессах, является фосфорилированное
соединение аденина и рибозы (нуклеотид)—
аденозинтрифосфат (АТР), образующееся
из дифосфатной формы (ADP) в реакциях,
связанных с дыханием. После выделения
энергии АТР вновь превращается в ADP,
и цикл повторяется. Процессы дыхания
обсуждались в гл. 11.
Фундаментальные физиологические
процессы обобщены на рис. 17.1. Отметим,
что поступление ресурсов в организм
ограничено затратами на их получение и
структурами, обеспечивающими питание.
Следовательно, даже когда пиша в окружающей
среде в избытке, ее количество, которое
может быть использовано для метаболизма,
лимитировано. Чем больше ресурсов
расходуется на одну метаболическую функцию,
тем меньше остается для других. Скорость
и пути использования ресурсов
контролируются ферментами и в конечном счете ге-
Еще раз об основных принципах
543
Рис. 17.1. Распределение
ресурсов (получаемых с пищей)
между анаболическими и ката-
болическими процессами —
основа физиологии животных
(по Calow, 1986)
Влияние на
приспособляемость
Выживание
Продолжительность
развития
--*-Плодовитость
нами. Способ распределения ограниченных
ресурсов решающим образом влияет на
биологию организма на нескольких
уровнях: от относительного вклада их в
анаболические и катаболические процессы
зависит физиология животного; от
распределения между различными видами
активности—его поведение', от использования
ограниченных продуктов анаболизма на
образование различных структур — форма тела, на
развитие соматических и репродуктивных
тканей — биология размножения и
жизненный цикл. Более того, степень
удовлетворения каждой из этих потребностей будет
существенно влиять на выживаемость и
плодовитость, а значит, на приспособленность
организма. Поэтому, согласно
дарвиновскому учению, отбор благоприятствует
таким генетически детерминированным
особенностям (стратегиям) распределения
и утилизации ресурсов, которые
максимизируют передачу в поколениях кодирующих
их генов. Значит, хотя в основе физиологии
всех организмов лежат общие принципы,
у каждой особи она «настроена»
естественным отбором на конкретные экологические
условия протекания перечисленных выше
процессов — это и называют адаптацией.
Отсюда вытекает, что в
функциональном отношении организм действует как
интегрированное целое. Компромиссное
обеспечение метаболических потребностей
может вести к компромиссу между
элементами приспособленности: так, увеличение
затрат на локомоцию обычно повышает
выживаемость, но снижает вклад ресурсов
в размножение. Принцип интеграции важен
также для морфологических структур,
поскольку развитие одной из них должно быть
совместимо с существованием других.
Отсюда следуют два вывода. Во-первых,
должен быть непосредственный контроль
формообразования, физиологических функций
и поведения. Он обеспечивается
химическими и электрическими сигналами (см. гл. 16).
Во-вторых, на интеграцию ориентирован
эволюционный отбор признаков: каждая
генетически детерминируемая черта должна
быть совместима с окружающей организм
средой, т. е., взаимодействуя с ней, должна
повышать шансы на выживание и
оставление потомства. Следовательно, гены
и определяемые ими признаки отбираются
как часть неделимого целого. Этого организ-
менного, или холистического, подхода, до
некоторой степени отличающегося от
«философии генного пула», которая
рассматривает организм как набор самостоятельных
«эгоистичных» генов, мы и придерживались
в предыдущих главах.
17.2. Главное—репликация
и размножение
Вклад в репродуктивные процессы —
очень важное, возможно, основное
направление использования ресурсов организмом.
Они идут на образование стадий
размножения (пропагул), которые являются
средством передачи в поколениях генетической
информации.
Иногда, как отмечалось выше (разд.
14.1.1), клетка или группа клеток,
отделяющаяся от родительской особи, полностью
544
Глава 17
Ж
!
Метасраза Телосраза
Постмитоги- Премитоти
ческая интер-ческая
фаза* интерфаза
Т Профаза
' Новый цикл
Рис. 17.2. Поведение пары хромосом при
митозе (по Paul, 1967)
воспроизводит геном последней. В основе
такого бесполого размножения лежит митоз
(рис. 17.2), за счет которого клетка или
клетки пропагулы образуются сами и
в свою очередь дают генетическую копию
родительского организма.
В противоположность этому отдельные
половые клетки (гаметы), возникающие
обычно в специализированных органах
(гонадах), для возникновения нового
организма должны, как правило, сливаться с
другими половыми клетками. Гаметы
образуются в процессе меиотического деления,
даюшего клетки с количеством ДНК и
хромосом вдвое меньшим, чем в обычных
соматических клетках (рис. 17.3). Полный на-
1 V J
эа
хэС
Предшествующая (Пахитена) (Диплотена)
телофаза митоза
/
(Диакинез)
1-я телофаза
\
2-я телофаза
Рис. 17.3. Поведение пары хромосом при
мейозе. Обратите внимание, что процесс крос-
синговера приводит к иному, чем у родителей,
сочетанию генов в хромосомах потомства (по
Paul, 1967)
бор хромосом восстанавливается при
слиянии гамет (сингамии, оплодотворении),
а поскольку они часто происходят от
разных родителей, потомство получает
смешанные генетические программы. Этот
процесс называют половым размножением
(см. гл. 14).
17.3. Онтогенез
Продукты размножения мельче и
обычно устроены проще, чем родители. При
половом процессе они одноклеточные, в то
время как у родителей много
функционально дифференцированных клеток.
Следовательно, для индивидуального развития, или
онтогенеза, необходимы клеточные
деления, увеличивающие размер (рост), и
клеточная специализация (дифференцировка).
Поскольку специфические клетки находятся
в строго определенных местах организма,
должен происходить гистогенез. Продукты
размножения обычно более или менее
сферические, а взрослый организм имеет
сложную форму, т. е. развитие включает
формообразование (морфогенез).
Деление зиготы (дробление
оплодотворенного яйца) происходит путем митоза.
Он точно воспроизводит ее геном и описан
выше. Некоторые из первых эмбриологов
(особенно Август Вейсман, 1834—1914)
считали, что дифференцировка включает
постепенное отделение в специфических
клеточных линиях ненужной части
наследственного материала. Однако изучение митоза и
открытие возможности полной регенерации
некоторых беспозвоночных из мелких
фрагментов их соматической ткани показали,
что каждая соматическая клетка содержит
более или менее полную копию исходного
генома. По мере дифференцировки в
различных клетках должны происходить
«включения» или «выключения»
специфических частей этой наследственной
программы. Таким образом, клетки, имея
одинаковый генотип, синтезируют различные белки
и, следовательно, функционируют по-
разному.
Хорошо известная модель такого
переключения основана на изучении бактерии
Escherichia coli. Одной бактериальной
клетке дают разделиться на две дочерние; затем
Еще раз об основных принципах
545
культивируют одну из образовавшихся
клеток в среде, содержащей как источник
энергии глюкозу, а вторую— в среде с другим
сахаром—лактозой. В присутствии
лактозы бактерии продуцируют фермент (3-
галактозидазу, который превращает ее
в глюкозу, поскольку непосредственно
лактоза использоваться не может. В среде
с глюкозой этот фермент не образуется.
У клеток в обеих средах один и тот же
геном, однако процессы синтеза различны,
т. е. можно сказать, что они
дифференцированы в отношении друг друга.
Клетки в глюкозной среде также
содержат гены, кодирующие Р-галактозидазу,
однако .обычно они «выключены» белком-
репрессором. Он препятствует
специфической генной транскрипции, связываясь с
соответствующим оператором. Лактоза,
попадая в клетку, превращается в аллолакто-
зу, которая взаимодействует с репрессором
таким образом, что он перестает ингибиро-
вать оператор и начинается синтез РНК,
служащей матрицей для синтеза фермента,
называемого в данном случае
индуцированным, т. е. запускаются метаболические
процессы, необходимые для утилизации
лактозы. Модель изображена на рис. 17.4.
По аналогии с такой регуляцией путем
включения в принципе возможна и
регуляция путем выключения, хотя понятно, что
она менее экономична. О молекулярной ос-
/' О G1 G2 G3
i i 1 1 1— Хромосома
JmPHK
Рис. 17.4. Модель генного контроля. Ген i
кодирует репрессор R, который связывается со
специфической частью хромосомы —
оператором (О) и ингибирует транскрипцию
генов G,, G2 и G3. Если для ферментов Р,, Р2 и Р3
имеется субстрат (Р), он, связываясь с
репрессором, изменяет его конфигурацию, не
позволяя ему блокировать оператор. При этом
транскрибируются G,, G2 и G3, синтезируются
Р,, Р2 и Р3 и метаболизируется Р (по Maynard
Smith, 1986)
нове генной регуляции при дифференциров-
ке эукариотических клеток известно мало,
однако похоже, что ее механизм близок
к рассмотренному выше.
Гистогенез требует упорядоченной
работы переключающих факторов. Он
обеспечивается либо детальным набором
инструкций о том, как клетка должна реагировать,
занимая определенное положение в
эмбрионе, либо менее детальной информацией,
позволяющей клеткам «узнавать» свое
местонахождение и действовать в соответствии
с ним. Поскольку генетические инструкции
находятся внутри клетки и крайне
маловероятно, что детальная информация может
поступать в нее извне или даже
передаваться из ее цитоплазмы к генетической
программе внутри ядра (запрет, известный как
«центральная догма молекулярной
биологии»), более правдоподобно предположение
о контроле, обеспечиваемом позиционной
информацией. Однако до сих пор остается
загадкой, на чем она основана, если все
клетки эмбриона генетически идентичны.
Возможно, это связано с цитоплазмой
яйцеклетки. Она часто даже визуально
неоднородна (например, пигмент и желток
распределены в ней неравномерно), и уже
продукты первого деления могут иметь разную
цитоплазму, а следовательно, и
неодинаковые условия для экспрессии своих
геномов.
Можно видеть, что этот процесс
соответствует предположению Вейсмана, за
исключением того, что в ходе дробления
клетки начинают различаться цитоплазматиче-
ским, а не ядерным материалом. Это делает
возможным материнское влияние на
развитие через цитоплазму яйца. В конечном
счете такой контроль, независимо от его
природы, бывает либо жестким,
определяющим судьбу клеток на ранних стадиях
онтогенеза {детерминированный процесс), либо
более гибким, позволяющим
экспериментально изменять судьбу отдельных клеток
даже на достаточно поздних стадиях
(недетерминированный процесс).
Морфогенез сочетает перемещения
клеток и дифференциальный рост. Первый
процесс хорошо описан для ранних стадий
развития прозрачного эмбриона морского ежа
(рис. 17.5, А и гл. 15): различные клетки вну-
35—1314
546
Глава 17
ггпп
Рис. 17.5. А. Раннее развитие морского ежа.
ПМ—клетки первичной мезенхимы —
мигрируют в «углы» эмбриона, используя фи-
лоподии; ГК—грушевидные клетки (по Gustaf-
son, Wolpert, 1967). Б. Изменение формы
клеток в зоне инвагинации. БМ—базальная
мембрана
три него можно не только наблюдать, но
и регистрировать с помощью замедленной
киносъемки. На ранней стадии эмбрион
образует состоящий из клеток полый
шар — бластулу. Затем некоторые клетки
должны переместиться внутрь, чтобы
сформировать внутренние органы, например
кишечник. Этот процесс, называемый гастру-
ляцией, происходит в два этапа. Сначала
внутрь проникают отдельные клетки, затем
происходит инвагинация всей стенки.
Миграция начинается утратой контакта клеток
друг с другом; они движутся в бластоцель
(внутреннюю полость бластулы) и затем
перемещаются вдоль его внутренней
поверхности, подтягиваясь филоподиями.
Инвагинация включает две стадии: медленное
прогибание стенки сменяется после
некоторого перерыва быстрым ее впячиванием.
Первая фаза, вероятно, связана с
изменением адгезии клеток: они слабее прилипают
друг к другу, но остаются прикрепленными
к базальной мембране. Это придает им
грушевидную форму и может вызывать
прогибание вовнутрь, как показано на рис. 17.5,
Б. Быстрая фаза, вероятно, обеспечивается
клетками, образующими в этой зоне фило-
подии, которые, установив контакт с
крышей бластоцеля, сокращаются. Эти
процессы, включающие межклеточные
мембранные взаимодействия и активную миграцию
клеток с помощью филоподий, вероятно,
довольно типичны для морфогенетических
изменений у животных. Основной вопрос,
который остается нерешенным,— как они
контролируются, обеспечивая
формирование полностью организованной особи.
Наконец, дифференциальный рост
внешних и внутренних органов может вести
к формообразованию. Джулиан Хаксли
(Julian Huxley) в своей книге «Problems of
Relative Growth» («Проблемы относительного
роста») в 1932 г. отмечал, что, если по-
Рис. 17.6. Пример аллометрического роста
насекомого Carausius morosus. Зависимость
длины задней части проторакса (А), ширины
головы (Б) и диаметра глаза (В) от общей длины
тела. Координаты логарифмические (по Wig-
glesworth, 1965)
Еще раз об основных принципах
547
строить в двойных логарифмических
координатах график зависимости размера одной
части организма от размера другой (или
всего организма), он часто будет
представлен прямой линией (рис. 17.6). Равные
интервалы на логарифмической шкале
соответствуют увеличению в одинаковое число
раз, т.е. эти прямые предполагают, что
каждый орган увеличивается (растет)
относительно другого с некоторой
специфической скоростью. Понятно, что аллометри-
ческие зависимости существенно влияют на
морфогенез организма.
В гл. 15 обсуждены детали основных
процессов развития, т. е. дифференцировки,
гистогенеза и морфогенеза, применительно
к беспозвоночным. Здесь мы затронули
лишь некоторые вопросы их регуляции.
17.4. Онтогенез и филогенез
Поскольку и онтогенез, и филогенез
внешне прогрессивны, заманчиво
предположить, что последний происходит путем
появления у первого дополнительных
стадий. В таком случае онтогенез
рекапитулирует филогенез. Эту точку зрения
сформулировал Эрнст Геккель в конце XIX в. В
соответствии с ней онтогенез «высшей»
формы повторяет взрослые формы более
низких уровней организации. Фон Бэр,
современник Геккеля, напротив, утверждал, что
ни один взрослый организм не повторяет
взрослых стадий проще организованных
животных, однако, поскольку развитие
всегда идет от недифференцированного
состояния к дифференцированному, его
начальные фазы у различных таксонов должны
быть одинаковы. Следовательно,
эмбриональные, а не взрослые формы, в различных
филетических линиях повторяются, что
вполне соответствует принципам развития,
изложенным в последнем разделе.
Эволюция путем появления
дополнительных стадий онтогенеза точно
соответствует ламаркистской теории
приобретенных признаков, которые обычно
развиваются в течение жизни и добавляются к
существующим структурам. Однако менделев-
ская генетика опровергает эту точку зрения,
поскольку мутация может привести к
изменению на любой стадии развития.
Действительно, сейчас известно, что скорость
развития контролируется генами. Изменения
последних могут либо остановить его
(устраняя «прошлый» взрослый организм),
либо продлить за нормальные пределы.
Вальтер Гарстанг (W. Garstang, 1868—1949)
был одним из первых зоологов, оценивших
важное значение генетики для развития
Процесс
Результат
Изменение скорости
Сдвиг конечной точки
Сдвиг начальной точки
Педоморфоз
U
X
/у
i
i
lm
1 т
1 Г)
1 X
11~
1с>
и
Замедление или неотения
Прогенез
Смещение назад
Пераморсроз
О
U
Т
_|_
/
/
/ у
/ /
//
//
4?
X.
ш
та
1 о
О
Акселерация
Гиперморфоз
Смещение вперед
Рис. 17.7. Классификация возможных эволюционных сдвигов в развитии По оси абсцисс—
продолжительность развития или размер; по оси ординат—морфологические изменения (по
Calow, 1983)
548
Глава 17
33-й день 45-и день 48-й день 62-й день
Рис. 17.8. Стадии развития многоножек. Ше-
стиногая стадия возрастом 33 дня иногда
используется как аргумент в пользу педоморфного
происхождения насекомых (см. также Gould,
1977)
и эволюционные возможности,
открываемые изменением его темпов.
На рис. 17.7 приведен полный спектр
эволюционных сдвигов, которые могли бы
возникнуть таким путем. Каждый график
включает траектории развития,
отражающие зависимость формы от размера или
возраста. Сплошные линии соответствуют
предковым траекториям, а прерывистые —
возникшим у потомков. Отмечены начало
развития и его прекращение. В верхнем
ряду графики отражают замедление
(неотения) или «усечение» (прогенез) развития;
такие его изменения называются педоморфо-
зом. При этом эмбриональные, личиночные
или ювенильные признаки сохраняются
у взрослой формы. Нижний ряд
иллюстрирует противоположную тенденцию (пера-
морфоз).
Вальтер Гарстанг особенно
интересовался личиночными формами и полагал,
что эволюция шла в основном за счет педо-
морфоза. Действительно, его роль в ряде
случаев могла быть велика (гл. 2).
Например, среди современных видов половозре-
лость у самцов некоторых эхиурид и
ракообразных (и рыб) наступает при мелких
размерах тела; такие самцы «паразитически»
прикрепляются к значительно более
крупным (иногда на несколько порядков)
самкам. Происхождение шестиногих
насекомых от многоножкоподобных предков
скорей всего также связано с педоморфозом
(рис. 17.8). Наконец, широко признанная
теория эволюции позвоночных
предполагает, что им дали начало личиночные
формы, напоминающие свободноплавающую
личинку-«головастика» современных
оболочников (см. рис. 7.30).
Возможен и филогенез за счет ускорения
развития. Рассмотрим эволюционные
тенденции лопастных линий на раковинах
аммонитов (рис. 17.9). Наклон графика у
потомков круче, чем у предков, т. е. имело
место именно ускорение (акселерация) их
формирования. К тому же рост лопастных
линий потомков идет дальше, чем у предков,
что предполагает гиперморфоз (рис. 17.7).
Наконец, графики у потомков проходят
«выше», т.е. потомки начинают развитие
раньше—происходит его смещение вперед
(рис. 17.7).
Теперь обобщим. Онтогенез не
рекапитулирует филогенез, но, поскольку
независимо от таксона он подчиняется ряду общих
принципов, различия, обусловленные диф-
ференцировкой, размерами, гисто- и
морфогенезом (см. последний раздел),
возникают на основе общих для всех животных
процессов—митоза, дифференциальной
экспрессии генома, движения клеток,
дифференциального роста. В связи с этим (как
предполагал фон Бэр) раннее развитие всех
2000
1000
о
х
ь
о
1
100 •
200 300
Диаметр раковины, мм
Рис. 17.9. Спектр изменений развития у
аммонитов: 1 —предок, 2—5—потомки. Более
старые части стадий — справа (по Newell, 1949;
см. также Calow, 1983)
Еще раз об основных принципах
549
организмов несколько сходно. Тем не менее
глубокое влияние на филогенез, по-
видимому, оказали небольшие изменения
темпов этих процессов, вызванные,
например, незначительными генетическими
отклонениями. Последние могли косвенно
привести к скачкам между уровнями
организации животных (гл. 1,2). Эту точку
зрения отстаивал Вальтер Гарстанг.
17.5. Дополнительная
литература
Calow Р. 1983. Evolutionary Principles. Blackie,
Glasgow and London.
Gould S. J. 1977. Ontogeny and Phylogeny. Harvard
University Press, Cambridge, Massachusets.
Huxley J. S. 1932. Problems of Relative Growth.
Methuen, London.
Maynard Smith J. 1986. The Problems of Biology.
Oxford University Press, Oxford.
Paul J. 1967. Cell Biology. Heineman, London.
СЛОВАРЬ ТЕРМИНОВ
Абдомен—задний отдел тела,
разделенного на три отдела, передний из которых —
голова (ср. торакс).
Аборальиый - относящийся к
поверхности тела, противоположной той, на которой
расположен рот, например у животных
с ртом в центре нижней или верхней
поверхности тела.
Автотомия—индуцированная самим
организмом ампутация конечности или части
тела, например как способ избегания
хищника, почкования (см.) или фрагментации
(см.).
Аденозинтрифосфат (АТР)— главная
энергосодержащая молекула организма.
Аддуктор—мышца, смыкающая и
удерживающая в сомкнутом состоянии створки
раковины.
Акклимация—изменение физиологии
в результате воздействия на организм
изменившихся окружающих условий.
Акрон—передний, не относящийся к
сегментам отдел тела членистоногих.
Акросома -волокнистая трубка (или
трубки) на переднем конце сперматозоида
(см.), вступающая в контакт и сливающаяся
с клеточной мембраной яйцеклетки (см.) во
время оплодотворения (см.).
Активный транспорт—движение
растворенных веществ против градиента
концентрации, требующее затрат энергии.
Амебоцит—клетка, способная к
амебоидному движению.
Анабиоз —см. криптобиоз.
Анаболизм—см. метаболизм.
Анаэробный—безвоздушный; термин,
обычно используемый для описания
дающих энергию метаболических (см.)
процессов, протекающих в отсутствие кислорода.
Аноксия —отсутствие свободного
кислорода.
Антенны—нитевидные и часто
удлиненные хемосенсорные придатки на голове
некоторых членистоногих, полихет, онихофор
и др.
Аподема—направленный внутрь тела
вырост экзоскелета членистоногих.
Апомиксис—см. бесполое размножение.
Арренотокия—форма партеногенеза
(см.), при которой неоплодотворенные
яйцеклетки развиваются в гаплоидных
самцов, а оплодотворенные — в диплоидных
самок.
Ацеломический—не имеющий полости
тела, если не считать просветов кишечника
и других органов.
Базальная мембрана — аморфный слой,
подстилающий эпителий (см.); образован
коллагеном (см.) и углеводом.
Бентосный —относящийся к дну или
субстрату в водных системах (ср.
пелагический).
Бесполое размножение—форма
размножения, не включающая редукционного
мейотического деления и слияния гамет
(см.) (ср. половое размножение).
Билатеральная симметрия—тип
симметрии, при котором тело можно разделить
только на две зеркально идентичные
половины.
Биотурбация—нарушение бентосных
(см.) осадков деятельностью животных.
Бирадиальная симметрия—тип
симметрии, при которой каждый из четырех
квадрантов сферического организма или
эмбриона такой же, как противоположный,
но отличается от соседних.
Бластема—группа
недифференцированных делящихся клеток, способных дать
начало дифференцированным клеткам (см.
дифференцировка).
Бластопор—отверстие, через которое
Словарь терминов
551
эмбриональная кишка гаструлы (см.)
сообщается с внешней средой.
Бластоцель—полость внутри бластулы
(см.).
Бластула—полый шар из клеток,
образующийся из зиготы (см.) в ходе дробления
(см.) при эмбриогенезе.
Брожение — ферментативное
разложение органических веществ с образованием
АТР в условиях аноксии (см.).
Брюшко—см. абдомен.
Вилочка—см. фурка.
Вторичнорогость—результат развития,
когда бластопор (см.) не становится ртом
взрослой особи, но может образовать ее
анус (ср. первичноротость).
Гамета—гаплоидная половая клетка,
способная сливаться с половой клеткой
противоположного пола, образуя зиготу
(см.); см. сперматозоид и яйцеклетка.
Ганглий—оформленное скопление
нервной ткани, содержащее нейроны (см.).
Гаплодиплоидия—см. арренотокия.
Гаплоидия—присутствие одинарного
хромосомного набора в соматических (см.)
клетках или гаметах (см.) (ср. диплоидия,
полиплоидия).
Гаструла—эмбриональная стадия,
следующая за бластулой (см.), на которой
однослойная структура превращается в
двуслойную за счет клеточной миграции,
инвагинации и т. д. и происходит обособление
клеток мезодермы (см.).
Гаструляция—процесс развития
гаструлы (см.) из бластулы (см.).
Гемму ла—многоклеточная пропагула
(см.) при бесполом размножении (см.)
некоторых губок, покрытая защитной
оболочкой.
Гемоцель — полость тела, образованная
кровеносными синусами; часто возникает
из бластоцеля (см.) (ср. целом и
псевдоцель).
Гермафродит—организм, способный
одновременно или в разные периоды
производить как яйцеклетки (см.), так и
сперматозоиды (см.).
Гетеротрофность — питание готовыми
органическими соединениями.
Гидравлический—приводимый в
действие давлением жидкости.
Гидростатический скелет—система
передачи силы мышечных сокращений
жидкостью в полостях тела или тканях.
Гиперполяризация—увеличение
разности электрических потенциалов,
существующих в состоянии покоя по обе стороны
клеточной мембраны (ср.
деполяризация).
Гипоксия—низкая доступность
кислорода.
Гипостом—возвышение, на котором
находится рот у книдарий.
Гистолиз—разрушение тканей.
Глазок—простой светочувствительный
орган (ср. сложный глаз).
Глия (нейроглия)—клетки, образующие
обкладку нейронов (см.).
Глотка—отдел передней кишки (см.),
находящийся позади ротовой полости
и перед пищеводом.
Головогрудь—часть тела некоторых
ракообразных, образованная слиянием
головы и груди (см.).
Гомойотермность — поддержание
относительно постоянной температуры тела (ср.
пойкилотермность).
Гоноподия—видоизмененная
конечность, служащая органом копуляции (см.)
(ср. циррус, пенис).
Гонопор— выводное отверстие для
гамет.
Грифельки—мелкие парные
нечленистые выросты, связанные с основаниями
конечностей у ряда многоножек и
присутствующие в эквивалентном положении на
некоторых абдоминальных (реже—и на
торакальных) сегментах большинства первич-
нобескрылых насекомых.
Грудь—см. торакс.
Двухлучевая симметрия—см. бирадиа-
льная симметрия.
Деполиризация—уменьшение разности
электрических потенциалов, существующих
в состоянии покоя по обе стороны
клеточной мембраны (ср. гиперполяризация).
Дерма—немышечный слой стенки тела
мезодермального (см.) происхождения,
лежащий под эпидермисом (см.).
Детерминированное развитие—см.
мозаичное развитие.
Детрит—частицы разложившегося или
разлагающегося органического вещества
вместе с колонизировавшими его микроор-
552
Словарь терминов
ганизмами в воде, на субстрате или в его
толще.
Диапауза — фаза покоя в жизненном
цикле, при которой интенсивность
метаболизма (см.) низкая, а организм устойчив к
неблагоприятным окружающим условиям.
Диморфизм—наличие у одного вида
двух различных
(обычно—морфологически) форм (ср. мономорфизм,
полиморфизм).
Диплоидия—присутствие двойного
хромосомного набора в соматических (см.)
клетках (ср. гаплоидия, полиплоидия).
Дифференцировка — процесс
специализации тотипотентных (см.) клеток эмбриона
(см.) для выполнения различных функций.
Дробление—митотические деления
зиготы (см.), формирующие многоклеточный,
но недифференцированный эмбрион (см.);
роста клеток при этом не происходит.
Дыхальце—выводное отверстие
трахейной (см.) системы.
Дыхательный пигмент—молекула,
обратимо соединяющаяся с кислородом
и служащая для его переноса или запасания.
Естественный отбор—эволюционный
механизм, предложенный Ч.Дарвином,
основанный на различиях в выживании
и оставлении потомства особями в среде
с ограниченными ресурсами.
Жгутики—выступающие из клетки ор-
ганеллы, совершающие круговые или
спиральные вращательные движения; обычно
относительно длинные и встречаются по
одному — по два на клетку; строение
жгутиков эукариот (см.) и бактерий
принципиально различное (ср. реснички).
Живорождение - развитие эмбриона
(см.) в теле родителя с частичным
использованием непосредственно поступающих от
последнего ресурсов.
Жизненная сила—мистическая
нефизическая сила, которая, как некогда считали,
обусловливает жизнедеятельность,
направляет развитие и эволюцию организмов.
Задняя кишка—последний отдел кишки
с эктодермальными (см.) происхождением
и выстилкой (ср. передняя кишка, средняя
кишка).
Зигота—клетка, возникающая при
слиянии сперматозоида (см.) с яйцеклеткой
(см.), т.е. при оплодотворении.
Зооид—особь модульной колонии (см.),
возникшая в ходе неполного почкования
(см.); термин обычно не применяется к кни-
дариям.
Зооксантеллы—симбиотические водо-
росли-динофлагеллаты в тканях различных,
главным образом морских,
беспозвоночных.
Зоохлореллы—симбиотические зеленые
водоросли (хлорофиты) в тканях
различных, главным образом пресноводных,
беспозвоночных.
Имагинальиый диск—группа
недифференцированных клеток внутри личинки
(см.), из которой развивается
специфическая система органов взрослого животного.
Интегумент—немышечные слои стенки
тела.
Интерстициальная— (а) фауна,
населяющая пространства (интерстициаль) между
частицами осадка; (б) тотипотентная эпи-
дермальная клетка у книдарий.
Интроверт—выворачивающийся и
втягивающийся передний отдел тела.
Инфауна—бентосные (см.) животные,
обитающие в толще субстрата или в норках
(ср. эпифауна).
Карапакс—защитный экзоскелетный
щит, покрывающий целиком или частично
дорсальную и боковые поверхности тела
членистоногого.
Карта зачатков — формализованное
описание пространственного расположения
в зиготе (см.), эмбрионе (см.) или имагина-
льном диске (см.) клеток или областей,
которые в норме дают начало тем или иным
частям организма.
Кладон—группа таксонов,
происходящих от общего предка.
Клейдоическое яйцо—яйцо (см.),
покрытое защитной оболочкой.
Клоака—расширенный последний
отдел кишечника, куда открываются протоки
других систем органов.
Книдоцит—клетка книдарий,
содержащая нематоцисты (см.).
Когорта—группа особей в популяции,
родившихся примерно в одно и то же
время.
Коксальные мешки—выворачивающиеся
тонкостенные пузыри у оснований ног
некоторых унирамий, служащие для поглоще-
Словарь терминов
553
ния окружающей влаги; сходные
образования известны и у онихофор.
Коллаген—волокнистый белок, обычно
связанный с соединительной тканью или ку-
тикулярными решетками.
Колония—группа организмов,
возникших бесполым путем и оставшихся
ассоциированными между собой; у многих
животных объединяющий тканевый контакт
с другими полипами (см.) или зооидами
(см.) является результатом неполного
почкования (см. также модульный организм);
особи, возникшие половым путем и
образующие полупостоянные скопления.
Кольчатость—разделение наружной
поверхности цилиндрического организма,
органа и т. п. бороздками на ряд колец,
внешне выглядящих как сегменты, но не
соответствующих им.
Комиссура—поперечное соединение
между нервными стволами или ганглиями.
Комменсал— организм, тесно
связанный с организмом другого вида (например,
живущий в его норке), но не приносящий
ему ни вреда, ни пользы.
Коннектива—см. комиссура.
Копуляция—перенос спермы органом
одной особи в тело другой; широко
распространенная, но не единственно возможная
стадия, предшествующая внутреннему
оплодотворению.
Критический облик—см.
покровительственный облик.
Криптобиоз —устойчивое состояние без
видимых проявлений жизнедеятельности во
время периодов с неблагоприятными
условиями среды (обычно с дефицитом влаги),
позволяющее, например, обитать в
пересыхающих водоемах.
Ктенидий—жабра моллюсков,
состоящая (исходно) из двух супротивных рядов
филаментов, отходящих от центральной
оси.
Куколка - неподвижная переходная
стадия развития некоторых насекомых
между личиночными стадиями и имаго.
Кутикула—неклеточный устойчивый
покров, секретируемый эпидермисом (см.)
и периодически линяющий; часто
орнаментирован и/или локально утолщен в
пластинки.
Лемниски —трубчатые мешки,
связанные с хоботком скребней и присутствующие
по крайней мере у некоторых бделлоидных
коловраток; в принципе — эпидермальные
.впячивания, функционирующие, вероятно,
как гидравлические резервуары.
Лецитотрофность — развитие за счет
внутренних ресурсов (желтка), полученных
от матери; обычно термин применяется
к морским личинкам (ср. планктотроф-
ность).
Литораль—приливно-отливная полоса
побережья.
Личинка—ювенильная стадия, заметно
отличающаяся по морфологии и экологии
от взрослого животного.
Ложная клешня—конечность
членистоногого, дистальный членик которой,
подгибаясь, прижимается к предпоследнему;
выполняет ловчую функцию.
Лофофор—система гидравлически
управляемых ресничных пищедобывательных
щупалец, образованных выростами стенки
тела и окружающих рот (но не анус).
Макромеры—крупные, богатые
желтком клетки раннего эмбриона (см.) (ср.
микромеры).
Макрофаг—организм, питающийся
относительно крупными пищевыми
частицами (ср. микрофаг).
Макроэволюция — становление
таксономического разнообразия, т.е.
эволюционные изменения, приводящие к появлению
новых видов и надвидовых таксонов (ср.
микроэволюция).
Максиллы—ротовые придатки
членистоногих, расположенные позади мандибул
(см.) и функционально их дополняющие.
Мальпигиевы сосуды—слепые
трубчатые экскреторные дивертикулы кишки.
Мандибулы—жвалы, челюсти.
Мантийная полость—полость,
окруженная мантией моллюсков и плеченогих, в
которой находятся соответственно органы
дыхания и захвата пищи.
Медуза—одна из двух форм тела у кни-
дарий: пульсирующая, обычно
пелагическая (см.), диско-, колоколо- или
зонтиковидная, часто студенистая (ср. полип).
Мезенхима—диффузные
соединительнотканные клетки, находящиеся в
студенистом матриксе.
Мезоглея—имеющий разную толщину
36—1314
554
Словарь терминов
клеточный или неклеточный слой
студенистого вещества между наружным и
внутренним клеточными слоями
кишечнополостных.
Мезодерма—зародышевый слой,
формирующийся между эктодермой (см.)
и энтодермой (см.).
Мезосома — второй отдел тела трехраз-
дельных олигомерных (см.) животных; его
целом (мезоцель) может гидравлически
поддерживать лофофор (см.) (ср. просома,
метасома).
Метаболизм—обмен веществ;
химические процессы в организме, включающие
разрушение структур и соединений
(катаболизм) и их образование (анаболизм).
Метамерия—тип строения тела,
характеризующийся линейной
последовательностью нескольких (многих) сегментов (см.)
(ср. мономерия, олигомерия).
Метаморфоз—резкое изменение
строения тела во время превращения личинки
(см.) во взрослое животное.
Метанефридий—открытый нефридий
(см.) с внеклеточным выводным протоком
(ср. протонефридий).
Метасома—третий отдел тела трехраз-
дельных олигомерных (см.) животных; его
целом (метацель) образует главную
полость тела (ср. просома, мезосома).
Метахрональный ритм—тип
синхронизированного движения ресничек (см.) или
конечностей, при котором движение
каждого элемента происходит с определенным
сдвигом по фазе относительно другого
элемента.
Микроворсинки—многочисленные
мелкие пальчатые выросты на свободной
поверхности клетки, участвующие в
абсорбции или (при соответствующей
специализации) в сенсорном восприятии; у цестод
называются микротрихами.
Микромеры—мелкие, бедные желтком
клетки раннего эмбриона (см.) (ср.
макромеры).
Микротрихи—см. микроворсинки.
Микрофаг—организм, питающийся
мелкими пищевыми частицами (ср.
макрофаг).
Микроэволюция—изменение частот
генов в популяции (ср. макроэволюция).
Миксонефридий—см. нефромиксий.
Мимикрия—сходство с неживым
объектом или с другим организмом,
потенциально позволяющее за счет «ложной
идентичности» ему избежать нападения.
Модульный организм—колония (см.) из
повторяющихся соединенных между собой
единиц («особей»), возникших бесполым
путем (см. полип, зооид).
Мозаичное развитие—тип развития, при
котором уже на ранней стадии онтогенеза
будущее каждой клетки эмбриона точно
определено (особенностями материнской
цитоплазмы), поэтому возможность
замещения утраченных частей зародыша очень
ограничена (ср. регуляционное развитие).
Мономерия—тип строения тела,
характеризующийся его неразделенностью на
сегменты (см.) (ср. олигомерия, метамерия).
Мономорфизм—наличие у вида
единственной формы тела (ср. диморфизм,
полиморфизм).
Моноцикличность—свойство
размножаться только один раз в жизни, после чего
погибать.
Нанопланктон—планктон (см.) с
наибольшим промером тела 2—20 мкм.
Наяда—водная нимфа (см.) некоторых
насекомых, сильнее, чем нимфа,
отличающаяся от имаго в результате специфических
адаптации к жизни в воде, например к ловле
водной добычи и/или потреблению
растворенного кислорода.
Недетерминированное развитие—см.
регуляционное развитие.
Нейрон—клетка, специализированная
для проведения электрических сигналов,
переносящих информацию.
Нейропиль — область нервной системы,
в которой сосредоточены синапсы (см.)
нервных волокон.
Нейросекреторная клетка—нейрон (см.)
с функциями железы, обычно секретирую-
щий гормоны.
Нектон—пелагические (см.) животные,
способные плыть против течения (ср.
планктон).
Нематоциста—внутриклеточная орга-
нелла книдарий с выворачивающейся
спирально закрученной нитью, используемая
для ловли добычи, защиты и т.д.;
находится в книдоцитах (см.).
Неотения—см. педоморфоз.
Словарь терминов
555
Непрямое развитие — развитие через
личиночную (см.) стадию (ср. прямое
развитие).
Нефридий—осморегуляторно-экскре-
торный орган эктодермального (см.)
происхождения (ср. целомодукт).
Нефромиксий—похожий на метанефри-
дий (см.) орган с участками
эктодермального (см.) и мезодермального (см.)
происхождения.
Нимфа—ювенильная стадия
насекомого, мало отличающаяся от взрослой особи,
если не считать размеров и развития
некоторых систем (например, крыльев, гонад);
обычно зачатки крыльев находятся снаружи
(в отличие от личинки насекомых).
Окончательный хозяин—хозяин
паразита (см.), в котором происходит его половое
размножение (см.).
Олигомерия—тип строения тела,
характеризующийся наличием двух или трех
сегментов (см.) (ср. мономерия, метамерия).
Онтогенез—индивидуальное развитие
от зиготы (см.) до взрослой особи.
Опистосома—задний отдел тела тех хе-
лицеровых, у которых оно четко разделено
на два отдела (ср. просома); иногда термин
применяется и к другим животным с дву-
раздельным телом.
Оплодотворение — процесс слияния
гамет (см.) с образованием зиготы (см.).
Оральный—ротовой.
Орган—структурно-функциональная
единица тела, образованная одной или
несколькими тканями (см.).
Остии—поры, например, для
прохождения воды в тело (у губок) или крови в сердце
(у животных с незамкнутой кровеносной
системой).
Осфрадии—хеморенепторные участки
или органы в мантийной полости (см.)
моллюсков.
Панцирь—наружный или почти
наружный защитный покров тела, обычно
состоящий из нескольких элементов.
Паразит—организм, постоянно или
временно обитающий внутри другого или
на нем и приносящий ему ущерб.
Паренхима—диффузная ткань (см.) из
вакуолизированных клеток, часто
заполняющая пространство между эпидермисом
и кишкой у ацеломических (см.) животных.
Партеногенез—форма размножения
(см.), при которой яйцеклетка (см.)
развивается в новую особь без оплодотворения
(см.).
Педицеллария—сложный подвижный
придаток некоторых иглокожих,
действующий как щипцы или пинцет.
Педоморфоз—процесс эволюционной
ювенилизации (см.), при котором либо
взрослая форма сохраняет ювенильные
признаки (неотения), либо половозрелость
наступает у животных ювенильного
возраста и ювенильной формы (прогенез).
Пелагический — обитающий в толще
воды (ср. бентосный).
Пенис—орган копуляции (см.),
увеличивающийся во время нее в размерах, но не
выворачивающийся (ср. циррус, гонопо-
дия).
Первичноротость—результат развития,
при котором бластопор (см.) образует рот
взрослого организма (ср. вторичноро-
тость).
Передняя кишка—передний отдел
кишки с эктодермальными (см.)
происхождением и выстилкой.
Перикардиальная полость — полость,
в которой расположено сердце.
Периостракум—белковый покров
раковины моллюсков и плеченогих.
Перистальтика—волны сокращений
кольцевых и продольных мышц, проходящие
по трубчатому органу или организму и
служащие для проталкивания его самого или
его содержимого.
Перитонеум—мезодермальная
выстилка целома (см.).
Пигидиум—задний, не относящийся
к сегментам отдел аннелид.
Пиннула—мелкая боковая ветвь
щупальца.
Пиноцитоз—поглощение клеткой
мелких капель жидкости.
Питание осадками—потребление
детрита (см.) и/или связанных с ним организмов
на поверхности и/или в толще субстрата.
Питание суспензиями—улавливание
и потребление частиц, взвешенных в воде,
обычно с помощью фильтра.
Плазмодий—много ядерная амебоидная
масса, окруженная общей клеточной
мембраной.
36*
556
Словарь терминов
Пламенные клетки—ресничные клетки
на проксимальном конце протонефридия
(см.).
Планктон—пелагические (см.)
организмы, взвешенные в воде и не способные
плыть против течения (ср. нектон).
Планктотрофность—питание (по
крайней мере частично) планктоном (см.);
термин обычно применяется к морским
личинкам (ср. лецитотрофность).
Плеоподы—абдоминальные (см.)
придатки многих ракообразных, часто
используемые для плавания.
Поверхностное дробление—тип
дробления (см.), при котором зигота (см.) дает
начало синцитию (см.); его ядра движутся
к поверхности эмбриона, где отделяются
друг от друга клеточными мембранами.
Пойкилотермность—зависимость
температуры тела от температуры внешней
среды (ср. гомойотермность).
Покровительственный облик — облик,
позволяющий скрываться от врагов за счет
сходства с окружающими объектами.
Полиморфизм—наличие у одного вида
более двух различных форм тела (ср.
мономорфизм, диморфизм).
Полип—одна из двух форм тела у кни-
дарий: сидячий или прикрепленный
цилиндр, обращенный к субстрату абораль-
ной (см.) поверхностью и несущий вокруг
рта кольцо щупалец; часто входит в состав
колонии (см.); иногда термин используется
как синоним зооида (см.).
Полиплоидия—наличие более двух
копий каждой хромосомы в соматических
(см.) клетках (ср. гаплоидия, диплоидия).
Полифиле гический—относящийся к
группе организмов, происходящих от
нескольких предковых форм.
Половая пора—см. го но пор.
Половое размножение—форма
размножения, при котором происходит обмен
генетическим материалом между
хромосомами во время мейоза и слияние гамет (см.)
в процессе оплодотворения (см.) (ср.
бесполое размножение).
Почкование—форма бесполого
размножения (см.) с образованием новых особей из
выростов на теле родителя; затем они
могут от него отделяться и жить
самостоятельно или оставаться с ним соединенными
(или связанными иным образом) в составе
колонии (см.).
Поясок—секреторный эпителий (см.),
образующий кокон у поясковых аннелид.
Предротовая полость— пространство
перед ртом, в котором работают ротовые
придатки или происходит наружное
пищеварение.
Предупреждающая окраска — особый
яркий, контрастный узор (например, черно-
желтый, красно-черный) потенциального
вида-жертвы, часто сочетающийся с его
несъедобностью или токсичностью.
Проглоттида—сериально
повторяющаяся единица строения тела у цестод.
Прокариоты—организмы с клетками
без окруженных мембранами органелл
и ядра (ср. эукариоты); бактерии.
Промежуточный хозяин —хозяин
паразита (см.), в котором он не размножается
или размножается только бесполым (см.)
путем.
Пропагула—репродуктивная структура,
отделяющаяся от родителя; может быть
многоклеточной (вегетативной) или
одноклеточной; в последнем случае возникает
путем мейоза (гаметы при половом
размножении) или же митоза, а также
аберрантного мейоза, не сопровождающегося
редукцией числа хромосом (при бесполом
размножении).
Просома—(а) передний отдел тела
(включающий голову) хелицеровых с
телом, явственно разделенным на два отдела
(ср. опистосома); иногда термин
применяется и к другим животным с двураздельным
телом (например, к веслоногим
ракообразным); (б) первый отдел тела трехраздель-
ных олигомерных (см.) животных; его
целом называется протоцелем (ср. мезосома,
метасома).
Простомиум—передний, не
относящийся к сегментам отдел аннелид.
Протонефридий—слепо начинающийся
нефридий (см.) с внутриклеточным
выводным протоком (ср. метанефридий).
Протоэукариоты—гипотетические,
возможно, способные к фагоцитозу (см.)
клетки, которые, став хозяевами различных
эндосимбиотических прокариот (см.), дали
начало первым эукариотам (см.).
Прядильные органы—сосочки с вывод-
Словарь терминов
557
ными отверстиями желез, образующих
паутинные нити.
Прямое развитие —развитие без
личиночных (см.) стадий (ср. непрямое
развитие).
Псевдокопуляция—тесное сближение
при выведении из тела гамет (см.)
спаривающихся животных с наружным
оплодотворением; при этом яйцеклетки (см.)
оплодотворяются (см.) сразу же после выхода из
гонопора (см.) самки.
Псевдоподия—временный лопастевид-
ный вырост протоплазмы, образуемый
амебоидными клетками при движении,
фагоцитозе (см.) и т. п.
Псевдоцель — любая полость тела, не
являющаяся целомом (см.).
Рабдиты —см. рабдоиды.
Рабдоиды— палочковидные
образования с неясной функцией в эпидермисе (см.)
плоских червей и сходных с ними животных;
некоторые образуются из железистых
клеток и называются рабдитами.
Рабдом—упорядоченное расположение
фоторецепторных микроворсинок (см.),
например, в сложном глазу.
Рабдоцель—слепая кишка турбелларий
без боковых ветвей и дивертикулов.
Радиальная симметрия—симметрия
относительно любой плоскости, проходящей
через орально-аборальную ось.
Радиальное дробление—тип дробления,
при котором плоскости клеточного деления
проходят параллельно и перпендикулярно
полярной оси бластулы (см.) (ср.
спиральное дробление).
Регенерация восстановление за счет
компенсационного роста и дифференциров-
ки (см.) утраченных частей организма.
Регуляционное развитие — тип развития,
при котором эмбрион (см.) способен
компенсировать утрату клеток и давать
нормальную личинку или взрослую особь,
поскольку будущее клеток опеределяется
только на поздних стадиях эмбриогенеза (ср.
мозаичное развитие).
Реснички — выступающие из клетки
двигательные органеллы, бьющие в одной
плоскости (жесткий удар и гибкое отведение
назад), обычно относительно короткие; могут
находиться на клетке в большом
количестве; специализированные неподвижные
реснички служат для сенсорного
восприятия; известны только у эукариот (см.) (ср.
жгутики).
Ресничное пламя—реснички пламенных
клеток (см.). ■
Самозарождение —ненаучная теория,
признающая возможность возникновения
живых организмов непосредственно из
неживого вещества (например, ила).
Сегмент—полуавтономная, сериально
повторяющаяся единица строения тела;
сегментация может затрагивать только его
стенку с ее структурами или помимо этого
все элементы внутреннего строения.
Семеприемник—мешок, в котором
животное накапливает полученные от
партнера по спариванию сперматозоиды (см.)
перед оплодотворением (см.) своих
яйцеклеток (см.).
Септа—мембрана, отделяющая один
отдел тела от другого.
Симбиоз—тесная ассоциация двух
различных организмов, взаимодействующих
между собой (один обычно зависит от
другого).
Синапс - контакт, служащий для
передачи информации между двумя клетками,
из которых по крайней мере одна — нейрон
(см.); передаюшая клетка называется преси-
наптической, воспринимающая—пост-
синаптической.
Синцитий—многоклеточная структура
с частично или полностью отсутствующими
границами между клетками; может
представлять собой и многоядерную цитоплаз-
матическую массу без заметного
разделения на клетки, и сеть клеток, цитоплазма
которых объединена межклеточными
мостиками.
Скалиды—кутикулярные или эпидер-
мальные выросты различной формы
(шиловидные, булавовидные, перистые,
чешуевидные), расположенные кольцами вокруг
интроверта (см.) киноринхов, лорицифер,
приапулид и личинок волосатиков; могут
выполнять сенсорную, локомоторную,
ловчую или прободающую функции.
Скелет—система, обеспечивающая
передачу силы мышечных сокращений
и/или опору частям тела.
Склерит—пластинка, составляющая
часть экзоскелета.
558
Словарь терминов
Склеротизация—химическое
упрочнение (с потемнением) участков кутикулы
(см.); результат процесса, аналогичного
дублению.
Слизь—смесь мукопротеинов
(соединений мукополисахаридов с белками), секре-
тируемая слизистыми клетками.
Сложный глаз—глаз, образованный
несколькими (многими) отдельными
зрительными единицами — омматидиями, каждая
из которых имеет собственные хрусталик,
поле зрения, рецепторные клетки и т. п. (ср.
глазок).
Сократительная вакуоль —
внутриклеточная окруженная мембраной вакуоль
с осморегуляторными функциями;
заполняется жидкостью и, резко сокращаясь,
выталкивает ее наружу из клетки.
Соматический—относящийся к клеткам
организма, не являющимся половыми.
Сперматозоид (спермий)—мужская
гамета (см.), обычно способная к активной
локомоции.
Сперматофор—пакет сперматозоидов
(см.), заключенный в защитную
оболочку.
Спиральное дробление—тип дробления,
при котором плоскости клеточного деления
проходят косо по отношению к полярной
оси бластулы, что приводит к
чередующимся смещениям вокруг этой оси по
часовой стрелке и против нее клеток,
образующихся при поперечных делениях после
стадии четырех бластомеров (ср. радиальное
дробление).
Средняя кишка—отдел кишки с энто-
дермальными (см.) происхождением и
выстилкой (ср. передняя кишка, задняя
кишка).
Старение —необратимые деградацион-
ные изменения особей с течением времени,
приводящие к повышенной уязвимости
и пониженной жизненности.
Статобласт—многоклеточная пропагу-
ла (см.) при бесполом размножении (см.)
некоторых мшанок, окруженная защитной
оболочкой.
Статоцист—орган, воспринимающий
силу тяжести и/или ускорение.
Стернит вентральный склерит (см.)
сегментарного экзоскелета членистоногих.
Стилет—твердое, заостренное, ноже-
видное образование, используемое для
протыкания клеток или тканей.
Столон—стебле- или корнеподобная
структура, соединяющая животных друг
с другом или с субстратом, иногда отпочко-
вывающая новые особи.
Субумбреллярный—связанный с
нижней, обычно вогнутой поверхностью
медузы (см.).
Схизоцель — целом (см.),
формирующийся в толще мезодермальной (см.) ткани
за счет образования пространства между ее
клетками (ср. энтероцель).
Тегумент—синцитиальный (см.)
наружный эпителий паразитических плоских
червей.
Телеология—учение об эволюционном
развитии, направленном к заранее
определенной цели.
Тельсон—задний, не относящийся к
сегментам отдел членистоногих.
Тергит—дорсальный склерит (см.)
сегментарного экзоскелета членистоногих.
Ткань—совокупность клеток одного
или нескольких типов, выполняющих
общую функцию и обычно соединенных
межклеточным материалом (ср. орган).
Тонический—устойчивый, длительный.
Торакс—средний отдел любого тела,
разделенного на три отдела, передний из
которых—голова (ср. абдомен).
Тотипотентность—способность клеток
многоклеточного организма к дифференци-
ровке (см.) в любую специализированную
клетку.
Трахеола—терминальная капилляропо-
добная часть трахеи (см.).
Трахея—трубка, проводящая воздух из
внешней среды непосредственно к тканям.
Трохофора—ранняя личиночная стадия
многих морских беспозвоночных,
характеризующаяся двумя околоротовыми
поясами ресничек.
Ультрафильтрация - прохождение
жидкости под давлением через
полупроницаемую мембрану.
Уроподы—последняя пара
абдоминальных (см.) придатков у десятиногих раков;
вместе с тельсоном (см.) образует
«хвостовой веер».
Уросома—опистосома (см.) веслоногих
ракообразных.
Словарь терминов
559
Фагоцитоз — процесс окружения
псевдоподиями (см.) амебоидной клетки пищевой
частицы и заключения ее в вакуоль.
Филогенез—эволюционное
происхождение таксонов.
Филоподии—нитевидные выросты
цитоплазмы.
Флоскулы—крошечные выступающие
сенсорные органы, оканчивающиеся
сосочками; присутствуют на туловище
некоторых приапулид, лорицифер, киноринхов
и коловраток.
Фотопериодизм—способность
физиологически реагировать на изменения
относительной долготы дня.
Фоторецептор—особая
светочувствительная клетка.
Фотосинтез—синтез органических
соединений с использованием энергии
солнечного света и участием хлорофилла по
общему уравнению
СО, + 2Н2Х -»(СН,0) + Н,_0 + 2Х.
При оксифотосинтезе у некоторых бактерий
и всех фотосинтезирующих эукариот X —
кислород; при аноксифотосинтезе у ряда
бактерий X—не кислород, а часто (но не
всегда) сера, т.е. Н,Х—сероводород.
Фрагментация - форма бесполого
размножения (см.), включающая разделение
тела на две и более частей, каждая из
которых способна вырасти в целую особь.
Фумарола—отверстие земной коры, из
которого выделяются горячая вода и
восстановленные соединения.
Фурка—пара варьирующих по форме
отростков на тельсоне (см.)
ракообразных.
Хемоавтотрофность—бактериальное
питание за счет хемосинтеза (см.).
Хемосинтез—синтез органических
соединений по общему уравнению
СО, + 2Н,Х -> (СН,0) + Н,0 + 2Х
с использованием энергии окисления (и
часто— восстановительных свойств)
неорганических субстратов типа Fe2+, CH4, NH3,
NO,-, H+, S и др. или органических
соединений (ацетата, формиата и др.);
осуществляется только бактериями в
анаэробных условиях (ср. фотосинтез).
Хитин— азотсодержащий полисахарид.
Хлоропласт—органелла эукариот—
место протекания фотосинтеза (см.);
встречается у некоторых протистов и
большинства растений.
Хоаноциты—характерные жгутико-
воротничковые клетки губок.
Хоботок — удлиненный вырост на
голове или передней части тела, используемый
для питания.
Хорда—упругий дорсальный скелетный
тяж, развивающийся из сильно вакуолизи-
рованных клеток, связанных между собой
внутри общей оболочки; характерный
признак хордовых.
Хроматофор—пигментная клетка.
Хромосома—нитевидная структура
клеточного ядра, несущая генетическую
информацию.
Целом—заполненная жидкостью
полость тела мезодермального (см.)
происхождения, окруженная мезодермальной
мембраной (ср. псевдоцель, гемоцель);
представлена как обширными
гидростатическими полостями, ограниченными перитонеу-
мом (см.) (см. схизоцель, энтероцель), так
и выстланным эпителием (см.)
пространством внутри мезодермальных органов.
Целомодукт—(а) слепо начинающаяся
осморегуляторно-экскреторная железа с
протоком мезодермального происхождения
(ср. нефридий); (б) открытый в целом (см.)
мезодермальный проток для выведения
наружу гамет или целомической жидкости.
Целомоцит — клетка, взвешенная в
целомической (см.) жидкости.
Цефализашш — развитие головы в ходе
филогенеза (см.) или онтогенеза (см.).
Церки—пара варьирующих по форме
придатков на последнем абдоминальном
сегменте многих насекомых.
Циррус—выворачивающийся копуля-
ционный орган (ср. пенис, гоноподия).
Циста—инкапсулированная,
устойчивая к высыханию стадия жизненного
цикла.
Цитохромы—дыхательные пигменты,
локализованные в митохондриях; по
структуре напоминают гемоглобин.
Чешуйчатый эпителий -эпителий (см.),
образованный уплощенными клетками.
Щетинки—мелкие жесткие выросты на
теле аннелид, погонофор, эхиур и др.; могут
содержать клеточный материал или иметь
кутикулярную природу.
560
Словарь терминов
Щупальце—тонкий гибкий придаток,
часто с сенсорной функцией; иногда
используется для улавливания пищи.
Эволюция—происхождение и изменение
в геологических масштабах времени.
Эвтелия—состояние, при котором
клетки взрослой особи не делятся, т. е. число их
постоянно (обычно невелико), а рост тела
идет за счет увеличения размеров клеток.
Экзоподит—наружная ветвь двуветви-
стой конечности членистоногих (ср. эндопо-
дит, эпиподит).
Эктодерма—наружный зародышевый
слой, покрывающий гаструлу (см.) (ср.
энтодерма, мезодерма).
Эктотермия—получение организмом
тепла из внешней среды, а не за счет
собственного метаболизма.
Эмбрион—зародыш, ранняя стадия
развития, выходящая из яйца (см.) или
остающаяся в нем и не способная к
самостоятельной жизни.
Эндоподит—внутренняя ветвь двувет-
вистой конечности членистоногих (ср.
экзоподит, эпиподит).
Энтероцель—целом (см.),
сформировавшийся путем отшнуровывания мезодер-
мальных карманов от эмбриональной
кишки (ср. схизоцель).
Энтодерма — внутренний зародышевый
слой, образующий эмбриональную кишку
гаструлы (см.) (ср. эктодерма, мезодерма).
Эпиболия—обрастание подвижными,
бедными желтком клетками клеток,
богатых желтком, во время гаструляции (см.).
Эпидермис — наружный клеточный
покров тела, формирующийся из эктодермы
(см.).
Эпиподит—отросток на базальном
членике конечностей членистоногих;
возможно, от эпиподита происходят грифельки
(см.) (ср. эндоподит, экзоподит).
Эпителий—слой ткани (см.),
покрывающий свободную поверхность, например
выстилающий полость.
Эпифауна—бентосные (см.) животные,
населяющие поверхность субстрата (ср. ин-
фауна).
Эукариоты — организмы, в клетках
которых находятся окруженные мембранами
ядро и органеллы (ср. прокариоты); все
организмы, кроме бактерий.
Ювенилизация—приобретение
взрослым организмом признаков
неполовозрелой особи.
Ювенильный—неполовозрелый.
Яйцеклад — трубчатый орган некоторых
насекомых, используемый для откладки яиц
в специфические места.
Яйцеклетка—женская гамета (см.).
Яйцо—неопределенный термин,
обозначающий исходную стадию развития
животного— яйцеклетку (см.), зиготу (см.) или
совокупность клеток развивающегося
эмбриона (см.), пищевых запасов и т.д.,
находящуюся внутри общей капсулы или
оболочки.
ИСТОЧНИКИ ИЛЛЮСТРАЦИЙ
Alexander R.McN. 1979. The Invertebrates.
Cambridge University Press, Cambridge.
Alldredge A. 1976. Sci. Am., 235(1), 94—102.
Anderson D. T. 1964. Embryology and Phylogeny in
Annelids and Arthropods. Pergamon Press, New
York.
Anderson P.A.V., Bone Q. 1980. Proc. R. Soc.
Lond(B), 210, 559- 574.
Atkins D. 1933. J. Mar. Biol. Ass., UK, 19, 233 -252.
Atwood H. L. 1973. Am. J. Zool., 13, 357—378.
Austin C.R. 1965. Fertilisation. Prentice Hall Inc.,
New Jersey.
Baehr J. C, Porcheron P., Dray F. 1978. С R. Acad.
Sci. (Paris), 278D, 253 -525.
Barnes R.D. 1980. Invertebrate Zoology, 4th edn.
Saunders, Philadelphia.
Barnes R.S.K., Hughes R.N. 1982. An Introduction
to Marine Ecology. Blackwell Scientific
Publications, Oxford.
Bastiani M. J., Doe C. Q., Helfand S. L., Goodman
C. S. 1985. Trends in Neurosciences, 8, 257—266.
Bayne B.L., Thompson R.J., Widdows J. 1976. In:
B. L. Bayne (Ed.) Marine Mussels: Their
Physiology and Ecology. Cambridge University Press.
Cambridge.
Besker G. 1937. Z. Morph. Okol. Tiere, 33, 72—127.
Belk D. 1982. In: S.P. Parker (Ed.) Synopsis and
Classification of Organisms, 2. 174 -180.
McGraw-Hill, New York.
Bergquist P. R. 1978. Sponges. Hutchinson, London.
Berrill N.J. 1950. The Tunicata. Ray Society,
London.
Biscardi H.M., Webster G.C. 1977. Exp. Geront.,
12, 201—205.
Blower J.G. 1985. Millipedes. Brill, Leiden.
Boeckh J.. Ernst K-D.. Sass H., Waldow U. 1975. In:
D.Denton (Ed.) Olfaction and Taste, V, 239 245.
Academic Press, New York.
Boss K.J. 1982: In: S. P. Parker, (Ed.) Synopsis and
Classification of Living Organisms, 1, 945—1166.
McGraw-Hill, New York.
Brill В. 1973. Z. Zellforsch., 144, 231—245.
Buchsbaum R. 1951. Animals Without Backbones,
Vol. 1. Pelican, Harmondsworth.
Bullough W.S. 1958. Practical Invertebrate
Anatomy, 2nd edn. Macmillan, London.
Cain A. J., Sheppard P.M. 1954. Genetics, 39, 89-
116.
Calkins G.N. 1926. The Biology of the Protozoa.
Bailliere Tindall and Cox, London.
Calow P. 1985. Causes de la mort i costos-d'autopro-
teccio. In: Biologia Avui. Fundacio Caixa de
Pensions, Barcelona.
Calow P. 1986. In: R. Peberdy and P. Gardner (Eds)
The Collins Encyclopedia of Animal Evolution,
pp. 90—91. Equinox, Oxford.
Calow P., ReadD. A. 1986. In: S.Tyler (Ed.)
Advances in the Biology of Turbellarians and Related
Platyhelminthes, pp. 263—272. D. W. Junk
Publishers, Dordrecht.
Carpenter W. B. 1866. Phil. Trans. Roy. Soc. Lond.,
156, 671—756.
Caullery M., Mesnil F. 1901. Arch. Anat. Micros., 4,
381—470.
Clark A. H. 1915. US Natn. Mus. Bull, 82, Vol. 1(1),
1—406.
Clark R. B. 1964. Dynamics in Metazoan Evolution
Clarendon Press, Oxford.
Clarke K. U. 1973. The Biology of the Arthropoda.
Arnold, London.
Clarkson E. N. K. 1986. Invertebrate Palaeontology
and Evolution, 2nd edn. Allen and Unwin,
London.
Clement A. C. 1962. J. Exp. Zool., 149, 193 215.
Cloudsley-Thompson J. 1958. Spiders. Scorpions.
Centipedes and Mites. Pergamon Press, London.
Cohen A. C. 1982. In: S. P. Parker (Ed.) Synopsis and
Classification of living Organisms, 2, 181- 202.
McGraw-Hill, New York.
Colwin L.H., Colwin A.L., 1961. J. Biophys. Bioc-
hem. Cytol., 10, 231 -254.
Conway Morris S. 1979. Ann. Rev. Ecol. Syst., 10,
327—349.
Conway Morris S. 1985. Phil. Trans. Roy. Soc. Lond.
(B), 307, 507—582.
Corliss J. O. 1979. The Ciliated Protozoa, 2nd edn.
Pergamon Press, Oxford.
Cuenot L. 1949. In: P-P. Grasse (Ed.) Traite de Zoo-
logie, VI, 3—75. Masson, Paris.
Danielsson D. 1982. Norw. N-Atlantic Exped.
(1876—1878) Rep. Zool., 21, 1 28.
Darnell J. E., Lodish H.F., Baltimore D. 1986. Mole-
562
Источники иллюстраций
cular Cell Biology. Scientific American Books Inc.,
New York.
Davies J. 1983. Ageing. Edward Arnold, London.
Dehorne A. 1933. Bull. Biol. Fr. Belgique, 67, 298—
326.
Dethier V.E. 1976. The Hungry Fly. Harvard
University Press, Cambridge, Mass.
Dixon A.F.G. 1973. Biology of Aphids. Studies in
Biology No. 44. Edward Arnold, London.
Durchon M. 1967. L'endocrinologie chez le Vers et les
Molluscs. Masson, Paris.
Eakin R. M. 1968. Evol. Biol., 2, 194—242.
Elner R. W., Hughes R.N. 1978. J. Anim. Ecol., 47,
103—116.
Epel D. 1977. Sci. Am., 237(5), 129 -138.
Fewkes J. 1883. Bull. Mus. Сотр. Zool., Harward,
11, 167—208.
Fingerman M. 1976. Animal Diversity, 2nd edn.
Holt, Rinehart and Winston, New York.
Fox H. M., Wingfield C. A., Simmonds B. G. 1937. J.
Exp. Biol., 14, 210—218.
Fraser J. H. 1982. British Pelagic Tunicates.
Cambridge University Press, Cambridge.
Fretter V., Graham A. 1976. A Functional Anatomy
of Invertebrates. Academic Press, London and
New York.
Friesen W. Q., Poon W., Stent G. S. 1976. Proc. Nat.
Acad. Sci. (USA), 73, 3734—3738.
George J.D., Southward E.C. 1973. J. Mar. Biol.
Ass. UK, 53, 403—424.
Gibson P.H., Clark R.B. 1976. J. Mar. Biol. Ass.
UK, 56, 649—674.
Gibson R. 1982. In: S.P. Parker (Ed.) Synopsis and
Classification of Living Organisms, 823—846.
McGraw-Hill, New York.
Gilbert L. E. 1982. Sci. Am., 247(2), 102—107B.
Glaessner M.F., Wade M. 1966. Palaeontology, 9,
599—628.
Gnaiger E. 1983. J. Exp. Zool., 228, 471 490.
Golding D. W., Pow D. V. 1987. In: M. С Thorndyke
and G.J. Goldsworthy (Eds) Neurohormones in
Invertebrates. Cambridge University Press,
Cambridge.
Golding D. W., White A. C. 1974. Tissue Cell, 6,
599—611.
Golding D. W., White A. C. 1977. Int. Rev. Cytol.
Suppl., 5, 189—302.
Goodrich E. S. 1945. Quart. J. Micros. Sci., 86, 113—
393.
Gordon D.P. 1975. Cah. Biol. Mar., 16, 367—382.
Grasse P.P. (Ed.). 1948. Traite de Zoologie, XI.
Masson, Paris.
Grasse P.-P. (Ed.). 1965. Traite de Zoologie, IV.
Masson, Paris.
Green J. 1961. A Biolody of Crustacea. Witherby,
London.
Gupta B.L., Berridge M.J. 1966. J. Morphol., 120,
23—82.
Gustafson Т., Wolpert L. 1967. Biol. Rev., 42, 442—
498.
Hackman R.H. 1971. In: M.Florkin, B.T. Scheer
(Eds) Chemical Zoology, 6, 1—62. Academic
Press, New York.
Hansen K. 1978. In: G. I. Hazelbauer (Ed.) Taxis and
Behaviour Receptors and Recognition, 5B, 231—
292. Chapman and Hall, London.
Hardy A. C. 1956. The Open Sea. The World of
Plankton. Collins, London.
Hedgpeth J. W. 1982. In: S. P. Parker (Ed.) Synopsis
and Classification of Living Organisms, 2, 169—
173. McGraw-Hill, New York.
Hermans С. О., Cloney R. A. 1966. Z. Zellforsch. 72,
583—596.
Hermans C. O., Eakin R. M. 1974. Z. Morph. Tiere,
79, 245—267.
Hescheler K. 1900. In: A.Lang (Ed.) Lehrbuch der
Vergleichenden Anatomie der Wirbellosen Thiere,
3rd edn. Fischer, Jena.
Hess R. 1987. Z. Wiss. Zool., 62, 247—283.
Higgins R. P. 1983. Smithsonian Contrib. Mar. $ci.,
18, 1 -131.
Hodgkin A.L., Huxley A.F. 1952. J. Physiol., 117,
500—544.
Holland N. D., Grimmer T. C, Kubota H. 1975. Biol.
Bull., 148, 219—242.
Holt C.S., Waters T.F. 1967. Ecology, 48, 225—
234.
Hughes Т. Е. 1959. Mites or the Acari. Athlone,
London.
Hummon W. D. 1982. In: S. P. Parker (Ed.) Synopsis
and Classification of Living Organisms, 1, 857 -
863. McGraw-Hill, New York.
Hyman L.H. 1940. The Invertebrates. Vol. I.
Protozoa through Ctenophora. McGraw-Hill, New
York.
Hyman L.H. 1951. The Invertebrates. Vol. II. Platy-
helminthes Phynchocoela. McGraw-Hill, New
York.
Imms A.D. 1964. A General Textbook of
Entomology, 9th edn., revised reprint. Methuen, London.
ho Y. 1980. Comparative Ecology. Cambridge
University Press, Cambridge.
Jagersten G. 1973. The Evolution of the Metazoan
Life Cycle. Academic Press, New York.
Jeannel R. 1960. Introduction to Entomology.
Hutchinson, London.
Jones A.M., Baxter J. M. 1987. Molluscs: Caudofo-
veata, Solenogastres. Polyplacophora and Scapho-
poda. Brill, Leiden.
Jones J.D. 1955. J. Exp. Biol., 32, 110—125.
Jones M.L. 1985. In: S. Conway Morris et al. (Ed.)
The Origin and Relationships of Lower
Invertebrates, 327—342. Clarendon Press, Oxford.
Joosse J., Geraerts W. P. M. 1983. In: A. S. M. Saleu-
din and К. М. Wilbur (Eds) The Mollusca, Vol. 5.
Academic Press, New York.
Источники иллюстраций
563
Kandel E. R., Schwartz J.H. 1982. Science, 218,
433—443.
Kennedy D. 1976. In: J. С Fentress (Ed.) Sinpler
Networks and Behaviour. Sinauer, Sunderland,
Massachusetts.
Kershaw D.R. 1983. Animal Diversity. University
Tutorial Press, Slough.
Krebs J. R., Erichsen J. Т., Webber M. I., Charnov
E. L. 1977. Anim. Behav., 25, 30—38.
Kudo R.R. 1946. Protozoology, 3rd edn. Thomas,
Springfield.
Lacaze-Duthiers F.J.H. de. 1861. Ann. Sci. Nat.
(Zool.), 15, 259—330.
Lamb M.J. 1977. Biology of Ageing. Blackie,
Glasgow.
Lemche #., Wingfield KG. 1959. Galathea Rep., 3,
9—71.
Lester S.M. 1985. Mar. Biol., 85, 263—268.
Lewis J.G.E. 1981. The Biology of Centipedes.
Cambridge University Press, Cambridge.
McFarland W.N., Pough F.N., Cade T.J., Heiser
J.B. 1979. Vertebrate Life. MacMillan, New
York.
MacKinnon D.L., Haws R.S.J. 1961. An
Introduction to the Study of Protozoa. Clarendon Press,
Oxford.
McLaughlin P. A. 1980. Comparative Morphology of
Recent Crustacea. Freeman, San Francisco.
Manton S.M. 1952. J. Linn. Soc. (Zool.), 42, 93—
117.
Manton S.M. 1965. J. Linn. Soc. (Zool.), 45, 251—
483.
Marcus E. 1929. Klassen und Ordnungen des Tier-
reichs, 5, 1—608.
Margulis L., Schwartz K. V. 1982. Five Kingdoms.
Freeman, San Francisco.
Marion M.A.-F. 1886. Arch. Zool. Exp. Gen. (2), 4,
304 -326.
Meglitsch P. A. 1972. Invertebrate Zoology, 2nd edn.
Oxford University Press, Oxford.
Millar R. H. 1970. British Ascidians. Academic Press,
London.
Miller R.L. 1966. J.Exp. Zool., 162, 23^4.
Miyan J. A., Ewing A. W. 1986. J. Exp. Biol., 116,
313- -322.
Moore R. C. (Ed.). 1957. Treatise on Invertebrate
Paleontology, Part L. Mollusca, 4, University of
Kansas Press, Lawrence.
Moore R. C. (Ed.) 1965. Treatise on Invertebrate
Paleontology, Part H. Brachiopoda. University of
Kansas Press, Lawrence.
Morgan C.I. 1982. In: Parker S.P. (Ed.) Synopsis
and Classification of Living Organisms, 2, 731—
739. McGraw-Hill, New York.
Morgan С I., King P.E. 1976. British Tardigrades.
Academic Press, London.
Mortensen T. 1928—1951. A Monograph of the Ec-
hinoidea. 5 vols. Reitsel, Copenhagen.
Muscatine L. et al., 1975. Symp. Soc. Exp. Biol., 29,
175—203.
Newell N. D. 1949. Evolution, 3, 103—240.
Nichols D. 1962. Echinoderms, 3rd edn. Hutchinson,
London.
Nichols D. 1969. Echinoderms, 4th edn. Hutchinson,
London.
Noble E. R., Noble G. A. 1976. Parasitology. Lea and
Febiger, Philadelphia.
Ohnishi Т., Sugiyama M. 1963. Embryologia, 8, 79—
88.
Olive P. J. W. 1980. In: D. С Rhoads and R. A. Lutz
(Eds), Skeletal Growth in Aquatic Organisms.
Plenum Press, New York.
Olive P.J. W. 1985. Symp. Soc. Exp. Biol., 39. 267 -
300.
Olive P.J. W. 1985b. In: Syst. Assosiation, series 28,
42—59. Oxford University Press, Oxford.
Oschman J.L., Berridge M.J. 1971. Federation
Proceedings, Federation of American Societies for
Experimental Biology, 30, 49—56.
Pashley H.E. 1985. The Foraging Behaviour of
Nereis diversicolor (Polychaeta). PhD Thesis,
University of Cambridge.
Pearson К G. 1983. J. Physiol. (Paris), 78, 765—771.
Pennak R. W. 1978. Fresh-water Invertebrates of the
United States, 2nd edn. Wiley. New York.
Phillipson J. 1981. In: С R. Townsend and P. Calow
(Eds) Physiological Ecology, 20—45. Blackwell
Scientific Publications, Oxford.
Pierrot-Bults A. C, Chidgey К. С 1987. Chaetognat-
ha. Brill, Leiden.
Ptingle J. W. S. 1975. Insect Flight. Oxford
University Press, Oxford.
Rice M. 1985. In: S. Conway Morris J. D. George R.
Gibson and H.M. Piatt (Eds) Origins and
Relationships of Lower Invertebratebrates. Clarendon
Press, Oxford.
Ritter-Zahony R. von. 1911. Das Tierreich, 29, 1—35.
Robbins Т.Е., Shick J.M. 1980. In: Nutrition in the
Lower Metazoa, Pergamon Press, Oxford.
Russell-Hunter W. D. 1979. A Life of Invertebrates.
MacMillan, New York.
Sanders D. S. 1982. Insect Clocks, 2nd edn.
Pergamon Press, Oxford.
Sanders H.L. 1957. Syst. Zool., 6, 112—128.
Savory Т. Н. 1935. The Arachnida. Edward Arnold,
London.
Schepotieff A. 1909. Zool. Jb. Syst., 28, 429-^48.
Schmidt-Nielsen K. 1984. Scaling. Cambridge
University Press, Cambridge.
SebensK.P., De Riemer K. 1977. Mar. Biol., 43,
247—256.
Sedgwick A. 1888. Quart. J. Micros. Sci., 28, 431 -
493.
Shelton G.A. B. 1975. Proc. R. Soc. Lond. B, 190,
239—256.
564
Источники иллюстраций
Shepherd G. M. 1983. Neurobiology. Oxford
University Press, Oxford.
Sheppard P. M. 1958. Natural Selection and
Heredity. Hutchinson, London.
Shick P. M., Dykens J. A. 1985. Oecologia, 66, 33—
41.
SihaP.H.D.H. de. 1962. J. Exp. Biol., 39, 483 -
490.
Singh C.L. 1975. Cell Tiss. Res., 158, 391—407.
Sleigh M. A., Dodge J. D., Patterson D.J. 1984. In:
R. S. K. Barnes (Ed.) A Synoptic Classification of
Living Organisms, 25—88. Blackwell Scientific
Publications, Oxford.
Smart P. 1976. The Illustrated Encyclopedia of the
Butterfly World. Hamlyn, London.
Smyth J.D. 1962. Introduction to Animal
Parasitology. The English Universities Press, London.
Smyth J.D., Hahon D. W. 1983. The Physiology of
Trematodes. Cambridge University Press,
Cambridge.
Snodgrass R. E. 1935. Principles of Insect
Morphology. McGraw-Hill, New York.
Snow K.R. 1970. The Arachnids: An Introduction.
Routledge and Kegan Paul, London.
Southward E. С 1980. Zool. Jb. Anat. Ontog., 103,
264—275.
Southward E. С 1982. J. Mar. Biol. Ass., UK, 62,
889—906.
Spengel J. W. 1932. Sci. Res. Michael Sars N.
Atlantic Deep Sea Exped., 5(5). 1—27.
Stein P.S.G. 1971. J. Neurophysiol., 34, 310—318.
Sterrer W.E. 1982. In: S.P. Parker (Ed.) Synopsis
and Classification of Living Organisms, 1, 847—
851. McGraw-Hill, New York.
Stiasny G. 1914. Z. Wiss. Zool., 110, 36—75.
Strausfeld N.J., N asset D.R. 1981. In: H. Autrum
(Ed.) Handbook of Sensory Physiology, Vol.
VII/6B. Springer-Verlag, Berlin.
Stretton A. O. W., Davis R. E., Angstadt J. D., Don-
moyer J.E., Johnson CD. 1985. Trends in Ne-
uroscienes, 8, 294—299.
Strumwasser F. 1974. Neurosciences Third Study
Program, 459—478.
Treherne J. E., Foster W. A. 1980. Anim. Behav., 28,
1119—1122.
Trench R.K. 1975. Symp. Soc. Exp. Biol., 29, 229—
265.
Trinkans J. P. 1969. Cells into Organs. Prentice Hall,
New Jersey.
Trueman E. R., Foster-Smith R. 1976. J. Zool.,
Lond., 179, 373—386.
Turner CD. 1966. General Endocrinology.
Saunders, Philadelphia.
Valentine J. W., Moores E.M. 1974. Sci. Am.,
230(4), 80 89.
Wallace M.M.H., Mackerras l.M. 1970. In:
C.S.I.R.O., The Insects of Australia, 205—216.
Melbourne University Press, Melbourne.
Warner G. F. \9П. The Biology of Crabs. Elek,
London.
Waterman Т.Н. 1960. The Physiology of Crustaces.
Academic Press, New York.
Welsch U., Storch V. 1976. Comparative Animal
Cytology. Sidgwick and Jackson, London.
Wenyon CM. 1926. Protozoology. Bailliere, Tindall
and Cox, London.
Whittington H. B. 1979. In: M. R. House (Ed.) The
Origin of Major Invertebrate Groups, 253—268.
Academic Press, London.
Widdows J., Bayne B.L. 1971. J. Mar. Biol. Ass.,
UK, 51, 827—843.
Wigglesworth V.B. 1940. J. Exp. Biol., 17, 201 -
222.
Wigglesworth V.B. 1972. Principles of Insect
Physiology, 7th edn. Chapman and Hall, London.
Wine J. J., KrasneF.B. [982. In: D.C. Sandeman
and H. L. Atwood (eds) The Biology of Crustacea,
Vol. 4, 241—292. Academic Press, New York.
Wright A. D. 1979. In: M. R. House (Ed.). The
Origin of Major Invertebrate Groups, 235—252.
Academic Press, London.
Wrona F.J., Davies R. W. 1984. Can J. Fish.
Aquatic Sci., 41, 380—385.
Yager J., Schram F. R. 1986. Proc. Biol. Soc. Wash.,
99(1), 65—70.
Young J. Z. 1939. Phil. Trans. Roy. Soc. Lond. В.,
229, 465—503.
Young J.Z. 1962. The Life of Vertebrates, 2nd edn.
Clarendon Press, Oxford.
Young J.Z. 1963. Nature (Lond), 198, 636—630.
Zullo V. Z. 1982. In: S. P. Parker (Ed.) Synopsis and
Classification of Living Organisms, 2, 220—228.
McGraw-Hill, New York.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
квогадро число 377
Агглютинины 391
Адаптация 543
Адаптивная радиация 42, 43, 45, 47. 242, 368
Аденозиндифосфат (ADP) 339
Аденозинтрифосфат (АТР) 309, 339—342, 542
Адипокинетический гормон 541
Акклимация 356—357
Акросомная нить 440—443
Акселерация 548
Аксоны 488. См. также Гигантский аксон
Активный транспорт и экскреция 374, 375
Актин 311
Актинии 62, 63, 72, 73, 289
— симбиоз 75
Актинотроха 172
Алломоны 385, 524
Амакриновые клетки 489
Амбулакральная система 318—319
Амбулакральные ножки 269, 318. 319
Амебоидные клетки 389—391
Амебоидный рост 533
Амебоциты 62
Аминокислоты 12
Аммиак 361, 362
Аммониты 43, 548
Аммонотелия 361, 362
Амфибластула 57
Амфиды 536
Амфиподы см. Бокоплавы
Анаболические процессы 542, 543
Анаэробный метаболизм 340, 341
Аннелиды (кольчатые черви, кольчецы) 31—32, 135—149
— гаметогенез 417
— нервная система 494, 495, 500
— питание 267, 270, 279. 498
— развитие 448
— размножение 403
— регенерация 481—483
— сегментация 136, 137
— сетчатка 507
— старение 394
— циркуляторная система 343, 345
— экскреция 369
Аноксия 340
Аноплиды 102
Антеннальные (зеленые) железы 371, 374, 529
Антенны 270, 462
Антигены 391
Антифертилизин 440
Анус 277, 278
Анхетрон 29
Аплизия (морской заяц) 532—533, 535
Аппендикуларии 209—211, 270
Аристотелев фонарь 201, 281
Артроподы см. Членистоногие
Археоциаты 42
Архианнелиды 141, 143, 144
Асконоидный тип организации 55
Аспартат 341
Асцидии 205—207, 384, 445
— развитие 452, 470, 471
Атриум 285
Ацетилхолинэстераза 471
Бабочки 241
— защита 389
— зрение 503
Бабочки-данаиды 385
Бактерии как пища 265, 266
размножение 396, 430, 431
— симбиотические 275
Бделлоидеи 123—124
Беззамковые 173 -175
Беззубки 410
Белки 12—13
Бензохиноны 387
Березовая пяденица 382—383
Бесполое размножение 85, 111, 544
Бессяжковые 234
Билатеральная симметрия 79
Биологические часы 501, 535
Биомеханика 308—338
Биотопы 50
Биохимия .14
Благородный коралл 74
Бластомеры 469
Бластула 24, 446
Блохи 330
Блюдечки 161, 353, 357, 405
— локомоция 315
Бокоплавы 260, 262, 270, 366, 367
— движение 353
Бомбикол 520, 524
Бора эффект 345
Бочоночник 209
Брюхоногие моллюски (гастроподы) 159—162, 364
дыхание 160
зрение 504, 507
питание 270, 275
Брюхоресничные черви 105—107
Брюшные сегменты 239
Бычий цепень 91
Велигер 312, 453
Вентральная мускулистая глотка 141
Вертикальные миграции 381
Веслоногие рачки 36, 253—255, 270
размножение 404
Ветвистоусые рачки 401
Вирусы 396, 429, 430
Витамин Е 379
Вителлогенины 526
Вихревое движение воздуха 337—338
Влагосберегаюгцие органы 215
Водители ритма 499, 501
566
Предметный указатель
Водоотталкивающий покров 238
Водоросли как пища 267, 274, 275, 284
Водяной ослик 367
Волосатики 114—115, 403
Восприятие 509—510
Вторичноротые 39—40, 45, 184—212
Выделительные системы 370—375
Выживание, выживаемость 43—47, 379, 425—428
Вымирание 18, 42—44, 47. 48
Р-Галактозидаза 545
Гаметы 396, 544
Гамма-облучение 378, 379
Гастродермис 61, 64
Гастроподы см. Брюхоногие моллюски
Гастротрихи 29—30, 39, 99, 403
Гаструляиия 23—24, 446—450
Гемато-энцефалический барьер 491
Геммула 57
Гемоглобин 345, 346
Гемоцель 29
Гемоцианин 345—348
Гемоциты 391
Гемэритрин 345—348
Генералисты 502
Генетическая программа 12—13, 465—466
Ген-оператор 545
Геофизические циклы 418
Гепатопанкреас 278
Гермафродитизм 404, 411
Гетеробранхии 161
Гетеротрофы 265, 542
Гигантский аксон 484, 487, 497
— лобулярный детектор движения (LGMD) 511—512,
515—516
Гидроидные полипы (гидроиды) 61—69, 440
движение 500
нейроны 514
регенерация 463
Гидромедузы 66—68
— движение и нервная система 494, 500
Гидростатический скелет 29, 316, 333—334
Гидры 61, 62, 66—69
— движение 500
— регенерация 493
Гимнолематы 180 -181
Гиперморфоз 548
Гиперосмотическая среда 367
Гипоневральная система 495
Глаза 504—509. См. также Рабдомерные глаза,
Сложные глаза
Глазки 506
Глазные железы 420
Глиальные клетки 489—499
Гликолиз 340
Глотка 141, 278, 280, 285, 286
Глоточный насос 109, 111
Гнатостимулиды 29—30, 103—105, 403
Головной активатор 523
Головоногие моллюски 165—169, 321, 337
размножение 403, 420
Головохордовые см. Ланцетники
Голодание 85—86, 355
Голожаберные моллюски 387
Голометаболические насекомые 458
Голотурии (морские огурцы) 201—204
— дыхание 345, 349
— питание 269
— размножение 403
Гомеобоксы 462
Гомеозисные гены 462, 479—481
Гомойотермные животные 355
Гонадостимулирующее вещество (GSS) 525
Гонангий 440
Гонозоиды, 67, 70
Гонопор 371
Горгонарии 389
Гормоны 419, 420, 520, 521, 524
Горячие линии 497—499
Градиентов теория (развития) 475—477
Гребешок 504
Гребневики 386
Гребные пластинки 76
Грибы как пища 276, 282
Гуанин 362
Губки 19, 22—23, 27, 51, 54—59
— защита 384
— нейроидная система 492—493
— питание 267—268, 278, 402
— реагрегация клеток 389
Губоногие многоножки 231—232
защита 387, 388
локомоция 324—325
Дактилозоиды 67, 69
Дафнии 507
Двигательная активность, генерирование программ
517—518
Двигательные центры 516
Движение 308—338, 500
— в толще субстрата и трубках 319—321
— клеточные механизмы 309—312
— нервные механизмы 516—520
Двукрылые насекомые, защита от хищников 386
оогенез 414, 415
— старение 393
Двупарноногие многоножки 232—233, 324
Двустворчатые моллюски 162—164
дыхание 345, 350
жизненный цикл 410
метаболизм 341
мыщцы и движение 519
размножение 403
экскреция 366, 367
Двухвостки 234, 235
Дезоксирибонуклеозиды 15
Демографическая теория жизненного цикла 425
Десятиногие раки 258—262
Детерминация 450—453, 460, 545
Диапауза 410, 421 424
Дивергенция 511
Динофлагеллаты 51
— как пища 289
— симбиоз 273
Диуретический гормон 541
Дифференцировка 16, 544
Диффузия 376—377
ДНК 12, 13, 15
Докембрийские остатки животных 18—19
Дорсальный непарно-срединный нейрон (DUM) 519
Древесина как пища 282
Дробление 445-^146, 466—477, 544
— радиальное 468—469
— спиральное 37, 355, 466—468
Дрозофила см. Drosophila melanogaster
Дыхание 342—355
— анаэробное 542
— аэробное 542
Дыхательные органы 348—350
— пигменты 345—346
Дыхательный ток воды 160
Естественный отбор 13—14, 16, 48, 426, 427
Жаброногие рачки 250—251, 404
Жабры 348, 349
Жабы 511
Жало 301, 386
Предметный указатель
567
Жгутики 309—311
Жгутиконосцы 22, 23, 51
— симбиоз 282
Жгучие (огненные) кораллы 69
Желточник 417
Желточные клетки 445
Желудок 278, 287
Жертвы 293, 294, 299—303, 307
— защитные приспособления 301
Жидкости тела см. Полостная жидкость
Жизненные циклы 263, 433—438, 543
и бесполое размножение 397—399
и половое размножение 407—412, 432
морских беспозвоночных 435—436, 471—479
насекомых 436—438, 479—481
Жужжальца 536
Жуки-бомбардиры 387
Жук-носорог 389
Загрязнители 392—393
Заднежаберники 384
Замковые 175
Запасные вещества 416
Зародышевые листки 24
Защита 378—395
Защитная окраска 381
Защитное вооружение 301
Зеленый краб 224, 294
Земляной червь 490, 497, 540
локомоция 316, 317
экскреция 374, 375
Зеркала 506—507
Змеехвостки см. Офиуры
Зоб 278
Зооиды 38, 178—181, 209, 515
Зооксантеллы 75, 289, 290
Зоохлореллы 75, 289
Зрение 503—509
Зрительные пигменты 503—504
Иглокожие 39, 40, 191—204
— нейрорегуляция 490
— нервная система 493. 495
— питание 275
— происхождение 19
— развитие 193, 446, 454, 469—470
— размножение 403
— циркуляторная система 345
Избегания реакция 380, 514, 519
Известковые губки 58
Изоосмотический раствор 376
Изотонический раствор 376
Имагинальные диски 458—462
Иммунные реакции 389—392
Интернейроны 488, 516, 517
Ионная регуляция 363—364, 376—377
Ископаемые остатки 39—43
История изучения беспозвоночных 10 -11
Кайромоны 385
Калиевые каналы 532
Кальмары 167, 168, 348, 484
— зрение 506
— нервная система 519, 530—531
— плавучесть 364
— феромоны 524
Каракатица 167, 168, 364, 365
— защита 381
Карапакс 259, 260
Карбоксилирование 341
Карденолиды см. Кайромоны
Кардиальные ганглии 501
Карповые вши (карпоеды) 252—253
Катаболизм 339—340, 542, 543
Кетокислоты 361
Кинетическая энергия 331—332
Киноринхи 31, 100, 116—117
— размножение 403
Кислород 357—359
— кривая диссоциации 345—347
— поглощение 342 -343, 351, 357—358
Киты, питание 304
Кишечник 277—279, 289
Кишечнодыщащие 39, 188—189, 372
Кишечнополостные 19, 23, 24, 77
— питание 27, 278
Кладистская школа 41
Кладоны 20, 46
Классификация, пятииарственная система 52
Клетки, гибель 533
— двигательные механизмы 309—312
— дифференцировка 16
— программа развития 533—535
— происхождение и эволюция 14—15
— ренетты 111
— сенсорные 503
Клеточная реагрегация 389
Клеточные линии 473—474
— мембраны 486
Клещи 224
Клоп-гребляк 366
Клопы 241, 299—300, 526
— питание 276
Книдарии 19, 24, 25, 268—269, 284, 363, 364
— нервная система 492, 493, 513
— строение 463
Книдоцисты 24, 62—64
Коксальные железы 371
Колеоидеи 167—168
Коллобласты 23, 76
Коловратки 31, 100, 121—125, 398
— размножение 403
Колониальность 21, 45
Кольчатые черви см. Аннелиды
Командные нейроны 516 517, 539
Комары 241, 367
Комменсализм 75, 80
Кампартменты 21
Компетенция 452
Конвергенция 511
Конечности червей 213
— членистоногих 321—326
Конкурентное исключение 47
Конкуренция за пищевые ресурсы 263
— межвидовая 48
Консументы 277, 283
Контралатеральные детекторы движения (DCMD) 516
Конформеры 357—358
Концентрациклоидеи 198, 200
Копеподы как пища 279
Кораллы, коралловые полипы 59, 61, 69, 73—75, 272,
290, 343
— рефлекторные действия 513
Корацидий 91—92
Кортикальная реакция 441, 443
Коэволюция 426
Крабы 258, 262
— зрение 507, 508
— конечности и локомоция 323—325
— нервная система и поведение 518. 539
— питание 306
— экскреция 367
Креветки 258, 367, 502 '
Кристаллический стебелек 287
Кровь 345—348
— концентрация ионов и осмотическое давление 363,
369
Круглые черви см. Нематоды
568
Предметный указатель
Крыло насекомых, жилкование 240, 241
защитные узоры 369
и полет 326—329, 337, 338
формы 326
Кры ложаберные 188—191
Ксенобиотики 392
Ктенидии 33, 268, 269, 347—349
Кубомедузы 69
Кузнечик, нервная система и поведение 533, 534
— питание 385
— прыжки 330
Куколки 245, 457, 458
Кутикула 213, 368, 529
— и экскреция 362
— нематод 108
— членистоногих и хождение 324
Лактатный путь (путь молочной кислоты) 340
Лактоза 545
Ланцетники (головохордовые) 211—216
Латеральное торможение 510
Легочная книжка 222, 349
Легочные моллюски 363
— слизни 384
Лейцинаминопептидаза 365—366
Лемниски 100
Ленточные черви 91—96
Лестница природы по Аристотелю 10, 11
Летательные мышцы 327, 520
Линзы 506—507
Линька 454
Липофусцин 393
Литораль 355, 356, 368
Личинки, личиночное развитие 403. 408. 411, 436
— дрозофилы 479
— морских беспозвоночных 408, 436, 437, 453—457,
477-^178
— морского желудя 478
— морских улиток 472
— насекомых 367, 457-^158, 461
— пелагические 454—455
Локомоция 308- -338
— животных с мягким телом 335—337
— ресничек 312
— членистоногих 321—326
Лопатоногие (скафоподы) 164—165
Лофофор 269
Лофофоровые (щупальцевые) 32, 37—39, 45, 170—183
— питание 267, 268, 287
Лоциферы 100. 117, 119
Лучистые 77
Магний 364
Макромеры 469
Макроэволюция 41—42, 47
Малощетинковые кольчецы (олигохеты) 144—148
Мальпигиевы сосуды (трубочки) 372, 373, 375
Маскировка 381, 383
Маточное молочко 524
Медоносная пчела 386—388, 400
— - биологические часы 501
феромоны 524
Медузы 61—65, 69—71
— нервная система 502
Мезентерий 72, 73
Мезоглея 23
Мезомеры 469
Мезохил 56
Мейоз 416, 430—432, 440, 525, 544
Меланизм 382—383
Мембраны 376, 486
Мерцательное пламя 371
Метаболизм 350—353, 362
Метаболическое окисление 368
Металлотионеины 392 393
Метаморфоз 420, 453—462, 477-^183
Метанефридии 370, 371, 374, 375
Метацель 37
Метахрональная ритмичность 312, 313, 317
1-Метиладенин 419, 520, 525
Механическая работа 309
Механические си.ш 309—312
Механорецепторы 536
Меченые линии 511
Мечехвосты 223, 345, 510
Мидии 294, 355, 365
— как пища 306
— развитие 457
— реакция на загрязнителя 392
Миноги 212
Микрокалориметр 352
Микромеры 469
Микроэволюция 41
Миксонефридии 371
Мимикрия 279, 301, 302, 385—386
Миозин 311
Мистакокариды 252
Митоз 544
Многоклеточность 21, 45, 51—98
Многоножки 230—234, 548
Многощетинковые кольчецы см. Полихеты
Многоядерные протисты 21
Модуль (патрон) нейронов 510
Мозаичное развитие 451—453, 471—474
Мозаичные глаза 507
Мозг 497-^199, 526, 541
— спонтанная активность 499
Мозговые волны 499
Мокрицы 345
Моллюски 33, 150—169
— гаметогенез 416, 417
— защита 383—384
— нервная система 495
— нога 315—316
— оогенез 419
— питание 267, 275, 287, 289
— развитие 453, 466, 469—474
— размножение 403, 420
— репродуктивные и выделительные органы 372
— экскреция 362
Моли 377
Моляльность 377
Молярность 377
Моноплакофоры 156—158
Морская среда 364. 418
Морские беспозвоночные 364, 418—420
жизненные циклы и развитие 435—436, 471—479
оплодотворение 397, 400
— водоросли как пища 275
— ежи 47, 200—206, 389
гаструляция 447
оплодотворение 441, 442
питание 275
развитие 452, 454, 455, 465, 475^77, 545, 546
размножение 403
— желуди 248, 255—257, 361
развитие 428, 457, 478
— звезды (астероидеи) \9& -198, 389, 403
гормоны 524—525
локомоция 318—319 ,
нервная система 495
развитие 441, 454
— лилии 195—196
нервная система 495
питание 273
размножение 403, 417, 418
— осадки 266
— пауки 224—227
Предметный указатель
569
— улитки 471—474
— уточки 256
Морфаллаксис 482
Морфогенез 544—546
Мотивация 530
Мотонейроны 488, 516, 518—519
Моча 367
Мочевая кислота 361—363
Мочевина 14, 361—363
Мочевой пузырь 374
Мукопротеины 287
Муравьи 283
Мутации 15
Мухи, зрение 537—538
— поведение 510
Мшанки 38, 39, 46, 47, 175—181
— питание 269, 287
— размножение 403
Мышечное сокращение 335
Мышечные волокна 311—315
Мышцы, работа 313—315
Мягкотелые животные 313—321, 370
Наземная среда 367—368, 421—424
Наземные беспозвоночные 276
Насекомые 234—245, 321 -323, 368, 370
— гаструляция 450
— гормоны 529
— жизненные циклы 411, 436—438
крылатые 237 245
— локомоция 325—329
— многоножкоподобные 236
— первичнобескрылые 234—237
— полет 520
— развитие 446, 456—462, 477—483
— ротовой аппарат 242
— сенсиллы 535—536
— сетчатка 537, 538
— эмбрионы 464
— эндокринная система 526—529
Натриевые каналы 531
Натриевый насос 486
Науплиус 247
Наутилоидеи 166
Наутилус 506
Научение 532—533
Независимые эффекторы 512—513
Нейрогемальный орган 522
Нейромедиаторы (химические медиаторы) 490—491,
Нейронные связи 533—535
— цепи 516—518
Нейроны 23, 488—489, 497, 516, 533—535
— командные 516—517, 539
— сенсорные 503
Нейропептиды 523
Нейропиль 495, 496
Нейросекреторные клетки 488, 521—522
Нейросекреция 521—522
Нейросуперпозиционные глаза 509
Нейроэндокринные системы 523—524, 541
Нектар 290
Нематоген 96, 97
Нематоды, нервная система 495, 518, 533
— плавание 518
— развитие 451
старение 111, 281
— экскреция 111
Нематофоры 100
Нематоцисты 23, 24, 63, 290
Немертины 29, 100—103, 403
— локомоция 314
— экскреция 369, 370
Необласты 464
Неомениоморфы 156
Непарный шелкопряд 526
Нервная система 484—485, 492—501, 533—535
автономные функции 500—501
спонтанная активность 499—501
Нервные волокна 502
— окончания 488
— сети 492—493
Нереиды 417, 420
Нерест 420
Нефридии 370, 371
Нефромиксий 372
Никотинамидадениндинуклеотид (NAD) 340
Ногохвостки 234
Норы, локомоция внутри них 319, 320
— роль в спасении от хищников 301, 302
Ньютона законы движения 308
Обгрызатели 281—284
Обезвоживание см. Потери воды
Облегченная реакция 513
Оболочники (личиночнохордовые) 205—211, 384
— нервная система 494, 502
— пелагические 207
— питание 270, 286
— размножение 403
Огнетелки 207
Одноветвистые рачки 45, 227—230, 239
локомоция 322
размножение 404
ротовой аппарат 229
экскреция 371
Окраска покровительственная (криптическая.) 381
— предостерегающая 385
Оксигеназы 392
Оксифотобактерии как симбионты 273
Октокораллы 74
Октопин 519, 520
Окукливание 527
Олигохеты [44—148,
Омар, локомоция 325
— нервная система 500
Омматидии 507, 508
Онихофоры 362, 371, 404
Онкосфера 91
Онтогенез 544—549
Опиновый путь 340
Оплодотворение 432, 439—445
Опсонины 391
Оптико-суперпозиционные глаза 509
Орган Y 529
Органические вещества 11, 14—15
— полимеры, добиологический синтез 14—15
Органогенез 544, 545
Ортонектиды 97—98
Осмоконформеры 365; 376—377
Осморегуляторы 365, 366
Осмос 376
Осмотическая регуляция 363—364
Осмотическое давление 376
Осьминоги 167
— защита от хищников 391
— Зрение 504
— мозг 499
— нервная система 495, 502
— пищедобывающее поведение 498—499
Отбор и защита от хишников 385. См. также
Естественный отбор, Стабилизирующий отбор
Офиуры (змеехвостки) 198, 199
— питание 269
— размножение 403
Охотники (преследователи) 279—281, 299
Оцеллоиды 505
Падальная муха 411, 498
37—1314
570
Предметный указатель
Палоло 417
Палочники 302
Память 498
— долговременная 533
Панцирные 403
Панцирь 301
Парабиоз 521
Паразитоиды 280—281
Паразиты 111—113, 277, 279—281
— защита от хозяина 391—392
Параподии 317
Паренхимула 57
Партеногенез 398
— и половое размножение 401
Парусник 67, 70
Патогены 389
Пауки 279
— зрение 507
— легочная книжка 349
— локомоция 325
— экскрепия 362, 367, 370—373
Паукообразные 222, 224, 225, 411
Пауроподы 233—234
Педипальпы 280
Педицелларии 196, 198, 389
Педоморфоз 41, 47, 548
Первичный бульон 15
Переваривание 278
Передняя кишка 277—278, 284
Перепончатокрылые 300
Перистальтика 319
Периферическая нервная система 493—495
Пескожил 319—320
Печеночная двуустка 90
Пигменты см. Дыхательные пигменты, Зрительные
пигменты
Пилидий 102
Пилильщики 300
Питание 265—307, 355, 498, 542
— взвесями и осадками 267—269, 272, 273, 284—289,
303—305
— слизисто-ресничное 272
— сосущий механизм 279—280
— фильтрационное 270, 271
— чистая прибыль 292—293
Пища, питательная ценность 293—299
Пищеварение 542
— внутриклеточное 389
Пищеварительные адаптации 267, 284—287
Пищеварительный тракт 277—278, 289
— термита 282
Пищевые предпочтения 293
— цепи 276—277
Пищедобывающее поведение 498
Пиявки 144, 147, 148
— гаструляция 448
— дыхание 350
— зрение 507
— локомоция 314, 317
— питание и переваривание пищи 279—280
— плавание 496, 497
— поведение 514
— реакция испуга 514
— сегментарное строение 481
— экскреция 374
Плавание 321, 323, 336—337, 496
Плавучесть У(Л
План строения 47, 48, 51, 248
Планарии 315, 464
Пластинчатые 59—61
Пластрон 350
Плеченогие 37—38, 43, 46, 172—175
— размножение 403
— экскреция 372
Плоские черви 19, 25—28, 78—96
локомоция 313, 316
нервная система 494
питание 267, 273, 498
развитие 417
размножение 402
регенерация 463
эволюция 27—28, 46
Плутеус 312, 454, 455
Поведение 485
— защитное 299, 300
— механизмы 512—516
— пишедобывающее 279, 299—303, 498
Погонофоры 130—135, 403
Позвоночные 370
Позиционная информация 482—483
Поисковики 279
Поисковое время 299—302, 307
Пойкилотермные животные 355
Пол, определение 432—433
Полет 326—329, 337—338, 393
Полимеры 14
Полинуклеотиды 14, 15
Полиплоидия 54
Полипы 61—66
Полисахариды 14
Полиспермия 440, 441
Полифосфаты 14
Полихеты (многощетинковые кольчецы) 139—144
- гаструляция 448, 449
— деление 399
— дыхание 348. 350
— защита 383
— зрение 507
— локомоция 317—320, 325
— нервная система 493^—496
- оогенез 414
— оплодотворение 443
— определение пола 432
— питание 269, 280
— плавание 332
— развитие 453—454, 457, 466, 477-^178
— размножение 403, 419
— регенерация 464, 482
— сегментное строение 481
— хеморецепторы 503
— циркуляторная система 343
Половое размножение 406, 544
Половые гормоны 529
Полости тела 29, 30
Полостная жидкость 363—367
Полярная плазма 452
Популяции 426—428
Порфирин 345
Потенциал действия 487—488, 531—532
Потенциалы 486—487
Потери воды 367
Потребители 299—305
Почвенные нематоды 529
— животные 369
Почки 374
Почкование 398
Поясковые кольчецы 144—148, 403
Пресноводная среда 365—367
Пресноводные беспозвоночные 45, 365—367, 410, 421—
424
#г<?рш«ге (герегеяевт в пресную воду Ш
Прерывистого равновесия гипотеза 16
Приапулиды 28, 31, 119—121, 403
Привыкание 532
Прилежащие тела 526. 528
Проглоттиды 91
Прогормоны 523
Происхождение жизни 14—15
Предметный указатель
571
Прокариоты 15
Проприорецепторы 501
Простейшие 52
Протесты 24—27, 52—54
— гетеротрофные 45
— как пища 265—267
— фотосинтезирующие 273
Протокишечнополостные 25
Протонефридии 79, 80, 370, 371, 374
Протонимфон 226, 227
Проторакальные железы 527
Проторакотропный гормон (РТТН) 526, 527, 541
Протоцель 29
Протоэукариоты 15
Прудовая улитка 280
Прыгающие насекомые 329—330
Прямая кишка (ректум) 278
Прямокрылые 436
Псевдоцель 29, 31
Птихоцисты 63
Пуринотелия 361, 363
Пурины 361, 362
Пчелы 509. См. также Медоносная пчела
Пятиустки 36, 217—218, 404
Рабдоиды 81—82, 385
Рабдомерные глаза 504—505
Рабдомы 504
Равноногие рачки 248, 260, 262, 362, 368
питание 282
Радиальная симметрия 23, 24
Радиальное дробление 468—469
Радула 33, 152, 267, 281
Развитие 439—483. См. также Метаморфоз, Эмбрис
нез
— гемиметаболическое 457
— генетический контроль 479—481
— голометаболическое 458
— градиентная теория 475—177
— и дифференцировка 544
— регуляционное 451—453
Размеры тела и дыхание 353—355
и интенсивность метаболизма 266—267
Размножение 391, 421—428, 543—544
— бесполое 397—399
— и регенерация 465
— половое 397, 399-^13
Раки 258
Раки-отшельники 385, 493
Раковина 33, 153, 364, 365
— маскировка 381—384
Ракообразные 36, 47, 246—262
— дыхание 350
— изменение окраски 525
— конечности и локомоция 321- 326, 539
— личинки 437
питание 270, 355
— плавучесть 364
— размножение 404
— экскреция 362, 366—368, 371, 374
Ракоскорпионы 44
Ракушковые рачки 267, 404
Распознавание 509—510
Растения как пища 276, 282, 283
Растительноядные (фитофаги) 26, 276, 282—284, 302
Реактивное движение 337
Регенерационная бластема 462—464
Регенерация 462-^165, 481—483
Регуляторы биологические 357—358
Рейнольдса число 309, 329
Ректальные жабры 367
Ремипедия (слепой рачок) 251, 252
Репарация 393—394
Репликаза 15
Репликация 13—15, 543
Репродуктивная ценность 425
— эффективность 425
Репродуктивные органы 372
— стратегии 425
Реснички 310—313
— в фоторецепторе 504
— роль в фильтрации пищи и экскреции 374—375
— у турбелларий 80, 81
Ресничные глаза 504—505
Ретиналь 503
Ретинотопия 511
Рефлексы 498, 513—516, 533
Рефлекторные действия 513 -515
— дуги 514—515
Рецепторные клетки 501
Рецепторы 501—503
— кодирование информации 502
— растяжения 514
Речной рак 258
поведение 514, 517 518, 538—539
экскреция 374
Рибосомы 13
РНК 13, 15
Родопсин 503
Рост 465, 544, 546
Рот, ротовое отверстие 277, 278
Ротовая полость 278
Ротовой аппарат 242, 270, 272. Си. также Радула
колюще-сосущий 276
Сальны 208, 209, 286
Самозарождение жизни 15
Саранча, нейроэндокринные системы 541
— нервная система 512, 515, 516
— поведение 498, 515—516
— полет 328
— развитие 457
Свайник 111, 113
Сверчки 529
Свет поляризованный 504
Сегменты 20, 36—37, 495-^197
Сенсибилизация 532—533
Сенсиллы 535 536
Сенсорная информация 509—512
Сенсорные клетки 501
— модальности 501, 502
— органы 497, 501
— поля 511
— пути 510
— системы 510—511
Сердце 501
Сетчатка 507, 537, 538
Сила 331—333
Симбиоз 15, 75, 273, 274, 283, 289—292
Симбиотические бактерии 279
Симбиотический фотосинтез 273—274
Симфилы 232
Синапсы 487, 490^92
Сипункулиды 31, 127—129, 348. 403'
— оогенез 414
— экскреция 372
Скелеты, скелетные системы 313—315. См. также
Гидростатический скелет
— жесткие 324, 334—335
Сколекс 91
Скорпион 224, 325
Скребни 31, 125—127, 403
Слизистые сети 271, 272
Сложные глаза 507—509, 525
Слух 536
Слюна 279
Слюнные железы 278
Смертность 378, 380
37*
572
Предметный указатель
Соленость 359, 365. 366
Соленоциты 371
Сосальщики 86—96, 279
— дигенетические 89
— моногенетические 86—89
Соскабливатели 281—284, 303
Сосущий механизм поглощения пищи 279—280
Спаривание 418, 419
Сперматозоиды 391, 440—441
Специалисты 502
Специфическое динамическое действие (СДД) 355
Спикулы 23, 54, 58
Спиральное дробление 37, 445, 466—468
Спироцист 63
Средняя кишка 278
Стабилизирующий отбор 44
Ставромедузы 69
Старение 378—379, 393—394
Статобласты 178
Статолиты 502
Статоцисты 502
Стеногалинные животные 377
Стерниты 116
Столоны 141
Стомодеум 65
Стратегии 543
Стрекающие 61—65, 402
Стрекозы 327, 367
Супероксиддисмутаза (СОД) 359
Суша, завоевание животными 44—45, 368—370
Схватывание добычи 279, 280 299
Схизоцель 29, 31. 100
Сцифомедузы 65, 75, 343, 493
Таннины 283
Таракан 510
Телифоны 224
Температура, влияние на дыхание 355—357
Тергиты 116
Термиты 282
Тип (таксономическая единица) 50
Тихоходки 372, 404
Токсины 384, 385
Тоническое сокращение 520
Тотипотентность 52
Трансдетерминация 479
Трансплантация 461, 479
Трахейные системы 344, 345, 349
Трематоды 86, 89—91
— питание 281
— размножение 402
Трение 331—332
Трикарбоновых кислот цикл (ТКК) 339, 340
Трилобиты 19, 36, 42, 44, 46
Трохофора 128, 448, 449
Туника 384
Турбелларии 80, 81, 343, 385, 394
Тутовый шелкопряд 421
Тяжелые металлы 392
Угри 321
Улитки, защита от хищников 381—382, 384
— локомоция 315, 316
— развитие 452
Ультрафильтрация 374—375
Уреотелия 361
Урикотелия 361, 362
Уровни организации 16
Урохордаты 211
Условные рефлексы 532
Усоногие рачки 255—257, 270, 404
защита от хищников 383
Устойчивый комплекс действия 513, 515
Фаги 429, 430
Фагоцитоз 25, 289, 389—391
Фазические рецепторы 501
Фазмиды 110
Фекальный шит 384
Фенотипы 542—543
Феромоны 502, 520, 521, 524
Фертилизин 440
Физиология 543
Филактолематы 178
Филогенез 18—49, 547
Филогенетическое древо 41, 43, 46
Фильтрация, фильтраторы 303—305, 374—375
Фитонематоды 111
Фитофаги см. Растительноядные
Форма тела 543
Форониды 37, 38, 171—172, 268
— выделительная система 372
— размножение 403
Фосфагены 341
Фосфоенолпируват (ФЕП) 341
Фосфорилирование 340
Фотопериодические реакции (фотопериодизм) 418,421—
424
Фоторецепторы 503—509
Фотосинтез 266
— морской 26—27
— симбиотический 273—274
Фрагментация 298
Функциональная биология беспозвоночных 263—264
Фуражирование 293—299
Хелицеровые 34, 36, 44, 45, 220—227
— локомоция 321, 322
— размножение 404
Хемокинез 440
Хеморецепторы 536
Хемодермоморфы 153— 155
Химическая коммуникация 521—524
Химические медиаторы см. Нейромедиаторы
— элементы 12
Химический состав живого 11, 12, 263
Хирономиды 348
Хитин 279, 324
Хитоны (панцирные) 157—159
Хищник—жертва, взаимосвязь 294—295
Хищники и защита против них 380—389
— их жертвы 293, 299—303, 307
Хлорагогенные клетки 374
Хлорокруорин 345, 346
Хлоропласты 289, 291
Хоаноциты 22, 23, 54—57, 267—268, 312
Хоботок 99, 100
Хождение 324—326
Хордовые 212
— беспозвоночные 267, 268, 285
Хроматофоры 525
Хроновиды 40, 41
Хрусталик 506, 507
Царства 45
Цветовое зрение 503—504
Целлюлазы 284
Целлюлоза как пища 275, 276
Целомодукты 370—371
Центральная нервная система 493—495
Церкарии 90
Цестоды 91—93, 402
Цефализация 83, 497
Цефалокариды 250
Цианобактерии 15, 21
— как пища 289
Циклический аденозинмонофосфат 491
Циклы размножения 421—424
Предметный указатель
573
Циркадный (циркадианный) ритм 421—424, 535
Циркуляторные системы 343—345
Цитоплазматическая локализация 470—471
Цитоплазматические органеллы 15
Челюсти насекомых 242, 281
— членистоногих 386
Червеобразное движение 314
Червеобразные 28—33
Черви 28—33, 99—149
— ископаемые 19, 28
— локомоция 316, 336
— паразитические 111—114, 281
— питание 292
— размножение 420
— регенерация 482
— старение 394
— схизоцеломические 100
Чернильный мешок 381
Чешуйницы 237
Членистоногие 34—37, 45, 213—264
— испарение воды 370
— локомоция 321—337
— нервная система 495
— питание и пищеварение 267—270
— развитие 448
— регенерация 462, 464
— циркуляторная система 343
Шестилучевые губки 58
Шипы (органы захвата пищи) 279, 280
Шистосомоз 90
Шистосомы 90—91, 391
Шмели 356
Щелевые контакты 491
Щетинкочелюстные (хетогнаты) 40, 185—186, 279,
Щетинохвостки 237
Щупальца 268, 269, 272, 273
Эвнициды 141
Эволюционная экология 16
Эволюция 11 -16. См. также Макроэволюция,
кроэволюция
— групп животных 40—45
— и филогения беспозвоночных 18—49
— первые животные 20—28
— постепенная 41, 42
— прерывистое равновесие 42
— способов питания 265—277
Эвригалинные животные 377
Эвтелия 31
Эвфаузиевый рачок 270
Экдизон 520, 529, 541
Экдистероиды 526, 527
Экзоптериготы 457
Экзоскелет 362, 366, 367
Экзоцитоз 490^91, 522, 540
Экскреторные системы 111
Экскреты 362
Экскреция 361—363, 542
Экспериментальная эмбриология 475—477
Эктодерма 61
Эктолецитальные яйца 417
Эктоневральная система 495
Эктопаразиты 86, 279
Эктотермные животные 355
Электронный транспорт 340
Электрохимическое равновесие 486
Эмбриогенез, эмбриональное развитие 469—477
Эмбриональные зачатки, карты 459, 470—474
Эндокринная система 521, 524—529
Эндокринный баланс 525
— пусковой механизм 524
Эндолецитальные яйца 417
Эндопаразиты 86, 90, 98, 280
Эндоптериготы 458
Эндостиль 285—286
Эноплиды 102—103
Энтодерма 61
Энтопрокты 181—183
Эпиболия 448
Эпигамия 141, 142
Эпидермис 61, 62
Эпитокия 141, 142
Эррантные виды 141
Этология 513
Эхиуриды 31, 129—130, 372, 403
— оогенез 414
Эхиуры 270
Ювенильные стадии 436—438
Ми- Ювенильный гормон 520. 528
Яйцеклетки, метаболизм 440—445
— эктолецитальные 417
— эндолецитальные 417
Янтарной кислоты путь (сукцинатный путь) 340—341
УКАЗАТЕЛЬ
Acariformes 225
Acoela 25, 26, 79, 84, 85
Acrothoracica 257
Acrotretida 175
Actinia 357
Aedes aegypti 367
Aeolosomata 148—149
Aglantha 519
Aglaura 69
Atcidis 386
Alcyonaria 74
Allogastropoda 161
Amblypygi 225
Amiskwia 42
Ampharetidae 346
Amphionidaceae 261
Amphipoda 260, 262
Amphiprion 75
Ancylostoma 111—113
Ancylus 357
Anemonia 357
Animalia 16
Annelida 32, 100, 135—149
Anodonta 366, 367, 410
Anomalocaris 35
Anomalodesmata 163
Anopla 102, 103
Anostraca 251
Antherea polyphenols 524
Anthozoa 65, 66, 72—75
Aphelocheirus 350
Aphragmophora 186
Apicomplexa 52
Apis melliphera 388
Aplousobranchia 206
Aplysia 523, 532, 533, 535
Apterygota 234—237
Arachnida 224
Araneae 225
Arbacia 441—443
Archaeocyatha 19
Archaeognathata 237
Architeuthis 506
Arenicola 319, 346, 347, 500
Arenicolidae 138
Argonemertes 370
Armandia 507
Anemia 346, 367
Articulata 175
Ascaris 112, 113, 346, 451, 495, 496, 518
Ascetospora 52
Aschelminthes 99, 100
Ascidiacea 205—207
Ascothoracica 255, 256
Aspidogastrea 90
Aspiraculata 205—206
Astacus 367
Astellus 367
Asterias 441
Asteroidea 196—198
Astropecten 346
Amelia 69, 71, 363, 364, 493
Bacillariophyta 22
Balanus 355, 428
Bdelloidea 100, 122—124, 403
Bdellonemertea 103
Beroe 78
Bilateria 79
Bistort betularia 382
Bivalvia 162—164
НАЗВАНИЙ
Bombyx 528
— moo 421, 524
Bonellia 404, 433
Bougainvillea 67, 506
Brachinus 387
Brachiopoda 37, 170, 172—175, 248, 250—251, 323
Brachyura 323
Branchiobdella 148
Branchiocaris 35
Branchionus 401
Branchiura 250, 252—253
Bryozoa 38, 170, 175—181
Bulimulus 363
Bullia 357
Burgessia 35
Bursovaginoida 104
Caenorhabditis 484, 485, 495, 529, 533
Calanus 381
Calcarea 58
Callinectes 324
Callipltora 540
Campanulariu 440
Carausius morosus 546
Carcinus 294, 306
— maenas 295, 363
Caryophyllida 93, 95
Cassida 384
Cassiopeia 69, 75
Cepaea nemoralis 381, 382
Cephalocarida 250, 323
Cephalochordata 211—212
Cephalodiscida 191
Cephalodiscus 187, 190
Cephalopoda 165—169
Cerastoderma 381
Ceriantharia 63, 66
Cestoda 91—96
Cestodaria 93—95
Chaetodermoinorpha 153, 155, 403
Chaetognatha 39, 185—186
Chaetonotida 107
Chaetoptcrus 270, 271, 481. 482
Cltarnia 19
Cheilostomata 181
Chelicerata 36, 213, 220—227
Chilopoda 231—232
Chirortomus 346
Chlorhaemidae 346
Chlorophyta 22
Choanoflagellata 22, 52, 54
Chordata 10, 204—212
Chrysophyta 22
Ciliophora 26, 52, 54
Cirripedia 248, 250, 255—257
Cladocera 250, 251
Clitellata 138, 144—149
Clymenella 481, 482
Cnidaria 19, 23, 51, 61—75
Coelenterata 51, 77
Coleoidea 167
Coleoptera 242
Collembola 234
Collothecida 124, 125
Comantlms 417—419
Concentricycloidea 198, 200
Conchostraca 250, 251, 258
Copepoda 248, 250, 253—255
Corallina 477
Corallium 74
Cordylophora 514
Указатель латинских названий
Corophium 293
Crepidula 405
Crinoidea 195—196
Crisialella 176
Crustacea 36, 213, 246—262
Ctenophora 23, 51, 75—78
Clenoplana 78
Ctenostomata 181
Cubomedusae 64, 69
Cubozoa 69
Cyclorhagida 116, 117
Cvclomedusa 19
Cumacea 260
Danaus plcxippus 386
Daplmia 346, 366, 367, 401, 404, 410
Decapoda 258, 262
Demospongiae 58
Dendroslomum 348
Deuterostomia 184—212
Dinomischus 42 ,
Dickinsonia 19
Dicyemida 96
Didenmum 359
Digenea 86, 89
Dinophilus 432
Dinophyta 23
Diopatia 357
Diopatra 414
Diphvlloboihrium 93
Diplopoda 232—233
Diplura 234
Diptera 242
Dirofilaria 113
Docoglossida 161
Dodecaceria 399
Doliolida 209
Dromia 363
Drosophila melanogaster 379, 414- 416, 433,
461, 479
Echinodermata 39, 191—204
Echimoidea 200—201
Echiura 31, 100, 129—130
Echiurus 130
Eledone 391
Enchytraeidae 144
Enopla 102—103
Enter obius 112, 114
— vermicularis 111
Enleromorpha 293
Enteropneusta 188—189
Emobdella 88
Entognatha 234
Entoprocta 39, 181—183
Erioptera 293
Escherichia coli 544
Eucarida 261, 262
Eucestoda 93
Euchlora rubra 77, 386
Eunice 417, 419
Eunicida 140
Eunicidae 138
Euphausiacea 261
Eurypterida 44
Euscorpius 346
Facetotecta 257
Filospermoidea 104, 105
Flosculariida 124
Gammarus 353, 366, 367
Gaslrophilus 346
Gastropoda 159—162
Gastrotricha 29, 99, 105—107
Glossiphonia 374
Gnathostomula 29, 100, 103—105
Golfingiu minulu 129
Gordioida 114, 115
Gymnolaenata 47, 180 181
Gymnomorpha 161
Gyrocoiyle 95
Gyrodaclylus 87
Haliclyslus 72
Halkryplus 346
Hallucigenia 42
Halobales 299, 300
Harmothoe 522
Heikonius 283, 386
Heliocranchia 364
Helix 315, 316
Hemichordata 39, 186—191
Hemilepislus 368
Heterobranchia 161
Heterocyemida 96
Heteroda 111
Heterodonta 163
Heteronemertea 102
Heteronereis 142
Heteroptera 242
Hexactinellida 58
Hexapoda 234—245
Hirudinea 137, 144—148
Hirudo 146, 147, 321, 507
Hohhuisana 369
Homalorhagida 117
Homalozoa 39, 40
Hoplocarida 259, 260
Hoplonemertea 103
Hyalinoecia 357
Hydra 66
452, 458, 459, Hydrobia 293
Hydroides 441, 443
Hydrozoa 62, 65—69
Hymenolepis nana 93
Hyolitha 33
Vyanassa 452
Inarticulata 173—175
Isopoda 248, 260, 262
iohoia 35
Kinetoplasta 52
Kinochulus 118
Kinorhyncha 31, 99, 116—117
Larvacea 209—211
Laukarliara 506
Lepidoptera 242
Leptograpsus 507
Limeniiis 386
Limnoria 282
Limulus 346, 510
Linens 102, 313
Lingula 346
Lingulida 173, 175
Liltorina 297, 375
Loligo 348, 484, 530
Lophophorata 170—183
Loricifera 31, 99, 116, 403
Luidia 346
Lumbricus 144—146, 346, 490, 522, 540
Lymantria 527
Lymnaea 363, 523
Macoma 293, 357
576
Указатель латинских названий
МасгоЬгасШит 355
Macrodasyida 106
Magelona 346
Maia squinado 363
Malacobdella 103
Malacostraca 47, 248, 250, 258—262, 323, 404
Manduca 526
Marrella 35
Marihasierias 363
Maxillopoda 250
Medusuniles 19
Megoura 424
Meloidogyne 111
Membranipora 400
Merostomata 223 -224
Mesozoa 27, 51, 96—98, 402, 446
Metamonada 52
Metapenaeus 365
Metazoa 96, 302, 446
Meiridium 73, 74
Microcoryphia 234
Microspore 23, 24, 52
Millepora 70
Milleporina 69
Mimetaster 35
Mnemiopsis 77
MoUusca 150—169
Moniliformis 126, 127
Monogenea 86—89, 403
Monogonata 124—125. 403
Monopisthocotylea 86, 87
Monoplacophora 156—157, 403
Musca domestica 538
Mya 515
Myriapoda 230—245
Myrientomata 234
Mysidacea 260
Mystacocarida 250, 252
Mytilus 294, 356, 358
— eduiis 295, 355, 356, 365, 457
Myxicola 487
M'yxozoa 23, 24, 52
fiamonia 493
Nasonia 379
Nassarius 452, 371. 474
Nautiloidea 166
Nautilus 365
Nebalia 125
Nectonematida 114, 115
Nematode 30, 99
Nematomorpha 30, 99, 100, 114—116
Nemertea 29, 99—103
Neomeniomorpha 156, 157, 403
Neoptera 245
Nephros 363
Nephlys 481, 503
— honbergii 347, 348
Neptunea 363
Nereidae 409
Nereis 136—142, 317, 325, 343, 503, 580, 525
— diversicolor 270. 292—294, 304
Neriia 506
Notoplana 498
Notostigmata 225
Notostraca 251
Nuculida 163
Nuda 77, 78
Obelia 66, 67
Obturata 134—135
Octopoda 167
Octopus 495
Odontogriphus 42
Oicopleura 494, 503
Olifochaeta 137
Oligoehtomota 234
Onchidiwn 504
Onchocerca 113
Onichophora 36, 213, 218—220
Opabinia 42
Opalinata 52
Ophiuroidea 198
Ophryotrocha 481, 483
Opiliones 225
Oryctes 389
Orthonecta 96—98
Orthonectida 96
Ostracoda 250, 257—258
Owenia 139
Oxyspirura 113
Vachygrupsus 363
Palaemonetes 355, 367
Palaeoheterodonta 163
Palaeonemertea 102
Palaeoptera 242
Palpigradi 225
Pancarida 259, 261
Papilio lag 386
Parabasalia 52
Parufilaria 113
Paramecium 346, 432
Parasitiformes 225
Parazoa 19
Parus 307, 383
Parvancorina 19
Patella 357, 367, 494, 506
— Pauropoda 233—234
Pecten 363, 504
Pentastoma 36, 213, 217
Peracarida 260, 261
Perviata 132—134
Phagocytellozoa 61
Phascolosoma 363
Philodinidae 124
Phlebobrahchia 206
Phorona 31, 37, 170—172
Phoronis 171
Phragmophora 186
Philactolaemata 178
Phyllocarida 259
Phyllodocida 140
Phyllodocidae 138
Phyllopoda 250
Physalia 66, 70, 364
Placozoa 27, 51, 59—61, 446
Planctosphaera 191
Planctosphaeroidea 191
Planorbis 346
Platasterias 197
Platyhelminthes 19, 25, 28, 29, 51, 78—96
Pleurobranchia 530
Ploima 124
Podotricha 386
Poekilocerus 385
Pogonophora 32, 100, 130—135
Polychaeta 137, 139—144
Polyclada 84, 313
Polynoidae 138
Polyopisthocotylea 86, 87
Polyplacophora 157—159
Polyorchis 500, 504
Polystoma 87
Porifera 19, 22, 51, 54—59
Porites 513
Poromyida 163
Portunidae 324
Указатель латинских названий
577
Poiamophilax 383
Praecambridium 19
Priapula 31, 99, 100, 119—121
Priaputus 120, 346, 370, 371
Prosobranchia 160
Protista 22
Protobranchia 163
Prltlzoa 51—54
Protura 234
Psammechinus 455
Pseudoceros 504
Pseudoscorpines 225
Pterygota 234, 239, 243, 244
Pteriomorpha 163
Pterobranchia 189—191
Pterygota 237—245
Pycnogona 224—227
Pyrosomida 207
Radiata 77
Ragnea 19
Remipedia 250—252
Rhabdilis 108
Rhabdocalyplus 492
Rhabdopleurida 191
Rhixocephala 255
Rhodnius 521, 527, 528
Rhombozoa 96—97
Ricinulei 255
Rotifera 31, 99, 121—125
Sabella 319, 320, 346, 347, 482
Sabellaria 139
Sabellidae 187, 346
Salpa 363, 515
Salpida 209
Sarcodina 22, 54
Sarcophaga 423
Sarotrocercus 35
Sarsia 493
Scaphopoda 164—165
Schistocephalus 95
Schistosoma mcmsoni 90
Schistosomatidae 90
Schixomida 225
Scieractinia 74
Sclerospongiae 59, 60
Scorpiones 225
Scutigerida 231
Scyphozoa 65, 66, 69—72
Seison 125
Seisonidae 124, 125, 403
Selenaria 176
Sepia 363
Sepiida 167
Serpula 346, 364
Serpulidae 346
Seticoronaria 121
Sigara 366
Sipuncula 31, 100, 127—129
Sipunculata 330—331, 415
Solpugida 225
Simdella 186
Spio 139
Spionidae 138
Spirorbis 346, 457, 477, 478
Spriggina 19
Stenoglossa 161
Stenolaemata 46, 178—180
Stephanqfilaria 113
Stephanopogonomorpha 26, 52
Stolidobranchia 206
Stomphia 73
Slrongylocentroms 357
Strongyloides 112
Styela 445, 452
Stylasterina 69
Stylommatophora 161
Siynapla 446
Sydnium 400
Symbiodinium 75
Symphyla 232
Synapia 446
Syncarida 259, 261
Taenia saginata 91, 93
Tamoya 503—506
Tanaidacea 260
Tardigrada 36, 213, 215—217
Teleogryilus 529
Temnocephalida 80, 81
Tenebrio molitor 383
Tentaculata 77—78
Tetrahymena 346
Teuthiida 167
Thaliacea 207—209
Thelazia 113
Theromyzon 374
Thoracica 255
Thysanura 234
Tivela 346
Trematoda 86, 89—91
Tribranchidium 19
Trichomonas 282
Trichonympha 282
Trichurus 114
Triclada 84, 85, 313
Tritonia 515
Tubifex 293, 343, 385
Tubularia 66, 68
Tullimonstrum 42
Turbellaria 80—86
Viva 293
Uniramia 36, 227, 245, 265, 457
Urechis 129
Urochordata 205—211
Uropygi 225
Valvata 506
Vampyromorpha 167
Vanadis 507
Velamen 11
Velella 67, 70
Vermetidae 271
Vertebrata 212
Veslimenifera 134
VJiwaxia 42
Wiwaxiida 33
Xanthophyta 22
Xenophyophora 54
Xenopneusta 130
Xiphosura 223
Xyloplax 198, 200
Zygoentomata 237
ОГЛАВЛЕНИЕ
Предисловие к русскому переводу 5
Предисловие (пер. М.А. Софронова) 7
2.5. Происхождение, радиация и
вымирание групп животных 40
2.6. Резюме и эволюционный обзор . . 45
2.7. Дополнительная литература . 48
ЧАСТЬ I. ЭВОЛЮЦИОННЫЕ
ВОПРОСЫ . . 9
Глава 1. Введение: основной подход
и принципы 10
1.1. Почему беспозвоночные? ... . 10
1.2. Свойства живых сушеств . 11
1.2.1. Введение II
1.2.2. Программа .... 12
1.2.3. Открытость 13
1.2.4. Эволюция путем
естественного отбора — неизбежное следствие
возобновления систем за счет ре- 13
пликашш
1.3. Происхождение жизни 14
1.3.1. Добиологический синтез
органических полимеров . . . . 14
1.3.2. Происхождение и эволюция
клеток . . . . 15
1.3.3. Почему самозарождение
жизни не происходит все время? . . 15
1.4. Уровни организации организмов . 16
1.5. План книги 16
1.6. Дополнительная литература ... 17
Глава 2. Эволюционная история
и филогения беспозвоночных .... 18
2.1. Введение 18
2.2. Первые животные .... 20
2.2.1. Губки 22
2.2.2. Кишечнополостные 23
2.2.3. Плоские черви 25
2.2.4. Placozoa и Mesozoa . . . . 27
2.3. Производные плоских червей . . 27
2.3.1. Группы червеобразных . . 28
2.3.2. Моллюски 33
2.4. Более отдаленные потомки плоских
червей 34
2.4.1. Членистоногие . . 34
2.4.2. Лофофоровые .... 37
2.4.3. Вторичноротые 39
ЧАСТЬ II. ТИПЫ
БЕСПОЗВОНОЧНЫХ 50
Глава 3. Параллельные подходы
к многоклеточное™ у животных . . 51
3.1. Protozoa (простейшие) 51
3.2. Тип Porifera (губки) 54
3.2.1. Этимология 54
3.2.2. Диагностические и
специфические признаки . . .... 54
3.2.3. Классификация 58
3.3. Тип Placozoa (пластинчатые) . 59
3.3.1. Этимология 59
3.3.2. Диагностические и
специфические признаки . . ... 59
3.3.3. Классификация 61
3.4. Тип Cnidaria (стрекающие:
гидроидные, сцифомедузы, актинии, кораллы) . 61
3.4.1. Этимология 61
3.4.2. Диагностические и
специфические признаки 61
3.4.3. Классификация 65
3.4.4. Симбиоз 75
3.5. Тип Ctenophora (гребневики) . . . . 75
3.5.1. Этимология ... 75
3.5.2. Диагностические и
специфические признаки 75
3.5.3. Классификация 77
3.6. Тип Plathyhelmintes (плоские черви) . 78
3.6.1. Этимология 78
3.6.2. Диагностические и
специфические признаки 78
3.6.3. Классификация 80
3.7. Тип Mesozoa 96
3.7.1. Этимология 96
3.7.2. Диагностические и
специфические признаки 96
3.7.3. Классификация 96
3.7.4. Родственные связи ... 98
3.8. Дополнительная литература .... 98
Оглавление
579
Глава 4. Черви 99
4.1. Тип Nemertea (немертины) . . . . 100
4.1.1. Этимология ... 100
4.1.2. Диагностические и
специфические признаки 100
4.1.3. Классификация 102
4.2. Тип Gnathostomula (гнатостомулиды) 103
4.2.1. Этимология 103
4.2.2. Диагностические и
специфические признаки .... 103
4.2.3. Классификация 105
4.3. Тип Gastrotricha (брюхоресничные
черви) 105
4.3.1. Этимология 105
4.3.2. Диагностические и
специфические признаки 105
4.3.3. Классификация 106
4.4. Тип Nematode (круглые черви) ... 107
4.4.1. Этимология 107
4.4.2. Диагностические и
специфические признаки 107
4.5. Тип Nematomorpha (волосатики) . 114
4.5.1. Этимология 114
4.5.2. Диагностические и
специфические признаки 114
4.5.3. Классификация 115
4.6. Тип Kinorhyncha 116
4.6.1. Этимология 116
4.6.2. Диагностические и
специфические признаки 116
4.6.3. Классификация . ... 117
4.7. Тип Loricifera ... 117
4.7.1. Этимология 117
4.7.2. Диагностические и
специфические признаки 117
4.7.3. Классификация 119
4.8. Тип Priapula (приапулиды) 119
4.8.1. Этимология 119
4.8.2. Диагностические и
специфические признаки . 119
4.8.3. Классификация 121
4.9. Тип Rotifera (коловратки) . . 121
4.9.1. Этимология 121
4.9.2. Диагностические и
специфические признаки 121
4.9.3. Классификация 123
4.10. Тип Acanthocephala (скребни) . 125
4.10.1. Этимология 125
4.10.2. Диагностические и
специфические признаки 125
4.10.3. Классификация . . 127
4.11. Тип Sipuncula (сипункулиды) . . 127
4.11.1. Этимология 127
4.11.2. Диагностические и
специфические признаки 127
4.11.3. Классификация. . . ... 129
4.12. Тип Echiura (эхиуриды) 129
4.12.1. Этимология 129
4.12.2. Диагностические и
специфические признаки 129
4.12.3. Классификация 130
4.13. Тип Pogonophora 130
4.13.1. Этимология ... 130
4.13.2. Диагностические и
специфические признаки 130
4.13.3. Классификация 134
4.14. Тип Annelida (кольчатые черви,
кольчецы) .135
4.14.1. Этимология ... . 135
4.14.2. Диагностические и
специфические признаки 135
4.14.3. Классификация 137
4.15. Дополнительная литература . . 149
Глава 5. Моллюски (пер. А.Б.Цет- 150
лин)
5.1. Тип Mollusca 150
5.1.1. Этимология 150
5.1.2. Диагностические и
специфические признаки 150
5.1.3. Классификация 153
5.2. Дополнительная литература . . . . 169
Глава 6. Lophophorata: лофофоро-
вые, или щупальцевые 170
6.1. Тип Phorona (форониды) . . 171
6.1.1. Этимология 171
6.1.2. Диагностические и
специфические признаки 171
6.1.3. Классификация 172
6.2. Тип Brachiopoda (плеченогие) ... 172
6.2.1. Этимология 172
6.2.2. Диагностические и
специфические признаки 172
6.2.3. Классификация . . . . 173
6.3. Тип Bryozoa (мшанки) . . 175
6.3.1. Этимология 175
6.3.2. Диагностические и
специфические признаки 175
6.3.3. Классификация . ... 177
6.4. Тип Entoprocta 181
6.4.1. Этимология 181
6.4.2. Диагностические и
специфические признаки 181
6.4.3. Классификация 183
6.5. Дополнительная литература . .183
Глава 7. Deuterostomia: вторичноро-
тые 184
7.1. Тип Chaetognatha (щетинкочелюст-
ные) 185
7.1.1. Этимология 185
7.1.2. Диагностические и
специфические признаки 185
580
Оглавление
7.1.3. Классификация 186
7.2. Тип Hemichordata (полухордовые) . 186
7.2.1. Этимология 186
7.2.2. Диагностические и
специфические признаки 186
7.2.3. Классификация 188
7.3. Тип Echinodermata (иглокожие) . . . 191
7.3.1. Этимология 191
7.3.2. Диагностические и
специфические признаки .191
7.3.3. Классификация . . . . 194
7.4. Тип Chordata (хордовые) 204
7.4.1. Этимология 204
7.4.2. Диагностические и специфи- 204
ческие признаки
7.4А. Подтип Urochordata (личиночнохор-
довые, или оболочники) 205
7.4А. 1. Диагностические и специфи- 205
ческие признаки ...
7.4А.2. Классификация 205
7.4Б. Подтип Cephalochordata (головохор-
довые, или ланцетники) 211
7.4Б. 1. Диагностические и
специфические признаки 211
7.4Б.2. Классификация 212
7.5. Дополнительная литература . . 212
Глава 8. Беспозвоночные с
конечностями: членистоногие и сходные
группы 213
8.1. Тип Tardigrada (тихоходки) 215
8.1.1. Этимология 215
8.1.2. Диагностические и
специфические признаки 215
8.1.3. Классификация 217
8.2. Тип Pentastoma (пятиустки) . . . . 217
8.2.1. Этимология 217
8.2.2. Диагностические и
специфические признаки 217
8.2.3. Классификация 218
8.3. Тип Onychophora 218
8.3.1. Этимология 218
8.3.2. Диагностические и спепифи-
ческие признаки 218
8.3.3. Классификация 220
8.4. Тип Chelicerata 220
8.4.1. Этимология 220
8.4.2. Диагностические и
специфические признаки 220
8.4.3. Классификация 222
8.5. Тип Uniramia (одноветвистые) .... 227
8.5.1. Этимология 227
8.5.2. Диагностические и
специфические признаки 227
8.5.3. Классификация 230
8.5.ЗА. Подтип Myriapoda
(многоножки) 230
8.5.3Б. Подтип Hexapoda
(насекомые) . 234
8.6. Тип Crustacea (ракообразные) . . 246
8.6.1. Этимология 246
8.6.2. Диагностические и
специфические признаки 246
8.6.3. Классификация 249
8.7. Дополнительная литература . . 262
ЧАСТЬ III. ФУНКЦИОНАЛЬНАЯ
БИОЛОГИЯ БЕСПОЗВОНОЧНЫХ
(пер. И. В. Успенский) 263
Глава 9. Питание 265
9.1. Введение: эволюция способов питания
животных 265
9.2. Типы питания животных: способы
захвата и переработки пищи 277
9.2.1. Классификация типов
питания 277
9.2.2. Общие черты 277
9.2.3. Охотники и паразиты .... 279
9.2.4. Обгрызатели и соскабливате-
ли 281
9.2.5. Питание взвесями 284
9.2.6. Питание осадками 287
9.2.7. Питание с участием
симбионтов 289
9.3. Затраты и выигрыш при различных
способах питания: оптимальное
фуражирование 292
9.3.1. Введение 292
9.3.2. Теория оптимального
фуражирования ... 293
9.3.3. Подвижные потребители и их
жертвы 299
9.3.4. Потребители суспензий
и осадков 303
9.4. Выводы 305
9.5. Дополнительная литература 306
Приложение 9.1. Выбор мидий крабами 306
Приложение 9.2. Поисковое время и
выбор жертвы: синица и мучные черви . . . 307
Глава 10. Биомеханика и движение
(локомоция) 308
10.1. Принципы локомоции 308
10.2. Генерирование механических сил
клетками животных 309
10.3. Движение с помощью ресничек . . 312
10.4. Работа мышц и скелетные системы 313
10.5. Локомоция мягкотелых животных . 315
10.5.1. Передвижение по твердой
поверхности 315
10.5.2. Движение в толще субстрата
и в трубках 319
10.5.3. Плавание 321
Оглавление
581
10.6. Локомоция беспозвоночных с
членистыми конечностями 321
10.6.1. Локомоция ракообразных . 323
10.6.2. Наземная локомоция:
хождение 324
10.6.3. Полет 326
10.6.4. Наземная локомоция:
прыгающие насекомые . . . . 329
10.7. Выводы 331
10.8. Дополнительная литература ... 331
Приложение 10.1. Термины и
определения механики . . .331
Приложение 10.2. Кинетическая энергия
и трение 331
Приложение 10.3. Разложение сил на
составляющие 332
Приложение 10.4. Гидростатический
скелет 333
Приложение 10.5. Жесткие скелеты -
принцип рычага 334
Приложение 10.6 335
Приложение 10.7. Плавание мягкотелых
червей 336
Приложение 10.8. Реактивное движение 337
Приложение 10.9. Вихревое движение
воздуха и полет 337
Глава 11. Дыхание 339
11.1. Центральная роль АТР в дыхании 339
11.2. Основной путь катаболизма .... 339
11.3. Получение АТР без кислорода . . 340
11.4. Поглощение кислорода 342
11.4.1. Главное — это диффузия . . 342
11.4.2. Циркуляторные системы . . 343
11.4.3. Кровь 345
11.4.4. Органы дыхания 348
11.4.5. Вентиляция . . . 350
11.5. Измерение метаболизма 350
11.6. Факторы, влияющие на дыхание . . 353
11.6.1. Размер тела 353
11.6.2. Активность ' 353
11.6.3. Питание 355
11.6.4. Температура . . 355
11.6.5. Концентрация кислорода
(Ро.) 357
11.6.6. Соленость 359
11.7. Выводы 359
11.8. Дополнительная литература . . 360
Глава 12. Экскреция, ионная и
осмотическая регуляция, плавучесть . . 361
12.1. Экскреция 361
12.2. Осмотическая и ионная регуляция 363
12.2.1. Морская среда 364
12.2.2. Отступление по поводу
плавучести 364
12.2.3. Пресноводная среда .... 365
12.2.4. Гиперосмотическая среда 367
12.2.5. Наземная среда 367
12.2.6. «Завоевание» суши и
пресных вод 368
12.3. Выделительные системы 370
12.3.1. Строение 370
12.3.2. Функция 374
12.4. Выводы 375
12.5. Дополнительная литература . . . 375
Приложение 12.1. Некоторые
определения 376
Глава 13. Защита 378
13.1. Классификация опасностей . . 378
13.1.1. Две основные группы . . . 378
13.1.2. Экологические (внешние)
факторы смертности . . . 378
13.1.3. Старение 378
13.1.4. Классификация 379
13.2. Защита 380
13.2.1. Против хишников 380
13.2.2. Против внутренних врагов 389
13.2.3. Реакция на загрязнители . . 392
13.2.4. Репарация —защита против
старения? 394
13.3. Выводы 395
13.4. Дополнительная литература . . 395
Глава 14. Размножение и жизненные
циклы 396
14.1. Введение 396
14.2. Значение полового и бесполого
размножения 397
14.2.1. Бесполое размножение в
жизненных циклах беспозвоночных . 397
14.2.2. Типы полового
размножения 399
14.2.3. Механизмы определения
пола 400
14.2.4. Адаптивное значение
различных способов полового
размножения 405
14.3. Разнообразие полового
размножения: жизненные циклы 407
14.3.1. Введение 407
14.3.2. Морские беспозвоночные . 407
14.3.3. Пресноводные и наземные
беспозвоночные 410
14.4. Контроль процессов размножения . 412
14.4.1. Первичные и
непосредственные контролирующие факторы . . 412
14.4.2. Гаметогенез 413
14.4.3. Синхронное размножение
беспозвоночных 417
14.4.4. Циклы размножения и диа-
пауза в наземной и пресноводной
средах 421
582
Оглавление
14.5. Размножение и распределение
ресурсов 424
14.6. Выводы 428
14.7. Дополнительная литература . . . 429
Приложение 14.1. Системы полового
размножения (миксиса) . . . 429
Приложение 14.2. Жизненные циклы
и половое размножение 432
Приложение 14.3. Системы определения
пола 432
Приложение 14.4. Жизненные циклы
беспозвоночных 433
Приложение 14.5. Жизненные циклы
морских беспозвоночных 435
Приложение 14.6. Функциональный
анализ жизненных циклов насекомых .... 436
Глава 15. Развитие 439
15.1. Оплодотворение и инициация
развития 439
15.2. Раннее развитие . .... 445
15.2.1. Дробление 445
15.2.2. Гаструляция 446
15.3. Экспериментальная эмбриология
беспозвоночных: детерминация судьбы
клетки 450
15.3.1. Введение 450
15.3.2. Мозаичное и регуляционное
развитие . 451
15.4. Личиночное развитие и метаморфоз 453
15.4.1. Метаморфоз морских
личиночных форм 453
15.4.2. Развитие и метаморфоз
насекомых 457
15.4.3. Экспериментальный анализ
развития имагинальных дисков Dro-
sophila melanogaster 459
15.5. Регенерация 462
15.5.1. Введение 462
15.5.2. Природа регенерационной
бластемы 462
15.5.3. Регенерация и регионарная
организация 463
15.5.4. Регенерация, рост и
размножение 465
15.6. Выводы: развитие беспозвоночных
и генетическая программа 465
15.7. Дополнительная литература .... 466
Приложение 15.1. Спиральное дробление 466
Приложение 15.2. Радиальное дробление 468
Приложение 15.3. Регуляционное и
мозаичное развитие I 469
Приложение 15.4. Судьба клеток, карты
зачатков и цитоплазматическая
локализация 470
Приложение 15.5. Регуляционное и
мозаичное развитие II 471
Приложение 15.6. Экспериментальный
анализ развития морского ежа 475
Приложение 15.7. Метаморфоз и выбор
субстрата у морских личинок 477
Приложение 15.8. Серийная
трансплантация имагинальных дисков и открытие
трансдетерминации 479
Приложение 15.9. Генетический контроль
и регионарная организация: гомеозисные
гены 479
Приложение 15.10. Позиционная
информация и каудальная регенерация у аннелид 481
Глава 16. Системы контроля . . 484
16.1. Введение 484
16.2. Потенциалы 486
16.2.1. Мембраны . . 486
16.2.2. Потенциалы 486
16.2.3. Проведение градуальных по- 487
тенциалов
16.2.4. Потенциал действия . 487
16.2.5. Спайковые и неспайковые 488
нейроны
16.3. Нейроны и их связи 488
16.3.1. Нейроны 488
16.3.2. Глиальные клетки . . . . 489
16.3.3. Химические синапсы .... 490
16.3.4. Электрические синапсы . . 491
16.4. Организация нервных систем .... 492
16.4.1. «Нейроидная» система губок 492
16.4.2. Нервные сети 492
16.4.3. Центральная и
периферическая нервные системы 493
16.4.4. Дифференцировка нейро-
пиля 495
16.4.5. Сегментарное строение 495
16.4.6. «Горячие линии» 497
16.4.7. Роль мозга .... ... 497
16.5. Спонтанная активность . 499
16.5.1. Нервная инициация . 499
16.5.2. Движение 500
16.5.3. Автономные функции . 500
16.5.4. Биологические часы . . 501
16.6. Рецепторы 501
16.6.1. Основная характеристика . 501
16.6.2. Классификация рецепторов 501
16.6.3. Специалисты и генералисты 502
16.6.4. Кодирование интенсивности 502
16.6.5. Сенсорные клетки и
сенсорные нейроны 503
16.7. Зрение 503
16.7.1. Зрительные пигменты . . . 503
16.7.2. Ресничные и рабдомерные
глаза 504
16.7.3. Глазки и глаза 506
16.7.4. Линзы и зеркала 506
16.7.5. Сложные глаза 507
Оглавление
583
16.8. Обработка сенсорной информации 509
16.8.1. Смысловое восприятие мира 509
16.8.2. Сенсорные системы .... 510
16.8.3. Конвергенция и дивергенция 511
16.8.4. Меченые линии . .511
16.8.5. Выделение признаков ... 511
16.8.6. Центробежный контроль 512
16.9. Нервные основы поведения ... 512
16.9.1. Независимые эффекторы . 512
16.9.2. Элементы поведения . . 513
16.9.3. Рефлекторные действия . . 513
16.9.4. Устойчивые комплексы
действий .... 515
16.9.5. Вариации на две темы ... 515
16.10. Организация двигательного
«выхода» 516
16.10.1. Аспекты организации . . 516
16.10.2. Командные нейроны . . . 516
16.10.3. Генерирование программы 517
16.10.4. Мотонейроны 518
16.10.5. Нервно-мышечная
иннервация 519
16.11. Химическая коммуникация . . 521
16.11.1. Спектр химической
коммуникации . . 521
16.11.2. Критерии эндокринной
активности .... . . . 521
16.11.3. Нейросекреция 521
16.11.4. Размытая граница . . . 522
16.11.5. Нейропептиды 523
16.11.6. Нейроэндокринные
системы ... 523
16.11.7. Феромоны 524
16.12. Роль эндокринной системы .... 524
16.12.1. Эндокринный пусковой
механизм 524
16.12.2. Эндокринный баланс . . . 525
16.12.3. Эндокринный контроль
развития у Nereis ... 525
16.12.4. Эндокринный контроль
развития насекомых 526
16.13. Выводы 529
16.14. Дополнительная литература . . . 530
Приложение 16.1. Система гигантских
волокон кальмара Loligo 530
Приложение 16.2. Потенциал действия . 531
Приложение 16.3. Клеточные механизмы
научения 532
Приложение 16.4. Становление
дефинитивных нейронных связей 533
Приложение 16.5. Клеточные механизмы
биологических часов 535
Приложение 16.6. Сенсиллы насекомых 535
Приложение 16.7. Нейросуперпозиционная
сетчатка мухи .... 537
Приложение 16.8. Оптическая доля
комнатной мухи (Musca domestiea) 538
Приложение 16.9. Удары брюшка у рака 538
Приложение 16.10. Контроль мышечных
сокращений у ракообразных 539
Приложение 16.11. Нейросекреция путем
экзоцитоза . 540
Приложение 16.12. Нейроэндокринные
системы саранчи . ... . . 541
Глава 17. Еще раз об основных
принципах 542
17.1. Основные физиологические черты
фенотипов . . 542
17.2. Главное— репликация и
размножение . . 543
17.3. Онтогенез 544
17.4. Онтогенез и филогенез . 547
17.5. Дополнительная литература . . . 549
Словарь терминов 550
Источники иллюстраций . . . 561
Предметный указатель . ... 565
Указатель латинских названий. . . . 574
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее
оформлении, качестве перевода и другие просим
присылать по адресу:
129820, Москва
1-й Рижский пер., д. 2,
издательство «Мир»
УЧЕБНОЕ ИЗДАНИЕ
Р. Варне, П. Кейлоу, П. Олив, Д. Голдинг
БЕСПОЗВОНОЧНЫЕ.
НОВЫЙ ОБОБЩЕННЫЙ ПОДХОД
Заведующая редакцией
канд. биол наук М.Д. Гроздова
Ведущий редактор Ю. И. Амченков
Мл. научи, редактор И. А. Ермакова
Художник Е. Г. Шведова
Художественные редакторы А. Я. Мусин. Л.М. Аленичева
Технический редактор М.А. Страшнова
Корректор Т. М. Подгорная
. ■ IVUUUCI
ИБ № 7237
Сдано в набор 04.10.90. Подписано к печати 07.04.92. Формат
70 х 100'/i6- Бумага офсетная № 1. Печа1ъ офсетная. Гарнитура
Тайме. Объем 18,25 бум. л. Усл.печ.л. 47,45. Усл.кр.-отт. 94,90.
Уч.-изд.л. 54,55. Изд № 4/6774. Тираж 3500 экз. Зак. 1314. С 190.
Издательство «Мир»
129820, ГСП, Москва, И-110, 1-й Рижский пер., 2
Можайский полиграфкомбинат Министерства печати и
информации Российской Федерации
143200, г. Можайск, ул. Мира, 93.