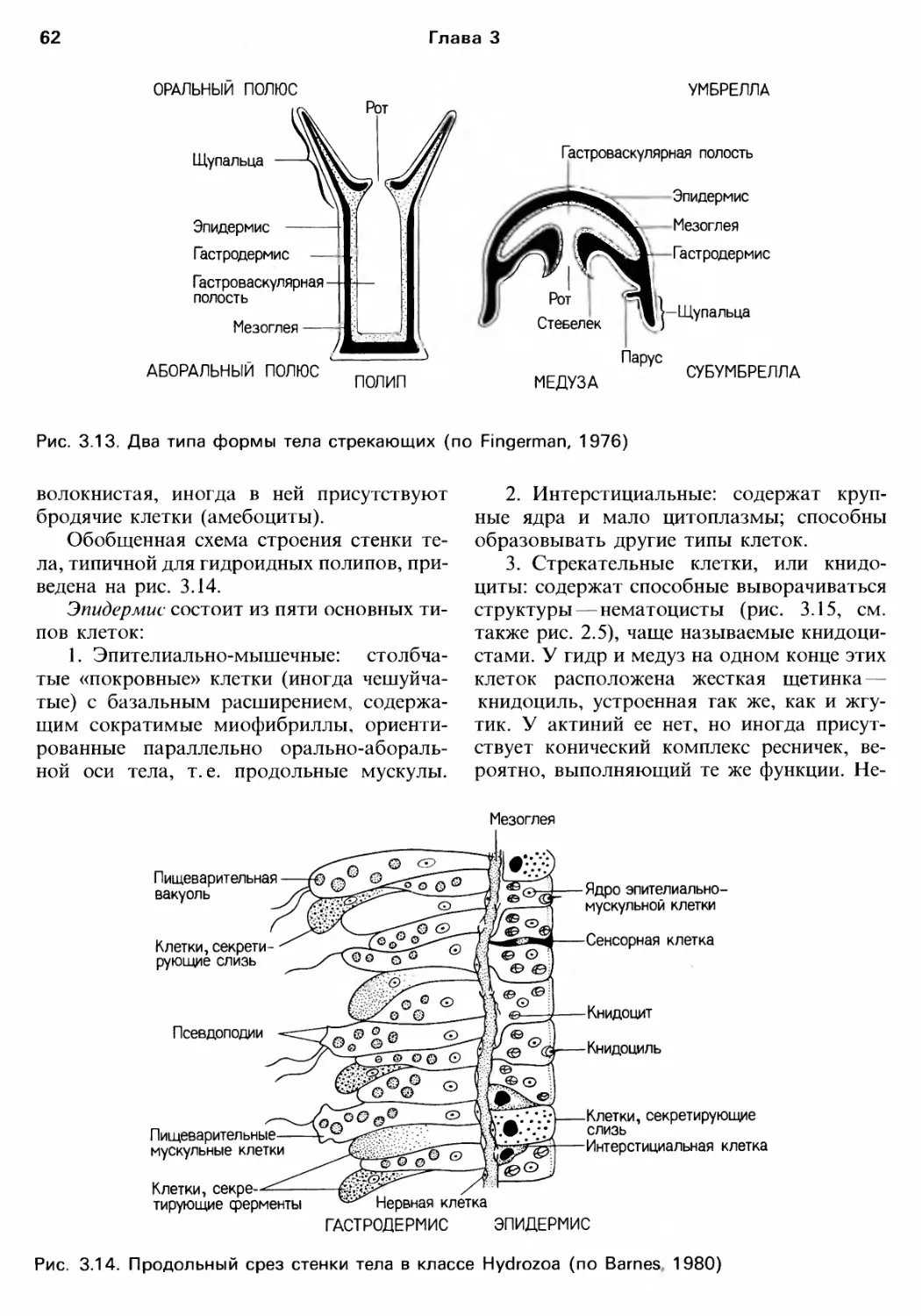
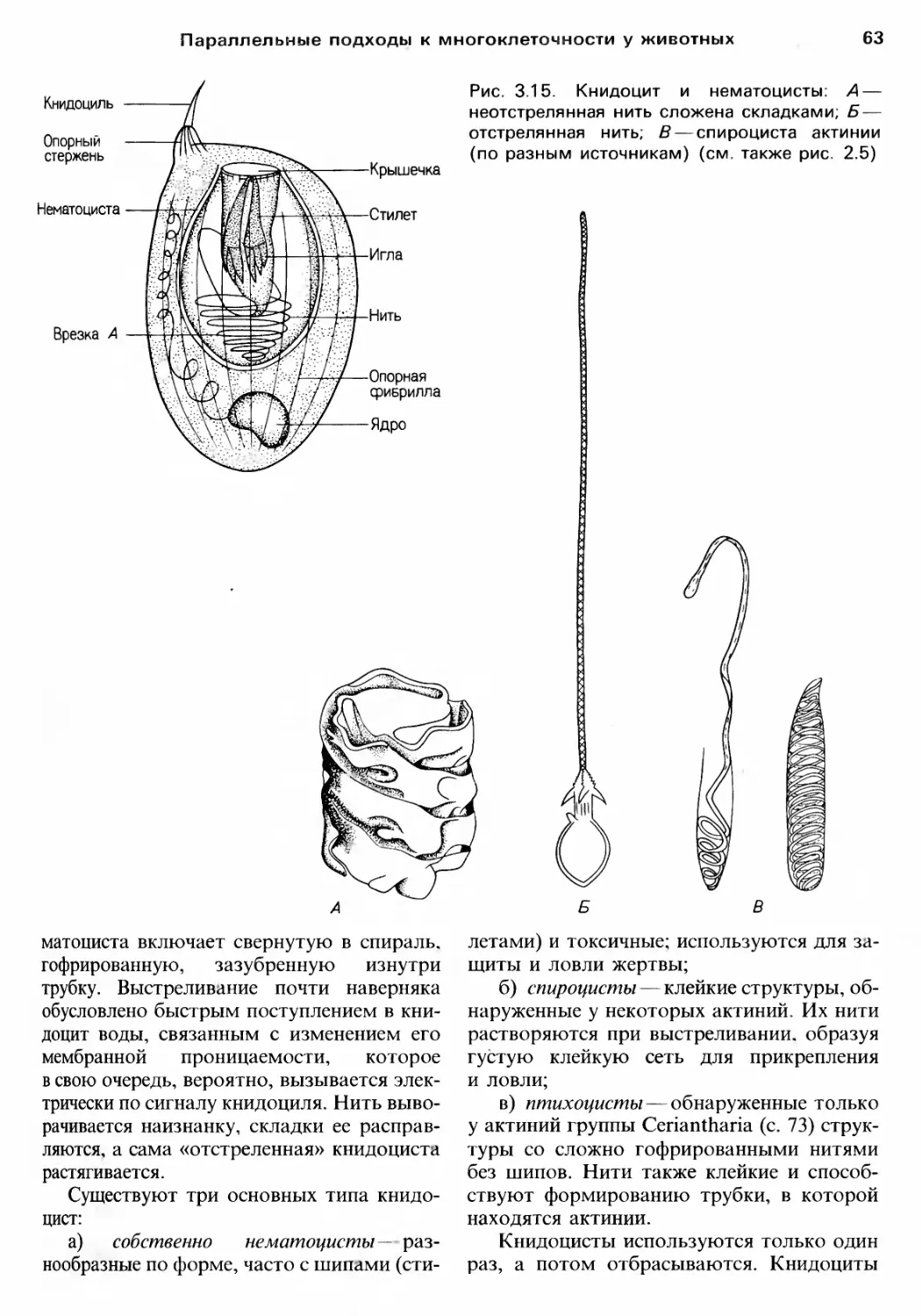
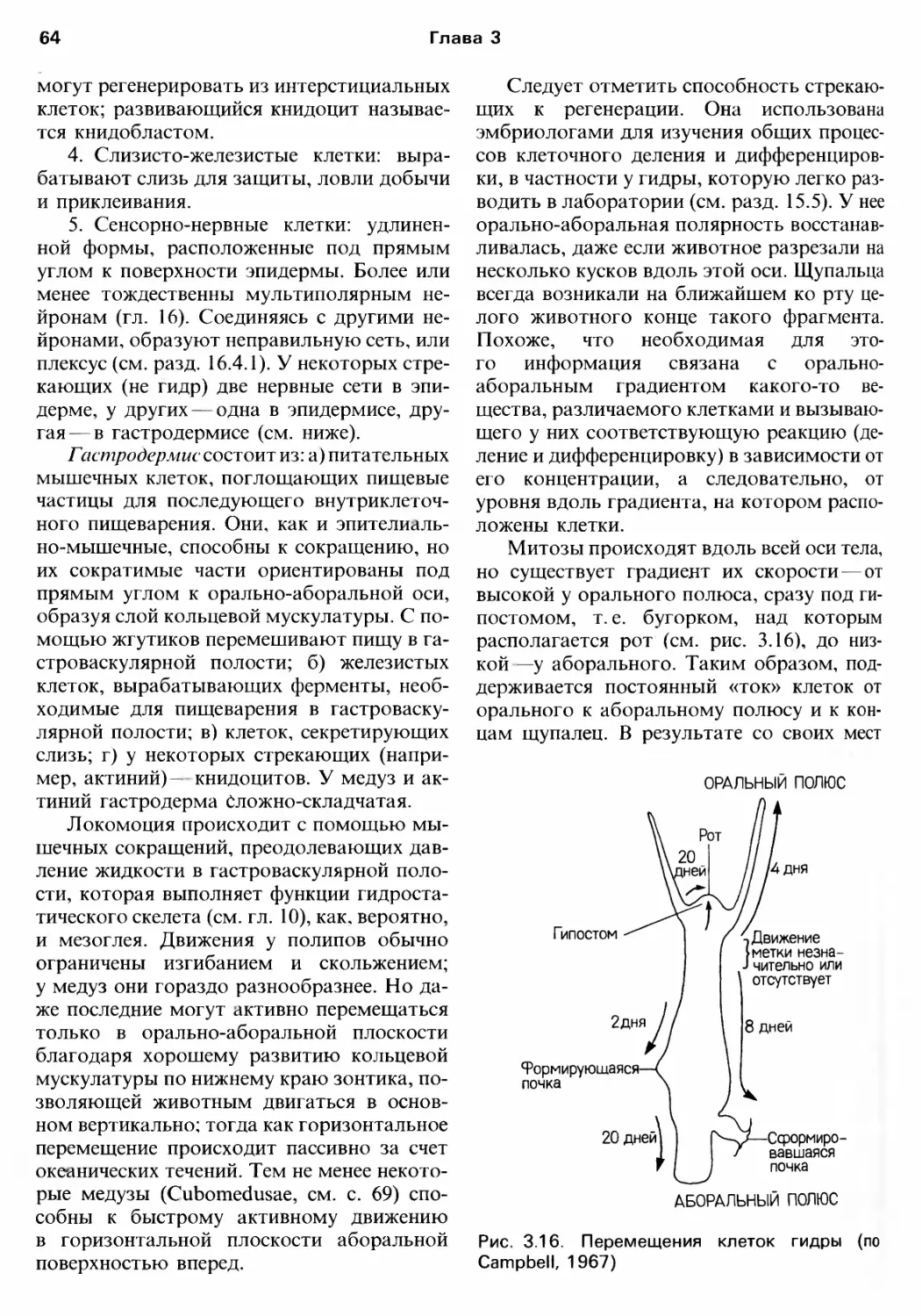
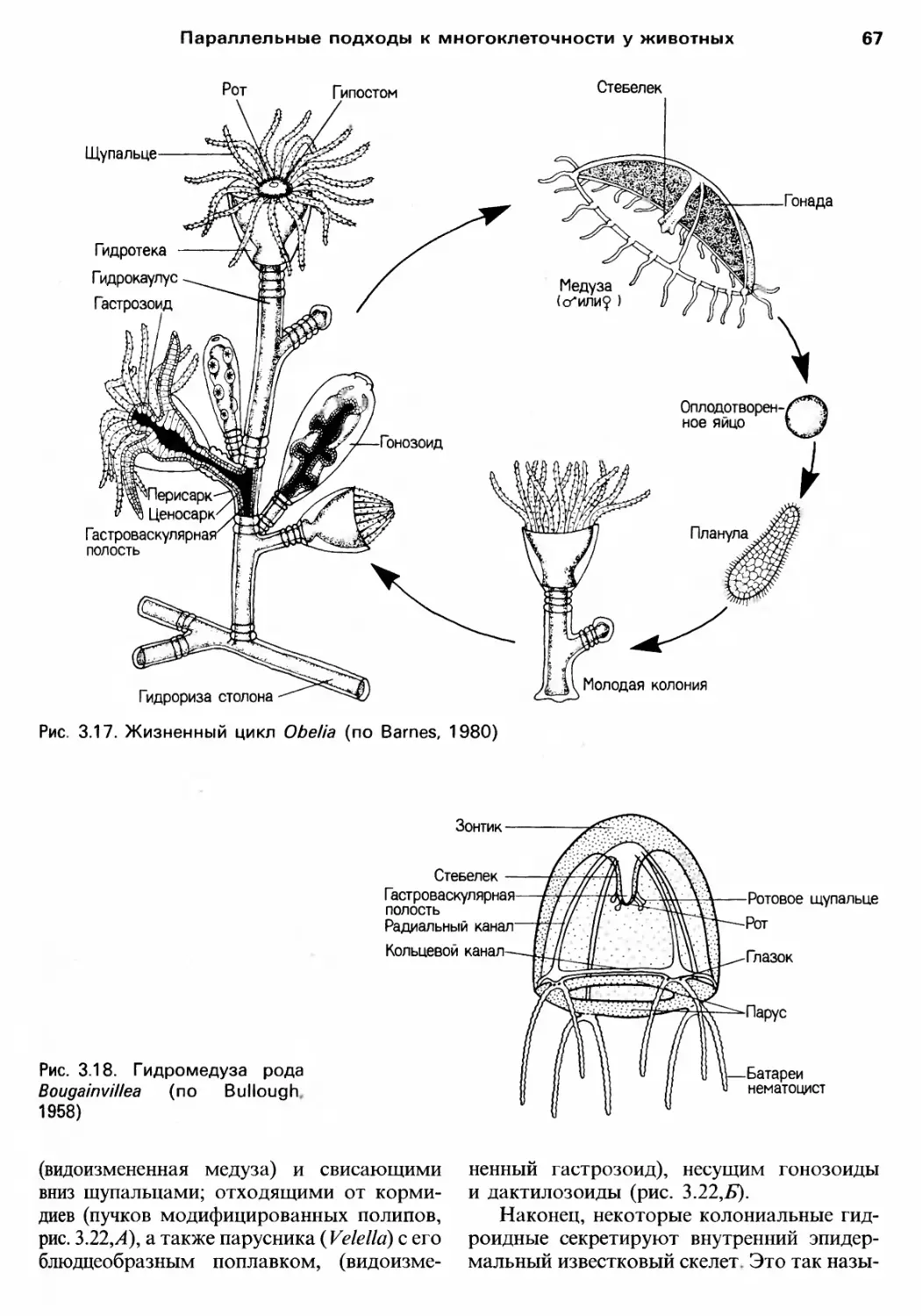
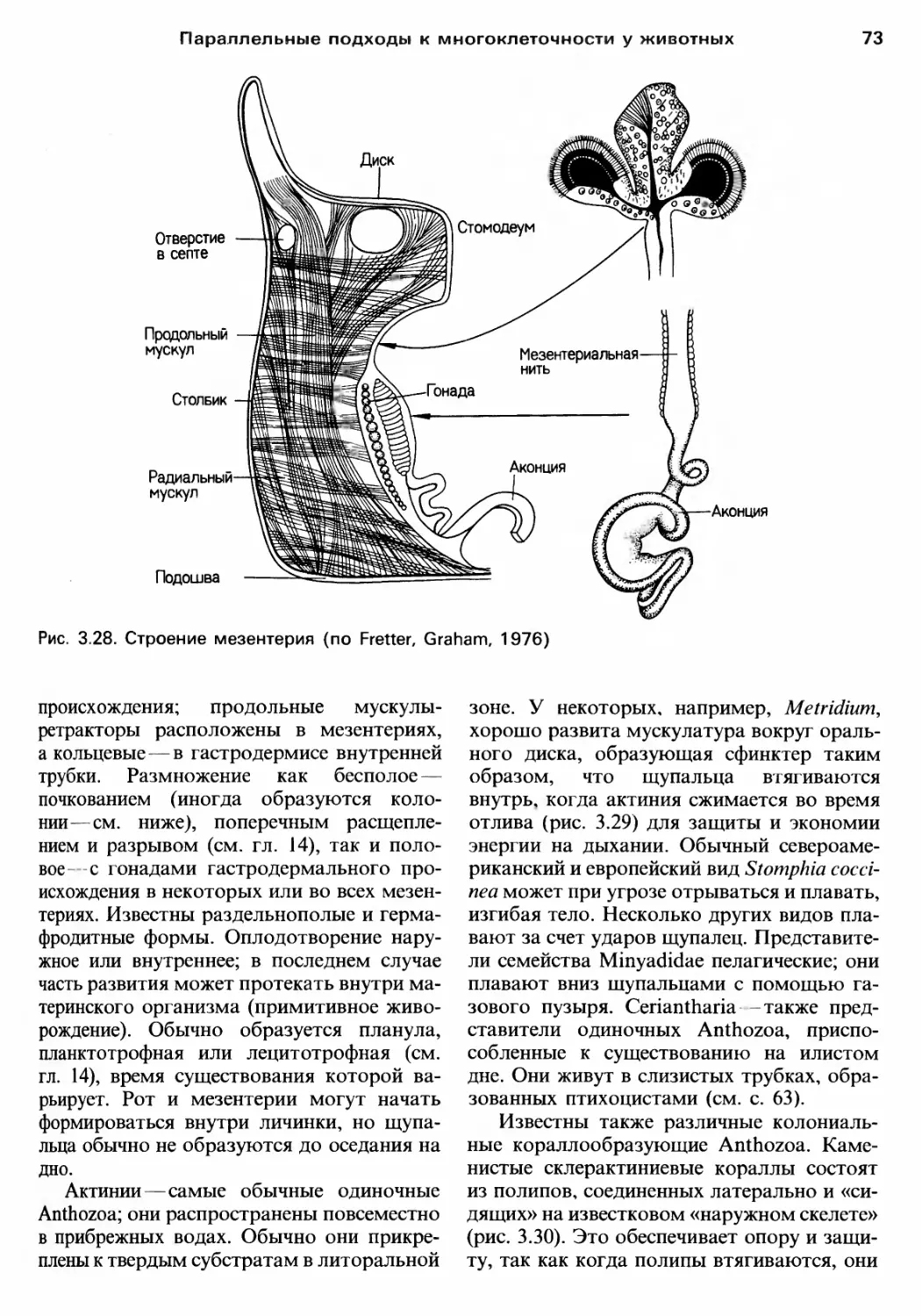
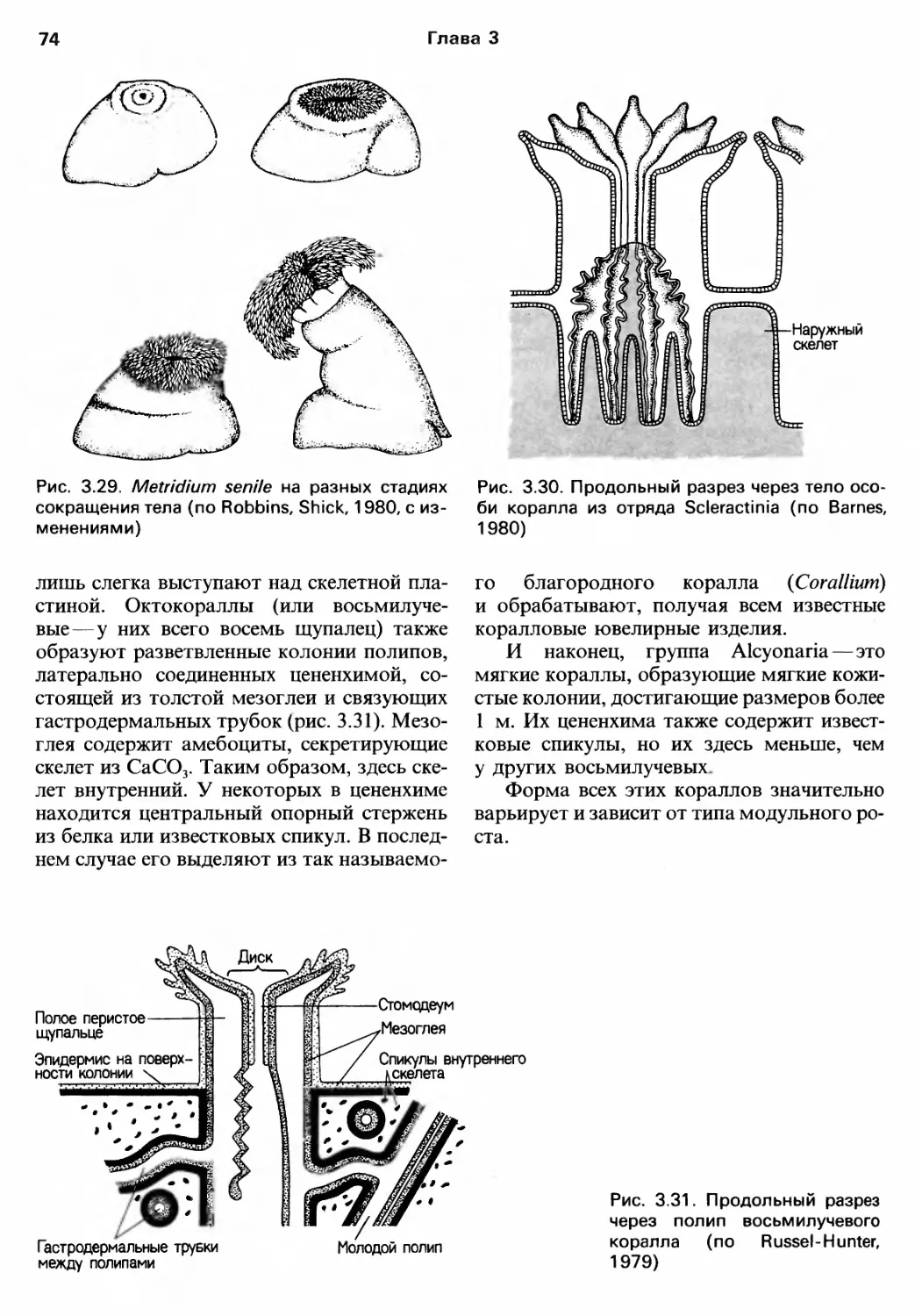
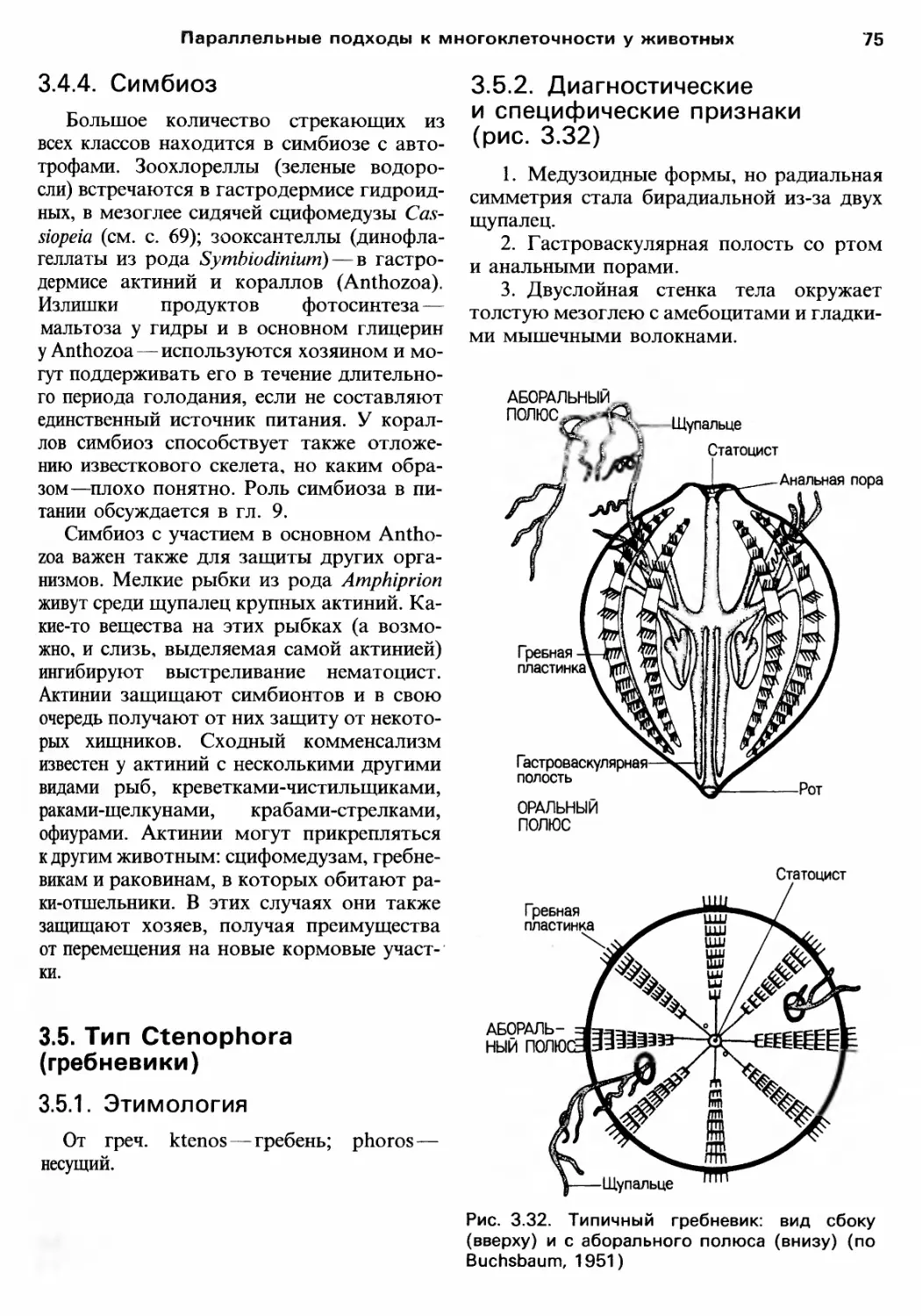
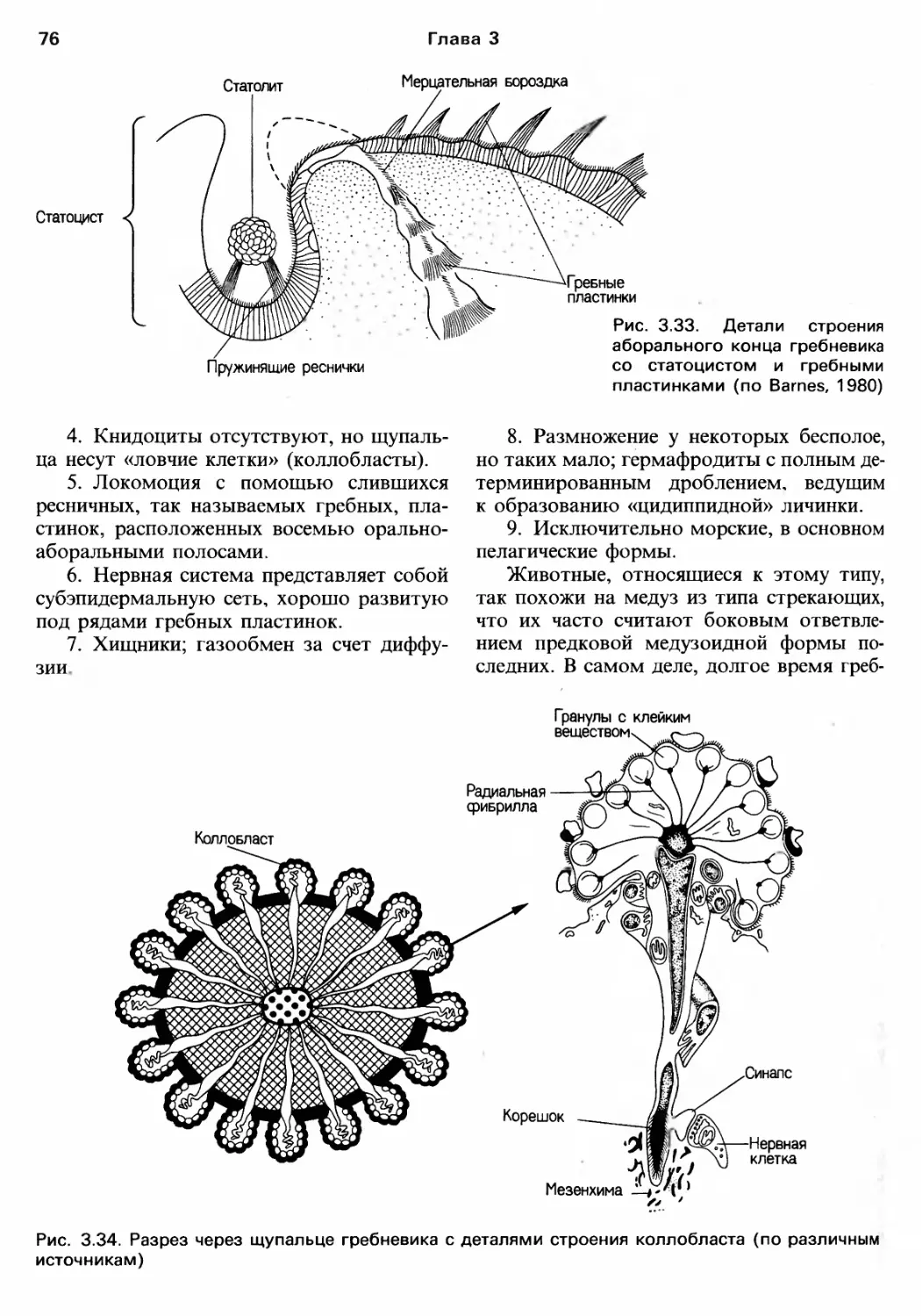
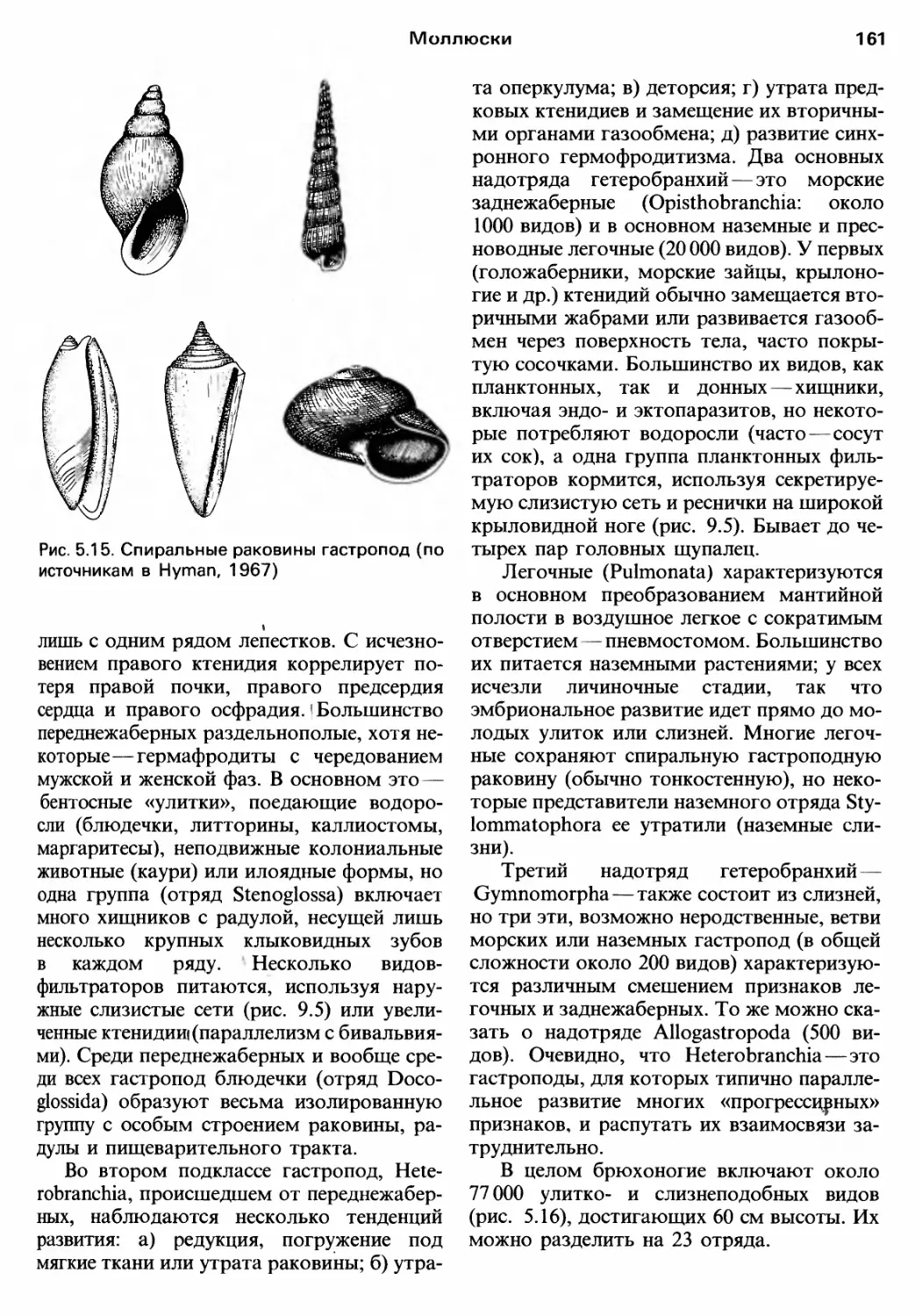
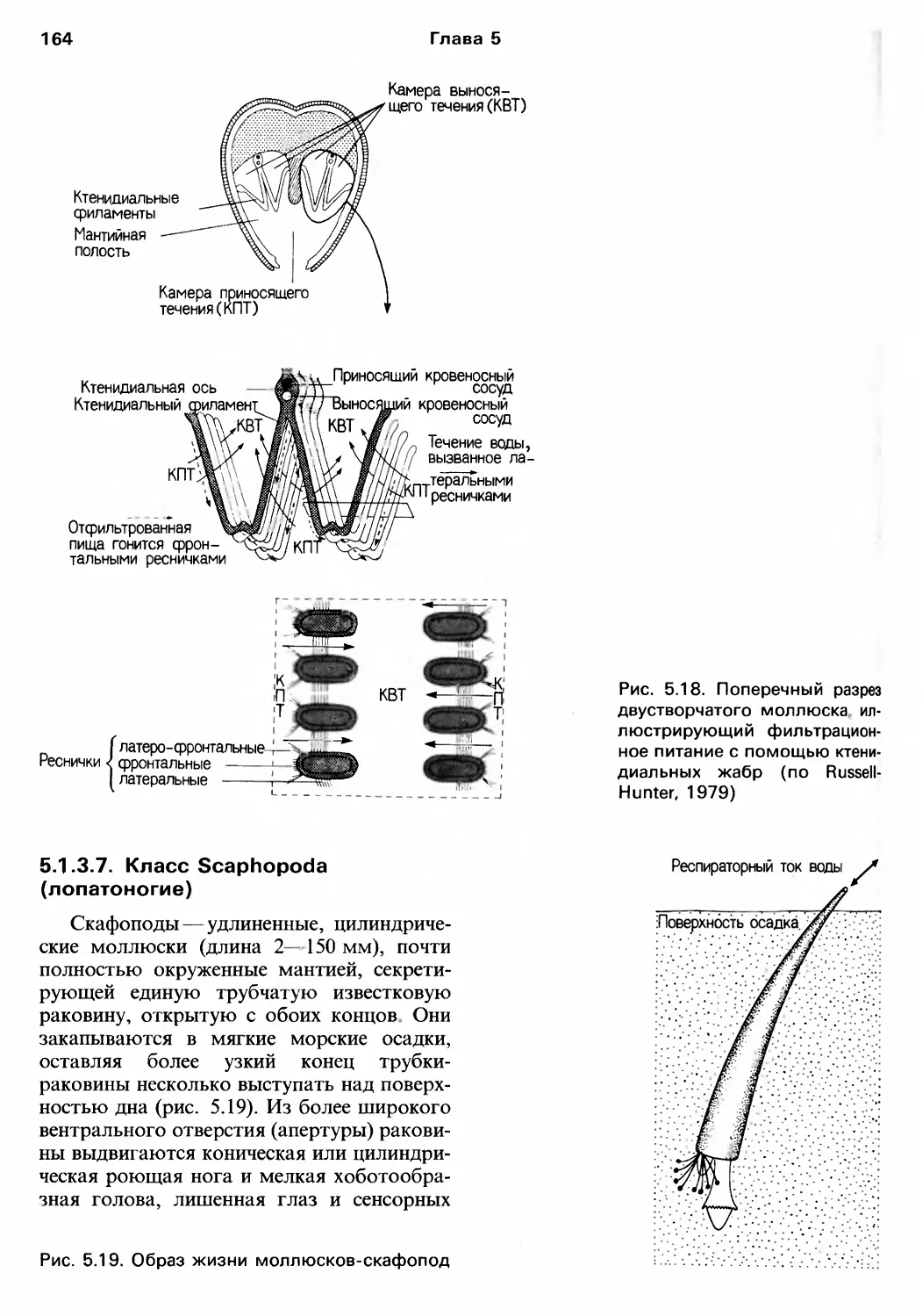
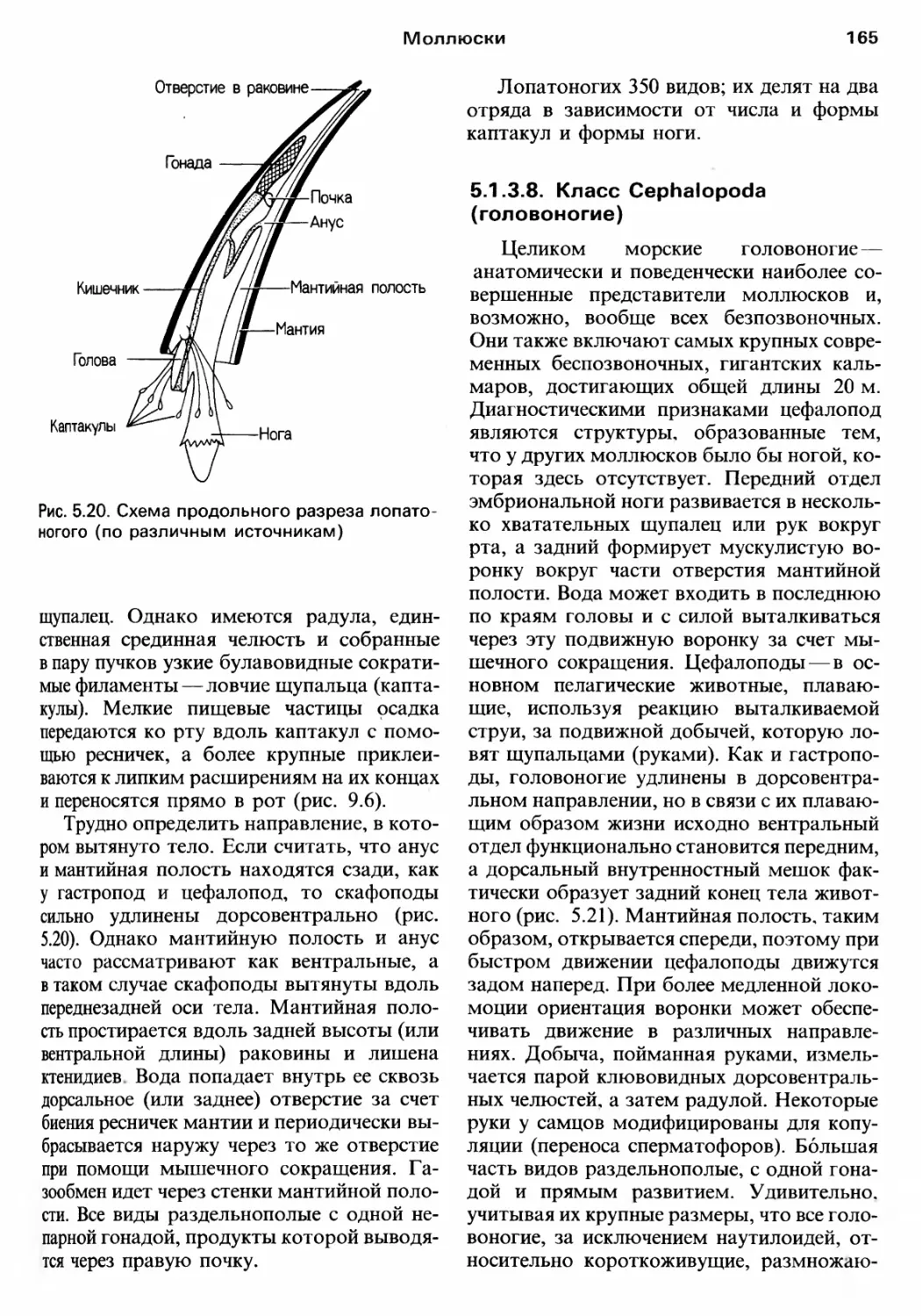
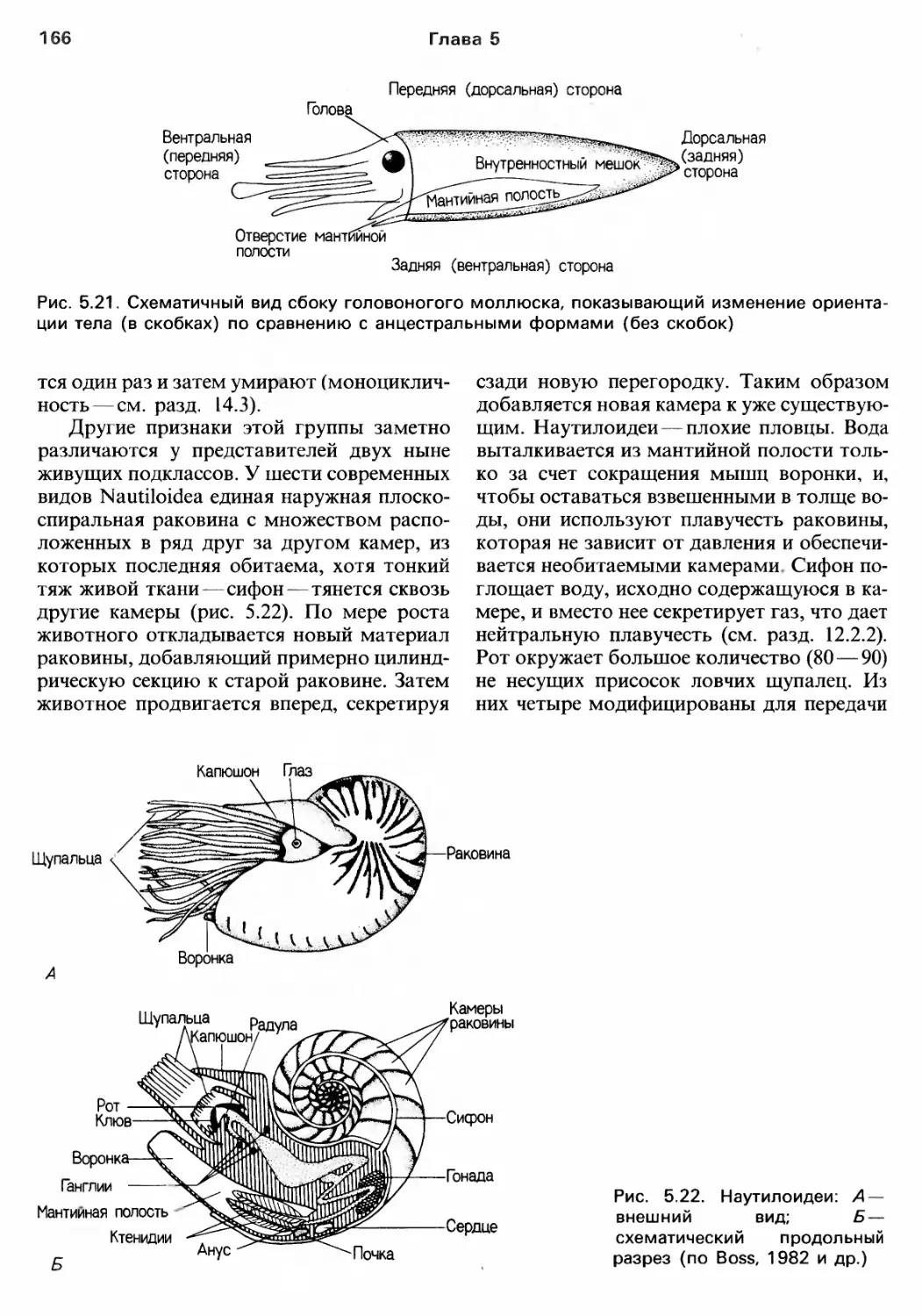
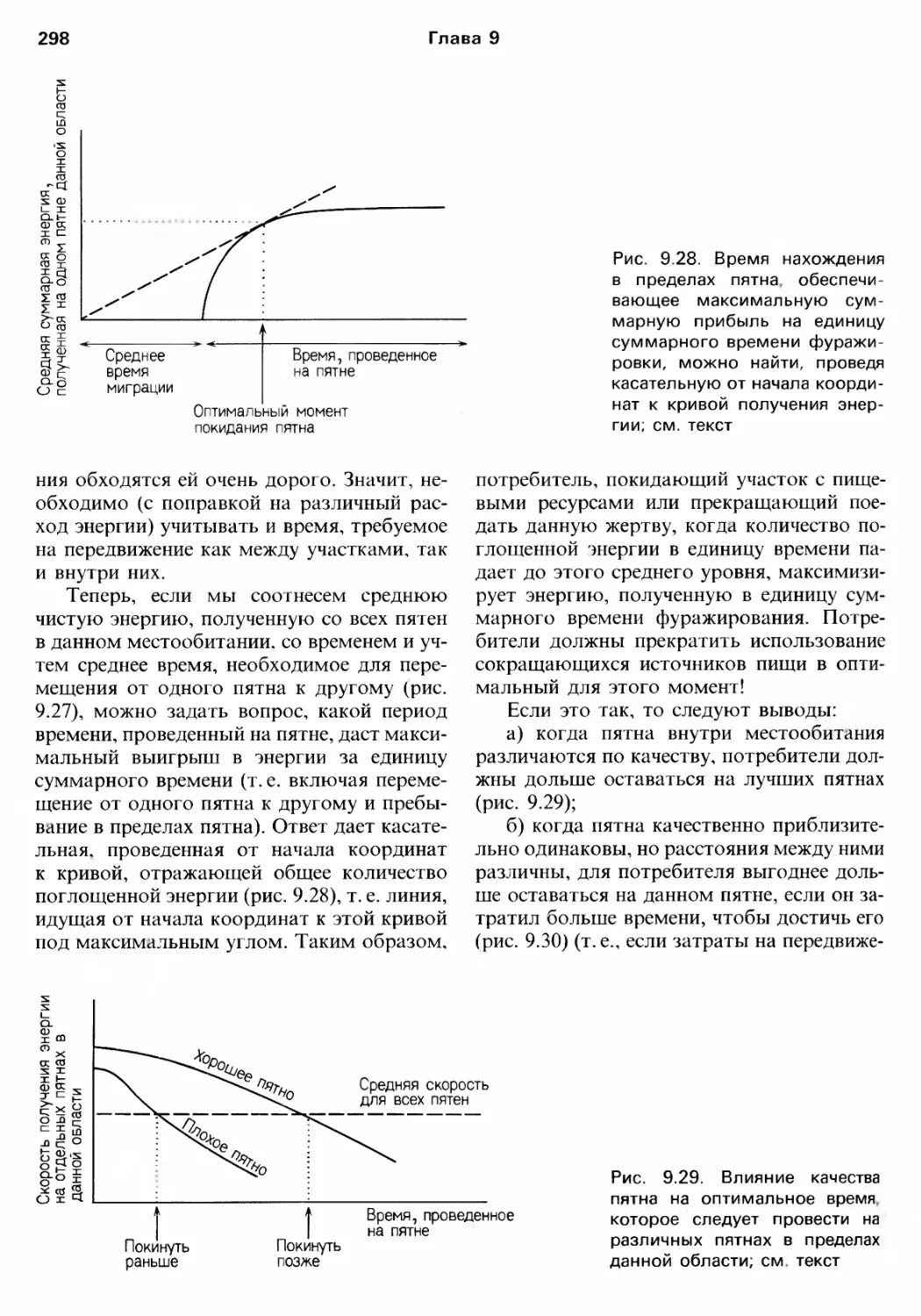
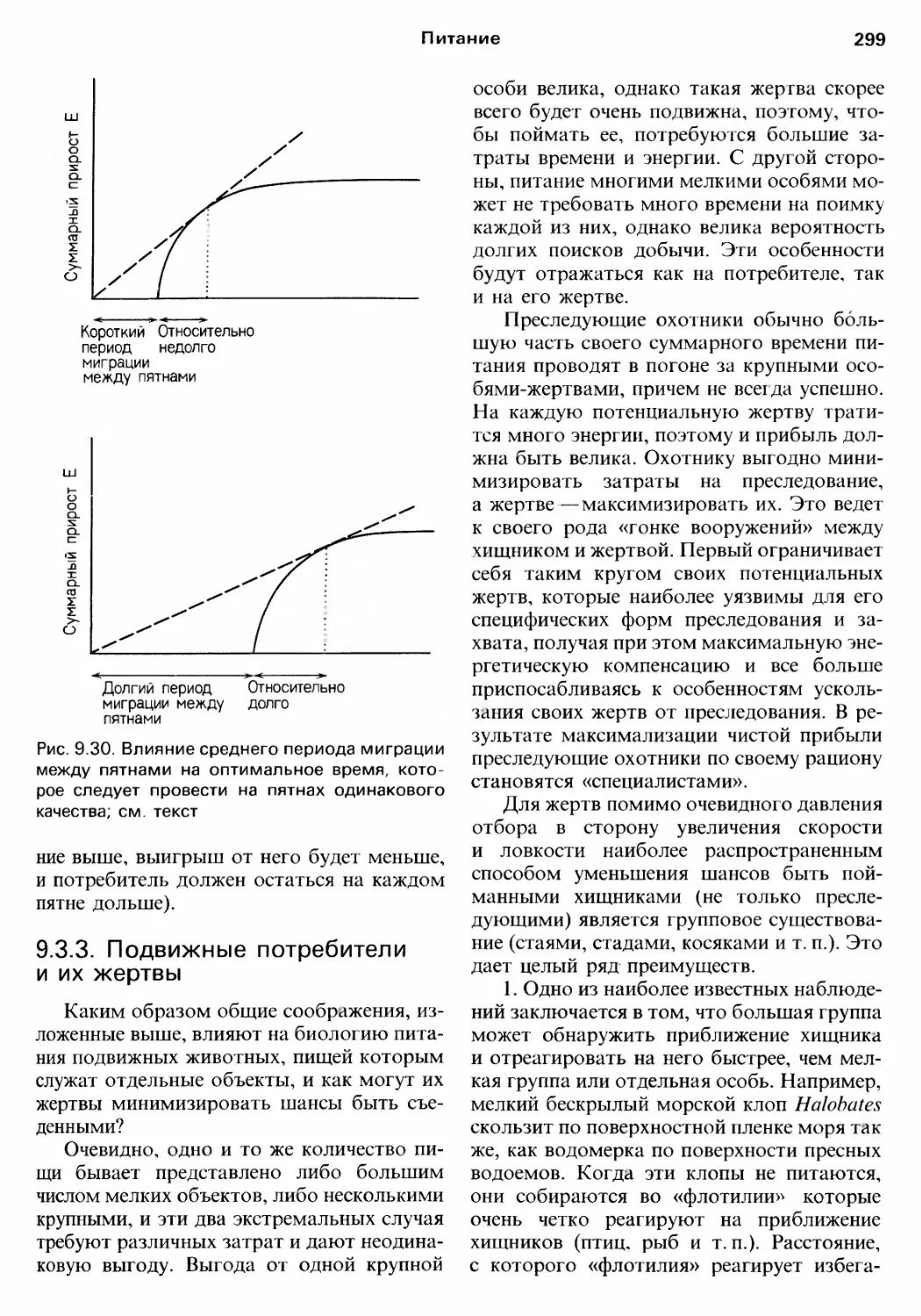
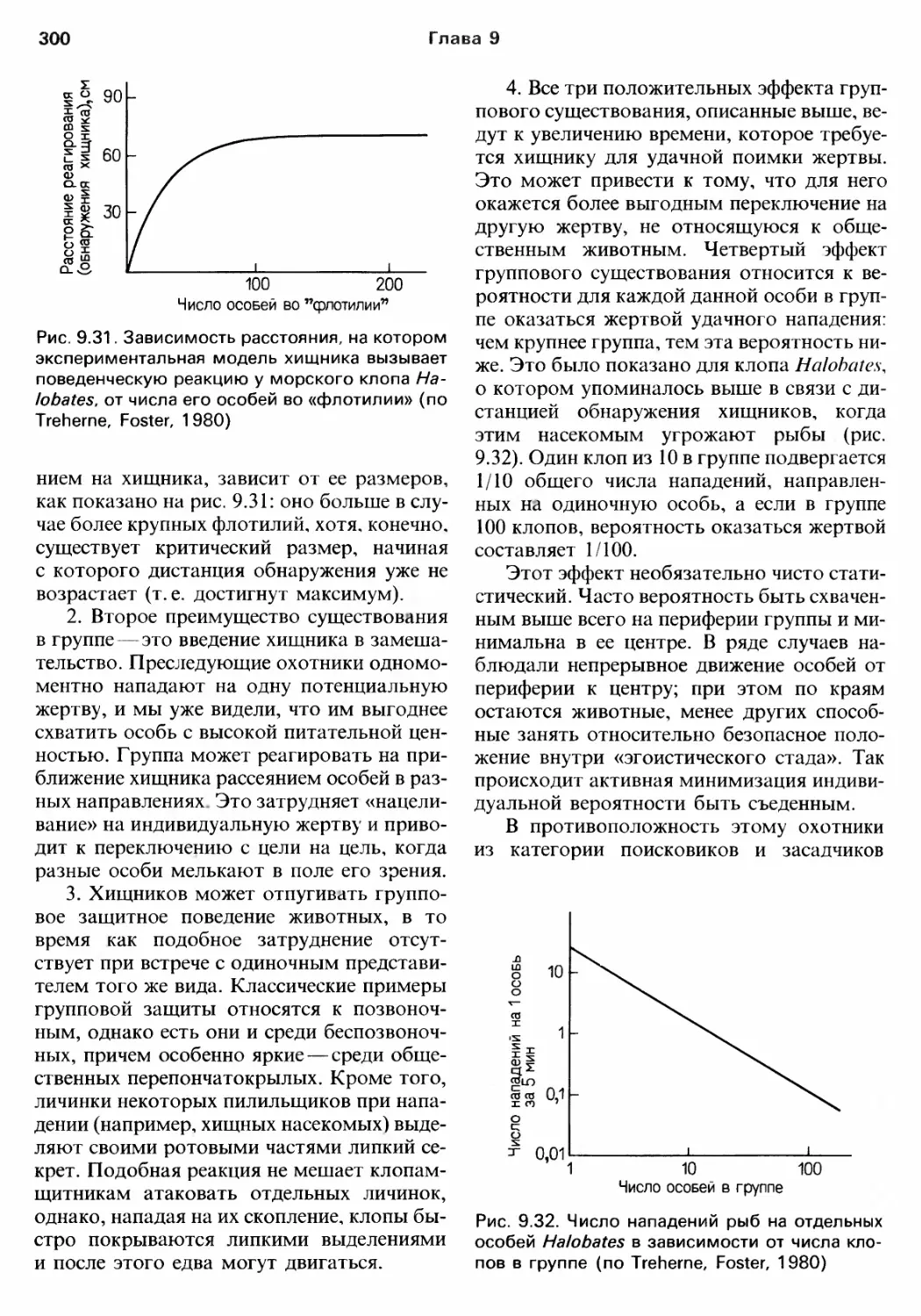
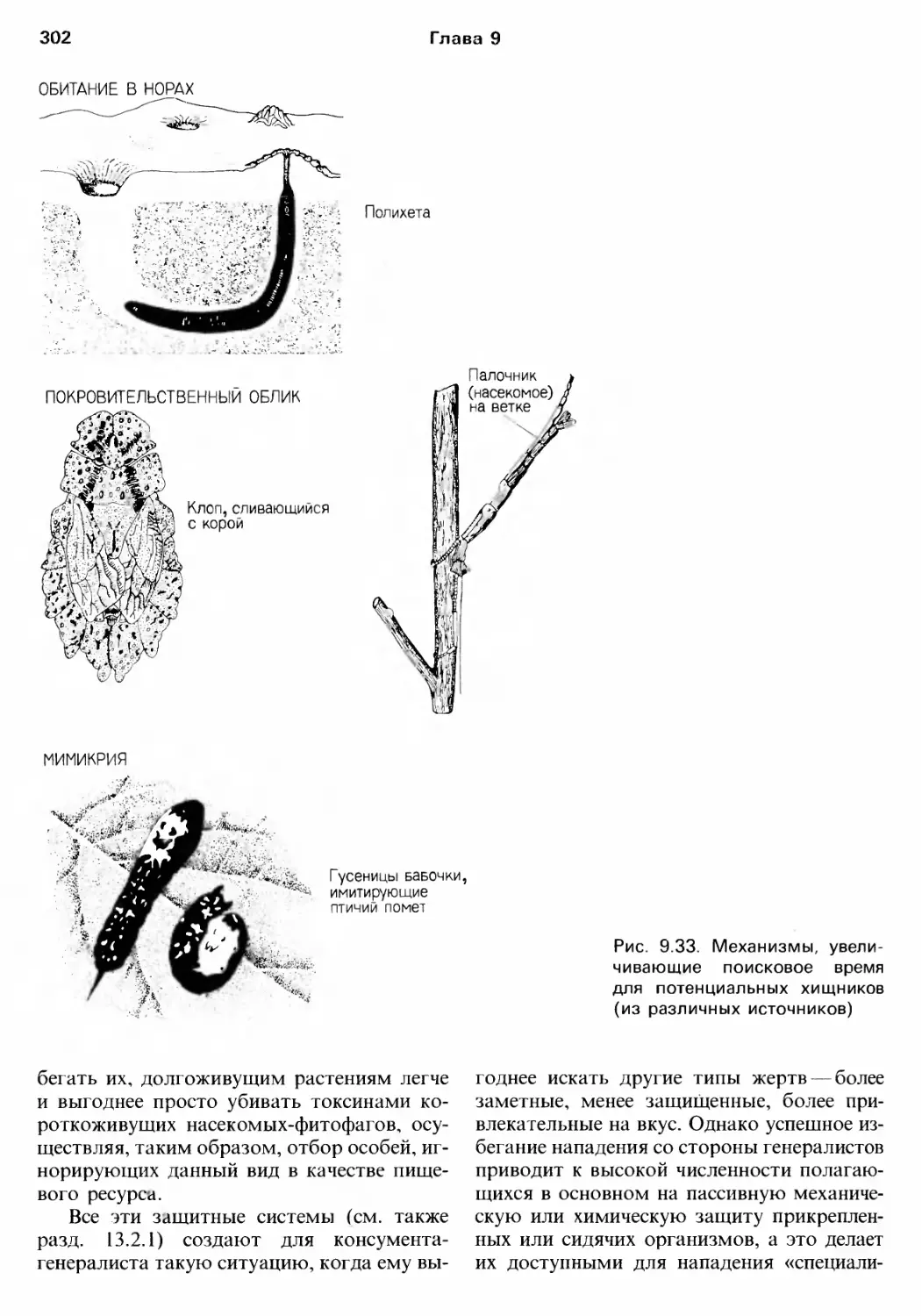
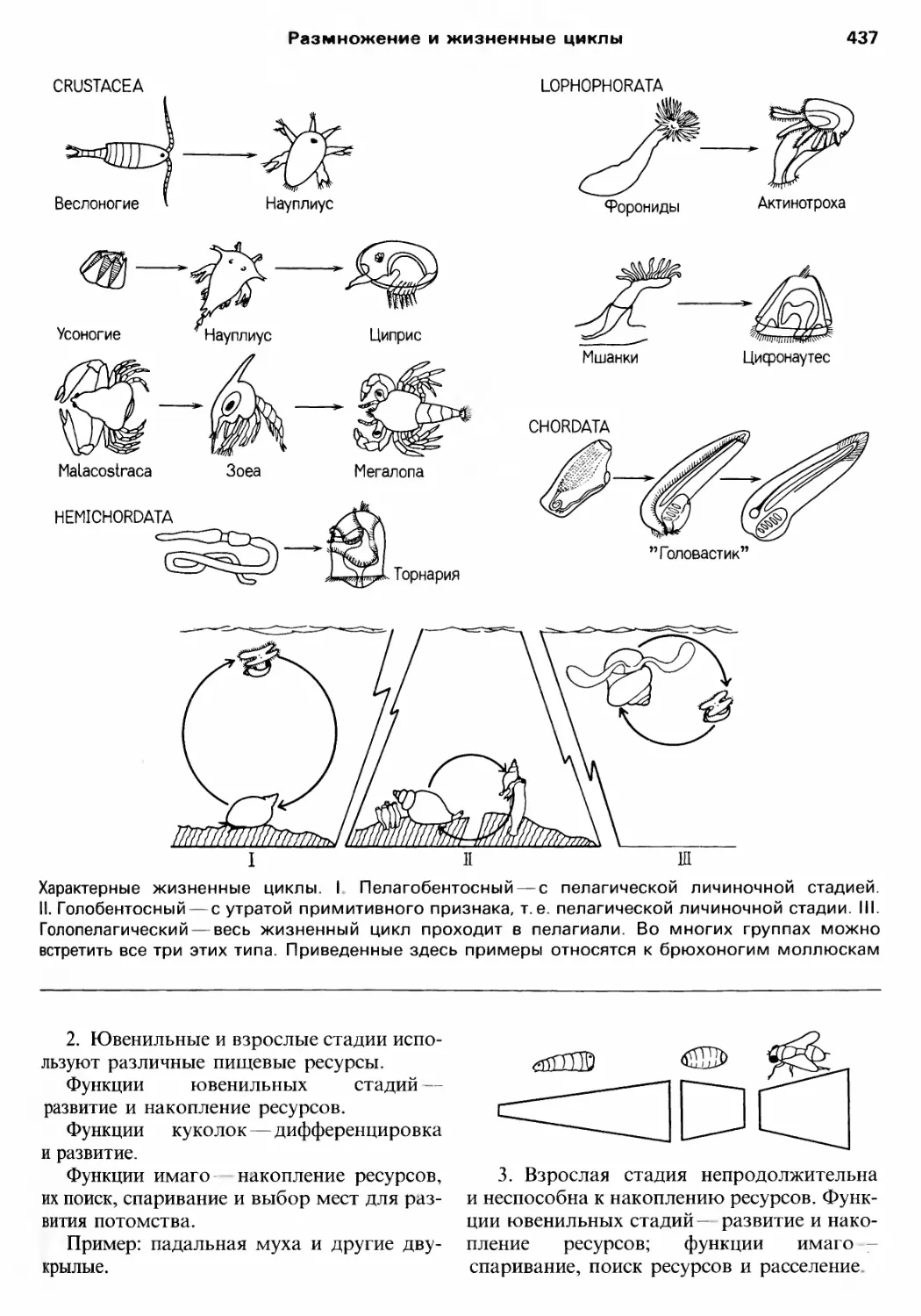
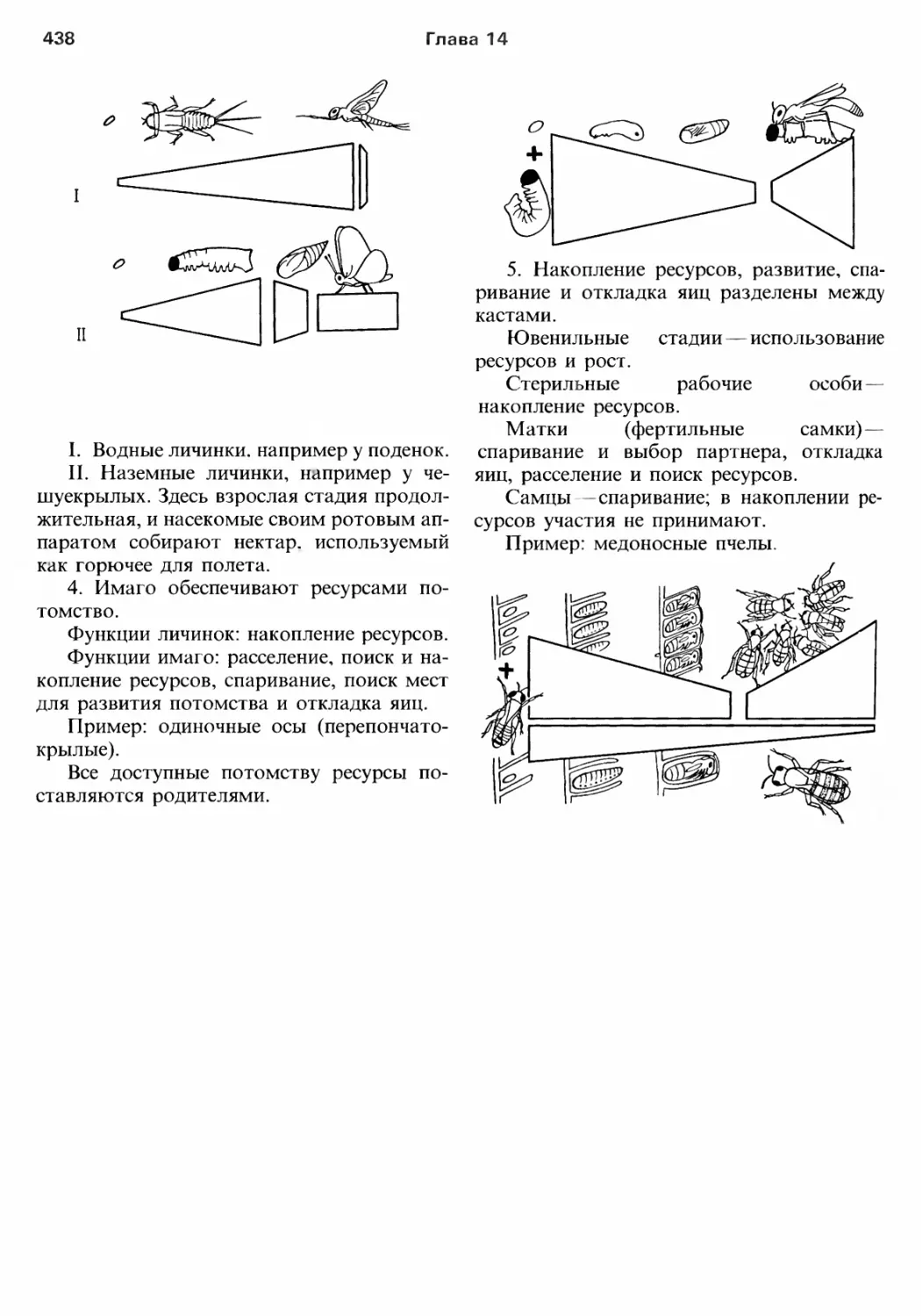
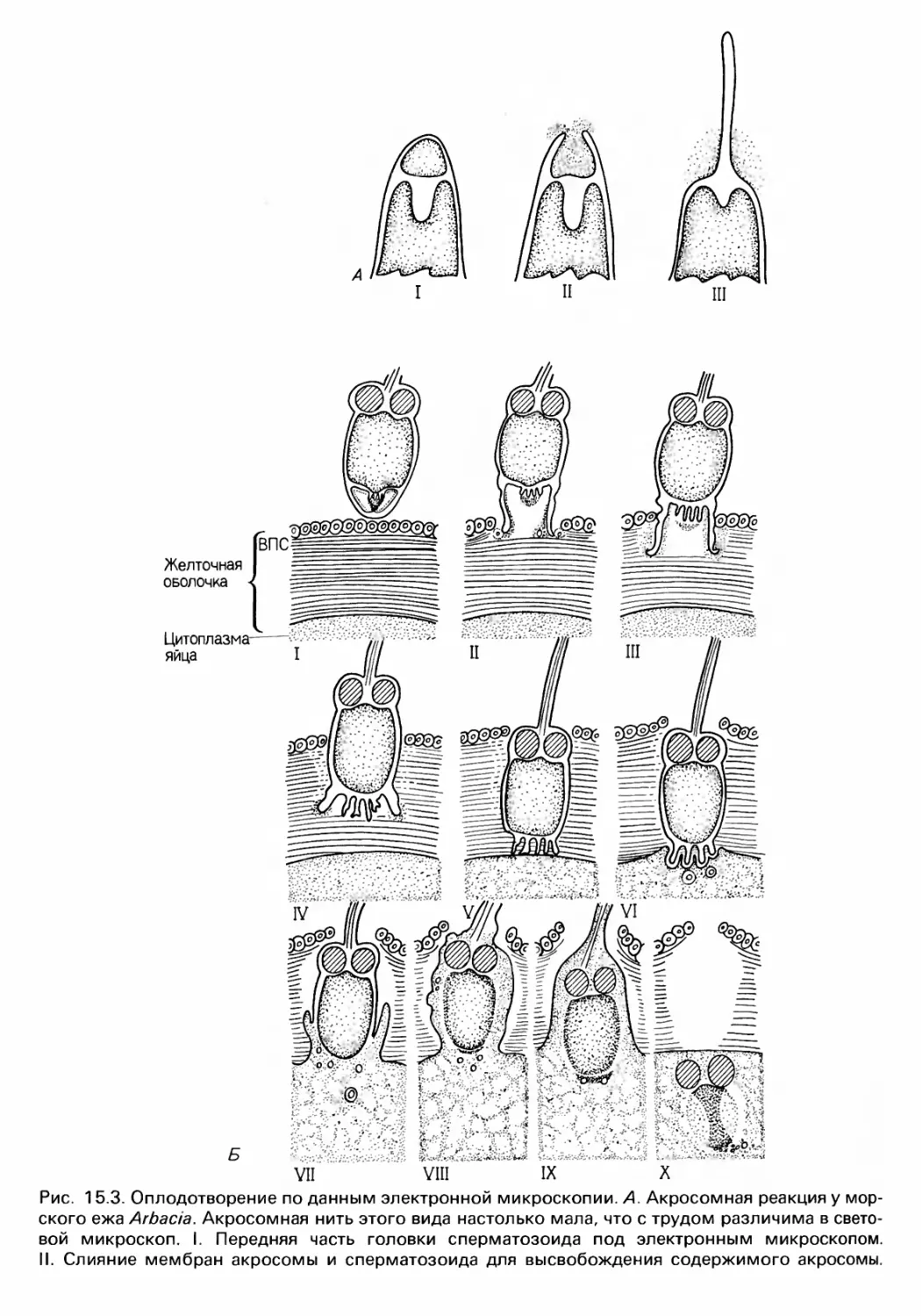
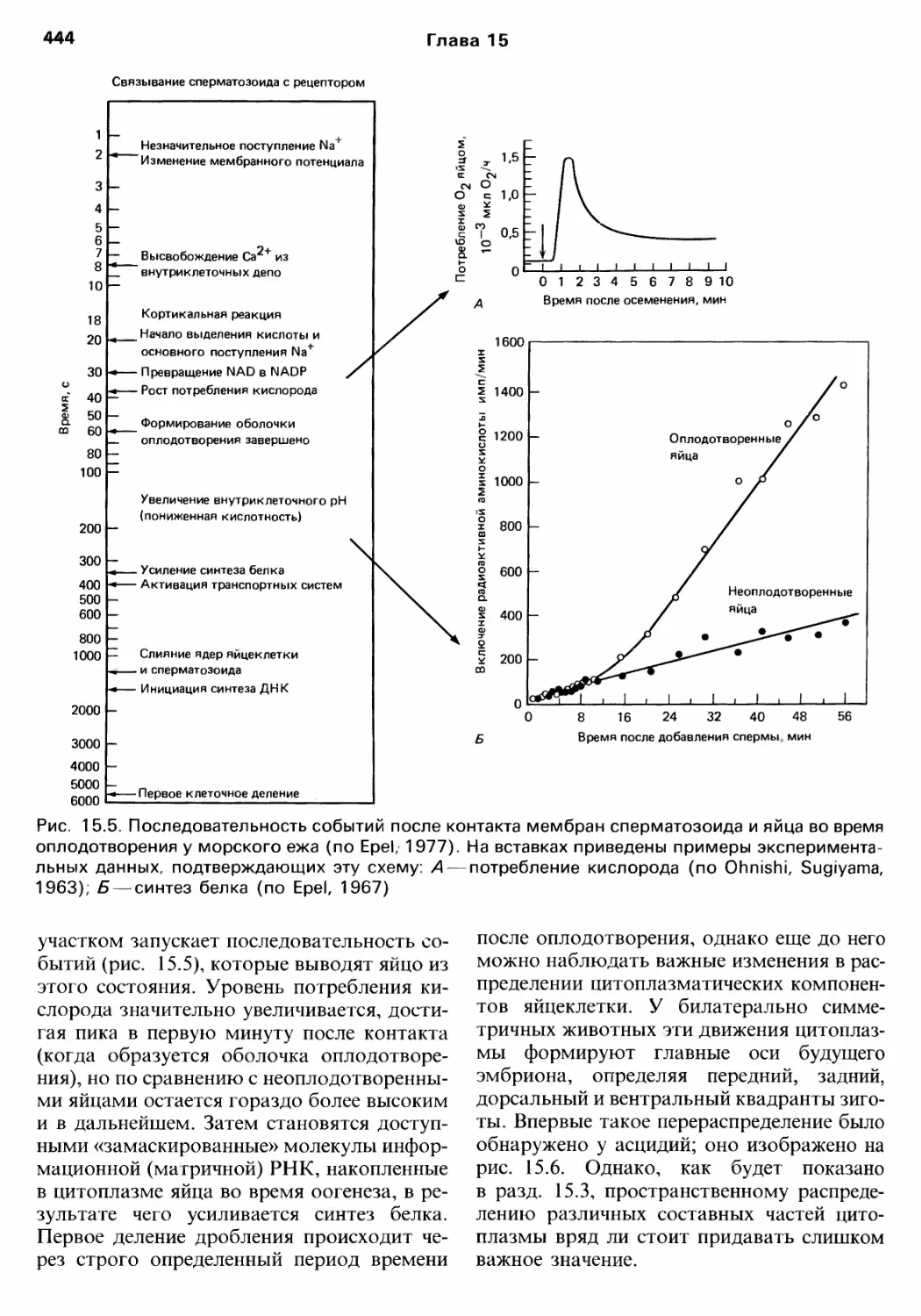
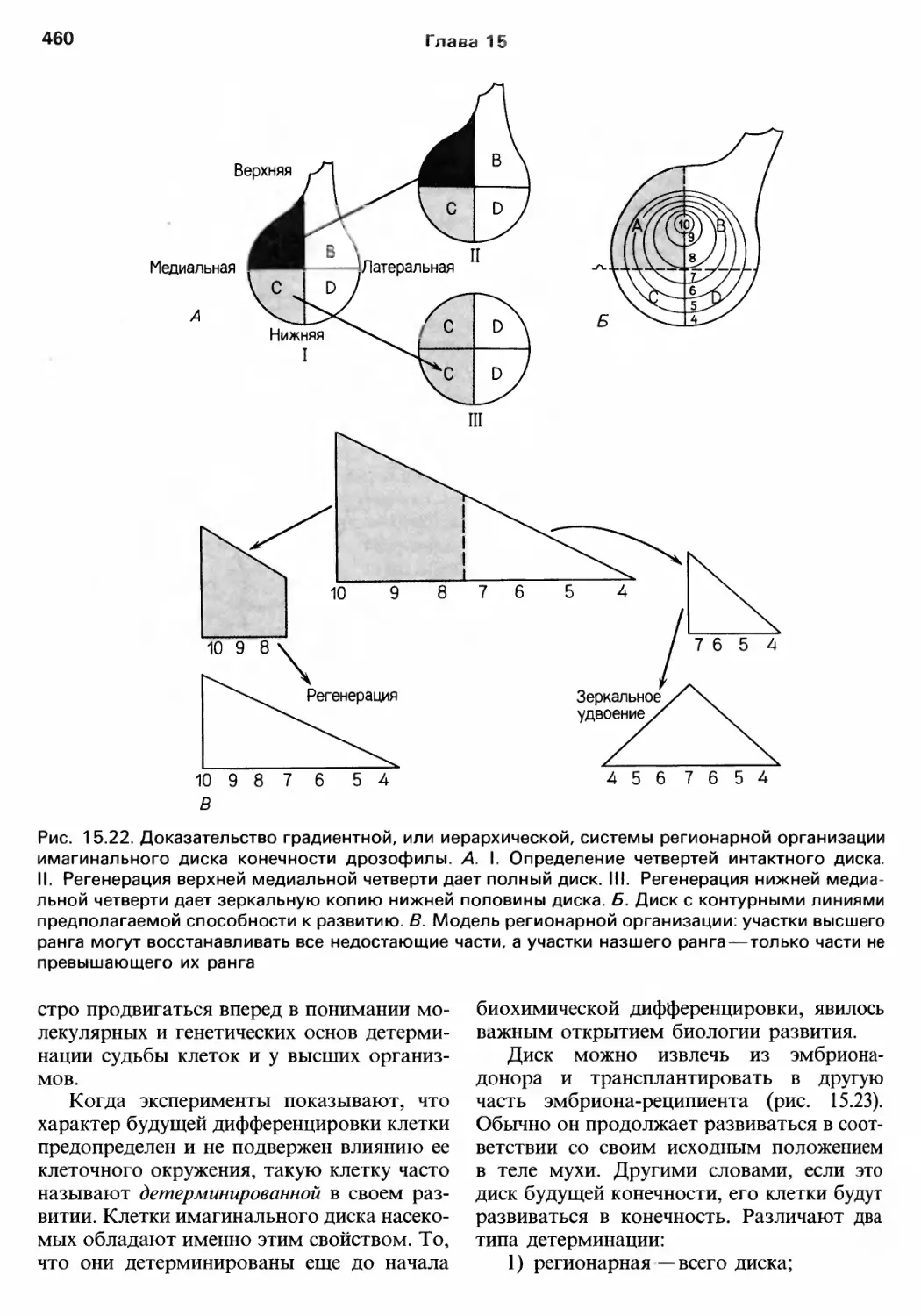
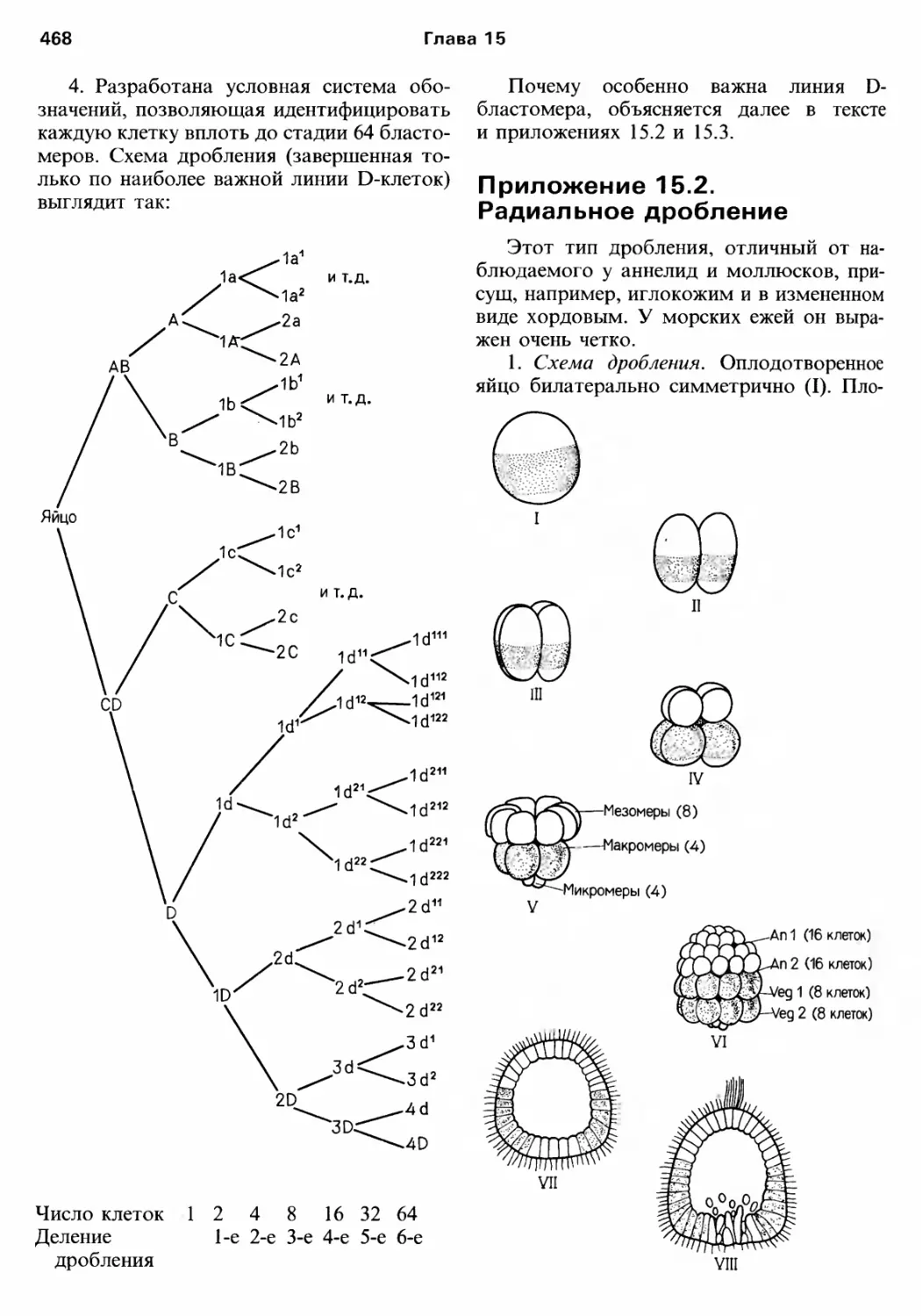
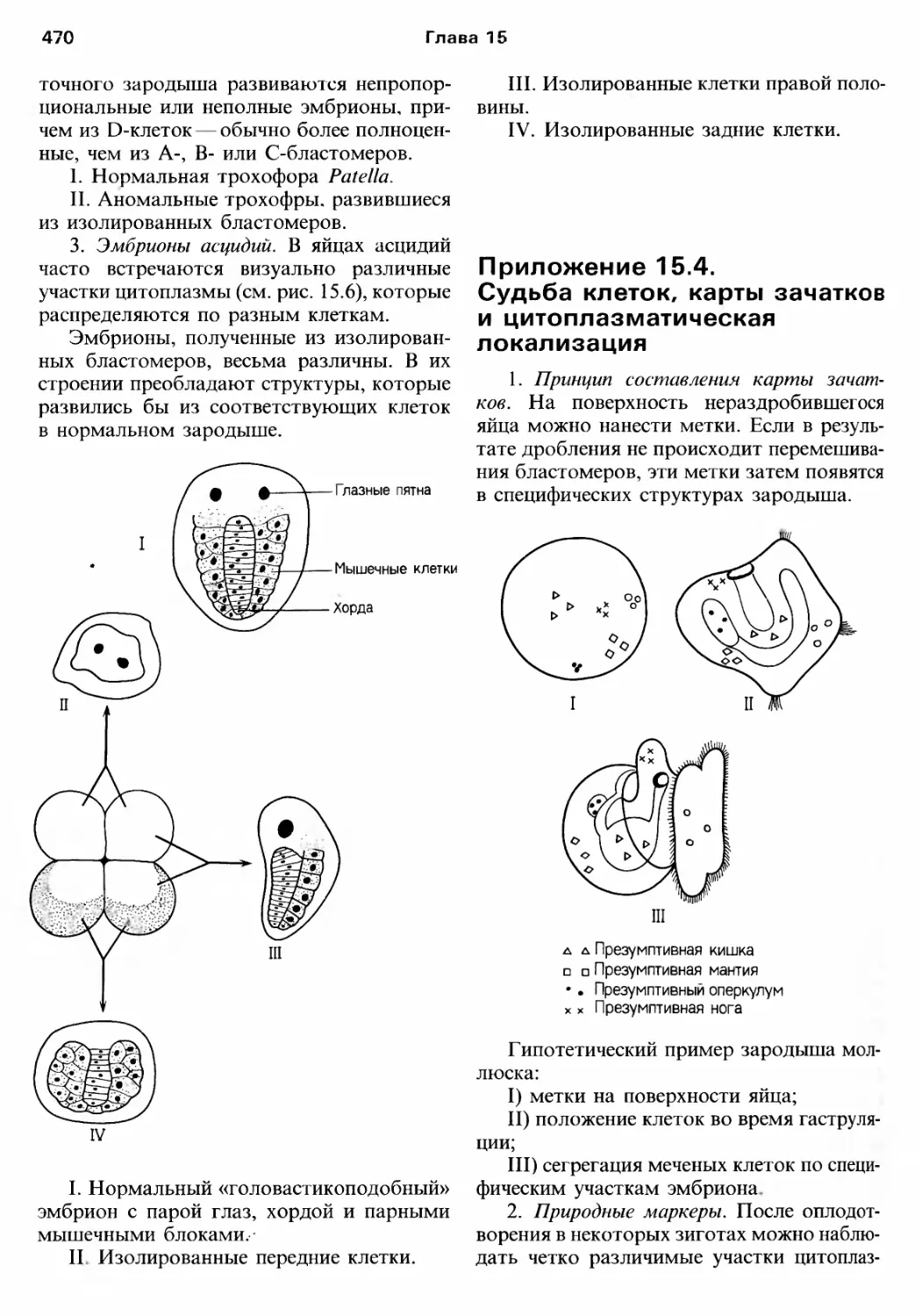
/












































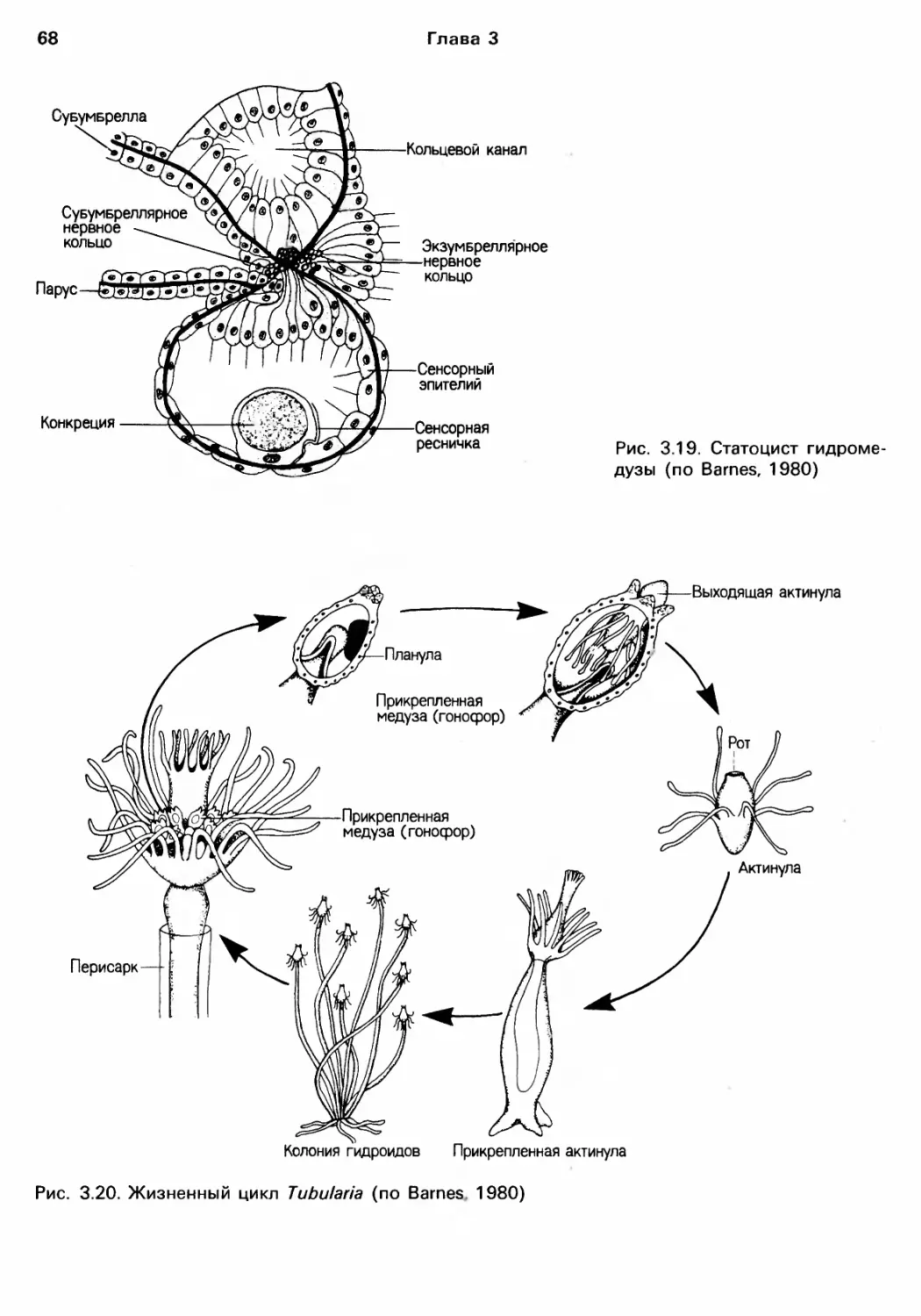
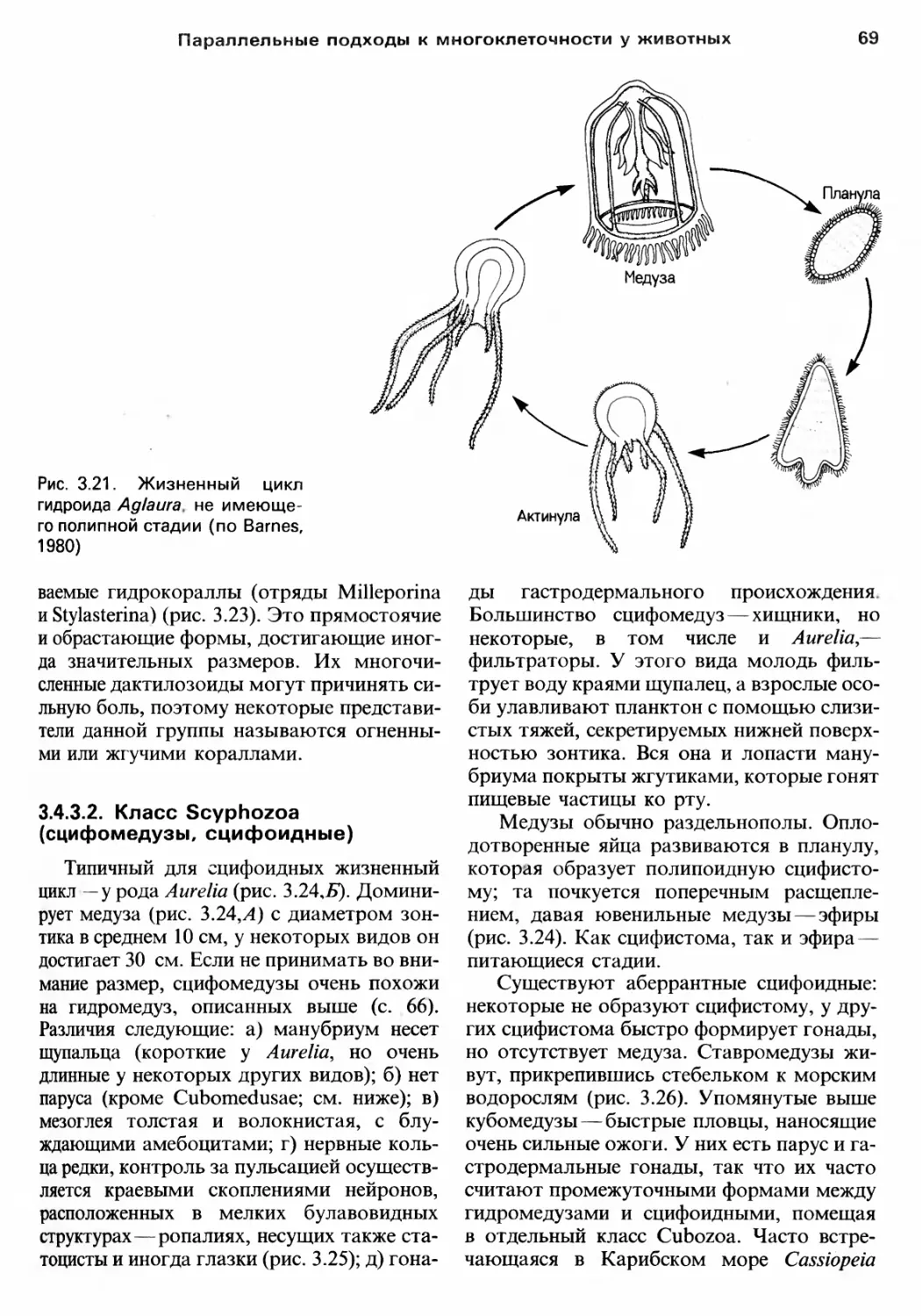
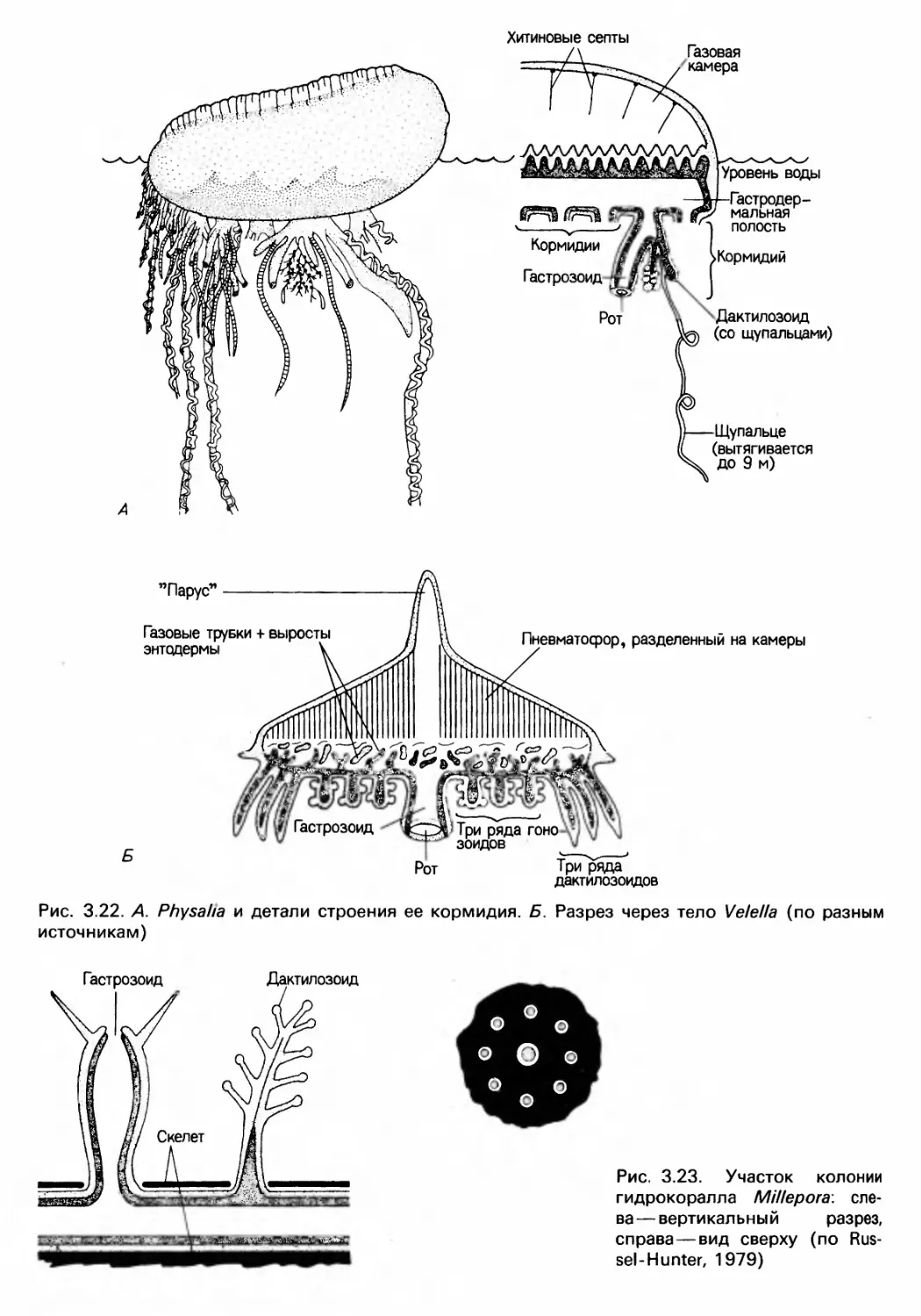
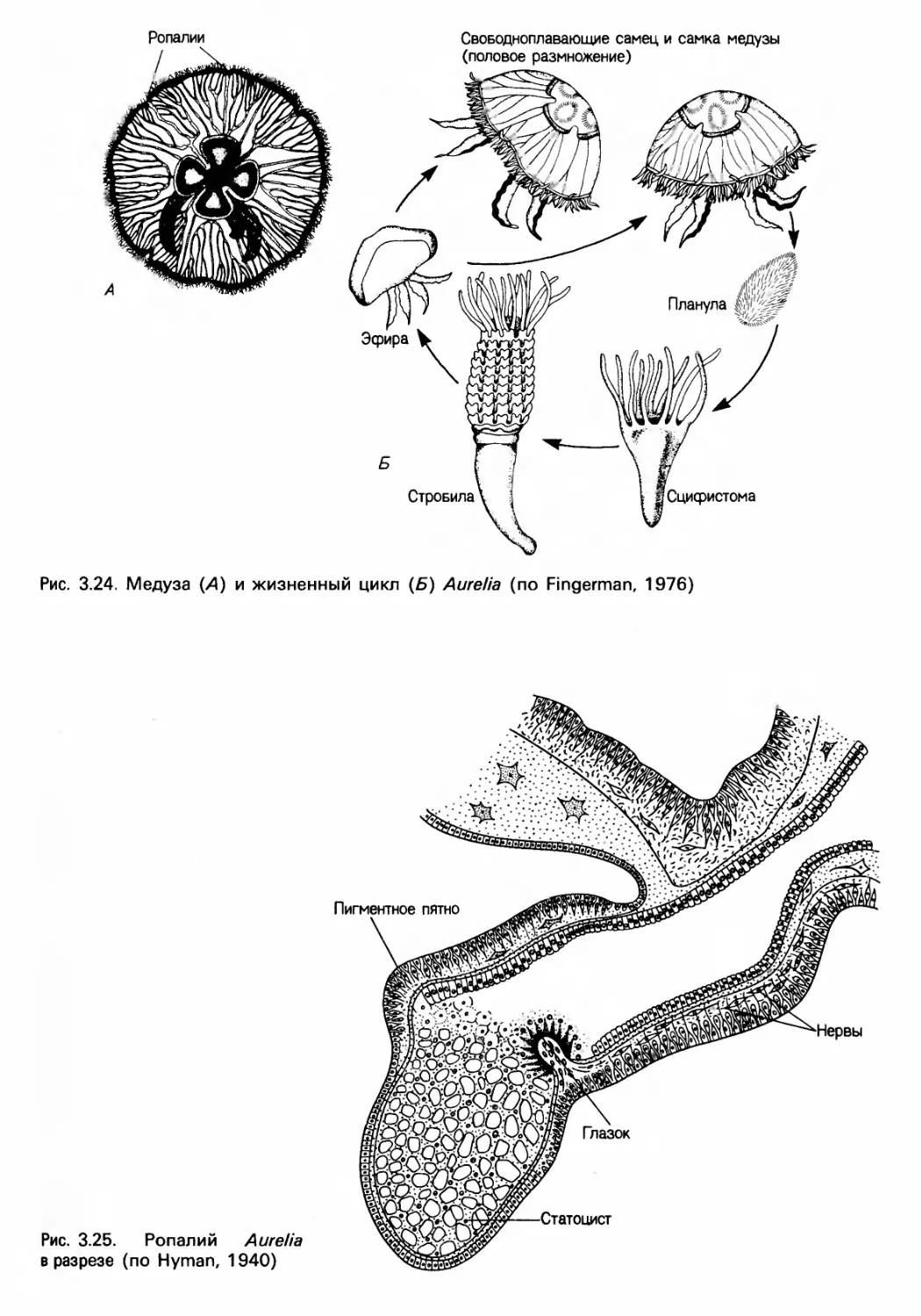

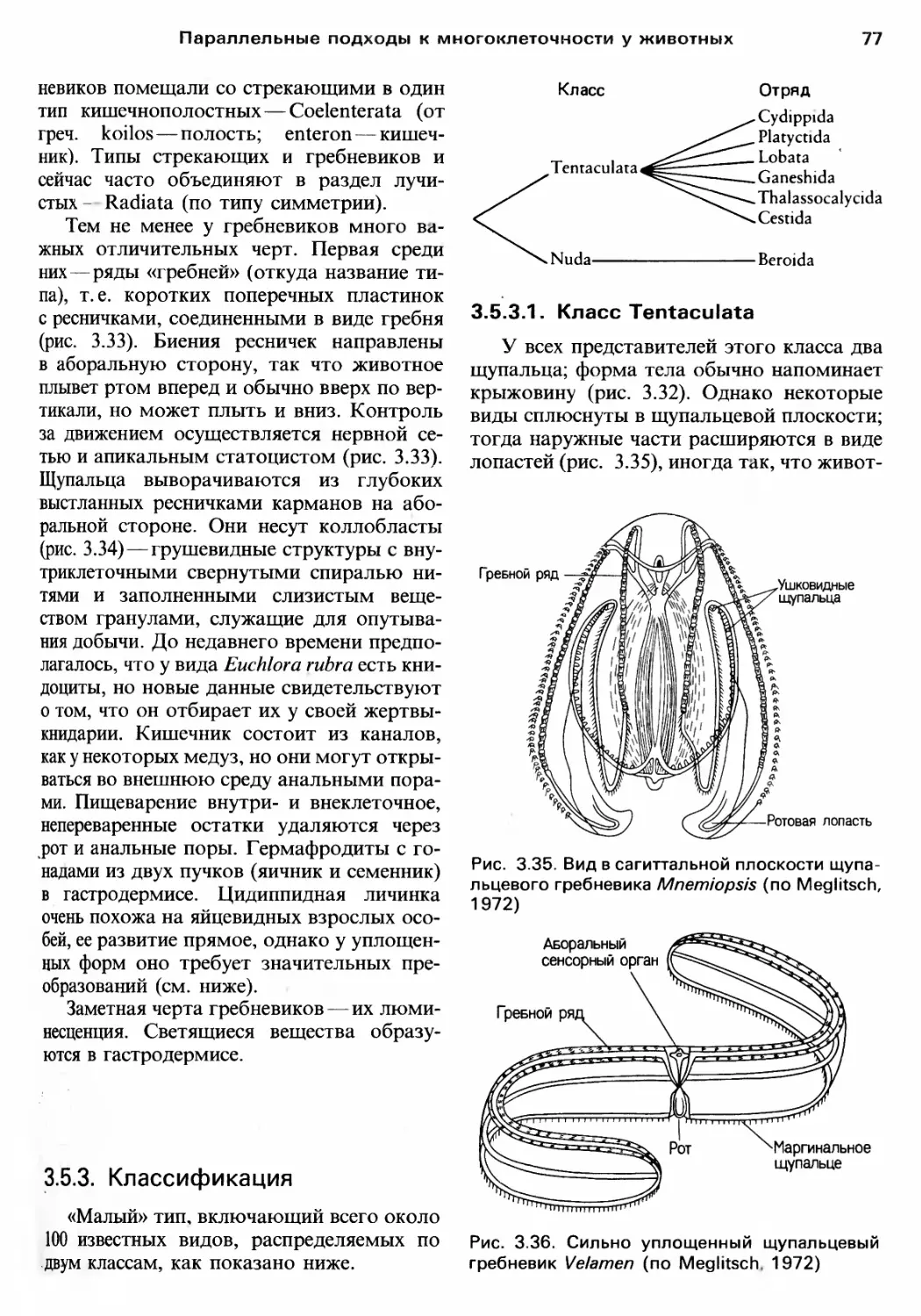
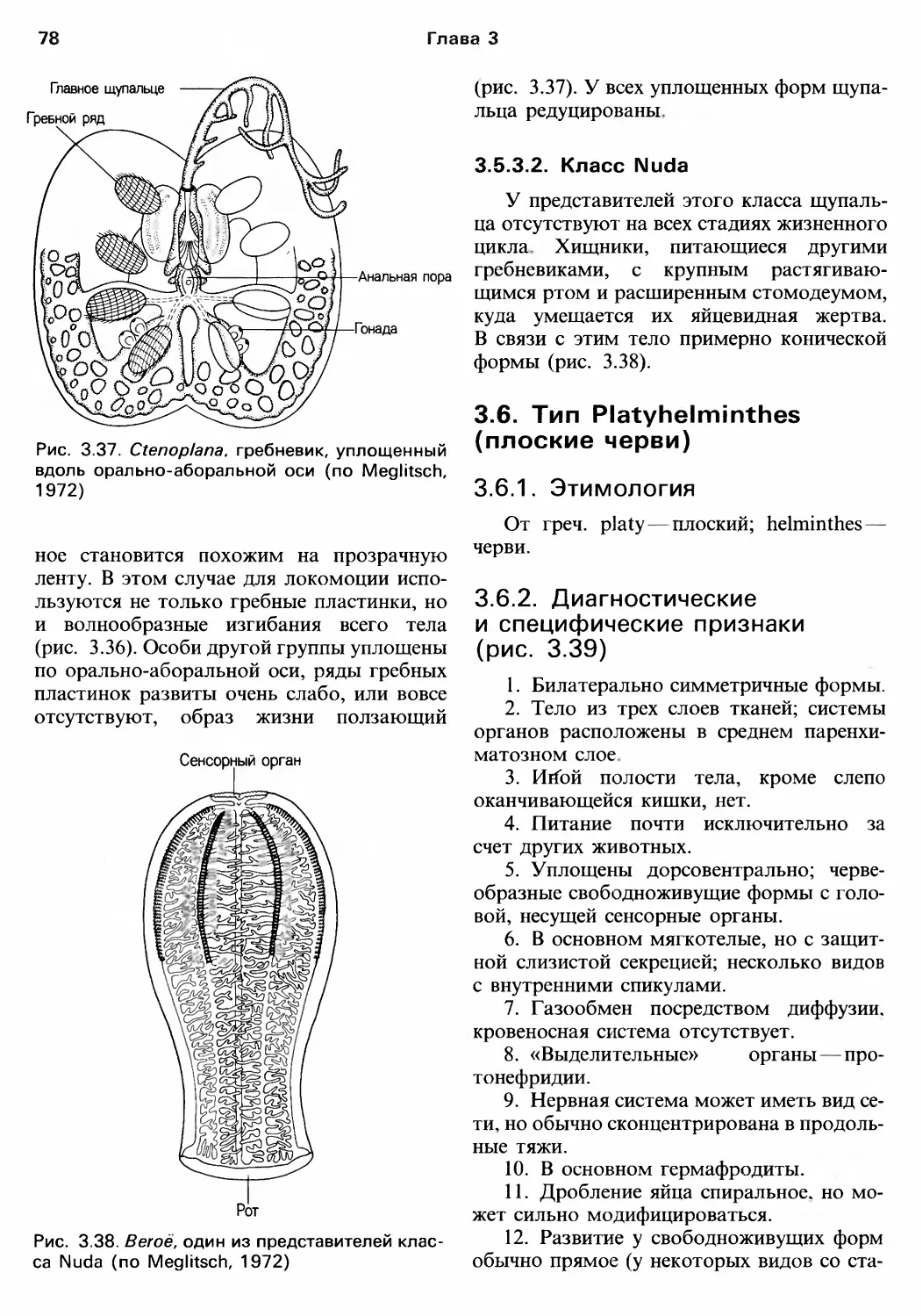
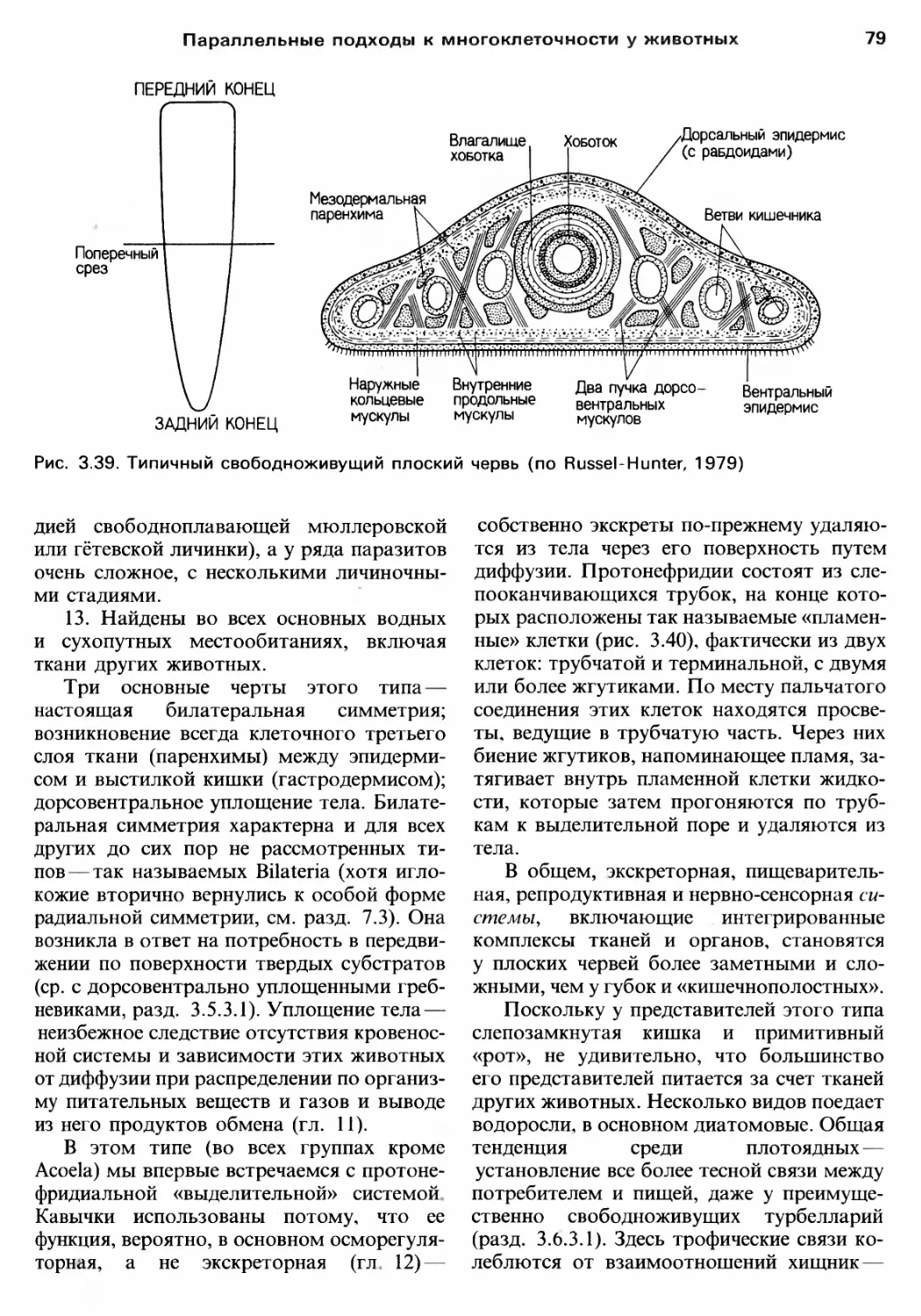
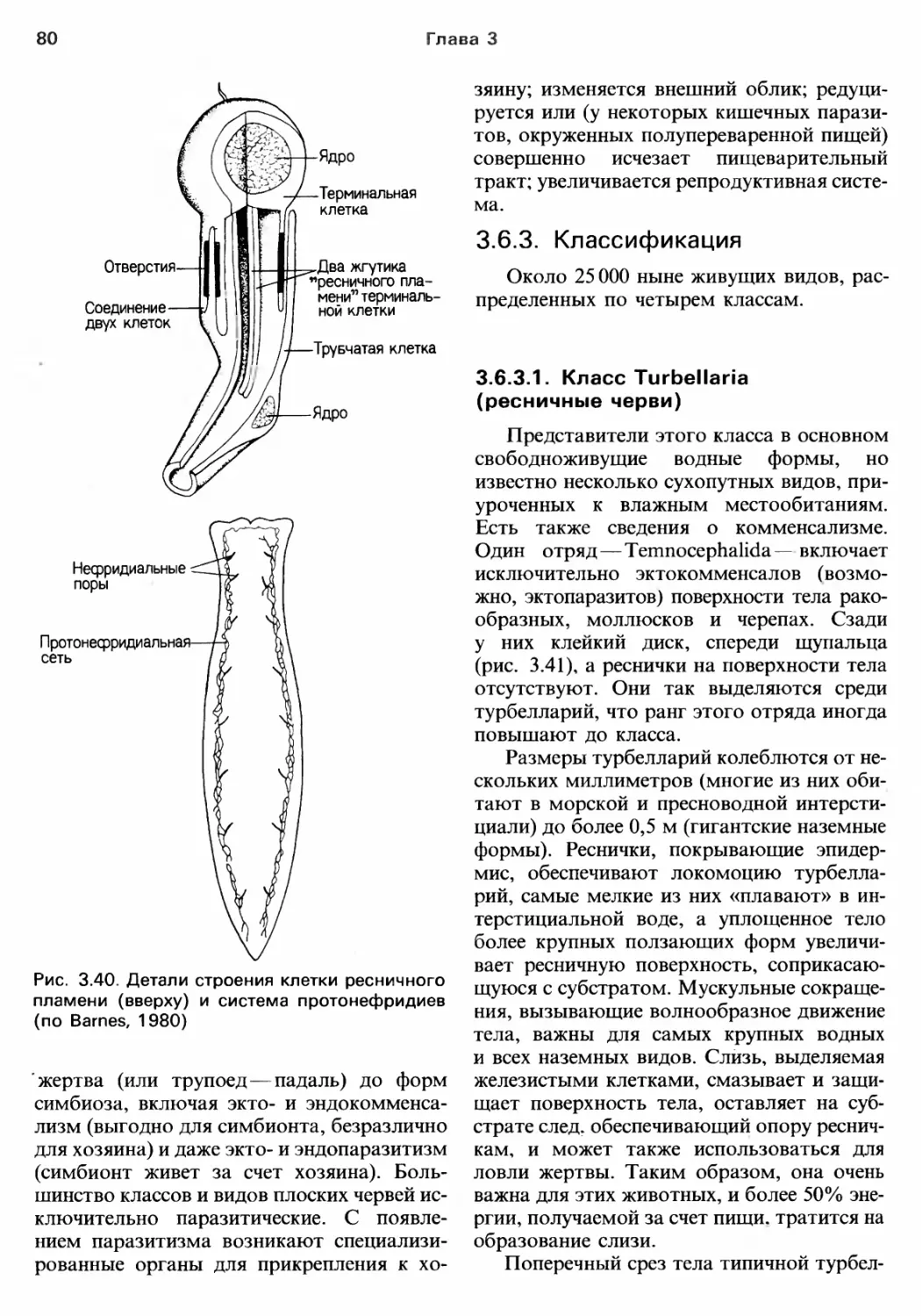






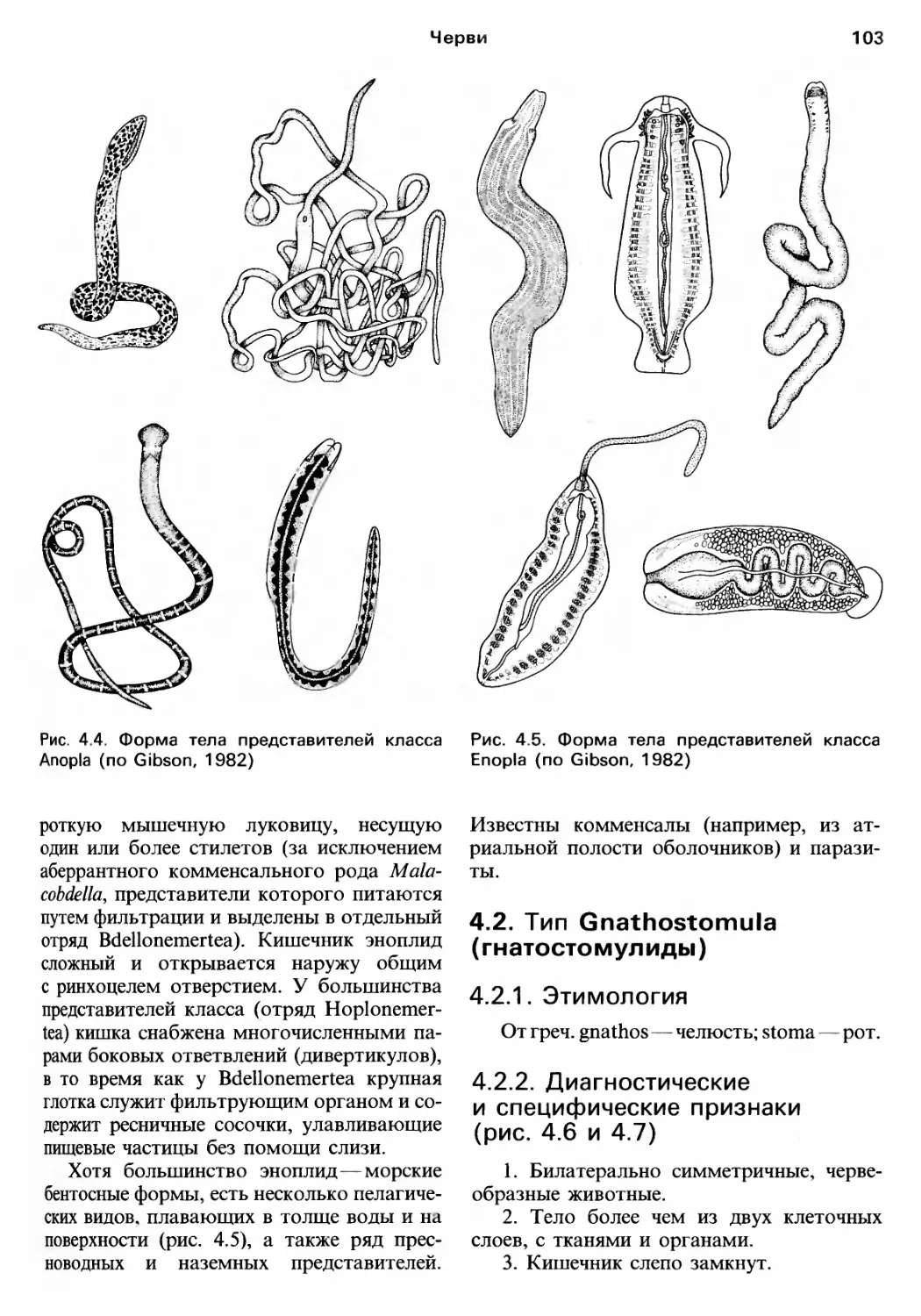
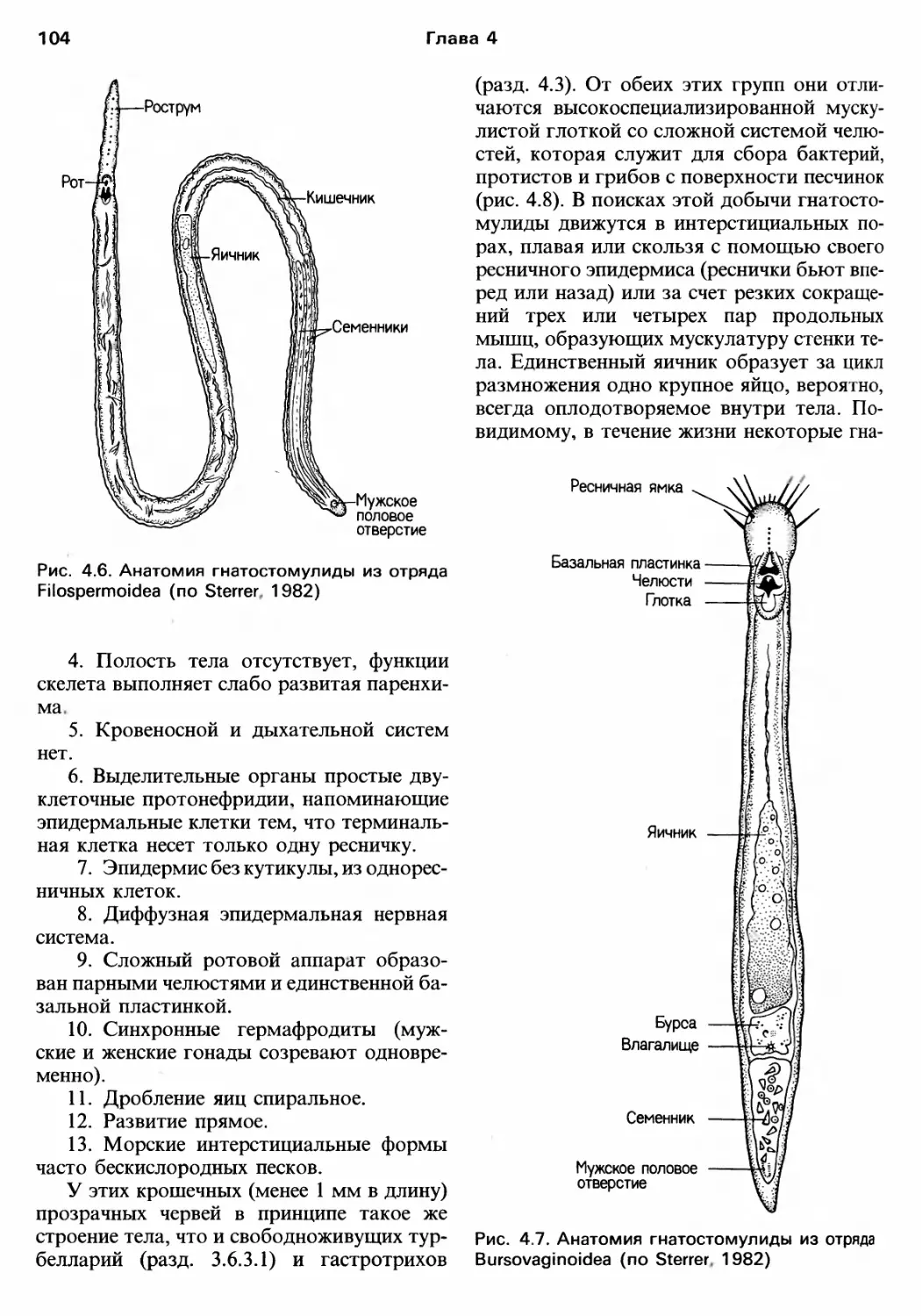
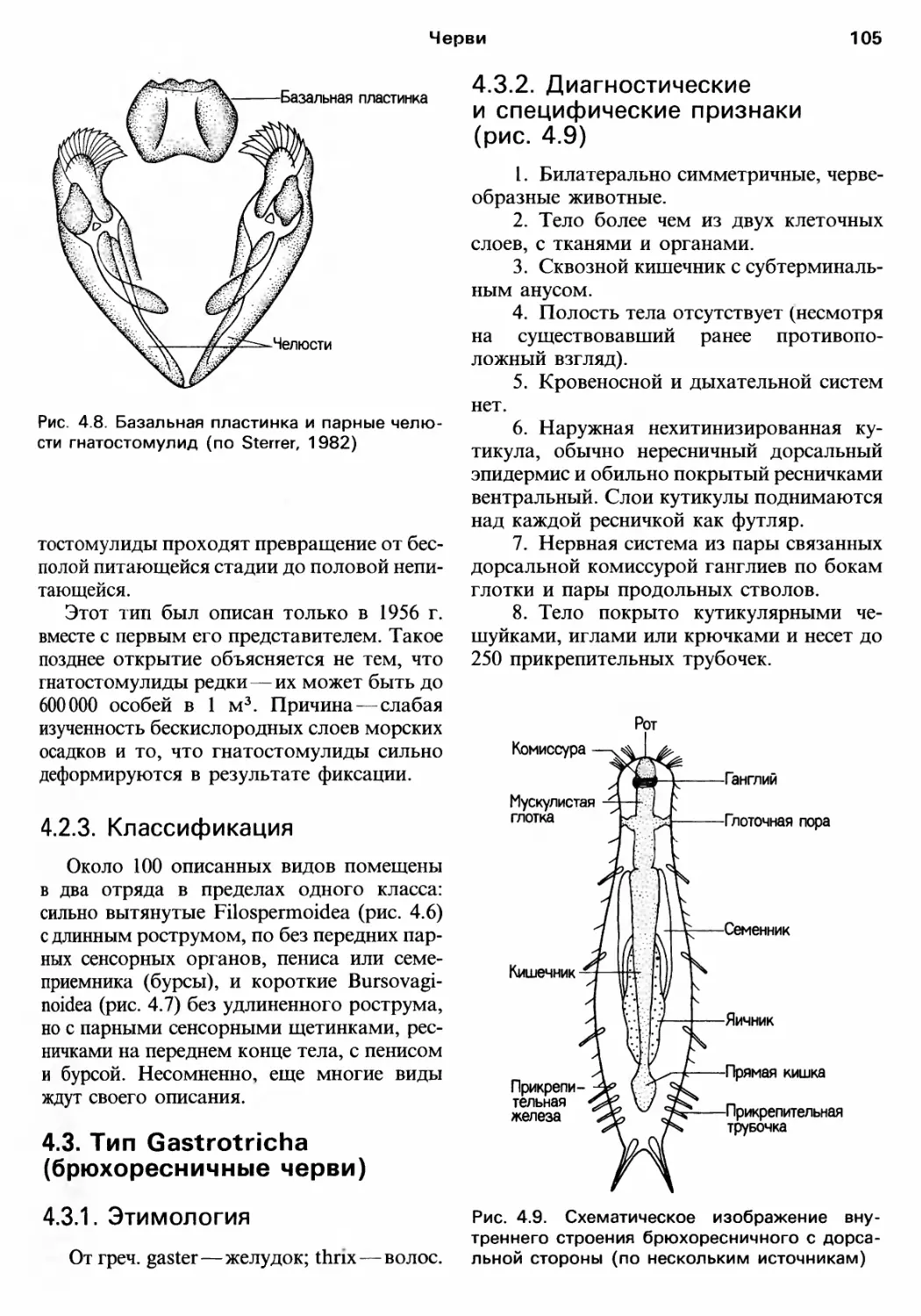
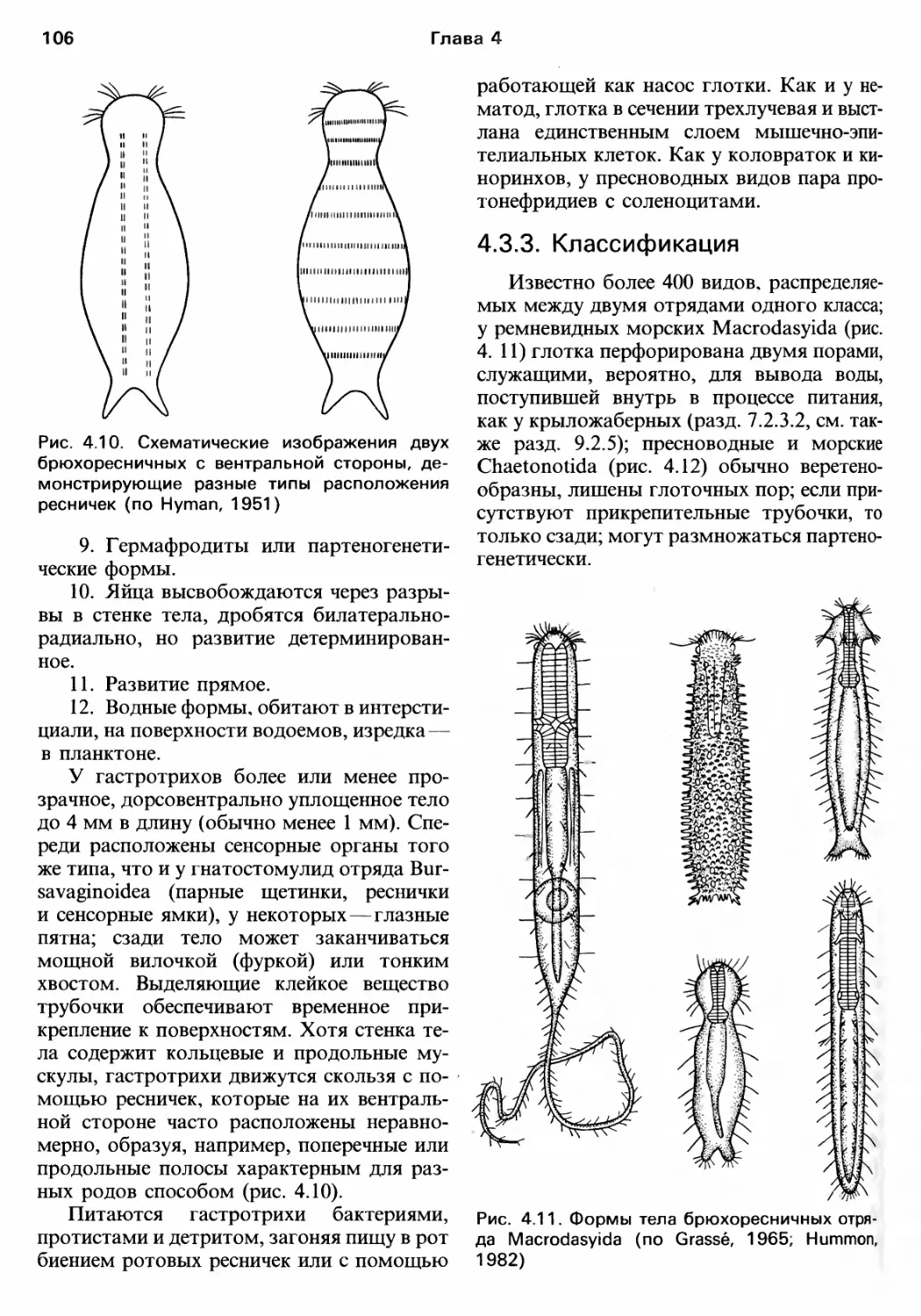










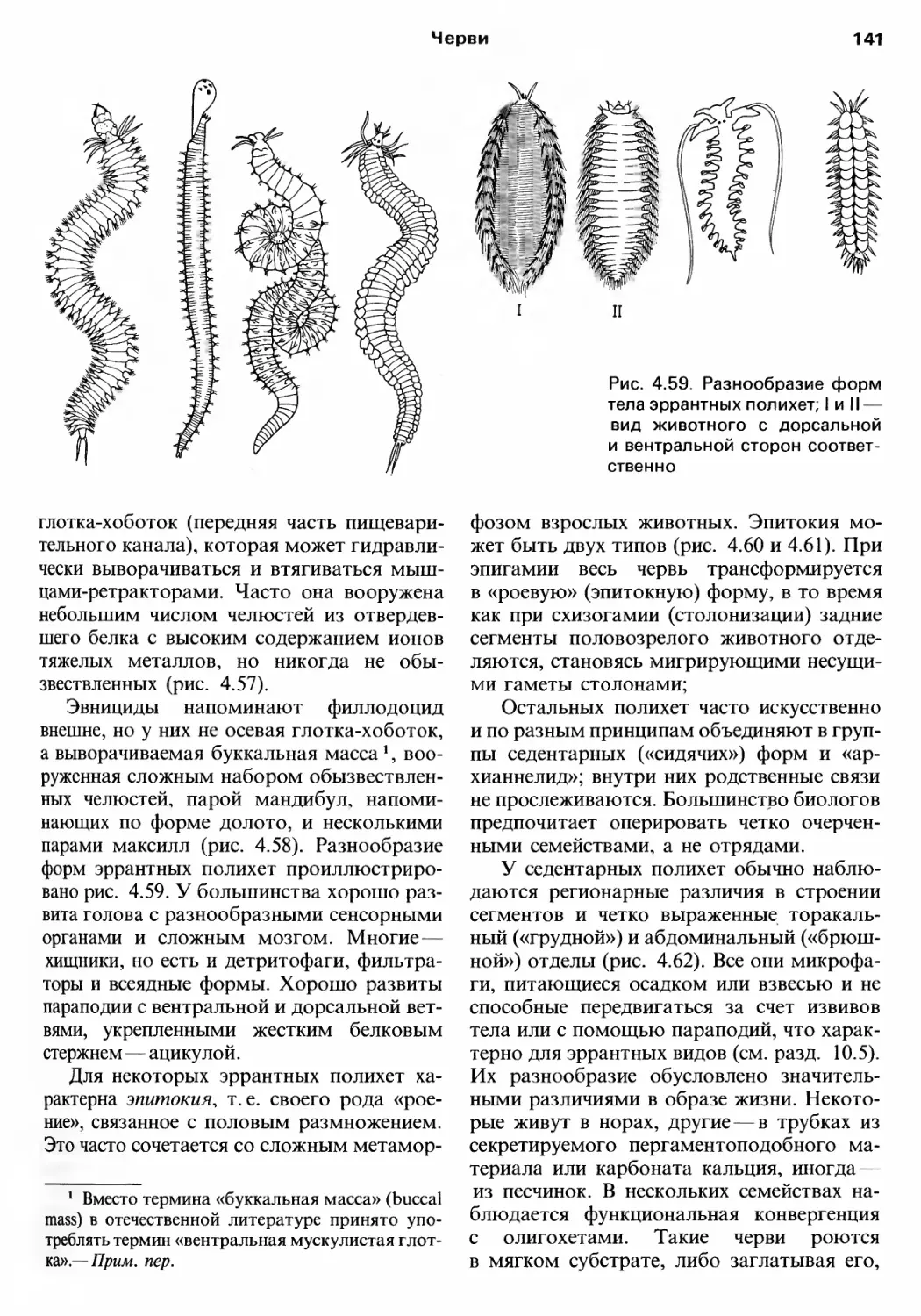
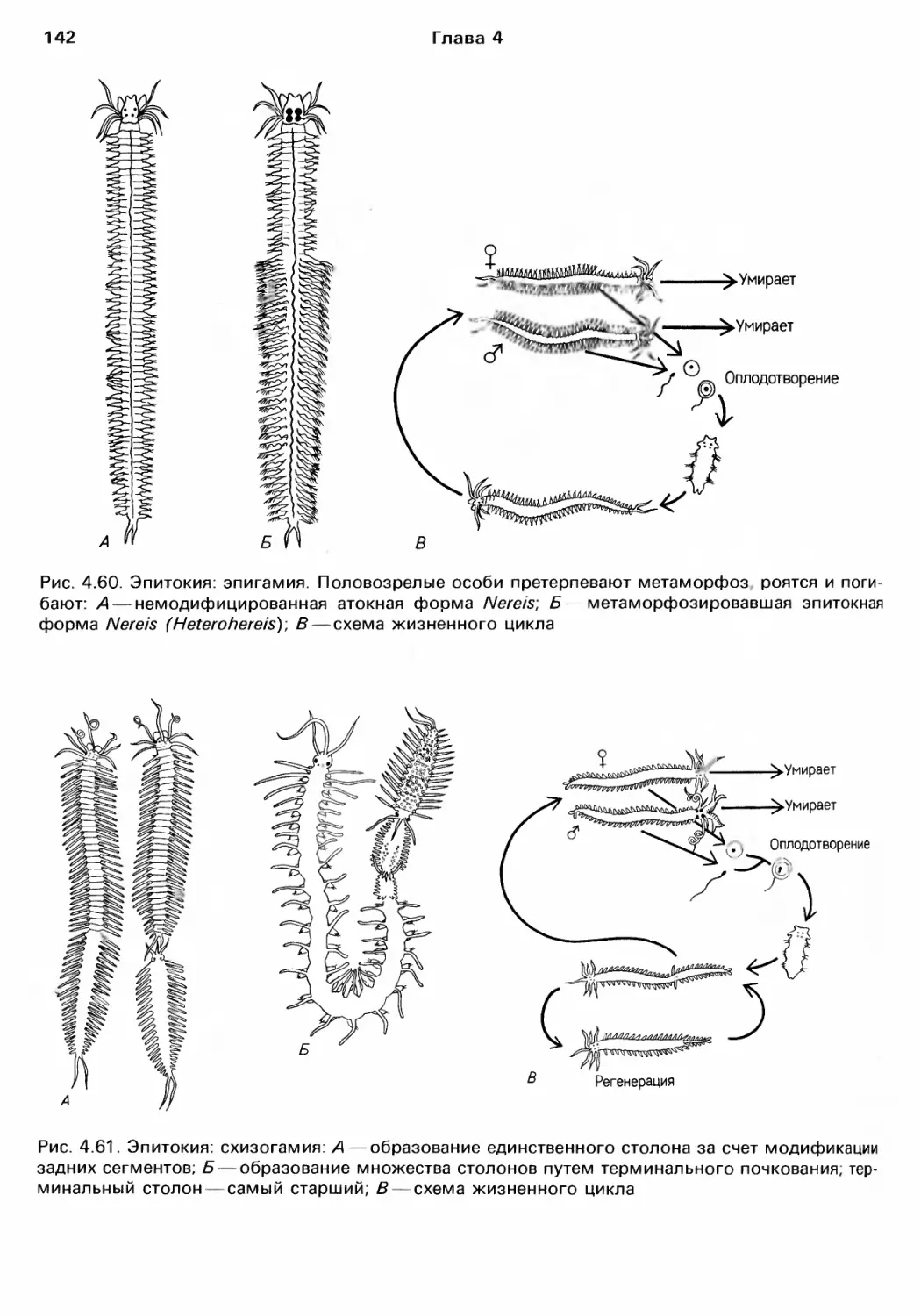
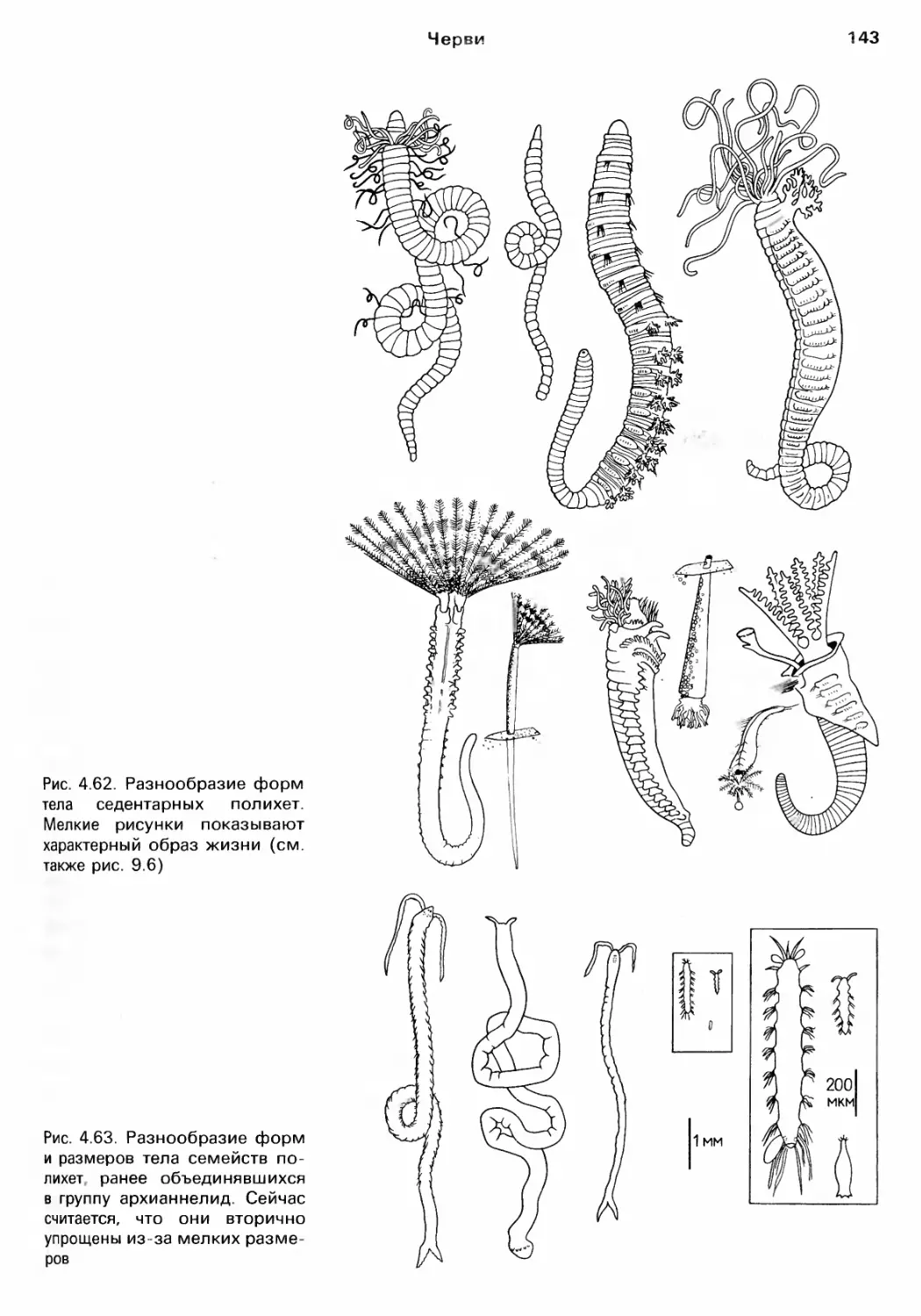

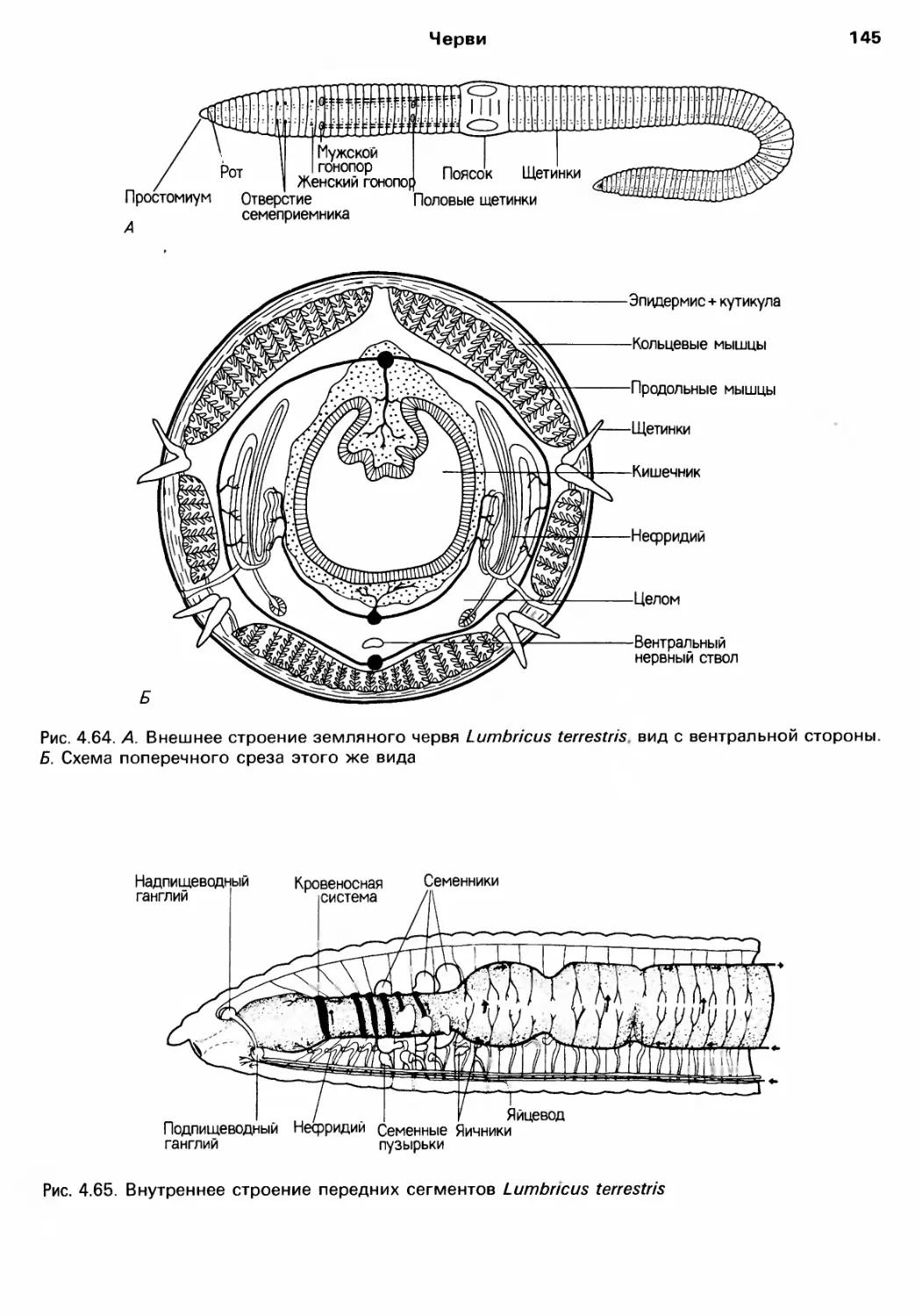


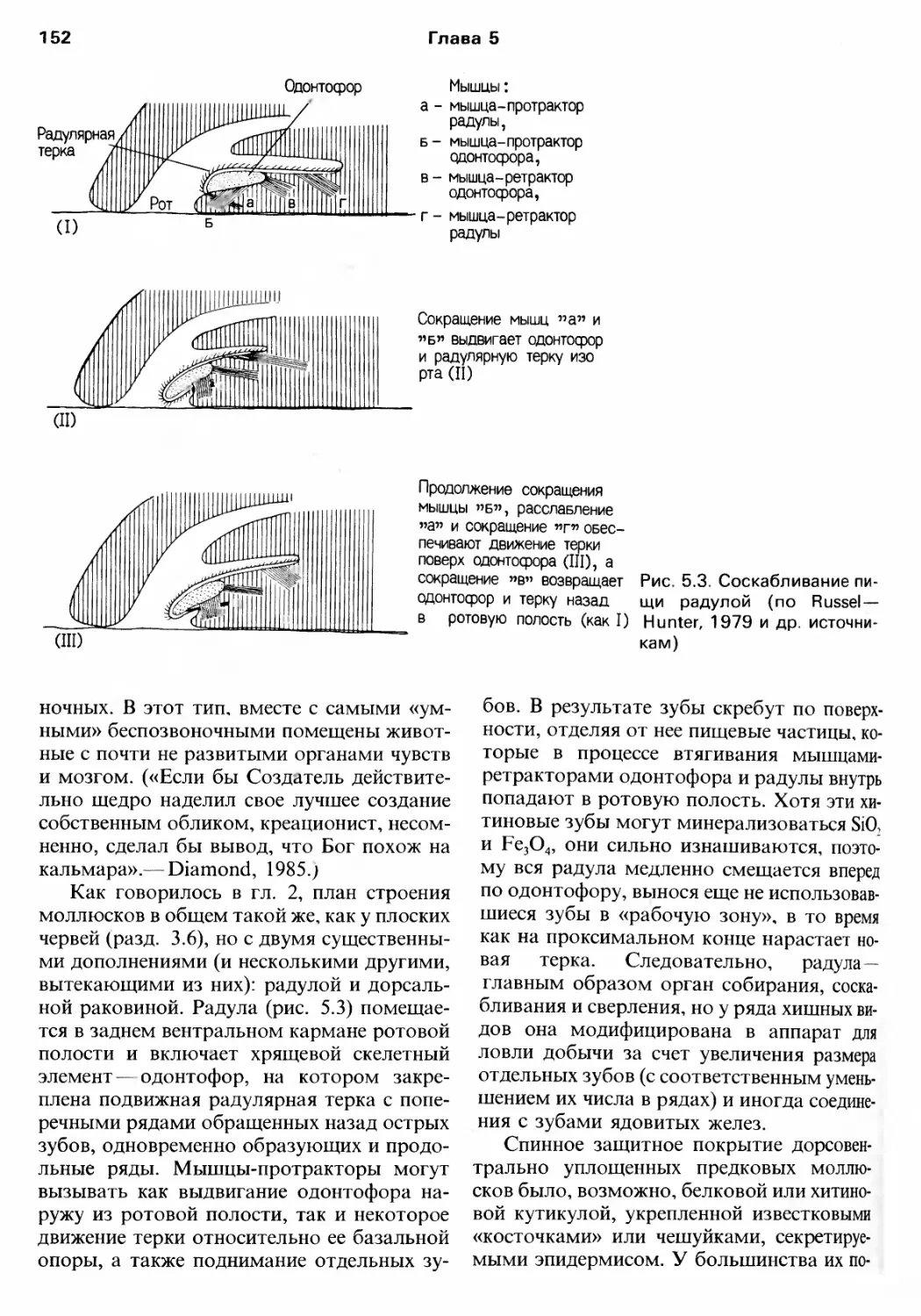

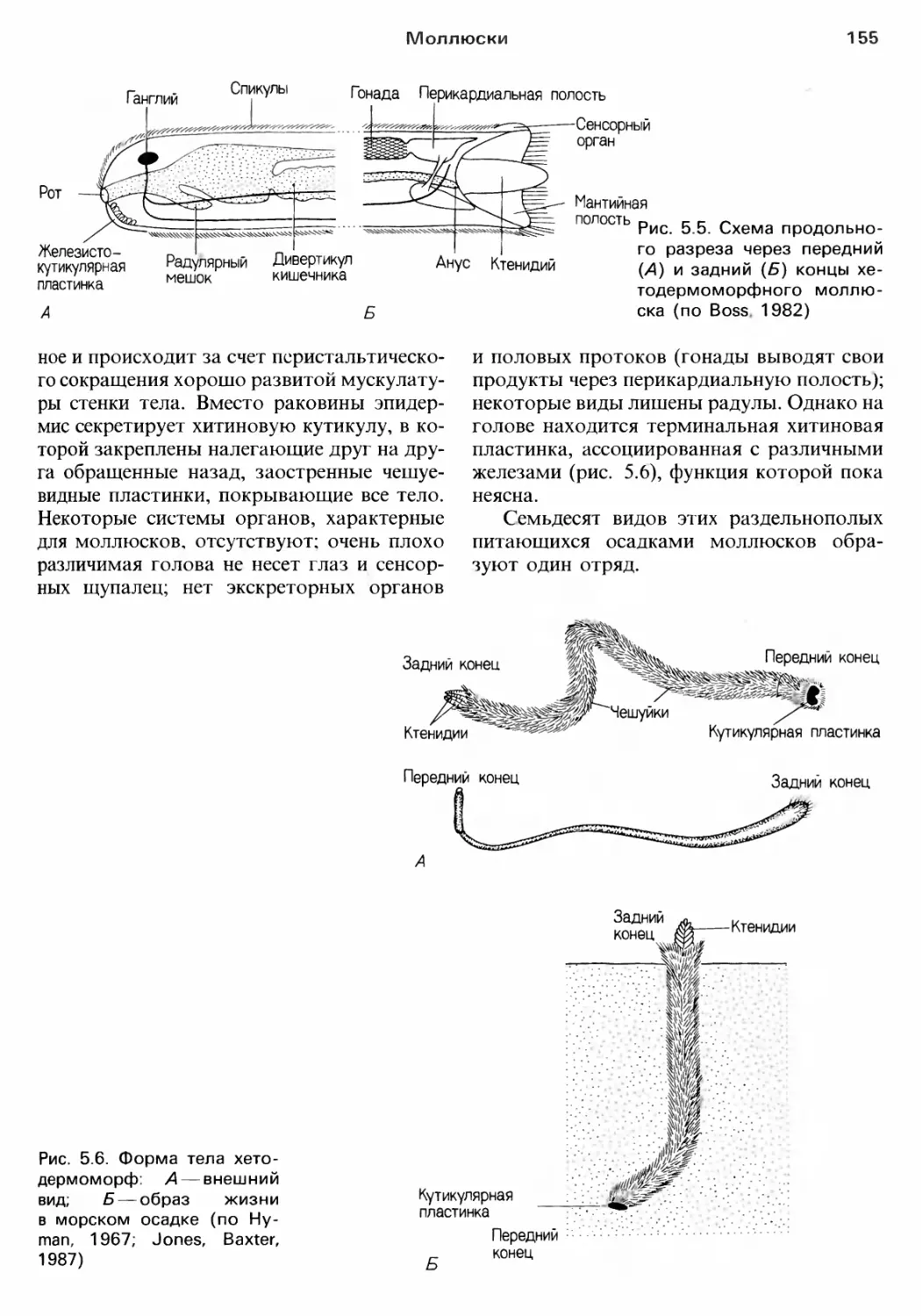
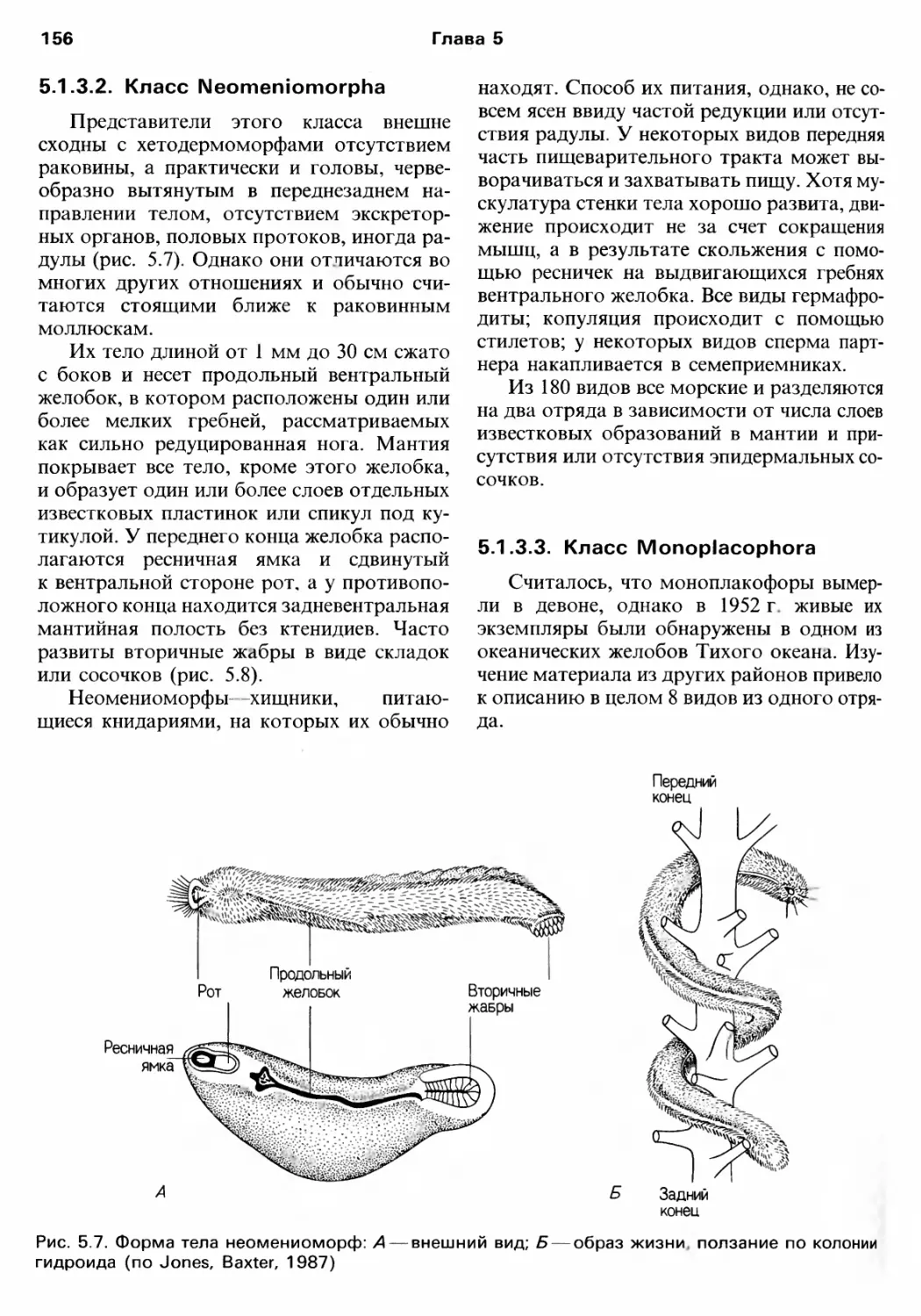


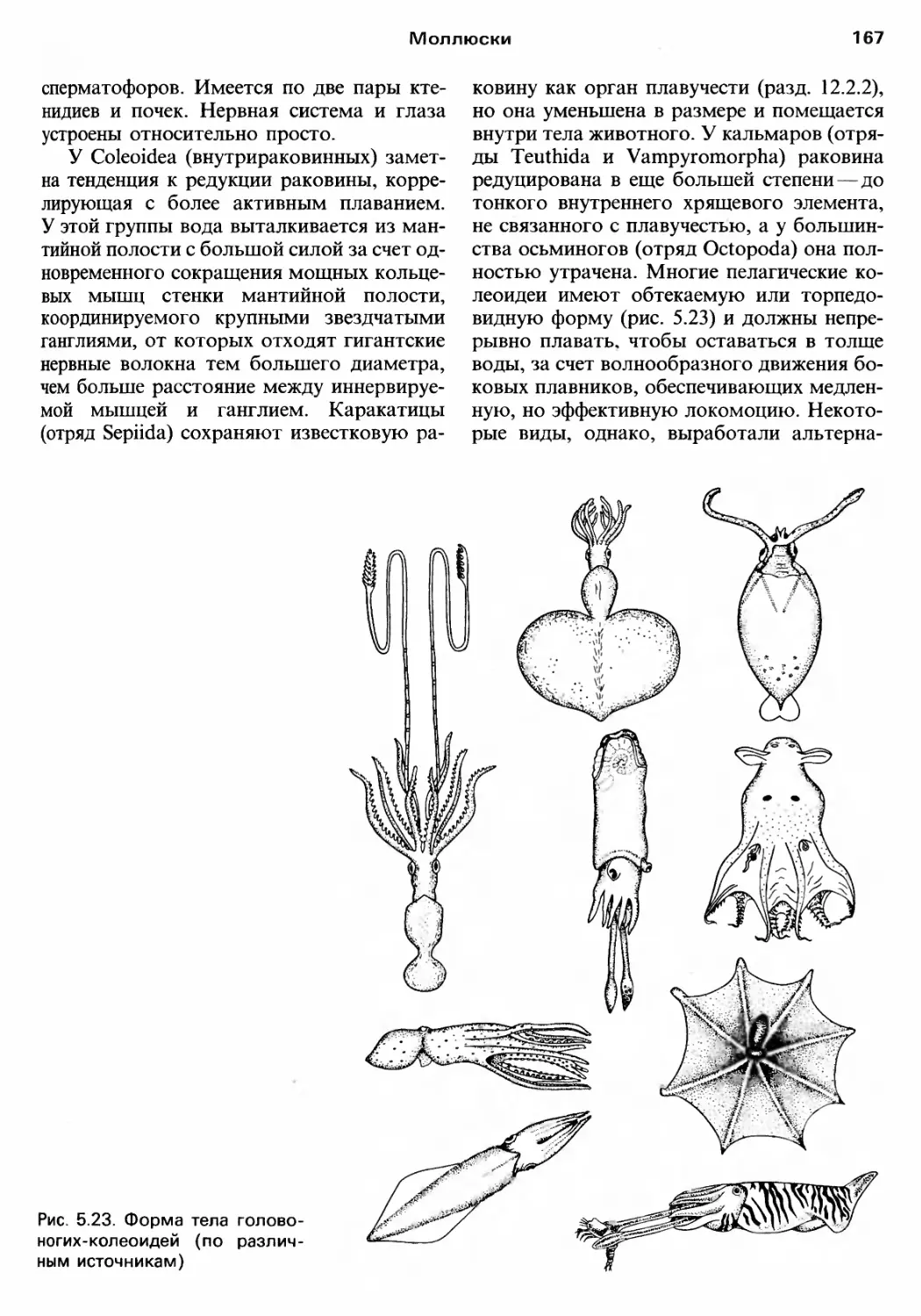
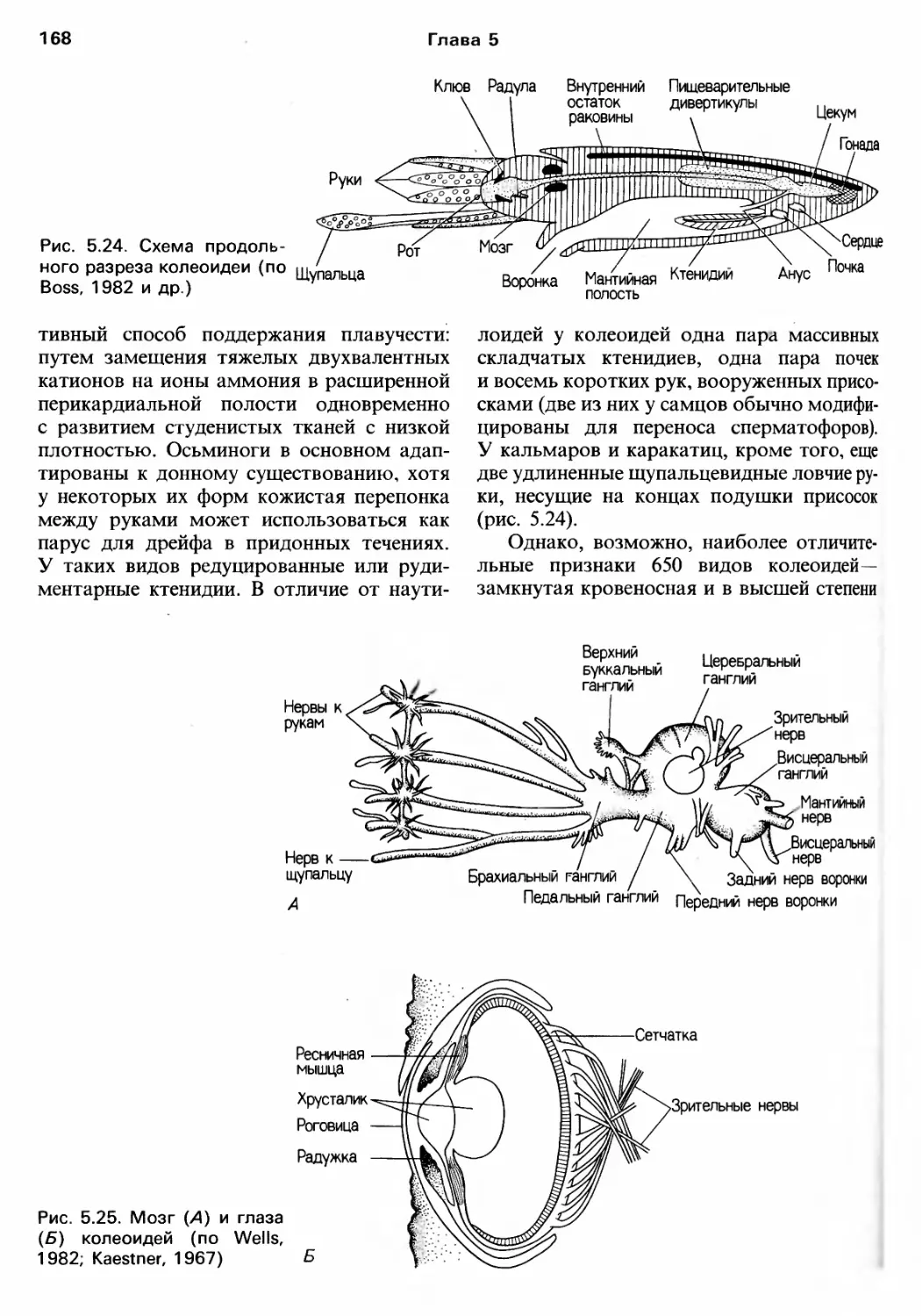


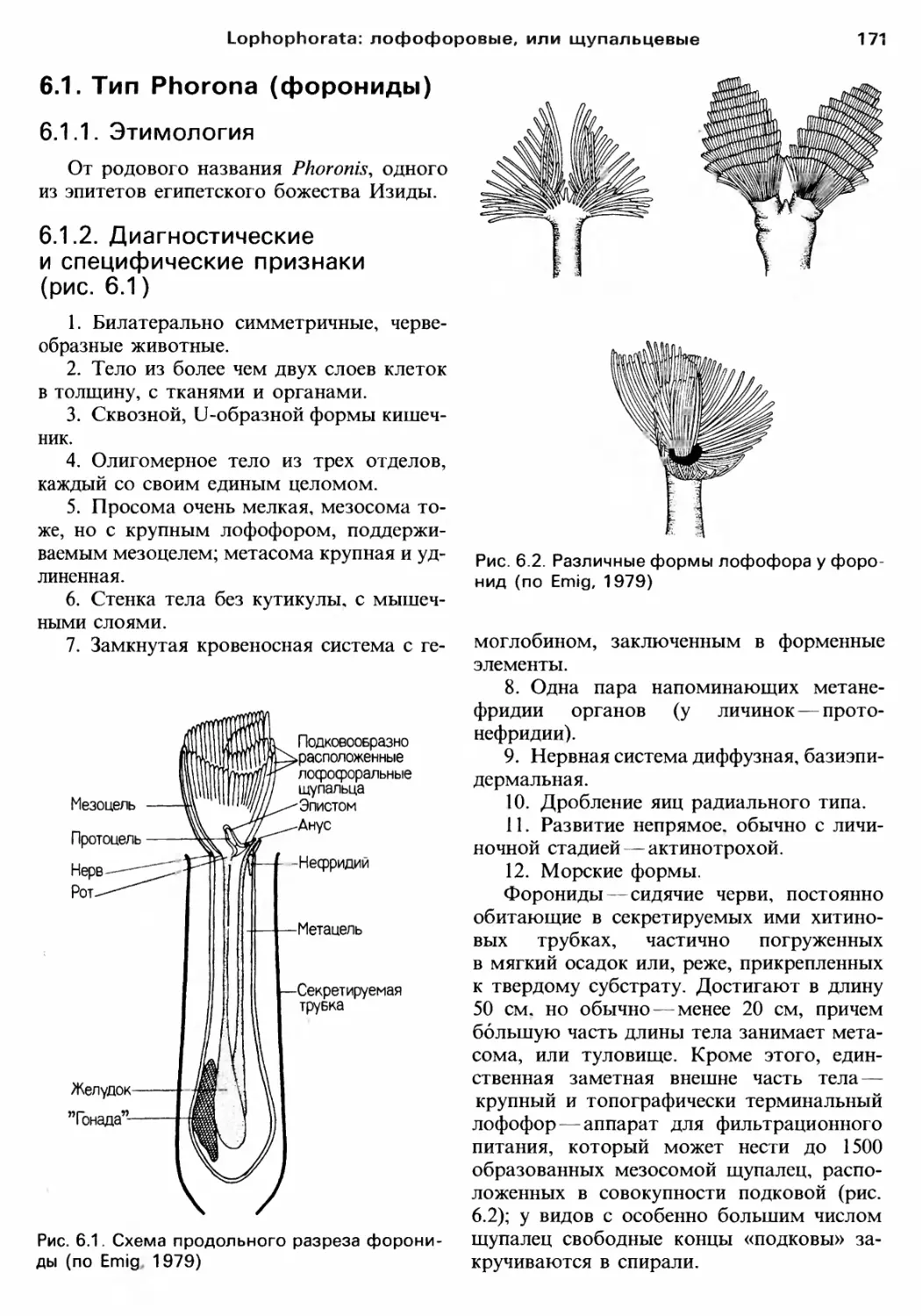


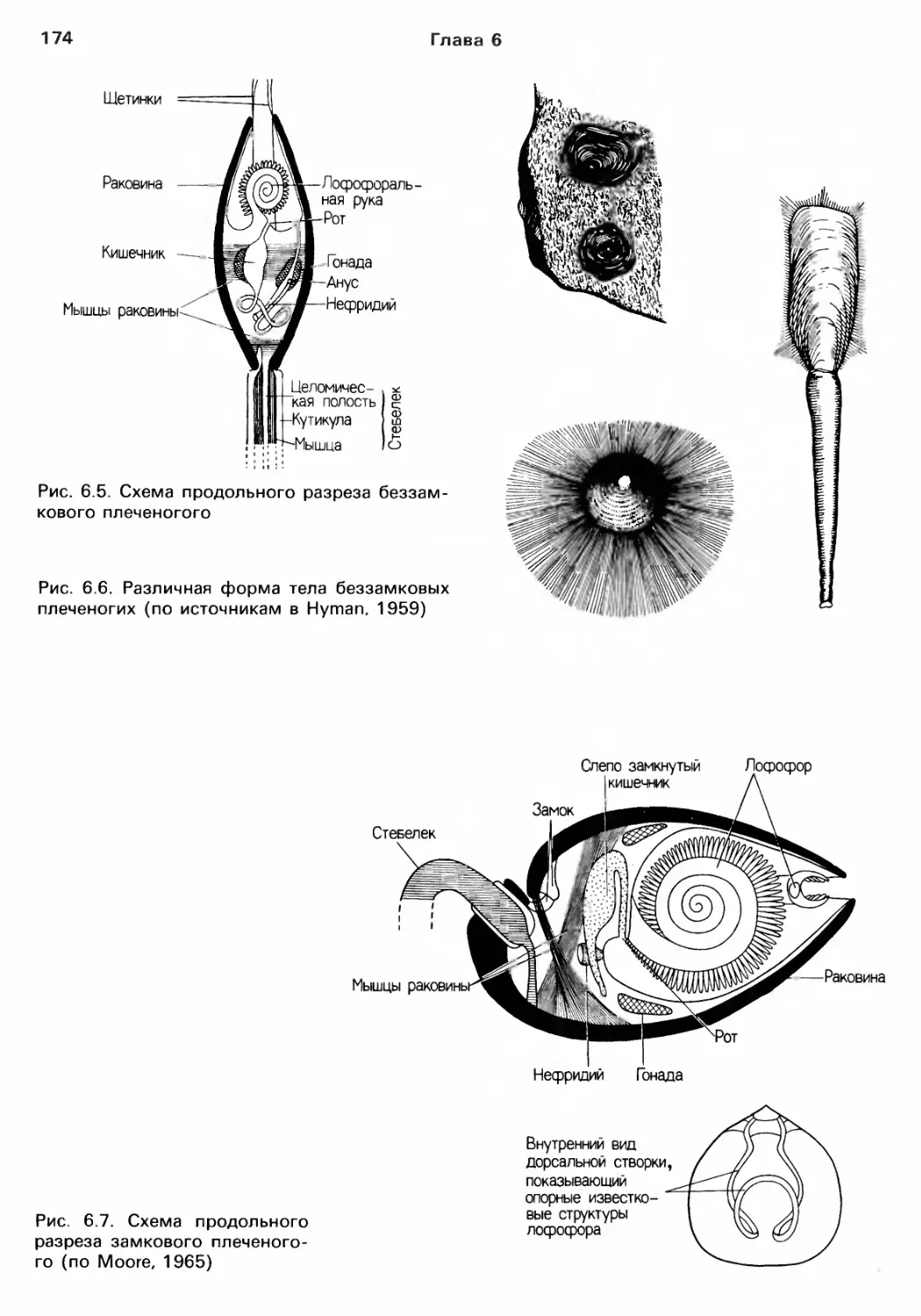






















































































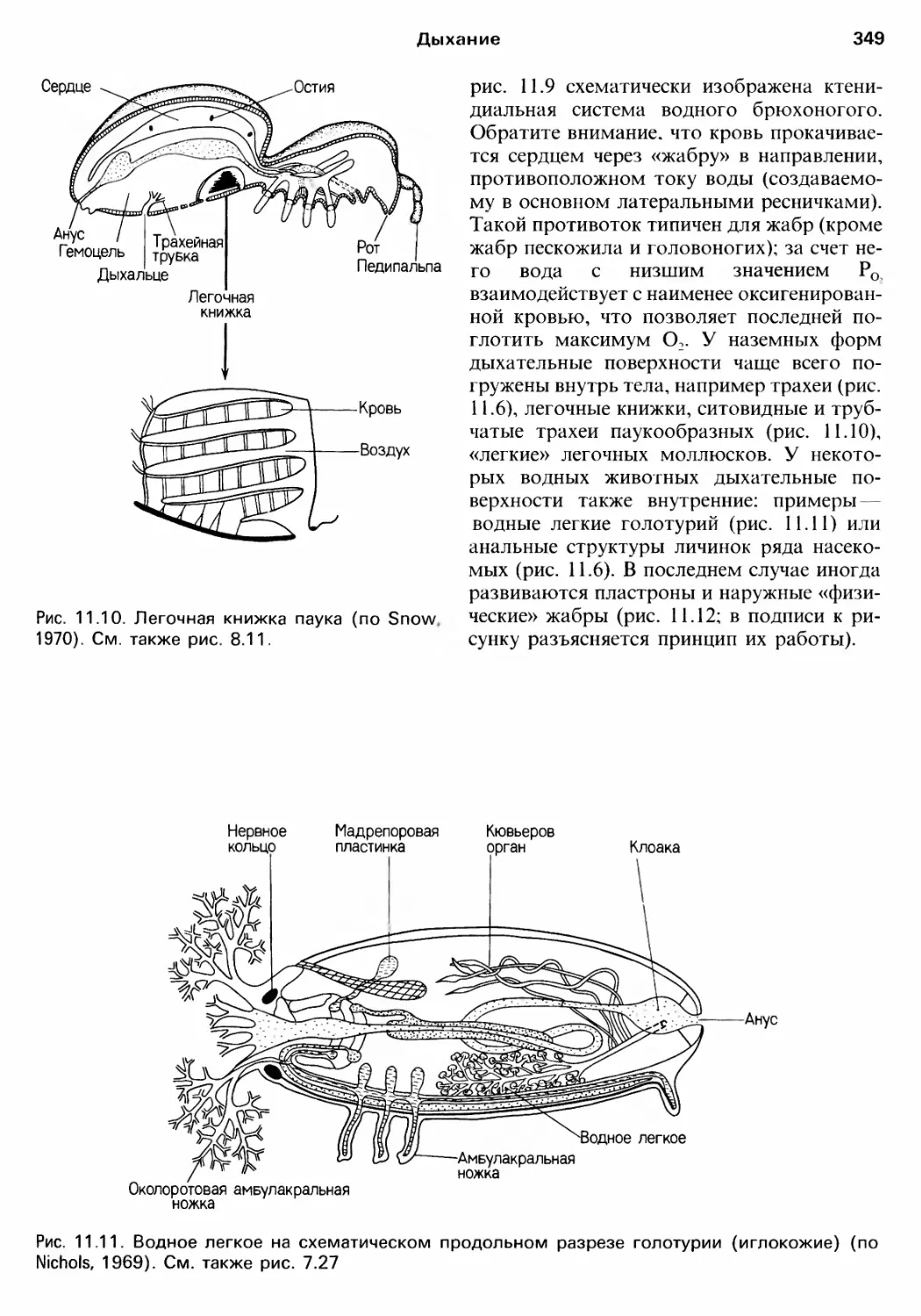


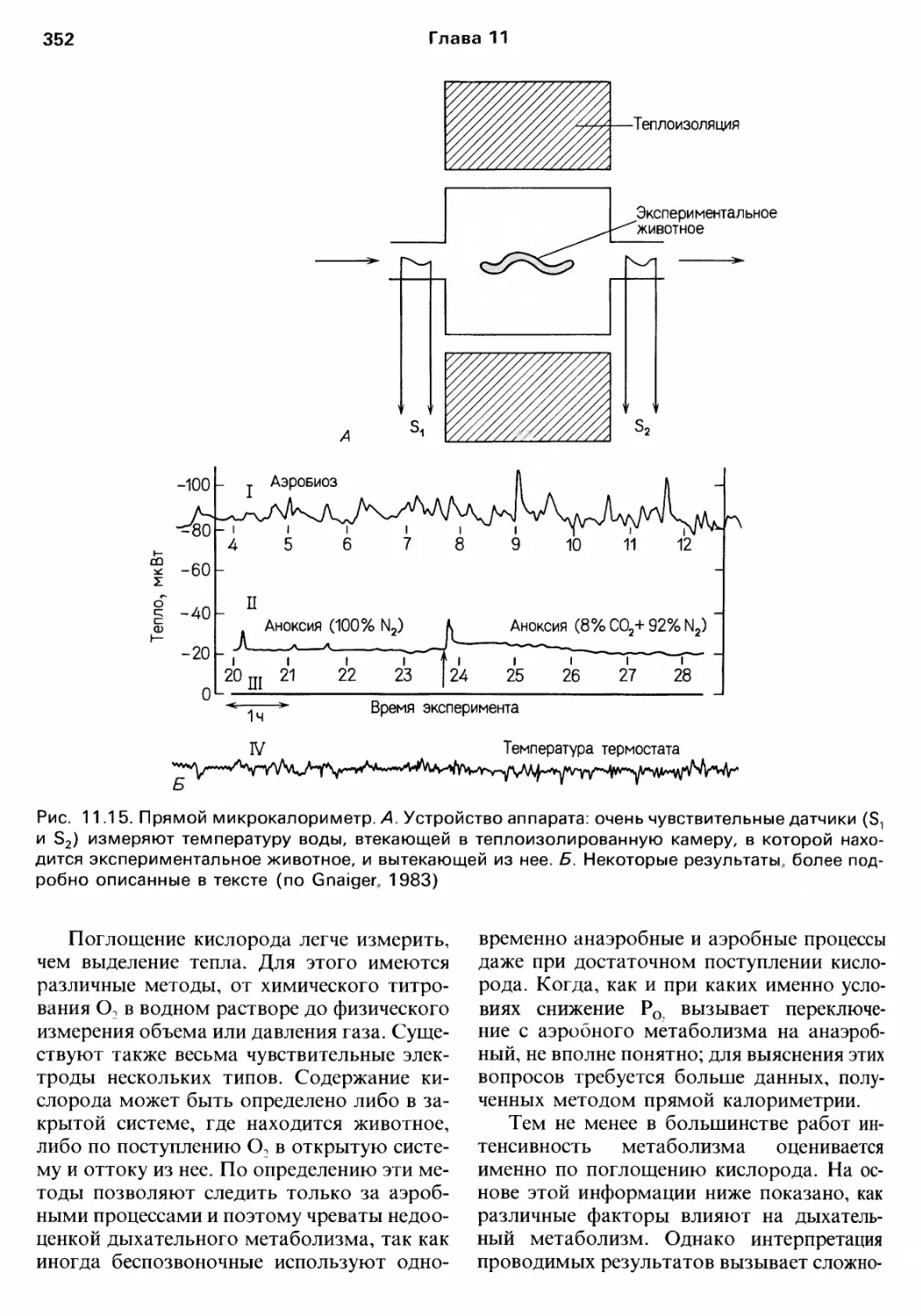

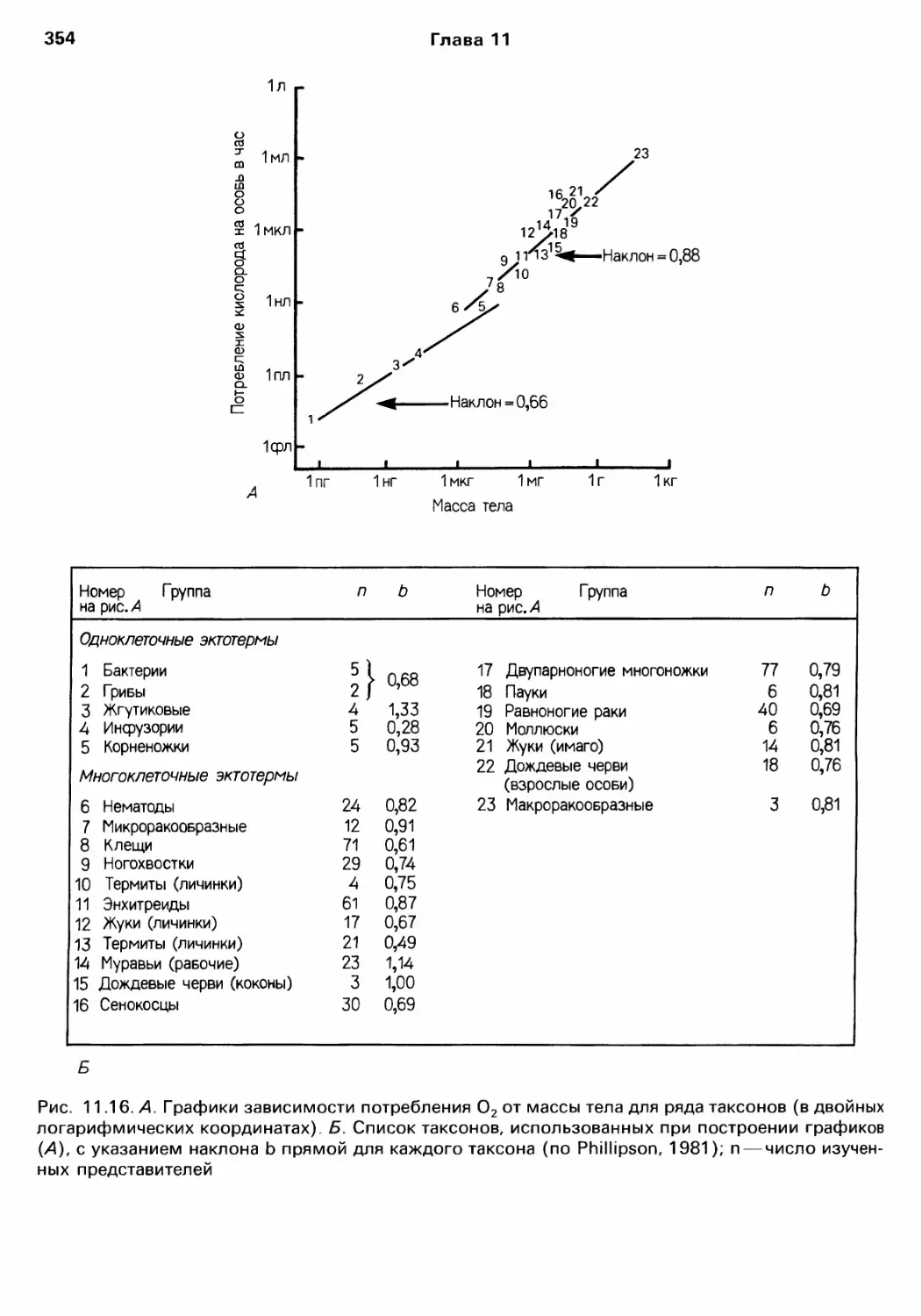

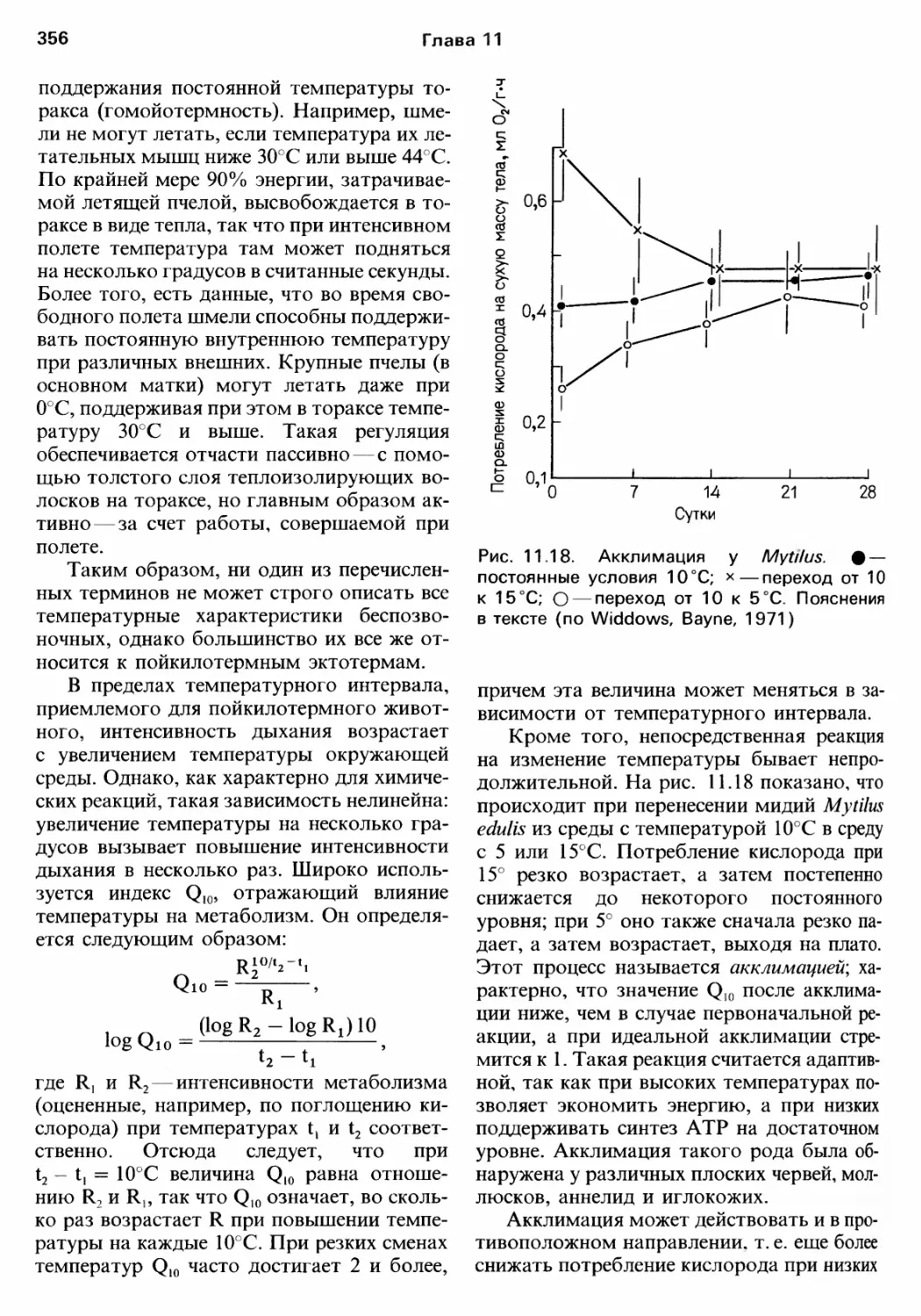
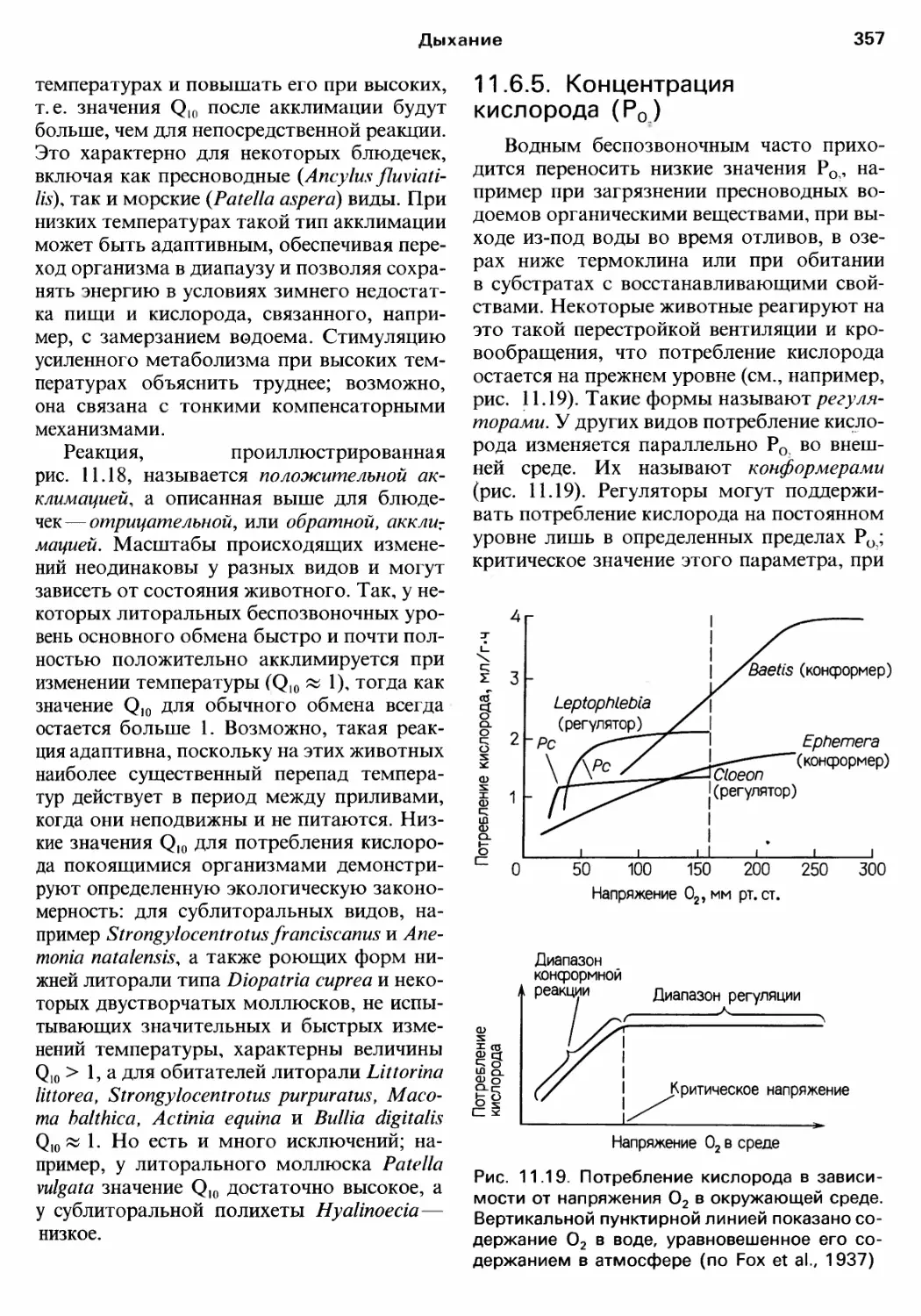
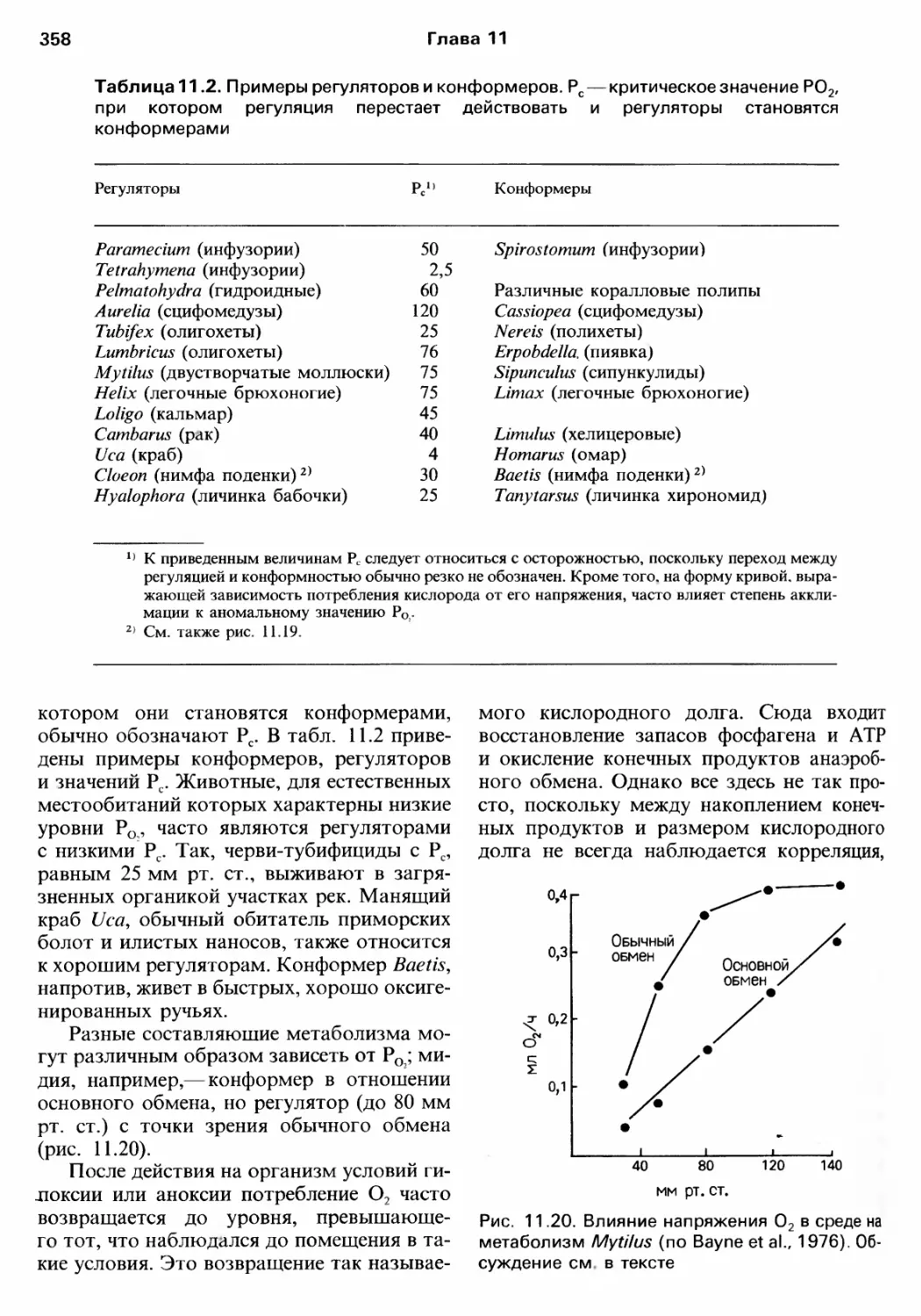
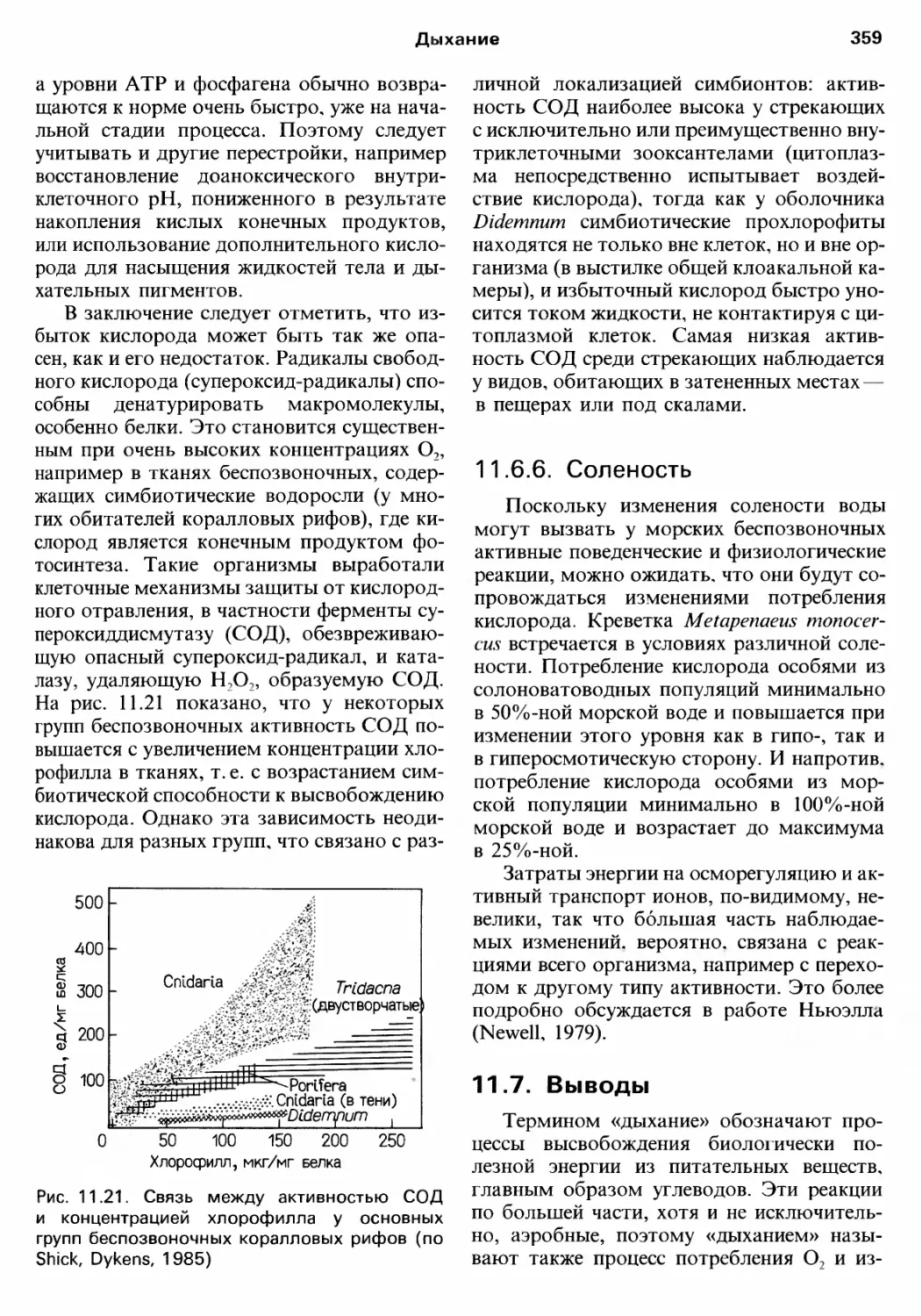






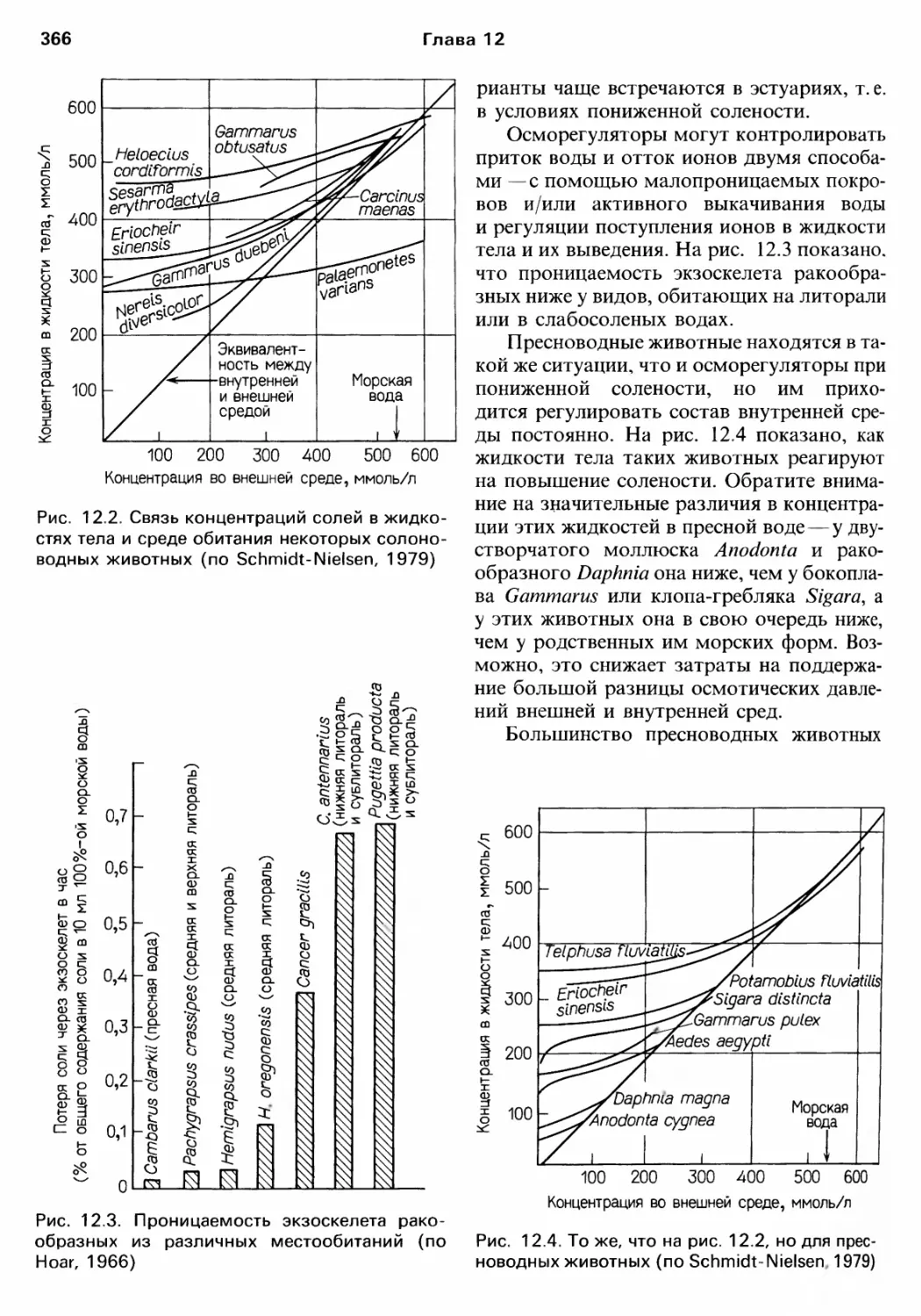









































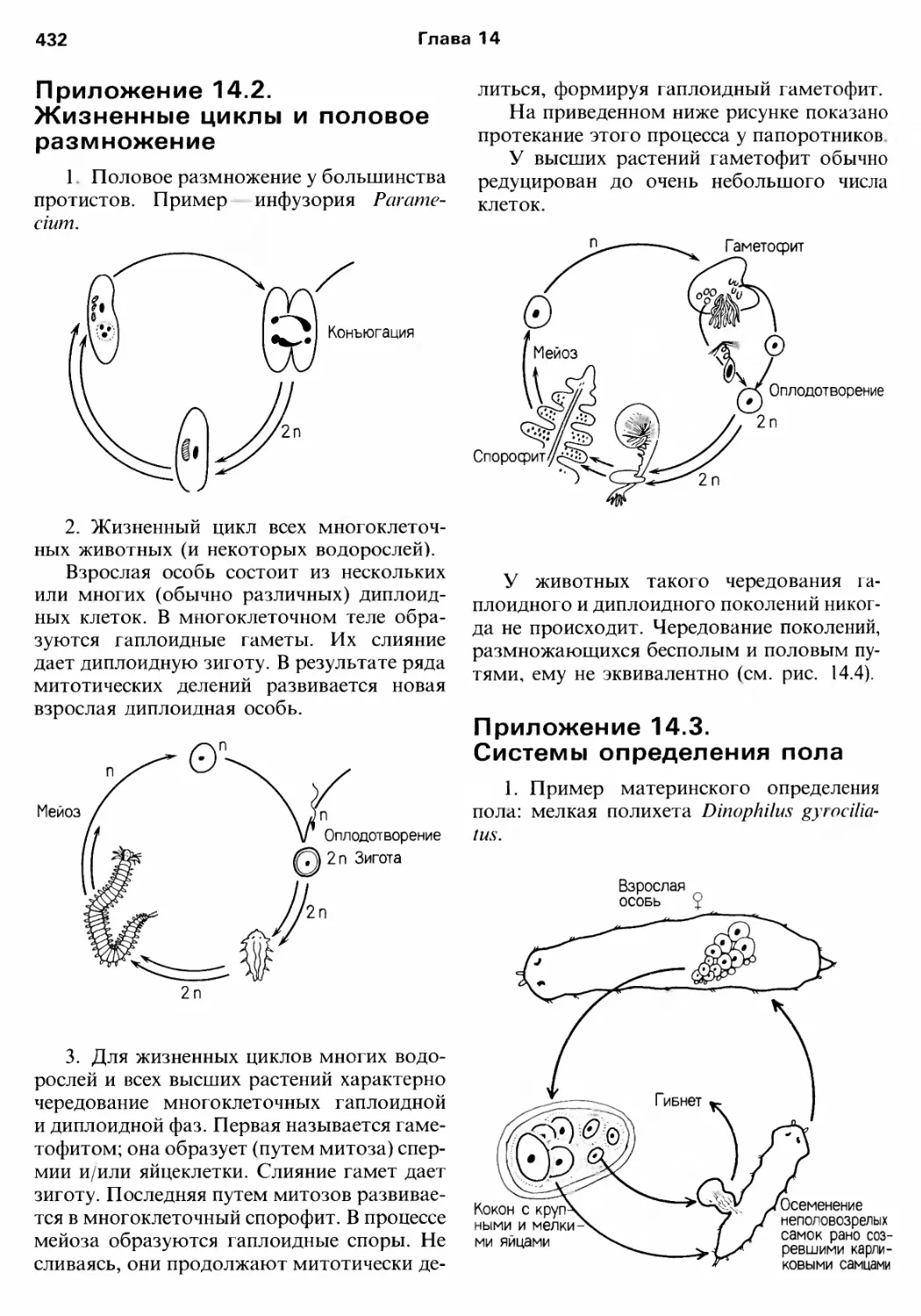
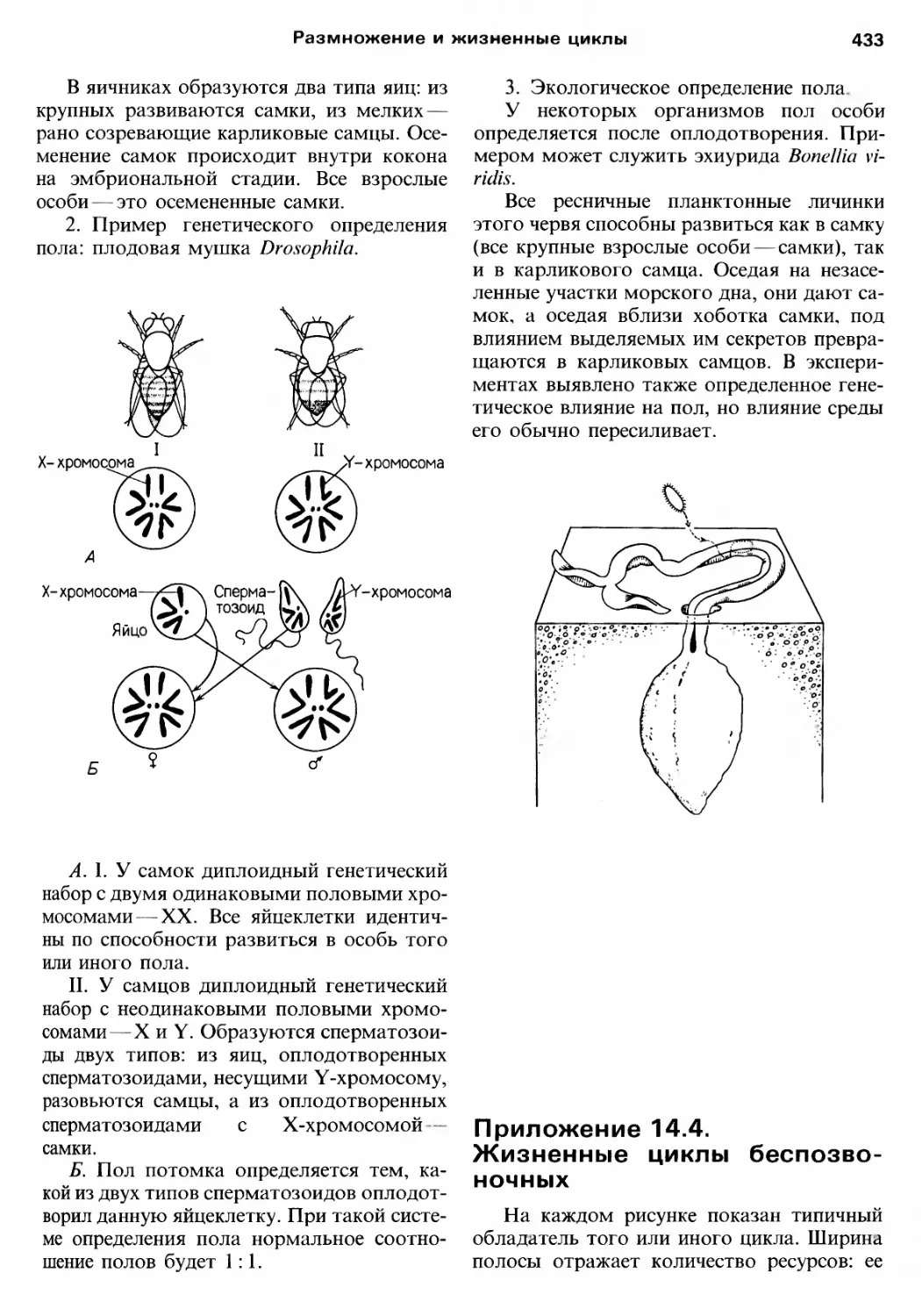
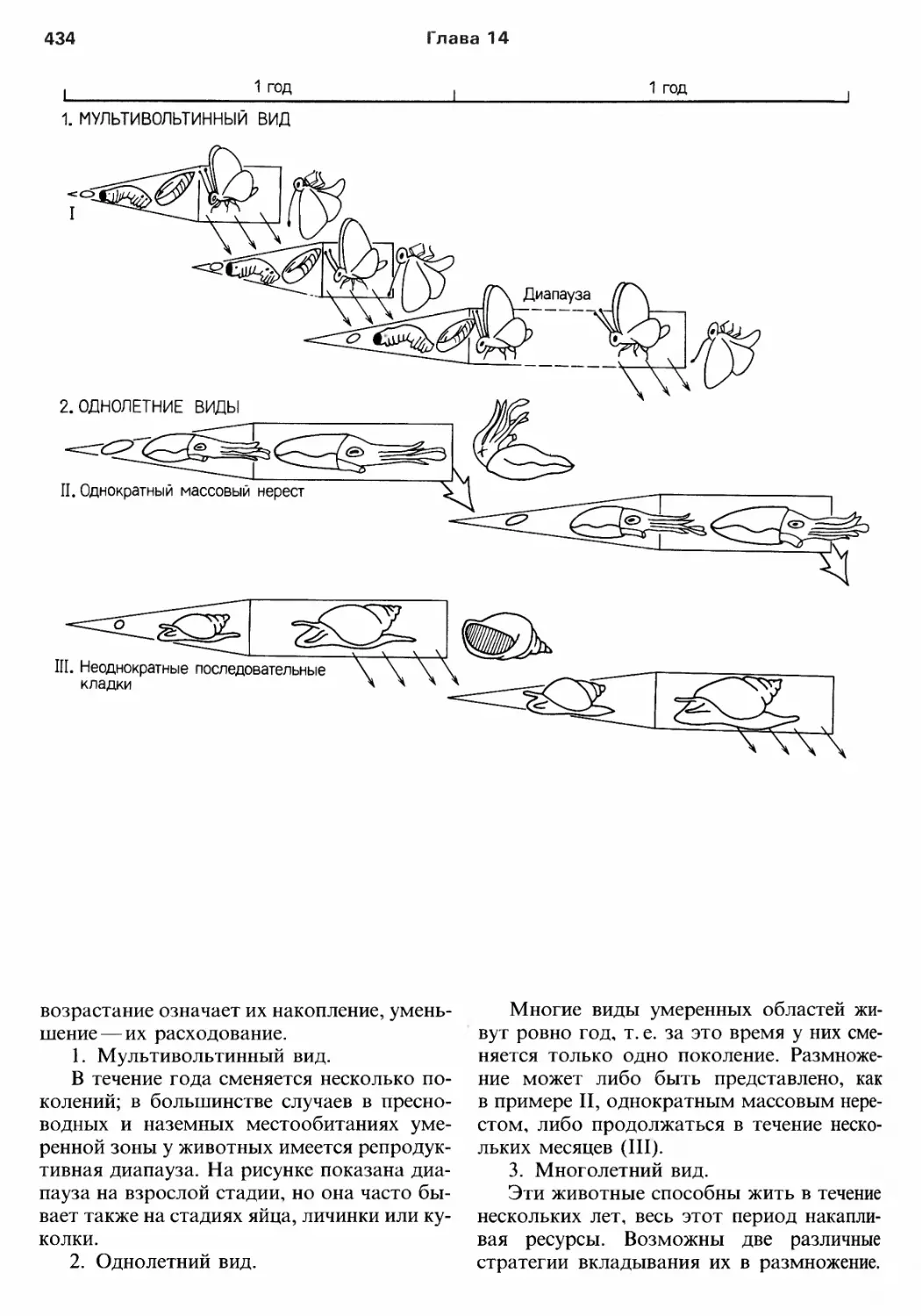
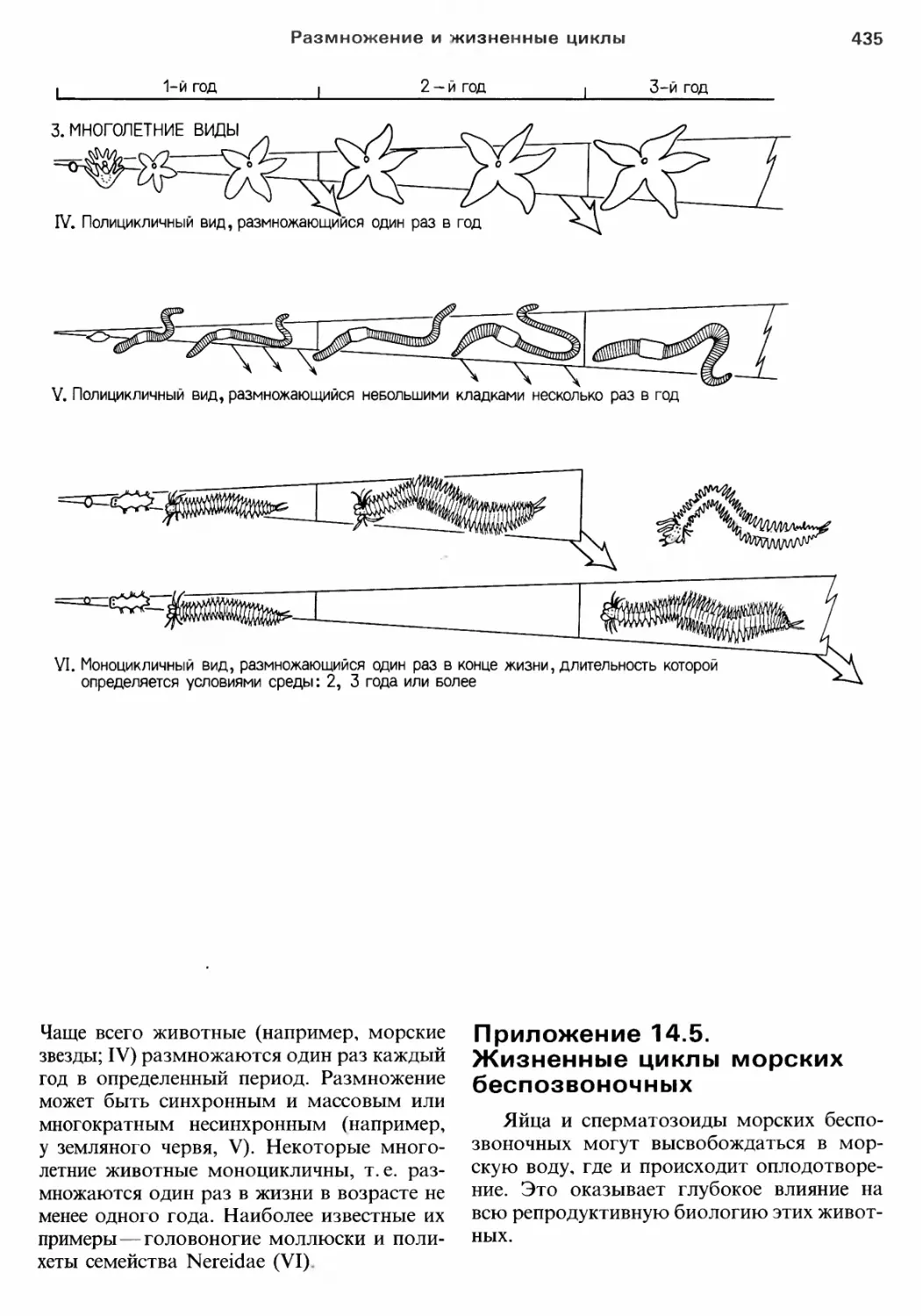




































































































Author: Барнс Р. Кейлоу П. Олив П. Голдинг Д.
Tags: invertebrata беспозвоночные зоология биология фауна
ISBN: 5-03-001496-9
Year: 1992




Text
ВЕ СПО3ВОНОЧ НЬIЕ
ТНЕ INVERTEBRATES
А N EW SYNTH ESIS
R.S.K. BARNES
Department 01 Zoology and 5t Catharine's Colledge,
U niversity 01 Cambridge, U К
Р. CALOW
Department 01 Zoology, U n iversity of 5heffield, U К
P.J.W. OLIVE
Department of Zoology,
University of Newcastle upon Тупе, U К
WITH А CHAPTER CONTRIBUTED ВУ
D.W. GOLDING
Department of Zoology,
University of Newcastle upon Тупе UK
,
BLACKWELL SCIENTIFIC PUBLICATIONS
Oxford London Edinburgh
Boston Palo Alto Melbourne
Е БАрне, п. КЕЙЛОУ, п. ОЛИВ, д. rолдинr
БЕспо3воночныIE
НОВЫЙ ОБОБЩЕННЫЙ
ПОДХОД
Перевол с анrлийскоrо
М. А. Сафроновой,
канд. биол. наук и. В. Успенскоrо,
канд. биол. наук А. Б. Цетлина
под редакцией
канд. биол. наук Б. я. Виленкина
МОСКВА «мир» 1992
ББК 28.6
Б53
УДК 592
Авторы: Барнс Р., Кейлоу П., Олив П., rолдинr Д.
Беспозвоночные: Новый обобщенный подход: Пер.
с анrл. М.: Мир, 1992. 583 с., ил.
ISBN 5 03 001496 9
в руководстве по зоолоrии беспозвоночных. написанном известными dнrлийски
ми учеными, впервые обобщены сведения по эволюции, систематике, морфолоrии
и физиолоrии беспозвоночных. Книrу характеризует полнота охвата предмета и Ha
r лядность изложения. Рассмотрены эволюция и систематика беспозвоночных, а также
их функциональная биолоrия: питание, дыхание, движение, выделение и осмореrуля
пия, репродукпия. онтоrенез, нервная и эндокринная системы. Приведен словарь Tep
минов.
Для сту дентов биолоrов, специалистов, интересующихся биолоrией беспозвоноч
ных, преподавателей биолоrии в школе.
Б 1907000000 190 101 92
041(01) 91
ББК 28.6
Редакцuя лuтературы по БUОЛО2UU
ISBN 5 03 001496 9 (русек.)
ISBN O 632 01638 8 (анrл)
(Q 1988 Ьу Blackwell Scientific Publications
(Q перевод на русский язык. Сафронова М. А.,
Успенский и. В., Цетлин А. Б., 1992
ПРЕДИСЛОВИЕ К РУССКОМУ ПЕРЕВОДУ
Задачи, построение и мноrие друrие oco
бенности предлаrаемой читателю книrи
вполне объяснены в предисловии ее aBTO
ров. Остается сказать лишь несколько слов
о том, какое, по нашему мнению, место MO
жет занять эта книrа в отечественной зооло
rической литературе и чем она отличается
от большинства наших руководств по зоо
лоrии и сравнительной физиолоrии беспо
звоночных.
Характерная ее особенность состоит
в том, что равное внимание уделено всем
rруппам беспозвоночных, независимо от
количества входящих в них видов, т. е. пла
козои, rнатостомулиды и форониды описа
ны так же подробно, как и моллюски или
членистоноrие. При этом вопросы филоrе
нии и происхождения тех или иных систем
opraHoB животных рассматриваются как
второстепенные для данной книrи, хотя
в ряде случаев они изложены достаточно
полно. Часто, rоворя о происхождении той
или иной rруппы, авторы приводят различ
ные rипотезы, обращая внимание читателя
на то, что эти проблемы еще далеки от
окончательноrо разрешения.
Там, [де это возможно, строение живот
ных описывается с функциональной точки
зрения, что, безусловно, помоrает читателю
разобраться в мноrообразии беспозвоноч
ных. Большая удача книrи рисунки и cxe
мы, которые не дублируют. а дополняют
текст.
И, пожалуй, основное отличие 01 pac
пространенных у нас руководств в том, что
описывается строение именно живых жи
вотных. Образно rоворя, эта книrа -
путеводитель скорее по зоопарку или aKBa
риуму, чем по анатомической коллекции.
Некоторые представления авторов MO
rYT показаться читателю непривычными.
Например, ими принимается так называе
мая архицеломатная теория происхождения
вторичноротых, описывается система брю
хоноrих моллюсков, считающаяся мноrими
архаичной, и т. п. Однако это не затрудняет
чтения книrи и делает ее, пожалуй, особо
интересной в качестве источника «альтерна
тивных» точек зрения на важные зоолоrиче
ские проблемы.
Как уже rоворилось, в предлаrаемой
книrе идет речь о живых животных, и в этом
отношении она уникальна среди распро
страненных у нас учебников и руководств
по зоолоrии. Сведения об орrанизации про
цессов жизнедеятельности у беспозвоноч
ных не столь обширны и не так равномерно
охватывают разные типы и классы, как дaH
ные об их анатомии и морфолоrии. В этом
отношении сравнительные эмбриолоrия,
физиолоrия, биохимия еще не достиrли Ta
кой цельности, как сравнительная анато"
мия, что отмечают и авторы руководства.
Соответствующие разделы (rл. 9 17) за
служивают внимания по двум причинам.
Во первых, здесь довольно полно собраны
современные сведения о механизме процес
сов жизнедеятельности у представителей
тех rрупп беспозвоночных. [де исследова..
ния TaKoro рода ДQстаточно продвинулись
вперед. BO BTOpЫX, что более важно для
зоолоrа, авторы сумели показать, что дале
кие от традиционных зоолоrических вопро
сов работы по физиолоrии, биохимии, MO
лекулярной rенетике, электрофизиолоrии
и т. п. имеют непосредственное отношение
к пониманию поведения, питания, адапта
ции видов к внешним условиям, их эволю
ции И прочим предметам, непосредственно
интересующим зоолоrов Можно надея
ться, что предложенная манера подачи и об
суждения нетрадиционноrо материала бу
6
Ilредисловие к русскому переводу
дет способствовать преодолению той не..
приязни к новейшим биолоrическим подхо..
дам" которую часто проявляют зоолоrи
классическоrо направления.
Эволюционный метод авторов четко
прослеживается в том, что они наряду
с устройством и возможностями эволю..
ционирующеrо объекта демонстрируют
и внешние силы, создающие давление отбо..
ра. Друrими словами, значительное внима..
ние уделено эколоrическим вопросам, т. е.
рассматривается большое количество эко"
лоrических классификаций, стратеrий и дру..
rих до некоторой степени умозрительных
построений. Нельзя, вероятно, утверждать,
что эколоrия в ее современном состоянии
вносит весомый вклад в целостное понима..
ние эволюционноrо процесса. Тем не менее
соответствующие разделы книrи способ..
ствуют развитию представления о зоолоrии
как о науке о живых орrанизмах, взаимо..
действующих с разнообразными биотиче..
скими и абиотическими факторами окру"
жающей их среды.
А. Б. Цеп1ЛUН
Б. Я. Вuленкuн
ПРЕДИСЛОВИЕ
Существует несколько вполне доступ
ных учебников, посвященных беспозвоноч
ным, поэтому появление еще одноrо Tpe
бует некоторых пояснений. Уже опублико
ванные книrи попадают в одну из двух кате..
rорий: либо это систематические обзоры,
последовательно рассматривающие различ
ные типы животных (например, R. D. Bar..
nes, "'lnvertebrate Zoology'\ Saunders, 1987)'1
либо функциональные исследования тех или
иных анатомических и физиолоrических
«систем» (дыхательной. двиrательной,
координационной и т. д.) В основном хоро..
шо изученных rрупп (например, E.J. W.
Barrington, "lnvertebrate Structure and Func..
tion", Nelson. 1979). Курсы беспозвоночных,
таким образом, требуют по крайней мере
по одной из книr обеих катеrорий.
Однако в общем за последние 25 лет
произошло сокращение учебноrо времени,
отведенноrо специально отдельным rруп..
пам, частично для Toro, чтобы высвободить
время для новых предметов, а также в связи
с потерей популярности систематических
обзоров разнообразия орrанизмов по cpaB
нению с временами классической зоолоrии.
В конечном итоrе и каждая пара суще..
ствующих учебников, и даже мноrие отпе..
льные работы содержат rораздо больше ин..
формации, чем это требуется для KpaTKoro
курса. Утопая в деталях, студенты не в со..
стоянии за деревьями увидеть леса.
Поэтому мы и предприняли попытку
объединить под одной обложкой основную
информацию о разнообразии беспозвоноч"
ных и различных их функциональных систе..
мах, информацию, которая, как нам ка..
жется, действительно требуется для универ..
ситетскоrо курса. Нашей rлавной пробле..
мой стал отбор Toro, что можно выкинуть,
а не Toro, что оставить, и здесь мы постара..
лись критически оценить существенные чер..
ты каждой rруппы или системы, обратив
внимание читателя именно на них. Далее.
мы твердо уверены, что эволюционный
подход стержень понимания всех аспек..
тов биолоrии, но, к сожалению, слишком
мало авторов смотрит на животных не как
на застывшие механические формы. Поэто..
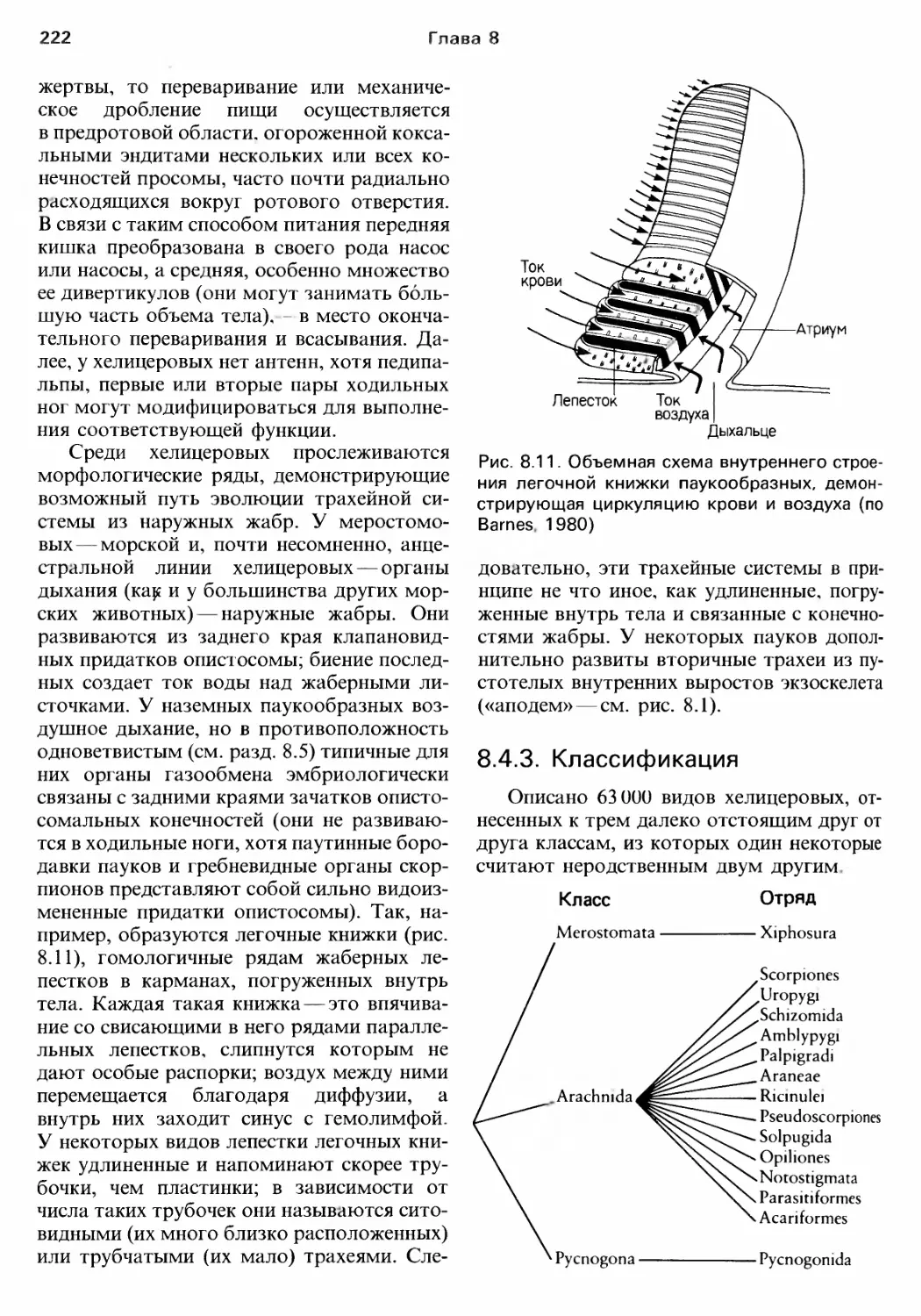
му везде, [де это возможно, мы вели речь об
эволюции, ставя перед собой цель охаракте..
ризовать разнообразие и функции беспозво..
ночных на фоне давления отбора и селе к..
тивных преимуществ rрупп в прошлом
и теперь. Это также повлияло на подбор
и обработку материала. Эта книrа, таким
образом, не обобшает уже опубликованные
сведения, а, как мы надеемся, является но..
вым критическим взrлядом на основные во..
пр осы биолоrии беспозвоночных.
Поскольку. как отмечалось выше, с каж..
дым десятилетием зоолоrические курсы все
меньше и меньше останавливаются на отде..
льных типах животных, мы считаем здесь
уместным выступить в защиту широкоrо
изучения беспозвоночных в высшей школе.
Множество наших современных представ..
лений о биолоrических процессах в целом
возникло в ходе исследований именно этих
животных; достаточно вспомнить плодо..
вую мушку В связи с rенетикой и кальмара
в связи с нейрофизиолоrией, чтобы все ста..
ло ясно. Однако до сих пор число детально
изученных типов животных, не rоворя уже
о видах, крайне мало и, конечно, не отра..
жает Bcero разнообразия структур и функ..
ций беспозвоночных. Мы верим, что в буду..
щем множество обобщений возникнет как
раз при изучении rрупп, не привлекающих
пока всеобщеrо внимания, и что без оценки
разнообразия, так же, как и единства жизни,
невозможно получить адекватноrо пред
8
Предисловие
ставления как о биолоrии в целом, так и
о том, в какой степени наши современные
знания опираются на крошечную необъек
тивную выборку.
За исключением rл. 16, эта книrа плод
совместных усилий трех авторов. Хотя
практически первые наброски разных rлав
или их частей были написаны каждым из
них поодиночке, весь текст затем перераба
тывался в ходе cOBMecTHoro критическоrо
обсуждения: мы все втроем принимаем на
себя ответственность за rл. 1 15, а также за
rл. 17. Однако нет книrи, которая была бы
произведением только ее авторов, и мы
очень признатеЛЬНЬJ множеству людей. KO
торые помоrали нам во время ее подrотов
ки. В частности, мы хотели бы выразить Ha
шу блаrодарность д. rолдинrу, взявшему
на себя rл. 16, и высоко опенить работу Xe
лен Крейтон, которая вместе с Б. Фостер
Смитом и п. Кинrстоном снабдили текст
рисунками. Несколько наших коллеr были
так любезны, что ознакомились с наброска
ми материала книrи: r. Беннет Кларк про
чел всю работу, а Б. Бейн, Дж. Коэн, с. Ko
нуэй..Моррис, п. KporaH, М. Джамrоз,
д. Джордж, п. rиббс, Р. Хьюс, П. Миллер,
Т. Ньюберри, д. Николс, дж. Райленд,
Р. Сид, С. Тайлор. и п. Уиллмер разные
ее части. Мноrие друrие предоставили нам
ту или иную информацию и поделились
своими мнениями. Их усилия спасли нас от
фактических ошибок и текстовых поrреш
ностей. Мы не рассчитываем на то, что
в книrе не осталось ни одной неточности
и ни одноrо отступления от общепринятых
положений, не в последнюю очередь
из за нашей неуступчивости перед лицом
справедливой критики. Р. Кемпбеллу
и с. Роллисону из издательства "Blackwell
Scientific Publications" мы весьма признате
льны за всестороннюю помощь. советы,
участие и административное содействие.
Наш долr перед собственными семьями MO
жет оценить только тот, кто коrда либо
посвяшал большую часть cBoero «свобод
Horo времени» TaKoro рода работе.
Большинство иллюстраций в книrе oc
новано на уже опубликованных в научной
литературе, хотя все они так или иначе пере
работаны. Ссылки на ориrинальные источ
ники даны в соответствующих подписях
к рисункам и в списке, прилаrаемом в конце
книrи.
Р.С.К.Б.
Л.К.
П.Дж.У.О.
ЧАСТЬ I
ЭВОЛЮЦИОННЫЕ ВОПРОСЫ
Основная идея, пронизывающая наш об
зор разнообразия беспозвоночных (часть 11)
и их функциональной биолоrии (часть
III), давление отбора и преимущества, OT
разившиеся на этих животных в прошлом
и продолжающие формировать их биоло
rию в настоящее время. В рамках вводной
части мы кратко описываем эти моrуще
ственные эволюционные факторы.
Слово «эволюция» означает просто «из
менение», а изменения можно анализиро
вать с двух различных точек зрения, KOTO
рые в общем относятся друr к друrу, как
причина к следствию или как механизм дей
ствия к ero проявлению: а) наличие пpoцec
сов, в конечном итоrе обусловливающих Ha
блюдаемые изменения; б) наличие общей
схемы или последовательности происшед
ших с течением времени изменений. Факти
чески хотя обычно считается, что Чарлз Дa
рвин продемонстрировал факт эволюции,
он лишь предложил ее правдоподобный Me
ханизм естественный отбор, который
Mor бы объяснить эволюционные измене
ния, признававшиеся и до Hero. Как уже ro
ворилось, эволюционное (или «филоrенети
ческое») древо типов беспозвоночных и про
цесс eCTecTBeHHoro отбора связаны друr
с друrом, но на практике можно видеть He
сопоставимость результатов и большое KO
личество разноrласий между, с одной CTO
роны, популяционными rенетиками, изу
чающими процессы отбора на живых opra
низмах, И, с друrой таксономистами,
строящими филоrенетические схемы и пы
тающимися понять причины возникновения
новых таксонов надвидовоrо уровня.
Здесь мы рассмотрим оба этих подхода
в основном раздельно; в этом смысле rл.
1 помимо Toro, что является введением
в книrу в иелом, описывает отбор как Mexa
низм изменений (этот аспект иноrда пазы..
вают «специальной теорией эволюции»),
в то время как в rл. 2 обсуждаются филоrе..
нетические связи rрупп беспозвоночных
(<<общая теория эволюции») и возникнове
ние разнообразия во времени. Тем не менее
в пределах каждой из этих rлав мы сочли
уместным представить часть материала
друrой rлавы, например комментируя Ta
кой противоречивый вопрос, как способ во..
зникновения классов и типов беспозвоноч"
ных
rЛАВА 1
ВВЕДЕНИЕ: ОСНОВНОЙ ПОДХОД И принципы
1.1. Почему беспозвоночные?
Эта книrа о беспозВ8ИОЧНЫХ, т. е. живот
ных без позвоночника. Такое определение.
основанное на отсуп1сп16ИИ, а не наличии xa
pdKTepHoro признака, необычно и подразу
мевает отклонение от стандартноrо типа,
у KOToporo этот признак имеется. Если бы
стандарта или нормы не было, подобноrо
рода определение вряд ли имело бы смысл.
Коrда Аристотель разделил животных
на полнокровных и бескровных, полразуме
валось, что присутствие крови норма. Он
верил в эволюцию живоrо к некой COBep
шенной форме, предполаrающей наличие
крови, и обобщил эту мысль в иерархиче
ской системе живых существ, называемой
«Лестница природы» (Scala naturae), OTpa
жающей развитие от бескровноrо состоя
ния к полнокровному, являюшемуся пелью
(табл. 1.1).
Аналоrичным образом, коrда Ламарк
(известный своим учением о блаrоприобре
тенных признаках) впервые отделил беспо
звоночных от позвоночных (в своей «Syste
те des Animaux sans Vertebres», Paris, 1801),
подразумевал ось, что последние норма.
Вероятно, это опять же вытекало из CBoe
образной эволюционной теории Ламарка,
которая предполаrала, что приобретенные
признаки наследуются в соответствии
с принципом не только выживаемости, но
и проrрессивноrо развития к некой высшей
форме, к которой ближе Bcero подошли по
звоночные и человек.
Современная зоолоrия отказалась от
концепций направленной эволюции (телео..
лоrии), однако деление на позвоночных
и беспозвоночных сохранил ось и оказало
влияние на мноrие поколения студентов.
Это удивительно, потому что проводимая
rРdница едва ли естественна и даже не впол..
не определенна; она отделяет rруппу, вклю"
чающую MHoro типов (беспозвоночные). от
части одноrо типа (у некоторых представи
телей типа Chordata нет настоящеrо позво
ночника!)
Тем не менее есть две друrие важные
причины для разделения учебноrо курса на
зоолоrию позвоночных и беспозвоночных.
Первая историческая; Ламарк создал
прецедент, от KOToporo, с тех пор как он YT
вердился как метод подхода к предмету,
оказалось трудно отказаться. Вторая, и, Be
роятно, более существенная
сохранившееся до сих пор ощущение, что,
поскольку у нас самих есть позвоночник,
животные с этим признаком достойны бо
льшеrо внимания, чем следует просто из их
таксономическоrо статуса.
Обращаясь здесь к биолоrии беспозво
ночных, мь] тем самым сохраняем это раз
деление, но не из за какой то философской
приверженности теории направленной эво
люпии или представлению о фунламеНТdЛЬ
ных биолоrических различиях между позво..
ночными и беспозвоночными. Наша пози
ция скорее праrматическая. Мы хотим по..
казать, что:
[) у всех живых орrанизмов есть общие
основные черты строения и функционирова
иия;
2) черты варьируют. и rруппы таксонов,
объединенные общими их «вариантами»,
называют типами;
3) варианты эволюционировали и, еле..
довательно, должны быть связаны обшим
происхождением;
4) в пределах каждой морфо
физиолоrической системы животные при..
способились к окружающим lколоrическим
условиям в ходе eCTecTBeHHoro отбора (в ка..
Введе"lие: ОСНОВНОЙ ПОДХОД И ПрИНЦIIJПЫ
11
Таблица 1.1. Лестница жизни, или Лестница ПРИРОДЫ (Scala naturae), по Ари
стотел ю
Живородящие
Яйцеродящие
с совершен..
ными
яйцами
Снесовер..
шенными
яйцами
ЧервеРОДЯll\ие
Возникают из зародышевой
слизи, путем почкования
или самозарождения из
неживоrо
Самозарождаются из нежи
Boro
Полнокровные
1. Человек
2. Волосатые четвероноrие (Ha
земные млекопитающие)
3. Cetacea (морские млекопитаю
щие)
4. Птицы
5. Чешуйчатые четвероноrие
и безноrие (рептилии и амфи
бии)
6. Рыбы
Бескрпвllые
7. Malacia (rоловоноrие)
8. Malacostraca (ракообразные)
9. Насекомые
10. Ostracoderma (моллюски кро"
ме rоловоноrих)
11. Зоофиты
кой степени эти микроэволюционные про..
цессы MorYT быть связаны с макроэволю"
ционными изменениями, отмеченными
в пп. 2 и 3, вопрос спорный, и мы вернемся
к нему позже).
При рассмотрении этих проблем было
БЬJ целесообразно каким..либо образом
оrраничить материал, и мы сделали это на
основе историческоrо прецедента. Более то..
ro, колоссальное разнообразие беспозво..
ночных позволяет полнее проанализиро..
вать пп. 2 . Однако прежде Bcero необхо..
ДИМО остановиться на основных чертах, об..
щих для всех живых орrанизмов (п. 1), их
отличиях от неживой природы. их возни..
кновении. Этому посвящено несколько сле..
дующих разделов.
1.2. Свойства живых существ
1.2.1. Введение
в химическом составе живых орrаниз--
мов обнаружено менее трети всех встречаю
щихся в природе элементов (табл. 1.2). Око..
ло 750/0 веса всех животных составляет во..
па. 500/0 сухой массы уrлерод снеболь..
шим количеством кремния или без Hero.
Земная кора, напротив, состоит более чем
на 200/0 из кремния и менее чем на 0,10/0 из
уrлерода.
Несмотря на оrраниченное количество
химических элементов, обнаруженных в жи..
вых орrанизмах, составляющие их молеку"
лы очень разнообразны по структуре
и функциям. Это объясняется тем, что ато"
мы уrлерода в отличие почти от всех прочих
элементов (только кремний несколько напо..
минает ero в этом отношении) способны со..
единяться друr с друrом в разнообразные
цепочки и кольца. Такими имеющими уrле..
родную основу молекулярными блоками
орrdНИЗМОВ являются: сахара, аминокисло..
ты, жирные кислоты и нуклеотиды; они
в свою очередь объединяются в макромоле..
кулы, образуя полисахариды, белки, липи..
ды и нуклеиновые кислоты. Свободные ор..
rанические вещества подобноrо уровня сло..
жности образуются внеживых систеМdХ
крайне редко (с. 14).
12
Таблица 1.2. Элементы обнаруженные в жи
вых орrанизмах
Элемент Символ Приблизи
тельная
атомная
масса, Д
Водород 1) Н 1
Уrлерод 1) С 12
Азот 1) N 14
Кислород 1) О 16
Фтор F 19
Натрий 2) Na 23
Маrний 2) Mg 24
Кремний Si 28
Фосфор 2) Р 31
Сера 2) S 32
Хлор 2) Сl 35
Калий 2) К 39
Кальций Са 40
Ванадий V 51
Хром Cr 52
Марrанец Мп 55
Железо 2) Fe 56
Кобальт Со 59
Медь Си 64
Цинк Zn 65
Селен Se 79
Олово Sn 119
Иод 1 127
Атомный
номер
1
6
7
8
9
11
12
14
15
16
17
19
20
23
24
25
26
27
29
30
34
50
53
1) Четыре первых по частоте встречаемости в жи
вых орrанизмах элемента.
2) Следующие по частоте встречаемости в живых
орrанизмах элементы. .
Еще более rлубокое различие между жи--
вым и неживым заключается в способе ор--
rанизации орrаническоrо вещества. В жи--
вых системах макромолекулы образуют
мембраны. оrраничивающие друrие неслу--
чайные скопления макромолекул, реаrи--
рующих друr с друrом в процессе упорядо--
ченноrо метаболизма. Эти «упаковки»
клетки. Множество клеток образует mhoro--
клеточный орrанизм, в котором они объ--
единены в хорошо управляемое и орrанизо--
ванное структурно--функциональное целое.
Само сушествование, устойчивость такой
упорядоченности и сложности долrое время
считались специфической и даже заzадочной
чертой орrанизмов, созданных и поддержи--
rлава I
ваемых мистической жизненной силой, по--
скольку неживая природа подчиняется фи--
зическому Второму закону термодинамики,
rласящему, что порядок и орrанизация
нестабильны. Энтропия, т. е. неупорядочен--
ность, должна постепенно возрастать во
всех реакциях и процессах.
Тем не менее сейчас признается, что
устойчивая орrанизация биолоrических си--
стем основана на двух отнюдь не таинствен--
ных чертах. присущих всему живому. Они
являются решающими для понимания основ--
ных принципов биолоzии, u мы постоянно бу--
дем к ним обращаться.
1. Орrанизмы запроrраммированы.
2. Этим проrраммам подчинены рабо--
чие системы и подсистемы, которые откры--
ты для поступления вещества и энерrии.
1.2.2. Проrрамма
Если взять самый фундаментальный
уровень, rенетическая проrрамма контро--
лирует свойства белков, определяя типы
и последовательность аМИНОКИСЛОТ, из KO
торых они образованы. Для животных ха..
рактерны только 20 аминокислот, но цепоч--
ка, включающая Bcero 100 из них (это не--
MHoro для белка), может иметь в принципе
20 100 различных конфиrураций! Это объяс--
няет большое разнообразие белков. Иеко--
торые из них относятся к ферментам.. кон--
тролирующим все процессы обмена ве..
ществ, друrие используются как строитель--
ный материал клеток орrанизма в целом.
Сама проrрамма закодирована в ну--
клеотидной последовательности ДИК. Че--
тыре типа нуклеотидов (аденин, тимин, [уа..
нин и цитозин) определяют 20 различных
аминокислот, так что однозначноrо соот..
ветствия первых последним быть не может.
Только комбинация как минимум трех ну..
клеотидов дает достаточное число альтер--
нативных сочетаний (43 == 64); такой три--
плетный код универсален для живоrо, при--
чем лишние альтернативы (около 40) обу..
словливают ero избыточность (одна амино..
кислота кодируется несколькими триплета..
ми) и наличие особой «пунктуации». Тем не
менее прямой передачи информации от
ДИК синтезирующимся белкам не происхо--
дит. Сначала она «переписывается» в виде
Введение: основной ПОДХОД и принципы
13
I ДНК I IPНКl
А
-\ Белок I
Ядерная
меМБрана
<f>ункция
мРНК
I rя РНК
ДНК
Б
Рис. 1.1. Схема синтеза белка на молекуляр
нам уровне (по Callow, 1976): rяРНК
rетероrенная ядерная РН К, впоследствии пре
абразующаяся в мРНК
молекул РНК (СХОДНЫ с ДИК, НО тимин за
менен урацилом), которые переносят ее
в цитоплазму, rде на содержащих особый
тип РНК частицах рибосомах служат
матрицами для упорядоченной сборки
определенных аминокислот. Аминокисло
ты переносятся и «вставляlOТСЯ» на место
дрyrим типом РНК транспортной. Вся
эта сложная система схематически изобра
жена на рис. 1.1.
1.2.3. Открытость
Упорядоченные системы орrанизмов
вплоть До их макромолекул постоянно под
верrаlOТСЯ «атакам энтропии», но их разру
шаlOщиеся компоненты MorYT замещаться
в соответствии с информацией, заКЛlOчен
ной в rенетических ПрОI'раммах. Это возмо
жно только В том случае.. если существуlOТ
постоянный приток упорядоченноrо «cы
рья» (в своей знаменитой книrе "What is Li
fe?", вышедшей в 1944 [., Эрвин Шрединrер
назвал это поrлощением не2энтропии!) и OT
ток утратившеrо орrанизациlO материала
(экскретов) и энерrии (в основном тепла).
Таким образом, орrанизмы и составляю
щие их клетки должны быть открытыми си
стемами, а это подразумевает, что даже
у нерастущих животных идет постоянная
замена клеток и/или молекул то, что Ше
нхеймер (Schoenheimer) в своей книrе "The
Dynamic State of Body Constituents" (1946)
определил как динамическое устойчивое co
стояние тела.
1 .2.4. Эволюция путем
eCTeCTBeHHoro отбора
неизбежное сл дствие
возобновления систем
за счет реп л и кац ии
Целые орrанизмы также MorYT заме
няться себе подобными, т. е. воспроизводи
ться путем репликации rенетической про
[раммы. Это предполаrает отделение MHO
rоклеточных или одноклеточных структур
(пропаrул), несущих полнуlO или неполнуlO
rенетическуlO проrрамму (reHoM) родите..
лей. Мноrоклеточные пропаrулы всеrда
имеlOТ более или менее полные копии роди
тельскоrо [енома; репродуктивные процес..
сы с их участием называются бесполыми
или веI'етативными. Одноклеточные пропа
rулы (rаметы), как правило, содержат лишь
часть родительской наследственной про
траммы и должны слиться с друrими raMe
тами, чтобы она стала полной (процесс
оплодотворения); только после этоrо начи
нается развитие. Это называют половым
размножением.
Таким образом, речь идет о системах,
реплицирующuх reHoM для воспроизведения
орrанизма. Однако reHoM не всеrда копи..
руется точно. Даже при бесполом размно"
жении мутации вызываlOТ ero изменения,
а при половом процессе дополнительная
«перетасовка» reHOB., связанная с мейозом.
и их «перемешивание» при оплодотворении
обусловливаlOТ значительные различия Me
жду родителями и потомством. Изменчи
вость rенетической проrраммы ведет к раз
нообразиlO формы и функций фенотипов,
что в CBOlO очередь влияет на способ их
взаимодействия с окружаlOщей средой,
а следовательно, определяет шансы особей
на выживание и скорость их воспроизведе..
ния. Из этоrо следует, что rенетические про
rраммы, лучше Bcero обеспечиваlOщие BЫ
живание и плодовитость в данной окружаlO
щей среде, т. е. наиболее адаптированные
к ней, станут чаще встречаться. Более Toro,
поскольку мир конечен и ресурсы необхо
14
димые для жизненных процессов, оrраниче..
ны, эти проrраММЬJ будут иметь тенденцию
замещать друrие менее выиrрышные. Так
схематически выrлядит процесс эволюции
путем еСnlественнос">о отбора, который впе..
рвые четко описан Чарлзом Дарвином в ero
книrе «Происхождение видов» (1859). Для
характеристики этоrо процесса он исполь..
зовал заимствованную у rерберта Спенсера
формулировку «выживание наиболее при..
способленных». Однако из сказанноrо выше
ясно, что приспособлеllносп1Ь nредставляеп1
собой свойсп1во одНО20 2еllеп1ически деп1еРJИИ"
llироваН1l020 признака расnросп1раllяп1ЬСЯ
в популяции за счеlп дРУ2ИХ а <-это требует
как выживания, так и nлодовип10сп1И.
1 .3. n роисхождение жизни
Самая фундаментальная черта живых
систем их способность сохранять упоря..
доченное и орrанизованное состояние за
счет процессов запроrраммированной ре..
пликации и воспроизведения. ЭвОЛlоция nу..
n1е./и еСn1ествеННО20 О/11бора являеnlСЯ авп10"
/Wап1ическuм следСn1вие.Atl ЭnlО20. Но каким
образом возникла такая система? Открытие
Toro как возникли орrанические молекулы,
составляющие живое (с. 11), дает только ча..
стичный ответ на этот вопрос. Мы должны
представить себе, как они ОР2анизовалuсь
в самореплицирующиеся системы.
Уrлеродсодержащие молекулы из кото..
рых состоят орrанизмы, коrда"то считали
настолько специфическими и уникальными,
что возможность их синтеза связывалась
только с живыми существами. Таким обра..
зом проводилось разrраничение между ор..
rdническими (== из живоrо) и неорrаниче..
скими веществами. Первая брешь вразде..
ляющей их стене была пробита Велером.
синтезировавшим в 1832 r. очень простую
орrаническую молекулу (мочевины) из циа..
ната аммония (неорrаническая молекула)
в результате простоrо ее наrревания. Это
положило начало рациональному и научно..
му подходу к химии живоrо, а тем самым
современной биохимии и молекулярной
биолоrии. Хотя, конечно. контролируемый
синтез мочевины это еще очень далеко от
спонтанноrо образования полисахаридов,
липидов, белков и нуклеиновых кислот, не..
rлава I
обходимоrо для возникновения живых си..
стем.
1 .3.1. Добиолоrический синтез
орrанических полимеров
О первоначальной земной атмосфере
известно очень мало, но, возможно, она
возникла в ходе «деrазации» планеты и бы..
ла очень похожа на rазовую смесь, выделяе..
мую ВУЛКdнами. Если это так, то в ней поч..
ти наверняка не было кислорода (см. rл. 11).
Эксперименты показали, что при этих усло..
виях ЛlOбое энерrетическое воздействие
освешение, ударные волны, ультрафиоле
товая радиация (поскольку отсутствовал
кислород, не было и озоновоrо экрана, от..
фильтровывающеrо волны этой длины из
солнечноrо спектра) или тепло вулканиче..
cKoro пепла привело бы к добиолоrическо"
му синтезу разнообразных «орrанических»
мономеров: сахаров, аминокислот и даже
нуклеотидов. При блаrоприятных обстояте..
льствах. например при высоких концентра..
циях неорrанических полифосфатов, они
MorYT объединяться в длинные цепочки,
образуя, в частности, полинуклеотиды и по..
липептиды. Все эти вещества, вероятно. на..
капливались в древнем океане, образуя так
называемый «первичный бульон».
В этом доБИОЛ6rическом мире шел свое..
ro рода отбор. поскольку молекулы. спо
собные полимеризоваться быстрее друrих
и/или оказавшиеся более стабильными, по..
лучали большее распространение. Но тем..
пы таких изменений были медленными
и вряд ли моrли привести к возникновению
принципиально новых структур, поскольку
образование каждоrо полимера происходи..
ло независимо и не закреплялось в rенетиче..
ской памяти. Следующий за этим шаr не
представлял особой трудности, поскольку
некоторые полимеры, однажды образовав..
шись, MorYT влиять на образование друrих
полимеров. Полинуклеотиды в частности
способны определять последователь..
ность нуклеотидов, действуя как матрица
для полимеризации. Если один полинуклео..
тид служит матрицей для комплементарно...
ro ему друrоrо полинуклеотида, который
в свою очередь является матрицей для пер..
Boro, можно rоворить о множестве моле
Введеl--lие: ОСНОВНОЙ ПОДХОД И принциnы
15
кул, связанных чем..то вроде rенетической
памяти. Полинуклеотиды, воспроизводя..
щиеся наиболее эффективно, увеличив а..
ются в количестве по сравнению с прочими.
т. е. имеют селективное преимущество.
Матричным системам свойственны
ошибки репликации, и новые полинуклеоти"
ды, образующиеся за счет «мутаций». ве..
роятно, конкурировали с друrими за orpa..
ниченное количество мономерных струк"
турных блоков. Поскольку дезоксирибону"
клеозиды (предшественники нуклеотидов)
синтезируются сложнее рибонуклеозидов
и поскольку РНК иrрает центральную роль
в современном белковом синтезе, широко
(но не всеми) признана rипотеза. соrласно
которой первыми самореплицирующимися
полимерами были короткие цепочки РНК.
1.2.3. Происхождение
и эволюция клеток
Следующий шаl по направлению к си..
стемам, иллюстрируемым рис. 1.1, предста..
вить труднее Bcero. При нормальной темпе..
ратуре спонтанная репликация., описанная
выше, протекала бы медленно и с высоким
процентом ошибок Включение в этот про..
цесс репликазы белка, способноrо ката..
лизировать репликацию, существенно бь]
ero ускорило. Каким образом это произо
шло не ясно, но, возникнув, такая систе..
ма получила преимушества. Более Toro,
должны быть определенные преимущества
в окружении матрицы и репликазы оболоч..
кой., чтобы выrоды от их взаимодействия не
моrли использоваться друrими., HeMHoro
отличными матрицами конкурентами. Так
появилась клетка и разница между rеноти"
пом и фенотипом. На эти примитивные
клетки отбор должен был действовать та..
ким образом. что те из них. у которых взаи..
модействие между rенотипом и фенотипом
стимулировало скорость репликации и ее
точность, распространялись быстрее дру"
rих. Хотя трудно точно представить себе
ход событий, но именно в ходе взаимодей..
ствия и отбора возникли и совершенствова..
лись сложные системы., включающие ДНК
и различные формы РНК.
Первые клетки были мелкими и просто
устроенными. Отчасти они напоминали со..
временных бактерий, так называемых про..
кариот. В некоторых из них шло дальней..
шее формирование мембран для отделения
rенетической информации от остальноrо
объема, что, вероятно., давало преимуще..
ства, поскольку обеспечивало лучшую за..
щиту rенетическоrо материала от повре..
ждений. Такие клетки, так называемые про..
тоэукариоты, возможно., позднее приобре..
ли цитоплазматические орrанеллы., из кото..
рых наибольшее значение имели митохонд"
рии. Последние очень сходны со свободно..
живущими прокариотами, напоминая их
размерами, формой, наличием собственной
ДИК и размножением путем деления над..
вое. Поэтому сейчас считается., что они во..
зникли за счет симбиоза между мелкими
прокариотами, похожими на ныне живу..
щую бактерию Pal"aCOCCUs, и более крупны..
ми.. содержашими ядра протоэукариотами.
Разрушая эукариотические клетки., можно
показать, что весь механизм аэробноrо ме..
таболизма связан именно с митохондрия"
ми, так что этот предполаrаемый симбиоз
развивалея параллельно накоплению в зем..
ной атмосфере кислорода вследствие фото..
синтетической активности древних циано..
бактерий.
1.3.3. Почему самозарождение
жизни не происходит все время?
Если крупные биомолекулы и даже клет..
ки возникли однажды., правомерен вопрос,
почему этоrо не происходит постоянно. Be
роятно, ответ заключается в том, что живые
существа сами создали условия. в которых
самозарождение невозможно. Например.,
кислород, продукт их обмена веществ.,
образовавшись, должен разрушать орrани"
ческие молекулы, из которых построено жи..
вое. В ero присутствии орrанические поли..
меры окисляются до простых неорrаниче..
ских составляющих. Следовательно., коrда
свободноrо кислорода накопилось доста..
точно MHoro. существование «первичноrо
бульона» стало невозможным. Кроме Toro,
сложные орrанические молекулы в ero со..
ставе, вероятно, были превосходным источ..
ником питания для первых орrdНИЗМОВ
и поедались или разрушались скорее, чем
синтезировались вновь
16
1.4. Уровни орrанизации
орrанизмов
ВРЯД ли вся совокупность физиолоrиче
ских процессов. коrда они «втиснуты» в oд
ну клетку (как у протистов; разд. 3.1), может
работать столь же эффективно, как в случае
их разделения между различными клетками
мноrоклеточноrо орrанизма. Мноrоклеточ"
ность обеспечивает болыпее пространство
для реакций, а разделение функций между
клетками означает сведение к минимуму
физиолоrических противоречий по крайней
мере в пределах каждой из них. Таким обра
зом, в сторону мноrоклеточности, т. е. про
исхождения царства Animalia, должно было
действовать значительное давление отбора.
Следующая rлава обобщает основные
черты беспозвоночных и соображения об их
родственных связях и эволюции. Она иллю
стрирует различные возможные тенденции
развития, постепенно раскрывающие фи
зиолоrический потенциал мноrоклеточных
животных, в частности:
1) ЭВОЛЮIIИЮ клеточной дифференци
ровки;
2) пространственное распределение кле
ток одноrо типа по тканям, а затем орrани"
зацию их в opraHbI (совокупности клеток,
обслуживающих общую функцию):
3) эволюцию сквозноrо кишечника, обе..
спечивающеrо большую специализацию
разных отделов тела;
4) развитие заполненной жидкостью по..
лости тела, которая позволяет кишке и дру..
rим opraHaM (например, сердцу) работать
независимо от мускулатуры стенки тела,
способствует диффузионному распределе
нию питательных веществ, И, действуя как
rидростатический скелет (разд. 10.5),
более эффективной локомоции;
5) эволюцию специфических систем
переноса питательных элементов идыха..
тельных rазов между тканями, позволяю..
щую избежать размерных оrраничений,
связанных с диффузионным распределе..
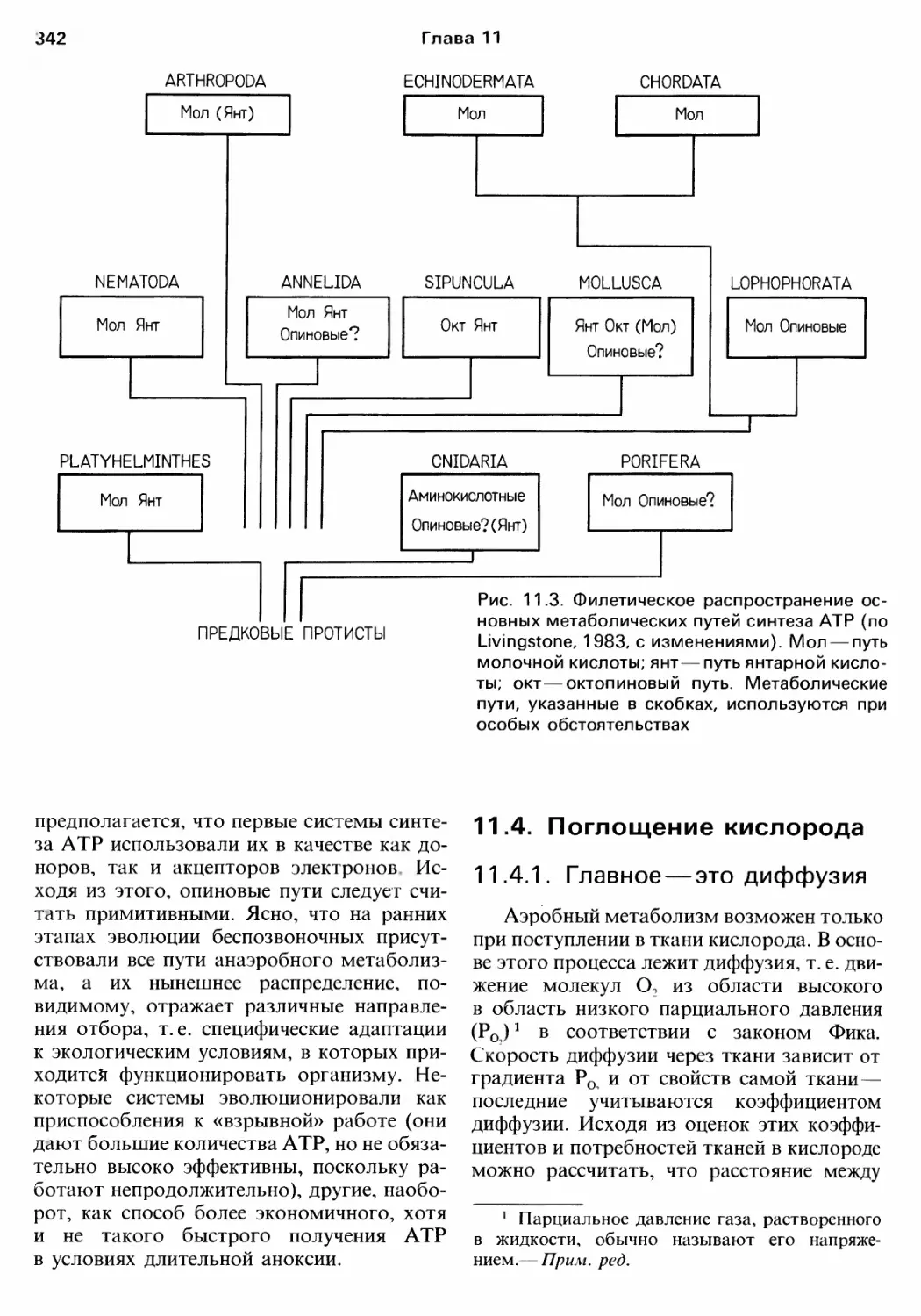
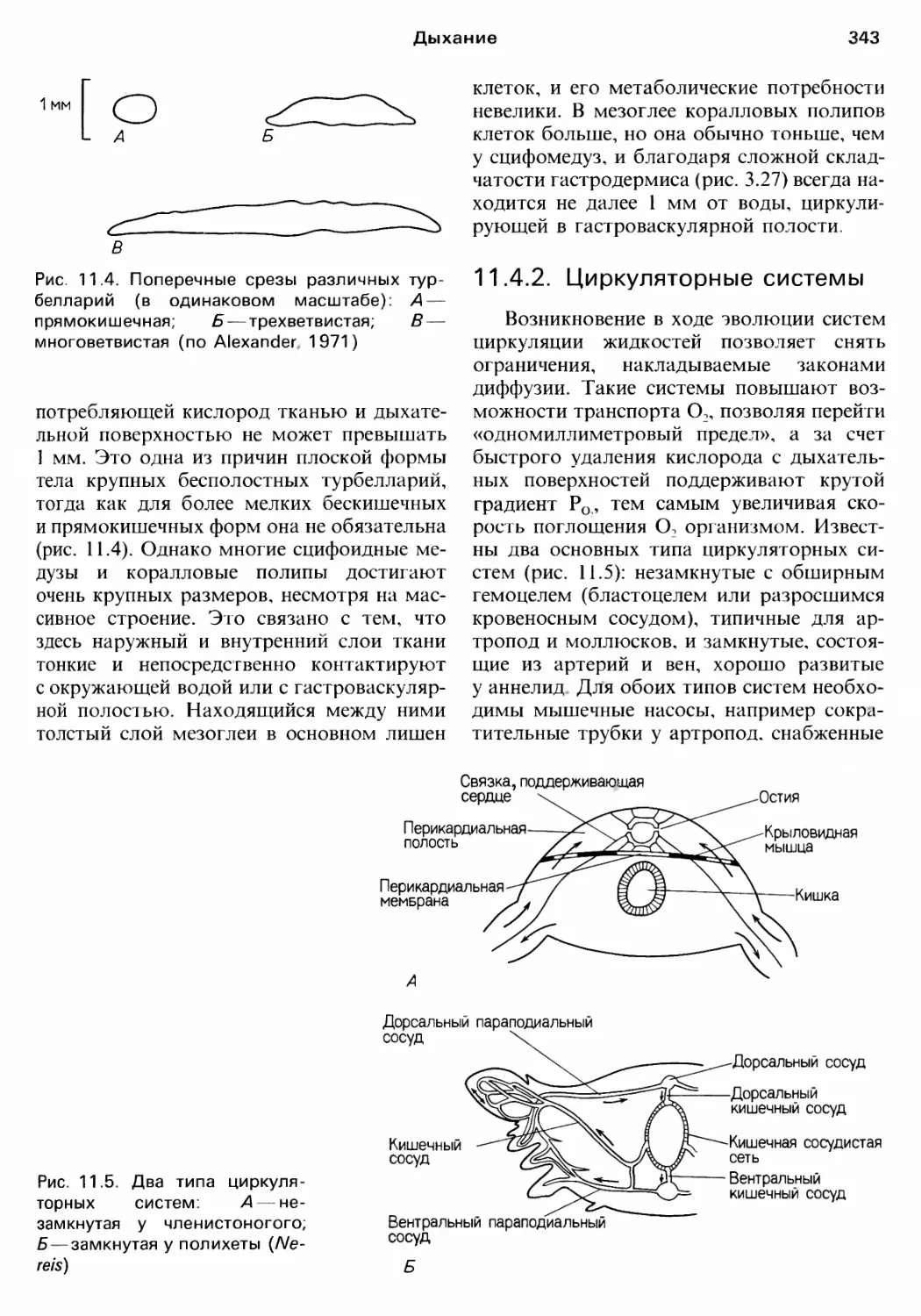
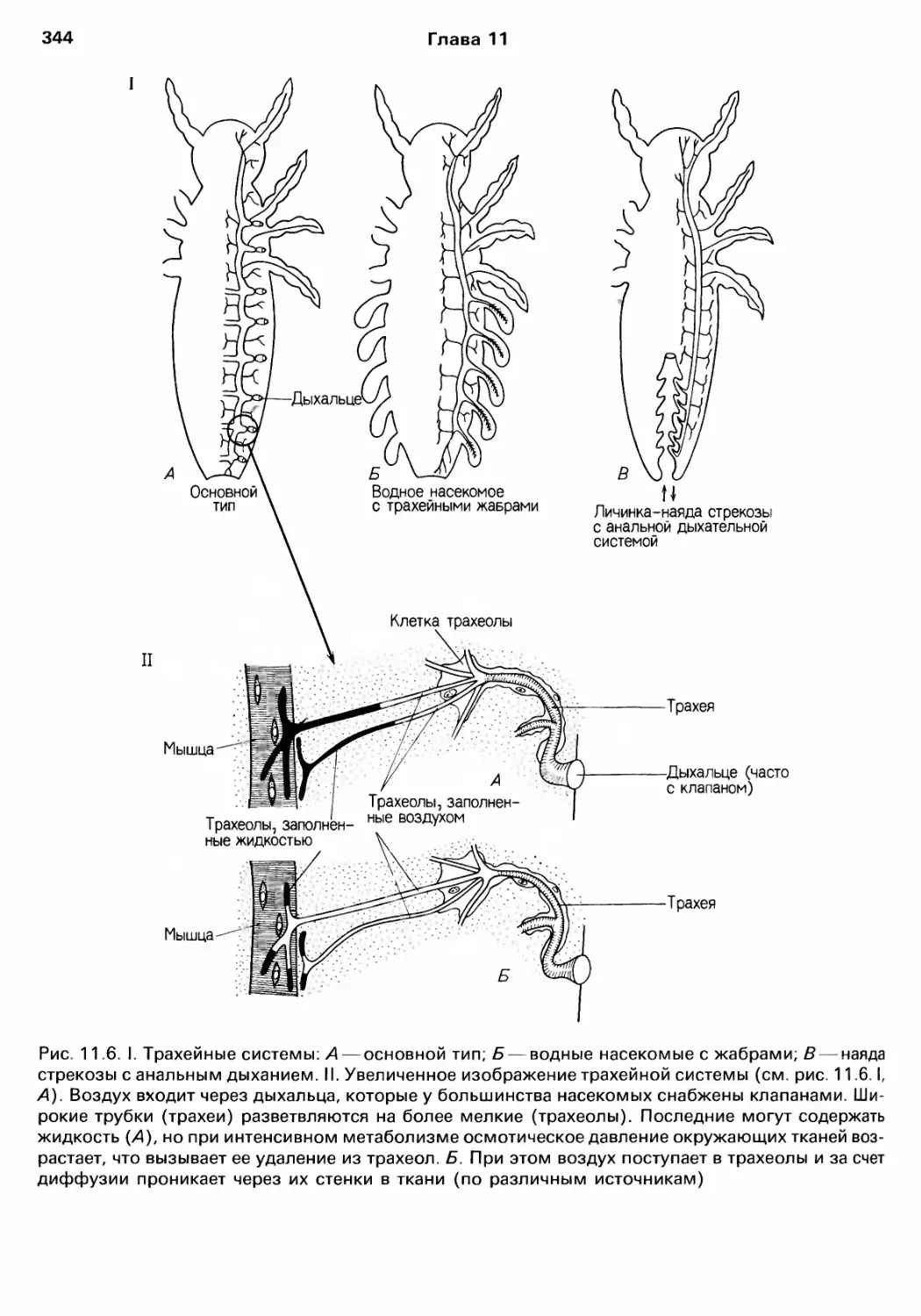
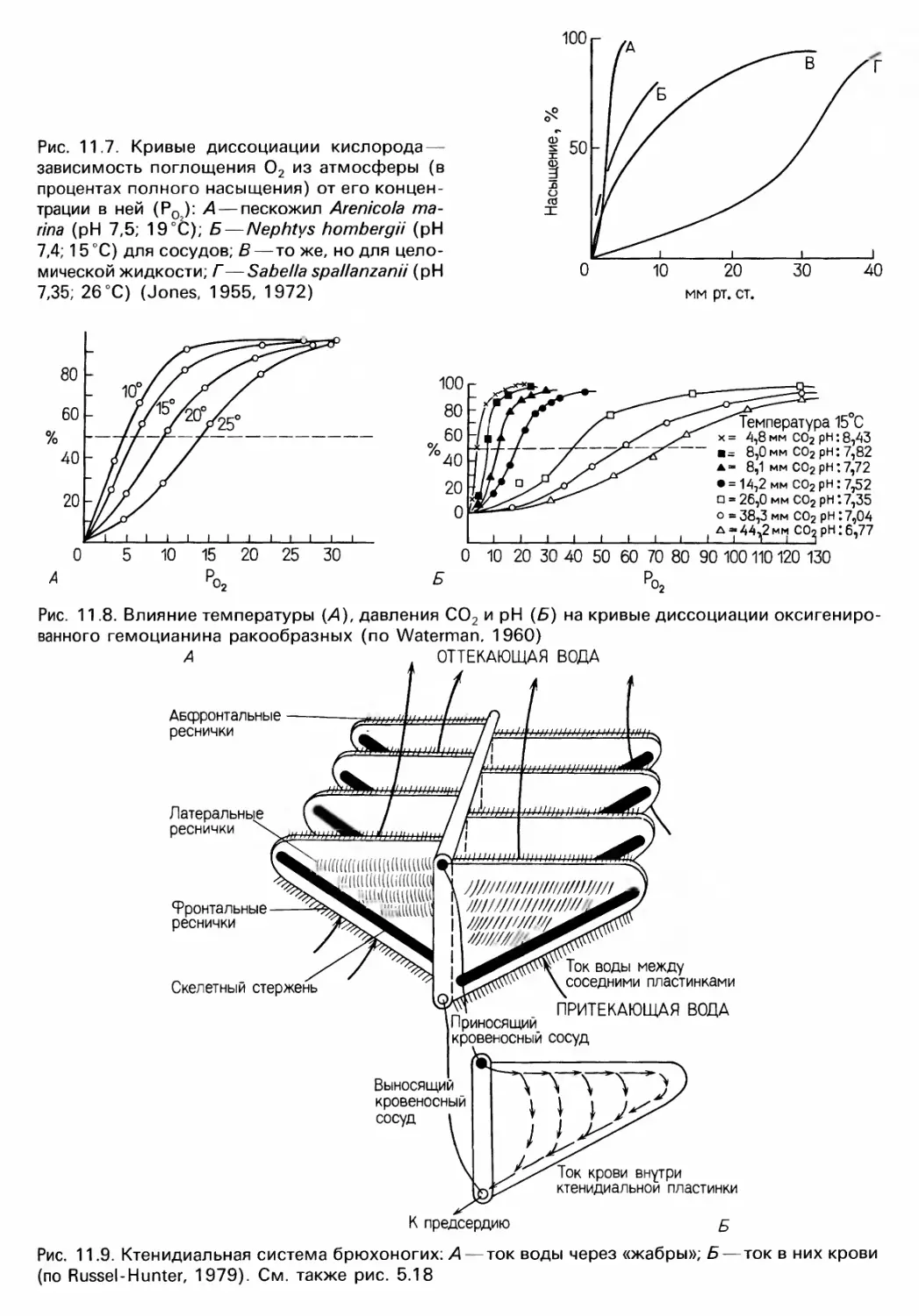
нием этих веществ (см. rл. 11);
6) эволюцию конечностей, значительно
расширяющую возможности локомоции,
особенно на суше и в воздухе.
Леrко показать. как естественный отбор
приводит к совершенствованию функций
rлава I
в пределах одноrо уровня орrанизации. Но
обеспечивает ли он переход с одноrо уровня
на друrой? Происходят ли эти переходы пo
степенно за счет непрерывных мелких изме
нений, улучшающих функционирование
и приспособленность орrанизмов, или пу..
тем скачков между уровнями, больше объ..
ясняющихся случайностью, чем естествен..
ным отбором? Эти альтернативы paCCMO
трены соответственно как rипотезы пocтe
пенности и прерывистО20 равновесия. Без co
мнения, эволюция беспозвоночных пред
ставляет собой отчасти прерывистый про..
цесс (rл. 2). Но поскольку такие скачки
обычно происходят в rеолоrических мас..
штабах времени, т. е. в течение нескольких
миллионов лет, они моrли произойти и под
действием eCTecTBeHHoro отбора. Похоже,
что ero давление время от времени сильно
менялось, что вызывало значительные KO
лебания темпов эволюции. Так что преры
вистый характер эволюции не исключает
действия дарвиновских механизмов. Этот
вопрос rорячо дебатировался в эволюцион
ной эколоrии (Calow, 1983) и мы позже Bep
немся к нему.
1 .5. План книrи
в этой rлаве кратко рассмотрено то, что
именно мы считаем основными признаками
живых систем.
Это ОР2анизованные систеJНЫ, пoддep
жание которых зависит от прО2рамм ре..
пликации и открып10сти.
Отсюда почти лоrически вытекают чер
ты животных.
Они потреБЛЯlот пищевые ресурсы из
окружающей среды и используют их так,
что это обеспечивает выживание и плодов и..
тость.
Различные уровни орrанизации живот..
ных эволюционировали, и в пределах типов
действовал естественный отбор, повышаю
щий адаптацию процессов получения и ути
лизации ресурсов. После предварительноrо
обзора разных уровней орrанизации в сле..
дующей rлаве мы более детально опишем
их в части 11. Это даст основу для более rлу
бокоrо рассмотрения поведения и физиоло..
rии беспозвоночных в части 111. [де мы co
средоточим внимание на отдельных аспек..
Введение: основной ПОДХОД и принципы
тах их функционирования. Таким образом,
в части 11 последовательно обсуждаются
разные типы, а в части 111 принят межтипо
БОЙ, функциональный подход к беспозво
ночным. Читатель, таким образом, может
выбирать, заняться ли ему в основном си
стематикой (часть 11), используя часть 111
как источник дополнительной информации,
или, наоборот, обратиться к Функциональ
ной биолоrии беспозвоночных (часть 111),
пользуясь частью 11 как «указателем» упо
минающихся таксонов. В любом случае обе
эти части составляют одно целое и призва
ны дать полное и всестороннее представле
ние об орrанизмах беспозвоночных.
17
1.6. Дополнительная
литература
Ca/ow Р. 1976. Biological Machines. Edward Ar..
nold, London.
Ca/ow Р. 1983. Evolutionary Principles. Blackie.
Glasgow.
Mayпard SmitJl J. 1986. The Problems of Biology.
Oxford University Press, Oxford.
Smith D. с., Doug/as А. Е. 1987. ТЬе Biology of
Symbiosis. Edward Arnold, London.
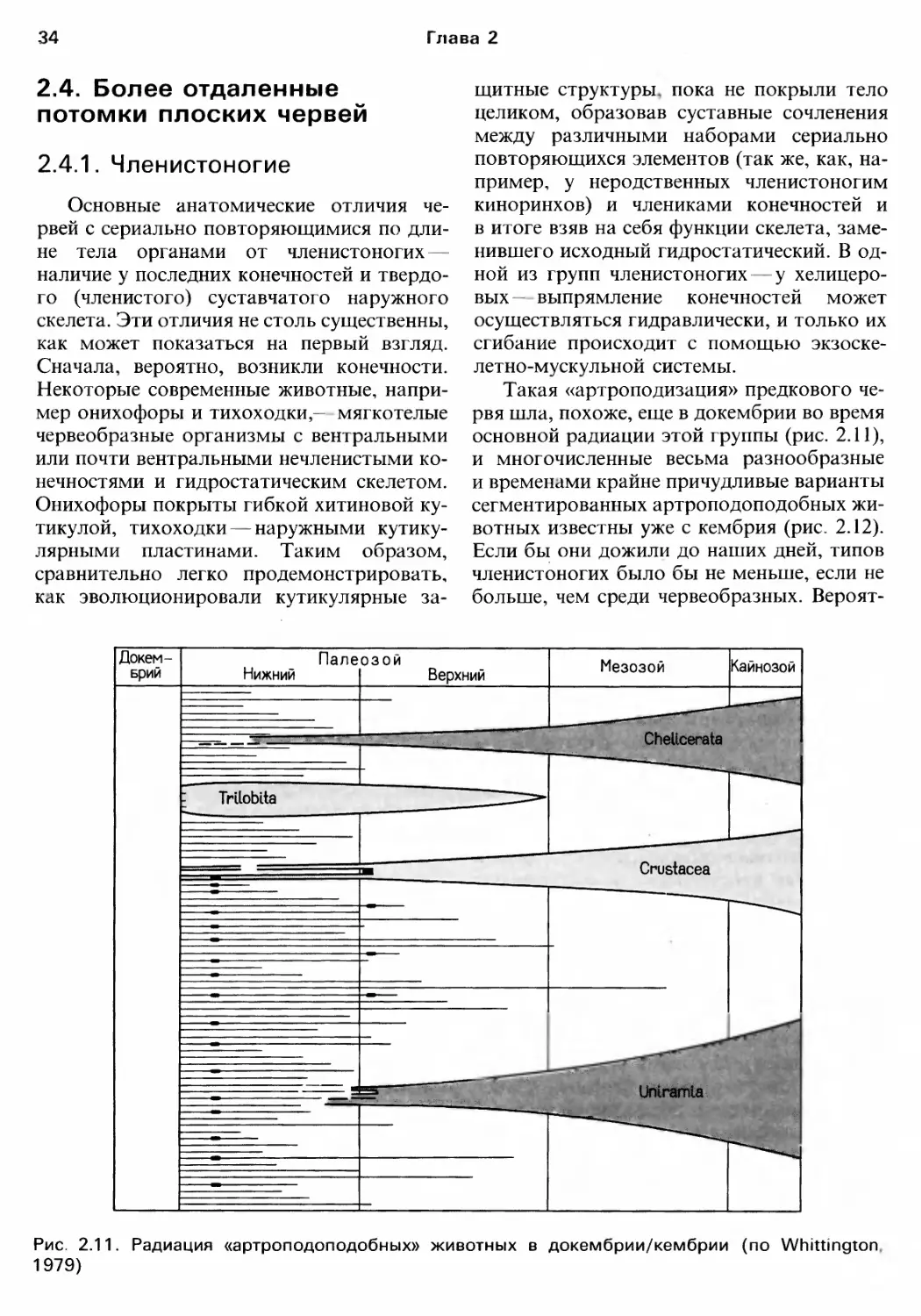
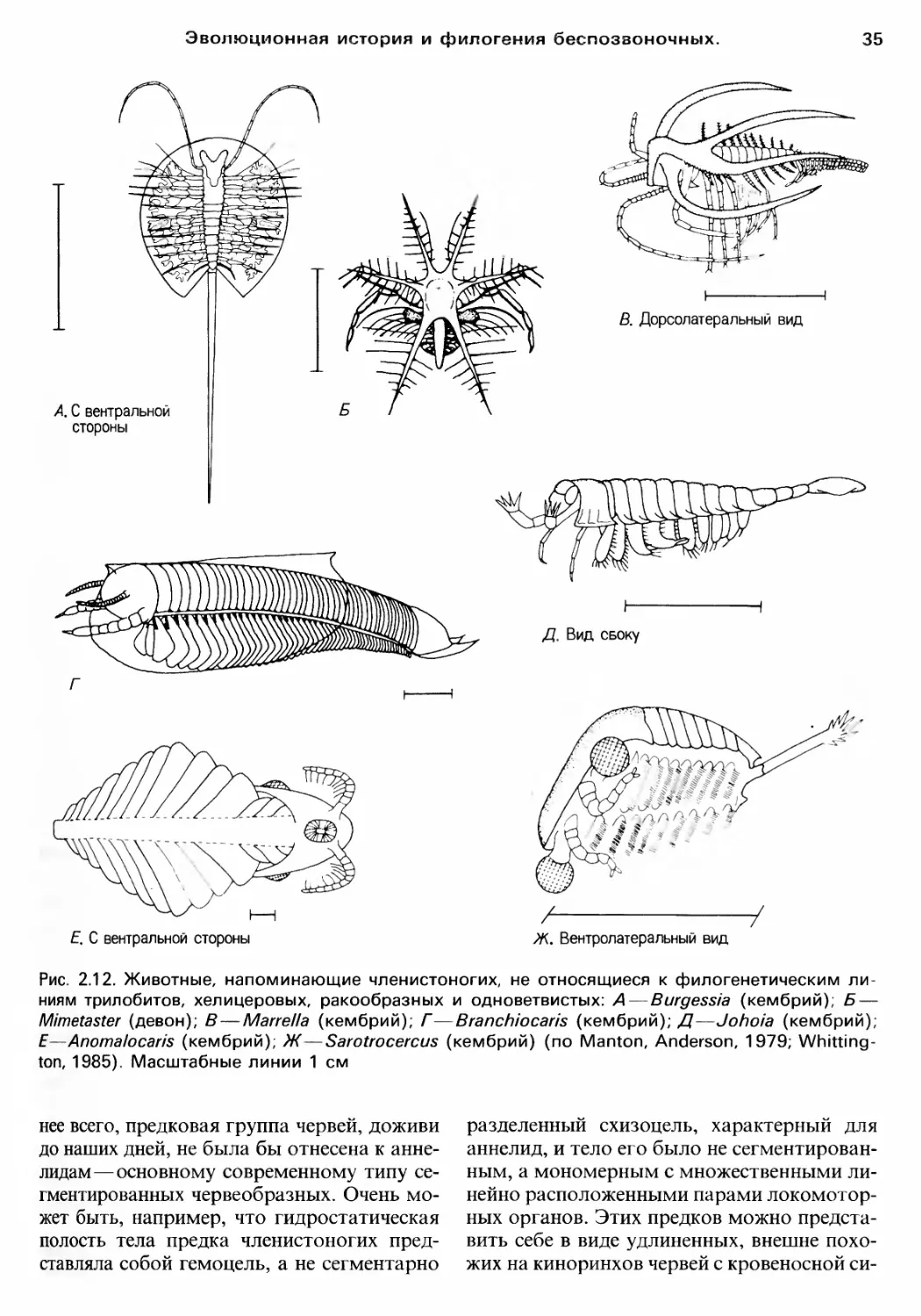
rЛАВА 2
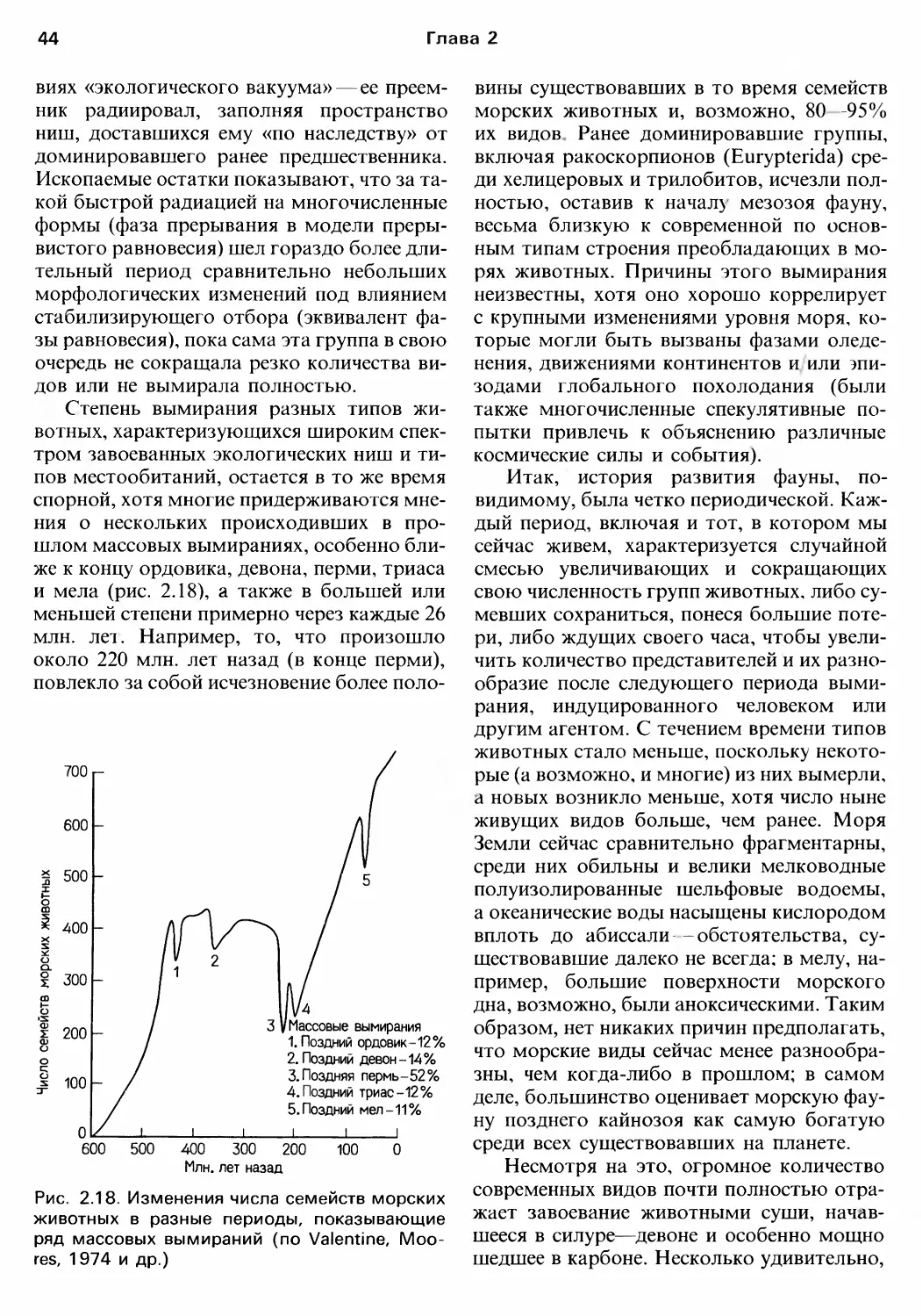
ЭВОЛЮЦИОННАЯ ИСТОРИЯ
И ФилоrЕНИЯ БЕспозвоночныx
Современные животные результат
эволюционной истории невозможно пол..
ностью понять современную биолоrию без
оценки ее прошлоrо и оrраничений, нало
женных им на возможности и образ жизни
орrанизмов. В этой rлаве мы опишем oc
новные черты ,волюционной истории жи
BOTHoro царства, в том числе ero происхо..
ждения.
Слишком часто филоrенетические pe
конструкции представляют собой довольно
бесплодное занятие остроумные. но ма..
лоперспективные попытки объединять пла..
нь] строения в разные комбинации с целью
получения удовлетворительной, т. е. более
или менее лоrичной картины что"то вро"
де составления картинки из кусочков, боль..
шинство которых утеряно. Для заполнения
пробелов в таких схемах предлаrались r и..
потетические формы. часто без учета Toro.
что такие постулируемые животные дол
жны были выживать, добывать ПИIЦУ, избе
rdTb хищников и размножаться. Причиной
бесплодности нескольких из этих попыток
было отчасти использование исключитель..
но анатомическоrо подхода, нескольких
«ключевых признаков» (часто эмбриолоrи
ческих по своей природе).. исследование ко..
торых на более широком спектре животных
выявило их меньшую определенность
и диаrностическую ценность, чем предпола
rалось изначально.
Здесь мы постарались дать современную
филоrенетическую оценку накопленных
данных, исходя из возможноrо действия OT
бора в разные периоды в прошлом и Be
роятных реакций на ero давление живших
тоrда орrанизмов. Мы также сосредоточи"
ли внимание на том, что rоворят ископае
мые находки о характере увеличения разно..
образия и ходе вымирания, продемонстри"
ровав, как наши знания о прошлом ВЛИЯЮl
на понимание эволюционноrо процесса.
Именно в этой области у палеОНТОЛОI ов BO
зобновились эволюционные споры, связан..
ные с широко известными rипотезами «(ма..
кроэволюции» и «прерывистоrо равнове..
сия»
Читатели должны обратить внимание
на две вещи. Во первых, в этой rлаве неиз
бежнь] ссылки на некоторые анатомические
черты, которые описаны в части [1 Мы счи..
таем более лоrичным дать общий обзор
перед детальным описанием отдельных
rрупп, даже если это подразумевает, что не..
которые о(;обенности строения и концепции
будут описаны позднее. Пока же характери
стики различных типов сведутся к необхо..
димому минимуму. Во"вторых, как подчер"
кивается ВJТОЙ rлаве, филоrенетические
связи животных остаются спорными вопро"
сами, а в нашей книrе не ставится задача об..
суждения всех противоположных точек зре..
ния. Мы просто приводим те представле..
ния.. которые кажутся нам наиболее правдо..
подобными, не вдаваясь в детальное обос..
нование всех принимаемых точек зрения.
rлавы части II т кже содержат ТОЛЬКQ кон..
кретную информапию (например.. относите..
льно (рундаментальных различий между ти"
пами членистоноrих), и те, кто интересуется
принципами таксономическоrо разделения,
должны обратиться к спискам «дополните
льной литературы», рекомендованной
в конце этой и друrих rлав.
2.1. Введение
Очевидно, что мноrоклеточные живот
ные, как и представители двух друrих
аарств мноrоклеточных rрибы и paCTe
ния, произошли от одноклеточных (или
Эволюционная история и филоrения беСПОЗВОI'IОЧНЫХ
19
w
Tribrachidiuт
Medusiпites
Parvaпcorlпa
Dickiпsoпia
Charпia
Spriggiпa
Cycloтedusa
Raпgea
Praecaтbridiuт
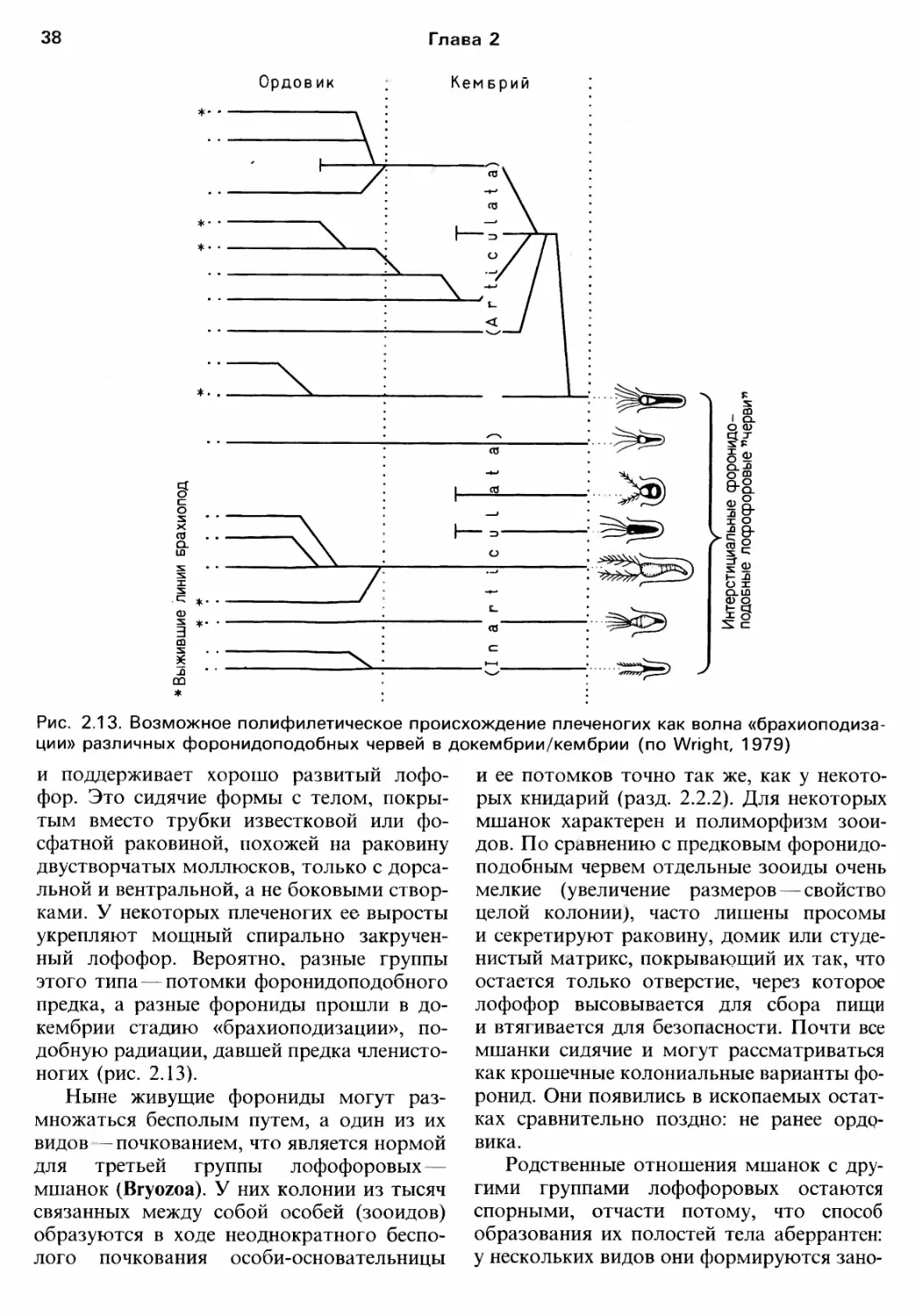
Рис. 2.1. Докембрийские (из отложений Эдиакары) животные (по Glaessner. Wade. 1966)
неклеточных) эукариотных протистов. Но
на этом ясность кончается. Большинство
типов животных, представленных виско..
паемых остатках, впервые появляются
«полностью сформированными» уже в кем..
брии. около 550 млн. лет назад. Это Kacae
тся и ТdКИХ продвинутых, анатомически
сложных типов, как трилобиты, иrлокожие,
плеченоrие и моллюски. Докембрийс.кие
остатки животных немноrочисленны. но
обычно считается, что книдарии и сеrменти
рованные черви тоrда уже существовали
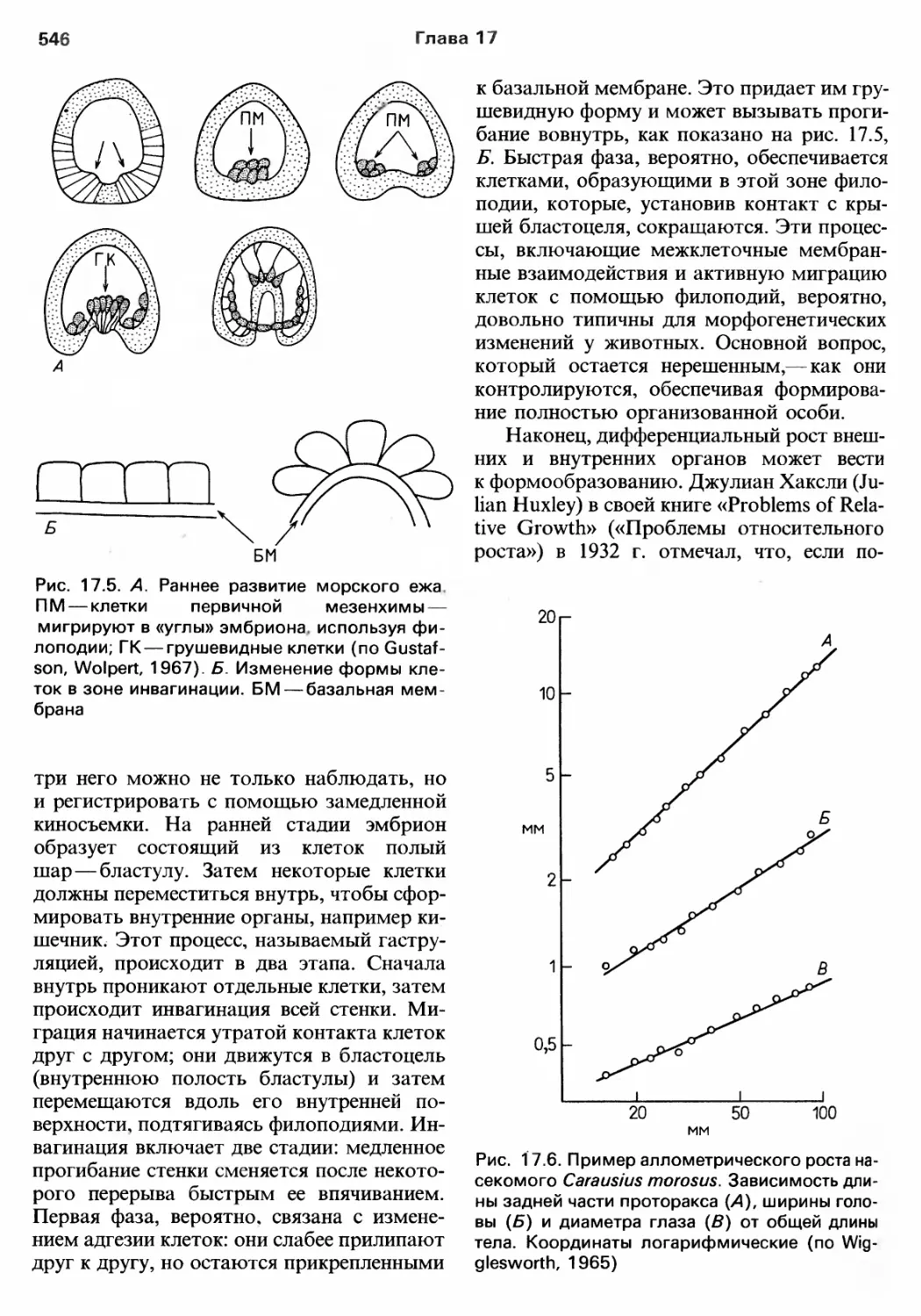
(рис. 2.1). Ископаемые находки, таким
образом. ничеrо не rоворят о возникнове..
нии и ранней диверI'енции разных типов, за
исключением Toro, что эти события должны
датироваться докембрием, вероятно, пе..
риодом 1200 900 млн. лет назад. Сама
жизнь, как считается, возникла 3500 млн.
лет назад.
Множество споров вызывает вопрос, ка..
кую rруппу животных считать наиболее
примитивной и от какой rруппы протистов
ее можно вывести. Если не учитывать сли..
шком фантастических rипотез, три типа
считались и считаются основными претен..
дентами на роль предков ой rруппы живот..
ных: rубки (Porifera), кишечнополостные (в
особенности Cnidaria) и плоские черви (Pla..
tyhelmlnthes). Ранние сторонники прими..
тивности той или иной из этих rрупп впо"
следствии стремились вывести из нее две
остальные, а в конечном итоrе и все прочие
известные типы. Сравнительно рано rубок
начали считать довольно специфической
изолированной rруппой. возможно Hepoд
ственной друrим животным, и поместили
в особое подцарство Parazoa'l отдельно от
всех остальных типов, кроме вымерших
Archaeocyatha. Вопрос тем самым был све..
ден к тому, произошли ли кишечнополост"
ные от плоских червей или наоборот?
Первые животные были, по видимому,
мелкими, состояли из относительно неболь..
шоrо числа клеток и не имели твердых Чd..
стей. Поскольку в подавляющем большин..
стве случаев сохраняются ископаемые
остатки орrанизмов именно с твердыми
структурами раковинами, пластинками
или скелетом, было бы нереальным ожи..
дать, что палеонтолоrические находки Kor..
да"нибудь помоrут определить предка жи..
вотных; в высшей степени невероятно, что
такая форма сохранилась. Таким образом,
зоолоrи вынуждены опираться лишь на
сравнение структуры и функций живых
представителей разных типов животных
и протистов И строить rипотезы о том, ка..
кие из существующих признаков живых ор..
rанизмов следует считать примитивными
20
чертами, дошедшими до наших дней. Здесь
надо помнить, что современные представи
тели всех rрупп орrанизмов отделены cpo
ком по крайней мере 600 млн. лет. а возмо
жно, И более, от момента возникновения
каждой из них, а значит, и всевозможными
изменениями в биохимии, физиолоrии,
эмбриолоrии. анатомии и т. д., которые MO
rли произойти за этот период.
Большинство ранних споров характери
зуется приверженностью к ныне непризна
ваемым «законам», например к формуле:
«онтоrенез повторяет филоrенез» (rл. 17),
выдумыванием чисто rипотетических про
межуточных стадий, отвечающих скорее
требованиям.. продиктованным выдвинутой
автором rипотезой, чем потребностям
успешноrо выживания, а также серией
остроумных кабинетных спекуляций, KOTO
рые, учитываяприродувопроса, впринципе
нельзя было ни проверить" ни опроверr
нуть. В качестве реакции на этот первонача
льный поток спекуляций и контрспекуляций
некоторые современные биолоrи считают
любую попытку реконструировать филоrе
нез пустой и ненаучной: если мы никоrда не
узнаем, правилен ли данный ответ, бесполе
зно ставить и сам вопрос! Хотя этот взrляд
в общем можно считать корректным, мало
вероятно, что пытливый человеческий pa
зум позволит сковать себя такими рамками.
Если cTporo придерживаться определения
биолоrии как науки, отверrа щей любую
область, в которой rипотезы нельзя прове
рить экспериментально, мноrие COBpeMeH
ные биолоrические исследования, в том
числе все, связанные с событиями прошло
ro" потеряли бы право на существование.
Свидетельства, полученные в результате
сравнительноrо изучения живых орrаниз
мов и ископаемых находок, можно исполь
зовать в качестве aprYMeHToB в пользу Toro
или иноrо пути филоrенеза, и именно на Ta
ких данных основан наш последующий об
зор. Тем не менее читатель должен пом
нить, что альтернативные интерпретации не
только возможны, но и фиrурируют в лите
ра туре.
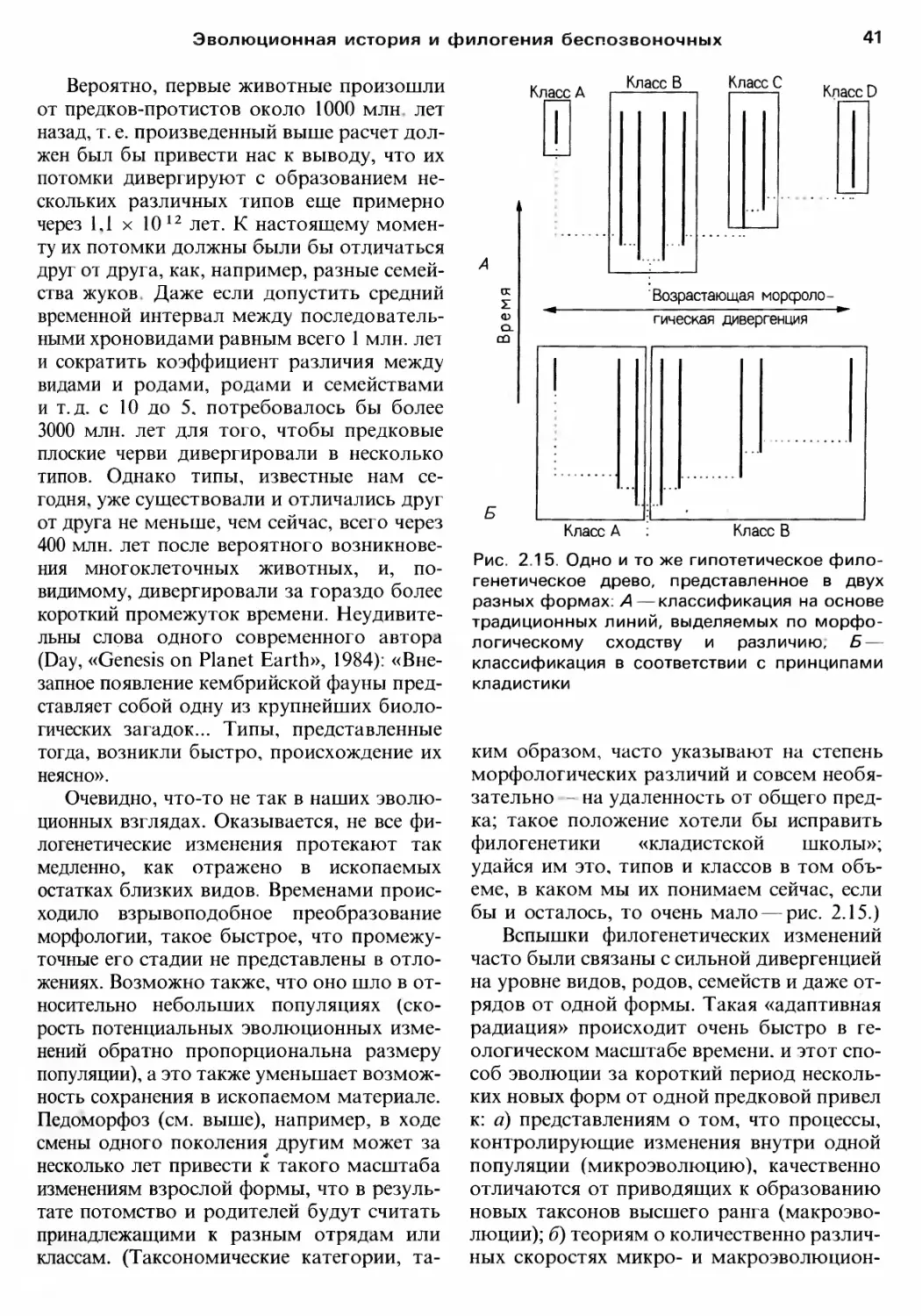
Одна из основных черт сравнительно HO
Boro подхода к разrадке тайн филоrенетиче
ской истории падение доверия к несколь
ким считавшимся решающими «ключевым
rлава 2
признакам», таким, как тип личинки или ce
rментация. В этой связи некоторые, возмо
жно, удивятся, прочтя на следующих CTpa
ницах. что. например. rлоточные щели
структура для удаления попадающей в пи..
щеварительный тракт воды, свойственная
не только хордовым и полухордовым, но
И неродственным им rастротрихам; что
«специфически нематодный» способ локо
моции с использованием продольно несжи
маемых скелетных элементов, продольной
мускулатуры и последовательных волно..
образных изrибов присущ и rоловохордо
БЫМ (и у тех и у друrих мышцы с OTpOCTKa
ми, идущими к нервному стволу, а не Hao
борот); что киноринхи сеrментированы так
же, как мноrие членистоноrие. Даже CTPYK
туры, подобные нематоцистам, характерны
не только для книдарий, но присутствуют
и У некоторых протистов.
Животные, сталкиваясь с одинаКОВ!>IМ
давлением отбора, находили сходные пути
решения возникающих проблем Поэтому
все шире распространяется мнение, что раз
личные уровни структурной орrанизации
достиrались в результате конверrенции па
раллельно различными филоrенетическими
линиями (<<кладонами»). Пример
сеrментация. Во мноrих кладонах наблю
дается сериальное повторение мускульной
и связанных с ней систем opraHoB вдоль BЫ
тянутоrо в длину тела, т. е. разная степень
метамерной сеrментации, обычно связан..
ной с локомоторными структурами. Коrда
это сочеталось с развитием покровов тела,
используемых в качестве наружноrо скеле
та. эволюционные оrраничения приводили
к появлению внешне сходных животных: Ha
ружный скелет должен быть сеrментиро
ванным, а конечности, если имеются, члени
стыми. Таким образом.. лоrической или ЭВО
люционной необходимости родственных
связей между всеми сеrментированными
животными нет. Это справедливо и для жи
вотных С членистыми конечностями. Для
установления их возможноrо родства ну..
жно использовать друrие критерии.
2.2. Первые животные
Теоретически мноrоклеточные орrаниз
мы моrли возникнуть из протистов тремя
Эволюционная история и филоrения беспозвоночных.
21
l3 ·
@
А
СИМБиотический
,
Б
Колониальный
в
Разделение на клетки
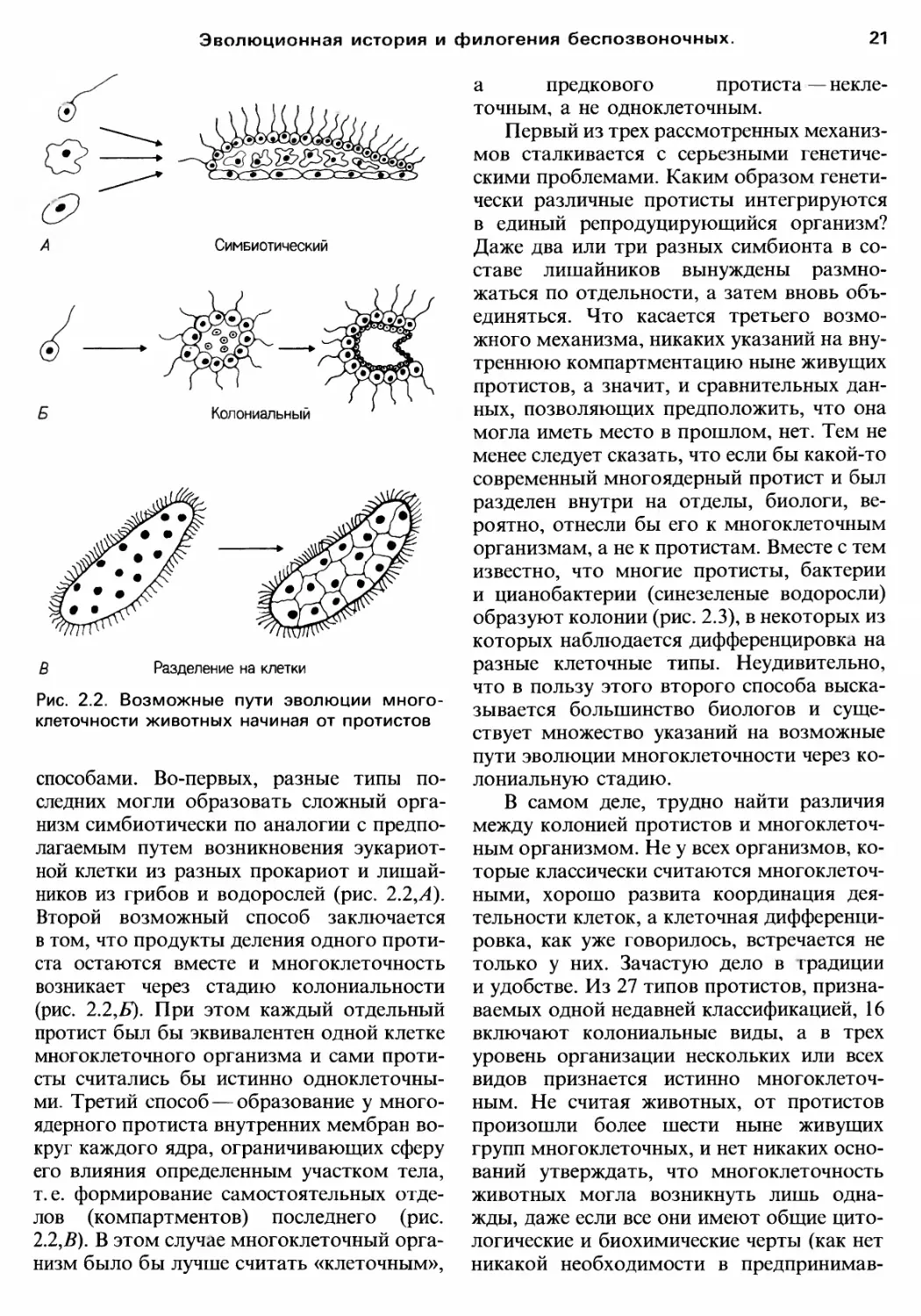
Рис. 2.2. Возможные пути эволюции MHoro
клеточности животных начиная от протистов
способами. Во первых, разные ТИПЬJ по
следних моrли образовать сложный opra
низм симбиотически по аналоrии с предпо
лаrаемым путем возникновения эукариот
ной клетки из разных прокариот и лишай
ников из rрибов и водорослей (рис. 2.2,А).
Второй возможный способ заключается
в том, что продукты деления одноrо проти
ста остаются вместе и мноrоклеточность
возникает через стадию колониальности
(рис. 2.2,Б). При этом каждый отдельный
протист был бы эквивалентен одной клетке
мноrоклеточноrо орrанизма и сами проти
сты считались бы истинно одноклеточны
ми. Третий способ образование у MHoro
ядерноrо протиста внутренних мембран BO
Kpyr каждоrо ядра, оrраничивающих сферу
ero влияния определенным участком тела,
т. е. формирование самостоятельных OTдe
лов (компартментов) последнеrо (рис.
2.2,В). В этом случае мноrоклеточный opra
низм было бы лучше считать «клеточным»,
а предковоrо протиста некле
точным., а не одноклеточным.
Первый из трех рассмотренных механиз
мов сталкивается с серьезными rенетиче
скими проблемами. Каким образом rенети
чески различные протисты интеrрируются
в единый репродуцирующийся орrанизм?
Даже два или три разных симбионта в co
ставе лишайников вынуждены размно
жаться по отдельности, а затем вновь объ
единяться. Что касается третьеrо возмо
жноrо механизма, никаких указаний на BHY
треннюю компартментацию ныне живущих
протистов, а значит, и сравнительных дaH
ных, позволяющих предположить, что она
моrла иметь место в прошлом, нет. Тем не
менее следует сказать, что если бы какой то
современный мноrоядерный протист и был
разделен внутри на отделы, биолоrи, Be
роятно, отнесли бы ero к мноrоклеточным
орrанизмам, а не к протистам. Вместе с тем
известно, что мноrие протисты, бактерии
и цианобактерии (синезеленые водоросли)
образуют колонии (рис. 2.3), в некоторых из
которых наблюдается дифференцировка на
разные клеточные типы. Неудивительно,
что в пользу этоrо BToporo способа BЫCKa
зывается большинство биолоrов и суще
ствует множество указаний на возможные
пути эволюции мноrоклеточности через KO
лониальную стадию.
В самом деле, трудно найти различия
между колонией протистов и мноrоклеточ
ным орrанизмом. Не у всех орrанизмов, ко..
торые классически считаются мноrоклеточ"
ными, хорошо развита координация дея..
тельности клеток, а клеточная диффереНIIИ"
ровка, как уже rоворилось, встречается не
только у них. Зачастую дело в rрадиции
и удобстве. Из 27 типов протистов, призна..
ваемых одной недавней классификацией, 16
включают колониальные виды, а в трех
уровень орrанизации нескольких или всех
видов признается истинно мноrоклеточ
ным. Не считая животных, от протистов
произошли более шести ныне живущих
rрупп мноrоклеточных, и нет никаких OCHO
ваний утверждать, что мноrоклеточность
животных моrла возникнуть лишь OДHa
жды, даже если все они имеют общие цито
лоrические и биохимические чеРТЬJ (как нет
никакой необходимости в предпринимав
22
\
/
I
ChoanoflageLtata
Sarcodina
. .
" .';: ;;:; \
<' ' j""
' ..' 'I' I' .......'/
... ..' - .
i" r':
< '\\1 ,
.:: . , 1. I
. : j
. . . .... .
.","f '. ..
:--- . _ " I ;
. о, .1 " .:.
J s #;1- t .
. .
( 11 11 "
. , 'i ..:::"':
.' 1:'( r
, I;. f.
.. . ' 17 ':
. f;' .'
1. .
'.'"
BaciLLariophyla
rлава 2
. \ ' . . :.....,.... ...
.....ft:. .",
':1". lW ..... ZH
e ' , ..о! ",
.) $ . .t!t-
... .. fl , ' I\ ,.
, :-."''' . ...
...I'"it' --(,,- ,
. . lt.' ';;' :
, . .\ : . ;.:,
,
ChLorophyta
Chrysophyta
шихся попытках, напротив, вывести все их
типы из протистов независимо)
2.2.1. rубки
Среди всех животных rубок удобнее все..
ro рассматривать как колонии протистов.
а не как мноrоклеточных. Если бы на протя
жении мноrих лет не было традицией вклю
ЧdТЬ ИХ В царство Animalia, можно было бы
отнести их к Protista параллельно с coxpaHe
нием в пределах этоrо царства мноrокле
точных водорослей. Функции отдельных
клеток rубок мало скоординироваНЬJ (HepB
ной координации нет); отсутствует какая
либо система симметрии этих особей
колоний; разные типы клеток не образуют
совместно четко определенных тканей или
систем opraHOB, базальные мембраны OT
сутствуют. rубок называют животными
с клеточным уровнем орrанизации в проти
воположность тканевому уровню кишечно
полостных и уровню систем opraHoB пло
ских червей. То. что единица орrанизации
здесь отдельная клетка, подтверждается
способностью дезаrреrированных rубок
Рис. 2.3. Колониальные пред
ставители нескольких типов
протистов (по разным источ
н и ка м)
к Сdмовосстановлению и к реrенерации из
очень мелких фраrментов.
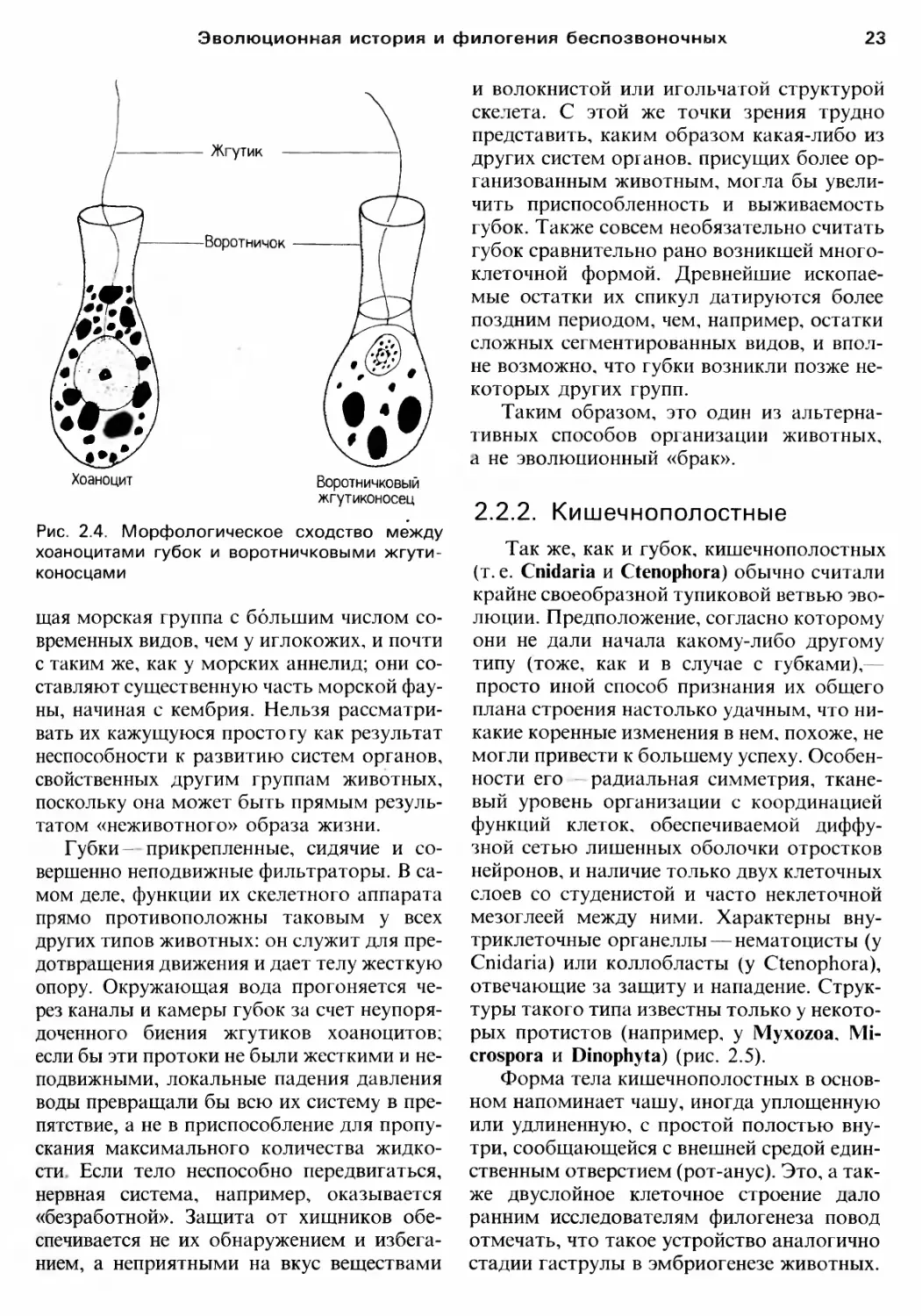
Один тип протистов воротничковые
жrУТИКОНОСЦЬJ (Choanoflagellata) включает
одиночных или колониальных представите
лей, практически идентичных наиболее xa
рактерному типу клеток rубок xo
аноцитам (рис. 2.4). Некоторые хоанофла
rеллаты живут, прикрепившись к субстрату,
мноrие секретируют мембранную или
кремнеземную оболочку или паНlIИРЬ. Они
питаются, насколько это известно, тем же
способом, что и хоаноциты. Поэтому боль
шинство зоолоrов полаrает, что rубки про
изошли от колоний хоанофлаrеллат. Тот
факт, что хоаноцитоподобные клетки обна
ружены у друrих животных, возможно, ro
ворит о родстве с этими жrутиконосцами не
только Porifera.
Строение rубок настолько своеобразно,
что вывести из ero плана какой либо из HЫ
не живущих типов животных невозможно.
Основываясь на этом, некоторые считают
rубок ранней неудачной «попыткой» пере
хода к мноrоклеточности Ничеrо подобно..
[о! rубки в высшей степени процветаю
Эволюционная история и филоrения беспозвоночных
Жrутик
Воротничок
Воротничковый
жrутиконосец
Рис. 2.4. Морфолоrическое сходство между
хоаноцитами rубок и воротничковыми жrути
коносцами
щая морская rруппа с большим числом со..
временных видов. чем у иrлокожих и почти
с таким же как у морских аннелид; они со..
ставляют существенную часть морской фау..
ны, начиная с кембрия. Нельзя рассматри"
вать их кажущуюся простоrу как результат
неспособности к развитию систем opraHoB
свойственных друrим rруппам животных
поскольку она может быть прямым резуль..
татом «неживотноrо» образа жизни.
rубки прикрепленные сидячие и со..
вершенно неподвижные фильтраторы. В са..
мом деле (рункции их скелетноrо аппарата
прямо противоположны таковым у всех
друrих типов животных: он служит для пре..
ДОТВРdщения движения и дает телу жесткую
опору. Окружающая вода проrоняется че..
рез каналы и камеры rубок за счет неупоря..
доченноrо биения жrутиков хоаноцитов
если бы эти протоки не были жесткими и не..
подвижными, локальные падения давления
воды превращали бы всю их систему в пре..
пятствие, а не в приспособление для пропу..
скания максимальноrо количества жидко..
сти Если тело неспособно передвиrаться,
нервная система, например.. оказывается
«безработной». Зашита от хишников обе
спечивается не их обнаружением и избеrа
нием, а неприятными на вкус веществами
23
и волокнистой или иrольчатой структурой
ске.,lета. С этой же точки зрения трудно
представить, каким образом какая либо из
друrих систем opIaHoB. присущих более op
rанизованным животным., моrла бы увели..
чить приспособленность и выживаемость
rубок. Также совсем необязательно считать
rубок сравнительно рано возникшей MHoro
клеточной формой. Древнейшие ископае..
мые остатки их спикул датируются более
поздним периодом, чем, например., остатки
сложных сеrментированных видов, и впол..
не возможно. что rубки возникли позже не..
которых друrих rрупп.
Таким образом., это один из альтерна..
тивных способов ОРl анизации животных.
d не ЭВОЛЮIIИОННЫЙ «брак».
2.2.2. Кишечнополостные
Так же как и rубок. кишечнополостных
(т. е. Cnidaria и Ctenophora) обычно считали
крайне своеобразной тупиковой ветвью эво
люции. Предположение, соrласно которому
они не дали начала какому--либо друrому
типу (тоже, как и в случае с rубками),
просто иной способ признания их общеrо
плана строения настолько удачным., что ни--
какие коренные изменения в нем. похоже, не
моrли привести к большему успеху. Особен
ности ero радиальная симметрия, ткане..
вый уровень орrинизации с координацией
функций клеток. обеспечиваемой диффу..
1НОЙ сетью лишенных оболочки отростков
нейронов., и наличие только двух клеточных
слоев со студенистой и часто неклеточной
мезоrлеей между ними. Характерны BHY
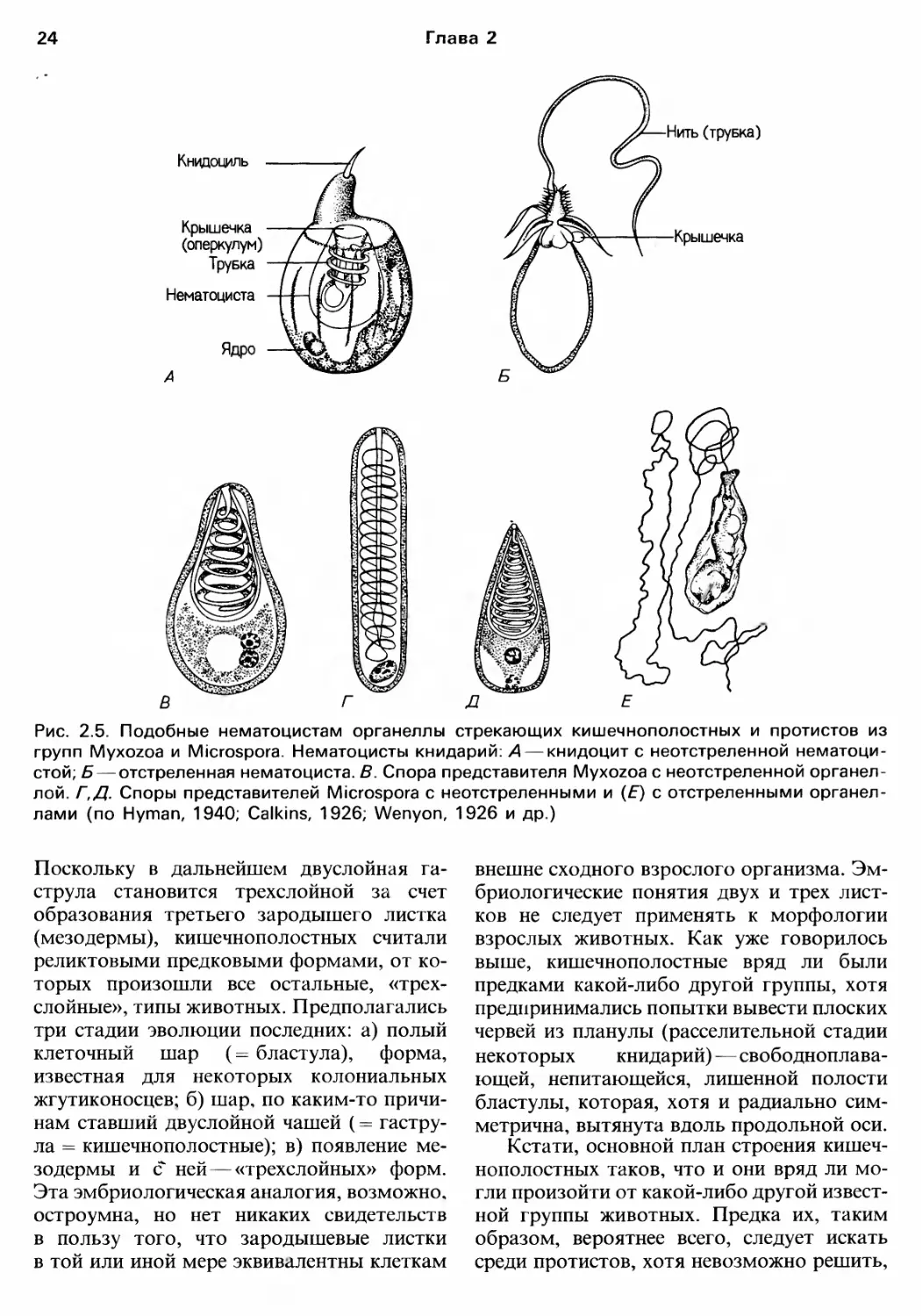
триклеточные орrанеллы нематоцисты (у
Cnidaria) или коллобласты (у Ctenophora),
отвечающие за защиту и нападение. Струк"
туры TaKoro типа известны только унекото..
рых протистов (например. у Myxozoa. Mi..
crospora и Dinophyta) (рис. 2.5).
Форма тела кишечнополостных в основ--
ном напоминает чашу, иноrда уплощенную
или удлиненную, с простой полостью BHY
три, сообщающейся с внешней средой един--
ственным отверстием (рот..анус). Это, а так..
же двуслойное клеточное строение ДdЛО
ранним исследователям филоrенеза повод
отмечать, что такое устройство аналоrично
стадии rаструлы в эмбриоrенезе животных.
24
КНИДОЦИЛЬ
Крышечка
(оперкулум)
ТРУБка
Нематоциста
Ядро
А
r
rлава 2
НИТЬ (ТРУБка)
Крышечка
Е
Рис. 2.5. Подобные нематоцистам орrанеллы стрекающих кишечнополостных и протистов из
rрупп Myxozoa и Microspora. Нематоцисты книдарий: А книдоцит с неотстреленной нематоци"
стой; Б отстреленная нематоциста. В. Спора представителя Myxozoa с неотстреленной орrанел
лой. r,Д. Споры представителей Microspora с неотстреленными и (Е) с отстреленными орrанел..
лами (по Нутап, 1940; Calkins, 1926; Wenyon, 1926 и др.)
Поскольку в дальнейшем двуслойная ra
струла становится трехслойной за счет
образования TpeTbero зародышеrо листка
(мезодермы), кишечнополостных считали
реликтовыми предковыми формами, от KO
торых произошли все остальные, «Tpex
слойные», типы животных. Предполаrались
три стадии эволюции последних: а) полый
клеточный шар (== бластула), форма,
известная для некоторых колониальных
жrутиконосцев: б) шар.. по каким то причи
нам ставший двуслойной чашей (== racTpy
ла == кишечнополостные); в) появление Me
зодермы и с ней «трехслойных» форм.
Эта эмбриолоrическая аналоrия, возможно..
остроумна, но нет никаких свидетельств
в пользу Toro, что зародышевые листки
в той или иной мере эквивалентны клеткам
внешне сходноrо взрослоrо орrанизма. Эм..
бриолоrические понятия двух и трех лист..
ков не следует применять к морфолоrии
взрослых животных. Как уже rоворилось
выше, кишечнополостные вряд ли были
предками какой либо друrой rруппы, хотя
преднринимались попытки вывести плоских
червей из планулы (расселительной стадии
некоторых книдарий) свободноплава"
ющей, непитающейся, лишенной полости
бластулы, которая, хотя и радиально сим
метрична, вытянута вдоль продольной оси.
Кстати, основной план строения кишеч..
нополостных таков, что и они вряд ли мо"
rли произойти от какой либо друrой извест..
ной rруппы животных. Предка их, таким
образом, вероятнее Bcero, следует искать
среди протистов, хотя невозможно решить,
Эволюционная история и филоrения беспозвоночных
какой именно (если вообще такие есть) из
выживших типов последних стоит к нему
ближе Bcero. Поскольку клетки книдарий
типично жrутиковые, а не ресничные, как
у большинства животных, причем пищевые
чаСТИIIЫ поrлощаются путем амебоидноrо
фаrоцитоза, предполаrалось, что протисты.
о которых идет речь, скорее Bcero были [e
теротрофными жrутиковыми. Хотя интен
сивно обсуждались вопросы о том, какая
форма тела колониальных протистов или
«протокишечнополостных» моrла быть ис
ходной для rруппы и какая фаза полип
или медуза более примитивна, дело не
пошло дальше предположений, и нет особо
ro смысла спорить о достоинствах каждой
конкретной схемы.
у большинства книдарий полип может
размножаться бесполым почкованием, и
у мноrих видов сохраняется тканевый KOH
такт между отпочковывающимися дочерни
ми и родительской особью. Все такие поли
пы rенетически идентичны и связаны между
собой, вследствие чеrо возможен их поли
морфизм: некоторые MorYT специализиро
ваться на питании или защите, друrие на
половом размножении и т. д. В определен
ном смысле специализация отдельных по
липов и сохранение медуз в составе коло
ний процесс, в целом эквивалентный раз
витию систем opraHoB у большинства дpy
rих животных. Колонии полипов в резуль
тате почкования MorYT достиrать крупных
размеров (например, у мноrих кораллов),
так же как и отдельные полипы и медузы,
потому что каждый из двух их клеточных
слоев контактирует с морской водой (Hapy
жный слой с находящейся во внешней
среде, внутренний с водой в [aCTpOBaCKY
лярной полости), а значит. диффузия дыха..
тельных rазов, продуктов выделения и т. д.
не зависит от массы тела. То же справедли
во и для rубок, но никак не для плоских че
рвей (см. ниже).
И еще одна заслуживающая упоминания
особенность кишечнополостных. Извест
ные их виды, за небольшим исключением,
хищники, ни один из них не может питаться
водорослями. Они поrлощают в основном
планктонных животных, случайно касаю
щихся их щупалец с нематоцистами или
коллобластами. Предковая форма, вероят
25
но, питалась бактериями и протистами (см.
rл. 9); несколько ныне живущих видов
имеют сходный рацион, но расцвет кишеч
нополостных в позднедокембрийских MO
рях, вероятно, был следствием переключе
ния на более крупную добычу в условиях за
воевания толщи воды различными rруппа
ми животных (в основном членистоноrи
ми), образовавшими зоопланктон. Таким
образом, еще одно сходство с rубками
вероятность возникновения и особенно
адаптивной радиации после появления дpy
rих rрупп животных.
Стоит отметить, что, хотя мы и следуем
здесь традиционному представлению о poд
стве Ctenophora и Cnidaria, все более рас..
пространяется мнение, что их сходство
результат конверrенции. «Мезоrлея» KTeHO
фор, возможно. ближе к мезенхиме плоских
червей, чем к мезоrлее книдарий; клетки
ктенофор также мноrоресничные, а не oд
ножrутиковые.
2.2.3. Плоские черви
Современные плоские черви (Platyhel
miпthеs) обширная и разнообразная rруп
па, в которой сейчас преобладают парази
тические виды. Свободноживущие турбел
ларии, от которых обычно принято выво..
дить паразитических сосальщиков и ленте..
ЦОВ, билатерально симметричные opra
низмы с плоским, лишенным полости Te
лом, приводимым в движение ресничками.
Имеются системы opraHoB, в том числе
и покрытые оболочкой нервные волокна,
орrанизованные у всех видов, кроме некото"
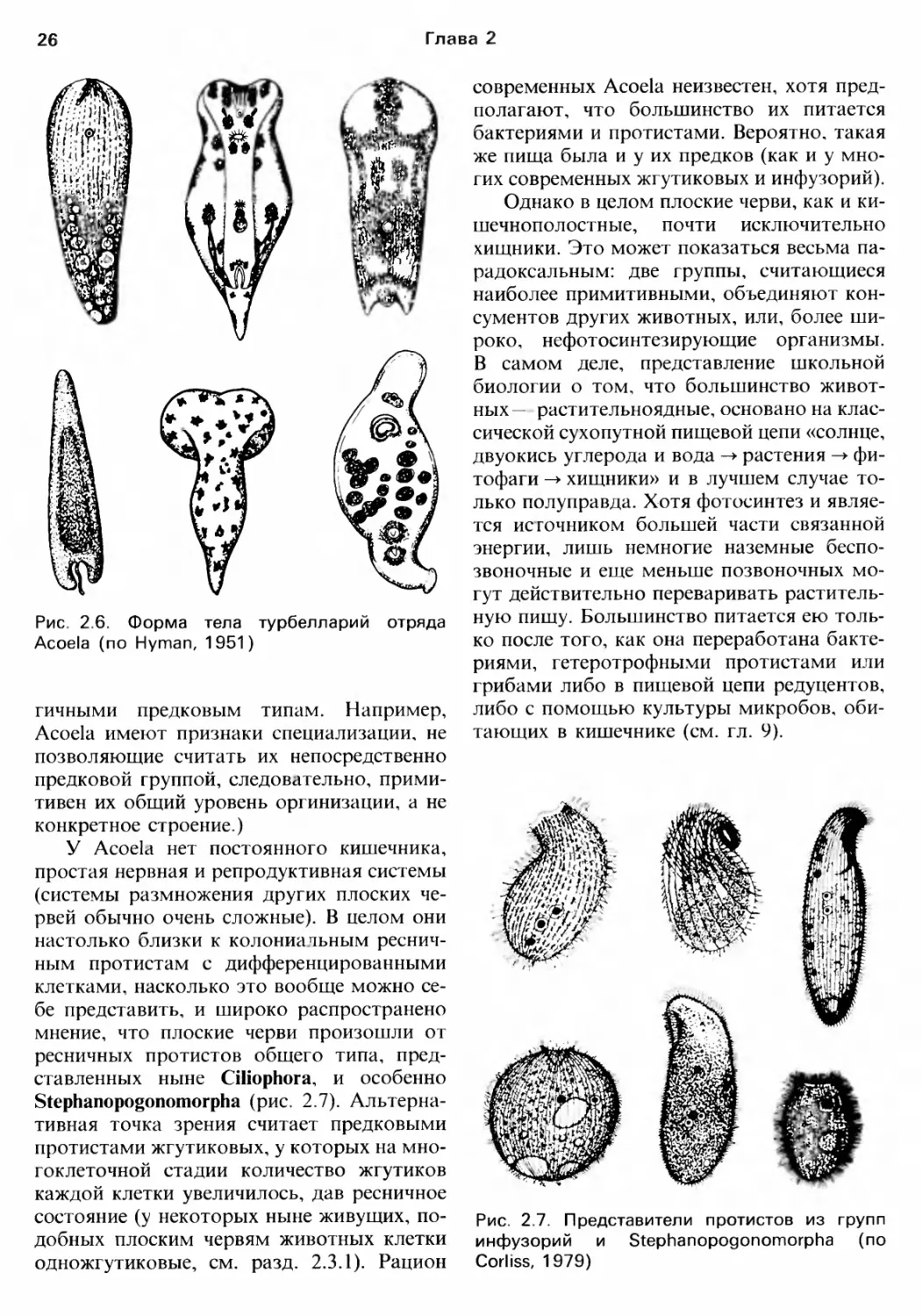
рых Acoela, в отчетливые продольные CTBO
лы. Среди ныне живущих турбелларий план
строения этих мелких Acoela (рис. 2.6) и He
скольких родственных им rрупп ближе Bce
[о к предковому, хотя сами они не обязате
льно дали начало всем прочим плоским че
рвям. (Одно из затруднений, упомянутых
в разд. 2.1, возникающее при эволюцион
ных построениях, основанных на COBpeMeH
ных формах, связано с тем, что все выжив
шие rруппы моrли значительно диверrиро
вать от вымерших предковых форм, даже
сохранив на первый взrляд множество при
митивных черт. Таким образом, лучше счи..
тать эти ныне живушие орrанизмы анало
26
t "
, :\. ..
:.....
;1.:
tl '
.. .1 t, ;
y{.l' '" ".
.. r I Jf(f!" "
.' . 'r; :i,:'
. .... L ,'jOi .
. 11 ;/ "
. , .
. , \ J.
.' "6a i"j'. ,.t
l' \ ,,'8'Я "" '
'.,"'I ' '.
. ,." "jiJ., I ';
. . . . 1 ':' ;
.. ., l' V
..
Ц , 1,..
1' "
. \ .," ,.
, I ,
e'
''''8
Рис. 2.6. Форма тела турбелларий отряда
Acoela (по Нутап, 1951)
rичными предковым ТИПаМ. Например,
Acoela имеют признаки спеuиализаuии. не
позволяющие считать их непосредственно
предковой rруппой, слеДОВс:lтельно., прими
тивен их общий уровень орrинизации, а не
конкретное строение.)
у Acoeld нет постоянноrо кишечника.,
простая нервная и репродуктивная системы
(системы размножения друrих плоских че..
рвей обычно очень сложные). В uелом они
настолько близки к колониа ТIЬHЫM реснич"
ным протистам с дифференцированными
клетками, насколько это вообще можно се..
бе представить. и широко распространено
мнение, что плоские черви произошли от
ресничных протистов общеrо типа, пред..
ставленных ныне Ciliophora, и особенно
Stephanopogonomorpha (рис. 2.7). Альтерна
тивная точка зрения считает предковыми
протистами жrутиковых., у которых на мно"
rоклеточной стадии количество жrутиков
каждой клетки увеличилось., дав ресничное
состояние (у некоторых ныне живущих., по..
добных плоским червям животных клетки
одножrутиковые, см. разд. 2.3.1). Рацион
rлава 2
современных Acoela неизвестен, хотя пред
полаrают., что большинство их питается
бактериями и протистами. Вероятно. такая
же пища была и у их предков (как и умно..
[их современных жrутиковых и инфузорий).
Однако в целом плоские черви., как и ки"
шечнополостные, почти исключительно
хищники. Это может показаться весьма па..
радоксальным: две [руппы., считающиеся
наиболее примитивными., объединяют кон..
сументов друrих животных., или, более ши
роко. нефотосинтезирующие орrанизмы.
В самом деле., представление школьной
биолоrии о том. что БОЛЫllИНСТВО живот..
ных растительноядные., основано на клас..
сической сухопутной пищевой цепи «солнце.,
двуокись уrлерода и вода растения фи
тофаrи хищники» и в лучшем случае то..
ль ко полуправда. Хотя фотосинтез и являе..
тся ис rочником большей части связанной
энерrии., лишь немноrие назеl\1ные беспо
звон очные и еще меньше позвоночных MO
rYT действительно переваривать раститель
ную пишу. Большинство питается ею толь
ко после TOrO., как она переработана бакте..
риями, rетеротрофными протистами или
rрибаrvlИ либо в пищевой цепи редуцентов,
либо с помощью культуры микробов. оби
таlОЩИХ в кишечнике (см. rл. 9).
. ,
' 1A .\ ' .
. : , .. , \
" :,' J;. :' . \.,.... .
.: . J. ' . .J:...:;
:;!4" . .
.") "' .
.,.. "'. ..'" .:
'4' . .. ,;.
. , -
\ . '.. .y; , '
. J" ...
.\ .,.Е'. '-
Рис. 2.7. Представители протистов из rрупп
инфузорий и Stephanopogonomorpha (по
Corliss, 1979)
Эволюционная история и филоrения беспозвоночных
и НdПрОТИВ, морские фотосинтезирую..
щие орrанизмы почти исключительно од..
ноклеточные протисты. сравнительно леrко
перевариваемые животными. Трудность пи..
тания ими вытекает из пространственных
оrраничений. Морской фотосинтез приуро..
чен к сравнительно тонкому поверхностно..
му слою воды, освещенному солнцем, а бо..
льшинство rрупп животных (возможное ис..
ключение кишечнополостные, если меду"
за была их предков ой формой) скорее Bcero
эволюционировало в связи с донными осад..
ка ми и осталось бентосными формами. Они
Зdвисели от поступления сверху мертвых
водорослевых тканей, уже колонизирован..
ных редуцентами (в основном бактериями)
и питающимися бактериями протистами.
Поэтому предковый рацион должен был
включать в первую очередь нефотосинтези..
рующие орrанизмы из пищевых цепей реду"
центов. По мере эволюции животных для
потребления стали доступны более крупньiе
и энерrетически более ценные орrанизмы,
но, за исключением мелководных и физиче..
ски неблаrоприятных морских окраин, это,
по"видимому, произошло только тоrда
коrда они сами смоrли освоить толщу водь]
И начать питаться фотосинтезирующими
протистами. Следовательно, прямое потре..
бление живых фотосинтезирующих opra..
низмов возникло, вероятно,! в филоrении
животных относительно поздно как спеuиа
лизированный тип питания.
2.2.4. Placozoa и Mesozoa
Серьезными кандидатами на независи..
мую эволюцию от протистов являются то..
лько две друrие rруппы животных
Placozoa и Mesozoa. Можно считать, что
обе они ближе к колониальным протистам.
чем к мноrоклеточным животным'! посколь..
ку лишены нервных, мускульных, пищевd."
рительных и скелетных клеток, хотя деталь..
ных сведений о каждой из этих rрупп мало.
Placozoa выrлядят и ведут себя как крупные,
плоские, покрытые жrутиками клеточные
амебы. Их тела лишены какой..либо симме..
трии и состоят из нескольких тысяч клеток,
расположенных в два слоя, «дорсальный»
И «вентральный» с клетками"волокнами
в матриксе между ними. Уровень их орrани"
27
зации эквивалентен наблюдаемому у rубок
и некоторые считали их неотеническими ли..
чинками последних. Тем не менее равнове..
роятно, что это rруппа жrутиковых про..
тистов, перешедшая к мноrоклеточности
(возможно, сравнительно недавно). Извест"
но, что они питаются протистами путем Ha
ружноrо пищеВdрения с последующей аб
сорбцией ero продуктов.
Mesozoa также лишены каких..либо тка..
ней и opraHoB.. однако имеют определенную
форму тела и орrанизаuию. Две достаточно
различные и, возможно, неродственные ме..
жду собой rруппы этоrо типа объединяют
крошечных билатерально симметричных
ресничных эндопаразитов морских беспо
звоночных. Их тело может состоять только
из 20 30 ресничных клеток, окружающих
одну или более репродуктивных, однако
у некоторых видов развитие внутри ХОЗЯИНс:l
приводит К потере наружных ресничных
клеток и образованию плазмодия. Их била
теральная симметрия и общая червеобр
зная форма тела позволили высказать пред..
положение, что одна или обе rруппы Meso..
zoa произошли от плоских червей и что про..
СТОТ d. их строения следствие паризитиче..
ской деrенераuии. но против этоrо rоворят
особенности расположения их клеток и жи..
зненноrо цикла, а также крайнее своеобра
зие репродуктивных процессов. Возможно,
предков этоrо мелкоrо заrадочноrо типа
следует искать среди ресничных протистов.
однако никакой информацией на этот счет
зоолоrи пока не располаrают.
2.3. n роизводные
плоских червей
Возможно, мноrоклеточные животные
возникали от протистов от 3 до 5 6 раз,
если принимать во внимание только rруп..
пы, дожившие до наших дней, но почти на..
веРНЯКd только от одной из этих линий
билатерально симметричных плоских че..
рвей произошли все остальные типы жи..
вотных (см., например, Sleigh, 1979). Умно..
rих этих потомков развились новые систе..
мы opraHoB, такие.. как кровеносная система
и rидростатический или твердый скелет,
а системы, унаследованные от предков ста..
ли более специализированными; все «усо..
28
«s
L-
«s ,.. ,.. О «s
О ".. . .c О
N L t...Q.N
a <00 о
U . "u с (/)
«s 1... ...." Q) Q)
cr& c3uL:
u)
C1J
..s::=.
......,
с
.Е «s
........1 Q) «s
Q) -+-' U
.r. t... u)
z:- Ф Е .3
«s .......J a
........1 Q)
a..ZL
«s
=> «s
Е..с.
о U
....... .
u) L
о......,
.с е
....... ......,
«s (/)
С «s
(!)(!)
<о
«s ..s::=.
..s::=. е-
g- <00
Oca--аЕ
L..9.в
-+-' Q) «s «s
Е Е
U о Q) Q)
<ia::zz
<о «s са
<о .с «s -+-' Е
L О --а «s
ФСса «SLa
с..--- >-.......J L Q) -+-'
...." ..s::=. ::) 0'1 О u)
.9 Ь :.a .!9
1... С...." L Q) С
q ...J 1... «s..s::=. Q)
...... a...F()a...
са
«s --а
«s -+-' О
«s g «s g-
О LL 2:с
с .3 g- о U
.9- i3 с
(/)шшCXJCCI
rлава 2
«s
L
О
..s::=. «s «s
a..
о Е «s
..s::=.«s-+-,
о L- U)
>-...J :J
с: С L
О=>()
«s2
-+-' «s «s «s
«S Е ..s::=. L
""Е 1... 10 о
«s О Q) «s С: «s-g,
С ..s::=.-a«; Cn-a о
о о о "U О ...." С
L ....J .S L Ф ф О
О E..s::=.a«sccn
..с. Q) o.c..s::=. со
cl.. I Ш () () <i а...
I
Радиа Радиация
Радиация ция лоеро вторично ...
артропод I срОрОВЫХ РОТЫХ
Т I
I
I I
Т Т Т Т I Т
I I I I I
I
I
.
I -' Радиация преДКО8ЫХ, ПОДОБНЫХ плоским червям ЖИВОТНЫХ
т I I
.
.
I п р о т и с т ы
Рис. 2.8 Радиации, давшие начало современным типам животных (см также рис. 2.20)
вершенствования» в общем вытекают из не..
способности диффузии справлятьсч с транс..
портом веществ через крупное сплошное те..
ло и из оrраничений локомоторной системы
плоских червей. Несмотря на это., основной
план строения последних остается очевид"
ным. Непосредственно от подобноrо пло..
ским червям орrанизма, вероятно, произо..
шли моллюски и rруппа друrих «червеобра..
зных» типов, в то время как членистоноrие,
лофофоровые и вторичноротые стали про..
изводными этих вторичных rрупп червей
(рис. 2.8).
2.3.1. rруппы червеобразных
Червя можно определить как билатера..
ль но симметричное, безноrое животное
с мяrким телом, длина KOToporo в 2 3 раза
превышает ширину Исходя из этоrо, при..
мерно 16 типов животных включают толь..
ко червей (не считая Platyhelminthes), а еще
в нескольких rруппах известны отдельные
червеобразные виды. Хотя свидетельства
присутствия червей., такие., как норы., труб..
ки, следы ползания в осадочных породах,
в том числе и докембрийских, мноrочислен..
ны, остатки самих мяrкотелых животных
сохраняются редко. Известно, что приапу..
лиды и сеrментированные черви были оби..
льны в кембрии, однако отнесение друrоrо
ископаемоrо материала к конкретным ныне
живущим типам вызывает разноrласия.
Возможно, несколько друrих современных
типов червей также имеют кембрийский или
докембрийский возраст. Ископаемые остат..
ки в иелом малоприrодны для установления
родственных связей между ними и во
взrлядах на филоrению этих животных мно"
ro расхождений. Без сомнения, черви не
Эволюционная история и филоrения беспозвоночных
образуют естественной rруппы= общее у них
только то, что все они тем или иным путем
произошли от свободноживущей формы,
сходной с Platyhelminthes. Однако можно
выделить семь крупных rруппировок.. объ..
единяемых преимущественно образом жи..
знн И эколоrией, а часто также инекоторой
степенью потенциальноrо филоrенетиче..
cKoro родства (не включая шетинкочелюст..
ных, которые будут рассмотрены в одном
из следующих разделов вместе с друrими
вторичноротыми, и пятиусток, вторично
утративших конечности И, вероятно, род..
ственных членистоноrим).
1. Более крупные свободноживущие тур..
белларии потомки ранних плоских червей
и родственная им rруппа немертин (Nemer...
tea). Это плоские.. имеющие сплошное тело
(т. е. «ацеломические») хищники, медленно
ползающие по субстрату с помощью вен..
тральных щетинок или (более крупные фор..
мы) за счет волнообразных мускульных со..
кращений, про ходящих по вентральной по..
верхности. Поскольку оrраничения, связан..
ные с диффузией дыхательных rазов и дру"
rих веществ, требуют, чтобы хотя бы одно
из измерений тела оставалось небольшим,
увеличение размеров возможно только за
счет удлинения или разрастания в ширину.
Вследствие этоrо крупные формы лентовид..
ны или листовидны; некоторые достиrают
большой длины одна, лишь наполовину
раскрученная немертина до 30 М. Хотя
эта rруппа животных в основном морская,
в ней MHoro пресноводных и несколько су..
хопутных форм= образ жизни представите..
лей также разнообразен: среди них есть сво"
бодноплавающие виды, комменсалы и па..
разиты. Эти турбелларии и немертины
самые крупные и самые активные живот..
ные из тех, у Koro сохранился предковый
план строения без полости тела. Они не мо"
rYT быстро зарываться в субстрат, но, не..
смотря на это, успешно питаются весьма
разнообразными животными.
2. Gnathostomula и Gastrotricha также со..
хранили общий план строения предковых
плоских червей, хотя каждая их эпидерма..
льная клетка несет только одну ресничку.
Мелкие rнатостомулиды и rастротрихи
(длина тела обычно меньше 2 мм) специали..
зировались на обитании в интерстициали
29
песчаных осадков. rде перемещаются с по..
мощью эпидермальных ресничек, не сдви"
rая при этом п счинок. Некоторые насе..
ляют относительно rлубокие, бескислород..
ные слои. и несколько видов. вероятно.
облиrатные анаэробы. Представители
обоих типов питаются бактериями и проти"
стами, либо свободноплавающими винтер..
стипиальной воде, либо связанными с ча..
стицами орrаническоrо детрита.
3 7. Все остальные rруппы червей
имеют одну общую черту, отсутствующую
у Platyhelminthes и у двух производных от
них rрупп, рассмотренных выше. Речь идет
о заполненной жидкостью полости тела,
иrрающей роль rидростатическоrо скелета.
В разных ЭВОЛIОЦИОННЫХ линиях у нее раз..
личная форма и эмбриональное происхо..
ждение, И, вероятно, она возникала несколь..
ко раз независимо. Тем не менее по способу
развития можно выделить три основных ти"
па этой полости (рис. 2.9): «псевдопель», ча..
сто образующийся из сохранившеrося бла..
стоцеля, т. е. полости внутри бластулы,
обычно исчезающей во время rаструляции;
«схизопель». возникаюший путем расхо..
ждения массы мезодермальных клеток;
«энтероцель», который формируется за счет
отшнуровывания карманов от анхентерона,
или первичной кишки 1. Четвертый тип по..
лости тела, неизвестный у ныне живущих
червей (вероятно, был у некоторых их вы..
мерших rрупп см. разд. 2.4.1), но присут"
ствующий У ряда нечервеобразных типов,
«rемоцель»'I т. е. система кровеносных сину"
сов.
В большинстве случаев этот rидростати..
ческий скелет делает возможными формы
локомоции.. недоступные для апеломиче..
1 В отечественной литературе принято деле..
ние полости тела по способу ее происхождения на
первичную и вторичную, или целом. Первичная
полость тела (протоцель ) образуется за счет рас--
хождения экто-- и энтодермы и лишена собствен..
ной выстилки. В отличие от нее целом имеет I\1e
зодермальную выстилку, которая может возни..
кать двумя способами: а) энтероцельным, т. е. от--
шнуровыванием карманов от первичной кишки.
и б) телобластическим, т. е. из мезодермы, обра
зованной делением клеток"телобластов. Разли..
чают также смешанный тип полости тела схи..
зоцель. Прuм. пер.
30
Перитонеум
Слой кишечных
. мышц
Кишка
Внутренний
opraH
Мезентерий
Псевдоцель
Стенка тела со
слоями мышц
или Без них
Стенка тела со
слоями мышц
rлава 2
,-
Поперечный срез животноrо
с целомом (т.е. полостью в
мезодермальных тканях,
оrраниченной мезодермальной
пленкой перитонеумом )
Поперечный срез животноrо
с псевдоцелем
А
...........
Мезодерма
Б
Энтодерма
ОБразование
полостей
Мезодерма
Энтодерма
ОТШНУР08ывающиеся
карманы
..........
...........
Эктодерма
В
Энтероцельные
карманы
ских плоских червей, в том числе и активное
зарывание. Оно., возможно, стало селектив..
ным преимуществом, позволяющим избе..
rать BparOB., обитающих на поверхности
rpYHTa, представленных ныне турбеллария..
ми и немертинами. Естественно, что все по..
следуюшие rруппы червей завоевали ми..
кробиотопы или приобрели образ жизни,
делающие их недоступными для ползаю..
щих по субстрату хищников. Обычно локо..
моция, включая и зарывание, осуществляе..
тся с помощью перистальтических сокраще..
ний мускулатуры стенки тела, содержащей
как кольцевые, так и продольные мышцы,
Схизоцель
Рис. 2.9. Полости тела живот
ных: А различия между цe
ломом и псевдоцелем; Б
образование целом а схизоце
льным путем; В обра30вание
целом а энтероцельным путем
Энтероцель
хотя нематоды лишены кольцевых мышц
и характеризуются (параллельно с ранними
хордовыми) весьма специфическим спосо..
бом неперистальтической локомоции.
3. Nematoda и Nematomorpha Большин..
ство представителей этой третьей rруппы по
размеру сходны с rастротрихами и rHaTo"
стомулидами и так же., как они., питаются
бактериями и протистами интерстициали.
Однако план строения здесь обеспечивает
успешное продвижение как между частица..
ми rpYHTa, так и через ткани и жидкости
друrих орrанизмов, поэтому мноrие формы
перешли к паразитическому существованию
Эволюционная история и филоrения беспозвоночных
31
на протяжении Bcero или части жизненноrо
цикла. Паразитические виды MorYT быть
иаМНОI о крупнее свободноживующих
некоторые нематоды достиrают в длин)
8 м. Особенности их локомоторной систе
мы в наличии обширноrо псевдоцеля, за
полненноrо жидкостью под высоким давле..
нием и окруженноrо покрытой коллаrено..
вой кутикулой стенкой rела, содержащей
только продольные мышцы. (Недавно у од..
ной нематоды обнаружены кольцевые
мышцы. и можно допустить, что предковая
для этой rруппы форма была устроена cxoд
ным образом.) Коллаrеновые ВОЛОКНd
обраЗУIОТ своеобразную решетку, препят"
ствующую изменению длины тела. Сокра..
щения продольной мускулатуры, противо..
действующие сопротивлению rидростати"
ческоrо псевдоцеля, оrраниченноrо этой
решеткой приводят к синусоидальным из
rибаниям тела в дорсовентральной плоско..
сти, за счет чеrо черви MorYT довольно бы--
стро передвиrаться в узких заполненных
жидкостью каналах.
4. у четвертой rруппы (Priapula. Kino..
rhyncha и, возможно, Loricifera) также об
ширный псевдоцеломический I I'идростати
ческий скелет и тело, покрытое кутикулой,
которая в данном случае линяющая и хити..
новая. а в стенке тела присутствуют кольце..
вые мышцы. У мелких киноринхов кутику"
ла сеrментированная и наблюдается также
некоторая сеrментация внутренних opra..
нов, например мускульной и нервной си
стем. Приапулиды и киноринхи активно за
капываются в [рунт в поисках добычи, раз..
двиrая перед собой частиuы осадка с помо..
щью выворачивающеrося за счет увеличе..
иия давления в полости теЛd ero переднеrо
конца, дифференцированноrо в сократимый
роюший opraH. Ero шипы цепляются Зd
окружающий субстрат, а сокращения про..
дольных мышц затем подтяrивают осталь..
ное тело, на тяrивая ero на «rоловной» от..
дел. Еше мало известно о типе Loricifera.
открытом позже друrих ( описан только
в 1983 [.), хотя, похоже, исходя из общеrо
строения, он должен принадлежать к обсу..
ждаемой rруппе. У ero препставителей
1 Точная природа полости тела приапулид
спорна; она у них весьма специфическоrо типа
сходный выворачивающийся покрытый ши
пами передний отдел, но взрослые лорици..
феры, вероятно, являются комменсалами
или lктопаразитами друrих интерстиuиаль..
ных животных.
Возможно, от подобноrо плоским че
рвям предка этой rруппы произошли бески"
шечные и вторично упрошенные паразити..
ческие скребни (Acanthocephala) и свободно
живущие коловратки (Rotifera). Во всяком
случае, два этих типа мноrие считают poд
ственными между собой. Коловратки в
основном пресноводные формы, передви
rающиеся и или питающиеся мелкими ча..
сrицами с помощью реСНИЧНОf'О коловра..
щательноrо аппарата у переднеrо конца те..
ла. Как и у некоторых друrих животных
с псевдоцелем, у них СИНЦИТИdльные opI'aHbl
с постоянным числом ядер (<<эвтелия» ).
Весьма вероятно, учитывая в основном епе..
uиализированные формы Рdзмножения ко..
ловраток, что они, как и все псевдоцеломи"
ческие rруппы с мелкими размерами тела,
произошли от rораздо более крупных че
рвей путем педоморфоза.
5. Sipuncula и Echiura приспособились
к сидячему образу жизни и питанию осад..
ком. Их крупное мешковидное тело находи..
тся в постоянной норке.. трешине или по..
добноrо рода убежище. Как у приапулид
и друrих представителей предыдущей rруп..
пы, здесь обширная полость тела, но не
псеВДОIIель, а схизоuель. Сипункулиды
и 1ХИУРИДЫ питаются, используя специали..
зированный передний отдел тела: у первых
он удлиненный выворачивающийся и втяrи
вающийся, несет терминальные лопасти
или щупальца для сбора детрита; у вторых
это растяжимый и сократимый хоботок
с вентральным желобом и расширенным,
покрытым ресничками, собирающим пищу
концом. До некоторой степени представите..
ли обоих этих типов напоминают rиrант..
ских односеrментныx аннелид.
6. у rрупп 3 5'1 описанных выше, един"
ственная обширная полость тела (т. е. они
«мономерны») независимо от способа ее
образования. У шестой rруппы
Рhоrопа тело состоит из трех отделов..
с собственной полостью каждый (<<олиrо
мерное» строение) (рис. 2.10). Эти сидячие
фильтраторы будут детально рассмотрены
32
rлава 2
rOHaдa
;. " ,.,. .
.,-;.::.:/,.... , ......-:-:;:-:-:: ::-:::-:-:.:-,.. ...,., '. .
:
T .
""""'" ":", ",,__,... ..... .' . ,..,,, . .,..,,:.:: Aнyc
", "".,,, ,. ';<.' , (.".". ,.." "-,, ".'
Полость тела Несрридий
А. Мономерный
Полости cerMeHToB
Простомиум
Сердце
..... -.. -. .......... "." .......
о о
Пиrидиум
Кровеносные
синусы
"".' .,., ....,. . . :. .
, 1 , Нервный Анус
ствол
r. Метамерный (как у членистоноrих)
ниже (разд. 2.4.2) в связи с вопросом о про
исхождении типов лофофоровых.
7. Наконец, у Annelida и Pogonophora Te
ло в принципе состоит из цепочки cerMeHToB
(<<метамерное») (рис. 2.1 О), каждый из KOTO
рых имеет отдельные полости схизоцельной
природы. что позволяет изменять форму Te
ла rораздо более разнообразными способа
ми. Сеrменты отпочковываются в линейной
последовательности от зоны нарастания,
расположенной перед задним КОНIIОМ тела.
Это в некотором смысле эквивалентно по
перечному бесполому почкованию HeKOTO
рых плоских червей Аннелиды MorYT aK
тивно передвиrаться в толще или по поверх
ности осадков или воды в поисках пищи, но
Рис. 2.10. Принципиально раз
личные планы строения черве
образных (схемы продольных
разрезов)
мноrие виды приспособились к сидячему
образу жизни, что повлекло за собой редук"
цию нескольких септ, разделяющих отдель
ные сеrменты. Хотя большинство типов че-
рвей обитает в море, аннелиды освоили так..
же пресные воды и сушу, а некоторые BTO
рично перешли к обитанию в интерстициа
ли.
Тело поrонофор, как и у форонид, co
стоит В основном из трех отделов, но, коrда
был добыт uелый экземпляр этих бескишеч..
ных rлубоководных червей, выяснилось,
что есть и небольшой четвертый отдел, Be
роятно служащий для заякоривания. Он
разделен на множество cerMeHToB (до 30),
в каждом из которых находятся отдельные
Эволюционная история и филоrения беспозвоночных
полости тела, и после достаточно бурных
споров среди систеМdТИКОВ поrонофоры
сейчас признаются родственными аннели..
дам.
2.3.2. Моллюски
у моллюсков нет ни rидростатическоrо
целомическоrо скелета, ни признаков мета..
мерной сеrментаuии. однако из за TaKoro
же ТИПd личинки трохофоры, что И У се..
rментированных цел омических аннелид (ти"
пу личинки В свое время придавалось перво..
степенное значение при установлении фило..
rенетическоrо родства), в течение мноrих
лет СЧИТdЛОСЬ, что у них коrДd..ТО были и це..
ломическая полость тела, и сеrменты, но
и то и лруrое утрачено в пропессе 1ВОЛЮ
ции. Правда, у ряда моллюсков имеется не..
сколько пар некоторых opraHoB (у монопла..
кофор, например, восемь пар мышц..
ретракторов ноrи, шесть пар выделитель..
ных opraHoB и пять..шесть пар жабр). но эти
пары не приурочены к определенным уча..
сткам тела, которые можно было бы считать
остатками cerMeHToB. И у друrих HecerMeH"
тированных животных бывают располо..
женные в линейной последовательности
множественные пары opraHoB, но при этом
никоrда не предполаrdЛОСЬ, что эти формы
произошли от метамерно сеrментирован"
ных предков (их иноrда называют «псевдо"
метамерными» ). У моллюсков целомиче..
екая полость окружает сердце. но более ве..
роятно, что она возникла в пределах данной
rруппы как пространство, в котором оно
может биться, и не является остатком Kor..
да"то более обширной полости, служившей
rидростатическим скелетом.
Таким образом, нет никаких убедитель..
ных доказательств изначальноrо присут"
етвия и последующей редукции или утраты
этоrо признака, и моллюсков. как правило.
уже не считают близкими родственниками
аннелид (см., например, Tl"ueman, Clal"ke,
1985) В самом деле, первые моллюски, по..
хоже, мало чем отличались от плоских че..
рвей с дорсальным защитным покровом
(белковым с рядом известковых чешуек или
пластин). Как и ныне живущие Platyhelmin..
thes, они аuеломические и несеrментирован
ные. Как и у крупных плоских червей и не..
мер тин (за исключением специализирован..
зз
ных видов), их движение осуществляется Зd
счет ресничек или волн мускульных сокра..
щений, про ходящих вдоль плоской вентра..
льной поверхности.
Помимо спинной защитной раковины
друrой основной чертой, отличающей пред..
KOBoro моллюска от плоских червей, было,
по"видимому, наличие в ротовой полости
зубчатоrо языковидноrо opraHa радулы.
Он позволял соскребать пленки и колонии
водорослей с твердых поверхностей, и ши
роко признается, что первые моллюски пи
тались именно водорослями"протистами
в мелководных местообитаниях. Из..за раз..
вития твердоrо покрова на дорсальной сто..
роне (единственной поверхности, открытой
воздействию токов воды. а также хищников
и волн) rазообмен через поверхность тела,
как и в случае плоских червей, сильно orpa
ничивается. Таким образом, параллельно
эволюции paKOB HЫ должно было идти раз..
витие специфических opraHoB дыхания
(жабр) и циркуляторной системы для транс..
порта rазов по телу. Парные жабры моллю"
сков, своеобразные и специфические для
)TOrO типа орrаны, называемые ктенидия"
ми, изначально располаrались сзади в ман..
тийной полости, скрытой раковиной. Две
друrие вымершие rруппы Hyolitha и Wi..
waxiida - похоже, реаrировали на давление
отбора, блаrоприятствующее появлению
Зdщитноrо покрова тела, сходным с моллю"
сками способом.
От TaKoro покрытоrо раКОВИНОЙ, напо..
минающеrо плоскоrо червя животноrо в хо..
де адаптивной радиации возникли весьма
разнообразные формы, описанные в rл. 5,
включая и наиболее сложно устроенные не
только среди потомков плоских червей. но
и среди всех беспозвоночных. Тем не менее
некоторые виды сохранили древний способ
питания за счет соскабливания водорослей.
Этот исходный образ жизни можеl быть
сравнительно леrко адаптирован к наземно..
му существованию, и моллюски COCT lB"
ляют одну из очень немноrих rрупп живот
ных, обладающих ферментами, способны..
ми разрушать стойкие структурные уrлево"
ды морских и наземных Рdстений.
34
2.4. Более отдаленные
потомки плоских червей
2.4.1. Членистоноrие
Основные анатомические отличия че
рвей с сериально повторяющимися по дли..
не тела орrанами от членистоноrих
наличие у последних конечностей и твердо..
то (членистоrо) суставчатоrо наружноrо
скелета. Эти отличия не столь существенны,
как может по казаться на первый взrляд.
Сначала, вероятно, возникли конечности.
Некоторые современные животные, напри..
мер онихофоры и тихоходки, мяrкотелые
червеобразные орrанизмы с вентральными
или почти вентральными нечленистыми ко..
нечностями и rидростатическим скелетом.
Онихофоры покрыты rибкой хитиновой ку"
тикулой, тихоходки наружными кутику"
лярными пластинами. Таким образом,
сравнительно леrко продемонстрировать.
как эволюционировали кутикулярные за..
rl1aBa 2
щитные структуры пока не покрыли тело
целиком, образовав суставные сочленения
между различными наборами сериально
повторяющихся элементов (так же, как, на..
пример, унеродственных членистоноrим
киноринхов) и члениками конечностей и
в итоrе взяв на себя функции скелета, заме..
нившеrо исходный rидростатический. В од..
ной из rрупп членистоноrих у хелиuеро..
вых выпрямление конечностей может
осуществляться rидравлически, и только их
сrибание происходит с помощью экзоске..
летно"мускульной системы.
Такая «артроподизация» предковоrо че..
рвя шла, похоже, еще в докембрии во время
основной радиации этой rруппы (рис. 2.11),
и мноrочисленные весьма разнообразные
и времеНdМИ крайне причудливые варианты
сеrментированных артроподоподобных жи..
вотных известны уже с кембрия (рис. 2.12).
Если бы они дожили до наII1ИХ дней, типов
членистоноrих было бы не меньше, если не
больше, чем среди червеобразных. Вероят"
ДOKeM Палеозой Мезозой Кайнозой
БрИЙ Нижний Верхний
........
. .,
''-i' ".'<
.... '5. ,.,...:. ,. "," ",,: ,',:, '1'. : ... CheLLderata
'" .... ('
,
. .....
т riLobita
.....
.............-
Crustacea
-
-----
..........;: . ' .,,--, ,:",./.';, ."Unttamta.;
, /', '..,".,3/"'" . ". .;:"..
.. <- -"':',;"".
..' ,"', ш
"i..
Рис. 2.11. Радиация «артроподоподобных» животных в докембрии/кембрии (по Whittington
1979)
Эволюционная история и филоrения беспозвоночных.
35
I
8. Дорсолатеральный вид
А. С вентральной
стороны
Д. Вид СБОКУ
r
r------1
Е. С вентральной стороны
/ /
/ /
Ж. Вентролатеральный вид
Рис. 2.12. Животные, напоминающие членистоноrих, не относящиеся к филоrенетическим ли
ниям трилобитов, хелицеровых, ракообразных и одноветвистых: А Burgessia (кембрий); Б
Miтetaster (девон); B Marrella (кембрий); r Braпchiocaris (кембрий); Д Jоhоiа (кембрий);
E Aпoтalocaris (кембрий); Ж Sarotrocercus (кембрий) (по Manton, Anderson, 1979; Whitting
ton, 1985). Масштабные линии 1 см
нее Bcero, предковая rруппа червей, доживи
ДО наших дней, не была бы отнесена к анне..
лидам основному современному типу се..
rментированных червеобразных. Очень мо"
жет быть, например, что rидростатическая
полость тела предка членистоноrих пред..
ставляла собой rемоцель, а не cerMeHTapHo
разпеленный схизоцель, характерный для
аннелид, и тело ero было не сеrментирован"
ным, а мономерным с множественными ли..
нейно расположенными парами локомотор"
ных opraHoB. Этих предков можно предста..
вить себе в виде удлиненных, внешне похо..
жих на киноринхов червей с кровеносной си..
36
стемой (возникшей по той же причине., что
и у моллюсков., см. разд. 2.3.1), сильно рас..
ширившейся для выполнения rидростатиче..
ских функций. Существуют также фунда..
ментальные эмбриолоrические различия
между аннелидами и разными rруппами
членистоноrих, которые также развиваются
по разным схемам (см.. например, Manton,
Anderson., 1979).
Лишь немноrие из большоrо числа
rрупп членистоноrих пережили кембрий,
а из них только четыре девон. Так. трило"
биты, существовавшие по КРайней мере 300
млн. лет, вымерли в конце перми. До наших
дней сохранились только три rруппы
хелиuеровые (Chelicerata), ракообразные
(Cl"ustacea) и одноветвистые (Unil"amia)., от..
личающиеся в основном строением и распо..
ложением конечностей и друrих придатков,
а также 1мбриолоrией и внутренним строе..
нием, причем до такой степени, что можно
rоворить об их неЗdВИСИМОМ происхожде..
нии от червеобразноrо предка. Похоже да..
же, что по крайней мере ракообразные и од..
но ветвистые потомки совершенно раз..
личных rрупп червей (АЬеlе, 1982). (Иноrда
Pycnogona считают четвертой rруппой та..
кото же статуса: в противоположном случае
их включают в Chelicel"ata.)
«Атроподизированными» предками всех
этих современных линий были.. ВОЗl'(10ЖНО,
удлиненные передвиrающиеся по субстрату
животные с большим числом сетментов, не..
сущих по паре придатков, морские в слу..
чае хелицеровых и ракообразных, и, вероят"
но., наземные в случае одно ветвистых.
Последняя rруппа до силура неизвестна.
Мноrие представители этих трех линий со..
хранили удлиненную предковую форму те..
ла в разной степени дифференцированноrо
на отделы (вместе с придатками), со ответ..
ствующие rолове и туловищ}, просоме
и опистосоме или rолове, rруди и брюшку
и т. д. Однако у друrих число сетментов ре..
дуuировано за счет утрать] или слияния
и еще более сократилось число пар Hor. Во..
зникновение мноrих линий., например мор..
ских и пресноводных планктонных рако..
образных и насекомых среди одноветви..
стых, предположительно связано с педо..
морфозом. Веслоноrие рачки, например.,
моrли произойти от планктонной личинки
rлава 2
крупноrо, удлиненноrо, бентосноrо предка.
Как уже rоворилось, обитание в толще во..
ды дает селективное преимущество тут
относительно мното корма (из..за близости
фотосинтезирующеrо слоя) по сравнению
с донными осадками, питательность кото..
рых зависит от оседающеrо неиспользован..
ното орrаническоrо материала.
Три ныне живущие rруппы считаются
родственными членистоноrим И, возможно,
представителями первой стадии «артропо..
дизации». Две из них онихофоры (Опу
chophora) и тихоходки (lardigrada) упомина
лись выше, третья пятиустки (Pentasto
та). Все три отличаются от членистоноrих
отсутствием их основных признаков
наружноrо скелета и членистых конечно
стей. Тело не сеrментировано, хотя пары
мясистых, заканчивающихся коrотками HO
жек расположены вдоль Bcero тела или ero
части, причем каждой паре соответствует
отдельный rанrлий HepBHoro ствола. У ТИ"
хоходок четыре пары таких ножек, у онихо..
фор 1 3, у пятиусток можно rоворить
о двух сильно редуцированных парах (три
пары у ранней личинки). У онихофор друrие
ортаны также сериально повторяются по
паре на каждый «сетмент» с ножками.
Пятиустки червеобразные эндопара..
зиты, и их предполаrаемое вторичное упро..
щение затрудняет филоrенетическую интер
претацию некоторые современные иесле..
дования наводят на мысль об их близости
к ракообразным. Хотя rидростатический
скелет всех трех rрупп rемоцель., харак"
терный., вероятно., для предковых червей,
и у пятиусток. И У тихоходок наблюдается
кратковременное присутствие энтероцель
ных карманов (три пары у первоrо типа,
две у BToporo), т. е. признака, характерно..
ro для лофофоровых и вторичноротых (см.
ниже). Эти «протоартроподы» являются
в сущности червями с мяrким телом и за..
кладывающимися чертами «членистоноrо
сти». Сейчас они обитают в основном на cy
ше (несколько видов тихоходок в море),
хотя известны кембрийские морские фор
мы., напоминающие онихофор. Мноrие ис
следователи считают онихофор близкими
по происхождению к одноветвистым.
Природа сеrментации членистоноrих,
протоартропод и киноринхов (и позвоноч
Эволюционная история и филоrения беспозвоночных
ных), таким образом, совершенно отлична
от таковой у аннелид. У первых она выте..
кает из MOHoMepHoro строения с множе..
ством пар конечностей (или у позвоноч..
ных мышечных блоков, обеспечивающих
плавание) и последующей сериализацией
связанных с движением скелетных, мышеч..
ных, нервных и сосудистых элементов, а
у вторых основана на линейной цепочке ча..
стично независимых единиц, каждая из ко..
торых имеет собственные полости тела
и наборы opraHoB, включая у примитивных
форм И rонады (рис. 2.1 О). Метамерные
rруппы членистоноrих, вероятно, возникли
независимо от предковой rруппы или rрупп
неметамерных червей. Тем не менее конеч..
ный результат эволюции членистоноrих
и аннелид внешне сходен: тело состоит из
пресеrментноrо переднеrо отдела (<<акрон»
у первых, «простомиум» у вторых), ряда ме..
тамерных cerMeHToB, добавляющихся сзади
по мере роста и/или развития, и конечноrо
постсеrментноrо отдела (<<тельсона» у чле..
нистоноrих, «пиrидиума» у аннелид). У ки"
норинхов новые сеrменты по мере развития
добавляются сходным образом.
2.4.2. Лофофоровые
Все потомки плоских червей. рассматри"
вавшиеся выше (за одним исключением),
имеют общие черты развития 1: а) дробле..
иие яйцеклеток спиральное (иноrда с мо"
дификациями): б) из бластопора возникает
рот, т.е. это первичноротые животные; в)
ход дальнейших преобразований бластоме
ров определяется на очень ранней эмбрио..
нальной стадии ( 16"клеточной бластулы).
т. е. развитие детерминированное; r) если
rидростатический скелет или полости вну"
три или BOKpyr opraHoB целомические
(оrраничены мезодермальной выстилкой),
то они образуются путем расхождения мас..
сы мезодермальных клеток, т. е. схизоцель..
ным путем (разд. 2.3.1, см. также разд.
15.2 15.4). Исключение составляет rруппа
червей. кратко упомянутая в разд. 2.3.1..
1 Из них первая спиральное дробление
наименее характерна; она присутствует только
в семи типах и не выражена, например, у брюхо..
ресничных, нематод и приапу лид.
37
с телом, разделенным на три отдела, со
своей собственной полостью каждый.,
Phorona.
Хотя они и первичноротые., дробление
яиц у них радиальное и недетерминирован
ное, и по крайней мере некоторые из поло..
стей и мезодеРМdЛЬНЫХ тканей образуются
так, что это можно считать модификацией
1нтероцельноrо способа (разд. 2.3.1; см.
разд. 15.2 15.4). Ныне живущие форони
ды мелкие сидячие черви, обитающие
в хитиновых трубках. Первый отдел их теЛd
(просома) очень мелкий., нависает над ртом.
Второй (мезосома) также небольшой, но
несет крупный лофофор и содержит полость
(мезоцель)., функционирующую как rидрав--
лическая система последнеrо. Лофофор
включает в себя полые, покрытые ресничка
ми щупальцевидные выросты стенки тела
BOKpyr рта., которые в первую очередь испо..
льзуются для питания взвесями. Самый
крупный отдел тела несеrментированная,
лишенная придатков метасома с поло..
стью метацелем, служащей основным
rидростатическим скелетом. Все три поло..
сти тела связаны между собой порами. По..
хоже, что от червей., подобных этим, про
изошли два друrих типа современных лофо..
форовых И., возможно., вторичноротые И
у лофофоровых, И У вторичноротых радиа..
льное недетерминированное дробление.,
тройная возникающая 1нтероцельным пу..
тем полость тела., причем некоторых лофо..
форовых можно считать вторичноротыми
(рот образуется не из бластопора, а путем
вторичноrо прорывания кишки). Друrие об..
щие чеРТ:РI включают личинок с системой
восходящих выстланных одно ресничными
клетками полосок для сбора пищи BOKPYI
рта. ТРdНСфОРМИРУЮЩИХСЯ при метаморфо..
зе в руки лофофора с такими же клетками,
мозr в мезосоме и мезодерму, возникаю--
щую из энтодермы или клеток веrетативно
ro полюса бластулы, что отличает ее от ме..
зодермы друrих билатерально симметрич
ных животных, развивающейся из rуб бла..
стопора.
Плеченоrие (Brachiopoda), появившиеся
в кембрии.. утратили мелкий передний от..
дел тела (просому); их метасома по cpaBHe
нию сметасомой форонид сильно редуци..
рована, однако мезосома очень крупная
38
rлава 2
Ордовик
КеМБрИЙ
*. .
\ .
/
*. .
"
,
"
*. .
.
:.... )
:....
...::@
:
i.
>
:. . . , . . :::::::
*. .
......."
со
g
с:
о
><
со
а.
ID
:s:
:s
::I:
.С:::: *. .
ф
:s: *. .
3
m
:S:
]i
со
*
цs
=>
U
L.
со
с:
"",:
""'"
J:
:S:
I
оф
c:tT
:S:I:
:1:ф
o
о...й
ош
&8-
фа
&
o
..й&
с:::о
c:::
::r ф
S;
..LI
U::I:
0.11)
8
:1:0
Sc:
Рис. 2.1 З. Возможное полифилетическое происхождение плеченоrих как волна «брахиоподиза
ции» различных форонидоподобных червей в докембрииjкембрии (по Wright, 1979)
и поддерживает хорошо развитый лофо-- и ее потомков точно так же, как унекото..
фор. Это сидячие формы с телом, покры-- рых книдарий (разд. 2.2.2). Для некоторых
тым вместо трубки известковой или фо-- мшанок характерен и полиморфизм зоои..
сфатной раковиной, похожей на раковину дов. По СРdвнению с предковым форонидо..
двустворчатых моллюсков, только с дорса-- подобным червем отдельные зооиды очень
льной и вентральной, а не боковыми створ-- мелкие (увеличение размеров свойство
ками. У некоторых плеченоrих ее. выросты целой колонии), часто лишены просомы
укрепляют мощный спирально закручен-- и секретируют раковину, домик или студе..
ный лофофор. Вероятно.. разные rруппы нистый МdТРИКС, покрываlQШИЙ их так, что
этоrо типа потомки форонидоподобноrо остается только отверстие, через которое
предка, а разные форониды прошли в до-- лофофор высовывается для сбора пищи
кембрии стадию «брахиоподизации», по-- и втяrивается для беЗОПdСНОСТИ. Почти все
добную радиации, давшей предка членисто-- мшанки сидячие и MorYT рассматриваться
ноrих (рис. 2.13). как крошечные колониальные варианты фо..
Ныне живущие форониды MorYT раз-- ронид. Они появились В ископаемых остат..
множаться бесполым путем, а один из их ках сравнительно поздно: не ранее ОРДQ"
видов почкованием, что является нормой вика.
для третьей rруппы лофофоровых Родственные отношения мшанок с дру"
мшанок (Bryozoa). У них колонии из тысяч rими rруппами лофофоровых остаются
связанных между собой особей (зооидов) спорными, отчасти потому, что способ
образуются в ходе неоднократноrо беспо-- образования их полостей тела аберрантен:
лоrо почкования особи--основательницы у нескольких видов они формируются зано"
Эволюционная история и филоrения беспозвоночных
во во время метаморфоза. Некоторые пред
полаrают, что мшанки близки к неясной
rруппе Entoprocta, с которыми их объеди--
няет высокоспециализированный тип фото--
рецепторов Эти спирально дробящиеся
крошечные сидячие животные несут распо--
ложенные по Kpyry шупальца, как и лофо--
форовые, однако строение их не соответ--
ствует структуре настоящеrо лофофора,
а полость тела здесь нецеломическая. за..
полнена мезенхимой Друrие предполаrают
родство с Entopl"octa коловраток, но свиде--
тельств близости этоrо типа с какой--либо
конкретной rруппой слишком мало.
2.4.3. Вторичноротые
Парадоксально, но хотя четыре ТИПd
вторичноротых полухордовые (Hemichor...
data), иrлокожие (Echinodermata) хордовые
(Chordata) и щетинкочелюстные (Chaetog...
паthа) образуют rруппу с БОЛЫI1ИМ коли--
чеством общих черт, об их связях между со--
бой почти ничеrо не известно. В целом мо--
жно считать, что различия между форони
допопобными лофофоровыми И полухордо--
выми крыложаберными выражены нечетко.
Тело последних разделено на три отдела,
каждый из которых содержит образовав--
шиеся энтероuельным путем полости (един
ственная в просоме, и парные в мезо-- и ме--
тасоме), а мезосома несет крупный rидрав"
лически управляемый лофофоральный ап--
парат. Единственные существенные разли--
чия заключаются в том, что просома кры--
ложаберных хорошо выражена и используе--
тся при движении, ее полость (протоцель)
образует rидростатический скелет, и разви--
тие животноrо происходит с образованием
вторичноrо рта. Из--за этоrо сходства широ..
ко распространено мнение о близости об..
щеrо плана строения крыложаберных
и предковой формы вторичноротых; при
этом друrая rруппа полухордовых черве..
образные кишечнодышащие, вероятно,
утратила лофофор в связи с возвращением
к свободноживущему роющему образу жи..
зни.
Хотя иrлокожие выrлядят совершенно
иначе из--за paHHero развития у них сфериче--
ской формы тела с почти радиальной СИМ--
метрией (в связи с сидячим существованием
39
и питанием за счет фильтрации), строение
их полостей тела ясно указывает на исход
ную тройственную их орrанизацию, сход--
ную с наблюдаемой у лофофоровых и полу..
хордовых Один протоцель образует осевой
синус, левые мезоцель и протоцель вме..
сте амбулакральную систему, правый ме..
зоцель сильно редуцирован у всех иr локо..
жих, за исключением аберрантных орrаниз"
мов.. а два метаuеля дают основные отделы
полости тела. Иrлокожие одна из мноrих
rрупп, появившихся впервые с уже харак--
терным для типа обликом, начиная с KeM
брия, хотя и не все ранние формы имеют
пятилучевую симметрию, свойственную со--
временным классам. О предковом плане
строения можно только доrадываться. Од--
на из наиболее правдоподобных rипотез
выводит иrлокожих из сидячих, близких
к крыложаберным животных, у которых ло..
. фофор с ero rидравлической системой мезо--
целя развился в систему радиальных кана--
лов амбулакральной системы; у некоторых
современных крыложаберных у лофофора
пять основных стволов с каждой стороны.
Помимо черт, характерных для всех BTO
ричноротых, общий признак иrлокожих
и хордовых система твердых защитных,
содержащих кальций пластинок мезодерма..
льноrо происхождения в коже; у некоторых
вымерших иrлокожих, похоже, были обра--
зования, ЭКВИВdлентные rлоточным щелям,
присутствующим также у ныне живущих
полухордовых (и ряда rастротрихов). Не..
удивительно поэтому, что большинство по..
пыток вывести хордовых из друrих rрупп
животных помещает иrлокожих неподалеку
от точки их происхождения, но, как и в слу
чае перехода полухордовые/иrлокожие, все
предложенные филоrенетические схемы
страдают недостатком потенциальных про--
межуточных форм; им нет подходящих ана..
лоrов среди ныне живущих и ископаемых
видов, если не считать своеобразных Нота..
lozoa кембрия дeBOHa (рис. 2.14), лишен..
ных какой бы то ни было симметрии, кото--
рых некоторые считают предками хордо--
вых. Друrие исследователи выводят послед..
них путем педоморфоза из личинок полу..
хордовых/иr локожих. Древнейший ископае..
мый материал, точно относящийся к хордо--
вым, датируется ордовиком, хотя из--за од..
40
n
/.'''\:' ,
;..,.:
:,..;...:. ... .' ......;:.:':.; . . :". :...
:' :. ....: .:" ":0"';:": ;: . ,. .,.'
,ц;' '; l : ::.. l : ;.r:i;;:..::
. ...-,........ ..........',. ., .
'1 . '. ...!.i.;' '.. ;'."'; "..'!:-"'.' .
' . ."' , :, ..- .': "":..: .
.' ( j,...J.....4,...4'. .. .:-..
1./"" .. , 8!;..,..,..-
...... ..
... .
I
=:
..
"
.,
.-
.-
Рис. 2.14. Иrлокожие rруппы Homalozoa
возможные предки хордовых (по Clarkson,
1 986)
Horo кембрийскоrо остатка и нескольких
позднекембрийских фосфатных ФРdrментов
время их возникновения не вполне ясно,
и если будет признано, что заrадочные ко..
нодонты это части скребущих opraHoB
бесчелюстных позвоночных (как сейчас ка..
жется), периодом возникновения хордовых
надо будет считать кембрий и даже докем..
брий.
Четвертый тип вторичноротых
щетинкочелюстные настолько же непоня"
тен с филоrенетической точки зрения, на..
сколько важен с эколоrической; возможно,
это rлавные беспозвоночные хищники мор..
cKoro планктона. Ранний эмбриоrенез этих
ПрОЗрdЧНЫХ мяrкотелых червей типичен для
вторичноротых, а их тело состоит из трех
отделов rоловы. туловища и постаналь..
Horo локомоторноrо хвоста, Соответ"
ствуют ли эти три отдела про.., мезо.. и ме..
тасомеполухордовых, проблематично, при..
чем некоторые черты щетинкочелюстных
явно сходны с таковыми у псевдоцеломиче..
ских червей. В настоящий момент об их
родственных связях с друrими вторичноро"
тыми ничеrо нельзя сказать. Мяrкотелые
планктонные виды хуже всех друrих живот..
ных представлены в ископаемых остатках,
rлава 2
и, возможно (и даже весьма вероятно),
в ПРОШЛОМ было MHoro типов таких живот..
ных (см., например, рис. 2.16, Т, Д), о суще..
ствовании которых мы совершенно не по..
дозреваем (СМ. разд. 2.5); предки щетинко..
челюстных моrли принадлежать к одной из
таких rрупп.
2.5. Происхождение, радиация
и вымирание rpynn ЖИВОТНblХ
Все современные животные MorYT бы 1 Ь
помещены в 35 типов: под типом riонимае..
тся rруппа кажущихся родственными друr
друrу орrанизмов, чьи родственные связи
с аналоrичными rруппами СПОРНЬJ или
чисто предположительны. Таким образом.
статус типа в принципе дань нашему не..
знанию, хотя в последующих разделах мы
попытаемся проследить вероятные poд
ственные связи. Эти 35 ныне живущих типов
тем не менее не являются результатом мед..
ленной эволюционной диверrенции, ведшей
к возрастанию разнообразия. Кажется ве..
роятным, что С очень небольшими ис..
ключениями все ныне живущие типы жи..
вотных уже существовали в кембрии. а ди..
верrенция подобных плоским червям пред..
шественников большинства rрупп произо..
шла еще в докембрии. Это cBoero рода за
[адка.
Ископаемые остатки демонстрируют
медленное изменение животных во време..
ни, так что формы., присутствующие в нача..
ле и в конце данной последовательности,
MorYT иметь достаточно различий для отне..
сения к разным видам (так называемым
«хроновидам»). Среднее время., которое
требуется для анатомических изменений та..
Koro масштаба, у самых разных беспозво..
ночных составляет примерно 10 млн. лет.
Если признать что роды животных отли"
ЧdЮТСЯ друr от друrа., скажем, в 1 О раз силь..
нее, чем виды одноrо рода, семейства в 1 О
раз сильнее, чем роды одноrо семейства,
и Т. д. вплоть до отрядов. классов и типов,
то на основе средней скорости изменений
хроновидов можно получить по крайней ме..
ре миллион миллионов лет для диверrенции
разных видов до такой степени, коrда МО"
жно будет признать их относящимися к раз..
ным типам.
Эволюционная история и филоrения беспозвоночных
41
Вероятно, первые животные произошли
от предков"протистов около 1 000 млн лет
назад, Т. е. произведенный выше расчет дол..
жен был бы привести нас к выводу, что их
потомки диверrируют с образованием не..
скольких различных типов еще примерно
через 1.1 х 1012 лет. К настоящему момен..
ту их потомки должны были бы отличаться
дрyr от друrа, как, например, разные семей..
ства жуков Даже если допустить средний
временной интервал между последователь..
ными хроновидами равным Bcero 1 млн. лет
и сократить коэффициент различия между
видами и родами, родами и семействами
и т. Д. С 10 до 5. потребовалось бы более
3000 млн. лет для Toro, чтобы предковые
плоские черви диверrировали в несколько
типов. Однако типы, известные нам се..
rодня. уже существовали и отличались друr
от друrа не меньше, чем сейчас, Bcero через
400 млн. лет после вероятноrо возникнове
ния мноrоклеточных животных, и, по..
видимому, диверrировали за rораздо более
короткий промежуток времени. Неу дивите--
льны слова одноrо cOBpeMeHHoro aBTopd
(Day, «Genesis оп Planet Earth», 1984): «Вне..
запное появление кембрийской фауны пред..
ставляет собой одну из крупнейших биоло..
rических заrадок... Типы, представленные
тоrда, возникли быстро, происхождение их
неясно» .
Очевидно, что..то не так в наших эволю..
ционных взrлядах. Оказывается, не все фи--
лоrенетические изменения протекают так
медленно, как отражено в ископаемых
остатках близких видов. Временами проис..
ходило взрывоподобное преобразование
морфолоrии, такое быстрое, что промежу..
точные ero стадии не представлены в отло..
жениях. Возможно также, что оно шло в от..
носительно небольших популяциях (ско"
рость потенциальных эволюционных изме..
нений обратно пропорциональна размеру
популяции), а это также уменьшает возмож--
ность сохранения в ископаемом материале.
Педоморфоз (см. выше), например, в ходе
смены одноrо поколения друrим может за
.,
несколько лет привести к TaKoro масштаба
изменениям взрослой формы, что в резуль..
тате потомство и родителей будут считать
принадлежащими I< разным ОТРЯДdМ или
классам. (Таксономические катеrории, та..
Кт А
Класс В
К(1асс D
Класс С
А
о:
L
Q)
а...
(()
...
Возрастающая морсроло
rическая верrенция
Б
Класс Д
Класс В
Рис. 2.15. ОДНО и то же rипотетическое фило
rенетическое древо, представленное в двух
разных формах: А классификация на основе
традиционНЫХ линий, выделяемых по морфо--
лоrическому сходству и различию, Б
классификация в соответствии с принципами
кладистики
ким образом, часто указывают На степень
морфолоrических различий и совсем необя..
зательно на удаленность от общеrо пред--
ка; такое положение хотели бы исправить
филоrенетики «кладистской школы»;
удайся им это. типов и классов в том объ..
еме, в каком мы их понимаем сейчас, если
бы и осталось, то очень мало рис. 2.15.)
Вспышки филоrенетических изменений
часто были связаны с сильной Диверrенцией
на уровне видов, родов., семейств и даже от--
рядов от одной формы. Такая «адаптивная
радиация» происходит очень быстро в re..
олоrическом масштабе времени. и -этот спо--
соб эволюции за короткий период несколь..
ких новых форм от одной предковой привел
к: а) представлениям о том, что процессы,
контролирующие изменения внутри одной
популяции (микроэволюцию), качественно
отличаются от приводящих к образованию
новых таксонов высшеrо paHra (макроэво--
люции); б) теориям о количественно различ--
ных скоростях микро" И макроэволюцион"
42
rлава 2
Б
А
r
в
II
I I
1
д
1
ж l
Рис. 2.16. Вымершие преимущественно мяrкотелые животные, которых нельзя поместить ни
в один из ныне известных типов. А Wiwaxia (кембрий); Б Opabiпla (кембрий); B
Dihoтischus (кембри й); r А тiskwia (кем бри й); д т u//iтoпstruт (карбон); Е Odoпtogriphus
(кембрий); Ж Ha//ucigeпia (кембрий) (по Clarkson, 1986; Conway, Morris, 1979; Whittington,
1985). Масштабные линии 1 см
ных пропессов; вторые идут быстрее (моде..
ли «постепенности» и <прерывистоrо paBHO
весия», rл. 17).
Не все типы, возникшие во время об
ширных докембрийской и кембрийской pa
диаций, представлены среди ныне живущих
беспозвоночных. Некоторые из них извест
ны только по ископаемым остаткам, напри
мер археОIIиаты и трилобиты, и В особенно
сти обычно не получающая формальноrо
статуса типа rруппа кембрийских членисто
ноrих (разд. 2.4.1). Однако их удивительно
lVlало, причем для большинства из них xa
рактерны тверлые части тела, т. е. признак.
похоже, возникавший в каждой линии срав"
нительно поздно. Таким образом, ископае..
мые остатки MorYT дать ложное представле..
ние о прошлом разнообразии животных. на
которое указывает и обилие неизвестных
типов мяrкотелых животных, обнаружен..
ных в тех редких осадках, rде их остатки со..
хранились (например, докембрийская фауна
Эдиакары и кембрийская Берджесских слан..
цев) (рис. 2.16).
Двадцать ныне живущих типов, часто
называемых «малыми», включают менее
Эволюционная история и филоrения беспозвоночных
500 видов каждый. За ОДНИМ исключением,
подтверждающим правило, это червеобра..
зные мяrкотелые формы, не сохранившиеся
в ископаемых остатках. Исключение
плеченоrие, насчитывающие Bcero 350 со..
временных видов и более 25 000 ископаемых
форм, известных по толстым наружным ра..
ковинам. Попытки объяснить тот факт, что
этим малым типам удалось дожить до на..
ших дней, имея в своем составе так мало ви"
дов, привели к предположению, что это ста..
тистически вероятно только в том случае,
если коrда"то существовало rораздо боль..
ше похожих (т. е. мяrкотелых) ВПОСЛ ,LIствии
вымерших типов (мы уже отметили такую
возможность в связи с предками щетинко..
челюстных; разд. 2.4.3). Ныне известные
ископаемые типы, таким образом, MorYT
представлять собой лишь небольшую и не..
типичную (из..за наличия твердых частеЙ)
выборку: возможно, членистоноrие всеrо
лишь окаменевшая «верхушка» докембрий..
CKoro и кембрийскоrо айсберrа вымерших
rрупп.
Итак, напрашивается мысль о широко..
масштабной радиации червеобразных жи..
вотных в кембрии и докембрии. давшей.
возможно, сотни эквивалентных типам так..
сонов. Большинство их скорее Bcero были
небольшими по количеству видов и вымер..
ли без следа. а ныне живущие черви лишь
напоминание об этом былом разнообразии.
В некоторых линиях возникли твердые за..
щитные или скелетные части, возможно,
в ответ на эволюционное давление хищни"
чества, и о них наши знания довольно зна..
чительны: считается, что сейчас найдено
и описано около 120/0 всех таких видов, Kor..
да..либо существовавших на планете. Не..
сколько rрупп, диверrировав, дали начало
большому количеству видов и высокому
внутритиповому разнообразию. В настоя..
щий момент 1 О типов включают более чем
по 10000 ныне живущих видов каждый, в то
время как плеченоrие и иrлокожие, коrда"то
достиrнув примерно TaKoro же разнообра..
ЗИЯ, потом сократили ero ниже этоrо
уровня. Эти типы называют «большими».
Хотя некоторые типы отличались высо"
ким видовым разнообразием в течение дли..
тельных rеолоrических периодов, обычно
оно не обеспечивал ось все это время одни..
43
Мел
Триас
Пермь
КарБОН
Девон
Рис. 2.17. Филоrенетическое древо аммони
тов, демонстрирующее радиацию вымирание
и вытеснение одной подrруппы друrой с дeBO
на до конца мела, коrда вся эта rpynna вымерла
(по данным Moore, 1957)
ми и теми же формами. В разные интервалы
времени преобладали разные представите..
ли отрядов и классов, причем несколько
коrда"то доминировавших классов к на..
стоящему моменту вымерли, уступив место
друrим. Подrруппы вступали в полосу ра..
диации по очереди. Это наиболее четко вид..
но на примере ископаемых аммонитов (рис.
2.17) и почти полностью вымерших наути"
лид среди rоловоноrих моллюсков, а еще
более на смене «эр» амфибий, рептилий
и млекопитающих в пределах позвоночных,
но характерно и для друrих типов, в том
числе книдарий, плеченоrих, иr локожих
и неrоловоноrих моллюсков. В целом заме..
щение одной подrруппы друrой, похоже,
происходило не за счет KOHKypeHTHoro ис..
ключения, которое, как можно ожидать,
должно последовать после возникновения
и эволюции у вытесняющей подrруппы ка..
коrо..либо морфолоrическоrо или иноrо
признака, получающеrо селективное преи..
мущество. Наоборот, сначала одна форма
вымирала, и лишь после видимо, в усло..
44
виях «эколоrическоrо вакуума» ее преем"
ник радиировал, заполняя пространство
ниш, доставшихея ему «по наследству» от
доминировавшеrо ранее предшественника.
Ископаемые остатки показывают, что за та..
кой быстрой радиацией на мноrочисленные
формы (фаза прерывания в модели преры..
вистоrо равновесия) шел rораздо более дли..
тельный период сравнительно небольших
морфолоrических изменений под влиянием
стабилизирующеrо отбора (эквивалент фа..
зы равновесия), пока сама эта rруппа в свою
очередь не сокращала резко количества ви"
дов или не вымирала полностью.
Степень вымирания разных типов жи..
вотных, характеризующихся широким спек..
тром 1авоеванных эколоrических ниш и ти"
пов местообитаний, остается в то же время
спорной, хотя мноrие придерживаются мне..
ния о нескольких происходивших в про..
шлом массовых вымираниях, особенно бли..
же к концу ордовика, девона, перми, триаса
и мела (рис. 2.18), а также в большей или
меньшей степени примерно через каждые 26
млн. леl. Например, то, что произошло
около 220 млн. лет назад (в конце перми),
повлекло за собой исчезновение более поло..
700
600
х 500 5
::о
I
О
CD
400
х
S
()
а.
о 300
CD
4
()
IS 3 Массовые вымирания
Q) 200
:I:
Q) 1. Поздний ордовик 12 %
()
о 2. Поздний девон 14 %
5 100 3. Поздняя пермь 52 %
S
:r 4. Поздний триас 12 %
5.Поздний мел 11%
О
600 500 400 300 200 100 О
Млн. лет назад
Рис. 2.18. Изменения числа семейств морских
животных в разные периоды, показывающие
ряд массовых вымираний (по Valentine, Moo
res, 1974 и др.)
rлава 2
вины существовавших в то время семейств
морских животных и, возможно, 80 950/0
их видов Ранее доминировавшие rруппы,
включая ракоскорпионов (Eurypterida) сре..
ди хелицеровых и трилобитов, исчезли пол..
ностью, оставив к начал) мезозоя фауну,
весьма близкую к современной по основ..
ным типам строения преобладающих в MO
рял животных. Причины 1Toro ВЫМИрdНИЯ
неизвестны, хотя оно хорошо коррелирует
с крупными изменениями уровня моря. ко..
торые Mor ли быть вызваны фазами оледе
нения, движениями континентов и или Jпи
зодами rлобальноrо похолодания (были
также мноrочисленные спекулятивные по..
пытки привлечь к объяснению различные
космические силь] и события).
Итак, история развития фауны, по..
видимому, была четко периодической. Каж..
дый период, включая и тот, в котором мы
сейчас живем, характеризуется случайной
смесью увеличивающих и сокращающих
свою численность rрупп животных. либо cy
мевших сохраниться, понеся большие поте..
ри, либо ждущих cBoero часа, чтобы увели..
чить количество представителей и их разно..
образие после следуюшеrо периода выми"
рания, индуцированноrо человеком или
друrим areHToM. С течением времени типов
животных стало меньше, поскольку некото"
рые (а возможно. и мноrие) из них вымерли.
d новых возникло меньше, хотя число ныне
живущих видов больше, чем ранее. Моря
Земли сейчас сравнительно фраrментарны,
среди них обильны и велики мелководные
полуизолированные шельфовые водоемы,
а океанические воды насыщены кислородом
вплоть до абиссали обстоятельства, су..
шествовавшие далеко не Bcerna: в мелу, Ha
пример, большие поверхности MopcKoro
дна, возможно, были аноксическими. Таким
образом, нет никаких причин предполаrать,
что морские виды сейчас менее разнообра
зны, чем коrда..либо в прошлом; в самом
деле, большинство оценивает морскую фау..
ну позднеrо кайнозоя как самую боrатую
среди всех существовавших на планете.
Несмотря на это, orpoMHoe количество
современных видов почти полностью отра..
жает завоевание животными суши, наЧdВ..
шееся в силуре девоне и особенно мощно
шедшее в карбоне. Несколько удивительно,
Эволюционная история и филоrения беспозвоночных
ЦАРСТВА
Monera
Protlsta
F,ungi.
Ptantae
AnimaLia
ЖИВОТНЫЕ
КишеЧНОПОЛОСТНblе Плоские черви
rУБК I ЧервеОБразные
Моллюски
Лосросроровые
Вторичноротые
РаКООБразные
Хелицеровые
Ри'с. 2.19. Число ныне живущих видов разных
царств и типов животных (по данным Barnes,
1984 )
что эта колонизация, похоже, осуществля"
лась в основном из литоральной зоны не..
посредственно на сушу., а не по воде из
моря в эстуарий, дальше в реку (озеро)
и т. д.; при этом большинство наземных жи..
вотных содержат MHoro жидкости и устой
чивы К высыханию, а не наоборот. (Основ..
ными rруппами, использовавшими менее
«популярный» водный путь., были позво
ночные и некоторые моллюски.) Пресные
воды заселялись rлавным образом форма..
ми, уже адаптированными к наземным
условиям. Мелкие интерстициальные про..
тисты, инфузории., нематоды, тихоходки
45
и т. д. никоrда не ПОКИДdЛИ водных пленок,
окружавших частицы осадка или почвы
и MorYT считаться наземными только в том
смысле, что их микроместообитание Haxo
дится на суше. Какой из двух путей ее засе..
ления они использовали, неизвестно и, воз..
можно. непознаваемо. На суше rораздо
больше, чем в воде, препятствий к распро
странению, а 1ТО привело к массовому ви"
дообразованию в пределах сравнительно
немноrих успешно освоивших сушу типов
(по той же причине rораздо больше бентос..
ных морских орrанизмов, чем пелаrиче
ских). Мноrие наземные виды представ
ляют собой rеоrрафически сменяющие друr
друrа варианты одноrо OCHoBHoro плана
строения. Сухопутные хелицеровые и одно..
ветвистые, особенно насекомые, сейчас дo
минируют среди беспозвоночных планеты
по числу видов (рис. 2.19) так что за послед..
ние 350 млн. лет центр разнообразия opra
низмов на видовом и родовом уровнях
переместился из моря, rде и произошло все
филоrенетическое развитие.. рассмотренное
в этой rлаве, на сушу.
2.6. Резюме
и эволюционный обзор
Около 1 I\1ЛРД. лет назад несколько ли
ний rетеротрофных протистов., обитавших
на морском дне.. вероятно.. пересекли крайне
плохо выраженную rраницу между коло..
ниальностью и мноrоклеточностью. дав на..
чало «животным». В течение следующих не..
скольких сотен миллионов лет потомки oд
ной из этих линий дали ряд крупномас
штабных радиаций., вызванных, по..
видимому., в основном наличием неисполь..
зованных источников пищи., давлением
хищничества, а также 1колоrическими
и эволюционными преимуществами, свя
занными с увеличением размеров тела (см.
разд. 9.1). Эти радиации привели к широко..
му разнообразию бентосных черве.. и мол..
люскообразных животных, от некоторых
червей эволюционировали членистоноrие.
лофофоровые и вторичноротые (рис. 2.20),
так что 600 млн. лет назад, вероятно.. уже
существовали все известные типы и множе..
ство неизвестных. С Toro времени разные
типы и входящие в них классы и отряды
Platyhelminthes Gastrotricha и Псевдо Arthropoda Nemertea
Gпаthоstоmulа целомические
rруппы I
(См. ниже)
1. Метамерия
2, Псевдоцель
становится
циркуляторной
3. Интерстициальные системой
формы . х ный
4, Внутреннее
ОП!10дотворение
5. Конечности
1. Линяющая 1.Циркуляторная 1.Циркуляторная 1.Схизоцель
кутикула система Система 2. Кишка
2.Специализирован 2. Сквозной кишеч
2. rермафродитизм 2,rермафродитизм 2.Псевдоцель ный хоботок ник с терминаль
С ным анусом
У 3. квознои
3. Утрата первичной з.Утрата первичной '-'о трата первичнои кишечник З. Ктенидии
личинки личинки личинки с терминаль
4.СКВОЗНОЙ кишечник ным анусом 4.Радула
с терминальным 5 Д о р салы-tая
I анусом .
5. МОАИwицированное раковина
дроолениl..
Предковые, подобные плоским червям животные
Мономерия Тело без полости
Раздельнополость Нет кровеносной системы
46
1, Эвтелия
2. Внутреннее
оплодотворение
1 Внутреннее
оплодотворение
1.Внутреннее
оплодотворение
4.Моноцилиарный
эпидермис
5.И терстициальные
формы
Трохофороподобная
личинка
rлава 2
Mollusca Annelida,
Echiura и
Pogonophora
I
1. Метамеr;>йя
2.. Циркуляторная
система
З. Прямой кишечник
с терминальным
анусом
4. Щетинки
Sipuncula Lophophorata Deuterostomia
I
1. LJ.образный
кишечник
(См ниже' I
1. Вторичноротые
1. Модифицированная
Личинка
2. Лофофор
З. Энтероцель
4. ЦиркуляторнаR
система
5. НедетеРМИНИрОванное
развитие
6. Радиальное дробление
7.0лиrомерия (три отдела)
8. Сквозной U-образный
кишечник
9. rермафродитизм, широко
распространено бесполое
почкование
Спиральное дробление
Наружное
оплодотворение
Слепо заканчивающая. Первичноротость
ся кишка
Псевдоцеломические и членистоноrие
ro ro
n;
ro
Q. .J::
Q) ro О (,) ro
(,) "о Е с ro "о
ro О З о > т ro
.J:: .... .J:: . ct
ё ro ro О (,) ::]
Е Е Q. "о .
О ro С i: т .........
tJ Q) Q) о ro
а: « z z ...J Q: S
Детерминированное
развитие
rpynna лофофоровыхjвторичноротых
ro ro
ro .... .J:: ro
т Е
"о "о ....
О ro Q:;
(,) <5 с ro
О ro а ro о') "о ....
Q. о о с .J:: О О ro
N .J:: О , ..... с "Е
о о tJ <5 Е Q)
ё ro О
.J:: Q) .J:: tJ
Ш Q) Q) Q.. I U ш U
I
Рис. 2.20. rипотетические схемы родственных отношений между потомками близких к плоским
червям форм с указанием предковых признаков (сохранившиеся у потомков, кроме особо oroBo
ренных случаев) и основных вторичных черт специализации (по крайней мере исходных) в разных
rруппах или кладонах. Подробно связи в пределах линий псевдоцеломических/членистоноrих
и лофофоровых/вторичноротых показаны отдельно
приобретали и теряли эколоrическое значе..
ние. Палеоэколоrи. например. на основе
форм с твердыми леrко переходящими
в ископаемое состояние остатками выде..
ляют три сменявшие друr друrа крупные
фауны., доминировавшие в морях: кембрий--
ско ордовикскую (трилобиты и друrие ран--
ние членистоноrие., беззамковые плеченоrие
и моноплакофоровые моллюски), позднепа--
леозойскую, или ордовикско--пермскую (ра..
ковинные rоловоноrие, морские лилии, зам--
ковые плеченоrие, мшанки класса Stenolae--
mata). и современную (мезозойско--кай--
нозойскую), характеризующуюся ведущей
Эволюционная история и ф 1лоrения беСПОЗВОI-IОЧНЫХ
ролью брюхоноrих и двустворчатых мол..
люскQв, ракообразных класса Malacostraca,
морских ежей, мшанок класса Gymnolaema..
ta и позвоночных. Каждая из этих фаун, по..
видимому. формировалась в мелководной
среде обитания, а позже перемещалась ('1)
в более rлубоководные зоны внешнеrо ше..
льфа и континентальноrо склона.
Одной из важнейших стадий в радиации
типов строения животных было завоевание
толщи воды, позволяющее использовать на
месте фитопланктонную продукцию по
верхностных слоев моря. Основным факто..
ром, позволившим животным оторваться
от MopcKoro дна, был педоморФоз (см. rл.
17), который дал врожденные репродуктив"
ные преимущества., заключающиеся в co
кращении времени rенерации. Изменения
в относительной хронолоrии процессов раз..
вития (т. е. мутации «реrуляторных [енов»),
сопровождающие педоморфоз., являются
мощным средством быстрых и крупных
морфолоrических преобразований: за не..
сколько поколеиий анатомия половозрелой
особи может измениться до такой степени.,
что предковую форму и ее потомка в систе..
мах классификации, основанных на морфо..
лоrических различиях, будут относить
к разным клаССdМ или даже типам. Извест"
но несколько современных морских личи..
нок, У которых, по"видимому, В связи С дли..
тельностью их периода оседания на дно
развитие rонал начинается еще до метамор..
фоза; недавно наблюдали, как личинки од..
Horo из видов rребневиков выметывали
вполне жизнеспособные [аметы (как иеко..
торые двустворчатые моллюски икру):
это, возможно, указывает на способ возни..
кновения педоморфических rрупп и на меха..
низм ряда крупных «макроэволюционных
событий» .
Последний этап в формировании разно..
образия животных имел место около зоо
400 млн. лет назад, коrда животные с конеч..
ностями начали колонизаuию суши. По
сравнению с морем, [де обитали их предки,
суша представляла собой в высшей степени
расчлененную с rеоrрафической точки зре..
ния среду с множеством препятствий для
расселения орrанизмов. Таким образом, ви"
дообразование у наземных форм стало идти
в неизвестных прежде масштабах: хотя к но..
47
вым окружающим условиям смоrло при
способиться очень мало крупных 2рупп, по..
давляющее большинство видон современ..
ных животных наземные.
Довольно любопытная черта истории
больших типов животных то, что, хотя
составляющие их подrруппы и имели тен..
денцию сменять друr друrа во времени, по..
хоже, ЭТQ очень редко происходило за счет
KOHKypeHTHoro исключения. Напротив,
предшествовавшая rруппа сначала вымира..
ла или резко сокрашала численность..
и лишь затем шла радиация последующей
для заполнения, очевидно, вакантных ниш.
Возможно, с этим перекликается то обстоя..
тельство, что, соrласно современным эко"
лоrическим данным. известно очень мало
фактов вытеснения анатомически сложно
устроенными животными морфолоrически
более простых из общих местообитаний:
членистоноrие и аннелиды. например.. и по
сей день сосуществуют в осадках с «прими..
тивными» приапулидами и сипункулидами
без каких..либо при знаков KOHKypeHTHoro
исключения.
Таким образом., следует избеrать собла..
зна считать, скажем., плоских червей и ки"
шечнополостных в каком..либо смысле «ни..
же» ракообразных или хордовых, отличаю..
щихся rораздо более сложным строением
и поведением. Разнообразные типы живот..
ных, возникшие в результате крупных до..
кембрийской и кембрийской радиаций., со..
ответствуют более или менее одно возраст..
ным альтернативным планам строения., не
демонстрируя постепенноrо возрастания
приспособленности и «совершенства» Дру"
rими СЛОВdМИ, вся жизнь эволюционирова..
ла от ранних докембрийских прокариот; He
которые из их потомков сейчас сложно
устроенные дубы, слоны и люди, но они ни
в коей мере не вытеснили прокариот, и бак..
терии до сих пор успешно доминируют
в большинстве типов местообитаний!
Вымирание тех или иных типов и их под..
rрупп, о котором упоминалось выше, по..
видимому., определялось в значительной
степени случайными причинами, как и то,
какие конкретно формы придут им нс! сме..
ну. Как уже rоворилось, rруппы не вымира..
ли из..за Toro. что их план строения в каком..
то смысле был хуже, чем у друrих; целые на..
48
боры неродственных между собой и хорошо
приспособленных животных, похоже, CTa
но вились жертвами радикальных измене
ний окружающей среды, природа которых
хотя и неизвестна, но хорошо коррелирует
с крупными изменениями уровня моря
и rлобальной температуры. Возможно.
мноrим типам с небольшим количеством
входящих в их COCTdB видов не. удалось
пережить эти критические периоды, а более
крупные rруппы. сохранившиеся, хотя и со
ЗНdчительными потерями в разнообразии,
исключительно блаrодаря большей числен
ности и более широкому rеоrрафическому
и эколоrическому распространению своих
представителей. получили в результате воз
можность новой радиации. Итак, хотя и не
возникает вопроса о том, является ли эко
лоrической реальностью межвидовая кон..
куренuия OHd как правило. тем интенсив
нее, чем больше сходства между конкури
рующими видами. Следовательно, ничеrо
удивительноrо, если ВЫЖИВdние в ходе [e
олоrической истории таксонов высокоrо
paHra, таких, как отряды, классы и типы,
было по большей части делом случая.
Е -. ...
'Раза paBHO
весия или
"стазис"
5.......
Радиация
"
}Прерывистость 2.7. Дополнительная
литература
Е. . . .. . . . . . . . . ., . . . . . . . . .. .
S.. . .
РадиаЦИ f
Рис. 2.21, Схематическое изображение двух
радиаций потомков одной филоrенетической
линии, демонстрирующее фазы прерывания
непрерывности и равновесия, а также YMeHb
шение разнообразия rруппы, вызванное Mac
совой смертностью из за эколоrических при..
чин (Е) и восстановления BbIcoKoro давления
отбора при насыщении местообитаний (5); см.
текст
rлава 2
Представители rрупп, переживших ос..
новные периоды вымирания, сразу после
этоrо должны были освоить сравнительно
свободные местообитания, в которых дaB
ление отбора, похоже, на время сильно сни
жалось. В ТdКИХ условиях последующая ра..
диаuия моrла идти без заметных оrраниче..
ний, приводя к широкому «эксперименталь
ному» разнообразию, сокращавшемуся
только позже в ходе вымирания, начинав
шеrося после HOBoro насыщения местооби..
таний с установлением относительно BЫCO
Koro давления отбора. Отбор в этом случае
можно рассматривать как оrраничитель
разнообразия, формирующеrося во время
каждой радиации после стадии непосред..
cTBeHHoro расселения (рис. 2.21, см. также
рис. 2.11). Именно пережившие отбор линии
в пределах каждой I руппы или подrруппы,
доминировавшей в период между зколоrи..
чески обусловленными фазами MaccoBoro
вымирания, в большей или меншей степени
давали начало новому эволюционному ЦИ..
клу.
Таким образом, случай решал, какая
пьеса будет поставлена, определял потен..
циальный состав исполнителей и открывал
занавес; отбор прослушивал кандидатов на
вакантные роли и следил за непрерывными
исправлениями, вносимыми в сценарий
и характеры персонажей.
;J.ля иллюстрации филоrенетической
точки зрения, противоположной высказан..
ной здесь, особенно в отношении родствен..
ных связей rребневиков, мшанок и щетин..
кочелюстных, а также возможноrо меСТа
возникновения основных rрупп животных
см. Nie/sel1 С. 1985. Animal phylogeny in the
ligl1t of the t[ochaea theory. Biol. J. Linn. Soc.
Lond., 25, 24J 299. (Этот автор предпола..
[ает, что предковые формы большинства
эволюционных линий животных были ра..
диально симметричными планктонными
орrанизмами, а билатеральная симметрия
сформировалась независимо у лофофоро..
ВЫХjВТОРИЧНОРОТЫХ И У первичноротых.)
Эволюционная история и филоrения беспозвоночных
Abele L. G. (Ed.) 1982. The Biology of Crustacea
(Vol. 2). Embryology, Morphology and Genetics.
Academic Press, New York.
Barпes R. S. К. (Ed.) 1984. А Synoptic Classification
of Living Organisms. Blackwel1 Scientific Publica
tions, Oxford.
Bryce о. 1986. Evolution and the New Phylogeny.
Llanerch, Dyfed,
Clarksol1 Е. N. К. 19S6. Invcrtebrate Palacontology
and Evolution, 2nd edn. Аllеп & Unwin. London.
Coheп J., Massey В. D. 1983. Larvae and the origins
of major phyl . Biol. J. Linn. Soc.. Lond., [9.
32] 8.
Сопи'ау Mon'is S., Geol'<. e J. D.. Gibsoп R.. Platt
Н. М. (Eds) 1985. The Origlns and Relationships
of Lower Invertebrates. Clarendon Press.
Oxford.
G/aessner М. F. 1984 The Dawn of Animal Life.
Cambridge University Press, Cambridge.
Goldsп1ith D. 1985. Nemesis. The Death Star and Ot
her Theories of Mass Extinction. Walker, New
York.
Heпsoп Е. D. ]977. The Origin and Early Evolution
49
of Anlmals. Wesley University Press. М iddleton,
Connecticut.
House М. R. (Ed.) 1979. The Origin of Major Inver
tebrate Groups. Academic Press, London.
Maпtoп S. М., Aпdersoп D. Т. 1979. Polyphyly and
the evolution of arth ropods. In: М. R. House
(Ed.) The Origin of Major Invertebrate Groups,
рр. 269. 321. Academic Press, London.
М cKerrOH' W. S. (Ed.) ] 978. The Ecology of F ossils.
Duckworth, LOl1dol1. Scientific Americal1 1982.
The Fossil Record al1d Evolutiol1, Freemal1.SaI1
FraI1cisco.
Sleig/1 М. А. 1979. RadiatioI1 of the eukaryote Protl
sta. Iп: М. R. House (Ed.) The Origil1 ofMajor In
vertebrate Groups. рр. 23 53. Academic Press,
London.
Trueтaп Е. R., C/arke М. R. (Ed.) 1985. The Mol1us
са (vol. 1 О). Evolutiol1. Academic Press. Orlando.
Valeпtlпe J. W. (Ed.) 1985. Phal1erozoic Diversity
Patterns. Princeton University Press. Pril1ceton.
New Jersey.
W/1ittiпgtoп Н. В. 1985. The Burgess Shale. У аlе
Ul1iversity Press, New Haven.
ЧАСТЬ 11
ТИПЫ БЕСПОЗВОНОЧНЫХ
в rлавах этой части мы проиллюстри"
руем и кратко охарактеризуем все извест..
ные современные типы беспозвоночных,
а также входящие в их состав классы.
Этот систематический обзор rораздо ме..
нее детален, чем в друrих учебниках по зоо..
лоrии беспозвоночных, так как вместо опи..
сания всех анатомических черт разных ти"
пов животных мы сосредоточили внимание
только на тех жизненно важных признаках
каждой rруппы, которые должен знать каж..
дый студент., т. е. на определяющих индиви"
дуальность каждоrо типа или класса, их
эволюционные преимущества или эколоrи..
ческую роль. Приводятся также списки
диаrностических черт. облеrчающие сравне..
ние отдельных типов.
В этой части мы постарались подчер"
кнуть разнообразие планов строения беспо..
звоночных, отраженное основным таксоно..
мическим делением животноrо царства.
При этом всем классам уделено одинаковое
внимание независимо от числа входящих
в них видов. Этот «уклон» исправлен тем,
что в части 111 преобладает материал обо..
лее крупных, хорошо известных и, без со..
мнения, более важных rруппах.
Деление на типы, классы и т. д.. приня"
тое в этой части, основано на работе Барнса
(Barnes, 1984, «А Synoptic Classification of
Living Organisms>)., Bldckwell Scientific Publi
cation. Oxford). Как известно. до сих пор
продолжается открытие новых таксонов
животных с 1980 r. описаны новый тип
Loricifera и два новых класса, предположи..
тельно из хорошо известных типов (Remipe..
dia из ракообразных и Concentricycloidea из
иrлокожих). Таким образом, хотя в книrе
полностью ОХВdчены все известные в 1987 [.
классы беспозвоночных, нет оснований счи..
тать. что человеку уже знакомы все rлавные
rруппы животных., живущих ныне на плане..
те. Похоже, что более детальное исследова
ние таких малоизученных биотопов, как
rлубоководная и интерстициальная зоны.
позволит обнаружить новые, совершенно не
похожие на известные типы существ. По..
следнее слово о разнообразии животных
еще долrо не будет сказано!
rЛАВА 3
пАрАллЕльныE подходы
К мноrОКЛЕТОЧНОСТИ у животныx
Protozoa
Ропfеrа
Placozoa
Coelenterata: Cnidaria и Ctenophora
Platyhelmin thes
Mesozoa
rруппы животных, рассматриваемые
в этой rлаве, в основном относятся к таким,
которые, соrласно предположению, выска--
занному в rл. 2, моrли возникнуть непос--
редственно в рамках царства Protista, по--
скольку они обладают принципиально раз..
личными чертами орrанизации. Таким
образом, их можно рассматривать как па..
раллельные эволюционные линии, достиr..
шие мноrоклеточности независимо друr от
друrа.
3.1. Protozoa (простейшие)
До начала 1970--х rr. было принято поме--
щать все орrанизмы в одно из двух царств:
фотосинтезирующие или растушие в земле
формы относили к растениям, а нефотосин--
тезирующие и способные к свободному
перемещению к животным. Является ор--
rанизм про-- или эукариотным. одно.. или
мноrоклеточным, считал ось при этом фун--
даментальном подходе к делению Bcero жи..
Boro неважным. В рамках подобной класси--
фикации мноrоклеточные растения потом
объединили в rруппу Metaphyta. в то время
как одноклеточные фотосинтезирующие
формы (включая rакже различных прока--
риот) бьши названы Protophyta или Algae.
Соответственно мноrоклеточные животные
составили rруппу Metazoa. а одноклеточ"
ные Protozoa.
Такая система из двух царств явно стра..
дала мноrими недостатками, два из кото--
рых важнейшие: во--первых, она не OTpa
жала реальную филоrенетическую дихото--
мию: нет никаких данных, подтверждаю--
щих, что цианобактерии, диатомеи, баrрян--
ки, мухоморы и хвойные, например, COCTaB
ляют одну естественную rруппу.. в то время
как амебы, кишечные жrутиконосцы, rубки
и иrлокожие друrую. Впрочем, такая
искусственность не была бы серьезной про..
блемой, если считать цели классификации
чисто праrматическими: два царства дол--
жны представлять собой лишь четко очер--
ченные, удобные, хотя и произвольные
взаимоисключающие подразделения opra..
низмов. Но второй недостаток как раз и за..
ключается в том, что катеrории растений
и животных реально не различаются. В пре..
делах rруппы явно близкородственных ар--
rанизмов (эвrленовые жrутиконосиы. на..
пример ) часть видов одноrо рода можно от--
нести к растениям, часть к животным,
а еще несколько форм приходится считать
и теми и друrими одновременно. Некото--
рые динофлаrеллаты удовлетворяют свои
пищевые потребности на 50/0 за счет фото..
синтеза и на 950/0 за счет rетеротрофноrо
питания. Более Toro, известны жrутиконос"
цы. способные менять царство. к которому
они были отнесены первоначально, на про--
тивоположное, просто. оставаясь сутки
в темноте! Таким образом, система из двух
царств не только не отражает филоrенети"
ческоrо родства, но и не является работаю--
щей схемой, поскольку, хотя множество ор--
rанизмов и можно с уверенностью назвать
«животными» или «растениями», суще..
52
ствует большое количество и таких, KOTO
рых не удается отнести ни к тем ни к дpy
rим.
В течение последних примерно 15 лет за
воевала приверженцев и сейчас шире Bcero
используется альтернативная классифика
ция, стремящаяся более точно отражать фи
лоrению, сохраняя при этом четкость под
разделения на rруппы. Она также постули..
рует фундаментальную дихотомию живых
орrанизмов, выделяя на этот раз Haдцap
ства прокариот и эукариот эукариоты деля
тся на 4 царства: одноклеточные (протисты)
и три царства мноrоклеточных: растения,
rрибы и животные (рис. 3.1). Протисты, Ta
ким образом, включают формы, ранее OT
носимые к водорослям и простейшим: при
этом ни те ни друrие уже не имеют форма
льноrо таксономическоrо статуса, оста..
ваясь лишь общими названиями, эквива..
лентными, например, термину «черви»; или.
друrими словами, Protozoa с некоторых пор
перестали считать валидной и практически
полезной систематической rруппой. Одна
из последних самых общих классификаций
Система из пяти царств
ф
:о
:J:
:r
О
Q.)
с:::
:!I:
е
о
:х:
JЗ L
J
>-
..D Q.)
Ь
s: о
fl
......... ... ....ш
Система из дJ3yx царств
D"Животные"
'''Животные О
и растения "'Растения"
одноеремен
HO'"
Рис. 3.1. Система классификации из пяти
царств и ее отношение к разделению всех opra..
низмов на растения и животных
rлава з
t
протистов делит это царство на 27 типов, из
которых более половины облиrатно нефо
тосинтезирующие, причем 13 из них коrда
то считались животными: Parabasalia *. Ki
netoplasta * , Opalinata * , Stephanopogono
morpha, Ciliophora, Apicomplexa *, Metamo
nada *, Sarcodina, Labyrinthomorpha *,
Choanophlagellata. Microspora *. Ascetospo
ra * и Myxozoa * (рис. 3.2) (звездочкой OTMe
чены типы, в основном или полностью со..
стоящие из эндосимбионтов или эндопара
зитов). Несколько друrих типов (например,
Dinophyta, Euglenophyta и Cryptophyta)
включают ряд форм, на которые претендо
вали как зоолоrи, так и ботаники.
Большинство «простейших протистов»
филоrенетически удалено от беспозвоноч-
ных и лишено черт, свойственных только
мноrоклеточным животным. Их общие
с ними отрицательные признаки, например
отсутствие .пластид, присуши также «rрибо-
подобным протистам» и собственно rри-
бам, а среди общих положительных можно
назвать только характерные для всех эука-
риот, включая и фотосинтезирующие rруп-
пы. Конечно, по определению разница ме-
жду нефотосинтезирующими протистами
и друrими эукариотными царствами, в том
числе и животными, заключается в том, что
последние мноrоклеточные, а первые
одноклеточные (одиночные, или колониаль
ные). Отдельная клетка протистов, таким
образом, способна выполнять все функции,
необходимые для выживания и размноже
ния, одновременно обеспечивая локомо
цию, питание, осмореrуляцию, воспроизве-
дение и т. Д., В то время как у животных эти
и друrие задачи осуществляются различны
ми специализированными rруппами клеток,
обычно объединенных в отдельные ткани
или орrаны. Тем не менее это различие
чисто фенотипическое. Все множество кле-
ток мноrоклеточноrо животноrо происх
дит от одной обычно диплоидной клетки и,
таким образом, rенетически тотипотентно,
как и отдельная особь протистов. Некото-
рые клетки, например амебоциты rубок,
MorYT даже оставаться недифференцирован-
ными и существовать в виде резерва, из KO
Toporo в течение всей жизни животноrо
дифференцируются по мере необходимости
специализированные клетки.
Рис. 3.2. Разнообразие форм тела протистов (по Sleigh et al., 1984)
54
ro
... 10 З
()
Ъ
N
s
а..
с: 10 6
ro
(f)
s
10 9
О
ll1
со
Q)
1:
.lI 10 12
()
о
а..
о
U
rлава з
.
.
.
.
e, ..
v.. '() ..
10 12
I
10 9 10 6 10 3 10
Масса тела, r
Рис. 3.3. Отношение между скоростью MeTa
болизма в покое и размерами тела для проти
стов И беспозвоночных, демонстрирующее бо
лее высокие метаболические потребности MHO
rоклеточных по сравнению с одноклеточными
тех же размеров (по Schmidt Nielsen, 1984)
(1 ккал == 4,2 кДж)
Неудивительно, что тело протистов
обычно HaMHoro мельче, чем у животных,
хотя это и не всеrда так. Размеры значите
льноrо числа сложно устроенных живот
ных., состоящих ИЗ тысяч клеток, Нd.пример
среди лофофоровых., членистоноrих или
хордовых, менее ] мм вдоль самой длинной
оси., а у большинства представителей псев..
доцеломических rрупп менее 2 мм. С дpy
rой стороны, клетки некоторых ресничных
протистов (тип Ciliophora) достиrают 5 СМ
в длину., а псевдоподии некоторых Хепо--
phyophora (тип Sarcodina) MorYT OXBaTЫ
вйть поверхность диаметром 25 см. Един
ственный способ., которым неколониальные
протисты отвечают на давление отбора.,
блаrоприятствующее увеличению размеров
(см., например., разд. 9.1), разрастание ca
мой клетки., часто сопровождающееся поли--
плоидией или появлением нескольких ядер;
при этом токи цитоплазмы формируют
впоследствии внутриклеточную систему
циркуляции. В известном смысле крупных
рdзмеров протистам достичь сравнительно
леrче., поскольку интенсивность метаболиз--
ма у одноклеточных ниже., чем у мноrокле..
точных (рис. 3.3).
Таким образом, орrанизмы, ранее отно--
симые к типу или подцарству Protozoa.
представляют собой неоднородный набор
протистов, ОТЧd.сти 'Эквивалентных одной из
клеток тела животных, отчасти целому
животному. Слово «эквивалентных» в пре..
дыдушем предложении очень важно., поско-
льку у ныне живущих протистов более
длинная самостоятельная эволюционная
история, чем у различных мноrоклеточных
орrанизмов., и только тип Choanof1agellata
сохранил некоторые признаки., позволяю-
щие rоворить о ero прямо м родстве с ж и..
вотными (rл. 2).
3.2. Тип Porifera (rубки)
3.2.1. 3тимолоrия
От лат. роrus пора; fеrrе нести
3.2.2. Диаrностические
и специфические признаки
(рис. 3.4)
1. Общей характерной симметрии нет
даже У видов, форма которых может иноrда
Зd.висеть от локальной 1колоrии.
2. Мноrоклеточные., но снебольшим
числом типов клеток; тканевая орrанизация
развита очень слабо; opraHbI отсутствуют
и координация функций клеток слабая: не-
рвной системы нет.
3. Характернь] хоаноциты., обеспечи-
вающие циркуляцию воды и ПИТd.ние.
4. Иноrда имеется сложный скелет либо
из известковых или кремнеземных спикул.
либо из белковых (коллаrеновых == спонrи-
новых) волокон, либо из элементов обоих
типов.
5. Клетки располаrаются BOKpyr БОДЯ-
ных камер или каналов разной степени сло--
жности; настоящая полость тела или кишка
отсутствуют.
6. Питание фильтрационное; rазообмен
путем диффузии.
7. Только сидячие (при крепленные) фор-
мы.
8. Размножение половое или бесполое:
известна раздельнополость, но обычен [е-
рмафродитизм; дробление полное; личинки
планктонные.
9. Исключительно водные, в основном
Параллельные подходы к мноrоклеточности у ЖИВОТНblХ
55
Спикула .. (врезка А)
Хоаноцит (врезка Б)
А ТРИУМ
ПОРОЦИТ
Мезохил
АмеБОЦИТ
Пинакоцит
Пинакодерма
Асконоидная орrанизация
Плоскость Биения
жrутика
Парус 'o
оротничковые "';
МИКрОВОрСИНКИ
Вода .
,...Пища / t I
: GPИБРИЛЛЯРНая <>
I сеть r "<:>";?:>=-d.,
,
Жrутик
Ядро
морские формы (примерно 10000 видов);
около 50 пресноводных видов.
Функционирование rубок определяется
ОДНОЙ чертой их строения: размещением
клеток BOKpyr системы камер и каналов, по
которым за счет биения жrутиков хоаноци--
тов циркулирует вода. Среда, таким обра--
зом, пропускается через орrанизм, а не
окружает ero, так что неизбежен фильтра..
ционный тип питания, как и размножение
А
Рис. 3.4. Асконоидный тип орrанизации: А
спикулы разных видов; Б срезы хоаноцита
(по Meglitsch, 1972; Alexander, R. McN., 1979;
см. также рис 2.4)
стадиями, выходящими из родительской
особи.
Простейшая асконоидная орrанизация
rубок изображена на рис. 3.4: в цeHTpe
единственная камера (атриум), окруженная
стенкой тела.. которая покрыта снаружи пи--
накоцитами (пинакодермой) и пронизана
порами (остиями), пропускающими внутрь
воду и образованными пороцитами; един--
ственное выводное отверстие оскулум.
56
,т1/ \_,111\.
. i . С .-
т 1 C: r :_
Jx' --.. С..
" . ..
Асконоидный
Сиконоидный
rлава з
t
Лейконоидный
Рис. 3.5. Разные типы складча
тости стенок тела у rубок (по
Buchsbaum, 1951)
t
Пинакоциты MorYT сокращаться. сжимая
rубку; некоторые из них секреТИРУIОТ ве..
щества для прикрепления к субстрату. Ме..
зохил 1, лежащий под пинакодермой, пред
ставляет собой студенистый белковый Ma
трикс с опорными скелетными элементами
и различными амебоцитами с широким
спектром функций от питания и секреции
коллаrена до преврашения в любой друrой
тип клеток (тотипотентность). Внекотором
смысле хоаноциты и пинакоциты образуют
1 Этот термин в отечественной литературе
употребляется пока редко, чаще вместо Hero
встречается термин «мезоrлея». Мезохил rубок
не rомолоrичен анатомически мезоrлее кишечно
полостных. Он отличается и по строению, имея
клеточную природу и представляя собой смешан
ную паренхиму с разнообразными функциями,
в то время как мезоr лея выполняет в основном
только скелетную функцию. ПРUJI\Л. пер.
Рис. 3.6. Два типа архитекто
ники rубок (по Barnes, 1980)
ткани (хоанодерму и пинакодерму), но база..
льные мембраны здесь отсутствуют.
Этот тип орrанизации оrраничивает уве..
личение размеров rубки, поскольку объем
воды в атриуме растет как куб длины тела,
в то время как жrутиковая поверхность
только как квадрат последней. Это оrрани
чение преодолено у некоторых видов за счет
складчатости стенки тела и уменьшения
атриума, что привело к образованию сико
ноидных и, наконец, лейконоидных форм
(рис. 3.5). Большинство лейконоидных [y
бок либо «сплошные» (смежные вводные
и выводные каналы проходят параллельно
друr друrу; поры, а иноrда и оскулумы раз..
бросаны по всей поверхности), либо полые
и трубчатые, с выводными каналами, ни
коrда не соединенными с поверхностью те..
ла, а открывающимися во внутреннюю по..
лость с дистальным оскулумом (рис. 3.6).
Параллельные ПОДХОДЫ К мноrоклеточности у животных
57
Это разделение по форме тела не отражает
систематики rубок (см. ниже). Наиболее
распространена лейконоидная орrанизация,
что свидетельствует о ее эффективности.
Она возникала в ходе эволюции rубок Heoд
нократно, иноrда., по видимому, через про
межуточноесиконоидноесостояние.Лейко
ноидные формы в своем развитии часто
сначала проходят асконоидную и сиконоид
ную стадии
Хоаноциты крайне важны для метабо
лизма rубок. Они проrоняют через них воду
за счет биения своих жrутиков, а также
улавливают мелкие пищевые частицы, зано"
симые внутрь тела токами воды., коrда вода
проходит через их «воротнички». представ
ляющие собой венчики из микроворсинок
(рис. 3.4). Более крупные пищевые частицы
MorYT поrлощаться амебоцитами. Как хоа..
ноциты, так и амебоциты передаI{)Т захва
ченные частицы пиши друrим клеткам. и.
похоже., основными пищеварительными
структурами служат амебоциты, а не хоано..
циты.
rубки размножаются бесполым путем.,
высвобождая небольшие фраrменты rела
или скопления необходимых для развития
клеток, в основном амебоцитов.Например.,
rеммулы (рис. 3.7, А), производимые прес
новодными rубками., состоят из археоцитов
(крупных специализированных амебоци
тов), покрытых прочной твердой оболочкой
из спонrина и спикул, позволяющей им зи..
мовать. Половое размножение. хотя обыч
но rермафродитное (черта, кореллирующая
с сидячим образом жизни), часто включает
перекрестное оплодотворение. При этом
спермий выходит через оскулум одной oco
би, проникает в пору друrой, улавливается
ее хоаноцитом и передается яйцеклетке.
Развитие до личиночной стадии чаще Bcero
протекает в материнском орrанизме. Ли
чинка обычно паренхимула со сплошным
телом., покрытым жrутиками, кроме одноrо
из полюсов (рис. 3.7, Б), но иноrда полая
амфибластула, одна из полусфер которой
состоит из мелких жrутиковых клеток.,
а друrая из макромеров без жrутиков
(рис. 3.7, В) покидает родительское тело
и короткий период времени перед оседа..
нием на дно ведет планктонный образ
жизни.
Микропиле
Защитная
ОБолочка
Внутренняя
меМБрана
Археоциты
А
Внутренняя клеточная
масса
Жrутиковый слой
Б
Макромер
8
Рис. 3.7. А. rеммула преСНО80ДНОЙ rубки. Б.
Паренхимула. В. Амфибластула (по Meglitsch,
1972)
58
rлава з
Неудивительна высокоразвитая способ
ность rубок к реrенерации (даже из одной
клетки).. обусловленная автономностью их
клеточных комплексов.
3.2.3. Классификация
Примерно 10000 описанных на сеrод
няшний день видов распределены по четы
рем классам.
3.2.3.1. Класс Hexactinellida
(шестилучевые rубки)
Представителей этоrо класса часто Ha
зывают стеклянными rубками. Их скелет
образован кремнеземными. преимуше..
ственно шестилучевыми спикулами (рис.
3.8), откуда их название; тело часто похоже
на вазу в среднем зо 40 см высотой (рис.
3.9). Преобладают сиконоидные формы.
Пинакоциты, присутствующие во всех дpy
rих классах, здесь отсутствуют, и наружные
покровы представляют собой сетевидный
синцитий, образованный соединенными
псевдоподиями амебоцитов. Исключитель
но морские виды, встречающиеся преиму
щественно на больших rлубинах. Следует
отметить, что некоторые авторы выделяют
их в отдельный подтип или даже тип (Sym
plaSlna).
3.2.3.2. Класс Calcarea (известковые
rубки)
Представители зтоrо класса отличаются
присутствием спикул из карбоната каль
Рис. 3.8. Кремнеземные спикулы rубок класса
Hexactinellida (по I:\lexander, 1979)
Рис. 3.9. Стеклянная rубка корзинка Венеры
(по Barnes, 1980)
ция кальцита или араrонита. Встречаю..
тся все три типа орrанизации ас..
коноидная, сиконоидная и лейконоидная.
Высота большинства видов менее 10 см. То..
лько морские формы.
3.2.3.3. Класс Demospongiae
(обыкновенные rубки)
Самый крупный класс, объединяющий
900/0 всех известных видов. Скелет состоит
из кремнеземных спикул и/или спонrиновых
волокон. Одно семейство (Spongiidae)
включает обычные туалетные rубки со CKe
летом только из спонrина. Орrанизация TO
лько лейконоидная, некоторые формы дo
стиrают значительных размеров (например,
тропические Splleciospoпgia более 1 м
в диаметре и в высоту). Окраска часто бле..
стящая.
Среди представителей этоrо класса так
называемые сверлящие rубки способны де..
лать отверстия в известняковых образова..
ниях кораллах или раковинах моллю"
сков. Есть и пресноводные виды, в основ..
ном из семейства Spongil1idae.
Параллельные ПОДХОДЫ К мноrоклеточности у животных
5S
3.2.3.4. Класс Sclerospongiae (корал
повыв rубки)
Класс содержит очень небольшое число
специализированных морских видов, оби..
тающих в [ротах и туннелях, связанных
с коралловыми рифами в разных частях CBe
та. Ero представители напоминают обыкно"
венных rубок, но отличаются сложным CKe
летом из кремнеземных спикул и спонrино
БЫХ волокон, внутри TOHKoro поверхностно
[о слоя живой ткани, лежащеrо на массив
НОМ основании из карбоната кальция (рис.
3.10). Все виды лейконоидные.
Класс
Отряд
Amphidiscosida
Hexactinelllda HexactlnoSIda
Lychniscosida
LуssаОПОSldа
Sclerospol1giL1t.>
Clathril1ida
LetIcettIda
L_etIcosolel1i ida
Syccttlda
Il1ozoida
Sphil1ctozoldL1
НОПl0 сlеrорhоrid;'1
Chorlstllia
SpirophoridL1
L_lthlsпdа
НаlirОlпсrlliа
Axil1cllilia
AgelasldL1
rI.1lichoJldrida
Poeci losctcrtda
Petrosiida
Н..1рlоsсlеПliа
Verol1gllda
DictyoceratilJa
DCl1droccratIdL1
--= Ccra roporellllia
т ahtllosp()l1gIJa
3.3. Тип Placozoa (пластинча...
TbI е )
3.3.1 3тимолоrия
От rреч. рlаkоs плоский; zoon
животное.
3.3.2. Диаrностические
и специфические признаки
(рис. 3.11)
1. Осей симметрии нет; форма может
меняться, как у амеб.
2. Нет отдельных тканей или opraHOB.
3. Нет полости тела или пищеваритель..
ной полости.
4. Отсутствует система нервной коорди"
напии.
5. Тело в форме толстой пластины, ко..
торая может двиrаться в любом направле..
нии в своей плоскости.
6. Единственный наружный слой жrути
ковых клеток окружает заполненный жид..
костью мезохил, содержащий сеть звездча
тых клеток"волокон.
7. Морские формы.
Верхняя поверхность плоскоrо диско..
видноrо тела образована тонким слоем че
шуевидных клеток, мноrие из которых He
сут жrутик. Более толстый нижний слой co
стоит из двух типов клеток столбчатых
жrутиковых, перемежающихся железисты..
ми безжrутиковыми (рис. 3.12). Последние
секретируют на добычу (разнообразных
протистов) ферменты, частично перевари..
вающие ее вне тела. Продукты переварива
ния поrлощаются теми же железистыми
клетками. Нижние жrутиковые клетки обе
спечивают скользящее движение животных:
изменения формы тела, делающие их похо..
жими на крупных амеб, обеспечиваются
координированными сокращениями и pac
слаблениями клеток волокон в мезохиле.
Размножение обычно бесполое как де..
лением, так и почкованием. Известно и по
ловое размножение, но оно и последующее
развитие зародыша очень плохо изучены.
Яйцеклетки, вероятно, образующиеся из
клеток нижнеrо слоя, по видимому, находя--
тся в мезохиле; в воде, [де находились жи..
вотные, обнаруживались t:пермии, но про..
цессы сперматоrенеза и оплодотворения ни
коrда не наблюдались.
Тело пластинчатых состоит из несколь..
ких тысяч клеток и достиrает ,J,иаметра
3 мм, хотя в нем представлены Bcero 4 кле..
точных типа, и эти животные относятся
к простейшим из известных представителей
мноrоклеточных. Клетки содержат меньше
60
rлава з
...
Оскулум
Известковый
скелет
Выводной
канал
Камера
\
Рис. 3,10. rубка класса Sclerospongiae с деталями строения верхней поверхности (по Bergquist,
1978)
Дорсальный (чешуйчатый)
клеточный слой
J)
Полость, заполненная
жидкостью
:::.:iЙЙ1!j 1i 1f:Ь I 1%t1: ;:, '
'z.:, '.J:.' ' ':,,: i: .. :" 1i(j.. -.d;. ' 0......\"'1. : C--':" i' :i, . . .:
: ':'\i: i *k ; l{ t: * If:,
: fI 'b. : ".' :'.) р :."д, j . r ;a G .. - .9. .::6. 9:':.
. .'. :, "'.,,'..A...{.:t.., ,\:l!:.::', .;. ':.;,n... fi1.;' . .. ,:. p.:.,. '; ;:?::
. .... .:..... .. . у. '1'1'-.) ,1J'!1,O:- ...... "'\. .<,.,q..: ,.;. , О.. ,......
'. :;:\t : t : !fj1{ :" ';:
100 мкм
I
Рис. 3.11. Общий вид Placozoa (по Barnes,
1 980)
Рис. 3.12. Схематический срез Placozoa (по
Margulis, Schwartz, 1982)
Параллельные подходы к мноrоклеточности у животных
ДНК, чем у всех прочих животных (количе
CTBd Toro же порядка, что у бактерий), xpo
мосомы очень мелкие, меньше 1 мкм В дли..
ну. Почти сто лет эти формы считали пла
нулоподобными личинками rубок или CTpe
кающих, но в конце 1960 x rr. было обнару
жено. что они MorYT достиrать половой зре
лости, и в 197] r, для них был описан новый
тип.
Пластинчатых чаще Bcero обнаруживали
в морских аКВ(lриумах, а в естественных
условиях находили на литорали. Вероятно.
они широко распространены, но их трудно
заметить.
3.3.3. Классификация
Описан только один вид (возможно, их
два). Этот тип часто помещают в отдельное
подцарство фаrоцителлообразных (Phago
cytellozoa), название KOToporo неудачно по..
скольку их способ питания обычно не фаrо
цитоз.
3.4. Тип Cnidaria (стрекающие:
rидроидные, сцифомедузы,
актинии, кораллы)
3.4.1. 3тимолоrия
От rреч. спidе крапива.
3.4.2. Диаrностические
и специфические признаки
(рис. 3.1 3)
1. Радиально симметричные животные
со щупальцами.. окружающими рот. на oд
ном из концов тела.
2. Есть внутренняя пищеварительная
(<<racTpoBacKY лярная») полость, открываю
щаяся во внешнюю среду только ртом.
3. Две основные формы тела: медуза,
приспособленная к пелаrическому образу
жизни, и бентосный полип. Полипы бывают
колониальными, а в пределах колонии по..
лиморфными.
4. Стенка тела образована двумя TKa
нями эпидермисом и rастродермисом],
1 В советской литера туре эти ТКdНИ принято
называть эктодермой и энтодермой соответ"
CTBeHHO. ПРUМ. ред.
61
состоящими из разноrо числа типов клеток.
Между этими двумя тканями находится
бедная клетками мезоrлея. Орrаны развиты
сла бо.
5. Во всех классах типа присутствуют
так называемые книдоциты со стрекатель..
ными орrанеллами (в основном не..
матоцистами), не встречающиеся больше
ни в одной из rрупп мноrоклеточных.
6. MorYT быть скелетные элементы из
хитина и/или карбоната кальция.
7. Нервная система в виде одной или
двух сетей.
8. Большинство хищники; rазооб
мен за счет диффузии.
9. Размножение бесполое и половое;
последнее часто включает фОРМИРОВdние
почти билатерально симметричной покры"
той реСНИЧКdМИ личинки планулы.
10. Только водные формы, большин--
ство морские.
В жизненных циклах стрекающих MorYT
участвовать две разные формы орrаниза
ции относительно подвижная медуза. не..
сущая rонады, и сидячий полип, который
исходно их, по"видимому, не имел (рис.
3.13). Полип трубчатый, на одном конце
у Hero рот, на друrом, базальный диск,
с помощью KOToporo животное приклепляе
тся к субстрату. У медуз рот расположен Hd
конце хоботка манубриума, прикреплен
Horo к нижней поверхности зонтика наподо
бие ручки. Полипы отпочковывают медуз
бесполым путем, а медузы образуют поли
пы в результате половоrо размножения, ча
сто с промежуточной стадией планулы
сплошной (без полости внутри) покрытой
ресничками личинки (см. rл. 14, приложение
] 4.5).
Это очень общее описание, и в преде
лах типа MHoro исключений: не все медузы
подвижны, не все полипы сидячие, HeKOTO
рые полипы несут rонады, некоторые Meдy
зы отпочковывают медуз, в ряде жизненных
циклов отсутствует стадия медузы, в дpy
rих стадия полипа, иноrда развитие про
текает без планулы.
Клеточная орrанизация полипа и Meдy
зы в основном одинакова, различия в ней
касаются частностей: у некоторых полипов
мезоrлея тонкий, в основном неклеточ
ный слой, а у медуз она толще, часто более
62
ОРАЛЬНЫЙ ПОЛЮС
Рот
Щупальца
Эпидермис
rастродермис
rастроваскулярная
полость
Мезоrлея J.
АБОРАЛЬНЫЙ ПОЛЮС
'.
,.
\
ПОЛИП
rлава з
УМБРЕЛЛА
rастроваскулярная полость
-
Рот }
С Щупальца
теБелек
Эпидермис
Мезоrлея
Парус
СУБУМБРЕЛЛА
МЕДУЗА
Рис. 3.13. Два типа формы тела стрекающих (по Fingerman, 1976)
волокнистая, иноrда в ней присутствуют
бродячие клетки (амебоциты).
Обобщенная схема строения стенки те--
ла, типичной для rидроидных полипов, при--
ведена на рис. 3.14.
Эпидермис состоит из пяти основных ти--
пов клеток:
1. Эпителиально--мышечные: столбча--
тые «покровные» клетки (иноrда чешуйча--
тые) с базальным расширением, содержа..
щим сократимые миофибриллы. ориенти--
рованные параллельно орально--абораль
ной оси тела, т. е. продольные мускулы.
ПИLЦеварительная
вакуоль
Клетки, секрети
руЮL.ЦИе слизь
Псевдоподии
Пищеварительные
мускульные клетки
Клетки, ceKpe
тирующие срерменты
2. Интерстициальные: содержат круп..
ные ядра и мало цитоплазмы; способны
образовывать друrие типы клеток.
3. Стрекательные клетки, или книдо--
циты: содержат способные выворачиваться
структуры нематоцисты (рис. 3.15, см.
также рис. 2.5), чаще называемые книДоци--
стами. У rидр и медуз на одном конце этих
клеток расположена жесткая щетинка
книдоциль, устроенная так же, как и жrу..
тик. У актиний ее нет. но иноrда присут--
ствует конический комплекс ресничек, ве..
роятно, выполняющий те же функции. Не--
Ядро эпителиально
мускульной клетки
Сенсорная клетка
Книдоцит
Книдоциль
Клетки, секретирующие
слизь
Интерстициальная клетка
Рис. 3.14. Продольный срез стенки тела в классе Hydrozoa (по Barnes 1980)
Книдоциль
Опорный
стержень
Нематоциста
Врезка А
Параллельные ПОДХОДЫ К мноrоклеточности у животных
Крышечка
Стилет
Иrла
Нить
Опорная
Cf)ИБрилла
Ядро
А
маТОIIиста включает свернутую в спираль..
rофрированную, зазубренную изнутри
трубку. ВыстреЛИВdние почти наверняка
обусловлено быстрым поступлением в кни"
доцит воды, связанным с изменением ero
мембранной проницаемости, которое
в свою очередь, вероятно, вызывается элек..
трически по сиrналу книдоциля. Нить выво..
рачивается наизнанку, складки ее расправ..
ляются" а сама «отстреленная» КНИДОЦИСТd
растяrивается.
Существуют три основных типа книдо"
цист:
а) собственно нематОЦUС111Ы раз..
нообразные по форме, часто с ШИПdМИ (сти"
63
Рис. 3.15. Книдоцит и нематоцисты: А
неотстрелянная нить сложена складками; Б
отстрелянная нить; В спироциста актинии
(по разным источникам) (см. также рис. 2.5)
Б
в
летами) и токсичные: используются для Зd..
щиты И ловли жертвы;
б) спuроцuсты клейкие структуры, об..
наруженные у некоторых актиний. Их нити
растворяются при выстреливании.. образуя
rустую клейкую сеть для прикрепления
и ловли;
в) пmuхоцuсmbl обнаруженные только
у актиний rруппы Ceriantharia (с. 73) струк"
туры со сложно rофрированными нитями
без шипов. Нити также клейкие и способ..
ствуют формированию трубки, в которой
находятся актинии.
Книдоцисты используются только один
раз, а потом отбрасываются. Книдоциты
64
MorYT реrенерировать из интерстициальных
клеток; развивающийся книдоцит называе..
тся книдобластом.
4. Слизисто..железистые клетки: выра..
батывают слизь для защиты, ловли добычи
и приклеивания.
5. Сенсорно"нервные клетки: удлинен..
ной формы, расположенные под прямым
уrлом к поверхности эпидермы. Более или
менее тождественны мультиполярным не..
йронам (rл. 16). Соединяясь с друrими не..
йронами, образуют неправильную сеть, или
плексус (см. разд. 16.4.1). У некоторых стре..
кающих (не rидр) две нервные сети в эпи..
дерме, у друrих одна в 1пидермисе, дру"
rая в rастродермисе (см. ниже).
ТаспlродерJ'VlUС состоит из: а) питательных
мышечных клеток, поrлощающих пищевые
частицы для последующеrо внутриклеточ"
Horo пищеварения. Они, как и эпитеЛИdЛЬ"
но"мышечные, способны к сокращению, но
их сократимые части ориентированы под
прямым уrлом к орально..аборальной оси,
образуя слой кольuевой мускулатуры. С по..
мощью жrутиков перемешивают пищу в ra..
строваскулярной полости; б) железистых
клеток, вырабатывающих ферменты, необ..
ходимые для пищеварения в [астроваску"
лярной полости В) клеток, секретирующих
слизь; [) у некоторых стрекающих (напри..
мер, актиний) книдоцитов. У медуз и ак"
тиний [астродерма сложно"складчатая.
Локомоция происходит с помощью мы..
шечных сокращений, преодолевающих дав..
ление жидкости в rастроваскулярной поло..
сти, которая выполняет функции rидроста..
тическоrо скелета (см. rл. 1 О), как. вероятно,
и мезоrлея. Движения у полипов обычно
оrраничены изrибанием и скольжением;
у медуз они rораздо разнообразнее. Но да..
же последние MorYT dКТИВНО перемещаться
только в орально..аборальной плоскости
блаrодаря хорошему развитию кольцевой
мускулатуры по нижнему краю зонтика, по..
зволяющей животным двиrаться в основ..
ном вертикально: Torna как rоризонтальное
перемещение происходит пассивно за счет
океdнических течений. Тем не менее некото"
рые медузы (Cubomedusae, см. с. 69) спо..
собны К быстрому активному движению
в rоризонтальной плоскости аборальной
поверхностью вперед.
rлава з
Следует отметить способность стрекаю-
щих к реrенерации. Она использована
эмбриолоrами для изучения общих пропес-
сов клеточноrо деления и дифференциров-
ки, в частности у rидры, которую леrко раз-
водить в лаборатории (см. разд. 15.5). У нее
орально..аборальная полярность восстанав-
ЛИВdлась, даже если животное разрезали на
несколько кусков вдоль этой оси. Щупальца
всеrда возникали на ближайшем ко рту це-
лоrо животноrо конце TaKoro фраrмента.
Похоже, что необходимая для это-
ro информация связана с орально-
аборальным rрадиентом KaKoro"To ве-
щества, различаемоrо клетками и вызываю-
шеrо у них соответствующую реакцию (де-
ление и дифференцировку) в зависимости от
ero концентрации, а следовательно, от
уровня вдоль rрадиента, на котором распо-
ложены клетки.
Митозы происходят вдоль всей оси тела,
но существует rрадиент их скорости от
высокой у оральноrо полюса, сразу под rи-
постомом, т. е. буrорком, над которым
располаrается рот (см. рис. 3.16)" до низ-
кой y аборальноrо. Таким образом, под..
держивается постоянный «ток» клеток от
оральноrо к аборальному полюсу и к КОН-
пам щупалеп. В результате со своих мест
ОРАЛЬНЫЙ ПОЛЮС
r ипостом
J Д8ижение
метки незна
чительно или
отсутствует
8 дней
СРормируюшаяся
почка
АБОРАЛЬНЫЙ ПОЛIOС
Рис. 3.16. Перемещения клеток rидры (по
Campbell, 1 967)
Параллельные ПОДХОДЫ К мноrоклеточности у животных
Класс Подкласс Отряд
Athecata
МilIероrmз
Stуlаstеrmз
Chondrophora
Hydrozoa Thecata
Limnomedusae
т rachylma
Slphonophora
Actinulida
Stauromedusae
Coronatae
Semaeostomeae
Rhlzostomeae
[Cubozoaj Cubomedusae
Cerlantlpatharla = Antipatharla
Ceriantharla
Protoalcyonarla
Stolonifera
т elestacea
Alcyonaria Gastraxonacea
Gorgonacea
Coenothecalia
Anthozoa Alcyonacea
Pennatulacea
Actiniaria
Zоапthшаrlа
Zoantharla Sсlеrасtlша
Corallimorpharla
Ptychodactiaria
вытесняются клетки которые отмирают
(происходит «линька») и используются для
формирования новых почек. Таким обра
зом идет непрерывное обновление клеток
в течение нескольких недель все они у oд
ной особи замещаются новыми что позво
ляет животным быть как бы бессмертными
(разд. 13.2.4).
3.4.3. Классификация
Описано около 10000 ныне живущих ви
дов. Их распределение по таксонам высше
ro ранта зависит от тото, какая стадия
полип или медуза доминирует в жизненном
цикле. В классе Hydrozoa (rидроидные) CTa
дия полипа обычно преобладает, в классе
Scyphozoa (сuифомедузы) все наоборот.
В третьем классе Anthozoa все формы
полипоиды, а медуз, вероятно, нет вовсе.
Однако термин «полипоид)) использован
в данном случае сознательно. поскольку эти
животные, хотя и сидячие, имеют мното об
65
щих черт со сцифомедузами: толстую кле
точную мезоrлею, ввернутый внутрь MaHY
бриум (так называемый стомодеум), ra..
стродермальные тонады. Известны и сидя
чие медузоидные формы (см. ниже). Так что
Anthozoa вполне можно считать сидячими
медузами без настоящей полипной стадии!
Тем не менее одна из существующих
классификаций рассматривает Anthozoa как
(коралловые) полипы, а отсутствие у них
медузной стадии как настолько cepьe
зный признак, что их помещают в отдель
ный от rидроидных и сuифомедуз подтип
(табл. 3.1). Те же артументы можно исполь
зовать, если считать Anthozoa медузами,
лишенными полипной стадии. Такой под
ход имеет свои преимущества: он позволяет
выделить у Anthozoa несколько классов, KO
торые, как мы увидим, сильно различаются.
Однако отсутствие в жизненном цикле Me
дузы или полипа не такая уже редкость и
у представителей друrих классов, следова
66
Таблица 3.1. Классификация стрекающих
Классическая
Альтернативная
Класс Hydrozoa
Класс Scyphozoa
Класс Anthozoa
Подкласс Alcyonaria
[Подкласс Zoantha..
ria 1 >]
Подтип Medиsozoa
Класс Hydrozoa
Класс Scyphozoa
Класс Cиbozoa
Подтип Anthozoa
[Класс Cerian ti patha..
ria 1 >]
Класс Alcyonaria
Класс Zoantharia
l)Включая Ceriantharia.
[ ] Некоторые считают
(Schmidt, 1972)
таксон искусственным
тельно, ему можно было бы не придавать
слишком большоrо значения и у Anthozoa.
Поэтому здесь мы используем более клас
сическую схему делящую стрекающих на
три крупных класса Hydrozoa, Scyphozoa
и Anthozoa, признавая ее, впрочем, не впол
не удовлетворительной.
3.4.3.1. Класс Hydrozoa
(rидроидные)
Нуdrа хорошо всем известная rидра
(рис. 3.16) не является типичным представи
телем этоrо класса, так как она: а) одиноч
ная; б) лишена медузной стадии. rораздо
более характерна Obelia (рис. 3.17). Три MO
мента характеризуют полипную стадию
этих животных: а) они состоят из несколь
ких полипов (модулей), связанных со столо
нами (стелющаяся rидрориза и восходящий
rидрокаулус), так что представляют собой
колониальные, или модульные, орrанизмы;
б) не все полипы одинаковы: некоторые, со
щупальuами, выполняют пищеварит-ель
ную функцию (rастрозоид, или троФозоид);
друrие (rонозоиды) репродуктивную, т. е.
отпочковывают медуз, третьи, отсутствую
щие у Obelia, но обычные у друrих rидроид
ных, булавовидные, несущие батареи кни
доцитов и служащие для защиты (дактило
зоиды, с. 70); в) присутствует опорный CKe
лет из хитина (иноrда обызвествленноrо),
называемый перисарком; ткань, которую
он окружает (ценосарк), охватывает общую
для всей колонии полость тела. У разных
rлава 3
таксонов MorYT быть значительные как ко..
личественные, так и качественные различия
в этих чертах.
у медузы Obelia MHoro характерных для
rидромедуз черт. Она мелкая (0,5 см), ее ro..
нады эпидермальноrо происхождения, ЯИЧ"
ники и семенники находятся на разных осо..
бях. Однако на внутреннем краю зонтика
присутствует только рудиментарный парус
(велум), rораздо лучше развитый у друrих
видов rидромедуз (ср. рис. 3.18). rастродер..
мис без мускулов. Вместо них специализи..
рованные мускулы по краю внутренней по..
верхности зонтика образуют кольцевые
слои, создающие движущую силу. Они со..
кращаются, выталкивая воду вниз, а меду"
зу вверх. Серии сокращений напоминают
пульсацию. Иноrда парус образует узкое
отверстие, создающее струю воды. Муску"
лы работают, преодолевая сопротивление
мезоrлеи, которая, хотя и неклеточная, од..
нако содержит белковые волокна, придаю..
щие ей упруrость. Движение вниз пассивно.
Нервная система также специализирова..
на плексус образует два нервных кольца
у нижнеrо края зонтика, иноrда даже с rаи..
rлиями. Последние связаны с сенсорными
орrанами, расположенными также по краю
зонтика: rлазками (реаrируют на свет)
истатоцистами (реаrируют на ускорение)
(рис. 3.19).
У некоторых видов медуза не плавает
свободно. а остается прикрепленной к поли..
пу, высвобождая только [аметы. rонозоид
в этом случае называется rонофором. Вслу..
чае Tubularia от rастрозоида не отделяется
не только медуза (rонофор), но и планула.
Вместо них высвобождается личинка..
актинула окруrлая пелаrическая поли..
поидная форма (рис. 3.20). У некоторых ви"
дов прикрепленная медуза редуuируется
и остаются только rонады (например, у rид..
ры). Прикрепленность и редукция медуз, ве..
роятно, возникали у rидроидных несколько
раз независимо. С друrой стороны, в ряде
СЛУЧаев из жизненноrо цикла исчезает по..
лип, а не медуза (рис. 3.21).
Колониальность и полиморфизм дости"
rли cBoero предельноrо выражения У неко"
торых своеобразных пелаrических видов:
знаменитоrо портyrальскоrо кораблика
(Physalia) с заполненным rазом поплавком
Параллельные подходы к мноrоклеточности у животных
67
РОТ
rипостом
СтеБелек
Щупальце
rидротека
r идрокаулус
rастрозоид
r ОН8да
Медуза
(d'или« )
оплод творен О :.
ное яицо . .
f
/
r идрориза столона
Молодая колониЯ
Рис. 3.17. Жизненный цикл Obelia (по Barnes, 1980)
Зонтик
Ротовое LЦупальце
Рот
СтеБелек
rастроваскулярная
полость
Радиальный канал
Кольцевой канал
rлазок
Парус
Рис. 3.18. rидромедуза рода
Bougaiпvi//ea (по Bullough
1958)
Батареи
нематоцист
(видоизмененная медуза) и свисающими
вниз шупальпами; отходящими от корми
диев (пучков модифицированных полипов.,
рис. 3.22,А), а также парусника (Velella) с ero
блюдцеобразным поплавком, (видоизме
ненный rастрозоид), несущим rонозоиды
и пактилозоиды (рис. З.22,Б).
Наконец., некоторые колониальные rид
роидные секретируют внутренний эпидер
мальный известковый скелет Это так назы
68
rлава з
Конкреция
Кольцевой канал
Парус
ЭКЗУМБреллярное
нервное
кольцо
Сенсорный
эпителий
Сенсорная
ресничка
Рис. 3.19. Статоцист rидроме
дузы (по Barnes, 1980)
Выходящая актинула
Прикрепленная
медуза (rOHOcpop)
ПРlAкрепленная
медуза (rOHOcpOp)
Актинула
Колония rидроидов
При крепленная аКТlAнула
Рис. 3.20. Жизненный ЦИКЛ Tubularia (по Barnes 1980)
Параллельные подходы к мноrоклеточности у животных
69
Рис. 3.21. Жизненный цикл
rидроида Ag/aura не имеюще
ro полипной стадии (по Barnes,
1980 )
ваемые rидрокораллы (отряды Milleporina
и Stylasterina) (рис. 3.23). Это прямостоячие
и обрастающие формы, достиrающие иноr
да значительных размеров. Их мноrочи
сленные дактилозоиды MorYT причинять си
льную боль, поэтому некоторые представи
тели данной rруппы называются оrненны
ми или жrучими кораллами.
3.4.3.2. Класс Scyphozoa
(сцифомедузы, сцифоидные)
Типичный для сцифоидных жизненный
цикл y рода Aurelia (рис. 3.24"Ь). Домини
рует медуза (рис. 3.24,А) с диаметром зон
тика в среднем 10 см, у некоторых видов он
достиrает 30 см. Если не принимать во вни"
мание размер, сцифомедузы очень похожи
на rидромедуз, описанных выше (с. 66).
Различия следующие: а) манубриум несет
щупальца (короткие у Aurelia, но очень
длинные у некоторых лруrих видов); б) нет
паруса (кроме Cubomedusae; см. ниже); в)
мезоrлея толстая и волокнистая, с блу..
ждающими амебоцитами; [) нервные коль..
ца редки, контроль за пульсапией осуществ"
ляется краевыми скоплениями нейронов,
расположенных в мелких булавовидных
структурах ропалиях, несущих также ста..
тоцисты и иноrда rлазки (рис. 3.25); д) [она..
/
План ла
'
Медуза
Актинула ,
ды rастродермальноrо происхождения
Большинство сцифомедуз хищники, но
некоторые, в том числе и Aurelia,
фильтраторы. У этоrо вида молодь филь..
трует воду краями щупалец, а взрослые осо..
би улавливают планктон с помощью слизи..
стых тяжей, секретируемых нижней поверх..
ностью зонтика. Вся она и лопасти ману"
бриума покрыты жrутиками, которые rонят
пищевые частицы ко рту.
Медузы обычно раздельнополы. Опло..
дотворенные яйца развиваются в планулу,
КОТОРdЯ образует полипоидную сцифисто"
му; та почкуется поперечным расщепле
нием, давая ювенильные медузы эфиры
(рис. 3.24). Как сцифистома, так и эфира
питающиеся стадии.
Существуют аберрантные сuифоидные:
некоторые не образуют сцифистому, у др у..
rих сцифистома быстро формирует [онады,
но отсутствует медуза. Ставромедузы жи..
вут, прикрепившись стебельком к морским
водорослям (рис. 3.26). Упомянутые выше
кубомедузы быстрые пловцы, наносящие
очень сильные ожоrи. У них есть парус и [а..
стродермальные rоналы, так что их часто
считают промежуточными формами между
rидромедузами и сцифоидными, помещая
в отдельный класс Cubozoa. Часто встре..
чающаяся в Карибском море Cassiopeia
Хитиновые септы
Рот
Уровень воды
rастродер
мальная
} полость
Кормидий
Дактилозоид
(со щупалы!Зми)
tf"'\1 rrr:;i ..." f" &;
'" v- J .:: 1\
Кормидии .' : ;\\
i j/ j.' :\
rастрозоид / J . ,)
I 1/1.
Щупальце
(вытяrивается
до 9 м)
А
"Парус"
Пневматоq:>ор, разделенный на камеры
rазовые ТруБКИ + выросты
энтодермы
. c:; C;::JJ ,/": р " r'[) ....... .......... C::J r со.
1: ." i' . У , .{ 1" & ;. D\ () tf). '0,.'
;,:, '!Щ Н .(' "'ij;;... '.: .2;. .,..;;:-...f,.....,!-« ';\,
i ./ j . j.) ,4 'f t O \"'., \,
.,' д' " r. t . 'iJ '
,'; ;' rастрозоид ( . Три РЯДа r Hol, '\ ;
З0ИДОВ
Рот Три ряда
дактилозоидов
Рис. 3.22. А. Physalla и детали строения ее кормидия. Б. Разрез через тело Velella (по раЗНblМ
источникам)
Б
r астрозоид Дактилозоид
I О о
о
о О о
о о
о
....."'...... .r . :. . . I .
Рис. 3.23. Участок колонии
rидрокоралла Mi//epora: еле-
ва вертикальный разрез,
справа вид сверху (по Rus-
sel.. Hunter, 1979)
..'W-:. '!' ""'." .. - ... .., ,
, ': " ":.".. L _ ;. ;.,.: _......:. (-.,? "'-: }.. . ; ;\"
о о
Ропалии
/
А
СВОБодноплавающие самец и самка медузы
(половое размножение)
Эсрира \
Б
Рис. 3.24. Медуза (А) и жизненный цикл (Б) Aurelia (по Fingerman, 1976)
Рис. 3.25. Ропалий Aurelia
в разрезе (по Hyman, 1940)
СТ8ТОUИСТ
72
Вторичные
щупальца
Рот
Отверстие
СУБУМБрел ...
лярной ямки
Ротовой
стеБелек
Положение
rOHaA внутри
СтеБелек
Пр крепитель",
ныи диск
Рис. 3.26. Haliclystus представитель отряда
Stauromedusae (по Bullough, 1958) .
прикрепляется куполом вниз на тихом мел..
ководье манrровых зарослей. У нее множе..
ство вторичных ртов, открывающихся не..
посредственно в rастроваскулярнуlO поло..
сть. Мелкие животные, уловленные поверх..
ностью оборчатых «рук», перемещаlOТСЯ
к этим ртам по слизистым тяжам.
3.4.3.3. Класс Anthozoa
Представители этоrо класса поли"
поиды, но (см. также выше" с. 059) крупнее
и сложнее по строению, чем полипы в дpy
rих классах (рис. 3.27). Их уплощенный ора..
льный диск несет щупальца и щелевидный
рот, ведущий в латерально уплощенный
стомодеум. В одном или в обоих «уrлах»
рта может присутствовать ресничный жело..
бок, сифоноrлиф, ведущий в стомодеум.
Сифоноrлифы, вероятно, направляlOТ воду
в rастроваскулярную полость" поддерживая
в ней rидростатическое давление. Вместе
с щелевидной формой рта наличие сифоно--
rлифов придает симметрии Anthozoa черты
бирадиальности. т. е. уже невозможно раз--
резать их тело на две эквивалентные поло--
вины вдоль ЛlOбоrо диаметра. Мезентерии
(или септы), т. е. выступающие складки ra..
rлава з
стродермиса с мезоrлеей внутри, разделяют
rастроваскулярнуlO полость. Некоторые из
них, так называемые полные, прикреплен..
ные к стомодеуму, но имеlOТ в районе рта
отверстия в стенках для облеrчения цирку..
ляции rастроваскулярной жидкости; друrие,
llеnОЛllые, к стомодеуму не прикреплены
(рис. 3.27).
Края мезентериев представляlOТ собой
трехлопастные тяжи (рис. 3.28). Наружные
лопасти покрыты ресничками, а uентраль..
ная несет книдоциты и клетки, вырабаты"
ваlOщие ферменты. Для Anthozoa характер..
но, что нематоцисты лишены крышечки
(нить часто вырывается из нематоцисты
просто через трехклапанное отверстие).
Книдоциль также отсутствует (см. с 62).
Септы MorYT нести и свободные нити
аКОНIIИИ. которые закручиваются BOKpyr до..
бычи и участвуlOТ в ее умерщвлении с помо..
щью мноrочисленных книдоцитов. Мезо..
rлея толстая, волокнистая, клеточная. Му..
скулатура в основном rастродермальноrо
ерфорация
в мезеkТерии
томодеум
Мезентериаль-
ная нить
Мускул ре ...
трактор
rOHaдa
Сисроноrлисры
Неполные
вторичные
мезентерии
Полные
первичные
мезентерии
Рис. 3.27. Схема строения типичной актинии,
Внизу поперечный разрез на уровне CTOMO
деума (по Alexander, 1979)
Неполный
третичный
мезентерий
Параллельные ПОДХОДЫ К мноrоклеточности у животных
73
Отверстие
в септе
ПРОДОЛЬНЫЙ
мускул
СТОЛБИК
Радиальный
МУСКУЛ
Подошва
Стомодеум
М езентериальная
нить
rонада
Аконция
Рис. 3.28. Строение мезентерия (по Fretter, Graham, 1976)
происхождения; продольные мускулы
ретракторы расположены в мезентериях,
а кольцевые в rастродермисе внутренней
трубки. Размножение как бесполое
почкованием (иноrда образуются коло
нии см. ниже), поперечным расщепле
нием и разрывом (см. rл. 14), так и поло
Boe c rонадами rастродермальноrо про..
исхождения в некоторых или во всех мезен
териях. Известны раздельнополые и repMa..
Фродитные формы. Оплодотворение Hapy
жное или внутреннее; в последнем случае
часть развития может протекать внутри ма..
теринскоrо орrанизма (примитивное живо..
рождение). Обычно образуется планула,
планктотрофная или лецитотрофная (см.
rл. 14), время существования которой Ba
рьирует. Рот и мезентерии MorYT начать
формироваться внутри личинки, но щупа
льца обычно не образуются до оседания на
дно.
Актинии самые обычные одиночные
Anthozoa; они распространены повсеместно
в прибрежных водах. Обычно они прикре..
плены к твердым субстратам в литоральной
зоне. У некоторых. например, Metridiuт,
хорошо развита мускулатура вокрут ораль..
Horo диска, образующая сфинктер таким
образом, что щупальца втяrиваются
внутрь. коrда актиния сжимается во время
отлива (рис. 3.29) для защиты и экономии
энерrии на дыхании. Обычный североаме..
риканский и европейский вид Stomphia cocci
пеа может при уrрозе отрываться и плавать,
изrибая тело. Несколько друrих видов пла..
вают за счет ударов щупалец. Представите
ли семейства Minyadidae пелаrические; они
плавают вниз шупальuами с помощью ra..
зовоrо пузыря. Ceriantharia также пред
ставители одиночных Anthozoa, приспо
собленные к существованию на илистом
дне. Они живут в слизистых трубках, обра
зованных птихоцистами (см. с. 63).
Известны также различные колониаль
ные кораллообразующие Anthozoa. Каме..
нистые склерактиниевые кораллы состоят
из полипов, соединенных латерально и «си..
дящих» на известковом «наружном скелете»
(рис. 3.30). Это обеспечивает опору и защи
ту, так как котда полипы втяrиваются, они
74
rлава з
.:'\' t/ '& I, ,",,
.'. ;;- }J..'t\, I,! l:=t
,1 J( :,;jl' :1 ;\
.' .( 1'1/;, 1 /i t,ч \ \\ ',\..'"
: :]"",II'..'..:" ' :, ','
Рис. 3.29. Metridiuт seпile на разных стадиях
сокращения тела (по Robbins, Shick, 1980, с из..
менениями)
лишь слеrка выступают над скелетной пла..
стиной. Октокораллы (или восьмилуче..
вые у них Bcero восемь щупалец) также
образуют разветвленные колонии полипов,
латерально соединенных uененхимой, со..
стоящей из толстой мезоrлеи и связующих
rастродермальных трубок (рис. 3.31). Мезо..
rлея содержит амебоциты, секретирующие
скелет из СаСО з . Таким образом, здесь ске..
лет внутренний. У некоторых в цененхиме
находится центральный опорный стержень
из белка или известковых спикул. В послед..
нем случае ero выделяют из так называемо..
Полое пери стае
щупальце
Эпидермис на поверх...
ности колонии
Стомодеум
Мезоrлея
rастродермальные ТруБКИ
между полипами
Наружный
скелет
.'
Рис. 3.30. Продольный разрез через тело oco
би коралла из отряда Scleractinia (по Barnes,
1980)
ro блаrородноrо коралла (Corallium)
и обрабатывают, получая всем известные
коралловые ювелирные изделия.
И наконец, rруппа Аlсуопаriа это
мяrкие кораллы, образуюшие мяrкие кожи..
стые колонии, достиrающие размеров более
1 м. Их цененхима также содержит из вест..
ковые спикулы, но их здесь меньше, чем
у дрyrих восьмилучевых
Форма всех этих кораллов значительно
варьирует и зависит от типа модульноrо ро..
СТа.
Рис. 3.31. Продольный разрез
через полип 80сьмилуче80rо
коралла (по Russel..Hunter,
1 979)
Параллельные ПОДХОДЫ К мноrоклеточности у животных
3.4.4. Симбиоз
Большое количество стрекающих из
всех классов находится в симбиозе с aBTO
трофами. Зоохлореллы (зеленые BOДOpO
сли) встречаются в rастродермисе rидроид
ных, в мезоrлее сидячей сцифомедузы Cas
siopeia (см. с. 69); зооксантеллы (динофла
rеллаты из рода Syтbiodiпiuт) B racTpo
дермисе актиний и кораллов (Anthozoa).
Излишки продуктов фотосинтеза
мальтоза у rидры и в основном rлицерин
у Anthozoa используются хозяином и MO
rYT поддерживать ero в течение длительно
ro периода rолодания, если не составляют
единственный источник питания. У корал
лов симбиоз способствует также отложе
пию известковоrо скелета.. но каким обра
зом плохо понятно. Роль симбиоза в пи
тании обсуждается в rл. 9.
Симбиоз с участием в основном Antho
zoa важен также для защиты друrих opra
низмов. Мелкие рыбки из рода Aтphiprioп
живут среди щупалец крупных актиний. Ka
кие то вещества на этих рыбках (а возмо
жпо" И слизь" выделяемая самой актинией)
инrибируют выстреливание нематоцист.
Актинии защищают симбионтов и в свою
очередь получают от них защиту от HeKOTO
рых хишников. Сходный комменсализм
известен у актиний с несколькими друrими
видами рыб, креветками чистильщиками,
раками щелкунами, крабами стрелками,
офиурами. Актинии MorYT прикрепляться
к друrим животным: сцифомедузам, rребне
викам и раковинам, в которых обитают pa
ки отшельники. В этих случаях они также
защищают хозяев, получая преимущества
от перемещения на новые кормовые участ '
ки.
3.5. Тип Ctenophora
(rребневики)
3.5.1. Этимолоrия
От rреч. ktеnоs rребень; phoros
несущий.
75
3.5.2. Диаrностические
и специфические признаки
(рис. 3.32)
1. Медузоидные формы, но радиальная
симметрия стала бирадиальной из за двух
щупалец.
2. rастроваскулярная полость со ртом
и анальными порами.
3. Двуслойная стенка тела окружает
толстую мезоrлею с амебоцитами и rладки
ми мышечными волокнами.
АБОРАЛЬНЫЙ _ .
ПОЛЮС .,,' - \
, ,r .
.......... /
/ / " 1
; \. - / .....,'
i . .
Шупальце
Статоцист
rреБная
пластинка
rастроваскулярная
полость
Рот
ОРАЛЬНЫЙ
ПОЛЮС
Статоцист
АБОРАЛЬ
НЫЙ ПОЛЮ
Рис. 3.32. Типичный rребневик: вид сбоку
(вверху) и с аборальноrо полюса (внизу) (по
Buchsbaum, 1951)
76
rлава з
Статолит
Мерцательная БОроздка
Статоцист
.' ""
'. "\..
" . ,.
. '.' t . .
"':l
Пру-жиняшие реснички
4. КНИДОЦИТЫ отсутствуют, но щупаль
ца несут «ловчие клетки» (коллобласты).
5. Локомоция с помошью слившихся
ресничных, так называемых rребных, пла
стинок, расположенных восемью орально
аборальными полосами.
6. Нервная система представляет собой
субэпидермальную сеть., хорошо развитую
под рядами rребных пластинок.
7. Хищники; rазообмен за счет диффу
зии
КолЛОБласт
Рис. З.33. Детали строения
аборальноrо конца rребневика
со статоцистом и rребными
пластинками (по Barnes, 1980)
8. Размножение у некоторых бесполое,
но таких мало; rермафродиты с полным де..
терминированным дроблением.. ведущим
к образованию «цидиппидной» личинки.
9. Исключительно морские, в основном
пелаrические формы.
Животные, относяшиеся к этому типу,
так похожи на медуз из типа стрекающих,
что их часто считают боковым ответвле..
нием предков ой медузоидной формы по-
следних. В самом деле, долrое время rреб..
Радиальная
ФИБрилла
Корешок
r ранулы с клейким
веществом
Рис. 3.34. Разрез через щупальце rребневика с деталями строения коллобласта (по различным
источникам)
Параллельные подходы к мноrОI<леТОЧI-iОСТИ у животных
невиков помещали со стрекающими 8 один
тип кишечнополостных Coelenterata (от
rреч. koilos полость; enteron кишеч
ник). Типы стрекающих и rребневиков и
сейчас часто объединяют в раздел лучи..
стых Radiata (по типу симметрии).
Тем не менее у rребневиков MHoro Ba
жных отличительных черт. Первая среди
них ряды «rребней» (откуда название ти
па), т. е. коротких поперечных пластинок
с ресничками, соединенными в виде rребня
(рис. 3.33). Биения ресничек направлены
в аборальную сторону, так что животное
плывет ртом вперед и обычно вверх по вер..
тикали, но может плыть и вниз. Контроль
за движением осуществляется нервной се..
тью и апикальным статоцистом (рис. 3.33).
Щупальца выворачиваются из rлубоких
выстланных ресничками карманов на або
ральной стороне. Они несут коллобласты
(рис. 3.34) rрушевидные структуры с BHY
триклеточными свернутыми спиралью ни..
тями и заполненными слизистым веще
ством rранулами, служащие для опутыва
пия добычи. До недавнеrо времени предпо
лаrалось, что у вида Euchlora rubra есть кни
доциты, но новые данные свидетельствуют
о том, что он отбирает их у своей жертвы..
книдарии. Кишечник состоит из каналов,
как у некоторых медуз, но они MorYT OTKpЫ
ваться во внешнюю среду анальными пора
ми. Пищеварение внутри и внеклеточное,
непереваренные остатки удаляются через
.рот и анальные поры. rермафродиты с ro
надами из двух пучков (яичник и семенник)
в rастродермисе. Цидиппидная личинка
очень похожа на яйцевидных взрослых oco
бей, ее развитие прямое, однако у уплощен
IIЫХ форм оно требует значительных пре
образований (см. ниже).
Заметная черта rребневиков их люми..
несценция. Светящиеся вещества образу
ются в rастродермисе.
.5.З. Классификация
«Малый» тип.. включающий Bcero около
100 известных видов, распределяемых по
.двум классам, как показано ниже.
77
Класс
ОТРЯД
CydipPlda
Platyctlda
Loba ta
Ganeshida
Thalassocalycida
Cestida
т enraculara
Nuda
Beroida
3.5.3.1. Класс Tentaculata
у всех представителей этоrо класса два
щупальца; форма тела обычно напоминает
крыжовину (рис. 3.32). Однако некоторые
виды сплюснуты в щупальцевой плоскости;
тоrда наружные части расширяются в виде
лопастей (рис. 3.35), иноrда так, что живот..
r реБНОЙ ряд ....
"'t
-'9
-q
/11
11
:;}
'1:1
..q
1}
,q
-JJ
Р",
ff.
p;
Р'р
Ушковидные
щупальца
Ротовая лопасть
Рис. 3.35. Вид в саrиттальной плоскости щупа
льцевоrо rребневика Mпeтiopsis (по Meglitsch,
1972)
rреБНОЙ ря
Марrинальное
щупальце
Рис. 3.36. Сильно уплощенный щупальцевый
rребневик Ve/aтeп (по Meglitsch 1972)
78
rлавное щупальце
Анальная пора
rOHaAa
Рис. 3.37. Cteпop/aпa. rребневик, уплощенный
ВДОЛЬ орально аборальной оси (по Meglitsch,
1972)
ное становится похожим на прозрачную
ленту. В этом случае для локомоции испо--
льзуются не только rребные пластинки, но
и волнообразные изrибания всето тела
(рис. 3.36). Особи друrой rруппы уплощены
по орально--аборальной оси, ряды rребных
пластинок развиты очень слабо, или вовсе
отсутствуют, образ жизни ползающий
Сенсорный opraH
Рис. 3.38. Веrоё, один из представителей клас..
са Nuda (по Meglitsch, 1972)
rлава з
(рис. 3.37). У всех уплощенных форм щупа--
льца редуцированы
3.5.3.2. Класс Nuda
У представителей этоrо класса щупаль--
ца отсутствуют на всех стадиях жизненноrо
цикла Хищники, питающиеся друrими
rребневиками, с крупным растяrиваю--
щимся ртом и расширенным стомодеумом,
куда умещается их яйцевидная жертва.
В связи с этим тело примерно конической
формы (рис. 3.38).
3.6. Тип Platyhelminthes
(плоские черви)
3.6.1. 3тимолоrия
От rреч. рlаtу плоский; helminthes
черви.
3.6.2. Диаrностические
и специфические признаки
(рис. 3.39)
1. Билатерально симметричные формы.
2. Тело из трех слоев тканей; системы
opraHoB расположены в среднем паренхи"
матозном слое
3. И:Ffой полости тела, кроме слепо
оканчивающейся кишки, нет.
4. Питание почти исключительно за
счет друrих животных.
5. Уплощены дорсовентрально; черве..
образные свободноживущие формы с rоло..
вой, несущей сенсорные орrаны.
6. В основном мяrкотелые, но с защит-
ной слизистой секрецией; несколько видов
с внутренними спикулами.
7. rазообмен посредством диффузии.
кровеносная система отсутствует.
8. «Выделительные» орrаны про"
тонефридии.
9. Нервная система может иметь вид се..
ти, но обычно сконцентрирована в продоль..
ные тяжи.
10. В основном rермафродиты.
11. Дробление яйца спиральное.. но мо"
жет сильно модифицироваться.
12. Развитие у свободноживущих форм
обычно прямое (у некоторых видов со ста..
Параллельные ПОДХОДЫ К мноrоклеточности у животных
79
ПЕРЕДНИЙ КОНЕЦ
Поперечный
срез
Наружные
кольцевые
м'ускулы
ЗАДНИЙ КОНЕЦ
Влarалише
ХОБотка
Дорсальный эпидермис
(с раБдоидами)
ХОБОТОК
Внутренние
продольные
мускулы
Два пучка дopco
вентральных
мускулов
Вентральный
эпидермис
Рис. 3.39. Типичный свободноживущий плоский червь (по Russel Hunter, 1979)
дией свободноплавающей мюллеровской
или rётевской личинки), а у ряда паразитов
очень сложное, с несколькими личиночны..
ми стадиями.
13. Найдены во всех основных водных
и сухопутных местообитаниях, включая
ткани друrих животных.
Три основные черты этоrо типа
настоящая билатеральная симметрия;
возникновение всеrда клеточноrо TpeTbero
слоя ткани (паренхимы) между эпидерми..
сом и выстилкой кишки (rастродермисом);
дорсовентральное уплощение тела. Билате..
ральная симметрия характерна и для всех
друrих до сих пор не рассмотренных ти"
пов так называемых Bilateria (хотя иrло..
кожие вторично вернулись к особой форме
радиальной симметрии, см. разд. 7.3). Она
возникла в ответ на потребность в передви--
жении по поверхности твердых субстратов
(ер. с дорсовентрально уплощенными rреб..
невиками, разд. 3.5.3.1). Уплощение тела
неизбежное следствие отсутствия кровенос..
ной системы и зависимости этих животных
от диффузии при распределении по орrаниз"
му питательных веществ и rазов и выводе
из Hero продуктов обмена (rл. 11).
В этом типе (во всех rруппах кроме
Acoela) мы впервые встречаемся с протоне--
фридиальной «выделительной» системой
Кавычки использованы потому. что ее
функция, вероятно, в основном осмореrуля..
торная, а не экскреторная (rл 12)
собственно экскреты по--прежнему удаляю..
тся из тела через ero поверхность путем
диффузии. Протонефридии состоят из сле..
пооканчивающихся трубок, на конце кот о..
рых расположены так называемые «пламен..
ные» клетки (рис. 3.40), фактически из двух
клеток: трубчатой и терминальной, с двумя
или более жrутиками. По месту пальчатоrо
соединения этих клеток находятся просве..
ты. ведушие в трубчатую часть. Через них
биение жrутиков, напоминающее пламя, за..
тяrивает внутрь пламенной клетки жидко..
сти, которые затем проrоняются по труб..
кам к выделительной поре и удаляются из
тела.
В общем, экскреторная, пищеваритель..
ная, репродуктивная и нервно..сенсорная си..
стемы, включающие интеrрированные
комплексы тканей и opraHoB, становятся
у плоских червей более заметными и сло..
жными, чем у rубок и «кишечнополостных».
Поскольку у представителей этоrо типа
слепозамкнутая кишка и примитивный
«рот», не удивительно, что большинство
ero представителей питается за счет тканей
друrих животных. Несколько видов поедает
водоросли, в основном диатомовые. Общая
тенденция среди плотоядных
установление все более тесной связи между
потребителем и пищей, даже у преимуще..
ственно свободноживущих турбелларий
(разд. 3.6.3.1). Здесь трофические связи ко..
леблются от взаимоотношений хищник
80
Ядро
Терминальная
клетка
Отверстия
Два жrутика
"ресничноrо пла
мени" терминаль
ной клетки
Соединение
двух клеток
ТРУБчатая клетка
Ядро
Несрридиальные
поры
Протонесрридиальная
сеть
Рис. 3.40. Детали строения клетки ресничноrо
пламени (вверху) и система протонефридиев
(по Barnes, 1 980)
.жертва (или трупоед падаль) до форм
симбиоза, включая экто и эндокомменса
лизм (выrодно для симбионта, безразлично
для хозяина) и даже экто и эндопаразитизм
(симбионт живет за счет хозяина). Боль
шинство классов и видов плоских червей ис
ключительно паразитические. С появле
нием паразитизма возникают специализи
рованные орrаны для прикрепления к xo
rлава 3
зяину; изменяется внешний облик; редуци
руется или (у некоторых кишечных парази..
тов, окруженных полупереваренной пищей)
совершенно исчезает пищеварительный
тракт; увеличивается репродуктивная систе..
ма.
3.6.3. Классификация
Около 25 000 ныне живущих видов, pac
пределенных по четырем классам.
3.6.3.1. Класс Turbellaria
(ресничные черви)
Представители этоrо класса в основном
свободноживущие водные формы, но
известно несколько сухопутных видов, при
уроченных к влажным местообитаниям.
Есть также сведения о комменсализме.
Один отряд Temnocephalida включает
исключительно эктокомменсалов (возмо
жно, эктопаразитов) поверхности тела paKO
образных, моллюсков и черепах. Сзади
у них клейкий диск, спереди щупальца
(рис. 3.41) а реснички на поверхности тела
отсутствуют. Они так выделяются среди
турбелларий, что paHr этоrо отряда иноrда
повышают до класса.
Размеры турбелларий колеблются от He
скольких миллиметров (мноrие из них оби
тают в морской и пресноводной интерсти"
циали) до более 0,5 м (rиrантские наземные
формы). Реснички, покрываюшие эпидер
мис, обеспечивают локомоцию турбелла..
рий, самые мелкие из них «плавают» в ин
терстициальной воде, а уплощенное тело
более крупных ползаюших форм увеличи..
вает ресничную поверхность, соприкасаю
щуюся с субстратом. Мускульные сокраще..
иия, вызывающие волнообразное движение
тела, важны для самых крупных водных
и всех наземных видов. Слизь, выделяемая
железистыми клетками, смазывает и защи-
щает поверхность тела, оставляет на суб-
страте след обеспечивающий опору реснич-
KaM и может также использоваться для
ловли жертвы. Таким образом, она очень
важна для этих животных, и более 500/0 эне..
рrии, получаемой за счет пищи.. тратится на
образование слизи.
Поперечный срез тела типичной турбел..
Параллельные ПОДХОДЫ К мноrоклеточности у ЖИВОТНblХ
81
Класс
Подкласс
т urbellaria
ОТРЯД
Nemertodermatida
Acoela
Catenulida
Macrostomida
Proplicastomata
Lecithoepitheliata
Prolecithophora
Neorhabdocoela
т emnocephalida
Proseriata
т ricladida
Polycladlda
Monogenea
MonoPlsthocotvlea
Polyopisthocotylea
<:::: Aspidogastrea
т rema toda
Digenea
Cestodaria
Eucestoda
ларии приведен на рис. 3.42 (см. также
рис. 3.39): видны реснички (у более крупных
форм они MorYT отсутствовать на спинной
поверхности) и несколько мускульных
слоев из кольцевых, продольных" косых
и поперечных волокон. Эти волокна не ис
черченные. Если учесть все сказанное о сли
зи, неудивительно, что железистые клетки
такие крупные. Они часто расположены
в паренхиме и имеют выводной проток, OT
крывающийся на внешнюю поверхность те..
ла. Заметны также различные палочковид
ные структуры рабдоипы. одни из них BO
Рис. 3.41. Представитель отряда Temnocephali..
da
Aspidohothria
StrIgeldida
Azygiida
Echinostomida
Plagiorchiida
Opisthorchiida
Amphllinida
:= Gyrocotylida
Caryophyllida
Spathehothriida
т repanorhynchlda
Pselldophyllida
l.ecanice(.")hali<:la
Aporlda
т etraphyltida
Dlphyllida
Litobothrida .
Nippotaenlda
Proteocepnalida
CyclophyHida
rлотка
Кишка
Репродуктивная
система
Прикрепительный
диск
82
rлава з
Ядро
IJ ' ."'.. = ый Э ИС
:.:.:..,',.,....'.'.:::::.:.:.:....... ,.,., .:................ Базальная меМБрана
Кольцевые мышцы
; ?<" -1.. 80t 8С..,.. оос' 1\.
"'I(.. .,..,. IJC. ос. :а.с о..с. Пиrмент
f f (? (? IJ О (J 17 Диaroнаlьные мышцы
о о о о о о О о о о О Продольные мышцы
00000000000
Эпидермис
Мышечный слой
ПареНХИМ8
rастродермис
Нервный
ствол
о о 000000000
000000000000
C)
.. .. ...о .. ..
:-:.: .......:.. о" .:.: ... '".. ........ :..:. :::.".:... ...... .....:..'" :..: ..... ...
., .
I
, , ' вентральный
эпидермис
Реснички
Просвет
Клетка фаrоцит
Дорсовентральная мышца
Тело мышечной клетки
,
"д \ \
I\
I! 111
1( 1 1 111
[1 !/J
\Jl t
Рис. 3.42. Поперечный разрез через турбелларию с деталями строения рабдоидов, дорсальной
и вентральной поверхностей и выстилки кишечника (по различным источникам)
зникают из эпидермальных, друrие (так Ha
зываемые рабдиты) из железистых кле
ток в паренхиме. Их функции окончательно
не ясны, но они выбрасываются, коrда жи
вотное потревожено, и служат для защиты
(см. разд. 13.2.1.2) и/или разрушаются,
образуя BOKpyr Hero толстый слой слизи.
Они есть почти у всех турбелларий. Про-
странство между внутренними орrанами за-
полняет паренхима скопление неправиль-
ной формы клеток. Среди более крупных
клеток встречаются помельче, с крупным
Параллельные подходы к мноrоклеточности у животных
83
Наземная срорма
т ricladida
Пиrментная
клетка
Пресновqдная срорма
т ricLadida
rолова CatenuUd&
Ресничная ямка
"Мозr"
ядром и тонким слоем цитоплазмы, похо
жие по структуре и функциям на интерсти
циальные клетки стрекающих. Они назы
ваются необластами и иrрают важную роль
в замене износившихся тканей, а также в pe
rенерации после бесполоrо размножения
или повреждений (см. разд. 15.5.2).
Подвижность турбелларий привела
к определенной цефализации, т. е. обо
соблению rоловноrо отдела мо
дифицированной передней части тела,
первой попадающей в новые условия при
"Булава"
сетчатки
Нерв
Клетка
сетчатки
Пиrментные
клетки
Рис. 3.43. Цефализация (специализация
rоловноrо конца) у турбелларий и детали
строения «rлазных пятен}) (по различным
источникам)
движении (рис. 3.43). Обычно здесь появ
ляются rлаза (два или MHoro), устроенные
по типу пиrментноrо бокала (разд. 16.7.3).
На rолове также заметны ресничные поло
ски, поrруженные в желобки. Вероятно,
это хеморецепторы, помоrающие Haxo
дить пищу. Для илобслуживания развилось
переднее скопление нервной ткани
«rоловной мозr». От Hero отходят пары
продольных нервных стволов, количество
которых сокращается у некоторых rрупп до
двух вентральных (рис. 3 44). Эпидермаль
84
rлава з
Статоцист
" Мозr"
Рис. 3.44. Нервная система турбелларий из отрядов Acoela (слева), Polycladida (в середине)
и Tricladida (справа) (по Barnes, 1980)
Простой мешковидный
кишечник
fJyковицевидная rлотка
Acoela
Большинство отрядов
Трехветвистый
кишечник
Мноrоветвистый
кишечник
ТРУБчатая выдвиrаюшаяся
rл от ка
т ricladida
PoL ycLadida
Рис. 3.45. Типы орrанизации кишечника и рта у турбелларий (по Russel.. Н unter, 1979)
ная нервная сеть может присутствовать или
отсутствовать, хотя некоторые Acoela
имеют только ее.
Кишка (рис. 3.45) варьирует от мощной
синцитиальной массы клеток (у бескишеч
ных турбелларий) и простоrо мешка (<<раб
доцель») до ряда латеральных выпячиваний
(три у Triclada, MHoro y Polyclada). Эти
варианты положены в основу классической
систематики турбелларий. С ними коррели..
руют способы соединения рта с кишечни-
ком (рис. 3.45).
Параллельные подходы к мноrоклеточности у животных
85
Семенники
Яичник
Архоосроры
А
Сперма
Яичник
Рис. 3.46. Системы размножения турбелларий
rонады хорошо выражены (за исключе
нием Acoela) и связаны с внешней средой
выводными трубками, на которых может
быть несколько расширений с различными
ФУНКIlИЯМИ (рис. 3.46). Половая система
самца всеrда состоит из мноrих семенни
КОВ, сперматозоиды обычно двужrутико
вые. у самки может быть либо MHoro яич
ников, rде образуются эндолецитальные
яйца, либо единственный яичник и множе
ство отдельных желточников, производя--
щих желточные клетки; в этом случае яйца
эктолецитальные (разд. 14.4).
А
Б
Семевыносящии
канал
Совокупительный
opraH
rOHonop
Неоосроры
" rоловка"
Б
Сперматоциты
Сперматоrонии
Турбелларий без желточников называют
архоофорными, с желт очниками Heo
офорными.
Друrая заметная черта турбелларий
их способность к бесполому размножению,
в основном путем поперечноrо деления,
коррелирующая с ярко выраженной способ
ностью к реrенерации. При этом, как и
у стрекающих, наблюдается полярность
(здесь переднезадняя), возможно, OCHO
ванная на осевом физиолоrически--
эмбриональном rрадиенте. И наконец, при
rолодании Triclada способны сильно YMeHЬ
86
шаться, иноrда от размеров взрослой особи
до размеров только что вылупившеrося жи
BOTHoro, а начав питаться, вырастают зано
во. Предполаrалось, что этот процесс OMO
лаживает ткани Triclada, делая их практиче
ски бессмертными.
3.6.3.2. Сосальщики
у следующих двух паразитических клас
сов настолько MHoro общих черт" что paHЬ
ше они считались одним классом (и paCCMa
триваются так до сих пор в некоторых учеб
никах). Отличие в том, что представители
класса Monogenea в основном эктопарази
ты с простым жизненным циклом и одним
хозяином (обычно рыбой), в то время как
в классе Trematoda почти все виды
эндопаразиты. Более Toro, у представите
лей OCHoBHoro подкласса трематод
Digenea сложный жизненный цикл со CMe
ной двух или более хозяев. Внешне по фор
ме тела сосальщики напоминают турбелла
рий, но есть и важные отличия: xapaKTep
ный для последних ресничный клеточный
эпидермис заменен без ресничным синци
тием (TerYMeHToM) (рис. 3.47); появилась
мускулистая rлотка" с помощью которой
пища активно вытяrивается из тканей xo
зяина; она открывается в двуветвистый ки
МИКРОТРУБОЧка rЛИКQКаликс
Апикальная
плазматическа
меМБрана
Цитоплазмати"
ческий мостик
Аппарат rольджи
ШЭР
Щелевой
контакт
А
Интерстициальное
вещество
rлава з
шечник (рис. 3.48); у взрослых особей OT
сутствуют специальные сенсорные орrаны.
Класс Moпogeпea
(МОНО2енетические сосальщики)
Представители этой rруппы как пра
вило, эктопаразиты рыб, хотя из этоrо пра
вила MHoro исключений: некоторые из них
обитают в полостях тела, открывающихся
наружу .во рту" на жабрах" в мочеполовой
системе, а несколько видов обнаружены дa
же в целоме; есть паразиты амфибий, репти
лий и rоловоноrих моллюсков.
Как и следует ожидать в случае эктопа
разитов, которым необходимо прикрепля-
ться к наружной поверхности тела часто
подвижноrо хозяина, как на переднем, так
и на заднем конце тела моноrеней заметны
орrаны прикрепления, включая присоски,
крючья и зажимы (рис. 3.49). Сзади они
особенно крупные, и их строение положено
в основу разделения этоrо класса на два от..
ряда: М oпophisthocotylea с цельным зад
ним фиксаторным аппаратом и парными
прикрепительными орrанами спереди, Ро..
lyophisthocotylea c расчлененным задним
фиксаторным аппаратом, но единственной
ротовой присоской или парой выворачи..
вающихся буккальных присосок.
r емидесмосома
П rликокаликс
_ оверхностное
впячивание
Шип
Интерсти альное
вещество
Клетка паренхимы
Тело клетки поrPi..
женноrо эпителия
с тельцами Т 2
Б
Рис. 3.47. Схема строения покровов моноrенетическоrо сосальщика (А) и трематоды (5): ШЭР
шероховатый эндоплазматический ретикулум
Семенник
Двуветвистый кишечник
Половое отверстие
Ротовая при соска
rлотка
Пищевод
Семенной пузырек
Сумка цирруса
Семепровод
Циррус
Предстательные железы
Присоска
Семевыносящии
канал
Выделительный
канал
Лауреров канал
Семеприемник ОРАЛЬНАЯ
Яичник СТОРОНА
елточный проток
Матка
Qотип
Скорлуповая железа
Желточные железы
Мочевой пузырь
Присоска
Рис, 3.48. Схема строения rенерализованной двуустки
Прикрепительный диск "Якорь" Прикрепительный диск
Б
Рис. 3.49. Моноrенетические сосальщики: А Gyrodacty/us из отряда Monopisthocotylea; Б
Po/ystoтa из отряда Polyopisthocotylea (по Meglitsch 1972)
Передний при крепительный
opraH
Прикрепительные железы
Рот
r лотка
Скорлуповая
железа
Рот
Несрридиальная пора
Копуляционное
отверстие
Семепроеод
Коnyляционное
отверстие
Яичник
Желточный проток
rенитально",кишечный
канал
Кишка
ЭМБрИОН
Яйцо
Кишка
Протонесрридий
Присоска
88
rлава з
Прикрепительная
поверхность
Рот
Железистые клетки rлотки
Выделительная пора
Внутренний
резервуар
Внешний
резервуар
Семеприемник
Яичник
Выносящий канал
сперматосрора
Кишечник
Внешний мускул
Прикрепительный
диск
rлазные
пятна
Рот
ресничноrо
пламени
Крючок
rлаз
r лотка
Пенис
Семеизверrательный
канал
Яйцо
Отверстие влаrалища
Влаrалище
Семенник
Краевой валик
Крючок
Краевой крючочек
Крючок
rлотка
Кишечник
Реснички
Прикрепительный
диск
Рис. 3.50. Типичный жизненный ЦИКЛ моноrенетическоrо сосальщика на примере паразита рыб
Eпtobde//a soleae (по различным источникам)
Параллельные подходы к мноrоклеточности у животных
89
Жизненный цикл моноrеней относитель
но прост. Яйца защищены скорлупой, при
вылуплении высвобождается свободнопла
вающая, покрытая ресничками личинка
онкомирацидий. Она служит для расселе
ния на новых хозяев (рис. 3.50).
Класс Treтatoda
Представители этоrо класса
эндопаразиты, мноrие из которых вызы
вают заболевания человека и домашних жи
вотных. Как и у моноrеней, есть орrаны
прикрепления, но у представителей крупно
ro подкласса Digenea (диrенетические coca
льщики) они расположены BOKpyr рта (po
товая присоска) и посередине брюшной по
верхности (брюшная присоска). В физиоло
rии этих животных TerYMeHT иrрает важную
роль. Он обеспечивает защиту, особенно от
ферментов у видов, обитающих в кишеч
нике, но также и от антител хозяина (см.
разд. 13.2.2.5). Через Hero непосредственно
от хозяина MorYT быть получены некоторые
питательные вещества, в частности амино
кислоты.
Жизненный цикл диrенетических сосаль
щиков сложен: хозяин взрослой (половозре
лой) особи называется окончательным; это
позвоночное. Хозяева, в которых развиваю
тся друrие стадии цикла, способные к беспо
лому размножению, называются промежу
точными; их может быть от одноrо до трех
(один обычно брюхоноrий моллюск).
Типичный жизненный цикл выrлядит
так (рис. 3.51):
1. Яйцо, покрытое скорлупой с крышеч
кой, попадает в кишечник хозяина и BЫBO
дится наружу с фекалиями (а у некоторых
диrенетических сосальщиков через моче
вой пузырь с мочой).
2. Из Hero вылупляется свободноживу
ВЗРОСЛАЯ ОСОБЬ
Сумка цирруса
ЦЕРКАРИЯ
Поведение
<Рототаксис
rеотаксис
Осмотическое давление
емпература
Свет РСО 2
l)t
МИРАЦИДИЙ
СПОРОЦИСТА
ОБнаружение улитки:
хе""ютаксис?
Миrрация и выход
церкарии наружу
Паразитическая кастра я,
ОБЩИЙ метаБОЛИЗМ,
Иммунитет Беспозвоночноro
Рис: 3.51. Типичный жизненный ЦИКЛ трематоды (по Smyth, Halton, 1983)
90
rлава з
щая, покрытая ресничками личинка
мирацидий.
3. Ее съедает моллюск, или она сама
внедряется в Hero через эпидермис. Затем"
потеряв реснички, паразит проникает в пи
щеварительную железу хозяина.
4. Там он превращается в спороцисту.
5. Спороциста содержит зародышевую
массу, из которой митотически возникает
несколько дочерних спороцист и/или редия.
6. Последняя содержит зародышевые
клетки, которые аналоrичным образом раз
виваются в личинки церкарии.
7. Они покидают хозяина, прободая ero
rкани, и становятся свободноживущими.
8. Церкарии проникают в окончатель..
Horo хозяина или съедаются им (жизненный
цикл завершен), или же находят промежу..
точноrо хозяина из позвоночных и там ин..
цистируются, образуя метацеркарию.
9. Если промежуточный хозяин" coдep
жащий метацеркарию, будет съеден оконча..
тельным хозяином, цикл завершен.
Половозрелые паразитические особи
производят rиrантское количество яиц (на..
пример, печеночная двуустка около
20000 вдень). Более Toro, каждая выходя
щая из яйца личинка размножается в ходе
дальнейших бесполых стадий. Такая BЫCO
кая репродуктивная способность (возмо
жно, связанная с тем, что эндопаразиты по
определению окружены сверхобильной пи
щей тканями хозяина) компенсирует Mac
совую rибель рассели тельных стадий при
случайной их передаче от одноrо хозяина
к друrому.
Schistosomatidae (шистосомовые)
семейство диrеней, выделяющееся по двум
причинам. Во..первых, оно включает виды,
вызывающие у человека опасное (иноrда
смертельное) заболевание шистосомоз.
В эндемичных областях им может страдать
большая часть населения; это один из са..
мых распространенных rельминтозов чело
века BO BTOpЫX, шистосомовые раздельно
полы, чем резко отличаются от друrих пло
ских червей. Почему раздельнополость во..
зникла у паразитов, [де шансы на встречу
особей разных полов проблематичны,
остается заrадкой. Тем не менее" коrда
встреча происходит, между ними возникает
тесная взаимосвязь: у самца по всей длине
Вентральная присоска самки
Ротовая присоска
самца
Вентральная присоска
caMua
Вентральный (rинекоq:)Qр
ный) жеЛОБ
Рис 3.52. Schistosoтa тaпsoпi (по Meglitsch,
1972)
тела проходит вентральный желоб" в кото..
рой помещается более тонкая самка. Взро..
слых животных часто обнаруживают имен..
но в такой ассоциации (рис. 3.52).
Представители подкласса Aspidogastrea
в чем..то сходны как с диrенетическими, так
и с моноrенетическими соеальщиками
(рис. 3.53). У них нет заднеrо прикрепите..
РОТ
Присоска
Ячейка
Рис. 3.53. Представитель подкласса Aspidoga
strea (по Meglitsch 1972)
Параллельные ПОДХОДЫ К мноrоклеточности у ЖИВОТНЫХ
91
льноrо аппарата, но присутствует либо
крупная присоска, покрывающая всю вен..
тральную поверхность тела, либо ряд вен..
тральных присосок. Это эндопаразиты.. об..
наруженные в кишечнике рыб и рептилий,
а также в перикардиальной и почечной по..
лостях моллюсков. Однако их жизненный
цикл прост; он включает двух" а иноrда все..
[о одноrо хозяина.
3.6.3.3. Класс Cestoda
Представители этоrо класса, обычно
известные под названием ленточных червей,
приспособились к паразитированию в ки"
шечнике позвоночных.. и.. таким образом,
стали самой специализированной rруппой
Platyhelminthes. Окруженные массой полу..
переваренной пищи хозяина, они утратили
и рот.. и кишечник. Вместо этоrо, как и у со..
сальщиков (и даже в большей степени), те..
rYMeHT (рис. 3.54) приобрел способность
к непосредственной абсорбции пищи. Он
покрыт микроскопическими выростами
/
(микротрихами), увеличивающими ero по..
верхность, а в своей толще содержит боль..
шое количество митохондрий, что свидете..
льствует об интенсивности происходящих
здесь процессов активноrо транспорта. На
поверхности TerYMeHTa также может проис..
ходить переваривание пищи с помощью
ферментов хозяина так называемое мем"
бранное пищеварение.
Сложно устроенный opraH прикрепле..
ния сколекс с присосками и крючьями
(рис. 3.55) удерживает червя в потоке пи..
ЩИ, создаваемом кишечной перистальти"
кой. Жизнь в длинном трубчатом простран..
стве с движущейся пищевой средой блаrо..
приятствовала эволюции особой формы те..
ла стробилы, состоящей из одинаковых
члеников (проrлоттид), отпочковываю..
щихся вблизи сколекса (стробилизация), со..
зревающих и постепенно отделяющихся на
противоположном конце тела в виде ме..
шочков, набитых яйцами. В средней части
этоrо цикла в них развивается половая си..
стема (рис. 3.55), очень похожая на поло..
вую систему диrеней. Если возможно, про..
исходит перекрестное оплодотворение. но
известно самооплодотворение между двумя
проrлоттидами в одной стробиле или даже
Микротрихи
Наружная
плазматическая
меМБрана
П иноцитозный
пузырек
Внутренняя
плазматическая
меМБрана
Базальная меМБрана
Соединительный
канал
Митохондрия
Аппарат rоль и
Эндоплазматический
ретикулум
Ядро
Рис. 3.54. Схематизированный разрез через
покровы цестоды (по Meglitsch, 1972)
в одной проrлоттиде. Характерно, что
образование и развитие cerMeHToB у кольча..
тых червей и членистоноrих происходит
в противоположном по сравнению с цесто"
дами направлении, т. е. с заднеrо конца тела
вперед, а не спереди назад (см. с. 37).
Оплодотворенные яйца заполняют матку,
и в зрелых проrлоттидах мало что остается,
кроме этой растянутой набитой яйцами
матки. Таkая «конвейерная» система раз..
множения обеспечивает по разительную
плодовитость. Например, взрослый бычий
цепень (Taeпia sagiпata) может состоять из
70 1000 проrлоттид, теряя от 3 до 10 из
них в день; по 100000 яиц в каждой.
Яйца из зрелой проrлоттиды вылуп"
ляются либо во внешней среде, либо после
проrлатывания этой проrлоттиды хозяи..
ном. Из них выходит личинка онкосфера;
иноrда она окружается ресничной мембра..
ной (эмбриофором) и В этом случае назы..
92
Матка
rЛ8ва з
Яичник
елточная железа
Половая пора
Половая пора
вается корацидием. Эта стадия в промежу
точном хозяине может развиваться в раз
ные типы личинок, основные из которых:
1. Процеркоид мелкая веретеновид
ная форма с массивным телом и крючьями
на заднем конце.
2. Плероцеркоид массивная форма
с дефинитивным сколексом, но без зароды
шевых крючьев процеркоида't из KOToporo
она развивается.
3. Цистицеркоид, состоящий из пере
днеrо пузыря с ввернутым внутрь сколек
Рис. 3.55, Форма тела ленточ
ных червей с деталями строе-
ния различных отделов и тремя
типами сколексов (по различ-
ным источникам)
сом и хвостовидноrо заднеrо отдела с личи-
ночными крючьями.
4. Цистицерк пузырь с единственным
ввернутым внутрь сколексом.
Друrие личиночные типы представляют
собой варианты этих четырех. Иноrда про-
исходит бесполое размножение сколексов
пузыревидной стадии в промежуточном хо-
зяине. Коrда ero съедает окончательный хо-
зяин, личинка попадает в кишечник и пре-
вращается во взрослоrо червя. На рис. 3.56,
A B показано несколько типичных жизнен-
ПараллеЛЬНblе ПОДХОДЫ К мноrоклеточности у ЖИВОТНblХ
93
А
Море
Прямой сток
в море
?
?
Взрослый червь
в кишечнике
Холодильник
(личинка поrИБает
через 10 дней
при goC)
' ; ; :: . I .
:'.,: ....:. ..,
::.. '."
. ,,-. .
..,. I '.
о'" ,., "
.... ... .
' I g заражение
:.. ЖизнеСПОСОБные
яйца проходят . # ..7.... яйца в С , токах , ;It i1t
сквозь кишечник Яица разносятся
пти ц ы .,\\11 питающимися Па .,\1/, * \\lц
)1I1Jl jli( млью Птwa ми "
)JlЦ о "0 о iIt о))щ o о о \14 jI( 02==+ve
..... ............... .. ...... .... .'. ......... ........ ....... ...... ..' о... .'...' .......... .....' ....... .......... ...... Онкоссрера ....' . '.' Цистицерк . . . . . . . . . . . . . .
Рис. 3.56. Жизненные циклы цестод: А Taeпia sagiпata: Б Diphyllobothriuт deпdriticuт
(с. 090); B Hyтeпolepsis папа (с. 091) (по. Smyth, 1962)
ных циклов, каждый из которых включает
некоторые из этих стадий.
Не следует думать, что ,раз ленточные
черви прочно пр икре плены сколексом, они
не MorYT передвиrаться внутри кишечника
хозяина. Их мускулатура под TerYMeHToM
состоит из кольцевых и продольных, а в па
ренхиме из продольных, поперечных
и дорсовентральных волокон. Более Toro't
в сколексе расположено переднее скопление
нервной ткани, от KOToporo отходят два ла
теральных ствола, тянущиеся назад вдоль
всей строБилыI. MorYT присутствовать дop
сальныI,, вентральные и дополнительные
латеральные стволы, а также связывающие
продольные стволы кольцевые комиссуры.
Карликовый цепень (Hyтeпo/epis) способен
миrрировать на крупные расстояния в ки
шечнике хозяина, вероятно, блаrодаря этой
сложной нервно мускульной системе.
Обычно класс ленточных червей делят
на два подкласса Eucestoda и Cestodaria.
В первый (собственно ленточные черви)
входят 12 отрядов, сильно отличающихся
друr от друrа формой тела и хозяевами.
К последним относится широкий спектр
теплокровных и холоднокровных позвоноч
ных. Специальноrо упоминания заслужи
вают представители отряда Caryophyllida
(рис. 3.57), паразитирующих на пресновод
Б
: I II ?? I II I I: : H ::::::::::::: ::::::: : ::::::::::::: ::: :;:::::::::: ::::::: ::::: :::::::: : : ЩI 1 1 I I I } ?:::::::::::::: ::::::: : I I j
................ ........... .. « Скрытыи период'" 7 днеи .. . . . . . . . ... . . .. . ...... к .......
III I?I? ? ::.." . . ......: .:.:. :.:.:. :. : ..: : Ж II III ? :
III II II {.. . . Ji!]Ш : : IМ!! ::::.. ":: II I{
::: :::::: ::::::::::: ::::::: ::::::::::. .':::: пеРВИЧНQ при... -:::::: ный плероцер ::::::: ':'. :::::::::::::::: ::::::: ::::::::
. . . . . . . . . . . . . . . . . . . . . . . . . ' . . ' . . крепляется К . . . ' . . ' . . "" ОИД . . . . . .. . .. . .. . .. . .. . \ . . . . . . . . . . . . . . .
.................. n ..............
.................. . ..... ...............
.:.:. :.:.:.:.:.:.:.:.:.:.:. :.: .:.: ........... П р ямой кишке ...... '.':''''''''''', :.:.:. :.:.:. :.:. :.:.:.:. :.:.
................................. ..:.:.:.:.:. :.:.:.................. .-.-.-.... .................. .........................:
........... ....... ....... ....... 8.8.....*.... ...........*.*.. ........... а.. ............ ..... а._...*. е. . ....... ........8...*.111. а..
:' . :' . : ' . : . . : . *: . . :' . : . . :' . : . ': . -:- . : . . :' . : ' . :' ..:.:.:.:.:.:.: . . . . . . . . . . . . . . . . . . .. :.:.:.:.:.: .:.:.:.:.:.:.:.:.:.:.:.:.:.: .:.:.:.:.:.:.:.:.: .: . . : . . : . . :. . :' . : ' . : ' . : . . :" . :' . : . . : . . :
....... ...... ........ ........... ..................... ...
................. ...................................... ........................................... .............
::::::: ::::: ::: ::: :::::: :::::: .::: :::::::::::::: ::::::::::::::::::::: Птица : :::: ::: :::::::: ::::::: ::::: ::::::::::::::: .:::::::::: ::::: :::::::::
Анус
,
Яйца в пропитанной
А хинонами ОБОЛОЧке
V с крышечкой выводятся
с срекалиями
, 8ыСВОБОЖдающиИся
А на C TY <:рермент вылупления
воздеиствует на крышечку .
\./ ....
......' .
k r 1": ,
(1
94
ЭМБриональное
развитие
8 дней при 2s 0 c
+02
Корацидий
Рис. 3.56 (продолжение)
ных рыбах (окончательный хозяин) и paKO
образных (промежуточный хозяин). У взро
слых форм сколекс слабо развит, а в нечле
нистой «стробиле» находится единственный
набор мужских и женских половых opraHoB
(один яичник, MHoro семенников, но один
копулятивный орrаи). Эти черви обитают
в полости тела а не в кишке хозяина. Их
Миrрация
rлавв з
Н 2 О
10°С
=Ovc
02-+ ve
рН= 7,0
Поедание
рЫБОЙ
Развитие до процеркоиnа
в reмоцеле
считают педоморфной личинкой (см.
rл. 17) неизвестной rруппы ленточных че.
рвей. У представителей Cestodaria есть об.
щие черты как с эвцестодами, так и с соса.
льщиками (рис. 3.58). У них отсутствуют
как рот и кишечник, так и стробилизация
и сколекс, зато присоски «моноrенетическо-
ro» типа. Несмотря на это, их личинка напо-
в
Анус
. Прямая инвазия яйцами
Прикрenительная
зона
Несрридиальная
пора
Нервный ствол
Рис. 3.57. Schistocepha/us sо/idus пред..
ставитель отряда Caryophyllida, встречающийся
в полости тела колюшек (по Smyth, 1962)
Семеприемник
Рис. 3.58. Gyrocoty/e представитель под..
класса Cestodaria (по Barnes 1980)
Трипсин и желчь
в двенадцатиперст
ной кишке активи...
руют яйца и стиму
лируют вылуnление
РОТ
Непрямая инвазия
цистицеркоидами
Инвазивность
цистицеркоида
96...144 ч
Влаrалище
Матка
96
минает онкосферу, но с 10 крючьями вместо
6. Они паразитируют в целомической и ки
шечной полостях пластиножаберных и при
митивных костистых рыб.
3.7. Тип Mesozoa
3.7.1. Этимолоrия
От rреч. mesos средний; zoon
животное (такое название предложено по
тому, что этих животных считали промежу
точными формами между мноrоклеточны
ми животными, т. е. Metazoa" и одноклеточ
ными Protozoa).
3.7.2. Диаrностические
и специфические признаки
1. Билатерально симметричные формы.
2. OpraHoB нет.
3. Полости тела, включая пищеварите
льную, отсутствуют.
4. Очень небольшое количество клеток
расположено Bcero в два «слоя»
наружный ресничный слой и внутренняя
клетка или клетки с репродуктивной функ
цией.
5. Некоторые клетки развиваются вHY
три друrих.
6. Эндопаразиты беспозвоночных на He
которых стадиях cBoero жизненноrо цикла.
7. Сложный жизненный цикл, включаю
щий половую и бесполую репродуктивные
фазы.
8. Морские животные.
3.7.3. Классификация
в этот тип входит 50 видов из двух ДOBO
льно различных rрупп RhоmЬоzоа и Or
thonecta, между которыми, вполне возмо
жно, нет родства.. поэтому ниже они paCCMa
триваются порознь.
Класс
ОТРЯД
== Dicyemlda
Heterocyemida
< Rhombozoa
Orthonecta
Orthonectida
rлава з
3.7.3.1. Класс Rhombozoa
Это паразиты почек rоловоноrих мол..
люсков. Их червеобразное тело длиной
0,5 7,5 мм состоит из 2 30 ресничных
соматических клеток (у каждоrо вила их
cTporo постоянное число), расположенных
по спирали BOKpyr одной удлиненной осе..
вой (аксиальной) клетки (рис. 3.59). Внутри
последней находятся от одной до мноrих
(более 100) клеток аксобластов.. дающих на..
чало половому и бесполому поколе..
ниям.
Жизненный цикл сложный и не изучен
до конца, но, по видимому, описанное вы..
ше строение свойственно двум совершенно
различным стадиям жизненноrо цикла, раз..
множающимся бесполым (HeMaToreH) и по..
ловым (ромбоrен) путями. Они различают..
ся дальнейшей судьбой аксобластов. Ром..
боrен стадия, характерная для взрослоrо
rоловоноrоrо моллюска; аксобласты ero
аксиальной клетки образуют «инфузориrе..
ны», эквиваленты [онад, высвобождаюшие
яйца и/или спермии. Оплодотворение, про
исходящее внутри осевой клетки, приводит
к инфузориевидной личинке, которая вскоре
уже содержит полный набор клеток, и даль..
нейшее развитие личинки происходит уже
за счет их роста и дифференцировки, без де..
лений. Внутриклеточно на этой стадии уже
заложены будущие rерминативные клетки.
Инфузориевидная личинка покидает ромбо..
[ен и выводится из моллюска с ero мочой.
Обладая отрицательной плавучестью (из..за
высокой концентрации инозитолrексафо..
сфата маrния в двух специальных клетках),
она опускается на морское дно. Затем не..
известным способом (возможно, на более
поздних стадиях собственноrо развития или
превратившись в друrую личиночную ста..
дию) она инфицирует молодоrо rоловоно..
roro моллюска и, проникнув в ero почку,
развивается в HeMaToreH. Внутри аксиаль..
Ресничная соматическая клетка
Аксиальная клетка
Рис. 3.59. План строения тела представителей
класса Rhombozoa (по Lapan, Morowitz, 1972)
Параллельные ПОДХОДЫ К мноrоклеточности у животных
Незрелая
червеОБразная
личинка
АКСОБласт
Рис. 3.60. Жизненный цикл
представителей класса R hom
bozoa (по Margulis, Schwartz,
1982 )
ной клетки последнеrо аксобласты через
стадию червеобразной личинки формируют
новые нематоrены; таким путем может Ha
копиться MHoro бесполых поколений. В KOH
це концов, коrда плотность HeMaToreHoB
в почках становится высокой и/или хозяин
достиrает половой зрелости, они либо
трансформируются в ромбоrены, либо про
изводят их новое поколение (рис. 3.60). Пи
тание всех эндопаразитических стадий про
исходит, вероятно за счет абсорбции сома..
тическими клетками питательных веществ,
растворенных в моче хозяина, однако ника
ких подробностей 3Toro неизвестно.
Описано только пять родов, входящих
в 2 отряда.
3.7.3.2. Класс Orthonecta
Ортонектиды эндопаразиты широко
ro Kpyra морских беспозвоночных: плоских
червей, немертин, аннелид, двустворчатых
моллюсков и иrлокожих. Для половой CTa
дии характерен половой диморфизм; это
свободноживущие мелкие формы (размер
самцов около 0,1 мм, самок чуть больше;
I
I
I
I
I
,
\
\
\
\
Взрослый
HeMaToreH
97
Ресничная
соматическая
клетка
Аксиальная
клетка
в
rоловоноrом
Незрелая
инсрузориевидная
личинка
ИНq:>узориевидная
личинка
рис. 3.61). И самцы и самки по строению
несколько напоминают взрослых Rhombo
zoa кольцевой наружный слой ресничных
соматических клеток окружает мноrочи
сленные яйца или спермии. Осевая клетка
отсутствует. Самцы выделяют сперму в MO
ре, она проникает в самок и оплодотворяет
содержашиеся в них яйца. Затем внутри Te
ла самки развивается инвазионная личинка,
(\
Сперма
... .._8
'1i .
. If..s oh"
.. :f (I\;V .
.\. -i ''1
:./'!q '
· .oi{ (" :
:.",\: O
.t 4 . ...,.
. й (p.. "
. aQ.!! '(ff.
"Z д' \
) fpf) "
q ar! I Яйцеклетки Б
. .' 11
А
Рис. З.61. Представители класса Orthonecta:
самка (А) и самец (5) (по Atkins 1933)
98
Развивающиеся ЭМБрИОНЫ
8еrетативные ядра
Рис. 3.62. Плазмодий ортонектид (по Caullery,
Mesnil, 1901)
состоящая из слоя ресничных клеток, OKPy
жающих несколько rерминативных клеток.
После высвобождения личинок и инва
зии тканей хозяина наружные ресничные
клетки утрачиваются, а rерминативные раз
множаются, образуя мноrоядерный плаз
модий (рис. 3.62). От Hero отделяются
фраrменты, образующие бесполым путем
новые плазмодии внутри хозяина. HeKOTO
рые клетки плазмодия затем дают особей
одноrо или обеих полов, которые покидают
хозяина и завершают жизненный цикл
Известны только 3 рода, образующие
один отряд.
3.7.4. Родственные связи
у некоторых эндопаразитов отсут"
ствуют различные системы opraHoB, xapaK
терные для свободноживуших форм, от ко..
торых они предположительно про изошли,
и простота строения Mesozoa раньше pac
сматривалась как результат такой связан..
ной с паразитизмом деrенерации. Однако
если это так, Mesozoa представляют собой
крайнюю стадию данноrо процесса они
не только лишены вообще каких либо opra..
нов, но и MorYT состоять менее чем из 30
клеток. Кроме Toro.. некоторые их особен..
ности, в частности внутриклеточное разви
тие половых и бесполых личиночных форм
Rhombozoa, не имеют аналоrов в животном
иарстве. Нет никаких свидетельств родства
Rhombozoa с какой..либо известной rруппой
rлава з
животных, т. е. они вполне моrли независи..
мо произойти от протистов (см. rл. 2), при..
чем rораздо позже, чем друrие rруппы мно"
rОК1.Iеточных, поскольку их хозяева относя..
тся К сравнительно продвинутым rруп..
пам.
Оrthоnесtа несколько менее аберрант
ные формы, и их происхождение от плоских
червей вполне возможно, хотя и далеко не
очевидно.
3.8. Дополнительная
литература
Bergquist Р. R. 1978. Sponges. Hutchinson. London
[Porifera ].
Fry W. G. (Ed.) 1970. ТЬе Biology of Porifera. Аса-
demic Press, New У ork [Porifera].
GreZl К. G. 1982. Placozoa. In: Parker S. Р. (Ed.) Sy-
nopsis and Classification of Living Organisms,
(Vol. 1)., р. 639. McGraw Hill. New У ork [Placo
zoa].
Нутап L. Н. 1940. ТЬе Invertebrates (У 01. 1). Pro
tozoa through Ctenophora. McGraw..Hill, New
у ork [Porifera, Cnidaria. Ctenophora, Mesozoa].
Нутап L. Н. 1951. the Invertebrates (Vol. 2). Platy-
helminthes and Rhynchocoela. McGraw..HiIl.
New У ork [Platyhelminthes].
Kaestпer А. 1967. Invertebrate Zoology (Vol. 1).
Wiley, New У ork [Porifera, Chidaria, Ctenopho-
ra, Platyhelminthes, Mesozoa].
Lapaп Е. А., М oro vitz Н. 1972. ТЬе Mesozoa.
Scient. Ат.,. 227, 9 101 [Mesozoa].
Mackie G. о. (Ed.) 1976. Nutritional Ecology and
Behavior. Рlепит, New У ork [Cnidaria].
MilZer R. L. 1971. Trichoplax adhaerens Schulze
1883: Return of ап Enigma. Biol. ВиН mar. Biol.
Lab., Woods Hole, 141, 374 [Placozoa].
Morris s. С., Geordge J. D., Gibsoп R., Platt Н.М.
1985. ТЬе Origins and Relationships of Lower In-
vertebrates. ТЬе Systematics Association, Special
V olume No. 28. Clarendon Press, Oxford.
Reeve М. R., Walker М. А. 1978. Nntritional ecolo-
gy of ctenophores а review of past research.
Adv. mar. Biol. 15., 24 287 [Ctenophora].
Schтidr Н. 1972. Die Nesselkapseln der Anthozoen
und ihre Bedeutung fiir die phylogenetische Syste
matik. HelgoHinder Wiss. Meeresunters., 23, 422
458 [Platyhelminthes].
SC/1ockaer l Е. R., Ball 1. R. 1981. ТЬе Biology of the
Turbellaria. Junk, ТЬе Hague [Platyhelminthes].
SmYlh Т. D. 1977. Introduction to Animal Parasito-
logy, 2nd edn. Wiley, New У ork [Platyhelminthes].
rЛАВА 4
ЧЕРВИ
Nemertea
Gnathostomula
Gastrotricha
Nematoda
Nematomorpha
Kinorhyncha
Loricifera
Priapula
Rotifera
Acanthocephala
Sipuncula
Echiura
Pogonophora
Annelida
Все включенные в эту rлаву 14 типов
первичноротые производные плоских че..
рвей, хотя общих специфических черт лише..
ны и не составляют естественную rруппу
родственных животных (см. rл. 2). Общим
для ее представителей, помимо червеобра..
зной формы тела, является скорее отсут"
ствие некоторых черт, характерных для дру"
rих rрупп или типов. Можно сказать, что
эти черви не имеют Hor, не покрыты ракови..
ной, не вторичноротые, не несут лофофора.
у червеобразных форонид есть лофофор
и они рассмотрены в rл. 6; кишечнодыша..
щие и щетинкочелюстные вторично"
ротые, поэтому речь о них пойдет в rл. 7;
пятиустки обсуждаются со своими предпо..
лаrаемыми сородичами..членистоноrими
ВrЛ. 8. Таким образом, практически 14 опи..
сываемых ниже rрупп простые животные,
сохранившие более или менее неизменной
червеобразную предковую форму тела
(длина в 2 3 1500 раз превышает шири..
. ну), крyrлые или уплощенные в сечении.
Хотя для этих «червей» не разработано
общепринятой системы родства.. широко
признается ее деление на четыре rруппы ти"
пов (см., например, рис. 2.20). У первой
и второй rрупп (Nemertea и Gnathostomu..
lajGastrotricha) идентичный с предками
плоскими червями план строения и отно"
сительно небольшое таксономическое раз..
нообразие в пределах каждой rруппы; для
обеих характерна высокая специализация,
которая делает невозможным их объедине
ние с Platyhelminthes.
rастротрихи, возможно, MorYT служить
связующим звеном с rораздо более крупной
и разнообразной третьей rруппой типов..
известной как Aschelminthes, или псевдоце..
ломические черви. В нее входит семь типов
,
распадающихся на две подrруппы. Kino..
rhyncha, Loricifera и Nematomorpha на неко"
торых стадиях жизненноrо цикла имеют
выворачивающийся интроверт" несущий
окруженный стилетами и круrами «скалид»
ротовой конус; у Priapula есть сходный ин..
троверт со скалидами, но без ротовых сти"
летов. Nematoda, похоже, также связань]
с этой подrруппой через недавно описанный
из Бразилии вид с аналоrичным интровер"
том и иrловидными скалидами; кроме Toro,
у Nematoda и Nematomorpha одинаковая
высокоспециализированная мускульно"
локомоторная система. И напротив, Rotife..
ra и Acanthocephala лишены интроверта.
] В '"
отечественнои литературе для интроверта
и хоботка обычно используется один термин
хоботок. Поскольку между интровертом и хобот
ком существуют выраженные функциональные
и морфолоrические различия, здесь и далее под
интровертом понимается передняя часть тела,
которая может выворачиваться и втяrиваться.
а под хоботком специальный, иноrда сложно
устроенный opraH. Прuм. пер.
.100
имеют внутриклеточную кутикулу, локомо
торный и/или клейкий «хоботок» и лемни
ски. У коловраток последние две CTPYKTY
ры только в отряде Bdelloidea. Если не
брать в расчет интроверт и паНIIИРНУЮ ли
чинку, Рriарulа довольно неТИПИЧН Iе
Aschelminthes; возможно, по форме тела
и репродуктивному типу они близки
к предкам. Друrие типы псевдоцеломиче
ских червей можно объединить по устрой
ству стенки тела, природе rлотки и полости
тела, феномену эвтелии (постоянство кле
точноrо состава), внутреннему оплодотво
рению с копуляцией, отсутствию способно
сти К реrенерации частей тела и бесполому
размножению. Для мноrих также xapaKTep
ны протонефридиальная система, линька
кутикулы. ее утолщение с образованием за
щитноrо панциря, мелкие размеры непара
зитических форм, сеrментация кутикулы (и
связанных с ней элементов стенки тела),
Флоскулы и дуоrландулярные прикрепите
льные железы.
Четвертая rруппа также включает раз
нообразных по признакам червей. Однако
Annelida, Pogonophora и Echiura объеди
няют такие черты, как схизоцельная поло
сть тела 1, хитиновые щетинки, замкнутая
кровеносная система, орrаны выделения ти
па метанефридиев; представители первых
двух типов метамерные, в то время как
эхиуриды явно мономерны. Также MOHO
мерны Sipuncula, у которых «метанефри
дии» И схизоцель часто считаются призна
ками. сближаюшими их с четвертой rруп
пой типов, хотя щетинок и замкнутой KpO
веносной системы нет. Только у Sipuncula
есть интроверт, но в отличие от KopoTKoro,
боченковидноrо, покрытоrо иrлами интро"
верта Aschelminthes, у них он представляет
собой длинную тонкую трубку, оканчиваю
щуюся лопастями или щупальцами для со..
бирания детрита.
Положение Sipuncula в rруппе схизоцело..
мических 2 червей, как и Priapula среди
1 ПОД схизоцелем здесь понимается вторич"
ная полость тела, образованная расхождением
мезодермальных клеток. см. рис. 2.9,Б. Прuм.
пер.
2 Схизоцеломические черви вторичнопо..
лостные черви, у которых целом образован схи..
зоцельным способом. Прuм. пер.
rлава 4
Aschelminthes" аберрантно (в отношении од..
них черт типичное, в отношении друrих
резко отличное от остальных типов).
В обеих этих rруппах одни анатомические
признаки MorYT быть истолкованы в пользу
параллельноrо приобретения одинаковоrо
уровня орrанизации, а друrие для доказа..
тельства филоrенетическоrо родства. То,
что один автор считает железой и ее прото..
ком, друrой рассматривает как rлотку и ро..
товой канал (этот пример две точки зре..
ния на строение личинок нематоморф).
В некоторых типах, в частности, Nemato..
morpha среди псевдоцеломических червей
и Pogonophora среди схизоцеломических
выделяют rруппы совершенно различноrо
происхождения. Одну rруппу нематоморф
можно вывести из мерметидных нематод,
в то время как друrую связывают с кино..
ринхами и лорициферами; не исключено,
что один класс (или подтип) поrонофор
близкородствен аннелидам, в то время как
друrой возник независимо, и схизоцель
в нем развился параллельно!
Ясно, что преждевременно объединять
какие..либо из этих rрупп червей в меньшее
число более крупных типов. Следует прове
сти еще MHoro исследований, прежде чем
можно будет сделать определенные филоrе..
нетические выводы, поэтому в данной rлаве
мы будем рассматривать все 14 rрупп как
независимые таксоны одноrо систематиче..
cKoro paHra.
4.1. Тип Nemertea (немертины)
4.1.1. Этимолоrия
От rреч. Nеmеrtеs средиземноморская
нимфа, дочь Иерея и Дориды
4.1 .2. Диаrностические
и специфические признаки
(рис. 4.1)
. 1. Билатерально симметричные, черве-
образные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и орrанами.
3. Сквозной кишечник с терминальным
анусом.
Отверстие ДЛЯ
выворачивания
ХОБотка
Латеральный
нерв
ХОБОТОК
rлаза
Мозr
Ринхоцель
rOHaAbI
Кишечные
дивертикулы
Мускул ретрактор
ХОБотка
Анус
Рис. 4.1. Схематическое изображение ВНУ"
TpeHHero строения немертины с дорсальной
стороны (по нескольким источникам, rлавным
образом по Pennak, 1978)
4. Полости тела нет, функции скелета
выполняет паренхима.
5. Наружный ресничный эпидермис ли..
шен кутикулы.
6. Тело дорсовентрально уплощено, ча
сто с сериально повторяющимися орrанами
.s:
о
сп
о
8.
s:
m о
() о х
о с:: :J:
0.0 s:
с: с: а..
а:
g
0.3
Q)s: о
C: Х
Черви
101
(например, кишечными карманами, прото
нефридиями, rонадами)
7. Есть кровеносная система. но дыха..
тельная система отсутствует.
8. Нервная система с rоловным мозrом
и обычно тремя продольными стволами
9. Имеется хоботок эктодермальноrо
происхождения, способный выворачиваться
и втяrиваться в продольную дорсальную
полость (ринхоцель), которая открывается
наружу вблизи рта.
10. Дробление яиц спиральное.
Немертины, длина тела которых коле..
блется от менее 0,5 мм до более 30 м, MorYT
быть названы «сверхплоскими червями»,
так как основные черты их строения в при..
нципе те же, что и у крупных турбелларий
(см. разд. 3.6.3.1). Тем не менее они отли"
чаются от этой rруппы тремя основными
признаками; во..первых, более эффективно
работающим сквозным кишечником; во..
вторых, замкнутой кровеносной системой,
ток крови (правда, нереrулярный) в которой
обеспечивается сокращениями стенок кро"
веносных сосудов и движениями тела;
в"третьих, характерным хоботком, совер..
шенно изолированным от кишечника и не
являющимся ero частью (рис. 4.2).
Этот хоботок помещается в трубчатой
полости ринхоцеле, проходящей почти
о.
<tS
О.
а.
'со
C::
.....
и О
>-IЛ
Ее
s:
:J:
:r
Q)
3
s:
Отверстие для '..:.': '::::. :":':::" ......:. .:.:...:.:.::: ........:. :.: ":" :..:. ....: .: "::'. :::'. '.' :'.' ::. .'::.: ::. :. '.; Ы:!.\ :,:,:.) .:.:
:gf;ивания ".:.'::':::. /:' - . ; , ; .. I , . " ;; . ; ' : 1, ' ;i _ ; :,,:, ;;! I :: ! : :::, ... :, .
. '.:.'. ". I .:.::;:... ::::::;':".:'..:',:: С О :'.:;;"-:;: ::". :'.:::; :.: ::.х.,::.:... :':':':'::-.-::
А Рот
Анус
":"" ": " : :':': " : . :: ,':';;:'. .: ! : -: " : " " : "" ..... ... : :;:;
,/':'::':.' .... ".:'.':. .... . ; ., ., : ." j . .:. : . '.... .:.. ..:: .... .: .. " ':' i . '. . ' .. :' »);.. .;. / ;.\\.\;
:' ::': : :)!.:; : ;., . · .. ,(.' ;,. ).:,: :;;:'.: ,,;:;;,; ;: ;:':. ;:: ::: \ ",:::": -\.;:, : ; ;: . :.\: ::,:ЕУ
Б
Рис, 4.2. Схема продольноrо разреза немертины, демонстрирующая ринхоцель и хоботок (А
втянутый; Б вывернутый) (по Gibson, 1982)
102
Рис. 4.3. Личинка немертин пилидий (по
Kershaw, 1 983)
вдоль Bcero тела и имеющей такое же CTpoe
ние, что и стенки последнеrо. Давление,
возникающее в ринхоцеле при сокращении
мышц ero стенок, выворачивает хоботок.
Втяrивается он с помощью продольных MY
скулов, тянущихся от ero заднеrо конца
к заднему краю ринхоцеля. Хоботок испо
льзуется в основном для ловли добычи, а
у наземных видов также для быстроrо пере
мещения: в вывернутом состоянии он при
крепляется к земле впереди животноrо, KO
торое затем как бы подтяrивается на нем
вперед.
Локомоция у немертин осуществляется
в целом так же, как у турбелларий: они
скользят за счет биения ресничек по Bыдe
ляемой слизи, а крупные формы переме
щаются блаrодаря волнообразным мышеч
ным сокращениям вентральной поверхно
сти тела. Некоторые виды MorYT плавать,
волнообразно извиваясь в дорсовентраль
ной плоскости.
Размножение происходит как бесполым
путем (фраrментацией), так и половым,
причем [онады являются временными
структурами, формирующимися из скопле
ний дедифференцированных мезенхимных
клеток, которые окружаются мембранами,
а в зрелом состоянии связываются с внеш
ней средой временными протоками. Боль
шинство видов раздельнополые, хотя ряд
наземных и пресноводных форм
rермафродиты, способные к самооплодо
творению. Развитие обычно прямое, хотя
у представителей одноrо отряда известна
rлава 4
личиночная стадия (рис. 4.3); оплодотворе
ние, как правило, наружное.
Почти все немертины хищники, пи-
тающиеся широким набором орrанизмов,
от простейших до моллюсков, членистоно
rих и рыб. К этому типу относится длинней-
шее из всех известных животных Lzпeus
loпgissiтus, обычно достиrающее 30 м, а
в некоторых случаях и вдвое больше в пол
ностью вытянутом состоянии.
4.1 .3. Классификация
Около 900 известных видов распределе..
ны по двум классам
Класс
ОТРЯД
PaJaeonemertea
Heteronemertea
< AnopJa
EnopJa
Hoplonemertea
--= Bdellonemertea
4.1 .3.1. Класс Anopla
у аноплид центральная нервная система
расположена в стенке тела, которая часто
включает три мускульных слоя: либо два
слоя кольцевых мышц и один про
дольных (между ними), как в отряде Palaeo-
nemertea, либо два слоя продольных и ме-
жду ними один кольцевых, как в отряде
Heteronemertea. Хоботок аноплид устроен
относительно просто, не имеет реrионарной
дифференцировки или стилетов, хотя несет
большое количество эпителиальных раб-
доидов Toro же типа, что и у турбелларий;
ринхоцель открывается совершенно изоли.
рованным от рта отверстием.
Большинство представите.цей этоrо
класса (рис. 4.4) морские бентосные жи-
вотные, но три вида пресноводные" а еще
несколько обитают в солоноватых Бодах.
Heteronemertea единственная rруппа не-
мертин, у которой есть личиночная стадия.
4.1 .3.2. Класс Enopla
у эноплид центральная нервная система
расположена изнутри относительно МУСКУ.
латуры стенки тела. Мускулатура двуслой-
ная; хоботок дифференци;рован на отделы;
центральный из них представляет собой ко.
Рис. 4.4. Форма тела представителей класса
Anopla (по Gibson, 1982)
роткую мышечную луковицу, несущую
один или более стилетов (за исключением
аберрантноrо комменсальноrо рода М а/а..
cobdella, представители KOToporo питаются
путем фильтрации и выделены в от дельный
отряд Bdel1onemertea). Кишечник эноплид
сложный и открывается наружу общим
с ринхоцелем отверстием. У большинства
представителей класса (отряд Hoplonemer..
tea) кишка снабжена мноrочисленными па..
рами боковых ответвлений (дивертикулов),
в то время как у Bdellonemertea крупная
rЛО1ка служит фильтрующим opraHoM и со..
держит ресничные сосочки, улавливающие
пищевые частицы без помощи слизи.
Хотя большинство эноплид морские
бентосные формы, есть несколько пелаrиче..
ских ВИlIОВ. плавающих в толще воды и на
поверхности (рис. 4.5), а также ряд прес..
новодных и наземных представителей.
Черви
103
Рис. 4.5. Форма тела представителей класса
Enopla (по Gibson, 1982)
Известнь] комменсалы (например, из ат..
риальной полости оболочников) и парази..
ты.
4.2. Тип Gnathostomula
(rнатостомулиды)
4.2.1. Этимолоrия
Отrреч. gпаthоs челюсть; stoma pOT.
4.2.2. Диаrностические
и специфические признаки
(рис. 4.6 и 4.7)
1. Билатерально симметричные, черве..
образные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и орrанами.
3. Кишечник слепо замкнут.
104
rлава 4
Рот
Рострум
Семенники
'1
,
'f .
!
t .
I .
I
,
Мужское
половое
отверстие
Рис. 4.6. Анатомия rнатостомулиды из отряда
Filospermoidea (по Sterrer 1982)
4. Полость тела отсутствует, функции
скелета выполняет слабо развитая паренхи--
ма
5. Кровеносной и дыхательной систем
нет.
6. Выделительные орrаны простые дву--
клеточные протонефридии.. напоминающие
эпидермальные клетки тем, что терминаль--
ная клетка несет только одну ресничку.
7. Эпидермис без кутикулы, из однорес--
ничных клеток.
8. Диффузная эпидермальная нервная
система.
9. Сложный ротовой аппарат образо--
ван парными челюстями и единственной ба--
зальной пластинкой.
10. Синхронные rермафродиты (муж--
ские и женские rонады созревают одновре--
менно).
11. Дробление яиц спиральное.
12. Развитие прямое.
13. Морские интерстициальные формы
часто бескислородных песков.
у этих крошечных (менее 1 мм в длину)
прозрачных червей в принципе такое же
строение тела, что и свободноживущих тур--
белларий (разд. 3.6.3.1) и rастротрихов
(разд. 4.3). От обеих этих rрупп они отли
чаются высокоспециализированной МУСКУ"
лист ой rлоткой со сложной системой челю
стей, которая служит для сбора бактерий,
протистов и rрибов с поверхности песчинок
(рис. 4.8). В поисках этой добычи rHaTocTo
мулиды движутся в интерстициальных по
рах, плавая или скользя с помощью CBoero
ресничноrо эпидермиса (реснички бьют впе..
ред или назад) или за счет резких сокраще
ний трех или четырех пар продольных
мышц, образующих мускулатуру стенки Te
ла. Единственный яичник образует за ЦИКЛ
размножения одно крупное яйцо, вероятно,
всеrда оплодотворяемое внутри тела. По..
видимому, в течение жизни некоторые [на..
Ресничная ямка
Базальная пластинка
Челюсти
rлотка
Яичник
Бурса
Влаrалище
Семенник
Мужское половое
отверстие
:....
,.
Рис. 4.7. Анатомия rнатостомулиды из отряда
Bursovaginoidea (по Sterrer 1982)
Базальная пластинка
Челюсти
Рис. 4.8. 6азальная пластинка и парные челю..
сти rнатостомулид (по Sterrer, 1982)
тостомулиды проходят превращение от бес
полой питающейся стадии до половой непи
тающейся.
Этот тип был описан только в 1956 [.
вместе с первым ero представителем. Такое
позднее открытие объясняется не тем, что
rнатостомулиды редки их может быть до
600000 особей в 1 м 3 . Причина слабая
изученность бескислородных слоев морских
осадков и то, что rнатостомулиды сильно
деформируются в результате фиксации.
4.2.3. Классификация
Около 100 описанных видов помещены
в два отряда в пределах одноrо класса:
сильно вытянутые Filospermoidea (рис. 4.6)
с длинным рострумом, по без передних пар
ных сенсорных opraHoB, пениса или ceMe
приемника (бурсы), и короткие Bursovagi
noidea (рис. 4.7) без удлиненноrо рострума,
но с парными сенсорными щетинками, pec
ничками на переднем конце тела, с пенисом
и бурсой. Несомненно, еще мноrие виды
ждут cBoero описания.
4.3. Тип Gastrotricha
(брюхоресничные черви)
Черви
105
4.3.2. Диаrностические
и специфические признаки
(рис. 4.9)
[. Билатерально симметричные, черве
образные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и орrанами.
3. Сквозной кишечник с субтерминаль
ным анусом.
4. Полость тела отсутствует (несмотря
на существовавший ранее противопо"
ложный взrляд).
5. Кровеносной и дыхательной систем
нет.
6. Наружная нехитинизированная KY
тикула, обычно нересничный дорсальный
эпидермис и обильно покрытый ресничками
вентральный. Слои кутикулы поднимаются
над каждой ресничкой как футляр.
7. Нервная система из пары связанных
дорсальной комиссурой rанrлиев по бокам
rлотки и пары продольных стволов.
8. Тело покрыто кутикулярными че
шуйками, иrлами или крючками и несет до
250 прикрепительных трубочек.
Рот
Кишечник
Прикрепительная
ТруБочка
Комиссура
rанrлий
Мускулистая
rлотка
r лоточная пора
Семенник
Яичник
Прямая кишка
Прикрепи ...
тельная
железа
4.3.1. Этимолоrия Рис. 4.9. Схематическое изображение BHY
TpeHHero строения брюхоресничноrо с дорса..
От rреч. gаstеr желудок; thrlХ ВОЛОС. льной стороны (по нескольким источникам)
106
Рис. 4.10. Схематические изображения двух
брюхоресничных с вентральной стороны, дe
монстрирующие разные типы расположения
ресничек (по Hyman, 1951)
9. rермафродиты или партеноrенети
ческие формы.
10. Яйца высвобождаются через разры
вы в стенке тела, дробятся билатерально
радиально, но развитие детерминирован
ное.
11. Развитие прямое.
12. Водные формы.. обитают в интерсти
циали, на поверхности водоемов, изредка
в планктоне.
у rастротрихов более или менее про
зрачное, дорсовентрально уплощенное тело
до 4 мм в длину (обычно менее 1 мм). Спе
реди расположены сенсорные орrаны Toro
же типа, что и у rнатостомулид отряда Bur
savaginoidea (парные щетинки, реснички
и сенсорные ямки), у некоторых rлазные
пятна; сзади тело может заканчиваться
мощной вилочкой (фуркой) или тонким
хвостом. Выделяющие клейкое вещество
трубочки обеспечивают временное при
крепление к поверхностям. Хотя стенка Te
ла содержит кольцевые и продольные MY
скулы, rастротрихи движутся скользя с по ·
мощью ресничек" которые на их вентраль--
ной стороне часто расположены HepaBHO
мерно, образуя, например, поперечные или
продольные полосы характерным для раз
ных родов способом (рис. 4.10).
Питаются rастротрихи бактериями,
протистами и детритом, заrоняя пищу в рот
биением ротовых ресничек или с помощью
rлава 4
работающей как насос rлотки. Как и у не..
матод, rлотка в сечении трехлучевая и выст-
лана единственным слоем мышечно эпи-
телиальных клеток. Как у коловраток и ки-
норинхов, У пресноводных видов пара про-
тонефридиев с соленоцитами.
4.3.3. Классификация
Известно более 400 видов. распределяе-
мых между двумя отрядами одноrо класса;
у ремневидных морских Macrodasyida (рис.
4. 11) rлотка перфорирована двумя порами,
служащими, вероятно, для вывода воды,
поступившей внутрь в процессе питания,
как укрыложаберных (разд. 7.2.3.2, см. так-
же разд. 9.2.5); пресноводные и морские
Chaetonotida (рис. 4.12) обычно веретено-
образны, лишены rлоточных пор; если при-
сутствуют прикрепительные трубочки, ТО
только сзади; MorYT размножаться партено..
rенетически.
Рис. 4.11. Формы тела брюхоресничных ОТрЯ
да Macrodasyida (по Grasse, 1965; Hummon,
1982)
Рис. 4.12. Формы тела брюхо
ресничных отряда Chaetonotida
(по Grasse, 1965; Hummon,
1982 )
4.4. Тип Nematoda
(круrлые черви)
4.4.1. Этимолоrия
От rреч. nema нить; eidos форма
4.4.2. Диаrностические
и специфические признаки
1. Билатерально симметричные, черве
образные формы с тенденцией к радиаль
ной симметрии BOKpyr продольной оси.
2. Тело из двух клеточных слоев, с TKa
иями и орrанами.
3. Сложная кутикула.
4. Кольцевые мышцы в стенке тела OT
сутствуют.
5. Полость тела псевдоцель, обычно
развивается из бластоцеля.
Черви
107
6. Мускулистая кишка проходит от рта
на переднем конце через мускулистую rлот..
ку к субтерминальному анусу.
7. Продольная мускулатура разделена
на четыре зоны вентральным, двумя лате
ральными и дорсальным эпидермальными
валиками.
8. Нервная система из продольных
тяжей, расположенных в вентральном
и дорсальном эпидермальных валиках и не..
посредственно контактирующих с мышеч
ными клетками.
9. Поперечный срез представляет со..
бой Kpyr; полостная жидкость всеrда нахо..
дится под большим давлением.
10. Кровеносной системы нет.
11. Выделительная система без клеток
с ресничным пламенем или нефридиев. Эк
скреторные трубочки в одной или несколь..
ких клетках ренеттах.
12 Дробление яйца не спиральное и не
108
rлава 4
Средний
слой
радиальное, но сильно детерминированное..
с Т образным расположением клеток на
стадии четырех бластомеров.
13. Развитие прямое, но у паразитиче
ских форм может быть стадия инвазионной
личинки.
Этот тип один из добившихся наи
большеrо успеха в животном царстве. Опи..
сано более 15 000 видов, но считают, что
сейчас их примерно 1 000000. в отличие от
дрyrих типов это не связано с морфолоrиче
ским разнообразием. Все нематоды устрое..
ны по одному общему плану, оказавшемуся
весьма «удачным». Он предполаrает спо..
собность противостоять потенциально
опасному окружению, и мноrие круrлые че
рви паразиты.
Основная черта их структурной орrаниза..
ции сложная кутикула (рис. 4.13). В ней
можно обнаружить до девяти слоев, но наи..
НеDвноеrлоточный БУЛЬБУС
КOJ'1bUO Кишечник
. ... .. ..
/
ВЬUlелительная пора
Клапан
Женское половое
отверстие
А
Базальныи
слой
I
Рис. 4.1 з. Строение кутикулы
нематод, состоящей из Hapy
жноrо исчерченноrо, BHYTpeH
Hero однородноrо и комплекса
волокнистых слоев (по Clark,
1 964)
более заметны среди них три.. состоящие из
перекрещивающихся волокон и образую-
щие BOKpyr тела спиральную сеть. Она вме..
сте с полостью тела, содержащей жидкость
под высоким давлением, служит основой
локомоторной системы нематод (см. разд.
10.4).
Нематода представляет собой «трубку
в трубке», заостренную с обоих КОНЦОВ
и круrлую в сечении (рис. 4.14). Терминаль-
ный рот открывается в мускулистую rлот..
ку важный признак орrанизации этоrо ти-
па. Поскольку полостная жидкость нахо..
дится под высоким давлением.. внутренние
трубчатые структуры нематод имеют тен-
денцию сплющиваться и для Toro, чтобы
было возможным питание, нужен механизм
введения пищи во внутреннюю трубку, Т. е.
пищеварительную систему. Переднее отвер-
стие пищеварительноrо тракта называется
Яичник
0000
. . . . .0 0О 0 е 0 .
.' . . : · : '.,: : : . : . . .....: Семen овод I КonyЛЯТИ8НЫЙ аппарат
Семенник р Клоака
Б
Рис. 4.14. Общее строение нематоды (за основу взят паразитический род Rhabditis): А caMKa;
Б самец
Передняя часть rлотки
няя часть rлотки
Стомв
с
/
)
\.
Рис. 4.15. Механизм работы rлоточноrо Haco
са нематод Диаметр переднеrо отдела rлотки
HaMHoro меньше, чем у заднеrо отдела. Клапан
сзади закрывается при расширении rлотки,
и жидкость втекает в ее переднюю часть. Коrда
rлотка сокращается, задний клапан OTKpЫBa
ется, и жидкость течет назад из за большеrо
диаметра заднеrо отдела (по Croll, Matthews,
1977)
стома. Оно ведет в мускулистую rлотку, на
друrом конце которой находится клапан.
[лотка закачивает пищу против rрадиента
давления причем однонаправленный поток
в сторону кишечника обеспечивается COOT
ношением объемов передней и задней ча
стей rлотки (рис. 4.15).
Мышечные клетки
rлотка
Б
Черви
109
Значительная часть полости тела занята
парными, сериально орrанизованными ре..
продуктивными орrанами, семенниками
или яичниками, которые часто спирально
закручены. Расположение их выводных от..
веретий показано на рис. 4.14.
Нематоды обычо раздельнополы, опло
дотворение внутреннее. «Яйца», отклады
ваемые самкой, фактически представляют
собой оплодотворенные зиrоты или ранние
эмбриональные стадии. Ооциты BToporo
порядка движутся по яйцеводам к семе..
приемнику, rде происходит оплодотворе..
ние. Затем они покрываются слоем белко
вой скорлупы, вырабатываемой маткой,
а сама зиrота секретирует две внутренние
яйцевые оболочки. «Яйuа» бывают крайне
устойчивы к внешним воздействиям и обра
зуются у некоторых паразитических видов
в оrромных количествах.
Строение стенки тела лучше Bcero видно
на поперечном срезе. Оно выделяет этот
тип среди всех прочих (рис. 4.16). Под слоем
сложной кутикулы лежит эктодермальный
эпидермис, включающий клетки нервной
и выделительной систем основные элемен..
ты которых расположены в четырех про
дольных валиках. Дорсальный и вентраль
ный валики содержат крупные дорсальный
и вентральный нервы. В промежутках Me
жду эпидермальными валиками лежат клет..
ки продольной мускулатуры; их небольшое
количество cTporo фиксировано... После ca
Латеральный
нервный ствол
Матка
Вентральный нервный
ствол
в
Рис. 4.16. Строение нематоды на поперечном срезе:А объемное изображение rоловы дeMOH
стрирующее треуrольную орrанизацию рта, rуб и связанных с ними сенсорных opraHoB; Б
поперечный срез в области rлотки; В поперечный срез в средней части тела
110
rлава 4
Тело мышечной клетки
Сократимая часть
мышечной клетки
Рис. 4.17. Объемное изображение
нервно мускульноrо комплекса
нематод. Показаны сократимые
части мышечных клеток, их тела
и иннервационные отростки, YCTa
навливающие прямые синаптиче
ские связи с нервными волокнами
в латеральных, дорсальном и BeH
тральном стволах
Нервные волокна
Кутикула
Эпидермис
r иподермальный валик
мых ранних стадий эмбриональноrо разви
тия рост тела происходит за счет увеличе
ния размеров клеток, а не их числа.
Мышечные клетки правильнее было бы
назвать нейромоторными комплексами
(рис. 4.17). Сократимые волокна каждой
клетки расположены дистально и приле
rают к эпидермису. Удлиненный иннерва
ционный отросток непосредственно KOHTaK
тирует с нервными тяжами дорсальноrо
или вентральноrо валиков в зависимости от
положения мышечной клетки. Каждая Ta
кая клетка дает несколько отростков, концы
которых образуют синапсы с друrими MЫ
шечными и нервными fJолокнами. Таким
образом, вероятно, формируется система,
обеспечивающая одновременное сокраще
ние всех мышечных клеток. Связь их OT
rоловная папилла
rоловная щетинка
Соматическая
щетинка
Спикулы
СРазмиды
Амсрида
rлазок
r еМИЗ0НИД
Внутренние
рецепторы
Соматическая
щетинка
Различные мужские
сенсорные opraHbI
Сосочки
Хвостовые
папиллы
Рис. 4.18. Обобщенная схема, показывающая
разные типы сенсорных opraHoB, обнаружен
ных у нематод. Ни у oAHoro вида нет их всех (по
Croll, Matthews, 1977)
ростков с нервами необычная черта, по..
скольку у большинства животных отростки
отходят наоборот от нервных к мотор..
ным клеткам (см. разд. 16.4.2).
У нематод известны различные сенсор..
ные приспособления. К ним относятся 20"
ЛО6ные папиллы (сосочки), амфиды (хемо"
чувствительные ямки, часто связанные
с передними амфидными железами), 20лов
ные щетинки, соматические щетинки, хво..
ст06ые папиллы и спикулы, 2лазки и фазмu
ды (рис. 4.18). Фазмиды представляют со..
бой расположенные в хвостовом отделе
сенсорные ямки, похожие на амфиды, при..
чем иноrда те и дрyrие связаны с железами.
Нематод делят на два класса в зависимости
от отсутствия или наличия фазмид.
Кл асс
ОТРЯД
Enoplida
Isolaimida
Mononchida
Dorylaimida
т rlChocephalida
1ermithida
М uspicelda
Araeolaimida
Chromadorida
Desmoscolecida
Desmodorida
Мопhуstеrldз
Adenophorea
Secernentea
Rhabditida
Strongylida
Ascaridida
Splrurlda
Camallanida
Diplogasterlda
т ylenchida
Aphelenchlda
Экскреторная система круrлых червей
пример исключительной экономии числа
клеток, характерной для типа. Исходно в ее
состав входят одна или две специализиро
ванные клеткu ренеттbl. В более продвину
том случае формируется система трубочек
внутри этих железистых клеток ренетт, KO
торая у некоторых форм утрачена (см. разд.
12.3.1 ).
Однотипный план строения нематод, OT
ражающийся на их внешнем облике, может
быть приспособлен к orpoMHoMY разно
образию внешних условий. Лучше Bcero это
иллюстрирует разнообразие их механизмов
питания или жизненных циклов паразитиче
ских форм. Мноrие свободноживущие He
матоды питаются бактериями" которых они
поrлощают в виде взвеси, или детритом.
Есть и хищники, поедающие в основном
друrих нематод или микроорrанизмы.
у них возникли разнообразные зубы или че
люстеподобные пластинки на передней по
верхности стомы. Эти «челюсти» проты
кают жертву, содержимое которой затем
высасывается с помощью работающей как
насос rлотки.
Ряд свободноживущих видов фи
тофаrи. Некоторые заrлатывают rрибы
и дрожжевые клетки целиком, но большин
СТВО протыкает клетки растений и Bыcacы
вает содержимое. Для Toro чтобы питаться
таким образом, мноrие нематоды фи
тофаrи располаrают в ротовой полости спе
циальным приспособлением: копьевидным
образованием для вскрытия клеток или по
лым стилетом, который в совокупности
с rлоточным насосом используется как Bca
сывающий шприц.
Эти механизмы питания свободноживу
щих видов MorYT быть использованы и па
разитическими формами. - Например, поло
сти кишеЧНИКd позвоночных наполнены бо
rатой смесью бактерий и разрушенных кле
ток, т. е. питательной средой для нематод,
заrлатывающих бактерий. Челюстные пла
стинки MorYT быть использованы для раз
рыва выстилки кишки и высвобождения
форменных элементов крови, которыми пи
таются нематоды паразиты.
Ослабление позвоночных, вызываемое
инвазией их пищеварительной системы He
матодами, частично зависит от биолоrии
Черви
111
питания последних. Детская острица (Eпte..
robius verтicu/aris) относительно безвредна,
пока питается бактериями в кишечнике,
и становится патоrенной" коrда ее популя..
ция возрастает настолько, что вызывает за
купоривание пищеварительноrо тракта.
Свайник двенадцатиперстной кишки (Aпky
/ostoтa duodeпa/e) более опасен.. так как BЫ
зывает кишечные кровотечения, питаясь
клетками крови. Человек, в котором живет
100 таких червей, может терять до 50 мл
крови В день.
Стома всех нематод фитопаразитов
вооружена стилетом или «копьем». Мноrие
из них образуют постоянные цисты, в KOTO
рых неподвижные самки питаются rиrант
скими клетками. Черви сами вызывают
образование таких клеток растением
хозяином, вероятно, воздействуя на проду
цирование им веществ, стимулирующих
рост, или их инrибиторов. Наиболее важ
ные роды фитонематод Неtеrоdа и Melol
dogyпe.
Паразитизм возникал в филоrенезе He
матод неоднократно. Этому способствова
ли некоторые основные черты орrанизации
типа, а именно:
1) сложная непроницаемая и прочная
кутикула;
2) внутреннее оплодотворение и способ
ность производить высокоустойчивые к
внешним воздействиям яйца;
3) микрофаrия, т. е. питание мелкоди
сперсным материалом;
4) мелкие размеры тела;
5) химическое разнообразие, включаю..
щее механизмы, необходимые для избеrа
ния защитных систем хозяина.
Паразитизм у мноrих беспозвоночных
ведет к сильному видоизменению взрослых
форм и развитию специализированных
жизненных пиклов со стадиями, способны
ми к бесполому размножению, как, напри..
мер, у мноrих паразитических плоских че..
рвей (разд. 3.6.3.2 и 3.6.3.3). Однако у нема..
тод этоrо не произошло. Взрослые особи
«преадаптированы» к паразитическому
образу жизни, и способ их развития остался
неизменным. Исходный жизненный цикл
изображен на рис. 4.19, А. Он включает
в себя спаривание самцов и самок и высво..
бождение хорошо защищенных «яиц». Раз
112
А nC!
L / [ e ОСОБИ
Я
В. Aпcylostoтa с!'
По кровеносной f взрослые ОСОБИ
системе к леrким шечнике
L Aepe Я й ц а
It трахею в
1' . . киш . еч . ник . . . ' П . .. ар . '. азитизм
l Выделение с
ПроникаёТ чер ез кожу ИЛ И, . ....... ': ейек алиями
заrлатыва .
через сли.
зистую
rлава 4
Б. Stroпgyloldes
4.Jf'
,
Жи ненный цикл
С8О6ОДнОЖИВУЩИХ
if!J .
взрослы8
ОСОБи'Qnлoдотворенные яйца
Д. Eпterobius
r. Ascarls C!
Через трахею Взрослые ОСОБИ
в кишечник в кишечнике
По рове
LIt ОСНОЙ Яйца
l систем Паразитизм
к леrки Выделение с среК8ЛИЯМИ
Про н кает через r ......... .
слиз тую ,- Почва и
L3 cpet<S11ИИ
вылп /
ление
в кишечнике/,
,
Поrлощ.ение.!' L 2 в яйце
с пищеи
Рис. 4.19. Жизненные циклы нематод: А основной цикл с четырьмя личиночными стадиями;
Б д модифицированные циклы видов, приспособившихся К эндопаразитизму (по Croll
и Matthews, 1977). L 1 L4 различные личиночные стадии
витие включает четыре личиночные стадии.
Из этоrо жизненноrо цикла МО)l(НО BЫBe
сти ряд более приспособленных к парази
тизму, которые тем не менее никоим обра
зом не следует считать последовательными
эволюционными стадиями.
Наименее модифицированный жизнен
ный цикл включает личинку, живущую CBO
бодно В почве. Станут взрослые формы па
разитами или нет, зависит от случая. На-
пример, особи Stroпgyloides на протяжении
Bcero цикла Moryт быть свободноживущи-
ми, но некоторые личинки третьей стадии
проникают в млекопитающее хозяина через
кожу и миrрируют через трахею к кишечни-
ку, rде живут размножающиеся партеноrе-
нетически самки (см. rл. 14). Яйца либо на-
чинают жизненный цикл во внешней среде,
А. Oxyspirura
Черви
113
Б. Thelazia
;рослые ОСОБИ в rлазнице
Яйца проходят через
слезные/носовые
протоки в кишечник
Птица
d'j ослые ОСОБИ В 1<01fЪ
тиваllbНbIХ мешках
Яй
1 Млeкonитающее !
вы ходи , т вс лезную \
жидкость \
L3 MYxa \ L.,
Дви "" ется К ЗаrлаТЫВ8:,ется
"" со слезнои
ротовым частяи L ,жидкостью, про
мухи 2 ходит из кишеч...
ника в яйцевые
сролликулы
8. Parafilaria r. StephanofilarlB
Взрослые ОСОБИ в подкожной dJ Взрослые ОСОБИ в
А ткани, миrрируют на поверхность кожи одкожной ткани
Микросриларии
LJt Яйца в кровоточащих LJt B изъязвлениях
ранках t Ii кожи
Лошадь Жвачное
Выходит вра , кorдa ""\ ...... ...... " ...... КроёОсосущее .....
насекомое П ж ется , \ L насекомое
Кровососушее L 3 'L
, Y' ./'
) \ 'Y
Е. Dlrofilarla
d"« rfрослые ОСОБИ в сердце
l. f)J .I1 ик роq>иларии
ев крови
J Хищник
Вых одит п ри следующем ........
КРОВОСОС8НИИ
Ш Комар
L 2
Рис. 4.20. Жизненные циклы эндопаразитических нематод, включающие смену хозяев: А
Oxyspirura; Б The/azia; B Parafi/aria; r Stephaпofi/aria; Д Опсhосеrса' E Dirofi/aria (по
Croll, Matthews, 1977)
Выходит в ЗОБУ.,
миrрирует в
rлазницу L
lIt
СЪеда тся С -<.t, "" кал:и ; ',
насекомым ------ 3arлатывание
1 , Tapa KaH L:;вnолости тела
инцистиру
в полости тела L;
ДОпсhосеrса
Взрослые ОСОБИ
дкожной ткани
j /; дМикросриларии
;! в дерме
Млекопитающее
Выходит пр и следующем ......
кровососании W ,
" {; Мошка
L;s
либо дают начало новым инвазионным ли
чинкам третьей стадии (рис. 4.19,Ь).
У свайника Aпkylostoтa сходный
жизненный цикл, но без свободноживущих
взрослых форм. Как самцы, так и самки
обитают в кишечнике (рис. 4.19,B). Первая
и вторая личиночные стадии типичные
рабдитовидные личинки микрофаrи, пи
тающиеся в почве. Третья, инвазионная,
стадия инцистируется и не питается.
у необычно крупной человеческой ки
шечной нематоды аскариды (Ascaris) CBO
бодноживущая личинка исчезает Яйца BЫ
деляются с фекалиями наружу, но личинки
первой и второй стадий остаются инцисти
рованными внутри яиц, которые заrлаты
ваются хозяином с заrрязненной пищей.
Личинки третьей стадии вылупляются
114
в просвете кишки, но не остаются там. Че..
рез кровеносную систему они миrрируют
к леrким, а оттуда обратно в кишечник (рис.
4.19,п. Такой сложный жизненный цикл
может отражать ero эволюционную исто..
рию, в которой был промежуточный хо..
зяин, утраченный впоследсrвии.
В сходных жизненных циклах паразитов
кишечника человека Trichurus и Eпterobius
(рис. 4. 19,Д) отсутствуют признаки двойной
миrрации перед поселением в кишечнике.
Самки Eпterobius MorYT вызывать очень си..
льный зуд BOKpyr анальноrо отверстия. Он
приводит к расчесыванию и в результате
к прямому повторному заражению и пере..
носу паразитов через пальцы к друrим по--
тенциальным хозяевам.
Некоторые круrлые черви, паразити..
рующие в позвоночных, имеют промежу..
точных хозяев и используют взаимоотно..
шения животных в пищевой цепи (рис. 4.20).
Друrой пример жизненные циклы нема..
тод--мермитид: личинка паразитирует в на..
секомых, а взрослые особи свободноживу..
щие (ср. тип Neтatoтorpha. разд. 4.5).
4.5. Тип Nematomorpha
(волосатики)
4.5.1. ЭТИМОPlоrия
От rреч. nematos нить; morpha
форма.
4.5.2. Диаrностические
и специфические признаки
(рис. 4.21)
1. Билатерально симметричные, тон--
кие удлиненные черви (диаметр менее 3 мм,
длина от 10 см до 1 м).
2. Тело более чем из двух' клеточных
слоев, с тканями и орrанами.
3. Кишечник сквозной прямой, часто
деrенерирует, вероятно, никоrда не функ--
ционирует
4. Тело мономерное с псевдоцеломиче..
ской полостью, часто заполненной мезенхи"
мой.
5. Стенка тела состоит из rибкой кол--
лаrеновой кутикулы, эпидермиса и слоя
rлава 4
Рис. 4.21. Форма тела взрослых волосатиков:
А Gordioidea, Б Nectonematida (по Margu-
lis Schwartz, 1982; Fewkes 1883)
ПРОДОЛЬНЫХ мышц; слой кольцевых мышц
отсутствует.
6. Кровеносная, выделительная и ды-
хательная системы отсутствуют.
7. Нервная система расположена в эпи-
дермисе, состоит из переднеrо кольца
и двух продольных стволов без rанrлиев.
8. Раздельнополые формы, с един-
ственной удлиненной rонадой или парой ro-
над; оплодотворение внутреннее с помо-
щью сперматофоров.
9. Взрослые особи свободноживущие,
живут недолrо.
Крючковидная
скалида
Ротовой канал
Кутикулярное
кольцо
Кишечник
Анус
А
Стилеты
Псевдоцель
Кутикулярно
кольцо С
епта
Б
Рис. 4.22. Личинка Gordioidea: А c BЫBepHY
тым интроверТQМ, Б c втянутым интровертом
(по Нутап, 1951; Pennak, 1978)
10. Личиночные стадии заражают чле
нистоноrих (или, редко, пиявок) и разви
ваются в их rемоцеле; у ювенильных особей
три ротовых стилета и три Kpyra крючко
видных скалид (рис. 4.22).
11. Обитатели пресных вод или влажной
почвы. Один род морской.
Строение тела взрослых волосатиков
и нематод (разд. 4.4) в принципе сходно.
Как и последние" они передвиrаются за счет
волн, проходящих вдоль тела в дopCOBeH
тральной плоскости. Морские виды и caM
цы пресноводных и наземных rрупп наибо
лее ПОlIВИЖНЫ. Более крупные пресновод
ные самки обычно ведут относительно oceд
лый образ жизни, они лежат, свернувшись
Черви
115
в спираль или в кольцо, на субстрате. Наи--
более очевидное отличие волосатиков от не..
матод деrенерация у первых кишечника
и/или ero отверстий, что связано с peДYK
цией взрослой стадии до короткоживущей
репродуктивной и расселительной фазы
жизненноrо цикла.
Питаюшаяся фаза паразитические
ювенильные особи, которые, хотя и имеют
кишечник, скорее Bcero получают большую
часть, а то и всю пищу за счет абсорбции че
рез поверхность тела из rемоцеля хозяина.
Личинки и ювенильные особи совершенно
непохожи на нематод; они напоминают
взрослых киноринхов (разд. 4.6) и лорици
фер (разд. 4.7) наличием ротовых стилетов,
скалид и строением передней части кишеч
ника (рис. 4.22). Коrда личиночный интро--
вер т вывернут (рис. 4.22,А) с помощью по
перечной септы, изолирующей от ero поло
сти переднюю часть псевдоцеля" эти стиле
ты и скалиды, по видимому, способствуют
проникновению через ткани хозяина в [eMO
цель. Происходит ли это через покровы или
через кишечник. пока не ясно.
4.5.3. Классификация
в настоящее время известно 250 видов"
помещаемых в два отряда одноrо класса.
Наиболее мноrочисленная rруппа прес
новодные Gordioidea (рис. 4.21,А), парази..
ты одноветвистых. Для них характерен
единственный (вентральный) нервный
ствол, парные [онады и полость тела, почти
целиком заполненная мезенхимными клет
ками, по крайней мере в пререпродуктив
ный период. Небольшое число известных
морских Nectonematida (рис. 4.21,Б)
паразиты десятиноrих раков отличается
вторым (дорсальным) нервным стволом,
единственной оформленной (у самцов) или
диффузной (у самок) rонадой и псевдоце..
лем, незаполненным мезенхимой. Они несут
также двойной ряд латеральных щетинок,
проходящий вдоль Bcero тела, что увеличи
вает ундулирующую поверхность во время
плавания. Некоторые зоолоrи считают poд
ство между этими двумя отрядами очень
отдаленным.. выводят нектонематид из He
матод и сближают rордиоидей с киноринха
ми и лорициферами.
116
rлава 4
4.6. Тип Kinorhyncha
4.6.1. Этимолоrия
От rреч. kinema движение; rynchos
рыло.
4.6.2. Диаrностические
и специфические признаки
(рис. 4.23)
1. Билатерально симметричные, черве
образные, но довольно короткие формы.
2. Тело более чем из двух клеточных
слоев, с тканями и орrанами.
3. Пищеварительная система трубча
тая. с задним анусом, мускулистой rлоткой .
и выступающим ротовым KOHYCOM't coдep
жащим ротовую полость.
4. Тело снаружи разделено на опреде..
ленное количество (13 или 14) cerMeHTOB,
или «зонитов» .
5. Эпидермис с иrольчатой кутикулой,
Ротовые стилеты
Ротовой конус
Скал иды
Шейная плас
тинка(плацида)
Сенсорное пятно о
(1)
@
(I!)
А
Б
Рис. 4.23. Наружное строение киноринхов: А
Cyclorhagida: (1) вид с вентральной стороны;
(11) вид спереди с втянутым интровертом; Б
Homalorhagida: (1) вид с вентральной стороны;
(11) вид спереди с втянутым интровертом за
щищенным пластинками TpeTbero зонита (по
Higgins, 1983)
включающей единственную дорсальную
и парные вентральные пластинки на каж
дом зоните.
6. Полость тела псевдоцель, проис
ходящий из сохраняющеrося бластоцеля 1.
7. Стенка тела с кольцевыми и диаrо..
нальными мышцами.
8. Выделительная система из протоне
фридиев в 11 M зоните соленоциты с пар
ными ресничками и мноrочисленными
ядрами.
9. Нервная система из переднеrо около
кишечноrо кольца и вентральноrо ствола,
на котором rруппы rанrлионарных клеток
расположены в соответствии с cerMeHTa
цией эпидермиса и мускулатуры.
10. Мелкие животные; почти всеrда BXO
дЯТ в состав морской мейофауны.
11. Наружных ресничек нет.
Киноринхи одна из наименее изучен
ных rрупп животных. Они явно близки
к друrим типам псевдоцеломических чер
вей, особенно к Loricifera и Nematomor-
pha.
Наиболее характерная черта типа
покрытая иrлами кутикула, отчетливо под-
разделяющаяся на определенное число по-
перечных зонитов. Сеrментация кутикулы
влияет Hd эпидермис, мускулатуру и нерв-
ную ткань, придавая им мета мерную струк-
туру, которая тем не менее не rомолоrична
наблюдаемой у целомических червей-
аннелид (разд. 4.14).
Все киноринхи крошечные формы
мейофауны морских илистых осадков. Опи-
сано около 100 видов, основное строение
которых сходное.
Ротовой конус со стилетами образует
выворачивающийся интроверт.. который
может втяrиваться во второй, «шейный»,
cerMeHT. Интроверт вооружен расположен-
ными по Kpyry заrнутыми скалидами
Туловишный отдел состоит из 10 одина-
ковых туловищных зонитов, каждый из ко-
торых несет единственную дорсальную пла-
стинку теР2uт, и пару вентральных
стеРllиты. Мускулатура стенки тела упло-
щает тулови ще TaK't что спинная и брюшные
1 Эмбриональное развитие этих животных
пока до КОнца не изучено, поэтому трудно ска-
зать, как образуется их полость тела. Прuм.
пер
Мозr
Слюнная железа
отовой конус
Скалида
Трихоскалида
rлотка
ПИLЦеваритenьная
железа
Средняя кишка
Половые opraHbI
Нефридий
Рис. 4.24. Внутреннее строение обобщенноrо
киноринха с вытянутым интровертом
пластинки сближаются, и давление в псев
доцеле выворачивает интроверт. Животные
передвиrаются, выталкивая ero вперед, зая..
кориваясь скалидами, а затем подтяrивая
все тело с помощью мышц ретракторов.
Некоторые черты BHYTpeHHero строения
киноринхов (рис. 4.24) также общие с друrи
ми типами псевдоцеломических червей.
Эпидермис частично синцитиальной CTPYK
туры образует продольные среднедорсаль
ный и латеральные валики (как у нематод)
и заходит в самые крупные иrлы кутикулы.
Ротовой конус
Рис. 4.25. Вид с дорсальной
стороны лорициферы (по Kri
stensen 1983 с упрощениями)
Черви
117
Ero утолщения «подушечки» вдаются
в псевдоцель. Хорошо развита мускулистая
rлотка, треуrольная в поперечном сечении,
что характерно для мноrих псевдоцеломи
ческих червей.
4.6.3. Классификация
в основе деления киноринхов на два OT
ряда лежит способ втяrивания интроверта.
у Cyclorhagida втяrивается только первый
зонит, который защищают крупные пласти
ны BToporo зонита (см. рис. 4.23,А). В OT
ряде Homalorhagida втяrиваются оба пер
вых зонита, обычно зашишаемые вентраль
ными пластинами TpeTbero (рис. 4.23,Ь).
4.7. Тип Loricifera
4.7.1. Этимолоrия
От лат. lоriса rрудной шит или латы:
fеrrе нести.
4.7.2. Диаrностические
и специфические признаки
(рис. 4.25)
1. Билатерально симметричные живот
ные.
2. Тело более чем из двух клеточных
слоев, с тканями и орrанами.
3. Прямой сквозной кишечник и терми
нальный анус.
4. Есть полость тела.
5. Тело делится на три отдела: пе
редняя выворачивающаяся rолова или ин;
Скал иды
Панцирь
.. . ,.... ..:...: ;.i.
Ротовой стилет Вытян ут ый инт р ове р т _
Шеиныи отдел
118
rлава 4
РОТОВОЙ конус
Скалиды
Сеrментированный
шейный отдел
Панцирь
А
Хвостовые ножки
Б
троверт с девятью круrами направленных
назад иrло или зубовидных скалид и вытя
rивающимся ротовым конусом. окружен
ным 8 9 твердыми ротовыми стилетами;
короткий наружно сеrментированный шей
ный отдел с рядами пластинок; туловище,
покрытое кутикулярным панцирем из ше
сти продольных пластин, несущим направ
ленные вперед полые иrлы. У взрослых oco
бей два передних отдела MorYT втяrиваться
в туловищный панцирь; пластинки шейноrо
отдела, по видимому, служат защитным
покровом после TaKoro втяrивания.
6. Пищеварительный тракт с крупным
мускулистым rлоточным бульбусом (луко--
вицей) и телескопически раздвиrающимся
ротовым каналом; весь тракт, вероятно,
выстлан кутикулой.
7. Одна пара rонад; раздельнополые
животные.
8. Хорошо развитый мозr и вентраль
ный нервный ствол с rанrлиями.
9. Личиночная стадия похожа на
уменьшенную взрослую особь (рис. 4.26),
если не считать отсутствия ротовых стиле
тов и наличия двух типов локомоторных
opraHoB: а) 2 3 пар иrл на передневентра
льной части панциря, с помощью которых
животное может ползать; б) пары напоми
нающих ножки подвижных хвостовых при
датков, используемых для плавания. HeCKO
лько возрастных стадий личинки разделено
линьками.
10. Интерстициальные формы морских
ракушечно rравийных осадков.
Строение этих крошечных (менее 0,3 мм)
Рис. 4.26. А. Вид с дорсальной
стороны личинки последней
стадии лорициферы. Б. Вид
с вентральной стороны ее ло
комоторных иrл (по Кristепsеп,
1983 с упрощениями)
животных известно пока лишь в общих чер-
тах; нет никакой информации об их эмбрио-
лоrии и образе жизни. Впервые они были
описаны в 1983 r. Одна из причин столь
позднеrо открытия, вероятно, то, что лори-
циферы прочно прикрепляются к частицам
осадка или дрyrим орrанизмам и не обнару-
живаются с помощью обычных для сбора
мейобентоса методик; друrая причина, по-
видимому, сходство животных во втянутом
состоянии с личинками коловраток или
приапу лид.
Наиболее характерная черта строения.
деление тела на передние интроверт и
шейный отдел и заднее туловище, покрытое
панцирем. У взрослоrо животноrо вся пере-
дняя часть может быть втянута в туловищ-
ный панцирь, в то время как личинка спо-
собна втянуть только интроверт в шейный
отдел. Эта особенность очень близка к то-
му, что наблюдается у приапулид, личинок
волосатиков, нематод рода Kiпochu/us
и особенно у киноринхов (разд. 4.6). Как
у последних, так и у лорицифер «rолова»
выворачивается за счет давления в полости
тела и втяrивается специфической мышцей-
ретрактором, в то время как ротовой конус
с ero стилетами втяrивается и вытяrивается
независимо с помощью мускулов Функции
выдвижения и втяrивания переднеrо конца
тела у лорицифер неизвестны; в друrих
rруппах это обеспечивает продвижение
в осадках или тканях животных хозяев, лов-
лю и поrлощение добычи. Возможно, лори-
циферы эктопаразиты.
Пищеварительный тракт по строению
Рот
Панцирь
Средняя
кишка
Мышца...
ретрактор
Ротовой стилет
Ротовой конус
Ротовой канал
Шея
Дополнительный
стилет
Мускулистаs:l
rлотка
Рис. 4.27. Передняя кишка лорицифер (pOTO
вой конус И шейный отдел соответственно вы..
тянуты и вы вернуты (по Kristensen, 1 983)
сильно напоминает таковой у тихоходок
(разд. 8.1). Ero передний отдел (рис. 4.27)
состоит из телескопически складывающе
rося pOTOBoro, или буккальноrо, канала,
мускулистоrо, несущеrо плакоиды rлоточ
Horo бульбуса, пары дополнительных сти
летав и пары крупных слюнных желез
(единственные железы, отходящие от ки
шечника). Пока еще не ясно, является ли это
сходство следствием конверrентной эволю
ции, хотя оно, по видимому, означает, что
лорициферы питаются, протыкая жертву
ротовыми стилетами и высасывая жидкость
с помощью rлоточноrо насоса.
4.7.3. Классификация
Хотя формально описан только один
вид, CBoero описания ждут еще несколько.
4.8. Тип Priapula (приапулиды)
4.8.1. 3тимолоrия
От rреч. Priapos фаллическое боже
СТБО, олицетворяющее мужскую дeTOpoд
ную силу.
4.8.2. Диаrностические
и специфические признаки
1. Билатерально симметричные живот
ные.
2. Тело более чем из двух клеточных
слоев, с тканями и орrанами.
3. Есть крупный втяrивающийся интро
Черви
119
верт, или просома, и покрытое иrлами или
чешуйками нерасчлененное туловище; иноr
Дd присутствуют ветвящиеся хвостовые
придатки.
4. Пищеварительная система с пере
дним ртом, окруженным иrло или крючко
видными скалидами, и задним анусом,
иноrда окруженным венчиком иrл.
5. Выделительная и половая системы
тесно связаны в уроrенитальный комплекс
с множеством соленоцитов.
6. Нервная система из околоротовоrо
кольца и вентральноrо ствола с rанrлиями.
7. Обширная полость тела возможно,
настоящий целом.
8. Раздельнополые формы; оплодотво
рение наружное.
9. Личинка без ресничек, заключена
в панцирь (т. е. покрыта пластинками).
10. Кровеносной системы нет, но в по
лости тела находятся форменные элементы,
содержащие rемэритрин.
Приапулиды небольшая, но четко
очерченная rруппа морских животных с He
ясным происхождением. Они внекоторой
степени сходны с типами псевдоцеломиче
ских червей, к которым их часто относят, но
функционально rораздо ближе к целомиче
ским червям, рассматриваемым в разд.
4.11 .14. Некоторые авторы считают их
полость тела целомом, поскольку она имеет
мезодермальную выстилку, но электронно
микроскопические исследования свидетель
ствуют о том, что она уникальна и непохо..
жа на присутствующую у друrих целомиче
ских форм. Требуется более детальное изу
чение происхождения этой полости. HeMHO
rие данные по эмбриолоrии приапулид ro
ворят о радиальном.. а не спиральном
дроблении их яйца.
Различные формы тела представителей
этоrо типа показаны на рис. 4.28. У боль
шинства видов оно состоит из крупной лу
ковицеобразной просомы, или интроверта,
и туловища. Интроверт оканчивается ртом,
окруженным кольцом из пяти иrловидных
скалид. Этот крупный бочковидный opraH
с продольными рядами скалид обычно вы..
вернут, но может втяrиваться в туловище,
от KOToporo обычно отделен хорошо Bыpa
женным шейным отделом. Часто кольчатое
туловище, как правило, покрыто шипами
120
"i4t 1
,,,.IIt.
. .. .,.L,
'." ."
(Щ1
." .....
....
Рис. 4.28. Разнообразные формы тела приапу
лид
Рот
Интроверт
Вторичные
кольца
Вентра,!ьный
нервныи ствол
Хвостовые
отростки
А
rлава 4
или чешуями и несет периодически линяю
щую кутикулу. На заднем конце тела Pr;..
apu/us расположена любопытная разветв..
ленная структура хвостовой придаток,
выполняющий, по видимому, респиратор..
ную функцию (рис. 4.29,А). Кутикула по..
крывает эпидермис, в котором находятся
заполненные жидкостью межклеточные
пространства; этим он отличается от эпи..
дермиса как целомических, так и псевдоце..
ломических червей. Стенка тела включает
и кольцевые, и продольные мышцы, со..
здающие давление в заполненной жидко
стью полости тела, приводящее к выворачи
ванию интроверта. Пищеварительный
тракт прямая трубка с ороrовевшим, зуб-
чатым ротовым отделом и мускулистой
rлоткой (рис. 4.29,Ь); нет ни pOToBoro KOHY
са, ни ротовых стилетов.
Уникальная черта приапулид объ-
единение половой системы и соленоцитов
протонефридиев с формированием парных
мочеполовых opraHoB (рис. 4.29,Б).
Панцирная личинка может быть пелаrи-
ческой, но покрыта кутикулярными пласти
нами (рис. 4.30). У нее уже развита втяrи
вающаяся чешуйчатая просома, и она леrко
зарывается в [рунт, в то время как взро-
слым формам зарываться трудно. Возмо-
жно, в норме личинка единственная ста-
дия жизненноrо цикла, предназначенная
Околоротовое
нервное кольцо
r лотка
Короткая мышца
ретрактор
Длинная мышца
ретрактор
Кишечник
Неq>ридий } Моче _
rOH a половои
opraH
Анус
Рис. 4.29. Внешнее (А) и схе-
матическое внутреннее (Б)
строение Priapu/us
Рис. 4.30. Панцирная личинка приапулид
для поrружения в субстрат. Внешнее cxoд
ство между ПОКРЫТЫМИ иrлами интроверта
ми приапулид и киноринхов.. а также между
панцирной ЛИЧИНКОЙ первых и коловратка..
ми не дает достаточно оснований для Toro,
чтобы считать ЭТИ типы близкородственны
МИ.
4.8.3. Классификация
Описано 16 видов приапулид, разделен..
ных на два отряда: Priapulida с более круп
пыми, более обычными хищными формами
(РИС. 4.29,А) и Seticoronaria, у представите
лей KOToporo тонкие скалиды образуют вен..
чик жестких «щупалец» (см. рис. 4.28, внизу
слева) и имеется околоанальное кольцо
крючьев. У приапулид долrая ЭВОЛЮЦИОН"
пая история; несколько их видов известны
еще с кембрия.
4.9. Тип Rotifera (коловратки)
4.9.1. 3тимолоrия
От лат. rоtа колесо; fеrrе нести.
4.9.2. Диаrностические
и специфические признаки
1. Билатерально симметричные живот
пые.
Черви
121
2. Тело более чем из двух клеточных
слоев, с тканями и орrанами.
3. Венчик ресничек в передней части Te
ла состоит из преоральноrо и посторально
ro поясков. Часто они образуют два колесо
видных ресничных opraHa, от чеrо и проис
ходит латинское название rруппы.
4. Пищеварительная система с пере
дним ртом, сложным челюстным аппара..
том, МУСКУЛИСТОЙ rлоткой и задним анусом,
открывающимся в общую с уриноrениталь
ной системой клоаку.
5. Эпидермис с небольшим фиксирован
ным числом ядер; в ero состав входит BHY
триклеточная кутик}'ла, часто утолщенная
и образующая футляр или панцирь.
6. Тело не сеrментировано.
7. Протонефридиальная выделительная
система.
8. Кровеносной системы и opraHoB ды..
хания нет.
9. Полость тела псевдоцель.
10. Раздельнополые животные, но во
мноrих случаях самцы редки или OTCYT
ствуют совсем, если есть, почти всеrда Kap
ликовые.
11. Очень мелкие формы, редко дости"
rают в длину 3 мм.
12. Развитие прямое, следует за МОДИ
фицированным спиральным дроблением.
После завершения эмбриоrенеза ядерных
делений не ПРОИСХОДИТ.
Коловратки одни из самых мелких
животных; размеры их взрослых особей Ta
кие же, как у ресничных простейших и личи
нок мноrих дрyrих типов. Их тело состоит
из небольшоrо.. cTporo определенноrо числа
клеток, а точнее сказать ядер, так как
мноrие ткани синцитиальные (общая черта
с некоторыми друrими типами псевдоцело
мических червей).
Взрослые коловратки обычно ведут при..
мерно такой же образ жизни, что и реснич
ные личинки более крупных беспозвоноч
ных; в самом деле, внешне они похожи на
трохоФор морских аннелид, сипункулид
и моллюсков. Характерная черта трохофо"
ры наличие эква ториальноrо ресничноrо
кольца прототроха, состоящеrо из двух
поясков преоральноrо и посторально"
rо, реснички которых бьют в противопо
ложных направлениях. Сходный аппарат
122
Рот
Предротовой
ресничный
венчик ==
ТРОХУС
Послеротовой
ресничный
венчик 1:
цинrулум
Коловра ...
щательный
аппарат
Кишка
Рис. 4.31. Схематический план строения коло..
враток. Показаны два венчика ресничек
имеет большинство планктонных коловра..
ток: реснички их венчика образуют два поя..
СК8 - преоральный ТРОХУС и посторальный
цинrулум (рис. 4.31).
Однако это сходство не подразумевает
rлава 4
близкоrо филоrенетическоrо родства,
и друrие черты свидетельствуют оболыпей
близости к псевдоцеломическим червям.
особенно скребням. Некоторые авторы счи-
тают коловраток неотенической rруппой,
но неизвестно, какой предковой линии, Ни
в одном из типов псевдоцеломических че-
рвей нет ресничной личинки, из которой мо-
жно было бы их вывести.
Снаружи тело коловраток часто покры-
то кутикулой, образующей чашевидный
панцирь открытый конец несет ресничный
венчик и рот. Коловращательный аппарат
венчика может быть высокоспеЦИdЛИЗИРО-
ванным, а у сидячих форм сильно редуциро-
ванным. В последнем случае (см. рис. 4.37)
реснички часто модифицируются в жесткие
сенсорные волоски. Обычно венчик может
втяrиваться в панцирь. Сзади тело суживае-
тся, образуя тонкую подвижную «Hory», ча-
сто кольчатую снаружи (псевдосеrментиро-
ванную). Как и венчик, Hora обычно может
втяrиваться в панцирь; при этом псевдосеr-
менты телескопически входят один в дру-
rой. Hora заканчивается двумя выроста-
ми «пальцами», служащими для постоян-
т росрусы
астакс
БУЛЬБУС с ресничным
пламенем
Протонефридий
Желудок
Желточник
Яйцо
Мочевой пузырь
Клоака
Педальная железа
А
Палец
Б
Рис. 4.32. Общие анатомические особенности коловраток класса Bdelloidea: А вид с вентраль-
ной стороны; Б вид сбоку
Маллеораматный
Рамус
СРулькрум
СРорципатный
Вирrатный
Рис. 4.33. Разные формы ротовых придатков
(трофусов) коловраток (по Donner, 1966)
Horo или BpeMeHHoro заякоривания живот--
Horo в субстрате; она может отсутствовать
или редуцироваться у планктонных форм.
Внутреннее устройство коловраток относи--
тельно простое (рис. 4.32) OДHO из след--
ствий мелких размеров. Тем не менее строе--
нне pOToBoro аппарата сложное и варьи--
рующее в зависимости от способа питания.
Основной план строения твердых частей
(трофусов) жевательноrо аппарата (м а..
стакеа) показан на рис. 4.33. Фулькрум, ле..
жащий ниже средней линии трофусов, под--
держивает два рамуса, расположенные от..
носительно Hero симмеrрично. Над ними
подвешены парные УНКУСЫ и манубрuu. Раз--
личные трофусы показаны на рис. 4.33. Они
MorYT быть приспособлены для прокалыва--
ния и высасывания клеточноrо содержимо..
ro, схватывания добычи (как пинцетом),
размалывания, раздавливания или разры--
вания ее.
у большинства коловраток два протоне--
фридия. состоящие из синцития с неболь--
шим количеством клеточных ядер. В прес--
новодной среде через нефридии с большой
скоростью выводится rипоосмотическая
жидкость.
, 4.9.3. Классификация
Тип подразделяют на три класса.
Черви
123
Класс
Отряд
Selsonidea
Selsonida
Bdelloidea
Bdel101da
Ploima
Monogonara Flosculariida
Collothecida
4.9.3.1. Класс Bdelloidea
у большинства представителей этоrо
класса характерный венчик из двух «колес»
(рис. 4.34) и тело, не покрытое панцирем;
эти виды ползают изrибаясь петлями, но
MorYT также хорошо плавать с помощью
нескладчатоrо коловращательноrо аппара--
та. Бделлоидеи приспособлены к жизни
в непостоянных и скрытых биотопах., обыч--
ны в интерстициали сыроrо песка и почвы
речных и озерных береrов, связаны с мхами
в периодически увлажняющихся наземных
биотопах. Они больше ползают, чем пла--
вают, хотя ресничные диски хорошо разви--
ты, а в Hore имеются массивные, крупные
цементные железы (рис. 4.35).
Биолоrия размножения очень необычна.
Похоже, вся эта rруппа амиктическая,
т. е. мейоза и спаривания здесь не происхо--
дит. Самцов не находили никоrда, известны
Рис. 4.34. Механизм питания пелаrических KO
ловраток. Крупные реснички переднеrо венчи
K (трохуса) создают ток воды в переднезад
нем направлении, обеспечивающий транспорт
пищевых частиц и сохранение положения тела
Реснички заднеrо венчика (цинrулума) бьют
в обратном направлении. Это создает BOДOBO
рот, затяrивающий пищевые частицы и Hecy
щий их ко рту
124
БУЛЬБУС С ресничным
пламенем
Протонесрридий
Парные яичники
с желточниками
Мочевой пузырь
Цементная железа
Палец
Рис. 4.35. Внутреннее строение коловраток
класса Bdelloidea (семейство Philodinidae)
Обратите внимание на парные яичники. Эти
животные исключительно амиктические; caM
цы никоrда не обнаруживались
r
д
ж
Рис. 4.36. Разнообразие форм тела коловраток
классов Monogonata и Seisonidea (по Donner,
1966)
rлава 4
лишь облиrатно партеноrенетические самки
(см. разд. 14.2.1). Характерна также способ
ность впадать при высыхании в анабиоз:
в таком высохшем состоянии они MorYT co
хранять жизнеспособность мноrие rоды,
выдерживая колебания температуры от
+ 40 до 200 0 С.
4.9.3.2. Класс Monogonata
В этот класс, для представителей KOTO
poro характерно наличие единственноrо
(непарноrо) яичника, входит большинство
коловраток. Выделяют несколько отрядов.
Самый крупный отряд Ploima
включает разнообразные сидячие и сво-
бодноплавающие формы (рис. 4.36,А,Б, Ж).
Hora, если она есть, несет два пальuа, а рот
лежит внутри, а не спереди цинrулума.
у этих коловраток часто есть жесткий паи-
цирь.
Отряд Flosculariida (рис. 4.36.B,Д)
вторая rруппа свободноплавающих или си-
дячих .коловраток, у которых ресничные
пояски коловращательноrо аппарата четко
разделены на трохус и цинrулум. Hora, если
она есть, не несет парных пальцев.
В отряде Collothecida коловращатель-
ный аппарат представляет собой крупную
воронку, часто несущую реснички, видоиз-
мененные в жесткие сенсорные волоски (см.
рис. 4,36,Т; 4.37). Самки всеrда сидячие,
прикрепленные к субстрату цементными
железами; их тело часто окружено студени-
стой массой.
В отличие от Bdelloidea коловратки
класса Monogonata размножаются поло-
вым путем. Хотя в популяциях большую ча-
сть rода преобладают самки, в определен-
ные периоды появляются относительно
просто устроенные, часто карликовые, но
свободноплавающие самцы.
Большую часть rода диплоидные амик-
тические самки производят яйца, развиваю-
щиеся без оплодотворения в молодых са-
мок. Время от времени (возможно, как ре-
акция на условия окружающей среды) появ-
ляются морфолоrически ОТЛИЧJIые миктиче-
ские самки, откладывающие rаплоидиые
яйца. Они либо быстро развиваются в ra-
плоидных самцов, либо, если происходит
оплодотворение, становятся зиrотами,
\
,
.
1
1
I
, I
, I
. ,
.1
1,
.,
.1
11
"
"
11
11
и
I
,
Рис. 4.37. Сидячая коловратка отряда Collothe
cida. Обратите внимание на модифицирован
ные реснички коловращательноrо аппарата
дающими начало амиктическим самкам.
Таким образом, миктические самки являю..
тся факультативно партеноrенетическими
(СМ. разд. 14.2.1).
4.9.3.3. Класс Seisonidea
Этот мелкий класс морских коловраток
обитает на жабрах ракообразных Nebalia
и некоторых равноноrих. Сюда входит то..
лько один род Seisoп (рис. 4.36,Ь). Ero пред..
ставители относительно крупны (несколько
миллиметров), у них сильно редуцирован..
ный коловращательный аппарат и высту"
пающий мастакс. Яичники, как и у видов
Bdelloidea, парные, но (что не типично для
коловраток) происходит обычное половое
размножение с участием нормально разви"
тых самок и самцов, встречающихся в попу..
ляциях с одинаковой частотой.
4.10. Тип Acanthocephala
(скребни)
4.10.1. 3тимолоrия
От rреч.
rолова
akantha шип.
,
kephale
Черви
125
4.1 0.2. Диаrностические
и специфические признаки
1. Билатерально симметричные, черве..
образные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и орrанами.
3. Пищеварительной системы нет
4. Сеrментация тела отсутствует, но
иноrда выражена внешняя поперечная коль..
чатость.
5. Выступающий хоботок вооружен
крючьями.
6. Полость теЛd псевдоцель.
7. Синцитиальный эпидермис снеболь..
шим фиксированным числом относительно
крупных ядер.
8. Нервная система с единственным
передневентральным rанrлием и парными
или одиночными нервами, отходящими
к opraHaM.
9. Иноrда присутствуют нефридии.
10. Кровеносной и дыхательной систем
нет.
11. Раздельнополые животные с вну"
тренним оплодотворением и живорожде..
нием.
12. Инвазионная личинка проникает
в промежуточноrо хозяина насекомое.
13. Взрослые особи паразитируют в пи..
щеварительном тракте позвоночных.
Общее строение тела скребней показано
на рис. 4.38. Большинство видов мелкие (от
1 мм до нескольких сантиметров длиной),
но некоторые достиrают длины 1 м. Наи..
более характерная черта хоботок с изо..
rнутыми шипами. Он вместе с шейным от..
делом может вворачиваться, но обычно
образует постоянный аппарат прикрепле..
ния к тканям хозяина. Как и у друrих псев..
до цел омических червей, некоторые ткани
состоят из четко фиксированноrо неболь..
шоrо количества клеток.
Стенка тела включает тонкую кутикулу,
покрывающую синцитиальный эпидермис,
содержащий внутриклеточную кутикулу
и небольшое количество четко расположен..
ных и довольно крупных ядер. Положение
этих rиrантских ядер хороший диаrно"
стический признак. Под эпидермисом нахо..
дится тонкий слой кольцевых и продольных
мышц, окружающих псевдоцель. В псевдо"
126
ХОБОТОК
Крючок
Семенник
r иrантское ядро
Цементная железа
Семенной пузырек
Клоака
Рис. 4.38. Анатомия типичноrо скребня на при
мере самца рода Neoechiпorhyпcus
цель вдаются две вертикальные складки
тканей (лемниски); в них от эпидермиса Be
дут заполненные жидкостью каналы. Знто--
дерма редуцирована до лиrамента, вдоль
KOToporo подвешены орrаны размножения.
Как и у мноrих паразитов, у взрослых
скребней наБЛIOдается сильное упроrцение
строения с утратой мноrих систем opraHoB,
характерных для свободноживущих живот..
ных, И rипертрофия репродуктивноrо аппа..
рата. Скребни раздельнополы, что необыч--
но для эндопаразитов (но см. разд. 4.4),
поэтому внутреннее оплодотворение обе..
спечивается сложным поведением при сп а..
ривании. Сперма вводится в половой тракт
самки., который после копуляции закупори--
вается с помощыо цементных желез. Опло--
дотворение происходит в псевдоцеле.
Рис. 4.40. Жизненный цикл Moпiliforтis
скребня, окончательный хозяин KOToporo
млекопитающее (по NobIe, NobIe, 1976)
rлава 4
ХОБОТОК
Моэr
Лемниск
Мускулатура
стенки тела
Скорлупа
Рис. 4.39. Стадии жизненноrо цикла скребней:
А яйцо с заключенной в нем личинкой; Б
вылупившаяся инвазионная стадия акантелла
из личинки жука
Молодая ОСОБЬ ВЫХОДИТ ИЗ цисты
Взрослый червь
Зараженный
таракан
съедается
КРЫСОЙ
ЯЙЦО , :.{.
съедается '.J:.
тараканом ..:
Акантор ПРОБодевает
стенку кишечника
у мноrих специализированных парази
тов строение личинки позволяет СУДИТЬ
о филоrенетическом родстве взрослых oco
бей, однако у скребней это не так. Их личин
ка также высокоспециализирована и сходна
с взрослыми животными. После BHYTpeHHe
ro оплодотворения ранние личиночные CTa
дии протекают в покрытом прочной CKOp
лупой «яйце» (рис. 4.39,А). Оно выводится
из хозяина с фекальными массами и, чтобы
происходило дальнейшее развитие, долж
но быть съедено насекомым про
межуточным хозяином. В нем личинка
вылупляется и проникает в rемоцель, rде
превращается в следующую личиночную
стадию акаНlпеллу (рис. 4.39..Ь). Такая ли
чинка может инцистироваться в насекомом,
и после Toro как ero проrлотит позвоноч
ное, развивается в этом окончательном xo
зяине ВО взрослоrо скребня. Рис. 4.40 иллю
стрирует жизненный цикл М oпiZifo"пis
паразита мышей, крыс, кошек и собак.
4.10.3. Классификация
Известно 1000 видов, распределяемых
по трем отрядам единственноrо класса. Два
отряда составляют паразиты пресноводных
рыб, а третий наземных четвероноrих.
4.11. Тип Sipuncula
(СИПУНКУЛИДЫ)
4.11 1. 3тимолоrия
От лат. sipunculus маленькая дудка.
4.11.2. Диаrностические
и специфические признаки
(рис. 4.41 и 4.42)
1. Билатерально симметричные, черве
образные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и орrанами.
3. Мускулистая U образная кишка со
ртом и анусом, последний раСПОЛQжен дop
сально на передней части тела.
4. Тело подразделяется на переднюю ча
сть интроверт, заканчивающийся pTOM,
и более толстое туловище сзади.
5. Тело не сеrментировано.
Черви
127
РОТ
Рис. 4.41. Внешний вид типичной сипункулиды
Мозr
Нервное кольцо
ВентраЛЬtiый
нервный СТВОЛ
Ретракторы
Задняя кишка и анус
Кишечник
Щупальцевый
мешок
Пищевод
Рис. 4.42. А. Внутренее строение сипункулиды;
левая сторона тела удалена. Б. Детали строе..
ния rидравлической системы щупалец и интро"
верта
6. Полость тела схизоцель, но без BHY
тренних септ.
7. Наружный эпителий покрыт кутику
лой, без волосков и щетинок.
128
rлава 4
8. Кровеносной или дифференцирован..
ной дыхательной систем нет.
9. Нервная система с передним мозrом,
околопищеводным нервным кольцом и BeH
тральным нервным стволом без rанrлиев.
10. Единственный нефридий или пара He
фридиев.
11. Развитие со спиральным дроблением
и обычно трохофорной личинкой, которая
у некоторых видов сменяется океанической
«пелаrосферой» личинкой характерной
только для данноrо типа.
Тело четко разделено на два отдела:
передний, интроверт, втяrивается в находя
щееся сзади туловище мощными МЫШIIами
ретракторами (см. рис. 4.42) и выворачивае
тся за счет rидравлическоrо давления, co
здаваемоrо туловищной мускулатурой. Ло
кальное расслабление кольцевых мышц MO
жет вызывать расширение тела, закрепляю
щее животное в субстрате при выворачива
нии интроверта.
Сипункулиды в целом неизбира
тельные детритофаrи. захватывающие пи
щу с помощью щупалец, окружающих рот
на вершине интроверта. Щупальца управ
ляются rидравлической системой, изолиро
ванной от общей полости тела. Из каждоrо
из них пара протоков ведет в кольцевой Ka
нал, от KOToporo отходят один или два Me
шочка, расположенных параллельно оси ИН
троверта. Эти дорсальный и вентральный
компенсационные мешочки MorYT сжима
ться и служат rидравлическими резервуара..
ми для системы щупальцевых каналов
(рис. 4.42, Б).
Мноrие виды живут во временных HOp
ках, прорытых в илистых субстратах, но He
которые заселяют пустые раковины моллю
сков или трубки червей. Сверлящие фОрf\.fЫ
проделывают ходы в известковых коралло
вых отложениях. Разнообразие форм иллю..
стрирует рис. 4.43.
Нервная система сипункулид напоминает
известную для аннелид и эхиурид: передний
rанrлий (мозr) в интроверте, околопище
водное нервное кольцо и вентральный HepB
ный ствол (однако без признаков cerMeH
тарных rанrлиев). .
Обширная rидравлическая полость тела
служит также для накопления rаметоцитов,
выпадающих в нее из просто устроенных
Рис. 4.43. Разнообразные формы тела СИПУН-
кулид
[онад на ранних стадиях дифференцировки
и хранящихся там до тех пор, пока количе..
ство зрелых rаметоцитов не станет доста..
точно большим Затем rаметоциты через
нефридии выводятся наружу. Эта схема ви"
&" /0 О
11
IV
0\ .'
t
#1
Рис. 4.44. Способы змбrиональноrо развития
сипункулид: (1) через трохофорную личинку;
(11) через непитающуюся трохофору и харак-
терную для данноrо типа пелаrосферу; (111) че-
рез крупную планктотрофную пелаrосферу,
характерную для OTKpbIToro океана; (IV) через
непитающуюся, боrатую желтком личинку
с подавлением пелаrических личиночных ста-
дий ('10 Rice, 1985 с изменениями)
доизменена у крошечной формы Golfiпgia
mzпutll протандрическоrо rермафродита,
производящеrо крупные яйца, развиваю..
щиеся без оплодотворения. Это
следствие мелких размеров животноrо, а
в целом способ размножения сипункулид
характерен для всех крупных морских бес..
позвоночных.
Из яйца у большинства сипункулид вы..
ходит пелаrическая ЛИЧИНКа, БЛИЗКdЯ к ли..
чинке некоторых аннелид. У некоторых ви"
дов эта трохофора прямо развивается в мо"
лодую особь, но чаще претерпевает мета..
морфоз до следующей личиночной ста..
ДИИ пелаrосферы, которая может быть
и бентосной, хотя обычно пелаrическая. Че..
тыре способа развития сипункулид про ил..
люстрированы рис. 4.44.
4.11.3. Классификация
Все 250 известных видов можно объеди..
нить в один класс и отряд.
4.12. Тип Echiura (эхиуриды)
4.12.1. 3тимолоrия
От rреч. echis змея; ura хвост.
4.12.2. Диаrностические
и специфические признаки
1. Билатерально симметричные, черве..
образные животные
2. Тело более чем из двух клеточных
слоев, с ткаНЯl\IИ и орrанами.
3. Мускулистая кишка с передним и зад..
НИМ отверстиями.
4. Тело с крупным вытяrИВdЮЩИМСЯ
и сократимым передним выростом, или
«хоботком», с латеральными складками. ве..
ДУЩИМИ ко рту.
5. Единственная неразделенная полость
тела схизоцель расположена между
мышuами стенки тела и кишкой
6. Тело несеrментировано.
7. Единственная пара щетинок располо..
жена спереди вентрально, иноrда в друrом
месте.
8. ЗаNfКнутая кровеносная система с дор..
сальным и вентральным сосудами; в одной
rруппе сосудистая система незамкнутая.
Черви
129
9. Нервная система соколопищеводным
колыIмM и вентральным нервным стволом
без выраженных rанrлиев.
10. Выделительная система включает до
400 неметамерно расположенных нефри
диев
11. Развитие со спиральным дроблением
и трохофороподобной личинкой.
Эхиуриды несеrментированные цело..
мические черви, родственные кольчецам
(разд. 4.14), к которым их прежде и относи
ли. Наиболее их характеРНdЯ черта
хоботок, rде находится передняя часть не..
рвной системы; возможно, он rомолоrичен
простомиуму аннелид (рис. 4.45). Парные
вентральные щетинки или крючья на пере..
днем отделе тела также черта, напоми..
нающая аннелид, как и нефридии с реснич--
ной воронкой, расположение мускульных
слоев в стенке тела и строение пищеварите
льной системы. Тем не менее эхиуриды от..
личаются от кольчецов полным отсут"
ствием сеrментации или метамерии. Можно
считать их близкими к аннелидной линии
филоrенеза. В самом деле, онтоrенетиче
ские данные rоворят о возможной метаме..
рии на ранних стадиях эволюции этой rруп..
пы.
Все эхиуриды морские животные, ве..
дущие сидячий образ жизни в мяrких осад--
ках. Большинство питается детритом и оби--
тает в постоянных норах; при этом хоботок
служит инструментом неизбирательноrо
сбора пищи; вдоль ero желобка она направ"
ляется ко рту (см. рис. 9.6). Разнообразие
форм тела иллюстрирует рис. 4.46. Для ро..
да Urechis характерен друrой способ пита..
ния. Эти эхиуриды живут в rлубоких U..
образных норках, через которые они проrо..
няют воду за счет волновых перистальтиче..
ских сокращений мышц.
Биолоrия размножения эхиурид
характерная для крупных целомических че..
рвей, обитающих в морской среде. Живот..
ные раздельнополы, и целом служит поло
стью, В которой rаметоциты развиваются
перед массовым выходом через нефридии
во время «нереста».
Один из родов эхиурид отличается спе..
IIиализированным типом половоrо размно"
жения, который обсуждается в разд.
14.2.3.3. Самцы здесь карликовые и живут
130
ЖеЛОБОК
Щетинка
"
...... ..
.."
Туловище
....'-".... .... J....
Вентральный
нервный
ствол
.. .
....
... . .......
......
... _ .loJ
... .,.,J
А
на хоботке самок. Наличие карликовых
самцов ранее считалось чертой, характер..
ной только для отряда Bonel1ida, но недав"
но этот феномен был обнаружен и у друrих
эхиурид; это, возможно, свидетельствует
rлаsа 4
Кишечный сосуд
Передняя кишка
Неq>ридий
Вентральный
нервный ствол
Вентральный KpOBe
носный сосуд
Средняя кишка
Анальный мешок
r он ада
Рис. 4.45. Внешнее и BHY
трен нее строение эхиурид:
А вид с вентральной CTO
раны Echiurus; Б вид
в разрезе с дорсал ьной CTO
раны
Прямая кишка
о том, что он возникал в пределах данноrо
типа неоднократно.
4.1 2.3. Классификация
Описано 150 видов, распределяемых по
трем отрядам в зависимости от расположе
ния мускульных слоев в стенке тела, количе
ства нефридиев, замкнутоrо или неЗdМКНУ
Toro характера кровеносной системы.
у представителей onHoro из отрядов X
nopneusta, задняя часть кишки ФУНКUИОНИ
рует как дыхательный opraH.
4.1 з. Тип Pogonophora
4.1 3.1. 3тимолоrия
От rреч. pogon борода p11ol"os
носитель.
4.1 3.2. Диаrностические
и специфические признаки
(рис. 4.47)
(Следует отметить, что интерпретация
различных анатомических черт поrонофор
Рис. 4.46. Разнообразные формы тела эхиурид все еще не УСТОЯЛdСЬ, как и их rомолоrиза
Рис. 4.47. Сильно схематизи
рованный вид с дорсальной
стороны обобщенных предста
вителей двух классов поrоно
фор, показывающий основные
отделы тела и ero полости (по
Southward, 1980; Jones, 1985)
Perviata
ЦИЯ, в частности.. структур недавно откры"
тых вестиментифер и друrих представите..
лей зтото типа. Например, до сих пор идут
споры о том, что считать вентральной,
а что дорсальной поверхностью.)
1. Билатерально симметричные. удли..
ненные, метамерно сеrментированные че..
рви (0,5 мм 3 см в диаметре, 5 см
3 м длиной), постоянно обитающие в хити..
новых или белковых трубках, внутри кото..
рых MorYT двиrаться.
2. Тело более чем из двух клеточных
слоев, с тканями и орrанами.
3. Рот и кишка отсутствуют на всех ста..
диях развития.
4. Одиночные и или парные полости те..
па неясноrо происхождения (хотя обычно
их и считают схизоцелями, перитонеальной
выстилки нет).
5. Тело состоит из четырех отделов:
передней «rоловной лопасти», несущей 1
более [000 щупальцевидных «жабр» с rид..
ростатическим скелетом: короткото «желе..
зистоrо отдела» с парными полостями,
иноrда в значительной степени заполненны..
ми мускульной тканью; крайне вытянутоrо
«туловища» с одной парой крупных rидро..
Черви
131
ЖаБрЫ
ОБтуракулум
елезистая ОБласть
r онадЫ
Туловище
Телосома
Щетинки
Obturata
статических полостей и часто с различными
клейкими или секреторными папиллами:
короткой прикрепительной части тело..
сомы, включающей до 30 сетментов (с
одиночной или парной полостью тела в
каждом), которые отпочковываются от тер..
минальной зоны роста
6. Стенка тела с кутикулой, эпидерми..
сом, слоями кольцевых и продольных
мышц; телосома и (у некоторых видов) ту..
ловище со щетинками; некоторые участки
тела с ресничными полосками.
7. Кровеносная система замкнутая..
с сердцем, окруженным перикардиальной
полостью. в rоловной лопасти.
8. Пара выделительных opraHoB, напо..
минающих метанефридии, в rоловной лопа..
сти.
9. Орrанами дыхания считаются жабры,
которые, возможно, участвуют и в питании.
10. Эпидермальная нервная система из
переднеrо кольца или скопления нейронов
и обычно единственноrо продольноrо ство"
ла без rанrлиев (ето обычно считают вен..
тральным); в некоторых частях тела может
быть несколько стволов.
11. Раздельнополые животные с парой
132
rлава 4
Кровеносные
СОСУДЫ пиннулы
Полость тела
Реснички
удлиненных [онад в туловище; мужские [о-
нады производят спеРМdТОфОРЫ.
12. Оплодотворение считается НdРУЖ-
ным, Рdзвитие непрямое, яйца и личинки не
покидают материнской трубки.
13. Морские, обычно rлубоководные
(100 4000 м) формы.
Хотя впервые эти обитатели морских
rлубин обнаружены в 1900 r., известно о них
мало. Например, только в 1964 [. получен
целый экземпляр поrонофоры (ранее мета-
мерная телосома была неизвестна)" а до
этоrо времени их обычно считали родствен-
ными лофофоровым и вторичноротым из..
за как будто олиrомерноrо строения тела,
состоящеrо из трех отделов. Обнаружение
сеrментированной, несущей щетинки тер-
минальной части и первых вестиментифер
в 1969 [. привело к изменению прежних
представлений и сближению их с кольчеца-
ми. В самом деле, некоторые авторы скло-
няются к тому, чтобы поместить один или
оба класса поrонофор в тип аннелид, но, как
Эпидермис
Пиннула
Рис. 4.48. Поперечный срез жабры Perviata, де..
монстрирующий две пиннулы и расположение
ресничек (по Ivanov 1963)
ЖаБрЫ
rоловная лопасть
Уздечка
Железистая
ОБласть
Пиннулы
Реснички
Увеличенные
папиллы
Постаннулярные
папиллы
Парные
папиллы
Непарные
папиллы
Туловище
А
Телосома/
Рис. 4.49. Поrонофоры класса Perviata: А схематическое изображение внешнеrо вида типичной
первиаты (сильно укорочено), показывающее ее основные черты (по George, Southward, 1973);
у живой поrонофоры тело ориентировано прямо и вертикально; Б r передние концы трех
первиат с разным количеством и расположением жабр (по Ivanov, 1963)
отмечено выше, этот и мноrие друrие во..
просы еще ждут cBoero разрешения
Поrонофоры обитают в прямых. плотно
их облеrающих трубках, образуемых желе..
зистым отделом и утолщаемых секретами
туловища. Внутри них они удерживаются
с помощью: а) туловищных и/или телосо..
мальных щетинок; б) некоторыми туловищ..
ными папиллами; в) валиками или крылья..
ми железистоrо отдела. Жабры выставлены
из трубки в окружающую воду и сильно ва..
рьируют по количеству и расположению.
Часто их основания слиты вместе или отхо..
дят от языковидной или rребенчатой струк"
туры. У представителей caMoro обширноrо
класса они расположены так, вероятно, для
Toro, чтобы образовать центральную труб..
чатую полость; у видов с единственным
толстым «щупальцем» тот же эффект до..
стиrается за счет спиральноrо скручивания
последнеrо (рис. 4.49,В). В эту полость
вдаются парные ряды «пиннул»
удлиненных выростов эпидермальных кле..
."
.'
Б
Рис. 4.49 (продолжение)
;..:- --
в ",
Черви
1ЗЗ
ток, сбоку от которых находятся длинные
эпидермальные реснички (рис. 4.48). Функ..
цией этих ресничек, вероятнее Bcero, явля
ется создание нисходящеrо тока воды в по..
лости, образованной жабрами, однако роль
пцннул неясна.
Отсутствие кишечника породило множе..
ство спекуляций относительно сцособа пи..
тания поrонофор. Наружное переваривание
пищи маловероятно, поскольку не обнару..
жено соответствующих секретирующих
ферменты клеток. Всасывание растворен--
ных орrанических веществ возможно, но Ha
личие в среде достаточных их концентраций
проблематично. Не исключено, что мелкие
частицы орrаники, приносимые током воды
в межжаберную полость, MorYT улавлива..
ться пиннулами и поrлощаться посред..
ством пиноцитоза; известно, что эпидермис
по крайней мере нескольких видов несет ми..
кроворсинки, сходные с присутствующими
на эпителии средней кишки друrих живот
ных. К настоящему времени доподлинно
'"
}
Уздечка
r
134
известно только то, что виды, живущие
вблизи подводных rидротермальных фума..
рол или участков просаЧИВdНИЯ холодных
вод, удовлетворяют свои потребности в ос..
новном (если не целиком) за счет внутри..
клеточных хемоавтотрофных бактерий, ме..
таболизирующих восстановленные соеди..
нения серы и MeTaH поступающие из жерл,
причем аналоrичные бактерии присут"
ствуют у изученных видов поrонофор клас..
са Perviata.
4.1 3.3. Классификация
Сто видов поrонофор делят на два клас..
са
Класс
Отряд
:= Аthесаnерhпа
Thecaneph ria
Perviata
< Obturata
Vestlmentifera
4.13.3.1. Класс Perviata
Эти типичные поrонофоры сравнитель..
но мелкие (диаметр менее 3 мм, длина ме..
нее 85 см); они живут в трубках, заякорен..
ных в мяrких осадках. У них HeMHoro (1
250) удлиненных жабр и выражен косой ва..
лик «уздечка)}; окружающий железистый
отдел (рис. 4.49). Возможно, он удерживает
переднюю часть тела в устье трубки. Описа..
но 95 видов.. разrеляемых на два отряда.
4.13.3.2. Класс Obturata
Единственный отряд Vestimentifera (рис.
4.50), составляющий этот класс, включает
червей rораздо больших размеров (1 3 см
в ДИdметре.. длина некоторых особей более
2 м) с более короткими и более мноrочи..
сленными жабрами (свыше 1000), располо..
женными на «обтуракулуме)} части тела,
прикрывающей отверстие трубки после то..
ro, как животное в нее втяrивается. В от ли..
чие от первиат у вестиментифер нет щети"
нок на туловище и уздечки, а последняя за..
менена двумя крупными крыльями, кото..
рые сходятся вдоль средней дорсальной ли..
нии и тянутся вперед над основанием обту"
ракулума.
По мере исслеДОВdНИЯ новых зон проса..
rлава 4
\S I II I IIII ,
' , '\\1\1111111111 1 '1
'I\I"'IIIIIIIЙ
1IЩ111II/Ir.r 111
1t\\ШI!ш;1 1:I 1I11 ОБтуракулум
11\111111"111/11
111111II1 1 111
IIIIIIIM' I
, 11111111111111
Реснички
Железный
отдел
: .
::. ;) .,
:..' :
( )}
:.-" 0.0:'
:: >';
. ....
:. .,: '
:. >:"
::;) : .
.' ;
'r
:' >:.
.". "..
?: .;.
: ':1
Туловище
t. > :' :
. .1"
i: : (i
' '. :
: > !.l
:: )..
{
: :>;;
) -:
? .
'. )
:. ."
. . :
Телосома
Рис. 4.50. Схематическое изображение внеш-
Hero вида типичной вестиментиферы (по Sout
hward, 1982)
чивания и rYlдротермальных жерл число от-
крываемых видов вестиментифер возра-
стает почти экспоненциально. О находке
первоrо вида было объявлено в 1969 [. ВТО-
poro B 1975 r., TpeTbero в 1981 [., еще
шесть видов описаны в 1986, вероятно, око-
ло дюжины новых видов описываются в на-
стоящее время. Хотя в одной из последних
публикаций таксономический paHr обтурат
поднят до уровня самостоятельноrо типа
с двумя классами и тремя отрядами ввиду
Toro, что сведения и суждения об этой rруп..
пе постоянно меняются и будут меняться
в дальнейшем по мере получения HOBoro
материала, мы предпочитаем сохранить
здесь традиционную систему из одноrо
класса и отряда, пока классификация обту"
рат как"то стабилизируется.
4.14. Тип Annelida
(кольчатые черви, кольчецы)
4.14.1 3тимолоrия
От лат. annel1us или annelus YMeHЬ
шительное от anulus. кольцо.
4.14.2. Диаrностические
и специфические признаки
1. Билатерально симметричные, черве..
образные животные.
2. Тело более чем из двух клеточных
слоев, с тканями и орrанами.
3. Мускулистый пищеВdрительный
тракт со ртом и анусом.
4. Тело сеrментировано (сеrментация
может быть внешне незаметна, но всеrда
видна по нервной системе).
5. Имеются пресеrментный простомиум
с нервным rанrлием и постсеrментный пи..
rидиум.
6. Полость тела представляет собой ряд
схизоцелей.. нечетко выражена у форм
с передней и задней присосками.
7. Полость тела часто разделена попе..
речными септами, но они часто неразвиты
или плохо различимы в нескольких или во
всех cerMeHTax.
8. Наружный эпителий покрыт кутику"
лой И несет эпидерм'!льные щетинки (еди
ничные или в пучках), за исключением
экземпляров с передней и задней присоска..
ми.
9. Стенка тела мускулистая, часто со
сплошными слоями кольцевых мышц и че..
тырьмя полосами продольных мыши.
10. Кровеносная система замкнутая.
11. Нервная система с пресеrментным
надпищеводным rанrлием, околопищевод..
ным кольцом и вентральным нервным ство"
лом с сеrментарными rанrлиями.
12. Сеrментарные выводные протоки
Черви
135
мезодермальноrо и эктодеРМdльноrо про..
исхождения MorYT быть объединены, orpa..
ничены одним или несколькими сеrментами
и/или частично редуцированы
13. В разной степени выражена цефали..
зация.
] 4. Развитие со спиральным дробле..
нием, но возможны ero модификации и ra..
струляция путем эпиболии у форм с боrаты"
ми желтком яйцами.
15. у морских форм планктонное разви"
тие со свободноживущей трохофорной ли..
чинкой. но эта стадия иноrда инкапсулируе..
тся; у пресноводных и наземных форм
внутри яиц, покрытых прочными оболочка..
ми.
В структуре аннелид сочетаются rидро..
статическая и функциональная особенности
целомическоrо уровня орrанизации с се..
rментированным строением тела. В самом
примитивном случае сеrментация отражае..
тся на эктодерме и мезодерме и включает
посеrментное разделение целома септами.
у мноrих аннелид тем не менее такой план
строения исчез или модифицирован. а септы
или целом вторично утрачены.
Сеrментация (рис. 4.51, см. также
рис. 4.54) возникает в онтоrенезе из двух
парных мезодермальных зон роста и соот..
ветствующей 'Эктодермальной полоски
перед пиrидиумом, т. е. задним отделом., че..
рез который энтодерма сообщается с внеш..
ней средой. Новые сеrменты образуются
спереди от пиrидиума так, что последний
сформировавшийся cerMeHT всеrда оказы..
вается задним (рис. 4.52,А). У некоторых
полихет первые три cerMeHTa образуются
очень рано и одновременно и MorYT специа..
лизироваться для жизни в планктоне, но по..
следующие возникают постепенно один за
друrим (рис. 4,52,Ь). Их число может уве..
личиваться в течение всей жизни, хотя
обычно достиrает определенноrо предела.
у взрослых полихет и олиrохет образова..
ние новых cerMeHToB можно индупировать
поперечным перерезанием, причем утрата
последних cerMeHToB приводит к формиро..
ванию HOBoro пиrидиума и препиrидиаль..
ной зоны роста (рис. 4.53) (см. разд. 15.5).
В наиболее общих чертах тело аннелид
можно представить себе состоящим из пре..
cerMeHTHoro простомиума с надпищевод..
136
rлава 4
Тело из множества щетинконосных cerMeHTOB
Перистомиум
Рот
Простомиум
Недавно ОБразовавшиес
cerMeHTbI Зона нарастания
(сеrментная Бластема)
Рис. 4.51. Основные отделы Te
ла аннелид на примере полихе
ты Nereis
Ресничный поясок
(прототрох)
Зона езодерма
нарастания
cerMeHTOB Эктодерма
А
Б
ным rанrлием, ряда более или менее иден
тичных структурных единиц cerMeHToB..
и задней постсеrментной части
пиrидиума, в передней вентральной обла
сти KOToporo находится сеrментная бласте
ма, или зона роста (рис. 4.54). Основные
черты такой сеrментации сводятся к сле
дующему:
1) эктодерма: cerMeHTapHoe расположе
ние щетинок; нервная система с cerMeHTap"
ными raHr лиями и связанными с ними He
рвами; эктодермальные нефридии;
2) мезодерма: сеrментация мускулату
ры и кровеносной системы, связанная с ще
тинковыми мешками и параподиями; разде
ление септами целома с образованием изо..
лированных полостей тела; cerMeHTaplloe
расположение rерминативных 1 эпителиев
и связанных с ними парных целомодуктов.
у предков аннелид, вероятно, были
сплошные септы и полные наборы эктодер"
мальных протонефридиев и мезодермаль
1 Термин rерминативный эпителий (germinal
epithelium) в отечественной литературе встреча..
ется очень редко; им обозначается часть перито..
ниальноrо эпителия, производящая клетки
предшественницы половых продуктов. Прuм.
пер.
Рис. 4.52. Образование cerMeH
тов в ходе paHHero эмбриона
льноrо развития А ресничная
поспрохофорная личинка; Б
метаморфозировавшая личин
ка
ных целомодуктов. Такая орrанизация не
сохранилась ни у одной из ныне живущих
форм. Протонефридии с rруппой клеток
ресничноrо пламени еще встречаются в не-
скольких семействах полихет, НО больше
распространены нефридии с открытой pec
ничной воронкой нефростомом. Эмбрио-
Нdльное происхождение нефридиев и цело-
модуктов разное (разд. 1.3.1), у олиrохет
они разделены. У полихет (морская в целом
Зона нарастания
(Бластема)
Рис. 4.53. Реrенерация у Nere/s Утрата cerMeH-
тов вызывает образование новой зоны их по-
очередноrо нарастания; задний cerMeHT.....
самый молодой
Рис. 4.54. Схематическое
изобра)f(ение компонентов
сеrментированноrо тела rи
потетической протоаннели
ды. Ныне живущие кольче
цы сохранили некоторые, но
не обязательно все эти чер
ты. А. Продольные блоки
мышц и кишка не сеrменти
рованы. Полость тела
настоящий целом с перито
неальной выстилкой. Б. Ce
rментация вентральной
нервной системы, лучше
Bcero сохранившаяся у co
временных аннелид предко
вая черта (см. раздел
о пиявках ниже): cerMeHTap
ные rанrлии и нервы. В. Раз
деление целома поперечны"
ми септами, сохранившееся
у большинства олиrохет
и некоторых полихет; может
сильно модифицироваться;'
утрачено у пиявок. r. Се..
rментация эктодермы и Me
зодермы, связанная с раз
витием параподий или ce
rMeHTapHo расположенных
щетинок. Хорошо выражена
у полихет, меньше у олиrо
хет, подавлена у пиявок. Д.
Сеrментация половых и вы..
делительных протоков и re..
рминативноrо эпителия.
Предполаrается, что у про
тоаннелид были полные Ha
боры мезодермальных це..
ломодуктов (для вывода ra
мет) и эктодермаllЬНЫХ He
фридиев, а также пара re..
рминативных эпителиев
в каждом отделе целома
Этоrо не обнаружено ни
у одной из современных aH
нелид; часты сильные OT
клонения от предложенной
схемы. Е. Сеrментация кро"
веносно сосудистой систе..
мы
rруппа) нефридий и целомодукт обычно
объединены в сложную структуру
нефромиксий. В тех семействах, rде сплош..
вые септы отсутствуют и целомические по..
лости сообщаются между собой, число re..
рминативных эпителиев и сеrментарных
выводных протоков сильно сокращено.
Черви
А
Б
r
Е
137
CerMeHTapHbIe нервы
Вентральный
нервный ствол
CerMeHTapHbIe rанrлии
Межсеrментная септа
Ацикула
Внутренние мыцыы
Пара подия
Внешние мышцы
Отверстие целомодукта ==
полова воронка и
половои проток
Несрридий
rонада
Дорсальный
кровеносный сосуд
CerMeHTapHbIe
кровеносные сосуды
Вентральны
кровеносныи сосуд
4.14.3. Классификация
Известно по крайней мере 75000 видов,
которых делят на три крупные rруппы: Ро..
lychaeta, Oligochaeta и Нirudiпеа и две
мелкие. Hirudinea рано диверrировали от
эволюционной линии олиrохет, и две эти
138
Класс
Polychaeta
Aeolosonlata
rлава 4
Подкласс
Отряд
Orbiniida
Сtепоdпlidа
Psammodrilida
СОSSl1пdа
Splonida
Qtlestilia
Capltclllda
Ophcliida
Phyllolioclda
АlllрhiПОПlidа
SРlпthсri<.lа
Еl1ПlCldа
Stсrпаsrid l
()v.:enlilla
fla hell igerida
Pocohllda
т erebcllida
Sahellilla
Nertll ili.l
Оiпорhilidа
Polygordillla
Protodri1ida
t\1 \,ZOStolllltl.l
Acolo OIllatilia
HirlIdinea
rруппы вместе с малочисленным подклас
сом Branchiobdella образуют класс Clitel1a
ta.
Рис. 4.55. Различные формы
параподий полихет. А. OCHOB
ной двуветвистый тип (как
у Nereis) с деталями морфоло--
rии некоторых щетинок. Б.
Примеры пара подий седентар--
ных семейств: I Spionidae;
11 Arenicolidae; 111
Sabellidae. В. Примеры парапо
дий и щетинок эррантных се..
мейств: I PQlynoidae; 11
Phyllodocidae, 111 Eunicidae
в
4.14.3.1. Класс Polychaeta
(мноrощетинковые кольчецы)
Обитающие в основном в море полихе..
ты замечательны своим морфолоrическим
и анатомическим разнообразием. Они полу..
чили свое название из..за мноrочисленных
щетинок (chaeta) обычно расположенных
двумя пучками на двуветвистых парапо..
9
! 1, I \ \
II 111 \'
А
Б
Черви
139
диях, попарно отходящих от каждоrо се..
rMeHTa (рис. 4.55). Развитие у мноrих не..
прямое (рис. 4.56).. хотя есть и исключения.
Примерно половину видов и семейств
полихет можно для удобства объединить
в rруппу эррантных (<<бродячих») форм
В основном сюла относятся два крупных
и четко очерченных отряда Phyllodocida
и Eunicida. У филлодоцид имеется осевая
8
Рис. 4.56. Примеры планктон
ных стадий развития полихет:
A Sabellaria; Б Spio; B
Nereis; r Oweпia
r
:--
140
rлава 4
Простомиум
rл от ка
(втянута)
Челюсти
Параrнаты
Рис. 4.57. Хоботок у отряда Phyllodocida мускулистая rлотка, которая выворачивается за счет
давления в целоме втяrивается при сокращении мышц ретракторов (А r). Она часто, как у Ne
reis, вооружена белковыми челюстями
Передний конец
Рукоятка
Передняя
поверхность
А
МаНДИБУЛЫ
1 мм
Максиллы(М)
Б
Рис. 4.58. Челюстной аппарат
у отряда Eunicida. А. Изолиро
ванный челюстной комплекс.
Б. I Челюсти втянуты в MYCKY
листую вентральную rлотку. 11.
Вывернутые челюсти (по Olive,
1980)
rлотка хоботок (передняя часть пищевари
тельноrо канала), которая может rидравли
чески выворачиваться и втяrиваться мыш--
цами"ретракторами. Часто она вооружена
небольшим числом челюстей из OTBepдeB
шеrо белка с высоким содержанием ионов
тяжелых металлов, но никоrда не обы--
звествленных (рис. 4.57).
Эвнициды напоминают филлодоцид
внешне, но у них не осевая rлотка--хоботок,
а выворачиваемая буккальная масса 1, BOO
руженная сложным набором обызвествлен
иых челюстей.. парой мандибул. напоми
иающих по форме долото, и несколькими
парами максилл (рис. 4.58). Разнообразие
форм эррантных полихет проиллюстриро--
вано рис. 4.59. У большинства хорошо раз..
вита rолова с разнообразными сенсорными
орrанами и сложным мозrом. Мноrие
хищники, но есть и детритофаrи, фильтра
торы и всеядные формы. Хорошо развиты
параподии с вентральной и дорсальной BeT
вями, укрепленными жестким белковым
стержнем ацикулой.
Для некоторых эррантных полихет xa
рактерна эпuтокuя.. т. е. cBoero рода «poe
иие», связанное с половым размножением.
Это часто сочетается со сложным MeTaMOp
1 Вместо термина «буккальная масса» (Ьиссаl
mass) в отечественной литературе принято упо..
треблять термин «вентральная мускулистая r лот..
ка». Прuм. пер.
Черви
141
II
Рис. 4.59, Разнообразие форм
тела эррантных полихет; 1 и 11
вид животноrо с дорсальной
и вентральной сторон COOTBeT
ственно
фозом взрослых животных. Эпитокия MO
жет быть двух типов (рис. 4.60 и 4.61). При
эпиrамии весь червь трансформируется
в «роевую» (эпитокную) форму, в то время
как при схизоrамии (столонизации) задние
cerMeHTbI половозрелоrо животноrо OTдe
ляются, становясь миrрирующими несущи--
ми raMeTbI столонами;
Остальных полихет часто искусственно
и по разным принципам объединяют в rруп
пы седентарных (<<сидячих») форм и «ap
хианнелид»; внутри них родственные связи
не прослеживаются. Большинстро биолоrов
предпочитает оперировать четко очерчен
ными семействами.. а не отрядами.
у седентарных полихет обычно наблю--
даются реrионарные различия в строении
cerMeHToB и четко выраженные. торакаль
ный (<<rрудной») и абдоминальный (<<брюш..
ной») отделы (рис. 4.62). Все они микрофа
rи, питающиеся осадком или взвесью и не
способные передвиrаться за счет извивов
тела или с помощью параподий, что xapaK
терно для эррантных видов (см. разд. 10.5).
Их разнообразие обусловлено значитель
ными различиями в образе жизни. Некото--
рые живут в норах, друrие в трубках из
секретируемоrо перrаментоподобноrо ма..
териала или карбоната кальция, иноrда
из песчинок. В нескольких семействах на..
блюдается функциональная конверrенция
с олиrохетами. Такие черви роются
в мяrком субстрате, либо заrлатывая ero,
142
rлава 4
) Умирает
в
1"'1 :;.
"1/11/,, 1'111/11"11/1'11 Умирает
,,11'" t ., ,
о' 11"1111/1\11"11/,,,,, ''''),
, Оплодотворение
{f
Рис. 4.60. Эпитокия: эпиrамия, Половозрелые особи претерпевают метаморфоз роятся и поrи
бают: А немодифицированная атокная форма Nereis; Б метаморфозировавшая эпитокная
форма Nereis (Heterohereis); B cxeMa жизненноrо цикла
) Умирает
) Умирает
Оплоnотво р ение
J .,....
/ :'>jb
a
( :::: )
в Реrенерация
Рис. 4.61. Эпитокия: схизоrамия: А образование единственноrо столона за счет модификации
задних cerMeHToB; Б образование множества столонов путем терминальноrо почкования; тер-
минальный столон самый старший; B cxeMa жизненноrо цикла
Рис. 4.62. Разнообразие форм
тела седентарных полихет.
Мелкие рисунки показывают
характерный образ жизни (см.
также рис. 9.6)
Рис. 4,63, Разнообразие форм
и размеров тела семейств по
лихет ранее объединявшихея
в rруппу архианнелид. Сейчас
считается, что они вторично
упрощены из за мелких разме
ров
Черви
143
o
о
1 мм
u
200
мкм
о
144
Аибо «облизывая» песчинки. У них неболь..
шой простомиум без выступающих сенсор..
ных opraHoB, простые шетинки на редуци..
рованных параподиях, хорошо развитая ко..
льцевая мускулатура и сплошные септы.
Представители нескольких семейств соби..
рают мелкие частицы или орrанический
осадок и доставляют их ко рту мяrкими
хватательными щупальцами (см. рис. 9.6),
которые часто отходят от простомиума.
Живущие в трубках черви некоторых отно"
сительно продвинутых семейств имеют вен..
чик жестких простомиальных щупалец,
функционирующий как настоящий филь..
трационный аппарат (рис. 9.3). Друrие ти"
пы пищедобывательной специализации об..
суждаются в rл. 9 (см., например, рис. 9.5).
Архианнелиды (рис. 4.63) представляют
собой rруппу неродственных друr друrу
крошечных полихет, которые независимо
приспособились к обитанию в интерстициа..
ли морских песков.
4.14.3.2. Класс Clitellata
(поясковые кольчецы)
Подкласс Oligochaeta
(малощетиНКОВblе кольчеЦbl)
Олиrохеты в основном наземные
и пресноводные аннелиды; некоторые виды
вторично перешли к оБИТаНИЮ в морской
среде, 'особенно в эстуариях и интерсти"
циали.
При локомоции олиrохеты используют
rидроста1ические свойства полностью се..
птированноrо целома и метамерной opra..
низации. Таким образом, у них сохранился
план строения, близкий к приписываемому
протоаннелидам (см. рис. 4.53), однако сам
подкласс не является примитивным. Для
олиrохет характерен синхронный repMa..
фродитизм, вынашивание небольшоrо ко..
личества крупных, боrатых желтком яиц
в защитной и питающей структуре
коконе, секретируемом пояском. и сильно
редуцированноечислоrонад.Расположение
rениталий используют для разделения оли..
roxeT на три отряда.
Функционально и эколоrически суще..
ствуют в принципе два типа олиrохет: в ос..
новном водные мuкродрuлы и сухопутные
меzадрuлы, или земляные черви.
rлава 4
Функциональная анатомия земляных че
рвей на удивление постоянна и может быть
проиллюстрирована на примере Luп1bricus
terrestris (рис. 4.64,А).
Поперечный разрез (рис. 4.64,Ь) дeMOH
стрирует слеДУЮlцие детали строения: BOДO
непронипаемая кутикула, смазываемая BЫ
делениями бокаловидных секреторных кле
ток эпидермиса; эпидермис; нервный слой;
сплошной слой кольцевой мускулатуры; по
лосы продольных мышц; одиночные или
расположенные небольшими rруппами (как
у Luтbricus) щетинки; целом, разделенный
сплошными септами и выстланный перито
неальным эпителием мускулатура кишеч
ной стенки; энтодермальная выстилка пи
щеварительноrо тракта.
План строения червя метамерный (cer
ментарный), но передний отдел сильно спе
циализирован, особенно строение пище
варительной и половой систем
(рис. 4.64,А,Б; рис. 4.65). У земляных че
рвей сложное псевдокопуляционное поведе
ние. при котором спариваюшиеся животные
покрываются общей слизью, а сперма пере
носится наружно в семеприемники или ce
менные мешки в области пояска (рис. 4.66).
Оплодотворение происходит внутри KOKO
на, причем после одноrо спаривания червь
может произвести MHoro яйцевых коконов.
Олиrохеты микродрилы мельче земля-
ных червей. К ним относят несколько се-
мейств, обитающих в основном в пресных
водах, хотя Enchytraeidae в основном су-
хопутная rруппа. Некоторые живут даже
в морских rлубинах. Форма тела здесь бо-
лее разнообразна, чем у земляных червей.
у некоторых имеются выступающие воло-
совидные щетинки, напоминающие щетин
ки полихет (рис. 4.67). Биолоrия размноже-
ния часто высокоспециализирована. Рас-
пространен бесполый процесс в форме
спонтанноrо разделения (схизоrенез); у не-
которых видов настоящее половое воспро-
изведение неизвестно совсем.
Подкласс Hirudinea (пиявки)
Специфические черты пиявок опреде.
ляются развитием присосок. позволяющих
им плотно прикрепляться обоими своими
концами. После этоrо тело функционирует
как единая rидростатическая система Соот-
Черви
145
/
П ростом иум
А
, Мужско.й
Рот rонопор П Ш
Женский rонопо оясок етинки
Отверстие Половые щетинки
семеприемника
Эпидермис + кутикула
Кольцевые мышцы
Продольные мышцы
Щетинки
Кишечник
Неq>ридий
Целом
Вентральный
нервный ствол
Рис. 4.64. А. Внешнее строение земляноrо червя Luтbricus terrestris вид с вентральной стороны.
Б. Схема поперечноrо среза этоrо же вида
Надпищеводный
rанrлий
Семенники
Кровеносная
система
Подпищеводныи
rанrлий
Яйцевод
Семенные Яичники
пузырьки
Рис. 4.65. Внутреннее строение передних cerMeHToB Luтbflcus terrestris
146
rлава 4
А. Копуляция
Б. Откладка яиц
8. Оплодотворение
ОБразование кокона
Рис. 4.67. Разнообразие форм тела олиrохет
микродрил
Рис. 4.66. Поведение Luтbricus
terrestris при спаривании. А.
Животные покидают свои HOp
ки И образуют покрытую сли
зью пару. Во время копуляции
происходит взаимный обмен
сперматозоидами, сперма
переходит из семенных пу
зырьков в семеприемники, как
показано стрелками, Б. Зрелое
яйцо выходит из женскоrо по
ловоrо протока и переносится
назад к пояску. В. Секретируе
мый пояском кокон BOKpyr He
большоrо количества яиц дви
жется вперед; яйца оплодотво-
ряются, коrда он проходит ми
мо отверстия семеприемника,
r Кокон с развивающимися
эмбрионами откладывается
в почву
Простомиум
POT
Поясок I
Задняя
присоска
Рис. 4.68. Внешнее строение пиявки Hurudo
Вторичная кольчатость показана слева, а на-
стоящая сеrментация справа
Черви
147
Дорсальный кровеносный сосуд
Кольцевая мышца
Косая мышца
Несрридий
Кутикула
Эпидермис
Ботриоидапьная ткань
Кишечник
Рис. 4.69, Поперечный срез Н,..
rudo. Целомическая полость
заполнена паренхиматозной
тканью. Пространство между
ее клетками может быть opra..
низовано в хорошо выражен..
ные синусы.
Семепровод
Семенник Вентрапьный нервный ствол
ветственно сеrментация целома, эктодермь]
и мезодермы по большей части утрачена.
Функционально пиявки напоминают пло..
ских червей.. но используют преимушества
целомических синусов (для создания HarHe..
тающеrо усилия), нервной системы с се..
rментарными raHr лиями (для координации
сложной локомоторной и поведенческой ак..
тивности) и замкнутой кровеносной систе..
мы с дыхательными пиrментами.
rлотка
Мышца...расширитель
rлотки
Пение
Яичник
Влаrалище
Семенник
Семепровод
Вентральный
нервный ствол
Сеrментарный
rанrлий
Рис. 4.70. Внутренее строение
HffUdo, вид с дорсальной CTO
роны. Кишка показана отдель
но
у всех пиявок ровно 33 сеrментарных
rанrлия. Передняя присоска образована
сеrментами I IV., а задняя
XXY XXXIII (рис. 4.68). Щетинки инару..
жная сеrментация не выражены, хотя тело
разделено вторичными кольцами, не совпа..
дающими с rраницами cerMeHToB. Попереч..
ный разрез показывает, что пространство
между мезодермой и мускулистой стенкой
кишки заполнено рыхлой «паренхимопо"
Слюнная железа
Дорсальный сосуд
Слепой вырост кишки
( цекум)
148
rлава 4
Рис. 4.71. Разнообразие форм тела пиявок;
правый рисунок иллюстрирует характерную
способность этих червей сильно изменять
форму cBoero тела
добной» ботриоидальной тканью 1
(рис. 4.69). Некоторые внутренние орrаны,
особенно rерминативные эпителии инефри..
дии, все же отражают предковую метаме..
рию (рис. 4.70).
Пиявки rермафродиты с несколькими
парам и семенников, парой яичников и един"
ственным половым отверстием. Оплодот..
ворение внутреннее, и при размножении
происходит перенос сперматозоидов между
партнерами, часто сопровождающийся сло..
жным копуляционным поведением. Умно..
rих видов сложно устроенные сперматозои..
ды вводятся «подкожно» и миrрируют
к яичникам между клетками животноrо..
реципиента. После спаривания взрослые
пиявки откладывают мноrочисленные кок о..
ны, с одним или несколькими яйцами каж..
дый. Некоторые крупные виды живут по не..
скольку лет, но большинство только
один rод, зимуя в эмбриональном состоя..
нии внутри кокона.
Разнообразие формы тела пиявок иллю..
стрирует рис. 4.71. Все они плотоядные, но
не все наружные паразиты или кровососу..
щие формы. Классификаuия основана на
строении ротовых частей и способе пита..
ния.
Acanthobdellida отряд своеобразных
1 Термином «ботриоидальная ткань» здесь
обозначена паренхима, происходящая из целоми..
ческой выстилки. ПРUМ. пер.
примитивных эктопаразитов лососевых
рыб. Передней присоски здесь нет, несколь..
ко cerMeHToB неСУI щетинки, целом cerMeH-
тарный. Таким образом. они представляют
собой связующее звено между уровнями ор-
rанизации олиrохет и пиявок.
Подкласс Braпchiobde//a
к этому подклассу относятся крошеч-
ные эктопаразиты пресноводных ракообра-
зных. Тело состоит из 15 или 16 cerMeHTOB,
из которых четыре первых слиты и обра-
зуют цилиндрическую rолову с присоской.
Эти малоизученные поясковые кольчецы,
похоже, представляют независимую эволю-
ционную линию, идущую от олиrохетопо-
добноrо предка и во мноrих отношениях па-
раллельную линии пиявок (рис. 4.72).
4.14.3.3. Класс Aeolosomata
Этих очень мелких аннелид ранее счита-
ли примитивными олиrохетами, ,но сейчас
их эволюцию признают независимой от ли-
нии поясковых. Некоторые крошечные фор-
мы обитают в интерстициали пресных и со-
лоноватых водоемов. rермафродиты; яич-
ник Bcero в одном cerMeHTe, а семенники
Рис. 4.72. Branchiobdella.
Внешнее строение ти"
пичноrо вида
Рис. 4.73. Aeoloso-
mata Типичный
вид
в cerMeHTax, расположенных спереди и сза--
ди от иеrо. «Поясок» образован вентраль--
ными железами и не rомолоrичен дорсаль--
ному пояску Clitel1ata.
Типичный представитель класса изобра--
жен на рис. 4.73. У иеrо покрытый реснич--
ками простомиум. образующий основной
локомоторный opraH, и довольно длинные
волосовидные щетинки. У Aeolosomata, как
и в семействах архианнелид, наблюдается
структурное упрощение. связанное с мелки..
ми размерами, а родственные связи с друrи..
ми аннелидами неизвестны. В класс входит
Bcero около 25 видов.
4.15. Дополнительная
литература
Bird А. F. 1971. ТЬе S tructure о f N еmа todes. Acade..
mic Press. New У ork [Nematoda].
Boadel1 Р. J. S. 1985. Why is а gastrotrich? In: Co
nway Morris S. et аl. (Ed.) ТЬе Origins and Rela..
tionships of Lower Invertebrates, рр. 248 260.
Clarendon Press, Oxford [Gastrotricha].
Cro/l N. А. 1976. ТЬе Organisation of Nematodes.
Academic Press. London [Nematoda].
Cro/l N. А., Маtllеи's В. G. 1977. Biology of Nema
todes. Blackie. London [Nematoda].
Da/es R. Р. /963. Annelids. Hutchinson, London
[Annelida] .
D'Hoпdt J. L. 1971. Gastrotricha. Oceanogr. mar.
Biol., Апп. Rev., 9, 141 -192 [Gastrotricha].
Doппel' J. 1966. Rotifers ltransl. Wright Н. G. S.).
Warne, London [Rotifera].
Edwards с. А., Lofty J. R. 1972. ТЬе Biology of
Earthworms. СЬарmап & НаН, London [Anneli..
da].
Черви
149
Glbsoп R. 1972. Nemerteans, Н utchinson, London
[N emertea].
Нуmап L. Н. 1951. ТЬе Invertebrates (У 01. 2). Pla
tyhelminthes and Rhynchocoela. McGraw HiH,
New У ork [Nemertea].
Нуmап L. Н. 1951. ТЬе [nvertebrates (Уоl. 3).
Acanthocephala, Aschelminthes and Entoprocta.
McGraw Hill, New У ork [Gastrotricha. Nemato..
da, Nematomorpha, Kinorhyncha, Priapula, Roti
fera, Acanthocephala].
lvaпov А. v. 1963. Pogonophora. Academic Press,
London.
Kaestпer А. 1967. [nvertebrate Zoology (У 01. 1).
Wiley, New У ork [Nemertea, Gastrotricha, Nema
toda, Nematomorpha, Kinorhyncha, Priapula,
Rotifera, Acanthocephala, Sipuncula, Echiura,
Annelida].
Kristeпseп R. М. 1983. Loricifera, а l1ew рЬуlит
with aschelminthes characlers from the meiobent
hos. Z. zool. Syst. Evolutionsforsch.. 21, 163 180
[Loricifera].
Mill Р. 1978. Physiology of the Annelids. Academic
Press, Londol1 [Annelida].
Norrevaпg А. (Ed.) 1975. ТЬе Phylogeny апd Syste
matic Position of Pogonophora. Parey, Hamburg
[Pogonophora].
Rice М. Е., Todoro\'lc М. 1975. Proceedings of the
International Symposium оп the Biology of Sipun..
сиlа and Echiura. Smithsonian Inst.. Washingtol1
[Sipuncula, Echiura].
Sterrer W. 1972. Systematics and evolution within
the Gnathostomulida. Syst. Zool., 21. 151 173
[Gnathostomula].
Stel'rer W., Maiпitz М., Reigel' R. М. 19 5. Gnatho..
stomulida: enigmatic as ever. In: Conway Моrпs
S. et аl. (Ed.). ТЬе Опgiпs and Relationships of
Lower Invertebrates. рр. 18] 199. Clarendon
Press, Oxford [Gnathostomula]
rЛАВА 5
моллюски
Моллюски, или мяrкотелые, ХОРОШО
очерченный и своеобразный тип, не имею
щий ни близкоrо сходства, ни (рилоrенети
ческоrо родства с какой либо из ныне живу
щих rрупп, если не считать сохранения ряда
признаков, указывающих на то, что их
предками были плоские черви. В rл. 2 они
и были описаны как плотные плоские черви
со спинными защитными щитками (панци
рем). Тем не менее большинство видов ЭТО
ro типа очень далеко диверrировало от фор
мы и образа жизни плоских червей. Дей
ствительно, представители в высшей степе
ни преуспевающеrо типа моллюсков дости
rли TaKoro же морфолоrическоrо разно
образия.. что и у целоrо набора типов, опи
санных в друrих rлавах, поэтому заслужи
ВdЮТ отдельной rлавы.
5.1. Тип Mollusca
5.1 .1. 3тимолоrия
От лат. mol1uscus мяrкий (орех или
rриб) .
5.1 .2. Диаrностические
и специфические признаки
(рис. 5.1)
1. Билатерально симметричные живот
ные.
2. Тело из более чем двух слоев клеток
в толщину, с тканями и орrанами.
3. Сквозной кишечник
4. Полости тела нет, за исключением
кровеносных синусов.
5. Тело несеrментировано и весьма из
менчиво по (рорме, но в основном Зdкручен
ное и ЧdСТО коническое, обычно вытянутое
в дорсовентральном направлении, так что
образуется «внутренностный мешок». Как
правило, спереди располаrается rолова, не..
сушая rлаза и сенсорные щупальпа. В боль
шинстве случаев с вентральной стороны
имеется массивная с уплощенной подошвой
Hora, а сзади мантийная полость, но все
это бывает сильно модифицировано.
6. Защитная наружная дорсальная рако--
вина из белка (конхиолина), упрочненноrо
известковыми спикулами, или из 1 8
известковых пластин, секретируемых эпи..
дермисом спинной и боковых сторон тела
(мантией) Иноrда раковина вторично реду"
цирована, покрыта мяrкими тканями, YTpa
чена вовсе или увеличена настолько, что
скрывает все тело.
7. Несущая зубы, хитиновая, языковид..
ная терка (радула), способная выдвиrаться
из ротовой полости наружу; соскобленные
ею частицы попадают на слизистый тяж.
который втяrивается в желудок с помощью
стебелька, помещающеrося в специальном
влаrалище (см. рис. 9.17 и разд. 9.2.5).
8. lазообмен осуществляется одной или
несколькими парами ктенидиальных жабр,
расположенных в мантийной полости (ино
rда они редуцированы).
9. Кровеносная система незамкнута:
сердце окружено перикардиальной поло..
стью мезодермальноrо происхождения,
сквозь которую проходит кишка.
10. Пара мешковидных «почек».. откры"
ваЮIЦИХСЯ проксимально в пеРИКdрдиаль..
ную полость и впадающих в мантийную по..
лость.
11. Нервная система с околопищевоД
ным нервным кольцом и двумя парами raH"
rлионизированных продольных стволов.
Иноrда rанrлии сильно сближены.
2. Типична одна пара rонад с протока..
ми, впадающими в мантийную полость.
Моллюски
151
Раковина
rOHaдa
Пищеварительный дивертикул
СтеБелек
Сенсорное
щупальце
Рис. 5.1, Схематичный про
дольны й разрез обобщен
Horo моллюска
Рот
Радула
у примитивных форм половые продукты
выводятся через перикардиальную полость
и почки.
13. Дробление яиц спиральное.
14. Развитие непрямое, с личиночными
стадиями трохофоры и велиrера (рис. 5.2).
Наблюдаются случаи вторичноrо прямоrо
развития.
Моллюски., насчитывающие почти
100000 видов. вероятно. второй после нема..
тод (тысячи видов которых еще ожидают
своето открытия и описания) тип животных
по численности Их преуспевание связано не
столько с какими..либо особыми анатомиче..
скими или эколоrическими свойствами, ско"
лько с исключительной пластичностью
и адаптационными возможностями основ..
Horo плана строения. Перечисленные выше
признаки типа модифицируются и очень
широко варьируют в разных eI'o классах.
Эту пластичность можно проиллюстриро"
вать, с одной стороны. изменчивостью
функций какой..либо одной структуры тела
моллюсков (например, раковина помимо
защиты может обеспечивать плавучесть,
Рис, 5.2. Личинки моллю
сков: А трохофора; Б
велиrер (по Нутап, 1967)
Сердце в перикардиальной
I полости
Почка
I
Продольные Hora
нервные стволы
Дыхательное
течение воды
быть роющим opraHoM или пластинкой
внутреннето скелета).. а с ДРУI'ОЙ стороны,
множеством структур, способных адапти"
роваться к выполнению какой..либо одной
функции (захватывать пищу мотут несущие
реснички щупальuа. пальпы. сильно увели..
ченные жабры, руки, вооруженные присо..
сками, радулярные зубы и т. д.).
По форме тела моллюски варьируют от
uилиндрических. сильно вытянутых вдоль
переднезадней оси, обитающих в вырытых
ими ходах «червей» без ноrи и раковины до
почти сферических, лишенных rоловы жи..
вотных. заключенных внутри крупной дву"
створчатой раковины. При этом длина ви"
дов составляет от 2 мм у планктонных и ин..
терстициальных форм до более 20 м у rи..
rантских кальмаров (включая руки). В по..
следнем случае это результат удлинения
дорсовентральной плоскости предков.
Моллюски занимают все основные типы
местообитаний и включают представителей
всех известных типов питания. К ним отно"
сятся как наиболее.. так и наименее подви"
жные формы свободноживущих беспозво..
152
Одонтосрор
rлава 5
Радулярная
терка
Мышцы:
а мышца...протрактор
радулы,
Б мышца протрактор
одонтосрора,
в... мышца... ретрактор
одонтосрора,
r мышца... ретрактор
радулы
(1)
Сокращение мышц "а" и
"Б" выдвиrает одонтосрор
и радулярную терку изо
рта (П)
(Н)
Продолжение сокращения
мышцы "Б", расслаБление
"а" и сокращение "р' ОБес...
печивают движение терки
поверх одонтосрора ан), а
сокращение "в" возвращает
одонтосрор и терку назаLl
в ротовую полость (как 1)
ОП)
ночных. В этот тип. вместе с самыми «ум--
ными» беспозвоночными помещены живот--
ные с почти не развитыми орrанами чувств
и мозrом. (<<Если бы Создатель действите..
льно щедро наделил свое лучшее создание
собственным обликом, креационист, несом--
ненно, сделал бы вывод, что Боr похож на
кальмара». Diamond, 1985.)
Как rоворилось в rл. 2, план строения
моллюсков в общем такой же.. как у плоских
червей (разд. 3.6), но с двумя существенны--
ми дополнениями (и несколькими друrими,
вытекающими из них): радулой и дорсаль--
ной раковиной. Радула (рис. 5.3) помещае..
тся в заднем вентральном кармане ротовой
полости и включает хрящевой скелетный
элемент одонтофор, на котором закре--
плена подвижная радулярная терка с попе..
речными рядами обращенных назад острых
зубов, одновременно образующих и продо--
льные ряды. Мышцы"протракторы MorYT
вызывать как выдвиrание одонтофора на..
ружу из ротовой полости, так и некоторое
движение терки относительно ее базальной
опоры, а также поднимание отдельных зу"
Рис. 5.3. Соскабливание пи
щи раду лай (по Russel
Нuпtеr, 1979 и др, источни
ка м)
бов. В результате зубы скребут по поверх-
ности, отделяя от нее пищевые частицы. ко-
торые в процессе втяrивания мышцами-
ретракторами одонтофора и радулы внутрь
попадают в ротовую полость. Хотя эти хи-
тиновые зубы MorYT минерализоваться SiO,
и FезО ф они сильно изнашиваются, поэто
му вся радула медленно смещается вперед
по одонтофору, вынося еще не использовав-
шиеся зубы в «рабочую зону».. в то время
как на проксимальном конце нарастает но-
вая терка. Следовательно.. радула
rлавным образом opraH собирания, соска-
бливания и сверления.. но у ряда хишных ви-
дов она модифицирована в аппарат для
ловли добычи за счет увеличения размера
отдельных зубов (с соответственным умень-
шением их числа в рядах) и иноrда соедине-
ния с зубами ядовитых желез.
Спинное защитное покрытие дорсовен-
трально уплощенных предковых моллю-
сков было, возможно.. белковой или хитино-
вой кутикулой, укрепленной известковыми
«косточками» или чешуйками, секретируе-
мыми эпидермисом. У большинства их по-
Моллюски
томков содержание карбоната кальция
в этой структуре сильно увеличено; она
представлена одной или несколькими круп..
ными известковыми пластинами, покрыты..
ми тонким слоем конхиолиновоrо пери о..
стракума. В типичном случае кальцит или
араrонит откладывается различными слоя..
ми (каждый в орrаническом матриксе).
Различают наружный призматический слой
и внутренние перламутровые. Появившись,
раковина эволюционировала, причем rлав..
ное направление роста моллюсков переста..
ло быть переднезадней осью, как обычно
у червеобразных животных, и сменилось
дорсовентральным, так что раковина и жи..
вотное, заключенное у большинства совре..
менных форм внутри нее, приняли более
или менее коническую форму, иноrда вытя..
нутую, и при этом закрученную в плоскую
или винтовую спираль. Мантия, секрети..
рующая раковину, обычно распространяе..
тся в виде складки поверх незащищенных
частей тела., поэтому раковина с помощью
мышц"ретракторов может надвиrаться на
rолову и веНТРdЛЬНУЮ локомоторную Hory,
обеспечивая безопасность животноrо. Од..
новременно Hora может прикрепляться
к субстрату, действуя как присоска.
Наличие спинноrо заIЦитноrо покрова
имеет прямые последствия для rазообмена
через поверхность тела. Развитию ракови..
ны должно сопутствовать преобразование
специализированной непокрытой ею обла..
сти в жабру одновременно с эволюцией си..
стемы циркуляции дыхательных rазов по
телу (если таковая не сформировалась ра..
нее). Охватывание раковиной и мантией те..
ла моллюска особенно заметно сбоку и
(или) сзади, [де мантийная полость замкну"
та (рис. 5.4). В ней расположены одна или
более пар ктенидиальных жабр. каждая из
которых состоит из уплощенной продоль..
ной оси, по обеим сторонам которой нахо..
дятся плоские треуrольные лепестки (фила..
менты). Каждый из них поддерживается
мелким хитиновым тяжем, идущим вдоль
фронтальной поверхности. Реснички на по..
верхности жабр rонят воду между отдель..
ными филаментами, через которые в проти"
вотоке движется кровь (см. рис. 11.9).
Обычно вода входит в мантийную полость
по бокам тела и выбрасывается наружу дор"
153
Прямая кишка
Приносящий Kp
веносный сосуд
Выносящий
кровеносный
сосуд
Опорный
хитиновыи
стержень
Рис. 5.4. Поперечный разрез мантийной поло
сти обобщенноrо моллюска; показаны ктени
диальные жабры и направления входящеrо
водноrо тока, образующеrося в результате
действия их ресничек
сально вдоль средней линии. Анус и почки
открываются в эту полость в зоне вынося..
щеrо течения.
Однако, как подчеркивал ось выше, при..
рода и развитие радулы, раковины и ктени"
диев, как и большинство друrих анатомиче..
ских признаков моллюсков, значительно ва..
рьируют в различных классах этоrо типа,
поэтому дальнейшая их характеристика бу..
дет дана ниже по отдельности. Поскольку
именно моллюски среди всех беспозвоноч"
ных составляют излюбленный объект ней..
робиолоrических исследований, сведения
о них приведены также в rл. [6.
5.1 .3. Классификация
Тип моллюсков состоит из 8 классов.
два из которых rастроподы (брюхоноrие)
и бивальвии (двустворчатые) включают бо..
лее 980/0 их современных известных видов.
5.1.3.1. Класс Chaetodermomorpha
Хетодермоморфы своеобразная I'рУП"
па лишенных раковины червеобразных мол..
люсков, сильно вытянутых вдоль передне..
задней оси. Они роют и заселяют веРТИКd"
льные ходы в мяrких морских осадках. Их
цилиндрическое тело длиной от 2 до ] 4 см
располаrается rоловным КОНIIОМ вниз.
в толщу осадка, который поrлощается тер..
минальным ртом. Мантийная полость, не..
сущая одну пару ктенидиев (с двумя рядами
филаментов каждый), расположена на зад..
нем конце тела (рис. 5.5), т. е. в устье хода.
Hora отсутствует, мантия покрывает все
тело; поэтому движения животных нетипич"
Класс
Подкласс
Надотряд
Отряд
Chaetodermomorpha
Caudofoveata
Neomeniomorpha
Aplotegmentarla
Pachytegmentaria
Monoplacophora
т rybIidiida
Polyplacophora
Lepidopleurida
Isсhпосhltoшdа
Aca.nthochitonida
Prosobranchia
Docog]ossida
Pleurotomariida
Anlsobranchlda
Lepetellida
NefltIda
Architaenioglossa
Ectobranchlda
Neotaenloglossa
Heteroglossa
Stenoglossa
Archaeopulmonata
Pulmonata < Basommatophora
Stylommatophora
onchidiida
Gyrnnomorpha SQleolifera
Rhodopida
Cephalasplda
Anaspida
Opisthobranchia Saccoglossa
Ntldibranchla
Pletlrobranchomorpha
UmbracuJomorpha
Allogastropoda Pyramidellomorpha
Gastropoda *
Protobranchla Ctenldiobranchia Nucl]lida
PaJaeobranchia Sol myida
Arcida
Pteriomorpha Mytilida
Pterllda
PaJaeoheterodonta<: т rigoniida
Lamellibrancllia UПlОПlldа
Vепепdа
Heterodonta -=::::::::::Myida
PhoJadomVlda
Anomalodesmata < Р " d '
oromYI а
Scaphopoda
Dentalida
Slphonodental]da
Natltiloidea
Cephalopoda <
CoJeoidea
N Зtltilidа
Sepiida
т etlthida
Octopoda
. Vampyromorpha
более фи lоrенетически обоснованным подходам.
При водимая нами cxeMa CKopee один из приме-
ров современных классификаций. ни одна из ко-
торых пока не получила общеrо признания.
* в настоящее время классификация брюхо
ноrих не устоялась. поскольку ста рая. разрабо
танная до 1930 [. система, основанная на разных
уровнях орrанизации постепенно уступает место
Моллюски
155
Рот
''' " ,'-" "',-.:... ......",-...::,
Железисто
J<yтикулярная
пластинка
Радулярный Дивертикул
мешок кишечника
А
Б
ное и ПРОИСХОДИТ за счет псристальтическо
ro сокращения хорошо развитой мускулату
ры стенки тела. Вместо раковины эпидер"
мие секретирует хитиновую кутикулу, в ко..
торой закреплены налеrающие друr на дpy
ra обращенные назад, заостренные чешуе
видные пластинки, покрывающие все тело.
Некоторые системы opraHoB, характерные
для моллюсков. отсутствуют очень плохо
различимая rолова не несет rлаз и ceHCOp
ных щупалец; нет экскреторных opraHoB
полость
Сенсорный
opraH
Анус
Мантийная
полость
Рис. 5.5. Схема продольно..
ro разреза через передний
(А) и задний (6) концы xe
тодермоморфноrо моллю
ска (по Boss 1982)
К тенидий
и половых протоков (rонады выводят свои
продукты через перикардиальную полость);
некоторые виды лишены радулы. Однако на
rолове находится терминальная хитиновая
пластинка, ассоциированная с различными
железами (рис. 5.6), функция которой пока
неясна.
Семьдесят видов этих раздельнополых
питаюшихся осадками моллюсков обра
зуют один отряд.
\\
Задний KOHeu \t '" Переднии конец
p,, \.. ....,' " "
fJP l '''' .,,:
a " iI
«' . Чешуйки / h
Ктен иди и ----=-< ;. Кутикулярная пластинка
Передний конец
Задний
конец . Ктенидии
\ !
\"\"l"
: . о' '. .. ..' 'If4.r .
о.. '.'::. :.: ...:.:,...:.: /Il: i :...:
. ': :.. : " '. .:,:. I l'f:lh ..
.' ......,... ' r
:- .' " ..,.:.. \ I .
. . "., ... '.
.' ::",: . : И' ' ::
: :: .. :-.' : /f "",.
'...,', ..., \ .....
: "о. .. : .. , , :.' : ::. ( ;, .:",
, '.' :, :. ' '.' . . \\ JI 11} ',..,.. . '
. . . " .I't'h . . . .
: . '. :'.. . . . :' : ' \ // : . : .
>:'... ...'..' ..... 1 ': '-:'.:,'
.', .,' . \lljll&,.' ',..
'. ," HI '.
'::",', :.- :: i : I 'II ;J , ':' ' .
. , . , J L k.. ' .
, .: .:. ':: . ':'v;lI/j;'1 :'" .....,:
", . ..... '1ll/ ".' : ,:: . ':
Кутикулярная ' ' '. .' . 'I'P" : ',. .. :.': ...' : '. ..'
пластинка '>'.:.: . :"::.::.. ,'> '. .:....:','.,.:
Передний : ,.':',' ..::'.. :': .... , '. : ': '.' .. .:.. . , : ' .... '. '. '. .: ....:...:
конец
А
Рис. 5.6. Форма тела xeTO
дермоморф: А внешний
вид; 6 образ жизни
в морском осадке (по Hy
тап, 1967; Jones, Baxter,
1987)
Задний конец
Б
156
5.1.3.2. Класс Neomeniomorpha
I1редставители этоrо класса внеlllне
сходны с хетодермоморфами отсутствием
раковины, а практически и rоловы, черве..
образно вытянутым в переднезаднем на..
правлении телом, отсутствием экскретор"
ных opraHoB, половых протоков, иноrда ра..
дулы (рис. 5.7). Однако они ОТТIичаются во
мноrих друrих ОТНОlllениях и обычно счи..
таются стоящими ближе к раковинным
моллюскам.
Их тело длиной от 1 мм до 30 см сжато
с боков и несет продольный вентральный
желобок, в котором расположены один или
более мелких rребней, рассматриваемых
как сильно редуцированная Hora. антия
покрывает все тело, кроме этоrо желобка,
и образует один или более слоев отдельных
известковых пластинок или спикул под КУ"
тикулой. У переднеrо конца желобка распо..
лаrаются ресничная ямка и сдвинутый
к вентральной стороне рот. а у противопо"
ложноrоконцанаходитсязадневентральная
мантийная полость без ктенидиев. Часто
развиты вторичные Жdбры в виде складок
или сосочков (рис. 5.8).
Неомениоморфы хищники., питаю..
щиеся книдариями, на которых их обычно
rлвва 5
находят. Способ их питания, однако, не со..
всем ясен ввиду частой редукции или отсут"
ствия радулы. У некоторых видов передняя
часть пищеварительноrо тракта может вы..
ворачиваться и захватывать пищу. Хотя МУ"
скулатура стенки тела ХОрОIllО развита, ДВИ"
жение происходит не за счет сокращения
МЫlllЦ, а в результате скольжения с помо..
щью ресничек на выДвиrающихся rребнях
вентральноrо желобка. Все виды rермафро..
диты; копуляция происходит С помощью
стилетов; у некоторых видов сперма парт..
нера накапливается в семеприемниках.
Из 180 видов все морские и разделяются
на два отряда в зависимости от числа слоев
известковых образований в мантии и ПрИ
сутствия или отсутствия эпидермальных со..
сочков.
5.1.3.3. Класс Monoplacophora
Считалось, что моноплакофоры вымер..
ли в девоне, однако в 1952 r живые ИХ
экземпляры были обнаружены в одном из
океанических желобов Тихоrо oKeaHd. Изу..
чение материала из друrих районов привело
к описанию в целом 8 видов из одноrо отря"
да.
Передний
конец
tf.<"(1'-;"' ", r(1." 6
'l .' 1, ",.....;r.; 9 ..;'/... /_?&'lр ,J;I-
: t* vщiЙ! ff{ ii i А \;
",, 1Q, . ' l&ff '
I<Щ ' "'''''''''' .
Продольный I
Рот жеЛОБОК Вторичные
жаБрЫ
А
-
A
i\ \\\\: . ; :.: :,......
"''\} :,...
t. ....\ ,
,.,\\. \
'\,\ , " "" , ,\
,\, t
Б
Задний
конец
Рис. 5.7. Форма тела неомениоморф: А внешний вид; Б образ жизни ползание по КОЛОНии
rидроида (по Jones, Baxter, 1987)
Моллюски
rанrлий
157
Кишка
rOHaдa
Перикардиальная полость
Рис. 5.8. Схема продольноrо
разреза неомениоморфноrо
молл юска (п о В 055, 1 982)
Ресничная Радулярный
ямка мешок
РОТ
Педальный
жеЛОБОК
Стилет
Мантийная
полость
Все это мелкие (3 мм 3 см) морские
раздельнополые потребители донных осад..
ков с телом, покрытым со спины единой ко..
нической или чашеобразной раковиной, ПОД
которой расположена окруrлая слабомуску"
листая Hora,! окруженная сзади и с боков
широкой мантийной полостью (рис. 5.9).
Внутри нее по бокам ноrи расположено
пять или шесть пар ктенидиев с одним
рядом филаментов каждый. Друrих opra..
нов также по нескольку пар: шесть пар до..
льчатых почек, открывающихся независимо
друr от друrа по бокам мантийной полости,
две парь] rонал и восемь пар мышц..
ретракторов ноrи (рис. 5.10). rолова разли..
Рот
аБрЫ
Hora
Раковина
Анус
Б
Рис. 5.9. Моноплакофорный моллюск с (А)
вентральной и (Б) боковой сторон (по Lemche
ing5trand, 1959)
чима, но плохо развита, не несет сенсорных
щупалец (за исключением расположенных
около рта) и rлаз Радула развита хорошо
Анус открывается в заднем конце мантий..
ной полости.
5.1.3.4. Класс Polyplacophora
(хитоны, или панцирные)
Овальные или более вытянутые, дорсо..
вентрально уплощенные хитоны отлича..
ются восемью расположенными в линию
и последовательно налеrающими друr на
друrа спинными пластинами панциря. Ман"
тия BOKpyr них развивается в широкий
«пояс», с кутикулой, часто несущей иrлы,
чешуйки или щетинки. У некоторых видов
он частично или полностью покрывает
спинные пластины (рис. 5.11). Разделение
дорсальноrо защитноrо покрытия на отде..
льные элементы позволяет хитонам свора..
чиваться в шар, как некоторым мноrонож
кам и мокрицам.
rолова плохо развита и спрятана под
передней частью мантийноrо пояса; rлаза
и сенсорные щупальца отсутствуют,! но ра..
дула крупная и несет в каждом поперечном
ряду помноrу зубов. Большая часть вентра..
льной поверхности тела занята мощной вы..
тянутой мускулистой ноrой'! с помощью ко..
торой хитоны медленно ползают по твердо..
му субстрату. Hora и пояс MorYT использо..
ваться как сильная присоска, плотно притя"
rивающая животных к субстрату. Узкая
мантийная полость окружает Hory со всех
сторон, кроме непосредственно передней,
и содержит 6 88 пар ктенидиев с двумя
рядами филаментов каждый; сзади в нее вы..
ходит анальное отверстие и протоки одной
пары почек. Последние крупные и, как
Рот
Околопищеводное
нервное кольцо
rонады
Почки
rонодукты
к тенидии
Сердце
Латеральныи
нервный ствол
Мантийная
полость
Анус
Мышцы ретракторы
ноrи
Мелкие непо
крытые участки
дорсальных
пластин
Рис. 5.10. Схема анатомическо
ro строения моноплакофор,
вид с вентральной стороны
с удаленными ноrой и стенкой
тела. Видны сериально повто
ряющиеся внутренние opraHbI
и ктенидии (по Lemche, Wing
stand, 1959)
Рис. 5.11. Вид с дорсальной
стороны четырех моллюсков-
полиплакофор, показывающий
различную степень покрытия
пластинок панциря мантией
(по источникам в Нуmап 1967)
Моллюски
Рот
Почки
rонодукт
Экскреторная пора
Анус
rолова
Ктенидии
Мантииная полость
,онада
,онопор
159
Сердце и перикардиальная
полость
Рис. 5.12. Схема анатомическоrо строения обобщенноrо панцирноrо моллюска; вид с вентраль
ной стороны с удаленными ноrой, стенкой тела и кишкой (по Hescheler, 1900)
и единственная (сросшаяся) [онада (с пар..
ными половыми протоками и половыми по..
рами), часто занимают всю длину тела (рис.
5.12). Нервная система устроена относите..
льно просто; ее стволы, к примеру, не raH..
rлионизированы.
Хитонов 550 видов, все они морские
и раздельнополые; большинство питается
соскабливая водоросли. Они MorYT быть от
3 мм до 40 см длиной. Имеются 3 отряда,
которые различаются по расположению
жабр в мантийной полости, присутствию
или отсутствию прикрепительных зубов на
пластинах панциря и степени покрытия пла..
стив панциря мантийным поясом.
5.1.3.5. Класс Gastropoda
(брюхоноrие)
rастроподь] обширная и разнообра..
зная rруппа, общий признак которой
ПОБОрОТ (торсия) BHYTpeHHoCTHoro мешка
в процессе развития примерно на 180 С про..
ТИБ часовой стрелки, так что мантийная по
лость занимает переднее положение, и впе..
ред соответственно открываются протоки
почек и анальное отверстие: брюхоноrие
как бы «скрученные» животные (рис. 5.13).
Это заметно по асимметричному развитию
двух мышц"ретракторов ноrи и, или нерав"
номерному росту (см. разд. 15.4.1). Возмо"
жность размещать хорошо развитую rоло..
Нервная
система
Кишка
Сердце
Почка
Мантийная
полость
К тенидий
Приносящее
течение
Выносящее течение
А
Б
Рис. 5.13. Процесс торсии у rастропод: A дo
торсии; Б после торсии
160
rлава 5
Отверстие в KpЫ
ше раковины
Жщы
/ "
А
.
Анус Экскреторные
поры
Б
Сисрон
Приносящее
течение \
ЖаБра
ву моллюска в мантийной полости при
втяrивании тела в рd.ковину, а также пере..
днее положение хеморецепторных opraHoB
мантийной полости (осфрадиев), позволяю..
щее воспринимать состав воды перед жи..
вотным, а не позади Hero.. по"видимому, да..
ли селективное преимущество этой rруппе.
Однако автоматическим следствием торсии
был бы поток воды из мантийной полости,
выносящий экскреты и экскременты.. непос..
редственно над rоловой животноrо. Заrря..
знение rоловных сенсорных opraHoB было
устранено за счет изменения траектории
этоrо потока, например развития щели или
одноrо или нескольких отверстий (аппертур )
в спинной части раковины, через которые
он может выходить из мантийной полости,
или же утраты ктенидия на правой стороне
тела и формирования тока воды с левой
стороны мантийной полости на правую
и далее во внешнюю среду (рис. 5.14). В по..
следнем случае левая (входная) сторона
мантийной полости часто вытяrивается
в подвижный сифон. Однако МНОI'ие фило..
rенетические линии rастропод вторично
подверrлись деторсии, в результате кото..
рой внутренностный мешок развернулся на..
зад на уrол около 900, так что мантийная
полость (если она сохранил ась) оказалась
Рис. 5.14. Два изменения
траектории дыхательноrо тока
воды у rастропод вызванные
торсией: А выносящий ток
выходит через дорсал ьное от-
верстие в раковине; Б потеря
правоrо ктенидия и образова
ние поперечноrо течения, по
направлению KOToporo ориен
тирован анус
с правой стороны тела, а анус обычно вновь
сзади.
Общий план строения rастропод (от ко-
Toporo мноrие их виды радикально откло-
нились) коренастое тело с хорошо разви-
той ноrой для ползания и хорошо диффе-
ренцированной rоловой, несущей радулу,
пару челюстей, пару rлаз и одну или более
пар сенсорных щупалец; как rолова. так
и Hora полностью втяrиваются в единую
толстостенную спирально закрученную ра-
ковину (рис. 5.15), отверстие которой ИНОf-
да закрывается при сокращении тела
известковой или орrанической крышкой
(оперкулумом), находящейся на заднедор-
сальной части ноrи. По крайней мере у при-
митивных форм мантийная полость содер-
жит одну пару ктенидиев с двумя рядами
лепестков каждый. Эти раздельнополые
моллюски развиваются с двумя личиночны-
ми стадиями трохофорой и велиrером.
Исходный план строения, включая рако-
вину, оперкулум и торсию большей частью
сохраняют 55000 видов в основном морско-
[о подкласса переднежаберных (Prosobran-
chia). При этом ктенидии имеют тенденцию
редуцироваться от примитивноrо парНОfО
состояния к сохранению только одноrо ле-
Boro элемента с двумя, а впоследствии
Моллюски
i
: :
Irl"
111'1'
' I'\"
. ,,..i
! ;',\I:
jr:Ij \ .
.![!!': \
; r!lf, \;"
i ' I :\\\ i\
1 I:';. \
:lii I!J!I,
;;!:!"i: : '\\'i
{ ( ,1.:IJI:lj ,
;;..
.......
;:ir. :i. \ _ :;{::
:i \\ /:: .:.
# :: :: '
Рис. 5.15. Спиральные раковины rастропод (по
источникам в Нутап, 1967)
,
лишь с одним рядом лепестков. С исчезно
вением правоrо ктенидия коррелирует по
теря правой почки, правоrо предсердия
сердца и правоrо осфрадия. I Большинство
переднежаберных раздельнополые, хотя He
которые rермафродиты с чередованием
мужской и женской фаз. В основном это
бентосные «улитки», поедающие BOДOpO
сли (блюдечки, литторины, каллиостомы,
марrаритесы), неподвижные колониальные
животные (каури) или илоядные формы, но
одна rруппа (отряд Stenoglossa) включает
MHoro хищников с радулой, несущей лишь
несколько крупных клыковидных зубов
в каждом ряду. Несколько видов
фильтраторов питаются, используя нару..
жные слизистые сети (рис. 9.5) или увели
ченные ктеНИДИИl(параллелизм с бивальвия
ми). Среди переднежаберных и вообще cpe
ди всех rастропод блюдечки (отряд Doco
glossida) образуют весьма изолированную
rруппу с особым строением раковины, pa
дулы и пищеварительноrо тракта.
Во втором подклассе rастропод, Hete..
robranchia, происшедшем от переднежабер
ных, наблюдаются несколько тенденций
развития: а) редукция, поrружение под
мяrкие ткани или утрата раковины; б) YTpa
161
та оперкулума; в) деторсия; r) утрата пред
ковых ктенидиев и замещение их вторичны
ми орrанами rазообмена; д) развитие синх
pOHHoro rермофродитизма. Два основных
надотряда rетеробранхий это морские
заднежаберные (Opisthobranchia: около
1000 видов) и в основном наземные и прес
но водные леrочные (20000 видов). У первых
(rоложаберники, морские зайцы, крылоно
rие и др.) ктенидий обычно замещается BTO
ричными жабрами или развивается rазооб
мен через поверхность тела, часто покры
тую сосочками. Большинство их видов, как
планктонных, так и донных хищники,
включая эндо и эктопаразитов, но HeKOTO
рые потребляют водоросли (часто сосут
их сок), а одна rруппа планктонных филь
траторов кормится, используя секретируе
мую слизистую сеть и реснички на широкой
крыловидной Hore (рис. 9.5). Бывает до че
тырех пар rоловных щупалец.
Леrочные (Pulmonata) характеризуются
в основном преобразованием мантийной
полости в воздушное леrкое с сократимым
отверстием пневмостомом. Большинство
их питается наземными растениями; у всех
исчезли личиночные стадии, так что
эмбриональное развитие идет прямо дО MO
лодых улиток или слизней. Мноrие леrоч
ные сохраняют спиральную rастроподную
раковину (обычно тонкостенную), но HeKO
торые представители наземноrо отряда Sty
lommatophora ее утратили (наземные сли
зни) .
Третий надотряд rетеробранхий
Gymnomorpha также состоит из слизней,
но три эти, возможно неродственные, ветви
морских или наземных rастропод (в общей
сложности около 200 видов) характеризую
тся различным смешением признаков ле
rочных и заднежаберных. То же можно CKa
зать о надотряде Allogastropoda (500 ви
дов). Очевидно, что НеtеrоЬrаnсhiа это
rастроподы, для которых типично паралле
льное развитие мноrих «проrрессц ных»
признаков" и распутать их взаимосвязи за
труднительно.
В целом брюхоноrие включают около
77000 улитко.. и слизнеподобных видов
(рис. 5.16), достиrающих 60 см высоты. Их
можно разделить на 23 отряда.
162
rлава 5
. t... " ,,"
.:"\"
..
/ ""-..>
," // ! , , : :.
Рис. 5.16. Форма тела различных rастропод (в основном по источникам в Нутап 1967)
5.1.3.6. Класс Bivalvia
(двустворчатые)
в сущности, бивальвии это сжатые
с боков моллюски, полностью помещаю--
щиеся между парой створок раковины. Бу--
дучи защищены таким образом, они мало..
подвижны или вообще неподвижны, иноr да
сцементированы с субстратом или прикре..
плены к нему друrим способом. Как и у дру..
rих животных, «спрятанных» внутри тол..
cToro защитноrо покрова, rолова этих мол--
люсков сильно редуцирована (рис. 5.17).
Радулы, rлаз и сенсорных щупалец нет, хотя
отсутствие rоловных opraHoB чувств может
быть эффективно восполнено щупальцами,
а иноrда и rлазами, располаrающимися по
краю мантии. Две створки раковины
являются боковыми, сочленены вдоль дор-
сальной средней линии и открываются пас-
сивно за счет действия упруrоrо дорсально-
ro лиrамента. Сохранение их в закрытом
состоянии требует сокращения двух (иноrда
их число редуцировано до одноrо) МУСКУ-
лов--замыкателей (аддукторов), которые
способны к так называемому запирательно-
му тонусу" т. е. MorYT длительное время
оставаться в сокращенном состоянии, не
требуя непрерывноrо расхода энерrии и че-
Моллюски
163
rонада Лиrамент Влаrалише
ЗУБЫ раковины стеБелька
Пищеварительный ди
Рот вертикул
Ротовая лопасть
Раковина
Мышuа аддуктор
Hora
А
Рис. 5.17. Схема продольноrо
разреза двустворчатоrо мол
люска: А c удаленной жа..
брей (ДЛЯ ясности); Б c жа..
брей в естественном положе
нии и указанием траектории
пищевых токов ВОДЫ (по неско..
льким источникам) Б
редующихся сокращений и расслаблений
отдельных волокон.
Собственно тело, у самых крупных ви
дов Достиrающее 1 м в длину, располаrае
тся внутри раковины, в дорсальной ее ча..
сти, а крупная мантийная полость занимает
оставшееся вентральное и латеральное про
странство. Коrда мускулы замыкатели pac
слаблены, створки несколько приоткры
ваются, позволяя току воды проходить че
рез полость и (у форм, закапывающихся
в мяrкий осадок) выдвиrаться наружу сжа
той с боков Hore. Часто вентральные и 60"
ковые края мантии срастаются вместе,
оставляя только отверстие для ноrи и мел..
кие отверстия для циркуляции воды. В
районе последних :rv1антия часто вытянута,
образуя вводной и выводной сифоны, иноr
да способные втяrиваться внутрь раковины.
На основе происхождения и функций
ктенидиев различают три крупные KaTero..
рии бивальвий. У примитивных форм (под..
класс J.>rotobranchia) парные ктенидии BЫ
полняют rлавным образом исходную функ
цию rазообмена, а питание, если оно не за
висит полностью от си:rv1биотических xe
моавтотрофов, осуществляется при помо
щи лопастей с обеих сторон рта. У одноrо
отряда (Nuculida) они образуют длинные
щупальца, которые, двиrаясь по поверхно
сти окружающеrо осадка, подrоняют пище..
вые частицы к ротовым лопастям, которые
сортируют их перед поеданием (см.
Почка
Сердце
/Мышuа аддуктор
Анус
Выводной
сисрон
Вводной сисрон
Место распо
ложения жаБрЫ
рис. 9.6). Эти виды питаются частицами
осадка с помощью слизисто..ресничноrо Me
ханизма. Однако У преобладающеrо боль
шинства видов (надотряды Pteriomorp a,
Palaeoheterodonta и Heterodonta) парныIe
ктенидии очень сильно увеличеНЬJ и сложе
ны в поперечном сечении W образно, обра..
зуя орrаны фильтрационноrо питания
(рис. 5.18). Пищевой материал в виде су..
спензии засасывается в мантийную полость
вместе с приносящим током воды, ero ча
СТИЦЬJ отфильтровываются, коrда этот ток
проходит сквозь жабры, а затем передаю
тся ко рту С участием ротовых лопастей,
т. е. (слизисто )ресничноrо механизма.
И наконец, у «переrородчатожаберных» ви
дов третьей rруппы (отряд Poromyida надо..
тряда Anomalodesmata) жабры сильно peдy
цированы и формируются мышечные «на..
сосные» переrородки, с помощью которых
и увеличенноrо вводноrо сифона мелкие
животные засасываются внутрь мантийной
полости, rде захватываются мускулистыми
ротовыми лопастями и поедаются.
Двустворчатые, насчитывающие 20000
ВИДОВ, это обычные бентосные животные
морей и пресных водоемов. Большинство
из них раздельнополые и характеризуются
непрямым развитием, хотя личинки HeKOTO
рых пресноводных бивальвий аберрантные,
поскольку приспособлены к паразитирова..
нию на рыбах. Различают 11 отрядов.
164
rлава 5
к тенидиальные
q:>иламенты
Мантийная
полость
Камера вынося
щеrо течения (КВТ)
Камера ПRиносяшеrо
течения (КПТ)
ОТСРИЛЫJ:юванная
пища rонится q:>pOH
тальными ресничками
i . .. ,: 1
1" I /" I
I I
'К '
:П.:." КВТ ... 1.:., П:
'Т :..-> Т'
J I
{ I J .:!I I
латеро срроНТалЬНы е ....... ... '11" I
Реснички срронтальные : VБ :
латеральные I \\. '!" 1 '. I
L
Рис. 5.18. Поперечный разрез
двуСТ80рчатоrо моллюска ил-
люстрирующий фильтрацион-
ное питание с помощью ктени-
диальных жабр (по Russell-
Hunter, 1979)
5.1.3.7. Класс Scaphopoda
(лопатоноrие)
Скафоподы удлиненные, цилиндриче
ские моллюски (длина 2 150 мм), почти
полностью окруженные мантией, секрети
рующей единую трубчатую известковую
раковину, открытую с обоих концов Они
закапываются в мяrкие морские осадки,
оставляя более узкий конец трубки..
раковины несколько выступать над поверх..
ностью дна (рис. 5.19). Из более широкоrо
вентральноrо отверстия (апертуры) ракови..
ны выдвиrаются коническая или цилиндри..
ческая роющая Hora и мелкая хоботообра..
зная rолова, лишенная rлаз и сенсорных
Респираторный ток воды /
:ПоверхнОСть осадка. :.',' .::' :-::.....::,:
: ..:..е :....* е. ..'. ... .. ",:.' .......... .:. .'.:
:: .:. '. ::.: .: '::':. .> /:.::i:', :;.::::.:: .:i:\':
: .. ... '...
. . .
.. ... .' .
. . .
.. .. .8..
.. ..."
.. .: : ." 8.: . е,
.. -, ..
, .... '. ':...:. :.'.......:::
Рис. 5.19. Образ жизни моллюсков"скафопод
.. .... . :.. .. .."'... ,"' ."' .. .. :
.. .." .... .. ........ ...... .. ..
". '." . ::" .../:.::.:.; : ..::.:-'::": :';: :' . ,,:
.. .. .. ".." ................ .. .... ...... '..
:...: ....:..:::.: .:. ::-.....::.: .: :....: ,...: :'; ':i ;::
Моллюски
165
rолова
Отверстие в раковине
rOHaдa
Кишечник
Мантийная полость
Мантия
Каптакулы
Hora
Рис. 5.20. Схема продольноrо разреза лопато
Hororo (по различным источникам)
щупалец. Однако имеются радула, един
ственная срединная челюсть и собранные
в пару пучков узкие булавовидные сократи
мые филаменты ловчие щупальца (капта
кулы). Мелкие пищевые частицы 9садка
передаются ко рту вдоль каптакул с помо
щью ресничек, а более крупные приклеи
ваются к липким расширениям на их концах
и переносятся прямо в рот (рис. 9.6).
Трудно определить направление, в KOTO
ром вытянуто тело. Если считать, что анус
и мантийная полость находятся сзади, как
у rастропод и цефалопод, то скафоподы
сильно удлинены дорсовентрально (рис.
5.20). Однако мантийную полость и анус
часто рассматривают как вентральные, а
в таком случае скафоподы вытянуты вдоль
переднезадней оси тела. Мантийная поло
сть простирается вдоль задней высоты (или
вентральной длины) раковины и лишена
ктенидиев Вода попадает внутрь ее сквозь
дорсальное (или заднее) отверстие за счет
биения ресничек мантии и периодически BЫ
брасывается наружу через то же отверстие
при помощи мышечноrо сокращения. ra
зообмен идет через стенки мантийной поло
СТИ. Все виды раздельнополые с одной He
парной rонадой, продукты которой выводя
тся через правую почку.
Лопатоноrих 350 видов; их делят на два
отряда в зависимости от числа и формы
каптакул и формы ноrи.
5.1.3.8. Класс Cephalopoda
(rоловоноrие)
Целиком морские rоловоноrие
анатомически и поведен чески наиболее co
вершенные представители моллюсков И,
возможно, вообще всех безпозвоночных.
Они также включают самых крупных COBpe
менных беспозвоночных, rиrантских каль
маров, достиrающих общей длины 20 м.
Диаrностическими признаками цефалопод
являются структуры. образованные тем,
что у друrих моллюсков было бы ноrой, KO
торая здесь отсутствует. Передний отдел
эмбриональной ноrи развивается в несколь
ко хватательных UIупалеп или рук BOKpyr
рта, а задний формирует мускулистую BO
ронку BOKpyr части отверстия мантийной
полости. Вода может входить в последнюю
по краям rоловы и с силой выталкиваться
через эту подвижную воронку за счет MЫ
шечноrо сокращения. Цефалоподы в oc
новном пелаrические животные, плаваю
щие, используя реакцию выталкиваемой
струи, за подвижной добычей, которую ло
вят щупальцами (руками). Как и rастропо
ды, rоловоноrие удлинены в дopCOBeHTpa
льном направлении, но в связи с их плаваю
щим образом жизни исходно вентральный
отдел функционально становится передним,
а дорсальный внутренностный мешок фак
тически образует задний конец тела живот
Horo (рис. 5.21). Мантийная полость. таким
образом, открывается спереди, поэтому при
быстром движении цефалоподы движутся
задом наперед. При более медленной локо
моции ориентация воронки может обеспе
чивать движение в различных направле
ниях. Добыча, пойманная руками, измель
чается парой клювовидных дорсовентраль--
ных челюстей. а затем радулой. Некоторые
руки у самцов модифицированы для копу
ляции (переноса сперматофоров). Большая
часть видов раздельнополые, с одной roha--
дой и прямым развитием. Удивительно.
учитывая их крупные размеры, что все rоло
воноrие, за исключением наутилоидей, OT
носительно короткоживущие, размножаю
166
rлава 5
Передняя (дорсальная) сторона
Вентральная
(передняя)
сторона
Отверстие мант й
полости
Задняя (вентральная) сторона
Рис. 5.21. Схематичный вид сбоку rоловоноrоrо моллюска, показывающий изменение ориента
ции тела (в скобках) по сравнению с анцестральными формами (без скобок)
тся один раз и затем умирают (моноциклич
HOCTЬ CM. разд. 14.3).
Друrие признаки этой rруппы заметно
различаются у представителей двух ныне
живущих подклассов. У шести современных
видов Nautiloidea единая наружная плоско
спиральная раковина с множеством распо
ложенных в ряд друr за друrом камер, из
которых последняя обитаема, хотя тонкий
тяж живой ткани сифон тянется сквозь
друrие камеры (рис. 5.22). По мере роста
животноrо откладывается новый материал
раковины, добавляющий примерно цилинд
рическую секцию к старой раковине. Затем
животное продвиrается вперед, секретируя
А
Б
Воронк
rанrлии
Мантииная полость
Ктенидии
Почка
сзади новую переrородку. Таким образом
добавляется новая камера к уже существую..
щим. Наутилоидеи плохие пловцы. Вода
выталкивается из мантийной полости толь
ко за счет сокращения мышц воронки.. и..
чтобы оставаться взвешенными в толще во..
ды, они используют плавучесть раковины,
которая не зависит от давления и обеспечи..
вается необитаемыми камерами Сифон по..
rлощает воду, исходно содержащуюся в ка..
мере, и вместо нее секретирует rаз, что дает
нейтральную плавучесть (см. разд. 12.2.2).
Рот окружает большое количество (80 90)
не несущих присосок ловчих щупалец. Из
них четыре модифицированы для передачи
Раковина
Камеры
раковины
Сифон
rOHaдa
Сердце
Рис. 5.22. Наутилоидеи: А
внешний вид; Б
схематический продольный
разрез (по Boss, 1982 и др.)
Моллюски
сперматофоров. Имеется по две пары KTe
иидиев и почек. Нервная система и rлаза
устроены относительно просто.
у Coleoidea (внутрираковинных) замет
иа тенденция к редукции раковины, Koppe
лирующая с более активным плаванием.
у этой rруппы вода выталкивается из MaH
тийной полости с большой силой за счет oд
HOBpeMeHHoro сокращения мощных кольце
БЫХ мышц стенки мантийной полости,
координируемоrо крупными звездчатыми
rанrлиями, от которых отходят rиrантские
нервные волокна тем большеrо диаметра,
чем больше расстояние между иннервируе
мой мышцей и rанrлием. Каракатицы
(отряд Sepiida) сохраняют известковую pa
167
ковину как opraH плавучести (разд. 12.2.2),
но она уменьшена в размере и помещается
внутри тела животноrо. У кальмаров (отря
ды Teuthida и Vampyromorpha) раковина
редуцирована в еще большей степени до
TOHKoro впутреннеrо хрящевоrо элемента,
не связанноrо с плавучестью, а у большин
ства осьминоrов (отряд Octopoda) она пол
ностью утрачена. Мноrие пелаrические KO
леоидеи имеют обтекаемую или торпедо
видную форму (рис. 5.23) и должны непре
рывно плавать.. чтобы оставаться в толще
воды, за счет волнообразноrо движения бо
ковых плавников, обеспечивающих медлен
ную, но эффективную локомоцию. HeKOTO
рые виды, однако, выработали альтерна
;' ,:: .""::>. :.: - :. : . ;.:. .
Рис. 5.23. Форма тела rолово
ноrих колеоидей (по различ..
ным источникам)
168
Руки
Рис. 5.24. Схема продоль
Horo разреза колеоидеи (по Шупальца
Boss, 1 982 и др.)
rлава 5
тивный способ поддержания плавучести:
путем замещения тяжелых двухвалентных
катионов на ионы аммония в расширенной
перикардиальной полости одновременно
с развитием студенистых тканей с низкой
плотностью. Осьминоrи в основном адап--
тированы к донному существованию, хотя
у некоторых их форм кожистая перепонка
между руками может использоваться как
парус для дрейфа в придонных течениях.
у таких видов редуцированные или руди--
ментарные ктенидии. В отличие от наути
Нервы к
рукам
Нерв к
щупальцу
А
Ресничная
мышца
Хрусталик
Роrовица
Радужка
Рис. 5.25. Мозr (А) и rлаза
(Б) колеоидей (по Wells,
1 982; Kaestner, 1 967) Б
Клюв Радула
Пищеварительные
дивертикулы
! КтенИДИЙ
полость
Анус
лоидей у колеоидей одна парd массивных
складчатых ктенидиев, одна пара почек
и восемь коротких рук, вооруженных присо-
сками (две из них у самцов обычно модифи-
цированы для пере носа сперматофоров).
у кальмаров и каракатиц, кроме Toro, еще
две удлиненные щупальцевидные ловчие ру-
ки, несущие на концах подушки присосок
(рис. 5.24).
Однако, возможно, наиболее отличите-
льные признаки 650 видов колеоидей
замкнутая кровеносная и в высшей степени
Верхний
6уккальный
rанrлий
Брахиальный Fанrлий
Педальный rанrлий
Зрительный
нерв
Висцеральный
raнrлий
Мантийный
:.:.... нерв
ВисцеральнbIЙ
нерв
Задний нерв воронки
Передний нерв воронки
Сетчатка
Зрительные нервы
Моллюски
169
концентрированная нервная системы, по
следняя с хорошо развитыми rлазами.
Ктенидии лишены ресничек (вода движется
через мантийную полость за счет мускуль
ното сокращения, обеспечивающеrо peaK
тивную локомоцию), но содержат капилля
ры, по которым кровь без системы противо
тока прокачивается жаберными сердцами.
Околопищеводное нервное кольцо расши..
рено в мозr, устроенный сложнее, чем у Ka
ких..либо иных беспозвоночных. Общее
строение парных rлаз, иннервирующихся от
мозrа, такое же, как у позвоночных: с poro
вицей, радужковой диафраrмой, хрустали
КОМ и сетчаткой (рис. 5.25). Однако в отли
чие от позвоночных фоторецепторы TaKoro
fлаза обращены в сторону падающеrо CBe
та. Нервная система контролирует также
мноrочисленные поверхностные xpOMaTO
форы. которые (что н.еобычно) управляются
мелкими мышцами и поэтому MorYT реаrи
ровать очень быстро, приводя к почти
MfHOBeHHoMY изменению окраски.
5.2. Дополнительная
литература
Frefter V., Peake J. (Eds) 1975 1978. Pulmonates (2
V 0Is). Academic Press. London.
Hughes R. N. 1986. А Functional Biology of Marine
Gastropods. Croom Helm. Beckenham.
Нутап L. Н. 1967. ТЬе Invertebrates (Vol. 6): Mol
lusca 1. McGraw.. Hill., New У ork.
Kaestпer А. 1967. Invertebrate Zoology (Vol. 1). Wi..
ley, New York.
М ortoп J. Е. [979. Moll'-:lscs, 5th edn. Н utchinson,
London.
Purchoп R. D. 1968. ТЬе Biology of the Mollusca.
Pergamon Press, Oxford.
Ruпhaт N. W., Huпter Р. J. 1970. Terrestrial Slugs.
Hutchinson, London.
SoZeт А. 1974. ТЬе Shell Makers. Wiley, Ncw York.
Wells М. J. 1962. Brain and Behaviour in Cephalo--
pods. Heinemann, London.
Wells М. J. 1978. Octopus. Chapman & НаН, Lon
don.
WiZbur К. М. (Ed.) 1983 1987. ТЬе Mollusca (12
V ols.). Academic Press, New У ork.
rЛАВА 6
LOPHOPHORATA: лофофоровы,,
ИЛИ щупАльцЕвыE
Phorona
Brachiopoda
Bryozoa
Entoprocta
Обычно к лофофоровым относят три ти
па: Phorona, Brachiopoda и Bryozoa. Общим
признаком этих rрупп, как указывает их Ha
звание, является лофофор, т. е. цикличе
ский, подковообразный, в виде сложноrо
,
завитка или складчатоrо кольuа opraH с по
крытыми ресничками полыми щупальцами,
функционирующий как аппарат для филь
трации пищевых частиц из толщи воды.
Этот лофофор, окружающий рот (но не
анус), rидростатически поддерживается co
бственной полостью тела.
В своей основе тело лофофоровых co
стоит из трех отделов с очень мелкой прео
ральной частью (просомой), небольшим
вторым отделом (мезосомой), несущим рот
и лофофор, и обширным третьим (MeTaco
мой), включающим остальные системы op
raHoB, а также анус у основания лофофора.
Далее, общий отличительный признак ло
фофоровых их «rонады», которые пред
ставляют собой не оформленные opraHbI,
а просто рыхлые перитонеальные скопле
иия половых клеток. У большинства лофо
форовых просома утрачена и, даже если
присутствует, крайне мала. Ее значение для
понимания родственных взаимоотношений
между типами этих животных rораздо Ba
жнее, чем в их жизни.
Хотя лофофоровые первичноротые
в том смысле, что их рот формируется из
бластопора, они имеют и ряд характерных
черт вторичноротых (rл. 7), включая план
строения с тремя отделами (у некоторых
вторичноротых просома хорошо раЗБита),
каждый из которых с собственной поло-
стью тела, в некоторых случаях образую-
щейся энтероuельно (разд. 15.2 15.4). Об-
щие признаки включают также радиальное
и недетерминированное дробление и ба-
зиэпидермальную нервную систему. У не-
которых вторичноротых известны даже вре-
менные перитонеальные «rонады» и мезо-
сомальный лофофор. Поэтому лофофоро-
вых рассматривают как rруппу, занимаю-
щую промежуточное положение между пер-
вично и вторичноротыми (см. разд. 2.4.3).
Лофофоровые при крепленные или си-
дячие животные, и их тело защищено секре-
тируемым наружным покровом Он бывает
в виде хитиновых трубок, внутри которых
животные свободно двиrаются, или же сту-
денистых, хитиновых или известковых ра--
ковин или домиков, постоянно соединенных
с эпидермисом.
Таким образом, лофофоровые представ-
ляются хорошо очерченной rруппой, и не-
которые считают, что всех их следует вклю-
чать в один тип Lophophorata. Эта точка
зрения не стала общепризнанной, с одной
стороны, из за продолжающихся споров по
поводу взаимоотношений мшанок с осталь-
ными rруппами. а с друrой из за особен-
ностей Entoprocta, которые остаются плохо
понятными. Служащие для питания щупа-
льца энтопроктов не подходят под опреде-
ление лофофора, но при этом сами ЖИВОТ-
ные отчасти сходны с мшанками, хотя у них
спиральное, детерминированное дробление,
явно не свойственное лофофоровым.
Поэтому включение энтопроктов в эту rла-
ву во MHoroM условно.
Lophophorata: лофофоровые, или щупальцевые
6.1. Тип Phorona (форониды)
6.1 .1. 3тимолоrия
От родовоrо названия Phoroпis, одноrо
из эпитетов еrипетскоrо божества Изиды.
6.1 .2. Диаrностические
и специфические признаки
(рис. 6.1)
1. Билатерально симметричные, черве
образные животные.
2. Тело из более чем двух слоев клеток
в толщину, с тканями и орrанами.
3. Сквозной, U образной формы кишеч
ник.
4. Олиrомерное тело из трех отделов,
каждый со своим единым целомом.
5. Просома очень мелкая, мезосома TO
же, но с крупным лофофором, поддержи
ваемым мезоцелем; метасома крупная и yд
линенная.
6. Стенка тела без кутикулы. с мышеч
ными слоями.
7. Замкнутая кровеносная система с re
Мезоцель
ПОДКОВООБРазно
расположенные
лосроq:юральные
щупальца
Эпистом
Анус
Протоцель
HepB
POT
Несрридий
Метацель
Секретируемая
труБка
Желудок
"roHaдa"
Рис. 6.1. Схема продольноrо разреза форони..
ды (по Emig 1979)
171
Рис. 6.2. Различные формы лофофора у форо
нид (по Emig, 1979)
моrлобином, заключенным в форменные
элементы.
8. Одна пара напоминающих MeTaHe
фридии opraHoB (у личинок прото
нефридии).
9. Нервная система диффузная, базиэпи
дермальная.
10. Дробление яиц радиальноrо типа.
11. Развитие непрямое.. обычно с личи
ночной стадией актинотрохой.
12. Морские формы.
Форониды сидячие черви, постоянно
обитающие в секретируемых ими хитино
вых трубках, частично поrруженных
в мяrкий осадок или, реже, прикрепленных
к твердому субстрату. Достиrают в длину
50 см.. но обычно менее 20 см, причем
большую часть длины тела занимает MeTa
сома, или туловище. Кроме этоrо, един
ственная заметная внешне часть тела
крупный и топоrрафически терминальный
лофофор аппарат для фильтрационноrо
питания, который может нести до 1500
образованных мезосомой щупалец, распо
ложенных в совокупности подковой (рис.
6.2); у видов с особенно большим числом
щупалец свободные концы «подковы» за
кручиваются в спирали.
172
Рис. 6.3. Личинка форонид актинотроха (по
Emig, 1979)
Большинство форонид rермафро
диты, некоторые раздельнополые. Поло
вые клетки образуются в перитонеуме, а за
тем попадают в метацель, откуда выводя
тся во внешнюю среду через нефридии.
Оплодотворение наружное, хотя яйца и ли
чинки (рис. 6.3) MorYT «инкубироваться»
в полости, образованной лофофором, или
внутри трубки в течение 40 75 0 А> периода
развития. Один вид способен размножаться
бесполым способом путем поперечноrо дe
ления и почкования, друrой только попе
речным делением.
у форонид целый ряд признаков, общих
как с остальными типами лофофоровых
(разд. 6.2 и 6.3), так и с вторичноротыми
(rл. 7), и их часто считают близкими по про
исхождению с обеими этими rруппами (rл.
2). Некоторые даже выступают за отнесение
их к собственно вторичноротым, несмотря
на развитие рта из бластопора.
6.1 .3. Классификация
Известно только десять видов, помещае
мых в два рода одноrо семейства.
rлава 6
6.2. Тип Brachiopoda
(плеченоrие)
6.2.1. 3тимолоrия
От rреч. brachion плечо; pous Hora.
6.2.2. Диаrностические
и специфические признаки
(рис. 6.4)
1. Билатерально симметричные живот
ные.
2. Тело из более чем двух слоев клеток
в толщину, с тканями и орrанами.
3. Кишечник либо сквозной, U
образной формы, либо вторично замкну..
тый на конце.
4. Олиrомерное, состоящее из двух OT
делов тело, каждый отдел с единым, в oc
новном энтероцельным по происхождению
целомом.
5. Просома отсутствует, мезосома мел
кая, но с крупным лофофоральным ком..
плексом, который поддерживается мезоце
лем (см. рис. 9.3), а у некоторых видов
известковыми выростами раковины; MeTa
сома мелкая.
6. Тело целиком, за исключением CTe
белька, заключено внутри двустворчатой
раковины, которая может быть сцементи"
рована с субстратом, при креплена к камню,
заякорена в мяrком осаДКе с помощью сте..
белька или вообще не прикреплена. Мелкое
тело животноrо располаrается в задней ча..
сти раковины, большая часть BHYTpeHHero
пространства которой занята лофофором.
7. Незамкнутая кровеносная система
с сердцем или сердцами.
8. Одна или две пары метанефридиепо..
добных opraHoB.
9. Нервная система из околопищеводно..
ro HepBHoro кольца с rанrлиями, от KOTOpO
ro отходят отдельные нервы.
10. Бесполоrо размножения нет: боль..
шей частью раздельнополые, без офор..
мленных rонад, но с четырьмя скоплениями
половых клеток, связанными с перитонеу..
мом. rаметы поступают в метацель ивыво..
дятся во внешнюю среду через нефридии.
11. Дробление яиц радиальное.
12. Морские формы.
Lophophorata: лофофоровые или щупальцевые
173
rOHaAa
Нервное кольцо
Желудок
СтеБелек
Пищевой жеЛОБОК
Боковая рука ло...
сросрора
Раковина
Мантия
Uентральная рука
лосросрора
Рот
rOHaдa
Несрридий
Полость тела
Мускулы раковины
Современные брахиоподы, являющиеся
остатком некоrда очень крупноrо типа (335
ныне живущих видов 26000 вымерших),
по своему внешнеl\1У виду и образу жизни
удивительно схожи с двустворчатыми мол..
люсками (разд. 5.1.3.6). Обе rруппы -
прикрепленные или в основном неподви
жные животные фильтраторы, помещаю
щиеся внутри двустворчатой раковины, ce
кретируемой эпидермальной мантией и по
крытой орrаническим периостракумом. Oд
нако фильтрационный аппарат плечено
rих циклический, петлевидный или спира
льный лофофор (а не ктенидиальные жа
бры), а две створки их раковины
дорсальная и вентральная (а не латераль
ные, как у бивальвий). Следующая точка
конверrенции редукция имевшеrося
у предков rоловноrо отдела (у БРdХИОПОД
утрата просомы) как следствие полноrо за
ключения тела внутрь толстой наружной
раковины.
Все ныне живущие брахиоподы
относительно мелкие (раковина менее 1 О см
в длину и ширину) бентосные животные.
В эволюционном смысле они, вероятно,
эквивалентны раковинным форонидам (rл.
2). Действительно, их план строения Mor BO
зникнуть от таких червеообразных орrаниз
мов полифилетически (рис. 2.13).
Рис. 6.4. Схема продольноrо
разреза обобщенноrо плечено
roro
6.2.3. Классификация
Различают два класса, диверrировавших
друr от друrа по мноrим признакам.
Класс
ОТРЯД
6.2.3.1. Класс Inarticulata
(беззамковые)
Беззамковые брахиоподы. как свиде
тельствует их название, характеризуются
створками раковины, состоящими из фо
сфата кальция и хитина и соединенными
друr с друrом только с помощью мышц.
В задней части тела между створками BЫXO
дИТ стебелек, Формирующийся как часть Te
ла животноrо из личиночной мантии. Он co
держит вырост метапеля и собственные
мышцы; в некоторых rруппах, rде стебелек
отсутствует, вентраЛЬНdЯ створка сцементи
рована с субстратом. В отличие от друrих
брахиопод анус у беззамковых есть (рис.
6.5), лофофор относительно простой, под
держивающих ero скелетных элементов
(выростов раковины) нет.
174
rлава 6
Шетинки
Лосросрораль
ная рука
РОТ
Кишечник ...............
'онада
Анус
Несрридий
Мышцы раковины<
Uеломичес х
кая полость
Кутикула ffi
Q)
Мышца ()
. ','U"IU I /
',,-
= . , t .,. - -
;': i$ > ' ii:
"'",'!i '
jJ"!\ '
II 11 . .
I I " · I
. . ,1 : I
. . .. . I
Рис. 6.5. Схема продольноrо разреза беззам
KOBoro плеченоrоrо
Рис. 6.6. Различная форма тела беззамковых
плеченоrих (по источникам в Нутап. 1959)
МЫШЦЫ раковинь
Раковина
Несрридий
rOHaдa
Рис. 6.7. Схема продольноrо
разреза замковоrо плеченоrо..
ro (по Moore, 1965)
Внутренний вид
дорсальной створки,
показывающий
опорные известко
вые СТРУКТУРЫ
лосросрора
Lophophorata: лофофоровые или щупальцевые
175
Личинка уменьшенное свободнопла..
вающее подобие взрослых животных с ра..
ковиной и лофофором, реснички KOToporo
обеспечивают движение в толще БОДЫ. Ме..
таморфоза при оседании на дно не происхо..
дит. В ходе развития мезодерма формируе..
тся из архентерона энтеропельным спосо..
бом а полости тела схизоцельно внутри
образовавшихся энтероцельно мезодер"
мальных клеточных масс.
К беззамковым относят 47 видов, разде..
ляемых на два отряда: роющие Lingulida
и сидячие, блюдечко видные Acrotretida
(рис. 6.6).
6.2.3.2. Класс Articulata (замковые)
у замковых раковины из карбоната
кальция створки которых сочленяются
с помощью зубов на вентральной створке,
входящих в уrлубление на дорсальной.
В вентральной створке находится также вы..
рез или отверстие, через которое выходит
стебелек (если он есть). Дорсальная створка
часто несет направленные внутрь выросты,
поддерживающие крупный, уложенный сло..
жными петлями или спиральный лофофор
(рис. 6.7). Ни собственных мышц, ни выро"
ста метацеля в стебельке нет. Он образуется
не из мантии, а за счет одноrо из трех отде..
лов тела личинки. Эта личиночная стадия
(рис. 6.8) сильно отличается от взрослых
животных и развивается с метаморфозом.
В отличие от беззаl\1КОВЫХ кишечник на кон..
це замкнут; как мезодерма. так и полости
тела образуются знтероцельно.
Замковые Формь] появились В процессе
эволюции брахиопод относительно поздно
Рис. 6.8. Личинка замковоrо плеченоrоrо (по
Lacaze Duthier, 1861)
(в ордовике); сохранилось почти 300 их ви"
дов, помещаемых в три отряда на основе
структуры лофофора и поддерживаюших
ero известковых образований.
6.3. Тип Bryozoa (мшанки)
6.3.1. Этимолоrия
От rреч. bryon мох; zoon животное.
6.3.2. Диаrностические
и специфические признаки
(рис. 6.9)
1. Модульные, колониальные живот..
ные; каждая колония состоит из отдельных
особей (зооидов), число которых может до..
ходить до миллиона. Колонии формирую..
тся за счет бесполоrо размножения (почко"
вания) особи..основательницы. Ткани каж..
доrо зооида контактируют с тканями ero
непосредственных соседей.
rлотка
Лосросрораль
ные щупальца
rанrлий
Анус
Мышца
ретрактор
Пилорическая
часть
Полость тела
Яичник
иeKYM
Защитное
покрытие
Семенник
;
Тканевый тя >к
Рис. 6.9. Схема продольноrо разреза обобщен
ной мшанки
176
rлава 6
2. Зооиды часто полиморфны; разли
чают их питающие, защитные, репродук
тивные и др. типы.
3. Колонии в целом rермафродитные,
хотя зооиды MorYT быть rермафродитными
или раздельнополыми.
риводимые ниже особенности относя
тся к отдельным зооидам.
4. Билатерально симметричные фОрl\1Ы.
5. Тело более чем из двух слоев клеток
в толщину, с тканями и орrанами.
6. Сквозной, U образной формы кишеч
ник.
7. Тело олиrомерное, из двух или трех
отделов. каждый со своей полостью тела,
формирующейся de novo в ходе метаморфо
за; некоторые с дополнительной MeTaco
мальной полостью тела.
8. Просома присутствует только у oд
ной rруппы" мелкая; мезосома тоже мелкая"
с относительно мелким циклическим или
подковообразным лофофором, который
поддерживается rидростатически мезоце
леl\1. Метасома крупная, мешковидная.
9. Тело замкнуто или поrружено в хити
новую, студенистую или известковую труб
ку, в домик или общий матрикс; сквозь OT
верстие в этой оболочке может выпви
rаться (за счет rидростатическоrо давления)
или втяrиваться (с помощью мышц
ретракторов) лофофор.
10. Циркуляторной И экскреторной си
стем нет.
11. Нервная система в виде rанrлия Me
жду ртом и анусом и отходящих от Hero
околопищеводноrо HepBHoro кольца и OT
дельных нервов.
12. rонады «перитонеальные», rаметы
попадают в метацель и выходят во внеш
нюю среду через «целомопоры», связанные
с лофофором.
13. Дробление яиц радиальное, обычно
они развиваются под защитой родительско
ro орrанизма.
14. Развитие обычно непрямое.
Мшанки похожи на крохотных (до 0,5
мм) форонид (разд. 6.1), формирующих за
счет бесполоrо размножения (почкования)
(рис. 6.1 О) обширные столонообразуюшие,
KOBpO ,! дpeBO или листовидные колонии.
Сходство с друrими типами лофофоровых
(разд. 6.1 и 6.2) в наличии лофофора для
фильтрационноrо питания, олиrомерноrо
плана строения тела, радиальноrо дробле..
ния, внешне аналоrичной rонадной системы
с половыми клетками, развивающимися из
клеток эпителиальной выстилки полости те..
ла. Вопрос о происхождении последней до
сих пор дебатируется, что связано с ОТСУТ"
ствием как ее эмбриолоrических предше-
ственников, так и прееl\1ственности между
зародышевыми листками и тканями взро"
слоrо животноrо. В ходе метаморфоза тка-
ни личинок подверrаются rистолизу, а за..
тем полностью реорrанизуются в тело
взрослой особи. Мезодерма и полость тела
остальных лофофоровых в основном знте..
роцельные и образуются из зародышевой
кишки; однако у мшанок, если она и имее-
rся, то исчезает в процессе метаморфоза.
Поэтому прямых свидетельств мезодерма-
льноrо происхождения полостей взрослых
животных нет и целомический статус, КОТО..
рый им обычно приписывают, базируется
в основном на аналоrии с форонидами
и брахиоподами. Единства взrлядов здесь
нет: альтернативная школа предлаrает
рассматривать полости тела мшанок как
эквивалент предположительно псевдоцело-
мической полости энтопроктов (разд. 6.4).
Как rруппа, мшанки BeCbl\1a преуспе..
вающая линия фильтраторов, причем на се-
rодняшний день более, чем какие либо иные
лофофоровые. За несколькими исключения..
ми, колонии неподвижные, покрывающие
относительно твердый субстрат или при-
крепленные к нему. Исключения
пресноводная форма CristateZZa, способная
медленно ползать (около 10 см в день) с по..
мощью мускулистой «ноrи», и морская
форма Seleпaria тaculata, передвиrающаяся
значительно быстрее (до 1 м/ч) на длинных
щетинках своих авикуларных зооидов (см.
разд. 6.3.3.3). S. тacu/ata обитает на мелко-
водьях коралловых рифов; показано, что
она движется к освещенным солнцем участ-
кам. Колония зеленоrо цвета, поэтому воз-
можно, что такое поведение связано с при-
сутствием в ее тканях симбиотических зоок-
сантелл, как и у мноrих друrих рифовых
беспозвоночных (разд. 9.1).
Lophopho."ata: лофофоровые, или щупальцевые
177
\\ 'lr. -
I t I t! J
. \ r '" ,f},!lfl
I I, \' '" J dffi';-.
' ,1 "\L,I \ ., J;;; /,.C! ,"
\1 , , ' , { , # !1
\ 1 Ije J'.}#
'tt.:. '1 "S,r'... if. 1I..JI.... .:fJ:.:
'" "»> '2 :""" t.f:
;, ..»»..:! y ,", ,:"
77 .... <""' !;.
__ X
Рис. 6.10. Форма колоний мшанок (по различным источникам)
6.3.3. Классификация
Большинство из 4000 описанных ВИ
дов морские формы, но один класс и OT
дельные виды в двух друrих классах
преСНО80дные.
Класс
Отряд
Phylactolaemata
Stenolaemata
Gymnolaemata
Plumatellida
Cyclostomata
= Ctenostomata
Cheilostomata
178
6.3.3.1. Класс Phylactolaemata
Филактолематы относительно неспе
циализированные мшанки. Тело зооидов
сохраняет предположительно анuестраль
ное разделение на три отдела. Просомаль
ная тканевая складка, эпистом, с rидроста"
тическим протоцелем нависает над рото..
вым отверстием, как у форонид. В стенке те..
ла ХОрОlllО развиты слои продольной
и кольцевой мускулатуры. Сокращение ко..
льцевых мышц создает давление, приводя..
щее к выдвиrанию крупноrо лофофора, He
сущеrо до 120 щупалец.. расположенных
в форме подковообразноrо rребня (рис.
6.11). Зооиды мономорфные, цилиндриче
ские, одни из наиболее крупных у мшанок.
Полости тела соседних особей часто связа
ны между собой.
Все виды пресноводные, эпидермис се..
кретирует необызвествленный хитиновый
или студенистый чехол (солей кальция
в пресной воде rораздо меньше, чем в мор..
ской) В процессе эволюции мшанки этоrо
класса выработали механизм образования
покоящихся стадий для распространения
и / или зимовки. Такие стадии весьма свое..
образны: на тяже ткани, идущем от кишки
к стенке тела, скопления эпидермальных
Пищевод
Яичник
Кишка
Желудок
Стенка тела
. ... ., A '
"'. I _... 1. '" . :'.: ..а.. _.. . - .L _.. L,#'L "":'&t.oL
Рис. 6.11. Расправленный зооид филактоле..
Ma изображенный прозрачным (по Pennak
1978)
rлава 6
Рис. 6.12. Статобласты филактолемат (по Hy
тап, 1959)
и «перитонеальных» клеток развиваются
в «статобласты» (рис. 6.12), Т. е. боrатые
желтком клетки.. обычно заключенные в хи..
тиновые дископодобные створки. ОНИ МО-
rYT оставаться прикрепленными к стенке те-
ла и обеспечивать восстановление колонии
in situ после ее отмирания или же высвобо-
ждаться при жизни или после rибели зооида
в окружающую среду. В последнем случае
они способны плавать и MorYT переноситься
на большие расстояния. Статобласты край-
не устойчивы к замерзанию и высыханию.
В умеренных широтах после зимовки они
«прорастают» в ЗООИДОВ"ОСНОВdтелей.
Один отряд этоrо класса включает око-
ло 50 видов.
6.3.3.2. Класс Stenolaemata
Зооиды стенолемат также цилиндриче-
ские, но лишены просомы и МЫlllечных
Рис 6.1 З. Зооиды стенолемат:
А сократившийся; Б рас..
правленный (по Ryland 1970)
Рис. 6.14. Срезы через камеры
зооидов различных rимноле..
матных мшанок, показываю..
щие как сокращение париета..
льных мышц приводит к де..
формации стенки и одновре"
менно к повышению давления
в полости тела: A r хей..
лостоматы; Д ктено"
стоматы (по Ryland, 1970)
Условные
ОБозначения:
Кишечник
Семенник
А
А
Б
в
r
д
', \ .f'
'J ? '/1
t ';/' .'" Лоqюqюр
,r,;.\' !
: i;
{,
Мышцы
дилататоры
Мышечный
ссринктор
.
Мышцы
ретракторы
Метацель
Б
..........
Париетальные
мышцы
Полость тела
.........
.........
Расположенный
под наружной
стенкои тела
мешок, содержа
ший морскую
воду и СООБща...
ЮЩИЙся с внеш...
ней средой
.........
.......
Поперечные
париетальные
мышцы
Продольные
париетальные
мышuы
оБывествленныыe части стеt-',(И тела
МеМБранные (десрормируемые) части
стенки тела
180
rлава 6
слоев в стенке тела. Их цилиндрические
трубки с окруrлым терминальным отвер'"
стием сильно обызвествлены под наружной
rиалиновой кутикулой или клеточным
слоем. Отверстие трубки после втяrивания
лофофора закрывается мембраной, но не
крышечкой (оперкулумом). Давление, тре...
буемое для выдвиrания IIиклическоrо, несу...
щеrо не более 30 щупалец лофофора, со--
здается мышцами--дилататорами (расшири--
телями), действующими на жидкость как
в метацеле, так и в экзосаккальной полости
тела, обычно считающейся псевдоцеломи--
ческой (рис. 6.13). Зооиды стен оле мат в не--
большой степени ПОЛИМОРфНЫ.
Репродуктивная система имеет ряд осо--
бенностей, rлавная из которых по--
ЛИЭl\1бриония. После оплодотворения дро...
бящаяся зиrота образует комок бластоме--
ров, затем распадаюшийся на несколько
вторичных эмбрионов, иноrда отпочковы--
вающих третичные эмбрионы; из одной зи--
rоты может образоваться более 100 бла--
стул.
Лосросрор
Мышечный
ссринктер
А
Большинство rрупп этоrо класса вымер..
ло; единственный современный отряд вклю
чает около 900 морских видов.
6.3.3.3. Класс Gymnolaemata
В настоящее время rимнолематы
наиболее мноrочисленные и преуспевающие
среди мшанок; их более 3000 видов, оби..
.....
тающих в основном в море, но также и в со..
лоноватых и пресных водоемах. Как и усте..
нолемат, их зооиды лишены просомы и мы..
шечных слоев в стенке тела и несут относи..
тельно мелкий циклический лофофор. Од..
нако эти обычно короткие, коренастые
зооиды создают давление, необходимое для
выдвиrания лофофора, за счет мускульной
деформации стенки тела с участием парие..
тальных мышц, действующих на особый
мембранный отдел (рис. 6.14). В связи
с этим стенка тела, если и обызвествлена, то
лишь частично. После втяrивания лофофо..
ра отверстие у большинства видов закры"
вается оперкулумом. Полиморфизм зоои
'::-
Париетальные
мышцы
Кишечник
Поперечные
париетальные
мышцы
Тканевые тяжи
Рис. 6.15 Зооиды ктеностомат:
А сократившийся; Б
расправленный (по Ryland,
1970)
lophophorata: ЛОфОфОРОВblе, или щупальцеВblе
181
А
Яичник Тканевый
тяж.
Лосросрор
Анус
Мышца, закрывающая
оперкулу м
Метацепь
Кишечник Латеральные
поры
Б
Рис. 6.16. Зооиды хейлостомат: А сократившийся; Б расправленный (по Ryland, 1970)
дов В пределах этой rруппы очень развит.
Различают питаЮlцие автозооиды, уплот--
няющие кенозооиды, захватывающие ави--
куларии, очищающие вибракуль] и оэции
выводковые камеры.
Выделяют два отряда: Ctenostornata
с необызвествленной стен ой, без оперкулу--
ма, часто цилиндрическими зооидами и тер..
минальным отверстием (рис. 6.15) и Cheilo--
stomata с уплощенными, коробообразными
зооидами, частично (часто сильно) обыз..
вествленными стенками, оперкулумом
и фронтальным отверстием (рис. 6.16).
6.4. Тип Entoprocta 1
6.4.1. 3тимолоrия
От rреч. entos внутри: proktos анус.
6.4.2. Диаrностические
и специфические признаки
(рис. 6.1 7)
1. Билатерально симметричные, бока..
ловидные животные.
2. Тело из более чем двух слоев клеток
в толщину, с тканями и орrанами.
3. Сквозной, U--образной формы кишеч--
НИК.
1 В русскоязычной литературе принят термин
«Kamptozoa». Прu/и. пер.
4. Пространство между стенкой тела
и кишкой, содержащее студенистую «мезен..
химу», интерпретируется некоторыми как
заполненная клетками псевдоцеломическая
полость тела.
5. Тело по форме представляет собой
полусферический или яйцевидный бокал,
несущий кольцо ЩУПалец BOKpyr рта и ан у..
Шупальца
Протонесрридий
rанrлий
Ресничный
кишечник
"Псевдоuель"
Мышцы стеБелька
I .
:
Рис. 6.17. Схема продольноrо разреза энто
прокта (по Becker 1937)
Анус
Ресничные
питаюшие
щупальца
А. Энтопрокт
I Направление
тока воды
Ресничные
питаюшие
щупальца
Анус
Рис. 6.18. Сравнение пищедобывательных тече
ний у энтопроктов (А) и мшанок (Б) относите
льно положения ануса
Б. Мшанка
./
..::::-
\
Вид СБОКУ
Вид с вентральной стороны Вид с дорсальной стороны
Рис. 6.19. Личинка энтопроктов (по Nielsen 1971)
Lophophorata: лофофоровые, или щупальцевые
183
са; прикрепляется к субстрату сократимым
стебельком.
6. Стенка тела с кутикулой, но без MЫ
шечных слоев.
7. Циркуляторной И rазообменной си
стем нет.
8. Одна пара или множество (у пресно
водных видов) протонефридиев.
9. Нервная система в виде rанrлия Me
жду ртом и анусом и отдельных отходящих
от Hero нервов.
10. Щупальца невтяrивающиеся" но MO
rYT сокращаться и заrибаться внутрь, за
полняя межщупальцевую полость.
11. Дробление яиц спиральное.
12. Развитие непрямое.
Энтопрокты (внутрипорошицевые)
мелкие (0,5 5,0 мм высоты), одиночные
или колониальные животные, временно или
чаще постоянно прикрепленные к субстра
ту, в том числе и к друrим орrанизмам. Все
они фильтраторы. От 6 до 36 выростов
стенки тела образуют щупальца с ресничка
ми, улавливающими частицы пищи из воды
и передающими их в слизи ко рту. У форм,
создающих собственные токи воды для пи
тания, она идет снаружи между щупальца
ми в окруженное ими пространство. а затем
из Hero вверх (рис. 6.18): это направление,
обратное наблюдаемому у остальных лофо
форовых. Анус на вершине коническоrо
возвышения выводит остатки пищи в зону
центральноrо выносящеrо течения.
Широко распространенное бесполое раз
множение почкованием может приводить
к возникновению модульных колоний. Be
роятно" все энтопрокты rермафродиты.
Внешне однополые особи соответствуют
различным половым стадиям rермафродит
Horo орrанизма Сперма выбрасывается
в воду, но оплодотворение" видимо" всеrда
внутреннее Личинка планктотрофная
или лецитотрофная трохофора (рис. 6.19),
в большинстве случаев метаморфозирую
щая во взрослое животное, но у некоторых
видов последнее развивается из образуемой
личинкой почки.
Взаимоотношения этоrо мелкоrо типа
с друrими спорны. Некоторые интерпрети
руют обширную «мезенхимную» ткань как
заполненный rемолимфой псевдоцель
и объединяют энтопроктов с друrими псев
доцеломическими типами (rл. 4); дрyrие
считают их близкими мшанкам на основе
сходства процессов развития" а также неяс
Horo происхождения полости тела (разд.
6.3.2).
6.4.3. Классификация
Описано 150 видов, помешенных в один
класс и один отряд. Все, за исключением oд
Horo пресноводноrо poдa, морские фор
мы.
6.5. Дополнительная
литература
Eтig С. С. 1979. British and Other Phoronids Аса..
demic Press, London [Phorona].
Нутап L. Н. 1951. ТЬе Invertebrates (У 01. 3).
Acanthocephala, Aschelminthes and Entoprocta.
McGraw..Hill, New York [Entoprocta].
Нутап L. Н. 1959. ТЬе Invertebrates (V 01. 5). Smal..
ler Coelomate Groups. McGraw..Hill, New У ork
[РЬотопа, Brachiopoda, Bryozoa].
Nie/seп С. 1971. Entoproct life..cycles and the ento..
proct/ectoproct relationship. ОрЬеНа 9, 209 341
[Entoprocta].
Rudwick М. J. S. 1970. Living and Fossil Brachio..
pods. Hutchinson, London [Brachiopoda].
Rylaпd J. S. 1970. Bryozoans. Н utchinson, London
[Bryozoa].
Wright А. п. 1979. Brachiopod radiation. [п: House
М. R. (Ed.) ТЬе Origin of Major Invertebrate
Groups. рр. 235 252. Ac",demic Press, London
[Brachiopoda ].
rЛАВА 7
DEUTEROSTOMIA: вторичноротыE
Chaetognatha
Hemichordata
Echinodermata
Chordata
о том, что вторичноротые
естественная rруппа родственных друr дpy
[у типов" rоворит наличие у них ряда общих
эмбриолоrических" структурных и биохими
ческих признаков, а также эволюционных
тенденций, неизвестных в дрyrих rруппах
Например, в ходе paHHero эмбриональноrо
развития большинства вторичноротых: а)
бластопор не образует рта, являющеrося
в результате вторичным отверстием кишки
(откуда термин «вторичноротые» ); б)
дробление идет по радиальному типу: в)
судьба каждой клетки в процессе развития
не определена окончательно до относитель
но поздних стадий морфоrенеза (HeдeTep
минированное развитие); [) полость тела
формируется путем отделения от эмбриона
льной кишки серии «энтероцельных» KapMa
нов (см., например, рис. 2.9). Все это в COBO
купности типично для вторичноротых, хотя
отдельные из названных признаков (особен
но б и [) известны и в друrих типах. Далее,
к характерным чертам вторичноротых OT
носятся фоторецепторы ресничноrо (ци
лиарноrо)типа(вотличиеотрабдомерных
фоторецепторов первичноротых), преобла
дани е одноресничных клеток в эпидермисе,
креатинфосфат как депо фосфатов (у пер
вичноротых более обычен в этой роли арrи
нинфосфат) и практически полное OTCYT
ствие хитина.
Предок вторичноротых был, вероятно,
похож на современных крыложаберных по
лухордовых (см. разд. 7.2.3.2), которые ca
ми напоминают форонид (см. разд. 6.1).
По видимому, он представлял собой KOpOT
кое сидячее червеобразное животное с оли
[омерным телом, состоявшим из трех OTдe
лов, второй из которых (мезосомальный)
нес лофофор" а третий (метасомальный) за..
ключал временные скопления перитонеаль
ных клеток, вероятно образующие [онады.
Их нервная система, по видимому, состоя
ла в основном из диффузноrо субэпидер..
маЛЬНОI О плексуса, с одним или более про..
дольными тяжевидными утолщениями.
В отличие от всех известных лофофорат
(однако по аналоrии с некоторыми [астро"
трихами, см. разд. 4.3.3) ток воды" транс..
портирующий захваченные лофофором пи..
щевые частицы в передний отдел кишечни..
ка.. выходит наружу через несколько боко
вых отверстий, ведущих из rлотки непос..
редственно на поверхность тела. Таким
образом кишка предков вторичноротых со..
общалась с внешней средой не только ртом
и анусом.
В большинстве эволюционных линий
вторичноротых лофофор утрачен или силь
но МОДИфИIlирован. В связи с развитием
в некоторых rруппах прикрепленноrо обра..
за жизни (иrлокожие, некоторые полухор..
довые) или личиночноrо свободноплаваю..
щеrо существования (большинство хордо..
БЫХ) исчезло исходное разделение тела на
три отдела. У двух rрупп, наконец, rлоточ..
ные перфорации модифицированы для
обеспечения фильтрационноrо питания
(ранние хордовые) или rазообмена (более
поздние хордовые и кишечнодышащие по..
лухордовые). К друrим эволюционным тен..
денциям, неизвестным у первичноротых,
относятся: сворачивание субэпидермально"
Deuterostomia: вторичноротые
185
ro HepBHoro плексуса в полую дорсальную
нервную трубку (хордовые и некоторые по
лухордовые): появление постанальноrо XBO
ста с локомоторной функцией, совершаю
щеrо с помощью продольных мышц волно
образные движения (щетинкочелюстные
и хордовые); появление защитных кожных
известковых пластин, которые в двух неза
висимых эволюционных линиях впослед
ствии начинают использоваться как часть
мощноrо BHYTpeHHero скелета, к которому
прикрепляется локомоторная мускулатура
(некоторые иrлокожие и хордовые).
7.1. Тип Chaetognatha (щетин--
кочелюстные)
7.1.1. Этимолоrия
От rреч. сhаitе длинный волос, gnat
hоs челюсть.
7.1 .2. Диаrностические
и специфические признаки
(рис. 7.1)
1. Билатерально симметричные черве..
образные животные.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и орrанами.
3. Прямой сквозной пищеварительный'
тракт инетерминальный (вентральный)
анус.
4. Олиrомерное тело разделено септами
на три отдела: rолову, туловище и поста..
нальный хвост. Туловище и хвост несут ла
теральные и каудальный немускулистые
плавники.
5. Каждый отдел тела с одним или
двумя целомами 1нтероцельноrо проис
хождения, однако постювенильные стадии
лишены перитонеальноrо эпителия.
6. Стенка тела снехитиновой кутикулой
и пучками продольных мышц.
7. Систем циркуляции, rазообмена
и экскреции нет.
8. Нервная система с околоrлоточным
кольцом, несушим дорсальный илатераль..
ные rанrлии, от которых отходят отдель
ные нервы.
9. rермафродиты с крупными парными
Щетинки
rлаз
r лотка
Яичник
Женский
rонопор
Туловищно...
хвостовая
септа
Хвостовой
плавник
Капюшон
rанrлий
Септа между rоловой
и туловишем
Кишка
Боковые плавники
Анус
Семенник
Семенной
пузырек
Рис. 7.1. Схематический вид щетинкочелюст
Horo с дорсальной стороны (по различным
источникам)
семенниками и яичниками (последние с
rонодуктами)
10. Дробление яйца радиальное.
11. Развитие, характерное для вторич
норотых, прямое.
12. Морские формы.
Хетоrнаты торпедообразные хищни
ки, плавающие с помощью быстрых биений
постанальноrо хвоста в дорсовентральной
плоскости Латеральные плавники выпол"
няют стабилизирующую функцию. Добыча
схватывается рядом подвижных нехитино
вых щетинок, расположенных спереди и
с боков вентральной камеры, которая ведет
в ротовую полость (рис. 7.2), однако, коrда
животное не питается, rолова вместе со ще
тинками закрывается складкой стенки тела.
Этот капюшон, вероятно, служит в OCHOB
ном для снижения лобовоrо сопротивления,
а также для защиты. На rолове пара rлаз,
186
Передние зуБЫ
РОТ
r раница капюшона
rлотка
Рис. 7.2. rолова щетинкочелюстноrо, вид с
вентральной стороны (по Ritter Zahony, 1911)
образованных за счет слияния отдельных
простых rлазков. Их фоторецепторы ци..
лиарноrо типа, как и у друrих rрупп вторич"
норотых. Но хетоrнаты не являются типич"
ными представителями Deuterostomia.
Планктонные ВИдЫ
il] "
о" .
': ';";i",""""Ш1!,,w.;&""*i;r,l.;'Ji!"":;,"!щ ; ;:: "":: :":,';; '
....
...........
.............
Бентосный вид
............
.... :
,
:
Рис. 7.3. Форма тела xeTorHaT (по Pierrot Bults,
Chidgey, 1987)
rлава 7
в частности, ни в одной из дефинитивных
полостей их тела нет перитонеальноrо эпи..
телия (диаrностический признак целома),
а хвостовые энтероцели здесь (один или
два) это вторичные производные парных
туловищных полостей, т. е. не образуются
в виде отдельных карманов из первичной
кишки. как у остальных олиrомерных rрупп
с тремя отделами тела. rоловная полость
непарная.
Хетоrнаты, ДОСТИI'ающие в длину 10 см,
MorYT быть доминантной rруппой планк..
тонных хищников в море; питаются различ..
ными орrанизмами от мальков рыб сход..
Horo с ними размера до простейших
7.1.3. Классификация
Известно 70 видов, помещаемых в один
класс с двумя отрядами, различающимися
присутствием (Phragmophora) или отсут"
ствием (Aphragmophora) поперечной вен..
тральной мускулатуры. Отряд Phragmopho..
ra включает среди прочих один бентосный
род Spadella (рис. 7.3).
7.2. Тип Hemichordata
(полухордовые)
7.2.1.3тимолоrия
От rреч. hеmi половина; Chordata
название родственноrо типа (разд. 7.3).
7.2.2. Диаrностические
и специфические признаки
1. Двусторонне симметричные живот..
ные.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и орrанами.
3. Сквозной прямой или U образно
изоrнутый пищеварительный тракт.
4. Олиrомерное тело, разделенное на
три отдела: крупный просомальный хобо..
ток, мелкий мезосомальный воротничок
и большое, вытянутое или мешковидное ту"
ловище.
5. Каждый отдел с одним (хоботок)
или двумя (воротничок и туловище) энтеро..
цельными полостями тела.
6. у некоторых на воротничке распо..
Oeuterostomia: вторичноротые
187
ложен лофофоральный opraH, rидростати"
чески поддерживаемый мезоцелем.
7. У всех, за исключением одной, rрупп
верхняя половина стенки rлотки несет от
одноrо до более 100 пар сквозных от вер..
стий, через которые выходит ток воды.
Только нижняя часть rлоточной трубки от..
носится к пищеварительному тракту.
8. Ресничный, лишенный кутикулы
эпидермис, у некоторых секретирующий
внешнюю нехитиновую трубку.
9. Циркуляторная система частично
незамкнутая.
10. Экскреторный орrан клубочек,
образованный выпячиваниями перитонеу..
ма в протоцель.
11. Нервная система диффузная, ба..
зиэпидермальная, у HeKOTOpыx c полым
дорсальным нервным тяжем в воротничке.
Базиэпидермальный плексус концентрируе..
тся в дорсальной и вентральной срединных
зонах.
12 Раздельнополые животные.
...-:::::
Лосросрораль
ные руки со
щупальцами
Ценоций
колонии
rлоточная щель
Рот
.. .,;'
"..-
"j"
Рис, 7.4. П ищедобывательные течения у крыло
жаберноrо полухордовоrо Cepha/odiscus Изо
rHYTbIe стрелки показывают движение воды
к лофофоральным рукам и сквозь них Стрел.
ки, направленные вверх обозначают вынося
щее течение в центре лофофоральноrо аппара..
та. Стрелки на теле соответствуют движению
частиц, направляемому эпидермальными pec
ничками: «наконечники» показывают движение
пищи ко рту (по Lester 1985)
Полоса OTBeprHYTbIX частиц
ХОБОТОК
Воротничок
Рис. 7.5. Пищедобывательные течения, вызы"
ваемые ресничками на хоботке и воротничке
кишечнодышащеrо полухордовоrо (по Barring
ton, 1886)
13. Дробление яиц радиальное.
14. Развитие, характерное для вторич"
норотых, непрямое.
15. Морские формы.
Полухордовые бывают двух основных
типов, которые значительно различаются
формой тела и образом жизни: с одной сто..
роны, червеобразные роющие кишечноды"
шащие, с друrой сидячие,! обитающие
в трубках крыложаберные. Несмотря на
это, в обоих ветвях сохраняется слизисто..
ресничный механизм питания. У крыложа..
берных лофофоральный opraH расположен
дорсально на воротничке. Ero руки расхо..
дятся конусом в сторону от вентрально
расположенноrо рта (у лофофоровых и ДРУ"
rих rрупп, питающихся с помошью кольuа
щупалец, последние окружают рот). Вода
заrоняется сквозь кольцо щупалец в интра..
лофофоральную полость (так же, как, на..
пример, у полихет из семейства Sabellidae
и у энтопроктов). Пищевые частицы затем
продвиrаются ресничками по внешней сто..
роне рук вниз, ко рту (рис 7.4). Кроме Toro,
они MorYT захватываться ресничками всей
поверхности тела (единственный способ
ресничноrо питания у лишенных лофофора
кишечнодышащих, ресничные дорожки ко..
торых особенно хорошо развиты на хобот..
ке; рис. 7.5).
У обеих форм потреблению пищевых ча..
стиц, возможно, помоrает (как и у друrих
микрофаrов) ток волы. заrоняемой в рото..
вую полость rлотки и, как у некоторых [а..
стротрихов (разд. 4.3.3), выходящей через
rлоточные отверстия во внешнюю среду.
188
Такой однонаправленный поток более
эффективная система, чем у лофофоровых,
которые, например, должны периодически
выбрасывать через рот избыток воды, за
хваченной вместе с пищевыми частицами.
Хотя обеспечение питания было, вероятно,
исходной функцией rлоточных перфораций
(см. разд. 9.2.5), сохраняющейся у крыло
жаберных, у кишечнодышащих (потре
бляющих также частицы осадка) ток воды
стал обслуживать rазообмен и количество
rлоточных пор сильно увеличено.
Как следует из названия типа, полухор
довые имеют ряд черт, общих с хордовыми.
В целом последние характеризуются че
тырьмя признаками: присутствием хорды,
rлоткой, перфорированной щелями или по
рами, постанальным хвостом и дорсальной
нервной трубкой (разд. 7.4). Из них у ки
шечнодышащих есть по крайней мере два
перфорированная rлотка и дорсальная He
рвная трубка, а возможно, и третий: у юве
нильных особей некоторых видов описан
постанальный хвост. Однако если у щетин
кочелюстных и хордовых хвост в принципе
локомоторная структура, о постанальном
отделе rемихордат этоrо сказать нельзя.
(Действительно, такой отдел свойствен
мноrим беСПQЗВОНОЧНЫМ, особенно с ки
шкой U образной формы, и ювенильный
«хвост» кишечнодышащих. вероятно, TOMO
лоrичен стебельку крыложаберных.) Хорда
у rемихордат определенно отсутствует,
хотя за нее в течение мноrих лет ошибочно
принимали направленное вперед выпячива
ние кишки, отходящее от ротовой полости.
Таким образом, некоторым полухордовым
все же свойствеННd половина специфических
ПРИЗНd.ков хордовых.. что оправдывает дaH
ное им название.
Как указывал ось выше, ряд признаков
объединяет крыложаберных с лофофоровы
ми, особенно с форонидами (разд. 6.1 ).
мезосомальный лофофор, поддерживаемый
rидростатическим давлением в мезоцеле,
трехраздельная система энтероцельных по
лостей тела, диффузный базиэпидермаль
ный нервный плексус и т. д. Единственные
важные отличия крыложаберных состоят
в развитии вторичноrо рта и парной приро
де мезо и метаuелей. Таким образом, две
rлавные rруппы полухордовых в анатоми
rлава 7
ческом отношении представляют собой зве
но, связующее первичноротых лофофоро
вых и друrие rруппы вторичноротых. Пред
полаrается, что именно крыложаберные,
а не форониды являются предковыми фор
мами для обеих филоrенетических линий
7.2.3. Классификация
Известно 100 видов полухордовых'l раз
деляемых на три класса.
Класс
Отряд
Helmlnthomorpha
Enteropneusta
Pterobranchia
Rhabdopleurida
Cephalodiscida
Planctosph-aeroidea
Planctosphaerida
7.2.3.1. Класс Enteropneusta
(кишечнодышащие)
Одиночные, подвижные, удлиненные че
рви длиной до 2.5 м: роют ходы в мяrких
субстратах или обитают под камнями, [де
питаются частицами осадка или взвешен..
ной в Боде пищей Тело состоит из (а) длин
Horo хоботка. реснички KOToporo создают
основную движущую силу при локомоции
животноrо и образуют транспортную си
стему для пищевых частиц; (6) KopoTKoro,
лишенноrо лофофора воротничка: (в) очень
длинноrо туловища с терминальным aHY
сом и мноrочисленными rлоточными пора
ми (рис. 7.6), количество которых увеличи
вается всю жизнь. В rлоточной стенке эти
перфорации имеют U образную форму
и поддерживаются системой скелетных
переrородок, но на поверхность тела BЫXO
дят в виде мелких, окруrлых'l дорсальных
отверстий. Нервный тяж и экскреторный
клубочек хорошо развиты, но полость тела
в основном заполнена мышечными воло
кнами и соединительной тканью
производными перитонеума, заменяющи
ми большую ч сть мускулаТУРЬJ стенки Te
ла
Имеется как половое, так и бесполое
размножение (фраrментация); MHoro пар
rонад, расположенных в передней части TY
ловища, открываются, каждая своим про..
током. Оплодотворение наружное. Разви
Deuterostomia: вторичноротые
ХОБОТОК
Стеsелек ХОБотка
Воротничок
Туловище
Анус
Рис. 7.6. Внешний вид кишечнодышащеrо (по
Marion, 1886)
тие идет через сталии, аналоrичные извест"
ным для мноrих иrлокожих, до у боль..
шинства видов личинки торнарии
(рис. 7.7).
Все 70 видов кишечнодышащих относят
к одному отряду.
Рис. 7.7. Личинка кишечнодышащих
торнария (по Stiasny, 1914)
189
7.2.3.2. Класс Pterobranchia
(крыложаберные )
Крыложаберные сидячие, обитающие
в трубках животные, достиrающие 12 мм
в длину, с коротким телом из (а) хоботка
в форме диска или щитка, обеспечивающеrо
как (ресничную) локомоцию внутри трубки,
так и секрецию этой коллаrеновой структу"
ры: (6) воротничка, несущеrо на вентраль..
ной стороне рот, а на дорсальной от од..
ной до девяти пар лофофоральных рук, от
каждой из которых отходят двойные ряды
ресничных, секретирующих слизь щупалец;
(в) KopoTKoro мешковидноrо туловища
с анальным буrорком около воротничка
и субтерминальным сократимым стебель..
ком (рис. 7.8). Этот стебелек может закан..
чиваться opraHoM BpeMeHHoro прикрепле..
ния, переходить в общий столон или закру"
чиваться BOKpyr опоры как цепкий хвост
мноrих древесных млекопитающих. в от ли..
чие от кишечнодышащих имеется не более
одной пары rлоточных перфораций, нерв..
ный тяж отсутствует, а экскреторный клу"
бочек развит очень плохо Целомические
полости не заполнены тканями: присут
ствует только одна пара [онад.
ll1ироко распространено бесполое поч..
кование, приводящее в одной rруппе к обра
rOHaдa
Лосросроральные
руки
Мезоцель
ХОБОТОК
Протоцель
Туловище
Желудок
Метацель
СтеБелек
Рис. 7.8. Схема продольноrо разреза крыложа..
берноrо (по McFarland et al, 1979 и др.)
Питаюшиеся ОСОБИ
Uеноций.
! Е...
. .
0.-. -J. ...
,-.
Рис. 7.9. Одиночные крыложаберные рода Се..
pha/odiscus питаются, цепляясь за выступы об
щеrо ценоция (по Lester, 1985)
Рис. 7.10. Трохофороподобная личинка крыло
жаберных (по Schepotieff 1909)
Лсхросроральные
щупальцз
Пищевой желоБОК
ХОБОТОК на стенке ТРУБКИ
Две лосросроральные
руки
Рот
ТРУБка
Туловище
СтеБелек
Б
I I
Рис. 7.11. Часть колонии рабдоплеврид (А) и увеличенный отдельный зооид (Б) (по Grasse 1948
и др.)
Deuterostomia: вторичноротые
зованию колоний с общим столоном из сте...
бельков, а в друrой к клонам отдельных
особей, живущих в общей оболочке
«ценоции» (рис. 7.9). О половом размноже..
нии известно мало, но личиночная стадия не
такая, как у кишечнодышащих (рис. 7.10).
Десять видов крыложаберных распреде...
лены между двумя отрядами. В отряд Се...
phalodiscida входят неколониальные фор...
мы, либо ползающие по поверхности коло...
ний rидроидов, либо живущие внутри обще...
ro «ценоция» и поднимающиеся к ero отвер'"
стиям для добывания пищи (рис. 7.9). У них
одна пара rлоточных перфораций, от четы...
рех до девяти пар лофофоральных рук и од...
на пара rонад. Отряд Rhabdopleurida объ..
единяет колониальные формы (рис. 7.11)
снепарной rонадой, одной парой лофофо...
ральных рук и без rлоточных щелей.
7.2.3.3. Класс Planctosphaeroidea
Этот класс выделен, чтобы поместить
в Hero (хочется надеяться, только временно)
rиrантские личинки (до 2,2 см в диаметре),
напоминающие своей общей формой ли..
чинки кишечнодышащих (рис. 7.12). Их на...
ходят в образцах планктона из Атлантиче...
CKoro океана с начала 1930...x rr. Поскольку
Рис. 7.12. P/aпctosphaera, вид с вентральной
стороны (по Spengel, 1932)
1q1
взрослая форма пока еще не известна, их
статус и особенности остаются неопреде...
ленными.
7.3. Тип Echinodermata
(иrлокожие)
7.3.1. 3тимолоrия
,
От rреч echinos еж; derma кожа
7.3.2. Диаrностические
и специфические признаки
(рис. 7.13)
1. Взрослые с пятилучевой симме...
трией, в большинстве случаев действитель...
но радиальные, некоторые билатеральные.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и орrанами.
3. Кишечник сквозной (у некоторых вто",
рично слепозамкнутый, у одной rруппы от...
сутствует ).
4. Форма тела в высшей степени разно...
образна. Анпестральные и современные си...
дячие формы со сферическим или чаше...
образным телом, прикрепленные абораль...
но к субстрату стебельком с обращенным
вверх ртом, окруженным пятью руками.
Производные свободноживущие формы без
стебелька, с телом, ориентированным так,
что рот обращен вниз, к субстрату или (ре...
же) вперед; с различимыми руками или без
них, иноrда билатерально симметричные.
5. Какой...либо дифференцированной ro..
ловы нет.
6. Исходно с олиrомерной, состоящей
из трех парных целомов полостью тела.
Метацели (<<соматоцели») образуют ее ос...
новную часть. У немноrих форм с прямым
развитием полость тела образуется схизо...
цельно.
7. Водно",сосудистая (амбулакральная)
система, формирующаяся в основном из ле...
Boro мезоцеля (левоrо «rидроцеля») и ча...
стично из левоrо протоцеля (левоrо «аксо"
целя»), содержит не связанную прямо
с внешней средой морскую воду и rидравли",
чески управляет движениями трубкообра...
зных локомоторных (амбулакральных) но...
жек и/или захватывающих пищу щупалец
( «подий»). Часть этой системы образует ко..
192
\
Околоротова
кольцевая
система
Кишечник
-В
Иrлокожее на стеБельке
rлава 7
х
Подии
АМБулакральные
ножки
Полость Ампулы
тела
у
rерминативный I
эпителий
Вторично
СВОБодноподвижное
иrлокожее
В
I
rонада
Мадрепоровая
пластинка
Условные ОБозначения:
. Нервы
.. .... .. АМБулакральная
система
8- - . -.... rемальная система
Каменистый
канал
х
ишечник
I > C '"-
РОТ
i Известковые структуры
Околоротовая
кольцевая
система
у
Рис. 7.1 з. Схемы разрезов двух обобщенных иrлокожих (по Nichols 1962)
льцевой канал BOKpyr пищевода, от которо"
ro начинаются радиальные каналы, идущие
вдоль каждой руки.
8. Имеется мезодермальная субэпидер..
мальная система известковых «косточек»
или пластинок, часто с выступающими на
поверхность буrОРКdМИ или шипами.
9. Экскреторных opraHoB нет.
10. Циркуляторная «rемальная» систе..
ма выражена слабо; ее функции в значитель..
ной мере выполняются целомическими
жидкостями.
11. Нервная система субэпидермальная,
в форме околопищеводноrо HepBHoro коль..
ца, от KOToporo начинаются диффузные не..
рвы, идущие вдоль каждоrо амбулакра.
12. Обычно раздельнополые
13. Дробление яиц радиальное.
14. Развитие типичное для вторичноро"
тых, как правило, непрямое, с ресничной,
билатерально. симметричной личинкой
(рис. 7.14).
15. Морские формы.
Иrлокожие в высшей степени свое..
образная rруппа с тремя весьма характер..
ными чертами: особой симметрией, из вест..
ковым мезодермальным скелетом, водно..
сосудистой (амбулакральной) и друrими це-
ломическими системами. Все эти три осо-
бенности MorYT быть прямо связаны с их
происхождением от сидячих фильтраторов,
которые, от каких БЬJ предков ни происхо-
дили, имеют тенденцию к развитию радиа-
льной симметрии ловчеrо аппарата (разд.
9.1). у иrлокожих этот процесс привел
к формированию Kpyra из пяти часто дихо-
томически ветвящихся рук. У предковых
форм это были эквиваленты и, возможно,
производные лофофоральных рук живот-
ных, подобных крыложаберным полухор..
довым (разд. 7.2.3.2). Как и у последних,
rидравлическая система рук и ресничных
щупалец на них поддерживается в основном
давлением в мезоцеле. Некоторые эволю-
ционные линии иrлокожих вторично стали
свободноживущими; большинство этих
форм как бы перевернулось «вверх ноrами»,
так что исходно обращенная вверх несущая
рот и руки поверхность стала контактиро-
вать с субстратом, а пищедобывающие щу-
пальца превратились в локомоторные. От..
носительно небольшие анатомические из-
менения сформировали из них амбулакра-
льные ножки (общая адаптация при этом
Deuterostomia: вторичноротые
Апикальный
ceHcopHЫ
хохолок
Ресничные
пояса
А. Виттелария морских лилий
Рот
Анус
в Осриоплутеус осриур
Ресничные
пояса
Анус
Д. Аурикулария rолотурий
Рис. 7.14. Личинки иrлокожих (по Barnes, 1980)
изменении Функций появление при каж
ДОЙ ножке rидравлическоrо резервуара" aM
пулы, позволяющей ей индивидуально co
кращаться и вытяrиваться). При этом руки
часто становились крупнее и шире (как
у морских звезд) или объединялись с телом"
заворачиваясь BOKpyr Hero (кйк у морских
ежей и rолотурий). В любом случае пять рук
все еще определяют план строения тела, так
что у большинства современных свободно
живущих форм радиальная симметрия co
храняется, и, следовательно, они MorYT дви
rаться в любом направлении (пятилучевая
193
Анус
Желудок
Б. Брахиолария морских звезд
Рот
Скелетные спикулы
Анус
r. Эхиноплутеус морских ежей
Кишечник
Ресничные
пояса
Рот
Анус
Е. Долиолария rолотурий
симметрия была не у всех ранних rрупп
иrлокожих известны спирально симме
тричные и даже асимметричные формы
см. рис. 2.14).
Сидячие животные нуждаются в механи
ческой опоре тела и защите ero от хищников
и действия волн У иrлокожих это обеспечи
вается отложением в дермисе системы «KO
сточек», каждая из которых представляет
собой пористую решетку из карбоната ка..
льция, ведущую себя как единый кристалл
кальцита Отдельные «косточки» часто уве..
личиваются в размерах, принимая форму
1Э4
r лана
пластинок, иноrда сцепляющихся с друrими
с образованием жесткоrо, почти наружноrо
панциря. Как и у друrих панцирных или ра..
ко винных животных (например" двуствор"
чатых моллюсков, плеченоrих) имевшаяся
у предков rолова утрачена. Покровный, как
правило ресничный, эпидермис иноrда ис
чезает. и части панциря оказываются внеш
ней rраницей тела. Известковые структуры
MorYT вторично выполнять локомоторную
функцию, замещая в этом отношении амбу..
лакральные ножки. МЫШIIЫ.. идушие от оп
них «косточек» к друrим, обеспечивают
движение по субстрату с помощью рук или
иrл (см. rл. 10).
Хотя тело иrлокожих мономерное, у них
развиваются три пары энтероцельных кар..
манов (иноrда в модифицированной фор..
ме)., как и у олиrомерных животных с тремя
отделами тела, Т. е. форм, которым" как
принято считать, родственны иrлокожие
(см. разд. 15.4.1). Как и у лофофоровых,
И полухордовых, метацели (<<соматоцели»)
формируют основную полость тела.. а мезо..
цели (у иrлокожих один из них), или «rид..
роцеЛИ», rидравлическую систему рук,
похожих на лофофоральные; правый мезо..
цель обычно редуцируется до мелкоrо co
кратимоrо мешочка. Правый протоцель
(<<аксоцель») образует аксиальный (осевой)
синус, т. е. пространство BOKpyr части [ема..
льной системы, функции которой неясны,
в то время как левый объединяется с мезо..
целем в амбулакральную систему. Прото
цели и мезоцели этих олиrомерных энтсро"
цельных животных связаны с внешней cpe
дой порой. Соrласно классическим
взrлядам, эта связь сильно развита у и[ло..
кожих как средство изменения объема жид..
кости в, амбулакральной системе. Считае..
тся. что через общий аксоrидропор в виде
пористой (мадрепоровой) пластинки проис..
ходит водообмен с внешней средой (факти..
чески в амбулакральной системе находится
не целомическая жидкость" а морская ВОДа
с небольшим количеством целомоцитов).
От этой пластинки так называемый «каме..
нистый канал» ведет к околопищеводному
кольцу. В действительности прохождение
воды через мадрепоровую пластинку ни
разу не наблюдалось, однако она, возмо"
жно, обеспечивает выравнивание rидроста..
тическоrо давления внутри и вне животно..
ro.
Таким образом, своеобразие иrлокожих
не столько в самом их анатомическом
строении, сколько в том, что это единствен..
ная процветающая rруппа свободноживу..
щих животных, происшедшая от прикре..
пленных сидячих предков. которые., по-
видимому, были способны двиrать лишь
несущими щупальца руками и изrибать сте..
белек.
7.3.3. Классификация
Хотя известно около 6250 современных
видов иrлокожих, нынешнее их разнообра-
зие значительно меньше. чем было в палео..
зое: сейчас осталось только 6 из 24 классов
этоrо типа.
Отряд
Isocrinida
Comatulida
МШеricriпidа
Вои rguеticrШldа
Суrtосrшidа
Класс
Crinoidea
Platyasterida
Paxillosida
Valvatida
Sр1ПulОSldа
Forcipulatida
Euclasterida
. Oegoph1Urida
Рhrупорhшr,dз
OphlUrida
Peripodida
Concentricycloidea
Cidar01da
Ech lПоth urOlda
Diadematoida
Pedinoida
Salenoida
Phymosomatoida
ArbaclOlda
т emnopleuroida
Echinoida
Holectypoida
Clypeasteroida
Cassiduloida
Spatang01da
Neolampad01da
Holasteroida
Dendrochirotida
Dactylochirotida
ASPldochlrotlda
Elaslpoda
Apodida
Molpadiida
Deuterostomia: вторичноротые
7.3.3.1. Класс Crinoidea
(морские лилии)
Криноидеи это единственные COBpe
менные иrлокожие, сохранившие aHцeCTpa
льную ориентаuию тела со ртом, обращен
ным вверх и окруженным слизисто
ресничными, несущими мелкие щупальца
(НОЖКИ, или подии) руками, фильтрующими
пищу из толщи воды. Хотя в принпипе рук
пять, они MorYT неоднократно разделяться,
давая от 10 до 200 ложных рук с мноrочи
сленными боковыми ответвлениями (пин
нулами) на каждой. По их обращенной
вверх поверхности проходит амбулакраль
ный желобок (рис. 7.15). Частицы пищи за
хватываются ресничными, секретирующи
ми слизь ножками и продвиrаются вдоль
ресничных амбулакральных желобков ко
рту и далее в U образный кишечник (рис.
195
7.16). Амбулакральная и друrие целоми
ческие системы простые, без ампул, управ
ляющих ножками, и мадрепоровой пла..
стинки. Каменистые каналы (часто MHoro
численные) открываются в соматоцели.
Ножки вытяrиваются за счет сокращения
снабженных сфинктерами каналов амбула..
кральной системы.
Шаровидное тело постоянно или Bpe
менно прикреплено к субстрату с помощью
несократимоrо аборальноrо стебелька дли
ной до 1 м, часто несущеrо мутовки так на..
зываемых цирр, с терминальным прикрепи..
тельным диском или системой цирр., спо
собных удерживаться за морское дно или
заякориваться в нем. У мноrих COBpeMeH
ных видов после KopoTKoro периода прикре
пленноrо существования животные CTaHO
вятся подвижными и плавают с помощью
Рис. 7.15. Форма тела кринои
дей (по Oanielsson, 1892; Car
penter, 1 866)
СВОБодноподвижная
СтеБельковая
196
rлава 7
Анус
Осевой opraH
и синус
Кишка
Uирры
перьевидных рук, лишь временно прикре..
пляясь к субстрату аборальными ЦИррdМИ.
Однако анцестральное положение обратен..
Horo вверх рта при этом сохраняется. Руки,
тело и стебелек несут сильно обызвествлен"
ные пластинки, занимающие большую ча..
сть их объема, так что пространство.. прихо..
дящееся на живые ткани, мало.
Ткань rонад распределена ДИФФУЗНО. r а..
меты образуются в области перитонеума
внутри рук, развиваются в мелких прокси"
мальных «rонадах» и высвобождаются че
рез разрывы в стенках пиннул. Характерная
личинка виттелария после метаморфоза
превращается в миниатюрное стебельковое
подобие взрослых животных.
В классе 625 видов. Различают пять от..
рядов, в основном по деталям строения CTe
бель ка и ero прикрепительной системы.
7.3.3.2. Класс Asteroidea
(морские звезды)
Астероидеи в целом характеризуются
уплощенным телом, постепенно переходя
щим в пять (иноrда до 40) рук. Они MorYT
быть короткими и широкими, придавая жи--
вотным пятиуrольные очертания, или же
тонкими и длинными (рис. 7.17). Животные
свободноподвижные, рот и амбулакраль--
ные борозды расположены на нижней по..
верхности тела, а мадрепоровая пластинка
и анус (если он есть) аборально. Амбула
кральные борозды хорошо развиты и защи
Нерв
АМБулакральная
::.>; система
. . ... -.' ..
rемальная
система
Рис. 7.16. Схематический раз
рез тела и части руки свобод
ноживущей морской лилии (по
Nichols. 1962)
щены с каждой стороны двойным рядом не
слившихся друr с друrом пластинок, через
вырезы в которых направляются протоки,
соединяющие ампулы с локомоторными
амбулакральными ножками, часто с присо..
сками на концах. В целом известковый ске..
лет развит относительно слабо, хотя несет
внешние буrорки и шипы, иноrда располо..
женные в определенном порядке. Некото"
рые из шипов модифицированы в педицел..
ЛdрИИ (рис. 7 .18) rруппы, обычно из трех
мелких «косточек», действующих как но..
жницы или щипцы и за счет этоrо очищаю..
щих животное от орrанизмов, пытающихся
осесть на ero поверхности.
Полости тела, включая амбулакральную
систему, обширны; соматоцели образуют
мелкие выросты (папулы), проходящие на
аборальной стороне сквозь стенку тела
и служащие для rазообмена (рис. 7.19). Не..
которые астероидеи rермафродиты;
известно несколько форм размножения
и онтоrенеза, включая бесполое почкова..
ние" своеобразную заботу о потомстве.
прямое (особенно в высоких широтах) и не..
прямое развитие с личиночными стадиями
бипиннария и брахиолария (рис. 7. 14,Ь).
Большинство видов трупоеды и хищники"
питающиеся сидячими животными, но
известны и детритофаrи, и фильтраторы.
При макрофаrии добыча заrлатывается це..
ликом или желудок, который очень круп..
ный, выворачивается через рот и охваты..
вает ткани жертвы. От желудка отходит па..
Oeuterostomia: вторичноротые
197
Рис 7.17. Форма тела астероидей (по источникам в Нутап, 1955)
ра слепых выростов в каждую руку.
Шесть отрядов, включающих L500 ви
дов, различают по типам педицелларий, pa
сположению амбулакральных пластин
и форме амбулакральных ножек Открытый
в 1962 r. род Platasterias сочли сохранив
шимся представителем давно вымерших
иrлокожих из класса Somasteroidea. Однако
сейчас ero принято считать специализиро
ванной астероидеей из отряда Platyastel"ida.
198
СтеБельковые Сидячая
Характерные типы
Мышцы
СтеБелек
Структура окончании стеБельковых педицелларий
Рис. 7.18. Педицелларии (по Mortensen, 1928
1951; Нутап, 1955)
Радиальный
вырост
кишки Папула
м адрепоровая ...............
пластинка (fj :
АМБулак
Нерв lемальная ральная
система система
Рис. 7.19. Схематический разрез тела и части
руки морской звезды (по Nichols, 1962)
7.3.3.3. Класс Ophiuroidea
(змеехвостки, или офиуры)
Как и морские звезды, которым они poд
ственны, офиуры плоские, свободноживу
щие иrлокожие со ртом на нижней стороне
тела. Однако у этой rруппы тело мелкое,
дискообразное и резко отrраниченное от
длинных узких рук (рис. 7.20). Известковый
rлава 1
скелет занимает большую часть их объема
и также хорошо развит в центральном ди-
ске. Пластинки рук слиты между собой,
образуя продольный ряд сочлененных дрyr
с друrом «позвонков», способных двиrаться
за счет «межпозвонковых» мышц. С помо"
щью TaKoro механизма офиуры MorYT ХО"
дить на своих руках; центральный диск при
этом часто приподнят над субстратом. Со..
ответственно амбулакральные ножки лише..
ны локомоторных функпий И только помо..
rают рукам опираться на субстрат; в основ..
ном они используются для захвата пищи.
Амбулакральная борозда поrружена
внутрь скелетных пластин. В отличие от
астероидей мадрепоровая пластинка
у офиур расположена на оральной стороне
тела.
Мноrие офиуры слизисто ресничные
фильтраторы, у некоторых из них пять ос-
новных рук дихотомически ветвятся. Одна..
ко среди змеехвосток обычны и детритофа-
rи., и трупоеды. У всех имеется крупный же-
лудок, но кишечник и анус отсутствуют.
Между желудком и оральной поверхностью
находятся 1 О впячиваний нижней поверхно-
сти тела бурс (рис. 7.21). Это орrаны ra..
зообмена. Вода попадает в них через распо-
ложенные BOKpyr рта щели за счет биения
ресничек или мышечноrо накачивания.
В бурсы открываются rонады и здесь же ча-
сто развиваются яйца. У некоторых видов
эмбрионы прикреплены к стенке бурс, так
что происходит cBoero рода «живорожде-
ние». Развитие прямое или с личиночной
стадией офиоплутеусом (рис. 7. 14.,В).
Около 2000 видов разделяют на три от-
ряда.
7.3.3.4. Класс Concentricycloidea
В 1986 r. по девяти экземплярам одноrо
HOBoro вида, найденным на затопленной
древесине на rлубине около 1000 м у побе-
режья Новой Зеландии, был описан новый
класс необычных медузоподобных иrлоко-
жих. Тело Xylop/ax тedus(forтis диаметром
8 мм имеет форму плоскоrо диска, ОКРУ-
женноrо шипами (рис. 7.22). Рук и каких-
либо признаков пищеварительноrо тракта
нет. Верхняя поверхность диска покрыта че-
шуевидными пластинками, а нижняя несет
два концентрических кольца амбулакраль-
Deuterostomia: i1торичноротые
199
Рис. 7.20. Форма тела различных ОФИVD (по источникам в Hvman. 1955)
Желудок
r онада
Бурса
Каменистый канал
и осевой opraH
Мадрепоровая
пластинка
Позвонки
Нерв rемальная АМБулакральная
система система
Рис. 7.21. Схематический разрез диска и части DVКИ ОФИVDЫ(ПО Nichols 1962)
200
А
rлава 7
ных каналов, от которых отходит един-
ственное краевое кольцо амбулакральных
ножек. Пять пар rонад; прямое развитие
в молодь, сходную по строению со взро-
слыми особями, идет, по видимому, в бур-
сах.
7.3.3.5. Класс Echinoidea
(морские ежи)
Эхиноидеи сферические или вторично
уплощенные свободноживущие иrлокожие
без рук (рис. 7.23) (руки в определенном
смысле включены в состав тела). Для них
характерен жесткий панцирь из соединен..
ных друr с друrом скелетных пластинок.
Каждая амбулакральная и интерамбула-
кральная зоны несут в принципе по две вер-
тикальные дуrовидные полосы таких пла-
стинок, идущих от аборальноrо полюса ко
рту. На панцире находятся педицелларии
(см. разд. 7.3.3.2) и подвижные иrлы, с по-
мощью которых некоторые виды MorYT
передвиrаться по поверхности субстрата
или зарываться в Hero. Помимо этоrо локо-
моция происходит за счет амбулакральных
ножек, выходящих сквозь поры в амбула-
кральных пластинках.
Рис. 7.22. Вид с дорсальной (А) и вентральной
(Б) сторон представителя концентрициклоидей
Ху/ор/ах (по Baker et al, 1986)
А Вид СБОКУ
Вид СБОКУ
r Вид С оральнои стороны
\,\ ./! I
,,\ 1\\ I! I
\I 11 t,l., /
'" . ,r ... t.) , .. /
> . :. :;i{' '\ : /
. . l: "- .
I ,. ,"'- "1
/" \
Б Вид СБОКУ
! (
, ,
':" : »,\\\"
:W- .. :
=', ' ':'"
," '
. ...; .=-..... --=- . .,
,...
' .........,
М"'JP /"'''"\' '"
в Вил СБОКУ
. А -,:, ,
' . :.:t . 'е7,
- ",,' ': :f. . ':','
" ,-. 'J. . ..tВ.
." 'Jl/J , '. . ;
, .-.. '-- 7 " :#1,: ;' ,'-
',) f(
Й:,' ,k:t
- ?Jfi .
". tl :O",
Вил СБОКУ
n Вид с аБоральной стороны
Рис. 7.23. Форма тела морских ежей: А, Б «правильные»; В Д «неправильные» (по несколь-
ким источникам, в основном в Нутап, 1955)
Deuterostomia: вторичноротые
201
Проксимальные
концы ЗУБОВ
МЫШЦЫ
Пирамиды, удерживающие
ЗУБЫ
Дистальные концы ЗУБОВ
Рис. 7.24. Аристотелев фонарь (по Нутап,
1955 )
Расположенный на нижней стороне тела
рот снабжен характерным rрызущим аппа..
ратом из пяти крупных и нескольких мелких
пластинок «аристотелевым фонарем»
(рис. 7.24), с помощью KOToporo пережевы..
ваются водоросли и сидячие животные. Для
этоrо «фонарь» выдвиrается изо рта.
у «правильных» видов эпифауны рот Haxo
дится в центре нижней поверхности тела,
а анус посередине аборальной. Некото"
рые виды эхиноидей роются в рыхлых осад"
ках и вторично стали билатерально симме..
тричными (<<неправильные» морские ежи).
у таких форм анус заметно смещен к «зад..
нему концу» тела, а рот иноrда отчасти
сдвинут к «переднему». Они детритофаrи,
в связи с чем аристотелев фонарь редуци..
руется или отсутствует, а питание осуществ"
rOHaдa
Кишка
ляется с помощью специализированных
слизисто..ресничных амбулакральных но..
жек.
Обширная полость тела (рис. 7.25)
образует выросты в виде сильно модифиuи--
рованных амбулакральных ножек (у «не..
прав ильных» ежей) или в виде околорото--
вых жабр (у «правильных» видов), служа..
щих для rазообмена. Четыре (у «неправиль--
ных») и пять (у «правильных» форм) круп..
ных rонад, вдающихся в полость тела, от..
крываются наружу аборальными rонопора..
ми. у некоторых наблюдается забота о по..
томстве, однако у большинства развитие
непрямое с личинкой офиоплутеусом
(рис. 7.14,1).
Классификация по отрядам базируется
в основном на вымерших формах; в 15 со..
временных отрядах около 950 ныне живу..
щих видов.
7.3.3.6. Класс Holothuroidea
(rолотурии, или морские оrурцы)
rолотурии лишены рук (как и у МОРСКИ)l
ежей они включены в тело), а их билатера.
льно симметричное тело, сильно ВЫТЯНУТОt
вдоль орально..аборальной оси, лежит Kal<
бы на боку. При этом одна ero сторона
с тремя рядами ножек, амбулакрами, (три-
виум) является «вентральной», а противо-
положная.. с двумя амбулакрами (биви-
ум) «дорсальной» (рис. 7.26). Стенка тела
также необычная, кожистая, с хорошо раз-
витыми кольцевыми и пятью продольными
Мадрепоровая
пластинка Анус
Каменистый канал
и осевой opraH
Нерв
АМБулакральная
система
r емальная
Аристотеле система
сронарь
Рис. 7.25. Схематический раз
рез MopcKoro ежа (по Nichols, ЖаБра
1962)
202
rлава 7
.f'. :.:....... . .
. .II;........Jt.'\t ....... .'....11
Рис. 7.26. Форма тела rолотурий (по нескольким источникам, в основном в Нутап, 1955)
мышечными пучками. Известковый скелеl
редуцирован до изолированных микроско
пических «косточек».
BOKpyr находящеrося «спереди» рта ам..
бу лакральная система поддерживает от 8 до
30 пальцевидных, разветвленных или ЩИТ-
ковидных ротовых щупалец, использую-
щихся для питания детритом или, реже, для
Deuterostom;a: вторичноротыр
Мадрепоровая rонада
пластин ка
Рот
203
Полость тела
Клоака
Ротовое
щупальце
Нерв rемальная Амбулакральная
система система
Водное леrкое
Рис. 7.27. Схематический разрез rолотурии (по Nichols, 1962)
фильтрования (рис. 9.3 и 9.6). Сквозной, ча..
сто очень длинный пищеварительный тракт
заканчивается «сзади» прямой кишкой или
клоакой и затем анусом. В клоаку у мноrих
видов открывается пара обширных впячи"
ваний, лежащих в полости тела; это раз..
ветвленные водные леrкие. Вода заполняет
их и с силой выталкивается обратно, обе..
спечивая rазообмен (рис. 7.27). Некоторые
rолотурии при раздражении выбрасывают
заднюю кишку включая водные леrкие че..
рез анус (эвисцерация). У нескольких видов
при раздражении выбрасываются специаль..
ные расположенные около леrких клейкие
или содержащие токсин кювьеровы трубоч..
ки. Мадрепоровая пластинка лежит свобод"
но в полости тела.
Локомоция очень медленная, у боль..
шинства видов с помощью амбулакраль--
ных ножек, которые часто разбросаны по
поверхности тела, а не оrраничены опреде...
ленными зонами (амбулакрами). Некото"
рые rруппы полностью лишены ножек
и движутся за счет перистальтических co
кращений мышц, отталкиваясь от осадка
заякоривающимися в нем, выступающими
из тела мелкими известковыми «косточка..
ми». Некоторые rолотурии сидячие жи..
вотные, использующие амбулакральные
ножки больше для прикрепления, чем для
локомоции. У ряда rлубоководных форм
сильно увеличенные и удлиненные амбула..
кральные ножки функционируют как ходу..
ли (см. рис. 7.28). Большинство видов
представители эпифауны, хотя некоторые
роются в рыхлых осадках, а одна rруппа
включает планктонные формы.
Обычно непарная [онада открывается
rонопором около ротовых щупалец. Рас..
пространена забота об эмбрионах, которые
у некоторых форм развиваются в полости
тела, а у одноrо вида в яичнике. Некото--
рые протандрические rермафродиты. Ес..
ли отсутствует забота о потомстве, разви"
тие непрямое с личиночными стадиями ви"
телларии (или аурикуларии) и затем доли о..
ларии (рис. 7.14,Д).
АМБулакральные ножки
Рис. 7.28. rолотурии, идущие по океаническо..
му дну с помощью крупных удлиненных амбу
лакральных ножек
204
По форме ротовых щупалец, характеру
амбулакральных ножек и форме тела Bыдe
ляют шесть отрядов.. объединяюших около
1150 ныне живущих видов.
7.4. Тип Chordata (хордовые)
7.4.1. 3тимолоrия
От лат. chorda CTpYHa (из кошачьей
кишки) .
7.4.2. Диаrностические
и специфические признаки
Тип хордовых включает три подтипа. из
которых два относятся к беспозвоночным,
поэтому обсуждаются в настоящей книrе.
Эти два подтипа достаточно сильно отли"
чаются друт от друта и будут рассмотрены
по отдельности. Тем не менее у них следую
щие общие признаки.
1. Билатерально симметричные живот..
ные.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и орrанами.
3. Кишечник сквозной, анус не терми
нальный.
4. Тело в принципе мономерное, без
rлотка Желудок
Нервная ТРУБка
Атриум
Эндостиль В
I Сердце
Атриальное отверстие
Срез на уровне А В
rлава 7
различимой rоловы, конечностей или челю..
стей.
5. Обширная rлотка со стенками, перфо..
рированными насквозь rлоточными щеля..
ми, число которых варьирует от нескольких
до очень мноrих.
6. Слизисто ресничные фИЛЬТрdТОрЫ
(см. разд. 9.2.5); ток воды, при носящий пи..
щевые частицы, покидает тело через rло..
точные щели.
7. Обычно rлоточная область окружена
вторичной стенкой тела, образующей
атриальную полость, связанную СОКРУ"
жающей средой единственной порой (СМ.
рис. 9.14).
8. Эпидермис не секретирует наружной
кутикулы и не несет ресничек.
9. На некоторых или на всех стадиях жи..
зненноrо цикла присутствует внутренний
дорсальный скелетный тяж хорда.
10. На некоторых или на всех стадиях
жизненноrо цикла дорсальнее хорды прохо..
дит нервная трубка.
11. На некоторых или на всех стадиях
жизненноrо цикла opraHoM активноrо пла..
вания служит мускулистый постанальный
хвост.
12. Циркуляторная система частично не
замкнута.
rлоточная и первичная
стенки тела
Атриум
Вторичная стенка тела
Рис. 7.29. План строения обоб
щенноrо беспозвоночноrо xop
довоrо
Deuterostomia: вторичноротые
13. Дробление яйца радиальное.
14. Развитие, характерное для вторич
норотых, обычно недотерминированное
инепрямое.
15. Морские формы.
7.4А. Подтип Urochordata
(личиночнохордовые,
или оболочники)
7.4А.1. Диаrностические
и специфические признаки
1. Хорда, нервная трубка и постаналь
ный хвост (если есть) только на личиночной
стадии и у одной неотенической rруппы.
2. Целомической полости нет
3. Экскреторных opraHoB нет.
4. Мышечные и друrие структуры не ce
rментированы.
5. Тело целиком заключено внутри ce
кретируемой туники, панциря или «доми
ка», обычно состоящих из целлюлозы и бел
ка и содержащих миrрировавшие из тела
клетки, а у некоторых форм
экстра корпоральные кровеносные сосуды.
6. Пищеварительный тракт U образной
формы с обширной rлоткой, обычно зани"
мающей большую часть объема тела.
7. Нервная система в виде rанrлия, pa
сположенноrо между ртом и атриальным
отверстием, с отходящими от Hero отдель..
ными нервами.
8. rермафродиты, обычно с непарными
семенником и яичником.
Оболочники сидячие или свободножи..
вущие, одиночные или колониальные филь..
траторы с признаками хордовых только на
стадии свободноплаваюшей личинки.
205
7.4А.2. Классификация
Около 1400 видов оболочников распре
делены между тремя классами.
Класс
ОТРЯД
Aplousobranchia
А d . Phlebobranchla
SCI lacea
Stolldobranchla
Asplraculata
Thaliacea
Larvacea
Copelata
7.4А.2.1 Класс Ascidiacea
(асцидии)
Этот класс включает около 1250 сидячих
бентосных видов. Обычно имеется свобод
ноживущая планктонная, похожая на rоло
вастика личинка (рис. 7.30), которая в KOH
це концов оседает на дно и прикрепляется
rоловным концом к субстрату. Затем HepaB
номерный рост в течение метаморфоза фак
тически приводит к повороту передне
задней оси тела на 1800, так что рот и OKPy
жающий ero жаберный сифон туники или
панциря оказываются на противоположном
точке прикрепления конце тела (рис. 7.31).
Этот жаберный сифон ведет через окружен
ное щупальцами отверстие в обширную
rлотку, обычно перфорированную боль
шим количеством мелких щелей (за исклю
чением rлубоководных видов из отряда
Aspiraculata, у которых rлотка реДУIlирова
на, щели отсутствуют, а жаберный сифон
модифицирован в ряд ловчих выростов).
Щупальца BOKpyr отверстия, ведущеrо
в rлотку.. используются для Toro. чтобы
Туника Нервная ТруБка
Рис. 7.30. Анатомия rоловасти
коподобной личинки асцидий
(по McFarland et al., 1979 и др.)
Хорда
Прикрепительные
opraHbI
Кишечник Сердце Эндостиль
206
rлава 7
предотвращать попадание в нее крупных ча..
стиц. Атриальное отверстие также офор..
млено в виде сифона, находящеrося
рядом с жаберным сифоном (рис. 7.32).
Сердце расположено базально и. что не..
обычно, способно выталкивать кровь в двух
периодически сменяемых направлениях.
Кровь интересна тем, что в некоторых ее
клетках высока конuентраuия связанных
с сульфат"ионами тяжелых металлов, ос о..
бенно ванадия (может быть в 105 106 раз
выше, чем в морской воде), железа, ниобия.
Распространено бесполое размножение
почкованием. Оно происходит несколькими
способами от различных частей тела асци..
дий. Как и у неродственных асцидиям кни"
дарий и мшанок, отпочковавшиеся особи
MorYT оставаться связанными друr с друrом
в колонии, однако полиморфизма зооидов
не наблюдается. Некоторые колониальные
формы окружены общим панцирем. умно..
rих общее атриальное отверстие, друrие
связаны базальными столонами; часто у ко..
лоний характерная форма. У некоторых ви"
дов почкование начинается еще на личиноч"
ной стадии. Однако большинство асцидий
одиночные; эти формы часто rораздо круп..
нее зооидов колониальных видов, достиrая
в высоту более 15 см (рис. 7.33).
Кроме аберрантных Aspiraculata выде..
ляют еще три отряда асцидий, в основном
по размещению rонад и структуре rлоточ"
ной стенки. У исключительно колониаль..
ных Aplousobranchia rонады окружены пет..
лей кишечника, стенка rлотки простая. У,
как правило, одиночных Stolidobranchia они
поrружены в стенку тела и лежат вдоль
rлотки с продольными складками и вну"
Рис. 7.31 . Метаморфоз rолова
стикоподобной личинки (по
Barnes, 1980)
тренними переrородками. У также большей
частью одиночных Phlebobranchia rонады
расположены, как у Aplousobranchia, rлотка
без складок, но с характерными продольны-
ми переrородками внутри, образованными
раздвоенными сосочками.
Приносящее
течение
Жа6ерн СИ
Ротовые
щупальца
rанrлии
Выносящее
/T
Дтриальный
сисрон
Д" . . .
rлотка с БОЛЬ
шим количест
вом персрора
ций
ЭНДОСТИЛЬ
Анус
Панцирь или туниra
Сердце
Желудок
rOHaдbI
/ СУБстрат
//
СРорма атриальной
полости
Срез на уровне Д в
Туника
д триальная
ПОлость
Просвет rлотки
ЭНДОСТИЛЬ
rлоточные
пер<:рорации
Рис. 7.32. Схематический разрез асцидии (по
нескольким источникам)
Ueuterostomi<=-: вторичноро-'-не
207
Колониальные q:юрмы
Одиночные срормы
Рис. 7.33. Форма тела и колоний асцидий (по Millar, 1970 и др.)
7.4А.2.2. Класс Thaliacea
(пелаrические оболочники)
Талиацей 70 видов, все они планктонные
и используют ток BOДЫ несущей пищевые
частицы, для реактивноrо движения. Жа
берное и атриальное отверстия расположе
ны на противоположных концах их BepeTe
новидноrо или боченкообразноrо тела.
у всех форм наблюдается бесполое почко
ванне от вентральноrо столона, отходящеrо
сразу за rлоточным эндостилем.
Наблюдаются два различных образа
жизни. Отряд Pyrosomida (оrнетелки) вклю
чает колониальных талиацей, которые по
мещаются в общем цилиндрическом доми..
ке с центральным просветом. открытым во
внешнюю среду только с одноrо конца.
Каждый зооид расположен так, что жабер
ное отверстие обращено вовне общеrо ци..
линдра. а атриальное в ero внутреннюю
полость, куда, таким образом, попадают
все пищевые токи воды, и вся трубчатая KO
лония длиной до нескольких метров движе
ОБщая полость
колонии
Отдельные зооиды
Отверстие
колонии
',А
Анус
Персрориро
ванная
rлотка
ОБЩИЙ
панцирь
rанrлий
Атриальная
полость
Атриальный
СИq>ОН
ОБщая полость колонии
Б
Выносяшее
течение
rлоточная
переrородка
Рис. 7.34. Схематический раз
рез I'<олонии (А) и отдельноrо
зооида (Б) пиросомы (по Gras-
5е, 1948; Fra5er, 1982)
Анастомозирующие инеполные
мышечные кольца
Атриальный
СИq>ОН
rанrлий
аБерный сиq>он
Семенник
А
Атриум
Одиночная Боковая
rлоточная перqpорация
Б
ЖаБерный Рис. 7.35. Схематичная анато-
сисрон мия двух стадий жизненноrо
цикла сальп: A
размножающееся половым пу
тем аrреrированное «поколе-
ние»; Б размножающееся
бесполым путем одиночное
«поколение» (по Beril', 1950;
Fra5er, 1982 и ДР.)
Атриальный
сиq>он'
Deuterostomia: вторичноротые
ЖаБерный
сисрон
209
rанrлий
Мышечные ОБОДЬЯ
Атриум
Атриальный
сифон
Анус
А
Эндостиль Семенник
Рис. 7.36, Схематичная aHaTO
мия двух стадий жизненноrо ЖаБерный
цикла боченочников: А сисрон
размножающееся половым пу
тем одиночное «поколение»;
Б размножающееся беспо
лым путем аrреrированное
«поколение» (по Berrill, 1950;
Fraser, 1982 и др.) Б
тся как единое пелое, напоминая крупную
морскую змею (рис. 7.34). У отдельных
зооидов множество rлоточных перфорациw,
внешне сходных с имеющимися у асцидий.
Отряды Salpida (сальпы) и Doliolida (бо..
ченочники) в основном одиночные жи..
вотные, хотя у них происходит чередование
одиночных форм и аrреrаций различных по..
ловых типов. У сальп (рис. 7.35) одиночная
стадия размножается бесполым путем, да..
вая цепочки особей, которые в конечном
итоrе отделяются в виде колонии половоrо
«поколения». у долиолид половым путем
размножается, напротив, одиночная стадия,
образующая бесполое аrреrированное «по..
коление» (рис. 7.36). В обоих отрядах на..
блюдается полиморфизм зооидов (на раз..
личных стадиях жизненноrо цикла), имею..
тся мощные ленты или обручи мышц, иду"
щие BOKpyr тела и служащие для выталкива..
Развиваюшиеся почки
r\
Атриальный
сисрон
Столон
Почки, миrриру
юшие от столона
к кадофору
ния воды (вместо ресничноrо механизма ее
продвижения), более или менее прозрачный
слизистый чехол и небольшое число круп..
ных rлоточных перфораций У сальп Bcero
две orpoMHbIe rлоточные щели (рис.
9.15).
Личиночные стадии есть только у боче..
ночников; от них, как обычно, считается,
произошли путем педоморфоза предст dВИ"
тели следующеrо класса.
7.4А.2.З. Класс Larvacea
(аппендикуларии)
Строение этих планктонных форм ха..
рактерно для личинок оболочников (рис.
7.37): мелкое тело с крупным хвостом, по
всей длине KOToporo проходят хорда и нерв..
ная трубка. Общая длина животных около
5 мм Эпидермальные железы секретируют
210
rлотка
Рот
Эндостиль
r лоточные щели
Анус
rлава 7
Яичник
Семенник
Кишечник
тонкий студенистый (нецеллюлозный) «до..
мик», образующий внешний фильтрацион"
ный аппарат (рис. 7.38). Вода поступает
в домик за счет биения хвоста через особые
сита, и пищевые частицы, в основном нана..
планктонные водоросли, отфильтровываю..
тся тонкой сетью внутри студенистой обо..
лочки (см. рис. 9.5), а затем 1аrлатывается
с помощью тока воды, создаваемоrо rло..
точными ресничками. Вода выходит из пи..
щеварительноrо тракта через пару rлоточ"
ных шелей, открываюшихся независимыми
отверстиями на поверхности тела. Роль
атриальной полости в результате иrрает
пространство внутри «домика». Коrда ero
сита и фильтры необратимо засоряются,
аппендикуларии MorYT покидать ero через
«дверь» (в норме закрытую) и «надувают»
новый уже заложенный к этому моменту
домик.
Хорда
<Рилырационная
система
Приносящее
течение
I
Рис. 7.37. Анатомия аппендикуларии (по Alld
redge. 1976)
Рис. 7.38. «Домик» аппендику
ларии и индуцируемые питаю
щие течения сквозь Hero (по
Hardy, 1956 и др.)
Сито
Передний
конец
>Кивотное внутри
домика
Отверстие юля выхода
из домика
Вид СБОКУ
Передний конец
Приносяuцее течение
Задний конец
Deuterostomia: вторичноротые
Все аппендику ларии одиночные фор--
мы, размножение только половое. Один
ВИД, что необычно для урохордат, раздель--
нополый. Bcero один отряд, включающий
70 видов.
7.4Б. ПОДТИП Cephalochordata
(rоловохордовые,
или ланцетники)
7.46.1. Диаrностические
и специфические признаки
(рис. 7.39)
1. Тело, сжатое с боков, рыбообразное.
2. Хорда проходит по всей ero длине.
3. Полый дорсальный нервный ствол
тянется почти вдоль Bcero тела. не расши--
ряется спереди в мозr.
4. Постоянный постанальный хвост.
5. Тело с сериально повторяющимися
мышечными блоками, нервами.. выдели
тельными орrанами и rонадами.
6. Полость тела формируется энтероце..
льно из множества сериально повторяю--
щихся карманов. Их вентральные части
Миомеры Хорда
Нервная ТРУБка
А
А триопор rонады
Х
Миомеры
Эндостиль
211
сливаются, а дорсальные вытесняются
мышцами.
7. Экскреторная система очень сходна
с протонефридиальной, но формируется
перитонеальными клетками.
8. rлоточный отдел покрыт вторичной
стенкой тела, образованной парой метапле..
вральных складок, растущих в вентральном
направлении и срастающихся по средней
вентральной линии тела.
9. rлотка обширная. 1анимающая поло..
вину длины тела.
10. Раздельнополые животные с MHoro..
численными парными rонадами.
rоловохордовые мелкие (до 10 см
длиной) свободноживущие животные, веду"
щие бентосный и малоподвижный образ
жизни, но способные плавать, меняя места
питания или спасаясь от хищников. При
этом хорда используется как несжимаемая,
но rибкая опора. Сокращение продольно
ориентированных мышц приводит к волно..
образному изrибанию тела, а не к ero уко"
рочению (эквивалентно системе движения,
наблюдаемой у xeTorHaT и нематод)
Пищедобывательная система rолово"
хордовых в сущности такая же. как у обо..
Х
у
Эндостиль
rлотка
Камера "плавниковоrо луча 1 '
Нервная ТРУБка
Хорда
rлоточные
щели
Просвет
rлотки
Атриум
Вторичная
стенка тела
у
..
Б
Рис. 7.39. rоловохордовые: А схематический продольный разрез; Б прижизненный внешний
облик (по Young, 1962)
212
rлава 7
Пиrментное
пятно
Хорда
Камеры
''плавниковых
лучей"
Мерцательный opraH
Парус
Ротовые цирры
Эндостиль
лочников. Ротовой отдел окружен рядом
щупалец (ротовыми циррами и веларными
щупаЛЬЦаМИ рис. 7.40), которые предот..
вращают попадание внутрь нежелательных
частиц. rлотка удлиненная, а не бочкообра..
зная и атриопор расположен на средней вен..
тральной линии тела сразу перед анусом.
Домика. окружающеrо тело и вытянутоrо
в сифоны, нет.
В некоторой степени rоловохордовые
являются связующим звеном между беспо..
звоночными и третьим подтипом хордо..
вых позвоночными (Vertebrata), особенно
с бесчелюстными, также лишенными конеч..
ностей и челюстей. У личиночной стадии
миноr, как и у ланцетника, слизисто..
ресничная фильтрующая rлотка, хотя в дан..
ном случае вода выталкивается из нее с по..
мощью мышечноrо сокращения. Однако
более очевидны отличия rоловохордовых
от позвоночных. Они состоят в отсутствии
дифференцированной rоловы с парными
сенсорными орrанами, мозrом и защищаю..
щим ero черепом, в особенностях строения
нервной трубки. к которой мышечные клет..
ки посылают свои отростки (формируя ло..
жные вентральные корешки), и перифериче..
ских нервных волокон, лишенных миелино..
вой оболочки, в очень тонком однослойном
эпидермисе. Тем не менее общие чеРТЬJ ro..
ловохордовых, вероятно, соответствуют
r лотка
Рис. 7.40. Передний конец ro
ловохордовоrо (по У oung,
1962)
орrанизации «протопозвоночных». Очевид-
но также сходство ланцетников с увеличен-
ным неотеническим вариантом личинки
урохордат, во всем прочем похожих на бес-
позвоночных.
7.46.2. Классификация
Известно 25 видов, помеЩаемых в один
класс и один отряд.
7.5. Дополнительная
литература
Alvarino А. 1965. Chaetognaths. Oceanogc mar.
Biol.. Апп. Rev.. 3, 115 194 [Chaetognatha].
Baker А. N., Rои,'е F. W.E., Clark H.E.S. 1986.
А new class of Echinodermata from New Zealand,
Nature, Lond., 321, 862 864.
Barrington Е. J. W. 1965, Ttle Biology of Hemichor-
data and Protochordata. 01iver & Boyd, Edin-
burgh [Hemichordata, Chordata].
Berri/Z N. J. 1950. The Tunicata. Ray Society, Lon.
don [Urochordata].
Нутап L. Н. 1955. The Invertebrales (V 01. 4) Ест.
nodermata. McGraw Hill, New У ork [Echinoder-
та ta].
Нуп1аn L. Н. 1959. The Invertebrates (Уоl 5) Smal.
ler Coelomate Groups. McGraw Hill, New York
[Hemichordata, Chaetognatha]
NicJzols D. 1969. Echinoderms, 4th edn. Hutchinson.
London [Echinodermata].
Youпg J. z. 1981. The Life of Vertebrates, 31'd edn,
Clarendon Press, Oxford [ChordataJ.
rЛАВА 8
БЕспозвоночныE С КОНЕЧНОСТЯМ-И:
ЧЛЕнистоноrИЕ И сходныE rруппы
Tardigrada
Pentastolna
Onychophora
Chelicera ta
U niramia
Crustacea
За исключением первичноротости, для
представителей шести рассматриваемых
в данной rлаве типов фактически общими
являются только два признака парные
конечности по всей длине тела или какой"то
ее части (каждая пара иннервируется обыч
но парны:rvfИ rанrлиями, расположенными
на продольных нервных тяжах) и псевдоце..
лом, часто заполненный rемолимфой и по..
тому называемый rемопелем.
у преДСТd.вителей трех типов (Tardigra
da, Pentastorna и Onychophora) жесткоrо Ha
ружноrо (экзо ) скелета нет; полость их тела
иrрает роль rидростатическоrо скелета;
в сущности эти животные черви с мяrки..
МИ, мясистыми, нечленистыми конечностя..
ми, вооруженными коrотками, т. е. с паль..
цевидными выростами тела, приводимыми
в движение внешними по отношению к ним
мышцами. В rл. 2 мы допустили, что таких
«червей с конечностями» можно было бы
считать «протоартроподами» и реликтами
исходной волны «артроподизации» (при..
обретения членистых конечностей) в ДOKeM
брии или в самом начале кембрия. Однако
если онихофоры и кажутся родственными
некоторым истинным членистоноrим (од..
новетвистым), особенности тихоходок так
же сильно, если не сильнее, сближают их
с (анцестральными?) псевдоцеломическими
животными, как и с друrими протоартропо"
дами, а истинные родственные связи пятиу..
сток по меньшей мере туманны, в значите..
льной степени из..за их специализированно..
[о паразитическоrо образа жизни.
у «истинных» членистоноrих (Crustacea,
Chelicerata и Unirarnia) жесткий, членистый,
склеротизированный кутикулярный экзо"
скелет из хитина и белка, иноrда пропитан..
ный карбонатом кальция (рис. 8.1). Он по..
крывает все тело, включая членистые конеч..
ности (слово артроподы, neреводимое как
«членистоноrие», происходит от rреч. arth..
ron cycTaB, роdоs ноrи) (рис. 8.2). Как
и у друrих животных, наличие кутикулы за..
трудняет рост, который требует ряда линек.
Особенно остро эта проблема встает имен..
но у артропод, rде кутикула выполняет
также роль скелета. Во время их линьки ста..
рый скелет частично резорбируется, затем
сбрасывается, а новый мяrкий скелет, обра..
зующийся под ним, растяrивается (блаrо
даря поступлению в тело воздуха или воды)
и отвердевает. В этот период животные осо..
бенно уязвимы, поэтому линька часто ocy
ществляется в убежищах. Изначально экзо"
скелет, вероятно, представлял собой ряды
защитных кутикулярных пластин или обру
чей, как, например, у киноринхов (разд. 4.6)
и некоторых тихоходок (разд. 8.1). Позднее
он адаптировался к выполнению скелетной
функции.. частично заменив анцестральный
rидростатический псевдоцель. В некоторых
rруппах эта замена так и осталась частич
ной, поскольку ряд членистоноrих все еще
использует rидростатическое давление для
выпрямления конечностей, а систему экзо"
скелет"мышцы только для сrибания их.
Исходно тело членистоноrоrо мономер..
но, хотя с каждой парой Hor связана замет..
ная метамерия стенки тела, экзоскелетd.
214
Эпипермис
rлава 8
\
\
\
\
\
\
\
\
\
\
,
\
\
\
\
\
\
\
\
\
\ .
\
\
Направленная
внутрь аподема
. .
т ОНОФИБРИЛЛЫ,
пронизывающие
клетки эпидермиса
\
\
\
Мышечный
пучок
Железистая
клетка
Эпидермис
Базальная меМБрана
Клетка, ОБразующая
щетинку
Рис. 8.1. Разрез кутикулярноrо экзоскелета членистоноrоrо, демонстрирующий различные слои
и внутренний вырост (а подем у), выполняющий функцию BHYTpeHHero скелета (по Hackman, 1971
и др.)
и некоторых внутренних структур (см., Ha
пример, рис. 2.1 О). В сущности артропод--
ный план строения следующий: небольшой
передний отдел (акрон) и эквивалентная ему
задняя часть (тельсон), не несущие конечно
стей, а между ними ряд промежуточных OT
резков (cerMeHToB).. каждый из которых
с парой конечностей и сериально повторяю
щимся набором обслуживающих их CTpYK
тур (мышечных, нервных, скелетных и т. д.).
При этом не связанные с конечностями ор-
rdНЬJ (например, выделительная и репро-
дуктивная системы) не метамеризуются. Во
мноrих линиях членистоноrих прослеживае-
тся значительное слияние несущих конечно-
сти cerMeHToB, утрата придатков и/или диф-
ференцировка различных отделов тела: осо-
бенно широко распространено преобразо-
вание передних конечностей в пищезахваты-
вающий аппарат. Однако кроме частичной
Терrит
r.1лейрит I
I
, """"" ,
" I
11' 'J,. /
I I .......... т "'у/
I I ... .1....'"
/\ I
1\ У
/)/
,
Рис. 8.2. Поперечный разрез
тела членистоноrоrо, демон-
стрирующий различные ЭКЗ0-
скелетные пластинки и члени-
стую конечность. Число отде-
льных члеников в конечности
в разных rруппах неодинаково;
в каждом типе они носят осо-
бые названия
Беспозвоночные с конечностями: членистоноrие и др.
215
ИЛИ полной МОДИфИКdЦИИ кутикулы В экзо
скелет (и вытекающих отсюда морфолоrи..
ческих особенностей, например, членистых
конечностей), отсутствия ресничек и TeHдeH
ции к развитию сложных rлаз, у артропод
мало общих особенностей. поэтому весьма
вероятно, что членистоноrость скорее
уровень орrанизации, а не отличительный
признак единой филоrенетической линии.
К артроподам относится подавляющее
большинство видов животных, rлавным
образом в результате успешноrо заселения
этой rруппой суши и той леrкости, с KOTO
рой может идти видообразование у мелких
наземных орrанизмов. Успех членистоно
rих как обитателей суши, резко отличаю
щий их от большинства rрупп беспозвоноч
ных, вероятно, в значительной степени обу
словлен эволюцией влаrосбереrающих op
[анов выделения и rазообмена, а также раз
витием препятствующей испарению непро
ницаемой эпикутикулы.
8.1. Тип Tardigrada
(тихоходки)
8.1.1.3тимолоrия
От лат. tardus медленный, gradu
шаr.
8.1 .2. Диаrностические
и специфические признаки
(рис. 8.3)
[. Мелкие, коренастые, билатерально
симметричные животные.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и орrанами.
3. Кишечник сквозной, прямой.
4. Тело мономерное, хотя с четырьмя
парами коротких нечленистых конечностей
с коrотками, служащих для ползания
(рис.8.3,А); конечности обслуживаются
внешними по отношению к ним мышцами
и иннервируются сериально повторяюши
мися парами нервных rанrлиев.
5. Скелет rидростатический, образован
хорошо развитым псевдоцелем.
6. Тело покрыто кутикулой. но мышеч
ных слоев под эпителием нет; в целом нехи
тинизированнаякутикуласбрасываетсяпри
линьке; ее поверхность часто скульптуриро
вана шипиками и/или чешуйками; тело про..
низано сетью отдельных rладких мышеч
ных волокон.
7. Число клеток тела cTporo фиксирова
но (эвтелия).
8. Мускулистая сосущая rлотка выстла
на хитиновыми чешуйками (плакоидами);
А
Рис, 8.3. Строение тихоходок: Мозr
А. Внешний облик животных, Рот .'-
ползающих по поверхностиСтенка тел
нитчатой водоросли (по Mar
cus, 1929). Б. Схема продоль
Horo разреза rенерал изован
ной тихоходки (по Cuenot,
1949) Б
rлотка
Мышечные
пучки
"Мальпиrиевы
сосуды"
Hora Псевдоцель
rанrлий
конечности
216
для поражения добычи из ротовой полости
может выдвиrаться пара стилетов.
9. OpraHoB кровообращения и rазооб
мена нет.
10. У ряда видов выделительная систе
ма состоит, вероятно, из трех «мальпиrие
вых сосудов».
11. Нервная система с rоловным моз"
rOM и вентральной парой стволов с rанrлия..
ми, иннервирующими конечности.
12. Раздельнополые животные, иноrда
наблюдается партеноrенез; rонада непар..
ная.
13. Развитие прямое.
14. Свободноживущие обитатели интер
стициали пресных и соленых вод, а также
водной пленки на наземных растениях: ве..
дут скрытный образ жизни.
Для тихоходок характерна специфиче..
ская смесь признаков: как и у друrих про
тоартропод_ у них парные конечности с KO
I
...:..:....:,
... . ". -..
.: + -' : .
.-: -=: . .:. . <:"
rлава 8
Вид с дорсолатеральной стороны
Вид СБОКУ
Рис. 8.4. Стадия «боченка» у тихоходок (по
Morgan, 1982)
rотками; как у лофофоровых и вторичноро"
тых, В их развитии, хотя и кратковременно,
прослеживается стадия парных (пять пар)
энтероцельных карманов; с псевдоцеломи"
чески:ми формами их сближает общий уро..
вень орrанизации и образ жизни.
Хотя ряд видов обитает в относительно
стабильных биотопах типа MopCKoro песка.
Вид с дорсальной
стороны
.:'f.
/J :: . 1
:. .-:, ...
. . . . :
.-,0.0 ".. .
I
Рис. 8.5. Разнообразие форм тела, скульптуры кутикулы, числа и формы KoroTKoB у тихоходок (по
Morgan, King, 1976 и др.)
Беспозвоночные с конечностями: членистоноrие и др
для мноrих характерна жизнь во временных
водных пленках и текучей воде. У послед
ни для переживания неблаrоприятных усло
вин раз вились различные устойчивые CTa
дии. Например, коrда водная пленка на ли
стьях мха высыхает, через проницаемую ку"
тикулу тихоходок также происходит испа
рение влаrи. Так они MorYT терять большую
часть жидкости тела, превращаясь в мелкие
сухие «бочоночки» (рис. 8.4), которые
остаются жизнеспособными до 10 лет (воз..
можно" и дольше). В этом состоянии их по..
требление кислорода в 600 раз меньше ис
ходноrо, они способны выдерживать более
8 ч температуру 2720 С и кратковремен..
ное наrревание до 150' С. Менее суровые
условия переносятся блаrодаря: (а) инци"
стированию (животное втяrивает конечно..
сти, сбрасывает кутикулу, как при линьке,
и с орачиваетсяl":" в шарик внутри кутикуляр
нои капсулы); (о) образованию толстостен..
ных покоящихся яиц. Возможно, что с уче..
том таких анабиотических фаз продолжите..
льность жизни тихоходок может превы"
шать 60 лет.
Все тихоходки сосущие орrанизмы,
хотя их кормовой спектр широк: от клеток
водорослей или наземных растений до Ta
ки: интерстициальных или ведущих скрыт..
'ныи образ жизни животных, как коловрат--
rки, нематоды или даже друrие тихоходки.
Несколько видов паразиты, один из
них в желудке брюхоноrих моллюсков.
Животное прикладывается ротовым отвер"
стием к пищевому объекту и прокалывает
ero двумя выдвиrающимися при помощи
мышц стилетами, после чеrо блаrодаря на--
сасывающему действию rлотки в желудок
перекачиваются жидкости и клеточные ор--
rанеллы. rлоточные плакоиды, вероятно..
служат для измельчения заrлатываемых
твердых частиц.
8.1.3. Классификация
400 современных видов (длина тела
O,05 1..2 мм) слабо различаются анатоми..
чески, если не считать орнаментации кути--
кулы, формы И числа KoroTKoB (рис. 8.5); все
виды относятся к одному классу.
217
8.2. Тип Pentastoma
(пятиустки)
8.2.1. 3тимолоrия
От rреч. pente пять; stoma poT.
8.2.2. Диаrностические
и специфические признаки
(рис. 8.6)
1. Билатерально симметричные, упло..
щенные, червеобразные животные.
2. Тело более чем из двух слоев клеток
в толщину" с тканями и орrанами.
3. Кишечник сквозной, прямой.
4. Тело мономерное, хотя внешне коль..
чатое; на переднем конце две пары «Hor»
с коrотками или просто две пары KoroTKoB
(рис. 8.7).
5. Псевдоцель в роли rидростатическо
ro скелета.
6. Тело покрыто хитинизированной ку"
тикулой, пронизанной порами и сбрасывае..
мой при линьке; под эпителием залеrают
слои кольцевой и продольной поперечнопо"
лосатой мускулатуры.
7. OpraHoB выделения, кровообращения
и rазообмена нет.
8. Нервная система с rоловным мозrом
,
rанrлиями. иннервируюшими каждый одну
«Hory» (иноrда все пять rанrлиев слиты вое..
дино), и вентральным нервным тяжем 1 .
9. Раздельнополые животные. одна или
две rонады.
10. Оплодотворение внутреннее путем
копуляции.
11. Развитие с тремя «личиночными CTd
диями»: у первой из них три пары лопаст..
ных, ноrоподобных, нечленистых придат--
ков.
12. Паразиты дыхательных путей и леr..
ких позвоночных; питаются кровью хозяев.
Как и у мноrих эндопаразитов, основ--
ную часть теЛd пятиусток занимает половая
система и образуется orpoMHoe количество
мелких яиц. Оплодотворение внутреннее;
оплодотворенные яйца (одновременно до
полумиллиона) до момента откладки
остаются в орrанизме самки. Матка" rде
1 Нервная цепочка лестничноrо типа с 4 5
парами rанrлиев. Прu.н. пер.
218
"Педальные железы"
Кишка
" Hora"
идет их накопление, фактически занимает
всю полость тела самки., увеличивающеrося
при этом более чем в 100 раз. Развитие
включает последовательно три «личиноч"
ные» или ювенильные стадии, первая из ко..
торых завершается еще до выхода из яйца.
Последнее выходит из тела хозяина через
ero пищеварительный тракт (все хозяева
наземные или пресноводные хищные позво..
ночные; 900/0 видов пятиусток паразиты
пресмыкаюшихся). Если личинка. еше Haxo
дящаяся внутри яйца, проrлатывается про
межуточным хозяином (всеядным или pa
стительноядным Насекомым., рыбой или
четвероноrим). она выходит из яйuа и aK
тивно движется через ткани позвоночноrо,
используя для этоrо три хитиновых стилета
1 мм
5 мм
1 см
Рис. 8.7. Разнообразная морфолоrия передне
ro конца тела пятиусток; видна степень разви
тия или редукции «Hor» (вид с вентральной CTO
роны ) (по Kaestner, 1968)
rлава 8
Анус
Рис. 8.6. Внутреннее строение
сам ки пятиустки на схематиче
ском продольном разрезе
(преимущественно по Cuenot,
1 949)
и короткие толстые конечности; достиrнув
специфической области тела (печени и Т. д.),
инцистируется и развивается во вторую ли
чиночную стадию. Если затем зараженный
промежуточный хозяин становится добы-
чей окончательноrо (рептилии, птицы или
млекопитающеrо ). появляется третья ли-
чинка, похожая на мелкий вариант взрослой
пятиустки, миrрирующая в леrкие или верх-
ние ДЫХd.тельные пути. У ряда видов во
время одной из последних личиночных ли..
нек происходит утрата конечностей, на ме..
сте которых остаются лишь терминальные
крючковидные коrотки. У личинок мноrих
видов на каждой Hore изначально пара ко-
[отков; на взрослой стадии их по одному
8.2.3. Классификация
Описано примерно 100 видов этих мел-
ких (2- 16 см длиной) «червей»; всех их мо-
жно отнести к одному классу.
8.3. Тип Onychophora
8.3.1. 3тимолоrия
От rреч. опусhоs коrотки; phoros
носитель.
8.3.2. Диаrностические
и специфические признаки
(рис. 8.8)
1. Билатерально симметричные живот-
ные с удлиненно цилиндрической., черве-
образной формой тела.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и орrанами.
3. Кишечник сквозной., прямой. Перед
ним Пd.ра ротовых придатков, каждый из
которых несет две коrтевидные мандибулы
(образуют внутреннюю и внешнюю че-
Беспозвоночные с конечностями: членистоноrие и др.
219
Клейкая
железа
rllаз
Ротовая
папилла
Выделительный
opraH
А Мозr
Рис. 8.8. Строение онихофор:
A cxeMa продольноrо разре..
за; Б внешний облик (по Se..
dawick, 1888; Cuenot, 1949)
Б
люстные пластинки). Передняя и задняя ки"
шка с кутикулярной выстилкой; выпячива..
пия (дивертикулы) пищеварительноrо трак..
та отсутствуют.
4. Тело с 14 43 парами коротких, не..
членистых, мясистых Hor по всей своей дли..
не. Каждая Hora полое выпячивание стен..
ки тела с терминальной подушечкой, парой
KoroTKOB и внутренней мускулатурой (хотя
движение ноrи вызывается сокращением
внешних, т. е. лежащих вне ее мышц); каж..
дой паре Hor соответствует пара остий cepд
ца и пара выделительных opraHoB.
5. В роли rидростатическоrо скелета хо..
рошо развитый rемоцель; сердце трубчатое,
друrих циркуляторных сосудов нет.
6. Тело покрыто очень тонкой, rибкой,
хитинизированной кутикулой. Под эпите..
лием расположены слои кольцевой, косой
и продольной rладкой мускулатуры.
7. OpraHbI выделения сериально пов..
торяющиеся пары мешковидных желез;
передняя пара образует слюнные железы,
задняя половые протоки.
8. OpraHbI rазообмена просто
устроенные трубчатые трахеи.. подходящие
пучками к мноrочисленным мелким дыха..
льцам.
9. Нервная система с rоловным моз"
rOM и парой очень широко расставленных
вентральных нервных тяжей, соединенных
9 10 КОМИССУРdМИ в каждом из несущих
конечности «cerMeHTOB», но без отчетливых
rанrлиев. Орrаны чувств пара кольчатых
антенн с небольшим простым rлазом в ос..
новании каждой.
\
Анус
10. Раздельнополые животные, rОНdДЫ
парные.. оплодотворение внутреннее с по..
мощью сперматофоров.
11. Развитие прямое.
12. Свободноживущие наземные фор..
мы.
Длительное время онихофоры вызывали
интерес исследователей rлавным образом
как живой пример промежуточной стадии
между уровнями орrанизации червей и чле..
нистоноrих. Как и у червей, у онихофор
мяrкое тело, rидростатический скелет, рес..
ничные выделительные каналы и слой rлад
кой мускулатурь] под эпителием. С члени..
стоноrими их сближает наличие конечно..
стей, трахей, сердца с остиями, продольно
разделенных кровеносных синусов и челю..
стей производных придатков, в данном
случае KoroTKOB, которыми оканчиваются
ходильные ноrи. Однако тонкая структура
их общих для артропод особенностей явно
указывает на независимую эволюцию они
хофор И всех известных rрупп членистоно..
rих. Если они и напоминают внешне, напри--
мер.. древних одноветвистых, то предками
их считаться никак не MorYT. Трахеи онихо..
фор, в частности, ЭТО просто устроенные
в основном неразветвленные трубочки, от..
крывающиеся пучками через мноrочислен..
ные (до 75) дыхальца (рис. 8.9), разбросан..
ные по поверхности каждоrо «cerMeHTa»
С конечностями. Челюсти онихофор, движу"
щиеся в переднезадней плоскости, дей..
ствуют независимо друr от друrа как раз
рывающие пищу приспособления (блаrо
даря своим заостренным концам), а не в ка..
220
Трахейные
ТРУБОЧКИ
Рис. 8.9. Дыхальце и пучок трахейных трубочек
(по Clarke, 1973)
честве жующих придатков. К друrим oco
бенностям относится строение вентральных
нервных тяжей с их мноrочисленными ко..
миссурами, но без «сеrментарных» [ан..
rлиев.
Онихофоры наземные животные,
связанные в основном с влажными микро"
местообитаниями. Толщина их проницае..
мой для воды кутикулы Bcero 1 мкм (MeHЬ
ше, чем у тонкой эпикутикулы членистоно..
rих, см. рис. 8.1); дыхальuа без замыкающе..
[о механизма. Однако потерянная влаrа мо"
жет быть восполнена блаrодаря выпячива..
нию через щели в кутикуле тонкостенных
пузырьков.. соприкасающихся с влажными
поверхностями. ОНИХОфОРЬJ в основном
ночные хищники, обнаруживающие добычу
с помощью антенн и охотящиеся даже на
достаточно подвижных животных типа ку"
знечиков, разбрызrивая на расстояние до
0,5 м слизеподобное вещество из желез, OT
крывающихся на парных ротовых сосочках
по бокам рта. Эта слизь почти MrHoBeHHo
застывает на воздухе, образуя очень клей
кую ловчую сеть, в которой запутывается
потенциальная добыча. Этот же прием
используется с целью защиты.
8.3.3. Классификация
Известно 70 видов онихофор длиной ДО
15 см, относящихся к одному классу и отря"
ду.
rлава 8
8.4. Тип Chelicerata
8.4.1. 3тимолоrия
От rреч. сhеlе клешня; cerata pora.
8.4.2. Диаrностические
и специфические признаки
(рис. 8.10)'
1. Билатерально симметричные артро-
поды длиной от менее 1 мм до 60 см; форма
тела от удлиненной до почти сферической.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и орrанами
3. Кишечник сквозной, прямой; от
среднеrо ero отдела отходят от двух до
множества пар слепых пищеварительных
выростов (дивертикулов), секретирующих
ферменты и внутриклеточно перевариваю-
щих и абсорбирующих пищу (они возни-
кают не как выпячивания эмбриональной
кишки.. а в результате разделения желтка за-
родыша до дифференцировки пищеварите-
льноrо тракта); р01 на переднем конце теЛd
с вентральной стороны.
4. Тело разделено на два отдела: пере-
дний (<<просома» ) образован акроном и ше-
стью сеrментами, несущими придатки, пол-
ностью или частично прикрыт карапаксом;
задний отдел (<<опистосома») без придатков
или же они сильно модифицированы.
5. Придатки одноветвистые; на просо-
ме это пара «хелицер» (клешневидных,
в форме ложных клешней или стилетов), па-
ра Horo.., щупальце.. или клешневидных «пе-
дипальп» и четыре пары ходильных Hor (все
крепятся вблизи вентральной срединной ли-
нии и у некоторых видов выпрямляются за
счет rидростатическоrо давления в rемоце-
ле); антенн и челюстей нет.
6. Ротовой аппарат образован только
одной парой придатков (хелицерами), хотя
в измельчении пищи и направлении ее в рот
MorYT также участвовать медиальные от-
ростки базальноrо членика друrих конечно-
стей (<<коксальные эндиты»).
7. Обычно (если не происходит вторич-
ной утраты ) на просоме находятся простые
1 Морские пауки сильно отличаются от дру-
rих хелицеровых, и к ним не относятся мноrие из
перечисленных ниже признаков.
Беспозвоночные с конечностями: членистоноrие и др
Клешня Срединные
педипальпы rлаза
.::J --
':..... i
".... ........ ........_.. \
" \
..... .......... :,.."...
rанrлиозная
масса
221
Пищеварительные
железы Мальпиrиев
сосуд
Сердце
rонада
Рис. 8.10. Схема продольноrо разреза тела обобщенноrо хелицеровоrо
срединные и боковые rлазки; в одной из
rрупп за счет скопления боковых rлазков
формируются сложные rлаза.
8. Опистосома иноrда BHellIHe разделе
па на cerMeHTbI (до 12), а у некоторых ви
ДOB Ha широкую переднюю «мезосому»
и узкую заднюю «MeTaCOMY» иноrда несет
постанальный шип, жало или жrутик.
9. Выделительная система из слепо
оканчивающихся коксальных желез в про
соме и/или разветвленных эндодермальных
мальпиrиевых сосудов опистосомы, OTXO
дящих от среднеrо отдела кишечника и BЫ
водящих rлавным образом rуанин.
10. Экзоскелет необызвествленный;
иноrда в просоме еще и пластинчатый мезо
деРМdЛЬНЫЙ эндоскелет.
11. Орrаны дыхания связаны с ПрИДdТ"
ками опистосомы или их эмбриональными
зачатками; у морских форм это наружные
жаберные «книжки», у наземных
внутренние леrочные «книжки» и производ"
пые от них ситовидные или трубчатые тра..
хеи.
12. В rазообмене участвует циркулятор
пая система; в rемолимфе обычно солержи
тся rемоцианин.
13. Нервная система с отдельными [ЙH
rлиями по всей длине теЛd или, чаще, слив
шимися в один просомальный узел.
14. Раздельнополые животные. В ис
ключительно морских классах оплодотво
рение наружное (хотя партнеры при СПdрИ"
вании объединяются друr с друrом, COBep
llIая псевдокопуляцию)., в rлавным образом
наземных rруппах внутреннее (копуляция
или с помощью сперматофоров); половые
поры расположены на втором cerMeHTe опи
стосомы.
15. Ювенильные особи внешне напоми
нают взрослых, обычно выходят из ЯЙЦd
С полным набором конечностей.
16. J:1сходно морские бентосные opra
низмы; один класс очень успешно заселил
сушу и пресные воды.
Хелицеровые отличаются от двух дpy
rих типов членистоноrих по ряду важней
llIИХ признаков, хотя эволюция мальпиrие
вых сосудов и трахей шла у них и у одновет..
вистых параллельно. Наиболее явная отли"
чительная черта их придатки. Ни у OДHO
[о из хелицеровых нет челюстей или друrих
конечностей, способных перемещаться друr
относительно друrа, пережевывая пищу.
Добыча схватывается и разрывается с по
мощью клешней или коrтей (на педипаль
пах и/или хелицерах).. но заrлатывается то..
ль ко в сильно размельченном или, как пра
вило, жидком виде. Чтобы в кишечник не
проникли крупные частицы, ротовое отвер"
стие обычно прикрыто щетинками. HeCMO
тря на это, хелицеровые почти исключи
тельно хищники. Процесс их питания co
стоит в удержании жертвы вблизи pOTOBO
[о отверстия.. впрыскивания в нее пищева
рительных ферментов и последующеrо вса..
сывания продуктов этоrо внешнеrо перева
ривания. Если ферменты не вводятся в тело
222
жертвы, то переваривание или механиче
ское дробление пищи осуществляется
в предротовой области.. оrороженной KOKca
льны ми эндитами нескольких или всех ко..
нечностей просомы, часто почти радиально
РdСХОДЯЩИХСЯ BOKpyr pOToBoro отверстия.
В связи с таким способом питания передняя
КИlllка преобразована в cBoero рода насос
или насосы, а средняя, особенно множество
ее дивертикулов (они MorYT 1анимать боль..
lllУЮ часть объема тела). в место оконча..
тельноrо переваривания и всасывания. Да..
лее, у хелицеровых нет антенн, хотя педипа..
льпы, первые или вторые пары ходильных
Hor MorYT модифицироваться для выполне..
ния соответствующей функции.
Среди хелицеровых прослеживаются
морфолоrические ряды, демонстрирующие
возможный путь эволюции трахейной си..
стемы из наружных жабр. У MepOCTOMO
вых морской и, почти несомненно, анце..
СТРdЛЬНОЙ линии хелицеровых орrаны
дыхания (Ka и у Болыllнстваa друrих MOp
ских животных) наружные жабры. Они
развиваются из заднеrо края клапановид..
ных придатков опистосомы; биение послед--
ных создает ток воды над жаберными ли..
сточками. У наземных паукообразных воз..
душное дыхание, но в противоположность
одноветвистым (см. разд. 8.5) типичные для
них орrаны rазообмена эмбриолоrически
связаны с задними краями зачатков описто..
сомальных конечностей (они не развиваю..
тся входильные ноrи, хотя паутинные боро..
давки пауков и rребневидные орrаны скор..
пионов представляют собой сильно видоиз"
мененные придатки опистосомы). Так, на..
пример, образуются леrочные книжки (рис.
8.11), rомолоrичные рядам жаберных ле..
пестков в карманах, поrруженных внутрь
тела. Каждая такая книжка это впячива..
ние со свисающими в Hero рядами паралле
льных лепестков. слипнутся которым не
дают особые распорки; воздух между ними
перемещается блаrодаря диффузии, а
внутрь них заходит синус с rемолимфой.
у некоторых видов лепестки леrочных кни
жек удлиненные и напоминают скорее тру..
бочки, чем пластинки; в зависимости от
числа таких трубочек они наЗЫВdЮТСЯ сито..
видными (их MHoro близко расположенных)
или трубчатыми (их мало) трахеями. Сле
rлава 8
А триум
Jg духаl
Дыхальце
Рис. 8.11. Объемная схема BHYTpeHHero CTpoe
ния леrочной книжки паукообразных, дeMOH
стрирующая циркуляцию крови и воздуха (по
Barnes 1980)
Лепесток
ДОВdтельно, эти трахейные системы в при..
нципе не что иное. как удлиненные. поrру-
женные внутрь тела и связанные с конечно..
стями жабры. У некоторых пауков допол-
нительно развиты вторичные трахеи из пу..
стотелых внутренних выростов 1кзоскелета
(<<аподем» см. рис. 8.1).
8.4.3. Классификация
Описано 63 ООО видов хелицеровых, от-
несенных к трем далеко отстоящим друr от
друrа классам, из которых один некоторые
считают неродственным двум друrим
Класс
Merostomata
Arachntda
ОТРЯД
Xiphosura
Scorpiones
Uropygt
Schizomida
AmbIypygt
Palpigradi
Araneae
RiClnulel
Pseudoscorpiones
Solpugida
Opiliones
Norostigmata
Parasiriformes
Acariformes
Pycnogonida
Беспозвоночные с конечностями: членистоноrие и др
223
8.4.3.1. Класс Merostomata
Хотя до пермскоrо периода это была до..
минирующая rруппа беспозвоночных, в на..
стоящее время меростомовые представле..
ны только четырьмя видами отряда Xipho..
sura (мечехвосты). Это крупные морские хе..
лицеровые с толстым карапаксом, напоми..
наЮIЦИМ очертаниями лошадиное копыто,
покрываюшим крупную про сому и нави..
сающим над ней спереди (ротовое отвер"
стие оказывается в центре вентральной сто..
роны) и С боков, прикрывая сверху все ко..
нечности. Небольшая подвижно причлен..
ная опистосома представляет собой пло..
скую пластину, частично входящую в вырез
карапакса и окаймленную с боков мощны..
ми шипами; терминально она несет длин..
ный постанальный хвостовой шип (рис.
8.12). Хелицеры, педипальпы и все ходиль..
ные ноrи, кроме последней пары, вооруже..
ны клешнями; последняя пара, несущая ряд
терминальных шипов или пластин, исполь..
зуется для рытья в [рунте (рис. 8.13).
В отличие от прочих современных хели..
церовых у мечехвостов почти полный Нdбор
Рис. 8.13. Полусхематический
вид мечехвоста с вентральной
стороны; показаны различные
придатки (по Savory, 1935)
Опистосома
Хвостовой шип
Рис. 8.12. Вид мечехвоста с дорсальной сторо"
ны (по Kaestner, 1968)
опистосомальных придатков: передняя их
пара неБОЛЫlIая и трубчатая, остальные
шесть пар плоские пластины" при помо..
щи которых животное плаВdет" перевернув..
шись вверх ноrами. Первая из этих пар пла..
стин служит также половой крышечкой,
Карапакс
Рот Хелицера
rнатоБЭза
Шипы на
ходильной
Hore пас
ледней
пары
Опистосома
Придатки
опистосомы
Хвостовой шип
224
rлава 8
а последние пять несут наружные жабры.
Друrие особенности, присущие морским
формам, но нетипичные для хелицеровых
в целом, отсутствие мальпиrиевых cocy
дов и внешнее оплодотворение. Среди всех
современных хелицеровых только у мече
хвостов имеются крупные, диффузные сло
жные rлаза достаточно своеобразноrо типа
по бокам карапакса. В отличие от пауко
образных rонады и пищеварительные ди--
вертикулы расположены в просоме.
Мечехвосты ночные, бентосные жи--
вотные прибрежноrо мелководья; они рою--
тся В рыхлых осадках и ползают по дну, пи--
таясь крупными моллюсками и полихета..
ми, раскусывая их коксальными эндитами
(<<rнатобазами») и размельчая в специали
зированном желудке 1.
8.4.3.2. Класс Arachnida
(паукообразные)
Почти целиком наземные формы, со..
ставляющие 980/0 современных видов хели..
церовых, с хорошо выраженными приспо..
соблениями к существованию на суше (на..
пример, выделительная система с мальпи..
rиевыми сосудами; орrаны дыхания, поrру..
женные внутрь тела; кутикула, непроницае..
мая для воды блаrодаря слою воска: вну"
треннее оплодотворение). ВнеllIНИЙ облик
паукообразных весьма разнообразен: от уд..
линенных, хорошо вооруженных артропод
с крупными хватательными педипальпами.
отчетливой наружной сеrментацией и опи..
стосомой, расчлененной на мезо.. и метасо..
му (скорпионы, телифоны и т. д.), ДО почти
сферических форм с тонким экзоскелетом.
незаметной наружной сеrментацией и менее
развитыми педипальпами (большинство
пауков, клещей и т. д.). Это разнообразие
находит отражение в делении класса на 13
отрядов (рис. 8.14). Скорпионовидная фор
ма тела, почти HeCOMHeHHO, примитивное
состояние, поскольку она слабо отличается
от облика некоторых (вымерших) paKOCKOp
пионов из меростомовых, которые вели
пресноводный, а возможно, и земноводный
1 Жевательный желудок лежит в передней ча
сти просомы И имеет кутикулярную выстилку
С rрубыми зубцами. При.'н. пер.
образ жизни. Некоторые современные ПdУ-
кообразные вновь вернулись в пресную во-
ду, а ряд клещей в море.
В отличие от мечехвостов у паукообра-
зных нет сложных rлаз и локомоторных
придатков на опистосоме; у них длинные
ходильные ноrи без клешней 1, а половые
железы и пищеварительные дивертикулы
расположены в опистосоме. В нескольких
rруппах паукообразных общая тенденция
к облеrчению экзоскелета привела к отделе-
нию двух последних cerMeHToB просомы от
карапакса (хотя на них и MorYT присутство-
вать самостоятельные дорсальные пластин-
ки) и к развитию мяrкой, rибкой опистосо-
мы. У большинства клещей эти два послед-
ных просомальных cerMeHTa слиты с опи-
стосомой, И тело состоит из двух отделов,
отличных от наблюдающихся у прочих хе-
лицеровых. Однако в общем случае сохра-
няется типичное деление на просому и опи-
стосому, разrраниченные внутренней диа-
фраrмой или узким стебельком. Такое раз-
деление позволяет повышать давление
в просоме (для выпрямления конечностей),
не изменяя ero вопистосоме.
Паукообразные один из наиболее
преуспевающих классов животных. Их
образ жизни связан rлавным образом с охо-
той на представителей друrой не менее
преуспевающей rруппы насекомых. К за-
мечательным приспособлениям для ловли
этих очень подвижных орrанизмов относя-
тся ядовитые железы, секрет которых мо-
жет впрыскиваться в жертву. а также обра-
зование паутинной нити - явление, дости-
rающее своей кульминации в постройке
ловчей сети пауками. Размеры паукообра-
зных колеблются от менее 0.1 мм до 18 см;
в классе свыше 62000 видов. Известны кле-
щи, питающиеся растительными тканями
и детритом; некоторые клещи паразиты,
8.4.3.3. Класс Pycnogona
(морские пауки)
Этих мелких бентосных морских беспо-
звоночных с длиной тела менее 6 см ряд
зоолоrов считает неродственными хелице-
ровым. а возможно. и всем прочим члени-
стоноrим. Первая пара конечностей этих
1 На просоме. При.и. пер.
Бее.l0ЗlВоночныв с конечностями: членистоноrие и др
225
Scorpiones
( скорпионы)
Palpigradi
Uropygi (телисроны., или
скорпионопауки)
SoLpugida
(сольпуrи,или сраланrи)
Pseudoscorpiones
(ложноскорпионы )
Schizomida
Amblypygt
(жrутоноrие)
Opiliones
( сенокосцы)
Ricinulei
Acariformes
Parasitiformes
Araneae (пауки)
Notostigmata
Рис. 8.14. Форма тела представителей различных отрядов паукообразных (по Savory, 1935; Huges,
1959 и др.)
226
rлава 8
Рис. 8.15. Форма тела морских пауков (преимущественно по Hedgpeth 1982)
животных, хелифоры, оканчивается. как и
у хелицеровых, клешнями; вторая, пальпы,
возможно, rомолоrична педипальпам; у бо..
льшинства видов присутствуют еще четыре
пары ходильных Hor. Однако у некоторых
видов ходильных Hor пять или шесть пар,
а между пальпами и первой парой ходиль..
ных Hor располаrаются дополнительные
придатки яйпеносы. В отличие от прочих..
латеральных, придатков (ходильные ноrи
причленены к мощным боковым выростам
тела) они крепятся вентрально (рис. 8.15).
У ряда видов хелифоры. пальпы и яйпеносы
рудиментарные или отсутствуют, но ходи..
льные ноrи всеrда хорошо развиты и бы..
вают очень длинными (до 75 см).
Цефалоторакс, или rоловоrрудь (просо..
ма?), не покрытая карапаксом, несет цен..
тральный выступ с двумя парами простых
rлаз и (спереди или вентрально) крупный
трубчатый невтяrивающийся хоботок с тер..
минальным ротовым отверстием. Абдомен,
или брюшко (ОПИСТОСОМd ?), крошечный
несеrментированный сосочек. лишенный
придатков. OpraHoB выделения и дыхания
нет. Необычно то, что rонады, как и пище-
Вdрительные дивертикулы, заходят внутрь
ходильных Hor. rде и созревают яйuа. В хо-
де псевдокопуляции [аметы выводятся из
множества половых пор (rонопоров), ра-
сположенных у основания каждой ходиль-
Рис. 8.16. Протонимфон морских пауков (по
Kaestner 1 968)
Беспозвоночные с конечностями: членистоноrие и др.
Рис. 8.17. Схема продольноrо
разреза тела и хоботка MOpCKO
ro паука (по King, 1973)
ной ноrи или их последних двух пар. После
оплодотворения самец собирает яйца и но..
сит в виде шаров., прикрепленных к яйцено..
сам. В итоrе вылупляется своеобразная ли..
чинка с тремя парами придатков
протонимфон (рис. 8.16), чаще Bcero веду..
щая полупаразитический образ жизни на
поверхности или внутри стрекающих или
моллюсков.
Взрослые морские пауки питаются в ос..
новном rубками., книдариями, мшанками.
Часть тела добычи захватывается хелифо..
рамп (если они имеются), а заrлатывание
тканей жертвы осуществляется за счет МОЩ"
Horo насасывающеrо действия rлотки. Пое..
данию пищи способствует rрызущее дей..
ствие трех зубов, расположенных на конце
хоботка В rлотке пища измельчается др у..
rими зубами или мошными щетинками;
у основания rлотки воротничок из щетинок
обеспечивает попадание в пищевод только
мельчайших пищевых частиц (рис. 8.17).
Описано более 100 видов, относимых
к одному отряду.
8.5. Тип Uniramia
(одноветвистые)
8.5.1. Этимолоrия
От лат. unus один и rarnus -ветвь.
8.5.2. Диаrностические
и специфические признаки
(рис. 8.18)
1 Двусторонне симметричные члени..
стоноrие с разнообразной формой тела (от
227
rлазной
Буrорок
I
Сердце
10 нада
АБдомен
Анус
Срезы диверти
кулов rонад и
кишечника в
конечностях
Брюшной нервный
ствол с rанrлиями
I
Рот
очень вытянутой ДО почти шаровидной)
длиной от менее 1 м до 35 см.
2. Тело более чем из двух слоев клеток
в толщину, с тканями и орrанами.
3. Кишечник сквозной, прямой, без пи..
щеварительных дивертикулов.
4. Тело разделено на два отпела
«rолову», образованную акроном и тремя
или четырьмя сеrментами с придатками,
и «туловище» с парами ходильных Hor.
В одном из подтипов в состав туловиша
входит до 350 относительно однородных се..
[ментов, подавляющее большинство из КО..
торых сходильными ноrами; в друrом
подтипе оно дифференцировано на «rрудь»
(торакс) с тремя парами Hor и «брюшко»
(абдомен), включающее до одиннадцати се..
[ментов с крайне видоизмененными при..
датками, если таковые вообще имеются.
5. Придатки одноветвистые; на rолове
находится по паре «антенн» (усиков)., «маи..
дибул» (жвал) и «максилл» (челюстей)., а
у ряда rрупп и вторая пара максилл: ту..
ловище несет только функциональные или
модифицированные ходильные ноrи; хели..
цер или клешненосных конечностей нет.
6. Ротовых придатков (мандибулы
и максиллы) две или три пары, причем эле..
менты каждой пары работают в фазе или
в противофазе с образующими друrую па..
ру: базальные членики максилл или вторая
пара максилл (в тех rруппах, [де их две па..
ры) срастаются в пластинку (нижнюю rубу,
или лабиум), оrраничивающую предрото..
вую полость снизу.
7. По бокам rоловы находятся rлазки.,
часто образующие сложные rлаза; иноrда
простые rлазки присутствуют и на средней
линии rоловы
228
мноrОНОЖКА
Антенна
Максиллярная
железа Кишечник
Мозr
rлава 8
r онада
Сердце
Ротовые Слюнная Дыхальца Мальпиrие... Нервный
пр атки железа вы сосуды тяж
rонопор
НАСЕКОМОЕ
Антенна
Сложный
rлаз
Церка
.......:. ":'..-:. ":" AHYC
Пенис
Класпер
Мальпиrиевы
сосуды
Дыхальца
Рис. 8.18. Схематические продольные разрезы rенерализованных представителей двух подтипов
одноветвистых
8. Туловище наружно сеrментировано,
rолова нет.
9. У большинства представителей oд
Horo из подтипов на rруди одна или две па
ры крыльев.
10. В rемоцеле находится жировое тело,
часто тесно связанное с кишечником.
11. Выделительная система представле
на 0 2 парами максиллярных желез в rоло
ве и 1 75 парами неветвящихся эктодерма
льных мальпиrиевых сосудов" отходящих
от задней кишки вблизи места ее соедине
ния со средней и выводящих rлавным обра
зом аммиак и/или мочевую кислоту.
12. Наружный скелет известковый или"
чаще, неизвестковый.
13. Орrаны rазообмена парные в виде
ветвистых трубчатых трахей, через которые
диффундирует воздух; исходно они OTKpЫ
ваются наружу парой дыхалец на каждом
cerMeHTe сходильными ноrами, а оканчи
ваются в теле трахеолами; эмбриолоrиче-
ски трахейная система не связана с придат-
ками.
14. rемолимфа не переносит дыхатель-
ных rазов, и дыхательные пиrменты в ней
отсутствуют (исключение некоторые ли-
чиночные стадии).
15. Раздельнополые формы; оплодотво-
рение внутреннее с помощью сперматофо-
ров или путем копуляции; исходно rонопо-
ры находятся на последнем туловищном се-
[менте, в ряде случаев это состояние пре-
терпело модификации.
16. В ряде rрупп выделяется четкая ли-
чиночная стадия" сильно отличающаяся от
взрослых форм; иноrда особь выходит из
яйца с неполным набором cerMeHToB (и Hor)
и они добавляются при линьках по одному,
причем в некоторых случаях и после дости-
жения половой зрелости.
17. В основном наземные формы, неко.
Беспозвоночные с конечностями: членистоноrие и др.
229
торые преСНОБодные, очень мало мор..
ских.
Возникшие, по"видимому, на суше одно..
ветвистые наряду с паукообразными, кото..
рых они превосходят и по видовому разно
образию, и по общей численности, сейчас
доминируюшая rруппа наземных беспозво..
ночных. Причины подобноrо успеха на су..
те отчасти те же, что и у паукообразных.
В обеих rруппах независимо появились си..
стемы rазообмена, основанные на диффу..
зии rазов из окружающей среды в ткани
и обратно, что rораздо эффективнее предот..
вращает потери влаrи, чем, к примеру, рес..
пираторная система четвероноrих позво
ночных, выдыхающих большое количество
водяных паров во время принудительной
вентиляции леrких. Характерная для арах..
нид и одноветвистых выделительная систе..
ма с мальпиrиевыми сосудами блаrодаря ее
связи с кишечником также, по крайней мере
потенциально, способствует экономии водь]
за счет реабсорбции ее из продуктов азотно..
ro обмена после их выведения в заднюю ки"
ШКУ. Наружный кутикулярный скелет в бо..
льшей степени препятствует потере влаrи,
нежели мяrкие влажные покровы кольчецов
и моллюсков. И все же в большинстве клас..
сов одноветвистых эти адаптации решают
проблему потери воды лишь частично, и их
представители связаны в основном с вла..
жными местообитаниями в почве или
А
Б
Мышца для ОТ
ведения назад
поблизости от нее. В частности, их дыхаль..
ца не способны закрываться, некоторое ко..
личество воды уходит с продуктами азотно..
ro обмена (представленных rлавным обра..
зом аммиаком), а кутикула относительно
проницаема. Лишь в одном классе потери
влаrи удалось сократить еще сильнее и
HaMHoro за счет появления механизмов
закрывания дыхалец, водонепроницаемой
-воскоподобной эпикутикулы и системы из в..
лечения воды из экскрементов в прямой ки"
шке. Именно этот класс, точнее крылатые
насекомые, стал самой крупной и разно..
образной rруппой одноветвистых.
Хотя перечисленные выше механизмы
экономии воды позволяют понять, почему
одноветвистые достиrли по крайней мере не
меньшеrо процветания, чем паукообразные,
все же для объяснения rораздо больших
масштабов их успеха этоrо мало. Не исклю"
чено, что значительную долю ero можно от..
нести на счет появления челюстей, способ..
ных кусать и жевать. Унирамии MorYT за..
хватывать в пищеварительный тракт твер..
дую пищу и, следовательно, не оrраничены
в своем рационе жидким кормом, а значит,
и добычей, приrодной для наружноrо пище..
варения. На деле это означает, что им до..
ступны растительные ткани, а суша как раз
и характеризуется прежде Bcero изобилием
этоrо относительно rрубоrо материала, ко..
торый приходится откусывать и разжевы"
в
.
#:;:.: . (. \
,...... I ......,..
":;' '?::: i: f.f)
. .
Верхняя ryБа
(лаБРУМ)
Мышца для BЫ
движения
вперед
Передний
аддуктор
МаНДИ6ула
МаНДИБула
Сросшиеся
максиллы
(лаБИУМ)
Верхняя ry6a
(лаБРУМ)
МаНДИБула
Максилла 1
-. .'<'. '..
Максилла 11
(лаБИУМ)
Рис. 8.19. РОТОВОЙ аппарат одноветвистых. А. Схема поперечноrо разреза через rолову, показы..
вающая примитивное состояние, при котором мандибулы расположены вертикально направлены
вниз и приводятся в движение примерно теми же мышцами, что и ходильные ноrи. Б. Вид спереди
и снизу на предротовую полость двупарноноrой мноrоножки: видна верхняя rуба (лабрум), обра..
зованная склеритами rоловной капсулы, и нижняя rуба (лабиум), образованная сросшимися мак..
силлами (заметно rоризонтальное положение мандибул). В. Вид сбоку rоловы HaceKOMoro: вид..
ны три пары ротовых придатков и причленение мандибулы к rоловной капсуле в двух точках (1
и 2) (по Kaestner, 1968)
230
вать, прежде чем проrлотить. В отличие от
почти исключительно плотоядных пауко"
образных здесь лишь ОДНd крупная rруп..
па rубоноrие полностью состоит из
хищников, хотя большинство представите..
лей типа потребляет все же мертвое иразла..
rающееся растительное вещество, а не
собственно живые растения (причины этоrо
рассмотрены в rл. 9).
Первоначально мандибула одноветви..
стых была ходильной конечностью первой
пары, у которой развивалась лишь базаль..
ная часть. Ее режущий край широкий, Ha
правленный внутрь ВЫРОСТ, неподвижно со..
единенный с остальной частью жвалы у всех
форм" кроме двупарноноrих и симфил. Ис..
ходно мандибула причленяется к телу так
же, как и прочие ходильные ноrи" т. е.
ориентирована перпендикулярно rоловной
капсуле (рис. 8.19.А). Однако у большин..
ства одноветвистых ее положение измени..
лось таким образом, что она располаrается
параллельно вентральной поверхности ro..
ловы, т. е. развернута вперед. Поэтому ее
первоначально дорсальное соединение с Te
лом стало задним, и у некоторых форм спе..
реди возникла еще одна точка причленения
(см. рис. 8.19.В). В итоrе движения жвал
оrраничиваются в основном раздвиrанием
и сдвиrанием Нижняя стенка или задний
край предротовоrо пространства, в кото..
ром работают ротовые придаТКИ, ре..
зультат слияния базальных частей максилл
или второй пары максилл (в rруппах" rде та..
ковые имеются), также раЗВИВllIИХСЯ из xo
дильных Hor (рис. 8.19.Б). Это совсем не по..
хоже на то, что мы видим в двух остальных
типах членистоноrих, нижняя rуба которых
представляет собой часть вентральноrо
экзоскелетноrо стернита. У :rvlноrоножек
строение ротовых частей в целом довольно
однообразно, но среди крылатых HaceKO
мых морфолоrия мандибул и максилл край..
не варьирует.. коррелируя с адаптивной ра..
диацией этих форм в широчайшем диапазо
не пищевых ниш {см. разд. 8.5.3Б.2).
8.5.3. Классификация
Представителей одноветвистых, кото..
рых известно свыше миллиона видов, мо"
rлава 8
жно разделить Hd два подтипа, один из них
с четырьмя классами, а друrой с шестью.
Однако общий план строения здесь относи
тельно постоянен, так что различия между
большинством классов выражены rораздо
слабее, чем во мноrих друrих крупных ти
пах, например среди моллюсков, хелиuеро..
вых и ракообразных.
8.5.3А. ПОДТИП Myriapoda
(мноrоножки)
Классы мноrоножек" вероятно" OTpa
жают первую крупную адаптивную радиа
цию внутри данноrо типа (или среди ero co
временных представителей); один из этих
классов дал затем второе важнейшее ОТ-
ветвление, положившее начало классам вто-
poro подтипа.
rлавная отличительная особенность
мноrоножек в том. что их туловише образо
вано рядом более или менее одинаковых ce
[ментов, каждый из которых, не считая oд
ноrо двух последних и иноrда первоrо, не-
сет пару ходильных Hor; [рудь и брюшко не
выражены. Можно назвать и друrие общие
признаки подтипа, хотя они присущи не TO
лько ему, но в некоторых случаях и первич-
нобескрылым насекомым (см. разд.
8.5.ЗБ.l). Так, имеется лишь одна пара Ma
льпиrиевых сосудов (как правило, вытяну-
тых); на rолове нет срединных rлазков, но
присутствуют «opraHbI Темешвари», ВОС-
принимающие, по"видимому, переносимые
воздухом химические соединения, а возмо
жно, и влажность; отдельные членики ан-
тенн обслуживаются собственной мускула-
турой; нервная система не сконцентрирова..
на, и rанrлии расположены во всех туло-
вищных cerMeHTax. Кроме Toro, как уже [о-
ворилось выше, механизмы экономии ВОДЫ
развиты относительно слабо. Классы раз-
личаются между собой в основном числом
пар максилл, расположением rонопоров,
числом терrитов и тем, какие сеrменты ли-
шены Hor.
Хотя в целом это суrубо наземная rруп-
па, в каждом классе по нескольку видов на-
селяет морскую литораль.
Беспозвоночные с конечностями; членистоноrие и др
Класс
ОТРЯД
Scutlgerlda
C h . 1 d LithObiid3
, I оро а S I
со opendrida
(;eophilida
Scolopendrellida
Polyxenida
C;lomeridesmida
Oniscomorph3
Polyzdnllda
temmlUllda
plrobolida
Iuliformida
т yphlogend
Chordeumatid3
Polydesmlda
Pauropodida
8.5.ЗА.1. Класс Chilopoda
(rубоноrие)
rубоноrие. . сильно или очень сильно
вытянутые, дорсовеНТРdЛЬНО уплощенные
мноrоножки с двумя парами максилл на [о..
лове и туловищем из 15. 181 и более се..
[ментов (их число всеrда непарное) с Hora
ми и двух хвостовых безноrих cerMeHTOB.
преrеНИТdльноrо и rенитальноrо (рис. 8.2О).
Все ноrи имеют сходное строение, хотя
иноrда длина их возрастает спереди назад;
только первая пара (это диаrностический
признак класса) видоизменена в хвататель..
ные opraHbI крупные ко тевидные крючья
с ядовитыми железами внутри (рис. 8.21).
Хотя у большинства видов количество
боковых rлазков достаточно велико, а в од..
ной rруппе (отряд Scutigerida един..
ственный случай среди мноrоножек) имею..
тся даже крупные сложные rлаза, эти хищ..
ные артроподы ведут ночной образ жизни
или обитают под поверхностью почвы, об..
наруживая добычу (олиrохет и членисто но..
fИХ) с помощью dHTeHH или, реже, Hor. Не..
которые rруппы rлаз лишены.
Как и у большинства мноrоножек, пере..
нос спермы происходит с помощью нару..
жноrо сперматофора, выдавливаемоrо из
XBoCToBoro rонопора лишь после сложноrо
ухаживания. Во всех rруппах, кроме одной,
у самца из Toro же отверстия выдвиrается
прядильный аппарат, выделяющий неско..
231
Рис. 8.20. Форма тела rубоноrих (по Lewis,
1981 )
лько паутинных нитей для подвешивания
сперматофора и направления к нему самки.
Некоторые виды охраняют яйца и даже мо"
лодь после вылупления.
Необычно для представителей этоrо
подтипа то, что у мноrих видов потомство
выходит из яиц с полным набором cerMeH..
тов и Hor (в том числе, как ни странно,
у форм с максимальным их числом), однако
в более типичном случае у свежевылупив..
шихся особей cerMeHToB меньше, чем
у взрослых, иноrда Bcero четыре.
Известно 3000 видов с длиной тела
до 27 см. Их разделяют на четыре отряда.
232
Видоизмененная
первая пара
туловищных HOr
Антенна
Максилла 1
Максилла II
Рис. 8.21. Сильно видоизмененная первая пара
туловищных конечностей rубоноrоrо (по Borror
et al., 1976)
8.5.ЗА.2. Класс Symphyla
Симфилы мелкие (менее 8 мм длиной)
мноrоножки; общий план их строения такой
же, как у rубоноrих, т. е. имеется rолова
с двумя парами максилл и туловище из две..
надuати (в данном случае) cerMeHToB с Hora..
ми и двух терминальных безноrих, послед..
ний из которых слит с тельсоном (рис. 8.22).
Прядильные opraHbI также находятся на по..
следнем cerMeHTe тела. Однако в отличие от
rубоноrих у симфил терrитов больше, чем
cerMeHToB (до 24), что делает тело более
rибким. rонопоры вторично открываются
в передней части тела (на третьем cerMeHTe),
а первая пара Hor не образует крючьев, хотя
Прядильный
аппарат
Рис. 8.22. Внешний вид симфил (по Kaestner,
1968). Обратите внимание, что терrитов боль
ше, чем пар Hor
rлава 8
эти конечности заметно мельче последую..
щих иноr да вдвое. Еще одна необычная
для мноrоножек черта единственная пара
коротких трахей, открывающихся дыхаль..
цами на rолове
Однако во мноrих друrих отношениях
этот класс напоминает представителей под..
типа Hexapoda, особенно первичнобескры
лых насекомых. В принципе сходны, напри
мер, их рртовые придатки, а рядом с Hora
ми на большинстве cerMeHToB развиты мел
кие rрифельки и коксальные мешочки, KOTO
рые способны выворачиваться наружу за
счет давления жидкости в rемоцеле и испо
льзуются для поrлошения влаrи из OKPY
жающей среды. Поскольку, как и большин
с [во мноrоножек, симфилы выходят из яиц
с неполным составом cerMeHToB, не исклю
чено. что Hexapoda произошли от симфило
подобных предков в результате педоморфо
за (некоторые двупарноноrие и пауроподы
также вылупляются лишь с тремя парами
Hor ).
Среди 160 известных видов этих мелких,
слепых, мяrкотелых и бледно окрашенных
мноrоножек большинство растительнояд
ные, питающиеся в основном мелкими жи..
выми корешками и обитающие в почве, ли
стовой подстилке, rниющей древесине, ПОД
камнями и т. п. Все виды относят к одному
отряду.
8.5.ЗА.З. Класс Diplopoda
(двупарноноrие)
Тело от KopoTKoro до очень сильно BЫ
тянутоrо (рис. 8.23). Наиболее характерный
и бросающийся в rлаза признак наличие
«сдвоенных» cerMeHTOB. Туловище двупар
ноноrих состоит из безноrоrо первоrо ce
[мента, следующих за ним трех cerMeHToB
с типичной для мноrоножек единственной
парой Hor на каждом, а затем ряда «KO
лец» (от пяти до 85 и более) с двумя парами
Hor, rанrлиев, остий сердца и т. д. В каждом,
после чеrо идет один или несколько безно
rих cerMeHToB, включая хвостовую зону их
прироста. Каждое сдвоенное cerMeHTHoe KO
льцо, из которых в основном и образовано
почти цилиндрическое или, реже, уплощен
ное туловище, возникает вследствие частич
Horo или полноrо попарноrо слияния туло
Беспозвоночные с конечностями: членистоноrие и др.
4
\ "!
. ...
"" _r: . .
, P'f ,
....,....,-.'1 "&'"
С':;:', ..-
, \. :' 41 /
:::.::::
/' 'O :
.
;, . :.! '
I ... ,
1"'....)..,.
.r::: :
. . , . 1.......
./ .(-......" :
Ittt_ r
ь. "';;;...#1 ....
, ..... ."
. r:';'", f I
r,: ; ., /
,,;:. {-": 1 z\ ..
'/ "'f :
J 1." '"
: _ , , . '> 11, . .
,."I, '.'.;1. . \' "... ,,,,,; .. ,J
, , ,,' ".' - " О 1\'",.'
. :1 ;. ." , "JI, :J, j"j,\ . 8.,' .
." . . 110 '" ,..". · ,,"".''''
'" , ,...... ,..
Рис. 8.23. Разнообразие двупарноноrих (по
разным авторам, rлавным образом по Blower,
1985 )
ВИЩНЫХ cerMeHTOB; у двух cerMeHToB caMO
меньшее может быть общий терrит. У сам..
цов мноrих видов одна две пары Hor (ко..
нечности VII сдвоенноrо cerMeHTa. или зад"
ние конечности VII и передние VIII) в раз..
личной степени видоизменены и образуют
копулятивные «rоноподии» (половые нож..
ки), забирающие сперму из rонопоров на 111
cerMeHTe и переносящие ее в соответствую..
щие отверстия самки. Друrие виды для
переноса спермы используют мандибулы;
иноrда самки и самцы тесно сближают свои
rонопоры., или же (классический для MHoro..
ножек способ) самец выделяет пакеты спер..
мы и паутинные направляющие нити. У бо..
льшинства диплопод в момент вылупления
2ЗЗ
Bcero семь туловищных «cerMeHTOB», а но..
вые добавляются всю жизнь даже спустя
долrое время после наступления половой
зрелости.
К друrим особенностям двупарноноrих
относится лишь одна пара максилл, боль..
шое число боковых простых rлазков., со..
бранных в rруппы., которые на первый
взrляд можно принять за сложные rлаза
(впрочем у ряда видов rлазки вовсе отсут"
ствуют)., а также обызвествленный нару..
жный скелет (исключая представителей од..
Horo отряда). Из всех одноветвистых по..
следний признак встреЧdется лишь у дипло..
под.
Класс выключает 1 О 000 видов и делится
на 10 отрядов. Это медленно передвиrаю..
щиеся потребители Рdстительных тканей
(обычно уже начавших разлаrаться) в тех же
местообитаниях, что и симфилы, т. е. rнию..
щих стволах, листовой подстилке и т. п. Са..
мые крупные представители достиrают
в длину 28 см.
8.5.ЗА.4. Класс Pauropoda
Эти крошечные существа (длиной менее
2 мм) обитают там же, rде симфилы иди..
плоподы; они надкусывают rифы rрибов
и высасывают их содержимое с помощью
передней кишки, работающей наподобие
насоса. Их трудно заметить, хотя числен..
ность их часто весьма высока В смысле
плана строения они примерно так же отно"
сятся к двупарноноrим., как симфилы к
rубоноrим. Так, у пауропод одна пара мак..
силл., первый и два последних cerMeHTa ту..
ловища лишены Hor, rонопор находится
спереди (на третьем cerMeHTe), у туловищ..
ных cerMeHToB наблюдаются начальные
этапы попарноrо слияния: общий терrит
полностью или отчасти покрывает cerMeH"
ты 1 и 11, 111 и IV, V и УI и т. д. (рис. 8.24).
CerMeHToB Bcero 11 12, последний сливае..
тся с тельсоном. У вылупившихся из яйца
особей cerMeHToB мало и в большинстве
случаев три пары Hor.
Однако пауропод нельзя считать просто
примитивными диплоподами., поскольку
у них есть и отличительные признаки. На..
пример, отсутствует сердце, а у большин..
ства и трахеи (если трахеи все же имеются,
234
ВИД СВЕРХУ
Ветвистая
антенна
ВИД СБОКУ
OpraH
Темешвари
Терrиты
Рис. 8.24. Внешний вид пауропод (по Kaestner.
1968 и Borror et al, 1976). Обратите внимание,
что терrитов меньше, чем пар Hor
они очень слабо развиты). Кроме Toro ан--
тенны ветвистые.
Все 500 видов класса слепые, мяrкотелые
и бесцветные. Их относят к единственному
отряду.
8.5.36. Подтип Hexapoda
(насекомые) 1
От мноrоножек представителей дан--
Horo подтипа отличает лишь один признак:
туловище разделено на rрудь из трех несу..
щих ноrи cerMeHToB и брюшко из одиннад..
1 В мировой И отечественной литера туре «Hace
комыми» в настоящее время чаl1 е Bcero назы
вают rруппу включающую отряды Thysanura
и Microcoryphia (объединяемые под названием
Apterygota. или первичнобескрылые насекомые),
а также всех Pterygota. В таком объеме этот TaK
сон принято именовать классом Insecta (иноrда
используют и название Hexapoda). Отряды Diplu..
ra, СоllетЬоlа и Protura либо противопоставляют
насекомым, объединяя их в класс Entognatha ли..
бо (как и здесь) считают самостоятельными клас..
сами Прu./и. пер.
rлава 8
цати cerMeHTOB без ходильных Hor, но иноr..
да с придатками различной формы, причем
абдоминальные сеrменты MorYT исчезать
или сливаться, так что их число бывает
меньше одиннадцати. Кроме Toro, у всех re..
ксапод две пары максилл., а rонопоры зани"
мают терминальное или субтерминальное
положение; в большинстве случаев имеются
срединные простые rлазки, а также простые
rлазки или сложные rлаза по бокам rоловы.
8.5.35.1. Классы первичнобескрылых
насекомых (Apterygota)
Выделяют пять rрупп первичнобескры"
лых насекомых. По всей видимости, каждая
из них отделилась от прочих на очень ран..
них стадиях эволюции насекомых (если они
вообще не произошли независимо от каких..
то предковых форм мноrоножек), поэтому
вполне можно rоворить о самостоятельных
классах. Лишь у одноrо из них ряд черт ука..
зывает на явное родство с крылатыми насе..
комыми, шестым классом rексапод. Все
классы первичнобескрылых очень невелики
по объему и включают Bcero по одному от..
ряду, поэтому удобнее рассматривать их
вместе.
Эти пять классов можно разбить на три
rруппы, первая из которых (двухвост..
ки == Diplurata; ноrохвостки == Oligoentorna--
td и бессяжковые == Myrientornata) характе..
ризуется частично поrруженными внутрь
rоловной капсулы ротовыми частями и еще
некоторыми особенностями, общими ско"
рее с мноrоножками, чем с прочими насеко..
мыми. По образу жизни, в том числе по
брачному поведению и местообитаниям,
эти формы также очень напоминают MHoro..
ножек, и неудивительно, что мноrие энто"
молоrи выступают 'Ja включение двухво..
сток, HoroxBocTOK и бессяжковых в подтип
Myriapoda. Среди сохранившихся у них
предковых черт сочленение мандибул
с rоловой в одной точке трехлопастный rи
пофаринкс (непарный, похожий на язычок
opraH, расположенный около слюнных же..
лез), выпячивающиеся коксальные мешоч..
ки. брюшные rрифельки.. собственная му"
скулатура в члениках антенн, орrаны Те..
мешвари (или очень близкие к ним образо--
вания), некоторые типы абдоминальных
Беспозвоночные с конечностями: членистоноrие и др.
Класс
Diplurata
235
Надотряд
Отряд
Diplura
Collembola
Prorur.l
Zygoentomata
Thysanura
Archaeognathata
Microcoryphia
Palaeoptera
-= Ephemeroptera
Odonata
Blattaria
Manrodea
I optera
Zoraptera
GrуllоЫаttап.l
Dermaptera
Orthoptera
Phasmida
Еm bloptera
Plecoptera
Psocoptera
Mallophaga
Anoplura
Thysanoprera
Homoptera
Heteroptera
Coleoptera
Strepsiptera
Hymenoptera
Raphidioida
Neuroptera
Megaloptera
Mecoptera
Diptera
Siphonaptera
т richoptera
Lepidoptera
Orthopteroidea
HemlPteroidea
Endopterygota
придатков. Все они малы по размерам (дли..
на тела, как правило, не превышает 7 мм) 1;
вероятно, по этой причине мальпиrиевых
сосудов нет, а продукты азотноrо обмена
удаляются через эпителий средней кишки;
rлаза и простые rлазки имеются лишь
у представителей одното класса из трех.
Двухвостки (рис. 8.25, А) лишены черт
специализации, которые столь ярко прояв--
ляются У остальных двух классов. У бессяж..
ковых (рис. 8.25, Б) антенны рудиментарны
(их функцию берут на себя трудные ноrи
1 Известны представители двухвосток дли
ной до 30 MM. Прuм. пер.
первой пары), а ман-дибулы превратились
в колющие стилеты. Как и у большинства
мноrоножек, число туловищных сетментов
после вылупления постепенно растет.. пока
не достиrнет полноrо взрослоrо набора (11
брюшных). Зато ноrохвостки (рис. 8.25, В)
сохранили лишь пять брюшных сетментов
(и тельсон), а две пары брюшных придатков
у них преобразованы в особое устройство,
за которое этот класс и получил свое назва..
ние. Конечности четвертоrо абдоминально"
то сетмента образуют отросток, способный
с силой распрямляться под действием дав..
ления полостной жидкости. В покое этот от..
росток удерживается в corHYTOM положении
236
rлава 8
.
А
Антенно
ОБразные
передние
ноrи
Вид сверху
Б
Вид СБОКу
Придаток
III cerMeHT8
Придаток
1 cerMeHTa
в
Вид СБОКУ
Рис. 8.25. Мноrоножкоподобные классы Apterygota: А ДBa представителя двухвосток; Б
бессяжковое; В различные ноrохвостки; показаны видоизмененные брюшные придатки (по
Imms, 1964; Wallace, Mackerras, 1970)
зацепкой, образованной придатком TpeTьe
ro cerMeHTa. Кроме Toro, конечности перво
ro cerMeHTa брюшка образуют крупную
вентральную трубку с выпячивающимися,
подобно коксальным мешочкам, пузырька
ми; назначение этой структуры окончатель
но не выяснено. Возможно, она используе-
тся для пополнения запасов влаrи из окру-
жающей среды. Ни в одном из трех классов
rонопоры не расположены на VIII (у самок)
или Х (у самцов) cerMeHTax брюшка, как
у всех прочих насекомых; отсутствуют так-
Беспозвоночные с конечностями: членистоноrие и др.
237
А
Б
Рис. 8.26. Насекомоподобные классы Aptery..
gota: A ДBa представителя чешуйниц; Б
представитель щетинохвосток (по Imms; 1964;
Daly et al., 1978)
же эффективные приспособления, предот--
вращающие потерю воды орrанизмом
Ко второй rруппе относятся чешуйницы
(класс Zygoentomata) (рис. 8.26. А), rораздо
больше напоминающие крылатых насеко--
мых, от которых их отличают помимо пер--
вичноrо отсутствия крыльев (а также сопут"
ствующих им структур rруди), по сути дела,
лишь три признака, сохранившиеся от пред--
ковых форм и присущие всем первичнобес--
крылым: наличие брюшных rрифельков, от..
сутствие копулятивноrо аппарата и перенос
спермы с помощью откладываемых 80
внешнюю среду сперматофоров, а также
продолжение линек после достижения поло--
вой зрелости. Поэтому их, как правило, счи..
тают близкими родственниками непосред--
ственных предков крылатых насекомых.
Щетинохвостки (класс Archaeognatha)
(рис. 8.26, Б), составляющие третью ветвь
первичнобескрылых, должны помещаться
rде--то между Zygoentomata и прочими
Apterygota, так как, обладая некоторыми
продвинутыми признаками чешуйниц, они
сохраняют множество предковых черт,
свойственных и «мноrоножкоподобным»
классам rексапод, причем часто в рамках
одних и тех же структур и opraHoB. Так,
хотя ротовые части и не поrружены в rолов..
ную капсулу. сочленение мандибул. особен--
ности rипофаринкса и максилл отражают
анцестральное состояние, присущее MHoro..
ножкам. Представлен также полный набор
примитивных признаков, сохранившихся
у Zygoentomata. Если крылатых насекомых
можно вывести из чешуйниц, последние,
возможно, происходят от щетинохвосток,
поэтому оба этих класса часто объединяют
в рамках одной rруппы «настоящих насеко..
мых». Надо сказать, что представители
обоих классов в среднем крупнее (до 2 см
длиной) и не так тесно связаны с влажной
подстилкой и почвенными микроэкотопа--
ми, как прочие аптериrоты.
Все пять рассмотренных классов вклю"
чают немноrим более 3100 видов, большая
часть которых потребители падали и раз..
лаrающихся растительных тканей, хотя не..
которые и нападают на друrих мелких re..
ксапод.
8.5.35.2. Класс Pterygota
(крылатые насекомые)
Третья крупнейшая вспышка разнообра..
зия внутри типа одноветвистых представле..
на 29 отрядами ныне живущих крылатых
насекомых (к которым относятся и вторич--
но бескрылые формы), включающими 98 0 А.
всех видов данноrо типа. Безусловно.. са..
мым важным обстоятельством, обеспечи--
вающим подобный успех, было то, что пте..
риrоты перестали зависеть от влажных ме..
стообитаний, с которыми тесно связаны все
прочие классы одноветвистых. Это в свою
очередь в значительной степени явилось
следствием приобретенной ими способно--
238
сти эффективно препятствовать потере BO
дЫ телом. Эволюция их cOBpeMeHHoro co
стояния, вероятно, включала целый ряд эта
пов, начиная с выработки особых свойств
кутикулы и трахей.
Возможно, восковой слой эпикутикулы
первоначально иrрал адаптивную роль BO
доотталкивающеrо rидрофобноrо покрова:
в местообитаниях, rде существует риск
переувлажнения, мелкие животные MorYT
захватываться водной пленкой, и поступле
иие в тело воды препставляет для них купа
более серьезную опасность, чем ее потеря;
максиллярные железы аптериrот, по
видимому, должны откачивать ее значите
льный избыточный объем. У некоторых HO
roxBocTOK возникли ряды покрытых воском
буrорков с водоотталкивающей функцией,
и, надо полаrать, предок крылатых HaceKO
мых имел аналоrичные приспособления.
у Hero также вполне моrла отсутствовать
rрахейная система, как и у большинства
мноrоножкоподобных аптериrот, что MO
жно приписать_ по видимому, прежде Bcero
мелким размерам любоrо орrанизма, BO
зникшеrо в результате педоморфоза
особенно если ero предок и сам по себе был
не особенно велик. Как только (вновь) раз
вились трахеи, rидрофобный восковой слой
cMor распространиться по всей поверхности
тела, никак не препятствуя rазообмену.
В то же время наземные членистоноrие_
по видимому, теряют через трахейную си
стему больше воды, чем через кутикулу,
rлава 8
поэтому сам по себе водонепроницаемый
покров не дал бы нужноrо эффекта.
Обычно у птериrот имеются мускулы,
закрывающие дыхальца (бывают и дру-
rие открывающие их), например, насеко-
мые сухих местообитаний MorYT открывать
их лишь на очень короткое время Иноrда
потери воды через дыхальца сокращаются
и друrими путями. Один из них
накопление в течение длительноrо времени
образовавшейся двуокиси уrлерода в Hera-
зообразной форме. Поскольку кислород из
пространства трахеи потребляется, не заме..
щаясь эквивалентным объемом уrлекисло-
[о rаза, возникает частичное разрежение,
обеспечивающее поступление новых порций
воздуха через дыхальца в виде однонаправ..
ленноrо потока, что противодействует диф..
фузии водяных паров наружу.
Последнее усовершенствование системы
сохранения воды связано с задней кишкой
и заключается в механизме обратноrо вса-
сывания воды из экскрементов и продуктов
выделения по мере их прохождения через
этот отдел пищеварительноrо тракта. Реаб-
сорбция может осуществляться разными
путями: среди них широко распространен
способ активной секреции неорrанических
и/или орrdнических растворимых веществ
в систему межклеточных пространств вну-
три буrорков или сосочков стенки прямой
кишки. В результате вода пассивно диффун-
дирует из просвета кишечника в эти про-
странства по осмотическому rрадиенту;
ПРОСВЕТ ПРЯМОЙ КИШКИ
Кути кула
Межклеточное
пространство
АБСОРБЦИЯ растворенных веществ
из жидкости в воронке
\ Поступление воды по
, \мо ическому rpaдиенту
,
Секреция растворенных
веществ в межклеточное
пространство
rЕМОЦЕЛЬ
Клапаны
Поступление воды после аБСОРБЦИИ
растворенных веществ в воронке
Рис. 8.27. Схематический разрез сосочка прямой кишки падальной мухи, показывающий систему
межклеточных полостей, в которые и из которых MorYT перемещаться растворенные вещества
и куда из просвета кишки вынуждена поступать вода, переходящая затем в rемоцель (с сильными
упрощениями по Gupta, Berridge, 1966)
Беспозвоночные с конечностями: членистоноrие и др.
коrда накапливающаяся жидкость перено
сится в rемоцель, растворимые вещества
абсорбируются из нее и вновь возвращаю
тся в KpyroBopoT (рис. 8.27).
Освоение местообитаний с недостатком
влаrи, ставшее возможным блаrодаря этим
усовершенствованным механизмам эконо
мии воды, должно было сильно сказаться
на биолоrии ранних птериrот, что и обусло
вило формирование мноrих характерных
черт этой rруппы. Откладывание капелек
спермы или сперматофоров, например,
в относительно сухих условиях было бы He
надежной стратеrией. и у крылатых HaceKO
мых сперма переносится от самца к самке
непосредственно путем копуляции, хотя
сперматозоиды по прежнему чаще Bcero за..
ключены в сперматофор. У примитивных
форм самки сохраняют унаследованные от
предков парные rонопоры, и самцы имеют
парные совокупительные орrаны, однако
в большинстве отрядов половое отверстие
самок непарное, а парные rениталии самца
соответственно срастаются в единую CTPYK
туру. У самца развита также пара класпе..
ров, т. е. захватов, которые удерживают
самку, в то время как выворачивающийся
дистальный отдел ero половоrо протока
(или обособленная rоноподия половая
ножка) вводится через половое отверстие
самки, чтобы отложить один или несколько
сперматофоров или пакетов спермы в ее со..
вокупительную сумку или друrой отдел по..
ловоrо тракта. При высвобождении из спер
матофора сперма, как правило, хранится
порой на протяжении длительноrо перио..
дa B семеприемнике (сперматеке), покуда
не оплодотворит все яйца по мере их OT
кладки, причем сперматозоиды попадают
в яйцо через крошечные поры в ero защит..
ной оболочке, характерной для птериrот.
у ряда насекомых самки спариваются Bcero
один раз в жизни, но полученная при этом
сперма может оплодотворить несколько
кладок яиц; ни один из видов не линяет по..
сле достижения половой зрелости. Унасе..
комых часто есть яйцеклад, что дает возмо"
жность откладывать яйца в особые, как
правило, укромные места; друrие способны
с этой же целью вытяrивать конец брюшка
в виде трубочки, тоrда как у некоторых
перепончатокрылых (пчел, ос и др.) яйце..
239
CerMeHT, несущий rонопор(ы)
о
Непарный пение
Класпер
Рис. 8.28. Модификации придатков брюшных
cerMeHToB предковых форм одноветвистых
способствующие откладке яиц и спариванию
крылатых насекомых (вид сбоку) (по Snod
grass, 1935). Яйцеклад самки представляет со..
бой базальные части придатков VIII и IX cer
ментов брюшка; rOHonopbI самца находятся на
Х cerMeHTe, однако у большинства насекомых
они смещаются вперед и кажутся структурой IX
cerMeHTa. Следует отметить, что у мноrих Pte
rygota сформировались вторичные совокупи
тельные opraHbI и ложные яйцеклады
клад видоизменился в жало. утратив тем ca
мым связь с размножением. Класперы и [о..
ноподия самца, как и яйцеклад самки
(рис. 8.28), произошли из анцестральных
придатков брюшных rенитальных cerMeH"
тов; кроме часто встречающихся парных
церков (придатки ХI cerMeHTa), эти орrаны
спаривания и откладки яиц представляют
собой единственные функционирующие
у взрослых птериrот остатки целой серии
сеrментарных придатков брюшка.
Однако больше, чем любой друrой чер..
той, крылатые насекомые выделяются сре..
ди всех членистоноrих наличием крыльев
еще одноrо приобретения, ставшеrо возмо
жным лишь после преодоления зависимо..
сти от влажных местообитаний. У взрослых
особей (имаrо) второй и третий [рудные ce
[менты несут кроме Hor по паре крыльев;
впрочем) одна или обе пары MorYT быть ре..
дуцированы или совсем утрачены. Происхо..
ждение этих крыльев, как и селективные
преимущества обладателей предшествую
240
r'лава 8
щих ИМ структур пока не известны, однако
скорее Bcero крылья развились из лопасте
видных боковых выростов rрудных терrи
тов с боrатой сетью трахей, встречаю
щихся, например, у некоторых чешуйниц
и мноrочисленных ископаемых rексапод
(СМ. разд. 10.6.3). Установлено. что у ныне
живущих птериrот крылья формируются из
выпячиваний стенки тела путем дорсолате
ральноrо разрастания эпидермальных за
чатков Верхняя и нижняя поверхности по
следних прирастают друr к друrу на всем
протяжении, кроме участков вдоль системы
каналов, в которых циркулирует rемолим
фа и расположены трахеи и сенсорные He
рвы. В конечном итоrе клетки эпидермиса
секретируют тонкую кутикулярную оболоч-
ку; коrда происходит последняя линька (на
имаrо) мелкие. несколько мясистые крыло-
вые зачатки расправляются под давлением
полостной жидкости, выстилка каналов
укрепляется КУТИКУЛОЙ (они становятся де-
финитивными «жилками крыла»), эпидер-
мис деrенерирует и крыло превращается
в тонкую двуслойную кутикулярную струк-
туру. Естественно поэтому, что ero движе..
пиями управляют мышцы, размещенные
в друrих местах; при этом они не всеrда кре-
пятся к основанию крыла. Кроме Toro, по..
следнее должно быть сочленено с rрудью
так, чтобы описывать необходимую для по..
лета траекторию в виде эллипса или вось"
...
... ,.) ...
Рис. 8.29. Разнообразное жил
кование крыльев насекомых
(по разным авторам). Прими-
тивные крылья С мноrочислен-
ными жилками показаны
в верхней части рисунка, а вни-
зу представлен ряд, демон-
стрирующий постепенное со-
кращение количества жилок
Беспозвоночные с конечностями: членистоноrие и др.
241
мерки. У мелких форм крылья MorYT рабо..
тать с частотой до тысячи взмахов всекун"
ду. Весьма необычно для животных то, что
летательные мышцы насекомых отвечают
на каждый полученный нервный им..
пульс несколькими сокращениями (см.
разд. 16.10.5). Крылья примитивных птери..
rOT, как правило, относительно велики
и снабжены мноrочисленными жилками, но
во мноrих эволюционных линиях их разме..
ры и количество жилок сильно сокрашаю..
тся (рис. 8.29); это соответствует эволюции
от длиннотелых животных с устойчивым,
но неманевренны!VI полетом к короткоте..
лым формам, у которых он неустойчив, за..
то высокоманеврен. В тех же эволюцион..
ных линиях возникла и способность полно..
стью складывать крыловую пластинку, Kor..
да она не используется тенденuия, до..
стиrшая cBoero наиболее полноrо развития
в видоизменении всех частей переднеrо кры"
ла в защитные покрышки для сложенных
задних крыльев. Полет насекомых рассма..
тривается в разд. 10.6.3.
Фактором, обеспечившим окончатель..
ный эволюционный успех крылатых насеко--
мых, оказалось, по всей видимости, разно..
образие доступных им источников пищи.
Это нашло свое отражение в крайне разно..
образном строении pOToBoro аппарата, обу..
словленном модификацией различных ero
придатков в разных отрядах, хотя в отличие
от большинства аптериrот (если не считать
чешуйниц) птериrоты и представляют со..
бой единую rруппу, поскольку у всех них
эти придатки не поrружены в rоловную кап..
сулу, мандибулы причленяются в двух точ"
ках, а rипофаринкс однолопастный. Пред..
ковой формой этих придатков был, без со..
мнения. стандартный ротовой аппарат мно"
rоножек, приспособленный для кусания
и пережевывания, причем такое ero строе..
ине сохранилось и у некоторых современ..
ных крылатых насекомых (рис. 8.30 А). Он
дал начало целому ряду направлений эво"
люции колю.щих И сосущих структур, рас..
смотрение которых мы оrраничим здесь
лишь немноrими примерами.
Так, клопы питаются в зависимости от
rруппы соками растений или животных
с помощью хоботка, образованноrо нижней
rубой (вторая пара максилл), внутри кото..
рой размещаются две пары развившихся из
мандибул и максилл стилетов (рис. 8.30, Б).
Мандибулярные стилеты прокалывают тка..
ни жертвы и позволяют максиллярным про..
никнуть в ранку. На внутренней поверхно..
сти последних находятся ДВа канала через
один из них выпускается слюна, через вто"
рой высасывается жидкость. У комаров
(рис. 8.30, В) есть два дополнительных сти"
лета, образованные верхней rубой (окру"
жаюшей пишевой канал) и rипофаринксом
(в ней находится проток для слюны). Одна..
ко в обоих случаях нижняя rуба иrрает ис..
ключительно защитную роль и склаДывае..
тся телескопически или отrибаясь локте..
образно назад по мере проникновения
остальных ротовых придатков в ткани.
Друrие насекомые сосут более леrкодо"
ступные жидкости такие, как нектар или
выделения из ран и трещин покровов. Ба..
бочки, например, пьют нектар через длин..
ный хоботок, способный расправляться Зd
счет наriIетания rемолимфы (коrда давле..
ние ее падает, он сам сворачивается в спи..
раль блаrодаря своим упруrим свойствам).
Хоботок образован частями максилл, все
же прочие ротовые придатки редуцированы
или бесследно исчезли (рис. 8.30, п. У ряда
ночных бабочек ero кончик заострен, несет
кисточку из длинных волосков и может
использоваться для прокалывания. У ком..
натной мухи для захвата пищи служит ни..
жняя rуба. Мандибулы здесь также отсут"
ствуют, но верхняя rуба и rипофаринкс по..
rружены в yrлубление разросшейся нижней
rубы несущей на конце большой.. мяrкий
двулопастный «лабеллум», действующий
подобно rубке (рис. 8.30, Д).
Есть насекомые с rиrантскими мандибу..
лами KOTopыe однако, не принимают ника..
Koro участия в питании, а служат для защи..
ты или для сражений между самцами (рис.
8.30,Е); кроме Toro, по меньшей мере в во..
сьми отрядах отмечены виды с рудимент ар..
ным ротовым аппаратом, вообще не пи..
тающиеся на имаrинальной стадии. у неко"
торых птериrот, особенно с полным мета..
морфозом или с водными ювенильными
стадиями, большая часть жизненноrо цикла
и вся активность, связанная с питанием, за..
вершается до наступления половой зрело..
сти, а взрослая особь представляет собой
242
r олова спереди
Lr
Li
А
rлава 8
rолова СБОКУ,
стилеты спрятаны
т.:х:
md
rолова СБОКУ Разрез в плоскости
x y
Lr тх Li
I
в
rолова СБОКУ
д
md h
Разрез в плоскости
x y
Li
При питании стилеты
выходят наружу
у
rолова СБОКУ
Разрез в
ПЛОСКОСТИ х...у
Li
Разрез в плоскости Х у
ОБЩИЙ ВИД тела сверху
md
Рис. 8.30. Примеры адаптивной радиации pOToBoro аппарата крылатых насекомых (см. текст):
А rрызущие мандибулы жука (Coleoptera); Б колюще сосущий хоботок клопа (Heteroptera);
В ротовой аппарат комара (Diptera); r хоботок нектароядной бабочки (Lepidoptera); д
лижущий ротовой аппарат комнатной мухи (Diptera); Е сильно разросшиеся мандибулы самца
жука роrача (Coleoptera), используемые в схватках самцов, а не для .питания (по ряду авторов,
rлавным образом по Borror et al., 1976) Lr верхняя rуба, md мандибула, тх максилла, Li
нижняя rуба, h rипофаринкс
y
ЛаБеллум
недолrовечный «автомат» для расселения
и размножения.
Миллион видов крылатых насекомых
можно разделить на четыре основные rруп..
пы отрядов (рис. 8.31). Paleoptera, или древ..
некрылые (два отряда)., отличаются rлав..
ным образом одним (примитивным) при..
знаком неспособностью укладывать сза..
Х
r
ХОБОТОК
rолова
rрудь
Е
ди на брюшке крылья, которые у них круп-
ные и несут rустую сеть продольных и попе-
речных жилок. Неполовозрелые стадии
(наяды, нимфы), живущие долrо и MHoro-
кратно линяющие, развиваются только
в воде, и., следовательно., их адаптации от-
личаются от имаrинальных. У мноrих, на-
пример, имеются наружные парные трахей-
Беспозвоночные с конечностями: членистоноrие и др
А
j \
I \
I \
I \
I I \
HOMOPTERA
(тли)
в
HETEROPTERA
(клопы)
':..
, . .....
PLECOPTERA
(веснянки)
243
DERMAPTERA
(уховертки)
J
r COLEOPTERA
(жуки)
STREPSIPTERA
(веерокрылые)
'" '-, \
7
. J
\
lJ.ерки \,
,
BLA ТТ ARIA
(тараканы)
Б
ORTHOPTERA
(прямокрылые)
ANOPLURA
(вши)
NEUROPTERA
(сетчатокрылые)
. .
"'f... ., ':;.:""
THYSANOPTERA
(трипсы)
Е
HYMENOPTERA
. 1/.. (перепончатокрылые)
::, (.1
& \ :,.';', " .iJ:
\ ': i.' .
" , '. --с.."' ' a "'1':.
\...... (. . . "," .а .;;.,: .
\ <.... ,. 1 " '\" <i -=:. ' ':':, . .
редуцирован I. · : .t:i;
ное заднее \ . .... .У'
крыло
LEPIDOPTERA
(БаБОЧКИ)
д
DIPTERA
(двукрылые)
(
Рис. 8.31. Разнообразие форм тела взрослых Pterygota: А древнекрылые; Б ортоптероиды;
В rемиптероиды; r нейроптероидные эндоптериrоты' Д мекоптероидные эндоптериrоты;
E rименоптероидные эндоптериrоты (по разным авторам)
244
rлава 8
ные жабры листовидные пластинки на
брюшке, напоминающие настоящие жабры,
за исключением тото" что вместо KpOBOC
набжающих структур (циркуляторная си
стема одноветвистых не участвует в перено
се дыхательных rазов) в них находится TY
стая сеть замкнутых трахей. Возможно" эти
аружные
зачатки
крыльев
Трахейные
жаБрЫ
А
EPHEMEROPTERA
,
, 1,
, I ""
I ,
1 ; .\
" .1 &.
i t......,.. !
,1F,
У,
HETEROPTERA
в ANOPLURA
ODONA ТА
THYSANOPTERД
HOMOPTERA
Е HYMENOPTERA
образования развились из предковых при
датков абдоминальных сетментов. Все про
чие птериrоты относятся к «новокрылым».
Т. е. в состоянии покоя способны плоско
укладывать крылья поверх брюшка.
Вторая rруппа (из 10 отрядов)
ортоптероиды. Для этих стоящих у основа..
BLATTДRIA
DERMAPTERA
Наружный зачаток крыла
PLECOPTERA
Б
ORTHOPTERA
COLEOPTERA
DIPTERA
Рис. 8.32. Неполовозрелые формы различных rpynn Pterygota, изображенных на рис. 8.31: A
наяды древнекрылых; 5, В ортоптероидные и rемиптероидные нимфы; r Е личинки эндоп-
териrот (по разным авторам)
r STREPSIPTERA
NEUROPTERA
д
Беспозвоночные с конечностями: членистоноrие и др.
ния линии новокрылых (Neoptera) форм ти"
пичны rрызущий ротовой аппарат.. нескон..
центрированная нервная система, неветвя"
щиеся, длинные мноrочлениковые антенны,
пара церков на конце брюшка, мноrочи..
сленные мальпиrиевы сосуды, выпячиваю
щиеся совокупительные орrаны. Нимфы не..
сколько раз линяют, постепенно меняясь
и приближаясь по облику к взрослому пасе..
комому (имаrо). Все орrаны взрослой осо..
би, включая крылья, сложные rлаза и т. п.,
развиваются по мере прохождения нимфа..
льных возрастов. Поэтому образ жизни
ювенильных особей примерно такой же, как
и у взрослых.
В шести отрядах следующей
rемиптероидной rруппы происходит та..
кое же постепенное изменение нимф до
взрослых форм, хотя в большинстве слу..
чаев наиболее серьезная перестройка opra..
низма приурочена к последней неполово"
зрелой стадии (или стадиям). В подавляю..
щем большинстве эти насекомые питаются
жидкой пищей, и определенные части мак..
силл превратились в склеротизированные
стилеты. Для них характерно небольшое
число (четыре и менее) мальпиrиевых сосу..
дов, отсутствие церков, сильно редуциро..
ванное жилкование крыла, короткие антен..
ны с небольшим числом члеников и хорошо
выраженная rоноподия. У нимф никоrда
нет простых rлазков.
Последняя rруппа эндоптериrоты 1
(1] отрядов) в чем..то сходна с предыдущей,
хотя у ее представителей имеются короткие
церки и может быть множество мальпиrие..
вых сосудов. Однако, если всем прочим но..
вокрылым свойственно постепенное пре..
вращение первой нимфальной стадии в има..
ro с развитием крыловых зачатков снаружи
тела, у эндоптериrот есть личиночная ста..
дия, резко отличающаяся от взрослой осо..
би по строению, питанию, а часто и по ме..
стообитанию уникальный случай среди
наземных членистоноrих. У мноrих мух
и перепончатокрылых, к примеру, личинки
безноrие, червеобразные, в то время как
у бабочек и друrих перепончатокрылых на
1 В отечественной литературе ее обычно Ha
зывают Holometabola, т. е. насекомые с полным
преврашением. Прu.М. пер.
245
Б
Diptera
Lepidoptera
CoLeoptera
Рис. 8.33. Вид куколок эндоптериrот с BeHT
ральной стороны; куколка двукрылоrо (в cepe
дине) заключена в «пупарий» оболочку из
шкурки, сброшенной личинкой последнеrо
возраста (по Jeannel, 1960; Wallace, Mackerras,
1970)
их брюшке находятся дополнительные вто"
ричные ходильные ноrи (<<ложные ножки»)
(рис. 8.32). При этом зачатки крыльев раз..
виваются под покровами и, следовательно,
снаружи неразличимы. Поскольку личиноч"
ные формы так существенно отличаются от
взрослых, переход между ними включает
так называемое полное превращение (см.
разд. 15.4.2). Этот процесс разворачивается
на особой неактивной стадии в «куколке»
(рис. 8.33); почти все ткани личинки разру..
шаются путем rистолиза, взрослый opra
низм формируется заново, и развиваю..
щиеся крылья впервые появляются на по..
верхности тела. Некоторые куколки сохра..
няют сброшенный последней личинкой по..
кров в качестве защитной оболочки (рис.
8.33, Б). В эволюционном отношении такая
стратеrия эколоrическоrо разделения личи..
нок И имаrо представляется весьма выи..
rрышной: 850/0 всех видов крылатых насеко..
мых относятся к эндоптериrотам. Личинки
ряда видов MorYT встречаться даже в таких
местообитаниях, которые для преимуще..
ственно сухопутных животных в высшей
степени неблаrоприятны, Нdпример в дон..
ных осадках приливной зоны и временных
водоемах; личинка одноrо африканскоrо
комарика способна после деrидратации
к криптобиозу, соперничая в этом отноше..
нии с тихоходками (см. с. 0222).
246
8.6. Тип Crustacea
(ракообразные)
8.6.1. 3тимолоrия
От лат. сrustа скорлупа, корка.
8.6.2. Диаrностические
и специфические признаки
(рис. 8.34)
1. Двусторонне симметричные членисто..
ноrие с разнообразной формой тела от
вытянутой до сферической; длина тела от
менее О., 1 мм до 60 см (размах Hor унекото..
рых форм достиrает 3.,5 м, масса друrих
более 20 Kr).
2. Тело из более чем двух слоев клеток
в толщину, с тканями и орrанами.
3. Кишечник сквозной, прямой; от ero
неБолыllrоo среднеrо отдела отходят два
пищеварительных дивертикула., в которых
происходит переваривание и всасывание пи..
щи; у видов., питающихся не только мель..
чайшими частицами или жидкой пищей.,
в переднем отделе кишечника находится из..
rлава 8
мельчающий аппарат; пищеварительные
дивертикулы возникают как выросты На за-
родышевой кишке.
4. Разные rруппы ракообразных очень
сильно отличаются друr от друrа xapaKTe
ром разделения тела на отделы и числом се..
[ментов.. составляющих каждый отдел. oд
нако, как правило, можно выделить rолову,
образованную акроном и пятью несущими
придатки сеrментами, туловище из 2 65
(и более) cerMeHTOB и тельсон, часто с парой
выростов «Фуркой». Первый и до семи (В
зависимости от rруппы) следующих туло"
вищных cerMeHToB часто срастаются с rоло..
БОЙ, образуя цефалоторакс (<<rолово"
rрудь»); туловище по типу придатков, как
правило, подразделено на торакс (rрудь)
и абдомен (брюшко), причем торакальные
cerMeHTbI.. не вошедшие в состав rоловоrру..
ди, составляют так называемый «переон»,
а абдоминальные «плеон». rоловоrрудь,
а в отдельных rруппах и значительная часть
Bcero тела или все оно целиком заключены
в особый вырост rоловы, так называемый
«карапакс», который разрастается в CTOpO
ны И закрывает бока тела.
А
Пищеварительный
дивертикул Сердце
rOHaдa
Задняя
кишка
rрудные
конечности
Рис. 8.34. Морфолоrия ракообразных. А. Схе-
ма продольноrо разреза через тело rенерали-
зованноrо представителя этоrо типа. Б r. Ха-
рактерные двуветвистые конечности (сильно
упрощено) (частично по McLaughlin, 1980):
Б с одной трубчатой и одной листовидной
ветвью; B c двумя листовидными ветвями
(типичная плавательная конечность); r c
обеими трубчатыми ветвями, одна из которых
сильно редуцирована (типичная ходильная но-
ra)
Б
в
Нервный тяж
с rанrлиями
Беспозвоночные с конечностями: членистоноrир и др
247
5. Цилиндрические или листовидные
придатки в основе своей двуветвистые., при
чем ветви, как правило, имеют разные раз
меры и форму, а часто несут и вторичные
ответвления; к придаткам rоловы относятся
две пары антенн (первую называют «антен..
нулы»)., одна пара мандибул и две пары
максилл; туловищные придатки широко
ВаРЬИРУЮТ в числе, строении и реrионарной
дифференцировке: исходно каждый сет'мент
несет пару конечностей, хотя на брюшке
они нередко отсутствуют; хелицер нет., но
отдельные конечности MorYT оканчиваться
клешнями.
6. Первичных ротовых придатков три
пары (мандибулы и две пары максилл), но
ВО всех rруппах бывает от одной до трех ДО"
полнительных пар «ноrочелюстей», отхо"
дящих от rpy дных cerMeHToB в составе rоло..
воrруди; элементы каждой пары MorYT дви
rаться в фазе или в противофазе с друrими
парами.
7. На rолове находятся срединные про
стые rлазки или сложные f'лаза по бокам
(часто на подвижных стебельках).
8. Обычно наружная сеI'ментация вы..
ражена на туловище (часто скрыта под ка..
рапаксом, иноrда исчезает) и отсутствует на
rолове
9. Выделительная система представле..
на слепо оканчивающимися антеннальными
и/или максиллярными железами.
10. Наружный скелет часто обызвеств"
лен.
11. rазообмен идет через внутреннюю
стенку карапакса, через всю поверхность те..
па или с помощью жабр.. развиваюшихся из
определенных частей [рудных или брюш..
ных конечностей.
12. Для циркуляторной системы харак"
терна наличие rемоцианина.. реже друrих
пиrментов.
13. Нервная система с парными raH..
rлиями в каждом cerMeHTe; вентральные
тяжи и rанrлии исходно раздельные, но ча..
ще слиты; иноrда все [рудные rанrлии сли..
ваются в единую массу.
14. Раздельнополые животные, редко
rермафродиты: оплодотворение внутреннее
путем копуляции с помощью rоноподий
или пенисов; расположение rонопора варьи..
рует, часто он расположен на rруди.
15 Яйпа самка обычно носит на себе
или держит в специальных карманах; у од..
них форм из яйца выходит особь с полным
набором cerMeHToB, у друrих личинка
«науплиус» лишь с тремя сеrментами (рис.
8.35) (ряд высокоспециализированных пара..
1итических форм удалось отнести к рако..
образным только блаrодаря наличию у них
на уплиуса).
16. По большей части морские формы.,
хотя встречаются и пресноводные (13 0/0 ),
реже наземные (30/0).
Ракообразные., по сути дела, препстав"
ляют членистоноrих в морских 1косисте..
мах: лишь 30/0 видов артропод мировоrо
океана не относится к этому типу Они пре..
обладают в планктоне (и в пресноводном
также) и входят в число трех..четырех ва..
жнейших бентосных rрупп, включая интер..
стициальный комплекс и макрофауну. Кро..
ме Toro, ряд видов ведет паразитический
образ жизни.
Процветание ракообразных можно
в определенной мере объяснить таким об..
щим для всех артропод признаком. как Ha
личие членистых конечностей. Их ноrи
и/или плавательные придатки позволяют
быстро передвиrаться и без труда осваиваrь
различные участки местообитания. Несом..
ненно, успеху способствует и друrая важная
черта артропод наружный скелет: круп..
ные, заключенные в крепкий панцирь
и мошно вооруженные формы rораздо ме..
ньше подвержены риску поедания хищника..
ми, чем мяrкотелые животные. Однако., мо"
жет быть, основная причина практически
повсеместноrо распространения водных ра..
кообразных разнообразие формы тела,
хотя внутренняя анатомия их достаточно
сходна
Рис. 8.35. Личинка науплиус (по Grееп, 1961)
248
Если строение хелицеровых (разд. 8.4)
и насекомых (разд. 8.5.3Б) соответствует He
кой общей схеме, а мноrоножки (разд.
8.5.3A) несмотря на все различия в числе ce
rMeHToB, неизменно сохраняют общую фор
му тела, у ракообразных дело обстоит со..
вершенно иначе; TaKoro понятия, как типич"
ный план строения тела ракообразных, по..
просту не существует. У одних есть rолова
и туловище, у друrих rолова, rрудь
и брюшко, у некоторых rоловоrрудь,
переон и брюшко а в одной крупной rруппе
лишь rоловоrрудь и брюшко; в немноrих
случаях брюшко отсутствует, а бывает, что,
по сути дела, нет и дифференцированной ro..
ловы или rруди. Кроме Toro, число cerMeH..
тов в каждом из этих отделов также варьи--
рует и зависит от rруппы иноrда внутри
одноrо класса. Аналоrичным образом, КО--
нечности бывают самой разнообразной
формы от ходильных Hor, практически
таких же, как у хелицеровых и одноветви..
стых, до листовидных плавательных при..
датков. Антенны MorYT иrрать роль сенсор--
ных opraHoB, служить для локомоции или
сбора корма, действовать как средство при..
крепления к хозяину, выполнять роль кла..
сперов.. удерживающих партнера при спари..
вании, и т. д. У MopcKoro желудя, паразити..
ческоrо веслоноrоrо, раковинноrо жабро..
Hororo краба и paBHoHororo из семейства
антурид (рис. 8.36) едва ли есть какие..либо
внешние черты, свидетельствующие об их
принадлежности к одной и той же rруппе
животных.
Такая пластичность строения позволяет
ракообразным плавать, закапываться
в rpYHT ползать, делать ходы в древесине,
прилепляться к скалам, охотиться, объ..
едать растения питаться взвешенной пищей
и донными осадками (см. рис. 9.4), парази--
тировать на животных различных типов (и
на друrих ракообразных) так что, вероят--
HO не будет преувеличением сказать, что
они занимают все возможные типы эколо..
rических ниш в морях. Ракообразные
прежде Bcero водные обитатели. Хотя пред..
ставители ряда их rрупп живут и на суше,
это все же не наземные животные в полном
смысле слова. В частности, у них сохраняе..
тся та же система rазообмена, что и у вод--
ных сородичей, а это оrраничивает распро..
rлава 8
д
Рис. 8.36. Разнообразие формы тела ракообра
зных: А морской желудь (Cirripedia) Б
паразитическое веслоноrое (Copepoda)' в
paBHOHoroe (Malacostraca); r раковинное
жаброноrое (Branchiopoda); д краб (Malaco-
straca) (по разным авторам)
странение лишь влажными место обитания-
ми. Более Toro, некоторые виды, например
наземные крабы, вынуждены возвращаться
в воду для размножения поскольку их цикл
развития по--прежнему включает фазу вод-
ной личинки. Только среди мокриц встре-
чаются формы с суrубо сухопутными при-
способлениями и среди наземных рако-
образных эта rруппа распространена наибо-
лее широко. В дополнение к жабрам на
плеоподах характерному признаку этоrо
отряда (Isopoda) у некоторых мокриц эти
придатки брюшка снабжены трахеевидны-
ми впячиваниями покровов, заходящими
в rемоцель конечностей. У друrих видов вы-
работались лишь механизмы позволяющие
постоянно поддерживать жабры в увла-
жненном состоянии, например направляя
на плеоподы капельки БОДЫ с поверхности
тела
Беспозвоночные с конечностями: членистоноrие и др
8.6.3. Классификация
249
ми. Все шире распространяется традиция
объединять эти девять таксонов в четыре
более крупные rруппы (их можно считать,
например, подтипами); при этом относите..
Почти 40000 видов, включаемых в этот
тип, распределены между девятью класса..
Класс
Cephalocarida
Надотряд
ОТРЯД
Brachypoda
Anostraca
Notostraca
Conchostraca
Cladocera
Necriopoda
Derocheilocarida
Arguloida
Calanoida
Cyclopolda
Misophrioida
Mormonllloida
Harpacricoida
Monstrilloida
Siphonostoma toida
Poecilostomatoida
?Т antulocaridida
Rhizocephala
Ascothoracica
Thorat:ica
Acrothoracica
Facetotecta
м yodocopida
Cladocopida
Podocopida
Platycopida
Palaeocopida
Phyl10carida Leptostraca
Hoplocarida Stomatopoda
Anaspidacea
Syncarida Stygocaridacea
Bathynellacea
Thermosbaenacea
Mysidacea
Сиmасеа
Spelaeogriphacea
т anaidacea
Mictacea
Isopoda
Amphlpoda
Euphausiacea
Amphlonidacea
Decapoda
Pancarlda
Peracarida
250
льно неспециализированные классь] Remi..
pedia и Malacostraca помещают в отдельные
монотипические подтипы, а остальные два
подтипа (возможно, обязанные своим во..
зникновением педоморфозу) включают со..
ответственно классы Cephalocari..
da + Branchiopoda (подтип Phyllopoda)
и Mystacocarida + Branchiura + Сореро..
da + Cirripedia + Ostracoda (подтип Maxil..
lopoda). Хотя Malacostraca часто называют
«высшими ракообразными», на самом деле
это довольно примитивная rруппа, находя..
ЩdЯСЯ rде"то у основания ствола ракообра..
зных; наиболее продвинутые черты орrани"
зации заметны у Phyl1opoda и Maxillopoda.
8.6.3.1. Класс Cephalocarida
Эти мелкие слепые рачки (длина их не
достиrает и 4 мм) обитают на морском дне
и питаются детритом. О них ничеrо не было
известно до 1955 r., несмотря на то что ме..
стами их численность может быть высокой.
Тело (рис. 8.37) делится на rолову. rрудь
и брюшко без каких..либо намеков на обра..
зование rоловоrруди или карапакса все во..
семь пар [рудных Hor похожи друr на друrа
и вполне идентичны 11 паре максилл, также
состоящих из листовидной и трубчатой ча..
сти. Брюшко включает 1] cerMeHTOB и ли..
шено придатков, если не считать сохранив..
шихся на первом cerMeHTe редупированных
конечностей, к которым прикрепляются
<Рурка
I
Рис. 8.37. Цефалокарида (вид сбоку) (по San
ders 1957)
rлава 8
яйцевые мешки. Цефалокариды
rермафродиты (что для ракообразных нети..
пично); у парных яичников и семенников об
щий проток; развитие протекает постепенно
и сопровождается мноrочисленными личи
ночными Iиньками.
Bcero известно десять видов; тело у всех
вытянутое, цилиндрическое, заканчиваю
щееся тельсоном с длинной фуркой.. каждая
ветвь которой несет щетинку. Все известные
формы относят к одному отряду.
8.6.3.2. Класс Branchiopoda
(жаброноrие)
Жаброноrие представляют собой дово"
льно разнообразную rруппу (рис. 8.38),
включающую rлавным образом пресновод"
ных рачков с мелкими или рудиментарны..
ми придатками rоловы (за исключением,
как правило, антенн), не приросшими к [о..
лове туловищными сеrментами и с тулови
щем. несущим ряд примерно одинаковых
конечностей.. которые обычно уменьшаю..
тся в размерах спереди назад (несколько по..
следних cerMeHToB вообще их лишены). Ko
нечности эти несут жабры. и в типичном
случае представляют собой листовидные
плавательные и/или фильтрующие орrаны,
форма которых поддерживается в основном
давлением полостной жидкости. а не кути
кулой. Мноrие виды размножаются парте..
ноrенетически и «вынашивают» на себе
яйца до выхода личинок. Отдельные пред..
ставители способны образовывать покоя
щиеся стадии.
Жаброноrих делят на четыре отряда.. си..
льно различающиеся по внешнему облику.
у раковинных (Conchostraca) и ветвисто..
усых.. или водяных блох (Cladocera). тело
короткое, иноrда почти окруrлое, антенны
выполняют локомоторные функции, фурка
в виде KoroTKoB, а выводковая камера поме
щается дорсально под сдавленным с боков
карапаксом. Однако если у раковинных до
более 30 туловищных cerMeHToB, а карапакс
покрывает все тело, включая rолову, у BeT
вистоусых последний, хотя часто велик, ни..
коrда не закрывает rоловы.. а иноrда реду"
цирован до небольшой выводковой камеры;
туловищных конечностей всеrда не более
шести пар (рис. 8.38, А, Б). Раковинные не
Беспозвоночные с конечностями: членистоноrие и АР.
ОБласть прикрепления
карапакса
Мышца...
аддуктор
МаНДИБула
Кольца
роста
Сложный
rлаз
Антенна
;арапакс
А
::\,
,
СРурка
Антеннула Туловищные
придатки
СРурка
Первая пара
туловищны
конечностеи.... &
J.
$
"
\ "...
...
Карапакс
т уловище
в
Ei
!
251
Сложный rлаз
Пищеварительный \
дивертикул
Антенна
Б
Сердце
Карапакс
Яйца в
выводковой
камере
Кишечник
СРурка
Антенна
I
Антеннула сложный rлаз
r
r енитальные
cerMeHTbI
Рис. 8.38. Морфолоrия жаброноrих. А. Раковинные (Conchostraca), вид сбоку, левая створка Kapa
пакса удалена (см. также рис. 8.36, r) (по Kaestner, 1970), Б. 8етвистоусые (Cladocera), вид сбоку
через прозрачный карапакс (по 8elk, 1982). В. Щитни (Notostraca), вид сверху (по Kaestner, 1970).
r. Жаброноrи (Anostraca), вид сбоку в естественном положении плавания дорсальной стороной
вниз (по McLaughlin, 1980)
сбрасывают карапакс в процессе линьки,
а наращивают ero концентрическими коль..
цами как двустворчатые моллюски; как и
у последних, створки «раковины» удержи..
ваются вместе сильной мышцей..
аддуктором, про rиводействующей упруrо..
му лиrаменту.
Зато у Notostraca (щитней) карапакс
представляет собой широкий дорсовентра..
льно уплощенный щит, по форме близкий
к отпечатку KOHcKoro копыта; из..под Hero
выступает назад узкий цилиндрический зад..
ний конец тела с двумя длинными MHoro..
членистыми ветвями фурки (рис. 8.38, В).
Интересно что эта часть туловища разделе..
на на сеrменты не полностью, так что на од..
ном видимом «cerMeHTe» может быть до
шести пар конечностей. Bcero туловищных
конечностей до 70 пар, причем одиннадца..
тая пара несет выводковые камеры, а пер..
вая крупнее остальных. У представителей
Anostraca (жаброноrов) никакоrо карапакса
нет (рис. 8.38, Т), а выводковая камера
образуется внутри тела из расширенноrо
влаrалища; коrда rенитальные cerMeHTbI на..
полнены яйцами, они образуют обширный,
выпирающий наружу яйцевой мешок. Как
щитни. так и жаброноrи обычно встречаю..
тся в водоемах с суровыми условиями
жизни (например, в пересыхающих лужах
и соленых озерах), поэтому у них развились
высокоспециализированные покоящиеся
формы. Яйца этих ракообразных хорошо
переносят сильные колебания температуры
и пересыхание, а в состоянии покоя способ
ны пребывать у некоторых форм по 10 лет.
Среди 850 известных современных видов
большинство имеет размеры до 3 см, лишь
отдельные достиrают 10 см в длину.
8.6.3.3. Класс Remipedia
Этот класс включает единственный
вид слепоrо рачка, описанноrо из одной
морской пещеры в 1980 r. (рис. 8.39); о ero
образе жизни до сих пор почти ничеrо не
252
Антенна rоловоrрудь
/
Боковые плавательные конечности
\
Ноrочелюсти
известно. Мелкое (меньше 3 см в длину) BЫ
тянутое прозрачное тело состоит из KOpOT
кой, лишенной карапакса rоловоrруди,
включающей rолову и первый туловищный
cerMeHT и длинноrо туловиша из более чем
30 похожих друr на друrа cerMeHToB, каж
дый из которых несет пару боковых листо
видных конечностей, служащих для плава
ния дорсальной стороной вниз. Среди при
датков rоловы необычны палочковидные
выросты спереди от антеннул и хвататель
ные ротовые орrаны, в том числе ноrочелю..
сти. Сейчас специалисты собираются опи
сать еще восемь видов; все они обнаружены
в морских пещерных местообитаниях в А т..
лантике.
8.6.3.4. Класс Mystacocarida
Мистакокариды (рис. 8.40) KpOXOTHыe
(менее 1 мм длиной) вытянутые, лишенные
пиrментации интерстициальные морские
рачки. Их характерные признаки прежде
Ноrочелюсrь
ЗУБчатая
Бороздка
Тельсон
<1'урка
Рис. 8.40. Мистакокарида, вид сверху (по
Kaestner 1970)
rлава 8
<1'урка
Рис. 8.39. Ремипедия, вид
с вентральной стороны (по Ya
ger, Schram, 1986)
Bcero разделение rоловы на небольшую
переднюю и обширную заднюю части,
а также туловище из 10 cerMeHToB, первый
из которых несет ноrочелюсти, хотя и не
срастается с rоловой. rоловные придатки
крупные (они используются для локомо"
ции), а придатки брюшка или редуцирова..
нь] до мелких одночленистых структур (на..
пример, на 11 V cerMeHTax).. или вовсе от..
сутствуют; на тельсоне мощная клещевид..
ная фурка. К примитивным признакам
rРУППЬJ относятся парные туловищные raH..
rлии, которые лишь соприкасаются между
собой, но не слиты. отсутствие сложных
rлаз, пищеварительных дивертикулов (ве..
роятно, по причине малых размеров тела).
Еще одна их любопытная черта уникальна
среди ракообразных: на каждом туловищ..
ном cerMeHTe находится пара расположен..
ных по бокам поперечных бороздок с зубча..
ты ми краями: назначение их не выяснено.
Все десять известных видов относят
к одному отряду.
8.6.3.5. Класс Branchiura
(карпоеды, или карповые вши)
Представители этоrо класса не..
большие (меньше 3 см длиной) формы,
часть жизненноrо цикла проводящие в каче..
стве эктопаразитов на морских и пресно"
водных рыбах; они прокалывают покровы
хозяина мелкими мандибулами и питаются
жидкой пищей чаще Bcero кровью, как ко..
мары. Их сильно уплощенное дорсовентра..
льно тело состоит из rоловоrруди (rолова
и приросший к ней первый rрудной cer..
мент), TpexcerMeHTHoro переона и двуло"
пастноrо нерасчлененноrо брюшка; rолово..
rрудь, а иноrда и большая часть пере она по..
крыты обширным плоским карапаксом
в виде круrлоrо, двудольчатоrо или напо..
Беспозвоночные с конечностями: членистоноrие и др.
Карапакс
253
Максиллярная присоска
Дыхательные участки
лавательные конечности
СРурка
А ВИД С ВЕНТРДЛЬНОЙ СТОРОНЫ
Максиллярная
присоска
Максилла II
Карапакс
Брюшко
Б ВИД С ВЕНТРАЛЬНОЙ СТОРОНЫ 8 ВИД С ДОРСАЛЬНОЙ СТОРОНЫ
Рис. 8.41. Морфолоrия и разнообразие карпоеДО8 (по Kaestner, 1970)
минающеrо наконечник стрелы щита, про..
стирающеrося в стороны или в стороны
и назад (рис. 8.41) и несущеrо пару сложных
rлаз.
Придатки rоловы MorYT быть либо силь
но уменьшены, либо превращены в орrаны
прикрепления к телу хозяина; они заканчи"
ваются крючьями, или же, как в случае мак..
силл 1. крупными стебельчатыми присоска..
ми. Все четыре пары rрудных придатков,
включая конечности cerMeHTa, полностью
или частично слитоrо с rоловой, представ..
ляют собой плавательные ноrи; на брюшке
придатки полностью отсутствуют. Неско..
лько необычно то, что самка не носит на се..
бе яйца до вылупления молоди, d прикре..
пляет их к субстрату или к донной растите..
льности.
Единственный отряд насчитывает по..
рядка 150 видов.
8.6.З.6. Класс Copepoda (веслоноrие)
Веслоноrие доминируют в морском и
в несколько меньшей степени в пресновод"
ном планктоне. К ним относятся и мноrочи..
сленные интерстициальные формы, а при..
мерно четверть всех известных видов
паразиты самых разных животных от ry..
бок до китов. Хотя в большинстве случаев
это мелкие рачки (меньше 2 мм длиной),
254
rоловоrрудь
Антеннула
МаНДИБула
Антенна
Просома
rлава 8
Максилла 1
Максилла П
Ноrочелюсть r рудные
плавательные
А конечности
Б
длинd одноrо свободноживущеrо вид 2 см,
а одноrо зктопаразита 0,3 м.
В типе тело веслоноrих ёостоит из rоло
вы с хорошо развитыми ротовыми придат
ками и антеннами, шестисеrментной rруди
с плавательными конечностями и брюшка
из пяти cerMeHToB без придатков. Однако
cerMeHTbI MorYT срастаться между собой са..
мыми различными способами (по меньшей
мере один rрудной cerMeHT всеrда слит с ro--
лов ой, а у мноrих форм в состав rоловоrру--
ди входит и второй), так что исходное раз
деление на rоловоrрудь, переов и брюшко
не обязательно отражает фактическое рас--
членение тела. если в нем вообще сохраняю
тся обособленные отделы. Так, у паразити
ческих видов можно видеть различные ста..
дии морфолоrической деrенерации, причем
в крайних случаях утрачивается всякая
внешняя сеrментация и исчезают придатки.
у свободноживущих и ряда паразитических
форм наблюдается функциональное разде
ление тела на заднюю область и сочленен
ную с ней переднюю. За исключением ин
терстициальных цилиндрических форм,
передний отдел «<просома») овальный,
иноrда удлинеННО ОВdЛЬНЫЙ, в то время как
Рис. 8.42. Морфолоrия весло
ноrих. А. Внешний вид свобод
ноживущей формы сбоку; по
казаны придатки. Б. Место co
членения просомы с уросомой
(показано стрелкой) у разных
веслоноrих; rрудные cerMeHTbI
пронумерованы, номера cer
ментов, слитых с друrими OT
делами, взяты в скобки (по
Kaestner, 1970)
задний (<уросома» ) тонко--трубчатый (рис.
8.42). Хотя в одной из rрупп rраница между
ними и в самом деле совпадает с делением
на rоловоrрудь и пере он + брюшко, 80
мноrих случаях она проходит между [11 и IV
сеrментами переона (соответствующими
V и VI туловищным cerMeHTaM), так что по-
следний cerMeHT пере она (и rруди) входит
в состав уросомы Карапакс и сложные rла-
за у веслоноrих всеrДd отсутствуют.
Паразитические формы MorYT иметь ca
мый разнообразный облик (рис. 8.43), при-
чем некоторые червеобразные или мешко-
видные виды можно отнести к ракообра-
зным лишь по признакам ранних стадий их
жизненноrо цикла. Они часто носят на себе
яйца в виде длинных тяжей в отличие от
свободноживущих веслоноrих, у которых
один два овальных яйцевых мешка.
Около 8400 видов мировой фауны rруп-
пируют в девять отрядов, из которых один
помещен в данный класс лишь условно. Эти
слабоизученные ра чки танту локариды
крошечные эктопаразиты морских рако-
образных, живущие постоянно на теле хо-
зяина, прикрепившись к нему ротовым ди-
ском; rоловные придатки у них ОТСУТ-
Беспозвоночные с конечностями: членистоноrие и др
255
Рис, 8.43. Разнообразие формы тела и ero деrенерация у паразитических веслоноrих (по разным
авторам)
ствуют, а из яйца выходит продвинутая ли
чинка танталус, осуществляющая зараже--
иие. Самка, по сути лела, представляет со--
бой мешок с яйцами, удерживаемый на теле
хозяина личиночной rоловой, а свободно--
живущий самец развивается из массы де..
диффереНllированной ткани внутри личин
КИ. Высказывалось предложение выделить
эти виды в самостоятельный класс Tantulo..
carida, возможно родственный Copepoda
или Cifl 4 ipedia.
8.6.З.7. Класс Cirripedia (усоноrие)
Усоноrие наиболее видоизмененная
rруппа ракообразных. представленная либо
сидячими формами, либо паразитами. Они
по сути лишены rоловы, у большинства OT
сутствует и брюшко, а от наружной cerMeH..
тации в лучшем случае сохраняются лишь
следы. Крайняя специализация наблюдае..
тся в 01ряде корнеrоловых (Rhizocephala),
представители KOToporo больше Bcero напо..
минают rриб"трутовик с сетью тонких тру--
бочек, распространяющейся по всему телу
хозяина (почти всеrда lесятиноr'оrо ра--
ка), и наружным мешком, [де располаrаю
тся [онады (рис. 8.44, А).
Слабее Bcero структурная специализация
в отряде Ascothoracica (рис. 8.44, 6). куда
относятся паразиты книдарий и иrлокожих.
у немноrих видов здесь сохраняются даже
рудименты rоловы, несущей антеннулы
с клешнями (что совершенно нетипично для
ракообразных). и [рудь с шестью парами
плавательных конечностей, покрытыми
двустворчатым карапаксом, из..под которо"
ro наружу выступает пятисеrментное брю--
шко. Наиболее известные представители
усоноrих морские желуди и морские
уточки (отряд Thoracica) прикрепленные
орrанизмы; шесть пар их [рудных Hor «(уси
ков») образуют фильтрующий аппарат лля
сбора пищи, а бочковидный карапакс уси--
лен известковыми пластинками (от несколь..
ких до множества) (рис. 8.44, В, Т), не сбра--
256
rлава 8
Внутренние ветвя иеся отростки
стеБелька (показаны лишь на
Одной стороне краБа)
Тело паразита
OTorHYToe БрЮШКО краба
А
Карапакс
Мышца аддуктор
Клешненосные
антеннулы
Б
Выводковая
камера
СтеБелек
Ротовой конус
Известковые
пластинки
домика
Мышца
ретрактор
; --,
I JJ -
\ ';..<:"'.. . I
:: " ;3!' I
' ,ry.. -
>-. ,,
Яйцевод
8
"'"
ело Антеннула
r
Рис. 8.44. Усоноrие. А. Вид с вентральной стороны краба пораженноrо представителем KopHero
ловых (покровы краба показаны прозрачными). Б. Схематический вид сбоку представителя Asco
thoracica (левая створка карапакса удалена). В. Морской желудь вид сбоку (часть карапакса и ero
пластинок удалены; см. также рис. 8.36). r. Внешний вид двух морских уточек прикрепившихея
к деревянному бруску (А и Б по Kaestner, 1970; В и r по Zullo, 1982)
Беспозвоночные с конечностями: членистоноrие и др.
257
еываемыми при линьке, а нарастающими
по краям. Как и у прочих усоноrих, перво..
начальное прикрепление к субстрату (ска..
лам, раковинам моллюсков или к водоро"
елям) осуществляется антеннулами; затем
предротовая область может увеличиться,
образуя вытянутый столбик (как у морских
уточек), или превратиться в тонкий прикре..
пительный диск, как у морских желудей.
Четвертая rруппа Acrothoracica в об..
щем похожа на предыдущую, но здесь от..
сутствуют известковые пластинки, и рачки
вбуравливаются в кораллы или, реже, в ра..
ковины моллюсков. У представителей двух
этих «желудеобразных» отрядов пенис, что..
бы достиrать соседних, также неподвижно
прикрепленных особей. бывает очень длин..
ным. У пятоrо отряда (Facetotecta) из вест..
ны только личинки, так называемые У..
науплиусы и У "циприды.
В этом классе заметна явная тенденпия
к rермафродитизму или к уменьшению
Рис. 8.45. Ракушковые рачки:
А внешний вид сбоку; Б то
же с удаленной левой створкой
карапакеа (по Cohen, 1982)
мужских особей до крошечных размеров,
а также к редукции кишечника: у мноrих ви"
дов он оканчивается слепо, а у корнеrоло..
вых и вовсе отсутствует. У всех усоноrих
имеется специфическая личиночная стадия
(после науплиуса) «циприсовидная», ко..
торая находит хозяина или место прикре..
пления на субстрате, а затем в ходе мета..
морфоза превращается в молодую взро--
слую особь.
Все известные виды (около 1000) оби..
тают в морях.
8.6.3.8. Класс Ostracoda
(ракушковые рачки)
Остракоды очень мелкие. большинство
их не достиrает в длину и миллиметра,
лишь немноrие приближаются к 2 см. У них
короткое овальное тело, заключенное в дву"
створчатую, часто известковую раковину,
образуемую карапаксом (рис. 8.45). Как и
Антенна
СРурка
А
ОБласть
прикрепления МаНДИБула Мышца аддуктор
карапакса
Б
Максилла 1
Максилла II
Первая пара
rрудных Hor
258
у раковинных рачков (Conchostraca) и ДBY
створчатых моллюсков, створки удержи..
ваются в сомкнутом состоянии поперечной
мышцей аддуктором, противодействующей
упруrому лиrаменту. Иноrда присутствуют
зубы замка. Однако в отличие от упомяну..
тых rрупп остракоды сбрасывают раковину
и образуют ее заново при каждой линьке.
На их мешковидном теле не заметно
сеrментации, но, если судить по придаткам.
половину ero объема занимает то, что paHЬ
ше было rоловой. Сохраняется Bcero от
пяти до семи пар придатков, причем первые
четыре из них несомненно принадлежат «ro
лове» (антеннулы, антенны, мандибулы
и первые максиллы), а две задние (одна или
обе MorYT отсутствовать) «rруди»; что же
касается лежащей между ними пятой пары,
взrляды специалистов на ее природу pacxo
дятся. Большинство считает эти придатки
второй парой максилл, некоторые
rрудными конечностями, т. е. ноrочелюстя
ми или первыми торакальными ноrами.
Если это, в самом деле, вторые максиллы,
ракушковые рачки оказываются единствен
ной rруппой ракообразных, в которой MO
rYT сохраняться только rоловные придатки.
Функции этих конечностей у разных форм
неодинаковы, но антеннь] обычно являются
основными плавательными орrанами,
а первые [рудные конечности.. если присут
ствуют, часто бывают ходильными. Яйца,
как правило, вынашиваются до вылупления
под карапаксом, хотя ряд видов просто при--
крепляет их к субстрату.. в том числе к под--
водным растениям.
Остракод делят на пять отрядов (из них
один описан лишь по пустым раковинам).
Bcero известно 5700 видов. по большей ча
rлава 8
сти морских. Есть и несколько наземных
форм" перешедших из пресных водоемов
к жизни в слое лесноrо rYMyca или подстил"
ки.
8.6.3.9. Класс Malacostraca
По объему этот класс HaMHoro превы"
шает все прочие и насчитывает примерно
23000 видов. По видимому. TaKoro разно..
образия форм тела нет больше ни в одном
классе животноrо мира; лишь один из 16
входящих сюда отрядов Decapoda
включает столь несхожие друr с друrом ор-
rанизмы, как крабы, речные раки, креветки
и раки отшельники. rлавный признак, обь..
единяющий представителей зтоrо класса
тело из rоловы, восьмисеrментной rруди
и брюшка из шести (редко семи) cerMeHToB,
причем все перечисленные отделы снабже-
ны полным набором придатков. Наблюдае-
мое разнообразие обусловлено тем, что
разное количество rрудных cerMeHToB (от
нуля ДО восьми) может объединяться с [о-
ловой в rоловоrрудь" О 3 пары rрудных
придатков бывают преобразованы в Horo..
челюсти" карапакс имеется или отсутствует
(как первично, так и вторично), а если он
есть, то покрывает от первых двух до всех
восьми [рудных cerMeHToB и до нескольких
брюшных.
К числу типичных черт специализации
Malacostraca относятся, во..первых, разви-
тие в передней кишке «желудка»" rде пища
тонко перемалывается" а все оставшиеся
крупные частицы отфильтровываются от
получающейся массы, которая затем посту-
пает в пищеварительные дивертикулы (рис
8.46). а во--вторых. преврашение послед-
Клапан
Пилорический
отдел желудка
--!адняя кишка
Кардиальный
отдел желудка
Пищевод
рительноrо
дивертикула
Рис. 8.46. Схематический про
дольный разрез через желудок
десятиноrоrо рака; видны же
вательные пластинки, измель
чающие проrлоченную пищу,
а также фильтрующая система
пилорическоrо отдела, при
крывающая отверстие одноrо
из пищеварительных диверти-
кулов (по Warner, 1977)
Беспозвоночные с конечностями: членистоноrие и др.
Мышца...
аддуктор
Карапакс
I
I
СРурка
А
r олова
259
Карапакс
Б
Хвостовой веер
Выв одковая камера
с яицами
r
Рис. 8.47. Разнообразие Malacostraca. А. Phyllocarida, вид сбоку (карапакс как бы прозрачен). Б.
Hoplocarida, вид сверху. В. Syncarida, вид сбоку r Pancarida, вид сбоку; карапакс изображенной
самки, используемый в качестве выводковой камеры, сильно увеличен и вздут (по разным aB
торам)
ней пары брюшных придатков (<<уропод»)
и тельсона в хвостовой веер, которым
и оканчивается тело вместо более обычной
для ракообразных фурки. Характерно TaK
же наличие пары хорошо развитых слож
ных rлаз, иноrда вторично утрачиваемых.
Мноrие виды крупные, с сильно обызвеств
ленными покровами, у некоторых наблю
дается значительная концентрация нервной
системы и сложное поведение. Malacostra
са существенный компонент MopcKoro He
ктон':! и бентоса; мноrие виды встречаются
в пресных ручьях, реках и озерах, друrие ве..
дут наземный образ жизни (сюда относятся
все ракообразные, способные выживать не
только в постоянно увлажненных местооби..
таниях).
Можно выделить шесть rрупп отрядов.
В двух из них сохранился исходный план
строения тела (т. е. rолова, [ру дь и брюшко)
без всякой тенденции к образованию rоло..
воrруди или добавочных ротовых частей.
В одной rруппе (Phyllocarida, рис. 8.47, А)
наблюдается нетипичный для этоrо класса
признак: VII брюшной cerMeHT и фурка,
а уроподы отсутствуют. Наиболее харак"
терный внешний признак филлокарид
крупный, сжатый с боков двустворчатый
карапакс. покрывающий [рудь и восемь пар
ее листовидных конечностей, щетинки кото..
260
А
Кар'апакс
Сидячие
сложные
rлаза
д
rлава 8
СтеБельчатые Карапакс
сложные rлаза
" .....,. :' :.;.,;::.... ;... . ,:.. ':'
,.' 'i'
Рис. 8.48. Разнообразие Malacostraca. Peracarida: A Tanaidacea; Б Mysidacea; B
Spelaeogriphacea; r Cumacea; д Isopoda (равноноrие раки; см. также рис. 8.36. В); E
Amphipoda (бокоплавы) (А и Д вид сверху, прочие сбоку) (по разным авторам). Обратите
внимание на неодинаковую степень развития карапакеа и разную форму придатков
рых окружают выводковую камеру. ВО BTO
рой rруппе Hoplocarida (раки боrомолы)
(рис. 8.47, Б) карапакс rораздо мельче
и закрывает лишь половину дорсовентраль
но сплюснутой rруди. Особенно xapaKTep
но, что первые пять пар rрудных конечно
стей здесь несут ложные клешни и не связа
ны с локомоцией; вторая пара очень круп
ная, хватательная, а последние три пары
у самок удерживают яйцевую массу У ряда
видов конечности второй пары превращены
в «кулаки», способные выбрасываться со
скоростью 1 CMjMC, нанося удар такой же
силы. что и пуля 22 ro калибра.
В отличие от двух этих rрупп следующие
три надотряда характеризуются слиянием
по меньшей мере одноrо (как правило, а са..
мое большее трех) rрудноrо cerMeHT3
с rоловой в rоловоrрудь, соответственно
переоном из семи cerMeHToB и брюшком из
Беспозвоночные с конечностями: членистоноrие и др.
в
261
Б
Вздутый карапакс,
служащий выводковой
камерой
Мяrкое,редуцированное,
асимметричное БрЮШКО
д
Е
Брюшко спрятано
ПОД карапаксом
Рис. 8.49. Разнообразие Malacostraca. Eu arida: А Euphausiacea; Б Amphionidacea; В
Ж десятиноrие раки (А В вид сбоку; r Ж вид сверху) (по разным авторам)
шести. Степень развития карапакса при
этом широко варьирует. Исключительно
пресноводные Syncarida (рис. 8.47, В) лише
ны ero вообще (изначально, как обычно по
лаrают) и демонстрируют тенденцию
к утрате придатков на достаточно OДHOpOД
ных туловищных cerMeHTax. Слепые Panca
rida (рис. 8.47, N, населяющие пещеры и ин
терстициальные местообитания (иноrда
в rорячих источниках с температурой воды
до 450 С), также в основном пресноводная
rруппа; они отличаются коротким карапак
сом, иrрающим роль выводковой камеры
и центра rазообмена. У третьей rораздо бо
лее обширной rруппы Peracarida (рис.
8.48) выводковая камера образуется OT
ростками конечностей переона. У предков
этоrо надотряда, вероятно. были карапакс
262
rлава 8
и сложные rлаза на стебельках, однако и то
и друrое претерпело сильную редукцию
в разных эволюционных линиях, так что
сложные rлаза стали по большей части си--
дячими, а в двух крупных отрядах
бокоплавов (Amphipoda) и равноноrих (Iso--
poda) исчез и карапакс. Перз.кариды
очень разнообразны по форме и местам
обитания: одни плавают. друrие роют нор--
ки, мноrие передвиrаются, используя при--
датки переона в качестве ходильных Hor,
есть и паразитические формы. Тело может
быть вытянутым или коротким. уплощен--
ным дорсовентрально, сжатым с боков или
«креветкоподобным». Мокрицы (из отряда
равноноrих) успешно заселили сушу (см.
разд. 8.6.2).
Последняя rруппа Eucarida (рис.
8.49) характеризуется включением всех
BOCЬM сросшихся между собой rрудных
cerMeHToB в rоловоrрудь, так что тело со--
стоит лишь из нее и брюшка, причем rоло--
воrрудь пеликом заключена в карапакс,
простирающийся обычно по бокам вниз
с образованием BOKpyr rрудных жабр за--
щитной камеры. Брюшные плеоподы, как
правило, используются для плавания, а
у самок десятиноrих раков для удержа--
ния яйцекладок, в то время как rрудные но--
rи служат для питания, ловли и удержива--
ния добычи и для локомоции. В наиболее
обширном отряде Decapoda (десятиноrие
раки) у особей три пары ноrочелюстей
и прослеживается тенденuия к репукции
брюшка, зашедшей дальше Bcero у крабов,
которые подrибают ero под крупный, часто
очень широкий карапакс, так что при
взrляде сверху оно совершенно незаметно.
Среди ракообразных только у крабов шири--
на тела подчас HaMHoro превышает ero
длину.
8.7. Дополнительная
литература
BOl ror D. J., De Long D. М., Trip/e/l01"n С. А. 1976.
Ап lntroduction to the Study of Insects. 4 th edn.
Holt, Reinhd.rt & Winston, New У ork.
Bouclreaux Н. В. 1979. Arthropod Phylogeny with
Special Reference to Insects. Wiley, New У ork.
С/lартаn R. F. 1969. ТЬе Insects: Structure and
Function. English Universities Press, London.
Clarke К. U. 1973. ТЬе Biology of the Arthropoda.
Edward Arnold. London.
Clouclsley T/loтpson J. L. 1968. Spiders, Scorpions,
Centipedes and Mites. Pergamon Prcss. Oxford
[Chelicerata, Myriapoda].
Da/y Н. V., Doyen J. Т., EhrZich Р. R. 1978. Intro
duction to Insect Biology and Diversity. McGraw
Hill, New У ork.
Gupla А. Р. (Ed.). 1979. Arthropod Phylogeny. Уап
Nostrand Reinhold, New У ork.
Kaeslner А. 1968. Invertebrate Zoology. (У 01. 2). Wi..
lеу, New York [Onychophora, Tardigrada, Penta
stoma, Chelicerata.. Myriapoda].
Kaeslner А. 1970. Invertebrate Zoology, tVo1. 3).
Wiley, New У ork [Crustacea].
King Р Е. 1973. Pycnogonids. Hutchinson. London.
Lеи'is J. с. Е. 1981. ТЬе Biology of Centipedes.
Cambridge University Press.. Cambridge.
Litt/e С. 1983. ТЬе Colonisation of Land. Cambrid
ge University Press. Cambridge [все наземные ap
троподы].
М anton S. М. 1977. ТЬе Arthropoda. Oxford Uni
versity Press, Oxford.
McLaughlin Р. А. 1980. Comparative Morphology
of Recent Crustacea, Freeman. San Francisco.
Raпlazzotti с. 1972. Il РЬуlиm Tardigrada, 2nd edn.
Istituto Italiano di Idrobiologia. Pallanza.
Savory Т. Н. 1977. Arachnida. Academic Press, New
у ork.
Schraт F. R. 1986. Crustacea. Oxtord U Пl vcrsity
Press, New У ork.
Sеdgи'iсk А. 1888. А Monograph of the Develop
ment of Peripatus capensis, and of the Species and
Oistribution of the genus Peripd.tus. Сlау, London.
ЧАСТЬ 111
ФУНКЦИОНАЛЬНАЯ Биолоrия
БЕСПОЗВОНОЧНЫХ
Если в предыдущей части было показано
морфолоrическое и биолоrическое MHoro..
образие беспозвоночных, то в этой основ..
ное внимание уделяется общим чертам их
функциональной анатомии, физиолоrии
и поведения. Совершенно ясно, что незави"
симо от строения, эволюции или эколоrии
все животные имеют некие общие потреб..
ности, обеспечивающие, по крайней мере
потенциально, выживание отдельных осо..
бей и сохранение их reHoB в следующих по..
колениях. В связи с этим животные распола..
rают эквивалентными наборами функцио"
нальных систем, позволяющих как полу..
чать необходимые ресурсы и информацию,
так и обрабатывать полученное, особым
образом упорядочивая er о. Возникшие
в процессе отбора селективные преимуше..
ства тех или иных планов строения, образов
жизни и местообитаний обусловливают не..
одинаковые способы решения этих обших
для всех проблем, поэтому, как и в преды..
дущей части, в rлавах этой функциональная
биолоrия беспозвоночных рассматривается
с учетом различных факторов отбора и оп..
тимальных вариантов взаимодействия с ни..
ми.
Некоторые потребности служат нуждам
непосредственноrо выживания, и в этих слу"
чаях отбор часто очень сильно действует на
особей. Например, животным необходимо
находить, потреблять и ассимилировать пи..
щу, содержащую необходимое вещество
и энерrию, нередко в условиях жесткой кон..
куренции за эти ресурсы (rл. 9) и в то же
время избеrать опасности стать пищей для
друrих животных (rл. 13). Друrие задачи,
обеспечивающие любому орrанизму саму
возможность существования, можно счи..
тать возникшими и разрешенными в 'Jволю--
ции большинства rрупп относительно рано,
поэтому сейчас они связаны в основном
с действием стабилизирующеrо отбора.
Так, большинство животных нуждается
в локомоторных системах, всем необходи--
мы подвижные части тела, чтобы захваты--
вать пищу, спасаться от хишников, избеrать
неблаrоприятные условия среды и Т. д. (rл.
10). Нужны также rазообмен с окружающей
средой и метаболические процессы, в ре..
зультате которых высвобождается энерrия
(rл. 11). Поскольку химический состав тела
не такой, как у окружающей средьУ. требуе..
тся реrулировать состав и/или концентра..
ции химических веществ в орrанизме, а зна
чит, и удалять «отходы» (rл. 12). Чтобы
мноrоклеточный орrанизм поведенчески
и физиолоrически проявлял себя как единое
целое (rл. (6), необходимо получать и оце..
нивать информацию о внутренней и внеш..
ней средах и, если нужно, реаrировать на
нее, координируя во времени и простран..
стве развитие и работу различных функцио--
нальных систем тела. Наконец (rл. 14 и 15),
животные должны обладать стратеrией вос"
производства и жизненным ииклом, позво..
ляющими передать максимум reHoB после
дующим поколениям, а их зиrоты или иные
пропаrулы должны в свою очередь развива--
ться в орrанизмы. которые сами способны
к воспроизводству, к потреблению и пере--
работке добываемых ресурсов; иноrда это
развитие включает своеобразные личиноч
ные стадии, приспособленные для распро--
264
Часть 111
странения" отыскания специфических хозяев
или питания до метаморфоза во взрослую
размножающуюся форму.
Поскольку исследованиями по всем
этим направлениям затронуто очень orpa..
ниченное число беспозвоночных, животные,
обсуждаемые ниже" составляют лишь He
большую часть от тех" что были описаны
в ч. 11. По вполне понятным причинам
экспериментальный материал относится
в основном к наиболее мноrочисленным
и относительно крупным среди беспозво..
ночных (членистоноrие, моллюски и др.).
Это, однако, послужит оправданием наше..
му кажущемуся невниманию к так называе..
мым «большим типам», обусловленному
подходом, принятым в ч. 11.
rЛАВА 9
ПИТАНИЕ
Питание животных представляет собой
до некоторой степени неразработанную
область исследования. Верно, что накоплен
оrромный материал.. например по механике
фильтрационноrо питания или о том, как
хищники охотятся за своими жертвами,
MHoro известно о строении пищеваритель..
ной системы и физиолоrии пищеварения, но
ответ на вопрос, почему животные питаю..
тся той или иной пищей, предполаrался оче..
видным поедается то, что можно добыть
и усвоить. Но почему большинство прими..
тивных типов животных почти исключите..
льно плотоядные? Почему так MHoro расти--
тельных тканей не потребляется? Почему
некоторые виды более «всеядны», чем дру"
rие? И почему некоторые болотные птицы
предпочитают мидий червям?
В этой rлаве мы рассматриваем филоrе..
нетические оrраничения, связанные с пита..
ни ем животных (эволюционное прошлое),
различные механизмы питания (наследие
прошлоrо), а также факторы, определяю..
щие выбор той или иной пищи в рамках ши..
pOKoro KopMoBoro спектра, и возможности
влияния жертвы на выбор хищника (эколо"
rическое настоящее).
9.1. Введение: эволюция
способов питания животных
Все типы животных (может быть, за ис..
ключением Uniramia) возникли в море,
очень СТ(lбильной и однородной среде. Если
не считать поrраничные приливно..
отливные и непосредственно прибрежные
зоны, такие физические факторы, как темпе..
ратура, соленость, ионный состав, насыще..
ние кислородом и т. Д., не представляют ни..
какой опасности для выживания морских
орrанизмов и не создают физиолоrических
оrраничений для их биолоrической активно..
сти. Поэтому жизнь обитателей моря лими..
тируется Bcero двумя, может быть, тремя
факторами, из которых большинство, если
не все, связаны с питанием. Животные ну..
ждаются в достаточном количестве пищи,
должны избеrать опасности стать пищей
для друrих и иметь определенное простран..
ство для удовлетворения первых двух по..
требностей. У спешное выживание особей
определяется только этим. Конечно, если их
[енам предписано существовать в течение
более длительноrо периода, чем жизнь од..
ной особи, необходимо также произвести
максимально возможное число потомков,
способных в свою очередь выживать и про..
изводить себе подобных (см. rл. 14). Все
анатомические, физиолоrические, биохими"
ческие или онтоrенетические особенности
лишь механистически способствуют повы..
шению шансов на решение этих rлавных
жизненных задач.
Что же моrло служить пищей для пер..
вых животных, населявших поверхность до..
кембрийскоrо MopcKoro дна (rл. 2)? Только
колониальные или одноклеточные бактерии
и протисты, которые во всех водоемах, кро"
ме самых мелководных, также были в ос..
новном rетеротрофами. Достаточно света
для фотосинтеза проникает в толщу воды
примерно на 100 м в открытом море, а на
мелководье обычно на меньшую rлубину
(20 30 м в береrовой зоне.. а иноrда лишь
несколько сантиметров в очень мутных во..
дах с илистыми взвесями). Большая часть
континентальноrо шельфа и все дно океана
вообще не знают солнечноrо света. Первич"
ная продукция орrаническоrо вещества фо..
тосинтетическими морскими протистами
266
образуется только в поверхностных слоях
воды, сильно удаленных от основных бен
тосных местообитаний. В наше время боль--
шинство донных животных почти полно--
стью зависят в конечном счете от оседания
сверху мертвых и уже частично разложив--
шихся тканей и от орrанизмов, обусловли
вающих их разложение как во время прохо
ждения через водную толщу, так и после их
попадания на морское дно.
Только в при6режных мелководных зо..
нах фотосинтезирующие орrанизмы были
непосредственно доступны животным, либо
отфильтровывающим их из взвешенноrо
в воде материала, либо заrлатывающим
прикрепленные формы. Если не считать фи--
лоrенетически изолированных rубок, ПИТй--
ние путем фильтрации было в эволюпии
животных rораздо более поздней специали..
зацией, чем обrладывание или ОЩИПЫВdние
(rл. 2). Вместе с тем хемосинтетическая дея--
тельность бактерий MopcKoro дна была, а
в некоторых местах все еще остается ва--
жным источником первичной продукции.
Бактериальный фотосинтез и хемосинтез
характерны для анаэробных или ми..
кроаэробных местообитаний, а также вос"
становленных субстратов.
Поэтому неудивительно, что наиболее
просто устроенные из доживших до наших
дней плоских червей и. по--видимому. их
предковые формы были и остаются потре--
бителями в основном бактерий и проти--
стов, связанных тем или иным образом
с донными осадками и камнями. Пожалуй.
более удивительно то, что явно и скрыто
ЭТОТ примитивный рацион доминировал
у всех форм, которым ДdЛИ начало плоские
черви, даже у пере шедших к наземному су--
ществованию. Численность бактерий и про--
тистов В морских осадках orpoMHa, но они
очень малы и часто относительно рассеяны
в пространстве.. редко встречаясь плотными
скоплениями, хотя бывают и исключения,
которые мы рассмотрим позже. Эти осо--
бенности имели два очень важных послед--
ствия для образа жизни животных
Во--первых, они должны обладать опре--
деленной подвижностью, чтобы отыскивать
новые источники пищи, коrда прежние ока--
зываются исчерпанными. Таким образом,
подвижность стала одним из условий жи..
rлава 9
BOTHoro образа жизни. Во--вторых.. потреби..
тель мелких, рассеянных в пространстве op
rанизмов и сам должен быть относительно
мелким. Чем крупнее животное, тем выше
ero метаболические потребности и тем бо
льше ему требуется пищи в единицу BpeMe
ни. Это можно видеть на rрафике зависимо
сти между размером тела и расходом эне
рrии, необходимой для ero существования
(рис. 9.1). Крупное животное не сможет про-
кормиться бактериями или мелкими проти-
стами, поскольку не сумеет потребить дo
статочное их количество в единицу времени.
Однако само увеличение размеров тела.
по--видимому., дает селективные преимуще
ства по трем причинам:
1) более крупные животные обычно спо
собны произвести больше потомства (Т. е.
повышается репродуктивный потенциал);
2) больший размер может повысить
шансы не быть съеденным друrими opra
низмами (т. е. повышается выживаемость);
3) более крупные животные., как прави
ло, MorYT оттеснить более мелких от orpa
ниченных источников общих пищевых рес-
урсов (т. е. опять же повышается выживае
мость).
Кроме Toro, при одинаковой усваивае..
мости 7нерrетически всеrда выrоднее поrло
щать крупные частицы данной пищи, чем
мелкие, а для этоrо тоже нужно обладать
большим размером.
Итак, с учетом селективных преиму-
ществ увеличения размеров неудивительно,
что большинство доживших до наших дней
(J
са
:т 1 мл
со
Q
..D
LD
О
()
О
N
О
cl) 1мкл
s:
:t:
Q)
с::
LO
cl)
а.
о
с:
1 нл
1 MKr
1r
1 Mr
Размер тела
Рис. 9.1. Связь между размером тела и интен-
сивностью метаболизма (по Philipson 1981)
(см. также рис. 11.16)
Питание
267
плоских червей крупнее видов, которые пи..
таются бактериями, и пищей им служат бо..
лее крупные объекты, т. е. животные. В на..
те время две наиболее значительные исход..
ные rруппы животных билатеральные
плоские черви и радиальные кишечнопо..
лостные представлены практически лишь
плотоядными формами. Тем не менее раз..
мер тела плоских червей оrраничен скоро..
стью диффузии (разд. 11.4.1), поэтому пре..
имущества, связанные с дальнейшим увели..
чением тела, делают выrодными любые
морфолоrические изменения, которые бы
сняли такое оrраничение. Это обстоятель..
ство и стремление избежать хищничества со
стороны плоских червей, возможно, и по..
служили двумя rлавными ТОЛЧКdМИ, при..
ведшими к широкому разнообразию черве..
образных форм, появившихся на ранних
этапах эволюции животных.
Однако в самых мелких морских водах
в изобилии присутствуют протисты в виде
нитчатых или слоевищных, колониальных
или мноrоклеточных водорослей. Хотя бо
лее крупные морские водоросли создают
ряд особых затруднений (см. ниже), нитча..
тые формы и ювенильные стадии макроско..
пических типов можно относительно леrко
отделить от субстрата, к которому они при
креплены. с помошью спепиальноrо соска..
бливающеrо opraHa (рис. 5.3), что делает
этот источник обильной и концентрирован..
ной фотосинтеЗИРУЮlцей ткани доступным
для потребления. Моллюски, одни из ран..
них эволюционных производных плоских
червей, использовали именно этот специа..
лизированный способ питания, обезопасив..
шись как от быстрых течений, характерн.ых
для мелких вод. так и от хищников дорсаль..
ной раковиной. Соскабливание с помощью
радулы является также эффективным сред..
ством добывания в качестве пищи сидячих
или прикрепленных животных.. защищен..
ных наружными раковинами, оболочками
или домиками, а также наземных растений,
и в течение своей последующей эволюции
моллюски распространили свой исходный
способ питания на большинство доступных
типов пищи.
Друrие рано диверrировавшие от пло..
ских червей животные либо сохранили пред..
ковый кормовой рацион, либо развили ме..
ханизмы для концентрации взвесей мелких
частиц, пр оплывающих мимо или осаждаю..
щихся из верхних слоев воды. Во мноrих
морских средах скорость осаждения и коли..
чество орrаническоrо материала.. взвешен..
Horo в воде, практически постоянны Зна..
чит, если консумент расположится так, что
обеспечит себе ero перехват и концентриро..
вание, он уже не будет нуждаться в большой
подвижности или даже в подвижности вооб..
ще, если не считать одной системы opraHoB,
и, став сидячим или прикрепленным, смо"
жет сократить энерrетические потребности
и. следовательно, необходимое количество
пищи на единицу веса тела. В результате от..
носительно крупные животные способны
существовать на рационе, состоящем в ос..
новном из бактерий. протистов и MepTBoro
орrаническоrо вещества.
Некоторые из этих ранних производных
плоских червей извлекали пищевые частипы
из водной взвеси. пропуская ее через некое
фильтрационное приспособление (потреби..
тели суспензий); друrие собирали 'Эти части..
цы и связанные с ними скопления микроор"
rанизмов уже после их оседания на поверх..
ность субстрата (потребители осадков); тре..
тьи перехватывали частицы во время их
пассивноrо оседания еще до Toro, как они
достиrали дна (потребители оседающих ча..
стиц) .
К потребителям суспензий относятся
rубки, лофофоровые, беспозвоночные хор..
довые, ряд представителей моллюсков,
кольчатых червей и членистоноrих, в том
числе некоторые формы, которые позже
в ходе филоrенеза перенесли с собой этот
способ питания в толщу воды вплоть до зо..
ны фотосинтеза. Пищеварительные адапта..
ции потребителей суспензий описаны в этой
rлаве ниже (разд. 9.2.5), а здесь мы остано..
вимся лишь на некоторых общих чертах их
питания. Всем им свойственны фильтр
определенноrо типа и либо ос060е положе..
ние участков захвата пищи, позволяющее
перехватывать естественный (и постоян..
ный) водный поток, либо приспособления.
создающие собственный ток воды через фи..
ль тр.
У rубок одни и те же клетки хоа..
ноциты создают ток воды за счет биения
своих жrутиков и улавливают из нее пище..
268
rлава 9
Пищеварительная
вакуоль
АмеБоидная клетка
мезохила
вые частицы, задерживающиеся на ворот--
ничке из микроворсинок BOKpyr жrутика
(рис. 9.2). Мелкий размер отверстий обра--
зуемоrо микроворсинками фильтра позво
ляет задерживать частицы, размер которых
меньше, чем у бактерий. Вполне очевидно,
что всем потребителям суспензий выrодно
фильтровать один и тот же объем воды не
более одноrо раза" и у rубок это обеспечи--
вается ее пропусканием через множество
мелких пор" распределенных по всей по--
верхности тела, и выведением через одно
или несколько крупных отверстий (см. разд.
3.2). Образующийся более сильный поток
ОТНОСИ1 профильтрованную воду от тела
rубки (у колониальных и планктонных orhe--
телок из оболочников с аналоrичной систе--
мой общий для всех зооидов выводимый
поток создает, кроме Toro, реактивную си--
лу, способствующую движению колонии
сквозь толщу воды; рис. 7.34).
В отличие от этоrо у разнообразных по--
требляющих суспензии потомков плоских
червей для фильтрационноrо питания адап--
тировались или образовались особые opra
ны щупальца на rолове или вблизи от нее
у аннелид, мезосомальный лофофор у фо--
ронид И их производных, сильно увеличен--
ные ктенидиальные жабры у двустворчатых
молюсков и т. д. (рис. 9.3). Однако принцип
действия здесь один и тот же. Биение ресни--
чек в определенных участках фильтрующе--
[о opraHa создает поток воды, в то время
Рис. 9.2. Хоаноциты rубок (по
Brill 1973)
кйк друrие ресничные поверхности осу-
ществляют захват и транспорт пищевых ча-
стиц ко рту с помощью слизи. Хотя захва-
тываются лишь чаСТИllЫ определенноrо
размера, имеются дополнительные реснич-
ные зоны либо на самом фильтре, либо на
каком--нибудь вспомоrательном opraHe,
в которых частицы сортируются на потре-
бляемые и отсеиваемые, часто в виде псев-
дофекальной массы. Эквивалентная систе..
ма есть и у беспозвоночных хордовых жи-
вотных (разд. 9.2.5).
Однако у членистоноrих фильтраторов
реснички отсутствуют в связи с наличием
экзоскелета, и вместо них функцию фильтра
выполняют щетинки на различных конечно-
стях (рис. 9.4). Поток воды, несущий пише-
вые частицы, создается как бы плаватель-
ными движениями фильтрующих или дру-
rих придатков. Фильтры из щетинок обыч..
но не такие мелкие, как ресничные, поэтому
захватывают только относительно крупные
частицы.
Все животные, использующие щупальце..
подобные структуры для питания суспен-
зиями (как и мноrие друrие фильтратары)"
являются прикрепленными или сидячими,
а их ловчие щупальца расположены в при-
нципе радиально--симметрично, обычно во..
Kpyr рта. Такая же функциональная система
наблюдается у первично радиальных книда-
рий (см. разд. 3.4). Тело обычно защищено
раковиной или трубкой или же помещается
Питание 269
Полихета (щупальца) Мшанка (лофофор)
Плеченоrое (лофофор) Двустворчатый моллюск
(ктенидиальные жаБРЫ)
Dq>иура (руки)
' , .' "
'. :с; '! ::.' ",:: : '::1. <, "::
. . "-' "o .... ... ). ....,:..
: ;,. :.!.;,;:.и.... .. " . .. ..:;." ....:. ,.
I 1 . ,,:, ;;r..f: ;' ','; ,f': "", ". '.
'.. .", ..:., .,." ,..: . . l' " ;,. .
':. ,..:.;.' rl '.. iJi:.'Y....'.,. ."',1
. . '.' ':0'. :.;:;;.;:.' ...:...;" :1 /' n'\ :'
'. ., ..:< <,....., j . '! ..',.1fJr.:.' "10;' о..,..
. .... /' ." " . " t;;:'. ,''>, \ ". ,:" .'i';.... :'; с.,
.." l',..!. .i:',," --'{,
. /. k):'.y,'.. " , .. ,
. 'f." , ' .';: ";;::. , . I .
С',' "::'i'i:C'
"; ,:' .,<;, . 1'" .
;. f\ .
' 1f t\If,
f; fj .
. \"',I'""f1
. rr f1f
.."
"\. \",р. ff
rолотурия (около ротовые
аМБулакральные ножки)
Рис. 9.3. Наружные opraHbI, используемые для слизисто ресничноrо типа питания суспензиями
(по различным источникам)
в норке, сделанной в субстрате. Это позво..
ляет животному и/или ero нежному филь..
трующему аппарату втяrиваться под обо..
лочку, обеспечивающую безопасность от
хищников. При таком полускрытом образе
жизни терминальный анус неизбежно при..
вел бы к прохождению фекальных масс
вдоль всей поверхности тела перед выносом
их во внешнюю среду. Поэтому у видов,
.
обитающих в трубках или эквивалентных
приспособлениях.. наблюдается заметная
тенденция к образованию U..образноrо ки..
шечника с расположением ануса, а часто
и выводных отверстий экскреторных opra..
нов вблизи переднеrо конца тела на пути
выходящеrо потока воды (см., например,
рис. 7.8). По тем же причинам сходной ана..
томической структурой обладают и мноrие
270
, I
'1 . f :
! -
l'
' t
Jf
(
Бокоплав (антенны)
(
'И
Веслоноrий рачок
(ротовой аппарат)
\\:),/ / ,
Неполнохвостыи рак
(ротовой аппарат)
rлава 9
(.
(,
о
о
... о
\
r
о
о
v
о
"
\.)
()
О. ')
J
Неполнохвостый рак
(антенны)
Эвсраузиевый рачок (конечности)
,/
Усоноrий рачок
(конечности)
Рис. 9.4. Системы щетинок, используемые различными ракообразными для фильтрации (по раз
личным источникам)
сидячие потребители осадков (см., напри--
мер. рис. 4.42). В друrих случаях у норки бы
вает по крайней мере два отверстия, и ее
обитатель создает через нее однонаправлен--
ный поток воды или же, что rораздо реже,
животное внутри трубки corHYTo пополам
так, что рот и анус оказываются у одноrо ее
выхода.
у некоторых аннелид, включая Nereis di-
versic%r и Chaetopterus. аппенликуларий из
оболочников, некоторых брюхоноrих мол-
люсков, эхиурид и т. д. фильтрующий аппа-
рат является не частью тела, d секретируе-
мой структурой в виде сети или мешка из
слизистых нитей, через которые пропускае-
тся поток воды, создаваемый движением
rолова
Рот
Секретируемыи
слизистыи мешок
Ресничная
дорсальная
БОрОЗДка
rлотка
Крыловидные
параподии
слизистый
мешок
ВИД СВЕРХУ
ПЕРЕДНЕЙ ЧАСТИ ТЕЛА
ВеерООБразные
выросты
....
....'.. ......
ВИД СБОКУ
А. ОБитающая В ТРУБках полихета Chaetopterus
Слизистый срилыр
Рот
Поток
воды
Крышечка
Раковина
Щупальца ноrи
f.; .Жf?"'i«\;\' ' t!6.i::::i; 'f'?< :iЙ
::J
:s:
-of
D,)
::t
:s:
CD
ХВОСТ
В. АппеНl1икулария
/
Б. ЧервеОБразный Брюхоноrий моллюск семейства Vermetidae
Секретируемая
слизистая ссрера
или воронка
.; ;.:::.: .:.: у.; . g; в и вырост
Раковина V
Д. Крылоноrий Брюхоноrий моллюск
"морская БаБочка"
r. Полихета Nereis diversicolor
Рис. 9.5. Фильтрационное питание с использованием секретируемых слизистых сетей (по различным источникам)
N
-....J
272
)КеЛОБОК ХОБОТОК
. :-...-....
......... .". ..........
fi :;: ;:е;.::.ц( tji t[ ;;f"v,rj,1! {j fi
rлава 9
Сифоны
.l; Ш[' ' ЙШN:;
; ;:«::: :..\::. \,} :;;./::
ХОБОТОК (эхиуриды)
Каптакула
ОБорчатые
rУБЫ
rолова
РОТ
Пища, скопившаяСЯ
в продольной Бороздке
Каптакулы (лопатоноrий МОЛЛЮСК)
:
Интроверт (сипункулиДы)
Ротовые
лопасти
.. .. .-. .. .
.- .. .. .." .. .... ..... .." ..: ".... .. .... ..: .'"...... .. ........ .... .'"
.'": :'". .. .. о" ........:.. :. .. .......: .. ..
.... '"....:.. ....:.......... ... . ... ...." ..
Ротовые лопасти (двустворчатый моллюск
подкласса Protobranchla)
специальных образований, служащих у дру"
rих животных локомоторными орrанами.
Через определенные периоды времени эта
сеть вместе с собранной на ней пищей съе..
дается, и животное секретирует новый
фильтр (рис. 9.5). У некоторых кораллов,
друrих брюхоноrих моллюсков и личинок
насекомых независимо образовались систе..
мы захвата пищи, сходные с этой, но испо..
льзующие в основном естественные, а не
специально создаваемые потоки воды.
Сбор пищи с поверхности субстрата по..
требителями осадков не требует таких сло..
жных приспособлений, как при питании су..
спензиями, и, действительно, некоторые че..
-.
.:.:
"!t!!i!!!ilti!1
Щупальца (полихеты)
Околоротовые щупальца
(модифицированные подии)
rолотурия (иrлокожие)
Рис. 9.6. OpraHbI обитающих в rpYHTe беспозво
ночных, используемые для слизисто рес
ничноrо питания осадками (по различным
источникам)
рви просто заrлатывают поверхностный
слой субстрата с помощью совершенно не..
специализированной ротовой области. Тем
не менее в ряде случаев, особенно у сипунку"
лид rолотурий и эхиурид. а также унекото..
рых аннелид и полухордовых около ротово..
[о отверстия развиты особые лопасти, щу"
пальца или растяrивающийся хоботок, по-
крытые ресничками и способные двиrаться
по субстрату или в нем, что позволяет более
эффективно собирать пищевые частицы
вблизи норок, [де живут такие формы (рис.
9.6). У потребителей осадков с лопастями
или щупальцами BOKpyr рта есть черты, об..
щие с потребителями суспензий (как отме..
Питание
чал ось выше в связи с U образным кишеч
НИКОМ)., и некоторые из них MorYT использо
вать оба этих способа питания. Однако две
проблемы касаются только потребителей
осадков: а) орrанический материал часто
составляет лишь малую часть последних;
б) основная часть самото орrаническоrо Ma
териала часто состоит из твердых и непере
вариваемых остатков, накапливающихся
именно из за использования их не животны
ми, а только бактериями (см. разд. 9.2.6).
В связи с этим, за исключением мелково
дий., тде на поверхности субстрата мотут
обитать бентосные фотосинтезирующие
протисты., потребители осадков., вероятно,
зависят от бактерий., способных превра
щать недоступную иначе орrанику в съедоб
ный материал, и от тех протистов и интер
стициальных животных, которые сами зави
сят от этих бактерий. Так или иначе. они BЫ
нуждены либо поrлощать большие количе
ства отложений (поэтому переоткладывают
бентосные осадки и являются мощными
аrентами биотурбаuии), либо проводить
большую работу по их сортировке и отбору
пищи. Щупальца мноrих потребляющих
осадки червей способны, например, вытяrи
ваТЬ9Я на большие расстояния от НОРКИ, [де
обитает животное. За исключением особо
«плодородных» участков, потребители
осадков не MorYT достиrать плотности Hace
ления, характерной для потребителей cy
спензий, у которых лимитирующим факто
ром часто является пространство, а не пи..
щевые ресурсы. Поскольку во мноrих слу..
чаях в конечном счете приходится полаrа
ться на продуктивность бактерий., а не на
скорость оседания детритных частиц, а про..
дуктивность бактерий в свою очередь MO
жет оrраничиваться не только поступле
нием уrлерода (например., дефицитом пита
тельных веществ в интерстициальных
водах), скорость роста потребителей ocaд
ков часто ниже, чем у потребителей суспен
зий.
Последняя rруппd. животных, питаю
щихся веществом, поступающим из поверх
ностных вод.. потребители оседающих ча
стиц, например некоторые стебельчатые
иrлокожие с телом., прикрепленным к MOp
скому дну, но значительно поднимаю
щимся над ним, чтобы перехватить частицы
273
СтеБелек
Система
прикрепле...
ния
СуБстрат
Рис. 9.7. Стебельковая морская лилия (иrлоко
жие), питающаяся оседающими частицами (по
Clark, 1915)
детрита до Toro, как они «разбавятся» дон..
ными отложениями (рис. 9.7). «Перехват»
производят несколько радиально располо..
женных рук, несущих rидравлически управ
ляемые и покрытые слизью сосочки; затем
по ресничным бороздкам пища направляе
тся к ротовому отверстию, расположенно
му в центре образованноrо основаниями
рук Kpyra.
Судя по выжившим представителям
древних rрупп, на очень ранних этапах исто
рии мноrоклеточных животных уже. Be
роятно, существовал еще один совершенно
особый способ питания. У некоторых MOp
ских обитателей мелководья в поверхност
ных тканях тела находятся симбиотические
оксифотобактерии (прохлорофиты и циано
бактерии) или фотосинтезирующие проти
сты (одноклеточные динофлаrеллаты или
хлорофиты). Питание некоторых плоских
червей и книдарий, по видимому, полно
стью зависит от этих симбионтов, хотя
в большинстве случаев они не способны их
переваривать. Такие бескишечные турбел
ларии., мяrкие кораллы и медузы постоянно
(сидячие формы) или временно (подвижные
формы) находятся на полном солнечном
свету. Их симбионты фотосинтезируют
274
: C#o
,,;; : , , ',
?' -- < - . .
rлава 9
,( ", . i > ," ' ' .'
-1>:.
.. ,
у. .
#Эяrодерм8' " .
Мезоrлея
J '
<if:
'.. .fo , '* .
, '"
'* !"," "" ..: l'
': 1__..
,'0 '
( :--
'"
.<>-;
.. : '
"i-
..
'<!о! Q ,:«
"" .
.: '-.. ..
*,
..
t ;,' t ..
t J«
i: t ;'
,. »
'... :
,
,.
J...
,&"
/1. .
ti. :,
,;; .с
"
j ,.
"':io' : .ttI»
'Ii':
..,: .
-i(>' ..
_ lt .
'{-'.:
. - )'
....f:,(?J?'
't:
;
./': .
.
': '\
;. а,.
'-" 'f ";t
о".." ' p
1' OiII
...+" "
.,,' r: .}; 4р .,!" .
4" ,. . "'J <C
,п.... '" ,.. ,
/. , "" "1" . ;; Jf
., ...Х7"" _' . '"
j :.. - "C _':
,.
;,.
. . ; f
... ....'!t'
:' )'
. t 'i"
.,.......' '. ,О,
</': " "
t
;
4. .."'
,#." "",
'1' :.'" '
.
#<' "
..а s:
. у
V ;.- :r:
"
.,
11
... , .i# <.:
J
. KTfJ,Aept1 '
. ,:>:' " '
. '. ..
,' -':.
O , ,.
. f*..
, .,.}
, '. .
, ,"')
. ".'
t
'..
<1" "//
;/
..' .t, :.'" ,;: ,.1
': ' ."-' .' .' .t.
! ".
.-- J ' --
6<
. f .',
/ " !t J. .
# '.
:>15'- i:
. .
..J..,( -
'J
.... " , 6If',
, '4 i,
.. % ..
.. '
t
...:
\(. :1
. , .
.". т:»
"";. '.. <'"
;j4' ,
it.
.' :.: /.If.:- f"
. "
Рис. 9.8. Электронная микрофотоrрафия водорослей симбионтов в тканях книдарии (Muscatine
et al., 1975)
обычным способом, но как источник мине..
ральноrо питания частично используют He
орrанические азот и фосфор, образующиеся
из орrанических соединений в ходе метабо..
лизма хозяина, а в качестве источника уrле
рода уrлекислый rаз, выделяющийся
в процессе ero дыхания. По--видимому, жи..
вотное..хозяин каким"то образом делает
клеrочные стенки симбионта более тонкими
и/или более проницаемыми, чем в норме,
и поэтому часть веществ, образующихся 3d
счет фотосинтеза (сахара, жиры и аминоки..
слоты), диффундирует в ткани хозяина. [де
dссимилируется продукты их распада за--
тем снова путем диффузии попадают
в клетки симбионта. Таким образом живот
ное «доит» этих автотрофов, плотность ко..
торых может достиrать 30000 в 1 мм 3 ткани
хозяина (рис. 9.8). Друrие виды морских жи
вотных хотя И частично, но также ЗdВИСЯТ
от фотосинтезирующих симбионтов.
Аналоrичным образом некоторые rлу--
боководные поrонофоры целиком или отча..
сти зависят от симбиотических хемоавто..
трофных бактерий, использующих восста..
новленные соединения серы и метан, выде..
ляющиеся из жерл и фумарол MopcKoro дна.
Недавно было обнаружено, что различные
олиrохеты и двустворчатые моллюски.
связанные с друrими, также боrатыми суль..
фидами, морскими местообитаниями, полу
чают около 500/0 фиксированноrо уrлерода
от бактерий. окисляющих серу. Тот факт.
что симбионты участвуют в питании пред
ставиrелей большинства типов животных,
причем особенно широко в rруппах, счи
тающихся наиболее близкими к предконым
формам. дает основание предполаrать, что,
хотя симбиотическое происхождение MHO
rоклеточности и спорно (1 л. 2), своему BO
зникновению и успеху древние rруппы жи
вотных моrли быть в значительной степени
обязаны симбиозу и, вероятно, особенно
тоrда, коrда пищевые ресурсы были оrрани"
чены (и в самом деле хлоропласты зеленых
водорослей, по видимому, произошли от
друrих, коrда то эндосимбиотических, эука..
риотных водорослей, а если это так, сим
биотическое объединение отдельных эука
риотных орrанизмов моrло дать начало не..
которым друrим эукариотам).
Способы питания и типы пищи, описан..
ные выше, были, по всей вероятности, в Te
чение сотен миллионов лет единственными
доступными свободно живущим животным,
и в наше время обитатели морей кормятся
Питание
в основном живыми и мертвыми бактерия..
ми, протистами и друr друrом. В конце кон..
цов возникшие в море формы 1аселили су..
ту, И, хотя наземные бактерии, протисты
и друrие животные моrли обеспечить им
практически такой же рацион (с поправкой
на неэффективность потребления суспензий
в наземных условиях), в новых местообита..
ниях они столкнулись С новым, обильным,
однако совершенно особым источником пи..
щи наземными бриофитами и сосудисты--
ми растениями. Серьезная проблема, встав..
тая перед их потенциальными потребите..
лями, заключалась в том. что большая
часть биомассы растительноrо орrаниз"
ма rрубые неперевариваемые клеточные
стенки из целлюлозы и почти инертных хи..
мически структурных полимеров rипа лиr..
нина. Предки большинства животных ни..
коrда не нуждались в целлюлазах или сход..
ных ферментах для усвоения пищи, и пер
вые наземные формы, во"видимому, оказа..
лись абсолютно неприспособленными для
использования этоrо обильноrо, но химиче..
СКИ стойкоrо источника пищи. Только в ме..
ловом периоде растительноядность широко
распространилась среди наземных живот..
НЫХ. Однако небольшое число морских ви"
дов уже до некоторой степени решило очень
близкую проблему. Крупные водоросли
морских мелководий также содержат ело..
жные уrлеводы, обсспечиваюшие устойчи"
вость к сильным движениям воды. Эти Ma
кроскопические формы так же потенциаль..
но приrодны в пищу, как и большинство
тканей наземных растений. но потреблять
этот в целом непривлекательный корм, по..
ка он живой, способны только две rруппы
морских животных.
Первая из них это некоторые морские
ежи. Большинство потребителей морских
водорослей нуждаются в обязательном
предварительном расщеплении их прочных
полисахаридов бактериями до усвояемой
формы. К таким животным относятся клас--
сические потребители осадков. Бактерии,
особенно анаэробы, способны разлаrать са..
мые разнообразные орrанические молеку"
лы, в том числе из нефти и мноrих пласт..
масс, однако, вместо Toro чтобы использо..
вать результаты lТИХ процессов, происходя
щих во внешней среде, некоторые морские
275
ежи перенесли их внутрь cBoero тела Спе..
циальный участок их кишечника содержит
культуру анаэробных бактерий. 3аrлатывая
rрубые ткани морских водорослей, морские
ежи обеспечивают бактерий пищей те раз--
лаrают эти ткани. и животные..хозяева пере--
варивают как продукты метаболизма бак..
терий, так и их самих. Следующая пробле--
ма, связанная с питанием морскими водо..
рослями, низкое содержание в них азота
на единицу веса, но в анаэробных условиях
некоторые бактерии способны фиксировать
атмосферный азот, растворенный в мор..
ской воде, т. е., живя в кишечнике иrлоко--
жих. MorYT повышать количество азота в их
преимущественно уrлеводной диете. Хотя
морские ежи как бы преадаптированы к на..
земной растительной пище. ни один из них
не перешел к жизни на с) II.tL . но мноrие из
наиболее успешных назеl\l11ЫХ фитофаrов
решили эту проблему сходным образом.
Друrая rруппа потребителей морских
водорослей зто БРI-Dхоноrие моллюски.
и неудивительно, что при их исходном
образе жизни на мелководье, включающем
соскабливание пищи с субстрата, они оказа..
лись среди немноrих rрупп животных. обла..
дающих ферментами. необходимыми для
разложения большоrо числа полимерных
уrлеводов, в том числе и целлюлазами.
В отличие от иrлокожих брюхоноrие стали
процветающей rруппой наземных фитофа
[ов, что может ldсвидетельствовать любой
садовод.
Хотя у некоторых сухопутных членисто..
ноrих есть целлюлазы, питание, основанное
на деятельности кишечных симбиотических
микроорrанизмов, характерно для мноrих
успешных наземных фитофа ов, включая
тараканов. термитов. некоторых жуков
и млеко питающих. Все они для переварива..
ния растительной пищи используют сим..
биотические бактерии и протистов, обеспе..
чивающих распад уrлевопных uепей и, или
поступление дополнительных количеств op
rаническоrо азота. Эти так называемые
«травоядные» животные столь же зависимы
от бактерий и простейших. как и бескишеч
ные плоские черви, сипункулиды и питаю..
щиеся осадками аннелиды
Однако не все части Растения одинакuво
химически устойчивы; например, молодые
276
фотосинтезирующие листья для животных
доступнее. И все же лишь немноrие из фито
фаrов MorYT разрушать стенки раститель..
ных клеток, чтобы добраться до их содер"
жимоrо, используя свой собственный фер..
ментный арсенал. Наземные беспозвоноч"
ные, не способные разлаrать целлюлозу
с помощью симбионтов или собственных
ферментов, выработали два особых метода
извлечения содержимоrо растительных кле..
ток или Рdстительных соков. rусеницы, ку"
знечики и друrие насекомые. питающиеся
листьями, откусывают их мелкие кусочки
или тонкие полоски и потребляют содержи..
мое клеток, разрушенных при разрывании
и последующем пережевывании тканей ра..
стения. Целые клетки при этом в пищу не..
приrодны, поэтому лишь относительно не..
большая доля (в среднем одна треть) отку..
санных частей усваивается и для возмеще..
ния низкой эффективности питания требуе..
тся переработать большое количество кор..
ма. Подобная система может существовать
только блаrодаря обильной биомассе ра..
стений. Второй способ основан на прони..
кновении в систему транспортировки жид..
костей внутри растения с помощью колю..
ще..сосущеrо pOTOBoro аппарата (рис. 9.9;
см. также рис. 8.30). Так питаются все рав"
нокрылые, например тли. Они избеrают
устойчивых химически структурных уrлево"
дов, однако иноrда нуждаются в симбиоти"
ческих кишечных бактериях для увеличения
I
j
1I
Стилеты 11
ti
l'
.f
U
Рис. 9.9. rолова клопа, питающеrося соками
растений; стилет введен в ткани растения (по
Barnes, 1980)
rлава 9
с их помощью количества азота в разбав"
ленной и бедной этим элементом кормовой
жидкости.
Растениям, используемым в пищу, оче
видно, были выrодны приспособления, пре
пятствующие потреблению фотосинтези
рующих тканей, и подобно друrим прикре
пленным орrанизмам (морским колониаль..
ным животным, например), мало приспое..
обленным к друrим способам обороны, они
развили сложную систему механической
и химической защиты (разд. 9.2.4). Как
обычно бывает в таких случаях, это поло..
жило начало «roHKe» наступательных и обо..
ронительных вооружений . образованию
токсинов у кормовых объектов и механиз..
мов детоксикации или устойчивости у их
потребителей.
Только одна катеrория фитофаrов
эффективно использует вещества растите..
льноrо происхождения без помощи симбио..
тических бактерий или протистов. Это те
виды, которые питаются веществами или
структурами, развившимися спепиально
для привлечения животных (нектаром, пло..
дами, семенами и т. д.), необходимых расте..
ниям для опыления и/или распространения.
Хотя плоды и семена в основном предна..
значены для позвоночных. членистоноrие
и моллюски также иноrда «приобщаются»
к этим ресурсам до Toro, как они съедены
птицами или млекопитающими.
[рибы с их хитиновыми клеточными
стенками и отсутствием прочных опорных
тканей не столь трудны для переваривания
животными и поэтому широко использую..
тся ими. Некоторые насекомые в этом пла..
не заходят столь далеко, что собирают ра..
стительные ткани, пережевывают их до
мяrкой массы и затем используют ее как
субстрат для выращивания rрибов, идущих
в конце концов в пищу. В какой"то степени
rрибы иrрают ту же роль, что и водоросли
в питании предков наземных животных.
Несмотря на широко распространенное
представление о преобладании на суше пи..
щевых цепей типа растение раститель..
ноядное животное плотоядное животное,
сложившееся в связи с большим внима..
нием. уделяемым в научных исследованиях
травоядным (особенно пастбищным млеко..
питающим), основу большинства наземных
Питание
277
пищевых сетей составляют не живые расти..
тельные ткани и не обrрызание их, аразла..
rающиеся орrанические остатки. Основные
потоки 1нерrии идут через животных, испо..
льзующих в пищу детрит и лесную подстил"
КУ С участием бактерий протистов и rрибов.
Таким образом наземные животные неда..
леко ушли от своих морских предков в смы"
сле способности «перерабатывать» различ..
ные типы корма, и даже сейчас фитофаrи
потребляют менее 3 О/о лесной ПрОДУКllИИ
в виде живоrо растительноrо материала.
9.2. Типы питания животных:
способы захвата
и переработки пищи
9.2.1. Классификация типов
питания
Влияние давних представлений о ва..
жней шей эколоrической роли цепей пита..
ния, включающих обrрызание растений, от..
ражено даже в названиях классических тро"
фических уровней. Мноrие тоды выделение
типов питания животных основывалось на
таких пищевых цепях и на систематической
принадлежности поедаемых орrанизмов;
отсюда, например, «растительноядные»,
«плотоядные» и (промежуточные ) «всеяд..
ные» формы. Подобная классификация по
рождает мното неудобств. Во..первых, ис..
ключительно растительноядных или пло
тоядных видов относительно мало, особен..
но если учитывать все стадии их жизненно..
ro цикла, т. е. большинство животных попа..
дают внеопределенную катеrорию «всеяд..
ных». Во"вторых, будучи основана на ста..
ром представлении о разделении живых ор..
rанизмов на два царства растений и жи..
вотных, она не решает вопроса о том, куда
отнести виды, питающиеся бактериями
и протистами и составляющие довольно об..
ширную rруппу. В"третьих филоrенетиче..
ские связи видов добычи необязательно ва..
жны для эколоrии питания ее потребителей.
Позже стало принято Рdзличать катето..
рии консументов по способам питания. Mo
жно rоворить о хищниках, паразитах, об
rрызающих и ощипывающих животных,
о потребителях суспензий и осадков, о пи..
тающихся с помощью симбионтов. Эта
классификация не учитывает систематиче..
ското положения орrанизмов, используе..
мых в пищу, например, «обrрызатели»
потребители суспензий и осадков мотут
поедать бактерий, протистов. rрибы. рас те..
ния и животных. Если способы питания жи..
вотных не делают различий между царства..
ми поедаемых орrанизмов, не следует де..
лать это и нам при анализе биолоrии пита..
ния Поэтому в следующем разделе мы от
них абстраrируемся и более детально рас..
смотрим особенности поrлощения и перева..
ривания пищи.
9.2.2. Общие черты
у большинства беспозвоночных «скво"
зной» кишечник с ротовым отверстием ра..
сположенным более или менее спереди
и служащим для захвата пищи, и анальным
отверстием, сдвинутым более или менее на..
зад и служащим для вывода из кишечника
во внешнюю среду непереваримых остатков
и экскретов. У некоторых животных кишеч..
ник заканчивается слепо, и единственное от..
верстие служит как для поrлощения пищи.
так и для освобождения от «отходов». Сю..
да относятся в основном книдарии и пло..
ские черви, но также и ряд друrих живот..
ных, у большинства из которых такое
строение носит вторичный характер. У дру"
rих форм (некоторые паразиты, но также
и свободноживущие виды) пищеваритель..
ный канал вообще отсутствует, и пища по..
ступает либо от внутренних симбионтов
либо через наружную поверхность тела.
Мното и отклонений от этих вариантов. На..
пример, у форм, ведущих прикрепленный
образ жизни, рот часто расположен в иен..
тре верхней стороны тела, а не в ето пере
дней части; у некоторых свободноживущих
форм, предки которых были прикрепленны"
ми например у свободно передвиrающихся
иrлокожих рот находится в центре нижней
поверхности тела, а у пищеварительноrо ка..
нала хордовых и ряда друrих животных на..
ружных отверстий множество (разд. 9.2.5).
С точки зрения развития пищеваритель..
ный канал животных включает три отдела:
эктодермальную переднюю кишку (обра..
278
Рот
/
Мышечные
стенки
Ротовая
полость
r лотка
Пищевод
Передняя
кишка
ЗОБ
Желудок
Пищеваритель... Средняя
кишка
ный дивертикул
Кишечник
Прямая
кишка
} Задняя
кишка
Анус
Рис. 9.10. Отделы пищеварительноrо тракта
обобщенноrо беспозвоночноrо (схема)
зуется при втяrивании части наружной по..
верхности тела), также эктодеРМdЛЬНУЮ
заднюю кишку и энтодермальную среднюю
кишку, rде происходят переваривание и Bca
сывание, часто снабженную слепыми дивер
lикулами. Каждый из этих основных OTдe
лов разделен на функционально различные
части (рис. 9.10). Передняя кишка может со..
стоять из а) ротовой полости, в которую Be
дет ротовое отверстие и открываются про..
токи «слюнных желез» (иноrда специализи..
рованных на образовании клейких секретов,
антикоаrулянтов или токсинов); б) мускули
стой rлотки, иноrда участвующей в поrло..
щении пиши.. либо действуя как насос, либо
образуя выворачиваемый opraH для CXBaTЫ
вания добычи, либо (как у хордовых) служа
фильтром при питании суспензией; в) KO
pOTKoro пищевода и/или ?) накапливающе..
ro opraHa зоба, особенно хорошо разви"
Toro у животных, поrлощающих через дли..
тельные интервалы времени большие коли
чества пищи, постепенно поступающей
в среднIOЮ кишку.
rлава 9
Средняя кишка обычно подразделяется
на а) мускулистый желудок, 6) различные
слепые секреторные и/или всасывающие от..
ростки и в) кишечник. В желудке происхо..
дит механическое измельчение и сортиров
ка проrлоченной пищи; иноrда в нем имее..
тся особый отдел для растирания пищевых
частиц. Иноrда здесь же начинается перева
ривание, но чаше переваривание и/или Bca
сывание протекают в различных дивертику"
лах слепых выростах, отходящих от ки
шки сразу за желудком. У моллюсков ира..
кообразных 1ТИ выросты преврашаются
в крупные сложные орrаны rепа
топанкреасы.
у примитивных форм переваривание
идет в основном внутриклеточно, коrда ча..
стиllы пищи фаrоцитируются клетками вы..
стилки абсорбирующеrо отдела пищевари..
тельной системы и расщепляются в их Ba
куолях Такой способ питания преобладает
у rубок. кишечнополостных, плоских червей
и ряда друrих животных, у которых пища
поступает в среднюю кишку в тонкоди..
сперсном состоянии. У более сложно opra
низованных животных, особенно у поrло
щающих пищу отдельными крупными Mae
сами, переваривание происходит в OCHOB
ном С помощью ферментов, секретируемых
в просвет кишки.. а ero продукты затем аб
сорбируются клетками ее стенки. У таких
видов дивертикулы имеют, как правило,
чисто секреторную функцию, и всасывание
происходит в кишечнике. Внеклеточное
переваривание способствует ферментной
специализации как отдельных клеток, так
и различных участков кишки, однако приво
дит к необходимости производить большее
общее количество ферментов. поскольку
просвет кишки крупная открытая сиете..
ма, в которой нелеrко поддерживать их ОП
тимальную для пищеварительных процее
сов концентрапию.
Наконец, задний отдел пищеварительно
ro тракта (если он есть) представлен прямой
кишкой (ректумом), [де может происходить
всасывание воды (у некоторых наземных
животных; см. рис. 8.27) и образование фе
КdЛЬНЫХ масс перед их выведением наружу
через анальное отверстие.
Все животные нуждаются в получении из
пищи энерrосодержащих веществ, исполь
Питание
зуемых немедленно или несколько позже,
аминокислот, необходимых для синтеза
структурных и метаболических белков,
и ряда друrих химических элементов и co
единений, например витаминов, участвую
щих в катализе биохимических реаКIlИЙ.
Друrими словами, животные должны пое..
дать уrлеводы, жиры, белки и витамины и,
если они не MorYT всасываться сразу, пере..
варивать их до усвояемых форм. Виды. пи..
ща которых бедна белком и/или витамина
ми, обычно нуждаются в симбиотических
кишечных бактериях, синтезирующих эти
вещества, а потребители сложных полимер..
ных yr леводов в сим биотических бакте..
риях и протистах, разлаrающих эти ве..
щества до более простых орrанических MO
лекул. Значительная часть фекалий состоит
из кишечных бактерий.
9.2.3. Охотники и паразиты
Охотники это подвижные животные.
которые атакуют, убивают и потребляют
одну особь..жертву (почти Bcer да также
подвижное животное) в данный момент вре..
мени. Среди охотников различают три
крупных типа: преследователи (например,
кальмар) доrоняют, хватают и обездвижи"
вают друrих быстро передвиrающихся жи..
BOTHЫX поисковики (мноrие rac1 роподы,
членистоноrие и т. д.) отыскивают менее
подвижную, чем они сами. добычу засадчи..
ки (такие, как пауки и боrомолы) большую
часть времени относительно неподвижны
и хватают жертву. совершая редкие рывки
или прыжки.
Преследователи и засадчики (в меньшей
степени поисковики) имеют специальное
вооружение для схватывания и обездвижи"
вания жертвы, Как правило, это либо opra..
ны, расположенные BOKpyr рта, например
клешневидные и ложноклешневидные при
датки у членистоноrих. руки rоловоноrих
с присосками или крючками, шипы у ще..
тинкочелюстных (рис. 9.11), либо сильные
челюсти, связанные с передней кишкой, как
у аннелид, rде они находятся на выворачи..
ваемой r лотке, которая с силой «выстрели..
вается» для захвата добычи. Некоторые за..
садчики привлекают жертв с помощью ми..
микрии. Например, у некоторых сифонофор
279
щупальца несут образования, напоминаю..
щие веслоноrих рачков. и их добычей стано..
вятся в основном животные, питающиеся
копеподами (обычно друrие ракообразные).
Пойманное животное либо заrлатывается
целиком. либо разрывается с помощью раз
личных придатков и заrлатывается по ча..
стям, либо из ero тела высасывается жид..
кость. Если жертва заI латывается целиком,
передняя часть пищеварительноrо тракта
способна растяrиваться в соответствии
с размерами пищи.
С друrой стороны, МНОI'ие поисковики
питаются относительно малоподвижными
животными. которые зашишаются от напа..
дения с помощью наружных покровов из
карбоната кальция, целлюлозы, хитина
и Т.д. (разд. 9.33 и 13.2.1). В связи с этим
у поисковиков иноrда развиваются специа
лизированные способы хищничества: про..
буравливание защитных покровов (напри..
мер, с помощью радулы у моллюсков) OT
ламывание их кусочков и выворачивание
в образовавшееся отверстие cBoero желуд..
ка, секреция на незащищенные ТКdНИ фер..
ментов и абсорбция продуктов наружноrо
пищеварения заrлатывание жертвы цели
ком и раздавливание ее раковины в желуд
Ke высасывание отдельных полипов или
100ИДОВ В колониях С помощью хоботков
или стилетов и rлоточноrо насоса (напри
мер. некоторые заднежаберные моллюски,
морские пауки и др.).
у некоторых охотников сосущий меха..
низм поrлощения пищи. В дополнение к уже
перечисленным rРУППdМ ряд хищных насе..
комых и прежде Bcero пауки высасывают
жидкость из своих жертв либо непосред"
ственно, либо сначала впрыскивая в пой..
манную добычу слюну. содержашую про..
теолитические ферменты (и парализующие
токсины), разжижающие ее ткани до TaKoro
состояния, что их можно всосать. Неболь..
шой переход отделяет описанный тип пита..
ния от эктопаразитизма, коrда животное
питается жидкостями хозяина, не убивая
ero caMoro. Катеrории охотников и парази..
тов различаются нечетко, и их разделение
в значительной мере опирается на относите..
льные размеры потребителя и объекта по..
требления. Пиявка, сосущая кровь крупноrо
млекопитающеrо, вряд ли может ero убить,
280
rлава 9
ноrи с ЛОЖНЫМИ
ПЕДИПДЛЬПЫ С КЛЕШНЯМИ ПЕДИПДЛЬПЫ С ЛОЖНЫМИ КЛЕШНЯМИ КЛЕШНЯМИ
Скорпион (хелицеровые)
Ложноскорпион (хелицеровые)
ВЫВОРАЧИВАЮЩАЯСЯ rЛОТКА
Жrутоноrое (хеЛlAцеровые)
Ложные
клешни
Рак "'Боrомол (раКООБразные)
Ложные
клешни
,,. . \ Ложные кпешн
"""
............... 't' ;.: .', .*'...........,.,.
.4".. --:-: ,.' : ;::;'.' ",
......,.,,- :. . ! :;. ' . "
:';' ..'""w!i..
Скорпионопаук (хелиuеровые)
Боrомол (одноветвистые )
I
Выворачивающаяся rлотка
Ядовитая железа
Проток ядовитой железы
Полихета (аннелиды)
РУКИ С ПРИСОСКАМИ
. - :; .. . ...:':: '.- :. /.!.; ..." ......
. ..
. .
rоловоноrий моллюск
ноrи с КЛЕШНЯМИ
Клешни
краб (раКООБразные)
rОЛОВНЫЕ ШИПЫ
Задние ЗУБЫ
Ловчие шипы
Щетинкочелюстное
Рис. 9.11. OpraHbI схватывания добычи (по различным источникам)
так как потеря крови будет для Hero незна..
чительна. Однако друrой вид пиявок, пи..
тающийся кровью мелкой прудовой улит..
ки, rубит последнюю. поэтому должен счи..
таться охотником. Даже в случае животных
одинаковоrо размера отнесение к той или
иной катеrории представляет трудности.
Некоторые планктонные полихеты напа..
дают на щетинкочелюстных и отъедают
у них передний конец тела. Жертвы от этоrо
не поrибают и способны реrенерировать He
достающую часть. Как квалифицировать
полихету как охотника или как паразита?
Столь же проблематична ситуация и со
мноrими эндопаразитами. Например, неко"
торые перепончатокрылые насекомые про..
водят часть жизненноrо цикла в теле друrих
животных, что приводит К rибели послед..
них. Эти так называемые «паразитоиды»
съедают свою жертву изнутри наружу, а не
наоборот, и не так быстро, как классические
охотники. Такие насекомые откладывают
Питание
яйцо или яйца в особь жертву (обычно дру"
roe насекомое), и развивающиеся личинки
питаются тканями хозяина до cBoero OKY
кливания и метаморфоза Имаrо здесь
типичные охотники, за исключением Toro,
что они только нападают на добычу, но не
поедают ее, а оставляют потомкам. И
в этом случае встает вопрос об относитель..
ных размерах животных. Личинка перепон"
чатокрылоrо или двукрылоrо HaceKoMoro
велика по отношению к размеру жертвы,
в которой она паразитирует, поэтому хо..
зяин может быть крупным лишь настолько,
чтобы обеспечить возможность развития до
окукливания одной или нескольким личин..
кам. В то же время взрослые формы, CKa
жем трематодь] или нематоды, хотя и пи..
таются тканями хозяина по существу таким
же образом. очень малы по сравнению.. на..
пример, с млекопитающим"хозяином и не
вызывают ero rибели. IJоскольку оба типа
питания, не считая их крайних форм, нераз..
личимы, неудивительно, что во мноrих
rруппах мелких хищных беспозвоночных
имеются и паразитические виды.
Более четкую rраницу можно провести,
по крайней мере в отношении биолоrии пи
тания. между охотниками и паразитами. пи..
тающимися жидкостями тела и/или ткан я..
ми своих жертв, с одной стороны, и бески"
шечными эндопаразитами, обитающими
в пищеварительном тракте хозяина, c
друrой, хотя опять..таки эволюционный
переход между эндопаразитическими само..
стоятельно переваривающими животными
и формами, всасывающими уже переварен..
ные хозяином продукты, не слишком велик.
Некоторые паразитические черви, особенно
цестоды и скребни, лишены кишечника, но
их наружная кутикула снабжена микровор"
синками (так называемыми микротриха..
ми), эквивалентными тем, которыми по..
крыты поrлощающие клетки кишечника
друrих животных. Внутри хозяина эти черви
абсорбируют продукты ero пищеварения.
а их кутикулы, кроме Toro, защищают их от
влияния ero расщепляющих ферментов.
В то время как друrие типы «паразитов»,
рассмотренные выше, фактически являются
микрохищниками, цестоды и скребни
истинные паразиты в обыденном понима..
нии этоrо слова, поскольку существуют за
281
счет ресурсов, добытых друrим видом Т о
же самое справедливо и в отношении дру"
rих кишечных паразитов (включая и HeKOTO
рых нематод с функционирующей пищева..
рительной системой), потребляющих толь
«о содержимое кишечника CBoero хозяина.
Однако друrие живущие в кишечнике жи..
вотные питаются тканями ero стенки и KpO
вью хозяина, поэтому относятся к хищни"
кам.
Виды, питающиеся тканями и жидкостя..
ми теЛа животных, потребляют леrко пере..
вариваемый, боrатый белком материал,
поэтому, как правило, секретируют протеа..
зы и имеют относительно короткий и про..
сто устроенный кишечник, специализиро..
ванный (если специализация вообще имеет
место) только в передней части с образова..
нием жевательноrо желудка (если жертва
заrлатывается целиком), зоба для накапли..
вания запасов или насасывающей rлотки.
Высокое содержание белка в животной пи
ще делает ее привлекательной и для ряда
неплотоядных животных на некоторых кри"
тических стадиях их жизненноrо цикла.
Иноrда самки двукрылых насекомых, на..
пример комаров, нуждаются в крови, чтобы
получить белок. необходимый для развития
яиц, хотя самцы и личинки MorYT обходи..
ться без животной пищи
9.2.4. Обrрызатели
и соскабливатели
Обrрызатели и соскабливатели это
подвижные потребители неподвижной дo
бычи, использующие в пищу доступные для
них ткани, но обычно не убивающие при
этом свои одиночные или колониальные
жертвы. На суше источниками питания для
них являются растения и rрибы, а в море
колонии животных (таких, как книдарии,
мшанки или оболочники), бактерий и мно"
rоклеточные водоросли. Добывание пищи
требует наличия твердых rрызущих или
скоблящих ротовых частей типа радуляр"
ной леНТЬJ моллюсков (рис. 5.3), аристоте..
лева фонаря морских ежей (рис. 7.24), скле
ротизованных челюстей насекомых (рис.
8.30) и т. Д., а поиск «орма здесь не состав..
ляет rлавной проблемы (сильное отличие их
от большинства типов охотников), посколь..
282
Задняя
кишка
А
ЗОБ
Средня
кишка
rлава 9
Анус
Б Trlcпoтoпas
Рис. 9.12. Пищеварительный тракт термита, питающеrося древесиной (А), и симбиотические жrу
тиконосцы (Б В) из ero задней кишки (А по Morton, 1979; Б, В по Mackinnon, Hawes, 1961)
ку съедобный материал, как правило, нахо..
дится в изобилии. Трудности этой KaTero..
рии животных в следующем: а) наличие си..
стем химической защиты у неподвижных
жертв: б) небольшая доля приrодной для ус..
воения пищи на единицу веса Bcero поrло..
щенноrо материала из..за большоrо количе..
СТВй в нем устойчивых структурных или за..
щитных элементов и часто низкое содержа--
r
ние оеЛКсl в усвоеннои opr d.нике.
. Проблема высокоrо содержания непере..
вариваемых уrлеводных полимеров в расте..
ниях и водорослях (arap, альrин, ламина..
рин, целлюлоза и др.) частично решается,
как уже rоворилось, блаrодаря специфиче..
ским фермеНТdМ и одновременному удлине..
нию пишеварительноrо тракта (особенно
средней кишки), увеличивающему размер
opraHoB, переваривающих устойчивые ма..
r
териалы, а частично за счет симоиотиче..
ских бактерий и протистов. обитающих
в особых частях кишки (часто в задней ки"
шке, но иноrда в зобе или желудке). Неред..
ко присутствует крупный зоб для накопле..
ния запасов.
Кишечные симбионты раЗЛсlrают поли
сахариды анаэробным путем с образова..
нием жирных кислот и простых уrлеводов,
приrодных для всасывания. Такой способ
достиrает cBoero наивысшеrо развития
у термитов, использующих в пищу наибо..
лее устойчивый из природных орrанических
материалов древесину. У этих насекомых
задняя КИIпка длиннее Bcero остальноrо пи
щеварительноrо тракта (рис. 9.12) и содер"
жит плотную популяцию жrутиковых Ну..
репnаstigiпа. Эти протисты Фdrоцитируют
частицы древеСИНЬJ и сами содержат сим..
биотические бактерии, которые, по..
видимому, в основном и отвечают за рас..
щепление целлюлозы. Друrой компонент
дерева, ЛИfНИН, вероятно, не переваривае..
тся. Равноноrий рачок LiпZ110J,'ia также пи..
тается древесиной, однако, по всей вероят"
ности, лишен кишечных симбионтов и пере..
варивает целлюлозу и rемицеллюлозы с по..
мошью собственных ферментов. Структур..
ные уrлеводы, если MorYT усваиваться, слу..
жат боrатым источником энерrии, но не
дают почти ничеrо, кроме нее. Liпl1un'ia по..
лучает необходимый белок из rрибов. живу..
ЩИХ в потребляемой этим ракообразным
древесине, которая без таких колонизиро"
вавших ее орrанизмов"редуцентов не может
r
удовлетворить потреоности животноrо.
Фактор размера также важен при потре..
блении растительноrо материала с относи..
тельно низкой питательной ценностью.
у крупных животных.. например позвоноч"
ных размером с кролика, низка удельная
Питание
скорость метаболизма, что снижает потреб
насть в знерrии на единицу веса тела Бла
rодаря большому объему они способны на..
капливать в переваривающем отделе кишки
MHoro растительноrо материала. Млекопи"
тающие, будучи rомойотермными живот
ными, поддерживают температуру, необхо
димую для l{рфективной раБОТЬJ пищевари
тельных ферментов. Поэтому они MorYT cy
ществовать за счет rрубых трав и подобных
им материалов, поrлощаемых в больших
количествах. Мелкие животные, например
растительноядные беспозвоночные. BЫHY
ждены питаться кормом более BbIcoKoro Ka
чества и обеспечивать себе доступ к coдep
жимому растительных клеток, протыкая их,
соскабливая целлюлозные стенки с помо
щью радулы или прокусывая их своими ро..
товыми частями. Поrлощение больших KO
личеств пищи здесь, как правило, невозмо"
жно просто из за мелких размеров тела. Да..
же мелкие птицы и млеКОПИТd.ющие не MO
[ут существовать за счет малопитательноrо
корма: если это растительноядные живот
ные, их рацион бrраничен боrатыми эне..
рrией семенами или сходными образова..
ниями.
у растений MHoro разных средств хими
ческой защиты., включая алкалоиды (нико
тин, кокаин хинин. морфин и кофеин).. rлю..
козиноляты., цианоrенные rликозиды и тан..
нины. Эти вещества., вероятно, специально
образуются в их тканях, чтобы воспрепят
ствовать потреблению последних, и иноrдd.
сильно токсичны, являясь естественными
инсектицидами например, у дерриса и пи..
ретрума. Таннины, высвобождаясь, связы
вают белки, делая их неусвояемыми и та..
ким образом уменьшая белковую ценность
поrлощенной пищи. Друrие соединения
имитируют собственные [ормоны и феро..
моны фитофаrа. отрицательно влияя на ero
рост, развитие, размножение или вызывая
неадекватные поведенческие реакции. У He
которых растений механизм химической за..
щиты включается только во время выеда..
ния, исключая не нужную трату ресурсов.
необходимых для роста и размножения; He
давно показано, что (по крайней мере в aд
нам случае) растения, даже не будучи объек
том нападения. MorYT реаrировать мобили
зацией своей химической защиты на пора
283
ОБразования" имитирующие яйца
J \
Рис. 9.1 З. На листьях и стеблях страстоцвета
находятся образования, имитирующие яйца ба
бочек Helicoпius. Это предотвращает откладку
яиц на растение (самки избеrают листьев, на
которые уже отложены яйца, возможно,
в связи с тем, что только что вылупившиеся ли
чинки способны поедать друr Apyra) (по Gil
bert, 1 982)
жение фитофаrами соседних экземпляров
Toro же вида.
Друrие защитные механизмы растений
бывают структурными. например волосо..
видные выступы, выделяющие при прикас..
новении липкие секреты или отпуrИВd.ющие
вещества., или анатомическое имитацион"
ное сходство здоровых листьев с листьями.
пораженными фитофаrами (рис. 9.13). а TaK
же симбиотическими., особенно заметными
в случае ассоциаций различных покрытосе
менных и друrих сосудистых растений с му"
равьями. Муравьи изrоняют насекомых.
обrрызающих растение, «в обмен» на eI о
нектар и место для устройства [незд. Сход..
ные структурные. химические и друrис си..
стемы защиты имеются и у морских живот..
ных, страдающих от rрызущих консумен..
тов, но они пока слабо изучены.
Тем не менее консументы коэволюцио..
нировали со своими неподвижными жертвd.--
ми в течение мноrих тысячелетий., и HeKOTO
рые соскабливатели и обrрызатели при..
обрели способность обезвреживать, избе
[ать. связывать или 1кскретировать спепи..
фические защитные метаболиты, обходить
284
структурные барьеры и ускользать от вни"
мания охраняющих растения муравьев пу..
тем химической мимикрии под этих насеко..
мых. И все же в большинстве Нd.земных ме..
стообитаний, блаrоприятных для развития
растений., накоплено orpoMHoe количество
биомассы живых растений так же, как
в океане водорослей. Это rоворит о том,
что большая часть тканей макрофитов на
сймом деле остается недоступной для rры..
зущих и скоблящих животных и/или не
используется ими и становится источником
пищи только после отмирания иразложе..
ния.
Поедание макрофитов ос) ществляется
не только путем обrрызания и соскаблива..
ния. Некоторые растительноядные живот..
ные как в море (пример заднежаберные
моллюски)., так и на суше (пример мноrие
клопы) обходят трудности. создаваемые
структурными уrлеводами, высасывая со..
держимое отдельных клеток или внедряясь
своими трубчатыми ротовыми частями
в ксилему и флоэму сосудистых растений
(рис. 9.9 и 8.30). В последнем случае rидрав"
лическое давление в про водящих элементах
растения бывает достаточным для закачи..
вания жидкости непосредственно в кишеч
ник, например, тли, которая паразитирует
на растении"хозяине примерно так же, как
клещи или самки комаров на животных.
Как отмечал ось выше в отношении пло..
тоядных охотников, лишь небольшая сту"
пень отделяет эктопаразитизм на крупном
хозяине от эндопаразитизма., и некоторые
нематоды паразитируют в растениях так
же, как друrие нематоды в животных" d. ли..
чинки ряда насекомых выrрызают ткани ра..
стений, обитая внутри последних.
Однако даже при наличии целлюлаз
и эквивалентных им ферментов или симбио
тических Мd.кроорrанизмов в кишечнике (й
тем более при отсутствии тех и друrих)
переваривание и ассимиляция тканей ма..
крофитов, как правило. идут очень неэффек"
тивно, а в случае питания жидкостями по..
ступление уrлеводов HaMHoro превышает
потребность в них. Поэтому фекальные
массы таких консументов обильны и содер"
жат MHoro неассимилированноrо орrаниче..
CKoro вещества. В связи с этим они являю..
тся важным эколоrическим путем ДОСТУПd.
rлава 9
к уrлероду., фотосинтетически фиксирован..
ному живыми маКРОфИТdМИ, для друrих ка..
теrорий животных консументов.. особенно
для потребителей осадков.
9.2.5. Питание взвесями
Основные механизмы, с помощью KOTO
рых животные, питающиеся суспензиями,
отфильтровывают пищевые частицы от во..
ды, были рассмотрены в разд. 9.1. У rрупп,
использующих для этоrо наружные секреты
неанатомическоrо характера (рис. 9.5), по..
видимому., отсутствуют друrие специаль..
ные приспособления для улавливания взве..
сей. Эти животные" по крайней мере по
строению cBoero кишечника, вероятно, наи..
более близки к охотникам..засадчикам,
улавливающим свои неБОЛЬШОI о размера
жертвы сетями. В самом деле, эти два спо
соба питания не имеют четкой rраницы:
паук, плетущий тенета., может рассматрива..
ться и кйк потребитель «воздушной суспен"
зии». и как охотник..засапчик. Такое же
двойственное положение у морских книда..
рий; BOKpyr их центральноrо pOToBoro от..
верстия расположены радиально симме..
тричные кольца щупалец, а в фазе полипов
эти животные ведут прикрепленный образ
жизни; оба этих признака характерны для
потребителей суспензий, и, действительно,
жертвами здесь являются в основном зоо..
планктонные виды" взвешенные в воде. Од..
нако их особи улавливаются не совсем пас..
сивно, а индивидуально поражаются нема..
тоцистами, пусть даже сама жертва способ..
ствует их выстреливанию" случайно коснув"
шись щупальца, и книдария не нападает «по
собственной инициативе».
В отличие от этоrо у rрупп со слизисто..
ресничной системой фильтрационноrо пи..
тания четко прослеживается специализация
пищеВd.рительной системы. У большинства
таких видов с фильтрационным аппаратом,
либо свободно омываемым водой. либо за..
щищенным снаружи раковиной (например,
у плеченоrих и двустворчатых моллюсков)
(рис. 9.3), передняя кишка представляет со..
бой короткую трубку, соединяющую на..
ружный фильтр с желудком. Однако у хор..
довых передняя кишка сама является ме..
стом, rде происходит фильтрация, и поэт о..
Питание
r иперБранхиальный жеЛОБОК (В котором
СОБирается слизистый тяж с пищевыми частицами)
I '/ТОК BO I
О r ПРОСВЕТ r АТРИУМ
t rлотки I О
му сильно специализирована. rлоточная
стенка у этих животных пронизана по бокам
мноrочисленными мелкими отверстиями
(<<стиrмами»).. которые.. прободая стенку те--
ла, открываются на ero поверхности. Вода
поступает в рот, оттуда в rлотку, затем про..
ходит через стиrмы и выводится обратно
в окружающую среду, т. е. ее поток однона--
правленный. Хотя сам по себе процесс фи--
'-" r
льтрации rлоткои и неооычен, аналоrичная
система известна и в друrих rруппах, что,
возможно, укаЗЫВает на ее эволюционное
происхождение. Животные" питающиеся
мелкими частицами, часто используют для
втяrивания их в рот поток воды., которая за--
тем должна как--то выводиться из орrаниз--
ма. У лофофоровых вода просто периопиче--
ски «выплевывается», и поrлощение частиц
в эти моменть] прекращается. У брюхорес--
ничных макродазиид (рис. 4. 9) и полухор
довых цефалодисцид (разд. 7.2.3.2) rлотка
несет пару перфораций" ведущих на поверх--
ность тела, через которые поrлощенная во--
да выводится наружу без перерывов в пита--
нии. Древнейшие хордовые, по--видимому,
специально приспособили такой поток BO
ДЫ для отфильтровывания пищевых частиц,
что избавило их от необходимости ВТЯI"И--
вать лофофороподобный opraH при уrрозе
со стороны хищников (что также прерывает
Отверстия
АТРИУМ
Ток слизи
Латеральные
реснички
285
питание). Кишечнодышащие полухордовые
используют этот же поток для rазообмена,
что свойственно и более поздним водным
хордовым.
rлотка питающихся суспензиями хордо--
вых очень велика и занимает большую
часть объема тела. Пронизанная тысячами
отверстий стенка тела в rлоточной области
у мноrих видов почти исчезает и поэтому
заменяется вторичной стенкой, образован--
ной у rоловохордовых складками ткани, а
у туникат секретируемой целлюлозной
оболочкой. В связи с этим собственно тело
целиком или частично окружено морфоло--
rически внешней полостью, или атриумом,
между истинной и ложной стенками тела,
Сюда поступает вода, проходящая через
стиrмы., и отсюда она выводится в окру--
жающую среду через единственное отвер--
стие, т. е. атриопор или атриальный сифон
(рис. 9.14). В сущности такая же система на..
блюдается у позвоночных. Однако у рыб
жаберных щелей (== стиrм), ведущих из ки"
шечника к поверхности тела, значительно
меньше, а сама стенка тела толще, поэтому
нет необходимости в дополнительной ло--
жной стенке, а поток воды используется
прежде Bcero для rазообмена, а не для по..
rлощения пищевых частиц.
Спеuиальная железа, энпостиль.. распо..
СРронтальные
реснички
Рис. 9.14. Схема поперечноrо среза rлотки беспозвоночноrо хордовоrо, показывающая действие
системы Фильтрационноrо питания (по различным источникам)
286
Ток
воды
Передний
конец
ЖаБерный
сисрон
ОБолочка
rанrлий
Слизистый мешок
rлава 9
rлоточная переrородка
(единственный остаток
дорсальной стенки rлотки)
Дтриальная
полость
А триальный
сисрон
Задний
конец
Вид сверху
Атриальная полость
Эндостиль
Ток воды
Задний конец
r лоточная
переrородка
ВИД СБОКУ
ложеННdЯ ВДОЛЬ вентральной средней линии
r
rлотки, выраоатывает слизь, которая TOH
кими слоями rонится вверх по перфориро
ванным боковым стенкам. Коrда Бода BЫ
текает наружу через стиrмы, 'Эти слои пере..
хватывают потенциальные пищевые час
тицы в некоторых случаях размером до
0,5 мкм. В конце концов потоки слизи с пи
щевыми частицами сходятся дорсально, [де
образуют продольный тяж в rипербранхиа
льном желобе. Движение в этой системе KdK
воды, так и слизи обеспечивается полоска
ми ресничек внутри rлотки (исключение
пелаrические сальпы).
у пелаrических оболочников поток воды
с пищевой взвесью обычно обеспечивает
и движение животноrо вперед, поскольку
ротовое и атриальное отверстия расположе
ны Hd противоположных концах тела.
В связи с этим реактивным типом движения
число стиrм уменьшается до двух (у сальп),
слизь образует одну внутреннюю сеть кони
ческой формы, подвешенную в полости
rлотки (рис. 9.15), а проталкивание воды
Рис. 9.15. У пелаrических обо
лочников (например, у сальп)
слизистый мешок перекрывает
оба крупных rлоточных OTBep
стия (по Berrill, 1950, и др,)
обеспечивается мышечными кольцами,
охватывающими тело (рис. 7.35 и 7.36).
Не только у 'Этих беспозвоночных xopдo
вых. но И У лофофоровых И моллюсков
фильтраторов пища поступает из филь
трующеrо opraHa (независимо от ero ТИПd)
в желудок в виде слизистоrо жrута, несуще
ro съедобные частицы. У всех животных со
слизисто ресничной системой питания в же..
лудке кислая реакция. Это снижает вязкость
слизистоrо жrута, обеспечивая выход из He
[о пищевых частиц. Переваривание и аб
сорбция питательных веществ происходят
в желудке и или в дивертикулах средней ки
шки, отходящих поблизости от Hero. Непе..
реваренные остатки попадают в обычно ко..
роткий кишечник.. [де шелочная реакпия
вновь увеollичивает вязкость слизи и способ..
ствует образованию фекальных комочков
или нитей.
Движущая сила, обеспечивающая до..
ставку слизистоrо жrута в желудок, также
обеспечивается ресничками, у хордовых
действующих на этот жrут непосредствен"
Питание 287
Пилорус Вращающийся стержень
Рис. 9.16, Желудок мшанок со
слизисто фекальным CTep
жнем, используемым для под
тяrивания пищи в пищеварите
льный тракт (по Gordon, 1975)
но, а у лофофоровых и моллюсков
косвенно. У двух последних rрупп в желуд..
ке находится вращающийся стержень, на..
матывающий на себя слизистый жrут. У ло..
фофоровых он состоит из слизи и фекаль..
ных масс, расположен в пилорической части
желудка и вращается расположенными там
ресничками (рис. 9.16). Время от времени
использованный стержень поступаеl в ки"
шечник, преврашается в фекальный KOMOK
выбрасывается наружу и заменяется новым.
у моллюсков этот стержень, известный
r
ПОД названием кристаллическоrо стеоелька,
rораздо крупнее, сохраняется значительно
дольше и состоит из rиалиновоrо мукопро"
теина. Он также вдвинут в желудок, но вы..
ходит из специальноrо дивертикула, так на..
зываеМОI о стебельковоrо мешка, и ero вра..
тают выстилающие этот мешок реснички.
Кончик стебелька не только работает как
лебедка, накручивая на себя слизистый жrут
пищево\
Зона
сортировки
Прямая кишка
с пищевыми частицами (рис. 9.1 7), но и по..
степенно растворяется в желудке, высвобо..
ждая фермент амилазу. Параллельно стебе..
лек медленно выдвиrается вперед в желудок
(опять"таки блаrодаря работе ресничек),
а секретируемый проксимально материал
поддерживает ero длину постоянной.
Поэтому кончик стебелька все время упи..
рается в стенку желудка, что создает эффект
вращающеrося жернова. и служит для от..
правки пищевых частиц из ставшеrо менее
вязким слизистоrо жrута в сортирующую
часть желудка, rде реснички продолжают их
разделение.
9.2.6. Питание осадками
Вероятно, большинство современных
животных пит dЮТСЯ детритом ( орrаниче..
ским материалом, состоящим из мелких ча..
стиц) или подстилкой (из частиц большеrо
Вход в пищеварительный
диве iJ'икул Кишечник
Слизистый
тяж с пищей
Кристаллический
стеБелек
СтеБельковый
мешок
Рис. 9.17. Желудок фильтрующеrо моллюска с кристаллическим стебельком используемым для
подтяrивания пищи в пищеварительный тракт (по Мопоп, 1979)
288
rлава 9
размера), но, несмотря Hd это, четкоrо пред
ставления о природе этоrо Рациона пока
нет. Лист, съедаемый, скажем, земляным
червем, ЭТО не просто мертвый лист, а цe
лая экосистема. На ero поверхности и в ero
ТКdНЯХ обитают бактерии и rрибы, обеспе
чивающие разложение орrаники, различные
протисты, потребляющие деструкторов (Ha
пример, амебы, ресничные, rетеротрофные
жrутиковые), а также лишь немноrим более
крупные нематоды и клещи. питающиеся
этими мелкими орrанизмами. На поверхно
сти листа иноrда оказываются фотосинте
зирующие водоросли и цианобактерии,
мертвые и разлаrающиеся остатки друrих
орrdНИЗМОВ (не в последнюю очередь из
фекалий животных). Все эти живые и MepT
вые компоненты MorYT быть поrлощены
животным вместе с листом, и задача co
стоит в том, чтобы определить, какие из них
перевариваются и ассимилируются и какие
в наибольшей степени удовлетворяют MeTa
болические запросы потребителя.
Это не обязательно означает.. что потре
битель просто заrлатывает основной MaTe
риал и переваривает все, что может. Теперь
известно, что некоторые потребители ocaд
ков способны к значительно более избира--
тельному поrлощению, чем представлялось
ранее (впрочем, лишь немноrие животные
были настолько детально изучены с этой
точки зрения).
Избирательное поrлощение. безусловно,
должно давать преимущества, поскольку
пищевая ценность различных компонентов
детрита весьма неодинакова. Сравнение op
rаники в осадке, заrлатываемом HeKOTOpы
ми морскими животными, не способными
к такому отбору, и их фекалий, а также
обработка известными ферментами из их
кишечника проб этоrо осадка показали, что
основная часть Toro небольшоrо количе..
ства орrаническоrо вещеСТВd, что содержи
тся в донных отложениях, остается таким
видам недоступной. Они усваивают менее
5 100/0 ero детритноrо «пула». (На чисто
песчаных пляжах вся орrаника иноrда co
ставляет менее 1 О/о заrлатываемоrо MaTe
риала. Слой лесной подстилки ею боrаче,
но все равно часто лишь небольшая ее доля
приrодна в пищу.)
К тому времени, коrда любой кусочек
орrанических остатков становится частью
поверхностноrо слоя подстилки или детри
та, он уже теряет большую часть своей пи..
тательной ценности. Если это лист, то перед
сбрасыванием все растворимые вещества
переходят из Hero в остающиеся на роди..
тельском растении ткани.. а выщелачивание
ПрИВОДИl к потере друrих орrанических
веществ уже в первые часы после падения на
землю. ОРI'анические частицы попадают на
субстрат только пройдя через кишечник
консумента, [де из них обычно извлекается
большинство питательных соединений,
а фекальные массы один из основных
источников орrаническоrо вещества в осад..
ках и почве. Поэтому орrаника в них это
чаще Bcero устойчивые структурные, скелет..
ные или защитные соединения, проходящие
без изменений через пищеварительные
тракты большинства животных (у детрито..
фаrов, как правило, отсутствуют целлюла..
зы).
Однако в случае свежих остатков друrих
орrанизмов потребитель осадка имеет шанс
усвоить некоторые содержащиеся в них ор..
rанические вещества, хотя и тоrда белка
скорее Bcero будет очень мало или он ока..
жется недоступен из за присутствия тании"
нов (см. рdЗП. 9.2.4). Вообще наиболее ве..
роятно, что потребители мертвой орrаники
(за исключением питающихся падалью) су..
ществуют за счет живых микроорrанизмов,
связанных с этими остатками.. служащими
лишь удобным транспортным средством
для ДОСТdВКИ пиши В кишечник. На суше и
в пресных водах rрибы, разлаrающие под..
стилку, возможно, являются основной пи..
щей для по rребителей осадков, а на при..
брежном морском мелководье наибольшую
питательную ценность, вероятно, имеют
обрастающие частицы rpYHTa фотосинтези..
рующие одноклеточные протисты. Показа..
но, что у животных, способных выборочно
питаться TO.lIbKO диатомовыми водоросля..
ми, эффективность ассимиляции достиrает
700/0, в то время как у неизбирательиых по..
требителей орrаническоrо материала она
менее 40/0. ТаК или иначе, на основной пло..
щади MopcKoro дна, [де фотосинтез невоз"
можен и частицы, осеДdющие из верхних
слоев воды" редки, к тому времени" коrда
они достиrают субстрата, в них остаются
Питание
289
только устойчивые к разложению вещества,
и для консументов источником как энерrии,
так и белка служат бактерии.
Пищеварительная система потребителей
осадков, как правило, не специализирована,
но кишечник часто удлинен (см. рис. 4.42
и 4.45), что обусловлено теми же причина..
ми, что и в случае обrрызающих и соска..
бливающих животных. Как и у последних,
обычно образуется очень MHoro фекальных
масс, питательная ценность которых для
друrих потребителей HaMHoro ниже. чем
у проrлоченноrо материала.
9.2.7. Питание с участием
симбионтов
Хотя питание относительно немноrих
видов животных полностью зависит от фо..
тосинтеза их эндосимбионтов, как это пока..
зано на рис. 9.18 (см. также разд. 9.1), неко"
торые представители таких филоrенетиче..
ски различных rрупп, как rубки, моллюски,
книдарии, хордовые и т. д.. В той или иной
степени используют в пищу продукты жиз..
недеятельности своих фотосинтезирующих
партнеров. Ими MorYT быть цианобактерии
(например, у некоторых rубок и эхиурид),
прохлорофиты (у оболочников). а чаще ди..
нофлаrеллаты (в виде зооксантелл во мно"
rих морских беспозвоночных) или зеленые
водоросли, представленные зоохлореллами
Рис. 9.18. Схема потоков уrле
рода (С) и питательных Be
щеСТ8 (N) при симбиотических
взаимодействиях (по Barnes,
Hughes 1982)
(обычно у пресноводных видов). Некоторые
Мdдрепоровые кораллы удовлетворяют 2/3
своих метаболических потребностей за счет
внутриклеточных зооксантелл, и только
1/3 за счет внешних источников питания,
включая типичное для кишечнополостных
улавливание 100планктона. У некоторых
актиний зооксантеллы и приспособления
для ловли добычи расположены в различ..
ных частях тела: «opraHbI» с симбионтами
расправляются в дневное время, а щупаль..
ца с нематоцистами ночью (рис. 9.19).
Симбиоз может дополнять растительнояд"
ность. Мешкоязычные моллюски высасы..
вают клетки макроскопических зеленых (и
друrих) водорослей: некоторые их виды мо"
[ут сохранять в дивертикулах средней
кишки целые функционирующие хлоропла..
сты своей добычи (рис. 9.20), которые захва..
тываются пищеварительными клетками..
фаrоцитами, но продолжают фотосинтези..
ровать в некоторых случаях еще более двух
месяцев. До половины фиксируемоrо уrле..
рода передается моллюску, чеrо вполне до..
статочно для удовлетворения ero энерrети"
ческих потребностей. (Возможно, хлоро--
пласты некоторых эвrлен имеют сходное
эволюционное происхождение.)
Несколько иные сим6иотические отно"
шения существуют между растениями. вы..
деляющими нектар, и насекомыми, кото..
рые посещают их для поедания этой боrа..
r етеротросрное животное
СРотосинтезирующий
СИМБИОНТ
Орrанические
С + N '..
Орrанические .....
с + N . . . . . . . .. :".
'.
Неорrанические
C+N
\
\
\ ".
,
'Opr анические
'C+N
Внешняя среда
290
rлава 9
ДЕНЬ
Lebrипia danae
Lebrunia coralligens
Bипodeopsis antillieпsis
НОЧЬ
Рис. 9.19. Использование различных частей тела тремя видами рифообразующих коралловых по
липов в разное время суток: opraHoB, содержащих З00ксантеллы при дневном освещении (слева)
и щупалец, несущих нематоцисты, ночью (справа) (по Sebens, Dе Riemer, 1977)
той сахарами жидкости и в результате обе..
спечиваlОТ опыление. У некоторых видов
имаrо потребляют только нектар (чтобы
обеспечить себя энерrией для полета), а пи
тающаяся фаза их жизненноrо цикла пред
ставлена личинками, накапливающими
достаточно ресурсов для формирования
взрослоrо орrанизма, который нуждается
Питание
291
"'
,.0
,* ...
41>.... 4 . . ' '
, .... 11 ' ,; . f 1> , J. .. ' . 1. ... ...." "'t ...'*'......:;.
.... , .... .. .. ... 4. ,.:,..... i
" '-' ....., ..,.. т !... ..... '" }<.,.
. 1\.,. \ :;... . '... .. Т:. .. ,... .;,
; N' ' "'... · ..' '., i" i!- · . ...;..
7" , . ",:: ' (' .. ...', .!)
/. ) :;. .' ." ,.. ' #,. "" А ,(..
. . \ '" ..t Il .
.. .. . >< .;. '". ...
{, е t -. .. f',,; ,
.. , " ' . ' \ 8. . ",. :.. .. .... ,. "'. '.. :}!J :
- .....,
.""
iJtt.' ;>," t .. , , I ; ',,\,
. ._ . ..- I '\ ..".., .. '.... .. '
. -.'. ... .'; -. ..\о<..... .#;.. .
: ,. . .. - -:- ' "... I . " _
'\" '/ \ ', ,,' , ':..." .<&. ,':/ ;.; ',;,\
'" ) I '. а" "J' t ... tf .. ) k,.
'(" ; ' ' : : '-':: ,;' 'i :: ,
" ..... ...,.. * . '-t . " *. , ,.. ., ,. r:.., .
... .... ..... , ... .) . ...
"",. ,. . " .. tiI" 4 -, *. .!
.,:..... t ...- :А, "f " '" ." . '. .., "
.., l 1 ..' . ..,' ....,. ... +. '" 1, J
'; . . :' 1 t,:, ' ';: ' ' M ' " ;<!i;f:,. .,' ',::J!: ,, ...
, ,. { ,. ", . .. ...
... "". ' ......,.fl. -" "'\;: ...., ,,: .'» ....' . ': ,...
.::. . J .. .. ... .."',. ,. ...
" .... .. .. " "" .. « '" -.. . (..
. .! .... М -., '\... ... . , .., ...' ." "', .J!t
-: ',. .': h 4' \: N. . =т: .. "''\. .... ).... ,'Ю.. 11> ,,1" ... . .'
t .P' ' ,"о.. .. <! .- .1' ',($ ,..; "" +. '('\ -:; . .
'1' ,"". .:..;. . ,':'. . ; {.,. .
. · .,.. 4.. r",.'tI'I """., , .....w:: "-"'" ') ""'''
..",. w t l 'f .. '..... . ... . tt -; ... 1 ..
...., . . : > "', 11 '*6. * . ...... .., .l", # f'"" "":J",,
..,. .. '.......4 w .. .,, .,. "
.- ... ф 1 " ... # , ... ktl<L>"" o/>J. --""{II w:: >< .\ '
.. d'" ..........,' "'.,.IOC .... ''''"3..]:,' ..
, ... . " ; ' 11'4..."'" .. ,. . ..... . '/ff.... ., 'Т'
7 .. -..;.. .. ........ .. ,j,.. ..:.. ,.. _;rc . .. .
.. .... . . ." ., ....... ;а:l\ "..r.. .. 1", :1.
#< , . .,.IIIf ( .ч. F ... ,, . '"i . .:'5
. ...... t- /« '. . , '1\... . . J:j-,,-, ,:" "N 1t-7"'\r- ... O..l-.i'''' ..:
, , . 110\. 4. "" ". r . + '(..,
· 7.. ......' "" 4J '" t: .-. ..... Jt .
.. . i :" ' '''.,' .: ; ' '.A
..... .... .........-: . .. :+,....... r.",' А -
...... . .. <... '''.' , :-, ... .. ..
.. - "\ ,,! .. ' :'"
... . .",. , <. . :--, " ".."', , .
fr ',;. .. 'у ". . .. .,.. , ....' '.
.. .,:- -: '..: f.1 /:' ",'
-". . ......., k 1; ,, " Jfo 'i
\ , .. ....,.', . ... .. 4..<>
: . . 1':';;," , "" 1., ... .. .- ':"
. , . N9<--:'::- . ...; .. "' ) , 5. :
, .' . -.: .... \ ',.,"\.
4 ......'- . .. 11/' \ ... .!t: ...,("'.."
· ,....... , . '1
.. ..1
'-' i' ...,, < 1f ·
Ф* *,,,, ф." I. : .". '1L.. '..... . » :;,. ...
" /,. " . * ::> !t. ....т .. . '" . __ .. .' . , --: . \ . t 'i.;..... ... . : , .. .
", "'" .. , . ... .. '" ''''' ':"Ir 4 ..., .-
ft..' .«'. ' :. ,.. ' . . JI' . ., '/' .. ",'" ,.." . -"It .t " \' .
'\О.... ., ",.f:; . '" ,'. ... ,... -» С
. ' . >;, 111 . "'., i ' .-- ....$'< ... · ""'':!''
::1. ,,' iJ lIIf1 ,. . . . ..... Jj ...
....,r
<'.
...
" .
,;", ,
...
.... ,1
" ,"
1s ""'
1.. ..,.
.
. .
...
, ..,.
..
.
<#
""
.. -
t
Рис, 9.20. Электронная микрофотоrрафия функциональных хлоропластов в пищеварительном
дивертикуле мешкоязычноrо моллюска (Trench, 1975)
в течение cBoero недолrоrо существования
лишь в источнике энерrии
Наконец, у ряда относительно крупных
свободноживущих животных кишечник от..
сутствует на всех стадиях жизнеННОI'О цикла
(некоторые двустворчатые моллюски, оли
[охеты, полихеты и все поrонофоры). Их
способ питания до сих пор остается заrад
292
кой хотя В тканях большинства таких ви
дов найдены симбиотические xeMoaBTO
трофные бактерии. Известно, однако, что
по крайней мере у некоторых поrонофор Hd
поверхности тела имеются микроворсинки,
эквивалентные покрывающим поверхность
тела у бескишечных паразитов обитающих
в пищеварительном тракте друrих живот
ных. Следовательно, возможно, что некото--
рые из этих свободноживущих червей спо--
собны извлекать орrанические соединения
из окружающей воды. Относительно друrих
мяrкотелых свободноживущих животных
предполаrается, что их пища частично со--
стоит из растворенной орrаники, хотя это
признается не всеми отчасти из за необхо--
димости при таком способе питания доста--
точно высокой, нечасто встречающейся
концентрации в окружающей среде орrdНИ--
ческих веществ. Друrая возможность
наружное пищеварение с последующим
всасыванием переваренных продуктов стен..
кой тела однако соответствующих желези..
стых клеток пока не обнаружено.
9.3. Затраты и выиrрыш
при различных способах
питания:
оптимальное фуражирование
9.3.1. Введение
Мы видим, что типы питания животных
cTporo оrРdничены их эволюционным про--
шлым: представителю линии потребителей
суспензий нелеrко стать охотником--пре--
следователем, и наоборот. Тем не менее
мноrие виды обладают некоторой пластич--
ностью, и некоторые «rенералисты» MorYT
даже переключаться с одноrо рациона на
совершенно иной и менять способы пита--
ния. Например. обитающая в эстуариях по--
лихета NeJ'eis diversic%r способна питаться
взвесью (рис. 9.5,п, осадком. быть охотни..
ком, поедать падаль и соскабливать фраr--
менты макроскопических водорослей (табл.
9.1).
Во время своей дневной активности жи--
вотные сталкиваются с различными потен--
циально съедобными объектами т. е. с
альтернативными пищевыми реСУРСdМИ.
rлава 9
Для большинства специализированных кон-
сументов это может означать выбор лишь
между различными особями одноrо и Toro
же вида жертвы, но обычно диапазон воз-
можных типов пищи шире. Таким образом,
в рамках оrраничений, наложенных их эво-
люционным прошлым животные в мае..
штабах CBoero эколоrическоrо времени (ка..
ждую минуту, каждый час, каждый день)
вынуждены выбирать, использовать тот
или иной объект в качестве пищи или отка..
заться от Hero в данных условиях в пользу
друrоrо. Такие «решения» имеют важные
последствия, поскольку выиrрыш от потре..
бления в пищу различных объектов неоди..
наков, т. е. одни из них будут «питательнее»
друrих. Однако наиболее питательная пища
предпочитается и б6льшим числом консу..
ментов, что ведет к более интенсивному ее
потреблению и в результате к истощению
этот'о ресурса. У друrих объектов питатель..
ная ценность ниже, поэтому они пользую..
тся меньшим вниманием KOHCYMeHTOB
а следовательно более обильны и более до..
ступны в любом местообитании.
Питательная ценность (на единицу по..
rлощенной массы) только один из факто..
ров. влияющих на выбор пищи. поскольку
кроме очевидной выrоды добывание ее вле..
чет Зd собой и определенные расходы. Неза..
висимо от способа питания на поиск за..
хват, переработку' и переваривание добычи
должны быть затрачены время и энерrия,
которые можно употребить на что--нибудь
друrое. Расходы на процесс питания неоди-
наковы при различных ero способах, однако
во всех случаях отбор блаrоприятствует
стратеrии, приносящей максимум «чистой
прибыли» (выиrрыш минус расходы). Ли..
ща одно из первых условий существова..
ния. поэтому любая особь. у которой эта
«чистая прибыль» наибольшая, будет иметь
селективное преимущество с точки зрения
и выживания, и воспроизведения перед дру"
rими, поскольку:
а) сможет тратить меньше cYMMapHoro
времени Hd поиск пищи (и таким образом
уменьшит для себя риск оказаться жертвой
хищников );
б) будет быстрее расти или дорастать
до больших размеров (с соответствующими
репродуктивными преимуществами);
ПитаНИд
293
Таблица 9.1. Пищевые предпочтения Nereis diversicolor в порядке убывания
количества усвояемой энерrии на пищевой объект
Тип питания и характер нищи Порядок пред
почтения
Порядковый номер
в ряду убывания yc
вояемой энерrии
Поедание мертвых двустворчатых моллю
сков Маео/па
Охота на живых олиrохет Tub t'ex 2
Охота на живых рачков бокоплавов Coro 3
p/1iulп
Охота на живых личинок двукрылоrо Hace 4
KOMoro E,'iopte,.a
Питание поверхностными частицами ДOH 5
ных осадков
Питание суспензиями 6
Обrрызание живых зеленых водорослей Ьп 7
(еl'ОI1101'р/1а
Обrрызание живых зеленых водорослей Ull'a 8
Охота на живых брюхоноrих моллюсков 9
Hytlrobia
2
4
3
5
6
7
8
9
в) будет «здоровее», а значит, устойчи--
вее к пара зитам и лучше Зdщищенной от
хищников и т. п.;
[) извлечет большую пользу из меньше--
[о количества ресурсов при дефиците пищи.
Каким же образом MorYT животные мак--
симизировать «чистую прибыль» в единицу
времени? Эта проблема изучалась на про--
стых моделях стратеrии оптимальноrо фу--
ражирования животных с последующей
проверкой проrнозов, полученных в этих
моделях, с помощью экспериментов и на--
блюдений. Обычно при постановке экспери--
ментов выделяют какую--нибудь одну пере..
менную, в данном случае выбор жертвы.
Следует помнить. однако. что в естествен..
ных условиях поведение животноrо в каж--
дый отрезок времени представляет собой
компромиссный ответ на множество разно..
направленных воздействий. Максимизапия
«чистой прибыли» от питания сама по себе
уже дает преимущества, но не менее выrод..
ны избеrание хищников, уход за потом..
ством И т. д. 'Эти друrие потребности часто
накладывают оrраничения на процесс пита..
ния, в результате чеrо потребление добычи
будет идти на уровне ниже оптимальноrо,
т. е. животное может иметь очень неболь--
шой выбор и поrлощать то, что удастся
найти за тот короткий период, который
останется ему для питания.
Кроме Toro, живая добыча отнюдь не
ждет, пока ее съест консумент! Фактически
стремление жертвы,не оказаться потреблен..
ной значительно больше, чем стремление
хищника поrлотить ее; это называют
принципом «жизнь/обед». Например, при
любой встрече между каракатицей икревет..
кой для первой это лишь проблематичная
возможность поесть, для второй вопрос
жизни и смерти, а значит, и давление отбо..
ра для этих партнеров различно.
Следующие разделы будут посвяще..
ны обсуждению способов максимизации
«чистой прибыли» консументами и ее мини--
мизации их жертвами" т. е. уменьшения ими
для себя опасности быть съеденными. Под..
вижные хищники, имеющие возможность
схватить или oTBeprHYTb ту или иную особь--
жертву, представляют собой простейший
случай, и мы особенно подробно разберем
именно ero. Потребители суспензий и осад--
ков сталкиваются с иными проблемами
и будут рассмотрены отдельно.
9.3.2. Теория оптимальноrо
фуражирования
Любой съедобный о"бъект имеет для по--
требителя свою особую «питательную цен--
ность». Потребляемые вещества служат для
294
живо I'HOrO источником энерrии и материа..
лом для роста соматических ирепродуктив..
ных клеток и тканей, значит, питательная
ценность должна учитывать обе эти потреб..
ности. К сожалению, они MorYT проявля"
ться в разное время rода или На различных
стадиях жизненноrо цикла, т. е. в какой"то
MOl\1eHT rлавной может быть потребность
в lнерrии, в друrой в орrаническом азоте,
а у некоторых видов (например, у наземных
брюхоноrих) бывают периоды, коrда боль..
ше Bcero прочеrо они нуждаются в Heopra
нических элементах или соединениях (на..
пример., кальция).
В большинстве исследований в качестве
условной меры питательной пенности пиши
используется ее энерrетическое содержание,
или калорийность; с помощью этоrо упро..
щенноrо термина питательная ценность мо"
жет быть выражена как энерrия.. полученная
при поедании данноrо объекта (Eg), минус
все энерrозатраты, связанные с ero поим..
кой., умерщвлением, обработкой и перева..
риванием (<<затраты на переработку».. E)J.
Помимо этоrо различные потенциально
съедобные объекты встречаются в данном
местообитании с различной частотой, и,
если потребитель предпочитает определен..
ный тип объекта., он может активно искать
именно ero., т. е. идти на «поисковые затра..
ты» (Es). Чистая энерrетическая прибыль
по,тому будет равна
Eg Е)l Es.
В свою очередь время., которое леrче из..
мерить, можно подставить в это выражение
вместо энерrии, и тоrдd. питательная цен..
ность будет измеряться энерrией, получен..
ной в единицу времени. Она составит E/T h ,
если встреча с жертвой уже произошла, или
Е/(Т'1 + Ts)" если требуется поиск жертвы
(Тh «время переработки», Ts
«поисковое время»., а Е доступное коли..
чество энерrии = Eg в выражении выше).
Возьмем сначала более простой случай.,
коrда отсутствуют затраты времени на пои..
ски. Потребитель случайно наталкивается
на различные потенциально съедобные объ..
екты в месте cBoero обитания. и возникает
вопрос: какой из них он должен съесть,
чтобы получить Мd.ксимальное количество
энерrии в единицу времени? По лоrике сле..
rлава 9
дует ожидать, что он предпочтет добыч)
с наибольшей питательной ценностью, вы..
раженной отношением Е/Т)l (или.. если
в данный l\10MeHT он нуждается в определен..
ном элементе N, с максимальным отноше..
нием N, T h ). Друrими словаl\1И, выбирается
жертва, из которой можно извлечь наиболь..
шую пользу.
Эта простая модель приводит к ряду
уже. проверенных проrнозов.
1 Потребитель будет поедать 1110.1bKO
жертв с наибольшей питательной пенно"
стью, если частота встреч с ними настолько
велика, что включение в Рd.пиов друrих
жертв (с более низкой питательной цен..
ностью) Уl\1еньшит среднюю скорость по
rлощения энерrии.
2. Если., однако, частота встречи с жертва..
ми с наивысшей питательной ценностью ни..
же, потребитель должен расширить диапа..
зон типов жертв, включив в их число сле..
дующие по питательной ценности объекты
и т. д.
3. Если на распознаВd.ние различных ти"
пов жертв требуется значительное время
(например, коrда это возможно только при
соприкосновении с ними), следует исполь..
зовать в пищу жертвы с более низкой пита..
тельной пенностью. но встречаюшиеся ча..
ще друrих, даже если более ценных жертв
остается MHoro. Коrда же идентификация
жертвы происходит MrHoBeHHo (например,
с помощью зрения), животное должно от..
BepraTb жертвы с меньшей питательной
ценностью, если более ценная пища вполне
доступна. (Установление пищевой ценности
жертвы просто дополнительный компо..
нент времени переработки. )
Все перечисленные проrнозы полностью
подтвердились, например, для зеленоrо
краба (Carciпus), питающеrося мидиями
(Mytilus) различноrо индивидуальноrо раз..
мера (рис. 9.21 9.23; приложение 9.1).
Опыты по изучению взаимодействия
хищника и жертв, относящихся к разным
видам. проводились реже. однако полихета
Nereis diversicolor как потребитель очень ши..
pOKoro диапазона пищи также служит при..
мером иерархическоrо подхода к выбору
жертвы., соответствующеrо количеству ас..
симилируемой энерrии., заключенной в каж..
дом типе добычи (табл. 9.1).
Питание
2
L
о
6 I.CI
«1
а.
х
]i
I
I
....4 Q.)
.д
()
Q
.... 2
..
.::s
::s:
L::s:
O
с::>.
()
Тт
/.......
/
/
/
/
/
/
/
..
1
'"
w
1
2
4
з
Длина мидии, СМ
Рис. 9.21. Связь между питательной ценностью
(Е/Т h) и выбором жертвы у краба Carciпus
maeпas (шириной 6,O 6,5 см), питающеrося
мидиями Mytilus edu/is; см. текст (по Elner,
Hughes, 1978)
'S
QJ
:r:
q: 20
Ll)
са
с')
><
::о
:r:
:r:
&
Q)
10
"-\Вторые 5 дней
\
\ ....
" ...(' .......! ретьи 5 дней
\ .... \ ....
". \ 1.. \ .'.
...... ,/...... .....
...
IS
S
q
S
1:
о
с::
()
:s
т
2 4
Длина мидии, СМ
Рис, 9,22. Выбор жертвы крабами Carciпus
тaeпas, коrда число мидий (Myti/us edulis)
предпочитаемоrо размера уменьшается в pe
зультате их выедания; см. текст (по данным EI
ner, Н ughes, 1 978)
6
Теперь можно усложнить базовую MO
де ль, включив в нее разное время, необхо
димое для поиска различных типов пищи.
Предположим, что T I1 постоянно для дaHHO
ro типа и данноrо размера жертв, а Ts
зависит от ЧdСТОТЫ различных потенциаль
ных особей жертв в данном местообитании.
Пусть будут два типа жертв х и у, при этом
питательная ценностью х больше, чем у у.
Тоrда
Е(х) Е(у)
> .
т h (х) Т h (у)
Очевидно, что, если потребитель обнаружи
вает жертву типа х, он всеrда ее съедает, по
295
%20 j
10
зо
% 20
10
50
40
% зо
20
10
60
50
.qo
о
iЪ зо
20
10
11
в наличии
в рационе
Рис. 9.23. Влияние доли менее питательных
мидий (Mytilus edulis) на выбор добычи краба
ми Carclпus тaeпas; см. текст (по данным Elner,
Н ughes, 1978)
скольку не получает н;икакоrо преимуще..
ства при отказе от нее в пользу жертвы у
(при прочих равных условиях). А если сна..
чала он встречается с жертвой у, съест он
ее или откажется, чтобы продолжать пои
ски более выrопной для Hero жертвы х?
С точки зрения максимизации чистой эне..
рrетической прибыли решение зависит TO
лько от частоты встречаемости х" т. е. от ве..
личины Т:,(Х). Особь типа у, встреченная жи..
вотным потребителем, должна быть съеде..
на, если
Е(у) Е(х)
> ,
Ть(у) Ть(Х) + Ts(X)
и OTBeprHYTa, если
Е(у) Е(х)
< .
Ть(у) Ть(х) + Ts(x)
Конвейерная лента
"Поисковое время"
для крупных
червей, с
ОБщее число
пищевых ОБЪ'"
ектов в минуту
Место
ночлеrа
Прозрачный
пластик
Выдвижная
крышка
Процент преДЪЯ8ЛЯ
емых крупных и
мелких червей
Крупные Мелкие
50 50
........
.......
......
......
Рис. 9.24. Аппарат, используе
мый для предъявления с Me
няющейся частотой различных
пищевых объектов (мучных
червей) неподвижному потре
бителю (большой синице); см,
текст (по Krebs et al., 1977)
Процент съеденных
крупных и мелких
червей
Крупные
53 Мелкие
147
.......
-:.:.;
б
50 750
><
х
><
49
1t 51
B
:-:.: 67
......
......
:-::::
........
......
.......
.:-: .:
.......
......
.:.:.:
......
75
А 40
з
50
18
27
33
87
13
Б 20
8
12
88
т. 12
E :: : ; :I
78
:iiШ 22
Рис. 9.25. Специализация на более крупных мучных червях при небольшом связанном с ними
поисковом времени, как в опытах В, r и Д:
Е(мелк)
Тh(мелк)
7
r
7
д
7
< Е(крупн) (по Krebs et al., 1977)
Th(KPYnH) + Ts(KPYnH)
Питание
297
Иными словами" данная встреченная особь
жертва должна съедаться" если за время" за..
траченное на ее переработку" найти и съесть
друrую" более блаrоприятную с точки зре..
ния полученной энерrии жертву невозмо"
жно (рис. 9.24 и 9.25" приложение 9.2).
Дальнейшее усложнение состоит в том"
что мноrие виды потенциальной пищи рас..
пределены в виде компактных скоплений.
т. е. отдельными пятнами" например opra..
низмы, обрастающие морские скалы, мура..
вьи в МУРCtвейниках, стаи мизид, заросли
крапивы и т. п. В этом случае консумент
сталкивается с проблемой друrоrо рода:
любое подобное скопление" участок или
область локальной концентрации содеРЖИ1
определенное количество пиши. поедание
которой может привести к уменьшению ее
обилия и снижению чистой прибыли для по..
требителя. Как же долrо следует эксплуати"
ровать данное пятно, прежде чем перейти
к следующему? Стоит ли сначала исчерпать
все ero ресурсы или выrоднее прекратить
использование раньше, а если да, то в какой
момент?
Хотя внешне иные. но по сути такие же
проблемы встают и перед друrими живот..
ными"потребителями. Например" в высоких
llIиротах количество доступной пищи зави..
сит от сезонных климатических условий.
и животные должны решать" коrда им поки..
дать rеоrрафическую область с уменьшаю..
щимися пищевыми ресурсами. Коrда дол..
жны улетать ласточки? Ряд особых взаи..
моотношений хищник жертва также от..
носится к этой катеrории" особенно если
хищникам требуется значительное время,
чтобы потребить все ткани пойманной
жертвы. в том числе высосать из нее все со..
ки. Пчелы" питающиеся нектаром из цветка,
водяные клопы" сосущие жидкость тела ко..
мариных личинок" и даже львы" пожираю..
щие антилопу. ВО всех этих случаях снача
ла скорость поrлощения пищи велика" но по
мере Toro" как основная часть более доступ
ных и более питательных ее компонентов
усваивается, процесс потребления замед..
ляется. В какой момент животному следует
оставить старую жертву и переключиться
на новую? После Toro как извлечен весь съе..
добный материал или не дожидаясь этоrо?
Коrда потребитель начинает эксплуати"
t-----
"
ш
т
:S:
:S:
L
а..
<1.)
:1:
(т)
':S:
О
:1:
:1:
Ф
Т
>-
а
с
со
1:
1:
>-
U
т
Рис. 9.26. Изменение со временем скорости
получения энерrии из оrраниченноrо источни
ка; см. текст
ровать еще нетронутое до Hero пятно" будь
то rеоrрафическая область или пойманная
особь жертвы, количество энерrии" извле
каемое в единицу времени, можно выразить
соотношением, представленным на рис.
9.26. Рано или поздно животное должно по..
кинуть этот источник пищи, перейдя на др у..
rой участок или поймав друrую жертву.
причем 11'01' переход может стоить ему мно"
[о времени и энерrии. Даже такое медленно
передвиrающееся животное, как береrовая
улитка Litto# il1a" тратит в единицу времени
в 12 раз больше энерrии на ползание" чем на
соскабливание водорослей на одном месте"
и, конечно" миrрации на большие расстоя..
I:S:
О
:1:
:1:
Q)
Т
>-
t::
О
Cs;
CO S
1: L
a..
>-
()(Т)
1Ii.
:J" 11(
Время Время, проввденное
миrрации на пятне
Рис. 9.27. Потребление энерrии при эксплуата..
ции пятна ресурса в зависимости от времени
проведенноrо в ero пределах и затраченноrо на
ero достижение; см. текст
298
:s:
Ь
с'о
t::
LO
О
.:s:
О
I
I
с'о
....t::[
О::
:s:Ф
LI
o..
Ф О:: .... . , .
IC
(Т)
O
C'OI
Iq
2"0
C'O
I
().
O:: -<
О::ф
I-;r
g>-
0..С:
ug
/..
.,/
.,/
",..,
.,/
",..,
",..,
.,/
",..,
=-
Время, проведенное
на пятне
Среднее
время
миrрации
Оптимальный момент
покидания пятна
ния обходятся ей очень дороrо. Значит, He
обходим о (с поправкой на различный pac
ход энерrии) учитывать и время" требуемое
на передвижение как между участками. так
и внутри них.
Теперь, если мы соотнесем среднюю
чистую энерrию, полученную со всех пятен
в данном местообитании. со временем и уч
тем среднее время, необходимое для пере
мещения от одноrо пятна к друrому (рис.
9.27)" можно задать вопрос, какой период
времени, проведенный на пятне, даст макси..
мальный выиrрыш в 1нерrии за единицу
cYMMapHoro времени (т. е. включая переме
щение от одноrо пятна к друrому и пребы
вание в пределах пятна). Ответ дает KacaTe
льная. проведенная от начала координат
к кривой, отражающей общее количество
поrлощенной энерrии (рис. 9.28), т. е. линия,
идущая от начала координат к этой кривой
под максимальным уrлом. Таким образом.
:s:
:s:
L
о..
(O
(Т)Х
0::<0
:s:I
I
фО::::s;
:r c .....
>-х о
t:: <o
0..Dt::
cJ5LO
..Dt:: 0
..... a.>,::s;
(Jc:Io
O.....I
g.OI
CU
UI
...ropo
'l.i..Je e
fJЯ'liО
Средняя скорость
для всех пятен
1
Покинуть
раньше
rлава 9
Рис. 9.28. Время нахождения
в пределах пятна обеспечи
вающее максимальную CYM
марную прибыль на единицу
cYMMapHoro времени Фуражи
ровки, можно найти, проведя
касательную от начала коорди
нат к кривой получения энер
rии; см. текст
потребитель, покидающий участок с пище
вы ми ресурсами или прекращающий пое
дать данную жертву, коrда количество по..
rлощенной 1нерrии в единицу времени па..
дает до этоrо среднеrо уровня" максимизи..
рует энерrию, полученную в единицу сум..
MapHoro времени фуражирования. Потре..
бители должны прекратить использование
сокращающихся источников пищи в опти..
мальный для этоrо момент!
Если это так" то следуют выводы:
а) коrда пятна внутри местообитания
различаются по качеству" потребители дол..
жны дольше оставаться на лучших пятнах
(рис. 9.29);
б) коrда пятна качественно приблизите..
льно одинаковы, но расстояния между ними
различны, для потребителя выrоднее доль..
ше оставаться на данном пятне, если он за..
тратил больше времени, чтобы достичь ero
(рис. 9.30) (т. е.. если затраты на передвиже..
Рис. 9.29. Влияние качества
пятна на оптимальное время
которое следует провести на
различных пятнах в пределах
данной области; см текст
Покинуть
позже
Время, проведенное
на пятне
Питание
ш
u
о
Q.
:S:
Q.
с
.:S:
])
I
Q.
Ф
L
>-
()
=- If:
Короткий Относительно
период недолrо
миrрации
между пятнами
w
u
о
Q.
s:
Q.
С
,.,.,,.,.,"'"
.:S:
::а
I
Q.
Ф
1:"
L
>-
()
,.,.,
,.,.,,.,.,,.,.,,.,.,
,.,.,
,.,.,"'"
,.,.,"'"
,.,.,
f..
Долrий период
миrраци между
пятнами
:J -=- ...
Относительно
долrо
Рис. 9.30, Влияние среднеrо периода миrрации
между пятнами на оптимальное время, KOTO
рое следует провести на пятнах одинаковоrо
качества; см. текст
пне выше, выиrрыш от Hero будет меньше,
и потребитель должен остаться на каждом
пятне дольше).
9.3.3. Подвижные потребители
и их жертвы
Каким образом общие сооБРdжения, из
ложенные выше, влияют на биолоrию пита..
пия подвижных животных, пищей которым
служат отдельные объекты, и как MorYT их
жертвы минимизировать шансы быть cъe
денными?
Очевидно, одно и то же количество пи
щи бывает представлено либо большим
числом мелких объектов, либо несколькими
крупными, и эти два экстремальных случая
требуют различных затрат и дают неодина
ковую выrоду. Выrода от одной крупной
299
особи велика, однако такая жерrва скорее
Bcero будет очень ПОДВИЖНd, поэтому, что
бы поймать ее, потребуются большие за
траты времени и энерrии. С друrой CTOpO
ны, питание мноrими мелкими особями MO
жет не требовать MHoro времени на поимку
каждой из них, однако велика вероятность
долrих поисков добычи. Эти особенности
будут отражаться как на потребителе, так
и на ero жертве.
Преследующие охотники обычно б6ль
шую часть cBoero cYMMapHoro времени пи..
тания проводят в поrоне за крупными oco
бями жертвами, причем не все! да успешно.
На каждую потенциальную жертву трати
тся MHoro энерrии, поэтому и прибыль дол
жна быть велика. Охотнику выrодно мини..
мизировать затраты на преследование,
а жертве. максимизировать их. Это ведет
к cBoero рода «roHKe вооружений» между
ХИЩНИКОl\1 и жертвой. Первый оrраничивает
себя таким KpyroM своих потенциальных
жертв, которые наиболее уязвимы для ero
специфических форм преследования и за
хвата, получая при этом максимальную 1не..
рrетическую компенсацию и все больше
приспосабливаясь к особенностям усколь"
1dНИЯ своих жертв от преследования. В pe
зультате максимализации чистой прибыли
преследуюшие охотники по своему раuиону
становятся «специалистами».
Для жертв помимо очевидноrо давления
отбора в сторону увеличения скорости
и ловкости наиболее распространенным
способом уменьшения шансов быть пой
манными хищниками (не только пресле
дующими) является rрупповое существова
ние (стаями, стадами, косяками и т. п.). Это
дает целый ряд' преимуществ.
1. Одно из наиболее известных наблюде
ний заключается в том, что большая rруппа
может обнаружить приближение хищника
и отреаrировать на Hero быстрее, чем мел
кая rруппа или отдельная особь. Например,
мелкий бескрылый морской клоп Ha/obates
скользит по поверхностной пленке моря т ак
же, как водомерка по поверхности пресных
водоемов. КоrДd эти клопы не питаются,
они собираются во «флотилии> которые
очень четко реаrируют на приближение
хишников (птиц.. рыб и т. п.). Расстояние,
с KOToporo «флотилия» реаrирует избеrа
300
о::() 90
S ...
:1:""""
<0<0
ro
0:1:
Q.:f 60
Ss
@х
(l)
Q.o::
Q)S
s:l: ЗА
:I:
g
......<0
():I:
()ш
<00
0............
100 200
Число ОСОБей ВО "флотилии"
Рис. 9.31. Зависимость расстояния, на котором
экспериментальная модель хищника вызывает
поведенческую реакцию у MopcKoro клопа Ha
lobates, от числа ero особей во «флотилии» (по
Treherne, Foster, 1980)
нием на хищника, зависит от ее размеров,
как показано Het рис. 9.31: оно больше в слу
чае более крупных флотилий. хотя. конечно..
существует критический размер, начиная
с KOToporo дистанция обнаружения уже не
возрастает (т. е. достиrнут максимум).
2. Второе преимущество существования
в rруппе это введение хищника в замеша
тельство. Преследующие охотники OДHOMO
ментно нападают на одну потенциальную
жертву, и мы уже видели, что им выrоднее
схватить особь с высокой питательной цeH
ностью. rруппа может реаrировать на при
ближение хищника рассеянием особей в раз
ных направлениях Это затрудняет «нацели
вание» на индивидуальную жертву и приво
дит К переключению с цели на цель, коrда
разные особи мелькают в поле ero зрения.
3. Хищников может отпуrИВdТЬ rруппо
вое защитное поведение животных, в то
время как подобное затруднение OTCYT
ствует при встрече с одиночным представи
телем Toro же вида. Классические примеры
rрупповой защиты относятся к позвоноч
ным, однако есть они и среди беспозвоноч
ных, причем особенно яркие среди обще
ственных перепончатокрылых. Кроме Toro,
личинки некоторых пилильщиков при напа
дении (например., хищных насекомых) Bыдe
ляют своими ротовыми частями липкий ce
крет. Подобная реакция не мешает клопам
щитникам атаковать отдельных личинок,
однако, нападая на их скопление. клопы бы
стро покрываются липкими выделениями
и после этоrо едва MorYT двиrаться.
rлава 9
4. Все три положительных эффекта rруп..
повоrо существования, описанные выше, ве..
дут к увеличению времени, которое требуе..
тся хищнику для удачной поимки жертвы.
Это может привести к тому, что для Hero
окажется более выrодным переключение на
друrую жертву, не относящуюся к обще
ственным животным. Четвертый эффект
rрупповоrо существования относится к ве..
роятности для каждой данной особи в rруп..
пе оказаться жертвой удачноrо нападения:
чем крупнее rруппа., тем эта вероятность ни
же. Это было показано для клопа Ha/o/Jates,
о котором упоминалось выше в связи с ди..
станцией обнаружения хищников, коrда
этим насекомым уrрожают рыбы (рис.
9.32). Один клоп из 10 в rруппе подверrается
1/10 общеrо числа нападений, направлен--
ных на одиночную особь, а если в rруппе
100 клопов, вероятность оказаться жертвой
составляет 1/100.
Этот эффект необязательно чисто стати..
стический. Часто вероятность быть схвачен..
ным выше Bcero на периферии rруппы и ми..
нимальна в ее центре. В ряде случаев на..
блюдали непрерывное движение особей от
периферии к центру; при этом по краям
остаются животные, менее друrих способ..
ные занять относительно безопасное поло
жение внутри «эrоистическоrо стада». Так
происходит активная минимизация индиви"
дуальной вероятности быть съеденным.
В противоположность этому охотники
из катеrории поисковиков и засапчиков
.1)
ш 10
о
()
о
<о
:1: 1
'5
S:I:
:l:s
Ф
L{)
с: 0,1
<0<0
:I:M
О
с:::
()
5
:r 0,01
1 10 100
Число ОСОБей в rруппе
Рис. 9.32. Число нападений рыб на отдельных
особей Halobates в зависимости от числа кло
пав в rpynne (по Treherne, Foster, 1980)
Питание
предпочитают добычу, которую леrко пой..
мать из..за небольшоrо (по сравнению
с хищником) размера. Любая с леrкостью
схваченная жертва тут же съедается, что
дает в сумме достаточно еды, и поискови..
кам леrче Bcero максимизировать свою
чистую прибыль, становясь по рациону re
нералистами. Затем, поскольку поисковое
время обратно пропорционально общему
обилию подходящей жертвы, вместообита..
ниях, r де мало добычи или в периоды сни"
жения ее количества, поисковики.. по..
видимому, наименее специализированы
в смысле питания. Зато в периоды пика оби..
лия определенноrо вида..жертвы они MorYT
тратить непропорuионально MHoro време..
НИ, потребляя именно ero (становясь вре..
менно «специалистами»), и переключаться
Hd друrие виды, только коrда их числен..
ность также достаточно возрастет.
Особям видов, выедаемых поисковика..
ми, выrодно максимизировать поисковое
время охотника. Этоrо можно добиться
Рd.зличными путями. увеличивающими
трудности обнаружения жертвы или распо..
знания в ней потенциальной пищи. Способы
следующие (рис. 9.33):
1) заселение таких микроместообитаний,
которые нелеrко обследовать большинству
потенци<tльных хищников. т. е. жизнь в ще..
лях, под камнями, в норах и т. д.;
2) незаметность, Т. е. особые (покровите..
льственные) форма тела. поза. окраска (или
запах), сливающиеся с фоном и таким обра..
зом увеличивающие время распознавания
(некоторые виды способны изменять цвет
и рисунок тела так. чтобы сливаться с не..
сколькими типами фона; друrие изменяют
свое окружение, чтобы оно более эффектив..
но скрывало их от хищника);
3) мимикрия. как и покровительствен"
ный облик, увеличивает время распознава
ния хищников за счет имитации малосъе..
добноrо или вовсе несъедобноrо объекта
(MepTBoro листа, сучка, кусочка фекалий
и т. д.. вида с плохим вкусом или способно..
ro к аrрессивной самозащите)
Увеличение необходимоrо поисковоrо
времени не единственный способ уменьше..
ния пищевой uенности отдельной особи для
хищника; к друrим относятся удлинение
периода переработки жертвы и уменьшение
301
энерrии, содержащейся в единице веса жерт"
вы.
Увеличение времени переработки часто
достиrается в ходе эволюции следующих
признаков (рис. 9.34):
1) защитноrо вооружения (жал.. челю..
стей и т. д.):
2) защитных покровов (раковин, извест..
ковых или хитиновых пластин и т. д.);
3) специальных образований, затрудняю..
щих захват жертвы, манипулирование ею
и/или заrлатывание (например., иrл). Иrлы
и подобные им выросты тела особенно
эффективны, поскольку увеличивают не то..
лько время переработки, но и видимый раз
мер тела жертвы (при небольшом приросте
тканей) и таким образом исключают живот..
ное из числа возможных объектов поедания
хотя бы некоторых хищников.
Содержание энерrии и или питательных
веществ на единицу веса или объема тела
может быть уменьшено путем «разбавле
ния» живых тканей инертными, мало пита
тельными или несъедобными веществами.
Вкусовая приrодность может быть снижеНd
присутствием особых веществ в некоторых
opraHax или во всем теле. В этом случае
хищнику необходим определенный уровень
научения методом «проб и ошибок».. и LIa
сто жертвы делают такое научение как мо"
жно более быстрым и эффективным, под
крепляя неприятный вкус яркой предупре
ждающей окраской.
Увеличение времени на переработку.
снижение содержания энерrии на единицу
веса и уменьшение вкусовой приrодности
(или явная токсичность) используются так..
же в качестве способов защиты пищевыми
объектами обrрызающих и соскабливаю..
щих животных, функционально эквивалент
ных охотникам"поисковикам. но потре
бляющих прикрепленные орrанизмы, часто
способные к фотосинтезу. Поисковики, как
правило, крупнее и живут дольше, чем их
жертвы"животные, а у соскабливателей
и обrрызателей обычно как раз наоборот
их добыча крупнее их самих и долrовечнее.
Таким образом, в то время как жертвам
охотников"поисковиков трудно использо..
вать какую..либо иную стратеrию. кроме
научения своих долrоживущих потребите..
лей с помощью отпуrивающих веществ из..
302
ОБИТАНИЕ В НОРАХ
' J;:::
... ,:,,,..'\',:,.' ,.
: ? ::<:?'
':':--<-;: ;-,
Q d. i!i _
..... ... ' .-.1',"
: .. , 1 .
Полихета
f .. : .\.. .:
.. ,-"
. r ...;Jfz -"
." l' . :..
.,. ..;;:
,', . .' '" ._ ;: i::-2i ;!"J.
ПОКРОВИТЕЛЬСТВЕННЫЙ ОБЛИК
Клоп, сливающийся
с корой
МИМИКРИЯ
v ::i ;:i:: . ,:!; _, ."
-,о,' ""W'f.""ft: '. """"' ,
:A(:; :t : . ", Б
- \.! ""А - .... .
.:t... ., · У- \.
..-: ; _. ,.' .. ..... /
' ';' - · ( . 4 " lt v J
.. ; .. -.
,.} ;
rусеницы БаБОЧКИ,
имитирующие
ПТИЧИЙ поме1
. 1:;.
"... : . .'.
' = :"
беrать их.. долrоживущим растениям леrче
и выrоднее просто убивать токсинами ко..
роткоживущих насекомых"фитофаrов. осу..
ществляя, таким образом, отбор особей.. иr..
НОРИРУЮIЦИХ данный вид в качестве пище..
Boro pecypcd.
Все эти Зd.щитные системь] (см. также
разд. L3.2.1) создают для KOHCYMeHTa
rенералиста такую ситуацию, коrда ему вы..
rлава 9
Палочник
(насекомое)
на ветке
Рис. 9.33. Механизмы, увели
чивающие поисковое время
для потенциальных хищников
(из различных источников)
rоднее искать друrие типы жертв более
заметные.. менее защищенные, более при..
влекательные на вкус. Однако успешное из..
беrание нападения со стороны rенералистов
приводит к высокой численности полаrаю..
щихся в основном на пассивную механиче..
скую или химическую защиту прикреплен..
ных или сидячих орrанизмов, а это делает
их доступными для нападения «специали..
Питание
заз
Карапакс
иrлы
(!! , \\\\\ ! I //
\\ : \, J 11 / /;, .
-'. '1 / >
... ,,/ ,/ /
. 1\f\---: :
.... 1/11/"
--; , f- v1 .'
',: ::'"ttj::1.'f (, .\. ','
Морской еж
\
- \
'-
РаКООБразное
Дорсальная створка
Раковина
f'
Личинка раКООБразноrо
Вентральная створка
Плеченоrое
ЖАЛА
r Жало
G;
Рис. 9,34. Структуры, увеличи
вающие время переработки для
потенциальных хищников (из }l
различных источников) Скорпион (хелицеРО8ые)
стов», ЭВОЛЮЦИОННО приспособленных для
преодоления таких барьеров. Обrрызатели
и соскабливатели, а также специалисrы..
потребители форl\t1. используюших покро"
вительственный облик, МИМИКРИIО и отпу"
rивающие вещества, принимали участие
в такой же эволюционной «roHKe вооруже..
ний» с поедаемыми видами, что и охотники..
преследователи с их чрезвычайно подви"
жными жертвами. В самом деле. специали..
сты"потребители, выработав мехаНИЗl\llЫ,
которые делают неэффективными химиче..
ские методы защиты их жертв" MorYT даже
накапливать токсичные вещества в со..
бетвенном теле, используя их для защиты
против своих BparoB.
ПереднежаБерный моллюск
9.3.4. Потребители суспензий
и осадков
Животные улавливающие пищу путем
процеживания потока воды, очевидно" TPa--
тят энерrию не на преследование или пои--
ски добычи, а на фильтрацию и отбраковку
ненужных частиц.
Рdзмеры отверстий в их фильтрах обыч..
но MorYT меняться лишь в масштабах ЭВО"
люционноrо в pel\ll ени " поэтому трудно
предполаrать возможность 01бора из воды
только частиц с наиболыпей питательной
ценностью: фильтр специализирован на
размер, а не на съедобность пищи. Прикре..
пленные фильтраторы MorYT максимизиро"
304
:s;: 20
:s;:
::f
са
а.
......
.D 15
с:::
:s;:
е
:s;:
()
о 10
а.
о
()
Q)
r.:1 5
I
.s:
о 2 4 б 8 10
Содержание усвояемой энерrии
во взвешенном материале, кДж/л
Рис. 9.35. Связь между концентрацией съедоб
ных частиц в воде и скоростью фильтрации
у полихеты Nereis diversicolor, питающейся cy
спензией (по Pashley, 1985) (см. рис. 9.5, r
и табл. 9.1)
вать чистую скорость потребления Jнерrии..
изменяя только следующие параметры:
1) скорость фильтрации в зависимости
от относительноrо обилия в воде частиц
разноrо типа;
2) скорость отбраковки в зависимости
от относительной питательной ценности ча..
стиц и затрат на отбраковку.
Скорость фильтрации должна увеличи..
ваться при повышении конпентрации ча..
стиц с высокой ПИТdтельной ценностью
(рис. 9.35), ОДНйКО здесь может быть верх..
ний предел, определяемый возможностями
пишеварительноrо тракта хищника перера..
батывать поступающий материал. При сли..
зисто"ресничном механизме это зависит от
движения слизистоrо тяжа внутри кишки,
обусловленноrо деятельностью ресничек.
у членистоноrих, просеивающих пищу с по..
мощью щетинок, может произойти полное
заполнение полости кишки заrлоченными
частицами. Как только это происходит, СКО"
рость поrлощения lнерrии оrраничивается
временем прохождения частиц через пище..
варительный тракт и интенсивностью их
переваривания. Следовательно, скорость
фильтрапии падает до величины, способ..
ствующей поддержанию кишечника в за..
полненном состоянии. Как и в ситуации
rлава 9
с хищниками, поедающими только наибо-
лее питательные части добычи, чтобы после
JTOrO перейти к друrой жертве (разд. 9.3.2),
потребителям суспензий, зависящим от ка..
чес [ва и обилия пищевых частиц, бывает
ВЫI'однее быстро пропускать пищу через ки"
шечник, усваивая лишь часть ее, чем мед..
ленно переваривать большую долю заrло..
ченноrо.
Затраты на отбраковку бывают высоки,
поэтому по мере роста затрат на отбраков"
ку частиц определенноrо типа можно ожи..
ДйТЬ увеличения скорости их заrлатывания.
КОI да эти затраты очень велики, бывает де..
шевле избавляться от даже полностью не..
съедобных частиц через кишечник, а не е по..
мощью сортирующих opraHoB. В этих уело..
виях потребитель суспензий может столкну..
ться с дефицитом энерrии, и ему иноrда вы..
rоднее полностыо прекратить питание до
изменения состава взвешенноrо в воде
вещества.
При подобных системах сбора пищи
возможность стать «спепиалистом» очень
мала (если вообще существует), разве толь..
ко в эволюционном масштабе времени и
в отношении определенноrо размерноrо
диапазона частиц. Например, у пелаrиче..
ских оболочников очень тонкоячеистые фи..
льтры, способные задерживать бактерии;
с друrой стороны, киты отцеживают круп..
ных планктонных эвфаузиид. Эти различия
связаны просто с величиной отверстий их
(рильтров и относительными размерами са-
мих фильтраторов. Киты не моrли бы испо..
льзовать qJИЛЬТр, способный собирать бак..
терий и протистов: им не удалось бы одно-
временно продвиrаться вперед и фильтро..
вать по той же самой причине, по которой
невозможно быстро протащить в воде мел-
кую планктонную сеть и что"нибудь в нее
поймать. Вода просто проталкивается впе-
ред, почти не проходя через ячеи. Такая
избирательность у потребителей суспензий,
естественно, не позволяет реrулировать пи-
тательную ценность частип. а проявляется
в основном у свободноживущих личинок,
имеющихся у мноrих видов, коrда они рас..
сеЛЯIОТСЯ в места, способные обеспечить
прикрепленные взрослые сталии необходи-
мыми взвешенными пищевыми частицами.
Потребители осадков представляют со..
Питание
З05
бой нечто среднее между потребителями су..
спензий и объединенной катеrорией охотни..
ков и обrрызателей"соскабливателей. Пи..
таясь неживым орrаническим веществом"
они потребляют пищу низкой питательной
ценности, которая соответственно очень
обильна и не может защищаться от поrло..
щения. Некоторые консументы этой KaTero..
рии просто пропускают LIерез кишечник не..
орrанический осадок, содержащий орrани"
ческий компонент" поэтому сталкиваю rся
с такими же проблемами переполнения ки"
шечника, как и потребители суспензий. Kor..
да содержание усвояемоrо вещества в осад..
ке ниже определенноrо предела. может
быть энерrетически выrоднее не поrлощать
такой материал, а перейти в друrое место.
Однако мноrие потребители осадков спо..
собны к избирательному поrлощеНИIО
(разд. 9.2.6) и MorYT вести себя как охотни
ки..поисковики С той разницей, что «поиск»
совершает не все животное, а очень подви
жные щупальца или подобные им образова..
иия.
В целом живые существа в Рdционе потре..
бителей суспензий и осадков очень малы по
сравнению с размером caMoro потребителя,
и, хотя некоторые из поrлощаемых видов
и располаrают химическими средствами за..
щиты, большинство" по"видимому" полно..
стью лишены препятствующих выеданию
систем. Вместо этоrо они наделены способ..
иостью быстро размножаться половым
и/или бесполым путем, которой более чем
достаточно для компенсации потерь от
хищничества. В этом они отчасти сходны
с добычей обrрызателей и соскабливателей,
обычно достаточно интенсивно растущей
или бесполо делящейся, чтобы возмещать
утраченные структурь] типа листьев" поли..
пов или зооидов. так что только при исклю..
чительно высокой скорости выедания на..
блюдается rибель отдельных особей или ко..
лоний.
9.4. ВЫВОДЫ
По всей видимости, первые животные
были мелкими потребителями бактерий..
протистов и друr друrа, причем эти исход..
ные рационы не только сохранились отно"
сительно неизменными в течение миллио..
нов лет, но и всеrда оказывали влияние Hd
питание животных. Селективные преимуще..
CTBd крупноrо размера способствовали раз..
витию механизмов, увеличивающих заrла..
тываемое количество этих мелких и pac
сеянных в среде объектов. но не привели
к появлению принципиально новых рацио..
нов. Потребление макрофитов, будь то во..
доросли или растения, возникло относите..
льно поздно и без участия кишечных ми
кроорrанизмов остается малоэффективным
не только из..за низкой питательности тка..
ней, боrатых структурными уrлеводами, но
и в связи с защитными приспособлениями
этих во всех друrих отношениях леrко дo
ступных объектов нападения.
Подразделение животных на «растите..
льноядных», «плотоядных>} и «всеядных>} не
способствует пониманию их биолоrии пита..
ния: с этой точки зрения большинство ви
дов относятся К всеядным. Более рациона..
льна классификация по типам питания
охотники/паразиты, соскабливатели/об..
rрызатели, потребители осадков. потреби..
тели суспензий, потребители продуктов жи..
знедеятельности эндосимбионтов и т. д. Т d"
кая система не считается с систематическим
положением поедаемых видов и ориенти"
руется скорее на потребителя, чем на пот ре..
бляемое.
Хотя и оrраниченные в разной степени
своим ЭВОЛЮIlИОННЫМ прошлым.. все живот..
ные тем не менее оказываются в масштабах
}колоrическоrо времени перед выбором: Ka
кую из особей..жертв предпочесть в данных
пространственно"временных условиях; про..
должать питаться на этом участке или пе..
рейти на друrой и т. д. По"видимому" потре..
бители не принимают случайных «реше..
НИЙ», и теория оптимизации дает модель,
довольно успешно предсказывающую пове..
дение животных в ходе фуражирования.
В рамках оrраничений создаваемых факто..
рами, не относящимися к качеству пищи
и обеспеченности ею (например. хищниче..
ством, биолоrией разможения), некоторые
животные, действительно, питаются таким
образом, чтобы максимизировать получае..
мую прибыль (выrода минус затраты).
С друrой стороны, отбор среди видов..
жертв ведет к максимизации относительных
расходов, связанных с ИСПОЛЬЗОВdнием их
З06
в пищу, за счет особенностей поведения,
морфолоrии или биохимии. Как и следова..
ло ожидать, различные типы питания под..
разумевают неодинаковые потенциальные
решения проблемы максимизации прибы..
ли, дающие селективное преимущество ли..
бо rенералистам" либо специалистам.
Пищевое поведение.. очевидно" ежеднев..
но находится ПОД давлением не менее ин..
тенсивноrо отбора" чем друrие" более изу..
ченные аспекты биолоrии животных.
9.5. Дополнительная
литература
Вап1аl"а С. J. (Ed.). 1985. Pl"oducers and Scroungel"s.
CI'oom НеlПl" Lопdоп.
Са/он' Р. ] 981. [nverrebI'ate Biology: А Functional
Approach. Croom Helm, London.
Сl'ан'/еу JvJ. J. 1983. Herbivory. Blackwe1J Scientific
Publications, Oxford.
HOllkiпsoп 1. п., Hu r;lles М. К. 1982. Insect Herbivo
ry. Chapman and НаН" New York.
Jeппiпgs п. Н., Lee D. L. (Ed.) 1975. SYnlbiosis.
Cambridge Unlversity Press, Cambridge.
Jeппiпgs J. W. 1972. Feeding" Digestion and Assimi
lation in Animals. 2nd edn. Macmillan, London.
JOl A geпsoп с. В. 1966, Biology of Suspension Fee
diпg. Pel"gamon Pl"ess, Oxford.
М asoп с. Е. 1977. Decomposition. Edward Arnold_
London.
M01'foп J. 1979. Guts, 2nd edn. EdwaI'd АП101d_
London.
Он'еп J. 1980. Feeding Strategy. OXfOI'd Uпivсrsitу
Press" Oxford.
SПlitll D. с.. Doиg/as А. Е. 1987. The Biology of
Symbiosis. Edward AI"nold" LondoI1.
ТаУ/Оl' R. J. 1984. PI'edation. Сhарmап ап(! Наll.
New YOI"k.
TOH'пseпll с. R., Са/он' Р. (Ed.) ] 981. Physiological
Ecology. Blackwel1 Scientific Publications, Ох..
f ord.
Vel A тe(j G. 1987. Evolution апd Еsсаlаtiоп. Ап Eco
10gical History of Life. Princeton U niversity Press,
Princeton.
Приложение 9.1.
Выбор мидий крабами
в результате наблюдений не представ
ляет большоrо труда определить время" KO
торое требуется крабу, чтобы разломать pa
ковину мидии и съесть ее содержимое (T h ).
Количество энерrии, полученное при поеда..
нии моллюска (Е), определяется с помощью
rлава 9
калориметрической бомбы. Краб ломает
раковину с помошью своих клешней, на что
уходит значительное «время переработки»,
которое более или менее экспоненциально
увеличивается с размером мидии. Таким
образом, для открывания крупных раковин
требуется очень MHoro времени, но им соот..
ветствует большое значение Е, а мелкие ми..
дии можно открыть быстро, НО они дают
и меньше энерrии. Для краба данноrо раз..
мера отношение Е/Т'l изменяется с измене..
нием размера мидий, как показано на рис.
9.21. На этом же rрафике указан размерный
диапазон мидий, поедаемых крабами рас..
сматриваемоrо размера. Очевидно. соотно"
шение между теми мидиями, которые съе..
дены, и теми, что обладаlОТ наибольшей
предсказанной питательной ценностью" т. е.
проrноз 1 (стр. 294), оказывается верным.
В этом опыте каждая съеденная мидия
тут же заменялась друrой TaKoro же разме..
ра, и таким образом частота мидий снаибо..
льшей питательной ценностью поддержива..
лась на уровне достаточно высоком для то..
ro, чтобы включение в рацион менее ценных
жертв привело к снижению средней скоро..
сти потребления энерrии. Во втором эк сп е..
рименте съеденные мидии не возмещались.
и частота наиболее питательных моллю
сков уменьшалась. Реакция на это крабов
показана на рис. 9.22: в соответствии с про..
rнозом 2 (с. 294) они расширили диапазон
своих жертв, включив в Hero следующие по
ценности размеры жертв.
Крабы распознают размеры мидий, при..
касаясь к ним.. и в третьем опыте изменяли
соотношение моллюсков разноrо размера.
Из рис. 9.23 следует" что в подтверждение
проrноза 3 (с. 294) в случае BbICOKoro оби..
лия менее пенных мидий они все же употре..
блялись в пищу, хотя особи оптимальноrо
размера были также мноrочисленны.
Таким образом, коrда Carciпus сталки..
вается с особями разноrо размера одноrо
типа жертвы, он способен выбрать тот раз..
мер, который даст ему максимальное коли..
чество энерrии за единицу времени. В ка..
кой то мере он, вероятно, оценивает питате..
льную ценность мидий перед тем, как пита..
ться ими.
Питание
307
Приложение 9.2. Поисковое
время и выбор жертвы:
синица и мучные черви
Одна из трудностей. связанных с провер..
кой вывода о том, что жертва будет съеде..
на, если искать друr)'ю менее ВЫl ОДНО, за..
ключается в проблеме правильной оценки
поисковоrо времени: Korna потребитель
передвиrается в предеЛаХ cBoero местооби..
тания, как установить, ищет ли он опреде..
Iенный тип жертвы или преследует иные це..
ли? Эта трудность была изящно обойдена
учеными из Оксфорда. Вместо Toro чтобы
дать потребителю возможность перемеща..
ться среди различных жертв, ero удержива..
ли на месте и предлаrали ему несколько 1И"
пов пищевых объектов снеодинаковой ча..
стотой. Эффект получался такой же, поско"
льку частота обнаружения пищи (эквива..
лент поисковоrо времени) реrулировалась
скоростью ее движения мимо ЖИВОТНОI О.
В этом опыте потребителеl\1 была птица
(большая синица, Parus), а жертвами
кусочки мучных червей (личинок жуков..
хрущаков) двух размеров, причем больший
по размеру кусочек был вдвое боrаче эне..
рrией, чем меньший.
Мучные черви продвиrались мимо сидя..
щей в клетке птицы на ленте конвейера
(рис. 9.24). и синица моrла либо съесть. ли..
бо проиrнорировать любой потенциальный
пищевой объект. Однако, склевывая одноrо
из червяков, она упускала возможность
схватить тех, которые моrли оказаться до..
ступны в течение следующих 5 15 с (время
переработки). Был проведен ряд опытов
с различными эквивалентами ПОИСКОВОI"О
времени для более ценных крупных червей
и различными частотами предъявления бо..
лее и менее выrодных пищевых объектов
(рис. 9.25).
Соrласно проrнозам модели максимиза..
ции энерrии в вариантах В. r и П (но не А
и Б), выбор падает на крупных червей.
В опытах это подтвердилось. Даже если
мелких жертв было в два раза больше, LleM
крупных (вариант Д). Korna поисковое
время для более выrодной пищи снижал ось.
потребитель иrнорировал менее выrодных
для Hero мелких жертв и склевывал в основ..
ном крупных, причем переКЛlочение на
избирательность происходило при близком
к предсказанному соотношении числовых
параметров.
rЛАВА 1 О
БИОМЕХАНИКА И ДВИЖЕНИЕ
(ЛОКОМОЦИЯ)
в этой rлаве мы познакомимся с тем, Ka
ким образом беспозвоночные MorYT Mexa
нически воздействовать на окружающую их
среду. В основном речь пойдет о типах ло
комоции, однако те же механические при--
нципы применимы и к при крепленным жи
вотным, которые тратят энерrию, переме
щая именно среду, а не самих себя.
Раздел, открывающий rлаву, посвящен
основным принципам механики и термино
лоrии. Затем обсуждаются биолоrические
аспекты rенерирования механических сил
клетками животных. Общая их черта
использование получаемой в ходе дыхания
энерrии (см. rл. 11) для образования пере
крестных связей между белковыми фибрил
лами.
Следующая тема сравнительное изу
чение локомоции животных. Обсуждается
влияние их относительноrо размера на ло
комоторные системы и дается обзор потен
циальных возможностей в этом смысле раз
личных уровней структурной орrанизации
беспозвоночных. Отдельно описываются
локомоторные системы мяrкотелых живот--
ных с raK называемым rидростатическим
скелетом и форм с твердыми скелетными
элементами, в частности ракообразных
и насекомых.
10.1. ПРИНЦИПЫ ЛQКQМQЦИИ
у беспозвоночных можно видеть широ
кое структурное разнообразие. Мноrие из
них мяrкотелые, у друrих развиты твердые
ткани или покровы. Фактически все они спо
собны совершать механическую работу,
в результате которой их тело перемещается
относительно окружающей ero среды или
среда относительно фиксированноrо тела.
Энерrия.. затраченная на эту работу, должна
принести выrоду. Последняя может Bыpa
жаться в повышении интенсивности пита..
ния или использования друrоrо ресурса,
в снижении опасности со стороны хитни..
ков, в избеrании неблаrоприятных абиоти"
ческих условий, в активизации расселения
и/или повышении вероятности контакта
с потенциальными партнерами для полово..
ro размножения.
Коrда животные передвиrаются, они
подчиняются тем же основным принципам,
что и все подвижные объекты, т. е. ньюто..
новским законам движения. Они rласят сле..
дующее:
1) если тело находится в состоянии по..
коя относительно cBoero окружения, оно
может быть приведено в движение только
за счет приложения внешней силы;
2) приложение неуравновешенной силы
к движущейся массе приводит либо куска..
рению. либо к замедлению движения дан..
ной массы в зависимости от направления
действия силы;
3) каждое действие вызывает против о..
действие. равное по силе и противополо"
жное по направлению;
4) энерrия внутри замкнутой системы
остается постоянной, хотя может перехо..
дить из одной формы в друrую.
Эти принципы универсальны, и мы про..
иллюстрируем их в общем виде до рассмо"
трения различных типов движения беспо..
звоночных. В приложении 10.1 приведены
основные законы, управляющие локомо..
цией животных, и определены некоторые
единицы измерения.
Коrда животное движется, оно должно
преодолевать силы. стремящиеся препят"
ствовать этому движению; их называlОТ си..
лами трения или лобовоrо сопротивления
(поскольку противодействие среды, через
которую движется объект, стремится за..
медлить ero движение, речь также идет о си..
БИQмехаНИI<а и движение (ЛQКQМОЦИЯ)
ле). Их принято выражать в единицах, при..
водимых в приложении 10.1. В любой систе..
ме имеется определенный запас энерrии, ко..
торая может быть превращена в кинетиче..
скую, связанную с движением массы, но
в конечном счете рассеиваемую в виде теп..
ла. Это показано в приложении 10.2.
Механическая работа, совершаемая при
локомоции, обеспечивается энеРI ией, выс--
вобождающейся при ХИl\Iических реакциях
в клетках тела животноrо (см. rл. 11), одна..
ко химическая работа необязательно равна
по величине механической.
Человек может очень устать.. стараясь
поднять слишком тяжелый для Hero пред--
мет, однако по определению он не совер..
шает при этом механической работы. Ра--
зумеется, в ero теле происходит метаболиче..
екая работа: мышцы используют А ТР, т. е.
энерrию, запасенную в ходе обмена ве..
ществ, и их усилия приводят К выделению
тепла. Поэтому при оценке эффективности
локомоции мы должны учитывать то. что
затраченная энерrия не всеrда вызывает
движение тела. Механическая работа опре--
деляется величиной массы и расстоянием,
на которое она переместилась. Это можно
назвать полезной рабоп10й. Общие затраты
энерrии при движении животноrо измерить
труднее, но их можно оценить по интенсив--
насти поrлощения кислорода. Тоrда зффек--
тивность проделанной животным работы
(как и к.п.д. любой машины) выразится от..
ношением полезная работа
затраченная энерrия
Мы увидим далее, что одни локомотор"
ные системы MorYT быть эффективнее, чем
друrие, но в то же время иметь оrраничен"
ную мощность, т. е. скорость выполнения
работы. Как и повсюду в биолоrии, здесь
наблюдается пелый ряд компромиссов. сре..
ди которых один из важнейших - между
эффективностью (к.п.д.) и абсолютной
мощностью.
Как показано в приложении 10.2. суще..
ствует инерция движения; кроме Toro, на
движущиеся орrанизмы влияют силы, дей..
ствующие в направлении, противополо"
жном направлеНИIО перемешения, обуслов"
ленные сопротивлением среды. Их роль
309
очень сильно зависит от вязкости (или тре..
ния) последней, относительноrо размера
орrанизма и ero абсолютной скорости. Эта
взаимозависимость выражается числом Рей
нольдса (Re):
Re == 1 V шах ,
V
rде ] длина движущейся системы; Vшах
ее максимальная скорость; v вязкость
среды (для воды v == 1 х 10 6 M2jc).
Число Рейнольдса это мера относите..
льноrо влияния инерции и трения. Для
очень мелких движущихся систем типа
сперматозоидов, жrутиковых, инфузорий,
по крытых ресничками личинок и крохот..
ных беспозвоночных вроде коловраток
(разд. 4.9) число Рейнольдса очень мало.
rде то между 10- 3 и 10 1. В этих случаях
движение почти целиком обусловлено сила
ми трения. Некоторые более крупные бес
позвоночные достиrают скоростей, при ко..
торых число Рейнольдса значительно боль
ше единицы; их движение уже нельзя объяс
нить лишь действием сил трения. Это отно"
сится прежде Bcero к насекомым и rолово..
ноrим моллюскам.
В приложениях 10.1 и 10.2 rенерируемые
силы считаются деЙСТВУIОШИМИ в направле--
нии движения, однако они MorYT приклады
ваться и в друrих направлениях. Такая си..
туация описана в приложении 10.3.
10.2. rенерирование
механических сил
клетками животных
Локомоция У всех животных обусловле..
на изменением формы клеток, противодей"
ствующим сопротивлению окружающей
среды, за счет использования энерrии, вые..
вобождающейся в ходе метаболизма. Локо"
моция обеспечивается тремя типами кле..
ток жrутиковыми, ресничными и мышеч..
ными (рис. 10.1). Несмотря на морфолоrи--
ческие различия, механизмы, приводящие
к изменению их формы, сходны; все они
ВКЛlочают зависимое от А ТР образование
поперечных мостиков между белковыми
фибриллами.
Жrутик это трубчатый вырост клеточ..
ной поверхности, окруженный мембраной,
310
rлава 1 О
II
А
Б
Рис. 10.1. Различные типы двиrательных клеток. А. Жrутиковые клетки: I свободноживущее
жrутиковое простейшее; II хоаноциты rубок. Б. Ресничные клетки: I свободноживушая инфу
зория (простейшее); 11 ресничные клетки животноrо. В. Мышечные клетки
Жrутик
1
А
Б
Рис. 10.2. Схематическое изо
бражение реснички: A по
перечный срез; Б продоль
ный срез; В сrибание
в
Ресничка
>-
Рис. 10.3. Направление силы,
создаваемой биением жrутика
(А) и реснички (Б)
Б
Биомеханика и движение (локомоция)
311
являющейся продолжением мембраны. по..
крывающей и всю клетку. Он содержит пу..
чок белковых фибрилл, так называемую ак..
сонему, которая, как показывает электрон..
ная микроскопия, состоит из лвух централь..
ных \1икротрубочек, oKpY)J(eHHbIX кольцом
из девяти пар микротрубочек (рис. 10.2).
Изменение формы жrутика вызывается ско"
льжением наружных дублетов ДРУI по друrу
вследствие образования поперечных мости..
ков между соседними микротрубочками.
Для образования каждоrо TaKoro мостика
требуется энерrия фосфатной связи, постав..
ляемая молекулой А тр (см. разд. 11). Тот
же самый механизм действует в ресничках.
Основное различие между этими структура..
ми касается их абсолютной длины, характе..
ра размешения и биения. Длина волны.. про..
беrающей по жrутику., обычно меньше ero
обrцей длины, поэтому он может rенериро..
вать более или менее постоянную силу, дей..
ствующую параллельно ero продольной
оси (рис. 1 0,3,А).
В отличие от жrутика ресничка относи..
тельно короткая, и цикл ее ДВИ)J(ений часто
ВКЛЮЧdет жесткий сильный удар и rибкое
возвращение назад (рис. 10.3,Б). Постоян..
ная движущая сила создается cTporo после..
довательным и координированным движе..
нием большоrо числа ресничек, располо..
женных на обширных участках поверхности
в виде поясков или полос из мноrих тысяч
этих орrанелл.
rенерирование сил мышечными воло--
кнами обусловлено их способностью стано..
виться короче., преодолевая сопротивление.
Они состоят из трубчатых сократимых эле..
ментов, миофибрилл, каждая из которых
диаметром примерно 1 мкм., В свою оче..
редь оБРdзованных белковыми филамента--
ми, расположенными в CTporo определен..
НОМ порядке. Во мноrих мышцах можно
выделить фУНКIIиональную единицу, так на..
зываемый саркомер, из совокупности тон--
ких И толстых филаментов, образованных
двумя разными белкаМИ .;ииозиНОАt и ак"
тинu и (рис. 10.4).
Между миозиновыми и актиновыми фи--
ламеНТdМИ MorYT возникать поперечные
мостики; их образование и изменения фор..
мы «rоловки» миозиновой молекулы при..
водят к скольжению тонких филаментов
А
Б
8
r
Рис, 10.4. Строение мышцы при различном
увеличении: А целая иннервируемая мышца;
Б отдельное мышечное волокно с нервным
окончанием (синапсом); В функциональная
единица мышцы саркомер; r поперечные
срезы саркомера под электронным микроско
пом
А
Е!; II""" '" l'
11111111111111
11111111\111111
11/111111111111\
j 1. . I 1" I . I I I 1. I
III" "''''''
111111111111111
IНlIIIIlII1III -
111111111111111
'1....1'1111"1 I
Б
" 1 1111\11111 1 1111111 1\ 1111111 1 1 1 !!..!..!..!..!l
111111111 111 111 ./1 " 11111 11 \11 1\ 1111. . .
............. /111111111111111111111.11\ 11111111\
11111111111111/111111 \11111111'111111
в
I 11,. ""iI I ,. 111111
1111111111 111 111111
1111111111 111111111
J 111 J 11111 11 111111 I
JII'II "'III
1 II
Рис. 10.5. Структура мышцы и rенерирование
силы мышечными клетками: A cxeMa CTpoe
ния поперечнополосатой мышцы; Б cxeMa
строения rладкой мышцы; В rенерирование
силы: 1 строение мышцы при различной CTe
пени сокращения; 11 зависимость развивае..
мой силы от относительной длины
312
rлава 1 О
ВДОЛЬ толстых, пока 'Этому движению не
воспрепятствует уравновешивающая сила,
действующая в противоположном направ
лении. Значительное сокращение мышечно..
ro волокна требует повторяющихся циклов
образования и разрыва поперечных мости..
ков, что обеспечивается энерrией фосфат
ных связей А ТР. Свойства мышечной систе..
мы определяются скоростью укорачивания
саркомеров и их числом в одном ряду; от
этоrо зависит скорость сокращения всей
мышцы. Сила.. действующая на единицу по
перечноrо сечения, зависит от числа парал..
лельно расположенных филаментов По ме..
ре увеличения степени их взаимоперекрыва..
ния число свободных мест для образования
поперечных связей снижается и мощность
мышцы падает до нуля (рис. 10.5).
10.3. Движение
с помощью ресничек
Беспозвоночные (за исключением очень
просто устроенных Placozoa; разд. 3.3) не
используют для локомоции жrутиков. хотя
движение с их помощью сохранилось почти
у всех сперматозоидов животных. Однако
у rубок жrутиковые клетки
хоаноциты проrоняют воду через камеры
и каналы тела.
Обычным локомоторным приспособле..
нием являются реснички, либо покрываю
щие всю поверхность тела, либо располо
женные в виде поясов или полос" но такой
способ движения характерен только для жи..
вотных длиной менее 1 мм. Это связано
с тем" что движение отдельной реснички
cTporo фиксированно" и единственный спо..
соб увеличения суммарной движущей силы
состоит в увеличении числа ресничек. Оно
оrраничено плошадью поверхности тела,
в то время как масса орrанизма увеличивае..
тся пропорционально ero объему. Более TO
ro, rидродинамический к. п. д. ресничных
орrанизмов уменьшается с увеличением их
размеров" следовательно, более крупным
орrанизмам, использующим этот тип дви"
жения" требуется rораздо больше энерrии
для достижения тех же относительных ско"
ростей.
у личинок мноrих беспозвоночных рес..
нички расположены в виде полос или поя..
А
Рис. 10.6. Личинки с полосами ресничек: A
велиrер брюхоноrоrо моллюска; Б плутеус
иrлокожеrо (офиуры)
сов, длина или число которых возрастают
по мере роста животноrо (рис. 10.6). Рес..
нички rребневиков образуют сложные rреб..
ные пластинки.. обеспечивающие скорость
до 15 мм/с.
Работа ресничек орrанизована таким
образом, что все они обычно имеют одина..
ковые характер и частоту биения. но движу"
тся В разных фазах. Координированное дей
ствие создает волну, пробеrающую от од"
ной реснички к друrой. Эта волна может ид..
ти в направлении движущей силы или в про..
тивоположном ему, так что каждая реснич"
ка совершает удар чуть позже той, которая
находится непосредственно перед ней. Во
время ударов отдельные реснички не rасят
усилие друr друrа. Такой тип координации
их активности называется меn1GХрОНG.1ЬНОЙ
РUn1Аiuчносп1ЬЮ (рис. 10.7).
Переход от ресничноrо к мышечному
движению произошел" по вилимому.
у близких к плоским червям предков била-
теральных форм, причем у некоторых СБО"
бодноживущих плоских червей инемертин
встречаются оба типа ЛОКОМОIIИИ.
Эпидермис свободноживущих турбелла-
рий и немертин обильно покрыт ресничка-
ми Самые мелкие их формы длиной около
Биомеханика и движение (локомоция)
З1З
А
W\I/ \lIt
Б
Внешний I
..
....... 1 ...... ..... II ...
Рис 10.7. Метахрональное биение ресничек. А.
Отдельные реснички в различных фазах цикла
биения. Б. Удар (1) и возвратное движение (11)
создают результирующую движущую силу во
внешнем поrраничном слое
1 мм находятся на верхнем пределе размер
ното диапазона животных, эффективно
использующих ресничную локомоцию. Бо
лее крупные плоские черви (TIiclada и Polyc..
lada), сохранившие реснички в качестве oc
новното средства передвижения (ползания),
способны к нему только блаrодаря плоской
форме тела площадь их поверхности уве..
личивается быстрее, чем квадрат линейных
размеров.
Самые крупные животные, ползаЮlцие
с помощью ресничек, ЭТО, вероятно, He
мертины, например вид Liпеиs /опgissiп1ИS,
длиной до нескольких метров при ширине
не более 1 2 мм, что обеспечивает ему Tpe
буемую площадь поверхности тела. Однако
такой выход из положения открыт только
для водных орrанизмов. На суше быстрая
потеря воды, неизбежная при такой боль
шой относительной поверхности, была бы
rубительна. Мышечные механизмы пере..
движения плоских червей и немертин разно
образны: сюда относятся локомоторные
волны. пробеrающие по нижней стороне те..
ла, перистальтические движения и «шаrа..
ния» с подтяrиванием заднеrо конца к пере
днему. Эти способы значительно сильнее
развиты в друrих типах животных и описа..
ны в следующих разделах.
10.4. Работа мышц
и скелетные системы
Химическая энерrия может использова..
ться клетками мышц таким образом, что
мышечные волокна укорачиваются. однако
эти изменения формы клеток лишь часть
локомоторной системы у орrанизмов со ске..
летными структурами, передающими тене..
рируемые силы.
Выделяют два основных типа скелетов
животных: .жидкие, свойственные широко..
му круту мяrкотелых беспозвоночных,
и жеС111кие, используемые членистоноrими
и некоторыми иrлокожими. Различия ме..
жду формами с жестким и жидким скелетом
далеко не абсолютны. У мноrих типов, кро"
ме членистоноrих, наблюдаются жесткие
скелетные элементы примерами MorYT слу..
жить иrлы морских ежей, «позвонки» офиур
и ацикулы в параподиях полихет.
Аналоrичным образом у членистоноrих
для передачи сил часто используется rидро..
статический скелет, например при движении
кишечника, а у паукообразных для рас..
прямления членистых конечностей. Жест
кие скелеты артропод являются наружными
(экзоскелеты), т. е. мяrкие ткани окружены
твердыми скелетными элементами. У xop
довых В качестве примитивной формы BHY
треннето скелета развился упруrий стер..
жень хорда. На ее основе сформирова..
лись хрящевые и костные скелеты высших
позвоночных.
Во всех локомоторных системах, ВКЛЮ"
чающих мышцы. .активность последних
оrраничивается сокращением; для восста..
новления их прежней длины требуется oco
бая сила. Чаще всето она обеспечивается ан..
таrонизмом между различными rруппами
мышечных волокон, однако ее источником
может быть и работа ресничек (например,
у некоторых стрекающих) или энерrия, Ha
копленная упруrими тканями. Жидкие CKe
леты мяrкотелых беспозвоночных мотут
использоваться по разному, но в основе их
функционирования лежат одни и те же при..
НlIИПЫ. рассмотренные в приложении 10.4
Мноrие мяrкотелые животные способны
значительно изменять свою форму. Этим
особенно отличаются немертины, однако
деформируемость имеет свои физические
оrраничения, поскольку объем тканей
и обычно полостей тела остается постоян..
ным. Унемертин нерастяrивающиеся воло
кна стенки тела расположены под уrлом
к ето продольной оси. Если их ориентиро..
rлава 1 О
314
4 А
3
2
1
10 20 зо 40 50 60 70 80 90
4 Б
. 3
c::r
ф
.
с::.
2
ф
,J)
u)
о 1
Geoпeтertes,
Rhyпchodemus
'Aтphlporus, L. gessereпsi
10 20 зо 40 50 60 70 80 90
4
3
2
1
о
10 20 30 40 50 60 70 80 90
Уrол между волокном и продопьной ОСЬЮ
Рис. 10,8. Физические свойства спиральной си
стемы неупруrих волокон и изменение формы
червеобразноrо животноrо. А. Связь между
внутренним объемом и наклоном неупруrих
волокон относительно оси цилиндра, BOKpyr
KOToporo они закручены. Б. Пределы деформи
руемости некоторых немертин, объем тела KO
торых меньше, чем теоретически допустимый
их системой волокон. В. Связь между уrлом
наклона волокон и объемом тела у нематод.
вать параллельно этой оси объем тела CTa
нет нулевым, то же самое произойдет если
расположить их к ней под уrлом 90< ). Между
этими двумя нереальными крайностями су..
ществует положение, при котором объем
тела максимален уrол волокон около 55'
01носительно продольной оси. Отношение
между объемом. оrраничиваемым этой си..
стемой волокон, и их уrлом с продольной
осью тела показано на рис. 10.8. По
рис. 10.8,A видно, что объем червя типа не..
мертины обычно меньше максимально воз
можноrо в теории. При максимальном co
крашении или максимальном растяжении
червь будет в поперечном сечении круrлым,
в прочих случаях "Эллиптическим. Изме
нения длины нескольких видов немертин
в pal\1KaX этих теоретических rраниц пред..
ставлены на рис. 10.8 Б. Механика движе..
ния нематод также определяется свойства..
ми волокон стенки тела. В отличие от не..
мертин эти черви относительно слабо под..
вержены деформации; орrанизованные
в сложную систему их волокна (СМ.
рис. 4.13) расположены под уrлом более 55
к продольной оси, а полостная жидкость на..
ходится под высоким давлением.
При сокращении продольных мышц
уrол прохождения волокон становится 60"
льше. и оrраниченный ими объем умень"
шается (рис. 10.8 B). Это в свою очередь
повьппает внутреннее давление противо..
действующее усилию продольных мышц
и возвращающее их к прежней длине.
Поэтому у нематод в отличие от модели,
представленной в приложении 10.4, нет ко..
льцевых мышll.
Мяrкотелые животные с косыми мышеч..
ными волокнами, например пи вки, способ..
ны устанавливать структурные волокна
Поскольку у этих животных высокое BHYTpeH
нее давление, их тело в поперечном сечении
круrлое. При сокращении продольных мышц
объем тела уменьшается, и вызываемое этим
увеличение давления противодействует MЫ
шечному сокращению. Эти животные обычно
перемещаются синусоидальными движениями,
при которых контралатеральные (т. е. лежащие
на противоположных сторонах тела) мышцы
сокращаются поочередно (из Clark, 1964)
Биомеханика и ДЕ ижение (ЛОКОМОЦI.1q)
стенки тела под уrлом 55 к продольной
оси, становясь фактически твердыми. Мыш..
цы, действующие на жесткие или членистые
скелеты, передают силы от места их [енера"
ции к точке их приложения к внешней среде.
Жесткий скелетный элемент работает как
рычаr, принципы KOToporo рассмотрены
в приложении 10.5. Следует отметить. одна..
ко, что в примере 7 приложения 10.4 речь
также идет о CBoero рода рычаrе, но осно"
ванном не на относительной длине, а на от..
носительной площапи.
10.5. ЛОКОМОЦИЯ мяrкотелых
животных
10.5.1. Передвижение
по твердой поверхности
Мноrие мяrкотелые беспозвоночные
способны передвиrаться по твердому суб..
страту, т. е. по такому, поверхность которо"
ro, соприкасающаяся с телом животноrо, не
сдвиrается по отношению к частям субстра
та, находящимся на расстоянии от этоrо те..
ла, коrда животное прикладывает к суб..
страту силу. Чтобы обеспечить локомоцию,
эта сила должна передаваться субстрату че..
рез фиксированную точку, часто называе..
МУЮ точкой опоры.
Во мноrих локомоторных системах
вдоль продольной оси тела Пdраллельно на..
правлению движения раСПРОСТРdНЯЮТСЯ
волны мышечноrо сокращения и расслабле..
иия. ПеремеIцение происходит либо в на..
правлении распространения jтих волн. либо
в противоположном ему. Это показано
в приложении 10.6. Плоские черви, некото"
рые книдарии и прежде Bcero брюхоноrие
Рис. 10.9. Строение ноrи мол
люска
315
моллюски движутся блаrодаря ВО..lнам мы..
шечных сокращений поверхности тела, при..
леrающей к субстрату (педальным локомо..
торным волнам).
Педальные локомоторные волны леrко
наблюдать на нижней поверхности теЛd
планарии или улитки, коrда они ползут по
стеклянной пластинке.. наклоненной поп не..
большим уrлом. У наземной улитки He/ir:
можно видеть одновременно несколько та..
ких волн, проходящих через всю Hory в на..
правлении движения животноrо.. но с боль..
шей, чем перемещение самой улитки, скоро..
стью. У друrих брюхоноrих, например
у MopcKoro БЛI-одечка (Palella), относитель..
но небольшое число волн проходит в на..
правлении, противоположном движению
животноrо. Более Toro, здесь, как правило,
наблюдается разница в половину длины
волны между правой и левой сторонами но..
rи. В результате различают педальные ло..
комоторные волны нескольких типов:
пРЯЛ1ые и обрапlllые, ,,\10НОl1zаксuчеС1\.uе иди..
п1Clксuческие.
Прямые монотаксические волны Неliх..
длина которых HaMHoro меньше длины но..
rи, ЭТО как бы двиrатель, работающий на
первой передаче: он обеспечивает относите..
льно низкую скорость движения и слабую
маневренность, ОДНаКО способен переме..
щать довольно большую l\1accy или преодо
левать значительные силы сопротивления.
Дитаксические системы с волнами такой же
длины, что и длина ноrи, или даже боль..
шей, позволяют двиrаться относительно
быстрее и более маневренно.
Hora моллюска весьма сложно
устроенный opraH Основной ero скелетный
элемент представлеп сложной системой
кровеносных синусов, Т. е., хотя сами жи..
316
rлава 1 О
вотные ацеломические. внутри их ноrи Ha
ходится rидростатическая скелетная систе
ма. rлавные мышечные волокна здесь не
продольные, а передние и задние косые
с противоположным действием (рис. 10.9).
Принято было считать, что при прохожде
нии локомоторной волны каждый отрезок
ноrи поочередно отделяется от поверхности
(как показано в приложении 10.6,А и Б), но
это маловероятно в связи с оrромным уси
лием, необходимым для разделения двух
липких влажных поверхностей. Образова..
ние точек опоры и зон движения зависит от
свойств слизистоrо слоя под ноrой. Слизь
ведет себя 'как упруrое твердое тело в усло
виях слабоrо сдвиrовоrо напряжения, а при
увеличении последнеrо как вязкая жидко
сть. В точках опоры она находится как раз
в первом состоянии, а под фронтом волны,
[де сдвиrовые силы больше, во втором.
Некоторые улитки, включая Неliх, MO
[ут передвиrаться и друrим способом
«rалопом», при котором rенерируются
обратные волны мышечных сокращений
с длиной волны, примерно равной длине HO
rи. Дальнейшая молификаuия этоrо типа
локомоции состоит в поочередном прикре
плении к субстрату переднеrо и заднеrо KOH
цОВ тела; примером может служить пере
движение с петлеобразным выrибанием Te
ла, характерное для пиявок (см. ниже).
у мноrих крупных плоских червей и бо--
льшинства немертин чередующиеся волны
сокращений кольuевых и продольных
мышц создают обратные перистальтиче
ские волны, расширяющие локомоторные
возможности, создаваемые работой поверх--
ностных ресничек. Такая система лучше Bce
[о развита у червей с септированным цело--
мом" особенно у земляных.
На рис. 10.1 О,А показан механизм дви
жения земляноrо червя. Обратим внимание"
что, Kor да кольцевые мышцы расслаблены"
а продольные сокращены, сеrменты тела не
перемещаются (см. приложение 10.6). Ce
[менты, находящиеся перед самым KOpOT
ким cerMeHToM. растяrиваются. отталки
ваясь от субстрата этой точкой опоры, Tor
да как задние сокращаются, обеспечивая
с ее помощью подтяrивание (рис. 10.10,Б).
Проскальзывание исключается за счет pac
ставления прочных щетинок.
А
(lIIIIIIIIIIIIIIIIIIJJ"IIID
с r I (IIlllIllIlllllllfllllD
с I I I I I (11111111 I 11111 11111 D
<1IfI1 III I]I/IIIIIIIIIII()
(11111 I I I I I (11111111111 D
(1111111111 II J/IIIIIID
( I I (111 I 1 I 1 I 1 J I I (1 I 11 D
с. J II rllllllllllJ II CID
спm
II fllllllllll'
Б
6
v
Рис. 10.10. А. Последовательные стадии дви
жения земляноrо червя. У каждоrо cerMeHTa
постоянный объем, как на рис. А. 11. приложе
ния 10.6. Коrда продольные мышцы сокраще
ны, а кольцевые расслаблены, cerMeHTbI непод
вижны относительно земли; объем cerMeHTa
фиксирован, и локомотарная волна распро
страняется назад. Б. Силы, действующие на He
подвижный cerMeHT, заякоренный щетинками
Волны изменений давления, возникаю..
щие при сокращении кольцевых и продоль..
ных мышц, независимы друr от друrа. так
как септыI эффективно изолируют cerMeHTbI
(рис. 10.11). Наивысшее давление отмечае..
тся в момент сокращения кольцевых мышц
и проталкивания cerMeHTa сквозь субстрат.
В фазе заякоривания давление ниже. Такая
изоляция давлений невозможна у животных
с неразделенной полостью тела. Полаrают,
что это одно из основных преимуществ се..
rментации. способствовавших эволюции
метамерии у аннелид. Каждый cerMeHT
представляет собой отдельный rидростати"
ческий элемент с постоянным объемом (см.
приложение 10.5). Хотя здесь рассмотрено
движение земляных червей по плоской по..
верхности, те же принципы лежат в основе
их перемещения в толще почвы к которому
они rлавным образом и адаптированы.
Совершенно по друrому обеспечивается
Биомеханика и движение (локомоция)
локомоция пиявок. Их передние и задние
присоски являются весьма эффективными
точками опоры. Все тело работает как един"
ственная rидростатическая система, причем
кишка единственная заполненная жидко..
стью полость. Прикрепление к субстрату
осуществляется поочередно переднеЙ и зад..
ней присосками (рис. 10.12,А).
Сходный способ передвижения встречае..
тся у личинок некоторых насекомых. На..
пример, у rусениц бабочек петлеообразные
выrибания тела эквивалентны сокращению
продольных мышц (рис. 10.12,Б). Полость
тела (целом) у пиявок фактически вытеснена
деформируемой ботриоидальной тканью.
так что с точки зрения архитектоники эти
А
д в
Б
. : .
::>
='
. .:
.
( .
( .
. =
. =
:
>
.
. :
.
"»
в
Рис. 10.11. Изучение локомоции земляноrо че
рвя. А. Система, позволяющая реrистрировать
давление внутри cerMeHTOB А и В с помощью
вставленных в них пластиковых трубок, непос
редственно связанных с датчиком давления. Б.
Последовательные положения cerMeHTOB А и
В при движении червя, В. Волнообразное из
менение давления (по Seymour 1969)
317
с
А
Б
Рис. 10.12. Шаrающие движения, при которых
точками опоры служат попеременно передний
и задний концы животноrо: А пиявка; Б
rусеница (личинка бабочки)
животные несеrментированные и ацеломи"
ческие, хотя, безусловно" относятся к анне..
лидам, и их происхождение от сеrментиро"
ванных целомических предков выдают не..
которые анатомические детали прежде все..
ro в строении нервной системы.
Эррантные полихеты используют способ
локомоции, отличающий их как от олиrо..
хет, так и от пиявок. Она обеспечивается
здесь мноrочисленными конечностями. кон..
цы которых движутся назад относительно
тела, перемещая при этом последнее вперед
по субстрату (см. приложение 10.5).
Коrда полихета. например Nereis. пол..
зет медленно (рис. 10.13,A), по ее парапо--
диям от хвоста к rолове проходит метахро..
нальная волна активности; при этом фазы
А
Б
Рис. 10.1 З. Схема локомоции полихеты Nereis:
А медленное ползание; Б быстрое полза
ние. Для простоты движение щетинок не пока
зано. Они MorYT выдвиrаться и втяrиваться,
увеличивая длину «шаrа»
318
rлава 1 О
движения параподий правой и левой сторон
тела различаются ровно на половину длины
волны. Поскольку волна направлена впе
ред, каждая конечность совершает сильное
отталкивание без риска интерференции
с толчком ближаЙIпей сзади параподии, так
как последняя оттолкнулась чуть рdньше.
При более быстром ПОЛЗdНИИ или при пла
вании Nereis к этой дитаксической прямой
локомоторной волне добавляется двиrате..
льная сила. rенерируемая СОКРdшением
продольной мускулатуры стенки тела
(рис. 10.12,Б). Активность продольных
мыиIЦ правой и левой сторон распростра..
няется так, что каждая пара подия совер..
шает мощное отталкивание, находясь на
[ре6не синусоидальной волны, пробеrаю
щей вперед по стенке тела синхронно с вол..
ной движения параподий
В этом случае силы, прикладываемые
к субстрату, rенерируются не только муску"
латурой параподий, но и контралатераль..
ными продольными мышцами. При таком
типе движения в соответствии с общим пра..
вилом прямые локомоторные волны дол
жны наблюдаться тоrда, коrда точки опоры
возникают в областях максимальной длины
продольных мышц.
У мноrих эррантных полихет продоль..
ные мышцы иrрdЮТ в процессе ползания He
А
в
'//////////
значительную роль, и движущая сила в ос--
новном создается мускулатурой параподий,
пересекаюшей rраницы cerMeHToB. Это oco
бенно характерно для червей, по крытых че..
шуйками, и близких к ним форм, у которых
межсеrментные септы сильно редуцирова
ны или утрачены. В локомоuии здесь уча..
ствуют как собственные мыиIЦЫ параподий,
так и мышцы, входящие в них извне. Ще..
тинки выталкиваются наружу и ВТЯI'иваю..
тся обратно собственными МЫIl1uами.
а движение параподий вверх--вниз и вперед
назад обеспечивается совместно собствен..
ными и входящими мышцами, что приво
дит К cBoero рода lпаrанию. и движение He
которых полихет напоминает сложную по..
ходку мноrоножек.
Уникальное локомоторное устройство
представляет собой амбулакральная систе
ма иrлокожих. Для морской звезды, напри--
мер, характерно наличие пяти лучей, в каж
дом из которых проходит амбулакральный
канал. начинающийся от околоротовоrо ко..
льца. Вдоль каждоrо радиальноrо канала
находятся ряд резервуаров..ампул и ам6ула..
кральные ножки (рис. lО.14,Б). МыиIЦЫ ам..
пул и амбулакральных ножек действуют ан..
Тdrонистически по отношению друr к друrу:
сокращение мышц, сжимающих ампулу,
выталкивает воду в амбулакральную нож..
*
r
Б
Ампула
'./ ////////// '
Рис. 10.14. Амбулакральные
ножки и ампулы амбулакраль
ной системы иrлокожих. А. Об
щая орrанизация. Б. Схема по
перечноrо среза луча морской
звезды, показывающая амбу
лакральную систему В. ЦИКЛ
шаrающеrо движения отдель
ной амбулакральной ножки (ее
мышцы ретракторы не показа
ны)
Биомеханика и движение (локомоция)
319
ку, а сокращение ножки вызывает отток во--
ды обратно в ампулы. При этом отдельная
ампула не служит резервуаром только од..
ной «своей» ножки, поскольку наблюдается
перемещение жидкости между частями всей
системы, и при полном растяжении ножки
она может достиrать большеrо объема, чем
соответствующая ей ампула в полностью
растянутом состоянии. Однако, как прави--
ло, мышцы, сокращающие ампулу, дей..
ствуют как антаrонисты тех, что укорачи--
вают ножку.
Амбулакральные ножки, вытяrиваясь
под действием rидравлическоrо давления,
MorYT совершать простые шаrающие дви--
жения; на их концах часто находятся липкие
присоски (рис. 10.14,B).
Особенность движения морских звезд
отсутствие какоrо--либо заметноrо метах--
рональноrо ритма в движении амбулакра--
льных ножек.
10.5.2. Движение в толще
субстрата и в трубках
Роющим червям свойственно наличие
крупных полостей тела и по крайней мере
частичное отсутствие септированности. Жи--
вотные с таким строением способны испо--
льзовать различные способы локомоции
внутри нор (рис. 10.15). Перисталыическое
движение бывает четырех типов, поскольк)
любая часть тела может одновременно уве--
личиваться и в длину, и в обхвате. Эти дви--
жения используются для прокачивания во--
ды через норку, для перемещения в ней или
для Toro и друrоrо вместе.
Несеrментированные и сеrментирован"
ные, но лишенные септ животные с крупны--
ми полостями тела MorYT существенным
образом менять форму последнеrо, что
используется для внедрения в субстрат.
Примером служат сипункулиды, способные
делать это чрезвычайно быстро; соответ--
ствуюшие движения показаны на рис. 10.16
вместе с rрафиком изменений. BHYTpeHHero
давления Эти животные MorYT развивать
высокую мощность, но обычно в течение
коротких периодов времени. При поддер--
жании BbIcoKoro давления в целоме все
мышцы тела должны расходовать метабо--
лическую энерrию для сохранения своей по--
А Б
в r
tшnru
II]ПIIПI
юrТПCШJ
EIIIIIIШ
Рис. 10.15. Четыре типа перистальтических
движений, используемые обитающими в труб..
ках червями. В случаях А, Б и r объем cerMeH
тов не фиксирован. В случае В (Sabe//a) ce
rMeHTbI разделены септами и имеют постоян
ный объем. Перемещение тела по трубке при
перестальтических движениях предотвраща..
ется за счет образования точек опоры cerMeH
там и промежуточной длины
стоянной длины. Механическая работа со..
вершается за счет контролируемоrо рассла--
бления определенных мышц, что позволяет
растяrиваться соответствующим участкам
тела (см. приложение 10.4). Мощность при
этом может быть очень высокой, но к.п.д.
будет низок, так как требуется непрерыв"
ный мышечный тонус для поддержания вы--
cOKoro давления в целоме. Полаrают, что
эволюционное возникновение поперечных
септ у аннелйд связано, по крайней мере ча..
стично, с необходимостью предотвращать
растяжение стенок тела без использования
мышечной работы, что увеличивает меха..
нический К.п.д. системы (ср. разд. 10.1).
Полихета Areпico/a пZllriпa (пескожил)
это сеrментированный червь, по--видимому,
вторично приспособленный к роюшему
образу жизни. Метамерное строение дает
ему ряд преимуществ. CerMeHTapHOe распо--
ложение rанrлиев улучшает нервную коор--
динапию; параподии со шетинками со..
здают точки опоры; имеется хорошо разви--
тая сосудистая система с сеrментарными
жабрами. С механической точки зрения ту..
320
rлава 10
А
(
с
{ (
с (:
с(
с
) 1
) 11
) ш
) IV
) v
) VI
) VII
Б
IП
Рис. 10.16. А. Схема последовател ьных движе..
ний сипункулиды при зарывании и перемеще
нии сквозь субстрат. Б. Изменение BHYTpeHHero
давления в различных фазах локомоторноrо
цикла. Обратите внимание, что максимальное
давление отмечено в момент проникновения
в субстрат (как и у земляных червей рис.
10.11 ), но при этом оно 80 всех частях тела
одинаково (по Trueman, Foster Smith, 1976)
А
Б
ЛОВИЩНЫЙ целом пескожила неразделен,
что позволяет ему COBepIl1aTb самые разно..
образные движения и развивать высокую
мощность. Это дает ему возможность внед..
ряться в субстрат путем попеременноrо тер..
минальноrо и пенетрационноrо заякорива..
ний (объяснение см. на рис. 10.17). Такой
же тип движения встречается у роющих кни
дарий и на поздних стадиях закапывания
у двустворчатых моллюсков (рис. 10.18).
Пескожилы обычно обитают в откры"
тых j--образных трубках, являющихся ча..
стью U..образной норовой систеl\1Ы
(рис. 9.32). Животное извлекает и поедает
песок в «забое» с помощью соскабливаю
щих и заrлатывающих движений хоботка.
Скорость этой работы невысока, и во время
ее выполнения передний целом изолируется
от остальной полости тела rлоточной ce
птой, клапаны которой позволяют затем
объединять ero с туловищным целомом.
Высокое Дdвление в последнем приводит
к вытяrиванию хоботка при более интенсив..
ном рытье.
Обитающие в трубках черви, сохранив..
шие межсеrментные септы (например, 5а..
Ьеllа), сталкиваются со специфической про..
блемой. Им необходимо прокачивать воду
через трубку с помощью перистальтических
движений. Имея постоянный объем сеrмеп..
тов, они t\tlorYT использовать только пе..
ристальтическую волну типа изображенной
на рис. lO.15,B; следовательно, их будет
сносить в направлении, противоположном
движению волны. Чтобы этоrо не произо..
шло" Sahella образует точки опоры сеrмеп
-
Рис. 10.17. Стадии движения пескожила в субстрате. А. Терминальное заякоривание: передняя
часть тела расширена, а задняя подтяrивается вперед. Б Пенетрационное заякоривание: cerMeHTbI
передней части тела образуют заякоривающие выступы, хоботок раскапывает песок и «rолова»
продвиrается вперед (по Trueman 1975)
Биомеханика и движение (локомоция)
... \ \ :., '
1 II ПI
Рис 10.18. Закапывание двустворчатоrо мол
люска. 1. Створки раковины раздвиrаются, co
здавая опору для проталкивания ноrи 11, 111
Конец ноrи расширяется, и раковина втяrива
ется в песок (по Trueman, 1975)
тами промежуточной длины, раздвиrая на
иих щетинки. Остальные сеrменты скользят
по очень rладкой выстилке трубки.
10.5.3. Плавание
Самые мелкие мяrкотелые беспозвоноч"
иые плавают за счет движущей силы ресни"
чек, как описано в разд. 10.3. Более круп..
иые животные используют работу мышц;
исключение составляют rребневики, несу..
щие ряды ресничных rребных пластинок.
Для плавания необходимо, чтобы сила, [е..
иерируемая мышцами, воздействовала на
среду (обычно lТО вода). сквозь которую
животное движется. Для относительно
крупных животных число Рейнольдса (см.
разд. 10.1) больше единицы, так что сле..
дует учитывать инерцию тела. Коrда плы..
вет крупное животное с Re > 1, масса окру"
жающей воды обычно приводится в вихре..
вое движение. Такой тип плавания особенно
характерен для позвоночных. но мноrие
мяrкотелые беспозвоночные. не обладаю..
щие в воде нейтральной плавучестью, также
способны l енерировать подъемную и дви
жущую силу за счет мышечной активности
Для lToro используются три основных ме..
ханизма: а) распространение назад волн по
телу животных с rладкой поверхностью б)
rребущие движения; в) реактивное движе..
иие.
Относительно мелкие беспозвоночные
с Re < 1 не MorYT использовать вихреобра..
зования, тем не менее также способны пла..
вать Их движение основано на различиях
321
в сопротивлении водь] при разных CKOpO
стях.
Основные работы по механике волно..
образноrо движения выполнены на уrрях.
но аналоrичные локомоторные волны reHe..
рируются и rладкотелыми червеобразными
беспозвоночными, например пиявкой Hil'и
llо. При плавании ее тело сплюшено дорсо..
вентрально и по нему пробеrают направ"
ленные назад волны. Волна движется назад
со скоростью U (относительно земли), ко..
торая выше скорости V перемещения тела
вперед. Принципы TaKoro типа плавания из..
ложены в приложении 10.7.
Механизм плавания полихет, у которых
тело шероховатое, иной. Волна движется
в том же направлении. что и тело. Частично
это связано со сложной динамикой жидко..
сти, омывающей колеблющееся тело с вы..
ступами, а частично с действием параподий,
совершаlОШИХ мах назад в тот момент. Kor..
да их достиrает rребень проходящей по те..
лу волны (см. рис. 10.13,Б).
Мноrие крупные позвоночные движутся
ТОЛЧКdМИ. в перерыве между которыми ско"
льзят по инерции; при этом энерrия толчка
сохраняется в виде кинетической энерrии те..
ла. rоловоноrие моллюски используют тот
же принцип в своем реактивном движении.
Вода с силой выталкивается из мантийной
полости при сокращении выстилающих ее
мышц. Поскольку вода имеет массу и при..
обретает ускорение, направленное назад,
должн,," возникнуть сила противодействия.
разrоняющая животное вперед. Этот при..
нцип более полно объясняется в приложе..
нии 10.8.
10.6. ЛОКОМОЦИЯ
беспозвоночных
с членистыми конечностями
Ракообразные, хелицеровые и одновет..
вистые rри разнообразные и процветаю..
щие rруппы беспозвоночных. Все они харак"
теризуются Нdличием твердоrо экзоскелета,
позволяющеrо иметь членистые конечно..
сти. Успех этих трех rрупп может быть ча..
стично связан с локомоторными преимуще..
ствами, которые дает такое строение по..
следних. Кроме Toro. насекомые это
единственные беспозвоночные, имеющие
322
rлава 1 О
Б
Рис. 10.19. А. Конечность типичноrо членистоноrоrо. Обратите внимание на мощные мышцы в ее
базальной части, сухожилия и суставы. Б. Характерное положение конечности членистоноrоrо.
При движении вперед происходит вращение проксимальноrо сустава в rоризонтальной плоско
сти и разrибание более дистальных суставов
крылья и способные к полету, чем отчасти
обусловлено их доминирование в наземных
местообитаниях. Несмотря на свое полифи..
летическое происхождение. ходильные KO
нечности наиболее высокоорrанизованных
ракообразных, хелицеровых и одноветви--
стых устроены весьма сходно в результате
конверrентной ЭВОЛЮlIИИ. Они состоят из
рЯДd. подвижно сочлененных элементов,
масса которых снижается по Нd.правлению
к дистальному концу (рис. 10.l9,A). Каж
дый сустав допускает движение только в од--
- -
ной плоскости; это обусловливает сrибание
и разrибание конечности" а также вращение
ее плоскости в суставе, rде происходит при--
членение к туловищу; последнее обычно
и определяет перемешение тела вперед. Ча
ще Bcero оно как бы подвешено между рас--
ставленными в стороны ноrами
(рис. 10.19,Ь), так что при ходьбе центр
тяжести движется rоризонтально.
За счет вращения конечности у основа--
ния и сrибания и разrибания в остальных
суставах животное может перемещать кон--
чик ноrи по прямой линии, параллельной
направлению движения. Конечность рабо--
тает как механический рычаr (см. приложе--
ние 10.5), и у длинноноrих артропод ero
устройство таково, что большая сила пере..
мещает относительно малый rруз на боль..
шое расстояние, обеспечивая тем самым
быструю ходьбу.
Наиболее массивные двиrательные
мышцы расположены не в конечностях,
а внутри туловища, и, поскольку животное
не подскакивает вверх--вниз при перемеще..
нии вперед, основные изменения момента
затраrивd.ЮТ только конечность. Они сводя..
тся К минимуму за счет общих для артропод
особенностей строения: масса конечности
убывает от основания к дистальному концу.
Боковые волнообразные колебания тела,
как у полихет (см. рис. 10.13) и некоторых
мноrоножек (см. ниже), конечно, приводят
к изменениям момента. и их подавление,
по--видимому, оказало существенное влия"
ние на эволюцию членистоноrих.
Хотя ходильные конечности насекомых
и продвинутых ракообразных, например
крабов, сходны по строению, их эволю..
ционная история существенно различается.
Конечности ракообразных происходят от
предковой структуры, выполнявшей поми..
мо локомоторной различные функции,
включая захват пищи и дыхание.
Биомеханика и движение (ЛОКОМQЦИЯ)
З23
10.6.1. ЛОКОМОЦИЯ ракообразных
Конечности ракообразных выводятся из
общеrо плана строения" включающеrо не..
сколько составных частей: два непарных ба..
зальных cerMeHTa, листовидный эпиподит
и пару дистальных частей эндоподит
И экзоподит (см. рис. 8.34.Б п. T совре..
менных листоноrих раков Cephalocarida
и Branchiopoda (см. рис. 8.37 8.39) подоб..
пые конечности имеются на каждом нецефа..
лизованном торакальном cerMeHTe и
участвуют не только в передвижении, но и
в питании.
Конечность ракообразных может дей..
ствовать как весло, поверхность KOToporo
максимальна во время rребка, а при воз..
вратном движении существенно уменьшае..
тся за счет складывания и изменения поло..
жени я щетинок (рис. 10.20).. что снижает со..
противление воды. Однако примитивные
ракообразные используют для плавания
также изменения объема пространства,
окружаемоrо конечностями. Вода затяrи..
вается внутрь Hero по средней линии
(рис. 10.21) и выталкивается назад. Однона..
правленное ее движение создается за счет
Toro, что экзоподит действует как клапан.
Пищевые частицы задерживаются щетинка..
ми эндоподита и передаlОТСЯ вперед со ще..
тинки на щетинку вдоль срединной вентра..
льпой борозды по направлению ко рту. Та..
ким образом, метахрональный ритм лвиже..
пия конечностей обеспечивает локомоцию,
питание, а возможно, и дыхание.
Отличительная черта локомоции рак о..
образных относительная леrкость пере..
f2I
А
Рис. 10.20, Роль щетинок при создании неоди
HaKoBoro сопротивления во время плавания pa
кообразных, для которых значения числа Ае
относительно низки: А положение при воз
вратном движении; Б положение во время
локомоторноrо rребка (по Hesseid, Fowtner,
1981 )
А
Б
Рис. 10.21. Конечности листоноrих ракообра
зных создают потоки воды, служащие для пла
вания и питания. А. Вид снаружи правой CTOpO
ны тела. Б. Вид изнутри левой стороны тела.
Показаны щетинки, фильтрующие пищевые ча
стицы, которые затем передаются вперед ко
рту. Вода затяrивается в срединную вентраль
ную полость, а затем проталкивается назад,
проходя через фильтры эндоподитов, т. е. BHY
тренних ветвей конечностей
хода от плавания к хождению. У мноrих
креветкоподобных Malacostraca торакаль..
ные конечности ходильные, а абдоми"
нальные (плеоподы) плавательные
(рис. 10.22). Во мноrих эволюционных ли..
ниях либо утрачивается способность к хо..
ждению (см. рис. 8.49,А), либо снижается
роль плавательных плеоподов, как у ома..
ров и крабов (см. рис. 8.49, r E). у омаров
хорошо развита реакция избеrания с уча..
стием уроподов последней пары абдоми"
нальных придатков. Мошные брюшные
мышцы сокращаlОТСЯ, вызывая резкое сrи
бание туловища, и в результате заrребания
воды уроподами животное отбрасывается
назад (см. разд. 16.10.4).
у крабов (Brachyura) брюшко редуциро..
вано, и большинство из них передвиrаются
Ходильные
ноrи
Уроподы
(для реакции
Плаватель ИЗБеrания)
ные ноrи
(плеоподы)
Рис. 10.22. Креветкоподобный представитель
Malacostraca с ходильными и плавательными
конечностями
324
rлава 1 О
Рис. 10.23. Плавающий краб Ca//iпectes. Ero
задние конечности при плавании действуют как
rребные лопасти
на пяти парах [рудных конечностей. HeCMO
тря на это, и здесь возможен переход к пла
ванию за счет уплощения и модификации
последних [рудных Hor" как у CaZliпectes из
семейства Portunidae (рис. 10.23). Краб пла..
вает за счет вращения этих конечностей" ко..
торые действуют как rребные лопасти.
Переходы между плаванием и хожде
ни ем в эволюции ракообразных следствие
обитания в водной среде" в основном orpa..
ничивающей распространение этих живот..
ных. В морской воде безпозвоночные MorYT
иметь почти нейтральную плавучесть" а BЫ
сокая (по сравнению с воздухом) вязкость
среды не позволяет развивать большую
скорость. Строение ракообразных позво..
ляет некоторым представителям этоrо типа
передвиrаться на суше" но друrие физиче..
ские особенности исключают широкую ра..
диаЦИIО в наземных местообитаниях. К ней
оказались спосоБНЬJ друrие членистоно
rие хелицеровые и одноветвистые.
10.6.2. Наземная локомоция
хождение
у животных, обитающих на суше, плот
ность значительно выше" чем у окружающе..
[о их воздуха, поэтому им необходимы
опорные структуры, и формы, которым
приходится часто или быстро передвиrа..
ться, используют для опоры о землю твер..
дые скелетные элементы. Последние дол
ЖНЬJ противостоять существенным дефор..
МИРУI{)ЩИМ силам: сrибающим, т. е. дей
ствующим перпендикулярно оси конечно..
сти, и скручивающим ее BOKpyr собственной
оси.
Структурой, устойчивой в таких усло..
виях и имеющей минимальную массу,
является полый цилиндр. Быстрое передви..
жениетребуеттакжеrибкихсочленений
ставов) между жесткими элементами, сухо..
жилий для передачи сил, rенерируемых
мышцами. и устройств для запасания эне..
рrии. Кутикула членистоноrих идеально
удовлетворяет всем этим требованиям
Она состоит в основном из уrлеводно
белковоrо комплекса хитина. Это про
чное и rибкое вещество" но само по себе не
приrодное для формирования жестких ске..
летных элементов. Жесткость может быть
повышена за счет образования связей ме..
жду составляющими ero белками. В резуль..
тате TaKoro «дубления» возникает белок,
называемый склеротином. Он может сооб..
щать кутикуле различную жесткость, иноr..
да сравнимую с жесткостью кости, в то
время как rибкий хитин образует суставы
конечностей. Как правило, движение в каж..
дом суставе ОI раничено только одной пло..
скостью, что обеспечивается развитием вну"
тренних выступов.
Обычно сrибание требует сокращения
мышц, а разrибание часто происходит за
счет упруrости мембраны сустава или с уча..
стием rидравлическоrо механизма.. коrда
сила, распрямляющая конечность, передае..
тся через заполняющую ее rемолимфу.
Шаrание наземных членистоноrих вклю"
чает вращение направленной вбок оси ноrи.
у двупарНОНОI'ИХ очень MHoro коротких ко..
нечностей (рис. 8.23), движение которых
следует направленному вперед метахро"
нальному ритму, так что каждая из них со..
вертает толчок чуть позже, чем находя..
ЩdЯСЯ непосредственно за ней. Это как бы
,...J:вижение «на малых оборотах», позволяю..
щее развивать относительно большое уси..
лие при низкой максимальной скорости.
Двупарноноrие фитофаrи" обитающие,
как правило, в rниющей древесине и анало..
rичных субстратах. так что такой способ
передвижения вполне подходит для их
образа жизни.
Мноrие rубоноrие мноrоножки, напро..
тив, активные хищники, способные передви..
rdТЬСЯ относительно быстро. На рис. 10.24
показано их движение с различными скоро..
стями. Характер локомоции здесь сходен
Биомеханика и движение (локомоция)
. . . <
):I-
Рис. 10.24. Движение rубоноrих мноrоножек
с различной скоростью. Черными точками обо
значены концы конечностей, контактирующие
с землей (по Manton, 1965)
с описанием для N e/.e;s (рис. 10.12). С po
стом скорости число конечностей, касаю..
щихся земли, уменьшается. а изrибания те..
ла усиливаются.
Боковые волнообразные изrибы тела
служат для увеличения длины шаrа у члени
стоноrих с короткими конечностями, но это
связано с существенными изменениями MO
мента. Их можно избежать путем удлине..
пия каждой конечности при снижении их
числа, и в различных эволюционных линиях
артропод прослеживается именно такая
тенденция. У десятиноrих раков их пять пар
(но для хождения часто используются толь
ко три или четыре пары), у паукооб
разных четыре. а у насекомых три. Хо..
ждение требует поднятия ноrи, ее движения
вперед, опускания и движения назад относи
тельно тела. Во время последней фазы KO
нец ноrи упирается в землю. а пентр тяже..
сти тела перемещается относительно Hero
вперед. Движения в различных суставах по
зволяют при этом удерживать тело на по
стоянной высоте над землей.
Рис. 10.25. Вид сбоку и траектории движения
конечностей скорпиона, Заметно, что TpaeKTO
рии отдельных конечностей не пересекаются
между собой (по Hesseid, Fowtner, 1981)
325
А
Б
в
r
Рис, 10.26. Траектории движения конечностей
различных членистоноrих: А краб; Б омар;
В паук; r насекомое (по Manton 1952)
На рис 10.25 показан вид сбоку и TpaeK
тории движения конечностей скорпиона при
ero движении вперед. Ноrи не мешают друr
друrу блаrодаря расположению их КОНЦОВ
на неодинаковых расстояниях от rела.
Траектории конечностей различны, что тре..
бует сложной координации движений в каж..
дом суставе. причем они обычно не пересе
каются друr с друrом, как показано на рис.
10.26. Большинство артропод движутся впе..
ред за счет вращения базальноrо членика
конечности относительно тела (рис. 10.26.Б..
N, однако крабы перемещаются вбок, сrи
бая и разrибая ходильные ноrи в дисталь..
ных суставах.
Если при малом числе конечностей часть
из них переставляется одновременно,
возникает проблема устойчивости. У Hace
комых с тремя парами Hor они должны
переставляться в такой последовательно..
сти, чтобы в любой момент времени по
крайней мере три ноrи опирались о землю,
а центр тяжести находился в пределах треу..
rольника, образуемоrо точками опоры.
Наиболее распространенные способы хо..
ждения насекомых удовлетворяют этим
условиям. Две rруппы конечностей пере..
ставляются поочередно, и каждая, CTaHO
вясь на землю, образует устойчивую опору.
етахрональная последовательность их
движений направлена вперед, и ноrи с двух
сторон одноrо cerMeHTa действуют в проти"
вофазе. То же самое наблюдается и у жи
вотных С большим числом конечностей Ha
пример у полихет"нереид (рис. 10.13) и rубо..
ноrих мноrоножек (рис. 10.24).
326
rлава 10
Такое «чередование треуrольников» мо"
жет обеспечивать различные скорости локо..
моции: чем больше отношение периодов
движения конечности вперед и назад, тем
быстрее перемещается животное.
у некоторых насекомых подобноrо «че
редования треуrольников» не наБЛIодается,
особенно если одна пара конечностей спе..
циализирована для какой..либо особой
функции, как, например'! у боrомолов, у ко..
торых первая их пара редко используется
....{ля опоры.
1 0.6.3. Полет
Физические свойства усиленной склеро
тином кутикулы позволили крылатым насе..
комым выработать способность к полету.
Кутикула обладает почти идеальными
свойствами для построения аэродинамиче..
ской структуры. Она может образовывать
тонкую'! леrкую'! rибкую мембрану. усилен..
ную склеротизованными жилками и ребра..
ми. Такая конструкция фактически является
rофрированной пластиной, но по своим
свойствам аналоrично значительно более
тяжелой сплошной конструкции аналоrич..
ных rабаритов. Кроме Toro, упруrие белки
способны запасать поставляемую мышца..
ми энерrию, чтобы использовать ее для re..
нерации быстрых движений крыла.
Первые летающие насекомые появились
около 200 млн. лет назад, и с тех пор меха..
низ м полета модифицировался и совершен..
ствовался. Поэтому у современных насеко..
мых структурные и аэродинамические при..
способления к полету значительно разно..
образнее, чем, скажем, у птиц и летучих мы..
шей. На рис. 10.27 показаны лишь некото"
рые формы крыльев насекомых; они изо..
бра жены одинаковоrо размера, но на самом
деле и по этому признаку существенно раз..
личаются (см. также рис. 8.29).
Структурные приспособления, сделав..
шие возможной эволюцию полета унасеко"
мых, изначально, по--видимому, служили
для термореrуляпии. Мноrие современные
насекомые до сих пор используют крылья
для этой цели: бабочки повышают темпера--
туру тела, поrлощая ими солнечную
энерrию.. а перепончатокрылые (например,
пчелы и осы) «разоrреваются» перед поле..
Рис. 10.27. Различные формы крыльев HaceKO
мых Изображения примерно одноrо размера,
но в действительности размеры этих видов си
льно различаются (см. также рис. 8.29)
том, быстро ими взмахивая. Возможно, из-
менение относительных размеров привело
к тому.. что крылья.. первоначально присло-
собленные для термореrуляции, приобрели
аэродинамические свойства.
В частности, для точной термореrуля-
пии необходимо пвижение крыловидных
придатков, так что их развитие должно бы..
ло бы сопровождаться появлением со ответ..
ствующей мускулатуры. Примитивные ле-
таюшие насекомые, по--випимому, были
способны лишь к планирующему полету,
А
п
Движение торакса
III
Биомеханика и движение (локомоция)
Б
11
Нервные импульсы
rnтmrrmmrrrm-
Движение торакса
ПI
Рис. 10.28. Структурные и нервно мышечные
приспособления к полету у насекомых А.
Прямые летательные мышцы, характерные Ha
пример, для стрекоз: 1 стрекоза; 11
поперечные срезы ее торакса с прямыми лета
тельными мышцами, прикрепленными непос
редственно к основания м крыл ьев; 111 зап ись
синхронных нервных импульсов и движений
торакса у таких насекомых. Б. Непрямая лета
тельная мускулатура, например, двукрылых:
1 муха; 11 попереч ные срезы торакса с oc
новными антаrонистическими летательными
мышцами; 111 запись асинхронных нервных
импульсов и движений торакса у насекомых
TaKoro типа (по Pringle, 1975)
при котором требуются относительно мед..
ленные движения крыльев для реrулирова..
ния стационарных аэродинамических сил.
Однако для понимания d-эродинамики мно"
rих ныне живущих насекомых необходимо
учитывать нестационарные инеустойчивые
услов:ця.
Современные виды используют два типа
структурных приспособлений для приведе..
пия крыльев в движение. Стрекозы счи..
таются одними из самых примитивных ны"
пе живущих крылатых насекомых. У них
крыло приводится В движение так называе..
327
мыми прямыми летательными МЫШЦd.ми
прикрепляющимися к ero основанию как
пока за но на рис. 10.28,А, причем наблюда..
ется однозначное соответствие между каж..
дым нервным импульсом, стимулирующим
эти мышцы, и соответствующим движе..
нием крыльев и торакса (рис. lO.28,A, 111).
Такую иннервацию летательных мышц на..
зывают синхронной (см. разд. 16.10.5).
Однако у большинства насекомых дви"
жение крыльев вызывается «непрямыми»
летательными мышцами. изменяющими
форму торакса, который посредством осо..
боrо сочленения с основаниями крыльев вы..
зывает их поднимание и опускание (рис.
10.28,Б). Нервные импульсы. стимулирую..
щие такие мышцы, не синхронизированы
с наблюдаемой частотой движений торакса
и соответственно крыльев (см. разд.
16.10.5). Это называется асинхронной инне..
рвацией летательных мышц. Нервные им..
пульсы, показанные на рис. 1 0.28,Б III под..
держивают эти мышцы в состоянии возбу..
ждения, характеризующемся высоким ВНУ"
триклеточным содержанием ионов Са 2+. В
таком состоянии сокращение вызывается
стимуляцией расположенных в мышцах ре..
цепторов растяжения. При этом частота
биения крыльев зависит от физических
свойств торакальных cerMeHToB. Сокраще..
ние одной rруппы летательных мышц при..
водит к растяжению мышц"антаrонистов
что в свою очередь вызывает их сократе..
ние, индуцируя каскад поочередных сокра..
щений мышц обеих rрупп, который не пре..
кратится, пока они находятся в возбужден..
ном состоянии.
Ранее считалось, что под действием не..
прямых летательных мышц торакс ДBYKPЫ
лых насекомых как бы «перескакивает» из
одноrо стабильноrо состояния в друrое с
поднятыми или опущенными крыльями.
Однако оказалось, что кинематика движе..
ния крыла не cor ласуется с этой моделью,
предскаЗЫВd-ющей замедление движения
крыла по мере приближения к среднему по..
ложению и роста напряжения в тораксе
с быстрым ускорением при переходе к CTa
бильным точкам (рис. 10.29,А).
Схема, иллюстрирующая альтернатив..
ную модель, изображена на рис. 10.29,Б.
Эта модель предполаrает, что движение
328
rлава 10
ь
ф о
sro
20
о ro 40
С:::о.
L
Ь ro 60
со 80
5 Ш 100
ro
о:: 120
140
т 160
00.
а 180
CI
2
6
4
1
А
о
А
Б /
/ / I ....
/ // :
// Акс 3 ::::.
/// .,::
(У/
В
Б
14
ДВМ Il
Рис. 10.29. А. I Изменение положения кончика крыла мухи, зафиксированное с помощью
сверхбыстрой киносъемки (4000 кадров в секунду); 11 кривая, предсказанная на основе просто
ro механизма «перескакивания». Б. Полусхематическая модель причленения крыла у двукрылоrо
HaceKoMoro. Опускание крыла из положения А в положение Б поднимает дорсальную пластинку
торакса (скутум) и тем самым активирует дорсовентральные мышцы (ДВМ 11), которые COKpa
щаются и поднимают крыло. Аксиллярные мышцы (Акс 3) MorYT увеличивать амплитуду движе
ния крыла (до положения В), что ведет к возрастанию энерrии, запасенной в упруrих белках TO
ракса при деформации. Это способствует быстрому подъему крыла, как показано на рис. 10,29,
А (по Miyan, Ewing, 1986)
мышц, опускающих крыло, вызывает под--
нятие сочлененной с ero основанием дор--
сальной стенки торакса. Это приводит к ра--
стяжению дорсовентральных мышц (стиму--
лирующему их сокращение) и накоплению
энерrии в упруrих компонентах системы,
высвобождающих ее при движении крыла
вверх.
Чтобы насекомое моrло лететь. на Hero
должна действовать сила противодействия,
одна из составляющих которой равна весу
иасекомоrо, а друrая не меньше, чем сила
сопротивления воздуха при движении
сквозь Hero. Обе эти составляющие
подъемная и тяrовая силы создаются
движением аэродинамической плоскости
в воздухе.
Результаты тщательных наблюдений за
,...:(вижениями крыльев саранчи, подтвер"
жденные затем на друrих насекомых, св иде..
тельствуют о том.. что крыло создает и по..
дъемную, и тяrовую силь] при движении как
вниз, так и вверх. Кончик крыла следует
почти по эллиптической траектории относи..
Биомеханика и движение (локомоция)
329
II
ПI
Рис. 1 0.30, Сlар/fliпg механизм rенерирования вихрей. 1. В верхнем положении крылья прижаты
друr к друrу. 11. Крылья начинают раздвиrаться, причем их более жесткие передние края расходя
тся первыми Это вызывает циркуляцию воздуха 80Kpyr крыла. 111. Движение крыла вниз сообщает
воздуху ускорение; совершается работа и возникает сила противодействия с направленными впе
ред и вверх составляющими (по Weis Fogh. 1975)
тельно тела HaceKOMoro, что означает зуб
чатую траекторию относительно непод..
вижной точки отсчета при движении живот..
Horo вперед.
Однако подъемый коэффициент у неко"
торых насекомых оказывается выше. чем
можно было бы ожидать от обычной аэро
динамической плоскости, исходя из изме..
реиныx значений числа Рейнольдса. Такое
несоответствие особенно велико для мух..
журчалок и друrих насекомых, способных
парить неподвижно. Для понимания этих
явлений неоБХОДИl\10 принять во ВНИl\1ание
нестационарные, неустойчивые силы. созда..
ваемые движением крыла. За летящим насе..
комым образуются вихревые потоки возду"
ха, и полет обусловлен силой противодей..
ствия, возникающей при rенеРИРОВdНИИ
вихревоrо движения BOKpyr поверхности
крыла. Более подробно этот принцип раз..
бирается в приложении 10.9.
Коrда крылья HaceKoMoro смыкаются
и затем РаСХОДЯТСЯ.. передние.. более жесткие
их края разъединяются первыми и воздух
устремляется в область низкоrо давления,
ВОЗНИКаЮЩУЮ между крыльями (рис. 10.30)
Работа затрачивается на ускорение массы
воздуха, закручивающеrося BOKpyr крыла,
а сила противодействия включает как по..
дъемную, так и тяrовую составляющие.
Как только движение воздуха ,J,остиrает
максимальной скорости, крылья перестают
совершать полезную работу до тех пор.. по..
Ка вихрь не будет «сброшен», что произой..
дет при смене крылом направления движе..
иия в нижней ero точке. В приложении 10.9
аналоrичным образом разбирается взлет
бабочки.
Насекомые настолько разнообразны,
что ко всем представителям этой rруппы
вряд ли может быть примеиена единая тео..
рия полета. Для самых мелких форм ТИПа
«мошкары» значение числа Рейнольдса на..
столько низко, что они MorYT разrребать
воздух, как будто это вязкая жидкость.
у таких насекомых крылья покрыты воло..
сками и утрачивают свойства аэродинами"
ческой поверхности (см. рис. 10.27). И на..
против, у крупных жуков Re достиrает
23000, и их жесткие надкрылья MorYT вести
себя как плоскости самолета.
1 0.6.4. Наземная локомоция:
прыrающие насекомые
Некоторые Нdсекомые способны пры..
[ать. Для большинства из них это важная
реакция избеrания; тот, кто хоть раз пы..
тался поймать блоху, знает, насколько
эффективно это непредсказуемое поведение
ззо
rлава 1 О
Способность к прыжкам наиболее развита
у блох, кузнечиков и цикадок.
Чтобы подпрыrнуть, насекомое должно
подействовать на землю с такой силой что..
бы сила противодействия сообщила старто--
вую скорость ero массе. Высота прыжка
определяется уравнением:
Кинетическая энерrия (1/2 тУ 2) == Потен--
IIиальная энерrия в высшей точке прыжка
(mgh),
rде т Macca HaceKoMoro;
V стартовая скорость;
g ускорение свободноrо падения;
h высота прыжка.
у 2
Следовательно, h == 2 g и
Кинетическая энерrия
Ь==
mg
Сила, с которой конец ноrи прыrdющеrо
HaceKoMoro воздействует на землю, имеет
вертикальную и rоризонтальную состав..
ляющие, как показано на рис. 10.31 для ку"
знечика. Вертикальная составляющая равна
F.sine.
у скорение продолжается только до тех
пор. пока ноrи HaceKoMoro касаются земли,
и стартовая скорость определяется силой
толчка и временем, в течение KOToporo она
действует (см. также приложение 10.2). Это
время зависит от длины Hor. и у всех пры--
rdЮЩИХ форм они относительно длинные.
Рис. 10.31. Кузнечик ДО и во время прыжка
При разrибании задней ноrи на землю дей
ствует сила. Чем длиннее ноrи тем дольше ее
действие, следовательно, больше ускорение
А
Б
в
Рис. 10.32. Прыжок блохи. А. Бедро поднимае
тся и в деформированных упруrих белках кути
кулы запасается энерrия. Б. Бедро «снимается
с предохранителя» при расслаблении мышцы.
Сила воздействия rолени на земл ю сообщает
блохе начальную скорость, от которой зависит
высота прыжка
в разд. 10.2 было показано, что скорость со..
кращения мышечных клеток оrраничена их
физиолоrическими особенностями: уле..
тающих насекомых это оrраничение прео..
долевается с помощью непрямых летатель..
ных мышц и способности запасать энерrию
в упруrих компонентах стенки тела при де..
формаuии (см. разд. 10.6.3). Аналоrичные
приспособления есть и у БОЛЬШИНСТВd пры..
rающих насекомых. Элементы ноrи 0606..
щенноrо HaceKOMoro показаны на рис.
10.19..A.. и мы будем ссылаться на этот рису..
нок при описании механизма прыжка. Нота
блохи характеризуется удлиненными тази..
ком и бедром. При ПОДrотовке к прыжку
бедро приподнимается в положение. пока..
занное на рис. IO.32,A. Затем в участке
очень упруrой кутикулы стенки торакса за..
пасается энерrия. Хотя блохи неспосо6ны
к полету. считается.. что этот участок про..
изошел от rомолоrичноrо ему места уле..
тающих насекомых, причем для деформа..
ции торакса используются прежние леТd"
тельные мышцы. Торакальные сеrменты
удерживаются в таком напряженном поло..
жении с помощью cBoero рода «предохра..
нителя)} из выступов и пазов в покровах,
и блоха находится как бы во «взведенном»
состоянии, rотовая к резкому высвобожде..
нию запасенной энерrии 80 время прыжка.
При расслаблении мышц, поднимающих
бедро, «предохранитель» снимается и ro..
лень с большой силой ударяет о землю при
возвращении cerMeHToB торакса впервона..
чальное положение.
Биомеханика и движение (локомоция)
10.7. ВЫВОДЫ
в этой rлаве дано общее введение в про
блему локомоuии у беспозвоночных Чита
тель должен уяснить себе значение разме..
ров: например, почему крупные животные
используют для движения мышечные клет"
ки, тоrда как более мелким достаточно pec
ничек или жrутиков.
Следует понимать, что двиrательные си..
стемы всех животных подчиняются одним
и тем же законам механики и что в передаче
силы Bcer да участвуют скелетные элементы.
Эта передача может осуществляться либо
посредством жидкости, заключенной в зам
кнутом объеме, либо с помощью системы
жестких рычаrов.
Некоторые беспозвоночные медлитель
ны и малоподвижны, друrие перемещаются
очень быстро и маневренно; такие живот
ные, как rоловоноrие моллюски и HaceKO
мые, располаrают весьма совершенными
локомоторными системами, одна из KOTO
рых основана на реактивном движении
а друrая -на использовании маховых KpЫ
льев. Здесь изложены лишь элементарные
принципы; читателей, желающих более под..
робно ознакомиться с проблемой, мы отсы"
лаем к приведенному ниже списку дополни
тельной литературы.
Понимание механизмов локомоции жи
вотных крайне важно для понимания эво"
люционноrо происхождения Тd.ксономиче
ских rрупп, поскольку любая предполаrае
мая предковая форма должна быть струк"
турно оправданной. Она должна «рабо
тать», подчиняясь основным законам физи..
ки точно так же, как и любой ныне живущий
орrанизм.
10.8. Дополнительная
литература
Alexaпdel' R. McN. 1982. Locomotion of Animals.
Tertiary Level Biology. Blackic Glasgow.
Clark R. В. 1964. Dynamics in Metazoan Evolution.
Clarendon Press. Oxford.
ElderH. У., Trueтaп Е. R. 1980. Aspects of Animal
Movement. Society for Experimental Biology Se
minar Series. Cambridge University Press, Cam
bridge.
Hesseid С. F.; FОИ'lпеr С. R. (Eds) 1981. Locomotion
ЗЗ1
and Energetics in Arthropods. РlепиПl Press, New
у ork.
KiпgsoZvel' J. С. 1985. Butterfly engineering. Scient.
Аm., 233 (2), 90 97.
Raiпey R. С. (Ed.) 1984 Insect Flight. Blackwell
Scientific Publications, Oxford.
Т,'иеп1ап Е. R. 1975. The Locomotion of Soft Bodied
Animals. Edward Arnold, London.
Weis FogJl Т. 1975. Unusual mechanisms for the ge
neration of lift in flying animals. Scient. Аm., 233
(5), 81 87.
Приложение 10.1.
Термины и определения меха...
ники
1. Сила про является в ее действии H l
массу.
Сила стремится изменить направление
или скорость движения массы. Сила есть
масса х ускорение: F == М . а.
В единицах системы СИ сила величиной
1 ньютон (Н), приложенная к массе 1 Kr, со..
общает ей ускорение 1 м/с.
2. Коrда одно тело прикладывает силу
к друrому. это равнозначно тому. что BTO
рое тело прикладывает к первому противо
положно направленную силу равной вели..
чины. Последняя называется силой противо
действия.
3. Коrда сила двиrает массу, соверша
ется механическая работа. Работа есть си
ла х расстояние:
W == F. d.
W == (М . a)d.
в системе СИ епиниuа измерения рабо..
ты джоуль (Дж). Скорость совершения
работы мощность, т.е. мощность это
сила х скорость ЕДИНИЦd СИ для
измерения мощности ватт (Вт): 1 Вт ==
== 1 Дж/с.
Приложение 10.2.
Кинетическая энерrия и трение
1. в приложении 10.1 сила F придает Ha
шей модели (осьминоr в лодке) такое YCKO
рение, что
F == м а.
Совершаемая работа будет зависеть от
расстояния, пройденноrо массой за время
действия данной силы: W == F. d.
ЗЗ2
rлава 1 О
2. Если бы лодка плыла без трения, ось..
миноr Mor бы двиrаться вечно за счет кине..
тической энерrии:
КЕ ==...lm' У 2
2 '
rде V CKOpOCTЬ, м/с.
Лодка останавливается., поскольку суще..
ствует сила трения, вызывающая замедле
нне. Эту силу называют также сопротнвле
ннем.
3. Кинетическая энерrия рассеивается
в виде тепловой энерrин. Чтобы продолжать
пвиrаться с постоянной скоростью., ось ми..
Hory нужно прикладывать силу., равную си..
ле трения.
.............. d
Иллюстрация принципа раБота == сила х расстояние
Иллюстрация концепции сил действия
и противодействия
1
r---d
2
<Q
о
.....,..
t I
< I ЛОБовое сопро
тивление,ИЛИ Tpe
з
Приложение 10.3.
Разложение сил на составляю..
щие
1. Любая сила действует в определен-
ном направлении. она является вектором.
2. Две силы, действующие на одну и ту
же массу., но в разных направлениях, можно
Силы, действующие в различных направле
ниях дают результирующую силу. действую
щую в KaKOM TO ином направлении
Биомеханика и движение (локомоция)
суммировать., получив результирующую си...
лу, действующую в своем собственном Ha
правлении.
3. а) Направления действия сил мо)кно
изображать линиями со стрелками., начи
нающимися от массыI.
б) Длина линии пропорциональна Be
"Iичине силы.
в) Направление и относительную ве...
личину результирующей силы можно опре
делить rеометрически.
F;
lO'
u
/5
g
с')
tf
rеометрическое разложение сил. Sin е == F 1 /F,
cos е == F 2 /F
4. Одна сила может быть разложена на
две составляющие., направленные перпенди
кулярно друr друrу. За направление одной
из составляющих обычно принимаЮl Ha
правление дви)кения вперед. Друrая COCTaB
ляющая., действующая перпендикулярно
этому направлению.. не имеет отношения
к движению., но TefvI не менее требует MeTa
болических за трат.
Выделение составляющей, действующей в Ha
правлении движения, если дана сила, дей
ствующая в ином направлении В направлении
движения действует сила, равная F cos е
ззз
Приложение 10.4.
rидростатический скелет
1. Сокращение мышц окружающих
ЗdМКНУТУЮ полость., заполненную )кидко"
стью. увеличивает давление этой жидкости:
Давление == Сила
Площадь .
В системе СИ давление измеряется
в ньютонах на квадратный метр: Н/м 2 .
2. Таким образом., сила., действующая
на любую поверхность. составит F == павле
ние х площадь.
3. Давление действует одинаково на все
(очки поверхности и направлено ПОД
прямым уrлом к ней (см. рисунок).
Давление в жидкости действует 80 всех направ
лениях
4. Внутреннему давлению в rидравличе..
ской системе противостоит сила противо..
действия оrраничивающей поверхности.
Если сопротивление этой поверхности
в каком"то участке меньше., чем то., которое
необходимо для сдерживания давления.,
движение жидкости приведет к изменению
формы поверхности. Это лежит в основе ло--
комоции всех животных с rидростатиче--
ским скелетом.
На приведенном ниже рисунке мышцы
в правой и левой ЧL1СТЯХ цилиндра дей..
сrвуют как взаимные антаrонисты.
OI1}ru )
t t t t
1) I1 wщ) ru r
+ t t tt J +
Антаrонизм между соединенными между co
бой правым (А) и левым (L) компартментами
системы
З34
rлава 1 О
5. Большая изменчивость системы обе
спечивается тем, что мышцы стенки тела
ВКЛЮЧd.ют продольные и кольцевые воло..
кна.
l b О
Антаrонистическая орrанизация
(снаружи) и продольных мышц
кольцевых
1 [IIJ ) 1]] II Ш)) ID
III ) IПIШ w QDJ
v QJ:CJJ
VI
I IV. Участки тела переменноrо объема: I
кольцевые мышцы сокращаются, жидкость BЫ
талкивается; 11 кольцевые мышцы рассла
бляются, жидкость входит; 111 кольцевые
и продольные мышцы сокращаются, жидкость
выталкивается; IV продольные и кольцевые
мышцы расслабляются, жидкость входит. V, VI.
Участки тела фиксированноrо объема: V
продольные мыщцы расслабляются, объем не
меняется; VI кольцевые мышцы расслабляю
тся, объем не меняется
6. Любой отрезок животноrо с разде..
ленным на изолированные отсеки телом ци..
линдрической формы может в результате
претерпевать разнообразные деформаuии.
7. При изменении формы может совер-
шаться механическая работа. Она равна
произведению действующей силы ( == давле-
ние х площадь) на пройденное расстояние.
В приведенном ниже примере хоботок
выталкивается наружу при сокращении 60"
льшей по объему части тела
Общая сила стенки тела
== Давление ==
Сокращающаяся площадь а 1
Сила, действующая на конец хоботка
Площадь хоботка а 2
F' F"
==p==
а 1 а 2
Это так называемый rидростатический ры-
чат (см, также приложение 10.5). Совершен
ная работа равна J == Р х а, х d'}.
d 1
82
d 2
81
Приложение 10.5. Жесткие
скелеты принцип рычаrа
1. Сила F может приводить в движение
rруз М, вращая ero BOKpyr точки опоры.
Если F приложенная сила, а F R сила
,"'\ @
F
Биомеханика и движение (локомоция)
З35
противодействия, они связаны равенством
F х d, == F R Х d 2 ,
[де d, путь, пройденный точкой приложе..
иия силы F, а d2 ПУТЬ' пройденный точкой
приложения силы F R 8
Обратим внимание, что в такой системе
работа совеРll1ается силой, противополо
жной по направлению приложенной.
2. неБолыl1яя приложенная сила может
перемещать крупный rруз, но количество
эиерrии должно остаться постоянным,
поэтому
F
== Выиrрыш в силе ==
F
d
== == ПРОИf'рыш В расстоянии.
d 2
3. Приложив БОЛЬ1l10е усилие, можно
передвинуть маленький rруз на значитель
ное расстояние.
Приложение 10.6
На рисунке изображено тело животноrо,
разделенное на пять отрезков (они MorYT
соответствовать или не соответствовать Ha
стоящим cerMeHTaM), прикрепленных к по..
верхности, а затем растяrивающихся (А)
или сокращающихся (Б). Волны МЫll1ечноrо
сокращения MorYT про ходить по мускули
стой Hore, как показано на рис. A(I) и Б(I),
или по цилиндрическому червеобразному
телу, как в случаях A(II) и Б(II).
\ '-
'l '
I bJ
V-.J
f;
"}J I
А (1) Опора на короткие cerMeHTbI Б (1). Опора на длинные cerMeHTbI
волна идет назад волна идет вперед
А (11) Опора на короткие
cerMeHTbI волна идет
назад (как например,
v дождевых червей)
Б (11). Опора на длинные cerMeHTbI
волна идет вперед. Для червя
цилиндрической формы такой
тип движения требует
ИСПользования щетинок
(однако см. также описание
движения по принципу
нереи д)
ЗЗ6
rлава 1 О
Результаты одинаковы. Если точки опо..
ры возникают в областях с максимальной
длиной отрезков. животное передвиrается
в одном направлении с волной мышечноrо
сокращения т. е. волна прямая. Если на..
против, точки опоры соответствуют участ
кам сокращения продольных мышп.. т. е. на
месте остаются самые короткие участки те..
ла животное перемещается в направлении,
противоположном движению волны мы..
шечноrо сокращения, которая в таком слу..
чае называется обратной.
Приложение 10 7.
Плавание мяrкотелых червей
1. Плавание мяrкотелых животных мо"
жно анализировать рассматривая движе..
ние отдельноrо продольноrо отрезка их те..
ла.
2. Такой отрезок описывает «восьмер..
ку» с уrлом е между ero продольной осью
и линией, перпендикулярной оси движения
Bcero тела. Мы рассмотрим только силы,
действующие на среднюю линию отрезка,
хотя 1TO конечно, упрощение.
ОСЬ движения
Для понимания действующих сил необ
ходимо знание правил разложения сил (см.
приложение 10.3).
3. Если отрезок тела находился в покое
а затем начал ДВИI аться под уrлом к оси
перемещения Bcero тела, он приложит силу
К массе окружающей воды и приведет ее
в движение (см. приложение 10.2).
F
Продольная ось
Одна из составляющих силы F, F", вызо"
вет движение воды вдоль поверхности тела,
а составляющая F" будет действовать ПОД
прямым уrлом к этой поверхности, сооб
щая воде ускорение.
F
Сила противодействия воды RF", равная
по величине и противоположная по направ
лению силе F".. может быть разложена на со..
ставляющие одна из которых параллельна,
а друrая перпендикулярна оси движения
Bcero животноrо.
RF'
F'
3а время про хождения полноrо ЦИКЛd
волнообразноrо движения составляющие,
перпендикулярные оси движения взаимоу-
ничтожаются, а составляющие. действую-
щие вдоль этой оси, складываlОТСЯ.
По мере Toro как животное набирает
скорость, уrол между направлениями дви-
жения отдельных отрезков ero тела и тела
целиком уменьшается. Этот уrол называют
уrлом атаки (а).
J]ействительное направление
движеНИR отрезка тела
с учетом скорости пере
мешения Bcero тела вперед
RF'
Биомеханика и движение (локомоция)
ЗЗ7
Это снижает силу противодействия., на..
правленную вдоль оси движения. Скорость
будет увеличиваться до тех пор. пока эта си..
ла не уравновесится силой лобовоrо сопро..
тивления
Приложение 10.8.
Реактивное движение
1. При реактивном движении сила, дей..
ствуюшая на воду. и сила противодействия
направлены вдоль линии движения живот..
Horo.
2. у rоловоноrих моллюсков сокраще..
ние мышц тела в стенке мантийной полости
повышает в последней давление. Таким
образом., в любой точке полости деЙствует
одинаковая сила., направленная перпендику"
лярно ее стенке.
Сила Давление х Площадь.
3. При открывании сифона эта сила обе..
спечивает ускорение массы воды., выходя..
щей из мантийной полости.
Произведенная работа == + m V 2
( == кинетическая энерrия).
Поскольку масса выталкиваемой воды
меньше, чем масса тела животноrо, ско"
рость этой волы выше. чем скорость лвиже..
нмя животноrо в противоположном направ"
лении. У реактивных двиrателей с такими
свойствами низкий к. п. д.
4. Часть работы, про изводимой мыш"
цами, сжимающими мантийную полость,
расходуется на деформацию упруrих стенок
тела; имеются также мышцы..антаrонисты.,
расширяющие полость до ее первоначаль..
Horo объема.
Приложение 10.9. Вихревое
движение воздуха и полет
1 Полет насекомых основывается на
нестационарных воздушных потоках, [ене..
рируемых движениями крыльев; важный
элемент этоrо процесса образование
м сбрасывание вихрей.
в составе вихря масса текучеrо вещества
вращается в виде кольца или эллипса во..
Kpyr центральной линии. как показано на
приведенном ниже рисунке. Вихревое коль..
цо обладает моментом и энерrией.
Индуцированный
поток
\ \\ \ и I f I
: vJl\1111 (/ f1,r/
"'( ('\] :'\
llltt (I'11 )1/ )2
h1 l ' I \ \' \ Направленныи
11 вниз момент
вихря
Схематическое изображение вихревоrо коль
ца Чтобы rенерировать вихрь в текучей среде,
необходимо затратить работу
2. При полете движение крыльев вызы
вает вращение воздуха BOKpyr их аэродина..
мической поверхности; этот воздух «сбра..
сывается» с крыла в виде вихревоrо кольца,
коrДа крыло изменяет направление Движе..
ния. наклон или форму.
Q
iC
" "-
(3 . '..уУ ) l
\\\ I
(\ \.. "" ((C
. .....'-' \\\\\ V
'-...:r \'
) ..' )}
' \'.. )
(:
Летящее насекомое поддерживается
в воздухе и движется вперед за счет силы
противодействия Q, возникающей в резуль..
тате «сбрасывания» вниз вихревых колец.
3. Вращение воздуха часто возникает
при разведении в стороны сомкнутых KpЫ
льев (см. также рис. 10.30). Это иноrда на..
зывают механизмом "'clapjfling".
3З8
rлава 1 О
Почти вертикальный взлет бабочки (см.
рисунок) возможен блаrодаря «сбрасыва..
нию» вниз воздушноrо вихря (по Kingsol..
ver, 1985).
1. Крылья соединены наверху.
11., 111. По мере Рdзведения крыльев во..
зникает циркуляция воздуха BOKpyr их пере..
дних краев (у большинства насекомых
более жестких см. характер жилкования
на рис. 10.27).
IY. Ускорение массы вращающеrося
воздуха rенерирует подъемную силу., и ба..
бочка отрывается от земли.
Y.Vl. Коrда крылья достиrают нижнеrо
положения., воздух «сбрасывается» с их по..
верхности в виде вихревоrо кольца, момент
KOToporo направлен вниз.
rЛАВА 11
ДЬIХАНИЕ
Потребность в кислороде СЧИТdется фун"
даментальным свойством Bcero живоrо. Он
поrлощается через дыхательные (респира..
торные) поверхности" такие" как жабры или
леrкие" и после поступления в орrанизм
используется для окисления орrанических
субстратов (в основном уrлеводов" хотя
и не только их) с выделением энерrии, необ..
ходимой для обеспечения процессов мета..
болизма. Однако жизнь зародил ась без ки"
слорода, поэтому аэробное дыхание свой..
ственно не всем орrанизмам, и некоторые
из них до сих пор функционируют по
анаэробному типу. В этой rлаве обсуждаю..
тся процессы аэробноrо и анаэробноrо ды..
ХdНИЯ у беспозвоночных. Мы начнем с раз..
бора биохимических основ дыхания, а за..
тем, обратившись к ero аэробному типу,
рассмотрим, как кислород извлекается из
окружающей среды и переносится в ткани
и как на ero поr лошение влияют факторы
окружающей среды.
11.1. Центральная роль АТР
в дыхании
Фосфорилированные нуклеотиды" и осо..
бенно адеНОЗll1-lпlрuфосфап1 (А ТР), иrрают
важную роль посредников впереносе эне..
рrии от «топлива» (питательных веществ)
к энерrопотребляющим процессам метабо..
лизма. Потенциальная энерrия пищи пере..
носится на так называемые Мdкроэрrиче..
ские (высокоэнерrетические) фосфатные
связи (обозначаемые "" Р) следующим обра..
зом:
Энерrия из потребленной пищи +
А Р "'" Р + р. А Р "'" Р '" Р"
I
rде А аденозин, а Pi неорrанический
фосфат Эта запасенная энерrия может за..
тем быть использована в ходе метаболизма
с образованием из А ТР аденозиндифосфата
(ADP) и Р,. Следует" однако" отметить" что
термин «макроэрrическая фосфатная связь»
не вполне точен. Энерrия не 1апасается в ко..
валентной связи между фосфатом и осталь..
ной частью молекулы. Правильнее [ово"
рить о том" что энерrия этой связи отражает
разницу между энерrетическим содержа..
нием молекул АТР и ADP.
11.2. Основной путь
катаболизма
rликолиз и цикл трикарбоновых кислот
(ТКК) являются «становым хребтом» ката..
болизма у всех типов беспозвоночных. Они
подробно изложены в друrих учебниках.
поэтому ниже дается лишь краткое их опи..
сание
На рис. 11.1 приведена очень Уl1роu/еJlная
схема этих двух путей. «Топливом» служит
rлюко3а
NA'j
lAD r
Лактат ///////////, Пируват
N
а
Q)
:s:
I
36Д тр
('t)
.D
а
с
()
's
U
Рис. 11.1. Очень упрощенная схема метаболи
ческих путей, связанных с rенерированием А тр
340
rлава 11
rлюкоза., получаемая непосредственно из
пищи или в ходе ферментативных превра..
щений друrих молекул пиши или резервов
тела.
ТЛUКОЛU3 происходит в цитоплазме и не
требует присутствия кислорода. Он дает
А ТР в ходе прямых превращений субстрата,
как бы продвиrающеrося по этому метабо..
лическому пути «вниз» в сторону упроще
ния молекул. Это фосфорuлuрованuе на
уровне с.J/бстрап1а.
ЦИКЛ ткк протекает в митохондриях
при участии кислорода. Он дает BOCCTaHOB
ленный нuкотинамидаденuндuн} клеотuд
(NAD r ), который в свою очередь использу
ется для образования А ТР. отдавая элек
троны (т. е. окисляясь с образованием N AD ())
в цитохромную транспортную систему, [де
в роли их конечноrо акцептора выступает
кислород. Это фосфорилuрование, сопря
:JICeH1l0e с lпранспорп10./И электронов. На рис.
11.1 отражено, что участие кислорода по
зволяет получить больше молекул АТР на
одну молекулу rлюкозы, чем бескислород
ная часть общеrо процесса.
11.3. Получение АТР
без кислорода
До появления в ходе эволюции фотоав
тотрофов орrанизмам приходилось обходи
ться без кислорода. Кроме Toro, эколоrиче
ская аноксия (отсутствие 02) может встреча
ться и В современных условиях; с ней стал
киваются литоральные животные, оказав
шись на воздухе во время отлива, формы,
роющие ходы в восстанавливающих суб
стратах, а также мноrие паразиты. Опреде
ленные ткани MorYT находиться в аноксиче
ских условиях, даже коrда орrанизм в цe
лом достаточно насыщен кислородом. На..
пример, в мышцах., участвующих в peaK
циях избеrания у двустворчатых моллю
сков, обычно протекает анаэробный MeTa
болизм.
Таким образом, «первые» орrанизмы
оыли анаэробами, а некоторые остаются
ими и по сей день. Теоретически возможно
большое число анаэробных метаболических
путей.. но лишь четыре из них были исполь
ЗОВd.ны беспозвоночными. Различные пути
возникли в ходе эволюции для удовлетворе
ния определенных энерrетических потреб
ностей.
Путь .А10,ЛОЧНОЙ КUlлопlЫ (или лактатный
путь) изучен лучше друrих. Он показан на
рис. 11.]; NADr' образующийся при rлико"
лизе, вновь окисляется при восстановлении
конечноrо продукта этоrо метаболическоrо
пути пирувата до лактата под дей..
ствием фермента лактатдеrидроrеназы. Как
уже отмечалось, это неэффективный способ
получения АТР, но он позволяет rенериро..
вать ero с высокой скоростью в отсутствие
кислорода и обычно используется для обе..
спечения интенсивной мышечной работы
у позвоночных, коrда мышца может вре..
менно оказаться в анаэробных условиях.
Лактатный путь встречается не во всех тка..
нях беспозвоночных. Он, по видимому,
присутствует у Насекомых в мышцах конеч..
ностей, но отсутствует в летательных мыш
цах, которые так обильно снабжены трахея..
ми и трахеолами (рис. 11.6,Б), что редко
оказываются в условиях аноксии.
ОпUНО6ЫЙ путь близок к лактатному
и приспособлен для «взрывной» работы.,
требующей быстроrо, но не обязательно
эффективноrо синтеза АТР. При этом уrле..
воды катаболизируются в ходе rликолиза,
но вместо восстановления пирувата проис..
ходит ero восстановительная конденсация
с аминокислотой, приводящая кобразова..
нию опина производноrо d-МИНОКИСЛОТЫ:
rлюкоза + 2 аминокислоты + 3 ADP +
3 p.
I
2Н 2 О + 2 опина + 3 АТР.
Можно выделить несколько опиновых
метаболических путей в зависимости от
участвующих аминокислот и ферментов.
Например, при резком сокращении мышц
мантии у rоловоноrих образуется вещество,
называемое октопином. Друrой опин
стромбин обнаружен у некоторых дву"
створчатых моллюсков.
Путь янтарной кислоты (сукцинатный
путь). у некоторых орrанизмов типа ДBY
створчатых моллюсков. обитаюших в анок..
сических илах, или паразитов, населяющих
анаэробные среды своих хозяев, например
кишечник позвоночных, в ходе эволюции
сформировались анаэробные метаболиче..
ские пути.. позволяющие получить на моле..
Дblхание
кулу rлюкозы больше А ТР, чем при опино"
вом И лактатном путях. но медленнее. При
этом энерrия NAD,., полученноrо на первых
этапах rликолиза, используется для фосфо..
рилирования, сопряженноrо с ТРdНСПОРТОМ
электронов., при котором конечным их aK
цептором вместо кислорода служит фума..
рат. Общая схема 1Toro процесса приведена
на рис. 11.2. Фосфоенолпируват (Ф ЕП)
предшественник пирувата при rликолизе
превраlllается в оксалоаuетат за счет при..
соединения СО 2 (карбоксилирования)., KaTa
лизируемоrо ФЕП карбоксикиназой, а за..
тем в фумарат. Фумарат окисляется до сук..
цината с участием электронотранспортной
системы, а сукцинат может метаболизиро..
ваться дальше до пропионата и друrих ле
тучих жирных кислот. Оксалоацетат, фума..
рат и сукцинат являются промежуточными
соединениями цикла ТКК.. однако там они
образуются в последовательности, проти"
воположной rолько что описанной. Поэто..
МУ путь янтарной кислоты является как бы
обращенным циклом ТКК. Часть пирувата
в такой сукцинатной системе может
превращаться в лактат, ацетат, аланин
и этанол, т. е. широкий спектр конечных
продуктов. В общей сложности этот про..
цесс позволяет синтезировать от 4 до 6 мо"
лекул А ТР на каждую молекулу rлюкозы.
r ликоrен
ADP+Pi
) с;.. .. ДТР
ADP+11 АТР
/ EO;' V
/ Пи ат Различные конечные
Оксалоацетат ............. продукты
..... ......
/............. .......................
I I
I I
I I
ADP+ I I
СРумарат I
I ТКК I
I I
I
t
I
I
I
J
............... ,..,.../
.............................",
NADo
NAD ..:
r
Система транспорта
электронов
341
у двустворчатых моллюсков описана
сходная последовательность реакций даю..
щая сукцинат в ходе восстановления окса..
лоацетата; последний, однако, образуется
не из ФЕП, а из аминокислоты (аспартата)
в реакции трансаминирования.
Фосфас?еIlЫ. Эти запасные соединения по..
лучают rруппу "'" Р от А ТР в периоды покоя
и отдают ее назад при интенсивной наrруз..
ке или аноксии:
Фосфоарrинин + АОР == Арrинин + АТР.
Такая система типична для бес..
позвоночных, тоrда как у позвоночных в ка..
честве донора используется фосфокреатин.
Исключениями являются иrлокожие, кото..
рые MorYT использовать оба фосфаrена.,
а также аннелиды, содержащие помимо фо..
сфоарrинина четыре друrих фосq)аrена.
Рис. ] 1.3 демонстрирует филоrенетиче..
ское распределение основных мета60личе..
ских путей (фосфаrены не указаны). Все они
распространены очень широко. Так. либо
путь янтарной кислоты Mor эволюциониро"
вать из системы rликолиз"цикл ТКК, либо
наоборот, т. е. один путь Mor возникнуть из
друrоrо путем обращения направления ре..
акций (см. выше). Принято СЧИТdТЬ" что
В ранней биотической среде были широко
представлены аминокислоты" поэтом)'
Рис. 11 .2. Вариант анаэробноrо метаболиче
cKoro пути, используемый эндопаразитами
и двустворчатыми моллюсками переживаю
щими длительные периоды rипоксии (по Ca
low, Townsend, 1981). ФЕП фосфоенол
пируват
J42
rлава 11
ARTHROPODA
Мол (Янт)
ECHINODER-MA ТА
Мол
NEMATODA
ANNELIDA
Мол Янт
Мол Янт
Опиновые '?
PLATYHELMINTHES
CHORDATA
Мол
SIPUNCULA
MOLLUSCA
LOPHOPHORATA
Окт Янт
Янт Окт (Мол)
Опиновые?
Мол Опиновые
CNIDARIA
PORIFERA
Мол Янт
Аминокислотные
Опиновые? (Янт)
Мол Опиновые?
ПРЕДКОВЫЕ ПРОТИСТЫ
предполаrается, что первые системы синте..
за А тр использовали их в качестве как ДО"
норов, так и акцепторов электронов Ис..
ходя из этоrо, опиновые пути следует счи..
TdTb примитивными. Ясно, что на ранних
этапах эволюции беспозвоночных присут"
ствовали все пути анаэробноrо метаболиз..
ма, а их нынешнее распределение.. по..
видимому, отражает различные направле..
ния отбора, т. е. специфические адаптации
к эколоrическим условиям, в которых при..
ходится функционировать орrанизму. Не..
которые системы эволюционировали как
приспособления к «взрывной» работе (они
ДdЮТ большие количества А ТР, но не обяза..
тельно высоко эффективны, поскольку ра..
ботают непродолжительно), друrие, наобо..
рот, как способ более экономичноrо, хотя
и не TaKoro быстроrо получения А ТР
в условиях длительной аноксии.
Рис. 11.3. Филетическое распространение oc
новных метаболических путей синтеза АТР (по
Livingstone, 1983, с изменениями). Мол путь
молочной кислоты; янт путь янтарной кисло
ты; OKT октопиновый путь. Метаболические
пути, указанные в скобках, используются при
особых обстоятельствах
11.4. Поrлощение кислорода
11.4.1. rлавное это диффузия
Аэробный метаболизм возможен только
при поступлении в ткани кислорода. В осно"
ве этоrо процесса лежит диффузия, т. е. дви"
жение молекул O из области высокоrо
в область низкоrо парциальноrо давления
(Р о) 1 в соответствии с законом Фика.
Скорость диффузии через ткани зависит от
rрадиента Р о. И от свойств самой ткани
последние учитываются коэффициентом
диффузии. Исходя из оценок этих коэффи"
циентов и потребностей тканей в кислороде
можно рассчитать, что расстояние между
I Парциальное давление rаза, paCTBopeHHoro
в жидкости. обычно называют ero напряже
нием. Прu/н. ред.
Дыхание
1 мм [
о
А
Б
в
==
Рис. 11.4. Поперечные срезы различных TYP
белларий (в одинаковом масштабе): A
прямокишечная; Б трехветвистая; В
мноrоветвистая (по Alexander 1971)
потребляющей кислород тканью идыхате..
лъной поверхностью не может превышать
] ММ. ЭТО одна из причин плоской q>ормы
тела крупных бесполостных турбелларий,
тоrда как для более мелких бескишечных
и прямокишечных форм она не обязательна
(рис. 11.4). Однако мноrие сuифоидные ме..
дузы и коралловые полипы достиrают
очень крупных размеров, несмотря на мас..
сивное строение. Это связано с тем, что
здесь наружный и внутренний слои ткани
тонкие и непосредсrвенно контактируют
с окружающей водой или с rастроваскуляр"
ной ПОЛОСl ью. Находящийся между ними
толстый слой мезоrлеи в основном лишен
34З
клеток, и ето метаболические потребности
невелики. В мезоrлее коралловых полипов
клеток больше, но она обычно ТОНЫllе, чем
у сuифомедуз. и блаrодаря сложной склад
чатости rастродермиса (рис. 3.27) всеrда на..
ходится не далее 1 мм ОТ воды, циркули
рующей в rастроваскулярной ПО.110СТИ.
11.4.2. Циркуляторные системы
Возникновение в ходе -эволюции систем
циркуляции жидкостей позволяет снять
оrраничения, накладываемые законами
диффузии. Такие системы повышают воз
можности транспорта 0')'1 позволяя перейти
«одномиллиметровый предел», а за счет
быстроrо удаления кислорода с дыхатель..
ных поверхностей поддерживают крутой
rрадиент Р о,, тем самым увеличивая CKO
рость поrлощения O орrанизмом. Извест--
ны два основных типа пиркуляторных си
стем (рис. 11.5): незамкнутые с обширным
rемоцелем (бластоцелем или разросшимся
кровеносным сосудом), типичные для ар..
тропод и моллюсков. И замкнутые. состоя
щие из артерий и вен, хорошо развитые
у аннелид Для обоих типов систем необхо..
димы мышечные насосы, например COKpd
тительные трубки у артропоп. снабженные
Связка, поддержи ваюuцая
сердце
Перикардиальная
полость
А
Кишка
Дорсальный параподиальный
сосуд
Кишечный
сосуд
Рис. 11.5. Два типа циркуля
торных систем: А He
замкнутая у членистоноrоrо;
Б замкнутая у полихеты (Ne..
reis)
Дорсальный сосуд
Дорсальный
кишечный сосуд
Кишечная сосудистая
сеть
Вентральный
кишечный сосуд
Вентральный параподиальный
сосуд
Б
344
rлава 11
1
Б
Водное насекомое
с трахейными жаБрами
Клетка трахеолы
II
.+ . ..
. .. .
, "
Мышца : =
.. А
Трахеолы, заполнен
Т рахеолы, заполнен ные воздухом
ные жидкостью
. .
. .
Мышца .
Б
t
Личинка наяда стрекозы
с анальной дыхательной
системой
Трахея
Дыхальце (часто
с клапаном)
Трахея
Рис. 11.6. 1. Трахейные системы: А основной тип; Б водные насекомые с жабрами; В наяда
стрекозы с анальным дыханием. 11. Увеличенное изображение трахейной системы (см. рис. 11.6.1,
А). Воздух входит через дыхальца, которые у большинства насекомых снабжены клапанами. Ши
рокие трубки (трахеи) разветвляются на более мелкие (трахеолы). Последние MorYT содержать
жидкость (А), но при интенсивном метаболизме осмотическое давление окружающих тканей B03
растает, что вызывает ее удаление из трахеол. Б. При этом воздух поступает в трахеолы и за счет
диффузии проникает через их стенки в ткани (по различным источникам)
Дыхание
парами отверстий (остий) с клапанами (рис.
11.5, А), или пульсирующие кровеносные
сосуды (латеральные «сердца») у аннелид.
у циркуляторных систем иrлокожих и полу..
хордовых строение промежуточное они
включают мелкие сосуды. впадающие в бо..
лее крупные синусы. Однако у rолотурий
(иrлокожие) хорошо развитая замкнутая си..
стема. У мокриц в связи с выходом на суш)
сформировались более крупные и более МУ"
скулистые. чем у друrих ракообразных.
сердца и сосудоподобные лакуны, образую..
щие почти замкнутую циркуляторную си..
стему. Незамкнутая система насекомых
практически не используется для транспор..
та кислорода. Вместо 3Toro у них сформи"
ровалась обширная и сложная система
стволов, трахей и трахеол (рис. 11.6), [аран"
тирующая почти прямую доставку rазо..
обраЗНОIО кислорода ко всем активно мета..
болизирующим тканям (как она работает,
объясняется в подписи к рис. 11.6).
11 .4.3. Кровь
Изначально кровь, по"видимому, была
бесцветной и близкой по составу к морской
воде, как у мноrих современных брюхо но..
rих и двустворчатых моллюсков. Такая со..
левая среда может переносить сравнитель..
НО небольшие количества кислорода, поэто..
му кровь мноrих беспозвоночных содержит
ero переносчики специализированные
белки, называемые дыхательными пиrмен..
тами, спо(.;обные обратимо связывать 02.
Например, в крови аннелид концентрация
pacTBopeHHoro кислорода составляет менее
1 О/О об. (мл 02/100 мл крови), тоrда как с ды..
хательными пиrментами связано 1 100/0
об. Структура этих пиrментов сходна: все
они состоят из белковой части, соединенной
спростетической rруппой, содержащей ме..
талл. Близкие по строению простетические
rруппы rемоrлобина и хлорокруорина на..
зывают [емом. Это порфирин, связанный
с одним атомом железа. У rемоцианина
простетическая rруппа образована не пор..
фирином, а полипептидом связанным с ме..
дью и серой. У rемэритрина ,то также не
[ем, хотя и включаеl железо. Некоторые
свойства и распределение в животном цар..
345
стве дыхательных пиrментов приведены
в табл. 11.1.
Количество переносимоrо кровью 02 (ее
кислородная емкость) зависит от содержа..
ния пиrментов и 01 их способности связы..
вать и отдавать 02 (сродства к кислороду).
Важный показатель отношение количеств
кислорода, способноrо связаться с пиrмен..
том и доступноrо в окружающей среде
(внешнее РО). Это отношение выражае..
тся так называемой кривой диссоциации ки"
слорода 1 , которую получают, насыщая
кровь кислородом при различных значе..
ниях Р 02 И измеряя связывание 02 по из..
менению оптических свойств пиrмента. Та..
кие кривые часто. хотя и не Bcer да (см. ни..
же), имеют S..образную форму (рис. ] 1.7).
Полезной характеристикой пиrмента являе..
тся величина Р О,, при которой он насы..
щается кислородом наполовину (Pso). Значе..
ния Pso ваРЬИРУIОТ в зависимости от темпе..
ратуры (ее снижение СДВИI'ает кривую дие..
социации влево рис. 11.8, А) и рН. У мень..
шение рН чаше Bcero понижает сродство
пиrмента к кислороду, т. е. сдвиrает кривую
вправо (рис. 11.8, Б). Это явление называют
эффектом Бора (нормальным, или отрица..
тельным). Рост конuентраuии CO
вызывает снижение рН, приводя к эффекту
Бора, а поскольку содержание СО') выше
Bcero в тканях с высокой интенсивностью
метаболизма, высвобождение кислорода
стимулируется там, [де он более Bcero необ..
ходим. у некоторых беспозвоночных. на..
пример у хелицеровоrо LiI11U/US (мечехво"
ста). обнаружен обратный (аномальный.
или положительный) эффект Бора. Мече..
хвост обитает на поверхности ила. l де пер..
иодическая нехватка 02 и повышенный уро..
вень С0 2 ' возможно, способствовали выра..
ботке такой реакции на рН. которая усили..
вает поrлощение кислорода из среды, хотя
при 1ТОМ затрудняется ero высвобождение
в тканях Не всем случаям обратноrо эффек"
та Бора можно дать такое раuиональное
объяснение, поэтому их и называют «ано..
мальными» .
1 Такое название неудачно, поскольку диссо
циирует не сам кислород. а ero комплекс с пиr
ментом, так что ПРdвильнее rоворить «кривая
диссоциации оксиrенированноrо пиrмент а»
При.н. ред.
346
rлава 11
Таблица 11.1. Дыхательные пиrменты (по Schmidt Nielsen, 1979)
Пиrмент
ОПИСШlие
Распространение в животном мире
Молекуляр
ная масса
rемоцианин
Мсдьсодержащий бсw'IОК. перено
СИl\AЫЙ в раствореН'НО\1 состоя
нии; синеrо цвета в оксиrени
рованной форме. бесцветный
R 1С10ксиrеНИРОВdННОЙ
rемэритрин
ЖелезосодеРЖаlНИЙ бслок; Haxo
дится внутри клеток; не сол.ер
жит порфирина: фиолетовоrо
цвета
Хлорокруорин
Белок. содержащий жеwlезопор
фириновый комплекс: перено..
сится в pdCTBOpCHHOM состоя
нии; красный в КОНllентриро
ванных растворах. зеленый
в разбавленных
r eMor лобин
Белок. содержаlНИИ жеwlезопор
фириновый комплекс: псрено"
сится в растворенном состоя
нии или внутри клеток; наибо
лее часто встречаюuJ.ИЙСЯ
у животных пиrмент; KpacHoro
цвета
300 000
9 000 000
Мо..1ЛЮСКИ: панцирные. rOwloBOHO
rие. переднежаберные и wlсrоч
ные брюхоноrие; отсутствует
у двустворчатых.
Ракообра1НЫС: крабы. 01VШРЫ.
Хелицеровые: Linпl/иS, Eиsco/,/JillS
Сипункулиды: ВСС изученные ви
ды.
Полихеты: Маgе/опа.
П риапуw'IИДЫ: H({/i(TJ,/Jtus. P/'iaplI
/lIs.
Плеченоrие: Li/ц(ulа
Распространение оrраНИLIСНО чс
тырьмя семействами ПО:lихет:
Sabellidae. Serpulidae. Chlorhae
midae. Ampharetidac.
П ростетическая rруппа u r JJe:IbHO
от белка обнаружена у морских
звезд (Lиidia. Astl'opecteп)
Позвоночныс.
Иr..lокожие: lОwIОТУРИИ.
Моллюски: P/ano/'his. Tit'e/a.
Н асекомые: C /li1"()пoпlUS. Gasf/'(}plli
/иs.
Ракообразные: Dap/111i({. А /.tc/llia.
Аннелиды: LU111hl'icus. Tи/Jf(ex. А/'е..
nico/a. Spi/"()1'bis (у некоторых ви"
дов rемоrлобин. у пруrих
ХwlОрОКрУОРИН. oCTawlbHbIe w1ише
ны дыхательных пиr:vtентов). Se
l'pu/a (rемоr.:lобин и хлорокруо
рин).
HCl\t1aTonbI: Ascal'is.
Плоские черви: паразитическис
трематол. ы .
Инфузории: Раl11Пlесill111, Tctl'allY
Пlепа
2750000
I 7 UUU
300 000
Различия по кривым диссоциации и по
значениям P o наблюдаются как внутри от..
дельных классов пиrментов.. так и между
ними, что., наверняка. отражает адаптацию
пиrментов к внутренней и внешней средам,
в которых им приходится функционировать
(см... например. рис. ] 1.7). Можно отметить
следующие корреляции.
1. Pso выше у животных. обитающих при
высоких внешних Po и имеющих дыхате..
льные поверхности. не представляющие су--
щественных препятствий для диф(рузии, T(tK
что Р O В их крови не очень отличается от
Р O В среде. Такая ситуация характерна
для полихеты Sa/Jclla (рис. 11.7) с дыхатеwlЬ"
ным пиrментом хлорокруорином (Р)О 30
мм рт. ст.). Это сублиторальное животное
обитает в трубке из ила и слизи. но способ..
но выставлять венчик из шупалеu в ОКРУ"
жающую хорошо оксиrенированную воду.
Кроме Toro, ею постоянно омывается по..
верхность тела за счет перистальrических
движений мускулатуры ero стенки. Друr-ой
пиrмент с низким сродством к КИСЛОРОДУ
Температура 15 0 с
)( = 4,8 мм СО2 рН : 8,43
. == 8,0 мм СО2 рн : 7,82
..=- 8,1 мм СО2 рН: 7,72
. = 14,2 мм СО2 рн: 7,52
0= 26,0 мм СО2 рН: 7,35
о = 38,3 мм СО2 рн : 7'104
А=-44 2мм со рН:б,77
О 5 10 15 20 25 ЗА О 10 20 ЗА 40 50 60 70 80 90 100 110 120 1ЗО
А РО2 Б РО2
Рис. 11.8. Влияние температуры (А), давления С0 2 и рН (Б) на кривые диссоциации оксиrениро
BaHHoro rемоцианина ракообразных (по Waterman. 1960)
А ОТТЕКАЮЩАЯ ВОДА
Рис. 11,7. Кривые диссоциации кислорода
зависимость поrлощения 02 из атмосферы (в
процентах полноrо насыщения) от ero KOHцeH
трации в ней (РО): А пескожил Areпico/a тa
riпa (рН 7,5; 19 ОС); Б Nephtys hoтbergii (рН
7,4; 15 ОС) дЛЯ сосудов; В TO же, но для цело
мической жидкости; r Sabe"a spa//aпzaпii (рН
7,35; 26 ОС) (Jones, 1955, 1972)
80
БО
%
40
20
АБсрронтальные
реснички
Латеральные
реснички
СРронтальные
реснички
100
*
...
50
:r:
1i
u
(tj
I
о
за
40
10 20
мм рТ. СТ.
100
80
60
%40
20
О
Ток воды между
соседними пластинками
Приносящий
кровеносный сосуд
Выносящий
кровеносный
сосуд
Ток крови внутри
ктенидиальнои пластинки
к предсердию
Б
Рис. 11.9. Ктенидиальная система брюхоноrих: А TOK воды через «жабры»; Б TOK в них крови
(по Russel Hunter, 1979). См. также рис. 5.18
348
rлава 11
rемоцианин кальмара Loligo (Р50 50 мм
рт. ст.). К этой же катеrории животных от..
носится эррантная роющая полихета Nep/l"
[у."; с rемоrлобином в крови Как видно из
рис. 11.7., у этоrо пиrмента довольно низкое
значение Р50( 5 7 мм рт. ст.), но следует
отметить, что кривая диссоциации здесь rи..
перболическая. поэтому по сравнению с S..
образной кривой со сходным Pso пиrмент
менее насыщен кислородом при более высо"
ких Р о,. Иными словами, пиrмент с S..
образной кривой полностью наСЫШClется
кислородом при более низких Р О,, чем
пиrмент с таким же Р 50' но С rипер6 оличе..
ской кривой диссоциации. Эта полихета
обитает в песке литоральной зоны во время
прилив а вода прокачивается через сделан..
ный ею ход с помощью мерцательноrо дви"
жения ресничек. В таких условиях пиrмент
должен быть способен как связывать. так
и отдавать 02 при высоких значениях РО:
Коrда наступает отлив, ход спадается
и происходит резкое снижение Р O по..
видимому, до TaKoro уровня, что поrлоще..
ние кислорода становится вообще невозмо"
жным., и животное скорее Bcero переходит
в этот период (около 6 ч) на анаэробный ме..
таболизм. Следует также отметить, что
у rемоrлобина сосудов NepJ1lys сродство
к кислороду выше, чем у целомическоrо [е..
моrл06ина. Возможно, это связано с посту..
плени ем 02 через всю поверхность тела,
сначала в целом. а уже оттуда в сосудистую
систему. ХОД кривых диссоциации сосуди..
cToro и целомическоrо rемэритринов сипун..
ку лиды DenC!,'OSlOпlUJ11 zosterico/uпl как раз
противоположный: у целомическоrо пиr..
мента сродство больше., чем у сосудистоrо.
у этоrо животноrо дыхательным opraHoM
служат щупальца. Кислород диффундирует
через них в сосудистую систему, а уже отту"
да в целомическую жидкость.
2. Р50 относительно низкое У животных
с затрудненной диффузией через дыхатель..
ные поверхности., обитающих в средах с вы..
соким Р О,, т. е. коrда значение Р О, в кро"
ви значительно ниже внешнеrо. К этой кате..
rории относятся мноrие десятиноrие раки
с Р S() В пределах от 1 до 20.
3. Р 'ю может быть очень низким « 1)
у животных., населяющих среду" 'с" низким
значением Р O . К этой катеrории относя..
тся, например, олиrохеты"тубифициды
и личинки комаров"хирономид (и те и дру"
rие с rемоrлобином в качестве дыхательно"
[о пиrмента), встречающиеся в местах. бо..
[йТЫХ (или заrрязненных) орrаническими
веществами. ХИрОНОМИДЫ живут в норках,
через которые активно прокачивают воду.
Однако они делают это не постоянно. так
что даже при насыщенности окружающей
воды кислородом внутри норки может на..
ступить rипоксия; в этих условиях пиrмент
с высоким сродством к кислороду важен
для ero поrлощения и, возможно, для запа..
сания на короткие периоды аноксии. Низкое
значение Р50 характерно также для MopcKoro
литоральноrо кольчеца пескожила, часто
роющеrо ходы в неаэрируемых илах (рис.
11.7). Как и хирономиды, он постоянно про..
rоняет воду через норку, и пиrмент с высо"
ким сродством К кислороду, возможно, ва..
жен для KpaTKoBpeMeHHoro запасания 02.
Тем не менее маловероятно, что запасен..
ный rаким образом кислород обеспеЧИВdет
пескожила В течение Bcero отливноrо перио..
да, так что в это время он. по"видимому,
способен частично переходить на воздуш..
ное дыхание. Наконец, следует отметить,
что кривая диссоциации у пескожила срав"
нительно мало чувствительна к низким зна..
чениям рН, что также может быть адапта..
цией к жизни в аноксических илах., часто бы..
вающих закисленными.
11 .4.4. OpraHbI дыхания
Еще одним фактором, оrраничивающим
поrлошение кислорода. является площадь
дыхательной поверхности. Для дыхания
плоских червей, длинных, тонких немертин,
нематод и некоторых аннелид достаточно
неспециализированной поверхности тела.
Однако дальнейшее увеличение размеров
или активности, а также эволюция относи..
тельно малопроницаемых защитных нару..
жных ПОКРОВОВ. в том числе раковины, по..
требовали возникновения васкуляризован"
ных дыхательных поверхностей. У водных
( OPM широко распространены выросты
стенки тела, например «жабры» некоторых
полихет и членистоноrих (рис. 11.6)., ктени"
дии моллюсков., жаберные книжки у мече..
ХВОСТа, ножки некоторых иrлокожих. На
Дыхание
349
Леrочная
книжка
1
Кровь
Воздух
Рис. 11.10. Леrочная книжка паука (по Snow
1970). См. также рис. 8.11.
Нервное
кольцо
Мадрепоровая
пластинка
u
1t
ji А afr
l' Р dd"'Ч
JFtIf
Околоротовая аМБулакральная
ножка
рис. 11. 9 схематически изображена ктени"
диальная система водното брюхоноrоrо.
Обратите внимание.. что кровь прокачивае..
тся сердцем через «жабру» в направлении.,
противоположном току воды (создаваемо..
му в основном латеральными ресничками).
Такой противоток типичен для жабр (кроме
жабр пескожила и rоловоноrих) за счет не..
то вода с низшим значением Р о,
взаимодействует с наименее оксиrенирован"
ной кровью., что позволяет последней по..
rлотить максимум О:!. У наземных форм
дыхательные поверхности чаще всето по..
rружены внутрь тела., например трахеи (рис.
11.6)., леrочные книжки, ситовидные и труб..
чатые трахеи паукообразных (рис. 11.1 0)'1
«леrкие» леrочных моллюсков. Унекото..
рых водных животных дыхательные по..
верхности также внутренние: примеры
водные леrкие rолотурий (рис. 11.11) или
анальные структуры личинок ряда насеко..
мых (рис. 11.6). В последнем случае иноrда
развиваются пластроны и наружные «физи..
ческие» жабры (рис. 11.12: в подписи к ри..
сунку разъясняется принцип их работы).
Кювьеров
opraH
Клоака
\
Анус
Водное леrкое
Амsулакральная
ножка
Рис. 11.11. Водное леrкое на схематическом продольном разрезе rолотурии (иrлокожие) (по
Nichols, 1969). СМ. также рис. 7.27
350
Вода
Воздух
Слой воздуха
Дыхальце
Трахея
r.naBa 11
Водоотталкивающие
б 2
волоски (10 /мм )
Кутикула
Ткань
Рис. 11 .12. Пластрон, Часть поверхности тела покрыта rидрофобными волосками, обеспечиваю
щими ее несмачиваемость и постоянное присутствие слоя воздуха. В результате она может дей
ствовать как несжимаемая жабра, в которую ИЗ воды диффундирует кислород. rустой слой воло
сков препятствует поступлению в воздушный слой воды по мере использования кислорода, а, так
как объем слоя остается постоянным, Р 02 В нем падает, вызывая диффузию 02 из окружающей
воды. У водноrо HaceKoMoro Aphe/ocheirus пластрон может выдерживать давление в несколько aT
мосфер (по Ramsay, 1962; Eckert, Randall, 1983)
11.4.5. Вентиляция
Еще ОДНИМ механизмом, способствую
щим поrлощению кислорода, является BeH
тиляuия дыхательных поверхностей. Она
обеспечивает постоянное поступление к ним
кислорода, т. е. поддержание на их уровне
как можно более KpYToro rрадиента KOHцeH
траций последнеrо. Вентиляция свойствен
на животным как со специализированными
дыхательными поверхностями, так и без
них, но отсутствует в трахейной системе Ha
секомых. Структура приспособлений, ocy
ществляющих вентиляцию. самая разная:
от ресничек на жаберных пластинках ДBY
створчатых моллюсков до мышечных Haco
сов, обслуживающих водные леrкие rолоту
рий или анальные структуры у стрекоз. Жи
вущие в трубках полихеты используют pec
нички и/или перистальтические движения
(см. выше). У ракообразных вентиляцию
обычно СОЗДают колебательные движения
придатков. При снижении РО! во внешней
среде вентиляция обычно усиливается. На
РИС. 11.13 показан аппарат, позволяющий
одновременно измерять характер активно..
сти (путем реrистрации изменений электри"
ческоrо импеданса на респирометре) и по..
rлощение кислорода (с помощью электро"
дов) у водных пиявок. При снижении
р O в среде усиление двиrательной актив..
ности увеличивает поr ЛОlцение 02 по срав"
нению с пиявками в состоянии покоя. При
этом упорядоченные движения MorYT так же
эффективно улучшать снабжение кислоро..
дом, как и реrулярная вентиляция.
11.5. Измерение метаболизма
Основная часть энерrии, получаемой за
счет дыхания. в конечном счете преВРdшае..
тся в тепло (рис. 11.14). Поэтому, если изме..
рить количество выделяемоrо животным
тепла, можно довольно точно оценить ин..
тенсивность метаболизма. Известны уст..
ройства с очень чувствительными термоэле..
J ыхание
351
А
Дыхательная камера
Пиявка в воде дышит через
поверхность тела
f
t<. кислородочувст
вительному электро
Ток воды
<
езервуар с реrуляцией
температуры и напряжения
кислорода
Б
в
Насыщение окой Неупорядочен Вентиляция
возд хом,% ное движение
Nephelopsis obscura
100 0,21:tO.,02 0,54:t0,09
50 0,04! 0,01 о,36:! 0,05
25 0,03! 0,01 0,18 :!: 0,01
О 0,02:!: 0,01 0,03:!: 0,01
Erpobdella punctata
100 0,271: 0,02 0,59 t 0,11
50 0,05:t0,01 0,35i: 0,07
25 0,03:t0,01 0,13::t0,04
О 0,02:!: 0,01 0,06:!: 0,01
D,52:t 0,10
0,32! 0,04
0,10!0,07
0,03:!: 0,02
Осциллоскоп
самописец
Вентиляция
Неупорядоченное движение
о,54:!: 0,09
0,55 :t 0,05
О,20:! 0,03
0,04 :t0,01
Покой
Время, мин
Рис. 11.1 З. Схема аппарата, описанноrо в тексте (А), и некоторые полученные с ero помощью pe
зультаты (5, В). 5. Типы движений, зареrистрированные путем измерения импеданса. В Интен
сивность потребления 02 (мкл/мr сухой массы в час :1:: среднеквадратичное отклонение) у двух ви
дав пиявок при различных концентрациях 02 в среде и разных типах активности. Можно видеть,
что движение связано с повышенным потреблением кислорода при любых ero концентрациях,
кроме самой низкой, причем существенной разницы в этом смысле между вентиляцией и неупо
рядоченным движением нет (по Wrona, Davies, 1984)
02.....
СО 2
Поступление
энерrии
Тепло
Тепло
Тепло
Тепло
Рис. 11.14. Схема использования энерrии, получаемой из пищи. Процессы дыхания rенерируют
АТР неэффективно, с потерей энерrии в виде тепла; большая часть запасенной в АТР энерrии TaK
же рассеивается в виде тепла после совершения работы
ментами способные зареrистрировать ма..
лые количества тепла, выделяемые беспо..
звоночными (их называют микрокалориме..
трами)" но используют их редко. На рис.
11.15 приведены результаты микрокалори..
метрическоrо эксперимента с водной олиrо
хетой Luп1briculus variegatus. На rрафике MO
жно различить: (1) аэробное состояние; (11)
аноксическое состояние; (111) состояние от..
равленноrо животноrо. Заметно что интен..
сивность аэробноrо метаболизма приблизи
тельно вчетверо выше, чем аноксическоrо.
352
rлава 11
>
А
51
Теплоизоляция
. . Экспериментальное
животное
>'
52
......
cl)
r-
О ...40
r::
с
Q)
r---
... 20
О
II
Аноксия (100% N 2 )
I I
20 111 21
'< >
1ч
N
I
22
I
23
Аноксия (8% СО 2 + 92% N 2 )
I
25
I
26
I
27
I
28
Время эксперимента
Температура термостата
Рис. 11.15. Прямой микрокалориметр. А. Устройство аппарата: очень чувствительные датчики (5,
и 52) измеряют температуру воды, втекающей в теплоизолированную камеру, в которой Haxo
дится экспериментальное животное, и вытекающей из нее. Б. Некоторые результаты более под
робно описанные в тексте (по Gnaiger 1983)
Поrлощение кислорода леrче измерить,
чем выделение теплd.. Для этоrо имеются
различные методы., от химическоrо титро..
Вd.ния O В водном растворе до физическоrо
измерения объема или давления rаза. Суще
ствуют также весьма чувствительные элек
троды нескольких типов. СодеРЖание ки
слорода может быть определено либо в за..
крытой системе, rде находится животное,
либо по поступлению 02 в открытую систе
му и оттоку из нее. По определению эти Me
ТОДЫ позволяют следить только за аэроб
ными процессами и поэтому чреВаТЫ Heдoo
ценкой дыхательноrо метаболизма, так как
иноrда беспозвоночные используют одно..
временно анаэробные и аэробные процессы
даже при достаточном поступлении кисло..
рода. Котда., как и при каких именно уело..
виях снижение Р о, вызывает пере ключе..
ние с аэрооноrо мётаболизма на анаэроб
ный., не вполне понятно; для выяснения этих
вопросов требуется больше данных., полу..
ченных методом прямой калориметрии.
Тем не менее в большинстве работ ин
тенсивность метаболизма оценивается
именно по поrлощению кислорода. На ос..
нове этой информации ниже показано, как
различные факторы влияют на дыхатель..
вый метаболизм. Однако интерпретация
проводимых результатов вызывает сложно
Дыхание
сти, так как метаболизм беспозвоночных
настолько зависим от состояния животноrо
и от условий, в которых оно находится, что
данные разных опытов MorYT быть несопо
ставимы npyr с друrом. Дыхательный MeTa
болизм подразделяют на основной обмен,
характеризующий орrанизм в состоянии
покоя, обычный обмен, измеряемый при
обычном для животноrо уровне активно
сти, пищевой обмен у только что питавше
rося животноrо и активный обмен при по
вышенной активности. Экспериментаторы
обычно стремятся определить основной об
мен, поскольку 'Это наиболее воспроизводи
мая оценка метаболизма.
11.6. Факторы, влияющие
на дыхание
Ниже перечисляются наиболее изучен
ные факторы, влияющие на дыхание у бес
позвоночных. Они расположены в порядке
снижения их связи с особенностями caMoro
орrанизма, т. е. начиная с неотъемлемых от
Hero свойств и кончая внешними влияния
ми.
11.6.1. Размер тела
Естественно ожидать, что крупные бес
позвоночные дышат более интенсивно, чем
мелкие, но, поскольку поrлощение кислоро
да оrраничивается поверхностью (см. BЫ
те), а поверхность тела пропорциональна
квадрату линейных размеров, тоrда как ero
масса их кубу.. следует также ожидать.
что зависимость между массой тела и по..
треблением кислорода нелинейна. CpaBHe
ине животных различноrо размера в преде..
лах данноrо вида или относяшихся к раз..
иым видам свидетельствует о том, что ин..
тенсивность дыхания в покое возрастает
с увеличением массы, но с убывающей СКО"
ростью. Это выражено следуюшим уравне..
иием:
Интенсивность дыхания == а(масса)Ь,
(11.1 )
rде d и b KOHCTaHTЫ, причем Ь обычно Me
иьше 1. Пролоrарифмировав.. получаем
log интенсивности дыхания ==
К + b(log массы),
(11.2)
З5З
Fде К == log а. Поэтому, если построить rpa..
фик зависимости лоrарифма поrлощения
кислорода от лоrарифма массы тела, полу
чится линейное отношение с наклоном, со..
ответствующим Ь (рис. 11.16).
Разделив- уравнения 11.1 и 11.2 на массу
тела, получим:
Интенсивность дыхания на единицу
массы == a(Macca)b 1,
log интенсивности дыхания на единицу
массы == К + (b 1) (log массы).
Поскольку Ь меньше 1, величина b 1 будет
отрицательной, так что rрафик зависимости
лоrарифма интенсивности дыхания на еди
ницу массы от лоrарифма массы будет
прямой линией с отрицательным наклоном
(рис. 11.17). Следовательно, как и ожида..
лось.. интенсивность дыхания на единицу
массы тела снижается с возрастанием по..
следней. Если бы это отношение определя
лось только увеличением поверхности тела
с возрастанием ero массы без учета друrих
факторов, Ь всеrда было бы 0,67, а b 1 co
ставляло бы 0,33. Объясняется это про
сто. Дых:ание, будучи зависимым от площа..
ди поверхности (см. выше), должно (для re
ометрически сходных тел) быть пропорцио
нально квадрату длины тела (Р). Масса про
порциональна объему и, следовательно
(опять же при rеометрическом подобии),
пропорциональна Р. Поэтому дыхание бы
ло БЬJ пропорционально кубическому кор..
3
ню из массы (.Ji3 == 1) в квадрате (== 12), т. е.
3
JМ2 == м О . б7 . Однако реальные значения
Ь редко точно соответствуют 0,67 и обычно
составляют от 0,67 до 1 (рис. 11.16). Считае
тся, что.. хотя rеометрические факторы
и вносят свой вклад в зависимость дыхания
от размера, они не являются единственны..
ми или хотя бы rлавными факторами, опре
деляюшими это соотношение. Остальные
предстоит выяснить.
11 .6.2. Активность
При движ нии l\10ллюск..блюдечко по
требляет приблизительно в 1,4 раза больше
кислорода, чем в покое; эта цифра состав..
ляет около 2 для бокоплава Gатп1аrиs, от
3 до 4 для обитающих в эстуариях креветок
354
rлава 11
1л
()
ro
"J" 1мл 23
m 16 21 /
.о
u)
о
() 20 22
о 171
CtS 1 мкл
:I: 121418
ro 9 1 5.... Наклон == 0,88
g
CL 7/'10
о
8
() 1нл ;7
:s:
:=='
Q)
:s:
:r:
Q) з,....4
r:::
u) 1пл
Q)
Q..
Наклон == 0,66
о ,..
С
1
1q:>л
1 пr 1 Hr 1 MKr 1 Mr 1r 1 Kr
А
Масса тела
Номер rpynna п Ь Номер rруппа п Ь
на рис. А на рис. А
Одноклеточные зктотермы
1 Бактерии п 0,68 17 Двупарноноrие мноrоножки 77 0,79
2 r рИБЫ 18 Пауки б 0,81
3 Жrутиковые 4 1,33 19 Равноноrие раки 40 0,69
4 И нq>узории 5 0,28 20 Моллюски 6 0,76
5 Корненожки 5 0,93 21 Жуки (имаrо) 14 0,81
Мноrоклеточные зктотермы 22 Дождевые черви 18 0,76
(взрослые ОСОБИ)
6 Нематоды 24 0,82 23 Макроракооsразные 3 0,81
7 Микроракооsразные 12 0,91
8 Клещи 71 0,61
9 Ноrохвостки 29 0,74
10 Термиты (личинки) 4 0,75
11 Энхитреиды 61 0,87
12 Жуки (личинки) 17 0,67
13 Термиты (личинки) 21 0,49
14 Муравьи (раsочие) 23 1,14
15 Дождевые черви (коконы) 3 1,00
16 Сенокосцы 30 0,69
Б
Рис. 11.16. А. rрафики зависимости потребления 02 от массы тела для ряда таксонов (в двойных
лоrарифмических координатах)_ Б. Список таксонов, использованных при построении rрафИКQВ
(А), с указанием наклона Ь прямой для каждоrо таксона (по Phillipson, 1981); п число изучен
ных представителей
Дыхание
Palaemoпetes, и может превышать 100 для
летящей саранчи. В этом нет ничеrо неожи
данноrо, так как движение требует биения
ресничек или жrутиков или же сокращения
мышц, что повышает интенсивность MeTa
болизма по сравнению с основным обме
ном.
11.6.3. Питание
Потребление кислорода обычно возра
стает сразу после приема пищи, но вскоре
опять снижается. Например.. у ракообразно
ro Macrobrachiuп1 roseпbergii эти колебdНИЯ
составляют 10 400/0. Такую дыхательную
реакцию на питание называют спецuфuче
ским дuнамическим действuем (СДД), специ
фическим дuнамическuм эффектом (СДЭ)
или калориrенным эффектом.
В основе СДЭ лежат следующие факто
ры:
1) энерrетические затраты на переработ
ку пищи в кишке;
2) энерrетические затраты на расщепле
иие и выделение лишних белков, поступив
ших с пищей;
3) энерrетические затраты на использо
вание веществ, полученных из пищи, для
синтеза компонентов новых тканей.
В исследовании, проведенном на мидии
Mytilus edu/is (Ваупе, Scullard, 1977), зареrи
стрировано возрастание на 250/0 потребле
иия кислорода после приема пиши; с энерrо
затратами на фильтрование пищи связано
более 80% этоrо прироста, на выделение
аммиака менее 200/0.
Длительное rолодание обычно сопрово
ждается снижением потребления кислоро
да. Это связано не только с отсутствием
СДЭ, но и с пониженной двиrательной aK
тивностью и даже с переходом на более эко"
иомичный «поддерживающий» метабо
лизм. Например, для разных видов морских
желудей, обитающих на различных уровнях
литорали и получающих неодинаковые KO
личества пищи, характерны разные интен
сивности потребления кислорода
(рис. 11.17). У Balaпus creпatus и В. rostratus,
которым ПИЩа доступна непрерывно в тече
иие большей части приливно отливноrо uи
кла, интенсивность метаболизма самая BЫ
сокая. У видов нижне и среднелиторальной
355
2,5
Balaпus glaпdula (сред)
Balaпus creпatus (СУБ)
8alaпus rostratus (СУБ)
Вalaпus cariosa
(НИЖ)
:r
.
L
"
ON 2,0
t::
L
CТI
..9 1,5
Chlhaтalus
fissus CtJttJamalus
(верх) dahi
(верх)
Polliclpes
polymerus
(сред)
0,5 1,0 1,5
Log массы (Mr)
Рис. 11 17. Связь между лоrарифмом потре
бления кислорода на единицу массы тела и ло
rарифмом этой массы для некоторых видов
морских желудей (по Newell, Branch, 1979);
верх, сред, ниж верхняя, средняя, нижняя ли
тораль; суб сублитораль
о
2,0
зон В. g/aпdula и В. cariosa уровень метабо
лизма промежуточный. тоrда как виды po
да Chthama/us, населяющие верхнюю лито..
раль и питающиеся лишь короткое время на
протяжении каждоrо приливно отливноrо
uикла.. а в периоды квадратурных приливов
вообще несколько дней не имеющие досту"
па к пище, отличаются очень низким ypOB
нем потребления кислорода.
11 .6.4. Температура
Изменения температуры среды MorYT
влиять на метаболизм, только если они
".,
влияют на температуру тела, что ООblЧНО
свойственно беспозвоночным, но не млеко
питающим или птицам, поэтому первых Ha
зывают пойкилотеРМНblМИ (от rреч. poiki
10 изменчивый), а вторых
20мойотеРМНblМИ (от rреч. homoio
остающийся таким же). Однако мноrие бес
позвоночные, например rлубоководные или
населяющие тропические районы океанов,
живут при постоянной температуре и, CTpO
ro rоворя, не являются пойкилотермными.
Более общие термины, учитывающие
источник тепла, определяют беспозвоноч
ных как эктотеРМНblХ, а млекопитающих
и птиц KaK эндотеРМНblХ животных. Но
и эта классификация не универсальна, так
как в летательных мышцах некоторых Hace
комых образуется так MHoro тепла (эндо
термия), что оно может использоваться для
356
rлава 11
поддержания постоянной температуры TO
ракса (rомойотермность). Например, шме..
ли не MorYT летать, если температура их ле
тательных мышц ниже ЗООС или выше 44 0 С.
По крайней мере 90% энерrии, затрачивае
мой летящей пчелой, высвобождается в TO
раксе в виде тепла, так что при интенсивном
полете температура там может подняться
на несколько традусов в считанные секунды.
Более Toro, есть данные, что во время CBO
бодноrо полета шмели способны поддержи
вать постоянную внутреннюю температуру
при различных внешних. Крупные пчелы (в
основном матки) MorYT летать даже при
ООС, поддерживая при этом в тораксе темпе
ратуру ЗООС и выше. Такая реrуляция
обеспечивается отчасти пассивно с помо
щью толстоrо слоя теплоизолирующих BO
лосков на тораксе, но rлавным образом aK
тивно за счет работы, совершаемой при
полете.
Таким образом, ни один из перечислен
ных терминов не может строто описать все
температурные характеристики беспозво
ночных, однако большинство их все же OT
носится к пойкилотермным эктотермам.
В пределах температурноrо интервала,
приемлемоrо для пойкилотермноrо живот
Horo, интенсивность дыхания возрастает
с увеличением температуры окружающей
среды. Однако, как характерно для химиче
ских реакций, такая зависимость нелинейна:
увеличение температуры на несколько Tpa
дусов вызывает повышение интенсивности
дыхания в несколько раз. Широко исполь
зуется индекс QlO) отражающий влияние
температуры на метаболизм. Он определя
ется следующим образом:
R 10/t 2 tl
QI0 == 2
R 1
10 (log R 2 10g R 1 ) 10
g QI0 t t '
2 1
[де R) и R2 интенсивности метаболизма
(оцененные, например, по поrлощению ки
слорода) при температурах t, и t 2 COOTBeT
ственно. Отсюда следует, что при
t 2 t) == 10 0 С величина QlO равна отноше
нию R 2 и R), так что QIO означает, во сколь
ко раз возрастает R при повышении темпе
ратуры на каждые 10 0 С. При резких сменах
температур QIO часто достиrает 2 и более,
;т
L.
"-
I
! 0,6 1'"
iд '"
X ,1
x IJ
' . I
04. · II O I
(1j , I " О
g o 1
1/1
о
ф I
:s:
:r:
Ф
LO
Ф
......
о
с:
I
'.х
ЛI
I
0,2
0,1
О
28
7
14
Сутки
21
Рис. 11 ,18. Акклимация у Mytl/us. .
постоянные условия 1 О ос; х переход от 1 О
к 15 ос; О переход от 1 О к 5 ос. Пояснения
в тексте (по Widdows, Ваупе, 1971)
причем эта величина может меняться в за..
висимости от температурноrо интервала.
Кроме тото, непосредственная реакция
на изменение температуры бывает непро
должительной. На рис. 11.18 показано, что
происходит при перенесении мидий Mytilus
edulis из среды с температурой 10 0 С в среду
с 5 или 15 0 С. Потребление кислорода при
150 резко возрастает.. а затем постепенно
снижается до HeKoToporo постоянноrо
уровня; при 50 оно также сначала резко па
дает, а затем возрастает, выходя на плато.
Этот процесс называется акклuмацuей; xa
рактерно, что значение Ql0 после акклима
ции ниже, чем в случае первоначальной pe
акции, а при идеальной акклимации CTpe
мится к 1. Такая реакция считается адаптив-
ной" так как при высоких температурах по-
зволяет экономить энерrию, а при низких
поддерживать синтез А ТР на достаточном
уровне. Акклимация TaKoro рода была об-
наружена у различных плоских червей, мол-
люсков, аннелид и иrлокожих.
Акклимация может действовать и в про-
тивоположном направлении.. Т. е. еше более
снижать потребление кислорода при низких
Дыхание
357
температурах и повышать ero при высоких,
т. е. значения QIO после акклимаuии будут
больше, чем для непосредственной реакции.
Это характерно для некоторых блюдечек,
включая как пресноводные (Aпcy/us fluviati
lis).. так и морские (PateZla aspera) виды. При
низких температурах такой тип акклимации
может быть адаптивным, обеспечивая пере
ход орrанизма в диапаузу и позволяя coxpa
нять энерrию в условиях зимнеrо HeДOCTaT
ка пищи и кислорода, связанноrо, напри
мер, с замерзанием ВGдоема. Стимуляцию
усиленноrо метаболизма при высоких TeM
пературах объяснить труднее; возможно,
она связана с тонкими компенсаторными
механизмами.
Реакция, проиллюстрированная
рис. 11.18, называется положuтельной aK
клuмацuей"! а описанная выше для блюде
чек отрuцательной, или обратной, акклu:
мацuей. Масштабы про исходящих измене
ний неодинаковы у разных видов и MOI YT
зависеть от состояния животноrо. Так.. у He
которых литоральных беспозвоночных ypo
вень OCHoBHoro обмена быстро и почти пол
ностью положительно акклимируется при
изменении температуры (QIO 1).. тоrда как
значение QIO дЛЯ обычноrо обмена всеrда
остается больше 1. Возможно, такая peaK
ция адаптивна, поскольку на этих животных
наиболее существенный перепад темпера
тур действует в период между приливами,
коrда они неподвижны и не питаются. Низ
кие значения QIO дЛЯ потребления кислоро
да покоящимися орrанизмами демонстри
руют определенную эколоrическую законо
мерность: для сублиторальных видов, Ha
пример Stroпgyloceпtrotus fraпciscaпus и Aпe
moпia пataleпsis, а также роющих форм ни
жней литорали типа Diopatria cuprea и HeKO
торых двустворчатых моллюсков, не испы
тывающих значительных и быстрых изме
нений температуры, характернь] веЛИЧИНЬJ
QIO > 1, а для обитателей литорали Littoriпa
littorea, Stroпgyloceпtrotus purpuratus, Масо..
та halthica, Actiпia equiпa и BuZlia digitalis
QIO 1. Но есть и MHoro исключений. на..
,
пример, у литоральноrо моллюска PateZla
vulgata значение QIO достаточно высокое, а
у сублиторальной полихеты HyaZiпoecia
низкое.
11 .6.5. Концентрация
кислорода (Р о )
Водным беспозвоночным часто прихо
дится переносить низкие значения Р O , на..
пример при заrрязнении пресноводных BO
доемов орrаническими веществами"! при BЫ
ходе из под воды во время отливов, возе..
рах ниже термоклина или при обитании
в субстратах с восстанавливающими свой..
ствами. Некоторые животные реаrируют на
это такой перестройкой вентиляции и кро"
вообращения, что потребление кислорода
остается на прежнем уровне (см., например,
рис. 1.19). Такие формы называют реzуля--
торами. У друrих видов потребление КИ !lО
рода изменяется параллельно Р о, во внеш..
ней среде. Их называют конформера.мu
(рис. 11.19). Реrуляторы MorYT поддержи
вать потребление кислорода на постоянном
уровне лишь в определенных пределах Р O ;
критическое значение этоrо параметра, при
4
:т
L..
"
С:::: 3
L
ctf Leptophlebia I
cl. (реrулятор) I
о 2
С:::: Ре Ephemera
о
:s: \ I (консрормер)
ф Cloeoп
::s:: I (реrулятор)
I 1
ф
С::::
LD I
ф
а.. I
ь
с О 100 150
200 250 300
Напряжение 02' мм РТ. СТ.
ф
:s:
Iro
фq
С::::О
Ша..
фо
а..с::::
ь()
c
Диапазон
консрормной
i ритическое напряжение
,
Напряжение 02 В среn.е
Рис. 11.19. Потребление кислорода в зависи..
мости от напряжения 02 в окружающей среде.
Вертикальной пунктирной линией показано co
держание 02 в воде, уравновешенное ero co
держанием в атмосфере (по Fox et al., 1937)
358
rлава 11
Таблица 11 .2. Примеры реrуляторов и конформеров. Ре критическое значение Р0 2 ,
при котором реrуляция перестает действовать и реrуляторы становятся
конформерами
Реrуляторы
Р 1)
с
Конформеры
Paraтeciuт (инфузории)
Tetrahyтeпa (инфузории)
Pe/тatohydra (rидроидные)
AureZia (сцифомедузы)
Tubifex (олиrохеты)
Luтbricus (олиrохеты)
MytiZus (двустворчатые моллюски)
He/ix (леrочные брюхоноrие)
LoZigo (кальмар)
Caтbarus (pdK)
и са (краб)
Cloeoп (нимфа поденки) 2)
Hya/ophora (личинка бабочки)
50
2,5
60
120
25
76
75
75
45
40
4
30
25
Spirostoтuт (инфузории)
Различные коралловые полипы
Cassiopea (сцифомедузы)
Nereis (полихеты)
Erpо bdeZZa, (пиявка)
Sipuпcu/us (сипунку лиды)
Liтax (леrочные брюхоноrие)
Liти1us (хелицеровые)
Hoтarus (омар)
Baetis (нимфа поденки) 2)
Taпytarsus (личинка хирономид)
1) К приведенным величинам Ре следует относиться с осторожностью, поскольку переход между
реrуляцией и конформностью обычно резко не обозначен. Кроме Toro. на форму кривой. Bыpa
жающей зависимость потребления кислорода от ero напряжения, часто влияет степень аккли
мации к аномальному значению Ро,-
2) См. также рис. 11.19.
котором они становятся конформерами,
обычно обозначают РС' В табл. 11.2 приве
дены примеры конформеров, реrуляторов
и значений РС' Животные, для естественных
местообитаний которых характерны низкие
уровни Р О,, часто являются реrуляторами
с низкими РС' Так, черви"туБИфИЦИДЬJ с РС,
равным 25 мм рт. ст., выживают в заrря..
зненных орrаникой участках рек. Манящий
краб Иеа, обычный обитатель приморских
болот и илистых наносов, также относится
к хорошим реrуляторам. Конформер Baetis,
напротив, живет в быстрых, хорошо оксиrе..
нированных ручьях.
Разные составляюшие метаболизма мо"
rYT различным образом зависеть от Р 02; ми..
дия, например, конформер в отношении
OCHoBHoro обмена, но реrулятор (до 80 мм
рт. ст.) С точки зрения обычноrо обмена
(рис. 11.20).
После действия на орrанизм условий rи..
локсии или аноксии потребление 02 часто
возвращается до уровня, превышаюше..
ro тот, что наблюдd.ЛСЯ до помещения в та..
кие условия. Это возвращение так называе..
Moro кислородноrо долrа. Сюда входит
восстановление запасов фосфаrена и А тр
и окисление конечных продуктов анаэроб..
Horo обмена. Однако все здесь не так про..
сто, поскольку между накоплением конеч..
ных продуктов и размером кислородноrо
долrа не всеrда наблюдается корреляция,
O 4
.
0,3
.
ОБЫЧНЫЙ /е
ОБмен / Основной
/е е/е
е:/
0,2
О
r::
L
0,1
.
40
80
120
140
мм рТ. ст.
Рис. 11.20. Влияние напряжения 02 в среде на
метаболизм Mytilus (по Ваупе et al., 1976). Об-
суждение см в тексте
Дыхание
359
а уровни А ТР и фосфаrена обычно возвра
щаются к норме очень быстро.. уже на нача
льной стадии процесса. Поэтому следует
учитывать и друrие перестройки, например
восстановление доаноксическоrо внутри
клеточноrо рН, пониженноrо в результате
накопления кислых конечных продуктов,
или использование дополнительноrо кисло
рода для насыщения жидкостей тела и ды
хательных пиrментов.
В заключение следует отметить, что из
быток кислорода может быть так же опа
сен, как и ero недостаток. Радикалы свобод
Horo кислорода (супероксид радикалы) спо
собны денатурировать макромолекулы,
особенно белки. Это становится существен
ным при очень высоких конпентрациях 02'
например в тканях беспозвоночных, содер"
жащих симбиотические водоросли (у MHO
rих обитателей коралловых рифов), rде ки
слород является конечным продуктом фо
тосинтеза. Такие орrанизмы выработали
клеточные механизмы защиты от кислород
иоrо отравления, в частности ферменты cy
пероксиддисмутазу (СОД), обезвреживаю
щую опасный супероксид радикал, и KaTa
лазу, удаляющую Н 2 О 2 , образуемую СОД.
На рис. 11.21 показано, что у некоторых
rрупп беспозвоночных активность СОД по
вышается с увеличением концентрации хло
рофилла в тканях, т. е. с возрастанием сим
биотической способности к высвобождеНИIО
кислорода. Однако эта зависимость неоди
иакова для разных rрупп.. что связано с раз
500
-.:
.:' :
400 . '-:iji. .
ffi 300 СПLdапа .;-j,f;fF ;. f ; :: Trldacпa
L ._: :,:<,,:: J':-: ", :;;..;:-'; ::(двустворчатые
200 ":J. ;' {j fJfifJ/f/i Ш
100 ЙШ :"' ' " orif
. :i" : : . ...........:.:.:.:..СПldаrLа (В тени)
.\" .;.;.:. .;.'. ..... .........:.:.:. ........ ......... Dlde ит
О 50 100 150 200 250
Хлорофилл, MKr /Mr Белка
Рис. 11.21. Связь между активностью СОД
и концентрацией хлорофилла у ОСНОВНЫХ
rрупп беспозвоночных коралловых рифов (по
Shick, Dykens, 1 985)
личной локализацией симбионтов: актив
ность СОД наиболее высока у стрекающих
с исключительно или преимущественно BHY
триклеточными зооксантелами (цитоплаз
ма непосредственно испытывает воздей
ствие кислорода). тоrда как у оболочника
Dideтпuт симбиотические прохлорофиты
находятся не только вне клеток, но и вне op
rанизма (в выстилке общей клоакальной Ka
меры), и избыточный кислород быстро уно"
сится током жидкости., не контактируя с ци..
топлазмой клеток. Самая низкая актив..
ность СОД среди стрекающих наблюдается
у видов., обитающих в затененных MeCTax
в пещерах или под скалами.
11 .6.6. Соленость
Поскольку изменения солености воды
MorYT вызвать у морских беспозвоночных
активные поведенческие и физиолоrические
реакпии, можно ожидать.. что они будут co
провождаться изменениями потребления
кислорода. Креветка Metapeпaeus тoпocer
cus встречается в условиях различной соле..
ности. Потребление кислорода особями из
солоноватоводных популяций минимально
в 500А> ной морской воде и повышается при
изменении этоrо уровня как в rипо , так и
в rиперосмотическую сторону. И напротив
потребление кислорода особями из мор..
ской популяции минимально в 1000/0 НОЙ
морской воде и возрастает до максимума
в 250/0 НОЙ.
Затраты энерrии на осмореrуляцию и aK
тивный транспорт ионов, по видимому, He
велики, так что большая часть наблюдае
мых изменений вероятно связана с реак"
циями Bcero орrанизма, например с перехо
дом к друrому типу активности. Это более
подробно обсуждается в работе НЬЮЭЛЛd
(Newell., 1979).
11.7. ВЫВОДЫ
Термином «дыхание» обозначают про
цессы высвобождения биолоrически по..
лезной энерrии из питательных веществ.
rлавным образом уrлеводов. Эти реакции
по большей части, хотя и не исключитель
но, аэробные, поэтому «дыханием» назы
вают также процесс потребления 02 и из
360
бавления от ненужноrо СО 2 . Ero биохими..
ческие аспекты иноrда называют «внутрен--
ним», или «тканевым дыханием»; а физио..
лоrические «внешним дыханием». В на..
стоящей rлаве рассмотрены механизмы,
действующие на обоих этих уровнях,
системы, обеспечивающие получение 02 из
окружающей среды и ero доставку к тка..
ням, а также внешние и внутренние факто..
ры, влияющие на интенсивность метаболиз..
ма.
11.8. Дополнительная
литература
A/exaпder R. McN. ] 971. Size and Shape. Edwards
Arnold, London.
Ваупе В. L., Scullard С. ]977. Ап apparent specific
dynamic action in Mytilus edu/is. J. mar. biol. As
soc.. UK, 57. 371 378.
Са/о"' Р., To"'пseпd С. R. ] 981. Resource utilization
in growth. ln: с. R. Townsend and Р. Calow (Eds)
Physiological Ecology: ап Evolutionary Approach
to Resoиrce Utilization. Blackwell Scientific PubIi..
cations, Oxford.
Eckert R., Raпdall D. 1983. Animal Physiology, 2nd
edn. W. Н. Freeman, New У ork.
Heiпric/l В. 1979. Bumble..bee Economics. Harvard
[пава 11
University Press, Cambridge. Massachusetts.
Joпes J. D. 1972. Comparative Physiology of Respi
ratory Mechanisms. U niversity of Pennsylvania
Press, Philadelphia.
Liviпgstoпe D. R. 1983. Invertebrate and vertebrate
.pathways of anaerobic metabolism: evolutionary
considerations. J. Geol. Soc. 140, 27 38.
Newell R. С. 1979. Biology of Intertidal Animals, 3rd
edn. Marine Ecological Sиrveys Ltd., Kent.
NeH'e/l R. С., B,'al1ch G. 1979. ТЬе influence of tem
perature оп the maintenance of energy balance lЛ
marine invertebrates. Adv. mar. Biol., 17, 329
346.
P/liZlipsuп J 1981. Bioenergetic options and phyloge
пу. In: С. R. Townsend, Р. Calow (Eds) Physiolo
gical Ecology: ап Evolиtionary Approach to Res
ource Utilization. Blackwell Scientific Pиblica
tions. Oxford.
Raтsay J. А. 1962. А Physiological Approach to the
Lower Animals. Cambridge University Press. Lon
don.
Schmidt Nie/seп К. 1979. Animal Physiology: Adap
tation and Environment. Cambridge U niversity
Press, Сат bridge.
Имеется перевод: Шмидт..Ниельсен К. Физио
лоrия животных: приспособление и среда: в 2 T.
М.: Мир, 1982.
rЛАВА 1 2
ЭКСКРЕЦИЯ, ИОННАЯ И ОСМОТИЧЕСКАЯ
РЕrУляция, ПЛАВУЧЕСТЬ
Вещества. количество которых превы"
шает метаболические потребности, должны
быть удалены из орrанизма ЖИВОТНОI'О.
К ним относятся непереваримые компонен..
ты пищи. выводимые с фекалиями (rл. 9),
и СО 2 , удаляемый в процессе дыхания
(rл. 11). Однако существуют и друrие избы..
точные вещества вода, различные ионы
и продукты распада лишних белков и ами"
нокислот. Именно последние азотсодержа..
щие соединения в физиолоrии обычно назы..
ваются продуктами выделения, или 1кскре..
ции (экскретами). Вместе с тем процессы
удаления лишних ионов, воды и азотсодер..
жащих веществ у беспозвоночных часто на..
столько тесно связаны, что лоrично рассма..
тривать их вместе. Поэтому в этой rлаве мы
сначала опишем истинную кскрецию, за..
тем разберем проблемы, связанные с pery..
ляцией водно"ионноrо баланса, и, наконец,
рассмотрим структуру и функuии выделите..
льных систем, связанных с ионной и осмо"
тической реrуляцией, но не обязательно
с удалением азота.
12.1. Экскреция
Ненужные аминокислоты MorYT появи..
ться в орrанизме при поступлении слишком
большоrо их количества с пищей или в резу..
льтате катаболизма белков. Обычно они
подверrаются дальнейшей окислительной
деrрадации с образованием кетокислот
и аммиака:
R
I
NH2 CHCOOH + 1/202 ==
Аминокислота
R
I
== о===ссоон + NН з .
Кетокислота Аммиак
Кетокислоты MorYT использоваться в дру"
rих метаболических превращениях, а ам..
миак, будучи крайне токсичным, должен
быть либо быстро удален из орrанизма, ли..
бо переведен в безвредное состояние (см.
ниже). Он леrко растворим и леrко выво..
дится в случае водных животных за счет
диффузии через покровы тела или дыхате..
льные поверхности. Если среди экскретов
преобладает аммиак, такой тип выделения
называют аммониотелией; он широко рас..
пространен у водных беспозвоночных,
в том числе у водных личинок насекомых (в
отличие от наземных имаrо; см. ниже).
у некоторых животных аммиак превра..
щается в менее токсичную мочевину:
NH 2
I
с===о
I
NH 2
Экскреция азота в виде мочевины известна
как уреотелия; такой тип выделения типи..
чен для млекопитающих. У беспозвоночных
он распространен слабо. Возможно, это
связано с тем, что, будучи еще токсичной,
мочевина требует для cBoero выведения по..
тери больших количеств воды. Тем не менее
уреотелия встречается у некоторых плоских
червей, аннелид и моллюсков.
Основным экскретом наземных беспоз..
воночных (а также наземных позвоночных,
за исключением млекопитающих) является
мочевая кислота, а основанный на ней тип
выделения называется урикотелией. По хи..
мическому строению она относится к клас..
су пуринов. Друrие вещества этоrо класса
также используются как экскреты некото"
рыми беспозвоночными (пуринотелия).
Появление в эволюции этих продуктов
выделения связано с их малотоксичностью
и низкой растворимостью, которая позво..
362
rлава 12
ляет удалять их в кристаллическом состоя
нии и тем самым сокращать потери воды.
Урикотелия характерна для онихофор од..
новетвистых и в меньшей степени для рако..
образных и моллюсков. У пауков основным
,кскретом является друrой пурин rуанин.
н н
N С """C,/N '"
I 11 СН
HC N/C"N/
Пурин
о
11 н
с N
HN/ "C '"
1 11 C O
O C"N/C"""N/
н н
Мочевая кислота
о
11
с N
HN/ "с./ '\
I 11 СН
./C N/C"N/
H 2 N н
rуанин
Исключение из указанной выше эволю"
ционной тенденции составляют ракоо6раз--
ные. Даже у наземных Рdвноноrих раков
преобладает аммониотелия. Кутикулярные
покровы этих животных леrко проницаемы,
поскольку почти лишены эпикутикулярноrо
воска, xapaKTepHoro для насекомых (разд.
12.2.5) и rазообразный аммиак может сво"
бодно удаляться через них путем диффузии.
Ткани ракообразных также, по"видимому,
менее чувствительны к действию аммиака,
чем у друrих животных. Кроме Toro, ero вы..
деление через покровы способствует фор..
мированию экзоскелета: щелочная среда,
возникающая на поверхности тела, необхо--
дима для образования карбонат--иона из ра..
cTBopeHHoro СО 2 и осаждения СаСО з . Мол--
люски также MorYT использовать' аммиак
для упрочнения раковины. Из друrих назем..
ных беспозвоночных непосредственное вы..
деление аммиака свойственно некоторым
олиrохетам и мноrоножкам, но они, как
и равноноrие раки, обитают во
влажной среде, rде опасность обеЗВОЖИВd"
ния очень мала.
Еще один фактор, сыrравший роль
в эволюции экскреторных систем,
экономия метаболических ресурсов. Так,
хотя пурины И менее токсичны, чем аммиак,
в их состав входит уrлерод
потенциальный источник энерrии. Мочеви"
на в этом смысле занимает промежуточное
положение. В табл. 12.] сопоставляются
токсичность и потери водь] и уrлерода
(энерrии), связанные с каждым из трех ос..
новных продуктов выделения. Однако. на..
сколько важны эти потери для «бюджета»
Bcero орrанизма, неясно; возможно, боль..
шее значение имеют энерrозатраты (выде..
ление тепла) в метаболических процессах.
ведущих к образованию ':)кскретов.
Важно понимать, что «продукты выде..
ления», особенно малотоксичные (напри..
мер. пурины). вовсе не обязательно должны
удаляться из орrанизма. Так, и у наземных,
и у водных леrочных моллюсков мочевая
Таблица 12.1. «Экономичность» экскретов. Число звездочек соответствует прибли..
зительной количественной оценке (Pilgrim, 1954)
Теплота сrорания
C/N
Т оксич Потребность
ность в Боде
кДж/моль кДж/моль N
Аммиак
Мочевина
Мочевая кислота
о
0,5
1,25
378
638
1932
378
319
483
***
***
**
*
**
*
Экскреция, ионная и осмотическая реrуляция
кислота накапливается в орrанизме по мере
ero роста и старения. Обнаружено также на..
копление мочевины в тканях тропическоrо
леrочноrо моллюска Buliтulus в период лет..
ней диапаузы. Значение этих процессов не..
известно, но помимо экскреторной функции
высокая концентрация мочевины в тканях
и жидкостях тела улитки может повышать
осмотическое давление, тем самым препят"
ствуя потере воды путем испарения. Не ис..
ключено, что это способствует пережива..
пию длительных засушливых периодов.
Мочевая кислота накапливается также в хо..
де эмбриональноrо развития клейдоических
ЯИЦ. Хорошим примером может служить
опять..таки леrочный моллюск: содержание
мочевой кислоты в яйцах Lyтпaea возра..
стает с 0,50/0 их свежеrо веса при дроблении
до 4,50/0 при выходе личинки. Есть мнение,
что именно возникновение клейдоических
ЯИЦ в ходе эволюции привело к появлению
363
пуринотелии. Как бы то ни было, оба этих
признака сыrрали важную роль в завоева..
нии суши.
12.2. Осмотическая и ионная
реrуляция
Жидкости тела животных предстаВЛЯЮ1
собой разбавленные солевые растворы, ос..
новным электролитом которых является
хлорид натрия; иными словами, они напо..
минают по составу морскую воду. Обычно
считается, что это следствие зарождения
жизни в море. Тем не менее между морской
водой и жидкостями тела имеются суще..
ственные различия (табл. 12.2). и Макал..
лум (Масаllит) в 1920..х rr. предположил,
что это отражает постепенное изменение со..
става океана с момента возникновения жи..
зни до наших дней. Однако результаты па..
леохимических исследований rоворят об от..
Таблица 12.2. Концентрации ионов в плазме крови или полостной жидкости, выраженные в про
центах от концентраций тех же жидкостей, диализованных против морской воды (по Barrington,
1979)
Na К Сй Mg Сl S04
Книдарии
Aurelia aurita 99 106 96 97 104 47
Иrлокожие
М arthasterias glacialis 100 111 101 98 101 100
Оболочники
Salpa тaxiтa 100 113 96 95 102 65
Аннелиды
Areпicola тariпa 100 104 100 100 100 92
Сипункулиды
Phascolosoтa vulgare 104 110 104 69 99 91
Ракообразные
М aja squiпado 100 125 122 81 102 66
Droтia vulgaris 97 120 84 99 103 53
Carciпus тaeпas 110 118 108 34 104 61
Pachygrapsus тarтoratus 1) 94 95 92 24 87 46
Nephrops пorvegicus 113 77 124 17 99 69
Моллюски
Pecteп тaxiтus 100 130 103 97 100 97
Neptuпea aпtiqua 101 114 102 101 101 98
Sepia officiпalis 93 205 91 98 105 22
1) Этот rрапсоидный краб единственное представленное в таблице животное с rипоосмотической rемолимфой
(концентрация ионов 860/0 их концентрации в морской воде).
364
rлава 12
сутствии существенных различий в составе
воды древних и современных океанов
Таким образом, из табл. 12.2 следует,
что даже морским животным приходится
реrулировать состав жидкостей cBoero тела.
У обитателей эстуариев и пресных вод эта
проблема стоит еше более остро, так как
они должны поддерживать в жидкостях Te
ла более высокие концентрации веществ,
чем во внешней среде. Обратная ситуация
наблюдается у животных. а) которые во
время жизни в пресной воде выработали
в своих жидкостях более низкую KOHцeHTpa
цию веществ, чем в морской воде, а затем
вернулись в море; б) обитающих в очень
концентрированной среде, например в соле
ных озерах. Для наземных форм, eCTeCTBeH
но, основная проблема сохранение воды.
Теперь давайте более детально paCCMO
три м вопросы, связанные с ионной и осмо"
тической реrуляцией в четырех перечислен..
ных эколоrических ситуациях. Основные
термины и понятия'! важные для понимания
этоrо материала, приведены в приложении
12.1.
12.2.1. Морская среда
У большинства морских беспозвоноч
ных жидкости тела изоосмотичны морской
воде, а сами животные являются OCMOKOH
формерами. Тем не менее, как уже rовори"
лось. ионный состав жидкостей их тела дo
вольно существенно отличается от состава
морской воды, так что необходима ионная
реrуляция. Это иллюстрируется данными
табл. 12.2. Стоит отметить следующее. а)
Aurelia реrулирует в основном KOHцeHTpa
цию ионов сульфата, поддерживая ее на бо..
лее низком уровне, чем в море. Возможно,
это связано с поддержанием плавучести: ио..
ны сульфата тяжелые, и их замена хлорид
ионами снижает плотность медузы. То же,
по видимому, справедливо для каракатицы
Sepia (см. также разл. 12.2.2). б) Единствен
ный ион, содержание KOToporo способны
реrулировать иrлокожие, ЭТО калий. Воз..
можно, это одна из причин, оrраничивших
их распространение морской средой. в)
Уровень маrния очень низок в жидкостях
тела членистоноrих. Это может быть связа
но с тем, что большинство членистоноrих
очень активны, а маrний имеет свойства
анестетика. Однако у весьма подвижной Ka
ракатицы Sepia концентрация маrния не по..
нижена.
12.2.2. Отступление по поводу
плавучести
Водные животные с плотностью выше,
чем у воды, должны опускаться на дно,
поэтому плавающим формам выrолно поп..
держивать свою плотность равной или ме..
ньшей, чем у воды, иначе им придется за..
трачивать дополнительную энерrию, чтобы
не поrружаться в rлубину. Уже rоворилось,
что этоrо можно достичь, по крайней мере
отчасти, реrулируя свой ионный состав. Ни
же мы проиллюстрируем это новыми при..
мерами, а также обратим внимание на дру"
rие, обеспечивающие плавучесть приспо..
собления.
У rлубоководиоrо кальмара Не liocraп..
cllia очень большая перикардиальная по..
лость. заполненная жидкостью. Она леrче
морской воды за счет rораздо более низко..
ro содержания ионов натрия и очень высо..
Koro ионов аммония. Последние, являясь
конечным продуктом катаболизма белков,
диффундируют в перекардиальную жид-
кость и удерживаются в ней за счет кислой
реакции. Кроме Toro, как и в случае Sepia
и Aurelia, анионы в этой жидкости представ..
лены почти исключительно хлоридом, за-
меняющим тяжелые ионы сульфата.
Существуют и друrие способы повыше-
ния плавучести: а) удаление ионов без их за..
мещения'! что, однако'! приводит к rипоос-
мотическому состоянию жидкостей тела,
поддержание KOToporo требует со ответ..
ствующих затрат; 6) устранение тяжелых
компонентов; например, у некоторых пела-
rических брюхоноrих моллюсков (В частно-
сти, птеропод и rетеропод) раковина реду-
цирована или вовсе отсутствует; в) увеличе-
ние солержания жиров (они леrче волы) рас-
пространено как у морских, так и у пресно-
водных планктонных ракообразных; 2)
использование заполненных rазом поплав-
ков с мяrкими стенками, как у книдарии
Physalia, или в виде твердостенных камер
раковины rоловоноrих наутилоидей и ее ру-
димента У каракатицы (рис. 12.1).
Экскреция ионная и осмотическая реrуляция
365
Отверстие
ТРУБКИ сифона
Жилая
камера
4% Заполненное rазом
t прос транство
,/ /
-
А "'
I 4% "....
I
......
Б
Рис. 12.1. А. Переrородчатая раковина моллюска Nautilus. По мере роста животноrо к ней при
бавляются одна за друrой новые заполненные rазом камеры. Сначала они заполнены жидкостью,
в которой растворен в основном NaCI. Затем Na t- удаляется посредством активноrо транспорта,
жидкость становится rипоосмотической по отношению к морской воде и удаляется, оставляя за
полненное rазом пространство. rаз поступает внутрь камеры путем диффузии, давление N 2
достиrает 0,8 атм, как в окружающей воде и тканях животноrо, а у 02 оно В камере t:lиже, чем BO
Kpyr нее. Поскольку у камеры жесткие стеlЧКИ, давление rаза остается около 0,9 атм, независимо
от rлубины поrружения животноrо. Б. Рудимент раковины у каракатицы имеет слоистую CTPYKTY
ру И заполнен rазом приблизительно TaKoro же состава и давления, что и у наутилуса. Однако
в наиболее старых, задних камерах (показанных черным) находится раствор NaCI. Количество ra
за можно изменять вместе с объемом жидкости, что позволяет каракатице реrулировать свою
плотность, а тем самым плавучесть и rлубину поrружения. Это обеспечивается за счет изменения
концентрации раствора NaCI в раковине: на небольшой rлубине он близок по осмотическим свой
ствам к морской воде, по мере поrружения концентрация соли снижается, жидкость становится
rиnоосмотичной морской воде и крови животноrо, а возникающее при этом осмотическое давле
ние уравновешивает rидростатическое давление, стремящееся заполнить «раковину», водой (по
Schmidt N ielsen, 1979)
12.2.3. Пресноводная среда
Если морских животных поместить
в разбавленную морскую воду, то либо oc
мотическое давление жидкостей их тела
уравняется с окружающим (осмоконформе--
рами), либо включатся механизмы, препят--
ствующие разбавлению их внутренней сре--
ДЫ (у осмореrуляторов). Примеры приведе..
ны на рис. 12.2. Обитатели эстуариев и ОТ..
части литорали в естественных условиях
периодически оказываются в разбавленной
воде, например во время отлива или после
сильноrо дождя. Среди таких животных
встречаются как осмоконформеры, так и ос--
мореrуляторы.
Не следует, однако, считать осмокон--
формеров обязательно более стеноrалин
ными, чем осмореrуляторы. Так, осмотиче--
ское давление крови мидии Mytilus edu/is со..
ответствует окружающему как в Северном
море (нормальная соленость), так и в Бал
тийском (менее половины нормальной со..
лености). Одна из причин такой толерант
ности в том, что эффективно реrулируется
не внеклеточная, а внутриклеточная среда.
По--видимому, важную роль в этом процес..
се иrрают аминокислоты: их концентрация
при разбавлении окружающей среды возра
стает, тем самым повышая осмотическое
давление внутри клеток. Выделение аммиа
ка мидиями усиливается при снижении
внешней солености. Это, вероятно, стиму--
лирует катаболизм белков и образование
свободных аминокислот под действием ос..
мотическоrо стресса. Подтверждением слу
жит тот факт, что у мидий обнаружена [eHe
тическая изменчивость по лейцинаминопеп
тидазе важному ферменту белковоrо ка..
таболизма, причем более активные ero ва-:-
366
500
с:::
о
со 400
с:::
ф
:s:
t; 300
о
c:t
:s::
ЭЕ
m 200
о::::
:S::
:::f
са
100
:r:
:r:
о
rлава 12
600
rflonetes
paL e a ns
va rL
Эквивалент
ность между
8нутренней
и внешней
средой
Морская
вода
100 200 300 400 500 600
Концентрация 80 внешней среде, ммоль/л
Рис. 12.2. Связь концентраций солей в жидко..
стях тела и среде обитания некоторых солоно..
водных животных (по Schmidt Nielsen, 1979)
ц)
.L1 13.й
,...... с::: :::> ,......
:ii CI) са""'" 1:) 0-.1)
g ::::> 0-.L1 а о С:::
...... о С::: L со
m L са Q..:S:: о-
ц) :s:: о- с::: о
':S:: ,....... с:: С::: о ц)
о с: ....... о:::: :s::
.L1 Q) о:::: :S: о:::: с:::
() С::: Q) Ш
о- са
о Q.. :S::
0,7 о :S:: Q. I
(jI ,,---,,:S::
':S:: :S:: .........,:S::
о с:::
,
о:::: ,.......
0,6 :r:
()О >< ,....... .L1
са О а.. .L1 CI)
ф с::: . --.J
'J" Q.. .......
m <о ........
CD с::: Q.. О ()
:S:: о
0,5 ...... :S::
ФО о:::: :S: С::: t::1)
C::: ,...... о:::: с:::
CD Qj I о:::: L
с:{ о:::: о:::: Q.)
() :S:: Q) о:: I ()
О с::: CD о- I g с::
('t) о 0,4 () с:{ <3
() о:::: ........., ф Q..
(') са а.. ()
м I: () .........,
() .о... .........,
Ф :r: Ф CI)
а..са (i) ........
Ф * 0,3 Q.. CI) CI)
'J"a.. с 1:) с::
........., Q)
:s:: ...... ::::>
() с: с::
с::: о а
8 () 0,2 ц)
.......
о:::: о () CI) (J) L
a.. L ft} а
2 t.:. С
о L1) .
СО 0,1 ц)
.Q .с::: Е
.... Е ()
о &
.......... о
Рис. 12.3. Проницаемость экзоскелета paKO
образных из различных местообитани й (по
Hoar, 1966)
рианты чаще встречаются в эстуариях, т. е.
в условиях пониженной солености.
Осмореrуляторы MorYT контролировать
приток воды и отток ионов двумя способа..
ми c помощью малопроницаемых покро"
вов и/или активноrо выкачивания воды
и реrуляции поступления ионов в жидкости
тела и их выведения. На рис. 12.3 показано.
что проницаемость экзоскелета ракообра..
зных ниже у видов, обитающих на литорали
или в слабосоленых водах.
Пресноводные животные находятся в та..
кой же ситуации.. что и осмореrуляторы при
пониженной солености, но им прихо..
дится реrулировать состав внутренней сре..
ды постоянно. На рис. 12.4 показано, как
жидкости тела таких животных реаrируют
на повышение солености. Обратите внима..
ние на Зt{ачительные различия в концентра..
ции этих жидкостей в пресной воде у дву"
створчатоrо моллюска Aпodoпta и рак о..
образноrо Daphпia она ниже, чем у бокопла..
ва Gaп1п1arus или клопа--rребляка Sigara, а
у этих животных она в свою очередь ниже,
чем у родственных им морских форм. Воз..
можно, это снижает затраты на поддержа..
ние большой разницы осмотических давле..
ний внешней и внутренней сред.
Большинство пресноводных животных
600
.L1
с:::
О
500
...
са
С:::
Ф
...... 400
:s::
()
о
300
ЭЕ
CD
о::::
3 200
са
о-
:r:
Ф
:::f
I 100
о
Pofaтoblus fluviatili
Sigara distiпcta
Gaттarus pulex
edes aegypti
Морская
вода
!
100 200 300 400 500 БОа
Концентрация во внешней среде, ммоль/л
Рис. 12.4. То же, что на рис. 12.2, но для прес
новодных животных (по Schmidt Nielsen 1979)
Экскреция, ионная и осмотическая реrуляция
367
реrулируют состав внутренней среды за
счет низкой проницаемости эпидермиса
(рис. 12.3) и выделения большоrо коw']иче..
ства мочи. Если ero выразить в процентах
к весу тела в сутки, эта величина будет [о..
раздо меньше 10% у большинства морских
беспозвоночных и HaMHoro больше у прес..
новодных. Например. для Astacus и Ga111тa..
ru\' она составляет около 400/0, для Dap/l"
l1iа более 2000/0, а для пресноводноrо дву"
створчатоrо моллюска Апоdопtа более
4000/0. Моча пресноводных беспозвоночных
обычно rипоосмотична жидкостям тела,
так как в соответствующих отделах «выде..
лительных» opraHoB (с. 375) из нее избира..
тельно извлекаются полезные ионы. Несмо"
тря на это, а также на низкую проницаемо"
сть эпидермиса, некоторые ионы все же те..
ряются, и их запас должен восполняться.
Некоторые из них поступают с пищей, но
мноrие пресноводные беспозвоночные спо..
собны и К прямому активному захвату ио..
нов из окружающей среды. В этом можно
убедиться, предварительно выдержав жи..
вотное в дистиллированной воде, чтобы
снизить концентрацию ионов в жидкостях
тела, а затем возвратив ero в обычную прес..
новодную воду. Очень скоро состав жидко..
стей тела возврашается к норме.. хотя в те..
чение Bcero эксперимента концентрация ио..
нов в среде ниже, чем в орrанизме. Следова..
тельно, они должны активно поrлощаться.
Эти процессы наиболее подробно изучены
у пресноводных ракообразных: речноrо ра..
ка Astacus, водноrо ослика Asellus и боко--
плава GaI11111al'us. Возможно, в активном
транспорте ионов какую"то роль иrрает вся
поверхность тела. но имеются убедитель..
ные данные, что он идет в основном через
жабры.
Если выращивать личинок комара Aedes
aegypti в дистиллированной воле.. уровень
натрия в их rемолимфе станет ниже 30%
нормы. При перенесении их в обычную
пресную воду концентрация натрия восста..
навливается за несколько часов. Экспери..
менты позволили установить.. что на 90%
поrлощение этоrо иона происходит через
анальные сосочки, а остальное в основ..
ном через кишечник. У личинок стрекоз
аналоrичную роль иrрают ректальные жа..
бры.
12.2.4. rиперосмотическая среда
Некоторые морские беспозвоночные, на..
пример креветка Pa/aeпl0пetes.. rипоосмо..
тичны морской воде (рис. 12.2). Обычно
считается, что такие животные происходят
от пресноводных форм, вернувшихся в MO
ре. Жидкости тела некоторых rрапсоидных
крабов также rипоосмотичны среде (см.
примечание к табл. 12.2), но, поскольку
возможность обитания их предков в прес
ных волах исключена. происхожление rи..
поосмотичности в этом случае неясно.
Основная проблема для орrанизма в rи
пер осмотических условиях состоит в том,
чтобы не про пускать внутрь соли и не выпу"
скать воду. Ее приходится решать и беспо..
звоночным, обитающим в более соленых
водах, чем морская. Ярким примером мо"
жет служить рачок Arte/11ia, живущий в co
леных озерах. Он поддерживает состйв
своей внутренней среды путем активной ре..
rуляции: «выпивает» большое количество
воды и выделяет избыток ионов через Жd..
бры во взрослом состоянии и через спеuиа
лизированный шейный opraH на личиноч..
ных стадиях развития.
12.2.5. Наземная среда
в принципе rлавная физиолоrическая
проблема для наземных животных
обезвоживание. Поверхность некоторых из
них.. например земляных червей. слизней
и улиток, влажная, а их распространение
оrраничено влажными местообитаниями
в почве и листовой подстилке, так что Ha
звать их истинно сухопутными нельзя. По..
сле дождя среда в почве может стать даже
rипоосмотической, и некоторые нематоды
в таких условиях активно удаляют воду из
тканей (возможно, через кишечник). Дру"
rую крайность представляют насекомые со
своим непроницаемым экзоскелетом, спо..
собные обитать в очень сухих условиях. Не..
проницаемость их покровов обусловлена
эпикутикулярным восковым слоем (рис.
12,5,А) (см. также разд. 8.5.3Б.2). Если этот
слой удалить, потери воды за счет испаре..
ния MHOrOKpd.THO возрастут. Значение эпи..
кутикулярноrо слоя для сохранения волы
показано также путем измерения скорости
368
rлава 12
А
Эпикути { Цемент
. кула Липид
Кути кули
Восковый
канал
3кзокутикула
Поровый
канал
Эн окутикула
70
60
50
-:i 40
с:{
о
m 30
:s:
g- 20
о
с: 10
24 26 28 30 32 34 36 38 40 42 44 46
Б Температура, ос
Рис. 12.5. А. Срез кутикулы HaceKOMoro с восковым эпикутикулярным слоем (по'Еdпеу, 1974). Б,
Потери воды через кутикулу таракана в зависимости от температуры воздуха (по Beament, 1958)
испарения при разных температурах. Она
резко увеличивается при достижении опре..
деленной температуры, соответствующей
точке плавления 1пикутикулярноrо воска
(рис. 12.5,Ь).
Как уже отмечалось (с. 362), у назем..
ных ракообразных BocKoBoro эпикутику
лярноrо слоя нет, и они обитают в OCHOB
ном во влажных условиях. Хотя МОКРИIIЫ
обычно встречаются в сырых местах, иноr
да им приходится переносить и воздействие
cyxoro воздуха. Неизбежные потери воды
при этом частично восполняются за счет
влажной пищи, которую потребляют эти
животные, разлаr d-ЮЩИХСЯ растительных
тканей; кроме Toro, некоторые их виды спо
собны пить воду И поrлощать ее через анус.
Пустынному paBHoHoroMY рачку Heпlilepi
stus избеrать высыхания позволяют пове
денческие приспособления: в жаркое ДHeB
ное время он прячется в норках. Их rлубина
достиrает 30 см. в них rораздо прохладнее..
чем на поверхности, а относительная влаж
ность может составлять 950/0 (подробнее
см. Edney, 1957).
Насекомые также способны пить воду,
получать ее из пищи или в результате мета..
болическоrо окисления орrанических моле
кул, например:
С<>Н ,2 О<> + 602 6C02 + 6Н20.
Более Toro, некоторые Нd-земные насекомые
и пауки поrлошают водяные пары непо..
средственно из атмосферы. Каким образом,
точно неизвестно; в различных rруппах
в этом процессе участвуют эпителии рото..
вой полости. прямой кишки и, возможно.
трахейная система.
12.2.6. «Завоевание» суши
и пресных вод
Никто не сомневался в том, что заро..
ждение жизни и адаптивная рd-диация opra..
низмов произошли в морской среде (rл. 2).
Завоевание друrих местообитаний шло как
прямо (из моря на сушу" из моря в пресные
воды), так и через промежуточные стадии
(из моря на сушу через пресные БОДЫ, из
моря В пресные воды через сушу). Кроме
Toro, выход из моря на сушу Mor происхо..
дить через литораль или интерстициаль.
Эти варианты схематически показаны на
рис. [2.6. Мы уже rоворили о несостоятель..
ности теории Макаллума (с. 363), но все же
разумно предполаrать, что предковые
образ жизни и место обитание отразились
на составе внутренней среды современных
Экскреция, ионная и осмотическая реrуляция
Суша
Эсту
арии
'!
Интер
стициаль
Море
Рис. 12.6. Пути «завоевания» суши ( ) и прес..
новодных местообитаний ( )
животных. Так, вполне вероятно, что OCMO
тическое давление жидкостей тела у назем
ных животных, происшедших непосред..
ственно от морских предков., выше., чем
у потомков пресноводных форм (с. 365).
Данные, приведенные в табл. 12.3, в основ..
НОМ подтверждают lТО предположение, но.,
369
как и следовало ожидать. не Bcerna. Напри..
мер, считается, что у сухопутноrо десятино..
roro рака Н olt/1uisaпa traпsversa пресновод"
ные предки, тем не менее осмотическое дав..
ление ero rемолимфы довольно высокое.
Хотя у мелких пресноводных ракообразных
осмотическое давление жидкостей тела низ..
ко « 300 мосмоль), пресноводные десяти..
ноrие раки сохранили более высокие ero
значения ( > 500 мосмоль), возможно.,
В связи с более крупными размерами. Та..
ким образом, высокое осмотическое давле..
ние у наземных представителей этой rруппы
не противоречит их происхождению от
пресноводных форм.
Животные, вышедшие на сушу через ин.
терстициаль., например некоторые немерти-
ны, нематоды и аннелиды, вероятно, пере-
жили в своей эволюционной истории уело..
вия, промежуточные между пресноводными
и наземными. Донные отложения забуфери..
вают соленость окружающей воды, однако
по мере продвижения из моря на сушу соле..
ность интерстициальной воды снижается,
так что почвенные животные., следовавшие
этим путем, вероятно, должны были выра..
ботать эффективные механизмы ионной ре..
rуляции. Поэтому вполне возможно, что
Таблица 12.3. Осмотическое давление крови некоторых наземных животных (Little, 1983, с из
менениями) 1)
ВИД
Таксономическое положе
ние
Осмотическое
давление, мос"
моль
Возможный путь заселения
суши
Немертины
Agroпeтertes deпdyi Hoplonemertea 145 Морская литораль
Аннелиды
Lumbricus terrestris Oligochaeta 165 Пресные воды
Моллюски
EutrochatelZa taпkervillei Prosobranchia 67 ..
Poteria liпeata " 74 "
PseudocycZotus laetus " 103 Солоноватые воды
He/ix poтatia Pulmonata 183 Соленые болота
Pomatias elegaпs Proso branchia 254 Морская литораль
AgrioZiтax reticuZatus Pulmonata 345 Соленые болота
Ракообразные
Talitrus sp. Amphipoda 400 Морская литораль
Holthuisaпa traпsversa Decapoda 517 Пресные воды
PorceZZio scaber Isopoda 700 Морская литораль
Cardisoтa arтatuт Decapoda 744 "
Coeпobita breviтaпus " 800 "
1) Приведены средние величины для активных животных во влажных условиях или в равновесии с пресной во..
дой.
370
rлава 12
концентрация жидкостей их тела низка, как
и у форм с пресноводными предками. При..
мером служит сухопутная немертина AI'go..
l1en1ertes c/eпd)'i. которая. как считается. про..
никла на сушу именно через интерстициаль
и характеризуется низким осмотическим
давлением внутренней среды (табл 12.3).
Даже из весьма оrраниченноrо набора
данных, приведенных в табл. 12.3, можно
сделать вывод, что большинство беспозво..
ночных вышли Hd сушу прямо из моря.
Хотя две важнейшие наземные rруппы
насекомые и паукообразные в этой та..
блице не отражены, они, судя по всему, так..
же происходят непосредственно от морских
предков. В отличие от беспозвоночных про..
исхождение наземных позвоночных исклю..
чительно пресноводное. Их покровы, кйк
И У наземных членистоноrих, особенно насе..
комых (разд. 8.5.3Б.2), эффективно проти"
востоят испарению. Мяrкотелые беспозво..
ночные избеrают высыхания, выбирая для
обитания влажные условия, однако иноrда
им, по"видимому. приходится переносить
частичные потери воды и повышение вну"
TpeHHero осмотическоrо давления. Живот..
ные с разбавленными жидкостями тела
(про исходящие от пресноводных предков)
обычно не способны выдержать такое повы..
шение; возможно, белки, ЭВОЛIОЦИОНИРО"
вавшие в разбавленных растворах, леI'че де..
натурируются в концентрированных.
Поэтому МЯI'котелые беспозвоночные
с пресноводным происхождением хуже при..
способлены к жизни на суше, чем непосред
ственные потомки морских форм.
Завоевав сушу, некоторые беспозвоноч"
ные (ряд немертин, мноrоножек, насеко..
мых, брюхоноrих моллюсков и Т. д.) вт о..
рично переселились в пресную воду. Насе--
комые в этом особенно преуспели: из при..
мерно 1 млн. их описанных современных
видов 25 000 35 000 про водят в пресной во..
де хотя бы одну стадию жизнеННОI'О цикла.
Так как кутикула взрослых особей непрони"
цаема, осмотические проблемы стоят для
них не так остро, как для личинок, покровы
которых rидрофильны и слабо хитинизиро"
ваны. Поэтому у них возникли сложные ме..
ханизмы реrуляции, в том числе ионные на..
сосы, локализованные в специальных струк--
турах (см. ниже). Эти личинки, как и мяrко"
телые беспозвоночные. вторично приспосо..
бившиеся к пресноводным условиям, испы..
тывают, по"видимому, такое же даВ,,'1ение
осмотических условий, Чl U И ) (рорм. пере..
шедших в пресные водоемы непосредс I'BeH
но из моря. В обоих случаях можно ожи..
дать снижения концентраций внутренней
среды OpI анизма
12.3. Выделительные системы
Спеuиализированные «выделительные
системы» встречаются во всех типах беспо
звоночных, за исключением стрекакяцих
и иrлокожих. У r-убок их тоже нет. но у прес
новодных видов имеются сократительные
вакуоли. «Выделительные системы» присут"
ствуют и У аммониотелических ЖИВОТНЫХ,
т. е. не всеr'ла участвуют в выведении азота.
Их основная функция почти всеrда заК.,1Ю"
чается в ионной и осмотической реrуляции
и лишь иноrда в истинной экскреции; то же
можно сказать и о сократительных BaKYO
лях rубок, поэтому сам термин «выделите..
льные системы» взят нами в кавычки.
В этом разделе мы сначала рассмотрим их
Сl роение. а затем (РУНКIlИЮ.
12.3.1. Строение
Исходя из особенностей развития. МО"
жно выделить две основные катеrории вы-
делительных систем: нефридии трубочки
эктодермальноrо происхождения, врастаю
щие внутрь тела, и целомодукты
мезодермальные протоки, развивающиеся
от внутренних opraHoB к поверхности.
Нефрuдuu бывают двух основных типов.
(1) Протонефридии это замкнутые систе..
мы трубочек, заканчивающиеся клетками
с мерцательным пламенем у плоских червей
(разд. 3.6.2) и немертин, луковицами (буль..
бусами) с мерцательным пламенем у коло..
враток и соленоцитами у PI"iapu/zl\\ некото"
рых rастротрихов, полихет и архианнелид
(рис. 12.7). (11) Метанефридии, оканчиваю-
щиеся ресничными воронками, характерны
для олиrохет (рис. 12.8) и, возможно, HeKO
торых друrих rрупп.
Ц е ЛО.l'ИодУКnlЫ это rрубча тые, иноrда
ресничные выделительные структуры, кото-
рые до Toro, как было изучено их эмбриона-
Экскреция, ионная и осмотическая реrуляция
1
А
Ядро
Жrутик
жrутик
/
I
Б
в
Рис. 12,7. А, Луковица с мерцательным пла
менем, замыкающая протонефридии коловра
ток: I продольный срез; 11 поперечный
срез. Б, В. Соленоциты приапулид и брюхорес
ничных соответственно (по Barrington, 1979)
льное развитие, часто не отличали от He
фридиев. Они встречаются у онихофор.. чле..
нистоноrих и моллюсков. У Peripatus по два
целомодукта (называемых коксальными
железами) почти в каждом cerMeHTe (рис.
12.9).. но у животных друrих rрупп их обыч--
но меньше. У ракообразных они открыв а..
ются в основании антенн (антеннальные же..
371
Рис. 12.8. Схема метанефридия аннелиды (см
также рис. 12.15)
лезы; рис. 12.9) и/или вторых максилл (MaK
силлярные железы; рис. 12.9) у некоторых
одно ветвистых они также связаны с макси л..
лами.. а у пауков протоки пары целомодук--
тов ведут на поверхность шестоrо cerMeHTa
(КОКСdльные железы., рис. 12.9). У моллю
сков «выделительные» целомодукты (<<поч..
ки») произошли из общеrо lачатка с серд--
цем и rонадами., но в ходе эволюции четко
обос06ились от этих opraHoB. как показано
на рис. 12.1 о. (Следует отметить., что., хотя
по определению эти целомодукты являются
«целомическими»., [. е. их полость окружена
мембраной мезодеРМdльноrо происхожде--
ния., они не связаны с целомом. На самом
деле животные с выделительными uеломо"
дуктами лишены целомической полости., а
у целомических беспозвоночных «uеломо"
дукты», связывающие целом с внешней сре..
дой, не используются для экскреции!)
В состав некоторых выделительных си
стем., Нdзываемых миксонефридиями или
нефромиксиями (рис. 12.11), входят как
эктодермальные, так и мезодермальные
компоненты. Они особенно распростране..
мж rп
Рис. 12.9. Выделительные и половые целомодукты онихофор и некоторых членистоноrих (по
Goodrich, 1945): аж антеннальная железа; кж коксальная железа; rп rонопор; мж
максиллярная железа
372
rлава 12
rонада
Перикардиальная
полость (П)
Сердце (С)
Почечно перикардиаль
ный канал
(Пч)
Кишка (К)
ПРИМИТИВНДЯ
<РОРМА
Мочеполовое (например, неоменио
отвер'стие орс:ры)
rOHonop (r) Мочевы r м
водящее
отверстие (м) ДВУСТВОРЧД ТЫЕ
)ДНОПРЕДСЕРДНЫЕ
БРюхоноrИЕ
м r
rоловоноrИЕ
ны У полихет, но «нефридии» форонид, си
пункулид, эхиурид И плеченоrих, по..
видимому, также относятся к этому типу.
«Выделительные системы» нематод
и крылатых насекомых совершенно иные.
у нематод это всеrда одноклеточная вен..
тральная железа, расположенная в псевдо"
целе, с терминальной ампулой, открываю..
rцейся порой на вентральной поверхности
тела. Она может быть связана с- каналами,
образуюrцими так называемую
Н"систему (рис. 12.12). Ее боковые каналы
внутриклеточные, а вся система одно ядер"
ная. У насекомых орrанами выделения слу..
жат мальпиrиевы сосуды, слепые трубочки
(от одной до мноrих), открывающиеся в пи..
rцеварительный тракт на rранице средней
и задней кишки (рис. 12.13). Сходные тру..
бочки, возможно, также с выделительными
функциями присутствуют у мноrоножек" не..
которых паукообразных и тихоходок, но
возникли они, по крайней мере у пауков
и тихоходок, независимо, так что представ--
Рис. 12.10. Реп родуктивные
и выделительные opraHbI мол
люсков (по Goodrich, 1945)
ляют собой результат конверrентной ЭБО"
люции. Наконец, erцe одной интересной БЫ"
делительной структурой является так назы"
ваемый клубочек кишечнодышаших полу..
(;)
r GJ
Q
Рис. 1 2.11 . Схема строения нефромиксия
(== миксонефридия): М мезодермальный
(== целомодуктовый) компонент; Э экто
дермальный (== нефридиальный) компонент;
r raMeTbI
3
А
Б
в
r
д
Е
ж
Рис. 12.12. Выделительные системы нематод. А и Б системы Н типа с двумя вентральными же
лезистыми клетками; В асимметричная система, состоящая из одной ветви Н системы; r
Н система без железистых клеток; Д и Е укороченные Н системы; Ж укороченная и редуци..
рованная Н система; З система из единственной вентральной железистой клетки (по Lee, Atkin
50П, 1976)
' .
I .
I .
I :
.......,. ,
. J
/
/
/
/
/
/
/
/
/
/
/
/
/
./
..... ........"
-+- .....
Рис. 12.1 з. Мальпиrиевы сосуды (М) и ректальная железа (РЖ) насекомых: движение пи
щи; ---+ движение воды и некоторых ионов (мальпиrиевы сосуды и ректальная железа распо
ложены так, что вода и ионы MorYT циркулировать по Kpyry); ... > мочевая кислота (по Potts, Parry,
1964). Показана также ультраструктура клетки мальпиrиевоrо сосуда (по Oschman, Berridge,
1971). Заметно большое количество митохондрий, что предполаrает участие этой клетки в актив
ном транспорте
374
rлава 12
хордовых. Он образован выпячиванием
перитонеума в пелом хоботка.
12.3.2. ФУНКЦИЯ
и так. существует множество типов раз--
личных по 1мбриональному происхожде..
нию «выде,,'1ительных систем». Несмотря на
такое разнообразие, принцип их действия
сходен. поскольку в образовании выделяе..
мой жидкости участвуют лишь два процес--
са.
1. Ультрафильтрация: жидкость под
давлением проходит через полупроницае..
мую мембрану, задерживающую белки
и друrие макромолекулы, но про пускаю..
щую воду и низкомолекулярные растворен--
ные вещества.
2. Активный транспорт: движение ве..
ществ против rрадиента их концентрации
с затратой энерrии. Если оно направлено
в выделяемую жидкость, процесс называют
активной секрецией, если в противополо--
жную СТОРОН), активной реабсорбцией.
12.3.2.1. Системы, участвующие
в ультра фильтрации
Считается, что давление, Нtобходимое
для ультрафильтрации, создается биением
200
..l)
о
L
L
... 100
Q
s
о...
о
х
о
км Л
м
к
Рис. 12.14. Антеннальная (зеленая) железа pa
ка и изменение содержания хлоридов в моче
по мере ее прохождения по ЭТОМУ opraHY:
КМ концевой мешочек; Л лабиринт; K
почечный каналец; М мочевой пузырь (по
Potts, Parry, 1964)
жrутиков или ресничек (мерцательноrо пла
мени) в протонефридиях и «давлением KpO
ви» в целомодуктах, например на уровне
KOHueBoro мешочка антеннальной (зеленой)
железы ракообразных (рис. 12.14) или
в сердце моллюсков. Из lТИХ структур уль
трафильтрат поступает в канальцы, а
у моллюсков через перикардиальную по..
лость в почки. Все низкомолекулярные KOr-.1
поненты жидкостей тела переходят в уль..
трафильтрат в концентрациях, пропорцио
нальных их содержанию в этих жидкостях.
Физиолоrически важные молекулы, в част
ности rЛIокоза, а у пресноводных 6еспозво
ночных также ионы Na +, К +, CI и Са 2+
В канальцах реабсорбируются (см., напри..
мер, рис. 12.14), а токсичные иненужные
вещества остаются в выделяемой жидкости.
Как уже отмечалось, этот избирательный
процесс связан с активным транспортом.
12.3.2.2. Системы, не участвующие
в ультрафильтрации
Фильтрация с помощью ресничек
Движение целомической жидкости
в «выделительные» каналы метанефридиев
и нефРОlV[иксиев обеспечивается биением
ресничек. По мере прохождения через эти
каналы состав жидкости изменяется за счет
активноrо транспорта (рис. 12.15).
'Этим путем MorYl выводиться и экскре
ты. Например, у земляных червей в выдели
тельных процессах участвует хлораl"оrенная
ткань эпителиальноrо происхождения, ле-
жащая в целоме на поверхности средней ки
шки. Хлораrоrенные клетки запасают жиры
и уrлеводыI' но, кроме Toro, в них происхо
дит дезаминирование, и образующиеся aM
миак и мочевина поступают в целом, а OT
туда в воронки нефридиев. Остатки, обра
1ующиеся при распаде хлораrоrенных кле
ток, также MorYT удаляться через нефридиа
льную систему. У пиявок функцию хлораrо
rенной ткани выполняет ботриоидальная
ткань. У некоторых rрупп, например в роде
T/1eroтyzoп, экскреты из нее удаляются че
рез открыIуюю систему нефридиев, однако
в друrих случаях, в частности у Glossiphoпia
и Hirudo, воронки метанефридиев отделены
от выводных протоков переrородкой.
В этих случаях биение ресничек направлено
Экскреция, ионная и осмотическая реrуляция
УЗКИЙ ШИрОКИЙ Мочевой
каналец каналец пузырь
200 I
I
о:: I
Ш 150 l
CL I
.....
I 100
g} . ..
I
О 50
о
Рис. 12.15. Осмотическое давление в различ
ных отделах метанефридия земляноrо червя
(по Potts, Рапу, 1964)
из воронок BHY111Pb цслома и служит для рас..
селения образующихся в них амебоцитов.
Протоки открываются наружу и, возможно,
служат для ионной и осмоrической реrуля..
ции.
МальпU2uевы сосуды насекомых
Прямое поступление O в ткани насеко..
мых через трахеи снизило роль находя..
щейся под давлением циркуляторной систе..
МЫ. Поэтому давление жидкости внутри
и снаружи маЛЬПИI ИСВЫХ сосудов примерно
одинаково что ИСКЛl0чает возможность
ультрафильтрации. BMecro этоrо ионы ка..
лия и" возможно, также друrие растворен..
ные вещества. активно секретирую rся
в просвет сосуда, d вода поступает туда за
счет осмоса. Секретируемые ураты (СО.,JИ
мочевой кислоты) поддерживаются в рас..
творенном состоянии блаI одаря высокому
значению рН в диста.'1ЬНОЙ части сосудов.
Из них жидкость поступаеI в заднюю ки"
шку, [де происходит реабсорбция воды
(особенно интенсивно ректальными железа
ми; см. рис. 8.27), значение рН снижается
а ураты осаждаются в виде мочевой кисло..
ты, которая выводится с фекалиями (см,
рис. 12.13).
12.3.2.3. Выделительная система
нематод
о принципах .LI:ействия этой системы
известно мало, однако функции каналов
и железистых клеток почти наверняка раз..
личны Первые, по видимому, участвуют
375
в ионной и осмотической реrуляции. хотя
и лишены ресничек для создания тока жил..
кости (обычно считается, что в процессе
участвуют ультра фильтрация и активный
транспорт). Вероятно, эта система не иrрает
сушественной роли в выведении азота. У не..
которых нематод ее секреты, по видимом},
обладаIОТ ферментативной активностью их
источником скорее Bcero является желези..
стая клетка.
12.4. ВЫВОДЫ
Теперь читателю должно быть понятно.
что у беспозвоночных не только rрудно" но
и бесполезно рассматривать экскрецию
(удаление азотистых отходов) отдельно от
ионной и осмотической реrуляuии и даже от
плавучести. Образование. транспорт и уда..
ление азотистых отходов на KaKOM TO этапе
обязательно сопряжены с током и )ильтра
цией жидкостей тела_ даже если экскреты
в конечном итоrе оказываются твердыми"
как, например, мочевая кислота. Поэrому
выделение тесно связано с движением жид
костей и ионов. а значит с проuессами ос..
моса, диффузии, активноrо транспорта
и ультрафильтрации. «Выделительные си
стемы» всеrда участвуют в ионной и осмо"
тической реrуляuии но не Bcerna
собственно в lкскреции. Несмотря на раз..
нообразие строения. их функционирование
основано на сходных процессах" и именно
на них сделан акиент в этой [лаве. И нако"
нец, замена ионов на ионы друr"ой массы
или жидкости на rаз влияет на п 1]0 IHOCTb
ткани и, как теперь должно быть понятно,
является важным фактором. контролирую..
щим плавучесть водных беспозвоночных.
12.5. Дополнительная
литература
B{l1Til1gtOI1 Е. J. W. 1979. 111\/errebrare Structllre al1d
Fllllctioll. 2nd еdп. Nеlsоп_ LопdОI1.
BealJ1ent J. W. L. 1958. The cffect of tcmperatllrc оп
the wаtсrрt'ооfiпg ll1echanis111 of аl1 Iпsесt. J. ехр.
Biol.. 35. 494 519,
Del1tol1 Ь . J.. Gilpiп B"OH'1l J. В. 1961. The distribu
tion оС gas alld liqllid within the cuttlebol1e. J. mar.
biol. Assoc., U К.. 41, 365 381.
Deп!on Е. J. Gilpiп BI'OH'11 J. В. 1966. Оп the
376
rлава 12
ыlуапсуy of the pearly Nautilиs. J. mar. biol. As..
soc., 46, 723 759.
Edпey Е. В. 1957. ТЬе Water Re1ations ofTerrestrial
Arthropods. Cambridge University Press. Сат..
bridge.
Edпey Е. В. 1974. Desert artropods. In: G. W. Brown
(Ed.) Desert Biology, (Vol. 2). Academic Press,
New У ork
Ноа, W. S. 1966. General and Comparative Physio
10gy. Prentice НаН, New Jersey.
НОl'пе } . R. 1971. Accumll1ation of urea Ьу а pulmo
nate snail during aestivation. Сотр. Biochem.
Physiol., 38А. 565 570.
Koe/lп R. К. 1983. Biochemica1 genetics and adapta
tions in molluscs. Iп: Р. W. Hochachka (Ed.) The
Mollusca (Уоl. 2), рр. 305 330. Academic Press,
New У ork.
Kl'Og/l А. 1939. Osmotic Regu1ation in Aqllatic Ani
ma1s Cambridge Univesity Press, Cambridge.
Lee D. L., Atkiпsoп Н. J. 1976. Physiology of Nema
todes, 2nd edn. Macmillan, London.
Litt/e с. 1983. ТЬе Colonisation of Land. Cambridge
Universlty Press. СаmЬпdgе.
Need/1aт J. 1931. Chemical Embryo10gy (3 vols).
Cambridge University Press, Cambridge.
Pilgriт R. L. с. 1954. Waste of carbon and energy in
nitrogel1 excretlOn. Nature (Londol1), 173, 491
492.
Potts W. F. W., Pal'ry G. 1964. Osmotic and lonic
Regulation in Al1ima1s. Oxford Ul1iversity Press,
London.
Schlпidr..Nielseп К. 1972. How Animals Work Сат..
bridge University Press, Cambridge.
SC/1п1idt..Nielseп К. 1979. Anima1 Physiology: Adap..
tation and Envirol1ment. Cambridge Uпivеrsitу
Press, Сат bridge.
Имеется перевод: Шмидт Ниельсен К. Физио..
лоrия животных: приспособление и среда: в
2 T. М.: Мир, 1982.
Приложение 12.1.
Некоторые определения
Если растворы разделены барьером
(мембраной), проницаемой как для раство"
рителя, так и для pacTBopeHHoro вещества,
то происходит следующее.
1. Растворенное вещество (ионы) пере..
ходит из более в менее концентрированный
раствор путем диффузии (движение ионов
подчиняется более сложным закономерно..
<;тям. если между разделенными мембраной
растворами существует разность электриче..
ских потенциалов; это более подробно обсу..
ждается в rл. 16).
2. Растворитель переходит из менее
в более концентрированный раствор Зd счет
OCJwoca; силу, обеспечивающую этот пере..
ход, называют осмотическим давление.м.
Оно зависит от числа молекул растворенно"
ro вешества.. но не от их природы
Процессы ] и 2 продолжаются до тех
пор, пока rрадиент концентраций не мече..
знет.
Мембраны живых орrанизмов обычно
лишь частично проницаемы для БОДЫ и рас..
творенных веществ, как показано на рису н..
ке, rде А ион; ......J> результирующее
движение ионов; движение воды.
r II
А А А А А А
+-
А А А А
.....
А А А А А А
Орrанизм (О) Внешняя О 8С
среда (ВС)
Ш
А А А А А
....
А ААА
А А ААА
О ВС
в случае 1 орrанизм изоосмотичен ОКРУ"
жающей среде;
в случае 11 он 2ипероемотичен ей, а
в случае 111 2ипоосмотичен по отноше..
нию к среде.
Термины изотонический и изооемотиче
ский (и по аналоrии остальные) часто иепо..
льзуют как синонимы, но это не вполне точ
но, так как тоничность описывает реакцию
объема клетки (орrанизма); если орrанизм
изоосмотичен внешней среде, он обычно
хотя И не всеrда, ей изотоничен.,Так, напри..
мер, яйца MopcKoro ежа изотоничны изоос..
мотическому раствору NaCl, но не изоосмо-
тическому раствору СаС1 2 (более детальное
объяснение см. Potts, Parry, 1964).
Экскреция, ионная и осмотическая реrуляция
377
Осмоконформерами называют живот
ных, у которых осмотическая -концентрация
жидкостей тела изменяется в зависимости
от среды, т. е. остается ей изоосмотич
ной.
Осмореzуляторами называют живот
ных, поддерживающих осмотическую KOH
центрацию внутренней среды отличной от
осмотической концентрации внешней cpe
ды.
Ионная реzуляция это поддержание
концентрации ионов в жидкостях тела на
уровне, существенно отличающемся от их
концентрации во внешней среде.
эвриzйлинныJvlиu называют животных,
жизнеспособных в широком диапазоне KOH
центрации солей в окружающей воде.
Стеноzалuнные животные MorYT перено
сить лишь оrраниченные колебания солено
сти воды.
Концентрации. Леrко понять, что такое
вес вешества. но число молей это более
сложное понятие. Моль {rpaMM
моль) это молекулярная масса, выражен
ная в rpaMMax, или масса вещества, в кото..
ром содержится число Авоrадро
(6,023 х 1023) ero молекул. Молярные кон..
центрации удобны, поскольку отражают
число молекул pacTBopeHHoro вещества,
а именно от Hero зависит осмотическое дaB
ление.
МОЛЯРl-lость число молей paCTBopeH
Horo вещества в литре раствора.
Моляльность число молей paCTBopeH
Horo вещества на килоrрамм чистоrо pac
творителя.
rЛАВА 1 3
ЗАЩИТА
в 1ТОЙ rлаве сначала дается класси( ика..
ция опасностей, которым подверrаlОТСЯ
беспозвоночные, а затем рассматриваются
способы защиты от них. Разнообразие воз
можных опасностей определяет широкий
крут затраrиваемых вопросов: от заЩИIЫ
против патоrенов до защиты от хищников
и даже от процессов старения, если, конеч..
но, такая защита возможна.
13.1. Классификация
опасностей
13.1 .1. Две основные rруппы
На рис. 13.1 представлены примеры кри--
вых выживаемости, т. е. численности (или
доли) особей, родившихся приблизительно
в одно и то же время (относяшихся к одной
KoropTe) и доживших до Toro или иноrо воз..
раста. Можно видеть, что в природных по..
пуляциях смертность либо наиболее высока
у молоди либо распределена довольно рав--
номерно по всем возрастным rруппам. rи
бель здесь, вероятно, вызывается rлавным
образом 1колоrическими, т. е. внешними по
отношению к орrанизму ( акторами типа
хищничества, несчастных случаев или бо..
лезней. Молодь обычно более уязвима
к ним. чем взрослые особи, но иноrда чув--
Сl вительность всех возрастных ['рупп сход--
на. В лабораторных культурах можно ис--
ключить большинство внешних факторов
смертности, и она наиболее высока у особей
самых старших возрастных rрупп. Это сви
детельствует об увеличении уязвимости ор..
rанизма с возрастом, вызванным, вероятно,
внутренними факторами, т. е. процессами
старения.
13.1.2. 3колоrические (внешние)
факторы смертности
Хотя существуеJ" множество разнооб..
разных 1колоrических причин смертности.
они MOI'YT быть объединены в четыре основ..
ные rруппы: несчастные случаи, болезни,
хищничество и CTpeccы вызываемые ОКРУ"
жающей средой. Первые три очевидны и не
нуждаю rся в специальном пояснении.
а с [рессы возникают либо при о [су [СТВИИ
какоrо--нибудь необходимоrо для орrаниз"
ма фактора, либо при наличии стрессора
природноrо токсина или аНТРОПОI'енноrо
заrрязнителя.
13.1 .3. Старение
С'тарение на первый взrляд идет при ие..
КЛlочении внеII1НИХ факторов Сl\лертноети
и может поэтому рассматриваться как про
иесс внутренней деrенераuии физиолоrиче..
ских систем, клеток и молекул. который, ве..
роятно, прослеживаеrся вплоть до денату"
рации биолоrически важных веществ (ну..
клеиновых кислот и белков) под влиянием
тепловоrо шума, ковалентной сшивки боко
вых цепей макромолекул, самоокисления
и т. д. Не следует, однако, полаrать, что на
эти внутренние пропессы не MorYT влиять
внешние q)акторы. На рис. 13.2 показано,
что облучение Bcero тела снижает продол..
жительность жизни дрозофилы, причем
э(рфект зависит от дозы радиации. Однако
форма кривых выживаемости при lТОМ не
меняется, и некоторые rеронтолоrи евязы..
вают укорочение жизни в данном случае
с ускорением старения возможно объяс..
няющимся в свою очередь повреждением
Рис. 13.1. А. Кривые выживае
мости полевых популяций бес
позвоночных; х' отклонение
(%) от средней продолжитель
насти жизни (по Ito, 1980). Б.
Кривая выживаемости и рас..
пределение по возрастам абсо
лютноrо числа поrибших oco
бей и их доли от общей числен
насти для лабораторной попу
ЛЯЦИИ Drosophi/a тe/aпogaster
(по Lamb, 1977); кривая выжи
ваемости лабораторной попу
ляции перепончатокрылоrо Ha
ceKOMoro Nasoпia vitripeппis
(по Davies, 1983)
За щита
А. Полевые популяции
о
90
99
99,9
*
....
.D
()
о
I
.....
cl.
Ф
L
()
о::
99.99
cl.
ro
L 99.999
L
>-
()
100
О
90
99
Б. ЛаБораторные ПОПУЛЯЦИИ
1000 .
S 900 Drosophlla
800 тelaпogaster
700
(l) 600
S 500
400
S 300
200
..а
(() 1 00
О
200
..а 150
100
50
О
100 l d
SL
с:: rcs 0.75
ф........
w () 0.50
O.2 = .,.,., " ,
а о 8 16 24 32 40 48 56 64 72 80
с:{ с Возраст, дни
Dlaptoтus
clavipes
50 О 50 100
Cerasfoderтa
edule
о 50 1 00 1 50
Пресноводные
пиявки
1 00 50 О 50
х
макромолекул. особенно ДHK высокоэнер..
rетическим облучением. С друrой стороны,
добавки некоторых веществ, Нdпример ви"
тамина Е, в культураЛЬНУIО среду нематод
или в пищу дрозофил удлиняют продолжи--
тельность их жизни. Эти вещества, вероят--
но, защищают макромолекулы от повре--
ждений (так, витамин Е, по--видимому, ОТ..
носится К антиоксидантам).
1 3.1 .4. Классификация
Как показано выше. смертность opra..
низмов всеrда в той или иной степени зави..
сит от условий окружаlощей среды, однако
некоторые ее причины более тесно связаны
с самим орrанизмом. чем друrие. В табл.
13.1 ее факторы классифицируются в со от..
ветствии со степенью такой связи: от напа..
дения хищников, с одной стороны, до про..
100
х
S
:3
т
S
11 50
т
.......
I
Ф
:::f
О
О
О
200
379
Nasoпia
vitripeппis
10 20
Возраст, дни
30
иессов старения. с друrой. Простота исклю--
чения Toro или ИНО1'О фактора в условиях
'3ксперимента уменьшается в обратном по..
рядке.
100
о:: 80
s
I
rcs
т 60
s
:а
т 40
.......
I
О 20
о-
с
О
О 10 20 30 40 50 60 70 80
Возраст, дни
Рис. 13.2. Влияние тотальноrо rамма об
лучения на самцов Drosophila тe/aпogaster (по
Lamb, 1977)
380
rлава 1 3
Таблица 13.1. Классификация факторов смертности
Фактор
Болезнь
Внутренний
стрессор (напри
мер, системные
нарушения)
Несчастный
случай
Хищниче
ство
Внешний
стрессор (Ha
пример, заrря
знитель)
Зависимость от поrиб..
шеrо животноrо
Леrкость исключения
данноrо фактора
в эксперименте
Реакция орrанизма
х о
х
хх
хх
ххх
ХХХ 1)
ххх
Защита
1) x низкая; ХХХ высокая.
ХХ
ХХ
х
Иммунная
защита
Устойчивость,
репарация
Репарация
1 3.2. Защита
13.2.1. Против хищников
Все животные MorYT быть съедены дpy
rими животными (см. rл. 9) и, чтобы избе
жать 3Toro, защищают себя разными спосо..
бами, объединяемыми в три основные rруп..
пы: избеrание потенциальных хищников,
пассивная защита и активный отпор.
13.2.1.1. Избеrа.ние
Этот способ включает либо избеrание
встреч с хищниками, либо незаметность для
них. Примером использования обеих CTpa
теrий служит массовая вертикальная миrра..
ция некоторых морских и пресноводных
планктонных животных. Хотя ее характер
обычно сложен, он часто включает уход
вниз от солнечноrо света в дневное время
и всплывание ночью. Тем самым животные
снижают вероятность быть замеченными
хищниками, руководствующимися при oxo
те зрением (рис. 13.3, А). Такое поведение
особенно выражено у наиболее уязвимых
для хищничества видов и возрастных клас
сов. Более Toro, в результате уникальноrо
исследования одноrо из видов веслоноrих
в нескольких изолированных [орных озерах
показано (Gliwicz, 1986), что вертикальные
миrрации этоrо рачка происходят только
там, [де обитают планктоноядные рыбы.
Удалось также пронаблюдать эту миrра
цию в одном из озер на разных стадиях ero
заселения такими рыбами. Через 12 лет по
сле первоrо их появления вертикальная ми..
rрация была выражена слабо, но еще через
23 rода четко прослеживалось поrружение
рачков в дневное время. Однако хищниче..
ство не служит единственным и исчерпы..
вающим объяснением эволюционноrо воз..
никновения вертикальных миrраций, ибо
некоторые беспозвоночные опускаются в их
процессе на rлубину, HaMHoro превышаю..
щую необходимую для Toro. чтобы скры"
ться от света. Кроме Toro, некоторые ми..
rрирующие зоопланктеры люминесцируют
по ночам, увеличивая этим свою ВИДИ..
мость. Друrие возможные объяснения вер..
тикальных миrраций включают оптимиза..
цию использования неравномерно распре..
деленной пищи (rл. 9), экономию энерrии
и связь с rоризонтальной миrраuией.
Сходные вертикальные миrрации, но
в меньшем масштабе, наблюдаются у прес-
но водных беспозвоночных, обитающих на
поrруженных камнях. Днем животные нахо..
дятся под ними В неактивном состоянии,
а ночью обычно перемещаются на поверх
ность валунов, [де ведут активный образ
жизни. Большое количество этих, как пр а..
вило, бентосных орrанизмов можно обна..
ружить пассивно плывущими в потоке во..
ды. Этот так называемый дрейф беспозво..
ночных особенно выражен в сумерках (рис.
13.3, Б), вероятно, потому, что именно в это
время они выползают на омываемую тече
нием верхнюю часть камней, откуда леrко
MorYT быть смыты.
Особые «реакции избеrания» представ
ляют собой крайнюю форму локомоторно"
Защита
ro ускользания от опасности. При них ис..
пользуются либо обычные двиrательные
стереотипы.. либо специализированные по..
веденческие ответы. Так, у каракатицы Se..
pia чернильный мешок содержит жидкость
с rранулами меланина. Будучи атакован,
моллюск выталкивает ее наружу't создавая
«чернильную завесу», тут же изменяет свой
цвет на очень бледный и уплывает под
прямым уrлом к предшествовавшему на..
правлению движения. Двустворчатые мол..
люски, например обычная прикрепленная
форма Cerasloderтa, MorYT избеrать хищни"
ка за счет внезапных и быстрых сокращений
мышц ноrи и аддукторов раковины. ИСКЛIО"
чительно мощную реакцию избеrания мол..
люсков этоrо рода вызывает прикосновение
амбулакральных ножек морской звезды; ве..
роятно.. она индуцируется какими"то веще..
ствами, выпускаемыми иrлокожим в воду.
Криптическая (маскирующая, или покро"
вительственная) окраска еще один шира..
381
ко распространенный метод избеrания хищ..
ников у беспозвоночных (см. рис. 9.33).
Примеры ее можно.. по"видимому.. обнару..
жить во всех типах животных, но наиболее
детально этот феномен изучен на некото"
рых улитках и насекомых.
1. Исчерченность раковины у Сераеа пе..
тoralis. У этоrо вида наземных улиток ши..
роко варьируют цвет и узор раковины за
счет изменения ее фоновоrо оттенка, а так..
же числа, ширины и яркости окраски полос.
Все эти варианты находятся под rенетиче..
ским контролем. Дрозды питаются особя..
ми Сераеа, находя раковины с помощью
зрения и разбивая их о камни., так называе..
мые дроздовые наковальни. В середине
1950..х rr. Кейн и Шеппард (А. J. Cain,
Р. М. Sheppard) показали, что окраска рако..
вин.. разбитых дроздами., в среднем лучше
заметна, чем у живых улиток, обитающих
в том же районе. Различные узоры по..
разному заметны в зависимости от место..
1: О 1600
{ 1 r Q
с
А
1900 0100 0700 1300
2200 0400 1000 1600 ч
VV 9QQ09
Ночь День
О
1:
... 50 lf
т
I
::s
LQ
:>о-
С
100
Б
2,0 0,2
Рис. 13.3. Вертикальные ми 1..;... 1 О 0,1
::s ,
rрации (А) веслоноrоrо рачка ::s
::1
Саюпus и (5) более rлубоко со О О
а.
L В
водной креветки ..акантефири.. ::s
ДЫ (по Barnes, Hughes, 1982). ..lI
Влияние света и темноты на () 0,04
8.. 0,4
миrрации некоторых пресно о
водных беспозвоночных (В) () 0,2 0,02
в открытом водоеме и (r)
в управляемой эксперимента О о о о о О о о о о
льной системе (по Holt, Waters, о
q о q о q q q о о
1967) r C'J со ф
C'J т"'" т"'" N О
382
Q-
Полосатая Светлая Темная
свет лая
ж
тТ Т
Ж Л
ТЖж ж тт
,ж Жж ж Ж
жжж ЖТТ
ЖтжтТ
Ж Т Т ж Т
ЖЖЛ ЖТ л
лл
Т л Л л л л
л л л л
л л
20 40 ЛtJ140
Процент Бесполосых
Рис. 13.4, Частоты встречаемости различных
морф Сераеа в разных местообитаниях. CBeT
лые раковины с полосами малозаметны в низ
кой растительности (Т TpaBa, Ж живые из
rороди), тоrда как темные однотонные ракови
ны труднее различимы в тенистых лесах (Л)
(по Calow, 1983; ориrинальные данные из pa
боты Cain, Sheppard, 1954)
100
80 Т
Ж
х 60 Т
:D
......
1::
Ф
40
......
I
О
Q. 20 Н
с:
о
о
rлава 1 3
обитания и времени [ода. Например, поло..
сатые светлые раковины трудно разrлядеть
в буйной растительности (луrа. живые изrо..
роди), [де взаимодействие проникающеrо
света и узких теней создает резко KOHTpa
стирующие темные и светлые участки. В Ie..
нистых лесах труднее Bcero РdЗЛИЧИТЬ TeM
ные раковины без полос (рис. 13 4),
2. Меланизм у березовой пяденицы (Bi
.\'10п he1и/aria). У мноrих бабочек и ДРУI"ИХ
насекомых сформировались на крыльях
и теле рисунки, делающие их незаметными
на деревьях, обросших лишайниками. Про..
мыIIленныыe заrрязнения rубят лишайники
и покрывают стволы копотью В таких рай
онах ранее адаптированные к фону свет..
лые бабочки типичной формы замещены
темными. Jwеланистuческu./wu формами. Это
явление известно как индустриальный мела
низм (рис. 13.5). В 1950..е rr. Кеттлуэлл
(Н. В. D. Kettlewell) показал ero связь у берс..
зов ой пяденицы с селективным выеданием
бабочек птицами. При прямом наблюдении
было установлено, что в заrрязненных Me
CTdX типичные формы более уязвимы для
насекомоядных птиц, чем меланистические.
Типичная
/
carboпaria
Рис. 13.5. Частоты трех морф березовой пяденицы (iпsu/aria и carboпaria меланистические) He
смотря на то что окраска iпsu/aria кажется промежуточной между типичной и carboпaria, она KOH
тролируется друrим reHHbIM локусом (по Sheppard 1958)
iпsu{ar{a
; '::\ i :': " ;:::, i.::8;Й;i /
. ., " ''''''' "', ; ;O:.
:::::. > . ) ;., . \\ :.? (
.!1 .
, .
и наоборот. Кеттлуэлл выпускал пядениц
в бочку, раскрашенную изнутри черными
и белыми вертикальными полосами. Оказа..
лось, что 650/0 бабочек предпочитали совпа..
ДdЮЩИЙ с ними по цвету фон (типичные
белый. меланистические черный), т. е. со..
ответствующий поведенческий механизм
позволяет им выбирать для отдыха такой
фон, на котором они наименее заметны.
(Как и следовало ожидать, эволюция по..
лосатости у улиток и меланизма у бабочек
протекала сложнее, чем представлено в дан..
ном здесь кратком описании. За более под..
робными сведениями можно обратиться
к rенетическим работам. например Веп.у,
1977. )
и наконец, маскировка не всеrда делаеl
животное незаметным, но и придает ему
иноrда сходство с несъедобными объекта..
ми. Мноrие насекомые напоминают внешне
сучки или листья растений. Личинки ранних
возрастов некоторых видов бабочек..
парусников хорошо 1аметны.. но избеrают
нападения, поскольку своей черной окрас..
кой с белым «седлом» на спине похожи на
птичий помет (см рис. 9.33)! Их узор резко
меняется. коrда они становятся чересчур
крупными для TaKoro способа маскировки.
Эриксен и др. (Еriсhsеп et аl., L 980) про..
вели ориrинальные опыты по выяснению
влияния покровительственной окраски на
выбор жертвы насекомоядными птицами.
Большой синице (Parus 111{ ior) предлаI'алось
выбрать между крупными и мелкими ли..
чинками мучноrо хрущака (Teпebrio /11o/i..
10r), помещенными в соломинки, причем
крупные особи находились в непрозрач..
ных соломинках, напоминающих веточки,
а мелкие в прозрачных. что делало их
леrко заметными. Знерrетическая ценность
крупных личинок примерно вдвое выше,
поэтому они были более выrодной пищей,
но их обнаружение требовало больше вре..
мени, потраченноrо птицей на рассматрива..
ние и перебирание «веточек». В эксперимен"
тах синицы выбирали мелких личинок, если
«веточек» было MHoro, и переключались на
крупных, если мало.
Друrие животные маскируются, прикре..
пляя к себе материал из окружающей ере..
ды. Например, крабы..пауки покрывают ку"
сочками водорослей и друrими предметами
Защита
З8З
острые выросты cBoero экзоскелета. Доми--
ки личинок ручейников, вероятно, rакже
используются для маскировки. Так. для ви"
да Potaп10p/ly/ax ciпgu/atus. у KOToporo они
построены из листьев, вероятность стать
жертвой {рорели меньше, коrда фоном слу..
жит дно, покрытое листьями, а не песком.
По мере роста личинки листовой домик
сменяется песчаным, но в этом случае ве..
роятность поедания уже не зависит от фон о..
Boro субстрата, что, возможно, объясняется
низкими вкусовыми качествами HOBoro ДО"
мика (см. ниже). (Более подробно см. Нап--
sell, 1984.)
13.2.1.2. Пассивная защита
Животные MOI-YT физическими или хи--
мическими методами «разубеждать» хищ..
ников нападать на них.
Широко используются для этоrо извест..
ковые защитные образования, например
спикулы rубок. скелеты кораллов. трубки
dннелид, раковины моллюсков и плечено..
rих, домики неко торых лофофоровых, пан..
цири иrлокожих. ХИТИНИЗИРОВdнные покро--
вы членистоноrих также MorYT использо--
ваться для защиты и в ряде случаев укрепле..
ны соединениями кальция. Это особенно за..
метно у сидячих усоноrих рачков, покрытых
толстыми известковыми пластинками (см.
рис. 8.36, А).
Иноrда неорrанические включения на..
столько снижают питательную ценность
тканей животноrо, что оно теряет свою при--
влекательность как источник пищи. Возмо--
жно, такова функция, по крайней мере ча..
стично.. известковых или кремнеземных ске..
летов rубок, известковых скелетов корал
лов, трубочек из rравия или песчинок, окру"
жающих тело некоторых полихет и личинок
ряда пресноводных насекомых (см. выше).
Удивительно, но лишь у небольшоrо числа
морских турбелларий в стенке тела обнару..
живаются известковые пластинки или шипы
(rл. 3). Их функuия. по"видимому, опорная.
но есть основания полаrать что они одно..
временно снижают питательную ценность
орrанизма. Более TOI'O, кажется вполне
правдоподобным.. что раковина моллюсков
произошла именно от таких образований.
Раковина наиболее очевидное сред"
384
rлава 13
Рис. 13.6. Защитная скульптурированность pa
ковин моллюсков
ство физической защиты (см. рис. 9.34). Ее
толстая стенка, сильная скульптурирован
ность поверхности, закрывающееся устье
и спиральная форма эффективно предохра
няют мноrих морских бентосных rастропод
от рыб, крабов, омаров и друrих хищников,
способных разламывать раковины друrих
форм (рис. 13.6). Крышка, прикрывающая
устье раковины наземных улиток, препят
ствует проникновению внутрь нее хищных
жуков. Сложная скульптура, включаюшая
разнообразные выросты, ребра, шишки
и шипы, служит не только для укрепления,
но и повышает эффективный размер, тем
самым снижая доступность животноrо для
хищников или увеличивая время переработ
ки жертвы. То, что у пресноводных моллю
сков нет толстостенных раковин со сложной
архитектурой, возможно, частично объяс
няется отсутствием в их местообитаниях
специализированных моллюскофаrов. Kpo
ме Toro, это может быть связано с меньшей
доступностью кальпия в пресной воде.
Необычные защитные покровы BCTpe
чаются у асцидий. Их наружный однослой
ный эпителий дополнительно окружен TY
никой (оболочники по латыни Tunicata;
см. разд. 7.4А), обычно достаточно тол..
стой, но сильно варьирующей по консистен"
пии от мяrкой и непрочной до rрубой
и хрящеподобной. Она состоит из волокни..
стото материала, основным компонентом
которото у мноrих, хотя и не у всех, асцидий
служит особая форма пеллюлозы. называе
мая туницином. Помимо этоrо в состав TY
ники входят белки и неорrанические вклю
чения, например кальций. Она бывает Ba
скуляризована и иноrда содержит амебоип
ные клетки, т. е. ее нельзя считать мертвым
покровом.
3ашитные структуры не всетда секрети..
руются самим обороняющимся орrаниз
мом. Хороший пример обратной ситуа
ции домики личинок ручейников. Они по
строень] из материалов, полученных из
окружающей среды. Сходным примером
может служить так называемый «фекаль
ный щит» личинок жука Cassida rubigiпosa ,
хотя в этом случае защита обеспечивается
отходами жизнедеятельности обороняюше
rося. Речь идет о спрессованной массе сбро
шенных покровов и фекалий, которую ли
чинка носит на прикрывающем ее спину
особом вилкообразном opraHe. Она может
маневрировать щитом, используя ето для
защиты от нападения друrих насекомых,
в частности муравьев. Раки отшельники за-
бираются в пустые раковины rастропод, из-
бавляясь таким образом от необходимости
создавать собственный экзоскелет. Их уро-
поды модифицированы, причем увеличен-
ная левая приспособлена для удерживания
за столбик раковины (см. рис. 8.49'1 Д).
Химические методы пассивной защиты
также широко распространены у беспозво-
ночных. у мноrих видов ткани тела содер-
жат токсины. Так, до 0,30/0 веса одной из не-
мертин составляют нейротоксины. Различ-
ные токсины, в том числе серную кислоту,
используют мноrочисленные безраковин-
ные брюхоноrие моллюски, например зад-
нежаберники и леrочные слизни. Некоторые
rубки выделяют раздражающие вещества;
было показано, что экстракт из пресновод-
ных rубок убивает мышей. Ряд животных
пользуется чужими токсинами. Так, некото-
рые крабы «украшают» себя rубками воз-
можно, для маскировки, а возможно, с це-
лью использования образуемых ими токси-
нов. Сходным образом определенные виды
раков--отшельников обеспечивают себе за
щиту, занимая раковины моллюсков с при..
крепленными к ним rубками и актиния--
ми.
Турбелларии, судя по всему, не выде..
ляют токсинов В ткани cBoero тела. но зато
образуют раБДОИДЬJ стержневидные
структуры, расположенные в эпидермисе
перпендикулярно поверхности тела и секре..
тируемые эпидермальными железистыми
клетками (разд. 3.6.3.1). При раздражении
червя они выбрасываются наружу. Защит..
ную роль рабдоидов леrко продемонстри"
ровать экспериментально. Колюшки ох от..
но питаются олиrохетой Tubifex tubifex
и MorYT выхватывать ее прямо из пинцета.
Однако, если предварительно покрыть че..
рвя слизью, выделяемой турбелларией при
раздражении, рыба отказывается от такой
пищи. Если же Tubifex покрыт слизью,
оставляемой на субстрате спокойно ползу..
щим плоским червем, приманка съедается.
Слизь раздраженных турбелларий содер--
жит MHoro рабдоидов, тоrда как в слизи не..
потревоженных животных их нет или очень
мало. Рабдоиды, вероятно, выполняют
и друrие функции, например участвуют
в быстром образовании самой слизи или
служат антимикробными аrентами.
Токсины обычны у насекомых, которые
в свою очередь MorYT «заимствовать» их от
растений, на которых питаются. Например"
кузнечик Poekilocerus Ьцfопius поедает моло..
чаи, содержащие сложные токсины (так
называемые карденолиды), способные
нарушать сердечную деятельность.
Кузнечик извлекает их из своей пищи, запа
сает в ядовитой железе, а при нападении
хищника выпускает содеРЖаЩУЮ эти ве..
щества жидкость. Если выдержать кузнечн"
ков на рационе без молочаев, содержание
карденолидов в выпускаемой жидкости сни
жается в 10 раз. Бабочки"данаиды также пи
таются молочаями, насыщая свои ткани
карденолидами, что делает их несъедобны..
ми для птиц. Если исключить эти растения
из рациона их личинок, данаиды оказываю..
тся безвредными для хищников. Токсичные
вещества типа карденолидов. не вырабаты"
ваемые в орrанизме насекомых, а получае..
мые ими из растений, иноrда называют кай..
Защита
385
pO tOHaJWU, противопоставляя их ал.лОJwона.\-t,
т. е. токсинам, используемым самими про..
изводящими их орrанизмами (в данном
случае растениями) (более детальное опи..
сание см. Nordlund" Lewis" 1976).
Присутствие токсинов часто сочетается
с предостереrающей окраской. Такую кор..
реляцию можно встретить во мноrих типах
беспозвоночных, от немертин и слизней до
насекомых. Окраска обычно связана с про..
стым узором.. часто включаюшим красный..
желтый, черный и белый цвета. Каждый хо..
рошо знаком с черными и желтыми полоса..
ми пчел и ос.
Возникновение токсинов и предостере..
rающей окраски в ходе эволюции
непростой процесс. Их преимущество про..
является только тоrда, коrда они защи..
щают от хищников, и в то же время един"
ственный способ для хищника научиться ра..
спознавать несъедобное животное - по..
пробовать ero на вкус. Одним из механиз..
мов может быть отбор сородичей" при ко..
тором rибель одной особи с rенами призна..
ков, предостереrающих о наличии токсина,
защищает особей, несущих такие же [ены.
rены MorYT распространяться аналоrичным
способом, если их носитель с трудом уя..
звим И способен выжить после обнаружения
ero хищником или если хищник отпуrивае..
тся каким..либо неприятным стимулом еще
до нападения. У большинства насекомых
с предостереrающей окраской прочные по..
кровы, а ярко окрашенные слизни часто
испускают сильные запахи. Немертины спо..
собны реrенерировать ткани, уничтоженные
хищниками.
Нетоксичные животные MorYT имитиро..
вать предостереrающие окраски; это свой..
ство называют бейтсовской мимикрией по
имени Toro, кто первым ero проанализиро..
вал. Поскольку предостереrающая окраска
у имитатора обман, а у животноrо..
модели правда, необходимо, чтобы а)
модель была несъедобной и ярко окрашен..
ной; б) модель встречалась чаще имитато
ра, ибо, если она встречается редко, хищник
не усвоит взаимосвязи между внешним
обликом и средствами защиты; в) имитато..
ры обитали рядом с моделями и были с тру..
дом от НИХ отличимы. Еще одна форма ми--
микрии называется мюллеровской; при ней
386
rлава 1 3
/ \ ,-,',
, f-"'--'
, 1 '-
,
'ii .
)
.'
t ;
А
;. \ / ...
0:."
'" ,
-.........,'. .....';....
д
"
,
.,..
'..
Ij
.. .
- -. / _.,
..-. I - ; ...
_.. ) _,,}:' .J<' 1\" - , ,
''I'/ ';..о '\ ,- ... .
. / ;, <:.. .:' ':'; ,' ': r:
." ..(:t ]! .' ."-" , .,. . t ""rr.
, ; : : , ': f.. ;;: .;;. . .
: Ff!; 1 -;: -' :
. a. ::. ,- . ,'
, :'f'- ',J:r. .. .;. p ·
. . .
,
, .1'
',1-
'1
Б
I
/ ..,;..1, .
.. '-
..;: ",' ,
..I,' ;' " " -: . ..-".-I' '
..j.'I'It!l>i-:SWii..... . ,. :"
.lr'4tI.... . -z:r..::.. IJ<: -
_..., .....
r
Е
P . 13.7, 6ейтсовская мимикрия между бабочками A/cidis agarthyrsus (А) и имитирующей ее Pa
plllo /ау (Б) и между североамериканским видом Liтeпitis archippus (В) и ее моделью данаидой
Daпaus p/exlppus (r). Мюллеровская мимикрия между Podotricha te/esiphe ( Д ) и He/icoпius te/esiп
he (Е) ,.,
несъедобные виды конверrируют морфоло--
rически, получая от этоrо ВЗd.имную BbIro..
ду. Критерий о здесь не существенен,
а CXOД TBO критерий в не обязательно
должно быть столь точным. У различных
видов ос и пчел одинаковый полосатый ри
сунок; это одно из проявлений мюллеров..
ской мимикрии. Мноrие двукрылые, в част..
ности мухи..журчалки и некоторые бабочки,
внешне походят на ос и пчел это бейтсов..
ская мимикрия. Примеры для чешуекрылых
можно видеть на рис. 13.7.
Ход 1ВОЛЮЦИИ мюллеровской мимикрии
понять нетрудно. Существенной проблемой
при бейтсовской мимикрии остается усло
вие В") т. е. достаточно близкое сходство ме--
жду имитатором и моделью, чтобы оно
обеспечило надежную защиту. Как в таком
случае моrли выжить формы, промежуточ--
ные между немимикрирующим предком
и современным имитатором? Одним из воз
можных путей может быть двухфазный 'JBO
люционный процесс: установление прибли
зительноrо, но достаточно адекваТНОI'О
сходства в результате одной крупной MYTa
ции с дальнейшей постепенной «подrонкой»
деталей путем отбора мелких rенетических
изменений.
13.2.1.3. Активный отпор
Орrаны, использующиеся для захвата
и умертвления побычи, часто MorYT приме
няться и для активной защиты от хищни
ков. Примером служат книдобласты CTpe
каюuJ,ИХ. Как это ни парадоксально.. книдо
бласты видов, которыми питаются rоложа
берные моллюски.. MorYT заимствоваться
последними для собственной защиты. He
разряженные нематоцисты съеденной кни
дарии транспортируются ресничками же
лудка не перевариваясь в выросты на ero
дорсальной поверхности (<<рожки», или цe
расы), дистальные концы которых.. книдоса
ки.. открываются наружу (рис. 13.8). OTCTpe
"'Iивание немаТОllИСТ, по--випимому, вызы"
вается сокращением кольцевых мыщц КНИ
досака. Запас нематоцист может быть BOC
становлен приблизительно за 1 О дней, при
чем большинство rоложаберников исполь
зуют лишь определенные их типы, присут"
ствующие у жертв. Некоторые турбелларии
аналоrичным путем пользуются нематоци
стами rидроидов.. а rребневик Eиcl1lo1 a ru
lJl"а нематоцистами медуз.
Хитинизированные челюсти и жала чле
нистоноrих также служат примером средств
нападения.. применяемых для защиты.
С друrой стороны.. жало иноrда иrрает TO
лько защитную роль. например у MeДOHOC
П щевод
Защита
387
Ядро клетки
книдосака
Слой кольцевых
мышц
А
Кишечник
Б
Рис. 13.8. Желудок rоложаберноrо моллюска со вскрытой дорсальной стенкой (А) и книдосаки
(Б) (по Barnes, 1980)
ной пчелы. Оно представляет собой моди..
фицированный яйцеклад, не используемый
для откладки яиц, состоит из парных зазу..
бренных ланцетов и непарноrо стилета (рис.
13.9) и в состоянии покоя находится в фут..
ля ре внутри седьмоrо абдоминальноrо се..
[мента. еханизм действия жала разобран
в подписи к рисунку. Ядовитая жидкость се..
кретируется парой продолrоватых желез
в брюшке и у медоносной пчелы содержит
помимо собственно яда ферменты, вызы"
вающие образование rистамина в тканях
жертвы.
Жуки рода Brachiпus, известные под на..
званием жуков..бомбардиров, разбрызrи..
вают особую жидкость для ОТРdжения ата..
ки хищников пауков. боrомолов и даже
ляrушек. Потревоженный жук выпускает ее
из пары желез, расположенных на кончике
брюшка. Последнее способно вращаться,
поэтому жидкость может быть направлена
точно в цель, фактически в любом направ
ленин. Ее действующие составляющие
бензохиноны, синтезируемые взрывооб
разно в момент разбрызrивания путем
окисления фенолов. Это сопровождается
хорошо слышимой детонацией, причем
жидкость изверrается при температуре
100 С!
Большинство двупарноноrих MHoroHo"
жек передвиrаются относительно медленно;
для своей защиты помимо толстоrо извест..
KOBoro экзоскелета они используют также
набор отпуrивающих желез (рис. 13.10). Их
выводные отверстия расположены по краям
терrитов. обычно по паре на cerMeHT хотя
на некоторых cerMeHTax полностью отсут"
ствуют. Состав секрета у разных видов не..
одинаков; в Hero MorYT входить альдеrиды,
хиноны, фенолы и синильная кислота. По..
следняя синтезируется непосредственно
перед ИСПОЛЬЗОВd.нием при смешивании ее
предшественника и фермента, образую..
шихся в разных долях железы. Эта жил..
кость токсична или репеллентна для друrих
мелких животных, а у крупных тропических
видов действует разъедающе на кожу чело..
века. Обычно она выделяется медленно.. но
у некоторых мноrоножек выпускается ПОД
большим давлением струей или в виде
брызr на расстоянии 10 зо см. Такое ее
извержение вызывается, по"видимому со..
кращением туловищных мышц, прилежа..
ЩИХ к секреторному мешку. Плотоядные
388
rлава 1 3
Ядовитый мешок
Квадратная
пластинка ер
утляр
.......&.
I
I
Рис. 13.9. Жало медоносной пчелы (Ap/s тe/l/fera). Стержень, состоящий из стилета и зазубрен
ных ланцетов опускается при сокращении мышцы м 1 , а затем сокращение мощных мышц м 2 , иду
щих от квадратной пластинки к передней части продолrоватой пластинки, приводит к повороту
треуrольной пластинки и к выталкиванию ланцета наружу. Втяrивание ланцета происходит при
сокращении мышц м з . Мышцы по обе стороны жала работают поочередно, так что за счет после
довательных выдвиrаний и втяrиваний ланцетов они все rлубже проталкиваются в тело жертвы.
Яд секретируется парой нитевидных кислых желез в брюшке. Их секрет накапливается в ядовитом
мешке, проток KOToporo открывается у основания жала в ядовитый канал. Функция так называе
мой щелочной железы точно неизвестна (по Imms, 1964)
t
в
I
I
rубоноrие мноrоножки передвиrаются бы
стрее, поэтому в меньшей степени оснаще
ны отпуrивающими механизмами, пола
rаясь в основном на спасение беrством и на
использование ядовитых KoroTKoB, приме
няемых также для захвата добычи (рис.
\
r
8.21). Тем не менее некоторые их виды обла..
дают отпуrивающими железами. Так, неко-
торые литобиоморфы несут большое число
одноклеточных желез на последних четырех
парах Hor, которыми они MorYT отшвыри-
вать Bpara, одновременно разбрасывая
Часть железис...
Toro мешка
Мышца,окружа...
ющая выводной
проток
Рис. 13.10. Отпуrивающие же
лезы двупарноноrих MHorOHO
жек (по Cloudsley.. Thompson,
1958)
клейкие капли секрета.
Педицелларии морских звезд и морских
ежей также относятся к opraHaM, сФормиро--
вавшимся специально для защиты. Они
представляют собой клешневидные придат--
ки, используемые в основном для борьбы
с личинками, которые MorYT поселяться на
поверхности тела (рис. 7.18). Выделяют три
основных типа таких структур: стебельча--
тые, сидячие и альвеолярные (поrружен--
ные). Один из нескольких типов педицелла--
рий морских ежей содержит железы, секре..
тирующие яд, способный быстро парализо--
вать мелких животных и отпуrнуть крупных
хищников. Аналоrичные функции у зоои..
дов..авикуларий некоторых мшанок (разд.
6.3.3.3).
Интересная форма защиты «обес..
кураживание» потенциальных хищников.
Узор на крыльях некоторых бабочек и дру"
rих насекомых напоминает rлаза позвоноч..
Horo. В состоянии покоя «rлаз» не видно, но
они MrHoBeHHo «открываются», стоит толь..
ко животное потревожить. Их присутствие,
возможно, связано и с отвлечением внима--
иия хищника на менее уязвимые части тела
или даже на opraHbI защиты; например
у некоторых ос белые абдоминальные пятна
находятся как раз около жала. Эксперимен--
ты с отловленными птицами продемон..
стрировали обе функции «rлазных пятен».
Резко меняющаяся окраска жертвы также
может обескураживать хищника; быстрое
изменение цвета потревоженной каракати--
цы яркий пример TaKoro рода.
13.2.2. Против внутренних BparOB
Некоторые патоrены MorYT проникать
через наружные защитные берьеры, но лю..
бой орrанизм располаrает внутренней ли..
пией обороны, способной противостоять
такому вторжению. У позвоночных это им..
мунная система с антителами, нейтрали--
зующими специфические чужеродные анти"
rепы у беспозвоночных такой системы нет,
по и они, конечно же, располаrают внутрен--
ними средствами защиты, в основном
фаrоцитарными клетками, способными
распознавать и уничтожать чужеродный
материал.
Защита
389
1 3.2.2.1. rлавное отличать
«свое» от «чужоrо»
Классическим примером распознавания
«CBoero» служит реаrреrация клеток у ry--
бок. Продавливая кусочки rубок через
ткань или помещая их в раствор этилендиа..
минтетрауксусной кислоты (ЭДТ А), можно
разделить весь орrанизм на отдельные
клетки. Смесь клеток разных видов или да..
же клонов одноrо и Toro же вида даст видо..
специфичные и клоноспецифичные pearpe..
[аты.
Опыты по пере садке тканей дают сход..
ные результаты. Например, rорrонарии (ко..
лониальные кораллы) отторrают ткани
друrих видов (ксенотрансплантаты) и даже
rенетически отличных особей cBoero вида
(аллотрансплантаты), но ткани друrих
частей той же колонии (автотранспланта..
ты) всеrда приживаются.
13.2.2.2. Роль фаrоцитов,
или амебоидных клеток,
в защите беспозвоночных
Все описанные выше реdКЦИИ характер--
ны для колониальных и обрастающих opra--
низмов, часто окаЗЫВdЮЩИХСЯ в условиях,
коrда их жизненное пространство оrраниче..
но и является объектом жесткой конкурен--
ции. Это.. по"видимому, блаrоприятствова..
ло эволюции способности распознавания
«cBoerO» как средства поддержания соб..
ственной целостности. Следовательно, та--
кие типы систем самораспознавания MorYT
быть результатом особоrо давления отбо..
ра, а вовсе не общей основой эволюции им..
мунных систем в царстве животных.
Иммунолоrические исследования были
начаты на заре ХХ в." коrда и. и. Мечников
ввел шипы розы под эпидермис личинки
бипиннарии морской звезды и вскоре обна--
ружил, что они «атакованы» амебоидными
клетками. Сходные результаты он получил
и при инъекции бацилл сибирской язвы ли..
чинкам жука"носороrа (Oryctes пasicorпis).
На основе этих наблюдений Мечников вы..
сказал предположение, что амебоидные
клетки, которые у мноrих примитивных
беспозвоночных участвуют в процессах вну--
триклеточноrо пищеварения (разд. 9.2.2),
390 rлава 1 з
Таблица 13.2. Типы беспозвоночных в которых зареrистрировано удаление чужеродных
материалов амебоидными клетками
Таксон животных
Реакция
Инъецированные частицы или вещество
Фаrоцитоз
Инкапсу лирова
иие
rубки
Индийская тушь, кармин
Эритроциты
Редии и церкарии трематод
Индийская тушь, кармин
Частицы железа
Эритроциты
Чужеродные сперматозоиды
Частицы латекса
Бактерии
Кармин, индийская тушь
Эритроциты. дрожжи, бактерии
Диоксид тория
Кармин
Бактерии
Бактерии
Частицы латекса
Железо, сахариды
Аралдитовые, имплантаты
BaciZlus thuriпgieпsis
Индийская тушь, кармин
Эритроциты, бактерии
Бычий сывороточный альбумин, бычий
rамма..rлобулин
Клетки MopcKoro ежа (в морской звезде)
Кармин
Частицы стекла
Трипановый синий
Диоксид тория
Аннелиды
Сипункулиды
Моллюски
Ракообразные
Одно ветвистые
Иrлокожие
Оболочники
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
сохранились у более высокоразвитых жи
вотных в качестве внутренней системы за
щиты. Действительно, фаI'ОЦИТЫ широко
распространены у беспозвоночных., и экспе
рименты по введению живым орrанизмам
чужеродноrо материала продемонстриро
вали их способность удалять различные чу
ждые орrанизму частииы (табл. 13.2).
13.2.2.3. Как же фаrоциты
отличают «свое» от «чужоrо»?
Фаrоциты должны одновременно «иrно..
рировать» нормальные ткани cBoero opra
низма и поrлощать чужеродные частицы.
По видимому, они участвуют и в удалении
«своей» ткани, поврежденной в результате
присутствия ксенобиотика, например за
rрязнителя.
о механизме этоrо процесса известно
сравнительно мало. Априорно можно пред..
положить, что распознавание происходит
при контакте фаrоцита с ero мишенью. По..
скольку маловероятно, что все чужеродные
частицы подают специфический сиrнал
«убей», разумно предположить подачу
своими клетками особоrо сиrнала «не уби..
вай». Нет данных о существовании у беспо..
звоночных такой же развитой, как у позво..
ночных, системы антител, служащих пр оме..
жуточным звеном при разпознавании анти"
reHoB, но есть указания на опсониновую ак..
тивность жидкостей их тела (опсонины
молекулы, связывающиеся с чужеродными
частииами, с одной стороны. и с Фаrоцита..
ми, с друrой, что облеrчает процесс фаrоци..
тоза). Так, амебоциты мечехвоста (хелице..
ровые) не обладают существенным бакте..
ОБозначения:
...... Бивалентный опсонин
. Опсониновый рецептор
. Н молекула ("своя")
v Анти Н рецептор
Сиrнал "не УБивай"
А
Рис. 13.11. Две модели фаrоцитарных иммун
ных систем беспозвоночных, см. объяснение
в тексте (по СоотЬе et el., 1984)
РИЦИДНЫМ действием без ero сыворотки, но
в ее присутствии убивают кишечную палоч..
ку. Сходным образом фаrоцитоз эритроци"
тов человека rемоцитами (амебоцитами
крови) осьминоrа Eledoпe cirrosa происхо..
дит только после Toro. как эритроциты
обработаны ero сывороткой. Экстракты из
мноrих беспозвоночных действуют как ar..
rлютинины, связывая друr с друrом различ..
ные клетки и бактерии in vitro (табл. 13.2).
Возможно, они обладают и опсониновыми
свойствами, связывая чужеродные частицы
с поверхностью фаrоцитов. (Более подроб..
ное обсуждение этих механизмов см. Соот..
Ье et al., 1984.)
Предполаrаемые механизмы распозна..
вания клеток фаrоцитами представлены на
рис. 13.11. Прямое узнавание (рис. 13.11,А),
вероятно, осуществляется фаrоцитами, «па..
трулирующими» ткани беспозвоночных
с массивным телом, Tor да как у животных
с заполненными жидкостями полостями
в качестве промежуточных факторов фаrо..
цитоза MorYT выступать опсонины (рис.
13.11,Ь).
Защита
391
13.2.2.4. Размножение связано
с определенными сложностями
Коrда происходит внутреннее оплодот"
ворение (rл. 14), в орrанизм животноrо по..
падает сперма с чужеродным rенотипом.
Оплодотворенные яйцеклетки и эмбрионы,
находящиеся в тканях матери (rл. 15), также
являются rенетически наполовину «чужи..
ми». Однако в норме они не должны разру..
шаться ее иммунной системой напротив,
материнский орrанизм должен служить им
надежным хозяином. Неизвестно, как кон..
кретно половые клетки избеrают разруше..
ния иммунной системой, но некоторое
представление об этом MorYT дать резуль..
тать] описанных ниже опытов. Спермато..
зоиды аллоrенных доноров не фаrоцити"
руются при введении в целомическую поло..
сть земляных червей, однако сперматозои..
ды млекопитающих или друrих видов до..
ждевых червей фаrоцитируются. Самцы си..
пункулид не MorYT инкапсулировать яйца
Toro же вида, введенные в их целом; это по..
зволяет предположить, что половые клетки
обладают каким..то универсальным меха..
низмом, позволяющим им избеrать атаки
иммунной системы орrанизма..хозяи..
на ведь в природе яйца в орrанизме сам..
цов не встречаются! Те же яйца, поврежден..
ные химической, тепловой или ультразвуко..
вой обработкой, быстро инкапсулируются,
в то время как предварительно заморожен..
ные яйца, хотя инежизнеспособны, инкап..
суляции не подверrаются. Интересующихся
мы вновь отсылаем к работе Кумба и др.
(СоотЬе et al., 1984).
13.2.2.5. Защита паразитов
от иммунноrо ответа хозяина
у спех пара зита зависит от ero способно..
сти защитить себя от иммунноrо ответа хо..
зяина. Этот вопрос был подробно исследо..
ван на трематодах, у которых открыты раз..
нообразные способы защиты. Так, шисто..
сомы окружают свое тело антиrенами,
близкими или идентичными антиrенам хо..
зяина. которые либо синтезируются самим
паразитом (молекулярная мимикрия), либо
берутся у хозяина и связываются с поверх..
ностью паразита Независимо от происхо..
392
rлава 13
ждения они маскируют собственные антиrе..
ны паразита, так что те более не вызывают
ответной реакции. Фасциолиды используют
иной механизм: они выделяют вещества..
токсичные для лимфоцитов и друrих клеток
иммунной системы хозяина. Кроме Toro,
rликокаликс покровов паразита с высокой
скоростью обновляется, что позволяет тре..
матодам «сбрасывать» антитела хозяина.
1 3.2.3. Реакция на заrрязнители
Стрессоры окружающей среды и вызы"
ваемые ими реакции столь разнообразны,
что невозможно обсудить все их достаточно
исчерпывающе. Некоторые способы «фи
зиолоrической защиты» от стресса, вызы"
BaeMoro кислородом, соленостью и т. п.,
уже обсуждались в предыдущих rлавах (на..
пример, разд. 11.6.5 и 12.2). Ниже мы рас..
смотрим некоторые общие реакции беспо..
звоночных на два типа заrрязнителей: ксе..
нобиотики (орrанические токсины) и тяже..
лые металлы.
13.2.3.1. Оксиrеназа с широкой
субстратной специфичностью
(ОШСС) и ксенобиотики
Орrанические заrрязнители типа уrлево"
дородов, попадающих в среду при утечках
нефти, MorYT проникать в ткани морских
беспозвоночных. Они липофильны, поэто..
му их метаболизация затруднена., и они мо"
[ут накапливаться в липидных отложениях
и липидных компонентах клеточных мем..
бран до такой концентрации, что это нару..
шает биохимические процессы. Однако
у ряда морских беспозвоночных, в частно..
сти у некоторых полихет, моллюсков ира..
кообразных, имеется ферментная система,
способная окислять такие токсины путем
присоединения атомов кислорода, что де..
лает эти вещества более rидрофильными и,
следовательно, леrче метаболизируемыми.
Эта система довольно неспецифична вотно..
шении субстратов, на которые воздей..
ствует, и состоит из нескольких ферментов
и цитохромов, связанных в основном с мем..
бранами микросомной фракции эндоплаз..
матическоrо ретикулума некоторых тканей
(например, пищеварительной железы или
ТОКСИН ОН
у
МетаБОЛИЗМ
И экскреция
Рис. 13.1 2. Упрощенная схема действия систе
мы ОШСС (по Calow 1985)
rепатопанкреаса). Процесс окисления тре..
бует больших затра1 }нерrии; в самом 06..
щем виде он представлен на рис. 13.12. Он
включает эпоксидирование, rидроксилиро"
вание и дезалкилирование. Работа системы
требует индукции, т. е. входящие в нее фер..
менты образуются только при наличии ксе..
нобиотика. Например. содержание соответ"
ствующеrо цитохрома в тканях мидий воз..
растает после суточноrо воздействия дизе..
льноrо топлива и возвращается к норме че..
рез 8 дней после снятия стресса.
Оксиrеназы с широкой субстратной спе..
цифичностью присутствуют и У раститель..
ноядных насекомых, [де они, вероятно, уча..
ствуют в обезвреживании природных opra..
нических токсинов, вырабатываемых расте..
ниями для защиты от фитофаrов. Напри..
мер, у насекомых..полифаrов активность
ОШСС обычно выше, чем у стенофаrов (60"
лее специализированных в смысле питания),
по"видимому, в связи С воздействием более
широкоrо спектра токсичных соединений,
включая фенолы. хиноны.. терпеноиды и ал..
калоиды.
13.2.3.2. Металлотионеины
Тяжелые металлы типа ртути, кадмия,
меди, серебра и олова MorYT быть исключи..
тельно токсичными для водных беспозво..
ночных. Например, они способны денатури"
ровать ферменты, так как, взаимодействуя
с ними, изменяют их третичную структуру.
Однако некоторые морские беспозвоноч"
ные, в том числе крабы двустворчатые
моллюски. блюдечки и некоторые аннели
ды, обезвреживают тяжелые металлы,
связывая их с особыми белками, называе..
мыми металлотионеинами. Это соединения
низкоrо молекулярноrо веса.. боrатые SH
rруппами из..за высокоrо содержания ами
нокислоты цистеина. SН rруппа может
связывать или хелатировать металлы, что
делает их менее токсичными.
13.2.4. Репарация защита
против старения?
в разд. 13.3.1 вскользь упоминалось,
что старение целоrо орrанизма вызывается
накоплением повреждений на суборrаниз"
менном уровне. Об этом свидетеЛЬСТВУЮ1
следуюшие данные.
Разрушение ПIканей. Старение у двукры"
лых насекомых сопровождается снижением
способности к полету, и оно в свою очередь
коррелирует с деrенеративными изменения..
ми в структуре летательных мышц. Показа..
но, что среднее число клеток в мозrе рабо
чих пчел падает с 522 при вылете из КУКО..IКИ
ДО 350 в возрасте 10 недель (рис. 13.13).
Лuпофусцин (пиrмент старения). Вероят
но, это продукт пероксидирования липидов,
550
500
450
о
ti>
с:::
:!IC:
О 400
с:::
()
s
:r
350
1 О 20 30 40 50 60 70
Возраст, ДНИ
Рис. 1 3.1 3. Снижение с возрастом числа клеток
МQзrа у пчел (по Rockstein, 1950)
Защита
З9З
0,6
t
0,5
. Самец
:s: О Самка
:s: 0,4
::1
I
()
Q)
о...
о
>-
7i
::1
:s: 0,2
I
:s:
q:
W
0,1
О
10 20
Возраст, ДНИ
40
За
Рис, 13.14. Накопление липофусцина (изме
ренное по ero флуоресценции) у Drosophila
тe/aпogaster (по Biscardi, Webster, 1977)
образующийся при разрушении клеточных
мембран. Показано, что липоФусцин HaKa
пливается с возрастом в тканях нематод
и насекомых (рис. 13.14).
Работа ферментов. Данные, получен..
ные в основном при изучении нематод, ro..
ворят о нарушении с возрастом структуры
и функции ферментов: они становятся более
чувствительными к тепловой денатурации
(что указывает на изменения в молекуляр..
ной орrанизации), проявляют аномальные
иммунолоrические свойства и снижают ка..
талитическую активность. Однако это не
относится к некоторым ферментам HeMa
тод, а также ко мноrим изученным фермен..
там друrих животных.
Основная причина всех этих нарушений
связана, вероятно, с молекулярными про..
цессами типа тепловоrо шума, вызывающи..
ми ошибки синтеза белков и друrих метабо
лических превращений. В то же время по..
врежденные белковые молекулы MorYT
в принципе в соответствии с rенетической
проrраммой заменяться новыми так же, как
394
t+
..1} 0::..L) ..1}
U O:: fO
fOroUIU U
o..I O та О
8 т@:s g.
т o. o..
.5: g >-М И
:о т gg
О О 0..0
о.. о.. с::. о с С::
с Фа
Фr::: с
то..
01:1
С:: Ф
00..
сс
Ф
I
Deпdrocoeluт
lacteum
(короткоживущий
моноцикличный )
з
2
1
о
е
О
I
Q:r
00
L.{)
ф
roС::
I
т
О
M ,"",
°IO::
:s:s
:s......... I
L ф
C::
0000
t::: 0
UФI
sс::ш
T O
rлава 13
t5 О:: о::..а ..а
rorou t;
o...IO та U
м тс:: sC:: О
О S Ф ......ф о.
т o. o..
. E:i ()
..а от от
с:: о.. О 0..0
Ф r::: с:: r::: с::
о..ФО Фа
м о..с c
OQ
CD ф
00..
ас
с
Ф
I
Polycelis feпi us
(долrоживущий
полицикличный)
Рис. 13.1 5. Скорость обновления клеток на
различных стадиях жизненноrо цикла двух ви
дав трехветвистых турбелларий долrо
живущеrо и короткоживущеrо. Последний
вкладывает rораздо больше энерrии в размно
жение, чем первый. В обоих случаях число кле
точных делений снижается с возрастом, но
и целые клетки (в данном случае путем
митоза) (rл. 1), Действительно, результаты
опытов на нематодах позволяют rоворить,
что белки с аномальной аминокислотной
последовательностью обновляются бы..
стрее нормальных, но этот процесс замед..
ляется по мере старения червей. Клеточные
деления у взрослых особей характерны для
книдарий, плоских червей, аннелид и мол..
люсков, но весьма оrраничены у нематод
и насекомых. У пресноводных трехветви--
стых турбелларий обновление клеток с воз--
растом замедляется (рис. 13.15), причем
особенно заметно с пере ход о м у моноци
кличных форм К размножению (см. разд.
14.4). Последнее связано с ускорением само..
ro процесса старения. Таким образом, био
лоrический возраст, или жизнеспособность,
орrанизма зависит от баланса между возни..
кновением дефектов и их заменой или ис..
правлением (репарацией). Неудивительно
поэтому, что процессы старения наименее
выражены у таких орrанизмов, как книда..
рии, у которых обновление тканей происхо..
дит на протяжении всей жизни, и наиболее
заметны у нематод и насекомых, rде оно
весьма оrраничено (табл. 13.3). А среди на..
секомых митотическое деление клеток за..
хватывает больше тканей и продолжается
дольше У видов с более продолжительной
жизнью, например у жуков.
у короткоживущей формы митотическая актив
ность в репродуктивном возрасте падает силь
нее. Считается, что замедление обновления
тканей вызывается ббльшим вкладом энерrии
в размножение и сокращает жизнь ускоряя
процессы старения (по Calow, Read, 1986)
Таблица 13.3. Старение у беспозвоночных
rруппа
Подозревается OT
сутствие старения
у некоторых видов
Подозревается cy
ществование CTa
рения у некоторых
видов
Доказано cтape
иие некоторых
видов
Книдарии
Плоские черви
Моллюски
Нематоды
Аннелиды
Коловратки
Членистоноrие
+
+
+
+
+
+
+
+
+
+
+
+
+
13.3. ВЫВОДЫ
Опасности, которым подверrаются ор..
rанизмы, весьма разнообразны. инеудиви..
тельно, что они вызывают широкий спектр
ответных реакций. Однако можно предло..
жить единую классификацию таких опасно..
стей в зависимости от их более или менее
тесной связи с орrанизмом, находящимся
под уrрозой. Сходным образом за внешним
разнообразием реакций можно различить
некоторые общие черты. Например.. все за..
щитные механизмы независимо от Toro, на..
правлены они против хищников, паразитов,
микроорrанизмов или изнашивания caMoro
орrанизма, требуют больших затрат ве..
щества и энерrии. Именно эти общие при..
нципы читатель должен постараться уяс..
нить из этой и друrих rлав данной книrи.
13.4. Дополнительная
литература
Berry R. J. 1977. Inherital1ce And Natural Selectiol1.
New Naturalist No. 61. William Collil1S and Со..
Glasgow.
.B/est А. п. 1957. ТЬе function of eyespot patterI1s in
the Lepidoptera. Behaviour, 11, 209 256.
Ca/ow Р. 1981. Invertebrate Biology. Croom Helm.
London.
Ca/ow Р. 1983. Evolutionary Pril1ciples. Blackie
Glasgow.
СоотЬе D. R., Еу Р. L., Jeпkiп С. R. 1984. Self/I1011
Защита
395
self recognition. Q. Rev. Biol.. 59, 231 255.
Davies /. 1983. AgeiI1g. Edward ArI101d LOl1doI1.
Eisпer Т., Vaп Tassell Е., Carre! J. Е. 1967. Defensi..
ve use of а "faecal shield" Ьу а beetle larva. Scien
се, N. У., 158, 1471 1473.
Erichseп J. Т.. K1 A ebs J. R., Houston А. /. 1980. Opti
mal foraging al1d cryptic prey. J. al1in1. Ecol.. 49,
271 276.
Gliи'iсz М. z. 1986. Predatiol1 and the evolutiol1 of
vertical n1igratio11 in zooplal1ktol1. Nature (Lon
dOI1), 320, 74б 748.
Haпsell М. Н. 1984. Al1imal Architecture and Buil
ding Behaviour. L0l1gn1al1, LondoI1.
Liviпgstoпe D. R., МООI'е М. N., LOH'e D. М., Nas..
ci с., Fan'ar S. V. 1985. Respol1ses of rhe cytoch
rome Р..450 mOI1oxygeI1ase system to diesel oil in
the соmmОI1 mussel. MytiZиs edulis L., and the pet"
iwil1kle, Littoriпa littol"ea L. Aquatic Toxicology.
7, 79 81.
N01'd/uпg D. А., Lеи'is w. J. 1976. Теrmiпоlоgу of
chemical releasing stimull il1 il1traspecific and iп
terspecific interactiol1s. J. сЬеm. Ecol., 2, 211
220.
Rocksteiп М. 1950. ТЬе relatiol1 of cholil1esterase ac
tivity to change in сеll l1umber with age il1 the
brail1 of the adult worker Ьее. J. сеН. соmр. РЬу..
siol., 35. 11 23.
Theodor J. L. 1976. Histo..incon1patibility in а l1atu..
ra] population of gorgonial1s. Zool. J. Lil1l1. Soc.,
58, 173 176.
Turпer J. R. G. 1984. Darwin's coffin and Dr. Рап..
gloss do adaptationist models explain mimicry?
111: В. Shorrocks (Ed.) Evolutiol1ary Ecology, рр.
313 361. Blackwell Scientific Publications, Ox
ford.
rЛАВА 1 4
РАЗМНОЖЕНИЕ И жизнЕнныE циклы
В принципе мноrоклеточные эукарио
ты это диплоидные орrанизмы, размно
Жdющиеся половым путем, образуя ra
плоидные [аметы, пары которых при слия..
нии дают диплоидную зиrоту. rенотип по
ловых клеток (raMeT) обычно отличается от
rенотипа родительскоrо орrанизма вслед
ствие перераспределения [енов в ходе мейо..
за. Более Toro, новый диплоидный opra..
низм обычно (хотя и не всеrда) образуется
при слиянии [амет различных rенотипов.
Таким образом у мноrоклеточных орrаниз
мов в результате половоrо размножения
и оплодотворения возникает клетка с но..
вым, уникальным набором [енов.
В этой rлаве рассмотрены процессы,
связанные с половым размножением, а да..
льнейшее развитие и дифференцировка зи..
[оты и продуктов ее деления описаны в rл.
15. Хотя жизненные циклы животных сле..
дуют одному основному плану, особенно..
сти половоrо и бесполоrо размножения
(также широко распространенноrо среди
животных) сильно варьируют. Мы рассмо"
трим это разнообразие, обусловливающие
ero факторы и адаптивное значение различ
ных вариантов, включая селективные преи..
мущества половоrо размножения перед бес..
полым. По ходу изложения станет ясно, что
различные особенности жизненных циклов
и размножения характерны для определен
ных филоrенетических линий, уровней opra
низации животных и эколоrических ситуа..
ций. Это позволит обсудить стратеrии раз..
множения беспозвоночных в свете совре..
менных теорий эволюции жизненных ци..
клов.
14.1. Введение
В rл. 1 высказывалась мысль о том, что
эволюция это естественное следствие са..
мовоспроизведения систем путем полукон..
сервативной репликации, причем у всех ор..
rdНИЗМОВ один и тот же носитель rенетиче..
ской информации ДИК, а наследственная
изменчивость возникает при неправильном
копировании последовательности азоти..
стых оснований в процессе репликации этой
молекулы. Однако эволюция мноrоклеточ"
ных существ требует полуконсервативноrо
воспроизведения не просто клеток, а opra..
низма в целом, т. е. процесса, называемоrо
размно.женuем. У подавляющеrо большин..
ства форм он сопровождается rенетической
рекомбинацией; в этом случае rоворят о по..
ловом размножении. Существуют различ..
ные ero системы, но их общей чертой остае..
тся обмен rенетическим материалом между
особями с разными rенотипами.
В приложении 14.1 приведены схемы
трех различных половых процессов. rенети"
ческий обмен у вирусов возможен при сме..
шанном "заражении одной клетки хозяина
вирусами разных штаммов. У бактерий
часть молекулы ДИК одноrо из партнеров
передается друrому в ходе конъюrации, по..
сле чеrо между rомолоrичными нитями
ДИК может происходить обмен. Все MHoro..
клеточные эукариоты используют третью
из представленных систем половоrо процес..
са.
у животных обычно образуются два си..
льно различающихся типа [aMeT
сперматозоид и яйцеклетка (яйцо) (разд.
14.4.2). Часто они происходят от специали..
зированных клеток, обособляющихся на
Размножение и жизнеННblе ЦИКЛbl
397
Таблица 14.1. Репродуктивные признаки морских беспозвоночных
Признак
Р,,-звитие
Пелаrическое
Непелаrическое
MHoro лет
Крупный:
длина> ] 000 мм
Простые
Н,,-ружное без хранения спермы
Размер яиц
Мелкий:
'" 50 мкм
Высокая:
106
Низкая:
] раз в rод
Одно
Плодовитость
Частота размножения
Число размножений за
жизнь
Продолжительность
жизни
(Время rенерации)
Размер тела
\1 ноrолетняя
Сперматозоиды
Оплодотворение
планкотрофное
1Jецитd'rрофное
смешанное
Крупный:
> 1000 мкм
Низкая:
1
Высокая:
MHoro раз в rод, почти непрерывно
Множество
Однолетняя
Меньшая
Несколько дней или недель
Мелкий:
< 1 ММ
Сложной формы
Внутреннее или вклю
чающее процессы пере
носа и хранения спермы
Небольшое; совершает..
ся р,,-ньше
Репродуктивное
усилие 1)
Большое' совершается позже
EG дЕ G
1) Репродуктивное усилие можно определить по формулам Е или дЕ ' rде EG энерrия, вло
s + EG s + EG
женная в половые структуры, Es энерrия, вложенная в СОМdтические ткани, .6.E G ежеrодный прирост половых
тканей, Es ежеrодный прирост соматических тканей
ранних стадиях эмБРИОI енеза, но внекото..
рых rруппах MorYT возникать в результате
дедифференцировки соматических клеток.
Жизненные циклы мноrоклеточных ор..
rанизмов разнообразны; например, умно..
rих растений чередуются rаплоидные иди..
плоидные поколения (приложение 14.2)
с одинаковой или различной морфолоrией.
у животных такой смены поколений не
встречается, но известно четкое чередова..
иие диплоидных фаз. размножающихся по..
ловым И бесполым путями (разд. 14.2).
Несмотря на общие черты половоrо
процесса у всех животных, условия ero осу..
ществления и детали жизненных uиклов
различаются очень сильно. Отличительные
признаки последних в каждой rруппе беспо..
звоночных часто в совокупности называют
несколько антропоморфным термином «ре..
продуктивная стратеrия» (табл. 14.1).
14.2. Значение половоrо
и бесполоrо размножения
14.2.1. Бесполое размножение
в жизненных циклах
беспозвоночных
Половое размножение распространено
очень широко, но мноrие орrанизмы MorYT
размножаться бесполым путем, т. е. без re..
нетической рекомбинации. rенотип потом..
ства при этом практически идентичен роди..
тельскому. Бесполое размножение может
идти как путем разделения тела животноrо
398
rлава 14
А
о
Б
Рис. 14,1. Схема двух типов
(амейотическоrо) размножения:
ментация; Б партеноrенез
бесполоrо
А фраr
на мноrоклеточные части (почкование
и фраz"неНI11ация), так и с образованием He
оплодотворенных яиц (парl11еНО2енез) 1. На
рис. 14.1 показаны оба этих механизма,
свойственные мноrим беспозвоночным.
Почкование и фраrментация особенно
распространены у мяrкотелых форм
rубок, книдарий, плоских червей, немертин,
аннелид и некоторых иrлокожих. Эти спо
собы размножения редко встречаются у жи
вотных С твердыми покровами и не обнару
жены у моллюсков и членистоноrих. Фраr
ментация может означать либо простое раз
деление орrанизма на две части, каждая из
которых реrенерирует недостающую (рис.
14.1,А), либо распад на множество пропа
rул, раЗВИВdЮЩИХСЯ впоследствии в целые
орrанизмы (рис. 14.2). Она обычно сочетае
1 Партеноrенез часто относят к половом)
(однополому) размножению. Прu..w. ред.
тся С половым размножением в сложных
жизненных циклах с чередованием половых
и бесполых «поколений» (рис. 14.4,А). За
труднения. вносимые почкованием и фраr..
ментацией в формулировку понятия
«особь», особенно выражены у тех орrаниз"
мов, [де эти процессы идут не до конца, так
что возникают колонии из большоrо числа
структурных единиц. Колониальность осо..
бенно распространена у книдарий, мшанок
и оболочников. Некоторые из таких беспо..
звоночных показаны на рис. 14.3. По
видимому, этих животных правильнее счи
тать состоящими не из особей, а из модуль..
ных единиц.
Ilартеноrенез у беспозвоночных встре..
чается также часто (нами под этим терми
ном понимается бесполое размножение по..
средством неоплодотворенных яиц.. но не..
которые авторы включают в 'Это понятие
и способы, описанные в предыдущем абза
це). Он включает либо модифицированный
мейоз с образованием диплоидноrо яйца,
для развития KOToporo не требуется слия..
ния с мужской rаметой, ..lибо развитие из
неоплодотворенных rаплоидных яиц сам..
цов. а из оплодотворенных диплоидных
самок (такой способ называется appeHOTO
кия). Облиrатный партеноrенез, т. е. полное
отсутствие половоrо размножения, встре..
чается редко. Он предполаrается, например,
у бделлоидных КОЛОВРdТОК (см. разд.
4.9.3.1), самць] которых не обнаружены. Ча
ще партеноrенез циклически чередуется
с половым размножением. После одноrо
или нескольких бесполо размножающихся
поколений образуются особи разноrо пола,
причем слияние их [амет обычно дает по
коящиеся, устойчивые к внешним воздей
ствиям яйца (рис. 14.4.А). Жизненные ци
клы TaKoro типа характерны для MOHoro
натных коловраток, мноrих мелких пресно-
водных ракообразных и тлей Диплоидная
особь. развивающаяся из оплодотворенно-
ro покоящеrося яйца, дает начало клону re-
нетически идентичных потомков. Скорость
роста популяции при этом максимальна,
и такой жизненный цикл можно рассматри"
вать как приспособление к максимальному
использованию непостоянных пищевых ре-
сурсов в условиях, коrда друrие биолоrиче-
ские факторы оrраничивают размеры тела.
РазмножеНI е и жизнеННblе ЦI КЛbl
А
1
ЯЙЦО (п)
Взрослый
орrанизм
(2п)
Сперматозоид (п
399
о f f
OOQ6Q6
д '-.... '-.... f t
Ы ",
п
Б
Яйцо зрослы й
или ]
орrанизм
Сперматозоид
Партеноrенез или
срраrментация
L Бесполое
размножение
в
Половое размножение
Рис, 14.2, Пример множественной фраrментации: полихета Dodecace"a. А. 1. Взрослая особь до
разделения на cerMeHTbI. 11. Стадия реrенерации: каждый cerMeHT формирует rолову и хвост путем
прироста новых cerMeHToB. Б. Образование первичных, а затем вторичных особей из одноrо ce
rMeHTa. В конечном счете этот cerMeHT оказывается в составе особи, прекратившей фраrмента
ЦИЮ, В. Схема жизненноrо цикла, включающеrо бесполое и половое размножение: бесполое CMe
няется половым, и все бесполые особи в конце концов поrибают (по Dehorne, 1933; Gibson, Clark,
1976)
На определенной стадии uикла ПОЯВЛЯIОТСЯ
особи, дающие потомков двух типов
самцов и самок, способных к половому раз..
множению. Переход к нему БЫВdет 'Эндоrен
но детерминирован, но чаше является peaK
цией на изменение окружающих условий,
например на перенаселение, изменение каче..
ства пищи или сокращение продолжитель..
ности дня (см. также разд. 14.4.4). Переход
к половому размножению может сопрово"
ждаться морфолоrическими изменениями
(рис. 14.4,Б,В), а у тлей эти изменения про..
исходят I-! на бесполой стадии (рис. 14.4,1).
14.2.2. Типы половоrо
размножения
Более 99% беспозвоночных размножаю..
тся половым путем на той или иной стадии
жизненноrо цикла, причем для подавляю..
щеrо большинства это единственный спо
соб воспроизведения. При нем всеrДd про..
исходит слияние относительно крупной и
неподвижной женской [аметы яйце..
клетки с более мелкой и подвижной муж..
ской сперматозоидом. Такое различие [а..
мет называют анuзоzамuей. Оно позволяет
различать мужские и женские функции, ко..
торые не обязательно выполняются разны"
ми особями. Виды, представленные самца..
400
rлава 14
А
II
в
Рис. 14.3. Примеры колониальных, или «моду..
льных}), беспозвоночных. А. rидроидный полип
Obe/ia: 1 ветвящаяся колония; 11 отдельный
полип. Б. Мшанка Meтbranipora: I часть ко..
лонии; lI отдельный зооид. В оболочник
Sydniuт. I три колонии с отдельными BXOД
ными и общим выводным сифонами; 11
отдельный зооид
ми и самками, называют раздельнополыми;
если одна и та же особь одновременно или
последовательно выполняет мужские и жен--
ские функции., животных называют серма--
фродUI11а/иu. Характеристики обоих типов
размножения представлены в табл. 14.2,
а их распространение в основных rруппах
беспозвоночных в табл. 14.3. При после--
довательном rермафродитизме смена пола
может наступать по достижении определен--
Horo, видоспецифичноrо размера или возра..
ста., так что самцов и самок одновременно
не встречается; в друrих случаях этот мо"
мент различен у разных особей в популя..
ЦИИ, что допускает наличие двух «чистых»
полов. Обе эти ситуации показаны на рис.
14.5.
14.2.3. Механизмы определения
пола
Эти механизмы у беспозвоночных
можно разделить на три основных типа:
а) материнский, б) rенетический и в) эколо..
rический.
14.2.3.1. Материнское
определение пола
В этом случае пол потомства детерми"
нируется самкой, образующей яйца двух ти"
пов. Пример приведен в приложении
14.3(1). При такой системе размножения не..
избежен инбридинr, и все особи зрелой по..
пуляции являются самками. Так, у пчел
матка способна контролировать оплодот"
ворение своих яиц; если оно произошло, ди..
плоидные яйца развиваются в стерильных
самок (рабочих особей)., но при определен..
ных условиях способны дать и плодовитых
маток; неоплодотворенные яйца партеноrе..
нетически развиваются в rаплоидных сам..
цов.
14.2.3.2. rенетическое
определение пола
у некоторых животных хромосомные
наборы самцов и самок 1аметно различаю..
тся: у самок две половые хромосомы иден..
тичны (так называемые Х--хромосомы),
у самцов различны (х-- и У"хромосомы).
Пол зиrоты определяется при оплодотворе..
нии половой хромосомой, оказавшейся
в сперматозоиде. Это явление, называемое
rетероrамией, распространено у беспозво..
ночных не так широко, как часто считают.
Несколько ero разновидностей встречаются
Покоящееся яйцо
2 n О результат половоrо размножения
1 (ДИПЛОИдНое, 2 п)
Вылупление партеноrенетической
О иплоrой самки
Диплоидные яйца
хНесколько поколений
1 Изменение условий:
« температура,
. освещенность,
If пищевые ресурсы,
стресс
Диплоидные самки дают
размножающееся половым путем
потомство
J l
Самцы Самки
! !
rаплоидные rаплоидные
сперматозоиды яйцеклетки
,.............. @
n
Оплодотворение
.......
1:
О
I
Ш
L
{
L
А
@ Покоящееся яйцо
о
Диплоидные
покоящиеся
-
яица /' 1 l
Сперматозои I
B о)
Неоплодотворенные
rаплоидные яйца
в
Вылупление
рачка из
эсриппия
Самка, ОБ
разующая
эсриппий
Б
ЛЕТО
крылатые Бесполые
с\сселительные срормы
,-.lj Бескрылые Бесполые
/ -: у I срормы, ОБеспечива
t Q J. ющие рост популяци
. Крылатые самки,
... дающие в потомст
J I t // ве яйцекладущих
I /!.... самок
......'
,,.,, "
".;",
ОСЕНЬ
Крылатые
самцы
покоящаяся"
стадия
r
Рис. 14.4. Чередование партеноrенеза и половоrо размножения в жизненных циклах беспозво
ночных. А. Элементы обобщенноrо жизненноrо цикла. Б. Жизненный цикл пресноводноrо ветви
cToycoro рачка Daphпia В. Жизненный цикл коловратки Braпch/oпus: за бесполой фазой следует
половая, коrда неоплодотворенные самки откладывают мелкие яйца, из которых развиваются
самцы, а оплодотворенные самки откладывают более крупные яйца, представляющие собой зи
мующую стадию жизненноrо цикла. r. Жизненный цикл вишневой тли. Для партеноrенетических
самок характерно живорождение и только осенние особи откладывают яйца. Хорошо выражен
полиморфизм, особенно в образовании крылатых и бескрылых форм (Б по Bell, 1982, r по
Dixon, 1973)
402
rлава 14
Таблица 14.2. Системы половоrо размножения животных
r ермафропитизм
Каждая особь в популяции образует либо мужские либо женские половые
клетки и имеет соответствующие вспомоrательны(' желсзы и opraHbI
Некоторые особи в популяпии образуют как мужские. так и женские поло
вые клетки и имеют вспомоr ательные железы и opraHbI связанные как
с мужской. так и с женской воспроизвопительными функuиями в тот и Т"IИ
иной период жизни
Синхрuнный zеРJ\1афродип1ИЗ.w
Взрослые животные одновременно образуют оба типа половых клеток. При
этом они MorYT либо обмениваться мужскими половыми клетками с дpy
rим орrанизмом (аутбридинr) либо размножаться са\100плодотворе
нием
Раздельнополость
Последоваlпельный zер.мдфродИlпuз.н
Каждое животное выполняет функции сначала ОДноrо затем друrоrо пола.
1. Протандрия. Мужские функции развиваются и выполняются раньше,
чем женскис. При активации женских функций мужские либо утрачива
ются, либо сохраняются (синхронный rермафродитизм)
2. Протоrиния. Женские фУНКIlИИ развиваются и выполняются pdHbIlJe
мужских. В конечном итоrе также может наблюдаться синхронный [ep
мафродитизм
3. Чередование. Протандрическое или протоrиническое. но с послеДУЮIЦИМ
обращением пола
Таблица 14.3. Типы половоrо размножения у беспозвоночных
Тип
Класс
Характер ПОЛОВОI о размножения
Плоские черви
Тур6елларии
Моноrенеи
Все способны к половому размножснию но MHO
rие также размножаются б сполым путем за
счет отделения фраrментов rеммул
Самооплодотворяющиеся rермафродиты с чере
дованием половоrо и бесполоrо поколений
Раздельнополые, но часто размножаются ПОЧКО
ванием или фраrментацией. Иноrда имеют
сложные жизненные циклы с пелаrическими
медузами образующими raMeTbI. и бентосны
ми rидроидами. размножающимися бесполым
путем
Синхронные rермафродиты по--видимому, пере
крестно"оплодотворяющиеся
Синхронные rермафродиты по видимому, пере
крестно"оплодотворяющиеся. Распространено
бесполое размножение фраrментацией. у неко"
торых форм половой процесс не обнаружен
Синхронные rермафропиты: при множественных
заражен иях, возможно, распространено пере
крестное оплодотворение. И Hor да самоопло
дотворяющиеся. На личиночных стадиях часто
размножаются множественной фраrментацией
Синхронные rермафродиты, обычно самоопло
дотворяющиеся. Один род раздельнополый
Синхронныеrермафродиты
rубки
Mesozoa
Стрекающие
rребневики
Трематоды
Цестоды
rнатостомулиды
Размножение и жизнеННblе ЦИКЛbl
403
ПродО.!l:нсеlluе lпабл. 14.3
Тип
Характер половоrо размножения
Класс
Немертины
r,,-стротрихи
Моллюски
Коловратки
Киноринхи
Скребни
Loricifera
Волосатики
Нематоды
Приапулиды
Сипункулиды
Эхиуриды
Аннелиды
Поrонофоры
Форониды L
Мшанки f
Плеченоrие
Внутрипорошицевые
Полухордовые
Иrлокожие
Хордовые
ChaetodcrmomOrPha }
М onoplacophora
Панцирные
Лопатоноrие
Брюхоноrие
Neomeniomorpha
Двустворчатые
rоловоноrие
Bdelloidea
Monogonata
Seisonidea
Полихеты
Поясковые
Морские ежи
IОЛОТУРИИ
Морские лилии
Морские звезды
Офиуры
Оболочники
Почти все раздельнополые; некоторые пресно
водные формы rермафродиты
Синхронные rермафродиты, но распространен
партеноrенез
Раздельнополые
Весьма различный характер размножения: пере
днежаберные в основном раздельнополые.
но иноrда последовательные (протандриче
ские) rермафродиты; заднежаберные в ОС..
новном синхронные rермафродиты; леrоч
ные синхронные rермафродиты с перекрест
ным оплодотворением
rермафродиты
Раздельнополые
,
Облиrатный партеноrенез; самцы неизвестны
Циклический партеноrенез; в половой фазе рdзде
льнополы (самцы карликовые)
Раздельнополые
"
.,
"
"
"
"
"
Раздельнополые: иноrда саМllЫ карликовые
Обычно раздельнополые; иноrда последоватсль
ные протандрические rермафродиты. изредка
синхронные rермафродиты. Некоторые виды
размножаются фраrментацией
Bcer да синхронные rермафродиты с перекрест..
ным оплодотворением
Раздельнополые
Синхронные rермафродиты; отдельные вилы
раздельнополые
В основном раздельнополые, изредка rермафро..
диты
Синхронные или последовательные rермафроди
ты
Раздельнополые
}
1
I
"
в основном раздельнополые. иноrда синхронные
rермафродиты. Некоторые виды размнож,,-
ются фраrментацией
Синхронные rермафродиты
404
rлава 14
Продол:женuе табл. 14.3
Тип
Класс
Характер половос'о размножения
Ракообразные
Жаброноrие
Рdкушковые
Весло ноrие
у соноrие
Malacostraca
Хелицеровые
Онихофоры
Тихоходки
Пятиустки
Одноветвистые
}
Раздельнополые, но распространен партеноrенез;
у некоторых видов самцы не обнаружены
Раздельнополые
Торациковые обычно rермафродиты. хотя HeKO
торые виды раздельнополые с карликовыми
самцами: остальные раздельнополые с КdрЛИ
ковыми самцами
Обычно раздельнополые. но иноr да последовате
льные (протандрические) rермафродиты
Почти всеrда раздельнополые
, ,
"
"
Почти всеrда раздельнополые, но иноrда встре-
чается партеноrенез или арренотокия (СdМЦЫ
обычно неизвестны)
у насекомых; оно подробно узучено на при
мере плодовой мушки Drosophi/a тe/aпoga
ster. Как показано в приложении 14.3, у ca
мок здесь две Х хромосомы, а у самцов Х
и У. В ходе мейоза все яйцеклетки получают
по одной Х"хромосоме, а ровно половина
сперматозоидов Y xpOMOCOMY. При слу
чайном слиянии raMeT устанавливается OT
ношение полов 1: 1. У мноrих беспозвоноч
ных rетероrамия не обнаружена, тем не Me
нее наблюдается фиксированное отношение
полов.
100
90
80
m 70
g 60
50
40
Q)
g ЗА
а.
с 20
10
А
Б
Размер тела
(длина, число cerMeHTOB и т.д.)
Рис. 14.5. Типы обращения пола при последо
вательном rермафродитизме: А ложная раз
дельнополость; Б несбалансированный re
рмафродитизм
14.2.3.3. Эколоrическое
определение пола
Пол особи не Bcerna определяется до
или в момент оплодотворения; иноrда он
зависит от окружающих условий во время
эмбриональноrо или личиночноrо разви"
тия. Одним из наиболее известных приме..
ров служит эхиурида Boпellia viridis; карли..
ковые самцы этоrо червя паразитируют на
самках (разд. 4.12). В начале нынешнеrо
столетия было показано, что пол свободно..
плавающих планктонных личинок не фикси..
рован оседая на дно, они становятся сам..
ками, а попадая на крупный хоботок самки
или оказываясь вблизи от Hero, превращаю..
тся в самцов (приложение 14.3).
Эксперименты показали, что самки вы..
деляют вещество, оказывающее сильное
маскулинизирующее действие на личинки:
в ero отсутствие почти все они развиваются
в самок. У большинства друrих эхиурид
определение пола rенетическое, но недавно
в семействах, не связанных близким род..
ством С Boпellia"l были обнаружены карлико..
вые самцы, что может отражать независи..
мую эволюцию сходноrо механизма в раз..
личных филетических линиях этой rруппы.
Эхиуриды раздельнополы.. но у мноrих по..
следовательных rермафродитов смена пола
также сильно зависит от внешних условий.
Считается, что у таких животных в процес..
Размножение и жизненные циклы
405
о
A
1
II
III
Рис. 14.6. Определение пола у блюдечка Crepi
dula forпicata: 1 ранняя стадия образование
пары; 11, III более поздние стадии
образование сложных комплексов, включаю
щих самцов, самок и интерсексов
сах определения пола участвуют феромоны.
Рис. 14.6 показывает это на примере блю
дечка Crepidula jorпicata. Животные обра
зуют плотные стопки, причем самые ни
жние особи всеrда превращаются в самок,
а верхние в саМIIОВ: находяшиеся между
ними животные обычно становятся repMa
фРОДИТdМИ.
14.2.4. Адаптивное значение
различных способов
половоrо размножения
14.2.4.1. Зачем нужно
половое размножение?
Одна из задач эволюционной биолоrии
состоит в том, чтобы понять. почему суще
ствуют различные типы размножения. По
чему, например, половое размножение Ha
столько преобладает над друrими типами?
Ответ не так самоочевиден. как может пока
заться. Половое размножение приводит
к разнообразию потомства и скрывает про
явление потенциально вредных мутаций, но
эти преимушества необходимо сравнить
с недостатками и «стоимостью» процесса.
Во..первых, снижается эффективность пере
дачи reHoB следующему поколению, поско
льку от каждоrо родителя потомку ДOCTae
тся лишь половина reHoMa. Поскольку при
способленность определяется именно спо
собностью к сохранению reHoB в ряду поко
лений, речь идет об основной «статье pacxo
дов)} на половое размножение. BO BTOpЫX
необходимо тратить ресурсы на развитие
половых признаков и на брачное поведение;
наконец, риск для жизни 80 время поиска
партнера часто повышается.
cf
Мноrие теории считают, что половое
размножение выrодно виду или rруппе
в эволюционном масштабе времени. а бес
полое дает кратковременное преимущество
на уровне отдельных особей (концепция
rрупповоrо отбора). В последнее время раз
работаны более приемлемые теории, при
знающие селективное преимущество поло
Boro размножения для особи, а не для rруп
пы или вида.
Предполаrалось. что у особей. размно
жающихся половым путем, разнообразное
rенетически потомство будет в среднем луч
ше приспособлено к изменившимся усло
виям среды в будушем. чем одинаковые по
томки бесполоrо орrанизма. Соrласно этой
теории, преимущество половоrо размноже..
ния максимально в нестабильной среде и те..
ряется в относительно устойчивых уело..
виях. Однако анализ распространения бес
полоrо размножения среди беспозвоноч
ных, обитающих в различных биотопах, не
подтверждает TaKoro проrноза. Оно особен
но часто встречается у орrанизмов завися
щих от изменчивых ресурсов, тоrда как по..
ловое размножение свойственно почти всем
животным самых стабильных местообита..
ний. Так. бесполое размножение более
обычно для пресноводных форм, чем для
морских представителей тех же таксономи
ческих rрупп.
Альтернативная теория предполаrает,
что преимущество размножающихся поло
вым путем орrанизмов обусловлено конку..
рентоспособностью их разнообразных по..
томков В сложно орrанизованной среде
с интенсивной конкуренцией: однообразное
потомство бесполоrо животноrо не сможет
вытеснить их из всех возможных местооби
таний. Ее создатель Белл (ВеН. 1982) счи..
тает, что половое размножение должно пре
обладать в стабильных, но сложных биото"
пах, а бесполое там, rде численность вида
ниже максимальной которую может под
держивать данная cpeдa и возможен 1КОЛО"
rический оппортунизм. В этих условиях
преимущество получают животные с макси
мальной скоростыо роста или увеличения
численности потомства.
Существуют и друrие теории, пытаю..
щиеся объяснить преимущество половоrо
размножения (рис. 14.7). Некоторые из них,
406
rлава 14
Нет
Половое размножение важно для сохранения видов
Половое размножение историческое наследие,
не связанное в настоящее время с отбором
Проzнозuруется ко еляция половоrо размножения с
таксономическим положением, но не с эколоrическими
пара метрами.
НЕ СООТВЕТСТВУЕТ НАБЛЮДЕНИЯМ отверzаеТСR
2. Половое размножение поддерживается кратковременным
отбором на адекватную изменчивость
I
З. Половое размножение
преадаптирует потомство
к новым условиям среды
в будущем
Проzнозuруется половое
размножение rлавным о разом
'в нестабильных средах,
а беспол Ое В стабильных
НЕ СООТВЕТСТВУЕТ
НАБЛЮДЕНИЯМ
отверzаеТСR как общая
теория
+
Половое размножение
следствие пространственной
и биотической HeOДHOpoД
ности среды. В таких усло
виях общая приспособленность
раЗi-lOобразноrо потомства
особей, размножающихся
половым путем, будет выше,
чем у однородноrо потомства
от бесполоrо размножения
т. е. половое размножение
дает селективное преиму
щество отдельным особям.
ПроzнозuруеТСR преоблада
ние половоrо размножения
в стабильных сложных
средах, а партеноrенеза
в простых и нарушенных
СООТВЕТСТВУЕТ НАБЛЮ
ДЕНИЯМ принимается как
основа для общей теории
Да
Отбор, действующий в настоящее время,
поддерживает половое размножение в связи с ero
преимуществами
ПрuзнаеТСR но каков механизм?
Половое размножение поддерживается долrовремен
ной эволюцией при неадекватной современной
изменчивости 1
Половое размножение поддерживается за счет
вымирания популяций, размножающихся бесполым
путем I
Вымирание обусловлено
неспособностью адапти
роваться к изменениям
занятых ниш
Преимущество только
на уровне популяции
ПроzнозuруеТСR быстрое
вымирание и преоблада...
ние половоrо размноже
ния внестабильных
средах
НЕ СООТВЕТСТВУЕТ
НАБЛЮДЕНИЯМ
О тв epzaeТСЯ
Вымирание обусловлено
накоплением вредных
аллелей в стабильных
средах
ПроzнозuруеТСR большая
стабильность популяций
с половым ра множе
нием в простых, быстро
меняющихся средах
НЕ СООТВЕТСТВУЕТ
НАБЛЮДЕНИЯМ
О тверzаеТСR
Рис. 14.7. Алrоритм выбора между альтернативными теориями селективноrо преимущества по
ловоrо размножения (по Bell 1982)
например предположение, что половое раз
множение является механизмом предотвра-
щения заражения Bcero потомства парази
тами, совпадают в своих проrнозах с более
общими теориями, в частности с теорией
Белла.
14.2.4.2. rермафродитизм
и раздельнополость
Задаваясь вопросом, каковы селектив..
ные преимущества половоrо размножения
по сравнению с бесполым, лоrично спро
сить, чем определяется распространение
раздельнополости и rермафродитизма. При
этом следует учитывать как таксономиче..
ские, так и эколоrические аспекты. Напри..
мер, из табл. 14.3 видно, что в некоторых
таксономических rруппах преобладают
rермафродиты, а друrие состоят почти ис-
ключительно из раздельнополых фОРМ.
Кроме Toro, rермафродитизм чаще встре-
чается в определенных условиях среды: на-
пример, пресноводные и наземные аннели-
ды и моллюски rермафродиты.. а их мор-
ские сородичи в основном раздельнополы.
Аналоrичным образом среди rлуБОКОБОД"
ных ракообразных rермафродитизм более
распространен, чем среди близких мелко-
водных форм. Разнообразие типов полов о..
ro процесса у беспозвоночных позволяет
предположить, что тот или иной из них во-
зник под влиянием eCTecTBeHHoro отбор
поэтому можно попытаться найти функцио-
нальное объяснение ситуаций, в которых
встречается rермафродитизм
Размножение и жизненные циклы
Предложено несколько моделей эволю..
ции, три из которых вкратце рассмотрены
ниже
Модель низкой плотности: если плотно..
сть популяции низка., или орrанизмы ведут
сидячий образ жизни, синхронный repMa..
фродитизм повышает вероятность опло..
дотворения при неизбежно редких встречах
между особями, а если встреч не происхо..
дит, деЛdет возможным самооплодотворе..
ние.
Модель преимущества размеров: если
успех спаривания у одноrо пола связан
с размерами тела, а у друrоrо нет, возни..
кает последовательный rермафропитизм.
Популяционно..rенетическая модель:
при низкой численности популяции повы..
шается вероятность инбридинrа и дрейфа
[енов; в 1ТИХ условиях rермафродитизм по..
вышает эффективную численность популя..
ции.
Была также предложена более общая
теория, включающая в себя все три описан..
ные модели. В ее основе лежит простое но
rлубокое наблюдение: при половом раз..
множении у любой особи только одна мать
и один отец. При этом ровно половина re..
HOMd зиrоты вносится каждым из родите..
лей, поэтому и мужская, и женская репро..
дуктивные функции обеспечивают одинако"
вый успех размножения.
Объем ресурсов. которые может исполь..
зовать орrанизм., оrраничен, и их следует
распределять на поддержание жизнедеяте..
льности и воспроизведение таким образом.,
чтобы суммарная приспособленность особи
была максимальной. Определенная часть
затрат на половое размножение связана
с развитием вспомоrательных CTPYKTYP
протоков и желез. 06леrчающих движение
[амет, а также с повышенным риском rибе..
ли при поисках партнера. rермафродитам
приходится «тратиться» И на мужскую., и на
женскую репродуктивные функции. поэто..
му У них общие расходы любоrо конечноrо
ресурса на размножение, по"видимому., вы..
ше. Следовательно, можно ожидать., что
давление отбора блаrоприятствует разпель..
нополости.
Однако ситуация меняется, если «от да..
ча» от затрат на какую то из репродуктив"
ных функций падает: можно полаrать, что
407
в таких условиях отбор будет блаrоприяr
ствовать rермаФродитизму. Например., по..
ниженная «отдача» от энерrозатрат на жен--
скую функцию., вероятно., свойствеННd жи..
вотным, выращивающим потомство в BЫ
водковой камере с оrраниченным внутрен--
ным пространством.
14.3. Разнообразие половоrо
размножения:
жизненные циклы
14.3.1. Введение
Существует MHoro типов половоru раз..
множения наиболее важные признаки.,
в сумме составляющие репродуктивную
стратеrию, следующие:
1) максимальная потенциальная продол..
жительность жизни;
2) число эпизодов размножения в тече..
ние жизни;
3) характер расходования [амет: нако"
плени е и массовое использование или по..
степенное расходование по мере образова..
ния;
4) степень синхронизации половых ии..
клов особей в популяции
5) избирательность спаривания и CTe
пень аутбридинrа в популяции:
6) относительный рdзмер и «стоимость»
[амет;
7) тип развития и степень подверженно--
сти ювенильных и взрослых особей давле
нию отбора:
8) относительная доля всех доступных
ресурсов, вкладываемых в размножение.
Жизненные циклы животных часто клас..
сифицируют в зависимости от числа эпизо--
ДО В размножения в течение жизни и от про
должительности половозрелой стадии.
Используется несколько невзаимоисклю..
чающих систем., некоторые из которых
СРdвниваются в таб Т"I. 14.4. Различные типы
размножения также схематически преДСТdВ
лены в прил:ожении 14.4.
14.3.2. Морские беспозвоночные
rлубокое влияние на тип размножения
животных оказывает среда их обитания
Морские беспозвоночные MorYT высвобо
408
rлава 14
Таблица 14.4. Системы классификации типов размножения
практически синонимы; второй термин используе
тся в основном исследователями аннелид
В 'Эту катеrорию попадают все насекомые; их жизненные циклы далее классифицируются следующим
образом:
униволь тинный (однолетний):
му ль тиволь тинный:
биволь тинный:
семиваль тинный
Размножение происходит несколько раз в жизни
Полицикличность (полителия)
Ежеrодная полицикличность (полителия):
Размножение происходит один раз в жизни
Моноцикличность 1) (монотелия):
Непрерывная полицикличность (полителия):
одно поколение в [од
MHoro поколений в [од
два поколения в [од
одно поколение каждые два rода
размножение в определенные периоды, Рdзделен--
ные обычно однолетними перерывами
размножение идет относительно непрерывно. Если
продолжительность жизни составляет не более [o
да, такой жизненный цикл бывает неотличим от
унивольrинноrо
1) В анrлоязычной литературе термину «МОНОЦИКЛИЧНОСТЬ» соответствует «semelpdrity», а «полициклично
сты> «iteroparity». но производные от них типа «семель"арносты> и «итеропарносты) в советской Jlитературе не
привились. Прuм. ред.
ждать rаметы прямо в окружающую .среду,
rде и происходит оплодотворение, но прес
новодные и наземные формы такой возмо"
жности лишены. После наружноrо оплодот"
ворения из яиц морских беспозвоночных ча..
сто развиваются подвижные планктонные
личинки (см. разделы по систематике и rл.
15). Это оказывает большое влияние на
стратеrию размножения.
Почти во всех типах животных с морски"
ми представителями большинство или хотя
бы некоторые виды характеризуются нару..
жным оплодотворением и наличием пелаrи..
ческих личинок. Такой жизненный цикл, на..
зываемый пелаrо..бентосным, проиллю..
стрирован в приложении 14.5. Однако во
всех этих типах с пелаrической личиночной
стадией встречаются также виды с непела
rическими, бентосными личинками, что
обычно считается проrрессивным призна..
ком, связанным с утратой или модифика-- '
цией личиночной стадии. В большинстве ти"
пов животных имеются также виды, пелаrи--
ческие личинки которых не питаются, а су--
ществуют за счет запаса желтка, полученно--
ro от родительскоrо орrанизма. Перечи
сленные типы личинок называют планкто..
трофными пелаrическими и лецитотрофны--
ми пелаrическими, а развитие без пелаrиче..
ской личиночной стадии прямым. Это
разделение не абсолютно, и развитие мно"
rих личинок смешанное. Наличие пелаrиче..
ской личинки обычно считают примитив..
ным признаком. У более 700/0 всех морских
беспозвоночных умеренных зон планкто..
трофные пелаrические личинки, что подра..
зумевает явные преимущества TaKoro спо..
соба развития в большинстве ситуаций, за-
ключающиеся, по--видимому, в следующем:
1) использование BpeMeHHoro источника
пищи во время «цветений» фитопланктона;
2) заселение новых местообитаний;
3) расширение rеоrрафическоrо ареала;
4) избеrание катастрофическоrо вымира..
ния при исчезновении локальных местооби..
таний.
,
5) избеrание локальной конкуренции
и конкуренции между сибсами;
6) предоставление разнообразному по..
томству максимальноrо разнообразия ме..
стообитаний.
Пелаrическое развитие часто (но не Bcer..
да) связано с наружным оплодотворением,
которое в свою очередь сочетается с еше од..
ним предположительно примитивным при..
знаком простой формой сперматозоида
с круrлой rоловкой вроде приведенноrо на
рис. 14.10. rаметы TaKoro типа встречаются
во всех rруппах животных, и две эти черты
характеризуют особый морской тип раз..
Размножение и жизненные циклы
409
Таблица 14.5. Примитивные и производные признаки морских беспозвоночных 1)
Исходные признаки
ПрОИЗБодные признаки
Яйца высвобождаются непосредственно в воду, Яйца не высвобождаются; часто вынашиваются
пелаrическое развитие
Наружное оплодотворение; форма сперматозои
дов примитивная
Небольшое количество желтка в яйце
Полное равномерное дробление
Бластула с б.;1астоцелем
rаструляция посредством инваrИНdЦИИ
Планктотрофная личинка
Автосинтетический ооrенез
Размножение один раз в [од в cTporo определен
ный период, с четкой синхронизацией raMeTore
неза у одной и у различных особей
Длительное хранение накопленных половых кле
ток
Крупный Рdзмер тела. часто обширная полость
тела или широкие межклеточные пространства
1) Некоторые данные из Jagersten, 1972.
Внутреннее оплодотворение, ЧdСТО СОПрОВОЖДdЮ
щееся хранением спермы; форма сперматозои
дав сложная
Большое количество желтка в яйце
Неравномерное дробление
Бластула более или менее заполнена
rаструляция друrоrо типа
Лецитотрофная личинка
Ооrенез друrоrо типа
MHoroKpaTHoe, часто относительно непрерывное
размножение: сезонность и синхронизация [а
метоrенеза не выражены
Частое использование небольших КОТIичеств поло..
вых клеток без их накопления
Тело небольшое, с частично заполненной поло
стью. Накопление [амет в межклеточных про
странствах невозможно
множения. С ними связаны и некоторые
друrие а все вместе они составляют ком--
плекс коррелирующих между собой особен--
ностей жизненных циклов морских беспо..
звоночных (табл. 14.5, «Исходные призна--
ки»). Производные.. или проrрессивные,
признаки, развившиеся на основе этих пред--
положительно примитивных, также перечи--
елены в табл. ]4.5. Они особенно характер..
ны для очень мелких морских беспозвоноч"
ных, обитающих, например, в интерстициа..
ли песчаноrо дна.
При эволюции относительно крупных раз..
меров тела оказалось возможным сохране--
ние типа размножения, считающеrося при..
митивным. В типах животных без крупных
форм, например у киноринхов, а также
у мелких моллюсков, аннелид и иrлокожих
(с крупными сородичами) наблюдаются
в основном продвинутые признаки. Они
встречаются также у морских беспозвоноч"
HЫX обитающих в экстремальных или не..
обычных условиях, например на большой
rлубине или в полярных районах, а также
в эстуариях.
Важным свойством, коррелируюшим
с «примитивным}) типом размножения,
является массовое высвобождение [амет во
время ежеrодных нерестов. Как правило,
морские беспозвоночные накапливают их
в полостях тела (часто в целоме), а затем
полностью расходуют в течение единствен
Horo HepecToBoro сезона. Одной из причин
'Эволюции продвинутоrо типа размножения
у мелких орrанизмов, возможно, была их
неспособность запасать достаточное коли--
чество raMeT. Два различных типа жизнен..
ных циклов С массовым и четко сезонным
нерестом рассмотрены в приложении
14.4(3). Более распространен ежеrодный не--
рест при продолжительности жизни два ro..
да или более. Однако некоторые виды. Ha
пример полихеты семейства Nereidae и все
rоловоноrие моллюски (за исключением
примитивных наутилоидей) нерестятся все..
[о раз в жизни. Их массовый нерест сопро--
вождается rенетически запроrраммирован
ной rибелью.
Для мяrкотелых пресноводных и назем--
ных беспозвоночных также характерны про--
двинутые репродуктивные признаки, приве..
денные в табл. 14.5. Они описаны в следую--
щем разделе.
410
rлава 14
14.3.3. Пресноводные
и наземные беспозвоночные
у животных, обитающих в пресных во--
дах или на суше, нет выбора между планк--
тотрофным и лецитотрофным пелаrиче--
ским или прямым развитием. В этих средах
осмотические и друrие оrраничеиия не до..
пускают выделения в среду незащищенных
сперматозоидов и яйцеклеток. так что опло..
дотворение должно быть внутренним. Раз..
вивающиеся эмбрионы также требуют за..
щиты от потерь водь] и осмотическоrо
стресса, например с помощью водонепро--
ницаемых покровов или внутри кокона.
Следствием этоrо явилась утрата прими--
тивной пелаrической личинки, и размноже..
иие крупных мяrкотелых неморских беспо--
звон очных характеризуется следующими
признаками:
1) живорождение или откладка яиц, за..
щищенных непроницаемыми мембранами
или коконами:
2) внутреннее оплодотворение, требую--
щее непосредственноrо спаривания партне--
pOB
3) структурно усложненные.. часто ните..
видные сперматозоиды;
4) относительно большой вклад мате..
ринских ресурсов в каждое яйцо, что сни"
жает плодовитость:
5) забота о потомстве или обеспечение
ero пищей;
6) мноrократная или эпизодическая от..
кладка яиц с использованием BHYTpeHHero
запаса спермы для их оплодотворения;
7) rермафродитизм (особенно характе..
рен для мяrкотель]х беспозвоночных, но не
свойствен членистоноrим).
Признаки некоторых пресноводных жи..
вотных выдают их недавнее происхождение
от морских предков. Так. пресноводные
двустворчатые моллюски. например без--
зубка {Aпodoпta), очень сходны со своими
морскими сородичами. Они., в tIастности,
образуют довольно мноrочисленные 2ЛОХU"
дии (МОПИфИIIированные личинки..
велиrеры), которые, однако, не являются
свободноживущими, а распространяются,
прикрепляясь к жабрам или коже рыб.
Жизненные IIИКЛЫ пресновопных пло..
ских червей., поясковых аннелид и леrочных
моллюсков очень сходны. Все они
синхронные rермафродиты (кроме утратив..
ших способность к половому размноже..
нию) и характеризуются сложным половым
поведением, высокоспециализированными
придаточными железами и способностью
зашишать развивающиеся эмбрионы. В их
жизненные циклы входят продолжительные
периоды откладки яиц (см. приложение
14.4). Основной компромисс, На который
они вынуждены идти. между плодовито
стью и продолжительностью жизни; неко"
торые виды живут Bcero [од или меньше,
друrие, обычно более крупные. несколько
лет.
Так как в пресноводной или наземной
средах колебания физических параметров
значительно сильнее. чем в море. большин..
ство обитающих в этих биотопах беспозво..
ночных способны переходить в состояние
физиолоrическоrо покоя, называемое диа..
паузой., при котором орrанизм пере носит
без вреда для себя потери воды и экстрема..
льные окружающие условия. Ero мета боли..
ческие потребности снижены, он может об..
ходиться без внешних источников энерrии
и за счет 1Toro переживать пеРИОДЬJ отсут"
ствия корма, характерные для умеренных
и полярных областей. У долrоживущих ви"
ДOB например некоторых леrочных моллю"
сков и поясковых кольчецов., диапауза Ha
ступает во взрослом состоянии, а у мелких
форм она часто соответствует стадии яйца.
Так, мноrие популяции пиявок в течение
зимних месяцев представлеНЬJ только
эмбрионами в коконах. Перезимовывание
в виде покоящихся яиц особенно характер..
но для таких крошечных форм, как коло..
вратки и некоторые rруппы ракообразных,
в частности для водяных блох (Dapll1lla).
у этих орrанизмов жизненные циклы вклю"
чают чередование половых и бесполых по..
колений (рис. 14.4): из диапаузирующеrо
яйца, образующеrося в результате полово..
[о размножения, весной вылупляются бес..
полые особи, которые дают начало летним
клонам.. используюшим обильные в это
время ресурсы. Крайняя форма диапаузы
наблюдается у бделлоидных коловраток: их
инцистированные яйца MorYT находиться
практически в состоянии анабиоза мноrие
[оды, пока не попадут в подходящие для ро..
Размножение и жизненные циклы
ста условия. Это свойство, возможно,
связано с отсутствием половоrо размноже..
ния: избеrая неблаrоприятных условий, они
живут только в оптимальной для себя
среде.
Н асекомые и па укообразные приспособи..
лись к наземной жизни особенно успешно
(см. разд. 8.4.3.2 и 8.5.3Б.2); кроме тото, не..
которые их личиночные, нимфальные или
половозрелые стадии (особенно унасеко"
мых) вторично заселили пресные воды. Ре..
продуктивная биолоrия этих rрупп отличае..
тся от свойственной мяrкотелым беспозво..
ночным одним очень важным признаком:
среди них очень редко встречаются терма..
фродиты; в подавляющем большинстве
случаев возможно только половое размно"
жение, причем всетда с раздельнополостью.
Эволюционный успех здесь в значительной
мере связан с появлением водонепроницае..
мых покровов. а также способностью
к образованию водонепроницаемых яиц.
Они должны быть оплодотворены до от..
кладки, поэтому у всех одно ветвистых вну"
треннее оплодотворение. часто связанное со
сложным брачным поведением. Преоблада..
ние раздельнополости в этих rруппах, ве..
роятно, объясняется высокой подвижно..
стью животных.
Жизненные циклы несекомых тради..
ционно классифицируют по типу разви..
тия c неполным (экзоптериrоты) или пол..
ным (эндоптериrоты) превращением (см,
rл. 15). Здесь, однако, будет использован
иной, функциональный подход. Жизненный
цикл насекомото можно рассматривать как
смену различных типов активности в юве
пильном и имаrинальном состояниях. По
скольку развитие включает несколько ди..
скретных стадий, разделенных линьками,
каждая стадия может выполнять специ(ри..
ческие фУНКIIИИ. основные из которых пере..
числены ниже:
1) развитие и ди(рференцировка;
2) накопление пищи и друrих ресурсов;
3) расселение и поиск ресурсов;
4) спаривание и выбор партнера;
5) обеспечение ресурсами потомства;
6) выбор места для развития потомства;
7) откладка яиц
Жизненные циклы некоторых насеко..
мых проанализированы с этой точки зрения
411
в приложении 14.6. Обратим внимание на
то, что накопление ресурсов и расселение
мотут происходить на различных стадиях
развития, а также на отличие по этим пара..
метрам большинства насекомых от мор..
ских беспозвоночных, у которых расселение
обычно происходит на ювенильной стадии,
а накопление pecypCOB BO взрослом со..
стоянии.
Самый простой тип развития такой.. при
котором переход от личинки к имаrо проис..
ходит постепенно, причем все стадии испо..
льзуют одни И те же пищевые ресурсы. На..
копление их также идет на всех стадиях. но
взрослые особи, кроме тото, выполняют
функции расселения, поиска ресурсов.. спа..
ривания и откладки яиц. Такой тип разви"
тия характерен для прямокрылых (напри
мер, саранчи) и про иллюстрирован в прило..
жении 14.6(1).
Однако довольно часто личинки и имаrо
различаются по характеру питания, и. сле..
довательно, давление отбора для них не..
одинаково. Например, личинки падальной
мухи (двукрылое) используют обильный, но
временный источник пищи мясо мертвых
животных. Найти ето задача родитель..
ской особи, которая часто питается иначе
[см. приложение 14.6(2)]. Личинки некото"
рых насекомых обитают в воде, так что от..
личие их Jколоrической ниши от имаrина..
льной еще более выражено.
На таком несходстве, вероятно, основа..
на эволюция жизненных циклов, в ко [орых
функции расселения и поиска ресурсов пол..
ностью отделены от функции их накопле..
ния, так что имаrо почти или совсем не пи..
таются. Это разделение в (рилоrенезе насе..
комых возникало неоднократно инезависи..
МО. Крайними ето примерами MorY1 слу..
жить поденки, веснянки и ручейники. Их ли..
чинки водные хищники. а взрослые особи
не питаются и живут Bcero несколько часов,
в течение которых должны спариться и от..
ложить яйца в места, подходящие для их
развития [см. приложение 14.6(3)].
у чешуекрылых жизненный цикл сход..
ный, хотя взрослые бабочки все же полу..
чают некоторое количество 1нерrии.. потре..
бляя нектар. Основная фУНКIIИЯ их личинок
(rусениц) накопление как можно больше..
то количества доступных ресурсов с макси..
412
rлава 14
мальной скоростью. Переход ко взрослому
состоянию, в задачи KOToporo входит спа..
ривание и поиск ресурсов, осуществляется
через непитающуюся стадию куколки [при..
ложение 14.6(3)]. Пищевые ресурсы бывают
доступны лишь в определенные сезоны, их
поиск может идти как в пространстве, так
и во времени" включая диапаузу на стадии
яйца, куколки или имаrо, а также миrра..
цию.
Некоторые взрослые насекомые кроме
расселения" спаривания и поиска ресурсов
занимаются также накоплением последних.
При этом они только находят их, чтобы
передать потомству. Такой жизненный цикл
известен у некоторых прямокрылых" жест..
кокрылых и двукрылых, но наиболее харак"
терен для перепончатокрылых [см. прило..
жение 14.6(4)]. Считается, что он сыrрал
rлавную роль в )волюции эвсоциальноrо
поведения пчел и ос [см. приложение
14.6( 5)].
14.4. Контроль процессов
размножения
14.4.1. Первичные
инепосредственные
контролирующие факторы
Жизненные циклы, обсуждавшиеся
в предшествующих разделах, включают
сложные переключения клеточных процес..
сов, которые должны быть скоординирова..
ны во времени и в пространстве как друr
с ДРУI ОМ, так с внешними факторами, в не..
которых случаях обеспечивая необходимый
уровень синхронизации жизненных отправ"
лений особей в популяции. Принято счи
тать.. что четкие IIИКЛЫ репродуктивной aK
тивности в популяциях животных связаны
с циклическими изменениями внешних
условий, в определенные периоды более
блаrоприятных для размножения. чем
в друrие. Это создает давление отбора, на..
1. Меняющиеся параметры внешней среды и
сиrналы времени
Долrота ОСОБые химические
Температура ДНЯ Пища вещества
\ \ I I
Восприятие сиrналов
Uентральная
нервная система
Ф длинные или короткие ДН
Температура
Приливы и сразы луны
Взаимоотношение сраз
2.
IСуммирование и интеrраЦИЯI
3.
Нейроэндокринная система:
ПРЕОБРАЗОВАНИЕ
4
Эндокринная реакция
Пролиq>ерация raMeT / /
Развитие raMeт /.
ВЫСВОБождение
raMeT
5. НаБлюдаемая цикличность ОБразования и ВЫСВОБождения raMeT
1. Изменение условий-
срепы
2. Восприятие этих
изменений
3. ПреОБразование
сиrнала
4. Прямое и косвенное
влияние на лимитиру...
ющие процессы
5. Внешний цикл raMe
тоrенеза и размно'"
жен ия
Рис. 14.8. Схематическое изображение цепочки контролирующих элементов участвующих в pe
rуляции репродуктивноrо цикла, синхронизированноrо с внешними событиями (по Olive 1985)
РазмножеНИЕ и жизненные циклы
правленноrо на синхронизацию репродук
тивной активности с такими оптимальными
периодами. Это давление обусловлено пep
вИЧНblМИ факторами, контролирующими
размножение Они не обязательно непос
редственно реrулируют репродуктивный
цикл. rаметоrенез (особенно ооrенез) иноr
да продолжается несколько месяцев, и для
правильноrо протекания этих клеточных
процессов, завершающихся размножением,
орrанизм должен ориентироваться по co
вершенно иным факторам внешней среды.
События, используемые для такой реrуля
дии, называют непосредствеННbl./ИИ факп1.0
рами Воспринятая информация об измене..
ЭМБРИОНАЛЬНОЕ
РАЗВИТИЕ
Сперматоrониальная
стволовая клетка
Сперматоrонии
(митотическо ) е
размножение
Сперматоцит
1 порядка
1\
Сперматоциты
II порядка
х2
? ерматиды @ @ @ @
СПЕРМАТО I I I I
rЕНЕЗ
i
МЕЙОЗ 1
413
ниях среды должна быть обработана в цeH
тральной нервной системе и передана
к ,ффекторам в виде нервных, нейроэндо
кринных или эндокринных сиrналов (разд.
16.12). Последовательность таких команд
определяет высокоупорядоченный видоспе
цифичный репродуктивный цикл (рис. 14.8).
Под их контролем находится дифференци
ровка половых клеток и распределение
энерrии между поддержанием жизни, poc
том и размножением.
14.4.2. rаметоrенез
Мужские половые клетки образуются
в результате сперматоrенеза, а женские
Ооrониальная стволовая
клетка
Ооrонии (митотическое
размножение)
Ооцит 1 поряпка
ВИТЕллоrЕНЕЗ
МЕЙОЗ 1
Ооцит П порядка +
полярное тельце
МЕЙОЗ II
(i) Яйцо + 3 полярных тельца
rАМЕтоrЕНЕЗ у САМКИ
rАМЕтоrЕНЕЗ у САМЦА
Рис. 14.9. Схематическое изображение основных клеточных процессов rаметоrенеза
414
rлава 14
А Б 8 r
Рис. 14.10. А. Примитивный сперматозоид
с круrлой rоловкой. Б r. Более продвинутые
нитевидные типы сперматозоидов которые
MorYT иметь различное внутреннее строение
ооrенеза; схемы этих весьма различных
пропессов показаны на рис. 14.9 и более де..
тально они рассмотрены ниже.
14.4.2.1. Сперматоrенез
у большинства беспозвоночных сперма
тоrенез протекает быстро; мейозу предше..
ствуют мноrочисленные митотические дe
ления.. что приводит К образованию очень
большоrо числа половых клеток. Каждый
сперматоrоний превращается в спермато
цит первоrо порядка, из KOToporo образую--
тся четыре сперматиды. Сперматозоиды
формируются в результате дифференциров
ки последних (рис. 14.9, А). У мноrих MOp
ских беспозвоночных сперматозоиды сход--
ные, с круrлой rоловкой (рис. 14.10, А), но
для некоторых характерно более сложное
строение (рис. 14.10, Б п; это обычно Ha
блюдается в жизненных циклах, включаю
щих хранение спермы, внутреннее оплодот
ворение., а также тоrда., коrда у яиц прочные
защитные покровы (см. выше).
14.4.2.2. Ооrенез
Ооrенез это продолжительный про--
цесс, в ходе KOToporo в развивающемся
яйце откладываются запасы питательных
веществ (обычно в течение длительной про
фазы мейоза), а мейоз завершается либо не..
посредственно перед оплодотворением, ли..
бо уже после Hero.
у некоторых морских беспозвоночных,
например сипункулид, эхиурид и некоторых
полихет, развивающиеся ооциты по отдель..
ности свободно плавают в uеломической
жидкости так называемый 0О2енез СОДИ"
f!)
v
А
в II
Рис. 14.11. Солитарный, фолликулярный и HY
триментарный типы ооrенеза. А. Солитарный
тип: ооциты не связаны с вспомоrательными
клетками, а свободно взвешены в жидкостях
тела. Характерен для эхиурид, сипункулид И He
которых (по видимому, немноrих) полихет. Б.
Фолликулярный тип: ооциты сильно зависят
от покрывающих их соматических клеток. В
Нутриментарный тип: ооциты связаны с ce
стринскими клетками, происходящими из Toro
же зачатка, что и сам ооцит. В результате He
полноrо цитокинеза часто возникает синцитиа
льный комплекс, в котором ооциты И питаю
щие клетки соединены цитоплазматическими
мостиками или канальцами. 1. Пример ооцита
без покрывающих фолликулярных клеток (по
лихета Diopatra). 11. Пример ооцита, окружен
Horo фолликулярными клетками, что xapaKTep
но для двукрылых насекомых, например дл.я
мухи Drosophila (см. также рис. 14.13)
Рис. 14.12. Синтез желтка в ооцитах. Автосин
тетический тип, характерный для солитарноrо
ооrенеза. Низкомолекулярные предшествен
ники поrлощаются микроворсиночной поверх
ностью ооцита, а синтез высокомолекулярных
запасных веществ происходит в ero цитоплаз
ме с участием информационных молекул
образующихся в ядре. ЭР эндоплазма
тический ретикулум
1
III
v
Низкомолеку
лярные предшествен
ники желтка
o
о
Рис. 14.1 З. Клеточные процессы при развитии яйца двукрылых насекомых (например, у Drosophi
'а), 1. При делении стволовой клетки образуется стволовая клетка следующеrо поколения и поло
вая клетка первоrо порядка. 11 V. Половая клетка первоrо порядка делится четыре раза, но за
счет неполноrо цитокинеза образуется комплекс из 16 клеток, соединенных цитоплазматически
ми мостиками. Обратим внимание, что у двух заштрихованных клеток на любом этапе деления
больше связей с соседними, чем у любых друrих. Одна из этих двух клеток превратится в ооцит,
остальные станут питающими. VI. Питающие клетки действуют как система rенной амплифика
ции: образующиеся в их ядрах информационные молекулы (мРНК) поступают в цитоплазму oo
цита. Весь комплекс окружается фолликулярным эпителием из соматических клеток. VII. Ооцит
растет за счет питающих клеток. VIII. Зрелое яйцо полностью заполняет фолликул. Оно rOToBo
к эмбриоrенезу после оплодотворения и откладки
416
rлава 14
Низкомолекулярные
преДUJественники
Высокомолекулярные
составляющие
желтка
3ндопиноцитоз
o
СРолликулярные
клетки
Рис. 14.14. rетеросинтетический тип синтеза запасных веществ, обычно (но не обязательно)
связанный с фолликулярным развитием ооцита. Сложные высокомолекулярные предшественни
ки желтка приносятся к ооциту жидкостями тела (кровью, rемолимфой, целомической жидко
стью и т. д.). Эти вещества синтезируются в неполовых клетках с использованием их информа
ционных молекул. Приведенная схема типична для некоторых насекомых
тарНО20 {пиnа (рис. 14.11, А). При этом бу
дущие запасные вещества поrлощаются 00"
цитом в виде низко молекулярных предше..
ственников (аминокислот, простых сахаров
и моноrлицеридов), из которых орrанеллы
ero цитоплазмы синтезируют высокомоле..
кулярные вещества, в совокупности назы..
ваемые «желтком» (как показано на рис.
14.12). Такой характер накопления послед..
Hero называют авrпОСUНJпеrпuчеСКUJИ; он
обычно (но не всеrда) связан с солитарным
типом ооrенеза (см. ниже).
Однако для большинства беспозвоноч"
ных характерны фоллuкулярный или нутри..
/vlенп1арный типы ооrенеза (рис. 14.11, Б, В),
при которых развивающийся ооцит тесно
связан с эпителиальным слоем соматиче..
ских клеток (называемых фолликулярными)
и/или со своими сестринскими клетками,
образующимися в результате неполноrо ци..
токинеза при митотических делениях мате-
ринской клетки (ооrония) до начала профа..
зы мейоза. Развитие сложноrо яичника та-
Koro типа детально рассматривается на рис.
14.13 на примере двукрылоrо HaceKOMoro
Dl"Osop/1ila. Накапливается все больше дан-
ных о том, что при более сложных типах
ооrенеза большинство высокомолекуляр-
ных веществ, накапливающихся в цитоплаз-
ме ооцита, синтезируется не им самим, а со-
матическими клетками в друrих частях ар-
rанизма и доставляется к ооциту жидкостя-
ми тела или кровью по сосудам (рис. 14.14).
Такой тип ооrенез(! называют ?етеросинте-
п1uческuм. Впервые он был обнаружен у на-
секомых, но затем оказался широко распро-
страненным, характерным, например, для
ракообразных и моллюсков.
Рис. 14.15. Схема строения турбелларии, показывающая обширную систему желез участвующих
в образовании сложноrо «яйца» состоящеrо из оплодотворенной яйцеклетки желточных клеток
и белковой оболочки. Стрелками показан путь движения яйца из яичника, в ходе KOToporo оно
оплодотворяется и окружается совместно с питающими клетками белком перед выведением на-
ружу (см. также рис. 3.46)
Размножение и жизненные циклы
417
Ранее считалось., что для полихет типи.. щий вещества, запасенные в ЦИТОПЛdзме
чен автосинтетический солитарный ооrенез, яйцеклетки.
но. как выяснилось, эта rруппа по разно--
образию и сложности вариантов ооrенеза 14.4.3. Синхронное размножение
и участвующих в нем структур не уступает морских беспозвоночных
насекомым. у некоторых видов обнаруже--
ны ero фолликулярный и нутrиментарный Наиболее распространенный тип раз..
типы, а у нереид (семейство Nereidae) он.. множения морских беспозвоночных (разд.
хотя и солитарный, но, вероятно, rетеросин-- 14.3.2) требует очень точной синхронизации
тетический. репродуктивных процессов как в одном ор..
«Яйцо» турбелларий обычно представ-- rанизме так и между особями одной попу..
ляет собой сложное образование.. состояшее ляпии. Эта точность бывает весьма впечат
из почти не содержащеrо желтка ооцита ляющей. В табл. 14.6 приведены даты нере--
и питающих клеток с цитоплазмой, запал-- ста тихоокеанскоrо червя палоло (Polychae
ненной желтком, синтеЗИРОВdННЫМ в осо-- ta, Eunicida) за последние 100 лет: ero пери..
бом opraHe - желточнике (рис. 14.15): все од жестко связан с третьей четвертью луны
ЭТО упаковано в плотную белковую оболоч-- наступающей после определенной даты
ку. Такие яйца называют эктолепитальны-- в начале октября. Сроки соблюдаются
ми; считается, что для плоских червей этот с точностью до одноrо дня, причем нерест
ПРИЗНdК продвинутый, а примитивное со.. каждый раз начинается в одно и то же время
стояние эндолецитальный тип с желтком, суток. Так же cTporo соблюдаются сроки
запасенным в цитоплазме ооцита; он встре-- размножения у японской морской лилии
чается у некоторых свободноживущих тур.. Coп1aпtl1us japoпicus; на рис. 14.16 показано,
белларий. У пресноводных и наземных ан.. что весь rодовой цикл rаметоrенеза у этих
нелид и моллюсков адаптапии сходные: от-- животных CTporo синхронизирован. Приве
кладываемые яйца окружены коконом, со.. денные примеры представляют собой, по..
держащим питательный белок, дополняю-- видимому, крайние случаи, но у большин..
Таблица 14.6. Подборка данных/ иллюстрирующая синхронность всплывания для нереста по
nQ8Qзрелых червей палоло (Euпice viridis) у островов Самоа
{ 1843
19 лет 1862
{ 1874
19 лет 1893
1926
]927
1928
19 лет 1929
1930
, 1943
1944
1945
Третья четверть луны Даты всплывания
Октябрь Ноябрь Октябрь Ноябрь
16 14 i 15 / 16
15 14 15/16 14/15
31 30 {Н I
31 29 31 I
27 26 28
17 15 17
5 4 4
24 23 25
14 13 14 15
20 19 20
8 7 7/9
27 26 28
[од
Эти данные свидетельствуют о том, что а) нерест никоrда не происходит ранее 8 октября б) он происходит
в октябре, если третья четверть луны наступает позднее 18 октября; в) каждые 19 лет нерест приходится на одно и то
же число. Кроме l'oro, время суток, коrда взрослые особи всплывают на поверхность моря также точно фиксирова
но.
418 rлвва 14
50% 13
250 I 43 3
L 4
L 200 4
m
о
150
:s: 6
=r 7
о
о
а. 100
ф
(о 50
:s:
с:[
Рис. 14.16. Синхронное развитие яиц у морской лилии Coтaпthus japoпicus. Цикл rаметоrенеза
занимает 12 месяцев и заканчивается синхронным спариванием всех особей популяции. Оно про
должается двое суток и cTporo коррелирует с фазами Луны. rоризонтальной полосой обозначено
время размножения rаметоцитов (по Holland et al., 1975)
ства морских беспозвоночных" относящихся
к любому типу, про является та или иная
степень синхронизации репродуктивных
процессов. Возникает вопрос чем же pery--
лируются эти циклы? Ранее считалось. что
основными сиrналами служат изменения
температуры окружающей среды, но для
обеспечения такой точности" как в двух при--
веденных выше примерах. этоrо. очевидно.
недостаточно, и вообще у температурных
циклов слишком велика случайная изменчи..
вость (т. е. у сиrнала слишком высок уро..
вень шума). чтобы они моrли служить един..
ственным реrулирующим фактором.
Таблица 14.7. rеофизические ЦИКЛЫ в морской среде
Море сложная среда с множеством
ритмических изменений (табл. 14.7), и ре..
продуктивные циклы морских беспозвоноч"
ных MorYT быть соrласованы по фазе с каж..
дым из средовых uиклов. Было показано.
что такие разные животные, как полихеты,
иrлокожие и ракообразные, способны четко
реаrировать на длину cBeToBoro дня, при..
чем иноrда не менее сложно.. чем в случае
описанных ниже фотопериодических реак"
ций наземных насекомых. Кроме Toro,
у них известны непосредственные реакции
Hd смены фаз луны. а также эндоrенные рит..
мы с примерно лунной или приливной пе..
Название
Периодичность
Повторение одинаковых сочетаний фаз СОЛНllа и Луны
Оборот Земли BOKpyr Солнца
Оборот Луны BOKpyr Земли
Повторение одинаковых сочетаний фаз приливноrо
и солнечноrо циклов (цикл квадратурноrо/сизиrийно"
ro приливав)
лунные сутки
1 солнечные сутки
Время между полной или малой водой и средней водой
Метонов
rодичный
Лунный
Полулунный
п риливный
Суточный
П ОЛУПРИ.,1ивный
19 лет
1 rод
29.5 сут
15 сут
24,8 ч
24 ч
12,,4 ч
Результаты наблюдений rоворят о том, что репродуктивная dКТИВНОСТЬ по крайней мере каких то морских бес-
позвоночных коррелирует с каждым из этих циклов. Это, однако, не позволяет выявить причинный фdКТОр (обсужде-
ние в тексте).
Размножение и жизненные циклы
III
Циркадианные
компоненты
419
Часы
О 10 20 30 40
I I I I I
60 70
I I
Свет/темнота
II
Лунны€
компоненты
'Разы о
Луны
Ir
1
r одовые
компоненты
Месяцы
Модуляция сракторами
внешней среды
Рис. 14.17. Схематическая модель высокосинхронизованноrо спаривания животных типа MOp
екай лилии Coтaпthus (рис. 14.16) или полихеты Euпice (табл. 14.6). Предполаrается, что раз
множение запускается взаимодействием эндоrенных ритмов, каждый из которых, возможно, Ha
страивается внешним времязадателем с rодовой, лунной или суточной периодичностью В резу
льтате размножение происходит в течение одноrо или двух месяцев в rод в дни, жестко связанные
с лунным циклом, и в CTporo определенное время суток (по Olive, 1985)
риодичностью, которые настраиваются
(<<захваТЫВdЮТСЯ») цикличностью соответ..
СТВУЮЩИХ внешних процессов (<<времязада..
телей»). причем эти внутренние «часы»
очень точно «показывают» лунное или при..
пивное время. Накапливаются сведения, [о..
ворящие о том, что в основе rодичных ре..
продуктивных uиклов MorYT также лежать
эндоrенные ритмы с периодом, близким [о--
ДУ (рис. 14.17).
Репродуктивные циклы морских беспо..
звоночных включают чередование двух ос..
новных процессов образования и высво..
бождения [амет. Эти циклы MorYT реrули..
роваться циклическими изменениями обра--
зования [ормонов. По характеру действия
можно выделить два основных типа послед..
них: rонадотропные и индуцирующие не..
реет или созревание и/или активацию [aMe
тоцитов. как показано на рис. 14.18. Такой
тип эндокринноrо контроля размножения
возникал неоднократно и независимо в ходе
эволюции беспозвоночных, поэтому ве..
щеетва, ero осуществляющие, MorYT иметь
у разных животных совершенно различную
молекулярную структуру. У иrлокожих [o
надотропная функция выполняется двумя
rормонами" типичными также для позво..
ночных,, проrестероном и эстроном, а не..
рест вызывается каскадом сиrналов, начи..
нающимся высвобождением нейросекре..
TopHoro пептида их радиальных нервов, LITO
в свою очередь индуцирует образование
в яичнике простоrо реrуляторноrо вещества
]..метиладенина. Последний стимулирует
сокращение мыши яичника, т. е. начало не..
реста, а помимо этоrо образование тре..
Tbero посредника на внутренней поверхно..
сти мембраны ооцита, что в конечном счете
ведет к исчезновению ero ядерной оболочки
и делает возможным оплодотворение.
Сходные функциональные взаимоотноше
ния обнаружены у полихет и моллюсков"
хотя УЧdствующие в них вешества имеют
совершенно иную молекулярную природу.
По"видимому, эти rормональные механиз..
мы 'Эволюционировали независимо.
Некоторые морские беспозвоночные
420
rлава 14
...... Активность rонадотропноrо ropMOHa
Активность ropMOHa нереста
....... ......... ....... ....... .............. ........"
/" \
I \
I \
I \
/ \
J
I
/
/
оматический рост МОБилизация запасов Q) Рассасывание яиц
:s:
I .....
ro ()
со ф Соматический рост
Накопление Накопление репродуктивных ф Q.
Q. Ф
М :r:
запасов клеток о Накопление запасо
u
:s:
t>
о
:r:
со
:s:
.....
со
()
Q)
с:[
:r:
S
А
Время
С
Б
в
Рис. 14.18. Контроль rодовоrо репродуктивноrо цикла rонадотропным и индуцирующим нерест
rормонами: А обобщенная схема; Б клеточные события связанные с различными фазами
rормональноrо цикла
размножаются не циклически, а лишь раз
в жизни (см. выше). Участие эндокринной
системы в контроле репродуктивных про..
цессов у этих животных, по--видимому, су..
щественно отличается от описанноrо вы--
ше здесь происходит обычно необрати--
мое переключение с соматическоrо роста на
раЗl\1ножение. Такой механизм наблюдае--
тся у нереид. Созревание и часто сопрово"
ждающий ero соматический метаморфоз за..
висят от постепенноrо снижения уровня
единственноrо [ормона (см. rл. 16), необхо--
димоrо для реrенерации и образования се--
[ментов, Т. е. по мере созревания животноrо
ero способность расходовать ресурсы на
данные процессы утрачивается. Это
адаптивный признак, так как после размно"
жения животное неизбежно умирает, новые
сеrменты все равно не моrли бь] повысить
ero будущую плодовитость И, таким обра--
зом. оказываются бесполезными.
у полителических червей (см. табл. 14.4)
ситуаuия иная.. поэтому у них способность
к реrенерационному росту не связана с раз..
множением так жестко. Характер эндокрин--
Horo контроля репродуктивных процессов
и общая репродуктивная стратеrия орrаниз
ма, по"видимому, тесно коррелируют, но
остается неясным, какой из этих двух фак
торов является причиной, а какой
следствием.
Сходная ситуация наблюдается у мол
люсков. Для большинства их видов xapaK
терна полицикличность, и считается, что их
эндокринные системы соответствуют об
щей схеме.. приведенной на рис. 14.18. Одна..
ко у большинства rоловоноrих при поло
вом размножении происходит быстрое Mac
совое перераспределение резервов: запас
ные метаболиты переходят в развиваю
щиеся половые клетки, и животные после
спариванияrибнут.Такоепереключениевы
зывается секретом rлазных желез, выпол
няющих таким образом rонадотропную
функцию; активность зрительноrо нерва по
давляет их секрецию. Изоляция желез или
перерезание зрительноrо нерва вызывает
преждевременное половое созревание и
в конечном итоrе смерть, а переключение,
о котором идет речь, обычно необратимо.
Размножение и жизненные циклы
421
14.4.4. ЦИКЛЫ размножения
и диапауза в наземной
и пресноводной средах
Репродуктивная биолоrия наземных
и пресноводных беспозвоночных в значите..
льной степени определяется их способно..
стью к внутреннему оплодотворению и хо..
рошей защищенностью откладываемых
яиц. С этим также связана способность са..
мок сохранять сперматозоиды и оплодот"
ворять яйца последовательно. Большин..
ство беспозвоночных, обитающих в умерен..
ных, бореальных и полярных широтах, ха..
рактеризуются сезонным размножением
с продолжительными периодами откладки
яиц, перемежающимися репродуктивным
покоем. Такой цикл характеризуется не сто..
лько синхронностью размножения.. сколько
закономерными переХОДdМИ между репро..
дуктивными активным и неактивным со..
стояниями Последнее может быть покое.И,
Т. е. непосредственной временной реакцией
на неблаrоприятные ОКРУЖdющие условия,
прекращающейся при их улучшении; факу"
/lbп1Gl11U61iOii диапау зой непосредственной
реакцией на неблаrоприятные условия сре..
ДЫ, которая не прекращается" пока не
пройдет определенный период времени;
облuzатной диапаузой прекращением ак..
тивности в определенное время [ода вне за..
висимости от неблаrоприятных условий.
для избежания которых оно бывает приспо..
соблен о.
Состояния покоя и факультативной диа..
паузы наступают на взрослой стадии леrоч..
НЫХ моллюсков, дождевых червей инекото..
рЫХ HaceKOMЫX облиrатная диапауза, на..
против, чаще связана с более ранними ста..
диями развития 'Эмбриональной, личи..
ночной или куколочной. Так, мноrие попу"
ляции наземных и пресноводных беспозво..
ночных в умеренных и бореальных областях
переживают зиму исключительно на стадии
яйца. Это явление особенно подробно изу..
чено на насекомых, причем установлены не..
которые факторы, ero вызывающие. Насе..
комых, которые активны летом и впадают
в диапаузу осенью, называют «длинноднев..
ными», а тех, что активны зимой, как, на..
пример, тутовый шелкопряд (ВОI11ЬУХ 1110"
ri), «короткодневными». Хотя эти терми
ны носят чисто описательный характер.. они
вполне отражают суть дела, так как именно
продолжительность cBeToBoro дня реrули
рует переход из одноrо физиолоrическоrо
состояния в друrое.
Циклические изменения долrоты CBeTO
Boro дня MorYT служить очень точным
источником информации о смене времен {'о..
да во всех областях, кроме тропических.
Ход этих изменений в течение [ода на раз..
личных широтах показан на рис. 14.19;
обратим ВНИМdние. что лва раза в [од 21
марта и 21 сентября продолжительность
и дня, и ночи составляет ровно 12 ч в любой
точке земной поверхности. Часто животные
реаrируют на изменение полrоты пня
и используют эту информацию для кон..
троля процессов, имеющих rодовую перио..
дичность. Это явление называется фотопе
риодизмом. Реакция МlIоrих животных Hd
относительную долrоту дня (или ночи)
обычно нелинейна, т. е. световой период"
меНЫllИЙ определенной критической вели
чины, воспринимается просто как короткий.
а превышаюший некую величину как
длинный. В этом можно убедиться. поме..
стив колонии насекомых в различные фото..
периодические условия и реrистрируя ча..
СТОТУ наступления у них диапаузы (рис.
14.20). Точность определения животными
времени находится при этом в пределах 30
мин, что привлекло значительное внимание
физиолоrов.. интересующихся природой
биолоrических часов.
Изучение различных насекомых показа..
ло, что фотопериодизм" вероятно, опреде
ляется различными процессаl\1И У HeKOTO
рых видов для оценки эффективной долrо..
ты дня, по"видимому, используется BHY
тренний циркадианный 1 осциллятор. Наи..
более отчетливо это демонстрируют опы..
ты, коrда насекомых содержат в условиях
с фиксированным световым днем и ночью
различной продолжительности (от 12 и Me
нее до 72 ч и более). При этом некоторые
«дни» животные воспринимают как длин..
ные, а друrие как короткие, причем таким
1 В советской литературе более употребим
термин «циркадный», однако rрамматически он
менее верен (от латинскоrо "dies" == «день»
остается только одна буква). ПрU.lи. ред.
24
700
700
650
18
16
14
'з"
..
о:: 12
I
q
т
10
о
L
t::
О
q
8
6
4
2
1
п III N V VI VH УН! IX
х XI ХН
Рис. 14.19. Сезонная изменчивость долrоты дня в зависимости от широты. Обратим внимание,
что в любой точке Земли дважды в rод день и ночь продолжаются по 12 ч. Период изменений
долrоты дня везде одинаков; от широты зависит только ero амплитуда (по Saunders, 1977)
Естественная долrота ДНЯ
r
А
"\
100
IS
Q)
LQ
О
()
О
20
:I:
О О
а.
С
I
/
Б ..-.---/
....... ........ ......
Q)
(")
:>о-
т
с
т
60
m
" А
;'
;'
;'
;'
;';'
;';'
6
СРотопериод
12
18
( Светлое время суток, ч)
Критическая долrота дня
Рис. 14.20 Схема фотопериодической реакции у (А) длиннодневных и (Б) короткодневных насе-
комых. Выраженность реакции оценивается по проценту особей, входящих в диапаузу при данном
фотопериоде
Размножение и жизненные циклы
423
А
L "L
8 24
11 8 36
III 8 48
IV 8 60
Б
4
8
50%
30
'j"
.. 10
g
s 12
Q.
ф
t:
О
Ь
&
14
16
20
24 36
'С,ч
48
60
72
образом., как будто есть осциллятор со
«свободнотекущим}) циркадианным (т. е.
примерно 24--часовым) ритмом, имеющим
светочувствительную фазу, определенным
образом сдвинутую по отношению к началу
Рис. 14.21. Данные, свидетельствующие об
участии свободнотекущеrо циркадианноrо рит..
ма в фотопериодических явлениях. А. Схема
резонансноrо опыта. Предполаrается суще..
ствование HeKoro эндоrенноrо осциллятора
(показан в виде синусоиды) с фоточувствите
льной фазой фl. Различные световые режимы
изображены чередованием заштрихованных
и светлых полос. В примерах 1 IV световой
период (С) фиксирован (8 ч), а темновой (Т)
варьирует; это дает различную продолжитель
ность циклов т. Обратим внимание, что в при
мерах 1 и 111 фl всеrда приходится на темновой
период, а в примерах 11 и IV иноrда на CBeTO
вой (*). Это может восприниматься как усло
вия длинноrо дня. Б. Экспериментальная дe
монстрация «резонанса», свидетельствующая
о существовании BHYTpeHHero осциллятора. Н а
rрафиках показан процент серых мясных мух
Sarcophaga, входящих в куколочную диапаузу
при выращивании в различных фотопериоди
ческих режимах с длиной cBeToBoro дня 4
20 ч и эффективной продолжительностью су..
ток 20 72 ч. Явно заметен резонанс (по Saun
ders, 1982)
цикла" устанавливаемоrо либо «рассветом»,
либо «закатом}). На рис. 14.21 показано., как
в таких «резонансных» опытах можно доби--
ться Toro, что эта фаза всеrда будет прихо--
диться на светлое время «суток» даже при
424
rлава 14
100
х
:s;
()
60
:S;X
а5ФО
бffi
0..5 U 20
C I
Q)
о..
со
с
24 36 48
Темновой период, ч
-
60 72
9,0
9 'i
9 'i
9 ji
1° i
12
А
100 1
х
90 1
Q)
:т
..... :s; 60 I
Iti)
g}Iб
о е 1: 20 I
0..0 со
Cr. U
5 1
о..
со
с
-
-
-
-
I I
13,5 1,5
Уасы
Б
4
о
__ AI
100
12
О
16
о
I: О
со
() 20
х О
:s;
24
U
Q) О
:::т
s:: 28
.....
Q) 0_ I
r.
Q)
L 32
о
I О
Q)
..... 36
о..
со О
с
40
О
::J:
Q) 44
::r
о О
о..
с 48
О
52
О
56
О
60
О
I I I
О 8 64
В Часы
Рис. 14.22. Данные, свидетельствующие о том,
что тли Megoura viciae измеряют продолжите
льность TeMHoBoro времени суток с помощью
механизма типа «песочных часов». А. Процент
бескрылых (летних) партеноrенетических ca
мок в колониях при различных фоторежимах
с фиксированной продолжительностью CBeTO
Boro времени (8 ч). Заметно отсутствие резо
нанса. Б. Процент бескрылых партеноrенетиче
ских самок, появляющихся, коrда после HaCTY
пления темноты возникает полуторачасовой
световой перерыв, а затем следует темновой
период различной длительности. В. Эффект
ПОСIUЯННОЙ продолжительности CBeToBoro
периода. Это возможно только при условии
«свободноrо течения» виутреииеrо пирка
дианноrо ритма в течение нескольких ци..
клов. Однако у некоторых животных таких
резонансных эффектов обнаружить не улае
тся.
Например, у тли .LИеgоиrа }liciae в опыте,
аналоrичном проиллюстрированному на
рис. 14.21, признаков резонанса не заметно.
Как следует из рис. 14.22, 1ТИ насекомые
используют для измерения времени Mexa
низм типа «песочных часов». Переход от
партеноrенетическоrо бесполоrо размноже
ния J( половому., характерный для жизнен
Horo цикла тлей, обычно индуцируется YKO
рочением cBeToBoro дня, что имеет явное
адаптивное значение. так как обильные Be
сенние и летние пищевые ресурсы к ЗТОl\1У
времени почти иссякают. Фактически этот
переход вызывается увеличением относите
льной продолжительности ночи., измеряе
мой внутренними «песочными часами» (т. е.
определяющими истекший период време..
ни). Как только длина TeMHoBoro периода
превысит 9,75 ч, возникает реакция на «KO
роткий лень» и ПОЯВЛЯIОТСЯ половые фор
мы, дающие на чало зимующим в состоянии
диапаузы ЯЙЦdМ.
14.5. Размножение
и распределение ресурсов
Образ жизни и циклы размножения бес
позвоночных весьма разнообразны. но при
этом видовые репродуктивные особенности
обычно жестко фиксированы. Одна из про..
блем современной биолоrии беспозвоно
ных заКЛЮЧdется в создании ЭВОЛЮIIИОННОЙ
одночасовоrо CBeTOBoro перерыва в различное
время очень длинной «ночи». Такой режим BOC
принимается как «длинный день» только тоrда,
KorAa свет подается через 8 ч после «заката».
Даже кратковременное прерывание темноты
в это время воспринимается как рассвет, насту-
пивший до истечения критическоrо периода,
что оказывает rлубокое влияние и физиолоrию
HaceKoMoro. Более ранний световой импульс
интерпретируется как новый «закат»; ero воз-
действие перекрывается следующей длинной
ночью (по Saunders, 1982)
Размножение и жизненные циклы
425
теории., объясняющей наблюдаемое в при
роде разнообразие жизненных циклов По
ка такой теории не существует. но уже MO
жно сформулировать некоторые ее общие
положения
Наиболее признана в настоящее время
так называемая «демоrрафическая теория
жизненноrо цикла». Она базируется на сле
дующих положениях:
1) естественный отбор действует Hё:J OT
дельные признаки жизненноrо цикла:
2) естественный отбор направлен на оп
тимизаuию адаптивных стратеrий (т. е. на
повышение приспособленности особей., r де
под приспособленностью понимается
эффективность передачи rенетическоrо ма..
териала последующим поколениям);
3) отдельные признаки жизненноrо uи
кла MorYT эволюционировать независимо.
Более Toro, поскольку доступные орrаниз
му ресурсы оrраничены., возникает потен
циальный конфликт между их использова
ни ем для выживания и плодовитости., KOTO
рый разрешается тем или иным компромис
сом.
Демоrрафическая теория полаrает., что:
1) увеличение репродуктивноrо усилия,
т. е. вклада ресурсов в размножение. ведет
к росту эффективности последнеrо., но в то
же время уменьшает 1атраты на поддержа
ние жизнеспособности caMoro орrанизма:
2) увеличение репродуктивноrо усилия
ведет к повышению собственной плодови..
тости и/или выживаемости потомства;
3) снижение вклада ресурсов в СОМdТИ
ческое развитие ведет либо к снижению BЫ
живаемости особи, либо к замедлению po
ста, а следовательно., к снижению эффектив
ности размножения в будущем;
4) возможен компромисс между СИЮМИ
путной репродУК111uвной эффективностью
и остаточной репродуктивной ценностью.,
определяемой KdK произведение выживае
мости и плодовитости в оставшийся период
жизни особи.
Там, rде возможны компромиссы., раз
личные решения будут оптимальными
в разных условиях, однако адаПТИВНdЯ из
менчивость орrанизма оrраничена ero эво
люционной историей. Например, высказы
валось предположение, что утрата стадии
личинки велиrера брюхоноrими моллюска
ми связана с очень rлубокими rенетически
ми изменениями., сделавшими этот шаr -эво..
ЛЮllИОННО необратимым. Если это так. сле
дует ожидать резкоrо таксономическоrо
расхождения rрупп rастропод, сохранив..
ших и утративших такую личиночную ста..
дию: это и подтверждается наблюдениями.
Важный для беспозвоночных компро..
мисс между численностью и выживаемо
стью потомства. Это иллюстрируется., на..
пример, выбором между планктотрофным
и лецитотрофным типами развития или ме..
жду стратеrиями заботы о потомстве или
отсутствия таковой. Одни и те же русурсы
можно вложить или в относительно боль
шое число «дешевых» потомков., или в не..
мноrих., но «дороrих»., каждый из которых
будет иметь более высокие шансы на выжи"
вание. Этот выбор совершается обычно не
на индивидуальном уровне., а в ходе эволю
ционной истории вида., так что вклад pecyp
сов в каждоrо потомка видоспецифичный
признак. Нередко даже близкородственные
виды по нему различаются.
Вторая важная переменная отно"
сительное количество доступных ресурсов.,
вкладываемых в каждый акт размножения.
Следует предполаrать., что любой вклад
в Hero снижает соматические возможности,
причем этот конфликт разрешается на ин..
дивидуальном уровне. Репродуктивные
стратеrии большинства беспозвоночных OT
нюдь не случайны., и между накоплением
ресурсов и их вкладом в размножение обыч
но проходит KaKoe TO время. У этой задерж
ки своя «стоимость». связанная с опреде
ленной вероятностью rибели особи до нача
ла размножения., а также с затратами на со..
хранение запасов. Затраты MorYT быть
уравновешены следующими преимушества
ми:
1) условия для выживания ПОТОМСТВd
бывают непостоянными. Это часто наблю
дается, если молодь использует временный
источник пищи или менее устойчива копре..
деленным условиям среды;
2) у видов с наружным оплодотворе
нием ero вероятность выше., если rаметы
выделяются всеми особями популяции од..
новременно;
3) размножение бывает сопряжено с ри
ском. Если, например, в этот период живот..
426
rлава 14
ное подверrается повышенной опасности
нападения хищника, может быть выrоднее
сначала накопить б6лыl1Io часть или все
доступные ресурсы, а затем вложить их
в размножение;
4) если накопление жизненноrо опыта
родительской особыо повышает ll1aHCbI по
томства на выживание, этоrо может оказа
ться достаточным для Toro, чтобы переве
сить стоимость позднеrо размножения.
Биолоrия популяций и взаимодействия
между членами биоценоза в значительной
мере определяIOТСЯ особенностями репро
дуктивной биолоrии составляIOЩИХ их oco
бей. Жизненные циклы выражаIOТ характер
распределения ресурсов между соматиче
скими и репродуктивными функциями. От
Hero зависят плодовитость и выживае
мость, т. е. в конечном счете возрастная
структура популяции. К сожалениIO, в на..
стоящее время не понятны ни функциональ
ное значение Bcero разнообразия жизнен
ных циклов и типов размножения беспозво
ночных, ни давления отбора, ответственные
за их формирование; однако в этой области
уже ПОЯВЛЯIOтся новые идеи.
Обычно считаIOТ, что признаки. xapaKTe
РИЗУIOщие жизненный цикл, подвержены
давлениIO отбора. С друrой стороны, необ
ходимо признать, что бываIOТ филоrенети
ческие и функциональные оrраничения, пре--
ПЯТСТВУIOщие оптимальному ответу на это
давление. Трудности возникаIOТ в связи
с возможностыо не одноrо, а нескольких
реll1ений. Например. иноrда близкород
ственные орrанизмы занимаIOТ сходные
эколоrические НИll1И, но существенно разли--
чаIOТСЯ по характеру распределения своих
ресурсов. Так, у морских беспозвоночных
часто встречается ситуация, коrда из двух
близких видов, обитаIOЩИХ в практически
идентичных условиях, один образует небо
льшое число крупных яиц с большим запа
сом желтка, а друrой множество мелких,
почти не содержащих желтка. Значит, MO
жно rоворить о компромиссе между числом
и размером яиц, который не обязательно
зависит от общеrо вклада ресурсов в раз
множение, причем не ИСКЛIOчено, что суще
ствует несколько компромиссных решений.
Вот примеры еще нескольких компромис
сов:
вклад в одноrо потомка - число по..
томков;
вклад в одноrо потомка
выживание потомства;
число потомков данноrо размера
выживание родительской особи;
число потомков данноrо размера
численность будущеrо потомства.
Один из подходов к пониманиIO разно
образия типов размножения выделение
совместно изменяIOЩИХСЯ признаков
и поиск теорий, которые бы предсказывали
их КОЭВОЛIOциIO. В качестве ИЛЛIOстрации
можно рассмотреть виды, обычно встре..
чаIOщиеся во временно «недоиспользуе..
мых>) эколоrических нишах. Условия суще..
ствования в них краткосрочны и непредска..
зуемы, и у видов, к ним приспособленных,
следовало бы ожидать появления призна..
ков, ПОЗВОЛЯIOщих производить максималь
но возможное количество потомства Им
MorYT быть противопоставлены виды кли ,
максовых сообществ, rде шансы на выжива..
ние отдельной молодой особи оrраничены,
а конкуренция за ресурсы сильна. Можно
предположить, что в этих двух противопо"
ложных ситуациях естественный отбор бу..
дет способствовать возникновениIO разных
наборов репродуктивных признаков, при
чем конкретные признаки вида станут отра..
жать относительнуIO интенсивность различ..
ных, действовавших на Hero давлений отбо
ра. Эта идея оказала очень большое влия
ние на развитие теории размножения, и,
хотя в настоящее время понятна ее недоста..
точность для полноrо объяснения эволю
ции репродуктивных признаков, она заслу
живает более детальноrо обсуждения.
Для описания ситуаций, в которых от..
бор идет преимущественно на образование
максимально возможноrо числа потомков,
используется термин «r отбор», rne r
константа, опредеЛЯIOщая скорость роста
популяции в уравнениях
dN rt
==rNилиNt==Nое .
dt
Отбор на максимальное значение r блаrо
приятствует раннему размножениIO, BЫCO
кой плодовитости И короткой продолжите
льности жизни. Можно предполаrать, что
«r стратеrи» характеризуIOТСЯ набором
Размножение и жизненные циклы
признаков, способствующих производству
максимальноrо числа потенциальных по..
томков, И что численность популяций таких
видов будет нестабильной, склонной к силь..
ным колебаниям. В условиях интенсивной
конкуренции. напротив, возможности для
роста численности невелики, и можно ожи..
дать корреляции максимальной приспо..
собленности с поздним размножением, низ..
кой плодовитостью и обеспечением каждо..
ro из немноrих потомков большим запасом
питательных веществ. Отбор TdKoro типа
называется К"отбором по константе К
в «лоrистическом» уравнении
dN (К N)
== rN .
dt К
Это уравнение описывает рост популяции,
плотность которой асимптотически стреми..
тся к предельной для данной среды. Одна из
трудностей TaKoro анализа связана с тем,
что величины r и К, cTporo rоворя, hecpdB--
нимы. Мальтузианский параметр , можно
измерить или оценить при исследовании ре..
альной популяции, тоrда как ее предельная
плотность для среды абстрактная безраз..
мерная величина, которую определить на
практике нельзя.
Параметры роста популяции часто вы--
водятся из исследований выживаемости
и плодовитости особей. Один из подходов
здесь использование демоrрафических
таблиц, отражающих среднюю выживае..
мость и среднюю плодовитость самок каж..
дой возрастной rруппы. Эти таблицы мо"
жно составить либо при наблюдении за по..
пуляцией в состоянии динамическоrо рав"
новесия, rде поддается определению воз..
раст любой особи, либо изучая выживаемо..
сть и плодовитость данноrо возрастноrо
класса популяции. Получение таких данных
о беспозвоночных сопряжено с особыми
трудностями, поскольку возраст обычно
трудно определить однако некоторые ме..
тоды все же дают нужную информацию; на..
пример, прикрепленные орrанизмы типа
морских желудей на каменистых береrах
можно метить и наблюдать в течение неско..
льких лет. Пример демоrрафической табли..
цы для одноrо из североамериканских ви"
дов этих ракообразных приведен
в табл. 14.8, rде УКdзаны средняя выживае--
427
мость l(x) и средняя плодовитость т(х) в зави..
симости от возраста (х).
Сумма произведений l(x) . т(х), I:l(x)m x '
представляет собой среднюю плодовитость
самки в течение всей ее жизни. Ее часто обо..
значают Ro. Время rенерации это средний
возраст особей в момент смерти; ero можно
вычислить из данных таблицы по формуле
1: х l(x) т(х)
Т== .
1: l(x) т(х)
Мальтузианский параметр часто прибли..
женно получают из уравнения
1nRo
r , rде Ro == L l(x) т(х).
т
Если значение ,. положительно (как в
табл. 14.8) ПОПУЛЯllИЯ paCTeT если ОТРИlld"
тельно она сокращается а если равно ну..
лю ее численность постоянна. Заметим,
что если r выводится из данных таблицы., то
величина К нет.
Условия среды всеrда изменчивы, но
в некоторых местообитаниях они стабиль--
нее, чем в друrих. В последнем случае мел--
кие и слабые молодые особи будут более
подвержены rибели, чем крупные и хорошо
приспособленные взрослые. Поэтому мо--
жно ожидать, что в стабильных условиях
отбор блаrоприятствует БОЛЫllОЙ продол--
жительности жизни с растянутым во време--
ни производством хорошо обеспеченноrо
запасом питательных веществ потомства.
И напротив во флуктуирующих условиях
среды все возраста будут примерно одина..
ково подвержены смертности. Эти доводы,
как и концепция r/ К--отбора, позволяют
предполаrать коэволюцию в стабильных
условиях таких признаков, как длительная
продолжительность жизни, низкая плодо..
витость и поздний переход к размножению,
а в неустойчивых условиях таких как ко..
роткая продолжительность жизни, высокая
плодовитость и раннее половое созревание.
Иными словами, можно ожидать что ста..
бильность среды обитания будет предска..
зуемым образом влиять на компромиссы
жизненноrо цикла, обсуждавшиеся выше.
Это не вполне подтверждается наблюдения..
ми. В рассмотренных в этой rлаве циклах
размножения морских беспозвоночных про--
428
rлава 14
Таблица 14.8. Демоrрафическая таблица для MopCKoro желудя Ba/aпus g/aпdu/a (Hines, 1979)
Возраст (х)
мес
Выживаемость к дaH
ному возрасту, 1 х
Среднее число яиц,
выметанное к возра
сту х, m х
1 х m\
хl (" ln (х)
Оценочн,,-я смертность О
до прикрепления 3
12
24
36
48
60
72
84
96
1.000
1,17хl0 4
2.04 х 1 О 5
3,84 х 1 О б
2,05 х 1 О б
1.28 х 1 О 6
7,33 х 10 "
4,18xIO 7
2 44 Х 10 7
1,42 х 1 О 7
о
о
20 504
66 814
113 125
140 742
159 435
1 70 S92
176 922
180 540
о
о
0.418
0,256
0,231
0.180
0,117
0,075
0.043
0,026
о
о
5.016
6,153
8.335
8.641
7.008
5.151
3 629
2,469
R о == r.l (х)т (х) == 1.346
Время rенерации Т == LX 1 (х)т (х) == 46.402 мес
111 Ro
Скорость при роста попу ляuии в месяu r == т == 0,006,
т. е., если ПОПУЛЯllИЯ сохранится, ее численность будет возрастать медленно
Выживаемость в данном случае оценивали по числу прикрепившихся особей на каждые 1000 выметанных яиц (на
более поздних стадиях путем прямоrо наблюдения). Эти данные состаВ lЯЮТ столбеu 1 х.
Плодовитость оценивали путем измерения базаЛЬНОl о диаметра (по отношению базальноrо диаметра же lУДЯ
naHHoro возраста и числа выметываемых яиu за один раз). Реrистрировали также число нерестов за [од. Произведе
ние этих величин дает опенку средней плодовитости в каждом возрасте (столбеu m х).
слеживается явный компромисс между раз
мером яиц и числом потомков. Однако в цe
лом орrанизмы с планктотрофным пелаrи
ческим развитием, очень высокой плодови
тостью И массовой смертностью молоди
образуют во взрослом состоянии довольно
стабильные популяции, живут относитель
но долrо и отличаются крупными размера
ми тела. И напротив, морские виды, адап..
тированные к использованию временных
эколоrических ниш, часто отличаются низ..
кой плодовитостью, а также леuитотроф
ным и прямым развитием, минующим пе..
лаrическую стадию.
Чтобы понять, почему каждый вид жи..
вотных размножается так. а не иначе, необ..
ходимо учесть оrраничения, накладывае
мые средой обитания, функциональной
морфолоrией орrанизма на различных ста..
диях развития и, самое rлавное, ero ЭВО
люционной историей, которая может су..
жать Kpyr доступных ему путей решения
этой проблемы.
14.6. ВЫВОДЫ
в последнее время интенсивно разраба..
тывается теория жизненноrо ЦИКЛd. стремя..
щаяся функционально объяснить разно..
образие типов размножения животных,
в частности беспозвоночных.
В этой rлаве мы кратко Рdссмотрели не..
которые данные, на которых базируются
эти теоретические построения. Очевидно,
что основной способ размножения Bcero
живоrо половой, хотя он может переме..
жаться периодами бесполоrо воспроизведе..
ния в сложных жизненных циклах мноrих
книдарий, плоских червей, аннелид, мелких
ракообразных.. насекомых и т. д.
Типы половоrо размножения и распре..
деления доступных ресурсов между ним
и соматическими нуждами поразительно
разнообразны. Это разнообразие рассмо--
трено с таксономической, эколоrической
и функциональной точек зрения. Ясно, что
далеко не все возможные комбинации ре--
Размножение и жизненные циклы
продуктивных признаков встречаются
в каждой среде обитания или таксономиче
ской rруппе. Так. развитие морских беспо
звоночных часто идет через стадию свобод--
ноживущей пеЛdrической личинки, и с этой
особенностью связаны мноrие друrие: нару..
жное оплодотворение,! массовые синхрон--
ные нересты, простое строение спермато..
зоидов,! яйца, бедные питательными ве--
ществами.
у всех пресноводных и наземных видов'!
а также в некоторых морских средах пелаrи..
ческое личиночное развитие отсутствует.
При 1ТОМ обычно наблюдаются внутреннее
оплодотворение., хранение спермы и нали..
чие большоrо запаса питательных веществ
в яйцах.
Любой тип размножения не СЛУЧdСН
и обусловлен контролируемым распределе
нием ресурсов между репродуктивными
процессами Если размножение животных
в популяции синхронизировано. должны
действовать механизмы восприятия и ней
роэндокринной передачи информации из
внешней среды. В данной l"лаве мы лишь
кратко коснулись этой проблемы: более
подробно она рассмотрена в rл. [6.
Приведенные здесь описания и анализ
репродуктивных процессов у беспозвоноч--
ных послужат основой для более детально--
[о разбора вопросов эколоrии и биолоrии
сообществ, а также индивидуальноrо разви--
тия (rл. 15).
14.7. Дополнительная
литература
ДеТdльные сведения по размножению беспо
звоночных можно получить из двух MHoroToM
ных трул.ов и следующей серии обзорных томов:
Adiyodi К. G., A(liyodi R. G. (Eds) 1983, Reproducti..
ve Biology of Invertebrates. Wiley, New У ork.
Уоl. 1. Oogenesis, oviposition and oosorption.
Vol. 2. Spermatogenesis and sperm function.
Следующие тома ждут выхода в свет
GieseA.G., Peal'seJ.S. (Eds) 1974 1977. Repr()
duction in Marine Invertebrates. Academic Press.
New У ork.
V ols 1 IV и следующие.
Advances in Invertebrate Reproduction. Elsevicr
SClence. Amsterdam.
Уоl. 1. Clark W.,! Adams Т. S. (Eds) 1981.
Уоl. 2. Engels W. (Ed.) 19S4.
429
Vol. 3. Porchet М. (Ed.) 1986.
В следуюших моноrрафиях рассмотрены
различные аспекты размножения беспозвоноч
ных:
Begoп М., Hal'pel' J. L., ТОИ'l1sепd С. R. I Y 6. Eco
10gy: Individuals, Populations and Communities.
Blackwel1 Scientific Publlcations Oxtord.
Имеется перевод: Биrон М.. Харпер Дж..
Таунсенд К. 3колоrия. Особи. ПОПу..1ЯЦИИ и сооб
щества: в 2 т.' М.: Мир. 1989.
Ве" С, 1982. The Masterpiece of Nature: The Evolu
tion and Genetics of Sexuality University of Cali
fornia Press,! Berkeley
B,'a(Z}' J. 1979. Biological Clocks (Studies in Biology.
104). Edward Arnold, London.
Са/он' Р. ] 978. Lifc Cycles. Chapman and НаН. Lon
don.
С/ипАnО\' Е. 1982. The Theory of Sex AHocation. Pri
11seton University Press, Princeton, New Jersey.
CO/len J. 1977. Reproduction. Butterworth, London.
Grеепи'о()(! Р. J., Allaпls J. 1987. The Ecology of Sex.
Edward Arnold, LOl1don.
М a,,'п!lI'd Sfпith J. 1978. The Evolutiol1 of Sex. Сат..
bridge University Press. Cambridge.
Sauпdel's D. S. 1977. Аl1 111trodllctiol1 to Biological
Rhythms. Blackie, Glasgow.
Приложение 14.1.
Системы половоrо
размножения (миксиса)
Половое размножение это систеМd
воспроизведения, обеспечиваюшая обмен
rенетической информацией (носителем ко..
торой является молекула ДИК) и появление
особи, чья rенетическая информация будет
результатом рекомбинации родительских
reHOB.
1. Вирусы. Бактериофаrи заражают бак--
териальные клетки. вводя в них свою Д IК.
При этом белковая оболочка вируса остае--
тся вне клетки. Вирусная ДНК внутри бак..
терии содержит информацию для синтеЗd
новых белковых оболочек фаrа и лизиса
бактерии--хозяина.
А. Одновременное заражение разными
мутантами по одному и тому же структур--
ному reHY не обеспечиваеl воспроизведение
фаrа, а одновременное заражение мутанта--
ми по разным структурным [енам дает ак--
тивные фаrи.
Б. Мутантные фаrи способны размно--
жаться в бактерии--хозяине.
В. При смешанном заражении фаrами
430
Белки, кодируемые
rенами Д и В
МУТ ДНТНЫЕ срормы
не СПОСОБНЫ синтези...
ровать ПОЛНЫЙ наБОр
reHHbIX продуктов
А
Мутант 1, reH Д
rлава 14
Белковая ОБолочка
6актериосраrа
Мутант II, reH А
Б Заражение некомпле Заражение компле
ментарными мутантами ментар'НЫМИ мутан
там и (инсрекция
"дикоrо типа")
. у .. :
Кроссинrовер
не дает срорм
дикоrо типа
в
с комплементарными мутациями кроссин
rOBep может привести к появлению фаrов,
способных заражать бактерии «в одиноч
ку», т. е. к половому размножению
2. Бактерии.
1. Разные штаммы бактерий способны
к конъюrации. При этом образуется при..
крепительный мостик (пилус), по которому
молекула дик (бактериальная хромосома)
передается от донора (<<самца») реципиенту.
11, 111. Длина переданноrо отрезка ДНК
зависит от времени конъюrации, которую
можно прервать путем интенсивноrо
встряхивания культуры.
: х :_
Кроссинrовер дает
вирусную частицу
дикоrо типа
III
асть ДНК
с rенами Д и В
_ i
J
Мутант III, reH В
Заражение некомпле
ментарными мутантами
9
э
: :
э
II
IV
Размножение и жизненные циклы
431
IV. Между ДУПЛИllированными участка
ми бактериальной хромосомы может про
изойти обмен rенами. При этом возникают
бактерии, совмещающие признаки обеих
родительских особей.
3. Половое размножение у высших op
rанизмов. ДИПЛОИДНЬiе клетки имеют два
набора хромосом, в каждом из которых co
держится rенетическая информация одноrо
из родителей. В ходе особоrо типа клеточ..
Horo деления, вызываемоrо мейозом, из ди
ПЛОИДных клеток образуются rаплоидные
[аметы. Их rенетическая информация MO
жет быть идентична родительской или OT
личаться от нее, если произошел кроссинrо..
вер.
rаплоидные половые клетки от двух ро..
пителей (или иноrда от ollHoro родителя)
сливаются (это называют оплодотворе..
нием) с образованием диплоидной зиrоты.
reHoM зиrоты отличается от [еномов ди..
плоидных клеток, павших начало [аметам.
В нем сочетаются rенетические признаки
обоих родителей, каждый из которых пред..
ставлен одним набором хромосом.
Диплоидная животная клетка.
reHbI последовательностей
а...е и A E находятся на
хромосомах, унаследованных
от ОБОИХ родителей.
Эта клетка rетерозиrотна
в ходе М Е ЙОЗД хромосомы
спариваются, и между их
хроматидами может
происходить rенетический
Обмен (кроссинrовер)
! МЕЙОЗ
В raMeTax может оказаться родительская или реКОМбинантная
последовательность reHoB
rAMETbI
Родительская РеКОМбинантная РеКОМбинантная Родительская
Частота кроссинrовера зависит от длины ДНК, разделяющей
reHbI на хромосоме
432
rлава 14
n риложение 14.2.
Жизненные циклы и половое
размножение
1 Половое размножение у большинства
протистов. Пример инфузория P{a aпle..
C1Uп1.
2. Жизненный цикл всех мноrоклеточ..
ных животных (и некоторых водорослей).
Взрослая особь состоит из нескольких
или мноrих (обычно различных) диплоид
ных клеток. В мноrоклеточном теле обра..
зуются rаплоидные [аметы. Их слияние
дает диплоидную зиrоту. В результате ряда
митотических делений развивается новая
взрослая диплоидная особь.
Мейоз
(:)n
v
n
Оплодотворение
o 2 n Зиrота
)J2n
fj{/
2n
3. Для жизненных циклов мноrих BOДO
рослей и всех высших растений характерно
чередование мноrоклеточных rаплоидной
и диплоидной фаз. Первая называется raMe..
тофитом; она образует (путем митоза) спер..
мии и/или яйцеклетки. Слияние raMeT дает
зиrоту. Последняя путем митозов развивае
тся в мноrоклеточный спорофит. В ПрОllессе
мейоза образуются rаплоидные споры. Не
сливаясь, они продолжают митотически де..
литься, формируя rаплоидный rаметофит.
На приведенном ниже рисунке показано
протекание этоrо процесса у папоротников
у высших растений rаметофит обычно
редуцирован до очень небольшоrо числа
клеток.
2n
у животных TaKoro чередования l a..
плоидноrо и ДИПЛОИДllоrо поколений никоr..
да не происходит. Чередование поколений,
размножающихся бесполым и половым пу..
тями ему не эквивалентно (см. рис. 14.4).
Приложение 14.3.
Системы определения пола
1. Пример материнскоrо определения
полd.: мелкая полихета Diпop/lilus gy' ()ci/ia..
tus.
Взрослая
ОСОБЬ 2
Кокон с круп
ными И мелки
ми яйцами
Осеменение
неполовозрелых
самок рано соз
ревшими карли..
ковыми самцами
Размножение и жизненные циклы
В яичниках образуются два типа яиц: из
крупных развиваются самки, из мелких
рано созревающие карликовые самцы. Осе--
менение самок происходит внутри кокона
на эмбриональной стадии. Все взрослые
особи это осемененные самки.
2. Пример rенетическоrо определения
пола: плодовая мушка DrosophiZa.
1
X XPOMOC OMa I
)..,
11'
А
XpOMOCOMa
Б
d'
А. 1. У самок диплоидный rенетический
набор с двумя одинаковыми половыми хро--
мосомами хх. Все яйuеклетки идентич--
ны по способности развиться в особь Toro
или иноrо пола.
11. у самцов диплоидный rенетический
набор с неодинаковыми половыми хромо..
сомами Х и У. Образуются сперматозои--
ды двух типов: из яиц, оплодотворенных
сперматозоидами, несущими У--хромосому,
разовьются самцы, а из оплодотворенных
сперматозоидами с Х--хромосомой
самки.
Б. Пол потомка определяется тем, ка--
кой из двух типов сперматозоидов оплодот
ворил данную яйцеклетку. При такой систе..
ме определения пола нормальное соотно--
теине полов будет 1 : 1.
4ЗЗ
3. Эколоrическое определение пола
у некоторых орrанизмов пол особи
определяется после оплодотворения. При..
мером может служить эхиурида Boпellia vi..
ridis.
Все ресничные планктонные личинки
этоrо червя способны развиться как в самку
(все крупные взрослые особи самки), так
и в карликовоrо самца. Оседая на незасе..
ленные участки MopcKoro дна, они дают са..
мок. а оседая вблизи хоботка самки. под
влиянием выделяемых им секретов превра..
щаются в карликовых самцов. В экспери..
ментах выявлено также определенное reHe..
тическое влияние на пол, но влияние среды
ero обычно пересиливает.
f}.;'.-;::;:D("::. ' :'"
o;.
." ' .
. '.
:. .
. .... :.':: .i: :{ o:;
О, 0.0."0
'.' (, '0' о;;
. . , .: ;i
.: 'О.:
.: :
Приложение 14.4.
Жизненные циклы беспозво
ночных
На каждом рисунке показан типичный
облаДатель Toro или иноrо цикла. ШИРИНа
полосы отражает количество ресурсов: ее
434
rлава 14
1 roA
1. МУЛЬТИВОЛЬТИННЫЙ ВИД
<О
I
2. ОДНОЛЕТНИЕ виды
:'3
II. Однократный массовый нерест
ПI. Неоднократные последовательные
кладки
возрастание означает их накопление, умень"
шение их расходование.
1. Мультивольтинный вид.
В течение [ода сменяется несколько по
колений; в большинстве случаев в пресно
водных и наземных местообитаниях YMe
ренной зоны у животных имеется репродук
тивная диапауза. На рисунке показана диа..
пауза на взрослой стадии" но она часто бы--
вает также на стадиях яйца, личинки или KY
колки.
2. Однолетний вид.
1 rод
1j5
qj \t
Мноrие виды умеренных областей жи..
вут ровно [од" т. е. за это время у них сме..
няется только одно поколение. Размноже..
ние может либо быть представлено, как
в примере 11, однократным массовым нере..
стом" либо продолжаться в течение неско..
льких месяцев (111).
3. Мноrолетний вид.
Эти животные способны жить в течение
нескольких лет" весь этот период накапли..
вая ресурсы. Возможны две различные
стратеrии вкладывания их в размножение.
Размножение и 'жизненные циклы
1...й roA
2 й roA
3 й roA
435
з.мноrОЛЕТНИ
. о о
IV. ПОЛИЦИКЛИЧНЫЙ ВИД, размножающийся ОДИН раз в roA
J l
\
v. ПОЛИЦИКЛИЧНЫЙ вид, размножающийся неБОЛЬШИМИ кладками несколько раз в roA
Q L
VI. МОНОЦИКЛИЧНЫЙ вид, размножающийся один раз в конце жизни, длительность которой
определяется условиями среды: 2, 3 roAa или Более
Чаще Bcero животные (например, морские
звезды; IV) размножаются один раз каждый
rод в определенный период. Размножение
может быть синхронным и массовым или
мноrократным несинхронным (например,
у земляноrо червя, У). Некоторые MHoro..
летние животные моно цикличны, т. е. раз..
множаются один раз в жизни в возрасте не
менее одноrо [ода. Наиболее известные их
примеры rоловоноrие моллюски и поли..
хеты семейства Nereidae (VI)
Приложение 14.5.
Жизненные циклы морских
беспозвоночных
Яйца и сперматозоиды морских беспо..
звоночных MorYT высвобождаться в мор..
скую воду.. [де и происходит оплодотворе..
ние. Это оказывает rлубокое влияние на
всю репродуктивную биолоrию этих живот..
ных.
436
CNIDARIA
iP
PLATYHELMINTHES
V
NEMERTEA
ANNELIDA
Полихета
MOLLUSCA
v
Брюхоноrие
>
rлава 14
. 1ft
ECHINODERMATA
Плутеус
.
Приложение 14.6.
Функциональный анализ
жизненных циклов насекомых
Планула
Мюллеровекая
личинка
Пилидий
т рохоqюра
т рохосрора
m
> М!
Нектохета
Велиrер
1. Ювенильные и взрослые сталии испо..
льзуют сходные пищевые ресурсы.
Функции ювенильных стадий
развитие и дифференцировка, накопление
ресурсов.
Функции имаrо накопление ресурсов,
расселение и поиск ресурсов, спаривание
и откладка яиц.
I
Морские звеЗ иплеврула
Бипиннария
Брахиолария
Осриуры !
Осро плутеус
Двустворчаты
Примеры взрослых и характерных личиночных форм основных rрупп морских беспозвоночных,
Взрослые животные изображены в значительно меньшем масштабе, чем личинки
c. .. = ... . t1t'll1тъ
,, ,v ,'
IОЛОТУРИИ Dолиолария
Пример: прямокрылые (саранча и кузне..
чики) .
Размножение и жизненные циклы 437
CRUSTДCEA LOPHOPHORATA
:.
,
8еслоноrие Науплиус СРорониды Актинотроха
)1
>'
Усоноrие Циприс t
Мшанки Цисронаутес
)1
...............
MaLacostraca
Зоеа
Меrалопа
HEMICHORDATA
Торнария
CHORDATA
" r оловастик"
1
1l
III
Характерные жизненные циклы. 1 Пелаrобентосный с пелаrической личиночной стадией,
11. rолобентосный c утратой примитивноrо признака, т. е. пелаrической личиночной стадии. 111.
rолоnелаrический весь жизненный цикл проходит в пелаrиали. Во мноrих rруппах можно
встретить все три этих типа. Приведенные здесь примеры относятся к брюхоноrим моллюскам
2. Ювенильные и взрослые стадии испо..
льзуют различные пищевые ресурсы.
Функции ювенильных стадий
развитие и накопление ресурсов.
Функции куколок дифференцировка
и развитие.
Функции имаrо накопление ресурсов,
их поиск, спаривание и выбор мест для раз
вития потомства.
Пример: падальная муха и друrие дву--
крылые.
oC]
3. Взрослая стадия непродолжительна
и неспособна к накоплению ресурсов. Функ--
ции ювенильных стадий . развитие и нако--
пление ресурсов; функции имаrо
спаривание, поиск ресурсов и расселение
438
rлава 14
t)
1
о
II
1. Водные личинки. например у поденок.
11. Наземные личинки, Нdпример у че..
шуекрылых. Здесь взрослая стадия продол..
жительная, и насекомые своим ротовым ап..
паратом собирают нектар. используемый
кйк rорючее для полета.
4. Имаrо обеспечивают ресурсами по..
томство.
Функции личинок: накопление ресурсов.
Функции имаrо: расселение" поиск и на..
копление ресурсов, спаривание, поиск мест
для развития потомства и откладка яиц.
Пример: одиночные осы (перепончато"
крылые).
Все доступные потомству ресурсы по..
ставляются родителями.
о
+
<IJ
5. Накопление ресурсов, развитие" спа..
ривание и откладка яиц разделены между
кастами.
Ювенильные стадии использование
ресурсов и рост.
Стерильные рабочие особи
накопление ресурсов.
Матки (фертильные самки)
спаривание и выбор партнера, откладка
ЯИII, расселение и поиск ресурсов.
Самцы спаривание; в накоплении ре..
сурсов участия не принимают.
Пример: медоносные пчелы.
\
rЛАВА 1 5
РАЗВИТИЕ
Большинство мноrоклеточных живот..
ных начинают жизнь с оплодотворенноrо
яйца, возникшеrо при слиянии двух очень
разных rаплоидных [амет. Слияние дает ди..
плоидную зиrоту, и именно эта довольно
специфическая клетка развивается во взро"
слый мноrоклеточный орrанизм. Последо..
вательность ее ранних, точно орrанизован"
ных делений называется дроблением; во
время Hero роста почерних клеток не проис
ходит. Зиrота особым способом расщепляе
тся на большое количество мелких клеток,
взаиморасположение которых в простран..
стве соответствует орrанизации оплопотво"
peHHoro яйца. Эта орrанизация может опре..
делить судьбу дочерней клетки на ранних
стадиях эмбриоrенеза. У беспозвоночных
обнаружено несколько различных типов
дробления яйца, среди которых наиболее
известны спиральное у большинства пер
вичноротых и радиальное у вторичноро"
тых.
Дальнейшее развитие эмбриона вклю"
чает ero пространственную реорrанизацию
в соответствии с планом строения взросло..
ro орrанизма, а на более поздних стадиях
дифференцировку клеток в зависимости от
их функций у личинки и взрослоrо орrаниз"
ма. Последовательность развития, таким
образом, включает следующие этапы:
1) оплодотворение;
2) активация метаболизма ЗИI'ОТЬJ
и трансляция материнской информацион"
ной РНК (мРНК);
3) дробление;
4) активация ядра зиrоты и транскрип"
ция новых специфичных для нее мРНК;
5) орrаноrенез;
6) дифференцировка.
Развитие часто включает метаморфоз.
коrда адаптированная к определенным
условиям окружающей среды личинка вне..
запно превращается в резко отличающееся
от нее взрослое животное, выполняющее
иноrда совсем друrие функции и адаптиро"
ванное к совершенно иным условиям (см.
rл. 14).
Мноrие беспозвоночные оказались весь..
ма ценными моделями для анализа биохи..
мических механизмов, участвующих в кле..
точной дифференцировке и орrаноrенезе.
Изучение развития важно и потому" что оно
проливает свет на филоrенетические взаи..
мосвязи между животными.
В этой rлаве развитие беспозвоночных
будет рассматриваться в основном на осно"
ве экспериментальных данных по механиз..
мам клеточной детерминаuии. Наш обзор
включает анализ событий, про исходящих
в момент оплодотворения яйца, и ero реак"
цию на это. Описание типов дробления дает
основу для экспериментальноrо изучения
клеточной детерминации, т. е. формирова..
ния проспективноrо значения клеток в ходе
онтоrенеза. Большое число описательноrо
материала по личиночным формам и мета..
морфозу закладывает фундамент экспери..
ментальных исследований, раздвиrающих
rраниuы нашеrо понимания развития
и пифференцировки. В этой rлаве обсужпае..
тся также реrенерация у беспозвоночных,
т. е. процесс восстановления целой структу"
ры из ее фраrмента. Участвующие в этом
механизмы сходны с обеспечиваюшими
нормальное развитие.
15.1. Оплодотворение
и инициация развития
Оплодотворение сложный процесс;
ero основные компонеНТЬJ таковы:
1) физический контакт [амет;
44е
rлава 15
2) взаимодействие мембранной поверх..
ности, ведущее к слиянию двух клеток;
3) физиолоrическая реаКlIИЯ оболочки
яйца, препятствующая дальнейшему прони..
кновению спермы и обычно называемая
блокадой полиспермии;
4) активация метаболизма ООlIита;
5) слияние пронуклеусов с формирова..
нием HOBoro диплоидноrо reHoMa зиrоты;
6) инициация paHHero дробления.
В понимании общих ПРИНIIИПОВ этоrо
процесса особую роль сыrрали морские бес..
позвоночные с наружным оплодотворе..
нием.
Точная последовательность событий за..
висит от степени зрелости «яйца» в момент
слияния raMeT. У некоторых беспозвоноч"
ных они сливаются до окончания мейоза
и отделения полярных телец. В друrих слу..
чаях мейоз заканчивается до оплодотворе..
ния. Например, у MopcKoro ежа он к этому
моменту завершен, а яйцеклетка морской
звезды вступает в метафазу 1 мейоза непос..
редственно перед выметыванием (см.
rл. 16) и, если не будет оплодотворена, он
дальше не пойдет, т. е. полярные тельца
появляются уже после взаимодействия ме..
жду яйцом и сперматозоидом.
Большинство морских беспозвоночных
выметывают яйца и сперму в воду, rде
и происходит оплодотворение. Можно бы..
ло бы ожидать" что между мужской и жен
ской rаметами существует KaKoe TO притя"
жение, но, по сути дела, это бывает доволь"
но редко. В морской воде сперматозоиды
становятся активными и совершают хаоти"
ческие движения, которые в случае до ста..
точно высокой концентрации яиц и сперма..
тозоидов приводят к физическому контакту
между rаметами. Лишь в некоторых слу..
чаях, например у оболочников, ряда мол..
люсков и колониальных rидроидов Сатра..
пuZaria наблюдается хемокинез. У упомяну..
тых rидроидов сперма должна проникнуть
в яйцо через сравнительно узкое отверстие
в rонанrии колбообразной структуре,
окружаюшей яйца Киносъемка показала,
что движения отдельных сперматозоидов
не хаотичны, а направлены к устью rOHaH..
rия (рис. 15.1). Вещество, экстраrированное
из этоrо устья" привлекает ИХ, будучи ра..
створенным в arape
Рис. 15.1. rонанrий rидроида Сатрапu/апа
и траектории движения сперматозоидов, заре
rистрированные на кинопленке. Это один из
первых случаев демонстрации хемотаксиса Me
жду яйцеклетками и сперматозоидами ДaH
ный вид, конечно, не исключение; друrие при
меры можно найти среди оболочников и мол
люсков, но насколько широко распространено
подобное явление, неизвестно (по Muller,
1966)
Механизм caMoro обычноrо оплодотво
рения яиu хаотически движущимися спер
матозоидами полностью еще не ясен В на..
чале века было обнаружено, что спермато-
зоиды MopcKoro ежа, помещенные в воду,
rде находились яйца. временно лишаются
подвижности, как бы аrrлютинируясь. Это
вызвано диффузией в воде высокомолеку-
лярноrо вещества (из студенистой оболочки
яиu) Ero назвали ФерпlUЛUЗUНОМ" а соответ..
ствующий белок--детектор на сперматозои-
де анпluфертuлu.зuном. На поверхности
мноrих яиц находится видоспеuифичный
коаrулянт спермы, но только в некоторых
случаях он диффундирует в окружающую
среду, как у MopcKoro ежа.
В световой микроскоп можно увидеть
физические изменения, происходящие со
сперматозоидом некоторых беспозвоноч-
ных при контакте с яйцом (рис. 15.2,А).
Сначала ero иrлообразный отросток сопри..
касается с цитоплазмой яйца. Этот ОТРО"
сток акросомная нить развивается из
самой передней части сперматозоида
акросомы. У друrих беспозвоночных он ОТ-
сутствует (рис. 15.2,Б).
Развитие
С;:П
:' ... .... .... .... ... ...... ... .... r .... .... ... ... .... ... "''"1 :
. '
. ,
: : : ; . . '
Б \ii:;.;:';:Sf: i.' !ff i; f}; ;:;'f { J;1Z ;i] - : I:.: ;i t : ;i \
---... "'"
:....""....
...
........ ... ......
'" "', ..",...-.""'"
. . .
. . . .
А ;; rСiI;Й:i::;!l. ;i.i
I
I
II
441
....
........
,.... ....
: {1J} :;,J: 1(; 5 j
III
IV
........
...........
IП
IV
Рис. 15.2. Изменения в ходе оплодотворения по данным световой микроскопии, А. Морская звез
да Asterias. 1. Приближение сперматозоида к яйцу, окруженному студенистой оболочкой. 11, AKpO
сомная реакция. Кажется, что тонкая нить выбрасывается из мелкой, напоминающей пузырек
акросомы спереди сперматозоида. 111. Акросомная нить соприкасается с мембраной яйцеклетки
и начинается кортикальная реакция, распространяющаяся от точки контакта. IV. Сперматозоид
поrлощается коническим выступом цитоплазмы ооцита. Б. Морской еж Arbacia. 1. Приближение
сперматозоида к студенистой оболочке. 11. Прохождение сперматозоида через студенистую обо
лочку. 111. Контакт rоловки сперматозоида с желточной оболочкой; иноrда, как показано на рисун
ке, заметна очень мелкая акросомная нить. IV. Кортикальная реакция и внедрение в ооцит ядра
сперматозоида (по Austin, 1965)
с помощью электронной микроскопии
установлено, что некоторые аспекты взаи..
модействия сперматозоид/яйцо сходны
у всех видов. поскольку все они включают
слияние мембран. На рис. 15.3 показаны
изменения, наБЛЮJ,аемые в момент контак..
та сперматозоида с яйцом у MopcKoro ежа
Arbacia и у полихеты Hyr1roides. В обоих
случаях сначала происходит слияние мем..
браны aKpOCOMHOI'O пузырька и плазмати..
ческой мембраны сперматозоида, в резуль..
тате чеrо высвобождается ферментное со..
держимое пузырька. Последующий лизис
обеспечивает расправление акросомных
трубочек или акросомной нити., их прони--
кновение через оболочки яйца и установле..
ние контакта с ero плазматической мембра..
ной. ДальнеЙIllее слияние мембран приво..
дит к образованию единой оболочки BOKpyr
яйца и сперматозоида
Начальные реакции., вызванные контак..
том, иноrда называют «блокадой полиспер..
мии» Существуют два компонента этоrо
явления. Первый невидимый., хотя у неко"
торых видов связан с изменением 'Электри"
ческоrо потенциаЛd у поверхности яйца.
Второй видимая кортикальная реакция,
которая через несколько секунд распростра..
няется радиально от места контакта спер..
матозоида с яйцом. Она представляет со..
бой слияние мембраны кортикальных rpa..
нул, формирующихся в позднем ооrенезе.,
с мембраной яйца и выход их содержимоrо
в пери вит ел ли новое (околожелточное) про..
странство (рис. 15.4).
Реакция TaKoro типа почти универсаль..
на; ее результат выделение студенистой
оболочки оплодотворения, которая., начи..
ная с этой стадии., препятствует СЛИЯНИIО
яйца с друrими сперматозоидами.
Теперь в яйце происходят rлу60кие ме..
таболические изменения. До оплодотворе..
ния оно как бы пребывает в анабиозе.
Связывание сперматозоида с рецепторным
А
I
Желточная
ОБолочка
Б
11
...............
IIJ
IП
УН VIII IX Х
Рис. 15.3. Оплодотворение по данным электронной микроскопии. А. Акросомная реакция у MOp
cKoro ежа Arbacia. Акросомная нить этоrо вида настолько мала, что с трудом различима в CBeTO
вой микроскоп. 1. Передняя часть rоловки сперматозоида под электронным микроскопом.
11. Слияние мембран акросомы и сперматозоида для высвобождения содержимоrо акросомы.
1
II
III
: ';':" ..., ,. .'_ " j, .,. ....... . ... --:-7":.-::' . '..' :- .--:- .::::,: ::. :::7 . ::. :
.: . : ' -!u \ '" :'.' " xi jf:" '.:" ," . .,::.. . ".;......."..... . ...:.......
:.':. :..\ (::ъ > .:>P:::;:::. .;:.; ::: '.:::': ):' "'.: {'::':':: <.:: ':': :'<: ."':::. :'.;':
. ...
А. Arbacia
IV
II
:
.,
..
.'
..
. .'
...... .......
Б. Nerels ПI
Рис. 15.4. Кортикальная реакция: отслаивание оболочки оплодотворения. А. Морской еж Arbacia.
1. Сперматозоид контактирует с плазматической мембраной яйца; кортикальные rранулы интакт
ные. 11. Кортикальные rранулы разрушаются, и отслаивается оболочка оплодотворения; реакция
постепенно распространяется от места контакта. 111. Оплодотворенное яйцо. Обратите внимание,
что студенистая оболочка присутствует еще до оплодотворения IV. Схематическое представление
цепи событий по данным электронной микроскопии. Б. Полихета Nereis. 1, Сперматозоид прибли
жается к яйцу; на этой стадии студенистоrо слоя нет. 11. Постепенное распространение кортикаль
ной реакции от точки контакта Наружная часть яйца состоит из крупных кортикальных ячеек, KO
торые разрушаются во время этоrо процесса. 111. Оплодотворенное яйцо. У большинства видов
содержимое кортикальных ячеек выделяется через оболочку оплодотворения (внежелточную
оболочку), формируя студенистую оболочку (по Austin 1965)
111. Удлинение акросомной нити. Б. Детали процесса оплодотворения у полихеты Hydroides. 1.
Приближение сперматозоида к внешнему поrраничному слою (ВПС) желточной оболочки. Этот
слой образуется из кончиков микроворсинок на поверхности ооцита. 11, 111. Слияние мембраны
aKpocoMHoro пузырька с ВПС, вызывающее высвобождение содержимоrо акросомы. Начало про
бодения желточной оболочки. IV. Продолжающееся проникновение rоловки сперматозоида и yд
линение мноrочисленных акросомных трубочек. V, VI. Контакт и окончательное слияние aKpOCOM
ных трубочек с плазматической мембраной ооцита. VII, VIII. Проникновение сперматозоида в
ооцит по мере Toro, как цитоплазма последнеrо смещается к месту слияния мембран ооцита
и сперматозоида. IX. Дальнейшее проникновение. Х. Ядро и митохондрии сперматозоида в цито
плазме яйца. Обратите внимание на отверстие, оставшееся в желточной оболочке (с электронных
микрофотоrрафий в кн. Colwin, Colwin, 1961)
444
1
2
3
4
5
6
7
8
10
18
20
30
(,J
rr. 40
50
Q)
а.
са 60
80
100
200
rлава 15
Связывание сперматозоида с рецептором
Незначительное поступление Na +
Изменение мембранноrо потенциала
Высвобождение Са 2 + ИЗ
внутриклеточных депо
Кортикальная реакция
Начало выделения кислоты и
OCHoBHoro поступления Na +
Превращение NAD в NADP
Рост потребления кислорода
g :r 1 ,5
):S: ""
rr. (".j
(".j о
о с:; 1,0
Q)
:s:
I: М
I 0,5
\D о
8
о
о 1 2 3 4 5 6 7 8 9 10
Время после осеменения, мин
300
400
500
600
800
1000
2000
Формирование оболочки
оплодотворения завершено
Увеличение внуТриклеточноrо рН
(пониженная кислотность)
Усиление синтеза белка
Активация транспортных систем
А
1600
I:
:s:
""
t:
1400
:s:
Слияние ядер яйцеклетки
и сперматозоида
Инициация синтеза днк
g 1200
(,J
:s:
о
1000
с'о
):S:
800
m
:s:
с'о
600
с:[
с'о
а.
400
I:
Q)
:r
2
200
са
о
Оплодотворенные
яйца
Неоплодотворенные
Яйца
о
о
24
40
48
56
32
8
16
Б
Время после добавления спермы мин
3000
4000
5000
6000
Рис. 15.5. Последовательность событий после контакта мембран сперматозоида и яйца во время
оплодотворения у MopCKoro ежа (по Epel; 1977). На вставках приведены примеры эксперимента
льных данных, подтверждающих эту схему: А потребление кислорода (по Ohnishi, Sugiyama,
1963); Б синтез белка (по Epel, 1967)
Первое клеточное деление
участком запускает последовательность со..
бытий (рис. 15.5), которые выводят яйцо из
этоrо состояния. Уровень потребления ки"
слорода значительно увеЛИЧИВается, дости"
rая пик а в пеРВУIО минуту после контакта
(коrда образуется оболочка оплодотворе..
ния), но по сравнению с неоплодотворенны"
ми яйцами остается rораздо более высоким
и в дальнейшем. Затем становятся доступ
ными «замаскированные» молекулы инфор"
маЦИОННОЙ (маrричной) РНК, накопленные
в цитоплазме яйца во время ооrенеза, в ре..
зультате чеrо усиливается синтез белка.
Первое деление дробления происходит че..
рез cTporo определенный период времени
после оплодотворения, однако еще до Hero
можно наблюдать важные изменения в рас..
пределении uитоплазматических компонен..
тов яйцеклетки. У билатерально симме..
тричных животных 'Эти движения цитоплаз..
мы формируют rлавные оси будущеrо
эмбриона, определяя передний, задний,
дорсальный и вентральный квадранты зиrо..
ты. Впервые такое перераспределение было
обнаружено у асцидий; оно изображено на
рис. 15.6. Однако, как будет показано
в разд. 15.3, пространственному распреде..
леНИIО различных составных частей цито"
плазмы вряд ли стоит придавать слишком
важное значение.
Развитие
::.: .: : .
::. . .::: :'.: "::\:\':
. ". . . . . :. .' . '..::.
. , . - · t ... .... l' '..
I . · · , , I' .. . ,
:..: # .' ' . · ' i!l :::' '. ".:
О О. .0 '. о . .:.
о Jo ,О to. 0 :"
. О О О '
00 о о о о () о
О о о О О 000 00
о О о о 00 о D
/ 000000
о о о
е.е.. .. .
. . . ' .' . II
i:? /- 0/ :: : ';. .:.:: ;:).
. . ; . : .. .. . .: . . .: 8.. :......
.:.......... ".
III
IV
v
Рис. 15.6. Движение визуально различимых
типов цитоплазмы, описанное в начале ХХ в,
у асцидии Stye/a partita. Эти наблюдения важны
для демонстрации потенциальной роли цито
плазматических факторов в детерминации cy
дьбы клеток. 1. Радиально симметричное не..
оплодотворенное яйцо. 11, 111. Миrрация «жел
той» цитоплазмы к веrетативному полюсу и ce
rреrация «серой» цитоплазмы в центральной
части яйца. IV. Сеrреrация и миrрация желтой
и прозрачной цитоплазмы к одной стороне
яйца. Яйцо становится билатерально симме
тричным. V. Билатерально симметричное яйцо
с локализацией цитоплазмы, соответствующей
первому делению дробления (см. приложение
15.3)
15.2. Раннее развитие
15.2.1. Дробление
Характер дробления зиrот большинства
животных, во время KOToporo относительно
445
крупное яйцо делится на все более мелкие
клетки, точно детерминирован с начала пер..
Boro митоза. Оно происходит таким обра
зом., что пространственная орrанизация
оплодотворенноrо яйца сохраняется в мно"
rоклеточном эмбрионе, причем у большин..
ства животных в результате возникает по
лый клеточный шар, называемый бласту"
лой.
Мноrим таксонам первичноротых (на..
пример немертинам.. аннеЛИДdМ. сипунку
лидам, зхиуридам, моллюскам и., возмо
жно, поrонофорам) присуще спиральное
дробление. Оно особенно хорошо известно
по работам, выполненным на аннелидах
и моллюсках, и более подробно описано
в приложении 15.1, поскольку имеет боль..
шое значение для экспериментальноrо aHa
лиза развития беспозвоночных (см. также
разд. 15.3).
Иноrда высказывается мнение, что спи
ральное дробление у названных таксонов
указывает Hd довольно близкую филоrене
тическую связь между ними. К этому пред"
положению, однако, следует относиться
с осторожностью. Считается, что указанные
таксоны берут начало от плоских червей,
причем некоторым примитивным видам по..
следних также свойственно спиральное
дробление. Особенно четко это заметно
у свободноживущих мноrоветвистых и бес..
кишечных турбелларий, хотя у последних
на веrетативном полюсе только два круп..
ных макромера, а не четыре, как обычно. От
плоских червей, возможно, произошли
и мноrие друrие таксономические rруппы.
включая псевдоцеломических червей., лофо..
форовых и членистоноrих, а также вторич"
норотые (rл. 2), хотя им и свойственны др у..
rие типы дробления. описанные ниже. Дей..
ствительно, для плоских червей спиральное
дробление необычно и известно лишь в не..
скольких rруппах, у которых желток запа..
сается в Ilитоплазме ООIlита.. а у видов со
специализированными желточными клетка..
ми (см. разд. 14.4.2.2) оно имеет аберрант"
ный характер.
Распространение спиральноrо дробле--
пия в нескольких типах rоворит лишь о том.,
что это консервативный признак, сохра..
нившийся в их эволюции, который нельзя
использовать как доказательство сравните..
446
1
п
IV
rлава 15
IП
V УI
Рис. 15.7. Дробление у иrлокожеrо Syпapta:
1 стадия двух бластомеров; 11. стадия четы
рех бластомеров; 111 стадия восьми бласто
меров; IV стадия 16 бластомеров; V
позднее дробление; VI стаДl1Я бластулы
льно недавней диверrеНIIИИ от общей пред
ковой ветви. ОДНdКО если у древних плоских
червей было спиральное дробление, то Ha
личие друrоrо ero типа у нескольких TaKCO
нов действительно может предполаrать бо
лее позднюю их эволюционную диверrен
цию. Это демонстрируют, например, BTO
ричноротые. У мноrих из них так называе
моерадиальноедробление.коrдапроизвод
ные поперечных делений лежат cTporo друr
над друrом. Сравнительно простой при
мер дробление у rолотурии Syпapta, по
казанное на рис. 15.7. Все дочерние клетки
. .
J"; ,L . . '
::.0 "
',. . -. . . . \
; ':-.: '. " :
p ;:.:...: :. i
I 11 ПI IV
Рис. 15.8. Поверхностное дробление яиц чле
нистоноrих (насекомых). 1. Единственное ядро
расположено внутри желтка в центральной ча..
сти яйца. 11. Деление ядра при водит к появле
нию нескольких ядер. но без клеточных rраниц
между ними. 111. Ядра движутся в перифериче
скую цитоплазму, относительно свободную от
желтка. IV. Между ядрами появляются радиа
льные клеточные стенки
в ходе постепенноrо развития до стадии по..
лой сферической бластулы здесь практиче..
ски одноrо размера Более сложное радий"
льное дробление у морских ежей, широко
использовавшихся в экспериментально..
эмбриолоrических исследованиях. На 64..
клеточной стадии отдельные бластомеры
здесь невозможно идентифицировать, так
как первые деления дробления дают один а..
ковые клетки, но зато у эмбриона появляю
тся различные по характеру делений слои
(см. приложение 15.2).
В яйцах членистоноrих., особенно HaceKO
мых, обильный желток занимает централь..
ное положение, поэтому свойственный им
тип дробления называют поверхностным,
или эндолецитальным. llитоплазма здесь
образует лишь поверхностный слой, а нача
льные деления ядра не сопровождаются ци..
токинезом клеток. В конечном счете ЯДрd
движутся к поверхности яйца. [де BOKpyr
них образуются клеточные мембраны (рис.
15.8).
15.2.2. rаструляция
Беспозвоночные., за исключением Placo..
zoa, Mesozoa, rубок и двух таксонов кишеч..
нополостных, ' трехслойные животные.
Друrими словами, их тело производное
трех эмбриолоrически различных клеточ..
ных слоев эктодермы, мезодермы и эито..
дермь] (у некоторых rрупп на взрослой ста..
дии следов энтодермы нет; например, у по..
rонофор). Бластула эмбриои. формирую..
щийся в результате paHHero дробления,
состоит только из одноrо слоя клеток. Если
в яйце не очень MHoro желтка, бластула
полая сфера и ПрОIIесс 2аСnlрУЛЯЦUU, даю..
щий в принципе трехслойный зародыш,
представляет собой инваrинацию клеток.
Этот способ хорошо заметен в случае про..
зрачных яиц иrлокожих (рис. 15.9).
Клетки первичной мезенхимы., про..
изводные микромеров зародыша на стадии
64 x бластомеров (см. приложение 15.2),
вторrаются в первичную полость тела
бластоцель. Затем веrетативный полюс за
счет изменения относительноrо слипания
своих клеток начинает впячиваться внутрь
с образованием уrлубления. У эмбриона
MopcKoro ежа этому движению внутрь МО"
1
Развитие
447
II
УТ
III
IV
Рис. 15.9. rаструляция у MopCKoro ежа. 1. Ранняя rаструляция, инваrинация первичной мезенхи
МЫ, 11, 111. Миrрация мезенхимы и начало инваrинации. IV. Образование контактов филоподий
вторичной мезенхимы. V. Поздняя rаструла, появление скелетных спикул, секретируемых первич
ной мезенхимой. VI. Сформированная rаструла (призматическая стадия зародыша). Закладка
оральноrо поля. VII. Вид paHHero плутеуса с оральной поверхности. VIII. Вид paHHero плутеуса
сбоку (по Trinkans, 1969)
жет содействовать прямой контакт филопо--
дий, протяrивающихся от клеток вторичной
мезенхимы к внутренней поверхности бла--
стоцеля (рис. 15.9.IV). Однако для rаструля--
ции такие филоподии не обязательны (на--
пример., у эмбриона морской звезды подоб
Horo рода контактов нет), поэтому нельзя
сказать, что инваrинация зависит от созда--
BaeMoro ими натяжения. Возможно, движе--
нне клеток результат сил, rенерируемых
их различным слипанием.
К концу rаСТРУЛЯIIИИ у эмбриона разли--
чают внешний слой эктодермы, открываю--
щуюся сзади внутреннюю трубку и мезен--
химные клетки между ними. Эта стадия Ha
зывается 2аспlРУЛОЙ. На ней дефинитивная
мезодерма еще не обособилась от первич
ной кишки (архентерона). Клетки мезенхи--
мы не принимаю т участия в образовании
дефинитивной мезодермы, но MorYT вклю--
чаться в скелетные и мышечные элементы
личинки, которые утрачиваются во время
метаморфоза. Формирование мезодермы
описано в разд. 15.4.
у вторичноротых бластопор становится
анусом функциональной личинки, а зача..
стую и взрослоrо орrанизма. Ротовое от..
верстие формируется путем инваrинации
эктодермы.
у первичноротых rdСТРУЛЯЦИЯ иде1 по--
иному. Она также включает внедрение
внутрь бластулы (интернализацию) пре..
зумптивной энтодермы и небольшоrо числа
клеток, дающих начало мезодерме взросло..
ro животноrо. У форм с малым количе..
ством желтка в яйце rаструляция осуществ--
ляется путем инваrинации., как показано на
рис. 15.10,1, а в случае яиц, боrатых желт..
ком, заключается в амебоидной миrрации
клеток в обхват относительно инертной за
полненной желтком энтодермы (рис. 15.10,
11, 111).
У форм с малым количеством зароды--
шевоrо желтка в состав дорсальной rубы
бластопора входят клетки, из которых фор--
мируется ротовое отверстие личинки. Не..
сколько таксонов объединены в rруппу пер..
вичноротых именно по этому., считающе..
муся фундаментальным признаку paHHero
развития.
В СЛУЧdе боrатых желтком яиц тип
дробления часто неясен, и rаструляпию
проследить трудно. Бластоцель обычно за..
полнен, а презумптивные клетки энтодермы
441
rлава 15
1
II
ПI
А
Б
Рис. 15.1 О, rаструляция у первичноротых
(примеры основаны на изучении полихет с раз
личным количеством желтка в яйцах).
А. Строение бластулы. Б. Средняя rаструла.
Морфолоrически важные клетки, берущие Ha
чало от 4d бластомеров и формирующи& Me
зодерму взрослоrо животноrо, обозначены
точками; клетки, образующие ротовое OTBep
стие, заштрихованы; клетки будущеrо кишеч
ника показаны мелкими кружками. 1. Относи
тельно бедное желтком яйцо. 11. Яйцо с проме
жуточным содержанием желтка. 111. Боrатое
желтком яйцо и rаструляция путем эпиболии
на веrетативном полюсе часто оказываются
внутри за счет Jпu60ЛUU, или обрастания их
клетками анимальноrо полюса, как показа..
но для пиявки на рис. 15.11. У некоторых
полихет ротовое отверстие формируется не..
зависимо от бластопора, хотя они и счи..
таются первичноротыми.
Таким образом, rаструляция это про..
цесс интернализации клеток с образованием
типичной трехслойной структуры. Можно
составить карту зачатков с точной локали..
зацией на поверхности бластулы будущих
внутренних тканей.
На рис. 15.12 показана такая карта для
бластулы полихеты; указаны также траек"
тории движения клеток, приводящие к фор..
МИРОВdНИЮ трохофоры. Нужно заметить,
А
Рис. 15.11. rаструляция в боrатых желтком
яйцах пиявки: А на поперечном срезе с левой
стороны; Б объемное изображение со CTOpO
ны веrетативноrо полюса, показывающее пло
скость среза
что клетки производные бластомеров 2d
и 4d иrрают в эмбриональном развитии
первичноротых очень важную роль. Про..
изводные 2d образуют эктодерму взрослоrо
животноrо, а производные 4d две парные
клетки мезоэнтодермы (мезентобласты),
дающие начало будущей мезодерме. Рис.
15.12 показывает относительное расположе..
ние этих клеток перед rаструляцией и их
движения, вызывающие инваrинацию пре..
зумптивной rэнтодермы И перемещение ме..
зентобластов и зктотелобластов на их дефи..
нитивное место в трохофорной личинке.
у личинки аннелид (рис. 15.12) все се..
rMeHTapHbIe структуры будут производны"
ми парных мезодермальных полосок
и эктотелобластическоrо кольца. Из друrих
частей трохофоры развиваются иесеrмен..
тарные структуры пиrидиум и просто..
миум (см. разд. 4.14).
у членистоноrих тип rаструляции со..
всем иной. «Бластопор» здесь представлен
бороздкой, через которую происходит ин..
тернализация вентральной полоски про..
спективных мезодермальных клеток, пред..
назначенных для формирования мускулату..
рь] тела, стенки кишечника, rонад и Т. д.
В обобщенном виде последовательность на..
блюдаемых при этом событий показана на
рис. 15.13. Внутренняя часть TaKoro эмбрио..
на не полая, d заполнена массой неклеточ..
Развитие
1
n
Бластопор
IП
Прототрох
Рот
Мезодермальные
полоски
ЭктотеЛОБластическое
кольцо
А
449
Б
Рис. 15.12. rаструляция у первичноротых на примере полихет: А вид с вентральной поверхно
сти; Б вид с левой стороны. 1. Схематическое изображение простой карты зачатков. 11. rаструля
ционные движения: сплошные стрелки показывают движение клеток к бластопору, пунктирные
миrрацию инваrинировавших клеток. Обратите внимание, что стомодеум, или рот, образован ин..
ваrинацией клеток через переднюю часть бластопора. Мезодермальные полоски берут начало от
парных мезентобластов, миrрирующих через задний край бластопора. 111. Сформированная TpO
хофора с парными мезодермальными полосками и эктотелобластическим кольцом. Эти два эле
мента образуют сеrментную бластему, которая сохраняется на передней стороне пиrидиума на
протяжении всей взрослой жизни. Простомиум, перистомиум и пиrидиум являются HecerMeHT..
ными структурами (частично по Anderson, 1964)
Horo желтка Одновременно инваrинируют
rанrлионарные клетки сеrментарной нерв--
ной системы.
Эмбрион HaceKOMoro окружен «внезаро--
дышевыми оболочками», позволяющими
провести ин ересные аналоrии между раз--
витием «высших» и «низших» беспозвоноч--
ных, с одной стороны, и «высших» и «низ..
ших» позвоночных с друrой.
450
I
II
III
IV
А
rлава 15
Б
Рис. 15,13. rаструляция у насекомых, А, Попе
речный разрез. Б. Вид с вентральной стороны.
1 Раннее деление ядра. 11. Поверхностное
дробление. 111, IV. Инваrинация мезодермы. V,
VI. Сеrментация нейроэктодермы образование
внезародышевых оболочек. VII. Сформирован
ная rаструла (по Slack, 1983)
15.3. Экспериментальная
эмбриолоrия беспозвоночных:
детерминация судьбы клетки
1 5.3.1. Введение
Сложное мноrоклеточное животное про-
исходит от одноклеточной, недиффереНIlИ-
рованной тотипотентной структуры
оплодотвореННQrо яйца. В процессе разви-
тия оно дает множество клеток. причем от-
дельные их rруппы различаюrся по химиче
скому составу, морфолоrии и функциям.
Экспериментальная эмбриолоrия это
наука о том. как в развивающемся эмбрио-
не возникаfОТ популяции дифференцирован-
ных клеток со специфическими функциями.
Эмбрионы и личинки беспозвоночных пред..
ставляют собой наиболее удобные модель-
ные системы для анализа участвующих
в этом процессов.
В ходе paHHero онтоrенеза жизнедея
тельность развивающеrося Эl\lбриона рету-
лируют материнские [ены. Однако затем
в действие вступает [еном собственно зиrо-
ты, а, поскольку со временем появляется
MHoro различных типов клеток, заключен-
ная в ядре информация должна в каждом
случае особым образом контролироваться.
Потенциальных rочек контроля l\1HOrO; они
включают один или несколько этапов такой
последовательности:
1) rенетическая информация в ядре зиrо-
ты;
2) распределение информации по дочер-
ним клеткам:
3) транскрипция rенетической информа-
ции;
4) ПОПУЛЯIlИЯ молекул мРНК:
5) перенос 1\IРНК из ядра в цито-
плазму;
6) популяция мРНК в цитоплазме дочер-
них клеток;
7) трансляция информации на '\10леку-
лах мРНК;
8) образование специфичных для клеток
белков;
9) функция специфических веществ.
В настоящее время общепринято, что
у большинства орrанизмов ядра дифферен-
цированных клеток содержат ту же rенети-
ческую информацию, что и ядро оплодот-
Развитие
1
451
Элиминируемый
хроматин
II
Примордиальные
репродуктивные
клетки
ПI
IV
Рис. 15.14. Пример диминуции хромосом во время paHHero эмбриональноrо развития (Ascaris).
1. Первое деление дробления; хромосомы не утрачиваются. 11. Элиминация XpOMocoMHoro MaTe
риала во время деления передней клетки. 111. Элиминация XpOMocoMHoro материала и neperpyn
пировка клеток на стадии четырех бластомеров. Только одна клетка сохраняет полный набор xpo
мосом и тотипотентность. IV. На стадии rаструлы полный набор хромосом остается в двух при
мордиальных половых клетках
BopeHHoro яйца. Доказательства этоrо ос--
нованы на результатах
1) экспериментальной трансплантаuии
ядер из дифференцированных клеток в ци--
топлазму энуклеированных яиц;
2) изучения цитолоrии хромосом во
время данноrо дробления;
3) сравнения характера исчерченности
rиrантских хромосом некоторых двукры--
лых и друrих насекомых;
4) изучения дифференцировки и редиф--
ференцировки во время реrенерации (см.
также разд. 15.5);
5) анализа «отпечатков пальцев» ДНК
из разных тканей.
Однако из этоrо обшеrо правила есть ис..
ключения. Например, у нематоды Ascaris
только примордиальные половые клетки
эмбриона получают полный набор хромо--
сом, а в друrих клеточных линиях часть их
утрачивается во время paHHero дробления
(рис. 15.14).
Если все ядра развивающеrося зародыI'
ша имеют одинаковый rенотип, дифферен"
цировка должна выражать взаимодействие
цитоплазматической информации клетки
и ее ядра.
15.3.2. Мозаичное
и реrуляционное развитие
в начале этоrо столетия эмбриолоrи..
экспериментаторы изучили возможности
нормальноrо развития изолированных кле..
ток зародышей беспозвоночных. В ряде слу..
чаев, например у MopcKoro ежа, каждый из
бластомеров на стадии четырех клеток да..
вал целую и совершенно нормальную ли..
чинку--плутеус (см. приложение 15.3,1). Та..
кое развитие назвали реrуляционным.
В друrих случаях эмбрионы, развиваю
щиеся из каждоrо бластомера на четырех--
клеточной стадии. были весьма неполно
ценными (см. приложения 15.3,2 и 15.3,3).
Такое развитие назвали мозаичным. Был
сделан вывод, TO отдельные клетки насле..
дуют особый тип цитоплазмы, определяю..
щий их дальнейшую судьбу. Более деталь..
ный анализ этих данных требует 1нания су"
дьбы различных блестомеров при нормаль..
ном развитии, которую можно отразить
картой зачатков на бластуле или нераздро--
бившейся цитоплазме яйца. Такую карту
составляют, маркируя специфические зоны
452
rлава 15
цитоплазмы и прослеживая судьбу меченых
клеток, а в случае асцидии Stye/a partita она
выявляется в ходе тшательных наблюдений
за яйцом после оплодотворения (см. рис.
15.6). Эти наблюдения rоворят, что блаrо
даря движениям цитоплазмы яйцо станови
тся явно билатерально симметричным"
а среди ero зон выделяется серповидный
участок цитоплазмы желтоватоrо цвета,
так называемый желтый серп. Ero можно
наблюдать от ранних стадий дробления до
формирования мышечных блоков у личин
ки «rоловастика». Все презумптивные мы..
шечные клетки наследуют некоторое коли..
чество желтой цитоплазмы, причем ни одна
из клеток, которая не станет мышечной, ее
не получает. Факторы, определяющие cyдь
бу клетки у беспозвоночных, более деталь
но обсуждаются в приложении 15.4.
Результаты экспериментов с асuидией
явились убедительным доказательством po
ли локализации цитоплазматических ком..
понентов в детерминации судьбы клетки.
Опыты с яйuами насекомых также показы
вают, что в процессе развития деятедьность
ядер может определяться цитоплазмой,
в которую они попали. На рис. 15.8 показан
ход дробления. На заднем полюсе яйца ча
сто находится хорошо различимая цито
плазма, так называемая полярная плазма.
Способностью формировать половые клет
ки (т. е. оставаться тотипотентными) обла
дают только ядра, оказавшиеся в клетках
с такой цитоплазмой. Эмбрионы с удален
ной полярной плазмой стерильны. Поляр..
ная плазма определенной rенетической ли
нии мухи DrosophiZa при введении в пере
днюю часть яйца мухи друrой линии вызы
вает формирование из клеток этой области
[амет.
Однако развитие нельзя рассматривать
исключительно как результат особенностей
распределения цитоплазматических дeTep
минантов в яйце. Оно также включает взаи
модействие между клетками. Эксперимен"
ты с улиткой Nassarius obso/etus (часто упо
требляют также родовое название /lyaпassa)
продемонстрировали взаимодействие цито"
плазматических детерминантов и межкле
точных влияний., определяющее судьбу
клетки в ходе эмбриоrенеза (более детально
см. приложение 15.5).
Nassarius obso/etus свойственно спираль-
ное дробление (см. приложение 15.1) с ПОЯВ-
лением при первом и втором клеточных де-
лениях выступающей полярной лопасти
Специфическая полярная плазма передается
только в D бластомер. Эмбрионы с удален-
ной полярной допастью становятся симме-
тричными, определенная хронолоrия
дробления в последовательности поколений
D клеток нарушается, и развитие приводит
к весьма неполноценной «безлопастной»
личинке (см. приложение 15.5). У таких ли-
чинок нет rлаз, статоцистов, ноrи, паруса,
раковины, сердца и оформленной кишки.
Однако некоторые продукты клеточной
дифференцировки можно распознать. Схе-
ма клеточных поколений в приложении 15.1
подтверждает эти наблюдения. Мноrие из
зависящих от полярной лопасти структур не
являются производными клеток, наследую-
щих ее материал. Ясно, что судьба клеток,
происходящих от lа и lс, дающих rлаза,
или от 2d, из которых формируется ракови-
на. определяется не только их цитоплазма-
тической наследственностью, но и взаимо-
действием с клетками D"линии во время
критическоrо периода, длящеrося от появ-
ления 2d до формирования 4d. Аналоrич-
ным образом клетки, компетентные дать
начало раковинной железе, являются про-
изводными единственноrо бластомера 2d,
однако они будут выполнять свои функции,
только если клетка 2d контактно взаимо-
связана с друrими клетками D"линии.
у эмбриона MopcKoro ежа судьба раз-
личных клеточных слоев еще в большей сте-
пени определяется их взаиморасположе-
нием и не закреплена цитоплазматической
наследственностью. Основные элементы
личинки здесь можно проследить вплоть ДО
бластомеров последних стадий дробления
в соответствии со следующей схемой:
Аn 1 апикальный ресничный султан;
Аn 2 оральное поле истомодеум;
Veg 1 эктодерма;
Veg 2 apxeHTepOH и целомические ме-
шки;
Микромеры первичная и вторичная
мезенхима.
Взаимосвязи слоев рассмотрены в при-
ложении 15.6. Поперечное разделение бла-
стулы четко выявляет ее части с разным по..
Развитие
тенциалом развития, однако rоворить
о простой мозаике этих частей нельзя. По..
видимому каждый клеточный слой раз..
вивается в соответствии с ero положением
относительно двух rрадиентов, направлен..
ных один от анимальноrо полюса к Bere..
тативному, а друrой наоборот. Некото"
рые эксперименты, приведшие к такой кон..
цепции., обсуждаются в приложении 15.6.
Различия между реrуляционным и мо"
заичным эмбрионами, видимо, заклады..'
ваются в момент первоrо разделения кле..
ток со специфическими цитоплазматически..
ми детерминантами.
15.4. Личиночное развитие
и метаморфоз
Жизненный цикл мноrих животных ха..
рактеризуется диверrенцией эколоrических
НИШ, занимаемых ювенильной и взрослой
стадиями. Особенно ярко это проявляется
у свободноживущих морских беспозвоноч"
ных (см. rл. 14). паразитических видов и не..
которых насекомых. Такая особенность
развития требует более или менее резкоrо
перехода между образом жизни, к которому
была адаптирована личинка, к новому., ха..
рактерному для взрослоrо орrанизма. Ча..
сто это сопровождается существенным из..
менением строения, так называемым MeTa
морфозом.
15.4.1. Метаморфоз морских
личиночных форм
Ресничные личинки r орских беспозво..
ночных адаптированы к пелаrическому
образу жизни. Полосы и пояса ресничек,
обеспечивающие их движение, не подходят
взрослым формам (см. rл. 10). Для таких
животных характерен метаморфоз с утра..
той ресничных поясов и переходом клоко..
моции С помощью мышечных клеток.
у rастропод метаморфоз личинки..
велиrера сопровождается постепенной ре..
дукцией паруса, который в качестве rлавно"
ro локомоторноrо opraHa в конечном итоrе
заменяется ноrой (см. рис. 15.15), поскольку
уже не в состоянии поддерживать разви"
вающуюся улитку. Метаморфоз двуствор"
453
Правый
ретрактор
Hora
1
,.
/
/
-
ПI
Рис. 15.15 Закручивание (торсия) во время
развития личинки моллюска на примере вели
repa брюхоноrих, К началу торсии мантийная
полость находится сзади, а после Hee
спереди, что соответствует тополоrии взро"
слой улитки. 1. Ранний велиrер с асимметрич..
ной правой мышцей..ретрактором. 11 Первая
фаза торсии быстрый поворот на 900 за счет
сокращения мышцы ретрактора. Это вызывает
частичное закручивание BHYTpeHHocTHoro ме..
шка относительно ноrи. 111. Вторая фаза тор..
сии закручивание BHYTpeHHocTHoro мешка
еще на 900 относительно ноrи за счет HepaBHO
MepHoro роста
чатых моллюсков часто происходит бы..
стрее, с резкой утратой паруса.
у полихет также существует противоре..
чие между характером локомоции личинки
454
и потребностями развития во взрослое жи..
вотное. Сеrменты тела добавляются на
эмбриональной или личиночной стадии по..
степенно (см. разд. 15.2.2) и берут начало от
парных вентральных тяжей в задней части
трохофоры. Мезодермальные клетки тяжей
являются производными парных мезен..
тобластов, образующихся из бластомера 4d
в ходе rипичноrо спиральноrо дробления
(см. разд. 15.2.] и приложение 15.1). У этих
зачатков cerMeHToB имеются мезодермаль..
ные и эктодермальные элементы. Два ТЯЖd
клеток, дающие мезодерму., разрастаются
в ряд тканевых масс, в которых формируе..
тся целом. В отличие от описанных ниже
иrлокожих у этих животных мезодерма об..
разуется не из полости архентерона. Такой
тип целома назван схизоцелем. CerMeHTbl
формируются путем координированноrо
орrаноrенеза мезодермальных и эктодерма..
льных тканей, в котором rлавную индук"
ционную И орrанизационную роль иrрает
вентральный нервный ствол. Каждый но..
вый cerMeHT развивается спереди от пиrи..
диума, увеличивая массу личинки. Часто
каждый cerMeHT во время пелаrической ста..
дии жизни снабжен локомоторными или
плавательными приспособлениями.
Развитие иrлокожих характеризуется од..
ной из наиболее впечатляющих форм мета..
морфоза в животном царстве. Полностью
сформировавшаяся их личинка билатераль..
но симметрична, в то время как у взрослоrо
животноrо доминирует пятилучевая симме
трия. хотя билатеральная иноrда наклады
Вdется на нее вторичцо (см. разд. 7.3.2). Це..
ломические мешки иrлокожих раЗБиваются
из боковых ВЫПЯЧИВdНИЙ вершины apxeHTe
рона через некоторое время после заверше..
ния rаструляции (см. рис. 15.9). Такой спо..
соб их формирования называется энтероце--
льным. Изучение ныне живущих иrлокожих
rоворит о том.. что изначально было три
парных целомических мешка: аксоцель, rид--
роцель и соматоцель. Их развитие и после..
дующий метаморфоз показаны на рис.
15.16. .У мноrих современных иrлокожих
правые dксоцель и rидроцель редуцированы
или полностью исчезли. Левый rидроцель
разделяется на rидроцель и вырост, обра..
зующий каменистый канал и rидропор (рис.
15.16, I IV).
rлава 15
Эти примитивные целомические зачатки
показаны у девятидневной личинки
плутеуса на рис. 15.16,V. Правый и левый
соматоцели распространяются BOKpyr же
лудка и формируют полости тела взрослоrо
орrанизма. Левый rидроцель преобразуется
в пятилучевой заполненный жидкостью за
ЧdТОК, из KOToporo развивается амбулакра
льная система взрослой особи. Дефинитив
ное ротовое отверстие, определяющее opa
льное поле MopcKoro ежа, будет формиро
ваться в центре кольца rидроцеля. Таким
образом. орально..аборальная ось взросло
[о орrанизма проходит примерно вдоль ле
во--правосторонней оси плутеуса (рис.
15.16, VI) Морской еж или морская звезда
развивается из имаrинальных дисков. раз
растающихся ВО время метаморфоза парал
лельно сморщиванию и сбрасыванию )KTO
дермальноrо эпителия личинки (рис.
15. 16..VII..VIII).
Животные с экзоскелетом не MorYT pa
сти и развиваться постепенно. Они должны
пройти через ряд линек, при каждой из KO
торых сбрасывается старый покров и opra
низм увеличивается в размерах вплоть до
затвердения HOBoro наружноrо скелета. Это
хорошо заметно у ракообразных. Их раЗБИ-
тие часто включает ряд морфолоrически
различных личиночных стадий. которые
в конечном итоrе линяют, давая взрослую
форму (см. приложение 14.5 и rл. 8). При
этом рост с линьками нередко продолжае-
тся и после Toro. как сформировалась мор-
фолоrия взрослоrо орrаНИЗМd.
У морских беспозвоночных личинка обе-
спечивает поселение орrанизма в приrод-
ных для дальнейшеrо развития условиях.
..Это очень важная стадия в жизненном ця-
. кле животноrо, особенно для видов, Beдy
щих во взрослом состоянии сидячий или
прикрепленный образ жизни, для которых
выбор подходящеrо субстрата бывает ре-
шающим для выживания фактором.
rидробиолоrи иноrда наблюдают тучи
только что прошедших метаморфоз личи-
нок, как будто случайно оседающих на не-
приrодные для жизни субстраты Однако
этот процесс не всеrда случаен. Мноrие ра-
боты rоворят о точности, с которой пелаrи-
qеская личинка способна «выбрать» блаrо-
приятный субстрат. При этом действуют
Целомичес
кие карманы
1
IV
Оральная
поверхность
Оральный
имаrинальный
диск
Развитие
II
V
АБоральный
имаrинальны
VH диск
455
ПI
VI
Рис. 15.16. Развитие целомической системы MopcKoro ежа и метаморфоз ero личинки во взро
слую форму. 1. Формирование целомических мешков из выпячиваний у вершины архентерона
после окончания rаструляции. 11. Парные целомические мешки, 111, IV. Разделение мешков.
V. Дифференцировка BToporo левоrо целома в rидроцель и комплекс каменистоrо канала
rидропора, рост третьих (левоrо и npaBoro) целомических мешков. У Psaттechiпus эта стадия ДО"
стиrается примерно на девятый день планктонноrо развития. VI. Формирование пятилучевоrо ам..
булакральноrо кольца, определяющеrо оральную поверхность развивающеrося иrлокожеrо,
и становление орально аборальной оси. VII. Постепенное развитие оральноrо и аборальноrо
имаrинальных дисков, сопровождающееся ростом ресничных локомоторных поясов плуте уса.
VIII. Эмбрион Psaттechiпus незадолrо до завершения метаморфоза. Личиночные ткани будут
разрушены, зачатки взрослой особи объединяются, сформировав уменьшенную копию MopcKoro
ежа
следующие механизмы:
1) последовательность поведенческих
актов, приводящая к контакту личинки
с блаrоприятным субстратом;
2) замедленный метаморфоз в ОТСУ1--
ствие TaKoI'o субстрата;
3) различение и выбор предпочтитель--
Horo субстрата;
4) аrреrационное поведение и хемосен--
сорное распознавание субстратов. уже засе--
ленных взрослыми и личиночными форма--
ми cBoero вида.
456
rлава 15
с::> #
а.
ф
м I
"'
а. I
I I
1i
::r:
..о
@ I
I
s
() I
о
::r:
О
ЭМБриональное Питание Питание Питание Питание Питание 1 e 2 p 3 e
1 й -личи-- 2 й личи- 3...й личи 4 й личи 5....й личи имаrи имаrи имаrи
развитие ночной ночной ночной ночной ночной нальное нальное нальное
стадии стадии стадии стадии стадии питание питание питание
ЯЙЦО Размнож.ение ,.
Неполовозрелые личинки
ИМАrо
А
ЯИUО
Питание
1 й личи
ночной
стадии
Зачатков
крыльев нет
НИМСРА
Питание
2 .й личи
ночной
стадии
Питание Питание
3...й личи 4.....й личи
ночной ночной
стадии стадии
Неполовозрелость
Наружные зачатки крыльев
НИМСРА (или ЛИЧИНКА)
Питание
5 й личи Имаrинальное питание,
ночной расселение
стадии
ЭМБриональное
развитие
Половозрелость
Крылья
ИМАrо
Б
Рис. 15.17. Схематическое представление роста и развития насеком ых. А. Аметаболические фор
мы: метаморфоза нет. В этом примере рост продолжается и после достижения половой зрелости.
Б. rемиметаболические формы: неполный или частичный метаморфоз. В этом примере крылья
появляются у личинки BToporo возраста в виде наружных зачатков. Полностью они формируются
после пятой (последней) линьки. Увеличение размера происходит в течение KopoTKoro периода
после каждой линьки. в. rолометаболические формы: полный метаморфоз. В этом примере че
тыре личиночные, куколочная и имаrинальная стадии. Рост происходит после каждой личиночной
линьки
Развитие
.
ЭМБриональное
развитие
Питание
1 й личи
ночной
стадии
Зачатков
крыльев не
ЛИЧИНКА
Пи ание Пi-!тание Пи ание Непитающаяся Имаrо
2 и л чи 3 и ли и и л чи 5 я личиночная Размножение и расселение
ночнои ночнои ночнои стадия или
стадии стадии стадии куколка Иноrда питание
Внутренние зачатки крыльев Наружные за
чатки крыльев
ЛИЧИНКА КУКОЛКА
ЯЙЦО
в
Рис. 15.17 (продолжение)
Например., сложная последовательность
поведенческих актов в процессе развития
наблюдается у личинки мидии MytiZus edu..
lis.
Установлено.. что личинки мноrих видов
высокоизбирательно относятся к заселяе..
мым субстратам. Это иллюстрируют опи..
санные в приложении 15.7 эксперимеНТЬJ
с выбором поверхностей заселения личин..
ка ми близкородственных видов живущих
в трубках полихет Spirorbis, взрослые особи
которых обычно встречаются на различных
субстратах.
Циприсовидные личинки морских желу..
дей также демонстрируют перед оседанием
на субстрат ярко выраженное и СЛО)I(ное по..
ведение. Это позволяет им выбрать удоб..
ные для метаморфоза места. Личинки ре..
аrируют не только на текстуру поверхности
(предпочитают rрубую и пористую ), но И на
присутствие друrих морских желудей, их
личинок, а также остатков взрослых особей
собственноrо вида.
15.4.2. Развитие и метаморфоз
насекомых
Тип Uniramia включает большую rруппу
животных, обычно называемых насекомы..
ми (подтип Hexapoda). У них три различ..
457
Крылья
ИМАrо
ных способа развития. Представители близ..
ких К мноrоножкам бескрылых rрупп, на..
пример ноrохвостки и щетинохвостки, раз..
виваются постепенно через ряд линек. Их
строение при этом не претерпевает каких..
либо сильных изменений, поэтому rоворить
о метаморфозе нельзя (рис. 15.17,А). Такие
формы называют аметаболическими, Пол..
ное дробление яип у мноrих видов этих жи
вотных примитивная черта, однако у не..
которых оно, хотя сначала и полное., в ко..
нечном счете приближается к типу, xapaK
терному для крылатых насекомых (рис.
15.18).
у друrих насекомых во время развития
наблюдается более или менее выраженный
метаморфоз и фиксированное число личи
ночных стадий, предшествующих взрослой
форме. В нескольких отрядах метаморфоз
не слишком резкий, поскольку преимаrина
льные формы, называемые «нимфами» или
«наядами» (если они водные), имеют Hapy
жные зачатки крыльев. Развитие этих насе..
комых (к ним относятся стрекозы и кузнечи..
КИ) называют rемиметаболическим, а их са..
мих экзоптериrотами. У кузнечиков и ca
ранчи., как показано на рис. 15.17.Б., нимфы
и имаrо занимают одну и ту же нишу,
поэтому при метаморфозе последней личи
ночной стадии в крылатое насекомое значи..
458
rлава 15
Рис. 15.18. Раннее развитие бескрылых Hace
комых. Можно видеть, что дробление сначала
полное (ер. рис. 15.8)
тельной перестройки строения не требуется.
Если же ювенильные стадии обитают в BO
де, изменение морфолоrии может быть зна
чительным, поскольку оно связано с дивер
rенцией неполовозрелых особей и имаrо по
нишам. Однако и такой тип метаМОРфОЗd
не столь впечатляющ, как у насекомых
с внутренними крыловыми зачатками
(эндоптериrот). Их развитие называют ro..
лометаболическим. Между последней личи..
ночной и имаrинальной стадиями здесь есть
промежуточная стадия куколки (рис. 15.17
и 8.33). Личинки l оломатаболических Hace
коrvlЫХ относятся к различным типам tрис.
15.19); им часто дают обобщенные назва..
ния -rусениuы, черви и т. д. Как показано
в rл. 14 и на рис. 15.17,В, это специализиро
ванные питающиеся стадии, в то время как
lr/rrJLl7
II
1
.....
. . .
. . .
IV
v
Ф111ПlIllJ
VI
Рис. 15.19. Примеры личинок rолометаболи..
ческих насекомых: I жужелицы (жуки); 11
жука щелкуна (<<проволочник»); 111
долrоносика (жуки); IV медоносной пчелы
(перепончатокрылые); V бабочки (rусеница);
VI падальной мухи (двукрылые). В каждом
случае морфолоrия личинки и имаrо COBep
шенно различна
rениталии
Рис. 15.20. Система имаrинальных дисков ли
чинки двукрылоrо (Drosophi/a). Обозначены
местоположение дисков в личинке и развиваю
...циеся из них имаrинальные структуры. Можно
видеть, что пищеварительная система при Me
таморфозе не заменяется
имаrо выполняют функuию расселения
и размножения. На стадии куколки движе..
ние и питание прекращаются, поскольку
происходит коренная структурная пере
стройка орrанизма.
у rолометаболических насекомых во
время paHHero эмбриоrенеза закладыва
ются rруппы клеток, которые дифференци"
руются только во время метаморфоза. Эти
имаrинальные К lетки. собранные довольно
рыхлыми rнездами или чаще оформленные
в виде компактных имаrинальных дисков,
дают начало всему внешнему покрову има..
ro с ero сложноЙ реrионарной структурой,
слюнным железам и друrим внутренним ор..
[анам. Схема расположения имаrинальных
дисков у личинки двукрылоrо и их соответ..
ствие имаrинальным структурам показаны
на рис. 15.20. В современной биолоrии ос О..
бый интерес приобрела система имаrиналь..
ных дисков плодовой мушки Dl"Osop/zila те..
laпogaster. В OCHOBHOl\1 это обусловлено 60"
raTCTBOM rенетической информаuии, нако"
пленной по этому орrанизму, однако связа..
но также и с преимуществами использова..
ния имаrинальных дисков в качестве моде..
ли для анализа судьбы клеток, пропессов
детерминации, дифференцировки и орrапо..
rенеза, которые обсуждались выше в разд.
15.3.
Развитие
15.4.3. Экспериментальный
анализ развития имаrинальных
дисков Drosophi/a тe/aпogaster
Имаrинальные диски rолометаболиче--
ских насекомых закладываются в раннем
эмбриоrенезе. Т. е. клетки должны рано при--
нять «решение», будут ли они функциониро--
вать в теле личинки или останутся недиффе--
ренцированными в составе имаrинальных
дисков. В последнем случае их дифференци--
ровка начнется под влиянием изменениЙ
rормональной среды во время окукливания.
Несмотря на отсутствие дифференци--
ровки, имаrинальные диски являются высо--
кодетерминированными структурами, по
свойствам больше напоминающими мо"
заичных, чем реrуляционных эмбрионов.
Однако, как показано ниже, имеются дан..
ные о том, что в основе этой детерминации
лежит система rрадиентов. Путем выбороч--
Horo удаления частей диска с последующеЙ
ero имплантацией в личинку последней ста--
дии можно составить карту ero зачатков. Во
время метаморфоза в соответствии с уда--
ленными частями будут развиваться непол--
ные имаrинальные структуры. Карта зачат--
ков дисков конечностей, например, имеет
концентрическое строение (рис. 15.21.А).
Из внешних зон развиваются проксималь--
ные элементы ноrи, а из внутренних
дистальные (рис. 15.21,Б,В). Диск конечно--
сти можн.о разрезать и имплантировать ero
фраrмент в молодую личинку. При этом
происходит компенсационный рост, и недо--
стающие части восстанавливаются. Однако
на морфолоrию получившейся конечности
влияет положение внутри диска исходных
фраrментов.
Если, например, диск конечности разде--
лен на секторы, как показано на рис.
15.22.А, то верхняя медиальная четверть
(А), реrенерируя, дает нормальный полныЙ
диск, а нижняя медиальная (С) зеркально
удвоенную нижнюю половину. Эти резуль--
таты сопоставимы с rралиентной теорией
орrанизации. Можно предположить суще--
ствование некой орrанизационной иерар--
хии, причем фраrменты, находящиеся на
высшей ее ступени, MorYT реrенерировать
стоящие ниже их по paHry, а в обратном на..
правлении реrенерация невозможна. Эта
459
А
Тазик
Вертлуr
.............. Бедро
r олень
Лапка
Б
в
Тазик
Вертлуr
Бедро
rолень
Рис. 15.21. Развитие конечности HaceKoMoro
из имаrинальноrо диска. А. Карта зачатков
имаrинальноrо диска, полученная путем селек
тивноrо удаления ero частей. Б. Постепенное
развитие конечности путем разрастания има
rинальноrо диска. В. Элементы сформирован
ной ноrи
модель представлена схематически на рис.
15.22,Б. Подобные модели ценны, но еле--
дует заметить, что, воспроизводя наблю--
даемое, они не дают никаких ключей к раз--
rадке биолоrической приролы действую--
щих механизмов. В то же время именно си--
стема имаrинальных дисков позволяет бы--
460
rлава 15
Верхняя 1\
6 II
Латеральная
D
Медиальная
А
10
9
10 9
8\
Реrенерация
10 9 8 7 6 5 4
В
..л..
Б
III
8
7 6
4567654
Рис. 15.22. Доказательство rрадиентной, или иерархической, системы реrионарной орrанизации
имаrинальноrо диска конечности дрозофилы. А. 1. Определение четвертей интактноrо диска.
11. Реrенерация верхней медиальной четверти дает полный диск. 111. Реrенерация нижней медиа
льной четверти дает зеркальную копию нижней половины диска. Б. Диск с контурными линиями
предполаrаемой способности к развитию. В. Модель реrионарной орrанизации: участки высшеrо
paHra MorYT восстанавливать все недостающие части, а участки назшеrо paHra только части не
превышающеrо их paHra
стро продвиrаться вперед в понимании МО--
лекулярных и rенетических основ детерми--
нации судьбы клеток и у высших орrаниз--
мов.
Коrда эксперименты показывают, что
характер будущей дифференцировки клетки
предопределен и не подвержен влиянию ее
клеточноrо окружения, такую клетку часто
называют детерминированной в своем раз--
витии. Клетки имаrинальноrо диска насеко--
мых обладают именно этим свойством. То,
что они детерминированы еще до начала
биохимической дифференцировки, явилось
важным открытием биолоrии развития.
Диск можно извлечь из эмбриона..
донора и трансплантировать в друrую
часть эмбриона--реципиента (рис. 15.23).
Обычно он продолжает развиваться в СООТ..
ветствии со своим исходным положением
в теле мухи. Друrими словами, если это
диск будущей конечности, ero клетки будут
развиваться в конечность. Различают два
типа детерминации:
1) реrи:онарная Bcero диска;
Развитие
Дополнительная
конечность
III
1 II
Рис. 15.23. Реrионарная орrанизация имаrина
льноrо диска фиксирована (см, также прило..
жение 15.8). Диск конечности личинки двукры
лоrо (1) удален хирурrически и (11) импланти
рован в брюшко эмбриона реципиента. После
завершения метаморфоза (111) у вышедшей из
куколки мухи на брюшке находится дополни
тельная конечность
2) локальная, т. е. отдельных клеток
внутри диска, обеспечивающая дифферен"
цировку орrанизованной интеrрированной
структуры.
Ниже будет показано, что эти два типа
детерминации MorYT быть разrраничены.
Клетки имаrинальноrо диска способны
к пролиферации (митотическому делению),
а ero рост является естественной состав..
ляющей развития. Диск способен также
к компенсационному росту, т. е. может
быть восстановлен из cBoero фраrмента (см.
рис. 15.22).
rормональная среда взрослоrо насеко..
Moro не препятствует пролиферации клеток
имаrинальноrо диска.. но подавляет или не
стимулирует их дифференцировку. Послед"
няя происходит только В том случае, если
диск поместить в rормональную среду ли..
чинки последней стадии или куколки.
Эти факты леrли в основу мноrих инте..
ресных экспериментов, результаты которых
часто оказывались совершенно неожидан..
ными.
Диск может оставаться недифференци"
рованным rораздо дольше, чем в норме,
если разводить ero серийную культуру вну"
три взрослой мухи. Низкий уровень линоч..
Horo ropMoHa в ее rемолимфе не тормозит
пролиферацию клеток диска, однако не по..
зволяет ему претерпевать метаморфоз.
Фраrменты диска можно последователь..
но трансплантировать в ряду мноrих поко..
лений, в любой момент проверяя состояние
461
их детерминации путем пересадки тестовых
участков в личинку последней стадии. Me
тод проиллюстрирован на рис. 15.24.
Тестируемые таким путем клетки имаrи
нальных дисков обычно сохраняют свою
исходную реrионарную детерминацию,
превращаясь во время метаморфоза
в структуру (rлаз, антенну, крыло и т. д.),
определяемую первоначальным местополо
жени ем диска. Однако это правило знает ис..
ключения: иноrда фраrмент диска дает на..
чало структуре, соответствующей друrой
части зародыша (приложение 15.8), т.е.
происходит трансдетерминация. Особенно
интересно, что различные клетки диска про..
должают при этом вести себя координиро
ванно, т. е. как составные части нормаль..
ной, но иной карты зачатков.
Большое число наблюдений свидетель..
ствует, что некоторые трансдетерминации
встречаются часто, друrие редко или вооб..
ще никоrда (см. приложение 15.8). Коrда все
клетки диска неожиданно изменяют свое со..
стояние детерминации, создается впечатле..
ние, что при этом действует хорошо отла..
женная система контроля Ее rенетическую
основу можно исследовать на нескольких
мутантах Drosophila (см. приложение 15.9).
Схема координированноrо развития диска,
соответствующая каждой специфической
I
Ф
.
ф
I Тестируемый
имплантат
I
.
Дальнейшее
культивирование
Рис. 15.24. Метод серийной трансплантации
имаrинальных дисков. Диск извлекают из ли
чинки донора, разрезают и помещают в имаrо,
rде он реrенерирует. Реrенерировавший диск
можно извлечь из хозяина, снова разрезать
и одну из ero частей протестировать на харак"
тер развития в личинке последнеrо возраста.
Друrой фраrмент переносят во взрослоrо хо..
зяина для дальнейшеrо культивирования (см.
также приложение 15.8)
462
rлава 15
области тела, контролируется rруппой re..
нов, называемых rомеозисными.
Например. rенетическая мутация Aпteп..
пapedia изменяет реrионарную орrаниза..
цию таким образом, что на месте антенны
развивается неполная конечность. Чтобы
предотвратить развитие антеннальноrо ди..
ска в конечность., необходим нормальный
(<<дикий») аллель reHa Aпteппapedia (см. при..
ложение 15.9).
rомеозисные rены контролируют также
дифференцировку друrих структур., отра..
жающих реrионарную орrанизацию. Неко"
торые из таких reHoB и свойственные им
эффекты приведены в приложении 15.9. Все
rомеозисные rены способны преврашать
один cerMeHT в друrой или одну часть се..
rMeHTa в друrую. Для исследования реrио..
нарной и сеrментарной орrанизаций живот..
ных эти rены являются крайне интересным
материалом. Так., известно., что некоторые
последовательности оснований у rомеозис..
ных reHoB Drosophila такие же, что и у др у..
rих reHoB животных самых различных си..
стематических rрупп. Эти последовательно..
сти называют «rомеобоксами»; их суще..
ствование свидетельствует о раннем эволю..
ционном происхождении таких reHoB реrио..
нарной орrанизапии.
Однако некоторые фундаментальные
вопросы до сих пор не получили объясне..
ния Реrионарная орrанизация paHHero
эмбриона несекомых запроrраммирована
в яйцеклетке, т. е. это результат процесса ra..
метоrенеза. В дальнейшем различные rруп..
пь] reHoB, по"видимому., транскрибируются
в имаrинальных дисках в соответствии с их
местоположением в раннем эмбрионе.
Установлено, что в этом участвуют rOMeo..
зисные I eHы, однако молекулярные основы
такой реrионарной дифференцировки и точ"
ная роль этих reHoB по сих пор не выяснены.
\
15.5. Реrенерация
15.5.1. Введение
Реrенерацию можно определить как спо..
собность к восстановлению утраченных
случайно или в результате автотомии ча..
стей тела путем компенсационноrо роста
и дифференцировки Способность к pereHe..
рации недостающих частей тела за..
мечательная особенность мноrих мяrкоте..
лых беспозвоночных. например rубок. стре..
кающих, плоских червей, немертин, анне..
лид и некоторых иrлокожих. Для них харак"
терно бесполое размножение путем фраr..
ментапии (см. rл. 14), очевидно тесно
связанной с реrенерацией. Беспозвоночные
с твердыми или высокоспециализированны..
ми покровами (например, членистоноrие,
псевдоцеломические черви, моллюски)
обычно слабо реrенерируют и часто не спо..
собны к размножению путем фРdrмента..
ции. у членистоноrих реrенерация происхо..
дит во время линьки и оrраничена, как пра..
вило, конечностями.
Процессы, происходящие во время pere..
нерации, сходны с происходящими при
нормальном развитии. К ним относятся
1) пролиферация недифференпирован"
ных клеток (как у бластулы) и формирова..
ние зачатка (бластемы);
2) мор(роrенез и орrанизация клеток
в пространственную иерархию;
3) диф(реренцировка и экспрессия специ..
фических r'eHOB.
В связи с этим явление реrенерации
представляет собой удобную модель для
изучения процессов развития. У некоторых
животных во время нее происходит дедиф..
ференцировка, не свойственная нормально..
му развитию.
Реrенерация означает реакцию орrаниз"
ма на утрату части тела, причем ответ дол..
жен включать как пролиферацию бластемы,
так и развитие соответствующей структуры
из пролиферировавших клеток.
15.5.2. Природа реrенерационной
бластемы
Новые клетки при реrенерационном ро..
сте MorYT происходить либо из резервной
популяции недифференцированных тотипо"
тентных клеток, либо из ранее дифференци"
рованных клеток путем их дедифференци"
ровки. Какой из этих альтернативных ва..
риантов осуществляется на самом деле, не
всеrпа ясно. Особенно хорошо выражена
способность к реrенерации у стрекающих.
Они располаrают запасом интерстициаль..
ных (I,,)клеток, которые в норме образуют
Развитие
Интерстициальные (I)-+<.летки
Рис. 15.25. Поперечный разрез книдариИ, по
казывающий интерстициальные клетки и их
пролиферацию перед реrенерацией
и постоянно замещаЮ1 различные лассы
дифференцированных клеток, например
киидобласты (рис. 15.25).
У rидры эти I клетки обычно скапли..
ваются в эктодерме перед бесполым раз..
миожением почкованием. Такую же их ре..
акцию вызывает повреждение тела. причем
рана становится местом их KOHKypeHTHoro
привлечения и на ее месте они формируют
реrенерационную бластему. У rидры в от..
личие от некоторых друrих книдарий 1..
клетки пролиферируют в течение всей жи..
зин, блаrодаря чему поддерживается по..
стоянный реrенерационный потенциал. Од..
нако rипотеза, рассматривающая их как са..
мовозобновляющуюся популяцию, обяза..
тельную для восстановительных процессов,
слишком упрощена. Если эти клетки разру..
шить химическим путем, орrанизм не утра..
чивает способности к реrенерации. PereHe..
рировать MorYT и фраrменты rидры., обыч..
но не содержащие I"клеток. Более Toro,
в блаrоприятной среде дифференцирован..
ные клетки тканевых эксплантатов rидрь]
дедиффереНIIИРУЮТСЯ в интерстициаЛЬН Iе
клетки, размножаются и редифференци
руются в клетки друrих типов. Это значит,
что обычно наблюдаемый путь к дифферен"
цировке через популяuию резервных клеток
не единственный.
Удивительная реI енерационная способ..
ность свободноживущих плоских червей
также связана снедифференцированными
463
резервными клетками, так называемыми
необластами. Этих животньiх используют
для изучения феномена реrенерации уже
в течение более 100 -чет. Из фраrментов., по..
лученных в результате поперечноrо перере..
зания планарии.. восстанавливаются две пе..
лые особи (рис. 15.26,А). Точно так же це..
лые черви реrенерируют из мелких фраr..
l\1eHTOB, включая и саrиттально отсеченные
(рис. 15.26,Б).
Первая стадия реrенерации фор..
1\1ирование раневой бластемы и ее после..
дующее заселение необластами. Роль этих
резервных клеток была продемонстрирова..
на экспериментами по облучению и транс..
плантации. Облучение рентrеновскими лу..
чами в дозе 3000 рад орrанизм не убивает.,
но препятствует пролиферации необластов.
flосле облучения животное утрачивает спо..
собность к реrенерации, однако она может
произойти, если в тело TaKoro червя им..
плантировать фраrмент необлученноrо до..
нора. Более Toro, если клетки имплантата
помечены., например по..друrому окрашены.
lVlОЖНО видеть, что цвет реrенерировавшеrо
фраrмента такой же, что у имплантирован"
ной ткани (рис. 15.27).
Тотипотентные резервные клетки не
всеrда участвуют в реrенерации., и их веду"
щая роль в этом процессе, доказанная для
планарий, может быть скорее исключением.
у аннелид реrенерационная бластема
6 OO ЭА
D
U \.CJ О lJ
а (l @ Б
Рис. 15.26. Реrенерация у плоских червей. А.
Поперечное разрезание и последующая pere
нерация передним или задним фраrментом
утраченной задней или передней части. Б. Дa
же мелкие кусочки MorYT восстановить целоrо
червя, причем исходные ткани сохраняют свою
реrионарную специфику
464
rлава 15
J L сз
!
с/
А 1 II III
G
:; I
;
,
1 II IV
Б
Рис. 15.27. Доказательство особой роли кле
ток «необластов» при реrенерации у планарии.
А. 1. Особь облучена жесткими рентrеновски
ми лучами. 11. Передняя часть удалена.
111. Задний фраrмент не способен к pereHepa
ции. Б. 1,11. См. А. 111. В среднюю часть тела им
плантирован небольшой фраrмент rенетически
отличной особи. IV. Теперь rолова может быть
реrенерирована; rенетически она идентична
имплантированному фраrменту
формируется по--иному: путем дедифферен--
цировки и переrруппировки дифференциро--
ванных клеток эктодермы и мезодермы.
Например, заживление раны при утрате
каудальных cerMeHToB идет за счет миrра--
ции целомоцитов к поврежденной поверх--
ности и восстановления зон роста" которые
появляются в виде тяжей характерных кле--
ток со сферическими ядрами и четко замет--
ными ядрышками. У нереид дифференци--
ровка этой области нарастания cerMeHToB
требует присутствия pocToBoro ropMoHa
мозrа, а формирование сеrментарных экто--
дермальных структур (сумок щетинок и па--
рапопий) идет под индукционным влиянием
вентральной нервной цепочки (см. ниже"
а также приложение 15.10).
15.5.3. Реrенерация
и реrионарная орrанизация
Мноrие мяrкотелые беспозвоночные
способны целиком восстанавливать реrио--
нарно орrанизованную структуру из очень
мелкоrо фраrмента тела. Это иллюстри"
руют рис. 15.26. а также рис. 14.2, на кото..
ром изображено бесполое размножение по-
лихеты с помощью спонтанно автотомиро"
ванных фраrментов.
Обшее для этих примеров восста-
новление целой структуры из ее части.
В каждом случае фраrмент сохраняет свою
исходную полярность, и компенсационный
рост возвращает ero на прежнее положение
в общем плане строения орrанизма. На рис.
15.26 фраrмент rоловы планарии восстанав-
ливает утраченную «хвостовую» область,
а «хвостовой» фраrмент утраченную [о-
лову. В составе целоrо каждой части прису-
ща реrионарная специфика, которая сохра..
няется при реrенерации. У сеrментирован"
ных животных эта специфика выражена еще
четче и закреплена в линейной иерархии се-
rMeHToB. У эмбриона насекомых число се-
rMeHToB фиксировано, и на каждом разви-
ваются свойственные только ему структу-
ры. Специфика этих cerMeHToB контроли..
руется или реrулируется особыми rенетиче-
скими локусами (см. приложение 15.9). Сле..
дует, однако, отметить, что rены bithorax
свидетельствуют о существовании морфо-
лоrических rраниц, дополняющих леrче ра..
спознаваемые сеrментарные rраницы.
Членистоноrие не MorYT восстанавли-
вать утраченные сеrменты, а мноrие анне-
лиды, строение которых также cerMeHTap-
ное, обладают такой способностью. Каж-
дый cerMeHT аннелид уникален и является
частью единоrо интеrрированноrо цело[о;
это выражается, в частности, в том, что
мноrим аннелидам свойственно фиксиро-
ванное их число. Позиционная информация,
по--видимому, поступает от вентральной
нервной цепочки (см. приложение 15.10).
у полихет семейства Nereidae для pereHepa-
ции «хвостовой» области необходим [ор-
мон мозrа, но это представляется вторич-
ной адаптацией, связанной с облиrатно МО-
ноцикличным размножением данных че-
рвей (см. разд. 14.3 и 14.4); тем не менее ско-
рость нарастания новых cerMeHToB остается
зависимой от позиционноrо контроля (при-
ложение 15.10).
Развитие
15.5.4. Реrенерация, рост
и размножение
Для реrенерации требуются ресурсы ор..
rанизма, и это, по видимому, должно при..
водить к их перераспределению в пользу со..
матических функций за счет уменьшения
вклада в половое размножение. Следова..
тельно, существует потенциальный конф"
ликт между реrенерацией и половым раз..
множением, и можно предполаrать наличие
реrуляторных механизмов, эффективно
контролирующих антаrонизм между этими
ПрОllессами Особенно это касается моноци"
кличных орrанизмов. Ресурсы, необходи..
мые для образования репродуктивных тка..
ней, должны расходоваться у них на pereHe
рационный рост только в том случае, если
он обеспечивает компенсационное увеличе..
иие выживаемости или жизнеспособности
потомства.
Подобный механизм следует предпола..
rать у МОНОIIИКЛИЧНЫХ нереид.. которые.. как
показано выше, способны к реrенерацион"
ному росту после утраты задних cerMeHToB.
Коrда эти полихеты достиrают половозре..
лости, все ресурсы полностью расходуются
на размножение, и после Hero черви поrи..
бают. Значит, реrенерация cerMeHToB в кон..
це репродуктивноrо периода теряет смысл.
На этой стадии жизненноrо цикла pereHepa..
ционный рост подавляется эндокринным
механизмом. Во время половоrо созревй--
иия постепенно снижается секреция [ормо"
на мозrа, и уменьшение ero концентрации
в циркуляторной системе способствует за..
вершению rаметоrенеза (см. разд. 14.4),
инициируя одновременно соматические из..
менения, связанные с размножением (см.
рис. 16.35). Этот же [ормон необходим для
каудальной реrенерации., поэтому половоз..
релые нереиды не способны восстанавли"
вать утраченные задние сеrменты. У поли..
хет, размножающихся несколько лет под..
ряд., подобный эндокринный механизм не
встречается. Реrенерационный рост здесь
идет и у половозрелых особей, поскольку
восстановленные сеrменты в последующие
rоды MorYT участвовать в размножении и их
репродуктивная ценность остается высо"
кой
465
15.6. Выводы: развитие
беспозвоночных и rенетиче..
екая nporpaMMa
3кспериментальное изучение развития
беспозвоночных началось в конце XIX в.
Оно развивал ось параллельно с rенетиче..
скими исследованиями, открытием молеку-
лярных основ клеточной наследственности
и быстрым расширением знаний в области
молекулярной биолоrии. На повестке дня
стоит выдающаяся по сложности задача
объединения этих дисциплин. Беспозвоноч"
ные являются кладезем удобных модель..
ных систем для ее решения. На этом пути
уже достиrнут значительный проrресс., и
большая часть материала, представленноrо
в настоящей rлаве, может быть переосмы"
слена с точки зрения молекулярной биоло..
rии.
Информационные молекулы животной
клетки это последовательности мРНК,
синтезированные на матрице ДНК в ядре.
В первых разделах этой rлавы rоворилось,
что почти у всех животных ядерная ДНК зи..
[оты полностью передается каждой дочер"
ней клетке во время paHHero дробления.
Эти молекулы мРНК можно изучать ме..
тодами современной биохимии. Во время
ооrенеза (см. rл. 14) молекулы материнской
мРНК накапливаются в цитоплазме ооци..
та. При оплодотворении они активируются
и обеспечивают ранний белковый синтез.
В конечном счете ядерный [еном зиrоты
становится источником rенетической ин..
формации Bcero орrанизма. Однако в про..
цессах дифференцировки и морфоrенеза
решаюшую роль сохраняет llитоплазма
ооцита. Мы видели, что у широкоrо Kpyra
беспозвоночных функции клеточноrо ядра
и информация, которая в конечном счете
транскрибируется или транслируется, де..
терминированы их предысторией в 1мбрио..
не.
Иноrда синтез тех или иных ферментов,
по"видимому, определяется специфически..
ми цитоплазматическими факторами (см..,
например, развитие оболочника Styela).
В друrих случаях функциональные реакции,
вероятно, вызываются межклеточными
контактами Роль таких взаимодействий
показана в экспериментах с эмбрионами
466
rлава 15
моллюсков и морских ежей. Ряд наиболее
удивительных открытий сделан при изуче..
нии эмбрионов насекомых.. особенно плодо..
вой мушки Drosophila тelaпogaster, в случае
которой обилие имеющихся rенетических
сведений сочетается с удобством постанов..
ки опытов. Вполне вероятно, что "Эl\tlбрионы
беспозвоночных еще долrо будут оста..
ваться в числе лучших модельных систем
для исследования сложных проблем разви"
тия животных.
15.7. Дополнительная
литература
Веn-и N. J. ] 97]. Developmental Biology. McGraw
Hill, New У ork.
Brookbaпk J. W. 1978. Developmental Biology: Em
bryos, Plants and Regeneration. Harper апd Row,
New У ork.
Вrои'dеr L. W. 19 4. Developmental Biology, 2nd
edn. Saunders, New У ork.
Ере! D. 1977. The program of fertilisation. Scient.
Аm. 237. 129 138.
Gehriпg W. J. 1985. The molecular basis of develop
ment. Scientific American. 253. 137 ]46.
Oppeпheimer S. В. 1980. Introduction to Embryonic
Development. Аllуп d-nd Васоп. New У ork.
Reverberi G. 197]. Experimental Embryology of
Marine and Freshwater Invertebrates. Amster
dam.
Slack J. М. W. ]983. From Egg to Embryo. Cam
bridge University Press, Cambridge.
Stearпs L. W. 1974. Sea Urchin Development: Cellи
lar and Molecular Aspects. Dowden, Hutchinson
and Ross, Pennsylvania.
Приложение 15.1.
Спиральное дробление
Этот тип дробления характерен для та..
ких таксонов первичноротых.. как немерти--
ны, аннелиды, сипункулиды, эхиуриды
и моллюски. Наиболее четко ero можно на..
блюдать у эмбрионов с относительно небо..
льшим количеством желтка, например
у морских полихет и моллюсков.
1. Неоплодотворенное яйцо радиально
симметрично относительно оси, соединяю..
щей анимальный (бедный желтком) и Bere..
ТdТИВНЫЙ полюса (1). При этом все cerMeH..
ТЫ, отсекаемые анимально"веrетативной
плоскостью, эквивалентны.
После оплодотворения яйцо билатера..
льно симметрично, и на нем можно просле..
дить основные плоскости эмбриона (II
IV):
11) поперечная разделяет переднюю
и заднюю части;
111) саrиттальная разделяет левую
и правую половины;
IV) фронтальная разделяет дорсаль
ную и вентральную стороны.
Развитие
2. Плоскости двух первых делений
являются продольными и делят пополам
уrлы между фронтальной и саrиттальной
плоскостями.
1
В результате образуются первые два
бластомера, АВ и CD (11), а затем первые
четыре бластомера А, В, С, D (111).
II
III
D..бластомер обычно крупнее трех OCTa
льных. Все последующие продукты дробле
ния MorYT быть идентифицированы относи
тельно этой клетки
3. Плоскость TpeTbero деления попереч
ная; она проходит над экватором и OTдe
ляет четыре меньшие клетки на анималь
НОМ полюсе от четырех более крупных на
веrетативном (1).
467
1С
10
II
1А
Четыре мелкие клетки называют пер..
вым квартетом микромеров. Они располо
жены над rраницами между макромерами,
поскольку плоскость дробления наклонена
относительно продольной оси эмбриона.
Если смотреть с анимальноrо полюса, Ka
жется, что микромеры сдвинуты по часовой
стрелке (11).
Восемь бластомеров эмбриона обозна
чают так:
la 1 Ь lс ld
lА lВ lС lD
Все последуюшие деления являются по
перечными и заключаются в разделении cy
ществующих микромеров и отделении но..
вых квартетов микромеров в результате He
paBHoro дробления макромеров. На рис, III
показано появление 16 клеточноrо эмбрио
на после четвертоrо деления дробления.
Обозначения бластомеров на этой стадии
следующие:
деления первоrо
квартета:
la 1 lb 1 lс 1 ld 1
, , ,
! ! ! !
1 а 2, 1 Ь 2 , 1 с 2 , 1 d 2
второй квартет: 2а, 2Ь, 2с, 2d
! ! ! !
макромеры: 2А, 2В, 2С, 2D
Плоскость каждоrо деления дробления
наклонена относительно продольной оси
таким образом. что направления сдвиrов
микромеров по часовой и против часовой
стрелки чередуются.
"
IП
468
rлава 15
4. Разработана условная система обо
значений, позволяющая идентифицировать
каждую клетку вплоть до стадии 64 бласто"
меров. Схема дробления (завершенная то..
лько по наиболее важной линии D"клеток)
выrлядит так:
1а 1
1а < и т.д.
1а 2
А" 2a
/ 1д
АВ \ '-- :1
1Ь/ ит.д.
/ "-1 Ь 2
В,-- 2b
1B
"""""'2 В
ЯЙЦО
1 с 1
1c
/ '-...1 с 2
С и Т. д.
" 2с
"1С / 1d 111
............... 2 С 1 d 11
/ "1 d 112
&1 d 12 1d 121
1d1V 1d122
/ 1d21 1d211
1d 1d212
1 ct 2
'" 1 d 221
""1 d 22
1 d 222
2 d 11
D \ 2d1
2d12
2d
/ 2d 21
1D/' 2d2
\ 2 d 22
3 d 1
.............3 d < 3 d 2
2О 4d
3D
40
CD
Число клеток
Деление
дробления
1 2 4 8 16 32 64
l..е 2..е 3..е 4..е 5..е 6..е
Почему особенно важна линия О..
бластомера, объясняется далее в тексте
и приложениях 15.2 и 15.3.
Приложение 15.2.
Радиальное дробление
Этот тип дробления, отличный от на..
блюдаемоrо у аннелид и моллюсков, при..
СУЩ, например, иrлокожим и в измененном
виде хордовым. У морских ежей он выра..
жен очень четко.
1. Схема дробления. Оплодотворенное
яйцо билатерально симметрично (1). Пло..
1
Qij .......
'. ...... .... .. ..-
: ./; ?' ::. ... :'
@ . .,
'.0 ." .........
... -.':: I t. ." :.;.....
'.t. . :: . f : ji
11
III
{' ' ":.::.:. :"" "}: ?i
..:.: ,.. :.'. .. ..,.
. " . ..... ..-......
'.. о.'
IV
Мезомеры (8)
Микромеры (4)
v
n 2 (16 клеток)
eg 1 (8 клеток)
eg 2 (8 клеток)
УI
УН
УIII
Развитие
скости двух первых делений дробления про
ходят вдоль анимально веrетативной оси
и делят яйцо сначала на два, а затем на че
тыре равных бластомера (11, III).
Третье деление дробления поперечное
и неравное, плоскость ero проходит непос
редственно над экватором (IV) и отделяет
четыре «меЗОJwера» от несколько более
крупных «макро,,«еров».
Четвертое деление дробления в нижнем
и верхнем клеточных слоях идет по
разному. В верхней половине оно продоль
ное и дает кольцо из восьми клеток, а в ни
жнем ярусе поперечное и неравное, в резу
льтате чеrо образуются четыре макромера
и четыре микромера. В этом примере ми
кромеры находятся на веrетативном полю
се (У).
Два следующих деления в верхней поло
вине поперечные, а в нижней продольное
и поперечное. В результате образуется
эмбриональная стадия из 64 бластомеров
(VI).
2. Идентификация бластомеров. Поско
льку первые четыре бластомера структурно
идентичны, идентифицировать отдельные
клетки невозможно. Однако MorYT быть
идентифицированы клеточные слои. На CTa
дии 64 бластомеров их условно обозначают
следующим образом:
первый анимальный слой == Anl == 16
верхних бластомеров;
второй анимальный слой == An2 == 16
нижних бластомеров анимальноrо полуша
рия;
первый веrетативный слой == Vegl == KO
льцо из восьми макромеров под экватором;
второй веrетативный слой == Veg2 == ни
жнее кольцо из восьми макромеров BereTa
тивноrо полушария;
микромеры == Mic == rруппа самых мел
ких клеток на веrетативном полюсе.
3. Бластула. В результате дробления
формируется бластула (VII) полый шар
из ресничных клеток.
rаструляция начинается инваrинацией
микромеРОБ (VIII). Она описана в разд.
15.2.2 и проиллюстрирована на рис. 15.9.
469
Приложение 15.3.
Реrуляционное и мозаичное
развитие I
в конце ХIХ начале ХХ вв. на
эмбрионах морских беспозвоночных стало
возможным изучать характер развития OT
дельных клеток, изолированных на ранних
стадиях дробления (двух или четырех бла..
стомеров). Результаты оказались на удивле
ние различными.
1. Эмбрионы И2локожих. Из каждой
изолированной на стадии четырех бласто
меров клетки может развиться нормальный
(но мелкий) плутеус. Эти мелкие эмбрионы
были названы реrуляционными.
Мелкий, но
)::;:/ ормальный
плутеус
1
t
п
I. Нормальный плутеус.
11. Результаты разделения бластомеров
Из бластомера, изолированноrо на че
тырехклеточной стадии, в результате деле
ния образуются два мезомера. макромер
и микромер, т. е. наблюдается схема
дробления нормальноrо эмбриона (см. при
ложение 15.2).
2. Эмбрионы моллюсков. Из изолиро
ванных бластомеров ДBYX и четырехкле
1
{д1
°6 U
II
470
rлава 15
точноrо Зародыша развиваются непропор
llиональные или неполные эмбрионы. при..
чем из D"клеток обычно более полноцен..
ные, чем из А.., В.. или С..бластомеров.
1. Нормальная трохофора Patella.
11. Аномальные трохофры. развившиеся
из изолированных бластомеров.
3. Эмбрионы асцидий. В яйцах асцидий
часто встречаются визуально различные
участки цитоплазмы (см. рис. 15.6), которые
распределяются по разным клеткам.
Эмбрионы, полученные из изолирован..
ных бластомеров, весьма различны. В их
строении преобладают структуры, которые
развились бы из соответствующих клеток
в нормальном зародыше.
r лазные пятна
1
.
Мышечные клетки
Хорда
111
IV
1. Нормальный «rоловастикоподобный»
эмбрион с парой rлаз, хордой и парными
мышечными блоками.,'
11 Изолированные передние клетки.
111. Изолированные клетки правой поло..
вины.
IV. Изолированные задние клетки.
Приложение 15.4.
Судьба клеток, карты зачатков
и цитоплазматическая
локализация
1. Прuнцип составления карты зачат-
ков. На поверхность нераздробившеrося
яйца можно нанести метки. Если в резуль..
тате дробления не происходит перемешива..
ния бластомеров, эти метки затем появятся
в специфических структурах зародыша.
1
III
А А Презумптивная кишка
о о Презумптивная мантия
. Презумптивный оперкулум
х х Презумптивная Hora
rипотетический пример зародыша мол..
люска:
1) метки на поверхности яйца;
11) положение клеток во время rаструля"
ции;
111) сеrреrация меченых клеток по специ..
фическим участкам эмбриона
2. Природные маркеры. После оплодот"
ворения в некоторых зиrотах можно наблю-
дать четко различимые участки цитоплаз..
1
III
V
Развитие
II
IV
VI
rаструляция и цитоплазматическая локализа
ЦИR у асцидий
мы и билатеральную симметрию. На рисун..
ке видно распределение различных цито"
плазматических участков яйца асцидии (1)
в клетках веrетативноrо полюса (11); во
время rаструляции эти клетки инваrини--
руют (111, IV). Из клеток, содержащих ви"
зуально различную цитоплазму, после [а..
струляuии разовьются различные эмбрио--
нальные структуры (У, У1). В связи с этим
возникает вопрос: «Определяют ли природ..
ные цитоплазматические маркеры судьбу
клеток?» .
3. У асцидии судьбу клеПIОК деЙСП16ИПlель..
11,0 определяют цитоплаз./wаПlические факпzо..
ры. Участок желтоватой цитоплазмы мо"
жно рассматривать как природный маркер
мышечных клеток rоловастикоподобной
личинки [см. приложение 15.3(3)].
471
4. Фермент ацеlпилхолинэсп1ераза спеuи..
фи чески стимулирует развитие мышечных
клеток; он может быть окрашен в цитоплаз..
ме.
Получены следуюшие эксперименталь..
ные данные:
а) обычно он появляется через 8 ч после
начала эмбриоrенеза;
б) фермент появляется в это время даже
при подавлении деления клеток химически..
ми препаратами;
в) при обработке актиномицином D в те..
чение 5 чили пуромицином В течение 7 ч по..
сле начала эмбриоrенеза фермент не появ..
ляется;
[) сжатие яйца во время дробления мо"
жет привести к перемещению желтой цито"
плазмы в те клетки, которые 6 нор./ие ее не
получают. После этоrо в них наблюдается
ацетилхолинэстеразная активность. Из 'Этих
экспериментов вытекает предположение,
что [eHЫ кодирующие ацетилхолинэстера..
зу, транскрибируются только в клетках на..
следующих некие факторы, обычно связан..
ные с желтой цитоплазмой.
Хронолоrия транскрипции и трансляции
этих [енов контролируются биолоrически"
ми часами, функционирующими независи--
мо от клеточных делений
Приложение 15.5.
Реrуляционное и мозаичное
развитие 11
Экспериментальное изучение морской
улитки Nаssаriиs оЬsо/еtиs
1. Нормальное развитие NаssаrlИS.
Улитке свойственно спиральное дробление
с появлением заметной полярной лопасти
во время первоrо и BToporo делений
дробления, что обеспечивает наследование
D"клеткой особой полярной плазмы (см.
приложение 15.1). Последовательность
дробления показана ниже.
472
rлава 15
1. Недробящееся яйцо.
11. Появление первой полярной лопасти
до первоrо деления дробления.
111. Образование «трилистника» во
время первоrо деления.
IV. Стадия двух бластомеров.
1
/
VIII
/
У, VI. Появление второй полярной лопа
сти во время BToporo деления дробления.
VII. Стадия четырех бластомеров (обра..
тим внимание на крупную D клетку).
VIII. Стадия восьми бластомеров вскоре
после появления первоrо квартета микро"
меров.
IX, х. Более позднии стадии дробления.
Восьмиклеточный эмбрион заметно
асимметричен, причем потомки D"клетки
подчиняются собственному ритму -деления,
отличному от ритма деления клеток друrих
линий.
2. Особая роль D клеток в развитии
N assarius
Изучение клеточных линий свидетель
ствует о развитии специфических личиноч
ных структур из производных отдельных
клеток, которые можно идентифицировать
и обозначить во время paHHero дробления.
С использованием обозначений из при-
ложения 15.1 ниже на схеме клеточных ли
ний показано экспериментально установ..
ленное происхождение личиночных CTPYK
тур из 29 бластомеров.
Обратим внимание, что клетки 2d и 4d
иrрают особенно важную роль в формиро..
вании велиrера. Бластомер 4d называется
мезентобластом.
11
} Парус
1 ............... r лаз
12
/21 } Раковинная
2............. железа и
2 2 мантия
2d/
3 abcd Hora
4 4 d Сердце
Орrанизация личинки"велиrера
Развитие
473
д/
/ ""1а
АВ 2B
" 18
" ,/ 2b
В
1b
3 А ....... ....... ....... ....... ....... Ки шка
2Д За
1A 2а 1
2a
2 а 2 .............. 1'1....... ....... ...... } Стомодеум
1 а 1 1 а 1 .............1 а 12 Левый rлаз
1 а ...............1 а
...............1 а 2 1 а 2
...... ....... ....... ....... ............ ...... П ару с
...... ....... ....... ...... ............. Киш ка
c
/ ""1 с
СО
20
1О
,/ 2d
p D ""-
""-1d
ЗВ
3Ь
2ь 1
2 ь 2 1 Ь 11 } Пространство
.,............. 1 Ь 1 ............... 1 Ь 1 < 1 ь 12 между r лазами
1 Ь ...............1 Ь 2 2
...............1 Ь ....... ....... ....... ....... ....... ...... П а рус
ЗС
2 С .............. ....... ....... .................... ....... ...... Ки шка
3 с ......................................................... J Правая сторона ноrи
1С 2 2с1 ОБласть мантии
с 2 2
С 1 11
1с 1 1c1 < С Правый rлаз
1 1с 12
С ...............1 с 2 1 с 2
4 D ...... ----. К
ЗD ...... :::: ишка
3 d d ........... Кишечник, сердце
1............. ....... ....... ....... ...... ....... ...... Левая стор она ноrи
2d }
:2 Раковинная железа и
2d 1 d 11 соматическая
1 d 1 ...............1 d 1 2 пластинка
1d1
1 d ...............1 d 2 1 d 2
Схема клеточных линий (Р полярные тельца)
{
Бластула
т рохосрора
Перемещение клеток 2d и 4d в специфическое для них положение во время rаструляции
474
rлава 15
3 Орrанизующая роль полярной плаз--
мы. Материал полярной лопасти во время
paHHero дробления можно удалить. микро--
хирурrическим путем. Результаты зтоrо
впечатляющие.
06разующаяся безлопастная личинка
утрачивает всю структурную орrанизацию,
сохраняя некоторые признаки клеточной
дифференцировки. Ниже изображены два
типичных примера таких личинок.
в классической серии экспериментов А.
Клемент изучал результаты последователь--
ности изъятия D клетки во время Jмбриоrе--
неза. Результаты сведены в приводимую ни..
же таблицу.
4. Интерпретация: доказательство ин..
дукционных взаимодействий. 11сходя из
схемы клеточных линий и таблицы, можно
отметить следующее: rлаза образуются из
производных клеток 1 а и 1 с, а раковинная
железа из производных клетки 2d. Клетки
1 а и 1 с есть у всех экспериментальных
эмбрионов. но rлаза формируются только
в опытах IV и У. Почему? Ответ: «rлаза
формируются только в присутствии клеток
D"линии, по крайней мере до образования
бластомера 3d».
Нормальная раковинная железа разви"
вается только в том случае, если клетка 2d
присутствует по крайней мере до образова..
ния бластомера 3d.
Доказательство мозаичноrо развития:
rлаза формируются из клеток la и lc. Раз..
рушение их препятствует развитию rлаз.
Вывод: только производные lа и lс ком..
петентны формировать rлаза, но для детер"
минации этоrо направления развития им
требуется индуктивное влияние клеток D..
линии.
Сердце обычно развивается из клетки
4d: только она компетентна обеспечить ero
дифференцировку.
Обобщение результатов последовательноrо удаления клеток D линии у Nassarius (по А. С. Cle
ment, 1962)
Удаленная клетка
Эмбрион
Обнаруженные дефекты
»
Раковина развит а в различной степени., УТР"- чены
кишка и сердце
Аве + ld + 2d + 3d + 4d Отсутствуют
I. D (СМ. приложение 15.1 )Аве
II. 1 D
III. 2О
IV. зо
Аве + ld
АВС + ld + 2d
Аве + ld + 2d + 3d
V.4D
Как и у безлопастноrо эмбриона, утрачены: КИШ
ка, сердце, раковина., HOra., статоцисты. rлаза
»
Развитие
Приложение 15.6.
Экспериментальный анализ
развития MopcKoro ежа
1. Клеточные слои. Дробление яйца
MopcKoro ежа описано в приложении 15.2.
Основные слои 64"клеточноrо зародыша
следующие:
Апl 16 клеток,
Ап2 16 клеток,
Veg 1 8 клеток,
Veg 2 8 клеток,
микромеры.
Клеточный слой Ап 1
Клеточный слой Ап 2
Клеточный слой Veg 1
Клеточный слой Veg 2
Микромеры
Бластула
rаструла
j
Развивающийся плутеус
(призматическая стадия)
Взаимное соответствие клеточных слоев бла..
ступы, rаструлы и paHHero плутеуса
2. Развитие изолированной половины
эмбриона. Изолированная анимальная по..
лавина эмбриона превращается в неразви"
вающуюся далее бластулу с аномально
крупным апикальным султаном ресничек.
Изолированная веrетативная половина дает
начало более или менее нормальному
эмбриону с чрезмерно развитым кишечни..
ком. Такой эмбрион называется веrетализи..
рованным.
475
Эти наблюдения можно было бы объяс..
нить самодифференцировкой первичных
клеточных слоев (<<мозаичная» теория раз..
вития), но дальнейшие эксперименты пока..
зывают'I что это не так.
Неразвивающаяся Бластула
(анимализированный ЭМБРИОН)
Анимальная поло
вина ЭМБриона
Веrетативная
половина ЭМБриона
Веrетализированный
ЭМБРИОН
3. Доказапlельство zрадиентной Т11еориu
развипlия .
Если изолированную анимальную поло..
вину эмбриона объединить с четырьмя ми..
кромерами, она развивается в нормальный
плутеус. На рисунке показано:
1) разделение эмбриона;
11) объединение анимальной полусферы
с четырьмя микромерами;
111) racTpy ляция;
IV) нормальный плутеус.
CimJ
CW
QlP
!
11(
IV
IH
476
rлава 15
в эксперименте, описанном в п. 3, о ми..
кромерах rоворят, что они «индуцируют»
архентеронформирующую способность
клеток слоя An..2. Клетки An..2 становятся
компетентными дифференцироваться в ар..
хентерон, хотя в норме они таковыми не
являются.
4. MHoro экспериментов было проведе..
но по развитию изолированных слоев
эмбриона и сочетаний различных слоев. Ре..
зультаты одноrо из них описаны ниже.
Нормальные эмбрионы получаются при
«равновесии» анимальной и веrетативной
тенденций развития:
An 1 + 4 микромера == нормальный плу
теус;
An 2 + 2 микромера == нормальный плу
теус;
Veg 1 + 1 микромер == веrетализирован
ный эмбрион
+4
микромера
+2 I
микромера
+1
микромер
+1
<микромер
Развитие изолированных
клеточных слоев
r радиент от анимальнorо полюса
5. Описанные выше реЗУЛЬТdТЫ можно
интерпретировать как наличие двухrра..
диентной системы. Предполаrается, что
в нормальном эмбрионе каждый клеточный
слой Зdнимает определенное положение от..
носительно двух rрадиентов.
r радиентная теория развития.
Нормальное развитие треБует
"равновесия" между двумя
rрадиентами
r радиент от веrетативноrо полюса
Развитие
6. Демонстрацuя системы zрадuентов.
Некоторые вещества вызывают развитие
аномальных анимализированных или Bere..
тализированных эмбрионов, которые похо
жи на развивающихся из веrетативной или
анимальной полусфер.
zп 2 +или CN О
ирующие
вещrт..в а @
l Ber
@) вещества lrJ
Ионы Li + обладают сильным веrетали--
зируюшим действием, а ионы Zn2+ и CN
анимализирующим.
Изолированная @
анимальная О
n
ализированный
с.::...:...........)
Нормальный
При этом ионы Li + MorYT вызвать нор--
мальную rаструляцию изолированной ани
мальной полусферы.
7. Демонстрация rрадиентов метаболи--
о мин
60 мин
90 мин 120 мин
Ход восстановления красителя в нормальных
эмбрионах. Можно видеть, что зоны реакции
распространяются от анимальноrо и веrетатив"
Horo полюсов
477
ческой активности. Прямое доказательство
этих rрадиентов у полюсов ранней ractpy--
лы получено с помощью прижизненноrо
окрашивания янусом зеленым. Этот краси..
тель становится розовым затем обесцвечи--
вается.
Соrласованность доказательств: ионы
Li + вызывают нормальное развитие ани--
мальной полусферы и подавляют подкисле--
иие януса зеленоrо на анимальном полюсе.
.
с Li ионо
8. rрадиентная система. по--видимому.
имеет физико--химическую основу, но ее мо--
лекулярные механизмы еще не выяснены.
Приложение 15.7.
Метаморфоз и выбор
субстрата у морских личинок
1. Мноrие бентосные орrанизмы оби..
тают на видоспецифичных субстратах.
Иноrда несколько близкородственных ви"
дов встречаются симпатрично, но на раз..
личных субстратах. Наблюдаемое распре..
деление взрослых особей связано с распо--
знающей способностью личинок. Для ряда
мелких, живуших в трубках полихет рола
Spirorbis это продемонстрировано экспери--
ментально. На рисунке показаны:
1) Spirorbis borealis на Fucus serratus;
11) S. trideпtatus на rолой скале;
111) s. coralliпae на Coralliпa officiпalis
111
. - . "' ;
_ .OI!! [,\
. ;, \
.,;; . \ \
/: - . l T"'"
-;' , I .п ,. .,,: .
1" .""
.. " .' ,L
...:-:-=-:,\ \\ /, :.
." . '.
: :t::.\;,.:: . .
-/ ,'
478
rлава 15
Эксперименты по выбору субстрата для посе
ления личинками различных видов полихет po
да Spirorbis (по Dе Silva, 1962 )
Вид
Субстрат
Общее число
личинок
Spirorbis borealis Fucus serratus 1297
Coralliпa o.fficiпalis 18
Fucus serratus 457
Камень 295
S. trideпtatus
F. serratus
Камень
с. officiпalis
Камень
с. ojficiпalis
Р. serratus
S. coralliпae
о
52
О
55
63
2
Коrда в эксперименте личинкам предла
rали «на выбор» два разных субстрата, у ви
дов обнаружились четкие различия в пред
почтениях (см. таблицу ниже)
2. Циприсовидная личинка MopcKoro
желудя (см. ниже) очень разборчива вотно..
шении места поселения.
Цементная железа
Прикрепительный Антеннула
и сенсорный Диски
Перед тем как окончательно выбрать
место для прикрепления, она передвиrается
по субстрату, демонстрируя исследователь
ское поведение, причем иноrда уплывет,
чтобы выбирать в друrом месте.
..(7
...
....(J
.... \qF'c:j5
3. Мноrие факторы MorYT увеличить Be
роятность поселения личинки MopcKoro же
лудя на специфическом субстрате.
Так, поселение и метаморфоз стимули
руют:
1) шероховатая поверхность;
2) ямчатая или бороздчатая поверх
ность;
3) остатки панцирей морских желудей;
4) присутствие поселившихся пиприсо
видных личинок. Наиболее важны среди
этих факторов те, что связаны с присут
ствием друrих морских желудей.
Непривлекательная поверхность может
стать ПРИВoIlекательной. если ее пропитать
экстрактом из тканей MopcKoro желудя. AT
трактантом служит белок, распознаваемый
циприсовидной личинкой уже на уровне MO
номолекулярноrо слоя.
Развитие
479
Приложение 15.8.
Серийная трансплантация
имаrинальных дисков
и открытие трансдетерминации
1. С помощью метода серийной транс..
плантации можно MHoroKpaTHo тестиро..
вать характер детерминации фраrмента до--
HopHoro имаrинальноrо диска.
с н орный диск конечности
о
ф
.
Тест 1
Лишняя Hora
ф
Тест 2 7/)'
Навсеrда ли
сохраняет диск
специсричность '?
ф
I
Т ест 3
и Т.Д.
Метод серийной трансплантации
Если характер детерминации донорноrо
диска сохраняется, каждый тестируемый
имплантат будет проявлять ero свойства.
Это обычно и наблюдается.
2. Однако в случае мноrократных тести..
рований имплантантов иноrда наблюда..
ются спонтанные изменения характера де..
терминации. При этом возможны не все
переходы. На рисунке про иллюстрированы
некоторые из них, встречающиеся у дрозо..
филы. Стрелки показывают направление
спонтанных изменений характера детерми--
нации.
rениталии
Ноrи
Антенны
рыль :/ rлаза
\ \. /
1'8/
' . '
i "
j
/ ,
,
Торакс /
Например, наблюдалось развитие из ди..
сков конечностей крыльев. антенн и pOTO
вых придатков, но из них никоrда не обра--
зуются rлаза, жужжальца или rениталии.
Следует отметить, что дальнейшая судь--
ба спонтанно изменяется сразу у всех кле..
ток диска., однако они должны сохранять
локальную специфику, поскольку из них
формируются реrионарно орrанизованные
структуры. Такое спонтанное изменение на--
зывается Пlрансдеn1ер/иuнацuей.
Приложение 15.9.
rенетический контроль
и реrионарная орrанизация:
rомеозисные reHbI
1. В культурах Drosophila иноrда BCTpe
чаются аномальные или уродливые мухи,
у которых из ИМdrинальноrо диска разви--
ваются структуры несоответствующеrо или
неправильноrо типа. Например, из имаrи..
нальноrо диска антенны может сФормиро--
ваться Hora
480
rлава 15
rенетический анализ показывает, что
причиной таких аномалий является мута..
ция по одному reHHoMY локусу. Иначе ro..
воря, существуют rены, каким то образом
поддерживающие или упорядочи ающие
характер детерминации отдельных имаrи..
нальных дисков. Эти КОНТРОЛИРУЮПlие re..
ны названы rомеозисными.
Название
мутанта
эффект
Символ rенный
локус
Aпteппapedia Abtp
3 48 ПРИВОДИТ к раз..
витию ноrи BMe
сто антенны (1)
2 68 ПРИВОДИТ к раз--
ВИТИЮ крыла из
rлазноrо диска
(11)
2 Приводит К раз..
ВИТИIО вместо
дорсальноrо про..
торакса крыла,
иноrда ноrи (111)
Ophthalтoptera Opt.
Hexaptera Нх
Эти мутапии доминантные, известны
и друrие рецессивные. В какой то степени
эффекты этих мутаций сходны со спонтан
ными изменениями при Пlрансдеп1ермина..
ции.
2. rомеозисные rены, по"видимому,
участвуют в процессах, контролирующих
реrионарную специфику. Это особенно на..
rлядно про является в случае reHoB комплек
са bithorax.
Комплекс ыlоrах этоo ряд тесно сце..
пленных reHOB третьей хромосомы дрозо..
филы, влияющих не некоторые особенности
реrионарной орrанизапии торакса или пере..
дней части абдомена. Фактически ero му"
танты демонстрируют ранее не различимые
морфолоrические rраницы.
Ниже на рисунке представлено схемати..
ческое изображение мухи, рассеченной на
основные структурные элементы rOMeo..
зисные мутанты по комплексу bithorax сви"
детельствуют о том, что существует реrио..
нарная rраница между передней и задней
частями мезоторакса (BToporo торакально..
ro cerMeHTa), мета торакса (третьеrо торака..
льноrо cerMeHTa) и первоrо абдоминально"
ro cerMeHTa.
: Дорсальная сторона
Вентральная сторона
'1 '"
Проторакс I Передний/задний
, метаторакс
Передний/задний
мезоторакс
Схема нормальной конфиrурации торакальных
структур дрозофилы
Мутант bitllorax (Ьх) это рецессивный
reH., приводящий к тому, что на переднем
метатораксе (rде в норме образуются жуж-
жальца) развивается передняя часть крыла,
как на переднем мезотораксе (1).
Развитие
481
rеи СИМВОЛ
1 bithorax Ьх
11. postbithorax рЬх
111. U/trabahorax Ubx
У, bithoraxoid bxd
Что меняется
Во что превращается
Передний метаторакс(перед
няя часть жужжальца)
Задний мета торакс (задняя
часть жужжальца)
Метаторакс (жужжальце)
Первый абдоминальный cer..
мент
Передний мезоторакс(перед..
няя часть крыла)
Задний мезоторакс (задняя
часть крыла)
Мезоторакс (крыло)
Метаторакс (жужжальце)
Обратим внимание, что двойное peцec
Ьх рЬх
сивное состояние Ь Ь (IV) cxoд
х р х
НО в ОТНОIllении реrионарной орrанизации
Ubx
с доминантным (111).
Ubx+
Приложение 15.10.
Позиционная информация
и каудальная реrенерация
у аннелид
1. Число ce Meптoв. Тело аннелид co
стоит из следующих отделов (см. также
рис. 4.51):
1) простомиум (rоловная лопасть);
2) специфическое число cerMeHTOB;
3) пиrидиум (постсеrментная лопасть).
у некоторых полихет, как и у пиявок,
число cerMeHToB невелико; например. у Cly
140
Десринитивное число
cerMeHTOB
.
m 120
о
.....
1:
Q)
1:
L
Ф
()
о
()
:s:
:r
Десринитив . о
ное чи сло. ....... ....... ...... .......
cerMeHTOB . ""g;;,.......-;; о о о
· .. <УО.......о о Nephtys
0/ longosetosa
/
/0
/
100
80
12345678
Длина тела, см
Взаимосвязь между числом cerMeHTOB и дли
ной тела у двух близкородственных видов Ne
phtys
тепеllа tоrqиаtа 22, у Ophryotroc/za риеri..
lis 25. Иноrда cerMeHToB значительно бо--
льше, но тем не менее их число может слу
жить видовым признаком.
у молодых животных скорость образо
вания новых cerMeHToB высокая, однако
у более старых особей рост в большей сте..
пени идет за счет увеличения размеров cy
ществующих cerMeHToB. При ампутации
«хвостовых» cerMeHToB восстанавливается
высокая скорость их формирования.. харак"
терная для молодых червей. После ампута
ции и заживления раны образуется новая
зона нарастания cerMeHToB У Nereis ero
скорость прямо пропорциональна числу
удаленных cerMeHToB.
m
о 7
....
:I:
Q)
6
L
Ф
()
Х 5
::о
:I: 4
:I:
со
m
о 3
а.
:s
а.
ф 2
:I:
Ф
L
Ф 1
а.
о
()
:s
:r
10
20
30
40
Число ампутированных cerMeHT08
2. Специфика сеzментов. Каждый ce
rMeHT аннелиды ведет себя как часть инте
rрированноrо целоrо, имеет особое CTpoe
ние и специфику. Это наиболее ясно Bыpa
жено у живущеrо в трубке червя Chaetopte
I"ИS vаriореdаtиs, изображенноrо на рис. 9.5.
Однако один..единственный cerMeHT этой
внешне сложно орrанизованной полихеты
способен реrенерировать части тела спере
ди и сзади от Hero, т. е. восстанавливать цe
лоrо червя!
482
rлава 15
Стадия реrенерации целоrо червя из изолиро
BaHHoro веероподобноrо cerMeHTa Chaetopte
rus variopedatus
Во время реrенерации фраrментов червя
Clyтeпella torquata, во взрослом состоянии
состоящеrо из 22 cerMeHToB, также можно
наблюдать реrионарную специфику каждо..
ro cerMeHTa.
c=cJ3 3 r e TO 15 nшпd
П
: 13 er ! OB : ' 8 [DJj
D1I[[s
IП
DIПIfIJ 9 '
... ... .. ::1 ...
N
}3 er o 21 JXJ
На рисунке изображены:
1) интактный червь;
II IV) компенсационная реrенерация
13 сеrментных фраrментов из разных участ
ков:
11) сеrменты 3 15;
111) сеrменты 6 18;
IV) сеrменты 9 21.
Обратим внимание, что в каждом случае pe
rенерирующие сеrменты сохраняют свое
изначальное положение в порядковой ие
рархии.
3. Морфаллаксuс. У некоторых полихет
утрата rоловных cerMeHToB вызывает мор..
фолоrическую перестройку оставшихся.
Это явление иноrда называют морфаллак--
сисом. Сеrменты как бы изменяют свое по..
ложение в иерархической орrанизации тела.
Это наблюдается, например, у червей
рода Sabella. По--видимому, для них xapaK
терна восстановление венца щупалец, время
от времени обкусываемоrо рыбами.
Тело у Sabella состоит из простомиума
со сложным венцом щупалец, перистомиу..
ма, фиксированноrо числа торакальных се..
rMeHToB с характерным расположением па..
раподий и мноrих идентичных абдоминаль..
ных cerMeHToB (1). Реrенерироваться может
не более трех передних cerMeHTOB. Если по..
лихета теряет их больше, задние сеrмеиты
утрачивают щетинки и превращаются в то..
ракальные (II IV).
'---"
.........
.......
II III N
На рисунке изображены:
1) интактная Sabella:
11) утрата простомиума, перистомиума
и всех торакальных cerMeHToB;
111) ранние стадии реrенерации и мор-
фаллаксиса;
IV) реrенерация венца щупалец, пер-
истомиума и торакальных cerMeHToB, в ко-
торые превращается соответствующее
число абдоминальных.
Такой путь реrенерации ловчих щупа-
лец самый быстрый. Очевидно также, что
кажДЫЙ cerMeHT имеет характерное строе-
ние блаrодаря определенному положению
в переднезаднем rрадиенте тела
Развитие
483
4. Модель реzенерацuu. Мноrие наблю..
дения можно описать следующей простой
моделью:
а) позиционная ценность простомиума
равна нулю:
б) позиционная ценность пиrидиума
равна 1 + видоспецифичное число cermeh--
тов;
в) зона нарастания (бластема) cerMeH
тов расположена у передней стороны пиrи..
диума;
r) скорость образования cerMeHToB про..
порциональна разности между позицион--
ными ценностями пиrидиума и последнеrо
cerMeHTa и равна нулю, коrда эта разность
составляет единицу;
д) образование cerMeHToB продолжается
до тех пор.. пока разность между позицион--
ными ценностями пиrидиума и caMoro мо"
лодоrо cerMeHTa не станет равна единице;
е) утрата каудальных cerMeHToB приво..
ДИТ К восстановлению пиrидиума с прису..
,
1
3
. . 4
25
.
А
.
.
щей ему высокой позиционной ценностью.
Эта модель проиллюстрирована на при..
мере полихеты OphryOlrocha. Она примени..
ма к нормальному эмбриональному (А)
и реrенераuионному (Б) росту.
Эксперимент (В) наводит на мысль, что
нервный ствол несет позиционную инфор
мацию. Отклонение ero в сторону, напри..
мер в 9 M cerMeHTe, вызывает формирова..
ние дополнительноrо пиrидиума. Этот
лишний «хвост» будет теперь образовывать
cerMeHTb] по описанным выше правилам.
Подобные двухвостые полихеты изредка
встречаются в природе и MorYT быть также
получены путем соединения фраrментов
двух особей. В любом случае скорость
образования cerMeHToB у каждоrо пиrидиу..
ма подчиняется законам нормальноrо ро--
ста и пропорциональна разности между по..
зиционными ценностями бластемы и по--
следнеrо cerMeHTa.
Б
в
.
111
»-
А. Нормальный рост Ophryotrocha Ценность простомиума О, пиrидиума 25. Последователь
НОСТЬ cerMeHToB во время роста устанавливается путем интеркаляции. Б. Реrенерационный рост
Ophryotrocha. После удаления каудальной части между последним cerMeHToM и восстановленным
пиrидиумом существует позиционный разрыв. В. Индукция дополнительноrо пиrидиума путем
хирурrическоrо нарушения вентральноrо HepBHoro тяжа
rЛАВА 1 6
СИСТЕМЬI КОНТРОЛЯ
Первые rлавы этой части книrи посвя
щены отдельным функциональным систе
мам питания, движения, дыхания и
T.Д., OДHaKO наш rлавный тезис заключа
ется в том, что отбор действует не на част
ные признаки, а на орrанизм в целом. Все
rены особи в совокупности обеспечивают ей
успех или rибель.
Уже подчеркивалось, что эволюционное
преимущество имеет две составляющие:
выживание caMoro индивида и более BЫCO
кую вероятность оставления им потомства.
Обе они требуют усвоения информации из
внутренней и внешней среды и успешноrо
осуществления необходимых реакций на
нее. Уже rоворилось, что на животных по
стоянно действует противоречивое давле
ние среды. Каждый вид активности влияет
на друrие и может, например. повышать Be
роятность rибели особи, которая вынужде--
на искать компромиссы и подстраховывать
свои действия. Поэтому для реrуляции раз--
вития от зиrоты до размножаюшейся взро
слой формы (см. rл. 15), для получения
и обработки требуемых данных, а затем для
инициации, контроля и видоизменения
координированных реакций (если нужно ре--
аrировать" чтобы вероятность непосред--
cTBeHHoro выживания была максимально
возможной) орrанизму необходимы особые
системы Они должны действовать на всех
стадиях жизненноrо цикла. В данной rлаве
как раз и рассматриваются такие нейробио--
лоrические и эндокринные системы кон--
троля.
16.1. Введение
Изучать системы контроля у беспозво
ночных практически начали по трем основ--
ным причинам. Во--первых, из чисто научно
ro любопытства, во--вторых, с целью
использования полученных данных в эк оно..
мике и медицине и, наконец, в связи с тем,
что эти системы представляют собой удоб..
ные «модели» для исследования научения,
обратной связи и т. д.
Ряд видов (<<лабораторные крысы» мира
беспозвоночных), rлавным образом насеко..
мые и моллюски, широко использовалиеь
для изучения фундаментальных проблем,
которое привело ко мноrим из наиболее
значительных достижений в области нейро..
биолоrии. Выдающуюся роль при этом
иrрали 2И2антскuе аксоны, или нервные БО"
локна" кальмара Loligo (приложение 16.1)"
однако крупные нейроны с диаметром тела
до 1 мм обычны для моллюсков, и в нейро..
биолоrии стало почти рутинным делом вво"
дить до четырех электродов в одну такую
клетку, реrистрируя ее импульсы, воздей..
ствуя на нее током и Т. д.
Нервные системы мноrих беспозвоноч"
ных отличаются простотой, которая позво..
лила исчерпывающе описать их строение
и функциональные взаимоотношения ме..
жду их частями. В отношении позвоночных
это пока лишь отдаленная мечта. Примера..
ми MorYT служить вся нервная система не..
матоды Caeпorhabditis e/egaпs, а также пе..
риферические кардиальный и cToMaToraCT"
ральный rанrлии ракообразных. Более то..
[о, в разных особях можно идентифициро"
вать одни и те же индивидуальные нейроны,
что позволяет исследовать их строение, фи..
зиолоrию и значение. Важна также неев ой..
ственная позвоночным устойчивость мно"
rих беспозвоночных к экспериментальным
воздействиям.
Изучение плодовой мушки дрозофилы
внесло исключительный вклад в понимание
rенетики нейробиолоrии и поведения. Ха..
Системы контроля
485
Тонкая стеклянная
нить
Капля раствора
электролита
Электронно ...
лучевой
осциллоскоп
Ми кропи пет ка
Кончик
микропипетки
}
()
L........J
1 мкм
о о
о о
о о
о о
Электрометр
(входной сиrнал
от микроэлектрод )
-=- Заземление
Аксон кальмара
)
Рис. 16.1. Микроэлектрод и установка для реrистрации внутриклеточных электрических потен
циалов (по Shepherd, 1983)
рактерно, что мноrие полученные мутанты
были прежде Bcero идентифицированы по
поведенческим отклонениям, что дало ма..
териал для исследования морфолоrических,
биохимических и rенетических основ пове..
дения. Для этих целей была также широко
использована нематода Caeпorhabditis.
у нее шесть хромосом (у дрозофилы
четыре) и только 3000 5000 reHoB. Мутан"
ты способны жить в питательной среде да..
же при наличии крупных дефектов (напри..
мер, будучи парализованными); возможно
и их размножение, поскольку один из полов
является самооплодотворяющимся repMa
фродитом. К 1984 r. было известно 228 му"
таций по 14 reHaM, в том числе делающие
животных нечувствительными к прикосно..
вению.
Для изучения нервных систем сейчас
используется множество методов Самые
передовые из них применяют внутриклеточ"
ные электроды и осциллоскопы, реrистри"
рующие электрические потенциалы (рис.
16.1). Недавно разработана методика
«patch--clamp» (локальной фиксации), с по..
мощью которой к отверстию микроэлек--
трода MorYT быть присоединены мельчай..
шие фраrменты клеточной мембраны для
изучения функций индивидуальных мем..
бранных каналов. Широко используются
также световая и электронная микроскопия
в сочетании с методами введения в отдель
ные нервные клетки красителей (например,
проционовоrо желтоrо) или электроноплот..
ных материалов, что позволяет проследить
распространение отростков нейронов и их
контакты с друrими клетками. В последние
rоды наблюдается настоящий информа..
ционный взрыв, связанный с применением
методов современной биохимии и молеку"
лярной биолоrии. Нейромедиаторы, ropMo"
НЫ. рецепторы и т. д. были идентифициро"
ваны химически, установлены кодирующие
их последовательности, к их молекулам по..
лучены антитела.
Нейробиолоrия особым образом связа..
на с поведением животных. Наблюдение за
поведением целоrо орrанизма можно срав"
нить с изучением «черноrо ящика» типа
калькулятора или компьютера. О мноrих
свойствах аппарата можно узнать, наблю..
дая ero в работе, сравнивая информацию на
ero входе и выходе и т. д. Однако для изуче..
ния ero внутренней структуры, свойств раз..
личных составных частей и механизмов их
функционирования необходимо «вскрытие
ящика», чем и занимается нейробиолоrия.
Как будет показано, вместе с эндокриноло"
rией она обеспечивает наиболее перспектив"
ный подход К всесторонней оценке поведе..
ния, физиолоrии и общей биолоrии opra..
низмов.
486
rлава 16
16.2. Потенциалы
16.2.1. Мембраны
Соrласно жидкостно"мозаичной моде..
ли, плазматическая мембрана клетки co
стоит rлавным образом из двойноrо слоя
(бислоя) фосфолипидных молекул, rидро..
фильные концы которых направлены нару..
жу. Она обычно отделена от мембраны"
оrраничивающей соседнюю клетку, элек
тронопрозрачным межклеточным про..
странством шириной около 20 нм. Бислой
образует rлавный барьер для диффузии ио..
нов и т. д. В Hero поrружены молекулы бел..
ков и rликопротеинов, мноrие из которых
пронизывают ero полностью, поэтому мо"
[ут функционировать в качестве «каналов»
и «насосов» для перемещения ионов через
мембрану. Мноrие компоненты мембраны
свободно плавают внутри нее, друrие про..
чно заякорены на специфических участках
клеточной поверхности, специализирован..
ных для восприятия стимулов, нервной
передачи и т. д.
Концентрация ионов в цитоплазме
и межклеточной жидкости зависит от пac
сивНО20 движения (диффузии) и активНО20
транспорта. В мембранах нервных волокон
находится «натриевый насос», отвечающий
за выведение из клетки ионов Na 1- и посту..
пление в нее ионов К +. Это движение про..
исходит против электрохимическоrо rpa..
диента (см. ниже) и зависит от метаболиче..
ской энерrии. Насос состоит из фермента,
Nа+jК+..АТРазы" названноrо так из..за
способности катализировать распад АТР
и использовать высвобождающуюся энер"
rию.
16.2.2. Потенциалы
Наличие у плазматической мембраны
разности электрических потенциалов
общая особенность живых клеток, основан..
ная на неодинаковом распределении ионов
и различной проницаемости мембраны.
Обычно наиболее важны при этом ионы на..
rрия и калия. Если мембрана проницаема
в основном для ионов К +, сконцентриро--
ванных в клетке, их диффузия из нее ведет
к появлению отрицательноrо заряда внутри
(анионы оказываются в избытке) и к поло..
к+
к'"
Na+
+
+
+
+
+
Na+
Рис. 16.2. Электрические потенциалы клеточ
ной мембраны, вызванные различной прони
цаем остью для ионов, особенно дЛЯ К+ (слева)
и Na+ (справа)
жительному заряду снаружи (рис. 16.2). По..
скольку противоположные заряды притяrи..
ВdЮТСЯ, а одноименные отталкиваются, во..
зникающий мембранный потенциал задер"
живает дальнейший отток К -t-, пока сила,
обусловленная разностью концентраций,
уравновешивается силой, создаваемой раз..
ностью потеНllИалов. Это состояние систе..
мы называется электрохимическим равнове..
сием. И напротив, если ион Na +, присут"
ствующий rлавным образом вне клетки,
способен проникать в нее.. то в момент рав"
новесия изнутри от мембраны установится
положительный заряд. Такие простые изме..
нения проницаемости являются основой
большинства сиrнальных процессов в нерв..
ной системе.
Потенциал, обычно свойственный неак..
тивной клетке, называется потенциалом по..
коя. В основном он определяется относи..
тельными концентрациями К +" поскольку
мембрана хорошо проницаема для этоrо
иона. Это леrко продемонстрировать, изме-
няя концентрацию К + В окружающей среде.
Оказалось также возможным выдавить из
аксона кальмара цитоплазму и заменить ее
экспериментальным солевым раствором.
В результате естественная разность концен..
трации ионов К + меняет знак, что в соот..
ветствии с ожиданиями изменяет на проти-
воположную полярность потенциала покоя.
Изменения мебранноrо потенциала мо-
rYT происходить спонтанно в связи с рабо..
той особых каналов, через которые в клетку
проникают ионы Са +, а в сенсорных клет..
ках из..за влияния стимулов окружающей
Системы контроля
487
среды. Они происходят также в местах спе..
цифическоrо контакта между нейронами,
называемых синапсами, [де обычно вызы"
ваются химическими веществами (нейроме..
диаторами), которые высвобождаются из
одной клетки и возбуждают или тормозят
активность друrой.
16.2.3. Проведение rрадуальных
потенциалов
rлавная функция нервных элементов
передача информации из одной точки тела
в друrую. Это обеспечивается проведением
изменений мембранноrо потенциала. Так
как нервное волокно обладает свойствами
электрическоrо кабеля, вдоль Hero распро..
страняются локальные изменения потен..
циала (рис. 16.3). Такое проведение назы..
вается пассивным" или электротоническим,
а происходящие изменения потенциалов
2радуальными и декрементными (затухаю..
щими) Их величина зависит от уровня на..
чальноrо потенциала и постепенно умень"
шается по мере распространения тока по
мембране. В rлазу мухи за время про веде..
пия по аксону фоторецепторной клетки та..
кой потенциал уменьшается примерно на..
половину. Аналоrичным был бы эффект от
броска камня в озеро. Высота расходя..
щихся волн зависит от размера камня и рас..
стояния от источника. Мноrие нервные
...40
...50
т
L
...60
"'Ю о 1 2 3
Расстояние от места деполяризации, мм
Рис. 16.3. Пассивное и декрементное распро
странение деполяризации по крупным и мел..
ким аксонам. «Утечка» потенциала зависит TaK
же от эффективности rлиальной изоляции (по
Oarnell et al., 1986)
клетки, «неспайковые нейроны», по..
видимому, фУНКIIИОНИРУЮТ только таким
образом, а все нейроны имеют аналоrично
действующие участки.
16.2.4. Потенциал действия
Частичная деполяризация мноrих аксо"
нов вызывает быстрое изменение их состоя..
ния, приводящее к исчезновению и перемене
знака разности потенциалов (приложение
16.2). Возникающий потенциал действия,
или «спайк», связан с пОР020вым эффектом,
т. е. наблюдается только тоrда" коrда сти"
мул достиrает определенноrо минимально..
[о уровня. Это ответ по типу «все или ниче..
20», поскольку дальнейшее увеличение ин..
тенсивности стимуляции ero не усиливает.
В орrанизме потенциалы действия вызы"
ваются спонтанными, рецепторными или
синаптическими потенциалами, которые
пассивно распространяются от мест возни..
кновения к способной активно реаrировать
области мембраны, откуда вдоль HepBHoro
волокна пойдет ток. Он деполяризует при..
лежащие области мембраны, стимулируя их
активность" т. е. обеспечение дальнейшей
деполяризации и т. д. Таким образом, по..
тенциал действия активно распространяе..
тся вдоль мембраны. Он не rрадуальный,
а соответствует принципу «все или ничеrо».
проводится с постоянной величиной, а не
декрементно. Аналоrией ему может слу..
жить поджиrание пороха запальным фити..
лем.
Скорость проведения сильно зависит от
диаметра волокна, поэтому передача им..
пульсов в реакциях избеrания и подобных
им у беспозвоночных обеспечивается 2И"
2антскими волокнами. Морская аннелида
Myxicola держит по их размеру первенетво:
у ее аксонов диаметром более 1 мм скоро..
сть проведения 20 м/с. в некоторых случаях
эта скорость сильно увеличивается за счет
изоляции участков волокна rлиальной обо..
лочкой (разд. 16.3.2). В результате проведе..
ние происходит сальтаторно (скачкообра..
зно ), т. е. от одноrо неизолированноrо
участка мембраны к следующему при вос"
становлении каждый раз потенциала дей..
ствия. У rиrантских волокон земляноrо че..
рвя по две такие «rорячие точки» на дорса..
488
rлава 16
льной поверхности в каждом cerMeHTe; в aK
сонах креветки «золотых медалистах»
животноrо мира по скорости проведения
(200 м/с при 20° С) эти точки располаrаю
тся в местах разветвления аксонов. Миели
новые оболочки, изолирующие нервное BO
локно У позвоночных, У беспозвоночных не
обнаружены.
16.2.5. Спайковые
и неспайковые нейроны
Неспайковые нейроны описаны в OCHOB
ном У членистоноrих и обычно COOTBeT
ствуют рецепторам (например, фоторецеп
торам rлаза насекомых) и интернейронам.
Неспайковые мотонейроны известны толь
ко в системе иннервации мышц стенки тела
у нематоды Ascaris. Мы просто не знаем,
почему по некоторым нейронам распро
страняются rрадуальные потенциалы, в то
время как по друrим потенциалы дей
ствия. Это нельзя объяснить только длиной
волокон. Если они иннервируют окончания
Hor у крупных беспозвоночных, потенциалы
действия им, несомненно, необходимы. Oд
нако в фоторецепторах ycoHororo рака по
волокнам длиной до 1 см распространяю
тся rрадуальные сиrналы" Torna как во MHO
rих нейронах с rораздо более короткими BO
локнами, включая некоторые амакриновые
клетки (разд. 16.3.1 ), потенциалы дей
ствия, как и в крошечных ооцитах. желези..
стых клетках и т. д.
Во мноrих клетках, rенерирующих rpa..
дуальные потенциалы, происходит почти
непрерывное высвобождение нейромедиа
тора, и незначительные изменения MeM
бранноrо потенциала (для фоторецептора
мухи Bcero 0,3 мВ) MorYT ero усилить или
ослабить. Даже присутствуя внебольшом
числе, что типично для систем беспозвоноч
ных, такие нейроны способны обеспечить
плавный rрадуальный контроль, который,
по видимому, требует rораздо большеrо
количества спайковых клеток с суммиро
ванным выходом. Информация, передавае
мая с помощью потенциалов действия, воз
можно, «HeHaMHoro сложнее чередования
точек и тире азбуки Морзе» (Adrian, 1932),
однако комбинирование частоты, последо
вательности импульсов и продолжительно
сти активности обеспечивает очень тонкую
систему кодирования. Обычны промежу..
точные варианты между двумя этими край..
ностями: небольшой участок клеточной
мембраны может rенерировать спайк, кото..
рый затем декрементно распространяется
к окончаниям нейрона. Расположенные
вдоль аксона и зависящие от мембранноrо
потенциала каналы для Са 2 + MorYT поддер..
живать этот потенциал, остающийся тем не
менее rрадуальным, сходные каналы в мем..
бране HepBHoro окончания способны усили..
вать поступающий rрадуальный сиrнал.
16.3. Нейроны и их связи
16.3.1. Нейроны
Нервные клетки (нейроны) характери..
зуются наличием удлиненных отростков
и способностью проводить электрические
потенциалы. Они традиционно подразде..
ляются на сенсорные (афферентные), пере..
дающие информацию в центральную нерв..
ную систему, мотонеЙРОНbl (эфферентные,
или двиrательные), передающие команды
из центра к эффекторам (мускулатуре, желе..
зам и т. д.), и инmернейроны, которые связы..
вают предыдущие два типа клеток (локаль..
ные интернейроны часто отличают от «ре..
лейных» клеток с более длинными отроет..
ками). Можно добавить еще нейросекретор"
ные клеп1КИ" выделяющие в кровь rормоны.
Сейчас уже обнаружены клетки с признака..
ми как сенсорных, так и двиrательных ней..
ронов, d также с друrими сочетаниями
свойств.
Нейроны часто описывают, используя
термины функциональной морфолоrии
(рис. 16.4). Входная (или дендритная) зо..
на это чувствительный рецепторный уча..
сток сенсорных и постсинаптическая об..
ласть друrих нейронов. Здесь возникают
и отсюда распространяются по мембране
rрадуальные рецепторные или синаптиче..
ские потенциалы. У спайковых нейронов
есть зона 2енерации импульсов, rде дости"
rающие пороrОБоrо значения rрадуальные
потенциалы запускают потенциалы дей..
ствия. Аксон проводит потенциалы к нерв..
ным окончаниям, rде осуществляется синап..
тическая передача (т. е. связь с друrой клет
СИСТ4 МЫ контроля
Аксон
Дендрит
ная зона
489
Передаю
щи е OKOH
чания
Рецептор
Хеморецептор растяжения
HaceKoMoro раКООБразноrо
Мотонейрон Нейросекреторная
HaceKoMoro клетка моллюска
Рис. 16.4. Нейроны с различными морфолоrией и функцией обладают сходной функциональной
орrанизацией с характерной динамической полярностью. Некоторые нейроны монополярные,
т. е. от их тела отходит единственный отросток (пример мотонейрон насекомых), друrие би или
мультиполярные
Аксон
r
А
Рис. 16.5. А. Дендриты мотонейронов CTOMa
тоrастральноrо rанrлия ракообразноrо (и MHO
rих друrих) одновременно получают и пере
дают информацию. Их выходные окончания
MorYT одновремена быть постсинаптическими
по отношению к друrим волокнам. Однако
естественное проведение импульса по аксону
кой). Заметим, что расположение тела
нейрона не важно для этих аспектов ero фи--
зиолоrии.
Поскольку мноrие дендриты, а также
пресинаптические окончания MorYT как по
лучать, так и передавать информацию (рис.
16.5), такая концеПlIИЯ динамической поляр..
ностu нейронов требует уточнения. Более
Toro, аксоны некоторых нервных клеток, ве..
роятно, проводят импульсы В обоих на..
правлениях, а у друrих (амйкрuновых) кле--
ток отростка, соответствующеrо аксону,
нет, причем их разветвления, вероятно, спо--
собны к независимой активности.
16.3.2. rлиальные клетки
Вторую катеrорию клеток, присут--
ствующих в нервной системе, называют ней--
РО2лuей. Их роль механическая (укрепле--
носит односторонний характер. Б. Изополяр
ный нейрон стрекающих. В. Нейрон rрибовид
Horo тела полихеты. В таких нейронах динами
ческая полярность, по видимому, отсутствует.
r у амакриновых клеток, например, из зрите
льной доли HaceKoMoro, отсутствует rлавный
отросток, соответствующий аксону
490
rлава 16
ние, разделение и защита нервных элемен
тов), электрическая (изоляция волокон
и увеличение скорости HepBHoro проведе
ния) и метаболическая (контроль ионной
средь] внутри нервной системы и разруше
ние нейромедиаторов после их выделения
из нейронов).
16.3.3. Химические синапсы
Мноrие нейроны передают информа
цию, секретируя химические вещества, Ha
зываемые нейромедиаторами и нейромоду
ляторами. В отличие от секретов обычных
железистых клеток, распространяющихся
на большие расстояния, эти продукты BЫC
вобождаются строто локализованно и ce
лективно в специальные места контакта
с друrими клетками. называемые синапса
ми.
Чаще всето химические синапсы обра
зуются между окончаниями нервных воло
кон. Они MorYT включать луковицеобраз
ные концевые расширения или вздутия (ва..
рикозы) по всей длине волокна. У мноrих
синапсов настолько характерная структура
(рис. 16.6), что существование функциональ
Horo контакта обычно предполаrается уже
на основе наблюдений в электронном ми
кроскопе, хотя подобная корреляция и BЫ
зывает споры. Заслуживает внимания и тот
факт.. что обнаруженный электрофизиоло
rическими методами «синаптический KOH
такт» между двумя клетками в действи
тельности может соответствовать тысячам
соединений (25000 в стоматоrастральном
rанrлии омара), различимых при помощи
электронной микроскопии.
Обычные нейромедиаторы, например
ацетилхолин, запасаются в синаптических
пузырьках, и принято считать.. что их BЫ
брос происходит за счет экзоцuтОЗQ, т. е.
слияния мембраны пузырька с мембраной
окончания и последующеrо выхода coдep
жимоrо в синаптическую щель. Слабое BЫ
деление медиаторов, по видимому, проис
ходит из окончаний даже в состоянии по
коя, но потенциал действия (или деполяри
зация не более чем на 0,3 мВ в неспайковом
нейроне. например фоторецептора мухи)
вызывает одновременный экзоцитоз MHO
жества пузырьков. Секреторные rранулы,
Секреторная rранула
Синаптические пузырьки
Пресинаптическая
""Т пластина'')
Синаптическая шель
Постсинаптическое
уплотнение
А
. ).' \Z: < :i:,:"
- -;:j" l "' r:,: ':',; :" ,
s ';,'i': , ,' ":' ::" , : " '
".. , '* . .\1',
, ' ",,';l" ,, ':. ,.";<,, . ;' ,,' " f, " ,,_.' ".
'; ."\"" >\>' : .
)\' J..-: '." '. ' t "' '
".
..
.;....
'. " 11:) . "',. -- s . . '
'. :"." :..,,,' ..; :!.. . , :< t. ( "/:.'. ,
',.. (J '. , 5' ;'" ·
,) ;ч, 7; ' ' : 1 ". '
1 ;,IЛ fr.: 1t ;; ,: ),: '
? . . . : ;'
*" <
'%,">
":t-" ......
;;,' . ; ,,: \(c "'.",
', :;;
. '" .:. "
....t-
."'Ч, -. ...
'.' .
,, "
-....OJO
.-1' . .
, .'"(.
. .- ..; . "". ,- .
I
, "", f',:,'
:Jj < ": ' .. :' :t ; :: ,- '"
. ;;::- ...:'
'\
$. . "," ".
'4 ' ,;1; ,1': ;
-,',," $
Б
: ,..':tk 11!11;;.
ф .,.jI'"
Рис. 16.6. А. Пресинаптическое окончание хи
мическоrо синапса у членистоноrих. Диаметр
синаптических пузырьков, rруппирующихся
у пресинаптическоrо утолщения, составляет
обычно ЗО 50 нм; секреторные rранулы, как
правило, находятся позади них и, возможно,
преобладают в объеме окончания. Мноrим си
напсам не свойственны некоторые из указан
ных признаков. Например, у иrлокожих ни спе
циализированных щелей, ни утолщений, ни BЫ
ростов не наблюдается. Б. Электронная микро
фотоrрафия синапса в церебральном rанrлии
земляноrо червя Luтbricus. Заметно диффу
зное пресинаптическое уплотнение. Стрелкой
показана синаптическая щель (х 40 000, Mac
штаб 100 нм)
Системы контроля
являясь местом запасания пептидных неЙ..
ромедиаторов и модуляторов, также высво..
бождают их путем экзоцитоза (см. прило..
жение 16.11). Фундаментальные механизмы
функционирования синапсов были изучены
на rиrантских волокнах кальмара, пре.. и
постсинаптические окончания которых до..
статочно велики для введения микроэлек"
тродов. Льинас (Llinas) с соавторами пока
зал, что потенциал действия открывает
в пресинаптической мембране зависящие от
б С 2+
мем paHHoro потенциала каналы для а ,
а выброс медиатора активируется повыше..
иием уровня Са 2 + внутри окончаний, а не
самой деполяризацией.
После высвобождения нейромедиатор
диффундирует через синаптическую щель
и связывается с белковыми рецепторными
молекулами постсинаптической мембраны.
После такой активации молекула..рецептор
может функционировать как селективный
ионный канал, причем крайне короткое вре..
мя не более нескольких миллисекунд. От..
вет рецептора может заключаться и в акти..
вации внутри клетки синтеза втОрО20 пос..
редника, например сАМР (циклический аде..
нозинмонофосфат), который обладает бо..
лее продолжительным метаболическим воз..
действием на ионные каналы
у отдельноrо нейрона обычно MHoro си..
наптических входов с различными медиато"
рами, вызывающими rенерирование возбу..
ждающих (деполяризующих) и тормозных
(обычно rиперполяризуюших) постсинапти"
ческих потенциалов. На нейроны действую r
также неЙРОМОДУЛЯТОРЬJ вещества, назы--
ваемые так из..за своих менее четких эффек"
тов (см., например, разд. 17.10.5). Действие
медиаторов быстро прекращается блаrо..
даря их ферментативной деrрадации (аце..
тилхолин, пептиды) или обратному поrло..
щению пресинаптическими окончаниями
(амины, аминокислоты).
Преимущество химических синапсов за..
ключается в их способности усиливать сиr..
нал. Например, нейромедиатор (возможно,
rистамин), выделяемый окончаниями фото--
рецепторных нейронов мухи, в ответ на
определенную деполяризацию вызывает
в 7 14 раз большее изменение мембранно..
ro потенциала (rиперполяризацию в дан..
ном случае) постсинаптических нейронов.
491
Друrая важнейшая для функционирования
нервной системы черта химических синап..
сов высокая пластичность их действия;
действительно. есть данные. что на этом oc
новань] различные типы научения (прило..
жение 16.3).
16.3.4. Электрические синапсы
Один из наиболее спорных вопросов
в истории нейробиолоrии касается механиз
ма синаптической передачи. По иронии
судьбы, у рака как раз к тому времени, Kor..
да вопрос наконец решался в пользу хими..
ческой rипотезы rлавным образом на oc
иове работ по периферическим нервным
контактам у позвоночных (СМ. приложение
16.9), B 1959 r. Фершпаном (Furshpan)
и Поттером (Potter) были открыты электри"
ческие синапсы. Они позволяют клеткам не..
посредственно стимулировать друr друrа
без участия медиаторов и их рецепторов.
1
Передача зависит от щелевых контактов ,
в которых белковые молекулы образуют
пересекающие межклеточное пространство
полые цилиндры, через которые может течь
ток, что обеспечивает распространение де..
поляризации непосредственно от одной
клетке к друrой. Передача происходит бы--
стро, исключая синаптическую задержку,
составляющую для химических стимулов
около 0,4 мс, и во мноrих случаях может
происходить в любом направлении. Однако
иноrда электрические синапсы действуют
как выпрямители, обеспечивая проведение
с низким сопротивлением только в одном
направлении.
Иноrда такие синапсы способствуют ин..
те2рации в единую функциональную единицу
нескольких клеток. Это может синхронизи"
ровать действие всей их rруппы (рис. 16.7),
например в случае рецепторных клеток от..
1 Щелевые контакты не следует путать
с плотными контактами, в которых происходит
слияние внешних листков мембран. Плотные
контакты препятствуют диффузии через межкле
точное пространство и являются структурной oc
новой 2емато"энцеФаJШчеСКО20 барьера. С помо
щью посл днеrо у насекомых, rоловоноrих и по
звоночных реrулируется химический состав cpe
ды, окружающей нервную систему.
492
rлава 16
ф
ЭС
эс
Рис, 16.7. Электрические синапсы придают
клеточной популяции свойства единоrо функ
циональноrо элемента. ХС химический си
напс; ЭС электрические синапсы
дельных элементов (омматидиев) сложноrо
rлаза саранчи и нейросекреторных клеток,
синтезирующих нерестовый ropMoH у мол..
люсков (как и в случае мноrих железистых
и мышечных клеток). Электрические синап..
сы обычны для нейронов с rиrантскими во..
локнами; они обеспечивают высокую ско"
рость функционирования их системы, рабо..
тающей в результате как одна оrромная
клетка (см. рис. 16.25).
Эмбриональные нейроны связаны мно"
жественными щелевыми контактами, мно"
rие из которых утрачиваются по мере раз..
вития ионных каналов и химических синап..
сов.
16.4. Орrанизация
нервных систем
16.4.1. «Нейроидная» система
rубок
у rубок не может сформироваться на..
стоящей нервной системы, поскольку у них
вообще нет фиксированных «тканей», одна..
ко эти орrанизмы способны сокращать
и отдельно оскулумы, и все тело. Таким
способом некоторые виды создают обрат..
ный ток воды, пр очищающий каналы,
а друrие используют мощные. кажущиеся
координированными сокращения для вы..
брасывания в окружающую среду личинок.
Мноrие реакции на стимуляцию, вероятно,
вызваны исключительно распространением
механических воздействий: миоциты, сокра..
щаясь, растяrивают соседние клетки и т. д.
Однако у RhabdocaZyptus не исключено элек..
трическое проведение возбуждения, по..
скольку электрический стимул rенерирует
ответ, распространяющийся через весь ор..
rанизм со скоростью 0,25 см/с и вызываю..
щий закрытие всех оскулумов. Эпителиаль..
ное проведение (см. ниже) представляет со..
бой друrой тип нейроидной функции.
16.4.2. Нервные сети
Нервная сеть это диффузное двумер"
ное сплетение би.. или мультиполярных
нейронов (рис. L 6.8); такая орrанизация ха..
рактерна для нервных систем книдарий.
Элементы сети расположены между основа..
ниями эпителиальных клеток поверх мезо..
rлеи. Они встречаются как в одном, так и
в обоих клеточных слоях и способны про во..
дить сиrналы со скоростью от 10 до 100
см / с.
Характерная черта нервной сети
распространение активности в любом на..
правлении из любой точки стимуляции.
Первые экспериментаторы вырезали из
стенки тела книдарий извилистые фиrуры
и показывали, что через них MorYT распро..
Эпидермис
Нейрон
Мезоrлея
rастродермис
Б
Рис. 16.8. А. Вид с поверхности нервной сети
книдарий, состоящей rлавным образом из изо
полярных и биполярных нейронов (детали си
напсов не показаны). Б. Поперечный разрез,
демонстрирующий нервную сеть в эпидерми
се. Коrда такие сети находятся в обоих эпите..
лиальных слоях, они MorYT контактировать че..
рез мезоrлею
СистеМЫI контроля
страняться импульсы, объясняя это симме..
тричностью (т. е. двусторонним проведе..
нием) синапсов. Однако были обнаружены
и обычные поляризованные контакты, и па..
ры взаимно противоположных поляризо..
ванных синапсов. Возможно, в стебельке
колониальной книдарии Naтoпia клетки со..
единены электрическими синапсами. В не..
которых сетях проведение активное, тоrда
как в друrих декрементное, т. е. сиrнал за..
тухает с увеличением расстояния от точки
ero возникновения (разд. 16.9.3).
Нервная сеть может быть реrионарно
дифференцирована. Например, в мезенте
риях актиний встречаются крупные ориен"
тированные по вертикали биполярные
нейроны. обеспечивающие быстрое актив..
ное проведение. Более Toro, в одном и том
же эпителии иноrда присутствуют две раз..
личные нервные сети. Для сцифомедуз (на..
пример.. Аиrеliа) характерен ряд rанrлиев,
расположенных по краю колокола. Их се..
нсорная стимуляция обеспечивается «диф..
Фузной нервной сетью», а двиrательные
сиrналы от них идут к плавательным мыш"
цам через быстро проводящую «нервную
reть из rиrантских волокон». Это первая
известная нервная сеть; ее описал в 1879 r.
Шафер (Schafer) Две сети покрывают при
мерно одну и ту же область на нижней по..
верхности колокола, но прямо между собой
не связаны.
Поведение книдарий, например актиний,
включает питание. плавание. вскарабкива..
ние на раковины комменсалов раков..
отшельников, «хождение» на щупальцах,
движение базальноrо диска и закапывание.
Сложность и интеrрапия этих действий.
если учесть крайне низкий уровень орrани"
зации нервной системы, поразительны.
у плоских червей присутствует ряд нерв..
ных сетей с теми же функциональными ха..
рактеристиками, что и у книдарий. Две из
них, сочетающие сенсорные и двиrательные
функции, иноrда расположены внутри вен..
тральноrо эпидермиса и непосредственно
под ним, а третья. исключительно двиrате..
льная, под лежащим rлубже мышечным
слоем. Описаны также хорошо различимые
нервные сети, связанные с дорсальной по..
верхностью. rлоткой и друrими частями пи..
щеварительноrо тракта. В rлубине тела они
493
соединяются с нервными стволами. Нерв..
ные сети, вероятно, иrрают важную роль
и в друrих rруппах животных: у иrлокожих,
в Hore моллюсков, в кишечнике (даже у по..
звоночных).
16.4.3. Центральная
и периферическая
нервные системы
Между центральной нервной системой,
состоящей из нервных стволов и rанrлиев,
и периферической нервной системой, вклю"
чающей нервы и рецепторы, существует ва..
жное отличие. Первая является «пунктом
назначения» сенсорных сиrналов. центром
синаптической интеrрации и источником
двиrательноrо контроля, тоrда как нервы
служат rлавным образом проводящими пу..
тями (нервные сети сочетают обе эти роли).
Функция миниатюрных периферических сан"
2 flиев промежуточная в том смысле.. что они
всеrда связаны с центральной нервной си..
стемой и находятся под ее влиянием, одна..
ко в значительной мере способны и к неза..
висимой активности. Примером MorYT слу..
жить rанrлии мелких клешнеобразных ор..
raHoB защиты (педицелларий) некоторых
иrлокожих (рис. 7.18).. кардиальные rанrлии
ракообразных и параподиальные rанrлии
полихет.
Считалось, что у беспозвоночных кле..
точные тела практически всех сенсорных
нейронов расположены на периферии. одна..
ко сейчас у мноrих rрупп они обнаружены
и в центральной нервной системе. Любо..
пытная черта нематод, некоторых плоских
червей и иrлокожих (рис. 16.9. I) заключае..
тся в наличии вместо нервных волокон, иду..
щих от мотонейронов к мускульным бло..
кам, длинных аксоноподобных о rpocTKoB
мышечных клеток. образующих постсинап
тические окончания на поверхности нервно..
ro ствола.
Эпителиальное проведение со скоростью
3 35 см/с иrрает важную роль у rидроме..
дуз и оболочников. В некоторых случаях
эпителий функционирует как единый рецеп..
торный opraH с оrромной поверхностью.
Так, у Sarsia механическая стимуляция лю..
бой точки внешней поверхности колокола
возбуждает потенциал действия, который
494
ЦереБральны
rанrлий
Нервный
ствол
Комиссуры
А
rлава 16
Надпищеводный
rанrлий
Б
ЦереБральный
rанrлий
Околопищеводные
коннективы
т оракальный
rанrлий
r
Подпищеводный
rанrлий
ЦереБральный
rанrлий
Плевральный
rанrлий
Висцеральные
rанrлии
Вентральный
нервный
ствол
8
Аникальная
r ипоневральная
Эктоневральная
д
Рис. 16.9. Разнообразие нервных систем беспозвоночных. А. Ортоrональная орrанизация парных
нервных стволов и сериально расположенных комиссур у плоских червей. Б. Сеrментированный
вентральный нервный ствол пиявок не медуллярный, как у друrих аннелид, а четко дифференци
рован на rанrлии, содержащие клеточные тела, и коннективы, образованные нервными волокна
ми. Подпищеводный rанrлий формируется несколькими сеrментарными rанrлиями. В. ДЛЯ мол
люска Patella показаны основные rанrлии и перекручивание висцеральных коннектив. Орrаниза
ция нервной системы самых примитивных моллюсков HeHaMHoro выше, чем у плоских червей,
в то время как у некоторых rоловоноrих моллюсков мозr и поведение такие же сложные, как и
у рыб. r. Высококонцентрированная нервная система десятиноrих раков. Д. Поперечный разрез
луча морской звезды. Нервные волокна эктоневральной сис:rемы слепо оканчиваются на поверх
ности rипоневральной системы. Выделяемое вещество, вероятно, действует как «локальный rop
мон» с диффузной активностью
распространяется через щелевые межкле
точные контакты по всему эпителию. Обыч..
но эпителиальное проведение дает входные
сиrналы нервной системе (например, у обо..
лочника Oikopleura через электрические
синапсы с эпидермальными сенсорными
нейронами). Оно также участвует в двиrате
льном контроле, передавая возбуждение,
возникающее в нервной системе, слою мы..
шечных волокон или ресничкам (например,
в rлотке оболочников).
у 2идромедуз вдоль края колокола про..
ходят два кольцевых нервных тяжа. В ну..
тренний, А1едуллярный, включает равномер..
но рассеянные тела нейронов (rлавным
образом биполярных) с вытянутыми парал..
лельно друr друrу отростками Медулляр..
ные стволы у плоских червей MorYT быть
Системы контроля
495
расположены по ортО20нальному плану
(рис. 16,9, А), который считается у первично"
ротых основным. Передние участки их
иноrда лишь слеrка утолщены, но уже
у высших плоских червей и нем ер тин обра..
зуется четко дифференцированный церебра..
льный rанrлий, или «мозr».
у большинства аннелид вдоль всей дли..
ны тела проходит единый вентральный нерв..
ный ствол, образованный путем попарноrо
объединения в каждом сетменте коннекти"
вами эктодермальных rанrлионарных руди..
ментов (рис. 16.9. Б). К периферии сетмен..
тов от них отходят пары нервов. Спереди
развивается надпищеводный rанrлий, а
у мноrих полихет хорошо развиты сенсор..
ные щупальца, rлаза и ольфакторные орта..
ны, отсылающие сиrналы различным ча..
стям этоrо «мозrа» (рис. 4.59). Членистоно..
2ИМ также свойственны сеrментированные
нервные стволы, но во мноrих rруппах еще
сильнее проявляется тендеНIIИЯ объедине..
ния сетментарных рудиментов в сложные
rанrлии (рис. 16,9, п. Обычно формируется
хорошо развитый мозr, состоящий из про..
то.., дейто" и тритоперебрума. Нервная си..
стема моллюсков состоит из ряда связанных
комиссурами и коннективами rанrлиев, от
которых к периферии отходят нервы. У не..
которых rастропод наблюдается интерес..
ное расположение коннектив в виде вось"
мерки (рис. 16.9, В) результат торсии. Во
мноrих rруппах rанrлии имеют тенденцию
концентрироваться, образуя околопище..
водное кольцо.
у нематод нервное кольцо вокрут пище..
вода соединяет несколько продольных ме..
дуллярных нервных стволов (например,
у Ascaris два rлавных и четыре более мел..
ких). У И2ЛОКОЖИХ, что хорошо заметно на
морских звездах, эктоневральная система
сохраняет примитивное эпидермальное по..
ложение (рис. 16.9, д) и совмещает сенсор..
ную и моторную функции. Радиальные не..
рвы простираются вдоль нижней поверхно..
сти каждоrо луча, отходя от околоротовоrо
нервното кольца. Для морских лилий харак"
терна rлубже залеrающая исключительно
моторная 2ипоневральная система, но наи..
более важна для них апикальная система на
аборальной стороне тела. У асцидий нерв..
ные волокна идут к периферии от един..
cTBeHHoro церебральноrо тант лия, располо
женноrо между сифонами, d. у их «rолова..
стикоподобной» личинки от этоrо rанrлия
вдоль хвоста тянется полая нервная трубка.
по"видимому. rомолоrичная таковой у по..
звоночных.
16.4.4. ДиффереНЦИРО8ка
нейропиля
Различают по крайней мере три типа rи..
столоrической орrанизации нервных ство"
лов и rанrлиев (рис. 16.1 О). Наиболее обыч..
ны участки дифференцироваННО20 нейропиля.
Тела нейронов, часто окруженные rлиаль..
ными клетками, оказываются изолирован..
ными в коре, или периферической зоне.
Сложное переплетение их разветвляющихся
отростков формирует сердцевину, или ней
ропиль, являющиЙся основной зоной синап..
тических контактов, хотя часть из них лока..
лизована на телах клеток (например. у МОЛ
ЛЮСКОВ).
В нервных стволах нематод и внутрен"
нем нервном кольце медуз можно видеть
параллельное расположение нейронов. Их
тела rистолоrически не изолированы, а си..
наптические контакты образуются между
соседними волокнами. У нейронов нематод
обычно по одному отростку (неразветвлен..
ному у Caeпorhabditis, с двумя точками
ветвления у большинства Ascaris), возмо"
жные синаптические контакты которото
оrраничены прилежащими волокнами
Наконец, наиболее проrрессивный
островковый тип орrанизации обнаружен
в мозrе осьминоrа (ОСlОРИS). Тела клеток
расположены не только в коре, но и рассея..
ны внутри нейропиля. Это напоминает
строение серото вещества у позвоночных.
16.4.5. CerMeHTapHoe строение
Передвижение мноrих удлиненных жи..
вотных происходит блаrодаря волнам со..
кращений.. распространяющимся в том или
ином направлении вдоль тела (например,
у аннелид; см. rл. 10). При этом мускулату..
ра представляет собой ряд блоков или со..
митов, способных к последовательному
возбуждению. Каждый cefMeHT снабжен
«стандартным набором» нервных структур,
496
rлава 16
Рис. 16.10. Характер дифференцировки ней
ропиля. А. Дифференцированный нейропиль.
Б. Параллельное расположение нейронов. В.
Нейропиль с островками Островковая орrанизация. r. Поперечный раз
тел клеток рез церебральноrо rанrлия полихеты Nereis.
т клеточные тела нейронов: o
соединительнотканная оболочка мозrа; Н
нейропиль; П пучки нейросекреторных aKCO
нов, идущих к нейроrемальным зонам 8 OCHO
вании мозrа (ер. разд. 16.11.3). х 125; масштаб
100 мкм (Golding, Whittle, 1974)
, . .:
ИЛИ коннектива O .'
Соединительнатканная
ОБолочка
А
Кора с клеточными
телами
Ядро
Синапс
Б
Кора с клеточными
телами
в
сериально повторяющихся вдоль всей дли
ны тела. У некоторых беспозвоночных
выявлены соответствующи.е структурные
единицы и их связи. «Принципиальная схе..
ма» системы, контролирующей плавание
пиявки, приведена на рис. 16.11 (см. также
приложение 16.4).
Обычно эта система включает мотоней..
роны с аксонами, идущими к мышцам, ло..
кальные интернейроны для rенерирования
соответствующеrо типа активности (разд.
16.10) и релейные интернейроны, координи"
рующие работу соседних cerMeHToB (у пияв..
ки С помощью тормозных волокон). На рис.
1 ф
>(
. ,':;. ,;;.1;,
-.
:, ,Ц
l' .
i
-# . ;;'
!;"j,' ,'С' '*
т ·
.: 'ii . "
. "',/J/J-
..
Jff .,.'
,
.. '! \-
"
-. .#
"
( .' ;, ", / ..' )';
...
. .
.
.' . ; .. ';'4 --.>
.,.,! '>; ..,:', , .' ""'; Н':
. . . ., '" ,', .' jl ,. .it'd.,. . '''>''' .,. ,;
.:. . _ . '" . ' . . , .... . : : . .. , , " . : . .: :'; '; ':' . '; :.>-:,," ..",. "",. iI '"
{ " " .. ., · ,:/:f .
."\': ...... " .. .
. t... .
. .... .. \, ,,'# .. *," ,
,
.-)'1'
) ..
,..,
..
;- ;'
.
.-Т
.,;,"" ',jj '>
" . . "
. .
-t'
. 11
"'"
'Н.
.
r
16.11 не показаны сенсорные клетки, ОДНИ
из которых обеспечивают проприорецеп..
торную обратную связь с мышцами, а дру..
rие, активируясь, ИНИIlИИРУЮТ плавание
Сериальное повторение нервных СТРУК"
тур встречается не только у сеrментирован"
ных животных. Аналоrично устроены ра..
диальные нервы иrлокожих и осевые ство"
лы в щупальцах rоловоноrих. Они вклю"
чают rруппы тел нейронов, зоны нейропиля
и билатеральные нервы, отходящие к пе..
риферии. У нематоды Ascaris пять rрупп
мотонейронов (из одиннадцати клеток каж..
дая) сериально расположены вдоль тела.
Системы контроля
497
Дорсальные
мышцы
I
I
I
I
I
@
/
/
/
ф
I
I
I
I
/
/ I
/ 6
I I
I I
C:J
Вентральные
мышцы
Рис. 16.11. CerMeHTapHo повторяющийся комплекс нейронов (обозначены одинаковыми цифра..
ми), контролирующих плавание пиявки. В передний rанrлий (слева) входят четыре интернейрона
с циклической активностью, обеспечивающие поочередную активацию возбуждающих мотоней
ронов 3 и 4, индуцирующих сокращение соответственно дорсальных и вентральных мышц. Справа
показаны синаптические связи внутри и между rанrлиями. Светлые утолщения возбуждающие,
темные тормозные химические синапсы; между клетками ЗЗ и 28 находится также выпрямляю
щий электрический синапс (по Friesen et al., 1976)
16.4.6. «rорячие линии»
Коrда земляной червь, выставив из нор..
ки передний конец тела, сталкивается с во..
шедшей в поrоворку «ранней пташкой»,
описанноrо выше межсеrментноrо распро
странения сиrналов со скоростью около
3 СМ/С дЛЯ спасения недостаточно
требуется сквозной ПРОБОДЯЩИЙ путь. Та..
кие пути часто состоят из «rиrантских» ак..
сонов (разд. 16.2.4), образованных, в част..
ности у земляных червей, рядом клеток (по
одной на rанrлий), некоторые из которых
продольно сливаются в мноrоклеточный
синцитий, а друrие соединяются электриче..
скими синапсами. У rидромедуз пучок
связанных электрически rиrантских воло..
кон обеспечивает очень быстрое проведение
возбуждения ко всем частям BHYTpeHHero
HepBHoro кольца, а тем самым
симметричные сокращения колокола, необ
ходимые для плавания. У друrих животных
с помощью таких путей осуществляется об..
щая стимуляция или торможение cerMeH..
тарных систем, конкретная работа которых
контролируется межсеrментными связями
(рис. 16.28). Мноrие волокна внутри HepB
ных стволов проходят через несколько се..
rMeHToB и являются промежуточными по
функции между двумя описанными выше
крайностями.
16.4.7. Роль мозrа
Передние rанrлии.. особенно у эв олю..
ционно продвинутых беспозвоночных, раз..
виты лучше друrих. Такая тенденция
часть более общеrо процесса цефализации,
т. е. дифференцировки rоловы.
Во первых, это явление, несомненно, во
MHoroM связано с неизбежной KOHцeHтpa
цией сенсорных opzaHOlJ на переднем конце
498
rлава 1 б
тела ПОДВИЖНЫХ билатерально симметрич
ных животных. Роль мозrа в обработке, ин
теrРdЦИИ сенсорных сиrналов и реаrирова
нии на них хорошо изучена у насекомых.
Протоцеребрум с крупными зрительными
долями получает информацию от rлаз, дей
тоцеребрум от антенн с хорошо развиты
ми хеморепепторами, а тритоперебрум OT
передней части пищеварительноrо тракта.
Выдающаяся черта протоцеребрума мноr'их
членистоноrих, как и церебральноrо [aH
rлия высших полихет и даже плоских че
рвеЙ, наличие «rрибовидных тел». Они co
стоят из большоrо количества крошечных
нейронов (у насекомых диаметром менее
З мкм), пучки аксонов которых образуют
«ножки». Эти клетки, по видимому., уча..
ствуют в интеrрации информации, посту..
паю щей от различных сенсорных opraHoB,
расположенных на rолове и друrих частях
тела.
Вторая функция передней части тела
нервный контроль пищедо6ывающеzо пoвeдe
НИЯ. У аннелид, например, оно подавляется
при удалении Haд и/или подпишеводноrо
rанrлия. Физиолоrия питания падальной
мухи была подробно описана Детье (De
thier, 1976) в ero книrе «The Hungry Fly»
(<<rолодная муха») Распознавание пищи
при помощи хеморецепторов на тарсаль
ных (дистальных) cerMeHTax Hor и на лабел..
луме (см. рис. 8.ЗО, д) вызывает прохожде
ние нервных импульсов к подпищеводному
rанrлию и активацию ero интернейронов.,
а. затем мотонейронов, что индуцирует BЫ
ворачивание хоботка и всасываюuдие дви
жения rлоточноrо насоса.
В третьих. передние rанrлии Функциони
руют как высшие центры 2лавНО20 контроля
рефлексов и спонтанной активности «НIIЗ
ших» уровней. У плоскоrо червя Notoplaпa
во время локомопии пиша попадает прямо
в рот, тоrда как неподвижное животное сна..
чала направляет ее к переднему концу тела
для идентификации. Децеребрированная
особь не способна к последней форме пове--
дения и продолжает питаться, даже если ки"
шечник заполнен до конца. Эти эсперимен
ты показывают, что мозr получает различ
ную сенсорную информацию, в данном слу
чае о состоянии кишечника и движении.
Он принимает «стратеrические решения»
и отдает «команды»., как бы выключая или
включая рефлекторные реакции низших
центров, обладающих собственным нерв..
ным аппаратом для орrанизации конкрет"
ной деятельности. Децеребрированные осо..
би у мноrих видов беспозвоночных обычно
характеризуются rиперактивностью.
В--четвертых, высшие центры контроли..
руют общее «возбуждение».. или «настрое-
ние», которое разносторонне влияет на всю
нервную систему животноrо. Например,
чувствительность к стимулам у нейронов
rлаза саранчи значительно увеличивается
при поступлении возбуждающих сиrналов
от МОЗI'а; одновременно стимулируются
мото.рные центры.
Наконец., мозr иrрает ключевую роль
в орrанизации ВЫСIllИХ нервных функций
и высокоразвитых форм поведения, напри..
мер у осьминоrа (рис. 16.12). Работы Янrа
(У oung) продемонстрировали. что в зависи..
мости от типа сенсорной информации
в процессе научения участвуют различные
части мозrа. Зрительные пути ведут от зри..
тельных к верхнефронтальным. а затем
к вертикальным долям, в то время как так..
тильное научение связано с нижнефронталь..
ными, субфронтальными и вертикальными
долями. Внутри каждоrо из этих путей ин..
формация переключается на ряд центров;
например, внутри вертикальных долей раз..
личаЮl четыре центра поступления зрите..
льной информации. Существуют также во..
локна., идущие в обратном направлении от
вертикальных к зрительным долям.
В любой нервной системе «следы памя"
ти» присутствуют более чем на одном уров"
не ее орrанизапии. Так., удаление вертикаль..
ных долей существенно ослабляет зритель..
ную память, хотя некоторые ее следы со..
храняются в друrих долях. Более Toro, ин..
формация, усвоенная одной стороной мозrа
при одностороннем предъявлении с тиму..
лов, через несколько часов закрепляется
друrой ero стороной.
Несмотря на выводы, которые можно
сделать из анализа механизма научения
в приложении 16 3, высшие нервные функ..
ции., по видимому, требуют orpoMHoro ко..
личества нейронов. Например, такие «паи..
высшие» центры., как вертикальные доли,
содержат 25 млн. крошечных «микронейро..
Системы контроля
СУБвертикальная
Дорсально
Базальная
Медиально
Базальная
Рис. 16.12. Надпищеводная
область мозrа осьминоrа на
продольном срезе (плоскость
сечения показана внизу) с He
которыми из rлавных долей
(по Young, 1963)
НОВ», мноrие из которых не имеЮl аксонов.
Эти доли облеrчают процесс научения, воз..
можно формируя кодированное отображе..
иие ситуации, что лежит в основе замечате..
льной способности осьминоrов к обобще..
пию.
16.5. Спонтанная активность
16.5.1. Нервная инициация
Важнейший вклад в нейробиолоrию,
внесенный изучением рефлекторных дуr,
имел прискорбный побочный эффект. Поя..
вилась тенденция считать, что нервная си..
стема зависит от внешнеI'О стимулирова..
ния'! как компьютер от источника тока.
Ничто не может быть дальше от истины в
этом убеждают любые, даже самые поверх..
ностные наблюдения за поведением opra..
низмов от амебы до осьминоrа. Способ..
насть нервной системы к спонтанной актив..
ности, ее роль в инициации событий являе..
тся такой же решающей, как и ее свойство
499
Вертикальная
Медиально верхне
срронтальная
реаrировать на изменение окружающих
условий.
Часто активность с эндоrенной инициа..
цией является ритмической, и, возможно'! ее
проявление в течение коротких BpeMeHHbIx
интервалов обусловлено спонтанно rенери"
руемыми потенциалами действия (разд.
16.2.2). Участвующие в этом клетки назы..
ваются водителями ритJwа. Подобная ак..
тивность может также возникать в резуль--
тате взаимноrо и чередуюшеrося торможе..
ния двух или более нейронов. Синхронные
ритмические вспышки активности «<мозrо..
вые волны»), охватывающие большое число
нервных клеток, хорошо известны у позво..
ночных'! включая и человека. У беспозво..
ночных они также широко распространены
и обычно отличаются высокой частотой
(около 50 rц), хотя у осьминоrа она ниже
(менее 25 rц) и удивительно близка к отме..
чаемой у позвоночных. Периоды друrих
ритмов значительно больше; как будет по..
казано, у них друrая клеточная основа.
500
rлава 16
16.5.2. Движение
Сжимание rидры в мелкий комок проис..
ходит блаrодаря последовательным спон..
танным сокращениям эпидермиса. Они вы..
зываются серией реrистрируемых на по..
верхности тела электрических «импульсов
сокращения», инициируемых в расположен..
ной под rипостомом области эпидермаль..
ной нервной сети. После двукратной обра..
ботки колхицином, избирательно токсич"
ным для нервных клеток, животное стано..
вится совершенно неподвижным. Возни..;.
кнув, импульсы распространяются во всех
направлениях, вероятно, как по нервной се..
ти, так и по эпидермису.
Спонтанная активность ряда краевых
rанrлиев сцифомедузы rенерирует ритмиче..
ские сокращения колокола, необходимые
для плавания. Такую активность прояв"
ляют одновременно все центры, но в тече..
ние HeKoToporo времени «задает темп» сна..
чала один rанrлий, затем друrой и Т. д. «Ли..
дер» rенерирует потенциалы действия с ин..
тервалом, скажем, 2 с у животноrо диаме..
тром 2 см и 20 с у 20..сантиметровоrо. По..
тенциалы распространяются по «нервной
сети rиrантских волокон» нижней поверхно..
сти колокола. Они стимулируют плаватель..
ные мышцы и реrулируют активность дру..
rих rанrлиев, чтобы она не препятствовала
установившемуся ритму.
у rидромедуз (например, Polyorchis)
спонтанная активность, задающая ритм
плавания, rенерируется rиrантскими нейро..
нами BHYTpeHHero HepBHoro кольца; по..
видимому, здесь присутствует единствен..
ный водитель ритма. Сокращение щупалец
происходит блаrодаря спонтанной активно..
сти rанrлиев внешнеrо HepBHoro кольца.
Добавляющиеся по мере роста новые щупа..
льца и rанrлии ведут себя независимо до тех
пор, пока не сформируются их нервные
связи. В дальнейшем их активность синхро"
низируется с активностью остальной систе..
мы.
16.5.3. Автономные функции
Аннелида Areпicola живет в песчаной
норке (рис. 9.32), которую она вентилирует,
обеспечивая дыхание (разд. 11.4.3). Перио..
дические насасывающие движения с ин те..
рвалом около 40 мин проrоняют через жа..
бры червя воду или пузырьки воздуха. Это
поведение включает сложную последовате..
льность движений: сначала сдвиrание к хво..
СТОВОМУ концу норки; затем перемещение
вперед в сочетании с волнообразными пе..
ристальтическими сокращениями стенки те..
ла; и наконец кратковременную смену на..
правления вентиляции на противополо"
жное. Эта активность не является рефлек"
торным ответом на недостаток кислорода,
а rенерируется находящимися в вентраль..
ном нервном стволе мноrочисленными
скоординированными водителями ритма.
Точно так же спонтанные пищедобываю..
щие движения с более коротким интерва..
лом (около 7 мин) rенерируются нервными
элементами в пищеводе.
Наиболее очевидная спонтанная ритми..
ческая активность животных наряду с дыха..
Перикардиальный opraH
Сенсорная
ОБратная
связь
Сердце
ВОЗБуждающее и 9
тормозящее
влияния из ЦНС
рис. 16.13. Кардиальный rанrлий омара про
являет спонтанную активность и rенерирует
сокращения сердца. 1 9 идентифици
рованные нейроны (по Shepherd, 1983)
Системы контроля
тельными движениями биение сердца.
у мноrих форм ее первичный источник ло
кализован в мышечных клетках.. и она толь
ко модулируется нервной системой. Однако
у ракообразных биение сердца невРО2енное
и контролируется кардиаЛЬНblМ 2аН2лием
(рис. 16.13). Все ero клетки про являют cпOH
танную активность., хотя координируются
одной клеткой, действующей как водитель
рипzма. rанrлий находится под влиянием
высших ценпzров (через СИНdптические связи
с центральной нервной системой), ceHCOp
ных нейронов сердца (по принципу обратной
связи) и эндокринной системы.
16.5.4. Биолоrические часы
Некоторые периодические явления дают
основание думать, что существует Mexa
низм узнавания времени суток или слеже .
ния за ходом времени. Он называется биоло..
2ическими часами (приложение 16.5). Такие
часы необходимы для животных, ориенти
рующихся по солнцу, например для пчел
и муравьев. Медоносная пчела не только
может сообщать друrим рабочим особям
направление на источник пищи и расстоя
ние до Hero, исполняя виляющий танец
перед входом в улей [как показал фон Фриш
(von Frisch) в 1950 r.], но и способна по про..
шествии времени корректировать этот зада
ваемый по солнцу азимут с учетом движе..
ния светила по небосводу.
Мноrие животные реаrируют на измене
ние долrоты дня, меняя, например, xapaK
тер развития или размножения с приходом
коротких осенних дней (см., например,
разд. 14.3.4), Это подразумевает способ..
ность «измерять» длину дня и/или ночи,
являющуюся meMnepamypHo KoMneHcaпzop"
НОЙ (т. е. независимой от температуры), что
необычно для эктотермных беспозвоноч
ных. В противном случае в измерении вре..
мени моrли бы быть большие ошибки.
16.6. Рецепторы
16.6.1. Основная характеристика
Чувствительность к воздействиям OKPy
жающей среды общая характеристика
живых клеток, наблюдаемая даже в отсут"
501
ствие явной структурной дифференцировки.
Например, протист амеба обладает фото
чувствительностью, а у насекомых восприя"
тие продолжительности cBeToBoro дня., по
видимому., происходит в мозrу и не зависит
от rлаз. Однако у большинства животных
сформировался ряд специализированных
рецепторных клеток., часто входящих в co
став мноrоклеточных сенсорных орсанов
(см. приложение 16.6). У подавляющеrо
большинства рецепторов чувствительная
часть клетки структурно дифференцирована
и несет реснички или микроворсинки (а в He
которых случаях и те и друrие) (см. рис.
16.16). Часто строение таких ресничек ви
доизменено, хотя это. по видимому, слабо
связано с их сенсорной модальностью. Зна
чение их, вероятно, заключается в увеличе
нии площади сенсорной клетки. В HeKOTO
рых случаях (например, в rлазу рака) на
электронно"микроскопическом уровне МО--
жно видеть внутримембранные частицы,
предположительно состоящие из зритель--
Horo пиrмента, которые концентрируются
в мембранах микроворсинок. являющихся.
по видимому, чувствительным участком.
у друrих видов (например, у дрозофилы)
чувствительность скорее Bcero более диффу
зная. так как распределение частиц внутри
микроворсинок И В окружающей их плазма..
тической мембране сходно.
16.6.2. Классификация рецепта..
ров
Аристотель различал так называемые
«пять чувств». которые затем были допол
нены друrими, иноrда довольно 'Экзотиче..
скими (например, чувствительностью к Mar..
нитным полям). Типы чувствительности на..
зываются сенсорными модальностями. Их
современная классификация основана на
физических свойствах соответствующих
стимулов (например, свет, механические,
химические воздействия и т. д.). Кроме то..
ro., принято различать экстерорецепторы.,
чувствительные к внешним раздражениям,
интерорецепторы, реаrирующие на BHY
тренние факторы, и проприорецепторы, сиr..
нализирующие о движении или положении
мышц, суставов и т. д. Выделяют также фа
зические рецепторы, отвечающие на измен
502
rлава 16
СУБУМБреллярное
нервное КОЛЬЦО
Сенсорная
ресничка
Се!:iСОрНЫЙ
неирон
Статолит
Рис. 16.14. Статоцист книдарии (по Siпglа,
1975) (см. также рис. 3.19 и 3.25)
чивость внешней среды, и тонические peцeп
торы, ак ивность которых зависит от абсо
лютноrо уровня стимуляции. Мноrие pe
цепторы являются сочетанием двух этих ти
пов.
Изучая какую либо сенсорную модаль
ность, можно получить информацию о дpy
rой. Так, статоцисты, чувствительные к си
ле тяжести, являются спепиализированны"
ми механорецепторами. Они широко pac
пространены у беспозвоночных, например
обнаружены у осьминоrа, а у медуз распо
ложены по краю колокола. Этот opraH co
стоит из пузырька, содержащеrо одно или
несколько твердых образований, спzатоли..
тов (рис. 16.14). Коrда статолит перемеlцае
ТСЯ под действием силы тяжести, он Kacae
тся сенсорных ресничек или изrибает их, ак..
тивируя тем самым рецепторные клетки.
у оболочника Оikорlеиrа статолит заменен
меланиновой каплей, которая, будучи леrче
воды, всплывает., указыIаяя таким образом
направление силы тяжести.
Роль статоцистов была продемонстри
рована Крайдлем (Kreidl) в 1893 r. на при
мере креветки. Эти животные во время ли
ньки заменяют песчинки, образующие их
статолиты, на новые. Крайдль подбросил
им вместо песчинок железные шарики, а за
тем с помощью маrнита заставлял креветок
плавать вверх ноrами, на боку и т. д., поско
льку маrнитное поле имитировало rравита
ционное. Хотя статоцисты чувствительны
к ускорению, у некоторых беспозвоночных
(например, у осьминоrа) имеется также си..
стема каналов.. аналоrичных полукружным
каналам позвоночных, в которых рецепт о..
ры активируются движением жидкости.
16.6.3. Специалисты
и rенералисты
Рецепторы данной сенсорной модально..
сти обычно обладают повышенной чувстви..
тельностью к определенному виду (или
диапазону) поступающей энерrии, что вы..
ражается в более низком пороrе раздраже..
ния и более интенсивном ответе на стимуля..
цию. Однако одни из них являются специа..
листами, а друrие zенералистами. У оль..
факторноrо специалиста спектр восприни"
маемых запахов крайне узкий; иноrда чет..
кую реакцию вызывает лишь одно соеди..
нение (/jepoMoH (разд. 16.10.6), секретируе..
мый как химический сиrнал друrой особью
Toro же вида. или только запахи. связанные
с пищей. Ольфакторные rенералисты реаrи-
руют на широкий диапазон стимулов одной
модальности, но, поскольку у каждоrо из
них свой собственный характер чувствите..
льности, животное способно распознавать
вещества за счет специфическоrо сочетания
активированных рецепторов.
16.6.4. Кодирование
интенсивности
Информация, поступающая от рецепто..
ров, обычно означает не только «да» или
«нет». но также и «сколько». Это закодиро"
вано в частоте импульсов, rенерируемых
в нервных волокнах, которая пропорциона..
ЛЬНd лоrарифму интенсивности стимула
или какой либо друrой аналоrичной функ..
ции от Hero. Диапазон интенсивности сти"
мула, к которому чувствителен орrанизм,
расширяется путем контроля чувствитель..
ности репептора центробежными волокна..
ми (проводящими импульсы К периферии),
иннервирующими, например, рецепторы
растяжения у ракообразных (см. рис. 16.24).
Это свойство особенно хорошо развито
у беспозвоночных. Наконец, различные
клетки MorYT работать в разных частях ши..
pOKoro диапазона. В классической работе
Системы контроля
Николлса (Nicholls) с соавторами показано,
что механорепепторы в стенке тела пиявки,
клеточные тела которых обнаружены в каж..
дом rанrлии вентральноrо HepBHoro ствола,
относятся к нескольким типам Клетки
«прикосновения» (Т) чувствительны даже
к току воды. Рецепторы «давления» (Р) на..
чинают реаrировать, коrда сила, приклады..
ваемая щупом, достиrает 7 r. Наконец,
клетки «ноцицептора» (N) активируются
только в ответ на сильные повреждающие
стимулы.
16.6.5. Сенсорные клетки
и сенсорные нейроны
Мноrие сенсорные орrаны позвоночных
состоят из сенсорных клеток без волокон.
Через химические синапсы они передают
информацию окончаниям сенсорных нейро..
нов, аксоны которых идут в центральную
нервную систему. Несмотря на свою пр оме..
жуточную функцию, такие нейроны (по не..
доразумению) раньше назывались первич"
ными, а периферические клетки
вторичными. У большинства беспозвоноч"
ных сам рецептор является нейроном. Ис--
ключение составляют рецептор Ланrерrан"
са у оболочника ОikорlеИl'а и фоторецепто--
ры у книдарии Taпzoya (рис. 16.15).
cЬ
11 i
к централь
ной нервной
системе
А
Б
в
r
Рис. 16.15. Сенсорные нейроны хеморецеп
торных нухальных opraHoB полихет (А) Nephtys
и (Б) Nereis. Сенсорные клетки Oikopleura (В)
и Татоуа (r) образуют с сенсорными нейрона
ми электрический и химический синапсы COOT
ветствен но
50З
16.7. Зрение
Зрение это сенсорная модальность,
дающая максимум информации о мире,
Это справедливо и для мноrих наиболее
сложно устроенных беспозвоночных, о чем
свидетельствует строение их rлаз.
16.7.1. Зрительные пиrменты
Чувствительность к свету., т. е. узкому
участку спектра 1лектромаI.нитноrо излуче..
ния, обусловлена зрительным пИ "_J\Ilенто.и
химическим веществом, поrлощающим лу..
чистую и выделяющим в ответ на эту сво--
бодную энерrию. Родопсин это соедине..
ние, состоящее из каротиноида ретиналя
(производноrо витамина А) и белка опсина.
Оно распространено почти у всех rрупп ор..
rанизмов и обнаружено, например, как у во..
доросли V olvox, так и у человека. Воздей..
ствие света превращает ретиналь из 11..цис
в 11..пlpaHc конфиrураuию, а затем диссо..
циация ретиналя и опсина приводит
к обесцвечиванию пиrмента. Эти изменения
влияют на ионные каналы фоторецептор..
ной мембраны. стимулируя синтез вещества
типа - сАМР (см. разд. 16.3.3), вероятно
cGMP (циклический rуанозинмонофосфат).
В ходе ферментативноrо синтеза пиrмент
реrенерируется.
Родопсин существует в различных фор..
мах, определяемых типами ретиналя и оп..
сина. Светляки, активные в сумерках, излу..
чают как сиrнал для друrих особей cBoero
вида желтый свет, в то время как их виды,
проявляющие активность в темноте, све..
тятся зеленым. Таким образом, максималь..
ная чувствительность ретиналевых пиrмен..
тов у различных видов точно отреrулирова..
на.
Цвеп10вое зрение ВdЖНО для мноrих рако..
образных. насекомых и паукообразных
(удивительно, что у осьминоrа ero нет) и ос..
новано на присутствии нескольких типов
рецепторов, пиrменты которых чувствите..
льны к различным частям спектра. Напри..
мер, различные рецепторы пчел восприни"
мают в желто..зеленой (540 нм), синей (440
нм) и ультрафиолетовой (340 нм) областях.
Бабочки чувствительны также к красному
свету. Большинство фоторецепторов мух
504
rлава 16
блаrодаря наличию родопсина и друrоrо
пиrмента обладают любопытной двойной
чувствuтельностью (в сине--зеленой u уль--
трафиолетовой областях). Диапазон спек--
тральной чувствительности друrих рецепто--
ров зависит от экранuрующuх пИ2ментов,
которые позволяют реаrировать только на
определенные длины волн., «отсекая» оста--
льные.
Некоторые животные способны разли--
чать плоскость полярuзацuu свепzа. У мух
эту функцию выполняют краевые элементы
сложноrо rлаза. Характер поляризации рас--
сеянноrо света неба позволяет пчелам опре--
делять положение солнца, даже если оно
скрыто облаками, и ориентироваться по не--
му. Осьминоr блаrодаря этой способности
«видит сквозь» маскирующую серебристую
чешую рыбы.
16.7.2. Ресничные
и рабдомерные rлаза
Как показал Икин (Eakin), подавляющее
большинство фоторецепторов принадлежат
к этим двум типам (рис. 16.16). У одноrо из
них фоторецепторные орrанеллы представ--
лены реснuчкамu или их модифицированны--
ми мембранами (например, у позвоноч--
ных). Набор микротрубочек в ресничке МО--
жет быть обычным или видоизмененным.
у BToporo типа фоторецепторные орrанел--
лы микроворсинки, образующие правиль--
ные ряды., называемые рабдома./wu. Следует
обратить внимание, что дендриты, несущие
микроворсинки, часто оканчиваются рес--
ничкой или ее рудиментом, т. е. можно
предполаrать., что ресничный комплекс
иrрает роль индуктора при развитии рабдо--
мов. Более Toro, у полихет семейства Serpu--
lidae на разных уровнях одной и той же
клетки обнаружены дефинитивные реснич--
ные и рабдомерные рецепторы. У книдарии
Polyorchis микроворсинки, образованные
клеточной и ресничной мембранами, распо--
ложены вперемешку; некоторые из них, по--
видимому, имеют смешанную природу.
Мноrие фоторецепторные мембраны
обоих типов ежесуточно в массе разрушаю--
тся и обновляются. У ночных пауков--
динопид рабдом занимает днем около 150/0
объема рецепторной клетки, а через час по--
сле захода солнца до 900/0. Этот избыток
разрушается в течение двух часов после ВОС"
хода. У плоских червей микроворсинки
в темноте деrенерируют и восстанавливаю..
тся между пятой и десятой минутами осве..
щения (по--видимому, из мембранноrо ма..
териала, накопленноrо в виде rотовых
к использованию внутриклеточных пузырь-
ков ).
Большинство ресничных элементов
является рецепторами «выключения». Тем--
нота приводит к открытию Nа+--каналов
(обычно), деполяризации и выделению ме-
диаторов из окончаний. Рабдомерные ре-
цепторы, напротив, обычно реаrируют на
«включение» (перечисленные эффекты вы-
зываются освещением). Эти различия леrко
наблюдать у rребешка Pecteп: верхний слой
рецепторов ero rлаза ресничный, а ни..
жний рабдомерный.
Ресничные рецепторы., например у поли-
хет--сабеллид и rребешка, по--видимому, хо-
рошо адаптированы к участию в «теневых
рефлексах» защитных реакциях при не..
ожиданном снижении освещенности. Одна..
ко у усоноrих рачков при этом использую..
тся рабдомерные рецепторы «включения}).
Уменьшение освещенности, вызывая их rи-
перполяризаuию, блокирует выделение
mОрМОЗНО20 медиатора и тем самым акти..
вирует нейроны мозrа. Однако из правила
всеrда есть исключения. Например, у брю-
xOHororo моллюска Oпchidiuт микроворсин-
ковые рецепторы, по--видимому, реаrируют
на «выключение». У сальп (оболочники) та-
кая реакция свойственна фоторецепторам
обоих типов. Принцип работы рецепторов
смешанноrо типа пока не выяснен.
Долrое время считал ось, что тип фото-
рецепторов имеет филоrенетическое значе-
ние: ресничный характерен для книдарий
и вторичноротых, а микроворсинковый
для первичноротых. Однако сейчас извест-
ны мноrочисленные исключения из этой
схемы. Например, у морской звезды рецеп-
торы микроворсинковые; в rлазах rребешка
представлены оба типа: правый личиноч..
ный rлазок плоскоrо червя Pseudoceros со-
стоит из нескольких микроворсинковых ре..
пепторов, а левый из трех микроворсин-
ковых и одноrо ресничноrо.
у мноrих жrутиковых протистов часть
СИСТtемы контроля 505
Ресничные сроторецепторы РаБдомерные (микроворсинковые)
сроторецепторы
Кольчецы
Членистоноrие
Щетинкочелюстные
Кольчецы /плоские черви
Стрекающие
1
Моллюски
Протисты
Ресничка
Коловратки
икроворсинки
Рис. 16.16. Разнообразие ресничных и рабдомерных фото рецепторов (в основном по Eakin
1968)
жrутика специализирована для выполнения
рецепторных функций, причем стиrма (пиr--
ментное пятно) обеспечивает дирекциона--
льную чувствительность (т. е. определение
направления на источник стимуляции).
В высшей степени удивительно наличие
у некоторых динофлаrеллат оцеллоuдов, т. е.
частей клетки, напоминающих простой rлаз
с роrовицей, хрусталиком и кристалличе..
ским сетчаткоподобным слоем, подстилае--
мым пиrментным бокалом, которые, как
предполаrается, способны формировать
изображения.
506
rлава 16
1 6.7.3. rлазки и rлаза
Уровень орrанизации фоторецепторных
opraHoB беспозвоночных весьма неодина--
ков (рис. 16.17). Простейшие из них, пазы--
ваемые rлазками дают информацию толь--
ко об интенсивности и направлении света.
При оценке эволюционноrо значения раз--
личных типов структур следует соблюдать
осторожность. поскольку все данные соот--
ветствуют ныне живущим видам. К тому же
последовательное усложнение может на--
блюдаться внутри мноrих крупных TaKCO
нов выделяют до 40 65 «филетических
линий сложности» (Salvini..Plawen, Mayr,
1977). Например, среди фоторецепторных
opraHoB книдарий встречаются и участки
пиrментированноrо эпителия, и rлаза с хру--
сталиком' (у У'атоуа): У моллюсков это
и простые открытые rлазные бокалы (у
блюдечек), и пятиклеточные rлазки (у неко--
торых rоложаберных), и rлаза rоловоноrих
с 20 млн. рецепторов, выдерживаюшие
сравнение с нашими собственными.
rлаза, формирующие хорошо сфокуси--
рованные изображения, называются камее--
1lыми. Во--первых.. они характеризуются oo
ширным слоем рецепторов, составляющих
сетчатку (ретину), которая может быть
прямой или инвертированной в зависимости
от направления рецепторных полюсов кле..
ток к источнику света или от Hero COOTBeT
ственно (рис. 16.18). Во--вторых, такие r лаза
снабжены усп1ройспzваJwи для фокусирования
света. хотя, если сетчатка толстая, а хруста--
А
лик прилеrает к ее поверхности, изображе..
иие будет нечетким.
Хорошо развитое зрение имеет решаю..
щее значение для хищников, например для
пауков"скакунов, которым необходимо оце..
НИВdТЬ скорость движения жертвы (у пауков
все rлаза состоят из пиrментноrо бокала
с хрусталиком). Камерные rлаза достиrают
наивысшеrо развития у осьминоrа и близ
ких К нему форм (рис. 5.25,Б). Совершен..
CfBO их выражается помимо прочеrо в появ--
лении век, подвижноrо зрачка, изменяюще
ro свою форму хрусталика, и внешних
мышц, KOTopbJe при участии сиrналов от
статоциста фиксируют зрение на интере..
сующем предмете независимо от движений
тела. rоловоноrие моллюски
рекордсмены по размеру rлаз. У rиrантско--
ro кальмара Аl"с/-zitеиthis их диаметр дости"
raeT 40 см, а сетчатка. по всей вероятности,
содержит до 1010 рецепторов (у человека
108). rлаз наутилуса представляет собой за..
rадку, поскольку лишен и хрусталика, и ро..
rОВИIIЫ. Он функционирует как камера
с крошечным входным отверстием; при
этом чувствительность и разрешение низки.
16.7.4. Линзы и зеркала
у наземных видов (например, пауков) за
прело мление .света отвечает rлавным обра..
зом выпуклая роrовица. У водных живот..
ных эту функцию выполняют линзы (хру"
сталики), даl0щие., вопреки ожиданиям,
лишь незначительную сферическую аберра..
сд
C 1 '
п
Рис. 16.17. Уровни орrаниза
ции rлазков с пиrментным бо
калом и rлаз книдарий (A
фоторецепторный эпителий
Laukartiara' В В о ugaiпvi//ia;
r Татоуа) и переднежабер
ных моллюсков (Б Раtе//а;
B Nerita; r Va/vata). ПК
пиrментная клетка, cд
сенсорный дендрит СН
сенсорный нейрон. Дендриты
несут реснички или микровор"
синки.
Системы контроля
507
Рис. 16 18. Сетчатка аннелид.
А. Прямая rлазной бока.м по
лихеты Vaпadis (по Hermans,
Eakin, 1974). Б. Инвертирован
ная rлазные бокалы в мозrу
rюлихеты Arтaпdia с един
ственным сенсорным нейро
ном и одноклеточным пиr
ментным бокалом (по Her
mans, Cloney, 1966). В. rлазной
бокал пиявки Hirudo с ceHCOp
ными нейронами, содержащи--
ми вакуолеподобную «фаосо--
МУ», нельзя отнести к KaKOMY
либо из этих типов Фаосомы
MorYT быть образованы и pec
ничными и микроворсинковы
ми фоторецепторами (по Hess,
1897)
цию. Маттиссен (Matthiessen) в 1886 r.
предположил, что у таких линз коэффи"
циент преломления изменяется от высоких
значений в центре до низких нс! периферии.
Это было подтверждено для rоловоноrих
и ряда брюхоноrих моллюсков., а также не..
которых веслоноrих и полихет..альциопид
(и рыб). Очевидно, «существует только
один путь создания хорошей линзы с испо..
льзованием биолоrическоrо материала»
(Land, 19 4).
Зеркала важны для фокусирования света
в большинстве rлаз и выполняют эту функ..
цию либо самостоятельно, либо в сочета..
нии с линзами. Отражая свет, они MorYT на..
правлять ero сквозь слой рецепторов
в обратном направлении, фактически уд..
ваивая эффективную освещенность (у неко"
торых пауков); иноrда на одном слое рецеп..
торов свет фокусируется линзой, а на вто"
ром перефокусируется зеркалом (у неко"
торых ракообразных).
16.7.5. Сложные rлаза
Сложные rлаза это орrаны, состоя..
щие из ряда практически идентичных эле..
ментов, ом матидиев, расположенных в [е..
ометрическом порядке. Их иноrда назы..
вают мозаичными 2лаЗQМИ, поскольку изо..
бражение формируется с участием всех ом..
матидиев, каждый из которых дает лишь
небольшую ero часть. По"видимому, такие
rлаза хорошо адаптированы к реrистрации
!
А
Б
в
движения., но их разрешающая способность
(за исключением мух) составляет в лучшем
случае 1 о, в то время как у позвоночных
и осьминоrа 1'. Если сетчатка пиrменти..
pOBaHHoro rлазноrо бокала воrнутая и дает
перевернутое изображение, сложные rлаза
и их сетчатки всеrда выпуклые, а получае..
мое изображение прямое (рис. 16.19). Такие
rлаза обнаружены у мноrих членистоноrих,
за исключением пауков. диплопод и т. д.
Они присутствуют также у двустворки Arca,
расположены на щупальцах полихет..
сабеллид, а у морских звезд образуют «зри..
тельные подуrпечки» на конпах лучей.
Острота зрения сложных rлаз зависит от
уrловоrо расхождения омматидиев. Напри..
мер, в rлазу дафнии 22 омматидия, направ"
пение каждоrо из которых отличается от на..
правления соседнеrо на 380, а у краба Lepto..
grapsиs их несколько тысяч с разницей ме..
жду соседними 1,5 . Разрешающая способ..
ность в последнем случае выше, а мозаич..
ное изображение из мелких кусочков четче,
чем из крупных, но это коррелирует с поте..
рей чувствительности. В rлазах некоторых
крабов наблюдаются мелкозернистые по..
лоски. образованные омматидиями с ма..
лым уrловым расхождением; они аналоrич..
ны желтому пятну в rлазах позвоночных.
у членистоноrих каждый омматидий
представляет собой удлиненный элемент,
обычно состоящий из роrовичной линзы,
кристаллическоrо конуса, ряда рецептор..
БЫХ (ретинулярных) клеток (у дафнии 4,
508
rлава 16
А
.,
" ....
.
' ).I;'щ'v . ..,
.../l)llJ j .... .':V
;j:!:E: ..iJ;;: i :Jt
.,;................4.'..:.::. .4.
,,#«;:.::. ::.:*IO:. JI .I:::::M .
· ;:.:: .,..:1I :..;.....:/::;:;
.. . l ';.....:..:..:..:.......:::i
'\. .... ",. . 45 «.фМ .- .
\ \ ....:...... .......:#::
.....:....../:::
f . 11 :.:....: .:.1i ........
. ... .....",. . It.*....
. '........: :....::
, J .'4 :.:.::.:«:.:.T.
, t ..........
. # fIJ::::,,:.:..:.:..:.: ....
.4. 1#,'#"....................
: -: '::*:.: ::=::;:::t\'
",:.4.._:. *. 8.. .... .",\\ 1W"
. .:.... -:.:- .......IJI:.. \ . .
:...:. ... *.... -..*-.\.
. .:.. .:.... . .\\\\ x;.
.. .:. .. ' t,.t.t..
". ... . \..\ ...
. .....*"'"
.:\., .
;'" .: :'f .:j..., \
х' ;,- \\\',.-. .......
1. .....;;*, Yo,,
. 4> ;". ;::; .*. 4' \ 't '
::' . ::" ':
· .'. \1
. I ..,.
...
,') 06- . .
{$1.::" ' , ,
1
1,
,"' .
10'
"
. 411,
" .
.
Б
f.
I
\,
'" .
Рис. 16.19. А. Разница в орrанизации rлазноrо
бокала (слева) и сложноrо rлаза (справа). Б.
rолова ктыря (двукрылые). Диаметр крупных
передних фасеток сложноrо rлаза около 60
мкм (с любезноrо разрешения профессора М.
Land, Университет Суссекса)
у крабов и Myx 8, у мечехвоста от 10 до
15) и примыкающих к ним пиrментных кле..
ток (рис. 16.20). Ретинулярные клетки pa
сположены как дольки в апельсине Упоря..
доченные наборы микроворсинок вдоль их
внутренних поверхностей образуIOТ рабдо
иеры, арабдомеры одноrо омматидия со..
СТd.вляют ero рабдом.
Омматидии некоторых членистоноrих
зависят от дифракции света. У эвфаузиевых
рачков и бабочек длинные, по форме напо..
минающие ПУЛI0, кристаллические конусы,
соrласно Экснеру (Exner, 1881), образуют
«линзовые цилиндры» с высоким значением
коэффициента преломления вдоль оси, по..
степенно снижающимся по направлению
к периферии. Однако сейчас известно. что
в друrих rлазах (например, у креветки
и омара) используется отражающая опти
ка; при этом пирамидальные конусы «посе..
ребрены» снаружи слоями кристаллов. По
верхность таких rлаз образована квадрат..
ными, а не шестиуrольными фасетками.
В аппозиционных 2лазах поля зрения со..
седних омматидиев обычно слеrка перекры"
ваются, хотя у ракообразноrо P/zroпiтa
каждый омматидий срединных rлаз воспри"
нимает свет от поля с уrлом 40, тоrда как
уrол расхождения омматидиев составляет
только 0,50. Возможно, наиболее замечате..
льные rлаза у раков..боrомолов. В то время
как у нас формируются два изображения
одноrо объекта, у этих животных шесть,
поскольку в каждом их rлазу находятся три
полосы омматидиев, ориентированных
Линза роrовицы
Кристаллический
конус
Пиrментные клетк
Ретинулярные
клетки
РаБДОМ
А
Дистальный пиrмент
Кристаллический
конус
Рис. 16.20. А. Продольный разрез омматидия
аппозиционноrо сложноrо rлаза (по Wiggles
worth, 1965). Б. Пути световых лучей в оптико
суперпозиционных rлазах при световой (ни
жняя полови на) и тем новой (верхняя полови
на) адаптации
Б
Системы контроля
в одном направлении! Два последних при..
мера свидетельствуют о том, что никакой
анализ возможностей сложных rлаз непра..
вомерен без учета «компьютерной» способ..
ности мозrа обрабатывать и интерпретиро"
вать информацию, получаемую от рецепто..
ров.
Типы сложных rлаз у членистоноrих
различаl{)ТСЯ по степени и природе функ..
циональной изоляции омматидиев. Аппози..
ционные rлаза (рис. 16.20,А), обнаружен..
ные, например, у крабов и пчел, адаптиро"
ваны к высокой освещенности. Пиrментные
клетки в них экранируют рабдом от любоrо
света, не проходящеrо через роrовичную
линзу данноrо омматидия.
Оптико--суперпозиционные 2лаза
(рис. 16.20,Ь), например, у креветок, ома..
ров и бабочек, имеют широкую «прозрач..
ную зону» между кристаллическим конусом
и рабдомом, причем последний короче пер..
Boro. При сильном освещении они функцио"
нируют как аппозиционные, обеспечивая
максимальную разрешающую способность.
Однако при слабом освещении рабдомы по..
лучают свет, проходящий через несколько
соседних омматидиев, блаrодаря измене..
нию формы пиrментных клеток или миrра..
ции в них пиrмента. если сами они остаются
неподвижными. Лучи от данноrо источни..
ка, падая на разные омматидии, изменяют
направление, тем самым увеличивая коли..
чество света.. возбуждающеrо один рабдом,
что максимизирует чувствительность rлаза
при тусклом освещени.и. Однако, вероятно,
в связи с рассеиванием света во время e] o
прохождения через мембраны разрешаю..
щая способность rлаза снижается.
В описанных выше типах rлаз невозмо--
жно разделить информацию, поступающу.ю
от отдельных ретинулярных клеток одноrо
омматидия. поскольку рабдом здесь «зам..
кнутый», т. е. различные рабдомеры как бы
вставлены друr в друrа микроворсинками.
Более Toro, каждая рецепторная клетка
образует синапсы на всех нейронах, объеди..
ненных в мелкую rруппу (<<патрон»), распо..
ложенную непосредственно под оммати--
дием. Совсем друrая картина наблюдается
в нейросуперпозиционных 2лазах мух.. пред..
ставляющих собой вершину развития зре..
ния у ченистоноrих (приложение 16.7).
509
16.8. Обработка сенсорной
информации
16.8.1. Смысловое восприятие
мира
Информация о внешней и внутренней
средах должна быть не только собрана и за..
кодирована в изменениях мембранноrо по
тенциала рецепторов, но и обработана, т. е.
видоизменена и преобразована так, чтобы
обеспечить адаптацию рассматриваемоrо
животноrо. Например, у саранчи простые
rлазки на вершине rоловы дают слабо сфо--
кусированное изображение и демон..
стрируют ,массовую конверrенцию
информация от приблизительно 1000 рецеп..
торов каждоrо rлазка передается сравните..
льно небольшим числом (25) нейронов вто"
poro порядка. Во время полета это обеспе..
чивает быструю обшую оценку положения
rоризонта, тоrда как сложные боковые rла..
за, как мы увидим ниже, MorYT различать
детали окружающеrо.
Совершенство ceHcopHoro и HepBHoro ап..
парата членистоноrих про является в распо..
'1навании ими признак а или сочетания при--
знаков среди множества простых альтерна
тивных вариантов. Пчела научается разли..
чать важные для нее участки, «как будто де..
лает их моментальные панорамные сним..
ки» (Collett, 1983). Некоторые южноамери..
канские пчелы, опыляющие орхидеи, ежед..
невно посещают в определенной последова..
тельности одни и те же цветы, положение
каждоrо из которых на постоянном марш..
руте протяженностью около 20 км они дол..
жны запомнить.
rлавным аспектом сенсорной обработки
является выделение признаков
распознавание или выбор в качестве объек"
та внимания особенно важной для животно..
ro информации. Так, у членистоноrих неко"
торые виды поведения стереотипны и ярко
выражены, например движение клешнями
при сексуальных демонстрациях у маняще..
ro краба. Они иrрают роль «сиrнальных
раздражителей», «высвобождающих» спе..
цифические поведенческие реакции у особей
cBoero вида, ничеrо не знача для друrих ви"
дов. По всей видимости, сенсорная и HepB
ная системы орrанизованы так, чтобы раз--
510
rлава 16
личать, проводить и усиливать ОДНИ сиrна..
лы, не реаrируя на друrие.
Интересно рассмотреть поведение CdM..
цов некоторых журчалок. Korna в поле зре..
ния появляется потенциальный партнер, са..
мец летит ему на перехват. Чтобы проло..
жить правильный курс, нужно знать рас..
стояние до объекта. ero скорость инаправ..
ление движения. Эти параметры, вероятно,
нельзя определить путем наблюдения, по..
скольку быстро передвиrающийся крупный
объект на большом расстоянии будет вы..
rлядеть почти так же, как медленно движу"
щийся мелкий объект, находящийся рядом
с самцом. По"видимому, муха «допускает»,
что объект примерно TaKoro же размера,
как она сама., и что он перемещается со ско"
ростью, близкой к ее собственной. Расстоя..
ние, с KOToporo он может быть замечен за..
,
висит от ero размера, следовательно, мо"
жно сделать вывод о расстоянии. Вместе
с направлением и скоростью движения объ..
екта в поле зрения оно позволяет (как в ком..
пьютере с соответствующей проrраммой)
проложить перехватывающий курс. Конеч
но, если объект представляет собой летя..
щий вдалеке реактивный авиалайнер, свадь..
ба не состоится, однако в запасе еще столь..
ко попыток!
r РИБовидные тела
16.8.2. Сенсорные системы
Информация, ПОСТУПdIOщая в цеНТРdЛЬ"
ную нервную систему по сенсорным путям.
проходя через центры синаптическоrо Бзаи..
модействия, видоизменяется (рис. 16.21).
На данном уровне передачи представлен
ряд модулей практически идентичных си..
наптических центров. Они часто связаны
локальными интернейронами и посред-
ством их способны влиять друr на друrа.
Среди TaKoI 1 o рода взаимодействий наибо..
лее изучено латеральное торможение, Бпе..
рвые описанное Хартлайном (Hartline)
в 1949 т. в ето классической работе по мече..
хвосту (Liтиlиs). Реакция омматидия на
данный уровень освещенности сильнее., если
он освещен один, поскольку одновременное
стимулирование нескольких соседних ом..
матидиев вызывает их взаимное торможе..
ние. Орrанизации сенсорных систем СБОЙ"
ственна также иерархия . информация по..
степенно передается от низших центров
к высшим.
Зрительный путь насекомых связан
с тремя участками мозrа ламинарной, ме..
дуллярной и лобулярной областями зритель..
ной доли, не считая протоцеребрума (при-
ложение 16.8). Каждому омматидию СООТ"
ветствует модуль, или патрон., нейронов
Местные
От антенны нейроны
Рис. 16.21. Ольфакторный
путь у таракана. Узелки (rломе
рулы) являются синаптически
ми центрами, состоящими из
триад, включаюших входные
окончания, местные нейроны
и релейные нейроны передаю-
щие обработанную информа
цию к высшим центрам (по
Boeckh et al., 1975)
Системы контроля
511
BToporo порядка в ламине и пучок клеток
TpeTbero порядка в медулле. Этот принцип
ретинопlопиu означает, что характер OCBe
щения rлаза неоднократно ОТРdжается
в морфолоrически упорядоченных формах
активности на четырех различных уровнях
орrdнизации зрительной системы.
16.8.3. Конверrенция
и диверrенция
Конвер2енция. свойственная rлазкам Ha
секомых, типична для мноrих сенсорных си
стем. С ней связана концепция сеНСОрНО20
поля набора рецепторов, передающих
ceHcopHYIO информацию одной клетке или
одному центру HepBHoro пути. Например,
у rиrантскоrо срединноrо волокна земляно
ro червя сенсорное поле переднее, т. е. rлав
ная информация поступает к нему от рецеп..
торов. расположенных в передней части Te
ла, тоrда как у пары боковых rиrантских BO
локон rлавным образом задние сенсор..
ные поля. Поля обоих типов волокон пере
крываются в средней части тела.
Для сенсорных путей характерна также
дивер2енция, поскольку информация от oд
поrо рецептора или их rруппы передается
в центральную нервную систему по множе
ственным, или параллельным., путя./н.. KOTO
рые MorYT быть использованы для выделе
пия и изоляции информации различноrо ти
па. Диверrенция также характерна для ce
псорных систем. каждая из которых хотя
обычно и соответствует только одной ce
псорной модальности, имеет выход на He
сколько моторных центров, влияя таким
образом на различные типы поведения.
16.8.4. Меченые линии
в отличие от эндокринных сиrналов, Ka
чественно отличающихся друr от друrа
в связи с химическими особенностями rop
монов, в нервной системе передача различ
пых элементов информации происходит
с помощью внешне идентичных последова..
тельностей электрических изменений и oд
поrо и Toro же химическоrо медиатора. Зна
чепие информации определяется участвую
щими в передаче нервными элементами,
каждый из которых как бы по своему «мече..
ный» .
Этот принцип можно наблюдать в слу..
чае реакции избеrания у тараканов и сверч"
ков. Стремительная атака хищника
жабы создает ток воздуха, который вос"
принимается сеНСОРНЫl\IИ волосками на
анальных церках HaceKoMoro (см., напри..
мер.. рис. 8.31..Ь).. удаление 'которых сни"
жает шансы на спасение. Волоски собраны
в несколько пучков, чувствительных к ве..
тру, дующему в разных направлениях. Каж..
дый пучок образует отдельный комплекс
соединений с rиrантскими интернейронами.
доходящими до торакса, а различные rи..
rdнтские интернейроны косвенно стимули
руют специфические мотонейроны. Напри..
мер.. интернейрон NQ 5 активируется только
струями воздуха от заднеrо квадранта этой
же стороны тела и возбуждает медленные
депрессорные мотонейроны (и мышцы)
этой стороны. что приводит К повороту от
источника ветра. Если церки в эксперименте
поменять местами, животное «rлупеет»
и реdrирует так, KdK будто поток воздуха
идет с друrой стороны.
16.8.5. Выделение признаков
Одноrо фотона достаточно для Toro..
чтобы вызвать заметный скачок мембран..
Horo потенциала фоторецептора. Внекото..
рых случаях единственная молекула способ
на возбудить хеморецептор. Поскольку
у одноrо орrанизма MorYT быть миллионы
рецепторов, понятно, что животное должно
отфильтровывать из поступающей сенсор..
ной информации значимые сиrналы. считая
остальные лишними.
Как показано выше, в зрительную долю
саранчи заходят все ретинотопные отрост..
ки. В конечном счете они образуют контакт
с нейроном, называемым 2И2антским лобу
лярным детектором движения (LGMD),
имеющим (рис. 16.22) множество вееро..
образно расходящихся дендритов, позво..
ляющих ему получать стимулы от элемен..
тов, соответствующих любой части поля
зрения. LGMD возбуждается только в том
случае, если объект в поле зрения, BO
первых, достаточно мал, так что саранча не
реаrирует, скажем, на движения раститель..
512
rлава 16
.
. s .:;:
: ,: ..
.-у. " -
./> ."",
, 1'.
А
.,. "f.'.
:.. <
а
f
.-.
r
,. ,
II
; - ::. :. :
т
Рис. 16.22. Нейрон LGMD в зрительной доле саранчи, заполненный раствором хлорида кобальта.
Д веерообразное расположение дендритов в зрительной доле; Т тело клетки; A aKCOH, иду
щий в мозr (слева). х 80; масштаб 100 мкм (с любезноrо разрешения доктора Claire Rind, УНИ8ер
ситет Ньюкасл апон Тайна)
ности. Информация о крупных объектах ча..
стично отфильтровывается блаrодаря лате..
ральному торможению (разд. 16.8.2), а ча..
стично за счет активации параллельноrо
пути, оказывающеrо на LGMD тормозное
воздействие. Во"вторых, объект должен дви--
zаться. Пресинаптические по отношению
к LGMD элементы являются фазически"
ми каждый из них, выпустив залп импу"
льсов, прекращает работу. Следовательно,
только движущийся объект, возбуждающий
целый ряд таких элементов, обеспечивает
последовательное поступление стимулов
к LGMD и ero активацию. Наконец, движе..
ние должно варьuровать неоднократная
стимуляция одних и тех же синаптических
входов LGMD вызывает нервную депрес--
сию и становится неэффективной.
16.8.6. Центробежный контроль
Информация передается по сенсорНЫI\-1
путям не в одном направлении; с любоrо
уровня нервные волокна MorYT направля"
ться назад, к более периферическим цен..
трам (т. е. центробежно), обеспечивая, на..
пример, контроль по принципу отрицатель..
ной обратной связи. Нейроны..детекторы
движения у саранчи находятся под влия"
нием как тормозных. так и стимулирующих
центробежных волокон, идущих от мозrа.
Коrда саранча шевелит rоловой, нейроны
LGMD тормозятся, поскольку иначе любой
мелкий стационарный объект двиrался бы
в поле зрения, активируя детекторную си..
стему. И напротив, коrда общее возбужде..
ние орrанизма центробежно повышается,
чувствительность этой системы к визуаль..
ной стимуляции может возрасти.
16.9. Нервные основы
поведения
16.9.1. Независимые эффекторы
Некоторые действия являются адаптив"
ной реакцией клеток на самостоятельно по..
лучаемые стимулы, т. е. клетки ведут себя
как незавuсuмые эффекторы. В эту KaTero..
рию входят некоторые железистые и мы..
шечные клетки хроматофоры иrлокожих,
непосредственно реаrирующие на свет; кни"
доциты (стрекательные клетки книдарий),
Системы контроля
разряжающиеся при контакте с чужерод
ным орrанизмом (хотя на них действует
и то, насколько животное rолодно). Анало
rичным образом координированное биение
ресничек обеспечивается механическими
взаимодействиями между смежными opra
неллами,
Ilротисты являются независимыми
эффекторами по определению, однако дe
монстрируют контрольные механизмы,
идентичные некоторым из действующих
в нервной системе. При встрече с препят
ствием при движении вперед в Paraтeciuт
возникаеl мембранная деполяризация. и
в клетку поступает Са 2 +, что изменяет на..
правление биения ресничек на противополо"
жное. Стимуляция сзади увеличивает про..
ницаемость мембраны для К +, повышает
потенциал покоя и ускоряет биение.
16.9.2. Элементы поведения
Большинство эффекторов не относятся
к независимым. Обычно поведение пред..
ставляет собой результат спонтанной ак..
тивности, rенерируемой в нервной системе,
или ответ на стимуляцию. Ero элементы
принято относить либо к рефлексаАt, либо
к устойчuвы./и ко./иплекса/vl дейспZ6UЙ. Ре..
флекс это стереотипный, относительно
простой двиrательный ответ на специфиче..
ский стимул. Ero сила и/или масштабы за..
висят от интенсивности стимуляции,
Концепция устойчивых комплексов дей..
ствий возникла в ходе развития Эl1'lОЛО?UU,
сравнительноrо изучения видоспецифично"
ro поведения, начатоrо Лоренцом и друrи..
ми. Такие комплексы представляют собой
стереотипные действия, часто весьма сло..
жные и не изменяющиеся в зависимости от
силы стимуляции. Они MorYT быть и спон"
танными, а если вызываются стимуляцией,
последняя действует только как пусковой
механизм. Один и тот же комплекс MorYT
вызывать совершенно различные стимулы;
например, рак принимает характерную позу
защиты в ответ на широкий диапазон сиrна..
лов уrрозы. Комплексы rенетически детер"
минированы и точное выполнение опреде..
ленной «проrраммы» обеспечивается opra..
низацией нервной системы. Обратные
связи, как таковые, их не контролируют.
513
Итак. две эти конuепuии различаются
rлавным образом ролью, которая отво"
дится стимулам. По отношению к рефлек"
сам она примерно такая же, как у человека,
иrраюшеrо на пианино, а в случае устойчи--
вых комплексов действий ближе всето будет
аналоrия с включением маrнитофонной за..
писи.
16.9.3. Рефлекторные действия
у мноrих книдарий в ответ на слабые
прикосновения можно наблюдать медлен--
ные локальные сокращения. распростране..
ние которых зависит от интенсивности, ко..
личества и частоты стимулов. Мы будем на..
зывать такой тип реакции «о6леzченным»
(употребляют также термины «декремент--
ный» и «инкрементный»), поскольку оди..
ночный стимул, даже если он не в состоянии
вызвать данный ответ, тем не менее облеr..
чает эту задачу следующему и т. д. (рис.
16.23). Классическое объяснение. пред..
ложенное Пантином (Pantin), основано на
явлении пресинаптическоrо облеrчения
в нервной сети, за счет которото второй
и следующие за ним импульсы, достиrая си..
напса, запускают постсинаптический ответ,
,
,
,
,
,
,
Рис. 16.23 Облеrченное сокращение полипов
у коралла Porites в ответ на пять последовате
льных идентичных электрических стимулов (по
Shelton, 1975)
514
rl1aBa 16
не возникающий при поступлении первоrо
импульса.
Примечательно, что облеrченные ре..
флексы, встречающиеся у rидроида Cordy..
lophora, опосредованы электрическими им..
пульсами в эпителии (разд. 16.4.3). Это сви"
детельствует о возможности имитации MHO
rих свойств химических синапсов щелевыми
контактами. Наконец, у некоторых корал..
лов (рис. 16.23) такие рефлексы обусловле..
ны потенциалами в нервной сети со скво"
зной проводимостью, т. е. распространяю..
щимися по всей колонии в ответ на каждый
стимул! Этот парадокс можно объяснить
постепенным снижением скорости проведе..
ния в случает серии импульсов. Любой сле..
дующий импульс будет неизбежно все боль..
ше и больше отставать от предыдущеrо..
временной интервал между поступлением
двух импульсов в данную точку
постепенно увеличиваться, а облеrчение в
ее нервно"мышечных синапсах умень"
шаться.
Рефлекс с участием рецептора растяже..
ния рака, как и коленный рефлекс человека,
является моносинаптическим, т. е. между
рецепторным и двиrательным нейронами
находится только один химический синапс.
Этот рефлекс отвечает за сохранение позы,
Командное волокно
Рис. 16.24. Нейронные цепи рака, связанные
с рецептором растяжения. Если внешние фак
торы препятствуют сокращению «рабочей»
и рецепторной мышц, вызванному активацией
командноrо волокна и мотонейрона, рецептор
активируется, Ч1"О ведет к дальнейшей стимуля--
ции мышцы. Рецептор участвует также в моно..
синаптическом рефлексе на растяжение (тор..
мозная иннервация рецепторной клетки цeH
тробежными волокнами не показана) (по Кеп..
nedy, 1975)
Мышца
Рис. 16.25. Нейронные цепи, участвующие
в реакции испуrа у пиявки. Передача от ceHCOp
ных Т клеток через соеиннерней
роны и ряд S"клеТQК к мотонейронам OCY
ществляется исключительно через электриче
ские синапсы. Только для нервно мышечных
синапсов характерна химическая передача. По
казана одна сторона двух смежных rанrлиев
поскольку, если соответствующая мышца
растяrивается, активация рецептора и затем
связанноrо с ним мотонейрона вызывает
противодействующее мышечное сокраще..
ние. Такая нервная цепь также контроли..
рует выполнение мышцами команд, посту..
пающих из центральной нервной системы
(рис. 16.24).
Во мноrих реакциях uзбеzанuя или uспу..
2а участвуют rиrантские аксоны (разд.
16.2.4). у пиявки их нет, однако по ее не..
рвному стволу проходит путь с быстрой
сквозной проводимостью, обеспечивающий
резкое сокращение Bcero тела. Он состоит
из ряда «S"клеток», по одной на cerMeHT,
продольно направленные аксоны которых
связаны с S"клетками соседних cerMeHTOB
электрическими синапсами. Эта система ил..
люстрирует важность электрических (т. е.
«быстрых») синапсов для реакции избеrа..
ния (рис. 16.25). Разные типы механорецеп..
торов стенки тела также образуют моноси..
наптические рефлекторные дуrи с мотоней..
ронами «cBoero» cerMeHTa.
Пожалуй, наиболее любопытный из всех
Системы контроля
515
Мозr
Пластинка
Сенсорное
волокно
Место стимуляции
Рис. 16.26. Рефлекторные дуrи внутри зооидов Sa/pa. Каждая дуrа состоит из ceHcopHoro пути,
идущеrо от эпидермальной пластинки в мозr, двиrательной иннервации эпидермиса и эпителиа
льноrо проведения к выходной пластинке (по Anderson, Вопе, 1980)
известных рефлексов наблюдается у arpera
ционной стадии некоторых сальп (оболоч
ники), которые образуют цепочки, вклю
чающие до 20 «зооидов». Каждый зооид
связан с соседним двумя эпидермальными
пластинками, проводящими в противопо
ложных направлениях (рис. 16.26). Клетки
на «выходе» каждой пластинки напоми
нают по ультраструктуре пресинаптические
окончания (но это эпителиальные клетки,
а не нейроны!). На «постсинаптической»
стороне, отделенной тонким слоем внекле
точноrо вещества, находятся 12 реснич
ных рецепторных нейронов. Их стимуляция
вызывает прохождение импульсов по aKCO
нам в мозr и возбуждение там мотонейро
нов, индуцирующих потенциал действия
в клетках эпидермиса, куда тянутся их OT
ростки. За счет эпителиальноrо проведения
возбуждение распространяется к «BЫXOД
ной» пластинке зооида и т. д.
16.9.4. Устойчивые комплексы
действий
Врожденная способность центральной
нервной системы rенерировать и координи
ровать упорядоченные комплексы двиrате
льных действий продемонстрирована
у мноrих rрупп животных. «Втяrивание
мантии и смыкание раковины у двустворча
Toro моллюска Муа,... копуляционные дви
жения ..., ритм полета, ходьбы ..., дыхатель
ные движения ..., биение сердца ..., плавате
льные сокращения у медузы показано,
что все это более или менее жестко (sic) KOH
тролируется центром» (Bullock, 1977).
Заднежаберный моллюск Tritoпia реаrи
рует на прикосновение хищной морской
звезды 30 ceKYHДHЫM «припадком» плава
ния. Это требует чередующихся сокраще
ний дорсальных и вентральных мышц с уча
стием либо сиrналов, rенерируемых центра--
льной нервной системой., либо рефлектор
ной цепи начальный стимул вызывает со..
кращение дорсальных мышц, это по при--
нципу сенсорной обратной связи через ре..
флекторную дуrу стимулирует вентральные
мышцы и т. д. Исследования показали, что
типичная картина наблюдаемой активности
мотонейронов может быть воспроизведена
в мозrу in vitro при стимуляции перерезан
ных сенсорных нервов. Это ясно показы
вает, что в данном случае для rенерирова
ния поведенческоrо ответа сенсорной
обратной связи, обеспечиваемой мышцами..
недостаточно.
16.9.5. Вариации на две темы
Мноrие элементы поведения заключают
в себе признаки как рефлексов, так и устой..
чивых комплексов действий (приложение
16.9). у саранчи единственная клетка рецеп--
тора растяжения, связанная с местами co
членения крыльев, активируется в конце
каждоrо взмаха. Здесь по крайней мере MO
жно предположить систему, функциони
рующую посредством рефлекторных дуr,
однако это не так, поскольку в изолирован
ных торакальных rанrлиях и без сенсорной
стимуляции наблюдается активность, xa
рактерная для полета. В то же время импу
льсы, rенерируемые рецепторами за каж
дый цикл, оказывают тоническое воздей..
ствие на скорость и силу взмаха крыла на
протяжении нескольких циклов.
Нейроны LGMD саранчи (разд. 16.8.5)
образуют синаптический выход на пару ней
5'16
rлава 16
Рис. 16.27. Нейронные цепи, контролирующие
прыжок саранчи, DСМ О нейрон детектор
движения; С С нейрон; М М нейрон; Е,
F мотонейроны соответственно разrибателя
и сrибателя; R 1 , R2 ceHcopHbIe нейроны, обе
спечивающие обратную связь при OДHOBpe
менном сокращении. Жирные линии
нервные пути, тормозящие сrибатели (по Pear
sоп, 1983)
ронов, называемых «контралатеральными
детекторами движения» (DCMD), отростки
которых спускаются в третий торакальный
rанrлий, тде каждый из них иннервирует два
нейрона «С» и «М» (рис. 16.27). Синапсы
каждоrо С"нейрона на мотонейронах двух
антаrонистических мышц разrибателя
и сrибателя rолени находятся в бедрах
третьей пары нот.
Сначала С"клетка вызывает как «взведе..
ние» ноrи с подrибанием rолени поп тело
насекомото, так и совместное сокращение
двух названных мышц"антаrонистов,
искривляющее эластичную кутикулу бедра.
Затем по принципу положuпzельной обрат..
ной связи кутикулярные рецепторы напря..
жения действуют на мотонейроны, усили--
вая сокращение мышцы..разrибателя. Нако"
нец, обратная связь, образуемая друrой
rруппой рецепторов, в ответ на одновре..
менное сокр щение стимулирует М"клетки.
Если это совпадает с продолжающимся по..
ступлением сиrналов от DCMD и т. д., М..
клетки активируются, тормозя мотонейро..
нь] сrибателя и тем самым индуцируют
прыжок с высвобождением запасенной
в искривленной кутикуле энерrии. Ясно, что
сенсорная обратная связь. являясь интеrра..
льной частью системы, контролирующей
прыжок, rарантирует неэффективность
внешней инициации последнеrо, пока ето
механизм полностью не подrотовлен (см.
разд. 10.6.4),
В заключение следует отметить «непо..
чтительность» природы к применяемым на..
ми катеrориям. Нервная система, безуслов..
но, обладает врожденной способностью те..
нерировать интеrрированные двиrательные
проrраммы. Однако некоторые устойчивые
комплексы действий оказываются «устой..
чивее» друrих. Например, одни быстрые ре..
акции сокращения тела у аннелид, опосре..
дованные rиrантскими волокнами, являю..
тся rрадуальными, в то же время как друrие
осуrцествляются по принципу «все или Биче..
ro». Исключение при их анализе эффектов
обратной связи дурная традиция. Они
так же заложены в rенетической проrрамме,
как и орrанизация центральной нервной си..
стемы, являясь одним из rлавных средств,
с помощью которых устойчивые формы по..
ведения достиrают поставленных природой
целей,
16.10. Орrанизация
двиrательноrо «выхода»
16.1 0.1. Аспекты орrанизации
Системы нейронов, контролирующие
двиrательный эффект (например. сенсор..
ные).. обычно орrанизованы иерархически.
Это хорошо заметно в цепях, управляющих
брюшными конечностями (плеоподами,
или плавательными ножками) рака (рис.
16.28). Различают нейроны трех уровней:
командные, uнпzернейроны 2енераторы
nрО2рамм и ./'vlотонейроны.
16.1 0.2. Командные нейроны
у рака к каждому из сетментарных rап..
rлиев подходят как стимулирующие, так
и тормозные нейроны. Они впервые были
открыты у ракообразных Вирсмой (Wiers..
та), который обнаружил, что активация
особых клеток запускает координирован..
ную поведенческую проrрамму. Команд..
ный нейрон отвечает за прuнятuе решения.
В то время как сенсорные пути диверrи..
руют, обеспечивая стимуляцию множества
двиrательных центров (разд. 16.8.3), ко..
мандные нейроны внутри двиrательных си..
стем являются местом конверrенции и иите..
rрации входных сиrналов от различных
сенсорных opraHoB. Их активация или от..
сутствие таковой определяют, начнется или
Системы контроля
517
Командный нейрон
reHepaTop nporpaMMbI
Межсеrментный
релейный
G) нейрон
Рис. 16.28. Основные компоненты нейронных цепей, КОНТРОЛИРУЮЩИХ движение плавательных
ножек рака (показаны клетки в ДВУХ смежных cerMeHTapHbIx rанrлиях). Обычно наблюдается бо
лее одноrо нейрона каЖдоrо типа (по Stеiп, 1971)
нет двиrательное действие. Следовательно,
для ero rенерирования работа таких клеток
необходима и достаточна.
rиrантские волокна, управляюшие уда..
рами брюшка у рака (приложение 16.9), счи..
тались моделью командных структур. Од..
HdKO более детальное исследование покаЗd"
ло, что ответ на их прямую эксперимента..
льную стимуляцию не совсем такой, KdK
в природе: естественные удары обычно бо..
лее вертикальные и продолжительные. Та..
ким образом, rиrантские волокна опреде..
ляют общий характер действия, но для ero
точноrо выполнения необходимы паралле..
льные неrиrантские нервные пути.
Командные клетки обычно не опреде..
ляют «деталей» двиrательной активности:
не устанавливают хронолоrии или порядка
прохождения импульсов в контролируемых
ими интер.. и мотонейронах. Исключение
составляют rиrантские волокна, располо..
женные по краю колокола медуз, а также
известные у аннелид и друrих удлиненных
животных и отвечающие за практически од..
новременные сокращения отдаленных друr
от друrа частей тела, характерные для реак"
ции избеrания.
Во мноrих двиrательных системах ни
одна клетка или rруппа клеток, по..
видимому, не соответствует командному
элементу (см., например, рис. 16.27). Более
Toro, хотя маутнеровские клетки у кости..
стых рыб и водных амфибий являются ко..
мандными нейронами, вся эта концепция
плохо приложима к позвоночным.
16.1 0.3. rенерирование
nporpaMMbI
rенерирование проrраммы двиrатель..
Horo ответа часто осуществляется не ко..
мандными и не двиrательными нейронами,
а расположенными между ними интерней..
ронами. Одна из наиболее обычных двиrа..
тельных проrрамм связана с двумя МЫШIIа
ми, работающими как антаrонисты. Нерв..
ная система должна rарантировать едино
временную стимуляцию только одной из
них Одним из примеров может служить
замкнутая петля из интернейронов, KOHTpO
лирующая плавание пиявки (рис. 16.11).
В каждом cerMeHTe рака одному неспай..
ковому интернейрону свойственны реrуляр..
ные спонтанные колебания мембранноrо
потенциала, происходящие в фазе с биением
плавательных ножек. Деполяризация и выI
ход медиатора стимулируют мотонеЙРОНbJ
МЫШII, отвечающие за rребущий удар.
и тормозят мотонейроны, обеспечивающие
возвратное движение конечности. Хотя от
командных клеток зависит, будет активен
этот интернейрон или нет, за rенерирование
nрО2раммы действий отвечает именно он.
Это можно продемонстрировать, ускоряя
или замедляя колебания ero потенциала
экспериментальным путем. Ритм оказыва..
518
rлава 16
Мышца депрессор
сокращается
Мышца леватор
расслаБляется
'
Мышца ле8атор
сокращается
Рис. 16.29. Нейронная цепь, контролирующая
вентиляцию жабр у краба. Два неспайковых ин
тернейрона (IN 11 IN 2 ), поочередно возбу
ждаясь, активируют мотонейроны мышцы
депрессора (DMN) и мышцы леватора (LMN)
соответственно. Прерывистые линии
неактивные волокна; светлые утолщения
возбуждающие синапсы, темные тормозные
синапсы
ется nеренастроенным он не COOTBeT
ствует BpeMeHHbIM характеристикам пред..
шествующеrо. Друrие интернейроны уча..
ствуют в межсеrментной координации, обе..
спечивающей метахрональный ритм биения
плавательных ножек.
Вторакальном rанrлии краба, контро..
лирующем вентиляцию жабр, обнаружена
сходная система: один неспайковый интер..
нейрон функционирует по описанной выше
схеме, а колебания потенциала BToporo не
совпадают с первым по фазе (рис. 16.29).
Работа обоих обеспечивает нормальные
респираторные движения.
Блаrодаря наличию нескольких интер..
нейронов, способных rенерировать различ..
ные проrраммы, индивидуальные мотоней..
роны MorYT быть использованы различным
образом, что делает ненужным дублирова
ние волокон, осуществляющих нервно..
мышечную иннервацию. Так, Берроус (Bur
rows) обнаружил. что у саранчи на един
ственный мотонейрон, идущий к мышце но..
rи, влияют по крайней мере двенадцать ин..
тернейронов.
16.10.4. Мотонейроны
Хотя за rенерирование двиrательной
проrраммы часто ответственны интерней
роны, мноrие мотонейроны в результате
своих взаимодействий также включены
в этот проиесс. У нематоды Ascaris вдоль
Bcero тела проходят пять крупных интер..
нейронов, обеспечивающие тоническую
стимуляцию плавания. Мотонейроны, воз..
буждающие дорсальную мышцу в каждом
Рис. 16.30. Нейронная цепь контролирующая плавание у Ascaris, обеспечивает поочередные co
кращения дорсальной (слева) и вентральной (справа) мышц. IN интернейроны; ОЕ, VE, DI, VI
дорсальные и вентральные возбуждающие и тормозные мотонейроны соответственно. Светлые
утолщения возбуждающие синапсы; темные утолщения тормозные синапсы (по Stretton et al.,
1 984)
Системы контроля
«cerMeHTe» тела, одновременно стимули
руют тормозные нейроны вентральной
мышцы и через них тормозят вентральные
возбуждающие нейроны (рис. 16.30). Акти
вация последних оказывает аналоrичное
влияние на дорсальные нейроны. Чередую
щаяся активность дорсальных и вентраль
ных мотонейронов, зависящая по частоте
и интенсивности от степени их стимуляции
интернейронами, вызывает попеременные
сокращения соответствующих мышц. Дpy
rие клетки образуют связи между соседни
ми участками тела и координируют их aK
тивность.
То, что rенерирование проrраммы пви
rательными элементами потребовало бы
дублирования последних, по видимому. BЫ
текает из приведенных наблюдений, поско
льку плавание вперед и назад у аскариды
контролируют различные типы мотонейро
нов. Возможно, в данном случае это HeBa
жно, так как набор совершаемых червем
движений оrраничен. Подобный вывод
справедлив в отношении мноrих двиrатель
ных систем беспозвоночных.
Нервные элементы, участвующие в peaK
циях избеrания, в большинстве случаев не
подтверждают правила. соrласно которому
один и тот же набор мотонейронов исполь
зуется в зависимости от входных сиrналов
в различных целях. У кальмара rиrантские
волокна участвуют в реактивном отплыва
нии при испуrе, тоrда как более мелкие
управляют сокращениями мышц, обеспечи
вающими дыхательные движения. У Meдy
зы Ag/aпtha реакuия избеrания выражается
в одном трех мощных сокращениях коло
кола, контролируемых восемью rиrантски
ми двиrательными волокнами под ero ни
жней поверхностью и боковыми мотоней
ронами меньшеrо диаметра. Обычные пла
вательные сокращения управляются только
последними элементами. Аналоrичным
образом у рака боковые rиrантские волокна
возбуждают не только (через пару цeHTpa
льных rиrантских волокон) от пяти до девя
ти быстрых «сrибающих» мотонейронов,
участвующих в пластичных реакциях избе
rания, но и пару rиrантских сетментарных
мотонейронов, ответственных исключите
льно за удары брюшка (приложение 16.9).
519
16.1 0.5. Нервно мышечная
иннервация
Контроль мышечной активности MOTO
нейронами связан с выделением химических
нейромедиаторов в нервно мышечных си
напсах между нервными окончаниями и MЫ
шечными волокнами (разд. 16.3.3 и прило
жение 16.10). Очень важно также наличие
или отсутствие распространяющихся потен
циалов действия внутри отдельных мышеч
ных волокон и/или их передача через щеле
вые контакты между ними. Способь] инне..
рвации мышц и желез чрезвычайно разно
образны. Так.. сердце асцидий (оболочники)
и некоторых насекомых ее лишено, и ero co
кращения миоrенные, возможно, с эндо
кринной модуляцией. И напротив, биение
сердпа у ракообразных невротенное и KOH
тролируется только возбуждающими Moтo
нейронами. Соматическая мышца аскариды
находится под двойным (возбуждающим
и тормозным) контролем.
у саранчи разrибатель rолени контроли
руется четырьмя нервНЫЛ1U волокнами. Воз
буждающие и тормозные волокна Bыдe
ляют медиаторы rлутамат и raMMa
аминомасляную кислоту соответственно.
Кроме Toro, окончания нейрона DUM (дop
сальный непарно срединный) секретируют
нейромодулятор октопамин (табл. 16.1).
Ero собственный эффект.. по видимому..
слаб, а основная роль заключается в прида
нии мышце способности отвечать на двиrа
тельное возбуждение более интенсивным
сокращением с быстрым расслаблением,
что важно для локомоции. В отсутствие OK
топамина сокращение становится более
длительным и способствует сохранению
позы.
у двустворчатых моллюсков мышцы
аддукторы очень выносливые они MorYT
держать створки раковины сомкнутыми
в течение нескольких часов или даже дней,
обеспечивая защиту от хищников. Сокраще
ние происходит в результате активации воз
буждающих мотонейронов с выделением
ацетилхолина. Коrда напряжение уже раз
вилось., мышца входит в состояние «замка»..
характеризующееся низким потреблением
кислорода и поддерживаемое независимо
от дальнейшей стимуляции Расслабление
520
rлава 16
Таблица .16:1. Химические медиаторы беспозвоночных
Нейромедиаторы
о
11
НЗС С О (СН2)2 N+ (СНз )3
О
11
rлутамат (аминокислота) H3N CH CH2 CH2 C O
I
с==о
I
о
HO ) fН СНl- NНЗ-
ОН
Ацетилхолин
Октопамин (амин)
rOpMOHbI
rоловной активатор rидры (пептид)
pGlu Pro Pro Gly Gly Ser l ys Val Ile Leu Phe
'.метиладенин NH
НЗС N 6 5 1
l з4 9/
N N
Н
Экдизон (стероид)
НО
НО
Ювенильный ropМOH I НзС
НзС
ОН
ОН
О
СООСН з
10
2
6
н н н н н н н н н н н н н н НН
H t t = = t t b d t OH
I ! I I I I I I I I I I
ННН ННННННННН
Фера мон
Бом6икол
индуцируется активацией тормозных воло
кон, медиатором которых является, по..
видимому, 5..rидрокситриптамин.
у насекомых с «прямыми}) летательны..
ми мышцами импульсы в нервных волокнах
«отбивают ритм» сокращений. В противо..
положность этому специализированные
«непрямые» летательные мышцы, напри..
мер у мух, сокращаются с частотой более
1000 раз в секунду, тоrда как число потен..
uиалов действия. прохопящих по нервным
и мышечным волокнам, составляет Bcero
5 10. Нервы обеспечивают лишь пl01iuче..
СКИЙ эффекп1, переводя мышцу в «активное
состояние». Сокрашение и расслабление
являются внутренними «рефлексами» мы..
шечных волокон которые индуцируются
воздействиями (растяжением и т. д.), сопро..
вождающими биение крыльев (см. разд.
10.6.3).
Системы контроля
521
16.11. Химическая
кация
коммуни
16.11 .1. Спектр химической
коммуникации
Нейромедиаторы это вещества, ceKpe
тируемые нейронами и обычно выделяемые
в непосредственной близости от друrих кле
ток, на чью активность они влияют
(табл. 16.1). В результате нервное воздей
ствие высокоспецифично в пространстве
и времени. В противоположность этому
20р./ИОНЫ ЭТО химические посредники. BЫ
рабатываемые клетками, которые обычно
объединены в :железы, и влияющие на дpy
rие, отдаленные клетки, переносясь к ним
kровОП10КО w или друrими внутренними жид..
костями. Железы, секретирующие rормоны,
называются эндокринными, поскольку BЫ
деляют свои продукты внутрь тела. Секре..
ты ЭКЗ0КРИННЫХ желез, например слизь или
ферменты. выводятся наружу, обычно через
протоки.
Если по функции обычный нейрон мо"
жно сравнить с телефоном, эндокринный
сиrнал напоминает команду, поданную че..
рез меrафон. rOpMOH часто действует сразу
по всему орrанизму, хотя это не означает
отсутствия у Hero высокоспецифичной aK
тивности. Эффект rормональноrо сиrнала
зависит от рецепторов на клетках мишенях.
Например, разные типы хроматофоров, да..
же с одним и тем же пиrментом, MorYT быть
чувствительными к разным ropMoHaM.
Друrие секретируемые вещества, назы..
ваемые феромона./ии, выделяясь в окружаю..
щую среду, имеют биолоrическое значение
для друrих особей. У протистов, например,
они обеспечивают аrреrаuию амебоидных
клеток слизевиков. Крайне uажна роль фе..
ромонов у общественных насекомы'\., кото..
рых можно сравнить с «подвижными бата
реями экзокринных желез» (Е. О. Wilson,
1975) .
16.11 .2. Критерии эндокринной
активности
Существует ряд критериев, позволяю..
щих судить об эндокринной активности ор..
rаиа. В частности, ero ХИРУР2ическое удале
Рис. 16.31. По две личинки пятоrо и четвертоrо
возрастов Rhodпius в состоянии парабиоза
Они соединены восковыми перемычками
с имаrо и «подключены» к ero кровообраще
нию. Личинки секретируют линочный и ювени
льный rapMoHbI, вызывая дополнительные ли
ньки имаrо, приводящие к ero ювенилизации
(по Wigglesworth, 1940)
ние должно прекращать действие предполо..
жительно вырабатываемоrо им ropMoHa,
а реи./иплантация данноrо opraHa или инъек..
ция экстрактов из Hero должны это действие
восстанавливать. Можно использовать пa
рабиоз или трансплантаuионный метод
(рис. 16.31). Альтернативный способ вклю
чает сохранение клеток или opraHoB in vitro.
Эффект внесения в культуральную среду
желез или их экстрактов можно использо..
вать для оценки предполаrаемой активно
сти.
16.11.3. Нейросекреция
в 1928 1. Эрнст Шаррер (Е. Scharrer) по..
стулировал, что некоторые клетки, назван..
ные нейросекреПl0рНЫ./ИИ, проявляют свой..
ства как нейронов. так и клеток эндокрин"
ных желез. Эта идея получила развитие
в исследованиях Берты Шаррер и друrих aB
торов на беспозвоночных. Было показано,
что такие клетки у них широко распростра..
нены. Особенно убедительные подтвержде
ния теории дали работы с насекомыми.
Тела нейросекреторных клеток обычно
собраны в центральной нервной системе
в rанrлионарные ядра (рис. 16.32). Их аксо"
ны образуют вздутые окончания, тесно
связанные с циркуляторной системой. Эти
окончания MorYT быть объединены в офор..
522
rлаВд 16
А
... .......
, .
.? .,
«.м: ...-:
..
.. ,.
.' "'.." '. '.
,
."" '" .
""
, . .4'
"
_ ,'\ !f> .'
..., ,.' .
. .,
"..,.. '
..."
,.,'
,. '*
. ", .'.
. '.r .
. ...(.. . .... : '
., · . 'J.': ,i :;
It: . t .'
" . '.
'<t.' 7>... :
. .. ff" ..
...." ;..\' ,." ..,.."
. .. ..'
'" < ""<'
'.' ,,., ,.-,.i
*'
'.' . <-=._-
.....'- .';'
"
t .;. 4:
. *" ,"," #- f
;":'-.(:t" ,-
1ft
%.
,::--'\: ,
. .. "
. A".. <..
, ...., :'\.
'.. '..
,
..
1'<
J<1fII
I
'.'1.,:;'. .:,'
." ....
I!W" .....
, ."'
,
..
# '.
. ......
1"
*
t:
JI! ''':..
,..."",. ,..
..
.'
..
*'"
Б
. .
); ;'
-)ф'
, 4'
_',.. 'vij
..,',? ."
";\, ... i'
,,,,"., -..,...
" ......
. .» .. ., .,"
'!"k.- : ".-, ". . "у 'O':- - -".
,,::';;,". ,.,...
. ,:,...'" " ' ... "''I-f.,. "
; <. ;''''Ч' " 'Ш Э t". '"
:: ;.."'::',Y" ' . ';"7
:,;: ш'. ",.
. i "..
". '; 'f :.
<f '
. . ." ,.- .
. ,: ': j -;::".
4'
:
., :
: '(",-'.
..
-
. "
. '
.
",t
. .,
.
. '..
")
"
.,
..
."'" ,,,
....
" .' ,"
- .
. ..... . '" .
, ,.
--.'. ....
..,? . ;
.....
'"
" .
'"
'.' .: "
......
;
. ':
'"
" H
..
. :. ..---.", >
fJ:- "'I''!'З
<;
.. ,
,..",-
: '."..
"
. ..
'. "' .:'''' ,".;:: цT/ .
,
.
",
--
..
AI! ..
.. :# ..,
, ' ..
f 1$,'
Рис. 16.32. А. Нейросекреторные клетки (на врезке) в церебральном rанrлии полихеты Harтot
hoe. Цитоплазма заполнена секреторным материалом, окрашенным «по rомори» в темный цвет
паральдеrид фуксином; ядра неокрашены; масштаб 100 мкм (Gоldiпg, Whittle, 1977). Б. Элек
тронная микрофотоrрафия нейросекреторной клетки земляноrо червя Luтbricus. Секрет состоит
из большоrо числа мелких rранул диаметром около 200 нм. Я ядро С ядрышком; ШЭР
шероховатый эндоплазматический ретикулум; х 6500, масштаб 1 мкм
мленное тело (неЙРО2еJиальный Ор2ан) или
рассеяны по неЙРО2еJиальной зоне на поверх
ности rанrлия или нерва (рис. 16.33).
Нейросекрет вырабатывается в телах кле
ток, переносится по аксонам течением аксо"
плазмы и накапливается в окончаниях. Под
электронным микроскопом видно" что он
состоит из мельчайших секреторных rранул
диаметром обычно 100 200 нм, которые
отпочковываются аппаратом rольджи.
Активность нейросекреторных клеток
возникает либо спонтанно, либо в результа
те синаптическоrо возбуждения. Она вызы
вает прохождение потенциалов действия по
аксонам, поступление ионов Са 2 + в оконча
ния и выделение ropMoHa в кровяное русло
за счет экзоцuтоза (приложение 16.11).
16.11.4. Размытая rраница
Недавние открытия сrладили различие
между нейросекреторными клетками
Системы контроля
523
Рис. 16.33. Церебральный raH
rлий моллюска Lyтпaea. Пока
заны нервные тракты от двух
типов нейросекреторных кле
ток к нейроrемальным зонам
поверхности комиссуры и rуб
ным нервам. Дорсальные Te
ла эндокринные opraHbI He
нервной природы (по Joosse
Geraerts, 1983)
и «обычными» нейронами. Нейроны, напо..
минающие нейросекреторные клетки, были
описаны у книдарий и плоских червей, ли..
шенных сосудистых систем. У rидры обна..
ружен 20ловной активатор (табл. 16.1),
очищенный в 1981 r. Шаллер (Schaller) с co
авторами. Это вещество в концентрации
1 «rидра"эквивалент»/мл, т. е. менее
10 1O моль/л, вызывает in vitro реrенерацию
щупалец фраrментами тела. Интересно, что
с помощью антител к этому пептиду в моз"
ry млеко питающих была обнаружена систе..
ма содержащих ero нейронов.
Более Toro, некоторые описанные на се..
rодняшний день нейронь] функционируют
как классические нейросекреторные клетки
(т. е. секретируют ropMoHbI), но лишены ха..
рактерной для них цитОЛО2UU. Друrие ней..
роны, напротив, имея цuтОЛО2UЮ типич"
ных нейросекреторных клеток, не выпол"
няют их функций: вместо выделения секрета
в циркуляторную систему они непосред..
ственно контактируют с клетками..
мишенями.
16.11 .5. Нейропептиды
Принципиальное сходство нейросекре..
торных и обычных нейронов доказывается
секрецией мноrими, а возможно, и всеми ней..
ронами (включая и те, что образуют обыч..
ные медиаторы) особых пептидов, хотя пер..
воначально это считали специфическим
свойством нейросекреторных клеток Более
r УБНОЙ нерв
Toro, типичные нейросекреторные клетки
MorYT оказывать координированное воздей..
ствие на физиолоrию и поведение животно..
ro. В «сумковых клетках» моллюска Aplysia
нейросекрет синтезируется в виде высоко..
молекулярноrо белка, или nрО20рмона. В се..
креторных rранулах он расщепляется на бо..
лее короткие фраrменты под действием
ферментов, так что выделяется не един..
ственный ropMoH, а «коктейль» пептидов.
Один из них «rормон откладки яиц»
через кровоток достиrает rонад и индуци"
рует икрометание. Друrой стимулирует не..
рвные окончания и нейроrемальном ком..
плексе, обеспечивая их максимальную ак"
тивность. Ряд ropMoHoB, включая ropMoH
откладки яиц, диффундирует по всему абдо..
минальному rанrлию и стимулирует осо..
бые нейроны, запускающие высокоспецифи..
ческий тип поведения, сопровождающий
икрометание.
16.11 .6. Нейроэндокринные
системы
Нервная, эндокринная и нейросекретор"
ная системы функционально не изолирова..
ны друr от друrа., а образуют интеrрирован"
ные (у эволюционно продвинутых беспозво..
ночных сложные) нейроэндокринные си..
стемы. В рефлекс, включающий сенсорное
восприятие., обработку и интеrрацию интер..
нейронами и контроль эффекторов, входит
эндокринный этап, опосредующий послед..
524
rлава 16
нюю ero часть. Таким образом, эндокрин
ная система обычно контролируется нерв--
ной, и наоборот, ее секреты часто оказы--
вают rлубокое влияние на нервную систему
(приложение 16.12).
16.11.7. Феромоны
Первым идентифицированным феромо--
ном был оомбuкол (см. табл. 16.1)
вещество. секретируемое самками TYToBoro
шелкопряда (ВотЬух тori). В 1959 [. куль--
минацией ero исследований стала работа
Бутенандта (Butenandt) с сотрудниками,
связанная с экстраrированием продуктов
полумиллиона абдоминальных желез. По..
трясает то, что единственная молекула бом..
бикола, по--видимому, оказывает заметное
воздействие на ольфакторный репептор ан..
тенны самца, а 200 молекул, вызывающих
200 импульсов, достаточно для запуска по--
веденческоrо ответа. Самец начинает харак--
терный зиrзаrообразный полет против ве--
тра.
Более позднее исследование продемон--
стрировало важность САlесей компонентов.
Сочетание химических веществ. возможно,
необходимо для возбуждения начальноrо
эффекта, например свойственноrо полово..
му аттрактанту. Не исключено также, что
один из компонентов оказывает первичное
длинно дистанционное воздействие (напри--
мер, аттрактивное), а друrой вторичное,
на близком расстоянии (например, иниции--
рует копуляцию). Даже бомбикол секрети..
руется в сочетании с друrим, сходным со..
единением БОАlБuкалем, тормозяще
влияющим на полет. Вблизи друr от друrа
самцы мноrих бабочек стимулируют друr
у друrа секрецию половоrо феромона. вы..
зывающеrо реакцию у самки. Изучение по..
добных веществ, очевидно, имеет orpoMHoe
потенциальное значение для борьбы снасе..
комыми--врепителями.
Феромоны очень важны для реrулирова..
ния кастовой структуры общественных на..
секомых Наиболее известный при мер
секрепия матОЧНО20 молочка мандибуляр--
ными железами матки медоносной пчелы.
Два ero компонента 9"оксодекановая и 9..
rидроксидекановая кислоты действуют
как синерrисты, тормозя развитие яичников
у рабочих особей и постройку ими специа..
льных ячеек, в которых развиваются матки.
Коrда матку удаляют из улья, это торможе..
ние снимается, и вместо нее развиваются
новые активные в половом отношении сам..
ки. Это же «молочко» важно как половой
аттрактант, а также во время роения и осно"
вания новой семьи.
Вещества, секретируемые особями одно..
ro вида, MorYT заметно влиять на друrой и
в этом случае называются алломона..НU. На
них основаны взаимоотношения мноrих
симбиотических орrанизмов. У некоторых
насекомых вещество, действующее как фе
ромон на особей cBoero вида. влияет как ал
ломон на представителей друrоrо, напри..
мер привлекая паразитов. У аннелиды Ne..
reis снижение концентрации ropMoHa МОЗfа
вызывает нерест (после KOToporo животное
поrибает), а паразиты..споровики (проти"
сты), «настраиваясь» на этот сиrнал, синх..
ронизируют свое собственное размножение
с размножением хозяина.
у бабочки дубовоrо шелкопряда Aпtherea
poZypheтus, личинки которой питаются ли..
стьями KpaCHoro дуба, к размножению при--
водит интересная последовательность хи..
мических сиrналов. Алломон, образуемый
листьями дуба, стимулирует у самки выде..
ление нейросекреторноrо 20рмона, вызы"
вающеrо выдвиrание rениталий и секрецию
феромона. Он привлекает самцов, и размно"
жение происходит в месте, боrатом пищей
для рdЗВИТИЯ следующеrо поколения.
16.12. Роль эндокринной
системы
у беспозвоночных ropMoHbI участвуют
в реrуляции широкоrо спектра процессов.
Ниже будут описаны лишь некоторые из
этих механизмов.
16.12.1. Эндокринный пусковой
механизм
Некоторые [ормоны выделяются в от..
вет на специфический стимул и действуют
как «пусковой механизм», вызывая бы..
стрый ответ по принципу «все или ничеrо».
у морской звезды выметывание половых
продуктов у особей каждоrо пола может
быть вызвано инъекцией экстрактов радиа..
Системы контроля
525
льных нервов в полость тела взрослоrо жи
BOTHoro. Странно, что участвующий в этом
пептидный ropMoH «<rонадостимулирую
щее вешество» или GSS) распространен
ный по всем радиальным нервам и в HepB
ном кольце, по видимому, секретируется
слоем субкутикулярных опорных клеток
(т. е. ненастоящими нейронами). Как он
транспортируется по нервным тяжам и уча
ствует ли в ero переносе к тканям целомиче
ская жидкость, вопрос спорный. С этой He
обычной системой связано еще MHoro Hepe
шенных проблем.
rонадостимулирующее вещество вызы
вает созревание raMeT в яичниках или в их
фрcirментах in vitro, что, казалось бы, свиде
тельствует о ero прямом действии. Однако
на самом деле GSS стимулирует синтез
и выделение BToporo вещества фолликуляр
ными клетками rонад. Это «индуцирующее
мейоз вешество» (MIS), как показал Кана..
тани (Kanatani) с сотрудниками в 1969 r.,
оказалось 1 метиладенином (табл. 16.1).
MIS любопытным образом сочетает
эндокринную и параэндокринную функции.
Во..первых оно влияет на фолликулярные
клетки, которые ero синтезируют, что при
водит к их отделению от raMeT и выделе
нию последних через половые протоки. Bo
вторых, MIS диффундирует через межкле
точное пространство, индуцируя мейоз
ооцитов. Наконец, оно диффундирует в цe
ломическую жидкость и стимулирует COKpa
щение мышll УЧd.ствуюших в выметывании
raMeT, а у некоторых видов вызывает cBoero
рода заботу о потомстве.
16.12.2. Эндокринный баланс
Некоторые функции контролируются не
единственным ropMoHoM, а балансом анта..
rонистических факторов. Уrлеводный. соле
вой и водный обмен ЧdСТО реrулируется
именно таким путем, обеспечивающим бы..
струю обратимость эндокринноrо эффекта.
у ракообразных изменение окраски HO
сит физиолоrический характер, т. е. не
связано с количеСlпво./w пиrмента, а обуслов"
лено движением (или «МИI рацией») ero [ра..
пул в клетках, называемых ХРОЛ1атофора ии
(рис. 16.34). Сами клетки при этом не изме
няют формы и положения.
,
;{
1
"
1
2
1:1
3
4
5
Рис, 16.34. Хроматофоры с различной степе
нью дисперсии пиrмента
rормоны, контролирующие хроматофо
ры, обычно выделяются в ответ на внешние
стимулы (освещение фона и т. д.) но в неко"
торых случаях их секреция подчиняется
циркадианному или приливно"отливному
ритму. Обычно наблюдаются два антаrони"
стических rOpMOHd.. один из которых вызы"
вает рассеивание пиrмента, а друrой ero
концентрирование. Однако в некоторых
случаях на хроматофоры ОДНОI'О цвета
влияет множество факторов. что приводит
к различным окраскам тела. rOpMOHbI воз..
действуют также на миrрацию ретинальных
пиrментов, связанную с изменением физио..
лоrии зрения сложноrо rлаза (рис. 16.20.Б).
16.12.3. Эндокринный контроль
развития у Nereis
Характер эндокринной реrуляции у по
лихет нереид сравнительно прост и похож
526
на известный для немертин и некоторых
друrих беспозвоночных (но не для боль
шинства друrих аннелид). Как впервые по
казал Деркон (Durchon) в 1948 [., дeцepe
брация этих червей приводит к преждевре
менному созреванию. На последней стадии
жизненноrо цикла большинство видов пре
вращаются в 2етеронереидную форму
(рис. 4.60), адаптированную к «роению»
и нересту в поверхностных морских водах.
Этот процесс также подавляется [ормоном
мозrа.
Децеребрация на ранних стадиях жи
зненноrо цикла приводит к аномалиям раз
вития ооцитов или их деrенерации, а также
к неполному метаморфозу. Это позволяет
предполаrать" что у [ормона сложное,
двойственное действие, включающее тpo
фический эффект, поддерживающий раннее
развитие [амет и тела, и тормозное влияние
на процессы, связанные с последними CTa
диями развития. У самок трофическое влия
ние на [аметы может быть косвенным, по
скольку синтез специфических белков желт
ка (витеЛЛО2енинов). необходимых для раз
вития ооцитов, происходит В целомоцитах.
rOpMOH необходим также для образования
новых cerMeHToB тела (см. разд. 14.4.3).
На ранних стадиях жизненноrо цикла
уровень [ормона высок, но впоследствии
интенсивность ero секреции снижается. Это,
вероятно, зависит от окружающих условий.
Однако вещество, продуцируемое созре
вающими rаметами, подавляет эндокрин
ную активность мозrа, что в свою очередь
ускоряет развитие [амет, приводя к Bыдe
лению еще б6льших количеств их инrиби
рующеrо вещества и т. д. (эффект положи
тельной обратной связи). Вопрос о том, Ka
кая здесь связь прямая, а какая обратная,
спорный.
Считалось" что множество процессов,
участвующих в образовании cerMeHToB, [a
метоrенезе и метаморфозе, контролируется
простыми изменениями секреции «мозrово
[о [ормона». Однако электронная микро
скопия позволяет предполаrать, что мозr
нереид является источником множества
[ормонов. Более Toro, недавно у этих и дpy
rих нечленистоноrих беспозвоночных обна
ружены экдuстероuды (разд. 16.12.4), а
у пиявок они, вероятно, контролируют ли
rlавэ 16
ньку. Несомненно, эндокринные механизмы
у нереид HaMHoro сложнее, чем представля
лось ранее.
16.12.4. Эндокринный контроль
развития насекомых
В течение последних 50 лет насекомые
привлекают большое внимание эндокрино
лоrов из за скачкообразноrо характера их
развития, включающеrо ряд линек, и в He
которых rруппах метаморфоз от личинки
ДО куколки и имаrо. Сложность этих про..
цессов у членистоноrих отражается на KOH
тролирующей их нейроэндокринной систе
ме (приложение 16.12).
Впервые значение МОЗ2а было продемон
стрировано Копецом (Корес) в классиче
ских экспериментах 1917 1923 rr. (рис.
16.35). rусеница непарноrо шелкопряда бу
дет окукливаться" только если испытывает
влияние мозrа в течение HeKoToporo мини
мальноrо (крuтuчеСКО20) периода. Уча
ствующий в этом проторакатропный 20p
мон (РТТН) необходим для линьки на лю
бой стадии жизненноrо цикла. У бабочки
Maпduca 1 источником этоrо [ормона, по
видимому, является единственная нейросе
креторная клетка с каждой стороны мозrа,
а местами ero хранения и выделения служат
нервные окончания внутри кардиальноrо
и прилежащеrо тел (rлавным образом в по
следнем) (приложение 16.12). Стимулы, ин
дуцирующие выделение [ормона у разных
насекомых, неодинаковы. У клопа Rhodпius
пусковым сиrналом служит растяжение
брюшка в результате питания кровью (жи..
вотное может быть «обмануто» заполне..
нием брюшка воздухом), а у Maпduca
достижение критической массы тела.
Целью недавней работы Исидзаки (Ishi..
] в настоящее время установлено, что у М ап..
duca источником РТТН являются четыре латера
льные клетки (по две с каждой стороны мозrа).
Их аксоны проходят через кардиальное тело. не
ветвясь, а затем по аллатальному нерву дости
rают прилежашеrо тела. rде дают мноrочислен
ные разветвления, характерные для нейроrемаль
Horo opraHa. Местом выделения и запасания rop
мона является только прилежащее тело (М. А
O'Brien et аl., J. Neural Sci., 1988, v. 8 (9), рр.
3247 3257). Прuм. пер.
Системы контроля
527
aIUlJJIlДD
aЦJJJJlIJJ}J
t Линька
t 7 дней
16 дней t
Рис. 16.35. Эксперименты, демонстрирующие роль мозrа в реrуляции окукливания у Lyтaпtra.
После критическоrо периода ero ropMoH уже выделился в достаточном количестве поэтому деце..
ребрация или лиrатура не MorYT блокировать развития
zaki) и rилберта (Gilbert) с соавторами бы
ла очистка РТТН. Из мозrа бабочки Boт
Ьух выделены два пептида с молекулярны
ми массами около 5000 и 22000, способные
стимулировать секрецию экдизона прото
ракальными железами. Меньший из них co
стоит из двух аминокислотных цепей,
связанных дисульфидными мостиками. Это
послужило основанием для rипотез относи
тельно филоrенетическоrо значения cxoд
ства этоrо соединения с инсулином и инсу
линоподобными факторами роста позво
ночных. Установление физиолоrической po
ли двух этих пептидов еще впереди.
Роль проторакальных желез продемон..
стрирована в экспериментах на куколках
бабочки цекропиевоrо шелкопряда (Saturпia
cecropia). Если в ИЗОЛИРОВdнное брюшко
имплантировать активный мозr, оно не
в состоянии линять. Если же для этой цели
использовать одновременно активный мозr
инеактивные проторакальные железы, раз
витие происходит. Это свидетельствует
о том, что [ормон мозrа стимулирует в про
.
торакальных железах или их rомолоrах ce
крецию BToporo фактора, стероида экдUЗ0на
(табл. 16.1). Карлсон (Karlson) с соавтора..
ми использовал 1000 Kr куколок шелкопря..
да для получения 200 Mr чистоrо вещества,
ставшеrо в 1963 r. первым химически иден..
тифицированным [ормоном беспозвоноч
ных. В тканях экдизон превращается в 20
rидроксиэкдизон, который и является со..
бственно [ормоном линьки.
Для подобных соединений, известных под
общим названием экдuстероuды, разрабо..
тан ряд чувствительных количественных
анализов. Каллифоровый тест, использо..
ванный Карлсоном, основан на способно
сти экдизона вызывать окукливание у пере..
вязанных личинок мухи. Сравнительно не..
давно стал широко применяться радUОUJИ
МУНОЛО2uческuй анализ. Антитела к экдизон..
белковому комплексу связывают радиоак..
тивный экдизон. Масштабы этоrо процесса
зависят от концентрации немеченоrо экди
зона., конкурирующеrо за места связыва..
ния, и, таким образом, позволяют судить
о ero количестве. У клопа Rhodпius един
ственный пик экдизона наблюдается перед
каждой линькой (рис. 16.36).
РТТН типичный пептидный [ормон
в том смысле, что ero воздействие на ре цеп..
торы плазматических мембран проторака
льных желез индуцирует внутриклеточный
синтез сАМР. Стероиды., напротив., прони
кают внутрь клетки и (предположитель..
но ),связываясь с цитоплазматическими бел..
ками, стимулируют rенетический аппарат
ядра. Для исследования этой проблемы иен..
ным материалом являются мухи, так как их
слюнные железы содержат «rиrантские»
528
rлава 16
Rhodпiиs
с;
........
L..
:I: 2000
:I:"
О
М
S
с:а:
с')
s
u
8. 1000
с:а:
s
L..
6
N
о
Дни О
9 12 О 3 6 9 12 15 18
А
4 я стадия
5 я стадия
50
с;
........
L..
:I:
:I:
О
а.
о
L..
'S
:ii
з:
с;
s
з:
Q)
ш
Q
Б
Rhodпius
Линька
о
Дни О
4 я стадия
5 я стадия
Рис. 16.36. Уровни в rемолимфе 20 rидроксиэкдизона (А) и ювенильноrо ropMOHa (Б) в течение
последних личиночных стадий Rhodпius (по Baehr et al., 1978)
хромосомы" которые в 1 О раз длиннее и
в 100 раз шире обычных. Во время развития
различные «диски» таких хромосом сильно
утолщаются, образуя «пуффы», rде повы--
шенная активность reHoB выражается в ин--
тенсивном синтезе РНК. Эксперименталь--
ным путем была показана индукция экди--
стероидами появления специфической по--
следовательности ранних и поздних хромо--
сомных пуффов, идентичной той, что обыч--
но наблюдается перед линькой.
Третий ropMoH ювеНUЛЫ-LЫЙ (JH)
(табл. 16.1) секретируется прилежащими
телами (приложение 16.12). Три основные
A
формы JH близкие по структуре молеку-
лы; каждое насекомое секретирует одну из
них или более. Роль JH наиболее четко про-
демонстрирована в экспериментах судале..
нием прилежащих тел и их трансплантапией
в особей, находящихся на разных стадиях
развития (рис. 16.37). На ранних этапах жи..
зненноrо цикла наблюдается высокий ура..
вень JH, который обеспечивает появление
после линьки (запускаемой экдизоном) еле..
дующей личиночной стадии. У Rhodпius во
время пятой (последней) личиночной ста-
дии уровень JH сильно снижаетс (рис.
16.36). и в результате индуцируемой
.
Б
.
в
.
. ............... '
.'
Рис. 16.37. Эксперименты, демонстрирующие роль прилежащих тел у ВотЬух. А. Нормальное
развитие: личинка пятоrо возраста, куколка и имаrо. Б. Ускоренное развитие личинки TpeTbero
возраста после удаления прилежащих тел. В. rиrантская личинка шестоrо возраста полученная
путем имплантации активноrо прилежащеrо тела в личинку пятоrо возраста развивается в rиrант
скую куколку и имаrо (по Turner, 1966)
Системы контроля
экдизоном линьки появляется имаrо (или
сначала куколка, а затем имаrо, как, напри
мер, у бабочек).
На ранних стадиях развития ювениль
ный ropMoH защищен от распада спеuифи
ческим белком rемолимфы, с которым он
связывается. Низкий уровень JH перед по
следней линькой обеспечивается снижением
скорости ero синтеза и образованием специ
фическоrо фермента, разрушающеrо rop
мон, несмотря на ero связь с белком.
у насекомых ювенильный ropMoH уча
ствует также в контроле размножения. BMe
сте с родственными химическими соедине
ниями он используется в борьбе с вредите
лями, обладая такими явными преимуще
ствами, как нетоксичность и способность
к биодеrрадаuии. Кроме Toro, насекомые
с трудом вырабатывают устойчивость
к веществу, от KOToporo зависит их HopMa
льное развитие.
rор./ион выхода секретируется мозrом
и запускает сложную последовательность
событий, про исходящих при сбрасывании
насекомым старой кутикулы (через HeKOTO
рое время после образования lпидермисом
новой кутикулы под влиянием экдизона).
у сверчка Те/еоgrуllиs различают четыре oc
новные стадии этоrо процесса. Во первых,
кутикула отслаивается и прикрепляется
к субстрату затем насекомое выбирается из
старой шкурки; имаrинальные крылья, ноrи
и т. д. расправляются и приобретают OKOH
чательную форму; наконец, старая кутику
ла съедается. Эти стадии включают стере о..
типную и координированную последовате
льность по крайней мере 48 специфических
движений, повторяющихся не менее 4 ч.
Понятно, что о нейросекреторных клетках,
вырабатывающих ropMoH выхода, можно
rоворить как о «командных нейронах»
(разд. 16.10.2).
Новая кутикула вскоре темнеет и OTBep
девает. Ее «дубление» стимулируется ней
росекреторным ropMoHoM бурсиконом, KO
торый секретируется после выхода из CTa
рой шкурки мозrом или абдоминальными
rанrлиями.
Интересно сравнить эндокринный кон..
троль развития у насекомых и друrих чле..
нистоноrих, особенно ракообразных. У них
такой же стероидный ropMoH линьки
529
20 rидроксиэкдизон, секретируемый эндо
кринными железами ненейронной приро
ды У орrанами. Секреuия стероида KOH
тролируется нейросекреторной системой
I'лазноrо стебелька. Однако" если у HaceKO
мых РТТН оказывает стимулирующее дей
ствие, у ракообразных соответствующий
I OpMOH Y2Hefпaem работу У opraHoB, поэто
му удаление rлазноrо стебелька ускоряет
.:Iиньку или даже вызывает серию быстрых
линек. И наконец, недавно показано, что
у ракообразных есть JH Он секретируется
аllтеннаЛЬ/--lЫМИ .железа 'Ии и ero роль сходна
с таковой у насекомых.
rлавное отличие заключается в присут
ствии У ракообразных половых 20р./ионов.
Как показал Шарнио Коттон (Charniaux
Cotton) в 1954 [., мужской ropMoH секрети
руется андроrенными железами, связанны
ми с семепроводами, а один или несколько
женских ropMoHoB яичником. Эти Be
щества стимулируют rаметоrенез и диффе
ренцировку вторичных половых признаков.
у большинства насекомых rенотип, по
видимому, влияет на половую дифференци
ровку непосредственно, без участия эндо
кринной системы.
16.13. ВЫВОДЫ
Мы завершим наш обзор реrуляторных
систем у беспозвоночных двумя примера
ми, свидетельствующими о проrрессе, дo
стиrнутом за последние rоды в объяснении
поведения и физиолоrии с точки зрения их
нервных механизмов.
Почвенная нематода Caeпorhabditis e/e
gaпs была детально изучена Бреннером
(Brenner) и ero коллеrами в Кембридже.
Этот червь почти прозрачен, длина ero Bce
[о 1 мм, а жизненный цикл в лабораторной
культуре при 20 0 С длится 3,5 дня. Строение
ero нервной системы было полностью pe
конструировано по электронным микрофо--
тоrрафиям серийных срезов.
у этоrо rермафродита ровно 302 нейро
на, каждый из которых характеризуется
определенной формой и местоположением.
Более Toro, сейчас известны синаптические
контакты каждой клетки, т. е. построена
«принципиальная схема» нервной системы.
Обнаружено около 600 щелевых контактов
530
rпаеа 16
и примерно 7000 химических синапсов" хотя
их число и положение MorYT варьировать.
Прослежены последовательность кле..
точных поколений, предшествующих
в эмбриоrенезе каждому нейрону, стадии
развития, На которых происходит их специ..
фическая дифференцировка, а также измене..
ния их синаптических связей.
Изучение MopcKoro заднежаберноrо
моллюска РZеиrоЬrапсhiа caZiforпica Дейви..
сом (Davis) с соавторами в Калифорнии
оказалось наиболее успешным с точки зре..
ния выяснения нервных механизмов, лежа..
щих в основе поведения почти у любоrо жи..
BOTHoro.
Различные формы поведения орrанизо"
ваны иерархически. Так, реакция избеrания
важнее пищевой, ее стимуляция активирует
тормозные синапсы единственноrо «ко..
мандноrо» нейрона, контролирующеrо пи..
тание. Мотивация питания зависит от пред"
шествующеrо потребления пищи" а актив и..
руемые им сенсорные пути тормозят этот
командный нейрон. Конкурирующие сти"
мулы вызывают различные ответы, завися..
щие от относительной силы стимуляции,
и животное должно обладать способностью
к принятию решения, поскольку в каждый
момент времени оно обычно занято только
одним делом. Основой этоrо является инте..
rрация сенсорной информации в нервной
системе. Например, если пищевой стимул
сопровождается стимулом для втяrивания
ротовой лопасти, предпочтение будет отда..
но первому, поскольку активируются два
нейрона, блокирующие двиrательный вы..
ход, ответственный за втяrивание. Если же
пищевой стимул слабый или животное на..
сытилось, активируются друrие цепочки
и доминирующим оказывается сиrнал на
втяrивание ротовой лопасти. В этих процес..
сах участвует и эндокринная система: пита..
ние тормозится до тех пор, пока на нейроны
буккальноrо rанrлия влияет ropMoH отклад..
ки яиц.
Поведение упорядоченно и адаптивно,
и этим оно четко отличается от разнообра..
зных конкурирующих влияний, С которыми
сталкивается животное. Изучение нервной
системы беспозвоночных открыло некото"
рые свойства нервной орrанизации, лежа..
щие в основе их поведения. Тем не менее.
несмотря на составление полных «принци..
пиальных схем» нейронных соединений, мы
до сих пор не знаем, что эти животные «чув..
ствуют» и чувствуют ли они вообще. Это
должно сделать нас осторожными в проrно"
зах при будущем исследовании более разви"
тых нервных систем.
16.14. Дополнительная
литература
Bullock Т. Н., Horridge G. А. [965. Strиctиre and
Function in the Nervous System of lnvertebrates.
Freetnan, San Francisco.
Eatoп R. с. (Ed.) 1984. Neиral Mechanisms ofStart-
le Behavioиr. Plenum Press, New У ork.
Ha//iday Т. R., Slater Р. J. В. (Eds) 1983. Animal Ве-
havioиr. Vol. 1, Causes and Effects. Blackwell
Scientific Pиblications, Oxford.
Mill Р. J. 1982. Comparative Neurobiology. Ed. Ar
nold, London.
Nighпaт К. L., Hill L. 1977. The Comparative En
docrinology of the Invertebrates, 2nd edn. Ed. Ar
nold, London.
Roberts А., Bush В. М. Н. (Eds) 1981. Neиrones
Without Impиlses: Their Significance for Verte-
brate and Invertebrate Systems. Cambridge Uni
versity Press, Cambridge.
Sheltoп G. А. В. (Ed) 1982. Electrical Condиction
and Behavioиr in "Simple" Invertebrates. Claren-
don Press, Oxford.
Приложение 16.1.
Система rиrантских волокон
кальмара Lo/igo
А. Аксоны двух rиrантских нейронов
первоrо порядка сливаются по средней ли..
нии тела и иннервируют несколько клеток
BToporo порядка в задней части мозrа. Бо..
льшинство последних являются мотонейро..
нами, непосредственно управляющими
мышцами rоловы и воронки, однако аксо"
ны двух клеток идут к звездчатому [ан"
rлию, rде образуют синапсы с rиrантскими
волокнами TpeTbero порядка (показана то..
лько одна пара) (по Joung, 1939)
Системы контроля
531
rиrантское
волокно 2 ro
порядка
А
Нервы от звездчатоrо
rанrлия
Б
Б. Звездчатый rанrлий. rиrантские воло--
кна BToporo порядка образуют «дисталь--
ные» синапсы с волокнами третьеrо поряд--
ка, а дрyrие rиrантские волокна образуют
с ними «проксимальные» контакты. Каждое
волокно третьеrо порядка возникает путем
слияния отростков нескольких клеток, и все
ОТХОДЯIЦие от звеЗДЧdтоrо rанrлия нервы
включают по одному такому волокну (по--
казано только пять нервов). Самый задний
и самый крупный нерв является излюблен--
+
rиrантское
волокно 3 ro
порядка
r иrантское
волокно 2 ro
порядка
в
ным объектом исследования нейробиоло--
rOB.
В Синаптические контакты между rи..
rантскими волокнами BToporo и третьеrо
порядков. Физиолоrическое соединение
двух этих элементов опосредуется множе..
ством контактов, в каждом из которых по..
стсинаптический отросток как бы зубцами
вставлен в пресинаптический участок.
Стрелки указывают направление потока
импульсов.
Приложение 16.2.
Потенциал действия
Открытие механизма возникновения по..
тенциала действия Ходжкином (Hodgkin)
и Хаксли (Нихlеу) из Плимута и Кембриджа
1 й
порядок
2 и
порядок
BopOHKa
Звездчатый
rанrлий
з ...й
порядок
и друrими привело к присуждению им Но--
белевской премии 1963 [. С помощью так
называемоrо метода «фиксации потенциа--
ла» и путем замешения одноrо из ионов
веществом, для KOToporo мембрана непро..
ницаема, были измерены токи определен..
ных ионов через мембрану при различных
потенциалах. Друrой метод заключался
в избирательной блокировке ионных кана--
лов ядами: натриевых тетродотоксином,
выделенным из иrлобрюха, калиевых
тетраэтиламмонием.
Возникновение потенциалов действия
обусловлено наличием в мембране ионсе..
лектuвНblХ каналов с воротным потенциа..
лом. Натриевые каналы, по"видимому, ме..
няют свою форму при деполяризации, от..
532
rлава 16
крывая «ворота», через которые ионы Na+
входят в аксон, приводя к еще большей де..
поляризации мембраны. Это открывание
явление кратковременное, вслед за ним бы..
стро наступает «натриевая инактивация».
Существуют также калиевые каналы
с воротным потенциалом. Они открыва..
ются после натриевых, и диффузия ионов К +
через них (достаточно одноrо из каждых 107
ионов К + на импульс) восстанавливает
потенциал покоя. За этим следует короткий
период «отдыха», В течение KOToporo мем"
брана рефрактерна_ т. е. не способна активи
роваться. В клетках мноrих моллюсков oc
новной вклад в возникновение потенциала
действия вносят Са 2 + ..каналы. Известен му"
тант дрозофилы s/laker (трясущийся), Ha
ЗВd.нный так из..за паралитических вздраrи..
ваний конечностей во время эфирной ане..
стезии. Калиевые каналы с воротным по..
тенциалом у Hero отсутствуют или дефект..
ны (затронутые этой мутаuией rены иденти"
фицированы). В результате реполяризация
Зdпаздывает, и потеНЦИdЛЫ действия растя..
rиваются во времени.
50
со
:L
...
с:
О
::r
:I:
ф
о
t:
.S
1i
:I:
:I: ...50
ro
CL
111
:L
ф
L:
2
Время., МС
4
На rрафике показаны изменения мем..
бранноrо потенциала Р (ось ординат) в лю..
бой точке аксона в зависимости от времени
( ось абсцисс) и обусловливающие их после..
довательные изменения ЧИСЛd. открытых
К+.. И Nа+"каналов (по Hodgkin, Huxly,
1952).
Приложение 16.3.
Клеточные механизмы науче...
ния
Одни и те же типы научения обнаружены
во всем животном царстве_ но только недав"
но Канделом (Kandel) и друrими были ис..
следованы их клеточные механизмы.
Мантийная полость моллюска Ap( 'sia
имеет сифон и защитную пластинку. Леrкое
прикосновение активирует сенсорный путь
от каждой из 'Этих структур, нейроны кото..
poro образуют синаптические контакты не..
посредственно с мотонейронами., управ..
ляющими защитной реакuией втяrивания
ктенидия (моносинаптическая рефлектор..
ная дуrа см. разд. 16.9.3). Сенсорные
нейроны, иннервирующие хвостовую зону,
стимулируют интернейроны, образуюшие
синапсы с сенсорными окончаниями.
В пресинаптических окончаниях сенсор..
ных нейронов происходят изменения, опре..
деляющие МОДИфИКd.цию поведения, т. е.
научение. Частая стимуляция дыхательных
структур, вызывая неоднократное проведе..
ние потенциалов действия в эти окончания,
приводит к rрадуальной инактивации Са 1 +..
каналов (разд. 16.3.3). уменьшению количе..
СТВа выделяемоrо в ответ на каждый сти"
мул медиатора и ослаблению поведенче..
ской реакции. Эта простейшая форма науче..
ния называется привыканиеЛIl.
Если на «хвостовую» зону действует
мощный., уrрожающий животному раздра..
житель, ответ на широкий Kpyr друrих сти"
мулов, включая и леrкое прикосновение
к дыхательным структурам. усиливается.
Такая форма неассоциативноrо научения,
называемая сенсибилизацией, опосредуется
медиатором 5..rидрокситриптамином (5..
НТ), выделяемым интернейронами и запу..
скающим целый каскад метаболических
процессов в сенсорных окончаниях:
5.. НТ Синтез сАМР Активация фер..
мента протеинкиназы Фосфорилирова..
ние специфических белков К + ..каналов мем..
браны --4 Инактивация К + ..
каналов Увеличение длительности потен..
циаловдействия УвеличениепритокаСа 2 +
внутрь клетки Усиление выброса
медиатора.
Системы контроля
5ЗЗ
Выработка условНО20 рефлекса это oд
на из форм ассоцuапluвНО20 наученuя (пом
ните собак Павлова?). Так. если за серией
слабых стимуляций жаберной пластинки
каждый раз немедленно следует уrрожаю
щее раздражение «хвоста», животное нач
нет реаrировать rораздо силы[еe на стиму
ляцию пластинки, чем, например, сифона.
Описанное сочетание стимулов приводит
к выделению из интернейронов 5 HT и син
тезу сАМР во всех сенсорных окончаниях
как раз тО2да. КО2да с приходом потенциа
лов действия повышается уровень ионов
Са 2 + в окончаниях только стимулированных
сенсорных нейронов пластинки. Считается,
что Са 2 + активирует сАМР, а тот в свою
очередь усиливает выделение медиатора,
KOToporo в результате высвобождается зна
чительно больше, чем во время сенсибили
зации.
Описанные выше формы научения
являются кратковременнымu они coxpa
няются в течение нескольких минут или,
в лучшем случае, часов. Однако применение
особых проrрамм привопит к формирова
АБдоминальный
rанrлий
Ктен иди й
А
А. Нервная система и дыхательная камера
Ap/ysia (вид сверху). Б. Синаптические связи
нейронов в абдоминальном rанrлии, исполь
зованные для изучения клеточных механизмов
процессов научения: C ceHcopHыe нейроны;
И интернейрон; М мотонейрон
нию dОЛ20вреNlенной паJ'v!яп1И (у Aplysia
более трех недель), обусловленной не не..
устойчивыми метаболическими сдвиrами_
а Cтp}'KlпypHblJ'v!U UЗJwенения.I'vlИ. Число и раз
мер синаптических участков, d также число
синаптических пузырьков существенно воз..
растают при долrовременной сенсибилиза
ции и снижаются в ходе привыкания.
Приложение 16.4.
Становление дефинитивных
нейронных связей
Хотя окончательное строение нервной
системы невероятно сложное, она развива
ется из простоrо эпителия. На конце OT
ростков эмбриональных клеток находятся
конусы аJ'v!е60uдНО20 роста, оканчиваю
щиеся псевдоподиями. Формирование «пра..
вильных» связей нейронов имеет решающее
значение для. дальнейшей работы нервной
системы (разд. 16.8.4). еханизмы этоrо
процесса изучены у насекомых, в частности,
у кузнечиков, поскольку их эмбриональные
rанrлии достаточно прозрачные, а число
клеток в них невелико. В результате отдель..
ные нейроны и паже их растушие аксоны
MorYT быть надежно идеНТИфИЦИРОВdНЫ.
В то время как одни клетки развива
ются, друrие становятся uзбыпl0ЧНЫ.. tu
и элиминируются. Хотя различные rанrлии
в нервном стволе эмбриона насекомых co
держат одинаковое число клеток, некото"
рые клетки абдоминальных rанrлиев утра..
чиваются, а их аналоrи в тораксе развиваю..
тся для управления конечностями (они со..
храняются, даже если конечности удалить).
Аналоrичным образом у rермафродитной
формы нематоды Caeпol'llahc!itis 302 нейро
на. а у самцов в связи с репродуктивными
функциями сохраняются еще 79 нейронов.
Такой процесс запроzраМ1vlированной 2u6ели
клепl0К детерминирован rенетически. «Про..
[рамма» развития деrенерирующих клеток
такая же, что и у их выживающих «сестер»,
однако у них включается «проrрамма ca
моубийства». Один из наиболее интересных
мутантов у Caeпorhabditis (по [ену ced 3)
в этом отношении дефектен. Клетки. KOTO
рые в норме должны поrибать, выживают
и формируют функциональные связи; по
видимому, на животном это не отражается!
534
rлава 16
.. 11
.. 11
U2
А
1 .
l'
А. Становление нейронных путей в cerMeHTap"
ном rанrлии кузнечика на седьмой день двад..
цатидневноrо развития. Б. После удаления
пионерных нейронов U 1 и U 2 нейрон аСС дез
ориентируется. В. При удалении пионерных
нейронов МР1 и dMP2 из этоrо же и соседнеrо
rанrлия дезориентируется нейрон рСС (по Ba
stiani et al., 1985)
Выросты различных клеток в rанrлиях
эмбриона кузнечика появляются в четкой
последовательности; отростки, заканчи
вающие развитие первыми" называются
пионерными волокнами. Прорастая от raH
rлия к rанrлию или от эпидермальных ce
нсорных клеток в центральную нервную си
стему, они следуют стереотипными путями.
Ориентация конусов роста, по видимому,
определяется различными факторами их
окружения, но rлавным образом, вероятно,
"---, 1
U2 ( ...) \.. ..... .....,
':= .,;:: ..... [ ..... E -тr .....,'
U1 ,( ..., '- ..... .LL.....--. 1 1
\,. .:::::" .... ..... ---- \, 1
аСС е P:I
1111
рСС tf-:J) 1; 11
1111
1111
" IIJ . ......... ... ..
, '" .,
\ '"' ..... ..... ........
"- ..... .......... '"
Б
i
1111
1111
1111
1111
........ ....... ....... ....... н 1....... ........ .......
1111
1111
уМР2 0 1111
1111
,...... vu
d М Р 2 ((.... ..... J \......... ..... ..... ..... ..... ...
..... / ..... ..... ..... "'" ,
r 11
МР1 \ (..... ) =-=--=.....---=-- "" 11
.............. )1 1
1111
1111
.1 1
в
1111
1111
1111
1111
1111
I111
1111
1111
,1 Р
.., IJ 1..... ..... ....... ........
.11
положением базальных мембран и rлиаль..
НЫХ клеток.
Волокна, вырастающие позднее, часто
следуют за пионерными и в их отсутствие
(при экспериментальном удалении отдель..
ных эмбриональных клеток методами ла..
зерной микрохирурrии) дезориентируются,
Если в эксперименте задержать развитие
клетки, ее аксон «отыщет» правильный
путь, несмотря на появление множества дo
полнительныx факторов.
(;истемы контроля
Использование моноклональных анти--
тел к центральной нервной системе дрозо--
филы показало.. что различные волокна
в составе HepBHoro пучка имеют одинако--
вую специфическую молекулу. По
видимому, она является «фактОрО.Nl УЗ1lава--
1lИЯ», а нейрон «запроrраммирован» следо
вать сначала одному химическому указате--
лю, затем друrому и Т. д. Более Toro, хотя
конкретный, окончательно сформирован--
ный нейрон может значительно различаться
своими дендритами у разных особей, он
в любом случае образует практически оди
наковые синаптические связи.
Приложение 16.5.
Клеточные механизмы
биолоrических часов
Среди наиболее известных ритмов био--
лоrической активности некоторые связаны
с реrулярным чередованием дня и ночи.
ЦuркадuаН1lЫЙ (circa dies около суток)
ритм у моллюска Aplysia захватывается (на..
страивается) изменениями в суточном ци--
кле освещенности, однако, если животное
содержать в неизменных условиях, он со..
храняется в течение нескольких дней, лишь
HeMHoro изменяя длину каждоrо цикла. Это
свойство циркадианноrо ритма дает opra
низму возможность реаrировать на факто..
ры окружающей среды еще до их наступле..
ния.
Ритмическая активность широко рас..
пространена среди нейронов центральной
нервной системы и сохраняется в течение
нескольких недель даже при извлечении raH..
rлиев из тела и содержании их в культураль
ной среде. Различные клетки имеют свои со--
бетвенные ритмы, однако в природе они
синхронизируются «rлавными часами», ло
кализованными в «D--клетках» rлаза (это не
фоторецепторы, а клетки, непосредственно
получающие от них информацию через си--
напсы). Циркадианный ритм может быть
зареrистрирован в D--клетках изолирован..
Horo rлаза, которые оказывают свое влия
ние не за счет секреции ropMoHoB, как коrда..
ТО считалось, а через аксоны, проходящие
почти по всей нервной системе.
Были исследованы фундаментальные
механизмы работы биолоrических часов.
535
5 До удаления rлаза
....
>-
1:
о
5 После удаления rлаза
....
>-
1:
о
Время
л окомоторная активность Ap/ysia в течение дня
и ночи до удаления обоих rлаз и через три дня
после Hero (по Strumwasser, 1974)
Обработка анизомизином, который блоки..
рует синтез белка в клетке, связываясь с од..
ной из субъединиц рибосом, уничтожает
ритм, а короткое воздействие этоrо ве..
щества «останавливает часы» на время, т. е.
заставляет их «отставать». Джаклет (Jac--
klet, 1981) следал вывод, что «суточный син..
тез белка основное условие хода цирка--
дианных часов». Соответствующие белки
MorYT быть ферментами протеинкиназами,
которые, как известно, влияют на ионные
каналы и, следовательно, на импульсную
работу друrих систем.
Приложение 16.6.
Сенсиллы насекомых
у насекомых некоторые рецепторные
нейроны связаны с кутикулярными волоска
ми, шипиками, пластинками, ямками и Т. д.
Такие сенсорные орrаны называются се..
1lсилламu; тело буквально изобилует этими
536
rлава 16
т ормоrенная
клетка
Материн ...
ская клетка
Дистальные cerMeHTbI
(сенсорные реснички)
ОБкладка
Волосковый
стержень
Развитие и строение сенсиллы HaceKoMoro. Тормоrенная клетка секретирует материал для сочле
новочной впадины, а трихоrенная клетка для волосковоrо стержня (06е они участвуют в обра
зовании лимфы); 06кладочная клетка формирует чехол BOKpyr ресничек (по Dethier, 1976; Нап
sen, 1978)
крошечными «антеннами». Все клетки, BXO
дящие в состав сенсиллы, образуются путем
деления одной материнской клетки У нема--
тод хеморецепторные орrаны" называемые
амфuдамu (рис. 4.18), развиваются сходным
образом.
Вкусовые или обонятельные хе/иорецеп--
торы у разных животных чрезвычайно схо--
жи. Дендрит (или «внутренний cerMeHT»)
каждоrо биполярноrо рецепторноrо нейро--
на направлен к основанию этоrо opraHa, rде
образует одну или более ресничек (<<нару..
жный cerMeHT»). Проходя через множество
пор (до 15000 на волосок), химические ве..
щества достиrают этих ресничек по особой
омывающей их жидкости (<<лимфе»).
Хорошо развитые механореце1ll110ры на..
секомых, как и у подавляющеrо большин--
ства друrих животных, ресничные. Движе..
ние щетинки opraHa осязания деформирует
ресничку и тем самым стимулирует рецеп..
торную клетку. КаJипанuфор./нные сенсuллы,
состоящие из единственноrо нейрона,
связанноrо с жестким кутикулярным кол--
пачком, воспринимают напряжение в кути--
куле. Хордоп10нальные сенсuллы дают ИН..
формацию о положении или движениях су..
ставов у насекомых и ракообразных. Они
состоят из рецепторов, каждых из которых
покрыт специализированной сколопоuдной
клеткой, связанной с тяжами соединитель..
ной ткани. Такие рецепторы служат, в част..
ности, сенсорными компонентами тимпа..
нальных Ор2аnов слуха. встречающихся
в разных частях тела насекомых. Эти opra..
ны состоят из модифицированных частей
трахейной системы и имеют воздушные ме..
шки с тимпанальными мембранами. вибра..
ция которых в ответ на звук стимулирует
связанные с ними рецепторы.
у насекомых, по--видимому, нет особых
структур, чувствительных к силе тяжести
и ускорению, и соответствующую инфорrvfа..
цию они получают от мноrочисленных се..
нсорных opraHoB, связанных с суставами,
и т. д. Однако у мух задние крылья превра..
щены в булавовидные :J1СУЖ:J1сальца. кото..
рые быстро вибрируют во время полета
и по принципу rироскопа, используемоrо
в навиrации, дают информацию о повара..
тах тела.
Системы контроля
537
Приложение 16.7.
Нейросуперпозиционная
сетчатка мухи
Разрешающая сила нейросуперпози--
ционноrо rлаза может быть в 100 раз выше,
чем у сложных rлаз друrих типов. Рабдоме..
ры омматидия здесь остаются разделенны"
ми, т. е. рабдом «открытый». Каждый раб..
домер получает свет, падающий под опре..
деленным уrлом, а ero аксон идет к патро"
ну" общему не для всех рабдомеров данноrо
омматидия, а для ориентированных вдоль
той же оптической оси рабдомеров сосед..
них омматидиев. В результате лучи света от
одноrо источника падают на шесть ретину..
JlЯРНЫХ клеток смежных омматидиев. Их
нервные волокна, спирально закручиваясь,
образуют синапсы на одном патроне в оп..
тическом rанrлии.
На рисунке показаны пути световых лу..
чей от двух точек в поле зрения (светлые
и темные линии), их проекции на ретинуляр"
ные клетки соседних омматидиев и схожпе..
ние (конверrенция) волокон активируемых
ими рецепторов на оптических патронах ла..
мины. Cпpaвa CBeTOBыe лучи от одной
точки попадают на различные ретинуляр--
ные клетки смежных омматидиев" но аксо"
ны последних сходятся на одном патроне.
Из восьми ретинулярных клеток каждоrо
омматидия в нервной суперпозиции обычно
участвуют только шесть (по Strausfeld, Nas..
se 1, 1 98 1 ).
\ '11' I " I "
\ I / J//,
\ I I rjlJ
\ ' /
, ' I
\ ' I
\ I I
, I
\ , I
I
О °CJ
О 8 I
I
,,\\
Ретинулярные
клетки сетчатки
Патрон ламины
Окончание аксона
ретинулярной клетки
Постсинаптическая
монополярная клетка 2 ro порядка
Ламина
538
rлава 16
Приложение 16.8.
Оптическая доля комнатной
мухи (Musca doтestica)
«Сложность сетчатки насекомых ошело
мляет, обескураживает и не имеет аналоrов
у друrих животных. Размышляя о запутан
ной системе сложных, или фасеточных,
rлаз, проникая в лабиринт нейронов и co
единяющихся волокон трех крупных ce
rMeHToB сетчатки,... исследователь COBep
шенно теряет rолову» (Ramon, Cajal, 1937).
'/
Все типы клеток ламины и большинство
образуемых ими связей сейчас идентифици
рованы, и мноrие из них изучены электро
физиолоrически Достиrнут также большой
проrресс в исследовании медуллы и лобулы
(рисунок из Strausfeld, Nassel'l 1981).
Приложение 16.9.
Удары брюшка у рака
Удары брюшка у рака считают типич
ными устойчивыми комплексами действий.
Мышцы'"
разrИБатели
Мышцы
сrИБатели
А
Б
А. Упрощенная схема нервных цепей из rиrант
ских нейронов и «неrиrантских» проводящих
путей, присутствующих в каждом абдоминаль
ном cerMeHTe и контролирующих удар брюшка
у рака. Эти системы активируются внешними
стимулами (стрелки). ur центральный rи
rантский нейрон; БС быстрый мотонейрон
мышцы сrибателя; r rиrантский KOMaHД
ный нейрон; ТМ тормозный мотонейрон;
ТВ тормозный вход рецептора растяжения;
rМ rиrантский мотонейрон; MP
мышечный рецептор растяжения; нп
неrиrантский нервный путь; C ceHcopHыe He
йроны; СМ стимулирующий мотонейрон.
rиrантское волокно и rиrантские мотонейроны
связаны первыми известными науке электри
ческими синапсами (см. разд. 16.3.4). Б. Удар
брюшка у рака (по Wine, Krasne, 1972)
Системы контроля
Неожиданный уrрожающий стимул, ДOCTa
точно интенсивный для возбуждения един
cTBeHHoro импульса в rиrантских волокнах,
инициирует быструю высокостереотипную
реакцию избеrания «удар брюшка» за
счет синхронноrо сокращения мышц
сrибателей абдоминальных cerMeHToB. Дей
ствие совершается настолько быстро, что не
может быть модифицирано обратной
связью.
Активность в rиrантских волокнах TOp
мозит как мотонейроны разrибателей (aH
таrонистов сrибателей), так и рецепторы
растяжения разrибателей (поскольку иначе
они помешали бы удару), пока сrибатели не
разовьют максимальную силу. Таким обра
зом, rиrантские волокна действуют как KO
мандные нейроны (разд. 16.1 0.2), запускаю
щие различные координированные элемен
ты двиrательноrо акта.
Однако после подrибания брюшка про
исходит быстрое распрямление тела за счет
сокращения мышц разrибателей. Этот
«возвратный удар», будучи неизменной ча
стью естественной последовательности, не
2енерuруется центральной нервной cиcтe
мой, а является рефлекторным. Как только
прекращается торможение мышечных pe
цепторов, они стимулируются растяжением
и активируют разrибатели через описанную
ранее рефлекторную дуrу (разд. 16.9.3). Ин
тересно, что торможение сrибателей, сопут
ствующее обусловленному обратной
связью сокращению разrибателей" стиму
лируется (хотя и с задержкой) rиrантскими
волокнами.
Приложение 16.10.
Контроль мышечных сокраще--
ний у ракообразных
у краба отдельные мышць] состоят из
волокон с различной структурой и ФУНКIIИО
нальными характеристиками. Некоторые из
них быстро rенерируют потенциалы дей
ствия и развивают напряжение, в то время
539
Быстрый !
аксон
! тормозный
аксон
меДFlеННЫИ J
аксон
1 1
}) п)
Иннервация мышц у краба (по Atwood, 1973)
как друrим свойственны в основном rрадуа
льные потенциалы и более медленное co
кращение. МЫШIIЫ имеют сложную полu
нейронную иннервацию и в отличие от COMa
тических мышц позвоночных «принятие
решения» о характере их работы часто ocy
ществляется периферией, rде, как и у MHO
rих друrих беспозвоночных, внутри инне
рвируемой структуры сталкиваются проти
воположные влияния.
Отростки единственноrо быстропрово
дящеrо аксона контактируют rлавным
образом с быстрыми мышечными волокна
ми; эти синапсы в ответ на повторную сти
муляцию вскоре «устают». Окончания HepB
ных волокон малоrо диаметра подходят
в основном к медленным . мышечным воло
кнам; в ответ на повторную стимуляцию
здесь наблюдается облеrчение. Следовате
льно, степень сокращения контролируется
не подключением дополнительных двиrате
льных единиц, как у позвоночных, а rрадуа..
льным управлением мышечными волокна..
ми. Кроме Toro, тормозные волокна Bыдe
ляют rамма аминомасляную кислоту, как
в синапсах, образуемых с возбуждающими
окончаниями (пресинаптическое торможе
иие), блокируя выброс возбуждающеrо Me
пиатора rлутамата, так и в синапсах на MЫ
шечных волокнах (постсинаптическое TOp
можение), что тормозит любое их возбу
ждение.
540
rлава 16
Приложение 16.11.
Нейросекреция путем экзоци
тоза
Секрет из нейросекреторных нейронов
выделяется, как и из большинства обычных
железистых клеток, путем ЭКЗОЦUJnоза. Во
время этоrо процесса мембрана rранулы
сливается с плазматической мембраной
окончания аксона, что приводит к диффу
зии ее сопержимоrо в кровь. Впервые это
было установлено при изучении беспозво
ночных, в частности в работах Норманна
(Normann, 1965) по падальной мухе Calli
pJl0ra.
Позже исследования на аннелидах. а за
тем на широком Kpyre друrих беспозвоноч--
ных показали, что секреторные rранулы из
«обычных» нервных окончаний (т. е. обра--
зующих синаптические контакты с друrими
клетками) также выделяются путем экзоци"
тоза. Это означает, что синаптическим
......
cr
.. ,.
А
А, Электронная микрофотоrрафия, дeMOH
стрирующая экзоцитоз секреторных rранул из
синаптическоrо окончания в церебральном
rанrлии земляноrо червя Luтbricus. Стрелкой
показано выделяемое содержимое rранулы.
cr ceKpeTopHыe rранулы (х 60000 масштаб
100 нм). Б. В синаптических окончаниях дей
ствуют два различных секреторных механизма
с участием пузырьков и rранул соответственно.
у л ьтраструктура ней росекреторных окончани й
идентична (Golding, Pow, 1987). rэР
rладкий эндоплазматический ретикулум
окончаниям свойственны два различных се..
креторных механизма.
Синаптические пузырьки. ПО ВИПИМОМУ.
формируются локально внутри окончаний
и захватывают обычные медиаторы, напри..
мер ацетилхолин, из клеточноrо сока, rде
эти вешества синтезируются. Пузырьки
обычно выделяют свое содержимое в ме..
стах синаптических утолщений (рис. 16.6).
Нейропептиды, напротив, синтезируются
и упаковываются в rранулы в теле клетки.
После переноса в нервное окончание rpaHY"
лы высвобождают свое содержимое путем
экзоцитоза, по видимому, через неспециа..
лизированные участки мембраны.
После экзоцитоза мембраны пузырьков
и rранул отделяются от плазматической
мембраны, переходя внутрь ОКОНЧdНИЯ
(эндоцuтоз). Однако в то время как пузырь..
ки осушествляют КРуzоворопl (т. е. вновь за
полняются и используются), мембраны rpa..
нул, вероятно, разрушаются.
, ,
Аппарат
rольдж
"
@
Секреторны
@ rранулы
@
..
rэр
О
,; .,
О
.
dO
'еrр ц
Лизосома О o
о (J ...... ?........ 0%
о:>
О \
.:L..
Рец кли О f
000
00 О
00 О О t
Б О о ..
Эндоцитоз
Системы контроля
Приложение 16.12.
Нейроэндокринные системы
саранчи
Разнообразие и сложность нейроэндо"
кринных систем некоторых беспозвоночных
хорошо иллюстрирует пример саранчи.
Систе./иа J. Диуретический 20рМОН син..
тезируется нейросекреторными клетками
мозrа и транспортируется по аксонам в за..
пасающую долю кардиальноrо тела (нейро..
rемальный opraH), из которой выделяется
в кровь. rOpMOH стимулирует образование
мочевины мальпиrиевыми сосудами. Это
эндокринная система первоrо порядка
только с одним rормональным этапом.
Система 2. Нервные волокна, идущие
к железистой доле кардиальноrо тела.. обра..
зуют там типичные синаптические контак..
ты с железистыми клетками, работой кото..
рых они управляют, выделяя нейромедиа..
тор октопамин. При стимуляции клетки вы..
деляют адипокинетический 20рМОН пептид"
ной природы, идентифицированный Мор..
дью (Mordue) с соавторами в 1976 r. Он мо"
билизирует липиды жировоrо тела для
использования в качестве источника эне..
рrии при полете.
Система З. Друrой ropMOH, секретируе..
мый нейросекреторными клетками ком..
плекса мозr/кардиальное тело, называется
проторакотропным (РТТН). Он стимули..
рует секрецию проторакальными железами
стероидноrо ropMOHa экдизона, который по..
сле модификации в тканях запускает линь..
ку. Это эндокринная система BToporo по..
541
Llиуретический,,'
ropMOH It
Мозr и эндокринные opraHbI саранчи (относи
тельные размеры кардиальноrо и прилежаще
ro тел сильно увеличены)
рядка с двумя эндокринными этапами.
Система 4. Секреция ювенильноrо rop
мона прилежащим телом, по"видимому.
контролируется как прямой иннервацией,
так и rуморальными факторами.
Обратная связь. Эндокринные систеМЬJ
обычно действуют по ПРИНIIИПУ обратной
связи. Например, высокий уровень цирку..
лирующеrо 20..rидроксиэкдизона, влияя на
нейросекреторные клетки мозrа, тормозит
синтез экдизона проторакальными желе..
зами.
rЛАВА
17
ЕЩЕ РАЗ ОБ OCHOBHblX ПРИНЦИПАХ
17.1. Основные физиолоrиче...
ские черты фенотипов
Для построения новых соматическиих
и репродуктивных тканей, замены отжив
ших соматических тканей и получения эне
рrии для -этих процессов животным требу
ются ресурсы. В отличие от автотрофных
орrанизмов, которые MorYT синтезировать
орrанические соединения из неорrаниче
ских, используя в качестве источника эне
рrии солнечный свет, животные должны пи
таться друrими орrанизмами (см. rл. 9).
Эта их 2етеротрофность требует развития
способности к передвижению для поиска
и добычи ресурсов, а также как средства
спасения от друrих животных. Появление
локомоции (см. rл. 10) обусловило коэво
люционный отбор хищников и их жертв на
эффективность передвижения. Локомоция
также требует энерrии (rл. 11). Кроме Toro,
ресурсы необходимо вкладывать в различ
ные процессы и структуры, обеспечиваю
щие защиту от друrих орrанизмов (см. rл.
13).
Поступившие с пищей макромолекулы
для их усвоения тканями расщепляются на
субъединицы в ходе ферментативноrо про
цесса, известноrо как пищеварение. Затем
эти субъединицы либо используются для
ресинтеза макромолекул (анаболизм), либо
распадаются с выделением необходимой
для жизнедеятельности энерrии (катабо
лизм). Излишек ресурсов, особенно амино
кислот, и «отработанные» тканевые белки
также катаболизируются и экскретируются
(см. rл. 12).
rлавным энерrетическим ресурсом для
метаболизма (анаболизма и катаболизма)
являются уrлеводы, обычно моносахариды,
которые MorYT запасаться в виде полисаха
ридов (часто rликоrена) и / или жира. Энер
rия высвобождается в ходе их катаболиче..
cKoro окисления. В период между возникно..
вением жизни и появлением фотосинтеза,
коrда атмосфера Земли была лишена кисло..
рода (rл. 1 и 2), оно шло путем переноса
электронов к орrаническим соединениям,
которые в восстановленной форме накапли..
вались как конечные продукты обмена.
С появлением кислородной атмосферы ста..
ло возможным более полное и более эффек"
тивное окислени.е.. потребовавшее поступле..
ния из внешней среды 02 и удаления из ор..
rанизма С0 2 в качестве одноrо из «ОТХО..
дов» жизнедеятельности. Аэробное дыхание
сейчас распространено в животном царстве
наиболее широко, хотя у некоторых видов
основу катаболизма все еще составляет
анаэробное дыхание. Универсальной фор..
мой KpaTKoBpeMeHHoro запасания и перено..
са энерrии" выделяемой в катаболических
процессах, является фосфорилированное со..
единение аденина и рибозы (нуклеотид)
аденозинтрифосфат (А ТР), образующееся
из дифосфатной формы (ADP) в реаКIIИЯХ,
связанных с дыханием. После выделения
энерrии АТР вновь превращается в ADP,
и цикл повторяется. Процессы дыхания об..
суждались в rл. 11.
Фундаментальные физиолоrические
процессы обобщены на рис. 17.1. Отметим,
что поступление ресурсов в орrанизм orpa-
ничено затратами на их получение и СТРУК-
турами, обеспечивающими питание. Следо..
вательно, даже коrда пища в окружающей
среде в избытке, ее количество, которое мо"
жет быть использовано для метаболизма,
лимитировано. Чем больше ресурсов расхо..
дуется на одну метаболическую функцию,
тем меньше остается для дрyrих Скорость
и пути использования ресурсов контроли..
руются ферментами и в конечном счете [е..
Еще раз об O(::HOBI-iЫХ принципах
543
Рис. 17.1. Распределение ре..
сурсов (получаемых с пищей)
между анаболическими и ката..
болическими процессами
основа физиолоrии животных
(по Calow, 1986)
нами. Способ распределения оrраниченных
ресурсов решаюшим образом влияет на
биолоrию орrанизма на нескольких ypOB
нях: от относительноrо вклада их в анабо
лические и катаболические процессы зави
сит фИЗИОЛО2ИЯ животноrо; от распределе
ния между различными видами активно
сти rо поведение; от использования orpa
ниченных продуктов анаболизма на образо
вание различных структур форма тела, на
развитие соматических и репродуктивных
тканей биолоrия размножения и жизнен
ный цикл. Более Toro, степень удовлетворе
ния каждой из этих потребностей будет cy
шественно влиять на выживаемость и пло
довитость, а значит, на приспособленность
орrанизма. Поэтому, соrласно дарвинов
скому учению, отбор блаrоприятствует Ta
ким rенетически детерминированным oco
бенностям (страте2ИЯМ) распределения
и утилизации ресурсов, которые максими
зируют передачу в поколениях кодирующих
их reHoB. Значит, хотя в основе физиолоrии
всех орrанизмов лежат общие принципы,
у каждой особи она «настроена» eCTeCTBeH
ным отбором на конкретные эколоrические
условия протекания перечисленных выше
процессов это и называют адаптацией.
Отсюда вытекает, что в функциональ
ном отношении орrанизм действует как ин
теrрированное целое. Компромиссное обе
спечение метаболических потребностей MO
жет вести к компромиссу между элемента
ми приспособленности: так, увеличение за
трат на локомоцию обычно повышает BЫ
живаемость, но снижает вклад ресурсов
в размножение Принцип интеrрации важен
.. :;ло
W Влияние на
ПРИСПОСОБляемость
.... Выживание
Продолжительность
развития
Плодовитость
также для морфолоrических структур, по
скольку развитие одной из них должно быть
совместимо с существованием друrих. OT
сюда следуют два вывода. Во первых, дол
жен быть непосредственный контроль фор
мообразования, физиолоrических функций
и поведения. Он обеспечивается химически
ми и электрическими сиrналами (см. rл. 16).
BO BTOpЫX, на интеrрацию ориентирован
эволюционный отбор признаков: каждая re
нетически детерминируемая черта должна
быть совместима с окружающей орrанизм
средой, т. е., взаимодействуя с ней, должна
повышать шансы на выживание и оставле
ние потомства. Следовательно, reHbJ
и определяемые ими признаки отбираются
как часть недеЛUМО20 цеЛО20. Этоrо Ор2аниз"
меННО20, или холистuчеСКО20, подхода, до
некоторой степени отличающеrося от «фи
лософии reHHoro пула», которая рассматри
вает орrанизм как набор самостоятельных
«эrоистичных» reHoB, мы и придерживались
в предыдущих rлавах.
17.2. rлавное репликация
и размножение
Вклад в репродуктивные процессы
очень важное, возможно, основное направ
ление использования ресурсов орrанизмом
Они идут на образование стадий размноже
ния (пропаrул), которые являются cpeд
ством передачи в поколениях rенетической
информации.
Иноrда, как отмечалось выше (разд.
14.1.1), клетка или rруппа клеток. отделяю
щаяся от родительской особи, полностью
544
rлава 17
I
rn
.....
I
.......
I I
Постмитоти Премитоти Метасраза
ческая интер ческая
сраза 1 интерсраза
Просраза
Новый цикл
Телосраза
Рис. 17.2. Поведение пары хромосом при ми
тозе (по Paul, 1967)
воспроизводит reHoM последней. В основе
TaKoro бе(nОЛО20 раЗJwно.жения лежит митоз
(рис. 17.2). за счет KOToporo клетка или
клетки пропаrулы образуются сами и
в свою очередь дают rенетическую копию
родительскоrо' орrанизма.
В противоположность этому отдельные
половые клетки (rаметы), возникающие
обычно в специализированных opraHax (ro
надах), для возникновения HOBoro орrаниз
ма должны, как правило, сливаться с друrи
ми половыми клетками. rаметы образу
ются в процессе мейотическоrо деления,
дающеrо клетки с количеством ДНК и xpo
мосом вдвое меньшим, чем в обычных co
матических клетках (рис. 17.3). Полный Ha
B ==
Предшествующая (Пахитена) (Диплотена)
телосраза митоза
I
/ I
= 1; J I / I \......J .............. ,
...
(Диакинез) f'---.J "'"
1 я телосраза I ,. I
2 я телосраза
"'1
I
"1
Рис. 17.3. Поведение пары хромосом при
мейозе. Обратите внимание, что процесс крос--
синrовера приводит к иному, чем у родителей,
сочетанию reHoB в хромосомах потомства (по
Paul 1 967)
бор хромосом восстанавливается при слия..
нии [амет (синrамии, оплодотворении),
d поскольку они часто происходят от раз..
ных родителей, потомство получает сме..
Шdнные rенетические проrраммы. Этот
процесс называют половы/vt раЗ.МНОJlсение/vt
(см. rл. 14).
17.3. Онтоrенез
Продукты Рdзмножения мельче и обыч..
но устроены проще. чем родители. При по..
ловом процессе они одноклеточные, в то
время КйК у родителей MHoro функциональ..
но дифференцированных клеток. Следова..
тельно.. для индивидуальноrо развипlИЯ, или
онтО2енеза, необходимы клеточные деле..
ния, увеличивающие размер (рост), и кле..
точная специализация (диффереllцировка).
Поскольку специфические клетки находятся
в cTporo определенных местах орrанизма,
должен происходить 2испl02енез. Продукты
размножения обычно более или менее сфе..
рические, а взрослый орrанизм имеет сло..
жную (рорму, т. е. развитие включает фор..
мообразование (МОрфО2енез).
Деление зиrоты (дробление оплодотво"
peHHoro яйца) происходит путем митоза.
Он точно воспроизводит ее reHoM и описан
выше. Некоторые из первых эмбриолоrов
(особенно ABrycT Вейсман, 1834 1914) счи..
тали, что дuфференцировка включает посте..
пенное отделение в специфических клеточ..
ных линиях ненужной части наследственно..
ro материала. Однако изучение митоза и от..
крытие возможности полной реrенерации
некоторых беспозвоночных из мелких фраr..
ментов их соматической ткани показали,
что каждая соматическая клетка содержит
более или менее полную копию исходноrо
reHoMa. По мере дифференцировки в раз..
личных клетках должны происходить
«включения» или «выключения» специфиче..
ских частей этой наследственной проrрам"
мы. Таким образом, клетки, имея одинако"
вый rенотип. синтезируют различные белки
и, следовательно, функционируют по..
разному.
Хорошо извеСТНdЯ модель TaKoro пере-
КЛlOчения основана на изучении бактерии
Escherichia сои. Одной бактериальной клет-
ке дают разделиться на две дочерние; затем
Еще раз об основных принципах
культивируют одну из образовавшихся кле
ток в среде, содержащей как источник энер
rии rлюкозу, а вторую в среде с друrим
сахаром лактозой. В присутствии лакто
зы бактерии продуцируют фермент
rалактозидазу, который превращает ее
в rлюкозу, поскольку непосредственно лак
тоза использоваться не может. В среде
с rлюкозой этот фермент не образуется.
у клеток в обеих средах один и тот же re
ном, однако процессы синтеза различны,
т. е. можно сказать, что они дифференциро
ваны в отношении npyr друrа.
Клетки в rлюкозной среде также coдep
жат rены, кодирующие В rалактозидазу, oд
нако обычно они «выключены» белком
репрессором. Он препятствует специфиче
ской rенной транскрипции, связываясь с co
ответствующим оператором. Лактоза, по
падая в клетку, превращается в аллолакто
зу, которая взаимодействует с репрессором
таким образом., что он перестает инrибиро
вать оператор и начин ется синтез РНК,
служащей матрицей для синтеза фермента,
называемоrо в данном случае индуцирован
ным, т. е. запускаются метаболические про
цессы, необходимые для утилизации лакто
зы. Модель изображена на рис. 17.4.
По аналоrии с такой реrуляцией путем
включения в принципе возможна и реrуля
ция путем выключения, хотя понятно, что
она менее экономична. О молекулярной oc
i О
Ф ...
. .
R
Хромосома
Рис. 17.4. Модель reHHoro контроля. reH i KO
дирует реп рессор R который связывается со
специфической частью хромосомы
оператором (О) и инrибирует транскрипцию
reHoB G" G 2 и G з . Если для ферментов Р" Р 2 И Р з
имеется субстрат (13), он, связываясь с репрес
сором, изменяет ero конфиrурацию, не позво
ляя ему блокировать оператор. При этом
транскрибируются G 1 , G 2 и G з , синтезируются
Р 1 . Р 2 И Р З и метаболизируется J3 (по Maynard
Smith, 1986)
545
нове rенной реrуляции при дифференциров
ке эукариотических клеток известно мало,
однако похоже, что ее механизм близок
к рассмотренному выше.
FистО2енез требует упорядоченной pa
боты переключающих факторов. Он обеспе
чивается либо детальным набором инструк
ций о том, как клетка должна реаrировать,
занимая определенное положение в эмбрио
не, либо менее детальной информацией, по
зволяющей клеткам «узнавать» свое MeCTO
нахождение и действовать в соответствии
с ним. Поскольку rенетические инструкции
находятся внутри клетки и крайне малове
роятно, что детальная информация может
поступать в нее и вне или даже передава
ться из ее uитоплазмы к rенетической про
rpaMMe внутри ядра (запрет, известный как
«центральная доrма молекулярной биоло
rии»), более правдоподобно предположение
о контроле, обеспечиваемом позиционной
информацией. Однако до сих пор остается
заrадкой, на чем она основана, если все
клетки эмбриона rенетически идентичны.
Возможно, это связано с цитоплазмой яйце
клетки. Она часто даже визуально HeOДHO
родна (например, пиrмент и желток распре
делены в ней неравномерно ), и уже продук
ты первоrо деления MorYT иметь разную ци
топлазму, а следовательно, и неодинако
вые условия для экспрессии своих reHo
мов.
Можно видеть, что этот процесс COOT
ветствует предположению Вейсмана, за ис
ключением Toro, что в ходе дробления клет
ки начинают различаться цитоплазматиче
ским, а не ядерным материалом. Это делает
возможным материнское влияние на разви
тие через цитоплазму яйца. В конечном сче
те такой контроль, независимо от ero при
роды, бывает либо жестким, определяю
щим судьбу клеток на ранних стадиях OHTO
rенеза (детерминированный пропесс), либо
более rибким, позволяющим эксперимента
льно изменять судьбу отдельных клеток дa
же на достаточно поздних стадиях (Heдeтep
минированный процесс).
МОрфО2енез сочетает перемещения кле
ток и дифференциальный рост Первый про
цесс хорошо описан для ранних стадий раз
вития прозрачноrо эмбриона MopCKoro ежа
(рис. 17.5, А и rл. 15): различные клетки BHY
546
rлава 17
А
Б
Рис. 17.5. А. Раннее развитие MopcKoro ежа
ПМ клетки первичной мезенхимы
миrрируют в «уrлы» эмбриона используя фи
лоподии; rК rрушевидные клетки (по Gustaf..
50П, Wolpert, 1967). Б. Изменение формы кле
ток в зоне инваrинации. БМ базальная MeM
брана
три Hero можно не только наблюдать, но
и реrистрировать с помощью замедленной
киносъемки. На ранней стадии эмбрион
образует состоящий из клеток полый
шар бластулу. Затем некоторые клетки
должны переместиться внутрь, чтобы сфор
мировать внутренние opraHbI, например ки
шечник. Этот процесс, называемый [aCTpy
ляцией, происходит в два этапа. Сначала
внутрь проникают отдельные клетки, затем
происходит инваrинация всей стенки. и
rрация начинается утратой контакта клеток
друr с друrом; они движутся в бластоцель
(внутреннюю полость бластулы) и затем
перемещаются вдоль ero внутренней по
верхности't подтяrиваясь филоподиями. Ин
ваrинация включает две стадии: медленное
проrибание стенки сменяется после HeKOTO
poro перерыва быстрым ее впячиванием.
Первая фаза, вероятно.. связана с измене
ни ем адrезии клеток: они слабее прилипают
друr к друrу, но остаются прикрепленными
к базальной мембране. Это придает им [py
шевидную форму и может вызывать проrи
бание вовнутрь, как показано на рис. 17.5,
Б. Быстрая фаза, вероятно, обеспечивается
клетками, образующими в этой зоне фило
подии, которые, установив контакт с кры"
шей бластоцеля, сокращаются. Эти процес
сы, включающие межклеточные мембран..
ные взаимодействия и активную миrрацию
клеток с помощью филоподий, вероятно,
довольно типичны для морфоrенетических
изменений у животных. Основной вопрос,
который остается нерешеННЫМ, как они
контролируются, обеспечивая формирова..
ние полностью орrанизованной особи.
Наконец, дифференциальный рост внеш
них и внутренних opraHoB может вести
к формообразованию. Джулиан Хаксли (Ju..
lian Huxley) в своей книrе «Problems of Rela..
tive GI'owth» (<<Проблемы относительноrо
роста») в 1932 r. отмечал, что, если по..
20
А
10
5
мм
2
1
0,5
20
50
100
мм
Рис. 1'7.6. Пример аллометрическоrо роста Ha
ceKoMoro Carausius тorosus. Зависимость ДЛИ
ны задней части проторакса (А), ширины rоло
вы (Б) и диаметра rлаза (В) от общей ДЛИНЫ
тела. Координаты лоrарифмические (по Wig
gle5worth, 1965)
Еще раз об ОСНОВНЫХ принципах
строить в двойных лоrарифмических коор"
динатах rрафик зависимости размера одной
части орrанизма от размера друrой (или
Bcero орrанизма).. он часто будет представ..
лен прямой линией (рис. 17.6). Равные ин..
тервалы на лоrарифмической шкале соот..
ветствуют увеличенйю в одинаковое число
раз, т. е. эти прямые предполаrают, что
каждый opraH увеличивается (растет) orHo"
сительно друrоrо снекоторой специфиче..
ской скоростью. Понятно, что аллометри..
ческие зависимости существенно влияют на
морфоrенез орrанизма.
В rл. 15 обсуждены детали основных
процессов развития, т. е. дифференцировки,
rистоrенеза и морфоrенеза, применительно
к беспозвоночным. Здесь мы затронули
лишь некоторые вопросы их реrуляции.
17.4. Онтоrенез и филоrенез
Поскольку и онтоrенез, и филоrенез
внешне проrрессивны, заманчиво предполо..
жить, что последний происходит путем
появления у первоrо дополнительных ста..
дий. В таком случае онтоrенез рекапитули..
руепl филоrенез. Эту точку зрения сформу"
лировал Эрнст rеккель в конце XIX в. В со..
ответствии с ней онтоrенез «высшей» фор..
мы повторяет взрослые формы более низ..
Проuесс
Результат
547
ких уровней орrанизации. Фон Бэр, совре..
менник rеккеля, напротив, утверждал, что
ни один взрослый орrанизм не повторяет
взрослых стадий проще орrанизованных
животных, однако, поскольку развитие
всеrда идет от недифференцированноrо со..
стояния к дифференцированному, ero нача..
льные фазы у различных таксонов должны
быть одинаковы. Следовательно" эмбрио..
нальные, а не взрослые формы, в различных
филетических линиях повторяются, что
вполне соответствует принципам развития,
изложенным в последнем разделе.
Эволюция путем появления дополни..
тельных стадий онтоrенеза точно со ответ..
ствует ламаркистской теории приобретен..
ных признаков, которые обычно развиваю..
тся в течение жизни и добавляются к суще..
ствующим структурам. Однако менделев..
ская rенетика опроверrает эту точку зрения,
поскольку мутация может привести к изме..
нению на любой стадии развития. Действ и..
тельно, сейчас известно, что скорость раз..
вития контролируется rенами. Изменения
последних MorYT либо остановить ero
(устраняя «прошлый» взрослый орrанизм),
либо продлить за нормальные пределы. Ва..
льтер rapcTaHr (W. Garstang, 1868 (949)
был одним из первых зоолоrов, оценивших
важное значение rенетики для развития
Изменение скорости Сдвиr конечной точки Сдвиr начальной точки
Педоморсроз
I
I
1
lco
I
о I
1 :I:
I
'8
Замедление или неотения
Пераморсроз
I
,
/1
/ I
/ I
о / I
/ 1:1:
I I
t 18
Акселерация
Смещение назад
,
1
.А
/1
,
1
1
I
1
1
r иперморсроз
Смещение вперед
Рис. 17.7, Классификация возможных эволюционных сдвиrов в развитии По оси абсцисс
продолжительность развития или размер; по оси ординат морфолоrические изменения (по
Calow, 1983)
548
rлава 17
33 й день 45...й день 48...й день 62...й день
Рис. 17.8. Стадии развития мноrоножек. Ше..
стиноrая стадия возрастом 33 дня иноrда испо..
льзуется как aprYMeHT в пользу педоморфноrо
происхождения насекомых (см. также Gould,
1977 )
и эволюционные возможности, открывае..
мые изменением ero темпов.
На рис. 17.7 приведен полный спектр
эволюционных сдвиrов, которые моrли бы
возникнуть таким путем. Каждый rрафик
включает траектории развития, отражаю..
щие зависимость формы от размера или
возраста. Сплошные линии соответствуют
предковым траекториям, а прерывистые
возникшим у потомков. Отмечены начало
развития и ero прекращение. В верхнем
ряду rрафики отражают замедление (неоте..
ния) или «усечение» (проrенез) развития; та..
кие ero изменения называются педоморфо"
зом. При этом эмбриональные, личиночные
или ювенильные признаки сохраняются
у взрослой формы. Нижний ряд иллюстри"
рует противоположную тенденцию (пера...
морфоз).
Вальтер rapcTaHr особенно интересо..
вался личиночными формами и полаrал,
что эволюция шла в основном за счет педо..
морфоза. Действительно, ero роль в ряде
случаев моrла быть велика (rл. 2). Напри..
мер, среди современных видов половозре..
лость у самцов некоторых эхиурид ирако",
образных (и рыб) наступает при мелких раз..
мерах тела; такие самцы «паразитически»
прикрепляются к значительно более круп..
ным (иноrда на несколько порядков) сам..
кам. Происхождение шестиноrих насеко..
мых от мноrоножкоподобных предков ско"
рей Bcero также связано с педоморфозом
(рис. 17.8). Наконец" широко признанная
теория эволюции позвоночных предпола..
raeT, что им дали начало личиночные фор..
мы, напоминающие свободноплавающую
личинку..«rоловастика» современных обо..
лочников (см. рис. 7.30).
Возможен и филоrенез за счет ускорения
развития. Рассмотрим эволюционные те н..
денции лопастных линий на раковинах ам..
монитов (рис. 17.9). Наклон rрафика у по..
томков круче, чем у предков, т. е. имело ме..
сто именно ускорение (акселерация) их фор
мирования. К тому же рост лопастных ли..
ний потомков идет дальше, чем у предков,
что предполаrает 2uперморфоз (рис. 17.7).
Наконец, rрафики у потомков проходят
«выше», т. е. потомки начинают развитие
раньше происходит ero смещение вперед
(рис. 17.7).
Теперь обобщим. Онтоrенез не рекапи
тулирует филоrенез, но, поскольку незави"
симо от таксона он подчиняется ряду общих
принципов, различия, обусловленные диф..
ференцировкой, размерами, rисто.. и мор..
фоrенезом (см. последний раздел), возни..
кают на основе общих для всех животных
процессов митоза, дифференциальной
экспрессии reHoMa, движения клеток, диф..
ференциальноrо роста. В связи с этим (как
предполаrал фон Бэр) раннее развитие всех
5
....
s:
s:
I
s:
с::
J
.s:
О
I
t>
(о
с
О 100
с::
J:i
I
s:
с::
q
со
I
s:
m
о
а
с
4l
3
2
1
10 100 200 300
Диаметр раковины, мм
Рис. 17.9. Спектр изменений развития у aMMO
нитов: 1 предок, 2 5 потомки. Более CTa
рые части стадий справа (по Newell, 1949;
СМ. также Calow, 1983)
Еще раз об основных принципах
549
орrанизмов несколько сходно. Тем не менее
rлубокое влияние на филоrенез, по
видимому, оказали небольшие изменения
темпов этих процессов, вызванные, напри
мер, незначительными rенетическими OT
клонениями. Последние моrли косвенно
привести к скачкам между уровнями opra
иизации животных (rл. 1, 2). Эту точку зре
иия отстаивал Вальтер rapcTaHr.
17.5. Дополнительная
литература
Ca!ow Р. 1983. Evolutionary Principles. Blackie,
Glasgow and London.
Gou!d S. J. 1977. Ontogeny and Phylogeny. Harvard
University Press, Cambridge, Massachusets.
Них/еу J. S. 1932. Problems of Relative Growth.
Methuen, London. О.
М ayпard Sтith J. 1986. The Problems of Biology.
Oxford University Press, Oxford.
Раи! J. 1967. СеВ Biology. Heineman, London.
СЛОВАРЬ ТЕРМИНОВ
Абдомен задний отдел тела, разделен
Horo на три отдела" передний из которых
rолова (ер. торакс).
Аборальныii относящийся к поверхно
сти тела, противоположной той, на которой
расположен рот., например у животных
с ртом в центре нижней или верхней поверх
ности тела.
Автотомия индуцированная самим op
rанизмом ампутация конечности или части
тела, например как способ избеrания хищ
ника, почкования (см.) или фраrментации
( см. ).
Аденозинтрифосфат (А ТР) r лавная
энерrосодержащая молекула орrанизма.
Аддуктор мышца, смыкающая и yдep
живающая в сомкнутом состоянии створки
раковины.
Акклимация изменение физиолоrии
в результате воздействия на орrанизм изме
нившихся окружающих условий.
Акрон передний, не относящийся к
cerMeHTaM отдел тела членистоноrих.
Акросома волокнистая трубка (или
трубки) на переднем конце сперматозоида
(см.), вступающая в контакт и сливающаяся
с клеточной мембраной яйцеклетки (см.) во
время оплодотворения (см.).
Активный транспорт движение paCTBO
ренных веществ против rрадиента KOHцeH
трации, требующее затрат энерrии.
Амебоцит клетка, способная к aMe
боидному движению.
Анабиоз CM. криптобиоз.
Анаболизм см. метаболизм.
Анаэробный безвоздушный термин,
обычно используемый для описания даю
щих энерrию метаболических (см.) процес
сов, протекающих в отсутствие кислорода.
Аноксия отсутствие свободноrо ки
слорода.
Антенны нитевидные и часто удлинен
ные хемосенсорные придатки на rолове не..
которыхчленистоноrих,полихет,онихофор
и др.
Аподеl\lа направленный внутрь тела
вырост экзоскелета членистоноrих
Апомиксис см. бесполое размножение.
Арренотокия форма партеноrенеза
(см.), при которой неоплодотворенные
яйпеклетки развиваются в rаплоидных caM
цов, а оплодотворенные в диплоидных
самок.
Ацеломический не имеющий полости
тела, если не считать просветов кишечника
и друrих opraHoB.
Базальная мембрана аморфный слой,
подстилающий эпителий (см.); образован
коллаrеном (см.) и уrлеводом.
Бентосный относящийся к дну или
субстрату в водных системах (ср. пелаrиче..
ский) .
Бесполое размножение форма размно
жения, не включаюшая реДУКIlионноrо
мейотическоrо деления и слияния raMeT
(см.) (ср. половое размножение).
Билатеральная симметрия тип симме..
трии, при котором тело можно разделить
только на две зеркально идентичные поло..
вины.
Биотурбация нарушение бентосных
(см.) осадков деятельностью животных.
Бирадиальная симметрия тип симме..
трии, при которой каждый из четырех квад..
рантов сферическоrо орrанизма или
эмбриона такой же, как противоположный,
но отличается от соседних.
Бластема rруппа недифференцирован"
ных делящихся клеток, способных дать Ha
чало дифференцированным клеткам (см.
дифференцировка ).
Бластопор отверстие, через которое
Словарь терминов
эмбриональная кишка rаструлы (см.) сооб
щается с внешней средой.
Бластоцель полость внутри бластулы
(см.).
Бластула полый шар из клеток, обра
зующийся из зиrоты (см.) в ходе дробления
(см.) при эмбриоrенезе.
Брожение ферментативное разложе
ние орrанических веществ с образованием
АТР в условиях аноксии (см.).
Брюшко см. абдомен.
Вилочка CM. фурка.
Вторичноротость резуль тат развития,
коrда бластопор (см.) не становится ртом
взрослой особи, но может образовать ее
анус (ср. первичноротость).
rамета rаплоидная половая клетка,
способная сливаться с половой клеткой
противоположноrо пола, образуя зиrоту
(см.); см. сперматозоид и яйцеклетка.
rанrлий оформленное скопление HepB
ной ткани, содержащее нейроны (см.).
rаплодиплондия CM. арренотокия.
rаплоидия присутствие одинарноrо
XpOMocoMHoro набора в соматических (СМ.)
клетках или raMeTax (см.) (ср. диплоидия,
полиплоидия).
rаструла эмбриональная стадия, сле
дующая за бластулой (см.), на которой oд
послойная структура превращается в ДBY
слойную за счет клеточной миrрации, инва
rинации и Т.д. и происходит обособление
клеток мезодермы (см.).
rаструляцня процесс развития racTpy
лы (см.) из бластулы (см.).
rеммула мноrоклеточная пропаrула
(см) при бесполом размножении (см.) HeKO
торых rубок, покрытая защитной оболоч
кой.
rемоцель полость тела, образованная
кровеносными синусами; часто возникает
из бластоцеля (см.) (ср. целом и псевдо"
цель) .
rермафродит орrанизм, способный
одновременно или в разные периоды про
изводить как яйцеклетки (см.), так и сперма
тозоиды (см.).
rетеротрофность питание rотовыми
орrаническими соединениями.
rидравлический приводимый в дей
ствие давлением жидкости.
rидростатический скелеl система
551
передачи силы мышечных сокращений жид..
костью в полостях тела или тканях.
rиперполяризация увеличение разно..
сти электрических потенциалов, существую
щих в состоянии покоя по обе стороны
клеточной мембраны (ср. деполяриза
иия)
rипоксия низкая доступность кисло..
рода.
rипостом возвышение, на котором Ha
ходится рот у книдарий.
rистолиз разрушение тканей.
r лазок простой светочувствительный
opraH (ср. сложный rлаз).
rлия (нейроrлия) клетки, образующие
обкладку нейронов (см.).
rлотка отдел передней кишки (см.),
находящийся позади ротовой полости
и перед пищеводом.
rоловоrрудь часть тела некоторых pa
кообразных, образованная слиянием rоло..
вы и rруди (см.).
rомойотермность поддержание отно"
сительно постоянной температуры тела (ср.
пойкилотермность ).
rоноподия видоизмененная конеч
ность, служащая opraHoM копуляции (см)
(ср. циррус, пенис).
rOHonop выводное отверстие для ra
мет.
rрифельки мелкие парные нечлени..
стые выросты, связанные с основаниями KO
нечностей у ряда мноrоножек и присут
ствующие в эквивалентном положении на
некоторых абдоминальных (реже и на TO
ракальных) cerMeHTax большинства первич
нобескрылых насекомых.
rрудь см. торакс.
Двухлучевая симметрия см. бирадиа..
льная симметрия.
Деполяризация уменьшение разности
электрических потенциалов, существующих
в состоянии покоя по обе стороны клеточ..
ной мембраны (ср. rиперполяризация).
Дерма немышечный слой стенки тела
мезодермальноrо (см.) происхождения, ле
жащий под эпидермисом (см.).
Детерминированное развитие см. мо"
заичное развитие.
Детрит частицы разложившеrося или
разлаrающеrося орrаническоrо вещества
вместе с колонизировавшими ero микроор
552
Словарь терминов
rанизмами в воде, на субстрате или в ero
толще.
Диапауза фаза покоя в жизненном ци--
кле. при которой интенсивность метаболиз--
ма (см.) низкая, а орrанизм устойчив к не--
блаrоприятным окружающим условиям.
Диморфизм наличие у одноrо вида
двух различных ( обычно морфоло--
rически) форм (ср. мономорфизм, полимор--
физм).
Диплоидия присутствие двойноrо хро--
MocoMHoro набора в соматических (см.)
клетках (ср. rаплоидия, полиплоидия).
Дифференцировка процесс специали--
зации тотипотентных (см.) клеток эмбриона
(см) для выполнения различных функций.
Дробление митотические деления зи--
rоты (см.), формирующие мноrоклеточный,
но недифференцированный эмбрион (см.);
роста клеток при этом не происходит.
Дыхальце выводное отверстие тра--
хейной (см.) системы.
Дыхательный пиrмент молекула,
обратимо соединяющаяся с кислородом
и служашая для ero пере носа или запасания.
Естественный отбор эволюционный
механизм, предложенный ч. Дарвином, ос--
нованный на различиях в выживании
и оставлении потомства особями в среде
с оrраниченными ресурсами.
Жrутики выступающие из клетки ор--
rанеллы, совершающие круrовые или спи--
ральные вращательные движения; обычно
относительно длинные и встречаются по
одному по два на клетку; строение жrути"
ков эукариот (см.) и бактерий принципиаль--
но различное (ср. реснички).
Живорождение развитие эмбриона
(см.) в теле родителя с частичным использо..
ванием непосредственно поступающих от
последнеrо ресурсов.
Жизненная сила мистическая нефизи--
ческая сила, которая, как некоrда считали,
обусловливает жизнедеятельность, направ--
ляет развитие и эволюцию орrанизмов.
Задняя кишка последний отдел кишки
с эктодермальными (см.) происхождением
и выстилкой (ср. передняя кишка, средняя
кишка) .
Зиrота клетка, возникающая при слия--
нии сперматозоида (см.) с яйцеклеткой
(см.), т. е. при оплодотворении.
Зооид особь модульной колонии (см.),
возникшая в ходе неполноrо почкования
(см.); термин обычно не применяется к кни--
дариям.
Зооксантеллы симбиотические водо--
росли--динофлаrеллаты в тканях различных,
rлавным образом морских, беспозвоноч--
ных.
Зоохлореллы сим биотические зеленые
водоросли (хлорофиты) в тканях различ--
ных, rлавным образом пресноводных, бес..
позвоночных.
Имаrинальный диск rруппа недиффе..
ренцированных клеток внутри личинки
(см.), из которой развивается специфиче..
ская система opraHoB взрослоrо животноrо.
Интеrумент немышечные слои стенки
тела.
Интерстициальная (а) фауна, населяю--
щая пространства (интерстициаль) между
частицами осадка; (6) тотипотентная эпи..
дермальная клетка у книдарий.
Интроверт выворачивающийся и втя"
rивающийся передний отдел тела
Инфауна бентосные (СМ.) животные.
обитающие в толще субстрата или в норках
(ср. эпифауна).
Карапакс защитный экзоскелетный
шит. покрывающий целиком или частично
дорсальную и боковые поверхности тела
членистоноrоrо.
Карта зачатков формализованное
описание пространственноrо расположения
в зиrоте (см.), эмбрионе (см.) или имаrина--
льном диске (см.) клеток или областей, КО--
торые в норме дают начало тем или иным
частям орrанизма.
КJIадон rруппа таксонов, происходя--
щих от общеrо предка.
Клейдоическое яйцо яйцо (см.), покры"
тое защитной оболочкой.
Клоака расширенный последний от..
дел кишечника, куда открываются протоки
друrих систем opraHoB.
Книдоцит клетка книдарий, содержа..
щая нематоцисты (см.).
Коrорта rруппа особей в популяции,
родившихся примерно в одно и то же
время.
коксальныe меllIКИ выворачиваюшиеся
тонкостенные пузыри у оснований Hor неко"
торых унирамий, служащие для поrлоще..
Словарь терминов
ния окружающей влаrи; сходные обраЗОВd
ния известны и у онихофор.
Коллаrен' волокнистый белок, обычно
связанный с соединительной тканью или KY
тикулярными решетками.
Колония rруппа орrанизмов, возник
ших бесполым путем и оставшихся ассоции
рованными между собой: у мноrих живот
ных объединяющий тканевый контакт
с друrими полипами (см.) или зооидами
(см.) является результатом неполноrо поч
кования (см. также модульный орrанизм);
особи, возникшие половым путем и обра
зующие полупостоянные скопления.
Кольчатость разделение наружной по
верхности цилиндрическоrо орrанизма.. op
raHa и т. п. бороздками на ряд колец, BHeIII
не выrлядящих кйк сеrменты, но не COOTBeT
ствующих им.
Комиссура поперечное соединение Me
жду нервными стволами или rанrлиями.
Комменсал орrанизм, тесно связан
ный с орrанизмом друrоrо вида (например,
живущий в ero норке), но не приносящий
ему ни вреда, ни пользы.
Коннектива см. комиссура.
Копуляция перенос спермы opraHoM
одной особи в тело друrой; широко распро
страненная., но не единственно возможная
стадия, предшествующая внутреннему
оплодотворению.
Криптический облик см. покровитель..
ственный облик.
Криптобиоз устойчивое состояние без
видимых проявлений жизнедеятельности во
время периодов с неблаrоприятными усло
виями среды (обычно с дефИIIИТОМ влаrи),
позволяющее, например, обитать в пересы
хающих водоемах.
Ктенидий жабра моллюсков, состоя..
щая (исходно) из двух супротивных рядов
филаментов., отходящих от центральной
оси.
Куколка неподвижная переходная
стадия развития некоторых насекомых Me
жду личиночными стадиями и имаrо.
Кутикула неклеточный устойчивый
покров, секретируемый эпидермисом (см.)
и периодически линяющий; часто opHaMeH
тирован и/или локально утолшен в пластин
ки.
Лемниски трубчатые мешки, связан
553
ные с хоботком скребней и присутствующие
по крайней мере у некоторых бделлоидных
коловраток; в принципе эпидермальные
.впячивания, функционирующие, вероятно,
как rидравлические резервуары
Лецитотрофность развитие за счет
внутренних ресурсов (желтка), полученных
от матери: обычно термин применяется
к морским личинкам (ср. планктотроф"
ность).
Литораль ПРИЛИВНО ОТЛИВНdЯ полоса
побережья.
Личинка ювенильная стадия, заметно
отличающаяся по морфолоrии и эколоrии
от взрослоrо животноrо.
Ложная клешня конечность членисто..
Hororo, дистальный членик которой, подrи
баясь, прижимается к предпоследнему; BЫ
полняет ловчую функцию.
Лофофор система rидравлически уп
равляемых ресничных пищедобывательных
щупалец, образованных выростами стенки
тела и окружающих рот (но не анус).
Vlакромеры крупные, боrатые желт..
ком клетки paHHero эмбриона (см.) (ср. ми
кромеры).
Макрофаr орrанизм, питающийся OT
носительно крупными пищевыми частица
ми (ср. микрофаr).
Макроэволюция становление TaKCOHO
мическоrо разнообразия, т. е. эволюцион
ные изменения, при водящие к появлению
новых видов и надвидовых таксонов (ср.
микроэволюция ).
Максиллы ротовые придатки члени
стоноrих, расположенные позади мандибул
(см.) и функционально их дополняющие.
Мальпиrиевы сосуды слепые трубча
тые 'Экскреторные дивертикулы кишки.
Мандибулы жвалы, челюсти.
Мантийная полость полость, окружен
ная мантией моллюсков и плеченоrих. в ко..
торой находятся соответственно орrаны
дыхания и захвата пищи.
Медуза одна из двух форм тела у кни
дарий: пульсирующая. обычно пелаrиче
ская (см.), диско , колоколо или зонтико
видная, часто студенистая (ср. полип).
Ме.зенхима диффузные соединитель
нотканныIe клетки, находяшиеся в студени
стом матриксе.
Мезоrлея имеющий разную толщину
554
Словарь терминов
клеточный или неклеточный слой студени
cToro вещества между наружным и BHYTpeH
ним клеточными слоями кишечнополост
ных.
Мезодерма зародышевый слой. фор
мирующийся между эктодермой (см.)
и энтодермой (см.).
Мезосома второй отдел тела трехраз
дельных олиrомерных (см.) животных; ero
целом (мезоцель) может rидравлически
поддерживать лофофор (см.) (ср. просома,
метасома).
Метаболизм обмен веществ; химиче
ские процессы в орrанизме't включающие
разрушение структур и соединений (катабо
лизм) и их образование (анаболизм).
Метамерия тип строения тела, xapaK
теризующийся линейной послеловательно
стью нескольких (мноrих) cerMeHToB (см.)
(ср. мономерия, олиrомерия).
Метаморфоз резкое изменение CTpoe
ния тела во время превращения личинки
(см.) во взрослое животное.
Метанефридий открытый нефридий
(см.) с внеклеточным выводным протоком
(ср. протонефридий)
Метасома третий отдел тела трехраз
дельных олиrомерных (см.) животных; ero
целом (метацель) образует rлавную по
лость тела (ср. ПрОСОМd, мезосома).
Метахрональный ритм тип синхрони
зированноrо движения ресничек (см.) или
конечностей, при котором движение каждо
ro элемента происходит с определенным
сдвиrом по фазе относительно друrоrо эле
мента.
Микроворсинки мноrочисленные мел
кие пальчатые выросты на свободной по
верхности клетки, участвующие в абсорб
ции или (при соответствующей специализа..
ции) в сенсорном восприятии; у цестод Ha
зываются микротрихами.
Микромеры мелкие, бедные желтком
клетки paHHero эмбриона (СМ.) (ср. MaKpo
меры>,.
Мнкротрихи см. микроворсинки.
Микрофаr орrанизм, питающийся
мелкими пищевыми частицами (ер. MaKpo
фаr) .
Микроэволюция изменение частот re
нов в популяции (ср. макроэволюция).
Миксонефридий см. нефромиксий.
Мимикрия сходство снеживым объек"
том или с друrим орrанизмом, потенциаль..
но позволяющее за счет «ложной идентич"
ности» ему избежать нападения.
Модульный орrанизм колония (см.) из
повторяющихся соединенных между собой
единиц «<особей»), возникших бесполым пу
тем (см. полип, зооид).
Мозаичное развитие тип развития, при
котором уже на ранней стадии онтоrенеза
будущее каждой клетки эмбриона точно
определено (особенностями материнской
цитоплазмы), поэтому возможность заме..
щения утраченных частей зародыша очень
оrраничена (ср. реrуляционное развитие).
Мономерия тип строения тела, харак"
теризующийся ero неразделенностью на
сеrменты (см.) (ср. олиrомерия. метамерия).
Мономорфизм наличие у вида един"
ственной формы тела (ср. диморфизм, по..
лиморфизм).
Моноцикличность свойство размножа..
ться только один раз в жизни, после чеrо
поrибать.
Нанопланктон планктон (см.) с наи..
большим промером тела 2 20 мкм.
Наяда водная нимфа (см.) некоторых
насекомых, сильнее, чем нимфа, отличаю
щаяся от имаrо в результате специфических
адаптаций к жизни в воде, например к ловле
водной добычи и/или потреблению paCTBO
peHHoro кислорода.
Недетерминированное развитие см. ре..
rуляционное развитие.
Нейрон клетка, специализированная
для проведения электрических сиrналов't
переносящих информацию.
Нейропиль область нервной системы,
в которой сосредоточены синапсы (см.)
нервных волокон.
Нейросекреторная клетка нейрон (см.)
с функциями железы, обычно секретирую"
щий ropMoHbI.
Нектон пелаrические (см.) животные,
способные плыть против течения (ср. планк
тон).
Нематоциста внутриклеточная opra..
пелла книдарий с выворачивающейся спи..
рально закрученной нитью, используемая
для ловли добычи, защиты и т. Д.; нахо..
дится в книдоцитах (см.).
Неотения см. педоморфоз.
Словарь терминов
555
Непрямое развитие развитие через ли..
чиночную (см.) стадию (ср. прямое разви"
тие) .
Нефридий осмореrуляторно экскре..
торный opraH эктодермальноrо (см.) проис..
хождения (ср целомодукт).
Нефромиксий похожий на метанефри..
дий (см.) opraH с участками эктодермально"
ro (см.) и мезодермальноrо (см.) происхо
ждения.
Нимфа ювенильная стадия насекомо"
ro, мало отличающаяся от взрослой особи,
если не считать размеров и развития HeKO
торых систем (например, крыльев, rонад);
обычно зачатки крыльев находятся снаружи
(в отличие от личинки насекомых).
Окончательный хозяин хозяин парази..
та (см), в котором происходит ero половое
размножение (см.).
Олиrомерия тип строения тела, хйрак"
теризующийся наличием двух или трех се..
rMeHToB (см.) (ср. мономерия, метамерия).
Онтоrенез индивидуальное развитие
от зиrоты (см.) до взрослой особи.
Опистосома задний отдел тела тех хе..
лицеровых, у которых оно четко разделено
на два отдеЛd (ср. просома); иноrда термин
применяется и к друrим животным с ДBY
раздельным телом.
Оплодотворение процесс слияния ra..
мет (см.) с образованием зиrоты (см.).
Оральный ротовой.
OpraH структурно функциональная
единица тела, образованная одной или не..
сколькими тканями (см.).
Остии поры, например, для прохожде..
ния воды в тело (у rубок) или крови в сердце
(у животных с незамкнутой кровеносной си..
стемой).
Осфрадии хеморецепторные участки
или opraHbI в мантийной полости (см.) мол..
люсков.
Панцирь наружный или почти нару..
жный защитный покров тела, обычно со..
стоящий из нескольких элементов.
Паразит орrанизм, постоянно или
временно обитающий внутри друrоrо или
на нем и приносящий ему ущерб.
Паренхима диффузная ткань (см.) из
вакуолизированных клеток, часто запол
няющая пространство между эпидер:t\1ИСОМ
и кишкой у ацеломических (см.) животных.
Партеноrенез форма размножения
(см.), при которой яйцеклетка (см.) разви"
вается в новую особь без оплодотворения
(см.).
Педицеллария сложный подвижный
придаток некоторых иrлокожих, действую..
ший как щипцы или пинцет.
Педоморфоз процесс эволюционной
ювенилизации (см.), при котором либо
взрослая форма сохраняет ювенильные
признаки (неотения), либо половозрелость
наступает у животных ювенильноrо возра..
ста и ювенильной формы (проrенез).
Пелаrнческий обитающий в толще во..
ды (ср. бентосный).
Пенис орrан копуляции (СМ.), увеличи..
вающийся во время нее в размерах, но не
выворачивающийся (ср. циррус, rонопо..
дия).
Первичноротость резу ль тат развития..
при котором бластопор (см.) образует рот
взрослоrо орrанизма (ср. вторичноро"
тость) .
Передняя кишка передний отдел ки"
шки с эктодермальными (СМ.) происхожде..
нием и выстилкой.
Перикардиальная полость полость,
в которой расположено сердце.
Периостракум белковый покров рако..
вины моллюсков и плеченоrих.
Перистальтика волны сокращений ко..
льцевых и продольных мышц, проходящие
по трубчатому opraHY или орrанизму и слу..
жащие для проталкивания ero caMoro или
ero содержимоrо.
Перитонеум мезодермальная выстил..
ка целом а (см.).
Пиrидиум задний, не относящийся
к cerMeHTaM отдел аннелид.
Пиннула мелкая боковая ветвь щу"
пальца.
Пиноцитоз поrлощение клеткой мел..
ких капель жидкости.
Питание осадками потребление детри"
та (см.) и/или связанных с ним орrанизмов
на поверхности и/или в толще субстрата.
Питание суспензиями улавливание
и потребление частиц, взвешенных в воде,
обычно с помощью фильтра.
Плазмодий мноrоядерная амебоидная
масса, окруженная общей клеточной мем..
браной
556
Словарь терминов
Пламенные клетки ресничные клетки
на проксимальном конце протонефридия
(см. ).
Планктон пелаrические (см.) орrаниз"
мы, взвешенные в воде и не способные
плыть против течения (ер. нектон).
Планктотрофность питание (по край..
ней мере частично) планктоном (см.); тер..
мин обычно применяется к морским личин..
кам (ер. лецитотрофность).
Плеоподы абдоминальные (см.) при..
да тки мноrих ракообразных, часто исполь..
зуемые для плавания.
Поверхностное дробление тип дробле..
ния (см.), при котором зиrота (см.) дает на..
чало синцитию (см.); ero ядра движутся
к поверхности эмбриона.. rде отделяются
друr от друrа клеточными мембранами.
Пойкилотермность зависимость тем--
пературы тела от температуры внешней
среды (ер. rомойотермность).
Покровительственный облик облик,
позволяющий скрываться от BparoB за счет
сходства с окружающими объектами.
Полиморфизм наличие у одноrо вида
более двух различных форм тела (ер. моно--
морфизм, диморфизм).
Полип одна из двух форм тела у кни--
дарий: сидячий или прикрепленный ци--
линдр. обращенный к субстрату абораль--
ной (см.) поверхностью и несущий вокрут
рта кольцо щупалец; часто входит в состав
,
колонии (см.); иноrда термин используется
как синоним зооида (см.).
Полиплоидия наличие более двух ко..
пий каждой хромосомы в соматических
(см.) клетках (ер. rаплоидия, диплоидия).
Полифилетический относящийся к
rруппе орrанизмов, происходящих от не..
скольких предковых форм.
Половая пора см. rонопор.
Половое размножение форма размно--
жения, при котором происходит обмен re..
нетическим материалом между хромосома--
ми во время мейоза и слияние raMeT (см.)
в процессе оплодотворения (см.) (ср. беспо--
лое размножение).
Почкование форма бесполоrо размно--
жения (см.) с образованием новых особей из
выростов на теле родителя; затем они мо"
rYT от Hero отделяться и жить самостоя..
тельпо или оставаться с ним соединенными
(или связанными иным образом) в составе
колонии (см.).
Поясок секреторный эпителий (см.),
образующий кокон у поясковых аннелид.
Предротовая полость пространство
перед ртом, в KOTOpO:t\1 работают ротовые
придатки или происходит наружное пище..
варение.
Предупреждающая окраска особый
яркий, контрастный узор (например, черно..
желтый, красно..черный) потенциальноrо
вида--жертвы, часто сочетающийся с ero не..
съедобностью или токсичностью.
Проr лоттида сериально повторяю
щаяся единица строения тела у цестод.
Прокариоты орrанизмы с клетками
без окруженных мембранами орrанелл
и ядра (ер. эукариоты); бактерии.
Промежуточный хозяин хозяин пара..
зита (см.), в котором он не размножается
или размножается только бесполым (см.)
путем.
Пропаrу ла репродуктивная структура,
отделяющаяся от родителя; может быть
мноrоклеточной (веrетативной) или одно..
клеточной; в последнем случае возникает
путем мейоза (rаметы при половом размно"
жении) или же митоза, а также аберрантно"
ro мейоза, не сопровождающеrося редук"
цией числа хромосом (при бесполом раз..
множении).
Просома (а) передний отдел тела
(включающий rолову) хелицеровых с те..
лом, явственно разделенным на два отдела
(ср. опистосома); иноrда термин применяе..
тся и К друrим животным с двураздельным
телом (например, к веслоноrим ракообра..
зным); (6) первый отдел тела трехраздель..
ных олиrомерных (см.) животных; ero це..
лом называется протоцелем (ср мезосома,
метасома).
Простомиум передний, не относя..
щийся к cerMeHTaM отдел аннелид.
Протонефридий слепо на чинающийся
нефридий (см.) с внутриклеточным вывод..
ным протоком (ер. метанефридий).
Протоэукариоты rипотетические, воз
можно, способные к фаrоцитозу (см.) клет
ки, которые, став хозяевами различных
эндосимбиотических прокариот (см.), дали
начало первым эукариотам (см.).
Прядильные орrаны сосочки с вывод..
Словарь терминов
ными отверстиями желез, образующих пау--
тинные нити.
Прямое развитие развитие без личи..
ночных (см.) стадий (ср. непрямое разви"
тие) .
Псевдокопу ляция тесное сближение
при выведении из тела raMeT (см.) спари--
вающихся животных с наружным оплодот--
ворением; при этом яйцеклетки (см.) опло--
дотворяются (см.) сразу же после выхода из
rонопора (см.) самки.
Псевдоподия временный лопастевид--
ный вырост протоплазмы, образуемый аме--
боидными клетками при движении, фаrоци--
тозе (см.) и т. п.
Псевдоцель любая полость тела., не
являющаяся целом ом (см.).
рабдитысм.. рабдоиды.
Рабдоиды палочковидные образова--
ния с неясной функцией в эпидермисе (см.)
плоских червей и сходных .с ними животных;
некоторые образуются из железистых кле
ток и называются рабдитами.
Рабдом упорядоченное расположение
фоторецепторных микроворсинок (см.), на..
пример, в сложном rлазу.
Рабдоцелъ слепая кишка турбелларий
без боковых ветвей и дивертикулов.
Радиальная симметрия симметрия OT
носительно любой плоскости, проходящей
через орально--аборальную ось.
Радиальное дробление тип дробления,
при котором плоскости клеточноrо деления
проходят параллельно и перпендикулярно
полярной оси бластулы (см.) (ср. спираль--
ное дробление).
Реrенерация восстановление за счет
компенсационноrо роста и дифференциров--
ки (см.) утраченных частей орrанизма.
Pery ляционное развитие тип развития,
при котором эмбрион (см.) способен ком--
пенсировать утрату клеток и давать нор--
мальную личинку или взрослую особь, по--
скольку будущее клеток опеределяется то--
лько на поздних стадиях эмбриоrенеза (ср.
мозаичное развитие).
Реснички выступающие из клетки дви--
rательные орrанеллы, бьющие в одной пло--
скости (жесткий удар и rибкое отведение на--
зад), обычно относительно короткие; MorYT
находиться на клетке в болыuом количе--
стве; специализированные неподвижные
557
реснички служат для ceHcopHoro восприя--
тия; известны только у эукариот (см.) (ср.
жrутики) .
Реснично. пламя реснички пламенных
клеток (см.).
Самозарождение ненаучная теория,
признающа . возможность возникновения
живых орrанизмов непосредственно из He
живоrо вещества (например, ила).
CerMeHT полуавтономная, сериально
повторяющаяся единица строения тела: ce
rментация может затраrивать только ero
стенку с ее структурами или помимо этоrо
все элементы BHYTpeHHero строения.
Семеприемник мешок, в котором жи--
вотное накапливает полученные от партне--
ра по спариванию сперматозоиды (см.)
перед оплодотворением (см.) своих яйце..
клеток (см.).
Септа мембрана., отделяющая один
отдел тела от друrоrо.
Симбиоз тесная ассоциация двух раз--
личных орrанизмов, взаимодействующих
между собой (один обычно зависит от др у..
roro) .
Синапс контакт, служащий для пере--
дачи информации между двумя клетками,
из которых по крайней мере одна нейрон
(см.); передающая клетка называется преси..
наптической, воспринимающая пост..
синаптической.
Синцитий мноrоклеточная структура
с частично или полностью отсутствующими
.
rраницами между клетками; может пред
ставлять собой и мноrоядерную цитоплаз--
.
матическую массу без заметноrо разделе--
ния на клетки, и сеть клеток. цитоплазма
которых объединена межклеточными мо--
стиками.
скалидыкутикулярныыe или эпидер"
мальные выросты различной формы (шипо--
видные, булавовидные, перистые, чешуе--
видные), расположенные кольцами BOKpyr
интроверта (см.) киноринхов, лорицифер"
приапулид и личинок волосатиков; MorYT
выполнять сенсорную, локомоторную, лов--
чую или прободающую функции.
Скелет система" обеспечивающая
передачу силы мышечных сокращений
и/или опору частям тела
Склерит пластинка, составляющая
часть экзоскелета.
558
Словарь терминов
Склеротизация химическое упрочне
ние (с потемнением) участков КУТИКУЛЬJ
(см.); результат процессd., аналоrичноrо дy
блению
Слизь смесь мукопротеинов (соедине
ний мукополисахаридов с белками), ceKpe
тируемая слизистыми клетками.
Сложный f'лаз rлаз, образованный He
сколькими (мноrими) отдельными зритель
ными единицами омматидиями, каждая
из которых имеет собственные хрусталик,
поле зрения, рецепторные клетки и т. п. (ср.
rлазок).
Сократительная вакуоль внутри--
клеточная окруженная мембраной вакуоль
с осмореrуляторными функциями; запол
няется жидкостью и, резко сокращаясь, BЫ
талкивает ее наружу из клетки.
Соматический относящийся к клеткам
орrанизма, не являющимся половыми.
Сперматозоид (спермий) мужская [a
мета (см.). обычно способная к активной
локомоции.
Сперматофор пакет сперматозоидов
(см.), заключенный в защитную обо
лочку.
Спиральное дробление тип дробления,
при котором плоскости клеточноrо деления
проходят косо по отношению к полярной
оси бластулы.. что привопит к чередую
щимся смещениям BOKpyr этой оси по часо
вой стрелке и против нее клеток, образую
щихся при поперечных делениях после CTa
дии четырех бластомеров (ср. радиальное
дробление).
.
Средняя кишка от дел кишки с энто
дермальными (см.) происхождением и BЫ
стилкой (ср. передняя кишка, задняя ки
шка).
(:тарение необратимые деrрадацион--
ные изменения особей с течением времени,
приводящие к повышенной уязвимости
и пониженной жизненности.
Статобласт мноrоклеточная пропаrу
ла (см.) при бесполом размножении (см.)
некоторых мшанок, окруженная защитной
оболочкой.
Статоцист орrаи, воспринимающий
силу тяжести и/или ускорение.
Стернит вентральный склерит (см.) ce
rMeHTapHoro экзоскелета членистоноrих.
Стилет твердое, заостренное, ноже
видное образование., используемое для про
тыкания клеток или тканей.
Столон стебле или корнеподобная
структура, соединяющая животных дрyr
с друrом или с субстратом, иноrда отпочко
вывающая новые особи.
Субумбреллярный связанный с ниж"
ней, обычно воrнутой поверхностью Meдy
зы (см.).
Схизоцель целом (см.)., формирую..
щийся в толще мезодермальной (см.) ткани
за счет образования пространства между ее
клетками (ср. энтероuель).
Теryмент синцитиальный (см.) наруж..
ный эпителий паразитических плоских
червей.
Телеолоf'ИЯ учение об эволюционном
развитии, направленном к заранее опреде..
ленной цели.
Тельсон задний, не относящийся к се..
[ментам отдел членистоноrих.
Терf'И'I дорсальный склерит (см.)
cerMeHTapHoro экзоскелета членистоноrих.
Ткань совокупность клеток одноrо
или нескольких типов, выполняющих об..
щую функuию и обычно соединенных меж..
клеточным материалом (ср. opraH).
Тонический устойчивый, длительный.
Торакс средний отдел любоrо тела,
разделенноrо на три отдела, передний из
которых rолова (ср. абдомен).
Т отипотентность способность клеток
мноrоклеточноrо орrанизма к дифференци"
ровке (см.) в любую специализированную
клетку
Трахеола терминальная капилляропо..
добная часть трахеи (см.).
Трахея трубка, проводящая воздух из
внешней среды непосредственно к тканям.
Трохофора ранняя личиночная стадия
мноrих морских беспозвоночных, характе..
ризующаяся двумя около ротовыми пояса..
ми ресничек.
Ультрафильтрация прохождение жид
кости под давлением через полупроницае..
мую мембрану.
Уроподы последняя пара абдоминаль..
ных (см.) придатков у десятиноrих раков;
вместе с тельсоном (см.) образует «хвосто..
вой eep».
Уросома опистосома (см.) веслоноrих
ракообразных.
Словарь терминов
Фаrоцитоз процесс окружения псевдо
подиями (см.) амебоидной клетки пищевой
частицы и заключения ее в вакуоль
Филоrенез ЗВОЛЮllионное происхо
ждение таксонов.
ФИЛОПОДИИ нитевидные выросты ци
топлазмы
Флоскулы крошечные выступающие
сенсорные орrаны. оканчивающиеся сосоч
ками; присутствуют на туловище HeKOTO
рых приапулид, лорицифер, киноринхов
и коловраток.
Фотопериодизм способность физиоло
rически реаrировать на изменения относи
тельной долrоты дня.
Фоторецептор особая светочувстви
тельная клетка.
Фотосинтез синтез орrанических co
единений с использованием энерrии солнеч
Horo света и участием хлорофилла по обще
му уравнению
СО 2 + 2Н 2 Х ----+ (СН 2 О) + Н 2 О + 2Х.
При оксифотосинтезе у некоторых бактерий
и всех фотосинтезирующих эукариот Х
кислород; при аноксифотосинтезе у ряда
бактерий Х не кислород, а часто (но не
всеrда) сера, т. е. Н 2 Х cepOBOДOpoд.
Фраrментация форма бесполоrо раз
множения (см.), включающая разделение
тела на две и более частей. каждая из KOTO
рых способна вырасти в целую особь.
Фумарола отверстие земной коры, из
KOToporo выделяются rорячая вода и BOC
становленные соединения.
Фурка пара варьирующих по форме
отростков на тельсоне (см.) ракообраз
ных.
Хемоавтотрофность бактериальное
питание за счет хемосинтеза (см.).
Хемосинтез синтез орrанических co
единений по общему уравнению
СО 2 + 2Н 2 Х (СН 2 О) + Н 2 О + 2Х
с использованием энерrии окисления (и ча
сто восстановительных свойств) Heopra
нических субстратов типа Fe 2 +, СН 4 , NH' H
N0 2 , Н+, S И др. или орrанических co
единений (апетата, формиата и др.); ocy
ществляется только бактериями в анаэроб
ных условиях (ср. фотосинтез).
Хитин азотсодержащий полисахарид.
Хлоропласт орrанелла эукариот
место протекания фотосинтеза (см.); BCTpe
559
чается у некоторых протистов и большин
ства растений.
Хоаноциты характерные жrутико
воротничковые клетки rубок.
Хоботок удлиненный вырост на rоло
ве или передней части тела, используемый
для питания.
Хорда упруrий дорсальный скелетный
тяж, развивающийся из сильно вакуолизи--
рованных клеток, связанных между собой
внутри общей оболочки; характерный при
знак хордовых.
Хроматофор пиrментная клетка.
Хромосома нитевидная структура кле
точноrо ядра, несущая rенетическую инфор
мацию.
Целом заполненная жидкостью по
лость тела мезодермальноrо (см.) происхо
ждения, окруженная мезодермальной мем--
браной (ср. псевдоцель, rемоцель); пред--
ставлена как обширными rидростатически
ми полостями, оrраниченными перитонеу
мом (см.) (см. схизоцель, энтероцель), так
и выстланным эпителием (см.) простран
ством внутри мезодермальных opraHoB
Целомодукт (а) слепо начинающаяся
осмореrуляторно экскреторная железа с
протоком мезодермальноrо происхождения
(ср. нефридий); (6) открытый в целом (см.)
мезодермальный проток для выведения
наружу raMeT или целомической жидкости.
Целомоцит клетка, взвешенная в цело..
мической (см.) жидкости.
Цефализация развитие rоловы в ходе
филоrенеза (см.) или онтоrенеза (см.).
Церки пара варьирующих по форме
придатков на последнем абдоминальном
cerMeHTe мноrих насекомых.
Циррус выворачивающийся копуля
ционный opraH (ер. пенис, rоноподия).
Циста. инкапсулированная., устойчи"
вая к высыханию стадия жизненноrо
цикла.
Цитохромы дыхательные пиrменты
локализованные в митохондриях; по CTPYK
туре напоминают rемоrлобин.
Чешуйчатый эпителий эпителий (см.),
образованный уплощенными клетками.
Щетинки мелкие жесткие выросты на
теле аннелид, поrонофор, эхиур и др.; MorYT
содержать клеточный материал или иметь
кутикулярную природу.
560
Словарь терминов
Щупальце тонкий rибкий придаток,
часто с сенсорной функцией; иноrда исполь
зуется для улавливания пищи.
Эволюция происхождение и изменение
в rеолоrических масштабах времени.
Эвтелия состояние, при котором клет
ки взрослой особи не делятся, т. е. число их
постоянно (обычно невелико), а рост тела
идет за счет увеличения размеров клеток.
Экзоподит наружная ветвь двуветви
стой конечности членистоноrих (ср. эндопо
дит, эпиподит).
')ктодерма наружный зародышевый
слой, покрывающий rаструлу (см.) (ер.
энтодерма, мезодерма).
Эктотермия получение орrанизмом
тепла из внешней среды, а не за счет соб
cTBeHHoro метаболизма.
Эмбрион зародыш, ранняя стадия раз..
вития, выходящая из яйца (см.) или остаю..
ЩdЯСЯ в нем и не способная к самостоятель..
ной жизни.
Эндоподит внутренняя ветвь двувет"
вистой конечности членистоноrих (ср. 1КЗО"
подит, эпиподит).
Энтероцель целом (см.), сформиро--
вавшийся путем отшнуровывания мезодер"
мальных карманов от эмбриональной
кишки (ср. схизоцель).
Энтодерма внутренний зародышевыIй
слой. образуюший эмбриональную кишку
rаструлы (СМ.) (ср. эктодерма, мезодерма).
Эпиболия. обрастание подвижными,
бедными желтком клетками клеток, боrа
тых желтком во время rаструляции (см.).
Эпидермис наружный клеточный по..
кров тела, формирующийся из эктодермы
( см. ).
Эпиподит отросток на базальном чле..
нике конечностей членистоноrих; возмо"
жно, от эпиподита происходят rрифельки
(см.) (ер. эндоподит, экзоподит).
Эпителий слой ткани (СМ.). покрываю..
щий свободную поверхность, например вы..
стилающий полость.
Эпифауна бентосные (см.) животные,
населяющие поверхность субстрата (ер. ин..
фауна).
Эукариоты орrанизмы, в клетках KO
торых находятся окруженные мембранами
ядро и орrанелль] (ср. прокариоты); все ор..
rанизмы. кроме бактерий.
Ювенилизация приобретение ВЗРО--
слым орrанизмом признаков неполовозре..
лой особи.
Ювенильный неполовозрелый.
Яйцеклад трубчатый opraH некоторых
насекомых, используемый для откладки яиц
в специфические места
Яйцеклетка женская raMeTa (см.).
Яйцо неопределенный термин. обозна--
чающий исходную стадию развития живот..
Horo яйцеклетку (см.), зиrоту (см.) или СО..
вокупность клеток развивающеrося
эмбриона (см.), пищевых запасов и т. д.. на..
ходящуюся внутри общей капсулы или 060"
лочки.
ИСТОЧНИКИ ИЛЛЮСТРАЦИЙ
Alexander R. McN. 1979. ТЬе Invertebrates. Caт
bridge University Press
Cambridge.
Alld/'eclge А. 1976. Sci. Ат., 235(1), 94
102.
Anderson D. Т. 1964. Embryology and Phylogeny in
Annelids and Arthropods. Pergamon Press, New
у ork.
Aпdel'son Р. А. V., Вопе Q. 1980. Proc. R. Soc.
Lond(B), 210, 559 574.
Atkiпs D. 1933. J. Mar. Biol. Ass.
UK, 19, 233
252.
АIИJfJпd H
L. 1973. Ат. J. Zool., 13, 357
378-
Aиstiп с. R. 1965. Fertilisation. Prentice Н,,-Н Inc.,
New Jersey.
Вае/l1' J. С., РОl'С/lеl'оп Р., пl'ау Р. 1978. С. R. Acad.
Sci. (Paris), 278О, 253 525.
Bal'nes R. п. 1980. Invertebrate Zoology, 4th edn.
Saunders, Philadelphia.
Bal'nes R. S. К., Hughes R. N. 1982. Ап [ntroduction
to Marine Ecology. Blackwell Scientific PubIica
tions, Oxford.
Bastiaпi М. J., Doe С. Q., He((aпd S. L., Goodтaп
С. S. 1985. Trends in Neurosciences, 8, 257
266.
Ваупе В. L.. Thoтpson R. J., Widdои's J. 1976. In:
В. L. Ваупе (Ed.) Marine Mussels: Their Physiolo
gy and Ecology. Cambridge University P)'ess.
Сат bridge.
Beskel' с. 1937. Z. Morph. Okol. Tiere, 33, 72
127.
Belk D. 1982. In: S. Р. Parker (Ed.) Synopsis and
Classification of Organisms. 2. 174
180.
McGraw
Hill, New У ork.
Bergquist Р. R. 1978. Sponges. Hutchinson, London.
Berrill N. J. 1950. ТЬе Tunicata. Ray Society, Lon
don.
Biscardi Н. М." Webstel' G. С. 1977. Ехр. Geront.,
12. 201
205.
Blowel' J. G. 1985. Millipedes. Brill" Leiden.
BoeckJl J.. El'пst K..D.. Sass Н.. Wаldои' и. 1975. In:
D.Denton (Ed.) 01faction and Taste, V" 239
245.
Academic Press. New У ork.
Boss К. J. 1982: In: S. Р. Parker, (Ed.) Synopsis and
Classification of Living Organisms, 1, 945
1166.
McGraw..Hill, New У ork.
Вl'П! В. 1973. Z. Zellforsch., 144, 231
245.
Buchsbauт R. 1951. Animals Without Backbones,
V 01. 1. Pelican, Harmondsworth.
Bullough W. S. 1958. Practical lnvertebrate Anato..
ту, 2nd edn. Macmillan. London.
Cain А. J., S/leppard Р. М. 1954. Genetics, 39, R9
116.
Ca/kins с. N. 1926. ТЬе Biology of the Protozoa.
Bailliere Tindall and Сох, London.
CalOH! Р. 1985. Causes de lа mort i costos -d'autopro
teccio. [п: Biologia А vui. Fundacio Caixa de Pen
sions, Barcelona
Са/о и' Р. 1986. In: R. Peberdy and Р. Gardner (Eds)
ТЬе Collins Encyclopedia of Anima] Evolution.
рр. 90
91. Equinox, Oxford.
Са/он' Р., Read п. А. 1986. In: S.Tyler (Ed.) Advan
ces in the Biology of Turbellarians and Related
Platyhelminthes, рр. 263
272. О. W. Junk Publis..
hers. Dordrecht.
C'al'pentel' W. В. 1866. Phil. Trans. Roy. Soc. Lond.
156, 671
756.
Caullery М., MesпU F. 1901. Arch. Anat. Micros., 4,
381
470.
C/al'k А. Н. 1915. US Natn. Mus. Виll." 82, Уоl. 1(1),
1
406.
Clal'k R. В. 1964. Dynamics in Metazoan Evolution
Clarendon Press, Oxford.
Clarke К. и. ] 973. ТЬе Biology of the Arthropoda.
Arnold, London.
C/arkson Е. N. К. 1986. Invertebrate Palaeontology
and Evolution. 2nd edn. Аllеп and Unwin, LOJ1
don.
Clement А.С'. 1962. J. Ехр. Zool., 149" 193 215.
Cloud
ley
T/10тpSOn J. 1958. Spiders. Scorpions.
Centipedes and Mites. Pergamon Press, London.
Coheп А. С. 1982. In: S. Р. Parker (Ed.) Synopsis and
Classification of living Organisms, 2, 181
202.
McGraw..Hill, New У ork.
Colwiп L. Н., Соlи'iп А. L.. 1961. J, Biophys. BIOC
hem. Cytol.. 10, 231
254.
Coпway MOl'ris S. 1979. Апп. Rev. Есоl. Syst.. 10
327
349.
Сопи'ау MOl'ris S. 1985. Phil. Trans. Roy. Soc. Lond.
(В), 307, 507
582.
C01'!iSS J. о. 1979. ТЬе Ciliated Protozoa, 2nd edn.
Pergamon Press. Oxford.
Cuenot L. 1949. In: Р..Р. Grasse (Ed.) Traite de Zoo..
10gie. VI. 3
75. Masson. Paris.
Danielssoп D. 1982. Norw. N
Atlantic Exped.
(1876
.1878) Rep. Zool., 21, 1 28.
Darпell J. Е., Lodish Н. F., Baltiтol.e п. 1986. Mole
562
Источники иллюстраций
cular СеВ Biology. Scientific American Books Inc.,
New York.
Davies 1. 1983. Ageing. Edward Arnold, London.
Dehorпe А. 1933. ВиН. Biol. Fr. Belgique, 67, 298
326.
Dethiel' V. Е. 1976. ТЬе Hungry Fly. Harvard Uni
versity Press. Cambridge, Mass.
Dixoп А. Е. с. 1973. Biology of Aphids. Studies in
Biology No. 44. Edward Arnold, London.
Durchoп М. 1967. L'endocrinologie chez le Vers et les
Molluscs. Masson. Paris.
Eakiп R. М. 1968. Еуоl. Biol., 2, 194
242.
Elпer R. W.. Hughes R. N. 1978. J. Anim. Есоl.. 47.
103
116.
Ере! D. 1977. Sci. Аm., 237(5). 129
138.
Fеи'kеs J. 1883. ВиН. М us. Соmр. Zool., Harward,
11, 167
208.
Fll1gel'тaп М. 1976. Animal Diversity, 2nd edn.
Holt, Rinehart and Winston. New У ork.
Fox Н. М.. Wiпgfield С. А., Siттoпds В. с. 1937. J.
Ехр. Biol., 14, 21Q
218.
F,'ase,' J. Н. 1982. British Pelagic Tunicates. Сат..
bridge University Press, Cambridge.
F,'ettel' V., GraJlaт А. 1976. А Functional Anatomy
of Invertebrates. Academic Press, London and
New У ork.
F,'ieseп W. Q., Рооп W., Steпt с. S. 1976. Proc. Nat.
Acad. Sci. (USA), 73, 3734- 3738.
Geol'ge J. D., Sоuthи'аrd Е. с. 1973. J. Mar. Biol.
Ass UK, 53, 403
24.
Gibsoп Р. Н., Clal'k R. В. 1976. J. Mar. Biol. Ass.
UK. 56, 649
674.
Gibsoll R. 1982. In: S. Р. Parker (Ed.) Synopsis and
Classification of Living Organisms, 823
846.
McGraw..Hlll, New У ork.
Gilbel.t L. Е. 1982. Sci. Аm., 247(2), I 02
1 07В.
Glaessпel' М. F., Wade М. 1966. Palaeontology, 9,
599
628.
Gпaigel" Е. 1983. J. Ехр. Zool., 228, 471
490.
Gиldiпg D. W., Рои; D. V. 1987. In: М. С. Thorndyke
and G. J. Goldsworthy (Eds) Neurohormones lП
Invertebrates. Cambridge University Press, Сат..
bridge.
Goldiпg D. W.,White А. с. 1974. Tissue СеН. 6,
599
611.
Go/diпg D. W., White А. С. 1977. Int. Rev. Cytol.
Suppl., 5, 189
302.
GOOd,"ich Е. S. 1945. Quart. J. Micros. Sci.. 86. 113
393.
Gordoп D. Р. 1975. СаЬ. Biol. Mar.. 16. 367
382.
Grasse Р...Р. (Ed.). 1948. Traite de Zoologie, XI.
Masson, Paris.
G,'asse Р... Р. (Ed.). 1965. Traite de Zoologie, IV.
Masson, Paris.
С,'ееп J. 1961. А Biolody of Crustacea. Witherby,
London.
Gupta В. L.. Bel'ndge М. J. 1966. J. Morphol., 120,
23
82.
Gustafsoп Т., Wolpel.t L. 1967. Biol. Rev., 42, 442
498.
Hackтaп R. Н. 197]. In: М. Florkln, В. Т. Scheer
(Eds) Chemical Zoology, 6, 1
62. Academic
Press, New York.
Haпseп К. 1978. In: G. 1. Hazelbauer (Ed.) Taxis and
Behaviour Receptors and Recognition. 58. 231
292. СЬарmап and НаН. London.
Hard.J;' А. С. 1956. ТЬе Ореп Sea. ТЬе World of
Plankton. CoHins, London.
Hedgpeth J. W. 1982. In: S. Р. Parker (Ed.) Synopsis
and Classification of Living Organisms, 2. 169
173. McGraw..HiH. New У ork.
Herтaпs С. О., Cloпey R. А. 1966. Z. Zel1forsch. 72,
583
596.
Hel'тaпs С. О., Eakiп R. М. 1974. Z. Morph. Tiere,
79, 245
267.
HescJzeler К. 1900. In: А. Lang (Ed.) Lehrbuch der
Vergleichenden Anatomie der Wirbellosen Thiere,
3rd edn. Fischer, Jena.
Hess R. 1987. Z. Wiss. Zool., 62, 247
283.
Higgiпs R. Р. 1983. Smithsonian Contrib. Mar.
ci.,
18, 1
131.
Hodgkiп А. L., Huxley А. F. 1952. J. Physiol., 117,
500
544
Hollaпd N. D., Griттer Т. с., Kubota Н. 1975. Biol.
ВиН., 148, 219
242.
Ho!t С. S., Watel'S Т. F.. 1967. Ecology, 48, 225
234.
Hughes Т. Е. 1959. Mites or the Acari. Athlone, Lon..
don.
Ниттоп W. D. 1982. In: S. Р. Parker (Ed.) Synopsis
and Classification of Living Organisms, 1, 857
863. McGraw
Hill, New У ork.
Нутап L. Н. 1940. ТЬе 111vertebrates. Yol. 1. Proto..
zoa through Ctenophora. McGraw..Hill, New
У ork.
Нутап L. Н. 1951. ТЬе lnvertebrates. У 01. 11. Platy
helminthes РЬупсЬосоеlа. McGraw
HiH, New
у ork.
1ттs А. D. 1964. А General Textbook of Entomolo..
gy, 9th edn., revised reprint. Methuen, London.
1to У. 1980. Comparative Ecology. Cambridge Uni..
versity Press, Cambridge.
Jiigersteп с. 1973. ТЬе Evolution of the Metazoan
Life Сусlе. Academic Press, New У ork.
Jeaппel R. 1960. Introduction to Entomology. Hutc..
hinson, London.
Joпes А. М., Baxtel' J. М. 1987. MoHuscs: Caudofo..
veata. Solenogastres. Polyplacophora and Scapho..
poda. Brill, Leiden.
Joпes J. D. 1955. J. Ехр. Biol.. 32, IIO
125.
Joпes М. L. 1985. In: S. Conway Morris et al. (Ed.)
ТЬе Origin and Relationships of Lower Inverte..
brates, 327
342. Clarendon Press, Oxford.
Joosse J., Geraerts W. Р. М. 1983. In: А. S. М. Saleu..
din and К. М. Wilbur (Eds) ТЬе Mol1usca, У 01. 5.
Academic Press, New York.
564
Источники иллюстраций
Shepherd G. М. 1983. Neurobiol0gy. Oxford Univer
sity Press, Oxford.
Sheppard Р. М. 1958. Natural Selection and Heredi..
ty. Hutchinson, London.
Shick Р. М., Dykeпs J. А. 1985. Oecologia, 66, 33
41.
Silva Р. Н. D. Н. de. 1962. J. Ехр. Biol., 39, 483
490.
Siпgla С. L. ]975. Сеll Tiss. Res., 158, 391
07.
SZeigh М. А.. Dodge J. D.. Pattersoп D. J. 1984. In:
R. S. К. Barnes (Ed.) А Synoptic Classification of
Living Organisms, 25
88. Blackwell Scientific Ри..
blications, Oxford.
Smart Р. 1976. ТЬе IIlustrated Encyclopedia of the
Butterf1y W orld Наmlуп, London.
Sтyth J. D. ] 962. lntroduction to Animal Parasito
10gy. ТЬе English Universities Press, London.
Smyth J. D., HaZtoп D. W. 1983. ТЬе Physiology of
Trematodes. Cambridge University Press, Сат..
bridge.
Sпodgrass R. Е. ] 935. Principles of Insect Morpholo
gy. McGraw..Hill, New У ork.
Sпow К. R. [970. ТЬе Arachnids: Ап Introduction.
Routledge and Kegan Ра иl, London.
Sоutlп't'аrd Е. с. 1980. Zool. Jb. Anat. Ontog., 103,
264
275.
Southward Е. С. 1982. J. Mar. Biol. Ass., UK, 62,
889
906.
Speпgel J. W. 1932. Sci. Res. Michael Sars N. Atlan..
tic Оеер Sea Exped., 5(5), 1
27.
Steiп Р. S. G. 1971. J. Neurophysiol., 34, 310
318.
Sterrer W. Е. 1982. In: S. Р. Parker (Ed.) Synopsis
and Classification of Living Organisms, 1, 847
851. McGraw--Нlll, New У ork.
Stiasпy G. 1914. Z. Wiss. Zool.. 110, 36
75.
StrausfeZd N. J., Nassel D. R. 1981. In: Н. Autrum
(Ed.) Handbook of Sensory Physiology, V 01.
VIIj6B. Springer.. V erlag, Berlin.
Strettoп А. о. W., Davis R. Е., Aпgstadt J. D., Doп
тoyer J. Е., Johпsoп с. D. 1985. Trends in Ne..
uroscienes, 8, 29
299.
Struтwasser F. 1974. Neurosciences Third Study
Program, 459
78.
Treherпe J. Е., Foster W. А. 1980. Anim. Behav., 28,
1119
1122.
Treпch R. К. 1975. Symp. Soc. Ехр. Biol., 29, 229
265.
Triпkaпs J. Р. 1969. Cells into Organs. Prentice Наll.
N ew J ersey
Truemaп Е. R., Foster..8mith R. 1976. J. Zool.,
Lond., 179. 373
386.
Turпer с. D. 1966. General Endocrinology. Saun
ders, Philadelphia.
Valeпtiпe J. W., М oores Е'. М. 1974. Sci. Аm..
230(4), 8
89.
Wallace М. М. Н., Mackerras 1. М. 1970. [п:
C.S.I.R.O., ТЬе Insects of Australia, 205
216.
Melbourne University Press. Melbourne,
Warпer G. F. 1977. ТЬе Biology ofCrabs. Elek. Lon..
don
Watermaп Т. Н. 1960. ТЬе Physiology of Crustaces,
Academic Press, New У ork.
We/sch И., Storch v. 1976. Comparative Animal Су-
tology. Sidgwick and Jackson. London.
Weпyoп С. М. 1926. Protozoology. Bailliere, Tindall
and Сох, London.
Whittlпgtoп Н. В. 1979. In: М. R. House (Ed.) ТЬе
Origin of Major Invertebrate Groups. 253
268.
Academic Press, London.
Widdows J., Ваупе В. L. 1971. J. Mar. Biol. Ass.,
UK, 51, 827
843.
WigK/esworth V. В. 1940. J. Ехр. Biol.. 17. 201
222.
Wiggles'l-vorth V. В. 1972. Principles of Insect Physio
logy, 7th edn. СЬартап and Наll, London.
Wiпe J. J., Krasпe F. В. 1982. (п: О. С. Sandeman
and Н. L. Atwood (eds) ТЬе Biology ofCrustacea,
V 01. 4, 241
292. Academic Press, New У ork.
Wright А. D. 1979. In: М. R. House (Ed.). ТЬе Ori..
gin of Major Invertebrate Groups, 235
252. Aca
demic Press, London.
Wroпa F. J., Davies R. W. 1984. Сап J. Fish. Aqua..
tic Sci., 41, 380
385.
Yager J., Sch,'am Р. R. 1986. Proc. Biol. Soc. Wash.,
99(1), 65 70.
Youпg J. Z. 1939. Phil. T,'aпs. Roy. 80с. Loпd. В.,
229. 465
503.
у ouпg J. z. 1962. ТЬе Life of Vertebrates, 2nd edn.
Clarendon Press, Oxford
у ouпg J. z. 1963, Nature (Lond), 198, 636
630.
ZulZo V. Z. 1982. In: S. Р. Parker (Ed.) Synopsis and
Classification of Living Organisms, 2, 220
228.
McGraw..Hill, New У ork.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Авоzадро число 377
Arr лютинины 391
Адаптация 543
Адаптивная радиация 42, 43, 45, 47, 242, 368
Аденозиндифосфат (АОР) 339
Аденозинтрифосфат (АТР) 309, 339 342, 542
Апипокинетический ropMoH 541
Акклимация 356 357
Акросомная нить 440 43
Акселерация 548
Аксоны 488. См. mаК.же rиrантский аксон
Активный транспорт и экскреция 374 375
Актин 311
Актинии 62, 63, 72, 73, 289
симбиоз 75
Актинотроха 172
Алломоны 385, 524
Амакриновые клетки 489
Амбу лакральная система 318 319
Амбу лакральные ножки 269, 318, 319
Амебоидные клетки 389 391
Амебоидный рост 533
Амебоциты 62
Аминокислоты 12
Аммиак 361, 362
Аммониты 43, 548
Аммонотелия 361, 362
Амфибластула 57
Амфиды 536
Амфиподы СМ. Бокоплавы
Анаболические процессы 542, 543
Анаэробный метаболизм 340, 341
Аннелиды (кольчатые черви, кольчецы) 3] 32, 135 149
rаметоrенез 417
нервная система 494, 495, 500
питание 267, 270, 279, 498
развитие 448
размножение 403
реrенерация 481 83
сеrментация 136, 137
сетчатка 507
старение 394
циркуляторная система 343, 345
экскреция 369
Аноксия 340
Аноплиды 102
Антеннальные (зеленые) железы 371, 374, 529
Антенны 270, 462
Антиrены 391
Антифертилизин 440
Анус 277, 278
Анхетрон 29
Аплизия (морской заяц) 532 533, 535
Аппендикуларии 209 211, 270
Аристотелев фонарь 201, 281
Артроподы см Членистоноrие
Археоциаты 42
Архианнелиды 141, 143, 144
Асконоидный тип орrанизации 55
Аспартат 341
Асцидии 205 207, 384, 445
развитие 452, 470, 471
А триум 285
Ацетилхолинэстераза 471
Бабочки 241
защита 389
зрение 503
Бабочки данаиды 385
Бактерии как пища 265 266
размножение 396, 430, 431
симбиотические 275
Бделлоидеи 123 124
Беззамковые 173 175
Беззубки 410
Белки 12 13
Бензохиноны 387
Березовая пяденица 382 383
Бесполое размножение 85, 111, 544
Бессяжковые 234
Билатеральная симметрия 79
Биолоrические часы 501, 535
Биомеханика 308 338
Биотопы 50
Биохимия .14
Блаrородный коралл 74
Бластомеры 469
Бластула 24, 446
Блохи 330
Блюдечки 161,353,357,405
локомоция 315
Бокоплавы 260, 262, 270, 366, 367
движение 353
Бомбикол 520, 524
Бора эффект 345
Бочоночник 2()9
Брюхоноrие моллюски (r dСТРОПОПЫ) 159 162, 364
дыхание 160
зрение 504, 507
питание 270, 275
Брюхоресничные черви 1 05 1 07
Брюшные cerMeHTbI 239
Бычий цепень 91
Велиrер 312, 453
Вентральная мускулистая rлотка 141
Вертикальные миrрации 381
Веслоноrие рачки 36, 253 255, 270
размножение 404
Ветвистоусые рачки 401
Вирусы 396, 429, 430
Витамин Е 379
Вителлоrенины 526
Вихревое движение воздуха 337 338
Влаrосбереrаюшие opraHbI 215
Водители ритма 499 501
566
Предметный указатель
Водоотталкивающий покров 238
Водоросли как пища 267, 274, 275, 284
Водяной ослик 367
Волосатики 11 115, 403
Восприятие 509 51 О
Вторичноротые 39 0, 45, 184 212
Выделительные системы 370 375
Выживание, выживаемость 43 7, 379, 425 428
Вымирание 18, 42 44, 47, 48
р rалактозидаза 545
raMeTbI 396. 544
rамма облучение 378, 379
rастродермис 61, 64
rастроподы С-,И. Брюхоноrие моллюски
rастротрихи 29 30, 39, 99, 403
rаСТРУЛЯl!ИЯ 23 24, 446 50
rемато энцефалический барьер 491
rеммула 57
rемоrлобин 345, 346
rемоцель 29
rемоцианин 345 348
rемоциты 391
rемэритрин 345 348
r енералисты 502
rенетическая проrрамма 12 13, 465 66
rен оператор 545
rеофизические циклы 418
rепатопанкреас 278
rермафродитизм 404, 411
r етеробранхии 161
fетеротрофы 265, 542
rиrантский аксон 484, 487, 497
лобулярный детектор движения (LGMD) 511 512.
515 516
fидроидные полипы (rидроиды) 61 69, 440
движение 500
нейроны 514
реrенерация 463
rидромедузы 6 68
движение и нервная система 494, 500
rидростатический скелет 29, 316, 333 334
rидры 61, 62, 66 69
движение 500
реrенерация 493
rимнолематы 180 181
rиперморфоз 548
rиперосмотическая среда 367
fипоневральная система 495
fлаза 50 509. См. тaK.JICe Рабдомерные rлаза, Сло
жные r лаза
fлазки 506
rлазные железы 420
r лиальные клетки 489 499
r ликолиз 340
rлотка 141, 278, 280, 285, 286
r лоточный насос 109, 111
fнатостимулиды 29 30, I03 105, 403
fоловной активатор 523
rоловоноrие моллюски 165 169, 321, 337
размножение 403,420
r оловохордовые ('.I'И. Ланцетники
fолодание 85 86, 355
fоложаберные моллюски 387
fолометаболические насекомые 458
rолотурии (морские orypUbl) 201 204
дыхание 345, 349
питание 269
размножение 403
r омеобоксы 462
r омеозисные [ены 462, 479 481
rомойотермные животные 355
rонадостимулирующее вещество (GSS) 525
r онанrий 440
rонозоиды, 67, 70
rонопор 371
f орrонарии 389
f ормоны 419, 420, 520, 521, 524
f орячие линии 497 99
fрадиентов теория (развития) 475 477
rребешок 504
fребневики 386
rребные пластинки 76
rрибы как пища 276, 282
rуанин 362
rубки 19, 22 23, 27, 51, 54 59
защита 384
нейроидная система 492 93
питание 267 268, 278, 402
реаrреrация клеток 389
rубоноrие мноrоножки 231 232
защита 387, 388
локомоция 32 325
ДdКТИЛОЗОИДЫ 67, 69
Дафнии 507
Двиrательная активность, rенерирование проrрамм
517 518
Двиrательные центры 516
Движение 308 338, 500
- в толще субстрата и трубках 319 321
клеточные механизмы 309 312
нервные механизмы 516 520
Двукрылые насекомые. зашита от хишников 386
ооrенез 414, 415
старение 393
Двупарноноrие мноrоножки 232 233, 324
Двустворчатые моллюски 162 164
дыхание 345, 350
жизненный цикл 410
метаболизм 341
мышцы и движение 519
размножение 403
экскреция 366, 367
Двухвостки 234, 235
Дезоксирибонуклеозиды 15
Демоrрафическая теория жизненноrо цикла 425
Десятиноrие раки 258 262
Детерминация 450 53, 460, 545
Диапауза 410, 421 424
Диверrенuия 511
Динофлаrеллаты 51
как пища 2 9
симбиоз 273
Диуретический [ормон 541
Дифференцировка 16, 544
Диффузия 376 377
ДНК 12, 13, 15
Докембрийские остатки животных 18 19
Дорсальный непарно срединный нейрон (DUM) 519
Древесина как пища 282
Дробление 445 446, 466 77, 544
радиальное 468 469
спиральное 37, 355, 466 68
Дрозофила ОН. Drosop/lila 11lelallogastel'
Дыхание 342 355
анаэробное 542
аэробное 542
Дыхательные opraHbI 348 350
пиrменты 345 346
Дыхательный ток воды 160
Естественный отбор 13 14. 16, 48, 426, 427
Жаброноrие рачки 25 251, 404
Жабры 348, 349
Жабы 511
Жало 30 1, 386
Предметный указатель
rутики 309 311
Жrутиконосцы 22, 23, 51
симбиоз 282
rучие (orHeHHbIe) кораллы 69
Желточник 417
Желточные клетки 445
Желудок 278, 287
Жертвы 293, 294, 299 303, 307
Зdщитные приспособления 301
Жидкости тела CJW. Полостная жидкость
Жизненные циклы 263, 433 38, 543
и бесполое размножение 397 399
и половое размножение 407 12, 432
морских беспозвоночных 435 36, 471 79
насекомых 436 38, 479 481
Жужжальца 536
Жуки бомбардиры 387
Жук носороr 389
Заrрязнители 392 393
Заднежаберники 384
Замковые 175
Запасные вещества 416
Зародышевые листки 24
Защита 378 395
Защитная окраска 381
Защитное вооружение 301
Зеленый краб 224, 294
Земляной червь 490, 497, 540
локомоция 316,317
экскреция 374, 375
Зеркала 50 507
Змеехвостки см. Офиуры
Зоб 278
Зооиды 38, 178 181, 209,515
Зооксантеллы 75, 289, 290
Зоохлореллы 75, 289
Зрение 503 509
Зрительные пиrменты 503 504
Иrлокожие 39, 40, 191 204
нейрореrу ляция 490
нервная система 493. 495
питание 275
происхождение 19
развитие 193, 446, 454, 469 70
размножение 403
циркуляторная система 345
Избеrания реакция 380, 5) 4, 519
Известковые rубки 58
Изоосмотический раствор 376
Изотонический раствор 376
Имаrинальные диски 458 462
Иммунные реакции 389 392
Интернейроны 488, 516, 517
Ионная реrуляция 363 364, 376 377
Ископаемые остатки 39 3
История изучения беспозвоночных 1 11
Кайромоны 385
Калиевые каналы 532
Кальмары 167, 168, 348, 484
зрение 506
нервная система 519, 530 531
плавучесть 364
феромоны 524
Каракатица 167, 168, 36 365
защита 381
Карапакс 259, 260
Карбоксилирование 341
Карденолиды см. Кайромоны
Кардиальные rанrлии 501
Карповые вши (карпоеды) 252 253
Катаболизм 339 340, 542. 543
567
Кетокислоты 361
Кинетическая энерrия 331 332
Киноринхи 31, 100, 116 117
размножение 403
Кислород 357 359
кривая диссоциации 345 347
поrлощение 342 343, 351, 357 358
Киты. питание 304
Кишечник 277 279, 289
Кишечнодышащие 39, 188 189, 372
Кишечнополостные 19. 23, 24. 77
питание 27, 278
Кладистская школа 41
Кладоны 20, 46
Классификация, пяти царственная система 52
Клетки, rибель 533
двиrательные механизмы 309 312
дифференцировка 16
проrрамма развития 533 535
происхождение и эволюция 14 15
ренетты 111
сенсорные 503
Клеточная реаrреrация 389
Клеточные линии 473 74
мембраны 486
Клещи 224
Клоп rребляк 366
Клопы 241, 299 300, 526
питание 276
Книдарии 19, 24, 25, 268 269, 284, 363, 364
нервная система 492. 493, 513
строение 463
Книдоцисты 24, 62 64
Коксальные железы 371
Колеоидеи 167 168
Коллобласты 23, 76
Коловратки 31, 100. [21 125, 398
размножение 403
Колониальность 21, 45
Кольчатые черви С.И. Аннелиды
Командные нейроны 516 517, 539
Комары 241. 367
Комменсализм 75. 80
Кампартменты 21
Компетенция 452
Конверrенция 511
Конечности червей 213
членистоноrих 321 326
Конкурентное исключение 47
Конкуренция за пищевые ресурсы 263
межвидовая 48
Консументы 277, 283
Контралатеральные детекторы движения (DCMD) 516
Конформеры 357 358
Концентрациклоидеи 198, 200
Копеподы как пища 279
Кораллы, коралловые полипы 59. 61. 69, 73 75. 272,
290. 343
рефлекторные действия 513
Корацидий 91 92
Кортикальная реакция 441, 443
Коэволюция 426
Крабы 258, 262
зрение 507. 508
конечности и локомоция 323 325
нервная система и поведение 518, 539
питание 306
экскреция 367
Креветки 258, 367. 502
Кристаллический стебелек 287
Кровь 345 348
концентраuия ионов и осмотическое давление 363,
369
Круrлые черви см. Нематоды
568
Предметный указатель
Крыло HaceKOMЫX жилкование 240, 241
защитные узоры 369
и полет 326 329. 337 338
формы 326
Крыложаберные 188 191
Ксенобиотики 392
Ктенидии 33 268 269, 347 349
Кубомедузы 69
Кузнечик, нервная система и поведение 533 534
питание 385
прыжки 330
Куколки 245, 457, 458
Кутикула 213 368, 529
и 'Jкскреция 362
нематод 108
членистоноrих и хождение 324
Лактатный путь (путь молочной кислоты) 340
Лактоза 545
Ланцетники (rоловохордовые) 211 216
Латеральное торможение 510
Леrочная книжка 222, 349
Леrочные моллюски 363
слизни 384
Лейцинаминопептидаза 365 166
Лемниски 100
Ленточные черви 91 96
Лестница природы по Аристотелю 10, 11
Летательные мышцы 327 520
Линзы 506 507
Линька 454
Липофусцин 393
Литораль 355, 356, 368
Личинки личиночное развитие 403, 408 411 436
дрозофилы 479
морских беспозвоночных 408. 436. 437. 453 457,
477 478
MopCKoro желудя 478
морских улиток 472
насекомых 367, 457 58, 461
пелаrические 45 55
Локомоция 308 -338
животных с мяrким телом 335 337
ресничек 312
членистоноrих 321 326
Лопатоноrие (скафоподы) 16 165
Лофофор 269
Лофофоровые (щупальцевые) 32, 37 39, 45, 170 183
питание 267, 268 287
Лоциферы 100, 117, 119
Лучистые 77
Маrний 364
Макромеры 469
Макроэволюция 41- 42 47
Малощетинковые кольчецы (олиrохеты) 14 148
Мальпиrиевы сосуды (трубочки) 372 373 375
Маскировка 381, 383
Маточное молочко 524
Медоносная пчела 386 388 400
биолоrические часы 501
феромоны 524
Медузы 61 65, 69 71
нервная система 502
Мезентерий 72, 73
Мезоrлея 23
Мезомеры 469
Мезохил 56
Мейоз 416, 430 32, 440 525, 544
Меланизм 382 383
Мембраны 376, 486
Мерцательное пламя 371
Метаболизм 350 353 362
Метаболическое окисление 368
МетаЛJIотионеины 392 393
Метаморфоз 420, 453 462 477 83
Метанефридии 370, 371 374, 375
Метацель 37
Метахрональная ритмичность 312. 313, 317
I Метиладенин 419,520, 525
Механическая работа 309
Механические СИ IЫ 309 312
Механорецепторы 536
Меченые линии 511
Мечехвосты 223, 345, 51 О
Мидии 294, 355, 365
как пища 306
развитие 457
реакция на заrрязнителя 392
Миноrи 212
Микрокалориметр 352
Микромеры 469
Микроэволюция 41
Миксонефридии 371
Мимикрия 279.301. 302. 385 38б
Миозин 311
Мистакокариды 252
Митоз 544
Мноrоклеточность 21, 45, 51 98
rvlноrоножки 230 234, 548
Мноrощетинковые кольчецы см Полихеты
Мноrоядерные протисты 21
Модуль (патрон) нейронов 510
Мозаичное развитие 451 53, 471 74
Мозаичные rлаза 507
МОЗI 497 499, 526, 541
спонтанная активность 499
Мозrовые волны 499
МОКРИllЫ 345
Моллюски 33 15 169
rаметоrенез 416, 417
защита 383 384
нервная система 495
Hora 315 316
ооrенез 419
питание 267 275, 287 289
развитие 453 466, 469 74
размножение 403, 420
репродуктивные и выделительные opraHbl 372
)кскреция 362
Моли 377
Моляльность 377
Молярность 377
Моноплакофоры 156 158
Морская среда 364. 418
Морские беспозвоночные 364, 418 420
- жизненные циклы и развитие 435 436. 47] 479
. оплодотворение 397 400
водоросли как пища 275
ежи 47, 200 206, 389
rаструляция 447
оплодотворение 441 442
питание 275
- развитие 452 454 455 465 475 77, 545, 546
. размножение 403
желуди 248, 255 257, 361
. развитие 428, 457, 478
звезды (астероидеи) 19 198, 389, 403
ropMOHbI 524 525
. локомоция 318 319
нервная система 495
развитие 441. 454
лилии 195 196
нервная система 495
питание 273
размножение 403, 417, 418
осадки 266
пауки 22 227
Предметный указатель
569
улитки 471 474
уточки 256
Морфаллаксис 482
Морфоrенез 5 546
Мотивация 530
Мотонейроны 488, 516, 518 519
Моча 367
Мочевая кислота 361 363
Мочевина 14, 361 363
Мочевой пузырь 374
Мукопротеины 287
Муравьи 283
Мутации 15
Мухи, зрение 537 538
поведение 51 О
Мшанки 38, 39,46, 47, 175 181
питание 269, 287
размножение 403
Мышечное сокрашение 335
Мышечные волокна 311 315
Мышцы, работа 313 315
Мяrкотелые животные 313 321, 370
Наземная среда 367 368, 421 424
Наземные беспозвоночные 276
Насекомые 234 245, 321 323, 368, 370
rаструляция 450
ropMOHbI 529
жизненные циклы 411, 436 38
крылатые 237 245
локомоция 325 329
мноrоножкоподобные 236
первичнобескрылые 23 237
полет 520
развитие 446, 45 62, 4 77 483
ротовой аппарат 242
сенсиллы 535 536
сетчатка 537, 538
эмбрионы 464
эндокринная система 526 529
Натриевые каналы 531
Натриевый насос 486
Науплиус 247
На утилоидеи 166
На утилус 506
Научение 532 533
Независимые эффекторы 512 513
Нейроrемальный opraH 522
Нейромедиаторы (химические медиаторы) 49 91, 520
Нейронные связи 533 535
цепи 51 518
Нейроны 23, 488 89, 497, 516, 533 535
командные 516 517, 539
сенсорные 503
Нейропептиды 523
Нейропиль 495, 496
Нейросекреторные клетки 488, 521 522
Нейросекреция 521 522
Нейросуперпозиционные r лаза 509
Нейроэндокринные системы 523 524. 541
Нектар 290
HeMaToreH 96, 97
Нематоды, нервная система 495, 518, 533
плавание 518
развитие 451
старение 111, 281
экскреция 111
Нематофоры 100
Нематоцисты 23, 24, 63, 290
Немертины 29, 10 103, 403
локомоция 314
экскреция 369, 370
Необласты 464
Неомениоморфы 156
Непарный шелкопряд 526
Нервная система 48 85, 492 501, S33 535
автономные функции 500 501
спонтанная активность 499 50 1
Нервные волокна 502
окончания 488
сети 492 93
Нереиды 417, 420
Нерест 420
Нефридии 370, 371
Нефромиксий 372
Никотинамидадениндинуклеотид (N АО) 340
Ноrохвостки 234
Норы, локомоция внутри них 319, 320
роль в спасении от хищников 301, 302
Ньютона законы движения 308
Обrрыза тели 281 284
Обезвоживание С.М. Потери воды
Облеrченная реакция 513
Оболочники (личиночнохордовые) 205 211. 384
нервная система 494, 502
пелаrические 207
питание 270, 286
размножение 403
Оrнетелки 207
Одноветвистые рачки 45, 227 230, 239
локомоция 322
размножение 404
ротовой аппарат 229
экскреция 371
UKpacKa покровительственная (криптическая..) 381
предостереrающая 385
Оксиrеназы 392
Оксифотобактерии как симбионты 273
Октокораллы 74
Октопин 519. 520
Окукливание 527
Олиrохеты 1 148,
Омар, локомоция 325
нервная система 500
Омматидии 507, 508
Онихофоры 362, 371, 404
Онкосфера 91
Онтоrенез 54 549
Опиновый путь 340
Оплодотворение 432, 439 45
Опсонины 391
Оптико суперпозиционные r fIaза 509
OpraH У 529
Орrанические вещества 11, 1 15
полимеры, добиолоrический синтез 14.-.......15
Орrаноrенез 544, 545
Ортонектиды 97 98
Осмоконформеры 365; 376 377
Осмореrуляторы 365, 366
Осмос 376
Осмотическая реrуляция 363 364
Осмотическое давление 376
Осьминоrи 167
защита от хищников 391
зрение 504
мозr 499
нервная система 495, 502
пищедобывающее поведение 498 99
Отбор и защита от хищников 385. См. также ECTeCTBeH
ный отбор, Стабилизирующий отбор
Офиуры (змеехвостки) 198, 199
питание 269
размножение 403
Охотники (преследователи) 279 281, 299
Оцеллоиды 505
Падальная муха 411, 498
570
Предметный указатель
Палоло 417
Палочники 302
Память 498
долrовременная 533
Панцирные 403
Панцирь 301
Парабиоз 521
Паразитоиды 280 281
Паразиты [11 113, 277, 279 281
защита от хозяина 39J 392
Параподии 317
Паренхимула 57
Партеноrенез 398
и половое размножение 401
Парусник 67, 70
Патоrены 389
Пауки 279
зрение 507
леrочная книжка 349
локомоция 325
экскреция 362, 367, 370 373
Паукообразные 222, 224, 225, 411
Пауроподы 233 234
Педипальпы 280
Педицелларии 196, [98, 389
Педоморфоз 41, 47, 548
Первичный бульон [5
Переваривание 278
Передняя кишка 277 278, 284
Перепончатокрылые 300
Перистальтика 319
Периферическая нервная система 493 95
Пескожил 319 320
Печеночная двуустка 90
Пиrменты СМ. Дыхательные пиrменты, Зрительные пиr
менты
Пилидий [02
Пилильщики 300
Питание 265 307, 355, 498, 542
взвесями и осадками 267 269, 272, 273, 28 289,
303 305
слизисто ресничное 272
сосущий механизм 279 280
фильтрационное 270, 271
чистая прибыль 292 293
Пища, питательная ценность 293 299
Пищеварение 542
внутриклеточное 389
Пищеварительные адаптации 267, 28 287
Пищеварительный тракт 277 278, 289
термита 282
Пищевые предпочтения 293
цепи 27 277
Пищедобывающее поведение 498
Пиявки 144, 147, 148
rастр,ляция 448
дыхание 350
зрение 507
локомоция 314, 317
питание и переваривание пищи 279 280
плавание 496, 497
поведение 514
реакция испуrа 514
cerMeHTapHoe строение 481
экскреция 374
Плавание 321, 323, 33 337, 496
Плавууесть 364
План.... строения 47, 48, 51, 248
Планарии 315, 464
Пластинчатые 59 61
Пластрон 350
Плеченоrие 37 38, 43, 46, 172 175
размножение 403
экскреция 372
Плоские черви 19, 25 28, 78 96
локомопия 313, 316
нервная система 494
питание 267, 273. 498
развитие 417
размножение 402
реrенерация 463
эволюция 27 28, 46
Плутеус 312. 454, 455
Поведение 485
защитное 299, 300
механизмы 512 516
пищедобывающее 279, 299 303, 498
Поrонофоры 130 135, 403
Позвоночные 370
Позиционная информация 482 83
Поисковики 279
Поисковое время 299 302, 307
Пойкилотермные животные 355
Пол, определение 432 33
Полет 32 329, 337 338. 393
Полимеры 14
Полинуклеотиды 14, [5
Полиплоидия 54
Полипы 61 66
Полисахариды 14
Полиспермия 440, 441
Полифосфаты 14
Полихеты (мноrощетинковые кольчецы) 139 144
rаструляция 448, 449
деление 399
дыхание 348, 350
защита 383
зрение 507
локомоция 317 320, 325
нервная система 493 96
ооrенез 414
оплодотворение 443
определение пола 432
питание 269, 280
плавание 332
развитие 453 454, 457, 466, 477 478
размножение 403, 419
реrенерация 464, 482
cerMeHTHoe строение 481
хеморецепторы 503
циркуляторная система 343
Половое размножение 406, 544
Половые [ормоны 529
Полости тела 29, 30
Полостная жидкость 363 367
Полярная плазма 452
Популяции 426 28
Порфирин 345
Потенциал действия 487 88. 53 1 532
Потенциалы 486--------487
Потери воды 367
Потребители 299 305
Почвенные нематоды 529
животные 369
Почки 374
Почкование 398
Поясковые кольчецы 14 148, 403
Пресноводная среда 365 367
Пресноводные беспозвоночные 45, 365 367, 41 о, 421
424
Kr<7t7K4f'fi<1e «еtтееелек«е в пресную воду {1(/
Прерывистоrо равновесия rипотеза 16
Приапулиды 28, 31, 119 121, 403
Привыкание 532
Прилежащие тела 526, 528
Проrлоттиды 91
Проrормоны 523
Происхождение жизни 1 15
Предмеl'НЫЙ указаl'ель
Прокариоты [5
Проприорецепторы 50 I
П ростейшие 52
Протисты 2 27, 52 54
rетеротрофные 45
как пища 265 267
фотосинтезирующие 273
Протокишечнополостные 25
Протонефридии 79, 80, 370, 371, 374
Протонимфон 226,227
Проторакальные железы 527
Проторакотропный [ормон (РТТН) 526, 527, 541
Протоцель 29
П ротоэукариоты 15
Прудовая улитка 280
Прыrающие насекомые 329 330
Прямая кишка (ректум) 278
Прямокрылые 436
Псевдоцель 29, 31
Птихоцисты 63
Пуринотелия 361, 363
Пурины 361, 362
Пчелы 509. С.М. такж:е Медоносная пчела
Пятиустки 36, 217 218, 404
Рабдоиды 81 82, 385
Рабдомерные r лаза 504 505
Рабдомы 504
Равноноrие рачки 248, 260, 262, 362, 368
питание 282
Радиальная симметрия 23 24
Радиальное дробление 468 69
Радула 33, 152, 267, 281
Развитие 439 83. С.И. J11аК.же Метаморфоз, Эмбриоrе
нез
rемиметаболическое 457
rенетический контроль 479 81
rолометаболическое 458
rрадиентная теория 475 77
и дифференцировка 544
реrуляционное 451 53
Размеры тела и дыхание 353 355
и интенсивность метаболизма 266 267
Размножение 391, 421 28, 543 544
бесполое 397 399
и реrенерация 465
половое 397, 399 13
Раки 258
Раки отшельники 385, 493
Раковина 33, 153, 364, 365
маскировка 381 384
Ракообразные 36, 47, 246 262
дыхание 350
изменение окраски 525
конечности и локомоция 321 326, 539
личинки 437
питание 270, 355
плавучесть 364
размножение 404
экскреция 362, 366 368, 371, 374
Ракоскорпионы 44
Ракушковые рачки 267, 404
Распознавание 509 51 О
Растения как пища 276, 282, 283
Растительноядные (фитофаrи) 26, 276, 282 284, 302
Реактивное движение 337
Реrенерационная бластема 462 64
Реrенерация 462 65, 481 83
Реrуляторы биолоrические 357 358
Рейнольдса число 309, 329
Ректальные жабры 367
Ремипедия (слепой рачок) 251, 252
Репарация 393 394
Репликаза 15
571
Репликация 13 15, 543
Репродуктивная ценность 425
эффективность 425
Репродуктивные opraHbI 372
стратеrии 425
Реснички 31 313
в фоторецепторе 504
роль в фильтрации пищи и экскреции 17 375
У турбелларий 80, 81
Ресничные rлаза 504 505
Ретиналь 503
Ретинотопия 511
Рефлексы 498. 513 516, 533
РефлеКIорные действия 5 13 51 5
дуrи 51 515
Рецепторные клетки 501
Рецепторы 501 503
кодирование информации 502
растяжения 514
Речной рак 258
поведение 514. 517 518, 538 539
экскреция 374
Рибосомы 13
РНК 13, 15
Родопсин 503
Рост 465, 544, 546
Рот, ротовое отверстие 277. 278
Ротовая полость 278
Ротовой аппарат 242, 270, 272. С.И. тaK.JK'e Радула
колюще сосущий 276
Сальпы 208, 209. 286
Самозарождение жизни 15
Саранча, нейроэндокринные системы 541
нервная система 512, 515, 516
поведение 498, 515 516
полет 328
развитие 457
Свайник 111, 113
Сверчки 529
Свет поляризованный 504
CerMeHTbI 20, 36 37, 495 97
Сенсибилизация 532 533
СеllСИЛЛЫ 535 536 .
Сенсорная информация 509 512
Сенсорные клетки 50 I
модальности 501. 502
opraHbI 497, 50 I
поля 511
пути 510
системы 510 511
Сердце 501
Сетчатка 507, 537. 538
Сила 331 333
Симбиоз [5, 75, 273, 274. 283, 289 .292
Симбиотические бактерии 279
Симбиотический фотосинтез 273 274
Симфилы 232
Синапсы 487, 490 92
Сипункулиды 31. 127 129, 348, 403
ооrенез 414
экскреция 372
Скелеты. скелетные системы 313 315. С.И. J1ШК.JI('е rид
ростатический скелет
жесткие 324, 334 335
Сколекс 91
Скорпион 224, 325
Скребни 31, 125 127, 403
Слизистые сети 27[, 272
Сложные rлаза 507 509, 525
Слух 536
Слюна 279
Слюнные железы 278
Смертность 378, 380
572
Предметный указатель
Соленость 359, 365, 366
Соленоциты 371
Сосальщики 8 96. 279
диrенетические 89
моноrенетические 8 89
Соскабливатели 281 284, 303
Сосущий механизм поrлощения пищи 279 280
Спаривание 418, 419
Сперматозоиды 391, 440 41
Специалисты 502
Специфическое динамическое действие (СДД) 355
Спикулы 23, 54, 58
Спиральное дробление 37, 445, 46 68
Спироцист 63
Средняя кишка 278
Стабилизирующий отбор 44
Ставромедузы 69
Старение 378 379, 393 394
Статобласты 178
Статолиты 502
Статоцисты 502
Стеноrалинные животные 377
Стерниты 116
Столоны 141
Стомодеум 65
Стратеrии 543
СтрекаIQщие 61 65, 402
Стрекозы 327, 367
Супероксиддисмутаза (СОД) 359
Суша, завоевание животными 4 5, 368 370
Схватывание добычи 279, 280, 299
Схизоцель 29, 31, 100
Сцифомедузы 65, 75, 343, 493
Таннины 283
Таракан 510
Телифоны 224
Температура, влияние на дыхание 355 357
Терrиты 116
Термиты 282
Тип (таксономическая единица) 50
Тихоходки 372, 404
Токсины 384, 385
Тоническое сокращение 520
Т отипотентность 52
Трансдетерминация 479
Трансплантация 461, 479
Трахейные системы 344, 345, 349
Трематоды 86, 89 91
питание 281
размножение 402
Трение 331 332
Трикарбоновых кислот цикл (ТКК) 339 340
Трилобиты 19, 36, 42, 44. 46
Трохофора 128. 448. 449
Туника 384
Турбелларии 80, 81, 343, 385, 394
Тутовый шелкопряд 421
Тяжелые металлы 392
Уrри 321
Улитки, защита от хищников 381 382, 384
локомоция 315, 316
развитие 452
Ультрафильтрация 37 375
Уреотелия 361
У рикотелия 361, 362
Уровни орrанизации 16
Урохордаты 211
Условные рефлексы 532
Усоноrие рачки 255 257, 270, 404
защита от хищников 383
Устойчивый комплекс действия 513, 515
Фаrи 429, 430
ФаrОllИТОЗ 25, 289, 389 39]
Фазические рецепторы 501
Фазмиды ] 1 О
Фекальный щит 384
Фенотипы 542 543
Феромоны 502, 520, 521, 524
Фертилизин 440
Физиолоrия 543
Филактолематы 178
Филоrенез 18 9, 547
Филоrенетическое древо 41, 43, 46
Фильтрация, фильтраторы 303 305, 374 375
Фитонематоды 111
Фитофаrи СМ. Растительноядные
Форма тела 543
Форониды 37, 38, 171 172, 268
выделительная система 372
размножение 403
Фосфаrены 341
Фосфоенолпируват (ФЕП) 341
Фосфорилирование 340
Фотопериодические реакции (фотопериодизм) 418, 421
424
Фоторецепторы 503 509
Фотосинтез 266
морской 26 27
симбиотический 273 274
Фраrментация 298
Функциональная биолоrия беспозвоночных 263 2б4
Фуражирование 293 299
Хелицеровые 34, 36, 44, 45. 22 227
локомоция 321, 322
размножение 404
Хемокинез 440
Хеморецепторы 536
Хемодермоморфы 153 155
Химическая коммуникация 521 524
Химические медиаторы см. Нейромедиаторы
элементы 12
Химический состав живоrо 11, 12, 263
Хирономиды 348
Хитин 279, 324
Хитоны (панцирные) 157 159
Хищник жертва, взаимосвязь 29 295
Хищники и защита против них 38 389
их жертвы 293, 299 303, 307
Хлораrоrенные клетки 374
Хлорокруорин 345, 346
Хлоропласты 289, 291
Хоаноциты 22, 23, 54 57, 267 268, 312
Хоботок 99, 100
Хождение 32 326
Хордовые 212
беспозвоночные 267, 268, 285
Хроматофоры 525
Хроновиды 40, 41
Хрусталик 506, 507
Царства 45
Цветовое зрение 503 504
Целлюлазы 284
Целлюлоза как пища 275, 276
Целомодукты 37 371
Центральная нервная система 493 495
Церкарии 90
Цестоды 91 93, 402
Цефализация 83, 497
Цефалокариды 250
Цианобактерии 15, 21
как пища 289
Циклический аденозинмонофосфат 491
Циклы размножения 421 24
Предметный указатель
573
Циркадный (циркадианный) ритм 421
24, 535
Циркуляторные системы 34J 345
Цитоплазма тическая локализация 470------471
Цитоплазматические орrанеллы 15
Челюсти насекомых 242, 281
членистоноrих 386
Червеобразное движение 314
Червеобразные 28
33
Черви 28 33, 99
149
ИСКОПdt:мые 19. 28
локомоция 316, 336
паразитические 111 114, 281
питание 292
размножение 420
реrенерация 482
старение 394
схизоцеломические 100
Чернильный мешок 381
Чешуйницы 237
Членистоноrие 3
37, 45, 213 264
испарение воды 370
локомоция 321
337
нервная система 495
питание и пищеварение 267
270
развитие 448
реrенерация 462, 464
циркуляторная система 343
Шестилучевые rубки 58
Шипы (opraHbI захвата пищи) 279, 280
Шистосомоз 90
Шистосомы 9
91, 391
Шмели 356
Щелевые контакты 491
Щетинкочелюстные (xeTorHaTbI) 40, 185
186, 279, 280
UЦетинохвостки 237
Щупальца 268, 269, 272, 273
Эвнициды 141
Эволюционная эколоrия 16
Эволюция 11
16. См. также Макроэволюция, Ми
кроэволюция
rрупп животных 40
5
и филоrения беспозвоночных 18
9
первые животные 20
28
постепенная 41, 42
прерывистое равновесие 42
способов питания 265
277
Эвриrалинные животные 377
Эвтелия 31
Эвфаузиевый рачок 270
Экдизон 520, 529, 541
Экдистероиды 526, 527
Экзоптериrоты 457
Экзоскелет 362, 366, 367
Экзоцитоз 490
91, 522, 540
Экскреторные системы 111
Экскреты 362
Экскреция 361
363, 542
Экспериментальная эмбриолоrия 4 7 5
77
Эктодерма 61
Эктолецитальные яйца 417
Эктоневральная система 495
Эктопаразиты 86, 279
Эктотермные животные 355
Электронный транспорт 340
Электрохимическое равновесие 486
Эмбриоrенез, эмбриональное развитие 4б9
77
Эмбриональные зачатки, карты 459, 470------474
Эндокринная система 521, 52
529
Эндокринный баланс 525
пусковой механизм 524
Эндолецитальные яйца 417
Эндопаразиты 86, 90, 98, 280
Эндоптериrоты 458
Эндостиль 285
286
Эноплиды 1 02
1 03
Энтодерма 61
Энтопрокты 181
183
Эпиболия 448
Эпиrамия 141, 142
Эпидермис 61, 62
Эпитокия 141, 142
Эррантные виды 141
Этолоrия 513
Эхиуриды 31, 129
130, 372, 403
ооrенез 414
Эхиуры 270
Ювенильные сталии 436
38
Ювенильный ropMOH 520. 528
Яйцеклетки, метаболизм 440
45
эктолецитальные 417
эндолецитальные 417
Янтарной кислоты путь ( сукцинатный путь) 340
341
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Acariformes 225
Acoela 25, 26, 79, 84, 85
ACl'othoracica 257
ACl'otl'etida 175
Actiпia 357
Ae(/es aegypti 367
Aeolosomata 148
149
Аg/апt/ш 519
Ag/alll-a 69
Alcidis 386
Alcyonaria 74
Al1ogastropoda 161
AmbIypygi 225
AтiskH'ia 42
Ampharetidae 346
Alnphiol1idaceae 261
Amphipoda 260, 262
Aпlp/lipl'iol1 75
AпCl'[ostoтa 111
113
Aпc
'/us 357
AпeтOl1ill 357
Animalia 16
Annelida 32, 100, 135
149
Anudonta 3-66, 367, 410
Aпoтa/ocaris 35
Anomalodesmata 163
Al10pla 102, 103
Al10straca 251
Ant/lel'ea pO/YP/leп1Us 524
Al1thozoa 65, 66, 72
75
Ap/le/ocheirus 350
Aphragmophora 186
ApicOlnplexa 52
Apis тellip/lera 388
Aplousobranchia 206
Ap(l'sia 523, 532, 533, 535
Apterygota 23
237
AI'achnida 224
Аt"Ю1еае 225
AI'bacia 441
3
Al'chaeocyatha 19
Archaeognathata 237
AI'c/liteut/lis 506
А ,'el1ico/ll 319, 346, 347, 500
Arenicolidae 138
Ai'goпeтenes 370
Arтaпdia 507
Arteтia 346, 367
Al'ticulata 175
Ascaris 112, 113, 346, 451, 495, 496, 518
Ascetospora 52
Aschelminthes 99, 100
Ascidiacea 205
207
Ascothoracica 255. 256
Aspidogastrea 90
Aspiraculata 205
206
Astacus 367
Astellus 367
Asterias 441
Asteroidea 19
198
Astropecteп 346
Aurelia 69, 71, 363, 364, 493
ВотЬух 528
mori 421, 524
Boпellia 404, 433
Bougainvillea 67, 506
Bral'Jliпus 387
Brachiopoda 37, 170, 172
175, 248, 25
251, 323
Brachyura 323
Branchiobdella 148
Branc/liocaris 35
Braпc/liol1US 401
Branchiura 250, 252
253
Bryozoa 38, 170, 175
181
Buliтu/us 363
Bullia 357
Blirgessia 35
Bursovaginoida 104
Bacillariophyta 22
Balal1us 355, 428
Bdelloidea ] 00, 122
124, 403
BdeIlonemertea 103
Веrоё 78
Bilateria 79
Biston betu/aria 382
Bivalvia 162
164
Cael10rhabditis 484, 485, 495, 529, 533
Clllanu.'i 381
Calcarea 58
Calliпectes 324
Callip/lOrll 540
Campaпu/aria 440
Caraиsius п10rosus 546
Carcinu.s 294, 306
тaenas 295, 363
Caryophyllida 93, 95
Cassida 384
Cassiopeia 69, 75
Сераеа neтoralis 381, 382
Серhаlосю"idа 250, 323
Cephalochordata 211
212
Cephalodiscida 191
С ep/lll/odiscus 187, 190
Cephalopoda 165
169
Cerasl0derтa 38]
Ceriantharia 63, 66
Cestoda 91
96
Cestodaria 93
95
ChaetodermOlnorpha 153, 155. 403
Chaetognatha 39, 185
186
Chaetonotida 107
CJlaetopterus 270, 271, 481, 482
C/larпia 19
Cheilostomata ] 81
Chelicerata 36, 213, 22
227
Chilopoda 231
232
CJliroпoтus 346
Chlorhaemidae 346
Chlorophyta 22
Choanoflagellata 22, 52, 54
Chordata ] О, 20
212
Chrysophyta 22
Ciliophora 26, 52, 54
Cirripedia 248, 250, 255
257
Cladocera 250. 251
Clitellata 138, 14
149
C/ymenella 481, 482
Cnidaria 19, 23, 51, 61
75
Coelenterata 51, 77
Co]eoidea 167
Coleoptera 242
Collembola 234
Collothecida 124, 125
ComantJ1Us 417
19
Concentricycloidea 198, 200
Conchostraca 250, 251, 258
Copepoda 248, 250, 253
255
Corallina 477
Coralliuт 74
Cordy/ophora 514
Указатель латинских названий
с orop/uuт 293
Crepitlllla 405
Crinoidea 195 196
Cristatella 176
Crustacea 36, 213, 24
262
Ctenophora 23, 51, 75
78
Cteпop/aпa 78
Ctel1ostomata 181
Cubomedusae 64, 69
Си bozoa 69
Cyclorhagida 116, 117
C
,tcloтedusa 19
с'иmасеа 260
Оапаи.'!" pZexippus 386
Dap/тia 346, 366, 367, 401, 404, 410
Decapoda 258, 262
Demospongiae 58
Deпd"ost0I11Ul11 348
Deuterostomia 184
212
Diпoтisc/IUS 42
Dickiпsoпia 19
Dicyemida 96
Dideтпunl 359
Digenea 86, 89
Diпop/lI"/us 432
Dinophyta 23
Diopatia 357
Diopatra 414
Dip/lyl/ohot/lriuт 93
Diplopoda 232
233
Diplura 234
Diptera 242
DiJ"().fiZaria 113
Docoglossida 161
Dodecaceria 399
Doliolida 209
Droтia 363
Drosop/lila me/anogaster 379, 41
.416, 433. 452, 458, 459,
461, 479
Echinodermata 39, 191
204
Echimoidea 20
20]
Echiura 31, 100, 129
130
Echiurus 130
Eledoпe 391
ЕпсЬ ytraeidae 144
Enopla 1 02
103
Enterobius 112, 114
verтicu/aris 111
Eпteroтorplla 293
Enteropneusta 188.
189
Entobt/ella 88
Entognatha 234
Entoprocta 39, 181
183
Erioptera 293
Esc/lericllia сои 544
Eucarida 261, 262
Eucestoda 93
Euc/llora rubra 77, 386
ЕипЁсе 417, 419
Eunicida 140
Eunicidae 138
Euphausiacea 261
Eurypterida 44
Euscorpius 346
Facetotecta 257
Filospermoidea 104. 105
Flosculariida 124
Gaттarus 353, 366. 367
Gastroplli/us 346
Gastropoda 159
162
575
Gastrotricha 29, 99. 1 05
I 07
Glossiphonia 374
Gnathostomula 29, [00, 103
105
Go/jil1gia miпuta 129
Gordioida 114, 115
Gymnolael1ata 47, 180
181
Gymnomorpha 161
Gyrocotyle 95
Gyrodactylus 87
Ha/ic/
,tstus 72
Ha/ic,=yptиs 346
Hallucigenia 42
Halobates 299, 300
Harтot/lOe 522
He/iconius 283, 386
He/iocraпc/lia 364
Helix 315, 316
Hemichordata 39. 186
191
Heпlilepistus 368
Heterobranchia 161
Heterocyemida 96
Heteroda 111
Heterodonta 163
Heteronemertea 102
Heteroпereis 142
Heteroptera 242
Hexactine11ida 58
Hexapoda 23
245
Hirudinea 137, 144
148
Hirudo 146, 147, 321, 507
Holt/lUisaпa 369
Homalorhagida 117
Homalozoa 39, 40
Hoplocarida 259, 260
Hoplonemertea 103
Hyaliпoecia 357
Hydra 66
H.pt/robia 293
Hyt/roit/es 441, 443
Hydrozoa 62, 65
69
Hyтel1o/epis папа 93
Hyolitha 33
Ilyal1assa 452
Inartictllata 173
175
Isopoda 248, 260, 262
JO/lOia 35
Kil1etoplasta 52
Kiпocl1U/иS 118
Kinorhyncha 31, 99. 11
117
Larvacea 209
21 1
Laukartiara 506
Lepidoptera 242
Leptograpsus 507
Liтel1itis 386
Liml10ria 282
Liтu/us 346, 510
Liпeus 102, 313
Liпgula 346
Lil1gulida 173, 175
Littorina 297, 375
Loligo 348, 484, 530
Lophophorata 17{},-",,-,183
Loricifera 31, 99, 116, 403
Luidia 346
Lumbricus 1
146, 346, 490, 522, 540
Lyтaпtria 527
Lymnaea 363, 523
MacOlпa 293, 357
576
Указатель латинских названий
Macrobrac/liuт 355
Macrodasyida 106
М ageloпa 346
М aia squiпado 363
Malacobdella 103
Malacostraca 47, 248, 250, 258
262, 323, 404
Maпduca 526
Маууеllа 35
М arthasterias 363
Maxillopoda 250
Medusuпites 19
М egoura 424
М eloidogyпe 111
Meтbraпipora 400
Merostomata 223
224
Mesozoa 27, 51, 9
98, 402, 446
Metamonada 52
М ecapeпaeus 365
Metazoa 96, 302, 446
Metridium 73, 74
Microcoryphia 234
Microspora 23, 24, 52
MilZepora 70
Milleporina 69
Mimetaster 35
Mпeтiopsis 77
Mollusca 150
169
Moпiliforтis 126, 127
Monogenea 8
89, 403
Mono
onata 124
125. 403
Monopisthocotylea 86,' 87
Monoplacophora 15
157, 403
Musca doтestica 538
Муа 515
Myriapoda 230
245
Myrientomata 234
Mysidacea 260
Mystacocarida 250, 252
Mytilus 294, 356, 358
edulis 295, 355. 356. 365. 457
Myxico/a 487
Myxozoa 23, 24, 52
OicopleUl'a 494, 503
Olifochaeta 137
Oligoehtomota 234
Oпchidium 504
ОпС/l0сеуса 113
Onichophora 36, 213, 218
220
Opabiпia 42
Opalinata 52
Ophiuroidea 198
Ophryotrocha 481, 483
Opiliones 225
Oryctes 389
Orthonecta 96
98
Orthonectida 96
Ostracoda 250, 257
258
Oweпia 139
OXY
1}irUl.a 113
Pachygrapsus 363
Palaemoпetes 355, 367
Palaeoheterodonta 163
Palaeonemertea 102
Palaeoptera 242
Palpigradi 225
Pancarida 259, 261
Papilio lag 386
Parabasalia 52
Parafilaria 113
Parameciuт 346, 432
Parasitiformes 225
Parazoa 19
Parus 307, 383
Parvaпcoriпa 19
Patella 357, 367, 494, 506
Pauropoda 233
234
Pecteп 363, 504
Pentastoma 36, 213, 217
Peracarida 260, 261
Perviata 132
134
Phagocytellozoa 61
P/lUSc%soma 363
Philodinidae 124
Phlebobrahchia 206
Phorona 31, 37, 17
172
Phoronis 1 71
Phragmophora 186
Philactolaemata 178
Phyllocarida 259
Phyllodocida 140
Phyllodocidae 138
Phyllopoda 250
Physalia 66, 70, 364
Placozoa 27, 51, 59
1, 446
Plaпctosp/1Qera 191
Planctosphaeroidea 191
Plaпorb;s 346
Platasterias 197
Platyhelminthes 19, 25, 28, 29, 51, 78
96
Pleurobraпchia 530
Ploima 124
Роdоtriс/ш 386
Poekilocerlls 385
Pogonophora 32, 1 00, 130
135
Polychaeta 137, 139
144
Polyclada 84, 313
Polynoidae 138
Polyopisthocotylea 86, 87
Polyplacophora 157
159
Polyorchis 500, 504
Polystoma 87
Porifera 19, 22, 51, 54----------59
Porites 513
Poromyida 163
Portunidae 324
Namoпia 493
Nasoпia 379
Nassarius 452, 371, 474
Nautiloidea 166
Nautilus 365
Nebalia 125
Nectonematida 114, 115
Nematoda 30, 99
Nematomorpha 30, 99, 100, 114
116
Nemertea 29, 99
103
Neomeniomorpha 156, 157, 403
Neoptera 245
Nephros 363
Nephtys 481, 503
hoпbergii 347. 348
Neptuпea 363
Nereidae 409
Nereis 1 36
142, 317, 325, 343, 503, 580, 525
t[ivers;color 270, 292
294, 304
Nerita 506
Notop/aпa 498
Notostigmata 225
Notostraca 251
N uculida 163
Nuda 77, 78
ОЬеиа 66, 67
Obturata 134----------135
Octopoda 167
Octopus 495
Odontogriphus 42
Potaтophilax 383
Praecaпlhritlium 19
Priapula 31, 99, [00, 119
121
Priapu/us 120, 346, 370, 371
Prosobranchia 160
Protista 22
Protobranchia 163
Prltlzoa 51
54
Protura 234
Psammec/liпus 455
Pseudoceros 504
Pseudoscorpines 225
Pterygota 234, 239, 243, 244
Pteriomorpha 163
Pterobral1chia 189
191
Pterygota 237
245
Pycnogona 224
227
Pyrosomida 207
Radiata 77
Ragпea 19
Remipedia 25
252
Rhabditis 108
Rhabdocalyptus 492
Rhabdopleurida 191
Rhixocephala 255
Rhodпius 521, 527, 528
Rhombozoa 9
97
Ricinulei 255
Rotifera 31, 99. 121
125
Sabella 319, 320. 346. 347. 482
Sabellaria 139
Sabellidae 187, 346
Sa/pa 363, 515
Salpida 209
Sarcodina 22, 54
Sarcophaga 423
Sarotrocercus 35
Sarsia 493
Scaphopoda 164
165
Schistocep/1QZus 95
Schistosoтa тaпsoпi 90
Schistosomatidae 90
Schixomida 225
Scieractinia 74
Sclerospongiae 59, 60
Scorpiones 225
Scutigerida 231
Scyphozoa 65. 66. 69
72
Se;soп 125
Seisonidae 124, 125, 403
Se/eпaria 176
Sep;a 363
Sepiida 167
Serpu/a 346, 364
Serpulidae 346
Seticoronaria 121
Sigara 366
Sipuncula 31, 1 00, 127
129
Sipunculata 330
331, 415
SoIpugida 225
Spade//a 186
Spio 139
Spionidae 138
Sp;rorb;s 346. 457. 477. 478
Spriggina 19
Stenoglossa 161
Stenolaemata 46, 178
180
Stephaпo.filar;a 113
Указатель латинских названий
577
Stephaпopogoпoтorpha 26. 52
Stolidobranchia 206
Stoтph;a 73
Stroпgy/oceпtrotus 357
StroпgyZo;des 112
Stye/a 445, 452
Stylasterina 69
Stylommatophora 161
Styпapta 446
Sydпium 400
Syпlb;od;пiuт 75
Symphyla 232
Syпapta 446
Syncarida 259, 261
т аеп;а sagiпata 91, 93
Татоуа 503
506
Tanaidacea 260
Tardigrada 36, 213, 215
217
Teleogryllus 529
Temnocephalida 80, 81
Teпebr;o тolitor 383
Tentaculata 77
78
Tetrahymena 346
Teuthiida 167
Thaliacea 207
209
T/leZaz;a 113
Theroтyzoп 374
Thoracica 255
Thysanura 234
T;vela 346
Trematoda 86, 89
91
Tribraпc/lidiuт 19
Tric/lomoпas 282
Tr;choпJ'mp/1Q 282
Tr;churus 114
Triclada 84, 85, 313
Tritoпia 515
Tubifex 293, 343, 385
Tubu/aria 66, 68
Tu!l;тoпstrum 42
Turbellaria 8
86
UZva 293
Uniramia 36, 227, 245, 265, 457
и rechis 129
Urochordata 205
211
U ropygi 225
Va/vata 506
Vampyromorpha [67
Vaпat!is 507
Ve/aтeп 77
Ve/ella 67, 70
Vermetidae 271
Vertebrata 212
Vestilnenifera 134
Wi
f,,'axia 42
Wiwaxiida 33
Xanthophyta 22
Xenophyophora 54
Xenopneusta 130
Xiphosura 223
XyZoplax 198, 200
Zygoentomata 237
оrЛАВЛЕНИЕ
Предисловие к русскому переводу
Предисловие (пер. М. А. Софронова)
ЧАСТЬ 1.
ПРОСЫ
ЭВОЛЮЦИОННЫЕ
ВО..
rлава 1. Введение: основной подход
и принципы .........
1.1. Почему беспозвоночные?
1.2. Свойства живых существ
1.2.1. Введение .
1.2.2. П porpaMMa
1.2.3. Открытость
1.2.4. Эволюция путем естественно..
[о отбора неизбежное следствие
возобновления систем за счет ре..
пликации ....
1.3. Происхождение жизни
1.3.1. Добиолоrический синтез ар..
rанических полимеров
1.3.2. Происхождение и эволюция
клеток . . .
1.3.3. Почему самозарождение жиз..
ни не происходит все время?
1.4. Уровни орrанизации орrанизмов
1.5. План книrи . . .
1.6. Дополнительная литература
rлава 2. Эволюционная история
и филоrения беспозвоночных . . . .
2.1. Введение . .
2.2. Первые животные
2.2.1. rубки . .
2.2.2. Кишечнополостные
2.2.3. Плоские черви
2.2.4. Placozoa и Mesozoa
2.3. Производные плоских червей
2.3.1. rруппы червеобразных
2.3.2. Моллюски . . . . . . . . .
2.4. Более отдаленные потомки плоских
червей . . . . . . . . . . . . . .
2.4.1. Членистоноrие
2.4.2. Лафофоровые . .
2.4.3. Вторичноротые . . . . . . . .
5
7
10
10
11
11
12
13
13
14
14
15
15
16
16
17
18
18
20
22
23
25
27
27
28
33
34
34
37
39
2.5. Происхождение, радиация ивымира..
ние rрупп животных ......... 40
2.6. Резюме и эволюционный обзор 45
2.7. Дополнительная литература . . . 48
9
ЧАСТЬ 11. ТИПЫ
НЫХ
БЕСПО3ВОНОЧ..
rлава з. Параллельные подходы
к мноrоклеточности у животных
3.1. Protozoa (простейшие) . . .
3.2. Тип Porifera (rубки)
3.2.1. Этимолоrия . .
3.2.2. Диаrностические и спеuифи..
ческие признаки . . .
3.2.3. Классификация . .
3.3. Тип Placozoa (пластинчатые) . .
3.3.1. Этимолоrия
3.3.2. Диаrностические и специфи..
ческие признаки .. .....
3.3.3. Классификация . . . . . . . .
3.4. Тип Cnidaria (стрекающие: rидроид..
ные., сцифомедузы, актинии, кораллы) . .
3.4.1. Этимолоrия . . . . . . .
3.4.2. Диаrностические и специфи..
ческие признаки . .
3.4.3. Классификация . .
3.4.4. Симбиоз . . . . . . .
3.5. Тип Ctenophora (rребневики)
3.5.1. Этимолоrия
3.5.2. Диаrностические и специфи..
ческие признаки . . .
3.5.3. Классификация
3.6. Тип Plathyhelmintes (плоские черви) .
3.6.1. Этимолоrия . . . . . . . .
3.6.2. Диаrностические и специфи..
ческие признаки . .. ....
3.6.3. Классификация
3.7. Тип Mesozoa
3.7.1. Этимолоrия . . . . . . . . . .
3.7.2. Диаrностические и специфи..
ческие признаки ....
3.7.3. Классификация .
3.7.4. Родственные связи
3.8. Дополнительная литература . . . . .
50
51
51
54
54
54
58
59
59
59
61
61
61
61
65
75
75
75
75
77
78
78
78
80
96
96
96
96
98
98
rлава 4. Черви
4.1. Тип Nemertea (немертины) .
4.1.1. Этимолоrия . . . . . . . . .. 100
4.1.2. Диаrностические и спепифи..
ческие признаки .. . 100
4.1.3. Классификация . . . . 102
4.2. Тип Gnathostomula (rнатостомулиды) ] 03
4.2.1. Этимолоrия . . 103
4.2.2. Диаrностические и специфи..
ческие признаки . . . ] 03
4.2.3. Классификация ........ 105
4.3. Тип Gastrotricha (брюхоресничные
черви) . . . . . . . . . . . . .
4.3.1. Этимолоrия . .
4.3.2. Диаrностические и специфи..
ческие признаки .. .
4.3.3. Классификация . . . . .
4.4. Тип Nematoda (круrлые черви)
4.4.1. Этимолоrия . . . . . . .
4.4.2. Диаrностические и специфи..
ческие признаки . . . . . . . . . .
4.5. Тип Nematomorpha (волосатики) .
4.5.1. Этимолоrия . . . . . . . . .
4.5.2. Диаrностические и специфи--
ческие признаки . . .
4.5.3. Классификация
4.6. Тип Kinorhyncha . . . .
4.6.1. Этимолоrия . .
4.6.2. Диаrностические и специфи
ческие признаки _ . .
4.6.3. Классификация
4.7. Тип Loricifera . .
4.7.1. Этимолоrия . . .
4.7.2. Диаrностические и специфи--
ческие признаки .
4.7.3. Классификация . . .
4.8. Тип Priapula (приапулиды) .
4.8.1. Этимолоrия . .
4.8.2. Диаrностические и специфи
ческие признаки . . .
4.8.3. Классификация .
4.9. Тип Rotifera (коловратки)
4.9.1. Этимолоrия
4.9.2. Диаrностические и специфи
ческие ПРИЗНdКИ . . . . 121
4.9.3. Классификация . . . . . . 123
4.10. Тип Acanthocephala (скребни) . . 125
4.10.1. Этимолоrия ....... 125
4.10.2. Диаrностические и специфи--
ческие признаки. . . . . . .
4.10.3. Классификация .....
4.11. Тип Sipuncula (сипункулиды) .
4.11.1. Этимолоrия .....
4.11.2. Диаrностические и специфи..
ческие признаки . . . . .
4.11.3. Классификация _ .
4.12. Тип Echiura (эхиуриды)
Оrлавление
99
100
105
105
105
106
]07
107
107
114
114
1 14
115
116
116
116
117
1 ] 7
1]7
] ] 7
119
. ] 19
119
119
121
121
121
125
127
127
127
127
129
]29
4.12.1. Этимолоrия
4.12.2. Диаrностические и специфи..
ческие признаки. .
4.12.3. Классификация
4.] 3. Тип Pogonophora . . . .
4. ] 3.1. Этимолоrия ..
4.13.2. Диаrностические и специфи--
чески е признаки . . . . . .
4.13.3. Классификация .. _ . .
4.14. Тип Annelida (кольчатые черви, коль
чецы) ......... . . . . .
4.14.1. Этимолоrия .........
4.14.2. Диаrностические и специфи--
ческие признаки . . . . . .
4.14.3. Классификация
4.15. Дополнительная литература
rлава 5. Моллюски (пер. А. Б. Цeт
лин) . . ... . . . . . . . . . . . . . .
5.1. Тип Mollusca . . . . .
5.1.1. Этимолоrия . .
5.1.2. Диаrностические и специфи--
ческие признаки ..
5.1.3. Классификация . . .
5.2. Дополнительная литература
rлава 6. Lophophorata: лофофоро..
вые, или щупальцевые. . . . . . .
6.1. Тип Phorona (фораниды)
6.1.1. Этимолоrия . . . . . . _ . _ .
6.1.2. Диаrностические и специфи..
ческие признаки
6.1.3. Классификация . . . . .
6.2. Тип Brachl0poda (плеченоrие)
6.2.1. Этимолоrия. .....
6.2.2. Диаrностические и специфи--
ческие признаки
6.2.3. Классификация
6.3. Тип Bryozoa (МI1Jанки)
6.3.1. Этимолоrия .
6.3.2. Диаrностические и специфи
ческие признаки
6.3.3. Классификация
6.4. Тип Entoprocta . . . .
6.4.1. Этимолоrия . .
6.4.2. Диаrностические и специфи--
ческие признаки . . . .
6.4.3. Классификация .
6.5. Дополнительная литература
rлава 7. Deuterostomia: вторичноро"
тые ..
7.1. Тип Chaetognatha (щетинкочелюст
ные) . . . . . . . . . . . _ . . .
7.1.1. Этимолоrия . . . . . .
7.1.2. Диаrностические и специфи..
ческие признаки . . . . . . . . . .. 185
579
129
129
130
130
130
130
134
135
135
135
137
149
150
150
]50
150
153
169
170
171
171
171
172
172
172
172
173
175
175
175
177
181
181
181
183
183
184
185
185
580
Оrлавление
7.1.3. КлассификаllИЯ . . . . 186
7.2. Тип Hemichordata (полухордовые) 186
7.2.1. Этимолоrия . . . . . . . . 186
7.2.2. Диаrностические и специфи--
ческие признаки . .... . 186
7.2.3. Классификация . . . . . 188
7.3. Тип Echinodermata (иrлокожие). . 19]
7.3.1. Этимолоrия . . . . . . . . 19]
7.3.2. Диаrностические и специфи
ческие признаки .. . 191
7.3.3. Классификация . 194
7.4. Тип Chordata (хордовые) 204
7.4.1. Этимолоrия . 204
7.4.2. Диаrностические и специфи 204
ческие признаки .. . . . .
7.4А. Подтип U rochordata (личиночнохор"
довые, или оболочники) ... 205
7.4А.l. Диаrностические и специфи.. 205
ческие признаки . . . .. .....
7.4А.2. Классификация ...... 205
7.4Б. Подтип Cephalochordata (rоловохор"
довые, или ланцетники) . 211
7.4Б.I. Диаrностические и специфи
ческие признаки 211
7.4Б.2. Классификация . . 212
7.5. Дополнительная литература 212
rлава 8. Беспозвоночные с конечно
стями: членистоноrие и сходные
rpYnnbI . . . . .. ..... 213
8.1. Тип Tardigrada (тихоходки) 215
8.1.1. Этимолоrия . . . . . . . . . . 215
8.1.2. Диаrностические и специфи..
ческие признаки ..... 215
8.1.3. Классификация. . . 217
8.2. Тип Pentastoma (пятиустки) 217
8.2.1. Этимолоrия . . . . . 217
8.2.2. Диаrностические и специфи..
ческие признаки ..... 217
8.2.3. Классификация 218
8.3. Тип Onychophora . . . 218
8.3.1. Этимолоrия . . 218
8.3.2. Диаrностические и специфи--
ческие признаки . . . . . 218
8.3.3. Классификация 220
8.4. Тип Chelicerata . . . . 220
8.4.1. Этимолоrия . . . . . . . 220
8.4.2. Диаrностические и специфи..
ческие признаки . . . . . 220
8.4.3. Классификация . . 222
8.5. Тип Uniramia (одно ветвистые) 227
8.5.1. Этимолоrия. .... 227
8.5.2. Диаrностические и специфи..
ческие признаки . . . . . . 227
8.5.3. Классификация . . . . . . . . 230
8.5.3А. Подтип Myriapoda (MHoro
ножки) . . . . . . . . . . . . . . . . . 230
8.5.3Б. Подтип Hexapoda (насеко..
мые) . . . . . . . . . . . . . . . 234
8.6. Тип Crust,,-cea (ракообразные) 246
8.6.1. Этимолоrия . . . . . . . 246
8.6.2. Диаrностические и специфи..
ческие признаки . . . . 246
8.6.3. Классификация . 249
8.7. Дополнительная литература . 262
ЧАСТЬ 111. ФУНКЦИОНАЛЬНАЯ
Биолоrия БЕСПОЗВОНОЧНЫХ
(пер. И. В. Успенский) 263
rлава 9. Питание . 265
9.1. Введение: эволюция способов питания
животных . . . . . . . . . . . . . . . 265
9.2. Типы питания животных: способы за
хвата и переработки пищи. . . . . 277
9.2.1. Классификация типов пита..
ния . . . . 277
9.2.2. Общие черты . . 277
9.2.3. Охотники и паразиты 279
9.2.4. Обrрызатели и соскабливате--
ли .... . . . . . 281
9.2.5. Питание взвесями 284
9.2.6. Питание осадками . . . . 287
9.2.7. Питание с участием симбион"
тов . . . . . . . 289
9.3. Затраты и выиrрыш при различных
способах питания: оптимальное фуражи..
рование . . . . . . . . . . 292
9.3.1. Введение . . . . . . . . . . . . 292
9.3.2. Теория оптимальноrо фура..
жирования .............. 293
9.3.3. Подвижные потребители и их
жертвы ....... . . . . . . . 299
9.3.4. Потребители суспензий
и осадков . . . . . 303
9.4. Выводы . ..... 305
9.5. Дополнительная литература 306
Приложение 9.1. Выбор мидий крабами 306
Приложение 9.2. Поисковое время и вы..
бор жертвы: синица и мучные черви . . . 307
rлава 10. Биомеханика и движение
(локомоция) . 308
10.1. П ринципы локомоции . . . . . 308
10.2. rенерирование механических сил
клетками животных . . . . . . . . . . 309
10.3. Движение с помощью ресничек 312
10.4. Работа мышц и скелетные системы 313
10.5. Локомоция мяrкотелых животных. 315
10.5.1. Передвижение по твердой
поверхности ...... ...... 315
10.5.2. Движение в толще субстрата
и в трубках . . . 319
10.5.3. Плавание . . . . . . . . . . . 321
Оrлавление
10.6. Локомоция беспозвоночных с члени..
стыми конечностями . . . . . . . . . . 321
10.6.1. Локомоция ракообразных . 323
10.6.2. Наземная локомоция: хо..
жление 324
10.6.3. Полет . . . . . . . . . . . . . 326
10.6.4. Наземная локомоция: пры..
rающие насекомые 329
10.7. Выводы. . . 331
10.8. Дополнительная литература .... 331
Приложение 10.1. Термины и определе..
ния механики ................ 331
Приложение 10.2. Кинетическая энерrия
и трение . . . . . . . . . . 331
Приложение 10.3. Разложение сил на со..
ставляющие - . . . . . . . . . . . 332
Приложение 10..4. rидростатический CKe
лет . . . . . 333
Приложение 10.5. Жесткие скелеты
принцип рычаrа . . . . 334
Приложение 10.6 ...... ....... 335
Приложение 10.7. Плавание мяrкотелых
червей . . . . . . . . . . . . . . . . . . .. 336
Приложение 10.8. Реактивное движение 337
Приложение 10.9. Вихревое движение
воздуха и полет . . . . . . . 337
rлава 11. Дыхание . . . . .. .....
11.1. Центральная роль А ТР в дыхании
11.2. Основной путь катаболизма .
11.3. Получение АТР без кислорода .
11.4. Поrлощение кислорода .....
11.4.1. r лавное это диффузия .
11.4.2. Циркуляторные системы .
11.4.3. Кровь . . . . . . . .
11.4.4. Орrаны дыхания
11.4.5. Вентиляция. . . . .
11.5. Измерение метаболизма . . .
11.6. Факторы? влияющие на дыхание.
11.6.1. Размер тела .
11.6.2. Активность .
11.6.3. Питание
11.6.4. Температура
11.6.5. Концентрация
(РО) ...
11.6.6. Соленость .
11.7. Выводы .
11.8. Дополнительная литература
339
339
339
340
342
342
343
345
348
350
350
353
353
353
355
. 355
кислорода
357
359
359
360
rлава 12. Экскреция, ионная и ОСМО"
тическая реrулЯЦИЯ, плавучесть 361
12.1. Экскреция . . .. ...... 361
12.2. Осмотическая и ионная реrуляция 363
12.2.1. Морская среда ...... 364
12.2.2. Отступление по поводу пла..
вучести 364
12.2.3. Пресноводная среда .... 365
581
12.2.4. rиперосмотическая среда
12.2.5. Наземная среда . .
12.2.6. «Завоевание» суши и прес..
ных вод . . . . . . .
12.3. Выделительные системы
12.3.1. Строение
12.3.2. Функция
12.4. Выводы . . . . . . .
12.5. Дополнительная литература
П риложение 12.1. Некоторые определе
ния .
367
367
368
370
370
374
375
375
376
rлава 1 З. Защита 378
13.1. Классификация опасностей 378
13.1.1. Две основные rруппы 378
13.1.2. Эколоrические (внешние)
факторы смертности 378
13.1.3. Старение . . . 378
13.1.4. Классификация 379
13.2. Защита . . . . . . . . . . 380
13.2.1. Против хищников. 380
13.2.2. Против внутренних BparoB 389
13.2.3. Реакция на заrрязнители 392
13.2.4. Репарация защита против
старения? . . . . . 394
13.3. Выводы . . . . . . . . . . . . . 395
13.4. Дополнительная литература 395
rлава 14. Размножение и жизненные
циклы . . . . . . 396
14.1. Введение . .. ..... .... 396
14.2. Значение половоrо и бесполоrо раз..
множения . . . . .. ..... 397
14.2.1. Бесполое размножение в жи..
зненных циклах беспозвоночных . 397
] 4.2.2. Типы половоrо размноже
ния . . . . .. ..... 399
14.2.3. Механизмы определения по..
ла ..... ..... 400
14.2.4. Адаптивное значение раз
личных способов половоrо размно"
жения ...... . . . . . . . . . . . 405
14.3. Разнообразие половоrо размноже
ния: жизненные циклы . . . 407
14.3.1. Введение . 407
14.3.2. Морские беспозвоночные 407
14.3.3. Пресноводные и наземные
беспозвоночные .......... 410
14.4. Контроль процессов размножения . 412
14.4.1. Первичные инепосредствен..
ные контролирующие факторы . . 412
14.4.2. rаметоrенез ........ 41 3
14.4.3. Синхронное размножение
беспозвоночных. . . . . . . . . . . . 417
14.4.4. Циклы размножения и диа
пауза в наземной и пресноводной
средах . . . . . . . . . . . . . . . . . 421
582
Оrлавление
14.5. Размножение и распределение pecyp
со в . . . . . . 424
14.6. Выводы. . . 428
14.7. Дополнительная литература .... 429
П риложение 14.1. Системы половоrо раз..
множения (миксиса) . . . . . . . . . . 429
Приложение 14.2. Жизненные циклы
и половое размножение . . . . 432
Приложение 14.3. Системы определения
пола . . . . . . . . . . . . . . . . . . . . . . 432
Приложение 14.4. Жизненные циклы бес
позвоночных . . . . . . . . . . . . . . . . . 433
Приложение 14.5. Жизненные циклы мор..
ских беспозвоночных . . . . . . . . . 435
Приложение 14.6. Функциональный ана..
лиз жизненных циклов Насекомых 436
rлава 15. Развитие
439
15.1. Оплодотворение и инициация разви"
тия . . . . . . . . . . . .
15.2. Раннее развитие.
15.2.1. Дробление
15.2.2. rаструляция .
15.3. Экспериментальная эмбриолоrия
беспозвоночных: детерминация судьбы
клетки . . . . . . . . .. ..... 450
15.3.1. Введение . . . . . . . . 450
15.3.2. Мозаичное и реrуляционное
развитие . . . . . 451
15.4. Личиночное развитие и метаморфоз 453
15.4.1. Метаморфоз морских личи
ночных форм . .. ......... 453
15.4.2. Развитие и метаморфоз Hace
комых . . . . . . . . . 457
15.4.3. Экспериментальный анализ
развития имаrинальных дисков Dro--
sop/ll/a тe/aпogastel . . . .
15.5. Реrенерация .. .
15.5.1. Введение
15.5.2. Природа реrенерационной
бластемы . . . . . . . 462
15.5.3. Реrенерация и реrионарная
орrанизация ........... . 463
15.5.4. Реrенерация, рост и размно"
жение .. .............. 465
15.6. Выводы: развитие беспоз оночных
и rенетическая проrрамма . . . . . 465
15.7. Дополнительная литература 466
П риложение 15.1. Спиральное дробление 466
Приложение 15.2. Радиальное дробление 468
Приложение 15.3. Реrуляционное и мо--
заичное развитие l ......
Приложение 15.4. Судьба клеток, карты
зачатков и цитоплазматическая локализа..
ция . . . . . . . . . . . . . . . . .. .... 470
Приложение 15.5. Реrуляционное и мо"
заичное Рdзвитие 11 . . . . . . . . . . . . . 471
439
445
445
446
459
. 462
462
469
Приложение 15.6. Экспериментальный
анализ развития MopcKoro ежа . 475
Приложение 15.7. Метаморфоз и выбор
субстрата у морских личинок . . . . . . . 477
Приложение 15.8. Серийная транспланта
ция имаrинальных дисков и открытие
трансдетерминации . 479
Приложение 15.9. rенетический контроль
и реrионарная орrанизация: rомеозисные
rены . . . . . . . . . . . . . . 47У
Приложение 15.10. Позиционная инфор"
мация и каудальная реrенерация у аннелид 481
rлава 16. Системы контроля . 484
16.1. Введение . . . 484
16.2. Потенциалы . . . . 486
16.2.1. Мембраны 486
16.2.2. Потенциалы . 486
16.2.3. Проведение rрадуальных по.. 487
тенциалов . . . . . . . .
16.2.4. Потенциал действия .... 487
16.2.5. Спайковые инеспайковые 488
нейроны . .
16.3. Нейроны и их связи 488
16.3.1. Нейроны . . . . 488
16.3.2. rлиальные клетки 489
16.3.3. Химические синапсы . 490
16.3.4. Электрические синапсы 491
16.4. Орrанизация нервных систем . . 492
16.4.1. «Нейроидная» система rубок 492
16.4.2. Нервные сети ........ 492
16.4.3. Центральная и перифериче..
ская нервные системы . . . . . 493
16.4.4. Дифференцировка нейро..
пиля . . . . . . . . . . . . .
16.4.5. CerMeHTapHoe строение
16.4.6. «rорячие линии»
16.4.7. Роль мозrа . . . . .
16.5. Спонтанная активность ...
16.5.1. Нервная инициация .
16.5.2. Движение . . . . . . .
16.5.3. Автономные функции
16.5.4. Биолоrические часы
16.6. Рецепторы . . . . . . . . . . . .
16.6.1. Основная характеристика
16.6.2. Классификация рецепторов
16.6.3. Специалисты и rенералисты
16.6.4. Кодирование интенсивности
16.6.5. Сенсорные клетки и сенсор"
ные нейроны . . . 503
16.7. Зрение ......... ...... 503
16.7.1. Зрительные пиrменты 503
16.7.2. Ресничные и рабдомерные
r лаза . . . . . . . . . . . .
16.7.3. rлазки и rлаза
16.7.4. Линзы и зеркала
16.7.5. Сложные rлаза
495
495
497
497
499
499
500
500
501
501
. 501
501
502
502
504
506
506
507
16.8. Обработка сенсорной информаuии 509
16.8.1. Смыслово.е восприятие мира 509
16.8.2. Сенсорные системы . 51 О
16.8.3. Конверrенция и диверrенция 511
16.8.4. Меченые линии . . . . 511
16.8.5. Выделение признаков . . 511
16.8.6. Центробежный контроль 512
16.9. Нервные основы поведения . . .. 512
16.9.1. Независимые эффектары . 512
16.9.2. Элементы поведения . .. 513
16.9.3. Рефлекторные действия .. 513
16.9.4. Устойчивые комплексы дей
ствий 515
16.9.5. Вариации на две темы . . . 515
16.10. Орrанизация двиrательноrо «выха..
Да>) . . . . . . . . . . . . . . . . . . . .
16.10.1. Аспекты орrан.изации . .
16.10.2. Командные нейроны
16.10.3. rенерирование проrраммы
16.10.4. Мотонейроны . . . . .
16.10.5. Нервно"мышечная инне..
рвация . . . . . . . . . . . . . ., .. .
Химическая коммуникация
16.11.1. Спектр химической комму..
никации . . . . . . . .
16.11.2. Критерии эндокринной ак..
тивности ........
16.11.3. Нейросекреция . . .
16.11.4. Размытая rраница
16.11.5. Нейропептиды . .
16.11.6. Нейроэндокринные систе..
мы . . . . . . . . .
16.11.7. Феромоны
Роль эндокринной системы
16.12.1. ЭНДОКРИННЫЙ пусковой ме..
ханизм . . . . . . . . . . . . .
16.12.2. ЭНДОКРИННЫЙ баланс . . .
16.12.3. Эндокринный контроль
развития у Nel
eis ..........
16.12.4. Эндокринный контроль
развития насекомых . . . . . . . . .
16.11.
16.12.
Оrлавление
583
516
516
516
517
518
519
521
521
521
521
522
523
523
524
524
524
525
525
526
16.13. ВЬIВОДЫ . . . . 529
16.14. Дополнительная литература 530
Приложение 16.1. Система rиrантских во..
локон кальмара Loligo . . . . . . . . .. 530
Приложение 16.2. ПотеНllиал действия 531
Приложение 16.3. Клеточные механизмы
научения ........ .... 532
Приложение 16.4. Становление дефини
тивных нейронных связей .. 533
Приложение 16.5. Клеточные механизмы
биолоrических часов . . . . 535
Приложение 16.6. Сенсиллы насекомых 535
П риложение 16.7. Нейросуперпозиционная
сетчатка мухи ............. 537
Приложение 16.8. Оптическая доля ком..
натной мухи (Musca d0l11estica) ...... 538
Приложение 16.9. Удары брюшка у рака 538
Приложение 16.10. Контроль мышечных
сокращений у ракообразных . . . . . . . . 539
Приложение 16.11. Нейросекреция путем
экзоцитоза ............. 540
Приложение 16.12. Нейроэндокринные
системы саранчи . . . .. ...... 541
rлава 17. Еще раз об основных
принципах .. 542
17.1. Основные физиолоrические черты
фенотипов . . . . . 542
17.2. rлавное
репликация и размноже
ние . . . . 543
17.3. Онтоrенез ....... 544
17.4. Онтоrенез и филоrенез . 547
17.5. Дополнительная литература 549
Словарь терминов 550
Источники иллюстраций . 561
Предметный указатель 565
Указатель латинских названий. . .. 574
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книrи, ее офор
млении, качестве перевода и друrие просим при
сыла ть по адресу:
129820, Москва
1
й Рижский пер., д. 2,
издательство «Мир»
УЧЕБНОЕ ИЗДАНИЕ
Р. Барнс. п. Кейлоу. п. Олив, д. rолдинr
БЕСПОЗВОНОЧfIЫЕ.
НОВЫЙ ОБОБЩЕННЫЙ ПОДХОД
Заведующая редакцией
канд. биол. наук М. Д. rроздова
Ведущий редактор Ю. И. Амченков
Мл. научн. редактор И. А. Ермакова
Художник Е. r. Шведова
Художественные редакторы А. я. Мусин, Л. М. Аленичева
Технический редактор М. А. Страшнова
Корректор Т. М. Подrорная
ИБ N!:! 7237
Сдано в набор 04.10.90. Подписано к печати 07.04.92. Формат
70 х 1001/16. Бумаrа офсетная N2 1. Печать офсетная. rарнитура
Тайме. Объем 18,25 бум. л. Уел. печ. л. 47,45. Уел. Kp.
oтт. 94.90.
Уч.
изд. л. 54,55. Изд. N2 4/6774. Тираж 3500 экз. Зак. 1314. С 190.
Издательство «М ир»
129820, rсп, Москва. И
110. l
й Рижский пер.. 2
Можайский полиrpафкомбинат Министерства печати и информа
ции Российской Федерации
143200, r. Можайск, ул. Мира, 93.