


































































































































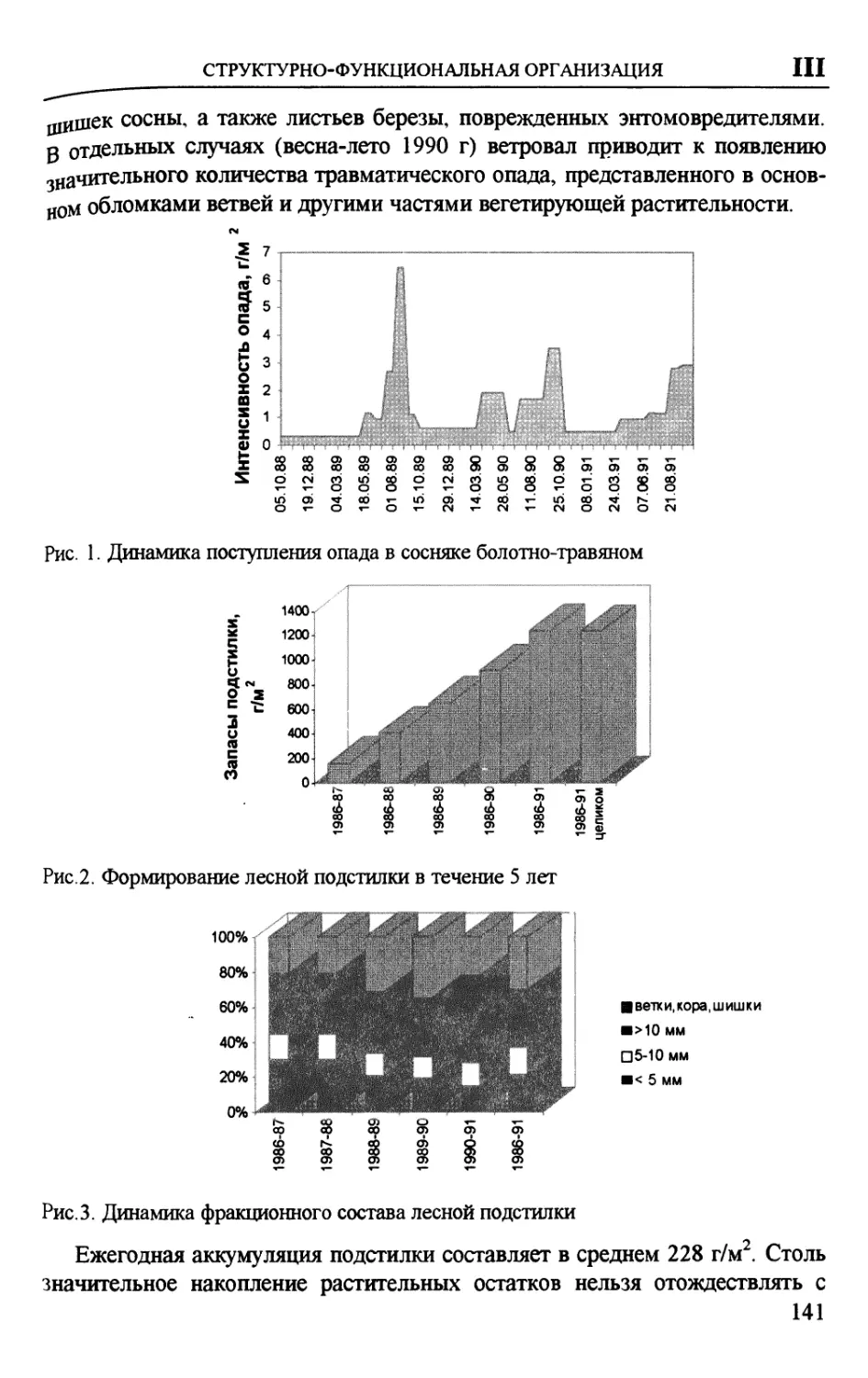

























































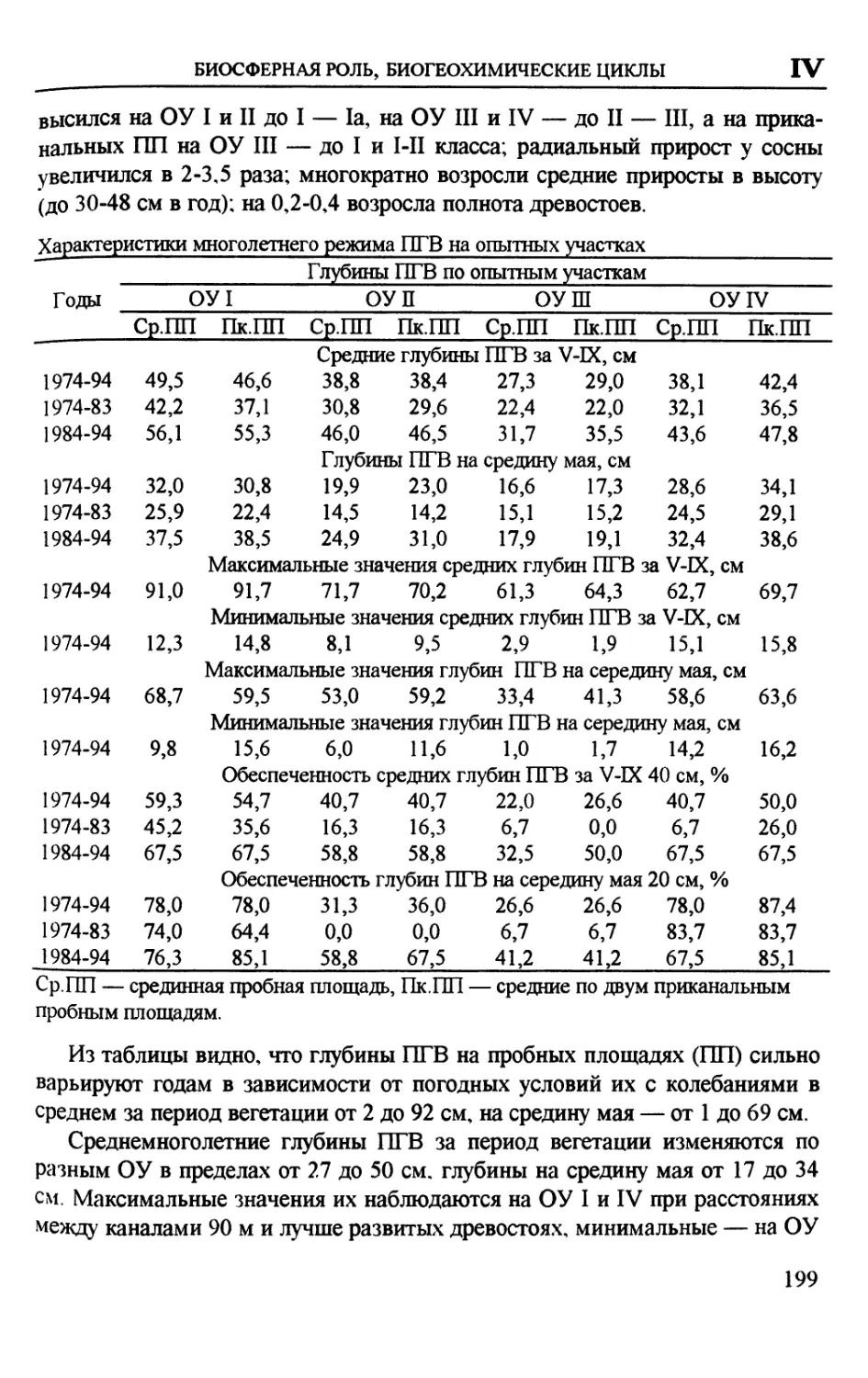


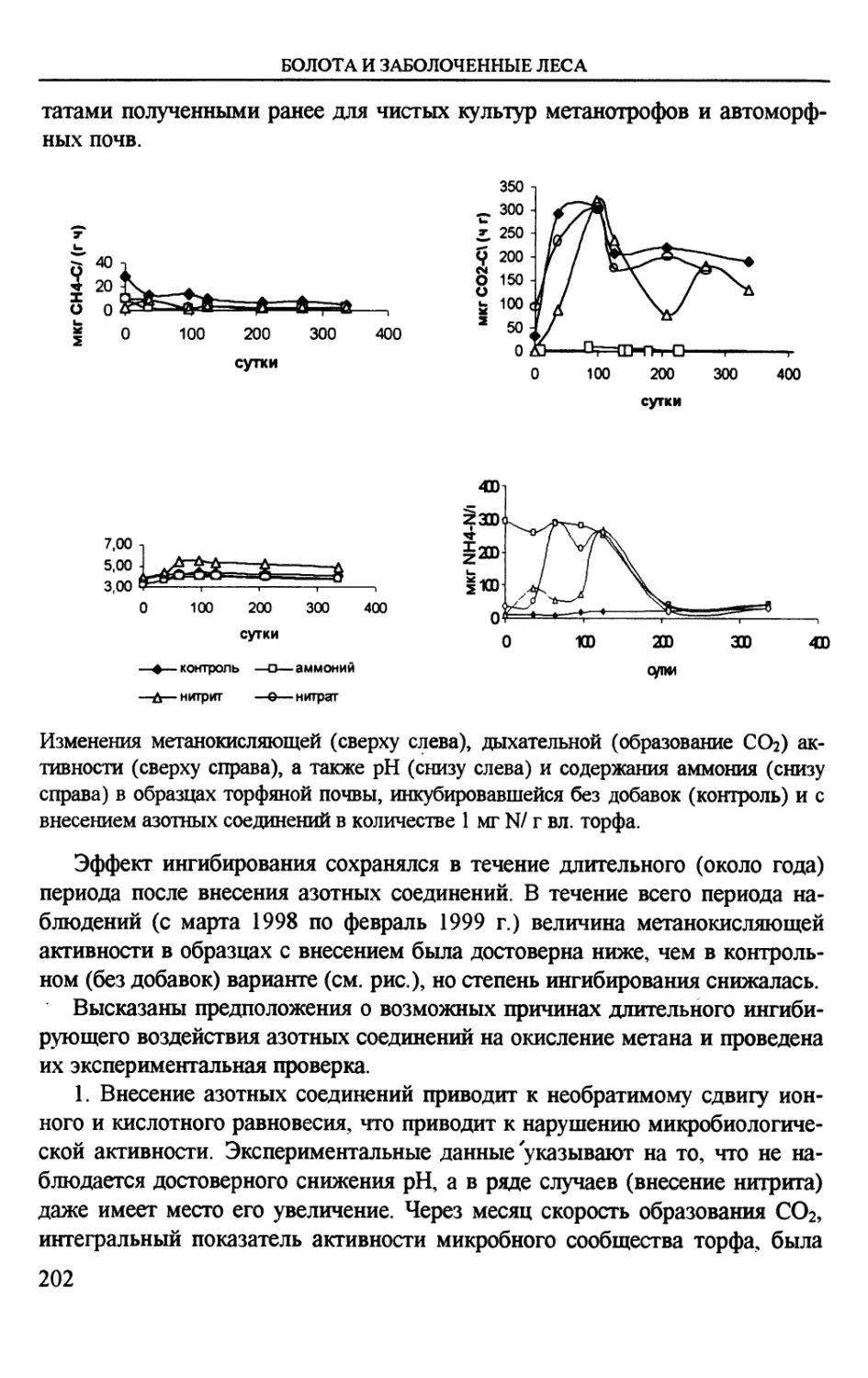

























































































































































































Author: Вомперский С.Э. Сирин А.А.
Tags: лесное хозяйство лесоводство религии месопотамии природа охрана окружающей среды гидрология суши гидрология рек, озер, водоемов гидрология болот болотоведение гидрометрия лиманы и так далее природоведение
ISBN: 5-89118-091-X
Year: 1999




БОЛОТА
И ЗАБОЛОЧЕННЫЕ
ЛЕСА
В СВЕТЕ ЗАДАЧ
УСТОЙЧИВОГО
ПРИРОДОПОЛЬЗОВАНИЯ
МАТЕРИАЛЫ
КОНФЕРЕНЦИИ
МОСКВА
1999
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ФЕДЕРАЛЬНАЯ СЛУЖБА
ЛЕСНОГО ХОЗЯЙСТВА РФ
275
лет
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ФЕДЕРАЛЬНАЯ СЛУЖБА ЛЕСНОГО ХОЗЯЙСТВА РФ
Научный совет РАН по экологии биологических систем
Научный совет РАН по лесу
Межведомственный совет по гидролесомелиорации
Институт лесоведения РАН
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
В СВЕТЕ ЗАДАЧ
УСТОЙЧИВОГО ПРИРОДОПОЛЬЗОВАНИЯ
МАТЕРИАЛЫ КОНФЕРЕНЦИИ
Москва
ГЕОС
1999
ББК 26.222.7
Б 79
УДК 630.182:252.6:502
Болота и заболоченные леса в свете задач устойчивого
природопользования. Материалы совещания. - М.: ГЕОС, 1999.-392 с.
ISBN 5-89118.091-Х
Представлены 151 статья, подготовленные к конференции «Болота и
заболоченные леса в свете задач устойчивого природопользования»
специалистами России, Беларуси и других стран. Отражен широкий круг знаний по
распространению, динамике болот и заболоченных лесов, их структурно-
функциональной организации, биосферной роли, биоразнообразии, методах
и последствиях хозяйственного использования, мониторингу и охране, как
научной основы для перехода к устойчивому пользованию этими угодьями
в современных экономических условиях.
Книга будет полезна для широкого круга специалистов в области экологии,
болотоведения, лесоводства, управления природопользованием, всех
интересующихся рациональном использованием и сохранением экосистем болот
и заболоченных лесов.
Табл. 60. Ил. 18. Библ. 329 назв.
Материалы совещания опубликованы при поддержке Западнодвин-
ского лесхоза Управления лесами Тверской области и Института
лесоведения Российской академии наук.
Ответственные редакторы
С.Э. Вомперский, А.А. Сирин
На обложке: Грядово-мочажинный комплекс на верховом болоте.
Высокопродуктивный сосняк на осушенном мелкооторфованном болоте. Черно-
ольшанник папортниковый. Фото С.Э. Вомперского
ISBN 5-89118-091-Х
©ИЛ АН, 1999 г.
©ГЕОС, 1999 г.
ПРЕДИСЛОВИЕ
ПРЕДИСЛОВИЕ
Данная книга — сборник статей авторов, откликнувшихся на участие в
Международной конференции «Болота и заболоченные леса в свете задач
устойчивого природопользования», приуроченной к 25-летию Западнодвин-
ского лесоболотного стационара Института лесоведения РАН (Тверская
область).
Начиная с Конференции ООН в Рио-де-Жанейро (1992 г.) и Генеральной
Ассамблеи ООН в Нью-Йорке (1997 г.), выработавших концепцию
«Устойчивого развития», ряда последующих конвенций и других международных
актов по борьбе с деградацией природной среды, сохранению
биоразнообразия и техногенному изменению климата, мировое сообщество стремится в
рамках каждой страны добиться «Устойчивого развития», т.е. найти в
природопользовании приемлемые компромиссы между экологическими и
социально-экономическими императивами. Хотя главным тормозом
экологизации методов и технологий выступает обычно их более высокая цена, тем не
менее, выяснилось, что ученые и специалисты из-за недостатка знаний часто
не готовы обоснованно сформулировать сами эти экологические требования
и реалистичные ограничения, отвечающие новым задачам.
В полной мере это относится и к болотам, и к заболоченным лесам,
которые широко используются в сельском, лесном хозяйстве, торфяной
промышленности, добыче лекарственных и пищевых продуктов, рекреации и
т.д. Значимость этой проблемы для России определяется масштабом
заболоченности ее земель: более 1/5 ее территории — оторфованные площади. В
торфе болот и заболоченных земель сосредоточен громадный пул
биогенного углерода — 113,5 млрд. тонн (Вомперский и др., 1994), свидетельствуя о
выдающейся биосферной роли болот России в глобальном цикле углерода.
В таежной зоне процессы болото- и лесообразования не только
соседствуют, но часто совмещены во времени и пространстве, с чем связано
образование заболоченных и болотных лесов. Подавляющее большинство всех
болот страны входит в состав земель Гослесфонда, а лесные болота наряду с
другими видами пользования подвергаются и лесоэксплуатации. Сказанное
в известной мере отражается заглавием книги — «Болота и заболоченные
леса...».
Многообразие природы болот в географически разных условиях
обусловливает неодинаковость в количественном отношении проявление ими
биосферных и средообразующих функций. С другой стороны, разная их
эксплуатация влечет разные следствия для природной среды и неодинаковую
хозяйственную и экономическую эффективность. Все это крайне важно
знать для выработки региональных критериев и индикаторов устойчивого
3
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
использования болот, сохранения их биоразнообразия и выполняемой ими
биосферной роли.
К сожалению, как за рубежом, так и особенно у нас недостаток знаний о
природе болот, низкая точность учета их площадей в России,
противоречивые оценки современной динамики торфонакопления и заболачивания
новых площадей, различие взглядов ученых на последствия эксплуатации
болот для окружающей среды и ряд других дискуссионных вопросов мешают
выработке согласованных рекомендаций по рациональной стратегии
использования и охраны болот, экологизации методов эксплуатации болотных
экосистем, оценки их биосферной роли. Внимательный читатель данной
книги найдет эти и другие остающиеся спорными или неясными вопросы в
статьях разных авторов.
Оргкомитет пытался акцентировать внимание участников конференции
на экосистемном анализе своих данных, так как устойчивое природопользоН
вание базируется именно на экосистемном уровне знаний. Поэтому сложивн
шаяся традиция научных и научно-практических совещаний по частньщ
разделам болотоведения и пользования болотами (по генезису болот, их|
классификации, истории в голоцене, гидролесомелиорации и т.д.) хотя sj
очень важна, но недостаточна для выработки новых концепций в природой
пользовании. Сознаем, что этого полностью нам не удалось достичь, но на-!
деемся что данная конференция — шаг в верном направлении. В данную
книгу помещено подавляющее большинство заявленных работ: хотелось
полнее выявить «географию» современных исследований болот. В ходе под^
готовки книги была сделана лишь необходимая техническая правка
представленных работ и за научное содержание их ответственны авторы.
Публикуемые материалы конференции — попытка отразить состояние
исследований по широкому кругу знаний о генезисе, динамике болот,
заболоченных и болотных лесов, их учету и распространению, разнообразному
использованию и охране в современных экономических условиях России,
Беларуси, Латвии и Литвы в надежде, что синтез этих сведений расширит
наши экосистемные представления о естественных и эксплуатируемых боло^
тах и поможет реализации идеи и методов устойчивого природопользования.
В организации конференции, публикации ее материалов, подготовке
натурных объектов, экскурсий и другую большую помощь оказала
Федеральная служба лесного хозяйства РФ и ее учреждения. Особая признательность
Оргкомитета конференции — Начальнику Тверского управления лесами
М.А. Федотову и директору Западнодвинского лесхоза В.М. Черникову без
личного энтузиазма которых, сотрудников управления и лесхоза
конференция была бы невозможной.
С.Э. Вомперский
. РАСПРОСТРАНЕНИЕ, ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ I
L РАСПРОСТРАНЕНИЕ, ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ
БОЛОТ, ЗАБОЛОЧЕННЫХ ЗЕМЕЛЬ И ЛЕСОВ
Г АГУЛЬСКАЯ КОТЛОВИНА — УНИКАЛЬНОЕ ВОДНО-БОЛОТНОЕ
УГОДЬЕ ЗАПАДНОГО САЯНА
А.Н. Байкалов, А.П. Савченко, В.В. Лаптенок, Н.А. Рябинина
Красноярский государственный университет, биологический факультет
Гагульская котловина — заболоченная долина рек Гагуль и Теплая,
находится в среднегорной тайге Западного Саяна у северного макросклона
Куртушибинского хребта на высоте около 1000 м над ур. м. Эта межгорная
котловина представляет собой сочетание различных ландшафтов —
степного, горнотаежного, пойменных зарослей, открытых кочкарниковых и
верховых болот, небольших озер, речек и ручьев. Благодаря высокой мозаичности
местообитаний здесь сложились уникальные условия для обитания
животных, обуславливающие значительное видовое разнообразие птиц разных
экологических групп [1,2,3,4,5,6]. Исследования проводились в 1990-1992
гг. по разработанному методическому комплексу [7]. ,
Орнитофауна котловины представлена гнездящимися и пролетными
видами птиц. В ходе работ установлено пребывание 133 видов, относящихся к
14 отрядам. Из них — 12 ввдов (черношейная поганка, красношейная
поганка, черный аист, касатка, орел-карлик, могильник, беркут, сапсан, серый
журавль, филин, черногорлая завирушка, полярная овсянка) занесены в
Красные книги России и Красноярского края. В миграционный период
характерны концентрации водоплавающих птиц (до 1,5-2 тыс.), здесь их
отмечается 16 видов из 3 отрядов (Podicipediformes, Anseriformes, Gruiformes),
суммарная интенсивность ночной миграции составляет 3392 особи/км.
Впервые в Западном Саяне на пролете отмечен белолобый гусь.
Значительна доля и пролетающих через котловину воробьиных птиц (суммарная
интенсивность ночной миграции — 2948 особей/км). Функционально-
значимой она является также и для хищных видов, общая численность
которых в регионе низка. Из млекопитающих - высокая численность отмечается
для зайцев (беляк и русак), косули, ондатры, встречаются также марал, лось,
кабан, медведь и лисица. Известны случаи встреч снежного барса [8].
Антропогенное влияние, не смотря на большую удаленность от
населенных пунктов и труднодоступность котловины, имеет свое действие из-за
привлекательности угодья. Сюда стремятся попасть как из Красноярского
края, так и из Тувы. Особенно значимыми являются браконьерство
(водоплавающие, копытные, редкие виды), выпас скота, приводящий к
вытаптыванию болотной и других видов растительности, фактор беспокойства.
5
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Ранее на территории Гагульской котловины действовал видовой заказник
для охраны и воспроизводства ондатры. В настоящее время территория
котловины с прилегающими участками Куртушибинского хребта вошла в
организуемый Гагульский заказник. Наибольшее внимание следует уделить
сохранению видов внесенных в Красные книги, а также охране пролетных
группировок водоплавающих и других видов птиц.
[1] Байкалов А.Н., Лаптенок В.В. Некоторые сведения об осенней миграции
водоплавающих птиц в Западном Саяне // Фауна и экология животных Средней Сибири.
Межвуз. сб. научн. тр. Красноярск: Изд-во КГПУ, 1996. С. 25-39. Щ Байкалов А.Н.
и др. Общая картина осенних миграций пеночек на юге Средней Сибири // Фауна и
экология наземных позвоночных Сибири. Красноярск: Изд-во Краснояр. гос. ун-та,
1997. С. 126-142. [3] Гаврилов И.К. Орнитофауна высокогорного пояса Саян
(состав, численность и размещение) // Фауна и экология наземных позвоночных
Сибири. Красноярск: Изд-во Краснояр. гос. ун-та, 1997. С. 72-94. [4] Коровицкий
Е.М. и др. Серая цапля Минусинской котловины и сопредельных территорий //
Вопросы орнитологии. Тез. докладов к V конф. орнитологов Сибири. Барнаул, 1995. С.
106-108. [5] Лаптенок В.В., Байкалов А.Н. Миграции трясогузок в Западном Саяне
// Вопросы орнитологии. Тез. докладов к V конф. орнитологов Сибири. Барнаул,
1995. С. 53-56. [6] Лаптенок В.В., Байкалов А.Н. Распространение некоторых
редких видов птиц в Западном Саяне // Состояние и пути сбережения генофонда диких
растений и животных в Алтайском крае. Тез. докладов к конф. Барнаул, 1992. С. 41-
43. [7] Савченко А.П. Методический комплекс по изучению миграций птиц: Метод,
рекомендации. Красноярск: Краснояр. гос. ун-т, 1991. 48 с. [8] Соколов Г.А.
Современное состояние популяций некоторых редких и исчезающих видов
млекопитающих юга Сибири // Редкие наземные позвоночные Сибири. Новосибирск: Наука,
1988. С. 212.
ОЛЬШАНИКИ НА ЗАБОЛОЧЕННЫХ МЕСТОПОЛОЖЕНИЯХ
ЦЕНТРАЛЬНОЙ ЛЕСОСТЕПИ РОССИИ
Т. А. Благодарова, А. И. Сиволапов
НИИ лесной генетики и селекции,хВоронеж;
Воронежская государственная лесотехническая академия
В Центрально-Черноземной полосе ольшаники занимают 48 тыс. га. На
всем этом пространстве ольха черная {Alnus glutinosa) растет, как правило,
на достаточно увлажненных местах с близким залеганием грунтовых вод, по
берегам рек, озер, ручьев на плодородных торфянистых, заболоченных,
дерново-луговых почвах. На специфических черноольховых трясинах в пойме
р. Хопер (Хоперский заповедник) ольха образует высокопродуктивные
чистые насаждения на огромной площади, что позволило выделить на площади
1118 га генетический резерват из чистой ольхи и провести селекционный
отбор насаждений. При отборе решающее значение придавалось продуктив-
6
РАСПРОСТРАНЕНИЕ, ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ I
ности, качеству и состоянию. Максимальный запас отобранных ольшаников
616-683 м3/га в сырых и мокрых местоположених. Эти уникальные
древостой отличаются высоким бонитетом и полнотой, они состоят из
цилиндрических стволов толщиною 30 см и более с высоко поднятой кроной. Средний
годичный прирост в средневозрастных и приспевающих насаждениях
составляет до 12-16 м3/га. Они образованы, в основном, здоровыми деревьями
по глазомерной оценке и пробам, взятым приростным буравом. В блюдце-
вых заболоченных понижениях на неглубоких торфянистых почвах Усман-
ского и Хреновского боров встречаются микропопуляции
высокопродуктивной ольхи, где единично произрастает осина и береза пушистая, на
повышениях - береза повислая.
Большой интерес представляет опыт создания быстрорастущих черно-
ольховых культур в Суджанском и Рыльском лесхозах Курской области. На
торфянистых почвах Суджанского лесхоза 18-летние культуры имели запас
288 м3/га. Культуры созданы сеянцами двухлетками рядовой посадкой в
гребень борозды. В зависимости от влажности почвы посадка сеянцев
проводилась осенью механизированным, конным и ручным способами. Почвы
под культурами торфяно-болотистые имеют нейтральную реакцию верхнего
торфяного горизонта, что соответствует оптимуму для ольхи (рН-6,6). Ход
роста по высоте модельных деревьев показывает, что на торфянистых
почвах интенсивный рост ольхи наблюдается до 14 лет, далее идет замедление
прироста по высоте.
Возобновление вырубок идет порослевым и семенным путем. Например,
возобновление 30-летней вырубки в Хоперском заповеднике представлено
почти на 70% семенными деревьями, общий запас по данным пробной
площади, 338 м3/га. В отношении преимуществ ольхи черной перед другими»
древесными породами надо отметить редкую возможность этой породы
занимать свою экологическую нишу, то есть самые низинные, сырые и
мокрые, заболоченные местоположения, там, где другие лесные породы расти
не могут. В настоящее время заметно повышается роль ольшаников как
стабилизирующего фактора в экосистеме Центральной лесостепи России.
Работа выполнена при поддержке ФЦП "Интеграция".
ГЕОГРАФИЧЕСКАЯ СПЕЦИФИКА БОЛОТНОГО ПОКРОВА В
ПРИЕНИСЕЙСКОЙ СИБИРИ
СМ. Горожанкина
Институт леса и древесины им. В.Н. Сукачева СО РАН, Красноярск
На современных схемах обзорного районирования болот приенисейская
Сибирь поделена по руслу Енисея между двумя зонами: выпуклых олиго-
7
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
трофных болот западно-сибирского типа на левобережье и мерзлых
крупнобугристых среднесибирского типа на правобережье. Изыскания показали
своеобразие болотного покрова этой территории, который сочетает как
западносибирские, так и среднесибирские черты и не может быть целиком
отнесен ни к одному из зональных типов. Специфику приенисейских болот
мы рассматриваем для участка с географическими координатами 59,5-62,9°
с.ш. и 88,9-91,7° в.д. Сравнительный анализ болотного покрова выполнен по
обзорным космическим теле- и фотоизображениям.
1. Особенности плановой структуры болотного покрова.
Физико-географические условия приенисейской полосы (всхолмленный рельеф и глубокий
эрозионный врез) не способствуют широкому развитию болотообразования.
Несмотря на тяжелый механический состав фоновых почвогрунтов, затор-
фованность в среднем составляет 10-15% (самый низкий показатель для
междуречья Обь-Енисей), благодаря чему приенисейская полоса резко
обособлена в составе Западно-Сибирскоо региона. Заболоченность выше
фоновой (местами до 80%) отмечается лишь на уплощенном песчаном
междуречье Сым-Дубчес, испытывающем слабые тектонические опускания. На
правобережье более всего заболочены верховья рек Вахта и Столбовая — 25-
30%. В целом болотный покров приенисейской части имеет мелкоконтурное
сложение и специфичную топографическую локализацию. Болотные очаги
сосредоточены на противоположных концах орографического профиля
местности: на поймах (и низких надпойменных террасах) и в истоково-приво-
дораздельных участках гидросети — водосборных «цирках». Междуречья
достаточно дренированы и сплошь залесены. Подобная структура болотного
покрова более характерна для среднесибирского типа заболоченности,
поскольку западно-сибирский тип отличается преимущественно
крупномассивным залеганием торфяников, покрывающих уплощенные междуречья.
2. Типологическая специфика приенисейских болот. По этому признаку
они не могут быть отнесены целиком ни & западно-сибирскому зональному
типу (с преобладанием выпуклых олиготрофных комплексов), ни
среднесибирскому (с преобладанием евтрофного компонента). Здесь содоминируют
плоские травяно-моховые экосистемы болот низинного и переходного
питания. Участками формируются миксотрофные комплексы, сочетающие
элементы разного типа питания (преимущественно низинного и переходного).
Олиготрофные экосистемы не имеют значимого распространения.
Локальные вариации типологического состава болот здесь обусловлены
палеогеографическими и литологическими факторами, высотной поясностью мезо-
климата. На общем фоне суглинистых почвообразующих пород резко
обособлено песчаное междуречье Сым-Дубчес, где развита обширная сеть
ложбин древнеледникового стока. Здесь болотный покров унаследовал днища
ложбин и носит западно-сибирский облик благодаря слитномассивному за-
8
РАСПРОСТРАНЕНИЕ, ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ I
леганию и широкому распространению олиготрофных мочажинных
комплексов. На правобережье при абсолютной высоте местоположения более
350 м характерны долинные ерниковые болота — явление довольно редкое
для относительно пониженного левобережья. На правобережье, помимо
того, болотные экосистемы отличаются сильной обводненностью поверхности,
что физиономично сближает их с болотами среднесибирского типа.
Заболачивание в приенисейской полосе протекает в гидротермических условиях,
которые не способствуют развитию мерзлоты среднесибирского типа.
Торфяники здесь преимущественно талые. Единичные местонахождения
крупнобугристых форм торфяного криогенеза зафиксированы на правобережье в
истоках реки Столбовой на высоте более 450 м.
3. Стратиграфическая специфика приенисейских торфяников.
Стратиграфия пойменных торфяников не обнаруживает существенных отличий от
аналогов западно-сибирского типа. Торфонакопление в поймах протекает по
низинному типу и пережило две основные фазы — лесо-топяную и
доминирующую топяную. Преобладающие виды торфа — осоковый, травяной, гип-
новый. В наложенных поймах, помимо того, фиксируются лесная фаза тор-
фонакопления и вступление процесса в переходную и даже верховую
стадии. Более значима специфика истоково-приводораздельных торфяников.
На суглинках их развитие изначально протекало по низинному типу с
последующим вступлением в переходную стадию. Удельная мощность
низинных торфов, фиксирующих продолжительность стадии, колеблется в
широких пределах — от 6 до 99%. В развитии процесса преобладает топяная
фаза при подчиненной значимости лесо-топяной. Основные виды торфа —
гипновый и осоковый низинный, сфагновый переходный. Для сравнения —
ландшафтные аналоги среднесибирского типа характеризуются отсутствием
переходной фазы развития и на всю глубину сложены низинными торфами.
Западно-сибирские аналоги имеют совершенно иную стратиграфию:
абсолютно преобладают верховые торфа (фускум, ангустифолиум, комплексный)
при подчиненной значимости переходных, а низинные часто вообще не
выявляются. На песчаных породах приенисейские торфяники эволюционируют
по другому пути. Залежь имеет трехслойное сложение, включая придонный
низинный слой, затем переходный и кроющий верховой. Удельная
мощность низинного слоя нарастает с продвижением на север: от 2% на широте
59,5° до 43% на широте 61,9°. Максимальная (до 91%) удельная мощность
переходного слоя отмечена на крайнем юге характеризуемой территории, а
верхового (68-72%) — в средней ее части, на междуречье Сым-Дубчес, где
болотный покров носит сходный с западно-сибирским облик
Таким образом, смена западно-сибирского зонального типа болот на
среднесибирский осуществляется в приенисейской Сибири через
формирование вдоль Енисея буферной переходной полосы со своеобразной структу-
9
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
рой болотного покрова, сочетающей черты обеих зон. Эту территорию вслед
за Е.А. Романовой предлагаем обособить в ранге самостоятельного
болотного выдела, по содержанию сопоставимого с зональными экотонами
растительности (лесотундрой, лесостепью). Накопленных к настоящему времени
материалов еще недостаточно для того, чтобы означить границы болотного
экотона на всем его протяжении. В этих целях необходимо расширить
географию болотоведческих исследований в приенисейской Сибири.
Исследования выполнены при поддержке РФФИ (проект 98-04-48385).
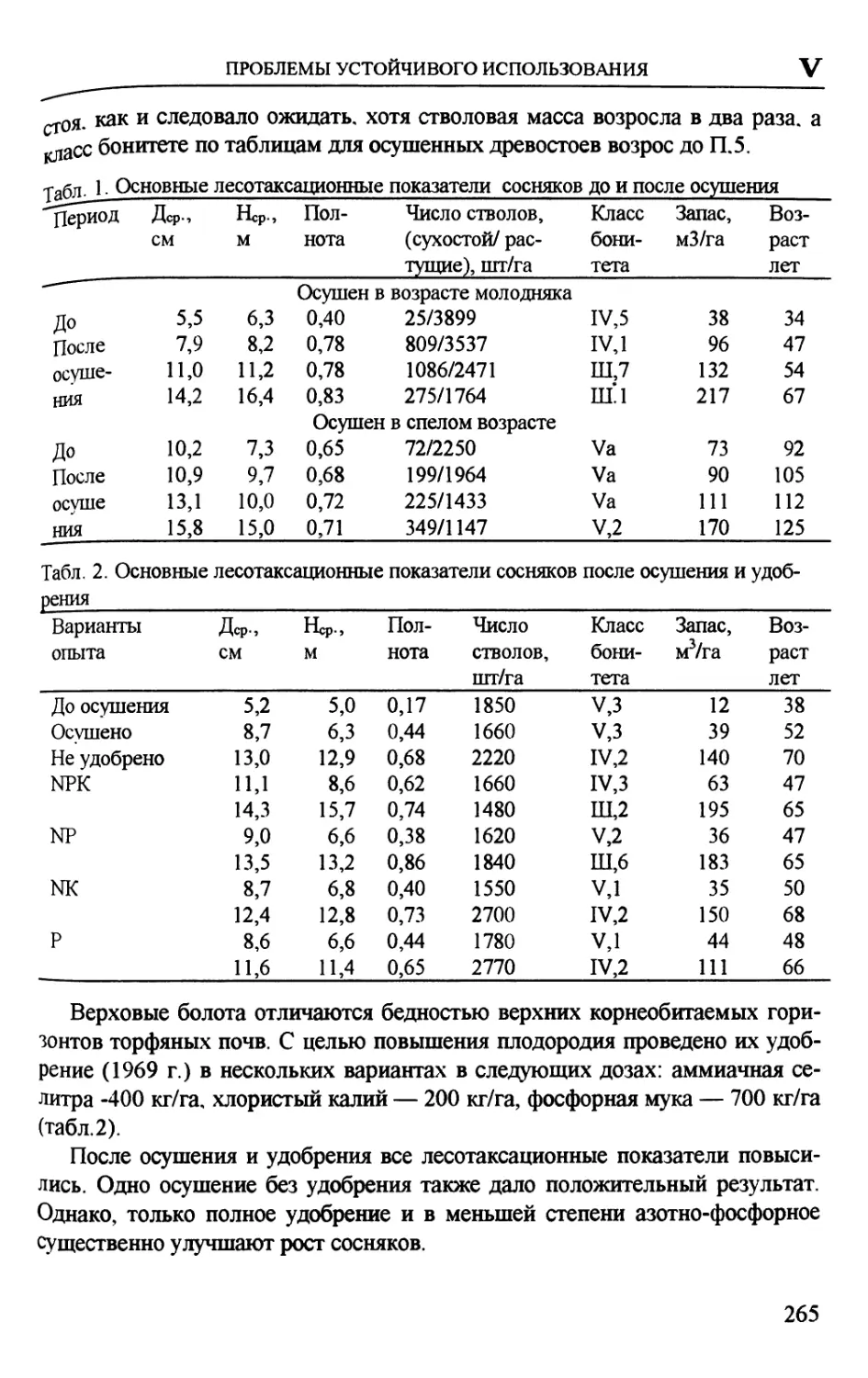
ТАКСАЦИОННАЯ ХАРАКТЕРИСТИКА ИЗБЫТОЧНО УВЛАЖНЕННЫХ
ЛЕСОВ БЕЛАРУСИ
Л.Д. Есимчик, Т.А. Колодий
Институт леса НАН Беларуси, Гомель, Беларусь
Общая площадь земель лесного фонда в Беларуси составляла на
1.01.1997 г. 9,09 млн. га. При этом, избыточно увлажненные земли (ИУЗ)
равны 2,06 млн. га или 22,6% общей площади лесного фонда (табл. 1).
Табл. 1. Общие сведения о лесах Беларуси
Годы Площадь, тыс. га: Запас древесины, млн.
учета общая / избыточно увлажненная м3: общий / ИУЗ
общая лесная лесопокрытая спелых всего спелых
1997 9088.6 8158.9 7849.7 385.0 1132.5 81.5
2055,9 1684,0 1624,7 104,3 144,5 18,5
Общий запас древесины в республике превысил миллиардный рубеж, а
запас спелых лесов достиг 81,5 млн. м3. Наибольшие запасы древесного
сырья сосредоточены на территории Гомельской и Минской областей,
традиционно наиболее многолесных. Более богаты спелыми лесами Гомельская и
Витебская области, в которых объем пригодной к потреблению древесины
достиг соответственно 19,6 и 17,1 млн. м3.
Табл. 2. Запасы древесины на 1 га избыточно увлажненных земель
Показатели
Беларусь -
всего
поМинлес-
хозу
С
157
73
160
74
Запас
Е
201
91
202
90
на 1 га по породам, м3:
Д
139
133
135
130
тв/л Б
143 Hi
108 84
137 ' 139
104 87
всего лесного
Ос
137
119
169
139
Ол(ч)
135
114
142
119
фонда
м/л
80
70
116
96
/ИУЗ
Итого
153
89
159
91
Лесной фонд Беларуси является собственностью государства и
сосредоточен в основном в Министерстве лесного хозяйства (74,2%). Значительная
часть его (13,7%) находится в ведении Министерства сельского хозяйства и
10
РАСПРОСТРАНЕНИЕ, ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ I
продовольствия. Большая часть (77,6%) ИУЗ находится в ведении Минлес-
хоза. Здесь же сосредоточено 80,6% запасов древесного сырья. Последние
составляют 12,8% от общего запаса всех лесов республики. Запас на 1 га
покрытых лесом избыточно увлажненных площадей значительно ниже
суходольных и составляет 61,8% (табл. 2).
По сосне и ели запасы насаждений на 1 га покрытой лесом площади ИУЗ
в 2,2 раза ниже по сравнению с этим показателем всего лесного фонда, а по
всем породам — в 1,7 раза. По мягколиственным породам отклонение
значительно меньше. Так, запасы березовых лесов на этих землях ниже в 1,6
ра-за, ольхи черной в 1,2 раза. Последняя в основном произрастает по
низменным берегам рек, ручьев, водоемов, на илистых почвах с проточной
водой, так называемых ольховых трясинах. Общий запас древесины на ИУЗ
достиг 144,5 млн. м3 (табл.3).
Табл. 3. Распределение покрытых лесом ИУЗ и запасов насаждений по сериям ти-
пов леса и преобладающим породам
Серия Итого По породам: площадь тыс. га / запас тыс. м3
типов
леса
Крапивная
Долго-
мошная
Багульни
-ковая
Сфагновая
Осоково-
сфагнов.
Осоковая
Таволго-
вая
Прируч.-
травян.
Папорот
никовая
Болотно-
папор
Всего
129.6
14303
350.6
32789
90.7
6028
169.2
11017
161.6
12280
256.4
22699
181.7
20361
122.8
9342
128.1
12275
34.0
3410
1624.7
144504
С
-
-
187.1
17765
78.0
4979
159.9
10235
97.5
6086
50.7
3319
-
-
46.8
3043
-
-
-
-
620.0
45427
Е
11
250
29.5
3750
CL7
41
-
-
02
41
11
152
-
-
10.1
510
12.5
633
-
-
58.9
5377
Д
и
225
-
-
-
-
-
-
-
-
-
-
-
-
-
-
8,3
1103
-
-
10.0
1328
тв/л
16
384
-
-
-
-
-
-
-
-
-
-
-
-
-
-
02
80
-
-
41
464
Б
13.3
1117
131.7
11063
12.0
1008
9Л
782
35.9
3018
126.3
10617
1Q
336
35.9
3018
50.5
4245
-
-
418.9
35204
Ос
гл
271
-
-
-
-
-
-
-
-
и
136
-
-
гл.
252
6,4
756
-
-
11.9
1415
Ол(ч)
104.6
11926
L2
133
-
-
-
-
27.5
3135
73.1
8328
172.4
19654
12.9
1466
45.0
5130
23.4
2668
460.1
52440
м/л
1Л
130
UL
78
-
-
-
-
-
-
2J,
147
13
371
15.0
1053
4J
328
10.6
742
40.6
2849
Наиболее распространенными породами, произрастающими на ИУЗ,
являются сосна (38,2%), ольха черная (28,3%) и береза (25,8%). Эти же поро-
11
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ды лидируют и по запасам древесины. Вместе с тем, ольха черная по этому
показателю выходит на первое (36,3%) место, сосна занимает
соответственно второе (31,4%) и береза третье (24,4%) место. Запасы твердолиственных
пород не превышают 1,2%, осины — 1,0%, а ольхи серой и других мягколи-
ственных пород - 2,0%. Наибольшие запасы древесины находятся в долго-
мошниковой (22,7%), осоковой (15,7%) и таволговой (14,1%) сериях типов
леса. Наименьший запас древесины сосредоточен в болотно-папоротниковой
серии типов леса. При этом наибольший удельный вес (78,2%) в этой серии
приходится на ольху черную. Она же доминирует и в таволговой серии.
Возрастная структура лесов на ИУЗ приведена в таблице 4. Возрастная
структура лесов на ИУЗ фактически мало отличается от этого показателя
всех лесов. Здесь наблюдается та же картина: преобладают по площади и
запасу средневозрастные древостой и дефицит спелых насаждений (табл.5.).
Табл. 4. Распределение ИУЗ и запасов насаждений по группам возраста
Группы
породы
Хвойные
в т.ч.
ель
Тверд.
листв.
в т.ч.
дуб
Мягк.
листв.
в т.ч.
береза
Всего
Итого
679.0
50804,0
59.1
5377,1
14.2
1792,7
10.0
1328,4
931.4
91907,5
418.9
35203,6
1624.6
144504,2
1кл.
59.5
674,8
10.5
81,9
L6
25,7
L2
20,9
96.7
1515,9
39.4
768,3
157.8
2216,4
в том числе по группам возраста:
площадь, тыс. га
молодняки
Пкл.
207.3
8770,2
12.1
493,7
12
238,5
Ш
192,6
133.5
5983,0
51.5
2178,5
344.7
14991,7
Всего
266.8
9445,0
22.6
575,6
йЛ
264,2
4,2
213,5
230.2
7498,9
90.9
2946,8
502.5
17208,1
t / запас, тыс. м3
ср. возр
273.8
23003,0
25.2
2767,0
£2
821,1
11
487,3
503.7
51699,3
267.3
24858,9
783.2
75523,4
присп.
99.7
12427,5
9£
1693,4
LS
401,9
L6
364,8
133.1
20472,1
46.5
5566,1
234.6
33301,5
спелые
38.7
5928,5
1Л
341,1
11
305,5
LQ
262,8
64.4
12237,2
14.2
1831,8
104.3
18471,2
Табл. 5. Возрастная
Показатели
Беларусь - всего
по Минлесхозу
структура лесов на избыточно
Группы возраста: %
молодняки
30.9
11,9
28.5
11,0
средневозр.
48.2
52,2
49.4
52,1
увлажненных землях
от площади / % от запаса
приспевающие
14.4
23,1
15.8
24,7
спелые
6,5
12,8
§Л
12,2
Процент спелых лесов на ИУЗ как в целом по республике, так и в Мин-
лесхозе выше на 1,6 процентных пункта, по сравнению со всеми лесами.
12
РАСПРОСТРАНЕНИЕ, ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ I
Краткая характеристика лесных ИУЗ позволяет выработать многоцелевой
подход к использованию их. Эти земли должны подлежать устойчивому
управлению и удовлетворять также социальные, экономические,
экологические, культурные и пространственные потребности человека нынешнего и
грядущего поколений.
с
ЗОНАЖНО-КЛИМАТИЧЕСКИЕ И ЛАНДШАФТНО-
ТИПОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РАСПРОСТРАНЕНИЯ
ЗАБОЛОЧЕННЫХ ЛЕСОВ КАРЕЛИИ
В.А. Коломыцев
Институт леса Карельского НЦРАН, Петрозаводск
Лесо- и болотообразовательный процессы находятся в стадии
динамического неравновесия, выражающееся в преимущественном развитии одного
из них в зависимости от типа ландшафта или тренда климатических
изменений. При исследовании заболоченности Карелии на зональной ландшафт-
но-типологической основе были выявлены три весьма существенных
обстоятельства с географической, лесоведческой и экологической позиций.
Большое внимание было уделено выяснению вопроса о влиянии подзо-
нальных (северо- и среднетаежных) условий тайги Восточной Фенноскандии
на заболоченность и потенциальные возможности болотообразовательного
процесса. Сделано заключение, что идентичные по морфогенезу группы и
типы ландшафтов в северотаежной подзоне заболочены в большей степени,
чем среднетаежные. Причем, соотношения открытых болот, с одной
стороны, и лесных болот и заболоченных лесов, с другой, в этом аспекте по
подзонам могут различаться в еще большей степени (см. табл.).
Соотношение общей заболоченности и открытых болот по подзонам и морфогене-
тическим группам ландшафта Восточной Фенноскандии (ср. взвеш.)
Группы типов ландшафтов Заболоченность: общая / открытых болот, %
_ северная тайга средняя тайга
Равнины 65/33 57/14
Водно-ледниковые 20/10 26/9
Денудационно-тектонические
Слаборасчлененные 57/26 40/13
Денудационно-тектонические
^Сильнорасчлененные 30/1 22/1
Средние взвешенные значения уклонов поверхности соответствующих
категорий переувлажненных земель в северотаежной подзоне в 1,5-2 раза
выше, чем в среднетаежной. При том, что характер кривых распределения
меящу категориями земель по уклонам поверхности в обеих подзонах иден-
13
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
тичен. Было выявлено сравнительно небольшое превышение средних
значений уклонов поверхности заболоченных лесов над лесными болотами (в 1,5
раза), тогда как градиенты уклонов поверхностей открытых болот гораздо
ниже (в 2,8-4,5 раза), чем в этих двух категориях земель.
Заболоченные (точнее, заболачивающиеся) леса в таежной зоне являются
^важным и весьма устойчивым компонентом ее структурной организации.
Они представляют большой теоретический интерес и практическую
значимость в силу их широкого распространения, типологического разнообразия
и необходимости особых режимов их эксплуатации. Они объективно служат
основным гидролесомелиоративным фондом.
Полученный эмпирический материал позволил затронуть лишь наиболее
общие моменты особенностей распространения заболоченных лесов на
уровне основных морфогенетических групп ландшафтов: 1) озерно-леднико-
вых и морских равнин, 2) ледниковых и водно-ледниковых холмистых и
грядово-холмистых и 3) денудационно-тектонических холмисто-грядовых и
грядовых (сельговых и скальных).
В группе равнинных ландшафтов озерно-ледникового и морского генези-1
са степень заболоченности варьирует в пределах 40-90%, характерно
преобладание заболоченных лесов над болотами. Различия между подзонами
проявляются в значительно меньшем распространении заболоченных лесов
в северотаежной подзоне, тогда как в средней тайге их преобладание на^
категорией болотных земель служит наиболее существенным элементом
структурной организации экосистем равнин. Данная особенность, выявлен^
ная в ходе ландшафтных исследований, не объяснима с геоморфологических"
позиций, поскольку морфометрические характеристики их практически;
идентичны. Определяющим фактором здесь служит климат, большая cypo-j
вость которого на севере способствует развитию моховых формаций за счет
снижения конкурентной способности древесной растительности в условиях|
избыточного увлажнения и быстрому прохождению сукцессионных фаз. |
Группа ландшафтов холмистых и грядово-холмистых водно-ледниковоп^
генезиса, которая от предыдущей отличается хорошо выраженными
скульптурными формами рельефа, так же характеризуется преобладанием рыхлых
четвертичных отложений в качестве почвообразующих пород. Степень
заболоченности варьирует в пределах 15-50%. Их особенностью, в отличие от
равнин, служит явное преобладание категории болотных (в том числе
лесных) земель над заболоченными в большинстве типов ландшафтов. Следует
отметить, что чем сильнее развиты холмистые формы рельефа, тем меньше^
заболоченность и участие заболоченных земель в структуре ландшафтов,
тогда как грядовый рельеф в большей степени способствует формированию
заболоченных лесов. Подзональные особенности распределения
заболоченных земель в водно-ледниковых ландшафтах проявляются в менее резкой
14
РАСПРОСТРАНЕНИЕ, ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ I
форме, чем на равнинах, но довольно однозначно: среднее взвешенное
значение доли заболоченных земель по типам ландшафтов свидетельствует о
большем распространении заболоченных лесов в среднетаежной подзоне.
группа холмисто-грядовых и грядовых ландшафтов денудационно-
т^утонического генезиса в Карелии является наиболее распространенной
(фоновой), занимая около 60% тещжтории. Почвообразующие породы —
преимущественно морены и, реже, скальные выходы. Степень
заболоченности ландшафтов варьирует от 15 до 60%, что обусловлено
морфологическими особенностями их рельефа и находится в обратно пропорциональной
зависимости от степени расчлененности последнего. Заболоченные леса в
зависимости от типа ландшафта формируют от 20 до 90% общей
заболоченности. Несмотря на морфологические различия внутри этой группы
ландшафтов и генетические — с озерно-ледниковыми равнинами, в них явно
прослеживается сходство в подзональных особенностях по степени
заболоченности и доле участия заболоченных земель. В северотаежной подзоне
степень заболоченности так же в целом выше, чем в среднетаежной. тогда
как заболоченные леса имеют меньшее распространение.
Полученные данные позволяют сделать ряд выводов: а) потенциальная
энергия процесса заболачивания в северотаежной подзоне выше, и оно
может происходить в менее благоприятных геоморфологических условиях, чем
в среднетаежной подзоне; б) лесные болота и заболоченные леса являются
довольно устойчивыми во времени экосистемами, благодаря особенностям
своего топо-эдафического распространения и сохранению естественного
дренажа; в) заболоченные леса являются важным структурным
компонентом таежных ландшафтов. Они формируют от 40 до 70% всех
переувлажненных местообитаний равнинных ландшафтов, до 30% в
водно-ледниковых и около 40% (в среднем) в денудационно- тектонических ландшафтах.
Доля их участия в формировании полугидроморфных и гидроморфных
экосистем при движении на север снижается, хотя общая степень
заболоченности в этом направлении увеличивается.
БОЛОТА ПРИОЗЕРНЫХ ЛАНДШАФТОВ СЕВЕРО-ДВИНСКОЙ
ВОДНОЙ СИСТЕМЫ
А.Н. Краснова
Институт биологии внутренних вод РАН, Ярославская обл.
При строительстве Северо-Двинского водного пути, эксплуатируемого с
1828 г., трасса, длиной 126 км., пролегла через болотные массивы юго-запа-
Да Вологодской области, в основном в полосе южной тайги. На шекснин-
15
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ском участке водораздела преобладают евтрофные болота, сухонском .—
мезотрофные.
Евтрофные болота, непосредственно прилегающие к каналам, оказались
осушенными вследствие понижения уровня грунтовых вод. Их торфяные
залежи разной мощности выступают по откосам каналов и сверху прикрыты
отсыпанным грунтом, создавая впечатление доголоценовых погребенных
торфяников. Современная растительность на месте существовавших болот
представлена луговыми и вторичными березово-осиновыми ценозами,
которые формируют антропогенный лесолуговой ландшафт. На сохранившишся
евтрофных болотах распространены осоковые и гипново-осоковые ценозы с
преобладанием Carex lasiocarpa, С limosa, С rostrata, Calamagrostis neg-
lectay гипновых мхов (преимущественно видов Calliergon\ некоторых
сфагновых — Sphagnum subsecundum, S. teres, S. warnstorfii.
В микропонижениях и элементах внутриболотной гидрографической
микросети характерны гипергигрофильные Menyanthes trifoliata, Comarum
palustre, Calla palustris, Phragmites australis, Carex acuta. Этот тип болот
занимает небольшие депрессии и послеледниковые котловины, часто с
остаточными незаторфованными водоемами.
Своеобразный тип представляют осоково-пушицево-гипновые болота,
приуроченные к долинам малых рек. В травяном покрове доминируют
Eriophorum vaginatum, E. latifolium, Carex acuta, C. aquatilis, Equisetum
fluviatile и гипновые мхи с отсутствием или слабым участием сфагновых.
Евтрофные лесные болота в районе Северо-Двинской водной системы
занимают ограниченные площади, хотя и распространены повсеместно.
Древостой сложен Betula pubescens, Populus tremula, в приручейных типах — с
примесью Alnus glutinosa. Травяной покров часто дифференцирован. На
пьедесталах вокруг стволов и разложившихся пней растут в основном
обычные лесные гигрофильные виды, в мочажинах — типичные болотные —
Carex acuta, С vesicaria, Equisetum fluviatile и другие.
Мезотрофные болота, больше распространенные на сухонском и слабее
на шекснинском водоразделах, по Т.К. Юрковской [1980] представлены
европейско-западносибирским вариантом сфагновых торфяников. Они
приурочены к значительным по площади приозерным послеледниковым
депрессиям, которые перерезает трасса канала. Пример — обширный
болотный массив, со всех сторон окружающий Кишемское озеро. Его
особенностью является обилие сплавин, надвигающихся на водоём. В растительном
покрове этого и других болот этого типа преобладают березово-сосновые и
берёзовые кустарничково^осоково-сфагновые олиго- и мезотрофные ценозы.
Разреженный древесный ярус состоит из Pinus sylvestris, Betula pubescens,
Picea abies. На кочках вокруг пьедесталов и пней растут Chamaedaphne
calyculata, Ledum palustre, Vaccinium uliginosum, Sphagnum magellanicum, S.
16
РАСПРОСТРАНЕНИЕ, ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ I
fuscum, S. centrale, S. teres. Между кочками развиваются Carex lasiocarpa,
Eriophorum vaginatum, Oxycoccus palustris. Меньшее распространение
имеют кустарниково-травяно-сфагновые и травяно-сфагновые мезотрофные
болота. В травяном покрове доминируют Carex lasiocarpa, С. rostrata, С.
dioica, Eriophorum vaginatum. Из мхов чаще встречаются Sphagnum fuscum,
S. magellanicum, S. warnstorfii. Обильно развивается Oxycoccus palustris.
Болота всех типов в районе Северо-Двинской водной системы за
длительный период её эксплуатации во многом утратили региональные
особенности. Это связано с их полным или частичным осушением, а на некоторых
участках, наоборот, обводнением. Другой вид антропогенного воздействия
представляет непосредственная хозяйственная деятельность —
осушительные мелиорации, рубки, выпас на пригодных участках.
БОЛОТА ЮЖНО-КАМЧАТСКОГО ФЕДЕРАЛЬНОГО ЗАКАЗНИКА
В.Ю. Нешатаева, В.Ю. Нешатаев
Ботанический институт РАН, Санкт-Петербург;
Санкт-Петербургская государственная лесотехническая академия
Растительность болот Камчатки исследована слабо. Для Восточнокам-
чатской болотной провинции характерны грядово-мочажинно-озерковые
травяно-сфагново-гипновые практически безлесные болотные массивы,
имеющие сходство с аапа-болотами северной Европы [2]. При этом вопрос о
южной границе распространения такого типа болот на Камчатке оставался
открытым. Исследования проведены в 1985-86 и 1990-91 гг. на территории
Южно-Камчатского заказника (ЮКЗ) площадью 247 тыс. га,
расположенного в зоне активного вулканизма.
Общая заболоченность территории ЮКЗ невелика в силу гористого
рельефа. Относительно крупные болотные массивы расположены в долинах рек
и на прибрежноморских низменных равнинах, сформировавшихся на месте
бьюших морских лагун в результате поднятия суши на последних этапах
четвертичной истории. Растительность болот охарактеризована по
материалам, собранным на профилях, пересекающих 5 крупных болотных массивов.
Болото в устье р. Ильинская (Восточное побережье Камчатки). От
Тихого океана оно отделено береговым валом с колосняковыми лугами. Имеет
простую структуру и сложено практически единым сообществом
ассоциации Uliginiherbetum caricoso-trichophorosum. Характерно преобладание Tri-
chophorum alpinum с согосподством других осоковых Carex cryptocarpa, С.
aPpendiculata9 С. gynocrates, Eriophorum polystachyon, и пгановым моховым
покровом из Нурпит lindbergii, Drepanocladus aduncus, Scorpidium scor-
Phides, постоянное участие Sanguisorba tenuifolia, Equisetum palustre, E.
17
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
variegatum, Calamagrostis neglecta, Trientalis europaea, Salix arctica, Par*
nassia palustris, Iris setosa, Drosera rotundifolia, Platanthera tipuloides, Ptar*
mica camtschatica. Микрорельеф слабокочковатый, мощность торфа 7-12 см.
Болота Западного побережья (по левому берегу р. Теплая, по левому и
правому берегам р. Камбальная) расположены на прибрежноморских низ^
менных равнинах, сформировавшихся на месте бывших морских лагун, и
отделены от Охотского моря узкой (50-100 м) полосой дюн, покрытых ко-
лосняковыми лугами. Небольшие реликтовые дюны встречаются и посреди
болотных массивов. Эоловые наносы песка нередко попадаются под мхами
и в торфе этих болот. Поверхность болот слабо наклонена в сторону мор*
На востоке болота граничат с суходолами, покрытыми ольховыми и
кедровыми стланиками. Мощность торфяной залежи не превысила по данным 140
замеров 100 см и варьировала от 10-20 см на окрайках до 60-100 см в
центральных частях болот. В ней отмечены прослойки вулканического пепла.
Окрайки болот заняты сообществами Herbosphagnetum empetroso sibirici -
caricosum cryptocarpae и Magnocaricetum caricosum cryptocarpae. Ассоциация
Herbosphagnetum empetroso sibirici - caricosum cryptocarpae отличается
преобладанием в моховом покрове Sphagnum warnstorjfii реже S. squamosum и
S. teres, в травяно-кустарничковом ярусе постоянно встречаются олиготроф-
ные кустарнички Empetrum sibiricum, Vaccinium uliginosum, Oxycoccus
microcarpus, Andromeda polifolia, Salix fuscescens, Betula exilis и
требовательные к условиям минерального питания травы Carex cryptocarpa,
Equisetum palustre, Comarum palustre, Sanguisorba tenuifolia, Polygonum
viviparum, Luzula capitata.
Для асе. Magnocaricetum caricosum cryptocarpae характерно наряду с
преобладанием Carex cryptocarpa, высокое обилие Sanguisorba tenuifolia,
развитый покров из гипновых мхов, постоянное присутствие болотных и
тундровых кустарничков Betula exilis, Salix fuscescens, Rubus arcticus,
Andromeda polifolia, Empetrum sibiricum, Oxycoccus microcarpus и трав
Calamagrostis neglecta, Polygonum viviparum, Comarum palustre, Equisetum
palustre, Trientalis europaea, Viola hultenii, Trichophorum alpinum, Luzula
capitata, Gentianella auriculata, Festuca rubra, Galium trifidum.
Центральные части болот заняты грядово-мочажинными и грядово-
озерково-мочажинными комплексами. На высоких грядах в этих комплесах
представлены сообщества Herbosphagnetum empetroso sibirici - caricosum
cryptocarpae и Empetretum sibirici sphagnosum. Для ассоциации Empetretum
sibirici sphagnosum характерно невысокое покрытие сфагнов (5-10%),
постоянное участие Calamagrostis neglecta, Comarum palustre, Andromeda
polifolia, Carex cryptocarpa, С rariflora, Oxycoccus microcarpus, O. palustris,
Menyanthes trifoliata. В ассоциации Empetretum sibirici sphagnosum нами
выделены следующие субассоциации, названные по основным доминантам:
18
РАСПРОСТРАНЕНИЕ, ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ I
1) caricosum rariflorae; 2) menyanthosum; 3) betulosum exilis; 4) vaccinietosum
uliginosii; 5) myricosum.
Топяные участки заняты сообществами Uliginiherbetum caricosum limo-
sae. В мочажинах и мочажинах-озерках распрстранены сообщества
Uliginiherbetum caricosum cryptocarpae и Uliginiherbetum menyanthosum. В травя-
но-кустарничковом ярусе сообществ асе. Uliginiherbetum caricosum limosae
преобладают мелкие длиннокорневищные гигрофильные осоки (Carex chor-
dorhiza, С rariflora, С limosa) и обильно болотное разнотравье (Сотагит
palustre, Menyanthes trifoliate, Equisetum palustre и др.). Моховой покров
слабо развит и состоит из гипновых мхов. Сообщества ассоциации описаны
на низких грядах (коврах) с уровнем вод от поверхности в 5-10 см. В
ассоциации по преобладающим и обильным видам для ЮКЗ нами выделены
следующие субассоциации: 1) menyanthosum; 2) myricosum; 3) dicranosum;
4) caricosum rariflorae.
Ассоциация Uliginiherbetum caricosum cryptocarpae отличается
преобладанием в травяно-кустарничковом ярусе Carex cryptocarpa (10-30%) и
высоким обилием Сотагит palustre (10-30%). В ассоциации выделено 3
субассоциации. Для субассоциации drepanocladetosum характерно наличие
развитого мохового покрова (70-95%) из Drepanocladus, Calliergon, Scorpidium
scorpioides. Сообщества ассоциации занимают обводненные мочажины, в
которых вода в начале августа 1990 г. стояла у поверхности (±5 см).
Мощность торфа составляет 40-70 см. Субассоциации typicum и menyanthosum
имеют менее развитый моховой покров из тех же видов в силу их большей
обводненности (уровень воды от 0 до + 30 см). При этом субассоциация с
обильным Menyanthes trifqliata занимает мочажины-озерки с глубиной воды
10-15 см и является переходной к следующей ассоциации.
Ассоциация Uliginiherbetum menyanthosum отличается слаборазвитым
травяным ярусом (8-30%) из Menyanthes trifoliata, приподнимающимся из
воды. Для многих сообществ характерно присутствие плавающих в воде
Vtricularia macrorhiza, Sparganium hyperboreum, Hippuris vulgaris. Кроме
сообществ, отнесенных к типичной субассоциации (субассоциация typicum),
встречаются сообщества с обильным Eleocharis palustris (субассоциация
eleocharidosum palustris) и Drepanocladus fluitans (субассоциация
drepanocladetosum). Крупный болотный массив с озерково-топяными комплексами
сформировался также в самой южной оконечности полуострова Лопатка,
сложенной морскими песчаными отложениями.
Таким образом, болота ЮКЗ могут быть отнесены к особому
южнокамчатскому типу аапа болот, отличающемуся мелкой торфяной залежью и
особенностями флоры. Они представляют собой ценные места гнездования
водоплавающих птиц: на болоте в устье р. Камбальная было учтено [1] 199
пар птиц на 1 кв. км. Болотные массивы имеют значение для охраны нерес-
19
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
тилищ лососевых. Изученные болотные системы ЮКЗ соответствуют крите-i
риям Рамсарской конвенции и заслуживают отнесения их к водно-болотньпц
угодьям международного значения.
[1] Лобков Е.Г. Гнездящиеся птицы Камчатки.Владивосток. АН СССР, 1986. 291 с.
[2] Нешатаев В.Ю. и др. Растительность болот. // Растительность Кроноцкого госу
дарственного заповедника (Восточная Камчатка). Тр. БИН РАН. Вып. 16, СПб,
1994. С. 167-196.
ЗАБОЛОЧЕННОСТЬ ТЕРРИТОРИИ НА ТОПОГРАФИЧЕСКИХ КАРТАХ
РАЗНОГО МАСШТАБА (НА ПРИМЕРЕ УЧАСТКА ЦЕНТРА ЕТР)
Сенникова М. М., Сирин А.А.
МГУ, географический факультет;
Институт лесоведения РАН, Московская обл.
Оценка заболоченности территории необходима для решения многих
научных и практических задач по рациональному использованию болот и
заболоченных лесов. При широком и постоянно расширяющемся (в том числе
благодаря развитию методов дистанционного зондирования) спектре
источников пространственной информации о заболоченности, топографические
карты (ТК) продолжают оставаться важным, и, вероятно, наиболее
унифицированным его составляющим. Обеспеченность ТК разного масштаба
существенно различается по территории России. Целью работы было оценить
заболоченность на примере участка (75 км2) таежной зоны центра ЕТР (юго-
запад Тверской области) с использованием ТК масштабов 1:10000, 1:25000,
1:100000, 1:200000, 1:300000. Отсутствовал масштаб 1:50000, а 1:200000
был представлен ТК Тверской области (1992).
При оцифровке ТК выделялись три категории объектов, отмеченные
условными знаками заболоченности, в том числе: очерченные точечными
контурами на бесцветном (а) и светло-зеленом (б) фоне, а также неоконтурен-
ные на темно-зеленом фоне (в). Им можно условно сопоставить открытые
болота (а), болотные мелколесья и редколесья (б), лесные болота и
заболоченные леса (в). Анализ проводился с использованием ГИС Maplnfo 4.O.
Хотя с уменьшением масштаба происходило снижение заболоченность
участка: 28,8% (1:10000), 28,3% (1:25000), 26,9% (1:100000), 23,3%
(1:300000), результаты показывают приемлемость использования ТК
масштаба 1:100000 и в определенной степени обзорно-топографических карт
масштаба 1:300000 для оценки заболоченности. На рассмотренных картах
масштаба 1:200000 заболоченность составила 15,4% подтверждая обзорный
характер этой серии. Потери заболоченности при переходе с масштабов
1:10000-1:100000 к 1:300000 происходили преимущественно за счет отбора
20
РАСПРОСТРАНЕНИЕ, ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ I
объектов, в меньшей степени за счет обобщения их очертаний. Не
изменялась категория (в), составляющая основную долю (70%) заболоченности на
ТК всех масштабов. На одну пятую сократилось присутствие категории (а)
при увеличении категории (б). Главные потери происходили за счет
мелкоконтурных открытых (низинных и переходных) болот, расположенные в
долинах рек и по окрайкам верховых болотных массивов.
БОЛОТНЫЕ ФАЦИИ ТУНКИНСКОЙ КОТЛОВИНЫ (ЮГО-ЗАПАДНОЕ
ПРИБАЙКАЛЬЕ): ВЛИЯНИЕ ПОТУХШИХ ВУЛКАНОВ
с В.А. Снытко
Институт географии СО РАН, Иркутск
Между Тункинскими гольцами и отрогами Хамар-Дабана в Тункинской
котловине располагается массив — Коймарские болота. Протекающая по
ним р.Тунка и ее притоки, с одной стороны, питают массив, а с другой —
его дренируют. Происхождение болотного массива обусловлено
тектонической деятельностью. Располагающийся западнее песчаный массив Бидар
испытывает воздымание, также как и восточнее находящаяся Еловская
перемычка между Тункинской и Торской котловинами. В центре массива
располагаются озера, в большинстве случаев проточные.
В непосредственной близости от болотных фаций располагаются широко
известные в Сибири потухшие Тункинские вулканы. Несомненно их влияние
на прилегающие территории в период активизации вулканической
деятельности. В современной обстановке это продолжается. Оно выразилось и в
поступлении в болотные фации вещества магматического происхождения,
переработанного процессами гипергенеза, и в тепловом обогащении.
Ряд вулканических гряд располагаются в непосредственной близости от
заболоченных фаций. В этом случае в явном виде можно говорить о
сопряжении хода ландшафтно-геохимических процессов. Аккумуляция вещества
обусловила формирование в болотных фациях сложного комплекса
болотных, в значительной мере солончаковатых почв. Характерна сравнительно
малая мощность торфяного горизонта. Торф представлен продуктами
разложения травянистых болотных растений.
По сравнению о почвами, формирующимися на вулканических
образованиях, почвы Коймарских болот имеют большие значения рН, в них
значительно больше содержание гумуса. Химический состав почв болотных
фаций характеризуется обогащением кальцием, стронцием, по сравнению с
почвами на вулканических массивах. В то же время в болотных почвах
понижено содержание железа, никеля, кобальта, меди, марганца, что свиде-
21
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
тельствует о повышении интенсивности миграции указанных элементов в
болотных фациях.
Болотный массив характеризуется широким распространением влажно-
луговых ассоциаций с высокой продуктивностью. Это одно из свидетельств
активного влияния на них сопряжения с фациями потухших вулканов. О
более благоприятных условиях на месте сопряжения этих фаций
свидетельствует хозяйственная деятельность издревле селившегося здесь человека.
БОЛОТНЫЕ ЗЕМЛИ КИРОВСКОЙ ОБЛАСТИ И
ИХ ХОЗЯЙСТВЕННОЕ ИСПОЛЬЗОВАНИЕ
А.Ф. Тимофеев
Вятская государственная сельскохозяйственная академия, Киров
В Кировской области более 2 млн. /а занимают болотные и
заболоченные земли, из них 1,8 млн. га расположены на землях лесного фонда.
Преобладают на территории области низинный процесс заболачивания:
низинные и близкие к ним переходные и смешанные болота занимают более 72%,
а верховые — менее 28%, причем, последние расположены в основном в
северных, более заболоченных районах области. Особенностями болот
области являются: высокая степень разложения торфа (в среднем 40-50% для
низинных и переходных и 30-40% для верховых болот) и высокая зольность
его: в среднем 8-15% для низинных и 3-5% для верховых болот [1]. 84%
болот области заняты древостоями сосны, ели и березы V-III классов
бонитета. По нашим измерениям, средний годичный прирост сосны в возрасте
10-20 лет равен от 6-7 см (в кустарничково-сфагновых типах леса) до 16-30
см в пушицево-сфагновых и осоково-сфагновых типах леса) [2].
Эти особенности болот можно объяснить условиями их образования и
расположения. Обычно образование болот связывают с большим
количеством осадков. Для Кировской области коэффициент прихода-расхода влаги
только для северных и центральных районов равен 1,2-1,0, а для более
южных районов он снижается до 0,8-0,7. Основную роль в процессах
заболачивания играют, во-первых, неравномерное выпадение осадков и резко
неравномерное их поступление в почву (особенно зимних); во-вторых,
перераспределение осадков, в связи с выраженным стоком, перераспределение
осадков, в связи с выраженным стоком, в соответствии с рельефом
поверхности. Поэтому заболачивание начинается обычно с отдельных котловин и
других понижений с малыми уклонами поверхности. Их наличие характерно
для Кировской области. Именно с этих понижений и начинались процессы
заболачивания. Далее, по мере роста болот в высоту (а он достигал, по
нашим измерениям по сосенкам, до 0,5-2 см в год), происходило слияние от-
22
РАСПРОСТРАНЕНИЕ, ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ I
дельных болотных массивов в один более крупный массив с выпуклой
поверхностью и ростом в стороны. В отдельных случаях образование болот
происходило путем зарастания водоемов, особенно стариц рек. В этих
случаях болота в плане имеют продолговатую, но извилистую форму.
Конечно, процессы переувлажнения и заболачивания протекали и
протекают независимо от деятельности человека. Но по мере освоения лесной
зоны человек способствовал процессам заболачивания в лесном фонде.
Здесь можно выделить два направления: 1) устройство дорог, особенно
железных и автомобильных, когда дороги обычно спрямляются и на
пониженных участках прокладываются по насыпям, но без достаточного количества
водоотводящих сооружений (в Кировской области таких «болотных
новообразований» достаточно); 2) как показано многими исследованиями,
пахотные угодья в нечерноземной зоне создавались, как правило, на более
повышенных и более крутых участках водосборов (как более дренированных),
причем, пашня часто располагается выше поверхности прилегающих
лесных земель до 10-20 м и более. Следовательно, сток воды с пахотных земель
повышал степень увлажнения лесных земель и способствовал их
заболачиванию. Оба указанных пути заболачивания можно назвать антропогенными.
Преимущественное расположение лесных земель на пониженных и
слабоуклонных участках водосборов и способствовало тому, что заболоченные и
болотные земли в большинстве случаев располагаются на лесных землях,
что касается как Кировской так и других областей нечерноземной зоны.
Болота Кировской области имеют большое гидрологическое и в целом
экологическое значение. Поэтому во многих случаях их надо сохранять и
использовать в естественном состоянии (для сбора ягод, прежде всего
клюквы, лекарственного сырья и пр.). Однако, часто наши хозяйственные
потребности приводят к необходимости их коренного изменения.
Применяются 2 пути: 1) осушение и использование осушаемых участков болот и
заболоченных земель для выращивания сельскохозяйственных культур и
лесонасаждений (или в совокупности тех и других), 2) осушение и добыча торфа
для различных целей (на топливо, удобрение и пр.), но обязательно с
последующим интенсивным освоением т.н. выработанных торфяников в
сельскохозяйственных и лесохозяйственных целях (а лучше комплексно). В
Кировской области распространены оба пути, но в основном осушение под добычу
торфа с последующим хозяйственным освоением.
Лесохозяйственное освоение земель после торфоразработок в Кировской
области проводится с 1963 г. С этого времени проводятся и наши
исследования на этих землях как по изучению свойств этих земель, так и по
особенностям создания здесь лесных культур. Результаты исследований
опубликованы, поэтому остановимся только на основных выводах.
23
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Выработанные торфяники представляют собой специфические
природные и антропогенные образования. На них создаются особые
гидрологические, температурные и почвенные условия, причем, несколько
разнообразные в зависимости от мощности остаточного слоя торфа, различного
высотного положения (несмотря на послойную, фрезерную добычу торфа),
состояния осушительной сети и пр. Надлежащие условия для роста лесных
культур создаются на повышенных и средних участках, плохие — на
пониженных, культуры погибают на котловинных местоположениях. Лучшей
породой для лесных культур является сосна, на повышенных
местоположениях с небольшой мощностью остаточного слоя торфа растут лиственница и
ель, а также некоторые другие. Естественно возобновляется береза, иногда
создаются естественные березовые насаждения невысокой продуктивности и
как примесь к посадкам сосны. На повышенных местоположениях посадки
сосны образуют насаждения I-II классов бонитета: в возрасте 25-26 лет
средняя высота достигает 10-13 м, средний диаметр — 10-12 см, запас 160-
225 м3/га при полноте 0,7-1,0. Развивается подлесок из малины, смородины,
рябины, образуется лесная подстилка. Непременное условие дальнейшего
интенсивного роста — исправное состояние осушительной сети.
[1] Тимофеев А.Ф. Болотные и заболоченные земли Кировской области и
эффективность их осушения. Сб. «Лес и почва», Красноярск, 1968. [2] Тимофеев А.Ф.
Повышение продуктивности лесов путем мелиорации. М.: Лес. пром-стъ, 1975. [3]
Тимофеев А.Ф. Использование мелиорируемых земель. Киров, 1980.
24
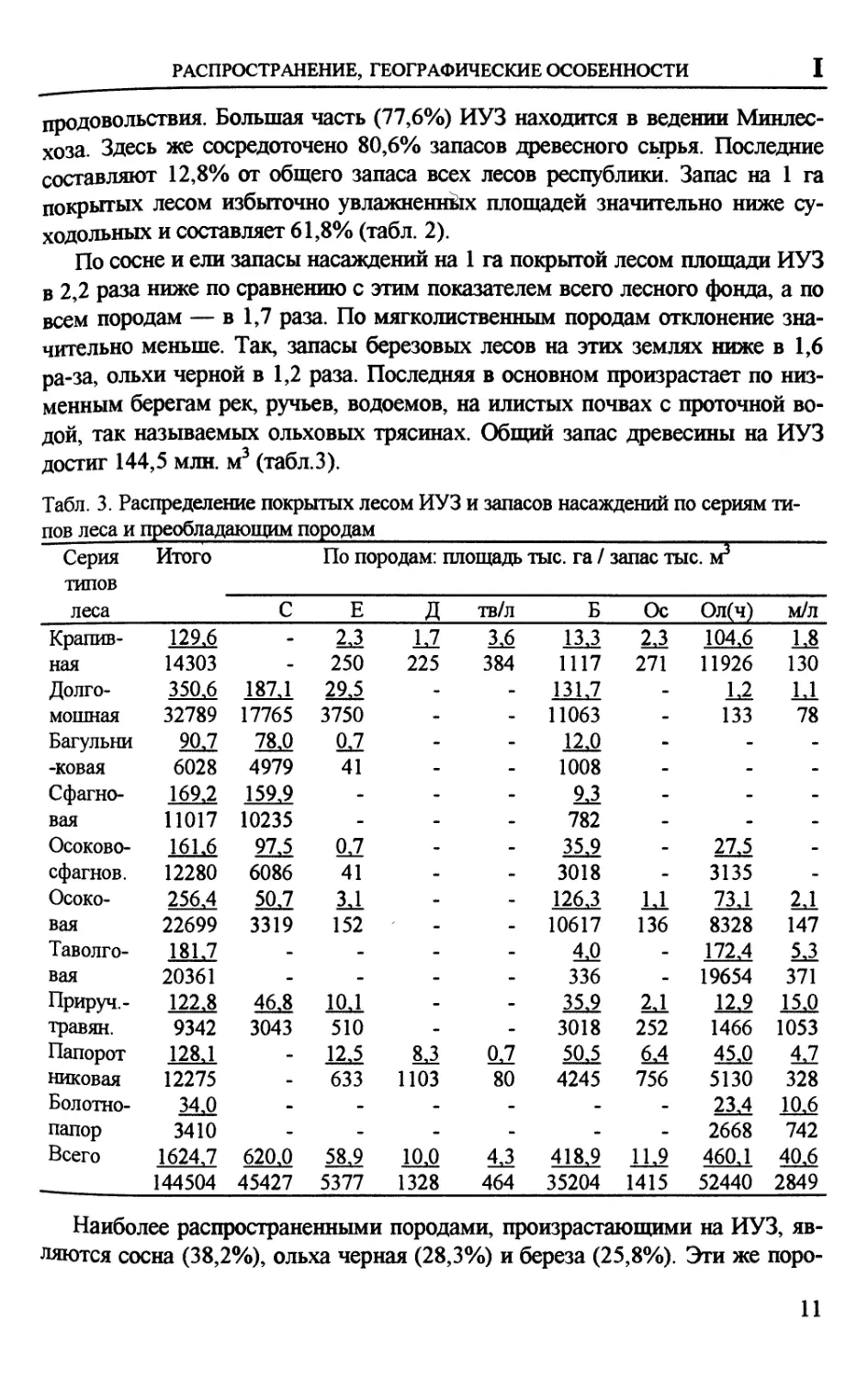
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
П. ГЕНЕЗИС, ДИНАМИКА БОЛОТООБРАЗОВАНИЯ
(В ПРОШЛОМ, НАСТОЯЩЕМ)
ОБ ИЗУЧЕНИИ ДИНАМИКИ БОЛОТООБРАЗОВАТЕЛЬНОГО
ПРОЦЕССА В ТАЕЖНОЙ ЗОНЕ ЗАПАДНОЙ СИБИРИ
В. А Базанов1, Н.Н. Пологова2, В. А. Льготин3
1 НИИ биологии и биофизики при Томском государственном университете;
2Филиал института леса им. В.Н. Сукачева СО РАН, Томск;
3ТЦ «Томскгеомониторинг», Томск
По масштабам и интенсивности развития процесс болотообразования
относят к одному из природных феноменов общепланетарного уровня.
История развития болот таежной зоны Западно-Сибирской равнины насчитывает
всего 8-10 тысяч ле£г За этот относительно короткий (в геологическом
отношении) срок экспансия болотного процесса достигла таких масштабов
влияния на все компоненты природной среды, что его справедливо
рассматривают в ранге сильно действующих экзогенных преобразующих процессов.
Для Томской области такой взгляд имеет особое значение: этим процессом в
разной степени охвачено более 50% территории.
Последовательность развития болот на территории реализуется в
постоянном и повсеместном воспроизводстве определенного типа структуры
связей и отношений под воздействием системообразующих потоков влаги.
Динамика болотного процесса наиболее полно и достаточно точно может быть
представлена с помощью ряда последовательно сменяющих друг друга во
времени и пространстве состояний. Действительно, природные единицы,
выступая одновременно в виде сложного природного процесса и его
результата, выстраиваются в последовательные этапы (фазы) своего развития. Под
этапом процесса мы понимаем индивидуальную степень интеграции
показателей, характеризующих его временные аспекты. Пространственные
особенности различных этапов — состояний — принято характеризовать
набором физиономических, структурно-вещественных и других показателей.
Для Томской области, где на большей части преобладают активно
развивающиеся преимущественно верховые болота, описаны 5 основных
пространственно-временных состояний, характеризующихся диагностическими
критериями и функциональными соотношениями компонентов, а также и
физиономическими признаками.
1. Состояние скрытого заболачивания. Характеризуется изменениями
водно-физических и химических свойств почвенно-грунтовой толщи| в
первичных очагах и по границе торфяного болота с минеральными
отложениями. В результате кольматации органо-минеральными веществами цллюви-
25
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
альных горизонтов происходит снижение водопроницаемости почв,
усиление увлажнения корнеобитаемого слоя на участках заболачивания Первым
результатом изменения условий увлажнения является частичная деградация
или структурная перестройка отдельных элементов растительности. Это
доминирование в древесных насаждениях хвойных пород (кедр, сосна), в
напочвенном покрове гипновых мхов, типичных представителей таежного
разнотравья и кустарничков. Этому состоянию соответствуют слабо
заболоченные леса, в таксационной оценке (для ранга высокобонитетных насаждений)
соответствующие показателям запаса <200 м3/га, сомкнутостью полога 0,6-
0,7, количеством подроста 2-3 тыс. шт. на га. Для сравнения — элитные
насаждения автоморфных местообитаний характеризуются запасами до 400
м3/га, сомкнутостью полога 0,8-1,0, количеством подроста >5 тыс. шт./га.
2. Состояние начального заболачивания характеризуется кардинальными
изменениями морфологии и водно-физических свойств приповерхностной
почвенно-грунтовой толщи, теряющей фильтрационные свойства. Из
напочвенного покрова исчезает часть видов мохово-кустарничкового яруса
типично лесной экологии, на смену приходят растения — обитатели болот.
Повышается степень увлажнения поверхности, вызывающая резкое снижение
скорости минерализации растительных остатков, появляются первые
признаки торфообразования. Изменения происходят и в строении рельефа
поверхности, появляются выраженные понижения и клумбообразные приком-
левые повышения вокруг деревьев. Химизм поверхностных вод
приближается к водам торфяных болот. Состоянию заболачивания соответствуют
мелкозалежные болотные массивы и их части. Запасы древесины (для той
же группы высокобонитетных насаждений) снижаются до 120 м3/га,
сомкнутость насаждений 0,4-0,5, количество подроста до 1 тыс. шт./га.
3. Состояние собственно болотного процесса является ключевым звеном
в экзогенном преобразовании территории, поскольку ведущим системообра-
зующми фактором становится торфообразование. Этот специфичный по
водно-физическим и геохимическим параметрам процесс трансформирует
при накоплении торфа в понижениях первичный рельеф и плоские
прилегающие пространства. Кроме того его воздействие проявляется и на
значительных расстояниях от массивов в изменении первичной геохимической
обстановки вмещающих литологических толщ. Свидетельством этому —
зоны геохимических аккумуляций, изменение реакции среды почв
приболотного пояса. Основными источниками водного питания торфяных болот
являются атмосферные осадки и грунтовые воды, которые различаются
между собой степенью минерализации, составом количеством химических
элементов, сезонной ритмикой уровней воды и т.д. По этому признаку все
болота принято делить на верховые, источником питания которых являются
только атмосферные осадки, и низинные, питающиеся преимущественно за
26
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
чет грунтовых вод. Растительность верховых болот образована очень
специфичным составом, принадлежащих к разным систематическим группам:
многочисленными видами сфагновых мхов (типичных болотных),
карликовыми формами древесных пород.
4. Состояние характеризуется снижением интенсивности торфонакопле-
дяя^ что индицируется образованием небольших по площади (1-20 м2)
постоянных водоемов в составе различных по строению и конфигурации
комплексов — грядово или клумбово-озерковых. Как весьма распространенная
на территории фаза болотного процесса, оно в целом свидетельствует о
смене прогрессивного периода развития на регрессивный. Смена этапов ведет
не только к снижению активности торфонакопления но и к кардинальным
изменениям и всех других составляющих процесса.
5. Активного развития расширения площади озер и снижения
интенсивности образования торфа в границах существующих болотных массивов. В
настоящее время площадь подавляющего большинство озер в области
составляет 1-10 га, и только на отдельных массивах она достигает нескольких
сотен гектар. Имеющиеся данные позволяют предположить, что площадной
рост озер происходит в значительной степени автономно. И определяется, на
наш взгляд, в основном волновой деятельностью воды, ведущей к
разрушению торфяных берегов. В условиях усиления активности озерообразования
торфяные болота теряют целый ряд функций и в первую очередь торфообра-
зуюшую. По мере расширения площади озер (с учетом открытости болотных
пространств) можно ожидать усиление волновой деятельности озер, что в
конечном счете повлечь усиление тенденции трансформации исходных
отложений торфа и преобразование их в донные отложения. Рост площади
озер на наш взгляд может привести к изменению в будущем природной
ситуации в регионе в сторону увеличения увлажненности.
Географический анализ используемых материалов позволил выявить
реальные генетически различные поверхности, в которых обеспечивается
формирование специфичных экологических условий под воздействием болото-
образования. При выделении ориентировались в основном на
физиономические характеристики с использованием тематических карт
(геоморфологических, топографических, почвенных, геологических, гидрогеологических,
интенсивности болотообразования, торфяных месторождений, планов
лесных насаждений), дистанционных материалов разных лет.
Выделенные состояния позволили провести районирование Томской
области, что представлено в обзорной карте М 1:1000000. Единицами
картографирования послужили этапы и их композиция, сочетания. Единицами
легенды стали отображаемые в соответствующем масштабе ареалы
собственно выделенных выше единиц, а также их сочетания (по признак
соотношения занимаемых ими площадей). Составленная карта позволяет на об-
27
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
зорном уровне проводить прогноз динамики ландшафтов, может быть ис^
пользована при решении разных задач регионального природопользования.
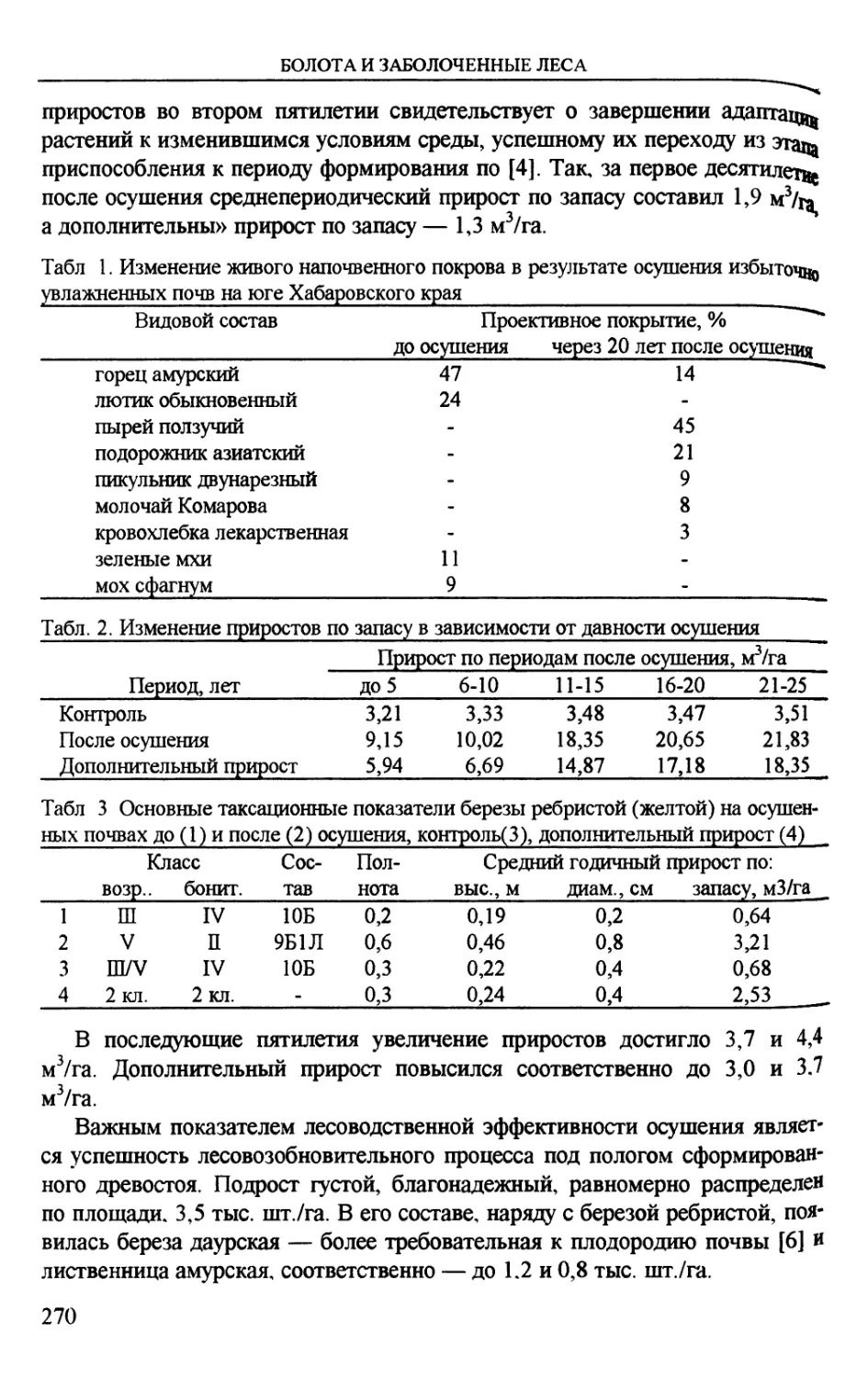
ДРЕВНЕЕ БОЛОТНОЕ ПОЧВООБРАЗОВАНИЕ
И ЕГО РОЛЬ В РАЗВИТИИ БИОСФЕРЫ
В.К. Бахнов
Институт почвоведения и агрохимии СО РАН, Новосибирск
Океан считают самой древней частью биосферы. Дальнейшее ее разви-
тие было направлено на освоение организмами воздушной среды и лито-
сферной оболочки суши. Данный процесс осуществлялся при активном
участии болотного почвообразования. Кроме утилизации органического
углерода и формировании кислородсодержащей атмосферы, древние болотные
почвы выполнили еще одну не менее важную функцию в истории биосферы.
Эти почвы оказались наиболее благоприятной экологической средой в пери-;
од выхода растений из воды и освоении ими суши. По крайней мере, предки
представителей зеленого ствола растительного мира экспансию суши
осуществили через болотное почвообразование [1].
Воздушная среда и литосферная оболочка суши различаются между
собой и отличаются от водной среды резкой экологической контрастностью.;
Процесс одновременного освоения организмами, прежде всего растениями,
этих новых сред обитания был бы непомерно сложным. Вероятно, он
совершался постепенно и характеризовался определенной
последовательностью. Болото можно рассматривать и как водоем, где вода связана
органикой, и как сушу, содержащую 80-90% воды и 20-10% сухого вещества.
Обладая таким двуединством, экологический контраст между водной средой и
сушей в болоте является наименьшим. Благодаря этому, болотное
почвообразование в истории биосферы выполнило своего рода роль переходного
моста, по которому растения вышли из воды на сушу.
Болотные почвы на Земле появились в конце силура — начале девона, то
есть примерно 400 млн. лет тому назад. Этим же временем специалисты
эволюционной фитоценологии датируют выход из водной среды предков
современных растений. На данном этапе растения во многих отношениях
были сходны с водными предками и длительное время сохраняли
потребность в постоянном обильном увлажнении среды обитания. Роль фактора,
стабилизировавшего водообеспечение, выполнило органическое вещество,
которое по своей природе обладает высокой способностью поглощать и
удерживать воду в больших количествах. Эта особенность органического
вещества послужила предпосылкой использования создаваемых самими же
растениями запасов мертвых остатков в качестве субстрата — почвы. Пер-
28
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
выми представителями растений суши (вернее сказать болот) считают пси-
лофиты. Тело этих растений представляло корневище, погруженное в воду,
й лишь на короткое время на поверхность воды выбрасывался небольшой,
высотой около 20 см, спорангий [3]. Псилофиты просуществовали недолго:
появились в конце силура и во второй половине девона вымерли. В карбоне
и перми огромную биомассу создавали представители голосеменных и
папоротникообразных. Остатками этих растений были сформированы
болотные почвы, представленные в нщпе время в виде залежей каменного угля.
В процессе эволюции растения не только приспосабливались к среде, но
одновременно становились и менее зависимыми от нее, приобретали
некоторую самостоятельность, которая достигалась и путем создания ими
субстрата — почвы. На примере относительно примитивных растений, какими
являются мхи, которые могли быть одними из первых растений суши,
можно видеть один из возможных вариантов кардинального приспособления
растений к условиям жизни вне водной среды. Уникальная способность
мхов состоит в том, что одновременно с вертикальным ростом верхушки
происходит отмирание нижних частей стебля, остатками которых
формируется исключительно влагоемкий субстрат, служащий одновременно
источником влаги и зольных элементов для вегетирующих частей растений.
Болотный период в развитии биосферы продолжался около 200 млн лет.
За это время организмы адаптировались к жизни в условиях воздушного
окружения. Произошли важные эволюционные преобразования
растительного мира: сформировался приспособленный к существованию и активному
функционированию вне водной среды фотосинтезирующий аппарат,
появилось большое генотипическое разнообразие, формирование которого
сопровождалось морфоанатомическим и физиологическим прогрессом,
осуществилась дифференциация и специализация тканей, выработалась способность
растений развиваться в вертикальной плоскости, широкое распространение
получили древовидные формы. Растения эволюционно были подготовлены к
переселению на литосферную оболочку суши. Однако, у них была
слаборазвитая корневая система, приспособленная к условиям влажных
органогенных почв. Дальнейшее совершенствование растений, прежде всего корневых
систем, происходило уже в период освоения литосферы суши.
Древнее болотное почвообразование отразилось не только на эволюции
растений, но и животных. Обилие влаги и органического вещества,
являющегося основным источником энергии для жизнедеятельности животных,
оказалось, очевидно, основной предпосылкой для освоения болотных почв
многими обитателями бентали. Дальнейшая эволюция животных следовала
за формированием первичного почвенного покрова на литосферной
оболочке суши. Было показано [2], что животные, прежде всего беспозвоночные,
29
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
населяющие почвы суши, являются выходцами из влажных местообитаний
— заболачивающихся водоемов и болот.
[1] Бахнов В.К. Биогеохимические аспекты болотообразовательного процесса.
Новосибирск: Наука, Сиб отд., 1986. 193 с. [2] Гиляров М.С. Особенности почвы как
среды обитания и ее значение в эволюции насекомых. М.-Л: Изд-во АН СССР,
1949 279 с. [3] Комаров В.Л. Происхождение растений. М.: Изд-во АН СССР, 1961.
190 с
К ВОПРОСУ О ПРОИСХОЖДЕНИИ ГРЯДОВО-МОЧАЖИННЫХ
КОМПЛЕКСОВ
В.К. Бахнов
Институт почвоведения и агрохимии СО РАН, Новосибирск
Происхождение грядово-мочажинных комплексов (ГМК) — давняя
проблема в болотоведении. Неоднократно предпринимались попытки объяснить
причины и механизмы их образования. Большинство гипотез [2,5,6] прямо
или косвенно связывают образование ГМК с влагой, выступающей в
качестве физической силы или источника элементов питания болотной
растительности. Предлагается также [7,8] рассматривать торфяный купол как
геологическую структуру, мочажины и озерки — как системы закономерно
ориентированных разломов, образовавшихся в результате выхода на
поверхность болота газов и внутризалежных вод. Главным критерием
признания гипотезы считается объяснение причин, вызывающих расположение
гряд и мочажин перпендикулярно уклону поверхности болота. Однако, в
действительности, по крайней мере на болотах Западной Сибири,
встречается и иное расположение данных элементов микрорельефа [1,4]. Ориентацию
гряд и мочажин поперек уклона поверхности, очевидно, следует
рассматривать как господствующее, но не строго закономерное явление на болотах.
Нерешенность проблемы происхождения.ГМК связываем с тем
обстоятельством, что болотоведы не учитывали роли минерального ложа в
образовании рельефа на современных болотах. Инструментальный профиль,
проложенный через ГМК на Бакчарском болоте, выявил четкую
закономерность расположения гряд и мочажин относительно положительных и
отрицательных форм микрорельефа минерального ложа. Обнаружен^
определенная зависимость некоторых параметров гряд и мочажин от фор^ы и
степени выраженности микрорельефа ложа [1]. Связь рельефа поверхности
болота с рельефом поверхности подстилающего его ложа свойственна не
только исследованному микроландшафту. Она имеет место и в грядово-
озерковом комплексе, где положение озерков и гряд достаточно строго
приурочено соответственно понижениям и повышениям рельефа ложа [3].
30
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
Основные свойства торфа, в том числе зольный состав, определяются
составом растений-торфообразователей. Проведенными ранее
исследованиями, основу которых составил биогеохимический подход [1], установлено,
что основное количество элементов минерального питания в органогенном
профиле накапливается на начальной стадии развития болота. Основным
источником является почва, подвергшаяся заболачиванию. Нарастание
почвенного профиля сопровождается «перекачкой» биогенных элементов снизу
вверх биологическим путем. Происходит постепенное обеднение верхней
части профиля элементами минерального питания растений, что приводит к
сукцессиям растительных ассоциаций, к смене одного вида торфа другим.
Пространственная и временная вариабельность растительного покрова,
динамический характер рельефа на поверхности болот в процессе их
развития — следствие отражения влияния рельефа и пестроты былого почвенного
покрова минерального ложа. Почвенный покров и рельеф предболотной
стадии — ведущие факторы, которые как бы «программируют» ход
развития болотного массива.
[1] Бахнов В.К. Биогеохимические аспекты болотообразовательного процесса.
Новосибирск: НаукаСиб. отд., 1986. 193 с. [2] Богдановская-Гиенэф ИД. Образование
и развитие гряд и мочажин на болотах // Сов. ботан. 1936. № 6. С. 35-52. [3] Болота
Западной Сибири, их строение и гидрологический режим / Под ред. К.Е. Иванова,
СМ. Новикова. Л.: Гидрометеоиздат, 1976. 447 с. [4] Бронзов А.Я. Гипновые болота
на южной окраине западно-сибирской равнинной тайги // Почвоведение. 1936. № 2.
С. 224-246. [5] Кац Н. и др. Движение поверхности сфагновых болот и
формирование их микрорельефа // Землеведение. 1936. Т. 38. Вып. 1. С. 1-34. [6] Ниценко А.
А. О происхождении грядово-мочажинного рельефа на болотах // Вестн. ЛГУ. Сер.
биол. 1964. Вып. 4. № 21. С. 75-87. [7] Фриш В.А. Торфяная тектоника // Изв. ВГО.
1978. № 2. С. 108-116. [8] Фриш В.А. Ландшафтный и структурно-геологический
анализ развития болот // Изв. ВГО. 1981, Т. 113. № 2. С. 122-129.
ТОРФЯНЫЕ БОЛОТА ЗАПАДНОЙ СИБИРИ (ПРОЕКТ РОССИЙСКО-
ГОЛЛАНДСКИХ ИССЛЕДОВАНИЙ)
ВВ. Блейтен1, Е.Д. Лапшина2, А.А. Величко3, И.М. Гаджиев, СВ.
Васильев4, СП. Ефремов5, К.В. Кременецкий, В.А. Климатов, Э.М. Зеликсон3
Университет Утрехт, Нидерланды; 2Томский государственный
университет; ^Институт географии РАН, Москва;АИнститут почвоведения и
агрохимии СО РАН, Новосибирск;5Институт леса и древесины СО РАН,
Красноярск
В 1998 г. начались работы по совместному российско-голландскому
проекту ? поддержанному научным фондом Нидерландов (NWO).
31
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
В Западной Сибири сосредоточено около половины мировых запасов
торфа. Исходя из общих оценок годовой прирост запасов углерода в болотах
Западной Сибири составляет около 500 кг/га. Однако до сих пор глобальные
модели цикла континентального углерода весьма в слабой мере учитывали
запасы углерода, сосредоточенные в болотах Западной Сибири. Если в
результате деятельности человека при наблюдаемом глобальном потеплении
начнется массовое высвобождение в атмосферу углерода, связанного в
торфах Западной Сибири, это может оказать серьезное воздействие на
глобальные климатические процессы.
Существующие модели климата исходят их важной климатической роли
парниковых газов (метана и углекислого газа), попадающих в атмосферу в
значительной мере в ходе хозяйственной деятельности. Для оценки
возможных климатических последствий важно учитывать буферную роль торфяных
болот, являющихся крупнейшим резервуаром стока углерода из атмосферы.
Прирост запасов углерода оценивается исходя из расчетов прироста запасов;
торфа за продолжительный период времени (голоцен). Эти данные нужда-*
ются в верификации, поскольку по палинологическим данным на
протяжении голоцена происходили неоднократные изменения климата. Таким
образом необходимо выяснение взаимоотношения между аккумуляцией углерода
и специфическими климатическими параметрами (осадки, температура, ра-
диация) и типом экосистем (растительность и гидрологические условия).
Эти закономерности необходимо знать для правильной оценки роли облас-1
тей торфонакопления в глобальном изменении климата.
Осуществление проекта предполагается в несколько этапов: 1. Подсчет
биологической продуктивности современных болотных экосистем. 2. Оценка
динамики аккумуляции углерода в голоцене исходя из имеющихся данных и
с помощью радиоуглеродного датирования. 3. Раззработка реалистической,
пространственно-временной модели распространения и эволюции болот]
Западной Сибири в голоцене. 4. Создание базы данных по распротсранению
болотных экосистем в центральнйо и южной частях Западно-Сибирской
низменности, о соотношении болот с рельефом, почвенным покровом, усло-1
виями дренажа. Сбор данных о возрасте торфов. 5. Создание ГИС по расН
пространению торфяных болот в Западной Сибири. 6. Изучение образцов
торфа в 3-4 ключевых участках для определения содержания углерода Щ
радиоуглеродного датирования. 7. Подсчет скоростей прироста запасов
углерода в торфяных болотах и возможной эмиссии углерода при осушений
торфяных болот. 8. Замеры эмиссии метана. 9. Подсчет баланса углерода
дал всей площади болот Западной Сибири.
В полевой сезон 1998 г. велись работы на трех ключевых участках в
южной части Западной Сибири (Томская область). В сезон 1999 г. работы про-
32
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
одились в северной части таежной зоны в районе Ноябрьска (Ханты-
Мансийский округ Тюменской области).
УСТОЙЧИВОСТЬ БОЛОТ К ИЗМЕНЕНИЮ КЛИМАТА:
ВОЗМОЖНЫЕ НАПРАВЛЕНИЯ АНАЛИЗА
К. Вайсман, А.А. Сирин, В.Н. Солнцев
Мюнхенский технический университет; Институт лесоведения РАН,
Московская обл.; МГУ, географический факультет;
Предполагаемые антропогенные изменения климата определяют
актуальность анализа устойчивости к ним болотных экосистем. В экологии и
географии рассматриваются разные концепции, выделяющие два аспекта
понятия «устойчивость» экосистем и природно-территориальных
комплексов [4-6 и др.]: а) способность поддержания определённого состояния или
последовательности состояний системы; б) способность системы
противостоять внешним возмущениям и воздействиям при сохранении ее
качественной целостности. В российской практике заметна тенденция называть
первый аспект «стабильностью», а второй — «устойчивостью» [4]. Для
оценки устойчивости используются два различных подхода: а) наблюдение
за реакцией системы (феноменологический подход), б) анализ ее
характеристик и свойств, потенциально влияющих на устойчивость (функциональный
подход). Наиболее эффективным представляется сочетание обоих подходов.
Анализ последствий климатических изменений остро ставит проблему
временного шага. Длительное саморазвитие болот происходило на фоне
естественной сезонной динамики и различных по длительности (внутриве-
ковых и многовековых) климатических флуктуации. В этих условиях
необходимо выявить наиболее отчетливо проявляющиеся устойчивые тенденции
развития систем, обусловленные изменением климата, а также, что более
сложно, признаки смены одной тенденции развития другой. При этом
следует учитывать, что по мере развития болотного массива его устойчивость к
однотипным внешним воздействиям меняется.
Ключевым моментом при исследовании устойчивости является выбор: а)
аспекта рассмотрения системы (состояние, структура, функция); б)
параметров системы, подлежащих рассмотрению. В общем случае этот выбор
зависит от научно-практической задачи анализа. При рассмотрении состояния
болота в целом, необходимо основываться на характеристиках,
определяющих болото как целостную особенную систему: степень увлажнения,
характер болотной растительности, специфика торфа [1].
Климат оказывает воздействие на разные стороны
структурно-функциональной организации болот, хотя наиболее часто его влияние оценивается по
33
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
изменению режима болотных вод. Поэтому делаются попытки
спрогнозировать последствия изменения климата по данным изменения болот при лесо-
осушении [8]. Нам представляется это малооправданным, так как, даже
предполагая близкое изменение водного режима (что дискуссионно),
скорость реализации этих изменений может иметь принципиально разные
последствия для болот. Известны попытки проведения активных
экспериментов с искусственным прогревом торфяных монолитов с болотной
растительностью (NRRI, University of Minnesota, Duluth), однако их эффективность
ограничена размером лизиметров и временными рамками исследования.
Феноменологический подход может быть осуществлен путем анализа
изменений растительного покрова (по ботаническому составу торфа),
интенсивности торфонакопления (по изменениям объемной массы и содержания
углерода в торфе), уровней боло!ных вод и их рН (по реконструированному
составу растительного покрова). Этот анализ должен быть привязан к
различным палеоклиматическим периодам последних 2-3 тысячелетий и,
особенно. Малому климатическому оптимуму (МКО) голоцена на рубеже
последних двух тысячелетий. Климат МКО наиболее близок предполагаемым
сценариям изменений современного климата [7]. В этот период времени
большинство бореальных болот находилось фактически в той же стадии
развития, что и в настоящее время. Это обстоятельство упрощает анализ.
На сформулированных выше подходах основаны исследования по оценке
устойчивости верховых болот, проводимые на Западнодвинском стационаре
Института лесоведения РАН (Тверская область) [2,9]. Рассматриваются три
расположенных рядом болотных массива одного возраста, которые a priori
развивались при одинаковых палеоклиматических изменениях, но в
различных геоморфологических и гидрогеологических условиях и, как следствие, в
разных условиях оттока болотных вод, определяющего структуру их водного
баланса. Хронологическое расчленение разрезов основывается на выделении
пиков палеоклиматических кривых [3], датированных комплексом палео-
хронологических данных и обеспечивающих стратиграфическую привязку
характерных палеоклиматических эпизодов с точностью до столетия.
Функциональный подход используется при анализе возможных причин
разной реакции изучаемых верховых болотных массивов на потепление в
МКО. и в последовавшее за ним похолодание Малого ледникового периода.
Он позволяет провести только относительную оценку общей устойчивости
болотных массивов ттем сравнения механизмов их частной устойчивости.
При этом анализируются особенности водного баланса разных болотных
массивов, и сравнивается высота «шапки» массива, а также характер
сложения залежи для оценки потенциала механизма «дыхания» болот, т.е.
отекания «шапки» в сухие годы, предохраняющего от катастрофических
понижений уровней болотных вод. Одновременно проводится анализ эколо-
34
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
гяческих диапазонов отдельных видов болотных растений
(реконструированных по ботаническому составу торфа) по фактору влажности. Состав
торфообразователей мбжет свидетельствовать об устойчивости торфяной
почвы к изменению водного режима. Присутствие остатков сосудистых
растений, обусловливающее повышение трофности, можно трактовать как
индикатор условий, способствующих при изменениях водного режима более
интенсивной деструкции органического материала.
Для проведения всесторонней оценки устойчивости болот, конечно,
необходимо рассматривать не один, а многие механизмы устойчивости,
которые на современном уровне знаний представляются существенными для
сохранения изучаемых характеристик.
[1] Гидрология суши. Термины и определения: ГОСТ 19179-73 Изд. офиц. М.: Гос.
комитет стандартов Совмина СССР, 1973. 34 с. [2] Гунова B.C., Сирин А.А. Палео-
геграфические условия развития верховых болот Западно-Двинской низины в
голоцене // Палинология в России. М.: Нац. ком. геологов России. 1995. Т. 2. С.27-36. [3]
Климанов В.А., Сирин А.А. Динамика торфонакопления болотами Северной
Евразии за последние 3000 лет // ДАН. 1997. Т.354. № 5. С. 683-686. [4] Механизмы
устойчивости геосистем /Под ред. Н.Ф. Глазовского, А.Д. Арманда. М.: Наука, 1992.
[5] Устойчивость геосистем. М: Наука, 1983. [6] Grimm V., Wissel С. Babel, or the
ecological stability discussions: An inventory and analysis of terminology and a guide for
avoiding confusion / Oecologia, 1997. V.109. P. 323-334. [7] Intergovernmental Panel on
Climate Change IPCC, Climate Change 1994, Radiative forcing of climate change and an
evaluation of the EPCC IS92 emission scenarios: Cambridge Univ. press, 1995. [8] Laine
J. et al. Effect of water-level drawdown on global climatic warming: northern peatlands //
Ambio. 1996. V.25. N 3. P. 179-184. [9] Sirin A.A. et al. Role of local hydrology in
raised bog development in the Central Russia // Dynamics of mire ecosystems of
Northern Eurasia in Holocene. 1998. Petrozavodsk: Karelian Scientific Center Russ.
Acad. Sci. 1998. P. 19-20.
О ПРИРОДЕ ГРЯДОВО-МОЧАЖИННЫХ КОМПЛЕКСОВ
С. В. Васильев
Институт почвоведения и агрохимии СО РАН, Новосибирск
Вопрос о генезисе грядово-мочажинных комплексов (ГМК) занимает
исследователей уже около века. В качестве причин их возникновения видят
явления солифлюкции, [1,12,16,22], неравномерного распределения
снежного покрова [8], сезонного промерзания и протаивания [1.13.18]. ГМК
рассматриваются как перигляциальные явления [19]. Формирование ГМК
объясняют биологическими причинами, связывая с постепенным обеднением
торфяной залежи минеральными элементами [7], с возрастающим
обводнением или, напротив, иссушением торфа [5]. На гетеротрофных болотах типа
35
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
аапа предполагают эффект подпруживания стока по мере их роста вверх по
склону [2,3]. Согласно [20,21] развитию мочажин способствует накопление
метана в торфяной залежи и выход его на поверхность. Предполагается [4]
наследование болотом рельефа минерального дна. В вопросах происхожде*
ния и развития ГМК нет единства взглядов и окончательных теорий [17].
Все это косвенным образом свидетельствует о множественности
механизмов их образования, однако распространенность явления и постоянство
таких свойств, как связь с определенными уклонами поверхности болота ц
параллельность гряд, вытянутых перпендикулярно уклону предполагает не«
которую общность условий в которых они формируются. ГМК развиты 1
различных про трофности болотах, с различным ботаническим составом
сообществ и торфа. Следует полагать, что ведущей причиной их
устойчивости является гидрология фильтрующегося потока воды. В этом аспекте
собран большой материал [6,9], позволяющий рассмотреть это подробней.
Согласно закону Дарси скорость фильтрации (v) жидкости в пористой
среде определяется пьезометрическим уклоном (/) и коэффициентом
фильтрации (к): v=ki. Установлено [6,9], что под грядами образуется торф,
коэффициент фильтрации которого во много раз (от 10 до 200) меньше, чем
мочажинах. Согласно условию неразрывности течения жидкости расход воды
по длине потока при отсутствии бокового притока и расхода остается
постоянным. Предположим в начале неизменность толщины слоя фильтрующейся
воды, тогда скорость ее фильтрации в грядах и в мочажинах должна
оставаться одинаковой, а именно v0=k2i2=kMiMi где /г, iM — частные уклоны по-,
верхности потока в гряде и мочажине, кг, км — средние по толщине
фильтрующего слоя коэффициенты фильтрации в торфе гряды и мочажины. Так
как коэффициенты фильтрации в грядах и в мочажинах различны,
равенство может быть достигнуто только при эквивалентной разнице в уклонах.
Иными словами, отношение уклона водного потока в гряде к уклону потока
в мочажине обратно пропорционально отношению коэффициентов
фильтрации: C=i/iM= кукг. Это означает, что уклоны потока воды в грядах в 10-200
раз больше, чем уклоны в мочажинах, а ГМК представляет собой каскад,
где мочажины — «террасы», а гряды — «уступы», что и установлено
нивелировкой [9].
ГМК развиваются при уклонах от 0,004 до 0,0008. При различных
уклонах соотношение гряд и мочажин по площади, а равно и по суммарной
длине в профиле стока, различно. Простые геометрические рассуждения
позволяют определить долю трядрг по длине профиля стока: рг= (Cio-iJ/fCi^iJ, a
учитывая, что C=i/iM, это можно записать и иначе pe=(io~&/(h-iJ-
Отсюда следует тривиальный вывод, что отсутствие гряд в ГМК
наблюдается тогда, когда общий уклон равен уклону в мочажинах, а отсутствие
мочажин — тогда, когда общий уклон равен уклону водного потока в гря-
36
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
дах. Используя величины /г=0,004 и /^=0,0008 в качестве предельных,
получаем простую линейную зависимость с помощью которой можно рассчитать
среднее соотношение гряд и мочажин по суммарной длине вдоль профиля
стока исходя из среднего уклона поверхности болота:
0^днийуклон(по[9]) : 0,0037 0,003 0,0025 0,0017 0,0009
Наблюдаемая доля гряд в площади ПИК, % 80 70 60 30 10
Расчетная доля гряд по длине профиля стока 91 69 53 28 3
При столь грубых рассуждениях, полученное соответствие эмпирических
и расчетных данных надо признать хорошим. Тем не менее, реальные
уклоны в ГМК существенно варьируют. Этому имеется объяснение.
Соотношение гряд и мочажин определяется водным балансом конкретного участка
болота. При одном и том же общем уклоне частный уклон водного потока в
грядах может быть больше или меньше. Больший уклон на грядах
соответствует большему уклону в мочажинах при сохранении отношения С, что
отвечает большей скорости потока фильтрующейся воды и большему
расходу. Пропорциональный рост частных уклонов при сохранении общего
уклона поверхности болота с увеличением стока влечет уменьшение доли гряд и
увеличение доли мочажин. Сам расход в конкретной точке профиля стока
определяется внешними факторами: количеством падающих осадков,
испарением, грунтовым стоком и площадью дренируемого бассейна. При одном
и том же среднем уклоне поверхности болота доля мочажин возрастает вниз
по профилю стока, так как увеличивается площадь водосбора, сток с
которого проходит через данный участок профиля. Такой же эффект имеет и гуми-
дизация климата: на болотах Эстонии [10,11] больше всего мочажин
сформировалось во влажные периоды голоцена. ГМК распространены в более
гумидных районах. На верховых болотах подтаежной зоны и северной
лесостепи Западной Сибири ГМК не развиты.
Равенство единичных расходов по длине стока при разных
коэффициентах фильтрации достигается не только за счет изменения частных уклонов,
но и за счет изменения толщины слоя фильтрации, согласно чему меняется
скорость водного потока. Толщина слоя фильтрующейся воды минимальна у
верхнего края мочажины. Здесь же максимальна скорость горизонтальной
фильтрации, за счет чего происходит насыщение текущей воды кислородом.
Согласно [14,15] на участке от верхнего края мочажины к середине
количество растворенного С02 снижается до 26 мг/л, а концентрация СЬ
увеличивается до 4 и 6 мг/л. Это способствует аэробному разложению торфа. К
нижнему краю мочажины за счет подпруживания стока грядой толщина
слоя фильтрации увеличивается, а скорость водного потока снижается. На
обращенный вверх по склону берег гряды со стороны мочажины в верхнем 5
см слое поступает вода с высоким содержанием углекислоты (80-90 мг/л), а
37
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
концентрация 02 снижается до 1-0,2 мг/л [15], что способствует автотроф-
ным процессам и снижает активность аэробных гетеротрофов. Можно
предполагать, что именно на этом участке стока происходит опережающий рост
торфяной залежи и формирование гряд, а с другой стороны, у верхнего края
мочажин, где скорость тока воды максимальна — их деградация. Это
подтверждает микростратиграфия: гряды и мочажины по мере роста
«смещаются» вверх по склону в сторону выпуклости болота [15,17].
Наблюдения [15] находятся в согласии с рассмотренными особенностями
стока болотных вод и свидетельствуют о наличии положительной обратной
связи между растительностью и торфом, который она формирует, с одной
стороны и скоростью водного потока — с другой. Любая неоднородность,
вызванная самыми разными причинами, приводит к дифференциации
уклонов потока фильтрующейся воды, а разница в скоростях фильтрации
провоцирует дальнейшую дифференциацию растительности гряд и мочажин и
рост контраста условий микроместообитаний. Каков конкретный механизм,
приводящий к возникновению первичной неоднородности, не столь важно.
[ 1 ] Auer V Uber die Entstehung der Strange auf der Torfmooren // Acta forestalia Fen-
nica 1920 Issue 12. P. 1-145. [2] Foster D.R , et al. Origin of string patterns in boreal
peatlands // Nature. 1983. V. 306. P. 256-258. [3] Foster D.R., et al Bog development
and landform dynamics in central Sweden and south-eastern Lamrador, Canada // J.
Ecology. 1988. V. 76. P. 1164-1185. [4] Бахнов В.К. Биогеохимические аспекты болото-
образовательного процесса. Новосибирск: Наука, 1986. 193 с. [5] Богдановская-Гие-
неф И Д Закономерности формирования сфагновых болот верхового типа на
примере Полистово-Ловатского массива Л: Наука, 1969. 188 с. [6] Болота Западной
Сибири их строение и гидрологический режим. Л.: Гидрометеоиздат, 1976. 448 с.
[7] Вильяме В Р Почвоведение. М.. Сельхозгиз, 1949. 471 с. [8] Гришин И.С
Влияние снежного покрова на формирование микрорельефа торфяных месторождений
верхового типа и их природных комплексов // Изучение состава и свойств торфа в
целях его использования в народном хозяйстве. М.: 1977. С 109-123. [9] Иванов
К Е. Основы гидрологии болот лесной зоны и расчеты водного режима болотных
массивов.- Л.: Гидрометеоиздат, 1957. 500 с. [10] Карофельд Э.К. О временной
динамике грядово-мочажинного комплекса на верховых болотах Эстонии // Ботан. ж.
1986 Т. 71. № И. С. 1535-1542. [И] Карофельд Э.К. Об образовании мочажин на
верховых болотах Эстонии // Структура и развитие болотных экосистем и
реконструкция палеогеографических условий. Тез. док . Таллинн: АН ЭССР, 1989. С. 70-74.
[12] Кац Н Я , Кириллович М М, Лебедева Н.В Движение поверхности сфагновых
болот и формирование их микрорельефа // Землеведение. 1936. Т 38. Вып. 1 С 1-
32 [13] Кац Н.Я Болота и торфяники. М • Учпедгиз, 1941. 400 с [14] Конойко МА
Условия развития верховых болот Белоруссии // Генезис и динамика болот. Вып.1.
М: МГУ, 1978. С. 157-162. [15] Конойко М.А. Методика и результаты
исследований грядово-мочажинного комплекса // Структура и развитие болотных экосистем и
реконструкция палеогеографических условий. Тез. док. Таллинн: АН ЭССР, 1989. С
75-80. [16] Краснов И.И. О болотной солифлюкции и современной трансгрессий
38
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
болот на равнинах в зоне тайги // Проблемы физической географии. 1941. Вып. 10.
С. 3-34. [17] Кузьмин Г.Ф. Болота и их использование. Спб.: ВНИИ торфяной
промети, 1993. 140 с. [18] Пьявченко Н.И. К вопросу познания природы грядово-моча-
жиНных комплексов карельского типа // Тр. Ин-та леса АН СССР. 1953. Т. 13. С.
130-147. [19] Уошборн А.Л. Мир холода. Геокриологические исследования. М.:
Мир, 1988. 384 с. [20] Фриш В.А. Торфяная тектоника // Изв. ВГО. 1978. Т. ПО.
Вып. 2. С. 108-112. [21] Фриш В.А. Торфяная тектоника и динамика ландшафтов //
Изв. РГО. 1993. Т.125. Вып. 2. С. 66-73. [22] Цинзерлинг Ю.Д. Растительность
болот// Растительность СССР. М.: АН СССР, 1938. Т. 1. С. 355-428.
МАСШТАБЫ СОВРЕМЕННОГО ЗАБОЛАЧИВАНИЯ ЗЕМЕЛЬ ЛЕСНОГО
ФОНДА ЛЕНИГРАДСКОЙ ОБЛАСТИ
Г.Б. Великанов
Комитет по лесу Ленинградской области
О заболачивании земель в таежной зоне России существуют различные
мнения (М.С. Боч, Г.А. Елина, Ф.З. Глебов, В.Н. Кирюшкин, Г.Ф. Кузьмин,
А.А. Корчагин и М.Н. Нейштадт, О.Л. Лисе, Н.И .Пьявченко и др.). Одни
считают, что этот процесс затухает, т.к. дальнейший рост болот,
контролируемый крупными формами рельефа, практически прекратился, и
взаимоотношения между лесом и болотом находятся в стадии, близкой к равновесию.
Другие убеждены, что масштабы заболачивания еще велики и даже не
компенсировались объемами выполненных лесоосушительных работ.
Анализ этих и других (А.А. Вейнерт, С.Э. Вомперский, К.Н Дьяконов,
К.Я. Казаков, М.В. Калинин, В.К. Константинов, Н.А. Красильников, Х.А.
Писарьков, М. А. Шапошников и др.) литературных источников и результатов
исследований автора дает основание экспертно считать (см. табл.), что среди
факторов, определяющих динамику наблюдаемого переувлажнения и
заболачивания земель в области, одним из ведущих и долговременным является
техногенный (строительство линейных сооружений; дорог, нефте-газопро-
водов и плотин). Ежегодный прирост земель постоянного переувлажнения в
лесном фонде может достигать 0,2-2 тыс. га за счет разрастания болот, 0,2-0,3
тыс. га только за счет дорог и на 1/3 площади сплошных вырубок.
Еще более быстрыми темпами происходит вторичное заболачивание ранее
осушенных земель (до 8 тыс. га в год) из-за плохого состояния осушительной
с^ти, недостаточной степени осушения, принятой в проектах, и из-за
строительной деятельности бобров. Прокладка линейных сооружений (в т.ч. и
кабелей связи вдоль дорог) без экологического обоснования и надлежащего
согласования с землепользователями существенно усложняет и даже
исключает дальнейшие осушительно-мелиоративные работы на вышележащих
водосборах. При проектировании этих сооружений необходимо учитывать
39
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
густоту гидрографической сети, которая (С.Х. Будыка) влияет на развитие бо«
лотообразовательных процессов.
Масштабы естественного и антропогенного заболачивания лесного фонда в Ленин-
градской области. ДПЗ — ежегодный прирост переувлажненных земель
Причина заболачивания ДПЗ, тыс. га Характер переувлажнения ]
1 Рост болот и ПЗ под влиянием 0,2-2 Необратимый и долговременный
естественных факторов
2. Заболачивание вырубок 1,5-2 На 1/3 площади — необратимый,
на 2/3 - кратковременный
3. Строительная деятельность до 2 Различный, в зависимости от ис-
бобров 1
4. Ухудшение работы
осушительной сети из-за плохого ее
содержания2
5. Недостаточналйроектная сте-
пень осушения
6. Водохранилища (Свирские и
Нарвское)3
7. Дорбги4
до 2
До 4
0,2-0,3
0,2-0,3
ходных лесорастительных условий,
пока площадь занята бобрами
Различный, в зависимости от
исходных лесорастительных условий
ее
-«.
Долговременный
1 - Затоплено до 50-60 тыс. га осушаемых и естественно дренированных земель.
Дальнейшее изменение площади зависит от численности бобров. 2 - Увеличения
общей площади переувлажненных земель не происходит, но сокращается площадь
осушаемых и эффективно осушаемых земель. За послевоенные годы вторично
заболотилось не менее 90 тыс. га и более чем на 150 тыс. га водный режим
недостаточен для нормального роста насаждений. 3 - С момента постройки водохранилищ
подтоплено 50 тыс. га земель, в т.ч. 4,5-7 тыс. га суходолов. 4 - Очаги подтопления
наблюдаются повсеместно, но данных по ним нет.
Суммарное воздействие всех факторов переувлажнения и заболачиваю»
земель лесного фонда области может оказать негативное воздействие на их
состояние, которое сейчас смягчается буферной рблью ранее выполненных
лесоосушительных работ.
РЕКОНСТРУКЦИЯ РАЗВИТИЯ КОТЛОВИННОГО БОЛОТА
ЮЖНОЙ ТАЙГИ
Н.В. Веселов, В.В. Панов
Тверской государственный университет;
Тверской государственный технический университет
Болото Локотенское расположено в Лихославльском районе Тверской
области. Площадь болота около 140 га, глубина — 5,5 м, из них 1,5 м озер-
40
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
^ie отложения. Болото находится на границе Ладожско-Ильменско-Запад-
нодвинской провинции широколиственно-хвойных лесов и выпуклых грядо-
во-мочажинных болот и Московско-Верхнеднепровской провинции хвойно-
щироколиственных лесов, евтрофных и олиготрофных сосново-сфагновых
болот [Кац, 1971], что совпадает с границей южной тайги и хвойно-широко-
лиственных лесов.
040 80% . 040»0% 040 80%
ф
ft
Т5
и
со
1
•о
с
Г-З
ф
л
-С
• гэ
СО <П
го
а
к
<ъ
<•"•
X О
а> —
я т
(Ъ <0
о *-\
lit
SA-3
SA 2
за-:
SB-2
SB-1
во-:
.ja-2
?в-:.
DR-3
Рис. 1. Распределение растительных остатков в торфах
Пертюлы I Глубина, j Йсеа ; "
aL..J_ .. . .
Рис.2. Спорово-пыльцевая диаграмма
На рис. 1 приведены данные ботанического анализа торфов, слагающих
залежь в генетическом центре болота, который соответствует первичному
очагу заболачивания. На 20% площади болота торфяная залежь подстилает-
41
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ся сапропелем, максимальной мощностью около 1,5 м.
Начальные условия заболачивания определялись природными условиями
приледниковой (перигляциальной) зоны. Первичное тундровое озеро
зарастало осоками, зелеными и сфагновыми мхами. На рубеже послеледникового
и позднеледникового времени — 10300-10500 лет назад на Локотенском
болоте формируются евтрофные торфа древесно-травяной группы (рис. 2). В
РВ-2 уменьшилась облесенность центра Русской равнины. Формируются
низинные осоково-гипновые торфа. В начале бореального периода Локотен-
ское болото переходит в стадию мезотрофного болота, а в середине
бореального периода болото перешло в олиготрофную стадию. Во второй
половине этого периода в растительном покрове болота господствовали фускум-
сообщества.
В начале атлантического периода торфяник представлял собой фускум-
залежь. заключительным этапом развития которой часто является
денудационный комплекс. С середины AT большую роль в растительном покрове
начинает играть сосна и пушица. Слой торфа, отложившийся в АТ-2, по
своим свойствам напоминает пограничный горизонт. Для суббореального
периода характерно появление Sphagnum magellanicum и S. angustifolium и
уменьшение роли сосны и пушицы. Субатлантический период
характеризуется увеличением роли S. magellanicum и S. angustifolium при почти полном
отсутствии сосны. На примере Локотенского болота видно, что для развития
болот большее значение имеют эндогенные факторы.
КОРОТКОПЕРИОДНЫЕ КОЛЕБАНИЯ ОТОРФОВАННОСТИ
ЗАБОЛОЧЕННЫХ ЛЕСОВ
С.Э. Вомперский, М.И. Вомперская, Н.А. Валяева, М.Ю. Зазнобин
Институт лесоведения РАН, Московская обл.
Мелкооторфованные земли (представленные преимущественно
заболоченными лесами) в избыточно увлажненной бореальной зоне
распространены значительно больше собственно болот (со слоем торфа более 30 см).
Оценка вклада заболоченных лесов в углеродный цикл биосферы, насколько
нам известно, не исследована. Между тем, принято считать [3,6,2], что в
подавляющем большинстве для таких экосистем характерна обратимость
процессов заболачивания — разболачивания, соответственно колебаниям
погоды или хозяйственным мерам (рубки, лесоводственные мероприятия).
Причем такой взгляд подкреплялся только наблюдениями за динамикой
травяно-мохового покрова, который отражает не только изменение условий
увлажненности, но главное, не позволяет количественно оценить
направленность и интенсивность болотообразовательного процесса.
42
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
В настоящей работе излагаются результаты прямого наблюдения за
запасом органического вещества в торфянистом горизонте заболоченных
лесов в интервале с 1963-65 по 1998 г. Увеличение его рассматривалось как
доказательство повышения увлажненности почвы и роста торфонакопления,
а изменение запасов органического вещества — как показатель разболачи-
вания и усиления лесообразовательных процессов.
Изучались производные от ельников березняки на
торфянисто-подзолистых суглинках в естественном состоянии и подвергнутые реконструкции
(рубка березы, поверхностная гидромелиорация, закультивирование елью), а
также сосняки на торфянисто-сильноподзолистых глеевых песчаных почвах.
В обоих случаях в профиле почвы на глубине 0,5-1,0 м имелся практически
водоупорный горизонт. Типы леса: березняки (ельники) чернично-сфагно-
вые. ивово-долгомошные и долгомошные, а также сосняки долгомошные.
Исследования велись на 4-х участках, удаленных друг от друга от
нескольких до десятков километров, на которых было 6 постоянных пробных
площадей. Более подробные сведения об этих объектах приводились ранее [1].
Методика работы предполагала обеспечение статистически достоверных
оценок разности запасов органического вещества в торфянистом слое в
начале и конце исследуемого периода на одних и тех же площадях. Это
достигалось: 1) определением мощности торфянистого горизонта, объемной
массы большим почвенным пробоотборником (стальной цилиндр 0 20 см) и
зольности подгоризонтов, 2) небольшим размером постоянных площадей
(0,05-0,1 га), с которых брались почвенные образцы с повторностью в
каждом сроке от 12-18 до нескольких десятков, 3) участием одних и тех же
исследователей в начальной и конечной серии работ (через 22-30 лет),
исключившим ошибки несовпадения изучаемых участков или влияния
неодинаковой техники определений.
Исследованный регион относится зоне избыточного увлажнения, но с
нередким наличием лет близких к нормальному или даже недостаточному. На
плакорах и слабых склонах мозаично размещенные мезо- и микроучастки
(разной формы и площадью от долей до нескольких га) оказываются
подверженными наиболее переменному режиму заболачивания — разболачива-
ния. Это обуславливается тем, что в дождливые годы водный баланс
каждого микроучастка в значительной мере определяется, помимо фсадков,
транзитным и боковым притоком-оттоком поверхностных и внутрипочвенных
вод, которые создают временный избыток воды, стимулирующий процессы
торфонакопления. Решающую роль здесь играют локальные микроводосбор-
нью уклоны поверхности, наличие слабопроточных понижений,
механический состав почв близость водоупора и его «рельеф» [4]. В годы
нормального или недостаточного увлажнения в таких микро- и мезоучастках
дополнительный источник увлажнения отсутствует, ранее накопленный торфяни-
43
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
стый горизонт, разлагаясь, становится источником эмиссии С02. Такой вод*
ный режим и проявления процессов заболачивания свойственны многие
лесам таежной зоны. В районе наших исследований в бассейне р.Нахта, ела*
гающие ее водосборы до 1/5 представлены торфянистыми почвами [5].
10 -,
8 i
СО
О
б н
СО 2
о х
» 3
s о
" g
5
3
5 2
'S
X
к 3
X О
' 2
е
§
с!
S
Уклон поверхности почвы:
0.0035 0.0050 0.0035 0.0050 0.0050 Q.0050
I 1 начало опыта; Доверительная вероятность разности запасов ОВ ** - 0.95;
■Вконец опыта; ***-0,99; I-IV - номера участков
Щ Запас в 1998 г. после пожара 1973 г.
Изменение запасов обеззоленного органического вещества (ОВ) в торфянистом
горизонте почв заболоченных лесов за 2-3 десятилетия в естественных условиях
Анализируя полученные данные, частично представленные на рисунке,
можно сделать следующие выводы:
1. За последние 2-3 десятилетия заболачивание ельника на тяжелых
суглинках при уклонах поверхности почвы 0,0035 и менее достоверно
увеличили запас органического (обеззоленного) вещества торфа со среднегодичны-
ми темпами 22-68 гм"2год-1. При больших уклонах (0,0050) достоверного
увеличения заболачивания и роста торфяного слоя не обнаружено (см. рис.).
2. Сосняки долгомошные за этот же период и при уклонах 0,0050
показали достоверное увеличение запаса органического вещества торфянистого
слоя (72 гм^год'^.Возможно, это связано с несколько меньшим
задержанием осадков сосняками в сравнении с ельниками, а также несколько более
влажным периодом наблюдений в первых.
44
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
3. Особенно большой прирост грубой органической массы — очеса из
^кушкина льна — 255 гм^тод'1 обнаружен после пожара (1973 г.) в забо-
доченных долгомошных сосняках. Это соответствует теоретическим
представлениям о повышенной скорости накопления органического вещества
при первичном болото- и почвообразовании. Имело значение также
отсутствие задержания осадков древесным пологом в первые годы, а возможно, и
экологическая приуроченность кукушкина льна к пожарищам.4.
Неизменность запаса органического вещества в лучше дренированных и менее отор-
фованных ельниках долгомошных по сравнению с ельниками чернично-
сфагновыми, ивово-долгомошными и сосняками долгомошными можно
объяснить двумя причинами. Во-первых, невысокой в среднем избыточной
влажностью климата последних двух-трех десятилетий района исследований
(за май — сентябрь коэффициент увлажнения 1,11-1,15); относительно
более дренированные ельники долгомошные на этом временном интервале с
таким избытком влаги вполне «справились» лучшим у них стоком или
меньшим притоком к ним поверхностных вод. Во-вторых, не исключено
небольшое изменение в запасах органического вещества за период опыта и в
этих типах леса, но принятой повторностью определений его нельзя было
доказать.
5. Можно полагать, что действительные флуктуации углеродного пула
заболоченных почв на более коротких (в несколько лет) фактически очень
влажных периодах и очень сухих — были существенно большими, чем
выявлено нами осреднением для 21 — 30 лет (по разным объектам). Это
позволяет рассматривать заболоченные леса как источник существенных
ошибок в оценке текущего вклада биоты в углеродный цикл, т.к. их роль
подобно другим лесам оценивается без короткопериодных флуктуации пула
углерода торфянистой почвы. Оказалось неожиданным, что торфяный горизонт
заболоченных почв характеризуется сравнительно пониженным, чем торф
болот, содержанием углерода в обеззоленном абсолютно сухом
органическом веществе: в слое АТ1Пот 29 до 40%, а в АТШ (лежащем на
микрорельефном горизонте) — от 24 до 46-51%.
[1] Вомперская М.И. Поверхностное осушение временно переувлажненных лесных
почв. М. Наука. 1989. 133 с. [2] Вомперский С.Э. Лес и болото: особенности
круговорота веществ и проявления биосферной роли // Лесоведение. 1991. № 6. С.54-64.
[3] Кощеев А.Л. Заболачивание вырубок и меры борьбы с ним. М. Изд-во АН СССР.
1955. 167 с. [4] Орлов А.Я. и др. Типы лесных биогеоценозов южной тайги. М.:
Наука. 1974. 226 с.[5] Осипов В.В. и др. Территориальное размещение типов леса в
связи с рельефом и гидрографической сетью // Лесоводственные исследования в
подзоне южной тайги. М. Наука, 1977. С.86-92. [6] Пьявченко Н.И. Лесное
болотоведение. М. Изд-во АН СССР. 1963 г. 192 с.
45
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
КАРТОГРАФИЧЕСКИЕ МОДЕЛИ СОСТОЯНИЯ РАСТИТЕЛЬНОГО
ПОКРОВА СФАГНОВЫХ МЕЗООЛИГОТРОФНЫХ БОЛОТ
О.В. Галанина, Т.К. Юрковская.
Ботанический институт РАН, Санкт-Петербург
В холмистых, преимущественно песчаных, ландшафтах краевых зон
последнего оледенения мезоолиготорофные сфагновые болота являются
постоянным компонентом наряду с озерами. В течение ряда лет мы изучали такие
болота в Архангельской, Ленинградской, Новгородской областях и Карелии.
Они располагаются в небольших, но довольно глубоких (4-8 м)
котловинах и имеют овальную форму; в центре или ближе к одному краю часто
сохраняется первичное озеро. Торфяная залежь сложена в основном травяно-
сфагновыми переходными торфами, которые подстилаются сапропелем. Во
время снеготаяния, они, подобно озерам, собирают весь сток с окружающих
холмов. Растительность образована преимущественно мезотрофными и ме-
зоолиготрофными сфагновыми сообществами, существенно участие в
сложении растительного покрова олиготрофных сообществ, а наименьшая доля
принадлежит сообществам из группы бедных низинных.
Девственные (ненарушенные) болота этого типа имеют четкую поясную
структуру. Их центральную часть (да 75% площади) занимает топь, по
которой иногда разбросаны редкие (5-15%) плоские сфагновые кочки. Здесь
преобладают Scheuchzeria palustris, Carex limosa или Rhynchospora alba, а в
моховом покрове — Gymnocolea inflata, Cladopodiella Jluitans, Sphagneta
cuspidati coll. Топь обычно окружена узкими полосами травяно-сфагновых
(Enophorum vaginatum — S. fallax, E. vaginatum — S. balticum, Trichopho-
rum cespitosum — S. balticum и т.п. с участием участием Andromeda polifo-
lia, Oxycocus palustris, иногда Carex rostrata, С pauciflora), кустарничково-
сфагновых (с Chamaedaphne calyculata, Calluna vulgaris, Sphagnum
angustifolium, S. magellanicum) и сосново-кустарничковых сообществ (с
Pinus sylvestris f. litvinowii и f. uliginosa, Ledum palustre, Vaccinium
ulitinosum, Be tula nana no S. fuscum, S. nemoreum. Вдоль кромки озера
тянется узкая (0,2-0.5 м) полоса разнотравно-осоковых сообществ (Carex
rostrata, реже — Carex lasiocarpa, Rhynchospora alba, Comarum palustre,
Galium uliginosum). На границе болота и леса в олиготрофных сосново-
кустарничково-сфагно-вых сообществах появляются минеротрофные
болотные и лесные виды, иногда заросли ивы.
В рассматриваемом типе ландшафта между лесом и болотом
установились довольно стабильные взаимоотношения. О стабильности в развитии
болот и структуре их растительного покрова свидетельствует и однородность
торфяной залежи. Однако в некоторых районах происходит самоосушение
таких болот. Это отмечается для территорий, испытывающих изостатиче-
46
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
ское поднятие суши (Балтийский щит). В результате самоосушения
окраинные сообщества постепенно надвигаются на центр. Это сопровождается
изменением видового состава с исчезновением топяных видов и появлением
лесных. В результате этого процесса упрощается структура растительного
покрова, исчезает центральная топь и ее место занимают пушицево-сфагно-
вые сообщества ковров, которые прежде представляли переходную полосу
между центром и окраиной. В результате самоосушения происходит резкое
обеднение биоразнообразия. Сфагновые мхи заменяются зелеными мхами.
Явления естественного самоосушения хорошо обнаруживаются при
сравнении разновременных аэрофотоснимков по увеличению количества и высоты
деревьев. Антропогенное воздействие, которое мы наблюдали, заключается
в осушении этих болот или, напротив, в их обводнении. В обоих случаях
усиливается евтрофикация и появляются несвойственные этим болотам
виды. В случае мелиорации нарушается прежде всего поясная структура
растительного покрова. От степени эффективности осушения зависит
изменение биоразнообразия. При недостаточном осушении, а также в первые годы
после осушения увеличивается разнообразие видов и сообществ, при
эффективной мелиорации через несколько лет оно резко снижается.
Обводнение происходит вследствие общего повышения уровня
грунтовых вод в зоне действия плотин вокруг искусственных водохранилищ. При
обводнении почти полностью исчезают олиготрофные сообщества.
Появляются большие и широкие полосы краевого заболоченного леса или краевой
лесной топи с обилием евтрофных болотных и лесо-болотных видов {Сагех
nigra, Calla palustris, Comarum palustre, Eriophorum polystachyon, Equisetum
fluviatile). Значительно увеличивается видовое разнообразие.
Особенность болот, расположенных в котловинах холмистого флювиог-
ляциального ландшафта, заключается в небольших размерах, четком
отграничении от окружающих лесов, малом количестве видов и сообществ,
хорошо выраженном поясном распределении сообществ. Вследствие простоты
структуры, небольшого набора сообществ и малого числа видов сфагновые
топяные болота — удобный объект для картографирования и
моделирования. Растительность всех изученных болот была закартирована и на
основании конкретных карт составлены 3 модели, соответствующие разным
состояниям растительного покрова этих болот — контроль (девственное
состояние) — спонтанное самоосушение — мелиорация. Такие
картографические модели позволяют установить характер и направленность изменений
структуры растительного покрова и прогнозировать эти процессы.
47
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ГОЛОЦЕНОВАЯ ИСТОРИЯ, ФАКТОРЫ И СКОРОСТИ
ЗАБОЛАЧИВАНИЯ СРЕДНЕТАЕЖНОГО СТРУКТУРНО-ЭРОЗИОННО-
ЛЕДНИКОВОГО ЛАНДШАФТА (ЮГ АРХАНГЕЛЬСКОЙ ОБЛАСТИ)
К.Н. Дьяконов, Ю.Г. Пузаченко, Т.А. Абрамова, А.В. Хорошев
МГУ, географический факультет
Цель исследования — решение фундаментальной проблемы ландшафго-
ведения — установление факторов саморазвития и развития локальных
геосистем среднетаежного структурно-эрозионно-ледникового ландшафта (юг
Архангельской области). При этом под саморазвитием понимались
необратимые, векторизованные изменения компонентов природно-территориаль-
ного комплекса (ПТК) в их взаимодействии. Векторизация подразумевает
существование генерального направления развития, скорость которого
может модифицироваться при изменении внешних климатических (во
времени) и геохимических (в пространстве) переменных.
Объектами полевых исследований были сопряженные комплексы
верховых болот, развивающихся в естественных суффозионнных понижениях в
приводораздельной части слабоволнистой равнины и лесных ПТК
окружающих склонов. Восстановлена история развития ландшафта за последние
10 тыс. лет, более детально — за 6500 лет — с начала развития процесса
заболачивания суходолов в первой половине атлантического периода.
Поскольку рельеф выступает как матрица, определяющая
перераспределение тепла и влаги, миграцию химических элементов и через это
опосредованно пространственную структуру и дифференциацию почв и
растительного покрова, то был выполнен сопряженный фрактальный и многомерный его
анализ. Основываясь на данных нивелировки профиля протяженностью
7400 м, с шагом нивелирования 12,5 м, топографических картах и АФС,
доказана разрывно-блочная схема организации структуры территории при
широком диапазоне масштабов. Рельеф в интервале от 25 м до 25 км имеет
строго фрактальную самоподобную структуру, определяемую механизмом
дробления коренных пород на блоки. Структура фундамента определяет
строение чехла многочленных четвертичных отложений. Последние, имея
небольшую мощность (5-30 м) почти не экранируют морфоскулыпуру
коренных пород.
Расчет регионального влагооборота в современной климатической
системе в условиях ослабленного дренажа создает предпосылки для развития
заболачивания по верховому типу. При норме годовых осадков 680-700
мм/год, слой стока — 270-290 мм/год; испарение, рассчитанное по методу
теплового баланса, составляет 385 мм/год.
На основе 14 датировок по радиоуглероду подтверждена гипотеза
зависимости вертикальной скорости накопления торфа от размера болота. Для
48
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
дзух изолированных болот с существенно различными размерами возраст
заболачивания установлен в 6500-6350 лет, при мощности торфа 96-100 см
(«большое» болото) и 85-88 см («среднее» болото).Скорость накопления
т0рфа скорее всего функция размера болот и варьирует от 0,14 до 0,5
мм/год. Наибольшие скорости роста отмечены у молодых лесных болот,
имеющих возраст 500-700 лет. Установлено, что «среднее» болото длиной
500 м и шириной 150 м растет в среднем со скоростью 10 м за 100 лет.
«Большое» болото — со скоростью около 6 м за 100 лет.
Анализ зависимости мощности торфа на болотах и органогенного
горизонта с учетом его возраста показывает, что торфянистые почвы являются
элементом единого генетического ряда общего заболачивания.
Распределение 23 видов сфагновых мхов согласуется со стадиями заболачивания и
гидрохимическими условиями в конкретной точке. Статистический анализ
связи размера торфяного и органогенного горизонта с формами микро-, ме-
зо- и макрорельефа, механическим составом почвообразующих пород и рН
почвы показывают, что эти переменные на 90% описывают все
пространственное варьирование органогенного горизонта. При этом наибольшее
влияние оказывают формы рельефа с линейными размерами около 500 м.
Большие значения рН замедляют развитие органогенного горизонта. Суглинкам,
при прочих равных условиях, соответствует большая мощность Ат, пескам
— меньшая.
Разнообразие типов болот от травянистых до грядовых верховых
определяется мозаикой щелочно-кислотных условий. Многократными
измерениями рН грунтовых вод и почв по трансету (1994-1998 гг., летние и осенние
серии), глубины вскипания почв и наличия щебня мергелей и карбонатных
новообразований установлены закономерности пространственной
изменчивости щелочно-кислотного фона и условий в почвах, которые
характеризуются высокой контрастностью как в латеральном, так и радиальном
направлениях. Амплитуда колебаний во времени достигает 2 единиц.
Скорость торфонакопления резко увеличивается при отрыве болота от
линзы минерализованных грунтовых вод.
Выявлены «правила» формирования пространственной структуры
современного растительного покрова. Главное: в' условиях близкой разгрузки
грунтовых вод и близком залегании карбонатов распространены сообщества
с неморальным типом травяного покрова.
Полученные материалы по спорово-пыльцевому анализу позволили
восстановить историю развития ландшафта с 9700 лет. В этот период, как и
всюду на Севере Русской равнины, существовали лесотундровые
ландшафты. Затем наступил криоксеротермический период, маркируемый слоистыми
лессовидными суглинками. Постепенно лесотундровые ландшафты
сменялись северотаежными. Более детальная история восстановлена начиная с
49
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
6500 лет. Опорный разрез, характеризующий динамику растительности в
окрестностях «среднего» переходного болота, находящегося в контакте с
зоной разгрузки сильно минерализованных вод, в субатлантике (SA-1, 2500*
2700 лет назад) выявляет небольшое участие широколиственных пород (дуб^
липа, лещина и даже граб). На «большом « болоте пыльца широколиствен^
ных пород не обнаружена. Спектр пыльцы более однообразен и характера
зует условия южнотаежных ландшафтов в атлантический период, среднего
ежные — в суббореале. Таким образом, в полном соответствие с современч
ной структурой растительности неморальные виды широколиственных лесо^
в AT и S А скорее всего как и в настоящее время в первую очередь связаны ^
наиболее богатыми карбонатными почвами.
Выявленная самоподобная иерархическая организация территории, фак*
торы, механизмы и скорости природных процессов в настоящее время и 4
голоцене, позволяют с достаточной детальностью восстановить прошлук*
структуру ПТК от уровня урочищ до региона и предсказать возможные
траектории развития ландшафта в будущем.
Гранты РФФИ 96-05-65495, 99-05-65097.
БОЛОТНЫЕ ГЕОСИСТЕМЫ ЦЕНТРАЛЬНОЙ МЕЩЕРЫ И ИХ
ВЗАИМООТНОШЕНИЯ С ПРИЛЕГАЮЩИМИ ТЕРРИТОРИЯМИ
А.И. Иванов
МГУ, географический факультет
Болотные геосистемы (ГС) являются неотъемлимой частью ландшафтов
Центральной (Озерной) Мещеры. На территории Центрально-Мещерского
полигона кафедры физической географии и ландшафтоведения, где с 1977 г.
проводятся полустационарные исследования, заболоченность достигает 40%.
На рассматриваемой территории выявлено семь видов болотных ГС: олиго-
трофные а) древнетермокарстовые котловины с редколесными сфагновыми
сосняками; мезотрофные б) древнетермокарстовые западины с сосново-бе-
резовыми редколесьями травяно-сфагновыми; в) западины в днищах
ложбин стока с сабельниково-осоково-сфагновыми сообществами, закустарен-
ными ивой; евтрофные г) западины с черноолынаниками крупнотравными;
д) с березово-ивняковыми болотнотравными мелколесьями; е) древнеозер-
ные западины с осоковыми сообществами; ж) озерные равнины (поймы) с
сабельниково-осоковыми и березово-ивняковыми сообществами.
Взаимодействие болотных ГС с прилегающими территориями
представляет одну из интереснейших и вместе с тем недостаточно разработанных
задач современной науки. Судя по имеющимся в литературе данным,
господствующей тенденцией в бореальном поясе в голоцене является прогрес-
50
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
сйрующее развитие болот и расширение их площадей. Наряду с этим
известны случаи регрессии болотных массивов вследствие изменений
климата, поднятия территории, усиления эрозионных процессов, самоосушения и
др. В наших исследованиях мы исходили из того, что рассматриваемый 20-
летний хроноинтервал недостаточен для выявления непосредственных
результатов взаимодействия болот с прилегающими территориями, но в ряде
случаев может выявить тенденции развития. Наряду со стационарными
наблюдениями в реальном времени, использовались и другие методы: эколого-
флористический анализ подроста и древостоя и их состояние, степень
разложения подстилки, соотношение процессов подзолообразования и торфо-
накопления, видовой состав фитоценозов и др. Исследования проводились в
контактных зонах разных видов болот, охарактеризованных ранее.
В контактной зоне «лес-болото» анализировалось 12 площадок. На пяти
из них за рассматриваемый хроноинтервал зафиксированы изменения,
которые можно интерпретировать как усиление процессов заболачивания. К
ним относятся выраженная многолетняя тенденция уменьшения разницы
между УГВ на болоте и в лесном приболотье (этот процесс особенно
выражен с начала 90-х гг. и связан с общим поднятием УГВ на полигоне в этот
период ), увеличение в составе фитоценозов доли сфагновых мхов и
специфических болотных видов трав (тростник, осока черная и др.), ухудшении
состояния подроста и т.д. На трех площадках изменения выражены слабее (в
пределах точности наблюдений). На четырех площадках изменения в
структуре ГС не зафиксированы. При этом, граница между лесом и болотом на
всех площадках оставалась неизменной.
В контактной зоне «луг-болото» анализировалось четыре площадки.
Исследовались луга низкого уровня, непосредственно окаймляющие болотные
массивы. В качестве основного индикатора процессов в контактной зоне
«луг-болото» использовалась экологическая шкала увлажнения Л.Г. Рамен-
ского. Все луга представляют собой щучники с различными содоминантами.
На трех луговых площадках, окаймляющих евтрофные осоковые болота,
динамика значений шкалы увлажнения носит флуктуационный характер, без
направленных изменений. На одной из площадок, находящейся в нижней
части древней ложбины стока, также начиная с 1990 г. прослеживается явно
выраженная тенденция увеличения влагообеспеченности (среднемноголет-
нее значение шкалы увлажнения 79,5, за 1991-97 гг. — 84,5). Одновременно
здесь произошло усложнение горизонтальной структуры растительности: на
месте исходной калганово-щучковой ассоциации образовалось четыре с
преобладанием осоки черной, сабельника болотного и таволги вязолистной.
Совокупность изменений свидетельствуют о тенденции к заболачиванию.
Специфика взаимоотношений болот с прилегающими территориями во
многом определяется ландшафтной структурой. Наиболее динамичны рас-
51
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
пространенные во всех ландшафтах неглубокие древнетермокарстовые
западины с сосново-березовыми травяно-сфагновыми редколесьями, где чаще
всего фиксировались случаи наступления болот на прилегающие террито*
рии. В зандровых ландшафтах, где олиготрофные болота находятся
преимущественно в древнетермокарстовых котловинах, процесс
горизонтального роста болот имеет место, но лимитируется относительно большим углом
падения болотных впадин и поэтому носит характер языков. В озерно-
зандровых ландшафтах с преобладанием евтрофных осоковых и березово-
ивняковых болот на древнеозерных и озерных равнинах границы между
болотными массивами и прилегающими территориями относительно
стабильны. В долинно-зандровых, где преобладают мезотрофные болота по
днищам ложбин стока, этот процесс, вероятно, носит колебательный
характер и определяется водоудерживающим потенциалом ландшафтов.
Главным фактором, определяющим многолетнюю динамику в условиях
Центральной Мещеры, является изменение влагообеспеченности ГС.
зависящее от количества осадков и водоудерживающего потенциала
ландшафтов. Последний в основном регулируется болотами. Основной природной
тенденцией в регионе является заболачивание территории, фиксируемое как
непосредственно стационарными исследованиями, так и косвенными
методами. В более длительном хроноинтервале основные природные тенденции
развития будут определяться изменением увлажненности территории
вследствие климатических изменений.
РЕКОНСТРУКЦИЯ ЛЕСНОЙ И БОЛОТНОЙ РАСТИТЕЛЬНОСТИ
ЗА ПОСЛЕДНИЕ 9500 ЛЕТ СРЕДНЕТАЕЖНОЙ ЧАСТИ
ПРИЕНИСЕЙСКОЙ СИБИРИ
Л.В. Карпенко
Институт леса им. В.Н. Сукачева СО РАН, Красноярск
Представлены результаты палинологического и стратиграфического
анализов 7 торфяных разрезов, мощность торфа которых 4,0-5,0 м, заложенных
в среднетаежной части Приенисейской Сибири (междуречье Сыма и Дубче-
са. левых притоков Енисея). Реконструированы сукцессии растительного
покрова болот и окружающих их суходолов в изученном регионе на
протяжении голоцена — 9500 лет назад.
Первые пять разрезов, условные названия которых «Хойбаж
«Лебединое-1», «Лебединое-2», «Никулино-1», «Никулино-2», находятся в
северной части исследованного района, в междуречье Нижней и Дубчеса,
левых притоков Енисея (в Тугуланской котловине). Болота образуют здесь
единую систему, покрывая сплошным плащом надпойменные террасы Ени-
52
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
сея. Центральная часть котловины занята в основном сильно обводненными
грядово-мочажинными и грядово-мочажинно-озерными комплексами.
Песчаные островки среди болот покрыты лишайниковыми борами олиготроф-
ного ряда. Следующие два разреза («Мочажина» и «Бугор») заложены на
верховом болотном массиве, находящемся в южной части района
исследований (в долине Енисея) у пос. Кривляк. Болото покрыто олиготрофными
кустарничково-сфагновыми рямами и грядово-мочажинными комплексами.
Благодаря комплексному анализу стратиграфии торфяной залежи, споро-
во-пыльцевым спектрам и датированию торфяников по 14С, удалось
реконструировать следующие сукцессии растительного покрова болот и
окружающих их суходолов в изученном регионе на протяжении голоцена.
Предбореальный период отмечен в разрезе «Хойба». В первой его
половине были развиты елово-лиственничные и березовые редколесья с полын-
но-разнотравными группировками растительности. Во второй — сосново-
березово-еловые леса с папоротниково-хвощово-разнотрав-ным
напочвенным покровом. Период формирования елово-лиственничных лесов
совпадает с большой обводненностью болотных массивов. Образование их началось
с евтрофной стадии. Ботанический состав торфа и спектр пыльцы трав
указывают на преобладание во флоре болот Equisetum palustre, Menyanthes
trifoliata, Car ex chordorrhiza, C. diandra, С caespitosa и др., а также Thy pa
latifolia, Calla palustris, Comarum palustre, Naumburgia thyrsiflora.
Бореальный период выделен в разрезах «Хойба», «Лебединое-2»,
«Бугор», «Никулино-2». В начале периода на суходолах, окружающих
болотные массивы, были развиты березовые и березово-еловые леса, в
напочвенном покрове которых господчгвовали осоки, вересковые кустарнички,
сфагновые и гипновые мхи. Еловые и пихтовые зеленомошные леса были
сконцентрированы, вероятно, в речных долинах и на склонах водоразделов,
преимущественно теплых экспозиций. Судя по возрастанию участия
древесной пыльцы, во второй половине периода увеличилась полнота автоморф-
ных лесов. Значительная доля появившегося в спектре кедра, а также ели и
сосны свидетельствует о начале формирования тайги. Болота этого периода
— преимущественно грунтового питания. Во болотной флоре первой
половины периода продолжали господствовать растительные группировки из
хвоща, вейника, осок, гипновых и евтрофных сфагновых мхов. В конце
периода почти повсеместно происходит смена этих группировок мезотрофны-
ми. Растительный покров болот образовывали Scheuchzeria palustris,
Eriophorum vaginatum, Carex lasiocarpa, C. inflata, C. limosa. а также
сфагновые мхи Sphagnum apiculatum, S. magellanicum, S. Dusenii и др.
В первой половине атлантического периода, как свидетельствует состав
спорово-пыльцевого спектра всех исследованных разрезов, доминировали
березовые леса с примесью ели, сосны, и незначительным участием пихты.
53
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Вторая половина периода характеризуется значительным сокращением
пыльцы березы в составе спектров и значительным увеличением
содержания пыльцы кедра и сосны. Содержание пыльцы пихты составляет ее
максимум. Следовательно, березовые лесные формации уступили место
соснякам и кедровникам. Происходит заметное увеличение роли пихты, которая,
возможно, образовывала чистые насаждения. Ель и пихта могли входить в
состав кедровых насаждений, которые были распространены не только в
речных долинах, но и на междуречьях. В напочвенном покрове —
господство осок, разнотравья, хвощей и папоротников. Гидрологический режим
болот был грунтово-атмосферным, что вызвало смену растительного
покрова на них. Доминирующими растительными группировками этого времени
были мезотрофные травяно-сфагновые, шейхцериево-сфагновые, сосново-
кустарничково-сфагновые и сосново-шейхцериево-пушицево-сфагновые.
Спорово-пыльцевые спектры суббореального периода всех разрезов
свидетельствуют о господстве на плакорах темнохвойной тайги с
преобладанием кедра. Ель, береза и пихта имеют второстепенное значение в лесообразо-
вании. В напочвенном покрове были развиты вересковые кустарнички,
злаки, разнотравье, сфагновые и гипновые мхи.
В субатлантическом периоде уже сформировались ландшафты, близкие
современным. Для пыльцевого спектра древесных характерно вытеснение
пыльцы березы с лидирующих позиций, преобладание пыльцы сосны и
кедра, что свидетельствует о господстве сосновых и кедровых суходольных и
заболоченных лесов. Темнохвойные леса из ели и пихты не имели прежнего
распространения. Повышенные участки рельефа с суглинистыми почвами
были заняты березовыми травяно-зеленомошными лесами. В понижениях
рельефа были распространены березовые заболоченные (осоковые и осоко-
во-сфагновые леса). Болота перешли в олиготрофную фазу своего развития
(свидетельством чему является стратиграфия торфяной залежи всех
разрезов), началось формирование бугорково-мочажинных и грядово-моча-
жинных комплексов. Растительность их была представлена Betula nana,
Chamaedaphne calyculata, Eriophorum vaginatum, Scheuchzeria palustris,
Carex limosa, а также сфагновых и зеленых мхов — Sphagnum fuscum, S.
cuspidatum, S. balticum, Pleurozium Schreberi, Polytrichum srtictum.
УСТОЙЧИВОСТЬ БОЛОТНЫХ СИСТЕМ
B.H. Кирюшкин
РГПУ, Санкт-Петербург
Структурно-антропогенные преобразования болотных систем тесно
связаны с их устойчивостью. В.В. Куликов [1976] рассматривает устойчивость
54
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
как свойство геосистемы активно поддерживать значение своих параметров
в пределах, не превышающих критических величин, и сохранять
определенный характер функционирования, проявляющийся при воздействии на них
возмущающих факторов. К.Е. Иванов [1975] устойчивость биофизической
системы определяет как степень сохранения ее структуры, если под
воздействием внешних и внутренних факторов система претерпела существенные
изменения и отклонения от нормального состояния.
Устойчивость болотных систем зависит от сложности и слитности
болотных систем и от величины соприкосновения с внешней средой. В результате
произведенных исследований установлено, что устойчивость компактных
(слитных) и рыхлых (слабокислых) болотных систем различна по
отношению к антропогенным и природным воздействиям.
Анализ аэрофотоснимков и фондовых материалов за 30-летний период
позволяет сделать вывод, что рыхлые слабослитные системы менее
устойчивы под воздействием природных возмущающих факторов. Это
объясняется тем, что величина контактной зоны болотных урочищ, входящих в
системы, незначительна. Чаще всего наблюдается точечный контакт, что
позволяет сохранять болотным урочищам структурную автономию. Контактные
(слитные) системы в аналогичных условиях могут длительное время
сохранять свое функционирование.
Противоположная картина наблюдается при антропогенных нагрузках,
где высокоорганизованные компактные болотные системы менее устойчивы,
чем рыхлые. Первые развиваются как одно общее целое и нагрузки,
приложенные в какой-то части болотной системы, передаются на значительные
расстояния. В рыхлых болотных системах составляющие их структурные
единицы могут сохранять длительное время свою индивидуальность.
Антропогенные нагрузки приводят лишь к локальным структурным
нарушениям, мало изменяя общую схему функционирования болотных образований.
При осушении какой-то части болотной системы в целом не происходит
заметных изменений в развитии ее динамичных структур.
ХАРАКТЕРИСТИКА ПРОЦЕССОВ ЗАБОЛАЧИВАНИЯ МЕЛКОВОДИЙ
НА ВОДОХРАНИЛИЩАХ ВЕРХНЕЙ ВОЛГИ "
О. И. Козловская
Институт биологии внутренних вод РАН, Ярославская обл.
Заболачивание мелководий водохранилищ — динамический процесс,
заключающийся в изменении во времени видовой структуры и биоценотиче-
ских процессов. Биоценоз и его отдельные компоненты формируют виды,
Дополняющие друг друга и соответствующие другим видам и условиям сре-
55
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ды сами по себе и в складывающейся совокупности. При этом происходит
выработка экологических ниш — функциональное размежевание в
возникающем множестве или его дополнение при малом разнообразии. На жиз-
непригодном субстрате биотические сообщества последовательно
формируют закономерный ряд биосистем, ведущий к наиболее устойчивой в данных
условиях фазе: климаксовой в чисто природных условиях или узловой при
природно-антропогенном режиме (принцип сукцессионного замещения).
В заболачивании и развитии растительного покрова искусственных
водоемов выделяется несколько этапов, плавно переходящих один в другой
[Калинина, 1945; Зеров, 1967; Довбня, Экзерцев, 1988; Корелякова, 1989;
Экзерцев и др., 1990; и др.]. На каждом этапе растительность, развиваясь,
готовит условия и среду для наступления следующего этапа. Характер этого
процесса искусственных водохранилищ определяется комплексом
природных и антропогенных факторов: уровенный режим, рельеф береговой линии
и морфологические особенности дна, трофические условия, грунтовые воды,
характер исходных биотопов и грунтов и др. [Калинина, 1945; Долгов, 1948;
Шмелева, 1954; Потапов, 1959, 1960; Экзерцев, 1963; Белавская, Кутова,
1966; Зеров, 1967; Корелякова, 1989]. Влияние данных факторов может
изменяться от года к году в зависимости от хозяйственной деятельности
человека, что сказывается на развитии растительного покрова искусственных
водоемов.
Первый этап характеризуется интенсивным зарастанием вновь
затопленных территорий, значительным участием кратковременных и пионерных
группировок, вспышками развития свободноплавающей и погруженной
растительности. Он охватывает период наполнения водохранилищ и первые 5-8
лет их существования. Для него типичны абразионные и эрозионные берега,
среди грунтов преобладают первичные, трансформированные песчаные или
глинистые. По мере заиления грунтов разреженные группировки растений
приобретают вид рассеянно-пятнистых зарослей, формирующиеся прежде
всего в заливах, затопленных устьях рек, на защищенных мелководьях. Для
фитоценозов характерна одноярусность, низкая флористическая
насыщенность, неустойчивый видовой состав. Чем сильнее изрезана береговая линия,
тем интенсивнее идет процесс зарастания. На верхневолжских
водохранилищах на данном этапе в первые 2-3 года благодаря наличию в водах
водохранилища большого количества биогенных элементов и обилию диаспор
наблюдалось развитие на защищенных мелководьях нитчатых водорослей,
свободноплавающих и погруженных растений Elodea canadensis, Lemna
trisulca, Ceratophyllum demersum, которые затем вытеснялись
группировками широколистного воздушно-водного разнотравья —Alisma plantago-aqua-
tica, Sagittaria sagittifolia, Oenanthe aquatica [Богачев, 1952; Экзерцев, 1960,
1963, 1972; Экзерцева, 1961; Экзерцев, Экзерцева, 1966].
56
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
Второй этап, продолжающийся 10-15 лет, характеризуется
формированием и распространением относительно устойчивых фитоценозов,
преимущественно зарослевого типа, образованных достаточно мощными в ценоти-
ческом отношении видами растений, но не достигшими однако своих
оптимальных показателей в продуцировании фитомассы; интенсивными
процессами смен, направленных в основном на вытеснение ценотически слабых
сообществ; более медленными темпами зарастания новых мелководий и
меньшим участием растений-временщиков. На верхневолжских
водохранилищах для второго этапа характерна смена широколистного
воздушно-водного разнотравья узколистным — Typha latifolia, Glyceric* maxima, Phragmi-
tes australis. Период характеризуется началом острых конкурентных
взаимоотношений, в результате которых из состава растительного покрова
выпадают «временные» виды и формируются устойчивые, ценотически
мощные сообщества. Растительность представлена, как правило, куртинными,
пятнистыми, реже поясовыми зарослями; четкой поясности не послеживает-
ся. Продолжаются достаточно интенсивно абразионные процессы берегов,
влияющие на развитии растительности. Интенсивно протекают процессы
заиливания мелководий, преимущественно заливов, устьев рек и отшнуро-
ванных мелководий и накопления биогенов в грунтах [Потапов, 1954;
Экзерцев, Экзерцева, 1963а, 1966; Белавская, Кутова, 1966; Экзерцев, 1972].
Третий этап характеризуется господством на значительных площадях
относительно устойчивых, ценотически мощных сообществ, замедлением
процессов смен, отсутствием в растительном покрове группировок
«временной» растительности, слабым зарастанием новых мелководий и
формированием растительного покрова зонального характера. Продолжительность
этого этапа зависит от экологических условий, создавшихся на водоеме. На
водохранилищах со значительным колебанием уровня и крутыми склонами
абразионные процессы берегов еще продолжаются, на водоемах с низкими
берегами и нерезкими колебаниями уровня воды эрозионная активность
прекращается. Данные процессы отражаются на развитии растительности.
На верхневолжских водохранилищах в этот период наблюдается
доминирование узколистной воздушно-водной растительности. Создается огромная
фитомасса, большая часть которой отмирая, остается в водоеме, приводя к
накоплению в литорали илов растительного происхождения. Накопление
большого количества органики подготавливает среду для эндогенных смен
одних сообществ другими [Экзерцев, 1963, 1973; Экзерцев, Довбня, 1974].
Четвертый этап проявляется в развитии процессов заболачивания и
образовании сплавин. Наиболее часто наблюдаются следующие варианты
сплавинообразования: отчленение отдельных кусков старых болотных
торфяников, отдельных участков тростниковых, рогозовых и др. торфяников,
образование сплавин за счет нарастания обильной, слабо разлагающейся
57
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
покрова дигрессивного характера. Биогеоценозы остаются по-прежнему бо-
лотными, т.е. относятся к категории переувлажненных земель, причем тщ.
рологический режим становится еще более контрастным из-за потери
сфагнового яруса, а из растительного покрова исчезают самые ценные в
хозяйственном отношении ресурсы — сфагновые мхи, а с ними и клюква и редкие
охраняемые и неохраняемые виды из семейства орхидных: хабенария, скру-
ченник китайский. С потерей хозяйственного значения эти экосистемы
утрачивают свою биосферную роль, как накопители углерода, поскольку
деградация сфагнового покрова приводит к прекращению торфообразования.
Между тем, биосферная роль приамурских коренных сфагновых болот
весьма велика и по вкладу не уступает лесным экосистемам. По данным
наших исследований общие запасы органического вещества в этих
экосистемах оцениваются в 150-170 т/га (фитомасса + фитодетрит), но это без
торфа. Если принять среднюю мощность торфа на этих болотах за
минимальную (0,63 см [4]), то запас углерода в торфе составит еще около 1 тыс.
т/га [5]. Для сравнения, запас органического вещества лесной экосистемы
Среднего Сихотэ-Алиня от кедровника багульникового на подзоле сухотор-
фянистом (олиготрофная система) до кедровника с липой и дубом на буро-
земо-дерновой почве (мезо-мегатрофная система) составляет от 186 до 485
т/га (фитомасса + фитодетрит + гумус) [6]. Запас органического вещества
кустарничково-травяного и кустарникового (ерник) ярусов мезотрофного
болота (пирогенная смена) колеблется от 27 т/га в условиях переувлажнения
до 56 т/га в сухих условиях (фитомасса + фитодетрит, без гумуса) [1].
Коренные гетеротрофно-мезотрофные кустарничково-сфаг-новые
сообщества очень хорошо адаптированы к чрезвычайно контрастным условиям
увлажнения Приамурья. Оптимумы продуктивности основных их
продуцентов — доминантов кустарничково-травяного и мохового ярусов находятся
на разных концах градиента увлажнения. При резкой смене
гидрологических условий продуценты меняются ролями и снижение продукции одного
компонента фитоценоза компенсируется увеличением продукции другого.
Что нельзя сказать про безмоховые осоково-кустарничковые ерники. В
контрастных условиях увлажнения Приамурья величины годичной продукции
сосудистых растений болот могут различаться в 3 раза в надземной части и
в 7-40 раз в подземной.
В ходе пирогенных смен трансформация растительности коренных гете-
ротрофно-мезотрофных кустарничково-сфагновых болот,Приамурья
приводит к уменьшению их продуктивности, значительному снижению
биоразнообразия и необратимой потере хозяйственной ценности. Экосистемы
фактически утрачивают свою уникальную роль накопителя углерода и
производителя кислорода в биосфере.
[1] Копотева Т.А. Экзогенные сукцессии на болотах Приамурья // Ботан. ж. 1995. Т.
60
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
ЯО № 5, С. 68-73. [2] Костырина Т.В. Динамика экстремумов пожароопасных
периодов (на примере Хабаровского края) // Геогр. и природа, .ресурсы. 1983, №3. С.
115-120. [3] Куренцова Г.Э. Растительный покров Приуссурийской части бассейна
Среднего Амура. Владивосток: Дальневост. книжн. изд., 1965. 72 с. [4] Прозоров
jo с. Закономерности развития, классификация и использование болотных
биогеоценозов. М.: Наука, 1985.208 с. [5] Трансформация мелиорированных почв торфя-
лых почв в Приамурье. Владивосток: Дальнаука, 1995. 125 с. [6] Утенкова А.П.,
Мотузова Г.В. Особенности биогеохимии природных экосистем Центр. Сихотэ-
длиня // Почвоведение. 1993. № 5. С. 5-13. [7] Шлотгауэр С.Д. Антропогенная
динамика растительности Хабаровского края // Вестн. дальневост. отд. РАН. 1993. №
6. С.84-91. [8] Grosvenier P., et al. Growth potential of three Sphagnum species in
relation to water table level and peat properties with implications for their restoration in
cut-over bogs // J. Appl. Ecology. 1997. V. 34. P. 471-483.
ЗАБОЛАЧИВАНИЕ ТЕХНОГЕННО ТРАНСФОРМИРОВАННЫХ
ПОЙМЕННЫХ ВОДОЕМОВ В ЮЖНОЙ ТАЙГЕ
Е.Г. Крылова
Институт биологии внутренних вод РАН, Ярославская обл.
Эволюция пойменных водоемов идет в сторону их прогрессирующего
заболачивания и заторфовывания, чему посвящена обширная литература. Эти
процессы ускоряются на техногенно трансформированных пойменных
водоемах. Один из типов последних представляют озера, оказавшиеся в зоне
инженерной защиты водохранилищ, созданных на реках. Таких водоемов в
европейской России немного, но они интересны в научном и практическом
отношении. Нами исследовались сукцессии растительности пойменных озер
Волги, расположенных в зоне инженерной защиты Горьковского
водохранилища. От последнего они отделены дамбой и другими гидротехническими
сооружениями с целью сохранения ценных в сельскохозяйственном и
историко-культурном отношении земель. До создания водохранилища вся пойма
с расположенными на ней озерами ежегодно находилась под влиянием
аллювиального режима. Фактор аллювиальности в значительной мере
ослаблял, а иногда и подавлял болотообразовательный процесс. После того, как
водоемы вышли из-под влияния аллювиального режима, болотообразование
активизировалось и стало протекать по ускоренной схеме. Однако скорости
зарастания водоемов различные и в этом отношении их можно разделить на
следующие группы:
1. Озера притеррасного и собственно пойменного залегания. Они
занимают почти 90% всей площади озер района. Зарастают по смешанном}' типу
с участием комплекса разных по экологии и сложению сообществ —
погруженных, плавающих и воздушно-водных: Ceratophylletum demersi, Elodee-
tiuv canadensis, Stratiotetum aloides, Nymphaetum candidae, Nupharetum lutei,
61
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Scirpetum lacustris, Typhetum latifoliae и др. Структура зарастания
отличается сложностью: пояса в прибрежьях, часто сложенные ценозами воздушно-
водной растительности; на остальной акватории — мозаика сообществ
самой разной физиономии — воздушно-водной, плавающей на поверхности
воды и погруженной растительности. Отличительной особенностью этих
озер является интенсивное развитие сплавин, что объясняется характером
грунтов, представленных сильно разжиженными сапропелями,
затрудняющими укоренение растений. В сложении сплавин, кроме типичных сплави-
нообразователей — Calla halustris, Comarum palustre, Menyanthes trifoliata,
участвуют также Турка latifolia, Carex pseudocyperus, Bidens cernua,
Phalaroides arundinacea и др. В этих озерах следует ожидать усиления
зарастания, тенденция к которому наметилась.
2. Малые по площади водоемы центральной части поймы с
незначительными глубинами. Озера окружены надвигающимися болотами с
преобладанием сообществ Caricetum acutae. Подвержены более интенсивному
зарастанию с преобладанием сообществ Stratiotetum aloides, Ceratophylletum de-
mersi, Nypharetum lutei, LemnoSpirodeletum polyrhizae, Elodeetum
canadensis. Следует ожидать почти полное зарастание водоемов этой группы. В
настоящее время степень их зарастания составляет 50-90%. Самые малые
водоемы площадью <1 га, расположенные возле защитной дамбы, полностью
заросли и перешли в стадию активного заболачивания и заторфовывания.
3. Озера речного типа, заходящие в пределы боровой террасы,
отличающиеся крутыми берегами, песчано-илистыми грунтами и глубинами до 5-7
м. Развитие растительности лимитируется глубинами, поэтому она
представлена узкими прибрежными полосами с преобладанием сообществ
Scirpetum lacustris, Phragmitetum communis, Typhetum latifoliae, Eguisetetum
fluviatilis. В озеровидных расширениях значительные площади заняты
ассоциациями Nupharetum lutei, Stratiotes aloides, Nymphaetum candidae,
Elodeetum cana-densis, Ceratophylletum demersi, Potametum pectinati.
Дальнейшее развитие растительности здесь будет лимитироваться и
максимальная степень зарастания не превысит 10-15%. Озерная стадия их развития
будет продолжаться неопределенно долгое время.
Активное зарастание техногенно трансформированных пойменных
водоемов изученного района сопровождается столь же активным их заторфовы-
ванием за счет ускоренного накопления биомассы неразложившихся
растительных остатков. Столь же интенсивно, вследствие отсутствия
аллювиального режима, происходит развитие древесно-кустарниковой растительности
побережий некоторых озер, в основном на данной стадии пока за счет
усиления ценотической роли Salix cinerea.
62
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
ДИНАМИКА БОЛОТООБРАЗОВАТЕЛЬНОГО ПРОЦЕССА В ГОЛОЦЕНЕ
НА СЕВЕРО-ЗАПАДЕ РОССИИ И НЕКОТОРЫЕ ЗАДАЧИ УПРАВЛЕНИЯ
ПРИРОДОПОЛЬЗОВАНИЕМ
Г.Ф. Кузьмин
НИИ торфяной промышленности, Санкт-Петербург
Краткая характеристика заболоченности и заторфованности Северо-За-
пяда России. На Северо-Западе России размещены крупнейшие торфяные
бассейны Европейской части России: Уломский, Полистовский, Волхов-
Тихвинский и др. Заболоченность колеблется от 5% (Приозерский р-н
Ленинградской области) до 60-70% (Поддорский р-н Новгородской области и
ряд граничных с ним районов Псковской области, а также вся западная
часть Вологодской. В целом по областям заболоченность составляет:
Ленинградская — 42%, Новгородская — 40%, Псковская — 46% и западная часть
Вологодской области — 60%. Из них 2/3 площади составляют заболоченные
земли, остальные — месторождения торфа.
Динамика торфообразовательного процесса в голоцене. Торфонакопле-
ние на Северо-Западе России началось примерно 300-ь2000 лет после дегля-
циации территории (-10-12 тыс. л.н.) с заболачивания безлесных
пространств. В бореале большинство первичных водоемов на болотах
прекратило свое существование. В атлантике наблюдалось активное торфонакоп-
ление в болотных мезоландшафтах и бурный процесс образования болотных
систем (БС), сопровождаемый заболачиванием лесов. В суббореале
продолжилось формирование БС, активное наступление олиготрофных
растительных комплексов на евтрофно-мезотрофные. Началось массовое накопление
сфагнового торфа и нормирование внутриболотной гидрографической сети
(мочажины, озерки, озера, ложбины стока и др.). В субатлантику БС
оформились до современного положения и состояния. Продолжается
формирование внутриболотной гидрографической сети и накопление сфагнового торфа,
трансгрессия БС на леса. Активизируется процесс олиготрофизации.
Количественная оценка процесса торфонакопления. Образование болот
на всей территории Северо-Запада началось одновременно 9-10 тысяч лет
назад. Выполнены исследования на ряде БС региона о накоплении торфа,
показавшие существенное отличие его в различные периоды голоцена и в
различных частях БС. Просматривается тенденция к выравниванию величин
накопления торфа по мере развития БС. Вертикальный прирост торфяной
залежи на БС во времени таков: 7,14 мм/год (0-100 лет), 2,29 мм (0-300
лет), 2,28 мм (0-500 лет), 0,97 мм (0-1000 лет), 0,77 мм (0-2500 лет), 0,89
мм (0-4500 лет), 0,62 мм (0-7000 лет), 0,59 мм/год (0-9500 лет).
Современные процессы заболачивания. В настоящее время новые очаги
заболачивания возникают, как правило, только при антропогенном воздей-
63
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ствии. Активен процесс заболачивания по периферии БС. Воды,
попадающие на окраины болот и окружающие леса, имеют кислую среду, низкую
минерализацию, повышенное содержание органики и дефицит кислорода
(весной до 98%, летом-осенью — 54-72%). Стекающие с БС талые воды
аккумулируются окраинами и окружающими лесами и приводят к
существенному уменьшению периода вегетации лесной растительности, тем самым
к существенному уменьшению как прироста биомассы, так и товарной
ценности продукции. В гумидной зоне естественный ход болотообразовательно-
го процесса направлен на максимальное покрытие равнинных территорий
болотами (до 70-80% площади ландшафта), чему активно мешает человек.
Экологическая роль болот. Болота - неотъемлемый элемент всех
ландшафтов лесной зоны, потому их роль в природе лесной зоны, развитии ее
производительных сил значительна. При торфообразовательном процессе
наряду с накоплением больших масс растительных и иных остатков,
происходит накопление многократно больших масс кислых вод, связанных с
этими остатками. Следствием этого является активная трансгрессия болот на
окружающие леса. Торфообразовательный процесс вызывает коренную
перестройку геохимической системы: резкое увеличение подвижности железа,
кремния, алюминия, марганца, цинка, меди, никеля, кобальта, свинца,
создает предпосылки увеличения концентрации этих элементов и в речном
стоке заболоченных водосборов. Меняется тип обмена веществ с кальциево-
азотного на кремниево-азотный; плоскостной смыв минеральных почв
заменяется смывом органики. Отрицательна роль болот и особенно верховых
в формировании речного стока, но в гумидных районах с молодым плоским
рельефом и слабо врезанной речной сетью проявляется определенная
положительная роль болот за счет перевода части атмосферных осадков в
прямой оборот: осадки-испарение+транспирация. В освоенных человеком
районах эту функцию успешно выполняет мелиорация. Необоснованно
завышается роль болот в биосфере, в частности в возможности утилизации С02 в
случае увеличения концентрации С02 в атмосфере.
Баланс накопления и добычи торфа. Проблема воспроизводства торфа.
Новые данные о приросте торфяной залежи [Кузьмин, 1993] позволяют
принять для болот Северо-Запада России среднегодовую величину
вертикального прироста болот в 3-4 мм. Исходя из этой величины прироста и
свободных площадей месторождений торфа (не подверженных осушением)
можно строить политику использования торфа как в отдельных
территориях, так и в регионе (стране) в целом. Баланс добычи и воспроизводства
торфа может быть сохранен при условии восстановления и интенсификации
торфонакопления на части выработанных площадей. Ключом решения
проблемы использования торфа как возобновляемого природного ресурса
является применение принципа компенсационного природопользования.
64
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
При решении задач оптимального природопользования нужно учитывать
двойственный характер месторождений торфа, являющихся одновременно
элементом ландшафта и месторождением полезного ископаемого.
Корректив решения получаются формированием целевых торфяных фондов в
пределах водосборного бассейна.
ГЕНЕЗИС И ФИТОГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ
ЧЕРНООЛЬХОВОГО КОМПЛЕКСА
А.И. Кузьмичёв
Институт биологии внутренних вод РАН, Ярославская обл.
Под черноольховым, или альнетальным комплексом, понимается ранее
выделенная автором [Кузьмичёв, 1992] группа видов, объединенных с
определенного момента их истории общими ценогенетическими связями.
Комплекс начал вычленяться в плиоцене из сборного прадолинного, давшего
впоследствии веер неокомплексов, в частности также и древненеморальный.
Формирование альнетального происходило на фоне эволюции видов ольх
систематического ряда Glutinosae в течение миоцен-плиоцена. К этому ряду,
кроме Alnus glutinosa, относятся географически замещающие A. barbata, A.
serrulata, A. serrulatoides. Ареалы первых двух видов частично
перекрываются и на Кавказе ольха клейкая ценотически замещается ольхой
бородавчатой. В Европе альнетальный комплекс из состава влажных долинных
лесов стал вычленяться вследствие распадения строя долинных лесов, прежде
всего выпадения термофильных древесных видов в ходе общего процесса
бореализации и континентализации климата под влиянием
прогрессирующего охлаждения Полярного бассейна. К началу плейстоцена он имел
современный состав и структуру ценоэлементов, продолжая однако терять в
периоды межледниковий некоторых теплолюбивых спутников.
Окончательно он оформился в голоцене, имея очерченный ареал, дифференцированный
на определенные хорионы. В структуре и эволюции растительного покрова
лесных болот этот флороценотический комплекс сыграл вьщающуюся роль.
На стратиграфических профилях юго-запада Русской равнины болото-
образование часто начиналось с лесной черноольховой стадии. На
отдельных массивах, почти без перерыва, она продолжается и поныне. Это связано
с климатическими и эколого-топографическими условиями данной
территории, благоприятствующими проявлению в наибольшей степени преадапта-
Ционных возможностей вида. Вообще ценотическая часть ареала A. glutinosa
охватывает Полесско-Припятскую и Польско-Германской низменностей.
В этих районах в естественных ненарушенных экотопах ольха черная
формирует высокопродуктивные древостой. Однако в большинстве сущест-
65
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
вующих насаждений вследствие неоднократных рубок она порослевого про.
исхождения и часто растет на пьедесталах, усиливая дифференциацию тра*
вяного покрова. По этой причине характерная ассоциация Alnetum carici
elongatum, связывающая восточноевропейские варианты черноолыпаников
со среднеевропейскими представлена производными ценозами.
В период атлантического оптимума голоцена A. glutinosa продвинулась в
северные районы восточной Европы. Миграционными путями послужила
долины рек, где этот вид был ассимилирован сборным и пока слабо
изученным комплексом уремы и в новых условиях вступил в контакт с
аборигенным A. incana, образовав веер переходных гибридных форм, впоследствии
почти полностью выпавших. Отголоском формообразовательных процессов
являются нетипичные, широко распространенные на севере европейской
России популяции ольхи черной с изрезанными краями листовых пластинок,
совершенно не характерные для Полесско-Припятской низменности. В
северной половине она редко встречается на лесных болотах, будучи больше
связанной с выходами вод в районах со сложно расчлененным рельефом.
В пределах юго-запада Русской равнины (Украина, Молдова, южная
Беларусь) черноолыпаники выполняют водоохранную и водорегулирующую
роль.
ВАЖНЕЙШИЕ НАПРАВЛЕНИЯ СОВРЕМЕННОГО БОЛОТОВЕДЕНИЯ
О. Л. Лисе
МГУ, биологический факультет
Как самостоятельная дисциплина болотоведение оформилось в начале 20
века. В его развитии выделяется два основных этапа: описательный и каузальный.
В настоящее время проблемы, стоящие перед болотоведами свидетельствуют о
необходимости перехода к функциональному этапу. Во-первых, огромный банк
данных, накопленный в течение 100 лет касающийся природы болот, требует
некоторого переосмысления с использованием современных подходов.
Во-вторых, познание значения болот в биосфере невозможно без детального изучения
их экологических функций в пространственно-временном аспекте. В-третьих,
для изучения роли болот в биосфере необходимо организовать подбор
региональных (зонально-подзональных) ключевых болотных систем по всей России.
Россия занимает одно из первых мест в мире по площадям болот и запасам
торфа, 30% площади болот планеты приходится на её долю. За последние
десятилетия в связи с интенсивным осушением болот и добычей торфа площади
болот значительно со1фатились и в России, и в зарубежных странах. В отдельных
странах Европы болота оказались полностью уничтоженными, что отрицательно
сказалось на экологической обстановке соответствующих территорий.
66
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
Болотные системы (БС) — уникальнейшие по своим функциям природные
^разования. С одной стороны, в них накапливается торф и аккумулируется уг-
дерод, с другой стороны, болота выполняют важные для биосферы
экологические функции: регулирование теплового баланса, гидрологического режима,
геохимических процессов, сохранение биологического разнообразия и др.
Ценность БС для человечества состоит и в том, что они являются источником
сырьевых (торф, депонированный в нём углерод) и растительных (пищевые,
кормовые, лекарственные растения) ресурсов.
Если к настоящему времени достаточно полно изучена природа БС, их
типология, районирование, различные качественные и количественные показатели,
то уточнение роли экологических функций этих образований требует
дальнейшей существенной проработки. Тем более, что некоторые из них до сих пор
остаются дискуссионными. Кроме того, взаимодействие конкретных БС и
окружающих их ландшафтов имеет региональные особенности. Этот аспект также до
сих пор достаточно не изучен.
Для получения исчерпывающей информации относительно экологической
роли БС в биосфере необходимо создание базовой геоинформационной системы
(ТИС) сбора данных о состоянии ключевых БС и окружающей их среды.
Разработка базовой гас должна базироваться на результатах инвентаризации и
круглогодичного разнопланового мониторинга. Она подразумевает в первую очередь
составление серии следующих региональных карт-схем:
— карты-схемы уровневого и теплового режимов, радиационного баланса и
испарения с БС. Положение уровня воды относительно поверхности болот
является одной из главных характеристик их обводнённости. Располагая данными о
режиме колебания уровней болотных вод, используя известные в настоящее
время связи уровней с водно-физическими характеристиками деятельного слоя
(водоотдачей, водопроводимостью, влажностью и др.) можно производить
расчёты водообмена и водного баланса болот, судить о свойствах торфяных
отложений, осуществлять регулирование водного режима заболоченных территорий;
— карты-схемы динамики гидрологических характеристик стока воды с БС.
Одним из проявлений гидрологической функции болотных систем является их
водоохранная роль, которая может быть как положительной, так и
отрицательной. Решение вопроса о водоохранном значении болот зависит от того, какому
конкретному звену влагооборота оно применяется. Известно, что болота на
водосборной площади несколько снижают объём речного стока в замыкающем
створе бассейна, но одновременно и консервируют значительные запасы влаги в
торфяных отложениях. Хорошо дренированные выпуклые олиготрофные
сфагновые болота осуществляют водосберегаюшую функцию в сухие сезоны года,
резко снижая испарения при обсыхании сфагновых мхов. Водообмен БС с
окружающими ландшафтами происходит посредством поверхностного и грунтового
стока. Поверхностный сток осуществляется через водотоки, озёра, топи (т. н.
67
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
гидрографическою сеть болотных систем) и путём фильтрации в деятельном
горизонте. Поверхностный сток, поступая на периферийные участки болот, их
переувлажняет и подпитывает верхние горизонты подземных вод. Влияние бо-
лог на сток с речных водосборов (уменьшение максимальных модулей
половодий и паводков) зависит от их регулирующей ёмкости. Наибольшее снижение
стока, а также более равномерное его распределение обеспечивают крупные оли-
готрофные системы таёжной зоны с господством грядово-мочажинноюзерковых
комплексов. Хорошей регулирующей способностью обладают мезотрофные и
евтрофные болота, которые, наряду с поверхностным, получают обильное
грунтовое питание и имеют благоприятные условия для формирования подземного
притока в реки. Болота в тундре и лесотундре (в условиях вечной мерзлоты) не
имеют значительной регулирующей ёмкости. Следует также обратить особое
внимание на роль поверхностного стока с БС в формировании речной сети.
Особенно это касается регионов с очень высокой заболоченностью (до 70-90%);
— карты-схемы формирования и динамики химического состава и качества
болотных, речных и подземных вод. БС способны аккумулировать большой
спектр загрязняющих веществ окружающей среды. Они являются
геохимическими барьерами с очень высокой адсорбционной ёмкостью, выполняют
функции биологических фильтров, накапливая токсичные техногенные элементы,
органические загрязнители, пестициды, консервируя их и выводя из круговорота
веществ в биосфере;
— карты-схемы структуры ключевых БС, динамики, прогноза торфонакоп-
ления и сукцессии растительного покрова. Зонально-подзональный подбор
ключевых болотных систем подразумевает выбор репрезентативных для
соответствующих регионов болот по условиям геоморфологического залегания, генезису,
стратиграфии, современным стадиям развития. Эти объекты должны быть
предназначены для проведения деятельной инвентаризации и организации
круглогодичного мониторинга, которые должны осуществляться по единой программе.
Программа должна быть ориентирована на разработку базовой ГИС;
— карты-схемы биологических ресурсов (торфяных и растительных, в том
числе редких и исчезающих видов) ключевых болотных систем. При
составлении этих карт особое внимание следует обратить на региональные
закономерности накопления углерода и эмиссии С02;
— карты-схемы ранжирования ключевых БС по порогам устойчивости к
антропогенному воздействию. Для создания серии таких карт необходимо
разработать критерии устойчивости БС.
Анализ базовой ГИС позволит разработать методику выявления
оптимальных для соответствующих регионов соотношений между осушаемыми землями
и не подлежащими мелиорации. Знание этих соотношений — необходимое
условие для составления региональных программ мелиоративных мероприятий.
68
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
В настоящее время в рамках программы как Международной организации
Wetlands International, так и аналогичной российской программы, особую
актуальность приобретает проблема сохранения БС, их биоразнообразия и
природных ресурсов. В первую очередь к охране следует представить региональные
ключевые БС, где планируется организация мониторинга.
БОЛОТНЫЕ ФОРМЫ СОСНЫ ОБЫКНОВЕННОЙ КАК ИНДИКАТОРЫ
УСЛОВИЙ ПРОИЗРАСТАНИЯ
О.Л. Лисе, Е.М. Волкова
МГУ, биологический факультет;
Тульский государственный педагогический университет
Существование возможности индицировать по растительному покрову
болот строение и глубину торфяной залежи известно давно [Романова, 1961;
Богдановская-Гиенэф, 1969; др.] Однако, многолетние исследования
показывают, что на болотах, различных по происхождению и закономерностям
развития, под одинаковыми растительными сообществами строение залежи
может быть различно. Только однотипные по происхождению и развитию
биогеоценозы имеют торфяные залежи сходного строения. Подобные
результаты указывают на необходимость учета размеров, географического и
геоморфологического положения болотной системы, местоположения в ней
индицирующих фитоценозов. При учете всех перечисленных признаков
можно в болотных биогеоценозах, одинаковых по характеру растительного
покрова, распознать различия в строении торфяных отложений. Поскольку в
большинстве сообществ древесный ярус образован сосной Pinus sylvestris L.,
представляет интерес существование связей между болотными формами
сосны, глубиной торфа и стратиграфией залежи.
По окрайкам болотных массивов или на склонах с уклоном поверхности
от 0,004 до 0,01 в сосново-кустарничково-сфагновых сообществах
преобладает сосна f. uliginosa. Она индицирует пушицево-сфагновую и фускум-за-
лежь, глубина которой варьирует на болотах разных зон, составляя в
среднем 1-2 м [Лисе, 1990].
В центральных частях болот и по окрайкам, на заболоченных террасах и
водораздельных равнинах от северной подзоны тайги до лесостепи
произрастает сосна f. litwinowii, являющаяся компонентом сосново-кустарничково-
сфагновых фитоценозов и грядово-мелко(средне)-мочажинных комплексов.
Доминирование деревьев данной формы слабо коррелирует с мощностью
торфяной залежи, которая колеблется в широких пределах: от 1-2 м [Лисе,
1990] до 1,5-5,5 м [Романова, 1961]. В составе залежи преобладают фускум,
69
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
а также ангустифолиум, магелланикум и пушицево-сфагновые виды торфа,
реже встречается комплексная мочажинная залежь.
Сокращение площади, занятой грядами, до 10-30% диагностирует грядо-
во-крупномочажинные комплексы. Обычно они приурочены к поверхностям
с уклоном 0,0008-0,001 и заняты пушицево-сфагновыми сообществами.
Сосна встречается единично и представлена f. willkommii. Значительная доля
мочажин объясняет присутствие сфагнового мочажинного и шейхцериевого
торфов в залежи. Мощность верховой толщи не более 2,0-4,0 м.
Сосна f. willkommii является неотъемлемым компонентом центральных
частей крупных болотных массивов, характеризующихся различными
вариантами озерковых комплексов (озерково-грядово-мочажинный, мочажинно-
озерковый). В таких комплексах биогеоценозов торфяные отложения
представлены комплексной, шейхцериево-сфагновой, мочажинной, фускум-
залежью. При этом мочажинный торф встречается преимущественно под
мочажинами, фускум — под грядами [Лисе, 1990]. Глубина торфяной толщи
достигает 4,0-6,0 м.
Таким образом, четко выраженная зависимость между формами сосны и
глубиной торфяной залежи отсутствует, однако в наиболее автономных —
центральных частях болот, где размеры залежи максимальны, показано
преобладание деревьев f. willkommii, тогда как по периферии болот
древесный ярус образован сосной f. uliginosa, индицирующей торфяную толщу
меньших размеров.
НАКОПЛЕНИЕ ХОРОШО- И СИЛЬНОРАЗЛОЖИВШЕГОСЯ
ВЕРХОВОГО ТОРФА В ЗАВИСИМОСТИ ОТ ВРЕМЕНИ НАЧАЛА
БОЛОТООБРАЗОВАНИЯ
Г.Л. Макаренко
Тверской государственный технический университет
К хррошо- и сильноразложившемуся верховому торфу относятся: пуши-
цевый, сосново-пушицевый и сосновый виды, образующиеся в условиях
обедненного водно-минерального и активного гидродинамического режима
с олиготрофным ходом развития залежи [1,2,3,4]. Это ценный источник
сырья для получения битумов, воска и сопутствующих продуктов.
Исследуемые болота в соответствии с началом борото- и торфообразова-
ния по климатическим периодам голоцена были разделены на четыре
группы: первая — аллерёд-дриас AL-DR, вторая — предбореальный период РВ,
третья — бореальный период ВО, четвертая — атлантический период AT
[2] (см. рис.). Сдои хорошо-\ й сильноразложившегося верхового торфа
начали формироваться в!юреаяъномлщриоде (около 9,4 тыс. лет назад), а за-
70
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
вершение этого процесса происходило в субатлантическое время (около 0,6
тыс. лет назад). Наиболее молодые по возрасту болота имеют наибольшую
продолжительность и более позднее время вступления и завершения
процесса накопления слоев хорошо- и сильноразложившегося верхового торфа.
Независимо от времени формирования болотных экосистем, слои
хорошо- и сильноразложившегося верхового торфа начинают накапливаться в
атлантическом периоде голоцена. Завершение этога процесса относится уже
к атлантическому, суббореальному и субатлантическому преиодам голоцена
(см. рис. и табл.). При этом стадии вступления и стадии завершения этого
процесса в составе болот не являются единовременными. Следовательно
здесь сказывается влияние локальных природных факторов, естественным
образом, регулирующих меняющуюся среду и условия произрастания
болотной растительности и соответствующее накопление слоев хорошо- и
сильноразложившегося верхового торфа [2]. Это, прежде всего,
гидрохимический состав и гидродинамический режим природных вод, питающих
болото в соответствии с геолого-геоморфологической обстановкой надболот-
ной части и геологическим окружением водосборной площади.
i?VI
\\\ \
i X Л 1
Yv
i
,
i i
1 < !
III
I Ь
i Pi i
1
i : '
LL_J
"Tj
s\*
1
i
i
i
i
. i
1; i
• i
•
i
i !
AL.DR PB BO AT
Возраст болот
ALDR PB BO AT
Возраст болот
Временные интервалы и средний возраст накопления слоев хорошо- и
сильноразложившегося верхового торфа в зависимости от возраста болот. Климатические
колебания в голоцене на Русской равнине; атлантико-континентальный тип (по
Н.А.Хотинскому, 1977): 1— температура, 2 — увлажненность.
Выявленные выше закономерности позволили проследить взаимосвязь
возраста болот с временными этапами накопления слоев хорошо- и
сильноразложившегося верхового торфа. Правильное понимание геологической
71
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
природы этого явления дает возможность не только прогнозировать, но и
управлять процессом развития болотных и лесоболотных экосистем.
Начало болотообразо- Временные этапы формирования слоев хорошо- и силь-
вания по периодам норазложившегося верхового торфа, тыс. лет назад
голоцена Начало Окончание Продолжительность
Аллерёд-дриас AL-DR 9,1...3,8 8,0...2,5
6,8 9,2 1,6
ПредбореалРВ 9,1...4,0 5,0...2,5
6,8 3,8 3,0
БореалВО 7,8...4,8 5,5...1,2
6,2 3,0 3,2
Атлантика AT 7,0...3,0 3,8...0,7
5,0 2А ЗА
[1] Макаренко Г. Л. Геологическая природа слоев хорошо- и сильноразложившегося
верхового торфа в залежном слое болот // Генезис, эволюция и роль болот в би-
сферных процессах: Тез. докл. Междунар. конф. Мн., 1994. С. 124-125. [2]
Макаренко Г.Л. О генезисе слоев хорошо- и сильноразложившегося верхового торфа в
залежном слое торфяных месторождений // Технология и комплексная механизация
торфяного производства. Тверь: ТГТУ, 1997. С. 115-121. [3] Макаренко Г.Л.
Генетическая классификация торфяных отложений на основе водного режима торфоген-
ного слоя болотного массива // Методы исследования торфяных и сапропелевых
отложений. Тверь: ТвеПИ, 1991. С. 28-43. [4] Makarenko G. Peat classification on the
basis of the water regime of deposit's dinamical path // Fen Restoration Workshop.
Central Forest Nature Reserve, 1997. P. 50-51.
ВОЗРАСТ И ДИНАМИКА БОЛОТ ЗАПАДНОЙ СИБИРИ
СМ. Новиков, Л.И. Усова, Е.С. Малясова
Государственный гидрологический институт, Соккт-Петербург
Исследованием вопросов образования и развития западно-сибирских
болот занимались многие ученые [Бронзов А.Я., Пьявченко НИ., Нейштадг
М.И., Тюремнов С.Н., Глебов Ф.З., Волков И.А., Лисе О.Л. и др.].
Результаты этих исследований, обычно базирующихся на ограниченном материале
радиоулеродных датировок и спорово-пыльцевых диаграмм опубликованы в
многочисленных работах, наиболее полный перечень которых приведен в
диссертационной работе О.Л. Лисе [ 2 ].
Тридцатилетние (1958-1990 гг.) экспедиционные исследования
заболоченных территорий Западно-Сибирской равнины, проведенные ГТИ,
позволили получить уникальный материал по типологии, строению и
водно-тепловому режиму болот этого обширного региона (от Барабы до северной
части полуострова Ямал). В процессе исследований изучалась также страти-
72
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
j-рафия торфяной залежи, определялся её возраст. За время работы Западно-
Сибирской экспедиции было отобрано в разных болотных зонах
(полигональных, бугристых, выпуклых олиготрофных и плоских евтрофных и мезо-
хрофных болот) 48 образцов торфа на радиоуглеродный возраст и 30
колонок на спорово-пыльцевой анализ. Сведения о районах отбора образцов
торфяной залежи и результатах их анализа приведены в табл. 1.
Данные по возрасту болот охватывают в меридиональном направлении
практически всю Западно-Сибирскую равнину (от полуострова Ямал до г,
Тюмень ), что позволяет при привлечении еще и литературных источников
по этому вопросу проследить процесс заболачивания столь обширного
региона. Ранее обобщенные нами материалы ГТИ по возрасту и генезису
бугристых болот опубликованы в работах [3,5,6]. Подробная характеристика
болотных зон и групп болотных микроландшафтов Западной Сибири
приведена в работах [1,4,7], поэтому здесь на рассмотрении этих вопросов
останавливаться не будем. Рассмотрим лишь два вопроса: вопрос образования,
точнее заболачивания территории Западной Сибири и вопрос интенсивности
торфонакопления в разные периоды голоцена. Анализ накопленных данных
по стратиграфии торфяной залежи и возрасту болот Западной Сибири
позволяет высказать следующие соображения по этим вопросам.
1. Процесс заболачивания Западно-Сибирской равнины начался в пред-
бореальный период (9200-9300 л.н.) практически одновременно, за
исключением района Крайнего Севера (северная половина полуострова Ямал,
полуостров Гыданский) и южной части равнины (лесо-степная зона), где
начало болотообразования приходится на суббориальный период.
2. В зоне бугристых болот прослеживается заметная разница в возрасте
залежи плоскобугристых и крупнобугристых болот, расположенных в
единых болотных системах. Первые примерно на 2500 лет старше вторых.
Столь значительную разницу в возрасте можно объяснить лишь
запозданием появления благоприятных условий для образования крупнобугристых
болот. Крупнобугристые болота образуются в условиях значительного
избытка влаги и поэтому приурочены к переувлажненным окрайкам
плоскобугристых болотных массивов, поймам рек и озерным котловинам. Возраст
крупнобугристых болот расположенных в поймах рек относится к предбори-
альному или бореальному периоду.
3. Возраст придонных слоев торфяной залежи на буграх (зона бугристых
болот) колеблется в пределах от 5110+100 до 9320±120 лет, в межбугорных
понижениях (топях, ложбинах, западинах) — от 8301160 до 4750±110 лет,
т.е. возраст бугров значительно древнее возраста пониженных элементов
мезорельефа. Последнее объясняется генезисом бугристых болот [6].
4. Абсолютный возраст верхних слоев торфяной залежи на буграх,
составляющий от 100 лет до 490±50 лет (табл. 1), и хорошо развитый на них
73
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
растительный покров свидетельствует о наличии торфообразовательноц)
процесса в современную эпоху. Наблюдения за развитием бугристых болот
показывают, что в настоящее время в динамике этих болот одновременно
прослеживаются два процесса: процесс разрушения торфяных бугров под
влиянием водно-тепловой эрозии и процесс образования новых бугров на
межбугорных топях под действием мерзлотного пучения. Процесс
зарождения новых бугров пучения является многофакторным.
Табл. 1. Толщина торфяных слоев (см) и интенсивность торфонакопления (мм/гоц)
в различные периоды ив среднем за голоцен на территории Западной Сибири
Типы
болот
Периоды голоцена (по Хотинскому, 1977)
SA SB AT2 ATI ВО
РВ
_£ESL
Полигональные
сев. тундра
южн. тундра
плоскобугристые
пологовыпукло-
бугристые
крупнобугристые
олиготрофные
мохово-лесные
евтрофные тра-
вяно-моховые
36
0.01
20-35
0Л1
22-70
0.11
-
20
0.08
55-125
4 0.30
-
20
0.01
10-38
0.34
15-55
0.13
20
0.24
20-120
0.31
55-110
0.42
-
-
75
0.20
20-50
0.16
10
0.04
•20-120
0.28
10-65
0.17
-
-
40-69
0.24
15-40
0.16
10
0.04
35-90
0.68
10-75
0.48
-
-
13-20
0.09
10
0.08
-
20-45
0.18
20-60
0.27
-
-
-
10-60
0.04
10
0.17
30-60
0.02
0-10
0.01
-
-
-
-
0.22
-
0.11
-
0.39
-
0.35
-
0.67
Средняя интенсивность торфонакопления в голоцене определена по
радиоуглеродным данным, прочерк в таблице - отсутствие данных.
Табл. 2. Радиоуглеродный возраст придонных слоев торфяной залежи различных
типов болот Западной Сибири (ЭМР — элемент мезорельефа, пов — в поверхност-
ном слое залежи, *— на заданной глубине залежи).
Район исследований
Тип болота ЭМР
Глубина,
м
Лабораторный индекс
Возраст
по
р.Яропенза
п.Новый Порт
р.Пангола
п.Пангода
р.Лонг-Юган
г.Надым
полигональ
ное
крупнобугристое
плоскобугристое
полигон
бугор
бугор
топь
бугор
бугор
бугор
топь
топь
0,10*
0,30*
2,80
3,25
0,70
пов
0,55*
1,40
0,20*
0,60
УПИ-301
УПИ-302
ЛУ-563
ТА-752
ТА-745
ЛУ-944
УПИ-303
ЛУ-942
УПИ-298
УПИ-300
2050±150
3170±170
51101100
5810170
750160
190140
64301190
8500150
50301130
73351110
74
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
^
р.Тыдыотта (место
слияния
рек Сев. и Юж. Тыдыот-
та
р.Пур-Пе
(Комсомольский БМ)
р.Тром-Еган (верховье,
Холмогорский БМ)
(Федоровский БМ)
р.Вах (Самотлорский
БМ)
р.Тура (Тарманский БМ)
_
полого-
выпукло-
бугристое
крупнобугристое
мелкобугри
стое
плоско-
бугристое
плоскобугристое
полого-
выпукло-
бугристое
плоскобугристое
бугор
бугор
топь
бугор
топь
бугор
бугор
бугор
топь
бугор
бугор
топь
бугор
бугор
бугор
бугор
топь
бугор
бугор
бугор
бугор
топь
бугор
бугор
бугор
топь
бугор
бугор
топь
бугор
бугор
олиготрофное
мохово-лесное
олиготрофное
мохово-лесное
евтрофное
травяно-моховое
0,40
0,50
0,65
1,20
0,50
0,60
0,35
0,30
0,40
пов
1,60
0,40
пов
0,60
0,25
0,90
0,65
3,80
1,80
1,15*
1,60
0,85
1,40
1,00
2,15
1,35
0,85*
1,60
0,60
1,50
2,80
1,00
2,30
3,00
3,50
3,90
1,60
3,00
ЛУ-943
УПИ-304
УПИ-296
УПИ-297
УПИ-295
УПИ-299
ЛУ-928
ЛУ-926
ЛУ-950
ЛУ-946
ЛУ-934
ЛУ-932
ЛУ-947
ЛУ-929
ЛУ-930
ЛУ-936
ЛУ-935
ЛУ-933
ЛУ-931
ЛУ-731
ЛУ-732
ЛУ-723
ЛУ-717
ЛУ-722
ЛУ-725
ЛУ-719
ЛУ-735
ЛУ-737
ЛУ-736
ЛУ-562
ЛУ-583
ЛУ-564
ЛУ-559
ЛУ-560
ЛУ-584
ЛУ-581
ЛУ-582
ЛУ-738
7150±80
8140±120
12301140
8660±100
370±90
6640±130
860±260
720±250
83Q1160
490150
9320±120
830±160
<100
2290+150
4070±110
8930±120
2900±120
8420±180
9320±320
5220160
6630150
47501110
7670190
3900170
52801240
4140180
58901110
88101100
2860190
6680180
92201100
31001140
8660180
8800180
8650150
9340180
4010180
31801110
Все образцы отобраны Л.И.Усовой. Лаборатории: ЛУ
УПИ — Уральский пед. ин-т, ТА — Институт зоологии и
- Ленинградский гос. ун-т,
ботаники АН ЭССР.
5. На основе датировок по 14С и палинологических данных оценена
интенсивность торфонакопления болотами разных типов в различные периоды
75
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
голоцена. Результаты представлены в табл. 2. Наибольшая интенсивность
торфонакопления наблюдалась в раннеатлантический период (0,16-0,68
мм/год), наименьшая (0,08-0,27 мм/год) — в предбореальный и бореальный
периоды. Относительная высокая интенсивность торфонакопления была и в
суббореальный период (0,13-0,42 мм/год). В субатлантический период
прослеживается снижение интенсивности торфонакопления (0,30-0,01 мм/год) в
северном направлении от олиготрофных до полигональных болот.
[1] Болота Западной Сибири, их строение и гидрологический режим / Под ред. К.Е.
Иванова, СМ. Новикова. Л.: Гидрометеоиздат, 1976. 446 с. [2] Лисе О.Л.
Закономерности развития болотных систем в голоцене и их рациональное использование
(на примере Западной Сибири) // Автореф. дис. докт. геогр. наук. М. 1990. 48 с. [3]
Малясова Е.С. и др. Динамика торфонакопления и процесс образования бугристых
болот Западной Сибири // Ботан. ж. 1991. Т. 76. № 9. С. 1227-1237. [4] Новиков
С,М., Усова Л.И. О природе и классификации бугристых болот // Тр. ГТИ. Л.:
Гидрометеоиздат, 1976. Вып. 261. С. 3-13. [5] Новиков СМ. и др. Генезис и возраст
бугристых болот Западно-Сибирской равнины // Изотопные и геохимические
методы в биологии, геологии и археологии. Тарту, 1981. С. 87-91. [6] Новиков СМ.,
Усова Л.И. Генезис бугристых болот Западной Сибири // Тр. ГГИ. Л.:
Гидрометеоиздат, 1983. Вып. 303. С 11-16. [7] Новиков СМ., Усова Л.И. О классификации и
дешифровочных признаках полигональных болот (на примере Западной Сибири) //
География и ресурсы. Новосибирск. Наука. 1987. С. 131-137. [8] Хотинский Н.А.
Голоцен северной Евразии// М.: Наука. 1977. 198 с.
ДИНАМИКА ЛЕСНОЙ РАСТИТЕЛЬНОСТИ И
БОЛОТООБРАЗОВАТЕЛЬНОГО ПРОЦЕССА В ГОЛОЦЕНЕ
В ГОРАХ СРЕДНЕГО УРАЛА
Н.К. Панова
Ботанический сад УрО РАН, Екатеринбург
Одной из специфических черт горных стран является значительное
разнообразие природных условий на сравнительно небольшой территории.
Заболоченность в горной полосе Свердловской области не превышает 2-3%.
Многие болота уникальны и нуждаются в охране в качестве ландшафтных
памятников и природных летописей истории развития растительности и
климата [1 и др.].
Для изучения особенностей истории формирования и развития лесной
растительности и болотообразовательного процесса в центральной горной
полосе Среднего Урала проанализирован методами спорово-пыльцевого и
ботанического анализа ряд разрезов торфяных болот, расположенных в при-
водораздельной части восточного (окрестности г. Екатеринбурга, подзона
южнотаежных сосновых лесов) и западного (район Висимского заповедни-
76
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
ка, примерно в 100 км к северо-западу от г. Екатеринбурга, подзона
южнотаежных пихтрво-еловых лесов) макросклонов. Выявлены существенные
различия и в истории лесов этих частей региона, обусловленные, очевидно,
субмеридиональной протяженностью горных хребтов и связанными с этим
особенностями циркуляции атмосферы в разные периоды голоцена.
СРЕДНИЙ УРАЛ
^ Э ЕЗз Ш* Ш* ЕИ« ЕЗгЕЗя ЩП>
fF№ Е№ HHS« IHf3 И'4 S!/5ИЗ" ЁПк (ИЗ'*
Сопоставление торфяных разрезов по высотному профилю: 1.Песчаное, 2.
Романовское, 3. Карасьеозерское, 4. Галашкинское, 5. Средне-Сулемское, 6. Шайтанское.
Условные обозначения: 1-12 - Растительные остатки в торфе: 1,2-сфагно-вые мхи
(1-низинные, 2-переходные), 3-гипновые мхи, 4-осоки, 5-пушица, 6-хвощ, 7-вахта,
8-разные травы, 9-древесный торф, 10-ель, 11-сосна, 12-береза, 13-сапропе-листый
торф, 14-сапропель, 15-глина, 16-суглинок, 17-песок, 18-галька, дресва.
В позднем дриасе и предбореальном периоде на всем Среднем Урале
господствовала травяно-кустарниковая растительность в сочетании с
редколесьями: елово-лиственничными с примесью сосны и березы на востоке и
77
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
елово-березовыми — на западе. На западном склоне, в пойме р. Сулем со„
хранились пихта, липа, ильм. В бореальном периоде формируются таежные
леса: на восточном склоне — березово-сосновые с примесью лиственницы,
ели, кедра сибирского; на западном — преимущественно еловые с примесью
пихты, сосны, кедра, березы.
Потепление способствовало распространению из убежищ
широколиственных видов, которые достигли наибольшего развития в конце
атлантического периода. В это время в обеих частях региона произрастали наиболее
разнообразные по составу лиственно-хвойные леса с примесью пихты и
широколиственных. В районе Висимского заповедника ильм и липа
поднимались до высоты 700 м, а в долине р. Сулем появились лещина и дуб. На
восточном склоне, кроме липы, ильма, дуба, лещины встречались клен и граб.
Последний распространялся из южноуральского рефугиума.
Похолодание в начале суббореального периода остановило дальнейшее
продвижение неморальной флоры. На восточном склоне резко возросла роль
сосны, участие широколиственных уменьшилось. На западном — в середине
суббореала преобладали пихтово-еловые леса с примесью
широколиственных, даже большей, чем в атлантическое время. В субатлантическом
периоде в обеих частях региона (на востоке — на фоне преобладания сосны)
увеличилась роль ели, пихты, сибирского кедра. На современном этапе
сохраняется природная тенденция к прогрессивному развитию сосны на
восточном склоне и темнохвойных видов — на западном. Однако, в результате
хозяйственной деятельности за пределами Висимского заповедника и в
окрестностях г. Екатеринбурга сокращается участие в лесах хвойных пород и
увеличивается распространение березы, что находит отражение в субре-
центных пыльцевых спектрах.
Судя по стратиграфии разрезов (см. рис.), все исследованные торфяники
на высотах 250-400 м образовались путем заболачивания водоемов. Начало
органогенных (сапропелевых) отложений относится в большинстве разрезов
к предбореальному времени. В бореальном периоде началось торфонакоп-
ление в мелких водоемах. Обмеление и заторфовывание более глубоких
озер произошло (с коррекцией местными гидрологическими условиями)
примерно в одно и то же время — в конце атлантического — первой
половине суббореального периодов. Тогда же на ранее сформировавшихся
торфяниках поселяется древесная растительность. Эти явления, очевидно,
связаны с общерегиональным повышением сухости климата.
Болото Шайтанское, расположенное на вершине горы на высоте 700 м
над у.м., образовалось путем суходольного заболачивания, и, по своему
местоположению, растительности и торфяной залежи, является уникальным
для Среднего Урала [2]. Несмотря на небольшую глубину, оно имеет,
согласно спорово-пыльцевым данным, достаточно древний возраст.
78
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
п] Маковский В.И. О рациональном использовании и охране торфяных болот
Свердловской области // Проблемы экологии, рационального использования и
охраны природных ресурсов на Урале. Свердловск, 1980. С.52-53. [2] Маковский В.И.
Итоги изучения заболоченных лесов и болот Висимского заповедника // 10 лет Ви-
симскому государственному заповеднику. Свердловск, 1981. С. 14-16.
АВТОНОМНОЕ ЗАБОЛАЧИВАНИЕ КЕДРОВЫХ ЛЕСОВ
ЗАПАДНОЙ СИБИРИ
Н.Н. Пологова, А.Г. Дюкарев
Филиал института леса им. В.Н. Сукачева СО РАН, Томск
Вопросы автономного заболачивания лесов вследствие эндогенеза
остаются дискуссионными, требующими в изучении системных подходов из-за
сложности взаимодействия ландпшфтоформируюпщх лесо- и болотообразо-
вательного процессов. Мы исходим из определения заболачивания как
этапного процесса перехода ландшафта из автоморфного состояния в гидро-
морфное, сопровождающегося участием и изменением в этом процессе всех
его компонентов. Ступенчатость развития процесса заболачивания ставит
задачу выявления критических состояний в лесных экосистемах, которые
соответствуют переходу их в качественно иное состояние — моменту
«выбора» направления их эволюции. Как для любого природного процесса, эти
состояния можно определить этапами зарождения, формирования, развития,
кульминации и последействия (распад и зарождение нового) процесса.
Лесным экосистемам присущи свойства, которые определить можно как
консервативность и стабильная динамичность компонентов.
Консервативность обязана длительному пребыванию дерева в одной и той же точке
биогеоценоза, в результате которой динамика компонентов вблизи дерева
приводит к необратимым изменениям в свойствах почв. С этой позиции в
полидоминантных среднетаежных лесах именно долгоживучесть кедра после
отпада менее долговечных сопутствующих лесообразователей обеспечивает
заметную трансформацию почвенного компонента в конечной стадии -
перестойных кедровых лесах. Для кедровников Западной Сибири,
формирующихся на слоистых почвообразующих породах, бесспорны факты усиления
степени гидроморфности местообитаний к этой стадии.
С точки зрения лесообразовательного процесса модель формирования
кедрового биогеоценоза можно рассмотреть как последовательность
функционирования, раскрывающуюся через его структуру и охватывающую
развитие и взаимодействие всех его компонентов (подсистем). В ряду
восстановления кедровников зеленомошных увеличение гвдроморфности
характеризуются следующими моментами изменения всех подсистем.
79
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Происходит изменение строения минерального профиля почв с заменой
гумусово-аккумулятивного горизонта тиксотропным глеевым и соответст-
венно увеличением плотности до предельных значений. Иллювиальные
горизонты трансформируются наиболее заметно по содержанию органо-мине-
ральных соединений, поскольку снижение поступления органического
вещества приводит к увеличению окристаллизованных форм соединений железа,
ортштейновых новообразований которые в данном случае можно
рассматривать как тенденцию превращения почвенного тела в безжизненную
породу. Тиксотропность верхних горизонтов становится причиной слабой
устойчивости древостоя к ветровалу.
В части биогеогоризонтов относящихся к органопрофилю почв
изменения наиболее существенны. Возрастает мощность подстилки, изменяется ее
структура, фракционный состав с заменой от моховой на мохово-кустарнич-
ковую и сфагновую, запасы ее увеличиваются от 20 до 120 т/га; как
показатель снижения качества органического вещества отношение углерода к азоту
в подстилках увеличивается от 20 до 30 единиц.
Изменяется архитектоника корневых систем возобновления кедра,
которые в соответствии с циклом оборота поколений подстилки реализуют
способность образовывать придаточные корни длительное время, а в целом
повторяют структуру основных компонентов слагающих этот биогеогори-
зонт. На конечной стадии проявления гидроморфизма корневая система
подроста и питающие корешки взрослых деревьев перемещаются в подстилку.
Становится иным характер распределения элементов питания в подстилке.
В процессе смен поколений древостоя мозаично-ярусная структура
кедрового биогеоценоза воспроизводится в соответствии с закономерностями
формирования GAP-струкгуры лесных биогеоценозов — чередования во
времени парцелл окон и возобновления в них новых поколений древостоя. С
возрастом между парцеллами окон и крон древостоя увеличивается
различие в увлажненности, особенно в органогенной части почв, усиливается
плотность верхних горизонтов, увеличивается кислотность, причем эти
различия затухают с глубиной. В соответствии с характерным временем
развития каждой из подсистем, мозаичность вызванная эдификаторным
воздействием, перерастает в комплексность.
Каждая из подсистем трансформирующегося кедрового БГЦ может
принимать на себя главную средообразуюшую функцию по отношению к лесо-
образующей породе: тиксостропность почвенного компонента снижает
возможность проникновения корней и ведет к неустойчивости древесного
яруса; замедленность разложения подстилки приводит к формированию
органического субстрата, куда смещается деятельность корневой системы
подроста. Показателем переходного состояния лесного биогеоценоза является
неустойчивость оборота поколений, на что указывает неполночленность воз-
80
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
растного состава, снижение плотности популяционных локусов
возобновления. В приближенных предварительных вычислениях эти значения
снижаются до 38% от исходных незаболоченных состояний.
Таким образом, устойчивость кедрового биогеоценоза, если ее
рассматривать как сохранение последовательности самовоспроизводства в рамках
цели — неизменности лесообразовательного процесса — переходит на
уровень изменчивости — при которой каждая из подсистем и система в целом
вырабатывает механизмы адаптации к изменяющимся внутренним
условиям среды. Прерывание процесса внешними причинами — пожар, вырубка,
возвращает систему в исходное состояние, но на некотором ином уровне
трансформированное™ компонентов. Восстановление почвенной системы
на стадии мелколиственных лесов сопровождается переходом тиксотропно-
сти в состояние тиксостабильности, изменяется органопрофиль. Однако
возможность бифуркации из состояний равновесия, как появление новой
«цели» системы, сохраняется. Различное характерное время трансформации
подсистем определяет цикличность сукцессии и растягивает время
окончательного перехода в гидроморфное состояние. Посколы^ наиболее
консервативен почвенно-грунтовый компонент, то от степени .накопления им
трансформационных свойств зависит и момент бифуркации.
Изменение всех подсистем приводит в конечном счете к формированию
нового свойства в одном из компонентов — подсистеме стоковой
составляющей ландшафта — результирующим гидрологическим условиям,
определяющим момент заболачивания — формированию экологически
избыточного увлажнения в лесном биогеоценозе как стадии поселения болотных
растений, способных сохранять и удерживать самостоятельный
гидрологический горизонт. На фоне усиления профильной дифференциации и тиксо-
тропности почв снижается вертикальная фильтрация, усиливается
аккумуляция влаги в органогенном слое.
Горизонт с повышенной влажностью чаще всего пространственно
локализован в микропонижениях рельефа. Внутри однородного по рельефу эко-
топа с кедровником зеленомошным, пятна сфагновых мхов приурочены к
понижениям с минимальными перепадами высот, которые при
внимательном рассмотрении оказываются участками поверхностного
перераспределения влаги. На пространственном уровне происходит перестройка
элементарной гидрологической ячейки стоковоформирующего комплекса экотопа.
Из имеющихся фактов и данных следует отметить, что транзитно-акку-
мулятивные позиции ландшафта на супесях и слоистых почвообразующих
породах предопределяют наиболее высокую вероятность заболачивания
кедровников. На плоских слабодренированных водоразделах в кедровниках
3-4 бонитетов формирования комплексности выражено еще более ярко и
81
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
момент перехода в заболоченные леса обширных площадей наступает при
слиянии очагов переувлажнения в микрозападинах.
Для равнинных территорий Западной Сибири по мере развития
растительного компонента, трансформации почвогрунтов и микрорельефа
достижимы конечные гидроморфные состояния кедровых биогеоценозов. Об
успешности завершения конечных стадий их трансформации в заболоченные
можно судить, приняв во внимание ландшафтные характеристики условий
местообитаний и выявив положение их в системе стока территории.
КОНКУРЕНЦИЯ ОЛИГОТРОФНОГО И ЕВТРОФНОГО
ЗАБОЛАЧИВАНИЯ В ТАЕЖНОМ ЛАНДШАФТЕ С БЛОКОВОЙ
СТРУКТУРОЙ
А.В. Хорошев
МГУ, географический факультет
Широко распространенные на моренных равнинах севера Европейской
России двучленные отложения обусловливают высокую радиальную
контрастность геохимической обстановки. Контакт супесчаной и суглинистой толщ
на глубине 40-50 см способствует поверхностному контактному оглеению и
существованию двух почти изолированных горизонтов почвенных вод с
разными свойствами. Кислая верховодка находится в пределах верхней
супесчаной толщи, а щелочные грунтовые воды — в пределах карбонатного
ледникового или озерно-ледникового суглинка. При близком залегании
пород, обогащенных карбонатами, в таежной зоне ход процессов
заболачивания определяется геохимической связью поверхности почвы с глубинными
карбонатными горизонтами. Как правило, верхняя супесчаная толща
относительно изолирована от влияния грунтовых вод, что благоприятствует
развитию ацидофильной растительности. Но при наличии даже мелких
разрывных нарушений в коренных породах грунтовые воды могут выходить на
поверхность и повышать реакцию в пределах всего почвенного профиля до
слабощелочных значений, что препятствует распространению ацидофилов.
Полигоном для изучения конкурентных взаимоотношений олиготрофно-
го и эвтрофного заболачивания в условиях близкого залегания карбонатных
пород и густой сети разрывных нарушений послужил трансект длиной 7350
м с шагом описания 25 м в среднетаежном моренно-эрозионном ландшафте
на юге Архангельской области (бассейн р.Кокшеньги). Коренные пермские
мергели здесь перекрыты московской карбонатной мореной и озерно-ледни-
ковыми отложениями мощностью от 0 м по склонам долин до 30 м на
водоразделах. Строго линейное простирание природно-территориальных
комплексов (ПТК) и резкие смены направления водотоков свидетельствуют о
82
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
блоковом строении территории. В котловинах и ложбинах, связанных с
разрывными нарушениями, развиваются гидроморфные ПТК.
На границе котловин и ложбин, связанных с разрывными нарушениями,
часто наблюдается контраст геохимических условий, связанный с
погружением щелочных грунтовых вод с минерализацией до 600 мг/л на глубину.
Их место в пределах почвенного профиля занимают сильнокислые
слабоминерализованные болотные воды. С такими контактами связаны узкие зоны
дренированное™, выраженные в формировании почв с ослабленными
признаками оглеения. Слабощелочная среда в почвах препятствует расширению
болот. При перепаде высот не более 0,5 м на расстоянии 25-50 м типичные
торфяно-глееземы сменяются типичными подзолистыми почвами без
признаков оглеения и даже с развитием гумусонакопления (при этом почвы
переходят в тип дерново-подзолистых). В пределах разрывного нарушения
щелочные грунтовые воды могут выходить на поверхность под напором,
исключая распространение сфагнума. При этом болото становится
осоковым, а местами формируются небольшие незарастающие озерки диаметром
10-15 м с рН 6,1.
Для болотных котловин свойственна асимметричность, что связано с
общим направлением уклона с северо-востока на юго-запад и
раздробленностью территории на блоки. Один склон котловины оказывается более
коротким и пологим, чем противоположный. У контакта крутого склона и днища
котловины разгружаются щелочные грунтовые воды, способствуя
формированию перегнойного горизонта. При этом конкурируют две траектории
развития, зависящие от мощности разгружающегося потока и характера днища:
подщелачивание среды с накоплением перегноя и подкисление с торфонако-
плением. Энергичный поток, разгружающийся в котловину (ложбину) при
значительном уклоне, приводит к накоплению мощного до 40 см слоя
перегноя; происходит эвтрофное заболачивание. При меньшей разгрузке и
наличии сфагнового покрова в днище перегной постепенно перекрывается
сфагновым торфом, что означает расширение площади олиготрофного болота и
смену щелочно-кислотного режима. На плоских водораздельных
поверхностях блоков рост гидроморфности провоцируется застоем верховодки над
суглинистой толщей и также обычно начинается с накопления перегноя. Из-
за отсутствия постоянной минеральной подпитки происходит
распространение сфагнума, приводя к олиготрофному заболачиванию. Перегной,
достигая мощности не более 5 см, перекрывается торфом.
Заболачивание в пределах плоских водораздельных участков, связанных
с поверхностями блоков, не является повсеместным. Разнообразие вносят
самые мелкие разрывные нарушения шириной не более 25-50 м,
выявляемые по увеличению мощности песчано-супесчаной толщи и «провалу»
уровня грунтовых вод. Это сопровождается аэрацией почвенного профиля, ха-
83
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
рактерной красноватой окраской, обилием охристых железистых
новообразований. Вместо перегнойного или торфянистого горизонта формируется
гумусовый. Другой вариант, при котором накопление слаборазложившейся
органики сменяется гумусонакоплением, связан с напорной разгрузкой
щелочных грунтовых вод на плоских участках без выхода их на поверхность. В
травянистом покрове доминируют Geum rivalis, Oxalis acetosa, Deschampsia
caespitosa, Filipendula ulmaria. На приводораздельных склонах при
глубоком погружении щелочных грунтовых вод переувлажнение может
создаваться верховодкой на глубине 25-30 см. В приповерхностном же слое
сохраняется окислительная среда. Устойчивый латеральный поток верховодки
способствует: 1) формированию резкого контраста
окислительно-восстановительной обстановки в верхней супесчаной толще с образованием
горизонта ортштейнов, 2) росту мощности подзолистого горизонта благодаря
свободному выносу продуктов разрушения почвенных минералов, 3)
сокращению или прекращению торфонакопления. Преобладают типичные таежные
виды Vaccinium vitisidaea, Rubus saxatilis, Majanthemum bifolium.
Классификация почвенных профилей по цветовой гамме с
использованием шкалы Манселла позволила получить информацию о соотношении
тенденций развития разных ПТК, о постепенном сближении или расхождении
свойств. Установлены наиболее отличные друг от друга по цветовой гамме
почвы («ядра типичности»). Они представляют различные типы
почвообразования. Выявлены и переходные варианты с большими дистанциями от
центра класса. Соседство ядер типичности, принадлежащих к разным
классам, свидетельствует об устойчивом расхождении почвенных признаков на
коротком расстоянии без каких-либо промежуточных вариантов, то есть
увеличении резкости границ. Это обычно связано с перегибами рельефа —
бровками склонов долины, локальными водоразделами. При компактном
расположении точек, принадлежащих к разным классам, но с большими
дистанциями от их центров, можно предполагать неустойчивое сочетание
почвенных процессов, что свидетельствует о сближении свойств почв.
Приуроченность таких зон к приболотьям позволяет выявить тенденцию
расширения болот и размыва границы между заболоченными и незаболоченными
ПТК. Неустойчивость цветовой гаммы почв, выявляемая кластерным
анализом, может предварять во времени появление морфологических признаков
заболачивания, таких как формирование торфянистого горизонта в
подзолистой почве. Почвы болот отнесены к трем разным классам цветовой гаммы,
но лишь для одного из них, в центральной части самого крупного верхового
болота, можно выделить ядро типичности. Два других класса представлены
в краевых зонах крупного верхового болота и в мелких переходных
шириной 100-200 м. Несмотря на возможность их выделения, разброс значений
цветовых характеристик велик, цветовая гамма не вполне самостоятельна я
84
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
тяготеет к другим классам. Это свидетельствует, о том, что размер болота
недостаточен для полного исключения процессов окисления, ожелезнения,
гумусонакопления, нарушающих «болотную» цветовую гамму.
ШДРОГЕОЛОГИЧЕСКИЕ УСЛОВИЯ ВОДНО-БОЛОТНОГО
КОМПЛЕКСА ВЕРХОВЬЕВ Р.ЯУЗЫ.
И.В. Цветкова, ГА. Гвоздикова, М.С. Орлов.
МГУ, геологический факультет
Водно-болотные комплексы (ВБК) можно отнести к очень хрупким и
сложным экосистемам. Их нормальное функционирование зависит, в
частности, от гидрогеологических условий. Подземные воды часто выпадают из
поля зрения гидрологов и экологов при рассмотрении ВБК, что, как
показала практика, является недопустимым [3]. В работе рассматривается влияние
гидрогеологических условий на состояние ВБК р.Яузы, находящегося на
территории Национального парка (НП) «Лосиный остров».
С 60-х годов происходит поступательное затопление поймы р. Яуза и
заболачивание прилегающих территорий. За период 1961-92 гг. уровень воды
в замыкающем створе ВБК повысился на 1,3-1,5 метра. Акватории ВБК
характеризуются интенсивными процессами эвтрофикации и заболачивания.
Это приводит к разрушению сложившейся экосистемы ВБК, негативно
влияет на состояние НП в целом. Основными возможными причинами
подтопления являются: а) гидрологическая, обусловленная возникновением
подпора в русле р.Яузы; б) гидрогеологическая, связанная с работой
водозабора «Главные Мытищи>>. От причины зависит метод борьбы с
подтоплением. Гидрологическая причина может быть обусловлена несколькими
факторами. В 50-е годы на территории ВБК велись торфоразработки изменившие
русло реки Яузы, была создана система каналов для осушения болот; в
нижней части ВБК имеется система дамб пешеходных и автомобильных дорог.
НЦ «Союзводпроект» был разработан рабочий проект «Восстановление
природной среды и организация функционального использования водно-
болотного комплекса верховьев р. Яузы от истоков до Ярославского шоссе».
Основные проектные мероприятия направлены на борьбу с гидрологической
причиной подтопления [1]. В рамках создания ОВОС к проекту и
проводилась данная работа. Целью было выяснение гидрогеологических условий
территории, для оценки возможности их влияния на развитие подтопления.
Подземные воды играют важную роль в формировании водного баланса
территорий и их эксплуатация может серьезно влиять на его изменение. На
территории ВБК находится старейший подмосковный водозабор «Главные
Мытищи», режим водоотбора которого изменялся. С пятидесятых годов
85
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
дебит водоотбора сокращался, что могло вызвать подъем уровня грунтовых
вод и способствовать подтоплению территории. Водозабор эксплуатирует
первый от поверхности водоносный горизонт, покрывающие отложения
имеют небольшую и невыдержанную мощность, что позволяет говорить о
несомненной связи с грунтовыми и болотными водами.
'Для изучения взаимосвязи водоносных горизонтов и оценки влияния
изменения водоотбора водозабором «Главные Мытищи» на положение их
уровней была создана гидрогеологическая модель данной территории.
Трехмерная стационарная модель описывает систему из трех водоносных
пластов (современных болотных, флювиогляциальных днепровско-москов-
ского межледниковья и карбонатных отложений карбона) и двух
разделяющих пластов (морены и верхней юры), имеющих гидрогеологические окна.
Для каждого пласта задавалась мощность, коэффициент фильтрации.
Граничные условия: сверху граница с постоянным расходом (инфильтрацион-
ное питание), снизу непроницаемая граница; для верхних двух водоносных
пластов по контуру модели задавалась непроницаемая граница
(водоразделы), для третьего водоносного пласта — боковая граница с постоянным
расходом (региональный поток подземных вод), в реке задавался расход,
связанный с положением уровня в ней. Водозабор моделировался одной
скважиной, фильтр которой располагался во втором водоносном горизонте.
Гидрогеологическая модель была реализована в компьютерной
программе MCG [2]. Прогнозная задача заключалась в выяснении взаимосвязи
первого от поверхности горизонта подземных вод и водно-болотного комплекса
р. Яузы. При решении прогнозной задачи были рассмотрены два вида
условий: состояние русел в настоящий момент времени и предполагаемое
состояние русел после осуществления следующих проектных мероприятий: их
чистки и снижения уреза на 1,2 м. В результате моделирования получилось,
что в настоящее время при дебите водоотбора 10 м3/сут, уровень во
флювиогляциальных отложениях на большей части территории располагается
выше уровня болотных вод и происходит перетекание из флювиогляциальных
отложений в болотные. При изменении дебита скважины водозабора
«Главные Мытищи» положение уровней водоносных горизонтов изменяется, при
уменьшении дебита усиливается питание болотных вод за счет первого
водоносного горизонта, а при увеличении дебита происходит инверсия
уровней и питания, так при уменьшении водоотбора в два раза уровень
грунтовых вод поднимается на 0,5-1 м. Взаимодействие каменноугольного и флю-
виогляциального горизонтов осуществляется в основном через
гидрогеологические окна, путем перетекания вниз (уровень в карбонатных отложениях
ниже за счет развития региональной воронки депрессии). Осуществление
чистки русел и снижения уреза реки будет в гидрогеологическом отношении
вызывает усиление разгрузки болотного и флювиогляциального горизонтов
86
ГЕНЕЗИС, ДИНАМИКА (В ПРОШЛОМ, НАСТОЯЩЕМ) II
3 реку, но это нельзя считать достоверным результатом поскольку
практически не учитывались гидрологические особенности территории. В результате
моделировании естественной ситуации получается, что на взаимодействие с
рекой идет около 3% всего расхода потока первого водоносного пласта.
Получены следующие выводы: — ВБК является областью питания
каменноугольного водоносного комплекса, что указывает на необходимость
охраны и защиты подземных и грунтовых вод; — основное взаимодействие
четвертичных и каменноугольных подземных вод происходит в нижней
части ВБК, где имеется большое гидрогеологическое окно (погребенная долина,
на отдельных участках размывшая слабопроницаемые отложения); —
выявлена тесная взаимосвязь уровней грунтовых вод и водно-болотного
комплекса; — доказано влияние объема водозабора на уровень грунтовых вод.
Для более точной оценки причин подтопления на территории ВБК
необходимо начать мониторинг объекта до начала реализации проектных
решений. В целом можно говорить о том, что развитие подтопления на
территории ВБК обусловлено как гидрологическими, так и гидрогеологическими
причинами. Это вызывает необходимость поиска варианта их совместного
изучения для решения вопроса об их взаимодействии и соотношении, что
необходимо для выбора эффективных методов борьбы с подтоплением.
[1] Гвоздикова Г.А., Цветкова И.В. Гидрогеоэкологические условия водно-болотного
комплекса р.Яуза // Физические проблемы экологии: Тез. докл. Москва, 1999. [2]
Гидрогеодинамические расчеты на ЭВМ. М.: Изд-во МГУ, 1994. 335 с. [3] Орлов
М.С. Гидрогеоэкология Москвы // Бюлл. МОИП. Отд. геол. 1997. Т.72. Вып.5. С.
18-25.
87
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Ш. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ,
ПЕРВИЧНАЯ ПРОДУКТИВНОСТЬ БОЛОТ
И ЗАБОЛОЧЕННЫХ ЛЕСОВ
СРАВНИТЕЛЬНАЯ ОЦЕНКА ВОДНОГО РЕЖИМА
ЗАБОЛОЧЕННЫХ ЕЛЬНИКОВ
В.И. Абражко
Ботанический институт РАН, Санкт-Петербург
Важнейшие черты струюурно-функциональной организации таежных
биогеоценозов теснейшим образом связаны и отражают в себе особенности
динамики водного баланса экотопа и водообмена в системе: почва —
фитоценоз — атмосфера. В этой связи оценка роли влаги должна включать в
себя и изучение эколого-физиологических процессов и реакций древостоев
на условия влагообеспеченности и главнейшие режимы литосферы. Такой
подход лучше всего согласуется с современными задачами лесоведения и
практическими нуждами лесного хозяйства, поскольку позволяет в
наибольшей степени выявить значение влаги в регуляции важнейших
процессов, протекающих в природных экосистемах. Он является научной основой
для разработки практических мероприятии, направленных на устранение
нежелательных эффектов избытка влаги, повышение продуктивности и
устойчивости древостоев.
Многолетние исследования осуществлялись на территории Центрально-
лесного биосферного государственного заповедника в относительно
разновозрастных субклимаксовых ельниках: липняково-ясменниковом, чернично-
кисличном, сфагново-черничном, чернично-пупвдцево-сфагновом (см.
табл.). В экологическом отношении они составляют ряд, отражающий их
реакции на увеличение избыточного увлажнения, ухудшение аэрации,
питательного режима почв.
В отличие от высокопродуктивных ельников, занимающих относительно
хорошо дренированные участки, ельники сфагновой группы приурочены к
слабопроточным понижениям и депрессиям мезорельефа. Они испытывают
постоянный приток воды с соседних территории, а её отток из почвы
протекает очень медленно и в течение всего года. Это приводит к накоплению
влаги, избыточному увлажнению почвенного профиля и постоянно
высокому уровню верховодки. Наиболее сильному и длительному затоплению
подвергается торфяная почва ельника чернично-пушицево-сфагнового, где ель
находится на пределе своего синэкологического ареала, и её эдификаторные
позиции сильно ослаблены. Содержание кислорода в почвенных водах этих
сообществ крайне низкое. Оно не превышает 0,5 мг/л даже после дождей.
88
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
Комплекс неблагоприятных условии, обусловленный анаэробиозом,
сокращает корнеобитаемый слой до 5-10 см.
Некоторые показатели водного режима ельников (числитель — средняя величина,
знаменатель — диапазон изменчивости)
Тип ельника
липняково-
ясменниковый
чернично-
кисличный
сфагново-
черничный
чернично-пуши-
цево-сфагновый
Класс
бонитета
1а
I
IV
Va
Запасы
влаги в слое
0-20 см, мм
67.7
27,5-104,6
65.3
27,5-101,7
115Л
16,7-178,7
159.8
69,3-184,9
Глубина
верховодки
см
70.2
+2,1-234,5
60.8
+2,4-215,5
21.7
+5,5-128,2
5,4
+18,0-69,4
Среднедневная
интенсивность транспи-
рации хвои, мг/г ч
80
36-146
78
23-130
98
20-152
115
32-193
Существенно менее сильное затопление испытывала торфяная почва
ельника сфагново-черничного, концентрация 02 в воде достигала 3,0-3,2 мг/л.
Но ограничения высокого уровня почвенно-грунтовых вод и анаэробиоза на
нормирование корнеобитаемого слоя и подбор видового состава сообщества
ещё очень велико. Отрицательное влияние кратковременных подтоплении
на жизнедеятельность корней деревьев в верхнем слое подзолистых почв
высокопродуктивных ельников невелико, а запасы влаги значительны.
Анализируя нормы реакций ели на факторы почвенной среды следует
отметить, что наиболее значительные различия между древостоями
наблюдаются в степени развития, структуре и характере распределения в почве
физиологически активной части корневых систем, играющих важную роль в
поглощении питательных веществ и воды. Несмотря на уменьшение
корнеобитаемого слоя для заболоченных ельников характерно развитие у
деревьев относительно более мощного всасывающего корневого аппарата, что
уменьшает у них коэффициент водного обслуживания хвои физиологически
активными корнями более, чем в 1,5-4 раза по сравнению с
высокопродуктивными системами. Наряду с этим в силу более слабого развития и
значительно меньших размеров деревьев в пологе заболоченных древостоев
создается более благоприятный режим освещённости и усиливается
проветривание крон. С уменьшением высоты деревьев сопротивление водному току
проводящей системы снижается, скорость поступления воды в кроны
возрастает. Эти факторы — важная предпосылка поглощения большего
количества почвенной влаги древостоями избыточно увлажненных фитоценозов,
они способствуют повышению их водного обмена, снижению водного
дефицита и водного потенциала в корнях и хвое. Интенсивность транспирации
деревьев в сфагновых ельниках обычно в 1,5 раза выше, чем в высокопро-
89
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
дуктивных сообществах, продуктивность транспирации значительно ниже.
В годы высокого количества осадков в результате сильного подтопления
корнеобитаемого слоя и отмирания корней происходит гибель деревьев й
распад древостоев ели. Цепь взаимообусловленных причинных связей,
характеризующих отмирание ели можно представить в следующем виде: про.
должительное затопление почвы — нарушение аэрации в корнеобитаемом
слое, анаэробиоз — отмирание всасывающих корневых окончаний —
уменьшение подачи воды в кроны — рост водного дефицита в хвое —
подавление и прекращение фотосинтеза, частичное осыпание хвои старшего
возраста — отсутствие пластических веществ для восстановления
всасывающего аппарата корней — исключение возможности улучшения
водоснабжения, дальнейшее развитие водного стресса -осыпание хвои последних
лет и отмирание деревьев. Главным звеном в цепи этих связей становится,
очевидно, нарушение продукционного процесса и тесно связанное с ним
отсутствие возможности регенерации корневых систем. Такое состояние
наступает гораздо раньше достижения сублетальных значении водного
дефицита в хвое и тем самым косвенно сокращает адаптационную возможность
ели выдерживать более высокий водный стресс в ассимиляционном
аппарате, как это наблюдается в засуху в высокопродуктивных древостоях.
Разрушение сфагновых ельников в результате сильного подтопления может
сопровождаться экзоэкодинамическими сменами сообществ. Сосна
обыкновенная и береза пушистая, обычно составляющие заметную часть в
ельниках, обладают более совершенными механизмами устойчивости к
затоплению и лучше переносят неблагоприятные условия аэрации почвы, чем ель
европейская. На первом этапе после распада древостоев ели часто
нормируются временные редкостойные насаждения с преобладанием этих пород.
ТОРФЯНИКИ ЮЖНОГО ПОБЕРЕЖЬЯ ОЗ.БАЙКАЛ: МИКРОФЛОРА И
БИОХИМИЧЕСКАЯ АКТИВНОСТЬ
A.M. Антоненко
Институт географии СО РАН, Иркутск
Проведены экспериментальные исследования функционирования
болотных фаций, окаймляющих оз.Байкал от п.Култук до п. Байкальск по р. Баб-
ха. Болотные ландшафты прибрежно равнинной полосы находятся в зоне
активного воздействия выбросов местных предприятий, автомобильного и
железнодорожного транспорта. Изучались верховые болота, расположенные
в 2,5 км восточное п. Утулик (фация 1), в 5 км юго-восточнее п.Слюдянка
(фация 2) и в 4 км южнее п.Култук в районе мыса Шаманский (фаций 3, 4).
Современная растительность представлена в основном древесно-
90
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
кустарничково-моховыми типами фитоценозов. Болота залесены: кедр, кедр
с сосной, реже береза. Древостой разреженный, подлеске.— кедровый стла-
дик (фация 2). Видимая мощность торфяного слоя — 0,8-1,3 м.
Как показали исследования в метаболизме органического вещества
торфяной толщи (до 30 см) велика роль микроскопических грибов (гифомице-
тов), представленных небольшим разнообразием. Выявлена тенденция
приуроченности гифомицетов к различным субстратам торфяных подстилок.
Наибольшая встречаемость (до 20%) микроскопических грибов отмечена в
подстилках, где повышены количественные параметры кедра (фация. 1),
усиливается участие березы, полнота и сомкнутость древостоя (фация 2).
Более стабильной и экологически приспособленной группой были
гетеротрофные микроорганизмы. Число бактерий, использующих минеральный
азот, в 3-5 раз ниже (для слоя 0-20 см) в зависимости от местоположения
фации, что свидетельствует о низкой минерализации органического
вещества и подтверждается накоплением его в виде торфяных отложений.
Функционирование специфического комплекса микроорганизмов,
обусловленного экологическими условиями почвенной среды, сопряжено с
низким уровнем активности отдельных биохимических процессов, связанных с
круговоротом азота, углерода и окислительными превращениями
органических остатков. Повышенная обводненность и невысокий уровень теплообес-
печенности торфяного субстрата препятствуют трансформации
азотсодержащего органического вещества. Гидролиз протекает очень слабо, о чем
свидетельствует низкий уровень активности уреазы. С глубиной она еще
уменьшается и составляет 7-25% ее величины в верхнем (0-10 см) слое
торфа. Такая же тенденция отмечена и в распределении подвижного азота.
Особенности превращения и накопления легкоусвояемых форм углерода
в торфяных почвах определяются качественным составом растительного
опада и гидротермическими условиями природной среды. Наилучшие
условия для круговорота углеводосодержащих соединений отмечаются в
болотных фациях с обильным поступлением лиственного и травянистого опада,
содержащего легкогидролизуемые углеводы. Максимум в активности инвер-
тазы, катализирующей превращения углеводосодержащего органического
вещества, отмечен в верхнем (20 см) слое торфяной залежи, где
сосредоточена основная масса корней и достаточная аэрация почвенной среды.
Активность инвертазы уменьшается вниз по профилю в 20-40 раз в
зависимости от местоположения фации. Тенденция уменьшения активности
гидролитических ферментов по профилю торфяного слоя отмечена при изучении
сосново-кустарниково-сфагновых болот Западной Сибири (южно-таежное
Прииртышье у с. Горно-Слинкино).
Динамика активности окислительных ферментов имеет свои
особенности. Увеличение в верхней части торфяной толщи (0-10 см) фенольных со-
91
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
единений типа танинов сопровождается относительным увеличением актив*
ности пероксидазы. Здесь же выявлены минимальные значения рН (4,2-4,0)
торфяного слоя. Недоокисленные формы фенольных соединений, митри*
рующие из верхней части профиля в нижележащие слои торфа (20-30 см),
подвергаются активной гумификации при участии полифенолоксидазы.
Низкий уровень биохимической трансформации остатков болотной
растительности приводит к медленным темпам обмена веществом и потери
продуктивности фитоценозов в верховых болотах исследуемого региона.
ЛИШАЙНИКИ ЗАБОЛОЧЕННЫХ ТЕРРИТОРИЙ ОКСКОГО
ЗАПОВЕДНИКА
B.C. Барсуков, Н.В. Пелепец, Т.Ю. Толпышева
МГУ, биологический факультет
Окский государственный биосферный заповедник (ОГБЗ) расположен в
юго-восточной части Окско-Клязьменского междуречья. Специфика
характерного для этой территории эолового ландшафта приводит к чередованию
ксероморфных сосновых лесов, формирующихся на гребнях дюн, и
многочисленных (зачастую и небольших по площади) болот в понижениях мевду
дюнами. При этом преимущественно пойменное расположение заповедника
создает условия для формирования также и значительных по площади мезо-
трофных болот осоко-вейникового типа («кочкарники») в поймах крупных
рек. Болота занимают не менее 40% площади основной территории ОГБЗ.
В ходе лихенологических работ 1997-98 гг. были обследованы 3
характерные для ОГБЗ болотные ассоциации: заболоченный лес по краю
переходного болота, заболоченное междюнное понижение и заболоченная гарь,
образовавшаяся после верхового пожара в сфагновом сосняке. Участок леса
по краю переходного болота сформировался на торфяной болотной почве
(кв. 130) в условиях постоянного переувлажнения. Древостой представлен
преимущественно березой IV бонитета, на повышениях изредка встречаются
сосна, осина и ольха. В травяном ярусе доминирует Calamagrostis canescens,
в моховом — различные виды Sphagnum. Заболоченное междюнное
понижение (кв. 102) занимает сравнительно небольшую площадь, но, в силу
особенностей микрорельефа, имеет сложную структуру. В древостое
преобладают береза и осина, в моховом ярусе — Pleurozium schrebeh и виды
Sphagnum. Для заболоченной гари в междюнном понижении (кв. 39)
характерно полное отсутствие древостоя, но много валежа сосны. Моховой ярус
развит очень хорошо и представлен преимущественно видами p. Sphagnum.
Общим для всех исследованных участков является отсутствие эпигейных
лишайников. Повышенная обводненность создает условия для пышного раз-
92
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
вития мохового покрова и препятствует развитию лишайников. В условиях
избыточного увлажнения они вымокают и не могут конкурировать со мха-
ин. Лишайники развиваются только на стволах деревьев (на коре и на эпи-
фитных мхах), а при их отсутствии (заболоченная гарь) на валеже.
Общее число видов невелико — 52, что обусловлено отсутствием эпи-
гейных лишайников. Среди лишайников преобладают виды p. Cladonia —
17. Видовое разнообразие других родов меньше: Physcia и Lecanora — по 4,
phaeophyscia — 3, Cladina, Physconia, Caloplaca, Candelariella, Lecidella,
Pertusaria - no 2, Anaptychia, Biatora, Evernia, Hypocenomyce, Hypogymnia,
Parmelia, Parmeliopsis, Tuckermanopsis, Usnea, Vulpicida, Xanthoria, — no 1
виду. Повышенная влажность воздуха способствует развитию соредиозных
лишайников: 50% макролишайников составляют соредиозные виды.
Видовой состав лишайников исследованных сообществ обусловлен как
количественным соотношением пород форофитов в древостое, так и
особенностями субстрата. Наибольшее число видов лишайников (33) найдено в
заболоченном междюнном понижении. В других сообществах их меньше: в
березняке, по краю переходного болота, — 22, на гари 20 видов.
В заболоченном березняке найдены практически все лишайники,
отмеченные для междюнного понижения. Исключение представляют Cladonia
macilenta var. bacillaris (Genth.) Schaerer, Physcia stellaris (L.) Nyl.,
Candelariella aurella (Hoffin.) Zalbr., Biatora helvola (Koerber) Hellbom,
Hypocenomyce scalaris (Ach.) Choisy. Последний вид тяготеет обычно к обгорелому
субстрату и является хорошим индикатором прошедших пожаров. Особенно
бурно он развивается сразу после пожаров, но постепенно вытесняется
другими видами. Особенно много его в настоящее время на гари.
На заболоченной гари, наряду с Я. scalaris, основной фон составляют
виды сем. Cladoniaceae, развивающиеся на валеже. Необычно нахождение в
этих условиях Cladonia bellidiflora (Ach.) Schaerer и С turgida Hoffm.
Первый вид - типичный арктоальпиец, встречающийся, как правило, на почвах
в тундре и лесотундре. Для валежа он не характерен. В московской области,
расположенной севернее, он не зарегистрирован. Не исключено, что он был
вытеснен в этот регион во время последнего оледенения, а своеобразные
условия Мещеры способствовали его выживанию. С. turgida на подобном
субстрате встречается очень редко. Вид обычно произрастает на сухих
песчаных почвах. Во влажных экотопах ранее отмечен не был. Для развития
обоих видов большое значение имеет повышенная освещенность, что,
вероятно, и определило их произрастание на гари, где древостой отсутствует.
В сообществе междюнного понижения найдены нитрофильные,
неморальные лишайники: Anaptychia ciliaris (L.) Koerber и Phaeophyscia
nigricans (Florke) Moberg. Они очень редко встречаются на территории ОГБЗ и
приурочены к осинам. Возможно они редки и на территории рязанской об-
93
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ласти. Ph. nigricans занесена в Красную Книгу Московской области [1998].
Лихенофлора осины заметно отличается от лихенофлоры других форо-
фитов как по числу видов лишайников (24), так и по их составу. По
сравнению с другими древесными породами, произрастающими в заболоченных
условиях, осина имеет субнейтральную кору, что и определяет эти
различия.. Все виды сем. Physciaceae, Lecanora allophana Nyl., L.argentata (Ach.)
Malme, Lecidella elaeochroma (Ach.) Hazsl., Xanthoha parietina (L.) Th. Fr.
найдены исключительно на осинах.
Только валеж несколько превосходит осину по числу видов лишайников
— 25. За исключением 5 видов, все они относятся к сем. Cladoniaceae. В
основном на валеже развиваются виды, либо перешедшие с почвы,
например, Cladonia furcata (Hudson) Schaerer, С crispata (L.) Flotow, С gracilis
(L.) WillcL а также виды р. Cladina, либо встречавшиеся ранее как эпифиты,
например, Hypogymnia physodes (L.) Nyl.. Lecanora populicola (DC.) Duby.
Интересно нахождение Pertusaria albescens (Huds.) M. Choisy et Werner.
Большинство эпифитных видов р. Pertusaria растет на лиственных породах
деревьев, нередко поверх мхов. Особенно обильны они в районах с высокой
влажностью воздуха. Вид широко распространен в мире, но на равнинах он
встречается реже, чем в предгорьях.
На эпифитных мхах найдено всего 6 видов лишайников; 5 из них
относятся к p. Cladonia. Преимущественно это виды, которые в лесах часто
растут на моховом покрове на комлях деревьев: Cladonia coniocraea (Florke)
Sprengel, С fimbriata (L.) Fr., С pyxidata (L.) Hoffm., С floerkeana (Fr.)
Florke, а также Hypogymnia physodes — эврисубстратный лишайник.
В целом лихенофлора болотных сообществ ОГБЗ довольно бедная и
представлена в основном широко распространенными видами. Однако
наличие здесь редких видов лишайников позволяет говорить об определенных
микроклиматических условиях, способствующих их развитию.
СООТНОШЕНИЕ ГУМУСООБРАЗОВАНИЯ И ТОРФОНАКОПЛЕНИЯ В
ЗАБОЛОЧЕННЫХ ЛЕСАХ СРЕДНЕЙ И СЕВЕРНОЙ ТАЙГИ
Л. Г. Богатырев, В.А. Сапожникова
МГУ, факультет почвоведения
Заболоченные леса являются одними из основных, и часто
преобладающими компонентами таежных биогеоценозов, особенно в средней и
северной тайге. Вместе с тем, своеобразие соотношения процессов
преобразования органического вещества на различных территориях, и прежде всего гу-
мусообразования и торфонакопления, обусловлено различными причинами.
Помимо гидротермических условий, которые являются общими для всех
территорий, региональная специфика обусловлена целым рядом других фак-
94
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
хоров. В их числе важнейшими являются характер почвообразующих пород,
особенности рельефа, история развития района в голоцене. Исследование
северотаежных экосистем севера Западной Сибири (бассейн реки Лонг-
Юган). лесов Архангельской области и республики Коми, включающее как
изучение морфологии органопрофилей, так и оценку гумусного состояния в
целом, позволило установить некоторые общие закономерности.
Особенности торфонакопления на севере Западной Сибири обусловлены
реликтовым характером торфяных бугров, находящихся на различных
стадиях деградации. Различная мощность торфа на фоне подстилания
тяжелосуглинистыми или песчаными отложениями обусловливает своеобразный
характер современного эволюционного развития. При незначительной
мощности торфа, подстилании песчаными отложениями и поселении леса
возможна трансформация торфа с его последующей дифференциацией и
образованием грубогумусных горизонтов. На тяжелосуглинистых отложениях и
при значительной толщине торфа, его последующая эволюция явно носит
замедленный характер. Существует и современная эволюция торфа,
обусловленная явлениями перехода болотных экосистем из гидроморфной
стадии в автоморфную с постепенной профильной дифференциацией в
зависимости от степени поднятия над уровнем болота, сопровождаемого
образованием мерзлотных форм мезорельефа, часто плоскобугристого характера.
Гумусообразование в этом случае также диагностируется по слабой прокра-
шенности горизонтов, залегающих под торфом. В низкобонитетных лесах на
песчаных подзолах гумусообразование выражено чрезвычайно слабо.
Отмечено своеобразие пространственного распределения торфянистых и
деструктивных подстилок. Если в лесах европейского севера современное торфона-
копление обусловлено характером микропонижений, то в лесах севера
Западной Сибири их генезис связан с характерными остаточными формами
мерзлотного комплекса, при котором торфянистый характер приобретают
подстилки при выходе к поверхности тяжелосуглинистых пород,
чередующихся в горизонтальном отношении с легкими песчаными породами.
Торфонакопление в заболоченных лесах европейского севера, например,
в республике Коми, также имеет своеобразный характер. Если
торфонакопление в сосновых экосистемах имеет часто локальный характер,
обусловленный особенностями рельефа, то торфонакопление в еловых лесах
характеризуется значительной пространственной протяженностью, например, в
пределах распространения суглинистых пород, характерных для территории
изученного Сосногорского лесхоза. Обращает на себя внимание, что на
большой территории мощность торфа все-таки ограничена, и часто не
превышает 60-70 см. что, вероятно, является своеобразным пределом для
данных гидротермических условий. Существенно, что на песчаных отложениях,
причем занимающих значительные территории, преимущественная роль
95
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
принадлежит процессам преобразования и минерализации органического
вещества на фоне отсутствия его закрепления в профиле почв. Исследования
территорий бассейна реки Айювы, Тобыси, Кедвы показали, что в этих ус-
ловиях, характерных для северотаежных и частично среднетаежных ланд,
шафтов, современные центры гумификации перемещаются от автоморфных,
часто занятых заболоченными лесами, ландшафтов, к транзитным формам
рельефа. Последние характеризуются приуроченностью к склонам, с
характерным проточным увлажнением, с повышенной трофностью, и
соответственно с более высоким бонитетом произрастающих лесных биоценозов.
Аналогичная ситуация описана нами и для лесов Архангельской области,
где накопление гумуса в перегнойных горизонтах также приурочено к лесам
с проточным характером увлажнения, тогда как в автоморфных позициях
интенсивность гумусообразования выражена незначительно, за
исключением случаев формирования лесов на карбонатных породах в условиях
хорошего дренажа. Следует отметить, что смещение максимумов
гумусообразования к полугидроморфным условиям на фоне преобладающего
распространения заболоченных лесов является подтверждением некоторых
предположений И.В. Тюрина о роли гидроморфизма в гумусообразовании.
Образовавшийся в таких условиях гумус может быть довольно устойчив во
времени. Некоторые изученные почвы сосновых лесов в пределах лесостепной
зоны (Присурский лесной массив), характеризуются не только высоким
содержанием гумуса, достигающим 10% при его гуматном составе (Сгк:Сфк
=1,7-2,3), но и его явно углефицированным характером, что подтверждается
микроморфологическими исследованиями. Можно предположить, что при
близком залегании тяжелосуглинистых грунтов, подстилающих песчаные
отложения, накопление гумуса проходило в предыдущую стадию развития
почв при гидроморфном режиме. В дальнейшем, по мере развития
дренирующей овражной сети, происходило постепенное обсыхание почв,
сопровождающееся конденсацией продуктов преобразования органического
вещества и образованием унифицированного органогенного материала. В виде
реликта он сохранился в составе гумуса до настоящего времени.
Таким образом, с учетом того, что гумусное состояние почв интегрально
отражает все стадии преобразования органического вещества, показано, что
как торфонакопление, так и в ряде случаев гумусонакопление носят
зачастую реликтовый характер, и связаны с предыдущими гидроморфными
стадиями эволюции почв. Направление современных стадий эволюции
заболоченных лесных почв связано с соотношением процессов гумусообразования
и торфообразования, которое, в свою очередь, определяется комплексом
биоклиматических, гидрологических, литологических условий и характером
рельефа.
96
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
КОМПЛЕКС МИКРОСКОПИЧЕСКИХ ГРИБОВ В БОЛОТАХ
СЕВЕРНОЙ ТАЙГИ
Г.И. Булавко
Институт почвоведения и агрохимии СО РАН, Новосибирск
Одной из важнейших проблем экологии является изучение процесса
распада органических соединений биологического происхождения, таких как
клетчатка и лигнин. Эти вещества — главные составные части
растительного опада и их разложение важно для поддержания круговорота соединений
углерода в каждой экосистеме. Характер и скорость разложения
растительных остатков определяются тремя основными факторами: составом
растительного материала, гидротермическими условиями, комплексом
микроорганизмов-деструкторов. Исследования проведены в нефтедобывающих
районах Тюменской области, расположенных в зоне северной тайги.
Особенностью экосистем северной тайги являются низкая температура, высокая
влажность и кислотность почв - факторы определяющие преимущественное
развитие микроскопических грибов среди почвенной микробиоты.
Микроскопические грибы — абсолютные гетеротрофы, их развитие
определяется наличием отмерших растительных остатков. Этим потребностям
в полной мере соответствуют торфяники. В живых растениях' содержание
углерода колеблется от 47 до 50%, в разложившихся торфах оно достигает
60% [1]. Лимитирующим фактором в процессе разложения растительных
остатков является содержание азота. Чем выше содержание азота и ниже
соотношение C:N, тем интенсивнее идет процесс разложения. Источником
азота в торфе являются торфообразующие растения, в которых содержание
азота колеблется от 0,5 до 3%. В разложившихся прослойках сфагнового
торфа было 0,95; 1,1; 1,5% N, в разложившемся торфе — 0,7; 0,9; 0,8%. В
связи с вышеизложенным именно комплекс микромицетов в данном случае
будет определять интенсивность деструкционных процессов. Для оценки
участия микроскопических грибов в деструкционных процессах необходима
оценка их содержания в почве. Самой общей характеристикой комплекса
микромицетов является величина их общей массы, а также соотношение
активной (мицелий) и неактивной (споры) частей.
Для определения величины параметров комплекса микроскопических
грибов (длина и масса мицелия, численность и масса спор с выделением
темно-окрашенных форм) использован метод мембранных фильтров
Хансена в модификации [2]. Величина грибной биомассы на болотных участках
сопоставима с верховыми торфяниками. Ее общие запасы и показатели
отдельных компонентов различалась в зависимости от типа болот и
положения участка.
Величина общей биомассы, длины и массы мицелия в 3-4 раза выше в
97
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
мочажинах относительно гряды или кочки (см. табл.). С глубиной
происходит изменение как величины общей биомассы, так и составляющих
компонентов: с глубиной общая биомасса в мочажине снижается, на гряде -
возрастает; численность спор, как правило, и на гряде и в мочажине возрастает
с глубиной. Изменение величины суммарной биомассы отражало
преобладание того или иного компонента в составе комплекса и имело разный
характер в каждом случае. Наиболее благоприятные условия для развития
грибов — в мочажине, пределы варьирования общих запасов биомассы
здесь составляли 6736-1006 мкг/г, тогда как на кочке — 5765-827 мкг/г.
Комплекс микроскопических грибов в болотах Тюменской области
Глу- Кочка, гряда Мочажина, межкочье
бина, Мице- Споры, Суммарная Мице- Споры, Суммарная
см лий, млн/г биомасса, лий, млн/г биомасса,
м/г мкг/г % спор м/г мкг/г %спор
Р.1 Грядово-мочажинное мезоолиготрофное болото
0-10
10-20
0-10
10-20
20-30
0-10
10-20
20-30
0-10
10-20
20-30
106
1506
3,5
6,3
827 47 711
5755 6 467
Р.2. Полигональное олиготрофное болото,
347
684
650
359
438
556
4,3
7,6
2,0
1899 24 1512
3557 21 187
2799 4 466
Р.З. Полигональное олиготрофное болоте
0,8
2,2
3,9
1650 10 1463
2015 10 216
2490 8
Р.4. Мезотрофное болото
1562
495
1096
6,0
8,3
3224
2855
загрязнение
4,7
4,3
9,5
>, контроль
7,0
2,1
23,2
7,4
13,4
6394
1006
2573
6736
1103
7565
2280
5329
9
33
3
23
26
И
13
15
10
15
Значительные различия выявлены при сравнении мочажин и межкочий
болотных участков разной трофности. Наиболее богато грибами
мезотрофное болото, незначительно от него отличается олиготрофное, величины
общей массы и массы мицелия в мезо-олиготрофном примерно в два раза
ниже. В целом, исследованные участки отличаются высоким содержанием
общей биомассы микроскопических грибов и активно развитым мицелием.
[1] Доктуровский B.C. Торфяные болота. ОНТИ НКТП СССР. 1935. 225 с. [2] Дем-
кина Т.С., Мирчинк Т.С. Определение грибной биомассы в почвах методом
мембранных фильтров // Микология и фитопатология. 1983. Т. 17. № 6. С.517-520.
98
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
РАДИАЛЬНЫЙ ПРИРОСТ СОСНЫ ОБЫКНОВЕННОЙ
НА БОЛОТАХ И СУХОДОЛАХ
С. В. Васильев, А. М. Перегон
Институт почвоведения и агрохимии СО РАН, Новосибирск
Реакции деревьев на изменения климата легче всего определяются
измерением радиального прироста древесных стволов. В общем случае эти
реакции в различных местообитаниях различны. Известно, что реакция деревьев
на климат наиболее выражена в неблагоприятных условиях. Там, где
условия теплообеспеченности и увлажнения близки к оптимальным, разногодич-
ные вариации температуры воздуха и количества осадков мало сказываются
на приросте деревьев. Представлены результаты сравнительного анализа
изменчивости прироста сосны обыкновенной, произрастающей на болотах и
на сухих песчаных почвах северотаежной подзоны Западной Сибири.
Предполагалось, что деревья, произрастающие на хорошо проницаемых для
влаги песчаных почвах, в жаркие годы могут страдать от засухи, тогда как на
болотных почвах они страдают в большей мере от избытка осадков.
Для анализа радиального прироста собраны поперечные спилы сосны в
60 км к северу от г. Сургут (Сургутское Полесье) и в районе г. Ноябрьск
(Сибирские Увалы). Образцы отбирались в лишайниковых и зеленомошных
лесах на автоморфных песчаных почвах (24 образца) и в древостоях сосно-
во-кустарничково-сфагновых сообществ олиготрофных болот (17 образцов).
При построении обобщенных рядов первоначально выделялся тренд
многократным повторным сглаживанием ряда диаметров простой средней
арифметической по трем точкам с фиксированными значениями крайних членов
ряда. После 128 итераций сглаживания выделенный тренд вычитался из
данных исходного ряда для получения индексов прироста. Независимо был
опробован и стандартный метод, используемый при дендроклиматологиче-
ских исследованиях и доступный, благодаря свободно распространяемым
программам типа ARSTAN (International Tree-Ring Data Bank Program
Library, 1997). Построено два обобщенных рада индексов прироста для
Сургутского района (SB — 9 моделей с болотных и SF — 6 моделей с
автоморфных местообитаний), два для района г. Ноябрьск (NB — 8 моделей с
болотных и SN — 18 моделей с автоморфных местообитаний) и кроме того
два сводных рада по болотным (АВ) и суходольным (AF) местообитаниям.
Данные по климату были получены в Центре глобальных климатических
исследований (National Climatic Data Center, Asheville, NC); данные
распространяются свободно. В анализ вошли ежемесячные данные метеостанции
Сургута по количеству осадков (1917-1983 гг.) и по максимальной и
минимальной температурам (1931-1983 гг.). Помимо этого проанализирован рад
чисел Вольфа (среднегодовое число солнечных пятен за период с 1917 по
99
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
1983 год), полученный по Интернету в Sunspot Index Data Center,
Королевской обсерватории Бельгии. Анализ связи приростов с климатом проводился
двумя методами: многомерный регрессионный анализ и вейвлет-анализ.
Вейвлет-анализ [1] является вариантом спектрального анализа и основан
на разложении исходного временного ряда на непрерывную сумму
гармонических составляющих с изменяющейся по времени амплитудой. При стан-
дартном Фурье преобразовании выделяется дискретная сумма гармоник с
постоянной по времени амплитудой. Преимуществом вейвлет-анализа
является получение локальных спектров на любом отрезке времени.
Для летних осадков выявлены циклы с периодом 25-30 лет, 44 года и
более слабые циклы с периодом 18-19 лет. По зимним осадкам выделены
циклы с периодом 10, 19-20 лет и 44 года. Колебания максимальной и
минимальной температур почти синхронны с периодами 14, 20 лет и 32 года. Ряд
чисел Вольфа имеет доминанту с периодом около 10 лет. С 1917 по 1950
годы циклы были чаще (с периодом 9-10 лет), в настоящее время, начиная с
1950-х годов частота уменьшилась и длительность циклов приближается к
11 годам. Вейвлет-анализ позволяет предполагать некоторую связь осадков
с солнечной активностью: по сумме выделенных гармоник в диапазоне
частот с периодами 6-16 лет, коэффициенты корреляции между числами
Вольфа и осадками (летними и зимними) равны +0.46 и +0.76 соответственно.
Колебания приростов сосны на болотах и на суходолах в целом
совпадают, но колебания на болотах часто запаздывают по фазе на 2-3 года
относительно лесных местообитаний. Для приростов сосны выделяются две
доминирующие гармоники с периодами 7 лет и 10-11 лет. В некоторых случаях
они расслоены и отделены друг от друга, как это наблюдается для приростов
на суходольных местообитаниях под Сургутом. В других случаях они могут
постепенно переходить друг в друга, другими словами, имеется некоторая
периодическая компонента, частота которой не остается постоянной с
течением времени, а изменяется. При этом до 1950 года доминируют циклы
продолжительностью около 7 лет, после 1950 года — циклы длительностью
10-11 лет. Существенных корреляций между климатическими переменными,
числами Вольфа и приростами деревьев на этом уровне не выявлено.
Связь между отдельными элементами климата и рядами прироста была
выявлена регрессионным анализом и расчетом частных корреляций. Как
независимые переменные регрессионной модели использованы
ежемесячные данные по сумме осадков, максимальным температурам и по амплитуде
температур (разность tmax и tmm за месяц). Данные с октября по декабрь
брались за год предшествующий приросту. Значимых корреляций было
выявлено немного. По сводным рядам высокие коэффициенты корреляции
обнаруживаются для осадков апреля, для максимальных температур июня и для
амплитуды температур за июнь и за октябрь предшествующего года; Боль-
100
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
me всего достоверных коэффициентов для болот района Сургута (см. табл.).
Наиболее интересным результатом проведенного анализа является то,
что сосновые древостой на болотах и суходолах на изменения климата
реагируют сходным образом. Это проявляется в совпадении частотного спектра
колебаний индексов радиального прироста и в совпадении регрессионных
моделей (по меньшей мере в знаках коэффициентов корреляций). Можно
полагать, что лимитируюпщм фактором роста леса на олиготрофных
болотах является не переувлажненность местообитаний, а иные факторы.
Наиболее вероятным является недостаток элементов минерального питания.
Полученные выводы могут быть интересны при объяснении появления прослоек
древесного торфа в олиготрофных торфах суббореального и атлантического
периодов голоцена. Возрастание теплообеспеченности и снижение
увлажненности климата непосредственно не способствуют росту древостоев на
болотах. Их действие проявляется опосредованно, однако приведенных
данных для обсуждения конкретного механизма действия пока недостаточно.
Частные коэффициенты корреляции между рядами индексов прироста и
климатическими переменными (подчеркнуты значимые на 95% уровне достоверности)
Ряды Осадки Температуры Амплитуды
приростов Апр. Янв. Фев. Март Июнь Сент. Окт.я Дек. Июнь Окт.
Nf 0.431 0.224 0.541 -0.387 -0.499 -0.104 -0.202 -0.447 0.639 -0.500"
Nb 0.305 -0.045 0.306 -0.276 -0.210 -0.021 -0.113 -0.181 0.349 -0.124
Sf 0.556 0.028 0.408 -0.169 -0.331 -0.192 -0.047 -0.157 0.459 -0.296
Sb 0.429 0.619 0.353 -0.555 -0.670 -0,582 -0.550 -0.539 0.686 -0.656
Af 0.574 0.211 0.603 -0.406 -0.549 -0.171 -0.199 -0.455 0.689 -0.540
Ab 0.538 0.535 0.486 -0.613 -0.680 -0.527 -0.540 -0.561 0.731 -0.637
Работа выполнена при частичной поддержке NWO, Netherlands по
совместному проекту «Carbon storage and atmospheric exchange within West
Siberian peat layers related to global climatic change process», 1998-2000 гг.
[1] Torrence Ch., Compo G.P. A Practical Guide to Wavelet Analysis // Bulletin of the
American Meteorological Society. 1998. V. 79. N. 1. P. 61-78.
ПСИХРОФИЛЬНЫЕ АЦИДОФИЛЬНЫЕ МЕТАНОТРОФЫ В
СФАГНОВЫХ БОЛОТАХ
Л.В. Васильева, Ю.Ю. Берестовская, Г.А. Заварзин
Институт микробиологии РАН, Москва
Глобальным источником метана — второго по значимости после
углекислоты парникового газа — являются болотистые ландшафты северного по-
101
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
лушария. Как и для других парниковых газов, главным источником метана
является деятельность бактерий. Выход метана в атмосферу обусловлен
дисбалансом между метаногенным и метанотрофным сообществами. В це.
лом метаногены и метанотрофы образуют взаимосвязанную систему
биологического круговорота метана, называемую циклом Зёнгена.
Кислые торфяные болота увлажнённых земель являются доминирующи,
ми наземными экосистемами бореальной лесной зоны Северной Америки и
Евразии. Болота покрывают 21% территории России. Значения рН водной
фазы в таких экосистемах колеблется от 3,5 до 5, а концентрация
минеральных солей для верховых болот находится в пределах 5-100 мг/л. Низовые
болота с высокой минерализацией и интенсивными процессами
биологического круговорота в общем балансе метана в России играют второстепенную
роль вследствие замкнутости цикла Зёнгена. Напротив, верховые болота с
малой интенсивностью обмена вследствие занимаемой ими площади
рассматриваются как наиболее мощный источник метана, образуемого по всему
профилю болота, включая его глубокие горизонты. Вероятной причиной
служит дисбаланс в биогенной продукции и окислении метана в кислых
условиях. Все ранее описанные метанотрофы из болот относятся к группе ней-
трофилов, развивающихся в низовых болотах с высоким содержанием
минеральных веществ, прежде всего кальция, и близь нейтральным значением
рН [1]. Организмы, ответственные за процесс окисления метана, в
сфагновых болотах с рН 3,5-4,0 и низкой минерализацией были неизвестны.
Неизвестны были и метанотрофы, развивающиеся при низкой температуре.
Изучалось поглощение метана в пробах, отобранных из сфагновых болот
разных мест северной части России — Чукотки, Западной Воркуты,
^Колымы, Камчатки, Архангельской области. Исследованный материал одоатил
север России с запада на восток, характеризующийся низкими
температурами, низкими значениями рН 3-5, а также малой минерализацией воды с
содержанием солей порядка 100 мг/л.
Было установлено, что в слое «белого мха»,, где происходит разрушение
хлорофилла, но растение ещё сохраняет свою анатомию и пустые клетки и
расположенного на расстоянии 5-10 см от поверхности болота, происходит
окисление метана. Во всех случаях потребление метана коррелировало с
развитием микроорганизмов с типичной морфологией метанотрофов,
характеризующимися большими тёмными клетками круглой и овальной формы,
начинёнными пакетами мембран.
Были получены накопительные метанокисляюпЫе культуры из проб
сфагновых болот Воркуты (ZV-1; ZV-2), Колымы (ZK-1; ZK-2), Камчатки
(ZU-1; ZU-4) и Чукотки (ZCH-2; ZCH-5). Накопительная культура ZV-1
окисляла метан одинаково при разных значениях рН. Напротив, скорость
окисления метана при рН 4.0 и рН 6,1 была различной для камчатских и
102
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
колымских культур: при рН 4,0 потребление метана в них начиналось
раньше и происходило с более высокой скоростью. Для образцов из Колымы и
Камчатки она была в 3-3,5 раза выше при рН 4,0, чем скорость при рН 6,1 и
с0Ставила значения 1,7 ммоль метана на литр культуры в сутки и 0,5
ммоль/л/сут для культуры ZU-1; 1,03 ммоль/л/сут и 0,34 ммоль/л/сут для
культуры ZK-1. Результаты указывают на существование ацидофильного
психрофильного метанокисляющего сообщества со скоростью окисления
при рН 4,0 превышающего скорость его окисления при рН 6Д в 3-3,5 раза,
что позволяет говорить о существовании метанотрофов, адаптированных не
только к низкотемпературным, но также к кислым условиям среды.
На накопительной культуре из проб Чукотки установлено, что
природным носителем бактериальных клеток является сфагнум. Основная масса
клеток обнаружена в его гиалиновых клетках, лишившихся внутренних
перегородок. Лишь незначительная часть клеток находилась вне сфагнума в
свободной жидкости. Потребление метана культурой ZCH-2 при рН 4,0
составило 103 ммоль/л, а для ZCH-5 — 94,8 ммоль/л в течение 3-х недель
роста. Основной организм, окисляющим метан в психрофильных и кислых
условиях, — метанотроф, морфологически сходный с Methylobacter [2].
Результаты показывают, что на всей территории севера России от
Чукотки, Камчатки до Полярного Урала, находящейся в зоне вечной мерзлоты,
работает психрофильный метанокисляющии бактериальный фильтр.
Метанокисляющий бактериальный фильтр тесно связан с растениями
сфагнума и представляет собой растительно-микробиальную систему —
физически организованную структуру, по-видимому, способную
контролировать поток метана в атмосферу.
[1] Заварзин Г.А. Биологический цикл метана на территории России // "Глобальные
изменения природной среды и климата" Москва, 1997. С. 347-373. [2] Васильева
Л.В. и др. Психрофильные ацидофильные метанотрофы из сфагнеты зоны вечной
мерзлоты // ДАН 1999 г.(в печати).
ОСОБЕННОСТИ ПРОСТРАНСТВЕННОГО РАСПРЕДЕЛЕНИЯ И
ТАКСОНОМИЧЕСКОГО СОСТАВА САПРОТРОФНОГО КОМПЛЕКСА
БАКТЕРИЙ В БОЛОТНО-ЛЕСНЫХ ЭКОСИСТЕМАХ
А.В. Головченко
Институт почвоведения МГУ-РАН,
Институт проблем экологии и эволюции РАН, Москва
Болотообразование — сложный биогеохимический процесс, изучение
которого требует комплексного подхода, предполагающего сопряженный
анализ водного режима, почвенных процессов, растительного покрова, жизне-
103
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
деятельности беспозвоночных животных и микроорганизмов. Наименее
изученным представляется бактериальный мир болот. Причем среди
значительного количества работ, посвященных микробоценозы торфяников
Белоруссии, Карелии, Западной Сибири и т.д., единичны исследования полного
профиля торфяников. Преобладает анализ только верхних горизонтов,
вовлекаемых в дальнейшем в сельскохозяйственные и лесохозяйственные
работы. Определение таксономического состава бактериальных комплексов
осуществляется большей частью на уровне выявления физиологических
групп бактерий, что не правомочно, в связи с полифункциональностью
микроорганизмов. Кроме того, приводимые в литературных источниках (50-80
гг.) перечни групп или родов бактерий, встречающихся в торфяниках,
получены с помощью старых определителей и в большинстве случаев не
соответствуют принятым в настоящее время названиям бактериальных таксонов.
Целью работы было изучение профильного распределения и
таксономической структуры бактериальных комплексов болотно-лесных экосистем.
Объекты исследования включали почвы болот: низинный нормально-
зольный торфяник под сосняком болотно-травяным, низинный
высокозольный торфяник под черноолыиаником крупнотравно-папоротниковым и
верховой торфяник под сосняком андромедово-пушицево-сфагновым (Западно-
двинский стационар Института лесоведения РАН, Тверская область).
Образцы (зелёные и сухие части растений, подстилки по слоям, почвенные
горизонты) анализировали по сезонам в течение двух лет. Почвенные пробы
отбирали через каждые 20-50 см. буром: вплоть до подстилающих пород (до
3-7 м — в низинных и 5,5 — в верховом торфяниках). Изучались бактерии
сапротрофного блока, включающего аэробные и факультативно-аэробные
формы, являющиеся характерными обитателями растений, подстилок, почв.
Основная тенденция в распределении бактерий по толще торфяников —
довольно плавное изменение их численности с глубиной с небольшими
увеличениями или уменьшениями плотности популяций (в пределах одного
порядка) в отдельных горизонтах, в отличие от уменьшения численности
бактерий по профилю, характерного для большинства типов почв.
Исследуемые торфяные почвы залегают на глубоких торфах, таким образом, весь
их профиль состоит из органогенных горизонтов — именно эта особенность
торфяных почв определяла равномерное, либо даже увеличивающееся с
глубиной распределение бактерий по вертикали. Следует также отметить,
что контуры распределения бактерий по профилю исследуемых почв,
полученные на основании прямого и чашечного методов, в большинстве случаев
коррелировали и не зависели от ботанического состава торфа [1,2].
В исследуемых образцах были обнаружены представители 22 родов
бактерий, среди них на долю грациликутов приходится 73 %, на долю фирма-
кутов — 27 % [1,2]. Список потенциальных доминантов является одним из
104
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
наиболее репрезентативных показателей таксономической структуры
микробных комплексов, тесно связанных с типом экосистемы {3,4]. В
сообществах бактерий болот в спектр потенциальных доминантов из протеобактерий
вошли представители семейств Enterobacteriaceae (роды: Enterobacter,
Klebsiella, Proteus), Vibrionaceae (роды: Aeromonas, Plesiomonas, Vibrio),
Spirillaceae (род: Aquaspirillum); из скользящих бактерий — бактерии
группы Flavobacterium-Cytophaga\ из актинобактерий — бактерии рода
Streptomyces. Спектр родов и видов бактерий, выделенных из торфяников,
отражает адаптацию прокариотных микроорганизмов к обитанию в
земноводных экосистемах с пониженным содержанием кислфода и низкими
значениями рН. В качестве доминирующих в болотно-лесных экосистемах
выявлялись специфические, характерные для водных сред организмы. Это
подвижные спириллы, вибрионы, энтеробактерии, способные к росту в
условиях, складывающихся при режиме затопления, характерном для болот.
Выявленные из торфяников виды цитофаг: Cytophaga aurantiaca, С. fleves-
cens, С. psychrophila и С. succinicans — согласно их первоначальному
описанию были выделены из воды, а вид С. succinicans является кроме того и
факультативно-анаэробным видом. Выделенные штаммы рода Beijerinckia
были отнесены к одному виду — В. mobilis, обладающему способностью
расти при низких значениях рН, поэтому понятна приуроченность этих
бактерий к исследуемым кислым почвам. Среди видов бацилл, обнаруженных в
торфяниках, кроме типичных для северных лесов видов, были обнаружены
— Вас. lentus и Вас. flrmus, характерные для мхов и торфов, а также Вас.
circulans, являющийся факультатавно-анаэробным видом. Таким образом,
особенностями типологической структуры прокариотных комплексов гид-
роморфных почв являются явное доминирование протеобактерий и особый
видовой состав цитофаг, спороцитофаг и бацилл, среди которых
значительную долю составляют факультативно-анаэробные виды [1,5].
Особенностью ярусной структуры бактериальных комплексов болотно-
лесных экосистем является континуальный характер распределения по
ярусам представителей одних и тех же групп, в основном спирилл и
факультативно-анаэробных бактерии семейств Enterobacteriaceae и Vibrionaceae.
Доминирование этой группы протеобактерий было подтверждено
сравнительным анализом структуры этих комплексов в 16 различных точках из
поверхностных горизонтов и в 25 точках на выборочных глубинах
почвенного профиля каждого из исследуемых торфяников. Во всех вариантах
процентное содержание этой группы гидробионтов колебалось от 44 до 90% по
горизонтальной структуре и 64-96% по вертикальной [1,5].
Экологическая структура бактериальных комплексов болот особенно
показательна: преобладание протеобактерий, относящихся к группам копио-
трофов и олиготрофов и низкая доля бактерий гидролитического комплекса,
105
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
особенно в верховых болотах; что коррелирует с замедленными процессами
деструкции органического материала в болотных экосистемах. Более
детально был изучен гидролитический комплекс бактерий с помощью метода
репликаторов. Основными агентами, способными осуществлять среди
прокариот деструкцию полимеров в верховом торфянике, оказались бациллы и
стрептомицеты; причём бациллы брали на себя роль полифункциональных
гидролитиков, разлагая карбоксиметилцеллюлозу, крахмал, лецитин, твин и
белки. В низинном нормальнозольном торфянике кроме бацилл и стрепго-
мицетов в разрушении полимеров в пределах метровой толщи принимали
участие цитофаги, целлюломонады и миксобактерии.
Проанализировав ярусную, типологическую и экологическую структуры
бактериальных комплексов сапротрофного блока торфяников удалось
выявить уровень их разнообразия, степень простанственной дифференциации и
выполняемые ими функции в болотно-лесных экосистемах.
Работа выполнена при поддержке ФЦНТП «Биоразнообразие».
[1] Головченко А.В. Особенности пространственного распределения и структуры
микробных комплексов болотно-лесных экосистем: Дис. канд. биол. наук. М.: МГУ.
1993. 156 с. [2] Головченко А.В. и др. Особенности пространственного
распределения и структуры микробных комплексов болотно-лесных экосистем. //
Почвоведение, 1993. № 10. С. 78-89. [3] Чернов И.Ю. География микроорганизмов и структура
экосистем. // Изв. РАН, сер. географ. 1993. № 6. С. 49-58. [4] Добровольская Т.Г. и
др. О показателях структуры бактериальных сообществ. // Микробиология, 1997. Т.
66. № 3. С. 408-414. [5] Добровольская Т.Г., Головченко А.В. Эколого-таксономи-
ческая структура бактериальных сообществ лесных почв гидроморфного ряда. //
Веста. МГУ, сер. 17. Почвоведение. 1999. № 1. С. 55-59.
ЖИЗНЕСПОСОБНОСТЬ МИЦЕЛИЯ И СПОР ГРИБОВ В ТОРФЯНИКАХ
А.В. Головченко, Л.М. Полянская
Институт Почвоведения МГУ-РАН, Институт проблем экологии и
эволюции РАН; МГУ, факультет почвоведения
Для оценки роли почвенных микроорганизмов в круговороте
органического вещества и энергии в болотных экосистемах необходимо иметь
репрезентативные данные относительно их запасов в торфяниках.
Почвенный аспект изучения микробных комплексов болотных экосистем
стал весьма актуальным благодаря исследованиям, проведенных нами на
объектах Западнодвинского стационара Института лесоведения РАН.
Показано, что торфяники представляют собой бесценные природные кладовые
микроорганизмов. Общая микробная биомасса в торфяниках в расчете на
всю толщу может достигать сотен тонн на гектар, причём биомасса эукари-
отных микроорганизмов (82-99%) значительно превышает биомассу прока-
106
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
ряотных; в биоморфологической структуре микромицетов мицелий (70-
99%) доминирует над спорами. В изученных торфяниках зафиксировано
равномерное вертикальное распределение микроорганизмов по всей толще ,
что связано с их органогенной природой. Выявлено достоверное
превышение уровня межсезонных колебаний численности микроорганизмов над
межгодовым, что соответствует представлениям о существовании здесь
годовой сукцессии комплекса почвенных микроорганизмов [1].
Наиболее дискуссионным и только частично разрешённым оставался
вопрос о состоянии, в котором находятся микроорганизмы в торфяниках,
особенно на глубине, где возраст слоев определяется тысячелетиями. О
жизнеспособности хотя бы части микробных сообществ косвенно
свидетельствовала способность к росту на питательных средах всех групп
микроорганизмов, высеваемых с разных глубин вплоть до подстилающей породы. На
реальное размножение хотя бы части микробного комплекса указывало
наличие сезонной динамики численности микроорганизмов, как прокариотных,
так и эукариотных по всему профилю торфяников. Особое внимание в
дальнейшей работе следовало уделить микромицетным комплексам торфяников,
так как на их долю в исследуемых почвах приходится более 90 % от
суммарной микробной биомассы.
Для оценки состояния, в котором находятся микромицеты в торфяниках,
применили разработанный метод определения жизнеспособности грибных
пропагул и спор по их прорастанию [2]. О прорастании спор судили по
выбрасыванию ростовой трубки, о прорастании дрожжеподобных клеток — по
почкованию, о прорастании мицелия — по удлинению гиф и образованию
микроколоний. Объектом исследования был выбран верховой торфяник под
сосняком андромедово-пушицево-сфагновым (мощность залежи — 5,5 м).
Процент прорастания мицелия грибов был максимальным в подстилке
(за подстилку принимали слой, включающий очёс живого сфагнового мха, в
который попадают опавшая хвоя сосны и другие остатки кустарничковых и
травянистых растений). Именно в этом слое были зарегистрированы
мощные микроколонии, образованные проросшими гифами и спорами. На
глубине 10-50 см количество проросших гиф сокращалось вдвое, а в
следующем полуметровом слое — вчетверо по сравнению с подстилкой. Глубже 1
метра гиф, способных к прорастанию, обнаружить не удалось, за
исключением слоя 4-5 м, где зафиксирована одна грибная микроколония (рис. 1).
Непроросшие гифы имели четкие контуры, хорошо выраженные
перегородки и септы, обладали ярким свечением, выглядели как живые.
Возможно, прорастание такого рода гиф могло бы иметь место, но в отличных от
условий опыта. Средние значения диаметров грибных гиф (проросших и не
проросших) не отличались от контрольных вариантов (рис. 2).
107
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
В отличие от гиф, способность к прорастанию у спор и дрожжеподобньос
клеток была отмечена по всему профилю торфяника. Процент их
прорастания варьировал в метровой толще от 46 до 87 %, в остальной части профиля
— от 6 до 40 % (рисЛ). При прорастании спор их диаметр увеличивался в
1,5 раза по сравнению с контролем и составлял в среднем 8,3 мкм (рис.2Х
Используемый нами метод определения жизнеспособности спор почвенных
грибов, позволил выявить ещё одну особенность исследуемой почвы —
обнаружение по всему профилю большого количества дрожжевых клеток,
благодаря их способности размножаться почкованием, которое фиксировали на
экспериментальных препаратах.
.ii.ii.Ji. п.
JL
_пц
|ТПрСП|
^. n.dl,
П
0 0- 0.02- 0.05- 0.08- 0.30-
0.02 0.05 0.08 0.30 0.50
3.5- 4.0-
2.0 2.5
Рис. 1. Прорастание гиф (ГП) и спор (СП) грибов на разных глубинах верхового
торфяника
0.00,02
■*
0,05-0.08 j
■
0,30-0,50
1.0-1.5
2,0-2,5
3.5-4,0
5,0-5,5
J
^^^^^^^^^^^^^^^^^^^^^^^^^
■■■^^^^^^^^
^^^^^^^^^^^^
ттшш^^^^ш
3
3"
[■2
|Р1
киЛ*
Рис. 2. Средние диаметры спор (СП) и
пропагул (ГП) грибов в образцах
верхового торфяника (1-контроль, 2-пророс-
шие, 3-непроросшие)
Рис. 3. Длина грибного мицелия в
контрольных (1) и экспериментальных (2)
образцах верхового торфяника.
Процент жизнеспособности спор и грибных гиф определяли по
соотношению числа фрагментов мицелия и спор в экспериментальных и
контрольных препаратах. Общей закономерностью является обнаружение высокого
содержания жизнеспособной грибной биомассы по всему профилю олиго-
трофного торфяника. Процент жизнеспособности для гиф составил 60-100%,
108
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
для спор - 83-100%. Максимальные значения этого показателя для гиф были
зарегистрированы в верхних горизонтах, где длина грибного мицелия в
опыте была значительно выше чем в контроле, и в нижних горизонтах (2-5,5
м) где плотность мицелия оставалась практически на одном уровне в
контрольных и экспериментальных образцах (рис. 3).
Грибы, являющиеся основными деструкторами органического вещества,
находятся в жизнеспособном состоянии по всему профилю торфяника. Но
лишь в пределах метровой толщи их гифы обладают способностью
прорастать. Глубже пропагулы грибов скорее всего «законсервированы» в
почвенном профиле, т.е. находятся в состоянии анабиоза, поддерживая высокие
запасы живого углерода в болотных экосистемах.
Работа выполнена при поддержке ФЦНТП "Биоразнообразие".
[1] Головченко А.В. и др. Особенности пространственного распределения и
структуры микробных комплексов болотно-лесных экосистем // Почвоведение. 1993. № 10.
С. 78-89. [2] Полянская Л.М. и др. Определение жизнеспособности спор и мицелия
почвенных грибов // Микробиология, 1998. Т. 67. № 6.
ОСОБЕННОСТИ ФОРМИРОВАНИЯ БЕРЕЗОВЫХ СООБЩЕСТВ
НА МЕЗОТРОФНЫХ ТРАВЯНО-СФАГНОВЫХ БОЛОТАХ
ПОД ВЛИЯНИЕМ ОСУШЕНИЯ
СИ. Грабовик
Институт биологии Карельского НЦРАН, Петрозаводск
В южной Карелии комплексное изучение болот и заболоченных лесов в
естественном состоянии и под влиянием осушения начато в 1970 г. на лесо-
болотном научном стационаре Киндасово, в подзоне средней тайги.
Болотные массивы мезотрофного сфагнового лесного типа не занимают в Карелии
больших площадей и до настоящего времени изучены слабо. Древесные и
древесно-сфагновые болотные участки встречаются также в составе других
типов болот, особенно часто на их окрайках. Данная работа посвящена
динамике биологической продуктивности мезотрофных травяно-сфагновых
болот в пространственно-временном аспекте под влиянием осушения.
Пробная полощадь 1 (пр.пл.) заложена в центральной открытой части
болота Близкое, относящегося к мезотрофному травяно-сфагновому типу,
где до осушения в центральной части доминировал кочковато-топяной
комплекс Sphagneta magellanici+Herbeto-Sphagneta subsecundi, занимая
половину его площади. Подавляющая часть комплекса (70%) приходилась на
сообщества мочажин: Carex lasiocarpa-Menyanthes trifoliata и Carex lasio-
carpa-Sphagnum subsecundum. Растительный покров кочек (30%) был
представлен фитоценозами ассоциации Chamaedaphne calyculata-Sphagnum
109
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
magellanium + S. angustifolium. Часть кочек была облесена Betula pubescens
или Pinus sylvestris высотой до 2 м. Торфяная залежь неглубокая — 0,8 ц
низинная, подстилается глиной.
Пробная площадь 2 расположена в периферическом облесенном кольце
вдоль линии стока, который обозначен небольшим ручьем, имеющим
местами выраженное русло на окрайке болота Койвусуо, относящегося к
южнокарельскому варианту кольцевого аапа типа в слабооблесенном кочковато-
западинном комплексе Sphagneta angustifolii+Heitoeta с редкой Betula
pubescens и занимала 34% площади. На топяном фоне, занятом ассоциацией
Phragmites australis-Carex lasiocarpa (56%), вдет образование сфагновых
кочек (44%): Betula pubescens-Chamaedaphne calyculata-Pleurozium schre-
beri и Andromeda polifolia-Carex lasiocarpa-Sphagnum angustifolium.
Деревья небольшие (2-4 м). древостой малосомкнутый 0,2. Кочки
сформировались недавно, о чем свидетельствует состав торфа под ними: ниже 25 см они
подстилаются осоковым торфом с небольшой примесью сфагновых мхов.
Глубина торфяной залежи 1,5 м, низинная.
До осушения на изучаемых болотных участках запасы надземной фито-
массы и ее фракционный состав на кочках и понижениях значительно
различались и зависели от видового состава и структуры растительного
покрова. На пр.пл. 1 запасы фигомассы кочек и топей близки, но структура фиго-
массы различна: на кочках она складывается из фигомассы кустарничков и
мхов, а в топях, в основном, за счет трав. Запасы фигомассы на кочках 25,2,
на понижениях 26,3 ц/га. На пр.пл.2 надземная фитомасса кочек была почти
в 3 раза больше за счет более густого растительного покрова. Запасы
фигомассы на кочках 79,3; на понижениях 30,3 ц/га. Запасы фигомассы
древесного яруса на 1 и 2 пр.пл. не определялись.
Повторные определения биологической продуктивности проводились
через 13 и 25 лет после осушения. Исследования показали существенные
изменения структуры растительного покрова и биологической продуктивности
болотных участков. В результате действия осушения через 13 лет на пр.плЛ
на месте болотного участка Sphagneta magellanici+Heibeto-Sphagneta subse-
cundi сформировался древесно-травяной комплекс Betuleto-Herbeta с
кочковато-равнинным микрорельефом. Состав древостоя 10Б20+С, относительная
полнота 0,3. На пр.пл.2 на месте кочковато-западинного слабооблесенного
болотного участка сформировался облесенный комплекс Betuleto-Sphagneta
angustifolii. Состав древостоя 8Б 302С+Е, относительная полнота 0,7.
Фигомассу напочвенного покрова определяли на кочках и на коврах.
Через 13 лет после осушения запасы надземной фигомассы на пр.пл. 1 и 2 на
кочках составили 36,0 и 25,5 ц/га, а на коврах — 252,6 и 466,6 ц/га
соответственно. Ее резкое увеличение произошло за счет древесного яруса на 1
пр.пл. — 218,5, а на запасы кустарничков и трав приходится — 12,7 и 21,4
ПО
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
ц/га. На пр.пл.2 запасы древесного яруса составляют 421,1 ц/га, а на кустар-
длчки, травы и мхи приходится 42,7, 0.9 и 1,9 ц/га соответственно.
Через 25 лет после осушения на пр.пл.1 на месте прежнего болотного
комплекса сформировался древесно-травяной Betuleto-Herbeta с кочковато-
равнинным микрорельефом. Состав древостоя 9Б351С50, запас — 78 м3/га,
полнота — 0,8. Запасы надземной фитомассы составили 543,8 ц/га. Резкое
ее увеличение произошло, в основном, за счет древесного яруса — 507,7
ц/га, а на запасы трав и мхов приходится 17,4 и 18,7 ц/га соответственно.
Доля участия древесного яруса, трав и мхов от общих запасов надземной
фитомассы составляет 93,4, 3,2 и 3,4%.
На 2 пр.пл.сформировался кочковато-равнинный облесенный комплекс
Betuleto-Sphagneta angustifolii. Состав древостоя 10Б 45 ед. Е, С, запас —
148 м3/га, относительная полнота — 1,3. Подрост ели — до 1000 шт/га.
Присутствие естественного возобновления ели под пологом короткопроиз-
водных лиственных насаждений будет способствовать переформированию
их в длительнопроизводные еловые насаждения. Запасы надземной
фитомассы древесного яруса составляют 968,7 ц/га, запасы кустарничков и мхов
— 42,7 и 15,5 ц/га соответственно, доля участия которых от общих запасов
надземной фитомассы составляет 94,3; 4,2 и 1,5% соответственно.
Таким образом под влиянием осушения и сукцессии растительного
покрова происходит выравнивание микрорельефа: от кочковато-топяного до
кочковато-равнинного. Через 25 лет после осушения существенно
изменяются структура растительного покрова и биологическая продуктивность.
Наблюдается снижение участия в надземной фитомассе основных растений-
торфо-образователей и возрастание роли в них древесных пород.
Происходит образование насаждений с преобладанием березы.
НАСЕЛЕНИЕ ПТИЦ ЗАБОЛОЧЕННЫХ ЛЕСОВ
КОСТОМУКШСКОГО ЗАПОВЕДНИКА
Е.А. Даниленко, Е.Н. Матюшкин
МосНПО «Радон», Москва; МГУ, географический факультет
Заболоченные леса в заповеднике — сосняки, насаждения сосны и ели,
небольшие участки ельников, часто с примесью березы, произрастающие в
небольших понижениях или на пологих склонах в местах выхода грунтовых
вод. Часто они бывают приурочены к основаниям склонов, граничащих с
болотами или с заболоченными поймами рек.
Характерной особенностью группировки птиц этого местообитания
является относительно высокая численность овсянки-ремеза 40,5 особей км"2 (см.
табл.), которая доминирует здесь наряду с чижом и пеночкой-весничкой
111
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
(16,3 и 10,2%). По сравнению с долинными и склоновыми ельниками час-
ленность веснички здесь заметно ниже (с 27,4 до 23,5 особей км"2), чижа ^
почти вдвое больше (с 19,0 до 37,5).
Характеристика населения птиц заболоченных лесов Костомукшского заповедника
(протяженность маршрутов 11,8 км, конец мая — июнь 1987-1989 гг.)
Вид
Овсянка-ремез
Чиж
Весничка
Юрок
Зяблик
Клест-сосновик
Пухляк
Горихвостка-лысушка
Пищуха
Лесной конек
Желтоголовый королек
Мухоловка-пеструшка
Кукша
Глухарь
Теньковка
Снегирь
Фифи
Трехпалый дятел
Большой пестрый дятел
Серая мухоловка
Лесная завирушка
Свиристель
Рябчик
Зарянка
Всего 24 вида:
Количество особей
На 1 км маршрута
1,9
3,9
3,4
3,4
2,9
1,8
1,2
2,0
0,3
1,4
0,5
0,7
0,4
0,3
0,5
0,3
0,7
0,2
0,2
0,2
0,2
0,3
ОД
0,2
26,8
на 1 км
40,5
37,5
23,5
17,9
16,5
15,6
13,5
11,7
9,2
7,8
5,9
5,2
4,2
2,9
2,8
2,3
2,1
2,1
1,5
1,8
1,5
1,4
1,4
1,2
230,0
Доля вида в
населении, %
17,6
16,3
10,2
7,8
7,2
6,8
5,9
5,1
4,0
3,4
2,6
2,3
1,8
1,3
1,2
1,0
0,9
0,9
0,7
0,8
0,7
0,6
0,6
0,5
100,0
Примерно такая же картина прослеживается и для 5 видов из группы со-
доминантов. Только у зяблика численность незначительно ниже, небольшое
ее увеличение отмечено для клеста-сосновика; у пухляка и юрка в
заболоченных лесах она выше почти вдвое, у горихвостки-лысушки - более чем в 6
раз. Напротив, показатели численности рябчика и зарянки ниже: в
долинных ельниках эти виды входили в категорию господствующих, здесь же
попали в разряд третьестепенных видов.
Преобладание в группах доминантов и содоминантов птиц,
предпочитающих разреженные леса и опушки, и заметное участие лесного конька,
характеризует это местообитание в целом как «опушечное». При этом чис-
112
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
ценность и участие в населении птиц, тяготеющих к сомкнутым ельникам —
желтоголового королька, зарянки и теньковки — сравнительно низки.
Разнообразие видов, обитающих в заболоченных лесах, свидетельствует
о широком спектре экологических условий характеризующих данное
местообитание. Так, присутствие здесь среди многочисленных видов птиц таких
как юрок, зяблик, горихвостка-лысушка, а среди обычных — мухоловки-
пеструшки, связывают его с боровой группой местообитаний. В то же время
с населением птиц еловых лесов эту группировку сближают: из
многочисленных видов — клест-сосновик, из второстепенных — королек, рябчик,
теньковка и трехпалый дятел. Два вида — чиж и овсянка-ремез, как бы
связывают в классификационном плане сообщество заболоченных лесов, с
одной стороны, с сосняками и ельниками (чиж), с другой — с ельниками и
открытыми и малооблесенными болотами (овсянка-ремез). Сходство с
населением переувлажненных местообитаний (болот) выражается в присутствии
в качестве обычных видов птиц, играющих в этой группировке различную
роль: это лесной конек (второстепенный вид) и фифи (третьестепенный).
Только в этом местообитании отмечены лесная завирушка и
обыкновенная пишуха, относительно редкие для заповедника виды птиц. Регистрация
здесь на учетах большого числа видов птиц (24), весьма разнообразных
экологически, объясняет, по-видимому, и более высокие коэффициенты
общности населения этого местообитания с таковыми «сосновой» и «еловой»
групп, чем, например, с открытыми и малооблесенными болотами или
хвойно-лиственными молодняками.
ПРОДУКТИВНОСТЬ ТРАВЯНОГО БОЛОТА
НАЗАРОВСКОЙ ЛЕСОСТЕПИ
С.С. Дубынина
Институт географии СО РАН, Иркутск
Изменение продуктивности представляет важную часть исследований,
главной целью которых является разработка мероприятий по оптимизации
природопользования в условиях Канско-Ачинского
топливно-энергетического комплекса (КАТЭК). Предусмотрено выяснение многих сторон поведения
органического вещества в спонтанных и антропогенно измененных
геосистемах. Наблюдения за биологической продуктивностью травяного болота
гидроаккумулятивной фации осуществлялись в естественных условиях
Березовского участка Назаровской лесостепи Красноярского края.
Пространственное изменение и динамическое состояние геосистем
определялись по комплексу признаков, причем основное внимание уделялось
структуре растительных сообществ, наиболее точно отражающей состояние
113
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
геосистемы в целом. Величина продуцируемой фитомассы зависит от
условий тепла и влаги не только в годы наблюдений, но и в предшествующие им
периоды, а также от местоположения каждой конкретной фации.
Болота на Березовском участке занимают сравнительно незначительную
площадь и по своему расположению большая часть травяных болот связана
с поймами рек и озер. Лабазниково-осоково-вейниковое закочкаренное,
высыхающее болото в лесном березовом колке занимает аккумулятивное
местоположение в днище неширокого лога поймы ручья Отножки. В зимний
период происходит перераспределение снега, в результате которого высота
снежного покрова достигает 100-140 см. Соответственно происходит
значительное переувлажнение верхних горизонтов почв в весенний период. В
профиле почвы влаги содержится обычно в 2 раза больше наименьшей вла-
гоемкости. поэтому характерной особенностью лугово-болотных почв
является их высокая гигроскопичность и водоудерживающая способность.
Увлажнение атмосферное, грунтовое, избыточное. Уровень фунтовых вод
подвержен сильным колебаниям. Необходимое для вегетации количество
тепла в корнеобитаемом слое накапливается поздно, этому способствует
большое количество мертвых растительных остатков, которые не только
препятствуют прогреванию верхнего почвенного слоя, но задерживают рост
молодых побегов растений. Поэтому развитие растений в болотных фациях
начинается несколько позже. Почвообразующая порода пролювиальные
отложения. Почва лугово-болотная иловато-карбонатная слоистая не вскипает
от соляной кислоты по всему профилю, оглеена с поверхности. Поверхность
неровная, закочкаренная. Средняя высота кочек 15-20 см, диаметр 30 см.
Травяной покров очень густой, довольно однороден: осока дернистая, вей-
ник Лангсдорфа. лабазник вязолистный, крапива жгучая, обыкновенная,
чемерица Лобеля. Проективное покрытие 95-105%. Средняя высота
травостоя 95-105 см. видовое разнообразие невелико 12 видов на 1 м2. Строение
2-х ярусное. Вегетационный период оцениваем (по переходу среднесуточных
температур через 0°С) с мая месяца по середину октября.
Структура и запасы фитомассы — результат взаимодействия двух
разнонаправленных процессов функционирования геосистем: образования и
расходования растительной массы. В лесных фациях основная часть
растительного вещества аккумулируется в запасах живой массы (надземной и
подземной), отношение отмершего вещества к живому изменяется от 0,1 до 0,5,
корневая масса в общей фитомассе составляет около 30%. Минимальные
запасы зеленой массы наблюдаются в травянистом покрове лесных фаций
под древесным пологом — 70 г/м2. Высокая изменчивость структуры
растительного компонента лесных фаций, обусловленная их разновозрастностью,
особенностями функционирования и мерой их использования. В болотных
геосистемах наблюдается выравнивание запасов вещества в надземной и
114
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
подземной частях, но заметное увеличение отмершего вещества - его
отношение к живому 1,8. Для динамики запасов мертвого вещества свойственен
-плавный", сглаженный ход кривой. Запас живых корней составляет 16-30%
от общего запаса корневой массы, но эта масса живых корней в травяном
болоте позволяет образовать наибольшую величину фитомассы. Средние
запасы надземной массы лабазниково-осоково-вейникового болота
достигают 1432 г/м2, подземной — 4677 г/м2, общее органическое вещество
составляет в среднем 5970 г/м2, с максимумом до 7578 г/м2.
Быстрое нарастание зеленой массы в мае месяце. Максимум
характеризуется двумя пиками — июль и сентябрь, особенно в периоды сухих лет. Во
влажные годы максимум зеленой массы приходится на август месяц.
Колебания условий тепла и влаги по годам не только отражаются на величине
фитомассы, но и перестраивают ее структуру, меняют соотношение живого
и отмершего вещества, их дальнейшее распределение по разным
трофическим уровням, обеспечивая тем самым устойчивость геосистемы.
Многолетний ряд наблюдений с 1981 по 1997 гг. дает возможность проследить
флуктуации величины запасов фитомассы болот. Процесс флуктуации
ритмичный, периоды повышенной продуктивности сменяются периодами
пониженной продуктивности. В цикле сухих и теплых лет происходит осушение
низинных форм рельефа. Засушливый период 1981 года сказался на запасах
надземной массы 1982 года, которая была ниже среднемноголетней в 1,7
раза. Очевидно, пересыхание верхнего слоя почвы тормозит разложение
мертвых подземных органов из-за недостаточности кислорода, а также рост
и развитие растительности надземной части. Особенно ярко это выражается
в ярусности фитоценоза. В засушливые годы высота растений 1-го яруса
снижается от 150 до 70 см, который хорошо выражен только в
благоприятные по увлажнению годы, его образуют генеративные побеги
доминирующих видов: из злаков — вейник, из разнотравья — лабазник, чемерица,
крапива и др. Не только засуха влияет на понижение запасов надземной и
подземной массы, а также сильное переувлажнение во влажные годы.
Экстремально низкие запасы надземной массы гидроаккумулятивной фации в
период влажных лет с 1987 по 1988 были ниже среднемноголетней — в 1,5
раза, а запасы корней — ниже в 2 раза. Наиболее флуктуирующими
величинами в болотных геосистемах являются запасы мортмассы надземных и
подземных органов и менее флуктуирующими — сумма запасов корневой
массы. Относительная устойчивость величины корневой массы связана с
совпадением во времени процессов прироста живых корней и разложения
мертвых. Замедленное разложение мертвых остатков болотных
биогеоценозов в момент пересыхания и переувлажнения происходит вследствие того,
что микробиологические процессы заторможены. Заторможенность
разложения подземных растительных остатков травяного болота связано с низки-
115
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ми температурам почвы и дефицитом кислорода.
Продукция органического вещества — основной показатель
функционирования геосистем. Во всех фациях продукция растительного вещества,
накапливается в течение теплого периода года, в надземной части прирост
происходит в основном в первую половину лета, в подземной наблюдается 2
пика прироста — весенний и позднелетний. Ежегодно в природных
геосистемах продуцируется в среднем 2500 г/м2'растительного вещества, причем в
надземной части лишь 1/4 этой величины. Процент продуцирования зеленой
массы в целом от общей фитомассы в разных геосистемах по среднемного-
летним данным составляет 32% в болотных фациях, в луговых — 20% и
самый низкий процент продуцирования зеленой массы в лесах всего 10%.
Установлено, что период обновления живых корней колеблется
незначительно от 0,4 до 0,8. Отношение величины живых корней к чистой
продукции корневой массы болотных биогеоценозов — 0,7 т.е. период обновления
в среднем близок одному году. Среднемноголетняя продукция зеленой
массы возрастает от лесных фаций к болотным от 279 до 1025 г/м2 в год,
соответственно, живых корней от 1937 до 2215 г/м2 в год. Период формирования
первичной продукции растительного вещества в естественных геосистемах
длиннее, чем в антропогенно измененных (агроценозы), которые по
продолжительности короче примерно на 1,5 месяца. Общая среднемноголетняя
продукция живого вещества изменяется от 950 г/м2 в год (антропогенно
нарушенных) до 3240 г/м2 в год, с максимумом до 3765 г/м2 в год для
болотной фации. Динамика и функционирование продукционных процессов в
болотных фациях зависит прежде всего от биологии самих растений, от
умения перестраивать структуру вещества путем урегулирования соотношения
живой и мертвой части фитомассы, обеспечивая тем самым устойчивость
геосистем в данной обстановке природопользования.
ВНУТРИВЕКОВАЯ ДИНАМИКА РАДИАЛЬНОГО ПРИРОСТА
ДЕРЕВЬЕВ В СВЯЗИ С ЦИКЛАМИ СОЛНЕЧНОЙ АКТИВНОСТИ
К.Н. Дьяконов, Ю.Н. Бочкарев
МГУ, географический факультет
Поиск пространственно-временных закономерностей фитопродукционно-
го процесса и геофизических факторов, его определяющих, представляет
собой многоплановую проблему, поставленную и разрабатываемую био- и
гелиобиологами, физико-географами, геофизиками, дендроклиматологами
многих поколений. Рассматриваются два вопроса: синхронность прироста в
различных подзонах лесной зоны Западно-Сибирской и Русской равнины и
возможная его связь с циклами солнечной активности, которые выражены
числами Вольфа. Известно, что в работах многих ученых [А.Э. Дуглас, В.Н.
116
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
ддаменко, Т.Т. Битвинскас, А. Звиедрис, СИ. Костин, Г.Е. Комин, Н.В.
довелиус и др.] отмечена прямая или опосредованная зависимость
радиального прироста от солнечной активности, причем понижение прироста
соответствует периодам ее повышенных значений.
Отбор спилов и кернов осуществлен в шести пунктах.
1. Северная тайга Западной Сибири. В 6 км на ЮВ от г. Ст. Надым.
Пологая аллювиальная верхнеплейстоценовая равнина, с развитием болотных
отложений. Сложена супесями и песками с линзами торфа. Относится к
южной субарктической зоне многолетней мерзлоты. Развиты долинные ело-
во-кедрово-лиственничные кустарничково-зеленомошно-лишайниковые
слабосомкнутые (0,2-0,3) леса на подзолах иллювиально-гумусовых. Возраст
деревьев — 105-135 лет.
2. Средняя тайга лесоболотной зоны. Междуречье Конды и Мал.Сосьвы.
Моренно-ледниковая пологая, слабопереработанная денудацией,
верхнеплейстоценовая возвышенность. Сложена средними, реже легкими
суглинками и супесями, в сочетании с торфяниками. На дренируемых участках
преобладают высокосомкнутые (0,9) березово-пихтово-еловые с кедром мел-
котравно-зеленомошные с пятнами долгомошно-сфагнового покрова леса на
торфянисто-подзолисто-элювиально-глееватых почвах. На плоских
междуречьях — багульниково-осоково-сфагновые сосновые редколесья на
торфяниках. По террасам — сосняки бруснично-зеленомошные на подзолах иллю-
виально-гумусово-железестых глееватых. Возраст деревьев — 120-185 лет.
3. Южная тайга лесоболотной зоны. Пос. Ягодное (учебно-научная база
Томского гос. ун-та), бассейн р. Кеть. Низменная аллювиальная среднеплей-
стоценовая песчаная и супесчаная равнина. Сосняк среднесомкнутый (0,5-
0,6) бруснично-лишайниково-зеленомошный на торфянисто-подзолистых
почвах. Возраст деревьев — 90-130 лет.
4. Подтайга Западно-Сибирской равнины. Керны отобраны в двух
районах на северо-востоке Омской области. Низменная озерно-аллювиальная
слаборасчленненная равнина, сложенная легкими лессовидными суглинкам
и супесью. Сосняк среднесомкнутый (0,7) бруснично-хвощево-разнотравный
папоротниковый на дерново-слабоподзолистых почвах. Возраст сосен — 80
лет. Керны елей взяты на высокой террасе Иртыша, в 12 км на ВЮВ от г.
Тара, сложенной супесью и легкими суглинками в сосново-еловом
можжевеловом кустарничково-зеленомошном среднесомкнутом (0,7) лесу. Возраст
елей — 102 года.
5. Средняя тайга восточноевропейского типа. В 25 км на юг от ст. Кос-
тылево (юг Архангельской области). Слабоволнистая структурно-]эрозион-
но-ледниковая равнина, сложенная маломощными моренными суглинками,
лимногляциальными супесями, реже песками. Отбор кернов и]спилов
осуществлен в сосновом кустарничково-багульниково-сфагновом редколесье на
117
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
мощных торфяниках; в заболоченном елово-березово-сосновом осоково*
багуниково-сфагновом и в ельнике разнотравно-зеленомошном на высокой
пойме р. Заячьей. Возраст 80-88 лет.
6. Южная тайга восточноевропейского типа. Молого-Шекснинское
междуречье, плоская песчаная озерно-аллювиальная равнина. Отбор кернов
осуществлен в двух комплексах — багульниково-кассандрово-осоково-сфаг-
новом сосновом редколесье на торфяниках; возраст —- 120-140 лет и в
сосняке чернично-зеленомошном на торфянисто-подзолстых почвах, где
возраст сосен 80 лет.
При проверке гипотезы о существовании статистической связи между
изменчивостью радиального прироста деревьев и циклами солнечной
активности для элиминирования фактора возраста, во-первых, из рассмотрения
исключались первые 15-20 лет жизни дерева, когда биологически
обусловленная тенденция к снижению проявляется в максимальной степени. Во-
вторых, путем использования методов статистики и частотного
фильтрования, основанного на спектральном разложении кривой исходного ряда,
находились тренды прироста. Отфильтровывались колебания с периодами
более 30-50 лет и менее лет, так как на таком уровне цикличность солнечной
активности практически не проявляется; что следует из периодограммы
чисел Вольфа.
Далее по исходным и преобразованным рядам с помощью кластерного
анализа в программе STATISTICA были выделены типы приростов
деревьев для выявлений районов синхронных (асинхронных) по динамике
прироста. При кластеризации первичных неотфильтрованных рядов хорошо
выделились два типа прироста (ранговый коэффициент корреляции Кэнделла
внутри групп — 0,5-0,8): первый — в северной, средней и южной тайге
Западной Сибири, а второй — в южной тайге Западной Сибири и в средней и
южной тайге Восточно-Европейской равнины. Внутри них хорошо
выявились подгруппы. В случае профильтрованных рядов ранговая корреляция
между ними в пределах одного района заметно ниже (0,2-0,6). Главное:
годичный радиальный прирост деревьев на региональном уровне имеет
некоторые общие факторы.
Далее была осуществлена кластеризация профильтрованных рядов
прироста с участием ряда чисел Вольфа. Лишь единичные деревья вошли в одну
группу с числами Вольфа, причем «представители» разных районов (часть
среднетаежных сосен и пихт Западной Сибири, сосны на болоте Молого-
Шекснинского междуречья). Была построена также линейная регрессионная
модель с циклами солнечной активности. Статистически значимая связь
оказалась только у некоторых сосен с Молого-Шекснинского междуречья, У
отдельных сосен в районе базы Томского ун-та и кедров Кондо-Сосьвия-
ского междуречья. У большинства деревьев непосредственная связь прирос-
118
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
^ с циклами солнечной активности не проявилась.
Вышеизложенное, однако, полностью не опровергает наличие
зависимости прироста от циклов солнечной активности. Возможно, деревья реагиру-
jQT с запозданием на изменение последней, что приводит к несовпадению
фаз солнечной активности и приростов. Поэтому при устранении фазового
сдвига ряды приростов могут коррелировать с числами Вольфа. Для
проверки этой гипотезы осуществлено сравнение периодлограмм приростных
рядов и рядов чисел Вольфа. У ряда деревьев оказались общие периоды, в
частности у елей (период 10,8-10,9 лет), некоторых сосен и кедров (период
11,5-11,7 лет) Кондо-Сосьвинского междуречья и у архангельских деревьев
(период 12,3-12,4 лет). При этом только у архангельских деревьев связь
статистически значимая, отрицательная с лагом (запаздыванием) на 4 года.
Результаты регрессии и кросскореляции не позволяют говорить о
солнечной активности как непосредственном факторе биологической
продуктивности природных комплексов. Однако там, где она все же прослеживается,
связь отрицательна. Существование совокупности общих факторов
динамики прироста на уровне регионов, действующих во всех местообитаниях, в
которой цикличность солнечной активности не играет главной роли,
позволяет предположить, что первостепенное значение имеет фактор изменения
климата как главнейший и всеобщий, действующий на уровне не только
физико-географических стран и зон, но подзон и провинций. Выводы
следует считать предварительными. Дальнейшее развитие исследования
предусматривает прежде всего существенное расширение пунктов опробования.
Грант РФФИ 99-05-65097.
СОСТОЯНИЕ И ДИНАМИКА ЛЕСНОЙ РАСТИТЕЛЬНОСТИ
НА ЕСТЕСТВЕННЫХ БОЛОТАХ БЕРЕЗИНСКОГО ЗАПОВЕДНИКА
B.C. Ивкович, Е.Н. Ивкович
Березинский биосферный заповедник Витебская обл., Беларусь
Болота и заболоченные леса являются основным элементом ландшафта
Березинского биосферного заповедника. Занимая более половины его
территории, они во многом определяют его общий облик, ландшафтные
особенности, биологическое разнообразие. В связи с трудной доступностью и из-за
относительно низкой хозяйственной ценности, болотные леса заповедника
не испытывали прямого антропогенного воздействия и сохранили
девственное строение с естественными процессами возобновления, роста и
отмирания деревьев. Они могут служить одним из основных модельных объектов
изучения динамики ненарушенных растительных сообществ.
Исследования состояния и динамики лесной растительности на различ-
119
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ных типах болот начаты с конца 70-х годов и проводятся на сети
постоянных пробных площадей (21 шт.), охватывающих основные типы болотных
лесов. С периодичностью в 5 лет производится таксация древостоя,
устанавливается его структура, количественная характеристика прироста и отпада
естественного возобновления, видовой состав и структура напочвенного но-
крова. Контроль за изменением уровня грунтовых вод ведется ежегодно.
Наиболее обширные площади на болотах заповедника занимают
сосновые леса (15,6 тыс. га). Они произрастают на всех типах болот и отличаются
значительным видовым разнообразием растительного покрова. В
типологическом отношении преобладают сосняки сфагновые и осоково-сфагновые. В
сосняках сфагновых, произрастающих на верховых болотах, динамика
растительного покрова определяется местоположением фитоценозов в пределах
болотного массива. Так, в сосняках пушицево-сфагновых, расположенных в
центре выпуклых верховых болот наблюдается процесс изреживания
древостоя с отмиранием наиболее старых и толстых деревьев, сопровождающийся
снижением запаса и числа стволов на единице площади.
В живом напочвенном покрове отмечается закономерный рост участия
подбела и олиготрофных сфагнумов: бурого и красного. Причина —
дальнейшее нарастание и олиготрофизация центральной выпуклой части болота.
В сосняках кустарничково-сфагновых на пологих склонах и окраинах
верховых болот, таксационные показатели практически не меняются, что
обусловлено постоянным пополнением древостоя за счет молодых особей.
Происходит закономерное увеличение статистических показателей рядов
распределения числа стволов по толщине (коэффициента вариации, эксцесса и
положительной асимметрии). Для напочвенного покрова характерно
высокое постоянство и обилие пушицы влагалищной и кустарничков семейства
вересковых.
Для переходных и низинных болот характерны смешанные березово-
сосновые древостой осоково-сфагнового и осокового типов леса. Древостой
сосны в этих условиях относительно разновозрастный и отличается весьма
высокой стабильностью, также благодаря постоянному пополнению за счет
подроста. Высоким постоянством отличается и напочвенный покров, основу
которого составляют осока и сфагновые мхи группы олигомезотрофов. В то
же время, на отдельных окраинах болотных массивов в сосняках осоково-
сфагновых отмечается массовое усыхание сосны вследствие резкого
изменения гидрологического режима. За исследуемый период (15 лет) отпад
деревьев сосны составил 3,9-4,5 % в год, запас снизился в 2,5-3,0 раза.
Явление вызвано усилением застойного увлажнения окраинных участков болот
вследствие развития верхового болотного массива.
Особенно уникальны в научном и природоохранном аспекте болотные
черноолыианики (площадь 10,4 тыс. га), представляющие собой девствен-
120
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
дий, не тронутый рубками, осушением и вообще редко посещаемый
человеком лес. В зависимости от их эдафических условий и степени
обводненности, естественную примесь составляют береза пушистая и ель, а по экото-
нам, примыкающим к минеральным почвам — дуб, ясень.
Динамика таксационных показателей и структуры древостоя исследована
в двух основных типах ольшаников: таволговом и осоковом. Доля пород-
примесей осталась без изменений, основные таксационные показатели ольхи
черной и сопутствующих пород характеризуются ростом значений.
Характерен естественный процесс смены поколений ольхи черной. Под пологом
девственного леса от высоковозрастных жи-вых стволов появляется
предварительное порослевое возобновление. Жизненность деревьев материнского
полога неодинакова, поэтому поросль появляется и получает возможность
успешно развиваться в разные годы. Это определяет значительную разно-
возрастность естественных черноольховых лесов на болоте.
Болотные леса Березинского заповедника характеризуются значительной
устойчивостью, определяющим фактором их естественной динамики
является водный режим экотопов. В настоящее время они не нуждаются в
активных формах управления, необходимы продолжение длительного
мониторинга их состояния и исключение возможности аномальных изменений
гидрологического режима территории заповедника.
К МЕТОДИКЕ ОПРЕДЕЛЕНИЯ ЛИНЕЙНОГО ПРИРОСТА И
ПРОДУКЦИИ СФАГНОВЫХ МХОВ НА МЕЗО-ОЛИГОТРОФНЫХ
БОЛОТАХ ЗАПАДНОЙ СИБИРИ
Н.П. Косых
Институт почвоведения и агрохимии СО РАН, Новосибирск
Сфагновые мхи — основные эдификаторы мезо-олиготрофных болот
южной тайги Западной Сибири. Их вклад в продукцию травяно-кустарнич-
кого-мохового яруса этих болот достигает 25-60%.
Для определения линейного прироста и продукции была применена
новая методика «индивидуальных меток», являющаяся модификацией метода
«перевязок» и позволяющая изучать прирост отдельных экземпляров
разных видов мхов. У растения каждого вида стебель окольцовывался
крючочками из тонкой проволоки под головкой мха. На другой конец проволочки,
расположенный на расстоянии 5-10 см от растения, прикрепляется бирка из
фольги с номером. Колечко из проволоки свободно лежит на веточках, не
травмируя стебель и не мешая его развитию даже при наклонном росте мха
в мочажинах. В каждой ассоциации было окольцовано 70-100 растений.
Отбор меченых растений проводился через год после кольцевания. В лаборато-
121
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
рии у каждого экземпляра измерялось расстояние от кольца до отметки под
головкой растения. Приросшая часть стебля отрезалась, высушивалась до
воздушно-сухого веса и взвешивалась. Полученный вес соответствовал
годичному приросту одного растения.
Масса годичного прироста каждого вида сфагновых мхов равнялась
произведению среднего прироста одного растения на количество растений на
единице площади. Во всех ассоциациях дополнительно отбирался мох с
небольших площадок (100 см2) и в каждой пробе подсчитывалось
количество стеблей. По количеству стеблей на единице площади и среднему приросту
одного стебля определялась годичная продукция выбранного вида мха.
В результате применения метода «индивидуальных меток» было
выделено 3 группы мхов по приросту и по плотности (количество стеблей мхов на
единице площади). Количество стеблей на единице площади у разных видов
мхов отличается очень сильно. К первой группе с наибольшей плотностью и
с наименьшим линейным приростом относится Sphagnum fuscum,
образующий кочки на грядах. Плотность этого вида достигает 700-960 стеблей
растений на 1 дм2. Линейный прирост его наименьший среди всех видов
изученных мхов. В разные годы линейный прирост колеблется от 5,0 до 30,0
мм в год, а продукция от 100 до 350 г/м2 в год.
Ко второй группе относятся Sphagnum angustifolium и S. magellanicum.
Плотность стеблей изменяется от 100 до 200 на 1 дм2. Линейный прирост
этих видов не превышает 70,0 мм, а продукция 300 г/м2.
Третью группу образуют Sphagnum balticum, S. majus, S. fallax, S. jense-
пищ которые преобладают на самых низких элементах микрорельефа, в
мочажинах. Эти мочажинные мхи растут очень рыхло, в среднем их плотность
не превышает 200 растений на 1 дм2. Прирост мочажинных мхов достигает
наибольшей величины (35,0-150,0 мм). Продукция этих мхов изменяется
очень широко от 100 до 500 г/м2.
Продукция мхов зависит как от линейного прироста, так и от количества
растений этого мха на единицу площади. Линейный прирост мхов в
мочажинах наибольший, при наименьшем количестве стеблей на единице
площади. За счет линейного прироста продукция мхов в мочажинах выше.
СОДЕРЖАНИЕ РАДИОНУКЛИДОВ В РАСТЕНИЯХ
НИЖНИХ ЯРУСОВ ЗАБОЛОЧЕННЫХ ЛЕСОВ
Г. А. Крючкова, Н.Н. Малышева
МосНПО «Радон», Москва
Исследования проводились в подзоне хвойно-широколиственных лесов,
в центральной части Клинско-Дмитровской гряды, на территории, находя-
122
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
щейся в сфере воздействия пункта захоронения радиоактивных отходов
(ПЗРО). Определялось поступление радионуклидов в лесные экосистемы,
окружающие ПЗРО.
Концентрация радионуклидов (238+239Pu/90Sr+90Y/90Sr/137Cs) в компонентах
напочвенного покрова заболоченных лесов, Бк/кг сухой массы. Обозначено: - ниже
чувствительности прибора. Ценозы: I — березняк хвощево- злаковый сфагновый, П —
березняк вейниковый сфагновый, Ш — березняк борово-хвощевый со сфагнумом в
западинах, IV — сосново- березовый пушицевый долгомошно-сфагновый.
Виды/ценозы
I
Ш
IV
Ajuga reptans
Calamagrostis
neglecta
Carex sp.
Comarum
palustre
Chamaedaphne
calyculata
Deschampsia
caespitosa
Dryopteris
carthusiana
Equisetum
sylvaticum
Eriophorum
vaginatum
Galeobdolon
luteum
Galium palustre
Impatiens
nolitangere
Lysimachia
vulgaris
Milium effusum
Oxycoccus
palustris
Pedicularis
palustris
Poa palustris
Vaccinium
myrtilus
Vaccinium
-/740/320/-
69/440/220/-
-/850/400/82,4
-/1600/620/52,6
-/1600/800/-
-/380/170/97,1
-/1400/700/210
260/930/160/85,2
-/780/-/140
-/530/-/77,8
62,2/780/-/96,6
-/1100/-/330
-/780/-/-
-/360/-/-
-/1300/220/
52,7
-/1100/-/-
-/1400/-/190
-/960/-/-
-/2300/350/-
-/1100/120/
28
-/1200-/66,8
-/320/-/35,7
-/620/-
/230
-/890/-
/210
-/350/-
/84
-/720/-
/360
-/450/
190/110
-/1200/
-/410
-/480/-
/150
-/250/-
123
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
vitisidea
Мхи зеленые
Мхи политри-
ховые
Мхи
сфагновые
Подстилка
-/780/340/160
-/760/300/110
-/630/140/-
50/710/-/110
130/720/-/-
-/590/250/-
1-
100/960
/-/180
60/770/
-/310
-/670/-
/150
Результаты определения содержания радионуклидов в напочвенном
покрове позволяют выявить следующие закономерности/см. табл.).
1. Типичные болотные виды: Comarum palustre, Poa palustris, Cala-
magrostis neglecta, содержат меньше радионуклидов, чем влаголюбивые
виды с более широкой экологической амплитудой: Eqisetum sylvaticum,
Impatiens nolitangere, и виды-мезофиты, встречающиеся в заболоченных
лесах на самых сухих участках: Milium effusum, Ajuga reptans, Galeobdolon
luteum. Это согласуется с данными о пониженной активности процессов
жизнедеятельности у болотных растений (в частности интенсивности
корневого всасывания), и замедленности их роста.
2. Кустарнички содержат радионуклиды в меньшем количестве, чем
травянистые растения. Это отмечается не только в заболоченных, но и в других
обследованных лесных сообществах.
3. Мхи накапливают несколько отличный от трав и кустарничков спектр
радионуклидов. Удельная активность мхов относительно невелика и
составляет от 700 до 1000 Бк/кг. Фитомасса мхов в обследованных ценозах в 2-4
раза превышает фитомассу трав и кустарничков, запас радионуклидов в
моховом ярусе намного больше, чем в травяно-кустарничковом.
4. Альфа-излучающйе радионуклиды отмечены только в некоторых
травянистых видах (сабельник и мятлик болотный) и во мхах.
ВЛИЯНИЕ СТЕПЕНИ ОСУШЕНИЯ ТОРФЯНЫХ ПОЧВ НА ФАУНУ
ПАНЦИРНЫХ КЛЕЩЕЙ
Л.М. Ласкова, Д. А. Криволуцкий
Институт леса Карельского НЦ РАН, Петрозаводск;
МГУ, географический факультет
В болотных почвах Карелии в комплексах почвенных животных
доминируют панцирные клещи — Oribatei [1]. Торф является специфической
средой их обитания. Высокая влажность, плохая аэрация приводят к
накоплению и консервации органического вещества в виде слаборазложившихся
растительных остатков. Гидролесомелиорация приводит к смене процессов
124
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
накопления торфа его минерализацией. В этом процессе большая роль
принадлежит и панцирным клещам.
Существуют данные о влиянии мелиорации на численность и состав мик-
роартропод в торфяных почвах. Благоприятные изменения в составе
почвенных сапрофагов отмечены в мелиорированных болотах Литвы [3].
Результаты [2] показали, что при осушении болот увеличиваются численность
и количество групп животных, минерализующих растительные остатки.
Исследования выполнены в сосняке травяно-сфагновом на переходной
торфяной почве. Болото осушено в 1972 году сетью открытых каналов при
расстоянии между ними 190 м. Материал собирался в 1982-84 г.г. в центре
межканавья на удалении 30 м от канала. Пробы отбирались до глубины 10
см. Вегетационные периоды годов существенно отличались по
метеорологическим условиям. Лето 1982 г. было относительно холодное и дождливое,
1983 — теплое и сухое, 1984 — теплое и дождливое. Независимо от
погодных условий в разные годы наблюдений высокий уровень стояния воды на
участках был отмечен в мае, низкий — в августе. На осушенных торфяниках
нами обнаружено 32 вида панцирных клещей (см. табл.).
Виды Расстояние от осушительного канала
30м 95м
Hypochthonius rufulus C.L.Koch
Nothrus pratensis Sellnick
N.borussicus Sellnick
Camisia borealis (Thorel)
C. biverrucata (C.L. Koch)
Platynothrus peltifer (C.L.Koch)
Malaconothrus punctulatus Hammer
Heminothrus longisetosus Willmann
Nanhermannia coronata Berlese
Ceratoppia quadridentata (Haller)
Carabodes marginatus Michael
Tectocepheus velatus Michael
Suctobelbella palustris (Forsslund)
S.hammeri (D.Kriv.)
S.acutidens (Forsslund)
S.forsslundi (Strenzke)
S.singularis (Strenzke)
Oppiella nova (Oudemans)
Oppia neerlandica (Oudemans)
0.minus (Paoli)
O.minutissima Sellnick
O.globosa (Michael)
Autogneta tragardhi Forsslund
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
125
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Scheloribates laevigatus (C.L.Koch)
Diapterobates oblongus (C.L.Koch)
Galumna obvia (Berlese)
Pilogalumna tenuiclava (Berlese)
Steganacarus striculus (C.L.Koch)
Phthiracarus ligneus Willmann
P.borealis (Tragardh)
P.nitens (Nicolet)
Euphthiracarus cribrarius (Berlese)
Всего видов: 32
+
+
+
+
+
+
+
-
-
30
Полученные данные о видовом и количественном составе орибатид на
опытных участках, расположенных на расстоянии 30 и 95 м от
осушительного канала, говорят о разнице между ними. На сильно осушенном участке
найдено 30 видов клещей, на слабоосушенном — 21, но в обоих случаях
доминируют одни и те же виды: Oppiella nova, Steganacarus striculus, Nan-
hermannia coronata, довольно многочисленна и Oppia neerlandica. На
интенсивно осушенном участке средняя плотность орибатид составляет
примерно 9000 экз./м2 начиная с мая и достигает максимума в октябре (11800
экз./м2). На слабо осушенном участке численность орибатид в течение
вегетационного сезона претерпевает сильные изменения. Плотность панцирных
клещей в мае-июне не превышает 4500 экз./м2, но в осенних образцах их
численность очень высока (в октябре — 16050 экз./м2).
Результаты можно объяснить различной степенью влажности торфяных
почв. Водный режим интенсивно осушенного участка более стабильный и
численность орибатид не колеблется сильно в течение вегетационного
сезона. На удаленном от осушителя участке уровень грунтовых вод сильно
меняется: в мае близок к поверхности почвы, к августу-сентябрю снижается.
Работа выполнена при поддержке РФФИ (проект № 97-05-65396).
[1] Ласкова Л.М. Состав и численность микрофауны беспозвоночных торфяных
почв в зависимости от степени осушения // Изменение лесоболотных биогеоценозов
под влиянием осушения. Петрозаводск, 1986. С. 193-204. [2] Козловская Л.С. Роль
беспозвоночных в трансформации органического вещества болотных почв. Л., 1976.
211 с. [3] Педобиологическая характеристика типичных болот Литовской ССР.
Вильнюс, 1972. С. 107-140.
СУБФОССИЛЬНЫЕ ПАНЦИРНЫЕ КЛЕЩИ КАЛУЖСКОЙ ОБЛАСТИ
Л.М. Ласкова, М.С. Соддатов, А.И. Карпов
МГУ, географический факультет
В 1998 г. исследованы панцирные клещи в торфяных отложениях из
Бутовского болота Боровского района Калужской области, расположенного на
126
+
+
+
+
+
21
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
еоритории учебного полигона географического факультета МГУ, на водо-
зделе рек Протва и Лужа. Минеральное ложе торфяной залежи выстлано
пылеватым суглинком на глубине от 1,3 до 2,2 м. В начальной стадии
формирования болото развивалось по низинному типу, потому нижняя часть
т0рфяной залежи представляет собой плохо разложившиеся остатки
крупных болотных растений, возрастом около 6000 лет. По мере нарастания
т0рфяной подушки центр болота превысил уровень грунтовых вод и
оказался на питании исключительно атмосферными осадками, иными словами,
развитие болота пошло по верховому типу. Именно поэтому в пробах с
глубины 1-1.8 м начинает попадаться сфагнум, из которого практически на
100% состоит торфяная толща от поверхности до глубины 0,5-1,5 м.
Скважина для отбора образцов была заложена в центральной части
болота, где торфяная залежь имеет максимальную толщу. Отбор образцов
проводился почвенным буром с глубины 1,8-2,0 м. Окружающая
растительность состоит преимущественно из Sphagnum medium, Eriophorum polysta-
chyon, Oxycoccus quadripetalys, Vaccinium uliginosum, Ledum palustris, Cha-
maedaphe calyculata. К центральной части болота приурочен разреженный
древостой из сосны и березы, высотой до 3 м, средний диаметр 12 см,
возраст 60-65 лет.
В образцах торфа найдено 14 видов орибатид, среди которых
доминировали обычные для европейской части России гидрофильные виды Hydro-
zetes lacusths (61 экз.), Я. lemnae (23 экз.), единично Limnozetes foveolatus,
L sphagni, гигрофилы Trimalaconothrus novus (62 экз.), Т. glabra (36 экз.),
Nothrus pratensis (4 экз.), единичны эврибионты Punctoribates frimaculatus.
Oppiella neerlandica, Ceratoppia quadridentata. В сборах впервые обнаружен
северный циркумполярный вид Melanozetes sellnicki и свойственный
широколиственным и смешанным лесам Pilogalumna tenuiclava.
ЦИКЛИЧЕСКИЕ СМЕНЫ ДРЕВОСТОЕВ НА ВЕРХОВОМ БОЛОТЕ:
АНАЛИЗ ПРИЧИН И ПОСЛЕДСТВИЙ ЧАСТИЧНОЙ ГЖЕЛИ СОСНЫ
А.А. Маслов, Ю.В. Петерсон
Институт лесоведения РАН, Московская обл.
Процессы динамики коренных сообществ олиготрофных сосновых болот,
а также природные факторы, влияющие на их состояние (в отдельные годы
и/или десятилетия) представляют большой интерес для теории
фитоценологии. Их исторический тренд на протяжении столетий может быть установлен
по данным ботанического анализа торфа. Но относительно краткосрочные
флуктуации, протекающие без существенной смены видового состава, могут
не находить должного отклика в составе залежи и выявляются только по
127
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
результатам наблюдений на постоянных пробных площадях. Цель работы
анализ причин и последствий массового усыхания сосны на болоте, про.
гноз динамики насаждения на основе гипотезы о цикличности изменений.
С 1979 г. изучался сосняк в краевой части верхового болота общей пло-
щадью 7 га у д. Волково (Звенигородский лесхоз Московской обл.).
Мощность залежи 80-100 см. Возрастная структура чисто соснового древостоя в
1981 г. — относительно разновозрастная, с пиком возобновления в 1871-
1900 годы. Бонитет Vе, полнота превышала 1. Подрост и подлесок не был
выражен. В травяно-кустарничковом ярусе доминировали пушица
влагалищная, клюква и хамедафна на почти 100%-ном покрове Sphagnum
magellanicum и S. angustifolium [2]. Регулярные наблюдения за древостоем,
подростом и нижними ярусами проводились в 1981-1996 гг.
Предполагалось, что сосняк на верховом болоте будет представлять
образец устойчивости и минимальной флуктуационной изменчивости. Однако,
в течение нескольких лет наблюдалось массовое усыхание болотных сосен.
Вне пробной площади этот процесс на болоте имел местами сплошной
характер. Одновременно с гибелью сосен появился и стал быстро развиваться
подрост березы пушистой. Драматические изменения были отмечены и в
нижних ярусах [3].
Большой сосновый лубоед как непосредственный главный фактор
смертности сосен. За 15 лет с 1981 по 1996 г. число живых стволов сосны на га
уменьшилось в 3 раза — с 1500 до 490. Массовый отпад деревьев был
непосредственно связан с тремя последовательными вспышками размножения
большого соснового лубоеда, максимумы которых пришлись на 1984, 1988 и
1992 гг. (рис. 1). Каждая последующая вспышка была сильнее предыдущей.
К 1996 г. усыхание деревьев из-за вспышек лубоеда прекратилось. Гибель
деревьев происходила в результате массового поражения стволов в их
нижней части, дополнительное питание жуков в кронах деревьев играло
меньшую роль. Попытка связать предпочтения лубоеда с возрастом или
размером деревьев, их состоянием, не получила статистического подтверждения.
Большой сосновый лубоед — довольно опасный вредитель сосны
обыкновенной: заселяет ствол в районе толстой коры, зимует на корневых лапах,
начало лета обычно в третьей декаде апреля - начале мая при
среднесуточной температуре около 5 ° С и освобождения от снега приствольных кругов
[4]. В районе исследований он был отмечен как фоновый фактор с 1979 г.
Динамика лубоеда в связи с климатическими флуктуациями. Для
выявления причин неожиданных вспышек лубоеда проведен анализ
климатических характеристик за 1971-1990 гг. по данным метеопоста «Звенигород»
(закрыт в 1991 г.). Показано, что вспышки четко приурочены к сериям
сухих (засушливых) лет и прекращались во влажные годы (см. рис.).
Дополнительно, на динамику популяции короеда большое влияние оказывают
128
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
температурные условия мая - первого месяца развития личинок. Май 1988 и
1992 гг. был аномально теплым и сухим, близким по своим параметрам к
маю 1988 и 1989 гг. [6]. Все указанные годы соответствуют периодам
вспышек. До 1988 г. подобные условия мая наблюдались только сто лет
назад — в 1886 г., причем именно в конце прошлого века наблюдалось
массовое возобновление сосен.
35 -,
25
15
-15
-25
-J-]—i—i—i—i—п—л—i—i—п—г
20 -
10 -
0 J
И
I Шр^.
Шт
лар-р
ср
Ljl
\sp
0>0*-C4CO^TlO<Or-.COO>QT-(NCO^lO«>N-
h»cococooooo<ocoooooooo)0><J>0)0)0)0>0)
Годовое количество осадков по метеопосту Звенигород, % отклонения от среднего
— 640 мм за 1971-90 гг. (вверху) и количество усохших сосен по годам
наблюдений (внизу). Затемнены серии сухих лет, совпадающие со вспышками лубоеда.
С другой стороны отмеченные сухие годы нельзя считать
неблагоприятными для соснового древостоя с точки зрения водного режима: показано [1],
что сухие годы благоприятно сказываются на росте поверхностных
корневых систем деревьев, способствуют деструкции торфа. В изученном объекте
не отмечено отрицательного влияния засух на рост деревьев по диаметру
даже в экстремальные 1972 и 1975 годы.
Возобновление березы как результат усыхания сосен. С начала массовой
129
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
гибели сосен (1983-85 гг.) наблюдалось массовое появление и быстрый рост
подроста березы пушистой. Через 10 лет, к 1996 г.. она сформировала
второй ярус высотой 3-6 м. и появление новых деревьев березы прекратилось.
В подросте отмечена также сосна и единичные экземпляры ели высотой до
70 см. Опубликованные свидетельства столь интенсивного внедрения
березы пушистой на верховом болоте нам неизвестны.
Предположительная причина внедрения и быстрого роста березы в
период массового отпада сосны — «залповое» поступление питательных веществ
в верхние слои почвы. Опад хвои, ветвей, коры и корней сосны на верховых
болотах разлагается на 40-50% в течение года [7] с высвобождением
зольных веществ и азота. С окончанием интенсивного поступления питательных
веществ возобновление березы прекратилось, однако рост ее продолжается
за счет второй, медленной фазы высвобождения.
Погребение корневой шейки как глубинный Фактор ослабления
древостоя. При поиске более общей причины ослабления древостоя сосны,
способствующей возникновению вспышек лубоеда, мы обратили внимание на
гипотезу В.Н. Сукачева [5], согласно которой нарастание слоя торфа на
верховых болотах приводит к постепенному погребению корневой шейки сосен
и корневых систем в целом. Погребение корневых шеек сосны на глубину
30-50 см является критическим. Предельным возрастом топяной формы
сосны болотной называется [5] 120-150 лет. Именно к этой форме относится
сосна на изученном болоте.
Для проверки этой гипотезы была измерена в трех разных выборках
сосен глубина залегания корневой шейки. У деревьев, выживших на пробной
площади к 1996 году, средняя глубина залегания составила 23,5 см, у
погибших во время третьей вспышки (1991-96 гг.) 29,9 см, а в зоне сплошной
гибели древостоя (вне пробной площади) 34,0 см. Различия средних для
всех трех выборок достоверны по критерию Стьюдента на 95% уровне.
Концептуальная модель циклической динамики древостоя. На основании
анализа данных предложена концептуальная модель динамики.
Возникновение относительно одновозрастного соснового древостоя и нарастание слоя
торфа через 100-120 лет приводит к серьезному ослаблению части деревьев.
Гибель сосен может происходить под действием разных факторов. Серия
лет, благоприятных климатически для развития короедов, приводит к
вспышкам и отмиранию массы деревьев. Высвобождение большого
количества питательных веществ за короткий период времени приводит к «взрыву
возобновления», возможно, со сменой на березу. Если модель верна, то
исчерпание дополнительного питания должно сравнительно скоро привести к
гибели подроста березы как более требовательного к условиям
минерального питания. Вероятно, что это произойдет во время серии слишком влажных
лет с высоким уровнем грунтовых вод на болоте. В дальнейшем возможна
130
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
серия флуктуации с возобновлением как сосны, так и березы, и с
постепенным возобновлением чисто соснового древостоя, замыкающим цикл.
Работа проводилась при поддержке РФФИ, грант № 98-04-48675.
[1] Вомперский С.Э. Биологические основы эффективности лесоосушения. М.:
Наука, 1968. 312 с. [2] Маслов А.А. Количественный анализ горизонтальной структуры
лесных сообществ. М.: Наука, 1990. 160 с. [3] Маслов А.А. Динамика фитоцено-
экологических групп видов и типов леса в ходе природных сукцессии заповедных
лесов центра Русской равнины // Бюл. МОИП. Отд. биол. 1998. Т. 103. № 2. С. 34-
43. [4] Маслов А.Д. и др. Защита леса от болезней и вредителей: Справочник. 2-е
изд. М.: Агропромиздат, 1988. 414 с. [5] Пьявченко Н.И. О приспособляемости
древесных растений таежной зоны к болотным условиям // Вопросы адаптации
растений к экстремальным условиям Севера. Петрозаводск: Карельский ф-л АН СССР,
1975. С. 52-63. [6] Рудичева Л.М. Обзор погоды в мае 1992 г. // Метеорол. и
гидрология. 1992. № 8. С. 121-123. [7] Смагина М.В. Результаты многолетнего опыта по
разложению растительных остатков на поверхности торфа и в его деятельном слое
// Гидротехническая мелиорация земель, ведение лесного хозяйства и вопросы
экологии. СПб: СПбНИИИЛХ, 1997. С. 142-145.
НАСЕЛЕНИЕ ПТИЦ ОТКРЫТЫХ И МАЛООБЛЕСЕННЫХ БОЛОТ
КОСТОМУКШСКОГО ЗАПОВЕДНИКА
Е.Н. Матюшкин, Е.А. Даниленко
МГУ, географический факультет; МосНПО «Радон», Москва
Открытые и малооблесенные болота (рямы), объединенные в группу
переувлажненных местообитаний птиц, занимают в заповеднике большую
площадь, сопоставимую с площадью сомкнутых лесов. Среди них
встречаются не только очень крупные, но также небольшие и даже очень мелкие по
размерам участки. Своеобразие этой группы местообитаний птиц состоит в
том, что открытые и малооблесенные болота часто соседствуют, поэтому на
всех топографических картах этот комплекс отображается одним контуром с
четкой общей границей. Их дифференциация проводится только на уровне
крупномасштабной лесотаксационной съемки при описании дробных
контуров. Хотя в типологическом отношении подразделения данной группы
местообитаний птиц значительно различаются между собой, по населению
птиц они довольно близки, что позволяет рассматривать их вместе.
Специфические черты группировки птиц этого местообитания сводятся к
следующему: наличие компактной группы видов-индикаторов; низкое в
целом видовое разнообразие (см. табл.). К числу видов-индикаторов из
господствующей группы можно отнести лесного конька, желтую трясогузку и
овсянку-ремеза, из второстепенных — фифи и лугового чекана, а также
черныша из третьестепенных.
131
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Эти птицы большей частью — наземногнездящиеся. Лесной конек хотя и
проводит большую часть времени на малооблесенных болотах, все же
гнезда устраивает вне их, хотя и в непосредственной близости от болотных
массивов. Для устройства гнезда этот вид выбирает сухие, хорошо
прогреваемые солнцем места на опушках или в разреженных участках леса под
прикрытием кустарников или камней. Из перечисленных видов, пожалуй,
только фифи, черныш и желтая трясогузка могут гнездиться среди сильно
переувлажненных участков болот, лишенных древесной растительности. Однако
и они чаще всего заселяют участки с наличием хотя бы очень разреженного
древостоя из низкорослых сосенок.
Характеристика населения птиц открытых и малооблесенных болот Костомукшско-
го заповедника (конец мая — июнь 1987-89 гг., длина маршрутов — 14,6 км,)
Вид
Лесной конек
Желтая трясогузка
Овсянка-ремез
Фифи
Пухляк
Луговой чекан
Глухарь
Черныш
Всего 8 видов:
Количество особей
на 1 км маршрута
8,4
1,8
0,8
1,4
0,3
0,3
0,1
0,1
12,1
на 1 км2
48,0
20,0
17,9
4,3
3,1
2,5
1,6
0,7
98,1
Доля вида, %
49,0
20,4
18,3
4,4
3,2
2,6
1,6
0,7
100,0
Так как открытые болота и рямы со всех сторон окружены лесом, вокруг
них формируются опушечные полосы большого протяжения, где
концентрируется значительное число лесных видов птиц. Привлекают их сюда более
благоприятные, чем внутри лесных массивов, условия обитания, что вообще
свойственно линиям контакта резко контрастных местообитаний (так
называемый опушечный эффект). Эти виды птиц часто вылетают далеко за
пределы своих основных местообитаний, что приходилось принимать во
внимание при обработке полевых материалов: некоторые встречи птиц, явно не
закономерные, в подсчет не включались.
В данном типе местообитаний, с учетом отсеивания случайных встреч,
зарегистрировано самое малое число видов — 8. Невысока и общая
плотность населения птиц (98,1 особей/км2), самая низкая из отмеченных где-
либо в заповеднике. Эта величина более чем наполовину ниже
установленных для остальных вариантов населения, за исключением лишь сосняков со
вторым ярусом ели.
Абсолютное господство здесь принадлежит лесному коньку, на долю
которого приходится почти половина (49%) всего населения птиц. Суммарная
доля участия его и двух других видов-доминантов — желтой трясогузки
132
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
(20,4%) и овсянки-ремеза (18,3%) - 87,7%. Четыре вида — пухляк, луговой
чекан, фифи и глухарь — второстепенные члены сообщества, хотя по
встречаемости фифи лишь немного уступает желтой трясогузке (1,4 против 1,8
особей на 1 км маршрута), занимающей второе место в ряду убывания
показателей численности. Коэффициенты общности видового разнообразия и
населения птиц этого местообитания с остальными — самые низкие, что
указывает на большое своеобразие данного типа сообществ птиц.
О СТРУКТУРЕ КОРЕННОГО ЕЛЬНИКА НА МЕЗООЛИГОТРОФНОМ
БОЛОТЕ ЮЖНОЙ ТАЙГИ
И.Б. Прокуронов
МосНПО «Радон», Москва
В ряду болотных экосистем умеренной зоны особое значение
принадлежит лесным насаждениям завершающих стадий олиготрофной гидросерии,
которые в настоящее время имеют узко локальное распространение. В ходе
исследований в резервате «Кологривский лес» (лесорастительный район
пихтово-еловых лесов южных отрогов Северных Увалов; Костромская
область) выявлен и описан уникальный тип леса — ельник морошково-
сфагновый. В пределах резервата известно только одно его местообитание.
Ельник (общая площадь около 3 га) занимает депрессию близ тылового
шва 1-ой надпойменной террасы, отделенной от водотока невысокой гривой.
Глубокие торфяные почвы мезоолиготрофного типа соответствуют субак-
вальному типу геохимического ландшафта. Мощность торфа более 1 м, в
центральной части не менее 3 метров.
Формула древостоя 9EnolBi2o+C2oo. Ярусная стратификация отсутствует.
Древостой абсолютно разновозрастный, сомкнутость 0,4-0,5; с большим
количеством окон и прогалин с тонкомером и подростом ели. Число стволов
около 900 шт./га; полнота 0,7. Бонитет IV-V; запас порядка 180 м3/га.
Средний возраст ели 170 лет, диапазон от 90 до более 350-ти лет. Высота 15 м,
иногда до 20 м; преобладающий диаметр 16-24 см. Крона узкая
конусовидно-цилиндрическая; высота прикрепления крон 1,5-5,0 м; очищенность от
сучьев плохая. Сосна (возраст 200 лет) высотой около 20 м, диаметр около
50 см. Крона распростертая, короткоконусовидная. Прикрепление кроны —
на высоте полога ели (15-18 м). Очищение от сучьев хорошее. Береза
высотой 15-18 м, диаметром 24-32 см; средний возраст превышает 100 лет.
Обильный подрост представлен елью (>19 тыс. шт./га) сомкнутостью 0,3-0,7
(sp-cop1); очень редко — сосна, пихта, береза. Виды подлеска отсутствуют.
В травяно-кустарничковом ярусе (проективное покрытие порядка 30%)
отмечено 14 видов растений, среди которых особенно заметно доминирова-
133
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ние морошки. Моховой покров с общим проективным покрытием 90%, в
том числе сфагнов — до 80%; из девяти видов мхов шесть — сфагновые.
Доминирует кустарничково-сфагновая парцелла (37% площади); участки
парцеллы, сливаясь, образуют общий фон. На сплошном покрове сфагновых
мхов (Sphagnum angustifolium, S. centrale, S. compactum, S. girgensohnii, S.
magellanicum, S. ripahum, S. russowii) обильна морошка, кустарнички:
Ledum palustre, Lyonia calyculata, Vaccinium myrtillus, V. uliginosum, V. vitis-
idaea с проективным покрытием 10-60% и попеременным доминированием;
изредка клюква (Oxycoccus microcarpus, О. palustris), осоки Carex canes-
cens, С. globularis. Парцелла елового подроста на ковре из сфагнов занимает
30% площади. Изредка черника и брусника (до 10% проективного
покрытия). Подрост высотой до 3 м и более имеет сомнутость до 1,0. Мелкотрав-
но-зеленомошная парцелла (27%) приурочена к подкроновому пространству.
Сплошной покров мхов с преобладанием Dicranum polisetum, Hylocomium
splendens, Pleurozium schreberi. Куртины брусники, черники; таежное
мелкотравье Linnaea borealis, Oxalis acetosella и др. Осоково-сфагновая
парцелла занимает 6% площади. На фоне сфагновых мхов — Sphagnum angusti-
folium, S. centrale, S. compactum, S. girgensohnii, S. magellanicum, S. ripahum
— осока Carex canescens: изредка примесь Eriophorum vaginatum, V. vitis-
idaea. Участки парцеллы крайне невелики и мозаично рассеяны по
площади. В ряде случаев границами парцелл служит значительное количество ва-
лежа разной степени разложения.
В отличие от более известных в подобных местообитаниях сериальных
сосновых насаждений, ельник морошково-сфагновый отвечает
завершающей стадии олиготрофно-гидросериальной сукцессии со сменой сосны на
ель. При этом высокий возраст эдификатора древесного яруса — ели,
обильное произрастание морошки, наличие отдельных сосен, в массе
отмечаемых только на значительном расстоянии, позволяет в то же время
высказать мнение о «реликтовом» характере экосистемы ельника морошково-
сфагнового. В пользу этого могут свидетельствовать также данные
исследования населения жужелиц [Еремин, 1986]: здесь впервые для региона
отмечены виды Bimbidion bruxellense и Agonum bogemanni; карабидофауна
ельника отличается высокой экологической специфичностью доминантов с
преобладанием лесо-болотных и болотных видов.
Исследование популяционной структуры еловых древостоев показало
следующее. Ельник морошково-сфагновый характеризуется абсолютно
разновозрастной структурой с отсутствием локализации возрастных структур (в
отличие от кисличных ельников климакса с циклично-, или ступенчато-
разновозрастной структурой). Вплоть до 200-летнего возраста преобладают
деревья подчиненного ранга — IV, V классов Крафта, тогда как в древосто-
ях климакса почти на столетие раньше начинают доминировать особи выс-
134
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
щего ранга. Это позволяет констатировать в данной ценопопуляции ели
преобладание К-стратегии, особенно заметно проявляющейся в сравнении с г-
стратегией ценопопуляции ели климакса. Меньшая в целом изменчивость
биометрических характеристик древостоя исследуемого типа леса по
сравнению с ельниками климакса свидетельствует о соответственно различной
степени внутрипопуляционной конкуренции.
Материалы исследований могут служить подтверждением известного
тезиса о видовой дифференциации ели сибирской (Picea obovata) и ели
европейской (P. abies) в связи с местоположением их популяций в зоне контакта.
В «Кологривском лесу», где ель в целом отнесена к гибридной форме Picea
х fennica (Regel) Kom., различие в структуре ценопопуляции может
расцениваться как свидетельство тяготения признаков древостоев олиготрофных
местообитаний к ели сибирской, климаксовых — к ели европейской.
НАСЕЛЕНИЕ ГЩЩ БОЛОТ ЮГА ЛЕСНОЙ ЗОНЫ
ВОСТОЧНО-ЕВРОПЕЙСКОЙ РАВНИНЫ
Е.С. Равкин, И.И. Бышнев
МосНПО «Радой», Москва;
Березинский биосферный заповедник, Витебская обл., Беларусь
Материалы собраны авторами в пределах южной тайги и подтаежных
лесов Европейской части России и Белоруси (Псковская, Новгородская,
Смоленская области, Березинский заповедник). Учеты птиц проводились в
разные годы на низинных, переходных и верховых болотах различной
степени облесенности. Птицы подсчитывались в первую половину лета на
постоянных маршрутах с -четырехкратной повторностью. Во время учетов
записывались все встреченные птицы, независимо от расстояния до них.
Видовая плотность населения рассчитывалась по средне групповым
дальностям обнаружения [Ю.С. Равкин, 1967]. Кроме того, использованы
результаты учетов птиц, предоставленные Ю. А. Буйволовым, проведенные на
верховых болотах Тверской области в Центрально-лесном биосферном
заповеднике, а также — опубликованные данные по населению птиц болот
Ярославской, Тамбовской областей и Березинского заповедника. Все
использованные материалы (22 варианта населения птиц) сгруппированы в
соответствии с природно-зональным делением территории и типами болот,
представляющих собой местообитания птиц. По усредненным данным в каждой
из двух групп местообитаний (верховые болота и низинные с переходными)
вычислены суммарные показатели орнитокомплексов (см. табл.). Расчеты
проводились по материалам и с помощью пакета прикладных программ
банка данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН.
135
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Сравнивая население птиц верховых и низинных с переходными болот
южной тайги можно отметить, что обилие особей и суммарная биомасса
птиц на низинных и переходных болотах почти втрое больше, чем на
верховых. Фоновых видов (обилие которых составляет от 1 и более особей на кв.
км) там в полтора раза больше. Но общее число зарегистрированных во
время учетов видов на верховых болотах южной тайги почти вдвое больше,
чем на низинных и переходных. Это скорее всего связано с наличием грядо-
во-мочажинно-озерного комплекса и облесенных участков болот (рямов). За
счет такого разнообразия происходит проникновение на верховые болота
водно-околоводных, лесных и луговых видов птиц. На более продуктивных,
но менее мозаичных низинных и переходных болотах меньше встречено
типично лесных видов, хотя остальные встречаются в большем количестве.
Для обеих групп орнитокомплексов южной тайги общим доминантом
является желтая трясогузка, более преобладающая в населении верховых
болот. Кроме нее на верховых болотах по численности лидируют лесной и
луговой коньки, а на низинных и переходных болотах бекас и чибис. Около
половины птичьего населения верховых болот приходится на
представителей европейского типа фауны, чуть более трети населения составляют транс-
палеаркты. Встречаются представители и сибирского типа, составляющие
лишь 6% орнитокомплекса по обилию особей. По видовому составу
преобладают транспалеаркты, доля числа видов европейского типа меньше, чем
доля их населения, а сибирского — наоборот больше. Последнее связано с
проникновением с севера и северо-востока на верховые болота южной тайги
видов сибирского типа фауны. Орнитокомплекс низинных и переходных
болот отличается повышенной долей транспалеарктов, меньшим участием
птиц сибирского типа фауны и появлением представителей китайского типа.
В подтаежных лесах прослеживаются аналогичные различия суммарных
показателей населения птиц разных групп болотных местообитаний, однако
превышения общего числа видов на верховых болотах не отмечено. Орни-
токомплексы разных групп болот сходны по двум лидирующим видам, но
на верховых болотах преобладает еще и лесной конек, отсутствующий на
низинных и переходных. По фаунистическому составу на верховых болотах
больше птиц европейского типа фауны, встречаются птицы сибирского типа
и меньше транспалеарктов (по обилию особей).
Орнитокомплексы верховых болот южной тайги и подтаежных лесов
различаются более высокими суммарными показателями, большим
участием птиц сибирского типа фауны и меньшим — птиц европейского типа в
южной тайге. Различия в населении птиц низинных и переходных болот
обеих подзон неоднозначны: видовое богатство больше в подтаежных лесах,
общее обилие особей и суммарная биомасса в южной тайге. Последнее
связано с преобладанием более крупных птиц (бекаса и чибиса), составляющих
136
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
20% населения. По фаунистическому составу в подтаежных лесах больше
птиц европейского типа (по обилию особей) и меньше транспалеарктов. В
южной тайге в число лидеров входят представители сибирского типа фауны.
Суммарные показатели населения птиц болот юга лесной зоны
Местообитания птиц
Верховые
болота
южной тайги
Низинные и
переходные
болота южной
тайги
Верховые
болота
подтаежных лесов
Низинные и
переходные
болота
подтаежных лесов
В среднем
Всего видов/
Фоновых
видов/Всего
особей на км2/
Сумм,
биомасса, кг/км2
57/17/105/16,4
32/25/390/42,1
38/11/85/10,5
72/37/300/31,1
103/32/238/26,
2
Лидеры по
обилию (%)
Лесной конек
(31), Желтая
трясогузка
(20), Луговой
конек (6)
Желтая
трясогузка (13),
Бекас (12),
Чибис (8)
Лесной конек
(60), Луговой
чекан (10),
Луговой конек
(6)
Желтая
трясогузка (18),
Луговой конек
(12), Луговой
чекан (10)
Желтая
трясогузка (16),
Лесной конек
(9), Луговой
конек (9)
Фаунистическ
ий состав по
обилию (тип
фауны, %)
Европейский
(48), Транс-
палеаркты
(39),
Сибирский (6)
Транспалеарк
ты (63),
Европейский
(36),
Китайский (0.8)
Европейский
(80), Транс-
палеаркты
(16),
Сибирский (3)
Европейский
(52), Танспа-
леаркты (46),
Китайский
(0.6)
Европейский
(50)
Транспалеарк
ты (48), Арк-
тический (0.9)
Фаунистическ
ий состав по
числу видов
(тип фауны,
%)
Транспалеарк
ты (46),
Европейский
(30),
Сибирский (17)
Транспалеарк
ты (47),
Европейский
(47),
Сибирский (3)
Транспалеарк
ты (53),
Европейский
(32),
Сибирский (13)
Транспалеарк
ты (49),
Европейский
(42),
Арктический (14)
Транспалеарк
ты (42),
Европейский
(41), Сибир-
ский(12)
Орнитокомплексы болот достаточно сходны между собой по суммарным
показателям. Различия связаны в основном с типом местообитания,
продуктивностью, облесенностью и мозаичностью болот, занимаемой ими
площадью, в меньшей степени с расположением в разных подзонах.
Коэффициенты сходства Жаккара-Наумова более высокие между сообществами птиц
однотипных болот в разных подзонах (39 и 40%), чем разнотипных — в
одной подзоне (по 11%). Сходство сообществ птиц верховых болот южной
тайги с низинными и переходными болотами подтаежных лесов несколько
137
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
выше (18%), за счет общих лидеров по обилию: желтой трясогузки и
лугового конька. Указанные различия подтверждают преобладающее влияние на
облик населения птиц болот типа местообитания, то есть высокую интразо-
нальность болотных сообществ птиц. По сравнению с населением птиц дру.
гих местообитаний они весьма специфичны и бедны. Они имеют лишь
небольшое сходство с лугово-полевыми и в еще меньшей степени с лесными.
СТРУКТУРА РАСТИТЕЛЬНОГО ПОКРОВА НА ГРАНИЦЕ
ЛЕС-ВОДОЕМ В ПОЙМАХ РЕК ОКСКО-ДОНСКОЙ РАВНИНЫ
А.В. Славгородский
Институт биологии внутренних вод РАН, Ярославская обл
Растительный покров на границе лес-водоем в поймах Окско-Донской
равнины представлен разнообразными сообществами, находящимися в
различных, иногда уникальных сочетаниях, испытывающих влияние
гидрологического режима рек. На его формирование оказали воздействие и
природные, и антропогенные факторы [СлавгородскийЛ 997]. Исследования 1995-
98 гг.. проведенные в поймах рек Битюг, Цна, Хопер и некоторых их
притоков, показали структурную неоднородность этого компонента биоты.
Используя теорию экотопологической дифференциации растительного
покрова [Камелин, 1973. 1979; Юрцев, Семкин, 1980; Шеляг-Сосонко, Ди-
дух. 1980; Клоков, 1980; Юрцев, 1987; Новосад, 1992; Кузьмичев, 1992] в
поймах рек выделены семь экотопологических комплексов. В них
объединяются виды растений на основе общности средообразующих факторов и
сходные экогенетически [Кузьмичев, 1992; Новосад, 1992] (см. табл.).
В результате обработки полученных данных все разнообразие границ
лес-водоем сведено к восьми типам. В формулах показана
последовательность смены экотопологических комплексов от центра водоема к
водоразделу в пределах поймы: Тип 1. Hyd-»Dr; Тип 2а. Hyd-»Hy->Dr; Тип 2b. Hyd
-»Ps-»Dr; Тип 2с. (Hyd-»Hy->Dr)n; Тип За. Hyd-»Hy->Dr-»Pr-»Dr; Тип ЗЬ.
Hyd->Ps Dr->Pr->Dr; Тип 4а. Hyd-»Hy->Pr ->Dr; Тип 4b. Hyd-»Ps->Pr->Dr;
Тип 5. Hyd->Hy<->Pl)n ->Dr; Тип 6a. Hyd-*Pr; Тип 6b. Hyd->Ps->Pr, Тип 7а.
Hyd->Hy->Ant; Тип 7b. Hyd-»Ps->Ant; Тип 8а. Hyd->Hy-»Pr ->Ant; Тип 8b.
Hyd->Ps ->Pr-»Ant.
Помимо буквенных обозначений каждый тип представлен в виде
рисунков-схем. Для каждого типа на местности выбран один модельный участок,
наиболее близкий рисунку-схеме. Схемы представляют структуру границы
на верхнем уровне иерархии. При детальном рассмотрении в каждом из
семи экотопологических комплексов может быть выделена своя структура.
138
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
Экотопологичекие
комплексы
Основной средообразующий
фактор
Экогенетические связи
Hydrophyton (Hyd)
Hygrophyton (Ну)
Paludophyton (PI)
Drymophyton (Dr)
Pratophyton (Pr)
Psammophyton (Ps)
Antropogenophyton
(Ant)
Водная среда
Колебание
уровня воды
Популяционная жизнь мхов,
болотный субстрат (торф)
Популяционная жизнь
древесных биоморф
Популяционная жизнь
крупных животных-фитофагов,
сенокошение
Песчаный, олиготрофный,
периодически увлажняемый
субстрат
Деятельность человека
Пресные и слабосоленые,
теплые воды
Побережья пресных и
слабосоленых, теплых водоемов
Выклинивающиеся воды
и мочажины
Неморальный, боровой,
таежный, мелколистный и
альнетальный комплексы
Лесолуговой доагрикультур-
ный макрокомплекс
Песчаные и супесчаные
почвы
Формирующийся комплекс со
слабыми связями
В поймах рек Окско-Донской равнины выделенные экотопологические
комплексы состоят из видов формирующих сообщества ассоциаций 15
классов. Hydrophyton формируется сообществами классов: Charetea Fukarek
1961; Lemnetea R. Tx. 1955; Potametea Klika in Klika et Novak 1941.
Hygrophyton - Phragmitetea R. Tx.et Preising 1942; Bolboschoenetea maritimi Viche-
rek et Tx.1969 ex Tx.et Hulb. 1971; Bidentetea tripartiti Tx. Lohm.et Prsg. in
Tx. 1950. Paludophyton - Scheuchzerio-Caricetea nigrae (Nordh.1936) Tx.1937.
Drymophyton - Alnetea glutinosae Br.-Bl. et Tx.1943 em. Muller et Gors 1958;
Salicetea purpureae Moor 1958; Querco-Fagetea Br. - Bl. et Vlieger in Vlieger
1937 em. Klika 1939. Pratophyton - Asteretea tripolium Westhoff et Beeftink
1962 ex Beeftink 1962; Chenopodietea Br.-Bl. 1951 em Lohm. J. et R.Tx. 1961
ex W. Matsz. 1962; Plantaginetea majoris Tx. et Prsg. in Tx. 1950; Galio-Urti-
cetea Pass. 1967; Urtico-Sambucetea Doing 1962 em Pass 1968. Psammophyton
- Juncetea maritimi Br.-Bl. et al. 1952 em. Beeftink 1965. Antropogenophyton -
культивируемыми и поддерживаемыми человеком сообществами
(сельскохозяйственные угодья).
В результате анализа структуры растительности полученных восьми
типов отмечены следующие закономерности. 1. Постоянным компонентом в
пойме являются сообщества гидрофитона и дримофитона. Последний при
антропогенном воздействии замещается сообществами антропогенофитона,
а при зоогенном — сообществами пратофитона. 2. Сообщества пратофито-
на. гигрофитона, палюдофитона, псаммофитона и антропогенофитона
присутствуют в пойме благодаря постоянно идущим нарушениям разного уров-
139
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ня иерархии (гидрологический режим, меандрирование, антропогенное, зоо-
генное и др. воздействие). 3. Общими для всех восьми типов границ явля*
ются сообщества классов Lemnetea и Potametea. Это указывает на
мозаичный характер размещения сообществ синтаксонов ранга классов в поймах
рек Окско-Донской равнины. Исключение составляют сообщества двух
вышеуказанных классов располагающиеся линейно, вдоль русел рек.
ЭКСПЕРИМЕНТАЛЬНАЯ ОЦЕНКА КИНЕТИКИ ФОРМИРОВАНИЯ
ЛЕСНОЙ ПОДСТИЛКИ В ОСУШЕННОМ СОСНЯКЕ
НИЗИННОГО ТИПА ЗАБОЛАЧИВАНИЯ
М.В. Смагина, А.В. Смагин
Институт лесоведения РАН, Московская обл.;
МГУ, факультет почвоведения
Количественная оценка динамики органических веществ (ОВ) в
болотных экосистемах — актуальная задача не только мелиорации, но и экологии
в связи с важнейшими проблемами современности — загрязнением
атмосферы и парниковым эффектом. Прямых экспериментальных данных для
решения данную задачу явно недостаточно. В первую очередь это касается
параметров аккумуляции и минерализации ОВ торфяной залежи, включая
наиболее активные верхние горизонты. Предпринята попытка определения
интенсивности аккумуляции лесной подстилки и ее отдельных фракций в
осушенном сосняке болотно-травяном низинного типа заболачивания
(Западнодвинский стационар ИЛАН, Тверская обл., 1986-1991 гг.).
В процессе эксперимента учитывалась динамика поступления
растительного опада и послойная аккумуляция подстилки на опытных площадках
(20x20 см2) в 10-ти кратной повторности. Слои подстилки отделялись друг
от друга посредством тонкой сетки (ячея 3 мм) из инертного синтетического
волокна. Анализ массы подстилки и ее фракций производился ежегодно
нарастающим итогом — на различных площадках материал накапливался
суммарно в течение 1, 2, 3, 4 и 5 лет. При этом оценивалась динамика массы
ОВ отдельных слоев подстилки (вариант опыта с сетчатыми прослойками), а
также общее количество ОВ, накопленное за 5 лет (вариант без послойного
дифференцирования). Результаты эксперимента приведены на рис. 1-3.
Общая масса ежегодного опада колеблется в небольших пределах (349-
430 г/м2) при среднем значении 388 г/м2. В течение года интенсивность
поступления опада неодинакова. Максимум, как правило, в октябре. При
значительном увеличении массы опада в конце августа 1989 г., основной
листопад наблюдался в последней декаде октября. Некоторое увеличение опада
в летние месяцы происходит за счет поступления соцветий, семян березы и
140
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
шишек сосны, а также листьев березы, поврежденных энтомовредителями.
5 отдельных случаях (весна-лето 1990 г) ветровал приводит к появлению
значительного количества травматического опада, представленного в
основном обломками ветвей и другими частями вегетирующей растительности.
2 7
i6
5 5
С
О 4
V
I 2
Ш
S л .
О 1
X л
0) 0
Ё 8
S с
и
5 8
> <N
) О)
S
8
^
S
8
об
8
8
*-
00
о
1Л
8
oi
я
8
8
^т
§
8
об
8
8
*-
S
о
ю
го 5
5 8
Й Я
5
8
г^
5>
8
•«-
СМ ч- (N
Рис. 1. Динамика поступления опада в сосняке болотно-травяном
Б
2
и
(Q
С
14001
12CO-J
woo]
8001
6004
400|
20о|
oi
Рис.2. Формирование лесной подстилки в течение 5 лет
■ ветх и, кора .шишки
■>10 мм
□5-10 мм
■< 5 мм
Рис.3. Динамика фракционного состава лесной подстилки
Ежегодная аккумуляция подстилки составляет в среднем 228 г/м2. Столь
значительное накопление растительных остатков нельзя отождествлять с
141
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
аккумулирующей способностью болотных экосистем, поскольку темпы их
минерализации также велики. Расчеты по линейной кинетической модели
[3] с использованием экспериментальных данных о поступлении опада и
динамики подстилки показывают, что в течение 40-60 лет формируется
стационарный запас подстилки, при котором поступление и минерализация ОВ
полностью сбалансированы и никакого депонирования Сорг не происходит.
Вместе с тем, ежегодно небольшая часть подстилки измельчается и
трансформируется, образуя специфичную для мелиорированных объектов форму
ОВ — торфогумус [2]. В эксперименте эта доля ОВ оценивалась фракцией
подстилки <5 мм. Последняя характеризовует количество трудноразлагае-
мого материала или потенциальный сток ОВ в торфяную залежь. Ее среднее
значение составило 50 г/м2год, что в пересчете на углерод соответствует
современным оценкам депонирования Сорг болотными экосистемами [1].
[1] Вомперский С.Э. Роль болот в круговороте углерода // Биогеоценологические
особенности болот и их рациональное использование. М.: Наука, 1994. С.5-38. [2]
Пальчински. А. Очерк фитоценологии торфяных болот Полыни и генетическая
классификация торфов, основанная на эколого-фитоценологическом принципе.//
Ботан. ж.. 1969. Т. 54. № 12. С.1921-1938. [3] Смагин А.В. К теории устойчивости
почв //Почвоведение. 1994. № 12. С.26-33.
ВЛИЯНИЕ ОСУШИТЕЛЬНОЙ МЕЛИОРАЦИИ
НА СТРУКТУРУ МИКОБИОТЫ В ПОЧВАХ
НИЗИННОГО И ВЕРХОВОГО ТИПОВ ЗАБОЛАЧИВАНИЯ
В. А. Терехова, А.В. Головченко1, М.В. Смагина2
1 Институт почвоведения МГУ-РАН, Институт проблем экологии и
эволюции РАН, Москва, 2Институт лесоведения РАН, Московская обл.
Изучение микобиоты в гидроморфной среде представляет интерес в
разных аспектах. Во-первых, не потеряла свою актуальность проблема
биологического разнообразия микроскопических грибов, сохранение его в ходе
естественной эволюции экосистем со специфическими для развития
микроорганизмов условиями и при вмешательстве человека. Во-вторых,
продолжается обсуждение эффективности грибной минерализации растительного
материала в переувлажненных почвах, в болотах разных типов, в водоемах
и т.п. Задача настоящей работы состояла в определении численности и
таксономического состава микромицетов в лесных торфяно-болотных почвах
разных типов, подвергшихся экспериментальному осушению. Для
выявления возможного влияния мелиорации проведено сравнение комплекса
микромицетов на одних и тех же объектах с интервалом более 20 лет.
Объектами исследования были почвы: торфяная низинная типичная
нормально-зольная и верховая торфяная обычная; обе залегающие на глубоких
142
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ
III
торфах (Западнодвинский стационар Института лесоведения РАН, Тверская
область). Они соответствуют двум типам болотных лесов: сосняк болотно-
травяной на низинном болоте с евтрофным типом водно-минераль-ного
питания (объект Грустинка, НТ) и сосняк андромедово-пушицево-сфаг-новый с
большой разреженностью древесного полога на верховом болоте централь-
но-олиготрофного типа развития с атмосферным питанием (объект Сосвят-
ское, ВТ). Экспериментальное осушение болот было проведено в 1972-73 гг.
сетью открытых канав. Микологические наблюдения проводили в два этапа:
I этап — в первые годы после осушительных мероприятий (1975-77 гг). II
этап — в более поздний период (1998 г.). Пробы отбирали буром с
определенной глубины. Микромицеты выделяли методом посева почвенной
суспензии из серийного разведения на подкисленный агар Чапека. Численность
грибных пропагул выражали в колониеобразующих единицах (КОЕ) на куб.
см исследуемого образца. Видовую идентификацию микромицетов
проводили по наиболее используемым современным определителям.
Полученные результаты свидетельствуют о том, что почвы обоих типов
обильно заселены грибами. В 1 куб. см отобранных образцов насчитывали
десятки и даже сотни тысяч грибных пропагул. На первом этапе более
богатым по численности грибов был верховой торфяник, тогда как в 1998 г.,
наоборот, в низинном более чем на порядок микромицетов было больше.
Это явление вполне объяснимо произошедшими, как показал видовой
анализ, изменениями в таксономическом составе, поскольку разные виды
обладают разной спорулирующей способностью.
В более ранние сроки наблюдений отмечалось более-менее равномерное
распределение грибов по профилю обоих объектов, чего никогда не
наблюдается в зональных типах почв [1] и, наоборот, характерно для микобиоты
водных экосистем [2]. Распределение микроскопических грибов по профилю
торфяно-болотных почв в 1998 г. характеризуется уменьшением
численности грибных пропагул с глубиной, эта закономерность оказалась общей вне
зависимости от их трофности. Более резкое падение численности в нижних
горизонтах было характерно для верхового торфяника, в котором уже с
глубины 50 см типичные колонии сапротрофного комплекса не выделялись.
Наибольшее значение при изучении устойчивости сообщества
организмов к внешним воздействиям и оценке биоразнообразия имеют
таксономические показатели его структуры. Исследуемые почвы имеют довольно
богатый видовой состав микромицетов. Всего идентифицировано 72 вида
микроскопических грибов, представляющих 27 родов. Основная часть относится
к несовершенным (класс Deuteromycetes). среди которых бесспорными
доминантами были грибы рода Penicillium (24 вида), несколькими видами
были представлены роды Trichoderma (5), Fusarhun (4), Aspergillus* Paeci-
lomyces и др. Класс Zygomycetes представляли 6 видов Mortierella. встре-
143
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
чался мицелий базидиальных грибов.
Почвы разной трофности существенно различаются по набору видов.
Верховой торфяник более чем в пять раз беднее низинного, причем, эта за*
кономерность обнаруживается на всех этапах . Соотношение числа видов в
разных типах торфяников в ранний период наблюдений выражалось как
4:23, а в более поздний — 8:45. Количество выявленных видов в 1998 году
увеличилось в два раза. Коэффициенты таксономического сходства
оказались очень низкими, наибольшее сходство (около 35%) — между образцами
из низинных торфяников, отобранными в разные сроки.
Таксономическое сходство комплексов микромицетов в торфяных почвах низинного
(НТ) и верхового (ВТ) типов заболачивания в разные сроки наблюдений (видовой
состав - справа вверху, родовой состав - слева внизу), %
НТ1975
ВТ 1975
НТ1998
ВТ 1998
НТ 1975
16,7
35,0
14,2
ВТ 1975
0
17,6
20,0
НТ 1998
6,56
2,17
25,0
ВТ 1998
0
10,0
5,66
Из результатов проведенного анализа следует, что в ходе почти
двадцатилетнего периода развития болотно-лесных почв обоих типов
заболачивания видовая структура микобиоты претерпела существенные изменения.
Различия по видовому составу грибов между контрастными биотопами не
сгладились. Наибольшим видовым разнообразием характеризовался
комплекс микромицетов, выделенный в 1998 г. на объекте Грустинка, в связи с
чем можно предполагать, что здесь сохранилась более высокая деструкци-
онная активность грибов. Как показано ранее, уже в первые годы после
осушения в почвах низинного типа заболачивания наблюдалась более
интенсивная деятельность целлюлозоразрушающих микроорганизмов [3]. В
почвах верхового типа заболачивания как через пять, так и через 20 лет
после осушения продолжают сохраняться условия, благоприятные для
аккумуляции и консервации органического вещества. Что касается изменений в
профильном распределении численности грибов, произошедших за два
десятилетия после осушения, то падение этого показателя с глубиной дает
некоторое основание говорить об эффекте мелиорации, поскольку
равномерное распределение больше характерно для типично водных экосистем.
[1] Звягинцев Д.Г. и др. Структура сапротрофного комплекса микроорганизмов в
торфяниках.// Микробиология. 1991. Т. 60, Вып. 6. С. 155-164. [2] Семенова ТА,
Терехова Т.А. Микромицеты Куйбышевского водохранилища Ш. Сезонные и
годичные изменения в структуре микобиоты // Микология и фитопатология. 1992. Т.
26. Вып.2. С. 112-118. [3] Смагина М.В. Активность целлюлозоразрушающих
микроорганизмов торфяно-болотных почв.// Лесоведение. 1984. №1, с. 52-58.
144
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
ЛИШАЙНИКИ ЗАПАДНОСИБИРСКИХ БОЛОТ
Т.Ю. Толпышева
МГУ, биологический факультет
В лихенологическом плане болота России изучены слабо. Лихенологи на
болотах, как правило, не работают, а болотоведы, при описании болотной
растительности, учитывают обычно наиболее распространенные эпигейные
лишайники. Эпифитные лишайники, при составлении подобных
геоботанических описаний, за редким исключением [2 и др.], вообще не учитывают.
Также выпадают из поля зрения болотоведов эпигейные лишайники,
которые встречаются эпизодически или как примесь среди фоновых видов.
Болота Западной Сибири — не исключение. На сегодняшний день здесь
зарегистрирован 81 вид лишайников, что, вероятно, составляет немногим более
половины видов, которые могут произрастать на болотах в этом регионе.
В зональном отношении болота изучены крайне неравномерно. Так в
тундровой зоне отмечен 31 вид лишайников, в лесотундре — 12, в таежной
зоне — 61, в лесостепи — 2. Увеличение числа видов в таежной зоне
связано как с появлением эпифитов, так и с лучшей изученностью лишайников
болот, особенно средней и южной тайги. Уменьшение видового богатства в
лесотундре, по сравнению с тундровой и таежными зонами, не соответствует
действительности и только подтверждает отсутствие полных данных о
лишайниках болот, поскольку сюда проникают арктоальпийские и бореальные
виды и появляются эпифиты.
В тундровой зоне на болотах преобладают арктоальпийские виды
лишайников, способные выносить суровые условия существования, криофиты,
такие как Alectoria ochroleuca (Hoffm.) A. Massal., Bryocaulon divergens
(Ach.) Karnefelt, Flavocetraria nivalis (L.) Karnefelr et Thell, F. cucullata
(Bellardi) Karnefelt et Thell, Asahinea chrysanta (Tuck.) W.L. Culb et C.F.
Culb. и психрофиты, например, Thamnoiia vermicularis (Sw.) Schaer.,
Nephroma arcticum (L.) Torss.
Только на болотах тундровой зоны, в ивово-осоком комплексе, отмечены
виды p. Peltigera. Несмотря на то, что эти лишайники широко
распространены в лесах, на болотах в таежной зоне они не найдены. В таежной зоне
они, как правило, отсутствуют. Основное ядро здесь составляют бореальные
мезофитные лишайники: большинство видов p. Cladonia, виды родов Вгуо-
ria, Usnea и др. В напочвенном покрове преобладают виды сем. Clado-
niaceae, причем виды p. Cladina в определенных ассоциациях достигают
пышного развития и нередко становятся фоновыми.
На болотах лесостепной зоны зарегистрировано всего 2 вида эпигейных
лишайников: Cladina rangiferina (L.) Nyl. и С. mitis (Sandst.) Hustich. Оба
вида широко распространены на болотах западносибирской равнины, встре-
145
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
чаются в различных растительных ассоциациях и в тундре и в тайге. Их
можно найти на олиготрофных, мезотрофных и евтрофных болотах.
В целом олиготрофные комплексы болот значительно богаче
лишайниками, чем евтрофные и мезотрофные. По мере продвижения к центру таких
болот увеличиваются как видовое разнообразие эпигейных видов, так и
площадь проективного покрытия лишайников. Некоторые эпигейные
лишайники, распространенные на болотах, к ним в частности относятся и
виды p. Cladina, имеют довольно широкую субстратную амплитуду. Они могут
произрастать на различных субстратах, как на сильно кислых, так и на
субнейтральных, способны развиваться как на оголенном торфе, так и, при
определенных условиях обводненности, успешно конкурировать со мхами.
Если на распространение эпигейных лишайников на болотах большое
влияние оказывает уровень болотных вод, то на распространение эпифитов
в большей степени влияет наличие подходящего субстрата — форофита.
Видовой состав эпифитных лишайников по окраинам и в центральной части
болот зависят только от присутствующих здесь пород деревьев, а их обилие
нередко остается, практически, на одном уровне. Некоторые виды
лишайников встречаются только на хвойных породах, другие только на
мелколиственных. На хвойных деревьях обычны кустистые лишайники из родов
Usnea, Bryoria, Evemia. На мелколиственных породах одним из фоновых
видов является Melanelia olivacea (L.) Essl. Часто на болотах
распространены лишайники, которые одинаково хорошо растут как на хвойных, так и на
мелколиственных породах, например Hypogymnia physodes (L.) Nyl.
Многие виды лишайников, встречающиеся обычно в лесах в нижней
части стволов, на болотах нередко переходят на ветви, например, Parmeliop-
sis ambigua (Wulfen) Nyl., P. hyperopta (Ach.) Arnold, Vulpicida pinasth
(Scop.) J. - E. Mattsson et M. J. Lai и др. Это связано с тем, что скорость
роста зеленых мхов, которые обычно окружают стволы деревьев на болотах,
довольно высокая и, постепенно, они затеняют светолюбивые лишайники,
произрастающие в нижней части стволов. В то же время относительная
влажность воздуха на болотах высокая, что и позволяет влаголюбивым
видам развиваться на более высоких уровнях, чем в лесных фитоценозах.
Тип фитоценоза на видовой состав эпифитных лишайников на болотах
влияет значительно меньше, чем в лесах. На болотах древостой
разреженный и условия освещенности и влажности в различных болотных
фитоценозах оказываются более выровненными, чем в лесных. Только наличие или
отсутствие определенной древесной породы в данном типе фитоценоза
нередко влияет на присутствие того или иного вида лишайника.
Огромные заболоченные пространства способствовали сохранению
лишайников прошлых климатических эпох [1]. На болотах Среднего Приобья
найдены лишайники, ближайшие местонахождения которых находятся за
146
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ II]
сотни километров. Большинство из них впервые отмечены для Западное
Сибири. К ним относятся: Cetrelia braunsiana (Mull. Arg.) Culb. et Culb. •
вид встречающийся на Дальнем Востоке, Chromatochlamys muscorum (Fr.)
H. Mayrhofer et Poelt - арктоалышйский вид с рассеянным распространением
на территории России (Чукотка, Карелия, Коми, Калининградская обл.,
Кавказ), Me lane lia lacinulatula (Flag., ex Oliv.) Essl. — в Азии отмечен
впервые, ближайшие местонахождения — Карпаты и юг Скандинавии,
Opegrapha rufescens Pers. и О. varia Pers.— ближайшие местонахождения в
Бурятии (Байкальский заповедник), Thelocarpon superellum Nyl. и
Protothelenella sphictinoides (Nyl.) H. Mayrth. et Poelt — виды известные
ранее с Кольского полуострова и из Карелии, Cladonia maxima (Asah.) Ahti
— ближайшее местонахождение Алтай.
Все эти виды, за исключением С. maxima, представлены крошечными
экземплярами, что свидетельствует о неблагоприятных условиях
существования. По всей видимости, они находятся на грани исчезновения, и только
климатические особенности западносибирской равнины помогли им дожить
до настоящего времени. Несомненно, что более целеноправленное изучение
лишайников болот Западной Сибири позволит расширить этот список и
поможет проследить историю развития и становления флоры этого региона.
[1] Толпышева Т.Ю. Об интересной лихенологической находке из Среднего При-
обья //Новости систематики низших растений. 1990. Т. 27. [2] Храмов А.А., Валуц-
кий В.И. Лесные и болотные фитоценозы Восточного Васюганья (структура и
биологическая продуктивность). Новосибирск. 1977.
РОЛЬ КЛИМАТА В СОВРЕМЕННОЙ СТРУКТУРНО-
ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ БОЛОТ ЗАПАДНОЙ СИБИРИ
И.Е. Трофимова
Институт географии СО РАН, Иркутск
Заболоченность Западно-Сибирской равнины М.И. Нейнггадг назвал
мировым феноменом, время зарождения которого относится к началу
голоцена. Первопричиной возникновения процесса заболачивания стало
переувлажнение множества бессточных депрессий рельефа, оставшихся после
отступления ледникового покрова. В силу плоской формы рельефа очаги
заболачивания со временем сливались между собой, образуя обширные
пространства, которые, благодаря механизму горизонтального саморазвития
болот за счет большой массы накопленной ими влаги, и в настоящее время
имеют тенденцию к росту. Поэтому изучению процессов заболачивания
территории Западной Сибири уделяется большое внимание.
147
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
В структурно-функциональной организации западно-сибирских болот
первостепенными остаются геоморфологический и климатический факторы,
а точнее интегральный эффект сочетания выровненное™ рельефа и слабой
дренированности с повышенным увлажнением и пониженной теплообеспе-
ченности территории, вполне достаточный для проявления процесса
заболачивания и последующего его развития. Климатическую составляющую
следует рассматривать: — с позиции природно-экологического фактора,
обусловливающего структуру и функционирование ныне существующих
болотных образований и определяющего направление дальнейшего процесса
заболачивания в региональной системе контакта лес-болото; — с точки зрения
трансформации климата деятельной поверхностью болот, значимой в энер-
пк тепло- и влагообменных процессах в системе атмосфера-суша
регионального и топологического среза лес-болото.
Первый аспект: климат — болотоформирующий, регулирующий и
контролирующий фактор. Основной климато-экологический критерий,
определяющий формирование, функционирование и структуру любых природных
образований — соотношение телоэнергетических ресурсов и влаги.
Процессы заболачивания лесов также подчиняются общим законам развития
природы. Умеренно гумидный климат западно-сибирской геосистемы
обусловливает равное развитие дренируемых лесных и заболоченных (болотных)
комплексов и создает при этом условия для проявления их зональности. По
существу произошла интеграция (или сбалансированность) компонентов
среды в рамках двух основных природных комплексов, в результате чего
стало возможным их сосуществование. Однако функционирование болот
возможно лишь при условии постоянного переувлажнения, которое
обеспечивается особыми взаимоотношениями в системе тепло - влага - деятельная
поверхность. И в этом случае через влагообмен в системе контакта
происходит взаимопроникновение деятельных поверхностей.
Каждой природной (болотной) зоне соответствует определенная
разновидность ведущего ландшафтообразующего процесса. Климато-экологичес-
кие условия создают фоновый эффект, который выражается в их плавном
переходе от одной зоны к другой от основного ядра. Причем выделяется три
типа соотношений тепла и влаги. Для самой обширной зоны выпуклых оли-
готрофных (сфагновых) болот эти соотношения (тип 1) оптимальны и
определяются максимальной для Западно-Сибирской равнины величиной
атмосферных осадков и достаточностью тепла. К югу (плоские евтрофные и ме-
зотрофные, вогнутые евтрофные и засоленные болота) ресурсы атмосферной
влаги уменьшаются, а тепла увеличиваются (тип 2), севернее
(крупнобугристые, плоскобугристые и полигональные болота) убывают как
ресурсы тепла, так и влаги (тип 3).
148
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
Второй аспект: деятельная поверхность болот — фактор климатообразо-
вания. Регионально-фоновый климат — это, в сущности, потенциальный
климат водораздельного (долинного) открытого пространства (стандартные
метеостанции), который в естественных условиях преобразуется под
влиянием деятельной поверхности леса и. болота. В зависимости от типа
поверхности дифференцируется радиационный баланс, а как следствие —
температура воздуха, поверхности почвы и верхних ее горизонтов, а также
испарение, являющееся связующим звеном в процессах теплового взаимодействия
и влагообмена деятельного слоя природных образований с атмосферой.
Для выявления закономерностей формирования климата в среднетаеж-
ной подзоне экспериментальные исследования проводились в системе
контакта лес - болото, а для сравнения и в опорной точке с мохово-травянистым
покровом водораздельного типа. Это позволило оценить влияние неодно-
родностей деятельной поверхности на перераспределение составляющих
радиационного и теплового баланса.
Радиационный баланс темнохвойного леса летом на 15-20%,а в целом за
год на 50% выше регионально-фонового. Соотношение между значениями
радиационного баланса болот и опорной точки в определенных ситуациях
противоположно по знаку и не превышает 12%.
В условиях заболоченных пространств затрата тепла на испарение
преобладает в структуре теплового баланса и составляет в среднем в течение
вегетационного периода около 70-80% от величины радиационного баланса.
Испарение с деятельной поверхности болот и темнохвойного леса
значительно выше регионально-фонового. Становиться очевидным, что в
формировании регионального климата западно-сибирской геосистемы особая роль
отводится характеру и состоянию ее деятельного слоя.
Общие климатические ресурсы территории обусловливают особенности
термического режима почвогрунтов. Однако в системе контакта лес-болото
топологического уровня температура минерального субстрата и мерзлотные
явления варьируют в зависимости от мохово-органогенной толщи.
Увеличение ее с 20 до 50 см снижает температуру нижележащего минерального слоя
в период максимальной аккумуляции тепла в почве на 8-10°С (по
отношению к опорной точке). В таком случае возможна тенденция к уменьшению
теплоаккумулирующей способности территории, так как минеральный
субстрат торфяно-глеевых почв становится инертным, отключенным от сферы
активного теплообмена. Аккумулятором тепла начинает служить
органогенная толща. Торфяные почвы в связи с постоянной их переувлажненностью
имеют повышенную теплоаккумулируюшую способность по сравнению с
торфяно-глеевыми. В целом для верховых болотных почв зависимость тем- ^
пературы минерального горизонта от органогенного носит криволинейный
характер со свойственным для логарифмической зависимости замедлением.
149
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
МЕЛИОРАТИВНАЯ ЭКОЛОГИЯ БОЛОТНЫХ ПОЧВ
ЗАПАДНО-СИБИРСКОЙ НИЗМЕННОСТИ
М.Т. Устинов, Т.Н. Елизарова, В.А. Казанцев, Л.А. Магаева
Институт ЗапСибгипроводхоз, Новосибирск;
Институт почвоведения и агрохимии СО РАН, Новосибирск
Западно-Сибирская низменность — уникальный полигон для изучения
болотного почвообразования. Здесь сформировался широкий генетический
спектр болот от полигональных на Крайнем Севере до травяных засоленных
и южных рямов в Барабе. Локальные болота, болотные массивы,
заболоченные земли составляют сельско-, лесохозяйственный и мелиоративный фонд
территории и являются основным объектом исследования мелиоративной
экологии. Ее необходимость и перспективность, как нового научного
направления, доказывает 100-летний мелиоративный опыт освоения болотных
почв в Барабе, начатый экспедицией И.И. Жилинского. Комплексное
решение вопросов водногообустройства этой территории заключалось в
органичном вплетении осушительных каналов в современную гидросеть (в
гидрографические бассейны — реки, озера, палеореки). Часть каналов без
восстановительных работ функционирует до настоящего времени.
Болотные ландшафты, как правило, имеют агрессивный, интенсивный
характер развития. В Западной Сибири сложность мелиорации болот
определяется следующими неблагоприятными природными особенностями:
значительной заболоченностью территории при слабой естественной дрениро-
ванности, ритмическими и резкими колебаниями гидротермических
показателей, 11-, 22-, 32-летними циклами увлажнения и иссушения (переменой
полюсов гидроморфизма); высоким уровнем грунтовых вод и их
значительным, увеличивающимся с севера на юг засолением; низкой
водопроницаемостью и сезонным промерзанием грунтов.
Мелиоративные мероприятия, особенно осушение, полностью меняют
эволюционно-генетическую природу болотной экосистемы, активизируют
биохимические процессы, усиливают миграцию веществ на всех природных
уровнях: почвенно-профильном, внутри ландшафтном, региональном.
Обширные массивы болотных засоленных почв (мелкоторфяные, а также
мощные торфяные почвы приболотного пояса) используются в
сельскохозяйственном производстве под сенокосы, что предполагает проведение
специальных природоохранных и мелиоративных мероприятий по оптимизации в
них и в смежных экосистемах водно-воздушного, теплового, солевого и
пищевого режимов. Для обоснования комплекса природоохранных и
мелиоративных мероприятий разработаны ландшафтно-экологический и
бассейновый принципы оценки территории, способы определения эколого-
мелиоративного потенциала почвенного покрова и устойчивости к мелиора-
150
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ II]
тивным воздействиям эколого-мелиоративных комплексов почв, пород зонь
аэрации, грунтовых и напорных вод.
Ландшафтно-экологический и бассейновый подходы к изучению
природных условий позволяют установить способность болот к
самовосстановлению; их экологическую нишу в структуре бассейна; степень утраты
генетической цельности ландшафта, произошедшей в результате
сельскохозяйственного использования земель; снижение или повышение природно-ресурс-
ного потенциала ландшафта или его природной буферности; угнетение,
деградацию или «процветание» биогеоценозов; неблагоприятные процессы и
явления — выгор, заростность, вторичное засоление и заболачивание;
направленность развития и степень трансформации болот; дифференциацию
на заболоченные, суходольные и болота озерного типа.
Критериями эколого-мелиоративного потенциала болотных почв служат:
высота стояния зеркала грунтовых и атмосферных вод, степень и тип их
засоления; степень минерализации органогенных поверхностных
(деятельных) горизонтов и активность жизнедеятельности микроорганизмов;
присутствие в торфяной залежи двух генетически разнородных горизонтов -
деятельного и инертного.
Природоохранное значение болот зависит от геохимических
особенностей территории и меняется по природным зонам. Болота тайги и подтайги,
способствующие сохранению режима рек и озер и не ослабляющие
эрозионную и дренирующую роль речной сети, не подлежат освоению. Их
необходимо сохранить. Болота лесостепи после проведения мелиорации,
соответствующих их эколого-мелиоративному потенциалу, могут использоваться под
сенокосы и пастбища. Болота степи должны оставаться в естественном
состоянии, поскольку служат регуляторами микроклимата территорий.
Эффективность осушения болотных почв, особенно торфяно-засоленных,
имеющих широкое использование в сельскохозяйственном производстве на
юге региона, находится в прямой зависимости от эффективности работы
двустороннего регулирования их водно-воздушным режимом (шлюзования).
Исследования, проведенные в районах нефтепромыслов Среднего При-
обья показали, что болотные биогеоценозы способны к биоаккумуляции и
адсорбции многих химических соединений. Они активно самоочищаются и
самовосстанавливаются, являются глобальным буфером, нейтрализующим
влияние шламовых амбаров и «выбросы» нефти в окружающую среду.
Разработка теоретических и практических основ мелиоративной
экологии болотных почв позволяет выявить их экологические функции,
организовать мониторинг мелиорированных болотных земель разработать
мелиоративные приемы по оптимизации природопользования.
Работа выполняется при поддержке РФФИ (грант № 99-04-49983).
151
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ТРАНСФОРМАЦИЯ ТРАВЯНОГО ЯРУСА ГИДРОМОРФНЫХ
СООБЩЕСТВ В СВЯЗИ С ОСУШЕНИЕМ И СЕЗОННЫМИ
КОЛЕБАНИЯМИ ПОГОДЫ
Ю.С. Федоров
РосНИИВХ, Екатеринбург
Проведение осушительных мелиорации в системе частного водосбора
вызывает различные по глубине трансформации экологических систем.
Растительные сообщества, попадающие в зону воздействия осушения,
претерпевают структурно-функциональные изменения по основным своим
характеристикам (видовой состав, продуктивность и другие).
Результаты исследования по влиянию гидрологического режима на фи-
тоценотическую составляющую болотных экосистем подтверждают, что
имеется определенная закономерность зависимости УПГВ и основных
характеристик фитоценозов, отражающаяся на продуктивности и
функциональной организации растительных сообществ вплоть до их трансформации
и деградации. В этой связи травянистый ярус выступает чувствительным
индикатором таких процессов.
Основной подход в изучении данного вопроса сводится к сравнению
состояния болотных экосистем до осушения и после осушения. На наш взгляд,
этого недостаточно, так как в этом случае не учитывается естественный
природно-климатический тренд. Нередко влияние ряда засушливых лет
ставит под сомнение выводы о роли осушения в снижении продуктивности
болотных растительных сообществ. Наоборот же, дождливые периоды могут
нивелировать влияние осушительных мероприятий.
В этой связи сделана попытка учесть естественный тренд влияния
природно-климатических факторов, выбрав в качестве контроля неосушенный
заболоченный водосбор - аналог, и на аналогичных рядах проследить
закономерность флуктуации и трансформаций типичных болотных экосистем в
условиях Среднего Урала. Для выявления особенностей фитоценотической
составляющей экосистем заболоченного водосбора были выбраны бассейны
-аналоги: существующий в естественном состоянии и подверженный
осушению. В пределах этих водосборов на основе геоботанических исследований
были выделены аналогичные эколого-фитоценотические ряды, включающие
по мере усиления гидроморфности следующие сообщества: 1. сосняк разно-
травно-вейниковый, 2. сосняк — березняк хвощево -разнотравный, 3.
березняк дернистоосоково- пурпурновейниковый, 4. березняк дернистоосоковый,
5. березняк дернистоосоково-тростниковый. Продуктивность травяно-кус-
тарничкового яруса гидроморфных сообществ определялась в конце
вегетационного периода (август) на учетных площадках.
Динамика продуктивности травяного яруса изучаемых сообществ являет-
152
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
ся достаточно интегральной характеристикой состояния данного
растительного сообщества, а также его ответной реакцией на условия внешней среды.
В связи с этим анализ и интерпретация данных требует привлечения
дополнительной информации по основным метеорологическим факторам среды.
Для изучения изменения продуктивности растительных сообществ и
исключения естественного влияния климатических факторов нами
рассматривались: сумма температур и средняя температура воздуха, количество
осадков за вегетационный период (май-август), гидротермический коэффициент
(ГТК), а так же обеспеченность по осадкам и сумме температур по годам
исследований.
Период проведения наблюдений за продуктивностью травяного яруса
гидроморфных сообществ характеризуется чередованием разнообразных по
обеспеченности как осадками, так и теплом вегетационных периодов.
Характеристика лет наблюдений по обеспеченности осадками не обнаруживает
достоверной связи с теплообеспеченностью (К-0,3).
Выявление коррелятивных отношений продуктивности травяного яруса
гидроморфных сообществ естественного болота и осушаемого показывает
тесную связь продукционных процессов гидроморфных сообществ
естественного болота со значениями суммы температур, и средними
температурами за вегетационный период (май-август) и, соответственно, отрицательную
корреляционную связь с обеспеченностью по осадкам за этот же период.
Совершенно другой характер коррелятивных отношений обнаруживается
при анализе продуктивности гидроморфных сообществ осушенного болота.
Наибольшая положительная зависимость продукционных процессов в
подверженных осушению растительных сообществах выявлена с параметрами
увлажнения (осадки за год). В этом случае осадки выступают как наиболее
значимый ( лимитирующий) фактор, определяющий продукционные
процессы травяного яруса гидроморфных сообществ, подвергающихся осушению.
У растительных сообществ на суходоле (№1) в любых вариантах в
большей или меньшей степени сохраняется лимитирующей доминантой
фактор теплообеспеченности. Такое положение, еще в большей степени
подтверждает довод в пользу смены лимитирующей доминанты
теплообеспеченности в условиях естественного состояния болотных экосистем на
увлажнение в условиях воздействия осушения.
Сложение гидроморфных сообществ болот низинного типа определяется
малым разнообразием доминантного состава травяного яруса. Изучение
изменения продукции каждой доминантной группы по годам как в
естественно- природных условиях, так и при осушении, выявляет противоречивую
реакцию участия каждого доминанта в формировании продукции травостоя.
Основные выводы: 1. Травяно-кустарничковый ярус гидроморфных
сообществ (продуктивность и ее состав) является чутким индикатором изме-
153
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
нения гидрологического режима как болотных растительных сообществ, так
и прилегающих к ним суходолов. 2. Среди изученных гидроморфных
сообществ наибольшей продуктивностью травянистой компоненты выделяется
березняк дернистоосоково- тростниковый. 3. Чувствительность
гидроморфных сообществ болотных экосистем к изменению гидрологических условий
их обитания проявляется уже на второй год, а сообществ прилегающих
суходолов на 3-4 годы после проведения осушительных работ. 4.
Направленность и глубина трансформации фитоценозов болотных экосистем зависит
от метеорологических условий (осадки, температура). 5. В условиях
осушения происходит смена доминанты экологического фактора с теплообеспе-
ченности на величину суммарного количества осадков.
ВИДОВОЙ СОСТАВ И СРУКТУРА МИКРОМИЦЕТОВ
БОЛОТНО-ПОДЗОЛИСТЫХ ПОЧВ
Ф.М. Хабибуллина
Институт биологии Коми НЦ УрО РАН, Сыктывкар
В умеренно-континентальных районах Европейского Северо-Востока
широко распространены почвы с торфяными горизонтами, формирующиеся
под сфагновым покровом в условиях затрудненного дренажа и
переувлажнения вследствие застоя атмосферными осадками. В подзоне средней тайги
(район исследований) ими заняты около 20% территории.
Почвенные микромицеты в сфагновых местообитаниях изучены неполно,
сведения о них фрагментарны [Зименко, 1965; Клевенская и др., 1970;
Гришкан, 1994]. Между тем в микологическом отношении почвы этих
местообитаний особенно интересны: деятельность микроорганизмов протекает
здесь в неблагоприятных условиях низкой тепообеспеченности,
сильнокислой реакции трудноминерализуемого опада растений и в подобной
обстановке именно грибы, благодаря ряду адаптивных особенностей, являются
основными деструкторами поступающей в почву органики [Мирчинк, 1988].
Изучены качественные характеристики микофлоры (видовой состав,
комплекс типичных видов) и ее количественные показатели (численность
различных групп, длина мицелия) в почвах сфагновых местообитаний в
окрестностях Максимовского почвенного стационара Института биологии
Коми научного центра УрО РАН. Работу проводили в еловом лесу с единичной
примесью осины, березы. Состав древостоя 6Е+2Б+20С. Возраст 110-130
лет. В подлеске рябина, жимолость, ольха, в подросте пихта, ель.
Напочвенный покров представлен сфагнумом, черникой, майником, хвощом.
Были выбраны 2 типа заболоченных почв: торфянисто-подзолисто-глее-
ватая (П61) и торфяно-подзолисто-глеевая (П62). В торфянисто-подзолисто-
глееватой почве органогенный горизонт имеет мощность 11 см, причем по
154
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ
III
степени разложенности торфа можно выделить слаборазложившуюся часть
(0-4 см) и среднеразложившуюся (4-11 см). В более гидроморфной торфяно-
подзолисто-глеевой почве мощность торфяного горизонта достигает 25 см,
который хорошо дифференцирован на следующие подгоризонты: слабораз-
ложившийся (0-7 см), среднеразложивпшйся (7-20 см) и хорошо
разложившийся (20-25 см). В направлении от Пб] к Пб2 происходит уменьшение
мощности элювиального и иллювиального горизонтов. Максимальное
содержание органического углерода приходится на органогенные горизонты и
составляет 38,9-33,8% для П62, 42,7-37,6% для П61 [Козачок, 1997].
Возрастание мощности торфяного горизонта ведет к менбе резкому снижению
содержания органического углерода вниз по профилю.
Образцы взяты в конце июня, июля и августа 1998 г. Анализировались
подстилки (по слоям) и почвенные горизонты до глубины 1 м. Для
установления набора видов использовался метод разведения почвенной суспензии с
последующим высевом на агаризованные питательные среды Чапека, Гет-
ченсона с порошковидной целлюлозой и агаризованную вытяжку из
субстрата, где отбирались пробы. Одновременно готовили препараты для
измерения длины мицелия по методу Шульгиной в модификации В.Е. Чайки
[1972]. Комплекс типичных видов определяли на основе пространственной и
временной встречаемости по методу Т.Г. Мирчинк с соавторами [1982] с
учетом данных по всем трем средам.
Видовой состав. В исследованных почвах выявлено 33 вида микромице-
тов, относящихся к 11 родам и 4 классам. Кл. Zygomycetes: Mucor rocemosus
Fres., Mucor sp., Mortierella ramanniana (Moller) Linnem., M. vinacea Dixon-
Stewart. Кл. Ascomycetes: Chaetomium globosum Kunze et Fr., Ch. spirale
Zopf, Ch. indicum Corda. Кл. Hyphomycetes: Acremonium strictum Gams,
Aspergillus sp., Alternaria tenuis Nees ex Fries, Paecilomyces sp., Penicillium
camemberti Thom, Penicillium sp., P. decumbens Thom, P. lividum Westl , P.
lanosum Westl, P. paxilli Bainier, P. restrictum Gil. et Abb, P. Rolfsii Thom, P.
spinulosum Thom, P. Thomi Maire , P. verrucosum var.cyclopium Samson et
all., Trichoderma album Preuss, Trichoderma sp., T. koningii Oudemans, T.
lignorum (Tode) Harz, T. sympodianum Kulik, T. viride Persoon ex Fries,
Cladosporium herbarum (Person) Link ex Fries, CI. potebniae Pidopliczko et
Deniak. Пор. Mycelia sterilia: Mycelia sterilia (Mucedin.), Mycelia sterilia
(Dematiac). Кл. Basidiomyces: Mycelia sterilia.
В почвах сфагновых местообитаний по численности и видовому
разнообразию преобладают пенициллы (10 видов), единичны аспергиллы,
темноцветные гифомицеты, многочисленны виды рода Trichoderma (5 видов),
светло- и темноокрашенные формы стерильного мицелия, а также мицелий
базидиальных грибов с пряжками.
Структура комплекса микромицетов. К доминирующим и частым видам
155
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
грибов в торфянисто-подзолйсто-глееватых почвах относятся Trichoderma
sympodianum, Tr. viride, Penicillium sp., Mortierella ramanniana, а также
мицелий базидиальных грибов. В торфяно-подзолисто-глеевой почвах к
доминирующим и частым относятся мицелий высших грибов и белая пушистая
форма стерильного мицелия. Количество и встречаемость видов в разное
время сезона были примерно одинаковыми; несколько разнообразнее —
августовская микофлора. Целлюлозоразрушающие грибы представлены
видами рода Trichoderma, Chaetomium, Paecilomyces sp. и Cladosporium
herb arum. Олиготрофная микофлора насчитывает всего 5 видов.
Стекла обрастания, установленные по почвенным горизонтам показали,
что грибы развивают вегетативный мицелий лишь в верхнем 30 см слое
почвы, основная же масса его размещена в подстилке. Длина мицелия в
подстилке за весь период наблюдений варьировала от 117 до 4055 м /г
сухого субстрата. По нашим данным длина мицелия в почве положительно
коррелирует с содержанием органического вещества и температурой. Запасы
биомассы мицелия и спор грибов составляют десятки т/га сухого вещества.
ОЦЕНКА СОСТОЯНИЯ РАСТИТЕЛЬНОГО ПОКРОВА ПОСЛЕ
СИЛЬНОГО ТОРФЯНОГО ПОЖАРА
СЮ. Цареградская, В.Н. Косицын
ВНИИЛМ, Московская обл.
В условиях сильных засух, когда сумма осадков за пожароопасный
период (конец весны — начало лета) оказывается значительно меньше (на 50-
80%) среднемноголетних данных, в заболоченных лесах олиготрофного ряда
развития и на верховых болотах может происходить углубление «огня»
низового пожара в более нижние слои почвы, вызывая тем самым
возникновение торфяного (подземного) пожара. Наиболее часто торфяные пожары
имеют место в зоне хвойно-широколиственных лесов. Отдельные торфяные
пожары носят катастрофический характер, вызывая существенные
изменения природной среды и сукцессионные смены растительного покрова. В
современных условиях глобального потепления климата и усиления
антропогенного воздействия на лесо-болотные экосистемы вероятность
возникновения торфяных пожаров возрастает. В связи с этим актуальной становится
проблема изучения динамики растительного покрова после торфяных
пожаров сильной интенсивности, важной частью которой является организация
системы мониторинга растительности на гарях с целью установления
тенденций развития отдельных компонентов фитоценозов. Это, в свою очередь,
позволит разработать перспективную стратегию хозяйственного освоения
участков лесного фонда, пройденных сильными торфяными пожарами.
156
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ
III
На территории Сергиево-Посадского опытного лесхоза Московской
области (северная подзона зоны хвойно-широколиственных лесов) нами
ведется мониторинг растительности на гари площадью около 260 га,
образовавшейся на месте сосняка сфагнового после сильного торфяного пожара в
1972 году. Постоянные круговые и ленточные пробные площади заложены в
центральной части гари и на ее окраинных участках, прилегающих к
верховому болоту, гидролесомелиоративной сети и озерной акватории. На
пробных площадях ежегодно в определенные фенологические периоды
производится учет видового состава растительного покрова и макромицетов,
биоморфологических и биопродукционных признаков видов. Ведется учет
основных параметров режимов освещенности и увлажнения, термического
режима и эдафических условий. При мониторинге растительного покрова на
гари используются и дистанционные методы, в частности метод
дешифрирования цветных спектрозональных среднемасштабных аэрофотоснимков.
При торфяном пожаре наблюдалось полное сгорание напочвенного
покрова, подстилки и толстого слоя аккумулирующего органического вещества
— торфа (глубина прогорания торфяной почвы достигала 50 см),
вываливание деревьев после сильного обгорания их корневых систем. В результате
гибели лесного насаждения на гари произошло уплотнение верхнего
торфяного слоя, его проседание и подъем уровня болотно-грунтовых вод. Это
привело к заболачиванию гари и формированию здесь кочкарно-моча-
жинного микрорельефа. В результате сгорания сфагнового покрова и
обогащения поверхностного торфяного слоя щелочными зольными элементами
после пожара на гари отмечалось хорошее развитие самосева сосны
обыкновенной. Через 25 лет после пожара на исследуемой гари произрастал
равномерный и густой молодняк сосны (12-15 лет); однако из-за избыточного
увлажнения и маломощности почвенного слоя он отличался крайне
замедленным темпом роста. Через 5 лет после пожара и в дальнейшем в составе
напочвенного покрова гари преобладали пушица влагалищная и сфагнумы.
Обследование гари, проведенное в последние годы, показало следующее.
В более заболоченной части (площадь мочажин составляла 25%), удаленной
от пушицево-сфагнового верхового болота на расстоянии 200-250 м,
молодой сосновый древостой имел густоту 7,8 тыс. шт./га, среднюю высоту 1,25
м и диаметр 2,3 см. Здесь же наблюдалось разрастание пушицы и клюквы
болотной. Встречаемость клюквы увеличилась по сравнению с контролем
(сосняк сфагновый в возрасте 65 лет) в 1,5 раза и составляла 71%.
Урожайность ягод клюквы в этой части гари равнялась 130,4 кг/га и была близка к
контролю (152,4 кг/га). Восстановление вегетативной и генеративной сферы
клюквы на гари до прежнего уровня (до пожара) произошло через 18 лет
после пожара. В менее заболоченной части гари (площадь мочажин — 15%)
молодой древостой сосны характеризовался большей густотой (15,2 тыс.
157
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
шт./га) и имел среднюю высоту 1,0 м и диаметр 1,5 см. По сравнению с
контролем (сосняк сфагновый в возрасте 75 лет) проективное покрытие
большинства видов травяно-кустарничкового яруса в этой части гари снизилось:
черники в 5,7 раза, морошки в 3,5, брусники в 2,3 и багульника в 2,0 раза.
Здесь же не было отмечено плодоношения черники и морошки при
урожайности их ягод на контроле соответственно 16,4 и 133,1 кг/га.
Таким образом, и через 25 лет после сильного торфяного пожара на
месте сосняка сфагнового растительный покров на гари находится все еще на
ранних стадиях формирования лесного фитоценоза. В настоящий момент
такая гарь представляет интерес для лесохозяйственной практики только в
качестве промыслового угодья для заготовки ягод клюквы.
[1] Иванов А.В. Лесовозобновление гарей в сосняках Республики Марий Эл: Авто-
реф. дис. канд. с.х. наук. Йошкар-Ола: МарГТУ, 1996. 20 с. [2] Косицын В.Н.
Влияние торфяного пожара на состояние и продуктивность некоторых видов пищевых и
лекарственных растений в северной подзоне зоны смешанных лесов // Растительные
ресурсы. 1998. Т. 34. Вып.2. С.30-35.
ОСОБЕННОСТИ ЕСТЕСТВЕННОГО ВОССТАНОВЛЕНИЯ
СФАГНОВОГО ПОКРОВА В ЗАБОЛОЧЕННЫХ ЛИСТВЕННИЧНИКАХ ПРИАМУРЬЯ
ПОСЛЕ ПРОВЕДЕНИЯ МЕЛИОРАТИВНЫХ РАБОТ
В.В. Чаков, В.А. Купцова
Институт водных и экологических проблем ДВО РАН, Хабаровск
На равнинах и низменностях Приамурья заболоченные лиственничники,
несмотря на их незначительные по площади размеры, встречаются
практически повсеместно. В большинстве своем они тяготеют к высоким
геоморфологическим поверхностям, испытывающим временное или постоянное
переувлажнение почвенного субстрата. Здесь, это могут быть и выположен-
ные участки делювиальных шлейфов, и места плавных переходов
минеральных останцов (релок) к заболоченным равнинам, среди которых они
присутствуют. Болотные экосистемы этого типа в регионе изучены
достаточно подробно [Прозоров, 1961; Анисимов,1973; Чаков, 1986 и др.].
Древесный ярус заболоченных лиственничников обычно сформирован
лиственницей даурской, к которой иногда может примешиваться береза
маньчжурская. Суммарное проективное покрытие названных видов
колеблется в пределах 5-30%, а средние показатели составляют 10-15%.
Вариабельность показателей проективного покрытия древесного яруса так же, как
и вариабельность таксационных показателей (IV-Va классы бонитета),
связана с экологией и разновозрастностью древесных насаждений. Эта разно-
возрастность на болотах Приамурья объясняется особенностями
возобновительных процессов, которые протекают здесь более равномерно по годам,
158
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ
III
нежели в суходольных лиственничных лесах [Колесников, 1947]. Одновоз-
растность древостоев здесь наблюдается крайне редко, преимущественно в
^олодняках. По всей видимости, это связано прежде всего, с
периодическими пожарами. Кустарниковый ярус обычно представлен березой овальноли-
стной с наиболее вероятным проективным покрытием 3-5%. Крайне редко в
8 описаниях из 100 этот показатель может достигать отметки 20%.
Относительным многообразием видового состава отличается
растительный покров кустарничково-травяного яруса характеризуемых болот.
Доминирующими видами здесь служат хамедафне болотная и багульник
болотный. В качестве сопутствующих растений чаще других выступают ива
черничная, голубика обыкновенная, клюква мелкоплодная и осока волосистоп-
лодная. Полный списочный состав растений яруса насчитывает 16 видов.
Проективное покрытие яруса колеблется от 45% до 80%.
Гораздо большим проективным покрытием почвы (65-95%) отличаются
растения мохового яруса. Формирующие его, сфагновые мхи подчас
образуют сплошной ковер. В большинстве описаний преобладает сфагнум ма-
гелланский, формирующий сфагновые подушки различной конфигурации и
размеров. Очень часто, но в меньшем количестве здесь встречаются сфаг-
нумы узколистный и ленский. Крайне редко только в двух описаниях из
одиннадцати на подушках можно встретить сфагнум красноватый с
проективным покрытием 20-40%. Кроме сфагновых мхов в сложении яруса
принимают участие зеленые мхи и лишайники, суммарное проективное
покрытие которых не превышает 15%. Их видовой состав крайне беден и в
основном представлен аулакомниумом болотным.
Среди мелиорированных болотных массивов на территории Приамурья
заболоченные лиственничники встречаются крайне редко. Исключение
составляют опытные лесхозы, где проводятся эксперименты по повышению
продуктивности древостоев на осушенных болотах, да фрагменты
лиственничников, вкрапленных в другие типы болот, подвергшихся масштабным
осушительным мероприятиям связанным с увеличением
сельскохозяйственных угодий или созданием торфокарьеров. Именно, на территории такого
заброшенного торфокарьера в междуречье рек Хор и Кия в 1997-98 гг.
авторами проводились исследования по естественному восстановлению
мохового покрова на осушенных картах через 15 лет после проведения на них куль-
туртехнических работ. Каждая из карт площадью по 2 га (400x50 метров)
оконтурена мелиоративными каналами глубиной от 1,5 до 2,5 метров.
Таким образом, торфяная залежь, с которой сняли сфагновый покров и
удалили древесную растительность 15 лет назад, все это время увлажнялась
исключительно за счет атмосферных осадков. В результате этого сфагновый
покров, в настоящий момент полностью восстановившийся, по видовому
составу значительно скуднее, а по экологии ближе к олиготрофным видам.
159
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Доминирующим видом здесь обычно выступает сфагнум магелланский
который успел за означенный период сформировать подушки высотой до 25-
30 см и диаметром от 1,5 до 2,5 метров. Этот вид встречается на всех 20-ти
площадках, а его проективное покрытие варьирует в широких пределах от
30% до 90%, но обычно в пределах 45-60%.
На склоновой части подушек и в понижениях между ними повсеместно
отмечен сфагнум балтийский с проективным покрытием в диапазоне 3-63%.
На 9-ти площадках этот вид имеет проективное покрытие от 3% до 10%. На
следующих 7-ми характеризуемый показатель составляет 10-40%. На
остальных площадках сфагнум балтийский выступает в качестве доминанта.
Третьим и последним в списке сфагновых мхов, восстановившихся
естественным путем на осушенных лиственничниках, является сфагнум бурый,
который обнаружен только на 11 площадках. Максимальный показатель
проективного покрытия у этого мха здесь составляет 56% и отмечен только
на одной площадке. На трех площадках этот показатель составляет
величину 30-40 %, на всех остальных 10-20%.
Существуют многочисленные примеры достаточно быстрого
самопроизвольного восстановления сфагновых мхов в болотных экосистемах,
подобных описываемой [Wheeler, Shaw, 1995; Wheeler, 1995; Joosten, 1995].
Большинство исследователей болот Сибири и европейской части страны
отмечают ту же тенденцию восстановления мохового покрова на массивах с
нарушенным растительным покровом. По данным Н. Г. Москаленко [1980]
любые болотные массивы после их освоения через 10 лет самопроизвольно
восстанавливают моховой покров, но доминантные виды при этом
меняются. По наблюдениям А. В. Прянишникова [1954] на отдельных видах торфа
полное восстановление мхов наблюдается вообще через 5-6 лет.
Наряду со сфагновыми мхами здесь на 16-ти площадках можно
встретить политрихум сжатый. Только на трех из них его проективное покрытие
может варьировать в диапазоне 24-44%. Во всех остальных случаях участие
этого вида в сложении яруса колеблется на уровне 4-10%.
Крайне беден здесь и списочный состав растений кустарничково-травя-
ного яруса, который формируют всего шесть видов. Ярко выраженного
доминанта среди них нет. Содоминантами обычно выступают багульник
болотный и хамедафне обыкновенная с проективным покрытием 15-30%.
Крайне редко этот показатель на описываемых площадках может достигать
отметки в 50%, еще реже 60%. Из других кустарничков здесь на двух
площадках выявлен подбел многолистный с проективным покрытием 10-20% и
на семи клюква мелкоплодная с низким (5%) проективным покрытием.
Травянистые растения представлены глицерией и касатиком гладким.
Первая присутствует повсеместно за исключением двух площадок,
последний наоборот отмечен только на одной. Проективное покрытие травянистых
160
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
видов не превышает 15% и обычно держится на минимальной отметке 5%.
Таким образом, подводя итог сказанному, следует отметить, что на
большинстве моховых болот лесотаежной зоны можно осуществлять
масштабные заготовки возобновляемых растительных ресурсов из числа
пищевых, кормовых и лекарственных без серьезных опасений за причинение
ощутимого ущерба болотным экосистемам.
ФЛОРИСТИЧЕСКИЙ СОСТАВ ЛИСТОСТЕБЕЛЬНЫХ МХОВ БОЛОТ
МЕЗЕНСКО-ВЫЧЕГО ДСКОЙ РАВНИНЫ
Т.П. Шубина, Г.В. Железнова
Институт биологии Коми НЦ УрО РАН, Сыктывкар
Мезенско-Вычегодская равнина располагается в северо-восточной части
европейской России от Северных Увалов на юге до Среднего Тимана на
севере (60° 10' и 64°30' с.ш., 46°50' и 52°50' в.д.). Площадь равнины
составляет немногим более 117000 км2. Изученный регион представляет собой
моренную равнину с преобладающими высотами около 150 м. Климат
умеренно-континентальный. Согласно геоботаническому районированию
Республики Коми [3] территория равнины относится к двум растительным
подзонам: северной и средней тайги. Мезенско-Вычегодская равнина лежит в
зоне избыточного увлажнения. Значительная часть равнины заболочена (50-
60%). Болота представлены низинным, переходным и верховым типами.
Флористический состав листостебельных мхов болотных ценозов
равнины более чем в 2 раза беднее лесных. На обследованных болотах
зарегистрировано 68 видов листостебельных мхов из 33 родов и 20 семейств, т.е. 1/3
видов региональной бриофлоры. Моховой покров на низинных болотах
представлен 50 видами листостебельных мхов, относящихся к 31 роду и 19
семействам. По числу видов лидируют семейства Amblystegiaceae (12
видов) и Sphagnaceae (9), а из родов — Sphagnum (9), Calliergon (4). Роль
сфагновых мхов в формировании растительного покрова низинных болот
Мезенско-Вычегодской равнины в сравнении с зелеными мхами невелика,
несмотря на то, что род Sphagnum обладает здесь наибольшим
разнообразием. Чаще всего в изученных болотных сообществах отмечались такие
представители этого рода, как Sphagnum warnstorfii, S. girgensohnii, S.
squarrosum, реже — Sphagnum riparium и единично S. inundatum.
Бриофлора низинных болот исследованного региона обогащена рядом
мезоэутрофных видов мохообразных из родов Plagiomnium, Pseudobryum,
Rhizomnium, Climacium, не отмеченных на верховых болотах. К наиболее
обычным видам мхов низинных болот Мезенско-Вычегодской равнины
относятся Pseudobryum cinclidioides, Plagiomnium ellipticum, Calliergon cordi-
161
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
folium, Hypnum lindbergii, Climacium dendroides, Bryum pseudotriquetrum,
Brachythecium mildeanum, Sanionia uncinata, Warnstorfia exannulata. В
примеси к ним встречаются Warnstorfia fluitans, Aulacomnium palustre,
Polytrichum commune, Calliergon giganteum, С stramineum, Calliergonella cuspi-
data, Campylium polygamum. По окраинам низинных болот на гниющей
древесине и приствольных повышениях деревьев, кроме доминантов
напочвенного покрова низинных болот, отмечены также лесные виды: Hylocomium
splendens, Rhytidiadelphus triquetrus, Fissidens bryoides, Homalia trichoma-
noides, Saelania glaucescens, Plagiomnium cuspidatum, Rhizomnium pseudopu-
nctatum, R. punctatum. Индикаторами ключевого питания болот являются
Paludella squarrosa, Helodium biandowii, Tomentypnum nitens,
произрастающие преимущественно на небольших повышениях микрорельефа среди
более сырых участков или на умеренно увлажняемых местах ключевых болот.
Флористическое разнообразие мхов, обнаруженных на переходных и
верховых болотах Мезенско-Вычегодской равнины, значительно ниже, чем
на низинных, и составляет 30 видов из 10 родов и 8 семейств. Аналогичные
сведения о крайней бедности видового состава мхов олиготрофных и мезо-
эутрофных болот приводятся как для европейской северной тайги [4], так и
для лесостепной и степной зон [2]. Господствующим семейством и по числу
видов, и по участию в образовании мохового покрова на изученных болотах,
является Sphagnaceae, представленное одним родом Sphagnum и 15 видами.
По сравнению с низинными болотами на переходных и верховых болотах
несколько возрастает роль семейства Dicranaceae (4 вида) и рода Dicranum
(4). На умеренно увлажняемых местах, а также на микроповышениях (часто
возле стволиков Betuia nana, Vaccinium uliginosum, Ledum palustre)
сплошной моховой ковер в рассматриваемых болотных сообществах образуют
Sphagnum angustifolium, S. fuscum, S. magellanicum, S. capillifolium, S. russo-
wii, реже — S. flexuosum, S. fallax. Наряду с указанными видами
мохообразных здесь произрастают и зеленые мхи: Aulacomnium palustre, Calliergon
stramineum, Polytrichum commune, P. strictum, Dicranum bergeri, D. poly-
setum. В ямках с водой регистрировались гидрофитные сфагновые Sphagnum
riparium, S. squarrosum, S. cuspidatum, а также Calliergon stramineum,
Warnstorfia exannulata, W. fluitans, Polytrichum commune. На гниющей
древесине на переходных и верховых болотах поселяются только Pleurozium
schreberi, Tetraphis pellucida, Polytrichum strictum, Pohlia nutans.
Анализ географических элементов флоры мхов низинных, переходных и
верховых болот выявил значительное преобладание бореальных видов (50
видов или 73,5%). Среди экологических групп бриофитов по отношению к
влажности субстрата лидируют гигрофиты (26 видов или 38,2%), мезофиты
(19 видов или 27,9%) и гидрофиты (13 ввдов или 19,1%).
162
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ III
На исследованных болотах произрастают виды листостебельных мхов
редкие как для территории равнины, так и для европейского Северо-Востока
— Sphagnum subsecundum, S. inundatum, Dicranum elongatum, Philonotis
marchica, Calliergon richardsonii, Pohlia sphagnicola. В «Красную Книгу
Республики Коми» [1] включено 4 вида листостебельных мхов (Philonotis
marchica, Calliergon richardsonii, Sphagnum quinquefarium, Bryum elegans)^
зарегистрированных на болотах Мезенско-Вычегодской равнины.
Полученные сведения о редких видах мхов болотных сообществ вошли в
компьютерную базу данных «Редкие растения Республики Коми».
[1] Красная Книга Республики Коми. М., 1998. 528 с. [2] Хмелев К.Ф., Попова Н.Н.
Бриофлора лесных и болотных экосистем бассейна Среднего Дона // Брио-
лихенологические исследования в СССР. Апатиты, 1986. С. 66-70. [3] Юдин Ю.П.
Геоботаническое районирование // Производительные силы Коми АССР. М.Д,
1954. Т. Ш. Ч. 3. С. 323-369. [4] Yurkovskaya Т.К. The regional and local features of
the moss flora in the Northern taiga mires of European Russia // Ботан. ж., 1998. Т. 83.
№ 6. C.46-50.
163
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
IV. БИОСФЕРНАЯ РОЛЬ БОЛОТ. БИОХИМИЧЕСКИЕ ЦИКЛЫ
ВЕЩЕСТВ ЕСТЕСТВЕННЫХ И ЭКСПЛУАТИРУЕМЫХ
БОЛОТНЫХ ЭКОСИСТЕМ
БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ АЗОТА И УГЛЕРОДА
В ЕСТЕСТВЕННЫХ И АНТРОПОГЕННО НАРУШЕННЫХ
БОЛОТНЫХ ЭКОСИСТЕМАХ
Н.Н. Бамбалов, В.А. Ракович, В.В. Марыганова, Т.Я. Беленькая,
А. А. Шишко
Институт проблем использования природных ресурсов и экологии
НАН Беларуси, Минск
Особенности биогеохимических циклов углерода и азота на
естественных болотах обусловлены их незамкнутостью и переходом гумифицирован-
ного материала в виде торфа из малого биогенного в великий геологический
круговорот веществ, где углерод и азот консервируются в торфяных залежах
на многие тысячелетия. Углекислый газ и азотистые соединения поступают
в растения из атмосферы и воды, трансформируются в органическое
вещество, которое после отмирания растения гумифицируется и вследствие
высокой обводненности верхнего слоя болотной почвы не успевает в течение
годового биоцикла полностью разложиться и аккумулируется в виде торфа.
В условиях Беларуси за тысячу лет может накопиться от 0,5 до 1,5 м торфа.
В незаболоченных лугах и лесах отмершие и гумифицированные
растения разлагаются почти полностью, поэтому даже через тысячу лет в лесах
образуется маломощный гумусный слой — не более 8-12 см с содержанием
органического вещества около 1,0-1,5%. На лугах за это же время
образуется гумусный слой не более 20 см с содержанием органического вещества
около 2,0-2,5%. В лесных и луговых ландшафтах практически весь углерод,
выведенный из атмосферы, снова возвращается в атмосферу вследствие
интенсивного разложения отмерших растений. В лесах атмосферный углерод
может задерживаться на десятки-сотни лет в древесине живых растений до
отмирания и разложения последних. Следовательно, в отличие от болот леса
и луга не переводят углерод и азот из биогенного круговорота в
геологический, либо этот процесс осуществляется в ничтожно малом масштабе.
Количество углерода и азота, ежегодно выводимого из биогенного
круговорота в геологический, зависит от условий обводнения болот,
минерального питания растений и продолжительности периода биологически активных
температур. В Беларуси один гектар естественного болота выводит из
атмосферы через биогенный круговорот в геологический в зависимости от фито-
164
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ ГУ
ценозов около 150-500 кг углерода, что составляет 550-1800 кг С02 в год.
Количество азота, ежегодно выводимого из атмосферы и почвенно-грун-
товых вод в геологический круговорот, составляет на низинных болотах 10-
30 кг/га, на верховых — около 10 кг/га.
При осушении болот увеличивается глубина зоны аэрации торфяных
залежей, и происходит интенсивное развитие аэробных микроорганизмов,
минерализующих органическое вещество торфа. По этой причине в результате
осушения болот прирост торфа сменяется его разрушением, и
аккумулятивный тип баланса вещества и энергии сменяется господством геохимического
выноса, величина которого зависит от интенсивности и характера
использования торфяной залежи. На осушенных болотах потоки углерода и азота
приобретают противоположную направленность по сравнению с неосушен-
ными болотами, а именно: в результате деятельности аэробных
микроорганизмов эти элементы переходят из геологического круговорота в
биогенный, в том числе в атмосферу и почвенно-грунтовые воды. Ежегодный
вынос углерода с осушенных для сельского хозяйства торфяных почв
составляет 1500-6000 кг/га (5500-22000 кг/га углекислого газа), азота — 100-300
кг/га. При этом минимальный вынос этих элементов наблюдается при
возделывании многолетних трав, максимальный — пропашных культур.
Промышленная разработка торфяных месторождений увеличивает скорость
выноса азота и углерода с болот в 300-500 раз по сравнению с осушением.
Таким образом, естественные болота очищают атмосферу и почвенно-
грунтовые воды от соединений углерода и азота, а осушенные болота,
наоборот, переводят углерод и азот прошлых эпох из торфяных залежей в
атмосферу и почвенно-грунтовые воды. Из этого следует, что современные
способы сельскохозяйственного и промышленного использования торфяных
месторождений противоречат законам развития нашей планеты и должны
быть заменены другими, более совершенными способами.
В настоящее время в Беларуси начали развиваться новые биосферно
совместимые методы использования природных ресурсов болот по четырем
направлениям: экологическому, культурно-рекреационному,
агрономическому и энерготехнологическому [1]. Экоболотоводство — сохранение в
естественном состоянии или восстановление антропогенно уничтоженных
болотных экосистем с целью поддержания экологического равновесия в
природно-территориальных комплексах: водного баланса, гидрологического
режима, качества поверхностных и грунтовых вод, микроклимата, состава
атмосферного воздуха, создания противоэрозионных, геохимических и
санитарно-гигиенических барьеров, сохранения генофонда растительного и
животного мира, а также процесса торфообразования.
Культурно-рекреационное болотоводство — сохранение в естественно состоянии или восста-
165
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
новление антропогенно уничтоженных болотных экосистем для
использования в качестве учебно-познавательных объектов, природных лабораторий,
научных полигонов и стационаров, памятников природы, историко-
археологических заказников, мест туризма, охоты, сбора ягод, грибов и т.д.
Агроболотоводство — управление естественным развитием болот или
восстановление антропогенно уничтоженных болот с целью получения урожаев
диких, полукультурных и культурных сортов болотных ягод, медоносных и
лекарственных растений. Энерготехнологическое болотоводство —
интенсификация фотосинтеза болотных фитоценозов для ежегодного
воспроизводства энергетического и органического сырья в виде биомассы болотных
растений, которая может перерабатываться в твердое, жидкое/газообразное
топливо, компосты, картон, бумагу, упаковочные материалы и т.п. [2].
Указанные направления предусматривают использование ресурсов болот
без прекращения болотообразовательного процесса. Кроме этого важной
задачей биосферно совместимого использования торфяных месторождений
является их повторное заболачивание после выработки торфа. Общая
площадь повторно заболоченных выработанных торфяных месторождений и
участков в Беларуси приближается к 50 тыс. га и в перспективе должна
составить около 120-150 тыс. га. Начинается повторное заболачивание многих
участков ранее мелиорированных торфяных почв в связи с выходом из
строя мелиоративных систем. В ближайшие 10-15 лет площадь таких
территорий может составить около 200-250 тыс. гектаров. Важно обеспечить
создание управляемых фитоценозов на таких площадях с формированием
насаждений ольхи, ивы, тростника и других ценных болотных растений, чтобы
максимально реализовать питательные вещества бывших пахотных
горизонтов, накопленные в процессе их сельскохозяйственного использования.
[1] Бамбалов Н.Н. Основные задачи болотоводства // Торфяная промышленность.
1991. № 12. С.25-26. [2] Бамбалов Н.Н. Космические и земные факторы торфообра-
зования // Торфяная промышленность. 1991. № 1. С.2-7.
БИОСФЕРНАЯ РОЛЬ БОЛОТ, ЗАБОЛОЧЕННЫХ ЛЕСОВ И
ПРОБЛЕМЫ ИХ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
С.Э. Вомперский
Институт лесоведения РАН, Московская обл.
Под биосферным значением экосистем обычно понимается их роль в
трансформации (поддержании) глобального круговорота веществ и энергии
на Земле. Эта роль — влияние выходит за «границы» конкретного
биогеоценоза, экосистемы (разного объема понимания) и в конечном итоге сказы-
166
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
вается на динамике состояния биосферы в целом или крупных ее
составляющих.
Устойчивое пользование болотами, как и другими экосистемами
предполагает, во-первых, воспроизводимость изымаемых ресурсов или, по крайней
мере «значительную» длительность (условную неистощимость) этого
пользования, а во-вторых, учет биосферных следствий антропогенного
вмешательства в природу. В частности, в наше время ряд Международных
форумов предложил сопоставлять выбросы С02 от сжигаемого топлива в стране
с размером связывания С02 ее биотой, а дисбаланс, т.е. загрязнение
атмосферы восстанавливать за счет государств — загрязнителей. Такое одно из
условий устойчивого развития, независимо от внедрения его в мировую
практику, потребовало от ученых, с одной стороны — лучшего понимания
механизма функционирования биогеоценозов и более точного измерения его
параметров, а с другой — перехода к другим пространственно-временным
масштабам. То есть от оценок «in situ» к ландшафтам, регионам и стране в
целом, от оценок за минуты, часы, сутки, сезон к прогнозным оценкам за
годы и десятилетия. Все это крайне непросто по существу, ограничено
материально-техническими возможностями научных исследований,
недостаточным учетом в масштабе страны типов экосистем, их площадей, состояния и
т.д. и вряд ли с желаемой точностью будет быстро разрешено.
В настоящей работе преимущественное внимание будет уделено влиянию
болот на связывание атмосферного углерода при торфонакоплении. Будучи
одним из самых главных углеродный цикл биогеоценоза во многом с
качественной стороны характеризует весь круговорот веществ и соответственно
биосферную роль экосистем. Кроме того, в течение многих лет водный цикл
болот, его трансформация при гидромелиорациях, использовании
торфяников в лесном и сельском хозяйстве интенсивно изучался [1, 3, 7-10].
Прежде всего как оценивается совокупная роль всех болот и оторфован-
ных земель в связывании углерода атмосферы? Как известно болота с при-
ростающими торфяниками обладают среди наземных экосистем
исключительным свойством — незамкнутостью круговорота вещества и энергии на
масштабе времени от года до возможно многих еще тысячелетий. То есть
приходящий из окружающей среды поток веществ и энергии в болото не
компенсируется его выходом. В результате накапливается торф, а вместе с
ним исключаются из глобального круговорота определенная часть углерода
атмосферы, воды других элементов геохимического цикла, а также энергии.
Обычным способом подсчета суммарной роли болот в связывании
углерода атмосферы сводится к оценке площадей болот их глубины и среднего
прироста их торфяников. Несколько более половины сухой массы ежегодно
наросшего торфа принимается за массу, аккумулированного в нем углерода.
167
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Однако сведения о площадях болот их глубине и интенсивности нарастания
в них торфа очень противоречивы. Исследователи прибегают к экспертным
оценкам многих показателей, что с одной стороны вело к внутренним
противоречиям самих этих работ, а с другой итоговые результаты оценок
площадей болот, их глубин и ежегодного связывания атмосферного углерода
получались конечно совершенно разными. Например, согласно [12] площадь
болот бывшего СССР принималась 164,8 млн. га (в более ранних
публикациях эти авторы исходили из вдвое меньшей площади болот). А по [6]
площадь только безлесных болот в России — 273 млн. га, если же учесть, что в
лесной зоне согласно [5] лесные болота и заболоченные леса составляют 114
млн. га, то количество болот России становится значительно более чем в 2
раза большим чем болот в прежнем СССР по [12]. Столько же
необъяснимого в упомянутых работах и относительно принимаемых возрастов болот
средних глубин торфа и прироста торфяников ...
Целью наших работ последних лет было объективизировать методы
оценки биосферной роли болот России в углеродном цикле, сделать эти
методы «прозрачными», а сами оценки более точными [2,4]. В качестве
исходных данных использовалась «Почвенная карта РСФСР», М 1:2,5 млн.,
данные Торфгеологии по запасам торфяного сырья, анализ всех доступных
абсолютных 14С датировок торфа болот России и литературные источники.
Установлено [2], что площадь оторфованных земель России — 369 млн.
га, в том числе болот (с глубиной торфа более 30 см) — 139 млн. га, а также
впервые была подготовлена серия карт размещения по трапециям (1°
широты и 2° долготы) торфяных отложений разной глубины. Запас биогенного
углерода в торфе болот и заболоченных земель России оказался — 113,5
млрд. т, что не менее 1/3 мировых запасов его в торфе, согласно оценке
Международного общества по торфу — 120-240 млрд. т.
Пожалуй наиболее интересные и неожиданные результаты дала новая
методика выявления репрезентативных средних по регионам приростов
торфяников. Оказалось, что в пределах относительно однородного региона
по природным условиям глубина торфяных отложений хорошо коррелирует
со средним за историю болота годичным вертикальным приростом
торфяника. Заключение сделано на основе обработки данных по 6 регионам. Для
центра Европейской части России, Западной Сибири, а также для 3-х
регионов Канады и Финляндии коэффициент корреляции этой связи колебался от
0.63 до 0,92, а в среднем для 6 регионов (с использованием преобразования
Z по Р.А.Фишеру) оказался — 0,77. Это означает, что для правильных
оценок среднего темпа связывания углерода болотами в голоцене надо знать
средние глубины болот по регионам.
168
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
Однако до сих пор исследователи всех стран располагали датировками
преимущественно самых глубоких болот в их генетических центрах.
Прирост торфа в этих точках нетипично высок даже для площади данного
болота, не говоря о репрезентативности таких оценок для всех болот в регионе.
Поэтому полученные таким путем разными авторами оценки связывания
углерода болотами, как по странам так и глобальные — завышены.
Анализ данных датировок базального торфа в болотах России и учет
резко различных условий болотообразования в стране позволил нам в
первом приближении разделить территорию России на 10 регионов, определить
в них среднюю глубину торфяников и соответственно ей среднеголоценовый
прирост торфа и связывании углерода атмосферы. В среднем для болот с
глубиной торфа более 0,5 м (97,5 млн. га) долговременное годичное
связывание оказалось равным лишь 13,53 г См'2год"1. С нашей точки зрения эта
цифра вполне объяснима, принимая во внимание, что 65% наших болот
расположены в зоне вечной мерзлоты, где прирост мал, а общая по стране
средняя глубина болот — 1,7 м. Между тем другие авторы, например [12],
исходили из самых высоких в мире расчетных величин годичного
накопления в торфе углерода — 31,56 г См"2 год*1 — средневзвешенное для всех
болот бывшего СССР. Суммарное годичное связывание углерода всеми
болотами России (включая с глубиной от 0,3 до 0,5 м) согласно нашей оценке
составляет — 17,7 млн. т С год"1. Более подробную информацию об этом
можно найти в работе [4].
Полученные оценки среднего годичного долговременного (за голоцен)
связывания углерода болотами России, представляются нам весьма близко
отражающими их современную способность накапливать углерод, так как
надежных непосредственных методов определения «актуальной»
(краткосрочной) аккумуляции углерода болотами для больших территорий нет,
либо они крайне дороги. Отсутствуют прямые методы измерения массы
растительных остатков ежегодно переходящих в инертную толщу (catotelm),
велико фактическое варьирование анаэробного потока углерода из толщи
торфяника в атмосферу, нет постоянства прихода и разложения растительного
материала в болото соответственно модельным расчетам [13,14],
противоречивы мнения о приросте бугристых торфяников (что можно найти даже в
данной книге), а также различны представления о прогрессивном или
остановившемся процессе расширения болот и заболачивания лесов в таежной
зоне и ряд других соображений не позволяют нам более или менее надежно
установить величину текущего связывания углерода болотами.
Существенным фактором, уменьшающим точность оценок современного
связывания ССЬ атмосферы является переменный баланс углеродного пула
заболоченных лесов, о чем говорится в статье С.Э. Вомперского и др. в дан-
169
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ной книге. Площадь заболоченных земель со слоем торфа менее 30 см в
России — 230 млн га в 1,6 раза больше болот и даже небольшие короткопе-
риодные колебания (в несколько лет) запаса углерода в них создают
большой источник ошибок текущих оценок баланса углерода всех болот,
годичное депонирование которых тоже колеблется по годам и периодам лет.
Наконец, нельзя не отметить неясность площадей лесных болот,
учитываемых как леса, но имеющих два пула биогенного углерода — в фитомассе
и в торфе. Частично эти площади входят в состав болот, а частично нет.
Можно лишь предполагать, что с этим связано некоторое занижение
фактического связывания углерода нашими лесоболотными экосистемами, но
какова величина его?
Другой вопрос на котором мы хотели бы остановиться: как
трансформируется углеродным баланс болот при их хозяйственном использовании. Если
не рассматривать наиболее экологически безвредное рекреационное
пользование болотами — сбор ягод, грибов, лекарственного сырья и охоты, то
наиболее массовым является лесоосушительная мелиорация. В бывшем
СССР за всю советскую и дореволюционную историю было осушено около 6
млн. га. которые однако примерно на половину из за отсутствия уходов за
сетью «вернулись» в исходное состояние. Лесоосушение и лесопользование
на болотах хотя и затрагивает всю систему взаимосвязей в биогеоценозе, но
сравнительно меньше, чем при сельхозпользовании и особенно в его
интенсивных вариантах. Наконец, наиболее радикальным является добыча
торфяного сырья. Восстановление торфа в выработанных торфяниках, если
возможно, то лишь за многие столетия.
В литературе по лесоосушению болот существует очень большой
разнобой мнений о деградации торфяников, либо даже об их нарастании или
суммарном (с фитомассой древостоя) росте запаса углерода в лесном
биогеоценозе на торфянике. Главное, что строгость методических требований в
такого рода экспериментальных исследованиях обычно недостаточна для
более или менее надежных балансовых расчетов всех потоков углерода.
Исходя из накопленного опыта в самом общем виде мы уже
высказывались [15] о том. что на малом масштабе времени (5-25 лет) после
эффективной мелиорации можно ожидать потери запаса углерода в торфе, а на
большом — спустя 4-6 десятилетий при исправном содержании осушительной
сети система переходит полностью на более высокий лесной уровень
сбалансированного (замкнутого) круговорота веществ.
В сущности всякое освоение естественных угодий в растениеводстве и
лесоводстве, в частности, направленное на повышение продуктивности
связано с увеличением мощности и скорости круговорота веществ в
экосистемах. Период перехода на более интенсивный круговорот согласно термоди-
170
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
намическим представлениям будет обязательно связан с некоторой потерей
или рассеиванием вещества и энергии в этих экосистемах. Если внутренний
потенциальный запас их как и внешний привнос искусственными методами
(агрономии, лесоводства) недостаточен, происходит падение биологической
продуктивности и деградация систем до уровней ниже исходных,
естественных. Устойчивая эксплуатация угодий предполагает по идее
сбалансированные с интенсивностью пользования адекватные методы хозяйства
индивидуальные для каждого участка (поля, водосбора), что с научной,
хозяйственной и экономической стороны представляет собой очень сложную,
фундаментальную проблему. По этому в конкретных природных, экономических и
социальных условиях что считать в методах хозяйствования «хорошо», а
что «плохо» вопрос не простой. Поэтому так называемые «критерии» и
«индикаторы» устойчивого природопользования по разным видам
природных ресурсов в мировом сообществе являются предметом дискуссий.
Хотя в современной экономической ситуации эксплуатация болот резко
упала, осушенные площади из-за отсутствия уходов вторично
заболачиваются, все же с методической стороны вопрос оценок следствий пользования
болотами остается крайне важным и спорным. Можно лишь отметить что по
исследованиям, касавшимся бывшего СССР, болота его имеют
отрицательный баланс (главным образом из-за торфодобычи) с общим излучением
углерода в атмосферу от более 40 млн. т С год"1 [11] до 70 млн. т С год"1 [12].
Все изложенное говорит об острой необходимости широких
исследований биосферной роли болот и ее трансформации при различном
использовании без чего невозможны осознанные действия по оптимизации
управления природопользования в целом.
[1] Булавко А.Г. Влияние осушения болот на элементы водного баланса рек
Белорусского Полесья. Л.: Гидрометеоиздат, 1961. 152 с. [2] Вомперский С.Э. и др.
Заболоченные органогенные почвы и болота России и запас углерода в их торфах //
Почвоведение. 1994. №12. С. 17-25. [3] Вомперский С.Э. и др. Формирование и
режим стока при гидролесомелиорации. М.: Наука, 1988. 167 с. [4] Вомперский С.Э. и
др. Заболоченность территории России как фактор связывания атмосферного
углерода// Избр. Научн. труды по проблеме «Глобальная эволюция биосферы.
Антропогенный вклад». М.: Научный Совет НТП «Глобальные изменения природной среды
и климата», 1998. С. 124-144. [5] Гидромелиоративный фонд СССР и основные
направления лесоосушения. М.: Гослесхоз СССР, «Союзгипролесхоз», 1975. 51 с. [6]
Ефремов СП. и др. Запасы углерода в экосистемах болот// Углерод в экосистемах
лесов и болот России. Под ред. В.А.Алексеева и Р.А.Бердси. Красноярск, 1995/1994.
С. 128-139. [7] Залитис П.П. Основы рационального лесоосушения в Латвийской
ССР. Рига, «Зинатне», 1983. 230 с. [8] Иванов К.Е. Основы гидрологии болот лесной
зоны и расчеты водного режима болотных массивов. Л.: Гидрометеоиздат, 1957. 500
с. [9] Киселев П.А. Влияние осушения на режим и баланс грунтовых вод при их
171
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
напорном питании. (На примере Полесья) М.: «Недра», 1980. 122 с. [10] Шебеко
В.Ф. Гидрологический режим осушенных территорий. Минск, «Урожай», 1970. 298
с. [11] Armentano T.V., Menges E.S. Patterns of change in the carbon balance of organic
soil wetlands of the temperate zone //J. Ecology. 1986. V. 74. P. 755-774. [12] Botch
M.S., Kobak K.I., Vinson T.S. and Kolchugina T.P. Carbon pools and accumulation in
peatlands of the former Soviet Union// Global Biogeochemical Cycles, 1995. Vol.9. №1.
P. 37-46. [13] Clymo R.S. The limits to peat bog growth // Phil. Trans. Royal Society of
London B. 19846. V. 303. P. 605-654. [14] Korhola A. et al. Estimating Long-term
Carbon Accumu-lation Rates in Boreal Peatlands by Radiocarbon Dating // Radiocarbon,
1995, Vol. 37 №2. P.575-584. [15] Vompersky S.E. Current status of forest drainage in
the USSR and problems in research // Peat and Peatlands — diversification and
innovation. Jeglum J.K., Overend R.P. (Eds.). Canad. Soc. for Peat and Peatlands, 1991.
V.l.P. 13-18.
РАЗЛОЖЕНИЕ ОПАДА И ЭФФЕКТИВНОСТЬ
ГИДРОЛЕСОМЕЛИОРАЦИИ В КАРЕЛИИ
Н.И. Германова, В.И. Саковец
Институт леса Карельского НЦ РАН, Петрозаводск
Исследование особенностей разложения торфообразующих растений в
Карелии было начато в районе Киндасовского стационара (средняя подзона
тайги) одновременно с проведением там лесоосушительных работ — в
конце 60-х — начале 70-х гг. и продолжается по сегодняшний день.
К 20-летнему рубежу лесомелиорации в Карелии четко обозначились ее
успехи и неудачи. Осушение верховых болот оказалось бесперспективным.
В составе их фитоценозов не происходит изменений, способных повлиять на
состав опада, жизнедеятельность почвенной биоты и плодородие почвы. Ле-
соводственный эффект здесь очень низок. Успешность осушения
переходных болот под лесными насаждениями очевидна: низкопродуктивные и
редкостойные до осушения V а класса бонитета березняки осоково-сфагновые и
разнотравно-хвощовые растут сейчас по II классу. Высокий уровень
микробиологических процессов в почвах под березняками сохраняется в течение
70 и более лет как при интенсивном, так и слабом осушении. Формирование
биологически активных лесных подстилок в березняках снижает угрозу
высокого расхода органического вещества торфа в результате минерализации.
Сосняки травяно-сфагновые имели до осушения запас 70 мЗ/га, через 20
лет он составляет 140 в середине и 170 мЗ/га с края межканавья. Величина
поступления органического вещества в почвенный блок до и после осушения
изменилась незначительно и размеры расхода его на минерализацию близки
и не превышают 32% (около 15 ц/га) исходной массы свежего опада [1].
172
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ 1\
Объекты исследований — сосняки травяно-сфагновые на переходны>
торфах с идентичными свойствами, расположенные r одном болотном
массиве: неосушенный участок и 3 пробных площади на осушенной 120-
метровой межканальной полосе, сосняк травяно-сфагновый, не затронутые
рубками (контроль к рубкам); участок рубки по улучшению породного
состава (через 3 года после осушения удалено 24% хвойных и 64%
лиственных деревьев); участок сплошной рубки с сохранением подроста и
тонкомера хвойных пород, проведенной одновременно с выборочной рубкой). В
опыте по разложению были использованы практически все виды опада на
каждом участке (92-94% общей его массы).
В результате проведения рубок в осушенном сосняке травяно-сфагновом
изменились масса и состав опада и интенсивность биологического
круговорота. Основная масса опада в сосняках — это древесные компоненты (хвоя,
листья, ветви), составляющие в неосушенных насаждениях 56% всей массы,
в осушенных 84-86%, что определяет своеобразный характер и замедленные
темпы его деструкции. На сфагновые мхи приходится от 8 до 12% массы
свежего опада, которые теряют за год 10-11% массы и в дальнейшем в
круговороте участия практически не принимают. Травы и кустарнички
составляют на осушенных объектах 3-6%. на неосушенном 34% массы опада. По
степени доступности микроорганизмам все компоненты опада можно
объединить в группы и расположить в ряд по убыванию: травы — листья березы
— хвоя сосны — ветви сосны и березы — сфагновые мхи. Пищевая
ценность опада для почвенной биоты зависит от соотношения масс опада
каждой группы. Особую роль в массе опада играют листья березы,
составляющие на пробных площадях от 28 до 45% общей массы опада. Масса опада
листьев березы на контроле, изреженном насаждении и молодняке в 2,7, 1,8
и 1,2 раза больше, чем опада хвои, а элементов питания с ним в почву
поступает на контроле в 4-5 раз, на участке выборочной рубки от 1,4 до 7 раз,
в молодняке до 4,5 раз больше, чем с хвоей. При изреживании древостоя
масса листового опада березы уменьшилась на 330 кг/га и увеличилась на
180 кг/га хвойного и на 310 кг/га веточного опада, т.е. состав опада, с точки
зрения пищевых качеств, заметно ухудшился. Через 20 лет после
проведения сплошной рубки общая масса опада в молодняке более чем на 1 т
меньше, чем на остальных участках и соотношение в нем легко- и трудноразла-
гаемых компонентов складывается не в пользу первых.
Осушенные участки близки по количеству опада трав, сфагновых мхов и
хвои. Представляет интерес соотношение масс опада — листья березы,
ветви березы и сосны, по которому можно сравнить питательную ценность
опада. В сосняках на неосушенном участке, контроле, изреженном древостое и
молодняке оно равно соответственно 70:30. 64:36, 54:46 и 45:55. Опад с до-
173
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
минированием в нем относительно легкоразлагаемых компонентов
разлагается быстрее — 35% убыль массы за год на неосушенном объекте, 31 — на
контроле. 27% в насаждениях, пройденных рубкой.
Со свежим опадом азота и зольных элементов в неосушенном сосняке
поступает 32 и 258, на контроле — 41 и 185. в изреженном насаждении 39 и
159, в молодняке - 28 и 101 кг/га. Темпы разложения опада не совпадают со
скоростью высвобождения из него элементов питания. На долю древесного
опада в осушенных насаждениях приходится от 82 до 91% азота, 61-86%
фосфора, 79-85% калия от их содержания во всем опаде. С удалением части
насаждения азота, фосфора и калия с опадом поступает на 7,23 и 16%
меньше, чем на контроле. В молодняке разрыв с контролем еще более
увеличивается, здесь соответствующих элементов в опаде меньше на 33, 53, 43%.
Высвобождение элементов из опада в сосновых лесах протекает в
соответствии с составом опада очень медленно. На осушенных пробных
площадях из опада, в том числе из сфагнума, высвобождается до 80% калия и от
16 до 44% зольных элементов (суммы). Азот и фосфор высвобождаются
мизерными порциями в основном из опада трав, интенсивность
высвобождения этих элементов снижается в ряду: контроль — изреженное
насаждение — молодняк.
Основные стадии высвобождения элементов из опада наступают через
год и более с начала его разложения на поверхности почвы. Интенсивность
процесса зависит от качества оставшейся массы. По сравнению с контролем
оно ухудшается в изреженном насаждении и еще более на участке сплошной
рубки, где соотношение травы: листья березы: хвоя сосны: ветви равно
3:31:22:44. На контроле, в изреженном насаждении и молодняке в остатке
опада содержится азота и зольных элементов соответственно 46 и 139, 46 и
131 и 28 и 81 кг/га. или в перерасчете на 1 ц годичного прироста на этих же
участках в остатке опада содержится соответственно 0,39, 0,32, и 0,26 кг/га
азота. 0,052, 0.024 и 0,018 фосфора и 0,069, 0,029 и 0,022 кг/га калия.
Текущий бонитет древостоев равен 11,8 — на участках выборочной и 11,5
— сплошной рубки (на контроле III, III.2). Это может свидетельствовать об
отсутствии напряженности пищевого режима древостоев. В результате
большой выборки деревьев на участках сократился общий объем выноса
элементов* на формирование годичного прироста, и, по всей вероятности,
запасы доступных элементов питания в почве к 20-летнему сроку
мероприятия еще не истощились, и активно используются насаждениями. С
увеличением возраста древостоя на участке сплошной рубки размеры фитомассы и
мортмассы и интенсивность круговорота элементов должны достигнуть
контрольных показателей или превысить их.
174
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
[1] Германова Н.И., Егорова Р.А. Баланс органического вещества при освоении
торфяных почв среднетаежной подзоны // Лесоведение. 1998. № 3. С. 12-17.
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ ЭМИССИИ МЕТАНА ИЗ
БОЛОТ В АТМОСФЕРУ
М.В. Глаголев
Институт микробиологии РАН, Москва
Метан является важным «парниковым» газом в климатической системе и
сильно влияет на фотохимию атмосферы [3]. Значительный источник
метана — анаэробное разложение в болотах. Около 60% эмиссии обусловлено
богатыми торфом болотами, находящимися в полосе от 50 до 70° с.ш. [11].
Задачи уменьшения неопределенностей в оценках текущей эмиссии метана и
предсказания ее изменения в будущем требуют (для расчета потока СН4,
исходя из данных об окружающей среде) построения математической
модели [3]. Существующие модели не в полной мере решают эти задачи в силу
ряда причин: 1) Эмпирические модели, например [6,7,9,14], связывающие
интенсивность эмиссии метана напрямую с температурой, уровнем
грунтовых вод и др. факторами внешней среды, успешно работают тогда, когда
значения входных переменных близки к тем значениям, при которых
коэффициенты модели были идентифицированы. Однако в задачах предсказания
эмиссии, приходится значительно отклонятся от вышеуказанных значений,
но в этом случае расхождение модельного расчета и последующего
эксперимента становится, как правило, катастрофически большим. 2) Модели,
основанные на подробном описании фундаментальных закономерностей
метаногенеза, как биолого-химических (например, трофические модели
[1,2,10]), так и биолого-физических [15], настолько сложны, что их
коэффициенты возможно идентифицировать для каждой экосистемы лишь при
помощи большого числа дорогостоящих лабораторных экспериментов. Таким
образом эти модели не могут быть в настоящее время рекомендованы для
предсказания эмиссии в глобальном масштабе. 3) Модели промежуточного
типа (основанные на грубом описании закономерностей метаногенеза и в
значительной степени эмпирические), например [4,5], будучи
идентифицированы по одним экосистемам и (в целях прогноза) применены к другим
экосистемам, дают пока не очень точные результаты, хотя и не столь
абсурдные, как чисто эмпирические модели. Целью работы была доработка и
проверка одной модели эмиссии СБЦ [8], на основании двух наборов
экспериментальных данных, не использовавшихся при создании модели (один из
этих наборов был вообще получен уже после создания модели).
175
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Исследования проводились в июле-августе 1995-98 гг. в районе
стационара «Плотниково» Института почвоведения и агрохимии СО РАН (Бакчар-
ский р-на Томской обл.: 56°5Г с.ш., 82°53' в.д.) на Бакчарском болоте.
Методы измерения потока метана, биомассы расгений, температуры почвы
приведены в [13]. Математическая модель была детально описана ранее [8].
Расчеты по модели для эмиссии метана из Бакчарского болота хорошо
соответствовали экспериментальным данным во временном масштабе
нескольких лет (измеренный с погрешностью 20% средний поток метана
варьировал от 4 до 19 г/м2/сезон в зависимости от температуры, уровня
грунтовых вод и типа растительной ассоциации, а рассчитанный по модели — от 3
до 14 г/м2/сезон), а также в сезонном масштабе времени для летних месяцев
(относительная погрешность около 20%). Особый интерес представляло
предсказание эмиссии в июле-августе 1998 г., т.к. модель была создана в
1997 г. и данные 1998 г. изначально в ней не могли быть учтены. Сравнение
теоретического предсказания и результатов полевых экспериментов выявило
незначительную погрешность около 10%. В меньшем (суточном) масштабе
времени предсказания модели и эксперимента не совпадали.
Попытка предсказания динамики осеннего уменьшения эмиссии не
увенчалась успехом: модель предсказывает значительно более быстрое падение
скорости эмиссии СНЦ, чем наблюдается в природе. Одно из возможных
объяснений: осенью температура верхнего слоя почвы значительно падает и
это сильно снижает потребление СН4 метанокисляющими
микроорганизмами, в то время как в глубине почвы температура падает не столь
значительно, поэтому производство СН4 метанобразующими бактериями,
уменьшается не сильно. Таким образом нарушается пропорция между образованием и
потреблением метана, в то время как в модели она принята постоянной.
Если это предположение правильно, то весной, после возможного выброса (во
время таяния снега) накопившегося за всю зиму метана, что было показано,
например, в [16], следует ожидать значительно более низких величин
эмиссии, чем осенью (при тех же средней температуре и уровне грунтовых вод).
Применение модели к 5 типам болотных экосистем на юго-западном
берегу залива Джемс (Онтарио, Канада) давало удовлетворительное
соответствие экспериментальным данным из работы [12] (измеренный поток СИ*
варьировал от 0,05 до 16,61 г/м2/сезон в зависимости от факторов среды, а
рассчитанный по модели — от 0,67 до 15 г/м2/сезон), хотя следует признать,
что для большинства экосистем модель слегка завышала результаты.
[1] Васильев В.Б. и др. 1994. Имитационная модель метаногенного сообщества. //
Журн. общ. биологии. 1994. Т. 55. № 1. С. 211-237. [2] Калюжный СВ. и др. Биогаз:
проблемы и решения // Биотехнология. Итоги науки и техники. М: ВИНИТИ, 1988.
Т. 21. С. 1-180. [3] Cao M. et al. Modeling methane emissions from rice paddies. Global
176
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
Biogeoch. Cycles, 1995. 9. P. 183-195. [4] Cao M. et al. Global carbon exchange and
methane emissions from natural wetlands: Application of a process-based model // J.
Geoph. Res. 1996. 101. 14399-14414. [5] Christensen T.R. et al. Methane flux from
northern wetlands and tundra. An ecosystem source modelling approach // Tellus, 1996.
48B. 652-651. [6] Dise N.B. et al. Environmental factors controlling methane emissions
from peatlands in Northern Minnesota // J. Geoph. Res. 1993. 98. 10583-10594. [7]
Frolking S., Crill P. Climate controls on temporal variability of methane flux from a poor
fen in southeastern New Hampshire: measurement and modeling // Global Biogeoch.
Cycles. 1994. 8. 385-397. [8] Glagolev M. et al. Modeling of methane emission to
atmosphere in West Siberian wetland (Bakchar Bog, Tomsk Area) // Proc. Sixth Symp.
on Joint Siberian Permafrost Studies. Tsukuba: «Isebu». 1998. 175-190. [9] Granberg G.
et al. Sources of spatial variation in methane emission from mires in northern Sweden: A
mechanistic approach in statistical modeling // Global Biogeoch. Cycles, 1997. 11. 135-
150. [10] James R.T. Sensitivity analysis of a simulation model of methane flux from the
Florida Everglades // Ecological Modelling, 1993. 68. 119-146. [11] Matthews E., Fung
I. Methane emission from natural wetlands: global distribution, area, and environmental
characteristics of sources // Global Biogeoch. Cycles, 1987, 1. 61-86. [12] Moore T.R. et
al. Methane emissions from wetlands, southern Hudson Bay lowland // J. Geoph. Res.
1994. 99. 1455-1467. [13] Panikov N.S. et al. Variability of methane emission from west-
siberian wetlands as related to vegetation type // Ecol. Chemistry. 1997. 6(1). P. 59-67.
[14] Shurpali H.J. et al. Seasonal distribution of methane flux in a Minnesota peatland
measured by Eddy correlation // J. Geoph. Research. 1993. 98. 20649-20655. [15] Walter
B.P. et al. A process-based model to derive methane emissions from natural wetlands //
Geoph. Res. Letters. 1996. 23. 3731-3734. [16] Windsor J. et al. Episodic fluxes of
methane from subarctic fens // Can. J. Soil Science. 1992. 72. 441-452.
ОЦЕНКА ПЕРЕНОСА МЕТАНА ИЗ ПОЧВЫ В АТМОСФЕРУ
БОЛОТНЫМИ РАСТЕНИЯМИ
М.В. Глаголев1, С.А. Голышев2, СЮ. Фирсов3
институт микробиологии РАН, Москва; 2МГУ, биологический
факультет; ^Московский городской дворец творчества детей и юношества
Введение. Среди экосистем одну из ведущих позиций по выделению
метана занимают болота. Наряду с исследованием процессов образования и
потреблением метана, необходимо изучение механизмов его транспорта от
мест синтеза к местам потребления и в атмосферу, т.к. некоторые
взаимосвязи между факторами среды и интенсивностью эмиссии газа
определяются именно механизмом транспорта. Например для чисто диффузионного
механизма не следует ожидать взаимосвязи между атмосферным давлением
и интенсивностью эмиссии. Кроме того, понимание механизма транспорта
важно и при разработке методов воздействия на метанобразующие
природные системы с целью снижения потока этого газа из почвы в атмосферу.
177
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Известны следующие механизмы транспорта: диффузия газа через воду и^
воздух, заполняющие поры в грунте, пузырьковый перенос и транспорт газа
посредством растений [7]. В работе рассматривается эксперимент, позвск
ляющий оценить интенсивность переноса СЬЦ через растения.
Объекты и методы исследования. Работы проводились в июле 1998 года
на Бакчарском болоте (Томская область, Бакчарский район, 152 км по
трассе Томск-Бакчар, 56°5Г с.ш., 82°53' в.д.) в районе почвенного стационара
«Плотниково» Института Почвоведения и Агрохимии СО РАН. Это
верховое сфагновое болото является частью Большого Васюганского болота,
расположенного в Западно-Сибирской низменности.
Для улавливания выделяющегося газа использовались стеклянные
воронки с площадью широкой части 172 см2. Узкий конец каждой воронки
герметично закрывался с помощью резиновой пробки, через которые
проходила металлическая трубка диаметром 2-3 мм, соединенная с резиновым
шлангом длинной 0,5 м. Шланг заканчивался малой стеклянной воронкой с
диаметром широкой части 1 см. Воронки укреплялись в деревянном
штативе на затопленном участке грунта узкой частью вверх, так, чтобы широкое
основание воронки оказывалось на несколько сантиметров ниже уровня
воды. После установки воронки заполнялись водой до контрольной отметки,
нанесенной на ножке воронки. Контролем служили воронки на участках,
покрытых только лишь Sphagnum fuscum (сосудистые растения
отсутствовали). В опытах воронки помещали на известное число растений
определенных видов. Исследовались: Carex rostrata, Eriophorum vaginatum, Menyan-
thes trifoliata, Equisetum palustre, Oxycoccus palustris, Sphagnum magellani-
cum. Sphagnum fuscum (мхи присутствовали и в воронках, содержащих
сосудистые растения). Регулярные измерения проводились в течение
9-дневного эксперимента с периодичностью в среднем 1 раз в 40 часов. Пробы газа
отбирались шприцем сквозь пробку в малой воронке на конце шланга.
Концентрация метана определялась с помощью газового хроматографа ХПМ-4 с
пламенно-ионизационным детектором. Для расчета массы метана в пробе
использовалось уравнение состояния идеального газа. По завершении
эксперимента растения были срезаны на уровне мха, высушены и взвешены.
Статистическую обработку результатов измерений и расчетов проводили при
помощи критерия Стьюдента [1].
Результаты и обсуждение. В большинстве опытов с высокой
надежностью было показано различие потоков метана в опыте (где присутствовали
сосудистые растения) и контроле (где сосудистые растения отсутствовали), а
также различие потоков метана в условиях затенения и при естественном
режиме освещения (табл, 1). Был вычислен поток метана в расчете на сухой
вес находившихся внутри воронки стеблей растений (табл. 2).
178
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
^ £Л 1; Надежность различия потоков метана по критерию Стьюдента
Вариант Надежность различия потока при расчете на
количество стеблей массу стеблей
Er. vaginatum/Kompojib
Er. vagina turn: свет/темнота
С. rasfrata/контроль
С. гоуГгаГахвет/темнота
М. trifoliata/KompoJib
Ы. гп/я/шгахвет/темнота
Е. palustre/коктрояъ
Е. palustrexBerr/TGMHOTa
О. palustris/конгрояъ
поток не рассчитывали <0,95
<0,95 <0,95
поток не рассчитывали 0,9999
0,96 <0,95
поток не рассчитывали 0,995
0,998 0,998
поток не рассчитывали 0,9998
0,997 0,997
поток не рассчитывали 0,9999999
Табл. 2. Поток метана через различные растения
Растение п™
Поток метана (мгС/г/час)
Среднее значение Абсолютная погрешность
Er. vaginatum
С. rostrata
М. trifoliata
О. palustris
Sph. sp.
Е. palustre
0,01
0,097
0,032
0,0042
0,0004
0,11
0,01
0,046
0,016
0,0012
0,0003
0,05
Полученные результаты в основном находятся в соответствии с
результатами других авторов. Так, в [2] было показано, что основную роль в
транспорте метана из затопленных почв рисовых полей играют растения риса, а в
[3], что некоторые водные растения (Avicennia nitida, Cladium jamaicense,
Eleocharis interstincta, Glyceria striata, Juncus effusus, Nymphaea odorata,
Naphar luteum, Peltandra virginica, Pontedeha cordata, Sagittaria graminea,
S. lancifolia, Typha latifolia, Zizaniopsis miliacea) способны обеспечить
скорость эмиссии метана от 0,1 до 14,8 мг/день/растение. В работе [8]
утверждается, что в болотах с доминированием Сагех (расположенных на
территории Квебека и Онтарио) более 90% эмиссии происходит за счет
транспорта, обусловленного растениями.
Несколько удивительно отсутствие в наших опытах значимого потока
через Eriophorum vaginatum, тогда как в [6] сообщалось о том, что удаление
этого растения (два участка в северных болотах Швеции) уменьшало поток
метана на 55-85%. Также наши результаты несколько расходятся с
результатами из [4], где было показано, что растения обеспечивают
доминирующий транспортный путь для метана, независимо от уровней освещенности
или фотосинтеза (правда, в условиях тундры). Мы не обсуждаем здесь
механизм транспорта, хотя в литературе высказывались по крайней мере 2
179
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
гипотезы: об активном [8] и пассивном переносе по корням и стеблям
растений за счет молекулярной диффузии [5,8].
Авторы выражают признательность администрации МГДТДиЮ за
финансовое содействие.
[1] Румшиский Л.З. Математическая обработка результатов эксперимента. М.:
Наука, 1971. 192 с. [2] Nouchi I., Mariko S. Mechanism of Methane Transport by Rice
Plants // Biogeochemistry of global change: radiatively active trace gases. (R.S. Orem-
land, ed.). New York; London: Chapman & Hall, 1993. P. 336-352. [3] Sebacher D.I., et
al. Methane emissions to the atmosphere through aquatic plants // J. Environ. Qual.,
1985. V. 14. P. 40-46. [4] Torn M.S., Chapin F.S. Environmental and biotic controls over
methane flux from arctic tundra // Chemosphere, 1993. V. 26. P. 357-368. [5] Van der
Gon H.A.C.D., van Breemen N. Diffusion-controlled transport of methane from soil to
atmosphere as mediated by rice plants // Biogeochemistry. 1993. V. 21. P. 177-190. [6]
Waddington J.M., et al. Water table control of CH4 emission enhancement by vascular
plants in boreal peatlands // J. Geoph. Res. 1996. V. 101. P. 22775-22785. [7] Walter
B.P., et al. A process-based model to derive methane emissions from natural wetlands //
Geoph. Res. Letters, 1996. V. 23. P. 3731-3734. [8] Whiting G.J., Chanton J.P. Plant-
dependent CH4 emission in a subarctic Canadian fen // Global Biogeoch. Cycles. 1992.
V. 6. P. 225-231.
ГОДИЧНАЯ ЦИКЛИЧНОСТЬ УГЛЕРОДА В БОЛОТНОМ ЛЕСУ КАК
ИНДИКАТОР КОРЕННОГО РАЗЛИЧИЯ МЕЖДУ ЛЕСОМ И БОЛОТОМ
Ф.З. Глебов, Л.В. Карпенко
Институт леса и древесины СО РАН, Красноярск
Рассмотрены особенности годичной цикличности углерода в разных
типах болотного леса, являющихся промежуточным природным образованием
между автоморфным лесом и болотом. В качестве моделей использованы
следующие типы: девственные сосняк чернично-зеленомопшо-сфагновый и
кедровник болотноразнотравный и осушенный березняк
малиново-крапивный. Уточнены понятия леса и болота.
Исследования проводились на Томском стационаре Института леса СО
РАН в биогеоценозах двух контрастных типов девственного болотного леса,
резко отличающихся характером фитоценоза и трофностью почвы — олиго-
трофного чернично-зеленомошно-сфагнового сосняка и евтрофного болот-
норазнотравного кедровника.
Соотношение между приходом и разложением торфа в верхнем
активном слое торфяной залежи было определено методом «черного ящика»: на
его входе — масса С в годичном опаде фитоценоза, на выходе — суммарное
количество азота, состоящее, во-первых, из количества валового азота, со-
180
БИОСФЕРНАЯ ЮЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
держащегося в годичной продукции фитоценоза (NPP), и, во-вторых, —
минерального азота, обнаруженного на различных глубинах подстилочно-поч-
венно-торфяного профиля. По этому суммарному количеству азота
рассчитано то количество торфа (и содержащегося в нем С), которое должно
разложиться, чтобы освободился из него азот.
Приводим результаты изучения биогеоценозов названных двух типов
девственного болотного леса, находящихся на мощной торфяной залежи.
Содержание С в общем количестве живой фитомассы в обоих типах леса
оказалось одинаковым (87 и 89 т/га) и на 66% и 68% приходящимся на
древесину. Содержание С во всей надземной фитомассе значительно
превышает таковое в подземной фитомассе и составляет в обоих типах леса 70%
общего содержания. Напротив, содержание С в подземной фитомассе травяно-
кустарничкового яруса (состоящего, главным образом, из болотных
растений) больше, чем в надземной фитомассе и составляет 52% и 68% от общего
ее количестве. Количество С в моховом покрове обратно пропорционально
трофности почвы. Содержание С в чистой первичной годичной продукции
NEP подсчитано по равенству: NPP - L = NEP, где NPP — первичная общая
продукция, L — опад и отпад.
Видно, что в евтрофных условиях в компонентах круговорота
преобладает древесная масса, в олиготрофных — масса травяно-кустарничково-
мохово-го покрова. Повышение трофности интенсифицирует круговорот С.
Из этого вытекают следующие положения:
1. Фитоценоз болотного леса образован деревьями, кустарничками, гиг-
ро- и гидрофильными травами, а также мхами. Торфообразование, т.е. дли-
тельновременное связывание С происходит за счет разложения отмерших
подземных частей растений.
2. Надземная часть деревьев намного больше подземной. Зольность
древесины незначительна и деревья долговечны: роль стволов и ветвей в
биологическом круговороте и торфообразовании невелика. Годичный приход в
почву зольных веществ и азота из быстро нейтрализующегося древесного
опада интенсифицирует процесс разложения торфа, концентрирует азот и
зольные вещества в поверхностном слое торфяного отложения, стимулирует
биологический круговорот и снижает скорость торфонакопления, т.е.
скорость связывания углерода.
3. Гигро- и гидрофиты состоят из травянистых растений, у которых
подземная часть превышает надземную, и мхов, у которых разложение
надземных частей очень незначительно, а рост происходит в верхушках стеблей
при одновременном отмирании нижних «подземных» частей. Эти растения,
таким образом, замедляют процесс торфоразложения (освобождения С); они
рассредоточивают химические элементы по вертикальному профилю торфя-
181
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ного отложения, сдерживают биологический круговорот и усиливают, таким
образом, торфообразование (связывание С).
4. Деревья стимулируют на болотах лесообразовательный процесс, а гиг-
ро- и гидрофиты усиливают болотообразовательный. В условиях высокой
трофности болот возникает обратная связь: древостой болотного леса
состоят из видов, опад которых обогащен зольными элементами (темнохвойные
породы, береза), что, в свою очередь, стимулирует биологический
круговорот и усиливает скорость разложения торфа. Аналогично снижению
трофности почвы действует повышение географической широты, т.е. продвижение
на север. Результаты исследования в березняке малиново-крапивном
показали, что масса С в общей годичной продукции фитоценоза (NPP) — 4.9 т/га
и в общем количестве разложившегося торфа — 5.5 т/га, так же как в
чистой годичной продукции фитоценоза (NEP) — 2.4 т/га и в чистом
количестве разложившегося торфа (разница между общим количеством
разложившегося торфа и опадом) — 2.9 т/га попарно весьма близки. Это может
свидетельствовать о вещественно-энергетическом балансе в биогеоценозе этого
осушенного леса: освобождение С из торфа в результате его разложения, т.е.
вынос С в атмосферу, компенсируется фитоценозом при фотосинтезе
атмосферного С. Уточним теперь определения понятий болота и автоморфного
леса. Природа болотного леса показывает, что болото, как таковое, —
единственная наземная гидроморфная экосистема, в которой происходит
непрерывное анаэробное накопление мортмассы, связывающей атмосферный С и
сохраняющей тем самым в атмосфере 02, пошедший бы на окисление
мортмассы. Тогда как автоморфный лес — экосистема, в которой механизм
непрерывного накопления мортмассы отсутствует, из-за чего
освобожденный при фотосинтезе 02 тратится на ее окисление, т.е. на освобождение из
нее С, что свидетельствует о меньшей по сравнению с болотом роли
автоморфного леса в смягчении парникового эффекта.
ОЦЕНКА НЕКОТОРЫХ БИОТИЧЕСКИХ КОМПОНЕНТОВ
УГЛЕРОДНОГО ЦИКЛА БОЛОТ И ЛЕСОВ.
Т.В. Глухова, А.Г. Ковалев, М.В. Смагина, С.Э. Вомперский
Институт лесоведения РАН, Московская обл.
Вопросы углеродного цикла болот от фундаментальных до прикладных
нельзя считать удовлетворительно разрешенными. Биогеоценология
вступила в стадию количественных исследований круговорота веществ и энергии,
знание которых пока недостаточно, чтобы делать хорошие прогнозы
естественной и антропогенной динамики болот, а тем более предлагать надежные
182
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
методы управления ими [1].
В связи с этим на Западнодвинском лесоболотном стационаре в Тверской
области на постоянных пробных площадях, заложенных в ряде болотных и
суходольных лесов, изучают компоненты углеродного цикла, такие как
ежегодное поступление древесно-кустарникового опада, почвенные потоки:
ССЬ. освобожденный из опада и отпада в аэробных условиях
гетеротрофными организмами и СН4 как результат анаэробного гетеротрофного дыхания.
Нетто-экосистемная продукция (NEP) оценивается накопленной массой
торфа и запасами органического углерода в нём. Поступление древесно-
кустарникового опада в различных типах леса фиксируется в течение 7 лет,
данные по четырем из которых приведены в табл. 1.
Табл. 1. Динамика поступления древесного опада, гм2 абс.сух.в-ва
Тип леса
Черноольшаник крупно-
травно-папоротниковый
Ельник кислично-зеленчу-
ковый
Сосняк чернично-зелено-
мошный (мелкооторф.)
Сосняк лишайниково-
зеленомошный
Сосняк кустарничково-
сфагновый
Грядово-мочажинный
комплекс с редкой внебо-
нитетной сосной
1993-94
269,08
243,39
200,98
172,01
115,83
20,03
1995-96
297,86
297,72
271,47
161,18
132,27
-
1996-97
342,06
329,37
294,90
222,37
153,88
-
1997-98
372,74
367,90
277,75
232,84
188,70
-
Сред.
320,43
309,59
261,27
197,10
147,67
20,03
Показано, что в высокопродуктивных болотных и суходольных лесах
поступает больше опада, чем в низкопродуктивных. Олиготрофные, почти
безлесные болота, по поступлению опада (20 гм'2, без продукции мхов) сильно
уступают мезотрофным — 148 и особенно евтрофным — 320 гм"2.
Существенным различием лесов и болот в фракционном составе формируемого
годичного прироста фитомассы является то, что главным продуцентом в лесу
выступают деревья, на болотах же. даже лесных, значительное место в
создании органического вещества принадлежит растениям нижних ярусов —
болотным мхам, травам, кустарничкам.
Исследования по учету и оценке сезонной динамики выделения С02 и
СН-1 с поверхности почвы проводят на стационаре в лесах разной
увлажненности и на болотах различной трофности. Оказалось, что процесс выделения
С02 с поверхности почвы очень динамичен, по отдельным срокам сезона его
интенсивность может не отличаться у болот разного генезиса. Однако, выяв-
183
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
лена отчетливая связь интенсивности почвенного дыхания (куда входит и
дыхание корней), прямая — с продуктивностью лесных биогеоценозов и
обратная — с развитием процессов заболачивания. Максимальная эмиссия
углерода (С02) характерна для зонально наиболее продуктивных ельников-
кисличников (727 г См'2 за период май-октябрь, табл.2).
Табл. 2. Эмиссия углерода (С02) за 184 дня (май-октябрь), г Ом"2
Тип леса
Ельник кислично-
зеленчуковый
Сосняк кустарничково-
сфагновый
Сосняк чернично-зелено-
мошный (мелкооторф.)
Сосняк лишайниково-
зеленомошный
Черноолыпаник крупно-
травно-папоротниковый
Грядово-мочажинный
комплекс с редкой внебонитет-
ной сосной
1993
913,61
493,65
705,88
448,49
340,52
321,33
1995
575,81
485,57
404,08
-
390,03
-
1996
959,25
710,63
655,21
-
441,33
-
1997
459,81
401,09
292,34
-
373,56
-
Сред.
727,12
522,73
514,38
448,49
386,36
321,33
Примерно в 1,5-2,0 раза меньшая эмиссия характерна по многолетним
данным для черноолыпаника крупнотравно-папоротникового и сосняка кус-
тарничково-сфагнового (386 и 523 г См"2 соответственно), что говорит о
неодинаковой скорости аккумуляции углерода в торфе и замедленной
деструкции годичного опада в бедной минеральными элементами среде.
Трудностью использования оценок почвенного дыхания в балансовых
расчетах является невозможность разделения регистрируемого потока С02
на составляющие части: обязанную деструкции органического вещества
(собственно гетеротрофному дыханию) и образуемую дыханием живых
корней.
Согласно современным представлениям, метан образуется только в
анаэробной среде. По некоторым оценкам [2,3] выделяемый в виде СКЦ углерод
составляет от 0,5 до 2-7% первичной нетто-продукции болот. Тем не менее
они рассматриваются как главный естественный биогенный поставщик СН*
в атмосферу. Эмиссия метана с поверхности болот дискретна, что не всегда
позволяет в срочных наблюдениях оценить сумму выделившегося СН4 за
сезон. Нередко разброс оценок потока СН4 отличается на два порядка и более.
Наши исследования характеризуют эмиссию метана с поверхности болот
резко различных по трофности (табл. 3). Из болотных черноольховых топей
она оказалась на порядок более низкой (0,3-0,6 гСм"2 за период май-ок-
184
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
тябрь), чем из олиготрофного грядово-мочажинного болота (3,6-4,6 гС-м"2).
Эти данные существенно влияют на использовавшиеся до сих пор оценки в
глобальных расчетах вклада болот в биосферный баланс метана и углерода.
Табл. 3. Эмиссия углерода (СНд) с поверхности болот за 184 дня (май-октябрь) раз-
ных лет наблюдений, г См"2
Тип болота 1993 1995 1996 1997 Сред.
Олиготрофное грядово-мочажинное 3,77 3,60 4,08 4,60 4,01
с редкой внебонитетной сосной
Евтрофное черноольховое 0,37 0,39 0,64 0,34 0,43
Результирующая баланса углерода оценивается накопленной массой
торфа. По данным ряда исследователей [1] величина долговременного
годичного NEP от первичной нетто-продукции (NPP) у болот находится чаще всего в
пределах 7-30%, а текущий(с учетом эмиссии СКО — на 1/3 меньше. О
накоплении торфа судят по скорости вертикального прироста поверхности
болот с учетом объемной массы торфа. Для определения средней скорости
прироста болот в высоту за все время их существования, т.е.
долговременного среднего прироста, необходимы датировки абсолютного возраста
торфяника и его глубины. Согласно имеющимся у нас датировкам по 14С
представленные здесь торфяники прирастают в среднем (со времени
возникновения) со скоростью 0,51, 0,31 и 0,46 мм год1 олиготрофные, мезотрофные
и евтрофные соответственно. По запасам органического углерода в торфе на
первое место выходят высокопродуктивные низинные болота, так черно-
олынаник крупнотравно-папоротниковый запасает в метровой толще около
68 кг м'2 органического углерода, переходная торфяная залежь сосняка кус-
тарничково сфагнового около 60, а верховая — грядово-мочажинного
болота лишь 27. В автоморфных почвах под ельником кислично-зеленчуковым
на дерново-подзолистой почве в метровом слое запас органического
углерода составляет 16 кг м"2, причем примерно половина углерода запасается в
подстилке, в сосняке лишайниково-зеленомошном на песчаных отложениях
запас углерода не превышает 8 кг м"2.
[1] Вомперский С.Э. Роль болот в круговороте углерода.// Биогеоценотические
особенности болот и их рациональное использование. XI чтения памяти академика В.Н.
Сукачева. М Наука, 1994, с. 5-37. [2] Aselman I., Crutsen P.J. Freshwater wetlands
and rice paddies, their net primary productivity, seasonality and possible methane
emissions // J. Atmos. Chem. 1989. N 8. P.307-358. [3] Brown A., Mathur S.P., Kushener
D.J. An ombrotrophic bog as a methane reservoir // Global Biogeochem. Cycles. 1989.
Vol.3, N3. P.205-213.
185
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
КИНЕТИКА РАЗЛОЖЕНИЯ ОРГАНИЧЕСКОГО ВЕЩЕСТВА ТОРФОВ В
ИСКУССТВЕННО АЭРОБНЫХ УСЛОВИЯХ
Л.И. Инишева, Е.В. Белова, Т.В. Дементьева1, Б.Г. Агеев2
1СибНИИТСО РАСХН, Томск;
2Институт оптики атмосферы СО РАН, Томск
Известна роль болот в круговороте углерода. Соотношение между интен-
сивностями потоков углерода (вход и выход) определяется продуктивностью
болотных экосистем, запасом депонированного углерода ж торфяной
залежи, биохимическими свойствами торфов, слагающих торфяную залежь и
скоростью их трансформации» гидротермическим режимом болот.
Целью работы было изучение динамики выделения СОг в процессе
микробиологической трансформации органического вещества торфов разного
ботанического состава, а следовательно, разного химического состава с
целью прогноза суммарной эмиссии С02 из торфяной залежи в условиях
антропогенного воздействия. При этом исходили из следующей гипотезы.
Наиболее интенсивно процесс минерализации протекает в аэробных
условиях, поэтому этот процесс, прежде всего, и подлежит моделированию. А
использование широкого круга модельных экспериментов позволяет
исследовать максимально возможное количество разновидностей торфа за короткий
промежуток времени. Подход даст возможность составить прогноз потока
С02 в атмосферу в целом из болот Западной Сибири.
Главной причиной, определяющей скорость распада торфов является
состав органического вещества, имеющий различную биохимическую
устойчивость. Микробиологами принят следующий ряд соединений по
устойчивости к разложению в природных условиях: сахара < гемицеллюлозы <
целлюлозы < лигнин. В естественных болотах преобладают анаэробные
микроорганизмы. Аэробные и анаэробные микроорганизмы резко различаются
между собой по своему каталитическому аппарату, термодинамике действия
и направлению разрушений и превращений, совершающихся при их
контактах с органическим веществом [1-3]. При смене
окислительно-восстановительных условий (например, осушении) происходит резкая смена вида
микробов. Микроорганизмы чрезвычайно чувствительны к внешним условиям.
Их развитие и рост при стимулировании могут возрастать на 300-400%.
Предлагается использовать эти свойства микроорганизмов и в процессе
лабораторных модельных экспериментов изучить кинетику трансформации
органического вещества торфов в аэробных условиях.
Для эксперимента были отобраны 24 образца торфа из репрезентативных
торфяных залежей в естественном состоянии в пределах наиболее заторфо-
ванных болотных областей Западной Сибири. Использовались два метода
186
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
измерения потока С02: хемосорбционный метод определения кинетики
минерализации органического вещества почв [4] и метод оптико-акустической
спектрометрии [5]. В последующем проведенные эксперименты по кинетике
выделения С(Э2 двумя независимыми методами доказали закономерность
полученных кривых по минерализации органического вещества торфов и
показали возможность использования любого из этих методов.
Исследования проводились в модельном опыте при температуре 20°С и
естественной влажности. Повторность — трехкратная. Длительность опыта
(16 суток) была принята исходя из следующих соображений. В торфяной
залежи in situ под действием внеклеточных ферментов, выделяемых
гетеротрофными организмами (прежде всего это относится к бактериям, актино-
мицетам и грибам), происходит распад торфов. Метаболитическая
активность этой микрофлоры определяет скорость минерализации органического
вещества торфов до С02 [6,7]. Поэтому было принято, что в условиях
оптимальных внешних условий (влажность, температура, свет) скорость
процесса минерализации будет значительно увеличена, что позволяет провести
модельный эксперимент за небольшой промежуток времени.
Результаты модельных экспериментов показали, что верховые и
низинные торфа разной степени разложения и зольности (и, следовательно,
разного химического состава) различаются по интенсивности минерализации.
По величине накопления С02 в процессе минерализации верховые
западносибирские торфа располагаются в следующий ряд: сфагново-
мочажинный > комплексный > фускум-торф > шейхцериево-сфагновый >
пушицево-сфа-гновый > шейхцериевый > пушицевый. Соответственно
кумулятивное накопление С-С02 за 16 суток по видам верхового торфа
составило 133-106-101-74-73-63-46 мг на 100 г сухого торфа.
Торфа низинного типа по величине накопления С02 располагаются в
ряд: гипиовый > осоковый > осоково-гипновый > вахтовый > древесный >
древесно-осоковый. Кумулятивное накопление С-С02 за 16 суток по видам
низинного торфа составило 198-144-125-107-76-64 мг на 100 г сухого торфа.
Получены зависимости скорости минерализации органического вещества,
рассчитано кумулятивное накопление С02 для 24 видов торфа. В отдельных
низинных торфах динамика процесса минерализации имеет вид простой
экспоненциальной функции, что являет собой классический пример
разложения органического вещества. В верховых и некоторых видах низинных
торфов динамика носит синусоидальный характер. Высказана гипотеза, что
синусоидальный вид кривой определяется сменой видов микроорганизмов и
их пулом. Проведены определения видового состава микрофлоры на
торфяном агаре, аэробных целлюлозоразрушающих микроорганизмов на твердой
среде Гетчинсона-Клейтона и гумусоразлагающих микроорганизмов на
187
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
жидкой среде Никитина. Эксперимент подтвердил наличие таких связей, что
предполагает введение в модель трансформации органического вещества
торфов поправки на интенсивность микробиологических процессов, степень
проявления которого определяется химическим составом торфов.
Работа выполнена при финансовой поддержке РФФИ (№ 99-04-48487).
[1] Богопольский ГД. Адсорбция бактерий низинным, торфом// Торфяное дело. 1933.
№ 1. С. 21-33. [2] Звягинцев Д.Г. Взаимодействие микроорганизмов с твердыми
поверхностями М. МГУ. 1973. 175 с [3] Инишева ЛИ., Васильева А.Н.
Микробиологические превращения азотсодержащего вещества в торфяных пойменных почвах
Томской области// Тр. Всес. НИИ с.-х. микробиологии. Л., 1986. Т. 56. С. 18-32. [4]
Иванникова ЛЛ. Способ определения кинетики- минерализации органического
вещества почвы. Патент СССР. № 1806375 от 08.04.1991. [5] Агеев Б.Г. и др.
Кинетика выделения СОг земной растительностью при стрессовых ситуациях// Оптика
атмосферы и океана. 1997. Т.10. № 4-5. С.437-448. [6] Clark F.E., Paul E.A. The
microflora of grassland // Adv. Agron. 1970. 22. P. 375-435. [7] Cambell C.W. Soil
organic carbon, nitrogen and fertility / Soil organic matter. N.Y.: Elsevier, 1978. P. 173-
271
ЭМИССИЯ C02 ИЗ БОЛОТ ЮЖНО-ТАЕЖНОЙ ПОДЗОНЫ
ЗАПАДНОЙ СИБИРИ
Л.И. Инишева, Т.В. Дементьева, Е.В. Белова1, Н.Г. Инишев2
1 СибНИИТ СО РАСХН, Томск;:'Томский государственный университет
Натурных оценок потока С02 с поверхности болот Западносибирского
региона очень мало [1-4]. Поэтому нами были проведены исследования
интенсивности продуцирования С02 на биогеоценозах олиготрофного ряда в
нативном состоянии: высокий рям (сосново-кустарничково-сфагновый
фитоценоз) — низкий рям — открытая топь (осоково-сфагновый фитоценоз), а
также на антропогенных болотных экосистемах: Сухое — Вавиловское (Бак-
чарский р-н) и Таган (Томский р-н). Суммарный поток С02 в атмосферу
определяли методом Штатнова [5].
Объекты Бакчарского района (56°03'-57' с.ш., 82°22'-42' в.д.) относятся к
Западносибирской таежной болотной области бореально-атлантических
выпуклых олиготрофных моховых болот активного заболачивания [6].
Подстилающие породы — лессовидные суглинки. Торфяная залежь ряма
достигает мощности 3 м и имеет смешанный топяной вид строения. В основании
залежи лежит слой хвощевого низинного торфа, далее — низинный
осоковый торф, который перекрывается мощным слоем верхового торфа двух
видов — магелланикум и фускум. Торфяная залежь осоково-сфагновой топи
достигает мощности 2,5 м. В основании залежи — низинный торф осокового
188
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
й древесно-осокового вида, далее переходный шейхцериево-сфагновый и с
поверхности до 60 см — верховой сфагново-мочажинный торф.
В связи с поздней весной и поздним сходом снега уровни болотных вод в
1998 г. установились ниже отметки средней поверхности только в середине
июля. Наиболее высокая амплитуда колебаний УБВ за вегетационный
период отмечалась на ряме — 41,5 см, на открытой топи — 14 см. Отсюда
окислительно-восстановительный потенциал (ОВП) в торфяной залежи олиго-
трофного ряда имел амплитуду колебаний от -226 до +882 мВ. В торфяной
залежи сосново-кустарничково-сфагнового фитоценоза до глубины 40 см
окислительно-восстановительные условия (ОВУ) характеризовались
значениями от -166 до + 857 мВ (при температуре слоя от 10°С и выше). Ниже 40
см находится горизонт с устойчивой восстановительной обстановкой
(температура слоя 15°С и ниже). В метровом слое торфяной залежи осоково-
сфагновой топи выделение такого слоя представляется затруднительным,
так как ОВУ имеют переменный характер, изменяясь от -186 до +785 мВ
(температурные условия аналогичны сосново-кустарничково-сфагновому
фитоценозу). Это свидетельствует об окислительном режиме внутрипочвен-
ного стока, что подтверждается флористическим составом фитоценоза и
результатами химического состава болотных вод.
Наиболее высокая величина эмиссии С02 (в среднем за вегетационный
период 216,2 мг м"2 час"1) получена в пределах высокого ряма. На осоково-
сфагновой топи выделение С02 снижается до 75,8 мг м"2 час"1, что связано с
менее значительной мощностью зоны аэрации и менее интенсивным
протеканием биохимических процессов в торфяной залежи за счет высоких
уровней болотных вод. В течение вегетационного периода в пределах высокого
ряма величина эмиссии С02 изменялась от 78,6 до 425,0 мг м"2 час"1; в
пределах низкого ряма — от 0,0 до 256,4 мг м"2 час'1 и соответствовала
динамике уровней болотных вод и температуре на поверхности торфяной залежи.
Болотная экосистема «Сухое Вавиловское»., относящаяся по болотному
районированию к той же болотной области, что и вышеописанный профиль
олиготрофного ряда, является осушенной и выработанной 20 лет назад. В
настоящее время остаточный слой торфяной залежи имеет мощность 2 м и
сложен осоковыми, осоково-гипновыми и древесными торфами высокой
степени разложения с зольностью 11-50%. По всему профилю отмечается
заиленность. Это является причиной отрицательных значений ОВП по всей
торфяной залежи, начиная с глубины 20 см. Только поверхностный
десятисантиметровый слой имеет положительные значения ОВП, изменяясь от
+550 до +685 мВ. Высокая заиленность профиля торфяной залежи позволяет
проникнуть активным температурам (более 10°С) глубже 1 м.
189
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Интенсивность выделения ССЬ в течение вегетационного периода здесь
колеблется от 0 до 263,1 мг м"2 час"1. Максимальное увеличение
интенсивности выделения С02 наблюдалось в период с последней декады июля по
первую декаду августа. К осени интенсивность выделения ССЬ снижается, а в
конце сентября наблюдается поглощение С02 почвой. В среднем за
вегетационный период интенсивность выделения С02 составила 144 мг м"2 час"1.
Болотная экосистема «Таган» (Томский район), в настоящее время
частично выработанная и рекультивированная, относится к Западносибирской
подтаежной болотной области атлантических плоских евтрофных травяно-
моховых болот умеренного заболачивания. Расположена на второй
надпойменной террасе р.Томи. Торфяная залежь низинного типа и сложена
древесным, осоково-древесным, древесно-гипновым торфом. Подстилающие
породы — пески. Низкая зольность и объемный вес торфов обусловливают
высокую увлажненность профиля торфяной залежи, отрицательные значения
ОВП и неглубокое проникновение активных температур в профиле.
Интенсивность выделения С02 в течение вегетационного периода
колеблется от 20,24 до 937,61 мг м"2 час1. В сезонной изменчивости выделения
С02 отмечаются два максимальных пика: в конце июня — 937,6 мг м"2 час"1
и в начале августа — 449,2 мг м"2 час"1. В это время в торфяной залежи
отмечается благоприятное сочетание тепла и влаги. К концу вегетационного
периода интенсивность выделения С02 составила 99 мг м"2 час'1.
Таким образом, среди исследуемых болотных экосистем максимальная
величина эмиссии С02 зарегистрирована на рекультивированном объекте
«Таган», что обусловлено повышенной биологической активностью торфов
данного объекта вследствие более благоприятных гидротермических
условий и сельскохозяйственного использования. Интенсивность выделения С02
почвами выработанной болотной экосистемой «Сухое Вавиловское» по
сравнению с объектом «Таган» ниже в 1,7 раза. Эмиссия С02 естественной
болотной экосистемой олиготрофного ряда зависела от вида фитоценоза и
снижалась в ряду: высокий рям — низкий рям — осоково-сфагновая топь.
[1] Ефремова Т.Т. и др. Биологическая продукпшностъ и почвы болот южного Ва-
сюганья// Сибирский экологический ж. 1994. № 3. С.253-267. [2] Титлянова А.А. и
др. Запасы органического углерода в почвах Сибири, эмиссия парниковых газов и
сток СО2 в почвы Западной Сибири // Тез. док. П съезда об-ва почвоведов. СПб,
1996, Кн. 1.С. 221-222. [3] Инишева Л.И. и др. Прогноз эмиссии С02 из болот //
Гидрологические исследования в Сибири // Мат-лы науч.-практ. конф., посвящ. 40-
летию кафедры гидрологии ТГУ. Томск, 1997. С32. [4] Дементьева Т.В., Инишева
ЛИ. Интенсивность трансформационных процессов в торфах разного
ботанического состава // Сибирский вести, сельскохоз. науки. 1998. № 3. [5] Агрохимические
методы исследования почв. М.: Наука, 1975. 656 с. [6] Лисе О.Л. Закономерности
190
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
развития болотных систем в голоцене и их рациональное использование (на
примере Западной Сибири). Автореф. дисс. докт. геогр. наук. М, 1990. 20 с.
ХИМИЧЕСКИЙ СОСТАВ, ВЫНОС МИНЕРАЛЬНЫХ И ОРГАНИЧЕСКИХ
КОМПОНЕНТОВ ВОДАМИ ОЛИГОТРОФНЫХ БОЛОТНЫХ МАССИВОВ
И.Л. Калюжный
Государственный гидрологический институт, Санкт-Петербург
Гидрохимический режим, химический состав органических и
минеральных компонентов, а также вынос их водами олиготорфных болотных
массивов является слабо изученной областью гидрохимии болот.
Рассматриваются результаты наблюдений за химическим составом болотных вод,
проводимые на сети специализированных болотных станций Росгидромета.
Наблюдения выполнялись по единым методам отбора проб воды и
производства химических анализов. Исследования велись на олиготрофных болотных
массивах основных болотных провинций России и сопредельных стран.
Комплекс гидрологических, метеорологических, гидрофизических и
гидрохимических наблюдений позволяет рассмотреть формирование
химического состава болотных вод и вынос органических и минеральных
компонентов в неразрывной связи с элементами гидрологического и
метеорологического режима болотного массива.
Основным источником поступления минеральных компонентов в воды
олиготрофных болот являются атмосферные осадки. Сезонное содержание
ионов в болотной воде близко к их содержанию в атмосферных осадках.
Однако ионный состав осадков отличается от болотных вод количеством
хлора и высоким содержанием сульфатов. Воды верховых болот имеют
хорошо вьфаженный сульфатный характер. На болотном массиве Ламмин-Суо
содержание сульфатов в атмосферных осадках весной в 3 раза, летом в 2
раза, а в осенний период в 9 раз больше, чем в болотных водах. Содержание
хлора имеет обратную тенденцию: в осадках его в среднем в 8 раз меньше,
чем в болотных водах. Ионов кальция в болотных водах в 2,5 раза больше,
чем в осадках. Содержание ионов аммония и магния имеет переменную
тенденцию в различные месяцы года.
Сравнения показывают, что на различных болотных массивах
наблюдаются близкие значения содержания сульфатного иона, кальция и магния.
Концентрация ионов хлора на различных болотных массивах различается
весьма существенно. Так на болоте Тирели она в среднем в 3 раза больше,
нем на Ламмин-Суо и других болотных массивах.
Содержащиеся в осадках ионы в порядке убывания концентрации распо-
191
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
латаются в следующей последовательности: анионы - S042>Cr>N03",
катионы — Са2+>Ыа+>К+>М^+. Такая же последовательность сохраняется для
вод болотных массивов с небольшим изменением для катионов: Ca2+>Mg2+>
Na+>K+.
Результаты сравнения химического состава вод олиготрофных болотных
массивов, их общей минерализации с атмосферными осадками показывает,
что зональное распределение минерализации осадков обуславливает и
зональное распределение минерализации болотных вод.
Наибольшая минерализация вод болотных микроландшафтов (до 60
мг/л) наблюдается в предпаводочный период при наступлении наибольшей
глубины промерзания. В среднем при увеличении глубины промерзания на 1
см минерализация воды возрастает на 0,6-0,8 мг/л. Увеличение
минерализации объясняется тем, что при промерзании залежи происходит
концентрация ионов в воде за счет вытеснения их из замерзшего раствора.
Наименьшая минерализация болотных вод (5-12 мг/л) наблюдается в
период весеннего снеготаяния, когда талые слабоминерализованные снеговые
воды разбавляют болотные. В этот период коэффициент вариации общей
минерализации по площади болотного массива принимает наименьшее
значение, не более 7-8%.
В начальный период вегетации и до наступления летней межени
минерализация болотных вод увеличивается равномерно, в основном под влиянием
испарения. В среднем она возрастает на 0,04-0,05 мг/л при испарении 1 мм
слоя воды. Общая минерализация в меженный период составляет на
различных болотных массивах от 12 до 50 мг/л.
Средняя годовая минерализация вод центральной части болотных
массивов не одинакова в течение ряда лет и в значительной мере зависит от
количества зольных элементов, выпавших на поверхность болота. Связь
минерализации болотных вод центральной части массива с водами периферийных
микроландшафтов определяются простыми эмпирическими зависимостями
с коэффициентами корреляции не ниже 0,75.
Установлены пределы колебания важнейших ионов в водах
олиготрофных болот. Воды олиготрофных болот характеризуются большим
содержанием водорастворимого органического вещества, которое в своем составе
содержит органические кислоты. Поэтому между окисляемостью и
водородным показателем существует определенная зависимость: рН уменьшается с
увеличением окисляемое™.
Перманганатная и бихроматная окисляемость является косвенным
показателем органического вещества в болотных водах. Используя результаты
работ П.П. Воронкова, построена расчетная зависимость между
содержанием органического вещества и бихроматной окисляемостью. Имея наблюден-
192
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
ные среднемесячные величины бихроматной окисляемости и наблюденные
объемы стока болотных вод по дренирующим болото ручьям, рассчитывают
ежемесячные величины выноса органического вещества за пределы
болотного массива. Анализ результатов расчета показывает, что наименьший
вынос органики наблюдается в зимний период — 0,41 г/м2 за сезон. В
весенний периодом составляет 31,2% от годового. Наибольший вынос его
наблюдается в апреле месяце и в осенний период — 56% от годового. В летний
период вынос органики не превышает 10%.
Общий вынос растворенного органического вещества поверхностным
стоком за год с болотного массива Ламмин-Суо составляет 14,14 г/м2
грунтовыми водами —7,57 г/м2, в сумме — 21,71 г/м2. Дана оценка величин
выноса легко и тяжело растворимых фракций органического вещества.
Приведены результаты расчета баланса минеральных компонентов в
водах болотного массива Ламмин-Суо за конкретный год. Показано, что
приходная часть минеральных компонентов за счет выпадающих осадков
составляет 9,78 г/м2 в год. Поверхностными водами выносится 3,58 г/м2, ин-
фильтрационным путем — 1,40 г/м2. В целом вынос составляет 4,98 г/м2 за
год или 50,9% от приходной части. Основной вынос минеральных
компонентов происходит в весеннее половодье. Аккумуляция в болотном массиве
составляет 4,80 г/м2 за год. В конкретный год наблюдений болотный массив
задержал 49.1% поступающих минеральных компонентов.
Комплексные гидрологические, гидрофизические и гидрохимические
исследования на болотных массивах в системе Росгидромета позволили
детально исследовать формирование химического состава болотных вод,
оценить диапазон изменения различных его компонентов и объем выноса
минерального и органического вещества за пределы болотного массива.
АККУМУЛЯЦИЯ УГЛЕРОДА В БОЛОТАХ
ЛЕНИНГРАДСКОЙ ОБЛАСТИ
К.И. Кобак, Н.Ю. Кондрашева, И.Е. Турчинович
Государственный гидрологический институт, Санкт-Петербург
Оценки площадей болот Ленинградской области, в последние годы
независимо выполненные двумя коллективами научных сотрудников
(Ботанический институт РАН и Государственный гидрологический институт),
различаются несущественно. Они свидетельствуют, что эта площадь составляет
1017 тыс. га, а площадь заболоченных земель — 434,8 тыс.га.
Заболоченность области — 12%, а с учетом заболоченных земель, площадь которых
определена с меньшей точностью, — 17%. В общем заболоченность области
193
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
значительно больше средней заболоченности России (8%). В данной работе
в основу классификации положены типы болотных местообитаний,
отражающие в основном различия в минеральном питании (олиготрофные, мезо-
и евтрофные). Верховые болота абсолютно доминируют, занимая более 75%
площади всех болот, на долю мезотрофных приходится 15%, а евтрофных
— 10%.
Запасы торфа составляют 2103,1 млн. т абсолютно сухой субстанции и в
основном сосредоточены в верховых болотах (75% общего запаса торфа
области), на долю мезотрофных болот приходится 14% , а евтрофных — 11%.
Общее содержание углерода оценено как 1150,4 млн. т. Распределение
содержания углерода в разных типах болотных местообитаний
приблизительно такое же. как и запасов торфа — 72,8% в олиготрофных, 15% в
мезотрофных и 12,2% в евтрофных. Некоторые расхождения в распределении
запасов торфа и углерода объясняются разной зольностью и концентрацией
углерода в торфах разных типов. Среднее содержание углерода составляет
1112.6 тС/га. что в общем соответствует среднему содержанию углерода в
болотах европейской части России — 1048,3 тС/га и больше среднего
содержания углерода в болотах всей России (871.6 тС/га).
На болотах Ленинградской области запасы фитомассы составляют от 8,3
т/га (грядово-мочажинные болота) до 21,4 т/га абсолютно сухого вещества
на облесенных верховых болотах. Пул фитомассы оценен как 6,77 млн. т С
(при 50%-ном содержании углерода в фитомассе), что значительно меньше
содержания углерода в торфе (0,6% от углеродного пула торфа).
Средняя за весь голоцен скорость нетто-аккумуляции углерода в торфе
была определена на основании данных о средней скорости нарастания
торфяной залежи в болотах разных типов, плотности торфа и данных
радиоуглеродного датирования возраста торфа в почвенном профиле. В болотах
всей Ленинградской области значение средней многолетней
нетто-аккумуляции углерода составляет 371 тыс. т углерода в год (0,359 тС/га в год), т.е.
всего 0,8% от общей нетто-аккумуляции углерода болотами России.
Скорость этой нетто-аккумуляции в области несколько выше, чем в среднем по
России (0,286 тС/га в год). Большая часть углерода депонируется в
верховых болотах (67,9%), в мезотрофных — 25,9%, а в евтрофных всего 6,2%.
Современное и предстоящее глобальное потепление климата может
оказать заметное влияние на болотные экосистемы. В северо-западном регионе
России возможно прогрессивное увеличение площади болот и нетто-
аккумуляции в них углерода, так как прогнозируется и рост температуры
воздуха, и увеличение количества осадков.
194
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
РОЛЬ БОЛОТНЫХ ЭКОСИСТЕМ В ДИНАМИКЕ ЛАНДШАФТОВ
УРБАНИЗИРОВАННЫХ РАЙОНОВ ПРИАНГАРЬЯ
Т.И. Коновалова, И.Б. Воробьева
Институт географии СО РАН, Иркутск
Регион, расположенный у берегов Байкала, отличается разнообразными
и уникальными природными комплексами, которые подвергаются в
большинстве своем активному антропогенному воздействию. Особенно
существенное влияние оказывают крупные промышленные центры региона —
города Иркутск, Ангарск, Усолье, Братск и т.д., промышленность и
коммунальное хозяйство которых перешли допустимые пределы по степени
антропогенной нагрузки на природу и человека. В связи с этим весьма актуально
решение задач оценки состояния окружающей среды, степени воздействия
источников вредных выбросов, устойчивости ландшафтов. Устойчивость
при этом можно рассматривать как особый природный ресурс, поскольку
ассимиляционная способность природной среды по отношению к выбросам
вещества и энергии в результате антропогенной деятельности может
служить одним из критериев устойчивого управления природопользованием [1].
Большинство крупнейших промышленных центров области расположено
в пределах подгорного подтаежного класса геомов, отличительной чертой
которого является сочетание гумидных и полуаридных систем. С ареалами
подтаежных ландшафтов «связаны основные проблемные ситуации в
бассейне оз. Байкал, влияющие на такие системообразующие отношения при
взаимодействиях, как естественное «загрязнение», пониженная
самоочищающая способность водных объектов, регуляция типа функционирования
таежных систем при антропогенных воздействиях» [2].
В пределах Приангарья отмечается изменение теплового режима
ландшафтов, связанное как с многолетней динамикой климата, так и с
антропогенным воздействием на отдельных участках, которое является критическим
фактором в динамике ландшафтов внутриконтинентальных секторов
умеренного пояса в целом и региона в частности. Отмечена тенденция к замене
подтаежных светлохвойных комплексов на более теплолюбивые и ксеро-
фитные. Эти выводы подтверждаются точкой зрения [3] об антропогенной
природе возникновения степей региона. Стабилизирующим фактором в
динамике геосистем Приангарья является наличие болот и заболоченных лесов
и лугов, которые воздействуют на метеоэнергетику и геохимию ландшафтов
региона. В условиях значительной засушливости климата и почв
возобновление растительности, которая является мобильным звеном в динамике
геосистем, стабильно в условиях оптимального сочетания тепла и влаги, т.е. в
195
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
условиях гумидного климата, который характерен для районов развития
гидроморфных ландшафтов.
В пределах района, в основном, развиты евтрофные и мезотрофные
болота, в силу климатических и геологических особенностей территории. Это
подгорно-долинные лугово-болотные гидроаккумулятивные комплексы
предгорной части Восточного Саяна, представленные кустарничково-осоко-
во-гипновыми болотами внутренних дельт, которые являются
экстраобластными, а значит и уникальными для региона, а также долинные
заболоченные кустарничково-осоково-моховые комплексы в сочетании с осоковыми
болотами и заболоченными лесами. Все болотные группы фаций относятся к
системе факторально-серийных, наименее устойчивых в системе фактораль-
но-динамических рядов ландшафтов. Колебания природных режимов и
сопутствующие катастрофическим разрушениям трансформации геосистем
наиболее выражены в серийных фациях, развитие которых определяется
усиленным проявлением различных процессов, таких как запасы почвенной
влаги, физико-механические свойства и трофность почвенного субстрата для
гидроморфного динамического ряда фаций [3].
С точки зрения [1] гидроморфные фации не имеют собственных
механизмов устойчивости, поскольку их состояние в наибольшей степени
зависит от внешних факторов, а отчасти — от других компонентов ландшафта.
Так, например, влагооборот содействует усилению как внутреннего оборота
вещества и энергии в ландшафте (укрепление устойчивости), так и внешних
потоков, которые приводят к потере субстанции и дестабилизации
ландшафта. Соотношение этих двух противоположных функций колеблется в весьма
широком диапазоне в зависимости от структуры ландшафта. Проведенные
исследования структуры и динамики всех типов ландшафтов региона
показали, что в условиях значительного техногенного воздействия на
экосистемы происходит снижение продуктивности видовых популяций, перестройка
структуры фитоценозов, уменьшение типологического разнообразия,
изменение гидротермического и геохимического режимов. Переменные
структуры, вызванные длительным техногенным воздействием городов, могут быть
отнесены к разряду устойчиво длительно производных, так как преобладают
процессы преобразования ландшафта. Преобразования ландшафтов под
воздействием крупных промышленных центров региона, по нашим
наблюдениям, прослеживаются в большинстве случаев на топологическом уровне -
- возникает новая интеграция природных режимов с формированием
своеобразного ландшафтообразующего эффекта, снижается роль саморегуляции
ландшафтов. Геосистемы с нарушенной структурой делятся на относительно
сохранившие свои спонтанные потенции и способные воспроизвести
первоначальную структуру за счет факторов саморегуляции топологического по-
196
БИОСФЕРНАЯ ЮЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ ГУ
рядка и коренным образом изменившие свою структуру, восстановление
которой допустимо в очень длительный срок под воздействием планетарно-
региональных движущих сил. Последняя категория геосистем характерна
для промышленных центров Приангарья, первая — их окружения.
Не смотря на это, гидроморфные ландшафты выполняют большую сре-
дообразуюшую функцию в регионе. С одной стороны, они регулируют
водно-тепловой баланс территории, с другой стороны — они служат
геохимическими барьерами.
Геохимически подчиненные супераквальные ландшафты низинных
болот, питаемые делювиальными и грунтовыми водами, выступают как глее-
вые, глеевые карбонатные, сорбционные, а в субаридных областях и как
испарительные и сероводородные барьеры для многих макро- и
микроэлементов. Микроэлементный состав низинных болот контролируется
гидрохимическим стоком из окружающих ландшафтов, глубиной залегания и
составом дочетвертичных пород. В низинных болотах кларки концентрации
почти всех элементов >1 и достигают максимальных величин для Мп (25,8),
Ti (35,0), Mo (17,0), Ni (14), Со (10), что свидетельствует об их аллохтонном
происхождении.
Были проведены исследования снежного и почвенного покрова осоково-
злакового заболоченного луга, расположенного в зоне техногенного
воздействия крупного промышленного центра — г. Иркутска. Изучены следующие
элементы — Си, Pb, Sr, Со, V, Сг, Mn, Ni, Ti. Исследования показали, что
количество тяжелых металлов, выявленных в твердой фазе снега, было
выше, чем в почве. Это объясняется тем, что при таянии снега не все вещества
попадают в почву, часть их выносится с талыми водами. В болотных почвах
аккумулируется больше свинца, чем в снеге, так как среди тяжелых
металлов РЬ наименее подвижен и при таянии снега все его количество оседает в
почве. Снеговые воды имеют слабощелочную реакцию среды (7,85 значений
рН). В заболоченном луге кларки концентраций элементов в твердой фазе
снега выше, чем в почве, кроме показателя свинца и составляют в твердой
фазе снега: РЬ — 4-6; Си — 2-3; Sr — 1-1,2; Сг — 1,2-1,8; Ni — 0,7-1,4; в
почве: РЬ — 6-9; Си — 0,8-1,5; Sr — 0,6-1,2; Сг — 0,8-1,3; Ni — 0,6-1,2.
Таким образом, на основании полученных данных можно сделать вывод,
что заболоченные и болотные экосистемы, являются геохимическими
барьерами, выполняют существенную средообразующую функцию в условиях
региона, но вместе с тем наименее устойчивы, что требует ограничения
природопользования с учетом факторов их дестабилизации .
[1 ] Исаченко А.Г. Широтная зональность и механизмы устоййчивости ландшафтов к
антропогенным воздействиям // Изв. РГО. 1997. Т. 129. Вып.З. [2] Михеев B.C.
197
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Ландшафтно-географическое обеспечение комплексных проблем Сибири.
Новосибирск: Наука, 1987. [3] Сочава В.Б. Географические аспекты сибирской тайги.
Новосибирск: Наука, Сиб отд., 1980. [4] Крауклис А.А. Проблемы экспериментального
ландшафтоведения. Новосибирск: Наука, 1978.
РЕЖИМ ПОЧВЕННО-ГРУНТОВЫХ ВОД ОСУШЕННЫХ
МЕЛКИХ ТОРФЯНИКОВ
В.П. Косарев, А.И. Тимофеев
Санкт-Петербургская лесотехническая академия
Приводятся некоторые результаты многолетних (1974-1994 гт.)
исследований режима почвенно-грунтовых вод (111 В) на мелких торфяниках поч-
венно-гидромелиоративного стационара кафедры почвоведения и
гидромелиорации СПб ЛТА «Малиновский». К таким торфяникам мы условно
относим болота с глубиной торфа менее глубины осушительных каналов.
Действие осушительной сети в таких условиях существенно отличается от
глубоких торфяников. В частности, при расстояниях между каналами более 90 м
кривые депрессии на межканальных полосах не формируются, а образуются
только кривые спада непосредственно у каналов, основная масса стока
поступает в каналы по верхнему хорошо водопроницаемому слою торфа и по
контакту его с минеральным слабоводопроницаемым грунтом.
Стационар заложен на территории Лисинского учебно-опытного лесхоза
ЛТА в 1973 (в год осушения). Подробная характеристика опытных объектов
стационара приведена нами в ранее опубликованных работах [Бабиков и
др., 1996]. поэтому ниже приводятся только основные сведения. Опытные
участки (ОУ) I и II заложены на переходном болоте, а ОУIII и IV — на
верховом болоте. Мощность торфа на ОУ I составляет 20-30 см, на остальных
участках она колеблется в пределах от 40 до 80 см. На всех ОУ торф
подстилается метаморфизированной практически водонепроницаемой
ленточной глиной. Расстояния между осушителями на ОУ I и IV около 90 м, на ОУ
И — 110 м и на ОУ III — 160 м. Проектная глубина осушителей около 1 м,
к настоящему времени глубины каналов уменьшились и колеблются в
пределах от 0,6 до 0,8 м. За каналами осуществлялся периодический уход.
На год осушения древостой на ОУ I был представлен редкостойной
средневозрастной сосной (полнота 0,1-0,2) со вторым ярусом из молодняка
березы (полнота 0,4-0,5); на ОУ II — приспевающей сосной (полнота 0,6-0,7) с
небольшим участием молодой березы во втором ярусе; на ОУ III и IV —
низкополнотной средневозрастной сосной с небольшой долей в составе
молодой сосны (полнота 0,1-0,3). Сразу же с первого года после осушения рост
древостоев на всех ОУ резко улучшился: текущий бонитет к 1994 году по-
198
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ ГУ
высился на ОУ I и II до I — 1а, на ОУ III и IV — до II — III, а на
приканальных ПП на ОУ III — до I и 1-Й класса; радиальный прирост у сосны
увеличился в 2-3,5 раза; многократно возросли средние приросты в высоту
(до 30-48 см в год); на 0,2-0,4 возросла полнота древостоев.
Характеристики многолетнего режима ПГВ на опытных участках
Глубины ПГВ по опытным участкам
Годы ОУ1 ОУП ОУШ ОУГУ
Ср.ПП Пк.ПП Ср.ПП Пк.ПП Ср.ПП Пк.ПП Ср.ПП Пк.ПП
Средние глубины ПГВ за V-ГХ, см
1974-94 49,5 46,6 38,8 38,4 27,3 29,0 38,1 42,4
1974-83 42,2 37,1 30,8 29,6 22,4 22,0 32,1 36,5
1984-94 56,1 55,3 46,0 46,5 31,7 35,5 43,6 47,8
Глубины ПГВ на средину мая, см
1974-94 32,0 30,8 19,9 23,0 16,6 17,3 28,6 34,1
1974-83 25,9 22,4 14,5 14,2 15,1 15,2 24,5 29,1
1984-94 37,5 38,5 24,9 31,0 17,9 19,1 32,4 38,6
Максимальные значения средних глубин ПГВ за V-ГХ, см
1974-94 91,0 91,7 71,7 70,2 61,3 64,3 62,7 69,7
Минимальные значения средних глубин ПГВ за V-ГХ, см
1974-94 12,3 14,8 8,1 9,5 2,9 1,9 15,1 15,8
Максимальные значения глубин ПГВ на середину мая, см
1974-94 68,7 59,5 53,0 59,2 33,4 41,3 58,6 63,6
Минимальные значения глубин ПГВ на середину мая, см
1974-94 9,8 15,6 6,0 11,6 1,0 1,7 14,2 16,2
Обеспеченность средних глубин ПГВ за V-EX 40 см, %
1974-94 59,3 54,7 40,7 40,7 22,0 26,6 40,7 50,0
1974-83 45,2 35,6 16,3 16,3 6,7 0,0 6,7 26,0
1984-94 67,5 67,5 58,8 58,8 32,5 50,0 67,5 67,5
Обеспеченность глубин ПГВ на середину мая 20 см, %
1974-94 78,0 78,0 31,3 36,0 26,6 26,6 78,0 87,4
1974-83 74,0 64,4 0,0 0,0 6,7 6,7 83,7 83,7
1984-94 76,3 85,1 58,8 67,5 41,2 41,2 67,5 85,1
Ср.ПП — срединная пробная площадь, Пк.ПП — средние по двум приканальным
пробным площадям.
Из таблицы видно, что глубины ПГВ на пробных площадях (ПП) сильно
варьируют годам в зависимости от погодных условий их с колебаниями в
среднем за период вегетации от 2 до 92 см, на средину мая — от 1 до 69 см.
Среднемноголетние глубины ПГВ за период вегетации изменяются по
разным ОУ в пределах от 27 до 50 см, глубины на средину мая от 17 до 34
см. Максимальные значения их наблюдаются на ОУ I и IV при расстояниях
межцу каналами 90 м и лучше развитых древостоях, минимальные — на ОУ
199
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
III при наибольшем расстоянии (более 150 м) и низкополнотном древостое.
Разница глубин при этом на срединных ПП достигает 11-22 см. Среднемно-
голетние глубины на средину мая на ОУ I и IV на всех ПП близки (29-34
см) и существенно больше глубин на ОУ II и III (17-23 см), где расстояния
между каналами 110 и 160 м.
Известно, что на осушенных торфяниках для обеспечения
удовлетворительного водно-воздушного режима необходимо понизить уровни ПГВ не
менее 20 см на начало периода вегетации (средина мая) и не менее 40 см в
среднем за период вегетации. Такие значения среднемноголетних глубин за
период вегетации наблюдаются у нас только на интенсивно осушенных ПП
(на ОУ I и ОУ 1У).Глубины ПГВ более 40 см даже на интенсивно
осушенных ОУ I и IV во многие годы не достигаются. Эмпирическая
обеспеченность такой глубины изменяется от 22-27% на ОУ III до 50-59% на ОУ I.
Даже на тех ПП, где текущий класс бонитета в настоящее время I — 1а,
средняя за период вегетации глубина ПГВ более 40 см наблюдается не чаще
чем в 60% лет. На средину мая по среднемноголетним данным понижение
ПГВ на глубину 20 и более см достигается на всех ПП (глубины изменяются
от 19,9 до 34,1 см), за исключением ОУ III. Обеспеченность такого
понижения за 21 год наблюдений на ОУ I и IV более 78 %, на ОУ II — 31-37%, а на
ОУ III — всего лишь 27%.
При сравнении средних глубин ПГВ за десятилетие (1974-1983 гг.) и за
последующие 11 лет (1984-1994 гг.) отчетливо выявляется усиливающееся
влияние на них улучшивших рост и состояние насаждений. По средним
метеорологическим данным эти периоды существенно не различаются. На ОУ
I, II и IV средние за 11-летние глубины ПГВ за V-IX увеличились на 11-18
см и на всех ПП превысили 40 см, а глубины на средину мая возросли на 8-
17 см, превысив на всех ПП 20 см. Если в первом десятилетии на ОУ II и IV
средние глубины ПГВ за период вегетации больше 40 см отмечались 1-3
года, а на ОУ I — 4-5 лет, то во второй, одиннадцатилетний период такие
глубины наблюдались на ОУ II в 7, а на ОУ I и IV в 8 годах.
Результаты показывают, что на мелких торфяниках по сравнению с
глубокими в начальном периоде осушения требуется несколько меньшее
понижение уровней ПГВ. Так, в первом десятилетии после осушения рост
насаждений по I классу бонитета на ПП стационара происходил при
среднемноголетних глубинах ПГВ за период вегетации соответственно 22-31 см и при
глубинах на средину мая около 15 см. По-видимому, глубины 30 см в
среднем за период вегетации и 15 см на средину мая с обеспеченностью их в
многолетнем ряду не менее 50% можно принять в качестве нормы осушения
для мелких торфяников. Расстояния между каналами более 90 м в подобных
условиях не обеспечивают требуемого понижения уровней ПГВ.
200
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
ВЛИЯНИЕ АЗОТНЫХ СОЕДИНЕНИЙ НА ОКИСЛЕНИЕ МЕТАНА В
ВЕРХОВОМ БОЛОТЕ, ТВЕРСКАЯ ОБЛАСТЬ
И.К. Кравченко, Е.В. Токарева
Институт микробиологии РАН, Москва
Одна из важнейших задач современной экологии состоит в объяснении и
прогнозировании климатических изменений, связанных с увеличением
содержания в атмосфере так называемых парниковых газов, которые
определяют современное глобальное потепление климата Земли. В последние годы
пристальное внимание исследователей привлекают переувлажненные почвы
северных широт, которые содержат до 20-30% мировых запасов углерода и
являются важнейшим источником поступления парниковых газов (в первую
очередь СОг и СН* в атмосферу.
Поступление в атмосферу метана, образующегося в болотных почвах,
регулируется деятельностью так называемого «метанового фильтра».
Микробное окисление метана является существенной частью его глобального
бюджета и составляет в почвах бореальных болот около 20% от его продукции.
В то же время, именно деятельность микробного блока в этих процессах и
регулирующая роль факторов внешней среды остается наименее изученной.
В настоящее время установлено, что снижение активности метанокисления в
аэробных почвах связано с поступлением в них азотных соединений в виде
удобрений и/или с атмосферными осадками. Эти закономерности были
подтверждены нами для болотных почвах Западной Сибири [1]. Установлено,
что в болотные почвы Западно-Двинского стационара поступает
значительное количество минеральных, в том числе азотных, соединений [2], что
может привести к нарушению деятельности «метанового фильтра».
Цель работы состояла в оценке влияния внесенных азотных соединений
на окисление метана в образцах торфяной почвы в длительных (в течение
года) инкубационных лабораторных экспериментах. Объектом
экспериментальных исследований были образцы торфяной почвы верхового болота
«Сосвятское» Западнодвинского стационара Института лесоведения РАН из
верхнего (0-20 см) слоя торфяной залежи. В течение всего периода
наблюдений оценивали в динамике потенциальную метанокисляющую активность,
дыхательную активность и изменения рН, азотных (NH/, NO2"' NO3") и
углеродных (РОВ, жирные кислоты) соединений.
Исследования подробной динамики потенциальной метанокисляющей
активности в образцах торфа выявили ее снижение в вариантах опыта с
внесением минеральных азотных соединений различной степени окисленности
в концентрациях, превышающих 0,1 MrN/r влажного торфа. Наибольшим
ингибирующим эффектом обладали соли аммония, что согласуется с резуль-
201
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
татами полученными ранее для чистых культур метанотрофов и автоморф-
ных почв.
100
4 Д П
200 300
сутки
400
400
7,00
5,00
3,00
100
200
сутки
300
400
- контроль
-нитрит
-аммонии
-нитрат
Изменения метанокисляющей (сверху слева), дыхательной (образование СОг)
активности (сверху справа), а также рН (снизу слева) и содержания аммония (снизу
справа) в образцах торфяной почвы, инкубировавшейся без добавок (контроль) и с
внесением азотных соединений в количестве 1 мг N/ г вл. торфа.
Эффект ингибирования сохранялся в течение длительного (около года)
периода после внесения азотных соединений. В течение всего периода
наблюдений (с марта 1998 по февраль 1999 г.) величина метанокисляющей
активности в образцах с внесением была достоверна ниже, чем в
контрольном (без добавок) варианте (см. рис.), но степень ингибирования снижалась.
Высказаны предположения о возможных причинах длительного ингиби-
рующего воздействия азотных соединений на окисление метана и проведена
их экспериментальная проверка.
1. Внесение азотных соединений приводит к необратимому сдвигу
ионного и кислотного равновесия, что приводит к нарушению
микробиологической активности. Экспериментальные данные 'указывают на то, что не
наблюдается достоверного снижения рН, а в ряде случаев (внесение нитрита)
даже имеет место его увеличение. Через месяц скорость образования С02,
интегральный показатель активности микробного сообщества торфа, была
202
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
на уровне контроля или несколько превышала ее.
2. Снижение метанокисляющей активности связано с конкурентным ин-
гибированием метанмонооксигеназы аммонием. Внесенные соединения
аммония в течение достаточно длительного (около 5 месяцев) периода
сохранялись в торфе на достаточно высоком уровне. Кроме того,
зарегистрировано образование высоких количеств аммония в вариантах с внесением
нитрита и нитрата. Это может быть результатом ассимиляции и реминерализации
азота в форме NH/ микробной биомассой. В пользу этого предположения
свидетельствует динамика потребления внесенных окисленных азотных
соединений.
3. Внесение азотных соединений приводит к образованию продуктов
микробного метаболизма, которые ингибируют активность метанокисляю-
щего сообщества. В вариантах с внесением нитрита и нитрата отмечено
возрастание по сравнению с контролем количества водорастворимых углерод-
содержащих соединений. Увеличивается количество алифатических жирных
кислот ацетата и формиата. Возможной причиной ингибирования при
внесении нитрита и нитрата может быть образование в кислых условиях среды
газообразных окислов азота за счет хемоденитрификации, а также за счет
процессов нитрификации и денитрификации. Это предположение требует
экспериментального подтверждения.
4. Изменения в активности окисления метана связаны с перестройками в
структуре метаноксиляющего сообщества торфяной почвы (изменение
соотношения активных и покоящихся форм в пользу последних, а также
сукцессия в ходе которого меняется и количество и/или таксономический состав
метанокисляющего сообщества). Это предположение пока не получило
экспериментального подтверждения для изучаемых почв, но основано на
полученных нами результатах изучения торфяных почв Западной Сибири и
выделенных из них метанокисляющих ассоциаций.
Исследования показали, что даже однократное поступление относительно
небольших количеств азотных соединений в верхние слои торфяной почвы
может привести к нарушению процесса потребления метана в течение
длительного периода времени. Механизм этого явления сложен и требует
дальнейших исследований. Ингибирование метанокисления связано с
комплексным воздействием внесенных соединений и вклад того или иного фактора
меняется в зависимости от формы, концентрации соединения и времени
прошедшего после его внесения.
[1] Кравченко И.К. 1999 Влияние азотных соединений на окисление метана в
верховом сфагновом болоте Западной Сибири // Микробиология. Т.68. № 2. С.254-258.
[2] Глухова Т.В. 1995. Влияние атмосферных осадков и пыли на питание болот // Ж.
экологической химии. № 4. С.282-287.
203
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ЗАПАСЫ УГЛЕРОДА В ТОРФЯНЫХ ЗАЛЕЖАХ ТОМСКОЙ ОБЛАСТИ
В.Н. Крештапова, П.И. Хорошев
Почвенный институт им, В.В. Докучаева, Москва
Проблема подсчета содержания и баланса углерода в различных
экосистемах становится все более актуальной в связи с распушим беспокойством
о возможности глобального изменения климата Земли, связанного с ростом
концентрации парниковых газов в атмосфере и, в первую очередь, углерода.
Педосфера планеты является крупнейшим резервуаром углерода
наземных экосистем. Очень важную роль в этой экосистеме играют торфяные
залежи, которые в естественном состоянии выполняют аккумулирующую
функцию, депонируя углерод атмосферы в виде торфа, тем самым выключая
его из «малого» круговорота. И наоборот, в случае добычи и использования
торфа, весь накопленный за 10-12 тыс. лет углерод тем или иным путем
попадает в атмосферу. Площадь торфяных месторождений в России составляет
56,8 млн. га, запасы торфа — 186 млрд. тонн (табл. 1).
Согласно [1] запасы углерода в торфяных болотах России оцениваются
величиной 101 млрд. т. По данным других авторов [Ефремов, 1994; Botch et
al. 1995: Бирюкова, Орлов, 1996] эта величина составляет 116,5-210 млрд.
т. Два основных фактора влияют на результат подсчета запасов углерода : 1)
оценка запасов торфа в торфяной залежи и 2) методика пересчета из
объемов торфяной залежи величину запаса углерода в ней. Не вдаваясь в анализ
точности результатов работ приводимых выше авторов, применяемых
методик подсчета запасов углерода, исходных материалов, используемых для
этих целей и т.п., авторы предлагают свой вариант подсчета запасов
углерода в торфяных залежах. Он состоит в том, что подсчет ведется по каждому
объекту — в данном случае по каждому торфяному месторождению. Ниже
приводятся сведения о первом этапе работы — подсчете запасов углерода в
торфяных залежах Томской области.
Табл. 1. Распределение выявленных и разведанных торфяных запасов в России [2]
Показатель Всего Европейская Запад, и Вост. Дальний
часть России Сибирь Восток
Количество месторождении
Площадь в границах торфяной
залежи 0,7 м, млн га
Верховой тип. млрд.т.
Смешанный, млрд.т.
Переходный, млрд.т..
Низинный, млрд.т.
Всего запасов, млрд.т.
44200
56,8
94,5
5,8
26,8
58,9
186,0
33900
10,2
18,7
1,2
5,3
13,2
38,4
5300
42,2
75,3
4,6
19,2
42,2
141,3
5000
4,4
0,5
-
2,3
3,5
6,3
204
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
Томская область — одна из наиболее заторфованных территорий
Российской Федерации. Заторфованность области (отношение площади
торфяных месторождений в границах промышленной залежи к общей площади
территории области) составляет более 25%. Специальные
геологоразведочные работы развернулись с 50-х годов, когда стал широко применяться ка-
мерально-экспедиционный метод выявления торфяных месторождений с
использованием материалов аэрофотосъемки, а позднее — аэрокосмической
съемки крупномасштабных карт в сочетании с аэровизуальными съемками и
последующим наземным обследованием [4]. В настоящее время все
торфяные месторождения Западной Сибири выявлены, а геологические запасы
торфа подсчитаны.
Торф представляет собой систему, состоящую из органической и
минеральной частей и воды. Органическая масса состоит из углерода, водорода,
серы, азота и кислорода. Содержание углерода в органической массе торфа
колеблется от 48,0 до 65,1%. В среднем по России для верхового типа
залежей содержание углерода составляет 56%, переходного 58,6%, низинного —
58%. Минимальное содержание отмечено у верхового фускум-торфа малой
степени разложения, максимальное характерно для верхового торфа
высокой степени разложения. По составу торфа, его агрохимическим и
техническим свойствам накоплен огромный статистический материал,
насчитывающий миллионы элементоопределений, который позволил провести
исследование связей между свойствами торфа.
Используя методы множественной корреляции, были выведены формулы
уравнения регрессии для определения элементного состава и других
технических характеристик торфа [3]. Уравнение множественной регрессии для
определения содержания углерода, предложенное этими авторами,
показывает, что С = f(R, A, Q, рН), где С — содержание углерода, % от
органического вещества торфа; R — степень разложения торфа, %; А — зольность
торфа, %; Q — теплота сгорания, ккал; pHkci — кислотность. Подставляя
значения R, A, Q, рН в эту формулу, определяем содержание углерода в
процентах от органического вещества на каждом торфяном месторождении
Томской области, которое затем пересчитываем на абсолютно сухое
вещество торфа. Общий список торфяных месторождений Томской области
составляет 990 объектов с запасами торфа 30500 млн. т., запасы углерода в
области составляют 9414 млн. т., в пересчете на абсолютно сухое вещество торфа.
В табл. 2 приводятся данные по запасам торфа и углерода в объектах
площадью более 40000 га, на которых сосредоточены основные запасы торфа в
области. Запасы торфа в торфяных месторождениях площадью до 500 га
составляют 47 млн. т, углерода — 14,5 млн. т. Запасы торфа в границах
торфяной залежи до 0,7 м составляют 305 млн. т, углерода 94 млн. т.
205
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Табл
. 2. Запасы торфа и
углерода в
площадью более 40000 га
№/
№'
!(Г~
28
57
79
227
268
288
380
397
405
433
449
477
480
520
523
546
547
555
565
574
599
650
653
866
Название
Киевское
Александровское-
Куликово
Шаровое-
Лым-Бель-
Карамовское
Компаское
Пульсецкое-
Ельцовское
Васюганское
Малое Васюганс.
Озерное Болыыое-
Кулай
Коноваловское-
Юголовское
Шубное-
Сазанье-
Каршаинское
Тайжольское
Комарное
Чур-Няр
Лягушка-
Лукьяново
Костяное-
Борковое-
Лотар
Площадь2,
га
50438
75023
65479
50466
42408
78884
52048
59443
2321
xlOOO
141717
583617
72004
197351
46484
102702
49828
98211
40987
432900
57780
112635
105059
59488
81225
51795
торфяных месторождениях Томской области,
Сред.
мощн.
торфа,
м
2,31
2,30
2,45
2,30
1,88
1,80
2,00
2,14
2,79
2,50
2,49
3,86
2,40
2,43
2,26
2,41
2,57
2,57
3,22
3,30
2,06
2,56
2,42
1,79
1,60
Запасы
торфа3,
млн. т
149135
219142
206574
147411
102050
186158
133243
179316
9082184
xlOOO
446408
1815812
364248
666197
149408
389483
178296
257450
142652
1677002
225434
329294
413446
234851
233328
118075
Запас4
с,
млн. т
45767
66514
63162
44742
31260
56920
41188
58739
277426
xlOOO
137994
583519
112734
214381
48969
126672
56912
83187
45694
508065
68297
96000
134465
76381
69459
36741
Тип
торфа5
"в
В
В,П
В
В
В,П
В
П,В
В,П,Н
В
В
П
Н
В,П
в,п,н
BJI,H
в,п,н
в,п
в,п
в,п,н
в,п
в,п
в,п
п,в
в,п,н
п,в,н
п,в,н
R,
%
26
23
25
23
24
26
30
25
20
21
12
23
25
-
-
-
30
20
10
15
14
14
21
30
30
27
30
~аГ*
%
"ТГ"
5,8
5,4
5,8
5,3
5,4
4,8
4,1
8,1
4,8
2,0
5,0
3,9
-
-
-
4,5
3,0
2,3
2,8
5,9
5,9
7,4
4,5
4,5
9,2
6,9
1 по карте торфяного фонда,2 в границе промышленной залежи (0,7 м),3 40%
влажности, 4 на абсолютно сухой вес. 5 В - верховой тип торфа, П - переходный, Н -
низинный
При продолжении исследований будут получены достоверные данные о
запасах углерода в торфяных залежах России. Высокая достоверность дан-
206
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
ных пообъектного подсчета будет характерна для подавляющей части
территории страны. Все геологические материалы по выявлению, оценке и
подсчету запасов торфа хранятся в Геологическом фонде Министерства
природных ресурсов РФ.
Работа проведена при финансовой поддержке NWO (Нидерланды) в
рамках проекта «Запасы углерода и атмосферный обмен в торфяных
залежах Западной Сибири в связи с процессами изменения климата», грант №
047-006-001.
И] Вомперский С.Э. и др. Заболоченные органогенные почвы и болота России и
запас углерода в их торфах // Почвоведение. 1994. № 12. С. 17-25. [2] Kosov V.I.,
V.N. Kreshtapova: The peat resources of Russia and their utilization // Global Peat
Resources, Ed. E. Lappalainen, UNESCO, 1996. С 127-131. [3] Лиштван Н.И., H.T.
Король. Основные свойства торфа и методы их определения. М.: Наука и техника,
1975. 320 с. [4] Логинов П.Е., Хорошев П.И. Торфяные ресурсы Западно-Сибирской
равнины. Москва, 1972. 148 с.
ХИМИЧЕСКИЙ СОСТАВ ТОРФЯНИКОВ И СНЕГА
ЮЖНОГО ПРИБАЙКАЛЬЯ КАК ПОКАЗАТЕЛЬ
ТЕХНОГЕННОГО ВОЗДЕЙСТВИЯ НА ОКРУЖАЮЩУЮ СРЕДУ
В.А. Кузьмин
Институт географии СО РАН, Иркутск
Торфяники являются сложными комплексными геохимическими
барьерами, способными удерживать влагу, газы и химические элементы. Это
свойство используется для выявления прошлого естественного
геохимического фона и интенсивности его изменения при техногенном загрязнении.
При исследовании верховых болот Русской равнины выявлено обогащение
верхних горизонтов свинцом, цинком, марганцем, кадмием, никелем и
хромом. Распределение свинца в торфяниках оказалось очень похожим на
показатели, отражающие нарастание этого элемента в разрезе ледникового щита
Гренландии. Различие состоит в неодинаковой концентрации его в торфе и
во льду. Судя по абсолютному возрасту слоев торфа, рост концентрации
свинца начался в нем около 1,5 тыс. лет назад. Загрязнение атмосферы
свинцом связывается с учащением лесных пожаров, вызываемых человеком
еще в неолите.
Указанные материалы послужили основанием для проведения подобных
исследований на юге Байкала, включенного в список объектов Всемирного
Наследия. Намечалось выявить характер и степень техногенного
химического воздействия на толщу торфяников, определить основные элементы, по-
207
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ступающие на подстилающую поверхность в составе аэрозолей локального,
регионального или глобального переноса.
В сентябре 1997 г. в пяти точках были отобраны пробы торфа на отрезке
протяженностью около 100 км от низовий р. Мишихи до г. Байкальска.
Пробы отбирались по глубине в зависимости от степени разложения торфа:
живой мох, отмерший, слабой (10-20%) и средней (30-40%) степени
разложения. Максимальная глубина взятых проб 2 м. После высушивания
образцов в них, а также в растениях определяли зольность рН; в пробах, озолен-
ных при 450° на спектрографе ДФС-8 выявлены 13 макро- и
микроэлементов. Средняя зольность торфа, представленного в основном сфагнумом
близка к 5%. Реакция торфяников кислая, рН изменяется в пределах 3,5-5,0.
Распределение свинца в профиле определяется степенью разложения торфа.
В очесе и отмершем мхе его в 3-4 раза больше чем в среднеразложившемся
торфе. Причем максимум уровня концентрации свинца чаще приходится не
на живой, а на отмерший неразложившийся мох. При разложении торфа
свинец как элемент, связывающийся с органическим веществом, выносится.
В живом мхе более всего аккумулируется марганец, коэффициент
концентрации которого (Кк) = 7-12. Даже при слабой степени разложения (10-
20%) его концентрация резко снижается, что может служить показателем
миграции элемента в двухвалентной форме в кислой восстановительной
среде. Кроме перечисленных элементов в живом мхе выявлено накопление
магния и меди с Кк 2-3. К элементам слабого захвата с Кк менее 1 относятся
железо никель кобальт хром и ванадий.
Полученные данные о химическом составе верховых торфяников
Южного Прибайкалья, удаленных от промышленных центров, не свидетельствуют
о влиянии техногенеза на их состав, как это выявлено на Русской равнине.
На территории, прилегающей к г. Байкальску, в живом мхе концентрация
почти всех исследованных элементов возросла, что можно объяснить
загрязнением выбросами Байкальского целлюлозно-бумажного комбината
(БЦБК).
Растения на торфяниках по сравнению со сфагнумом обеднены
химическими элементами. В широко распространенных здесь багульнике, голубике,
кассандре, морошке и хвое кедра концентрация большинства элементов
ниже чем в сфагнуме.
Чтобы выяснить поступление химических элементов из атмосферы за
зимний период в марте 1998 г. на этих же участках были отобраны пробы
снега и проведен гидрохимический анализ, а спектральным методом в
твердой и растворимой фазе определены те же химические элементы, что и в
торфе. рН снеговых вод в четырех исследованных пунктах колеблется в
пределах 4,8-5,7, что близко к фоновым показателям, а в окрестностях
208
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
г.Байкальска (вблизи его западной окраины, в 5 км от БЦБК) достигает 6,2.
Минерализация снега в местах удаленных от промышленных центров,
составляет 7-11 мг/л, а вблизи Байкальска — вдвое больше. Твердая фаза
снега (или труднорастворимые частицы) изменяется соответственно от 13-80
мг/л. По минерализации и составу снеговые воды близки к данным осадков
на. озерных и прибрежных станциях Байкала.
В растворимой фазе находятся преимущественно кальций и стронций, а
другие элементы — в твердой, где доля большинства элементов в 4-10 раз
выше. Различие в химическом составе снега вблизи БЦБК и на расстоянии
от него состоит в относительном обогащении снега около комбината
кальцием, стронцием и обеднении никелем, хромом и свинцом. Содержание
большинства элементов в снеге вблизи Байкальска по сравнению с пробами,
отобранными вдали от промышленных центров, выше в 2-5 раз. Различия в
химическом составе снега наиболее четко выявляется по соотношению
элементов в твердой и растворимой фазах. Если в пробах снега, взятых в
удаленных от Байкальска местах, концентрация их в твердой фазе по
сравнению с растворимой изменяется от 0,5 до 12, то вблизи Байкальска это
соотношение для ряда элементов увеличивается до 40-47. Можно полагать, что
элементы с высоким соотношением концентраций содержаться в выбросах
промышленного производства, т.е. обогащение элементами твердой фазы
является показателем антропогенного воздействия.
ОСОБЕННОСТИ ФОРМИРОВАНИЯ ОРГАНИЧЕСКОГО ВЕЩЕСТВА В
БОЛОТНО-ПОДЗОЛИСТЫХ ПОЧВАХ
Е.М. Лаптева, Г.Я. Елькина, Е.В. Шамрикова, Ж.Н. Козачок
Институт биологии Коми НЦ УрО РАН, Сыктывкар
В среднетаежной зоне Республики Коми широко распространены болот-
но-подзолистые почвы (БПп), которые в комплексе с подзолистыми (Пп)
образуют почвенный покров водоразделов. Болотно-подзолистые почвы
формируются в микропонижениях (торфянисто-подзолисто-глееватые П61)
и западинах (торфяно-подзолисто-глеевые П62) в условиях избыточного
увлажнения под долгомошными и сфагново-дол-гомошными лесами.
Морфологически БПп отличаются от Пп наличием мощной двух-,
трехслойной оторфованной подстилки, более значительным, чем в автоморфных
подзолистых почвах потечно-гумусовым горизонтом A2h, уменьшением
мощности элювиального горизонта и существенным оглеением профиля.
Эти особенности морфологического строения профиля П61 и П62
обусловлены застойно-инфлюкционным и застойным водным режимом почв за счет
209
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
латерального, поверхностного и внутрипочвенного стока атмосферной влаги
с более высоких элементов рельефа, где залегают подзолистые почвы.
Застойный режим увлажнения обусловил более низкий уровень
окислительно-восстановительного потенциала (ОВП) в П61 и П62, особенно ярко
выраженный во влажные годы, благодаря сохранению верховодки в течение
всего вегетационного периода. В такие годы в потечно-гумусовом горизонте,
ниже уровня верховодки, были зафиксированы восстановительные (390 мВ)
значения ОВП при средней слабоокислительной обстановке (430-440 мВ).
Резкое снижение ОВП в этих горизонтах отмечалось в теплое время года
при возрастании микробиологической активности. Однако торфяные
органогенные горизонты БПп даже в условиях избытка влаги, благодаря их
хорошей водо- и воздухопроницаемости характеризовались интенсивно-
окислительной обстановкой. В целом уровень ОВП в БПп был более низким,
чем в автоморфной Пп. В тоже время при длительной «сухой» погоде
окислительно-восстановительные условия в них приближались к Пп.
С характером увлажнения и окислительно-восстановительными
условиями тесно связаны жизнедеятельность микробиоты, протекание процессов
минерализации, гумификации, торфообразования и торфонакопления.
Исследования Ф.М. Хабибуллиной показали, что в почвах болотно-подзолис-
того ряда (Пп — П61 — П62) по мере усиления степени гидроморфизма
наблюдается резкое снижение доли несовершенных грибов. Уменьшение
численности и видового состава микромицетов, принимающих активное
участие в деструкции растительного опада, сопровождается его накоплением
в виде оторфованной лесной подстилки мощностью от 10-15 см (П61) до 25-
30 см (П62) по сравнению с гор. АО типичной Пп (5-7 см).
При разложении растительных остатков в условиях контрастного
окислительно-восстановительного режима (АО подзолистой и слой свежего опада
01 болотно-подзолистых почв) образуется значительное количество
водорастворимых органических соединений преимущественно неспецифической
природы. Их продуцирование резко возрастает во влажные годы (720-820
мгС/100 г), а в сухие доля водорастворимой фракции органического
углерода снижается до 300-400 мгС/100 г. Это может быть связано, с одной
стороны, с относительным накоплением в органогенных горизонтах
водорастворимых органических соединений благодаря резкому замедлению процессов
внутрипочвенной нисходящей миграции веществ при застое атмосферной
влаги в годы с большим количеством осадков. С другой стороны, возможна
активизация минерализации и включения в процессы гумификации
водорастворимых соединений микробиотой в годы с благоприятным окислительно-
восстановительным режимом (интенсивно-окислительная обстановка).
210
БИОСФЕРНАЯ ЮЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ ГУ
Групповой и фракционный состав органического вещества
свидетельствует о значительном влиянии гидроморфизма на протекание процессов
гумификации в БПп. Подавление жизнедеятельности микробиоты, накопление
слабо- и среднеразложенного растительного материала бедного зольными
элементами обусловили образование высокодисперсных, подвижных,
«агрессивных» гумусовых веществ, представленных преимущественно
фракциями ГК-1 и ФК-1. Фракция ГК-2 в составе гумуса БПп, равно как и в
типичных Пп, полностью отсутствует.
В оторфованных горизонтах при переходе от Пп к П61 и П62
наблюдается возрастание суммы гуминовых кислот (ГК) и образование гумуса гумат-
но-фульватного типа (Сгк:Сфк=0,62-1,21). Наиболее высокие величины
Сгк:Сфк отмечены для горизонтов 02. В оглееном потечно-гумусовом
горизонте A2hg болотно-подзолистых почв сохраняется гуматно-фульватный тип
гумуса (Сгк:Сфк=0,62-0,80), в нижележащих горизонтах величина СпсСфк
сужается до 0,32-0,54.
Изучение препаратов гуминовых кислот, выделенных из генетических
горизонтов (02, 03, A2hg, Big) болотно-подзолистых почв, показало, что
наиболее обуглерожены ГК, выделенные из хорошо разложенного горизонта
03. Содержание С в них составляет 61,0%. Они же характеризуются
максимальной степенью бензоидности (59,7%). В горизонтах 02 и A2hg степень
бензоидности значительно меньше и составляет соответственно 30,4 и
28,0%, что указывает на менее выраженный характер ароматичности
молекул ГК этих горизонтов и на увеличение в них доли периферической части с
кислородсодержащими функциональными группами. Об этом
свидетельствует расширение соотношения 0:С в ГК горизонтов 02 (0,52) и A2hg (0,46)
по сравнению с горизонтом 03 (0,34).
По всей видимости, более выраженная гуматность гумуса органогенных
горизонтов БПп и, в первую очередь, горизонта 02 связана не столько с
интенсификацией процессов гумификации в торфяных горизонтах при
активизации жизнедеятельности микрофлоры в условиях умеренно-окислительной
обстановки («сухие» годы), сколько с недостатками метода анализа и
возможной «искусственной гумификацией» полуразложенных растительных
остатков при обработке образцов кислотами и щелочами в ходе фракцион-
но-группового анализа.
Формирование органопрофиля БПп в условиях среднетаежной зоны
определяется в первую очередь степенью гидроморфизма, которая оказывает
непосредственное влияние на окислительно-восстановительный режим
изученных почв, жизнедеятельность микробиоты и процессы минерализации и
гумификации почвенного органического вещества. С характером
увлажнения и окислительными условиями тесно связан тип биоценоза. На авто-
211
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
морфной подзолистой и на подзолистой поверхностно-глееватой почве
произрастает осиново-березово-еловый лес чернично-бруснично-зеленомошный
на подзолистой и чернично-зеленомошный на более увлажненной
подзолистой поверхностно-глееватой почве. В древостое на торфянисто-подзолисто-
глееватой почве и торфяно-подзолисто-глеевой присутствуют ель и береза, в
напочвенном покрове преобладают сфагнум и кукушкин лен.
ВЗАИМОДЕЙСТВИЕ МЕТАЛЛОВ СТОЧНЫХ ВОД
С ТОРФЯНЫМИ ПОЧВАМИ ЕСТЕСТВЕННЫХ БОЛОТ
Л.К. Мамаева, О.А. Милицына
РосНИИВХ, Екатеринбург
Одним из источников поступления металлов в поверхностные воды
являются болотные экосистемы. Естественные болота в силу особенностей
условий формирования сами регулируют баланс металлов. Все металлы,
поступившие на поверхность болота с пылью, с осадками, с отмершими
растительными остатками частично используются развивающимися
растениями, частично поглощаются верхним деятельным горизонтом торфяной
почвы, частично мигрируют с болотными водами. Контролируют миграцию
металлов органические вещества: гуминовые и фульвокислоты, с которыми
металлы могут образовывать малоподвижные соединения.
При антропогенном воздействии на болотные экосистемы, в частности
при сбросе на них промышленных сточных вод увеличивается поступление
металлов на болотные экосистемы. Для оценки взаимодействия металлов с
торфяными почвами использован балансовый метод. Суммированием всех
поступлений определяется приходная часть металлов, поступивших на
болотную экосистему, которая принимается за 100%. Оценка поступления
металлов из болотной экосистемы (в процентах) производится путем
сопоставления количества металлов, сбрасываемых с болотной экосистемы с
количеством металлов, поступивших на болото.
Болотную воду, которая является неизменным компонентом любой
торфяной почвы, рассматривали как водную вытяжку, в которую из торфа
переходят элементы, находящиеся в растворенном состоянии. Химический
состав болотных вод зависит от растворимости соединений, образующихся
при минерализации растительных остатков и от загрязняющих веществ,
поступающих извне.
Для изучения взаимодействия металлов с торфяной низинной почвой
проведены лабораторный и полевой эксперименты. При составлении
баланса металлов в торфяных почвах использовали статьи прихода: в лаборатор-
212
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
ном эксперименте поступление металлов со сточными водами, в полевых
условиях — с атмосферными осадками, с фоновым поступлением и со
сточными водами; статьи расхода: вынос металлов с фильтратом и с урожаем
растений. При расчете баланса использованы достоверные данные
качественного состава воды. Для проверки достоверности использован пакет
прикладных программ «Statsgraphics». До начала исследований и в конце
вегетационного периода были взяты пробы исходной почвы на глубину 30 см,
проведен учет урожая и отбор растительных проб. Содержание металлов в
растениях, почве и воде определено методом атомно-абсорбционной
спектрографии (с графитовым атомизатором — AAS).
При сбросе сточной воды объемом 45 литров, фильтрат составил 10,6
литров. Со стоками в почву поступило 616,4 г/га хрома. Вынос хрома с
фильтратом составил 25,5%, вынос с урожаем — 2,6%. Незначительный
вынос хрома с фильтратом обусловлен малоподвижностью и инертностью
этого элемента при рН среды до 7,85. Поступление меди со сточными
водами составило 1,47 мг. Вынос с фильтратом — 60,2%, вынос с урожаем
растений — 56% от поступившего. Высокая подвижность меди обусловлена
близкой к нейтральной реакцией водно-почвенной среды. Со сточными
водами поступило 1,72 мг марганца. Вынос с фильтратом марганца составил
53% от поступившего. С урожаем растений вынесено 172%. Высокий
процент выноса марганца обусловлен, вероятно, тем, что марганец относится к
подвижным элементам: он не образует прочных соединений с
органическими веществами. Со сточными водами поступило небольшое количество
никеля — 0,28 мг. Содержание никеля в фильтрате составило 26,6%. Больше
вынесено никеля с урожаем растений — 77,6%. В составе сточных вод,
сбрасываемых на торфяную низинную почву, поступило 4,22 мг цинка.
Вынос цинка с фильтратом составил 134%. С урожаем растений вынесено
163%. Кроме цинка, поступившего со сточными водами, в фильтрате,
вероятно, присутствует цинк, вымытый из торфяной низинной почвы. В сточных
водах самое низкое содержание кадмия: на торфяную низинную почву
поступило всего 0,226 мг. Вынос кадмия с фильтратом 18%. Больше кадмия
вынесено с урожаем растений — 58%. Остальной кадмий, вероятно,
поглощен торфяной почвой, чему способствовали связь кадмия с органическим
веществом при благоприятных условиях водно-почвенной среды. На
торфяную низинную почву со сточными водами поступило 13,9 мг железа. В
фильтрате оказалось 35% железа от внесенного со сточными водами. Вынос
с урожаем железа составил 48%. Остальное железо, возможно, закрепилось
в торфяной почве за счет образования прочных хелатов.
В естественных условиях с осадками поступило металлов (в % от общего
поступления): 3,98 марганца, 3,47 цинка, 2,28 меди, 1,5 железа, 0,25 хрома.
213
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Фоновое поступление металлов незначительное и составило, в %: марганца
8.6, железа 0.86, никеля 0.48, меди 0.4, кадмия 0.2, цинка 0,14, хрома 0,06.
Наибольшее количество металлов поступило с промышленными стоками,
что в процентном отношении составило: хрома, кадмия, никеля 99,7%-99,5
%, железа, меди — 97,6-97,3%, цинка 96,4%, марганца — 91,2%. Вынос
металлов с болотной экосистемы распределяется следующим образом: с
болотной водой вынос составил, в %: Мп — 144, Fe — 114, Ni — 108, Zn —
43, Си — 34, Cd — 27, Сг — 13; с урожаем болотных растений, %: Мп —
48,7, Си— 18,3, Fe— 18, Zn — 8,5, Ni —4,9, Cd —2,9, Сг —0,11.
Установлено, что вынос марганца, никеля и железа составляет свыше
100% по сравнению с поступившим на болото. Поступление марганца
отмечено с осадками, со сточными водами и фоновым стоком. Условия водно-
почвенной среды способствуют подвижности марганца, вероятно, этим и
обусловлено самое высокое содержание марганца в болотной воде и в
урожае болотных трав. Никель в условиях среды, близкой к нейтральной не
связывается гуминовыми кислотами и потому может мигрировать, чем и
обусловлено высокое его содержание в болотных водах. Незначительную
часть никеля (16%)составил вынос с урожаем. Отмечено присутствие железа
в осадках, поступление с фоном и со сточными водами. Достаточно высокое
количество железа присутствует в торфяных почвах и природных болотных
водах. Благодаря способности железа мигрировать с коллоидными
частицами железо преобладает в расходной части баланса. Около 18% железа
составил вынос его с урожаем.
Низкое содержание (%) в расходной части баланса металлов: хрома —
13,6, кадмия — 30,1, цинка — 51,6 и меди — 52,8 позволяет предположить,
что большая часть металлов, поступивших на торфяную почву, поглощена
ею или растительным покровом, сформировавшимся по ходу потока
сточных вод. Хром, кадмий, медь, цинк при взаимодействии с торфяными
почвами становятся малоподвижными или инертными, так как способны
образовывать малоподвижные органо-минеральные комплексы, чему
способствуют условия водно-почвенной среды.
Таким образом в экспериментальных условиях, где рассматривается
один антропогенный фактор — поступление со сточными водами для таких
металлов как хром, кадмий и железа преобладающей является приходная
часть баланса. При этом поступление металлов (мг) распределяется
следующим образом: Fe — 13,9, Сг — 7,5 , Zn — 4,2, Мп — 1,7, Си — 1,47, Ni
— 0,29, Cd — 0,23, ; вынос, мг, (%): Zn — 12,54 (297%), Мп — 3,9 (225%),
Си — 1,7 (116%), Ni — 0,3 (104%), Fe — 11,6 (84%), Cd — 0,17%), Сг —
2,12(28,1%).
214
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
В естественных условиях в приходной части баланса металлы (кг/га)
распределяются следующим образом: Сг — 479,5, Zn — 350,4, Fe — 161,3,
^[n — 51, Си — 24,56, Ni — 14,4, Cd — 4,8, в расходной части баланса, кг,
(%): Мп — 98,4 (192%), Fe — 213 (132%), Ni — 16 (112%), Си — 13 (53%),
Zn — 181 (52%), Cd — 1,4 (30%), Сг — 63 (13,6%).
ФОТОСИНТЕТИЧЕСКАЯ ПРОДУКТИВНОСТЬ ЗАБОЛОЧЕННОГО И
СУХОДОЛЬНОГО СОСНОВЫХ ДРЕВОСТОЕВ
А.Г. Молчанов
Институт лесоведения РАН, Московская обл.
Исследования проводили в Ярославской области. Получены
сравнительные зависимости фотосинтеза от солнечной радиации у сосны,
произрастающей в заболоченном и суходольном сосняках. В суходольном, в чер-
нично-кисличном типе леса 1а бонитета при оптимальных условиях
почвенного увлажнения, и в заболоченном типе, в сосняке пушицево-сфагновом, в
летний период, когда уровень грунтовых вод стоял почти у поверхности
почвы (0-10 см от поверхности) [2] и когда неблагоприятный водный режим
болота практически не сказываются, а уровень грунтовых вод был
относительно глубоко, 20 см.
Методы оценки газообмена растений — традиционны, основаны на
использовании инфракрасного газоанализатора. Оценку фотосинтетической
продуктивности проводили по нашей оригинальной методике [1] по
зависимости использования поглощенной солнечной энергии охвоенными или
облиственными побегами на фотосинтез от падающей ФАР и поглощению
ФАР пологом насаждения. Для рассчета пользовались программами:
EXCEL, STATGRAF, SUPERCALC.
Интенсивность фотосинтеза при световом насыщении в сосняке 1а
бонитета была 2,87±0,41, в заболоченнм сосняке, когда грунтовые воды
находятся на глубине 20 см итенсивность фотосинтеза была 2,18±0.42, а когда
грунтовые воды подходят ближе к поверхности интенсивность фотосинтеза
становится 1,74±0,2 мгС02/г час. Рассчитаны уравнения регрессии,
представленные в табл.1.
Различие между фотосинтезом при световом насыщении у сосны,
произрастающей на дренированной почве и в заболоченном типе леса, когда
уровень грунтовых вод находится глубже 20 см составляют около 24% , Это
различие можно объяснить по-видимому недостаточным уровнем
минерального питания в заболоченном насаждении и разбросом данных из-за
методических погрешностей. Зависимость фотоситнтеза от света в этих условиях
215
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
различается незначительно, насыщение фотосинтеза светом происходит
примерно при одной и той же интенсивности радиации 400 Вт, тогда как
при высоком уровне грунтовых вод (0-10 см) насыщение было при 250 Вт.
При высоком уровне грунтовых вод (0-10см) интенсивность фотосинтеза за
день сосны в заболоченном типе леса в два раза меньше, чем в насаждении I
бонитета [2]. Исходя из этого, мы уменьшили величины фотосинтеза у
сосны 1б и рассчитали уравнения зависимости использования поглощенной
ФАР на фотосинтез от падающей ФАР (табл. 2).
Табл. 1. Уравнения зависимости интенсивности фотосинтеза от света в разных ус-
ловиях водообеспеченности.
Тип леса и уровень Уравнение Число на- R-squ- Стандартная
грунтовых вод регрессии блюдений varel ошибка
Сосняк чернично- у=0,03+0,013х- 21 0,90 0,30
кисличный, (200см) 0,00001х2
Сосняк пушицево- у=0,13-Ю,009- 41 0,74 0,46
сфагновый, (20см) 8,6Ех-6х2
Сосняк пушицево- у=0,16+0,013х- 61 0,65 0,23
_сфагаовый, (0-10см) 0,000032х2+2,35Ех3
Табл. 2. Уравнения зависимости использования поглощенной ФАР на фотосинтез от
падающей ФАР в сосняке кислично-черничном и сосняке пушицево-сфагновом.
Тип леса и возраст хвои Уравнение Квадратичное Стандартная
отклонение ошибка
Сосняк кислично-черничник; у=1,19+1,45х- 0,87 0,4
хвоя текущего года 0,19х2 +0,006х3
Сосняк кислично-черничник; у=4,54- 0,98 0,1
хвоя первого года 0,37х+0,018х2
Сосняк пушицево-сфагновый; у=0,59+0,72х- 0,87 0,2
хвоя текущего года 0,09х2+0,003х3
Сосняк пушицево-сфагновый; у=2,27-
хвоя первого года 0,19x^-0,009x2
Используя нашу методику, рассчитали фотосинтетическую
продуктивность соснового насаждения 1а бонитета за 1995г. [1]. По полученными
ранее данным поглощения ФАР по глубине полога, ее сезонной изменчивости
и взятых в обсерватории МГУ данных солнечной радиации за 1995 годы,
рассчитали поглощение ФАР в течение вегетационного периода в трех слоях
полога за каждый день (табл.3). По этим же данным и ранее полученным
уравнениям зависимости коэффициента использования поглощенной ФАР
на фотосинтез от падающей радиации по слоям полога рассчитали
фотосинтетическую продуктивность насаждения за 1995 год.
216
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ ГУ
Табл. 3. Фотосинтетическая продуктивность и поглощение ФАР пологом заболо-
<Tffmoro и суходольного насаждений в течение вегетационного сезона 1995 г
-"""" Параметры Месяцы* Сум-
04 05 06 07 08 09 10 ма
*—" Сосняк кислично-черничный 1а бонитета
Поглощ. ФАРв1995г,Мджм-2 172 237 293 252 235 131 59 1379
Поглощ. С02в1995,гм-2 411 557 681 685 739 428 184 3685
Сосняк пушицево-сфагновый V6 бонитета
Поглощ. ФАРв1995г,Мджм-2 32 44 56 53 53 32 21 291
Поглощ. С02 в 1995г, г м2 39,3 54,7 63,9 53,1 47,1 31,7 27,4 317,2
Рассчитанное нами поглощение ФАР пологом соснового насаждения 1а
бонитета было 1379 Мдж м"2 год. Фотосинтетическая продуктивность была в
1995 — 3685 г С02 м"2. По месяцам колебание фотосинтетической
продуктивности довольно близко следует колебаниям поглощенной ФАР пологом
насаждения, максимальные различия составили 4% (в июне), в другие
месяцы точно соответствовали колебаниям поглощенной ФАР.
Рассчитали поглощение ФАР и С02 в сосняке пушицево-сфагновом. В
условиях, когда уровень грунтовых вод был на глубине 0-10 см,
интенсивность фотосинтеза за день равна 0.5 на единицу массы хвои интенсивности
фотосинтеза в сосняке кислотно-черничном [2]. По этим данным рассчитали
эффективность использования поглощенной ФАР охвоенными побегами на
фотосинтез и далее рассчитали уравнение зависимости использования
поглощенной ФАР на фотосинтез от падающей ФАР. В табл.2 показано эти
уранения, по ним, по падающей ФАР на насаждение и по поглощению ФАР
насаждением рассчитали фоситетическую продуктивность пушицево-
сфагнового насаждения, в условиях, когда весь год уровень грунтовых вод
стоял на глубине 0-10 см. Полученные данные представлены в табл.3. Из
этих данных видно, что в заболоченном сосняке фотосинтетическая
продуктивность меньше, чем в суходольном почти в 12 раз, хотя поглощение ФАР,
из-за слабой сомкнутости насаждения, меньше в 5 раз. Однако, несмотря на
такое снижение интенсивности фотосинтеза, говорить об отмирании
взрослых деревья сосны еще рано, для этого необходимо еще рассчитать затраты
на дыхание органов и величину отмирания корневых систем. В
оптимальных условиях увлажнения в осоко-сфагновом насаждении в связи с тем, что
интенсивность фотосинтеза выше почти в два раза, фотосинтетическая
продуктивность будет также больше в два раза, но все же по сравнению с
суходольным насаждением ниже в 6 раз.
[1] Молчанов А.Г. Экофизиологическое изучение продуктивности древостоев М:
Наука, 1983. 136 с. [2] Молчанов А.Г. Сравнение интенсивности фотосинтеза сосны
в разных эдафических условиях // Лесоведение. 1993. № 6. С. 76-80.
217
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ПРОДУЦИРОВАНИЕ С02 ТОРФЯНОЙ ПОЧВОЙ
СЛАБО ОСУШЕННОГО МЕЗОТРОФНОГО БОЛОТА
В СВЯЗИ С ГТ1ДРОТЕРМИЧЕСКИМИ УСЛОВИЯМИ СЕЗОНА
А.В. Наумов1, Т.Т. Ефремова, СП. Ефремов2
институт почвоведения и агрохимии СО РАН, Новосибирск;
2Институт леса СО РАН, Красноярск
Охрана и рациональное использование болотных экосистем представдя*
ют сегодня важный этап на пути к устойчивому управлению природным*
ресурсами. Масштабы территории заболоченных лесов и болот России <иц
ределяют биосферную роль этих объектов как участников глобального угле*
родного цикла. Депонирование углерода в форме торфяных отложенн|
представляет собой, как известно, результирующую нескольких процессор
Для болотных экосистем почвенное дыхание, или выделение углекислой*
газа с поверхности почвы, мало изучено [3]. Оценки годового потока угле»
рода в атмосферу для сфагновых болот, широко распространенных в наше!
стране, единичны и не дают надежной базы для масштабных (региональ
ных) балансовых расчетов.
Почвенное дыхание, как и любой процесс в природной экосистеме,
подвержено влиянию разных факторов. Однако температура и влажность окру*
жающей среды при мало меняющихся прочих условиях имеют первостепец»
ное значение для биологических ритмов живых организмов, выделяющих
углекислоту в процессе жизнедеятельности.
Наши исследования эмиссии С02 в нескольких болотных экосистемах
выявили заметную инерционность этого показателя по отношению к темпе*
ратуре почвы в суточном цикле. Запаздывание отклика экосистемы на
изменение температурного фактора может иметь двоякую природу: биологиче*
скую (инерционность продуцирования С02) и физическую (диффузия угле»
кислого газа из глубины к поверхности почвы и выделение его в атмосферу)*
На отдельных образцах в контролируемых условиях было установлено, что
продуцирование углекислоты есть процесс безинерционньга, т.е. реакция
дыхательного газообмена биологического компонента фактически адекватна
изменениям температуры окружающей среды [1]. Таким образом, фазовый
сдвиг эмиссии углекислого газа из болотной почвы по отношению к темпе»-
ратурному фактору имеет, по-видимому, диффузионную природу и оиреде*
ляется физическими свойствами почвенного субстрата.
Изучение продуцирования СОг в контролируемых условиях также
позволило выявить функциональную зависимость этого показателя от температу*
ры и влажности торфа [2]. Эта взаимосвязь была использована для
реконструкции сезонного хода дыхания почвы и оценки суммарного потока СОг *
218
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
оСферу по гидротермическим характеристикам слабо осушенного сфаг-
ового болота переходного типа. Наблюдения за гвдротермическим режи-
ом велись в течение трех сезонов, что позволило оценить диапазон измен-
ивости изучаемого показателя и характер его сезонной динамики. При этом
полагали, что экосистема находится в стационарном состоянии и
коэффициенты регрессионной модели, характеризующие дыхательную активность
биоты, мало меняются от сезона к сезону.
Расчеты показали, что за период с мая по октябрь болотная почва
продуцирует от 142 до 204 г С/м2 со средней интенсивностью 1,1-1,6 г С/м2-сутки.
Сезонная динамика почвенного дыхания имеет вид параболы с вершиной
(максимум) в конце июля — августе. Амплитуда сезонных изменений
скорости продуцирования С02 составляла 6,3-6,9 г С02 /м2-сутки.
Уровень стояния почвенно-грунтовых вод оказывает заметное влияние
на величину потока С02. Закономерное понижение его к концу сезона
увеличивает мощность слоя интенсивного газообмена. Это, в свою очередь,
приводит к сдвигу максимальных значений скорости дыхания на конец
августа — начало сентября.
Функциональный подход к оценке состояния природных экосистем
представляется перспективным. Выявляемые функциональные зависимости
важнейших интегральных параметров объекта от факторов среды дают
исследователю некоторый стандарт, отклонение от которого расценивается как
переход к нестационарному режиму функционирования с определенной
вероятностью деградационных явлений.
[1] Наумов А.В. Дыхание растений и эмиссия углекислого газа в болотной
экосистеме // Сиб. экол. журнал. 1997. № 4. С. 385-391. [2] Наумов А.В. Функционально-
экологический аспект изучения СОг-газообмена болотной почвы // Системный
подход в почвоведении. Препринт. Новосибирск, 1997. С. 44-48. [3] Raich J.W., Schle-
singer W.H. The global carbon dioxide flux in soil respiration and its relation to
vegetation and climate // Tellus, 1992. V. 44B. P. 81-99.
ВОДООБМЕН БОЛОТ С ПОДСТИЛАЮЩИМИ
МИНЕРАЛЬНЫМИ ГРУНТАМИ
СМ. Новиков
Государственный гидрологический институт, Санкт-Петербург
Слабой стороной воднобалансовых исследований болот является
отсутствие относительно доступных и надежных методов определения
вертикально водообмена болотных массивов с подстилающими минеральными
^Унтами. Это обстоятельство не позволяет «замкнуть» водный баланс бо-
219
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
лотных массивов и в чистом виде получить его невязку. Обычно в невязщ,
расчета водного баланса входит и величина водообмена. Сложность щ^
ректной оценки точности расчета водного баланса заключается и в том, 1ц*
водообмен может иметь не только различную величину, но и разный знак.
Водный баланс как метод исследования водообмена болот был использо»
ван [1,6] при изучении низинных болотных массивов, в [3,4] и — при ^
следовании верховых болот. Расчеты водообмена базировались на данных
измерений составляющих водного баланса конкретных болотных массивов
Иной метод оценки водообмена болот использован в работе [5], где скоросгъ
водообмена определялась по распределению трития в болотных водах.
Уникальные экспериментальные исследования по изучению водообмена
низинных болот с подстилающими минеральными грунтами выполнены
ГТИ на Старосельской осушительной системе в 1976-1979 г. г. Специально
для проведения этих исследований были оборудованы кусты наблюдатель-
ных болотных и гидрогеологических скважин с фильтрами, заложенными в
торфяную залежь и минеральный грунт непосредственно под разделяющий
слой, и определены коэффициенты фильтрации этого слоя. По данным на*
блюдений за уровнем болотных и грунтовых вод были вычислены декадные
и месячные значения слоев водообмена [2]. Анализ результатов этих
исследований показал, что величина и знак декадных значений водообмена изме-
няются во времени и по площади. Причем диапазон изменений величин
водообмена довольно значительный. Весьма большие изменения водообмена
по площади, соизмеримые с величинами осадков и испарения, показывают
на исключительную важность определения этой составляющей водного
баланса, особенно для низинных болот.
Результаты исследований водного баланса дают основания считать, что
ввиду сложных и весьма разнообразных гидрогеологических условий
залегания низинных болот водообмен их с подстилающими минеральными
грунтами в каждом конкретном случае имеет свой сугубо специфический
характер.
Касаясь вопроса водообмена верховых болот с подстилающими
минеральными грунтами, следует отметить, что в работах К.Е. Иванова и В.В.
Романова высказывается мнение о незначительном вертикальном оттоке
болотных вод. Количественная оценка величины потерь воды в
подстилающие минеральные грунты получена [3] для одного из верховых болотных
массивов Севера-Запада. Средняя величина вертикального оттока оказалась
равной 27 мм/год.
Методом водного баланса был рассчитан [4] водообмен верхового болота
Ламмин-Суо (Карельский перешеек) за 20-ти летний период и сделан вывод
о значительной величине потерь стока в подстилающие минеральные грУ**-
220
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
лрилегающие суходолы. Средняя величина водообмена составила
120
** /год при колебаниях от — 29 до 244 мм/год..Столь большую вариант10
М ичйн водообмена при относительно малых амплитудах колебания уров-
х болотных вод трудно объяснить только вертикальным оттоком воды в
лстилающие минеральные грунты, особенно, если учесть что водообмен
ттределялся как остаточный член уравнения водного баланса, куда вопДО
помимо невязки и величина неучтенного стока с болота. Иными словами
получить в чистом виде величину вертикального водообмена верхового
болота в работе [4] не удалось.
В связи с этим возникла необходимость сделать попытку оценки величи-
ны вертикального водообмена этого массива с подстилающими
минеральными грунтами, учитывая, что по данному болоту накоплен уникальной
экспериментальный материал по водному режиму, водному балансу и воД-
но-физическим свойствам торфной залежи. Чтобы исключить влияние
прилегающих суходолов и окраек болота на корректность расчета
вертикального водообмена (В) величина его определялась не для всего массива Ламмян-
Суо, а лишь для его центральной части. Расчет проведен для разных по
водности лет по уравнению: В = О - Е - Сф ± W, где О — осадки, Е — испаре-
ние, Сф — фильтрационный сток через деятельный горизонт, W —
изменение влагозапасов за расчетний период.
Величины О и Е получены по данным наблюдений, величина С
рассчитана по методу фильтрационных характеристик на основе ежедневных
наблюдений за уровнем воды и экспериментальных определений
коэффициента фильтрации деятельного слоя, W — определено по данным наблюдении
за уровнем болотных вод и водоотдачей торфяной залежи. Результаты
расчета представлены в таблице.
Водный баланс центральной части болота Ламмин-Суо за годы различной водности
год О, мм Е, мм AW, мм Сф, мм В, мм ^
1969
1972
1977
1979
1981
1984
реднее
659
627
823
764
1129
1245
874
505
491
383
400
385
404
428
+49
+42
-30
+9
-16
-9
+8
135
158
334
317
730
804
413
+68
+20
+76
+56
-3
+28
+41
Как видно из таблицы диапазон изменения годовых осадков за
выбранные годы довольно велик (от 627 до 1245 мм). Однако, и еще больший
диакон изменений у фильтрационного стока (от 135 до 804 мм). Вариация
годовых величин испарения относительно мала (от 383 до 505 мм). ВеличЯ-
221
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
на водообмена колеблется от — 3 мм (1981 г.) до 76 мм (1977 г.), причем в
5 из 6 случаев имеет положительный знак, что указывает на отток водц
Средняя величина вертикального водообмена за 6-ти летний период
оказалась равной 41 мм. Следует отметить, что уровень грунтовых вод под це&.
тральной части болота Ламмин-Суо находится ниже его дна, что и обусловь
ливает потенциальную возможность вертикальной фильтрации болотных
вод в подстилающие минеральные грунты. Колебания годовых величин во.
дообмена можно объяснить определенным влиянием режима уровней бо*
лотных и грунтовых вод, а также режима промерзания болот. Необходимо
заметить,что до сих пор не выявлен механизм процесса водообмена
верховых болот с подстилающим минеральным грунтом при наличии закольма,
тированного придонного слоя. Необходимо было бы провести эксперимент
тальные работы по определению толщины и коэффициента фильтрации
этого слоя на верховых болотах, различающихся по геоморфологическим уело*
виям залегания. Это бы послужило началом и хорошей основой для
проведения широких исследований водообмена верховых болот.
Следует отметить, что полученная нами средняя величина вертикального
водообмена верховых болот очень близка к величине, определенной К.Е.
Ивановым [3]. Более того достоверность этой величины подтверждается
результатами неводнобалансового подхода к оценке водообмена болот,
приведенными в работе [5]. Согласно этой работе возраст болотных вод
придонных горизонтов верховых массивов составляет 100-150 лет . Если
принять среднюю величину возраста этих вод равной 125 годам, а мощность,
залежи исследуемого массива 4,5 м , то средняя скорость фильтрации будет
равна 1,14 х 10*7 см/с, величина же вертикального оттока болотных вод
составит 36 мм/год (в наших расчетах 41 мм/год).
Таким образом полученные результаты подтверждают ранее
высказанное К.Е. Ивановым и В.В. Романовым положение о малой величине
вертикального водообмена верховых болот с подстилающими минеральными
грунтами.
[1] Бавина Л Г. Водный баланс низинных болот Полесской низменности // Тр. ГТИ.
Л- Гидрометеоиздат\1966. Вып. 135. С. 181-196. [2] Воробьев П.К. Исследование
водообмена торфяной залежи с нижележащим водоносным горизонтом // Тр. ГТИ.
Л.: Гидрометеоиздат. 1981. Вып.,281. С. 81-89. [3] Иванов К.Е. Водообмен в
болотных ландшафтов. Л.: Гидрометеоиздат. 1975. 280 с. [4] Рождественская В.Г.
Водный баланс болота Ламмин-Суо // Сб. работ по гидрологии. 1973. № 11. С. 129-139.
[5] Сирин А.А. и др.. Изучение водообмена в болотных водах с помощью
результатов анализа 3Н // Водные ресурсы. 1997. Т. 24. № 6. С. 679-687. [6] Шебеко В.Ф.
Гидрологический режим осушаемых территорий // Минск: Урожай, 1970. 298 с.
222
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
ПРИБЛИЖЕННАЯ ОЦЕНКА САМООЧИЩАЮЩЕЙ СПОСОБНОСТИ
БОЛОТ В ОТНОШЕНИИ СТОЧНЫХ ВОД
А.П. Носаль
РосНИИВХ, Екатеринбург
Общепринято считать болота естественными геохимическими
барьерами способными аккумулировать в торфяной толще значительные массы
поллютантов. Замедленный водообмен, преобладание анаэробных условий
позволяют выводить из природного круговорота многие загрязняющие
вещества. Например, при сбросе сточных вод в процессе прохождения по
болотному массиву концентрации загрязняющих веществ в транзитном потоке
претерпевают значительные трансформации до поступления в дренирующий
болото водоток. Эта трансформация содержания поллютантов по длине
потока, происходит как в результате прямого разбавления болотными водами,
так и в результате сорбции, седиментации, смены биохимических барьеров
и ассимиляции болотным биоценозом.
В ряде регионов России, характеризующихся относительно высокой
заболоченностью, в частности в Свердловской области, использование болот в
качестве водоприемника недостаточно очищенных и неочищенных сточных
вод получило широкое распространение. Болотный участок при этом
рассматривается с утилитарной точки зрения как природный буфер,
дополнительный блок доочистки, гаситель залповых аварийных сбросов сточных
вод. В подавляющем большинстве случаев сброс в болото организовывался
стихийно из-за удаленности водотоков и имевшихся послаблений в
требованиях к нормирование сбросов сточных вод на болота.
Принципы нормирования сброса сточных вод на болота до сих пор в
действующем федеральном природоохранном законодательстве четко не
обозначены, что обуславливает вольную трактовку подходов к данному
вопросу. Наиболее корректной, на наш взгляд, является практика сложившаяся
в Свердловской области : при сбросе сточных вод на болото нормирование
ПДС производиться с использованием эмпирических показателей
фактической самоочищающей способности конкретного болота. Такой подход
позволяет учитывать специфику болота как водного объекта и использовать
эти особенности для рационального природопользования.
Самоочищающая способность болота оценивается по коэффициенту
самоочищения или иначе трансформации (Кс) отдельных загрязняющих
веществ. Коэффициент самоочищения отдельных загрязняющих веществ
определяется как отношение начальной концентрации ингредиента на входе в
колото (сброс сточных вод) к концентрации на выходе или в промежуточной
точке без рассмотрения внутриболотных процессов.
223
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Исследования связанные с оценкой самоочищающей способности болот
проводились специалистами РосНИИВХ на болотах Урала и Западной
Сибири. Все имеющиеся исследования производились на действующих
выпусках сточных вод и по сути полученные показатели самоочищающей
способности Кс отражают реакцию адаптированной болотной экосистемы,
испытывающей антропогенную нагрузку в течение длительного времени.
Исследования на конкретных болотах, с одной стороны, позволили определить
самоочищающую способность с учетом индивидуальных особенностей, с
другой стороны, учет самоочищения происходит пост фактум.
При планировании новых выпусков сточных вод на болота для оценки
риска загрязнения водотока необходимо иметь хотя бы ориентировочное
представление о самоочищающей способности транзитного болота.
Автоматически переносить величину Кс с исследованного объекта на неизученный
некорректно. Необходимо критерии, позволяющих обосновать величину Кс
по отдельным поллютантам для неизученных болот в зависимости от
параметров, определение которых возможно на этапе проектирования.
Опыт исследований показал, что одним из наиболее важных факторов,
определяющих значение Кс, являются фоновые показатели болотных вод
Эта величина служит своего рода нижней границей возможного снижения
концентраций загрязняющих веществ. При сбросе стоков с концентрациями
меньше фоновых наблюдается их рост по длине транзитного потока.
На данном этапе исследований именно фон, а точнее разность
концентраций загрязняющих веществ в сточных и болотных водах,
рассматривались в качестве фактора, определяющего величину Кс. В качестве базовых
ингредиентов рассматривались показатели характерные для хозяйственно-
бытовых стоков: биохимическое потребление кислорода (БПК), азотная
группа, хлориды, нефтепродукты, фосфор, бихроматная окисляемость
(ХПК) и т.д.. Сбросной расход на исследованных болотах колеблется в
пределах от 0,5 л/с до 10 л/с.
На основании натурных гидрохимических наблюдений на низинных
болотах таежной зоны для каждого ингредиента построены графики связи
величины коэффициента самоочищающей способности болота Кс от разности
концентраций поллютантов в сточных водах и фона для отдельных болот. В
основном графики связи имеет линейный характер, но по некоторым
ингредиентам (сульфаты, др.) возможно наличие степенной или логарифмической
зависимости. Для большинства исследованных болот отмечается наличие
тесной связи по азотной группе, ХПК, БПК.
Осредненные показатели Кс и средняя разность концентраций
загрязняющих веществ в сточных и болотных водах по исследованным болотам
использовались для построения обобщенных региональных зависимостей.
224
БИОСФЕРНАЯ ЮЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
Обобщенные зависимости достаточной степени надежности получены для
большинства базовых ингредиентов (коэффициент корреляции превышают
О 7). кроме нефтепродуктов, хлоридов и фосфора фосфатов.
Сброс на болото обычно производится или планируется с очистных
сооружений небольших поселков. Требования соблюдения рыбохозяйствен-
яых ПДК на выпуске связано с необходимостью больших капвложений. Это
редко возможно и учет самоочищающей способности болот при
регламентировании работы реконструируемых или вновь строящихся очистных
сооружений может помочь снизить капитальные и эксплуатационные затраты без
ущерба для водного объекта.
Предлагается следующая схема оценки Кс для неизученных болот. 1.
Определяются фоновые показатели для неизученных в отношении
самоочищения болот: — на основании натурных данных при выполнении
инженерных изысканий на стадии проектирования, по данным Росгидромета, других
организаций; — по аналогии с болотами, находящихся в близких ланд-
шафтно-геологических условиях по справочной литературе. 2.
Концентрация конкретного поллютанта принимается либо по факту на действующих
очистных сооружениях, либо по проектным данным для строящихся. 3. По
обобщенным региональным зависимостям коэффициента самоочищения Кс
по отдельным поллютантам с разностью их концентраций в сточных и
болотных водах методом подбора: — проверяется соблюдение
природоохранных требований в контрольном створе водотока при выходных
концентрациях на современное состояние или по предварительному проектному
решению; — определяются предельно допустимые концентрации на выходе с
очистных сооружений и соответствующий коэффициент самоочищения Кс.
ОСОБЕННОСТИ ОРГАНИЧЕСКОГО ВЕЩЕСТВА
ОСУШЕННЫХ ТОРФЯНИСТО-ПОДЗОЛИСТО-ГЛЕЕВЫХ ПОЧВ
ЯРОСЛАВСКОЙ ОБЛАСТИ
Д.С. Орлов, Л.К. Садовникова, И.В. Сухова
МГУ, факультет почвоведения
В современных природных условиях торфянистые почвы являются
важнейшими аккумуляторами и консерваторами огромных запасов связанной
воды, органического вещества и потенциальной солнечной энергии.
Несмотря на обширные исследования состава органического вещества почв торфя-
но-болотного типа, до сих пор малоизученным остается вопрос об
особенностях процесса трансформации органического вещества в процессе осушения
болотных почв.
225
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Целью настоящей работы стало изучение возможных изменений, проис*
ходящих в составе органического вещества торфянисто-подзолисто-глеевь^
почв при их осушении. 30 проб торфяных горизонтов исследуемых почь
были отобраны на Северной лесной опытной станции Института лесоведе,
ния РАН (Рыбинский р-н, Ярославская обл.). В четырех лесничествах Р^
бинского лесокомбината были заложены пять опытных участка и проведено
разновариантное осушение мелкой сетью канав и борозд. В отобранных
пробах торфяных горизонтов торфянисто-подзолисто-глеевых почв были
изучены: а) общая химическая характеристика; б) особенности группового к
фракционного состава органического вещества; в) неспецифические соеда*
нения (липиды, углеводы, хлорофилл); г) показатели гумусного состояния.
Исследованные торфянисто-подзолисто-глеевые почвы характеризуютс*
кислой реакцией среды (рН водной вытяжки колеблется от 3,5 до 3,9),
типичной для торфяных почв верхового типа. Общее содержание углерода
составляет от 18 до 25%, общее содержание азота постоянно, — около 1%, %
практически не зависит от степени осушения. Обогащенность гумуса азотом
по отношению C:N очень низкая (36) и постоянна по всем образцам, незави-
симо от осушения. Содержание липидов сравнительно невысокое и состав*
ляет от 2 до 6 %. Содержание хлорофилла составляет в среднем от 30 до 60
мкг/г и в процессе осушения происходит незначительное снижение
содержание хлорофилла, связанное с понижением влажности. Тип гумуса — гу-
матно-фульватный, в процессе осушения изменяясь на фульватно-гуматный.
В составе гумуса в осушенных торфяных горизонтах происходит небольшое
возрастание фракции гуминовых кислот, связанное, очевидно, с трансфор*
мацией органического вещества. Наблюдается повышенное содержание
углеводов во фракциях фульвокислот. В результате осушения происходит
уменьшение содержания углеводов во фракциях гуминовых веществ, в связи
с их свойством легко разлагаться под действием некоторых
микроорганизмов. Показатель гумифицированности торфа (ГИТ) более ярко выражает
степень гумифицированности торфа, чем отношение Сгк/Собщ., поэтому его
следует использовать для оценки гумусного состояния торфянистых почв.
Наибольшие превращения органической массы происходит с торфами
моховой и травяной групп с низкой степенью разложения, и степень разложения
торфа резко увеличивается в результате осушения и других агрохимических
приемов за период в 30-40 лет. Очевидно, осушение торфянисто-подзолисто-
глеевых почв Ярославской области улучшает качественные характеристики
органического вещества и способствует увеличению биопродуктивностй
лесов.
Работа выполнена при финансовой поддержке РФФИ, проект 96-15-
97791 научная школа «Химические основы почвообразования» и 99-04-
226
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
i8007 «Формирование и типизация гумусных профилей главнейших почв
России».
rii Бамбалов Н.Н., Беленькая Т.Я. Фракционно-групповой состав органического
ешества целинных и мелиорированных торфяных почв // Почвоведение. 1997. №
12 С. 15-19. [2] Вомперская МИ. Поверхностное осушение временно
переувлажненных лесных почв М., Наука. 1982. 134 с. [3] Горелова Т.А. Особенности
органического вещества торфяных, торфяно-глеевых и торфянисто-подзолисто-глеевых
почв. Дисс. канд. биол. наук. М, МГУ, 1982. 160 с. [4] Ефимов В.Н. Торфяные
почвы и их плодородие. Л., Агропромиздат, 1986, 264 с. [5] Орлов Д.С., Бирюкова О.Н.
Запасы органического вещества // Почвоведение. 1993. № 10. С.45-50. [6] Садовни-
кова Л.К. Углеводные компоненты гумусовых веществ почвы. Дисс. канд. биол.
наук, М.: МГУ, 1976, 190 с. [7] Фридланд Е.В. Методика изучения липидной
(растворимой в спиртобензоле) фракции почвенного гумуса // Биологические науки.
1978. №5. С. 75-81.
ОСОБЕННОСТИ ХИМИЧЕСКОГО СОСТАВА ПРИРОДНЫХ ВОД
ЛЕСОБОЛОТНЫХ КОМПЛЕКСОВ СИБИРСКИХ УВАЛОВ,
ЗАПАДНАЯ СИБИРЬ
А.А. Сеньков
Институт почвоведения и агрохимии СО РАН, Новосибирск
Проведено исследование химического состава следующих категорий
природных вод: атмосферные осадки, торфяные воды гряд, бугров и
мочажин, речные и озерные воды, грунтовые воды почв. Все разновидности
природных вод исследуемой территории, не испытывающих техногенного
загрязнения, имеют очень низкую минерализацию и относятся к категории
ультрапресных. Общая концентрация солей в большинстве случаев
колеблется в пределах 5-20 мг/л. Случаи более высокой концентрации
обусловлены особыми условиями формирования природных вод. Различия в
минерализации связаны в основном с наличием или отсутствием
гидрокарбонатного иона в зависимости от величины рН.
В связи с бедностью минерального состава песчаных осадочных пород
основным источником поступления солей в природные воды на исследуемой
территории являются атмосферные осадки. Химический состав атмосфер-
ных осадков в районе исследования и среднемесячный состав осадков на
метеостанции г. Туруханск за 1975-80 гг. не имеет существенных различий.
Содержание солей в осадках обычно не превышает 10 мг/л. В составе
анионов преобладают сульфаты, а катионов кальций и натрий. Концентрация
солей в атмосферных осадках данной территории в 2-3 раза меньше чем в
Районах Северного Предуралья и таежной зоне Западной Сибири. Средние
227
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
значения химического состава атмосферных осадков г. Туруханск были цп
пользованы для расчета коэффициентов метаморфизации ионного состав
природных вод по формуле h=CiB/RCia, где Gb и Cia — концентрация но-
нов в природных водах и атмосферных осадках; R — степень упаривания
равная отношению содержания хлора в природных водах к содержанию eix>
в атмосферных осадках.
Химический состав природных вод, мг/л
ЕС рН Сорг. НСОз CI SQ4 Са Mg Na К jT*
Бугры
Гряды
Моча
жины
Окр
Омл
Реки
ГВ
Ос
М
CV
м
CV
м
CV
м
CV
м
CV
м
CV
м
CV
м
CV
122,7
0,37
42,07
0,36
31,95
0,51
32,17
0,48
19,01
0,53
26,02
0,53
22,39
0,70
12,86
0,56
5,78
0,16
4,63
0,14
4,93
0,15
6,28
0,14
5,02
0,11
6,16
0,15
5,19
0,16
5,64
0,09
132,5
0,41
35,73
0,39
21,79
0,46
10,45
0,09
12,87
0,60
10,21
0,37
11,05
0,92
19,91
0,63
3,95
1,07
3,99
0,80
13,18
0,65
4,88
0,46
10,36
0,89
4,27
0,58
1,29
1,15
5,16
0,35
2,62
0,36
2,39
0,35
1,65
0,27
1,62
0,48
1,66
0,42
1,90
0,40
1,08
0,67
7,44
0,53
2,59
0,38
2,67
0,39
2,68
0,53
2,20
0,34
1,95
0,51
2,69
0,48
2,32
0,86
1,28
0,80
0,83
0,56
0,78
0,71
3,15
0,70
0,46
0,47
1,77
0,74
0,79
0,52
0,50
0,90
0,19
1,04
0,19
0,87
0,11
0,68
0,63
1,39
0,15
0,90
0,58
0,88
0,11
0,90
0,40
0,62
1,00
1,63
0,79
0,81
0,62
0,73
1,38
0,59
0,37
0,41
0,99
0,65
0,78
0,88
0,53
0,98
0,54
1,39
0,64
0,86
0,47
0,82
0,55
0,43
0,21
0,56
0,31
0,79
0,61
0,56
0,22
0,96
35,53
0,35
11,59
0,40
11,02
0,39
23,22
0,54
9,89
0,30
17,62
0,67
11,15
0,27
6,24
0,64
М — среднее значение; CV — коэффициент вариации; ЕС — электропроводность,
цСм/см; £ — сумма; Окр — озера крупные; Омл — озера мелкие; ГВ — грунтовые
воды; Ос — атмосферные осадки.
Ионный состав торфяных вод олиготрофных болот формируется только
за счет атмосферных осадков. Основными факторами, определяющими их
химический состав, являются испарительное концентрирование и
биохимические процессы. Наилучшие условия для проявления процесса
концентрирования имеются на крупных буграх пучения. Этому способствуют хорошая
продуваемость и наличие многолетней мерзлоты, ухудшающей дренирован-
ность торфяной толщи.. Воды, выделенные из оттаявших образцов торфа из
мерзлого слоя крупных бугров пучения, имеют самую высокую
концентрацию ионов хлора (5-10 мг/л). Другой отличительной особенностью этих вод
является очень высокая концентрация органического вещества, многократно
превышающая общую концентрацию минеральных веществ. Содержание
органического углерода колеблется в пределах 50-215 мг/л. В торфяных
водах гряд и мочажин средняя концентрация хлора в два раза больше чем в
атмосферных осадках. Эти разновидности болотных вод не имеют сущест-
228
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
- различий по составу минеральных веществ. Различие наблюдается
06101 о в содержании органических веществ. Средняя концентрация органи-
^^ го углерода в торфяной воде гряд составляет 36 мг/л, а в воде мочажин
^уменьшается до 22 мг/л.
Величины коэффициентов метаморфизации сульфата, кальция, магния и
тоия во всех торфяных водах значительно меньше единицы. Это свиде-
льствует об относительном уменьшении их содержания в торфяных водах
сравнению с атмосферными осадками в результате биологического
голошения, закрепления в клеточных структурах и поглощающем комплексе
торфа Наибольший дефицит наблюдается для магния. Его концентрация
составляет всего 10-20% от поступившего с атмосферными осадками.
Видимо магний является наиболее дефицитным элементом питания для
растительности верховых болот.
В районе исследования озера по условиям питания, своим размерам,
химическому составу воды разделяются на две группы. К первой группе
относятся крупные озера с минеральным дном, имеющие размеры сотен и тысяч
метров. Уровень воды в них находится на 1-3 метра ниже, чем в
окружающих болотных массивах. Эти озера, как правило, имеют речной сток.
Помимо атмосферных осадков, поверхностного и дренажного стока с
окружающих ландшафтов в их солевом балансе большую роль играют
подземные воды. В химическом составе воды крупных озер отмечается
повышенная концентрация всех катионов и гидрокарбонатов. Особенно велико
относительное содержание кальция. Оно в четыре раза больше, чем это может
быть обеспечено поступлением с атмосферными осадками. Ко второй группе
относятся небольшие и неглубокие озера грядово-мочажинных болот,
плоско - бугристых торфяников. Размер их обычно не превышает десятков
метров. Концентрация солей в воде этих озер меньше чем в торфяных водах
гряд и мочажин. Также в них меньше в 2-3 раза концентрация
органического углерода. Это видимо связано с процессом вымораживания и
отжиманием солей и органических веществ в торфяную толщу при промерзании озер.
Болотные речки и их неширокие долины дренируют окружающие их
болотные массивы. Следовало бы ожидать большого сходства состава речных
и торфяных вод. Однако явное сходство по всем параметрам наблюдается с
водой озер первой группы. Речные воды имеют близкую к нейтральной
реакцию, повышенное содержание катионов кальция, магния и натрия,
небольшую концентрацию органического вещества. Видимо, как и в крупных
°зерах в питании болотных рек большая pojjb принадлежит подземным
водам. Более богатый минеральный состав речных вод обеспечивает
произрастание мезотрофной растительности в их поймах даже посреди окружающих
их олиготрофных болот. Воды болотных ручьев, русло которых неглубоко
229
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
врезано в торфяную толщу, имеет близкий с торфяными водами ионный
состав. Однако они имеют более высокую рН и небольшую концентрации
органического вещества.
Песчаные почвообразующие и подстилающие породы, состоящие более
чем на 95% из кварца, не оказывают существенного влияния на формирова*
ние химического состава грунтовых вод почв. Концентрация кальция и на*
трия фактически не изменилась по сравнению с атмосферными осадками
Видимо только повышенное содержание калия (в 1,6 раза) может быть обу.
словлено дополнительным его поступлением из почвообразующих пород, в
грунтовых водах, также как и в торфяных наблюдается крайне низкое
содержание магния, в большинстве случаев не превышающее сотых долей
миллиграмма в литре. В грунтовых водах иллювиально-железистых
подзолов концентрация органического углерода не превышаетЮ мг/л. В торфяно-
болотных почвах и иллювиально-гумусовых подзолах его содержание
увеличивается до 15-30 мг/л.
В водах с одинаковой минерализацией электропроводность может
отличаться в несколько раз. Это связано с различным содержанием ионов
органических веществ, которые оказывают сильное влияние на величину
электропроводности. Между содержанием органических веществ и
электропроводностью имеется тесная взаимосвязь. В торфяных и грунтовых водах
коэффициент корреляции между этими показателями составляет 0,72-0,88.
ПОТОКИ, ГЕНЕРИРОВАНИЕ И ЭМИССИЯ ПАРНИКОВЫХ ГАЗОВ
В ЗАБОЛОЧЕННЫХ ПОЧВАХ
А.В. Смагин1, М.В. Смагина, ТВ. Глухова2
!МГУ, факультет почвоведения;
2Институт лесоведения РАН, Московская обл.
Болотные экосистемы являются мощным резервуаром органического
углерода на планете. По данным [7] в составе торфяной залежи
аккумулируется не менее 15% глобальных запасов углерода почвенного покрова, что
соответствует 180-260 млрд. тС. При этом 114-118 млрд. тС [1,2], то есть как
минимум 40% мирового запаса сосредоточено в торфяниках России.
Учитывая существенное значение болот в проблеме парникового эффекта и
распространенность этих экосистем на территории нашей страны, следует
признать изучение газовой функции болот одной из наиболее приоритетных
задач отечественной экологии и смежных дисциплин. Очевидно, что
нерациональная хозяйственная деятельность по освоению болотных экосистем
может привести к их разрушению и мобилизации значительных количеств
230
БИОСФЕРНАЯ ЮЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
апливаемого веками углерода в виде С02. Вместе с тем природные забо-
**eHHbie земли выступают в качестве мощного источника другого парнико-
Л о газа — метана. По современным оценкам от 1/4 до 1/3 глобального
^оизводства CbL» (560 млн. т/год) приходится на долю переувлажненных
мель, в состав которых в первую очередь входят болотные массивы [7].
Умеренное осушение способствует формированию метанотрофного фильтра
поверхностных горизонтах торфяной залежи и резкому сокращению
выделения СН4 в атмосферу. Таким образом, возможность управления газовой
функцией заболоченных земель заключается в поддержании оптимального
уровня почвенно-грунтовых вод, при котором с одной стороны не
происходит значительного разрушения торфяника, а с другой — минимизирована
эмиссия метана. Очевидно, что реакция болотных экосистем на изменение
водного режима, а, значит, и параметры оптимального функционирования
этих объектов не могут быть достоверно определены без предварительного
изучения внутрипочвенных механизмов генерирования, переноса и
выделения газообразных веществ. Последняя задача составила предмет данного
исследования, где указанные процессы оценивались количественно по
данным о профильном распределении, эмиссии и диффузии парниковых газов.
Экспериментальный материал был собран в летние полевые сезоны
1997-98 г. на стационарах ф-та почвоведения МГУ (Московская обл.,
Солнечногорский р-н, п. Чашниково), Ин-та лесоведения РАН (Тверская обл.,
Западнодвинский р-н, д. Сосвятское) и Ин-та микробиологии РАН (Томская
обл., Бакчарский р-н, п. Плотниково). Участок №1 находится в
притеррасном понижении поймы р. Клязьма на заболоченной территории грунтового
питания, подверженной осушительной мелиорации (УОПЭЦ Чашниково).
Две следующих опытных площади расположены на территории низинного
болота смешанного типа увлажнения под черноольшаником крупнотравно-
папоротниковым (№2) и черноольшаником болотно-травяным (№3)
(Западнодвинский стационар, объект «Грустинка»). Участок №2 не подвержен
осушению и, УПГВ на нем за период наблюдений (июль-октябрь 97)
находился в пределах 10-17 см. На дренированном участке №3 почвенно-грун-
товые воды залегали существенно глубже (60-80 см) и лишь осенью после
обильных осадков поднимались до отметок 30-40 см. Последний объект
(№4) является элементом типичного западносибирского верхового болота
атмосферного питания под травяно-сфагновой растительностью. Дренажная
сеть оказывает слабое воздействие на исследуемую территорию, в
результате чего УПГВ в течение полевого сезона редко снижается до глубин 10-12
см и в основном варьирует в пределах верхних 5 см торфяной залежи.
Результаты полевых наблюдений за профилем концентраций
парниковых газов (С02, СКЦ) с использованием стационарных мембранных пробо-
231
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
отборников [4] выявили наличие экстремума в их вертикальном pacim*^
лении (рис.1). Это означает наличие двустороннего градиента концентрации
и, соответственно, противоположно направленных вертикальных диффу^
онных потоков газов. Для их оценки были определены эффективные код.
фициенты диффузии газов в зависимости от пористости аэрации (D(Ilg)) №
(рис.2). В отличие от работы [3] функция D(IIg) впервые изучалась во всей
диапазоне естественного варьирования, включая состояния предельно^
влагонасыщения. Расчет потоков показал, что геотропический вертикальны!
перенос в неосушенных торфяниках (q= 4-30 мг/м2час для С02 и 0,Ь2
мг/м2час для СНЦ) часто превьшшет восходящий (q= 2-6 и 0,06-1 мг/м2час)
что связано с наличием воздухоносной пористости и значительными града,
ентами концентраций в нижних слоях.
h,CM
35
30
25
Э
20
2, г/и
СО15
10
5
О1
LJ_h „ , , ,
-*-т.2.07.в7|
-*-т.2.08.971
-*- т.2.00.971
-^-т.2.10.в7|
И,см
h, см
Ь,см
Рис. 1 Профильные распределения парниковых газов в болотных почвах
В осушенных объектах доминирует восходящая диффузия,
обеспечивающая на 80-100% эмиссию газов с поверхности. Определение
гросс-продукции СОг почвой (V) с помощью балансовой кинетической модели [4]
выявило, что этот показатель превышает на 13-23 % эмиссию газа. Так для
объекта №2 суммарная за июль-октябрь величина V составила 108 г/м', а эмиссия
— 84 г/м2; для объекта №3 — 577 г/м2 при эмиссии 500 г/м^ Разница
обусловлена аккумуляцией, геотропическим переносом и последующим
латеральным стоком растворенного газа. Сопоставление данных о поступлении
232
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
етятельных остатков [6] и его минерализации в виде парниковых газов
С Р ^^кой на дыхание корней на объектах 2 и 3 показало, что депонирова-
С П тпег^па/ям&я место только в неосушенных болотных экосистемах.
Объект №1
у = 0008Эк+0 9153х + 012
Rg=0 9999
Объект№4
у = О 004бк+ 0 047%- 0 0878х +
XUfiu-
Рис.2. Эффективные коэффициенты диффузии СОг (объект №1) и СН4 (объект №4)
в болотных почвах, см2 час"1
[1] Вомперский С.Э. и др. Заболоченные органические почвы и болота России и
запасы в их торфах// Почвоведение. 1994. № 12. С. 17-25. [2] Ефремова Т.Т. и др.
Запасы и содержание соединений углерода в болотных экосистемах России//
Почвоведение. 1997. № 12. С. 1470-1477. [3] Икконен Е.Н., Толстогузов О.В Диффузия
углекислого газа в торфяной почве верхового болота // Почвоведение. 1996. № 7. С.
868-872. [4] Смагин А.В. Анализ поведения углекислого газа в почве // Вести. МГУ
сер 17. 1998. № 4. С. 28-35. [5] Смагин А.В., Смирнов Г.В. Методы определения
эффективных коэффициентов диффузии СО2 в почве // Вести. МГУ. сер 17. 1996. №
2 С 3-10 [6] Смагина М.В. Микроорганизмы и экологические особенности
трансформации органического вещества в осушаемых болотных лесах. Автореф. дисс.
канд биол. наук. Красноярск, 1988. [7] Bridges E.H., Batjes N.H. Soil gaseous
emission and global climatic change // Geography. 1996. V. 81 (2). P.155-169.
ПОЧВЕННО-ГЕОХИМИЧЕСКИЕ ПРОЦЕССЫ В
СОПРЯЖЕННЫХ ФАЦИЯХ ЗАБОЛОЧЕННОГО ЛЕСА
В. А. Снытко, А. И. Щетников
Институт географии СО РАН, Иркутск
Важное значение в диагностике параметров функционирования лесных
геосистем повышенной гидроморфности имеет подвижное вещество.
Высокое содержание свободной влаги обеспечивает возможность нахождения
химических соединений и элементов в относительно мобильном состоянии,
что в значительной мере определяет их системоорганизуюшую роль. На
основе знаний пространственной и временной динамики активно мигрирую-
233
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
щего в органо-литогенном субстрате геосистем вещества определяю^*
форма и характер, протекающих процессов, устанавливаются их интенсв&„
ность, мощность и направленность как в пространстве, так и во времен»
Для заболоченных лесов представляется возможным наметить тренды ра^
вития доминирующего процесса, его устойчивость и предсказуемость.
Анализ практически любой геосистемы, а заболоченного леса в особен*
ности, следует проводить в тесной связи с окружающим ее пространством
Нами было осуществлено исследование не только фаций собственно заболо»
ченного южно-таежного леса, но и связанных с ней потоком вещества фацд|
с убывающей гидроморфностью. Натурные стационарные ландшафтно-гео-
химические наблюдения показали, что в заболоченном лесу в течение всеп)
вегетационного периода наименее изменчивы такие почвенно-геохимичес*
кие показатели как окислительно-восстановительный потенциал (ОВП), со-
став почвенного воздуха (С02) и влага. По сравнению с транзитными и
автономными местоположениями здесь весьма высокую временную
активность проявляет водно-растворимое органическое вещество (ВОВ).
Достаточно чувствительны к сезонным флуктуациям природных параметров
соединения подвижного железа и фосфора. Если в местоположениях
автономного ряда среди таких показателей как влага, рН, ОВП, ВОВ, железо,
фосфор, С02, общий углерод, подвижные кальций и магний преобладают
достаточно высокие прямые корреляционные связи, то в условиях аккумулягив-}
ных супераквальных с доминированием процессов заболачивания
соотношение высоких прямых и обратных зависимостей уравновешивается.
Повышается контрастность процессов, усиливается антогонизм почвенно-гео-
химических показателей. Ужесточение внутрифациальной структуры и
взаимообусловленности параметров заболоченного леса по сравнению со
смежными менее гидроморфными местоположениями, свидетельствует о
меньшем потенциале устойчивости системы и ее пониженной толерантности по
отношению к внешним природным и антропогенным воздействиям.
НЕКОТОРЫЕ ОСОБЕННОСТИ ОРГАНИЧЕСКОГО ВЕЩЕСТВА
ПОЧВ РАЗНОГО ТИПА ЗАБОЛАЧИВАНИЯ
С.Я. Трофимов, Е.И. Дорофеева
МГУ, факультет почвоведения
Изучалось органическое вещество заболоченных почв верхового и
низинного типов заболачивания на территории Центрально-лесного
государственного биосферного заповедника (Тверская обл.). Объектами исследования
были верховая торфяно-глеевая почва под ельником сфагновым, болотно-
234
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
золистая профильно-глееватая почва под ельником черничным, низин-
П торфяная почва под ельником черноольховым травяно-болотным, а
кясе незаболоченные почвы — палево-подзолистая под ельником кислич-
тм и бурозем под ельником неморальным. В лабораторных условиях опре-
елялись следующие характеристики: скорость разложения органического
вешества почв в условиях оптимальной влажности и температуры
(влажность 80% от предельной влагоемкости, температура 20°) по интенсивности
вЫделения углекислого газа с титрометрическим окончанием; содержание
углерода (на анализаторе углерода), азота по Къельдалю, зольность.
Скорость минерализации органического вещества низинной торфяной почвы
100
90
о
X
S 70
5 60-
О 50-
40-]
30-
¥ -X
I
\. ^**^>»*^1
1 ♦
♦ L
■ F
дН
х 80-70 см
ж 140-150 см
"^^.^^ ■
♦ ^ч^чч
1
■ ***^
^4vSs^ ♦
I 1
л
R*=0,9092
^ = 0,9548
R* = 0,9245 >ч
10
20
30
ДНИ
40
50
60
Скорость минерализации органического вещества весьма
удовлетворительно описывалась линейной функцией (см. рис.), что позволило условно
отнести органическое вещество изученных почв к четырем классам
устойчивости к разложению, выбрав при этом в качестве критерия величину
коэффициента разложения Км): I — активное органическое вещество (Км>10%
Сорг/месяц); II — лабильное органическое вещество (Км 2,5-10%СОрГ/месяц);
III — устойчивое органическое вещество (Км 1-2,5%Сорг/месяц); IV —
стабильное органическое вещество (Км<1%Сорг/месяц).
Изучение скорости разложения образцов органогенных горизонтов
показало значительные различия в степени устойчивости их к разложению —
образцы из торфяной низинной и незаболоченных почв разлагались со ско-
235
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ростью, на два порядка превышающей скорость разложения торфяных гори*
зонтов почв верхового типа заболачивания. Наибольшая скорость разложе*
ния была свойственна верхним органогенным горизонтам торфяной низин*
ной почвы, бурозема и палево-подзолистой почвы; наименьшая — для орга*
ногенных горизонтов торфяно-глеевой почвы. Практически во всех почвах
наблюдалось убывание скорости разложения сверху вниз по профилю,
исключение — почва верхового типа заболачивания, где максимальная
скорость разложения несколько нарастала с глубиной. Вероятно, более свежие
остатки сфагновых мхов содержат больше физиологически активных
компонентов, ингибирующих развитие организмов-деструкторов.
Коэффициенты минерализации органического вещества
Объект Коэффициент минерализации, Км Класс устойчивости к
%С0рг/сутки %Сорг/месяц разложению
Болотно-подзолистая («оторфованная белоподзолистая» почва)
гор. L
гор. F
гор. Н
гор. A2h
Болотно-подзолистая
гор. Т1
гор. Т2
гор. ТЗ
гор. T3/A2g
0,427
0,116
0,067
< 0,008
: («торфянисто
0,009
0,024
0,011
0,019
12,81
3,48
2,01
<0,25
-белоподзолистая» почва)
0,27
0,72
0,33
0,57
Палевоподзолистая почва (на карбонатной морене)
гор. L
гор. F
гор. Н
Бурозем
гор. L
гор. F
гор. АО
гор. Ahf
0,817
0,444
0,138
0,562
0,221
0,203
<0,008
Верховая торфяная почва
гор. Т1
гор. Т2
гор. ТЗ
0,006
0,007
0,009
Низинная торфяная почва
L
F
Н
70-80 см
140-150 см
1,085
0,718
0,048
0,023
0,086
24,51
13,32
4,14
16,86
6,63
6,09
<0,25
0,18
0,21
0,27
32,55
21,54
1,44
0,69
2,58
I
П
Ш
IV
IV
IV
IV
IV
I
I
П
I
П
П
IV
IV
IV
Г/
I
I
ш
IY
П
236
БИОСФЕРНАЯ РОЛЬ, БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ IV
различия в скоростях минерализации как в пределах органопрофилей,
таК и между разными объектами согласуются с результатами определения
и зольности, углерода и азота. Максимальные скорости разложения ха-
оактерны для почв с близкой к нейтральной реакцией среды и с
относительно высоким содержанием азота и зольных элементов. К таковым относятся
т0рфяная низинная и палево-подзолистая почвы и бурозем.
Несмотря на колоссальные различия в скорости минерализации нативно-
го органического вещества, различия в скорости минерализации
унифицированного внесенного субстрата как в природных, так и в лабораторных
условиях оказались несущественными [1,2], что свидетельствует о высокой
потенциальной биологической активности почв верхового типа
заболачивания. Это согласуется также с результатами определения содержания
микробной биомассы в исследованных почвах, которое оказалось наибольшим в
верховой торфяно-глеевой почве (Л.М. Полянская и А.В. Головченко —
перс, сообщ.). При этом наименьшее содержание микробной биомассы было
отмечено в торфяной низинной почве, вероятно, вследствие
антагонистических отношений микроорганизмов с почвенной фауной, широко
представленной в данной почве. По-видимому, низкая скорость минерализации
органического вещества верховой торфяно-глеевой почвы при высоком
содержании микробной биомассы обусловлено спецификой состава растений-
торфообразователей, а не является следствием низкой потенциальной
биологической активности.
[1] Трофимов С.Я. и др. Запасы органического вещества и скорость его
минерализации в заболоченных почвах ельников Центрально-лесного государственного
биосферного заповедника // Почвоведение. 1998. № 4. С. 416-422. [2] Трофимов С.Я.,
Дорофеева Е.И. О разложении хвойного опада в южнотаежных почвах разной
степени гидроморфизма//Вестн. МГУ. сер. 17, почвоведение. 1999. № 1. 1999. С. 3-11.
237
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
V. ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ БОЛОТ
V. 1. ГИДРОМЕЛИОРАЦИЯ И ЛЕСНОЕ ХОЗЯЙСТВО
ГЛАВНОЕ И ПРОМЕЖУТОЧНОЕ ПОЛЬЗОВАНИЕ
В ОСУШЕННЫХ ЛЕСАХ КАРЕЛИИ
В. А. Ананьев
Институт леса Карельского научного центра РАН, Петрозаводск
В Карелии осушено 760 тыс. га избыточно увлажненных земель. На
осушенных землях произрастают насаждения различного породного состава,
возрастной структуры и строения. Для получения максимального эффекта
наряду с лесоосушительной мелиорацией необходимо проведение
различных лесохозяйственных мероприятий (рубки главного пользования, ухода и
реконструкции), способствующих формированию древостоев максимальной
продуктивности и улучшению товарной структуры. При определении форм
хозяйства по способам рубок в осушенных лесах следует учитывать качество
и состояние естественного возобновления, строение, возрастную структуру,
состав и особенности хода роста древостоев.
Наибольшей энергией роста после осушения обладают молодняки и
средневозрастные насаждения, на долю которых приходится 29% лесопо-
крытой площади. Молодняки и средневозрастные насаждения через 20 — 25
лет после осушения имеют высокую полноту (0,9-1,2) и густоту (2,5-6,5 тыс.
шт/га) и наращивают запас, позволяющий окупить затраты на разреживание
при проведении рубок ухода. Изменение густоты и смена пород на
осушенных землях зависят от потенциального почвенного плодородия, исходной
полноты и возраста. В травяно-сфагновых типах леса, отличающихся
высоким плодородием, рубки ухода должны быть направлены на уборку
лиственных пород. В более бедных условиях, где преобладают чистые, с
незначительной примесью березы сосняки, необходимо только регулирование
густоты. Первоочередными объектами рубок ухода являются сосново-
лиственные молодняки (3 и более единиц лиственных в составе) и
средневозрастные насаждения с примесью до 5 единиц лиственных и перестойной
сосны с целью ухода за молодым поколением. В средневозрастных
насаждениях разреживание может быть назначено в целях ухода за подростом
хвойных пород, если его численность составляет не менее 1 тыс. шт/га, при
этом достаточно, чтобы запас верхнего яруса обеспечивал окупаемость
заготовки, а интенсивность разреживания не превышала 30%.
Значительная часть осушенных лесов представлена спелыми и
перестойными насаждениями (по сосне — 60%, по ели — 71%). Характерной осо-
238
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
бенностью спелых и перестойных осушенных насаждений, произрастающих
на торфяных почвах, является их разновозрастность и низкополнотность.
Анализ лесоустроительных данных (возраст, состав, полнота) показал, что
хвойные древостой с полнотой 0,6 и менее занимают 73% лесопокрытой
осушенной площади. Под пологом вышеуказанных древостоев имеется
достаточное количество жизнеспособного подроста и тонкомера, сохранение
которых в процессе лесозаготовок обеспечило бы лесовозобновление и
формирование насаждений с преобладанием хвойных пород на значительной
площади вырубок на осушенных землях. Изучение естественного
возобновления на сплошных концентрированных вырубках на торфяных почвах,
разработанных по технологии с сохранением подроста показало, что на
лесосеках 6-8 летней давности с учетом последующего возобновления
насчитывается от 5,1 до 6,5 тыс. шт/га жизнеспособного подроста, которого
вполне достаточно для формирования, при условии проведения ухода, хвойных
насаждений. По результатам обследования насаждений, сформировавшихся
из соснового подроста на вырубках через 20 лет после узколесосечных
сплошных рубок, установлены нормативы подроста, подлежащего
сохранению при проведении рубок главного пользования. Сохранение
жизнеспособного хвойного подроста при сплошных рубках на торфяных почвах в
количестве 3-3,5 тыс. шт/га с последующим двукратным (через 5-10 лет)
уходом в молодняках обеспечивает формирование хвойных насаждений с
полнотой 0,7 и выше.
В спелых разновозрастных ельниках наряду с повышением
продуктивности лесов после осушения отмечена повышенная фаутность. Число деревьев
ели, у которых гниль обнаружена на высоте 1,3 м, достигает 23%.
Значительное распространение фаутности в осушенных разновозрастных
ельниках, указывает на необходимость проведения рубок по улучшению
санитарного состояния и товарной структуры насаждений. Результаты опытных
рубок с сохранением на корню молодых тонкомерных деревьев и
жизнеспособного подроста показали относительно хорошую устойчивость их к изре-
живанию и достаточно высокий текущий прирост — 3,0 м3/га.
В богатых условиях (на торфяных почвах низинного типа) произрастают
в основном лиственно-еловые насаждения. Для улучшения условий роста
наиболее ценных хвойных пород и изменение состава древостоя в
желательном для хозяйства направлении необходима реконструкция лиственно-
еловых насаждений. По данным исследования рубок 15-летней давности (с
полной выборкой лиственного яруса), густота ели увеличилась в двое за счет
подроста, достигшего перечетных размеров и насчитывает 1000 деревьев на
1 га. Текущий годичный прирост запаса равен 3,5 м3/га и весь приходится
на наиболее ценную еловую часть насаждения, а на контрольных участках
До 50% прироста составляет низкотоварная береза.
239
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Проведение рубок ухода и главного пользования будут способствовать ве
только улучшению лесоводственного состояния и товарной структуры наса»
ждений, но и повышению эффективности мелиорации.
ВЛИЯНИЕ ТЕМПЕРАТУРЫ И ОСАДКОВ ЗА ВЕГЕТАЦИОННЫЙ СЕЗОН
НА СТОК С ОСУШЕННЫХ ЛЕСНЫХ ЗЕМЕЛЬ
Б.В. Бабиков, А.С. Алексеев
Санкт-Петербургская государственная лесотехническая академия
Важнейшими климатическими факторами, определяющими сток в реки,
являются осадки и температура воздуха. Осадки увеличивают сток,
температура, воздействуя на испарение, должна снижать его. Для анализа
влияния этих факторов использованы результаты 15-ти летних наблюдений,
проведенных на небольших водосборах осушенных лесных болот.
Использованы данные за май-сентябрь каждого из 15 лет. Краткая характеристика
объектов й статистические среднемесячные за вегетационный сезон (май-
сентябрь) характеристики температуры и осадков приведены в табл. 1 и 2.
Табл 1. Краткая характеристика объектов исследований
№/№ Расстояние между
участков каналами, м
1 65
2 130
3 205
4 128
Площадь
водосбора, га
7,1
20,4
7,5
8,1
УГВ за май-
Сентябрь, см
27
19
14
49
Класс бони-
[ тега древостоя
Ш
Ш-IV
IV
1-П
Табл. 2. Среднемесячные статистические характеристики температуры и осадков за
вегетационный сезон
Показатель
Среднее значение
Осадки, мм
Среднее квадратическое отклонение
Min
Мах
63,4
27,0
5,2
149,7
Температура, °С
15,2
3,6
6,8
22,0
Для изучения влияния температуры и осадков на сток с осушенных
лесных земель и р. Тосно применялся метод множественного регрессионного
анализа. Определено, что зависимость стока от осадков лучше всего
описывается следующей зависимостью: у = ai х\ + а2 х2, где, у, xi их2 — сток
(мм), температура (t °С) и осадки соответственно, ai и а2 — коэффициенты
модели, которые показывают изменение стока при увеличении температуры
или осадков на единицу. Для определения относительного влияния
температуры и осадков на сток рассчитывались их нормы замещения, dxi/dx2-
240
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
-a?/ai Норма замещения показывает, какое изменение температуры
необходимо для компенсации влияния на сток 1-го мм осадков.
Результаты регрессионного анализа влияния температуры и осадков на
сток приведены в табл. 3, в которой осушенные лесные участки различаются
расстоянием между осушительными канавами.
Табл. 3. Результаты регрессионного анализа влияния температуры и осадков на сток
с осушенных лесных земель и р. Тосно (единица измерения стока для р. Тосно —
м3/сек, уровень статистической значимости для всех уравнений — более 99%).
Объект Коэффициенты уравнения Норма Коэффициент
аь mm/i°C А2, мм/мм замещения детерминации, %
Река Тосно - 0,039 - 44,3
65 -0,182 0,197 1,08 73,5
130 -0,274 0,193 0,70 74,4
205 -0,211 0,138 0,65 63,4
128 - ОД75 - 55,7
Из табл. 3 следует, что для р. Тосно и участка с расстоянием между
каналами 128 м не установлено влияния температуры за вегетационный сезон на
сток. Бассейн р. Тосно составляет 130 тыс. га и имеет лесистость 75%,
заболоченность 12%, озерность 1%. Для анализа влияния температуры и осадков
на сток использовались данные только одной метеостанции, поэтому
результаты, относящиеся к р. Тосно, следует рассматривать как предварительные.
Для участков с различной интенсивностью осушения при ее снижении
имеет место падение нормы замещения климатических факторов,
определяющих сток. При увеличении расстояния между каналами для
компенсации увеличения осадков на 1 мм требуется все меньшее увеличение
температуры, что свидетельствует об относительном снижении влияния
температуры на сток.
Исследования также показали, что по мере повышения уровня грунтовых
вод (УГВ) (участки 1-3) влияние осадков снижается. Это объясняется тем,
что поступление воды в каналы в основном определяется величиной напора.
При высоком положении фунтовых вод (ГВ) в почве достаточно много
гравитационной воды и осадки мало меняют ее. При понижении ГВ степень
влияния осадков возрастает. При значительном понижении ГВ выше их
уровня создается значительная емкость аккумуляции и осадки
задерживаются не пополняя ГВ.
Температура, воздействуя на транспирацию, усиливает ее, но в
определенных пределах. При относительно высоких УГВ (участки 1-3), когда
корни растений в процессе десукции влаги получают ее из зоны капиллярной
каймы влияние температуры усматривается. При низком положении УГВ
(участок 4) растения потребляют влагу верхних горизонтов почвы и величи-
241
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
на транспирации, зависящая от температуры воздух, не влияет на сток, щ*
этому и зависимость стока от температуры не усматривается.
Проведенный анализ позволяет составить прогноз возможных изменений
стока с осушенных земель в зависимости от ожидаемых вариаций темпера*
туры и осадков. Учитывая тот факт, что в результате вероятных изменений
климата возможно увеличение и температуры и осадков одновременно, а их
влияние на сток противоположно, пока нет оснований ожидать
существенных изменений последнего.
ОСНОВНЫЕ ТРЕБОВАНИЯ К ТЕХНОЛОГИИ РУБОК ГЛАВНОГО
ПОЛЬЗОВАНИЯ В ДРЕВОСТОЯХ, ПРОИЗРАСТАЮЩИХ НА
ИЗБЫТОЧНО УВЛАЖНЕННЫХ ЗЕМЛЯХ БЕЛАРУСИ
В.Ф. Багинский
Институт леса НАН Беларуси, Гомель, Беларусь
Избыточно увлажненные земли (ИУЗ) Беларуси имеют громадную
экологическую ценность как место депонирования значительного количества
углерода, регулятор водного режима больших территорий и среда обитания
многих специфических видов животных и растений. На фоне повсеместного
осушения ИУЗ в Западной и Центральной Европе и перевода их в другие
категории земель болота Беларуси привлекают особое внимание мировой
научной общественности [9]. Представители Европы настойчиво советуют
нам сохранить белорусские ИУЗ.
Перед учеными республики стоит трудная задача сбережения болот и в
то же время их разумной и интенсивной эксплуатации. Какой-то
компенсации за выведение ИУЗ из хозяйственного оборота Запад нам пока не
обещает, а наше государство тоже не может себе позволить исключить из
эксплуатации около 2,2 млн. га древостоев на ИУЗ [9]. Но и сбрасывать со счетов
проблемы экологии, возникающие при лесоэксплуатации в болотных лесах,
не представляется возможным [8].
Лесные ИУЗ могут дать разнообразную продукцию. Но главным ее
видом, как по объемам использования, так и по суммарной стоимости,
является древесина, получаемая при рубках главного пользования [9].
Значительную добавь прироста древесины и упрощение технологий лесозаготовок и
лесовосстановления дает осушение [2-6], но в лесах Беларуси оно
практически прекращено по экономическим и экологическим соображениям [9].
При разработке технологий главного пользования в любых экотопах
неизбежно возникает проблема лесовосстановления, что отмечал еще Г.Ф.
Морозов: «Рубка и лесовосстановление — синонимы». На ИУЗ эта пробле-
242
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
обостряется из-за ограниченных возможностей провести здесь искусст-
енное восстановление вырубок.
Как правило, в болотных лесах ориентируются на естественное
возобновление. В сосняках, произрастающих на верховых болотах, этот процесс
затягивается на 8-12 лет [7]. Ускорить его можно путем минерализации
почвы. Восстановление вырубок в мягколиственных древостоях идет
преимущественно порослевым путем [1,7]. В ольховых древостоях это стало
серьезной проблемой, т.к. на больших площадях возобновились порослевые
черноольшатники 2, 3 и даже 4 генерации, что ведет к их постепенной
деградации [1]. Поэтому технологии рубок главного пользования на ИУЗ
должны предусматривать возможности для семенного возобновления
мягколиственных пород.
До недавнего времени эта проблема выпадала из поля зрения ученых
Беларуси, но сегодня ее решением занимаются в Институте леса в рамках
выполнения проекта развития лесного хозяйства Беларуси до 2015 года,
реализуемого с помощью кредита Всемирного банка. Основные подходы здесь
сводятся к уменьшению ширины лесосек и оставлению семенных куртин
(или полос). Правда, пока неясно, будут ли оставляемые деревья устойчивы
против ветровала.
В дополнение к описанным технологическим приемам желательно
создание на лесосеках минерализованных полос. Лучше это делать плугом
ПКЛ-70. но на ИУЗ такое возможно не всегда и не везде. Как альтернатива
здесь рассматривается удаление растительного покрова фрезами в ранневе-
сенний период, когда почва только начинает оттаивать. В дальнейшем
желательно организовать специфический уход за древостоем, сохраняя
семенные деревья и удаляя порослевые.
Проведение лесосечных работ на болотах, как и в других местах, не
должно вести к сокращению биологического разнообразия и другим
тяжелым экологическим последствиям. С этой целью в Беларуси силами
Института леса (В.И. Мироненко) в рамках работ по экологической сертификации
разработан проект стандарта, которым определены экологические
требования к лесосечным работам, учитывающие внутриреспубликанские и
международные подходы в этой области.
Таким образом, основными требованиями, выдвигаемыми к технологиям
рубок главного пользования на лесных ИУЗ, являются следующие: —
минимизация затрат на заготовку, трелевку и вывозку кубометра древесины;
— соблюдение лесоводственных правил проведения рубок с целью
обеспечения надежного естественного семенного возобновления, что потребует
изменения правил рубок главного пользования; — обеспечение норм,
предусмотренных правилами по экологической сертификации лесосечных работ
на лесных ИУЗ; — одним из эффективных путей рационализации лесополь-
243
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
зования и лесовосстановления является осушение низинных и переходных
работ, но это мероприятие в современных кризисных условиях пока не
рассматривается из-за невозможности осуществить большие капитальные
вложения.
[1] Багинский В.Ф. Проблемы лесовосстановления в Беларуси // Природные ресур.
сы. Межвед. бюлл. Науч изд.. Минск: НАН РБ. 1997. № 2. С. 64-72. [2] Биогеоце-
нотическое изучение болотных лесов в связи с опытной мелиорацией. М: Наука,
1982 207 с. [3] Буш К.К Взаимосвязь между продуктивностью древостоев и
интенсивностью осушения // Вопросы гидролесомелиорации. Рига: Зинатне, 1968. С. 5-50.
[4] Вомперский С.Э. Научное обоснование оптимального лесоосушения // Лесное
хозяйство. 1972. № 6. С. 28-33. [5] Ипатьев В.А. Повышение продуктивности лесов
на осушенных землях Автореф. дис. докт с.-х наук. Л.: ЛТА. 1985. 33 с. [6] Смоляк
Л.П. Болотные леса и их мелиорация. Минск: Наука и техника. 1969. 208 с. [7] Юр-
кевич И.Д., Гельтман B.C. География, типология и районирование лесной
растительности Белоруссии. Минск: Наука и техника 1965. 288 с. [8] Юшкевич Н.Т.,
Рожков Л Н. Лесное хозяйство Беларуси на пути к устойчивому управлению лесами
// Устойчивое управление лесами. Мат-лы межд. науч.-практ. семинара. Минск:
БГТУ 1998 С. 3-9. [9] Янушко А Д и др Стратегический план развития лесного
хозяйства Беларуси Минск* МЛХ РБ. 1997. 178 с.
ВЛИЯНИЕ ОСУШИТЕЛЬНОЙ МЕЛИОРАЦИИ НА
БИОПРОДУКТИВНОСТЬ СОСНОВЫХ ОЛИГОТРОФНЫХ БОЛОТ
СЕВЕРА БЕЛАРУСИ
В.В. Валетов
Государственный педагогический институт, Мозырь, Беларусь
Оценка биопродукционной перестройки болотных лесов в результате
осушения довольна сложна. Совершенно очевидно, что степень
информативности различных показателей продуктивности (как прямых, так и
косвенных) неоднозначна. Наиболее приемлемы сравнительные данные
продукции и особенности её структуры. Такой подход особенно эффективен по
отношению к напочвенному покрову, который подвержен наиболее быстрой
сукцессии. Однако нельзя игнорировать анализом изменений фитомассы, в
особенности напочвенного покрова, где не учитываются возрастные
различия растений. Этот тезис относится и к древостою при условии
сопоставимости его возраста.
По нашим данным в результате осушения наименее продуктивных пу-
шицево-сфагновых сосняков формируются пушицево-кустарниково-сфаг-
новые сосновые леса. В напочвенном покрове происходит лишь
незначительное увеличение числа видов сосудистых растений (с 8 до 10). В таксо-
244
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
омическом аспекте на уровне семейств изменений не отмечено. Спектр
ялов представлен до и после осушения пятью семействами.
Осушение сосняка пушицево-сфагнового приводит к увеличению
суммарной фитомассы на 30-47%. Продукция (2,1-2,8 до осушения и 2,3-2,4 т/га
после осушения) не претерпевает значительных изменений. Однако её
структура имеет принципиальные различия. При осушении происходит
интенсификация роста древесных растений, которые в конечном итоге
занимают доминирующее положение в составе продукции (34-46, против 11-13%
д0 осушения). Данную тенденцию подтверждает информация об изменении
ширины годичных слоев сосны. Этот показатель возрастает вдвое и
достигает 1,0±0,2 мм (различия статистически достоверны, t факт.>3).
О положительном влиянии осушения на древостой сосны
свидетельствуют биометрические показатели хвои. Сравнительный анализ указывает на
то, что в результате мелиорации длина, масса и площадь поверхности хвои
возрастают на 27-48%. Длина верхушечного и боковых побегов сосны
увеличивается на 54-77%.
Фитомасса напочвенного покрова осушенного пушицево-кустарниково-
сфагнового сосняка остаётся на уровне (9,9-11,2 т/га) его естественного
аналога (10,5-11,2 т/га, с. пушицево-сфагновый). Изменения касаются
величины суммарной продукции, которая уменьшилась в результате осушения на
35-40% (с 2,0-2,5 до 1,3-1,5 т/га год). В составе напочвенного покрова
наблюдается перераспределение продукционной значимости между
ботаническими группами растений. Фитомасса сфагновых мхов снижается на 25-
40%, в наибольшей степени аналогичные изменения присущи кустарничкам
и травам. В противоположность отмеченным особенностям доля
кустарников увеличивается на 90-95%, что в конечном итоге определяет
практическое равенство суммарной фитомассы напочвенного покрова сравниваемых
сосняков. Что касается отмеченного уменьшения продукции, то
закономерность обусловлена существенным сокращением годичного прироста сфагнов
и трав (более чем вдвое), что не компенсируют кустарники. В результате
осушения в напочвенном покрове отсутствует шейхцерия, более чем на
порядок снижается фитомасса подбела и клюквы. В покрове отмечено
появление бриевых мхов.
Сосняк пушицево-кустарниково-сфагновый при осушении
трансформируется в кустарниково-сфагновый. Принадлежность к одному типу болота с
рассмотренным выше сообществом предопределяет определённую степень
общности биопродукционных изменений. Отличительным в данном случае
является более высокая трофность почвы и не столь выраженное обводнение
естественного пушицево-кустарниково-сфагнового сосняка. Этот факт
значительно отражается на роли древостоя в составе фитомассы и продукции.
245
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Суммарная фитомасса сосняка кустарниково-сфагнового возрастает ъ»
35-46%. Продукция характеризуется стабильной величиной (2,6-3,5 до осу*
шения и 2,3-3,1 т/га год после осушения). Роль древостоя в её сложении я&,
ляется определяющей (1,0-2,4 т/га год), в то время как до осушения прирос*
не превышал 0,6 т/га год. Продукция напочвенного покрова сокращается о»
2,4-2,9 до 0,7-1,3 т/га год.
В результате осушения пушицево-кустарниково-сфагнового сосняка про*
исходит сравнительно интенсивная биопродукционная деградация напоч*
венного покрова. Фитомасса сфагновых мхов снижается на 31-48%, продув
ция более чем вдвое. Наблюдается также уменьшение фитомассы и продув
ции кустарничков и трав (в среднем на 80-86%).
Активизация роста древостоя в конечном итоге определяет то, что про»
дукция сообщества (древостой + напочвенный покров) остаётся практическ*
такой же, как до осушения).
Третью группу ассоциаций сосновых неосушенных болот олиготрофноп*
типа представляет сосняк багульниково-сфагновый. В результате
осушительной мелиорации формируются багульниково-чернично-, багульниково-
бруснично-сфагновые и мшисто-кустарниковые сосняки. В видовой насьн
щенности травяно-кустарничкового яруса изменений не установлено. В per
зультате осушения общий годичный прирост возрос с 2,5-3,1 до 3,6-4,1 т/щ
год. Основное увеличение связано с древесным ярусом. В среднем
продукция древостоя увеличилась на 50% и достигла 2,6-3,2 т/га год величина при*
роста напочвенного покрова сократилась на 29-76%. На порядок снижаете!
фитомасса пушицы, в 4-6 раз клюквы. В результате осушения происходи!
существенная перестройка яруса кустарников. К примеру, мирт может во*
обще не встречаться в напочвенном покрове. Фитомасса багульника и
голубики уменьшается на 48-97%. Что касается сфагновых мхов, то они могут
отсутствовать в составе покрова осушенных сосняков. В противоположность
сфагнам фитомасса бриевых мхов возрастает более чем в 10 раз.
О ПЛАНИРОВАНИИ И ПРОИЗВОДСТВЕ
ГИДРОЛЕСОМЕЛИОРАТИВНЫХ РАБОТ В ЛЕНИНГРАДСКОЙ ОБЛАСТИ
Г.Б. Великанов
Комитет по лесу Ленинградской области
Необходимость продолжения гидролесомелиоративных мероприятий я
обязательного сохранения эффективно осушенных земель (как наиболее
ценного лесного фонда) в области обусловлена следующими факторами: —-
большой заболоченностью ее территории, что является сдерживающим
фактором интенсификации лесного хозяйства, особенно при плохом развитии й
246
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
стоянии лесных дорог; — большими объемами ранее выполненных лесо-
лгущительных работ, что обязывает поддерживать осушительные системы в
абочем состоянии и увеличивать отдачу осушаемого гектара; — новыми
кономическими условиями, при которых, как отмечает [1], из-за высоких
транспортных расходов следует ориентироваться на обеспечение
потребности в древесном сырье преимущественно за счет собственных ресурсов.
Рекомендуемый подход к планированию и проектированию
гидролесомелиоративных работ основан на учете различий ландшафтов, которые
могут определять приоритеты при определении масштабов, методов, способов
осушения и величину оставляемой площади переувлажненных земель и
направлен на обеспечение устойчивого управления их ресурсами,
контролируемого экологического равновесия, биоразнообразия и улучшения
эстетических свойств лесоболотных ландшафтов.
Прогноз баланса переувлажненных и осушаемых земель в лесном фонде Ленин-
градской области на 20-летие до 2015 года
Площадь земель лесного
фонда, тыс. га
Земли лесного фонда Комитета по в целом по
лесу области
1995 2015 1995 2015
Переувлажненные, в т.ч. вновь заболоченные 1830 1870 2300 2350
под влиянием разрастания болот, строительной дея- - 8 10
тельности бобров и др.
в результате строительства линейных и подпорных - 8 10
сооружений;
в результате временного заболачивания вырубок
Осушаемые (учтенные) на объектах коренной
гидролесомелиорации, в т.ч.
с обеспеченной нормой осушения*
с необеспеченной нормой осушения
перейдут в разряд вторично заболоченных
Неучтенные из числа осушенных с 1951 по 1985 гг.
Лт.ч. вторично заболоченные
* 60% от площади с давностью осушения не менее максимального нормативного
срока работы осушительной сети без капитального ремонта
Прогноз баланса переувлажненных и осушаемых земель в лесном фонде
области на 20-летие до 2015 года (см. табл.) указывает на ожидаемое
увеличение площади земель постоянного переувлажнения на 20 тыс. га и
временного заболачивания вырубок на 30 тыс. га. Если не организовать
техническую эксплуатацию лесоосушительных систем, не провести необходимые
работы по их реконструкции, к концу периода площадь земель с
обеспеченной нормой осушения на объектах коренной гидролесомелиорации сокра-
247
-
453
272
181
-
133
74
24
390
160
230
60
-
-
-
480
288
192
-
147
88
30
410
170
240
70
-
-
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
тится на 112 тыс. га и в разряд вторично заболоченных перейдут 70 тыс ^
Произойдет катастрофическое ухудшение эколого-лесоводственного coc&w
ния осушаемых земель.
Возрождению гидролесомелиоративных работ может способствовать в*
редача в долгосрочную аренду эксплуатируемых лесов крупным аренда^
рам: например, объединениям леспромхозов с деревообрабатьюающим^
целлюлозно-бумажными предприятиями, экономически заинтересованны^
в повышении производительности и продуктивности лесных земель.
К числу самых неотложных работ по гидролесомелиорации следует оц»,
сти реконструкцию и капитальный ремонт осушительных систем. Главны^
критериями выбора объектов для этих работ предложены давность осуц*
ния и качество осушаемых земель. В неотложной реконструкции и ремонт
нуждается до 35% площади лесоосушительных систем, что близко (36%) $
рекомендациям [2] по Беларуси. До 2015 г. рекомендуется их выполнить щ
170 тыс. га и осушить вновь 20 тыс. га переувлажненных земель.
С этой целью нами было выполнено специальное гидролесомелиорапщ.
ное районирование Ленинградской области [3] путем объединения ранее
выделенных СПбГУ и СПбНИИЛХ [4] ландшафтов с близкими природны*
ми характеристиками. Первоначально было выделено три группы ландшаф.
тов с девятью гидролесомелиоративными районами (ГЛМР) по три в
каждой группе, которые в дальнейшем были дополнены четвертой (см. статью
Г.Б. Великанова и др. в настоящем сборнике).
При установлении приоритетов гидролесомелиорации по ГЛМР для ила»
нирования, привязки объектов работ и принятия проектных решений следу*
ет учитывать характеристику и размещение следующих территорий: —
переувлажненных и осушаемых земель; — разрабатываемых торфяных
месторождений и выработанных торфяников; — охраняемых территорий; —
линейных сооружений и крупных водохранилищ; — наиболее горимых
территорий; — районов с концентрацией сплошно-лесосечных вырубок и
бобровых поселений; — территорий, загрязненных радионуклидами; —
ландшафтов с напряженным меженным стоком лесных рек и ручьев.
В первой группе ГЛМР особое внимание нужно обратить на ландшафты
12 и 31, где расположены Свирские и Нарвское водохранилища, и где может
потребоваться польдерное осушение. В этой группе находятся земли,
загрязненные «чернобыльским следом», реабилитация которых возможна с
применением методов гидротехнической мелиорации [5]. Во второй группе
гидролесомелиорация в целом менее эффективна в лесоводственном
отношении, а в восточных районах желательны осушение и канализация
верховых болот, что может, как вытекает из исследований ГТИ, СПбНИИЛХ [6,7]
и СПбЛТА [8,9], улучшить напряженную ситуацию с меженным стоком
лесных речек и ручьев. В третьей группе, куда входит и Карельский переше-
248
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
и назначении гидролесомелиоративных мероприятий следует в боль-
pit ПР** „
" мере обращать внимание на улучшение условии.отдыха населения и на
01 -панение биоразнообразия.
° проекты осушения и реконструкции лесоосушительных систем должны
' ггь комплексными, предусматривающими мероприятия по освоению и
гашению лесов и экологичными, отвечающими требованиям рациональ-
ого природопользования. Их необходимо увязывать с целым комплексом
Факторов: — с проектированием и эксплуатацией инженерных линейных и
подпорных сооружений; — с регулированием расселения бобров; — с
противопожарной профилактикой земель лесного фонда, которой уделяется
большое внимание; — с освоением выработанных торфяников; — с
проектами осушения смежных объектов сельскохозяйственной мелиорации и
разработки торфяных месторождений, что отвечает требованиям учета
межбассейнового влияния мелиорации.
[1] Ильин В.А. К проблеме финансирования лесоосушения // Гидротехническая
мелиорация земель, ведение лесного хозяйства и вопросы экологии: СПб.:
СПбНИИЛХ, 1997. С. 7-9. [2] Шараг Е.И. Формирование высокопродуктивных насаждений
на мелиорированных торфяных почвах: Автореф. дисс. канд. с.-х. наук. Гомель: ИЛ
НАН Беларуси, 1996. 20 с. [3] Великанов Г.Б. Ландшафтно-экологическое
обоснование гидролесомелиорации в Ленинградской области: Автореф. дисс. канд. с.-х. наук.
- СПб: СПбНИИЛХ, 1998. 24 с. [4] Общие принципы стратегии лесопользования и
лесовыращивания на ландшафтно-типологической основе: Сб. науч. тр./
СПбНИИЛХ. СПб, 1994. 135 с. [5] Ипатьев В.А. и др. Лес и Чернобыль (лесные экосистемы
после аварии на Чернбыльской АЭС, 1986-1994 гг.). Минск, ИЛ НАН Беларуси,
1994. 252 с. [6] Крестовский О.И., Книзе А.А. Общие закономерности стока воды в
реки с малых площадей в Северо-Западном регионе России// Гидролесомелиорация:
задачи и координация исследований. СПб: СПбНИИЛХ, 1994. С. 85-86. [7] Книзе
А.А., Бурневский Ю.И. Гидрологические функции лесов Ленинградской области в
структуре лесов географических ландшафтов// Гидролесомелиорация: наука -
производству: СПб.: СПбНИИЛХ, 1996. С. 64-66. [8] Бабиков Б.В. Использование
болотных вод// Лесн. хоз-во. 1985. № 3. С. 30-31. [9] Бабиков Б.В. Гидротехнические
мелиорации лесных земель. М.: Экология, 1993. 224 с.
СОСТОЯНИЕ И РОСТ КУЛЬТУРЫ СОСНЫ НА ОСУШЕННОМ
ПЕРЕХОДНОМ БОЛОТЕ В ЮЖНОЙ КАРЕЛИИ
В.Н. Гаврилов
Институт леса Карельского НЦРАН, Петрозаводск
Осушение болот и формирование на них высокопродуктивных
насаждений является одним из путей увеличения покрытой лесом площади лесфон-
Да. Для условий Карелии этот вопрос весьма актуален, так как
гидролесомелиоративный фонд республики составляет 37% площади гослесфонда. По
249
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
состоянию на январь 1990 года в Карелии осушенный фонд, после отчу*^
ния части земель под охранные зоны, составлял 651 тыс. га, из которь*
44,2% представлено болотами. К лесокультурному фонду отнесено 105 тъ*
га, из которых на 72 тыс. га созданы лесные культуры.
Эффективность лесоосушения болот напрямую зависит от темпов щ*
цесса их облесения, которая во многом определяется типологическими »
широтно-зональными условиями произрастания, интенсивностью осушень
и способом облесения. Согласно исследованиям в Карелии в 80-ых гг. [\\
наиболее успешно процесс облесения происходил в условиях мезотрофао|
группы болот на юге республики (среднетаежная подзона), как при ескэ^
венном, так и при искусственном лесовозобновлении. Однако, при лесокуд|.
турном освоении осушенных болот в большей мере гарантируется
образование ценных хвойных и хвойно-лиственных молодняков.
Основной породой в условиях Карелии, используемой при создание
культур на осушаемых торфяных почвах, являются сосна, так как ель неред.
ко повреждается в массовом количестве поздневесенними и раннелетнщц
заморозками на открытых пространствах. Предыдущие исследования
позволили разработать основные методы агротехники выращивания хвойных по*
род при искусственном облесении болот. Эти рекомендации касаются лищь
первого этапа лесообразовательного процесса — от момента создания куль-
тур до образования сомкнутого полога из древесной растительности. В про»
веденных нами в последнее десятилетие исследованиях основной упор
делался на особенности процесса формирования молодых древостоев.
Исследования по изучению состояния, роста и строения лесных культур
сосны обыкновенной проводились в Южной Карелии на территории
научного стационара «Киндасово» Карельского НЦ РАН. Опытный участок
расположен в пределах крупного болотного массива, осушенного в 1970 г.
(расстояние между осушителями — 160 м), где в 1972 г. на площади 17 га были
созданы культуры сосны обыкновенной по микроповышениям (пластам).
Подготовка почва — бороздование, расстояние между бороздами сильно
колеблется, при среднем — 8 м. Торфяная залежь переходного типа,
перекрытая небольшим слоем (6-10 см) верхового торфа. Зольность торфа до
глубины 70 см — 3-4%, кислотность — 3,4-3,8. Первоначальная густота
культур — 4,3 тыс. шт./га. Сохранность — 60-70%.
Результаты проведенных измерений показывают, что, при сохранности
культур сосны более 60%, через 20 лет после осушения на осоково-сфагно-
вом болоте образуется сложный по составу (по запасу) древостой — 4,5 Слк
(20) 1,6 С(25) 2,7 Б(25) 1,5 Б(15). Это обусловлено тем, что в данных
условиях произрастания до осушения имеется подрост сосны и березы, а также
активно идет возобновление березы. С удалением от осушения процент
березы уменьшается. Общая густота формирующегося молодняка — 6-8 тыс.
250
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
/гя относительная полнота — 0,8-0,9. На лесные культуры приходится
40-45% запаса стволовой древесины.
~"~ Анализ результатов свидетельствует о начале естественного изреживания
древостое, начиная с 15-летнего возраста, которое вызвано как отставани-
в росте и гибелью культур при угнетении рядом стоящими деревьями,
и механическими повреждениями при объедании побегов и сломе
стволов лосем. Отпад идет за счет деревьев низших ступеней толщины.
Продуктивность молодняков искусственного происхождения во многом
зависит от интенсивности осушения. В приканальной зоне (30 м) рост куль-
jyp сосны в период от 20 до 25 лет усиливается. Средний периодический
ярирост в высоту здесь в 1,5 раза выше, чем на удалении от осушения 50-80
м. Среднепериодическое изменение запаса (без учета отпада) составляет в
приканальной зоне 2-3 м3 /га в год. Проведение раздельного учета деревьев
по классам роста показало, что деревья-лидеры (высота выше средней
высоты древостоя на 15%) продуцируют 35-37% запаса стволовой древесины,
хотя их количество составляет лишь 15-16% от общего числа деревьев. Доля
запаса подчиненных и отставших в росте деревьев, при их числе 30-35% от
общего количества, не превышает 15%.
Таким образом, при искусственном облесении мезотрофных осушенных
болот южной Карелии образование молодых древостоев заканчивается к 15-
20-летнему возрасту. К этому моменту увеличивается количество
угнетенных деревьев, начинается процесс естественного изреживания.
Продуктивность молодняков довольно высокая (И-Ш класс бонитета), однако для ее
поддержания необходимо проведение комплексного ухода — осветление
культур и разреживание в рядах. Кроме этого, для сохранения лесных
культур в дальнейшем и активизации их роста требуется ремонт осушительной
сети и ее реконструкция с уменьшением межканального пространства.
[1] Саковец В.И., Гаврилов В.Н. Лесообразовательные процессы на осушенных
болотах Карелии // Петрозаводск, 1994,102 с.
гаДРОЛЕСОМЕЛИОРАЦИЯ КАК МЕТОД РЕГУЛИРОВАНИЯ
НАКОПЛЕНИЯ РАДИОНУКЛИДОВ ДРЕВЕСНЫМИ РАСТЕНИЯМИ
В.А. Ипатьев
Институт леса НАН Беларуси, Гомель, Беларусь
Одним из факторов, влияющих на миграцию радионуклидов как вглубь
почвенного профиля, так и в надземную часть растений, является влажность
почвы [1]. Нам представляется весьма важным использовать этот фактор
^я лесных насаждений, произрастающих в условиях крупномасштабного
Радиоактивного загрязнения после аварии на Чернобыльской АЭС в апреле
251
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
1986 г. В почвенном покрове Белорусского Полесья, по территории которое
пролегает северный след Чернобыльских выпадений, как известно [2], пр^,
валируют почвы с избыточным увлажнением, достаточно развита искусе**
венная дренирующая сеть, в т.ч. и на лесных землях.
Наши исследования в середине 80-х годов с искусственными радиоа*»
тивными источниками позволили выявить довольно тесную связь между
уровнем влагообеспеченности торфяно-болотных почв и интенсивность^
потребления из них древесными растениями радиоактивных веществ [3]. j
конце 80-х годов, после аварии на ЧАЭС, исследования в данном направлю
нии были продолжены, но уже непосредственно на радиоактивно загрязнен-
ных лесных почвах. Целый ряд поставленных вегетационных опытов с ра«
диоактивно загрязненными гидроморфными почвами, для которых создав*
лись строго регулируемые условия водно-воздушного режима в течение все*
го вегетационного периода, в целом подтвердил данные, полученные с и<х
кусственными радиоактивными источниками [4]. Следовательно, существу*
ет реальная возможность направленного воздействия на водно-воздушный
режим почвы с целью снижения корневого потребления радионуклидов
древесными растениями.
В процессе исследования было установлено, что на торфяно-болотных
почвах существенное снижение накопления 137Cs и ^Sr различными
компонентами (древесина, побеги, хвоя) древесных растений определяется строго
определенными параметрами уровня грунтовых вод (УГВ). Следовательно,
при гидролесомелиорации загрязненных радиоактивными веществами
лесных почв необходимо ориентироваться на такой средневегетационный УГВ,
который обеспечивает сбалансированность двух процессов: скорости
увеличения прироста мелиорированных лесных насаждений и интенсивности
потребления ими радионуклидов из почвы.
С начала 90-х годов в данном направлении проводится цикл
исследований в естественных условиях роста лесных насаждений на мезотрофных и
евтрофных болотах. Остановимся на некоторых результатах исследования в
сосняке болотно-травяном IV класса возраста, произрастающего при
плотности загрязнения 137Cs 20-25 104 Бк/м2. Лесной массив был осушен в 1965-
1966 гг. сетью открытых каналов (среднее расстояние между осушителями
180 м). Исследования проводятся в течение 8-ми лет при различном
удалении, а следовательно, и УГВ, от мелиоративного канала (15 и 145 м) и
идентичных ботаническом составе торфа и его агрохимических показателях [4].
Проведенные исследования позволили выявить, во-первых, устойчивое
снижение накопления 137Cs в корнеобитаемом слое торфа и увеличение
накопления в нижележащих (до 0,5 м) горизонтах. Это явление мы связываем
не только с повышением влажности в близлежащих к зеркалу грунтовых вод
слоях торфа, но и с более выраженной амплитудой колебания УГВ, возрас-
252
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
шего после проведения гидромелиоративных работ [3]. Во-вторых, мно-
летние радиометрические исследования в исследуемых насаждениях пока-
и на четко выраженное различие в скорости накопления l37Cs при сред-
невегетационных УГВ в данном типе леса в 27 и 56 см — величина
коэффициентов накопления данного радионуклида в древесину, однолетнюю
хвою и побеги — снижается в 2-2,5 раза быстрее при последнем значении
уГВ. который обеспечил к 1999 году 3-4-х кратное снижение содержание
радиоцезия в древесине сосны по сравнению с контролем (УГВ 27 см).
Полученные результаты по снижению интенсивности накопления 137Cs
древесным ярусом сосняка болотно-травяного при средневегетационном
уГВ 56 см подтверждаются данными 8-летних радиометрических
измерений при УГВ 56 и 27 см опада, формирующего лесную подстилку в данном
типе леса. Установлено математически достоверное увеличение различий в
содержании радиоцезия составляющими опада во времени — если в 1991
году оно составляло всего 20%, то к 1999 году возросло до 240%.
Проведенные исследования в вегетационных и естественных условиях
роста древесных растений позволили выявить весьма важную роль
гидролесомелиорации как метода, позволяющего выращивать лесные насаждения
на торфяно-болотных почвах с пониженным содержанием радионуклидов,
что весьма важно для ускорения реабилитации огромной части лесных
массивов, подвергнутых крупномасштабному радиоактивному загрязнению.
[1] Алексахин P.M. и др. Сельскохозяйственная радиология. М.: Экология, 1991. 215
с. [2] Дементьев А.В. и др. География Беларуси. Мн.: Высшая школа, 1965. С. 141-
158. [3] Ипатьев В.А. Водный режим и потребление элементов питания в
мелиорированных сосновых фитоценозах. Мн.: Наука и техника, 1990. 127 с. [4] Ипатьев
В.А. и др. Лес и Чернобыль // Лесные экосистемы после аварии на Чернобыльской
АЭС. Мн.: МНПП «Стенер», 1994. С.177-192.
СПОСОБ ОПРЕДЕЛЕНИЯ ОПТИМАЛЬНОГО РАССТОЯНИЯ
МЕЖДУ ОСУШИТЕЛЯМИ
А.Е. Касьянов
Московский государственный университет леса, Московская обл.
Известен метод оценки влияния расстояния между осушителями на
продуктивность лесных культур. Он включает устройство двух осушителей в
плане в виде клина со схождением от устьев к истокам. По оси клина между
осушителями расстояние изменяется от минимального в истоках до
максимального в устьях. Поперек осушителей разбивают не менее трех
наблюдательных створов: истоковый, устьевой и центральный. В створах
устраивает пробные площадки и фиксируют параметры продуктивности лесных
253
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
культур. Недостаток известного метода — низкая точность, обусловленная
отсутствием повторное™ на одном клине.
Нами предложено не менее четырех соседних осушителей выполнять со
схождением от устьев к истокам, создавая на участке контроля не менее
трех последовательных клиньев. Поперек осей всех осушителей разбивают
не менее трех наблюдательных створов и вдоль створов на каждом клине
устраивают пробные площадки. Расстояния между осушителями в цен*
тральном створе равно расчетному расстоянию между осушителями на ме*
лиоративной системе. Трехкратная повторность пробных площадок
обеспечивает повышение точности результатов оценки.
Параметры планового размещения осушительной сети на участке
контроля определяются следующими зависимостями:
/, = {s +/ + d)/cos2^a/,aJ+l = 2arctg((e(A0 -l)cos£ay)/(5 + (E-e)sin2^ajyi
0 0 0
J J
У j =9°-Хау;Ву+1 =/](sinay+i/sin^+i)^]+i =(0-^Zay)sina^i/sin^M
о о
J j J
l\ =((r + 5)/cos^ai/) + (j + (£-e)sinay+1/(i4o "l)cos]Ta7)-e/g£a,;
0 0 0
e = £(l-cr):A0 = E/e:a = elE\
j j
rj = (((s I cos £a,) + (£ - e)tgaj) /(A " 1))" d I cos £ a,;
о о
где e — половина расчетного междренного состояния, /, — длина дрены, %
— угол примыкания осушителя к собирателю, В^ — расстояние между
устьями осушителей. b}j — расстояние между истоками осушителей, ц — угол
схождения дрен.
В наблюдательных створах устраивают, также, воднобалансовые
площадки. Число контрольных участков на осушительной системе определяется
изменчивостью почвенно-мелиоративных условий по площади системы.
При их существенной вариации устраивают не менее одного участка
контроля на 100 га. Если в центральном наблюдательном створе контрольного
участка формируется оптимальный водный режим, то расчетные расстояния
на мелиоративной системе выбраны правильно, если ли такие условия
отмечаются в устьевом наблюдательном створе, то расчетные расстояния
занижены.
254
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
ВЛИЯНИЕ ЛЕСООСУШИТЕЛЬНОЙ МЕЛИОРАЦИИ
НА БИОРАЗНООБРАЗИЕ.
А. А. Книзе
Санкт-Петербургский НИИ лесного хозяйства
Такое лесохозяйственное мероприятие, как осушение, изменяя водный
0еясим осушенных площадей и за счет этого производительность
лесорастительных условий, рост леса, типологическую структуру лесов и другие
характеристики насаждений, оказывает разноплановое влияние на состояние
биоразнообразия. В результате осушения лесов происходит изменение
типологической структуры лесного фонда — сокращаются площади с
насаждениями на недостаточно дренированных условиях местообитания и
появляются насаждения с антропогенными типами леса.
После осушения происходит отмеченная еще К. К. Бушем [1970]
конвергенция типов леса, т.е. число исходных типов леса и болот обычно
превышает число типов осушенных лесов. На осушенных площадях
увеличивается производительность лесорастительных условий, увеличиваются приросты
и запасы насаждений, усложняется структура насаждений. В результате
повышения производительности лесорастительных условий у значительной
части ранее чистых насаждений усложняется состав, появляется второй
ярус. Увеличивается число видов и сомкнутость подлеска. Происходят
сложные изменения в напочвенном покрове. В первое время после осушения
увеличивается число видов напочвенного покрова, поскольку одновременно
существуют, как представители напочвенного покрова, занимавшие данную
площадь до осушения, так и растения, характерные для условий,
образовавшихся после осушения. По мере стабилизации условий и выпадения трав,
кустарничков и мхов, "характерных до осушения, число видов напочвенного
покрова снижается. Разнообразие живого напочвенного покрова на
осушенных площадях несколько выше, чем на неосушенных [Jansons, 1997].
Экологическое разнообразие избыточно увлажненных земель возрастает в
результате их частичного осушения [Jansons, 1997; Zalitis, 1996]. Наибольшего
значения индекс биоразнообразия достигает там, где осушено 50-70%
избыточно увлажненных лесов [Zalitis, 1996].
Одновременное наличие на осушенных площадях лесных насаждений
разного возраста или покрытой и не покрытой лесом площади (насаждения
и трассы каналов) и водных площадей (русла каналов) улучшает кормовые
Условия обитания зверей и птиц [Вейнерт, 1989].
Таким образом, после осушения в результате повышения
производительности лесорастительных условий возможно как сокращение
биоразнообразия при полном осушении всех или почти всех избыточно увлажненных
255
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
площадей, так и увеличение биоразнообразия при осушении части переу*.
лажненных земель.
Для выявления степени осушения территорий выбрана Ленинградская
область, где из 4,5 млн. га гослесфонда (ГЛФ) осушено около 600 тыс.га^
где доля осушенных лесных площадей (около 14%) наиболее высокая од
Северо-Западе России. В границах области по ландшафтам устанавливалась
площадь болот и площадь избыточно увлажненных покрытых лесом земель
в ГЛФ области. Это позволило определить площадь избыточно
увлажненных земель по ландшафтам области. Площади осушенных лесов брались по
данным Г.В. Великанова [1998], который определял их по проектам Союз-
гипролесхоза. По его данным за послевоенное время в ГЛФ Ленинградской
области осушено 581 тыс. га лесного фонда. По проектам лесоосушения уда-
лось установить размещение по ландшафтам осушенных лесов на площади
453 тыс. га, что составляет 78% от всех осушенных площадей ГЛФ области.
Остальные проекты утрачены или площади перешли к другим
землепользователям. Распределение площадей избыточно увлажненных и осушенных
земель в ГЛФ показаны в таблице. В данной таблице из 36 ландшафтов
Ленинградской области для экономии места приведены только те ландшафты,
где проводилось осушение ГЛФ.
Как видно из таблицы доля избыточно увлажненных земель в ГЛФ
области составляет 45% и по ландшафтам колеблется в пределах 13-65% Доля
осушенных площадей ГЛФ от избыточно увлажненных по выявленным
проектам лесоосушения составляет 22%, а с учетом не полного выявления
осушенных земель — около четверти от всех избыточно увлажненных. По
ландшафтам доля выявленных осушенных площадей колеблется от 0 до
43%, а с учетом не полного выявления осушенных площадей — до 50%.
Лишь в одном случае в Нижнесвирском ландшафте доля осушенных
площадей от избыточно увлажненных составляет 60%. Однако надо учитывать,
что в этом ландшафте, кроме лесов ГЛФ, имеется около 20 тыс. га лесов
Нижнесвирского заповедника Комэкологии. в которых осушения не
проводилось. Это снижает долю осушенных лесных площадей в целом по
ландшафту до 50%. Наиболее высокая доля осушенных площадей в тех
ландшафтах, где доля избыточно увлажненных земель в ГЛФ составляет 40 -
65%. Осушение в этих условиях даже половины избыточно увлажненных
площадей не может отрицательно повлиять на биоразнообразие, а только
увеличивает его. Однако для сохранения этого увеличившегося
биоразнообразия необходимо поддерживать мелиоративную сеть канав в рабочем
состоянии. Выход канав из строя приводит к снижению производительности
лесорастительных условий, ухудшению роста осушенных насаждений
вплоть до их гибели. Первыми страдают от плохого состояния канав
насаждения в тех местах, где мелиорация дала наибольший эффект, осушенные
256
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
насажу
ения имеют высокую производительность и высокий потенциал
биоразнообразия.
определение по ландшафтам площадей избыточно увлажненных (ИУЗ) и осушен-
ОЗ) земель в гослесфонде Ленинградской области. ПЛ — площадь ландшафта.
нь1£
Х°
ландшафта.
2
3
9
11
12
13
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
33
34
Итого
ПЛ,
га
ГЛФ в ПЛ,
га
ИУЗ в ГЛФ,
га
ИУЗ/
ГЛФ, %
оз*
га
ОЗ/ИУЗ
%
230000
240000
90000
150000
190000
140000
430000
160000
360000
630000
240000
130000
480000
80000
180000
350000
380000
110000
100000
120000
480000
270000
120000
110000
140000
90000
7324000
177160,1
177673,8
49202,4
49564,0
116799,1
70679,1
219043,4
122417,6
83115,9
473445,1
72040,4
16979,0
300812,1
61474,2
92763,0
276233,3
262501,5
76011,3
57106,0
83788,1
28887,7
25455,2
9747,7
74890,8
110038,6
49550,2
4569451,8
23562,3
46017,5
14071,9
11895,4
52326,0
36399,7
137340,2
79693,9
17786,8
253293,1
28095,8
9117,7
180788,1
31782,2
53709,8
172645,8
150675,9
47507,1
14847,6
25387,8
122772,7
145858,3
38405,9
26286,7
22337,4
12338,0
2061581,5
13
26
29
24
45
51
63
65
21
53
39
54
60
52
58
62
57
63
26
30
42
57
39
35
20
25
45
2174
6607
4200
3475
21415
9134
47297
33224
1068
103332
11386
500
77573
11479
20139
12263
26118
6101
1700
3097
5912
17433
23111
2002
1035
1506
453281
9
14
30
39
41
25
34
42
6
41
41
5
43
36
37
7
17
13
И
12
5
12
60
8
5
12
22
* - выявленные объемы лесоосушения. Ландшафты: 2. Северо-Западное Приладо-
ясье, 3. Приморский Северобережный, 6. Центрально-водораздельный, 7. Лемболов-
ский, 8. Нижнелужский, 9. Приморский Южнобережный, И. Южное Приладожье,
12. Нарвско-Лужский, 13. Ижорский, 15. Лужско-Оредежский, 16. Лужский, 17.
Верхнелужский, 18. Тосненско-Волховский, 19. Мгинский, 20. Путиловский, 21.
Пащско-Сясьский, 22. Тихвинский, 23. Вишерский, 24. Среднеоятский, 25. Капшин-
ский, 26. Тихвинская гряда, 27. Судско-Чагодский, 28. Нижнесвирский, 29. Свирско-
Олонецкий, 33 Свирско-Оятский Западный, 34. Свирско-Оятский Центральный.
257
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
РУБКИ ГЛАВНОГО ПОЛЬЗОВАНИЯ В ЗАБОЛОЧЕННЫХ
ЛЕСАХ БЕЛАРУСИ: ПРОБЛЕМЫ И ПУТИ РЕШЕНИЯ
П.В. Колодий, В.Ф. Багинский
Институт леса НАН Беларуси, Гомель, Беларусь
В Беларуси на долю избыточно увлажненных земель (ИУЗ) приходщ*
18,3% лесной площади Гослесфонда. Наиболее распространенными порола
ми. произрастающими на этих землях, являются сосна (38,2%), ольха чеп!
ная (28.3%) и береза (25,8%). Возрастная структура древостоев на ИУЗ мало
отличается от этого показателя для всех лесов республики.
К объектам наших исследований относятся насаждения тех типов леса
которые в соответствии с классификацией [10] произрастают на низинных
переходных и верховых болотах, а также примыкающих к ним минерала
ных полугидроморфных и гидроморфных почвах.
Хотя структура лесов на ИУЗ складывалась веками, она довольно про-
стая. Здесь ограничен маневр в части формирования породного состава, т.к.
последний в основном определяется биологическими особенностями
древесных пород, их приспособлением к условиям местопроизрастания [3].
Основным способом повышения рентабельности лесного хозяйства на ИУЗ
длительное время считалось осушение этих земель [3,8]. Для осушенных
территорий разработаны рекомендации по ведению лесного хозяйства, описаны
мероприятия, повышающие продуктивность осушенных древостоев и т.д.
Лесные насаждения на ИУЗ, где не проведено осушение, изучены хуже.
В определенной мере известна их биология и продуктивность, описаны типы
леса и условия местопроизрастания [11]. Что касается вопросов рубок, то
сведений об их проведении в этих условиях практически нет.
Нетрадиционные методы выполнения лесохозяйственных мероприятий на избыточно
увлажненных землях в литературе освещены недостаточно. Лесовосстанови-
тельные и несплошные рубки на ИУЗ практически не применялись даже в
опытном порядке.
Технологический процесс лесосечных работ во всех экотопах состоит из
подготовительных, вспомогательных, основных лесосечных работ и очистки
лесосек [4,6]. Подготовительные и вспомогательные работы на ИУЗ почти
не отличаются от суходольных лесов. Что касается основных лесосечных
работ, то здесь необходимы специальные подходы. В ряде научных работ
приведены конкретные способы разработки лесосек на ИУЗ. Предлагаются
соответствующие системы машин [1,4,5]. Большинство исследователей
единодушны в том, что в переувлажненных лесах бензомоторные пилы не
имеют пока конкуренции, так как. сырые почвы являются неблагоприятными
для лесоэксплуатации [1]. В этих условиях очень низкая проходимость
машин. В периоды распутицы лесосеки становятся совершенно не проезжими,
258
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
\& заполнены жидкой грязью даже в сухую погоду. В условиях Белару-
*°Л чень часто средняя температура в зимнее время остается положительной
сИ не превышает — 3-5°С. что не позволяет серийным лесозаготовитель-
^ машинам осуществлять трелевку и вывозку древесины на ИУЗ.
НЬ 'невозможность вырубить спелые древостой на ИУЗ приводит к система-
вескому ежегодному недорубу расчетной лесосеки в размере 0,4-0,6 млн.
Тз йЛи Ю-15% от всей расчетной лесосеки в республике. Этот недоруб
лбычно компенсируют заменой лесосечного фонда, что ведет к перерубам в
уходольных насаждениях. Поэтому необходимо разработать технологию
внесезонной заготовки древесины на ИУЗ. Частично решение этой
проблемы возможно при использовании различного рода канатных установок /4/
или специальной техники с высокой проходимостью. Но опыт их
применения в Беларуси пока ограничен работами в Пружанском лесхозе
(австрийская канатная установка стоимостью 120 тыс. долларов США) и Лиознен-
ском лесхозе («железный конь»).
В действующих в республике нормативных документах [2,6,7,9]
специальных разделов по проведению рубок на лесных избыточно увлажненных
землях не имеется. В отдельных нормативах перечисляются серии типов
лесов на ИУЗ, служащих объектом сплошных или несплошных видов рубок
главного пользования, приводятся некоторые организационно-технические
элементы на разработку лесосек [2,9], даны требования и ограничения,
предъявляемые к трелевке древесины при разработке лесосек [7],
предложены способы очистки и использования порубочных остатков [9].
Все документы рекомендуют вести лесозаготовительные работы на ИУЗ
только в зимний период. При этом отсутствуют предложения по технологии
разработки переувлажненных лесосек, в т.ч. с сохранением подроста
хозяйственно-ценных пород или обеспечивающих их надежное естественное
семенное возобновление.
Практический опыт показывает, что чаще всего главное пользование на
ИУЗ осуществляется путем сплошной рубки. При этом придержки по
ширине лесосек, срокам примыкания и т.п. достаточно велики [6]. Рубка
разрешена, если запас товарной древесины на 1 га составляет не менее 50 м3. По
этой причине, а также по экологическим соображениям в последние годы в
большинстве сосняков по болоту рубки главного пользования не назначают.
Волотные сосняки V бонитета и ниже, приуроченные к ягодным местам и не
имеющие подроста, исключены из расчета лесопользования. В то же время
низкобонитетные перестойные древостой желательно вырубить для
предотвращения потерь древесины.
Постепенные (несплошные) рубки главного пользования возможны
только на минеральных ИУЗ с глубиной торфа до 30 см. Эти земли
характеризуются временным избыточным увлажнением в весенний или в аномально
259
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
дождливый летний периоды. Здесь технология главного пользования и дп*
гая лесохозяйственная деятельность мало отличается от таковой в суходолу
ных лесах, поскольку приурочивается к сухому периоду, когда возмоисао
свободное передвижение людей и техники.
Основные проблемы возникают на ИУЗ, где глубина торфа значитель
ная. Трудности в лесозаготовках здесь связаны с сезонностью работ. Прц
современных технологиях проход лесозаготовительной и транспортной тех*
ники возможен при сильных морозах — не менее 10-15 градусов в течение
нескольких недель. Исключением могут быть аномально сухие годы (раз *
7-10 лет), когда болота пересыхают.
Недостаток конкретных рекомендаций вызывает значительные трудности
при организации и выполнении лесосечных работ в рассматриваемых
насаждениях. Это часто ведет к неоправданным затратам труда, увеличению
себестоимости заготавливаемой древесины, снижению рентабельности всего
лесозаготовительного процесса лесхозов и леспромхозов.
Помощь в организации рубок главного и промежуточного пользования
на ИУЗ может оказать четкое прогнозирование периодичности пересыхания,
промерзания болот, сроков затопления водоохранных лесов и т.д. Это
позволило бы более рационально планировать проведение лесохозяйственных
мероприятий. При этом необходимо заложить в правила порядок
компенсации недорубов прошлых лет на ИУЗ только вырубкой их в благоприятные
годы, не допуская замены на суходольные лесосеки.
При организации лесного хозяйства в таких насаждениях требуется
разработка специальных способов, методов и технологий рубок, так как
традиционные не всегда приемлемы из-за особых условий произрастания лесов.
Поэтому вначале надо четко разделить древостой на ИУЗ по типам леса,
лесорастительным условиям, породному составу, возрасту,
производительности и т.д. На основании такого разделения, по различным лесоводственно-
таксационным признакам следует установить хозяйственную значимость
насаждений: выращивание товарной древесины, формирование кормовой
базы, получение недревесной продукции, сохранение биологического
разнообразия флоры и фауны и т.п. Это позволит определить основные способы
рубок главного пользования: сплошные, постепенные, выборочные.
Исследуемые насаждения необходимо классифицировать по степени
увлажнения и несущей способности почв, а также изучить обеспеченность
древостоев достаточным естественным возобновлением и подростом, что
даст возможность установить организационно-технические элементы рубок
главного пользования и предложить технологи разработки лесосек, системы
машин и способы очистки мест рубок.
В настоящее время в Институте леса проводится комплексное изучение
затронутых вопросов, что позволит разработать рекомендации по проведе-
260
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
jjjo рубок главного пользования на лесных избыточно увлажненных зем-
дях.
Ш Жуков А.В. и др. Заготовка сортиментов на лесосеке. Технология и машины. М.:
Экология, 1993. 311 с. [2] Инструкция по организации проведения несплошных
рубок главного пользования в лесах Республики Беларусь. Минск: Минлесхоз РБ,
1997. 72 с. [3] Ипатьев В.А. и др. Ведение лесного хозяйства на осушенных землях.
и ■ Лесн. пром-ть, 1984. 144 с. [4] Матвейко А.П. Технология и машины
лесосечных работ. Минск: Высшая школа, 1984. 334 с. [5] Матвейко А.П. и др. Справочник
мастера лесозаготовок. М.: Экология, 1993. 286 с. [6] Правила рубок главного
пользования и лесовосстановительных рубок в лесах Белорусской ССР. Минск: Ураджай,
1979. 19 с. [7] Рубки главного пользования. Экологические требования к
технологиям Проект СТБ. Минск: Минлесхоз РБ. 23 с. [8] Смоляк Л.П. Болотные леса и их
мелиорация. Минск: Наука и техника, 1969. 208 с. [9] Технологический регламент
на организацию и проведение постепенных и выборочных рубок в лесах главного
пользования. Минск: Минлесхоз РБ, 1998. 40 с. [10] Юркевич И.Д. Выделение
типов леса при лесоустроительных работах. Минск: Наука и техника, 1980. 120 с. [11]
Юркевич И.Д., Гельтман B.C. География, типология и районирование лесной
растительности Белоруссии. Минск: Наука и техника, 1965. 288 с.
СОВРЕМЕННОЕ СОСТОЯНИЕ ГИДРОЛЕСОМЕЛИОРАЦИИ:
АКТУАЛЬНЫЕ ПРАКТИЧЕСКИЕ И НАУЧНЫЕ ЗАДАЧИ
В.К. Константинов1, Н.А. Красильников2, С.В.Хитрин3
^ПбНИИЛХ; 2Ассоциация «ГИЛМ», Санкт-Петербург;
3Федеральная служба лесного хозяйства РФ
1. В 2000 году исполняется 225 лет, когда в России было начато
осушение болота площадью около 970 га, лежащего близ столицы вдоль р. Охты и
покрытого густым ельником и ольшаником. Хотя, большую его часть
предполагалось использовать не для лесовыращивания, а под луга и выгоны. С
тех пор общая площадь осушенных в разное время переувлажненных земель
(лесов и болот) в лесном фонде нашей страны превысила 5 млн. гектаров.
Из этой площади 3,5 млн. га было введено в эксплуатацию в советский
период в 1954-1988 годах. Причем, по данным инвентаризации 1987-88 гг, на
учете состояло только 2 млн. гектаров. Следовательно, потери ранее
осушенных земель превысили 3 млн. га, и об их лесоводственном состоянии
нет ясного представления. Несомненно, что на части этих земель (не менее 1
млн. га) осушительные каналы давно не работают, а сами земли вновь
заболотились. Многие участки осушенных земель были преданы другим
землепользователям, а также сняты с учета и необоснованно списаны. Такое
положение с учетом объектов гидролесомелиорации — земель, в которые
были вложены значительные средства, — является ненормальным и должно
261
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
быть исправлено по итогам единовременной инвентаризации лесоосуцщ„
тельных систем в лесном фонде, проводимой в 1999 году.
2. Лесоводственная эффективность гидротехнической мелиорации пере*
увлажненных земель изучена достаточно детально, но только в отношений
сосны и ели и не в таком объеме, чтобы на этой основе можно было соста*
вить региональные таблицы хода роста насаждений, мелиорированных в
разном возрасте с учетом обеспеченности нормы осушения и его давности. В
зависимости от условий дополнительный прирост древесины от осушения
достигает 1-10 м3/га в год. Однако, реально при отсутствии промежуточной)
пользования лесом и надлежащего ухода за осушительной сетью среднее
дополнительное накопление запаса за многолетний период для больших
площадей не превышает 0,7-2 м3/га в год. Улучшение породного состава и
возрастной структуры насаждений на осушаемых землях будет
способствовать повышению отдачи мелиорируемого гектара. Методы современной)
лесоводства разрабатывались преимущественно на суходолах и тем более не
относятся к осушаемым землям. В настоящее время закладываются основы
«мелиоративного лесоводства», но масштабы этих исследований не
позволяют быстро решить эту проблему. За счет осушения безлесных болот
можно существенно увеличить покрытую лесом площадь. Как и в случае с
лесами, необходимо ограничить выбор объектов землями, по условиям
плодородия позволяющими выращивать на них надлежащие по продуктивности
насаждения без негативного нарушения гидрологического режима водосборов.
3. В современных экономических условиях, когда осушение новых
земель практически прекратилось, а возможность поддержания лесоосуши-
тельных систем в рабочем состоянии из-за отсутствия госбюджетного
финансирования и разрушения материально-технической базы ЛММС и их
ликвидации весьма ограничена, главной задачей гидролесомелиорации
является сохранение наиболее эффективно осушенных земель с хорошими
лесоводственными результатами. Эта задача может решаться путем
реконструкции и капитального ремонта лесоосушительных систем на наиболее
ценных объектах, которые следует выявить при их инвентаризации. В
практическом плане указанную работу можно профинансировать в некоторой
степени за счет передачи эксплуатируемых лесов в долгосрочную аренду
крупным арендаторам - например, объединениям леспромхозов с
деревообрабатывающими и целлюлозно-бумажными предприятиями, экономически
заинтересованными, при соблюдении определенных гарантий,
закрепленных механизмом передачи, в повышении производительности и
продуктивности лесных земель.
4. Особую тревогу вызывает заболачивание осушаемых и дренированных
земель вследствие строительной деятельности бобров и подпора
поверхностных и грунтовых вод инженерными сооружениями (дорогами, плотинами,
262
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
фте-газопроводами и др.). Масштабы этих процессов велики. Прокладка
ттинейных сооружений (в т.ч. кабелей связи вдоль дорог) без экологического
обоснования и надлежащего согласования с землепользователями может
усложнить и даже исключить дальнейшие осушительные работы на
вышележащих водосборах. Высокими темпами происходит вторичное
заболачивание осушенных земель из-за плохого состояния каналов. Эти
обстоятельства необходимо учитывать при определении объектов и объемов
неотложной реконструкции лесоосушительных систем, при разработке методов
лесоводства на осушаемых землях, где уход за каналами не осуществляется.
5. Планирование и проектирование гидролесомелиоративных
мероприятий должно строиться на ландшафтной основе и направлено на обеспечение
устойчивого управления ресурсами переувлажненных земель,
контролируемого равновесия и биоразнообразия на экосистемном и ландшафтном
уровнях и улучшения эстетических свойств лесоболотных ландшафтов. Для
отдельных групп схожих ландшафтов должны быть установлены свои
приоритеты, подходы к определению масштабов, методов и способов осушения,
величины оставляемой площади переувлажненных земель.
6. Потребность в гидролесомелиорации, как показывает исторический
опыт развития лесного хозяйства России, безусловно будет востребована
уже в недалеком обозримом будущем, по мере улучшения экономики
страны. Альтернативы этому мероприятию для существенного повышения
производительности лесов в таежной зоне, особенно в европейской части, где во
многих районах доля переувлажненных земель в лесном фонде достигает
40-50% и более, нет. Без осушения земель и дорожного освоения лесов,
проводимого в тесной увязке со строительством осушительных систем, здесь
будет невозможно обеспечивать потребности в древесине народного
хозяйства и населения, не нарушая принцип неистощительного лесопользования и
не ухудшая сложившуюся структуру лесов. В связи с изложенным, следует
быть готовым в научном и практическом плане к возрождению
гидролесомелиоративных работ. Необходимо развивать и углублять теоретические
исследования, используя, прежде всего, стационары НИИ и ВУЗов, а также
производственные объекты длительного лесоосушения, проводя на них
мониторинговые наблюдения за лесоводственным и техническим состоянием
гидролесомелиоративных систем для совершенствования технических
условий и нормативных руководств. Необходимо совершенствовать подготовку
инженеров лесного хозяйства, усиливая ее экологическую составляющую, с
тем, чтобы обеспечить возможность их использования при проектировании
и производстве гидролесомелиоративных работ, а также обучить методам
ведения лесного хозяйства на осушаемых землях. Необходимо возобновить
°пытно-конструкторские работы по созданию новой более
производительной мелиоративной техники - ресурсосберегающей и экологически безопас-
263
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ной, базирующейся на использовании новых базовых тракторов, мацщц
(шасси) повышенной проходимости на гусеничном и колесном ходу.
ЛЕСОВОДСТВЕННАЯ ЭФФЕКТИВНОСТЬ ОСУШЕНИЯ СОСНЯКОВ
БЫСТРИЦКОГО ЛЕСОБОЛОТНОГО МАССИВА
А.А. Корепанов, С.А. Корепанов
Нижегородская государственная сельскохозяйственная академия
Быстринкий лесоболотный массив расположен в 25 км юго-западнее г.
Кирова, входит в его зеленую зону и занимает 3200 га. В
геоморфологическом отношении он относится к типу месторождений второй надпойменной
террасы реки Быстрицы. Лесоболотный массив является Участком гослес-
фонда Быстрицкого лесничества Кировского лесхоза . Мелиоративный фоцд
занимает 2094 га, т.е. 65% от общей площади участка. В мелфонде
преобладают сосновые насаждения — 53,4%, затем березовые — 37,3% и еловые —
9,3%. Основные площади занимают травяно-болотные, осоково-сфагновые и
сфагновые типы леса.
Болото осушено в 1967 году с целью повышения производительности
лесов. Лесоводственная эффективность осушения насаждений на низинных
и переходных болотах не вызывает сомнения. Осушение верховых (олиго-
трофных) болот не дает лесоводственного эффекта. Однако для Быстрицкого
лесоболотного массива характерно преобладание сосняков кустарничково-
сфагновых на мезоолиготрофной торфяной залежи, эффективность
осушения которых в нашем регионе мало изучена.
Для определения ледсоводственной эффективности осушения и
состояния осушенных древоствоев важное значение имеет изучение изменения их
лесотаксационных показателей в течение нескольких десятилетий после
мелиорации. С этой целью нами были заложены постоя иные пробные
площади до осушения в 1964-65 годах, а в 1978, 1985 и 1997 гг. проводилась
повторная перечислительная таксация с рубкой модельных деревьев и их
обработкой. Кроме того, на верховом болоте в четырех вариантах в 1969 году
проведено удобрение молодняков с учетами в 1965, 1977 и 1997 гг.
Некоторые результаты этих исследований приводятся в табл. 1 и 2.
Как видно из табл. 1, осушение существенно повлияло на все лесотакса-
ционные показатели молодняков сосны. Они за 30-летний период после
осушения увеличили класс бонитета с V до III, а по таблицам для
осушенных древостоев до 1,5. Возросла стволовая масса, идет усиленный процесс
изреживания, появилось возобновление березы. Однако осушение спелых
сосняков существенно не повлияло на лесотаксационные показатели древо-
264
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
стоя, как и следовало ожидать, хотя стволовая масса возросла в два раза, а
класс бонитете по таблицам для осушенных древостоев возрос до П. 5.
тябл. 1 Основные лесотаксационные показатели сосняков до и после осушения
Период
Дер.,
см
Нср.,
м
Полнота
Число стволов,
(сухостой/ рас-
тущие), шт/га
Класс
бонитета
Запас,
мЗ/га
Возраст
лет
Осушен в возрасте молодняка
До
После
осушения
До
После
осуше
ния
5,5
7,9
11,0
14,2
10,2
10,9
13,1
15,8
6,3
8,2
11,2
16,4
7,3
9,7
10,0
15,0
0,40
0,78
0,78
0,83
25/3899
809/3537
1086/2471
275/1764
Осушен в спелом возрасте
0,65
0,68
0,72
0,71
72/2250
199/1964
225/1433
349/1147
IV,5
IV,1
Ш,7
Ш.1
Va
Va
Va
V,2
38
96
132
217
73
90
111
170
34
47
54
67
92
105
112
125
Табл. 2. Основные лесотаксационные показатели сосняков после осушения и удоб-
рения
Варианты
опыта
Дер-,
см
Нср.,
м
Полнота
Число
стволов,
шт/га
Класс
бонитета
Запас,
м3/га
Возраст
лет
До осушения
Осушено
Не удобрено
NPK
NP
NK
Р
5,2
8,7
13,0
11,1
14,3
9,0
13,5
8,7
12,4
8,6
11,6
5,0
6,3
12,9
8,6
15,7
6,6
13,2
6,8
12,8
6,6
11,4
0,17
0,44
0,68
0,62
0,74
0,38
0,86
0,40
0,73
0,44
0,65
1850
1660
2220
1660
1480
1620
1840
1550
2700
1780
2770
V,3
V,3
IV,2
IV,3
Ш,2
V,2
Ш,6
V,l
IV,2
V,l
IV,2
12
39
140
63
195
36
183
35
150
44
111
38
52
70
47
65
47
65
50
68
48
66
Верховые болота отличаются бедностью верхних корнеобитаемых
горизонтов торфяных почв. С целью повышения плодородия проведено их
удобрение (1969 г.) в нескольких вариантах в следующих дозах: аммиачная
селитра -400 кг/га, хлористый калий — 200 кг/га, фосфорная мука — 700 кг/га
(табл.2).
После осушения и удобрения все лесотаксационные показатели
повысились. Одно осушение без удобрения также дало положительный результат.
Однако, только полное удобрение и в меньшей степени азотно-фосфорное
существенно улучшают рост сосняков.
265
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Осушение хмезолиготрофных болот существенно повышает производи
тельность произрастающих на них сосняков кукстарничково-сфагновых
Положительную роль выполняет удобрение осушенных сосняков. Однако
спелые древостой реагируют на осушение слабо. Поэтому при проведение
гидролесомелиорации на мезоолиготрофных болотах спелые сосняки необ*
ходимо вырубать, оставляя вырубки под естественное заращивание, а в мо-
лодняках проводить рубки ухода с первого десятилетия после осушения.
ТИПЫ ЛЕСА НА ОСУШЕННЫХ ТОРФЯНЫХ ПОЧВАХ
СЕВЕРНОЙ БЕЛАРУСИ
Н.А. Красильников, В.Ю. Нешатаев
СПбНИИЛХ;
Санкт-Петербургская государственная лесотехническая академия
К настоящему времени в Беларуси леса на осушенных землях занимают
более 230 тыс. га. Леса на осушенных торфах характеризуются
существенными экологическими, лесоводственными и технологическими
особенностями, поэтому целесообразно отнесение их к особым типам леса.
В основу разработки типологической классификации лесов на
осушенных местообитаниях принято определение типа леса в понимании В.Н.
Сукачева, что соответствует традициям лесотипологической школы Беларуси.
Конкретизировав цели классификации биогеоценозов осушенных
местообитаний, мы пришли к выводу о том, что выделяемые типы должны
отличаться в пределах гидролесомелиоративных районов по хотя бы одному из
следующих показателей: 1) преобладающая древесная порода, 2)
потенциальная продуктивность основных лесообразователей: ели, сосны, березы, ольхи
черной; 3) вероятность смены древесных пород после рубки и без нее; 4)
требуемые параметры осушительной сети при ее реконструкции.
Единицей, раскрывающей варьирование биогеоценозов в пределах типа
леса, является его вариант (подтип, разновидность), традиционно
описываемый в белорусской лесотипологической системе соответствующей
растительной ассоциацией. Единицей, объединяющей корреспондирующие типы
леса и типы вырубок на их месте является серия типов леса. С учетом
особенностей постмелиоративной динамики серия типов биогеоценозов
объединяет относительно устойчивый (условнокоренной) тип биогеоценоза,
формирующийся при длительном осушении, а также начальные стадии
сукцессии под влиянием осушения (серийные мелиоративно-производные типы
леса), которые, при сохранении действия осушительной сети, переходят в него.
Полевые работы в 1998 г. были проведены в Витебской обл. на
территории Полоцкого и Глубокского лесхозов, на объектах осушенных в 1962-1971
266
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
г пробные площади размером 20650 м закладывали сериями по 2: вблизи
канала и на середине межканального пространства. На 60 пробных
площадях определяли высоту, диаметр, возраст, полноту, запас, сомкнутость крон
деревьев по элементам леса, высоту в момент осушения, сомкнутость под-
деска по видам, проективное покрытие видов и ярусов живого напочвенного
покрова, мощность, ботанический состав, степень разложения торфа по
горизонтам в разрезе глубиной 50 см, общую мощность торфа.
Проанализированы закономерности изменения состава и
производительности лесов северной части Беларуси под влиянием осушения. При
длительном оптимальном осушении формируются типы леса близкие по структуре
фитоценоза, характеру лесной подстилки и производительности древостоев
к влажным вариантам коренных типов леса суходолов. Оптимальное
осушение приводит к выравниванию условий увлажнения различавшихся по
степени увлажнения исходных биогеоценозов, поэтому различия между
формирующимися типами леса обусловлены различиями в трофности торфов.
Выделено 4 серии типов леса на торфяных почвах, формирующихся при
оптимальном осушении (см. приложение), рекомендуемые к использованию
в лесоустройстве. Дана характеристика выделенных типологических единиц
и разработан определитель типов леса и растительных ассоциаций на
осушенных местообитаниях. Для типов леса определены классы бонитета
соответствующие бонитету насаждений, которые осушены в молодом возрасте.
На бедных торфах после осушения верховых болот формируются
сосняки с покровом из олиготрофных кустарничков и зеленых мхов,
преимущественно рода Dicranum — Сосняк кустарничковый торфяной осушенный.
На менее бедных питательными веществами верховых торфах
небольшой мощности или подстилаемых на небольшой глубине переходными
торфами сукцессия идет также в сторону формирования зеленомошного
покрова с участием Dicranum и преобладанием Pleurozium schrebeh. В травяно-
кустарничковом ярусе преобладающим видом на стадии относительной
стабилизации Сосняк черничный торфяной осушенный становится черника.
Для этого типа леса характерно незначительное участие таежного
мелкотравья (седмичник, майник), приспособленного к наличию грубогумусной
подстилки. Смена после осушения березняков пушицево-сфагновых идет
сходным образом, завершаясь типом Березняк черничный торфяной осушенный.
При осушении в течение 20-30 лет травяно-сфагновых болот и лесов
биогеоценоз переходит в стадию, характеризующуюся преобладанием в
живом напочвенном покрове таежного мелкотравья, преимущественно
кислицы, и мхов-мезофитов. В результате формируются Сосняки, березняки
ельники кисличные торфяные осушенные. Леса, формирующиеся при
длительном и интенсивном осушении лесов на низинных торфах отнесены к
Ельникам, соснякам, березнякам, черноольшанникам разнотравно-кисличным.
267
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Приложение. Схема типов леса и растительных ассоциаций на осушенных тодф»
ных почвах северной части Республики Беларусь. Для каждой из 4 серий типов леса
указаны: характеристика условий местопроизрастания; преобладающая порода и
класс бонитета; сопутствующие породы; индикаторные виды растений; преобла.
дающие виды живого напочвенного покрова; ассоциации. Кустарничковая торфяная
осушенная: верховые торфяники мощностью >1м; сосна — IV-V; редко — береза-
багульник, голубика, мирт болотный, пушица, плевроций Шребера, сфагн узколист*
ный; багульник, голубика, мирт болотный, брусника, плевроций Шребера, дикраи
сфагн узколистный; багульниково-брусничная / голубично-брусничная / кустарнич-
ково-брусничная. Черничная торфяная осушенная: верховые торфяники мощностью
<1 м/ переходные и низинные торфяники со слоем верхового торфа <30 см; сосна
— П-Ш / береза — П-Ш / ель — П-Ш; ель; багульник, голубика, мирт болотный,
щитовники, майник, седмичник, рябина, крушина, черника, малина; черника,
плевроций Шребера, дикран, сфагн узколистный и магелланский; зеленомошно-черничная
/ сфагново-черничная. Кисличная торфяная осушенная: переходные и низинные
торфяники со слоем переходного торфа >30 см; сосна — I / ель — I / береза — I;
редко — ольха черная; кислица, майник, седмичник, цирцея альпийская,
щитовники, рябина, крушина, малина, крапива; кислица, малина, плевроций Шребера,
дикран, брахитеции, мнии; зеленомошно-кисличная / тростниково-кисличная. Разно*
травнокисличная торфяная осушенная: низинные торфяники, мощность
органических горизонтов почвы >20 см; сосна — 1а / ель — 1а / береза — 1а / ольха черная —
1а; ясень, дуб, вяз; крапива, недотрога, кочедыжник, зеленчук, звездчатка дубравная,
лютик ползучий, фиалка сверхуголая; крапива, малина, кислица, зеленчук,
кочедыжник женский, вейник сереющий, тростник, мнии, брахитеции; разнотравно-
кисличная / крапивная / тростниково-вейниковая / тростниковая. Примечание:
указаны классы бонитета лесорастительных условий.
ЛЕСОВОДСТВЕННАЯ ЭФФЕКТИВНОСТЬ ОСУШЕНИЯ
ЗАБОЛОЧЕННЫХ БЕРЕЗНЯКОВ НА ДАЛЬНЕМ ВОСТОКЕ
В.В. Острошенко
Чумиканский лесхоз, Хабаровский край
В начале 70-х годов впервые на Дальнем Востоке в Хехцирском опытно-
показательном лесхозе ДальНИИЛХа начато осушение заболоченных почв
для дальнейшего использования их в лесном хозяйстве. Однако динамика
роста растений на осушенных почвах в условиях Дальнего Востока не
известна. Наши данные о ходе роста лесных культур хвойных на осушенных
почвах показывают возможность перевода их в лесокультурный фонд с
последующим проведением лесовосстановительных работ [2].
Представлены результаты наблюдений за ходом роста древостоя березы
ребристой (желтой) и естественного лесовосстановления на осушенных
почвах. Участок — вырубка 1962 г., количество пней — 400 шт./га, класс
бонитета IV. Древесная растительность представлена березой ребристой, полнота
268
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
^ 0.2. возраст — 25 лет; высота — 4,8 м; диаметр — 5 см, запас древесины
^ 16 м/га. Возобновление березой ребристой, осиной Давида — около 3
тыс шт./га, средняя высота 1,35 м. В подлеске жимолость съедобная, спирея
сСурийская, голубика, багульник болотный. В напочвенном покрове
господствует горец амурский, встречаются лютик обыкновенный, осоки,
зеленые и сфагновые мхи; проективное покрытие 94%. Почва — избыточно
влажный буро-подзолистый, местами оглеенный, крупнопылеватый средний
суглинок.
По [1] лесорастительные условия участка относятся к четвертой
категории, где после удаления древостоя требуется поверхностное осушение
временными каналами и проточными бороздами [1]. Поверхностное осушение
проводили в 1970 г. Плугом ПКЛН-500, агрегатируемым с трактором ЛХТ-
55. на расстоянии 8-10 м друг от друга прокладывали канавы глубиной 0,7-
0.3 м и шириной по дну 0,4-0,5 м.
Влияние осушения на формирование подроста и подлеска, нарастание
древесины и изменение запаса древостоя изучали на прямоугольных
пробных площадях размером по 0,5 га каждая, заложенных в наиболее типичных
местах участка в соответствии с общепринятыми методами по определению
лесоводственной эффективности лесоосушения [4]. Контролем служили
площади аналогичного размера, примыкающие к пробным, на которых
осушение не проводили.
На пробных и контрольных площадях определяли видовой состав,
количество экземпляров и сомкнутость подлеска, видовой состав, высоту и
диаметр стволов подроста, их текущие приросты по высоте и диаметру.
Производили перечет деревьев по ступеням толщины, замеряли их высоту и
диаметр, определяли приросты по всем таксационным показателям.
Видовой состав, высоту и проективное покрытие напочвенного покрова
определяли на учетных площадках размером 2 х 2 м, заложенных в
трехкратной повторности в наиболее типичных местах по диагонали пробной и
контрольной площадей. Результаты исследований показали, что через 10 лет
после осушения общее проективное покрытие напочвенного покрова
возросло до 98 %. Изменившийся видовой состав напочвенного покрова
представлен пыреем, подорожником азиатским, горецом амурским, пикульником
Двунадрезным, молочаем Комарова, кровохлебкой лекарственной (табл. 1).
Появление на участке растений, предпочитающих плодородные почвы,
указывает на улучшение почвенных условий.
Изменившиеся условия среды благотворно отразились на повышении
энергии роста растений на осушенных площадях. До осушения насаждение
находилось в фазе снижения ростовой активности. Осушение обусловило
значительное увеличение приростов уже через 3-4 года после проведения
Работ и продолжалось в последующие 25 лет (табл. 2). Резкое увеличение
269
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
приростов во втором пятилетии свидетельствует о завершении адаптации
растений к изменившимся условиям среды, успешному их переходу из этап*
приспособления к периоду формирования по [4]. Так, за первое десятилетие
после осушения среднепериодический прирост по запасу составил 1,9 м3/га
а дополнительны» прирост по запасу — 1,3 м3/га.
Табл 1. Изменение живого напочвенного покрова в результате осушения избыток
увлажненных почв на юге Хабаровского края
Видовой состав
горец амурский
лютик обыкновенный
пырей ползучий
подорожник азиатский
пикульник двунарезный
молочай Комарова
кровохлебка лекарственная
зеленые мхи
мох сфагнум
Табл. 2. Изменение приростов по
Период, лет
Контроль
После осушения
Дополнительный прирост
ДО
запасу
Проективное покрытие, %
осушения
47
24
-
-
-
-
-
11
9
через 20
лет после осушения
14 *
-
45
21
9
8
3
-
-
в зависимости от давности осушения
Прирост по периодам после
до 5
3,21
9,15
5,94
6-10
3,33
10,02
6,69
11-15
3,48
18,35
14,87
осушения, м^га
16-20 21-25
3,47 3,51
20,65 21,83
17,18 18,35
Табл 3 Основные таксационные показатели березы ребристой (желтой) на
осушенных почвах до (1) и после (2) осушения, контроль(З),
Класс Сос-
возр.. бонит. тав
1 Ш IV 10Б
2 V П 9Б1Л
3 Ш/V IV 10Б
4 2 кл. 2 кл.
Полнота
0,2
0,6
0,3
0,3
дополнительный прирост (4)
Средний годичный прирост по:
вые, м
0,19
0,46
0,22
0,24
диам., см запасу, мЗ/га
0,2
0,8
0,4
0,4
0,64
3,21
0,68
2,53
В последующие пятилетия увеличение приростов достигло 3,7 и 4,4
м3/га. Дополнительный прирост повысился соответственно до 3,0 и 3.7
м3/га.
Важным показателем лесоводственной эффективности осушения
является успешность лесовозобновительного процесса под пологом
сформированного древостоя. Подрост густой, благонадежный, равномерно распределен
по площади. 3,5 тыс. шт./га. В его составе, наряду с березой ребристой,
появилась береза даурская — более требовательная к плодородию почвы [6] я
лиственница амурская, соответственно — до К2 и 0,8 тыс. шт./га.
270
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
Энергичный рост лиственницы амурской позволил ее небольшой части
гги в древесный полог насаждения. В целом за 25 лет после осушения
омкнутость полога древостоя повысилась до 0,6, а бонитет повысился на 2
класса (табл. 3). Таким образом, осушение избыточно увлажненных почв в
условиях юга Хабаровского края являются эффективным лесоводственным
мероприятием.
п] Основные положения по гидролесомелиорации / Под общ. ред. В.К.
Константинова. СПб.: СПбНИИЛХ, 1995. 59 с. [2] Острошенко В.В. Рост лесных культур на
осушенных почвах Дальнего Востока // Гидротехническая мелиорация земель.
Веление лесного хозяйства и вопросы экологии. СПб. 1997. С. 92-97. [3] Корякин В.Н.,
Грек B.C. Сортиментные и товарные таблицы древесных пород Дальнего Востока.
до.: Госкомлесхоз, 1988. 164 с. [4] Рубцов ВТ., Книзе А.А. Закладка и обработка
пробных площадей в осушенных насаждениях. Л., 1977. 44 с. [5] Справочник
таксатора / Сост. Н.В. Ефремов. Хабаровск: ВО Леспроект, 1955. 85 с. [6] Усенко Н.В.
Деревья, кустарники и лианы Дальнего Востока. Хабаровск.: Кн. изд-во, 1969. 416 с.
РОСТ ЛЕСНЫХ КУЛЬТУР НА ПЕРИОДИЧЕСКИ ИЗБЫТОЧНО
УВЛАЖНЕННЫХ ПОЧВАХ ПРИОХОТЬЯ
В.В. Острошенко
Чумикаиский лесхоз, Хабаровский край
В северном территориально-природном дальневосточном комплексе —
Приохотье [1] площадь заболоченных лесов и марей, вследствие общего
гидрологического режима территории, составляет около 20%.
В середине 50-х годов в регионе начато проведение лесо-культурных
работ на периодически избыточно-увлажненных почвах. На невозобновив-
шихся естественным путем заболоченных гарях подготавливали в ручную
микроповышения в виде площадок размером 1 кв. м. Размещение площадок
5 х 5 м. На 1 га подготавливали по 400 площадок. 2-х летние сеянцы
лиственницы даурской и сосны обыкновенной, выращенные в питомнике из
семян местного сбора, высаживали в площадки конвертным способом, по 5
штук в площадку. На 1 га высаживали по 2,0 тыс. шт. сеянцев.
Через несколько лет лесокультурные работы были свернуты и
возобновились с конца 80-х годов. В первые два года за высаженными растениями
велся двукратный уход: в площадках пололи сорняки, рыхлили почву. С
этого же времени начаты регулярные наблюдения за ростом и развитием
Осаженных растений. Работы проводили по методикам [2,3].
Таксационная характеристика и биомасса модельного дерева лиственницы
даурской, растущего на периодически избыточно увлажненных почвах Приохотья,
подстилаемых вечной мерзлотой (Н — высота; Д — диаметр на высоте 1,3 м; ДПв —
Текуцщй прирост по высоте; Дп — диаметр проекции кроны)
271
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Tai
пока
ксационные
затели за по-
следний год
н.
м
11,4
6,3
44,7
1,6
0,8
50
д,
см
8
6
25
0,5
-
-
роста
ДПв,
см
11
5
54,5
16
5
320
Крона
Протяжен- вес (хвои
Дп, ность, м /живых
м (% от длины ветвей/все-
ствола) го), кг
в насаждениях 40-летнего возраста
3,2 8,3(75,5) 5,3/7,6/12,0
Контроль
2,1 2,8(44,4) 2,9/4,3/7,2
превышение над контролем, %
34,4 76,3(31,1) 45,3/43,4/44,2
в лесных культурах 10-летнего возраста
0,8 0,9(62,7) 0,1/0,2/0,3
Контроль
0,6 0,5(62,5) 0,1/0,1/0,2
превышение над контролем, %
33,3 80(-) -/50/33,3
Вес, кг *
ствола/сухих отГ
мерших сучьев
/корневой
системы/всего дерева
17,1/0,6/4,2/34,8
9,3/2,2/2,4/21,1
45,6/-/42,9/39,4
0,2/70,2/0,7
0,1/-/0,1/0,4
50/-/50/42,9
Анализ хода роста проведенный по срединным сечениям модельных
деревьев высаженных в 50-е годы показал, что с первых лет роста сеянцев в
культурах наблюдается превышение их таксационных показателей по срав»
нению с контролем. Через 40 лет
средняя высота лиственницы
составляла 11,4 м, а сосны 9,6 м.
На контроле (растения,
произрастающие естественно рядом с
лесокультурной площадью) эти
показатели были равны,
соответственно 6,3 и 5,2 м. Диаметр
ствола на высоте 1,3 м составил
в лесных культурах
лиственницы 8 см, сосны — 10 см; на
контроле, соответственно: 6 и 10 см.
Развитие кроны, нарастание фи-
томассы в искусственно
созданных насаждениях превысило
контроль на 31,1-76,3% (см.
табл.). Более энергичный рост
растений в культурах обусловил
повышение продуктивности
насаждений. За 40 лет объем
древесины увеличился до 12 куб. м на 1 гектаре, против 5 куб. м — на
контроле.
Начало образования ярусности корневой
системы у лиственницы даурской 10-
летнего возраста высаженной 2-летними
сеянцами в микроповышения
272
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ V
регулярные 10-летние наблюдения за ростом лесных культур заложен-
bix в 80-е годы показали, что приживаемость высаженных сеянцев
колебать у лиственницы от 66,0 до 97,1%, у сосны — от 62,6 до 84,8%. В
последующее десятилетие отпад растений не превысил 6%. Отмеченный нами
оанее энергичный рост сеянцев на микроповышениях подтвердился и в
кульТУРах заложенных в 80-е годы. Основные таксационные и
биометрические показатели превышали контроль в пределах 33,3-80,0%.
Указанная дальневосточным дендрологом Н.В.Усенко [4] способность
лиственницы к образованию придаточных корней на влажных и мокрых
почвах наблюдалась и нами (см. рис.). Масса корневой системы повышается
в среднем на 50%. Наблюдения доказывают возможность повышения
продуктивности периодически избыточно увлажненных лесных земель за счет
лесовосстановления по ми-кроповышениям. Начало форми-рования ярусно-
сти корневой сис-темы показывает, что несмотря на наблюдаемую лесово-
дственную эффективность, рекомендация устройства микроповышений
размером 1 кв. м для условий региона преждевременна. Необходимы
дополнительные исследования.
[1] Вишневский Д.С. и др. География Хабаровского края. Хабаровск: Кн. изд-во,
1976. 123 с. [2] Захаров В.К. Лесная таксация. Изд. 2 М.: Лесн. пром-сть, 1967. 408
с. [3] Огиевский В.В., Рубцов НИ. Лесные культуры и лесные мелиорации. М:
Высшая школа, 1960. 451 с. [4] Усенко Н.В. Деревья, кустарники и лианы Дальнего
Востока. Хабаровск: Кн. изд-во,. 1984. 270 с.
ОПЫТ КОЛИЧЕСТВЕННОЙ ОЦЕНКИ ДИНАМИЧЕСКИХ
СОСТОЯНИЙ И УСТОЙЧИВОСТИ СОСНОВЫХ НАСАЖДЕНИЙ НА
ОБЪЕКТАХ ГИДРОМЕЛИОРАЦИИ
В.В. Пахучий
Институт биологии Коми НЦ УрО РАН; Сыктывкар
Предпринята попытка количественного анализа динамических состояний
насаждений с преобладанием в их составе сосны на объектах, осушенных в
40-х годах прошлого столетия (Лисинский учебно-опытный лесхоз
Ленинградской области). Использованы материалы лесоустройства на осушенных
площадях 1841. 1891. 1928. 1953-1954. 1961 и 1973 гг.. фрагменты таблиц
хода роста нормальных сосновых насаждений V класса бонитета для
Ленинградской области [8]. а также собственные таксационные описания
насаждений на пробных площадях, заложенных в 1976 г. в выделах, для которых
сохранились лесоустроительные материалы.
Для комплексной оценки динамических процессов был использован
факторный анализ. Его основные положения в связи с проведением лесово-
273
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
дственно-таксационных исследований на осушенных лесных землях изла^
лись ранее [5]. В массив исходных данных было включено 33 таксационц^
описания, характеристика насаждений выполнена по 25 параметрам (при*,
накам). В результате выполненных исследований установлено, что о6о£
щенными факторами Ф1, Ф2 может быть учтено 77% суммарной дисперсщ*
изучаемых признаков. Ф1 интерпретирован как обобщенный фактор, оцр^
деляющий форму (ярусность) насаждений и ее изменение со временем щ
сохранении доминирующей роли сосны. Второй обобщенный фактор инщ,
претирован как обобщенный фактор, связанный с лесоводственно-такеа-
ционными характеристиками ели.
Анализ размещения опытных объектов в факторном пространстве ф^
Ф2 показывает, что при движении из отрицательной в положительную об?
ласть Ф2 наблюдается переход от простых одноярусных к сложным 2-3 -*
ярусным древостоям, от чистых сосновых к смешанным по составу насаж*
дениям. При этом доминантом во всех случаях является сосна. Особый
интерес представляет анализ распределения в факторном пространстве Ф1, Ф2
таксационных описаний по материалам лесоустройства 1841-1973 гг. Такса*
ционное описание 1841 г., которое может быть принято в качестве лщ
трольного, занимает в данной группе опытных объектов крайнее положение
в отрицательной области Ф1. По мере повышения продуктивности,
усложнения формы и состава насаждений (объектов) после осушения, последние
перемещаются в положительную область Ф1 факторного пространства.
Под устойчивостью фитоценоза понимается его способность
противостоять внешним изменениям среды для сохранения числа видов, их
количественных соотношений в рассматриваемом растительном сообществе и
взаимосвязей в течение некоторого интервала времени [7]. Согласно лесово-
дственным оценкам изменение продуктивности насаждений может привести
к изменению (потере) их устойчивости [3,6], которая рассматривается как
один из аспектов динамического поведения сообщества [1,7]. Это позволяет
допустить использование траектории движения лесных сообществ в
факторном пространстве для оценки их устойчивости и описания динамических
состояний. Опорные точки, через которые пройдет траектория, будут
отражать динамическое состояние изучаемого сообщества в рассматриваемые
периоды времени [1]. Дополнительные возможности для решения
аналогичных задач представляет сочетание факторного и регрессионного анализа.
Анализ характера размещения опытных объектов в факторном
пространстве и относительно аппроксимирующей их положение кривой показывает,
что наибольшее отклонение от сглаженной кривой приходится на таксаця*
онное описание 1953-1954 гг., когда согласно материалам лесоустройства
впервые для древостоев на осушенной территории был выделен второй ярус-
Корреляционный анализ показал наличие тесной связи между обобщенным
274
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
Фактором Ф1 и средним диаметром и высотой сосны (R=0,80-0,89). Близкая
функциональной связь наблюдается между Ф1.и возрастом сосны (R=
О 95) Последнее подтверждает динамический характер Ф1 и согласуется с
лредложенным вариантом интерпретации этого фактора. Несмотря на
доминирующее положение сосны, в сложном по форме и составу фитоценозе на-
блюДается возрастание участия ели. Анализ зависимости доли запаса сосны
й еЛЙ (Р,%) в общем запасе древостоя от обобщенного фактора Ф1
показывает, что для сосны эта связь отрицательная (R=-0,88), а для ели
положительная (R=0,92). Зависимость между рассматриваемыми показателями мо-
жет быть выражена уравнениями: Р=-20,2Ф1+65>6 (для сосны); Р=
21.5Ф1+ЗОД (для ели). Уравнивая левые и правые части уравнений, можем
рассчитать, что участие сосны и ели в формировании общего запаса будет
одинаковым при Ф1, соответствующим 0,83. В соответствии с уравнением
д=44,8Ф1+109,1 возраст сосны (А), по достижении которого она может
уступить доминирующую роль ели, соответствует приблизительно 150 годам.
Таким образом на потенциально богатых почвах лесоосушение стимулирует
серию смен, продолжающуюся столетия. Это свидетельствует о том, что
полные оценки лесоводственных и экологических последствий гидромели-
рации могут быть только гипотетическими. При отсутствии
катастрофических природных явлений (пожар, ветровал, бурелом) или рубок, спустя
период, близкий к 1,5 векам, сосна уступит свое доминирующее положение
ели, и ель сохранит положение доминанта и эдификатора неопределенно
длительный период. По нашим данным [4] на участках с высоким эффектом
осушения смена доминантов древесных ярусов может сопровождаться
вторичным увеличением прироста, связанным с влиянием на водный режим
самих формирующихся насаждений. Т.е. в определенных условиях может
наблюдаться явление, противоположное описанному в специальной
литературе как «вторичное заболачивание» на объектах гидролесомелиорации [2].
[1] Буш К.К. Отражение динамики осушенных лесов в типологии//Динамическая
типология лесов. М.: Агропромиздат, 1989. С. 168-177. [2] Вомперский С.Э.
Биологические основы эффективности лесоосушения. М.: Наука, 1968. 310 с. [3] Дырен-
ков С.А. Структура и динамика таежных ельников. Л.: Наука, 1984. 174 с. [4]
Пахучий В.В. Лесоводственно-гидрологическое обоснование гидромелиорации лесных
земель (на примере Республики Коми). Автореф. дисс. докт. с.-х. наук. Санкт-
Петербург: СПбГЛТА, 1993. 30 с. [5] Пахучий В.В., Урнышев А.П. Факторы
производительности осушенных хвойных насаждений на слаботорфованных лесных
землях // Эффективность и организация работ по осушению лесных земель в Коми
АССР'\ Тез. док. совещ. Сыктывкар, 1988. С.91-92. [6] Романовский М.Г.
Биоразнообразие, устойчивость, продуктивность // Биологическое разнообразие лесных
экосистем. М., 1995. С.37-40. [7] Свирежев Ю.М., Логофет Д.О. Устойчивость
биологических сообществ. М.: Наука, 1978. 352 с. [8] Третьяков Н.В. и др. Справочник
таксатора. М; Л.: Гослесбумиздат, 1952. 853 с.
275
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ПОВЫШЕНИЕ ПРОДУКТИВНОСТИ ЛЕСОВ НА ПОЧВАХ
ИЗБЫТОЧНОГО УВЛАЖНЕНИЯ
А. И. Русаленко
Белорусский государственный технологический университет, Беларусь
В лесном фонде Беларуси избыточно увлажненные земли (ИУЗ)
занимают около 2,5 млн. га, из которых на долю верховых болот приходится 440
тыс. га, переходных — 560 и низинных — 660 тыс. га, а остальное
составляют земли временного избыточного увлажнения (черничные, долгомошные
и пойменные серии типов леса). На ИУЗ наиболее распространенными
древесными породами являются ольха черная (540 тыс. га), береза (500 тыс.
га), сосна (80 тыс. га) и ель (50 тыс. га).
До настоящего времени осушено открытыми гидромелиоративными сие-
темами одностороннего действия 271 тыс. га лесов. За последние 25 лет в
результате осушения дополнительный прирост древесины составил 8 млн.
м3. Окупаемость осушения лесов на избыточно увлажненных землях
составляет по Беларуси около 10-11 лет. Значительные площади лесов с
избыточным увлажнением, расположенные возле осушенных для
сельскохозяйственного использования болотных массивов, подверглись пассивному
осушению, что особенно проявилось на территории Полесья.
Гидромелиоративные осушительные системы требуют значительных
капвложений при строительстве. Нормальное их функционирование
невозможно без текущих затрат и капремонта, необходимость в котором
возникает, как правило, через 25-30 лет. К тому же, при осушении болот
наблюдаются негативные экологические изменения, которые во многих случаях не
учитываются при подсчете эффективности осушения из-за отсутствия
методик расчета. Наиболее распространенным негативным явлением при
осушении болот является нарушение гидрологической ситуации на прилегающей
территории, то есть в пределах водосборной площади болот. На территории,
которая непосредственно прилегает к осушенному болоту, наблюдается
пассивное осушение и улучшение водно-воздушного режима почв. На
остальной, значительно большей части водосборной площади снижение уровня
почвенно-грунтовых вод ухудшает влагообеспеченность растений и ведет к
падению продуктивности фитоценозов. Как установлено нами, при
снижении уровня почвенно-грунтовых вод на 1,0 м продуктивность сосняков
мшистых уменьшается более, чем на один класс бонитета, а потери древесины за
период возраста главной рубки составляют около 140 м3/га.
В связи с негативными экологическими последствиями в последние годы
на территории Беларуси осушение лесов, произрастающих на почвах с
избыточным увлажнением, не производится. Однако актуальность повышения
их продуктивности не вызывает сомнений.
276
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
В большинстве случаев при избыточном увлажнении, особенно на чер-
оольховых низинных болотах, деревья произрастают на кочках (микропо-
ышениях), имея ограниченный объем ризосферы. Черноольховые вырубки
озобновляются преимущественно порослевым путем. При этом на одной
кочке могут произрастать не одно, а два дерева и более, в результате чего
оНи имеют малый диаметр, флагообразную крону и искривленные стволы,
то есть формируются в основном древостой Ш-го класса товарности.
Одним из путей повышения продуктивности черноольшаников являются
рубки ухода. Достаточна двухприемная рубка. Первый прием целесообразен
через 3-5 лет после вырубки древостоя с оставлением на каждой кочке 2-3
деревьев, отдавая предпочтение семенным экземплярам и наиболее
развитым порослевинам с прямыми стволиками. Второй прием (окончательный)
проводится примерно через 8-10 лет после первого с оставлением на каждой
кочке только одного дерева. По предварительным расчетам проведение
рубок ухода повышает продуктивность черноольшаников примерно на 15%.
Более радикальным мероприятием повышения продуктивности
черноольшаников (на 70-110%) следует считать создание искусственных
микроповышений в дополнение к естественным, занимающим только около 20%
общей площади. Повышение продуктивности обеспечивается не только
изменением класса товарности, но и в результате увеличения полноты и запаса
древесного яруса. Искусственные микроповышения могут также
использоваться для посадки сосны и формирования смешанного хвойно-лиственного
насаждения, что особенно важно в лесах 1-й группы.
Для устройства искусственных микроповышений лучше использовать
песок рыхлый, в котором в меньшей степени будет проявляться избыточное
увлажнение. Размеры их и количество определяются конкретными почвен-
но-грунтовы-ми условиями. Предварительными расчетами установлено, что
при доставке грунта на расстояние около 5 км затраты по строительству
искусственных микроповышений не превышают затрат на строительство
осушительных систем.
Создание искусственных микроповышений возможно в лесах всех
формаций, но только на тех болотных массивах, на которых наблюдается
временный или постоянный поверхностный сток воды. Иначе, особенно при
малой площади болот, будет наблюдаться подъем уровня почвенно-грунто-
вых вод на болотах, что повлечет подтопление и затопление
произрастающих древостоев.
В отличие от лесоосушения повышение продуктивности лесов,
произрастающих на избыточно увлажненных землях, посредством устройства мик-
Роповышений имеет ряд преимуществ, основными из которых являются
следующие: не нарушается гидрологический режим как на болотах, так и
277
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
прилегающих суходолах; микроповышения функционируют длительное
время и не требуют ухода, текущего и капитального ремонта.
ВЛИЯНИЕ РАСТИТЕЛЬНОСТИ БЕРМ И ОТКОСОВ КАНАВ НА
СОСТОЯНИЕ ЛЕСНОЙ ОСУШИТЕЛЬНОЙ СЕТИ
Ю. Русецкас
Литовский институт леса, Гирионис, Литва
Работы по содержанию лесоосушительных систем в новых условиях
ведения лесного хозяйства в Литве проводятся экстенсивно, а в отдельных
лесхозах некоторое время вообще не проводились. Происходит зарастание
осушительных канав кустарниками и деревьями, т.е. идет ренатурализация
искусственной гидрографической сети. Последствия этого на техническое
состояние осушительной сети и, тем же самым, на рост осушенного леса,
неизвестно. Целью работы было определение влияния вида и
продуктивности растительности, произрастающей на бермах и откосах канав, на
среднегодовое уменьшение их глубины.
В 9-ти лесхозах Литвы исследовалось около 30 км канав, построенных
15-35 лет тому назад, которые или практически не ремонтировались или по
которым известны даты ремонта. Среднегодовое уменьшение глубины канав
определяли по методике В.К. Константинова [1979], а распределение
доминирующей травяной растительности на 75 учетных площадках (1 м2),
случайно ориентированных относительно дна и откосов канав. Биологическую
продуктивность травяной растительности определяли по методике Л.Е.
Родина, Н.П. Ремезова, Н.И. Базилевича [1968]. Типы леса описаны по
методике [Karazija, 1988], а таксация деревьев и кустарников, произрастающих в
откосах и бермах канав, — по общепринятым в лесной таксации и
лесоводстве методам [Kairiukstis, 1961]. Освещенность в зоне канав в зависимости
от их положения относительно стран света и среднего угла закрытости
поверхности канав подсчитана по методике [Junghaus, 1966], средний угол
закрытости канав установлен по методике С.Ф. Федорова [1981].
Исследования показали, что если не ремонтировать канавы, не удалять
деревья, растущие на бермах и откосах канав и не косить траву, то
создаются все условия для ренатурализации осушительных систем. Как показывают
наши данные и данные других исследователей [Константинов, 1979]
основные факторы, определяющие интенсивность зарастания канав травяной
растительностью, это интенсивность освещения местности, трофность почвы и
продолжительность стояния (течения) воды в канавах. Установлено, что
абсолютное большинство видов травяной растительности не переносят
затенения, вызванного кронами деревьев и при обрастании канав древесной
растительностью (в том числе и кустарниками), погибают. Это влияет на тех-
278
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
ническое состояние осушительной сети. Показано (см. табл.), что среднее
годовое уменьшение глубины канав под пологом древесной и кустарниковой
растительности составляет в среднем 0,133-0,640 см/год, а без древесной
растительности 0,173-0,773 см/год, т.е. на 19% больше. Канавы, обросшие
древесной и кустарниковой растительностью мелеют медленнее.
Средняя интенсивность зарастания (ИЗ) откосов канав деревьями и кустарниками и
средняя интенсивность уменьшения глубины (ИУГ) осушителей, построенных 15-25
лет назад в зависимости, от их местоположения: а) под пологом древесной и
кустарниковой растительности, б) в окнах и прогалинах леса и поросших травяной
растительностью
Тип лесорастительных условий/Типы почв
Сосняк сфагновый/торфяная (торф верховой,
глубокий)
Сосняк багульниковый/торфяная (торф
верховой, глубокий)
Сосняк осоково-сфагновый/торфяная (торф
переходный, глубокий)
Черноольшаник осоковый/торфяная и
торфянистая (торф низинный)
Черноольшаник крапивный/торфяная
(низинный торф, глубокий)
Сосняк черничный/песчанная на супесчанике
Ельник чернично-кисличный/супесчанная на
суглинке
Ельник кислично-
широкотравный/суглинистая на глине
Продуктивность травяной растительности, произрастающей в канавах,
зависит и от продолжительности стояния (течения) воды в канаве.
Установлено, что застойный режим в канавах благоприятствует растительности ок-
раек озер (тростник обыкновенный, хвощ топяной, рогоз
широколиственный, частуха подорожниковая, осока, аир, кипрейник болотный). Буйная
травяная растительность, достигающая 1,6-2,8 кг/м2 образуется на открытых
участках канав с отрицательными уклонами дна в типах леса с богатыми
почвами (например, в ельнике кислично-широкотравном). В типах леса с
бедными почвами (например, в сосняке сфагновом) при любых
экологических условиях формируется травяная растительность только умеренной
продуктивности. Максимальная отмеченная в этих типах леса продуктивность
травяной растительности не превышает 0,46-0,83 кг/ м2.
Полученные результаты позволяют сделать следующие выводы. 1. Если
канавы исправно функционируют (канавы высшей категории не подпирают
воду в канавах низшей категории, а осушители весною не подпирают грун-
279
ИЗ, % от
длины канав
2,0
4,0
19,0
23,0
22,0
20,0
23,0
22,0
ИУГ, см/год
а)
0,460
0,450
0,563
0,640
0,613
0,133
0,250
0,397
б)
0,510
0,530
0,667
0,773
0,730
0,173
0,323
0,473
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
товые воды выше норм осушения обеспечивая максимальный рост осуще^
ных древостоев) обрастание деревьями или кустарниками берм, даже отко*
сов канав не является критерием плохого состояния осушительной сети. 2
Наилучшим образом канавы выполняют свою гидрологическую функцию ц
подлежат механизированному ремонту, когда их бермы покрыты только
высокими кустарниками или деревья растут только на одной стороне канав.
МЕТОДИКА ОЦЕНКИ ВЛИЯНИЯ ГИДРОМЕЛИОРАЦИИ
НА СУХОДОЛЬНЫЕ НАСАЖДЕНИЯ
Е.Д Сабо
Московский государственный университет леса, Московская обл.
Вкратце эта методика заключается в следующем. Задаваясь
определенной точностью работ и зная вариабельность прироста, находят необходимую
повторность взятия образцов. Ряды данных подвергают на ПЭВМ
статистической обработке. Затем находят закономерность изменения исследуемого
признака во времени (уравнение) с аппроксимацией прямой или кривой по
методу наименьших квадратов. Определяют коэффициент корреляции (или
корреляционное отношение) и уровень его значимости для двух (или более)
периодов: до и после начала воздействия исследуемого фактора. Затем
определяют первую производную функций, характеризующих изменение
исследуемого признака (например, прироста по диаметру) до и после начала
воздействия на насаждение. Затем определяют разность первых
производных путем вычитания последующей из предыдущей. Положительное
значение разности в этом случае свидетельствует об уменьшении величины
исследуемого признака, а отрицательное — об увеличении.
Такие же расчеты производят и для контрольных пробных площадей, не
подвергшихся воздействию исследуемого фактора. Затем из разности
первых производных на контроле вычитают разность первых производных на
исследуемом объекте. Полученные значения являются конечным
результатом, характеризующим качественное значение влияния исследуемого
фактора. Положительный результат говорит о возрастании исследуемого
признака, нулевой — о его неизменности и отрицательный — о его убывании.
При этом сравнении результаты расчета сопоставляют с суммарной
ошибкой расчетов. Окончательный вывод должен быть подтвержден
необходимым превышением полученного результата по сравнению с ошибкой.
Затем на основе конечных значений исследуемого признака и ошибок
разности его конечных значений за период исследований на контрольных и
основных пробных площадях определяют коэффициенты достоверности
различия, сравнивают их с табличными, а при необходимости определяют и
280
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
вероятность различия в %. Надежное использование этой методики
основано на том, что при выборе контроля особое внимание следует обращать на
идентичность типа леса, возраста и полноты. Последнее особенно важно
вследствие большой вариабельности показателей при изменении полноты.
В качестве наглядного примера рассмотрим результаты применения этой
методики для определения влияния осушения на прилегающие к осушаемым
массивам суходольные насаждения Рязанской Мещеры. Основным
элементом измерения явился годичный прирост по радиусу средних деревьев на
высоте 1,3 м. Для определения статистических уравнений хода роста
средних деревьев по радиусу до и после осушения были взяты одинаковые
периоды роста продолжительностью 6, 7 лет. Затем были найдены линейные
уравнения, характеризующие ход прироста по радиусу, коэффициенты
корреляции и их уровень значимости. Весь пример построен на анализе
расчетов по сосняку долгомошному 24 лет, сосняку орляковому 74 лет и сосняку
зеленомошному 46 лет. Результаты расчетов приведены ниже.
До осушения После осушения
Уравнения хода прироста
С дм Р = 20,42999-0,30285 Т, Р = -2,71714+0,06285 Т,
С орл Р = 2,88278-0,02623 Т, Р = 2,66384-0,02414 Т,
С зм Р = 7,89892-0,10821 Т, Р = 4,17990-0,05228 Т,
где Р - прирост по радиусу, мм/год; Т - время, лет /год минус 1900/.
Коэффициенты корреляции и уровень значимости
С дм - 0,98*** + 0,86*
С орл + 0,87* - 0,94**
С зм - 0,98*** - 0,98***
где * - уровень значимости 0,05; ** - уровень значимости 0,01; *** - уровень
значимости 0,001.
После этого были получены первые производные прироста по времени и
их разности до и после проведения осушительных работ.
Первые производные [Р/Т]1, мм/год2
До осушения После осушения Разность
С дм -0,30285 +0,06285 -0,36570
С орл -0,02623 -0,02414 -0,00209
Сзм -0,10821 -0,05228 -0,05593
После проведения аналогичных расчетов на контрольных пробных
площадях были получены разности разностей первых производных, а также их
ошибки, при уровнях значимости 0,32 и 0,05. Результаты окончательных
расчетов сведены в таблицу.
Из таблицы (с учетом величины ошибки) видно, что сосняк долгомош-
ный показал явно положительную реакцию на изменение водного режима,
281
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
сосняк орляковый — слабо отрицательную и сосняк зеленомошный — щ^
тральную.
Влияние мелиорации на ход прироста по радиусу суходольных сосняков в Рязанской
Мещере (данные получены при 800-кратной повторности взятия образцов).
Тип Возраст, Разность разностей первых Ошибка разности при уровГ*
леса лет производных, мм/год2 не значимости
0,32 0,05^"
Сдм ~24 +0,245 0,043 (ЩГ""^
Сорл 74 -0,095 0,033 0,064
Сзм 46 +0,035 0,035 0,069
ЭФФЕКТИВНОСТЬ ЛЕСООСУШЕНИЯ В КАРЕЛИИ
ПО МАТЕРИАЛАМ ЛЕСОУСТРОЙСТВА
В.И. Саковец
Институт леса Карельского НЦРАН, Петрозаводск
Данных о лесоводственной эффективности осушения в условиях Карелии
много, но большинство их них получены при исследованиях на отдельных
пробных площадях. Для оценки результативности гидролесомелиорации,
организации мониторинга гораздо важнее определить лесоводственную
эффективность осушения на больших территориях — на объектах осушения,
площади лесничеств, лесхозов. Это возможно сделать путем сопоставления
разновременных материалов лесоустройства. Такой анализ материалов
лесоустройства позволяет показать пространственно-временную динамику
состояния осушенных земель за длительный период.
В Карелии были проведены исследования лесоводственной
эффективности гидролесомелиорации по материалам лесоустройства на территории
трех лесничеств: Киндасовском Пряжинского лесхоза, Заозерском
Петрозаводского лесхоза, Юркостровском Кондопожского лесхоза. В Заозерском
лесничестве давность осушения к 1992-1994.гг. составила в среднем 24 года,
Киндасовском — 22 и Юркостровском — 19 лет. На всех объектах наряду с
лесопокрытой площадью было вовлечено в осушение много болот. Особенно
неблагоприятное соотношение леса и болота в Юркостровском лесничестве,
где болот было осушено в 3,5 раза больше, чем лесной площади. В
Киндасовском соотношение лесной и нелесной площади 1:1, в Заозерском — 1:0,5.
Почвенные условия на объектах Заозерского и Киндасовского лесничеств
характеризуются более высоким потенциальным плодородием, чем в
Юркостровском. На территории первых преобладали травяно-сфагновые типы
леса, а среди болот — переходные, на территории второго — сфагновые и
багульниковые занимали около 10% площади, а среди болот преобладали
282
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
верховые и верхово-переходные типы. В связи с такими природными
особенностями и результативность гидролесомелиорации оказалась разной.
Анализ распределения осушенных площадей по категориям земель
показал. чт0 в Заозерском и Киндасовском лесничествах покрытая лесом
площадь увеличилась, а болот сократилась соответственно на 33,5 и 33,6%.
Плошадь болот на этих объектах через 20-24 года после осушения
составляет всего лишь 0,3-4,4%. хотя до осушения на их долю приходилось 34-50%.
В Юркостровском лесничестве за 19-летний период после осушения
покрытая лесом площадь увеличилась, а площадь болот сократилась лишь на
25,1%, нелесные площади по-прежнему занимают значительную часть в
осушенном фонде (53,5%) из-за бедности торфяных почв. Облесение
осушенных болот происходило в основном с преобладанием сосны. Однако
значительная часть сформировавшихся после осушения молодняков в
богатых условиях местопроизрастания имеют большую долю примеси
лиственных пород (3-6 единиц).
В распределении осушенных площадей по группам возраста также
произошли существенные изменения: значительно возросла площадь
молодняков и средневозрастных насаждений. В Заозерском лесничестве площадь
молодняков увеличилась в первые 10-15 лет с 3 до 214 га, Киндасовском —
с 344 до 2284 га, Юркостровском — с 46 до 944 га. Часть болот, на которых
имелось естественное возобновление, в связи с преобладанием крупного
подроста после осушения, минуя стадию молодняков, лесоустройством
переведена в средневозрастные насаждения. Под влиянием осушения на всех
объектах произошло и увеличение полноты древостоев.
Наиболее полное представление о лесоводственной эффективности
гидролесомелиорации дает величина текущего прироста древостоев по запасу и
дополнительный прирост, получаемый за счет осушения. Дополнительный
прирост в Заозерском лесничестве в первое десятилетие составил в среднем
1,7, а в второе — ЗД м3/га в год, в Киндасовском соответственно — 0,66 и
3,1. а в Юркостровском спустя 11-19 лет после осушения — 1,8 м3/га в год.
Таким образом, данные лесоустройства показывают, что худшие результаты
получены в Юркостровском лесничестве, где в осушение в значительных
объемах были вовлечены насаждения и болота с верховыми торфяниками.
Анализ материалов лесоустройства позволяет вести мониторинг
осушенных площадей на больших территориях, проследить временную динамику
трансформации болот и заболоченных лесов. Обобщая результаты
исследований, следует отметить, что при преобладании на объектах осушения
переходных и низинных болот и насаждений травяно-сфагновой группы типов
леса гидролесомелиорация является наиболее перспективным средством
повышения продуктивности лесов. Однако для сохранения достигнутого
эффекта необходимо проведение дополнительных лесохозяйственных меро-
283
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
приятии по улучшению породного состава, возрастной структуры и своевр^
менного ухода за осушительной сетью.
ИМИТАЦИОННАЯ МОДЕЛЬ ДЛЯ ПРОГНОЗИРОВАНИЯ
ТЕХНИЧЕСКОГО СОСТОЯНИЯ ОСУШИТЕЛЬНОЙ СЕТИ
И ПРОДУКТИВНОСТИ ДРЕВОСТОЕВ
A.M. Тараканов
Северный НИИ лесного хозяйства, Архангельск
Для прогнозирования результатов гидролесомелиорации и выбора нан-
более рациональных способов ведения хозяйства разработана имитационная
модель функционирования гидролесомелиоративных систем. С помощью
модели, построенной из последовательно связанных блоков, можно
прогнозировать динамику технического состояния каналов, производительность
лесорастительных условий и ход роста древостоев как в проектируемых, так
и функционирующих гидролесомелиоративных системах. Наименование и
величина исходных данных задаются в зависимости от цели прогноза. Для
проектируемых систем достаточно задать параметры осушительной сети,
тип грунта, тип леса, средние высоту, диаметр, запас и возраст древостоев
перед прогнозом. На выходе модели можно получить вероятные
среднестатистические величины динамики технического состояния мелиоративной
сети, роста и продуктивности древостоев. В функционирующих системах
дополнительно задаются давность осушения до начала прогноза и величина
уменьшения глубины каналов за предшествующий период. Вначале
проводится расчёт прогнозных показателей без проведения каких-либо
мероприятий. После просмотра полученных данных при необходимости задаются
сроки проведения лесохозяйственных мероприятий и новые входные
параметры. Далее расчёт прогнозных величин повторяется при новых заданных
входных данных. Перечень исходных данных, необходимых для прогноза
по всем вариантам расчёта, приведён в табл.
Для получения прогнозных показателей лесоводственной эффективности
лесоосушения на стадии проектирования гидролесомелиоративных систем
алгоритм имитационной модели её функционирования является следующим.
1. Определение прогнозной величины уменьшения глубины каналов (AhJ
в метрах на конец каждого шага прогноза (п,) в зависимости от типа поч-
венно-грунтовых условий! у п
Ah, = 1—
284
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
2 Определение прогнозной глубины каналов на конец каждого шага
„рогноза: h, = ho - Ah,, м
3 Определение средней прогнозной глубины каналов, соответствующей
средине каждого временного шага: _ /А +А\
2
4. Расчёт класса бонитета лесорастительных условий (КБЛУ) (Б,) для
середины каждого шага прогноза в зависимости от глубины каналов (h ) и
расстояния между ними на этот момент и исходного типа леса:
Б, = В0 + Bi h, + В2 e"h + В3 h, L + В4 L
-
м
м
лет
Тг
L
ho
а
Исходные данные для прогноза технического состояния каналов и продуктивности
осушаемых сосняков и ельников
Наименование показателей Ед. Сим-
^ вол
1. Порода С,Е
2. Тип леса и его шифр (С.тр-сф-1, С.вах-сф-2, С. ос-сф-3, С.сф-4, - Тл
Елр.-сф. на низ. Ср. мощн. торфах-5, Е.ос-сф,тр-сф на переход.
ср.мощн.торфах-6, Е.ос.-сф. на низ. Мал омощн. торфах,
подстил, песком-7, Е.тр-сф на торфянисто- и торфяно-перегн.,
подстил, глиной и тяж.суглинком-8, Е.тр-сф на перегн.-торфяно-
глеевых тяжелосугл., на глине -9)
3. Тип грунта и его шифр
4. Расстояние между каналами
5. Глубина каналов проектная или перед началом прогноза
6. Давность осушения перед началом прогноза (для
функционирующих систем)
7. Величина уменьшения глубины каналов перед прогнозом (для м дП()
функционирующих систем)
8. Средняя высота древостоя
9. Средний диаметр древостоя
Ю. Запас древесины на 1 га
11 Средний возраст древостоя
12. Продолжительность щага прогноза
13. Период от начала прогноза до ремонта мелиоративной сети
14. Глубина каналов после ремонта сети
,Jj- Расстояние между каналами после их сгущения
5.Определение среднепериодического изменения высоты (ДНО, диаметра
(AD,) и запас (ДМ*) для каждого шага прогноза в зависимости от КБЛУ для
этого шага (Б1). средних высоты (Н1М), диаметра (Dm) и возраста (Ам) дре-
востоев на начало каждого шага прогноза:
АН, = Bo+Bi Б|+В2 Ам+Вз Нм+В4 Б,2 +В5 Аи2 +В6 Нм2
AD, = В0 +Bi Bi+B2 Аи +В3 DM+B4 Bf+B5 Ая2 +В6 DM2
м
см
м3/га
лет
лет
лет
м
м
Н
д
м
А
Щ
ар
hp
_Lp_
285
КОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
AMi = Во +В] Б|+В2 Ам+В3 Ни+В4 DlA+B5 Б,2 +В6 Аь12 +В7 Ни2 +В8 DM2
6. Определение на конец каждого шага прогноза средних высоты (Н{\
диаметра (Dj), возраста (А,) и видовой высоты древостоя (FH0, запаса (Ц)
суммы площадей сечения (Gi) и числа стволов на 1 га (Н):
Н, = Нм + АН, п;
Д = Dm+АЦ п;
Mi = Mi_i + AMi n; Ai = Аи+п;
ВД = 0,8768 + 0,4410Hi - для сосняков;
FHi = 1,3216 + 0,4183Hi - для ельников;
Gr=Mi/FHi N1=4-104 Gi/ITD,2
Все расчеты по пунктам 1-6 повторяются на каждом шаге прогноза. При
этом данные предыдущего шага используются в расчетах последующего.
Расчет прекращается, когда давность осушения (а; ) равна 100. Просмотр
данных на выходе позвляет проанализировать изменения технического
состояния каналов и ход роста древостоев, установить сроки проведения
ремонта или реконструкции мелиоративной сети, выявить отзывчивость
различных возрастных поколений на осушение и при необходимости назначить
рубку отдельных элементов леса.
7. В функционирующих гидролесомелиоративных системах определение
прогнозной величины уменьшения глубины каналов (Ahi) на конец каждого
шага прогноза производится в зависимости от давности осушения (а) и
величины уменьшения глубины каналов (Aho) перед началом прогноза:
Дальнейший расчет прогнозных величин проводится согласно пунктов 2-6.
8. В тех случаях, когда требуется получить прогнозные данные о росте и
продуктивности древостоев после ремонта или реконструкции
мелиоративной сети, установить целесообразность их проведения или оптимальные
варианты, на входе программы задаются сроки проведения этих мероприятий
(ар ), глубина каналов (hp ) и расстояние между ними (Lp ). Расчет вероятной
величины уменьшения глубины каналов после их углубления проводится по
уравнению п.1. Исходные таксационные показатели принимаются те,
которые получены на конец периода до ремонта.
9. Для прогнозирования роста и продуктивности древостоев после
проведения в них несплошных рубок на входе программы задаются измененные
рубкой таксационные показатели древостоя (средние высота, диаметр,
возраст, запас) и давность осушения в момент рубки. Все остальные входные
данные остаются такими же, как в варианте без рубки. В том случае, если в
одном насаждении имеется несколько поколений древостоев, прогнозирова-
286
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
яле проводится по каждому из них и прогнозная величина запаса каждого
лемента леса умножается на долю участия его в. составе насаждения.
Обилий запас насаждения равен суммам редуцированных через состав запасов
элементов леса. Сравнение результатов различных вариантов рубок позво-
доет установить их эффективность и выбрать оптимальный вариант.
10. Аналогичным образом проводится прогнозирование при выполнении
комплексов мероприятий на различных временных этапах. Изменение
одного из факторов условий местопроизрастания или состояния древостоев
вызывает ответную реакцию древостоев через изменение их текущего
прироста, направленную на восстановление соответствия его таксационной
характеристики условиям роста.
11. Имея данные о распределении осушенных площадей по типам леса и
классам возраста, а также средние таксационные показатели древостоев
согласно этому распределению, с помощью модели можно прогнозировать
продуктивность не только отдельных древостоев, но и всей
гидролесомелиоративной системы. Для этого необходимо спрогнозировать продуктивность
в однородных стратах при хорошем, удовлетворительном и
неудовлетворительном состоянии сети и на основе процентного соотношения каналов
различного состояния на начало прогноза в зависимости от давности осушения
объекта установить вероятную продуктивность на всей площади.
Представленная модель является одним из возможных вариантов
прогнозирования технического состояния гидромелиоративной сети, роста и
продуктивности осушаемых сосняков и ельников Европейского Севера.
Основана на закономерностях роста древостоев в зависимости от различных
факторов, функциональных связях между таксационными показателями
древостоев и вероятной природе изменений в функционировании
мелиоративной системы. Работа с ней довольно легко осуществляется на
персональном компьютере и позволяет решать оптимизационные задачи по ведению
хозяйства в осушаемых лесах и своевременному предупреждению
нежелательных последствий. Проверка её на адекватность показала, что расчётные
данные достаточно точно отражают фактический ход изменений в
мелиоративной системе под действием приведённых выше факторов.
ЛЕСОХОЗЯЙСТВЕННАЯ ЭФФЕКТИВНОСТЬ
ГИДРОЛЕСОМЕЛИОРАЦИИ НА СРЕДНЕМ УРАЛЕ
А.С. Чиндяев, А.Р. Иматов, И.А. Иматова, М.А. Матвеева
Уральская государственная лесотехническая академия, Екатеринбург
На Среднем Урале, где болотные леса только в ГЛФ занимают более 3,5
Млн. га [3], имеет место лишь экспериментальное их осушение, проведенное
287
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
на стационарах «Песчаный», «Северный», «Мостовое», и продолжитель»
ность которого не превышает 20 лет. Осушены были как чистые по составу
сосновые, еловые, березовые, так и смешанные древостой V-IV класса во^
раста IV-Va класса бонитета, произрастающие на низинных болотах («Пес»
чаный». «Мостовое») и чистые сосновые древостой IV-VI класса возрасту
V-Va классов бонитета — на глубоких сфагновых торфах («Северный»).
Важными являются данные, характеризующие влияние осушения на poet
леса для новой отрасли науки — лесного болотоведения [по 1], развивающе.
гося на стыке лесоведения и болотоведения и сочетающего основы веденщ
лесного хозяйства на осушаемых землях.
Известно, что скорость изменения процессов и взаимосвязей, сущест-
вующих в лесоболотных биогеоценозах и степень их проявления во многом
зависят от фактора времени, а также от того, в какой мере новые экологиче-
ские условия соответствуют требованиям компонентов биогеоценоза.
Впервые в регионе полученные материалы характеризуют самый важный —
начальный этап (период перестройки и приспособления [2]) роста
мелиорируемых древостоев, поэтому они представляют значительный и
теоретический . и практический интерес.
Установлено, что на низинных болотах сосновые древостой
адаптируются к изменившимся экологическим условиям среды в течение первых 5-7
лет. В дальнейшем их рост улучшается: прирост по радиусу увеличивается в
3-5 раза, текущий прирост по запасу достигает 4 м3, а дополнительный —
2.9 м3/га. Активнее на осушение реагируют еловые древостой. Они с
первого-второго годк осушения увеличивают прирост по радиусу в 5-7 раза, их
текущий прирост по запасу достигает 6 м3. а дополнительный — 3,6 м3/га.
Важным является и тот факт, что деревья сосны и ели разного диаметра
неодинаково реагируют на осушение. Лучшей реакцией характеризуются у
сосны средние по крупности деревья, а у ели — тонкие и средние.
Осушение положительно сказалось и на лесовозобновительных
процессах. Имеющийся до осушения подрост ели быстро адаптируется к
изменившимся условиям среды и существенно улучшает свой рост. Численность его
в первые 6-10 лет осушения увеличивается в 3-5 раз.
С целью улучшения породного состава, структуры древостоев и
обеспечения устойчивого функционирования лесоболотных биогеоценозов после
осушения часто возникает необходимость проведения выборочных рубок.
Их интенсивность в хвойно-лиственных высокополнотных древостоях
должна составлять по запасу 45-50 %. а в среднеполнотных — 30-35 %.
Весьма важным для условий Среднего Урала является и 10-летний
положительный опыт лесокультурного освоения интенсивно осушенных
низинных болот. Здесь культуры лиственницы, кедра и ели рядовые и
биогруппами были созданы посадкой крупномерного посадочного материала
288
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
без подготовки почвы как на безлесных участках и вырубках, так и под по-
10ГОм изреженных до полноты 0,4-0,5 березовых древостоев. В настоящее
время они характеризуются хорошим ростом. Их сохранность составляет не
менее 70 %. Прирост по высоте у культур ели превышает 10-15, кедра — 13-
16. лиственницы — 30-35 см в год.
Хорошим ростом характеризуются 8-летние рядовые культуры кедра и
ели на площадях реконструкции березовых молодняков (стационар
«Мостовое») Они также созданы крупномерным посадочным материалом без
подготовки почвы в 5-метровых коридорах, подготовленных трактором зимой.
Культуры созданы на 3-й год после осушения. Сохранность культур кедра
здесь составляет 70 %, средняя высота превышает 79 см, а текущие
приросты по высоте составляют 15-20 см в год. Культуры ели имеют сохранность
77 %. среднюю высоту — 110 см, приросты по высоте 20-25 см в год.
Определенный интерес представляет и 10-летний опыт осушения
сфагновых болот с сосновыми древостоями («Северный»). Здесь изучается
комплексное влияние осушения и вырубки материнского древостоя на состояние
и рост подроста сосны предварительной генерации, как основы
формирования нового поколения леса. С этой целью на 84-метровой межканальной
полосе был сплошь вырублен сосняк кустарничково-сфагновый Va класса
бонитета, имеющий Дер. — 13 см, Нср. — 9 м, Аср. — 105 лет, полноту —
0.59. число стволов — 1988 шт/га, запас — 83 м3/га, подроста — 8,6 тыс.
шт/га. мощность сфагнового торфа составляет 5 м.
Выявлено, что проведение осушения и вырубки материнского древостоя
в 3 раза увеличило численность подроста, улучшило его качественное
состояние и в 2,5 раза увеличило прирост по высоте. Экспериментально
выявлено и раздельное влияние осушения и удаления материнского полога на
рост подроста сосны предварительной генерации. Установлено, что 10-
летний период после удаления материнского древостоя обусловил
увеличение прироста по высоте подроста сосны на 94 %, а последующее 10-летнее
осушение — еще на 94 %. В настоящее время на экспериментальном
участке сформировались молодняки сосны (возраст 38 лет), произрастающие по
Н-Ш классу бонитета.
Проведенный эксперимент позволяет констатировать, что осушение и
сплошные рубки в спелых сосняках с полнотой 0,5-0,6 являются вполне
приемлемым и высокоэффективным лесохозяйственных мероприятием по
ускоренному формированию высокопродуктивных сосновых древостоев на
сфагновых болотах Среднего Урала. Исследования убеждают в лесоэколо-
гической целесообразности осушения болотных лесов, возможности
повышения их эффективности и отвечают задаче устойчивого управления
осушенными биогеоценозами.
289
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
[1] Мелехов И.С. Предисловие // Гидролесомелиоративные исследования. Рига:
Зинатне, 1970. С. 3-5. [2] Рубцов В.Г. , Книзе А.А. Ведение хозяйства в
мелиорируемых лесах. М: Лесн. пром-ть, 1981. 100 с. [3] Сабо Е.Д. и др. Справочник гид.
ролесомелиоратора. М.: Лесн. пром-ть, 1981. 200 с.
СТОК С ОСУШЕННЫХ ЛЕСНЫХ ЗЕМЕЛЬ
С.Г. Шурыгин
Санкт-Петербургская государственная лесотехническая академия
Влияние лесоосушения на сток малых рек — водоприемников может
быть неоднозначным и зависит от физико-географических условий
конкретного бассейна, метеорологических показателей, размещения осушенных
площадей в пределах водосбора и других факторов. Известные нам данные
о влиянии осушения на годовые значения речного стока [Бабиков, 1976;
Новиков, 1980; Лайне, 1980; Чиндяев, 1995] свидетельствуют о выравнивании
внутригодового распределения стока, увеличении продолжительности
летнего стока и его объема, улучшая питание рек в меженный период. Некоторое
увеличение годового стока при лесоосушении отмечено в работах [Чесноков,
1980; Вомперский, 1988].
Исследования проводились в Ленинградской области на осушенных
торфяных и минеральных землях, на гидрометрических постах с самописцами
в течение двух гидрологических лет. Метеорологические условия были
близки к норме. Анализировались годовые гидрографы стока с осушенных
лесных водосборов торфяных (площадь 33 га) и минеральных (10,2 га) землях
и реки-водоприемника Лустовки (водосбор осушен, площадь 21 тыс. га, 20%
занимают торфяные почвы).
Модули стока р.Лустовки в осенний период не превышали 0,15 л/с га,
что в 2 раза ниже, чем на торфянике и минеральных землях. Зимой на реке
и торфянике отмечается устойчивый сток воды. Весеннее половодье на реке
начинается на 2 недели раньше, чем на торфяных и минеральных землях: на
водосборе р.Лустовки много нелесных площадей. Более раннее начало
половодья на реках-водоприемниках отмечалось ранее [Аире, 1977].
Максимальные суточные модули стока в период половодья на реке не превышают
0,42 л/с га, что 1,6 и 2 раза ниже, чем на минеральных и торфяных землях
соответственно. В вегетационный период по р.Лустовке наблюдался
стабильный сток воды, но максимальные модули стока также в 1,5-2 раза ниже,
чем на торфяных и минеральных землях. Сток воды с осушенного водосбора
р.Лустовки обладает меньшей изменчивостью, чем с осушенных торфяных и
минеральных почв: коэффициент вариации стока на речном водосборе равен
1.15, на торфянике — 1,75, а на минеральных землях — 1,83.
290
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
1994-95 гидрологический год характеризовался частыми и сильными
зимними оттепелями и засушливым летним периодом. В целом осадков
выпало на 7% больше нормы, а температура воздуха была на 2°С выше сред-
немноголетней. Слой стока с речного водосбора в осенний период был в 1,1
и 1,3 раза ниже, зимой (при частых оттепелях) в 3-4 раза выше, а за
вегетационный период в 1,2 и 1.4 раза ниже, чем на минеральных и торфяных
землях соответственно (см. табл.). Наибольший годовой сток наблюдается
по реке за счет интенсивного снеготаяния на нелесных площадях и быстрого
поступления талой воды в реку в период зимних оттепелей, и
соответственно меньшего суммарного испарения.
1995-96 гидрологический год характеризовался устойчивой зимой без
оттепелей и засушливым периодом вегетации. Осадков выпало на 18%
меньше нормы, температура воздуха была на 2°С ниже среднемноголетней.
В осенний период сток по реке был в 4 и 2 раза выше стока с минеральных и
торфяных земель. Зимой наблюдался устойчивый сток, в то время как на
минеральных и торфяных почвах он прекращался вследствие малых
напоров грунтовых вод и промерзания откосов и дна каналов. Весенний сток на
реке был близок к стоку с минеральных земель, что в 1,6 раза ниже стока с
торфяника. За период вегетации речной сток был в 1,2 и 1,3 раза меньше
стока с минеральных и торфяных почв соответственно. Это объясняется
несколько большими потерями влаги с речного водосбора глубокие слои
почвы и на эвапотранспирацию. Годовая величина стока была близка к стоку с
минеральных почв, но ниже стока с осушенных торфяных почв.
Слой стока с осушенных лесных водосборов, мм
Периоды р.Лустовка Минеральные почвы Торфяные почвы
Осень (X-XI)
Зима (ХП-П)
Весна (Ш-IV)
V-DC
Год
Осень (X-XI)
Зима (ХП-П)
Весна (Ш-IV)
V-DC
Год
мм
47
87
194
36
364
4
5
35
67
111
%
1994-1995
12,8
23,8
53,4
10,0
100,0
1995-1996
4,0
4,2
31,8
60,0
100,0
мм
%
гидрологический год
51
22
172
44
289
17,5
7,7
59,6
15,2
100,0
гидрологический год
1
0
33
80
114
0,9
0,0
28,9
70,2
100,0
мм
59
29
203
51
342
2
0
56
88
146
%
17,1
8,4
59,4
15,1
100,0
1,5
0,1
38,4
60,0
100,0
Сток в реках — водоприемниках характеризуется меньшей
изменчивостью (ниже максимальные модули стока, не наблюдается «нулевого стока»),
291
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
чем сток с осушенных лесных водосборов на торфяных и минеральных
почвах. Основные различия в стоке связаны с зимними сезонами. В годы с
частыми и сильными оттепелями отмечается несколько больший годовой сток с
речного бассейна, чем на малых по площади осушенных лесных водосборах,
а в годы с холодной и устойчивой зимой наоборот. Полученные выводы
следует рассматривать как предварительные, их проверка требует
дополнительных более длительных наблюдений.
ИСПОЛЬЗОВАНИЕ БОЛОТ И ТОРФА В СЕЛЬСКОМ ХОЗЯЙСТВЕ
ОЦЕНКА ЦЕЛЕСООБРАЗНОСТИ СЕЛЬСКОХОЗЯЙСТВЕННОГО
ОСВОЕНИЯ ВЫРАБОТАННЫХ ТОРФЯНИКОВ
В ОРЕХОВО-ЗУЕВСКОМ РАЙОНЕ МОСКОВСКОЙ ОБЛАСТИ
A.M. Гребенников, И.И. Лыткин
Почвенный институт им. В.В. Докучаева, Москва
Целесообразность сельскохозяйственного освоения выработанных
торфяников определяется уровнем затрат материальных и финансовых ресурсов
на проведение мероприятий, связанных с их окультуриванием,
регулированием водно-воздушного режима, внесением удобрений и мелиорантов.
Объем необходимых работ по освоению территорий торфяных выработок
зависит от ряда факторов. К их числу следует отнести способ выработки торфа,
мощность остаточного слоя, время, прошедшее после прекращения
торфоразработок, геолого-геоморфологические особенности участков с
выработанными торфяниками, их химические, физико-химические, физические
свойства, засоренность пнями и т.д.[1,2,3].
В ходе исследований выработанных торфяников Орехово-Зуевского
района Московской области было установлено, что на свойства остаточных
слоев торфа большое влияние оказала вышележащая (до момента выработки)
торфяная толща. Поскольку выработанная толща в исследуемом районе
представлена в основном верховым и переходным торфом, и, несмотря но
тск что остаточные слои торфа по ботаническому составу соответствуют
преимущественно низинному типу, их нельзя отождествлять с низинными
торфяниками, на которых добыча торфа не проводилась. По
физико-химическим свойствам остаточная мощность низинного торфа оказалась очень
похожей на выработанную толщу переходного или верхового типа.
Учитывая это обстоятельство, тип выработанных торфяников устанавливался нами
не по ботаническому составу остаточных слоев торфа, а по типу
выработанной торфяной толщи, идентификация которого проводилась по останцам,
где торфяная залежь оказалась ненарушенной.
292
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
Обобщение материалов исследования выработанных торфяников Орехо-
в0-3уевского района позволило провести их типизацию по целесообразности
сельскохозяйственного освоения. Было выделено три основных типа. К
первому- наиболее распространенному типу, принадлежат выработанные
верховые и переходные торфяники, занимающие водоразделы, вторые
надпойменные террасы и озерные котловины, расположенные на возвышенной
равнине. Выделять в отдельные типы переходные и верховые выработанные
торфа не имело смысла в связи с тем, что остаточные слои торфа и
подстилающий минеральный материал обладают примерно одинаковым
комплексом свойств на обоих типах торфяников. К второму, менее
распространенному типу, были отнесены выработанные низинные торфяники на первой и
второй надпойменных террасах. Третий, относительно редко встречаемый
тип. был представлен пойменными низинными торфяниками.
Многолетние исследования свойств выработанных торфяников Орехово-
Зуевского района проводились на отдельных ключевых участках,
отражающих основные типы торфяников. Выработанная толща торфяников первого
типа в основном сложена из сосново-сфагнового и пушицевого верхового
торфа со степенью разложения 20-40%, Остаточные слои представлены дре-
весно-осоковыми и пушицево-сфагновыми торфами. Реакция среды очень
сильно кислая — рН солевой составляет 2.3-3.2, гидролитическая
кислотность варьирует в пределах от 153,3 до 227.0 мг-экв. на 100 г почвы,
содержание подвижных форм алюминия велико — 10,6-14.5 мг/100 г почвы.
Количество общего азота очень низкое — 0,35-0,56%. Содержание подвижных
форм фосфора и калия низкое по всей толще. Крайне неблагоприятны
водно-физические свойства.
Выработанная толща и остаточные слои торфяников второго типа
состоят из древесно-осокового торфа со степенью разложения 25-50%.
Остаточные слои характеризуются сильнокислой реакцией — рН солевой равен 3,2-
3.5. высокой гидролитической кислотностью — 126,0-145,1 мг-экв. на 100 г
почвы, большим содержанием подвижного алюминия — 46,2-67,1 мг на 100
г почвы. Содержание общего азота несколько ниже среднего — 1.14-1.31%.
подвижного фосфора — очень высокое по всей остаточной толще — 90.6-
160.5 мг на 100 г почвы, подвижного калия — ничтожно мало. Водно-
физические свойства неблагоприятны.
Третий тип торфяников представлен остаточными слоями
древесно-осокового и тростниково-осокового торфов со степенью разложения 35-55%.
Величина рН солевой составляет 4,5-5,0. Гидролитическая кислотность
равна 20,0-80,1 мг-экв. на 100 г почвы, степень насыщенности основаниями -
3.0-80,6%. Содержание подвижного алюминия незначительно — 2.7-3.3 мг
на 100 г почвы. Высокое содержание подвижного фосфора — 96,1-118,0 мг
на 100 г почвы. Количество общего азота среднее — 1,65-1,74%, подвижно-
293
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
го калия низкое — 7,5-17,1 мг на 100 г почвы. Водно-физические свойства
близки к оптимуму для культурной растительности.
Освоение выработанных торфяников первого типа для их использования
в растениеводстве нецелесообразно в связи с большими затратами на их
рекультивацию, окультуривание и регулирование водно-воздушного
режима. Как при освоении, так и при выращивании сельскохозяйственных куль-
тур на этих землях, по сравнению с другими типами торфяников, приходит,
ся применять значительно больше удобрений, чтобы компенсировать
высокий геохимический вынос. Для поддержания реакции среды в оптимальном
диапазоне требуется вносить очень высокие дозы извести. Они в среднем в 2
раза превышают аналогичные величины для торфяников второго типа и в 4
раза — для торфяников третьего типа. Экстенсивно в качестве естественных
сенокосов могут использоваться лишь наиболее ровные и наименее
засоренные древесными остатками площади, выработанные фрезерным способом.
При условии благоприятного водного режима, на торфяниках данного типа
можно успешно выращивать лесные культуры, а в естественных условиях -
клюкву, голубику, багульник и другие ягодные и лекарственные растения.
Выработанные торфяники второго типа в зависимости от мощности
остаточного слоя на полях разлива гидроторфа и площадях, выработанных
фрезерным способом, целесообразно использовать не только под
естественные, но и под культурные сенокосы, а после проведения известкования, на
этих участках в севооборотах с большой долей трав можно выращивать
многие полевые культуры. Аналогичным образом, после проведения
соответствующей планировки, затраты на которую для рассматриваемого типа
торфяников вполне оправданы, можно использовать выработки, где торф
добывался резным и машинно-формовочным карьерными способами. Такие
участки, после проведения планировочных работ, становятся даже более
привлекательными для использования (вследствие большего плодородия),
чем фрезерные поля и поля разлива гидроторфа. Их увеличение плодородия
происходит в результате разравнивания по всей поверхности участка
межкарьерных бровок, состоящих из невыработанного, плодородного торфа.
Неокультуренные торфяники третьего типа в районе встречаются редко.
Из всех, представленных типов торфяников, естественное плодородие их
самое высокое. Поэтому, если мощность остаточной залежи более 50 см,
после очистки участков от древесных остатков и планировки, их можно ис-
пользовать под выращивание в севооборотах интенсивных
сельскохозяйственных культур при минимальных затратах на удобрения и известь. Таким
же образом, после проведения соответствующей планировки, целесообразно
использовать торфяные выработки, где торф добывался резным и машинно-
формовочным способами.
294
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
п] Бознюк СТ. и др. Генетические особенности и агромелиративная
характеристика выработанных торфяников западных районов УССР // Почвоведение. 1976. № 7.
q 94-103. [2] Лыткин И.И., Скрынникова И.Н. Изменение физико-химических
свойств торфяных почв при сельскохозяйственном использовании // Факторы и кри-
терии оценки плодородия почв. М, 1986. С. 12-20. [3] Марков В.Д. Оценка и
перспектива использования торфяно-карьерных площадей на территории РСФСР //
Природа болот и методы их использования. 1976. С. 233-236.
ПЕРСПЕКТИВЫ ИСПОЛЬЗОВАНИЯ БОЛОТ И ЗАБОЛОЧЕННЫХ
ЛЕСОВ РЕСПУБЛИКИ КОМИ ДЛЯ РАЗВИТИЯ'ОЛЕНЕВОДСТВА
Т.В. Евдокимова, В.А. Иванов
Институт биологии Коми НЦ УрО РАН; Институт социально-
экономических и энергетических проблем Севера УрО РАН, Сыктывкар
Система землепользования в значительной мере определяет характер и
остроту экологических проблем, степень нарушения территории ландшафтов
и их компонентов, состояния окружающей среды в целом, и в конечном
итоге — возможности и эффективность развития экономики. Среди природных
ландшафтов (геосистем) в этом отношении особое положение занимают
болота и заболоченные земли. Это связано с тем, что эти территории и их
ресурсы используются для добычи торфа, после проведения мелиорации - как
сельскохозяйственные угодья, как низкопродуктивные леса, охотничьи
угодья, для сбора ягод, лекарственных растений. Известна особая биосферная
роль данных геосистем, поэтому болота и заболоченные земли составляют
важную часть системы охраняемых территорий.
Специфичным для северных районов Республики Коми и Ненецкого
автономного округа (НАО) Архангельской области является использование
территории, в том числе — болот и заболоченных земель, в качестве
пастбищных угодий для выпаса домашних северных оленей. Оленеводство в
регионе является жизнеобеспечивающей отраслью хозяйства и основой
традиционного образа жизни этнических групп коренного населения: ненцев и
коми-ижемцев. На территории региона выпасается около 230 тыс. домашних
северных оленей (1998 г.), принадлежащих 6 хозяйствам и оленеводам
восточной части территории НАО (правобережье бассейна р. Печора) и 7
хозяйствам и оленеводам Республики Коми. Выпас ведется практически на
всей территории типичной и южной тундры, лесотундры, крайнесеверной
тайги и большой части северной тайги (за исключением территорий
осваиваемых месторождений, населенных пунктов, транспортных трасс и
нарушенных земель). В соответствии с характером преобладающей
растительности выделено 20 типов пастбищ. Для каждого из 6-и сезонов выпаса
наибольшую ценность представляют определенные типы угодий. Северотаеж-
295
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ные леса используются в качестве зимних пастбищ, а лесотундра - для вы-
паса в периоды поздней осени и ранней весны. Во все сезоны выпаса важное
значение имеют кормовые ресурсы болот и заболоченных земель: травяно-
моховые заболоченные тундры, низинные травяно-моховые болота, осоково-
моховые ивняки, полигональные, плоско- бугристые, крупно-бугристые в
грядово-мочажинные болота, различные типы заболоченных лесов.
В настоящее время в связи с активным освоением минеральных ресурсов
в тундре и лесотундре на оленьих пастбищах сформировались локальные
очаги с неблагополучной экологической ситуацией, в пределах которых
выпас оленей становится невозможным, а на прилегающих к ним участках
может производиться лишь с ограничениями. Это ведет к эскалации
увеличения нагрузки на пастбища, остающиеся в распоряжении оленеводов, и к
снижению их оленеемкости. нарушению баланса кормов на сезонных
пастбищах в целом. При этом ландшафты с наиболее ценными кормовыми
ресурсами (тундровые и лесные урочища) наиболее часто нарушаются.
В качестве одного из альтернативных решений, направленных на
сохранение истощенных тундровых и лесотундровых пастбищ от необратимой
деградации, а крайнесеверных лесов - от уничтожения, может быть
рассмотрено освоение территорий в северной тайге Республики Коми для
развития лесного оленеводства. Имеющиеся данные свидетельствуют о
возможности развития этой отрасли на территории, включающей бассейны рек
Цильмы. Пижмы, Заижемской, Ижмы, Кедвы, Сэбыси, верховьев Ухты,
Лыжи, Кожвы в пределах Ижемского, Усть-Цилемского, Ухтинского, Со-
сногорского, Печорского и севера Удорского районов. Здесь имеются
крупные массивы земель, общей площадью около 400 тыс. га,
характеризующиеся сочетанием разнообразных типов угодий. Широко распространены
еловые, березово-еловые, сосново-еловые и лиственично-сосновые кустарнич-
ково-лишайниковые, кустарничково-долгомошные леса, которые могут быть
использованы как ценные зимние пастбища с достаточным запасов
лишайниковых кормов. В то же время заболоченность данной территории
составляет более 30%. здесь представлены низинные травяно-моховые болота,
грядово-мочажинные болота, различные типы заболоченных лесов, которые
могут быть использованы для выпаса во все сезоны. Применяя известные
приемы ведения лесного оленеводства (управляемый выпас зимой и
регулируемый выпас на огороженных пастбищах в другие сезоны) можно создать
для оленей наиболее благоприятные условия, при которых они достигают
высокой упитанности и большого веса. Предлагаемая технология
содержания оленей в таежной зоне позволяет получить от каждых 100 январсих
оленей 28-30 ц мяса и решить многие социально-экономические проблемы.
Одним из преимуществ в этом случае является также выгодность
расположения оленеводческих бригад вблизи от рынка сбыта продукции. В соответст-
296
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
йй с расчетами здесь в течение года может выпасаться более 5000 оленей,
принадлежащих как крупным оленеводческим хозяйствам, так и фермерам-
оленеводам. Таким образом, освоение практически неиспользуемых сегодня
болот и низкопродуктивных в лесохозяйственном отношении заболоченных
1есов может служить одной из мер, направленных на поддержание
перспектив устойчивого развития оленеводства в данном регионе.
ОСНОВНЫЕ НАПРАВЛЕНИЯ РЕСТРУКТУРИЗАЦИИ
СЕЛЬСКОХОЗЯЙСТВЕННОГО ТОРФОПОЛЬЗОВАНИЯ РФ
В СВЯЗИ ЗАДАЧАМИ УСТОЙЧИВОГО РАЗВИТИЯ
А.И. Еськов. НЮ. Гармаш
ВНИПТИОУ, Владимирская область
1. Россия не имеет равных в мире по запасам и площади торфяных
залежей, разнообразию и ценности их ресурсов, по уникальности
информативной базы географической сети торфяных опытных полей, универсальности и
полноте научно-технологического потенциала, разнообразию перспективных
направлений торфопользования.
2. На обширной площади болота обеспечивают экологическое
равновесие в биосфере, накапливают биологически связанный углерод и азот,
регулируют водный режим местности и истоков рек, аккумулируют вредные
вещества атмосферных осадков и стоковых вод. являются продуктивными
ягодниками, местами обитания редких растений и животных. Торфяники
ограничивают начавшееся глобальное потепление климата. Уничтожение
лесов и торфяников способствует опустыниванию территории, подкислению
почв и загрязнению вод. сокращению жизненных ресурсов.
3. Учеными РАСХН установлено, что за 1975-96 гг. преобладающим
фактором риска и ущерба от чрезвычайных ситуаций в
сельскохозяйственном производстве России была засуха, охватывающая ежегодно 16-36
регионов с населением 30-75 млн. человек и приводящая к ежегодному убытку
порядка 9-12 млрд. рублей в ценах 1999 г. Решение комплексной проблемы
компенсации вреда от засухи предусматривает более эффективное
использование 80 млн. га природных торфяников в нулевой границе, 2 млн. га
выработанных торфяников и более 1 млн. га окультуренных и используемых в
сельском хозяйстве органогенных торфяных почв. Последние гарантируют
устойчивость высоких урожаев по годам.
4. В дореформенный период сельское хозяйство страны осваивало
торфяные залежи болот и заболоченных лесов, включая их в
антропогенно-биологический круговорот веществ в земледелии и животноводстве, в основном
через низкорентабельное рудно-сырьевое направление. При этом годовая
297
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
добыча торфа для сельского хозяйства к 90-м годам превысила 150 млн*
Участие торфа в органических удобрениях РФ достигло 12-15%, а в неко^
рых областях и автономных республиках Нечерноземья — до 50-60%.
5. Столь интенсивное использование торфа было обусловлено, %&*
первых, стремлением восполнить потери гумуса в почвах за счет традицн.
онных органических удобрений и торфа при недооценке биологическое
фактора и многолетних трав. Во-вторых, интенсивным строительством
животноводческих комплексов и птицефабрик с гидравлической системой на.
возоудаления. При отсутствии машин для использования бесподстилочнопэ
навоза и помета на удобрение их компостировали с торфом с последующим
применением машин для твердых органических удобрений.
6. В результате экономического кризиса в стране наиболее сильно
пострадал агропромышленный комплекс, что пагубно сказалось на
использовании торфа в земледелии и животноводстве. Потребление торфа
многократно сократилось. Мелиорированные торфяные почвы снизили
продуктивность и во многих хозяйствах оказались в запущенном состоянии.
7. В начале столетия предстоит возрождение АПК и торфопользования
страны на эколого-экономической основе при более полном освоении
научного потенциала. Предстоит реструктуризация всех направлений
торфопользования с оптимизацией режима гумуса в торфяных почвах в основном за
счет самих агроценозов, всеми средствами систем земледелия и
мелиорации, в севооборотах с многолетними травами. При этом исключается
уничтожение маломощных торфяников и ускоренная сработка торфяных почв,
как было в дореформенный период.
8. Использование торфяных залежей должно обеспечивать
экологическую устойчивость мелиорируемых систем, экономическую выгоду и
ресурсосбережение. Расход торфа в крае, республике и области не должен
превышать его естественный прирост на нетронутых площадях. Освоение залежи
для торфодобычи целесообразно при комплексном пролонгированном
использовании с обязательным сочетанием сырьевой выработки с
последующим освоением под агроценозы, лесокультуры, продуктивные водоемы.
9. Системы мелиорации, земледелия и удобрений должны быть
экологически безопасными в природопользовании не только для осваиваемых, но и
смежных территорий. В каждом конкретном случае нельзя допускать
излишних потерь органического вещества торфа и элементов питания
растений. В условиях жесткого температурного режима торфяных почв
многолетним травам принадлежит ведущая роль в комплексном
ресурсосбережении и продленном, устойчивом торфопользовании.
10. Приоритетна и наиболее выгодна глубокая технологическая
переработка торфа для сельского хозяйства, промышленности и бытового
потребления, обеспечивающая многократное повышение ценности новой продук-
298
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
йй в сравнении с исходным сырьем. В сельском хозяйстве предстоит
уверить потребление торфа с получением гидролизных Сахаров, кормовых
зоояокей и добавок в корма, получения стимуляторов роста растений и
жирных, прессованной и формованной торфопродукции, улучшенных торфо-
грунтов, биоконверсируемых удобрений и других продуктов.
11. Новое освоение торфяников под агроценозы вполне целесообразно,
так как после мелиорации и окультуривания агроценозы используют ФАР
для формирования органического вещества в 5-10 раз интенсивнее, чем
растительность природных болот. По нашим обобщенным расчетам почвенное
торфопользование под агроценозы в 22-30 раз сокращает расход торфа на
каждую зерновую единицу и экономичнее энегетически в сравнении с
использованием торфа в компостах.
12. Возрождение торфяной отрасли возможно на основе более
совершенных рекомендаций производству, учитывающих новые организационно-
экономические условия. Решение проблемы подъема торфопользования
требует совершенствования и укрепления технологического обеспечения
сельскохозяйственных, мелиоративных и торфяных предприятий.
13. Торфяной фонд России, как национальное богатство, должен
использоваться в интересах рационального природопользования регионов
предприятий и граждан. Исходя из этого, необходимо принять Федеральный
закон, учитывающий интересы всех торфопользователей. Контроль за
соблюдением его на местах позволит избежать многих ошибок и злоупотреблений
прежних лет. Для ускорения возрождения торфопользования необходимо
творческое объединение усилий науки, производства и административной
власти на региональном и федеральном уровнях.
ПОСЛЕДСТВИЯ ОСУШИТЕЛЬНОЙ МЕЛИОРАЦИИ ЛУГОВО-
ГЛЕЕВЫХ И ТОРФЯНЫХ ПОЧВ СРЕДНЕАМУРСКОЙ НИЗМЕННОСТИ
О.А. Кириенко
Институт водных и экологических проблем ДВО РАН, Хабаровск
Мелиорация земель является сильнейшим антропогенным фактором,
вызывающим изменения в биосфере. В связи с этим возникают проблемы не
только хозяйственного, агрономического, но и социально-экологического
характера. Возрастание объемов мелиоративных работ требует оценки
уровня антропогенной нагрузки на почвы с целью сохранения и повышения их
производительности, рационального использования и охраны [1].
Природно-климатические условия Дальнего Востока, в которых
формируются почвы, своеобразны и не имеют аналогов. Большая их часть
переувлажнена и нуждается в мелиоративных мероприятиях.
299
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Почвы Хабаровского края характеризуются высокой эрозионной уязви*
мостью в связи с разрушающим действием муссонно-континентальногх)
климата, пульсационного и крайне неустойчивого гидротермического
режима. Разрушение почв в условиях Дальнего Востока происходит в несколько
раз быстрее, чем в других регионах России и Европейских странах [2].
Микробные комплексы тесно связаны с направленностью почвообра-
зующих процессов, играют важнейшую роль в формировании почвенного
плодородия и являются надежными индикаторами состояния почвенной)
покрова. Осушительная мелиорация переувлажненных почв, с одной
стороны, интенсифицирует микробиологические процессы, приводит к активной
минерализации органического вещества, определяющего плодородие. С
другой стороны, активизация жизнедеятельности микроорганизмов
способствует усилению процессов деструкции органического вещества и деградации
плодородного почвенного слоя. Сработка органогенного слоя торфяных
почв в условиях Дальнего Востока происходит за 2-3 десятилетия [3,4].
Особую актуальность в условиях сельскохозяйственного освоения почв
приобретает проблема сохранения органического вещества в
мелиорируемых почвах на длительный срок. Важнейшей задачей современной
мелиорации является разработка способов воздействия на активность микробоце-
нозов. обеспечивающих высокий уровень плодородия почв в процессе
длительной эксплуатации и сельскохозяйственного использования.
Осушительная мелиорация, улучшая гидрологический режим тяжелых
переувлажненных лугово-глеевых и торфяных почв Приамурья,
положительно влияет на биологическую активность и микробиологические
процессы. Влажность и температура являются важнейшими факторами,
определяющими функционирование микроорганизмов и активность
биохимических процессов. Эффективность мелиоративных мероприятий связана с
понижением влажности почв и улучшением температурного режима.
Влажность 55-70% от полной влагоемкости и температура почвы 14-30°С
являются оптимальными для развития основных групп микроорганизмов и
активизации почвенных ферментов.
Микробиологические пейзажи служат качественной характеристикой
степени антропогенной трансформации почв. Пейзажи стекол обрастания
неосушенной лугово-глеевой почвы более богатые и разнообразные по
сравнению с пашней. Видовой состав микробоценозов осушенной целинной
почвы близок к микробном}' пейзажу неосушенной почвы.
Мероприятия, связанные с усилением антропогенной нагрузки на
мелиорированные почвы, коренным образом нарушают равновесное состояние
микробоценоза и приводят к необратимому формированию качественно
нового сообщества. В результате осушения целлюлолитические и протеолити-
ческие микроорганизмы, присутствовавшие в неосушенной почве, уступают
300
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
место бактериям и актиномицетам. потребляющим минерализованные
низкомолекулярные соединения азота и углерода.
По мере усиления минерализационного процесса в лугово-глеевой и
торфяных почвах под действием осушительной мелиорации происходит
обеднение почвы питательными элементами, сработка торфяного горизонта.
Соответственно происходит снижение биологической активности почв, падает
активность ферментов, преимущество получают олиготрофные
микроорганизмы, что свидетельствует о последней стадии сукцессии.
Для предотвращения выноса питательных легкоподвижных компонентов
из верхних пахотных горизонтов необходимо внесение органического
удобрения, закрепляющего минерализованные соединения в почве и
поддерживающие почвенное плодородие на высоком уровне. Наиболее эффективным
удобрением в условиях Хабаровского края являются корокомпосты.
изготовленные из отходов деревоперерабатывающей промышленности.
Внесение корокомпостов в лугово-глеевую почву повышает биохимическую и
микробиологическую активность, благоприятно сказывается на плодородии,
увеличивая содержание гумуса и азота, повышает урожайность растений.
[1] Ивлев A.M. и др. Экологическая оценка мелиорации земель. Учеб. пособие.
Владивосток: Изд. Дальневост. ун-та. 1995. 80 с. [2] Зархина Е.С. и др. Земельные
ресурсы Хабаровского края и пути их рационального использования // Рациональное
использование почв Приамурья. Владивосток: ДВНЦ АН СССР. 1983. С. 4-14. [3]
Климин М.А. Изменение структуры почвенного покрова мелиорированных
торфяных массивов Среднеамурской низменности при различной интенсивности
хозяйственного использования // География и природные ресурсы. 1988. № 3. С. 75-80. [4]
Матрошилов Ю.А. и др. Изменение гумусного состояния и буферных свойств
осушенных маломощных торфяных почв Приамурья при сработке торфа // Тез. док. IV
конф. молодых ученых. Пущине 1989. С. 79-81.
СОВРЕМЕННЫЕ ВОПРОСЫ МЕЛИОРАЦИИ БОЛОТ
ДЛЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЦЕЛЕЙ
Б.С. Маслов
РАСХН, Москва
1. Сельское хозяйство использует болота и торфяные месторождения в
земледелии для возделывания после осушения сельскохозяйственных
культур, для добычи торфа с целью приготовления органических удобрений, в
тепличном хозяйстве, для получения бытового топлива. Основной вид
использования наиболее ценных, с плодородными торфяными почвами
низинных и частично переходных болот, — земледелие. Еще Петром 1 в указе
1723 г. дозволялась торфодобыча на топливо только на болотах, которые
«на пашню и на сенные покосы не годятся». При определении целесообраз-
301
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ных направлений хозяйственной деятельности и для оценки эффективности
использования учитываются особенности болот, их строение и взаимосвязь с
окружающими территориями. В недавнем прошлом торф в огромном
объеме вывозился на удобрения (в 1985 г. — 160 млн. т), для чего ежегодно
осушали 1,0-1,5 тыс. га болот. Исследования и опыт показали, что
удобрительная ценность торфа в чистом виде, не пропущенного через животновод,
ческую ферму, ничтожна, а вывозка торфа на поля — бесполезное (даже
вредное) и дорогостоящее мероприятие.
2. Возделывание сельскохозяйственных культур на осушаемых торфяных
почвах обеспечивает получение урожая в 6-10 раз и более по сравнению с
естественным использованием болот. Болота представляют огромный резерв
в решении продовольственной проблемы, особенно в регионах, страдающих
от малоземелья. Согласно опыта западных стран (Англия, США, Германия,
Нидерланды, Бельгия и др.), где осушено 70-90% всех болот и заболоченных
земель, торфяные почвы являются лучшими, наиболее плодородными
землями, а площадь мелиорированных земель представляет важный показатель
возможностей и технического уровня земледелия [5,8 ].
3. В России площадь осушенных земель по состоянию на 1990 г.
составляла 5Л млн. га, из них 1,3 млн. га торфяных почв. В отличие от западных
стран Россия еще находится в начале пути мелиоративного освоения земель.
В последние годы, из-за бесхозяйственного отношения к ранее
мелиорированным землям (выделяется не более 1/3 необходимых средств на
эксплуатацию осушительных систем), идет непрерывный процесс выхода их из
строя и вторичного заболачивания земель [2].
4. Разработаны режимы (нормы осушения, допустимые сроки отвода
воды и пр.) и техника мелиорации болот, включающая открытые каналы,
пластмассовый дренаж, искусственные ложбины, агромелиоративные
мероприятия, определены оптимальные расчетные схемы для определения
расстояний между каналами (дренами), их параметров, условия применения и
конструкции осушительно-увлажнительных и осушительных систем поль-
дерного типа и регулирования рек-водоприемников, критерии экологически
допустимого осушения, нормативы по разработке раздела «Охрана
природы» в проектах мелиорации [3]. Имеется достаточное научное обеспечение
проектов осушения с учетом как положительных, так и возможных
отрицательных последствий на прилегающие территории и природную среду в
целом.
5. Болота характеризуются спецификой флоры и фауны. Они тесно
связаны с окружающим ландшафтом и выполняют определенные
природоохранные функции. Болота — важнейшее природное богатство с
экономической и экологической точек зрения. Оценка социально-экономической и
экологической эффективности осушения болот производится с учетом при-
302
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
родных и хозяйственно-экономических факторов. Их учет заключается в
детальном изучении почвенного покрова, климата, водных ресурсов,
рельефа, растительности, то есть всех компонентов, на которые направлено
мелиоративное воздействие и состояние которых обусловливает мелиоратив-
лую неустроенность всей территории.
6. В результате осушения изменяются экологические условия
заболоченных территорий, их ландшафты превращаются из аккумулятивных в
элювиальные и аккумулятивно-элювиальные. Уничтожаются естественные
многовидовые биогеоценозы, увеличивается дренированность территории,
понижается уровень грунтовых вод на прилегающих территориях.
Исследованиями последних лет доказано, что положительная гидрологическая роль
неосушенных болот преувеличена. Признано, что гидрологические функции
неосушенных болот в основном отрицательные. Необоснованной оказалась
их положительная роль в питании рек. Вследствие высокой влагоемкости
болота не только не отдают рекам в меженный период воду, но, наоборот,
нередко уменьшают их меженное питание, перехватывая, поглощая и
удерживая поток поверхностных и грунтовых вод, направленный к рекам.
Поглощенная влага расходуется не на питание рек, а на испарение в
атмосферу. Положительная роль болот заключается лишь в более равномерном
внутригодовом и внутрисезонном распределении стока.
При крупномасштабном осушении болот северных районов есть
основания ожидать ухудшения теплового режима и увеличения площади
многолетне-мерзлотных почв в пределах болотного массива и улучшения теплового
режима на прилегающих территориях.
С социальной точки зрения заболоченные и болотные территории
являются малоблагоприятными для жизни людей и их производственной
деятельности. В таких районах осушение болот необходимо как с
хозяйственной точки зрения, так и для улучшения условий жизни людей.
Проблема осушения особенно актуальна при освоении огромных затор-
фованных пространств Западной Сибири, крупных торфяных
месторождений Северо-Запада, Урала и Камчатки. Высокая заболоченность и заторфо-
ванность таких территорий (20-30, в отдельных регионах 50-60% и более)
создает неблагоприятные условия для жизни людей. Например, обилие
кровососущих насекомых сдерживает социально-бытовое устройство населения.
Осушение торфяных территорий в этих регионах является не только
необходимым хозяйственным мероприятием, но и улучшает биосферу.
Осушение болот является кардинальным средством улучшения
природной среды. Образовавшиеся после мелиорации ландшафты с обилием
лесных насаждений, животных, птиц и рыб весьма убедительно характеризуют
их как продуктивные угодья с благоприятными условиями для земледелия,
лесовозобновления и организации охотничье-звероводческих хозяйств, а
303
БОЛОТА И 'ЗАБОЛОЧЕННЫЕ ЛЕСА
также парковой зоны для отдыха населения крупных городских и про^ы
ленных центров. Сохранение редкой болотной флоры и фауны peiuaJyJ*
путем создания заповедников, заказников, резерватов, охотничьих хозяйств
7. Торфяные почвы после осушения и культуртехники (сведение леса
кустарника, удаление кочек и пр.) подвергаются окультуриванию, цедил
которого является создание в почве свойств, обеспечивающих наряду с вы
соким плодородием и продуктивностью сельскохозяйственных культуп
уменьшение минерализации торфяной залежи. Окультуривание включает в
себя систему мероприятий по повышению плодородия почвы путем улучще.
ния водно-физических, тепловых, агрохимических и биологических свойств
почвы. В комплекс приемов окультуривания почвы входят: первичная обра-
ботка почвы, известкование, планировка поверхности, внесение удобрений,
создание глубокого однородного пахотного слоя, добавка минеральной)
грунта (землевание). Период первичного окультуривания,
продолжительность которого составляет, в зависимости от уровня естественного
плодородия и интенсивности применяемых мелиоративных приемов, 3-5 лет.
8. Торфяные почвы, особенно окультуренные и старопахотные (начиная с
6-10-го годов введения в культуру) целесообразно использовать
преимущественно под сеяные злаковые травы (одно-двух-многокомпонентные).
Приоритет в использовании торфяных почв в Нечерноземной зоне должен быть
отдан луговодческому направлению. Это направление в использовании
торфяных почв соответствует свойствам и особенностям этих почв, позволяет
получать в хозяйствах разнообразные корма — сено, сенаж, силос,
травяную муку, зеленую массу. Высокий урожай трав (120-150 ц/га сена или 480-
600 ц/га зеленой массы) возможен только при строгом соблюдении всех
элементов технологии их возделывания, которая достаточно разработана.
Культурные пастбища на торфяных почвах обеспечивают высокую
продуктивность в течение 10 и более лет, что не наблюдается на других типах
почв. Наиболее пригодными для пастбищ следует считать низинные
торфяные почвы с разной мощностью торфяной залежи, если уровни грунтовых
вод за вегетационный период и по площади массива поддерживаются в
пределах не выше 60-100 см от поверхности.
Пропашные культуры (картофель, кормовая свекла, морковь, турнепс,
брюква, кукуруза и др.) более продуктивны на торфяных почвах по
сравнению с многолетними травами и другими культурами сплошного сева.
Однако, они способствуют (благодаря интенсивной минерализации
органического вещества) избыточному накоплению питательных элементов в почве в
подвижной форме и их потере в связи с этим с дренажными и грунтовыми
водами, ухудшают водно-физические и агрохимические свойства торфяных
почв и снижают их эффективное «долголетие»; плохо борются с сорняками
и вредителями, приводят к распылению верхнего слоя почвы, увеличивают
304
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
ку торфа. Системы земледелия для осушаемых торфяных почв, вклю-
^ сев0обороты? достаточно разработаны [6].
^9 долговечность сельскохозяйственного использования торфяных почв в
леделии зависит от интенсивности минерализации торфа и потери орга-
зе* еСкого вещества, прямо связанной с величиной осадки и сработки торфа,
п новным фактором, вызывающим изменение рельефа поверхности торфя-
дка и свойств торфяной почвы после осушения, является осадка торфа.
Интенсивность осадки почвы в первые годы после осушения и сработки торфа
процессе сельскохозяйственного использования, .зависят от исходных
свойств торфяной залежи, глубины понижения уровней грунтовых вод, ха-
оакгера сельскохозяйственного использования торфяных почв,
климатических условий местности. Оказывают воздействие и другие факторы:
уплотнение почвы сельскохозяйственными машинами и скотом, распыление и
дефляция торфа при обработках и под воздействием ветра, механическая и
химическая суффозия. Все они связаны с сельскохозяйственным
использованием осушенного торфяника. Среднегодовая величина понижения
поверхности болота достигает максимума в первые месяцы и резко уменьшается в
первые 5-8 лет после осушения. Затем следует переходная стадия (до 15-18
лет) постепенного ее уменьшения; в последующем изменение носит
стабильный характер. Для оценки осадки торфяной залежи и понижения
поверхности торфяника за время Т после осушения можно использовать
зависимость S = а 0,9 Г+1,2, где S — интенсивность осадки (включая сработку)
торфа, см/год, а — коэффициент, учитывающий вид торфа, интенсивность
осушения и характер сельскохозяйственного использования (для низинных
торфяников мощностью 2.5-3 м в Центральном районе Нечерноземной зоны
5,9). т — продолжительность осушения, лет. Формула справедлива при
выращивании пропашных и зерновых культур; при возделывании многолетних
трав коэффициент а значительно меньше. Осадка тор-фа по глубине
торфяной залежи изменяется в зависимости от величины понижения поверхности
почвы Sn по зависимости: S=Sn[l,0065-l,7575(h/m) + 2,4206(h/m)2 -
1.6813(h/m)3], где h — глубина слоя, m — мощность торфа [7,9 ].
10. После осушения и в процессе сельскохозяйственного использования
болот за счет минерализации торфа изменяются все водно-физические
свойства торфа: повышается зольность и объемная масса, плотность и
наименьшая влагоемкость, уменьшается пористость, полная влагоемкость и
водоотдача. Меняются химические свойства торфяной почвы: увеличивается
кислотность, сумма поглощенных оснований, содержание CaO, Si02 и Si03,
уменьшается содержание железа и алюминия. Изменение запасов
питательных веществ (азот, фосфор, калий) находится в прямой зависимости от
количества вносимых удобрений. Потери органического вещества составляют
2,0-5,4 т/га и больше, минимальные значения наблюдаются при возделыва-
305
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
нии трав. Уменьшить потери и добиться положительного баланса органнча.
ского вещества можно применением пожнивных и повторных посевов [1 ^
11. Для решения вопросов изучения, использования и охраны болот в у^
ловиях активной антропогенной деятельности, целесообразно использовал
материалы долговременных полевых опытов. В последние 90 лет, со време»
ни организации первых болотных и опытно-мелиоративных станций д^
изучения вопросов сельскохозяйственного использования болот (Архангель»
ской. Новгородской областей. Барабинской степи и др.), создана широкая
географическая сеть стационаров и опытных участков. Накоплена ценная
информация по природе болот и изменению их характеристик и свойств
торфа под влияние осушения, освоения и сельскохозяйственного
использования торфяных почв и прилегающих к болотам земель. По материалам
исследований на мелиоративно-болотных стационарах были разработаны
нормы и режимы осушения почв и увлажнения сельскохозяйственных
культур, рационального использования земель, конструкции осушительно-увлаж-
нительных систем, нормативы для проектирования экологически безопасной
мелиорации. В 1997 г. собраны материалы по мелиоративным стационарам
на болотах, характеристика 52 из них опубликована [4]. Идет подготовка
второго издания книги (более 70 стационаров).
К числу наиболее обстоятельно изученных болотных стационаров
относятся: «Щеголево» в Смоленской, «Дальний» и др. в Московской, «Каль-
ское». «Олех». «Вожа» в Рязанской, «Гадово болото» в Кировской,
«Решетникове» и «Усалка» в Тюменской, «Убинское» в Новосибирской областях,
«Корзинская низина» в Карелии и др. Результаты исследований на
долговременных стационарах являются незаменимым средством проверки
теоретических моделей, для развития теории мелиорации и природообустройства
в условиях устойчивого развития. Анализ их материалов показывает, что
объем, программы и методы исследований на них неодинаковые, что было
обусловлено как кругом поставленных задач, так и уровнем развития науки
на период закладки стационаров и компетенцией исследователей.
Приходится отмечать на ряде стационаров недостаточную изученность
гидрогеологических условий (водоносные горизонты, их участие в
водно-минеральном питании болот и др.). гидрохимического режима и солевого баланса
почв, водно-физических свойств торфяной залежи на всю глубину и др.
Стационары — богатство страны. К сожалению, некоторые из них в
последние годы были заброшены. Среди ученых воспитывается
отрицательное, пренебрежительное отношение к полевому опыту, его недооценка.
Многие стационары располагают информацией не только о мелиорации, но и об
эволюции почв (включая прилегающие территории), подземных вод и озер-
но-болотных комплексов, динамике растительности, об использовании
мелиорированных земель для целей охотничьего хозяйства, рыбоводства, рек-
306
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
аЦйи. Их информация многоплановая, использование которой позволяет
овершенствовать мелиорацию в системе адаптивно-ландшафтно-го земле-
еЛИя. инженерно-мелиоративного и агро-экологического мониторинга.
.и 5елковский В.И. Использование торфяных почв в системе зернотравя-ных сево-
<0ротов // Мелиорация переувлажненных земель. Минск: БелНИИМиЛ, 1996. [2]
Колганов А.В. Дренаж переувлажненных и засоленных земель в России: состояние,
яроблемы, решения // Мелиорация и водное хозяйство. 1998. № 1. С. 2-6. [3] Мас-
10В Б.С. Комплексная мелиорация: становление и развитие. М.: РАСХН. 1998. 280
с [4] Маслов Б.С. Мелиоративно-болотные стационары России. М.: РАСХН. 1997.
150 с. [5] Маслов Б.С. Россия еще в начале пути улучшения земель // Вопросы ме-
;1ИОра-ции. 1994. ? 3-4. С. 30-36. [6] Маслов Б.С. и др. Руководство по длительному
использованию осушаемых торфяных почв Нечерноземной зоны Российской
Федерации. М.: РАСХН. 1999. 105 с. (в печати). [7] Маслов Б.С., Лысенок А.П.
Изменение осадки торфа после осушения по глубине торфяной залежи // Доклады РАСХН.
1999. ? 1. С. 27-28. [8] Сохранять и приумножать плодородную силу гектара //
Вопросы мелиорации. 1994. ? 1. С. 7-13. [9] Maslov B.S. et al. Peat soils and their change
under amelioration. M: Rosselkhozizdat. 1996. 148 p.
ВОЗДЕЙСТВИЕ СТОКА ОТ СВИНОФЕРМЫ НА
РАСТИТЕЛЬНОСТЬ ВЕРХОВОГО БОЛОТА
В. А. Смагин
Ботанический институт РАН, Санкт-Петербург
Рассматривается динамика развития болотной экосистемы,
подвергшейся поливу стоками с животноводческих ферм и комплексов. После
прекращения воздействия развитие растительности, на одном из таких болот,
пошло не по болотному, а по лесному ряду.
Во второй половине 1970-х годов для Ленинградской области острой
экологической проблемой явилась утилизация и очистка сточных вод
интенсивно строящихся крупных животноводческих комплексов. Удаление навоза
проводилось путем гидросмыва и объем стоков достигал десятков
миллионов кубических метров в год. Стоки очищались на очистных сооружениях
(от твердой фракции), а доочистка проходила на полях орошения, полях
запахивания и биологических прудах. Все эти способы доочистки были
очень дороги и не всегда эффективны. Поэтому шли поиски других, менее
затратных путей решения проблемы. Наиболее дешевым и эффективным
методом оказалось использование фильтрационных свойств болот.
Очистные способности болотных экосистем довольно давно учитываются в США
и Канаде. В нашей стране на возможность использования этих свойств
болот обратила внимание М.С. Боч, благодаря энтузиазму которой опытно-
промышленная проверка очистки стоков была проведена на 2-х болотах Ле-
307
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
нинградской области: Гуриновском (900 га), рядом с которым расположен
откормочный комплекс «Пашский» (22 тыс. бычков) и небольшом болот»
вблизи пос. «Петровское», где была свиноферма (20 тыс. свиней), с 1995 г
не функционирует. Именно об этом болоте проводились наблюдения за вое*
становительными сукцессиями.
В отличие от болота «Гуриновского», обширного массива, состоящего аз
мезотрофных осоково-сфагновых и евтрофных осоковых, березово-осоко-
вых. березово-тростниковых, черноольховых болот, нарушения
растительности которого были минимальны (на болото поступали исключительно
осветленные стоки), и проявлялись только непосредственно в узких «рукавах»
стока, болото у пос. Петровское пострадало существенно. Оно представляло
собой небольшое (50 га) верховое болото покрытое однообразной сосново-
кустарнггчково-пушицево-сфагновой растительностью, с маломощной
торфяной залежью (1-2 м). Стоки подаваемые в центр болота по трубопроводу,
не всегда являлись «осветленкой», зачастую содержали твердую фракцию,
смена места выпуска стоков проводилась не так часто как следовало бы
(сказывалась экономическая ситуация — работы начались в 1988 г.). Но не
смотря на это, а также на малые размеры, в течение первых 4-х лет болото
успешно справлялось с очисткой стоков, очищая их в 100-200 раз по
содержанию аммиака и в 10-20 раз — фосфора. В последующие годы (1992-94
г.), когда уже почти вся площадь болота подверглась воздействию стоков,
качество стекающей с него воды ухудшилось. Тем не менее болото
справилось с ролью фильтра — в протекающую вблизи реку Черную, загрязненные
стоки не попали, но как показали дальнейшие наблюдения, ценой своей
гибели. С самого начала полива растительность болота претерпела
существенные изменения. Непосредственно вблизи выпуска стоков у сосны сразу же
началось пожелтение и опад хвои, через 3-5 месяцев она погибла. Сразу же
гибнет сфагновый мох. Быстро происходит гибель кустарничков, они
сохраняются лишь на высоких кочках в зоне удаленной от водовыпуска на 50 м
(при условии продолжения полива в этом месте не более полугода).
Наиболее стойким оказался — Eriophorum vaginatum. Более того, на удалении 100
м. от места разлива стоков, в зоне умеренной подпитки биогенными
веществами (NH4-IOO мг/л, Р2О5-40 мг/л) наблюдается резкое увеличение обилия
пушицы, ее проективное покрытие достигает 100%. Здесь происходит
увеличение прироста побегов (вдвое) и размеров хвои у сосны, увеличение
прироста и продуктивности у кустарничков, кроме низкорослых,
вытесняемых разрастающейся пушицей. У сосны наблюдается здесь интенсивное
возобновление. Но к сожалению малые размеры болота, и невозможность
(по экономическим причинам) переброски трубопровода на соседнее болото,
с целью дать время на восстановление экосистеме поливаемого болота,
предопределили перегрузку его стоками, повлекшую полную гибель естествен-
308
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
ной растительности на большей части его площади. Болото оказалось
помытым густыми зарослями сорняков, перемежаемых зарослями Турка
latifolia и Phragmites australis. На третий год после прекращения полива,
появляется обильный подрост березы (ранее произрастающие на болоте
немногочисленные и угнетенные деревца березы при поливе погибают). Рост
березы очень интенсивен. Подача стоков на болото прекращена к началу
1995 г., к этому же время прекращаются регулярные ежемесячные
наблюдения за качеством стекающей с болота воды, фенологические наблюдения,
ежегодное изучение продуктивности и динамики растительности.
Переописание и составление плана-карты растительности болота проведено спустя 3
года, в августе 1998 г., и результаты его были удивительными. Вместо
ожидаемой растительности березово-тростникового и рогозового болота, мы
увидели молодой густой березняк, поражающий, на фоне залитых водой в
условиях очень влажного лета 1998 г. окружающих болота лесов, своей
сухостью. Он образовался в центральной части болота, первой принявшей
стоки, и куда, с середины 1989 г. они более не подавались. В этой части
болота можно проследить весь временной ряд развития растительности за
период 10 лет. До полива произрастала растительность которую, по составу
доминантов, можно охарактеризовать как Pinus sylvestris - Chamaedaphne
calyculata + Eriophorum vaginatum - Rubus chamaemorus - Sphagnum angu-
stifolium + S. magellanicum. Полив начат в самом конце 1988 г., в начале
мая 1989 г. растительность выглядела так — Pinus sylvestris -
Chamaedaphne calyculata + Eriophorum vaginatum, причем покрытие и жизненность
оставшихся расте-ний снизились. Мхи сохранились лишь на высокой кочке
{Sphagnum magellanicum — 10%, Pleurozium schreberi — 10%). В конце мая
погиб мох и на кочках. В июле у сосны пожелтела вся хвоя, в нижних
ярусах все погибло (пушица — 1%). В конце августа сосна потеряла всю хвою и
погибла, несколько разрослась пушица (5%), появились сорняки: Taraxacum
officinale (5%), Tussilago farfara (3%), Epilobium adenocaulis (1%), и ряд
других. К концу сентября все эти виды, включая пушицу, увеличили
покрытие в полтора — три раза, a Epilobium adenocaulis — в 8 раз. В мае 1990 г.
растительность была очень разреженной, преобладали пушица и кипрей (по
8%)., к концу июля исчезла пушица и разрослись рогоз, иван чай и марь,
растительность вновь приобрела подобие ярусного строения — Typha
latifolia - Chenopodium alba + Chamenerion angustifolium - Epilobium
adenocaulis + Tusselago farfara. Через год, в июле 1991 г. Epilobium
adenocaulis образовал сплошные заросли (покрытие 100%), появилась
крапива (8%). Такое строение сообщество сохраняло на протяжении 3 лет, по
1993г включительно, на протяжении этого времени в его состав
добавлялись, в небольшом количестве, щучка дернистая, ежа сборная, канареечник,
паслен, череда. В 1992г появилась поросль березы. В 1993 г. подрост березы
309
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
достигает высоты 1,5-2 м.. но покрытие его не велико — 3%. такое же по»
крытие имела и Salix aurita (высота 1м). В 1994 г. происходит смена дома*
нанта, им становится Bidens tripartita (85%). В 1995 г растительность насчи»
тывала 11 видов, доминировали череда (65%), крапива (35%), кипрей
(10%). Береза достигала высоты 2-3 м, а ее покрытие 8-10%. Через 3 года
береза достигла высоты 3,5-4 м и образовала густой полог сомкнутостью
0.6; остальной состав был следующим: ива (5%), крапива (5), череда (2),
вейник (5). иван чай (3). щучка дернистая (8), луговик извилистый (8). ря.
дом располагаются густые заросли вейника (80-98%). над которым высится
подрост березы (вые. 3 м, покр. 10%), далее по направлению участков более
позднего полива, находятся заросли крапивы. В местах последнего,
наиболее продолжительного полива, возвышаются густые моновидовые заросли
рогоза и тростника, перемежаемые «полянами» из кипрея. Со всех сторон на
эти участки надвигается поросль березы. Болотная растительность сохрани-
лась лишь на одном участке болота 60x150 м., здесь произошел
существенный отпад древостоя, но он компенсируется интенсивным развитием
подроста (покр. 15%), сохранились все виды первоначальной болотной
растительности, более обильными, чем на обычном болоте, стали пушица и
клюква. Есть инородные включения: иван чай (1%), кипрей (1), вейник (+),
обилен подрост березы. Тем не менее, это фрагмент настоящего болота. И
представляется интересным проследить за его взаимоотношением с
образующимся вокруг него березовым лесом. Наблюдения за дальнейшей
динамикой растительности будут продолжены. В заключение отметим еще раз, что
наблюдать столь сильное осушение болота ранее не доводилось, ибо
образовавшиеся здесь растительность, да и ландшафт в целом напоминают то, что
видим на месте болот выработанных фрезерным способом и заброшенных,
— густой березняк шучковый или мертвопокровный. С помощью
гидроосушительной лесомелиорации такого результата достичь едва ли возможно.
ПИЩЕВЫЕ И ЛЕКАРСТВЕННЫЕ РЕСУРСЫ БОЛОТ
ЗАПАСЫ СЫРЬЯ БАГУЛЬНИКА БОЛОТНОГО НА ТЕРРИТОРИИ
ГАССИНСКОГО МОДЕЛЬНОГО ЛЕСА (ХАБАРОВСКИЙ КРАЙ)
А. А. Бабурин
Институт водных и экологических проблем ДВО РАН. Хабаровск
Багульник болотный {Ledum palustre L.) имеет обширный
голарктический ареал. Произрастает в лесной и тундровой зонах, а также в гольцовом
поясе гор. В связи с широким распространением представлен несколькими
жизненными формами (кустарник или кустарничек) и разновидностями (var.
310
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
vlilgare Ledeb,. var. dilatatum Wahlehb. var. angustum E.Busch., var. labratum
jCryl) имеющими определенную фитоценотическую приуроченность
[Гришин. 1986]. Является гигрофильным олиготрофным мохово-болотным видом
[КрыловЛ 984] и поэтому лучше всего чувствует себя на холодных и влаж-
нЫх кислых и очень кислых почвах торфяного ряда (заболоченные леса,
лесные болота, заросли кедрового стланика и т.д.).
На территории Гассинского модельного леса он образует заросли,
имеющие промысловое значение на лесных типах болот (верховых и переходных)
с изреженным лиственничным (или лиственнично-березовым) древостоем.
В кустарниковом ярусе этих фитоценозов, как правило, присутствуют хаме-
дафне, андромеда, береза Миддендорфа, голубика, ивы черничная и корот-
коножковая. Описания таких болот и геоботаническая характеристика их
приводятся в работах Ю.С. Прозорова [1961.1974]. Наибольшее обилие
багульника (Copi-Cop2) отмечено в следующих группах фаций (табл.1).
Табл. 1. Расчет запасов сырья и объемов ежегодной заготовки (03) багульника
болотного (кг/га) по группам болотных фаций. ОД — обилие по Друде, ПП —
проективное покрытие, СФ — свежая фитомасса (Об — общая, П — облиственных
побегов), ЗС — запас сырья (воздушно сухая масса), ПР — период ротации.
Группы болотных фаций ОД ПП, СФ, кг/га ЗС ПР, 03,
% Об П кг/г лет кг/г
Лиственничная кустарничко- Sp-Cop2
во-сфагновая
Кустарничково-осоково-
сфагновая
Кустарничково-сфагновая
(подушки)
Пушицево-кустарничково-
сфагновая
Сфагновая
Гидрофильно-сфагновая гря- Sol-Сорг 23
дово-мочажинная
Гидрофильно-сфагновая озер- Sol-Copi 11 2400 494 250 5 50
jcoBo-грядово-мочажинная
При определении запасов сырья и расчета безущербных объемов
заготовок использовались полученные на ключевых участках связи проективного
обилия яруса багульника с сырьевыми характеристиками его (табл. 2)
Приведем средние (выборка из 60 особей) данные по определению фи-
томассы багульника болотного. Масса одной оси в свежем состоянии равна
34,2 г, в том числе листьев — 5,4 г; стеблей и ветвей — 28,8 г. Соотношение
сырого веса листьев и стеблей у нас составляет 15,8 % к 84?2%.Таким
образом наши данные не подтверждают тезис Л.К. Позднякова [1973] об относи-
Sp-Cop2
Sol-Copi
Sol-Cop2
Sp-Cop2
Sol-Copi
Sol-Cop2
22
5
11
26
10
23
9000
700
2400
10050
1700
9300
1845
144
494
2060
365
1900
920
70
250
1030
180
950
5
5
5
5
5
5
184
14
50
206
36
190
311
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
тельном постоянстве соотношения масс стеблей и листьев в различных v
ловиях существования. В Якутии и Западном Саяне это соотношение 6toJ
соответственно 34:66 и 36:64, т.е. действительно близко. Листья сосредо»*
чены в основном на приросте текущего года. Средний вес облиственное
прироста текущего года равен 7,3 г, что составляет 20,5% общей надземной
фитомассы. Масса листьев на годичном побеге в среднем равнялась 74%.
Обследованная нами заросль багульника находилась в лиственничнике
сфагново-багульниковом. Мощность торфа 0,7-0,8 м. Высота кустов дост*.
гала 0,5-0,7 м (длина осей 0,8-1,1 м). Диаметр основания от 0,5-0,7см (у
более молодых) до 1,0 см у осей 12-13 летних. Средний возраст заросли
около 10 лет. Сомкнутость древесного яруса составляла 0,3-0,4. При повьь
шении сомкнутости древесного полога следует ожидать уменьшение
среднего веса особи и индивидуального годичного прироста, хотя общая
урожайность может и возрасти за счет увеличения обилия. Так, например, в
лиственничнике ольховниково-багульниковом, сомкнутость крон в котором
достигала 0,5, свежая фитомасса одного среднего стебля багульника составляла
17,1 г, в том числе листьев — 3,1 г. На долю однолетних облиственных
побегов приходилось 22,0% общей надземной фитомассы.
Табл. 2. Корреляция ресурсных параметров багульника болотного
Параметры
10 20
Проективное покрытие, %
30 40 50 60 70
80
90
Численность, пгг/м2 5 25 32 38 43 47 51 55 58
Сырье свеж., г/м2 36,5 182 233 277 313 342 372 400 423
Сырье воз.сух, кг/га 182 910 1165 1385 1565 1710 1860 2000 2115
Табл. 3. Возможные объемы ежегодных заготовок сырья багульника болотного на
территории Гассинского Модельного леса (ГМЛ) и их экономическая оценка
Типы и
группы болотных
фаций
Общ.
пло-
щадь,
Под ба-
гульни-
ком,
Запасы
сырья
воз.сух
Объемы ежегодных загото- Валовая
вок, т выручка
типы труп. га
га
Возможные Рентабельные тыс. руб
2 6
2 7
3 9
4 9
5 9
Итого:
6570
6570
37314
9288
21114
80856
6570
6570
37314 .
6968
10557
67979
6750
6050
2650
486
737
16673
1355
1210
522
97
148
3332
678
605
261
49
74
1666
13560
12100
5220
980
1480
33340
Типы фаций: 2. Мезотрофный древесно-сфагновый смешанного питания, 3.
Гетеротрофный сфагновый смешанного питания, 4. То же в сочетании с суходольными
мелколиственными колками и заболоченными лиственничниками. 5. То же в
сочетании с мелколиственными лесами на минеральных останцах. Группы видов болот;
ных фаций: 6. лиственничная сфагново-багульниковая надпойменных террас,7.
лиственничная кустарничково-сфагновая, 9. кустарничково-осоково-сфагновая.
312
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
ресурсно-экологический потенциал болотных экосистем богат и разнооб-
азен. Его всесторонняя комплексная экономическая оценка — интересная в
яучном плане и трудновыполнимая практически задача, которая еще ждет
своего решения [Петров,1993; Косицын, 1997,1998; Кулишкина, Косицын,
1995 и др] Мы сделали попытку оценить лишь один и далеко не главный
компонент этих биогеоценозов, а именно перспективы использования
багульника болотного как продуцента лекарственного сырья (табл.3).
Площадь болот, целесообразных для заготовки багульника, составляет в
{У[Л около 68 тысяч гектар. Подсчитанные на них запасы сырья — 16673 т
в воздушно-сухом состоянии. Возможные объемы ежегодных заготовок
определены в 3332 т, а «рентабельный» объем заготовок, составляющий 50%
от возможного (эксплуатационного), — 1666 т воздушно-сухой массы.
Считаем, что при вовлечении в хозяйственный оборот этих болотных массивов
рассчитанные нагрузки не окажут существенного негативного влияния на
состояние ресурсов багульника и на экологию болотных экосистем.
ИСПОЛЬЗОВАНИЕ РЕСУРСНОГО ПОТЕНЦИАЛА МОРОШКИ
НА ЕВРОПЕЙСКОМ СЕВЕРЕ РОССИИ
В.Н. Косицын
ВНИИЛМ, Московская обл.
Устойчивое управление природопользованием на заболоченных землях
лесного фонда предполагает неистощительное и непрерывное использование
всех видов биологических ресурсов, в том числе и пищевых, среди которых
одним из наиболее хозяйственно-ценных видов является морошка
приземистая (Rubus chamaemorus L.). Она широко распространена в заболоченных
лесах и на болотных массивах таежной зоны России. Плоды (ягоды)
морошки отличаются высокими пищевыми, вкусовыми и витаминными
свойствами. В связи с этим они имеют большую коммерческую ценность в странах
Северной Европы. Так, в Финляндии средняя закупочная цена за 1 кг
свежих ягод морошки составляла за последние пять лет 7,7 долларов США.
Общие биологические запасы морошки в России значительны; только в ее
европейской части они составляют 62,2 тыс. т [4]. Однако степень их
использования находится все еще на довольно низком уровне, что во многом
обусловлено недостаточной изученностью ресурсного потенциала вида.
В течение последних 10 лет велись исследования по оценке
продуктивности морошки в зонах тайги и хвойно-широколиственных лесов европейской
части России. Установлено, что одним из главных факторов, определяющих
распространение и продуктивность зарослей морошки, является тип
растительного сообщества. Получены среднемноголетние значения степени по-
313
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
крытия плодоносящими зарослями ягодника площади фитоценоза и биолл.
гической урожайности ягод по типам леса (болота): сосняк сфагновый - q*
ответственно 15% и 75 кг/га. ельник сфагновый — 10 и 20, березняк сфаг„
новый — 5 и 15. ельник долгомошный — 10 и 30, березняк долгомоцщы*
— 5 и 10. мезотрофное болото — 5 и 10. олиготрофное болото — 10% и 40
кг/га. Для оценки урожайности морошки в текущем году за 2-1,5 месяца до
начала периода массовых заготовок сырья предложена «Методика прогноза
биологического урожая ягод морошки в подзоне южной тайги» [1995]. В ее
основу был положен учет обилия цветков, изменчивость доли женских
цветков вида от возраста древостоя и коэффициент выхода ягод из цветков с
учетом степени выраженности заморозков в период цветения ягодника.
Установлены регрессионные зависимости урожайности морошки от
производительности труда сборщиков, себестоимости заготовки и величины ренты
от использования ресурсов. При биологической урожайности морошки 35
кг/га и выше величина ренты больше нуля, то есть ресурсы доступны для
освоения и пользователь ресурсами способен получить чистый доход от их
эксплуатации [1]. По нашим оценкам, доход от использования ресурсов
морошки в сосняке сфагновом более чем в 11 раз превышает доход от
заготовки древесины за период оборота рубки древостоя [3]. На основе
предложенных нами наземных и дистанционных методов оценки ресурсов морошки с
учетом эколого-экономических факторов во ВНИИЛМе разработаны
«Методика учета и оценки ресурсов морошки в подзоне южной тайги» [1997] и
«Методика оценки недревесных растительных ресурсов на типологической
основе при сдаче лесов в аренду» [1997].
На примере Вологодской области с учетом материалов лесорастительно-
го районирования нами выполнено районирование ресурсов морошки. В
лесном фонде области плодоносящие заросли морошки занимают 185.3 тыс.
га. а биологический запас ягод равен 8.3 тыс. т. Наиболее высокие запасы
ягод морошки сосредоточены в юго-западных, северо-западных и южных
районах области, составляя соответственно 32,6; 17,0 и 16,4% общего запаса
сырья. Следует отметить, что ресурсный потенциал ягодника
характеризуется не только природно-сырьевой функцией — биологическим запасом, но и
хозяйственно-экономической и социальной функциями. В связи с этим были
рассчитаны и другие категории запаса морошки. Эксплуатационный запас
морошки (с учетом технологических возможностей заготовителей и
пищевых потребностей лесной фауны) в Вологодской области равен 6,25 тыс. т,
экономически доступный запас (с учетом плотности дорожно-транспортной
сети) — 4.81 тыс. т и реально возможный для промышленной заготовки
запас ( с учетом объемов сбора местным населением) — 2,98 тыс. т. Таким
образом, ресурсы морошки в Вологодской области являются достаточными
как для использования сырья в местной пищевой индустрии, так и для по-
314
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
ав0к его на мировой рынок. В настоящее время Россия ежегодно экспор-
друет. главным образом в Финляндию, только 4-15 т свежемороженных
ягод морошки, в то время как, например. Швеция — 200-220 т.
В последнее время в связи с усилением антропогенного воздействия на
лесо-болотные экосистемы наметилась тенденция сокращения запасов
морошки. Проведенные нами по данной проблеме исследования показали
следующее. Долгосрочная осушительная мелиорация (со сроком действия
осушительной сети более 20 лет) вызывает резкое уменьшение плодоношения
морошки и ее продуктивные угодья должны быть исключены из
гидролесомелиоративного фонда. После сплошнолесосечных "рубок урожайность
снижается в 3-8 раз. Установлено, что. чем больше ширина лесосеки и
расстояние от стены леса, тем хуже плодоносит морошка на вырубках. После
сильного торфяного пожара морошка не плодоносит более 25 лет; в тоже время
торфяные пожары слабой интенсивности способствуют разрастанию
зарослей ягодника. Результаты химического анализа ягод морошки, собранных в
окрестностях металлургического комбината «Северсталь» (Вологодская
область), показали, что они отличаются избирательной способностью в
отличие от других дикорастущих ягод накапливать тяжелые металлы Ni, Cr, a
также Cd [2]. Существенное снижение урожайности морошки наблюдается с
увеличением рекреакционных нагрузок. Если на I стадии рекреационной
дигрессии перестойного сосняка сфагнового урожайность составляла 65
кг/га. то на II стадии она равнялась 13 кг/га. а на III стадии — 5 кг/га. Нами
разработаны «Эколого-лесоводственные требования к использованию в
лесном фонде ресурсов дикорастущих ягодных растений» (1998), в которых
обосновываются рациональный режим использования запасов морошки и
мероприятия по охране и воспроизводству ее ресурсов.
Полученные результаты свидетельствуют об экономической
целесообразности промышленной эксплуатации ресурсов морошки на заболоченных
участках лесного фонда европейского Севера России.
[1] Косицын В.Н. Создание нормативной базы для экономической оценки ресурсов
даорастущих ягодников // Проблемы организации многоцелевого лесопользования.
М.БШ'ШЛМ, 1997. С. 130-136. [2] Косицын В.Н. Накопление тяжелых металлов в
плодах морошки в условиях техногенного загрязнения на территории Вологодской
области // Освоение Севера и проблемы природовосстановления. Сыктывкар: Ин-т
биологии, 1998. С.53-54. [3] Кулишкина С.Г., Косицын В.Н. Сравнительная
стоимостная оценка древесных ресурсов и дикорастущих ягодников // Лесн. хоз. 1996. № 6.
С.23-24. [4] Страхов В.В. и др. Недревесные лесные ресурсы Российской Федерации
//Лесохоз. информ.. 1997. Вып.З. С.32-47.
315
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
СОЗДАНИЕ РЕГИОНАЛЬНОГО БАНКА ДАННЫХ РЕСУРСОВ
ТОРФЯНО-БОЛОТНЫХ ЭКОСИСТЕМ
В.И. Косов, Ю.Н. Женихов, В.В. Кузовлев
Тверской государственный технический университет
В основе создания компьютерного банка данных «Торфяно-болотные
экосистемы региона» лежит географическая эколого-экономическая оценка
ресурсов болот, которая предусматривает комплексное использование
извлеченного сырья торфяных месторождений, использование мелиорируемых
торфяных площадей под посевы сельскохозяйственных культур,
лесопосадки, а также применение растительных и других ресурсов сохраняемых в
естественном состоянии торфяно-болотных экосистем. Актуальность создания
такого банка данных возрастает для заторфованных территорий, к которым
относится и Тверская область. Он реализует фондовую,
нормативно-информационную и функцию управления ресурсами болот, включая
геоэкологическую и экономическую оценку и оптимизацию разных вариантов
использования ресурсов торфяно-болотных экосистем (ТБЭ) [1].
Первая функция банка данных фондовая. Необходимость ее реализации
связана с большим количеством источников информации и их держателей.
Возрастает число организаций, фирм, акционерных обществ, имеющих
лицензию на производство геологоразведочных работ на торф. Сработка
торфяных запасов осуществляется помимо промышленных предприятий
землепользователями, предпринимателями. В большей мере это относится к
мелким низинного типа торфяным месторождениям. Фондовая функция банка
данных решает задачу создания обобщающего обзора и средства
наблюдения за изменением торфяного фонда региона. Вторая функция банка данных
нормативно-информационная. Здесь размещены документы об охране ТБЭ,
ГОСТы, ТУ, ТО различных видов торфяной продукции, источники
информации, расшифровка категорий сырья и вспомогательная информация.
Основная функция банка данных — обеспечение охраны и
рационального использования ресурсов болот. Банк данных построен по иерархическому
принципу. Вертикальная иерархия учитывает административное и торфяно-
болотное районирование региона. Каждому уровню иерархии соответствуют
свои задачи по охране и рациональному использованию ресурсов болот. На
верхнем зфовне иерархии находится экономический район, далее следуют
административная область (край, автономная республика), торфяно-болот-
ный район и административный район.
В пределах экономических районов целесообразно решать
стратегические вопросы использования торфяного сырья, какими являются, например
вопросы глубокой физико-химической переработки торфа с получением
ценных продуктов: торфяного воска, активированных углей и других. Эф-
316
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
активность использования ресурсов ТБЭ на местном уровне связана с при-
язкой к административному делению территории на области и районы. В
же время есть необходимость учета торфяно-болотного районирования
j^ic в связи с охраной, так и рациональным использованием торфяных ре-
сурсов на основе их качественной дифференциации.
Горизонтальная иерархия структуры банка данных связана с
формированием фондов целевого назначения. Анализ направлений использования
ресурсов ТБЭ Тверской области показал целесообразность выделения четырех
целевых фондов: охраняемого, мелиоративного (земельного),
разрабатываемого и фонда выработанных, застроенных и затопленных торфяных
месторождений.
Формирование фондов целевого назначения не позволяет полностью
решить проблему рационального использования ресурсов торфяных болот. Это
связано с тем. что существует довольно широкий спектр возможных
направлений использования болот и их ресурсов в пределах каждого фонда в
отдельности, особенно разрабатываемого. Структура регионального банка
данных должна позволить установить пути оптимального использования
запасов конкретных ТБЭ. Другими словами, речь идет о
дифференцированной оценке свойств торфяных ресурсов каждого целевого фонда. Для этого
составлены так называемые паспорта-анкеты болотных систем каждого
фонда, содержащие подробные сведения о торфяном месторождении.
На территории области сохраняются 153 торфяных болота в
соответствии с рекомендациями ПГО «Торфгеология» и местных ведомственных
организаций. Помимо этого, 23 экосистемы сохраняются на территории
Завидовского госкомплекса. На территории заказников в разных
административных районах сохраняются 164 торфяных болота. Общая площадь всех
торфяных месторождений охраняемого фонда области равна 390,6 тыс. га в
нулевой границе и 271,0 тыс. га в границе промышленной глубины
торфяной залежи. Запас торфа 40 % влаги на сохраняемых месторождениях
составляет 1157,4 млн. т.
Объекты охраны расположены в 33 районах области. Наибольшее их
количество сосредоточено в Андреапольском (27), Калининском (26),
Кимрском (25), Кувшиновском (22), Удомельском (20), Вышневолоцком (16) и
Западнодвинском (15) районах. По размерам площадей основная часть
сохраняемых торфяных месторождений имеет площадь в нулевой границе от
100 до 1000 га. Площадь менее 10 га имеют 32 торфяных болота, большей
частью заказники. Заказниками, в основном, представлены и охраняемые
болота площадью от 10 до 100 га (88 из 95). Крупных (>1000 га) экосистем
на территории области насчитывается 72 (40,5 % всех объектов охраны).
Значительные по площади болота объявлены памятниками природы,
сохраняются как ценные клюквенники и объекты водоохранного значения.
317
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Кафедрой природообустройства и экологии Тверского гос. техн. ун-^
ведутся работы по изучению урожая, запасов ягодных и лекарственных ра^
тений. С 1995 года изучается временная динамика растительных ресурсу
на полевом стационаре «Жердовское» охранной зоны Центрально-Лесногх)
государственного биосферного заповедника. Средневзвешенное значение
биологического урожая клюквы с учетом площади выделов составило на
месторождении-ключе «Жердовское» 266 кг/га, а биологический запас --
18.1 т. Расчетная биологическая продуктивность 1 гектара площади болота,
определенная как отношение биологического запаса к общей площади,
оказалась равной 21.7 кг. Эти данные позволили дать прогнозную оценку
запасов клюквы в области в 7,3 тыс. т (см. табл.).
Прогнозные запасы клюквы на ТБЭ Тверской области
Торфяные болота Общая площадь, Биологический
тыс. га запас, т
Олиготрофные, мезотрофные болота разра- 68,0 1475,6
батываемого фонда
Олиготрофные и мезотрофные участки раз- 64,7 1404,0
нотипных болот разрабатываемого фонда
Болота-ягодники охраняемого фонда 205,5 4459,9
[1] Геоинформационные технологии для эколого-экономического анализа
рационального использования торфяных ресурсов / Косов В.И., Женихов Ю.Н., Кузовлев
ВВ., Масленников А.В. // Экология и жизнь (наука, образование, культура): Межд.
сб. статей. Новгород, 1998. Вып.З. С. 70-71.
ПРОДУКТИВНОСТЬ ЯГОДНЫХ И ЛЕКАРСТВЕННЫХ РАСТЕНИИ
НА ОСУШЕННЫХ ЛЕСНЫХ БОЛОТАХ В ТВЕРСКОЙ ОБЛАСТИ
Л.Е. Курлович, В.Н. Косицын
ВНИИЦлесресурс, Москва; ВНИИЛМ, Московская обл.
В результате проведения гидролесомелиоративных работ происходит
изменение водно-воздушного режима торфяной почвы и её
физико-химических свойств. Это вызывает долговременные сукцессионные смены
растительного покрова, проявляющиеся прежде всего в изменении видового
состава и структуры фитоценозов. В связи с этим важное значение
приобретают вопросы динамики продуктивности хозяйственно-ценных видов
растений на различных объектах осушения.
В 80- и 90-х годах изучалось состояние и продуктивность некоторых
дикорастущих ягодных и лекарственных растений (клюква болотная, брусника
обыкновенная, черника обыкновенная, голубика, багульник болотный) на
объектах гидролесомелиорации Западнодвинского лесоболотного
стационара Института лесоведения РАН (Тверская обл.), где густота осушительной
318
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
сети, глубина каналов проектировались с учетом широкого охвата
вариантов. Одним из таких объектов являлось верховое болото, осушенное в 1974-
1975 гг. каналами глубиной 0,8-1,2 м, расположенными через 30, 60 и 100
и В качестве контроля был взят неосушенный участок. Он
характеризовался слабокочковатым микрорельефом, имел мощность торфяной толщи 2,5 м
й более и уровень болотно-грунтовых вод 10-20 см. В травяно-кустарничко-
вом ярусе господствовала пушица влагалищная (проективное покрытие -ПП
30%). мохово-лишайниковом — сфагновые мхи (ПП 100%). Сильно
разрешенный древостой состоял из сосны 40-80-летнего возраста высотой 2 м.
На участках с расстоянием между осушителями в 30 м общее
проективное покрытие видов живого напочвенного покрова уменьшилось по
сравнению с контролем, в 1,5-2 раза. Практически выпали из него сфагновые мхи
(ПП уменьшается со 100 до 0-7%), их место частично заняли лишайники
(ПП достигает 30%) и зеленые мхи (ПП до 15%). Участие клюквы болотной
в растительном покрове также значительно сократилось (ПП составило 0,5-
2% встречаемость — 32-54%; на контроле, соответственно, 3,1-4,5% и
100%) На участках, где расстояние между осушителями равнялось 60 и 100
м. снижение общего проективного покрытия живого напочвенного покрова
было менее заметным. Оно равнялось, соответственно, 60-75,4 и 84-96%.
ПП сфагнумов уменьшилось, соответственно, на 30 и 25%. Лишайники (ПП
0,6-3.0%) и зеленые мхи (ПП 1-2%) только начали внедряться в состав мо-
хово-лишайникового яруса. Проективное покрытие и встречаемость клюквы
болотной остались близки к контрольным.
В годы наблюдений в изучаемом районе не было высоких урожаев
клюквы болотной. Однако очевидно, что на участках с расстоянием между
каналами 30 м клюква вообще практически не плодоносила (урожай не
превышал 0.4-5,0 кг/га). При расстоянии между каналами 60 м урожай был близок
к контрольному или превышал его в 2-3 раза (колеблясь в различные годы
от 6 до 92 кг/га на осушенном участке и от 3,5 до 35 на контрольном).
Наиболее высок урожай ягод клюквы был при расстоянии между каналами 100
м (колеблясь от 53 до 2000 кг/га и значительно превосходя контрольный).
На осушенном участке наблюдалось разрастание зарослей багульника (в
контроле отсутствовал). Наиболее густые заросли багульник образовывал на
участках осушенного болота с расстоянием между каналами 30 м. где его
ПП составляло 6,6%. На участках болота с расстоянием между каналами 60
и 100 м ПП багульника равнялось 2.0-2,1 %. Урожайность лекарственного
сырья багульника (облиственных молодых побегов) на осушенном участке
болота составляла 189-273 кг/га в воздушно-сухом состоянии.
Другим гидролесомелиоративным объектом на территории Западнодвин-
ского стационара, где проводились исследования, было пушицево-сфагновое
переходное болото, осушенное в 1980 г. неглубокими (0,3-0,5 м) и частыми
319
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
(через 5-7 м) бороздами-канавами. Такое поверхностное осушение слузкщ.
первой мерой при подготовке почвы для посадки лесных культур и заключ»
ется в создании условий для быстрого отвода избытка воды, накопившейся ь
верхних слоях торфяной залежи, в горизонтальном направлении. На иссль.
дуемом болоте культуры сосны были посажены в отвалы борозд.
На болоте с поверхностным осушением в результате зарастания межка.
навных участков самосевом березы и осины наблюдалось резкое снижение
поступления фотосинтетически активной радиации к напочвенному покрову
Общее проективное покрытие напочвенного покрова уменьшилось по срав-
нению с контролем на приканавном участке в 1,4 раза (69,3 %, при 100% на
контроле), а на середине межканавного пространства — в 2,1 раза (48%).
Сфагновые мхи полностью сменились зелеными мхами, ПП которых в при-
канавной полосе составляло 60,7%, а на середине межканавий — 29,3%,
На осушенном участке болота отмечалось некоторое снижение
встречаемости клюквы (87%) по сравнению с контролем (100%). В тоже время, из-за
повышения качества условий местопроизрастания ягодника наблюдалась
интенсификация побегообразовательного процесса у особей клюквы, что
нашло отражение в увеличение её ПП (7,2% при 3, 8% в контроле).
Урожайность ягод на осушенном участке была значительно ниже чем в
контроле, соответственно 40 и 125 кг/га. Следует отметить, что величины
проективного покрытия и урожайности ягод клюквы в приканавной полосе были
выше чем подобные показатели на середине межканавий соответственно в 3
и 7 раз. Это связано с тем, что в результате осадки уплотненного слоя торфа
произошли процессы деформации поперечного профиля канав, разрушения
их откосов, заиления и зарастания русла канав, которые привели местами к
вторичному заболачиванию приканавных участков.
Изучалось также влияние различных технологий осушения сосняков
сфагновой группы типов леса на состояние и плодоношение дикорастущих
ягодников (глубина каналов 0,8-1,2 м, расстояние между ними -30, 60 и 100
м). Установлено, что при любой технологии осушения из напочвенного
покрова практически исчезает клюква болотная, зато появляются и
разрастаются брусника, черника и голубика. Исключение составляют участки с
расстоянием между каналами 30 м. На таких участках, вследствие
равномерности и высокой интенсивности мелиорации, более чем в два раза
уменьшается проективное покрытие мохово-лишайникового и травяно-кустарничково-
го ярусов. На остальных участках брусника, черника и голубика интенсивно
разрастаются (ПП — 17-25%), особенно в непосредственной близости от
осушительных каналов (в полосе шириной 15-20 м их ПП достигает 45-
50%). Однако плодоношения их часто не наблюдается, т.к. лимитирующим
фактором являются условия освещенности.
320
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
Таким образом, анализ результатов многолетних исследований на За-
^днодвинском лесоболотном стационаре показал, что при давности
осушительной мелиорации до 20 лет продуктивность ягодных и лекарственных
«астений во многом зависит от параметров гидролесомелиоративной сети,
расстояния от осушителей и в ряде случаев она значительно превышает
таковую на неосушенных болотных массивах.
[11 Курлович Л.Е., Косицын В.Н. Продуктивность дикорастущих клюквенников на
объектах лесоосушительной мелиорации различной интенсивности // Лесн. хоз.
1995.№1.С.29-30.
ТИПОЛОГИЯ И РЕСУРСНЫЙ ПОТЕНЦИАЛ
БОЛОТНЫХ ЭКОСИСТЕМ ПРИАМУРЬЯ
В.В. Чаков
Институт водных и экологических проблем ДВО РАН, Хабаровск
Термин «устойчивое развитие» (sustainable development) подразумевает
такую модель поступательного движения вперед, при котором «достигается
удовлетворение жизненных потребностей нынешнего поколения людей без
лишения такой возможности будущих поколений» [Коптюг и др., 1997]. В
современном мире удовлетворение жизненных потребностей человека
немыслимо без добычи и переработки природных ресурсов и особенно
органического происхождения. В этой связи роль болот и заболоченных земель
на планете с их колоссальными запасами органики переоценить
невозможно. Вместе с тем масштабное освоение ресурсов болот в СССР не дало
ожидаемого эффекта. Более того, в Белоруссии это привело к обмелению
большинства водотоков, а в Приамурье усугубило «фенольную» проблему. В
первом случае проблема связана с понижением уровня грунтовых вод,
вызванным интенсивной сработкой осушенных торфяников, во втором — с
интенсивным поступлением концентрированной органики содержащейся в
дренажных водах мелиорированных болот и заболоченных земель в р.
Амур. Сейчас можно с уверенностью сказать, что работы по освоению
такого уникального органического сырья как торф в Советском Союзе
осуществлялись без должного учета экологических особенностей территорий и
серьезных проработок в области закономерностей формирования и размещения
болотных экосистем. Особенно остро это проявилось в Приамурье на
территории Хабаровского и Приморского краев. Невостребованность здесь
мелиорированных угодий из-за развала крупных коллективных и
государственных сельхозпредприятий сегодня оборачивается экологической
катастрофой, повлекшей к существенному увеличению поступлений различных
форм углерода в водотоки и атмосферу. Вместе с тем, несмотря на значи-
321
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
тельную удаленность региона от научных центров России, уже к тридцати,
годам текущего столетия имелись сведения об особенностях размещения и
развития целого ряда достаточно крупных болотных массивов в на террит^
рии Хабаровского и Приморского краев и предпринимались попытки ^
классификации [Минкина, Федорова; 1936, Эндельман, 1936 и др.]. К
началу 60-х годов, связанных с пиком активности в решении вопросов по
мелиорации болот и заболоченных земель Приамурья, уже существовала фитоце-
нотическая классификация маревых болот Среднеамурской низменности
[Прозоров, 1961]. Несколько позже она была преобразована в
классификацию болотных биогеоценозов [Прозоров, 1974]. В основу каждой из них
автором закладывался типологический принцип построения. Всего на
территории Приамурья выделено восемь типов болот: травяной эвтрофный
грунтового питания; древесно-травяной эвтрофный грунтового питания; древес-
но-сфагновый мезотрофный смешанного питания; мезотрофный травяно-
сфагновый смешанного питания; гетеротрофный сфагновый смешанного
питания; сфагновый гетеротрофный атмосферного питания; сфагновый оли-
готрофный атмосферного питания и древесный естественно дренированный
атмосферного питания. Каждый из перечисленных типов болот имеет свою
специфику строения торфяной залежи, структуру растительного покрова и
особенности размещения в пределах равнин и низменностей Приамурья.
Экосистемы первых четырех типов несмотря на незначительные по
площади территории представлены здесь повсеместно от Приханкайской
равнины до Нижнеамурских низменностей. При этом если мелкозалежные эв-
трофные болота грунтового питания доминируют только в Приморском
крае, то эвтрофные древесно-травяные и мезотрофные травяно-сфагновые,
равно как и древесно-сфагновые. тяготеют преимущественно к Верхнезей-
ской и Зее-Селемджинской равнинам.
Два последних типа из представленного списка (сфагновый олиготроф-
ный и древесный естественно дренированный атмосферного питания)
наибольшее распространение получили на Нижнеамурских низменностях. Чаще
всего для таких болот характерна мощная (до 6,0-8,0 м) залежь верхового
торфа и преобладание в растительном покрове сфагновых мхов.
Господство сфагновой растительности отмечается и в биогеоценозах
двух гетеротрофных типов болот смешанного и атмосферного питания,
которые доминируют преимущественно в границах Среднеамурской
депрессии. Здесь для экосистем этих типов болот мощность торфяной залежи
очень редко превышает отметку 4 метра и обычно варьирует на уровне
значений 1,5-2,0 м. За пределами депрессии на Нижнеамурских низменностях
или Верхнезейской равнине показатели мощности торфа на таких болотах в
3,0-4.0 м являются нормой.
322
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
Основная часть болотных экосистем Приамурья, за исключением При-
чанкайских. имеет достаточно представительный спектр типологического
состава, что определяет здесь не только наличие колоссальных объемов
ресурсного потенциала, но и их разнообразие. В большинстве своем все
ресурсы болот, за исключением водных, имеют растительное происхождение. По
значимости сегодня на первое место можно поставить торфяные и
земельные. На территории Приморского края основная масса болот и
заболоченных земель используется для пополнения сельскохозяйственных угодий, в то
время как в ЕАО разработана и реализуется программа по использованию
торфа в качестве топлива. Повсеместно в Приамурье в той или иной степени
осуществляется заготовка торфа для производства торфокомпостов.
На наш взгляд, если с пополнением возделываемого земельного фонда за
счет болот еще в какой то мере можно согласиться, то с производством
топлива и низкоэффективных торфокомпостов из торфа, при избытке отходов
дерево-переработки, следует основательно подумать, т.к. он относится к
разряд> невозобновляемых природных ресурсов. В качестве исключения для
этих целей можно развивать производства только на уже мелиорированных
массивах для предотвращения здесь процессов окисления органики.
С нашей точки зрения целесообразнее всего в регионе осваивать
возобновляемые ресурсы болот, основываясь на принципах «болотоводства»
[Бамбалов, 1991]. Здесь, прежде всего, следует обратить внимание на
жидкую фазу торфа, избыток которой проявляется повсеместно, т.к. территория
Приамурья относится к зоне избыточного увлажнения. Добывая и
перерабатывая ее по технологии, разработанной специалистами ИВЭП ДВО РАН,
можно получать такие биостимуляторы, как вода торфяная природная
«Приамурская» (ТУ 9154-002-00336415-95. «Реликт-05» (ТУ 9158-001-
22156174-95) и «Реликт концентрат» (ТУ 9158-008-2698312-97). Названные
препараты используются для производства широкого спектра парфюмерно-
косметической продукции (шампуни, гели, маски, кремы, регенераторы
кожи и т.д.), а также профилактического лечения дерматозов и заболеваний
волос. О высоких потребительских свойствах товаров парфюмерно-космети-
ческой группы, вырабатываемых на основе жидкой фазы торфа,
свидетельствует большая золотая медаль полученная пено-моющим средством
«ТЕЛЬМА» на международной выставке «INHEBA* 90» в Братиславе.
Следующим по значимости из ресурсов болот в регионе сегодня следует
признать сфагновые мхи — уникальный природный материал растительного
происхождения. Сфера применения его в народном хозяйстве чрезвычайно
разнообразна — от использования в производстве гигиенических средств до
выпуска всевозможных субстратов и питательных сред для выращивания
сеянцев и рассады лесных, садовых, овощных и цветочных культур. Сегодня
в Приамурье сфагновый мох используется преимущественно в производстве
323
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
субстратов для выращивания сеянцев древесных пород в закрытом гру^
«ИДЕАЛ» (ТУ 0391-005-02698312-96) и «ИДЕАЛ ПЛЮС» (ТУ 0391-0о£
02698312-96). Кроме того, отдельные виды мхов заготавливаются здесь дд.
экспортных нужд в Японию и страны Юго-Восточной Азии.
Списочный состав и продуктивность пищевых, кормовых и лекарствен»
ных растений на болотах Приамурья не уступают по объемам и параметра^
проявляющимся на болотах европейской части страны [Елина, Кузнецов
1973]. Вместе с тем, серьезный интерес к этим ресурсам в регионе наблюди
ется только со стороны иностранных партнеров Российско-Канадского
объединения «Модельный лес «Гассинский», функционирующего на территории
Нанайского района Хабаровского края.
КЛАССИФИКАЦИЯ И СОВРЕМЕННОЕ ИСПОЛЬЗОВАНИЕ
РЕСУРСОВ БОЛОТНЫХ ЭКОСИСТЕМ
ЕВРЕЙСКОЙ АВТОНОМНОЙ ОБЛАСТИ
Е. Яковлева
ИКАРПДВОРАН, Биробиджан
Болотами в Еврейской автономной области (ЕАО) занято 1,025 тыс. га
что составляет почти 27 % территории [Отчет Комитета по земельным ...,
1997]. Они входят в зону гетеротрофных сфагновых болот хвойно-широ-
колиственных и южно-таежных лесов [Прозоров, 1985], для которой
характерно преобладание низинных болот, различающихся по составу
растительности: вейниково-пушицево-осоковые. тростниковые (монодоминантные),
моховые (сфагновые) [Куренцова, 1967]. Наибольшее количество болот
расположено в Смидовичском, Ленинском и Биробиджанском районах.
Изучение использования ресурсного потенциала болот является, на наш
взгляд, достаточно актуальным, не только с точки зрения активного
вовлечения в хозяйственный оборот новых компонентов экосистемы, но более в
свете задач устойчивого управления природопользованием. Комплексная
оценка ресурсов экосистемы является главной отправной точкой при
выработке режима природопользования на данной территории, т.к. содержит
сведения о количестве и взаимодействии компонентов, входящих в перечень
ресурсов. Оценка ресурсного потенциала болот ЕАО может быть
произведена по уже имеющейся классификации [Боч, Мазинг, 1986] с небольшими
дополнениями. Начальным этапом комплексной оценки является
инвентаризация и состояние использования ресурсов болотных экосистем.
1. Материальные ресурсы.
1.1. Торфяные ресурсы. 1.1.1. Топливно-энергетические. Ежегодная
добыча торфа в области составляет 0,6 млн. т.. при общем суммарном запасе
324
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
18632 тыс. т. Известно 55 месторождений торфа с запасами свыше 100 тыс.
крупнейшие месторождения Джаварга и Киргинское. На месторождениях
а3вит низинный тип залежей. Их средняя мощность от 0,57 до 2,1 м. [Ря-
мов. Кондратьева 1990]. До недавнего времени торф как топливо не
использовался, но в виду сложной экономической ситуацией, сложившейся в
топливно-энергетическом комплексе области, на базе ИКАРП ДВО РАН
успешно проведены научно-производственные исследования по
использованию торфа в качестве топлива, в некоторых районах области уже построены
малые котельные, работающие на местных торфах. 1.1.2.
Сельскохозяйственные. Основное направление использования торфа в сельском хозяйстве в
виде удобрений, традиционно — компостирование его с навозом. Получение
и использование высокоэффективных торфяных удобрений предполагается
начать на территории ЕАО в ближайшее время; качественные
характеристики торфов позволяют наладить производство микроудобрения «Экстра
Пит» (концентрированный водный раствор гумусовых веществ и
микроэлементов) широкого спектра действия. Использование торфа в качестве
удобрения гораздо предпочтительнее чем топлива, с точки зрения сохранности
экосистем и рентабельности производства. 1.1.3. Химические.
Использование торфа в качестве сырья для получения химических веществ
сравнительно новое и перспективное направление, но технологически сложное и
дорогостоящее, поэтому в области еще не получило распространения. 1.1.4.
Бальнеологические. Лечебные свойства торфов известны с незапамятных
времен. По имеющимся данным [Чаков, Гуцева, 1981]. Кульдурское болото,
входящее в состав курортной зоны области, насыщенное минеральными
водами и газами обладает высокими лечебно-минеральными свойствами.
Использование в бальнеологии компонентов торфа расширит спектр
санаторно-курортных услуг населению курорта федерального значения «Куль-
дур». 1.1.5. Торфо-минеральные. Наличие в болотах растворов
минеральных веществ, а также скопления некоторых химических элементов
позволяют рассматривать болота, как источник торфо-минеральных ресурсов
[Боч. Мазинг. 1986]. С конца прошлого века до середины 60-х годов нашего
времени на территории области велась эксплуатации Сутарского золоторас-
сыпного месторождения [Журнист. УсикОв. 1998], расположенного в сильно
заболоченной пойме реки Сутара. По прогнозным данным возможные
потери золота при добыче составили 40% и находятся в отработанных
(техногенных) образованиях, которые в настоящее время представляют
интерес для вторичной переработки по более сложной технологии.
Представляет интерес возможность извлечения золота биомассой микроскопических
водорослей и грибов [Куимова и др., 1998]. Освоение и внедрение таких
технологий будет способствовать основному принципу рационального
природопользования — минимальной нарушенности экосистем.
325
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
1.2. Водные ресурсы болот. Кроме органического и минерального вещ*
ства болота являются естественными аккумуляторами и пресной воды. Бда.
годаря особому гидрологическому режиму ненарушенные болотные экоси*
стемы способны снижать уровень паводков, впитывая часть воды [Прозо*
ров, 1980]. Октябрьский, Смидовичский, Ленинский районы области частое
сильно страдают от наводнений в период муссонных дождей, поэтому ис-
пользование ресурсов болот на территории этих районов нужно проводить с
учетом особенностей их гидрологического режима. Вода болот может слу.
жить источником питьевой воды, а сама поверхность болот служить
естественным фильтром бытовых и сельскохозяйственных стоков.
1.3. Земельные ресурсы. Осушенные территории болот могут
использоваться как сельскохозяйственные угодья. Всего в области мелиорированных
земель 86,9 га они использовались для выращивания овощных культур и
кормовых трав, в настоящее время 10,4 га выведены из
сельскохозяйственного оборота, в связи с сокращением использования с/х земель.
Строительство и эксплуатация мелиоративных систем должны сочетать в себе
экономическую эффективность и экологическую целостность, обеспечивающую
ненарушенное функционирование болотных экосистем.
1.4. Биологические ресурсы. 1.4.1. Пищевые растения. Среди других
видов растений, произрастающих на болотах ЕАО особое место занимают
ягоды - клюква и голубика, запасы последней позволяют производить
промышленные заготовки. По данным Облученского Госпромхоза, крупнейшей
заготовительной организации области в 1988 г. голубики было заготовлено
7132,5 кг. В настоящее время промышленных заготовок ягод по причине
низкой рентабельности не ведется, зато возросло количество стихийных
индивидуальных заготовок, которые регулярно довольно интенсивно
проводятся вблизи населенных пунктов и наносят значительный ущерб не только
ягодникам, но и всей поверхности болот. 1.4.2. Лекарственные растения.
Лекарственная флора болотных растений в области достаточно
представительна, по предварительным данным, только 15 видов растений обладают
широко известными целебными свойствами — аир болотный, багульник
болотный, вахта трехлистная, касатик гладкий, пузырчатка средняя, вех
ядовитый, подбел многолистный и др. [Растительные ресурсы..., Л., 1986.,
Атлас ареалов и ресурсов...М, 1976., Атлас лекарственных и
родственных... Л., 1990] после уточнения ресурсных характеристик эти виды могут
быть рекомендованы для промышленных заготовок. Промышленными
заготовками из числа болотных, до недавнего времени было охвачено ничтожно
малое количество (череда трехраздельная, горец, мох сфагнум). Заготовки
лекарственного сырья должны проводиться в соответствии с ресурсно-
производственными характеристиками, с учетом оборота пользования
каждого вида растения. 1.4.3. Медоносные растения. Пчеловодство — традици-
326
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
яная отрасль автономии. Разнообразие и насыщенность растительных
• ормаиии растениями — медоносами (250 видов [Пельменев, 1964], внут-
0йсезонная периодичность их цветения позволяют получать мед с ранней
весны до поздней осени. Доля болотных растений — медоносов в общем
списке медоносной флоры, в сравнении с другими растительными
формациями несколько ниже — приблизительно 20 видов (Carex schmidtii,
Saussurea amvrensis, Angelica maximowiczii, Menyanthes trifoliata и др.)
[Пельменев, 1969] и обеспечить получение товарного меда не могут,
поэтому могут использованы для поддерживающего взятка; увеличить их
медовый запас можно путем посева бобовых и других нектароносов на площадях
бывших торфоразработок, мелиорированных землях. 1.4.4. Кормовые
растения. Некоторые виды болотных растений являются естественными
накопителями питательных веществ (белка, крахмала, витаминов и т.д.),
большинство из них съедобны и могут использоваться, как в пищу так и на корм
животным: рогоз широколиственный, ряска малая, частуха обыкновенная,
чистец шероховатый, тростник и др. Особое место занимают сенокосные
угодья, наиболее урожайными (до 4-5 т/га сена) оказываются чистые вейни-
ковые луга на низкой пойме [Ахтямов, 1988], расположенные в основном на
территории Ленинского, Октябрьского районов. 1.4.5. Древесные растения.
Древесная растительность болот, как правило, находится в угнетенном
состоянии и особого ресурсного значения не имеет. Ее значение велико в
поддержании экологического баланса экосистемы в целом. 1.4.6. Растения
различного хозяйственного использования. К таковым можно отнести мох
сфагнум, который издавна используется, как теплоизоляционный материал в
строительстве деревянных зданий, ивы используются местным населением
для изготовления плетеных изделий. 1.4.7. Декоративные. Рододендрон
даурский, мытник крупноцветковый, герань Власова, скрученник китайский и
др. обладают определенными эстетическими свойствами и могут быть
рекомендованы в качестве декоративных экземпляров потенциального фонда
культурных растений. 1.4.8. Грибы и бактерии. Микроорганизмы,
населяющие болота обладают широкой экологической амплитудой, их роль в
формировании и функционировании биоценозов является исключительно
важной. Об использовании микроорганизмов болот было сказано выше. На
окраинах болот встречаются некоторые виды шляпочных грибов — волнушки,
сыроежки и др.. промышленного значения они не имеют. 1.4.9. Животные.
Позвоночных животных, занимающих болотную экологическую нишу
насчитывается немного. Из промысловых животных области к таковым
относятся: утки — серая, кряква, чирок; енотовидная собака, ондатра.
2. Нематериальные ресурсы. 2.1 Информационные. Болотные
экосистемы — уникальные хранилища редких и охраняемых видов растений,
нашедшие здесь благоприятную, или по крайней мере, слабо нарушенную сре-
327
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ду обитания. Среди них касатик гладкий, лапчатка кустарниковая, mohoxiw
рия Корсакова, осока Миддендорфа и др.. — всего 13 видов растений вне.
сенных в Красную Книгу ЕАО, 2 из которых имеют статус охраняемы*
краснокнижных растений РСФСР — бородатка японская и осока рыхла»
[Красная Книга Еврейской...,1997]. 2.2 Культурно-исторические и рекреа.
ционные. В настоящее время такие виды ресурсов болот являются невостре-
бованными.
Комплексная оценка ресурсов болот должна включать несколько этапов-
1 Инвентаризация всех видов ресурсов; 2. Экономическая оценка матери-
альных ресурсов; 3. Определение Потенциальной природной уязвимости
ландшафтов (ППУ), образующих болотные экосистемы, по предложенной
методике [Нарбут. Мирзеханова, 1993]. 4. Научно-обоснованные
рекомендации рационального использования всех видов ресурсов.
РЕКУЛЬ ТИВАЦИЯ И ИСПОЛЬЗОВАНИЕ
ВЫРАБОТАННЫХ ТОРФЯНИКОВ
КРИТЕРИИ ВЫДЕЛЕНИЯ ВЫРАБОТАННЫХ ТОРФЯНЫХ
МЕСТОРОЖДЕНИЙ ДЛЯ ПОВТОРНОГО ЗАБОЛАЧИВАНИЯ
С.Г. Беленький
JlHcmumym проблем использования природных ресурсов и экологии НАН
Беларуси, Минск, Беларусь
В настоящее время в республике Беларусь площадь выработанных
торфяных месторождений составляет 209,5 тыс. га и еще 101,0 тыс. га
находится в стадии разработки. Таким образом, общая площадь нарушенных болот
в связи с добычей торфа оценивается в 310,5 тыс. га. В перспективе
площади выработанных торфяников будут увеличиваться по мере завершения
добычи торфа на месторождениях разрабатываемого и запасного фондов.
Существующая в республике практика предусматривает рекультивацию
выработанных торфяных месторождений под сельскохозяйственное
использование, реже — под строительство водохранилищ или под посадки леса. Из
выработанных площадей торфяных месторождений более 70 %
рекультивировано. Однако, часто капиталовложения на рекультивацию не дают
экономического эффекта вследствие того, что многие торфяные месторождения
по своим характеристикам (геоморфологическим, геологическим,
гидрологическим, агрохимическим и др.) вообще не пригодны для создания на них
сельскохозяйственных угодий.
Несмотря на большие материальные и финансовые затраты такие
площади остаются в деградированном состоянии, дестабилизируют биосферные
328
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
^цессы и являются местами повышенной пожароопасное™. Новая
стратегия использования выработанных торфяных месторождений исходит из то-
^ что они являются не только значительным территориальным ресурсом,
но могут быть весьма эффективным средством стабилизации и поддержания
благоприятной природной среды после их повторного заболачивания.
(Обязательным условием биосферно-совместимого использования после
разработки торфяного месторождения должно быть возобновление болото-
образовательного процесса с целью восстановления всех биосферных
функций болота, существовавшего до начала разработки. Принципы биосферно-
совместимого природопользования на болотах не запрещают проводить
промышленную разработку торфяных месторождений, а лишь
ограничивают направления последующего их использования после выработки торфа и
обязывают после завершения разработки провести восстановление болото-
образовательного процесса и биосферных функций болот. Всякое другое
использование выработанных торфяных месторождений, например,
сельскохозяйственное, лесохозяйственное или водохозяйственное будет
находиться в противоречии с биосферными процессами и дестабилизировать
равновесие в окружающей среде.
Анализ литературных данных, кадастровых и фондовых материалов по
выработанным и неиспользуемым торфяным месторождениям, изучение их
в полевых условиях позволили сделать следующие выводы. Выработанные
торфяники, будучи антропогенно уничтоженными болотными экосистемами,
без повторного заболачивания не восстанавливают свои первоначальные
биосферные функции даже спустя много лет после выработки торфа и
продолжают дестабилизировать биосферные процессы. Повторное
заболачивание можно проводить на всех без исключения выработанных
месторождениях независимо от их природных характеристик (генетических,
геоморфологических, геологических, гидрогеологических, агрохимических и др.).
С учетом генетических особенностей, способов добычи торфа,
экологической ситуации в зонах влияния и районах расположения выработанных
торфяных месторождений, а также экологической целесообразности все
многообразие выработанных торфяных месторождений подразделяется на 3
группы по очередности повторного заболачивания.
В первую группу входят выработанные торфяные месторождения, на
которых проведение повторного заболачивания должно быть обязательным, а
именно: — месторождения, в районах расположения которых сложилась
неудовлетворительная экологическая ситуация, произошло обмеление,
загрязнение или исчезновение озер и малых рек, уничтожение редких и
находящихся под угрозой исчезновения видов болотной флоры и фауны,
деградация ландшафтов; — месторождения, разработка которых проводилась
багерно-экскаваторным способом или на которых добывался гидроторф; —
329
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
месторождения, имеющие под торфяной залежью отложения сапроцел4
мергеля или торфотуфа; - месторождения, расположенные у истоков рек „
ручьев, в зонах водосборов озер; — месторождения, загрязненные радио,
нуклидами свыше 15 Ки/км~.
Ко второй группе относятся выработанные месторождения, которые цл
своим природно-генетическим особенностям можно заболотить наиболее
простыми техническими мероприятиями, наиболее экономически целесооб-
разно. К ним относят торфяные месторождения пойменного залегания,
поименно-притеррасные, бессточных котловин, пониженные участки
месторождений сточных котловин, месторождения староречий и проточных котловин.
В третью группу входят выработанные торфяники, на которых
регулирование водного режима для повторного заболачивания связано с
определенными трудностями, т.е. экономически менее целесообразно, чем для
месторождений первой группы. К ним относятся: месторождения первых
надпойменных террас, месторождения склонов надпойменных террас,
месторождения водораздельных пологоволнистых абляционных равнин и др.
Если в зоне влияния 2 и 3 групп произойдет резкое ухудшение
экологической ситуации, то торфяные месторождения подлежат обязательному
повторному заболачиванию. Основным техническим мероприятием при
повторном заболачивании выработанных торфяных месторождений является
подъем уровня грунтовых вод. Осуществляется он запруживанием водотоков
и созданием регулируемых плотин для поддержания уровня грунтовых вод,
а также сброса паводковых и дождевых вод.
К ВОПРОСУ РЕКУЛЬТИВАЦИИ ЗЕМЕЛЬ НА БОЛОТАХ,
НАРУШЕННЫХ ПРИ БУРОВЫХ РАБОТАХ
А.Е. Березин
НИИ биологии и биофизики Томского университета
При бурении нефтегазовых скважин в Западной Сибири широко
используется «амбарный» метод, при котором выбуренная горная порода (шлам) и
остатки бурового раствора сбрасываются в котлован. Котлован или
шламовой амбар (ША) капается в непосредственной близости от буровой
установки. Размер амбара зависит от количества скважин и глубины бурения. В
течении одного года после окончания буровых работ ША должны быть
рекультивированы. Это требование долгое время не выполнялось: только у
предприятия ОАО «Томскнефть» на начало 1998 г. имелось около 300 ША
нормативный срок рекультивации которых истек, в том числе старые ША
сброс отходов бурения в которые проводился более 10 лет назад. В 1995-97
330
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
было обследовано 30 старых ША с целью оценки их потенциальной
экологической опасности и разработка предложений по рекультивации.
Воздействие на окружающую природную среду старых ША, с одной сто-
ооны. определяется количеством и составом загрязняющих веществ (ЗВ)
находящихся в амбаре, а с другой, физико-географическими условиями
местоположения. Объем и состав ЗВ определяет потенциальную опасность
(виды и масштабы отрицательного воздействия) ША для окружающей сре-
д!,!. Ландшафтные особенности (климат, состав грунтов и рельеф, почвенно-
растительный покров и др.) определяют возможности распространения ЗВ
за пределы ША, а так же направление и скорость само очистки экосистем,
восстановления в них естественных природных режимов развития и
функционирования. Основной целью рекультивации, по нашему мнению,
является скорейшее восстановление этих естественных режимов.
Существующие требования к проведению рекультивации нарушенных
земель разрабатывались, как правило, в европейской части России с
использованием зарубежного опыта там, где высока доступность и степень
хозяйственно использования территории, а значит и стоимость земель. В этих
условиях оправданно требование восстановления в исходное состояние
нарушенных земель не смотря на высокую стоимость этих работ. Здесь на
первом месте находится экономическая целесообразность.
В условиях зоны деятельности ОАО «Томскнефть» нарушенные земли
имеют, преимущественно, потенциально низкую экономическую ценность
(высокая заболоченность, огромные расстояния и слаборазвитая
транспортная инфраструктура), здесь при рекультивации земель имеет значение в
первую очередь экологическая безопасность. Поэтому мы считаем в этих
условиях основным принципом при рекультивации земель — «не
нанесение» природе большего вреда в процессе рекультивации, чем уже было
нанесено (и компенсировано на основе действующего законодательства) в
процессе производства. Выполнить данный принцип возможно только при
дифференцированном подходе к проведению рекультивации при строгом
учете ландшафтных особенностей территории.
Приведем два примера различных действий по рекультивации ША в
условиях водораздельного болота и поймы реки. На верховом болоте буровой
шлам осаждается на дне амбара, дно заиливается, растворенные в воде
минеральные элементы (в том числе химреагенты) сорбируются верхним
слоем торфа и не распространяются на значительные расстояния. Через
несколько лет котлован под ША превращается в водоем несколько
отличающийся от множества озер на болоте только составом растительности (более
евтрофные виды). Никакой экологической опасности от данного объекта нет.
Проведение рекультивации ограничивается выравниванием отвала (если
таковой был). Засыпка в этих условиях ША минеральным грунтом приведет
331
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
к значительно большему вредному воздействию на природу, чем уже имеет*
ся. т.к. привнос минерального загрязнения только увеличит площадь нар^
шенных земель, без всякого положительного эффекта.
На территории поймы содержимое ША в период паводка попадает в р^
ку. что является наиболее вредным воздействием с экологической точка
зрения. Здесь рекультивация ША необходима в кратчайшие сроки и с при.
менением методов препятствующих как размыву рекультивируемой поверх*
ности. так и предотвращению загрязнения грунтовых вод. Эти примеры
наиболее показательны не только с точки зрения различного подхода к
рекультивации, но и по тому, что при обследовании старых ША нами
выявлено важная особенность. В обеих случаях через несколько лет ША могут не
представлять угрозы окружающей среде, но в первом случае (болото) ее и не
было, а во втором отрицательные последствия были максимальными.
Основной задачей рекультивации нарушенных земель является
содействие естественному восстановлению природных режимов и биоты.
Собственно биологический этап рекультивации на болотах необходимо применять
только для ускоренного разложения нефтяного загрязнения. Использование
микробиологических методов разложения нефти проводится до начала
горнотехнического этапа рекультивации. Искусственное занесение семян или
корневищ болотных растений в обводненные ША ускоряет период за
который нарушенные земли приобретают вид близкий к естественным участкам
и процесс восстановления растительности в целом.
Проведенные нами работы позволяют сделать предварительные выводы
об экологической опасности старых ША и предложить рекомендации по
использованию более обоснованных, с учетом фактического состояния
нарушенных земель и местных условий, подходов к рекультивации (в том
числе мотивированного отказа на болотах от наиболее затратных методов зем-
левания). Так в большинстве случаев старые ША на болотах (при
отсутствии свежего нефтяного загрязнения), имеющих торфяную залежь более трех
метров (не зависимо от типа подстилающих грунтов) или при меньшей
мощности торфа (минеральное ложе торфяника сложено грунтами тяжелого
механического состава) горнотехнический этап рекультивации
целесообразно ограничить выравниванием внешней обваловки путем сталкивания
грунта внутрь ША. Засыпка привозным минеральным грунтом ША в этих
условиях не только не принесет пользы природе, а наоборот усилит
отрицательное воздействие предшествующей хозяйственной деятельности.
На заболоченных участках с почвами тяжелого механического состава
старые ША обсохшие естественным путем либо после откачки воды и
жидкой фракции отработанного бурового раствора, во многих случаях
(отсутствие значительного нефтяного загрязнения и невозможности
распространения ЗВ за пределы нарушенных земель в результате размыва поверхности
332
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
текучими водами или действия других факторов) нет необходимости
полностью засыпать амбары привозным грунтом. Это не ускорит восстановление
естественной растительности на рекультивируемой поверхности, но
потребует значительных материальных затрат и приведет к необходимости
дополнительного нарушения земель (карьеры минерального фунта). В этих
условиях достаточно разрушить образовавшуюся корку на поверхности (при
небольшом слое отходов бурения) или насыпать 20-30см слой привозного
фунта (желательно плодородного почвенного слоя или смеси торфа с
песком), частично для этих целей используется грунт старой обваловки амбара.
ОСОБЕННОСТИ ЛЕСОРАЗВЕДЕНИЯ НА ВЫРАБОТАННЫХ
ТОРФЯНЫХ МЕСТОРОЖДЕНИЯХ ОЗЕРНОГО ГЕНЕЗИСА
Р. А. Крупнов, М.В. Попов
Тверской государственный технический университет
Приживаемость и сохранность лесных культур при рекультивации
выработанных торфяных месторождений зависит от ряда природных и
технологических факторов. К природным факторам относятся агрохимические и
агрофизические свойства остаточного слоя торфа и подстилающих грунтов.
Из технологических факторов наибольшее значение имеют микрорельеф
выработанных площадей, уровень стояния грунтовых вод, вид и сроки
обработки почвы, применение удобрений, способ посадки саженцев.
Особенностью торфяных месторождений озерного генезиса является
наличие под остаточным слоем торфа сапропеля, который существенно влияет
не только на природные, но и технологические факторы естественного
облесения и искусственного лесоразведения.
Внесение сапропеля в почву повышает агрохимические свойства,
улучшает ее физические, теплофизические и водно-воздушные свойства.
Увеличивается выход стандартных саженцев хвойных пород на 10-23%, а
накопление сухой массы повышается на 60-98%. В связи с этим, при выборе вида
механической обработки почвы предпочтение следует отдать способам,
обеспечивающим перемешивание торфяного пласта с подстилающим
сапропелем. Оптимальное соотношение перемешивания торфа с сапропелем
составляет (70-80%):(30-20%).
Одним из важнейших технологических показателей при лесоразведении
является уровень стояния грунтовых вод. Влажность торфяного горизонта на
выработанных площадях, подстилаемых сапропелем, выше на 2-6% чем на
аналогичных площадях подстилаемых суглинками и глинами и на 5-11%
при подстилающих грунтах легкого механического состава. Наличие
сапропеля под слоем торфа значительно затрудняет проведение осушительных
333
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
мелиорации, особенно при мощности сапропелевого пласта более 0,5 м. п
этому на таких выработанных площадях уровень грунтовых вод будет вса
гда высоким. В весеннее время эти площади могут подтапливаться, ц^
снижает приживаемость хвойных пород до 30-ти и даже 10-ти %.
Грунтовые воды на выработанных площадях, подстилаемых сапропелеы
необходимо поддерживать на глубине не менее 0,4 м. и исключить подгон
ление корней растений в период вегетации более чес на 4-5 дней. Для дос-
тижения указанной цели осушительная сеть прокладывается через 60-120 ц
в зависимости от мощности остаточного стоя торфа и сапропелевого пласта
Обработка почвы должна предусматривать создание микроповышений
что осуществимо при плужной пластовой обработки в свал и бороздовании с
одновременным формированием пластов. Расстояние между бороздами 4-6
м. При этом, борозды выводятся в осушительную сеть, глубина борозд 0,3-
0,5 м. ширина по верху 1.1-1,3 м. Приемы обработки и сроки проведения
работ в зависимости от степени осушенности и мощности сапропелевого
пласта на выработанных площадях приведены в таблице.
Обработка почв выработанных торфяников подстилаемых сапропелем
УГВ,
м
до 0,4
до 0,4
0,4-
0,7
0,4-
0,7
Мощность, м
торфа
ДО
0,2
0,2-
0,5
ДО
0,2
0,2-
0,5
Сапро
-пеля
до 0,5
0,5-
0,7
до 0,5
0,5-
0,7
Способ обработки
Пластовая вспашка (в свал) с созданием
микроповышений и припахивании сапропеля
Бороздование с формированием
микроповышений -пластов, обогащенных сапропелем
Глубокое рыхление перемешивание торфа и
сапропеля вспашкой и дискованием.
Глубокое фрезерование с последующим
бороздованием и перемешиванием торфа с
сапропелем
Сроки
работ
июль-
август.
июль-
август
июль-
сентябрь
июль-
сентябрь
Лучшими сроками подготовки поверхности выработанных площадей,
подстилаемых сапропелем, является ранняя осень года, предшествующей
посадке лесных культур. При таких сроках обработки почвы
интенсифицируются окислительные процессы, повышается биологическая активность
почвы и создаются условия для промораживания извлеченного на
поверхность сапропеля. В отношении способа и срока посадки наиболее
приемлемыми следует признать вертикальный весной с использование посадочного
материала 1-2-летнего возраста для сосны и 2-5-летнего — для ели.
334
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
ОТНОСИТЕЛЬНАЯ УСТОЙЧИВОСТЬ К ДЕГРАДАЦИИ
АНТРОПОГЕННО-НАРУШЕННЫХ БОЛОТНЫХ ЭКОСИСТЕМ
ПРИ ИХ ОКУЛЬТУРИВАНИИ
И.И. Лыткин, A.M. Гребенников,
Почвенный институт им. В. В. Докучаева, Москва
Объектами исследований были в разной степени окультуренные
торфяные почвы Озерецко-Никольского месторождения, развитые на остаточной
залежи выработанного верхового и переходного торфа (Московская обл.,
Орехово-Зуевский р-н) после окончания его добычи в 1930 г. Исследовались
используемые в сельскохозяйственном производстве рекультивированные
хорошо окультуренные и слаборекультивированные слабоокультуренные
почвы, а также неосвоенные для целей земледелия нерекультивированные
слаборазвитые почвы с произрастающей на них болотной растительностью.
Изучались температурный, водный и пищевой режимы в подекадных
наблюдениях в течение вегетационных периодов 1982-1992 годов.
Непосредственно в полевых условиях ионометрическим методом определялась
динамика напряженности окислительно-восстановительных процессов, реакции
среды, активности ионов К+. Са2+. СГ и N03" в генетических горизонтах
почв и в почвенно-грунтовых водах. В аналогичные сроки отбирались
пробы почвенно-грунтовых вод для химического анализа в лаборатории.
Участки с нерекультивированными слаборазвитыми торфяными
почвами, при повсеместной выраженности вторичного заболачивания,
характеризовались весьма высокой устойчивостью по почвенному профилю многих
определяемых показателей. Среди них — высокая водонасыщенность
торфов, очень кислая реакция среды, низкая активность ионов К+, Са2+, С1\
Шз" по всему почвенному профилю, что является характерным для
ненарушенной верховой торфяной толщи. Наибольшая динамичность свойств в
этих почвах происходит в самых верхних слоях 0-15 см, что связано, в
основном, с изменением гидротермического фактора. Вторичная болотная
растительность (березовый подрост, клюква, росянка, пушица, сфагнум и
др.) соответствует растительному покрову ненарушенных болот верхового
или переходного типа. Почвенно-грунтовые воды, как и вышележащая
торфяная толща, отличаются высокой кислотностью, слабой минерализацией,
высоким содержанием водорастворимых органических веществ. За 11 лет
наблюдений, ионная сила последних изменялась весьма незначительно: m =
1.29 mM, Cv = 8,8 %, п = 70, где т — средняя ионная сила. Cv —
коэффициент вариации, п — число определений. В почве преобладают процессы
торфообразования. Среди растений прогрессируют в развитии пушица,
сфагновые мхи и клюква.
335
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Высокая динамичность всех определяемых показателей за исследован*
ный период наблюдалась в слаборекультивированной перегнойно-торфяной
почве, интенсивно осваиваемой в сельскохозяйственном производстве лищь
с 1982 года. При мелиорации и интенсивном сельскохозяйственном
освоении данные почвы развиваются по элювиальному типу, что постепенно при*
водит к их деградации, так как вместо аккумуляции веществ до осушения
происходит их разрушение и вынос. Высокие активные концентрации
кальция, хлора, нитрат-ионов в почвенном профиле и почвенно-грунтовых водах
связаны как с внесением минеральных и органических удобрений под
возделываемые в севооборотах культуры, так и с биохимической сработкой
торфа. С почвенно-грунтовыми водами происходит большой вынос всех
водорастворимых химических элементов и соединений, образующихся в
результате ионообменных процессов ППК с удобрениями и под
воздействием растений. Наибольшие качественные и количественные изменения
происходили в почвах и водах в первые 2-3 года после осушения и начала
окультуривания. Внесение невысоких доз извести в кислые верховые и
переходные торфяные почвы, с высокой потечностью органического вещества,
в начальный период освоения, как правило, малоэффективно, вследствие
значительного выноса Са в составе органо-минеральных комплексов за
пределы почвенного профиля. Изменения условий гидромелиорации в
значительной степени определяют дальнейшее развитие этих почв. При
соблюдении оптимальных параметров увлажнения выработанных торфяных
массивов создаются условия для их прогрессирующего окультуривания, при
избыточном осушении — развитие получают деградационные процессы,
протекание которых связано с биохимической сработкой торфа, при
переувлажнении — происходит вторичное заболачивание. За время наблюдений, поч-
венно-грунтовые воды слаборекультивированной слабоокультуренной
торфяной почвы характеризовались значительной изменчивостью ионного
состава и высокой ионной силой: m = 27,78 mM, Cv = 21,3 %, п = 77.
Рекультивированные окультуренные перегнойно-торфяные почвы,
сельскохозяйственное освоение которых началось в 1971 году,
характеризовались стабильным выносом минеральных компонентов и слабым выносом
водорастворимых органических веществ за пределы почвенного профиля.
Повышенная активность ионов Са2+, СГ, N03" и слабокислая реакция среды
явились следствием окультуривания перегнойно-торфяной почвы при
длительном возделывании многолетних трав в составе севооборотов,
известковании высокими дозами СаСОз и ежегодном внесении минеральных
удобрений. Рекультивированная торфяная почва обладает относительно высокой
буферностью к подкислению. внесению минеральных удобрений и другим
химическим воздействиям, влияние которых слабо сказывалось на
плодородии и свойствах почвы. Почвенно-грунтовые воды этих почв за последние
336
ПРОБЛЕМЫ УСТОЙЧИВОГО ИСПОЛЬЗОВАНИЯ
V
\\ лет наблюдений характеризовались относительно стабильным ионным
составом: m = 5,86 mM, Cv = 9,9 %, п = 83.
Наибольшей относительной устойчивостью к развитию деградационных
процессов обладают почвы выработанных торфяников, которые не были
освоены для вьфащивания сельскохозяйственных кулыур. Процессы,
происходящие в этих почвах, в основном направлены на восстановление
выработанной торфяной толщи и растительного покрова, которые
соответствовали облику болот до начала торфоразработок. В большей степени
подвержены деградации агроэкосистемы, развитые на рекультивированных хорошо
окультуренных почвах. Условиями их существования в таком виде является
своевременная компенсация биологического и геохимического выносов
элементов питания внесением органических и минеральных удобрений, а также
постояннный контроль за работой мелиоративных систем. При нарушении
этих условий рассматриваемые агроэкосистемы относительно быстро
деградируют. Самыми неустойчивыми являются агроэкосистемы, развитые на
слаборекультивированных слабоокультуренных торфяных почвах.
Интенсивность использования их в сельском хозяйстве связана прямой
зависимостью со скоростью протекания деградационных процессов.
Учитывая очень низкую устойчивость к деградации слабоокультуренных
агроэкосистем и большие затраты на окультуривание и поддержания этого
уровня для хорошо окультуренных агроэкосистем, сельскохозяйственное
освоение выработанных торфяников верхового и переходного типов является
нецелесообразным и с экологических, и с экономич)еских позиций.
Предпочтительны: естественные ценозы, при выращивании клюквы, голубики и
лекарственных растений; залужение многолетними травами; лесовыращива-
ние. При этих условиях, вследствие малой емкости биологического
круговорота веществ, почти не происходит биохимической сработки торфа.
Коэффициент вариации ионной силы почвенно-грунтовых вод тесно
коррелирует со степенью устойчивости почв выработанных торфяников к
деградации и при соответствующей доработке может быть использован в
качестве критерия выраженности деградационных процессов в
рассматриваемых почвах.
337
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
VI. БИОРАЗНООБРАЗИЕ ЕСТЕСТВЕННЫХ И
МЕЛИОРИРОВАННЫХ БОЛОТ И ЗАБОЛОЧЕННЫХ ЛЕСОВ,
МОНИТОРИНГ И ОХРАНА
БИОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ ПИЩЕВЫХ И ЛЕКАРСТВЕННЫХ
РАСТЕНИЙ БОЛОТ БЕЛАРУСИ
В.В. Гримашевич
Институт леса НАН Беларуси, Гомель, Беларусь
Основу экологического капитала Беларуси определяют лесные и водно-
болотные системы. Данный экологический ресурс имеет практическое
значение не только для белорусской нации. «Лесо-водно-болотная» специфика
природы нашей страны имеет огромное значение для экологически
устойчивого развития Европейского континента [1].
Болота — исключительно сложные природные образования, которые
больше всего подверглись антропогенным преобразованиям.
Крупномасштабное и интенсивное осушение болот и заболоченных земель
осуществлялось с 1965 по 1975 г., менее интенсивное — в последующем до 1985 г. и
даже позже. В результате мелиоративного преобразования 1,775 млн. га
(42,4%) болот и заболоченных земель трансформировано в другие категории
угодий (в основном, сельскохозяйственные). В естественном состоянии ныне
осталось около 2.3 млн. га болот и заболоченных земель, из них открытых
— лишь 797,5 тыс. га (33,5%), или 3,8% площади Беларуси. Около 1,15
млн. га (48.3%) занимают лесные болота, что составляет 5,5% территории
страны [2]. Значительные территории заняты прилегающими к болотам
лесами с избыточным увлажнением (типы леса с гигротопом 4).
В структуре растительного покрова экосистемы естественных болот
занимают 2379 тыс. га, или 17,1%, из них на долю верховых приходится 433,9
тыс. га [3]. Верховые болота в основном сконцентрированы в Северной
подзоне — 79,1%, около 16,4% находится в Центральной и 4,5% — в Южной
подзоне. Существует две группы типов этих болот — сфагновые открытые
(35,7%) и сфагновые лесные (64,3%). Главная их особенность в том. что это
— автономные, в значительной степени саморазвивающиеся образования,
испытывающие влияние только климата и солнечной радиации и
реагирующие в основном только на их изменения.
Переходные болота развиваются в основном в сопряжении с крупными
массивами верховых болот. Они представлены двумя группами типов —
осоково-сфагновыми открытыми (25,2%) и осоково-сфагновыми лесными
(74,8%). Режим водного питания — смешанный. По геоботаническим под-
338
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
зонам занимают сходный удельный вес: в Северной — 37,5%, в
Центральной — 27,6 и Южной — 34,9% [3].
На верховых и переходных болотах встречается до 95 видов сосудистых
растений (5,7% флоры сосудистых растений Беларуси). Основную роль на
этих болотах играют мохообразные, преимущественно сфагновые ыхи.
Сфагновые мхи в прошлом использовались в Беларуси в качестве
дезинфицирующего перевязочного материала. В последнее время эти мхи находят
применение в медицинской промышленности стран дальнего зарубежья.
Верховые болота в основном сохраняются в естественном состоянии. На них
сосредоточены запасы клюквы и голубики, в значительной степени
багульника, в очень небольших объемах — ресурсы черники и брусники (листья и
ягоды), трава вереска, кора крушины.
Переходные болота имеют ограниченное использование для выпаса
сельскохозяйственных животных и заготовки сена. Они отличаются
довольно низким кормовым достоинством трав и малой продуктивностью фитоце-
нозов. На этих болотах сосредоточены ресурсы голубики, клюквы,
багульника, а также имеются незначительные запасы черники, брусники (листья и
ягоды), рябины обыкновенной (плоды), травы вереска и вахты трехлистной,
коры крушины. На переходных и верховых болотах очень редко встречается
морошка и этот вид требует особой охраны.
Низинные торфяные, или эвтрофные, болота - самый распространенный
и представительный класс болот в Беларуси. Они занимают 1453,8 тыс. га
(61,1% заболоченных территорий), развиваются в условиях богатого водно-
минерального питания, обильного увлажнения, достаточной аэрации и про-
точности. Размещены они неравномерно: в Южной подзоне (Полесье —
51,0%), в Центральной — 27,6% и Северной подзоне — 21,4% [3].
Травяные лесные низинные болота занимают 502,8 тыс. га (34,6%), или 43,7%
всех лесных болот страны. В напочвенном покрове основной фон образуют
осоки, тростник обыкновенный, болотное широкотравье. На низинных
болотах важную роль играют мохообразные, которые представлены 76 видами.
Лекарственные растения на этих болотах составляют 11,6% состава флоры
всех сосудистых растений. Из них 15 видов применяется в научной и 6 — в
народной медицине. Здесь значительны запасы вахты трехлистной, аира,
ольхи черной, в небольшом количестве встречаются валерьяна, синюха,
лабазник, паслен сладко-горький, крушина и калина. Из
пряно-ароматических растений здесь представлены аир и гравилат речной, из технических
(дубильные) — ивы, которые широко встречаются на более дренированных
болотах. На низинных болотах широко распространены осоки, злаки и
разнотравье, которые представляют грубые витаминизированные корма для
домашнего скота. Здесь имеются значительные запасы волокнистых
растений — тростника, вейника, крупных осок, ситников.
339
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
В заболоченных лесах (гигротоп — 4) также сосредоточены значите^
ные запасы пищевых и лекарственных растений. Кроме перечисленных вы*
ше растений к ним относятся: дуб, липа, береза, осина, калина, черемух»
лещина, смородина черная, малина, ежевика, крапива, сныть, водяной п&
рец, недотрога обыкновенная, копытень.
Ресурсы плодово-ягодных, пищевых и лекарственных растений на
избыточно увлажненных землях подвержены сильному антропогенному
воздействию. Так, исследованиями, проведенными нами на сети постоянных и
временных пробных площадей в Полесье в 1979-1997 гг. установлено, что в
результате осушительной мелиорации ресурсы голубики сократились за эти
годы в 7 раз [4]. Мелиорация повлияла и на ресурсный потенциал других
ягодных, пищевых и лекарственных растений, в первую очередь, на клюкву
болотную. На запасы дикорастущих ягодников в значительной степени
отрицательно повлияло в последние годы усиление массовых и хищнических
заготовок местным населением в связи с резким падением уровня жизни. В
результате этого нанесен непоправимый урон угодьям клюквы болотной,
черники, брусники и других плодово-ягодных растений. Кроме того, в связи
с массовым посещением болот и заболоченных лесов участились случаи
лесных пожаров, сокращающих ресурсный потенциал пищевых и
лекарственных растений. Так, после устойчивых лесных пожаров сильной
интенсивности заросли черники восстанавливаются лишь через 28, клюквы — 24,
голубики — 18 и брусники — 17 лет. Наибольший вред лесные пожары
нанесли в 1993, 1994 и 1995 годах [5]. В значительной степени на ресурсный
потенциал пищевых и лекарственных растений влияет косьба трав и выпас
скота. И, наконец, значительная часть этих растений выпадает из
хозяйственного оборота из-за радиоактивного загрязнения.
На запасы изучаемых растений косвенное воздействие оказывает
прогрессирующий парниковый эффект планетарного масштаба. Учеными
отмечено, что в последние годы происходит аридизация климата Полесья и
увеличение осадков на севере Беларуси, что приводит к трансформации
болотных и лесных фитоценозов.
[1] Первый Национальный доклад по выполнению Конвенции о биологическом
разнообразии в Беларуси. Мн.: Изд-во «БЕЛСЭНС», 1998. 86 с. [2] Национальная
стратегия и План действий по сохранению и устойчивому использованию
биологического разнообразия Республики Беларусь. Мн.: «Конкордия», 1997. 43 с. [3] Состояние
и использование биологического разнообразия Республики Беларусь: Аналитический
обзор. Мн.: «Конкордия», 1998. 112 с. [4] Гримашевич В.В. Рациональное
использование и охрана угодий дикорастущих ягодников и грибов на Полесье //Тез. док.
межд. науч. конф. «Современные проблемы изучения, использования и охраны
природных комплексов Полесья». Мн.: БЕЛСЭНС, 1998, С. 190. [5] Гримашевич В.В.
Влияние антропогенных факторов на продуктивность дикорастущих плодово-ягод-
340
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
1Х растений и грибов // Состояние и мониторинг лесов на рубеже XXI века. Мат-
ы Межд. науч.-практ. конф. Мн.: Ин-т. эксп. бот, 1998. С.194-196.
РЕДКИЕ РАСТЕНИЯ ЗАБОЛОЧЕННЫХ БЕРЕЗНЯКОВ
РЕСПУБЛИКИ КОМИ
СВ. Дегтева, Т.Н. Пыстина
Институт биологии Коми НЦ УрО РАН, Сыктывкар
Республика Коми относится к числу многолесных районов России. Лесо-
покрытая площадь составляет 29.8 млн. га. В процессе исторического
развития на территории республики сформировались коренные хвойные фитоце-
нозы, сложенные преимущественно породами сибирской полидоминантной
тайги, в основном Picea obovata, г также Abies sibirica, Larix sibirica. По
боровым террасам в долинах рек и на заболоченных участках водоразделов
распространены древостой Pinus sylvestris. В Приуралье и горах Урала в
насаждениях обычна примесь Pinus sibirica. Лиственные леса представлены
насаждениями Be tula pubescens, В. pendula, В. tortuosa, Populus tremula,
Alms incana. На их долю приходится около 20% лесопокрытой площади.
Насаждения лиственных пород долгое время оставались менее изученными,
чем хвойные. С 1995 г. специалисты Института биологии Коми НЦ УрО
РАН начали комплексное исследование березняков, осинников и серооль-
шаников подзон средней и южной тайги с целью разработки их типологии,
выявления флористического разнообразия [1,2].
Наиболее распространенная на исследуемой территории формация
лиственных лесов — березняки. Березовые леса встречаются в широком спектре
экотопов, характеризуются наибольшим флористическим разнообразием
среди формаций лиственных и хвойных лесов. В подзонах южной и средней
тайги распространены березняки, относящиеся к шести группам типов:
травянистые, зеленомошные, долгомошные, травяно-сфагновые, зеленомошно-
сфагновые и сфагновые. Три последние группы относятся к заболоченным
лесам. Они приурочены к притеррасным частям пойм и на слабо
дренированным участкам пойменных речных террас, реже отмечены в нижней части
пологих склонов, у истоков и в верховьях небольших рек и ручьев.
Формируются в процессе заболачивания зеленомошных, долгомошных и
травянистых березняков и ельников. Экотопические условия сдерживают развитие
древостоев. Сомкнутость крон не превышает 0,6, преобладающий бонитет
V-Va. Заболоченные березняки характеризуются значительной пестротой
экотопических условий, разнообразием состава и строения нижних ярусов
сообществ, поскольку представляют собой различные стадии временного
ряда болотообразовательного процесса. Наиболее широко распространены
341
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
на изученной территории березняки вейниково-осоково-сфагновые, б. осп*
ково-сфагновые, б. вахтово-сфагновые, б. разнотравно-сфагновые, б. хвл.
щево-сфагновые, б. пушицево-сфагновые.
Как показали проведенные исследования, заболоченные березняки,
особенно сформировавшиеся в экотопах с ключевым питанием, отличаю^*
наиболее своеобразными флористическими комплексами и являются
местообитаниями редких видов сосудистых растений и лишайников. В таких
лесах обитает Cyphpedium calceolus — вид, охраняемый на локальном уровне
[3] и включенный в «Красную книгу РСФСР» [1983], «Красную книгу
СССР» [1984], списки редких растений МСОП. В напочвенном покрове
здесь преобладают влаголюбивые виды зеленых мхов — Plagiomnium ellu
pticum, Tomentypnum nitens, Helodium blandowii, Rhizomnium pseudopun-
ctatum, Paludella squarrosa и др. Отмечены также сфагны {Sphagnum warns-
torfii, S. girgensohnii, S. squarrosum), но они часто играют подчиненную
роль. В березняках этой группы типов отмечены и другие редкие для
Республики Коми растения. Это прежде всего представители сем. Orchidaceae
— Dactylorhiza cruenta, Epipactis helleborine, Platanthera bifolia, Malaxis
monophyllos, а также Ranunculus lingua, Theliptehs palustris.
Флора лишайников заболоченных березовых лесов отличается
богатством видового состава (75 видов), что составляет 72% от всего разнообразия
лишайников березовых лесов таежной зоны республики (104 вида).
Представлены главным образом эпифитные виды (64), доля эпиксильных
лишайников незначительна (11), а в напочвенном покрове лишайники не
отмечены.
Наибольшим разнообразием отличается лихенофлора березы (39 видов).
Характерными и обильными видами стволовых сообществ березы являются
Melanelia olivacea, Hypogymnia physodes u Parmelia sulcata, покрывающие
значительные участки стволов и ветвей. Обычны Parmeliopsis ambigua, Par-
meliopsis hyperopta, Evernia mesomorpha, реже и в меньшем обилии
встречаются Vulpicida pinastri, Tuckermanniopsis chlorophylla, Platismatia glauca,
Hypogymnia tubelosa, Imshaugia aleurites и др. В комлевой части среди
мохообразных отмечены немногочисленные представители p.Cladonia.
Tuckermanniopsis sepincola массово заселяет тонкие веточки березы. Из кустистых
лишайников чаще других встречаются Bryoria furcellata, Bryoria fuscescens,
Usnea filipendula, Usnea subfloridana и др. Разнообразие лишайников
увеличивается за счет эпиксилов, поселяющихся на 'гниющей коре и обнаженной
древесине (виды pp. Chaenotheca, Chaenothecopsis, Micarea). На стволах
березы отмечено два вида, включенных в Красную Книгу Республики Коми:
Hypogymnia bitteri — категория 3(R) и Tuckermannia laureri — 5 (Cd).
Возможно, что в условиях равнины в заболоченных березняках, расположенных
на притеррасных участках поймы больших рек или на водоразделах в доли-
342
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
нах мелких речек и ручьев, складываются наиболее оптимальные условия
ддя роста и размножения Hypogymnia bitter7. В более чем половине
обследованных березовых лесов данного типа найден этот редкий вид.
Среди других деревьев и кустарников, встречающихся в заболоченных
березняках, богатую эпифитную лихенофлору имеют ива (31 вид), ель (30),
сосна (28), меньшее число отмечено на можжевельнике (15) и березе низкой
(2). Только на стволах древовидной ивы козьей, изредка встречающейся в
виде примеси в древостое, обитает Ramalina thrausta, редкий для
республики вид (категория 5(Cd). Особенностью лихенофлоры ели является
значительное обилие кустистых видов (рода Usnea, Bryoha, реже A lectori a),
сплошь увешивающих ветви деревьев.
Необходимо дальнейшее изучение ценотического и флористического
разнообразия заболоченных березняков Республики Коми и сохранение их
массивов, являющихся местами обитания редких видов.
[1] Дегтева СВ. К проблеме изучения биологического разнообразия вторичных
мелколиственных лесов среднетаежной подзоны Республики Коми // Биологическое
разнообразие антропогенно трансформированных ландшафтов Европейского
Северо-Востока России. Тр. Коми НЦ УрО РАН. № 149. Сыктывкар, 1996. С. 90-101. [2]
Дегтева СВ. Флористический состав среднетаежных осинников Республики Коми.
Науч. докл. Коми НЦ УрО РАН. Вып. 404. Сыктывкар, 1998. 28 с. [3] Красная книга
Республики Коми. М.: Д и К, 1998. 528 с.
О РАЦИОНАЛЬНОМ ИСПОЛЬЗОВАНИИ И СОХРАНЕНИИ БОЛОТ
И ЗАБОЛОЧЕННЫХ ЛЕСОВ КАМЧАТКИ В XXI ВЕКЕ
В. Д. Дмитриев, Н.В. Ловелиус
Ботанический институт РАН, Санкт-Петербург
Большую часть равнин и речных долин Камчатского полуострова
занимают болотные массивы. Они широко развиты на Западно-Камчатской
равнине, на севере в Парапольском доле, частично на юге полуострова и вдоль
восточного побережья по днищам долин, в дельтах рек и на морских
террасах. Локальные участки болот внутри полуострова возникли в результате
лавовых подпруд речных долин (или схода грязе-каменных потоков) с
последующим отмиранием таких подпрудных озер и их заболачиванием.
Особо следует выделить небольшие «болотца» вокруг или вблизи
выходов многочисленных горячих источников со специфическими, пока еще
слабо изученными, растительными сообществами и заболачивание термальных
озер расположенных внутри термальных полей. Они характерны, например,
для кальдеры вулкана Узон, где наблюдается активная газо-гидротермаль-
ная деятельность и сосредоточено более тысячи термальных источников с
343
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
различными гидрохимическими типами термальных вод, с колониями сине*
зеленых водорослей, термофильных бактерий и растений.
Растительность болот, строение торфяных залежей верховых, переход,
ных и низинных типов, их мощностей и минерального дна слабо изучены, за
исключением площадей небольших месторождений торфа. Болота с гипно.
во-травянистыми сообществами развиты на юге полуострова, травяно-сфаг-
ново-гипновые — на западе и востоке, гипново-бугристые — на севере, в
зоне развития криогенных процессов. Болотные массивы сильно
обводненные с грядово-мочажинным и грядово-озерковым микрорельефом. Их вод-
ное питание — атмосферные осадки и мелкие водотоки. Безлесные
болотные массивы чередуются с заболоченным редколесьем, представленным
ольховниками, каменноберезняками и частично лиственничниками с
кочковатым микрорельефом.
Динамика современных процессов заболачивания лесов и болотообразо-
вания происходит в условиях активных вертикальных и горизонтальных
тектонических движений, зачастую унаследованных от голоценовых, что
влияет на мощности и состав торфяных залежей. Перекосы блоков земной
коры предопределяют динамику водного режима и болотную солифлюкцию.
Это видно в бассейнах рек Авачи, Камчатки, Быстрой, где были проведены
повторные инструментальные замеры современных движений блоков и
разломов земной коры. Сквозной характер ряда разломов, их активизация в
голоцене (в т.ч. в историческое время) предопределяют вертикальную
миграцию геохимических элементов с формированием площадных и линейных
(приразломных) аномалий в торфяниках, например, на Западной Камчатке.
В 1960-80-ые годы широкомасштабный размах мелиоративных работ
привел к осушению крупных болотных массивов и заболоченных лесов в
долинах главных нерестовых рек: Авачи, Камчатки, Большой Быстрой, Па-
ратунки и др. и их сельскохозяйственному освоению. Свой след оставили и
многолетние нефтегазопоисковые работы на болотах Западной Камчатки.
В 90-е годы практически завершено осушение естественных болот с
одновременной вырубкой заболоченных лесных массивов под дачное
строительство, имевших непосредственное влияние на режимы водного питания и
водный баланс в низовьях реки Авачи, в бассейне р. Паратунки. Негативные
последствия для экосистем этих рек видны уже сегодня. Это ухудшение
климатических условий, усыхание на корню остатков спелых лесов,
браконьерская рубка леса на дрова, лесные пожары, сокращение лососевых
нерестилищ, общее захламление территории и еще сохранившихся
фрагментов естественных болот и заболоченных лесов.
Одновременное бесконтрольное хозяйственное освоение, особенно Пе-
тропавловско-Елизовской агломерации (по водотокам, впадающим в Ава-
чинскую губу), приводит к подтоплению, в том числе лесных земель, и их
344
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
вторичному заболачиванию. Эти процессы ухудшают физико-механические
свойства грунтов и снижают их сейсмоустойчивость, что представляет опас-
йость для городов Петропавловска-Камчатского, Вилючинска, Елизова.
При варварском уничтожении болот их продуктивность для питания
населения, в том числе для коренных народов, как и их биосферная роль, а тем
более — последствия изменения водного и углеродного режимов, не
учитывались. Это вызывает сожаление, ибо Камчатка является своеобразны
природным полигоном для изучения глобального изменения климата Земли, в
том числе углеродного баланса, процессов миграции геохимических
элементов и создания геохимических барьеров на основе взаимосвязей лесных,
водных и болотных экосистем с геологической средой и атмосферой.
Как известно, полуостров — это «родильный дом» для 1/3 части
мирового лососевого тихоокеанского стада. Для естественных нерестилищ
решающее значение имеет сохранение существующих водных, геохимических
связей между болотными массивами и руслами нерестовых рек с их подпиткой
родниками. Сегодня эти связи нарушаются (или уже нарушены) для многих
нерестовых рек, которые имеют национальное значение, но не обладают
экологическим статусом на федеральном уровне.
К сожалению, болотные массивы пока занимают подчиненное место в
системе охраняемых территорий. А при включении уникальных природных
объектов полуострова в Список Всемирного природного наследия ЮНЕСКО
основные болотные массивы, за исключением болот на территории Кроноц-
кого государственного заповедника, этого статуса не получили.
Дальнейшая деградация болот (вслед за мелиорацией земель) будет
связана с созданием инфраструктуры для добычи рудного золота, с добычей
углеводородного сырья и его транспортировкой по газопроводам, с
созданием единой энергетической сети полуострова, со строительством фанерного
комбината и сплошными рубками каменноберезняков (в т.ч. заболоченных),
без достаточной экологической безопасности при реализации этих планов.
Для устойчивого управления природными ресурсами и
социально-экономического развития Камчатки в XXI веке предлагается: — составление
кадастра болот на стоимостной основе с учетом их биоразнообразия и
социально-экономических показателей для рационального природопользования;
— включение болотных массивов разных типов в систему особо
охраняемых территорий и в охраняемый экологический каркас полуострова; —
придание ряду болотных массивов: в дельте р. Авачи, в низовьях
нерестовых рек Западной Камчатки и на ее равнине статуса Всемирного природного
наследия; — проведение мониторинга болотных систем и заболоченных
лесов с изучением их биосферной роли по программе Глобального
изменения климата Земли; — создание международных научных стационаров в
кальдере вулкана Узон и на локальных участках внутри Западно- и Восточ-
345
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
но-Камчатской болотных провинций; — геоэкологическое картирован»*
болотных экосистем Западной Камчатки при поисках и добыче углевода
родного сырья. При реализации этих предложений возможно управление
оптимальным использованием природных ресурсов болот и заболоченных
лесов Камчатки в ближайшей и в отдаленной перспективе.
ВОССТАНОВЛЕНИЕ БИОРАЗНООБРАЗИЯ РАСТИТЕЛЬНОСТИ НА
НАРУШЕННЫХ НЕФТЕДОБЫЧЕЙ ЗАБОЛОЧЕННЫХ
ТЕРРИТОРИЯХ РЕСПУБЛИКИ КОМИ
Т.В. Евдокимова, Г.В. Железнова, Е.Г. Кузнецова
Институт биологии Коми НЦ УрО РАН, Сыктывкар
В последние десятилетия на Европейском северо-востоке России активно
ведется освоение природных ресурсов, что привело к увеличению площади
территорий с нарушенными природными экосистемами. Практическую и
теоретическую значимость приобретает поэтому изучение процесса
формирования биогенных компонентов нарушенных экосистем на первых стадиях
самовосстановительной сукцессии, и, в частности, восстановления почв и
биоразнообразия растительности на техногенных грунтах нарушенных
участков в различных экотопах. В целях изучения процесса восстановления
почв и растительности на пост-техногенных участках нами в 1992-97 гг.
выполнялись исследования на территории Усинского района (Усинское
месторождение нефти), где фон техногенных нарушений на экосистемы
формировали воздействия, связанные с добычей и транспортом нефти.
Ландшафты территории отличаются чертами переходности, так как
район расположен в подзоне, пограничной с южной лесотундрой. Это
сказывается на переходности состава растительных сообществ, участии в
формировании ландшафтной структуры тундровых урочищ, более широком
распространении сочетаний болотных урочищ с березово-еловыми и елово-березо-
выми сфагновыми и кустарничково-долгомошными лесами. Доминантными
в пределах ландшафтов моренных равнин являются лесные урочища — 38%
(от общей площади), тундровые — 31,4%, болотные — 14,3%. Среди
урочищ ландшафтов группы озерно-ледниковых равнин преобладают лесные —
42,8% и болотные — 19,8%. Тундровые урочища занимают здесь 7,3%
площади, аквальные геосистемы — 5,9%. Особенностью ландшафтной
структуры территории является также значительное распространение
техногенных геосистем, занимающих соответственно, в пределах низменных
моренных и озерно-ледниковых равнин — 16% и 24% территории.
Наибольшая степень антропогенной трансформации характерна для лесных и
тундровых урочищ. Почвообразующими породами в исследованном районе яв-
346
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
дяются озерно-ледниковые пески и супеси или двучлены. Наиболее
распространенные типы почв — иллювиально-гумусовые и гумусово-железистые
подзолы, в сочетании с болотно-подзолистыми и болотными почвами.
Было установлено, что скорость и направление самовосстановления
растительного покрова связаны с типом исходного ландшафта. Ведущими
факторами успешного восстановления растительного покрова являются степень
дренированности исходного типа урочища и микроклиматические условия,
основные показатели которых также связаны с приуроченностью
техногенных площадок (верхняя, средняя, нижняя часть) и экспозиция склона (в том
числе — ветровая), контакт различных типов урочищ, наличие фрагментов
лесной растительности и их расположение по отношению к техногенным
площадкам. Самые неблагоприятные условия для самовосстановления
растительного покрова на площадках, расположенных в пределах болотных
урочищ. Антропогенные нарушения связаны здесь с уничтожением леса в
прилегающей к промысловым площадкам 60-метровой полосе, с
изменением микрорельефа, с разъездами, захламлением, формированием зон
подтопления и осушки, загрязнением поверхности нефтью и пластовыми водами.
Естественная флора Усинского района Республики Коми насчитывает в
целом 456 видов из 66-ти семейств и 162 родов [Акулынина, Шушпаннико-
ва. 1989]. Результаты пятилетнего мониторинга свидетельствуют о том. что
на поверхности технически рекультивированных промысловых площадок
формируются производные сообщества, заметно отличающиеся и по
структуре и по составу растительности от коренных фитоценозов. В зарастании
всех обследованных площадок принимают участие 70 видов из 19-ти
семейств и 47 родов сосудистых растений и 11 видов мохообразных, а в
пределах болотных участков — 33 вида сосудистых растений из 12 семейств и
25 родов и 3 вида мохообразных. Таким образом, на пионерной стадии
зарастания промысловых площадок, отсыпанных на этапе технической
рекультивации однотипным песчаным грунтом, принимает участие 15% видов
флоры всего района, а на площадках, расположенных в пределах
переувлажненных экотопов всего 7,2%. Ведущее положение в синантропной флоре
принадлежит уже на данной стадии процесса самовосстановления
биоразнообразия растительности семействам Asteraceae, Poaceae и Сурегасеае.
На первых стадиях зарастание площадок, расположенных в пределах
болотных урочищ, наиболее активно также происходит по периферии.
Постоянными оставались некоторые виды, характерные для окружающих
площадки природных ландшафтов (хвощ полевой, ива шерстистопобеговая), а
также пионерные виды (иван-чай узколистный, трехреберник непахучий,
кипрей болотный). Однако встречаемость даже активных видов
относительно невысока, и лишь у иван-чая достигла 13,8%. Хотя мохообразные
предпочитают переувлажненные местообитания, но в зарастании песчаного
347
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
грунта даже в пределах болотных урочищ принимают меньшее участие, ней
сосудистые высшие растения. Отмечалась зеленая сеть протонемы мхов
понижениях микрорельефа. По краям площадок в непосредственной
близости от сохранившихся фрагментов естественной растительности разраоа.
ются фунария гигроскопическая, бриум. Характерно, что на первых стадиях
самозарастания техногенных площадок, расположенных в пределах
болотных экотопов, практически не принимают участия типичные болотные
кустарнички. Общее проективное покрытие растительного покрова за годы
наблюдений изменилось незначительно, и составляло не более 9,8%.
Первый этап восстановления растительности характеризуется
господством трав-многолетников и, в меньшей степени, однолетников.
Соотношение жизненных форм растений на обследованных нами площадках довольно
резко меняется по сравнению с ненарушенными сообществами. Древесные,
кустарниковые и кустарничковые растения занимают чаще всего участки
близ окраин промысловых площадок, где сохранились фрагменты
естественной растительности; их доля составляет 30% от числа видов по всем
площадкам: от 34-35% в лесных урочищах до 13% — в болотных.
Основную ценотическую роль в зарастании площадок имеют корневищные и рых-
лодерновинные многолетники (28 и 12%, соответственно); на болотных
участках их доля достигает в целом 60%. На всех площадках отмечено восемь
видов, относящихся к одно-двулетникам, которые составляют в нарушенных
тундровых и болотных сообществах от 20 до 27%.
Основную долю (в среднем 62%) в составе флоры всех техногенных
площадок представляют мезофиты, которые составляют от общего числа
видов от 67 до 54,5%. Виды, предпочитающие переувлажненные
местообитания — гигрогидрофиты, гигрофиты, мезогигрофиты — меньшую часть
составляют на площадках, расположенных в лесных урочищах (11,5%) и
наибольшую — на болотном участке (26,7%).
Работа выполнялась в качестве разделов исследовательских ФЦНТ11 по
фантам «Биоразнообразие»: «Возобновление биоразнообразия на
техногенных территориях Севера в процессе ускоренного природовосстановления» и
«Арктические экосистемы».
СОСТОЯНИЕ И ЗАДАЧИ ОХРАНЫ ЕСТЕСТВЕННЫХ БОЛОТ В СВЯЗИ
С ПРОБЛЕМОЙ УСТОЙЧИВОГО РАЗВИТИЯ ТВЕРСКОЙ ОБЛАСТИ
Ю. Н. Женихов, В.В. Кузовлев
Тверской государственный технический университет
В настоящее время выявлены и достаточно апробированы на разных
регионах основные принципы отбора болотных массивов для охраны. Разра-
348
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
ботана система качественных критериев и методика оценки значимости объ-
еКГов отбора. Оценка природоохранной ценности объекта осуществляется с
использованием системы критериев отбора. Хотя система качественных
критериев достаточно разработана, требует совершенствования методика
оценки природоохранной ценности. Это касается методов определения
экологической, ресурсной, научной, рекреационной значимости, а также
способов получения необходимой для оценки информации. Необходимо четко
определить и увязать состав работ, исполнителей, форму представления
результатов исследований. При выделении болотного массива для охраны
необходимо его всестороннее изучение, проведение гидрологических,
метеорологических, ботанических, зоологических, ресурсоведческих, торфоведче-
ских, экономических и других исследований. Выбор наиболее значимых
объектов для охраны производится в том случае, когда общая площадь
объектов, потенциально требующих охраны, превышает площадь,
соответствующую оптимальной заболоченности. В основе выбора лежит система
бальных или экспертных оценок.
Торфяные месторождения, сохраняемые на территории Тверской
области, имеют разные статус, цели и режим охраны. На территории Центрально-
Лесного биосферного государственного заповедника и госкомплекса
«Завидово» располагаются 25 торфяно-болотных экосистем (ТБЭ). Статус
памятника природы присвоен 28 ТБЭ. В режиме заказников сохраняются 164
торфяных месторождения. Помимо этого, принято решение о сохранении в
естественном состоянии 95 экосистем, имеющих водоохранное значение и
29 экосистем, отнесенных к ценным ягодникам или имеющих
промышленные запасы лекарственных растений. Объекты водоохранного значения и
ценные ягодники долгое время не имели статуса охраны, но были
исключены из планов хозяйственного использования.. Сейчас такие объекты
объявлены к охране в режиме заказников.
Целостной системы организации особоохраняемых природных
территорий (ООПТ) в отношении торфяно-болотных экосистем в Тверской области
нет. На это указывает существование большого количества ТБЭ на
территориях, объявленных заказниками. То есть ТБЭ автоматически, а не в силу
своего особого значения, попали в число заказников. Основные же цели
объявления отдельных территорий заказниками в пределах каждого
административного района связаны с задачами воспроизводства охотничье-
промысловых видов животных. Между тем, отдельные ТБЭ, числящиеся
сейчас памятниками природы, объектами водоохранного значения или
ценными ягодниками, могли бы объективно стать ландшафтными,
биологическими, гидрологическими или геологическими заказниками, выполняя не
только хозяйственные, но и научно-информационные функции. Совершенно
не используется форма внедрения заказников в биосферные полигоны запо-
349
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ведников. Не определен режим охраны больших по площади памятников
природы. Еще в большей степени обоснованы опасения о потере
устойчивости болот - мест произрастания ягодников и лекарственных растений.
Охраняемый фонд области представлен 341 полностью охраняемой ТБЭ
и 11 объектами, где в естественном состоянии сохраняется лишь часть
площади. Объектами охраны природы являются 11,0 % общего числа торфяных
месторождений области. Общая площадь объектов охраны в нулевых
границах составляет 390,6 тыс. га (48,3 %) (см. табл.). На долю забалансовых
запасов торфа охраняемого фонда приходится уже 57,1% — 1157,4 млн.т.
Характеристика охраняемого фонда ТБЭ Тверской области
Состав охраняемого фонда Количество Площадь, Процент от площади
ТБЭ га всех ТБЭ области
Заповедники
Памятники природы
Ягодники
Водоохранное значение
Заказники
Всего
25
26
25
100
176
352
14438,9
158214
32076
124235
61636,7
390600,6
1,8
19,5
4,0
15,4
7,6
48,3
Наибольшее число объектов охраны и наибольшие охраняемые площади
приходятся на Западнодвинскую (24,1% всех охраняемых площадей), Мо-
логскую (19,7%), Верхневолжскую (19,0%), Калининскую (17,8%) группы
районов. Минимальное количество объектов охраны природы — 9
площадью 8,87 тыс. га — имеет Краснохолмская. Еще меньше охраняемой
площади — 5,7 тыс. га (1,5%) — приходится на Ржевскую группу.
Доля охраняемых площадей болот в общей их площади в среднем по
области составляет 47%. Нет объектов охраны в Молоковском, Кесовогорском,
Ржевском районах, низкая доля охраняемых болотных площадей в Калязин-
ском (2,6%) районе. Наивысшего значения доля охраняемых площадей
болот достигает в Жарковском (91,2%) районе. Более 50% всех торфяных
площадей сохраняется еще в 8 административных районах. Однако
соотношение сохраняемых и общих площадей болот, учитывая их влияние на
экологическое равновесие территорий, целесообразно рассматривать в пределах
торфяно-болотных районов. При таком подходе даже в Ржевской группе
районов охраняется каждый четвертый гектар болот. В остальных группах
районов процент охраняемых площадей гораздо выше. Для региона
Тверской области для сохранения экологического равновесия рекомендуется
сохранять около 15% всех площадей болот. Анализ показывает, что для
любого из 8 торфяно-болотных районов эта цифра значительно перекрывается.
Если из числа охраняемых площадей условно исключить площадь
ягодников (хозяйственные цели) и площадь заказников (считая, что охрана их не
вполне обоснована), то картина может измениться. Действительно, в этом
350
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
случае недопустимо низкий процент охраняемых площадей болот (8,2%)
отмечается для Лихославльской группы районов. Близка к пределу (15,1%)
доля в Ржевской группе районов. В остальных торфяно-болотных районах
доля охраняемых площадей болот находится в пределах от 22,4 до 42,9%.
Таким образом, в Тверской области выделено большое количество ТБЭ -
объектов охраны природы. Площадь охраняемых болот и их доля в общей
площади болот обеспечивают сохранение экологического равновесия
области в целом и отдельных ее регионов в отдельности. Проблема заключается в
создании научно-обоснованной системы охраняемых торфяно-болотных
экосистем, в обеспечении режима охраны. Следует пересмотреть статус
отдельных памятников природы, обосновать объявление болот заказниками с
разными целями охраны. Важно установить пути рационального использования
растительных ресурсов сохраняемых без статуса охраны торфяно-болотных
экосистем. Эта форма хозяйственного использования объектов охраны
природы в наибольшей мере соответствует активным методам их охраны на
основе сбалансированного взаимодействия человека с окружающей средой.
МНОГООБРАЗИЕ ОСУ1ШННЫХ ЛЕСОВ ЛАТВИИ
П. Залитис
Латвийский институт лесоведения «Силава», Саласпилс, Латвия
В течении последнего десятилетия лавинообразно выросло число таких
публикации, в которых значительная часть текста отведена рассуждениям о
биологическом многообразии. Это напоминает ту азартную страсть к
математизации биологических и экологических явлений, вспышка которой
отмечалась лет тридцать тому назад. В результате сложных и обширных
математических изложений тогда нередко были сформулированы лишь
тривиальные и до того уже известные закономерности. Сегодня в публикациях о
биологическом многообразии подобным же образом удивительно мало
встречается деловой информации как о конкретных величинах показателей
многообразия в сопоставимых совокупностях леса, так и об изменениях этих
показателей в результате лесохозяйственной деятельности. Доминируют
абстрактные рассуждения о 4-6 уровнях оценки многообразия, которые порой
иллюстрируются ошибочными примерами, в т.ч. о сокращении численности
козодоя в связи с обширным осушением верховых болот в Латвии, не
обращая внимания на то, что в течении последнего полвека верховые болота в
Латвии не были осушены вообще.
При оценке лесов как неотъемлемого компонента биосферы в целом их
заболачивание безусловно рассматривается как деградация лесной
экосистемы. Но в локальном масштабе частичное заболачивание лесных площадей
351
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
так же как частичная мелиорация повышают биологическое многообразие
сравнительно небольших территорий, хотя и такая лишь «частичная
мелиорация» связана с экономическим и социальным уроном.
Лесам Латвии свойственна весьма мозаичная структура: в пределах
одного квартала средней площадью около 40 га выделяются в среднем 20
отдельных участков леса, принципы разграничения которых соответствуют
принципам выделения отдельных биотопов. На каждые 100 га суходольных
лесных земель встречаются в среднем 54 выдела, в избыточно увлажненных
неосушенных — 46, но в избыточно увлажненных осушенных лесах — 49
выделов. Эти статистические величины ориентировочно свидетельствуют
как о сходной мозаичности насаждений во всех лесных совокупностях, так и
о сохранении высокого экологического многообразия в осушенных лесах.
Для взаимного сопоставления внутреннего многообразия отдельных
совокупностей леса целесообразно пользоваться индикатором Шенона Н =
т
V р i0g2 р , где / - индекс одной определенной группы выделов со сход-
ными насаждениями; pi — относительное количество выделов в одной
группе (0,0< р <1,0); т — общее число групп в пределах выбранной
совокупности лесов. Принадлежность отдельных выделов леса к одной из групп / в
пределах лесхоза определилась по 7 параметрам: тип условий
произрастания (5 позиций), класс бонитета древостоя (5 позиций), доминирующая
порода древесных (8 позиций) и удельный вес ее в составе древостоя (2
позиции), соучастие ели во втором ярусе древостоя (2 позиции), возраст
древостоя (3 позиции) и полнота древостоя (3 позиции). Максимально возможное
количество групп mmax=7200, но по отдельным лесхозам фактическое их
количество варьирует в пределах от 150 до 690; это при колебании числа
отдельных выделов леса от 1750 до 37020 штук. В среднем по выборке с 5
лесхозами Латвии получены следующие величины индикатора Н: в лесах на
суходоле Н=6,44; в избыточно увлажненных лесах с гидроморфными и
торфяными почвами соответственно Н=6,23 и 5,65; в осушенных лесах с
гидроморфными минеральными и торфяными почвами Н=6,20 и 6,17.
Располагая информацией накопленной в базе данных «Лесной фонд
Латвии», на компьютере сыгран сценарий выявления изменений индикатора Н
как функции относительной величины осушенных лесов. Показатели
экологического многообразия переувлажненных лесов кульминируют, если
осушено 40-70% от ими занятой территории. Сохраняя максимальное
экологическое многообразие в лесах Латвии целесообразно дополнительное
осушение еще 300 тыс. га избыточно увлажненных лесов.
Прогнозируя экологические последствия гидротехнической мелиорации
избыточно увлажненных лесов необходимо разобраться в сходстве осушен-
352
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
ных и неосушенных лесов. Например, в Вентспилском лесхозе в неосушен-
яых лесах индикатор Н=6,55; такую же величину индикатора получим если
будет осушено 90% всех избыточно увлажненных лесов. Вполне допустимо,
что при одинаковом Н структура осушенных лесов будет совершенно иной.
Сходство неосушенных и осушенных лесов характеризовано с помощью
коэффициента Чекановского
т ( т т \
К= £2min(4,B.): £4+ХД,]> ™e А, и В, — удельный вес сходных
/=1 ^ i=i /=1 '
групп насаждений в совокупностях А и В, т.е. в выборках неосушенных и
лет 20-50 тому назад частично осушенных избыточно увлажненных лесов.
Полученные данные свидетельствуют о значительном сходстве изученных
лесов в пределах одного лесхоза.
Сходство двух совокупностей, в одной из которых осушено 70% всех
избыточно увлажненных лесов, а во второй представлены только неосушенные
леса, характеризуется высоким коэффициентом сходства К в пределах 0,80-
0,88; если осушены все избыточно увлажненные леса, то 0,69<К<0,85. В то
же время сходство между совокупностями исключительно неосушенных
лесов в двух на расстоянии 350 км территориально отдаленных лесхозах —
Вентспилс и Цесвайне — характеризуется К=0,23. Это подтверждает, что
экологическое многообразие избыточно увлажненных лесов Латвии
определяется естественными геоклиматическими условиями в гораздо большей
степени, чем антропогенным воздействием, в т.ч. гидротехнической
мелиорацией.
РОЛЬ ЗАБОЛОЧЕННЫХ СОСНЯКОВ В СОХРАНЕНИИ
БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ БЕСПОЗВОНОЧНЫХ
Г. А. Козулько
ГНП «Беловежская пуща», Брестская обл., Беларусь
Многочисленными исследованиями установлено, что заболоченные леса
играют важную роль в сохранении биологического разнообразия растений и
животных [Козловская, 1959, Почвенные беспозвоночные Кандалашского
заповедника, 1986]. В 1988-89 гг. изучали почвенных беспозвоночных в
сосняке долгомошном (кв. 942 Б). Исследования проводили методом поч-
венно-зоологических раскопок по стандартной методике с размером пробы
25 см х 25 см х 40 см [Гиляров, 1975]. Разборку подстилки осуществляли
несколько усовершенствованной методикой: просеянную мелкую фракцию
подстилки собирали в мешочки, доставляли их в лабораторию и разбирали
под электрической лампой. Это дало возможность наиболее полно учесть
мелких беспозвоночных. Пробы отбирали 4 раза в год (апрель, июнь, ав-
353
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
густ, октябрь). За 1 раз (в течение месяца) брали 16 проб, за сезон — 64 (4
м2). Всего взято 128 проб и собрано 4 199 экземпляров беспозвоночных.
Сосняк долгомошный. I ярус — 9С1Б. Возраст — 70 лет. Бонитет Щ
Полнота - 0.7. Запас древесины - 220 м3/га. В подлеске крушина, ива. В под-
росте единично ель. В живом напочвенном покрове - мхи кукушкин лен и
сфагнум, осока, черника и др. Проективное покрытие — 97%. Общая фвдхк
масса — 2348 кг/га. Запас подстилки — 160 т/га. Почва торфяно-глеевая на
оглеенном песке рыхлом водно-ледниковом.
В заболоченном сосняке долгомошном зарегистрирована самая высокая
плотность беспозвоночных среди хвойных типов леса (464-585 экз/м2) (см.
табл.). Значительные колебания плотности беспозвоночных в разные годы
исследований связаны с погодными условиями сезонов — холодным и
дождливым 1988 г. и теплым и сухим 1989 г. Общая зоомасса
беспозвоночных в этом типе леса оказалась также выше, чем в других зеленомошных
сосняках. В сосняке долгомошщном среди беспозвоночных массовыми
являлись паукообразные (275,2-330,5 экз/м2; 56,5-59,3%), плотность
насекомых была несколько ниже (111,0-130,8 экз/м2; 22,3-23,9%). Из насекомых
наиболее многочисленны жесткокрылые, из жесткокрылых — стафилиниды.
Для сосняка долгомошного характерна низкая плотность дождевых
червей, играющих огромное значение в переработке растительного опада и
формировании гумусового слоя почвы. Они представлены единственным
видом Dendrobaena octaedra Sav. Однако здесь очень большой плотности
достигают некоторые группы подстилочных сапрофагов. Так, плотность
Diplopoda составила 49,8-92,5 экз/м2. В литературе не отмечено подобных
высоких значений плотности Diplopoda в сосняках с полным преобладанием
в составе древостоя сосны. Значительна здесь плотность моллюсков (9,5-
11.2 экз/м2), постоянными обитателями являются мокрицы (2,0- 3,5 экз/м2).
Объяснение этого феномена, по-видимому, заключается в особенностях
формирования почвообразующих пород на территории Беловежской пущи и
связанной с этим спецификой почвообразовательных процессов. Неглубокое
залегание моренных суглинков обогащает грунтовые воды кальцием и
магнием. В результате этого количество СаО в глубинных почвенных слоях
сосняка долгомошного не уступает таковому в других типах сосняков.
Количество же ионов магния в почве сосняка долгомошного может даже
превосходить количество этого элемента в почвах данных сосняков. Распределение
кислотности по почвенному профилю также имеет интересную
закономерность: крайне низкое значение рН в торфяном слое (3,08) уже на глубине 1 м
в горизонте CgO повышается до величины 5,19, что превосходит
аналогичные показатели во всех обследованных сосняках.
Применение усовершенствованной методики выборки мелких животных
дало возможность установить высокие значения плотности для хищных под-
354
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
стилочных беспозвоночных — паукообразных (34,0-330,5 экз/м2) и стафи-
динид (40,6-104,6 экз/м2). В сосновых биогеоценозах моховой слой играет
важную роль в распространении этих групп беспозвоночных. С. Копонен
[1987] указывает, что при ручной разборке плотность пауков была самой
высокой в мощном моховом слое субарктического березового леса — 400
экз/м". Показательно то, что в сосняке долгомошном Беловежской пущи
численность паукообразных значительно превышает численность их
потенциальных жертв. Такой избыток хищников поддерживается, по-видимому,
за счет псевдоканнибалистических отношений (поедание хищников
хищниками одного и того же трофического уровня) и способцрстью их к
длительному голоданию [Берман, 1970].
Основу фаунистических группировок сосняка долгомошного составляют
гигрофитные виды (Actenicerus sjaelandicus Mull., Tipula signata Staeg.,
Smittia sp.* Rhamphomyia sp., Dolichopus sp. - из индикаторных групп). В
сосняке долгомошном практически все почвенное население сосредоточено
в подстилочном ярусе.
Средняя плотность (экз/м2) почвенной мезофауны в сосняке долгомошном
Группы мезофауны
Lumbricidae
Mollusca
Oniscoidea
Arachnida
Chilopoda
Diplopoda
Insecta
Hemiptera
Dermaptera
Neuroptera
Homoptera
Blattoptera
Coleoptera
Carabidae
Staphylinidae
Elateridae
Curculionidae
Проч. Coleoptera
Lepidoptera
Hymenoptera
Diptera
Общая плотность
1988 г.
5,2±1,0
9,5±1,8
2,0±1,0
275,2±19,9
11,5±1,7
49,8±6,9
111,0±8,0
2,5±0,8
-
+
18,8±4,6
2,0±0,7
66,8±5,2
8,8±1,5
42,5±4,5
7,2±1,7
2,0±0,8
6,3±1,6
6,2±1,4
1,3±0,5
13,2±1,9
464,2±27,2
1989r.
5,8±1,1
11,2=1=1.9
3,5±1,1
330,5±15,4
11,2±1,8
92,5±8,2
130,8±7,3
4,2±1,4
+
+
33,5±4,6
+
72,8=fc4,l
8,8±1,5
43,5±3,4
9,2±1,7
2,3±0,7
9,0±1,4
7,2±1,3
3,5±0,8
8,5±1,3
585,5±22,2
Примечание: + единично встречаемые особи; — отсутствие особей
355
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ОХРАНА БОЛОТ В РОССИИ: СОСТОЯНИЕ И ПЕРСПЕКТИВЫ
Т.Ю. Минаева1, А.А. Сирин2
Центрально-Лесной биосферный государственный заповедник, Тверская
область, Российское представительство Всемирного фонда дикой
природы (WWF), Москва;2Институт лесоведения РАН, Московская область
Болота и заболоченные земли как отдельные объекты охраны
практически не упоминаются до 60-х годов нашего столетия. С древних времен на
Руси охранялись преимущественно леса и места обитания охотничье-про-
мысловых видов животных [10,15]. За редким исключением [11,12] даже в
болотоведческой литературе уже нашего столетия не отмечалась
необходимость защиты болот. Но косвенно они сохранялись. В силу обширности
территорий в природоохранном планировании в России преобладал
ландшафтный подход [4], нашедший отражение в таких формах как священные земли
(рощи), охотничьи заказники (зверинцы, местности), засеки, в которые
топологически входили болота и заболоченные земли.
После 60-х годов интенсивное использование ресурсов, связанных
происхождением или механизмом восстановления с болотными экосистемами,
а также политическая поддержка идеологии рационального
природопользования способствует появлению разработок, направленных на обеспечение
возобновимости ресурсов болот. Указывается на необходимость сохранения
естественного разнообразия болот [14]. В 70-90-е годы развиваются научные
основы территориальной охраны болотных экосистем [1-3,5-7 и др.]. В
последнее пятилетие широкое внедрение через политику доноров «западной»
природоохранной культуры в российскую практику формирует устойчивое
восприятие болотных экосистем как наиболее ценных и угрожаемых
природных объектов.
Современный региональный анализ показывает наибольшую активность
охраны болот в регионах, где они занимают значительную долю площади,
играют заметную роль в социально-экономической структуре региона,
являются частым объектом научных исследованиях. Эти регионы (Северо-
запад ЕТР, Приветлужье, Поволжье, Коми, Западная Сибирь и др.)
отличаются высоким уровнем сохранности болот. В тоже время в районах, где
болота уже находятся под угрозой исчезновения (Центрально-Черноземные и
южные области ЕТР, степные районы Предуралья и Зауралья и пр.), роль
специальных проектов сохранения болотных экосистем невелика.
Отраслевой анализ природоохранного планирования в отношении болот
демонстрирует приоритет специалистов по мелиорации земель.
Мелиоративные задачи стимулировали развитие современных представлений о
структурно-функциональной организации болот. Практически во всех
работах по мелиорации, начиная с 70-х годов, рассматриваются подходы для
356
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА VI
рационального использования ресурсов болотных экосистем. Следующей
группой являются геологи: например, ПГО «Торфгеология» сделана оценка
природоохранной ценности торфяных месторождений Европейской части
России [9]. Роль ботанической науки выразилась через деятельность секции
болотоведения Ботанического общества и группы «Телма», авторитет
руководителей которой, В.В. Мазинга и М.С. Боч, способствовал изменению
ментальности многих региональных администраторов, земле- и ресурсо-
пользователей. В основу выделения болот под охрану Государственной
лесной службой лежало выведение низкопродуктивных древостоев из состава
расчетной лесосеки, однако именно их площади составляют основу
современной сети охраняемых болот.
Территориальная охрана природы предполагает создание определенных
типов особо охраняемых природных территорий (ООПТ), установленных в
законах об ООПТ РФ или субъектов Федерации. В границах этих
территорий наряду с другими ландшафтами сохраняются и болотные экосистемы.
На территории заказников и заповедников ЕТР сохраняется 1265 торфяных
болот общей площадью 1066,6 тыс. га. На территории таких заповедников
как Юганский, Полистовский, Рдейский, Дарвинский болота преобладают.
Большие площади занимают болота, имеющие статус «Памятник природы»
[9]. Специальной категории ООПТ для охраны болот на федеральном
уровне нет, хотя администрациями ряда областей ЕТР в 80-е годы была введена
категория ООПТ «Болото, сохраняемое в естественном состоянии». Их
площади не отчуждались у землепользователя, но изымались из планов рубок и
торфодобычи.
Болота, выделенные под охрану отраслевыми структурами с
закреплением режима землепользования в хозяйственных распоряжениях (материалы
лесоустройства, планы разработки торфяных месторождений и т.д.), реально
охраняются. На землях Гослесфонда болота часто отнесены к особо
защитным участкам (ОЗУ) в ранге водоохранных зон. Но законодательный статус
этих болот расплывчат и нарушение режима сложно оспорить в судебном
порядке. Опубликованные списки ценных болотных систем и массивов
(например, список «Телма») не имеют правового статуса и носят
рекомендательный характер. Исключение — болота из списка водно-болотных угодий
(ВБУ) России, определенного постановлением Правительства РФ (№1050 от
13.09.94 г.) и признанного странами-участницами Рамсарской конвенции
[8]. Эти болота подпадают под нормы международного права,
превалирующего над внутренним законодательством. Но механизм пользования этой
нормой в национальных судах не ясен. В целом, правовые основы охраны
болот в РФ практически не разработаны.
Вхождение России в международные проекты по охране болот началась
в 1967 году, когда в рамках ЮНЕСКО, Международного союза охраны при-
357
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
роды (МСОП) и Международной биологической программы (МБП) был
организован проект «Телма» (20 стран-участников). В 1968 г. создана
советская группа при Национальном комитете МБП. В результате 10-летней
работы были сформулированы цели охраны болот и выделены факторы,
неблагоприятные для их сохранения, составлен список охраняемых и
намеченных к охране болот [9].
Важную роль в наднациональной охране торфяных болот является
Международное Общество по торфу (IPS), в рамках которой действует группа по
рациональному использованию торфяников, их охране и восстановлению, а
также неформальная Международная группа по охране болот (IMCG),
объединяющая болотоведов многих стран, включая российских. К сожалению
активность Общества болотоведов (Society of Wetland Scientists)
практически ограничена Северной Америкой.
Наиболее значимая международная программа по сохранению болот
очерчена Рамсарской конвенцией (Тегеран, 1971), изначально направленной
на сохранение ВБУ в основном, как мест гнездования и миграции
водоплавающих птиц. В 90-х годах в ее рамках стала формироваться идеология
сохранения биологического разнообразия на ландшафтном уровне,
биосферных функций ВБУ. Но собственно.болота и заболоченные земли до сих пор
являются второстепенными по сравнению с другими ВБУ (см. табл.), на
которые нацелены национальные и международные программы [16].
Представленность торфяных болот в Рамсарской конвенции [по 17]
Рамсарские Страны - Общая площадь,
ВБУ участницы тыс.га
ВБУ, где торфяные болота 75 9,6% 21 22,8% 3179 6,1%
- доминирующий компонент
- минимальный компонент 155 19,9% 31 33,7% 7592 14,6%
Всего 778 100% 92 100% 52000 100%
В ходе рабочего совещания «Global Mire and Peatland Conservation»
(1996 г.) были разработаны рекомендации по охране торфяных болот в
добавление к Стратегическому плану 1997-2002 гг., принятому Шестым
Совещанием Конференции Сторон (Брисбен, Австралия, 1996 г.) [13]. В 1999
году стратегия по сохранению торфяных болот дополнительно поддержана
решениями Седьмого Совещания. Однако недоучет их особого значения
остается. В связи с этим роль России в международных программах по
сохранению болот представляется неадекватной. Необходима поддержка
усилий Канады в отношении уточнения критериев и других положений
Рамсарской конвенции касательно особой значимости торфяных болот.
Целесообразно активно сотрудничать, прежде всего, с Канадой и Индонезией (как
358
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
«держателей» наряду с РФ наиболее крупных пулов торфяников), в
выдвижении соответствующих инициатив на международном уровне.
Одной из организаций, активно реализующей решения стран-участниц
Рамсарской конвенции является Wetlands International, российское
представительство которого действует с 1997 года, и важнейшим направлением
деятельности которого в настоящий период является участие в разработке
Стратегии сохранения водно-болотных угодий РФ. Однако, посвященная ей
Всероссийская конференция (Москва, 24-26 февраля 1999 г.)
свидетельствовала о недоучете значения торфяных болот и на национальном уровне.
Перенос в российскую практику опыта стран с невысокой степенью
сохранности естественных болот представляется мало оправданным, учитывая
высокую заболоченность территории страны и значение болот для разных
аспектов хозяйственной жизни населения.
Сохранение болотных экосистем на национальном уровне должно занять
адекватное место в природоохранной политике и практике. Болота и
заболоченные земли занимают, по меньшей мере, 20 % территории России
составляя большую часть общей площади ВБУ страны. Они обладают отличными
функциями, свойствами и пользой, включающими: ресурсные (топливо,
сырье, пищевые продукты и др.), регулирующие (климатические,
гидрологические, геохимические и пр.), т.н. «несущую» (местообитания и др.) и
нематериальные (биоразноообразие, научная, образовательная и др.). Их
количество и относительное значение, постоянно возрастало и изменялось по мере
развития общества, что будет происходить и в дальнейшем. Они
представляют интерес с различных социально-экономических точек зрения, могут
изменяться в пространстве и времени, способствовать возникновению
противоречий интересов различных социальных групп, интересами местного,
регионального, национального и международного уровня, интересами
настоящего и будущего поколений. Это должно быть учтено в Стратегии.
Стратегия должна учитывать особое значение болот в России, не
ограничиваясь принятыми в настоящее время Рамсарскими определениями и
дефинициями. Цели конвенции существенно расширились в сторону
сохранения функций ВБУ, в т.ч. регулирующих. На большинстве пространственных
уровней (водосбора, местном, региональном, глобальном) они не могут быть
сохранены путем охраны конкретных болот, — только посредством
регулирования использования всей их совокупности («популяции»)
рассматриваемой территории. Это может быть осуществлено лишь посредством их
рационального использования с многоуровенной системой охраны болот, в
том числе строго заповедания отдельных из них..
Авторы благодарны Dr. Hans Joosten (Университет Грайфсвальд,
Германия) за дискуссии по обсуждаемой проблеме.
359
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
[1] Антипин В.К., Кузнецов О.Л. Охрана разнообразия болот Карелии // Биоразно.
образие, динамика и охрана болотных экосистем восточной Фенноскандии.
Петрозаводск: КНЦ РАН, 1998. С. 10-13. [2] Антропогенные изменения, охрана
растительности болот и прилегающих территорий, Минск, 1981. 280 с. [3] Болота охраняемых
территорий: проблемы охраны и мониторинга. Л.: ВБО, 1991. 145 с.[4] Бородин
И.П. Речь делегата от России на Бернской конференции 1913 г. // Мировая охрана
природы, № 2. Петроград, 1915. С.94-96. [5] Боч М.С., Мазинг В.В. Экосистемы
болот СССР. Л., 1979. 188 с. [6] Боч М.С., Ниценко А.А. Об охране болот в СССР //
Вопросы охраны ботанических объектов. Л., 1971. С.36-42. [7] Боч М.С., Смагин
В.А. Флора и растительность болот Северо-Запада России и принципы их охраны.
СПб: Гидрометеоиздат, 1993. 225 с. [8] Водно-болотные угодья России. Т.1. Водно-
болотные угодья международного значения. М.: Wetlands Intern. Publ. N. 47. 1998.
256 с. [9] Водно-болотные угодья России. Т.2. Ценные болота. М.: Wetlands Intern.
Publ. N. 49. 1999. 88 с.[10] Граве Н.П. Охрана природы в Древней Руси // Лесное
хозяйство, 1993, № 4, с.23-24. [11] Доктуровский B.C. О торфяниках Пензенской
губ. // Тр. по изучен, заповеди., 3, 1925. [12] Кац СВ. Два замечательных болота на
севере Московской губ. // Московский краевед, М., 1928. № 4. [13] Конвенция о
водно-болотных угодьях // Документы 6-го совещания Конференции Сторон, т.4.12.
Резолюции и рекомендации. Рамсарская Конвенция. Брисбен, Австралия, 19-27
марта 1996 г. 95 с. [14] Ниценко А.А. Об охране природы на Северо-Западе СССР //
Научн. докл. высш. шк. Биол.науки, 1962, № 2. С.23-27. [15] Реймерс Н.Ф., Штиль-
марк Ф.Р. Особо охраняемые территории. М.: Мысль, 1978. 295 с. [16] Clayton R.
Introduction to the Workshop and overview of the global peat resource // Proc. Intern.
Workshop "Global Mire and Peatland Conservation", Brisbane, Australia, March 18,
1996. [17] Frazier, S. (a) Major peatland sites from the List of Wetlands of International
Importance and (b) All sites recording peatland components from the List of Wetlands of
International Importance // Updated lists to December 31, 1995 prepared by Wetlands
International for the Ramsar Secretariat. Slimbridge, United Kingdom. Unpublished.
РОЛЬ НАЦИОНАЛЬНОГО ПАРКА «СЕБЕЖСКИЙ» В СОХРАНЕНИИ
ТОРФЯНИКОВ ПСКОВСКОГО ПООЗЕРЬЯ
В.Ю. Мусатов
Государственный комитет по охране окружающей среды
Псковской области,
Южная часть Псковской области расположена в пределах участка
полосы Белорусско-Валдайских гряд, включающего Себежскую и
Великолукскую гряды, которые соответствуют длительной задержке края валдайского
ледника [4]. Сильно пересеченный холмисто-котловинный рельеф полосы
гряд является одним из факторов определяющим широкое развитие озерных
седиментационных обстановок после дегляциации территории, в следствии
чего она получила название Белорусско-Валдайского Поозерья, а часть тер-
360
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
ритории Себежской гряды, характеризующаяся максимальным показателем
озерности (более 10%), соответственно — Псковского Поозерья.
В настоящее время значительную площадь в пределах полосы
Белорусско-Валдайских гряд, в том числе и Себежской гряды, занимают болота , в
которых происходит накопление торфов. В Псковском Поозерье болотные
(фитогенные) отложения представляют собой, обычно, конечную фазу
заполнения многочисленных ледниковых водоемов кластическим материалом.
Это подтверждается наличием озерных отложений, подстилающих
торфяные залежи. Мощность погребенных сапропелей нередко достигает 6,0 м.
Начало образования современных торфяников в области валдайского
оледенения колеблется в широких пределах от пребореального до
субатлантического времени [2].
Несмотря на то, что торфяники являются одним из доминирующих
компонентов Псковского Поозерья и выполняют важнейшие функции по
регулированию гидрологического режима и геохимических процессов в регионе
представленность их в системе особо охраняемых природных территорий
области до недавнего времени была явно недостаточной . В качестве
памятников природы в 1970-е годы были взяты под охрану только несколько
торфяных болот — Бронское I, Ирдынец, Копот, Красная Лука и Ямница [3].
Ситуация с охраной торфяников в Псковском Поозерье существенно
изменилась после создания в 1996 году национального парка «Себежский»
Федеральной службы лесного хозяйства России (площадь 50021 га).
Себежский национальный парк — наиболее типичная и хорошо сохранившаяся
часть Псковского Поозерья, имеющая важное природно-историческое,
ресурсно-сырьевое, природоохранное и культурно-рекреационное значение.
Основу ресурсоохранного потенциала парка составляют запасы торфа и
аккумулированной в торфяных залежах воды, общее количество которой
значительно превосходит запасы депонированного органического вещества.
В границах национального парка с различной степенью детальности
оценено и разведано 69 торфяников [8,9]. Наиболее крупные торфяники —
Большой Мох I (1021 га), Выдрино (573 га), Красная Лука(534 га) и Старина
(397 га). Как в целом в Псковском Поозерье, так и на территории
национального парка в частности, преобладают месторождения низинного типа с
торфяной залежью, сложенной главным образом торфами топяного подтипа.
Суммарная площадь всех торфяников в пределах национального парка
составляет 7370 га (14,7% от общей площади парка), а геологические запасы
торфа в пределах залежей, мощность которых позволяет осуществлять
промышленную разработку — 26604 тыс.т при 40%-ой влажности. Средние
мощности торфяных залежей изменяются в широком диапазоне от 1,09 м
(Имшки) до 6,01 м (Ровы-Мох), а максимальные мощности соответственно
— от 2,10 м (Большая Лужа, Мшара) до 8,30 м (Нивьевское).
361
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
На территории национального парка разведано месторождение торфяных
лечебных грязей Горелое П. Лечебный торф месторождения практически
полностью состоит из органического вещества и характеризуется высокой
степенью разложения и содержания битумов (18%) [6]. Данное
месторождение может быть оперативно востребовано как в рекреационных целях, так и
для обеспечения потребности в лечебных грязях населения постоянно
проживающего на территории парка, в том числе и жителей г. Себеж [7].
По своему функциональному назначению торфяники национального
парка разделяются на две группы. Первую группу составляют 37 торфяников с
общей площадью 5066 га, которые расположены на болотах, включенных в
состав парка с изъятием их у землепользователей. В соответствии с
утвержденным Положением о национальном парке разработка этих торфяников
запрещена. Потенциальная стоимость изъятых из хозяйственного
использования запасов торфа по ним, даже при значительно заниженной
современной оценке, составляет более 20 млн. долл. США. Вторая группа включает
32 торфяника, расположенных на болотах, включенных в состав парка без
изъятия у землепользователей (сельскохозяйственных формирований).
Разработка торфяников, относящихся ко второй группе, может осуществляться
по согласованию с администрацией национального парка.
Проведенный анализ свидетельствует о чрезвычайно большой роли
национального парка «Себежский» в сохранении торфяников Псковского
Поозерья. Полученные данные в соответствии с Рекомендацией 6.1
Конференции Сторон Рамсарской конвенции {5] должны учитываться в качестве
дополнительного критерия при обосновании включения национального парка
в перспективный Список Рамсарских угодий [1].
[1] Водно-болотные угодья России, рекомендованные для включения в Список
водно-болотных угодий, охраняемых в соответствии с Рамсарской конвенцией
(«теневой» Список водно-болотных угодий, имеющих международное значение).
Предварительные материалы. 1999. М.: 29-31. [2] Геология СССР. Том I.
Ленинградская, Псковская и Новгородская области. Геологическое описание. 1971. М.:
356-358. [3] Государственные памятники природы Псковской области и правила их
охраны. 1984. Псков: 46-49. [4] Исаченков В.А. 1988. Проблемы морфоструктуры и
древнеледниковой морфоскульптуры. Л.: 101-105. [5] Конвенция о водно-болотных
угодьях .Стратегический план. Задачи и действия 1997-2002 гг. Резолюции и
рекомендации. Документы 6-го совещания Конференции Сторон, том 4.12. 1997. М: 42-
43. [6] Корнилова Р.П. и др. 1968 . Отчет о климатических, гидрогеологических и
грязеразведочных работах, проведенных в 1968 г. в Себежском районе Псковской
области для выявления возможности обеспечения будущей здравницы лечебными
природными ресурсами. М.:181 с. [7] Мусатов В.Ю. 1998. О возможности
использования минерально-сырьевых ресурсов национальных парков для развития
рекреационной инфраструктуры (применительно к условиям Себежского национального
парка) // Проблемы и перспективы сбалансированного развития в бассейне Псков-
362
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
ско-Чудского озера. Мат-лы межд. обществ.-науч. конф. 4.1. Тез. и статьи. Псков:
92-97. [8] Торфяной фонд Псковской области по состоянию разведанности на 1
января 1969 года. 1970. М: 400-467. [9] Список торфяных месторождений Псковской
области, разведанных в 1969-1977 годах. 1978. М.:30-32.
ПРОБЛЕМА ИНВЕНТАРИЗАЦИИ И МОНИТОРИНГА РАМСАРСКОГО
УГОДЬЯ «ПСКОВСКО-ЧУДСКАЯ ПРИОЗЕРНАЯ НИЗМЕННОСТЬ»
В.Ю. Мусатов, В.И. Черевичко1, Л.С. Щеблыкина,
Л.П. Урядова. В.В. Борисов2
Государственный комитет по охране окружающей среды Псковской
области; 2Псковский государственный педагогический университет
Необходимым условием эффективного функционирования объявленных
Российской Федерацией 35 Рамсарских угодий и основой для планирования
в них природоохранных мероприятий является мониторинг динамики
процессов в их экосистемах. Наиболее остро в настоящее время вопрос
сохранения и развития системы мониторинга стоит в тех угодьях, где юрисдикция
Конвенции распространяется не на территории существующих
государственных природных заповедников и национальных парков. Одна из причин
этого — ведомственная разобщенность систем мониторинга.
Такая децентрализованная инфраструктура мониторинга
сформировалась, в частности, в водно-болотном угодье «Псковско-Чудская приозерная
низменность» (площадь 92,4 тыс. га), представляющем собой интеграцию
особо охраняемых природных территорий (ООПТ) федерального и
регионального значения [1,2]. На территории угодья с 1956 г. существует
орнитологическая станция Псковского государственного педагогического
университета, где осуществляется учет и кольцевание птиц. Многолетние
наблюдения на станции позволили установить видовой состав мигрирующих птиц,
закономерности их пролета, выявить редкие и охраняемые виды [5,6,7,9,10,
и др.]. С 1991 г. проводится также комплексный геоморфологический,
гидрологический, флористический и фаунистический мониторинг дельты р.
Великой [3,4]. Псковским отделением ГосНИОРХ более 30 лет ведется
гидрологический, гидрохимический и биологический (фитопланктон,
зоопланктон, зообентос и ихтиофауна) мониторинг Псковско-Чудского озера. В
дополнение к ранее осуществляемым Управлением охотничьего хозяйства
области учетам численности охотничьих видов животных Балтийским фондом
природы Санкт-Петербургского общества естествоиспытателей с 1997 г.
проводится инвентаризация флоры и фауны угодья: исследования
приоритетно ориентированы на выявление мест обитания видов, включенных в
Красные книги России и Балтийского региона.
363
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
СТРУКТУРА МОНИТОРИНГА ВОДНО-БОЛОТНОГО УГОДЬЯ
«ПСКОВСКО-ЧУДСКАЯ ПРИОЗЕРНАЯ НИЗМЕННОСТЬ»
Гос. комитет РФ по охране окружающей среды Администрация Псковской обл.
Государственный комитет по охране окружающей среды Псковской области
(Обобщение представляемой информации; Мониторинг источников
антропогенного воздействия; Эколого-геохимический мониторинг)
• Северо-Западный региональный геологический центр (Локальный мониторинг
подземных вод)
• Псковский центр по гидрометеорологии и мониторингу окружающей среды
(Гидрологический мониторинг)
• Управление лесами Псковской области (Мониторинг лесов)
• Управление охотничьего хозяйства Псковской области (Мониторинг
охотничьих видов животных)
• Псковское отделение ГосНИОРХ (Гидрологический, гидрохимический и
биологический мониторинг Псковско-Чудского озера)
• Псковский гос. пед. ун-т (Комплексный мониторинг дельты р. Великой, Учет
птиц в период миграции)
• Балтийский фонд природы Санкт-Петербургского общества
естествоиспытателей (Инвентаризация флоры и фауны)
Госкомэкологии области осуществляет мониторинг источников
антропогенного воздействия, а также с учетом рекомендаций 6.1 и 6.14
Конференции Сторон Рамсарской конвенции совместно с Северо-Западным
региональным геологическим центром в 1998 г. проведена инвентаризация
торфяников, являющихся доминирующим компонентом угодья [8], а в 1999 г.
планируются эколого-геохимические исследования по оценке степени
загрязнения почв тяжелыми металлами и нефтепродуктами. Вместе с тем
современное финансовое положение государственных служб мониторинга, а
также указанных научных и учебных учреждений, общественных
организаций не гарантирует сохранение и тем более развитие существующей
системы мониторинга.
Ряд вопросов, связанных с организацией и осуществлением мониторинга,
может решаться и решается на уровне субъектов РФ. Так, Госкомэкологии
Псковской области разрабатывается соглашение о создании общего банка
данных о состоянии экосистемы угодья «Псковско-Чудская приозерная
низменность». Но обозначенная проблема не может быть решена без единой
государственной политики на уровне Федерации. Прежде всего, необходимо
решить вопрос сохранения мониторинга федерального уровня (Рослесхоза,
Росгидромета и др.) в уже объявленных угодьях, а также в тех угодьях,
планируемых в ближайшее время включить в Рамсарский Список .
[1] Иванов В.Н., Мусатов В.Ю. Опыт организации особо охраняемых природных
территорий в рамках реализации международных конвенций // Охраняемые
природные территории и объекты Белорусского Поозерья: современное состояние, пер-
364
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА VI
спективы развития. Витебск, 1997. С. 11-12. [2] Иванов А.В., Мусатов В.Ю. Об
эффективности функционирования водно-болотных угодий международного значения
// Проблемы природопользования, экологического воспитания и образования.
Материалы Российско-Белорусской экологической конф. Великие Луки, 1998. С. 55-60.
[3] Лебедева О.А. и др. Состояние экосистемы Псковско-Чудского озера в условиях
прогрессирующего антропогенного воздействия // Краеведение и охрана природы.
Псков, 1993. С. 15-26. [4] Лебедева О.А. и др. Экосистема устья реки Великой //
Проблемы экологии и рационального природопользования Северо-Запада России и
Псковской области. Матер, научно-практической конф. Псков, 1995. С. 52-69. [5]
Мешков М.М. Псковско-Чудской микрорайон на Беломоро-Балтийской пролетной
трассе // Сообщения Прибалтийской комиссии по изучению-миграции птиц 11.1978.
С. 3-11. [6] Мешков М.М., Урядова Л.П. О зависимости пролета птиц от внешних
условий // Сообщения Прибалтийской комиссии по изучению миграций птиц 4.
1967. С. 3-18. [7] Мешков М.М. и др. Инвентаризационные виды птиц на
территории Псковской области // Материалы 9-й Прибалтийской орнитологической конф.
Вильнюс, 1976. С. 152-156. [8] Мусатов В.Ю., Филатов А.Т. Характеристика
торфяников водно-болотного угодья "Псковско-Чудская приозерная низменность" //
Проблемы и перспективы сбалансированного развития в бассейне Псковско-Чудского
озера. Материалы международной общественно-научн. конф. Ч. 1. Тезисы и статьи.
Псков, 1998. С. 89-92. [9] Урядова Л.П. Некоторые особенности миграций
вьюрковых и овсянковых в Псковско-Чудском микрорайоне // Сообщ. Прибалтийской
комиссии по изучению миграций птиц И. 1978. С. 12-23. [10] Щеблыкина Л.С.,
Урядова Л.П. Птицы северной части водно-болотного угодья «Псковско-Чудская
приозерная низменность» // Природа Псковского края 2. 1998. С. 19-21.
БИОРАЗНООБРАЗИЕ ФЛОРЫ БОЛОТ ВОЛОГОДСКОЙ ОБЛАСТИ
А.В. Папанов, Т.А. Суслова, В.И. Антонова,
Вологодский государственный педагогический университет
В Вологодской области болота занимают обширные площади, составляя
10-12 % ее территории. Они располагаются неравномерно в связи с
различиями в рельефе, литологическом составе четвертичных отложений и
гидрологических условий различных ее частей. Сильнее заболочена западная
часть (Молого-Судское междуречье), а восточная — в меньшей степени —
1-2% [1]. В систему охраняемых территорий области входит 260 болот,
общая площадь которых более 361 тыс. га, что составляет 20 % от площади
болот и 2,5 % от всей площади области [2].
Сотрудниками кафедры ботаники Вологодского государственного
педагогического университета определены продуктивность и запасы сырья для
двух видов болотных растений — Ledum palustre и Menyanthes trifoliata,
выявлены редкие растения болот, изучены онтогенез и структура популяций
некоторых видов.
365
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Ledum palustre растет в сосняках багульниково-сфагновых и на верховых
болотах. Проективное покрытие багульника в сосняках колеблется от 5,8±
0,4% до 32,1±4,2%, урожайность воздушно-сухого сырья изменяется от
115,6+10,2 кг/га до 326,4±36,5 кг/га; на верховых болотах эти показатели
составляют 12,7±1,6% и 168,4±15,6 кг/га соответственно. Максимальные
запасы сырья сосредоточены в западных районах — Вытегорском, Устюжен-
ском, Бабаевском, Череповецком, Кадуйском, значительно меньше в
восточных — Никольском, Бабушкинском, Кич-Городецком районах. Как
показали ресурсоведческие исследования биологический запас сырья багульника
болотного в Вологодской области в целом составляет 1240 тонн,
эксплуатационный 310 тонн. Ежегодный возможный объем заготовки сырья, исходя
из периодичности эксплуатации зарослей этого вида, равен 40 тоннам (в
сухом весе). На основании полученных данных Ledum palustre отнесен нами
к первой сырьевой группе лекарственных растений, имеющих в области
обеспеченную сырьевую базу.
Menyanthes trifoliata растет на болотах и заболоченных берегах озер,
входит в состав травяно-болотной группы типов леса. Наиболее
перспективными для заготовки сырья являются низинные и переходные болота. На
низинных болотах в осоково-вахтовых и хвощово-вахтовых ассоциациях
проективное покрытие Menyanthes trifoliata изменяется от 17,5±2,3% до
30,2±4,1 %, а урожайность воздушно-сухого сырья — от 206,2±18,5 кг/га до
411,8±40,5 кг/га; на переходных болотах в сосняках осоково-вахтово-сфаг-
новых эти показатели изменяются от 9,6±1,2% до 45±3,5% и от 85,0+10,0
кг/га до 374,0±28,6 кг/га соответственно. Основные заросли вахты (от 50 до
100 т) выявлены в западных районах области. Чуть беднее сырьевая база
(10-40 т) в Харовском, Тарногском, Бабушкинском районах. В целом в
Вологодской области биологический запас сырья вахты трехлистной
составляет 690 тонн, эксплуатационный — 340 т. Ежегодный возможный объем
заготовки лекарственного сырья, с учетом периодичности эксплуатации
популяций этого вида, составляет 115 т (в сухом весе). На основании этих
данных Menyanthes trifoliata отнесена также к первой сырьевой группе
лекарственных растений.
На болотах зарегистрировано 27 видов редких растений, из них 6 видов
растет на верховых болотах (в), 2 — на верховых и переходных (в, п), 3 —
на переходных (п), 1 — верховых и низинных (в, н), 7 — низинных и
переходных (н, п), 8 — низинных (н).
Для каждого вида определен статус редкости. Три вида Cypripedium cal-
ceolus, Dactylorhiza traunsteineri, Ophrys insectifera — это растения,
занесенные в Красную книгу (!). Dactylorhiza traunsteineri обнаружен на
сфагновых болотах Грязовецкого, Вашкинского районов и на территории
Национального парка (НП) «Русский Север». Ophrys insectifera известна только из
366
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
двух местообитаний. Первое — Бабаевский р-н — окраина топкого
ключевого болота близ д. Конецкий (сведения 1902 года, не подтвержденные
современными находками). Второе — НП «Русский Север» — низинное
болото, юго-восточнее горы Мауры, где сформировалась нормальная
полночленная популяция. Для Cypripedium calceolus зарегистрированы 31
местонахождение, в том числе в заболоченных лесах и на переходных болотах, где он
встречается единично и больших популяций не образует. Остальные 24 вида
имеют областные статусы редкости: вид подлежит охране (ПО), очень
редкий вид (ОР), редкий вид (Р), довольно редкий (ДР). Видовой состав редких
растений болот Вологодской области приведен в табл.
Редкие растения болот Вологодской области
Название вида
Baeothryon alpinum (L.) Egor.
Be tula nana L.
Betulan humilis Schranc.
Blysmus compressus (L.) Panz. ex Line.
Carex pseudocyperus L.
Caltha minor Mill.
Cicuta virosa L.
Cypripedium calceolus L.
Corallorrhiza bifida Chatel.
Dactylorhiza curvifolia (Nyl.) Czer.
D. traunsteineri (Saut.) Soo.
Drosera anglica Huds.
Epipactis palustris (L.) Crantz.
Gymnadenia conopsea (L.) R. Br.
Hammarbya paludosa (L.) O.Kuntze.
Herminium monorchis (L.) R.Br.
Ligularia sibirica (L.) Cass.
Listera cordata (L.) R.Br.
Malaxis monophyllos (L.) Sw.
Ophrys insectifera L.
Pedicularis sceptrum-carolinum L.
Petasites frigidus (L.) Fries.
Rhynchospora alba (L.) Vahl.
Saxifraga hirculus L.
Thelypteris palustris Schott.
Utricularia intermedia Hayne.
U. minor L.
Местообитание
ВД1
в
н,п
н
н
н,п
н,п
п
в,н
п,в
в
в
н,п
н
в
н
н,п
в
н
н
н,п
п
в
н
п
н,п
н
Статус
ПО
ДР
Р
Р
Р
Р
ДР
1
р
ОР
!
Р
ДР
ДР
р
по
по
ОР
р
!
ОР
Р
ОР
ПО
ДР
ДР
ДР
[1] Абрамова Т.Г., Козлова Г.И. Геоботаническое районирование Вологодской
области. // Природное районирование Вологодской области для целей сельского
хозяйства. Вологда, 1970. С. 169-238. [2] Уханов В.П., Сакович А.В. Охраняемые болта. //
367
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Особо охраняемые территории, растения и животные Вологодской области.
Вологда, 1993. С 172-179.
ВЛИЯНИЕ ФРАГМЕНТАЦИИ БОЛОТ НА БИОРАЗНООБРАЗИЕ
(ПТИЦЫ И БАБОЧКИ): ГОЛЛАНДСКИЙ ПОДХОД
М.С. Райнен1, Г.С. Еремкин, P.O. Бутовский, Г.М. Алещенко,
И.М. Есенова, А. Мишустин, Д.М. Очагов2
институт лесного хозяйства и природных исследований, Нидерланды;
2ВНИИприроды, Москва;
Группа российских и голландских специалистов в течение полевого
сезона 1998 г. изучала распределение так называемых ключевых видов,
выбранных из редких охраняемых птиц (11 видов: Tringa glareola, Tringa
nebularia, Larus canus, Lams ridibundus, Grus grus, Tetrao tetrix, Tetrao
urogallus, Picoides trydactyla, Motacilla citreola, Emberiza rustica, Lanius
excubitor) и дневных бабочек (10 видов: Argynnis laodice, Ahchana
melanaria, Boloria' aquilonaris, Boloria euphrosyne, Coenonympha hero,
Coenonympha tullia, Colias palaeno, Euphydryas maturna, Plebejus idas,
Polyommatus optilete) на всех 100 торфяных болотах Петушинского р-на
Владимирской области. Составлялись геоботанические описания.
Использованы также материалы фаунистических исследований и
физико-географических описаний болот, выполненные авторами ранее, топографические и
торфяные карты, материалы космической съемки.
Распределение «жизнеспособных» видов бабочек на торфяных болотах Петушихин-
ского р-на Владимирской области
368
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
Для анализа влияния фрагментации на птиц и бабочек, тесно связанных
с торфяно-болотными экосистемами, использована специальная программа
kARCH, разработанная в Институте лесного хозяйства и природных
исследований. Расчеты, в частности, показали, что среди бабочек максимальное
число жизнеспособных видов, отмеченных в одном биотопе достигает 6, из
них максимум 3 вида имеют высокую «степень жизнеспособности». Обилие
«жизнеспособных» видов бабочек на болотах района приведено на рис.
Для птиц картина несколько иная: максимальное число
«жизнеспособных» видов в одном биотопе составляет 3, из них только 2 вида обладают
высокой «степенью жизнеспособности». Однако необходимо учитывать
«пограничный эффект», связанный с тем, что характерный размер района
исследований сравним со средней дистанцией ежедневных кормовых разле-
тов птиц. Это должно внести коррективы в расчеты.
По результатам исследований подготовлены природоохранные
рекомендации для наиболее ценных торфяно-болотных биотопов северной Мещеры,
направленные на сохранение редких видов животных торфяных болот путем
создания ООПТ.
Проект осуществлен при финансовой поддержке фонда RIN-MATRA.
ВЛИЯНИЕ ХОЗЯЙСТВЕННОЙ ДЕЯТЕЛЬНОСТИ НА ВИДОВОЕ
РАЗНООБРАЗИЕ ЧЕРНООЛЬХОВЫХ СООБЩЕСТВ
ЗАПОВЕДНИКА «ВОРОНИНСКИЙ», ТАМБОВСКАЯ ОБЛАСТЬ
Е.П. Сарычева
Пущинский государственный университет, Московская область
Черноольховые леса Европы во второй половине 20 века претерпели
значительную деградацию вследствие проведения на их территории ряда
хозяйственных мероприятий (осушение, рубки) [3-5]. Это вызывает
некоторые опасения, поскольку черноолынаники относятся к лесным территориям,
характеризующимся значительным видовым разнообразием [1]. Известно,
что черноольховые леса относятся к достаточно устойчивым, климаксным
сообществам и при благоприятных условиях могут существовать на одной и
той же территории неограниченное время, вплоть до тысячелетий (доказано
на торфяных отложениях) [2]. Черноольховые леса Европы в настоящее
время сильно фрагментированы. Литературных данных о современном
состоянии черноольшаников Европейской части России в общем и Тамбовской
области в частности практически нет. Целью работы было оценить
современное состояние черноольховых сообществ заповедника «Воронинский»
(Тамбовская область).
369
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Для изучения флористического богатства черноолыпаников в последних
проводились геоботанические описания. Кроме того, при описании
черноолыпаников изучались особенности семенного и вегетативного
возобновления ольхи. Проводилась экспертная оценка возрастной структуры популяций
других древесных видов: вяза шершавого, дуба черешчатого, осины, березы.
Это было сделано с целью получения предварительной оценки сукцессион-
ного статуса исследованных сообществ.
Все исследованные черноолынаники заповедника относятся к одной
ассоциации — Carici elongatae-Alnetum (по флористической классификации).
Но поскольку ольшаники сильно отличались друг от друга по степени
увлажнения, они все-таки отличались друг от друга и по совокупному набору
видов. В процессе исследования все черноолыпаники были разделены на
три группы в зависимости от степени увлажнения: сухие, сырые и
обводненные черноолынаники. Разделение на эти три группы не случайно,
поскольку именно степень увлажнения, по нашему мнению, определяет
различия в видовом составе травяного яруса черноолыпаников. Кроме
увлажнения сильно влияет на видовой состав хозяйственная деятельность человека,
прежде всего рубки, выпас и сенокошение. Поэтому каждая группа
черноолыпаников была разделена на две подгруппы: сильнонарушенные и слабо-
нарушенные. Но в результате выяснилось, что среди сухих
черноолыпаников встречаются исключительно сильнонарушенные, а слабонарушеннные
не встречаются вообще, поскольку все сухие черноолыпаники интенсивно
используются (или использовались) в хозяйстве. Среди обводненных
ольшаников, напротив, сильнонарушенные не встречаются, поскольку
хозяйственное их использование затруднено по причине высокого уровня воды в
них. Таким образом, были получены следующие группы черноолыпаников:
сухие сильнонарушенные черноолыпаники, сырые сильнонарушенные,
сырые слабонарушенные и обводненные слабонарушенные черноолыпаники.
Для каждой группы ольшаников характерен свой набор видов, поэтому
каждую группу ольшаников можно причислить к определенной ассоциации
(по доминантной классификации). Так, все сухие сильнонарушенные
черноолыпаники можно отнести к ассоциации Alnetum urticosum (ольшаник
крапивный), которая включает следующие субассоциации: Alnetum impatiense-
urticosum (ольшаник недотрогово-крапивный), Alnetum ruboso-urticosum
(ольшаник малиново-крапивный) и Alnetum filipenduloso-urticosum
(ольшаник таволгово-крапивный). Сырые сильнонарушенные ольшаники также
относятся к ассоциации Alnetum urticosum, но поскольку влажность здесь
более высокая, увеличивается обилие водно-болотных видов. Сырые слабо-
нарушенные ольшаники относятся к ассоциации Alnetum varioherbosum
(ольшаники разнотравные). В отличие от сухих и сырых сильнонарушенных
ольшаников в травяном покрове доминирует не один вид, а несколько. 06-
370
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
водненные слабонарушенные ольшаники относятся к ассоциациям Alnetum
varioherbosum (ольшаники разнотравные) и Alnetum caricosum (ольшаники
осоковые). В ольшаниках осоковых, как уже можно судить по названию,
обильны осоки. Часто встречаются Carex vesicaria, Carex pseudocyperus.
Carex elongata встречается реже.
Видовое разнообразие напрямую связано с разнообразием
микроместообитаний [1,4,5]. Важным моментом исследований было следующее: при
проведении хозяйственных мероприятий значительно уменьшается число
микроместообитаний, а, следовательно, уменьшается число
произрастающих на них видов растений. Частые рубки в прошлом привели к тому, что
во многих ольшаниках отсутствовал валеж как удобный субстрат для
семенного возобновления ольхи и многих других видов. Практически во всех
ольшаниках отсутствовали папоротники, хотя, как показали наблюдения в
других заповедниках средней полосы, ольшаники являются обычным
местообитанием для Dryopteris carthusiana, Athyhum filix-femina, Thelipteris pa-
lustris. Значительно редки или совсем не встречались некоторые водно-
болотные, нитрофильные, неморальные виды, обычные для ольшаников.
Мероприятия по изменению гидрологического режима привели к тому,
что многие ольховые местообитания стали более сухими, а, следовательно,
менее подходящими для ольхи. Вместе с ольхой исчезают и пристволовые
повышения ольхи — местообитание многих неморальных и бореальных
видов. Во многих ольшаниках существует тенденция к смене последних
другими типами сообществ. Активно возобновляются вяз шершавый, затем
идут клен американский, дуб черешчатый; клен татарский, осина.
Таким образом, хозяйственная деятельность человека, на территории
заповедника «Воронинский» (Тамбовская область) привела к существенному
уменьшению площадей, занятых черноолыыаниками, а также негативно
сказалась на разнообразии видов растений, характерных для этих лесов.
[1 ] Сарычева Е.П. Пространственная структура и видовое разнообразие черноольхо-
вых сообществ Неруссо-Деснянского Полесья // Ботан. журн. 1998. Т.83. №10. С 66-
71. [2] Marek, S. (1965) Biologia i stratigrafia torfowisk olszynowych w Polsce (Biology
and stratification of alder peatlands in Poland) // Zeszpoly Problemow Postepow Nauk
Roln 57, 5-158. [3] Prieditis, N.(1993) Swamp forests of Latvia: status and conservation.
WWW, Riga. [4] Prieditis, N.( 1997a) Alnus glutinosa - dominated wetland forests of the
Baltic Region: community structure, syntaxonomy and conservation // Plant Ecology 129,
49-94. [5] Prieditis, N.( 1997b) Vegetation of wetland forests in Latvia: a synopsis //
Annales Botanici Fennici 34, 91-108.
371
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ОХРАНЯЕМЫЕ БОЛОТНЫЕ ТЕРРИТОРИИ И БИОРАЗНООБРАЗИЕ
БОЛОТ БЕЛАРУСИ
В.В. Смирнова, А.В. Козулин
Институт проблем использования природных ресурсов и экологии НЛН
Беларуси, Минск; Институт зоологии НАН Беларуси, Минск
Болотные экосистемы являются важным звеном в цепи взаимосвязанных
и взаимодействующих компонентов природы. Болота* как экологические
системы характеризуются специфической флорой и фауной и, как правило,
тесно связаны с прилегающими территориями, поэтому играют важную
роль в водном режиме сопредельных территорий, поддерживая уровни
грунтовых вод. Болота и торфяные месторождения, выполняющие важные
природоохранные функции, включены в заповедники, заказники, национальные
парки и другие охраняемые природные территории.
В Беларуси созданы крупные особо охраняемые территории: —
Государственный национальный парк «Беловежская Пуща», главной целью
которого является сохранение биологического разнообразия, углубленное изучение
лесных растительных сообществ, закономерностей их произрастания; —
Березинский биосферный заповедник, на болотах которого проводятся
работы по изучению и сохранению биогеоценозов Белорусского Поозерья,
исследования по гидрологии, ботанике, зоологии, лесоведению; — Припятский
ландшафтно-гидрологический заповедник организован для изучения
изменений гидрологически замкнутых ландшафтов Полесской низменности.
В Республике Беларусь созданы ботанические заказники, имеющие
статус охраняемых территорий. Общая площадь их составляет 142,7 тыс. га, в
том числе 2,4 тыс. га занимают 33 торфяные месторождения. В основном
это небольшие по размерам мелкозалежные месторождения, на которых
произрастают лекарственные или редкие растения.
На болотах и торфяных месторождениях, выполняющих водоохранную
роль, созданы гидрологические заказники общей площадью 83,0 тыс. га.
Наиболее крупными из них являются Ельня, Дикое, Выгонощанское и др.
Для сохранения дикорастущих ягод созданы ягодные заказники. В
настоящее время в республике имеется 20 заказников-клюквенников общей
площадью 26,5 тыс. га, в том числе 23,5 тыс. га составляет площадь болот.
В Беларуси впервые составлен Красный список болот, куда вошло более
200 объектов. В него включены болота: соответствующие критериям
международной значимости и представляющие собой специфический тип водно-
болотного угодья, редкого или необычного (уникального) для данного
биогеографического региона; входящие в состав заповедников, национальных
парков, заказников различного назначения (гидрологические,
зоологические, ботанические, клюквенники и др.), а также болота и торфяные место-
372
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА
VI
рождения, являющиеся частями озерно-болотных комплексов, и на которых
запрещено изменение водного режима.
Болота и болотные комплексы представляют собой весьма специфичные
и ничем незаменимые природные местообитания. Разнообразие видового
состава растений и животных, обитающих на болотах, невелико, однако
почти все они являются редкими или уязвимыми, так как могут обитать
только на болотах, и сокращение площади болот неизбежно ведет к
сокращению численности, а иногда и к полному исчезновению многих
уникальных видов. В настоящее время в результате мелиорации и торфоразработок
болота сохранились лишь островными участками, поэтому большинство
болотных видов обитает локальными, изолированными друг от друга
группировками.
Растительность белорусских болот имеет богатый флористический
состав, охватывающий 267 видов цветковых и высших споровых растений, в
том числе 37 видов древесных и кустарниковых растений, 167 видов
травяных, 31 вид сфагновых мхов и 32 вида зеленых мхов. На белорусских
болотах произрастает более 50 видов ценнейших лекарственных растений, среди
которых валериана, багульник, подбел и др., а также ягодные растения:
клюква, брусника, голубика, черника. Это разнообразие видов растений в
зависимости от степени обводненности, кислотности среды и других
факторов создает на болотах большое количество фитоценозов - тростниковых,
осоковых, моховых, лесных и т.д.
Растительный покров болот в сочетании с водными окнами создает
благоприятные условия для обитания журавлей, бобров, лосей, кабанов,
боровой и водоплавающей дичи. Состояние значительного числа видов птиц
целиком зависит от состояния болотных экосистем, поскольку большая часть
их популяций обитает именно на болотах. Так, на верховых и переходных
типах болот обитает большая часть белорусской популяции следующих
видов: скопа (Pandion haliaetus). змееяд (Circaetus gallicus), беркут (A quilla
chrysaetos), дербник (Falco columbahus), тетерев (Lyrurus tetrix), белая
куропатка (Lagopus lagopus), золотистая ржанка (Pluvialis aprikaria), фифи
(Tringa glareola), большой улит (Tringa nedularia), средний кроншнеп
(Numenius phaeopus).
Наибольшее значение для сохранения биоразнообразия Европы имеют
открытые низинные болота Полесья. В недавнем прошлом эти болота были
широко распространены в Европе, однако в результате осушительной
мелиорации они сохранились только в регионе Полесья. Большая часть
естественных низинных болот расположена в Беларуси. Одновременно с
сокращением площади болот сокращалась и численность видов, обитавших на этих
болотах. Численность одного из таких видов — вертлявой камышовки
(Acro-cephalus paludicola) снизилась до угрожаемых пределов, и этот вид
373
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
был объявлен глобально угрожаемым в Европе. Более чем 50% европейской
популяции этого вида обитает в Беларуси. Из других видов, популяции
которых зависят от состояния низинных болот, следует отметить болотную
сову (Asio flammeus) и серого журавля (Grus grus). В большой степени от
состояния болот зависят популяции волка, лося и енотовидной собаки.
Значительная часть растений, произрастающих на болотах, также
являются редкими и угрожаемыми видами: это хищные растения росянка,
сфагновые мхи, пушица изящная, клюква болотная, брусника, черника,
голубика, вахта трехлистная.
Особую ценность для сохранения общеевропейских и общепланетарных
популяций болотных видов имеют болотные ландшафты Беларуси. Здесь, в
центре Европы, в естественном состоянии сохранились крупные евтрофные
болота, сильно заболоченные обводненные поймы рек и озерно-болотные
комплексы. Большинство из них расположено на путях миграции водно-
болотных птиц из Европы и Африки на север России и обратно.
Девственные белорусские болота дают пищу и отдых мигрирующим птицам, часть их
остается здесь на гнездование. Только на болотах Полесья гнездится 15%
европейской популяции черного аиста {Ciconia nigra), 9% белого аиста
(Ciconia ciconia). 45% малого подорлика (Aquiila pomarina), 46% погоныша
(Porsana porsana), 59% коростеля (Crex crex), 7% дупеля (Callinago media),
23% травника (Tringa totanus), 57% вертлявой камышовки (Acrocephalus
paludicola) и других.
Согласно международным критериям Рамсарской конвенции и
территорий, особо важных для птиц (IBA), ряд болот Беларуси имеет
международное значение для сохранения биоразнообразия, например, Дикое, Выгоно-
щанское, Ельня и др. Несомненно, что многие болотные угодья Беларуси
представляют собой общеевропейскую и общемировую ценность и должны
быть включены в состав Европейского и Всемирного природного наследия.
ОРГАНИЗАЦИЯ И ВЕДЕНИЕ ЭКОЛОГО-МЕЛИОРАТИВНОГО
МОНИТОРИНГА ОСУШЕННЫХ ЗЕМЕЛЬ
ГОСЛЕСФОНДА БЕЛАРУСИ
Е.И. Шараг
Государственное предприятие «Беллесинвест», Минск, Беларусь
Важнейшая задача лесоводов Беларуси в настоящее время - увеличить
удельный вес лесного сектора в экономике республики, для чего необходимо
добиться максимальной отдачи от каждого гектара лесной площади,
обеспечив при этом непрерывность, неистощительность и относительную
равномерность лесопользования, а также сочетание эколого-природоохранных и
374
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА VI
хозяйственно - экономических функций леса при приоритете первых. В
значительной мере эта задача относится к мелиорированным лесам, площадь
которых в лесном фонде Беларуси составляет около 270 тыс. гектаров. Наши
расчеты показывают, что только за последние 20 лет дополнительное
накопление запаса в этих лесах составило более 8 млн. куб. м древесины.
Приведенная цифра еще раз убедительно подтверждает выводы о том, что
проведение гидролесомелиоративных работ с целью повышения продуктивности
лесных насаждений на избыточно увлажненных землях было обоснованным
мероприятием и в настоящее время имеется объективная необходимость
сохранить освоенный лесомелиоративный фонд республики.
Вместе с тем нельзя отрицать тот факт, что проведение
гидролесомелиоративных работ связано с изменениями в природных процессах лесных
экосистем, которые имеют различные направления. В первую очередь это
изменения в водообменных и почвообразовательных процессах,
отражающиеся на количественных и качественных показателях как водной, так и поч-
венно-грунтовой составляющих. В последующем эти изменения оказали
влияние на состояние древостоев, их продуктивность и товарность, состав
напочвенного покрова и подлеска.
Значительное влияние на состояние лесных экосистем оказала
сельскохозяйственная мелиорация. Безусловно, экологические последствия
сельскохозяйственной и лесной гидромелиорации несопоставимы. При проведении
гидролесомелиорации исключаются существенная сработка органического
вещества, механическое разрушение почвенного профиля в результате куль-
туртехнических и планировочных работ, эрозия почв. Из-за относительно
неглубокой осушительной сети не возникает проблема сбрасывания вековых
запасов природных вод
Особое место в решении вопросов гидролесомелиорации отводится
организации эколого-мелиоративного мониторинга мелиорированных земель в
гослесфонде республики. Совет Министров Республики Беларусь своим
постановлением «О реализации программы Национальной системы
мониторинга окружающем среды в Республике Беларусь» в августе 1998 года
одобрил Технический проект Национальной системы мониторинга окружающей
среды в Беларуси, составной частью которого является эколого-мелиоратив-
ный мониторинг. Его организация позволит проводить регулярные
наблюдения за изменениями избыточно увлажненных почв под влиянием
мелиорации и объективными проявлениями этих процессов в лесных экосистемах.
Основной целью создания системы эколого-мелиоративного
мониторинга является обеспечение информацией соответствующих служб Минлесхоза
и Минприроды о состоянии лесных экосистем и их изменении под влиянием
активной и пассивной лесомелиорации для обеспечения рационального ве-
375
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
дения лесного хозяйства и минимизации отрицательных последствий
мелиоративного воздействия на биогеоценозы.
Основными задачами эколого-мелиоративного мониторинга
мелиорированных земель являются: — организация регулярных наблюдений за
мелиорированными землями и гидролесомелиоративными системами; — изучение
влияния мелиорации на состояние гидрологического режима, почвенного
покрова и лесной растительности; — оценка и прогноз изменения лесорас-
тительных условий в зависимости от параметров осушительной сети и
длительности осушения; — разработка рекомендаций по ведению лесного
хозяйства на мелиорированных землях и контроль их выполнения; —
представление мониторинговой информации по установленным формам и
регламентам в информационно — аналитический центр мониторинга лесной
растительности.
Принципиальное отличие разрабатываемой системы мониторинга от
лесного мониторинга, проводимого в настоящее время по общеевропейской
методике UNEP/ECE, состоит в отсутствии регулярной сети (16 х 16 км; 8 х
8 км) и придании главенствующей роли при размещении сети наблюдений
следующим факторам: — разнообразие природных условий территории
Республики Беларусь; — современное состояние мелиоративных объектов в
гослесфонде республики; — влияние крупномасштабной
сельскохозяйственной мелиорации на лесорастительные условия.
Функционирование мониторинга будет осуществляться при выполнении
следующих требований: — сбор мониторинговой информации
осуществляется на постах, включенных в Государственный реестр пунктов наблюдений
Национальной системы мониторинга окружающей среды (НСМОС),
имеющих реестровый номер, адресную и географическую привязку; —
наблюдения проводятся по обязательному (приоритетному) перечню
контролируемых показателей и дополнительному списку, состав которого подлежит
периодической корректировке; — методическое и методологическое единство
информации, получаемой при ведении мониторинга лесной растительности
в рамках НСМОС; — сбор, хранение и представление информации по
единым формам с соблюдением минимальной избыточности; — применение
единой для мониторинга растительности в составе НСМОС системы
нормативов качества элементов окружающей среды; — выполнение анализа и
обобщений мониторинговой информации, разработка прогнозов с
применением современных методов и технологий; — научное сопровождение работ
осуществляется через государственные и ведомственные научные и научно -
технические программы.
Система мониторинга организуется на 3-х уровнях. 1-уровень —
организация наблюдений за уровнями грунтовых вод, ходом роста
мелиорированных насаждений, состоянием гидролесомелиоративных систем на наиболее
376
БИОРАЗНООБРАЗИЕ, МОНИТОРИНГ, ОХРАНА VI
значимых объектах активной и пассивной гидролесомелиорации. 2-уровень
— организация системы стационаров на мелиорированных землях и неосу-
шенных аналогах для глубокого и всестороннего изучения
причинно-следственных связей между длительностью осушения, параметрами
гидролесомелиоративных систем и состоянием лесорастительных условий. При этом
важной проблемой является разработка методики определения «порога»
экологической надежности гидролесомелиоративной системы, т.е. черты, за
которой ни при каком ожидаемом экономическом эффекте нельзя проводить
гидролесомелиоративные работы, включая реконструкцию осушительных
систем. 3-уровень предполагает решение поставленных задач посредством
применения дистанционных методов съемки, что позволит осуществлять
оперативный контроль за состоянием избыточно увлажненных земель
лесного фонда, включая мелиорированные.
Результаты наблюдений эколого-мелиоративного мониторинга будут
накапливаться в головной организации Минлесхоза, а затем передаваться в
Главный информационный центр Национальной системы мониторинга
окружающей среды Республики Беларусь (БелНИЦ «Экология») для сведения
воедино всей мониторинговой информации. Обмен данными будет
осуществляться средствами связи, используемыми в ведомственных
автоматизированных системах.
377
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Абражко В. И. — 88
Абрамова Т.А. — 48
Агеев Б.А. — 186
Алексеев А.С. — 240
Алещенко Г.М. — 368
Ананьев В.А. — 238
Антоненко A.M. — 90
Антонова В.И. — 365
Бабиков Б.В. — 240
Бабурин А.А. — 310
Багинский В.Ф. — 242, 258
Базанов В.А. — 25
Байкалов А.Н. — 5
Бамбалов Н.Н. — 164
Барсуков B.C. — 92
Бахнов В.К. — 28, 30
Беленькая Т.Я. — 164
Беленький С.Г. — 328
Белова Е.В. — 186, 188
Березин А.Е. — 330
Берестовская Ю.Ю. — 101
Благодарова Т.А. 6
БлейтенВ.В. — 31
Богатырев Л.Г. — 94
Борисов В.В. — 363
Бочкарев Ю.Н. — 116
Булавко Г.И. — 97
Бутовский P.O. — 368
Бышнев И.И. — 135
Вайсман К. — 33
Валетов В.В. — 244
Валяева Н.А. — 42
Васильев СВ. — 31, 35, 99
Васильева Л.В. — 101
Великанов Г.Б. — 39, 246
Величко А.А. — 31
Веселов Н.В. — 40
Волкова Е.М. — 69
Вомперская М.И. — 42
Вомперский С.Э. — 3,42, 166, 182
Воробьева И.Б. — 195
Гаврилов В.Н. — 249
378
АВТОРОВ
Гаджиев И.М. — 31
Галанина О.В. — 46
Гармаш Н.Ю. — 297
Гвоздикова Г.А. — 85
Германова Н.И. — 172
Глаголев М.В. — 175,177
Глебов Ф.З. — 180
ГлуховаТ.В. — 182,230
Головченко А.В. — 103, 106, 142
Голышев С.А. — 177
Горожанкина СМ. — 7
ГрабовикСН. — 109
Гребенников A.M. — 292, 335
Гримашевич В.В. — 338
Даниленко Е.А, — 111, 131
ДегтеваСВ. — 341
Дементьева Т.В. — 186,188
Дмитриев В.Д. — 343
Дорофеева Е.И. — 234
Дубинина СС — 113
Дьяконов К.Н. — 48, 116
Дюкарев А. Г. — 79
Евдокимова Т.В. — 295, 346
Елизарова Т.Н. — 150
Елькина Г.Я. — 209
Еремкин Г. С. — 368
Есенова И.М. — 368
Есимчик Л. Д. — 10
Еськов А.И. — 297
Ефремов СП. — 31,218
Ефремова Т.Т. — 218
Железнова Г.В. — 161, 346
Женихов Ю.Н.--316,348
Заварзин Г.А. — 101
Зазнобин М.Ю. — 42
ЗалитисП.П. — 351
Зеликсон Э.М. — 31
Иванов А.Н. — 50
Иванов В.А. — 295
ИвковичВ.С — 119
ИвковичЕ.Н. —-119
Иматов А.Р. — 287
УКАЗАТЕЛЬ АВТОРОВ
Иматова И.А. — 287
ИнишевН.Г. — 188
Инишева ЛИ. — 186, 188
Ипатьев В.А. — 251
Казанцев В.А. — 150
Калюжный И.Л. — 191
Карпенко Л.В. — 52, 180
Карпов А.И. — 126
Касьянов А.Е. —253
Кириенко О.А. — 299
Кирюшкин В.Н. — 54
Климанов В.А. — 31
КнизеА.А.—255
КобакК.И. — 193
Ковалев А.Г. — 182
КозачокЖ.Н.—209
Козловская О.И. — 55
Козулин А.В. — 372
Козулько Г.А. —353
Колодий П.В. — 258
Колодий Т. А. — 10
Коломыцев В.А. — 13
Кондрашева Н.Ю. — 193
Коновалова Т.И. — 195
Константинов В.К. — 261
Копотева Т.А. — 58
Корепанов А. А. — 264
Корепанов С.А. — 264
Косарев В.П. — 198
Косицын В.Н. — 156, 313, 318
КосовВ.И. —316
Косых Н.П. — 121
Кравченко И.К. — 201
Красильников Н.А. — 261, 266
Краснова А.Н. — 15
Кременецкий К.В. — 331
Крештапова В.Н. — 204
Криволуцкий Д. А. — 124
Крупное Р.А. — 333
Крылова Е. Г. —61
Крючкова Г.А. — 122
Кузнецова Е.Г. — 346
КузовлевВ.В. — 316, 348
Кузьмин В.А. — 207
Кузьмин Г.Ф. — &3
Кузьмичев А.И. — 65
Купцова В.А. — 158
КурловичЛ.Е. — 318
Лаптева Е.М. — 209
Лаптенок В.В. — 5
Лапшина Е.Д. — 31
Ласкова Л.М. — 124, 126
Лисе О.Л. — 66, 69
Ловелиус Н.В. — 343
ЛыткинИ.И.—292,335
Льготин В.А. — 25
Магаева Л. А. — 150
Макаренко Г.Л. — 70
Малышева Н.Н. — 122
Малясова Е.С. — 72
Мамаева Л.К. — 212
Марыганова В.В. — 164
Маслов А. А. — 127
Маслов Б.С. — 301
Матвеева М.А. — 287
Матюшкин Е.Н. — И, 131
Милицына О. А. — 212
Минаева Т.Ю. — 356
Мишустин А. — 368
Молчанов А.Г. — 215
Мусатов В.Ю. — 360, 363
Наумов А.В. — 218
Нешатаев В.Ю. — 17. 226
Нешатаева В.Ю. — 17
Новиков СМ. — 72. 219
Носаль А.П. — 223
Орлов Д.С. — 225
Орлов М.С. — 85
Острошенко В.В. — 268, 271
Очагов Д.М. — 368
Папанов А.В. — 365
Панов В.В. — 40
Панова Н.К. — 76
Пахучий В.В. — 273
ПелепецН.В. — 92
Перегон A.M. — 99
ПетерсонЮ.В. — 127
Пологова Н.Н. — 25, 79
Полянская Л.М. — 106
Попов М.В. — 333
Прокуронов И.Б. —133
Пузаченко Ю.Г. — 48
379
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
ПыстинаТ.Н. — 341
РавкинЕ.С. — 135
Райнен М.С — 368
РаковичВ.А. — 164
Русаленко А.И. — 276
Русецкас Ю. — 278
Рябинина Н.А. — 5
Сабо Е.Д.—280
Савченко А.П. — 5
Садовникова Л.К. — 225
СаковецВ.И. — 172,282
Сапожникова В. А. — 94
Сарычева Е.П. — 369
Сенникова М.М. — 20
Сеньков А.А. — 227
Сиволапов А.И. — 6
Сирин А.А. — 20, 33. 356
Славгородский А. В. — 138
СмагинА.В. — 140,230
Смагин В.А. — 307
Смагина М.В. — 140, 142, 182, 230
Смирнова В.В. — 372
СныткоВ.А. — 21,233
Солдатов М.С. — 126
Солнцев В.Н. — 33
Суслова Т.А. — 365
Сухова И.В. — 225
Тараканов A.M. — 284
Терехова В.А. — 142
Тимофеев А.И. — 198
Тимофеев А.Ф. — 22
Токарева Е.В. — 201
Толпышева Т.Ю. — 92. 141
Трофимов С.Я. — 234
Трофимова И.Е. — 147
Турчинович И.Е. — 193
Урядова Л.П. — 363
Усова Л.И. — 72
Устинов М.Т. — 150
Федоров Ю.С. — 152
Фирсов СЮ. —177
Хабибуллина Ф.М. — 154
ХитринС.В. — 261
Хорошев А.В. — 48, 82
Хорошев П.И. — 204
Цареградская СЮ. — 156
Цветкова И.В. — 85
ЧаковВ.В. —158,321
Черевичко В.И. — 363
ЧиндяевА.С—287
Шамрикова Е.В. — 209
Шараг Е.И. — 374
ТТТитттко А.А. — 164
Шубина Т.П. —161
Шурыгин СГ. — 290
Щеблыкина Л.С — 363
Щетников А.И. — 233
Юрковская Т.К. — 46
Яковлева Е. — 324
380
АДРЕСА ОРГАНИЗАЦИЙ
АДРЕСА ОРГАНИЗАЦИЙ
АООТ НИИ торфяной промышленности (НИИТП) — 191041, Санкт-Петербург,
Марсово поле, 5
Ассоциация гидролесомелиораторов «ГИЛМ» — 194021, Санкт-Петербург,
Институтский просп., 21
Белорусский государственный технологический университет — 220680, Беларусь,
Минск, ул. Свердлова, 13-а
Белорусский государственный университет — 220733, Беларусь, Минск, ул.
Академическая, 27
Березинский биосферный заповедник — 211188, Беларусь, Витебская обл., Лепель-
ский р-н, п. Домжерицы
Ботанический институт им.В.Л.Комарова РАН — 197376, Санкт-Петербург, ул.
Проф. Попова, 2
Ботанический сад УрО РАН — 620134, Екатеринбург, ул. Билимбаевская, 32а
Вологодский государственный педагогический университет — 166000, Вологда, ул.
С. Орлова, 6
Воронежская государственная лесотехническая академия — 394613, просп.
Тимирязева, 8
Всемирный фонд дикой природы (WWF), Российское представительство — 125319,
Москва, п/я 55
Всероссийский научно-исследовательский и информационный центр по лесным
ресурсам (ВНИИЦлесресурс) — 117418, Москва, ул. Новочеремушкинская, 69-а
Всероссийский научно-исследовательский конструкторский и проектно-техноло-
гический институт органических удобрений и торфа (ВНИПТИОУ) — 601242,
Владимирская обл., Судогодский р-н, и/о Вяткино
Всероссийский НИИ лесоводства и механизации лесного хозяйства (ВНИИЛМ) —
141200, Московская обл., г. Пушкино, ул. Институтская, 15
Всероссийский НИИ охраны природы и заповедного дела (ВНИИприроды) —
113628, Москва, Знаменское—Садки
Вятская государственная сельскохозяйственная академия — 610000, Киров,
Октябрьский просп., 133
Государственный гидрологический институт (ГГИ) — 199053, Санкт-Петербург,
В.О., 2-ая линия, 23
381
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Государственный комитет по охране окружающей среды Псковской области —.
180001, Псков, ул. Некрасова, 23
Государственный национальный парк (ГНП) «Беловежская Пуща» — 225063,
Беларусь, Брестская обл., Каменецкий р-н, п/о Каменки
Государственное предприятие «Беллесинвест» — 220002, Беларусь, Минск, ул. Вар-
вашени, 77
Государственный педагогический институт им. Н.К. Крупской — Беларусь,
Гомельская обл., г. Мозырь, ул. Студенческая, 28
Институт биологии внутренних вод им. И.Д.Папанина РАН — 152742, Ярославская
обл., Некоузский р-н, пос.Борок
Институт биологии Карельского НЦ РАН — 183610, Петрозаводск, ул. Пушкинская,
11
Институт биологии Коми НЦ УрО РАН — 167610, Сыктывкар, ул.
Коммунистическая, 28
Институт водных и экологических проблем (ИВЭП) ДВО РАН — 680000,
Хабаровск, ул. Ким-Ю-Чена, 65
Институт географии РАН — 109017, Москва, Старомонетный пер., 29
Институт географии СО РАН — 664033, Иркутск, ул. Уланбаторская, 1
Институт ЗапСибгипроводхоз — 630087, Новосибирск, ул. Немировича-Данченко,
167
Институт зоологии НАН Беларуси — 220072, Беларусь, Минск, Академическая, 27
Институт комплексного анализа региональных проблем (ИКАРП) ДВО РАН —
682200, Биробиджан, ул. Шелом Алейхема, 4
Институт леса НАН Беларуси — 246654, Беларусь, Гомель, ул. Пролетарская, 71
Институт леса Карельского НЦ РАН — 185610, Петрозаводск, ул. Пушкинская, 11
Институт леса и древесины им. В.Н. Сукачева СО РАН — 660036, Красноярск,
Академгородок
Институт лесного хозяйства и природных исследований — Institute for forestry and
nature research (IBN-DLO) — Postbus 23, 6700 AA, Wageningen, The Netherlands
Институт лесоведения РАН — 143030, Московская обл., Одинцовский р-н, п/о
Успенское
Институт микробиологии РАН — 117811, Москва, просп. 60-летия Октября, 7, к. 2
Институт оптики атмосферы СО РАН — 634055, Томск, просп. Академический, 1
Институт почвоведения МГУ-РАН — 119899, Москва, Воробьевы Горы, МГУ
382
АДРЕСА ОРГАНИЗАЦИЙ
Институт почвоведения и агрохимии СО РАН — 630099, Новосибирск, ул.
Советская, 18
Институт проблем использования природных ресурсов и экологии НАН Беларуси —
220114, Беларусь, Минск, Староборисовский тракт, 10
Институт проблем экологии и эволюции РАН — 117071, Москва, Ленинский пр., 33
Институт социально-экономических и энергетических проблем Севера УрО РАН —
167610, Сыктывкар, ул. Коммунистическая, 26
Институт устойчивых сообществ — 117312, Москва, ул. Губкина, 14, оф.75-76
Комитет по лесу Ленинградской области — 197046, Санкт-Петербург, ул.
Куйбышева, 21
Красноярский государственный университет — 660041 Красноярск, просп.
Свободный, 79
Латвийский институт лесоведения «Силава» — Latvian Forestry Research Institute
«Silava» — LV-2169, Latvia, Salaspils, Rigas iela, 111.
Литовский институт леса — Lithuanian Forest Research Institute — LT-4312,
Lithuania, Kaunas-Girionys
Московский городской дворец творчества детей и юношества — 117978, Москва,
ул. Косыгина, 17
Московский государственный университет (МГУ) им. М.В. Ломоносова — 119899,
Москва, Воробьевы горы
Московский государственный университет леса — 141001, Московская обл., г.
Мытищи, Институтская ул., 1
Мюнхенский технический университет — Technische Universitat Mtinchen, Lehrstuhl
fur Vegetationsokologie — D 85350 Freising-Weihenstephan, Am Hochanger 6,
Germany
НИИ биологии и биофизики при Томском государственном университете — 634050,
Томск, просп. Ленина, 36
НИИ лесной генетики и селекции — 394087, Воронеж, ул. Ломоносова, 105
Нижегородская государственная сельскохозяйственная академия — 603107, Нижний
Новгород, просп. Гагарина, 78.
Объединенный эколого-технологический НИЦ по обезвреживанию радиоцонных
отходов и охране окружающей среды (МосНПО «Радон») — 119121, Москва,
Ростовский 7-й пер., 2/14
Почвенный институт им. В.В.Докучаева — 109017, Москва, Пыжевский пер., 7
Псковский государственный педагогический университет — 180000, Псков, ул.
Советская, 21
383
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Путинский государственный университет — 142292, Московская область, г. Пущи-
но, просп. Науки, 3
Российская академия сельскохозяйственных наук (РАСХН) — 11J218, Москва, ул.
Кржижановского, 15, кор.2
Российский государственный педагогический университет (РГПУ) им. А. И. Герцена
— 190000, Санкт-Петербург, наб. Мойки, 48
Российский НИИ комплексного использования и охраны водных ресурсов
(РОСНИИВХ) — 620049, Екатеринбург, ул. Мира, 23
Санкт-Петербургская государственная лесотехническая академия — 194021, Санкт-
Петербург, Институтский просп., 5
Санкт-Петербургский НИИ лесного хозяйства (СПбНИИЛХ) — 194021, Санкт-
Петербург, Институтский просп., 21
Северный НИИ лесного хозяйства — 163062, Архангельск, ул. Никитова, 13
Сибирский научно-исследовательский проектно-технологический институт
комплексного использования торфа (СибНИИТ) СО РАСХН — 634050, Томск, ул.
Гагарина, 3
Тверской государственный университет — 170000, Тверь, ул. Желябова, 33
Тверской государственный технический университет — 170026, Тверь, наб.
Афанасия Никитина, 22
Томский государственный университет — 634050, Томск, просп. Ленина, 36
Тульский государственный педагогический университет им. Л.Н.Толстого —
300026, Тула, просп. Ленина, 125
ТЦ «Томскгеомониторинг» — 634000, Томск, пер. Совпартшкольный, 13
Университет Утрехт — University Utrecht — P.O. box 80.115, 3508 ТС Utrecht, The
Netherlands
Уральская государственная лесотехническая академия — 620067, Екатеринбург, ул.
Сибирский тракт, 37
Федеральная служба лесного хозяйства Российской Федерации — 113184, Москва,
Пятницкая ул., 5 9/19
Филиал института леса им. В.Н. Сукачева СО РАН — 634021, Томск, пр.
Академический, 2
Центрально-Лесной государственный биосферный заповедник — 172513, Тверская
обл., Нелидовский район, и/о Заповедник
Чумиканский лесхоз — 682460, Хабаровский край, Тугуро—Чумиканский р-н, с.
Чумикан, ул. Северная, 5
384
СОДЕРЖАНИЕ
СОДЕРЖАНИЕ
Предисловие 3
L Распространение, географические особенности болот, заболоченных
земель и лесов 5
Байкалов А.Н., Савченко А.П., ЛаптенокВ.В., РябининаНА. Гагульская
котловина — уникальное водно-болотное угодье Западного Саяна 5
Благодарова ТА, Сиволапов АИ. Ольшаники на заболоченных
местоположениях Центральной лесостепи России 6
Горожанкына СМ. Географическая специфика болотного покрова в приени-
сейской Сибири 7
Есимчик Л.Д., Колодий ТА. Таксационная характеристика избыточно
увлажненных лесов Беларуси 10
Коломыцев В А. Зонально-климатические и ландшафтно-типологические
особенности распространения заболоченных лесов Карелии 13
Краснова АН. Болота приозерных ландшафтов Северо-Двинской водной
системы 15
Нешатаева В.Ю., Нешатаев В.Ю. Болота Южнокамчатского федерального
заказника -,т 17
Сенникова М.М., Сирин А.А. Заболоченность территории на
топографических картах разного масштаба (на примере участка центра ЕТР) 20
Снытко В А. Болотные фации Тункинской котловины (юго-западное
Прибайкалье): влияние потухших вулканов 21
Тимофеев А. Ф. Болотные земли Кировской области и их хозяйственное
использование 22
Ц. Генезис, динамика болотообразования (в прошлом, настоящем) 25
Базанов В А., Полозова Н.Н., Льготин В А. Об изучении динамики болото-
образовательного процесса в таежной зоне Западной Сибири 25
Бахнов В.К. Древнее болотное почвообразование и его роль в развитии
биосферы 28
Бахнов В.К. К вопросу о происхождении грядово-мочажинных комплексов.... 30
Блейтен В.В., Лапшина Е.Д., Величко А А., Гаджиев И.М., Васильев СВ.,
Ефремов СП., Кременецкий КВ., Климате В А., Зеликсон Э.М.
Торфяные болота Западной Сибири (проект российско-голландского
сотрудничества) 31
Вайсман К, Сирин АА, Солнцев В.К Устойчивость болот к изменению
климата: возможные напрвления анализа 33
Васильев СВ. О природе грядово-мочажинных комплексов 35
Великанов Г.Б. Масштабы современного заболачивания земель лесного
фонда Ленинградской области 39
Веселое Н.В., Панов В.В. Реконструкция развития котловинного болота
южной тайги 40
Вомперский С.Э., Вомперская М.И., Валяева НА., Зазнобин М.Ю. Коротко-
периодные колебания оторфованности заболоченных лесов 42
385
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Галанина О.В., Юрковская Т.К. Картографические модели состояния
растительного покрова сфагновых мезоолиготрофных болот 46
Дьяконов К.Н., Пузаченко Ю.Г. Абрамова Т.А., Хорошев А.В. Голоценовая
история, факторы и скорости заболачивания среднетаежного структурно-
эрозионно-ледникового ландшафта (юг Архангельской области) 48
Иванов АЛ. Болотные геосистемы Центральной Мещеры и их
взаимоотношения с прилегающими территориями 50
Карпенко Л.В. Реконструкция лесной и болотной растительности за
последние 9500 лет среднетаежной части Приенисейской Сибири 52
Кирюшкин В.Н. Устойчивость болотных систем 54
Козловская СИ. Характеристика процессов заболачивания мелководий на
водохранилищах верхней Волги 55
Копотева ТА. Пирогенный фактор и динамика растительного покрова болот
Приамурья 58
Крылова Е.Г. Заболачивание техногенно трансформированных пойменных
водоемов в южной тайге 61
Кузьмин Г. Ф. Динамика болотообразовательного процесса в голоцене на
Северо-Западе России и некоторые задачи управления
природопользованием 63
Кузьмичев А.И. Генезис и фитогеографические особенности черноольхового
комплекса 65
Лисе О.Л. Важнейшие направления современного болотоведения 66
Лисе О.Л., Волкова Е.М. Болотные формы сосны обыкновенной как
индикаторы условий произрастания 69
Макаренко Г.Л. Накопление хорошо- и сильноразложившегося верхового
торфа в зависимости от времени начала болотообразования 70
Новиков СМ., Усова Л.И., Малясова Е.С. Возраст и динамика болот
Западной Сибири 72
Панова Н.К Динамика лесной растительности и болотообразовательного
процесса в голоцене в горах Среднего Урала 76
Полозова Н.Н., Дюкарев А.Г. Автономное заболачивание кедровых лесов
Западной Сибири 79
Хорошев А.В. Конкуренция олиготрофного и эвтрофного заболачивания в
таежном ландшафте с блоковой структурой 82
Цветкова И.В., Гвоздикова Г.А., Орлов М.С. Гидрогеологические условия
водно-болотного комплекса верховьев р.Яузы 85
III. Структурно-функциональная организация, первичная
продуктивность болот и заболоченных лесов 88
Абражко В. И. Сравнительная оценка водного режима заболоченных
ельников 88
Антоненко A.M. Торфяники Южного побережья оз.Байкал: микрофлора и
биохомическая активность 90
Барсуков B.C., ПелепецН.В., Толпышева ТЮ. Лишайники заболоченных
территорий Окского заповедника 92
386
СОДЕРЖАНИЕ
Богатырев Л.Г., Сапожникова В. А. Соотношение гумусообразования и тор-
фонакопления в заболоченных лесах средней и северной тайги 94
Булавко Г.И. Комплекс микроскопических грибов в болотах Северной тайги. 97
Васильев СВ., Перегон А.М. Радиальный прирост сосны обыкновенной на
болотах и суходолах 99
Васильева Л.В., Берестовская Ю.Ю., Заварзин ГА. Психрофильные
ацидофильные метанотрофы в сфагновых болотах 101
Головченко А.В. Особенности пространственного распределения и таксоно-
метрического состава сапротрофного комплекса бактерий в болотно-
лесных экосистемах 103
Головченко А.В., Полянская Л.М. Жинзеспособность мицелия и спор грибов
в торфяниках 106
Грабовик С.Н. Особенности формирования березовых сообществ на мезо-
трофных травяно-сфагновых болотах под влиянием осушения 109
Даниленко Е.А., Матюшкин Е.Н. Население птиц заболоченных лесов Кос-
томукшского заповедника 111
Дубинина С. С. Продуктивность травяного болота Назаровской лесостепи 113
Дьяконов КН., Бочкарев ЮЛ. Внутривековая динамика радиального
прироста деревьев в связи с циклами солнечной активности 116
Ивкович B.C., Ивкович Е.Н. Состояние и динамика лесной растительности на
естественных болотах Березинского заповедника 119
Косых Н.П. К методике определения линейного прироста и продукции
сфагновых мхов на мезо-олиготрофных болотах Западной Сибири 121
Крючкова ГА., Малышева Е.Н. Содержание радионуклидов в растениях
нижних ярусов заболоченных лесов 122
Ласкова Л.М., Криволуцкий Д.А. Влияние степени осушения торфянистых
почв на фауну панцирных клещей 124
Ласкова Л.М., СолдатовМ.С, /^«овЛ.Я/Субфоссильныепанцирнью
клещи Калужской области 126
Маслов А.А., Петерсон Ю.В. Циклические смены древостоя на верховом
болоте: анализ причин и последствий частичной гибели сосны 127
Матюшкин Е.Н., Даниленко Е.А. Население птиц открытых и малооблесен-
ных болот Костомукшского заповедника 131
Прокуронов И.Б. О структуре коренного ельника на мезоолиготрофном
болоте южной тайги 133
Равкин Е.С., Бышнев И.И. Население птиц болот юга лесной зоны Восточно-
Европейской равнины 135
Славгородский А.В. Структура растительного покрова на границе лес-водоем
в поймах рек Океко-Донской равнины 138
Смагина М.В., Смагин А.В. Экспериментальная оценка кинетики
формирования лесной подстилки в осушенном сосняке низинного типа
заболачивания 1^0
Терехова В.А., Головченко А.В., Смагина М.В. Влияние осушительной
мелиорации на структуру микобиоты в почвах низинного и верхового типов
заболачивания 142
387
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Толпышева Т.Ю. Лишайники западносибирских болот 145
Трофимова И.Е. Роль климата в современной структурно-функциональной
организации болот Западной Сибири 147
Устинов М.Т., Елизарова Т.Н., Казанцев В.А., МагаеваЛА. Мелиоративная
экология болотных почв Западно-Сибирской низменности 150
Федоров Ю.С Трансформация травяного яруса гидроморфных сообществ в
связи с осушением и сезонными колебаниями погоды 152
Хабибуллина Ф.М. Видовой состав и структура микромицетов болотно-под-
золистыхпочв 154
Цареградская С.Ю., Косицын В.Н. Оценка состояния растительного покрова
после сильного торфяного пожара 156
Чаков В.В., Купцова В.А. Особенности естественного восстановления
сфагнового покрова в заболоченных лиственничниках Приамурья после
проведения мелиоративных работ 158
Шубина ТИ.^Железнова Г.В. Флористический состав листостебельных мхов
болот Мезенеко-Вычегодской равнины 161
IV. Биосферная роль болот. Биогеохимические циклы веществ
естественных и эксплуатируемых болотных экосистем..» 164
Бамбалов Н.Н., Раковин В.А. Марыганова В.В., Беленькая Т.Я., ШишкоАА.
Биохимичесике циклы азота и углерода в естественных и антропогенно
нарушенных болотных экосистем 164
Вомперский С.Э. Биосферная роль болот, заболоченных лесов и проблемы
их устойчивого использования 166
Германова Н.И., Саковец В.И. Разложение опада и эффективность шдроле-
сомелиорации в Карелии 172
Глаголев М.В. Математическое моделирование эмиссии метана из болот в
атмосферу 175
Глаголев М.В., Голышев С.А., Фирсов СЮ. Оценка переноса метана из
почвы в атмосферу болотными растениями.., 177
Глебов Ф.З., Карпенко Л.В. Годичная цикличность углерода в болотном лесу
как индикатор коренного различия между лесом и болотом 180
Глухова Т.В., Ковалев А.Г., СмагинаМ.В., Вомперский С.Э. Оценка
некоторых биотических компонентов углеродного цикла болот и лесов 182
Инишева Л.И., Белова Е.В., Дементьева ТВ., Агеев Б.Г. Кинетика
разложения органического вещества торфов в искусственных анаэробных
условиях 186
Инишева Л.И., Дементьева ТВ., Белова Е.В., Инишев Н.Г. Эмиссия СОг из
болот южно-таежной подзоны Западной Сибири 188
Калюжный И.Л. Химический состав, вынос минеральных и органических
компонентов водами олиготрофных болотных массивов 191
Кобак К.И., Кондрашева Н.Ю., Турчинович И.Е. Аккумуляция углерода в
болотах Ленинградской области 193
Коновалова Т.Н., Воробьева И.Б. Роль болотных экосистем в динамике
ландшафтов урбанизированных районов Приангарья 195
388
СОДЕРЖАНИЕ
Косарев В.П., Тимофеев А.И. Режим почвенно-грунтовых вод осушенных
мелких торфяников стационара Малиновский 198
Кравченко И.К., Токарева Е.В. Влияние азотных соединений на окисление
метана в верховом болоте, Тверская область 2W
Крештапова В.К, Хорошев П.И. Запасы углерода в торфяных залежах
Томской области 204
Кузьмин В.А. Химический состав торфяников и снега Южного Прибайкалья
как показатель техногенного воздействия на окружающую среду 207
Лаптева ЕМ., Елькина Г.Я., ШамриковаЕ.В., КозачокЖ.Н. Особенности
формирования органического вещества болотно-подзолистых почв 209
Мамаева Л.К., Милицына О.А. Взаимодействие металлов сточных вод с
торфяными почвами естественных болот 212
Молчанов AT. Фотосинтетическая продуктивность заболоченного и
суходольного сосновых древостоев 215
Наумов А.В., Ефремова Т.Т., Ефремов СП. Продуцирование СОгторфяной
повой слабо осушенного мезотрофного болота в связи с
гидротермическими условиями сезона 218
Новиков СМ. Водообмен болот с подстилающими минеральными грунтами.. 219
Носаль АЛ. Приближенная оценка самоочищающей способности болот в
отношении сточных вод 223
Орлов Д.С, Садовникова Л.К., Сухова И.В. Особенности органического
вещества осушенных торфянисто-подзолисто-глеевых почв Ярославской
области 225
Сеньков А.А. Особенности химического состава природных вод лесоболот- 227
ных комплексов Сибирских Увалов, Западная Сибирь
Смагин А.В., Смагина М.В., Глухова ТВ. Потоки, эмиссия и генерирование
парниковых газов в заболоченных почвах 230
СныткоВА., Щетников А.И. Почвенно-геохимические процессы в
сопряженных фациях заболоченного леса 233
Трофимов С.Я., Дорофеева Е.И. Некоторые особенности органического
вещества почв разного типа заболачивания 234
V. Проблемы устойчивого использования болот. 238
V. 1. Гидромелиорация и лесное хозяйство. 238
Ананьев В.А. Главное и промежуточное пользование в осушенных лесах
Карелии , 238
Бабиков Б.В., Алексеев А.С Влияние температуры и осадков за
вегетационный сезон на сток с осушенных лесных земель 240
Багинский В.Ф. Основные требования к технологии рубок главного
пользования в древостоях, проюрастающих на избыточно увлажненных землях
Беларуси 242
Валетов В.В. Влияние осушительной мелиорации на биопродуктивность
сосновых олиготрофных болот севера Беларуси 244
Великанов Г.Б. О планировании и производстве гидролесомелиоративных
работ в Ленинградской области... 246
389
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Гаврилов В.Н. Состояние и рост культур сосны на осушенном переходном
болоте в южной Карелии 249
Ипатьев В А. Гидролесомелиорация как метод регулирования накопления
радионуклидов древесными растениями 251
Касьянов А.Е. Способ определения оптимального расстояния между
осушителями 253
Книзе АА. Влияние лесоосушительной мелиорации на биоразнообразие 255
Колодий П.В., Багинский В.Ф. Рубки главного пользования в заболоченных
лесах Беларуси: проблемы и пути решения 258
Константинов В.К, Красильников Н.А., Хитрин СВ. Современное
состояние гидролесомелиорации: актуальные практические и научные задачи... 261
Корепанов А.А., Корепанов С А. Лесоводственная эффективность осушения
сосняков Быстрицкого лесоболотного массива 264
Красильников НА., Нешатаев В.Ю. Типы леса на осушенных торфяных
почвах северной Беларуси 266
Острошенко В.В. Лесоводственная эффективность осушения заболоченных
березняков на Дальнем Востоке 268
Острошенко В.В. Рост лесных культур на периодически избыточно
увлажненных почвах Приохотья 271
Пахучий В.В. Опыт количественной оценки динамических состояний и
устойчивости сосновых насаждений на объектах гидромелиорации 273
Русаленко А.И. Повышение продуктивности лесов на почвах избыточного
увлажнения -.-.■ 276
Русецкас Ю. Влияние растительности берм и откосов канав на состояние
лесной осушительной сети 278
Сабо Е.Д. Методика оценки влияния гидромелиорации на суходольные
насаждения 280
Саковец В.И. Эффективность Десоосушения в Карелии по материалам
лесоустройства 282
Тараканов AM. Имитационная модель для прогнозирования технического
состояния осушительной сети и продуктивности древостоев 284
Чиндяев А.С, Ищтов А.Р., Иматова И.А., Матвеева МА. Лесохозяйствен-
ная эффективность гидролесомелиорации на Среднем Урале 287
ШурыгинСГ. Сток с осушенных лесных земель 290
V.2. Использование болот и торфа в сельском хозяйстве. 292
Гребенников AM., Лыткин И.И. Оценка целесообразности
сельскохозяйственного освоения выработанных торфяников в Орехово-Зуевском
районе Московской области 292
Евдокимова Т.В., Иванов В А. Перспективы использования болот и
заболоченных лесов Республики Коми для развития оленеводства 295
Еськов А.И., Гармаш Н.Ю. Основные направления реструктуризации
сельскохозяйственного торфопользования РФ в связи с задачами устойчивого
развития 297
Кириенко О А. Последствия осушительной мелиорации лугово-глеевых и
торфяных почв Среднеамурской низменности 299
390
СОДЕРЖАНИЕ
Ыаслое Б.С Современные вопросы мелиорации болот для
сельскохозяйственных целей 301
Смагин В.А. Воздействие стока от свинофермы на растительность верхового
болота 307
V.3. Пищевые и лекарственные ресурсы болот. 310
Бабурин А А. Запасы сырья багульника болотного на территории Гассинско-
го Модельного Леса (Хабаровский край) 310
Косицын В.Н. Использование ресурсного потенциала морошки на
европейском Севере России 313
Косое В.И., Женихов Ю.Н., Кузовлев В.В. Создание регионального банка
данных ресурсов торфяно-болотных экосистем 316
Курлович Л.Е., Косицын В.Н. Продуктивность ягодных и лекарственных
растений на осушенных лесных болотах в Тверской области 318
Чаков В.В. Типология и ресурсный потенциал болотных экосистем
Приамурья 321
Яковлева Е. Классификация и современное использование ресурсов
болотных экосистем Еврейской автономной области 324
V.4. Рекультивация и использование выработанных торфяников. 328
Беленький СГ. Критерии выделения выработанных торфяных
месторождений для повторного заболачивания 328
Березин А.Е. К вопросу о рекультивации земель на болотах, нарушенных при
буровых работах 330
Крупное Р.А., Попов М.В. Особенности лесоразведения на выработанных
торфяных месторождениях озерного генезиса 333
Лыткин И.И., Гребенников AM. Относительная устойчивость к деградации
антропогенно-нарушенных болотных экосистем при их окультуривании... 335
VL Биоразнообразие естественных и мелирированных болот и
заболоченных лесов, мониторинг и охрана 338
Гримашевич В.В. Биологическое разнообразие пищевых и лекарственных
растений болот Беларуси 338
Дегтева СВ., Пыстина Т.Н. Редкие растения заболоченных березняков
Республики Коми 341
Дмитриев В.Д., Ловелиус Н.В. О рациональном использовании и сохранении
болот и заболоченных лесов Камчатки в XXI веке 343
Евдокимова Т.В., Железнова Г.В., Кузнецова Е.Г. Восстановление
биоразнообразия растительности на нарушенных нефтедобычей заболоченных
территориях Республики Коми 346
Женихов Ю.Н., Кузовлев В.В. Состояние и задачи охраны естественных
болот в связи с проблемой устойчивого развития Тверской области 348
Залитис П.П. Многообразие осушенных лесов Латвии .. 351
Козулько Г.А. Роль заболоченных сосняков в сохранении биологического
разнообразия беспозвоночных 353
Минаева Т.Ю., Сирин А.А. Охрана болот в России: состояние и перспективы. 356
Мусатов В.Ю. Роль национального парка «Себежский» в сохранении торфяг
ников Псковского Поозерья 360
391
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
Мусатов В.Ю., Черевичко В.И., Щеблыкина Л.С., Урядова Л.П., Борисов
В.В. Проблема инвентаризации и мониторинга Рамсарского угодья
«Псковско-Чудская приозерная низменность» 363
Папанов А.В., Суслова Т.А., Антонова В.И. Биоразнообразие флоры болот
Вологодской области 365
Райнен М.С., Еремкин Г.С., Бутовский P.O., Алещенко Г.М., Есенова ИМ.,
Мишустин А., Очагов ЦМ. Влияние фрагментации болот на
биоразнообразие (птицы и бабочки): голландский подход 368
Сарычева ЕЛ. Влияние хозяйственной деятельности человека на видовое
разнообразие черноольховых сообществ заповедника «Воронинский»,
Тамбовская область 369
Смирнова В.В., Козулин А.В. Охраняемые болотные территории и
биоразнообразие болот Беларуси 372
Шараг Е.И. Организация и ведение эколого-мелиоративного мониторинга
мелиорированных земель Гослесфонда Беларуси 374
Указатель авторов 378
Адреса организаций 381
392
Научное издание
БОЛОТА И ЗАБОЛОЧЕННЫЕ ЛЕСА
В СВЕТЕ ЗАДАЧ
УСТОЙЧИВОГО ПРИРОДОПОЛЬЗОВАНИЯ
МАТЕРИАЛЫ КОНФЕРЕНЦИИ
Подписано к печати 20.07.1999 г.
Формат 62x94 1/16. Бумага офсет № 1,80 г/м2
Гарнитура Тайме. Печать офсетная.
Усл. печ. л. 24,5. Уч.-изд. л. 26,0.
Тираж 400 экз.
Тип. зак. № 336 С , Москва
Издательство ГЕОС
Изд. лицензия Л.Р. № 050112 от 09.03.95
109017, Москва, Пыжевский пер., 7.
Тел.:(095)230-80-92
Факс:(095)951-04-43
Болота и заболоченные леса в свете задач устойчивого
природопользования. Материалы совещания. - М.: ГЕОС, 1999. - 392 с.
ISBN 5-89118-091-Х
Оторфованные земли в России занимают более 1/5 площади
страны. Болота являются важным объектом различного хозяйственного
использования (в лесоводстве, сельском хозяйстве, добыче
торфяного, пищевого, лекарственного сырья). Одновременно болота
выполняют разнообразную средообразующую роль и особенно в
регулировании круговорота углерода в биосфере.
В книге представлены более 150 статей, подготовленных к
конференции «Болота и заболоченные леса в свете задач устойчивого
природопользования» специалистами России, Беларуси и других стран.
Отражен широкий круг знаний по распространению, динамике болот
и заболоченных лесов, их структурно-функциональной организации,
биосферной роли, биоразнообразии, методах и последствиях
хозяйственного использования, мониторингу и охране, как научной
основы для перехода к устойчивому пользованию этими угодьями в
современных экономических условиях.