/











































































































































































































































































Text
АТЛАС
ПОРОДООБРАЗУЮЩИХ
ОРГАНИЗМОВ
АТЛАС
ПОРОДООБРАЗУЮЩИХ
ОРГАНИЗМОВ
На соеремеьисм уровне геологических знаний при изучении осадочных пород боль-
шое значение имеют ископаемые организмы. В настоящем Атласе рассмотрены
микроструктуры ископаемых известковых и крем-
невых организмов, встречающихся в отложе-
ниях, начиная с докембрийских и кончая чет-
вертичными. Приводятся данные по условиям их
жизни, указывается фациальное, стратиграфи-
ческое и породообразующее значение.
В Атласе рассмотрено большинство групп орга-
низмов, которые могут быть обнаружены в шли-
фах и препаратах.
Атлас состоит из трех частей: 1 — известковые
организмы (водоросли, животные): 2—кремне-
вые организмы (водоросли, животные); 3 — следы
жизнедеятельности организмов.
Многие материалы, вошедшие в Атлас, публи-
куются впервые.
АКАДЕМИЯ НАУК СССР
КОМИССИЯ ПО ОСАДОЧНЫМ ПОРОДАМ
ПРИ ОТДЕЛЕНИИ ГЕОЛОГИИ, ГЕОФИЗИКИ И ГЕОХИМИИ
ОРДЕНА ТРУДОВОГО КРАСНОГО ЗНАМЕНИ
ГЕОЛОГИЧЕСКИЙ ИНСТИТУТ
ACADEMY OF SCIENCES OF THE USSR
COMMISSION ON SEDIMENTARY ROCKS
DEPARTMENT OF GEOLOGY, GEOPHYSICS AND GEOCHEMISTRY
ORDER OF THE RED BANNER OF LABOUR
GEOLOGICAL INSTITUTE
ATLAS
OF ROCK-BUILDING
ORGANISMS
(CALCAREOUS AND SILICEOUS
ORGANISMS)
COMP1LATOR V. P. Maslov
e
PUBLISHING OFFICE «NAUKA»
MOSCOW
1973
АТЛАС
ПОРОДООБРАЗУЮЩИХ
ОРГАНИЗМОВ
(ИЗВЕСТКОВЫХ
И КРЕМНЕВЫХ)
СОСТАВИТЕЛЬ В. П. Маслов
в
ИЗДАТЕЛЬСТВО «НАУКА»
МОСКВА
1973
УДК 56(084.4) : 552.54 + 552.55
Атлас породообразующих организмов (известковых и кремневых) М.,
«Наука», 1973
Настоящий атлас — руководство для широкого круга геологов
и микропалеонтологов при изучении осадочных пород под микро-
скопом. Он охватывает большинство групп организмов, которые
могут встретиться в шлифах и препаратах. В нем рассматриваются
микроструктуры ископаемых организмов, условия их жизни, пока-
зано их стратиграфическое, фациальное и породообразующее зна-
чение, а также некоторые спорные организмы, следы их жизнедея-
тельности и микрокопролиты. Часть вошедших в Атлас материалов
публикуется впервые.
Табл. 6. Илл. 128 + 76 фототаблиц. Библ. 282 назв.
Редакционная коллегия:
академик В. В. Меннер, С. В. Максимова,
В. С. Яблоков (ответственный редактор)
Editorial Board:
Academician V. V. Menrter, S. V. Maksimova,
V. S. Yablokov (Responsible Editor)
02101; 0293-0366
A-----------------212-71(11) Издательство «Наука», 1973
042(02)-73
ПРЕДИСЛОВИЕ
Широкие геологические, литологические и геологоразведочные
работы, проводящиеся на огромной территории СССР, вызывают
необходимость определения остатков организмов из образцов по-
род и кернов скважин под микроскопом. Такому изучению в свое
время помогали отдельные работы и атласы микроскопических
обломков организмов Л. Кайё (Сауеих, 1931) и В. П. Маслова (1937а),
ставшие библиографической редкостью.
Комиссия по осадочным породам при Отделении геологии, геофи-
зики и геохимии АН СССР, учитывая создавшееся положение и неод-
нократные запросы от разных геологических учреждений, решила ор-
ганизовать совместно с Геологическим институтом АН СССР и с
привлечением специалистов из ряда организаций составление нового
Атласа породообразующих (известковых и кремневых) организмов.
Общее руководство работой, начатой в 1964 г., принадлежит
В. П. Маслову, им же написано большинство разделов Атласа.
Авторами остальных разделов являются: Д. М. Раузер-Черноусова,
В. В. Миссаржевский, А. Ю. Розанов (Геологический институт
АН СССР), Г. Г. Астрова, Г. А. Афанасьева, Т. Г. Ильина, И. И. Чу-
динова (Палеонтологический институт АН СССР), С. В. Максимова
(Институт геологии и разработки горючих ископаемых Министерства
нефтяной промышленности СССР), И. Т. Журавлева (Институт
геологии и геофизики СО АН СССР), В. И. Яворский, 3. И. Глезер
(Всесоюзный научно-исследовательский геологический институт
Министерства геологии СССР), А. П. Жузе, С. Б. Кругликова (Ин-
ститут океанологии АН СССР), Б. А. Поярков (Геологическое
управление Киргизской ССР), Л. В. Липецкая (Институт геологии
и геохимии горючих ископаемых АН УССР, Львов), С. В. Шумейко
(Харьковский госуниверситет), В. Покорны (Карлов университет,
Прага, Чехословакия), Ф. Ю. Адамчак (Геологический институт.
Стокгольм, Швеция), Г. А. Бачинский (Институт зоологии АН
УССР), Э. М. Юргенсон, В. Я. Вийра (Геологический институт
АН Эстонской ССР).
Работа по составлению Атласа была в основном закончена в
1967 г.
После кончины В. П. Маслова (12 сентября 1968 г.) перед сда-
чей в печать было проведено некоторое сокращение текста и иллю-
страций, сделаны небольшие добавления и ссылки на работы послед-
них лет, работа была окончательно отредактирована.
Атлас содержит громадное количество иллюстраций (рисунки в
тексте и отдельные таблицы) с изображением в шлифах и аншлифах
и зарисовках известковых и кремневых остатков различных орга-
низмов и большой пояснительный текст.
Методика пользования Атласом как определителем может быть
разной. Если исследователь уже имеет представление, к какой груп-
пе организмов относятся изучаемые обломки, следует по оглавле-
нию найти соответствующий раздел и внимательно ознакомиться
с текстом и иллюстрациями, чтобы либо уточнить свои предполо-
жения, либо их отвергнуть. Если принадлежность обломков неясна
или неизвестна, необходимо после изучения и зарисовки остатков
в шлифе внимательно просмотреть все таблицы и рисунки, подо-
брать наиболее подходящие и детально ознакомиться с соответ-
ствующими текстами, после чего будет найдено наиболее правиль-
ное решение.
В Атласе упомянута только литература, непосредственно связан-
ная с разбираемыми вопросами. Палеоонтологическая и страти-
графическая литература (общая и по отдельным группам организ-
мов), как советская, так и зарубежная, не включена в списки. Необ-
ходимые сведения желающие могут найти в многотомных изданиях
«Основы палеонтологии», «Стратиграфия СССР» и «Геология
СССР».
В. П. Маслов в течение многих лет неустанно трудился над изу-
чением осадочных, главным образом карбонатных, пород и остат-
ков организмов, слагающих эти породы. Его многочисленные ра-
боты широко известны в СССР и за рубежом. Издание Атласа
породообразующих известковых и кремневых организмов, состав-
ление которого было возможно только при громадных знаниях,
трудолюбии и энтузиазме Владимира Петровича, является вечным
памятником ученому.
В. В. Меннер, С. В. Максимова, В. С. Яблоков
ВВЕДЕНИЕ
Организмы играют огромную роль в образовании осадочных по-
род. Концентрируя определенные соли, животное или растение вы-
деляет особый твердый «скелет»1, сохраняющийся в ископаемом
виде, который и изучается в шлифах.
Некоторые исследователи считают породообразователями те
организмы, остатки которых составляют преобладающую часть
породы. Таких организмов не так много, и мы не считаем себя вправе
ограничиваться столь узким понятием. Многие спорадически встре-
чающиеся остатки животных и растений имеют не только породо-
образующее значение, но могут играть роль указателей геологи-
ческого возраста и условий накопления включающих их осадков.
Поэтому мы поместили в настоящем Атласе такие организмы, как,
например, голотурии, конодонты и др. Некоторые из этих животных
получили важное стратиграфическое значение. Правда, часто их
невозможно определить в шлифе более подробно, чем тип, порядок,
семейство, а их исследование производится микропалеонтологами
в дезинтегрированном виде (выделенными из породы: конодонты,
голотурии, кокколиты, харофиты и др.). Этим организмам также
посвящены специальные очерки в надежде на то, что литологи и
геологи ими заинтересуются и обратят внимание при изучении
легкой фракции пород.
Единственными относительно точными указателями глубин
могут служить остатки донных водорослей. Поэтому им уделено
наибольшее внимание.
Основной методикой изучения остатков организмов в данном
Атласе принято микроскопическое исследование обломка или скеле-
та их в шлифах для определения по маленькому обломку под микро-
скопом типа, класса или более узкой систематической единицы, к
которым принадлежит организм.
Минеральный состав обломка дает некоторую грубую ориенти-
ровку при определении. К организмам с кремневыми остатками
принадлежат часть губок, радиолярии и диатомовые водоросли.
Фосфатные скелеты образуют позвоночные и два вида брахиопод.
Вся остальная преобладающая масса организмов образует карбо-
натные скелеты, чехлы или раковины.
Необходимо заметить, что часто обломки организмов, пере-
кристаллизовываясь, меняют свой минеральный состав: кремневые
1 Здесь и в дальнейшем под скелетом будет подразумеваться твердое минеральное образо-
вание. оставшееся после организма, вне зависимости от его принадлежности к животному
или растительному миру. Термин этот употребляется и в применении к породам (скелет
породы).
губки, например, замещаются кальцитом и, наоборот, известковые
обломки окремневают.
Среди таких процессов замещения необходимо различать окрем-
нение, кальцитизацию, доломитизацию, образование кристаллов
гипса и ангидрита, выпадение соединений железа (в том числе их
пиритизацию) и глауконитизацию. При замещении кремнеземом
известковых обломков микроструктура скелетной ткани организмов
нередко хорошо сохраняется. То же можно сказать о кальцитизации
кремневых организмов. В противоположность этому при доломити-
зации микроструктура скелетных обломков быстро исчезает, а при
общей доломитизации породы расплываются их очертания, так что
в шлифе различаются только неясные реликты органических ос-
татков.
Таким образом, при определении остатков организмов особенно
большое значение имеют форма и микростроение обломков. Форма
особенно важна при определении водорослей, фораминифер и таких
организмов, как трилобиты, гастроподы, мшанки и др. Из них
гастроподы, перекристаллизовывающиеся нацело, теряют обычно
свою микроструктуру, но легко узнаются по своей спирально сверну-
той раковине. Поэтому литологу нужно обладать хорошим про-
странственным представлением, чтобы восстанавливать по сечению
спираль, цилиндр, конус и т. п. Для определения формы обломка
существенную помощь оказывает более темный или, наоборот,
более светлый цементирующий материал, который подчеркивает
границы обломка и внутренние пустоты.
Наконец, одним из важных признаков при определении обломков
организмов является микроструктура данного обломка.
Кроме остатков фауны и флоры в Атласе рассмотрены такие
часто встречающиеся в шлифах образования, как капролиты и сле-
ды жизнедеятельности сверлящих водорослей.
В ряде очерков Атласа приведены еще не опубликованные сведе-
ния и новейшие исследования. В некоторых сомнительных случаях
приводятся разные, иногда противоположные мнения о данном
организме, особенно в разделе о спорных и сомнительных орга-
низмах.
Вопросы классификации ископаемых организмов и методы их
палеонтологического, т. е. родового и видового, определения с
большой полнотой освещены в обстоятельном многотомном изда-
нии «Основы палеонтологии», к которому мы и отсылаем всех, кто
пожелал бы заняться органическими остатками не как составной
частью осадочных пород, а как объектом самостоятельного изу-
чения.
Часть I. ИЗВЕСТКОВЫЕ ОРГАНИЗМЫ
ОБЩИЕ ВОПРОСЫ
Табл. 1—6
Под известковыми организмами мы будем подразумевать те
организмы, которые выделяют арагонитовый, или кальцитовый,
или магнезиально-карбонатный, или фосфористо-карбонатный ске-
лет, панцирь или чехол. Кратко коснемся общих вопросов, связанных
со способом отложения извести вообще и ролью организмов в обра-
зовании некоторых биогенных пород.
Способы выделения извести организмами. Выделение извести
организмами — разнообразный и большей частью сложный про-
цесс, еще недостаточно изученный. Животные отлагают известь
особыми клетками или органами, образуя наружный панцирь или
внутренний скелет. Этот процесс относится к «органическому»
выделению извести, так же как и у некоторых водорослей, и связан
с жизнедеятельностью организмов, которые, усваивая известь из
воды, выделяют известковые образования разной микроструктуры
и разного химического состава, в зависимости от способов связи
кальция с органическими молекулами у различных групп организмов.
Иной процесс карбонатообразования, известный только у рас-
тений, называете < «физиологическим отложением извести». Он выра-
жается в том, то на поверхности клеток выпадают неправильно
расположенные кристаллы карбоната. Такое выпадение карбоната
обусловлено фс 'осинтетической деятельностью растения в водной
среде. Для это) о процесса необходимо, чтобы растение усвоило
бикарбонат кал >ция, Н2СО3, НСО3, СО2 во время фотосинтеза.
Все эти компоненты воспринимаются нижней частью листа. Известь
выделяется верхней частью листа и образует твердый осадок. Такая
«физиологическая полярность» осуществляется обычно в тех расте-
ниях, которые имеют билатеральное строение.
Таким образом, различие между «органическим» и «физиоло-
гическим» отложением извести заключается в следующем. При
«органическом» процессе известь выделяется из клеточного сока
внутри клетки или в ее стенках, при «физиологическом» известь
выпадает снаружи клеточных стенок. При «органическом» процессе
выделенный минеральный осадок состоит из мелких, закономерно
расположенных кристалликов кальцита, которые при хорошей
сохранности дают определенные оптические эффекты (затухание
волной и т. п.). при этом наблюдается тонкое чередование их с орга-
ническим веществом. Микроскопическое строение выпавшего каль-
цита специфично для отдельных групп водорослей. При «физио-
логическом» процессе выпадающий кальцит обычно чист от орга-
нического вещества, крупнозернист и его кристаллы не дают законо-
мерного затухания. Если при «органическом» процессе консерви-
руется анатомическое строение водорослевых клеток, то при «физио-
логическом» отложении извести образуется известковый чехол
вокруг слоевища, фоссилизируется его внешняя форма и реже
внешняя форма клетки.
«Органическое» отложение извести свойственно всем животным
и некоторым водорослям. Именно такой процесс, по-видимому,
наблюдается у некоторых багрянок из семейства Corallinaceae, в
результате чего слоевище водорослей становится твердым, известь
консервирует внутреннюю прижизненную анатомическую струк-
туру, которую можно поэтому изучать в ископаемом состоянии.
К «органическому» процессу отложения извести можно отнести
известковые выделения кокколитофорид. Эти ничтожно малые
планктонные микроорганизмы образуют вокруг своей клетки из-
вестковый «панцирь» из мелких известковых частиц разной формы
(кокколиты). Процесс образования кокколитов еще недостаточно
ясен, но изучение их под электронным микроскопом показало, что
они состоят из очень мелких (единицы и доли микрона) плоских
кристалликов кальцита, закономерно расположенных: обычно ра-
диально и одновременно черепицеобразно. При скрещенных нико-
лях кокколиты хорошей сохранности дают эффект сферолитового
характера.
«Физиологическое» выделение извести наблюдается у зеленых
водорослей (сифоновых), где голые ассимилирующие части слое-
вища усваивают кальци; из раствора и отлагают его в некоторых
местах на наружной поверхности клетки также в виде известкового
чехла. Эти известковые оболочки у сифоней чрезвычайно характер-
ны и позволяют реконструировать почти все растение и изучать его
в ископаемом виде. По-видимому, к этому же процессу нужно от-
нести отложение извести некоторыми зелеными (например, Chae-
tophora) и синезелеными водорослями, образующими локальные
известковые футляры или чехлы.
У харовых водорослей в одном и том же экземпляре известь мо-
жет отлагаться физиологическим путем в вегетативных частях
слоевища и органическим способом в оболочках женских органов
плодоношения (ооспорангиев).
Вследствие «органического» процесса выделения извести извест-
ковая оболочка ооспорангия харовых водорослей приобретает осо-
бую слоистую текстуру как у современных, так и у ископаемых
представителей этих растений. Рентгено-кристаллографический ана-
лиз показал, что спелые известковые оболочки ныне живущей Chara
globularis состоят из кальцита. По петрографическим исследованиям
нескольких авторов чисто кальцитовые, очень тонкие слои (2—3 д)
переслаиваются со слоями кальцита, обогащенными органическим
материалом, в результате чего получается тонкая штриховатость
внутреннего слоя известковой оболочки. Вегетативные части хар
отлагают известь физиологическим способом. У современных ви-
дов Nitella flexilis, Chara baueri, Ch. braunii, Tolypella nitricata Аренс
нашел «физиологическую мультиполярность». Это выражалось в
кольцевых инкрустациях извести вокруг стержней, которая выделя-
лась кольцевыми зонами клеточных оболочек. Одна часть зон усваи-
вала гидроокись кальция, а другая, чередующаяся с ними, его выде-
ляла. В результате получается сплошной известковый чехол, ко-
торый покрывает все вегетативные части харовых водорослей.
Строение такого футляра, или чехла, резко отличается от строе-
ния известковых оболочек ооспорангиев того же самого растения,
так как чехол слагается мелко- и крупнокристаллическим кальцитом.
Существует и еще один, третий процесс отложения карбоната,
который можно назвать «биохимическим». Водные растения при
этом процессе играют косвенную роль, меняя pH среды при усвое-
нии ими углерода из воды и способствуя химическому выпадению
карбонатного осадка. Этот осадок не отличим от кристаллов карбо-
ната, образованных при чисто хемогенной садке, но, располагаясь
между нитями водорослей, кристаллики карбоната могут заполнять
пространства между нитями и образовывать, таким образом, ло-
кальную структуру, отличающуюся от обычного осадка.
8
Известковые организмы
Биохимический процесс в сочетании с химическим и физиоло-
гическими процессами участвует в образовании карбонатных пост-
роек строматолитов, причем относительное значение каждого из
них очень непостоянно. Биохимическое или смешанное отложение
извести свойственно также онколитам, т. е. желвакам, образован-
ным, вероятно, синезелеными водорослями и бактериями.
Рассмотренные нами процессы образования карбонатного осад-
ка интересны также с точки зрения связи этого осадка с условиями
окружающей среды. Если «биохимический» и «смешанный» процес-
сы тесно связаны с достаточным количеством карбонатов в растворе
и в некотором смысле отражают солевой карбонатный состав этого
раствора, то «органический» процесс не зависит от концентрации рас-
твора и его состава, необходимо лишь присутствие в растворе со-
лей кальция. «Физиологический» процесс, по-видимому, занимает
промежуточное положение, так как интенсивность обызвествления
может увеличиться с увеличением концентрации солей кальция в
растворе (Маслов, 1961 б), но все же этот процесс специфичен только
для определенных форм водорослей. «Органический» и «физиоло-
гический» процессы отложения извести создают такие структуры,
которые могут быть выявлены под микроскопом. При «органи-
ческом» процессе получаются известковые органогенные остатки
разнообразного вида, определяющие группу животных или расте-
ний. При «физиологическом» процессе получаются карбонатные
корки, позволяющие восстановить внешнюю морфологию растения.
«Биологический» и «смешанный» процессы отложения извести соз-
дают причудливые и трудно дешифрируемые карбонатные (иногда
доломитовые, иногда известковые) структуры. Ниже мы обратимся
к некоторым сооружениям, образованным при помощи известковых
выделений организмов, и приведем несколько примеров биогермных
текстур1.
Водорослевые биогермные текстуры. Самыми показательными и
наглядными являются биогермные известняки, образованные кор-
ковыми багряными водорослями. Они широко распространены в
современных тропических морях, и если кораллы составляют глав-
ную, легко отличимую часть современных рифов, то корковые
багрянки скрепляют их и наращивают волноупорную зону рифа.
Поскольку у нас нет образцов современных рифовых образований,
мы приводим пример из третичных тортонских биогермов Медо-
бор (Толтр). Биогермы здесь сложены, как это указал еще Ласкарев
в 1914 г., «литотамниями» и серпулами. Более детально они были
изучены И. К. Коро л юк (1952) и В. П. Масловым (1962). Биогермные
известняки сложены слоистыми корковыми багряными водорослями,
преимущественно из рода Lithothamnium, которые образуют нерезко
выраженные купола диаметром в 5—10 см. Порода в целом плот-
ная, массивная, но пористая. Естественно, что биогермные извест-
няки следует изучать в пришлифовках и крупных шлифах, чтобы
охватить достаточное пространство иногда очень неоднородной
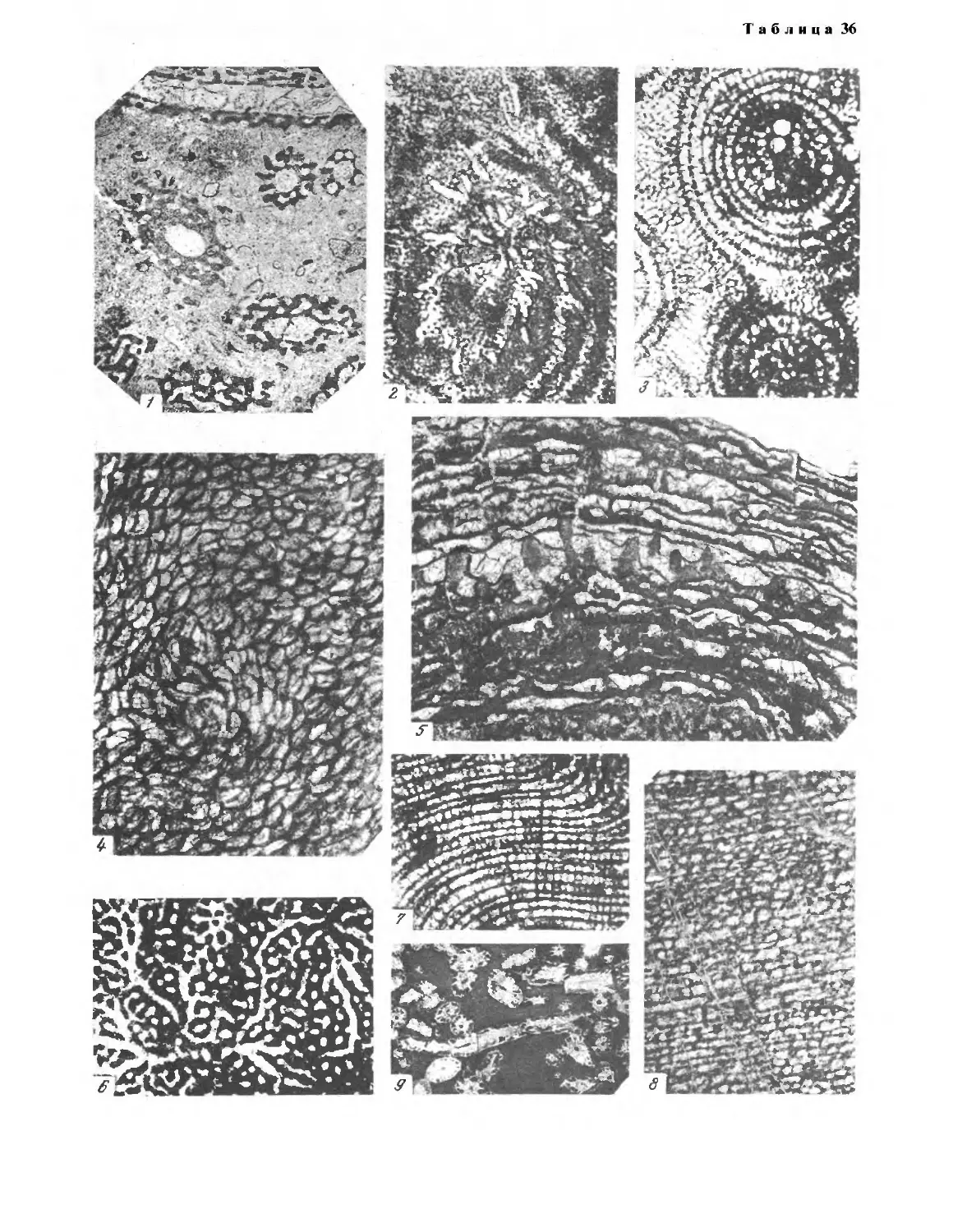
породы. Такая пришлифовка изображена на табл. 1, фиг. 1.
Биогермные известняки, образованные остатками животных. Из
одноклеточных животных активно создают биогермные структуры
некоторые прикрепленные фораминиферы — нубекулярии. В СССР в
среднем сармате Молдавии известны мощные и эффектные извест-
няковые массивы (разрабатываемые промышленностью), состоящие
из таких биогермов, в которых главным образователем являются
прикрепляющиеся фораминиферы — нубекулярии совместно с не-
которыми водорослями. В великолепно обнаженных массивах в
карьерах около г. Кишинева отчетливо видно, что по краям массивы
распадаются на ряд бугров, столбов и т. п., сливающихся в центре
В результате работы двух Всесоюзных палеоэколого-литологических сессий (1965, 1966 гг.).
посвященных ископаемым рифогенным образованиям, биогерм определяется как мас-
сивное карбонатное тело, возвышающееся над прилегающими синхронными отложения-
ми иного литологического состава и образованное остатками прикрепленных животных
и растительных организмов, захороненных преимущественно в прижизненном поло-
жении (прим. ред.).
массива (табл. 1, фиг. 2, 4, 5). Каждый биогерм в свою очередь сла-
гается грубыми, выпуклыми вверх слоями желваковатого строения.
Макроскопически видно, что большинство слоев сложено мелкими
столбиками нубекулярий с пустыми пространствами между столби-
ками. В эти промежутки часто попадает органогенный детритус
(табл. 1, фиг. 2). Эти слои чередуются со слоями из уплощенных ко-
лоний нубекулярий вместе с синезелеными и корковыми багряными
водорослями. Строение массива неоднородно. В некоторых местах
(Оргеевский массив) встречались линзы, сложенные кораллиновыми
водорослями (членистые багрянки) (Маслов, 1963г).
Похожие биогермные известняки описаны И. В. Хворовой (1951)
в пермских осадках Приуралья.
Мшанки являются другим образователем биогермов, встречаю-
щихся от силура до кайнозоя. Широко известны третичные керчен-
ские «рифы», построенные мшанками, по-видимому, с участием си-
незеленых водорослей. Особенно часто мшанки образуют биогерм-
ные текстуры в верхнем палеозое Приуралья. Такие текстуры были
описаны не раз (Маслов, 1945, 1950; Сермягин, 1938) и характерны
пятнистым распределением разных компонентов. Приведенные на
табл. 2—4 рисунки и фотографии пришлифовок керна буровых
скважин из ишимбаевских нижнеартинских погребенных массивов
публикуются впервые. Биогермные структуры образованы тонкими
сеточками мшанок, покрытыми корочкой вторичного карбоната
и образующими замкнутые полости, которые заняты химическим
осадком в виде вторичного карбоната или ангидрита. Открытые
полости заполнены детритусовым или пелитоморфным карбона-
том. Поскольку разные полости заполнялись в разное время, полу-
чалась довольно пестрая картина, а в случае битуминизации тонко-
зернистых разностей — разноцветная. Совершенно ясно, что от-
дельные мелкие шлифы не дадут полной картины биогермной струк-
туры и могут ввести в заблуждение исследователя.
Как в пермских (Приуралье), так и в юрских (Западная Украина)
отложениях известны биогермные структуры, образованные гид-
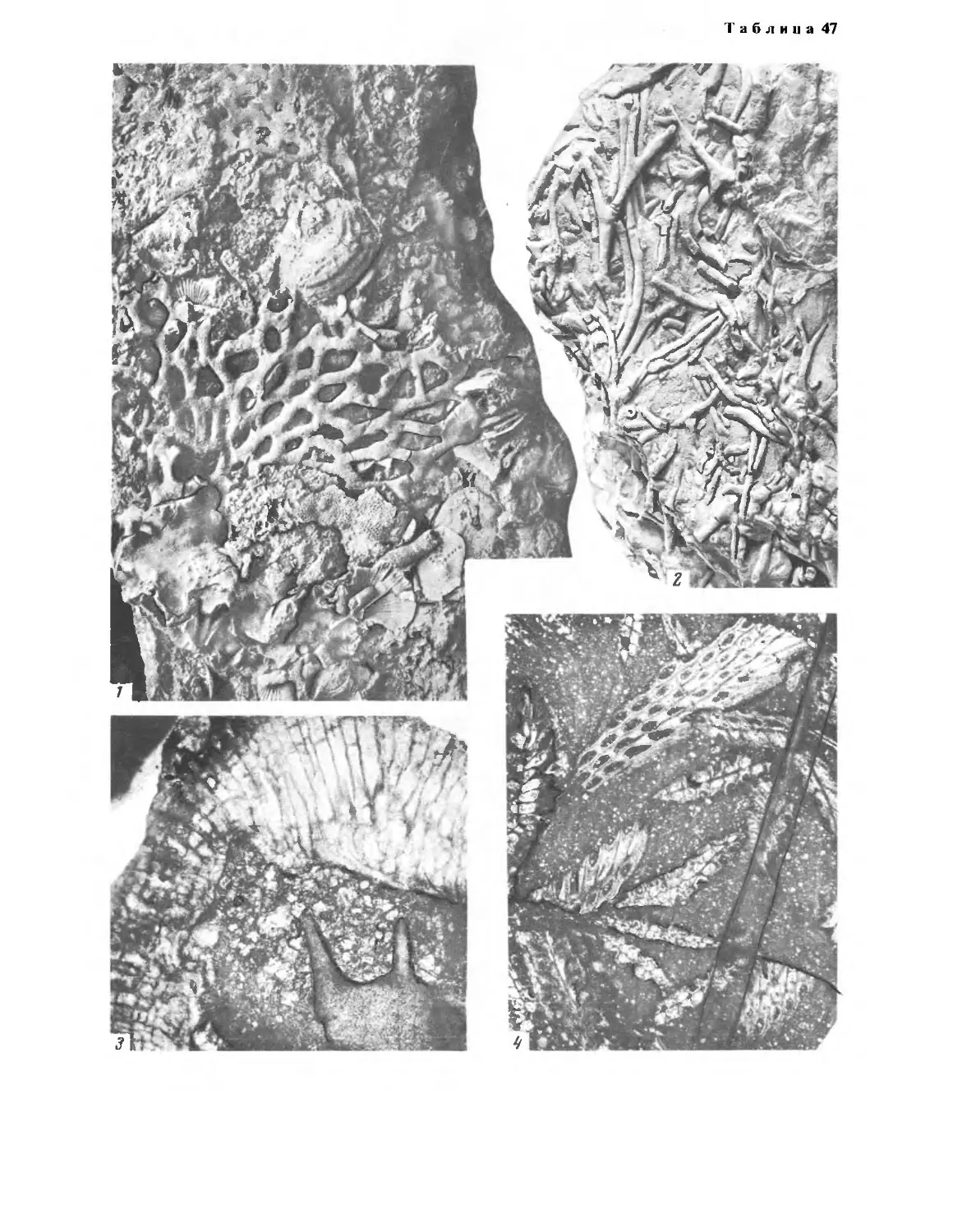
роидными полипами (табл. 3, фиг. 1).
Эти биогермные известняки характерны еще более трубой макро-
скопической структурой. Благодаря толстым пластинам полипня-
ков и легкой их перекристаллизации обнаружить их в шлифах иногда
невозможно. В то же время они бывают отчетливо видны на при-
шлифовках. На табл. 3 изображен исключительный случай запол-
нения ангидритом полостей между полипняками, причем полипняки
относятся к раннеартинекому времени, а вторичный по отношению
к ним ангидрит — к кунгурскому.
Бывают случаи, когда не удается обнаружить главного биогермо-
образователя, между тем как текстура и форма тела определенно
указывают на наличие биогерма. Такие случаи нередки, примером
могут служить биогермы вазалеммского горизонта ордовика на
ст. Вазалемма в Эстонии. Хотя некоторые геологи утверждают, что
эти биогермы образованы водорослями, автору не удалось обнару-
жить растений в биогермах. Корковые формы мшанок и прикреп-
ленные иглокожие иногда покрывали внешнюю поверхность био-
герма. Пример биогермных текстур с неопределимым биогермооб-
разователем приведен на табл. 2, фиг. 4 из нижнеартинских осадков
Ишимбая.
Биогермные известняки, образованные проблематическим орга-
низмом спорного происхождения. В перми Приуралья широко извест-
ны «червяковые известняки» раннеартинского и позднекаменно-
угольного времени, которые образованы главным образом извест-
ковым организмом Tubiphytes (= Schamovella). В. П. Маслов, описав-
ший этот организм, отнес его к синезеленым водорослям (Маслов,
1956). Но некоторые зарубежные исследователи сомневаются в во-
дорослевом происхождении этой формы, поэтому мы и не причле-
няем образуемые этим организмом текстуры к водорослевым.
Тонкие (в 1—2 мм) извивающиеся стерженьки тубифитес, пере-
плетаясь между собой, создают основной каркас биогерма, в кото-
Общий очерк. Синезеленые водоросли
9
ром живут и застревают другие организмы (мшанки, брахиоподы,
иглокожие и т. п.). В результате создается биогерм, возвышающийся
над дном и часто разрушающийся движением воды (прибоем ?),
благодаря чему у его подножия скапливаются обломки тубифитес,
образуя целые пласты. Биогермный известняк, образованный жизне-
деятельностью этого организма, в пришлифовке имеет пятнистый
вид (табл. 4, фиг. 3), но количество самих тубифитес невелико. Про-
странство между ними прихотливо распределяется между светло-
серым и темно-серым известняком. Это распределение, по-видимому,
сингенетичное и обязано расположению разных организмов и нерас-
шифрованных пленок (может быть, исчезнувших водорослей).
ВОДОРОСЛИ
ОБЩИЙ ОЧЕРК
Под термином «водоросли» подразумевается обширное царство
растительных организмов, живущих преимущественно в воде и об-
ладающих относительно примитивным строением. Но необходимо
сказать, что в состав водорослей входят очень различно построен-
ные организмы и что многие из них живут на суше. Это необходимо
помнить, даже если считать существование водорослей на суше
исключением.
Водоросли, живущие на суше, могут быть разделены на две ка-
тегории:
1. Эндолиты, или сверлящие водоросли, живущие внутри (близ
поверхности) известковых скал, камней, раковин и т. п. Растворяя
известь субстрата, они внедряются под поверхность и переносят
засушливый сезон, задерживая влагу внутри камня.
2. Водоросли почв и поверхностей предметов на суше (стен,
стволов деревьев и т. п.). Эти водоросли включают целый ряд групп,
не родственных между собой. Так, Э. А. Штина (1964), останавли-
ваясь на участии водорослей в процессах почвообразования, замечает,
что таких водорослей, обитающих на суше, в почвах и на их по-
верхности, насчитывается более 1000 видов (из них синезеленых —
320, диатомовых — 273, желтозеленых — 75, эвгленовых—18, пир-
рофитовых — 4, золотистых — 3 вида).
Известны еще наземные водоросли (некоторые виды синезеленых
и зеленых), которые живут на влажных скалах, стенах, стеллажах
оранжерей, используя воду, орошающую основную посадку (Елен-
кин, 1936). Некоторые из этих водорослей при жизни отлагают из-
весть в форме трубочек, облекающих нити (Scyionema drilosyphon).
Как известно, водоросли нуждаются в свете для фотосинтеза.
За редкими исключениями (водоросли пещер) они живут в хорошо
освещенных мелких водоемах, и лишь некоторые группы опускают-
ся до 100—180 м глубины. Таким образом, для геолога остатки
водорослей являются довольно четкими показателями мелководных
фаций (Маслов, 1935).
Большинство водорослей интенсивно окрашено особыми пиг-
ментами, которые поглощают солнечную энергию для синтеза орга-
нических веществ. Тип пигмента или комбинация нескольких пиг-
ментов придают водорослям определенный цвет, который послужил
для разделения современных водорослей на: 1) синезеленые, 2) зеле-
ные, 3) бурые, 4) красные, 5) желтозеленые (гетерогенная группа,
классифицируемая различно разными авторами). Особенности
структуры клеток, органов размножения и т. п. являются основными
признаками для деления на группы. Как видим, для палеонтолога
бблыпая часть признаков современных водорослей является недо-
ступной, так как ни цвет, ни особенности строения живой клетки
не сохраняются в ископаемом состоянии. Но все же некоторые,
обычно внешние признаки позволяют с разной степенью точности
относить фоссилизированные остатки к той или иной группе водо-
рослей, тем более что цвет имеет лишь относительное значение для
классификации.
В Атласе мы рассмотрим породообразующие водоросли, облада-
ющие известковым и кремневым скелетами1.
Царство водорослей необходимо разделить на две обширные
группы: 1) синезеленые водоросли (Cyanophyta) и 2) все остальные
типы водорослей. Синезеленые водоросли — очень примитивные
организмы, легко приспособляющиеся к разным условиям, не имеют
ядра, чем отличаются от других водорослей. Но некоторые другие
примитивные одноклеточные или нитчатые водоросли имеют морфо-
логическое сходство с синезелеными водорослями, и отличить эти
формы можно только в живом виде.
Таким образом, в ископаемом состоянии хорошо определяются
лишь высокоорганизованные водоросли.
СИНЕЗЕЛЕНЫЕ ВОДОРОСЛИ (CYANOPHYTA)
Табл. 5—7
Эти примитивного строения, исключительно микроскопические
водоросли не только очень многочисленны, но и широко распро-
странены на Земле, произрастая в самых разнообразных условиях,
как водных, так и вневодных, как в планктоне, так и на дне.
Химический состав. Химический состав карбонатных отложений
синезеленых водорослей наиболее тесно связан с химическим соста-
вом окружающего осадка, что доказывается анализом стромато-
литов и биохимическим способом отложения карбоната.
Современные пресноводные синезеленые водоросли характери-
зуются повышенным содержанием азота и фосфора (Виноградов,
1935). Вопрос об известковых отложениях этих водорослей и соот-
ношении в них солей кальция и магния остается во многом неясным,
но известны формы, выделяющие кальций в виде инкрустаций.
Анализы строматолитов (МаслЬв, 19606), в построении которых
участвовали синезеленые водоросли, показали, что содержание
углекислого магния в них мало чем отличается от содержания маг-
ния породы, окружающей строматолит, или, иными словами, если
осадок, окружающий строматолит, кальцитовый, то и сам стромато-
лит беден магнием, если же осадок — доломит, то строматолит
сильно магнезиален. Здесь сказывается биохимический способ от-
ложения извести. При анализе современной синезеленой водоросли
ривулярии, отлагавшей известь физиологическим способом, карбо-
нат оказался чистым кальцитом с содержанием 84.88% СаСОэ и
0,69% MgCO3.
Экология. Представители синезеленых водорослей, за редкими
малоизученными исключениями, нуждаются в солнечном свете. По-
этому они произрастают в пресных водоемах на незначительных
глубинах, а в морях — в области литорали и сублиторали (до глу-
бины 60 м). Они приспосабливаются к воде различной степени со-
лености: от сильносоленой до совершенно пресной, а также к разной
температуре: от —1,8° до +80° и более. Таким образом, ни одна
группа организмов не сравнима с этими водорослями по широте
условий обитания. Многие синезеленые водоросли приспосабли-
ваются также к смене соленых и пресных вод, которую другие орга-
низмы вынести не могут.
Синезеленые водоросли устойчивы при высыхании водоемов и
могут переживать засушливые сезоны. Они также могут жить на
поверхности суши при условии периодического увлажнения. Неко-
торые синезеленые водоросли являются «сверлильщиками», иста-
чивающими известковый субстрат, который предохраняет их от
гибели при неблагоприятных условиях.
Другие формы часто являются «кальцитофикаторами», т. е. в ре-
зультате их деятельности отлагаются известковые туфы. Еще реже
в горячих источниках и гейзерах они участвуют в отложении крем-
1 Литературу по водорослям см.: В. П. Маслов, 1956. I960, 1962.
10
Известковые организмы
невых туфов и являются одним из основных образователей стромато-
литов как в современных морях, так и в ископаемых породах. Благо-
даря нахождению строматолитов и онколитов в древних осадках
мы можем утверждать, что синезеленые водоросли существова-
ли в нижнем протерозое и, вероятно, были первыми организмами,
появившимися на Земле. В систематическом отношении синезеленые
водоросли-кальцификаторы принадлежат к разным классам и по-
рядкам.
Геологическое значение. Синезеленые водоросли имеют большое
значение как породообразователи в докембрии и нижнем палеозое
(как строматолитообразователи). В более поздние периоды большое
значение у них приобретает физиологический способ отложения из-
вести (см. стр. 7), благодаря чему мы с несколько большей долей
уверенности можем определять эти организмы. Современные сине-
зеленые водоросли хорошо изучены, но их карбонатные выделения
совсем не исследованы, так как альгологи для определения водо-
росли растворяют известь и имеют дело с органической частью
клеток.
Анатомия. Синезеленые водоросли бывают одноклеточными, ко-
лониальными и нитчатыми. Одноклеточные формы имеют клетки
разнообразного вида, но большей частью шаровидные и овальные.
В ископаемом виде они обнаруживаются очень редко. Объединение
нескольких живых клеток в общей слизи создает колонию в форме
шара, эллипсоида и т. п. Эти водоросли в ископаемом виде трудно
определимы. Чаще всего встречаются остатки от нитевидных много-
клеточных синезеленых водорослей. Они бывают в виде изолирован-
ных нитей или (посредством слизи) объединенных в шаровидные
или полушаровидные колонии, дерновинки и т. п. Нить состоит из
трихома — собственно ряда клеток разной формы (шаровидных,
эллипсовидных, бочковидных, цилиндрических и т. п.) — и слизис-
того влагалища, которое может быть одним для нескольких три-
хомов.
Трихомы могут ветвиться разными способами: 1) настоящее
ветвление (рис. 1,о); 2) ложное ветвление (при нем трихом разры-
вается внутри влагалища и, прорывая его, выходит наружу — оди-
ночное ветвление; если оба конца выходят наружу — двойное вет-
вление) (рис. 1,6 — г); 3) V-образное ветвление (трихом образует
изгиб в виде латинской буквы V) (рис. 1, д); 4) петлеобразное ветвле-
ние (ложное ветвление, образующее петлю) (рис. 1, е). В основании
ложного ветвления иногда присутствует гетероциста — погранич-
ная клетка — важный систематический признак, редко сохраняю-
щийся в ископаемом состоянии.
Влагалище для понимания ископаемых форм имеет большое
значение. Оно может быть очень тонким и очень толстым. Форма его
бывает от цилиндрической до воронковидной, в виде насаженных
друг на друга усеченных конусов. Иногда влагалище бывает во мно-
го раз толще трихома и имеет иную форму: трихом утоняется к
концу, а его влагалище расширяется (рис. 1, ж).
Мы остановились на этих особенностях современных водорос-
лей подробнее потому, что известковые инкрустации при биохими-
ческом способе отложения карбоната образуются в слизи между
нитями, т. е. снаружи влагалища, а при тонком влагалище или при
физиологическом способе отложения извести — снаружи трихома
в толще влагалища. Соответственные слепки зависят не только от
Рис. 1. Строение слоевища у синезеленых водорослей
а— Haplosiphon Jontunalis. настоящее ветвление. по Фремн; о — г — схема ложною ве>в-
ления: б — у Plectonema (одиночное и двойное), в — у Polypothrix (одиночное у гетероцисты).
— у Scytonema (двойное между двумя гетероцистами), по Еленкицу (1936): б — различные
стадии возникновения V-образного ветвления у Kyrtuthrix dalntatica. по Эрцеювичу (Егсе-
govic. 1927): е—Scytonema crustaceum f. intrusions, петлевидное ветвление, по Косинской
(1948): ж — схематическое изображение слоистого «воронкообразного» влагалища, по
Еленкину (1936); з — вертикальное сечение известкового нароста Rivularialithus herbidus
Masi.: наверху располагаются обволакивающие форамнниферы Nubecularia со светлыми
полостями камер; чокрак; Западная Грузия, р. Рнон, дер. Баджи
Синезеленые водоросли
11
формы нити, но и от ее расположения, переплетения нитей, плот-
ности колонии, способа ветвления и т. п. Легче всего расшифровы-
ваются случаи с физиологическим отложением извести, когда каль-
цитовые трубки (Girvanella) или удлиненные воротнички [(современ-
ная Scytonema drilosiphon (Klitz)] охватывают трихом. Вообще же
определение остатков синезеленых водорослей является наиболее
трудной задачей ввиду малого количества признаков, специфических
для них. В этих случаях исследователь часто увлекается предполо-
жительными реконструкциями и дает волю своей фантазии. Между
тем определение ископаемого остатка как синезеленой водоросли
несет в себе ответственность в отношении условий произрастания
организма и условий осадконакопления. Когда кроме данного иско-
паемого существует фауна или другие водоросли, вопрос является
менее острым, чем в том случае, когда сохранился только данный
организм. Поэтому 6oJiee осторожное систематическое определе-
ние (как, например, до раздела «Синезеленые или зеленые водо-
росли») явится более научным, чем безапелляционное отнесение
к определенной группе современных водорослей.
Мы рассмотрим ниже формы ископаемых синезеленых водорос-
лей, имеющих породообразующее значение и сравнимых с современ-
ными известьотлагающими синезелеными водорослями, а также
спорные синезеленые водоросли и некоторые карбонатные струк-
туры строматолитов. Формы же, которые могут быть отнесены или
относятся некоторыми исследователями к синезеленым или к ко-
диевым (зеленым) водорослям, рассмотрим после характеристики
кодиевых.
Ниже помещена таблица современных синезеленых водорослей,
отлагающих известь, составленная по определителям синезеленых
водорослей СССР. При сравнении максимальной ширины нити
с шириной каналов некоторых ископаемых кодиевых мы видим,
что диаметр последних не превышает ширины нитей синезеленых
водорослей. Значит, цифровой признак для отнесения ископаемого
остатка к кодиевым или синезеленым водорослям отпадает. Опре-
деление должно быть основано на анализе строения канала, формы
его стенок, наличия дополнительных структур и т. п.
Классификационные вопросы ископаемых водорослей кроме чис-
то палеонтологического имеют и геологическое значение. Кодиевые
водоросли — жители морей нормального солевого режима, а сине-
зеленые могут переносить опреснение. Как видим из табл. I, в совре-
менное нам время часть их живет в море, а часть (при этом наиболь-
шая) — в пресных водоемах.
Это далеко не полный список известьотлагающих форм, так как
он составлен по определителям синезеленых водорослей СССР,
где обызвесткование («инкрустация извести») входит как видовой
признак при определении. Среди современных известьотлагающих
синезеленых водорослей широко известен род Rivularia, предста-
вители которого (морские и пресноводные) образуют сферические
и полусферические колонии, инкрустированные известью и обладаю-
щие радиально расположенными нитями. Трихомы ривулярий сужи-
ваются к концу, в то время как влагалища ведут себя различно у
разных видов, часто расширяясь к периферии и превосходя трихомы
по ширине. Похожие на ривулярий образования встречены в верх-
ней юре запада Украины и в нижнем мелу Далмации—род
Ukrainotrichium. Водоросль образовывала сферические и полусфе-
рические желвачки, в сечении которых отчетливо видны каналы,
расположенные радиально (табл. 5, фиг. 9). В центре желвачка иног-
да различимы отдельные «клетки», частично связанные с каналами-
нитями. Может быть, эти «клетки» являются гетероцистами? Харак-
терно ветвление нитей этого рода: каналы-нити не ветвятся, а но-
вый канал возникает сбоку, тесно прилегая своим основанием. Он
как бы «приклеен» своей стенкой к другому каналу. В этом можно
видеть ложное ветвление трихомов. Эта водоросль очень обильна
в комковатых известняках с большим количеством пелитоморфного
карбоната.
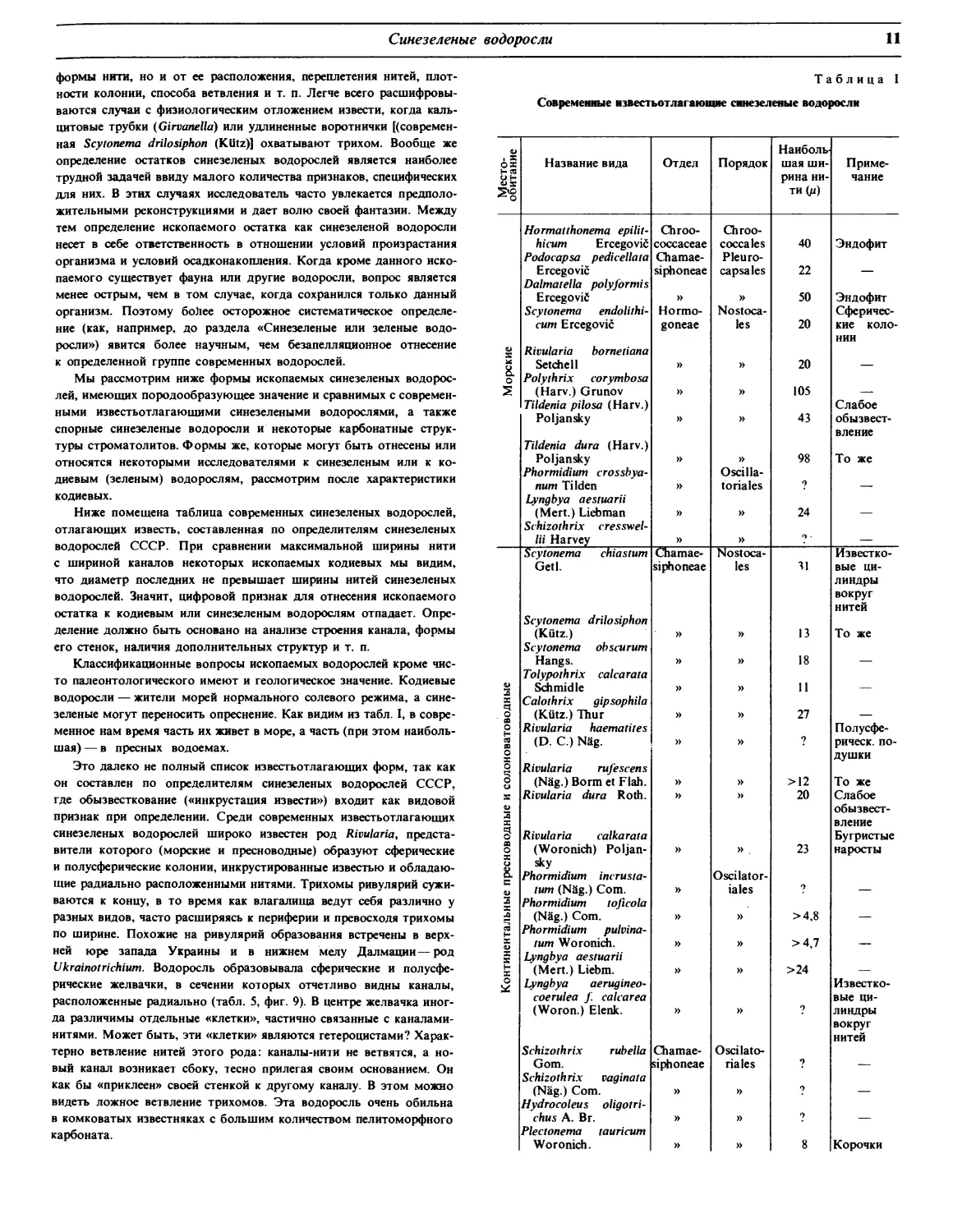
Таблица I
Современные нзвестьотлагающне синезеленые водоросли
Место- обитание Название вида Отдел Порядок Наиболь- шая ши- рина ни- ти (д) Приме- чание
Hormatthonema epilit- hicum Ercegovic Chroo- coccaceae Chroo- coccales 40 Эндофит
Podocapsa pedicellata Ercegovic Chamae- siphoneae Pleuro- capsales 22 —
Dalmatella polyformis Ercegovic » » 50 Эндофит
Scytonema endolithi- cum Ercegovic Hormo- goneae Nostoca- les 20 Сферичес- кие коло-
и S ж о Rivularia bornetiana Setchell » » 20 НИИ
tx о S Polythrix corymbosa (Harv.) Grunov » » 105 —
Tildenia pilosa (Harv.) Poljansky » » 43 Слабое обызвест-
Tildenia dura (Harv.) Poljansky » » 98 вление То же
Phormidium crossbya- num Tilden » Oscilla- toriales ? —
Lyngbya aestuarii (Mert.) Liebman » » 24
Schizoihrix cresswel- lii Harvey » » О • —
Scytonema chiastum Getl. Chamae- siphoneae Nostoca- les 31 Известко- вые ЦИ-
Scytonema drilosiphon (Kutz.) » » 13 линдры вокруг нитей То же
Scytonema obscurum Hangs. » » 18 —
и Tolypothrix calcarata Schmid le » » 11
X ч о Calothrix gipsophila (Kutz.) Thur » » 27
о н е« (В Rivularia haematites (D. C.) Nag. » » ? Полусфе- рическ. по-
о X о ч о Rivularia rufescens (Nag.) Borm et Fl ah. » » >12 душки То же
X Rivularia dura Roth. » » 20 Слабое
и 3 X О X О Rivularia calkarata (Woronich) Poljan- » » л 23 обызвест- вление Бугристые наросты
X о 8. X и sky Phormidium incrusta- tum (Nag.) Com. » Oscilator- iales 9
3 X л Phormidium tojicola (Nag.) Com. » » >4,8 —
X н X о Phormidium pulvina- tum Woronich. » » >4,7 —
X X н X Lyngbya aestuarii (Mert.) Liebm. » » >24
о Lyngbya aerugineo- coerulea f. calcarea (Woron.) Elenk. » » 9 Известко- вые ци- линдры
Schizoihrix rubella Gom. Chamae- siphoneae Oscilato- riales 7 вокруг нитей
Schizoihrix vaginata (Nag.) Com. » » 9 —
Hydrocoleus oligotri- chus A. Br. » » 9
Plectonema tauricum Woronich. » » 8 Корочки
12
Известковые организмы
Другой род — Rivularialithus, менее четкий в смысле сохран-
ности, встречен в третичных осадках Западной Грузии (чокрак)
и Молдавии (сармат) в виде известковых подушковидных наростов
1 мм толщиной. Нити росли вертикальной щеткой, имели ширину
4—5 д, суживались у начала и ветвились под очень острым углом.
В шлифе остатки от нитей имеют вид просветов на темном зернис-
том фоне. Водоросль ассоциировалась с обволакивающими фора-
миниферами — нубекуляриями (рис. 1,з; табл. 5, фиг. 8) и скреп-
ляла осадок на дне водоема.
Очень распространенная ископаемая синезеленая водоросль
Girvanella известна от нижнего кембрия до верхней юры. Она рас-
пространена по всему миру и встречается вместе с морской фауной.
Ее строение весьма простое: известковые трубки с одинаковым на
всем протяжении внутренним диаметром переплетаются друг с
другом, редко разветвляясь. В одних случаях наблюдается направ-
ленность трубок в одну сторону (табл. 5, фиг. 5), а в других — лежа-
щие трубки сменяются торчащими (табл. 5, фиг. 4). Иногда гирва-
неллы образуют желвачки диаметром до 1 см и более, навиваясь
вокруг обломка (табл. 6, фиг. 1,2). Виды гирванелл различаются
по величине диаметра трубок. В стратиграфическом разрезе эти
виды имеют широкое распространение и не могут служить возраст-
ными указателями. Неоднократно внутри трубочек гирванелл встре-
чались поперечные перегородки, что позволило Фреми и Донжару
сравнить одну юрскую форму из Францйи с современным родом
Symploca. Но относятся ли остальные гирванеллы к этому послед-
нему роду синезеленых водорослей, сказать нельзя. Гирванеллы
иногда играют значительную роль в образовании известняков. Так,
в палеозойских (девон-карбон) известняках Южного Урала встре-
чаются желвачки, образованные трубочками гирванелл, закручен-
ными вокруг какого-нибудь обломка (табл. 6, фиг. 1, 2), достигаю-
щие диаметра 2 см и слагающие породу на 50%. Аналогичные жел-
вачки наблюдались в девоне Тувы. Хотя в литературе высказыва-
лось мнение, что гирванеллы образуют оолиты, но исследователи
XX в. не подтвердили этого, так же как и мнение Кайе о том, что
гирванеллы бывают сверлящими организмами.
Другой палеозойский род синезеленых водорослей — Coactilum
резко отличается от предыдущих наличием поперечных перегородок
и вздутыми «клетками» обычно в виде переплетающихся цепочек.
Некоторые изгибы этих нитей напоминают V-образные и ложные
ветвления, что заставляет относить этот организм к синезеленым
водорослям. Но некоторые исследователи стремятся отнести Coa-
ctilum к кодиевым водорослям (табл. 5, фиг. 6, 7). Этот организм
встречается вместе с обломками раковин морских животных от
ордовика до карбона.
Органические спорные образования
и микроструктуры, условно отнесенные
к жизнедеятельности синезеленых водорослей
В этот раздел мы помещаем спорные организмы, трактуемые
разными авторами по-разному, но имеющие больше признаков
синезеленых водорослей. Ниже будут приведены микроструктуры
строматолитов — неясные, но, по-видимому, относящиеся к жизне-
деятельности синезеленых водорослей, а возможно, и бактерий.
Многие из этих образований играют важную породообразующую
роль, а большинство связывается с очень мелководными и часто
опресненными бассейнами и поэтому является показателем условий
осадкообразования. Из описанных ниже организмов Epiphy ton.
Tubiphytes и Nuia жили в морских, иногда слабо опресненных бас-
сейнах. Остальные относятся к пресноводным условиям, в которых
с успехом могли существовать синезеленые водоросли.
Под названием Tubiphytes obscurus Maslov в 1956 г. был описан
организм в виде известковых червеобразных цилиндров и обвола-
кивающих масс, состоящих из темного, мало прозрачного карбо-
ната. При хорошей сохранности и тонком шлифе видно, что темная
масса состоит из переплетающихся очень тонких нитей пелитоморф-
ного карбоната. Внутри этих тонких масс и цилиндров, достигаю-
щих 1 мм толщины и 6 мм длины, часто наблюдаются каналы, во-
круг которых происходит нарастание темной массы, а внутри встре-
чаются включения спикул губок, обломков мшанок и других орга-
низмов (табл. 7, фиг. 9, 10). Тубифитес близок к описанному Джон-
соном и Копией (Johnson a. Konishi. 1956) Retephycus globosus из
миссисипских отложений США. Отличие заключается в закономер-
ном расположении ветвящихся нитей (3—5 д толщиной) у Retephycus.
Два автора отнесли этот организм к зеленым (?) водорослям, для
чего, по нашему мнению, очень мало оснований. Tubiphytes = Nig-
riporella Rigby = Schamovella Rauser имеет большое значение как
породообразователь в верхнекаменноугольных и пермских осадках
Приуралья, слагая рифогенные массивы в районе рек Уфы и Сылвы.
Он также часто попадается в шлифах пластовых известняков. Джон-
сон (Johnson, 1960) указывает на его значение в образовании уголь-
ных пластов, что совершенно не отвечает действительности и описа-
ниям осадков СССР (благодаря ошибке в переводе?). Распростра-
нен этот организм в перми Ирака, п-ова Аравия, Австрии, Афгани-
стана, Бирмы, Южного Китая, Японии, в верхнем карбоне и перми
СССР и США.
Микрокодии известны с 1906 г., когда Глюк описал эти об-
разования как кодиевые водоросли. Microcodium elegans Gliick обра-
зует известковые желвачки, цилиндры и корочки, состоящие из па-
лисадно расположенных вытянутых или уплощенных известковых
пирамид («члеников»), иногда полых в середине (рис. 2, 3).
Рис. 2. Mi crocodium из палеогена Ферганы
Слева — подушковидная колония, ciciikh клеюк темные, iio.iocdi светлые: справа — дру)ая
сохранность удлиненной колонии: оенки клеток светлые, а полости темные (материал
А. Н. Шевнина)
Рис. 3. Microcodium sepimentoforme Masi.
а — отдельный спораший ('.’). встреченный в виде изолированною образования, со светлы-
ми стенками и перегородками; палеоген. Фергана: 6 — поперечное сечение группы члени-
ков микрокодия с темными полостями и тонкими стенками: такое сеченне легко смешать
с группой призм иноцерамов; палеоген: Фергана
Синезеленые водоросли
13
В 1956 г. В. П. Маслов описал третичные микрокодам и выделил
условное семейство Microcodiaceae (из синезеленых водорослей)
с тремя родами: Microcodicum (пермь-третичные), Nuia (ордовик)
и Nannoconus (мел). Найдя нечто вроде спорангиев (рис. 3) среди
пирамид «члеников» и сравнивая способ роста и форму клеток-
полостей в члениках с клетками современной синезеленой водоросли
Dermocarpa, В. П. Маслов отнес семейство Microcodiaceae к Dermo-
carpales (?). В дальнейшем исследования французских авторов пока-
зали, что разнообразные пресноводные, лагунные и морские усло-
вия жизни микрокодий скорее всего подтверждают их принадлеж-
ность к синезеленым водорослям.
Микрокодии часто встречаются начиная с перми в пресноводных
или лагунных осадках. Они или нарастали на неизвестковых водо-
рослях, образуя в ископаемом виде цилиндры, или произрастали
на дне в виде подушек или корок, которые состояли из палисадно
расположенных клеток («члеников»). Пермские формы этих орга-
низмов характеризуются более или менее округлыми пустотелыми
известковыми члениками, иногда тесно прижатыми друг к другу
(рис. 4). Иногда же они произрастают одиночно, причем некоторые
из них росли на растительных стержнях (обычно не сохраняющихся)
(табл. 6, фиг. 7). Для сравнения на рис. 5 приведен рисунок совре-
менной водоросли Dermocarpa, растущей на растительном субстра-
те. Третичные микрокодии отличаются от пермских вытянутостью
тесноприжатых члеников и часто полной перекристаллизацией кар-
бонатных выделений клеток и их полостей (табл. 7, фиг. 2—4). Ордо-
викский род Nuia по характеру своих члеников имеет больше сход-
ства с третичными формами, чем с пермскими. Это цилиндры с
осевой полостью, со стенками, состоящими из более или менее оди-
наковых, палисадно расположенных кристаллов-члеников (рис. 6;
Рис. 6. Nuia sihirica Masi, (схема)
Продольное сечение: ордовик: р. Ангара
Рис. 4. Microcodium permicum Masi.
( гучайпые сечения колоний и отдельных члеников: пермь: Северное Приуралье, с. Ху-
доельское (материал Р. П. Сливковой)
Рис. 5. Клетки современной синезеленой водоросли Dermocarpa
versicolor var. subsalina Рг.-Lavr., нарастающие на другой водоросли
Из работы Прошкиной-Лавренко. 1951. стр. 71. фиг. 9. х 400
а
Рис. 7. Epiphyton
(I - «кустик» эпифит они. pacmniiii над кубком археоциаты: б— поперечное сечение ра-
диально расходящихся «веточек»: кембрий: Сибирь
табл. 7, фиг. 5, 6). В отличие от рода Microcodium нюя встречается
только в морских осадках, отсутствуя там, где растут строматолиты.
Ее остатки иногда составляют в известняках более чем 50% (Мас-
лов, 1967).
Очень важным породообразователем в кембрийских отложениях
является Epiphyton, органическая структура которого до сих пор
остается загадочной. Epiphyton обычно представляет собой нежные
кустики, состоящие из тонких известковых «нитей», разветвляющих-
ся вверх, находящихся в положении роста и сложенных пелитоморф-
ным карбонатом (рис. 7, о — 6). Внутренняя структура нитей не-
различима, но иногда заметны горизонтальные «щели» в виде
светлого карбоната. К. Б. Кордэ (1954) наблюдала ряды «клеток»
неопределенной формы, также расположенных горизонтально или
вертикально, и на основании этого признака, а также на основании
проблематических органов размножения (без анатомической струк-
туры) отнесла этот организм к багряным водорослям. Ранее же он
относился к синезеленым водорослям. Исходя из того, что этот
мелководный организм, участвующий в образовании кембрийских
биогермных массивов в Туве, не разрушался, несмотря на свою
хрупкость, В. П. Маслов (1962, стр. 14—15) выдвинул гипотезу захо-
ронения каждой нити по мере ее роста, т. е. так, как это происходит
со строматолитами. Иными словами, эпифитоны являются своеоб-
разными микростроматолитами, выступавшими над осадком на
незначительную высоту и очень быстро засыпавшимися осадком.
На табл. 6 эпифитоны изображены вместе с археоциатами (фиг. 5)
и гирванеллами (фиг. 6) в положении роста, что у относительно
14
Известковые организмы
глубоководных ветвистых багряных водорослей бывает редко.
Epiphyton встречается главным образом в кембрии, но его предста-
вители найдены и в девоне Урала и Киргизии.
Целая группа карбонатных сгустковидных образований, описан-
ная под разными названиями: Renalcis, Nubecularites, Chabacovia,
Izhella, Schuguria и др., имеет, вероятно, один и тот же принцип
построения: тонкие нити синезеленых водорослей, разрастаясь в
одну или во все стороны, окружались пелитоморфным карбонатом.
При фоссилизации нити сгнивали, а их полости заполнялись тонко-
зернистым же карбонатом. Поскольку полости, оставшиеся от ни-
тей водоросли, измеряются единицами микрон, их очень редко мож-
но увидеть, а остатки от обызвествленных колоний описывались
под разными названиями, в зависимости от их морфологии: Renalcis
(кембрий — девон) — пузыревидные пустотелые образования
(табл. 7, фиг. 8); Nubecularites (кембрий — ордовик) — пузыревид-
ные облакоподобные образования, разрастающиеся периодически
или с заполненной серединой (табл. 7, фиг. 7); Chabacovia (кембрий) —
пузыревидные пустотелые образования, разрастающиеся кустисты-
ми нитями (в поперечном сечении такие нити дадут образования,
неотличимые от Renalcis); Izhella (девон) — пузыревидные образо-
вания с острыми окончаниями оболочки во внутренней полости,
и др.
Все эти образования в значительной мере проблематичны и мо-
гут быть экологическими формами одного исчезнувшего рода сине-
зеленых водорослей. Поэтому следует с осторожностью пользо-
ваться ими для стратиграфических целей, обязательно связывая их
изучение с фациальным анализом.
ЗОЛОТИСТЫЕ ВОДОРОСЛИ (CHRYSOPHYTA)
Табл. 61, фиг. 1—4
Обширная группа золотистых водорослей (большей частью од-
ноклеточных) мало известна в ископаемом состоянии и еще реже
образует известковые выделения. Однако их группа, известная под
названием кокколитофорид, и близкие к ним ископаемые организмы
являются породообразователями и описаны ниже в разделе «Кокко-
литофориды ...» (автор С. И. Шумейко).
В последние годы В. П. Масловым и Н. В. Ренгартен (1964) в чет-
вертичных отложениях (покровные лёссовидные породы Юга СССР)
найдены известковые образования, условно отнесенные к золотис-
тым водорослям, похожим на современные роды Phacodermatium
и Chrysothallus. Эти остатки названы Lithochrysiies calcareus Maslov.
Они участвуют в образовании лёссовых пород и представляют со-
бой известковые наросты — трубчатые корки вокруг центральной
полости, связывающиеся с исчезнувшим обрывком стебля растения,
на котором эпифитизировала водоросль. Стенки этих трубочек
образованы плотно прижатыми друг к другу нитями клеток, чаще
всего целиком замещенных кальцитом. Реже встречаются клетки
с центральной полостью, выполненной темным непрозрачным ма-
териалом. Нити или цепочки клеток обычно располагаются парал-
лельно оси центральной полости корки. Форма клеток в продольном
сечении удлиненная, а контуры их бывают или бочковидные (если
уровни поперечных перегородок данной клетки и клеток соседних
нитей не совпадают), или параллелепипедальные (если уровни пере-
городок совпадают). В поперечных к нитям сечениях клетки изомет-
ричны и обычно имеют шестигранные очертания. Размеры клеток
сильно изменяются по длине и меньше по ширине. Под микроско-
пом (при скрещенных николях) кальцит, заполняющий клетку, или
ведет себя как монокристалл, или распадается на различно гасну-
щие полигоны, или проявляет радиально-лучистое строение. В пос-
леднем случае наблюдается резкое несовмещение центров клетки
и сферолита.
Корковые наросты — трубки имеют разную форму: прямую,
изогнутую, раздувающуюся к одному концу и т. д. Нити клеток
обычно вытянуты параллельно поверхности предмета, к которому
прикреплялась живая водоросль. В одной и той же колонии встре-
чены клетки двух типов: 1) субкубической формы, дающей в разрезе
бочковидные (шестигранные) или широко параллелепипедальные
(иногда слегка овальные) очертания; размеры: длина 60—116 д,
ширина 50—80 д; 2) вытянутой формы; размеры: длина 35—170 д,
ширина 20—76 д (преобладают клетки длиной 170 д и шириной
35 д) (рис. 8).
Рис. 8. Lithochrysiies calcarea Masi.
а — два ряда клеток с темным содержимым: б — клетки первого типа разной формы:
в. г — продольные сечения клеток второго типа: д — клетки первого типа неправильной
формы
Корки часто загрязнены органическим веществом; местами они
гранулированы и тогда плохо различимы (табл. 61, фиг. 1—4). •
Восстанавливая условия образования водорослевых инкруста-
ций, необходимо заметить, что водоросли и другие растения, на
стеблях которых селились инкрустирующие водоросли, являлись
эфемерами. Их произрастание связывается с временными увлажне-
ниями, с эпизодическими появлениями в областях накопления оса-
дочных образований покровной толщи луж, степных блюдец и т. п.
Такие кратковременные увлажнения субаэральных осадков быст-
ро сменялись их осушениями, приводящими к гибели растительных
эфемер. По-видимому, общий засушливый климатический режим
обусловливал сильную насыщенность временных луж солями каль-
ция. В силу этого водоросли принуждены были откладывать из-
весть внутри своих клеток. В периоды общего повышения увлаж-
ненности климата, когда приобретали господство почвенные про-
цессы, условия существования водорослей становились менее благо-
приятными. Увеличение количества гумусовой органики способ-
ствовало быстрейшему растворению карбонатов.
Описываемые остатки распространены (по данным Н. В. Ренгар-
тен) в антропогеновых покровных осадках бассейнов рек Прута,
Дуная, Днестра, Днепра, а также в Средней Азии, на Алтае и в Китае.
В этих отложениях они играют породообразующую роль, концен-
трируя известь в ощутимых количествах. Эти органические остатки
легко обогащаются при отмывке.
Золотистые водоросли
15
Кокколитофориды (Coccolithophoridae)
и близкие к ним организмы
Табл. 8—10
Кокколитофориды — одноклеточные микроскопические планк-
тонные водоросли, выделяемые у нас в порядок Coccolithophora-
les, класса хризомонад (Chrysomonadineae), который относится к
типу золотистых водорослей (Chrysophyta) (Маслов, 19636).
Кокколитофориды — в основном морские организмы, однако
некоторые их виды, живущие в настоящее время, обитают и в прес-
ных водах (Курсанов и др., 1953). Ископаемые кокколитофориды,
описанные в литературе, являются микропланктоном экваториаль-
ных и теплых морей, а также морей умеренного пояса1.
Достоверные остатки кокколитофорид описаны с юры до нашего
времени, но есть основания предполагать, что кокколитофориды
существовали уже в палеозое. Максимума развития кокколитофо-
риды достигли в позднемеловое время, весьма распространены
они также в третичных и некоторых четвертичных отложениях.
В верхнемеловых и третичных морских отложениях кокколито-
фориды играют часто роль породообразующих организмов. Коли-
чество их фрагментов в 1 мм3 достигает нескольких миллионов; Не
исключено, что роль кокколитофорид как породообразователей
сейчас недооценивается вследствие того, что они еще слабо изучены
палеонтологами, а из-за своих малых размеров выпадают из поля
зрения литологов. Характерным примером в этом отношении мо-
жет служить установление только в послевоенные годы породооб-
разующей роли кокколитофорид в так называемой фораминиферо-
вой свите Северного Кавказа (Малышек, 1948).
В последнее время для изучения кокколитофорид с успехом при-
менен метод электронной микроскопии (Deflandre et Fert, 1954;
Hay a. Towe, 1962 а, б; Шумейко, 1962; Nofe’l, 1964).
Морфология и классификация. Протоплазма кокколитофорид с
поверхности покрыта защитным панцирем, состоящим из несколь-
ких известковых [кальцитовых или арагонитовых, иногда кремне-
вых и витеритовых (?)] щитков — кокколитов.
В ископаемом состоянии весь панцирь сохраняется лишь в мо-
лодых (четвертичных, иногда третичных и очень редко в более древ-
них) отложениях. В подавляющем же большинстве случаев прихо-
дится иметь дело с отдельными разрозненными фрагментами пан-
циря — кокколитами, размер которых колеблется от 1 до 20—30 ц
(обычно от 2—3 до 10—15 д). Морфология кокколитов и их очерта-
ния чрезвычайно разнообразны, о чем особенно убедительно свиде-
тельствует электронная микроскопия (табл. 8—10). При этом пан-
цирь может состоять из кокколитов различной морфологии, а один
и тот же кокколит может иметь различное строение дистальной и
проксимальной сторон (табл. 10, фиг. 3, 4). На рис. 9—12 изобра-
жены различные формы кокколитов, которые обозначаются осо-
быми названиями (циклолиты, зиголиты, сфенолиты и т. д.).
Изучение кокколитов с помощью поляризационного микроскопа
позволяет разделить их на два подпорядка (в зарубежных работах
порядка): гелиолитов и ортолитов. Гелиолиты сложены из кальци-
товых элементов — кристаллов, подобно сферолитам (рис. 9—11),
и в скрещенных николях дают либо крест, либо изогиры. Ортолиты
обладают мозаичным угасанием и состоят из беспорядочно ориенти-
рованных кристаллов. Следует отметить, что такое разделение-
весьма искусственно, так как довольно большое количество кокко-
литов имеет периферическую часть, построенную по типу гелиоли-
та, а внутреннюю — по типу ортолита.
' Для существования и нормального развития современных кокколитофорид необходимы:
водная среда, богатая кислородом с высоким pH (8—8,5), постоянная температура +10 —
----1-26 , чистая вода. Они живут на небольшой глубине, но их цисты, споры и пальмел-
лоидные стадии опускаются на большие глубины. Распределение кокколитофорид свя-
зано также с морскими течениями, ветрами, удаленностью от берега (большинство их
составляет прибрежный планктон), количеством растворенных в воде нитратов и фос-
фатов. Некоторые коккосферы живут в солоноватоводных бассейнах, а некоторые при-
крепляются к диатомовым водорослям (прим. В. П. Маслова).
Рис. 9. Кокколиты
а — коккосфера: б — дискосфера: а. б— по Дефляндру (Deflandre. 1936): в — з— различ-
ные типы дисколитов: в. г — по Камптнеру. вид сбоку и сверху, д — з— по Дефляндру,
и — Calcidiscus medusoides Kamptner: к — м — типы калиптролитов: к. м — вид сбоку.
л — вид сверху, по Дефляндру (Deflandre. 1936): н. о — цнклолнт: н — боковое сечение.
о — вид сверху, по Камптнеру: п — с — типы зиголитов: п. р — вид сверху. < — вид сбоку,
по Дефляндру
16
Известковые организмы
Рис. 10. Кокколиты.
а— стефанолит. по Дефляндру (Deflandre, 1952); б — г — сфенолиты: в—Micula deyussa-
ta Veksch.. по Векшиной (1959). б, г— Sphenolithus radians Deflandre: д—к—типы лопадо-
.штов: д — и — вид сбоку, к — вид сверху, по Дефляндру (Deflandre. 1936. 1952): .» — р—
типы тремалитов: л. м—по Камптнеру. и — р — по Дефляндру (Deflandre. 1936): с—г—
типы крибролнтов: с — по Архангельскому (1912), т, у — по Векшиной (1959); ф — ч—
типы рабдолитов^ по Дефляндру
Рис. 11. Кокколиты
а— Braarudosphaera bii/clari Gran et Braarud: 6—Teiralithus pyramidus Gardet.: a — e—
типы мнкрантолитов. по Дефляндру: ж. з—Scapholilhus. по электроиномнкроскопиче-
скому снимку (Deflandre. Fert. 1954): и — о — дискоастеры: и. к — по Тан-син-хок (Тап-
Sin-Hok. 1927). л — по Шамрай и Лазаревой (1956). .и— по Клумпп. н. о— Heliadiscoasier.
по Клумпп; и — < —фрагменты микрорабдолитов, по Дефляндру: т. у — нстмолиты. по
Дефляндру: ф — ч — Nannacanus calami (Lapp.), по Дефляндру (Deflandre. 1952)
Золотистые водоросли. Зеленые води/гт ли
17
Рис. 12. Кокколит в известняке
3 >e‘<p<>ii|b<M>ik|>.»'L»ii>i4ccM<ii снимок iHonjo. Ficher. 1464с х 551*' юра: A iwiu
Методика исследования. Кокколитофориды можно наблюдать
в петрографических шлифах и порошковых препаратах как под обыч-
ным поляризационным микроскопом, так и при помощи электрон-
ного микроскопа. При наблюдении под обычным микроскопом
желательно использовать увеличения 600—1000, применяя иммер-
сионные объективы. .
В петро! рафических шлифах кокколиты и их фрагменты обычно
различимы лишь на краях и в тонких местах шлифа. Возможности
для диагностики кокколитов в шлифах весьма ограниченны, однако
внимательное исследование шлифа дает возможность установить
наличие целых кокколитов или их крупных фрагментов в породе
(табл. 8, фиг. 1—3) н тем самым ориентировать исследователя
на более детальное изучение образца в порошковых препаратах.
В шлифах легко можно определить такие характерные группы, как
днскоастеры (табл. 8, фиг. I), рабдолиты, стефанолиты, причем
дискоастеры могут быть диагностированы более точно (до рода).
Значительно большие возможности для исследования и диагно-
стики кокколитофорид дают порошковые препараты. Они изготов-
ляются путем размачивания кусочка породы в пробирке или путем
отмучивания определенных размерных фракций. Препараты могут
приготавливаться иа воде, глицерине, желатин-глицерине, бальзаме
и т. д. Особенно подходят для этой цели вязкие среды, позволяющие
путем осторожной притирки покровного стекла получить гонкий
слой препарата и при этом менять ориентировку кокколитов (табл. 8,
фиг. 4—8). Для диагностики кокколитофорид желательно приме-
нение увеличений около 1000, хорошо отрегулированной освети-
тельной системы и фазово-контрастных устройств.
Современная классификация кокколитофорид основана на данных
обычной микроскопии, полученных при изучении порошковых
препаратов. Однако предел разрешения световых оптических систем
(примерно 0.25 д) сильно ограничивает возможности обычной оп-
тики, для которой детали строения кокколитов в I д и менее практи-
чески недоступны. Зарисовки кокколитов под микроскопом обычно
идеализированы и ие лишены субъективизма.
С 50-х годов для изучения кокколитов стали уверенно применять
электронную микроскопию. При изучении кокколитов «иа просвет»
получается теневое изображение (табл. 9, фиг. I), так как кокколиты
«непробиваемы» электронным пучком. Такой метод, естественно,
далеко ие исчерпывает возможностей электронной оптики.
Для изучения поверхности кокколитов применяется метод элек-
тронномикроскопических углеродных реплик (отпечатков). Сам кок-
колит растворяется слабой кислотой, а непосредственно под микро-
скопом исследуется тончайший углеродный «футляр» — реплика,
проницаемая для электронного пучка (Шумейко, 1962).
Электронная микроскопия, помимо исследования тончайших де-
талей морфологии кокколитов в порошковых препаратах, позволяет
исследовать их, а также вмещающую породу на сколах и пришли-
фовках (табл. 10, фиг. 1, 2, 5). При этом становится возможным об-
наружение кокколитов или их остатков даже в некоторых крепко
сцементированных породах (известняках) (рис. 12) (Honjo, Fischer,
1964). При изучении же наиболее тонкодисперсных компонентов
породы, как. например, «порошкового кальцита», мела и мергелей
(табл. 10, фиг. 7,8), только электронная микроскопия дает возмож-
ность сделать правильный вывод об органогенном или неоргано-
гениом генезисе породы.
ЗЕЛЕНЫЕ ВОДОРОСЛИ (CHLOROPHYTA)
Эта обширная группа водорослей характеризуется зеленым цве-
том пигмента и болыйим разнообразием своего строения. Но в
ископаемом состоянии чаше всего встречаются обызвествляющиеся
лонные прикрепленные неклеточные водоросли или сифоновые.
Тело этих водорослей, при значительных размерах и сложном внеш-
нем строении, не разделено на отдельные клетки; если же клетки
существуют, то они очень крупные н обладают многими ядрами
(разделение на клетки происходит в позднем онтогенезе). Обычно
сифоновые выделяются в порядок Siphonales, но некоторые бота-
ники выделяют особый порядок Siphonocladiales для водорослей, об-
ладающих делением клетки. Так как в ископаемом состоянии трудно
судить об одноклеточном или многоклеточном строении слоевища,
удобнее относить все сифоновые к одному порядку Siphonales.
Строение сифоновых бывает двух типов: 1) единая клетка обра-
зует несколько центральных сифонов и отходящих от них ответ-
влений — семейство Codiaceae; 2) центральный сифон один и от
него отходят более или менее закономерно ответвления к перифе-
рии— семейство Dasycladaceae, или мутовчатые сифонеи.
Кодиевые сифонеи (Codiaceae)
Табл. 11.12; табл. 13. фиг. 1—3
Эта группа водорослей характеризуется слоевищем в виде ог-
ромной сильно разветвленной клетки, очень разнообразной внешней
формы: в виде тонковетвистых кустиков, членистых кустиков, лис-
товидных пластин и т. п. Из них представители подсемейства Udo-
toideae отлагают известь на поверхности клетки и сохраняются в
ископаемом состоянии. Из современных представителей известко-
вых кодиевых широко известен и лучше изучен род членистых ко-
диевых Halimeda.
Химический состав. Изучен только у нынеживущих представи-
телей. Известь отлагается физиологическим путем в виде арагонита,
кристаллы которого не превышают 2—9 д в длину и 0,5 ц в ширину.
Естественно, что в ископаемом виде мы встречаем перекристалли-
зованный кальцит. Углекислый магний содержится в небольших
количествах — от 0,44 до 5,5%. У современного рода Halimeda
констатировано 1,3% и даже до 2,3% SrCO3, что является повышен-
ным содержанием по сравнению с известковыми багряными водо-
рослями, у которых количество углекислого стронция не превы-
шает 0,35%. Сводные таблицы некоторых химических анализов
современных кодиевых, так же как и сведения по современной лите-
ратуре. приведены у Кониси (Konishi. 1961).
Экология. Кодиевые являются теплолюбивыми растениями и
занимают тропическую и субтропическую зоны. Так, род Halimeda
встречается между 45° с. ш. и 34° ю. ш.
Глубины, на которых произрастают кодиевые, обычно ие превы-
шают 50 м, но зависят от прозрачности воды; как исключение,
они встречаются до глубины 120 м. Обломки кодиевых. обнаружен-
ие?
2
18
Известковые организмы
ные на значительных глубинах (до 240 м), вероятно, занесены
ураганами, цунами, подводными оплываниями и т. п. Лучше всего
изучено батиметрическое положение разных видов современного
рода Halimeda. По Тейлору, намечается два интервала их оптималь-
ного произрастания: одни виды живут на глубинах от 0 до 10—15 м,
главным образом в лагунах рифов и в лужах, не просыхающих при
отливе, а другие виды произрастают на глубинах от 40 до 60 м
(Н. opuntia до 90 л<). Особенно богаты этими водорослями лагуны
атоллов, защищенные от волн. На Багамской банке большинство
кодиевых произрастает на глубинах от 2,5 до 36 м на скальном
основании.
Нынеживущие кодиевые, обладая кустистым или листовидным
слоевищем, чувствительны к сильному движению воды. Поэтому
они селятся там, где движение воды слабое, благодаря чему кустики
не разрываются и не раздавливаются перемещающейся по дну галь-
кой.
Геологическое значение. Прежде чем рассматривать значение
кодиевых водорослей, необходимо отделить истинных кодиевых от
сомнительных. Истинные кодиевые могут сравниваться по своей
морфологии и анатомии с современными известковыми кодиевыми
водорослями. Сомнительные же* кодиевые, выделенные в особое
подсемейство Garwoodioideae, среди современных известьотлага-
ющих форм аналогов не имеют, анатомическое их строение сходно
со строением некоторых синезеленых водорослей, эволюция их не
может быть прослежена и экологические условия их жизни ближе к
синезеленым водорослям, чем к истинным кодиевым.
Стратиграфическое значение немногочисленных родов истинных
кодиевых бесспорно, но нужно оговориться, что некоторые кемб-
рийские формы, как, например, Ascosoma, Milscherlichia, для кото-
рых имеются лишь первичные описания, требуют дополнительного
изучения, так как их анатомия вызывает ряд сомнений.
Истинные кодиевые известны с ордовика, когда появляются
Dimorphosiphon и Palaeoporella.
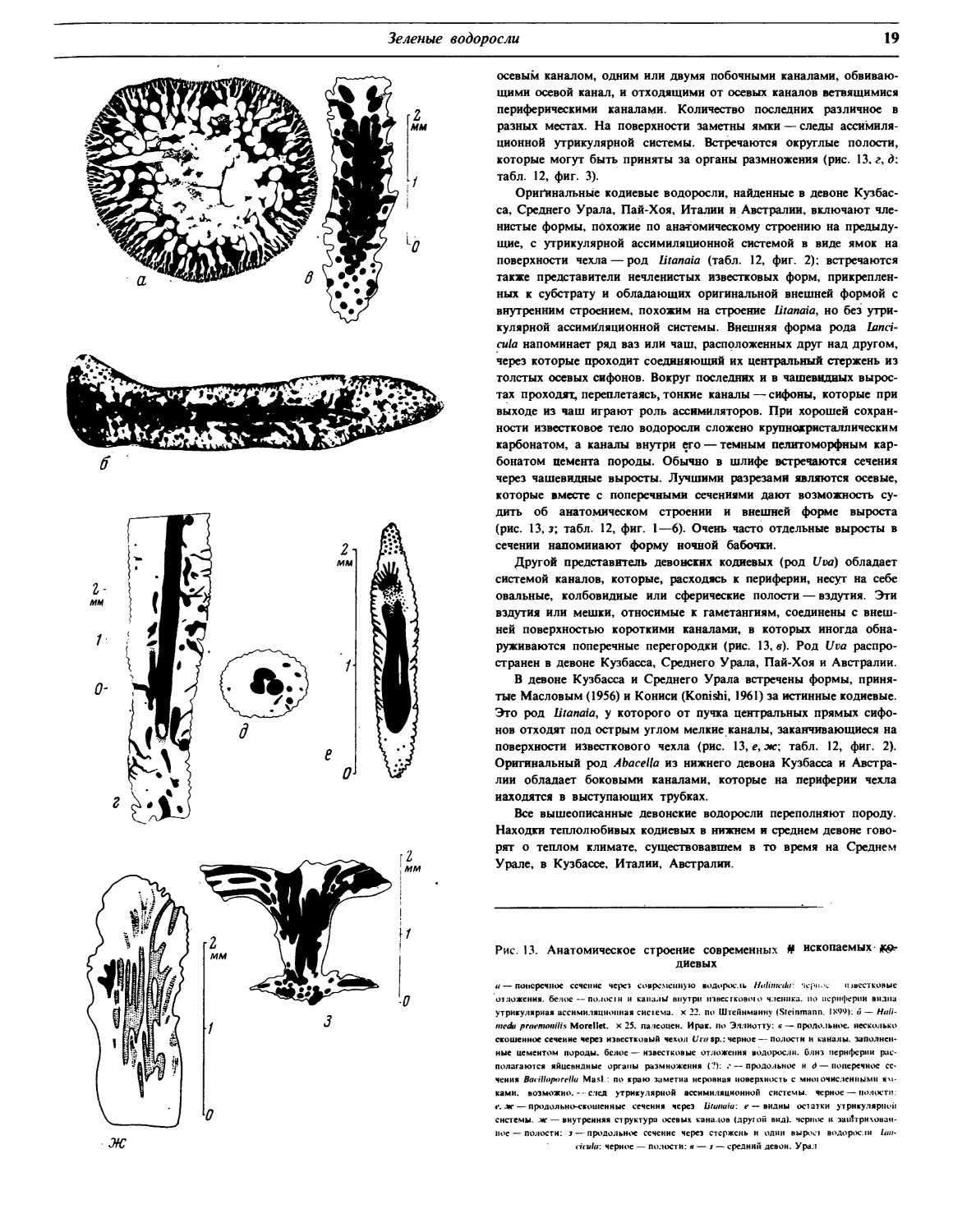
В девоне известны lancicula, Abacella, litanaia, Uva.
В карбоне распространены Calcifolium и Ivanovia.
Со среднего карбона до нижней перми распространены описан-
ные японцами Eugonophyllum и Anchicodium.
В перми известны роды Cyninocodium (некоторыми авторами
относимый к багряным водорослям) и Penicilloides. От перми до
мела распространен род Permocalculus.
В мелу известен род Boueina, близкий к третичному и современ-
ному Halimeda.
В третичных осадках кроме этого последнего рода встречаются
Ovulites и Penicillus.
Некоторые виды этих родов характеризуют ярусы.
Подсемейство Garwoodioideae (сомнительных кодиевых) вклю-
чает до 10 родов, среди которых Headstroemia распространен от
ордовика до нижнего карбона, a Orionella — от силура до верхнего
карбона. Если принять во внимание, что юрско-меловая форма
Cayeu.xia сходна с предыдущим родом и по существу от него не от-
личается, эволюция этих родов весьма медленная, что говорит об
их примитивности и, может быть, о принадлежности к синезеленым
водорослям.
Современные известковые кодиевые бывают породообразующи-
ми. В лагунах рифов и атоллов количество обломков Halimeda бы-
вает настолько велико, что они образуют известковые пески, а чет-
вертичные детритусовые известняки переполнены обломками их
члеников. В более отдаленные времена они не были столь обильны и
в отложениях обычно составляют небольшой процент органических
обломков. Но некоторые пластинчатые формы (Ivanovia в карбоне)
создавали нечто вроде биогермных известняков.
Истинные кодиевые водоросли, по-видимому, как и современные,
являлись теплолюбивыми.
Морфология и аъатомия. Современные кодиевые прикрепляются
к субстрату небольшой частью своего слоевища и возвышаются над
дном, колеблемые движением воды (табл. 11. фиг. 1). Их анатомия
иногда довольно сложна, несмотря на одноклеточное строение.
Многочисленные ветвления гигантской клетки создают перепутан-
ную сеть ветвящихся сифонов (разную у разных родов). Обычно
имеется система более крупных центральных сифонов, от которых
отходят более мелкие ответвления. Концы этих ответвлений играют
роль ассимиляторов и устроены различно: у Halimeda и родствен-
ных форм они образуют у поверхности членика так называемую
утрикулярную систему — ряды мелких мешков, соединенных с под-
водящими сифонами (рис. 13. а: табл. II. фиг. 7). У ископаемых
форм эта система имеет вид мелких Ямок, расположенных на по-
верхности известковых члеников. У пластинчатых форм, как, на-
пример, у современного рода Udoiea, мелкие ответвления, прохо-
дящие параллельно поверхности «листа», обладают короткими
пенькообразными отростками (папиллами), которые своими кон-
цами выступают на поверхности слоевища. В ископаемом состоя-
нии можно уловить только следы папилл: у рода Ivanovia, возможно,
им отвечают затемненные каналы или «клетки» у самой поверх-
ности пластины (табл. 11, фиг. 5).
Органы размножения современных кодиевых (гаметангии и спо-
рангии) обычно в виде мешков грушевидной или колбовидной формы
располагаются на поверхности или близ поверхности слоевища.
В ископаемом виде в известковых чехлах водоросли Uva были встре-
чены сферические и грушевидные (иногда колбовидные) полости,
принимаемые за гаметангии (см. рис. 13. в). Образования несколько
иной формы, в виде вытянутых вздутых окончаний центральных
сифонов, встречены у рода lancicula. Полости сферической формы
у Gymnocodium, Permocalculus1, Orlhriosiphon. Eugenophyllum также
принимаются за гаметангии кодиевых.
Перейдем теперь к характеристике некоторых ископаемых ро-
дов. Роды Calcifolium, Ivanovia, Anchicodium, Eugonophyllum имеют
слоевища в форме известковых пластин. По краю этих пластин у
Ivanovia и Anchicodium и Eugonophyllum расположен ряд затемнений
и каналов, принимаемых за кортикальную систему ассимиляторов.
Середина пластин перекристаллизована.
Calcifolium — лучше изученный и более четкий в смысле сохран-
ности, он существенно отличается от этих трех родов: тонкие плас-
тинки имеют один или два слоя, проходящих параллельно поверх-
ности ветвящихся тонких канальцев. При этом пластинки часто
образованы темным пелитоморфным карбонатом, а каналы —
светлым (соотношение, обратное всем истинным кодиевым иско-
паемым водорослям). Форма этих пластин различная — от почти
плоской, до вогнутой в виде буквы S (табл. 12, фиг. 7 — 9).
Триба Halimedeae характеризуется членистым слоевищем, у
которого уплощенные или цилиндрические членики образованы
толстыми осевыми сифонами, окруженными ответвляющимися от
них тонкими сифонами. На самой периферии образуется утрику-
лярная ассимиляционная система в виде ряда мелких вздутий, сое-
диненных с основными сифонами (рис. 13. а: табл. 12, фиг. 4). От-
дельные вздутия утрикулярной системы у современных галимед
значительно меньше внутренних каналов. Кодиевые с таким строе-
нием появились в ордовике (Papaeoporella), известны в девоне и
выше, а с мела (род Boueina) они очень близки к современному
роду Halimeda.
Ордовикские формы кодиевых водорослей характерны прямыми
известковыми члениками, внутренняя структура которых состоит
из более толстых осевых каналов, от которых отходят ветвящиеся и
неветвящиеся, более тонкие каналы. Dimorphosiphon имеет от 7
до 25 осевых каналов и отходящие под прямым углом периферичес-
кие, более тонкие каналы. В девоне Среднего Урала встречен род
Bacilloporella в виде длинных прямых члеников с одним основным
Некоторые авторы относят пермские роды Gymnocodium н Permocalculus к багряным
водорослям (Пиа. Джонсон, Эндо), а другие считают их кодиевыми (Маслов, Кониси).
Зеленые водоросли
19
о
осевым каналом, одним или двумя побочными каналами, обвиваю-
щими осевой канал, и отходящими от осевых каналов ветвящимися
периферическими каналами. Количество последних различное в
разных местах. На поверхности заметны ямки — следы ассимиля-
ционной утрикулярной системы. Встречаются округлые полости,
которые могут быть приняты за органы размножения (рис. 13, г, д',
табл. 12, фиг. 3).
Оригинальные кодиевые водоросли, найденные в девоне Кузбас-
са, Среднего Урала, Пай-Хоя, Италии и Австралии, включают чле-
нистые формы, похожие по анатомическому строению на предыду-
щие, с утрикулярной ассимиляционной системой в виде ямок на
поверхности чехла — род litanaia (табл. 12, фиг. 2); встречаются
также представители нечленистых известковых форм, прикреплен-
ных к субстрату и обладающих оригинальной внешней формой с
внутренним строением, похожим на строение litanaia, но без утри-
кулярной ассимиляционной системы. Внешняя форма рода Lanci-
cula напоминает ряд ваз или чаш, расположенных друг над другом,
через которые проходит соединяющий их центральный стержень из
толстых осевых сифонов. Вокруг последних и в чашевидных вырос-
тах проходят, переплетаясь, тонкие каналы — сифоны, которые при
выходе из чаш играют роль ассимиляторов. При хорошей сохран-
ности известковое тело водоросли сложено крупнокристаллическим
карбонатом, а каналы внутри его — темным пелитоморфным кар-
бонатом цемента породы. Обычно в шлифе встречаются сечения
через чашевидные выросты. Лучшими разрезами являются осевые,
которые вместе с поперечными сечениями дают возможность су-
дить об анатомическом строении и внешней форме выроста
(рис. 13, з; табл. 12, фиг. 1—6). Очень часто отдельные выросты в
сечении напоминают форму ночной бабочки.
Другой представитель девонских кодиевых (род Uva) обладает
системой каналов, которые, расходясь к периферии, несут на себе
овальные, колбовидиые или сферические полости — вздутия. Эти
вздутия или мешки, относимые к гаметангиям, соединены с внеш-
ней поверхностью короткими каналами, в которых иногда обна-
руживаются поперечные перегородки (рис. 13, в). Род Uva распро-
странен в девоне Кузбасса, Среднего Урала, Пай-Хоя и Австралии.
В девоне Кузбасса и Среднего Урала встречены формы, приня-
тые Масловым (1956) и Кониси (Konishi, 1961) за истинные кодиевые.
Это род litanaia, у которого от пучка центральных прямых сифо-
нов отходят под острым углом мелкие каналы, заканчивающиеся на
поверхности известкового чехла (рис. 13, е, ж; табл. 12, фиг. 2).
Оригинальный род Abacella из нижнего девона Кузбасса и Австра-
лии обладает боковыми каналами, которые на периферии чехла
находятся в выступающих трубках.
Все вышеописанные девонские водоросли переполняют породу.
Находки теплолюбивых кодиевых в нижнем и среднем девоне гово-
рят о теплом климате, существовавшем в то время на Среднем
Урале, в Кузбассе, Италии, Австралии.
Рис. 13. Анатомическое строение современных # ископаемых
диевых
и— поперечное сечение через современную водорос.н. l/uliincdu черное швестковые
отложения, белое — полос!» и капали внутри известковою членика, но периферии видна
утрикуляриая ассимиляционная система, х 22. по Штейнманну (Steinmann. 1899); о — Huli-
medti praemonilis Morellet. х 25. палеоцен. Ирак, по Эллиотту: « — продольное, несколько
скошенное сечеиие через известковый чехол 17 гн sp.: черное— полости и каналы, заполнен-
ные цементом породы, белое — известковые отложения водоросли, близ периферии рас-
полагаются яйцевидные органы размножения ('.’): — продольное и д—поперечное се-
чения Bacilloporella Masi : по краю заметна неровная поверхность с mhoi очисленными ям-
ками. возможно.- след утрикулярной ассимиляционной системы, черное — полости:
е. ж— продольно-скошенные сечення через Liuniaia е — видны остатки утрикулярной
системы, ж — внутренняя структура осевых каналов (другой вид), черное и заштрихован-
ное— полости: з— продольное сеченне через стержень и один вырост водоросли Ь/п-
cicula: черное — полости: в — з — средний девон. Урал
20
Известковые организмы
Синезеленые или кодиевые водоросли
Сомнительные, или спорные, кодиевые водоросли объединены
Кониси в подсемейство Garwoodioideae. Сам Кониси считает, что
это подсемейство, вероятно, относится к зеленым водорослям.
Наиболее характерные и часто встречающиеся роды Ortonella
и Cayeuxia образуют известковые желвачки с пронизывающими их
радиальными светлыми, более или менее прямыми каналами с
близкими по величине диаметрами. Ветвление каналов, угол ветвле-
ния и характер его считались родовыми признаками, по которым
выделялись роды: Ortonella с симметричным ветвлением в виде
буквы V (рис. 14), Cayeuxia с ветвлением в виде буквы U или пере-
вернутого П, Garwoodia и Hedstroemia с ветвлением в виде остроги
или буквы Ш (с несколькими ответвлениями). Но встречены пред-
ставители рода Ortonella, у которых в основании ветвление под
Рис. 14. Спорная кодиевая или синезеленая водоросль
Ortonella Jurcata Garwood, х 20
I o.ioihii: нижний карбон: Ашдия: по Гарвуд} (Garwood. 1914)
одним углом, а на периферии при плотно расположенных и парал-
лельных каналах угол ветвления, сильно уменьшаясь, такой же,
что и у рода Cayeuxia. С другой стороны, и у последнего (мезозой-
ского) рода были встречены ветвления в виде буквы V (табл. 11,
фиг. 3). Таким образом, определение геологического возраста по
ортонеллам и кайексиям затруднено. Эти организмы часто встре-
чаются вместе с морскими животными, но иногда изобилуют в ком-
коватых известняках с остатками синезеленых водорослей, т. е.,
возможно, живут в опресненных бассейнах (нижний мел Далмации,
верхняя юра Западной Украины и др.). Роль желвачков этих водо-
рослей как породообразователей иногда значительная.
Мутовчатые сифонеи (Dasycladaceae)
Табл. 13, фиг. 4—10; табл. 14, фиг. 4—7; табл. 15, 16
Это семейство исключительно донных водорослей обладает
большим количеством ископаемых представителей начиная с кемб-
рия и до современного времени. Разнообразная структура известко-
вого чехла, быстрое вымирание одних и появление других форм,
значительная роль в породообразовании некоторых известняков
делают их важными стратиграфическими указателями.
Химический состав. Химический состав известкового чехла у
живущих форм близок к чистому кальциту. Отложение извести про-
исходит физиологическим путем и обычно представлено прозрач-
ным кристаллическим карбонатом, за исключением рода Konin-
ckopora (карбон), который характеризуется тонким известковым
чехлом, сложенным пелитоморфным кальцитом.
Экология. Современные представители мутовчатых сифоней
встречаются в морях океанической солености в субтропической и
тропической зонах с температурой воды не ниже +20° С и отсутст-
вуют в умеренном климате1. В ископаемом состоянии в третичное
и меловое время мутовчатые сифонеи хотя и располагались поло-
сой вдоль экватора, но занимали значительно более широкую зону,
чем теперь.
Глубина обитания мутовчатых сифоней не превышает 60 .и. Не-
которые из них, например представители современного рода Neome-
ris, живут главным образом на литорали в лужах, не пересыхающих
при отливе, а некоторые виды этого рода (Nannulata и N. van bosseae)
обитают также на глубине до 10 м (Konishi, Epis, 1962) и даже до
50 м. Род Acicularia обычен на глубине 30 м (Hamme г ling, 1944).
Геологическое значение. В ископаемом состоянии мутовчатые
сифонеи часто слагают известняки и являются важными породо-
образователями. Особенно обильны ими мелководные фации. Встре-
чаются они в ископаемом состоянии в обломках в виде детрита и
редко в неразрушенном виде. В ордовике и силуре ими слагаются
иногда пропластки известняков. В карбоне СССР мелкие формы из
группы березелл иногда почти нацело слагают известняки. В западно-
европейском триасе известны известняки, переполненные обломками
мутовчатых сифоней. Довольно часто они встречаются в мелковод-
ных известняках юры Крыма, Югославии, Италии и в мелу Средней
и Малой Азии (Carozzi, 1955; Elliott, 1958). Встречены они также
в третичных породах Украины (Маслов, 1956; Маслов, Ярцева, 1969).
Стратиграфическое значение мутовчатых сифоней велико. Кем-
брийские примитивные и относительно проблематичные и редкие
мутовчатые сифонеи, которых здесь можно не касаться, начиная с
ордовика, быстро эволюционируя, дают ряд групп и форм, позво-
ляющих пользоваться ими как руководящими ископаемыми.
Количество родов в разных системах колеблется: в кембрии 5
родов, ордовике — 8, силуре — 3, девоне — 2, карбоне—15, пер-
ми— 21, триасе—12, юре — 22, мелу—14, третичной системе —
311 2. Первые четыре системы характеризуются присутствием отно-
сительно примитивных крупных форм, в карбоне появляются мел-
кие и крупные усложненные формы, а также формы с обызвествлен-
ными органами размножения. В перми появляются представители
групп, широко развитых в мезозое, многие из которых достигают
большой сложности слоевища уже в триасе. Ранее считалось, что
наибольшего развития мутовчатые сифонеи достигали в ордовике,
триасе и третичном периоде. Но, по-видимому, такое суждение было
связано с низкой степенью изученности этих водорослей. Микро-
палеонтологические исследования последнего десятилетия показали,
что начиная с карбона и до третичного периода происходит отно-
сительно равномерное развитие этих водорослей.
Определение мутовчатых сифоней возможно в шлифах и в об-
ломках в неориентированных сечениях, но при наличии разрезов,
пересекающих ось центральной полости (или ячеи).
Сечения через известковый чехол (продольные, поперечные и
тангенциальные) в совокупности часто позволяют реконструировать
известковый чехол. Рекомендуется давать ряд изображений сечений
и реконструкцию чехла. Обилие признаков у сифоней позволяет
в шлифах выделять роды и виды, быстро сменяющиеся в страти-
графическом разрезе.
Анатомия и методика изучения. Эти очень интересные водоросли
имеют закономерно и сложно разветвленную систему «сифонов»
одной гигантской клетки. Отложение извести происходит в опре-
деленных местах на поверхности этой клетки в виде известковой
корки, или чехла, определенной для данного вида толщины. Чехлы
1 За некоторыми редкими исключениями, во внутриконтинентальных бассейнах.
2 В этом списке некоторые роды переходят из одной системы в более молодую систему,
поэтому количество новых родов с трнаса несколько меньше: триас — 10, юра — 19,
мел — 11, третичные — 24. Сомнительные роды не вошлн в список.
Зеленые водоросли
21
бывают с неразветвленной и разветвленной осевой частью: палочко-
видные, грушевидные, шаровидные на ножке, четковидные (членис-
тые), зонтиковидные и т. п. В шлифах их внешняя форма не всегда
легко расшифровывается, и большое значение имеют ответвления
от центральной осевой части, которые после гибели живого орга-
низма оставляют в известковом чехле канальчики или поры.
Ответвления от центральной полости или образуют сквозные
поры или замкнуты внутри стенки известкового чехла.
Открытые поры (пересекающие стенку чехла) могут быть:
I) простыми (неветвяшимися): 2) ветвящимися на ответвления вто-
рого, третьего и т. д. порядков: 3) расширяющимися к поверхности;
4) сужающимися к поверхности.
Поры замкнутые могут заканчиваться вздутием или сужением.
Кроме того, в порах или рядом с ними образуются округлые или
продолговатые полости для спор.
Расположение пор бывает: 1) беспорядочное, 2) в мутовках,
3) в мутовках пучками.
Ответвления, выходя из пор известкового чехла, играют роль
ассимиляторов, как у кодиевых водорослей. Ассимиляторы бывают
двух типов: в виде волосков или в виде лепешек. Род Turkmeniaria
Masi, обладает обоими типами ассимиляторов, что легко распоз-
нается по его известковому чехлу (рис. 15, а, б).
Упомянутые сферические или овальные полости, которые при
жизни служили вместилищем для спорангиев или гаметангиев, не-
редко обызвествляются у сифоней, а некоторые роды только и оп-
ределяются по этим органам плодоношения, как, например, род
Acicularia. Особые известковые спорангиофоры (иглы или палочки)
этого растения, собранные в плодоносные мутовки или «зонты»
(похожие на цветы ромашки), несут ряды сферических углублений
или полостей по краю и внутри известковой палочки (рис. 16; табл. 13,
фиг. 8; табл. 16, фиг. 10—12).
Рис. 15. Turkmeniaria adduct a Masi, с двумя типами ассимиляционных
окончаний; в виде нитей и в виде полусфер, по В. П. Маслову (1963а)
косое сечение чехла: г> реконструкция чехла: в — продольное и поперечное сечения:
внешняя поверхпосiь: <>—реконструированные предположительные ассимиляторы
Рис. 16. Внешний вид
зонта Acicularia persica
L. et. J. Morellet
Средний сармат: Иран
Начиная с ордовика развиваются несомненные мутовчатые си-
фонеи, вначале представленные двумя типами слоевищ: 1) цилин-
дрической формы с деветвящимися или ветвящимися порами (ка-
налами от боковых ответвлений); 2) сферическими головками на
цилиндрической ножке. Сферическая головка образована радиально
расположенными ответвлениями с расширением на внешнем конце.
Эти расширения покрываются тонкой известковой корочкой, кото-
рая в совокупности с соседними образует сферический ячеистый
известковый чехол.
Типичным представителем первого типа является род Rhabdo-
porella, распространенный от ордовика (кембрия?) до карбона.
Построение известкового чехла простое: он цилиндрической фор-
мы и пронизан прямыми неветвящимися порами с небольшим рас-
ширением на внешнем конце. Другой ордовикско-силурийский род —
Vermiporella, с цилиндрическим ветвящимся известковым чехлом,
пронизанным прямыми или несколько изогнутыми порами. Работа
Пратурлона (Praturlon. 1963, стр. 123) ставит под сомнение принад-
лежность этого рода к мутовчатым сифонеям. Вермипореллы часто
являются породообразующими и в шлифах дают причудливые пет-
ли сечений известкового чехла, при ветвлении образующего тре-
угольные расширения и неправильные раздувы, что не характерно
для истинных мутовчатых сифоней (табл. 14, фиг. 5—7).
Потомком рабдопорелл, по-видимому, является род Масгоро-
rella, появившийся в перми и развитый в мезозое. Это прямые
или слабо изогнутые известковые цилиндры, пронизанные простыми
расширяющимися кнаружи порами (табл. 15, фиг. 3). Близкие по
цилиндрической форме (иногда со слабыми искривлениями) каменно-
угольные роды Beresella и Dvinella имеют мутовчатые пояски, чере-
дующиеся с безмутовчатыми поясками. Чаще всего поры сливаются
в темную полоску, опоясывающую известковый чехол, а промежу-
точные беспоровые пространства образуют светлые пояски1. Род
Samarella не имеет мутовчатых и беспоровых поясков, а поры у
него располагаются сплошь. Таким образом, определение этих
форм при их хорошей сохранности возможно в тонких шлифах
и при значительном увеличении. Но вся триба (или группа) Beresel-
leae, к которой принадлежат все эти роды, распространена только
в каменноугольной системе, легко определима в шлифах и имеет
следующий общий, часто присутствующий признак: тонкая светлая
карбонатная корочка снаружи закрывает наружные отверстия пор.
Присутствие такой корочки можно объяснить как обызвествление
старых, неассимилирующих частей растений, не пользующихся
солнечным светом для фотосинтеза (табл. 15, фиг. 4). Значение
березелл как породообразователей мелководных известняков иног-
да очень большое: по-видимому, временами они образовывали
настоящие заросли. Встречаются они всегда в обломках.
Род Antracoporella характерен ветвящимся слоевищем, у кото-
рого ответвления также ветвятся. Обычно это тонкие правильные
известковые цилиндры, стенки которых пронизаны тонкими, вет-
вящимися к поверхности порами. Местами эти водоросли имеют
существенное породообразующее значение (карбон Донбасса, си-
лур Казахстана).
Перейдем теперь к сферической форме чехлов. В палеозое триба
(группа) Cyclocrineae включает несколько родов. Из них ордовикско-
силурийский род Cyclocrinus (наиболее типичный) состоит из не-
обызвествляющейся ножки, раздутой центральной части и радиаль-
но отходящих ответвлений, на концах которых образуются чаше-
видные раздувы. Эти раздувы, сильно обызвествляясь и сливаясь
в наружную корку, образуют сферу, снаружи пронизанную порами.
Род Mastopora принципиально не отличается от предыдущего рода,
но центральная часть его сферического чехла не сохраняется и по-
род Beresella встречен в карбоне Русской платформы. Урала н Турции. Род Dvinella рас-
пространен в среднем и верхнем карбоне Русской платформы, Урала. Центральной Азии.
Японии. Туниса, Турции. Род Samarella встречен в карбоне Русской платформы и Турции.
22
Известковые организмы
чти не изучена. Известковый же чехол состоит из радиально распо-
ложенных призматических известковых ячей, имеющих внизу одну
пору, а на поверхности несколько пор (табл. 15, фиг. 7).
Следующий пермский род — Mizzia, часто играющий значитель-
ную породообразующую роль, надо считать условно отнесенным
к трибе Cyclocrineae. Его известковые «сферы» имеют овальную и
несколько вытянутую форму и два крупных отверстия. Водоросль
была членистой, имела вид «ожерелья». Поры, пронизывающие
известковую оболочку, расширены к внешней поверхности и отхо-
дят от бочковидной центральной ячеи. Несколько более сходен
с раннепалеозойскими представителями трибы Cyclocrineae род
Koninckopora. У этого рода вытянутый булавовидный, иногда близ-
кий к сферическому, тонкостенный известковый чехол составлен
из ряда призматических ячей с тонкими (обычно темными) стенка-
ми. Род известен только в карбоне и легко определим в шлифах.
Известковый чехол достигает иногда больших размеров — 50 мм
(табл. 15, фиг. 1,2)*.
Переходя к мезозойским сифонеям, нужно сказать, что неко-
торые роды принципиально устроены так же, как и палеозойские,
но в большинстве случаев значительно усложнены. Одним из харак-
терных признаков является наличие индивидуализированных чехлов
вокруг ответвлений первого порядка, а также обызвествленные ор-
ганы плодоношения. В поздней юре и ранеем мелу встречаются
известковые чехлы сферической формы (род Kopetdagaria), сходные
с Cyclocrinus. Сходство с этим палеозойским родом подчеркивает
иногда сохраняющаяся ножка, расположенная ниже сферического
чехла. Но ячеи, образующие сферу, имеют принципиальное отличие
от циклокринусов: они не являются ассимиляторами, а предназна-
чены для органов размножения. Поры, вернее, внешние отверстия
являются одиночными и временными (открываются после созрева-
ния). По-видимому, и отверстие, соединяющее центральную ячею
с полостью периферической ячеи, при созревании закрывается. Та-
ким образом, чехлы представителей рода Kopetdagaria образованы
рядом камер, похожих на камеры рода Koninckopora, но менее пра-
вильной формы (табл. 16, фиг. 3,4, 13). Иногда известковые головки
копетдагарий переполняют породу (табл. 16, фиг. 8).
В мезозое продолжают встречаться просто построенные цилин-
дрические известковые чехлы, как, например, Macroporella и Gripho-
porella. В последнем роде объединены виды с тонким известковым
чехлом, по которому трудно (большей частью невозможно) судить
о форме ответвлений и положении места обызвесткования на слое-
вище (табл. 16, фиг. 5, 6).
Наиболее интересными и важными породообразователями яв-
ляются водоросли из группы клипеин (роды Munieria, Clypeina,
Actinoporella и др.), которые имеют ответвления с индивидуализи-
рованными известковыми чехлами, -сливающимися у своего основа-
ния или в цилиндр, или в мутовчатое кольцо. Фигуры, которые
получаются в шлифах при случайных сечениях, приведены в табл. 13,
фиг. 10; табл. 16, фиг. 1, 7, 9. Водоросли эти очень ломкие, поэтому
их фрагменты чаще не определимы, хотя иногда переполняют по-
роду (табл. 16, фиг. 1).
Более простыми в отношении обызвествления, но также слож-
ными в отношении анатомического строения являются роды мезо-
зойских сифоней Triploporella, Turkmeniaria и др. У рода Triplo-
porella ответвления первого порядка, раздуваясь, играют роль спо-
ровместилищ, а ответвления второго порядка размещены пучками
по четыре вместе (табл. 13, фиг. 5, 6). У туркмениарий ответвления
чередуются в мутовке: одна простая с одной раздувающейся в ле-
пешку (рис. 15, а, б).
Дальнейшие усложнения приводят нас к родам, дожившим до
третичного времени и даже до современного периода. В триасе
появляется род Acicularia, который обызвествляет свой споронос-
ный зонтик, состоящий из отдельных радиально расположенных
известковых игл — спорангиофоров, со сферическими полостями
для спор. В ископаемом виде спорангиофоры залегают отдельно
друг от друга (табл. 13, фиг. 8; табл. 16, фиг. 10—12). Более пробле-
матичными третичными мутовчатыми сифонеями являются шарико-
образные известковые тельца с округлыми мелкими полостями для
спор (табл. 13, фиг. 8,9), которые рассматриваются как отдельные
обызвествленные спорангиофоры (роды Oilaria, Catellaria, Terque-
mella). В литературе известны похожие третичные’ спорангиофоры,
сидящие на стержне (как початок кукурузы) и описанные под родо-
вым названием Chalmasia. К сожалению, они не изучены в шлифах.
В девоне встречаются спорные организмы в виде мелких извест-
ковых полых телец, стенки которых пронизаны простыми тонкими
порами и двумя полярными отверстиями. Форма телец разнооб-
разна и изменчива, приближаясь к округлой. Были встречены груп-
пы индивидов, у которых наблюдалось соприкосновение отверстий
двух соседних тел. Поэтому они были приняты за известковые чле-
ники мутовчатых сифоней рода Cateniphycus (Маслов, 1956), близ-
кого к роду Mizzia (табл. 15, фиг. 6). По-видимому, Cateniphycus,
как и пермский род Mizzia, образовывал четковидную, но ветвя-
щуюся нить с известковыми члениками. Этот род иногда обилен в
известняках девона Кузбасса.
БАГРЯНЫЕ ВОДОРОСЛИ (RHODOPHYTA)
Табл. 17—22
Багряные, или красные, водоросли — группа организмов, оби-
тающих исключительно в водной среде (главным образом в море),
обладающих усложненными органами размножения, многоклеточ-
ным строением слоевища и красным (или оттенками красного) цве-
том. В ископаемом состоянии сохраняются представители лишь
некоторых семейств, а для литолога важны те формы, которые
отлагают известь в стенках своих клеток. Особенно интересны ши-
роко известные «литотамнии» или «нуллипоры» — багряные водо-
росли из семейства Corallinaceae (кораллинациевые), живущие в
настоящее время. Описание их породообразующей роли и связи с
фациями, а также деталей анатомического строения и т. п. можно
найти в работе В. П. Маслова (1962) главным образом для третич-
ных кораллинациевых.
Химический и минеральный состав. А. П. Виноградов (1935,
стр. 125) делит современные багряные водоросли по отношению
накопления солей кальция на три группы; 1) багрянки, содержащие
кальций в обычном количестве, как у всех водорослей: 2) багрянки,
концентрирующие известь в виде инкрустаций (арагонита ?); 3) маг-
незиально-известковые багрянки семейства Corallinaceae, обызвест-
вляющие все слоевище1. Если первая группа почти не фоссилизи-
руется, то вторая содержит ощутимые количества извести и может
сохранить форму слоевища в виде известкового футляра. Химичес-
кие анализы золы представителей этих родов содержали от 63 до
88% СаО. К сожалению, в ископаемом виде эти роды неизвестны.
А. П. Виноградов приводит большую таблицу содержания MgCO3
и СаСО3 у современных кораллинациевых из разных географических
пунктов, а также таблицу полных анализов. Приведена также таб-
личка количества магния и кальция в молодых и старых частях
слоевища. Количество углекислого магния сильно колеблется. У lit-
hophyllum pachidermum молодые части .содержат 24,95% MgCO3,
а старые части—15,43%, у Gonioliton strictum соотношение обрат-
ное: 22,98 и 23,74% MgCO3. Однако в анализах ископаемых остатков
* Род распространен в среднем н верхнем карбоне Русской платформы. Урала. Централь-
ной Азин, Япоиин, Туниса, Турции и в перми Армении.
1 К этому семейству необходимо прибавить некоторые роды семейства Squamariaceae.
Багряные водоросли
23
из семейства Corallinaceae (в последнее входит род Lithotamnium)
было обнаружено очень небольшое количество MgCO3 (до 1,65%).
Карбонат в стенках клеток известковых багрянок в ископаемом
виде встречается в двух разновидностях: 1) в виде пелитоморфного
очень тонкозернистого слабопроэрачного (в шлифе) темного каль-
цита и 2) в виде тонкозернистого прозрачного кальцита, часто
плеохроирующего в параллельных николях. Поскольку у современ-
ных багрянок кальцит в стенках клеток связан пектиновым вещест-
вом. а кристаллики его расположены закономерно, грануляция кар-
боната. вероятно, вторичная, а волнообразное затухание стенок
клеток некоторых палеозойских багрянок (унгдарелл) — по-види-
мому, первичный признак. Перекристаллизация стенок клеток у
палеозойских соленопор приводит к посветлению ткани, в то время
как вторичные эпигенетические изменения мезо-кайнозойских ко-
раллинациевых приводят к полной грануляции, затемнению и ис-
чезновению клеточной структуры. Все эти изменения зависят, воз-
можно, от величины первичных кристаллов карбоната и количества
органического вещества между ними в стенках клеток (табл. 76).
Экология. Экологические условия произрастания багряных водо-
рослей изучены на современных растениях и частично у каменных
багрянок (Corallinaceae) кайнозоя. Багрянки — морские растения,
обитающие на глубинах от 0 до 150 м. Отдельные виды, особенно
корковые формы, приспосабливаются к жизни в литоральной зоне
в лужах и гротах, смачиваемых прибоем. В сублиторальной зоне
нижняя граница жизни багрянок колеблется в зависимости от мест-
ных условий: от 17 .и у берегов Шотландии до 156 .м у о. Фунафути.
Необходимо заметить, что глубже 100 м живут только представи-
тели рода Lithothamnium. Некоторые кустистые формы нечленистых
багрянок (Melobesioideae и Solenopora) не прикрепляются к суб-
страту, образуя желвачки, шевелящиеся движением воды и даже
перекатывающиеся по дну (рис. 17, а — г; табл. 21). Такие желвачки
встречаются в изобилии, образуют пласты и линзы известняков в
третичных осадках западной части Украины (Маслов, 1962).
В некоторых случаях желваки, достигая определенных размеров,
теряют подвижность, а продолжающееся нарастание организмов
заполняет пространство между желваками, создавая сплошную
породу (табл. 21, фиг. 3). Обычно в таком нарастании участвует
несколько организмов, например корковые багрянки, черви и обво-
лакивающие фораминиферы. В третичных осадках нередки биоцено-
тические желваки, образованные мшанками и корковыми багрян-
ками (табл. 20, фиг. 4).
Большинство багряных водорослей живет в воде океанической
солености, но некоторые виды приспосабливаются к небольшому
опреснению. Эти виды обычно относятся к родам Melobesia, Тепагеа
и корковым Lithothamnium. Временное опреснение выдерживают
также членистые багрянки.
Современные багряные водоросли* по отношению к освещен-
ности ведут себя различно: одни являются солнцелюбивыми, дру-
гие— тенелюбивыми и третьи индифферентны к освещенности. Так-
же различно ведут себя разные виды по отношению к температуре.
В современных морях каменные багряные водоросли произрастают
от экватора до 80сЗГ с. ш. и 73° ю. ш. Но некоторые «роды, например
Archaeolithothamnium, Mesophyllum и Amphiron, исключительно тепло-
любивы.
При решении вопросов, связанных с биогермами и рифогенными
фациями, особое внимание приобретает скорость роста мелобе-
зиевых. Известно обрастание подводного кабеля коркой багрянок
в 10 с.и толщиной в течение 20—30 лет. что дает прирост корки в
год максимум на 5 мм.
Геологическое значение. Багряные водоросли могут быть ин-
тересны геологу и литологу как определители геологического воз-
раста и породообразователи.
Многие формы багряных водорослей являются руководящими
для систем, отделов, ярусов. Широко распространенные по верти-
кали от ордовика до палеогена два рода, Solenopora и Parachaetetes,
могут быть руководящими только при условии определения вида.
Но зато Donezella и Ungdarella являются каменноугольными ис-
копаемыми, широко распространенными от Донбасса до Урала.
Современные роды кораллинациевых известны с мела, и некоторые
их вилы являются руководящими для отдельных ярусов. В работе
В. П. Маслова (1962) приведена таблица распространения отдельных
видов кораллинациевых для третичных отложений СССР.
Как интенсивные породообразователи багряные водоросли при-
обрели особое значение лишь в третичное время, слагая рифоген-
ные массивы и биостромы из желваков на банках длительного суще-
ствования (Маслов, 1962, стр. 163—182).
Анатомия. Классификация современных форм багрянок бази-
руется преимущественно на органах размножения. Ископаемые
багряные водоросли в большинстве случаев лишены органов раз-
множения. Однако водоросли из подсемейства Melobesioideae (се-
мейство Corallinaceae). фоссилизирующие свою анатомическую
структуру в ископаемом состоянии, делятся на роды главным обра-
зом по структуре их органов размножения. Последние чаще обна-
руживаются в виде пустот в слоевище, а половые органы сохраня-
ются как редкое исключение.
Каменное багряные водоросли образуют корки, бугры или жел-
ваки различной величины, а также ветвящиеся, бугристые, сталак-
титовые наросты на донных предметах или, наконец, членистые
гибкие кустики (собственно кораллины). Размеры их слоевищ ко-
леблются от микроскопически малых до величины человеческой
головы.
Анатомические различия между родами семейства Corallinaceae
заключаются не только в различии органов размножения, но и в
различии структуры и взаимного расположения клеточных нитей.
Слоевище кораллинациевых водорослей производит впечатление
многоклеточной ткани. Последняя возникает в результате тесного
соприкосновения параллельно залегающих клеточных нитей. Взаим-
ное расположение нитей и положение клеток в соседних нитях опре-
деляют ту или иную структуру «ткани», которая бывает двух видов:
1) базальная или срединная — так называемый гипоталлий (hypo-
thallium) и 2) внешняя или периферическая, или периталлий (peri-
thallium), включающая корковый слой у поверхности слоевища
(рис. 17, е, ж). У некоторых родов мелобезиевых иногда присут-
ствует особая переходная ткань — так называемый мезоталлий, рас-
полагающийся между периталлием и гипоталлием (mesothallium).
В ветвях обычно наблюдаются «срединный гипоталлий» (или сре-
динная ткань ветвей) и периталлий.
Кораллинациевые можно разбить на ряд групп по анатомичес-
кому строению слоевища.
В первой группе клетки в соседних нитях располагаются бес-
порядочно («ткань в нитях») (рис. 17, е): гипоталлий образован
ползущими по субстрату нитями, периталлий в принципе образован
также неориентированными в ряды клетками (род lithothamnium).
Вторая группа отличается тем, что клетки соседних нитей пе-
риталлия и гипоталлия находятся на одном уровне, а поперечные
перегородки нитей сливаются как бы в одну линию («ткань в се-
риях») (рис. 17, ж), вследствие чего слоевище кажется состоящим
из клеток, расположенных рядами, в виде правильной сети или
концентрически. Таким образом, в одном случае в вертикальном
сечении получается более или менее правильная сетка из четырех-
угольных ячей. В другом случае в вертикальном сечении получается
структура в виде располагающихся друг над другом веерообразных
рядов (серий) клеток (рис. 17, ж). Для Lithophyllum характерны и
тот и другой случаи, при этом резкой разницы в величинах клеток
периталлия и гипоталлия не наблюдается. Вторая группа («ткань
в сериях») является единственной структурой для всех членистых
кораллин (Corallinae), причем клетки гипоталлия (всегда средин-
ного, базального не бывает) во много раз длиннее клеток перитал-
лия (до 100 р длины при ширине 5—20 р).
24
Известковые организмы
Багряные водоросли
25
Третья группа имеет кроме гипоталлия и периталлия с ориенти-
рованными в ряды клетками еще промежуточную, или переходную
(по Конти), ткань — мезоталлий, отличающийся от первых харак-
тером и размером клеток, но также обладающий серийным их рас-
положением. К этой группе принадлежит Mesophyllum (табл. 19,
фиг. 1).
К четвертой группе относится род Archaeolithothamnium, в корко-
вых формах которого гипоталлий не имеет серийного расположе-
ния клеток, как у lithophyllum, тогда как клетки периталлия распо-
ложены в ряды, как у lithophyllum. Ветвистые же формы Archaeo-
lithothamnium по анатомической структуре одинаковы с Lithophyllum
и отличаются от него лишь органами размножения.
Наконец, вымершая группа соленопор по структуре ткани напо-
минает первую и вторую группы, но отличается от них большими
размерами клеток и более толстыми, чем у мелобезиевых, клеточ-
ными нитями (рис. 17, з).
Для ткани соленопор и мелобезиевых характерен еще один приз-
нак: размеры клеток и толщина их стенок в толстом периталлии
иногда периодически меняются. Благодаря утолщению известко-
вых стенок и поперечных перегородок и уменьшению величины кле-
ток получаются «сгущения» или полосовидные затемнения тканей
периталлия. В этом случае говорят о «зонах роста».
Вообще для диагностики родов мелобезий надежным признаком
являются органы размножения, но половые органы размножения
после выполнения их функций зарастают тканью и в ископаемом
состоянии не сохраняются. При изучении в шлифах мы наблюдаем
сечения через эти пустоты и должны по их сечениям реконструиро-
вать форму данного органа (рис. 17, к — и). Ниже мы остановимся
вкратце на этом вопросе.
Бесполое размножение происходит при помощи спор и вегета-
тивным путем. Последний способ, как и половой, в ископаемом виде
устанавливается с трудом, в то время как споровый способ имеет
большое значение. У ископаемых представителей багрянок обыч-
но образуются тетраспоры, которые располагаются по четыре спо-
ры в спорангии. Способ расположения спор в спорангии служит сис-
тематическим признаком для классификации современных предста-
вителей багрянок, но в ископаемом состоянии споры сохраняются
редко. Поэтому практически важными для изучающего ископаемые
багрянки являются органы размножения (спорангии, сорусы и кон-
цептакли).
Отдельные спорангии в виде вытянутых в вертикальном направ-
лении пустот наблюдаются у рода Archaeolithothamnium (рис. 17,.»).
Они обычно группируются рядами, отделены друг от друга стериль-
ными нитями, которые частично как бы огибают спорангии (табл. 18,
фиг. 7; табл. 22, фиг. 2). У рода Palaeothamnium спорангии более
вытянуты, тесно прижаты друг к другу и сгруппированы в неясные
Рис. 17. Строение слоевища у багряных водорослей
ч-• внешний вид желвачков Salenopora spoiKpiides (Dyb.). ордовик. Эсюння. лер. Сакку:
о—сечение через желвачок 1ермофнзного типа багрянки, верхний торгов, с. Микулницы
близ I. Тернополя (ма!ериал В. Г. Юрковой): в- сечение монофнтною желвака эпифн|-
пою шла (темные пятна — полости — проделаны сверлильщиками), нижний тортоп.
I . Львов; г—сечение через желвачок солспопоры i ермофитно) о типа, ордовик, вазалсм-
ский горизонт. Эстония: <) часть слоевища современной Corullinu. состоящею из извсС1-
ковых члеников разной величины, х 3.6: е блок-диаграмма слоевища типа лгпогампия.
по Конги (Conti. 1950): ж— блок-диаграмма слоевища типа лнтофиллума. по Конти
(там же): з — радиальное сечение через ткань Solenopora цеха Masi., нижний карбон. Казах-
стан. Каратау: и—концептакль типа литотамния, различные сечення и очертания его полости
(схема): к — концептакль типа лнтофн.глума. различные сечения и очертания ею полости
(схема); л — очертания полости концептакля lilhophyllum duplex Masi, в трех сечениях
(схема): .» — очертания спорангия в трех сечениях (схема): и — вертикальное сечение через
группу зарождающихся спорашнев I’alaeolluunniuin kossorense Masi., палеоцен. Северные
Карпаты, дер. Старый Самбор: о— вертикальное сече пне через предполагаемый спорангий
Solenopora spoiuioldes (Dyb.). ордовик. Эстония: н— участок сечения корочки Ar< haeoli-
iluiphyllum missourieitse Johnson с распределением клеток гипоталлия (слева) и пери таллия
(справа), х 67.5. по Рею (Wray. 1964)
сорусы (рис. 17, я; табл. 18, фиг. 6). lithothamnium, Mesophyllum и
некоторые Solenopora обладают удлиненными отчетливыми сору-
сами, в которых иногда встречаются спорангии. Каждому споран-
гию, так же как и у Palaeothamnium, в крыше соруса отвечает от-
верстие или канал, не всегда зарастающий. У родов Lithophyllum,
Melobesia и у членистых багрянок развивается концептакли, или уг-
лубления, имеющие формы перевернутого сердца с одной порой на-
верху. Тетраспоры располагаются по периферии нижней части кон-
цептакля, который имеет отчетливую оболочку из выстилающих
полость особых нитей клеток. В ископаемом виде клетки исчезают,
а оболочки замещаются темной пленкой (табл. 18, фиг. 8—И).
Самая простая внешняя форма у спорангиев археолитотамния —
эллипсовидная или бутылковидная в продольном осевом сечении
(рис. 17. и). В редких случаях она приближается к сфере. Концеп-
такли (рис. 17, к, л) обычны в виде перевернутого сердца, в разных
сечениях выглядят различно. Только в осевом сечении можно полу-
чить истинную форму концептакля. Сложнее всего дело обстоит
с сорусом, который может и не иметь оси, например при вытянутой
форме. В этом случае приходится руководствоваться вертикальными
сечениями, судить по нитям клеток, окружающих сорус. Для видо-
вых определений имеют значение величина и форма клеток гипотал-
лия и периталлия, особенности строения и размеры органов раз-
множения и т. п. (Маслов, 1962), но и форма органов размножения
может являться видовым признаком (табл. 18, фиг. 8—10).
Семейство Corallinaceae делится на два подсемейства по форме
своего слоевища: 1) подсемейство Melobesioideae имеет корковые,
бугристые и ветвистые сплошные слоевища, 2) подсемейство Coral-
linoideae — кустистые слоевища, разделенные на обызвествленные
членики, соединенные друг с другом необызвествленными гибкими
нитями. Кораллины имеют всегда серийное расположение клеток в
виде дуг в срединной ткани (или срединном гипоталли) и серийное
решетковидное расположение клеток в окружающем периталлии
(табл. 19, фиг. 7; табл. 20, фиг. 5—7). Их органы размножения —
концептакли — такие же, как у лнтофиллума (встречаются очень
редко).
У древних багрянок органы размножения встречаются также
редко (рис. 17, б), поэтому определения основываются на анатоми-
ческом строении ткани. Два главных и наиболее распространенных
рода, Solenopora и Parachaetetes, отличаются тем, что их клетки
значительно крупнее (шире), чем у литотамний. и тем, что у Soleno-
pora поперечные перегородки расположены беспорядочно (табл. 19,
фиг. 5,6: табл. 22, фиг. 1,6), а у парахететес они слиты вместе и
клетки находятся в горизонтальных сериях (табл. 22, фиг. 4; табл. 21,
фиг. 5).
Одним из любопытных родов является открытый в карбоне США
Джонсоном (Johnson, 1956) род Archaeolithophyllum (встреченный
также в Донбассе). Корковое и ветвистое слоевище этого рода сло-
жено гипоталлием, или срединной тканью, из крупных неправиль-
ных клеток, ориентированных в выпуклые дуги — серии, и внешней
мелкоклеточной тканью, или периталлием, у которого клетки также
имеют слитые поперечные перегородки, но нити здесь расположены
радиально, а серии — параллельно поверхности корки (рис. 17, л).
Овальные концептакли на периферии, по-видимому, имели одну
пору. Этот род отнесен к семейству Corallinaceae (?).
Похожий по анатомическому строению третичный род Suhter-
raniphyllum Elliott отнесен к членистым кораллинам. Цилиндричес-
кие членики этого рода достигают 1,5 .и,и длины и 0,75 мм ширины.
Они сложены грубой срединной тканью из неправильных удлинен-
ных клеток длиной до 117 р и шириной до 78 р, расположенных в
горизонтальной серии, которые у краев слоевища заворачиваются
вниз. Форма клеток в осевом сечении неправильно трапециевидная,
в горизонтальном разрезе — неправильно многоугольная. Пери-
таллий тонкий, из сливающихся темных мелких клеток размером
10—15 р. Этот род распространен от верхнего эоцена до основания
миоцена в Средиземноморье (Сицилия), Иране, Ираке, Омане.
26
Известковые организмы
За последние годы из порядка Cryptonemiales кроме семейства
Corallinaceae в ископаемом виде были обнаружены представители
семейства Squamariaceae. Сквамариевые по своему анатомическому
строению, обызвествлению и образу жизни близки к корковым
мелобезиевым (подсемейству Melobesioideae) и легко могут быть
смешаны с литотамниями. Они отличаются присутствием крупных,
более или менее округлых клеток в мезоталлии и часто плохим
обызвествлением поверхности корочки. В ископаемом виде известны
роды Karpathia Maslov (ископаемый род), Pseudolithothaninium Pfen-
der (ископаемый = Ethelia— современный род) и Peyssonella De-
caisme (современный род).
Палеогеновый представитель этого семейства — Karpathia sphae-
rocellulosa Maslov образует тонкие (0.20—0,25 и.и) корочки, сло-
женные крупными стелющимися удлиненными клетками базаль-
ного гипоталлия, переходящими в округло-полигональные крупные
клетки мезоталлия, или промежуточные ткани, которые к поверх-
ности переходят в мелкие клетки периталлия (табл. 19. фиг. 2).
Другой ископаемый представитель — Pseudolithothamnium al-
bum Pfender распространен от баррема до эоцена. По-видимому,
этот же вид встречен в датском ярусе Закарпатья и Грузии (Маслов.
1956, 1962). Характерной его чертой являются корочки с веерной,
напоминающей литотамнии структурой гипоталлия, в котором час-
ты нити крупных удлиненных клеток, нередко с черным внутренним
содержимым. Ближе к верхней и нижней поверхностям клетки ста-
новятся мелкими и в верхней поверхности местами ориентированы
в серии с толстыми поперечными перегородками, слитыми в линии.
Эти последние идут под острым углом к внешней поверхности
(табл. 17, фиг. 3).
Семейство Ungdarellaceae, которое теперь относят к багряным
водорослям, распространено только в карбоне и перми и отличается
строением слоевища и обызвествлением. Ветви и стержни унгдарел-
ловых в осевом сечении обладают «фонтанной» структурой: от
центральной нити, или от пучка нитей, или от внутренней полости
(которая получается благодаря разрушению осевой части) под ост-
рым углом к оси отходят нити клеток, обрываясь у внешней поверх-
ности (табл. 21, фиг. 3). При хорошей сохранности карбонат стенок
клеток в скрещенных николях под микроскопом затухает волной.
Отдельные роды унгдарелл отличаются количеством нитей в осевой
части, мощностью периферической ткани и формой клеток*. Опреде-
ление возможно только при хорошей сохранности. Принадлежность
рода Komia к водорослям подверглась сомнению. Унгдарелловые
жили вместе с морской фауной. Под названием Foliophycus Джонсон
(Johnson, 1951) описал из пенсильванских осадков Техаса водоросль,
близкую к унгдареллам.
Другие каменноугольные водоросли, известные под названием
Donezella и отнесенные к багряным водорослям (семейство Rhodo-
mellaceae ?), состоят из кустистых многонитчатых слоевищ несим-
метричной формы.
Слоевище Donezella lutugini Masi, в целом виде кустистое, много-
клеточное, рассеченное. Оно состоит из обызвествленных нитей кле-
ток цилиндрической или бочковидной формы (по преимуществу),
которые отделены друг от друга (в одной и той же нити) попереч-
ными перегородками (стенками) с одной порой посередине. В по-
перечном разрезе многоклеточное слоевище не имеет симметрии
и сложено разной величины клетками, так же часто соединенными
с соседними клетками, как это наблюдается у современных Rhodo-
mellaceae (рис. 18). В шлифах стенки клеток светлые, но в некоторых
случаях цемент внутри клеток и снаружи бывает магнезиальным
(Ирменский рудник Донбасса; рис. 18, а-—е) и удается окрасить
стенки клеток с помощью реакций Лемберга, благодаря чему полу-
1 Род Unydarella встречается в карбоне и перми Урала. Донбасса. Армении. Турции; рол
Knmia встречен в карбоне Урала, перми Турции. Японии. США (Техас. Невада).
Рис. 18. Donezella
а молодой проросюк Donevella в вер,пкальном ссчсппп в извешняке Ц, Донбасс Винту
видна подошва прикрепления с ризоидами (?) Шлиф окрашен (реакция Лемберт). цсмсп-
I ируюшнй доломит — белый. X 100: о. « — продольное сечение через iihiii Donezella. X 60:
<>- водоросль окрашена, в — обычный неокрашенный шлиф: г - проблемагическнй спо-
рокарпий у конца слоевища Donezella. водоросль окрашена (реакция Лемберш. х 100:
<>. е — поперечные срезы слоевища Donezella lutut/ini Masi.: д — молодые, е - взрос 1ыс
стосвища
чается четкая картина сечений слоевища. В результате наблюдались:
молодой проросток (рис. 18, а), проблематический спорокарпий
(рис. 18, г), проблематические антеридии на концах слоевища с
пузыреобразными пустотами в стенках, точки роста (?) в виде
заостренных концов и в виде трехлопастных окончаний (табл. 19,
фиг. 4), орган прикрепления (?) к субстрату (рис. 18, а), тетраспоран-
гий (?) и т. п. Размеры нитей и клеток различны в одном и том же
слоевище и зависят от положения нити. Боковые нити значительно
тоньше, а клетки меньше центральных. Единственный вид Donezella
lutugini Masi. (Маслов, 1929) широко распространен в морских
осадках карбона европейской части СССР (особенно в среднем
отделе) (табл. 17, фиг. 2; табл. 21, фиг. 2,4).
Ложные багряные водоросли
Как это часто бывает, некоторые организмы не сразу расшифро-
вываются в отношении их принадлежности к той или иной группе.
Так, Джонсон (Johnson, 1960, 1963 а. Ь) описал под названием Со-
neiphyeus texana Johnson организм из карбона, который давно был
мне известен и даже назван (Cribroporidium Masi.), но не опублико-
ван, так как не было уверенности в принадлежности его к гидроидам
или губкам. В продольном сечении узких пластин этого организма
получается ложное впечатление клеток, ориентированных в серии.
Отчетливые дугообразные линии (в вертикальном сечении) идут
от одного крыла пластины до другого. При вертикальном сечении
видны черточки, как бы отвечающие стенкам клеток. Но при не-
сколько скошенном, косом и поперечном сечениях эти «стенки»
выклиниваются и исчезают (рис. 19, а — в). При детальном иссле-
довании можно увидеть, что это не что иное, как столбики, отве-
чающие или столбикам строматопор, или вертикальным элементам
шестилучевых спикул губок. Похожие организмы описывались как
Багряные водоросли. Харофиты. или харовые водоросли
Z1
Рис, 19. Cribroporidiuni-Coneiphycus в известняках
про.к» и-ныс и скошенные сечения, клрооп: Депоасс
губки. Поэтому согласиться с предположением Джонсона, что это
членистые кораллины, нельзя. Этот организм (Cribroporidium)
обилен в некоторых относительно глубоководных фациях карбона
Донбасса (рис. 19; табл. 21, фиг. 9, 10) и еще недостаточно изучен.
В США (Coneiphyeus) известен из пенсильванских осадков Техаса
и Нью-Мексико.
ХАРОФИТЫ, ИЛИ ХАРОВЫЕ ВОДОРОСЛИ (CHAROPHYTA)
Табл. 23, 24
Эта группа высокоорганизованных исключительно водных водо-
рослей имеет кустистое слоевище, по внешнему облику напоминаю-
щее хвощи. Многие из представителей харовых обызвествляют
вегетативные части и органы размножения (ооспорангии), почему
и сохраняются в ископаемом состоянии.
Харофиты — зеленые растения, которые долгое время относи-
лись к зеленым водорослям (Chlorophyta). В последние годы боль-
шинством альгологов они выделяются в особый тип (Charophyta
или Charophycophyta).
Методике изучения ископаемых харофитов посвящена работа
В. П. Маслова (1963 а), включающая также историю их изучения
и обширную литературу.
Химический состав. Обызвествленные остатки харофитов нередко
встречаются в ископаемом состоянии. Характер их обызвествления
изучался рядом исследователей, в результате чего отчетливо выяви-
лась разница в структуре карбоната в вегетативных частях и обо-
лочках ооспорангия. В вегетативных частях кальцит зернисто-крис-
таллический отложился физиологическим способом, а в оболочках
ооспорангия он представлен тонкослоистым и очень мелкозернис-
тым карбонатом (чередующимся с тонкими слоями, окрашенными
органическим веществом), отложившимся органическим способом
(см. стр. 7).
Согласно рентгено-кристаллографическому анализу Карльтро-
ма, известковые оболочки ооспорангия Chara globuraris состоят из
кальцита.
Карбонат, выделенный вегетативными частями, обычно также
представляет собой довольно чистую известь — 94% СаСО3 (Твен-
хофел, 1936, стр. 281; Grabau, 1932, стр. 471).
Дэвис приводит анализ минеральной части слоевища современ-
ной Chara sp. (в %): СаСО3— 93,76; MgCO3 — 2,93; SiO2— 2.4:
R2O3— 0,89. Главную часть кремнекислоты нужно отнести за счет
диатомовых водорослей, живущих на харофитах. Шуэтт и Альдер
(Schuette, Alder. 1929) приводят полный анализ минеральной части
современной хары из Зеленого озера (в %): SiO2—0,83: Fe2O3 —
0,06; А12О3 —0,81: Мп3О4 — 0.08; СаО — 37,82; MgO-1,19;
Na2O — 0,35; К2О — 0,58; Cl — 0,29; СО2 — 39.00; S — 0,27; Р —0.06.
Экология. Современные харофиты живут на малой глубине в
пресных водах, прудах, каналах, ямах, затопленных карьерах, озе-
рах и реках и считаются типичными пресноводными водорослями.
Однако существуют отдельные виды, приспособившиеся к солоно-
ватоводным и даже морским прибрежным условиям. Олсен конста-
тировал, что Chara bahica живет на юге Балтийского моря в воде,
содержащей 18% хлора, т. е. в воде, по солености близкой к океа-
нической. Клауд сообщает, что харофиты пышно произрастают
вместе с морской сифоновой (Dasycladaceae) водорослью Ваго-
phora corstedi в солоноватоводных условиях у берегов п-ова Флорида.
По данным Фромана, pH солоноватоводных бассейнов, в кото-
рых живут современные Chara polyeanla A. Br., Ch. hipsiodia L..
Ch. aspera Willd., колеблется от 7 до 8. Ch. crinila Wallr. встречается
в соленых прибрежных водах Средиземного моря. Ла-Манша, Ат-
лантического океана; в пресных водах она встречается редко, а в
солоноватых водах каналов, вырытых за зоной галечника у берега
моря, обильна.
Корийон указывает на lamprothamnium papulosum как на вид,
приспособившийся к соленым лагунам, которые заливаются при-
ливом; pH этих вод близок к 8. Распространение этого вида — Бал-
тийское и Северное моря, Ла-Манш, Атлантическое и Средиземно-
морское побережья. Амберже дал краткую характеристику условий
жизни таких «полуморских» харофитов.
В литературе неоднократно дискутировался вопрос о место-
обитании ископаемых харофитов, так как они встречались как в
морских, так и в континентальных осадках. В -частности, Крофт,
обсуждая вопрос о солености воды, в которой жили трохилиски,
стремится доказать, что гирогониты, встреченные с морской приб-
режной фауной, являлись или перемытыми, или принесенными с
суши речными водами. Действительно, плоды харофитов могли
легко заноситься течениями воды в морской бассейн, так как их
содержимое было легче воды. Но вряд ли вегетативные части харо-
фитов могли переноситься на значительное расстояние, так как,
будучи ломкими, они теряли свое внутреннее содержимое, а удель-
ный вес кальцитовой оболочки значительно больше 1. Между тем
вегетативные части харофитов встречены в известняках морского
происхождения в карбоне Коми АССР (р. Углар) и верхней юры
Западной Украины. Кроме того, массовое нахождение гирогонитов
харофитов в среднем и верхнем сармате Одесского района, Север-
ного Кавказа и Грузии вместе с морской фауной трудно объяснить
одним механическим переносом. Более вероятно, что в это время
харофиты жили по берегам моря с не очень соленой водой. В извест-
няках палеогена Таджикской депрессии, а также во многих местах
в мезозое разных стран гирогониты в массовом количестве встре-
чены вместе с морской фауной.
По-видимому, нельзя категорически отрицать прибрежные мор-
ские условия как местообитание некоторых ископаемых харофитов.
А. П. Карпинский (1906) считал трохилиски прибрежно-морски-
ми растениями. Действительно, американские трохилиски и сици-
дии жили вместе с кораллами и другими морскими животными.
Пэк прямо помещает их в морские фации в противоположность
другим харофитам, отнесенным к пресноводным фациям.
Таким образом, сами по себе харофиты не могут служить ука-
зателями солености бассейна и условия их обитания должны быть
доказаны в каждом случае (Маслов, 1963а). Однако преобладающая
часть ископаемых и современных харофитов встречается в конти-
нентальных водоемах.
28
Известковые организмы
Геологическое значение. Отложения, образуемые современными
харами, эксплуатируются в некоторых озерах. Так, в Зеленом озере
в Висконсине (США) вес сухого вещества водорослей, добываемых
со дна, дает 2260 т с 1 га, что отвечает 40 т карбоната кальция на
1 /си2.
Встречающиеся иногда породы, образованные главным образом
известковыми остатками харофитов, называются харацитом. В пре-
делах СССР известен такой харанит из плиоцена Южной Тувы
(р. Хоолу), образующий метровый пласт писчего мела, эксплуати-
руемого для нужд местных школ. Сложена эта порода мелкоистер-
тым детритом вегетативных частей харофитов с редкими известко-
выми оболочками ооспор орган-рода Tectochara. Выше- и нижеле-
жащие мергели и известковистые глины и алевриты включают
гирогониты того же орган-рода. Вегетативные части здесь не сохра-
няются, быстро размалываясь движениями воды. Сохранность же
гирогонитов объясняется их плавучестью благодаря наличию внут-
ренней полости, более легкой, чем вода. Такие отдельные гирого-
ниты встречались в значительном количестве в палеогеновых мор-
ских известняках Таджикской депрессии (р. Вахш).
Мэгдефрау приводит данные Зёргеля и Вюста о хароците в меж-
ледниковых отложениях окрестностей Веймара, который залегает
среди травертинов и мергелей. Мощность «харового туфа», как на-
зывает харацит Мэгдефрау, достигает 1 м, слагается он главным
образом обломками вегетативных частей харофитов, более круп-
ными, чем в тувинском тонкоизмельченном известковом осадке.
Аналогами осадка тувинского типа в Веймаре, возможно, являются
мергели с гирогонитами харофитов, залегающие выше и ниже «харо-
вого туфа».
В последние годы на стратиграфическом распространении харо-
фитов и на их гипотетических филогенетических связях останавли-
вался ряд исследователей. Длительные исследования и многочислен-
ная литература по сицидиям и трохилискам убедительно доказали,
что эти формы встречаются только в девоне и нижнем карбоне.
Хотя еще не была опубликована сводка, показывающая распределе-
ние отдельных видов по разрезу, но из работ Пэка вытекает, что
для ярусов девона и карбона, вероятно, будут иметься свои руко-
водящие виды.
Исследования мелких харофитов в последние годы убедительно
показали руководящее значение Atopochara trivolvis Peck, которая
очень широко распространена в апте Венгрии, США, Сирии, Африки,
Узбекистана. Нижнетриасовые отложения (от Белоруссии через
Большой Донбасс до Волги) расчленены Л. Я. Сайдаковским (1960)
на ряд горизонтов с помощью харофитов.
Анатомия. Лучше всего изучены современные хары (или лучицы).
Это мелкие многоклеточные кустики, сложенные «стержнями» или
«стебельками», достигающими нескольких сантиметров в длину
и разделенными узлами на междоузлия. От узловых частей расхо-
дятся мутовками ответвления, или «листья». Листья в свою очередь
имеют вторичные мутовки ответвлений. Рост верхушечный. Нижняя
часть слоевища прикреплена к субстрату ризоидами. Размножение
вегетативное и половое. Вегетативное размножение напоминает
образование корневищ. Из нижних узлов отходят «побеги», кото-
рые, выпуская ризоиды, образуют новые «стебли». На ризоидах
и нижних узлах могут образоваться «клубеньки» из разросшейся
клетки, заполненной крахмалом. Эти «клубеньки» способны также
давать новые «побеги».
Как указывалось выше, вегетативная часть слоевища состоит
из кустистой системы стержней, образованных междоузлиями и уз-
лами. Междоузлия состоят из одной или нескольких длинных кле-
ток. Узлы всегда имеют многоклеточное строение и состоят из ко-
ротких клеток, к которым примыкают клетки междоузлий, ответ-
влений и органов размножения. Вегетативные клетки с поверхности
часто обызвествляются и сохраняются в ископаемом состоянии в
виде известковых трубок (табл. 23, фиг. II).
Междоузлия, состоящие из многих клеток, имеют следующее
строение: вокруг центральной трубки располагается ряд более тон-
ких периферических трубок (род Chara), завивающихся спирально.
После отмирания растения его органические части сгнивают,
а известковые дробятся из-за хрупкости трубочек и в ископаемом
виде встречаются в виде обломков.
Органы размножения харофитов и в ископаемом состоянии обыч-
но сохраняются благодаря известковой оболочке, называемой гиро-
гонитом (термин исключительно палеонтологический). Гирогонит
образован известковой и органическими оболочками. Первая состо-
ит из спиральных полос, называемых партекальцинами, и базальной
пластинки (рис. 20). Часто встречаются гирогониты с двумя поляр-
ными отверстиями.
У некоторых ископаемых харофитов (Clavatorales) кроме гиро-
гонита сохраняется еще наружная известковая оболочка вегетатив-
ного происхождения. Это так называемый утрикул, который обра-
зуется смыкающимися вокруг плода листочками или веточками.
Утрикул состоит из полых трубочек или из одного слоя (внутрен-
него) стенок этих трубок, тесно прижатых — слитых друг с другом.
Таким образом, утрикул состоит из кристаллического кальцита,
в то время как гирогонит состоит из спиралей с типичной микро-
структурой: она характеризуется тонкой «штриховкой» или. иначе,
чередованием очень тонких, загрязненных органикой темных и
светлых слойков (I—3 /( толщиной), параллельных внутренней и
наружной поверхностям спиралей (рис. 20: табл. 23. фиг. 2—4. 6—8).
У утрикулов мезо-кайнозойских харофитов такой штриховки
на наблюдалось.
Норинни
Рис. 20. Схематический вертикальный
разрез через четвертичный гирогонит
орган-рода Tectochara
Буквы на рисунке: к,. к2 — известковые отложения
клеток: sp. — оболочки клеток: st. — общая органи-
ческая оболочка: б. п. — базальная пластина
Поскольку внешняя форма и степень обызвесткования гирого-
нитов различны у разных представителей, то и в шлифе мы наблю-
даем различный облик известковых выделений в продольном се-
чении.
Девонские трохилиски, сицидии, хованеллы и умбеллы сильно
отличаются от остальных харофитов. Широко известные и давно
изучавшиеся трохилиски и сицидии определяются, как и большин-
ство харофитов, после предварительного их выделения из породы
(Маслов. 1962). Главным признаком их является поверхностная
скульптура: у трохилисков — спирали, завернутые по часовой стрел-
ке (с правым завиванием), а у сицидии — разной формы много-
угольники. расположенные по меридианам. Сейчас делаются по-
пытки определения трохилисков в шлифах, для чего необходимо
знать не только внешнюю форму, но и количество спиралей, нак-
лон их к экватору, их ширину и т. п. Сицидии в шлифах не опреде-
лялись (более подробно см. ниже, раздел «Методика определения
ооспорангиев...», автор Б. В. Поярков).
Хованеллы характерны своей внешней структурой и формой в
виде головки мака. Как трохилиски и сицидии, они обладают дву-
мя отверстиями на противоположных концах, но в отличие от вто-
рых в них найдены базальные пластинки, прикрывающие нижнее от-
Харофиты, или харовые водоросли
29
верстие изнутри. Внешняя скульптура, двухслойное строение обо-
лочки и наличие базальной пластинки позволяют считать, что у
хованелл мы имеем дело с гирогонитом, частично перекрытым сни-
зу утрикулом.
Трохилиски и сицидии имеют мировое распространение. Они
известны в СССР, Америке, Китае, Европе, Африке и т. д. Трохи-
лиски известны от нижнего девона до карбона включительно. Сици-
дии находились только в среднем и верхнем девоне. Хованеллы еще
сравнительно мало изучены, но, по-видимому, это только девонские
организмы.
В последние годы к харофитам отнесены организмы, известные
под названием умбелл (Umbella Maslov). Это небольшие округлые
пустотелые образования с характерной микроструктурой оболочки.
Орган-род Umbella представляет собой орган плодоношения харо-
фитов, который состоит из гирогонита и облекающего его утри-
кула. Обычно обе оболочки слиты в единый оптический слой, но
встречаются экземпляры с двумя и тремя оболочками. Умбеллы из-
вестны в девоне и в основании карбона и по современным данным
имеют почти планетарное распространение.
Необходимо отметить, что кроме этих харофитов в девоне по-
явились представители харовых с левым завиванием спиралей, ко-
торые дожили до наших дней (орган-род Eochara), найденные вне
пределов СССР.
Методика определения ооспорангиев девонских харофитов в шли-
фах и их значение1. В средне- и верхнедевонских отложениях СССР
часто встречаются остатки харовых водорослей. Они представлены
главным образом остатками ооспорангиев (органов плодоношения),
хотя иногда попадаются и вегетативные части растений. Тонкий
темный мелкозернистый слой кальцита, окружающий внутренюю
полость гирогонита (рис. 21, в, г; табл. 24, фиг. 17), возможно, соот-
ветствует мембране ооспоры. Гирогонит — известковой чехол, ок-
ружающий ооспору, — состоит из партекальцин, каждая из которых
является известковым отложением одной спиральной клетки. Пар-
текальцина слагается концентрически слоистым кальцитом. Однако
у ископаемых гирогонитов вследствие перекристаллизации сохра-
няются лишь реликты концентрически слоистой структуры. Основная
масса кальцита партекальцин приобретает радиально-лучистое
строение (табл. 24, фиг. 15, 16). Между смежными партекальцинами
иногда наблюдаются четкие швы, сложенные темным тонкозер-
нистым кальцитом (см. табл. 24, фиг. 15, 16). В нижней части гиро-
гонита расположена базальная пора, прикрытая базальной плас-
тинкой. У некоторых сохраняется также вершинная (апикальная)
крышечка, прикрывающая апикальную пору (табл. 24, фиг. 17).
Наличие утрикула у древних харофитов доказывается несом-
ненно, если под этой «вторичной» оболочкой находится гирогонит
(табл. 24, фиг. 4, 14), что бывает крайне редко. Чаще для отличия
гирогонита от утрикула могут быть использованы следующие приз- ,
наки.
Во-первых, стенка утрикула отличается от стенки гирогонита
грубостью и слитностью и отсутствием швов, в то время как гиро-
гонит имеет обычно швы между известковыми спиралями (Маслов,
1961а).
Во-вторых, гирогонит орнаментирован спиральными ребра-
ми (рис. 21, а, б), закручивающимися слева направо или справа
налево; скульптура же утрикула состоит из продольных и попереч-
ных ребер, валиков и других элементов, являющихся отпечатками
вегетативных частей (рис. 21, б, е, и, к, х, ц). Иногда поверхность
утрикула может быть гладкой (рис. 21, я, о, с, т). Если выделенные
из породы гирогониты при известном навыке более или менее легко
отличаются от утрикулов, то значительно труднее это сделать в
шлифах. В последнем случае необходимо иметь массовый материал
1 Раздел написан Б. В. Поярковым.
для того, чтобы подобрать более или менее ориентированные се-
чения.
Сечения могут быть продольные, поперечные и косые. За про-
дольное сечение гирогонита и утрикула принимается такое, которое
проходит через базальную и апикальную поры (рис. 21, в, ж, п, у, ч).
Поперечное сечение располагается перпендикулярно продольному
и проходит через экваториальную часть внутренней полости ооспо-
рангия. Косые сечения располагаются под углом к продольному
(табл. 24, фиг. 14). Абсолютно надежным критерием продольных
сечений является наличие в срезе апикальной и базальной пор
(табл. 24, фиг. 1) или хотя бы одной из них (табл. 24, фиг. 2—4,
6—9, 11 —13,17). Иногда базальная пора может зарастать.
Для определения поперечных сечений нет такого критерия. По-
этому могут быть 'использованы следующие косвенные признаки.
В поперечном сечении гирогонита будет наблюдаться, как явствует
из рис. 21, в, г, наименьшее количество ребер (сечений партекаль-
цин). У утрикулов с гладкой внешней поверхностью и с базальными
выростами в поперечном сечении толщина стенки утрикула будет
более или менее одинакова (рис. 21, р). У утрикулов, орнаментиро-
ванных продольными ребрами, толщина последних в поперечном
сечении будет наименьшая. Определить же характер скульптуры
внешней поверхности можно в большинстве случаев по продольным
сечениям (табл. II). Отличия между сечениями гирогонитов и утри-
кулов в шлифах приведены в табл. III. Из этой таблицы видно, что
наиболее «универсальным» критерием различия во всех сечениях
является микроструктура стенки гирогонита и утрикула. Необхо-
димо только иметь в виду, что швы между партекальцинами вслед-
ствие перекристаллизации могут исчезать (табл. 24. фиг. 17).
Таблица II
Зависимость внешней орнаментации продольных и поперечных
сечений гирогонитов и утрикулов от типа скульптуры последних
Тип скульпту- ры Продольное сечение Поперечное сече- ние Роды
Скульптура отсутствует Внешняя поверхность гладкая U mbella Quasium- bella
Спиральные ребра Орнаментировано реб- рами, количество ко- торых зависит от чис- ла спиральных ребер и числа оборотов пос- ледних Орнаментирова- но ребрами,ко- личество кото- рых равно чис- лу спиральных или продоль- ных ребер Trochillis- cus
Продольные ребра Внешняя поверхность гладкая Chovanella
Частые про- дольные и поперечные ребра Орнаментировано реб- рами, количество ко- торых зависит от чис- ла поперечных ребер, иногда из-за малых размеров последних внешняя поверхность почти гладкая Орнаментирова- но ребрами, количество ко- торых зависит от числа про- дольных ребер Sycidium
Редкие про- дольные, иногда по- перечные ребра, вре- менами лишь на нижней по- ловине ут- рикула Внешняя поверхность гладкая или с редки- ми ребрами Орнаментирова- но редкими ребрами Elenia
30
Известковые организмы
Рис. 21. Девонские харофиты
ч . Ir<><hiliscus nuishni SamoUova из озсрско-ховапскпх c iuch фаменско! о яруса Рязан-
ской области: а — вид сбоку, о вид сверху (</.<< из рабо|Ы Самойловой. 1955). х 54.
поперечное сечение. . — продольное сечение: <1 — з — Sn uliuni mein Sanbf. iiraliensis
KarpinskyiH3 фраиских от ложений Урала: <> — вид сбоку, е — вил сверху (из работы Кар-
IIIIHCKOI о. 1906). х 16.2. ж — продольное сечение, з — поперечное сечение: и - - .и — С1н>-
iiinellu koialeii Reitlinger et Jarzcva из данково-лебедяпских слоев фаменскою яруса. По-
1есье. район с. Давидовка: и — вид сбоку, к — вид сверху, х 54. л продольное сечение.
' поперечное сечение; н — р — Umhella belki Maslov из седанских слоев фрапскою яруса
Воронежской области: н — вид сбоку, о—вил сверху, х 90. и -продольное сечение.
р - поперечное сечение: < ф - - QiiitsiuniMki roumdu (1 Bykoxa) из данково-лебедяпских
слоев фаменскою яруса Во.п отрадской области: <•— внд сбоку, т - вид сверху, х 45.
г - продольное сечение, видна апикальная крышечка, ф — поперечное сечение: v — ш —
Eleiiia Idnieiui (Е. BykoVa) нз данково-лебедянских слоев фаменскою яруса Вол1 от радской
области: v — вид сбоку, ч — вид сверху, х 45. ч продольное сечение, ш — поперечное
сечение [и - ш нз работы Быковой. 1955).
Буквы на рисунке: а. п. — апикальная пора: б. п. - базальная пора: а. к. - апикальная
крышечка: б. в. - базальные выросты: с. -- споростииа: б. пл. — базальная пластинка
Харофиты, или. харовые водоросли
31
Таблица III
Сравнение продольных н поперечных сечений гирогонитов и утрикулов и схема определения родов девонских харофитов
У девонских харофитов голые гирогониты имеются у представи-
телей рода Trochiliscus. Гирогониты трохилисков различаются меж-
ду собой по общей форме и форме внутренней полости, по разме-
рам. количеству партекальцин, их ширине и числу их витков, но
углам наклона спиралей, форме скульптурных элементов спираль-
ных ребер, строению базальной и апикальной частей, форме базаль-
ной пластинки. Швы между партекальцинами при полном обыз-
вествлении вещества спиральных клеток располагаются между реб-
рами. при частичном — в центре ребер. Все перечисленные призна-
ки более или менее легко наблюдаются у выделенных из породы
гирогонитов. Однако часто девонские известняки, содержащие тро-
хилиски. сильно метаморфизованы. Поэтому изучение последних
возможно производить только в шлифах — в случайных срезах
гирогонита. Для облегчения диагностики трохилисков в шлифах
в табл. IV показано, какие признаки наблюдаются в тех или иных
сечениях.
Кроме представителей рода Trochiliscus в девонских отложениях
СССР широко распространены харофиты, обладающие утрикулами
(Маслов. 1961а, 1963а; Поярков, 1965). Это роды Sycidium, Chova-
nella. Unibella, Quasiumbella и Elenia. Утрикулы этих харофитов раз-
личаются по наличию и характеру скульптуры внешней поверх-
ности и присутствию базальных выростов (рис. 21. д, и. н, с. л).
Внешняя поверхность этих девонских харофитов может быть глад-
кой, покрытой только продольными или продольными и попереч-
ными ребрами.
Иногда скульптура может присутствовать только на нижней по-
верхности утрикула. В. П. Маслов (1961а) считает, что характер
рисунка скульптуры утрикула может несколько варьировать в пре-
делах одного вида. Поэтому в целях систематики им надо пользо-
ваться крайне осторожно, обращая внимание лишь на общий тип
рисунка скульптуры. Среди последней выделяются следующие типы:
частые продольные ребра через весь утрикул (рис. 21, и), сочетание
частых продольных и поперечных ребер (рис. 21, д), редкие про-
дольные и поперечные ребра, иногда расположенные только на ниж-
ней части внешней поверхности утрикула (рис. 21, л), полное отсут-
ствие скульптуры (рис. 21,//, <•).
Светлый кальцитовый слой утрикула у некоторых харофитов
образует у базального отверстия горбообразные выросты самой раз-
личной формы (базальные выросты), между которыми располагает-
ся базальная пора (рис. 21, и, р; табл. 24, фиг. 1,2, 4, 5). Правда,
последняя наблюдается в шлифах редко, так как она может быть
видна лишь в точно ориентированных продольных сечениях (табл. 24,
Таблица IV
Сравнение возможности установления отдельных морфологических
признаков в различных сечениях гирогонитов рода Trochiliscus
Признак Продольное сечение Поперечное сече- ние Косое сече- ние
Форма гиро- гонита или его внут- ренней по- лости Видно продольное очертание гирого- нита (внутренней полости) Видно поперечное сечение гирого- нита (внутренней полости) Видно ко- сое сече- ние гиро- гонита (внутрен- ней по- лости)
Количество партекаль- цин Точно определить нельзя Определяется по количеству види- мых «ребер» в поперечном сече- нии гирогонита Точно оп- ределить нельзя
Число вит- ков парте- кальцин Определяется путем деления количества «ребер», видимых в продольном сече- нии, на количество «ребер», видимых в поперечном сечении Определить нельзя
Угол накло- на спира- ли (а) а — видимая ширина партекальцин в продольном сече- чении tg ас b — видимая шири- на партекальцин в поперечном се- чении b Определить нельзя
Ширина пар- текальцин (/п) т = a sina т = b cos a Определить нельзя
Базальная пластинка Наблюдается Не наблюдается Наблю- дается не всегда
Строение ба- зальной или апи- кальной части Устанавливается Не наблюдается Наблю- дается не всегда
32
Известковые организмы
фиг. 1). Сочетание наличия и отсутствия базальных выростов с ти-
пом скульптуры служит для различия утрикулов родов девонских
харофитов.
Строение апикальной части утрикула В. П. Маслов (1963а) счи-
тает у ископаемых харофитов родовым признаком. Действительно,
строение апикальной крышечки весьма разнообразно (табл. 24,
фиг. 2, 6—9, 11 —13,17). Однако таксономическое значение этого
признака у девонских харовых водорослей в должной мере еще не
выяснено. Е. В. Быкова (1955) полагала, что у представителей умбел-
ловых строение вершины может служить видовым различием. К ви-
довым признакам также относится форма утрикула и его внутрен-
ней полости. Определение выделенных из породы утрикулов не
представляет особых затруднений, значительно сложнее диагности-
ка их в шлифах. Для облегчения последней составлена табл. V.
Таблица V
Сравнение возможности установления отдельных морфологических
признаков в различных сечениях утрикула умбелловых
Признак Продольное се- чение Поперечное се- чение Косое сечение
Ф орма утрику- ла и его внут- ренней полос- ти Видно продоль- ное сечение утрикула и его внутрен- ней полости Видно попереч- ное сечение утрикула и его внутрен- ней полости Видно косое се- чение утрикула и его внутрен- ней полости
Продольные ребра Не видны Видны в виде шипов—ребер
Поперечные ребра Видны в виде шипов—ребер Не видны Видны в виде шипов—ребер
Базальные вы- росты Виден продоль- ный срез в ви- де горбооб- разного выс- тупа у нижне- го полюса Не видны Иногда виден ко- сой срез в виде горбообразно- го выступа у нижнего выс- тупа
Базальная плас- тинка Наблюдается Не видна
Строение апи- кальной и ба- зальной час- тей Видно Не видно
Как видно из этой таблицы, у умбелл и квазиумбелл наибольшее
количество признаков видно в продольных сечениях. Поэтому при
диагностике и описании последних необходимо в основном опирать-
ся на продольные или близкие к ним сечения. Вместе с тем полное
представление о сицидиях, хованеллах и елениях нельзя составить
только по продольным сечениям, так как характер скульптуры ви-
ден у них на поперечных и близких к поперечным сечениях.
Исследования последних десяти — пятнадцати лет (Рейтлингер,
1954; Быкова, 1955; Рейтлингер, Ярцева, 1958; Поярков, 1966) пока-
зали, что представители харовых водорослей в девоне имели широ-
кое географическое и узкое вертикальное распространение, т. е.
время существования отдельных видов имеет строго ограниченные
рамки. Вместе с тем в течение периода существования происходило
массовое развитие представителей рода, который в этот отрезок
времени занимал доминирующее положение. Это позволяет в нас-
тоящее время устанавливать по данным изучения харофитов воз-
раст с точностью до яруса и коррелировать разрезы между собой.
Девонские харофиты не только хорошие указатели возраста
вмещающих пород. Они также являются индикаторами определен-
ных фаций. Анализ распределения харофитов в девонских отложе-
ниях (Поярков, 1966) показал, что они принадлежат к двум боль-
шим группам. Первая включает представителей родов Umbella,
Quasiumbella, Elenia, которые встречаются исключительно в морских
отложениях. При этом харофиты приурочены в основном к неболь-
шим слойкам, лишенным остатков макрофауны. Прослои, содер-
жащие утрикулы, заключены между пластами с явно морской фау-
ной (брахиоподами, тамнопоридами и мшанками). Следовательно,
умбелловые хотя и обитали в морском бассейне, но массовое их
развитие было приурочено к определенным микрофациям. Весьма
характерно, что в Тянь-Шане в девонское время районы развития
харофитов располагались вдоль областей суши, т. е. умбеллы,
квазиумбеллы и елении явно были обитателями прибрежных мелко-
водных частей моря. Вторая группа объединяет трохилисков и си-
цидий. Они, как правило, в массовых количествах не встречаются
совместно с представителями первой группы. Эти харофиты. по-
видимому, обитали в более опресненных условиях (лагунно-дельто-
вых). хотя встречаются и в прибрежно-морских.
Обычно в девонских известняках редко встречаются отдельные
экземпляры харофитов. Как правило, наблюдаются скопления гиро-
гонитов или утрикулов. Это свидетельствует о некогда пышном
развитии харовых водорослей на отдельных прибрежных частях
морских бассейнов. Ежегодно отмирание огромной массы вегета-
тивных частей могло оказывать влияние на ход седиментогенеза,
периодически создавая аномальный газовый режим придонных вод
(дефицит О2 и появление сероводородного заражения).
ЖИВОТНЫЕ ОРГАНИЗМЫ
ФОРАМИНИФЕРЫ (FORAMINIFERA)
Табл. 25—29
Фораминиферы — одноклеточные животные, относящиеся к типу
простейших, или Protozoa, и состоящие из раковины и протоплазмы.
Последняя находится как внутри раковины, так и снаружи. Особен-
но характерны для фораминифер псевдоподии — протоплазмати-
ческие тонкие выросты, разветвляющиеся и анастомизирующие
(рис. 22). Они являются органами захвата пищи и передвижения,
участвуют и в построении раковины.
У очень небольшого числа представителей фораминифер рако-
вина пленчатая, из псевдохитина — сложного вещества, состоящего
из гликопротеинов и углеводов. У остальных раковина твердая,
минерализованная, различного химического и минералогического
состава. Состав раковин определяется прежде всего типом ее по-
строения. Различают раковины агглютинированные (час-
то называемые песчаными) исекреционные (микрозернистые,
стекловатые или гиалиновые и фарфоровидные) (рис. 23, см. рис. 24:
табл. 25, фиг. 1 —12; табл. 26. фиг. 1).
Агглютинированные раковины состоят из песчинок, реже — ске-
летных частей организмов, извлеченных животным извне при помощи
псевдоподий и скрепленных цементом, выделяемым прото-
плазмой. Состав агглютинированных частиц бывает очень разно-
образным, обычно специфичным для разных видов, реже — родов;
некоторые виды обладают избирательной способностью. Реже сос-
тав зерен обусловлен природой окружающей среды. Минералоги-
ческий состав раковин может быть моно- или полиминеральным.
Чаще в стенках раковин наблюдаются кварцевые зерна, реже — из-
вестковые (у многих палеозойских родов и у текстуляриид разного
возраста). Цемент агглютинированных раковин бывает или органи-
ческий, содержащий мукополисахаридовые кислоты, и мягкий, так
что раковины с таким цементом легко деформируются, или мине-
Фораминиферы
33
Рис. 22. Общий вид Allogroniia ovoidea Rumb. (Schultze, 1854)
а. псенлохшиновая раковина: б эндон.шзма. в. --эктоплазма
Рис. 23. Агглютинированные раковины
»/ Cliiiulimi paiisieiiaix d’Orb., х 23. датский ярус: б — Guudryina sp.. х 20 Маастрихт
рализованный и твердый, с органическим веществом, соединенным
с кальцием и железом, иногда с кремнием.
Химический состав. Известковые секренионные раковины обычно
слагаются карбонатом кальция (кальцит, реже арагонит) и органи-
ческим веществом — псевдохитином. Последний бывает или в виде
пленок и прослоев в стенках раковин и выстилки полостей, или в
тесной связи с кристаллической частью стенки. Кроме того, отме-
чаются примеси различных элементов, из которых важнейшие Mg,
Sr и Ва. Следует отметить, что фораминиферы обладают наивысшей
способностью накопления стронция в раковинах (до 1—5%) по
1102 3
сравнению с другими морскими беспозвоночными. Иногда в сек-
реционных стенках находятся и агглютинированные частицы раз-
личного химического и минералогического состава (табл. 25. фиг. 2).
Арагонитовые раковины наблюдаются только у видов несколь-
ких семейств. Содержание Mg в известковых раковинах колеблется
в широких пределах.
Кремневые раковины состоят из халцедона и кварца. Халцедон
в раковинах многих одно- и двухкамерных видов, относимых обычно
к агглютинирующим формам, установлен А. К. Богдановичем и
Р. Г. Дмитриевой (1956). Отсутствие цемента в. таких раковинах,
весьма мелкая размерность зерен халцедона и их тесное расположе-
ние указывают на возможно секреционное происхождение халцедо-
на в раковинах.
Экология. В результате многочисленных исследований, особен-
но американских ученых, установлена определенная экологическая
смена придонных комплексов современных фораминифер по зонам
разных глубин морских бассейнов. В самых общих чертах в мелко-
водных условиях (верхняя часть сублиторали) преобладают ми-
лиолиды, эльфидии и цибецидесы. в нижней части сублиторали
наблюдается как более разнообразный родовой состав, так и доми-
нирование секреционных раковин, на большей глубине и в батиаль-
ной области преобладают булиминиды. боливины. увигерины. ла-
гениды и другие в более обедненном сообществе, а еще глубже —
агглютинирующие формы. Максимальная глубина, на которой были
найдены живые фораминиферы, равна 10 687 л/, но в зависимости
от местных условий нижняя граница распространения бентосных
фораминифер колеблется в широких пределах. Факторами, влияю-
щими на распределение фораминифер по дну морей и океанов, явля-
ются температура, соленость, пищевой режим, биохимический сос-
тав водных масс, свет и т. п.
Установлено, что в более холодных водах преобладают фора-
миниферы с агглютинированной раковиной, а в теплых — с рако-
винами известковыми и секреционными. В области приливов, в
прибрежных осолоняющихся и опресняющихся водоемах обитает
своеобразный комплекс фораминифер, среди них — виды с псевдо-
хитиновой или с агглютинированной стенкой раковин и мягким
псевдохитиновым цементом или с кремневыми раковинами (тро-
хаммины, «Rotalia» beccarii и др.).
В распределении планктонных фораминифер ярче выражены кли-
матическая зональность и резкое их уменьшение с приближением к
береговой линии. Сводка по экологии современных фораминифер
дана Фледжером (Ph lege г, 1964).
Многими исследователями установлено, что раковины живых и
мертвых фораминифер чаще занимают приблизительно одни и те же
участки дна водоемов. Выяснено также, что смена комплексов фора-
минифер в последовательных слоях какого-либо разреза соответ-
ствует изменениям в сообществах фораминифер. существовавших
во время образования данного слоя (Раузер-Черноусова. 1953;
Крашенинников, 1960). Следовательно, ориктоценоз, т. е.
захороненный комплекс фораминифер, отражает в большинстве
случаев в значительной мере характер прижизненного сообщества
фораминифер какого-то биоценоза. Это дает возможность восста-
новления по ископаемым фораминиферам внешних условий с ис-
пользованием данных по экологии современных фораминифер. Так.
палеоэкологические и фациальные исследования подтвердили общее
сходство в распределении комплексов фораминифер в отложениях
третичного и мелового времени и в донных осадках современных
бассейнов (Морозова и др., 1966, и др.).
Фораминиферы отличаются большой изменчивостью и способ-
ностью приспособления к окружающей среде. Морфологические
признаки, возникшие в процессе индивидуальной, внутривидовой
и видовой приспособительной изменчивости, могут дать указания
как на условия осадконакопления, так и на другие физико-географи-
ческие факторы внешней среды. Так, например, килеватые двусто-
ронне симметричные раковины могут указывать на быстрое осадко-
34
Известковые организмы
накопление, так как экспериментально у таких раковин была дока-
зана возможность выбираться на поверхность дна водоема при за-
сыпании их песком, тогда как виды с более округлыми раковинами
погибали при этих экспериментах. Для участков со значительной
скоростью осадконакопления характерны более удлиненные формы,
развертывание у спирально свернутых, появление горлышек у устьев
и пр. Но те же признаки могут быть вызваны большой плотностью
поселений фораминифер и конкуренцией за обладание пищей. Под-
вижная водная среда вызывает более укороченные раковины с более
крепким скелетом у фузулинид, тогда как при более спокойном
гидродинамическом режиме те же виды представлены формами
более удлиненными и более тонкостенными. В большинстве слу-
чаев размеры особей, толщина стенок раковин и численность особей
видов также являются хорошими показателями биономических ус-
ловий и характеризуются более высокими значениями при более
высокой температуре, нормальной солености и лучших пищевых
условиях.
Для планктонных или пелагических фораминифер существенное
значение имеют температура и плотность воды, а также пищевые
ресурсы. Отмечено, что правозавитые спирально-свернутые рако-
вины фораминифер чаще приурочены к более теплым водам, а лево-
завитые — к более холодным. Бесспорным приспособительным
признаком к парению в толще воды является шарообразная форма
раковины с пустотелыми вздутыми камерами и с разнообразными
выростами, увеличивающими поверхность раковин. Другие скульп-
турные украшения или дополнительные отложения извести (напри-
мер, ребра, кили, бугорки на поверхности раковины и отложения
извести по основанию оборотов раковин) способствовали укрепле-
нию (или утяжелению) раковин, обитающих в зоне подвижного
гидродинамического режима. Такое же значение имеет сильная
складчатость перегородок у фузулинид. Все же следует отметить,
что в отношении функционального анализа морфологических при-
знаков фораминифер сделано до сих пор еще очень мало. Наблюде-
ния литологов могут содействовать освещению этой важной для
фациального анализа области.
Геологическое и стратиграфическое значение. Фораминиферы
населяли моря и океаны начиная с кембрийского периода и доныне,
а с каменноугольного периода составляли значительную часть мор-
ской фауны. Раковины фораминифер накапливались на дне водое-
мов, и в силу быстрого размножения и многочисленности особей
число оседающих ежегодно раковин (жизненный цикл фораминифер
обычно ограничен одним годом) бывает весьма значительно. Рако-
вины фораминифер нередко почти полностью образуют донные
осадки современных и древних бассейнов (глобигериновые илы,
писчий мел, нуммулитовые и фузулиновые известняки и т. п.), участ-
вуют в образовании биогермов (нубекуляриевые известняки). Разви-
тие фораминифер с палеозойской эры протекало неравномерно,
в зависимости от цикличности геологических процессов, и эти осо-
бенности их эволюции используются для освещения хода геологи-
ческой истории. Существенный интерес для геолога представляют
и различия развития фораминифер в разных геотектонических и
палеогеографических областях. Так, для геосинклинальных областей
отмечается более раннее появление видов и родов и большая дли-
тельность их существования, но разорванность ареалов их обита-
ния. В разных биогеографических областях и провинциях система-
тический состав фораминифер обычно резко отличается. Стенофа-
циальность фораминифер также имеет существенное геологическое
значение.
Но, бесспорно, наиболее широко используются фораминиферы
со стратиграфическими целями. Исключительно быстрая эволюция
во времени, большая экологическая изменчивость, широкое и быст-
рое (в геологическом смысле) распространение фораминифер в бас-
сейнах прошлого Земли — эти три свойства фораминифер обеспе-
чили им признание как одной из основных групп, четко характери-
зующих стратиграфические подразделения, и послужили основанием
для выделения микропалеонтологии как особой области палеонто-
логии, имеющей наибольшее практическое значение.
Породообразующее значение. Раковины как бентосных (например,
фузулиниды, нуммулиты), так и пелагических (глобигерины, глобо-
трунканы и др.) фораминифер могут целиком, почти без цемента,
слагать породы (табл. 27, фиг. 2, 4, 8). Чаще породообразователями
бывают раковины не одного семейства или рода, а представители
нескольких семейств (табл. 28, фиг. 1; табл. 29, фиг. 2), но обычно
с преобладанием одного таксона. Таковы эндотировые известняки
нижнего карбона, лагенидовые породы верхней юры или верхнего
мела, третичные милиолидовые, нуммулитовые и нубекуляриевые
слои (табл. 27, фиг. 1,5: табл. 28, фиг. 1. 3, 4; табл. 29, фиг. 1). В по-
роде раковины породообразователей обычно бывают скреплены
цементом, содержание которого может колебаться от нескольких
процентов до 50% (табл. 28, фиг. 2, 5, 6).
Раковины бывают или заполнены осадком, или (реже) пустые.
Внутренние полости раковин обычно заполняются вторичным про-
зрачным кальцитом, иногда — железистыми соединениями (пири-
том и др.). В наружном обороте, или последних камерах, раковины
обычно наблюдается та же порода, что и вмещающая, йо нередко
более мелкозернистая.
В расположении в породе раковин удлиненной формы нередко
наблюдается параллельность плоскостям напластования (табл. 27,
фиг. 3), а также определенная ориентировка. Последняя указывает
на направление движения водных масс. Скопления раковин в опреде-
ленных тонких прослойках могут быть объяснены помимо других
причин также и сезонностью в репродукционных процессах.
Сортировка раковин проявляется чаще в соответствии размеров
раковин гранулометрическому составу пород. В тонких породах
наблюдаются очень мелкие раковины или молодые особи1, что,
по-видимому, следует отнести к переносу этих раковин в более глу-
бокие части бассейна. Переносом пустых, легких раковин в обрат-
ном направлении, в область более мелководную, объясняется на-
хождение видов, обычно обитающих в полосе илистых осадков,
среди прибрежных, более грубых пород. В этих случаях наблюдается
заполнение наружного оборота более грубозернистым осадком и
нередко окатанность раковин.
В породах нередко наблюдаются деформированные раковины.
В агглютинированных раковинах особенно часто обнаруживается
посмертное сдавливание при сохранении всей раковины целой (тро-
хаммины, гипераммины, реофаксы). Цемент стенок таких форм,
в состав которого входит органическое вещество, обладает некото-
рой пластичностью. Известковые раковины реагируют на давление
разломом раковины. И по характеру разломов можно судить о
направлении сил, деформировавших раковину. Следует указать, что
случаи посмертных поломов раковин легко отличимы от прижиз-
ненных, так как последние обычно залечиваются животными. Такая
регенерация раковин особенно часто наблюдается в области сильно-
го движения воды, например в полосе рифов.
Следует еще отметить, что при опускании пустых раковин пела-
гических фораминифер на дно океана происходит или их полное (на
глубинах в 3000—4800 .и), или частичное растворение (с образова-
нием изъеденных поверхностей раковин).
Приведенные случаи переноса раковин фораминифер и их захоро-
нения в одновозрастных, но инакофациальных отложениях следует
четко разграничивать от вторичного переотложения в породах
более молодого возраста в результате размыва вмещавшей форами-
ниферы породы. Переотложение их раковин наблюдается нередко
(нижнепермские отложения Урала, третичный флиш Кавказа, Альп
и др.). Чаще такие явления отмечаются в грубозернистых породах,
' Не следует смешивать это явление с наблюдаемой иногда «детской смертностью» фора-
минифер. когда одновременно с взрослыми раковинами отмечается большое число мо-
лодых, что указывает на неблагоприятные для организма прижизненные условия (напри-
мер, в гипсово-доломитовых отложениях Донбасса). В современных осадках раковины
молодых особей обычно составляют небольшой процент.
Фораминиферы
35
в начальных трансгрессивных фациях седиментационных циклов.
Установить случаи вторичного залегания не всегда легко удается,
и это вызывает большие затруднения и ошибки в стратиграфических
исследованиях. Твердые критерии переотложения указать трудно.
Разный характер вмещающей и заполняющей раковину породы,
а также окатанность раковин могут быть признаком переотложения,
но также наблюдаются и при захоронении в песчаных донных осад-
ках с небольшим переносом к берегу в пределах того же водоема.
Бесспорным показателем переотложения является сохранение вместе
с раковиной породы более древнего типа (табл. 27, фиг. 6). Другим
прямым доказательством переотложения является смесь фаун раз-
ного возраста. Но такие случаи иногда толкуются как переживание
видов (если возраст разных фаун близок) и не дают полной уверен-
ности в возрасте пород, устанавливаемом лишь по присутствию эле-
ментов самой молодой фауны.
Морфология. Раковины фораминифер бывают одно-, двух- и
многокамерными (рис. 24). Однокамерные состоят из одной рако-
вины, реже — из скопления однокамерных особей. Двухкамерные
раковины образованы начальной камерой и второй трубчатой ка-
мерой, не подразделенной на части перегородками (септами). По
характеру расположения камер среди двух- и многокамерных рако-
вин различают однорядные формы с прямолинейно расположенны-
ми камерами, формы с беспорядочно расположенными камерами и
Структура стенки раковин. Под строением стенки раковины
обычно понимают: 1) способ построения (образования) раковин;
2) химический и минеральный состав; 3) микроструктуру (характер
зерен и кристаллов в стенке, их форму, положение, размеры); 4) по-
ристость стенки; 5) характер слоев, слагающих стенку; 6) характер
соединений (швов) последовательных камер; 7) скульптуру рако-
вины.
Общий вид раковины как с поверхности, так и в шлифах опре-
деляется прежде всего микроструктурой и пористостью стенки. Спо-
соб построения (образования) раковин, а также химический и мине-
ралогический состав стенок рассмотрены в начале очерка. Микро-
структуре раковин уделяется наибольшее внимание в исследованиях
фораминифер.
Микроструктура стенки агглютинированных раковин обычно ха-
рактеризуется размерами зерен, реже — их формой и положением
(табл. 25, фиг. 1—3). Положение и размеры зерен внутренней и на-
ружной частей стенки бывают различными. Поверхность стенки
может быть шероховатой или гладкой, в зависимости от размера
частиц и плотности их кладки. Микроструктура цемента обычно
микрозернистая. Агглютинированные частицы в секреционных стен-
ках некоторых родов представляют особый интерес. Так, например,
у эндотиранопсид крупные кристаллы кальцита, резко отличные от
основной секреционной части стенки, хорошо окристаллизованы
Рис. 24. Различные типы раковин, по Покорны (Pokorny, 1958)
°—г — однокамерные; д — двухкамерные; е — с — многокамерные; а — шарообразная, б — лопастная, в — трубчатая, г — древовидная, д — двухкамерная, трубчатая спирально-
плоскостная. е — однорядная, ж — то же, с изогнутой осью, з — то же, с седловидными камерами, и — спирально-винтовая двурядная, к — то же, трехрядная, л — спирально-плоскост-
ная, зволютная, .и — то же, ииволютиая, н — то же, веретеновидная, о — спирально-коническая, л — трохоидная, р, с — милиолидовая
с навитыми вокруг оси. Навивание бывает клубкообразным, сигмои-
дальным, спиральным и пр. В последнем могут быть спирально-
плоскостные, спирально-конические (трохоидные, или низкокони-
ческие, и высококонические) и спирально-винтовые. Камеры бы-
вают расположены в один ряд или многорядно. Нередко раковины
на разных стадиях роста имеют разное строение (табл. 27, фиг. 8).
К существенным признакам кроме типа строения раковин относятся
строение их стенок (см. следующий раздел), типы устьев (отверстий
в последней камере для выхода наружу псевдоподий — протоплаз-
матических тяжей) (рис. 25), отверстий, соединяющих соседние ка-
меры и служащих для сообщения с внешней средой (форамены,
поры и др.), система каналов в стенках, дополнительные скелетные
образования, наружные (скульптура) и внутренние, строение пере-
городок и др. Таксономическое значение морфологических призна-
ков фораминифер может быть различным для разных системати-
ческих категорий и изменяется в процессе исторического развития
отдельных групп фораминифер.
(табл. 29, фиг. 8), что вызывает сомнение в их инородном происхож-
дении.
Микроструктура секреционных раковин лучше изучена у извест-
ковых кальцитовых стенок. Выделяются три основных типа микро-
структуры: микрогранулярная, фарфоровидная и стекловатая (или
гиалиновая), отличающиеся размерами, расположением и формой
зерен — кристалликов и положением их оптических осей (Wood,
1949; Крашенинников, 1956).
У стенки с микрогранулярной микроструктурой (табл. 25, фиг. 2,
4—6; табл. 26, фиг. 7) преобладают зерна размером около 3 д со зна-
чительными колебаниями в обе стороны. Зерна обычно равновеликие,
и их оптические оси расположены беспорядочно, вследствие чего
стенка в проходящем свете под микроскопом темная. Отдельные
зерна — кристаллы отчетливо видны в микроскопе при больших
увеличениях. Форма зерен обычно с округлыми контурами или не-
сколько вытянутыми, реже с угловатыми (лапчатыми). Иногда зер-
на располагаются рядами, создавая впечатление «волокнистой»
36
Известковые организмы
микроструктуры (табл. 25, фиг. 5), но никогда не дают оптических
явлений, свойственных последней. У зернистых стенок с очень мел-
кими зернами наблюдается пятнистость в результате обособления
отдельных участков. У некоторых видов отмечено как бы «расслое-
ние» стенки по слоям с разными размерами зерен (табл. 25, фиг. 6).
Для микрогранулярных раковин характерна непостоянная примесь
агглютинированных частиц. Этот тип стенки свойствен большин-
ству палеозойских форм и некоторым текстуляриидам и орбитоли-
нидам.
Фарфоровидная стенка раковин (табл. 25, фиг. 11; табл. 26, фиг. I)
слагается мельчайшими удлиненными кристаллами (в поперечнике
0.2—0.4 д, в длину 2—5 мм) с оптическими осями, расположенными
беспорядочно, чем объясняется низкая интерференционная окраска
(серые и желтые тона первого и второго порядка) при скрещенных
николях, почти не изменяющаяся при вращении столика. Иногда
на участках в наружной части стенок наблюдается расположение
оси кристалликов параллельно поверхности раковины, что выра-
жается в поляризованном свете лимонно-желтым цветом ободка
раковины. В отраженном свете фарфоровидные раковины белые.
Фарфоровидной микроструктурой обладают милиолиды.
Рис. 25. Типы устьев раковин
</ конечное крукюе: о — основное (база ните) ч ycibe с зубом: г — спювнлное:
d — дендритовидное: с ус»ье па конце шейки
Стекловатая (или гиалиновая) микроструктура наблюдается у
большинства мезозойских и кайнозойских известковых секреционных
раковин. Стенка их раковин отличается обычно некоторой прозрач-
ностью. что объясняется характером пористости и иным расположе-
нием кристаллов (кристаллических отдельностей) и их оптических
осей в стенке. Различаются две основные разновидности стеклова-
той микроструктуры: стекловатая зернистая и стекловатая радиаль-
но-лучистая.
Стекловатая зернистая микроструктура характеризуется округ-
лой или пластинчатой формой зерен — кристаллов (кристаллических
отдельностей), преобладающими их размерами в 0,5—10 д, бес-
порядочным расположением зерен, но, возможно, с кристаллогра-
фической ориентировкой ромбоэдра (1014) параллельно поверхности
стенки; в поляризованном свете окраска высоких тонов (табл. 25,
фиг. 7).
Стекловатая радиально-лучистая микроструктура стенок (табл. 25,
фиг. 8—10) образована удлиненными кристаллами кальцита с оп-
тическими осями, расположенными нормально к поверхности стенки
раковины. В тангенциальных сечениях стенок с такой микрострукту-
рой при скрещенных николях в сходящемся свете появляются от-
рицательные одноосные интерференционные фигуры — черный крест
с концентрическими цветными кольцами. В осевых и поперечных
сечениях раковин при скрещенных николях в параллельном свете
наблюдается четырехкратное погасание каждого из участков стенки
при вращении столика. Явления креста и погасания отмеча-
ются также у шарообразных камер, ведущих себя как монокри-
сталл.
По размерам кристаллов и их расположению различаются три
разновидности этой микроструктуры: 1) тонко радиально-лучистая
(табл. 25. фиг. 8) — преобладает толщина кристалликов в 0.5—1.5 д
и кристаллики проходят через всю стенку строго перпендикулярно
ее поверхности; 2) грубо радиально-лучистая (табл. 25, фиг. 10) —
толщина кристаллов 2—3 д. расположение то же, радиальная полос-
чатость лучше видна, чем в стенках с более тонкими кристаллами;
3) неясно радиально-лучистая (табл. 25, фиг. 9) — с кристалликами
(толщиной 1—2 д) изогнутой червеобразной формы и в длину не
проходящими через всю стенку (за исключением отдельных кри-
сталлов — волоконец), но с общим радиальным расположе-
нием.
Такие же микроструктуры, как у кальцитовых, наблюдаются и у
арагонитовых стенок. Их различают дифракционными методами
при помощи икс-лучей, методами химическими и окраски.
Микроструктура халцедоновых секреционных раковин крипто-
кристаллическая. При скрещенных николях обнаруживаются тесно
прилегающие друг к другу мельчайшие зерна, которые дают агре-
гатную поляризацию, волнистое погасание и беспорядочно меняю-
щуюся конфигурацию при вращении столика микроскопа. По раз-
меру зерен можно различать раковины с микрозернистой микро-
структурой (размеры зерен обычно не более 5 д) и с тонкозернистой
микроструктурой (5—15 д). Иногда наблюдаются отдельные более
крупные зерна. В сечениях при одном николе халцедон почти бес-
цветен, иногда слегка желтоватый. В поляризованном свете окраска
стенки серая (цвета интерференции первого порядка). Халцедон
исследованных раковин оптически отрицательный, одноосный,
реже — двухосный. Поверхность халцедоновых раковин обычно
шероховатая, матовая, иногда приближающаяся к виду раковин
с фарфоровидной микроструктурой и иногда даже со стекло-
ватой.
В последние годы применяется еще метод изучения микрострук-
тур стенок раковин при помощи реплик под электронными микро-
скопами (обычным и сканирующим). Получены очень разнообразные
и интересные изображения (табл. 26, фиг. 5, 6). Наиболее существен-
ным является показ тесной связи органической матрицы и мине-
ральной части раковины. Органическое вещество образует пленки
и выстилки между последовательными слоями кристаллов в стенке,
обволакивает кристаллы и кристаллические отдельности и даже
находится внутри единичных кристаллов.
Перекристаллизация стенок и замещение одних минералов други-
ми наблюдаются при любых микроструктурах стенок. При пере-
кристаллизации секреционного кальцита происходит увеличение
размеров зерен и кристаллов, но их оптические свойства сохра-
няются.
Пористость стенки (табл. 25, фиг. 5; табл. 26, фиг. 2, 3. 7, 9) яв-
ляется ее существенным признаком и находится в определенной за-
висимости от микроструктуры. Так, все раковины со стекловатой
радиально-лучистой стенкой обладают порами, размеры которых
колеблются от долей микрона до 5—10 д. Поры, отверстия на по-
верхности стенки и поровые канальцы, пронизывающие стенку
раковин, бывают различных размеров и формы: простые, в виде
цилиндрических или изогнутых трубочек различных диаметров, или
сложные, в результате слияния или разветвления простых поровых
каналов. При правильном строении сложных пор образуется ячеис-
тая структура стенки (табл. 26, фиг. 7; рис. 26.д). Беспорядочное рас-
положение пронизывающих стенку каналов и пустот характеризует
губчатую и лабиринтовую структуры стенок.
Фораминиферы
37
Стенки бывают сложены одним или несколькими слоями. Разли-
чают первичное и вторичное сложение стенки. Первичное может
быть однослойным, без ясно различимых слоев, или первично много-
слойным. с несколькими разноструктурными слоями. Например,
у первично двухслойных раковин сохраняется внутренний псевдо-
хитиновый слой в виде выстилки полости камер. При растворении
камер в соляной кислоте этот слой сохраняет форму камер и всей
раковины. У палеозойских форм стенка раковины иногда состоит
из двух слоев различного происхождения: внутреннего секрецион-
ного известкового стекловатого радиально-лучистого и наружно-
го агглютинированного (табл. 29, фиг. 9). Для многих форамини-
фер. особенно палеозойских (фузулиниды, эндотириды), характер-
ны первично многослойные стенки со слоями различного строения,
отличающиеся по микроструктуре, окраске, толщине (рис. 26;
табл. 25, фиг. 6, 12).
Вторичная многослойность стенок раковины является результа-
том нарастания новых слоев стенки при формировании новой ка-
меры на наружные части всей раковины с ее последующим утолще-
нием и облеканием. Слои нарастания разделены гранями нарастания
утолщений (табл. 26. фиг. 2—4). Число вторичных слоев в стенках
может быть различным в зависимости от характера перекрытия и
облекания числа камер и частей раковины при образовании новой
камеры (рис. 27). У вторично многослойных стенок ориентировка
а б
Рис. 26. Типы строения
стенок раковин фузулинид
и о.тиос тойттая стенка (приматекаI: б двухс.тойпая протека (i тектум. пр iipii-
матека). « i рехс.тонная (и. I. наружным тскторий: т. - тектухт: пр. ттриматека):
чстырехслойная стенка (н. т. — наружный тскторий: д. - диафаиотека: в. т.— внут-
ренний текторнй): д - кериотекальное строение (а. — норовые каналы керпотеки: б. —
межтюровая часть сiенки)
Рис. 27. Вторично-многослойная стенка
у лагенид
а — простое сочленение смежных камер: наружная
стенка всех камер от начальной до последней одно-
слойная: б — черепицеобразное нарастание: стенка
каждой последующей камеры перекрываез стенку
предыдущей камеры; в — облекающее, многослойное
нарастанне: стенка каждой последующей камеры
полностью облекает все предыдущие камеры: у всех
камер, кроме последней, стенка многослойная
Рис. 28. Предшовные утолщения
у Nodosaria sp.
Рис. 29. Виды родов фораминифер
Отряд Astrorhizida: a I’summo\pinu-rci /тт.мтт Schuh/c. -. 22.5. о.титонсн (по. О. К. Каи-
iapeiiKO-Чсриоусовой): б. в — Siucuiiiiiiina iik/luria Г. Bykoxa. х 37.5. фамсттский ярус (по
I В. Быковой): г — Hyperanunina elonyaHi Brady, х 30. современный (по Бреди); <) — Etulaii-
diu dcipins (Rauser cl Reitlinger). x 75. живсгский ярус (no E. В. Быковой)
Отряд Endothyrida: с. ж -- Glid,t>eudi>thyra f/hihulus (Eichw). х 30. виде, г Гула: з. и —
Cribroipira panderi Moeller: з — х 30. и— х 26. виде, т. Венев (по Е \. Рейглингер):
к — Ijidiiihvru oinphaloiii Rauser et Reiilinger. x 26. визе. Подмосковный бассейн (по
Е. А. Рсйплитн'чз)
Отряд Fusulinida: / схема строения фузулинид (и. к. — начальная камера, с. а. --
ссттгалытыс арки: с. — септы: с.6. — септальные борозды: х. — хоматы: у. — устье: т
туннель): .и - схема внутреннего строения вербеекины
38
Известковые организмы
кристаллов каждого слоя сохраняется одинаковой через всю стенку,
что иногда создает ложное впечатление пронизывания кристаллами
линий (граней) нарастания. Поровые каналы проходят насквозь
через все слои вторично многослойной стенки.
Линии сочленения камер обычно выражены на поверхности рако-
вин в виде швов, иногда сложного слоения, а в шлифах — в виде
граней и темных линий1. Иногда образуются предшовные утолще-
ния (рис. 28), которые, разрастаясь, дают многослойную стенку.
Скульптура раковин в виде различных ребрышек, струек, килей,
бугорков, шипов, игол и т. п. выростов и наростов (рис. 35, а — е)
может быть результатом как дополнительных образований прото-
плазмой скелетного вещества, так и утолщения на определенных
участках последовательных слоев нарастания стенки раковины
(табл. 26, фиг. 2). Скульптурные части раковин обычно не пористые,
и их микроструктура в таких случаях зернистая. Скульптура появи-
лась на раковинах лишь в самом конце палеозоя. В значительной
степени скульптура раковин, зависящая от активности секреционной
деятельности протоплазмы, служит показателем палеоэкологических
условий.
На рис. 29—35 даны изображения различных палеозойских и
мезозойских фораминифер (внешний вид, разрезы, схемы строения
сложных спирально свернутых раковинок).
Иногда впечатление темной линии на границе стенки и внутренней полости камер, за-
полненных крупнозернистым кальцитом, является лишь результатом оптических явлений.
Рис. 30. Схема строения неошвагерин
Буквы на рисунке: с.—септы; с. б.—септальные борозды; с.п,— септулы аксиальные;
с.п2— септулы спиральные; п. — парохоматы; у. — устье; б. о.—боковые отверстия
(по Г. А. Дуткевичу)
Рис. 31. Виды родов фораминифер
Отряд Ataxophragmiida; о, б — Verneuilina sp., х 30; в, г — Clavulinoides szaboi (Hantken), х 30, эоцен. Каракумы (по В. T. Балахматовой); д, е — Valvulammina ralnovskajae Kusina.
х 100. готерив-баррем. Западная Сибирь (по В. И. Кузиной); ж, з—Telrataxis conica Ehrenberg, х 30, визе, г. Тула; и — л — Glohivalvulina graeca Reichel, х 40, пермь, Греция (по
Рейчелу)
Отряд Rota Hida, надсемейство Discorbidea: ,w — о — Discorhis dampelae Mjatljuk, x 90, апт. p. Эльба (по E. В. Мятлюк); n—c—Eponides sibiricus Neckaja. x 72, сенон (по H. H. Суб-
ботиной). Надсемейство Ceratobuliminidea; m, у— Epistomina regular) s Terquem, x 40, байос (по Терквему)
Фораминиферы
39
Рис. 32. Виды родов отряда Rotaliida
Надсемейство Nonionidea: о — в — Anomalina ajfinis (Hantken). х 50, эоцен, п-ов Мангышлак (no Н К. Быковой); г. д — Nonion incrassatus (Fichtel et Moll), x 50, плиоцен, Албания
(по E. А. Храмой). Надсемейство Rotaliidea: e. ж — Elphidium macelluni (Fichtel et Moll), x 102, миоцен, Украина (по А. К. Богдановичу). Надсемейство Globigerinidea: з — Globige-
rina hulioides d’Orb., x 102, современная, экспедиция «Витязь»: u — л — G. aperture Cushman, x 47, эоцен, Кавказ; .и, и — Hantkenina mexicana Cushman, x 33, эоцен. Кавказ; о — p —
Globorotaiia ntembranacea (Ehrenberg), x 47, датский ярус. Крым: < — у — Rotalipora reicheli Mornod. x 47. сеноман, Кавказ; u — у — по H. H. Субботиной
Рис. 33. Схематический вид нуммулитов (а), ассилин (б) и оперкулин (<?)
Верхний ряд — продольные сечения; нижний ряд — экваториальные сечения
Рис. 34. Схемы, показывающие соотношение между экваториаль-
ными и боковыми слоями и боковыми камерами
а — Diseocyclina: 6 — Asterocyelina (не заштрихованная часть поперечного сечения соответ-
ствует слоям боковых камер)
40
Известковые организмы
Рис. 35. Строение фораминифер
Отряд Buliminida: а — Uiegerina costellata Moroz., х 72, эоцен. Волгоградская область. Тнпы скульптуры раковнн у представителей семейства Buliminidae: б — тонкие ребрышки
(Crammostomunt): в — радиальные ребра (Bulimina): г — складчатые ребра (Bulimina praeinjlata N. Bykova), x 75. эоцен, Туркмения: д — сосочковидные шипы (Uiegerina): е — шипо-
ватая скульптура (Uvegerina)
Отряд Heterohelicida: ж. з — Bolivina mississipiensis Cushman, х 102, нижннй олнгоцеи, Туркмения; и, к — В. aragonensis (Nutall ), х 77. средний эоцен, Приааралье: г — н — Hete-
rohelix americanus (Ehrenberg), х 102, сеномаи. Кавказ; о — р — Ciimhelina globulosa (Ehrenberg), х 72, Маастрихт. Кавказ: с — у — Pseudotextularia varians Rzehak. х 47. масстрихт.
Кавказ
Формы неясного систематического положения: ф. х — Archaediscus karrery Brady (ф— внешний вид, визе. Англия, по Бреди; ,v — продольное сечение, х 100. визе. Колво-Вишер-
ский край, по А. П. Гроздиловой и Н. С. Лебедевой); у— Propermodiscus ulmeri (Mikh.). продольное сечение. х!20. визе. Ленинградская область (по А. В. Михайлову)
>« — к — но Н. К. Быковой .1 — г -- по Н. Н. Субботиной
ТИНТИННИДЫ (TINTINNOIDEA)
Тинтинниды — это свободноплавающие и способные к флотации
микроскопические организмы, относимые к наннопланктону. Они
населяют океаны и окраинные моря (известно около 800 видов).
Незначительное число видов (25—30) живет во внутриконтиненталь-
ных морях, пресноводных водоемах и реках, причем часть из них
является морскими реликтами (Гаевская, 1933). Современные тин-
тинниды обитают преимущественно в верхних слоях воды, но на
больших глубинах известны придонные формы (Тагац, 1933).
Геологическое значение. Ископаемые тинтинниды широко рас-
пространены в тонких известняках и мергелях альпийской геосин-
клинальной зоны в титоне и неокоме. Отдельные виды обладают
еще более узкой стратиграфической приуроченностью. Тинтинниды
определяются в шлифах при увеличениях более ста. Поперечное
сечение их всегда круглое, продольное — разнообразной формы
(кувшинчика, колокола, цилиндра и пр.), замкнутое внизу и откры-
тое в верхней части. Характер продольного сечения — основной
систематический признак. Распространение на большой площади
при узкой стратиграфической приуроченности, оригинальная форма,
более или менее устойчивые размеры, однотипность химического
состава скелета, а также присутствие в породах, где другие опреде-
лимые остатки практически отсутствуют, дают право считать их
руководящими ископаемыми.
Систематическое положение ископаемых тинтиннид было проб-
лематичным до 1934 г. Вначале их относили к фораминиферам, из-
вестковым водорослям, радиоляриям или считали Incertae sedis.
К настоящему времени известно более 10 родов и около 30 видов
тинтиннид, описанных различными исследователями, и отмечено
их присутствие в титоне и неокоме не только европейской части
Тетиса, но и в Северной Африке, Гималаях, на Кубе, в Мексике
и в других местах. В Советском Союзе тинтинниды обнаружены
в 1935 г (Vassoevich, 1936), затем в 1955 г. в титоне и неокоме Кавказа
и Крыма, а также в Советских Карпатах и в Предкарпатском про-
гибе (Вялов, Липецкая, 1960).
Ископаемые тинтинниды распространены в различных известня-
ках и мергелях. Высокие концентрации тинтиннид свойственны чис-
тым тонкозернистым известнякам глубоководного облика, которые,
по представлениям многих исследователей, образовались в гео-
синклинальных депрессиях (Colom, 1948; и др.). В известняках с
примесью терригенного или глинистого материала, а также с появ-
лением другой фауны и флоры концентрация тинтиннид резко
уменьшается. В окраинных бассейнах во всех перечисленных раз-
ностях пород концентрация тинтиннид очень мала.
Анатомия. Современные тинтинниды — одноклеточные животные,
обладающие студневидной или псевдохитиновой, однослойной или
многослойной оболочкой (наружным скелетом), на поверхности
которой могут аккумулироваться минеральные или органические
частицы. Форма протоплазматической клетки колоколовидная или
коническая. Задний конец заострен (рис. 36). С его помощью клетка
Тинтинниды
41
прикрепляется к основанию или к стенке оболочки (домика, ракови-
ны). На переднем конце клетки расположена хорошо сократимая
дисковидная перистома, в углублении которой расположено ротовое
отверстие. Перистома окружена воротничком, несущим мембра-
неллы. При помощи спирали мембранелл тинтинниды оживленно
плавают в воде. Оболочка (раковинка) снабжена скульптурой ске-
летных килей, узких балочек и перекладин, повышающих ее проч-
ность. Химический состав, форма и структура оболочек современных
тинтиннид являются важным систематическим признаком. Предпо-
лагается, что ячеистые стенки раковинок некоторых видов тинтин-
а.
Рис. 36. Общий вид ныне живущего
вида Tintinnopsis campanula (Ehrenb.)
и его домика
Буквы на рисунке: а. — адоральная зона: 6. або-
ральная зона: в. — воротничок: к. — каудальный
отросток: но Фор-Фремье (Основы палеонтологии.
1962)
— В-
5Z7L
нид содержат внутри ячеек вещества более легкого удельного веса,
облегчающие флотирование тинтиннид в водной толще. Частым
остановкам в наиболее подходящем слое воды способствует отогну-
тый назад передний край раковины (воротничок; рис. 36, в).
У ископаемых тинтиннид раковинки известковые, преимущест-
венно кальцитовые, редко со следами окремнения (Colom, 1948;
и др.). Нет единого мнения о том, первичны ли оболочки ископае-
мых тинтиннид, или они претерпели изменения химического соста-
ва при фоссилизации. Одни исследователи считают, что первоначаль-
но раковинка была агглютинированной, а попав в осадок, была
замещена кальцитом (Deflandre, 1936; и др.). По представлениям
других, первоначально раковинка была органической (подобно
Рис. 37. Основные виды ископаемых Tintinnoidae юры и мела
« — Calpionella alpina Lorenz, титон; б — Calpionella elliptiea Cadisch. титон; в — Calpionella
undelloides Colom, титон; г, д — Calpionellites darderi (Colom), неоком; e —- Calpionellites
neocomiensis Colom, неоком; ж — Calpionellopsis thalmanni (Colom), валанжин; з — Favelloi-
des pseudoserrata Colom, неоком; и — Calpionellopsis simplex (Colom), неоком; к. t—Tin-
tinnopsella earpathiea (Murgeanu et Filipescu). титон и неоком: .« — Tintinnopsella oblonga
(Cadisch), титон и валанжин; и — Tintinnopsella longa (Colom), неоком; о. n — Tintinnop-
sella cadischiana Colom, неоком; p — Tintinnopsella doliphormis (Colom), неоком: c—Tin-
tinnopsella batalleri Colom, неоком; tn — Tintinnopsella maxima Colom, неоком; у—Tin-
tinnopsella romaniea Boiler, валанжин; ф — Favelloides bolearica Colom, неоком; r — Rhabdo-
nelloides inesperatta Colom, валанжин: у — Amphorellina subacuta Colom, неоком; ч — Ampho-
rellina lanceolata Colom, неоком: ш — Stenosemellopsis hispanica (Colom), неоком: ut— Cras-
sicolaria brevis Remane, верхний титон; з — Crassieolaria parvula Remane, верхний ти-
тон; ю—Crassieolaria massutiniana (Colom), титон; я — Crassieolaria intermedia (Durand
Delga). верхний титон; ы—Coxliellina berriasiensis Colom, бериас: Salpingellina levan-
tina Colom, валанжин—готерив.
a — m. ф— ш.ю.ы. ь— по Колому (Colom, 1948). x 220,5; r— no Болерру (Boiler, 1963).
x 216; щ.э. я — по Ремен (Remane. 1963): щ.э — x 238.5. _ч — x 247.5
42
Известковые организмы
псевдохитиновым оболочкам современных тинтиннид), затем в
осадке заместилась кальцитом (Colom, 1948; и др.). Некоторые пред-
полагают, что раковинки ископаемых тинтиннид были первично
кальцитовыми.
Наружная часть раковинки четко очерчена, внутренняя часто
имеет неясный изломанный контур. Стенка сложена волокнистым
кальцитом (Andrusov, 1950; и др.).
При диагнозе ископаемых тинтиннид употребляется термино-
логия, заимствованная из описаний современных форм. Открытый
конец раковинки называется оральной, или устьевой, частью. Про-
тивоположный конец — аборальным, у многих форм он заканчи-
вается тонким остроконечным отростком, названным каудальным.
В самом верху раковинки, над устьем, различается воротничок.
Последний может быть параллельным стенкам раковинки, направ-
ленным во внутрь и наружу. В систематике ископаемых тинтиннид
большое значение имеют форма раковинки, размеры и расположе-
ние воротничка (рис. 37). Учитывается длина раковинки, ширина,
ширина ее аборальной части (рис. 36, б), если раковинка расширена
книзу, ширина устья, присутствие каудального отростка (рис. 36, к),
который при захронении не всегда сохраняется.
Определение ископаемых тинтиннид производится по вертикаль-
ному или близкому к вертикальному сечению. В косых сечениях
происходит изменение основных параметров вида. Исчезает кау-
дальный отросток. В сечениях, параллельных вертикальному, форма
сильно сужается. В сечениях, прошедших через верхнюю часть рако-
винки, резко сокращается ее длина и ширина, но сохраняются очер-
тания воротничка. Длина раковинок ископаемых тинтиннид колеб-
лется в пределах 50—200 д, ширина — от 25—30 д и больше. Длина
каудального отростка у некоторых видов достигает двойной длины
раковинки (Rhabdonelloides inesperalla Colom).
ИЗВЕСТКОВЫЕ ГУБКИ (CALCAREA)
Табл. 30, фиг. 1—9
Известковые губки составляют один из отрядов (Calcarea) в типе
Porifera (губки). Известковые губки — исключительно морские дон-
ные неподвижные организмы. Скелет их внутренний, состоит из
спикул — разрозненных или спаянных вместе (рис. 38, а, б). В иско-
паемом состоянии известны преимущественно спаянные известко-
вые спикулы, называемые обычно фибрами. В шлифе фибры имеют
очень причудливые, извилистые очертания, что позволяет называть
скелетную ткань «узорчатой». Породообразующее значение извест-
ковые губки приобретают в том случае, когда они начинают участво-
вать в сооружении органогенных построек.
Химический и минеральный состав скелета. Химический состав
спикул известковых губок, независимо от того, разрознены ли они
или спаяны воедино, постоянен и определяется как углекислый каль-
ций (СаСО3). Минералогически — это кальцит, реже, при минера-
логической перестройке, — коллофанит.
Экология. Наиболее многочисленны известковые губки в зоне
литорали и сублиторали, с увеличением глубины бассейна они встре-
чаются все реже и почти исчезают на глубинах в 150—300 м. Извест-
ковые губки достаточно выносливы к колебаниям температуры,
встречаясь как в высоких широтах, так и в тропических морях нор-
мальной солености. Известковые губки распространены меньше,
чем кремневые. Часто ископаемые известковые губки встречаются
совместно с водорослями (литотамниевыми и др.), что определенно
указываез на небольшие глубины, на которых они селились. Так,
среди рифовых коралловых известняков верхнего мальма Лайза-
кера Вагнером (Wagner, 1964) был встречен пласт мощностью 0.5 л/,
переполненный остатками' известковых губок без примеси кремне-
вых. Наряду с известковыми губками в этом же пласте были обна-
Рис. 38. Спикулы и внутреннее строение известковых губок
«— разрозненные спикулы (грирадиата. тетрарадиата. диапозонная спикула): б — спаян-
ные спикулы современных фаретронных губок: в — хоаноцит (жгутиковая воротничковая
клетка) современной губки, х 1000: г. д — схематические поперечные разрезы известковых
губок с асконоидной (г), сииоидиой (d) ирригационными системами
ружены многочисленные остатки литотамниевых водорослей. В верх-
неюрских отложениях Горного Крыма М. В. Михайлова обнару-
жила наряду с коралловыми биогермами также биогермы губково-
кораллово-водорослевые. губ ково-водорослевые и криноидно-ко-
раллово-губковые (Михайлова, 1968).
Геологическое значение. Биостратиграфическое значение извест-
ковых губок до сих пор очень невелико. Это объясняется в первую
очередь слабой изученностью группы. Подавляющее большинство
родов ископаемых известковых губок приурочено к мезозою (юра —
мел, реже — триас), когда они переживали расцвет, и к кайнозою;
в палеозое они встречаются не так часто. Отдельные роды извест-
ковых губок имеют значительное вертикальное распространение,
вплоть до целых геологических систем. Так, Peronidella характерна
для мела Крыма и юга европейской части СССР, Elasmostoma ха-
рактеризует юру и мел Крыма и т. д.
Участие известковых губок в сооружении органогенных построек
отмечается с конца палеозоя и в мезозое.
Анатомия. Губки — это довольно нежные организмы, одиночные
или колониальные, редко превышающие 7 см в высоту. Тело их
часто имеет трубчатую, бочонковидную или цилиндрическую фор-
му. Иногда образуется ножка, при помощи которой губка при-
крепляется к субстрату. Поверхность известковых губок может быть
Известковые губки. Археоциаты
43
гладкой или покрытой торчащими из нее пучками игл. Разломив
губку, можно видеть, что она состоит из неровной, ноздреватой
массы, пронизанной идущими в разных направлениях полостями и
каналами (Колтун. 1968).
Скелет известковых губок строится спикулами, выделяемыми
специальными клетками — склеробластами. В связи с тем что скле-
робласты выделяют у известковых губок кальцитовый скелет, их
называют калькобластами.
По своему строению известковые спикулы довольно однооб-
разны: различают три основных типа спикул: трирадиаты (спикулы
с тремя лучами не в одной плоскости), тетрарадиаты (спикулы с
четырьмя лучами), рабды (одноосные спикулы) (рис. 38. л, б). Длина
лучей трех- и четырехлучевых игл редко превышает 0.3 мм: длина
одноосных игл может достигать 3 см. Спикулы могут быть рас-
сеяны в теле губки или соединены, спаяны, образуя так называемый
фаретронный скелет1. Отдельные спикулы в фаретронном скелете
преобразованы в волокна-фибры. У некоторых губок с фаретрон-
ным (спаянным, фибровым) скелетом покровные слои укреплены
еще обильными разрозненными спикулами, трехлучевыми и одно-
осными (рис. 39).
В осадочных породах рассеянные спикулы известковых губок
отмечаются редко. Вероятно, это связано с общей слабой изучен-
ностью группы, так как в среднем карбоне Московской синеклизы,
откуда органические остатки изучены очень полно, довольно часто
встречаются рассеянные трехлучевые спикулы с толстыми лучами
примерно одинаковой длины (Иванова. 1958).
Рис. 39. Современная известковая губка Minchinella lamellosa Kirkpat-
rik, X180
a — дермальная мембрана; 6 — дермальные спикулы; в — спикулы кортекса; г — фибры
хоаносомы (Ziegler. 1964)
Вторичные изменения скелета известковых губок. В процессе диа-
генеза и эпигенеза кальцит фибр у фаретронных губок может быть
замещен кремнеземом, железом, глауконитом: кальцит может пере-
строиться в коллофанит.
В процессе перекристаллизации и вторичного замещения скелета
первичная структура фибр исчезает, и тогда скелет известковых гу-
бок внешним рисунком (в шлифах) может напоминать скелет неко-
торых строматопороидей, гидроидов и т. д.
Термин «фаретронный» дан по наименованию Pharetrones— группы известковых губок
с массивным скелетом.
АРХЕОЦИАТЫ (ARCHAEOCYATHI)
Табл. 31. 32
Экология. Археоциаты — исключительно морские донные орга-
низмы. Как правило, они прикреплялись к дну с помощью каблучка
прирастания, реже, по-видимому, просто лежали или пассивно пе-
ремешались. Среди последних типичны либо дискоидальные, та-
рельчатые формы, либо мешковидные.
Глубины обитания археоциат, вероятно, не превышали 100 м.
а обычно, по-видимому, даже 20—30 м. о чем свидетельствует их
совместное нахождение с синезелеными водорослями (Renalcis, Epi-
phyton, Girvanella и т. п.). Температурные условия могут быть, с
известной долей вероятности, восстановлены из фактов широкого
развития археоциат в красноцветных карбонатных фациях Сибир-
ской платформы (Зеленое. 1957). Для археоциат наиболее благо-
приятна нормальная соленость бассейна, однако исследования
И. Т. Журавлевой (1960) показали возможность существования ар-
хеоциат при несколько повышенной концентрации окиси магния
в породе (до 8%. т. е. MgCO3 — до 35%). В этих породах состав
археоциат чрезвычайно однообразен и чаще всего они представле-
ны очень мелкими экземплярами. Формы с морфологически слож-
ными стенками практически не переносят изменения солености и
поэтому в первично доломитовых породах не встречаются. Архео-
циаты приспособлены к жизни в условиях биогермов, биостромов
и отдельными особями. Настоящих рифовых образований, построен-
ных археоциатами, в настоящее время неизвестно (Журавлева. Ро-
занов. 1962). Биогермы, как правило, имеют небольшие размеры,
обычно не более 2—4 м в высоту. В самых нижних горизонтах кем-
брия встречаются биогермы, размер которых не превышает 50—70 см
в поперечнике при высоте 10—30 см.
Форма биогермов несколько изменчива во времени (Журавлева,
1960). Состав комплексов в зависимости от нахождения в биогер-
мах или в слоистых породах бывает резко различным. Одни формы
распространены обычно только в биогермах, другие — в межбио-
гермовых фациях (Журавлева, 1960: Розанов. Миссаржевский. 1966).
Геологическое значение. Археоциаты — одна из ведущих групп
для разработки стратиграфии нижнего кембрия. За редким исключе-
нием (начало среднего кембрия), археоциаты приурочены к отложе-
ниям нижнего кембрия. Многие виды археоциат являются прекрас-
ными зональными формами. Наиболее подробные сведения о стра-
тиграфическом распространении археоциат приведены в следующих
работах: Журавлева (1960, 1963); Журавлева. Конюшков. Розанов
(1964); Репина, Хоментовский. Журавлева. Розанов (1964): Розанов.
Миссаржевский (1966): Розанов, Миссаржевский и др. (1969): Жу-
равлева. Коршунов. Розанов (1969) и др. Большое значение имеют
археоциаты для корреляции удаленных друг от друга разрезов, та-
ких, как, например, разрезы Сибири. Северной Америки и Австра-
лии (Журавлева, 1960: Debrenne, 1964. 1969; Розанов, Миссаржев-
ский. 1966).
Несомненно большое значение археоциаты имели как породооб-
разователи. Как уже говорилось, они нередко образуют биогермы,
причем, как правило, совместно с водорослями. Пласты с архео-
циатово-водорослевыми биогермами на площади могут занимать
десятки квадратных километров. Нередко кубки археоциат обра-
зуют огромные скопления, слагая породу почти нацело.
Морфология. Большинство археоциат представляет собой кубки
конической или субцилиндрической формы. Лишь небольшая груп-
па имеет мешковидную неправильную форму. Еще реже встречают-
ся археоциаты тарельчатой формы. Размеры археоциат обычно
колеблются в пределах 5—20 мм в диаметре и 5—80 мм по высоте.
Однако диаметр их может достигать 50 см, а высота 1.5 м. Скелет
археоциат состоит из следующих основных элементов: обычно
44
Известковые организмы
Рис. 40. Элементы скелета археоциат
а—синаптикулы, соединяющие перегородки у рода Archaei>liinyi<i: и — строение решетчатой стенки (реконструкция); в — гребенчатые днища (реконструкция); г — дополнительная
оболочка у Rabdocyathella (подотряд Monocyafhina): д — Chabakovicyathus tumulatus Коп. с двумя стенками и днищами (нет перегородок)
двух, реже одной стенки, перегородок или тений, и иногда днищ.
Кроме указанных основных элементов встречаются стержневидные
элементы, соединяющие стенки (табл. 31. фиг. 3. б. 8: табл. 32. фиг. 8)
и перегородки или тении (рис. 40. а), пузырчатая ткань (табл. 31.
фиг. 8. 13).
Наружные стенки археоциат могут быть: 1) с простой
пористостью (табл. 31. фиг. 2.6.8,9,(5.10.13: табл. 32. фиг. 3.4.
7—9. 11): 2) с дополнительными образованиями около устьев (ко-
зырьками или тумулами) (табл. 31. фиг. 3. а. 4. 5. 11: табл. 32. фиг. 5);
3) с дополнительной тонкопористой оболочкой, наложенной на
основной грубопористый каркас (табл. 31. фиг. 7. 12; табл. 32. фиг. 1,
4. 6; рис. 40. г: 4) решетчатые (табл. 32. фиг. 2: рис. 40. б), также
состоящие из двух элементов — горизонтальных пластин и верти-
кальных тонких стерженьков.
Внутренние стенки бывают: I) с простой пористостью
(табл. 31. фиг. 3. 8. 9. 13; табл. 32. фиг. 3. 6. 8. 10): 2) с дополнитель-
ной оболочкой (табл. 32. фиг. 3. 5); 3) с поровыми каналами различ-
ной формы (табл. 32, фиг. 2); 4) чешуйчатые (табл. 31. фиг. 11):
5) кольцевые, состоящие из кольцевых пластин, расположенных
горизонтально друг под другом (табл. 31, фиг. 10).
Элементы интервал л ума (межстенкового
пространства). Перегородки и тении. Пластинча-
тые образования, расположенные радиально в интерваллуме, носят
название перегородок или тений, в зависимости от формы и проис-
хождения. Перегородки и тении делят интерваллум на камеры.
Перегородки и тении характеризуются или простыми порами, или
их отсутствием. Днища — выпуклые или плоские горизонтальные
пластинчатые образования, как правило, с простыми округлыми
или угловатыми порами (табл. 32, фиг. 1. 3—6,9. 10: рис. 40.0).
Особняком стоят гребенчатые днища (рис. 40, в), представляющие
собой самостоятельные для каждого интерсептума сочетания коль-
цевого валика и перпендикулярных к нему стерженьков (табл. 31.
фиг. 10, 11; табл. 32, фиг. 2). Гребенчатые днища, как правило, рас-
положены очень неравномерно, в отличие от обычных днищ, распо-
ло'женных через определенные промежутки (ср. табл. 31. фиг. 10, II
и табл. 32. фиг. 1,4,6).
ТАБУЛЯТЫ (TABULATA)
Табл. 33. 34
Табуляты принимают большое участие в образовании карбонат-
ных пород, они широко распространены глобально и стратиграфи-
чески. Целые колонии и особенно части разрушенных (раздроблен-
ных) колоний табулят очень часто встречаются в шлифах.
Для определения табулят под микроскопом необходимо два
строго ориентированных шлифа через колонию: поперечный и про-
дольный. Случайные косые срезы табулят в шлифах, особенно в
тонких петрографических, трудны для определения.
Условия обитания палеозойских табулят и особенности их захоро-
нения и сохранности. Табуляты. как и все другие кораллы, являются
обитателями морей. В бассейнах с повышенной соленостью морской
воды табуляты отсутствовали, опреснение было для них также ги-
бельным. Табуляты обитали в водах нормальной солености, на глу-
бинах от 10 до 100 л/. но редко встречались формы, обитавшие в бо-
лее глубоких участках моря. В литоральной зоне табуляты тоже были
редки.
В зоне литорали и на мелководных участках с очень подвижной
водой обитали в основном стелящиеся на твердом субстрате упло-
щенные колонии с большой площадью прирастания (представители
отряда Auloporida). В зоне мелководья с неустойчивым режимом,
т. е. в зоне волнений, но достаточно удаленной от берега и лишен-
ной грубого терригенного материала, табуляты расселялись свое-
образными лагунами — зарослями. В более неспокойных участках
встречаются колонии табулят сферической и полусферической фор-
мы и особенно многочисленны массивные колонии (представители
подотрядов Favositina и Alveolitina).На соседних более тихих участ-
ках обитали табуляты с ветвисто-массивной и пластинчатой формой
колоний (представители подотрядов Thamnoporina и Alveolitina).
Табуляты очень часто встречаются в карбонатных породах всех
систем палеозоя, а особенно часто — в карбонатных осадках, сильно
обогащенных терригенным материалом. Массивные полипняки та-
булят или их обломки наиболее часты в известняках различного
типа: органогенных, органогенно-детритовых, детритово-шламмо-
вых и в различной степени глинистых. Кустистые, ветвисто-массив-
ные полипняки встречаются в сильноглинистых и битуминозных
известняках. В аргиллитах и алевролитах преобладают в основном
ветвисто-массивные полипняки табулят. В грубых терригенных по-
родах (песчаниках, конгломератах) очень редко встречаются лишь
обломки колоний табулят.
Сохранность полипняков табулят обычно очень хорошая как в
карбонатных, так и в глинистых породах. В большинстве случаев
табуляты бывают захоронены на месте обитания или вблизи, а по-
этому хорошо сохраняются все детали наружного и внутреннего
строения полипняков.
Фоссилизация табулят может быть сведена к трем основным
типам: 1) известковый скелет сохраняется хорошо, а все проме-
жутки между скелетом заполнены вторичным кальцитом (табл. 33.
фиг. 4. 6; табл. 34. фиг. 6): при этом скелет рельефно выступает на
светлом фоне благодаря своей более темной окраске и тонкокристал-
лической структуре: 2) скелет в какой-то степени перекристалли-
Табуляты
45
зован, а промежутки заполнены каким-нибудь осадком (глинистым,
железистым); в таком случае скелет выступает в виде светлой сетки
на темном фоне породы (табл. 34. фиг. 7); 3) скелет в какой-то сте-
пени подвергся окремнению, которое может затронуть или только
скелет, или скелет и породу; при этом стенки кораллитов в полип-
няке пятнистые: то светлоокрашенные, то темноокрашенные.
Геологическое значение. Табуляты в настоящее время широко
используются в СССР для определения возраста (яруса) всех систем
палеозоя. Исключительное богатство и разнообразие табулят в
палеозойских отложениях земного шара позволяет использовать
их для целей стратиграфической корреляции и для разработки дроб-
ных стратиграфических схем.
Роль табулят в породообразовании велика. Они принимают
большое участие в образовании карбонатных пород от конца кемб-
рия и до раннего триаса включительно. Табуляты являются также
одним из компонентов палеозойских биогермов. В образовании био-
гермов первая роль принадлежит строматопороидеям и водорос-
лям. на втором месте стоят кораллы (ругозы и табуляты).
Морфология. Типы полипняков. Скелетная постройка
колониального коралла называется полипняком. Полный же ске-
лет отдельного полипа (зооида) носит название кораллита. У табу-
лят в зависимости от характера соотношения кораллитов выделяют
три типа полипняков: массивный, кустистый и стелющийся (рис. 41;
табл. 33. фиг. 1—3).
Рис. 41. Кустистая колония Syringopora (отряд Syringoporida),
состоящая из цилиндрических кораллитов с соединительными
трубками
Нал. вел.; нижний карбон
В массивных полипняках кораллиты плотно прилегают друг к
другу всей своей поверхностью. Кораллиты — призматические с
полигональным, полигонально-округлым и сдавленно-полулунным
очертанием поперечного сечения (рис. 42. а — г. з: табл. 33. фиг. 1.
4. 5).
В кустистых полипняках кораллиты всегда отделены друг от
друга свободным пространством, но иногда они могут соприкасать-
ся друг с другом оковыми сторонами. Кораллиты — цилиндри-
ческой формы с округлым, эллиптическим, реже — тетра- или гек-
сагональным поперечным сечением (рис. 41: 42. е. ж: табл. 34.
фиг. 7).
В стелющихся полипняках мелкие рожкообразной формы корал-
литы образуют вытянутые по субстрату цепочки или сеточки
(табл. 33. фиг. 2).
Кроме трех основных типов полипняков встречаются ветвисто-
массивные. кустисто-массивные и цепочечные (табл. 33, фиг. 3).
Во многих случаях нижняя и боковая поверхности колоний имеют
общую стенку — тонкий морщинистый покров, т. е. базальную
эпитеку.
К о р а л л и т ы. Они всегда ограничены стенкой (текой). тол-
щина которой сильно колеблется у различных родов. Снаружи стен-
ка одета тонким покровом — эпитекой. По окраске эпитека сильно
отличается от собственно стенки и всегда несет следы нарастания
в виде тонких концентрических струек. На поперечных разрезах
кораллитов четко наблюдается следующая картина. В смежной паре
кораллитов различают несколько зон (рис. 43. а. б). В середине
обычно видна непрозрачная темная линия, которая называется
срединным, межстенным, швом. Это эпитека смежных кораллитов.
От срединного шва прослеживаются две периферические широкие
зоны, которые обладают фиброзной структурой и буровато-желтой
окраской. Эти зоны Составляют собственно стенку кораллитов и
имеют микрокристалл ческое строение. Скелетное вещество стенки
называется склеренхимой или стереоплазмой [последний термин
чаще употребляется для вторичных скелетных структур (у подотряда
Thamnoporina)].
По очертанию поперечного сечения внутренняя полость может
сильно отличаться от очертания наружного края стенки коралли-
тов. Это особенно четко выражено у тех родов, которые имеют на
периферии сильно развитую стереоплазму. Кораллиты в колониях
табулят характеризуются обычно постоянным поперечником.
Соедините льные образования. Все скелетные об-
разования. при помощи которых внутренние полости кораллитов
сообщаются друг с другом, а сами кораллиты как бы-скрепляются
ими, называются соединительными. Соединительные образования у
табулят бывают в виде соединительных — стенных пор
(рис. 42. а, б: табл. 33. фиг. 4,6). хорошо развитых на стенках и
ребрах кораллитов (табл. 33. фиг. 5,а. б). Стенные поры могут пере-
ходить в соединительные каналы в периферических частях полип-
няков с обильной стереоплазмой. Они могут быть в виде соедини-
тельных трубок различной длины и в виде горизонтальных вырос-
тов — пластин. Все соединительные образования могут быть рас-
положены ориентированно или беспорядочно.
Септальные образования. Вертикальные скелетные
элементы, связанные своим происхождением с радиальными склад-
ками тела зооида, называются септальными образованиями. Сеп-
тальные образования у табулят в основном двух типов. Это сплош-
ные или шиповатые септальные пластины (табл. 34. фиг. 1,«. б).
септальные ребра (рис. 42. к, л). или обособленные конические шипы
(рис. 42. е) или чешуйки (табл. 33. фиг. 4. 5). Табуляты со сплош-
ными септами обычно редки и типичны для раннего палеозоя. Они
являются исходной структурой по отношению к более молодым,
у которых сплошные септы распадаются на свободные трабекулы
(шипы или чешуйки).
Днища. Внутренняя полость кораллитов обычно пересечена
горизонтальными или наклонными скелетными элементами, т. е.
днищами (рис. 42. б. г. и. л). Днища прикрепляются к стенкам ко-
раллитов и фиксируют передвижение зооида по мере роста полип-
няка. Днища являются одним из простейших скелетных элементов
у табулят. они сложены короткими параллельными фибрами
(табл. 34, фиг. 4). Большинство табулят характеризуется простыми
горизонтальными днищами, которые могут несколько наклоняться,
прогибаться. Кроме полных днищ у табулят наблюдаются непол-
ные днища, изогнутые, пузыревидные, воронковидные с осевой
трубкой или без нее (рис. 42. е. ж: табл. 34. фиг. 5. 7).
Микроструктура скелетных образований.
Склеренхима образована мелкими кристаллическими волокнами —
фибрами, толщина которых не превышает 2 ц. У табулят. как и у
других кораллов, фибры всегда ориентированы перпендикулярно
поверхности эктодермы (рис. 43. а. б: табл. 34. фиг. 2. 3).
46
Известковые организмы
Рис. 42. Строение табулят
и. о. — Palaeofavosites corrugatus Socolov, х 4: а — поперечный разрез, о — продольный
разрез (к. — кораллиты с полигональным поперечным сечением: п. — поры: с. ш.—сеп-
тальные шипикн: д.—днища), нижний ллаидоверн, Прибалтика (Соколов, 1955): в. г —
Squameofavosites: х 6: в — поперечный разрез, г — продольный разрез (к. — кораллиты
с полигональным поперечным сечением: с. —стенка с фиброзной микроструктурой: п. —
поры: с. ч. — септальные чешуйки; д.—днища), девон, Австралия (Hill. Stumm. 1956);
д— Alveolites suborbicularis Lamarck, поперечный разрез, х 10 (к. — кораллиты с полу-
лунным очертанием поперечного сечения; п. — поры: с. ш.—септальные шипикн), фран-
ский ярус, Русская платформа (Соколов. 1955); е, ж — Syringopora ramulosa Goldfuss, х4:
е — поперечный разрез, ж — продольный разрез (к. — кораллиты цилиндрические с округ-
лым поперечным сечением: с. т.—соединительные трубки; д. — воронковидные дннща).
нижний карбон. Новая Земля (Горский, 1935): з, и— Ijchenaria e.xpressa Socolov, х5:
з — поперечный разрез, и — продольный разрез (к. — кораллиты с полигональным по-
перечным сечением; с. — стенки тонкие, слитные; д. — днища), средний ордовик. Север-
ный Урал (Соколов, 1955): к, л — Billingsaria lepida Socolov. х 5: к — поперечный разрез,
л — продольный разрез (к. — кораллиты с массивными стенками, образованные септаль-
ными трабекулами: с. р. — септальные ребра; с. — столбик; д. — днища), средний ордовик.
Сибирская платформа (Соколов. 1955)
Горизонтальные (днища) и вертикальные (септы, шипы, чешуйки)
скелетные образования различаются по расположению фибр. В го-
ризонтальных — короткие фибры расположены параллельно друг
другу, следуя изгибам базальной эктодермы (табл. 34, фиг. 4). В вер-
тикальных — фибры соединяются в пучки, последние образуют
сложные структуры, известные под названием трабекул. Под микро-
скопом в поперечном сечении пучки фибр в трабекулах представ-
ляют собой концентрически слоистые и радиально-лучистые агре-
гаты округлой или овальной формы. Трабекулы — это столбики,
состоящие из пучков фибр: они могут быть простые и сложные по
своему строению. Когда трабекулы тесно примыкают друг к другу,
они образуют сплошную септу (рис. 42. к. л; табл. 34. фиг. !.«. б),
при разобщении — разобщенную, т. е. септальные шипики или
чешуи (рис. 42. а — д: 43. а. в. табл. 33. фиг. 4. 5: табл. 34, фиг. 3. 5).
Полипняки табулят с трабекулярной структурой септ встречаются
в основном в раннем палеозое — в ордовике и силуре. Чаще сеп-
Табуляты. Четырех- и шестилучевые кораллы
47
Рис. 43. Микроструктура стенок кораллитов
а — в — фиброзная микроструктура стеиок кораллитов у табулят подотряда Thamnoporina: а — Thamnopora. х 40 (с.—средний шов—эпитека—смежных кораллитов; с. т.—в стерео-
плазме стенки наблюдаются четкие фибры, расположенные перпендикулярно срединному шву. слои нарастания слабо выражены), средний девон; 6 — Striatopora, х 40 (слои нарастания
стереоплазмы четко выражены; фибры тонкие и короткие; буквенные обозначения те же. что и иа а), нижний девон; в — ламеллярная (волокнистая) микроструктура стенок корал-
литов у табулят отряда Syringoporida (род Syringopora). х 20 (с. — склеренхима стенки с волокнистым строением, покрытая снаружи темной эпитекой — э.; с. ш. — септальные ши-
пики погружены в склеренхиму стенки; д. —днища), инжний карбон
тальный аппарат у табулят выражен в виде шипов, чешуек и бугор-
ков. и расположение фибр в них лишь незначительно отличается от
их расположения в склеренхиме стенки (рис. 43, а; табл. 34. фиг. 2. 3).
Стенки у большинства табулят образованы простой фиброзной
склеренхимой, с более или менее ясно выраженным наслоением
фибр, перпендикулярных эпитеке и параллельных друг другу. В зави-
симости от степени проявления тех или иных элементов в склерен-
химе стенки выделяют два типа микроструктуры: фиброзный
и ламеллярный (волокнистый). Ламеллярный (волок-
нистый) тип микроструктуры, как и фиброзный, сложен фибрами,
но при этом фибры очень короткие и как бы сплюснутые параллель-
но эпитеке; они расположены часто, в обшей массе склеренхимы
видны в виде струек, слабо заметных волокон на фоне четких слоев
нарастания (рис. 43, в; табл. 34. фиг. 5). Как фиброзный тип микро-
структуры, так и ламеллярный (волокнистый) встречается у табу-
лят на протяжении всего палеозоя, с разным их проявлением у тех
или иных систематических единиц.
Почти у всех девонских табулят есть один признак, который
еще не объяснен. Это — пигментация скелета, в основном стенок
кораллитов. У табулят девона Советского Союза. Европы, Север-
ной Америки стенки кораллитов окрашены в коричневый цвет с
различными оттенками, от светлых до темных тонов; стенки окра-
шены ровно или участками (табл. 33, фиг. 6—9). Пигментация ске-
лета, вероятнее всего, связана с какими-то определенными экологи-
ческими особенностями.
Использование данных о микроструктуре скелетных элементов
в сочетании с другими морфологическими особенностями имеет
большое значение и оказывает существенное влияние на точность
определения табулят и на биостратиграфические выводы.
ЧЕТЫРЕХ- И ШЕСТИЛУЧЕВЫЕ КОРАЛЛЫ
(TETRACORALLA, HEXACORALLA)
Табл. 35
Кораллы — это группа исключительно морских прикрепленных
организмов, обитающих в открытых морях с нормальной соле-
ностью, на различных глубинах и широтах. Большинство из них
обладает наружным известковым скелетом. Среди них встречаются
одиночные формы, ветвистые и массивные колонии.
Четырехлучевые кораллы существовали в течение почти всего
палеозоя начиная со среднего ордовика и до раннего триаса. Шести-
лучевые скелетообразующие кораллы надотряда Scleractinia из-
вестны со среднего триаса доныне.
Химический н минеральный состав. Кораллы обладают арагони-
товым скелетом. Определенные клетки эктодермы выделяют эле-
ментарные частицы скелета — волокна (фибры) арагонита. В про-
дольном сечении они кажутся уплощенными, пластинчатыми, в по-
перечном имеют вид зерен. Для большинства четырех- и шести-
лучевых кораллов характерна фиброзная структура. Фибры бывают
различной длины, группируются в агрегаты — игольчатые или
пластинкообразные. Существует три типа сочетаний фибр: склеро-
дермитно-трабекулярный, фиброзный и ламеллярный (волокнис-
тый). Расположение и сочетание фибр составляют микроструктуру
скелетных элементов.
В продольном сечении в поляризованном свете фибры прояв-
ляют двойное лучепреломление с яркими цветами, аналогично каль-
циту. Продольная ось совпадает с Np, биссектриса отрицательная.
В поперечном сечении, перпендикулярном этой биссектрисе, двой-
ное лучепреломление очень слабое. Гранулированные участки соот-
ветствуют темным пятнам.
При фоссилизации нередко арагонит замещается кальцитом, и
фиброзная структура при этом может полностью исчезнуть. Кайё
(Сауеих, 1931) различает четыре типа фоссилизации:
1. Единственное изменение — появление вторичного кальцита
в пустотах, когда мелкие кристаллы кальцита выстилают стенки
полостей.
2. Арагонит замещен кальцитом без нарушения структуры, что
обычно наблюдается у палеозойских и нередко мезозойских корал-
лов.
3. Арагонит замещается очень тонкозернистым кальцитом.
4. Крайняя степень минералогического изменения — арагонит
перекристаллизован в общую недифференцированную кальцитовую
массу, часто стекловидную. Такое глубокое изменение видно не-
вооруженным глазом по сахаровидному излому. Внешняя поверх-
ность кораллов может оставаться ненарушенной при полной внут-
ренней перекристаллизации с исчезновением структуры.
Кроме того, арагонитовый или кальцитовый скелет ископаемых
кораллов в процессе фоссилизации может замещаться железом, фос-
фатом, кремнеземом.
Экология. Экологические условия существования кораллов изу-
чены на современных склерактиниях. Кораллы обитают на различ-
ных глубинах (от литорали до абиссали), при температуре от +2
до +36° С, питаются они исключительно зоопланктоном.
48
Известковые организмы
За время личиночной стадии, которое длится до 23—30 суток,
кораллы могут расселяться очень быстро и разноситься течениями
на большие расстояния (более 3000 км).
Рифостроящие кораллы характеризуются симбиозом с однокле-
точными водорослями — зооксантеллами, изобилующими в их экто-
дерме. Эти кораллы обитают на глубинах от 0 до 46—90 м (опти-
мальная глубина 4—7 м\ температура от +18,5 до +36° С (опти-
мальная от +25 до +29° С); соленость 27—40%о (оптимальная —
36%о). Свет необходим для зооксантелл. В строительстве рифов
принимают участие главным образом колониальные прикрепленные
массивные и ветвистые кораллы.
Нерифостроящие кораллы известны на различных глубинах, до
10 710 м, но оптимальная глубина их от 180 до 550 .м; температура
+ 2° С и выше (оптимальная от +8 до +20° С); соленость 36%,, и
немного ниже. Это одиночные или слабоветвистые формы.
В условиях подвижной воды, прибоя или течений, в том числе
и на рифах, растут массивные или крупные ветвистые колонии со
сближенными ветвями и утолщенными внешними стенками, с не-
сколькими порядками септ. Большинство кораллов предпочитает
чистые и мелкие воды. Массивные колонии, как правило, могли
существовать только в чистой воде. Однако кораллы не боятся се-
литься в местах, куда приносятся обломочные частицы, так как
полипы могут активно бороться с осадками энергичными движе-
ниями ресничек — щупалец. Известны одиночные кораллы и, в
частности, плерофиллиды у четырехлучевых, которые приспосабли-
вались к существованию в условиях приноса илистого материала.
Некоторые ветвистые колонии и иногда одиночные кораллы могут
существовать и при условии приноса песчаного материала ( Waage-
nophyllum, Thecosmilia и др.). Современные кораллы, будучи засыпан-
ными песком, могут подниматься из-под него. Кораллы, живущие
на скалистом грунте в зоне волнений, нередко переворачиваются
волной или течением, но при этом они могут сами возвращаться в
прежнее положение. У тех одиночных форм, которые селились на
тонком илистом грунте, часто вырастали корневидные отростки,
поддерживавшие коралл в вертикальном положении. В том случае,
когда коралл лежал, частично погружаясь в ил, верхний конец отги-
бался кверху, в результате чего образовывалась рогообразная форма.
Сезонные изменения отражались на толщине и частоте распо-
ложения скелетных элементов, а иногда и на внешней форме корал-
лов (перижимы и вздутия), что позволило вычислить среднюю ве-
личину годичного роста у четырехлучевых кораллов — 5—12 мм.
Геологическое, стратиграфическое и породообразующее значение.
Географическое распространение как четырех-, так и шестилучевых
ископаемых кораллов не ограничено. Они найдены во всех широ-
тах, на всех континентах.
На территории СССР четырехлучевые кораллы широко распро-
странены в палеозойских отложениях на Русской платформе, в За-
полярье, Донбассе, на Кавказе и в Закавказье, в Крыму, Средней
Азии, на Сибирской платформе, в Приморье, в горах Алтая, Урала,
Тянь-Шаня, Салаира, в Саянах, Забайкалье, Кузнецкой и Минусин-
ской котловинах, а также в нижнетриасовых отложениях в Закав-
казье.
Ископаемые склерактинии известны на Кавказе, в Крыму, За-
кавказье, на Мангышлаке, Памире, в Приморье.
Как четырех-, так и шестилучевые кораллы имеют большое
стратиграфическое и палеоэкологическое значение. Будучи донными
организмами, кораллы обнаруживают тесную связь с фациями и
быстро реагируют на изменение экологических условий. Многие
виды и даже роды кораллов, характеризующиеся кратковременным
существованием, являются руководящими для отдельных систем,
отделов и ярусов.
Кораллы встречаются в осадочных породах различного соста-
ва, наиболее часто — в известняковых фациях, но известны также
в мергелях и аргиллитах.
Кораллы встречаются вместе с брахиоподами, фораминиферами,
головоногими моллюсками, реже — с мшанками, двустворчатыми
моллюсками и водорослями.
Четырехлучевые кораллы в палеозое и шестилучевые в мезозое
и кайнозое участвовали в образовании биогермов и биостромов
совместно с другими организмами, нередко являясь породообра-
зующими, переполняя отдельные пласты и линзы. Начиная с позд-
него триаса склерактинии принимают участие в строительстве ри-
фов наряду с гидроидными и водорослями, а с поздней юры стано-
вятся одними из активных рифостроителей.
Строение скелета. У всех вымерших кораллов, так же как и у
современных, мягкое тело полипа помещалось на верхнем конце
одиночного коралла или кораллита колонии в углублении — чашке.
Скелет отлагался эктотермальным слоем базальной части полипа
и состоит из вертикальных и горизонтальных элементов. К верти-
кальным относятся септы, внешняя и внутренние стенки, осевые
структуры — столбик или сложная осевая колонна. К горизонталь-
ным элементам относятся днища, диссепименты, иногда функцию
горизонтальной опоры выполняли отложения стереоплазмы, утол-
щавшей септы.
Одиночные кораллы и кораллиты большинства колоний огра-
ничены внешней стенкой и покрыты тонким слоем эпитеки. По
микроструктуре эпитека не отличается от днищ и диссепиментов
и состоит из коротких волокон, перпендикулярных ее поверхности
нарастания.
На поверхности коралла обычно выступают продольные ребра,
концентрические линии нарастания, реже — бугорки или шипы.
В нижней части одиночных кораллов и массивных колоний иногда
имеются рубцы прикрепления или корневидные выросты. Посколь-
ку эпитека очень тонкая, она не всегда сохраняется. Аналогично
эпитеке одиночных кораллов, нижняя поверхность массивных ко-
лоний нередко покрыта общим покровом, имеющим такое же строе-
ние.
Внешняя стенка может быть образована слиянием перифери-
ческих концов септ (септотека; рис. 44, б); накладывающимися
друг на друга краями днищ или диссепиментов (рис. 44, г); стерео-
плазмой, покрывающей непосредственно эпитеку (рис. 44, в); си-
наптикулами у шестилучевых (синаптикулотека; рис. 44, а) или
сочетанием тех или иных скелетных элементов.
У ряда четырехлучевых кораллов на границе зоны диссепиментов
и днищ возникает внутренняя стенка, образованная сливающимися
стенками диссепиментов (рис. 45, а, б) и нередко утолщенная сте-
реоплазмой. Кроме того, у некоторых кораллов возникает стенка
в области днищ, разделяя их на осевые и периферические. Это так
называемая трубка, или аулос (рис. 45, г, д), образованная отги-
бающимися концами септ или вертикальными выростами на дни-
щах.
Важнейшей частью скелета является септальный аппарат. У боль-
шинства кораллов он представлен вертикальными перегородками —
септами. Последние могут быть шиповидными (рис. 46, в, г; табл. 35)
или пластинчатыми (см. рис. 49, е); у некоторых верхнесилурий-
ских — пластинчато-шиповидными (рис. 46, а, б); у девонских че-
тырехлучевых кораллов развивались своеобразные образования —
септальные конусы (рис. 46, д — ж), в которых слои трабекул, или
фиброзной ткани, отлагались с перерывами, переслаиваясь с дис-
сепиментами. Боковые поверхности септ иноыа орнаментированы
гранулами, шипами или каринами (струйками, (ребнями;
см. рис. 49, а, б). У шестилучевых кораллов шиповидные выросты
соседних септ могут сливаться между собой, образуя поперечные
балочки — синаптикулы (рис. 44; см. рис. 51, а, б).
Микроструктура септ различна, наиболее часто встречается
трабекулярная (рис. 47, в, г), реже — фиброзная (волокна распо-
лагаются в направлении, перпендикулярном плоскости нарастания;
Четырех- и шестилучевые кораллы
49
Рис. 44. Структура внешней стенки
и синаптикулотека: б —септотека; в — эпитекальная стенка, утолщенная стереоплазмой
(э. - эпнтека: с. 1. — стереоплазма): г — днссепиментотека
Рис. 46. Структура септального аппарата
а. о — Cystiphyllum— коралл с шиловидными септами, х 1.6: «—поперечный разрез,
о -продольный разрез, силур. Готланд: в. г—Tryplasma— коралл с пластинчато-шило-
видными септами, х 2.4: в — поперечный разрез, г — продольный разрез, силур. Готланд:
д — ж — Nartlophyllum — коралл с септальными конусами, х 1.6: д.ж — поперечные
разрезы, е — продольный разрез, средний девон. Северный Урал
Рис. 45. Структура внутренних стенок
и — « — Carcinophyllum. х 2.4: а — поперечный разрез, о — продольный ра трез. « — внут-
ренняя cieiika. нижний карбон. Новая Земля (Сошкина. 1952): г. л Aceriiiluitti (а\. - труб-
ки. и nt avioc»: — поперечный разрез, х 2.4: г) — продольный разрез, х 1.9. верхний
снлур, Готланд (Сошкина. 1952)
Рис. 47. Микроструктура септ
« — ламеллярная: б — фиброзная; в. г — трабекулярная.
Буквы на рисунке: п. т. — простая трабекула: г. т. — голакантная трабекула: р. т. — раб-
докантная трабекула (Wang. 1950)
1102 4
50
Известковые организмы
Рис. 48. Расположение трабекул в септах
и — Масдееа, продольный разрез коралла с чашкой, верхний девон. Русская платформа (Сошкина. 1952): б — Neosiringophyllum. продольный разрез, х 2. септы iiocipoenbi из парал-
лельных наклонных трабекул, средний девон. Урал (Сошкина. 1952): н — ж— структура столбиков: в. г — Lithosrroiion. столбик образован уюпшенным конном одной септы («
поперечный разрез, г — продольный разрез), х 4, нижний карбон. Русская платформа (Сошкииа. 1952). б— Bophophytlidium. столбик образован с ihbiiiicm оптировавшихся концов
нескольких септ, поперечный разрез, х 3. верхний карбон, США (Hill. 1956). е, ж — Giganloslilix. столбик построен наложенными друт на друт а днищами [»< внешний вид коралла
с выступающим столбиком (с. т.) в чашке. Х1.5. е — структура столбика, х 7.5]. верхний триас, Альпы (Cuif, 1965); з—и - структура осевых ко тонн: т.н Iimsduhia. сложная
осевая колонна (о. к.) со срединной пластинкой (с. п.) септы на периферии коралла прерываются лонсдалеоидными диссепиментами (л. д.) (т поперечный разрет. и продольный
разрез). х2, нижний карбон. Русская платформа (Добролюбова и др.. 1964). к. л — Aulophyllum. сложная осевая колонна (о. к.) без срединной пластинки, септы т тайных квадраюв
в зоне дннщ утолщены (к — поперечный разрез, л — продольный разрез, х 1.25). нижний карбон. Западная Европа (Wells. 1956): и — Zaphreniiiea. около т тавной септы видна фос-
сула (ф ). х 4. нижний карбон. Кузнецкая котловина (коллекция Н. В. Кабакович)
рис. 47, в. г) и ламеллярная или волокнистая (волокна распола-
гаются параллельно плоскости нарастания; рис. 47, а). У четырех-
лучевых кораллов известны трабекулы голакантные (рис. 47, г),
рабдокантные (рис. 47, г) и монокантные (рис. 47, г). Последние
аналогичны простым трабекулам, состоящим из отдельных пучков
фибр.
У большинства четырехлучевых кораллов септы построены из
простых трабекул, но у девонских родов встречаются как слож-
ные трабекулы, так и сочетание тех и других. У шестилучевых ко-
раллов септы слагаются как простыми или сложными трабекулами
(рис. 50, ж, з), так и сочетанием тех и других (рис. 49, а — в).
Трабекулы могут быть мелкими и крупными, расположены в
септах горизонтально (рис. 48, 6), наклонно и веерообразно, с одной
(рис. 48, а; рис. 49, а — в) или несколькими осями дивергенции
(у шестилучевых; рис. 50, в).
Столбик у четырехлучевых образуется либо утолщенным концом
одной септы (рис. 48, в, г), либо отшнуровавшимися концами одной
или нескольких септ (рис. 49, д), или вертикальными выростами
на днищах. У шестилучевых столбик мог возникать из усиленного
конца одной или нескольких септ либо отшнуровавшимися от септ
трабекулами (рис. 48, в — д), либо наложенными друг на друга
днищами (рис. 48, е, ж).
Сложная осевая колонна встречается только у четырехлучевых
кораллов. Она состоит из радиальных пластинок иногда из сре-
динной, и осевых днищ. Радиальные пластинки обособлены от
септ или соединены с ними. Осевые днища имеют разную форму,
но чаще выпуклые и приподняты к срединной пластинке. Форма
и величина осевой колонны у разных родов сильно варьируют
(рис. 48, з — л).
Днища и диссепименты отражают остановки в росте полипа.
Они представляют собой тонкие изогнутые пластинки и имеют
такую же микроструктуру, как и эпитека. Для многих четырехлу-
чевых кораллов характерно наличие от одной до четырех фоссул
около отдельных протосепт. Фоссула представляет собой ямку,
образованную в результате прогибания днища и расширения меж-
септального промежутка. Наиболее часто она развивается около
главной септы (рис. 48, лг). У некоторых четырехлучевых наблю-
даются лонсдалеоидные диссепименты, прерывающие септы по
периферии коралла (рис. 50, а, 51).
У шестилучевых кораллов иногда развивается перитека, свя-
зывающая отдельные кораллиты в колонию. Она состоит из днищ,
на которых могут развиваться вертикальные трабекулярные шипы
(рис. 49, г), из диссепиментов или из склеродермитов, соединенных
разрозненными синаптикулами как в вертикальном, так и в гори-
зонтальном направлениях.
Все скелетные элементы как четырех-, так и шестилучевых ко-
раллов могут быть утолщены ламеллярной или фиброзной стерео-
плазмой в один или несколько слоев.
Четырех- и шестилучевые кораллы
51
п
Рис. 49. Расположение трабекул в септах
и « — Monilhallia, боковая поверхность септы, построенной чередованием простых
(н г) и сложных (с. т.) трабекул, расположенных веерообразно, с одной осью дивергенции.
Сложные трабекулы выступают на боковой поверхности септ в виде карин: а — боковая
поверхность септы, х 2.7. б — вид сверху, в — общий вид септы (Alloiteau. 1952); г—Cyph-
astraea, перитека состоит из днища (д.) с вертикальными трабекулярными выростами. х 21.
современный вид (Alloiteau. 1952); д. е— Caryophyllin, столбик (с.) окружен полями (п).
септы пластинчатые (с. п.) (Alloiteau. 1952); ж. з— Tahularia. сепгы гребневидные, высту-
пающие над днищами (д.): ж— поперечный разрез, i продольный разрез, х 2. нижний
силур. Средний Урал (Сошкина. 1952); и. к -- Calostylis. септы пористые, нижний силур.
Западная Европа; л, и — Naayenophylluni. септы на периферии сливаются в ободок (об ),
в центре коралла осевая колонна (о. к ): л — поперечный разрез, п продольный раз-
рез. х 4.5. пермь. Армения (Сошкина. 1952); н. о -- Aulina. астреевидная колония, состоя-
щая из кораллитов с осевыми трубками — аулос (ау ): и - поперечный разрез, о — про-
дольный разрез, х 4. нижний карбон. Русская платфор (Добролюбова и др.. 1964);
п — Aeiinusrruea. септы построены из простых трабекул (п. т.). расположенных в один
или несколько рядов, между септами видны лиссепименты (д ). х 48 (Alloiteau. 1952):
р — Slilina. в центральной части кораллитов виден i рифслевидный столбик (ст ), х I. верх-
няя юра. Швейцария (Wells. 1956)
52
Известковые организмы
r\]J\
и
Рис. 50. Расположение трабекул в септах
и — Isastrnea, поперечный разрез септы в осевой части кораллита: септа построена нз
простых трабекул, располагающихся в одни-два ряда, х 60; б — Cladocora. поперечный
разрез септы, построенной из простых трабекул. х48: в — Missu. септа состоит из мелких
простых трабекул, расположенных в несколько дивергентных систем: на боковой поверх-
ности септ видны зерна-гранулы (з. г.): г — Meandroria. поперечный разрез через два смеж-
ных ряда чашек: стенки образованы периферическими концами септ септотеки (ст ). х!5:
д — Amphiastraea. септа состоит из мелких простых трабекул, расположенных в два ряда,
х 48; е — Turhinolia. септы состоят из очень мелких трабекул, расположенных в несколько
рядов; стенка — синаптикулотека (с.—синаптикулы). х 54; ж — Cunnoliies. септа пост-
роена из сложных трабекул (с. т). х 48; з— Plesincunnoliies. септы построены из простых
(п. т.) и сложных (с. т.) трабекул, х 36
а, б, г — з по Аллуато (Alloiteau, 1952); в — по Уэлсу (Wells, 1956)
Четырех- и шестилучевые кораллы. Гидроидные полипы, ближе неопределимые
53
а
б
Рис. 51. Расположение трабекул в септах
а — Siderasiraea, септы построены из крупных простых трабекул, соединенных синаптикулами (с.), х 50; б — Siereocaenia, септы построены из мелких простых трабекул, расположен-
ных в несколько рядов (с. — синаптикулы: д. — диссептименты; ст. — столбик), х 50; в — Cyclastraea. волокнисто-зернистая структура септы, х 60. мел: г — Corbariastraea. зериисто-
ламеллярная структура септ (с. — синаптикулы). х 43; б — г — по Аллоито (Alloiteau, 1952)
ГИДРОИДНЫЕ ПОЛИПЫ, БЛИЖЕ НЕОПРЕДЕЛИМЫЕ
(HYDROZOA)
Табл. 30, фиг. 10, 11;
табл. 61, фиг. 10. 11
Некоторые ископаемые остатки относят к гидромедузам, в си-
стематическом отношении близко стоящим к строматопорам,
гидрокораллинам и табуляриям. В карбоне, перми и юре СССР
встречаются грубые пластины неправильной формы, образующие
биогермные известняки. Материал, слагающий пластины, обычно
нацело перекристаллизован, но в поперечных сечениях в шлифах
часто обнаруживает систему полостей и каналов. При наблюдениях
в обнажениях нижнепермских представителей, относимых к Ura-
lotimania Riabinin (то же — Paleoaplysina Krotov), можно видеть,
что пластины гидроида имеют толщину от нескольких миллимет-
ров до 3—4 см и даже до нескольких десятков сантиметров в
длину. Расположение их самое прихотливое; они обычно стелются
и, одновременно изгибаясь, возвышаются над дном (рис. 52). Сово-
купность пластин создает сильнопористый, кавернозный, массив-
ный известняк, участвующий в образовании погребенных рифоген-
ных массивов Ишимбая и Стерлитамака (табл. 3, фиг. 1) и мощных
нижнепермских биостромов Уфимского плато, карбона Урала и
Тимана. Похожие гидроиды были встречены в буровых скважинах
в юрских породах западной части Украины, где они образовывали
массивные перекристаллизованные известняки мощностью
10 — 20 м. Изученность остатков этих ископаемых организмов
очень слабая.
Под микроскопом при малых увеличениях видна упоминав-
шаяся выше система ветвящихся полостей. Внизу или в середине
пластины эти полости широкие и щелевидные; к поверхности они,
изгибаясь, уменьшают свою ширину, ветвятся и образуют систему
каналов. Все полости соединяются между собой в центральной части
пластины, где у уралотиманий они располагаются параллельно
Друг Другу. Пространства между каналами в результате перекри-
сталлизации заняты вторичным карбонатом (табл. 30, фиг. 10, 11).
При хорошей сохранности остатков организма между каналами
наблюдалась тонкая крупноячеистая сетка, сохраняющаяся чрез-
вычайно редко. Можно высказать предположение, что известковые
остатки уралотиманий первоначально были арагонитовыми и с
палеозоя не сохранили своей ячеистой структуры (табл. 61,
фиг. 10, 11). В. Н. Рябинин (1915) несколько иначе описывает ура-
лотиманий. Он берет за основу описания не полости, а известковые
промежутки между ними, считая, что эти последние образованы
пластинками. Может быть, это и так, но слившиеся толстые пла-
стинки неразличимы в шлифах, они образуют фон, на котором вы-
деляются полости, соединенные между собой. Пластины же в этом
случае нужно воображать, и обнаруживаются они только в неко-
торых сечениях нижней части ценостеума.
Рис. 52. Сечения через пластины уралотиманий в обнажениях ниж-
непермских известняков Уфимского плато
Преобладающая часть их (а. г5) имеет горизонтальный рост, но встречаются экземпляры
с вертикальным ростом (а). Зарисовки В. П. Маслова
54
Известковые организмы
СТРОМАТОПОРЫ (STROMATOPOROIDEA)
Табл. 36—38
В число породообразующих организмов входят также строма-
топороидеи, образующие одну из наиболее распространенных вы-
мерших групп кишечнополостных, которые до сих пор не имеют
вполне определенного таксономического ранга.
Наиболее древние представители строматопороидей известны
из среднекембрийских отложений СССР. В ордовике, за исключе-
нием верхнего его отдела, они довольно редки, но в силуре и девоне
пользуются широким развитием. В верхнем палеозое распростране-
ние их значительно сокращается: в мезозое наблюдается их расцвет
и существенное обновление состава. В конце мела они выми-
рают.
Экология. Строматопороидей были обитателями теплой морской
среды, при нормальной ее солености, и наиболее благоприятные
условия для своего развития находили в пределах накопления чисто
карбонатных фаций. Они развивались в открытом море на неболь-
шой I дубине и обычно поблизости от береговой линии. Наиболее
благоприятные условия приводили к развитию пластообразных
форм, которые лучше обеспечивались питанием и не подвергались
сильному влиянию подводных течений. Последние, так же как и
привнос терригенного материала и характер субстрата, сильно влия-
ли на изменчивость ценостеума (скелета) строматопороидей.
Остатки строматопороидей обычно встречаются совместно
с остатками кораллов, в частности с табулятами. и вместе с ними
принимают участие в рифообразных постройках, в биогермах и
биостромах палеозойских и мезозойских морей. Очень часто они
встречаются в большом числе и вне рифовых образований — само-
стоятельно в пластах известняков. Некоторые из представителей
этой группы (Amphipora. Paramphipora. Indiostroma. Stachyodes)
являются важнейшими породообразователями.
Встречаются слои и линзы известняков различной мощности
(от нескольких десятков сантиметров до 1,5 м), целиком сложенные
остатками Amphipora и Paramphipora.
Внутри ценостеума строматопороидей часто встречаются ос-
татки других организмов — Rugosa. Tabu lata и др. Стромато-
пороидеям. больше чем каким-либо другим организмам, свойст-
венно сожительство (комменсализм) с различными организмами.
К этим формам сожительства относятся нациопоровые трубки.
Имеются географически широко распространенные виды, которые
всегда встречаются в сожительстве с Syringopora (см. рис. 55).
Вполне правильное развитие роста обоих организмов указывает
на полную их взаимосвязь. Такой способ больше всего наблюдает-
ся у Clathrodictyon. Stromatopora и Stromatoporella.
Геологическое значение. . Помимо важного участия стромато-
пороидей в мелководных биоценозах палеозойских и мезозойских
морей и участия в породо- и рифообразовании, они очень важны
и как индикаторы возраста соответствующих отложений. В настоя-
щее время строматопороидей благодаря их достаточной изучен-
ности нашими палеонтологами широко используются в СССР
для целей стратш рафии включающих их отложений.
Морфология и анатомия. Значительное разнообразие внешней
формы строматопороидей. не только различных родов, но и видов,
не может служить основой для их изучения. Последнее базируется
на микроскопическом изучении внутреннего строения ценостеума.
для чего необходимо изготовление шлифов, сделанных в двух на-
правлениях в зависимости от формы ценостеума: вертикального
или радиального и поперечного или тангенциального. Для цилинд-
рических форм ценостеума кроме поперечного и продольного шли-
фов необходимо И31 отовление тангенциального шлифа.
По внешней форме колонии строматопороидей весьма разно-
образны: инкрус!ирующие (коркообразные), желвакообразные, сфе-
роидальные, полусфероидальные, массивные, куполообразные, дис-
кообразные. пластинчатые, куносовидные. цилиндрические, сте-
бельчатые. дендроидальные. Колонии — различной величины, и
некоторые из них достигают значительных размеров. Так. наблю-
даются пластинчатые формы до 2—3 м2 при толщине 10—20 см.
Разнообразие форм и размеров колоний строматопороидей рас-
сматривается как следствие влияния среды (субстрата, течений
и т. д.).
Многие из строматопороидей имели в своем основании
бесструктурную, концентрически морщинистую базальную эпитеку
(рис. 53). которой они прикреплялись к субстрату. Сфероидальные
формы либо прирастали к субстрату короткой ножкой, либо имели
на очень незначительной части нижней поверхности базальную
эпитеку. Наряду с этим встречается много форм, субстратом для
развития которых послужили отмершие формы тех же стромато-
пороидей (см. рис. 55). Нередко встречаются формы, развитие кото-
рых началось с оплетения одиночных кораллов Rugosa или Tabulata
(рис. 54). При этом, как показывает изучение таких форм, оплетание
кораллов начиналось при их жизни, после чего они погибали.
Не все представители этой группы животных прикреплялись
к субстрату. Встречаются формы, свободно лежавшие на дне во-
доема и перекатывавшиеся при волнении.
Имеющийся материал в отношении Amphipora и Paramphipora
не позволяет определить способ их прикрепления. Стебельки их
при диаметре 1—3 мм и длине до 10—13 мм встречаются в переотло-
женном состоянии, обломанными.
Рис. 53. Ценостеум. вид снизу с поверхности; нат. вел.
Рис. 54. Последовательное оплетение — обволакивание коралла
Rugosa строматопореллами и Alveolites sp.; хЗ
Строматопоры
55
Ценостеум строматопороидей обычно известковый, иногда
окремненный. В целом он образован параллельными прямыми
или в различной степени волнистыми тонкими концентрическими
пластинками (ламинами), отстоящими друг от друга на близком
(но разном для различных видов одного и того же рода) расстоянии,
именуемом межламинарным промежутком. Соседние пластинки
у одних форм соединены между собой столбиками, развитие ко-
торых ограничено двумя соседними пластинками (Clathrodiciron
и др.; рис. 55); у других форм такие столбики в своем развитии
ограничены двумя, тремя, четырьмя, другие — большим числом
пластинок (Atelodictyon и др.; табл. 36. фиг. 8); наконец, у третьих
столбики проходят непрерывно через всю толщину ценостеума
(Actinostroma и др.; рис. 56). Толщина столбиков либо равна тол-
щине ламин, либо различная. Такая структура ценостеума в про-
дольном или радиальном его сечениях, а при хорошей сохранности
образца — на выветрелой боковой его поверхности представляется
в виде прямоугольной сетки.
У многих форм среди концентрических пластинок через неболь-
шие промежутки (1 — 3 л/.»х) четко выделяются на боковой поверх-
ности несколько более толстые пластинки (латиламины). отвечаю-
щие периодичности роста ценостеума.
Имеются формы, у которых ламины замещены пузырчатой
тканью, пересекаемой столбиками ( labechia. Pseudolabechia:
рис. 57. 58). формы с рудиментарными столбиками (Rosenella:
рис. 59) или целиком лишенные столбиков (Cystostroma: табл. 36.
фиг. 4). Встречаются также формы, у которых ггри отчетливом выде-
лении радиальных (продольных) столбиков концентрические пла-
стинки могут выделяться неотчетливо (Stromatopora: рис. 61).
От этого общего плана строения ценостеума строматопороидей
наблюдается значительное отступление у родов Amphipora, Beatri-
геа и Cryptophragmus (рис. 60. 62—64).
Волокна ткани у различных форм строматопороидей различные.
У представителей гидроактиноидной группы они плотные
(рис. 54—56); у представителей миллепороидной группы волокна
ткани пористые (рис. 61). Поры микроскопические, круглые или
овальные. У Stromaloporella пористость нитевидного (струйчатого)
вида (рис. 65—66).
Рис. 57. Схема строения Libechia sibirica Yavorsky
lh «арчшая исань пересечена стишками: х9
Рпс. 55. Сожительство строматопоры Clathrodiction comic turn Yavors-
ky c Syringopora (трубки с днищами)
Впп i\ видно. чю HapaciaHiie верхней чаои происходи ю на oixtepiiicn форме icx же jkiiboi-
НЫХ. .IX л. КП
Рис. 56. Схема строения Actimstronia
Рис. 58. Вертикальное сечение Pseudolabechia tubercilata Yavorsky. >;3
mamontori Yavorskv
Cio.ioiikm проходят через весь цсносчеум: на верхней часгй видны аезрорпзы: х >.5
56
Известковые организмы
Рис. 59. Вертикальное сечение Rosenella lahechioides Yavorsky с руди-
ментарными столбиками; х3,3
Рис. 61. Схема строения Siromatopora typica Rosen
На верхней поверхности видна астрориза: х 8.3; силур
Рис. 60. Продольные и поперечное сечения Amphipora ramosa
Phill.; х9,8
Поверхность ценостеума разных форм строматопороидей раз-
личная. Для одних из них характерны выступающие на поверх-
ности бугорки, либо микроскопические, либо более крупные, за-
метные и простым глазом. Многие формы строматопороидей
имеют на верхней поверхности возвышения конической формы —
сосочки (mamelons), через вершину которых проходит цилиндри-
ческий канал. При этом на поверхности часто наблюдаются тонкие
каналы, как звезды, отходящие от общего центра и дальше более
или менее древовидно ветвящиеся,— это астроризы (см. рис. 56 и 61).
При наличии сосочков они располагаются по их поверхности.
Рис. 62. Продольное и поперечное сечения Paramphipora tschusso-
viensis Yavorsky; хб.8
Рис. 63. Продольное и поперечное Рис. 64. Продольное и поперечное
сечения Beatricae tenuipunctata сечения Cryptophragnus elegans
Yavorsky; x 5,4 Yavorsky; x 5,4
Микроструктуры стенок ценостеума строматопор1. Выше
В. И. Яворским были описаны типы структур скелета строматопор.
которые можно назвать мезоструктурами (различимыми при уве-
личении в 5—10 раз).
Микроструктуры можно увидеть при увеличении в 30—50 раз
в стенках этих мезоструктур.
Автор использовал небольшое количество литературы и имел
в своем распоряжении несколько шлифов строматопор из мезозоя
Крыма, в результате изучения которых сделано нижеследующее
дополнение к очерку строматопор.
Раздел написан В. П. Масловым.
Строматопоры. Иглокожие
57
Рис. 65. Схема строения Stroniatoporella sniatkovi Yavorsky: х9
Рис. 66. Схема строения Stroniatoporella schelgutaniensis Yovorsky; х8
Никольсон высказал предположение о первичном арагонитовом
составе ценостеума строматопор, но присутствие хорошей сохран-
ности строматопор в палеозойских осадках скорее говорит о пер-
вичном кальцитовом составе скелета этих животных. Обычно це-
ностеум представлен как бы загрязненным кальцитом, более тем-
ным, чем вторичный карбонат, отложившийся в полостях (табл. 38,
фиг. 1). Во встречающихся полностью перекристаллизованных
остатках строматопор различимы только границы между телом
скелета и вторичным карбонатом, расплывающиеся при значи-
тельных увеличениях. Часть представителей строматопор имеет
темные, полностью гранулированные элементы скелета (табл. 38.
фиг. 2).
При хорошей сохранности ценостеума в стенках иногда наблю-
даются так называемые трабекулы (темные полосы в середине:
табл. 38, фиг. 6), но многие строматопоры их лишены. Среди тем-
ного, очень тонкозернистого материала иногда сохраняются мелкие
неправильные пятна со светлым центром в середине. При малом
увеличении создается впечатление облаковидной структуры. Эту
структуру Никольсон и Кайё трактовади как ячеистую. В других
строматопорах наблюдаются округлые или овальные светлые
пятна — «поры» — полости, занятые вторичным карбонатом
(табл. 38, фиг. 4) и окруженные темным карбонатом. При хорошей
сохранности некоторых мезозойских строматопор в темном мате-
риале отчетливо заметны «фибры» или «волокна», расположенные
в виде пучков (табл. 38, фиг. 2, 7) или перпендикулярно трабекулам
(табл. 38, фиг. 5, 6).
У некоторых строматопор вертикальные столбики, соединяющие
ламины, обладают тонким светлым каналом. Мелкие округлые
посветления при малых увеличениях часто не улавливаются, а во-
локна или фибры — расплываются.
В породах иногда встречаются мелкие обломки строматопор.
Их можно определить по «загрязненному» облику кальцита, со-
ставляющего петли грубой сетки, желтоватому или буроватому
цвету или облаковидной внутренней структуре стенок сетки. Форма
обломков обычно неправильная. Определение их зависит от вели-
чины обломка и сохранности их мезо- и микроструктуры.
ИГЛОКОЖИЕ (ECHINODERMATA)
Обший очерк
Табл. 39—42
Иглокожие — самостоятельный тип животного мира, в который
входит пять классов: морские лилии (криноидеи), морские ежи,
голотурии, морские звезды и офиуры. Представители этих классов
весьма отличаются друг от друга по форме тела и скелета. Образ
жизни иглокожих также очень разнообразен: среди них есть непод-
вижно прикрепленные, свободно передвигающиеся, зарывающиеся
в грунт и плавающие животные, активные хищники, илоядные
и растительноядные формы и формы, питающиеся планктоном
и органическим детритом, взвешенными в воде.
Известковый скелет иглокожих, хотя и внутренний, большей
частью лежит под тонкой оболочкой у поверхности тела и слагается
из разной формы пластинок, игл, шипов, выростов, спикул, скле-
ритов и т. п., которые образуют сплошной панцирь или включены
в тело животного. После смерти животного скелет в большинстве
случаев рассыпается на составляющие его элементы.
Химический и минеральный состав. Скелеты иглокожих всегда
слагаются кальцитом с солями магния и примесью фосфора, а также
сульфата (CaSO4?). Количество углекислого магния у современных
видов колеблется от 5 до 15% на зольный остаток (Виноградов,
1937, стр. 37). Необходимо заметить, что процентное содержание
углекислого магния в скелетах ископаемых форм значительно
меньше, чем у современных, так как полости ячей, при жизни за-
полненные органическим веществом, при фоссилизации выполняют-
ся кальцитом. Такая замена полостей вторичным карбонатом уве-
личивает количество извести вдвое.
Анализы современных иглокожих с очевидностью показали за-
висимость количества углекислого магния в скелете от температуры
воды. Чем севернее, тем меньше наблюдается солей магния.
Минералогически можно считать, что скелетные элементы игло-
кожих образованы кальцитом. При этом каждый отдельный эле-
мент (шип, членик, пластинка), за исключением голотурий, отве-
чает минералогически и оптически одному целому кристаллу каль-
цита. Установлено, что оптическая ось проходит параллельно оси
шипа, иглы или членика и перпендикулярно плоскости (или средней
тангенциальной плоскости у кривой поверхности) пластинки.
Каждый элемент скелета (пластинка, членик, игла и т. п.) или
всякий кристалл кальцита в ископаемом виде обладает спайностью
по ромбоэдру р (1011), причем все три плоскости спайности довольно
ясные. На табл. 39, фиг. 1,2 показано сечение через несколько чле-
ников ископаемой морской лилии в прижизненном положении.
58
Известковые организмы
При изготовлении шлифа произошло растрескивание кальцита
по спайности, благодаря которому отчетливо видно, что три чле-
ника имеют общую спайность или, иначе, они сложены одним кри-
сталлом кальцита.
Другой примечательный факт говорит о том. что вторичный
кальцит, заполняющий ячеи решетки членика, иглы или пластины,
принимает ту же ориентировку кристалла кальцита, которую имеет
элемент скелета.
Третье явление, наблюдаемое на ископаемых объектах, это обра-
зование двойников на плоскости р(— 10Т1) в виде полосок, которые
не нарушают сохранность ячеистой решетки иглокожего. Такие
«двойниковые» остатки иглокожих более многочисленны в пере-
кристаллизованных или в затронутых метаморфизмом породах.
Несомненно, что монолитность кристалла в отдельном элементе
скелета иглокожего (игле, шипе, пластинке и т.п.) заложена еще
при жизни животного, но заполнение пустот, возникновение полно-
ценного кристалла кальцита (иногда общего для нескольких чле-
ников) и появление двойников, по-видимому, связаны с вторичными
процессами. Во всяком случае, такая особенность иглокожих обу-
словливает большую их сопротивляемость механическому раздроб-
лению. о чем упоминает Кайе (Сауеих. 1931). Благодаря этому
породы, слагаемые ими. обычно грубозернисты.
Экология. Все иглокожие обитают исключительно в морях и
очень чувствительны к понижению солености. Лишь немногие
виды способны переносить довольно значительное опреснение
(до 14—20"„о). В то же время повышенная соленость переносится
ими достаточно легко. Например. Красное море с соленостью
43—47%О изобилует самыми разнообразными иглокожими. Игло-
кожие живут в морях и океанах всех широт земного шара, на самых
разнообразных грунтах и глубинах: от приливо-отливной зоны до
максимальных глубин океанских впадин (более 10 000 .и). Значи-
тельное количество видов оказывается эврибатными формами,
диапазон вертикального распространения которых иногда превы-
шает 5000—7000 м.
Геологическое значение. Достоверные обломки скелетов игло-
кожих встречаются, начиная со среднего кембрия (табл. 42.
фиг. 12—14), а важными породообразователями они являются
начиная с ордовика и до настоящего времени.
Микроструктура. Микроструктура иглокожих, за исключением
голотурий, имеет много общего. Она представляет собой ячеисто-
сетчатую «ткань». Правильная микроструктура иглокожих построена
по принципу прямоугольных координат, так что ее можно предста-
вить себе как результат пересечения трех взаимно пересекающихся
решеток. Разрез такой микроструктуры дает решетку или сетку
в любом направлении. Ячеи или полости такой микроструктуры
всегда сообщаются друг с другом, но окна, прямоугольные или
округлые, могут быть изолированы от соседних. Таким образом,
ячеи — это мелкие полости, заключенные между известковыми
стенками или решетками, а окна являются отверстиями в решетке.
Рис. 67. Типы микроструктур морских лилий и морских ежей
и. б окрут ло-рядовая микроструктура иыы морскою ежа RhulHfocidariх хр (верхняя
юра. Подмосковье): а близ оси тплы. б у периферии пиша (ш.) (о. осевая ч.кн..
кр. поверхностный сдой): в извилисто-ячеистая сiрукiура осевой части шлы lohuvnci-
(kids sp. (средний карбон. Подмосковье): г- с мнот оуюлыю-ячсистая с i рук i у ра: .» -
пластинки морс koi о ежа из т руппы Cidaridae (верхняя тора. Крым), г плас i инки мориkoi о
ежа 1л Ииюсвпь sp. (верхний мел. Крым): эн миоюуюлыю-ячеистая струю ура со ci ру fi-
nal ым расположением ячей н поперечном к оси сечении итлы морскою ежа из i руины
Cidaridae (нижний мел, Крым): j - прямоугольно-ячеистая структура пластинки морской
лилии HcHiisrimis sp. (баррсм. Крым) со светлой решеткой (слева), переходящей (справа)
в темную решетку: и. к неясно-ячеистая структура: и мнот oyi о льнам (?) морскою ежа
нз труппы Cidaridae. к микроструктура осевой части иглы морскою ежа .-1r< haem idtiris
sp. (средний карбон. Подмосковье): ./ вторично измененная (ожс.тезнение) пеправилыто-
ячеисгая (типа с) структура пласишкн морского ежа Echinoctwys sp. (верхний мел. окрест-
ности т. Актюбинска), местами ячеи с зазубренными краями, черное железистые таио.т-
нсния
Иглокожие
59
В шлифе мы видели и то и другое, но обычно отчетлива решетка
и ее окна, параллельные плоскости шлифов.
Различные типы микроструктуры иглокожих изображены ниже
(см. рис. 67 76).
В шлифах решетка современных иглокожих имеет вид прозрач-
ной известковой сетки. У ископаемых иглокожих чаще всего решетка
темная, а ячеи заполняются вторичным карбонатом — светлым
(табл. 39, фиг. 6—7). Оба случая показаны на рис. 67, з. Дальней-
шая перекристаллизация приводит к посветлению решетки и ниве-
лировке цвета ячей и их стенок. При оруденении железо часто под-
черкивает микроструктуру, но часто и искажает ее. Когда железо
выпадает в диагенезе, заполняя ячеи решетки иглокожего, оно мо-
жет занять всю полость, подчеркивая эту структуру. Но когда по
стенке ячеи частично выпал вторичный карбонат в виде корочки
или щетки, железо, занимая оставшуюся полость, не принимает
первоначальную форму ячеи. В таких случаях ячеи имеют непра-
вильную форму с зазубренными краями (рис. 67, л).
Исследование железных руд показало, что многие «бобовины»
в действительности являются оруденелыми члениками криноидсй.
в месторождениях как девона, так и юры (см. рис. 70). Процесс
оруденения обломков иглокожих широко распространен в природе
благодаря их тонкопористой или ячеистой микроструктуре.
Строение скелетных элементов. Состоящий из множества различ-
ных пластинок скелет криноидей после смерти животного легко
рассыпается на отдельные элементы, так что в шлифах обычно
встречаются разрозненные членики стебля и пластинки чашечки.
Значительно реже можно увидеть обломок стебля, состоящий из
нескольких члеников, сохранивших связь.
Членики морских лилий в центре имеют осевой канал, незапол-
ненный или заполненный ячеистой микроструктурой иного вида,
чем окружающая его структура (рис. 68, 69, 70). В осевых или близ-
ких к осевому продольных сечениях члеников канал хорошо выде-
ляется и позволяет уверенно относить такие фрагменты к кринои-
идеям. В продольных сечениях, проходящих ближе к краю, членики
криноидей имеют вид однородных по структуре прямоугольников
или узких трапеций.
Морские лилии, или криноидеи
(Crinoidea)
Табл. 39, фиг. 1. 2, 5
Криноидеи являются организмами, наиболее часто встречаю-
щимися в известняках. Химический состав известковых остатков
современных криноидей магнезиально-кальцитовый.
Экология. Современные морские лилии живут исключительно
в морях и океанах и распространены во всех широтах и на всех глу-
бинах — от приливно-отливной зоны до ультраабиссали (9700 м).
При этом стебельчатые лилии обитают главным образом на боль-
ших глубинах, а лилии, лишенные стебля, предпочитают более
мелкие воды. Криноидеи бывают неподвижно прикрепленными в
течение всей жизни (стебельчатые формы), свободно лежащими на
грунте и даже подвижными (бесстебельчатые формы). Лилии, жи-
вущие на мягких тонкозернистых илах, прикрепляются тонкими,
сильно разветвленными корешками, которые играют роль
якоря.
Большинство ископаемых криноидей обитало в мелком море
с нормальной соленостью и вело прикрепленный образ жизни.
В частности, палеозойские морские лилии в большинстве были длин-
ностебельчатыми формами и жили па мелководье, образуя
заросли.
В мезозое стебельчатые криноидеи постепенно стали пересе-
ляться в глубокие участки моря.
Геологическое значение. Кайе считает, что морские лилии по-
являются в докембрии, но достоверных фактов о существовании
криноидей в докембрии не проводилось. Обычно к морским лилиям
в докембрии относят крупные обломки кальцита округлого сечения,
часто с двойниками, но без центрального канала. Затухание этих
кальцитовых обломков такое же, как у иглокожих, однако уверен-
ности в принадлежности их к этим животным нет.
Начиная с ордовика криноидеи и близкие им Peimatozoa иногда
образуют известняки, переполняя породу. Классический пример
таких эхинодерматовых известняков ордовика дает вазалемский
известняк в Эстонии, широко применявшийся как облицовочный
и декоративный «мрамор». В силуре и девоне криноидеи достигают
массового развития и большого разнообразия. В карбоне крино-
идные известняки также широко развиты, но в перми их количество
резко сокращается. В юре их значение еще велико, а в мелу и в тре-
тичное время они становятся более редкими. По мнению Кайё,
значение криноидей как петрографического инградиента приумень-
шено.
Рис. 68. Сечения через обломки стебля ископаемых морских лилий
it — поперечное, членик разделен на десять секторов, образованных двумя различными
ячеистыми сетками; б — продольное, видны различные микрос!руктуры по обе стропы
о! осевого канала. Рисунки В. П. Маслова
Рис. 69.Морская лилия Pentacrinus suhieres Goldf.
ч - срединный вертикальный разрез стебля в направленнн. указанном на г. к IX: б юрп-
зошальный разрез стебля в направленнн. указанном на в. х IX: в. г — стебель в наг вс '. .
белая юра. Вюртенберг. Рейхенбер) (по Цнттелю. 1934) (« — различные микроструктуры
по обе стороны от осевою канала: б — чередующиеся микроструктуры)
60
Известковые организмы
Рис. 70. Поперечное сечение оруденелых члеников юрских морских
лилий
и — скелет имеет кальцитовую сетку и темные ячеи, заполненные железом, х 67: 6. в —
имеют обратную псевдоморфозу с железистой решеткой н кальцитовыми ячеями, но и
в центре, и в полостях сплошь отложились окислы железа: о— х 59. в— х 46 (Сауеих.
1931)
Некоторые криноидеи имеют на поверхностях сочленения своих
члеников особую «скульптуру», или «рельеф», которая препятствует
горизонтальному вращению членика вокруг оси. Наиболее часто
встречающаяся форма такой скульптуры — зто радиальные валики
и желобки, которые при сочленении двух соседних члеников входят
друг в друга (табл. 39, фиг. 1,2). При косом или тангенциальном
к поверхности срезе могут получиться радиальные полоски-секторы,
как результат проявления поверхностей «скульптуры». В продоль-
ных сечениях при хорошей сохранности скелетных элементов такие
валики проявляются в виде зубчиков, окаймляющих прямоугольный
или трапецеидальный разрез членика с одной либо с двух сторон
(см. рис. 77, е).
В поперечном сечении членики криноидей чаще всего бывают
круглыми, но могут иметь также прямоугольные, треугольные и
шестиугольные очертания. Сечение осевого канала у каменноуголь-
ных и пермских линий всегда круглое, а у девонских отличается
разнообразием форм, т. е. может быть трехлопастным (см. рис. 77, б),
пятилопастным (см. рис. 77, г), звездчатым (см. рис. 77, в), пяти-
угольным (см. рис. 77, а), в виде восьмерки (см. рис. 77, д), тре-
угольным и, конечно, круглым.
Пластинки чашечки не имеют сколько-нибудь надежных при-
знаков, отличающих их от пластинок морских ежей, морских звезд
и офиур, и зто не дает возможности определять их в шлифе как кри-
ноидей. С уверенностью определение может быть доведено только
до типа иглокожих.
Морские ежи (Echinoidea)
Табл.40
Морские ежи — свободно передвигающиеся по дну животные,
обладающие амбулакральной системой, известковым панцирем,
состоящим из пластин и известковых подвижных придатков (игл,
педицеллярий и т. п.). Соответственно назначению форма пластинок
и придатков различная. Так, для выхода амбулакров наружу суще-
ствуют особые округлые отверстия или амбулакральные пластинки
с отверстиями, обычно расположенные радиальными рядами. После
гибели животного скелет обычно рассыпается на составные части,
но у плоских Clypeaster и Scutella имеются внутренние столбики,
палочки и перегородки, укрепляющие периферические части изнутри,
что создает условия для лучшего захоронения целого скелета.
Химическим состав. Скелетные части современных морских ежей
мало чем отличаются от скелетов морских лилий, так что в общем
скелет современных морских ежей можно считать магнезиально-
кальцитовым. Необходимо отметить, что разные части скелета
содержат разное количество MgCO3.
Экология. Морские ежи — донные животные, живущие обычно
близко друг к другу (для обеспечения оплодотворения яиц). Поэтому
в сезоны икрометания наблюдаются скопления особей одного вида.
Морские ежи живут на разных глубинах — от сублиторальной зоны
до абиссальных глубин. Некоторые морские ежи высверливают
ямки и норки в твердом субстрате, например в рифовом известняке,
другие — закапываются в осадок и передвигаются в нем, являясь
илоядами. Такие морские ежи обычно обладают тонким панцирем.
Геологическое зиачеиие. На территории СССР остатки морских
ежей распространены по геологическому разрезу и географически
очень неравномерно. В отложениях нижнего и среднего палеозоя
они редки, в карбоне довольно многочисленны, в перми, триасе,
нижней и средней юре почти не встречаются. В верхней юре и осо-
бенно в мелу морские ежи обильны и являются существенным эле-
ментом морских фаун, а в палеогене и неогене значение их снова
заметно уменьшается. Территориально ископаемые морские ежи
тяготеют преимущественно к западным и южным областям Совет-
ского Союза.
Для юры и мела морские ежи имеют большое стратиграфическое
значение, но определение их требует сохранности целого скелета.
В некоторых случаях остатки ежей бывают породообразующими.
Так, например, в известняках среднего и верхнего карбона Русской
платформы, Донецкого бассейна и Урала скопления игл и обломков
панцирей морского ежа Archaeocidaris иногда образуют целые
прослои.
Микроструктура. Иглы морских ежей могут быть разделены на-
три группы по своей внутренней микроструктуре в поперечном к оси
сечении: 1) равномерно ячеистая микроструктура без резкого ра-
диального расположения сетки; 2) радиальная сетчатая микро-
структура с ячеями, увеличивающими свой размер от центра к пе-
риферии (рис. 71); 3) радиальная микроструктура с септами-радиу-
сами сплошных или пронизанных порами утолщенных пластин,
расходящихся от оси иглы (рис. 72—76; табл. 40, фиг. 3, 6, 7).
Первые две группы игл часто встречаются у палеозойских пред-
ставителей морских ежей.
Из пластинок панциря морских ежей характерные признаки
имеют только амбулакральные пластинки, которые пронизаны
Рис. 71. Часть поперечного к оси сечения шлы современного морско-
го ежа Phyllocanthus duhia
Буквы на рисунке: аг. — осевой комплекс ячей: s. — радиальная ткань (по Гессе. — пер-
форированные септы); d. — периферический слой с канальчиками (Hesse. 1X99—I90I)
Иглокожие
61
отверстиями для амбулакральных ножек. Отверстия эти всегда рас-
полагаются у внешнего края пластинки, причем на простых пла-
стинках имеется только одна пара отверстий, а на сложных пла-
стинках, образованных слиянием простых, число пар отверстий
может доходить до 50. Когда в шлифах попадаются целые амбула-
кральные пластинки или их обломки с сохранившимися отверстия-
ми, их можно уверенно определять как остатки морских ежей. Для
других пластинок отличительные признаки не установлены. В ряде
случаев наблюдалось, что на краях пластинки в шовных участках
.появляется рядовое расположение мелких ячей перпендикулярно
границам пластинки (рис. 72, б; 73, б; табл. 40, фиг. 4, 5).
Рис. 75.
Сектор поперечного сечения иглы
Cidaris flexuosa Miinst.
Буквы на рисунке: а. — осевой канал; as. — осевой цилиндр; s.—септы; d. — границы
несохранившегося периферическо!о слоя (Hesse. 1899 —1901)
Рис.72. Сечения через остатки
современного морского ежа
•cl.
S’.
Рис. 76. Сектор поперечного сече-
ния иглы Cidaris hausmanni Wissm.
из кассианских сланцев
и — горизонтальное сечение через иглу морского ежа. о-ва Фиджи; б — сечение параллельно
верхней поверхности пластинки морского ежа из рода Spharrechinus (по Циттелю. 1934)
Рис. 73. Сечения через остатки морского ежа
Буквы иа рисунке: аг. — осевой комплекс:
s.—«сеты»: s'. — «перьевидные» септы:
d. — границы несохранившегося периферичес-
кою слоя (Hesse. 1899—1901)
а — поперечное сечение иглы с перфорированными септами и темными межсептальными
полостями; б — плоскостное сечеиие пластинки с разными микроструктурами (по В. П. Мас-
лову)
Рис. 74. Сечения оруденелых игл юрских морских ежей; х 35
п. б. г.д — поперечные и косые сечения, в — продольное сечение. Гематит заполняет по-
лости и подчеркивает сплошные белые септы, расположенные радиально. Межсептальныс
решетки частично или полностью разрушены (Сауеих. 1931)
Рис. 77. Схематизированные зарисовки С. В. Максимовой члеников
криноидей в шлифах
а — д — поперечные сечения члеников девонских лилий с разной формой центрального
канала: а — пятиугольный канал, нижний и средний девон. Кузбасс, б — сложный трех-
лопастной канал, средний девон. Кузбасс, в — звездчатый канал, средний девон. Дальний
Восток, г — пятилопастной канал, средний девон. Горный Алтай, д — канал «восьмерка»,
нижний девон. Кузбасс: е— продольный разрез членнка морской лилии с зубчиками,
отражающими ребрышки на поверхностях сочленения, нижний карбон. Кузбасс
Морские звезды (Asteroidea) и офиуры (Ophiuroidea)
Табл.41
Морские звезды — это свободноподвижные плоские иглокожие
со звездообразным или многолучевым телом. Офиуры в известной
мере сходны с морскими звездами по форме тела, но в отличие от
последних лучи у них сильно обособлены и очень подвижны. Целые
остатки морских звезд встречаются в отложениях всех геологических
систем начиная с нижнего ордовика, но находки их редки, и, сле-
довательно, стратиграфического значения они не имеют. Разроз-
а
62
Известковые организмы
Иглокожие
63
Рис. 78. Пластинки морских зьезд из меловых пород о-ва Рим сна
(ГДР)
ч. <» амбулакральная плас) инка Pydnusier rasiiiusseni Muller, х 15: ч - вид снизу. <i вид
сверху (ар. апофиз соч.тснення с нижними амбулакральными и.тастнкамк: с. тело
членика: da. краевой крыдовидный вырос!: de. краевая впадина: та. срединная
поверхность сочленения: ml. - место прикрепления спинных продольных мускулов: mli. —
место прикрепления ннжннх поперечных мускулов: mis. место прикрепления верхних
поперечных мускулов: р отросток пластинки: ра. — верхний крыловидный вырост:
рс верхняя впадина): к. г амбулакральная пластинка Mclnpuster poulsenii Br. Niel-
sen. x 13: ь вид сверху, г вид снизу (ag. - впадина для вырхкта амбулакральной
пластики; ар. адрпанальный вырост: fa. -- зона борозды: fe. свободное ребро; ia -
место прикрепления мускулов, соединяющих пластики: г. радиальные эубча!ыс ребра,
несущие иодамбулякральные шины): ч -- яс - Tsnrius? sp. ind. конечный (глазной)
членик, х 10: Ч вид сверху. <• - вид снизу, ж - вид с конца луча (In. - локомоторная
впадина: sn. впадина для ортанов чувств): > - к cyuepsiapi ина.тьная пластика Metn-
риысг аГГ. poiilfeilii Br. Nielsen, х 4: г- вид снаружи, и ттазная конечная сторона, к
вид изнутри (ds. спинная боковая поверхность: h. высота: im. межч.теннковая по-
верхность. is. внутренняя поверхность: о. внешняя поверхность: w. цитрина):
I ч иифромарт тотальная (нижняя краевая) пластинка Mdopaslcr а(Г. рчч1г~ечП Вт. Niel-
sen. 5: л вид снаружи, п вид с конца, и вид изнутри (vs. брюшная боковая
иоверхиоеть: остальные обозначения см. < к): ч. п спинная пластика Pydiiusier гч\-
inusseiii Muller, х 9: ч -- вид снаружи, ч -вид изнутри: р.с - пластинка ротовою диска
Meiopusier aff. poulsenii Вт. Nielsen. > 9: р - вил от центра диска. < вид снаружи лиска
(ро. - первая амбулакральная пора: рр. первый вырост: ps. - второй вырост: остальные
обозначения ем. «I: m ф - узловая пласт инка со ст иба /?<< ш itisier railidius (Spencer), х 10:
in вид со стороны рта. г- межрадиальный вид (сутурный). ф - вид с брюшной сто-
роны (а. — амбулакральная сторона; d. -- конечная сторона: ia. впадина для прикреп-
ления мускулов, соединяющих с соседней амбулакральной пластинкой, ip. - внаднна для
ттрикреп.тения поперечных мускулов, соединяющих с парной пластинкой: р. — выступ:
г. - ребра, несущие шипы: s. - сутурная сторона: sp. - свободный край, направленный
ко рту и несущий места прикрепления ротовых шипов: ss. сутурная сторона тела пла-
стинки; тг. — брюшная кромка тела—пластинки, соединяющиеся с соседней пластинкой):
т спинная пластинка Meiopuster iihihIus Spencer, вид сверху, х 6.
По Мюллеру (Muller. 1953), терминология частично изменена
Рис. 79. Позвонок офиуры зи! оспонди.ювого типа
" брюшной, б спинной, в * проксимальный. .> дистальный. ч боковой виды:
ютммерндж; Чехословакия. Южная Моравия. Павловские верхи (ортн шта т пзвтечстт Л. Лбсо-
. тоном)
ненные скелетные элементы звезд изучены в общем слабо, и изу-
чают их выделенными из породы, так как по микроскопической
структуре они ничем не отличаются от остатков других иглокожих.
Офиуры в большинстве случаев сохраняются в виде рассеянных
склеритов, которые, как и остатки звезд, для исследования выделяют
из породы. В некоторых осадках они встречаются часто, особенно
в юрских отложениях.
Строение скелета. Основной опорный скелет морских звезд
состоит из известковых пластинок различной формы, связанных
между собой соединительной тканью и мускулами (табл. 41, фиг. 1:
рис. 78). Кроме опорного скелета у морских звезд есть разнообраз-
ные и многочисленные скелетные придатки, чаще всего в виде игл
и шипов, имеющих защитное значение. Под микроскопом пла-
стинки. так же как и у других иглокожих, имеют ячеистую структуру
и оптические свойства монокристалла кальцита (табл. 41, фиг. 2, 10).
У офиур скелет также состоит из пластинок, часть которых
орнаментирована различными иголочками и бугорками. Своеоб-
разны два скелетных элемента офиур: 1) так называемые позвонки
(табл. 41,-фиг. 7; рис. 79), представляющие собой сросшиеся и ви-
доизмененные амбулякральные пластинки, 2) челюсти (рис. 80. в).
образованные видоизменением ближайших к ротовому отверстию
пластинок. Выделенные из породы пластинки и иглы офиур изобра-
жены на рис. 80. Под микроскопом пластинки дают обычную кар-
тину сетчатой, ячеистой ткани (табл. 41, фиг. 3—11), свойственной
всем иглокожим.
Рис. 80. Скелетные элементы офиур; х16
ч боковая пластинка Ophiacaniha’’ iloretki Hess (Ip. насечка для поры щупальца):
ч неон редел н мая радиальная 1ени)а.н>ная пластинка: я половина челюсти неопре-
делимой офиуры (слева направо: вид со спннной. брюшной, межра.'шалт.ной и радна тьнон
сторон): .• межра.тиатытая тсниталытая пластика неопределимой офнуры: г) ж-
шлы офиур: > брюшная пласт инка Palucocoma f/чгеп (Wright) (ip. насечка для тю-
диальной поры), и — спинная пластина неопределимой офиуры
а — и — нз осадков от плиоцена до тоара: Швейцария, Зеевен (Hess, 1962)
64
Известковые организмы
Таким образом, в шлифах скелетные обломки морских звезд
и офиур могут быть определены только до типа. Другими словами,
можно установить, что они принадлежат иглокожим, но нельзя
сказать, к какому их классу.
Более подробные сведения о морских звездах и офиурах имеются
во многих работах (Berry, 1957; Fraas, 1885—1886; Spencer, 1914—
1940, 1951; Cooper, 1957; Hyman, 1955; Welburg, 1939;Cuenolt 1948a;
Matsumoto, 1917; Cuenot, 1948a, b).
Голотурии, или морские огурцы
(Holothuroidea)
Табл. 42
Голотурии сильно отличаются по своей форме от остальных
иглокожих. Они обычно обладают удлиненным цилиндрическим
телом с ротовым и анальным отверстиями, находящимися на про-
тивоположных конца-х. Вокруг ротового отверстия располагаются
щупальца. Одни голотурии напоминают огурцы, другие — червей.
Некоторые редкие формы напоминают медуз (Pelagothuria), дру-
гие— сферы (Sphaerothuria). Их размеры колеблются от несколь-
ких сантиметров почти до 2 м. В большинстве случаев преобладает
пятилучевая симметрия, которая подчеркивается пятью продоль-
ными зонами амбулякр с амбулакральными ножками.
У голотурий стенки тела кожистые, несущие мелкие известковые
тела (склериты), погруженные в стенку. Немногие роды имеют
заметную чешуйчатую броню.
Экология. Голотурии—донные морские животные, они обычно
медленно ползают по дну с помощью амбулакральных ножек,
щупалец или мускульных сокращений тела, реже закапываются
в грунт. Несколько видов плавают в толще воды и являются настоя-
щими пелагическими формами. Из всех классов иглокожих голо-
турии наименее чувствительны к опреснению воды. Так, в мало-
соленом Черном море иглокожие представлены главным образом
голотуриями, а некоторые виды могут жить даже в слабосоленой
воде мангровых болот. Голотурии распространены во всех широтах
и на всех глубинах — от приливно-отливной зоны до дна океани-
ческих впадин (более 10 000 л/). Главным фактором, определяющим
распределение современных голотурий, является температура.
Геологическое значение. Отпечатки целого тела ископаемых
голотурий исключительно редки. Многие из них, считавшиеся го-
лотуриями, сомнительны. До сих пор только единственный остаток,
известный из литографских известняков знаменитого Золенгофен-
ского месторождения (верхняя юра), может рассматриваться как
голотурия. С другой стороны, изолированные известковые тела,
часто встречающиеся в некоторых осадках, описывались неодно-
кратно. Лучше документированные остатки относятся к карбону
и юре.
По Дефляндр-Риго (Deflandre-Rigaud, 1962). первые несомнен-
ные остатки голотурий известны из девона.
Строение скелетных частей. Величина микроскопических извест-
ковых частиц внутреннего скелета колеблется от сотых до десятых
долей миллиметра, и только редкие из них достигают нескольких
миллиметров. Эти частицы располагаются не только в оболочке
тела, но и в амбулакральных ножках, щупальцах, в стенках вторич-
ной полости тела и других внутренних органах. Известковые кольца
(«косточки», склериты, спикулы) имеют очень разнообразную
форму и первостепенное значение для определения современных
видов. Некоторые из склеритов очень специфичны и встречаются
только у голотурий. Другие имеют простую форму без особых
признаков, так что в ископаемом виде они могут быть определены
лишь как остатки иглокожих вообще.
Наипростейшая форма склеритов называется милиарные (про-
сяные) гранулы. Это — мелкие известковые тела овальной, сфери-
ческой, эллипсоидальной или палочковидной формы, которые не-
возможно определить в ископаемом виде. Некоторые склериты
имеют форму стержней, простых или ветвящихся, с шипами, или
бородавками, или без них (рис. 81, а. б. с). Очень характерны тела,
похожие на очки (рис. 81, и) и овальные пластинки с четырьмя,
шестью и более отверстиями в двух рядах, называемые «пуговками»
(по-английски) или «застежками» (по-немецки) (рис. 81. в). Очень
своеобразны склериты в форме якорьков (рис. 81..и; рис. 82. к),
характерные для безногих голотурий (Apoda). Так называемые
столики (башневидные склериты, башенки, табуретки) имеют пер-
Рис. 81. Примеры внешнего вида склероций ископаемых голотурий
а— Ornaticannula bonhourei Defl.-Rig., x 128: 5— О. solida (Defl.-Rig.). x 200: в — Calc-
latnnella elliptic» (Defl.-Rig.), x 200: г — C. transverse Defl.-Rig., x 200: d — Cucumarites
speciosus Defl.-Rig., x 80; e — C. compactus Defl.-Rig., x 200: ж. з — Priscapedatus afjinis
Defl.-Rig., x 200: u — Prisculatrites dejlandreae (Frizzel et Exline) x 200: к — Calclingula
elgeri Defl.-Rig., x 80: л. .ti — Synaptellus cocoenus (Schlumberger): .1 — x 62,5. .u — x 165:
и — Dictyothurites cabisenta Defl.-Rig., x 80: о — D. spatiligerus Defl.-Rig., x 80: n. p —
Chiridotites speciosus (Defl.-Rig.), x 200: n — стадия развития без периферического круга,
р — полностью развитое тело: с—Stichopites ntortenceni Defl.-Rig., x 22.5: m — Chirido-
tes atavus (Waagen), x 200; ,v — Myriotrochites elegans (Schlumberger), x 200: ф — Proto-
caudina ntortenseni Defl.-Rig., x 270; .v — Staurocuniites barteinsteini Defl.-Rig., x 90: ц —
Binoculites terquemi Defl.-Rig., x 40; ч. ш — Achistrum brevis Gutschik, x 20; щ. з — неопре-
делимое тело и его поперечное сечение, х 36: ю, я — Microanty.x boani Gutschik, известковое
тело и его сечение; х 16
а — г, е и, п, р, т, ф — Оксфорд. Ф ранция, Кальвадос, Вийе-сюр-Мер: д. к. о — сред-
ний олигоцен, инский ярус, ФРГ, Гольштейн; л, .и. у — лютеций. Франция. Сэн-э-Уаз,
Шосси: с — лейас, ФРГ. Вюртенберг, Гейнинген; .v. у— лейас, ФРГ. Верхний Эсслинген;
ч— з—нижний миссисипий. США. Индиана.
а—у — по Дефляндр-Риго (Deflandre-Rigaud, 1962): ч — я — по Гучику (Gutschick.
1959)
Иглокожие. Черви
65
форированный диск, который несет выступающую дырчатую ба-
шенку (рис. 82, а, б. рис. 81, лс). Очень характерными для порядка
Apoda (Synapsida) являются колесики, колесики со спицами, коле-
совидные склериты, т. е. округлые тельца, у которых от внешнего
ободка отходит от 6 до 24 спиц (лучей), соединенных в центре
(рис. 81. т). Дисковидные склериты также имеют вид колеса
(рис. 81. ю), но в онтогенезе они развиваются иначе, чем колесики
Apoda. Склериты, имеющие форму перфорированных пластинок
(рис. 81. д'). иногда трудно классифицировать, и известны случаи,
когда перфорированные пластинки личинок других иглокожих
принимались за элементы голотурий.
Оптические свойства скелетных элементов голотурий изучались
в скрещенных николях Шмидтом (Schmidt. 1924. 1925) и Дефляндр-
Риго (Deflandre-Rigaud. 1962). При этом выяснилось, что иногда
весь склерит является единым кристаллом, а иногда его части имеют
разную оптическую ориентировку. Согласно Дефляндр-Риго. не-
которые из описанных ею склеритов обладают очень характерной
фибровой структурой.
ЧЕРВИ (VERMES)
Табл. 43. 44
К червям относится несколько больших групп животных в ранге
типа, которые в большинстве случаев не обладают твердыми ске-
летными частями и потому не сохраняются в ископаемом состоя-
нии. Так называемые следы или ходы червей, как показали новые
исследования, далеко не всегда действительно принадлежат червям.
Определение группы животных, с которой связаны те или иные
следы, вообще «представляет большие трудности и в большинстве
случаев на современном уровне знаний невозможно» (Геккер, Уша-
ков, 1962, стр. 457). Поэтому при обычном литологическом изучении
пород такие образования лучше не приписывать червям, а называть
менее определенно: следами или ходами илоядных животных.
Достоверные остатки червей, которые мы встречаем в породах
и шлифах, принадлежат полихетам, так что все дальнейшее изло-
жение будет посвящено именно этому классу.
Распространение и экология современных полнхет. Полихеты,
или многощетинковые черви.— животные в основном морские,
но среди них есть и солоноватоводные формы и редкие пресновод-
ные. В морях и океанах они распространены очень широко: от при-
ливно-отливной зоны до глубины почти 11 000 м, от приполярных
областей до тропиков. Естественно, что при повсеместном распро-
странении группы в целом среди полихет встречаются формы,
приспособленные к самым разнообразным обстановкам обитания
и ведущие самый разнообразный образ жизни. Есть виды, связан-
ные с зарослями тех или иных водорослей (например, ламинарий
или известковых литотамниев). Есть глубоководные формы, оби-
тающие на тонких илах, и есть обитатели мелководья, тоже при-
уроченные к илистым грунтам. Среди этих последних некоторые
виды населяют жидкий черный ил с запахом сероводорода, т. е.
обитают в среде, почти бескислородной, точнее, с очень низким
содержанием О2. В экспериментальных условиях полихеты из био-
ценоза черного ила жили без кислорода 7 суток и от 3 до 6 суток —
при значительном содержании в воде сероводорода (Якубова.
Мальм, 1930). Иногда полихеты являются массовыми руководя-
щими формами в биоценозах опресненных заливов. Другие поли-
хеты представляют характерный и существенный компонент сооб-
ществ прирастающих организмов, обитающих на твердом субст-
рате в местах с сильными придонными течениями (например, в
серпулово-водорослевых рифах Багамской банки). Среди полихет
довольно много также видов, способных переносить значительные
изменения условий внешней среды.
В основном полихеты являются донными животными, и лишь
немногие виды ведут планктонный образ жизни. Донные формы
в свою очередь могут быть подвижными (бродячими) и неподвиж-
но прикрепленными (сидячими), зарывающимися в грунт или свер-
лящими твердые предметы. Характерной особенностью полихет,
как сидячих, так и многих свободноподвижных, является образо-
вание подвижных трубок разного типа: известковых, хитиновых,
кожистых и т. д. Многие Terebelloidea и Serpuloidea образуют очень
толстую трубку из ила, сравнительно слабо скрепленного слизисты-
ми выделениями. У некоторых форм трубка снаружи покрывается
крупными песчинками, камешками, обломками раковин и даже
Рис. 82. Типы склероций некоторых современных видов голотурий,
по Хикману (Hickman, 1962) из моря близ Тасмании
а —ж— Stuhopus mollis (Hutton): а, б — таблички с башенками, вид сверху и сбоку, в —
табличка из первичного круга, г — розетка из кожи около анального отверстия, д — па-
лочка из средней части кишки, е — из щупальца, ж — три частицы из известкового кольца;
з — м — Leptosynapta dolabrifera (Stimson): з — склериты из оболочки тела, и — из щупалец,
к — якорек из оболочки тела, л — пластинка для якорьков нз оболочки тела, м — «прося-
ные гранулы» над мускулами титегумента; н — т — Trochodola allani (Joshuo): н — коле-
сико нз оболочки тела, о — реснитчатая-воронка, и — сигмоидальная частица нз оболочки
тела, р — три частицы нз известкового кольца, с, т — частицы из щупалец
1102 5
66
Известковые организмы
целыми небольшими створками. В мелководной зоне полихеты
нередко образуют массовые поселения. Так, на литорали Коль-
ского полуострова густота поселений Amphicora fabricii достигает
150—175 тыс. экземпляров на 1 м2 (Пригоровский, 1948). Этот вид
строит трубки из ила длиной 1 —1,5 см и диаметром около
0,5—1,0 мм. Трубки, пронизывающие самый верхний слой грунта,
ориентированы более или менее перпендикулярно его поверхности
и частично возвышаются над осадком, образуя нечто вроде густого
ковра или мха. Во время прилива и отлива они ложатся на дно,
покрывая его сплошным войлоком и препятствуя взмучиванию
и размыванию осадка (Максимова, 1959). В Северном море песчаные
трубки Sabellaria, вплотную примыкающие друг к другу, образуют
низкие, плоские острова до 10 км длиной и до 6 м высотой (т. е. мощ-
ностью). Трубки расположены вертикально, и в каждой из них живет
один червь, не связанный с соседями (Richter, 1927).
Геологическое значение. Из всех полихет только серпулиды с
известковыми трубками принимают участие в создании новых
осадков, и в шлифах также почти исключительно встречаются се-
чения и обломки известковых трубок. Лишь очень редко в шлифах
обнаруживаются иловые трубки, склеенные из материала окружаю-
щего осадка (табл. 43, фиг. 4; табл. 44, фиг. 5). Однако группа эта
изучена очень плохо: палеонтологи полихет вообще и серпулид
в частности почти не изучают и остатки червей, встречающиеся
в осадочных породах, как правило, не определяются. Обычно все
мелкие спирально свернутые трубки называют просто Spirorbis,
а более крупные трубки, длинные и неправильно изогнутые, —
Serpula, т. е. обозначают родовыми названиями современных форм.
Конечно, названия эти совершенно условны и не имеют таксоно-
мического значения.
Полихеты представляют интерес как породообразователи и
существенный компонент карбонатных осадков некоторых фаций.
Достаточно мощные (до 2—3 м) слои известняка, почти целиком
сложенного трубочками серпулид, известны в мезозое ГДР, ФРГ,
в нижнем карбоне Англии, в третичных отложениях Юго-Запада
СССР. В том или ином количестве серпулиды встречаются в раз-
нообразных типах известняков, но для некоторых они очень харак-
терны, а в других редки и играют сугубо подчиненную роль. Так,
например, серпулиды характерны для определенного типа водо-
рослевых известняков, которые распространены в девоне и карбоне.
В этом типе, сложенном в значительной степени желвачками сине-
зеленых водорослей (иногда неопределимых, чаще таких, как Orto-
nella, Bevocastria или Mitcheldeania), фауна представлена главным
образом серпулидами и остракодами.
Скопления трубок серпулид (спирорбисов) иногда составляют
ядро, на которое нарастают концентрические слои, образованные
остатками водорослей. Чаще спирорбисы прирастают к поверх-
ности водорослевых желвачков или свободно лежат в тонком кар-
бонатном цементе между желвачками. Такие полихетово-водо-
рослевые известняки описаны из турнейских и визейских отложений
Кузнецкого бассейна (Максимова, 1956, 1961), из турне и визе Англии
(Garwood, 1931), из бейского горизонта (D|) Южно-Минусинской
котловины (Иванова и др., 1964). В северо-западной Англии эти
известняки, так же как известняки собственно серпуловые, входят
в комплекс осадков опресненных лагун или заливов, в котором
породы, сопровождающие водорослевые и серпуловые пласты,
содержат обильные остатки наземной растительности и специфи-
ческую фауну, состоящую из эвригалинных форм. В Минусинской
котловине желваковатые водорослевые известняки со спирор-
бисами представляют собой мелководные отложения полузамкну-
того бассейна, также опресненного (Иванова и др., 1964). В Куз-
бассе этот тип известняков формировался в заливах и лагунах с
особым солевым составом, отличающимся от нормально-морского
вследствие воздействия вулканогенного материала на гидрохими-
ческие особенности водоема (Максимова, 1961). Наконец, в юго-
западной Англии образование спирорбисово-водорослевых извест-
няков происходило в очень мелкой осолоненной лагуне, в которой
периодически шла интенсивная химическая садка доломита (Rey-
nolds, 1921).
Таким образом, рассматриваемый тип известняков во всех слу-
чаях является образованием лагун и заливов с аномальным гидро-
химическим режимом, хотя характер этой аномалии может быть
различным. Конечно, спирорбисы встречаются и в нормально-
морских фаунистических комплексах (табл. 44, фиг. 6), но тут их
совершенно оттесняют обильные и разнообразные представители
типичных морских групп. Зато в специфических условиях лагун
и полузамкнутых бассейнов спирорбисы, приспособленные к ши-
рокому диапазону условий обитания, развиваются беспрепятст-
венно, иногда в массовом количестве.
В мезозойских и третичных отложениях широкое распространение
получают серпулиды, образующие прочные сростки из многих тру-
бок, спаянных известковыми же соединительными перемычками
(см. ниже описание известняковых трубок второго типа). Благодаря
своей способности прирастать не только друг к другу, но и к любому
твердому предмету (кораллы, известковые водоросли, раковины
моллюсков, камни) трубки этого типа становятся характерным ком-
понентом рифовых построек и биогермов, и в ряде случаев сами
трубки образуют небольшие биогермы мощностью до 20 м. Хоро-
ший пример биогермов, в которых серпулиды являются главными
породообразователями. представляют сарматские «онкоиды» По-
долии, описанные И. К. Королюк. Эти небольшие массивы слагают-
ся частично собственно серпуловыми известняками, частично —
известняками серпулово-ракушечными, в которых огромное коли-
чество трубок серпул переплетается с многочисленными раковинами
моллюсков (Королюк, 1952) (рис. 83, 84).
Следует отметить, что сарматский бассейн, в котором форми-
ровались серпуловые биогермы, был опресненным.
Небольшие сростки трубок серпул довольно часто встречаются
в мелководной карбонатной толще верхней юры Узбекистана,
в известняках с грубой песчаной примесью. Вместе с полихетами
здесь в шлифах присутствуют остатки моллюсков и иногда мшанок
(обычно в небольшом количестве). В эоцене Ферганы серпулы встре-
чаются в известняках, нередко тоже песчанистых, вместе с комплек-
сом эвригалинных моллюсков, которые существовали при соле-
ности, близкой к нормальной, но легко переносили небольшое
опреснение (Осипова, 1954).
Таким образом, в геологическом материале остатки серпулид
приурочены к различным мелководным фациям. Такое ограничен-
а.
Рис. 83. Сечение стенки третичной серпулы
Буквы иа рисунке: а. — пластинчатый внутренний слой, переходящий в неправильно пу-
зыристый слой — б. (s.—внутренняя полость трубки червя). Внешний слой служит для
прикрепления и обволакивания других предметов. На данном рисунке таким захваченным
предметом служит маленькая раковина червя слева. Рисунок по шлифу и объяснение
В. П. Маслова
Черви
67
ное распространение эврибионтной группы объясняется двумя при-
чинами: во-первых, значительным преобладанием в разрезах мел-
ководных отложений, а во-вторых, тем, что глубоководные формы
лишены твердых скелетных частей и не сохраняются в ископаемом
состоянии. В то же время виды с прочными известковыми трубками,
прирастающими к субстрату, приспособлены к обитанию именно
в неспокойной обстановке мелководной зоны.
Строение скелетной ткани. В имеющемся у нас материале пред-
ставлены известковые трубки трех типов и один образец иловых
трубок, построенных из окружающего осадка. У известковых тру-
бок первого типа стенки состоят из двух или четырех концентри-
ческих слоев, параллельных длине трубки. Если слоев два, то один
из них светлый, а другой темный, сложенный микрозернистым каль-
цитом (табл. 43, фиг. 1—3). Если слоев четыре, то наблюдается
чередование темных и светлых слоев, совершенно таких же, как
в первом случае. Тонкие полоски светлых слойков при скрещенных
николях почти не затухают. Все наблюдавшиеся нами трубки этого
типа очень мелкие (0,9—2,0 мм длиной и 0,2—0,5 мм в диаметре),
обычно подковообразно изогнутые или свернутые в неправильную,
почти плоскую спираль, т. е. по существующей терминологии они
называются спирорбисами. Для них характерны сечения овальные,
округлые, бобовидные, в форме восьмерки и в виде трех-четырех
примыкающих друг к другу овалов с разным направлением длин-
ных осей (табл. 43, фиг. 3). Реже встречаются продольные сечения,
спиральные и изогнутые полукругом (подковообразные).
Известковые трубки второго типа известны у ряда современных
форм, а в ископаемом состоянии найдены пока только в мезозой-
ских и третичных отложениях. Трубки эти двухслойные. Внутренний
слой светлый и слагается слабо различимыми фибрами или пла-
стинками. расположенными под углом, обычно очень небольшим,
к поверхности трубки. Наружный слой темный, отчетливо волок-
нистый, с волокнами, изогнутыми пологой дугой или крутой пара-
болой (табл. 44, фиг. 3). Из этого наружного слоя выступают от-
дельные, сравнительно толстые волокна, которые прирастают к
находящимся рядом трубкам или другим твердым предметам
и образуют пузырчатую, ячеистую ткань срастания (табл. 44,
фиг. 1,4; особенно табл. 43, фиг. 5). Граница между слоями выглядит
резкой только при малом увеличении, когда различается преиму-
щественно разница в окраске слоев. При большом увеличении вид-
но, что более грубые волокна наружного слоя в своей пологой части
переходят во внутренний слой, разделяясь при этом на тонкие фибры
а
Рис. 84. Третичные серпулы
а — сечение через трубки третичных серпул (я.): видно, что только один червь (s,) образовал
полностью внутренний и внешний слой. Другие раковины или более поздние участки той же
раковины, примкнувшие позже (s.). пользуются более ранней поверхностью, как стенкой.
Зарисовка по шлифу и объяснение В. П. Маслова; 6 — нарастание червя на багряных во-
дорослях литотаминн (Lth.) и литофиллуме (Lph.) (s. — раковина червя). Третичные отло-
жения Медобор, Подолия (по В. П. Маслову)
и становясь более прозрачными и менее отчетливыми. Только что
описанная картина наблюдается в продольных и близких к про-
дольному сечениях. В некоторых косых сечениях-срезах изгиб воло-
кон наружного слоя не виден и вся стенка трубки выглядит косо-
волокнистой, с более резко выраженной структурой вблизи ее внеш-
ней поверхности (табл. 43, фиг. 5). В тангенциальных сечениях по-
лучается то более, то менее широкая поперечно-штриховатая полоса,
которая на краях расщепляется на отдельные волокна, разделенные
пустыми промежутками (табл. 44, фиг. 3). Размер трубок этого
типа различный: от 0,5—1 мм до I см в диаметре и до 10—12 см
длиной.
Для сравнения с ископаемым материалом были расшлифованы
трубки современных Serpula sp. и Spirorbis sp., ближе не определен-
ных. Во всех расшлифованных образцах, как у серпул. так и у спи-
рорбисов. стенки были построены одинаково, а именно так, как
у ископаемых известковых трубок второго типа. Спирорбисы от-
личаются по этим шлифам от серпул только тем. что у них слабо
развиты внутренний слой и пузырчатая ткань. Трубки такого строе-
ния при скрещенных николях слабо ирризируют (в основном внут-
ренний слой), но почти не затухают.
Известковые трубки третьего типа также двухслойные, но у них
внутренний слой темный, сложенный пелитоморфным кальцитом,
а наружный слой светлый и состоит из очень тонких фибр, распо-
ложенных перпендикулярно поверхности трубки. Благодаря такой
структуре для наружного слоя характерно продольное перебегаю-
щее погасание, сходное с погасанием остракод и трилобитов. Кроме
того, этот тип трубок отличается от первых двух более толстыми
стенками. В шлифах наблюдались сечения круглой, овальной,
цилиндрической и конической формы, последние — в скошенном
продольном разрезе.
Здесь следует сказать, что принадлежность трубок этого типа
к серпулидам вызывает некоторые сомнения, хотя именно они чаще
всего описываются и изображаются в литологической литературе
(Хворова, 1957; Швецов, 1958). Дело в том, что на серпулид очень
похожи гастроподы — верметиды, обладающие неправильно за-
крученной трубчатой раковинкой, состоящей из двух слоев. Внут-
ренний слой у трубок верметид слагается тонкими концентри-
ческими пластинками или фибрами, расположенными параллельно
оси трубки, а наружный слой — поперечно-призматический. И как
раз этот призматический слой рассматривается как признак, отли-
чающий гастропод от серпулид, для которых характерен «параболи-
ческий» наружный слой (Пастернак, 1955). Трудно сказать, насколько
это отличие, наблюдавшееся у меловых представителей обеих
групп, универсально и постоянно и насколько однотипным является
строение призматического слоя у разных верметид. Сейчас можно
только отметить, что у описанных в литературе трубок гастропод
призмы наружного слоя достаточно грубые и хорошо различаются
под микроскопом, а у трубок третьего типа, относимых к серпули-
дам, наружный слой выглядит однородным и только характер
погасания указывает на наличие тонких поперечных фибр в этом
слое.
Что же касается мезо-кайнозойских серпулид, обладающих
«параболическим слоем» (см. трубки второго типа), то у них есть
еще одно очень заметное отличие от гастропод — пузырчатая ткань
в местах срастания, которая сразу бросается в глаза при просмотре
шлифов. Чтобы закончить сопоставление серпулид (черви) и вер-
метид (гастроподы), остается упомянуть, что в карбоне встречают-
ся мелкие плоскоспиральные трубки с отчетливым поперечно-приз-
матическим строением стенки, но однослойные (табл. 43,
фиг. 2). Может быть, это древние, более примитивные верметиды,
но, поскольку трубки очень мелки и наблюдались только в виде
разрезов в шлифах, утверждать это с уверенностью нельзя.
Иловые трубки полихет, найденные в визейских известняках
Кузбасса, сложены однородным темным пелитоморфным кальци-
том и отличаются по материалу от вмещающего микрозернистого,
68
Известковые организмы
слабоалевритистого известняка только тем, что в них отсутствуют
обломочные кварцевые зерна. Внешние контуры трубок ясно выри-
совываются только при некоторой перекристаллизации окружаю-
щей породы. В совершенно свежих участках известняка трубки
видны благодаря тому, что кальцит, заполняющий их полости,
всегда в большей или меньшей степени перекристаллизован и про-
зрачен, так что они выделяются в виде светлых пятен на темном
фоне. Трубки прямые, цилиндрические, возможно, закрытые на
заднем конце, в породе располагаются более или менее перпенди-
кулярно плоскостям напластования. Длина замеренных продоль-
ных сечений колеблется от I до 2,5—3 .и.м, внешний диаметр попе-
речных сечений — от 0,3 до 0,7 мм, толщина стенки — от 0,06
до 0.17.мм. Такие трубки из ила, скрепленного слизистыми выде-
лениями, встречаются у представителей нескольких семейств по-
лихет: у сабеллид, серпулид и теребеллид, так что точно определить,
к какому именно семейству принадлежат наши ископаемые трубки,
невозможно. Полихеты, которым принадлежали новые трубки,
входят вместе со спирорбисами в сообщество прибрежно-лагунных
водорослевых известняков.
При макроскопическом изучении осадочных пород могут быть
найдены и неизвестковые трубки полихет. Так, например, в кембрии
Русской платформы, Англии и США встречаются черные хитиновые
трубки Sabellidites, круглые в сечении, с поперечно-бороздчатой
или поперечно-штриховатой поверхностью, в породе обычно сплюс-
нутые. В меловых и третичных отложениях встречаются, обычно
на раковинах моллюсков, песчаные трубки, состоящие из сцементи-
рованных песчинок.
БРАХИОПОДЫ (BRACHIOPODA)
Табл. 45, 46
Брахиоподы существуют с кембрия по настоящее время и де-
лятся на два класса — замковых и беззамковых. Они особенно
характерны для среднего и позднего палеозоя, где представлены
огромным числом разнообразных форм (главным образом замко-
вых), чрезвычайно многочисленны и являются одним из основных
компонентов морских фаун. В мезозое брахиоподы становятся го-
раздо однообразней, встречаются лишь в немногих фациях и те-
ряют свое значение, уступая ведущую роль моллюскам. В совре-
менных морях это совершенно второстепенная группа, не играющая
сколько-нибудь существенной роли среди донного населения. Если
учесть к тому же, что в палеозое было девять отрядов замковых
брахиопод, а в современных морских бассейнах их только два,
то станет очевидным, что данные по экологии современных форм
ничего не могут дать для понимания условий обитания разнооб-
разных древних представителей этого класса. Наоборот, для без-
замковых, таких, как лингула, просуществовавшая от начала ордо-
вика до наших дней практически без изменений, изучение экологии
современных видов имеет большое значение.
Экология и фации. Современные лингулы живут в прибрежной
полосе на глубине от 0 до 30—40 м, изредка до 100 м, при нормаль-
ной солености и в дельтах и эстуариях — в условиях сильного опрес-
нения. На литорали во время отлива они по нескольку часов остают-
ся без воды, а местами омываются пресной водой стекающих с
берега ручьев и речек и заносятся выносимым этими ручьями пес-
ком или илом. Через свежие наносы лингулы прокапывают ходы
длиной от 5 до 30 см к поверхности и хорошо переносят такую
резко изменчивую и неблагоприятную обстановку, хотя другие
беспозвоночные с твердым скелетом существовать здесь не могут.
Лингулы хорошо переносят также сильное загрязнение воды раз-
лагающимся органическим веществом, т. е. могут жить при очень
низком содержании кислорода у дна водоема.
Такими же экологическими особенностями обладали и ископаемые
лингулы. Именно поэтому они являются, например, характерным
компонентом фауны угленосных толщ, в состав коюрых входят
солоноватоводные отложения разной степени опреснения (нижняя
пермь Воркутского бассейна, средний карбон Донецкого). Здесь
лингулы часто встречаются в большом количестве в углистых аргил-
литах кровли угольных пластов — или одни, или вместе с пелеци-
подами, связанными с обстановкой значительного опреснения.
Изредка остатки лингул встречаются в породах на одной плоскости
наслоения с настоящими пресноводными моллюсками (Дембс-
кая, 1959). В верхнем девоне Главного поля лингулы входят вместе
с пресноводными рыбами и пресноводными водорослями (тро-
хилиски) в комплекс форм дельтовых песков (Геккер, 1957).
Наоборот, в отложениях морей нормальной солености и даже
их прибрежной полосы лингулы встречаются редко. Так, например,
в среднем карбоне Московской синеклизы лингулы вообще не най-
дены, несмотря на многолетние детальные исследования, а в верх-
нем карбоне представлены всего пятью экземплярами (Иванова,
1949). В нижнем карбоне Кузнецкого бассейна среди многих тысяч
собранных брахиопод оказалось всего 3 экземпляра лингул (Сары-
чева и др., 1963). В общем обильные остатки лингул являются
хорошим показателем крайнего мелководья и опреснения, а лин-
гулы при отсутствии в слое другой фауны — опреснения в сочетании
с пониженным содержанием кислорода.
Палеозойские замковые брахиоподы, как показывают их остатки,
захороненные в породах весьма различного петрографического
состава и фациально-генетического типа, обитали в достаточно
разнообразных условиях. Наиболее благоприятной для этих бра-
хиопод была обстановка неглубокого нормальносоленого моря,
с хорошей аэрацией и умеренной подвижностью воды. Типичными
осадками такой обстановки являются детритусовые известняки,
в которых брахиоподы богато представлены и по количеству экземп-
ляров, и по числу видов, которых насчитывается до 30—40 в одном
слое. Большого разнообразия достигают брахиоподы и в других
нормально-морских фациях: в детритусовых известняках рифового
комплекса, в мелководных глинистых известняках, в фациях пере-
слаивания известняков и глин, нередко в алевритистых аргиллитах
и алевролитах.
Однако распространение не всех брахиопод ограничивалось
этими благоприятными обстановками. Многие формы, способные
существовать в гораздо более широком диапазоне условий, насе-
ляли также фации, отличающиеся от оптимальных по солености,
гидродинамическому или газовому режиму. Так, замковые брахио-
поды известны из отложений бассейнов не только нормальной со-
лености, но также осолоненных и опресненных (солоноватоводных).
Бесспорными осадками осолоненных водоемов, в которых были
найдены брахиоподы, захороненные на месте обитания, являются
первичные седиментационные доломиты. Фауна этих доломитов
количественно небогата, но достаточно разнообразна и включает
не только брахиопод, но также мшанок, пелеципод, гастропод,
фораминифер и редких иглокожих. Брахиоподы в свою очередь
представлены здесь сравнительно большим числом видов. Напри-
мер, в первичных доломитах каширского горизонта Московской
синеклизы брахиоподы принадлежат к семи родам, из которых три
(linoproductus, Diclyoclostus, Cancrinella) представлены несколькими
видами каждый (Иванова, 1949). Терригенные отложения осоло-
ненных бассейнов выявляются, естественно, гораздо труднее, но
при геохимическом определении солености ранневизейского водоема
в районе г. Сызрани был обнаружен слой черной углистой глины
с брахиоподами, сформировавшийся в водоеме с повышенной со-
леностью (Максимова, 1955).
Замковые брахиоподы обитали также в опресненных участках
и водоемах, но такого сильного опреснения, как лингулы, они не
переносили. Типичные солоноватоводные комплексы брахиопод.
в противоположность комплексам первичных доломитов, харак-
Брахиоподы
69
теризуются очень небольшим числом видов (1—4), но зато массо-
вым развитием одного или двух из них. Так, например, в верхнем
девоне Кузнецкого бассейна выделяется комплекс Athyris, приуро-
ченный к плохо отсортированным известковистым песчаникам
с гравием и галькой и к известнякам с обильной грубой терригенной
примесью. В этот комплекс часто входят массивные мшанки, а из
брахиопод — только атйрисы, но эти последние часто образуют
массовые поселения. Песчаники и известняки с Athyris слагают
слои небольшой мощности в морских клиньях среди континенталь-
ных красноцветных отложений и на переходах от континентальных
условий к морским (Бельская, 1960). Из рудницкой подсвиты вор-
кутской угленосной серии (PJ описаны два ярко выраженных соло-
новатоводных сообщества с преобладанием замковых брахиопод:
хонетесовое и неофрикадотирисовое (Дембская, 1965). Сообщест-
ва эти замещают друг друга по простиранию одного и того же слоя
мелко- или тонкозернистого песчаника мощностью 0,3—5 .м и
отличаются следующими особенностями. Хонетесовый комплекс
слагается из массовых скоплений одного вида брахиопод — lisso-
chonetes rotundatus Toula, к которому примешиваются единичные
представители еще одного вида брахиопод (Rhynchopora nikitini
Tsch.) и редкие двустворки и гастроподы. Сообщество с неофрика-
дотирис несколько более разнообразно: в нем кроме тех же хонете-
сов, что и в предыдущем комплексе, в значительном количестве
встречаются гладкие спирифериды (Neophricadothyris asiatica Chao)
и двустворки Aviculopecten, а также слегка увеличивается частота
встречаемости ринхопор. Характерно, что и хонетиды и гладкие
спирифериды являются формами широкого фациального распро-
странения, встречающимися в самых разнообразных породах и
обстановках.
В ранневизейском бассейне Поволжья, о котором упоминалось
выше и который был весьма изменчив по всем гидрологическим
показателям, как во времени, так и в пространстве, наряду с осад-
ками осолоненных водоемов или участков были установлены отло-
жения опресненных вод, содержащие многочисленных хонетесов
и камаротехий вместе с отпечатками наземных растений (Макси-
мова, 1955).
Таким образом, широко распространенное представление о том,
что все брахиоподы, кроме лингул, являются обитателями морей
нормальной солености, не соответствует дейст-
вительности. Брахиоподы, как группа в целом (и в том числе
замковые), были приспособлены к достаточно широкому диапазону
содержания солей в морской воде, хотя существенно уступали в
этом отношении моллюскам, которые не только заселяли водоемы
совершенно пресные и сильцоопресненные, но и переносили и более
высокую степень осолонения. Так, в кунгурских отложениях перм-
ского Прикамья, при переходе от содержащих фауну первичных
доломитов к гипсам, сначала исчезают брахиоподы, затем пеле-
циподы, а за слоями с одними пелециподами следуют уже немые
загипсованные доломиты (Герасимов. 1953). Однако специальных
солоноватоводных форм, способных существовать только в
водах пониженной солености, среди замковых брахиопод не было
и солоноватоводные комплексы слагались просто эвригалинными
видами или родами, встречающимися и в нормально-морских
сообществах.
Влияние гидродинамического режима и химизма водоема на
комплексы брахиопод может проиллюстрировать табл. VI, состав-
ленная в результате детального литологического и палеонтоло-
гического изучения нижнего карбона Кузнецкого бассейна (Сары-
чева и др., 1962).
Следовательно, в наиболее благоприятных условиях обитают
все брахиоподы. и те. которые привязаны только к этой нормально-
морской обстановке, и те. которые живут во всех остальных фациях,
чем-либо отклоняющихся от нормы. В неблагоприятных фациях,
наоборот, процветают или формы, способные суи^ивовать в весьма
разнообразных условиях (Camaratoechia. теребратулиды), или формы.
Таблица VI
Влияние различных условий на развитие комплексов брахиопод
Оптималь- ные условия Неблагоприятные условия
Аномальная подвижность воды Аномальный химизм (примесь вулканогенных продуктов)
очень слабая (затишная область) очень сильная (область волнового поля) слабоано- мальный сильно- аномальный
Все группы брахиопод. Господству- ют крупные продукти- ды, Spirifer, Fusella Camara- toechia Chonetidae Tomipro- ductus elegantulus Camara- toechia Terehratu- lida Rhipido- mella N eo spirifer Composita Eumetria Camara- toechia Те rebratu- lida Neospirifer Chonetidae Rhipidomella Camara- toechia Terebratu- lida
приспособленные к каким-то определенным особенностям внешней
среды (Eumetria, Composita). Так, в обсуждаемом примере все оби-
татели области волнового поля отличались крепким замыканием
раковины благодаря наличию хорошо развитого замка или очень
сильной мускулатуры, что и позволяло животным противостоять
вредному механическому воздействию волновых токов воды.
Меньше всего можно сказать о связи брахиопод с глубиной
бассейна. Судя по фациальному облику вмещающих пород, огромное
большинство брахиопод палеозоя и мезозоя обитало в неглубоких
морях (вероятно, до 200 м). Есть основания думать, что только в
третичное время они продвинулись на ббльшие глубины, в том
числе абиссальные.
Таковы в общих чертах особенности фациального распростра-
нения брахиопод. Стратиграфическое значение этой группы обще-
известно. В палеозое, где брахиоподы представлены многими ро-
дами и видами различного вертикального распространения и раз-
личной фациальной приуроченности, их остатки обычно дают
возможность довольно детально расчленять вмещающие отложения.
Разрезы раковин и обломки брахиопод в том или ином количестве
могут быть встречены в самых разнообразных породах — песча-
никах, алевролитах, аргиллитах, доломитах, известняках и даже
в туфогенных отложениях, накапливающихся в некотором отдалении
от вулканического очага. Но существенную породообразующую
роль они играют только в известняках, как правило, вместе с остат-
ками других организмов (особенно иглокожих), а иногда являясь
основным, господствующим компонентом породы.
Строение раковин. У беззамковых брахиопод раковины могут
быть или кальцитовыми, или неоднородными, состоящими из
чередующихся слоев, иногда кальцита и органического вещества,
чаще — хитинового вещества и фосфорнокислого кальция (lingula,
Obolus, Discina). У ископаемых форм хитиновое вещество обычно
замещается кальцитом, фосфатом, пиритом или другими минера-
лами. Фосфатные слои не изменяются. В шлифах, при увеличении
в 100 раз и больше, многослойное строение хитиново-фосфатных
раковин хорошо видно (табл. 45, фиг. 1), а при небольшом уве-
личении их обломки отличаются характерной резкой штриховато-
стью. Беззамковые редко бывают породообразующими, но в от-
дельных случаях скапливаются в таком количестве, что образуют
фосфатную руду (верхнекембрийские оболовые фосфориты При-
балтики).
70
Известковые организмы
У замковых брахиопод раковина всегда кальцитовая (с неболь-
шой примесью углекислого магния) и характеризуется несколькими
типами строения скелетной ткани. Известковая раковина современ-
ных форм сверху покрыта тонким хитиновым слоем, но в ископае-
мом состоянии этот слой никогда не сохраняется. Раковина состоит
из двух створок и целого ряда внутренних структур, служащих для
прикрепления мускулов, поддержки внутренних органов животного,
сцепления створок и укрепления самой раковины (рис. 85). Микро-
скопическое изучение показало, что известковое вещество створок
слагается из двух слоев: первичного и вторичного. Первичный слой
имеет почти одинаковую, обычно незначительную толщину на всем
протяжении от макушки раковины до лобного края, а толщина
вторичного слоя заметно увеличивается по направлению к макушке
(рис. 86), и этот слой может быть развит в очень разной степени
у различных замковых брахиопод. За счет сильного развития вто-
ричного слоя увеличивается толщина створок раковины и многих
внутренних структур, осевую часть которых образует первичный
слой. Сам первичный слой всегда состоит из налегающих друг
на друга пластинок, в разрезе шлифа имеющих вид длинных косых
волоконец, направленных под острым углом к поверхности ра-
ковины. Вторичный слой может быть или волокнистым, или приз-
матическим.
В первом случае волокна одного слоя направлены под углом
к волокнам другого или наклонены в разные стороны, подобно
слойкам косослоистых пород в соседних косых сериях (рис. 87).
При тонкой волокнистости такое строение проявляется только при
включении анализатора, тогда как при параллельных николях обло-
мок выглядит однородно светлым (белым или желтоватым). Сле-
дует отметить, что в обломках раковин такого типа оба слоя сохра-
няются не часто. Обычно первичный слой благодаря своей незна-
чительной толщине при захоронении и фоссилизации разрушается
и остается один слой (вторичный) с однонаправленными волокнами.
Именно этим объясняется, что в шлифах чаще всего встречаются
однослойные обломки брахиопод с волокнистым строением
(табл. 45, фиг. 2). Если волокна не очень тонки, то при скрещенных
николях видно, что они угасают попеременно. Вдоль обломков
сильноскладчатых волокнистых раковин при вращении столика
микроскопа пробегает волна погасания, как у трилобитов, с кото-
рыми брахиопод не позволяет спутать как раз наличие волокни-
стости. Значительно реже (например, у некоторых спириферид
и пентамерид) вторичный слой состоит из довольно грубых и ко-
ротких призм, направленных почти перпендикулярно первичному
слою (табл. 46, фиг. 1).
Двуслойные раковины брахиопод могут быть пористыми или
непористыми. У пористых раковин тонкие канальцы, направленные
почти под прямым углом к поверхности, пронизывают или оба
слоя раковины (например, у ринхонеллид, теребратулид и некото-
рых спириферид), или только первичный, внешний слой (у многих
ортид). Если поры проходят через оба слоя, то в скошенном попе-
речном сечении обломки такой структуры выглядят как изгородь
с толстыми столбиками (табл. 45, фиг. 7), в тангенциальном сечении
напоминают сито (табл. 45, фиг. 8). Конечно, и в том и в другом
разрезах видна обычная волокнистость, но прежде всего бросаются
в глаза многочисленные отверстия. У тонкоскладчатых ортид в
тангенциальном сечении видно чередование полос пористого пер-
вичного слоя и волокнистого — вторичного (табл. 45, фиг. 9). В по-
перечном сечении обломки ортид показывают волокнистую ткань,
с одной стороны рассеченную канальцами или покрытую темными
штрихами (если канальцы заполнены минеральным веществом).
У непористых раковин (главным образом в тангенциальных сре-
зах) наблюдаются дополнительные структуры, отражающие скульп-
турные образования на поверхности створок. Так, в шлифах довольно
часто встречаются неправильной формы обломки волокнистых ра-
ковин, усеянные мелкими кружками или овалами, обрисованными
множеством тонких концентрических линий (табл. 45, фиг. 4). Эти
Рис. 85. Раковина брахиоподы отряда Spiriferida
Видны спирали ручного аппарата (по Давидсону)
Рис. 86. Схематический поперечный разрез двуслойной известковой
раковины брахиоподы
а — первичный слой: б — вторичный слон
Рис. 87. Поперечный разрез пористой раковины брахиоподы отряда
Terebratulida
а — первичный слой; б — вторичный слой; в— поры. Видно направление волокон первич-
ного н вторичного слоев, сильно увеличено
образования представляют собой срезы через основания тонких и
коротких ребрышек, покрывающих поверхность раковины, чаще
всего так называемых пустул у продуктид. Волокна раковины из-
гибаются и обтекают срезы ребрышек, образуя прихотливый ри-
сунок, хорошо видимый при скрещенных николях.
Существуют еще раковины ложнопористые, у которых волок-
нистую скелетную ткань пронизывают тонкие иглоподобные стер-
Брахиоподы. Мшанки
71
женьки из микрозернистого кальцита, расположенные под большим
или меньшим углом к поверхности и нередко изогнутые (рис. 88;
табл. 46, фиг. 8).
Наконец, у многих брахиопод раковины снабжены более или
менее длинными выростами — иглами, которые бывают очень
разнообразны по форме, количеству и расположению. У продуктид
цилиндрические иглы могут быть очень длинными (до 5—7 с.и)
и неправильно изогнутыми. В некоторых шлифах разрезы таких
игл встречаются в большом количестве. В поперечном разрезе они
имеют вид колец с широким каналом (табл. 45, фиг. 6), в скошен-
ном поперечном — толстостенных эллипсов, а в сечениях, близких
к продольному, — закругленных на одном конце цилиндров. Ха-
рактерная для брахиопод волокнистая структура в кольцах и эллип-
сах проявляется в виде многочисленных концентрических струек,
а в субпродольных сечениях — в виде линий, параллельных длине
иглы. Очень своеобразны двуствольные иглы некоторых спирифе-
рид (рис. 89), которые в шлифах дают трехлопастные фигуры, на-
поминающие птичьи следы (табл. 46, фиг. 3, 4).
Внутренние скелетные образования брахиопод в шлифах, как
правило, не распознаются, так как их обломки по своему микроско-
пическому строению ничем не отличаются от обломков створок.
Исключение представляют изредка попадающиеся в шлифах раз-
резы через конический спиральный ручной аппарат спириферид и
атрипид, который на плоскости отражается в виде нескольких тон-
ких палочек, расположенных параллельно друг другу (табл. 46.
фиг, 2). При большом увеличении видно, что эти палочки имеют
тонкое косоволокнистое строение.
Рис,. 88. Строение ложнопористой раковины брахиоподы отряда
Strifbmenida
Видны стерженьки микрозернистого кальцита, пронизывающие волокнистое раковинное
вещество (по Вильямсу)
Рис. 89. Строение игл брахиопод
рода Phricadothyris (отряд Spiri-
ferida, семейство Reticularidae)
Видны крупные и мелкие двуствольные иглы.
>с.1ожненные боковыми отростками, и их
основания на поверхности раковины (по Коз-
ловскому)
Совершенно особым типом раковины отличаются распростра-
ненные в пермских отложениях области Тетиса брахиоподы семей-
ства Richthofenniidae, по внешнему виду чрезвычайно похожие на
одиночных кораллов. В шлифах это сходство с кораллами прояв-
ляется. пожалуй, не менее ярко, чем в общей форме раковины, так
как мы видим здесь образования, совершенно подобные пузырчатой
ткани и днищам кораллов (табл. 46, фиг. 6, 7), которые у рихтго-
фенид называются ячеистым слоем и поперечными пластинами.
Поперечные пластины прекрасно представлены на фиг. 6 табл. 46.
а на фиг. 7 той же таблицы хорошо видны в поперечном сечении
два слоя этой своеобразной раковины: тонкий наружный (на снимке
черный) и толстый ячеистый. Рассмотрение при большом увеличении
этого плотного слоя или тонких стеночек отдельных ячеек обнару-
живает типичную для брахиопод волокнистую структуру ткани,
которая и позволяет отличить рихтгофенид от кораллов. Более
того, многочисленные тонкие стерженьки, внедряющиеся в волок-
нистый слой (табл. 46. фиг. 8), показывают, что рихтгофениды
принадлежат к типу ложнопористых раковин, как и многие другие
представители отряда Productida. В пределах Советского Союза
рихтгофениды распространены на Северном Кавказе, в Армении,
в Дарвазе и Приморье.
Форма обломков брахиопод в шлифах достаточно разнообразна
и особенно неправильной бывает в тех случаях, когда разрез захва-
тывает примакушечную часть раковины (табл. 46, фиг. 9). Но чаще
всего они имеют вид изогнутых палочек или длинных полосок,
нередко волнистых. Довольно обычны также разрезы через целые
раковины, с сохранившими связь двумя створками. Два таких раз-
реза изображены на табл. 46, фиг. 10. Один из них, с редко волни-
стыми линиями створок, представляет поперечное сечение камаро-
техии, но вообще контур такого типа характерен для всех складчатых
ринхонеллид. Принадлежность обломков любой формы к брахио-
подам устанавливается по наличию описанных выше микроструктур
и особенно по волокнистому строению хотя бы одного слоя, кото-
рое прекрасно сохраняется даже в палеозойских породах сильно-
складчатых областей. Нередко волокнистость различается в облом-
ках шламмового размера (0,1 .и.и) и вообще уничтожается только
при чрезвычайно сильной перекристаллизации породы в целом,
стирающей все признаки ее первичной структуры. В этом состоит
существенная разница между брахиоподами и пелециподами. ко-
торые дают сходные по форме обломки, так как пелециподы в па-
леозойских отложениях не сохраняют никаких следов первоначаль-
ного строения и их остатки слагаются довольно крупными светлыми
зернами раскристаллизованного кальцита.
Однако в мезозое, особенно в мелу, нередко встречаются пеле-
циподы. у которых створки имеют волокнистую структуру, очень
сходную со структурой раковин брахиопод. Поэтому волокнистые
обломки, встреченные в мезозойских породах, надо определять
с осторожностью, сообразуясь с составом целых фаунистических
остатков, найденных в данном слое. Можно только отметить, что
если в шлифе таких обломков много и они толстые, то более ве-
роятно, что это пелициподы.
МШАНКИ (BRYOZOA)
Табл.47—50
Остатки скелетов мшанок широко распространены в осадочных
породах различного состава и возраста. Эти организмы имеют боль-
шое породообразующее, стратиграфическое и экологическое зна-
чение, особенно для отложений палеозоя (табл. 47, фиг. 1—4).
Мшанки представляют собой самостоятельный тип колониаль-
ных прикрепленных организмов, обитающих главным образом в
морях и в значительно меньшем количестве в пресных водах. Их
колонии отличаются большим разнообразием размеров и формы.
72
Известковые организмы
Индивидуумы мшанок, составляющие колонии (зооиды или
автозооид ы), отличаются очень маленькими размерами, не
превышающими 1 мм. Их передний отдел с венцом щупалец и ки-
шечником представляет собой полипид, а задний отдел в виде ци-
линдрического мешка, тесно связанный со всей колонией, является
цистидом или ячейкой. Ячейка имеет отверстие, через
которое может вытягиваться передняя часть особи.
Химический состав. Наружный слой ячейки выделяет студнеоб-
разную или рогоподобную оболочку, которая у многих морских
мшанок пропитывается карбонатами Са и Mg. Содержание карбо-
натов в стенке ячейки и ее общий химический состав у современных
мшанок различны. Иногда даже одни и те же виды отличаются раз-
личным содержанием рогового вещества, СаСО3 и MgCO3. в за-
висимости от того, живут ли они в прибрежной зоне моря или в
открытом море.
У некоторых мшанок стенки ячейки покрыты только рогоподоб-
ной оболочкой и не содержат известковых выделений, в связи с чем
представители этой группы очень редко встречаются в ископаемом
состоянии.
Экология. Морские мшанки известны в настоящее время во всех
морях мира и на всех широтах. Более всего они распространены
в мелком море от прибрежной зоны и до глубины 400—500 м. На
больших глубинах мшанки встречаются реже.
В приливно-отливной полосе моря, где господствуют сильные
течения, мшанки прикрепляются к твердым скалистым и грубо-
обломочным субстратам. Здесь обычны уплощенные колонии,
прикрепляющиеся всей нижней поверхностью, обрастающие во-
доросли, раковины (рис. 90). В сублиторальной зоне, в более спо-
койных водах, например на коралловых и раковинных песках, раз-
виваются двухслойно-симметричные, листообразные и сетчатые
колонии, прикрепляющиеся или непосредственно нижней частью
колонии, или корневыми выростами.
Морские мшанки лучше переносят колебания солености, чем
коралловые полипы, но количество их в бассейнах с пониженной
соленостью значительно меньше, чем в бассейнах с нормальной
соленостью. Все же известно довольно большое количество видов,
живущих в сильно опресненных водах, например поблизости от
устьев больших рек (главным образом мшанки Ctenostomata).
Геологическое значение. Ископаемые остатки мшанок встре-
чаются среди различных осадочных пород, преимущественно среди
таких морских пород, как известняки разного типа, глины и гли-
нистые сланцы, аргиллиты, алевролиты, песчаники, гравелиты
и конгломераты.
Рис. 90. Мшанки
а — современная мшанка Hornera lichenoides Linnaeus, прикрепившаяся на гальке (Барен-
цево море); 6 — пластинчатая колония ордовикской мшанки Spatiopora, обрастающая
гальку
Особенно обильны остатки мшанок в глинистых и аргиллитовых
породах, переслаивающихся с детритовыми известняками (табл. 48.
фиг. 4, 6, 7). Большое обилие их здесь и хорошая сохранность обус-
ловлены захоронением на месте обитания в условиях спокойных
вод (табл. 49, фиг. I: табл. 50, фиг. 2, 4). Скопления мшанок, свя-
занные. очевидно, с их массовыми поселениями на обширных про-
странствах морского дна в различные моменты истории Земли,
дают начало сплошным «мшанковым слоям» разной мощности.
Примером их могут служить мшанковые известняки среднего ордо-
вика на о. Вайгач, состоящие из крупных ветвистых колоний тер-
постомат (табл. 47, фиг. 4; табл. 48, фиг. 1). среднеордовикские
мшанковые аргиллиты, переслаивающиеся с детритусовыми извест-
няками на Сибирской платформе, сложенные тонкими двухслойно-
симметричными лентовидными и ветвистыми колониями крипто-
стомат, мшанковые известняки верхнего карбона и нижней перми
Урала и Приуралья, состоящие в основном из сетчатых колоний
криптостомат (табл. 49, фиг. 2—7), и, наконец, верхнемеловые
мшанковые слои Мангышлака и других регионов, сложенные скоп-
лениями разнообразных колоний циклостомат и хейлостомат
(табл. 50. фиг. 1).
В тонкозернистых, часто темных и даже черных битуминозных
известняках остатки мшанок сравнительно редки и отличаются
специфическим характером. Здесь преобладают массивные полу-
сферические колонии, свободно лежащие на морском дне. и тонкие
ветвистые колонии, прикрепляющиеся в одной точке.
В кварцевых песчаниках скелеты сохраняются хуже всего, в
песках почти совсем не сохраняются, что определяется не условиями
обитания (многие современные мшанки живут на песчаном грунте),
а последующими условиями захоронения. Исключением являются
третичные мшанки, остатки скелетов которых нередко весьма обиль-
ны в глауконитовых песках (например, в эоцене Мангышлака).
В грубообломочных породах — гравелитах, конгломератах —
встречаются плоские колонии, обрастающие гальки, или очень круп-
ные ветвистые и неправильно массивные формы, часто состоящие
из многих, нарастающих друг на друга слоев. Обычно в этих по-
родах встречаются комплексы бедного видового состава, но в боль-
шом количестве экземпляров. Мшанки здесь после гибели могут
быть перемыты и переотложены (табл. 48, фиг. 2). Нередко их ко-
лонии несут следы обтирания и окатывания, а изучение их внутрен-
него строения обнаруживает мелкий обломочный материал, заби-
вающий устья ячеек.
В отдельные моменты геологической истории и в разных райо-
нах земного шара мшанки принимали участие в образовании рифов
или в качестве главных строителей, или вместе с другими организ-
мами. На территории нашей страны примером мшанковых рифов
являются верхнекарбоновые и нижнепермские рифы западного
склона Урала и Приуралья и неогеновые керченские рифы Крыма
(табл. 50, фиг. 3, 5). Главными рифообразователями верхнепалео-
зойских рифов являются сетчатые мшанки отряда Cryptostomata
и только в небольшом количестве ветвистые мшанки Treposto-
mata. Неогеновые рифы в основном слагаются представителями
отряда Cheilostomata, при этом керченские — почти исключительно
многочисленными видами рода Membranipora, благодаря чему они
получили название мембранипоровых.
Состав скелета и морфология. Детали строения скелета различных
мшанок и особенности их морфологии требуют изучения под микро-
скопом в прозрачных шлифах. Для определения видов необходимы
определенным образом ориентированные шлифы. Но знание глав-
нейших особенностей строения колоний, морфологии ячеек, их
составляющих, и микроскопической структуры их стенок позволяет
в обычных неориентированных шлифах устанавливать принадлеж-
ность каждой данной мшанки к определенному отряду, а также
условия ее обитания и захоронения (рис. 91 —101).
Ячейки, составляющие скелет колоний, отличаются разной фор-
мой у разных групп. У представителей отрядов Cyclostomata. Cys-
Мшанки
73
Рис. 91. Продольные сечения мшанок
а — продольное сеченне трубчатой ячейки мшанки отряда Trepostomata с диафрагмами
и цистифрагмами: б — продольное сечение двуслойно-снмметричной колонии мшанки
отряда Cryptostomata (я. —удлиненная ячейка, п. — псевдопора. н. г. — нижняя гемисепта,
в. г. — верхняя гемисепта, с. п. —срединная пластина)
Рис. 92. Лунарии в устьях
ячеек у рода Fistulipora (от-
ряд Cystoporata)
Буквы на рисунке: л. лунарии:
ц. — цистопоры
Рис. 93. Мезопоры и аканто-
поры на поверхности колонии
у мшанок отряда Treposto-
mata
Буквы на рисунке: у. я. —устье ячейки:
м. — мсзопоры: а. — акантогюры
Рис. 94.Различные типы цистопор у родов отряда Cystoporata
ч— Ceramopora. Dianulites', б — Favositella: в — Xenotrypa. г—Stellipora: д — Fistulipora:
е — Eridopora
Рис. 95. Схематическое строение ветвистой колонии мшанки отряда
Trepostomata
Буквы на рисунке: я. — ячейки: м. — мезопоры. а. — акантопоры, д. —диафрагмы, з. з. —
зрелая зона колонии, н. з. — незрелая зона колонии
Рис. 96. Типы структуры стенок ячеек у мшанок отряда Trepostomata
а — продольно-волокнистые стенки (род Diplot гура)', б — поперечно-пластннчатые стенки
(род Lioclema): в — косо-пластинчатые стенки (род Hallopora)
toporata и Trepostomata они имеют характер длинных цилиндри-
ческих и призматических трубок (см. табл. 47, фиг. 3; табл. 48.
фиг. 3. 5: рис. 91, г/, см. рис. 95): у большинства представителей
отряда Cryptostomata — грушевидную форму с расширенным
основанием и вытянутым трубчатым вестибюлем
(рис. 91,6; см. рис. 99). У мшанок отряда Cheilostomata—ячейки
ящикообразной формы с четырехугольным основанием. Устья
ячеек имеют различную форму. В области устьев ячеек часто
развит приподнятый валик, гладкий или несущий бугорки и шипы,
или полулунный выступ — лунарий (рис. 92). вдающийся в
устье своими острыми углами. У многих мшанок отрядов Cryp-
tostomata и Cheilostomata в устье имеется также известковая к р ы-
ш е ч к а, отличающаяся различной формой и строением.
74
Известковые организмы
Рис. 98. Пятно на поверхности
колонии у мшанок отряда Trepos-
tomata, состоящее из скоплений
мезопор и крупных ячеек
Рис. 97. Вторичные известковые
отложения в области стенок ячеек
у мшанок отряда Trepostomata
Рис. 'О'. Структура стенок ячеек у мшанок отряда Cryptostomata
а — Stictoporella; б — Pachydictya
Рис. 99. Схема строения сетчатой колонии мшанки отряда Сгур
tostomata
Буквы на рисунке: пр. — прутья; п. — перекладины; у. я. — устья ячеек; к. — киль; к. 6. -
килевые бугорки; я. — ячейки
Рис. 100. Схема поперечного сечения прута сетчатой колонии мшанки
отряда Cryptostomata (род Fenestella)
Буквы на рисунке: я. — ячейки в поперечном разрезе, к. б. — килевой бугорок: в. с. — внут-
ренние стенки, н. с. — наружные стеикн, к. — капилляры
В ячейках часто присутствуют горизонтальные перегородки —
диафрагмы (рис. 91, а), сплошные или неполные (отряды Tre-
postomata и Cystoporata), или полуперегородки — гемисепты,
отделяющие вестибюль от основания у ячеек грушевидной и удли-
ненной формы (рис. 91,6). Кроме диафрагм у некоторых мшанок
отряда Treptostomata имеются особые пузыреобразные структуры —
цистифрагмы (рис. 91, а).
Известковые стенки ячеек, отличающиеся у ряда мшанок сложной
микроскопической структурой (см., например, рис. 96, 97, 100, 101),
пронизаны различными типами пор. Соединительные
поры осуществляли сообщение между полипидами, псевдо-
поры и другие типы пор — газообмен.
Колонии мшанок в преобладающем числе полиморфны.
Кроме нормальных ячеек, вмещавших самостоятельно питавшихся
и размножавшихся особей, в колониях имеются особые ячейки,
которые представляли собой вместилища для видоизмененных по-
липоидов или были совсем лишены последних. Из их числа у па-
леозойских мшанок наиболее распространены м е з о п о р ы,
цистопоры и акантопоры (рис. 93—95).
Me зо поры — трубчатые полости меньшего диаметра, чем
нормальные ячейки, нередко более толстостенные, округлого или
угловатого сечения, с частыми прямыми диафрагмами (отряд
Т repostomata) (табл. 48, фиг. 3). Псевдомезопоры — сход-
ные образования, лишенные диафрагм у мшанок отряда Cryp-
tostomata) (рис. 91,6).
Цистопоры — неправильные трубчатые полости с варьи-
рующей формой сечения, с косыми и выгнутыми перегородками,
редкими или очень частыми (рис. 94).
Акантопоры — мелкие трубчатые образования с узкой
центральной полостью малого диаметра, с толстыми концентри-
чески пластинчатыми стенками. Акантопоры погружены в стенки
ячеек и поднимаются над поверхностью колонии в виде шипов
(табл. 48, фиг. 5; табл. 49, фиг. 7; рис. 93, 98). Они преимущественно
развиты у мшанок Trepostomata и у ветвистых представителей
Cryptostomata. Они выполняли защитную функцию в колонии.
Сходны с ними по строению килевые и устьевые бугор-
к и некоторых представителей отряда Cryptostomata (рис. 99).
Многочисленные опорные и механические элементы колоний
(например, корневые выросты) мшанок отрядов Cyclostomata.
Cryptostomata и Chelostomata являются также скелетными обра-
зованиями, служившими вместилищами для особых специализи-
рованных особей.
Моллюски
75
МОЛЛЮСКИ (MOLLUSCA)
Двустворчатые, или пластинчатожаберные, моллюски
(Bivalvia)
Табл. 51, 52
Моллюски — билатерально-симметричные, водные, донные жи-
вотные с раковиной, состоящей из двух створок, обычно с симмет-
ричными правой и левой створками.
Химический и минеральный состав. Раковина пластинчатожа-
берных моллюсков — пелеципод обычно состоит из трех слоев.
Наружный слой образован конхиолином (рогоподобное органи-
ческое вещество). Средний слой слагается призматическими кристал-
лами кальцита, одетыми в тончайшие конхиолиновые чехлы, а
внутренний, форфоровидный или перламутровый, слой состоит
из арагонита. Таким образом, по химическому составу в раковинах
двустворок преобладает СаСО3. К нему добавляется в большем
или меньшем количестве органическое вещество (конхиолин), и
совсем незначительную примесь составляют углекислый магний
(до 1%) и фосфорнокислый кальций (до 1,5%).
Экология. Большая часть двустворчатых моллюсков обитает
в морских и солоноватоводных бассейнах, и лишь около 20% обще-
го числа их видов населяет пресные воды. Для группы в целом
характерно приспособление к чрезвычайно широкому диапазону
условий обитания. Вследствие этого пелециподы распространены
во всех климатических зонах; от теплых вод тропических морей до
Арктики и Антарктики, от приливно-отливной зоны и прибрежных
мелководий до дна океанических впадин (более 10 000 м); в водоемах
самого различного типа: в океанах, окраинных и внутренних морях,
лагунах, эстуариях, реках, озерах и даже в прудах. Соответственно
наблюдается большое разнообразие размеров и строения раковин,
а также образа жизни отдельных видов. Самой крупной раковиной,
до 1,4 м в длину, обладает тропическая тридакна, тогда как размеры
некоторых глубоководных видов не превышают 2—3 мм. Одни
двустворки закапываются в донный осадок, другие свободно лежат
или ползают по его поверхности, третьи прирастают створками
к субстрату или прикрепляются к различным предметам пучком
прочных биссусных нитей. Некоторые свободно лежащие двуст-
ворки (например пектиниды) могут плавать, хлопая двустворками
и передвигаясь в воде короткими скачками. Существует также до-
вольно значительное количество двустворок, сверлящих дерево,
плотные илисто-глинистые грунты и известковые породы. При этом
представители семейства фоладида приспособлены к механическому
сверлению, а так называемые морские финики (Lithophaga) одно-
временно и точат и растворяют известняки при помощи кислых вы-
делений мантии.
Многие пелециподы могут довольно долгое время жить при
очень малом содержании кислорода в воде или даже при его отсут-
ствии. Так, Муа arenaria может жить в бескислородных условиях
при 14° С до 8 дней, а при 0° даже несколько недель. Виргинская
устрица также выносит такие условия в течение недели и более.
Жизнедеятельность моллюсков при отсутствии кислорода в воде
обеспечивается анаэробным процессом расщепления их запасных
веществ. Очень многие двустворки — активные фильтраторы,
засасывающие и пропускающие через свое тело значительные массы
воды, из которой они извлекают пищевые частицы. При этом не-
годные для питания минеральные частицы, пройдя через кишечный
тракт животного или через мантийную полость, выбрасываются
наружу, склеенные слизью, в виде комочков или псевдофекалий
и отлагаются на дне. «Мидии во всех морях, кроме тропических,
образуют, — пишет Л. А. Зенкевич (1955, стр. 146), — мощный при-
брежный бордюр, нередко давая скопления по нескольку десятков
килограммов на один квадратный метр. . .» В течение часа крупная
мидия пропускает через мантийную полость около 3 л воды. В те-
чение суток плотное поселение мидий на банке профильтровывает
от 50 до 280 м3 воды. На мелководье вся вода проходит через био-
фильтраторы. Этот процесс является мощным фактором обра-
зования грунта. Под поселениями мидий иногда наблюдаются
вязкие илистые грунты, образованные этими биофильтраторами.
По берегам Северного моря, по Л. А. Зенкевичу, илы, образован-
ные мидиями, имеют мощность 8 м. За один год может отложиться
слой мидиевого ила мощностью в 30 см. Эти илы склеены слизью
и сильно противостоят размыву и сносу в более глубокие участки
моря. Несомненно, такие осадки существовали и в прежние эпохи,
но мы плохо умеем их обнаруживать в ископаемом состоянии (см.
раздел «Микрокопролиты»).
Геологическое значение. Двустворчатые моллюски известны
по всему стратиграфическому разрезу начиная с кембрия. В кемб-
рийских отложениях имеют плохую сохранность и не играют важной
стратиграфической роли. В девоне, карбоне и перми в нормально-
морских отложениях двустворки представляют совершенно второ-
степенный компонент фаунистических комплексов, но широко
распространены в толщах, обогащенных органическим веществом
(битуминозных и особенно угленосных), для которых очевидно их
большое стратиграфическое значение. Такое распределение обус-
ловлено эврибионтностью пелеципод как группы в целом. В нор-
мально-морских условиях их оттесняли на задний план стеногалин-
ные морские формы, а в опресненных и совсем пресных водоемах,
где накоплялись угленосные осадки, пышно развивались эврига-
линные двустворки. Вероятно, имела значение и способность пеле-
ципод жить при пониженном содержании кислорода, временами
падавшем до нуля, так как в осадках с высоким содержанием разла-
гающихся растительных остатков и вообще органических веществ
кислород придонной воды в большом количестве расходуется на
окисление органики.
Для мезозойских и кайнозойских отложений пелециподы имеют
как большое стратиграфическое значение, так, временами, и поро-
дообразующее. На мелководье иногда они образуют ракушняки,
например третичные на юге Русской платформы, эксплуати-
руемые как строительный камень и балласт. В других случаях дву-
створчатые участвуют в образовании массивных биогермных из-
вестняков. Так, некоторые представители отряда рудистов, похожие
по внешней форме на массивные одиночные кораллы, в меловую
эпоху были активными биогермообразователями.
Микроструктура. В палеозойских отложениях раковины двуст-
ворчатых моллюсков обычно перекристаллизованы до полного
исчезновения первичной структуры. В третичных и мезозойских
осадках наружный конхиолиновый слой раковины сохраняется
очень редко, так что, как правило, наблюдаются два слоя: внеш-
ний— призматический, развитый различно у разных групп, и внут-
ренний — пластинчатый. У некоторых форм (Veneridae, Cardiidae)
призматический слой вообще не развит. Микроструктуры призма-
тического слоя встречаются в трех разновидностях: 1) собственно
призматической; 2) ячеисто-призматической, 3) пучковидной.
Собственно призматическая структура слагается из отдельных
призм, образованных одним или несколькими полигональными
кристаллами кальцита. Ограничивающие грани призм обычно
плоские, бывают несколько искривлены. В продольном сечении
такая структура в поляризованном свете выглядит полосчатой,
в тангенциальном разрезе дает картину брусчатки или сетки с более
или менее правильными ячеями. У иноцерамов, например, призмы
обычно очень правильные и равны между собой как по диаметру,
так и по длине (табл. 52, фиг. 1). В других группах, например у Pinna,
ширина призм варьирует значительно, по-видимому, в результате
слияния нескольких призм. Длина призм бывает также различна,
у иноцерамов иногда их можно видеть невооруженным глазом.
Все призмы с точки зрения оптической ориентированы независимо
друг от друга. Одни затухают параллельно длинной оси, другие —
косо, благодаря чему и получается пестрополосатая структура в
поляризованном свете.
76
Известковые организмы
В сечении, поперечном к призмам, в скрещенных николях полу-
чается картина брусчатки благодаря разновременному затуха-
нию призм, оптические оси которых расположены различно по от-
ношению к их длине. Таким образом, в одной ракушке при скрещен-
ных николях можно видеть в одно и то же время темные призмы,
затухающие и яркополяризующие. В ископаемых призмах часто
видна спайность. При дроблении раковин призматический слой
может распадаться на отдельные призмы, которые сохраняются
в осадке в виде определимых, более или менее закономерных облом-
ков. У Pinna призмы заканчиваются, заостряясь, у иноцерамов они
резко обрезаются.
Ячеисто-призматическая структура характерна для рудистов
и для некоторых устриц. Эта структура резко отличается от преды-
дущей тем, что если в простом свете видны грани между призмами,
то в поляризованном затухание происходит вне зависимости от
этих граней. Таким образом, «призм» не наблюдается. Ячеи полу-
чаются благодаря пересекающимся границам по слоям роста, и.
таким образом, эти ячеи обычно удлинены (табл. 52. фиг. 2—5).
Несмотря на то что оптические признаки и ячеи не выражены, гра-
ницы ячей-призм иногда не только отчетливы, но состоят из серого
кальцита и образуют как бы стенки этих ячей-призм.
Пучковидная структура образуется пучками пластин, состоящих
из параллельных тонких листочков, совершенно не расщепляемых.
Структура очень сложная и не выражается в шлифе в виде опре-
деленных геометрически правильных фигур.
Наиболее простая из таких структур в поперечном сечении к
раковине имеет вид призм. Эти «призмы» различно окрашены:
в серый, бурый и более светлые тона. Светлые чередуются с темными
«призмами», имеют одну толщину, прямы и расположены перпенди-
кулярно к поверхности раковины. На первый взгляд их нельзя отли-
чить от настоящих призм, но при большем увеличении заметна ко-
сая штриховатость. При прослеживании таких пучков-пластин видно,
что некоторые из них утончаются и сливаются с соседним пучком
или, наоборот, дихотомически разветвляются. В некоторых же мес-
тах эти псевдопризмы ветвятся и вновь соединяются много раз в
толщине слоя (табл. 51. фиг. 6, 7). В то же время толщина их начи-
нает меняться и они приобретают вид неправильных, меняющих
направление, изгибающихся и выклинивающихся пучков, состоящих
из тонких листочков (табл. 52, фиг. 10). В сечениях, направленных
косо к пучкам, изменение их видно еще резче.Тангенциальный раз-
рез пучков показывает сложность и изменчивость их рисунка. Полу-
чаются угловатые, зубчатые, ветвистые, различной длины пятна
на светлом фоне (табл. 51, фиг. 2).
Обычно в бесцветных пучках листочки расположены продольно,
в темных они лежат наискось к пучку-пластине.
Пучковидная структура имеет некоторую особенность при се-
диментации: в то время как настоящие призматические слои пла-
стинчатожаберных дробятся на отдельные призмы и, таким образом,
попадают в осадок, пучковидные слои никогда не расщепляются на
отдельные пучки или листики. Слои ломаются, образуя отдельные
осколки, в которых можно увидеть несколько случайно обломан-
ных пучков.
Так называемый пластинчатый слой может быть в трех разно-
видностях:
1. Листоватый параллельно поверхности раковины:
а. Параллельная слоистость может быть плохо заметна и видна
только в поляризованном свете. В простом же свете слой кажется
однородным.
6. Листочки (пластинки) хорошо видны в простом свете (табл. 51.
фиг. 3. 4). Ограничивающие их плоскости выделяются в поперечном
сечении раковины в виде четких линий. У некоторых устриц эти
плоскости так близко отстоят друг от друга, что при малых увели-
чениях сливаются и дают серое однородное поле.
в. Плоскости, разделяющие пластинки, могут не совпадать с
плоскостями нарастания. В этом случае пластинки выделяются
оптически в скрещенных николях.
г. Пластинчатая структура может быть только кажущейся и
обусловленной отложением окрашенных, возможно, органических
веществ, образующих параллельные линии.
д. Слой может иметь двоякое деление: параллельно поверхности
(в виде очень правильных и толстых пластин) и перпендикулярно
к ней. Получается своеобразная решетка с квадратными или пря-
моугольными ячейками.
2. Решетчато расположенный, плетенчатый.
«Плетенчатую» структуру дают две системы пластинок, пере-
секающихся друг с другом под различными углами и образующих
своеобразную правильную фигуру, похожую на стенку плетеной
корзинки. Иногда одна система пластинок, обрываясь, заменяется
другой с некоторым перерывом. Все эти пластинки могут пересе-
каться грубыми линиями роста параллельно поверхности раковины.
3. Волнистый или смятый.
Смятая пластинчатая структура очень характерна для ископаемых
устриц (Gryphea, Еходуга). Она отличается тем. что пластинки или
тонкие листочки располагаются более или менее беспорядочно,
изгибаясь в различных направлениях и образуя дуги, веера и даже
сферолитоподобные фигуры (табл. 52. фиг. 11).
Гастроподы, или брюхоногие моллюски (Gastropoda)
Табл. 53. фиг. 1^-9; табл. 54
Широко распространенные моллюски с обособленной головой,
уплощенной подошвообразной ногой и блюдцеобразной или спи-
рально завитой раковиной.
Химический состав. Раковины гастропод состоят из арагонита,
кальцита и конхиолина и легко перекристаллизовываются с исчез-
новением характерных структур. Они почти не содержат углекислого
магния (от следов до 1.78%).
Экология. Экологические условия жизни гастропод чрезвычайно
разнообразны: это и морские, и пресноводные, и наземные живот-
ные. В водной среде это в большинстве случаев донные, свободно
передвигающиеся организмы, но среди морских форм часть при-
надлежит к настоящему пелагическом планктону (крылоногие,
киленогие). Из бетонных представителей гастропод некоторые за-
рываются в песок и ил. другие прикрепляются, а иногда и прирастают
к камням (червеобразные Vermelus). третьи просверливают ходы
в известковых породах или в остатках организмов. Глубже 100 м
количество гастропод уменьшается, но многие формы живут и на
значительных глубинах, вплоть до дна океанических впадин
(10 687.м). Гастроподы живут как в полярных морях, так и в тро-
пиках и даже в горячих источниках с температурой воды до 53°.
Среди гастропод известны травоядные и хищные представители.
Примитивные формы гастропод появляются в раннем кембрии.
В силуре число групп гастропод сильно увеличивается, но в течение
всего палеозоя сохраняется примитивность строения раковины.
В мезозое происходит увеличение числа сифоностомных и легоч-
ников. В поздней юре появились первые представители пресноводных
брюхоногих, которые в третичное время достигли большого разви-
тия.
Как породообразователи гастроподы играют иногда значитель-
ную роль, образуя гастроподовые известняки в результате скопления
целых и ломаных раковин. Обычно это мелководные прибрежные
скопления. Часто ископаемые раковины бывают нацело перекристал-
лизованы и лишь темные оторочки налипшего пелитоморфного
карбоната или иного илового материала, заполнившего внутренние
полости и промежутки между раковинами, позволяют определить
их форму и принадлежность к классу гастропод.
По микроструктуре раковины гастропод имеют много общего
с раковинами двустворок. Но при определении гастроподы имеют
Моллюски
77
то преимущество, что спирально завитая, большей частью конусо-
видная раковина брюхоногих имеет в разрезе характерную форму
(табл. 53. фиг. 1).
Стенки раковин гастропод обычно многослойны. В принципе
они трехслойны, но внешний роговой слой обычно исчезает, а в
результате навивания раковины вокруг оси и охвата в той или иной
степени предыдущих слоев получается неодинаковое количество
слоев в разных участках раковины. Так, у некоторых ископаемых
мелких раковин наблюдается два слоя (табл. 53, фиг. 4, 5). У других
в участках навивания — более трех слоев (табл. 53, фиг. 6, 9), а при
перекристаллизации нарастание раковины создает неправильно-
многослойную структуру (табл. 53, фиг. 7).
В раковинах современных гастропод все слои состоят из листо-
вых сложных пластинок. Эти последние различно ориентированы
в разных слоях и никогда не встречаются в изолированном виде.
Пластинки представляют собой одну оптически ориентированную
единицу (кристалл) и в поперечном сечении напоминают призму
двустворчатых моллюсков. Пластинки параллельны друг другу,
иногда несколько изогнуты. По толщине они приблизительно равны
друг другу, но часто выклиниваются и утоняются, не достигая края
слоя. Иногда они дихотомически ветвятся (табл. 53, фиг. 3; табл. 54,
фиг. 1—4, 8). Пластинки различаются между собой по цвету: одни
бесцветные, кажутся сплошными, гомогенными или с тонкой штри-
ховатостью, идущей поперек пластинки: другие, перемежающиеся
с первыми, окрашены в серый или буроватый цвет, имеют отчетли-
вую продольную штриховатость (иногда наискось к границам пла-
стинки). Иногда же обе системы пластинок тонко штриховаты наис-
кось в противоположные стороны, что, несмотря на общую серую
окраску, позволяет отличить их друг от друга (табл. 54. фиг. 1. 2).
Поляризованный свет резче выявляет структуру пластинок.
Те же пластинки, пересеченные параллельно плоскостями, их
ограничивающими, обнаруживают тонкоштриховатую структуру
(табл. 54, фиг. 6, 7). Ъта пересекающая штриховатость аналогична
«плетеночной» структуре пластинчатожаберных. Она выражается
в очень правильно расположенных зигзагообразных линиях.
Необходимо заметить, что все описанные микроструктуры в ис-
копаемом материале встречаются редко или же они не ясны.. На
табл. 53 и 54 приведены фотографии шлифов главным образом сов-
ременных и третичных гастропод.
Аммониты (Ammonoidea)
Табл. 53, фиг. 10, 11; табл. 55. фиг. 1—3
Эта вымершая группа головоногих моллюсков известна с ранне-
го девона до мела включительно. Их спирально свернутые раковины
состоят из большого количества камер, разделенных перегородками,
обычно сложно изогнутыми, и связанных сифоном. В последней
камере, лишенной перегородок, помешалось тело животного. Осталь-
ные камеры были наполнены газом. Стенки раковин аммоноидей
слагаются арагонитом, кальцитом и конхиолином.
Палеоэкология. Аммоноидеи — животные исключительно мор-
ские. Анализ их распространения показывает, что в палеозое они
обитали в водах нормальной солености и населяли неритовую зону
морей, в особенности се прибрежные участки. В каменноугольных
и пермских морях особенно благоприятна для аммоноидей была
обстановка заливов и бухт со спокойным гидродинамическим ре-
жимом. Открытых пространств эпиконтинентальных морей аммо-
ноидеи. наоборот, избегали. Именно поэтому они чрезвычайно
редки в девоне и карбоне Русской платформы за пределами специ-
фических, так называемых доманиковых, фаций. В мощных девон-
ских известняках центральной части США и других отложениях
подобного типа аммоноидеи также или отсутствуют, или очень
редки.
Аммоноидеи не встречаются или представляют большую ред-
кость в известняках биоморфных и органогенно-детритусовых.
а также в черных сланцах, формировавшихся в условиях низкого
содержания кислорода. Вместе с аммоноидеями чаше всего мы на-
ходим радиолярий (иногда в массовом количестве), остатки кремне-
вых губок, гастропод и двустворчатых моллюсков. Фузулиниды.
кораллы, мшанки и брахиоподы в одном слое с аммоноидеями
находятся редко.
Современные палеонтологические данные показывают, что
палеозойские аммоноидеи при жизни держались в пределах опре-
деленных частей морского бассейна, в определенных фациальных
условиях и не плавали на большие расстояния: после смерти живот-
ных их раковины тоже далеко не разносились.
В мезозое область обитания аммоноидей резко расширилась.
С одной стороны, они заселили обширные открытые эпиконтинен-
тальные моря, а с другой — приспособились к жизни на значительной
глубине в осевых частях геосинклинальных бассейнов. Соответст-
венно изменились и стали более разнообразными их экологические
требования и их сочетания с другими группами беспозвоночных.
Об образе жизни аммоноидей почти ничего не известно. На
основании экспериментальных работ по определению обтекаемости
раковин различной формы и их плавучести и сопоставления экспе-
риментальных данных с геологическими материалами по распро-
странению этой группы специалисты пришли к выводу, что боль-
шинство аммоноидей принадлежало к категории некто-бентических
организмов. Другими словами, это были животные,свободно пла-
вавшие. но державшиеся у дна. где они добывали пищу.
Геологическое значение. Аммоноидеи имеют важное стратигра-
фическое значение, особенно для мезозоя. В стратиграфии палео-
зойских отложений они играют меньшую роль вследствие своего
локальною распространения. В точном смысле слова аммоноидеи
породообразователями не являются, но в особо благоприятных
условиях (например, в нижней перми Южного Урала) слагают круп-
ные линзы ракушечника, состоящего из смеси целых раковин и об-
ломков.
Микроструктура. Стенки раковин аммоноидей состоят, как и у
других моллюсков, из трех слоев: наружного тонкого — кинхио-
линового и двух арагонитовых (среднего форфоровидного и внут-
реннего пластинчатого). Однако обычно раковины аммоноидей
бывают нацело перекристаллизованы, а все слои замещены одно-
родным зернистым кальцитом. Поэтому для определения этой
группы в шлифах важна не столько структура стенки, сколько кон-
туры сечения раковины. Характерные для аммоноидей фигуры плос-
кой спирали отличаются от гастропод наличием поперечных пере-
городок (табл. 55, фиг. 1). Косые сечения перегородок дают сложные
извилистые фигуры благодаря гофрировке и сложному сочленению
со стенкой раковины (табл. 55, фиг.З).
Парные пластинки — крышечки, закрывающие устье раковин
аммоноидей. называются аптихами. Аптихи имеют треугольно-
полулунные очертания и слагаются тремя слоями кальцита. Внут-
Рис. >02. Аптихи аммоноидей
а — 6 — аптихи, вертикальные сечения: а — через Aplychus projundus. 6 — через Aplychus
puciaius (и. с. — наружный слой с порами; с. с. — грубояченстый средний слой; в. с.—
плотный внутренний слой)
78
Известковые организмы
ренний тонкий слой покрывается толстым ячеистым слоем. Внеш-
ний плотный слой иногда несет ребра или другую скульптуру или
обладает порами (рис. 102). Определение аптихов производится,
как и аммоноидей. по внешней форме. В шлифах обычно получается
ячеистая структура благодаря сечению ячеистою слоя, который
очень похож на ячеистый слой некоторых устриц и иногда неотли-
чим от нею.
Белемниты (Belemnoidea)
Табл. 55. фи!. 4—8
Вымершие головоногие моллюски с внутренней раковиной,
известные с триаса и до конца меловою времени, имели некоторое
сходство с современными кальмарами. Остатки белемнитов встре-
чаются обычно в виде ростров — цилиндровидных или сигарооб-
разных сплошных массивных известковых тел. заостренных с одного
конца и имеющих конусовидную впадину с другого В этой впадине
при исключительно хорошей сохранности находится фрагмакси —
собственно конусовидная раковина животного с камерами и си-
фоном.
Ростр белемнита слагается кальцитом, редко распадается и обыч-
но встречается в породах в целом виде или в крупных кусках. Так как
белемниты жили в открытых морях и. вероятно, хорошо плавали,
их скопления встречаются довольно редко (в большинстве случаев
переотложенными) и значение их как породообразователей невелико.
Но как руководящие ископаемые они играют большую роль при
изучении стратиграфии мезозойских осадков (юра — мел).
В нашем материале были шлифы ростров белемнитов. Ростры
белемнитов слаг аются радиально расположенными конусами —
призмами кальцита, расходящимися от центральной оси. Эти ко-
нусы могут быть различной толщины: от близкой к фибре до гру-
бой неправильной призмы (табл. 55, фиг. 4—8). В поперечном к оси
сечении благодаря затуханию конусов в продольном направлении
в скрещенных николях получается черный крест, как у сферолита
(табл. 55. фиг. 4. 7). У некоторых форм этот крест имеет несколько
изогнутые или веерообразно расходящиеся концы (табл. 55. фиг. 5).
В вертикальном сечении ростра некоторые формы обнаруживают
призматическую структуру, некоторые же в поляризованном свете
образуют две зоны: осевую, равномерно поляризующую (скрыто-
фибровую). и яснофибровую периферическую (табл. 55, фиг. 6).
В изолированном виде конусы и призмы не наблюдались.
РАКООБРАЗНЫЕ (CRUSTACEA)
Общие замечании
Ракообразные относятся к типу членистоногих и включают ряд
отрядов, сильно отличающихся друг от друга строением тела.
Для геолога являются интересными те из них. которые имеют твер-
дый известковый панцирь, способный сохраняться в ископаемом
виде. Ракообразные известны с кембрия, и некоторые широко рас-
пространенные формы служат для стратиграфических целей (остра-
коды и трилобиты, см. соответствующие разделы).
Рис. 103. Раковины современного оалануса
а —О —современный баланус на Каспийского мора в сечениях, перпендикулярных к стенкам пластин: а — целые «домики», прикрепленные к створке раковины моллюска, вправо —
тонкая «лесенка» —сечение через крышечку, прикрывающую подвижную пластину, х II. 6 — сечение через крышечку, изображенную на с. в скрашенных николях. х 73. в - сечение
через стенку н основание «домика» оалануса. видно ячеистое строение пластин и основания. х 11, г — сечение пластин в скрещенных николях. видно волнистое затухание пластинчато-
фибровой структуры, х 73. д — участок пластины с «шестоватой» линзой, похожей на линзы у крабов, простой свет, х 73
Ракообразные
79
Обычно это водные животные, свободно передвигающиеся по
дну. Многие из них обладают известковым панцирем с примесью
фосфата и хитина. Строение панциря различно и еще слабо изучено
у разных групп.
Изучались преимущественно панцири современных раков. Автор
настоящего раздела изучал в шлифах раковину современного бала-
нуса (морского желудя). Этот рачок, обитающий в морях, — типич-
ный представитель усоногих, единственных ракообразных, ведущих
прикрепленный и неподвижный образ жизни. Баланус прикрепляется
к донным и плавающим предметам и создает раковину — кони-
ческий домик, из которого высовываются лишь конечности. Сов-
ременное значение балануса большое. Иногда он дает биомассу
до 30 кг на 1 м1 дна (Зенкевич, 1955). Баланус известен с третичного
периода.
Раковина баланусов коническая или полусферическая, на широком
основании в виде венца и состоит из нескольких пар сросшихся бо-
ковых пластин и двух пар подвижных свободных пластин, закрываю-
щих отверстие.
По шлифам, сделанным перпендикулярно основанию, видно,
что пластины сложены ячеями, стенки которых образованы тонкими
листовидными пластинами, огибающими полости ячей, как у мша-
нок. Ячеи неодинаковой толщины и формы, но в сечении они пре-
имущественно четырехугольные. Ячеи крышечек более правильные
и в поперечном к пластине сечении почти квадратные (рис. 103, а—д).
Такие сечения крышечек при изменении структуры стенок ячей
легко смешать с корочкой однослойной мелобезии (багряные во-
доросли). От мшанок баланус отличается меньшей выраженностью
листоватых пластинок, которые в простом свете не видны; в скре-
щенных николях в плоскостном срезе затухают волнисто, а в по-
перечном не затухают.
Остракоды (Ostracoda)
Табл. 56
Остракоды — это мелкие (0,5—50 мм) водные ракообразные,
встречающиеся как в пресных водах, так и в сильно засолоненных
водоемах. Встречаются они в породах всех морских формаций и
фаций начиная с раннего ордовика. Впервые пресноводные формы
начали появляться, очевидно, в девоне.
Тело остракоды заключено в двустворчатый панцирь, состоя-
щий из углекислого кальция или хитина у планктонных форм. По-
верхность створок панциря часто бывает богато скульптирована
и является дополнительным признаком, облегчающим их опреде-
ление. Значение остракод для практических целей стратиграфии
широко известно. Они часто встречаются в массовом количестве.
Экология. Современные остракоды являются формами бентон-
ными и планктонными. Раковины планктонных форм в ископаемом
состоянии сохраняются редко, так как их панцирь очень слабо обыз-
вествлен. Бентонные формы ползают по растениям, по дну или же
живут в донном иле. Некоторые из бентонных форм могут плавать,
поднимаясь на короткое время над дном. Интерпретация палео-
экологии ископаемых форм является результатом их сравнения
г современными формами, экология которых известна. Панцирь
эстракоды, а в особенности его свободный край, видоизменяются
э зависимости от образа жизни животного. Для многих форм,
имеющих выпуклый свободный край панциря, принимается, что
эни приспособились к плавающему образу жизни, во всяком случае
1ля свободного приподнимания над дном на определенный отрезок
<ремени. Виды, имеющие вогнутый или прямой свободный край
эаковины, ползают по дну или живут, зарывшись в ил.
Температура, по-видимому, не оказывает большого влияния
ia размещение остракод. Многие роды выдерживают иногда зна-
штельные скачки температуры (2—30° С). Это касается также и
олености. Определенные морские формы могут жить в водоемах
с соленостью менее чем 17%о. Верхней границей солености для мор-
ских форм считается 55%О.
Очень существенным фактором размещения остракод является
тип донного осадка.
Строение нанцнрей. Раковины остракод состоят из углекислого
кальция (СаСО3) — разновидности кальцита с небольшой при-
месью MgCO3. Кальцит может быть аморфным или кристалли-
ческим.
Форма панциря остракод очень разнообразна. Есть раковины
округлые, вздутые, уплощенные с боков, удлиненные и т. п. На
поверхности часто имеется разнообразная скульптура в виде мелких
сеток, зернистости, бугорков и ямок. Одним из определяющих
элементов скульптуры является медиальная борозда, расположен-
ная в центральной либо ближе к передней части раковины; она кос-
венно показывает место прикрепления замыкающих мускулов,
функция которых сводится к закрыванию раковины. Медиальная
борозда имеет также определенное значение для ориентации рако-
вины; это значит, что с ее помощью можно определить как переднюю
или заднюю часть раковины, так и правую или левую створку.
Панцирь остракод состоит из нескольких слоев, часто разно-
окрашенных. Линии, тянущиеся под прямым углом к поверхности
раковины (Leperditia), интерпретируются как призматические струк-
туры или как слоистое строение панциря. Относительно наиболее
ясную структуру раковины имеют Leperditia. Их крупные (30—40 мм)
раковины — хороший материал для исследования. На поперечном
разрезе (табл. 56, фиг. 1—3) в вентральной части панциря явно видны
линии, тянущиеся под прямым углом к поверхности. Проявляется
также определенная слоистость раковины, выражающаяся в том,
что «поперечные линии» не доходят до поверхности раковины
(табл. 56, фиг. 1). Упомянутые линии можно интерпретировать с
известной степенью допущения как поровые канальчики.
Петрографические признаки нрн определении остатков остракод1.
Остракоды в шлифе можно узнать по общей форме створок и по
внутренней структуре стенок панциря. Обычно остракоды представ-
лены в виде отдельных целых створок или двух створок, сочленен-
ных вместе, и реже — обломками раковин. Целый панцирь, состоя-
щий из двух одинаковых правильных створок, в шлифе бывает
эллиптическим, овальным (табл. 56, фиг. 4—7), миндалевидным,
субквадратным и т. п.
Внутренняя структура фоссилизованных панцирей остракод
чрезвычайно характерна оптически единообразной ориентировкой
кальцита. Эта ориентировка очень тонких, при малых увеличениях
невидимых фибр кальцита обнаруживается при скрещенных николях
благодаря затуханию, которое происходит перпендикулярно по-
верхности раковины. При вращении столика микроскопа полоса
затемнения пробегает вдоль створки, однако этот эффект хорошо
виден только у толстостворчатых раковин.
Встречается пять форм сохранности остракод.
1. Толстые створки сохранили свою первоначальную структуру.
В этом случае в них можно различить несколько слоев (два и более).
Отдельные слои имеют ту же структуру, которая в скрещенных ни-
колях дает затухание перпендикулярно поверхности. Эту форму
сохранности легко узнать в обломках.
2. Очень тонкие створки без видимой фибровой структуры, но
с ясным затуханием перпендикулярно поверхности.
3. Весь панцирь перекристаллизован, но кальцит, заполняющий
его внутреннюю полость, резко обособляется от створок.
4. Весь панцирь и полость, образованная им, замещены вторич-
ным кальцитом и представляют собой кальцитовую «миндалину»
без признаков створок. Кальцит ® призмах, ориентированных пер-
пендикулярно внешней поверхности миндалины, не относится к тон-
кой и нежной структуре створок остракод, а является вторичным.
1 Раздел написан В. П. Масловым.
80
Известковые организмы
5. Как и в предыдущем случае, от панциря осталась лишь запол-
ненная вторичным кальцитом миндалина, но крупнозернистый каль-
цит в ней расположен беспорядочно. При дальнейшей перекристал-
лизации все отдельные кристаллы сливаются в один.
Такие четко ограниченные микроскопические миндалевидные
линзы, если они встречаются группами, без сомнения относятся
к остракодам.
Так как остракоды при линьке сбрасывают старый панцирь
и заменяют его новым, обычно он состоит из одного или двух слоев.
Но существуют формы, сохраняющие старый панцирь и наращиваю-
щие новый панцирь с внутренней стороны старой створки. Полу-
чаются многослойные (многопанцирные) створки, у которых крае-
вые части утолщены на толщину края предыдущего, более старого
панциря (табл. 56, фиг. 8).
Трилобиты (Trilobita)
Табл. 57, 58
Трилобиты — исключительно морские членистоногие вымершие
животные, покрытые снаружи известковым панцирем. Предпола-
гают, что кроме известковых солей панцирь включал тонкие слои
хитина, исчезнувшего при фоссилизации. Некоторые авторы счи-
тают, что в состав панциря трилобитов наряду с кальцитом входит
фосфорнокислая известь (19—36%). Но другие авторы (Boggild, 1930)
считают эти данные о содержании фосфора преувеличенными, и,
по их мнению, панцири трилобитов состоят преимущественно из
кальцита.
Геологическое значение. Трилобиты часто являются важной
составной частью пород, иногда образуя трилобитовые известняки
(табл. 58. фиг. 2, 3. 5, 6). Трилобиты известны с кембрия до перми,
но уже с девона их значение сильно падает. Обычно их остатки
встречаются среди других типично морских организмов: иглокожих,
брахиопод, археоциат и т. п. Некоторые из трилобитов, по-видимому,
жили на значительных глубинах, другие — в мелком море. Некото-
рые трилобиты жили в илу. зарываясь в него и питаясь илом.
Для раннего палеозоя трилобиты имеют большое стратигра-
фическое значение.
Строение панцирей. Обломки панцирей трилобитов имеют в
шлифах разнообразную форму. Это могут быть длинные, по-раз-
ному изогнутые или волнистые ленты (табл. 57. фиг. 3—6; табл. 58.
фиг. 1, 4), фигуры, напоминающие петлю от крючка (табл. 57,
фиг. 9). толстые кольца, дуги и т. д. В простом свете обломки три-
лобитов светлые, почти всегда однородные. При хорошей сохран-
ности они выглядят шагреневыми, шероховатыми, как бы мелко-
губчатыми. В поляризованном свете при вращении столика микро-
скопа вдоль «ленты» панциря пробегает волна затухания. Это зату-
хание сходно с затуханием панцирей остракод и. возможно, обу-
словлено расположением тонких кристалликов (фибр?) кальцита
перпендикулярно поверхности панциря. От остракод панцирь трило-
битов отличается размерами, резкими изменениями толщины в од-
ном и том же сечении и сложными контурами сечений (табл. 58,
фиг. 6).
Не менее распространен другой тип сохранности, при котором
затухание волной наблюдается, но не ярко и не отчетливо. В про-
стом свете видно, что панцирь как бы рассечен трещинами, прохо-
дящими параллельно внешней поверхности и соединяющимися друг
с другом под острым или прямым углом. Ясно, что здесь приме-
шались вторичные процессы, изменившие микроструктуру. Такие
«растресканные» панцири, связанные с характерным внешним кон-
туром сечений, не оставляют сомнения в принадлежности облом-
ков к остаткам трилобитов.
Кайё описал несколько разновидностей тонкой микроструктуры
панцирей, но на имеющемся у нас материале удалось наблюдать
только две разновидности.
1. Панцирь однородный с каналами, пересекающими его пер-
пендикулярно поверхности. В тангенциальном разрезе каналы пред-
ставляют собой систему темных точек. Обломки кембрийских три-
лобитов такого строения можно смешать с обломками перфориро-
ванных брахиопод, от которых они отличаются другими контурами
сечений и затуханием в скрещенных николях.
2. Тонкослоистый панцирь со следами тонких каналов (?) в
виде темных линий, расположенных перпендикулярно слоям и по-
верхности панциря. Этот тип встречается редко, и. по-видимому,
ему отвечает тип микроструктуры, отмеченный Циттелем как сло-
женный десятком очень тонких слоев карбоната и фосфата извести,
пересеченных тонкими поровыми каналами. Обломки такой микро-
структуры мне приходилось встречать в кембрийских осадках Си-
бири. где фосфатные слои и каналы были заполнены низкополяри-
зующим материалом.
Многослойные панцири значительно толще, чем однослойные,
и также пронизаны порами. Но эти поры в разных слоях не всегда
совпадают и слои отделены друг от друга тонкими прослойками
некарбонатного вещества. Может быть, здесь мы сталкиваемся
с явлением, аналогичным смене панциря некоторых остракод.
когда линька при смене панциря трилобита происходит до пре-
дельного роста, после чего панцирь не сбрасывается, а нарастает
периодически изнутри. Новые слои наращиваются, будучи связан-
ными со старым слоем.
КОСТИ ПОЗВОНОЧНЫХ (VERTEBRATA)
Табл. 59, 60
Строение костных остатков! В состав костных элементов входят
четыре твердые ткани, хорошо сохраняющиеся в ископаемом со-
стоянии: кость, дентин, эмаль и аспидин. Эти ткани в различных
комбинациях образуют структуры скелетных частей позвоночных,
давая возможность специалисту в общих чертах определить систе-
матическое положение ископаемого животного даже тогда, когда
от его скелета сохранились только очень маленькие обломки.
Кость при исследовании ее в шлифе под микроскопом харак-
теризуется наличием в ней многослойных полостей. В этих поло-
стях при жизни животного находились живые костные клетки —
остеоциты (рис. 104). Полости остеоцитов очень разнообразны по
своей форме. Если кость имеет пластинчатое строение, то костные
клетки (остеоциты) оказываются сильно вытянутыми в плоскости
костных пластинок.
От полостей остеоцитов во все стороны отходят ветвящиеся
канальцы, концы которых соединяются с концами таких же каналь-
цев соседних полостей. У живого животного в этих канальцах на-
ходились тонкие протоплазменные отростки костных клеток.
Полости остеоцитов и отходящие от них канальцы в ископаемом
состоянии нередко бывают заполнены породой, окрашенной в более
или менее темный цвет. Это делает и полости костных клеток,
и все их канальцы очень хорошо видимыми. В некоторых случаях
и полости, и их отростки оказываются ничем не заполненными,
но и тогда структура кости может быть легко изучена, если само
костное вещество окрашено в коричневые тона. Такая окраска у ис-
копаемых костей наблюдается очень часто. Если костное вещество
совершенно не окрашено, а полости остеоцитов ничем не заполнены,
то изучение структуры такой кости бывает очень затруднительным.
Раздел написан В. П. Масловым по А. П. Быстрову. Взято нз книги «Условия образова-
ния. свойства и минералы осадочных пород», т. I (Л.. Изд-во АН СССР. 1958.
стр. 460—461).
Кости позвоночных
81
Дентином называется твердое вещество, из которого пост-
роены стенки зубов всех позвоночных, где бы эти зубы ни находи-
лись и какую бы форму ни имели.
В дентине никогда не встречается полостей клеток, но все вещест-
во дентина всегда бывает пронизано более или менее параллельными
канальцами, носящими название дентиновых. Эти канальцы по
ходу обычно разветвляются. Кроме того, в стороны от них отходят
многочисленные тонкие веточки (рис. 105). В дентиновых канальцах
Рис. 104. Вертикальный разрез кости Glvpiolepis sp.: х50: средний
девон
Буквы на рисунке: к. — кость; д. — дентин: э. — эмаль
Рис. 105. Вертикальный разрез панциря Psamniolepis parado.xu Agas-
siz; х50; средний девон
Буквы па рисунке: л. — дентин: э. — эмаль: а. — аспидин
у живых животных находятся тонкие протоплазменные отростки
особых клеток — одонтобластов, которые помещаются на стенке
зуба в его пульпарной полости. Микроскопическое строение дентина
настолько характерно, что распознавание его в шлифе не представ-
ляет никаких трудностей.
Наружная поверхность зубов позвоночных обычно покрыта
слоем эмали. Эмаль является самой твердой тканью позвоноч-
ных и состоит из многочисленных, плотно прижатых друг к другу
и немного изогнутых многогранных призм. В шлифах окаменев-
шая эмаль обычно кажется совершенно прозрачным белым вещест-
вом. Толщина эмалевого слоя на зубах бывает различной. В одних
случаях этот слой бывает очень толст, в других — настолько тонок,
что с трудом обнаруживается под микроскопом даже при больших
увеличениях.
Четвертым видом твердой ткани скелетных элементов ископае-
мых позвоночных является так называемый ас пид ин. По своему
строению аспидин отличается от кости тем. что в нем никогда не
встречается никаких полостей клеток, и от дентина тем. что в нем
не бывает канальцев, подобным дентинным. Но в твердом веществе
аспидина можно видеть беспорядочно разбросанные группы каналов.
У живого животного в этих каналах помещались колагенные (даю-
щие клей) волокна соединительной ткани. Такие каналы с находя-
щимися в них соединительнотканными волокнами иногда встречают-
ся в кости. Аспидин встречается только в панцирных элементах ордо-
викских. силурийских и девонских бесчелюстных позвоночных.
Дентинно-эмалевые образования, т. е. зубы, встречаются с на-
чала ордовика. Кость у позвоночных впервые появилась только
в конце силурийского периода.
Химический состав и оптические свойства1 (табл. 59. 60). У позво-
ночных животных в ископаемое состояние переходят только кост-
ные части скелетов, если не учитывать слепков и отпечатков мягких
частей организмов, которые встречаются в породах очень редко.
Породообразующее значение позвоночных гораздо меньшее, чем
беспозвоночных. В природе не существует пород, сложенных пол-
ностью или в значительной степени ископаемыми костями и их
обломками. Однако костные остатки как водных, так и сухопутных
позвоночных довольно часто встречаются в морских и континен-
тальных отложениях.
Для правильного понимания физико-химических свойств иско-
паемой костной ткани необходимо учитывать основные закономер-
ности процесса фоссилизации костей. Захороненная в породе кость
медленно утрачивает свой основной органический компонент —
коллаген. В костях среднеплиоценового и более древненго возраста
коллаген уже практически не содержится, и, следовательно, они мо-
гут считаться полностью фоссилизированными. Такие кости сохра-
няют первоначальную структуру и первичный неорганический ком-
понент. представленный скрытокристаллической разновидностью
карбонатапатита. близкого к франколиту, который в ходе фоссили-
зации лучше раскристаллизовывается. Удаленный из кости колла-
ген лишь в незначительной степени замещается вторичными, при-
несенными извне в ходе фоссилизации минеральными соединениями:
карбонатами, глиноземом, гидроокислами железа и т. д. Процентное
содержание этого вторичного вещества в нормально фоссилизи-
рованиых костях достигает обычно 2 и не превышает 5. Лишь
в минерализованных костях, т. е. костях, у которых полости гавер-
совой системы и поры губчатой ткани заполнены тем или иным
минералом, процентное содержание вторичных минеральных сое-
динений резко повышается. Однако минерализация ископаемых
костей довольно мало распространенное явление, так как оно про-
исходит в условиях перенасыщенности окружавшего кость раствора.
Нормально фоссилизированные и минерализованные кости легко
распознаются в шлифах под микроскопом. Гораздо труднее, а под-
час и совершенно невозможно распознать обломок кости, под-
вергшийся вместе с вмещающей породой процессам катагенеза
или метаморфизма, в результате чего первичная структура костной
ткани может полностью исчезнуть, а составляющий первичный
минеральный компонент кости частично или полностью замес-
титься другими минералами.
Как свидетельствуют многочисленные химические и спектраль-
ные анализы, процентное содержание СаО в ископаемых костях
по мере утраты ими коллагена увеличивается от 28 до 50%, P2OS —
от 20 до 36%. Содержание СО2 достигает нескольких процентов.
Остальные компоненты в нормально фоссилизированных костях
имеют подчиненное значение. Содержание SiO2, Fe2O3. А12<%,
MnO. MgO. Na2O. К2О. F. Cl. SO3 в различных костях без видимой
закономерности колеблется в пределах от сотых долей до 2%.
В некоторых костях присутствуют также Be, Ti, Ni, Sn, Y, Ce, Cr,
1 Раздел написан Г. А. Бачинским.
ног 6
82
Известковые организмы
Си. La. Pb, Zr. содержание которых колеблется от следов до сотых
долей процента.
Неорганический компонент как современных, так и фоссилизи-
рованных костей представлен скрытокристаллической разновид-
ностью карбонатапатита. содержащего гидроксильную группу и
фтор в различных пропорциях. В шлифах видно, что костная ткань
сложена однородным, белым или слегка желтоватым, скрытокри-
сталлическим мономинеральным агрегатом с показателями прелом-
ления четвертой группы (как и у апатита). Иногда он плеохроирует
от синевато-сиреневого или бурого до белого. В минерализованных
костях в шлифах легко определяются вторичные, привнесенные ми-
нералы (карбонаты, гидроокислы железа, гипс, и т. д.). заполняющие
полости гаверсовой системы. Минерализация гиДроокислами же-
леза иногда окрашивает всю костную ткань или ее отдельные участ-
ки в желтый цвет.
Показатели преломления костной ткани, определяющиеся при
помощи иммерсионных жидкостей у полностью фоссилизированных
костей среднеплиоценового и более древнего возраста, колеблются
в интервале 1,606—1,609 (показатели подолита колеблются в пре-
делах 1.598—1.634). В ожелезненных участках показатели прелом-
ления повышаются и могут достигать 1,619. С другой стороны,
содержание в костях коллагена понижает показатели преломления.
Данная закономерность довольно хорошо выдерживается: чем
кость геологически моложе, тем коллагена в ней больше и соответ-
ственно показатель преломления меньше. Для современных и голо-
ценовых костей он уже колеблется в пределах 1,560—1.575.
Удельный вес полностью фоссилизированных. а также минера-
лизованных костей, определяемый пикнометрическим способом,
колеблется в пределах 2,85—3,00 г/см3 и чаще всего равен 2.90,
т. е. соответствует удельному весу скрытокристаллической разно-
видности карбонатапатита (курскиту). Содержание в костях колла-
гена также понижает их удельный вес, который, таким образом,
является довольно тонким показателем геологического возраста
ископаемых костей. Удельный вес почти фоссилизированных костей
колеблется в пределах 2,85—2.70 г/с.н3, полуфоссилизированных —
2.70—2.40 и слабофоссилизированных — 2,40—2,00 г!см\
У нормально фоссилизированных костей полностью сохраняется
первичная структура костной ткани. В шлифах, изготовленных
из поперечных срезов пластинчатой плотной костной ткани, видны
остеоны, сложенные серией концентрических пластинок, располо-
женных вокруг круглого поперечного среза гаверсового канала
(табл. 59 и 60). В каждой кольцевой пластинке видны также концент-
рически расположенные многочисленные маленькие полости —
лакуны костных клеток (остеоцитов). соединенные между собой
системой Тончайших канальцев. В шлифах, изготовленных из про-
дольных срезов, видны вытянутые, слегка изогнутые линзовидные
костные пластинки с лакунами остеоцитов и продольные разрезы
гаверсовых каналов (табл. 59. фиг. 7). Вследствие этого строение
кости кажется волокнистым.
В поляризованном свете в шлифах из поперечных срезов остеоны
наблюдаются в виде чередования концентрически расположенных
светлых и темных колец различного диаметра, а из продольных сре-
зов — в виде темных и светлых вытянутых линз и полос. Это. оче-
видно. объясняется тем. что в свежих костях пучки коллагеновых
фибрилл в смежных пластинках располагаются в различных направ-
лениях взаимно под углом, близким к прямому. Соответственно
с этим кристаллики карбонатапатита в скрытокристаллическом агре-
гате, составляющем пластинки, в различных пластинках ориенти-
рованы по-разному: в одних пластинках — приблизительно парал-
лельно плоскости среза, в других — приблизительно перпендику-
лярно. В первом случае пластинки проявляют двупреломление,
во втором — изотропны.
Цвета интерференции довольно низкие (от серого к белому)
и соответствуют двупреломлению апатита. Погасание прямое.
Вследствие скрытокристаллического строения оно проявляется
в поперечных сечениях в виде темных неподвижных крестов, нало-
женных на «коломорфные» остеоны, а в продольных сечениях —
волнистое.
СПОРНЫЕ И ПРОБЛЕМАТИЧЕСКИЕ ИЗВЕСТКОВЫЕ
МИКРООРГАНИЗМЫ
Табл. 61. фиг. 5—9: табл. 62: табл. 63. фиг. 16—18:
табл. 76, фиг. 3, 4, 6
Очень часто при микроскопических исследованиях карбонатных
пород встречаются остатки породообразующих организмов, кото-
рые даже при детальном и тщательном изучении не дают возмож-
ности отнести их с уверенностью к той или иной группе организмов.
Там. где эти остатки с большей или меньшей долей вероятности
можно отнести к водорослям, они рассматривались в соответствую-
щем разделе. Здесь мы коснемся тех остатков, которые пока не
могут быть отнесены ни к животному, ни к растительному царству
или положение которых разными исследователями трактуется
по-разному.
Род Pseudovermiporelki — это иногда ветвящиеся, сильно изо-
гнутые трубки со стенками, состоящими или из сетки с массой тесно
расположенных каналов, или же стенка трубки внутри сплошная
и сетка располагается на наружной ее поверхности. Детальное изу-
чение материала хорошей сохранности из перми Армении показы-
вает. что мы имеем дело с различными стадиями обызвествления
наружного чехла организма. Различная прозрачность стенок трубок
показывает, что первоначальная стенка, пронизанная каналами,
во внутренней части переходит в сплошной неперфорированный
слой (рис. 106. а, б). Диаметр трубок неравномерный и неодинаковый
даже у одного и того же экземпляра (рис. 106. в; табл. 62. фиг. 6).
Изгибы трубок разнообразны, и встречаются спирально свер-
нутые экземпляры (табл. 62. фиг. 4). Часто к трубке с большим диа-
метром примыкает дочерняя трубка с малым диаметром, причем
у нее образуется лишь внешняя стенка, а стенка, примыкающая к
материнской, отсутствует (табл. 62, фиг. 1,6). Встречаются экземп-
ляры со слабым обызвествлением (рис. 106. г. д). При этом чехол
постоянно сходит на нет. Ветвление также незакономерно и под
разными углами. Внутри стенок иногда наблюдаются округлые
полости разного размера и загадочного характера (табл. 62. фиг. 3, 5).
Вся совокупность наблюдений не может свидетельствовать о при-
надлежности Pseudovermiporelki к мутовчатым водорослям. С боль-
шей вероятностью можно предположить, что это остатки животных.
Остатки этого организма легко окремневают (табл. 76, фиг. 3.
4, 6) и становятся прозрачными или оруденевают и окрашиваются
в желтый и бурый цвета. Распространен род Pseudovermiporelki
в перми п-ова Аравия, Югославии, Армении, Японии и Турции.
Организм, близкий к предыдущему, но без каналов в стенках,
был описан Флюгелем (Fliigel. 1964) под названием Microtuhus
с типом рода Microtuhus communis Fliig. из триаса Северных Альп.
Остатки организма представляют собой более или менее прямые
или слабо изгибающиеся одиночные трубочки с темными мало-
прозрачными стенками и более или менее постоянным диаметром
просвета. Аналогичные организмы были описаны как водоросли
из кембрия Сибири П. С. Краснопеевой (1955) под названием Tubo-
phyllum, а В. П. Масловым (19376)—под названием Epiphytonl
jacutii Masi, и под названием Palaeonites (1956). Флюгель, сравни-
вая Microtubus с известковыми чехлами разных организмов, прихо-
дит к выводу, что больше всего эти трубочки похожи на трубочки
червей. Возможно, что к ним же относятся трубочки, описанные
П. С. Краснопеевой (1937) под названием Proaulopora glabra Krasn.
из кембрия Западной Сибири. С. В. Максимова (1959) похожие ило-
Спорные и проблематические микроорганизмы
83
вые трубочки из карбона отнесла к домикам червей-полихет. с чем
можно согласиться, так же как с мнением Флюгеля (последний
сравнивал этот организм с серпулидами) (табл. 62. фиг. 10. 11).
Похожие трубочки более крупного размера (до 1 .и.и в диаметре)
были описаны Бэн (Bein. 1932) под родовым названием Tuhulites.
Стонели (Stoneley, 1958). описывая их в верхней перми Англии,
относит этот род к водорослям без особых на то оснований. Стенки
как пермских и каменноугольных, так и кембрийских форм этих
остатков организмов в шлифах очень похожи.
Среди юрских организмов выделяется один, описанный под
названием Coscinoconus Leupold. имеющий вид светлого извест-
кового конуса со спиральным желобом — каналом на поверхности
(табл. 62. фиг. 8. 9). Многими исследователями он относится к фо-
раминиферам. В. П. Маслов (1958) относит Coscinoconus к органам
размножения мутовчатых сифоней. так как известковое тело орга-
низма по структуре аналогично известковому чехлу этих водорослей.
Рис. 106. Pseudoverniiporella Elliott ( - Vermiporella niponica Endo)
из пермских отложений Армении
I/. <i - сечения перпендикулярно известковой стенке и оси трубки (видны поры, заполненные
во вторую стадию образования известковою чехла, и сферические полости разного раз-
мера); н — разломанная трубка без пор; г. Э — ветвление трубки
плоские тела (рис. 107) длиной в несколько миллиметров. Внутри
такой пластины находятся полости, располагающиеся горизонталь-
ными рядами (рис. 107, д). Иногда можно увидеть нечто вроде ка-
налов, связывающих полости между собой. В полостях часто наблю-
даются «колонки» или столбики, связывающие верхнюю и нижнюю
стенки полости. В поперечном сечении (рис. 107, ж) такие столбики
выглядят в виде белых точек на темном фоне полости (табл. 63,
фиг. 16. 17). В. П. Маслов считает, что контортопоридии близки
Рис. 107.Contortoporidium из карбона Донбасса
и — d — продольные сечения пластин при разных увеличениях: и. г - черное — отложения
скелета, в — виден продольный центральный канал, соединяющий полости, «’- полости
пересечения в разных направлениях; d. е — черное—полости, белое—отложения скелета:
е — плоскостное, ж — поперечное сечения пластин
Крупные формы Coscinoconus встречаются в грубодетритусовых
известняках, а мелкие — Coscinoconus minutus Masi., С. oblongus
Masi., C. pagodaeformis Masi.. C. Jusiformis Masi, обычны в пели-
томорфных или шламмовых известняках. Эти организмы легко
определяются в шлифах и встречаются в верхней юре Крыма, Кар-
пат. Югославии, Швейцарии. ГДР, ФРГ, Франции, Испании, Сирии.
В некоторых глубоководных фациях юры встречаются извест-
ковые прямые или слабоизогнутые, тонкие и узкие пластинки и
палочки, которые были описаны под родовым названием Lom-
bardia. Систематическое положение этих микрообломков не ясно:
их относят к пластинчатым формам бурых водорослей, но основа-
ний для этого слишком мало. Их примитивная структура при от-
сутствии анатомических признаков заставляет воздерживаться от
какого-нибудь определенного мнения (табл. 62. фиг. 12).
Остатки одного организма, довольно распространенного в кар-
боне Донбасса, названы В. П. Масловым Contortoporidium. но опи-
сание его, как и Cribroporidium (Cuneiphycus Johnson), не было опуб-
ликовано. Морфология этих двух организмов очень похожа. Соп-
tortoporidium образует неправильно разрастающиеся ветвящиеся
к крибропоридиям (Cuneiphycus Johnson, 1963) и относятся не к
водорослям, а к животным, имеющим признаки или губок, или
гидроидных полипов.
Род Distichoplax Pia широко распространен в нижнетретичных
отложениях (преимущественно в эоцене) Средиземноморья и тро-
пиков Азии. Известковые остатки дистихоплакса изучались только
в шлифах. Для его сечения характерна центральная темная полоса,
от которой отходят под прямым или косым углом симметричные
ячеи, образованные прямыми стенками или перегородками. Эти
ячеи открыты снаружи, но бывают прикрыты темной пленкой,
происхождение которой не ясно (табл. 61, фиг. 6). Альгологи от-
носят дистихоплакс к багряным водорослям, обладающим пластин-
чатой структурой, у которых неизвестны органы размножения
(табл. 61, фиг. 5—9).
Последние исследования Кейдж (Keij, 1964) и Денизо и Массиё
(Denisot. Massieux, 1965) доказывают принадлежность дистихоп-
лакса к группе мелобезий.
Этот род распространен в следующих регионах: палеоцен Испа-
нии, Пиренеев. Пакистана, Бангладеш, Чехословакии; палеоцен
84
Известковые организмы
эоцен Ирака и о-ва Мадагаскара; эоцен Чехословакии. Марокко.
Альп (австрийских и итальянских), Индии (Ассам); олигоцен Ал-
жира. Род представлен типовым видом Distichoplax biserialis
(Dietrich) Pia и сомнительным видом D. uniserialis Pfender.
КОНОДОНТЫ (CONODONTA) И
СКОЛЕКОДОНТЫ (SCOLECODONTA)
Табл. 63, фиг. 1 —15
Конодонты — это маленькие (от долей миллиметра до 2—3 лсм)
клыкоподобные или зубчатые ископаемые неизвестного происхож-
дения (рис. 107—109). По общему облику они напоминают сколе-
кодонты— так называются в палеонтологии челюсти многощетин-
ковых кольчатых червей. Но сколекодонты легко отличимы от
конодонтов главным образом по своему составу (см. ниже) и по
своему характерному черному цвету. Конодонты имеют окраску
в коричневых тонах; от светлых янтарных до более темных, иногда
с сероватым оттенком, часто они прозрачны. Конодонты быстро
разлагаются в азотной и серной кислотах (на сколекодонты эти
кислоты не действуют) и не подвергаются действию уксусной и
муравьиной кислот, при помощи которых их легко можно выделить
из известняков и других известковистых пород. Наиболее широко
распространенным методом выделения конодонтов является раст-
ворение названных пород в 10—25%-ной уксусной кислоте.
Химический состав. Химический состав конодонтов похож на
состав костей и зубов. Они состоят из фосфорнокислого кальция,
СО2 и органического вещества. Содержание фтора может быть
разное. Химико-минеральный состав сколекодонтов менее изучен.
По всей вероятности, в их состав кроме фосфорнокислого кальция
входит больше органического вещества.
Геологическое и стратиграфическое значение. Конодонты рас-
пространены от кембрия до мела включительно (Сергеева, 1962;
Lindstrom, 1964; Muller, 1960). В ордовике конодонты становятся
довольно обычными ископаемыми, встречаясь местами даже в
большом количестве. Вместе с простыми формами уже в начале
ордовика распространяются сложные конодонты.
Силур характеризуется сравнительно редкими конодонтами по
сравнению с предыдущим ордовикским и последующим девонским
периодами. Здесь встречаются известные уже в позднем ордовике
рогообразные простые формы и листообразные многозубчатые
сложные конодонты.
В девоне появляются новые формы и господство получают почти
исключительно плоские конодонты, очень многочисленные и раз-
нообразные. Развитие и образование новых видов конодонтов про-
исходит такими быстрыми темпами, что в некоторых местах (в част-
ности, в Северной Америке) они успешно используются в целях
детальной стратиграфии.
Конодонты карбона характеризуются в основном также плоскими
формами. Начиная с верхнего карбона развитие постепенно замед-
ляется, и конодонты в позднем палеозое становятся все более ред-
кими. До сих пор в перми известны немногочисленные конодонты,
очень близкие к карбоновым формам. Конодонты триаса очень
маленькие. Хотя здесь встречаются роды, известные в палеозое,
они не имеют типичных палеозойских представителей. В осадках
выше триаса мы ничего не знаем о конодонтах, за исключением
верхнего мела Камеруна, где найдены последние в геологической
истории конодонты.
Сколекодонты имеют более широкое стратиграфическое распро-
странение; они известны от кембрия до нынешнего времени. На
современном уровне знаний ни один из родов сколенодонтов не
характеризует определенный геологический период. Большинство
родов распространяется начиная с ордовика. Некоторую страти-
графическую ценность имеют сколекодонты в среднем палеозое.
Лнтофацналыюе распределение. Конодонты, как и сколекодонты,
встречаются в разных типах морских отложений. Как известно,
первые конодонты были найдены в глауконитовом песчанике (Pan-
der, 1856). Последующие исследователи обнаружили их в глинистых
сланцах, известняках, песчаниках, а также в кремнистых сланцах.
Попытки установить (статистически) частоту встречаемости коно-
донтов в разных литологических типах пород не дали удовлетвори-
тельных результатов. Многие авторы до сих пор считали характер-
ной для конодонтов фацию глинистых сланцев. По М. Линдстрему,
в материале из нижнего палеозоя Скандинавии более богатыми
являются карбонатные отложения. По материалам нижнего палео-
зоя Эстонии (результаты растворения) можно сказать, что коно-
донты больше связаны с глауконитовыми песчаниками и извест-
няками. В шлифах они встречаются больше в глауконитовом пес-
чанике и гораздо меньше — в известняках.
Одной из характерных особенностей для всех содержащих коно-
донты пород является присутствие соединений фосфора. В отло-
жениях разного возраста (в ордовике Скандинавии и Прибалтики,
в ордовике, девоне, перми США и т. д.) вместе с конодонтами часто
встречались фосфоритовые желваки или в самих породах отмеча-
лось повышенное содержание фосфора.
Отношение сколекодонтов к определенному литофациальному
типу пород пока не установлено. В Эстонии они были найдены как
в песчаниках, так и в карбонатных породах.
Морфология и внутреннее строение. Внешний вид конодонтов
бывает очень разнообразным. По морфологическим признакам они
разделяются на три большие группы: простые, сложные и плоские
(плитчатые). Конодонты первой группы — клыкообразные конусо-
видные формы и имеют лишь один главный зубец (или острие).
Сложные конодонты имеют кроме главного зубца еще 1 —4 отростка
(Ь разных плоскостях), которые могут быть или не быть зазубрены.
Плоские конодонты с широкими низкими краями на верхней по-
верхности могут иметь ряды маленьких зубчиков или бугорков.
Конодонты могут быть билатерально симметричными и несиммет-
ричными, причем последние бывают как левыми, так и правыми.
Общая терминология конодонтов приведена на рис. 108.
Каждый конодонт имеет центр роста, который окружен после-
довательно налегающими концентрическими ламеллами (lamella)
(рис. 109, а, г), крайняя из которых определяет внешнюю форму
конодонта. Рост происходит радиально от центра так, что каждая
более молодая ламелла окружает предыдущую, т. е. наружные слои
нарастают на внутоенние. Концентрические ламеллы прерваны в
базальной части конодонта, где образуется базальная полость.
Глубина и форма этой полости обусловливаются характером на-
слоения. Главный зубец и некоторые зубцы большинства конодонтов
содержат так называемое белое вещество (рис. 109, б — г). В то
время как базальная часть хорошо сохранившихся конодонтов имеет
светло-коричневую или желтую окраску и прозрачна, зубчатая часть
в отраженном свете является белой (или сероватой) и непрозрачной.
Брансоном и Мелом (Branson, Mehl, 1933, 1934) было установ-
лено существование волокнистых конодонтов в среднем ордовике
Северной Америки, которые не имеют никаких ламелл, а состоят
из волокон, расположенных параллельно длинной оси. Хорошо
сохранившиеся волокнистые конодонты имеют янтарную окраску
и прозрачны, не содержат участков белого вещества. Но многие
исследователи считают, что волокнистые конодонты являются ре-
зультатом вторичных изменений.
У некоторых конодонтов, особенно у простых форм, имеются
каналы в базальной части, большинство из которых соединяет
базальную полость с наружной поверхностью конодонта. Каналы
могут быть ветвистыми, но всегда имеют одинаковую толщину.
Базальная полость конодонта заполнена базальным наполнением
(каллусом — по Сергеевой, 1964), которое может и не сохраниться.
Рост наполнения происходил, по-видимому, путем отложения но-
вых пластин на нижележащие. Направление роста было вниз.
Конодонты и сколекодонты
85
В шлифах конодонты могут выглядеть по-разному в зависимости
от места сечения (табл. 63, фиг. 3—9). Чаще в шлифах пород встре-
чаются базальные части, главные зубцы, главные лопасти, плитки.
Более хрупкие части, например мелкие зубцы, в шлифах встречаются
реже. Цвет конодонтов в шлифах коричневатый, желтоватый, иногда
они бесцветны. Цвет сколекодонтов темно-коричневый. У коно-
донтов довольно часто отмечается более темный внешний покров,
состав которого пока не ясен.
Внутренняя структура самого конодонта зависит в первую оче-
редь от его сохранности, а также от его сечения. В хорошо сохра-
нившихся конодонтах отмечается ламелларная внутренняя струк-
тура. Как показали более детальные микроскопические и рентгено-
метрические исследования (Lindstrom, 1964), апатитовый материал
в конодонтах является микрокристаллическим (диаметр кристаллов
меньше 0,01 мм). Кристаллы имеют удлиненную форму, параллель-
ны главному направлению роста и располагаются поперек ламелл.
I
Рис. 108. Морфологические признаки конодонтов
а, б — простые конодонты: в — е — сложные конодонты; ж — к — плоские конодонты
(все виды сбоку, кроме з — к, на которых з — вид сбоку, и — вид сверху, к — внд снизу):
л—схема роста конодонта, по Гроссу (Gross, 1957). Буквы на рисунке: с. — главный зу-
бец; d. — зу цы; Ь.—базальная часть: bf. — базальное наполнение; Ьс. — базальная по-
лость; рг. — отросток; ррг. — задний отросток; арг. — передний отросток; 1рг. — боковой
отросток; Ы. — лист; pl. — плитка: са. — карина; ibc. — перевернутая полость; I. — ламел-
лы роста; bt. — базальная воронка; tf. — накопление воронки
Рис. 109. Вертикальные разрезы конодонтов
а -ламеллярная структура конодонта с пустым пространством вдоль осей роста, х 97
(Lindstrdm. 1964); 6 —в главном зубце ось роста с пустыми пространствами и лучи белого
вещества, х134 (Lindstrom. 1964); в — белое вещество в главном зубце, х 57 (Lindstrom.
1964); е — конодонт с базальным наполнением (bf.) (Gross. 1957); d — плоский конодонт
с базальным наполнением, где видны ламеллы накопления (Ы ), х 84 (Gross, I960).
Линиями показаны видимые границы ламелл (1.). межламеллярные пространства —
темные, белое вещество обозначено точками
Такая структура хорошо выявляется при скрещенных николях
под микроскопом (табл. 63, фиг. 13). Некоторые части в конодонтах,
например в районе оси роста, могут оказаться под микроскопом
почти аморфными. Тело конодонта имеет более высокий показатель
преломления по сравнению с канадским бальзамом. В известняках
и доломитах разница между показателями преломления относи-
тельно небольшая. В порах и каналах конодонтов, особенно в формах,
встречаемых в глинистых сланцах, отмечаются кристаллики марка-
зита и пирита. Конодонты, подвергшиеся вторичным изменениям,
могут постепенно потерять свою ламеллярную внутреннюю струк-
туру.
В течение последних 30 лет найдены так называемые естественные
скопления конодонтов (рис. ПО, г), которые можно рассматривать
как остатки организма, носящего конодонты-элементы. Такие
cvieci венные, ненарушенные скопления состоят из 10—20 экземпля-
ров конодонтов, которые представлены четырьмя-пятью различ-
ными морфологическими типами конодонтов, раньше считавши-
мися неродственными, но теперь оказавшимися элементами одного
животного.
Челюсти ископаемых аннелид — сколекодонты разбросаны в
отложениях беспорядочно и имеют вид маленьких гребешков.
86
Ил.сстковые организмы
Рис. ПО. Типы конодонтов
а -г- простые конодонты: а. в- Stolo-odus. о—Drcpanodus. г - Oistodua: <) -ж - простые
1ппы сложных конодонтов: г), е — Fatodus. ж Selodina 1 — <> -- сложные н примитивные
плоские коиодоиты: з— Paracordylodus. и Roundya. к Cordylodus. .1 — Haddingodus.
и. а - Antbalodus. 11 — Polyplacogmitus: л. р — листообразные и плоские конодошы: п Spa-
thognathodus. р — Polygnathus: с — естественное скопление конодонтов (Rhodes. 1952):
т— ф—верхние челюсти сколекодонтов (Silvester. 1959)
ХИОЛИТЫ (HYOLITHA)
Табл. 64
Хиолиты впервые описаны Э. И. Эйхвальдом в 1840 г. из ордо-
вика Эстонии. Выделенный род Hyolithes этот автор отнес к пробле-
матичным ископаемым (Eichwald, 1840). И на сегодня систематиче-
ское положение хиолитов остается не вполне ясным, хотя большин-
ство исследователей склонны считать их особым классом в типе
Mollusca (Розанов, Миссаржевский, 1969). Нет также общеприня-
той систематики для этой группы окаменелостей. На рис. 111 при-
водится схема одной из предложенных в последнее время систематик
(Розанов, Миссаржевский. 1969). В ее основу положен принцип раз-
деления форм по степени и характеру дифференциации раковины
на брюшную и спинную стороны.
Химический и минеральный состав. Скелетные части хиолитов
состоят исключительно из углекислого кальция. Как правило,
раковина перекристаллизована и слагается изометричными. разно-
образно ориентированными кристаллами кальцита (B^ggild, 1930).
Редко сохраняются относительно слабо измененные участки стенок
раковин, строение которых в этом случае напоминает строение приз-
матического слоя раковин моллюсков (табл. 64, фиг. 1, 12,в).
Геологическое значение. Хиолиты встречаются в осадочных по-
родах исключительно морского происхождения (аргиллиты, пес-
чанники, известняки), а также во всех промежуточных разностях
этих пород.
В кембрийских отложениях Сибири хиолиты особенно часто
встречаются в пестроцветных глинисто-карбонатных породах плат-
формы. реже — в чистых разностях известняков, водорослевых и
археоциато-водорослевых известняках. В области геосинклинали,
где нижнекембрийские отложения представлены в основном мощ-
ными толщами известняков (обычно археоциато-водорослевые
известняки), хиолиты очень редки, бедны в видовом отношении и.
как правило, приурочены к прослоям, обогащенным глинисто-пес-
чанистым материалом. В нижних горизонтах кембрия Сибирской
платформы хиолиты обычно встречаются в массовом количестве
и ассоциируются с остатками археоциат, трилобитов, гастропод
и брахиопод, спикулами губок, трубками червей (которые часто
путают с раковинами хиолитов из семейства Circothecidae) и различ-
ными известковыми водорослями: Epiphyton. Girvanella. Renalcis.
Proaulopora. Razuniovskia и др. Иногда отдельные прослои, особенно
в самых нижних горизонтах нижнего кембрия, почти нацело состоят
из обломков и целых раковин хиолитов. Чаще же они образую!
локальные скопления в виде линз или выполняют карманы на по-
верхности отдельных пластов. Одной из интересных особенностей
нахождения раковин хиолитов в породе является то. что они бывают
забиты друг в друга до пяти и даже более раковин. Это происходит,
вероятно, в процессе перемещения пустых раковин течениями
(табл. 64, фиг. 10. 12, б). Подобные образования в шлифе можно
принять за многослойную раковину какого-либо животного.
Хиолиты существовали от кембрия до триаса. Согласно подсче-
там Г. В. Синклера (Sinclair. 1946), больше всего видов описано
из кембрия — 179. затем из ордовика — 111, силура — 29. девона —
51, карбона — 7, перми — 3, триаса — 1. В настоящее время коли-
чество видов достигло 500—550. главным образом за счет ранне-
кембрийских форм.
В стратиграфии ордовикских и более молодых отложений хио-
литы практически не используются. Пока мало они используются
и для расчленения кембрийских отложений. Это происходит потому,
что кембрийские хиолиты еще очень слабо изучены. Но можно ду-
мать, что роль их в стратиграфии кембрия, особенно его нижних
частей, и в установлении нижней границы кембрия будет достаточно
велика (Розанов, Миссаржевский, 1969).
Строение раковины. Остатками наружного скелета хиолитов
являются удлиненно-пирамидальные или пирамидальные раковины
(от 1—2 мм до 17—20 см длиной), часто с перегородками (сплош-
ными) в ее начальной части и крышечками, обычно попадающимися
отдельно от раковин. Некоторые исследователи относят к скелету
и тонкие изогнутые пластины—так называемые парные придатки,
находимые иногда вблизи устья раковины (Marek, 1963). Раковина
(и крышечка) билатерально симметричная, дифференцированная
на брюшную и спинную стороны. За брюшную (чисто условно) при-
нимают уплощенную, слабовыгнутую или вогнутую сторону (обычно
раковина захороняется лежа на этой стороне). Спинная часть резко
выгнутая, аркообразная и т. д. (рис. 112). Кроме того, у большинства
видов скульптура наружной поверхности раковины резко отлична
от скульптуры спинной и брюшной сторон. Характер строения
сторон очень хорошо передается на поперечных сечениях раковины,
которые исключительно разнообразны — особенно в ордовике —
силуре (рис. 113). Кроме того, у ряда форм брюшная сторона длин-
нее спинной и выступает в виде так называемой губы (рис. 112).
В случае отсутствия последней устье бывает перпендикулярно пло-
скости симметрии раковины или чуть скошено. Существуют также
формы, имеющие раковину, слабо дифференцированную на спин-
ную и брюшную части (Circothecidae). Это трубчато-пирамидальные
образования с округлым, эллиптическим, округленно-треугольным,
четырехугольным (?) сечениями. Скульптура одинакова на всех
сторонах раковины.
Хиолиты. Тентакулиты
87
класс Hyolitha
<-------------------------------
off? Orthothecida otr Hyolithida
Рис. 111. Схема классификации хиолитов
Приводятся схематизированные изображения раковин хиолитов по семействам
/
Сведения о микроструктуре раковин хиолитид в литературе
практически отсутствуют. В. А. Сысоев (1960) приводит фотографии
участка раковины Costatheca clinisepta (Syss.) с сохранившейся,
по его мнению, первичной структурой, со строением, сходным со
строением призматического слоя раковин моллюсков. Аналогичное
строение раковины у хиолитид наблюдалось и автором (табл. 64,
фиг. 1).
Рис. 112. Основные элементы морфологии раковины хиолитов (на
. примере семейства Sulcavitidae)
а — губа: б — брюшная часть раковины; в — спинная часть раковины: г — устье: с* —
перегородки: е — «мускульные впечатления»
Некоторые исследователи отмечали, что при механическом воз-
действии на раковину ее стенки расщепляются на несколько слоев.
Нами приводится фотография косопоперечного сечения экземпляра,
относящегося к семейству Sulcavitidae (табл. 64. рис. 4), у которого
видно расслаивание раковины на несколько слоев.
Большинство хиолитов довольно легко отличается от других
окаменелостей по поперечным или слабо скошенным сечениям ра-
ковин. которые нередко встречаются в шлифах. Все основные типы
поперечных сечений приведены на рис. 113 и табл. 64. Следует
отметить, что большинство видов хиолитов имеет довольно круп-
ные раковины, поперечные сечения которых превышают размеры
петрографического шлифа. Поэтому уверенная диагностика хиоли-
тов возможна лишь в палеонтологических шлифах. В то же время
Circothecidae имеют сечения, сходные с сечениями трубок червей,
спикул ханцеллорий и с различными тубулярными проблематич-
ными образованиями типа Coleoulus, Cambrotubulus, Hyolithellus
и др. Последние очень часто встречаются совместно с хиолитидами,
особенно в древнейших горизонтах кембрия. От хиолитов их можно
отличить по слоистому строению стенки раковины (табл. 64, фиг. 17).
Спикулы Chancelloria отличаются от раковин хиолитид значительно
более тонкими стенками скелета, не имеющими призматического
строения, которое наблюдается у ряда форм семейства Circothecidae
(табл. 64, фиг. 3. б, фиг. 12, в).
Рис. 113. Контуры наиболее характерных поперечных сечений ра-
ковин хиолитов
и — крышечка хиолитов семейства ADathecidae
ТЕНТАКУЛИТЫ (TENTACULITOIDEA)
Табл. 65. фиг. 14—17
Тентакулиты — это вымершие животные, обладавшие узко-
конической прямой или слабоизогнутой известковой раковиной
с округлым сечением, заканчивающейся на остром конце вздутой,
заостренной или каплевидной начальной камерой. Суженная часть
88
Известковые организмы
раковины обычно разделена поперечными перегородками иа ряд
изолированных камер. Раковины тентакулитов имеют внешнее
сходство с хиолитами, а структура стенок и поперечное сечение
раковин у них иные (см. раздел «Хиолиты»).
Систематическое положение тентакулитов до сих пор остается
не ясным.
Химический состав. Химический состав раковин тентакулитов
не изучен. В основном они состоят, по-видимому, из кальцита.
Экология. Тентакулиты встречаются вместе с морскими живот-
ными в различных фациальных условиях. Но необходимо заметить,
что раковины с замкнутыми камерами на конце легко могли пере-
носиться током воды, почему и встречаются танатоценозы с рако-
винами. ориентированными в одном направлении. Такие скопления
раковин Г. П. Ляшенко отмечает для силура и девона. Для дома-
никовых фаций, которые отлагались в спокойной воде, характерно
беспорядочное расположение раковин тентакулитов.
Приуроченность тентакулитов к различным фациям привела
исследователей к мысли о их планктонном образе жизни, что остает-
ся пока лишь предположением.
Геологическое значение. Тентакулиты широко распространены
в осадках силура и девона. Имеются редкие находки в ордовике.
Микроскопические исследования пород карбона показали, что мел-
кие раковины тентакулитов. с характерной только для них структу-
рой, встречаются и в известняках этого возраста. Г. П. Ляшенко
(1959) считает, что тентакулиты имеют большое стратиграфическое
значение для отложений силурийского и девонского возраста.
В девоне (например в доманике) раковинки тентакулитов нередко
слагают целые пласты известняка (так называемый птероподовый
или кониконховый известняк).
Строение раковин. Наружные макроскопические особенности
раковины тентакулитов позволяют выделить среди них ряд семейств,
родов и видов. Эти особенности заключаются в наружной скульп-
туре, выраженной в чередовании поперечных кольцевых ребер или
валиков и борозд между ними. Кольца (по терминологии Г. П. Ля-
шенко) бывают острыми, затупленными, асимметричными, вали-
кообразными и т. п. Имеют значение также форма начальной «эмб-
риональной» камеры, характер начальных перегородок, внутренняя
поверхность и т. д. При рассмотрении тентакулитов в сечениях в
шлифе играет роль микроскопическое строение стенок. Стенки в осе-
вом сечении раковины дают несколько характерных микроструктур.
Гладкие однослойные стенки без наружного орнамента характерны
для стилиолин (род Styliolina Karpinsky): однослойные волнистые
стенки — для вириателл (род Иriatella Ljaschenko). Многослойные
стенки относятся к ряду родов и обладают разнообразной микро-
структурой (рис. 114, а—е). Как указывалось выше, кольцевые ребра
бывают разной внешней формы, но и внутренняя микроструктура
их сильно варьирует. Стенки многослойных тентакулитов могут быть
разделены на ряд групп: стенки без каналов (рис. 114, а. б. з) и
стенки с каналами (рис. 114, в, д, е). И те и другие могут обладать
кольцевыми ребрами без темных дисков и кольцевыми ребрами
с «темными дисками» (рис. 114, д). Я называю «темными дисками»
Рис. 114. Схематические изображения сечений через кольцевые ребра
и стенки раковин тентакулитов
и — г.е — осевые сечения раковин:а. 6, г — стенки без каналов, в. е — стенки с каналами
(к.); — стенка с каналами и темным диском (т. д.); ж — тангенциальное сечение раковины
(полоски — т. д. — темные диски, точки — поперечные сечення каналов): з — косое про-
дольное сечение раковины у суженного конца с внутренними поперечными перегородками
затемненную середину кольцевого ребра, которая образуется в
результате слияния в середине ребра утолщений темных слоев, обо-
гащенных органическим веществом (рис. 114, г. д). Такие слои в
многослойной стенке тентакулита обычно чередуются с прозрачны-
ми кальцитовыми слоями. Темные слои очень неоднородной тол-
щины, легко раздуваются и выклиниваются, исчезая совсем.
Каналы в осевом сечении в одной и той же стенке бывают пря-
мыми или изогнутыми, в зависимости от внешней поверхности,
к которой они всегда расположены перпендикулярно. Каналы ни-
когда не открываются во внутреннюю полость и прикрыты изнутри
первым от оси слоем стенки (рис. 114, в, г). Комбинация всех этих
микроэлементов создает ряд типов структур, схематически изобра-
женных на рис. 114.
Лнтолог обычно имеет дело с неориентированными сечениями
организмов. Несмотря на это, тентакулиты с кольцевыми ребрами
легко узнаются в шлифе: в косом сечении большинство тентаку-
литов имеет зубчатые или извилистые (по краю) стенки, на конце
же раковины благодаря скошенному сечению кольцевые ребра
образуют острые или тупые линзы (рис. 114. а: табл. 65. фиг. 14. 15).
В тангенциальном срезе темные диски имеют вид темных попереч-
ных полос с расширениями на краях. Каналы в поперечном сечении
образуют темные точки (рис. 114. ж; табл. 65. фиг. 14). Обычно
каналы часты в промежутках между большими кольцевыми
ребрами.
Часть II КРЕМНЕВЫЕ ОРГАНИЗМЫ
Общие замечания
Некоторые низшие растительные и животные организмы ис-
пользуют водный кремнезем для построения твердого панциря или
опорного скелета. Кремневая функция, как назвал А. П. Виноградов
способность организмов отлагать соли кремнезема при жизни,
не развилась далее кремневых губок у животных и злаков у растений.
Перечислим группы кремневых организмов: водоросли диатомовые,
силикофлягелляты, перидинеи (частично), синезеленые водоросли
(частично): животные: радиолярии, кремневые губки.
Скелет современных кремневых организмов обычно состоит из
опала и гидрата окиси кремнезема с различным содержанием воды.
Минералогически состав скелета отвечает опалу. В ископаемом со-
стоянии часто происходит замещение кремневых выделений орга-
низмов другими минералами.
Остатки организмов с кремневым скелетом играют важную роль
как породообразователи. Кремневые организмы в ископаемом виде
встречаются в разных породах. Спикулы губок и панцири радиоля-
рий известны с докембрия и изобилуют в палеозое. Диатомовые
водоросли известны как редкое исключение в юрских осадках и
лишь в меловых отложениях начинают играть некоторую роль.
В третичное время диатомовые приобретают важное породообра-
зующее значение.
ДИАТОМОВЫЕ ВОДОРОСЛИ (BACILLARIOPHYTA)
Диатомовые водоросли — микроскопические одноклеточные рас-
тения. одиночные или колониальные, величиной от 0.005 до 0.6 мм.
изредка — более крупные. Среди них имеются планктонные формы,
донные и прикрепленные к субстрату. Протопласт клетки покрыт
пектиновой оболочкой, которую тесно облекает снаружи кремне-
вый панцирь, состоящий из двух отдельных створок, пронизанных
порами. Диатомеи распространены повсеместно не только в водной
среде, но и в почвах, на поверхности снежного и ледового покрова,
на скалах, в верховых торфянниках.
Химический и минеральный состав. Панцирь диатомей образован
опалом (SiO2H2O). Удельный вес кремнезема панциря 2,07, пока-
затель преломления 1.43. За время длительного захоронения опал
постепенно превращается в халцедон, а затем — в кварц. В меловых
и палеогеновых диатомитах кремнезем нередко замещается пири-
том.
Известно, что современные формы растворяются быстрее, чем
ископаемые. Причем среди современных форм существуют много-
численные градации по степени сохраняемости. До 50% известных
ныне видов вообще не сохраняется в ископаемом состоянии. Их
растворение происходит, как только клетка начинает отмирать.
Другая часть диатомей отличается хорошей сохраняемостью. Ско-
рость растворения первой группы форм сходна со скоростью раст-
ворения аморфного кремнезема, вторая группа более трудно раст-
ворима (Жузе, 1966).
Содержание кремнезема в панцирях различных видов диатомей
сильно колеблется. В тонкоокремненных стенках панциря коли-
чество кремнезема не превышает 10% (Jorgensen, 1955). В стенках
панциря таких видов, как Coscinodiscus. содержание кремнезема
колеблется от 40 до 60%. а иногда до 75%. От толщины стенок
панциря в первую очередь зависит сохраняемость диатомей.
Экология. По отношению к солености диатомеи делятся на три
основные группы: морские, пресноводные и солоноватоводные.
Они являются прекрасными индикаторами солевых условий место-
обитания, нередко более тонкими, чем химические, так как видовой
состав диатомей коренным образом изменяется в условиях раз-
личной солености. Однако наряду со стеногалинными формами
имеются многочисленные диатомеи, способные существовать при
широких колебаниях солености.
Географическая зональность ярко выражена в распределении
диатомей в морях и океанах там, где солевые условия более или
менее постоянны. В континентальных водоемах зональность в рас-
пределении диатомей маскируется многими другими факторами
среды.
Следует сказать, что представление о холодолюбивой природе
всех вообще диатомовых водорослей возникло в результате того,
что массовое их развитие приурочено к северным и южным окраинам
обоих полушарий. Объясняется этот факт, однако, прежде всего
тем. что в субарктических и бореальных областях и в приантаркти-
ческих районах Мирового океана поверхностные воды особенно
богаты минеральными солями (фосфаты, кремнезем, железо, нит-
раты и разнообразные микроэлементы), которые идут на построение
клеток диатомей. Поэтому так бедны диатомеями тропические райо-
ны всех океанов, поверхностные воды которых не могут обеспечить
богатое развитие фитопланктона. С другой стороны, экваториальная
зона, где к поверхности океана поднимаются глубиные воды, бо-
гатые питательными веществами, характеризуется высоким раз-
витием диатомей и всех других планктонных организмов.
Геологическое значение. Диатомовым водорослям принадлежит
важная роль в процессе накопления кремнистых осадков. О г ран-
диозности этого процесса можно судить по диатомовым илам,
образующимся в современных водоемах, и по диатомитам мелового,
третичного и четвертичного возраста, широко распространенным
по всему земному шару. Диатомовые илы образуются в настоящее
время как в пресных, так и в морских водоемах. В морях в образо-
вании кремнистых осадков принимают участие, кроме того, радио-
лярии и силикофлягеллаты. но в континентальных водах эта роль
принадлежит только диатомеям. Для третичных и четвертичных
отложений диатомеи имеют также большое стратиграфическое зна-
чение (Жузе, 1968: Жузе. Козлова. Мухина, 1969).
Многочисленные указания на то, что в палеозое существовали
диатомеи, не подтвердились на фактическом материале. Было много
и просто ошибок, когда третичные и даже современные виды, по-
павшие в палеозойские породы в процессе заноса водами, рассматри-
вались в первичном залегании. О позднем возникновении диатомей
говорят очень скудные находки их остатков в юрских отложениях и
довольно редкие находки в нижнемеловых породах. С поздним появ-
лением этого типа водорослей хорошо согласуются и такие факты,
90
Кремневые организмы
как их бурный расцвет в настоящее время, геологически недавнее
появление новых таксономических единиц ранга класса (эоцен —
олигоцен) и быстрое выделение в пределах этого класса последо-
вательных крупных подразделений.
Диатомеи четвертичного возраста в большинстве своем сохра-
нились до настоящею времени, гак что для четвертичных бассейнов
есть все основания ожидать близкою к современному распределения
этих водорослей по климатическим зонам. В этом заключается воз-
можность использования диатомей при выяснении палеогеографи-
ческой обстановки прошлого.
Морфология и структура панциря. Панцирь диатомей представляет
из себя коробку, состоящую из двух створок, которые вдвинуты
краями одна в другую. Наружная, большая, створка (эпитека)
соответствует крышке, а внутренняя, меньшая, створка (гипотека) —
коробке. Таким образом, эпитека надвинута на гипотеку, как крышка
на коробку. Створки бывают дисковидные, чашевидные, цилиндри-
ческие. линейные и т. д. (рис. 115—118).
’«СоО».
ю J с о о
Рис. 115.
Комплекс
. по»
Melosira. образующий
озерные диатомиты
а--г— Melosira praebaicalensis Kutz. f. ntiocenica Jouse. x 540. нижний миоцен. Примор-
ский край: а — М. italica Kutz. Г. niioeenica Jouse: е. ж — М. antiqua Mois. миоцен
Наиболее сложным морфологическим элементом панциря являет-
ся его структура, т. е. тонкий рисунок, образованный порами и аре-
олами. пронизывающими его стенку. Структура панциря служит
наиболее точным диагностическим признаком при определении
видов. Многие диатомеи, имея панцирь одинаковой формы, отли-
чаются лишь по его структуре. Поры на створках диатомей являются
обычно простыми отверстиями, большей частью круглыми, иногда
продолговатыми. Ареолы (loculi) — пяти-шестисторонние полые
камеры в стенке панциря. Обычно ареолы построены таким образом,
что боковые стенки их гладкие, непористые, внутренняя стенка,
обращенная в полость клетки, и наружная стенка снабжены порами.
К элементам структуры относятся также различные выросты на
створке: «глазки», «выпуклости», «рога», «ножки», сосковидные
выросты, шипы и т.п. (рис. 116).
У многих диатомей на створке панциря имеется шов. т. е. узкая
и глубокая щель, пронизывающая стенку створки. Строение и раз-
меры шва очень разнообразны. Он служит для сообщения живой
клетки с внешней средой и. кроме того, является органом движения.
По морфологическим и структурным особенностям панциря диа-
томовые водоросли делятся на два класса: Centricae (центрические)
и Pennatae (пеннатные). Centricae — древняя группа, известная с
юры доныне, характеризуется преимущественно круглыми, оваль-
ными и треугольными створками, на которых структурные элементы
располагаются радиально, концентрически или беспорядочно
(рис. 116). Шов у центрических диатомей отсутствует. Класс Penna-
tae достоверно известен с среднего эоцена до наших дней, его рас-
Рис. 116. Различные представители
центрических форм
а — Actinoptychus splendcns Ralfs, х 1000. сармат. Молдавия: б - Asteroniphalus robustus
Castr.. х 600. плиоцен. Сахалин: в — Aslerolampra inarylandica Ehrenb.. х 600. миоцен.
Сахалин, г - Auliscus schliptus Ralfs, х 600. четвертичные. Японское море: <* Corona
magnijlt a Let et Chen, x 300. верхний эоцен. Поволжье: e - - Monopsis inarginata Chen, x 300.
олигоцен. Украина: ж - Aulacodiscus septus A. S.. x 333, нижний сенон. Свердловская об-
ласть
цвет начинается с миоцена. Эта группа характеризуется большей
частью удлиненными створками самой разнообразной формы:
ланцентной. линейной. S-образной и т. д. (рис. 117). Структурные
элементы располагаются на створке перисто, по обе стороны глад-
кого осевого поля. У всех пеннатных диатомей, кроме самых при-
митивных и наиболее древних форм, имеется шов (рис. 118).
luamo.новые водорос.ш
91
Рис. 117. Бесшовные Araphiiialex и примитивношовные (Raphi-
dioineae)
а. б — Diatoma hiemale Heib.. х 1000. четвертичные. Кольский полуостров: а — колония.
б — створка: в. г — Plagiogramnia staurophorum Heib., х 1000. четвертичные. Баренцево
море: в — створка, г — внл с пояска: б. е — Plagiogramnia pulchelluni Grev.. х 1000. чет-
вертичные. Дальний Восток: б— створка. <• - вил с пояска: ж. з—Dimeropgramma minor
Ralfs, х 1000. верхний плиоцен. Молдавия: ж — створка, з— вид с пояска: и — Оре-
phora martyi var. amphio.xys Poret/ky. x 1500. плиоцен. Грузия: к. .1— Fragilaria bicapiiata
A. Mayer, x 1000, верхний миоиен. Западная Сибирь, р. Тым: .и — Ceratoneis arcus Kutz,
х 1000. четвертичные. Северная Европа; н — Synedra pulchella Kutz, х 500. литорниовые
отложения. Латвия; о, п — Thalassionema mitzshioides Grun.. х 1000. современная, Берин-
гово море: о— вид с пояска, л — створка: р. с—Thalassionema nitsschioides Grun. f. cur-
vata Jouse. x 1000. инжиин миоцен. Таманский полуостров: т. г - Thalassiolhrix longissinia
Cl. et Grun.. x 1000. четвертичные. Охотское море: г — часть створки: ф — Eunolia ра-
pilio Hust.. х 1500. четвертичные. Гренландия: х — Eunolia sparsimpunctaia Jouse. х 1000.
верхний миоцен, оз. Ханка: д — Eunolia nikolskiae Jouse. х 1000. там же
Многие диатомеи образуют колонии, которые возникают в про-
цессе последовательного деления одной клетки. Колонии образуют-
ся у тех диатомей, которые не способны двигаться. Таким образом,
колониальные формы наиболее часты у центрических диатомей,
а также у диатомей без шва.
Изучение диатомей производится под микроскопом при больших
увеличениях в специальных препаратах, предварительно отмучен-
ных и обработанных химически (удаление карбонатов, обугливание
органического остатка и т. д.). Для изучения тонких структур пан-
циря применяется электронный микроскоп.
92
Кремневые организмы
Рис. 118. Шовные диатомовые (Raphinales)
а — Amphipleura pellucida Kutz., х 1400, неоген. Приморский край: б — Frustulia rhom-
boides de Toni, x 1000, четвертичные. Кольский полуостров, в. г — Diatomella balfouriana
Greville, x 1000, четвертичные: в—створка, г — вид с пояска: д — Mastogloia smith'd van
lacustris Grun., x 1000, четвертичные.Чекалии: e— Diplonois didyma Cl., x 1000. четвертич-
ные, Ленинградская обл.: >к Amxiieaiieis serians Cl., x 1500. четвертичные. Кольский
полуостров: з— Stauroneis acuta W. Smith., x 500, четвертичные. Ярославская обл.; и —
Navicula digitoradiata A. Smith., x 500. четвертичные, Кольский полуостров; к — N.amphi-
bola Cl.. xl500, современные, Гренландия: л — N. lundstromii Cl., x 1500, современные.
Гренландия; .и — Pinnularia meisleri f. armenica Poretzky. плиоцен, Армения; н — Comp-
honemia lingulatum Husted, x 1000. плиоцен, Приморский край
КРЕМНЕВЫЕ ЖГУТИКОВЫЕ ВОДОРОСЛИ
(SILICOFLAGELLATA)
Табл. 65, фиг. 1 —13
К классу Silicoflagellatophyceae (кремневые жгутиковые водо-
росли) относятся микроскопические одноклеточные морские во-
доросли, имеющие внутриклеточный скелет и один жгутик. Размеры
водорослей колеблются от 5 до 80 д.
Скелет силикофлагеллат, как правило, состоит из полых крем-
неземных (опаловых) трубок, реже — из нерасчлененных пластинок.
Дифференцированной оболочки у клеток нет, поэтому форма клеток
целиком повторяет конфигурацию скелета (рис. 119, а).
Экология. Экология силикофлагеллат изучена слабо. Поскольку
они встречаются вместе с морскими диатомовыми водорослями,
можно предполагать, что для их развития требуются сходные усло-
вия. Одним из основных факторов, необходимых для развития си-
ликофлагеллат, является достаточное содержание в воде растворен-
ного кремнезема, который используется ими для построения ске-
лета. В течение всего времени существования эти планктонные во-
доросли населяли только поверхностный, хорошо освещаемый слой
воды морей и океанов. Некоторые особенности в морфологии ске-
лета (его ажурность, трубковидные перекладины, выросты) являются
приспособлением к парению.
Геологическое значение. Силикофлагеллаты никогда не бывают
породообразователями. Они встречаются вместе с диатомовыми
водорослями, участвуя в качестве второстепенного компонента в
образовании диатомитов, диатомовых трепеловидных и опоко-
Кремневые жгутиковые водоросли
93
Рис. 119. Кремневые жгутиковые водоросли, х 1000
а — Dictyocha fibula Ehr., клетка (Marshall. 1934); 6— Vallacerra tumidula Gies., саитои-
кампан, славгородская свнта, восточный склон Урала; в— Lyramula simplex. Hanna, воз-
раст и местонахождение те же; г—L. furgula Hanna, возраст и местонахождение тс же;
d — Cornua trifurcata Schulz, (скелет снизу), возраст и местонахождение те же; е — С. ро-
retzkiae Gies, (скелет снизу), возраст и местонахождение те же; ж— Dictyocha navicula
Ehr. (скелет снизу), нижний эоцен, серовская свита, восточный склон Урала; з — D. fibula
van pentagona Schulz, (скелет снизу), верхний миоцен, маямрафская свнта. Северный Саха-
лин, п-ов Шмидта; и—D. elata var. media Gies. f. media (скелет снизу), нижняя половина
верхнего эоцена, верхняя подсвита люлинворской свиты. Западно-Сибирская низменность,
п-ов Тазовскнй: к, л — D. rotundata van secta Gies.: к — скелет сверху, л — скелет сбоку,
нижняя половина' верхнего эоцена, верхняя подсвита люлинворской свиты. Западно-Сибир-
ская низменность. Среднее Прнобье; .«, и — D. frenguellii van carentis Gies., f. carentis:.u —
скелет сбоку, н —скелет сверху, верхний эоцен, ирбитская свита, восточный склон Урала;
о — D. triacantha var. fiexuosa St rad пег (скелет снизу), верхний олигоцен. карагиннская
свита, п-ов Мангышлак; п — D. hexacai.'ha Schulz (скелет снизу), верхняя половина верх-
него эоцена, верхняя подевнта киевской евнты. левый берег р. Днепр, севернее г. Днепро-
петровска; р — D. spinosa (Dcfl.) Gies, (скелет сверху), нижняя половина верхнего эоцена,
верхняя подсвита люлинворской свиты. Западно-Сибирская низменность, п-ов Тазовскнй;
с — D. transitoria Defl. (скелет сверху), верхний эоцен, ирбитская свита, восточный склон
Урала; т — Distephanus crux (Ehr.) Hack var. < rux (скелет сверху), верхний олигоцеи, кара-
гиинская свнта, п-ов Мангышлак; г — D. speculum var. septenarius (Ehr.) Jorg, (скелет сверху),
неоген, Камчатка, Усть-Камчатский район; ф — Mcsocena muticata Gies., верхняя половина
верхнего эоцена, верхняя подевнта киевской свиты, левый берег р. Днепр, севернее г. Днеп-
ропетровска; .г — Cannopilus sphaericus Gem. (скелет сбоку), миоцен, Венгрия (Frenguelli,
1940)
Буквы на рисунке; f. — зачатки базального кольца; gb. — базальное кольцо; h. — апи-
кальное кольцо; i. — апикальное окно; kl. — апикальные перекладины; т. — радиальные
рога; п.—дополнительные шипы; os.—опорные шипы; pl. — латеральные перекладины
94
Кремневые организмы
видных глин. По мнению ряда исследователей, силикофлагсллаты
могут быть с успехом использованы для расчленения и корреляции
разрезов морских верхнемеловых и кайнозойских отложений (Def-
landre, 1950; Жузе, 1951. 1955; Ichikava, 1956; Глезер, 1966; и др.).
Строение скелета. У большинства видов силикофлагеллат ске-
лет при боковом положении имеет форму усеченной пирамиды.
Основание пирамиды образовано так называемым базальным коль-
цом. Форма базального кольца различна у разных видов: она можег
быть округлой, овальной, треугольной или многоугольной. От
полюсов или углов базального кольца отходят выросты — радиаль-
ные рога или радиальные шипы, которые могут быть едва замет-
ными или очень грубыми и длинными. Верхушка пирамиды пред-
ставляет собой так называемый апикальный аппарат. Апикальный
аппарат соединяется с базальным кольцом при помоши боковых
(латеральных) перекладин, которые делят базальное кольцо на
несколько базальных окон. В самом простом случае апикальный
аппарат представлен одной трубчатой перекладиной или. сильно
уплощенной апикальной пластинкой. Более сложный апикальный
аппарат состоит из нескольких перекладин, образуюгцих апикальное
кольцо с одним окном. Дальнейшее усложнение апикального аппа-
рата состоит в том, что совокупность большого количества апикаль-
ных перекладин образует много аггикальных окон. У некоторых
форм отдельные элементы скелета могут отсутствовать.
Способы выделения из породы скелетов силикофлагеллат те же,
что и панцирей диатомовых водорослей. В соответствии со своими
размерами силикофлагеллаты в большом числе содержатся во фрак-
ции осадка с величиной частиц от 0,005 до 0.25 мм.
РАДИОЛЯРИИ (RADIOLARIA)
Современные радиолярии
Радиолярии — одноклеточные морские животные, населяющие
все океаны и многие моря. Они распространены во всех климати-
ческих зонах и встречаются во всей толще океанских вод — от по-
верхности до абиссальных глубин. Размеры радиолярий от десятков
микрон до 1—3 .w.w, глубоководных феодарий — до 10 мм и более.
Колониальные формы достигают иногда 4—6 см.
Химический состав. Скелет у представил елей различных отрядов
отличается по своему химическому составу, что и определяет их
различную сохраняемость в ископаемом состоянии. Однако относи-
тельно химического состава пока известно немногое. Из пяти отря-
дов радиолярий представители Spumellaria и Nassellaria имеют
почти чисто кремневый скелет и поэтому хорошо сохраняются в
осадках. Представители отряда Phaeodaria очень редко встречаются
в современных осадках, а достоверные сведения о нахождении их
в древних отложениях немногочисленны (Dumitrica. 1964, 1965).
В скелете феодарий кремневая кислота находится в соединении
с органическим веществом. Однако неизвестно, в каких именно
соединениях находится кремнезем и что определяет различную
сохраняемость феодарий, с одной стороны, и спумеллярий и нассел-
лярий — с другой.
Два отряда радиолярий — Acantharia и Sticholonchea — совсем
не сохраняются в донных осадках. Скелез акантарий состоит из це-
стина (SrSO<i). Как предполагав! Хабаков (1934), SrSO4 в скелетах
акантарий связан с органическим веществом. Шевяков (Schewia-
kofT, 1926) обнаружил в скелете акантарий до 2i,6"<} SiO, (аморфн.)
в составе сложного алюмокальциевого силиката.
Соотношение между различными группами спумеллярий и нас-
селлярий несколько изменяется по мере осаждения на дно скелетов
отмерших радиолярий. До дна не доходят наиболее тонкоокремнен-
ные виды радиолярий и значительная часть ювенильных форм.
В толще воды процессы растворения кремнезема скелетов радио-
лярий наиболее интенсивны в верхних слоях; максимальная потеря
веса скелетов радиолярий отмечается на глубине 250 м. Ниже 3000 н
скорость растворения скелетов радиолярий уменьшается, что свя-
зано с низкими темпера гурами глубинных слоев воды и повышен-
ным содержанием в них кремния.
Решающее влияние на сохраняемость радиолярий в донных от-
ложениях оказывает вещественный состав вмещающих осадков.
В кремнистых и терригенных осадках процент корродированных
радиолярий наименьший. Наибольшая степень коррозии и макси-
мальное число корродированных скелетов радиолярий — в карбо-
натных осадках.
Сохранность радиолярий в осадках холодноводных районов
намного лучшая, чем в тропических.
Экология. Радиолярии — исключительно морские животные,
обитающие в водах океанической солености (32—38 /оо) и ведущие
пелагический планктонный образ жизни. Радиолярии не живут
в морях с соленостью ниже 22%о(Черное. Каспийское, Белое и Азов-
ское моря).
Спумеллярий и насселлярии в тропических районах Тихого океа-
на встречаются во всех горизонтах от 0 до 5000 м, однако макси-
мальная численность радиолярий наблюдается на горизонте 100 м.
Количество радиолярий с глубиной постепенно уменьшается, и
на глубинах более 1000 м радиолярии исчисляются единичными
экземплярами в 1 м3. Видовой состав спумеллярий и насселлярий
довольно однороден от поверхности до глубин 100—300 .и; на боль-
ших глубинах при общем обеднении фауны появляются виды, не
встреченные у поверхности (Петрушевская, 1966, 1969).
В антарктических водах спумеллярий и насселлярии встречаются
начиная с горизонтов 30—50 л/-(Петрушевская. 1966), в поверхност-
ных водах радиолярии встречены не были. Максимальное количество
и наибольшее видовое разнообразие отмечаются в слое воды 200—
400 м. На больших глубинах количество радиолярий снижается.
Преимущественно на основании изучения фауны феодарий вы-
деляются две экологические категории радиолярий (Решетняк. 1955);
стенобатные. обитающие на строго определенных глубинах, и эври-
батные, обитающие на разных глубинах. Эврибатные радиолярии
обладают различной степенью эврибатности. Выделяются две груп-
пы видов, одна из которых встречается почти во всей толще воды
от 200 до 8000 и, вторая группа в своем распространении ог рани-
чена зоной от 200 до 2000 м.
Фауна спумеллярий и насселлярий пока изучена в очень малой
степени; характерные глубины и температуры обитания известны
лишь для очень небольшого числа видов.
Географическое распространение. На распространение радиолярий,
как и всех других групп микрофлоры и микрофауны, решающее
влияние оказывает климатическая зональность океана. Среди ра-
диолярий выделяются группы видов, связанные в своем распрост-
ранении с водами определенных биогеографических областей либо
широко распространенные в водах всего Мирового океана.
Основная масса радиолярий связана с теплыми водами. Из
шести тысяч ныне живущих радиолярий большая часть обитает
в тропической области Мирового океана.
В водах умеренных широт (как в бореальной, так и в нотальной)
фауна радиолярий много беднее, чем в тропической области.
Помимо радиолярий умеренных и тропических широт известно
небольшое число видов, приуроченных к арктическим и антаркти-
ческим районам. В значительной мере фауна радиолярий холодно-
водных районов Атлантического. Тихого океанов и Антарктики
сходна по видовому составу. Возможно также, что в водах Мирового
океана существует единый циркумтропический комплекс радиоля-
рий (речь идет о спумелляриях и насселляриях, скелеты которых
сохраняются в донных осадках, так как фауна радиолярий в планк-
тоне изучена в очень малой стеггени).
Радиолярии
95
Таким образом, имеющиеся в настоящее время сведения свиде-
тельствуют о зональном характере распределения фауны радио-
лярий в водах Мирового океана. \
Распределение в осадках. Скелеты радиолярий в значительных
количествах входят в состав пелагических осадков современных
океанов. Особенно велика их роль в областях распространения ра-
диоляриевых и радиоляриево-фораминиферовых илов. Типичные
радиоляриевые илы распространены исключительно в тропических
районах, занимая наиболее значительные площади в экваториаль-
ных зонах Тихого и Индийского океанов. Кроме радиоляриевых
илов радиолярии встречаются в большем или меньшем количестве
во всех типах современных пелагических осадков.
Наиболее высокие содержания радиолярий встречены в алеври-
тово-пелитовых и пелитовых илах, минимальная численность ха-
рактерна для более грубых осадков — крупных алевритов и песков.
Приуроченность радиолярий к определенным вещественно-гене-
тическим типам осадков определяется зональным характером раз-
вития планктона. Максимальные концентрации скелетов радиолярий
в северной половине Тихого океана (Кругликова. 1969) встречены
в областях распространения радиоляриевых. радиоляриево-фора-
миниферовых и слабокремнистых диатомовых илов, минимальные —
в терригенных, карбонатных осадках и в красных глинах.
Радиолярии чаще всего сосредоточиваются в пределах фракций
0,1—0,01 мм, т. е. в крупном и мелком алеврите, а в зависимости
от видового состава — преимущественно в одной из этих фракций.
В количественном распределении радиолярий в осадках выражены
черты зональности, присущие распределению других планктонных
организмов. В северной половине Тихого океана по содержанию ра-
диолярий в осадках можно выделить три зоны: бореальную (к се-
веру от 40° с. ш.), тропическую (40—15° с. ш.) и экваториальную
(15° с. ш. — 0°).
Для бореальной зоны наиболее характерны осадки с содержанием
радиолярий менее 50 тыс. экз/г сухого натурального осадка, но встре-
чаются и более высокие концентрации радиолярий, приуроченные
в основном к слабокремнистым диатомовым илам северного пояса
кремненакопления.
Большую часть тропической зоны занимают красные глины и
фораминиферовые осадки с минимальными для северной части
Тихого океана концентрациями радиолярий: менее 1 тыс. экз!г
сухого осадка.
Экваториальная зона характеризуется наиболее высоким содер-
жанием радиолярий в осадках. Именно к распространенным в этой
зоне радиоляриевым и радиоляриево-фораминиферовым илам приу-
урочены максимальные концентрации радиолярий, достигающие
500 000 экз/г и более.
Распределение радиолярий по глубинам не носит характера пря-
мой зависимости, а проявляется через накопление терригенного
и карбонатного материала. Типичные радиоляриевые илы в совре-
менную эпоху не образуются на глубинах менее 4600 м, т. е. являют-
ся типичными океаническими илами.
Зональный характер количественного распределения радиолярий
отвечает зональному характеру распространения ecei о зоо- и фито-
планктона, связанного в свою очередь с зонами подъема глубин-
ных вод, где в поверхностные слои воды поступает большое коли-
чество важнейших биогенных элементов: фосфатов, нитратов,
кремнезема, железа и ряда микроэлементов, обеспечивающих раз-
витие фитопланктона и других планктонных организмов.
Зональный характер распределения фаунистических комплексов
и численности радиолярий, изменения состава и численности этих
комплексов в толще четвертичных отложений позволяет использо-
вать радиолярии для выяснений вопросов палеогеографии и страти-
графии плейстоцена.
Строение скелета. Для каждого отряда радиолярий характерны
свои типичные формы скелета. У спумеллярий наиболее примитивная
форма его — отдельные, рассеянные в эктоплазме микроскопические
иглы. Наиболее типичны для спумеллярий скелеты в форме сферы
(одной или нескольких, вложенных друг в друга и соединенных ра-
диальными иглами), от которой часто отходят радиальные шипы.
Для ряда групп спумеллярий характерны формы скелета, в
основе своей производные от шара, — дисковидные и эллип-
соидальные. Другой ряд форм составляют скелеты, представляющие
собой различного рода спирали — одиночные, двойные, свернутые
в одной плоскости или в нескольких и т. д. (рис. 120, а—г).
Очень разнообразны скелеты насселлярий. Исходным элементом
здесь, по-видимому, является четырехлучевая спикула. Три луча
Рис. 120. Радиолярии
а семейство Astrosphaeridae: на рисунке представлен скелет, состоящий из трех сфер:
от наружной раковинки отходят тданные (более длинные) радиальные шлы. соединяющие
между собой внутренние шары, и дополнительные (более короткие), отходящие только от
наружной сферы: б--семейство Ellipsidae: простая эллипсоидальная раковинка с отходя-
щими от полюсов шипами: х - семейство Druppullidae: показан скелет, состоящий из
четырех сфер: видны отходящие от полюсов наружной раковинки шнпы и дополнительные
балки, соединяющие наружную раковинку с внутренними: г семейство Porodiscidae:
иа рисунке видна маленькая центральная сфера, ячеи камерных кругов
ее образуют треножник, поддерживающий центральную капсулу
(в ископаемом состоянии не сохраняется), четвертый направлен
вверх, образуя апикальную иглу (рис. 121, а — г). От основной спи-
кулы у форм, имеющих сложно устроенный скелет, отходят отростки,
дистальные концы которых, сочленяясь, образуют разнообразной
формы скелеты: ажурные кольца, шлемы, колоколы и т. д.
96
Кремневые организмы
Рис. 121. Радиолярии
а семснсгяо Plecloidne: oimciiihbo видна основная часть скелета — трехосная игла («ба-
зальная rpenoi а») с отходящими oi нее ответвлениями: <> — семейство Stephoidae: в сагит-
тальной плоскости расположена основная часть скелета — сагиттальный круг: в — семей-
ство Cyrtoidac. рол Cornutclla: раковинка состоит из лвух сегментов, виден апикальный
pAi (ш ла), отходящий от цсфалиса: г — семейство Cyrtoidac, род Theocalypla (?): видны
апикальный pot и «крылья» (лучи треножника), отходящие с трех сторон от раковинки:
<• семейство Cyrtoidac. род Liiluimiira: раковинка поделена на множество сегментов:
видны элементы внутреннею строения скелета; с — семейство Spyroidae: на рисунке пред-
ставлен однокамерный скелет (имеется только цефалне); сагиттальный пережим делит
цефалис на две доли
Древние радиолярии
Табл. 66. 76
Радиолярии известны с кембрия и с этого времени являются
основным или существенным породообразующим компонентом
таких пород, как яшмы и фтаниты. Эти породы широко распрост-
ранены в геосинклинальных областях палеозоя и мезозоя, где они
ассоциируются с вулканическими, вулканокластическими и терри-
генными отложениями, а также с редкими биогенными известня-
ками. Как показала И. В. Хворова (1968), условия образования
и особенности размещения геосинклинальных кремнистых осадков
прошлого и современных океанических осадков различны. Области
современного кремненакопления имеют широтную ориентировку
кремнистые пояса прошлого прослеживаются в соответствии с про-
стиранием (еосинклинальных систем, а не в соответствии с основной
планетарной климатической зональностью, как бы ее ни реконструи-
ровать. Так, например, в нижнем палеозое Северной Америки
меридиональный Кордильерский пояс кремненакопления располо-
жен под углом около 60 ‘ к Аппалачскому поясу, а в среднем палео-
зое Уральский кремнистый пояс лежит почти под прямым углом
к Среднеазиатскому. Внутри геосинклинальных систем радиоляриты
(яшмы, фтаниты) накапливались в узких трогах, к которым часто
были приурочены скопления основных магматических пород.
По составу древние радиоляриты отличаются от современных
радиоляриевых илов значительно более высоким содержанием
кремнезема. Если в современных радиоляриевых илах SiO2 (аморфн.)
составляет от 5 до 22% осадка (см. выше), то во фтанитах и яшмах
минимальное содержание свободного кремнезема равно 29%.
обычно оно заметно больше 50% и нередко превышает 90%. Кроме
того, яшмы и фтаниты почти не содержат карбоната кальция, тогда
как в радиоляриевых илах его содержание достигает 20% и довольно
обычны смешанные радиоляриево-фораминиферовые осадки.
В сумме перечисленные отличия показывают, «что геосинкли-
нальные существенно радиоляриевые породы (яшмы, фтаниты)
не являются гомологом современных океанических радиоляриевых
илов» (Хворова. 1968. стр. 116).
Глубина, на которой формировались яшмы и фтаниты. оцени-
вается разными исследователями по-разному. Однако анализ обшир-
ного фактического материала, накопившегося к настоящему времени,
показывает, что среди геосинклинальных кремнистых радиоляри-
товых осадков есть и глубоководные и мелководные разности (Хво-
рова. 1968). Несомненно мелководными являются радиоляриты,
чередующиеся в разрезах с грубыми песчаниками, и яшмовые пачки,
залегающие на подстилающих породах несогласно, с размывом.
Наоборот, терригенно-кремнистые формации, в составе которых
твердо установлены пелагические терригенные и карбонатные по-
роды. при достоверном отсутствии мелководных осадков можно
рассматривать как глубоководные, скорее всего батиальные образо-
вания.
В палеозое высококремнистые радиоляриты и породы, в которых
радиолярии обильны, слагают не менее 50% осадка, нередко встре-
чаются и вне геосинклинальных областей, в платформенных про-
гибах. На Русской платформе, например, радиоляриевые силициты
с содержанием свободной SiO2 до 77% распространены в верхне-
девонских и нижнекаменноугольных отложениях ряда прогибов
Камско-Кинельской системы (Трохова. 1968). В отличие от яшм и
фтанитов. практически бескарбонатных. платформенные радиоля-
риты всегда содержа/ то или иное количество карбоната кальция
и входят в состав кремнисто-карбонатных толщ. С ними ассоции-
руются радиоляриевые известняки, в которых все или почти все
раковинки радиолярий замещены кальцитом, а также известняки
микрозернистые, в разной степени кремнистые и битуминозные
карбонатно-кремнистые и глинисто-кремнистые сланцы, реже —
птсроподовые и сферово-сгустковые известняки.
Кремнистые радиоляриевые осадки Камско-Кинельской системы
прогибов распределены неравномерно и возраст кремнистых толщ
в пределах разных прогибов неодинаков. Естественно, что платфор-
менные радиоляриты и радиоляриевые известняки никак не могут
приравниваться к океаническим радиоляриевым илам.
Микроструктура. При хорошей сохранности радиолярии легко
определяются по характерной общей форме раковинки и ее ажур-
ному сетчатому строению (табл. 66. фиг. 13). От диатомовых во-
дорослей хорошо сохранившиеся радиолярии отличаются более
крупными размерами, более грубой крупнопористой сеткой и не-
редко наличием шипов. Чаще, однако, встречаются раковинки в той
или иной степени измененные: перекристаллизованные, кальцити-
зированные или пиритизированные, с частично утраченной струк-
турой (табл. 76). В тангенциальных сечениях может при этом (в боль-
шей или меньшей степени) сохраниться сетчатое строение, но ис-
чезнуть шипы (табл. 66, фиг. 4, 5). а могут, наоборот, сохраниться
шипы у раковинки, превратившейся в сплошной кальцитовый шарик
(табл. 66. фиг. 11).
Пиритизация часто фиксирует структуру раковинок даже в силь-
но перекристаллизованных радиоляриевых известняках, как бы про-
рисовывая сетку на тангенциальных сечениях (табл. 66: фиг. 8)
и контуры вложенных друг в друга шаров с соединяющими их игла-
Радиолярии. Кремневые губки
97
ми на поперечных разрезах (табл. 66, фиг. 7, 10). Раковинки, со-
стоящие из нескольких шаров, вложенных один в другой, в попереч-
ном сечении дают рисунок концентрически расположенных окруж-
ностей (табл. 66, фиг. 1). Иногда такие окружности бывают пунк-
тирными, что отражает пористое, решетчатое строение сферических
скелетных оболочек.
Наиболее трудны для определения разрезы раковинок, состоя-
щих из двух шаров, если у них не сохранились иглы и шипы, так как
они очень похожи на поперечные сечения спикул кремневых губок.
Иногда их истинная природа выясняется благодаря тому, что хотя
бы один шар представлен на разрезе прерывистой точечной или
пунктирной окружностью (табл. 66, фиг. 9). Как правило, в таких
сомнительных случаях надо внимательно просмотреть весь шлиф
и в нем всегда найдутся экземпляры с более или менее отчетливыми
следами первичной структуры. При этом следует помнить, что в
скоплении спикул губок всегда найдутся иглоподобные продольные
сечения, даже если в шлифе резко преобладают поперечники, а шаро-
образные раковинки радиолярий в любом разрезе дают только
круги.
В скелетах палеозойских радиолярий опал почти всегда превра-
тен в халцедон. При крайней степени изменения их раковинки мотут
стать сплошными однородными халцедоновыми шариками (табл. 66.
фиг. 2). В этом случае, так же как в предыдущем, установить радио-
ляриевое происхождение шариков помогают экземпляры с более
или менее уцелевшей структурой раковинки. Кое-какие следы струк-
турных особенностей радиолярий различаются даже на упомянутой
выше фиг. 2: внизу, в правой стороне снимка, виден тонкий
шип. отходящий от белого кружка. Но. пожалуй, более типична
такая сохранность материала, как на табл. 66. фиг. 1. где среди
совершенно бесструктурных кружочков попадаются сечения с ха-
рактерным для радиолярий строением.
КРЕМНЕВЫЕ ГУБКИ (S1LIC1SPONGIA)
Табл. 67. 68
Кремневые губки принадлежат к одному типу животных с извест-
ковыми губками и достоверно существуют с кембрия по настоящее
время. Есть указания, что они встречаются и в позднем докембрии
(Dubigner. Eller. 1963). Общее наименование «кремневые» относится
ко всем губкам, обладающим опаловыми спикулами, и охватывает
представителей двух классов: стеклянных губок (Hyalospongia)
и обыкновенных губок (Demospongia). К этим последним относятся
губки четырехлучевые и кремнероговые.
Экология. Кремневые губки — донные неподвижные животные,
колониальные, реже одиночные, в громадном большинстве обитаю-
щие в морях. К жизни в пресных водах приспособились лишь не-
многие кремнероговые формы. Некоторые губки, обитающие в
местах с сильными придонными течениями, плотно прирастают
к субстрату. Другие живут на рыхлых грунтах (песок, ил) при срав-
нительно слабом движении воды и либо закрепляются в осадке при
помощи корневого пучка спикул, либо свободно лежат на его по-
верхности. Современные кремневые губки встречаются во всех кли-
матических зонах, в теплых и холодных морях, и при подходящих
условиях образуют массовые поселения. Такие поселения известны
в морях Баренцевом. Охотском. Средиземном. Карибском. в районе
островов Индонезии в Антарктике и т. д. На дне морей кремневые
губки могут встречаться на различной глубине начиная от приливно-
отливной зоны вплоть до больших глубин океана. Обыкновенные
губки — четырехлучевые и кремнероговые — имеют широкий диа-
пазон вертикального распространения, встречаясь как на мелко-
водье. так и на больших глубинах. Стеклянные губки можно считать
препму щественно глубоководными организмами.
11С2 7
С точки зрения седиментации очень важна способность гу-
бок образовывать густые заросли, где. отмирая, они оставляют на
дне целый слой, сплошь состоящий из спикул, слагающих своеоб-
разную «стеклянную вату» с порами, заполненными минеральным
осадком (табл. 68. фиг. 8). В юго-западной части Баренцева моря
на глубинах 150—400.и развит биоценоз, в котором кремневые губ-
ки составляют 95—98% общей биомассы, почти полностью вытесняя
другие группы фауны с площади, занятой их зарослями. Губково-
спикуловый осадок в районе этих зарослей имеет могцность
30—60 с.ц (Филатова, 1938). Почти полное отсутствие других дон-
ных животных на площади, занятой губками, объясняется двумя
причинами: 1) чисто механическим вредным воздействием находя-
щихся в грунте спикул на донные, особенно зарывающиеся формы:
2) тем. что губки как мощные фильтраторы в первую очередь извле-
кают из воды всякую пищу.
Кроме однородных губковых поселений, дающих прктически
чистые спикуловые осадки, губки играют заметную роль в некоторых
более разнородных группировках сидячих форм. Так. например,
в Индийском секторе Антарктики на глубине 130—160 м был обна-
ружен биоценоз, в который входили в большом количестве крупные
кремневые губки, а также мшанки, серпулиды. брахиоподы и круп-
ные фораминиферы. В осадке спикулы губок составляли здесь более
50% всей его массы, остатки известковых организмов — около
30—40%. а на минеральный материал приходилось всего 10—20%
(Лисицын. 1966). Такие смешанные детритусово-спикуловые осадки
широко распространены. Они известны в Антарктике, в Баренце-
вом море и в дальневосточных морях (Петелин, 1954; Лисицын, 1966).
на глубинах от 150 до 600 м.
Определенную роль в осадкообразовании играют также свер-
лящие четырехлучевые губки — клионы. обладающие способностью
проделывать ходы в твердом известковом субстрате. Обычно таким
субстратом являются раковины моллюсков, известковые водоросли,
твердые части кораллов и известковые породы, обнажающиеся
на берегу или на дне моря. На поверхности раковины или известня-
ковой глыбы при поселении губки образуется небольшое углубле-
ние. а затем, по мере ее роста, просверливаются многочисленные
неглубокие каналы, сообщающиеся между собой и имеющие выход
наружу в виде округлых отверстий с диаметром в среднем около
1 ,и.и. Появление густой сети каналов приводит к разрушению суб-
страта. От него отпадают куски разного размера, образующие но-
вый осадочный материал — органогенный детрит, если разрушают-
ся скелетные части животных и известняковые песчинки и щебень
при распадении карбонатных пород. Так, на коралловых рифах
клионы вместе с другими сверлящими организмами активно ис-
сверливают отмершие скелеты кораллов, способствуя образованию
так называемых коралловых песков.
Геологическое значение. Уже в кембрийских морях существовала
довольно богатая фауна кремневых губок, представленных почти
всеми классами и отрядами, известными в настоящее время. Лишь
очень немногие ископаемые формы принадлежат к вымершему
отряду разнолучевых губок. Кремневые губки находят некоторое
применение в стратиграфии, в частности, юры и мела. Так, напри-
мер. юрские и меловые отложения ГДР, ФРГ подразделяются по
комплексам губок на отделы и ярусы, а иногда и на более мелкие
стратиграфические единицы.
Однако губки имеют гораздо большее значение как породообра-
зователи, так как во многих системах палеозоя и мезозоя спикуло-
вые накопления, чистые или с примесью другого органогенного дет-
рита. слагают пласты и пачки пород мощностью до 10—15 л/. Такие
породы в том случае, когда спикулы сохранили свой первичный
кремневый материал, называются спонгалитами. Несравненно чаще
все спикулы или большая их часть бывают кальцитизированы.
и тогда порода превращается в спикуловый известняк или спикуло-
вый мергель, в зависимости от состава минерального цемета.
Чистые спикуловые известняки, в которых спикулы составляют
98
Кремневые организмы
85 -95",, породы, очень характерны по внешнему виду: обычно они
серые, синевато-серые или темно-серые, очень плотные, бесструк-
турные (сливные), в большей или меньшей степени окремнелые.
с многочисленными стяжениями черною кремня и без фауны. Спи-
куловые известняки со значительной примесью обломков извест-
ковых организмов (до 20% породы) отличаются от чисто спику-
ловых разностей тем. что в них на поверхности выветривания видна
редкая фауна.
Спикуловые известняки широко распространены. В Советском
Союзе они известны в верхнем девоне и нижнем карбоне Урало-
Волжской области (Сюндюков, 1961; Теодорович и др., 1962), в
нижнем карбоне пермского Приуралья (Липина, 1960), Донецкого
бассейна (Айзенверг, 1958), Кузнецкого бассейна (Максимова.
1961), района верхнего течения р. Печоры, в среднем карбоне Вол-
гоградской области и Донецкого бассейна (Киреева, Максимова,
1959), в верхнем карбоне бассейна р. Печоры. Мощные пласты спи-
куловых мергелей (по 6—7 .и мощностью) встречаются в артинских
отложениях (PJ западного склона Среднего Урала (Осипова, 1950).
Спикуловые известняки прослеживаются в ряде скважин в основании
верхней юры Предкарпатского прогиба.
Пластовые спонголиты, или, по другой терминологии, спонгие-
вые силициты, также известны в разных геологических системах:
в верхнем девоне Латвии (Сорокин, Гравитис, 1964), в нижнем кар-
боне Урало-Волжской области и Кузнецкого бассейна, в верхнем
карбоне Горной Башкирии (Теодорович, 1949), в верхней юре Арме-
нии (Мандалян, 1964), в олигоцене Западного Кавказа и Грузии.
Приведенные примеры ни в какой мере не являются исчерпы-
вающими, но достаточно ясно показывают, насколько часто встре-
чаются спикуловые породы в морских отложениях самого различ-
ного возраста и самых различных регионов.
Строение скелета. Скелет кремневых губок состоит из множества
опаловых игл или спикул, имеющих разнообразную форму. Среди
спикул обычно различают макросклеры, составляющие
основную массу скелета, и более мелкие и иначе устроенные мик-
росклеры. В образовании скелета кроме игл нередко принимает
участие органическое вещество — спонгин, при помощи которого
спикулы склеиваются друг с другом. У стеклянных губок спикулы
шестилучевые (трехосные). Часто один или несколько лучей реду-
цируются, и тогда образуются пятилучевые, четырехлучевые и даже
одноосные иглы. Лучи этих игл могут быть гладкими или шипо-
ватыми, прямыми или изогнутыми, и длина их измеряется обычно
десятыми долями миллиметра. Микросклеры стеклянных губок
имеют довольно причудливую форму (рис. 122) и размер не более
0,1 мм. У некоторых стеклянных губок спикулы срастаются своими
концами так. что образуется правильный решетчатый каркас
(табл. 68, фиг. 6).
У четырехлучевых губок в типичном случае скелет образован
четырехлучевыми иглами — макросклерами с примесью одноос-
ных игл. гораздо реже — только одноосными спикулами. Спонгин,
как правило, отсутствует. Макросклеры крупные, нередко дости-
гают в длину 1,5—2,5 мм. Микросклеры часто звездчатые и шаро-
образные (рис. 123). У многих форм они тесно и в несколько слоев
лежат у поверхности тела, образуя жесткий панцирь. Видоизме-
нением обычных четырехлучевых спикул являются неправильной
формы ветвистые образования, покрытые неровностями и буграми
(табл. 67. фиг. 2; табл. 68, фиг. 3). Плотно соединяясь, спикулы
такой формы строят связный, так называемый каменистый, скелет.
Скелет кремнероговых губок состоит только из одноосных спи-
кул, которые могут быть гладкими или шиповатыми, прямыми,
цилиндрическими, веретеновидными, изогнутыми или змеевидно
искривленными, головчатыми или булавовидными. Всегда имеется
в том или ином количестве спонгин, с помощью которого спикулы
склеиваются в пучки или волокна. Длина спикул измеряется сотыми
или десятыми долями миллиметра. Для микросклер характерна
форма маленьких якорьков, шпилек или сигм (рис. 124).
Рис. 122. Спикулы кремневых (стеклянных) губок
Слева — макросклеры: шестилучевые, пятилучевые и одноосные иглы. Справа - мнкро-
склеры
Рис. 123. Спикулы кремневых четырехлучевых губок
Слева — макросклеры: различные четырехлучевые иглы и одноосная головчатая hi ла.
Справа — микросклеры: звездчатые иглы и сигмы
о
Рис. 124. Спикулы кремпероговых губок
Слева — макросклеры: разной формы одноосные иглы. Справа — микросклсры: хелы.
якорьки (вверху), шпильки, сигмы, дужки (внизу)
Кремневые губки
99
Рис. 125. Спикулы разнолучевых губок
а — в — спикулы Chancelloria. х 8,75. нижний кембрий. Сибирская платформа: г. д -
спикулы Astraeospongium (Основы палеонтологии. Губки. 1962. рис. 65. 66)
Спикулы вымерших разнолучевых губок (рис. 125) представляют
собой звездчатые тела. В простейших случаях спикула состоит из
плоской правильной шестилучевой звезды с двумя перпендикуляр-
ными к ней, торчащими в обе стороны осевыми лучами. Эти по-
следние могут редуцироваться. В других случаях лучей может быть
значительно больше, а расположение их менее правильным.
В геологическом материале, естественно, спонгин не сохраняется
и даже опал, слагающий спикулы в мезозойских породах, обычно
превращается в халцедон или замещается кальцитом. В палеозой-
ских осадках спикулы бывают или халцедоновыми, или кварц-
халцедоновыми, или вторично-кальцитовыми. Так называемые
каменистые губки со спикулами, спаянными в решетчатый скелет,,
в ископаемом состоянии сохраняются в виде обособленных кремне-
вых желваков и в шлифах встречаются сравнительно редко. Чаще
всего в шлифах наблюдаются разрозненные одноосные и четырех-
лучевые спикулы, которые как раз и слагают пласты спонголитов
и спикуловых известняков. Одноосные спикулы в продольных или
слегка скошенных сечениях выглядят как палочки или иголки с
каналом посередине, в поперечном сечении — как круги опять-таки
с каналом в центре (табл. 67, фиг. 1, 5). Четырехлучевые спикулы
в сечениях, приближающихся к продольному, выглядят как три-
листники (табл. 67, фиг. 4), рогульки и стрелки (табл. 68, фиг. 9).
В поперечных сечениях они обычно дают круги с осевым каналом,
такие же, как у одноосных спикул. Гораздо реже попадаются об-
ломки спикул, срезанные в месте разветвления лучей. В этом слу-
чае, как у четырехлучевых, так и у шестилучевых (трехосных) спи-
кул, в двух-трех лучах могут быть видны продольные сечения ка-
налов, а в месте их соединения — поперечный разрез канала основ-
ного длинного луча (табл. 67, фиг. 7).
Наличие канала весьма важно для уверенного определения спи-
кул губок, и поэтому всегда следует внимательно искать его следы.
У халцедоновых спикул канал иногда заполняется кальцитом,
чаще же халцедоном, но обычно его можно различить, так как в
стенках спикулы халцедон, как правило, бывает более крупнокри-
сталлическим, чем в канале. Даже в участках сплошного окремне-
ния породы при параллельных николях нередко прорисовываются
контуры индивидуальных спикул, в которых можно заметить следы
канала благодаря окаймляющей его тончайшей пленке органиче-
ского вещества (табл. 67, фиг. 3). У кальцитизированных спикул
стенки слагаются светлым прозрачным кальцитом, а канал запол-
няется обычно более мелкокристаллическим и более темным СаСО3
или точечными скоплениями органического вещества. В одних шли-
фах могут преобладать поперечные разрезы спикул (табл. 67, фиг. 8),
в других — продольные (табл. 67, фиг. 6), но к кругам всегда в ка-
ком-то количестве примешиваются палочки или рогульки, а к па-
лочкам или рогулькам — круги, так что характерное сочетание этих
двух форм позволяет узнавать спикулы даже при плохой сохран-
ности, т. е. при отсутствии следов каналов. Кроме того, при внима-
тельном просмотре шлифа в массовом скоплении спикул почти
всегда найдется хотя бы несколько экземпляров с достаточно хо-
рошо сохранившимися каналами.
Встреченные в шлифах единичные кальцитовые или халцедо-
новые «палочки» и «иголки», похожие по общей форме на спикулы,
но однородные, без канала, не следует относить к кремневым губ-
кам, так как подобные образования, лишенные внутренней струк-
туры, могут иметь весьма различное происхождение.
Часть Ш. СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ
ОРГАНИЗМОВ
В этом разделе помещены образования, которые не могут счи-
таться скелетом, оболочкой или специфическим минеральным вы-
делением какого-либо определенного организма. Сюда отнесены
строматолиты, онколиты, следы сверления и копролиты. По этим
остаткам жизнедеятельности, при всей неопределенности организ-
ма. можно все же в той или иной степени судить о режиме осадко-
накопления.
В силу этого в описываемых образованиях могут быть заинтере-
сованы исследователи, изучающие осадочные породы.
МИКРОСТРУКТУРА СТРОМАТОЛИТОВ
Табл. 70—73
Строматолиты — это карбонатные постройки, образованные
сериями слоев, в которых располагались предполагаемые синезе-
леные водоросли и, может быть, бактерии. Внешние формы стро-
матолитов очень разнообразны. Слоистые текстуры строматолитов
имеют много общего с так называемыми коломорфными текстурами
чисто минерального происхождения (марганцовыми, кремневыми,
малахитовыми и т. п.), поэтому нужно быть осторожным при опре-
делении строматолитов при отсутствии специфических микрострук-
тур.
По микроструктурам строматолиты делятся на две большие
группы: 1) с зернистокристаллическими микроструктурами, 2) с раз-
личными типами микроструктур.
Первая группа характерна главным образом для доломитовых
фаций с обильным выпадением карбоната из осадка. Микрострук-
туры этой группы состоят из слойков более крупнозернистого и
пелитоморфного карбоната (табл. 71, фиг. 1). Строматолиты этой
группы с трудом различаются по микроструктурам или совсем не
могут различаться. Но текстуры, т. е. сочетание слоев и их распо-
ложение, могут быть разными, что пытаются использовать геоло-
ги для классификации строматолитов. Так, например, на табл. 71.
фиг. 2 изображена часть строматолита, описанного под названием
Macronubecularites (Маслов, 1960 6), где при зернистой микрострук-
туре толстые слои нарастают не закономерно, а наподобие
облаков.
Вторая группа микроструктур характеризуется: 1) радиальным
распределением элементов; 2) слоями с местными раздувами;
3) сгустковой структурой; 4) наличием тонких каналов или про-
светов, относимых к полостям от нитей водорослей.
1. Радиальное расположение элементов^ отчетливо видно в
стержневой микроструктуре, которая может при пышном ее раз-
витии превратиться в макроструктуру. Эта микроструктура опи-
сывалась К. Б. Кордэ как особая водоросль (Actinophycus). Но
в такой же стержневой микроструктуре были встречены остатки
клеток и нитей водорослей (Hydrocorynites), не похожие на описан-
ные К. Б. Кордэ. Таким образом, стержневую микроструктуру
нужно считать способом обызвествления разных водорослей. Она
состоит из кристаллических столбиков, между которыми засыпался
терригенный осадок (рис. 126, б; табл. 70, фиг. 3). Иногда похожая
структура появляется на краю строматолита (табл. 70, фиг. 1)
под влиянием увеличения терригенного материала, скатывавшегося
с вершины строматолита. Стержневая структура обычно пересекает
ряд слойков строматолита, что подчеркивается ожелезненным
или пелитовым материалом (табл. 70, фиг. 3). Иногда же стержне-
вая микроструктура развивается на небольшом количестве слой-
ков в виде небольших перевернутых конусов (табл. 70, фиг. 2).
В строматолитах и онколитах встречаются также радиальные микро-
структуры, которые проявляются главным образом под влиянием
вторичных процессов (ожелезнение, выветривание и т. п.). На
табл. 72, фиг. 1 изображено сечение через онколит с таким радиаль-
ным расположением вторичного карбоната, выявление которого
может объясняться неорганическими процессами. Между тем в
темных неперекристаллизованных участках заметны прерываю-
щиеся тонкие светлые канальчики, которые можно отнести к сле-
дам от нитей водорослей1. В образце строматолита с радиальной
ориентировкой вторичного светлого и первичного темного карбо-
натов (табл. 72, фиг. 3) также можно видеть светлые тонкие черточки
внутри темных конусов — вероятные пустоты от нитей водорослей,
что позволяет сделать вывод о первичной радиальной ориентиров-
ке составляющих элементов2.
2. Слои с местными раздувами или почковидная микрострук-
тура характерны для слоев некоторых строматолитов типа Сопо-
phyton. Слои его имеют форму острых конусов, и в сечении, нормаль-
ном к оси, конусы дают концентрическую конфигурацию (табл. 71,
фиг. 3). Слои бывают неоднородной толщины и слагаются светлы-
ми почковидными мелкими желвачками, часто соединенными в
группы. Между этими светлыми и перекристаллизованными слоями
располагается темный пелитоморфный материал, часто засорен-
ный глинисто-железистым осадком. Почковидные желвачки состоят
из зерен с радиальным расположением и могут трактоваться как
остатки от структур колоний каких-то (синезеленых?) водорослей
(табл. 71. фиг. 4).
Некоторое отдаленное сходство с этой структурой приобретают
перекристаллизованные микроструктуры некоторых обычных ку-
половидных строматолитов. При этом разрастание вторичного
1 Не надо смешивать канальчики с трещинами. характеризующимися непрерывностью
на значительном расстоянии, в то время как канальчики. будучи цилиндрическими и не
абсолютно прямыми, секутся плоскостью шлифа и таким образом прерываются.
2 Эта первичность подчеркивается ветвлением темных радиальных элементов. б.татодаря
чему образуются «кустики» и «метелочки», похожие на ветвящиеся кустики эпифитонов
(см. раздел «Синезе.теныс водоросли»). Принимая во внимание радиальное расположение
некоторых синезеленых известьотлагающих водорослей, например ривулярий, многие
исследователи относят такие ископаемые радиальные т.темснты к несомненным сине-
зеленым водорослям. Но нужно заметить, что у современных ривулярий участки обыз-
вествления всегда светлые, довольно крупнокристаллические, близкие к вторичному
карбонату. Гстественно ожидать, что в ископаемом виде эти светлые участки не превра-
тятся в пелитоморфные. поскольку вторичный карбонат не претерпевает этих изменений.
Поэтому надо относить эти темные радиальные структуры лишь к остаткам от возмож-
ных синезеленых водорослей.
Микроструктура строматолитов. Желваки органического происхождения
101
Рис. 126. Микроструктура онколитов и строматолитов
а се|чато-с1устковая микроструктура одного из онкодиюв. х 25. пермь. Пермская область, ст. Верещагине: б — червячная микроструктура, состоящая из ясных и расплывчатых
темных ветвящихся нитей, протягивающихся перпендикулярно с юпсюсти. х 10. неоген,Керчь;« — стержневая микроструктура строматолита, х 10. ордовик, р. Ангара
карбоната приводит к похожей «узловатой» микроструктуре
(табл. 71. фиг. 5. 6).
3. Сгустковая микроструктура описывалась Гюрихом (Gurich,
1906) под родовым названием Malacosiroma1 и относилась им к жи-
вотному миру каменноугольного периода. В дальнейшем оказалось,
чю эта микроструктура обычна для многих строматолитов оз
докембрия до миоцена. Кроме тою. эга микроструктура встречает-
ся и не в строматолитах. Ее происхождение не веема может быть
связано с жизнедеятельностью водорослей, и каждый раз необхо-
димо доказывать ее органогенный генезис изучением карбонатного
стяжения пли породы. Элементы этой микроструктуры — сгустки
могут располагаться (в строматолите) беспорядочно и ничем не
отличаться от сгусткового известняка (тортонские строматолиты
нараевского горизонта Западной Украины), группироваться в нити
(рис. 126г/. б), образующие своеобразную сетчатую микрострук-
туру и сгущенные слои (табл. 72. фиг. 4. 5) или чередоваться с пе-
литоморфными или микрозернистыми слоями (табл. 72. фиг. 6).
4. Выше мы касались частично канальчиков. относимых к по-
лостям от нитей или трихомов водорослей, в толще строматоли-
товых слоев при описании радиальных структур. Эти канальчики
имели более или менее прямую форму. Чаще же нити синезеленых
водорослей имеют спутанную извивающуюся форму, поэтому и
канальчики. остающиеся от этой нити, имеют прихотливо изогну-
тый вид. Отчетливые канальчики встречаются редко. На табл. 70,
фиг. 6—8 изображены разные слои одного и того же строматолита,
описанного В. П. Масловым под названием Collenia uraliea Masi,
из перми Южного Урала (Маслов. 1936).
Характер и расположение этих полостей с очевидностью дока-
зывают участие разных водорослей в образовании одного и того же
строматолита. Кроме более или менее радиально расположенных
каналов (табл. 70. фиг. 6) встречаются и горизонтально расположен-
ные канальчики (табл. 70. фиг. 8 вверху), при этом в непосредствен-
ной близости к другим пустотам. Часты также канальчики. которые
в основании имеют горизонтальное направление, а от них отходят
ответвления вверх и в стороны (табл. 70. фиг. 7. 8 внизу), подобно
современной водоросли Haplosiphon, изображенной на рис. 1,«.
или Scvfoneina (см. рис. 1, е).
Нскоюрыс пнюры. например Джонсон, продолжаю! выделять этот «род» как синезеле-
ную водоросль.
Некоторые канальчики и полости в трубкообразных вертикаль-
ных строматолитах не могут объясняться жизнедеятельностью одних
водорослей. На табл. 70. фиг. 9 изображено вертикальное сечение,
которое предположительно отнесено к симбиозу известковой губки
и синезеленых водорослей и описано под названием Phytospongia
cylindrica Maslov (1960 6). Наконец, иногда тонкие канальчики про-
низывают мелкие подушечки разной формы, состоящие из темного
карбоната и располагающиеся между слоями крупного стромато-
лита. Такие подушки, возможно, целиком относятся к жизнедея-
тельности одной синезеленой водоросли (табл. 70. фиг. 4. 5).
ЖЕЛВАКИ ОРГАНИЧЕСКОГО ПРОИСХОЖДЕНИЯ
Табл. 69
Под гермином «онколиты», введенным Пиа (Pia. 1927). под-
разумеваются мелкие и крупные желваки, образованные облекаю-
щими обломок слоями разной структуры, которые условно считают-
ся постройками синезеленых водорослей. Обычно в онколитах на-
блюдается чередование темных и светлых слойков без следов орга-
низмов. Но встречаются экземпляры или участки в онколитах,
в которых можно заметить или тонкие светлые канальчики. или
сгустковую структуру, обычную для строматолитов. В тех случаях,
когда желвачок образован четкими трубками гирванелл. его не
причисляют к онколитам. Более детально онколиты описаны
В. П. Масловым. По происхождению и структурам онколиты близки
к строматолитам, характеризуют очень большое мелководье, обра-
зуются как в нормально-морской, так и в пресной воде, а также в
солоноватоводных и лагунных условиях. Онколиты известны с про-
терозоя до настоящего времени. Часто мелкие онколиты похожи
на оолиты и пизолиты неорганического происхождения (с которыми
могут быть смешаны) и даже на бобовины. являющиеся результа-
том химического выпадения карбоната. Многие онколиты описы-
вались под латинскими бинарными названиями, и им придавалось
стратиграфическое значение. Однако многие онколиты с примитив-
ным строением из разных геологических систем похожи друг на
друга. Некоторые онколиты, описанные под «родовым» названием
Osagia и не обладающие концентрической текстурой, похожи на
102
Следы жизнедеятельности организмов
О 1 2 см 0 1 2см
-J —' * 1
Рис. 127. Онколиты
а — онколит, росший вначале как неподвижный нарост, затем переворачивающийся на обратную сторону, пришлифовка. ордовик, усть-кутская свита. Восточная Сибирь, р. Илим;
о — онколит морфологического типа Ot/onosia. разросшийся вокруг гастроподы, шлиф, юра, Дальний Восток, р. Конгауз, район Шмакова; в — онколит-лепешка, вид снаружи снизу,
рост вокруг плоской раковины; г. д — бугристые онколиты, нараставшие вокруг пелеципод: наружный вид снизу и сверху; е, ж — лодочковидиые онколиты с гладкой поверхностью,
наросшие вокруг целых пелеципод; в — ж — пермь, татарский ярус, Пермская область, ст. Верешагнио
образования, относимые к копролитам (см. раздел «Микрокопро-
литы»).
В основе всякого органогенного желвака в центральной его зоне
лежит обломок породы или какого-нибудь организма, вокруг кото-
рого происходит нарастание слоев при медленном вращении жел-
вака. Как характерные примеры приведены схематизированные
рисунки (рис. 127) желваков, обладающих различным строением
слоев. Внешняя форма желваков в значительной мере зависит от
формы обломка и характера движения его по дну (рис. 127. в — ж).
Часто сильным движением (волновым) желвак может быть рас-
колот. и затем этот осколок вновь становится центром концентри-
ческого нарастания. Иногда желвак в конце своего роста становится
неподвижным, тогда нарастание слоев происходит вверху и с боков.
Все высказанные наблюдения относятся и к онколитам.
СЛЕДЫ СВЕРЛЯЩИХ ИЛИ ПЕРФОРИРУЮЩИХ
ОРГАНИЗМОВ
Табл. 5, фиг. 1—3; табл. 6, фиг. 8
Многие организмы внедряются в известковый субстрат, в том
числе в обломки животных и растений, и образуют «ходы» различ-
ного вида. Большинство ходов — это микроскопические полости,
оставшиеся от жизнедеятельности синезеленых водорослей или
грибов. Но известны также сверлящие или перфорирующие живот-
ные: губки, черви и моллюски.
Термин «сверлящие» обычно не отвечает сущности процесса,
потому что как у растений, так и у животных способ внедрения осно-
ван на растворении субстрата кислотами, а механическое сверление
часто отсутствует, особенно у растений. Термин этот, однако, яв-
ляется общепринятым, и мы будем им пользоваться, не забывая
сделанную выше оговорку.
В ископаемом состоянии чаще всего встречаются следы сверля-
щих синезеленых водорослей, называемые сборным термином
Palaeachlya, и следы грибов (?), также имеющие свое название —
Mycelites. К Palaeachlya Пиа (Pia, 19376) относит изгибающиеся
и иногда разветвляющиеся канальцы внутри раковин, не имеющие
пережимов и концевых вздутий. Диаметр таких канальцев равен
2—5 д. Они встречаются от ордовика до современного периода на
остатках фораминифер, известковых губок, кораллов, брахиопод,
мшанок, криноидей, моллюсков и на чехлах мутовчатых сифоней.
Одновременно со следами сверлений практически всегда на поверх-
ности иссверленных обломков наблюдается кайма из темного пели-
томорфного кальцита. Во многих случаях видно, что сеть темных
нитей канальцев становится все гуще к периферии обломка и, нако-
нец, сливается в сплошную темную массу, иногда образующую
бугристые выступы. Появление этой каймы и выступов объясняется
тем, что водоросли параллельно с разрушением пораженного ими
предмета отлагают на его поверхности микрокристаллический
кальцит (Надсон, 1932). Другими словами, они не просто иссверли-
вают остатки известковых организмов, но перерабатывают их,
уничтожая форменные скелетные фршмешы и от.ипая бесструктур-
ный пелитоморфный карбонат.
Следы сверлящих или перфорирующих организмов. Микрокопролиты
103
Грануляция краев скелетного обломка может быть и не связана
с жизнедеятельностью водорослей, и в общем случае только при-
сутствие ясно различимых отдельных канальцев позволяет говорить
о былом существовании сверлящих организмов. Лишь в некоторых
типах сгустковых известняков, где значительная часть органоген-
ного детрита в той или иной степени источена и переработана свер-
лящими водорослями так, что в шлифе наблюдаются все стадии
этой переработки, даже совершенно бесструктурные сгустки можно
считать результатом работы сверлильщиков.
В. П. Маслов (1937, 1950, 1956), С. В. Максимова (1960) и другие
в своих работах придавали большое значение перфорирующим
(сверлящим) водорослям как показателю мелководности бас-
сейна.
Название Micelites Пиа (1937) применяет к канальчикам в чешуе,
зубах и костях позвоночных животных. Канальчики Micelites имеют
диаметр 1—Зд и сферические вздутия. Они известны с девона по
плиоцен и, по-видимому, относятся к грибам.
Иногда в известковых остатках организмов встречаются
каналы, диаметр которых измеряется не микронами, а деся-
тыми долями миллиметра (0,1 -0,2 м.м). Они, вероятно, при-
надлежат уже не растениям, а животным, скорее всего свер-
лящим губкам.
МИКРОКОПРОЛИТЫ
Табл. 74, 75
Копролитами называют ископаемые фекалии или экскременты
животных. В породе они встречаются в виде известковых телец,
которые получились в результате переработки ила животными
илоядами. Фекалии современных беспозвоночных животных были
известны давно (с ХУЛ в), но изучены они слабо в силу малого
количества отличительных признаков, которыми они обладают.
Мур посвятил фекалиям беспозвоночных животных современных
морей ряд статей, из них последняя (Мооге, 1955) является сводкой
материала по этому вопросу.
Ископаемые извест ковые копролиты отмечались исследователями
начиная с Гюриха (Giirich. 1906). Этот автор принимал цилиндри-
ческие с округлыми концами или овальные тела, состоящие из тем-
ною пелитоморфного карбоната, за фекалии кишечнополостных.
Некоторое внимание копролитовым известнякам и копролитам
уделял М. С. Швецов, приведя их микрофотографии (Швецов,
1958. фото 171, 172, 173) без указания признаков, по которым дан-
ное образование можно было бы с несомненностью отнести к коп-
ролитам. Копрогенные известняки и копролиты как породообразо-
ватели описаны И. В. Хворовой (1953) из среднею карбона Москов-
ской синеклизы и С. В. Максимовой (1961) из нижнего карбона
КузнецкЬго бассейна. Однако специально микрокопролитам посвя-
щено очень мало работ (Маслов, 1960а; Parejas, 1948; Cuviller,
1954; Elliot, 1960).
Анализ и сопоставление всех имеющихся данных по современным
и ископаемым копролитам позволяют выделить ряд достаточно
достоверных сведений об особенностях самих копролитов и их гео-
логическом значении. В донных отложениях современных водоемов
([текальные комочки беспозвоночных распространены очень широко
и нередко являются основной составной частью осадка. В илах
Севастопольской бухты они слагают от 12 до 25% осадка. В заливе
Ферт-оф-Клайд местами грунт целиком состоит из копролитов.
без всякой примеси илистого материала, а, как правило, они со-
ставляют до 40% всей массы осадка (Мооге. 1931b). На дне Боль-
шого Соленого озера (США) большие площади покрыты чисням
копролитовым песком мощностью около 60 см (Eardley, 1938).
В Крыму в дошл \ > щках соленых озер копролиты целиком сла-
1ают многочисленные тонкие прослои (Первольф, 1953). Установ-
лено. что копролиты обладают большой прочностью: они не
разрушаются, когда их довольно грубо трогают руками, и даже
бывают совершенно твердыми в несцементированном осадке
на дне водоема. Распространенность массовых фекальных на-
коплений и прочность копролитов, способствующая их пере-
ходу в ископаемое состояние, заставляют ожидать и в гео-
логическом материале достаточно широкое развитие копроген-
ны.х пород, в частности известняков.
Форма фекальных комочков, как правило, имеет мало харак-
терных признаков, но некоторые простые формы копролитов свя-
заны в своем распространении с определенными фациальными усло-
виями (рис. 128). В морях нормальной и пониженной солености
наиболее распространены более или менее элипсоидальные копро-
литы с округлым поперечным сечением. Такая форма свойственна
представителям разных таксономических групп: очень многим
червям, целому ряду моллюсков и некоторым иглокожим. При
этом грунтоядные черви производят огромное количество копро-
литов — до нескольких тысяч на 1 с.»3 осадка (Мооге. 1931b). Внутри
копролитов, среди более тонкой основной массы, нередко попа-
даются мельчайшие минеральные зерна, обломочки известковых
раковин, панцири диатомей и спикулы губок. Если донный осадок
содержит много спикул, то можно встретить копролиты. почти
целиком состоящие из них. В противоположность этому в водоемах
с заметно повышенной соленостью встречаются только фекалии
в форме цилиндрических палочек, иногда изогнутых, с длиной,
превышающей диаметр в 3—10 раз. Эти цилиндрические копролиты
принадлежат мелким ракообразным Artemia. которые являются
единственными или резко доминирующими представителями фауны
в ультрагалинных бассейнах. В соленых озерах Крыма и в Большом
Соленом озере (США) донные отложения слагаются именно шигинд-
рическими копролитами артемий.
Ископаемые копролиты встречаются в весьма различных типах
известняков, но в известняках открыто-морских фаций, изобилую-
щих остатками разнообразной фауны, они всегда являются второ-
степенным компонентом породы, образуя либо небольшие скопления
в ходах илоядов, в норках роющих животных, либо характерные
«дорожки», вероятно тоже представляющие следы ползания илояд-
ных форм (табл. 75, фиг. 9). Зато в мелководных условиях, особенно
в сравнительно затишных зонах, копролиты нередко слагают целые
пласты известняка мощностью 0,5—3,0 .и, изредка до 6 м (Мос-
ковская синеклиза. Кузнецкий бассейн, юго-западная Англия).
Макроскопически такие копрогенные известняки сливные или мел-
козернистые. серые, голубовато-серые или белые, обычно образую-
щие в обнажении монолитные плиты. При микроскопическом изу-
чении они распадаются на две разновидности.
В известняках первой разновидности основным компонентом
породы являются копролиты Coprolites communis Masi. (sp. n.) —
темные комочки пелитоморфного карбоната, чаще однородные,
но нередко включающие органогенный шлам или единичные мелкие
минеральные зернышки. Комочки в сечении имеют круглую, оваль-
ную или яйцевидную форму. Их внешний контур четкий, гладкий
или слабобугристый. Копролиты погружены в микрозернистый
кальцит, в котором они «плавают» не соприкасаясь и от которого
отличаются более темным цветом и еще более мелким размером
слагающих их кристалликов кальцита (табл. 75, фиг. 8). Органи-
ческих остатков в породе немного, и среди них наиболее характерны
гастроподы и остракоды.
Во второй разновидности копрогенных известняков копролиты
имеют более однообразную форму, всегда приближающуюся в се-
чении к кругу, и сцементированы они разнозернистым прозрачным
кальцитом. Здесь обычно довольно много мелкого окатанного орга-
ногенного детрита и нередко присутствует в заметном количестве
алевритовая или песчаная примесь. Во многих слоях встречаются
немногочисленные зачаточные оолиты с копролитовым или ока-
104
Следы жизнедеятельности организмов
Рис. 128. Копролиты
а — и — наружный вид конроднiob современных морских беспозвоночных, по Эджу (Edge. 1934): а— г — иглокожих: а — голотурий Sliehopus caiifornicus. xl.8.<> — офиуры Ophio-
derma panuniensis. х 4.5. в — морской звезды Asterina miniata х 2.7. г — морского ежа Lptmhinus amtnesis. х 3.6: д — з — червей: <> — Nereis cexilvsa. х 18. е - Amphitrite sp.. х 12.6.
*< Lamhrinereis sp. ? х 4.5. J--серпулы. х 9: и — членистоио!ого lepidora myttps, X 10.8: к— .» — моллюсков: к— Tethis ealifornita. х 3.6. t — Hermissenda crassicornis. /6.3.
Cardium quadraigenarium. x 14.4; и— Polypus dintaculatus. xl.8 (кружочки сбоку — поперечные сечения): о — ц—наружный вид и поперечные сечения (кроме <>. т. ф) копроли-
юв современных и ископаемых животных, по Муру (Moore. 1955): о — ископаемые тела из калифорнийских сланцев, п — Leda minuia. р - Chlamis сапа. < •- Numla tenuis, m Mvu
truttcuta. ,v — Cibhula magus, ф — Emarginula fissura. v — Mytilus edulis. Ц— Galathea squamifera: ч. ш— продольное (ч) и поперечное (ш) сечения через копролиты Parreina salerensis
(Parejas). х около 26. пурбек, Франция, Верхняя Савойя (Parejas, 1948); u/.э — продольное (ш) и поперечное (?) сечения через Palaxieus hahanensis Bronn. et Norton, x около 20. мио-
цен. Куба. Гаванна
тайным детритом в ядре. Очень характерно для этой разновидности
наличие тонкой косой или косоволнистой слоистости, реже встре-
чаются знаки ряби. Ясные следы воздействия на осадок волнистых
токов воды позволяют считать, что в этой разновидности копро-
литы, как и обломки фауны, окатаны, чем и объясняется их одно-
образная, приблизительно сферическая форма.
Прямым доказательством копрогенной природы этих извест-
няков являются постоянно наблюдающиеся переходы по горизон-
тали. в пределах одного пласта, копрогенных известняков второй
разновидности в известняки первой разновидности с сохранившейся
первичной формой копролитов и микрозернистым цементом. Иногда
такие переходы можно наблюдать прямо в шлифе, где среди пе-
ремытого осадка местами встречаются небольшие участки копро-
генного известняка первой разновидности.
В этих древних известняках, так же как в современных морских
осадках, преобладают копролиты более или менее эллипсоидальной
формы. Вс зечаются в геологическом материале и скопления ци-
линдрических копролитов, ничем не отличающихся по форме от
копролитов современных артемий, формировавшиеся, как и совре-
менные, в ультрагалинных водоемах. На табл. 75, фиг. 3 изображен
шлиф третичного копрогенного известняка. Известняк слабо сце-
ментированный, в разрезе ассоциируется с гипсами и загипсован-
ными породами. Благодаря беспорядочному, неориентированному
расположению копролитов в породе в плоскости шлифа мы видим
Микрокопролиты
105
различные их сечения: продольные (длинные палочки), поперечные
(небольшие кружки) и скошенные под различными углами. Эти
последние в зависимости от крутизны скоса выглядят как овалы,
то короткие, то удлиненные. Сравнительно короткие овалы могут
также давать обломанные цилиндры в продольном или почти про-
дольном разрезе. Решающими для определения истинной формы
и природы всей массы комочков разных очертаний являются длин-
ные палочки, т. е. продольники хорошо сохранившихся копролитов,
которые всегда обнаруживаются в шлифе, хотя бы в небольшом
количестве.
Копролиты мелких ракообразных благодаря их цилиндрической
(палочковидной) форме легко отличаются как от обломочных из-
вестняковых песчинок, так и от всех частиц в известняках микро-
комковатой структуры. Отличия обычных морских копрогенных
известняков от других микрокомковатых пород не столь очевидны.
По структурным особенностям к копрогенным наиболее близки
так называемые микросгустковые известняки, сложенные мелкими
округлыми карбонатными комочками размером, как правило,
не больше 0,10 мм. Однако эти типы обладают вполне определен-
ными и постоянными различиями.
Прежде всего в микросгустковых известняках самого различного
возраста и из весьма удаленных друг от друга районов размер ос-
новной массы комочков остается постоянным (до 0,10 мм), тогда
как величина копролитов в разных местах и даже в разных слоях
одного разреза бывает неодинаковой и либо все они, либо значитель-
ная часть их оказывается крупнее 0,10 мм. У микросгустков контур
нерезкий, у копролитов — совершенно четкий, ясно выделяющийся
на фоне микрозернистого цемента. Микросгустки часто сливаются,
образуя пятна сплошной пелитоморфной массы; копролиты даже
при самом тесном расположении сохраняют индивидуальные очер-
тания. Наконец, копролиты нередко включают органогенный шлам
или мелкие минеральные зернышки, чего никогда не бывает в ко-
мочках микросгусткового известняка. Возможно, что микросгуст-
ковые известняки являются хемогенным осадком, а сами сгусточки
представляют образование, подобное современным дрюитовым
комочкам, которые тоже местами сливаются в более крупные участ-
ки однородной пелитоморфной структуры.
Oi снетков водорослево!о происхождения копролиты также
отличаются рядом признаков.
1. Размер водорослевых сгустков в каждом образце и в каждом
шлифе колеблется от сотых или десятых долей миллиметров до
нескольких миллиметров, иногда до сантиметра, а величина копро-
литов из одного образца варьируег в узких пределах (например,
от 0.05 до 0.20 .и.н, от 0.10 до 0,25 мм. от 0.20 до 0.50 мм).
2. Копролиты отличаются достаточно правильной, обычно
округленной формой, сгустки имеют самую разнообразную и не-
правильную форму, нередко с резко фестончатым контуром.
3. Внутри водорослевых сгустков наблюдаются пустоты, ка-
нальцы (губчатая структура) или обломки фауны, в разной степени
источенные сверлящими водорослями. Размер этих обломков раз-
личный и часто крупный. Обыкновенные широко распространенные
копролиты всегда плотные, однородные и мо1ут включать лишь
мелкий органогенный шлам.
Обломочные известняковые песчаники обнаруживают известное
сходство с копрогенными известняками только в том случае, когда
все карбонатные песчинки состоят из одинакового пели гоморфног о
однородного известняка. Однако даже хорошо окатанные песчинки
сохраняют следы огранки обломочных частиц, вследствие чего их
форма значительно более разнообразна, чем форма копролитов.
Копролиты. включающие посторонние предметы (фораминиферы.
обломки иглокожих, спикулы губок, минеральные зерна и т. д.).
отличаются от известняковых песчинок и галечек тем. что у галек
включения срезаются их поверхностью (галька как бы вырезана из
породы), а у копролитов включения всегда отделены от внешней
поверхности тонким слоем пелитоморфного карбоната, одинакового
с основной массой копролита.
В последние годы обнаружены копролиты. резко отличающиеся
своей внутренней структурой от других похожих образований. Так.
копролиты с продольными просветами — каналами уже имеют
свою историю исследования. Впервые их описали как «организм В»
Жуковский и Фавр (Joukovsky. Favre, 1913). В дальнейшем Пере-
жас (Parejas, 1948) отнес их к копролитам — фекалиям ракообраз-
ных. назвав их Coprolitus salevensis Par. К нему присоединились
Кароцци (Carozzi. 1948). Кювийе (Cuviller. 1954) и Маслов (1960а).
Эта же форма копролитов описана также под названием Finreina
и Palaxius.
Распространен этот тип копролитов от триаса до миоцена вклю-
чительно и особенно часто встречается в верхней юре запада Украи-
ны и в бухарском и алайском «ярусах» района р. Вахша и г. Душан-
бе. Зарегистрированы они в триасе Ирака, в юре и мелу Франции,
н-ова Аравия. Швейцарии и Северной Африки, известны в олигоцене
Анатолии, Марокко и Франции, в эоцене Гватемалы и в миоцене
Ирака. Форма этих копролитов неправильно-цилиндрическая или
усеченно-коническая, несколько сужающаяся к концам (табл. 74.
фиг. 2). В продольном к оси сечении тонкие, прямые просветы имеют
неодинаковую ширину. В поперечном к оси сечении копролиты не-
правильно округлены, с неровным контуром и светлыми точками
просветов, расположенных беспорядочно, иногда с некоторым отно-
сительным порядком. Эти просветы в поперечном сечении бывают
округлыми и угловатыми. При увеличении в 90 раз видно, что коп-
ролиты состоят из неоднородной массы пелитоморфного карбоната,
уплотненной близ светлых просветов и включающей иногда более
крупные зерна — песчинки карбоната. Концы копролитов или сре-
заны. или закруглены, но просветы — каналы не открываются на-
ружу, а закрыты тонким слоем пелитоморфной массы.
В последние годы Эллиот (Elliot, 1962), присоединившись к
мнению. что эти остатки являются копролитами. оставил родовое
название Faireina. По количеству продольных каналов, их форме
и размерам как каналов, так и самих телец Эллиот описал несколько
видов. Он объясняет каналы этих телец как результат воздействия
особых складок в желудочном тракте некоторых ракообразных,
например современного рачка Galathea, что не очень убедительно.
Различное количество каналов, размеры и т. п. принимаются Эллио-
том за родовые и видовые признаки. Им описаны восемь видов
фаврейн, два вида Prothocoprolites (трубка с толстыми стенками —
может быть, и не копролит). один вид Agregatella — бесструктурные
пелитоморфные цилиндры. Дальнейшие исследования покажут,
насколько признаки, выбранные Эллиотом, специфичны, так как
копролит одного и того же животного может несколько измениться
при движениях хозяина.
Другого типа копролиты встречены в среднем карбоне Куйбы-
шевской области. Они имеют внутренние светлые просветы, но при-
хотливо закручены наподобие кишок (табл. 74. фиг. 1). Форма всего
копролита неправильно овальная, иногда изогнутая, внешняя гра-
ница иногда неровная, с выступами, отвечающими выступам, обра-
зуемым петлями заворачивающихся каналов. Диаметр внутренних
просветов каналов непостоянный, поперечное сечение их более или
менее округлое. Закономерности в закручивании каналов подметить
не удается. Мы предлагаем назвать этот копролит Coprolitus ver-
micularis Masi. Внутренние каналы не всегда отчетливы, чаще их
нет совсем и копролит выглядит однородным, но размер и внешняя
форма сохраняются. Происхождение кишкообразных каналов
скорее всего объясняется выбросом червевидных кишечных пара-
зитов.
Наконец, необходимо отметить интересные копролиты, имеющие
небольшое заострение на конце, иногда выражающееся в маленьком
хвостике из пелитоморфного карбоната. Такие «хвостатые» коп-
ролиты часто относят или к водорослям, или к проблематикам,
или к образованиям механического происхождения (Carozzi, 1961,
106
Следы жизнедеятельности организмов
фиг. 1). Действительно, в образовании копролитов участвует и
механическое формирование при выходе его из кишечника илояда.
Крупные илояды могут деформировать оолиты, и тогда получаются
оолито-копролиты, раздавленные половинки которых разрушены
по краям и вытягиваются иногда цепочкой. Встречаемость отдельных
«хвостатых» половинок среди ненарушенных оолитов (оворит о коп-
ролитовом происхождении первых (Маслов, 1960 а). На табл. 74,
фиг. 5—8 изображены такие хвостатые копролиты из разных систем.
Некоторые авторы (Carozzi, 1961) трактуют такие хвостатые копро-
литы как псевдооолиты1, раздавленные и смятые механическими
агентами. Кароцци, изучая дробленые или, вероятнее, раздавленные
оолиты, приводит разные случаи. Соглашаясь с ним в части объяс-
нения участия механического дробления, нельзя согласиться с его
мнением, что причиной раздавливания служили только механические
усилия на осадок. Приведенный пример (табл. 75, фиг. 7), взятый
из работы Кароцци, говорит о том, что одни механические усилия
на осадок не могли образовать таких причудливых форм и отбор
по внутреннему содержимому «псевдооолитов». Рассмотрение того
же материала как продукта механической и, одновременно, хими-
Происхождение «псевдооолитов» не всегда ясно. По-видимому, часть из них ниже оь
носится к копролитам.
ческой переработки ила в кишечниках разных илоядов легко объяс-
няет разнообразную форму и внутреннее содержание этих образо-
ваний, а также цепочкообразное их расположение с причудливым
изгибом цепочки.
По-видимому, часть микропроблематик и озагий (онколитов)
относится к копролитам и требуется критический их пересмотр.
Два случая приведены на табл. 74, фиг. 4,5.
Копролиты, как результат жизнедеятельности илоядов, не могут
служить показателями глубины бассейна, так как илоядные животные
обитают на разных глубинах (0—4224 ж). Но, являясь породообра-
зующими. копролиты характеризуют определенные фации, иногда
лишенные других остатков организмов, и могут служить местными
корреляторами. С другой стороны, и илояды в течение геологиче-
ской истории появлялись и исчезали, заменяясь новыми, поэтому
могут быть и такие копролиты, которые окажутся возрастными ука-
зателями. Но для этого необходимы признаки, которые позволяли
бы не смешивать их с другими копролитами.
Копролиты могут переноситься и вторично образовывать осадок
механического происхождения, но обычно они встречаются вместе
с детритусом известковых организмов и произошли на месте. Есте-
ственно, что «хвостатые» копролиты при перемещении теряют свои
тоненькие окончания. Поэтому все такие хвостатые копролиты
нужно считать автохтонными.
ЛИТЕРАТУРА
А й з е н в е р г Д. Е. 1958. Стратиграфия и палеогеография нижнего
карбона Западного сектора Большого Донбасса. Киев. Изд-во
АН УССР.
АрхангельскийА. Д. 1912. Верхнемеловые отложения Вос-
тока Европейской России. — Материалы по геологии Рос-
сии, 25.
Б е л ь с к а я Т. Н. 1960. Позднедевонское море Кузнецкой котло-
вины, история его развития, население и осадки. М., Изд-во
АН СССР
Богданович А. К.. Дмитриева Р. Г. 1956. О халцедоне
в стенках фораминифер. — Докл. АН СССР, 107, № 6.
Б ы к о в а Е. В. 1955. Фораминиферы и радиолярии девона Волго-
Уральской области и Центрального девонского поля и их
значение для стратиграфии. — Труды ВНИГРИ, нов. серия,
вып. 87.
ВассоевичН. Б. 1950. О распространении Calpionella Lorenz
в Азербайджане. — Бюлл. МОИП. отд. геол.. 25, № 6.
Векшина В. Н. 1959. Кокколитофориды маастрихтских отло-
жений Западно-Сибирской низменности.—Труды
СНИИГГИМС. вып. 2.
Виноградов А. П. 1935. Химический элементарный состав
организмов моря. ч. I. М.. Изд-во АН СССР.
Виноградов А. П. 1937. Химический элементарный состав
организмов моря.—Труды Биохим. лаборат. АН СССР. 4.
Виноградов А. П. 1944. Химический элементарный состав
организмов моря, ч. III. — Труды Биохим. лаборат. АН СССР, 6.
В я л о в О. С., ЛинецкаяЛ. В. 1960. Об ископаемых инфузо-
риях из мезозоя Карпат.—Докл. АН СССР. 131. № 6.
ГеккерР. Ф. 1957. Введение в палеоэкологию. М. Госгеолтех-
издат.
ГеккерР. Ф., Ушаков Б. В. 1962. Черви. В кн.: Основы палеон-
тологии. Губки, археоциаты, кишечнополостные, черви. М..
Изд-во АН СССР.
Г ерасимовН. П. 1953. Солоноватоводные лагунные формации
пермской системы востока Русской платформы. — Уч. зап.
Пермского гос. ун-та, 7, вып. 1.
Г л е з е р 3. И. 1966. Кремневые жгутиковые водоросли (силико-
флягелляты). — В кн.: Флора споровых растений СССР. 7.
М.—Л., «Наука».
Горский И. И. 1935. Некоторые Coelenterata из нижнекамен-
ноугольных отложений Новой Земли. — Труды Аркт. ин-та,
28, серия геол.
Горюнов С. В. 1958. Некоторые закономерности развития и
распада планктонных форм водорослей в дальневосточных
морях.—Труды Океанограф, комиссии, 3. М.. Изд-во АН
СССР.
ДембскаяГ. И. 1959. Закономерности распределения биофа-
циальных комплексов в некоторых морских горизонтах вор-
кутской свиты (нижняя пермь Печорского бассейна). — Труды
II сессии Всесоюзного палеонтол. об-ва. М., Госгеолиздат.
ДембскаяГ. И., ДембскаяВ. И. 1965. Биофациальные комп-
лексы фаунистического горизонта рудницкой подсвиты вор-
кутской серии Печорского бассейна. — Материалы по геол,
и полезн. ископаемым северо-востока Европейской части СССР,
вып. 4. М., «Недра».
ДобролюбоваТ. А., КабаковичН. В.. Чудинова
И. И. 1964. Наставления по сбору и изучению палеозойских
коралов. М., «Наука».
Еленкин А. А. 1936. Синезеленые водоросли СССР, вып. I. М.,
Изд-во АН СССР.
Ж у зе А. П. 1951. Диатомовые и кремневые жгутиковые водоросли
верхнемелового возраста из северного Урала. — Ботанические
материалы Отдела споровых растений Ботанич. ин-та АН
СССР, 7. М —Л., Изд-во АН СССР.
Ж узе А. П. 1955. Кремневые жгутиковые водоросли палеогено-
вого возраста. — Ботанические материалы Отдела споровых
растений Ботанич. ин-та АН СССР, 10, М. — Л.. Изд-во АН
СССР.
Жузе А. П. 1966. Кремнистые осадки в современных и древних
озерах. — В сб.: Геохимия кремнезема. М.. «Наука».
Ж у з е А. П. 1968. Древние диатомеи и диатомовые породы Тихо-
океанского бассейна. —Литология и полезные ископаемые. № 1.
Ж у з е А. П., К о з л о в а О. Г., М у х и н а В. В. 1969. Диатомеи
и силикофлягелляты в поверхностном слое осадков Тихого
океана. — В кн.: Тихий океан. Микрофлора и микрофауна
в современных осадках океанов. М., «Наука».
Ж у р а в л е в а И. Т. 1960. Археоциаты Сибирской платформы.
М„ Изд-во АН СССР.
Ж у р а в л е в а И. Т. 1963. Археоциаты Сибири (одпос генные архео-
циаты). М., Изд-во АН СССР.
Ж у р а в л е в а И. Т. 1965. Класс Sphinotozoa. — В кн.: Развитие
и смена морских организмов на рубеже палеозоя и мезозоя.
Труды ПИН АН СССР, 108.
Ж у р а в л е в а И. Т., К о н ю ш к о в К. Н„ Р о з а н о в А. Ю.
1964. Археоциаты Сибири (двустенные археоциаты). М„ «Наука».
Ж у р а в л е в а И. Т., К о р ш у н о в В. И., Р о з а и о в А. Ю. 1969.
Атдабанский ярус и его палеонтологическое обоснование в
стратотипе.—В сб.: Биостратиграфия и палеонтология ниж-
него кембрия Сибири и Дальнего Востока. М., «Наука».
Ж у р а в л е в а И. Т.. Р о з а н о в А. Ю. 1962. Возраст и условия
образования археоциатовых известняков бассейна р. Енисей. —
Геология и геофизика, № 3.
3 е л е и о в К. К. 1957. Литология кембрийских отложений север-
ного склона Алданского массива. — Труды ГИН АН СССР,
вып. 8.
Зенкевич Л. А. 1955. Моря СССР, их фауна и флора. М.. Изд-во
АН СССР.
Иванова Е. А. 1949. Условия существования, образ жизни и
история развития некоторых брахиопод среднего и верхнего
карбона Подмосковной котловины. — Труды ПИН АН СССР,
21.
Иванова Е. А. 1958. Развитие фауны в связи с условиями сущест-
вования.— Труды ПИН АН СССР. 69.
И в а и о в а Е. А.. Б е л ь с к а я Т. Н., Ч у д и н о в а И. И. 1964.
Условия обитания морской фауны силура и девона Кузнецкого,
Минусинского и Тувинского бассейнов.—Труды ПИН АН
СССР, 102.
К а п т а р е н к о-Ч е р н о у с о в а О. К. 1954. Об остатках голо-
турий в юрских отложениях Днепровско-Донецкой впадины. —
Изв. АН СССР, серия геол., № 3.
Карпинский А. П. 1906. О трохилисках. — Труды Геол, коми-
тета, нов. серия, вып. 27.
К и р.е е в а Г. Д.. М а к с и м о в а С. В. 1959. Фациальные изме-
нения известняков свит С2. Cf, С2 Донецкого бассейна. —
Труды ВНИИГаз, вып. 4.
Колтун В. М. 1968. Тип губки (Porifera или Spongia). — В кн.:
Жизнь животных. 1 М., «Просвещение».
Кор д э К. Б. 1954. Кембрийские водоросли из окрестностей о. Бо-
гучаны на р. Ангаре. — В кн.: Вопросы геологии Азии, 1.
М., Изд-во АН СССР.
Королюк И. К. 1952. Подольские Толтры и условия их обра-
зования. — Труды ИГН АН СССР, вып. ПО, серия геол. (№ 56).
Косинская Е. К. 1948. Определитель морских синезеленых
водорослей. М.-Л., Изд-во АН СССР.
КраснопееваП. С. 1937. Водоросли и археоциаты древней-
ших толщ Потехинского планшета Хакассии. — Материалы
по геол. Красноярского края, № 3.
Крашенинникове. А. 1956. Микроструктура стенки неко-
торых кайнозойских фораминифер и методика ее изучения
в поляризационном свете. — В кн.: Вопросы микропалеонто-
логии, № I. М., Изд-во АН СССР.
Крашенинников В. А. 1960. Изменение комплексов форами-
нифер в ритмах осадконакопления миоценовых отложений
юго-запада Русской платформы. — В кн.: Вопросы микро-
палеонтологии, № 4. М., Изд-во АН СССР.
К р и в и н А. Л., М а с л о в В. П. 1962. Новые данные по страти-
графии и водорослям верхнего мела нижнего палеоцена Мар-
марошского массива. — Изв. АН СССР, серия геол., № 12.
Кругликова С. Б. 1966. Количественное распределение радио-
лярий в поверхностном слое донных осадков северной поло-
108
Литература
вины Тихого океана. — В кн.: Геохимия кремнезема. М..
«Наука».
К р у г л и к о в а С. Б. 1969. Радиолярии в поверхностном слое
осадков северной половины Тихого океана. — В кн.: Тихий
океан, 8, М., «Наука».
КурсановЛ.И. и др. 1953. Определитель низших растений.
1. — «Водоросли». М., «Советская наука».
Л и п и н а О. А. I960. Пограничные слои девона и карбона и нижне-
турнейские отложения Среднего Урала. — Докл. АН СССР.
133, № 5.
Л и с и ц ы н А. П. 1966. Основные закономерности распределения
современных кремнистых осадков и их связь с климатической
зональностью. — В кн.: Геохимия кремнезема. М.. «Наука».
Ляшенко Г. П. 1959. Кониконхии девона центральных и восточ-
ных областей Русской платформы. Л„ Гостехиздат.
Максимова С. В. 1955. Фациально-экологическая характерис-
тика продуктивной толщи Сызранского района. — Труды
Ин-та нефти АН СССР, 5.
М а к с и м о в а С. В. 1956. О некоторых мелководных известняках
нижнего карбона Кузнецкого бассейна. — Труды Ин-та нефти
АН СССР. 7.
Максимова С. В. 1959. Иловые трубки полихет из нижнего
карбона Кузнецкою бассейна. — Палеонтол. ж.. № 1.
Максимова С. В. I960. Фациально-экологическая характерис-
тика нижпедевонских отложений р-на г. Гурьевска (Кузнецкий
бассейн). — Материалы по геол, и нефти Кузбасса. Л.. Изд-во
АН СССР
М а к с и м о в а С. В. 1961. Литология и условия образования биту-
минозной известняковой толщи нижнего карбона Кузнецкого
бассейна. М.. Изд-во АН СССР.
М а л ы ш е к В. Т. 1948. Кокколиты — породообразующие фора-
миниферовой свиты Северного Кавказа. - Докл. АН СССР.
59. № 2
Манда л я н Р. А. 1964. О силицитах из верхнеюрских от ложений
северо-восточной части Армянской ССР. — Изв. АН АрмССР.
науки о Земле. 17. № 3—4.
МасловВ. П. 1929. Микроскопические водоросли каменноуголь-
ных известняков Донецкою бассейна. — Изв. Геол, комитела.
48. № 10.
МасловВ. П. 1935. Карбонатные водоросли как геолол ический
фактор. — Проблемы советской геологии. № 5.
М а с л о в В. П. 1936. О стромаголиговых постройках на Южном
Урале. — Труды ГИН АН СССР. вып. 5.
МасловВ. П. 1937а. Атлас карбонатных пород. ОНТИ. НКТП
СССР. ч. I. М.-Л.
Маслов В. П. 19376. О распространении карбонатных водорослей
в Восточной Сибири. — Проблемы палеонтол.. 2 —3. М.. Изд-во
МГУ.
МасловВ. П. 1945. К вопросу о фазах седиментации и карсто-
образовании погребенных массивов Ишимбая. - Изв. АН
СССР, серия геол., № I.
М а с л о в В. П. 1949. Происхождение кембрийских известняков
Тувы. — Изв. АН СССР, серия геол., № 2.
МасловВ. П. 1950. Геолого-литололические исследования рифо-
вых фаций Уфимского плато.—Труды ИГН АН СССР, вып. 118.
серия леол. (№42).
МасловВ. П. 1952. Карбонатные желваки орланическоло проис-
хождения. — Бюлл. МОИП, отд. геол.. 27. № 4.
Маслов В. П. 1956. Ископаемые известковые водоросли СССР. —
Труды ИГН АН СССР. вып. 160.
МасловВ. П. 1958. Находка в юре Крыма рода Coscinoconus
Leupold и его истинная природа. — Докл. АН СССР. 121, № 3.
МасловВ. П. 1960а. Копролиты и следы сверлящих орланнз-
мов — их значение для лнтолола.— Изв. АН СССР, серия
геол.. № 10.
МасловВ. П. 19606. Строматолиты. — Труды ГИН АН СССР,
вып. 41.
М а ел о в В. П. 1961а. Не являются ли сицидии и ховапеллы утри-
кулами харофитов? — Докл. АН СССР. 138. № 3.
М а с л о в В. П. 19616. Водоросли и карбонатоосаждеине. — Изв.
АН СССР, серия леол. № 12.
М а с л о в В. П. 1962. Ископаемые балряные водоросли СССР
и их связь с фациями. - Труды ГИН АН СССР. вып. 53.
МасловВ. П. 1963а. Введение в изучение ископаемых харовых
водорослей. — Труды ГИН АН СССР. вып. 82.
М а с л о в В. П. 19636. Класс хризомопад. — В кн.: Основы па-
леонтологии (водоросли, мхи. папоротники). М.. Изд-во АН
СССР.
МасловВ. П. 19бЗв. Красные или балряные водоросллл. В кн.:
Основы палеонтологии. Низшие растения. М.. Изд-во АН СССР.
Маслов В. П. 1963л. Членистые балряные водоросли Junta ллз
среднесарматских биолермов Молдавии. — Палеонтол. ж. №1.
МасловВ. П. 1967. Микрокодии. - Палеонто л. ж. №1.
М а с л о в В. П.. Р е п г а р л е н Н. В. 1964. Находка ископаемых
известковых водорослей в лёссах. - Док л. АН СССР. 159. № 3
М а с л о в В. П.. Я р ц е в а М. В. 1969. Мутовчатые сифоллеи
(Dasycladiaceae) из отложений палеоцена Украинского щита.
Украинский бол. жури.. 26. № 5.
М и х а й л о в а М. В. 1968. Биолермные массивы в верхнсюрских
отложениях Горною Крыма и Северною Кавказа. - В сб.:
Ископаемые рифы и методика их изучения. Свердловск, изд.
Уральск, фил. АН СССР.
М о и с е с в А. С. 1944. Водоросли, лубки, лидроидные полипы
и кораллы верхнего л риаса лл Кавказскою хребта.— Уч. зап.
ЛГУ. № 70. серия леол.-почв., вып 11.
М о р о з о в а В. Г.. Кожевникова Г. Е.. К у р ы л е в а А. М.
1966. Датско-палеоценовые разнофациалылые отложения Конет-
Дала и методы их корреляции ио фораминиферам. — Труды
ГИН АН СССР, вып. 157.
Н а д с о и Г. А. 1932. Малериалы к изучению сверлящих водорос-
лей. - Изв. АН СССР. № 7.
Ос и нов а А. И. 1950. Литология лл фациальный анализ верхне-
палеозойских отложений среднего течения р. Юрезани. — Труды
ПИН АН СССР. 30
Осипова А. И. 1954. Ферганский анализ палеогеновою моря,
история ею развития и условия обитания населявшей ело фауны
и флоры М.. Изд-во АН СССР.
Основы палеонтологии. 1958. Моллюски, л оловоногие. 2. М., Гос-
научтехиздат.
Основы палеонтологии. 1959. Общая часть. Простейшие. М.. Изд-во
АН СССР.
Основы палеонгололии. 1960. Миланки, брахиоподы. М.. Изд-во
АН СССР
Основы палеонгололии. I960. Моллюски панцирные, двустворчатые,
.лопатонолне. М.. Изд-во АН СССР.
Основы палеонтололии. I960. Моллюски брюхоногие. М.. Госнауч-
техиздат.
Основы палеонтололии. 1962. Губки, археоциаты, кишечнополост-
ные. черви. М.. Изд-во АН СССР.
Основы палеонтолол ии. 1962. Моллюски головоногие. 1. М.. Изд-во
АН СССР
Основы палеонгололии. 1963. Членистоногие — трилобитообразные
и ракообразные. М.. «Недра».
Основы палеонгололии. 1963. Водоросли, мохообразные. М.. Изд-во
АН СССР.
Основы палеонтолол ин. 1964. Ил локожие. гемихородовые, полоно-
форы гг лцегипко-челюст ные. М.. «Недра».
Пастернак I. С. 1955. Серпуллди крейдяних вщклад1в Волино-
Подгн>ско1' плити i ix значения для стратиграфа. — Наук. зап.
Природозн. муз. Льв1вск. Фи. АН УССР. 4.
П е р в о л ь ф Ю. В. 1953. Илы и условия их образования в соляных
озерах Крыма. Труды Лаборат. озеровед. АН СССР. 2.
П е г е л и и В. П. 1954. О современных кремнегубковых морских
осадках. — Бюлл. МОИП. серия леол.. 29. № 1.
Пел р у ш е в с к а я М. Г. 1966. Радиолярии в планктоне и в дон-
ных осадках. — В кн.: Геохимия кремнезема. М.. «Наука».
Пет р у ш е в с к а я М. Г. 1969. Радиолярии Spumcllaria и Nas-
sellaria в донных осадках как индикаторы лидро.логических
условий. — В сб.: Основные проблемы микропалеонтологии
и орл аногелнлол о осадконакопления в океанах и морях. М..
«Наука».
П о я р к о в В. В. 1965. О систематическом положении умбелл.—
Докл. АН СССР. 163. № 3.
П о я р к о в В. В. 1966. Девонские харофиты Тянь-Шаня.—Труды
ГИН АН СССР. вып. 143.
П р и г о р о в с к и й В. Г. 1948. Фауна мягких грунтов литорали
губы Дальне-Зе.ленецкой. — Труды Мурманской биостанции, 1.
Р а у з е р -Ч е р и о у с о в а Д. М. 1953. Периодичность в развит ии
фораминиг|)ер верхнего палеозоя и ее значение для расчленения
и сопоставления разрезов. — Материалы палеонтологическою
совещания по палеозою. 1951 л.. М.. Изд-во АН СССР.
Р е й л л и и л е р Г. А. 1954. Девонские фораминиферы некоторых
разрезов восточной части Русской платформы. — Палеонтол.
сб. ВНИГРИ. вып. 1. М.. Гостоптехиздат.
Р е и т л и н л е р Г. А.. Я р н е в а М. В. 1958. Новые харофил ы
верхиефаменских отложений Русской платформы. Докл. АН
СССР. 123. № 6.
Р е п и н а Л. Н.. Хоме и т о веки л“л В. В. Ж у р а в л е в а И. Г..
Р о з а и о в А. Ю. 1964. Биостра л ш рафия нижнею кембрия
Саяпо-Алтайской складчатой области. М.. «Наука».
Литература
109
Решения третьей палеэколою-литологической сессии. 8—22 июня
1965 I. Сверл, ювек. 1966.
Решения четвертой па.icojko.toio-.ihiojoi ической сессии, проходив-
шей в Крыму и Молдавии в сентябре 1966 i. М.. 1968.
Р е ш е 1 н и к В. В. 1955. Вертикальное распределение радиолярий
Курило-Камчатской впадины.—Труды Збол. ин-та АН СССР.
21
Р о ж к о в а Е. В.. Г о р е ц к и й Ю. К. 1945. Современные кремне-
вые отложения. — Труды ВИМС. вып. 117.
Розанов А. Ю.. М и с с а р ж е в с к и й В. В. 1966. Биостра ги-
I рафия и фауна нижних юризонтов кембрия.—Труды ГИН
АН СССР, вып. 148.
Роза и о в А. Ю.. М и с с а р ж е в с к и й В. В. 1969. Томмотский
ярус и проблема нижней 1 раницы кембрия. - Труды ГИН
АН СССР. вып. 206.
Р я б и н и н В. Н. 1915. Рифообразователи каменноу) ольны.х отло-
жений Урала и Тимана. - Труды Геол, комитета, нов. серия,
вып. 123.
С а й д а к о в с к и й Л. Я. 1960. Биострати> рафическая схема ниж-
нею триаса Диеп ровско-Донецкой впадины. — Геол. ж.
Укр. ССР. 20. вып. 6.
С а м о й л о в а Р. Б. 1955. Об озерско-хованских т рохи.тисках. —
Докл. АН СССР. 103. № 5.
Сары ч е в а Т. Г.. Соколье к а я А. Н., Безносов а Г. А..
М а к с и м о в а С. В.. 1963. Брахиоподы и палеогеография
карбона Кузнецкой котловины. — Труды ПИН АН СССР. 95.
С а р ы ч е в а Т. Г., С о к о л ь с к а я А. Н., М а к с и м о в а С. В..
Б е з н о с о в а Г. А. 1962. Фациальная зональность брахиопод
в каменноут ольных морях Кузнецкой котловины. — Палеон-
тол. ж.. № 4.
Сергеева С. П. 1962. Общее стратиграфическое значение коно-
донтов. — Вестник ЛГУ. № 12. серия геол, и географии, вып. 2.
Сергеева С. П. 1964. Конодонты. — В кн.: Основы палеонто-
логии. Бесчелюстные. Рыбы. М. «Наука».
С е р м я г и н В. А. 1938. Литологическая характеристика извест-
няков Ишимбаевского месторождения нефти.—Труды Нефт.
геол.-развел, ин-та. серия. А., вып. 101.
С о к о л о в Б. С. 1955. Табуляты палеозоя Европейской части
СССР. Введение. Общие вопросы систематики и история разви-
тия табулят.—Труды ВНИГРИ, нов. серия, вып. 85.
С о к о л о в Б. С. 1962. Подкласс Tabulata. Табуляты. — В кн.:
Основы палеонтологии. 2. М. Изд-во АН ССР
С о р о к и н В. С.. Г р а в и т и с В. А. 1964. Некоторые закономер-
ности распределения аутигенного кремнезема в отложениях
даугавской свиты. — Изв. высш, учебн. завед.. геол, и развед-
ка. № 8.
С о ш к и н а Е. Д. 1952. Определитель девонских четырехлучевых
кораллов. — Труды ПИН АН СССР. 39.
С ы с о е в В. В. 1960. Микроструктура раковины хиолитов и их
систематическое положение. — Докл. АН СССР, 131. № 5.
С ы с о е в В. В. 1968. Стратиграфия и хиолиты древнейших слоев
нижнего кембрия Сибирской платформы. Якутск. Якутск,
филиал АН СССР. Ротапринт.
Сюндюков А. 3. 1961. Карбонатные отложения верхнего девона
Западной Башкирии. Уфа. изд. Башкирск. фил-ла АН СССР.
Т а г а ц В. 1933. Некоторые данные по зоопланктону бухты Пат-
рокл. (Залив Петр Великий) за лето 1926 г. — Труды Гос. гид-
рол. ин-та исслед. морей СССР. вып. 19.
Т в е н х о в е л В. X. 1936. Учение об образовании осадков. Перев.
с англ. МЛ.. ОНТИ.
Теодор о вич Г. И. 1949. Карбонатные фации нижней перми —
верхнего карбона Урало-Волжской области. М.. Изд-во МОИП.
Т е о д о р о в и ч Г. И., Соколова Н. Н., Розанова Е. Д..
БагдасароваМ. В. 1962. Минералого-геохимические фа-
ции терригенных отложений нижнего карбона основной части
У рало-Волжской области. М.. Изд-во АН СССР.
ТроховаА. А. 1968. Отложения доманиковых фаций в связи
с их нефтегазоносностью. — В сб.: Тектоника и размещение
нефтегазовых месторождений востока Русской платформы.
М.. «Наука».
Т р о ш к и н а-Л а в р е н к о А. И. 1951. Новые виды водорослей
из соленых водоемов СССР. — Ботанические материалы От-
дела споровых растений Ботанич. Ин-та АН СССР. 7. М.—Л..
Изд-во АН СССР.
Ф илатоваЗ. А. 1938. Количественный учет донной фауны юго-
западной части Баренцева моря. - Труды ПИНРО. вып. 2.
X а б а к о в А. В. 1934. Отряд Radiolaria. — В кн.: К. Ц и т т е л ь.
Основы палеонтологии. М., Гос. научи.-тех. горно-геол. нефт.
изд-во.
ХвороваИ. В. 1951. Фораминиферовые биогермы в рифовых
массивах Ишимбаевского Приуралья.—Докл. АН СССР.
78. № 6.
ХвороваИ. В. 1953. История развития средне- и верхнекамен-
ноугольного моря западной части Московской синеклизы. —
Труды ПИН АН СССР. 43.
ХвороваИ. В. 1957. Изучение размера и формы органических
остатков и его значение. — В кн.: Методы изучения осадочных
пород, т. I. М.. Госгеолтехиздат.
ХвороваИ. В. 1968. Кремненакопление в геосинклинальных
областях. — В сб.: Осадконакопление и полезные ископаемые
вулканических областей прошлого, т. I. М.. «Наука».
Ц и т т е л ь К. 1934. Основы палеонтологии (Палеозоология), ч. 1.
Беспозвоночные. М., Гос. научно.-техн. горно-геол. нефт.
Изд-во.
Шам рай И. А., Л аза рева Е. П. 1956. Палеогеновые Сос-
colithophorideae и их стратиграфическое значение. — Докл.
АН СССР. 108, № 4.
Ш в е ц о в М. С. 1958. Петрография осадочных пород. Изд. 3.
М., Госгеолтехиздат.
Ш т и н а Э. А. 1964. Участие водорослей в процессах почвообразо-
вания. — Изв. АН СССР, серия биол.. № 1.
Шумейко С. И. 1962. Применение метода электронномикроско-
пических реплик к изучению верхнемеловых Coccolithophoridae.—
Докл. АН СССР. 147, вып. 2.
Яворе к ий В. И. 1947. Некоторые палеозойские и мезозойские
Hydrozoa, Tabulata и Algae.—В кн.: Монография по палеонтоло-
гии СССР. — Труды ВСЕГЕИ, 20, вып. I.
Якубова Л. И.. МальмЕ. Н. 1930. Явление временного ана-
эробиоза у некоторых представителей бентоса Черного моря. —
Докл. АН СССР, № 4.
ЯнетФ.Е. 1965. Микроструктурные особенности стенок эйфель-
ских и живетских табулят и хетитид Урала. — В сб.; Табуля-
томорфные кораллы девона и карбона СССР, Труды I Всесоюз-
ного симпозиума по изучению ископаемых кораллов. Труды
Ин-та геологии и геофизики СО АН СССР, вып. 2.
А I 1 о i t е a u J. 1952. Madreporaires post-paleosoiques. — Traite de
paleontologie, t. I.
A n d r u s о v D. 1950. Skamenliny karpatskich druhohor. I. Rastliny
a prvoky. — Prace St. GeoL list., 25.
Bein G. 1932. Die Stellung des Richelsdorfer Gebirges etc. — Z. dtsch.
Geol. Ges.. 84.
Berry С. T. 1934. Miocene and recent Ophiura skeletons. — Stu-
dies Geol. John Hopkins Univ., v. II.
Berry С. T. 1957. Asterozoa of the Post-Paleozoic. — In «Treatise
on marine ecology and paleoecology», v. 2. Paleoecology. — Mem.
Geol. Soc. America, 67.
Bpggild О. B. 1930. The shell structure of the mollusk. — Acad.
Roy. Sci. Lettres Danemark. Mem., ser. G., 2.
Boiler K. 1963. Stratigraphische und micropalaontologische Un-
tersuchungen im Neocom der Klippendecke (ostlich der Rhone). —
Eclog. Geol. Helvet., 56, N 1.
В г a n s о n E. В.. M e h 1 M. G. 1933—1934. Conodont studies 1 —4. —
Univ. Missouri Studies, 8.
Carozzi A. V. 1948. Etude stratigraphique et micrographique du
Purbeckien du Jura suisse. These N 1122, Geneve.
Carozzi A. V. 1955. Dasycladacees du Jurassique superior du bas-
sin de Geneve. — Eclog. Geol. Helvet., 48. N 1.
Carozzi A. V. 1961. Distorted oolites and pseudoolites. — J. Se-
diment. Petrol., 31. N 2.
С a у e u x L. 1916. Introduction a 1’etude petrographique des roches
sedimentaires. Paris.
Cayeux L. 1931. Introduction a 1’etude petrographique des roches
sedimentaires. Paris.
Chave К. E. 1952. A solid solution between calcite and dolomite.
J. Geol., 60.
С 1 a r k H. L. 1912. Fossil Holothurians. — Science, 35, N 3.
Cloud P. E. 1959. Geology of Saipan Mariana islands. Pan 4. Sub-
marine topography. — Geol. Surv. Profess. Paper., N 280-K.
Colom G. 1948. Fossil Tindnnids: loricated Infusoria of the order
of the Oligotricha. — J. Paleont., 22.
Conti S. 1950. Alghe Corallinaceae fossili. — Publ. Inst. geol. Univ.
Genova, ser. A, Paleontol.
С о о p e r G. A. 1957. Asterozoa of the Paleozoic. In: «Treatise on
marine ecology and paleoecology». 2. Paleoecology. — Mem.
Geol. Soc. America. 67.
С г о n e i s C.. Me Cormack J. 1932. Fossil Hololhuroides.—
J. Paleontol., 6. N 2.
по
Литература
Cue not L. 1948a. Classe des Asterides. — Traite de Ecologic. 11.
Paris.
Cuenot L. 1948b. Classe des Ophiurides. Traite de Ecologie. 11.
Paris.
C u v i 1 1 e r J. 1954. Nouveavx a coprolithes de crustaces. — Bull.
Soc. Geol. France, 4, fasc. 1 —3.
Debrenne F. 1969. Lower Cambrian Archaeocyatha from the Ajax
mine Beltans South Australia. — Bull. Brit. Mus. (Natur. Hist.),
Geology, 17, N 7.
Debrenne F. 1964. Archaeocyatha. Contribution a l-etude des faunes
cambriennes du Maroc. de Sardaine et de France. — Not. et Mem.
Service Geol.. N 179. Rabat.
Deflandre G. 1936. Tintinnoideiens et Calpionelles.Comparaison
entre les Tintinnoidiens, Infusoires loriques pelagiques des mers
actuelles, et les Calpionelles, microfossiles de I'epoque secondai-
re. — Bull. Soc. France, Microsc., 5.
Deflandre G. 1950. Contribution a 1’etude des Silikoflagellides
actuels et fossiles. — Microscopic, 2.
Deflandre G. 1952. Classe des Coccolithophorides.— Traite de
Paleontologie, 2. Paris.
DeflandreG., Fert Ch. 1954. Observations sur les Coccolitho-
phorides aclueles el fossiles en microscopie ordinaire et electro-
nique. — Ann. paleontol., 40.
Deflandre-Rigaud M. 1962. Contributions a la connaissance
des sclerites d’Holothurides fossiles. — Mem. Mus. Nat. Hist. Natur.
ser. C. Sciences de la Terre. 11, fasc. 1.
DenizotM., Massieux M. 1965. Observations sur le genre
Distichoplax. — Bull. Soc. geol. France, 7. N 3.
D о u b i g n e r J., E 11 e r J. P. 1963. Presence de spongiaires dans
chistes precambriens metamorphiques des Vosges. — Bull. Serv.
Carte Geol. Alsace et Lorraine, 16, N 3.
D u m i t r i с a P. 1964. Asupra prezentei unor Radiolari din Familia
Challengeridae (ord. Phaedaria) in Tortonianul din Subcarpati. —
Studii Cercet. Geol., Geofiz., Geogr. Acad. Rep. Pop. Rom.
D u m i t r i с a P. 1965. Sur la presence de Phaeodaires fossiles dans
le Tortonien des Subcarpathes Roumaines. — C. r. Acad, sci.,
Gr. 9, Miner., Geol., Paleont., PedoL, 260, N 1.
E a rd ley A. J. 1938. The sediments of Great Salt Lake. — Bull.
Amer. Assoc. Petrol. Geologists, 22, N 10.
E i c h w a 1 d E. 1840. Lieber des silurische Schichterisystem in Eusl-
land. — Z. Natur und Heilkunde, 1—2.
E d g e E. R. 1934. Faecal pellets of some marine invertebrates. —
Amer. Midland Naturalist., 15, N 1.
El 1 i о 11 G. F. 1958. Algal debris-facies in the Cretaceous of the Middle
East. — Paleontology, 1, pt 3.
E 1 1 i о 11 G. F. 1960. Further records of fossil calcareous algae from
the Middle East. — Micropaleontology, 2, N 4.
E 11 i о 11 G. F. 1960. Fossil calcareous algal flores of the Middle East. —
Quart. J. Geol. Soc. London, 65, 3.
E 1 I i о 11 G. F. 1962. More microproblematica from the Middle East. —
Micropaleontology. 8, N I.
E n d о R. 1961. Phylogenetic relationship among the calcareous algae. —
Sci. Rept. Saitama Univ., ser. B.
Ercegovic A. 1927. Trois nouveaux genres des Cyanophycees litho-
phytes.— Acta bot. Inst. bot. R. Univ. Zagreb. 3.
Fliigel E. 1964. Microproblematica aus den Rhatischen Riffkalken
der Nord Alpen. — Paleontd. Z., 38, N 1—2.
F r a a s E. 1885—1886. Die Asterien des Weissen Jura von Schwabe
und Franken etc. — Paleontographica, 32.
F r e m у P. 1924. Cyanophycees perforantes. — Mem. Soc. Linn. Nor-
mandie, Mem. spec., 1, fasc. 1.
F r i z z e 1 D о n L., E x 1 i n e H. 1957. Holothurians. — Mem. Geol.
Soc. America, 67.
Gaewskaja N. S. 1933. Zur Okologie. Morphologic und Syslematik
der Infusorien des Baikalsees.—Zoologica, 32.
Garwood E. 1914. Some new rock-building organisms from Lower
Carboniferous beds of Westmorland. — Geol. Mag., n. ser., 1, N 6.
Garwood E. J. 1931. The Tuodian beds of Northern Cumberland
and Roxburgshire. — Quart. J. Geol. Soc. London, 87.
G r a b a u A. M. 1932. Pripciples of slratigraphu. — N. Y.
G г о s s W. 1957. Uber die Basis der Conodonten. — Paleontol. Z., 31.
G г о s s W. 1960. Uber die Basis bei den Gattungen Pahnatolepis und
Polygnathus (Conodontidae). — Paleontol. Z., 34.
G u r i c h G. 1906. Les Spongiostromides du viseen de la province de
Namur. — Mem. Musee hist, natur. Belg., pt. 3.
Gutschick R. C. 1959. Lower Mississippian sclerites from the
Rockford Limestone of Northern Indiana. — J. Paleontol., 33.
HammerlingJ. 1944. Zur Lebensweise, Fortpflanzung und Ent-
wicklung verschiedener Dasycladaceen. — Arch. Protistenk., 97.
Hansen H. J. 1970. Electron-microscopical studies on the ultrastru-
ctures of some perforate calcitic radiate and granulate foraminifera.
Copenhagen.
H a у W. W.. Towe К. M. 1962a. Electronmicroscope studies of
Braarudosphaera higelowi and some related coccophorides. —
Science. 137. N 3528.
Hay W. W.. Towe К. M. 1962b. Electronmicroscopic examination
of some coccolithes. — Eclog. geol. Helvet.. 55. N 2.
H e s s H. 1960. Ophiurenrcste aus Malm des Schweizer Juras und des
Departaments Haut-Rhein. — Eclog. geol. Helvet.. 53, 1.
HessH. 1962, 1963. Mikropalaontologische Untersuchungen an
Ophiuren. 1. Einleitung. — Eclog. geol. Helvet., 55. 2.
Hesse E. 1899—1901. Die Microstructur der fossilen Echinoideen
Stacheln und deren systematische Bedeutung. — N. Jb. Min., Geol..
Paleontol.. XIII. BeiL-Bd.
H i с к m a n V. V. 1962. Tasmanian sea-cucumbers (Holothuroidea). —
Pap. and Proc. R. Soc. Tasmania, 96.
Hill D. 1956. In: M о о r e R. C. Treatise on Paleontology. Pt F. Coe-
lenterata. Rugosa.
H i 11 D.. S t u m m E. C. 1956. Tabulate. Treatise on invertebrate
paleontology. — Geol. Soc. Am.. Univ. Kansas Press.
H о n j о S., F i s c h e r A. 1964. Fossil coccoliths in limestone exami-
ned by electron microscopy. — Science, 144. N 3620.
HoweH.V. 1942. Neglected Gulf Coast Tertiary microfossils.—
Bull. Amer. Assoc. Petrol. Geologists, 26. 7.
H u d s о n R. G. S. 1959. A revision of the Jurassic stromatoporids
Actinostroinina. Astrostilopsis and Trupetostromaria. — Paleonto-
logy. 2, pt. 1.
H у m a n L. N. H. 1955. The Invertebrate: Echinodermata the Coe-
lomata Bilateria. N. Y. Toronto - London.
1 c h i к a v a W. 1956. Preliminary report on Silicoeflagellates from Neo-
gena Tertiary of the Hokuriku district, Japan. — Sci. Rept. Ka-
nazawa Univ., 5. N .
Johnson J. H. 1951. Permian calcareous algae from the Arache
Mountains. Texas. — J. Paleontol., 24, N 1.
JohnsonJ. H. 1956. Archaeolithophyllum, a new genus of Paleo-
zoic Coralline algae. — J. Paleontol.. 30, N I.
JohnsonJ. H. I960. Paleozoic Solenoporaceae and related red al-
gae. — Quart. Colorado School Mines. 55.
JohnsonJ. H. 1962. The algae genus Lithothanutium and its fossil
representatives. — Quart. Colorado School Mines, 57. 1.
Johnson J. H. 1963a. The algae genus Archaeolithothantnium and
its fossil representatives.—, J. Paleontol., 37, N 1.
Johnson J. H. 1963b. Pennsylvanian and Permian algae. — Quatr.
Colorado School Mines, 58, N 3.
J о h n s о n J. H., KonishiK. 1956. Mississippian algae from the
Western Canada basin and Montana. — Quart. Colorando School
Mines. 51, N 4.
J о r g e n s о n E. G. 1955. Solubility of the silica in diatoms. -Phy-
siol. plantarum, 8.
JoukovskyE., Favre J. 1913. Monographic geologique et pale-
ontologique de Saleve. — Mem. Soc. phys. hist, natur. Geneve, 37.
К e i j A. J. 1964. Distichoplax from Kudat peninsula and Banggi island,
Sabah, Borneo. — Bev. micropaleontol., 7, N 2.
К о n i s h i K. 1961. Studies of Paleozoic Codiaceae and allied algae.
Part I. Codiaceae. — Sci. Rept. Kanazawa Univ.. 7.
К о n i s h i К., E p i s R. C. 1962. Some early Cretaceous calcareous
algae from Cochise county. Arisona. — Micropaleontology, 8, N 1.
L e m о i n e M. 1960. Comparaison de Distichoplax beserialis et des
Rhabdopleura fossiles et actuelles. — Rev. micropaleontol., N 2.
Lindstrom M. 1964. Conodonts. Amsterdam — London — N. Y..
Elsevier Publ. Co.
Marek L. 1963. New knowledge on the morphology of Hyolithes.—
Sbornik geol. ved. Paleontol., sv. 1.
Matsumoto H. 1917. A monograph of Japanese Ophiuroidea.—
J. College Sci. Tokyo, 38, 2.
MooreH. B. 1931a. The specific identification of faecal pelletes.—
J. Marine Biol. Assoc., n. ser., 17, N 2.
MooreH. B. 1931b. Muds of Clyde Sea area III.—J. Marine Biol.
Assoc., n. ser., 17, N 2.
MooreH. B. 1955. Faecal pellets in relation to marine deposite.—
In: «Recent marine sediments». Tulusa.
M ii 11 e r A. N. 1950. Die Ophiuroideenresle aus Mucronalensenon
von Riigen. — Geologica. 5.
Mii Iler A. N. 1953. Die isolierten Skelettelemente der Asteroidea
(Asterozoa) aus der obersenonen Schreibkreide von Riigen. —
Beih. Geologic. N 8.
M ii 11 e r K. J. 1960. Wert und Grenzen der Conodonten-Stratigraphie.—
Geol. Rundschau, 49. Hl.
N i e 1 se n К. B. 1943. The asteroids of the Senonian and Danian depo-
sits of Denmark. — KgL Danske vid. selskab. Biol, skr., 11, 5.
Литература
111
N о е 1 D. 1964. Modalitee d’ulilisation du . microscope electronique
pour 1’etude des coccolithes fossiles. — C. r. Acad. Sci.. 259.
P a n d e r C. 1856. Monographic des fossillen Fische des silurischen
Systems des Russisch-Bahischen Gouvernements. St-Petersb..
Akad. Wiss. X.
ParejasE. 1948. Sur quelques coprolithes de crustaces. — Arch.
Sci. Ceneve, 1, fasc. 3.
P h I e g e r F. B. 1964. Foraminiferal ecology and marine geology.—
Marine geology. 1, N 1. Amsterdam.
P i a J. 1927. Thallophyta. In: Hirmer «Handbuch der Paleobo-
tanik».
P i a J. 1937. Die wichtigsten Kalkalgen des Jungpalaozoikums und
ihre geologische Bedeutung. — Compt. rend. Congr. avanc. etude
stratigr. Carbon. 2.
P i a J. 1937. Die kalklosenden Thallophyten. — Arch. Hydrobiol.. 31.
Pokorny V. 1958. Grundziige der zoologischen Mikropalaontologie.
Bd. II. Berlin, VEB Deutsch. Verl. Wiss.
PraturlonA. 1963. Dasycladaceae from Upper Permian of the
Dolomites. — Geol. Romana, 2.
R a s m u s s e n H. W. 1950. Cretaceous Asteroidea and Ophiuroidea
with special reference to the species found in Denmark. — Dan-
marks Geol. Undersogels. X Raekke. N 77.
RasmussenH. W. 1951. Cretaceous Ophiuroidea from Germany.
Sweden, Spain and New Jercey. — Medd. Dansk. Geol. Foren.. 12.
R e m a n e J. 1963. Les Calpionelles dans les couches de passage juras-
sique-cretace de la fosse vocontienne. — Lab. geol. Fac. sci. Gre-
noble.
R e у n о 1 d s S. H. 1921. The lithological succession of the carbonife-
rous limestones. — Quart. J. Geol. Soc. London. N 77.
R h od e s F. H. 1952. Classification of Pensylvanian Conodont Assem-
blages. — J. Paleontol., 26, N 6.
R i c h t e r R. 1927. Sandkorallen-Riffe in der Nordsee. — Natur und
Mus., 57, N 2.
RiedelW. R. 1951. Number of Radiolaria in Sediments. — Nature,
167.
SchewiakoffW. T. 1926. Acantharia Fauna e flora del Colfo di
Napoli. London.
SchnorfA. 1960. Parastromatoporidae nouveaux du Jurassic su-
perieur et du Valanginien inferieur du Jura. — Eclog. Geol. hel-
vet., 53, N 2.
S c h u e t i e H. A., A d 1 e r H. A. 1929. A note on the chemical com-
position of Chara from Green Lake, Wisconsin. — Trans. Wis-
consin Acad. Sci., 24.
S i e v e r t s-D о г e с к H. 1943. Ubersicht uber stratigraphische und
regionale Verbreitung fossiler Holothurien. — Z. Disch, geol.
Ges.. 95, N 1- 2.
S i e v e r t s-D о г e с к H. 1958. Echinodermen. In: «Handbuch der
Mikroskopie in der Technik», H. Freud (Ed.). Bd. II. 3. Frank-
furt a. M.
S i I v e s t e r R. K. 1959. Scolecodonts from Central Missouri. — J.
Paleontol., 33, N 1.
S i n с 1 a i r G. W. 1946. Notes on nomenclature of Hyolithes. — J.
Paleontol., 20.
SpencerW. K. 1914—1940. A monograph of the British Paleo-
zoic Asterozoa. — Paleontogr. Soc. London.
SpencerW.K. 1951. Early Paleozoic Starphish. - Philos. Trans.
Roy. Soc. London, ser. B, 235.
SteinmannG. 1899. Uber Bouenia eine fossile Alge aus der Familie
der Codiaceen. — Ber. naturforsch. Ges. Freibourg, 11.
S l о n e I e у H. N. 1958. The Upper Permian flora of England. — Bull.
Brit. Mus. (Natur. Hist.). Geol., 3, N 9.
V a s s о e v i t c h N. B. 1936. Les roches a Calpionella Lorenz du Cauca-
se et de la Crimee. — Compt. Rend. Soc. geol. France, 10.
W a g n e r W. 1964. Zum Skeletenbau oberjurassischer Kalksch-
wamme.—Mitt. Bayer. Staatssamt. Palaontol., Hist., Geol., N4.
W a n g H. G. 1950. A revision of the Zoantharia Rugosa in the light
of their minute skeletal structures. — Philos. Trans. Roy. Soc.
London.
WeissM. P. 1954. Notes on some Middle Ordovician fossils form
Minnesota. — J. Paleontol.. 28, N 4.
Wei burg J. 1939. Skelettreste von Ophiuren aus dem deutschen
Lias, ihre systematische Zugehorigkeit und Bedeutung als Mikro-
fossilien. — Palaontol. Z.. 21.
W e I 1 s J. 1956. n: M о о r e R. C. Treatise on paleontology. Pt F, Coe-
lenterata. Scleractinia.
Wood A. 1949. The structure of the wall of the test in the Forami-
nifera, its value in classification. — Quart. J. Geol. Soc., 104, pt. 2.
W г a у J. 1964. Archaelithophyllum, an abundant calcareous alga in
limestones of the Lansing Group (Pennsylvania), Southweastern
Kansas. — Bull. Kansas, 170, p. 1.
Z i e g 1 e r B. 1964. Bewuchs auf Spongien. — Palaontol. Z., 38.
ТАБЛИЦЫ
1—76
1—4
5—24
25—61
61—63 и 63—65
65
66—68
5, 6, 69—75
76
Биогермные известняки
Известковые организмы
Водоросли
Животные организмы
Спорные микроорганизмы
Кремневые организмы
Водоросли
Животные организмы
Следы жизнедеятельности
организмов
Вторичные минералы
в остатках организмов
114
Биогермные известняки
Таблица 1
Строение биогермов
Structure of Bioherms
Фиг. 1. Водорослевый мелобезиевый биогермный известняк. На фотогра-
фии различимы тонкие корочки багряных водорослей и многочисленные
первичные незаполненные каверны. Пришлифовка. Верхний тортон,
тарнопольский горизонт. Подолия, Медоборы, дер. Сбараж.
Фиг. 2. Нубекуляриевые биогермы. Видны грубые слои, образованные
мелкими столбиками нубекулярий с темными кавернами между ними.
Пришлифовка перпендикулярно слоистости биогерма; нат. вел. Средний
сармат. Молдавия, г. Кишинев (окраина города — карьер известняка).
Фиг. 3. Вертикальное сечение мшанкового биогермного известняка. Свет-
лые пятна — инкрустации вторичного карбоната в замкнутых порах,
черное — пелитоморфный карбонат открытых сингенетичных пор. При-
шлифовка; нат. вел. Раннеартинский возраст. Ишимбаевский погребен-
ный массив, глуб. 535 м.
Фиг. 4. Нубекуляриевый биогерм, высота купола 0,5 м. Молдавия,
г. Кишинев (окраина города—карьер известняка).
Фиг. 5. Стенка карьера с биогермами параллелепипедальной формы,
с вертикальными трещинами, заполненными детритусом. Ширина 1 м.
Внизу видна структура сплошного биогермного нубекуляриевого извест-
няка. Молдавия, г. Кишинев (окраина города)
Та б л и ц а I
116
Биогермные известняки
Таблица 2
Пришлифовки биогермных известняков пермских погребенных массивов*
Polished sections of bioherm limestones of Permian buried massifs
Фиг. 1. Косой срез мшанкового биогермного нижнеартинского известняка.
Темный пелитоморфный известняк заполняет пространства между замкну-
тыми полостями мшанковых «фонарей», расположенных косо и заполнен-
ных вторичным карбонатом. Вверху направо — слоистый осадок, пропи-
танный битумом; нат. вел. Артинский ярус. Ишимбай, с. Термень-Елга,
скв. 17/20, глуб. 1210 м.
Фиг. 2. То же, что и на фиг. 1, но полости вторичного карбоната ожелез-
нены. Видно, что эти полости заполнялись зонально; нат. вел. Ишимбай,
Западный массив, скв. 89/2, глуб. 529,5 м.
Фиг. 3. Вертикальный разрез мшанкового биогермного нижнеартинского
известняка. Светлые и желтоватые полоски — сеточки мшанок, обросшие
вторичной светлой коркой. Внутренние полосы (серые) заняты микро-
зернистым карбонатным материалом, черные полости — пелитоморфный
битуминозный карбонат, проникший снаружи (вероятно, иозднеартин-
ского времени); нат. вел. Артинский ярус. Ишимбай, Западный массив,
скв. 89/2, глуб. 627,5 лс.
Фиг. 4. Биогермный нижнеартипский известняк с невыясненным био-
гермообразователем. Светлые пятна — органогенные участки, занятые
детритусом и измененными организмами. Все сильно битуминизировано.
Темные участки — каверны, заполненные пелптоморфным битуминизи-
рованным материалом, по-видимому, более позднего, чем поздиеартипское,
времени; нат. вел. Артинский ярус. Ишимбай, дер. Кусянкулово,
скв. № 325, глуб. 715,5—734,7 м.
Рисунки художника В. В. Ивенсена.
Таблица 2
118
Биогермные известняки
Таблица 3
Пришлифовки биогермных известняков пермских погребенных массивов*
Polished sections of bioherm limestones of Permian buried massifs
Фиг. 1. Гидроактиноидный биогермный нижнеартинский известняк с ан-
гидридом. Серые неоднородные полосы — гидроактиноиды типа Paleo-
plaesina, голубое — ангидрид кунгурского времени, белое — гипс, се-
точка — мшанка; нат. вел. Артинский ярус. Ишимбай, Восточный мас-
сив, глуб. 652—658 м (№ 117).♦♦
Фиг. 2. Вертикальный разрез мшанкового биогермного нижнеартинского
известняка. Бурые полоски — сеточки мшанок, серые и желтые пятна —
полости, заполненные пелитоморфным и шламовым карбонатом во время
осадкообразования, фиолетовые полости — ангидрид кунгурского времени;
нат. вел. Артинский ярус. Ишимбай, Восточный массив, глуб. 706,8—
726,3 .и (№ 133).**
Рисунки художника В. В. Ивеисена.
Номера образцов керна музея г. Ишимбая.
Т яблицаЗ
120
Биогермные известняки
Таблица 4
Строение биогермов и строматолитов
Structure of Bioherms and Slromalolithi
Фиг. 1. 2. Мшанковые биогермные известняки раннеартинского возраста
погребенных массивов Ишимбая, пришлифопки; пат. вел. Фиг. 1 — верх-
няя часть керна, вверху с первичными кавернами, заполненными ангид-
ридом кунгурского времени. Фиг. 2 -*- нижняя часть керна на детритусо-
вом основании. Белые пятна в нижней части — вторичные полости, за-
полненные карбонатом.
Фиг. 3. Биогермный известняк погребенных массивов Уфимского плато,
образованный проблематической водорослью Tu.biph.ytes (светлые черве-
образные пятнышки) и другими организмами. Темные пятна — пелито-
морфный материал, заполнивший полости в биогерме при его росте.
Нришлифовка; пат. вел. Пермь.
Фиг. 4. Столбиковидно-веерпый строматолит па слоисто-бугристом осно-
вании кремового цвета. В промежутках между буграми — полости, в кото-
рых иногда встречаются мелкие гастроподы. Внутри светлых слоев встре-
чены кустиковидные пустоты, отнесенные к третичной (сипезеленой?)
водоросли Dendractis Reis. Нришлифовка, вид сбоку. Олигоцен. Алтай
(сборы Е. Н. Щукиной).
Таблица 4
122
Известковые водоросли. Следы жизнедеятельности организмов
Таблица 5
Синезеленые водоросли
Cyanophyta
Фиг. 1—3. Сверлящие водоросли. Фиг. 1 — полости внутри обломка
брахиоподы; X 10. Верхний карбон. Окрестности г. Куйбышева. Фиг. 2 —
полости, просверленные в обломке криноиды (может быть, и не водо-
рослью); X 14. Известняк F*/2, Карбон. Донбасс. Фиг. 3 — обломок гастро-
поды, просверленный, возможно, водорослями; Х73. Карбон. Донбасс,
балка Железная.
Фиг. 4. Girranella sibirica Masi, в виде перепутанных и «растрепанных»
трубочек; ХбО. Кембрий. Тува.
Фиг. 5. Giruanella со стелющимися трубочками; X 90. Девон. Армения
U\?2Ua, материал А. А. Аракеляна).
Фиг. б. 7. Coaclilum. Фиг. 6 — X 45; верхний девон — нижний карбоп.
Северный Кавказ, р. Маруха. бассейн р. Зеленчук (материалы С. М. Кро-
пачева). Фиг. 7. — С. straeleni (Lecompte); X 30. Верхний девон. Москва.
Фиг. 8. Itiiulurialithus herbidus Masi. — вертикальное сечение желвачка
с продольным разрезом ветвящихся канальцев; <90. Чокрак. Грузия,
р. Риони.
Фиг. 9. Ukrainolrichium в сечении через центр желвачка, где обособились
отдельные «начальные» клетки; Х45. Верхняя юра. Западная Украина,
г. Стрый.
Таблица 5
124
Известковые водоросли. Следы жизнедеятельности организмов
Таблица 6
Синезеленые водоросли и спорные организмы
Cyanophyta and controversial Organisms
<1>пг. 1, 2. Желвачки, образованные главным образом водорослью Girua-
nella, обволакивающей различные обломки, в том числе коралл Aulo-
рога. Фиг. 1 — пришлифовка; нат. вел. Фиг. 2 — шлиф, Х2. Граница
турне и этрепь. Южный Урал, р. Зиган.
Фиг. 3, 4. Спорный организм, похожий на Coactilum и на Rothpletzella,
обволакивающий обломки раковин; ?< 10 (материал М. II. Александровой).
Фиг. 3 — ордовик. Казахстан, Бетнак-Дала. Фиг. 4 — нижний девон?
Казахстан, Кызыл-Шоны.
Фиг. 5, 6. Кустики Epiphyton в продольных и косых сечениях, X 20.
Кембрий. Тува, р. Баин-гол, правый приток р. Каа-хема. Фиг. 5 — за-
росли между кубками археоциат. Фиг. 6 — кустик эппфитона, располо-
женный на корочке гирвапеллы, которая заметна внизу в виде серой
полоски.
Фиг. 7. Microcodium permicum Masi, в виде агрегата полых члеников
Х45. Пермь. Печора.
Фиг. 8. Ходы сверлящих организмов внутри раковины моллюска; Х45.
Пермь. Уфимское плата.
Фиг. 9. Проблематические сферулы в доломите. Происхождение этих сфе-
рул спорное. Возможно, что они являются результатом жизнедеятельности
бактерии, которые изменяли кислотность окружающей среды в’водоеме
и карбопат осаждался биохимическим путем в виде сферул между бакте-
риями. Но возможны и другие, такие же проблематические объяснения;
Х45. Карбон. Окрестности г. Куйбышева.
Таблица 6
126
Известковые водоросли
Таблица 7
Спорные синезеленые водоросли
Controversial Cyanophyta
Фиг. 1. Epiphyton в косом сечении; Х20. Кембрии. Тува.
Фиг. 2—4. Microcodiuni в виде пустотелых агрегатов радиально и «пали-
садно» расположенных, иногда также в виде пустотелых кристаллов.
Палеоген. Фергана. Фиг. 2 — в правой части фотографии чехлы — призмы
и конусы в дезинтегрированном виде, лежат в беспорядке, в отдельных
призмах заметны внутренние полости, заполненные пелитоморфным мате-
риалом; X 30. Фиг. 3 — сечение через «колонию» с правильным располо-
жением чехлов; Х46.- Фиг. 4 — продольные и поперечные сечения целых
агрегатов; Х20.
Фиг. 5, 6. Nuia sibirica Masi, в известняках. Ордовик, усть-кутская свита,
р. Ангара (низовья, Овсянкинская шивера). Фиг. 5— Х46. Фиг. 6 —
Х20.
Фиг. 7, 8. Nubecularites — Renalcis — Schuguria — сгустковидные обра-
зования различной формы и разрастания с внутренней неправильной
полостью. Фиг. 7 — X 18. Нижний кембрий. Восточная Сибирь, р. Иркут.
Фиг. 8 — Х20. Девон. Средний Урал.
Фиг. 9, 10. Tubiphytes obscurus Masi.; Х20. Нижняя пермь. Уфимское
плато. Фиг. 9 — косое и продольное сечения окончания цилиндра.
Фиг. 10 — разрастание периодически наслаивающихся корочек с внутрен-
ней спутанно-волокнистой микроструктурой.
Таблица 7
128
Известковые водоросли
Таблица 8
Кокколиты
Coccolithophoridae
Фиг. 1—8. Кокколиты под обычным микроскопом, ХЮ26
Coccolithophoridae under the Microscope; Xi026
Фиг. 1. Дискоастер и многочисленные фрагменты кокколитов в галифе
мергеля «форамипиферовой свиты». Эоцен. Г. Черкесск.
Фиг. 2. Фрагменты кокколитов в шлифе из тех же пород, в центре виден
целый тремалит.
Фиг. 3. Зиголпты и мелкие фрагменты кокколитов в шлифе. Мел, коньяк.
КМА, Лебединский карьер.
Фиг. 4. Порошковый препарат мергеля «форамипиферовой свиты». Видны
звездообразный дискоастер и круглые тремалпты, а также мелкие обломки
кокколитов. Водный препарат.
Фиг. 5. Тот же образец с овальными и круглыми тремалитами (циклокок*
колитами).
Фиг. 6. Рабдолиты и их фрагменты в порошковом водном препарате лан-
цеолятового мела. В центре — целый рабдолит тина знграблита. Харь-
ковская область.
Фиг. 7. Крибросферолит (крибролит) в том же образце. Видны мелкие
фрагменты кокколитов и рабдолитов.
Фиг. 8. Порошковый водный препарат четвертичного ила с звездообраз-
ными дискоастерами, дисколитами и циклолитами (?), а также их фраг-
менты. Тихий океан, экватор (образец из коллекции М. Г. Ушаковой).
Фиг. 9—16. Кокколитофориды под электронным микроскопом
Coccolithophoridae under the Electronic Microscope
Четвертичные отложения. Экваториальная часть Тихого океана. Об-
разцы из коллекции М. Г. Ушаковой. Фото автора
Фиг. 9, 10. Циклококколиты (эллипсоплаколиты, по Камптнеру). Угле-
родные реплики; Х5500 и 6800.
Фиг. 11. Тип дискоастера (Discoaster aff. broweri Tan Sin Hok). Углеродная
реплика; X 5800.
Фиг. 12. Тот же тип дискоастера, вид сбоку. Снимок «на просвет»; Х4100.
Фиг. 13. Тип дискоастера Discoaster aff. crassus Martini. Углеродная реп-
лика; Х6500.
Фиг. 14. Тип дискоастера. Discoaster aff. crassus Martini. Углеродная
реплика; X 6500.
Фиг. 15. Тип дискоастера, вид сбоку. Углеродная реплика; Х4000.
Фиг. 16. Тип дискоастера трехлучевого (рода Marthasterites Deflandre).
Углеродная реплика; Х6000.
Таблица 8
9
130
Известковые водоросли
Таблица 9
Кокколиты под электронным микроскопом
Coccolithophoridae under the Electronic Microscope
Фиг. 1. Тремалит. Coccolithus huxleyi (Lohman). Снимок «на просвет» не-
посредственно кокколита; ,'<23 000. Ил. Средиземное море, у Монако
(Deflandre, Fert, 1954).
Фиг. 2. Тип крибролита. Cyath.osph.aera duponyi (Deflandre et Fert). Угле-
родная реплика; Х21 ООО. Третичные отложения. Франция, Донзак (Неу,
Towe, 1962).
Фиг. 3. То же, целый панцирь; X 15 000 (Неу, Towe, 1962).
Фиг. 4. Циклолит. Cyclolithus bramletti (Неу et Towe). Углеродная реплика;
х 18 300. Третичные отложения. Франция, Донзак (Неу, Towe, 1962).
Фиг. 5. Стефанолит. Cyathosphaera crux (Defl. et Fort). Углеродная реплика;
X5000. Третичные отложения. Франция, Донзак (Неу, Towe, 1962).
Фиг. 6. Дисколит. Discolithus ocellatus (Bramlette et Sull.). Углеродная
реплика; X 14 500. Третичные отложения. Франция, Донзак (Неу, Towe,
1962).
Фиг. 7. Дискоастер. Discoaster lodoensis (Braml. et Riedel). Углеродная
реплика; X3200. Третичные отложения. Франция, Донзак (Неу, Towe,
1962).
Фиг. 8. Дискоастер. Discoaster kueppeni (Strand.). Углеродная реплика;
Х5000. Третичные отложения. Франция, Донзак (Неу, Towe, 1962).
Фиг. 9. Пенталит. Braarudosphaera bigelowi (Gran et Braar.) Defl. Углерод-
ная реплика; X7500. Лютецкий ярус. Франция, Донзак (Неу, Towe,
1962).
Фиг. 10. Tetralithus sp., скрещенные николи; X около 800. Оксфорд. Запад-
ный Казахстан, Шунгай (коллекция Б. К. Прошлякова).
Таблица 9
132
Известковые водоросли
Таблица 10
Кокколиты под электронным микроскопом
Coccolithophoridae under the Electronic Microscope
Фиг. 1. Крибролит. Cribrosphaerella archangelskii Choumenko. Углеродная
реплика скола породы; X10 500. Лянцеолятовый мел. Харьковская
область. Фото Н. Д. Самотоина (коллекция автора).
Фиг. 2. Тремалит. Coccolithus tenuicostatus cent. nov. Углеродная реплика
скола породы; X10 000. Сантонский мергель Донбасса.
Фиг. 3, 4. Тремалит. Tremalithus barnesae (Black et Barnes). Вид с прокси-
мальной и дистальной сторон. Углеродная реплика; Х8000. Туронский
мел КМ А.
Фиг. 5. Зиголит. Zygolithus derculinus cent. nov. Реплика co скола маастрихт-
ского мергеля; X 15 000. Донбасс.
Фиг. 6. Микрорабдолит. М icrorhabdulus decoratus (Deflandre). Углеродная
реплика; X 95 000. Сантопский мел Донбасса.
Фиг. 7, 8. «Порошковатый кальцит» — фрагменты кокколитов и рабдо-
литов. Углеродная реплика; X 10 500. Ланцеолятовый мел. Харьковская
область.
Таблица W
134
Известковые водоросли
Таблица И
Кодиевые водоросли (истинные и спорные)
Codiaceae (true and controversial)
Фиг. 1. Ныне живущий род Halimeda с о. Сайпан. Наружный вид. Кустик
Halimeda opuntia на основании (с крупными члениками) и с ним вместе
Halimeda tuna (мелкие «резные» членики); х0,8 (Cloud, 1959).
Фиг. 2. Ortonella из каменноугольных известняков Кузбасса. Род спорный
(синезеленые или кодиевые?). Сечение желвачка близ центра; х22 (мате-
риал С. В. Максимовой).
Фиг. 3. Cayeuiia из верхпеюрских известняков Крыма. Сечение через
желвачок, который оброс каким-то животным (губка?) с крупными по-
лостями (налево вверху снимка); Х20 (материал Л. П. Задорожной)
Фиг. 4. Permocalculus digitatus Elliott. Осевое сечение. Водоросль от-
носится к Gymnocodiaceae или к кодиевым или багряным водорослям;
Х28. Верхнепермские известняки Ирака (Elliott, 1955).
Фиг. 5. Ivanovia. Вероятная кодпевая водоросль. Косое и тангенциальное
сечение пластинки; Х20. Каменноугольные известняки Подмосковья.
Фиг. 6. Обломок чехла, близкого по строению к роду Orthriosiphon (John-
son, Konishi, 1956). Пересекающие чехол каналы неоднократно ветвятся.
Род относится Джонсоном к кодиевым (Johnson, 1956) и к спорным зеле-
ным (Johnson, 1961); Х20. Каменноугольные известняки.
Фиг. 7. Отдельный членик современной кодиевой водоросли Halimeda
cryptica Colinvaux et Graham с растворенным известковым чехлом. Вверху
и внизу сочленения — «узлы» с соседними члениками. Темные ветвящиеся
нити в ископаемом состоянии выглядят в известковом чехле в виде каналь-
чиков; Х9 (Colinvaux, Graham, 1964). Данная водоросль произрастает
у берегов Ямайки на глубине 25—65 м (чаще 30—50 .и) на скальном осно-
вании рифового склона.
Таблица 11
136
Известковые водоросли
Таблица 12
Водоросли. Кодиевые сифонеи
Codiaceae
Фиг. 1, 5, 6. Lancicula Masi. Продольные и поперечные сечения чашевид-
ных выростов. Фиг. 1 — в нижнем обломке вверху центральные каналы
заканчиваются овальными полостями, которые, возможно, относятся
к органам размножения; Х20. Фиг. 5 — несколько чашевидных выступов,
расположенных друг над другом; X 10. Фиг. 6 — поперечные и косые се-
чения через центральный стержень или промежутки между чашевидными
выступами; X 10. Все экземпляры из нижнего девона Кузбасса.
Фиг. 2. Litanaia rnira Masi, в косых, близких к оси сечениях; X 20. Нижний
девон. Кузбасс.
Фиг. 3. Bacilloporella uralica Masi., продольное осевое и поперечное сече-
ния. В центре видны основной сифон и более тонкие побочные сифоны, но
краю заметны мелкие воронки, открывающиеся наружу, — остатки от
утрикулярной ассимиляционной системы; X 13. Верхи нижнего — низы
среднего девона. Средний Урал, Ивдельский район (Юртищенское место-
рождение).
Фиг. 4. Boueina в осевом сечении с хорошо различимыми окончаниями кана-
лов у поверхности; Х20. Пермь. Армения.
Фиг. 7—9. Calctfolium', X 50. Фиг. 7, 8 — С. okense Schvez. et Bir. в сече-
нии, поперечном к стержню и пластинам (фиг. 8 — несколько скошенное).
Заметны светлые точки близ верхней поверхности, отвечающие попереч-
ным сечениям мелких канальцев. Нижний карбон. Подмосковье, Ерино.
Фиг. 9 — плоскостное сечение пластинки С. punctatum Masi. Подмосковье,
Баховский карьер.
Таблица 12
138
Известковые водоросли
Таблица 13
Ложные кодиевые сифонеи и мутовчатые сифонеи
Pseudocodiaceae and Dasycladaceae
Фиг. 1. Гидроидный полни Cladocoropis в продольном сечении, похожий на
фиг. 2. Шлиф; ;<2. Юра (лейас). Италия, Монте Латтари (Castro, 1962,
табл. XV).
Фиг. 2. Мicorocodium fertilis Endo, отнесенный к кодпевым водорослям,
но но структуре аналогичный фиг. 1; ,7. Юра. Япония (известняк Тори-
позу) (Endo, 1961, табл. 14).
Фиг. 3. Clypeina jurassica Favre в разных сечениях: звездочки — сечения
мутовок перпендикулярно оси чехла, ряды колец — тангенциальные
сечения к чехлу через несколько мутовок; х 8, 7 (Castro, 1962, табл. 18).
Фиг. 4. Ортонелла. Видны ветвления трубок и намеки па поперечные пере-
городки; X 90. Нижний карбон. Кузбасс.
Фиг. 5, 6. Triploporella karabiensis Maslov. Фиг. 5 — поперечное, фиг. 6 —
продольное сечения. На скошенных сечениях чехла видно групповое рас-
положение каналов чехла, а па близких к осп чехла — ветвление каналов;
X 19. Юра. Крым.
Фиг. 7. Ортонелла из корки, обволакивающей обломок. Каналы распо-
лагаются поперек корки; х 10. Нижний карбон. Кузбасс.
Фиг. 8. Acicularia (палочка внизу) и Catellaria (вверху — конус) с поло-
стями в стенках; X 50. Палеоген, бухарский ярус. 1’. Вахш (близ Тут-
каула).
Фиг. 9. Oilaria cava Maslov; X 50. Местонахождение то же, что и на фиг. 8.
Фиг. 10. Munieria baconica Hantken, продольное и тангенциальное сече-
ния. Виден ряд мутовок с неясными и сливающимися порами; X 12. Апт.
Венгрия, горы Бакони.
Таблица 13
140
Ишестков ые водоросли
Таблица 14
Кодиевые и мутовчатые сифоней (палеозойские)
Codiaceac and Dasycladaceac (Palaeozoic)
Фиг. 1—3. Iranoria tenuissima Chvorova. Голотип, поперечное и косое се-
чения пластины. Фиг. 1,2 — сечение на сгибе пластины, где видны поры
в поперечном и косом разрезах (фиг. 1 — X 22; фиг. 2 — Х43). Фиг. 3 —
окончание пластины, отчетливо виден наружный известковый чехол, про-
низанный широкими порами, различимыми лишь местами; X 10. Карбон.
Подмосковье (материалы II. В. Хворовой).
Фиг. 4. Известняк, переполненный мутовчатыми сифоиеями (тонкие тру-
бочки); X 20. Нпжпий карбон. Казахстан (Семпзбугу).
Фиг. 5. Vermiporella icestbergensis Moskalenko в темном иелитоморфпом
известняке; X10. Силур. Центральный Казахстан, водораздел Сары-су-
Тепиз (№ 3025, материал Т. Т. Шарковой).
Фиг. 6. 7. Vermiporella sp. и неопределимые сифопеи в разных сечениях из
того же образца, что и на фиг. 5; X 10.
Таблица 14
142
Известковые водоросли
Таблица 15
Мутовчатые сифонеи (палеозойские)
Dasycladaceae (Palaeozoic)
Фиг. 1, 2. Koninckopora sp. Фиг. 1 — косое продольное сечение, фиг. 2 —
поперечное сечение; Х20. Известняк. Средний карбон, башкирский ярус.
Киргизия (коллекция Н. А. Аносовой).
Фиг. 3. Мутовчатая сифоиея типа Macroporella. Осевое сечение ; X 20.
Известняк. Пермь. Армения (коллекция А. А. Аракеляна).
Фиг. 4. Продольное (близкое к осевому) сечение сифонеи Beresella erecta
Masi, et Kul. Внизу сечение проходит тангенциально, благодаря чему бес-
поровые пояски выглядят в виде светлых поперечных полосок. Известко-
вые отложения водоросли светлые, мутовки тонких пор образуют темные
кольца. Заметна светлая корочка, покрывающая весь обломок, — харак-
терная черта старых частей слоевища этих водорослей; X 20. Средний
карбон. Ишимбай, скв. 101/1, глуб. 1977.
Фиг. 5. Продольное сечение Amicus fortunatus Masi, (в горизонтальном
положении). Типичная сохранность для мутовчатых сифоней. Полости
известкового чехла водоросли заняты темным пелитоморфным карбо-
натом, или границы их подчеркнуты налипшим темным материалом; Х46.
Девон. Кузбасс.
Фиг. 6, 8. Членики Cateniphycus в разных сечениях. Некоторые сечения
прошли через несколько члеников в прижизненном положении (фиг. 8
и справа на фиг. 6). На фиг. 8 заметны тонкие поры и отверстия, соединяю-
щие членики друг с другом; Х46. Девон. Кузбасс.
Фиг. 7. Участок известкового чехла сифонеи сферической формы из трибы
Cyclocrinaea типа Mastopora. Светлый известковый чехол образует «ка-
меры» с одной порой внизу и несколькими наверху. С наружной стороны
(вверху) на водоросль нарастают слоистые синезеленые водоросли; Х45.
Силур. Казахстан (материалы Б. М. Келлера).
Таблица 15
144
Известковые водоросли
Таблица 16
Мутовчатые сифонеи (мезо-кайнозойские)
Dasycladaceae (Mezo-Cainozoic)
Фиг. 1. Различные сечения Munieria, Clypeina и Coscinoconus в верхнеюр-
ском известняке Яйлы Крыма; <20.
Фиг. 2. Поперечное сечение юрской мутовчатой сифонеи тина клипеин.
Концы ответвлений с индивидуальными чехлами загнуты вверх, поэтому
на периферии видны их поперечные сечения; 1 \. Юра, титон. Югославия,
Далмация (материалы Д. Анича).
Фиг. 3, 4. Сечение «головок» Кopeldagaria sphaerica Maslov в детритусовом
меловом известняке. Полости спороносных камер заполнены темным
пелитоморфпым карбонатом; X 15. Нижний баррем. Туркменская ССР.
Копетдаг.
Фиг. 5, 6. Поперечное и продольное сечения Griphoporella sp.; У. 17,5.
Баррем-аптские известняки. Югославия, Далмация (материалы Д. Анича).
Фиг. 7. Поперечное сечение четырех ответвлений актинопореллы. Разрез
прошел параллельно оси чехла. Рядом сечение косцппоконуса; 30.
Верхняя юра. Крым, Яйла.
Фиг. 8. Различные сечения спороносных головок Kopeldagaria iailensis
Maslov; X 20. Титов. Крым, Яйла.
Фиг. 9. Тангенциальное сечение известкового чехла водоросли типа ура-
гиелла; Х15. Титопские известняки. Югославия, Далмация (материалы
Д. Анича).
Фиг. 10, 11. Acicularia. Спороносные камеры светлые, известковые отло-
жения водоросли— темные. Такая сохранность для мутовчатых водорослей
необычна; ХЗО. Известняк. Сармат. Молдавия.
Фиг. 12. Внизу — продольное сечение через b'erganella, вверху — через
Acicularia. Обычная сохранность мутовчатых сифоней с полостями, запол-
ненными темным материалом; .47. Палеоген. Таджикская депрессия.
Фиг. 13. Сечение через спороносную головку Kopetdagaria dalmatiensis
Maslov. Спороносные камеры разной формы и разной степени зрелости;
Х20. Титон. Югославия, Далмация (материалы Д. Анича).
Таблица 16
1102 10
146
И шестков ые водоросли
Таблица 17
Багряные и проблематичные водоросли
Rhodophyta and Problematic Algae
Фиг. 1. Монофитный желвачок Solenopora sp. в радиальном сечении. За-
метны «зоны роста» в виде более темных слоев и радиальные нити. Шлиф;
X10. Ордовик. Казахстан, Бетпак-Дала.
Фиг. 2. Ветвление нити Donezella lutugini Masi. Шлиф; X 50. Известняк
К4. Карбон. Донбасс.
Фиг. 3. Pseudolithothamnium album I’fender [—Erbia? alba (Pfender)];
сечение корочки в продольном к нитям сечении; X 45. Даний — нижний
палеоцен. Закарпатье, Раковский р-п (Кобылецкая Поляна).
Фиг. 4. Сечение через желвачок Parachaetetes ( Toniilithon) johnsoni Masi.;
15. Нпжпий карбон. Западная Сибирь, р. Томь.
Фиг. 5, 10. Solenomeris afonensis Masi. Фиг. Ю — корочка, пересеченная на
участке с крупными камерами. Шлиф; . 90. Пижний эоцен. Причерно-
морье, Новый Афон.
•Риг. 6. Желвачок моиофитного типа Solenopora spongoides var. iuchuii
Masi.; пришлифовка; 2,5. Ордовик, горизонт Пыхви. Эстония.
Фиг. 7—9. Монофитные желвачки багряных водорослей — мелобезий
в радиальных сечениях. Светлые веточки — водоросли, между ними —
мергелистый пелитоморфный материал (черное). Пришлифовка, иат. вел.
Третичные отложения. Сирия (коллекция В. А. Крашенинникова). Фиг. 7,
8 — Пальмира, фиг. 9 — Кариатейп.
Таблица 17
148
Известковые водоросли
Таблица 18
Багряные водоросли
Rhodophyta
Фиг. 1. Гипоталлий и периталлий. Lythophyllum carpathicum Lem. Видна
корочка с дугообразными сериями гипоталлия, постепенно переходящими
в горизонтальные серии периталлия; X145. Нижний эоцен. Абхазия,
Новый Афон.
Фиг. 2. Часть «веточки» литофиллума с дуговидными сериями срединной
ткани и (слева) тонкоклеточным периталлием; X 150. Эоцен. Крым, сел. На-
сыпное.
Фиг. 3. Ткань гипоталлия Lithothamnium tcheruomoricum Masi, с почковид-
ными и разной формы клетками, расположенными в виде наклонных ни-
тей. Серии отсутствуют; X145. Нижний эоцен. Абхазия, Новый Афон.
Фпг. 4. Дуговидные серии гипоталлия литофиллума резко сменяются
мелкими клетками периталлия; Х90. Верхний тортон. Крым, п-ов Тархап-
кут.
Фиг. 5. Ассоциация червей, фораминифер и корковидных багрянок, обра-
зующих биогермного типа известняк. Направо видно сечение форамини-
феры, извивающиеся нити — Melobesia (Lithoporella) parasitica Masi.,
веерообразные наросты — Lithophyllum (Dermatolithon) nataltae Masi.;
X55. Верхний тортон. Крым, п-ов Тарханкут.
Фиг. 6. Сорусы Palaeothamnium kossovense Masi, в начальной стадии обра-
зования спорангиев; Х114. Палеоген. Северный склон Карпат, р. Старый
Самбор, сел. Стрельчиби.
Фиг. 7. Спорангии в ткани периталлия Archaeolithothamnium keenano
var. Ivovicum Masi., отделенные друг от друга стерильными нитями кле-
ток. Вертикальное сечение; Х90. Верхний тортон. Подолия, Медоборы.
Фиг. 8—10. Вертикальные (осевые) сечения через концептакли литофил-
лума. Фиг. 8 — Lithophyllum duplex Masi.; на месте пор находится камера,
что характерно для данного вида; Х114. Верхний тортон. Близ сел. Ол-
ховцы, запад Украины. Фиг. 9 — Lythophyllum corculumis Masi.; Х114.
Нижний тортон. Западная Украина, р. Гнила. Фиг. 10 — Lp. platicarpum
Masi.; Х55. Верхний тортон. Крым, п-ов Тарханкут.
Фиг. И. Косой срез слоевища с концептаклями Lithophyllum corcolumis
Masi.; Х55. Верхний тортон. Крым, п-ов Тарханкут.
81 в И и ir g * j
150
Известковые водоросли
Таблица 19
Багряные водоросли
Rhodophyta
Фиг. 1. Вертикальное сечение через корочку Mesophyllum schenkii var.
corticesum Masi. Внизу виден дуговидный гипоталлий, сменяющийся вверху
серийным периталлием, в котором расположен овальный сорус со многими
отверстиями в крышке; X 150. Нижний тортон. Западная Украина, Ополье,
гора Высокая Тура.
Фиг. 2. Корка Karpathia sphaerocellulosa Masi.; Х114. Палеоген. Северные
Карпаты, Старый Самбор.
Фиг. 3. Lithothamnium praefruticulosum Masi. Видны сорусы с отверстиями
в крышке и вторичный гипоталлий над нижним сорусом; Х90. Верхний
тортон. Подолия, дер. Романьевка, Медоборы.
Фиг. 4. Трехлопастное окончание нити Donezella lutugini Masi, в продоль-
ном сечении; Х55. Известняк К4. Карбон. Донбасс.
Фиг. 5. Solenopora spongoides var. iuchvii Masi.; срезы продольный и косой
к нитям; X 15. Нижний силур, горизонт Иыхви. Эстония, сел. Мадизе.
Фиг. 6. Сечение продольное к нитям гипоталлия Solenopora sudakensis
Masi, в зоне перехода к периталлию. Видны перегородки и вздутые бочко-
видные клетки; Х45. Оксфорд. Крым, р-н г. Судака.
Фиг. 7. Продольное сечение членика Jania dniestrovica Masi. (Corallinoi-
deae). В осевой зоне располагаются дуговидные серии срединного гипо-
таллия, резко сменяющиеся по краям клетками периталлия. Концы чле-
ника ограничены вогнутыми чашевидными поверхностями; Х114. Верх-
ний тортон. Дер. Старый Сбараж (западнее г. Львова).
Фиг. 8. Сечение через центр желвачка Solenopora concentrica Masi, в поля-
ризованном свете. Видны концентрические непостоянные серии клеток,
что сближает этот вид с родом Parachaetetes-, X 20. Девон. Москва.
Таблица 19
152
Известковые водоросли
Таблица 20
Багряные водоросли
Rhodophyta
Фиг. 1—3. Solenopora multiformis Belokryss. Фиг. 1 —сечение корки внизу
с косорасположеннымп нитями гипоталлия. Фиг. 2 — радиальное сечение
желвака с сорусами, расположенными по концентрам и иногда пересечен-
ными стерильными нитями (направо вверху). Фиг. 3 — поперечное сече-
ние нитей; X10. Верхний эоцен. УССР, Днепропетровская область,
г. Ипгулец (коллекция Л. Белокрыса).
Фиг. 4. Сечение через центр биоценотического желвака, состоящего из
концентрических слоев мшанок (белое) и багрянок (черное); ХЗ. Нижний
тортон. Запад Украины.
Фиг. 5—7. Членистые багрянки Corallinoideae, род Jania. Средний сар-
мат. Молдавия, сел. Оргеев. Фиг. 5 — вид в изломе породы нескольких
члеников вместе; Х5. Фиг. 6 — то же, шлиф; X 10. Фиг. 7 — то же; X 45.
Таблица 20
154
Известковые водоросли
Таблица 21
Багряные водоросли
Rhodophyta
Фиг. 1. Parachaetates {Tomilithon) johnsonii Masi. — радиальное сечение
периталлпя бугра. Видны погнутые вниз поперечные перегородки; Х80.
Нижний карбон. Р. Томь, ниже г. Кемерово (сборы С. В. Максимовой).
Фиг. 2, 4. Donezella lutugini Masi., продольное и поперечное сечения нитей
(фиг.2 — Х46).
Фиг. 3. U ngdarella uralica Masi. Продольное сечение веточки с ветвлением
в верхнем конце. Белые прямоугольные пятна — гипс.
Фиг. 5. Parachaetetes palaeozoicus (Masi.), продольное сечение через пери-
таллий; X 20. Карбон. Южный Урал, р. Зиган.
Фиг. 6. Два членика Jania, охваченные эпифитами — корочками Litho-
phulluni (Dennatholithon) ucrainiciun Masi.; Z 40. Верхний тортон. Крым,
и-ов Тарханкут.
Фиг. 7. Желвак монофитного типа из бугристых корочек литофиллума;
пат. вел.; наружный вид. Нижний тортон .Г. Львов.
Фиг. 8. Ассоциация червей и корковых мелобезий; Х5. Верхний тортон.
Крым, п-ов Тарханкут.
Фиг. 9, 10. Ложные багрянки (по-видимому, губки?) Cribroporidium.
Видно, что вертикальные стенки «клеток» не сплошные, а образованы стол-
биками, не выраженными в косом сечении (фиг. 9 — ХЗЗ; фиг. 10 — Х50).
Карбон. Донбасс, балка Кричик (коллекция Г. Д. Киреевой, № 197).
Таблица 21
156
Известковые водоросли
Таблица 22
Багряные водоросли
Rhodophyta
Фиг. 1,4. Solenopora spongoides var. iiiclioii Masi. Продольное и косое сече-
ния. На фиг. 1 различимы поперечные перегородки, па фиг. 4 видны по-
перечные сечения нитей и зона извилистых нитей вторичного гипоталлия
(слепа); X 15. Ордовик, горизонт Пыхви. Эстония, сел. Мадизе.
Фиг. 2. Сечение параллельно нитям через плодоносный бугорок Archa-
eolithothamnium keenenii var. Irovicum Masi. Многочисленные вытянутые
спорангии располагаются концентрически параллельно сериям клеток;
х46. Тортон. Львовский район.
Фиг. 3. Parachaetetes palaeozoicus (Masi.), продольное сечение периталлия.
Хорошо видно серийное расположение клеток в виде концентрических
рядов; X 12. Карбон (этрень). Южный Урал, р. Зиган.
Фиг. 5. Сечение через биоценотический желвак, образованный мшанками
(светлое) и Lithothamnium magnum Сар. (черное); X 10. Тортон. Львовский
район (дер. Лесники).
Фиг. 6. Продольное сечение ветви современного литотамния. Слева —
вертикальные нити срединной ткани, переходящие (справа) в периталлий
и кортикальную ткань у поверхности; Х46. Баренцево море.
T a б .1 и ц a 22
158
Известковые водоросли
Таблица 23
Харофиты, или харовые водоросли
Charophyta
Фиг. 1. Орган плодоношения харофита предположительно из сем. Clavato-
гасеае с гранулированными измененными стенками в шлифе известняка;
ХЗО. Палеоцен. Таджикская депрессия, сел. Туткаул.
Фиг. 2. Осевые сечения через гирогонит Croftiella escheri (Unger) Horn of
Rantzien. Внизу у базального отверстия с внутренней поверхности рас-
полагается базальная пластинка. Стенки вторично раскристаллизованы.
Шлиф; X 60. Олигоцен. ФРГ.
Фиг. 3. Осевое сечение через гирогонит Tectochara inerianii L. et N. Gram-
bast. Базальная пластинка отсутствует. Видно двухслойное строение
стенки гирогонита. Шлиф; X 50. Плиоцен. Австрия.
Фиг. 4. Осевое сечение гирогонита Rhabdochara langeri (Ettingsshausen)
Madler. Внизу гирогонита видна пирамидальная базальная пластинка,
отделившаяся от оболочки. В последней заметны границы между извест-
ковыми спиралями, которые снаружи имеют резко вогнутую форму.в виде
желобов. Шлиф; ХбО. О.тигоцеп. Швейцария.
Фиг. 5. Осевое сечение гирогонита Nodosochara cli.vu.lata (Peck et Reker)
Madler с оторвавшейся вершиной (проросший экземпляр). Внизу видна
призматическая базальная пластинка, отделившаяся от оболочки во время
захоронения. Гирогонит перекристаллизован. Шлиф; Хб5. Средний эоцен.
США, Флорида.
Фиг. 6. Деталь фиг. 3 — базальная пора Tectochara merianii L. et X.
Grambast в шлифе. Базальная пластинка отсутствует. Видна «штрихова-
тость» внутреннего слоя, обусловленная чередованием слоев кальцита,
обогащенных органическим веществом, <200.
Фиг. 7. Шлиф через два органа плодоношения Lagynophora sp.. который
прошел также через окружающие их вегетативные части (кружки и
трубки). Органы, в составе гирогонитов и утрикулов, прикреплены к цен-
тральному стержню, расположенному внизу снимка; <35. Верхний мел —
палеоцен. Италия.
Фиг. 8. Осевой шлиф через гирогонит Rhadochara krauselii (Rasky) Gram-
bast. Вершина оторвана, базальная пластинка отсутствует, спирали
вогнуты или разделены четкими «щелями» (органическими оболочками
ранее существовавших спиральных клеток); х 35. Верхний олигоцен.
Венгрия.
Фиг. 9. Деталь фиг. 2 — базальная пластинка; , 150 (фиг. 2—9 — по
Horn of Rantzien, 1959).
Фиг. 10. Осевое сечение гирогонита неизвестного харофита в известняке
(алайский ярус) Таджикской депрессии. Фотография слабо ретуширована.
В верхней части гирогонита оттянутая «шейка», может быть, говорит
о принадлежности этого органа к роду Lagynophora (?); <40 (материал
Зелеиовой, № 2785).
Фиг. И. Поперечное сечение через вегетативную часть харофита (междо-
узлие) с двумя рядами коровых трубок Clavatoraxis sp. Слева харофпт
соприкасается с обломком раковины, изъеденной сверлящими водоро-
слями Palaeachlya. Заметно кристаллическнзернпстое строение вегета-
тивных частей харофита, резко отличающееся от строения стенок гиро-.
гонптов. Шлиф; ;<40. Кимеридж. Западная Украина, с. Подлупы.
Фиг. 12. Смыкание концов спирален йершины гирогонита Tectochara
diluviana (Madler) Grambast, видимых изнутри гирогонита; <50. Четвер-
тичные отложения. Низовья р. Амударьи.
Фиг. 13—15. Внешний вид белого гирогонита Т ectoohara diluviana; >'ЗО.
Фиг. 13 — вид сбоку. Фиг. 14 — вид снизу на базальную нору. Фиг. 15 —
вид сверху па вершинные бугры и пазы, утоняющие спирали в месте из-
лома при прорастании ооспоры. Верхний миоцен — нижний плиоцен.
Северный Кавказ, Сунженский хребет (материалы А. А. Стеклова).
Фиг. 16. Гирогонит Tectochara diluviana, протравленный уксусной кислотой
и снятый под водой. Известковая часть удалена, блестящая органическая
пленка — стенки спиральных клеток — сохранилась под светлым наруж-
ным кальцитовым слоем. Вид сбоку; <30. Четвертичные террасы. Ни-
зовья р. Амударьи (материалы Е. К. Обонпцкой).
Фиг. 17. Вид сбоку гирогонита Charites sp. п. В желобках спиральных
клеток находится налипший известковый осадок. Сам гирогонит имеет
бурый цвет, что видно на межспиральпых выступающих ребрах; X 47.
Майкоп. Северный Кавказ, окрестности г. Майкопа.
Таблица 23
160
Известковые eodopoc.iu
Т а б .1 и и а 24
Девонские харофиты
Devonian Charopliyla
Фиг. I. Umbella bella Maslov. Продольное* сечение утрикула. Хорошо
видны базальные выросты утрикула. между которыми располагается
базальная пора, в верхней части видна апикальная пора; ХбО. Фраиские
отложения. Северная Фергана, р. Сумсар (коллекция Б. В. Пояркова).
Фиг. 2. Umbella rei'ularis Skvorzov. Продольное сечение утрикула; хорошо
видны базальные* выросты и апикальная пора, прикрытая апикальной кры-
шечкой; ХбО. Иижнефаменские отложения. Юго-восточный склеен Чат-
кальского хребта, (бассейн р. Падшаата (коллекция В. II. Скворцова).
<1>иг. 3. Quasiиmbella rotunda (Е. Bykova). Продольное сечение утрикула.
Виднее апикальное отверстие и шейка, на которой располагалась апикаль-
ная крышечка; X 60. Нижнсфаменские отложения. Северная Фергана,
г. Лкташ (коллекция Б. В. Пояркова).
Фиг. Umbella mica Pojarkov. Продольное* сечение* утрикула. Видны
небольшие базальные выросты, между которыми располагается базальная
шера, прикрытая изнутри базальной пластинкой. В левой половине под
утрпкулом видна тонкая полоска кальцита, которая, возможно, является
сечением гирогонита; ХбО. Верхнефаменские отложения, куркебайская
свита. Хр. Казахский Каратау, бассейн р. Ханташ (коллекция Б. В. По-
яркова).
Фиг. 5. Umbella bashkirica Е. Bykova. Продольное сечение утрикула.
Видны массивные базальные выросты, ХбО. Живетские еетложения. Се-
верная Фергана, верховья р. Сумсар (коллекция Б. В. Пояркова).
Фиг. (>. Quasiumbella папа (Reitlinger). Продольное* сечение утрикула.
Видна апикальная крышечка; X60. Фаменские отложения. Хр. Боркол-
дой, Кок-шаал (коллекция М. М. Цуркина).
Фиг. 7. Quasiumbella sp. Продольное сечение утрикула. Видна апикальная
пора и сдвинутая с места (наклоненная под углом) апикальная крышечка;
ХбО. Франские отложения. Северная Фергана, бассейн р. Сумсар (кол-
лекция Б. В. Пояркова).
Фиг. 8. Quasiumbella pararotunda Skvorzov. Продольное сечение утрикула,
в апикальной поре видна повернутая апикальная крышечка; ?<60. Нижпе-
фаменские отложения. Северная Фергана, верховья р. Сумсар (коллекция
Б. В. Пояркова).
Фиг. 9, 11, 13. Quasiumbella rotunda (Е. Bykova). Продольное сечение утри-
кула; видно, как апикальное отверстие прикрыто апикальной крышечкой;
ХбО. Иижнефаменские отложения. Юго-восточный склон Чаткальского
хребта, бассейн р. Падшаата (коллекция В. П. Скворцова).
Фиг. 10. Известняк, переполненный утрикулами умбелловых. Шлиф;
Х12. Нижнефамснскнс отложения. Юго-восточный склон Чаткальского
хребта, бассейн р. Падшаата (коллекция В. П. Скворцова).
Фиг. 12. Quasiumbella pararotunda Skvorzov. Продольное сечение утрикула,
в котором видно апикальное отверстие; ХбО. Нижнефамепские отложения.
Юго-восточный склон Чаткальского хребта, бассейн р. Падшаата (кол-
лекция В. П. Скворцова).
Фиг. 14. Quasiumbella sp. Косое сечение утрикула; видно, как под утри-
кудом располагается не полностью обызвествленный гирогонит; ХбО.
Нижнефамепские отложения. Юго-восточный склон Чаткальского хребта,
бассейн р. Падшаата (коллекция В. П. Скворцова).
Фиг. 15, 16. Trochiliscus surmilovae Pojarkov. Фиг. 15 — поперечное сече-
ние гирогонита, видны' четкие швы между партекальцинамп (темные по-
лоски внутри ребер),-Фиг. 16 — близкое к продольному сечение гирого-
нита, видны швы между партскальцинами; <60. Нижнефаменские отло-
жения. Восточные отроги хр. Джетым-тау (коллекция В. М. Рожанца).
Фиг. 17. Trochiliscus. Продольное сечение гирогонита. Видно апикальное
отверстие, прикрытое крышечкой, швы между партекальцинамп не наблю-
даются из-за перекристаллизации (?); ХбО. Иижнефаменские отложения.
Юго-восточный склон Чаткальского хребта, бассейн р. Падшаата (кол-
лекция В. II. Скворцова).
Таблица 24
1102 И
162
Известковые животные организмы
Таблица 25
Фораминиферы
Foraminifera
Фиг. 1. Rhabdammina sp. Продольное сечение раковины с агглютиниро-
ванной стенкой; X 20.
Фиг. 2. llaplophragmina sp. Продольное сечение раковины, стенка из-
вестковая, секрецпонная микрогранулярная, с агглютинированными ча-
стицами (спикула губок, фораминиферы); ;,75. Московский ярус. Верх-
няя Волга.
Фиг. 3. Cyclammina cancellata Brady. Сечение раковины с агглютинирован-
ной стенкой; Х45. Современная форма. Тихий океан.
Фиг. 4. Earlandia sp. Продольное сечение раковины с известковой секрс-
цнонпой стенкой; Х60. Визейский ярус. Московская область.
Фиг. 5. Endothyra ontphalota Raus. et Reithl. Поперечное и тангенциаль-
ное (в левой нижней части раковины) сечения стенок части раковины;
видны волокнистая микрогранулярная микроструктура и поровые каналы
в стенке; X 100. Визейский ярус, Московская область.
Фиг. 6. Globoendothyra globulus (Eichw.). Поперечное сечение части рако-
вины, секрециопиая стенка с микрограиулярной микроструктурой и с диф-
ференциацией на слои с зернами различного размера (а, 6) и с внутренним
светлым слоем стекловатой радиально-лучистой микроструктуры (в);
X100. Визейский ярус. Московская область.
Фиг. 7. Nonion soldani (d’Orb.). Стенка раковины со стекловатой зерни-
стой микроструктурой; николи скрещены; X 600. Средний миоцен. Украина.
Фиг. 8. Cassidulina sp. Степка раковины с тонкой радиально-лучистой
микроструктурой, николи скрещены; X 600. Современная форма. Барен-
цево море.
Фиг. 9. Elphidium latusovum Krash. Стенка с неясной радиально-лучистой
микроструктурой, онколи скрещены; Х600. Средний миоцен. Украина.
Фиг. 10. Rotalia beccarii (Linne). Стенка с грубой радиально-лучистой
микроструктурой; николи скрещены; X 600. Средний миоцен. Украина.
Фиг. 11. Quinqueloculina akneriana d’Orb. Стенка с фарфоровндпой крипто-
кристаллической микроструктурой; николи параллельные; Х570. Палео-
ген. Арало-Тургай.
Фиг. 12. Eogeinitzina devonica Lip. Продольное сечение, первично двух-
слойное строение стенки, внутренний слой микрограиулярный и наруж-
ный— стекловатый радиально-лучистой микроструктуры; X 140. Девон.
Подмосковный бассейн.
Таблица 25
164
Известковые животные организмы
Таблица 26
Фораминиферы
Foraininifera
Фиг. 1. То же, что на таблице 25, фиг. 11, но николи скрещены.
Фиг. 2. Thaltnaniella apenninica (Renz.). Поперечное сечение, вторично
многослойная стенка, с поровыми каналами; X144. Сеноман. Северный
Кавказ.
Фиг. 3. Rugoglobigerina kelleri (Subb.). Вторично многослойная пористая
стенка с шероховатой поверхностью; Х600. Маастрихт. Крым.
Фпг. 4. Lenticulina sp. Вторично многослойная стенка раковины; ХбО.
Киевский ярус. Украина.
Фиг. 5. Стенка раковины Heterolepa, реплика; органические пленки (тем-
ное) обволакивают кристаллы и кристаллические отдельности; электрон-
ный микроскоп (Hansen, 1970).
Фиг. 6. Стенка раковины Bulimina, реплика; органическое вещество
(темное) образует прослой между слоями стенки и норовые трубочки, вы-
стилающие поровые каналы; электронный микроскоп (Hansen, 1970).
Фиг. 7. Bradyina sp. Сложная пористость стенки раковины; Х50. Визсй-
ский ярус. Куйбышевская обл.
Фиг. 8. Lenticulina sp. Внешний вид стекловатой раковины, хорошо виден
киль; Х72. Палеоген. Лрало-Тургай.
Фиг. 9. Cibicidea sp. Внешний вид раковины с грубопористой поверх-
ностью; х72. Палеоген, Арало-Тургай.
Фиг. 10. Uvegerina sp. Стекловатая ребристая раковина; Х72. Палеоген,
Лрало-Тургай.
Таблица 26
166
Известковые животные организмы
Т а б л и ц а 27
Фораминиферы
Foraminifera
Фиг. 1. Лагснидовый мелководный палеоценоз: нодозарии, лептикулипы,
денталины, робулусы; Х40. Маастрихт. Пенза.
Фиг. 2. Псевдоштаффелловый известняк; X15. Московский ярус. Под-
московный бассейн.
Фиг. 3. Ориентированное положение фузулииелл с длинной осью парал-
лельно напластованию; Х7. Московский ярус. Среднее Поволжье.
Фиг. 4. Паратурамминовый известняк. X 20. Фамснскпй ярус. Р. Печора.
Фиг. 5. Пенероплисовый известняк. Пришлифовка; X 5. Миоцен. Подолия.
Фиг. 6. Обломочный известняк, песчаники из фузулинового известняка;
Х15. Московский ярус. Среднее Поволжье.
Фиг. 7. Мелкофорамииифсровый известняк; Х20. Московский ярус.
Среднее Поволжье.
Фиг. 8. Текстуляриевый известняк; X15. Московский ярус. Среднее
Поволжье.
Таблица 27
168
Известковые животные организмы
Таблица 28
Фораминиферы
Foraminifera
Фиг. 1. Форамипиферовый известняк: Endothyranopsis crassa (справа),
Lituotubella (внизу), Cribrospira sp. (слева вверху), Palaeoteitularia sp.,
Ilyperantmina sp. и др.; ?<20. Нижний карбон. Среднее Поволжье.
Фиг. 2. Порода с альвеолинами; Х5,5. Эоцен. Италия.
Фиг. 3. Милиолидовый известняк; X 14. Мел. Италия.
Фиг. 4. Нуммулитовый известняк; X15. Эоцен. Армения.
Фиг. 5. Dicyclina schlumbergerl Mun. Shalm. Xll. Верхний мел. Италия
Фиг. 6. Дискоциклииы в песчанике; X 15. Эоцен, Крым.
Фиг. 7. Kilianina sp.
Таблица 28
170
Известковые животные организмы
Таблица 29
Ф ора ми нифсры
Foraminifera
Фиг. 1. Пубекуляриевый известняк, нубекулярии обрастают мшанками,
мшанковые рифы; \ 15. Третичные отложения. Мангышлак.
«Риг. 2. «Рузулинидовый известняк; X 5. Верхняя пермь. Лаос.
«Риг. 3. Parathurammina paradagmarae Lip.; 60. «Раменский ярус.
Р. Печора.
«Риг. 4, 5. Tournayella costata Lip.; Х70. Поперечное и продольное сече-
ния. Верхний турне. Западный склон Среднего Урала.
«Риг. 6. Chernyschinella gio mi for mis Lip.; X 75. Турпейскпй ярус, Среднее
Поволжье.
«Риг. 7. liradyina rotula Moell.-, X30. Впзейский ярус. Среднее Поволжье.
«Риг. 8. Endothyranopsis crassus Brady; . 30. Впзейский ярус. Среднее
Поволжье.
«Риг. 9. Palaeotextularia ponderosa Beitl.; <35. Московский ярус. Среднее
Поволжье.
«Риг. 10. Spiroplectammina ex gr. carinata d’Orb., внешний вид; X72.
Палеоген. Лрало-Тургай.
«Риг. 11. Eostafella parastruvei Bans., осевое сечение; Х45. Пижнпй кар-
бон. Подмосковный бассейн.
«Риг. 12. Pseudostafella kremsi Bans.; X 35. Московский ярус. Южное
Притимапье.
«Риг. 13. Fusulinella bocki Moell., осевое сечение; <20. Московский ярус.
Среднее Поволжье.
«Риг. 14. Fusnlina cylindrica Fisch., осевое сечение; ;<20. Московски!!
ярус. Г. Касимов.
«Риг. 15. Pseudofusiilina jaroslarkensis Viss.. осевое сечение; X 7. Тастуб-
ский горизонт. Южный Урал.
«Риг. 16. N eoschwagerina ex gr. craticulifera (Scliwag.), осевое сечение; X15.
Верхняя пермь. Крым.
Таблица 29
172
Известковые животные организмы
Таблица 30
Известковые губки u Uralotimania
Calcarea and Uralotimania
Фиг. 1. Внешний вид Siphonococlia expansa Loroil (Pharetroncs); X2.
Нижний мел. Крым (коллекция А. Л. Эрлангера).
Фиг. 2. Поперечное сечение Siphonococlia expansa Loriol. Шлиф; Хб.
Нижний мел. Крым (коллекция Л. А. Эрлангера).
Фиг. 3. Продольное сечение Amblysiphonella sp. (Sphinctozoa). Шлиф;
Х5. Пермь. Фергана (коллекция Б. К. Лихарева).
Фиг. 4. Проблематичная известковая губка. Шлиф; Х28. Юра. Крым
(коллекция В. II. Маслова).
Фиг. 5, 6. Поперечное сечение Amblysiphonella sarytchevae Zhur. Шлиф.
Фиг. 5— <3. Верхняя пермь. Армения (коллекция Т. Г. Сарычевой).
Фиг. 6 — часть поперечного сечения в шлифе; Хб. Верхняя пермь. Арме-
ния (коллекция Т. Г. Сарычевой).
Фиг. 7. Часть продольного сечения Sahraja triassica Moisseiew. Шлиф;
ХЗ. Верхний триас. Северный Кавказ (Моисеев, 1944).
Фиг. 8. Внешний вид A mblysiphonella sarytchevae Zhur.; Х2. Верхняя
пермь. Армения (коллекция Т. Г. Сарычевой).
Фиг. 9. Колония Amblysiphonella sarytchevae Zhur. в поперечном сеЧепии
на выветрелой поверхности; Х1. Верхняя пермь. Армения (коллекция
Т. Г. Сарычевой).
Фиг. 10, 11. Uralotimania reticulata Biab.; X 5. Урал, дер. Луньевка
(по Рябинину, 1915). Фиг. 10— горизонтальное сечение. Фиг. И — вер-
тикальное сечение.
Таблица 30
174
Известковые животные организмы
Таблица 31
Археоциаты
Archaeocyathi
Фиг. 1. Выветрелая поверхность археоцпатового известняка. Видны
поперечные сечения кубков; ват. вел. Пижнпй кембрий, санаштыкголь-
скпй горизонт. Тува.
Фиг. 2. Продольное сечение Archaeolynthus sibiricus (Toll.); Х-3. Нижний
кембрий, камешковский горизонт. Кузнецкий Алатау.
Фиг. 3. Косопоперечные сечения: а — Ttiuitilocyallius pustulatus Vol.,
видны тумулы на наружной стенке; б — Dokidocyathus missarzaerskii
Roz.; Хб. Нижний кембр.пй, камешковский горизонт. Кузнецкий Алатау.
Фиг. 4. Косопоперечное сечение Tumuliolynthus sp.; X 3. Нижний кембрий,
камешковский горизонт. Кузнецкий Алатау.
Фиг. 3. Продольное сечение l-’ransuasaecyathus sp., с крупными тумулами
па наружной стенке; <6. Нижний кембрий, камешковский горизонт.
Кузнецкий Алатау.
Фиг. 6. Поперечное сечение Capsulucyathus irregularis Zhnr.; Х7. Нижний
кембрий, сапаштыкгольский горизонт. Кузнецкий Алатау.
Фиг. 7. Тангенциальное сечение наружной стенки Ladaecyathus sp.
Хорошо видны крупные поры каркаса и мелкие — дополнительные обо-
лочки; Х20. Ленский ярус. Кузнецкий Алатау.
Фиг. 8. Косопродольпое сечение Dictyocyathus sp. со стержнями в интер-
валуме; .<5. Нижний кембрий, камешковский горизонт. Кузнецкий
Алатау.
Фиг. 9. Поперечное сечение Ajacicyathus tkalschenkovi (Vol.); .< 3. Ниж-
ний кембрий, базапхекпй горизонт. Восточный Саян.
Фиг. 10. Косоноперечное сечение Thalamocyathus sp. с кольцевой’внутрен-
ней стенкой; в верхней части видно гребенчатое днище; Хб. Нижний кемб-
рий, сапаштыкгольский горизонт. Кузнецкий Алатау.
Фиг. 11. Косопоперечное сечение Jakutocyathus bugradi Roz., хорошо
видны вздутия на стенках и гребенчатое днище в верхней части снимка;
X 3. Нижний кембрий, камешковский горизонт. Кузнецкий Алатау.
Фиг. 12. Строение наружной стейки у llupecyathellus sc/tuberfi Boz.:
:<20. Тарынскпй горизонт. Сибирская платформа. Игарский район.
Фиг. 13. Косопродольпое сечение Loculicyathus tolli Vol., в нижней части
кубка видна обильная пузырчатая ткань; х 3. Нижний кембрий, саиаштык-
гольскнй горизонт. Восточный Саян.
Таблица 31
176
Известковые животные организмы
Таблица 32
Археоциаты
Archaeocyathi
Фиг. 1. Косопоперечное сечение Clathricoscinus inopinatus Roz., Х5.
Нижний кембрий, камешковский горизонт. Горная Шория.
Фиг. 2. Косопродолыюе сечение Piamaecyathus sajanicus Zhur. с решетча-
той наружной и губчатой внутренней стенками; Х5. Нижний кембрий,
санаштыкгольский горизонт. Западный Саян.
Фиг. 3. Косопродолыюе сечение Coscinocyathus bedfordi Vol.; X 10. Ниж-
ний кембрий, базаихский горизонт. Восточный Саян.
Фиг. 4. Косопродольное сечение раздробленного кубка Coscinocyathus sp.,
внизу поперечное сечение Docidocyathus sp.; Хо. Нижний кембрий, ка-
мешковский горизонт. Кузнецкий Алатау.
Фиг. 5. Поперечное сечение Alataucyathus exelenthis Roz. с бугорчатыми
тумулами наружной стенки; X 5. Нижний кембрий, камешковский гори-
зонт. Кузнецкий Алатау.
Фиг. 6. Косопродольное сечение Tomocyathus gini Miss et Roz., черное —
вторичные утолщения; Х5. Камешковский горизонт. Кузнецкий Алатау.
Фиг. 7. Тангенциальное сечение наружной стенки Dictyocyathus sp.;
Х20. Камешковский горизонт. Горная Шорня.
Фиг. 8. Поперечное сечение Dictuocyathus sp.; X 5. Камешковский гори-
зонт. Кузнецкий Алатау, р. Кия.
Фиг. 9. Косопродолыюе сечение Claruscyathus billingsi (Vol.); ,< 1,5.
Нижний кембрий, солонцовский горизонт. Алтай.
Фиг. 10. Косопоперечное сечение Tabulaecyathellus bidzhaensis Miss., Х5.
Камешковский горизонт. Кузнецкий Алатау.
Фиг. 11. Косопоперечное сечение Archaeocyathus yavorskii (Vol.) с сильно
изогнутыми тениями; X 3. Солонцовский горизонт. Алтай.
Фиг. 12. Косопродольное сечение представителя отряда Cribrocyathida
с пластинчатым строением стенок; х 20. Нижний кембрий. Кузнецкий
Алатау.
Таблица 32
1102 12
178
Известковые животные организмы
Таблица 33
Табуляты
Tabulata
Фиг. 1. Массивная колония Favorites полушаровидной формы (подотряд
Favositina); вид сверху; Х1. Нижний силур. Сибирь, р. Подкаменная
Тунгуска.
Фиг. 2. Стелющаяся колония Aulopora (отряд Auloporida); мелкие корал-
литы вытянуты в цепочку; х2. Нижний девон. Кузбасс.
Фиг. 3. Хали.зитоидная цепочная колония (отряд Halysitida); вид сверху;
Х1. Нижний силур. Сибирь, р. Подкаменная Тунгуска.
Фиг. 4. Squameofavosites dirissimus Dubatolov (подотряд Favositina).
Поперечный разрез, кораллиты с полигональным поперечным сечением.
Стенки кораллитов толстые, с фиброзной микроструктурой. Септальные
чешуйки крупные и темноокрашенные. Внутренние полости кораллитов
выполнены вторичным кальцитом, X 10. Средний девон, эйфельский ярус.
Кузбасс.
Фиг. 5. Squameofavosites sp. (подотряд Favositina). Фиг. 5а — поперечный
разрез. Фиг. 56 — продольный разрез. Кораллиты с полигональным по-
перечным сечением. Степки кораллитов тонкие, с фиброзной микрострук-
турой. Поры расположены па ребрах и гранях кораллитов. Септальные
чешуйки мелкие. Днища горизонтальные. Внутренние полости кораллитов
выполнены вторичным кальцитом; Х4. Верхний силур. Северный Урал.
Фиг. 6. Natalophyllum rarus Tchudinova (подотряд Aveolitina). Фиг. 6а —
поперечный разрез, фиг. 66 — продольный разрез. Кораллиты много-
угольно-округлые по очертаниям. Степки кораллитов топкие, темпоокра-
шениые в осевой зоне, сильно утолщенные на периферии, со слабым пиг-
ментом. Поры чистые. Внутренние полости кораллитов выполнены каль-
цитом; х4. Средний девон, живетский ярус, Кузбасс.
Фиг. 7. Thamnopora porosa Tchudinova (подотряд Thamnoporina).
Поперечный разрез ветвисто-массивной колонии. Кораллиты многоугольно-
округлые по очертаниям. Стенки кораллитов утолщены фиброзной стерео-
плазмой, слабо в осевой зоне и сильно в периферической. Пигмент сохра-
нился в осевой зоне колонии, на периферии частично растворен. Внутрен-
ние полости кораллитов в осевой зоне выполнены вторичным кальцитом,
с периферии они заполнены глинисто-известковистым цементом; х4.
Средний девон, живетский ярус. Кузбасс.
Фиг. 8. Ветвисто-массивная колония Thamnopora (подотряд Thamnopo-
rina) в косом поперечно-продольном разрезе, в темно-сером детритовом
известняке, с обломками створок раковин брахиопод, неполных ветвистых
колоний мшанок; пустоты выполнены вторичным кальцитом; Х4. Верх-
ний девон, франский ярус. Кузбасс.
Фиг. 9. Неполные ветвисто-массивные колонии Cladopora и Striatopora
(подотряд Thaninoporina) в темно-сером, сильноглинистом известняке;
косой поперечный разрез; Х4. Нижний девон. Кузбасс.
Таблица 33
180
Известковые животные организмы
Таблица 34
Табуляты
Tabulata
Фиг. 1. Parastriatopora rhizoides Sokolov (подотряд Thamnoporina). Фиг. la
и 16 — продольные разрезы. В периферической зоне ветвисто-массивной
колонии стенки кораллитов утолщены стереоплазмой. Септальные пла-
стины с сильно зазубренным внутренним краем и трабекулярной микро-
структурой; Х40. Нижний силур. Сибирь, р. Подкамепная Тунгуска.
Фиг. 2. Thamnopora proba Dubatolov (подотряд Thamnoporina), попереч-
ный разрез. Стенки кораллитов с фиброзной микроструктурой, фибры
расположены перпендикулярно средней темной линии (срединный шов,
т. е. эпитека двух смежных кораллитов). Внутренние полости кораллитов
выполнены вторичным кальцитом; Х40. Средний девон, живетский ярус.
Минусинская котловина.
Фиг. 3. Pseudojavosites jinilimus Tchudinova (подотряд Favositina), попе-
речный разрез. Стенки кораллитов с фиброзной микроструктурой. Фибры
расположены перпендикулярно срединному шву и продолжаются в сеп-
тальные шипики. Внутренние полости кораллитов выполнены вторичным
кальцитом; Х40. Нижний триас, индский ярус. Армения.
Фиг. 4. Khmeria pumila Tchudinova (отряд Auloporida), продольный раз-
рез. Небольшой участок стенки кораллита и днища с фиброзной микро-
структурой. Внутренняя полость кораллита выполнена вторичным каль-
цитом; Х40. Нижний триас, индский ярус. Армения.
Фиг. 5. Armalites novellas Tchudinova (отряд Syringoporida). Фиг. 5а —
поперечный разрез. Фиг. 56 — продольный разрез. Отдельный кораллит
кустистой колонии в темпом, сильно глинистом известняке. Стенки корал-
лита толстые, с четкой ламеллярной (волокнистой) микроструктурой.
Септальные шипики иногда имеют осевую полость и погружены в склерен-
химу стенки. Внутри кораллита видны воронковидные днища и вторичный
кальцит; Хб. Средний девон, эйфельский ярус. Кузбасс.
Фиг. 6. Michelinopora allata Tchudiniva (подотряд Favositina). Фиг. 6а —
поперечный разрез. Фиг. 66 — продольный разрез. Кораллиты с полиго-
нальным поперечным сечением, тонкими стенками с фиброзной микро-
структурой. Внутренние полости кораллитов выполнены вторичным каль-
цитом; Х4. Верхняя пермь, джульфинский ярус. Армения.
Фиг. 7. Syringopora bella Tchudinova (отряд Syringoporida). Фиг. 7а —
поперечный разрез. Фиг. 76 — продольный разрез. Кустистая колония
в черном глинистом битуминозном известняке. Стенки кораллитов очень
тонкие. Внутренние полости кораллитов выполнены вторичным кальцитом,
там же видны воронковидные днища; х4. Нижний карбон, турнейский
ярус. Сибирь, устье р. Лены.
Таблица 34
182
Известковые животные организмы
Таблица 35
Кораллы
Zoantharia
Фиг. 1, 2. Holacantia. Септы построены голаканатными трабекулами,
погруженными в ‘ламеллярную ткань: фиг. 1 — поперечный разрез;
фиг. 2 — продольный разрез; Х50. Верхний силур. Подолия (коллекция
В. А. Сытовой).
Фиг. 3, 4. Digonophyllum. Септальные конусы состоят из слоев трабекул:
фиг. 3 — поперечный разрез; фиг. 4 — продольный разрез; X 2. Средний
девон. Урал (коллекция Е.’ Д. Сошкиной).
Фиг. 5. Zonophyllum. Трабекулярное строение септального конуса, по-
перечный разрез; X 50. Средний девон. Закавказье.
Фиг. 6, 7. Margarosmilia. Септы построены из крупных простых трабекул,
иногда соединенных синаптикулами. Поперечные разрезы (фиг. 6— X 10;
фиг. 7 — X 15). Триас. Альпы (Volz, 1897).
Фиг. 8. Siderastrea. Септы построены крупными простыми трабекулами,
соединенными синаптикулами; Х25. Верхний мел. Франция (Alloiteau,
1952).
Фиг. 9. Stypophyllopsis. Септы пшповидные, состоят из простых трабекул,
соединяющихся в септу стереоплазмой. Поперечный разрез; X 15. Верх-
ний триас. Кавказ (коллекция Т. Г. Ильиной).
Фиг. 10. Stylosmilla. Септы построены из мелких простых трабекул.
Поперечный разрез; Х50. Верхняя юра. Кавказ (коллекция Т. Г. Ильиной).
Фиг. И. Calamophyllia. Септы соединены трабекулами. Поперечное се-
чение; X 50. Верхняя юра. Кавказ.
Фиг. 12. Confusastrea. Чередование простых и сложных трабекул в сеп-
тах. Поперечное сечение; X 50. Вёрхняя юра. Кавказ (коллекция
Т. Г. Ильиной).
Фиг. 13. Thamnasteria, Септы построены из крупных простых трабекул.
Поперечный разрез; Х50. Верхний триас. Кавказ (коллекция Т. Г. Ильи-
ной).
Фиг. 14, 15. Колония кораллитов без внешней стенки. Поперечные
сечения; X12. Палеоген. Таджикская депрессия, Устын-тау (коллекция
И. М. Архангельской).
Таблица 35
184
Известковые животные организмы
Таблица 36*
Строматопоры (палеозойские)
Sfromatoporoidea (Palaeozoic)
Фиг. 1. Амфипоры в поперечном и наверху в продольном сечениях;
X 10. Верхний девон. Южный Урал, р. Сим (коллекция В. Н. Крестовпи-
кова).
Фиг. 2, 7. Actinostroma stellulatum Nichols: фиг. 2 — тангенциальное
сечение; светлые ветвящиеся каналы — астрормзы, черные точки — се-
чения столбиков (Х10); фиг. 7 — вертикальное сечение (Х5). Средний
девон. Кузбасс, р. Кара-Чумыш (по В. И. Яворскому).
Фиг. 3. Actinostroma verrucosuin Goldf. в горизонтальном сечении. Кон-
центрические круги являются сечениями пластинок вокруг астроризаль-
ных цилиндров с полным осевым каналом; X 5. Местонахождение и возраст
те же, что на фиг. 2.
Фиг. 4. Cistostoma simplex Yavor. в горизонтальном сечении; X 10 (ма-
териалы В. И. Яворского).
Фиг. 5. Rosenella memorabilis Yavor. в вертикальном сечении; X 10 (ма-
териалы В. И. Яворского).
Фиг. 6. Stromatopora typica Rosen в горизонтальном сечении. Длинные
светлые каналы — астроризы; X10. Силур. Каменец-Подольский (по
В. Н. Яворскому)
Фиг. 8. Atelodictyon sp. в вертикальном сеченип; X10 (материалы
В. И. Яворского).
Фиг. 9. А мф и поры в натуральную величину. Поперечные сечения в при-
шлифовке. Средний девон. Кузбасс (материалы В. И. Яворского).
• Составил В. II. Маслов
Таблица 36
186
Известковые животные организмы
Таблица 37*
Строматопоры (мезозойские)
Stromatoporoidea (Mesozoic)
Фиг. 1, 2. Stromatopora (Parastromatopora) renngarteni Yavor.: фиг. 1 —
продольный, фиг. 2 — поперечный разрезы (по В. И. Яворскому, 1947);
X 10. Средний валанжин. Северный Кавказ, р. Баксан.
Фиг. 3, 4. Actinostromaria taurica Yavor.: фиг. 3 — поперечный, фиг. 4 —
продольный разрезы; х 10 (Яворский, 1947). Титон. Караби-Яйла, район
Симферополя.
Фиг. 5, 6. Tauripora astroites Yavor.: фиг. 5 — вертикальный разрез;
X10; фиг. 6 — тангенциальный разрез; Х5 (Яворский, 1947). Верхняя
юра. Крым, р. Кучук-Карасу.
Фиг. 7, 8. Milleporidium kaberdinense Yavor.: фиг. 7 — тангенциальный,
фиг. 8 — продольный разрезы; х 10 (Яворский, 1947). Средний валанжин.
Северный Кавказ, р. Баксан.
Фиг. 9, 10. Actinostromina grossa (Germ.): фит. 9 — вертикальное сече-
ние, Хб,7; фиг. 10 — поперечное сечение, Х8 (Hudson, 1959). Титон.
Югославия.
Фиг. И, 12. Astroporina siellifera Hudson: фиг. 11 —тангенциальное
сечение; х4; фиг. 12 — радиальное сечение; Х4 (Hudson, 1960). Верхняя
юра. Сомали.
Фиг. 13, 14. Astrostylopsis slouenica Germ.: фиг. 13 — вертикальное,
фиг. 14 — поперечное сечения; Х8 (Hudson, 1959). Верхняя юра. Юго-
славия.
* Составил- В. П. Маслов
Таблица 37
188
Известковые животные организмы
Т а б л и ц а 38*
Микроструктуры стенок ценостеума строматопор
Microstructures of Stromatoporoidea Cenosteum Walls
Фиг. 1, 2. Пучковато-волокнистые стенки амфипор. Черное — пелито-
морфный карбонат цемента породы. Известковые отложения организма
светлее основной массы породы и темнее вторичного карбоната, заполняю-
щего внутренние полости. Фиг. 1 — Х15; фиг. 2 — Х15. Титов. Крым,
массив Аргамыш (коллекция Л. П. Задорожной, № 577).
Фиг. 3. Пятнисто-волокнистая микроструктура верхнеюрской стромато-
поры со светлым вторичным карбонатом в полостях; Х51. Секван. Крым,
северный склон ялтинской Яйлы, р. Биюк-Узенбаш (коллекция Л. П. За-
дорожной, № 1018).
Фиг. 4. Косой срез Astroporina valanginensis Schnorf с пористой структу-
рой стенок; Х90. Юра. Швейцария (Schnorf, 1960).
Фиг. 5, 6. Поперечные сечения трабекул с волокнистой структурой юр-
ской строматопоры Actinostromina g г ossa (Germ.); X 100 (Hudson, 1959).
Фиг. 7. Волокнистая пучковидная структура строматопоры Milleporidium
kabardinense var. barremicurn Yavorsky, x80. Баррем. Крым, дер. Кучки
(Яворский, 1947).
• Составил В. ПГ Маслов.
Таблица 38
190
Известковые животные организмы
Таблица 39
Иглокожие. Морские лилии и ежи
Echinodermata. Crinoidea and Echinoidea
Фиг. 1, 2. Морская лилия Forbesiocrinus sp. Фиг. 1 — продольное сече-
ние стебля; фиг. 2 — поперечное к оси, несколько косое сечение. На фиг. 1
швы между члениками заполнены вторичным карбонатом и видно, что
выступы одного членика входят в бороздки соседнего. Весь шлиф пересечен
системой трещин, которые прошли по спайности одного кристалла под
углом 45° к оси стебля. На фиг. 2 темные радиальные полоски отвечают
выступам поверхности нижнего членика, захваченного плоскостью шлифа
только частично, а темные зубцы и светлые заливчики отвечают выступам
и бороздкам верхнего членика, который охватывает верхнюю часть фи-
гуры; х 20. Карбон. Подмосковье, сел. Мячково.
Фиг. 3. Поперечное сечение через иглу морского ежа Archaeocidaris
rossica. В середине располагается осевая полость. Справа видны сечения
продольных ребер в виде бугорков и пористый слой. Микроструктура
радиально-струйчатая при полном отсутствии септ; Х24. Карбон. Под-
московье, сел. Мячково (материалы В. Н. Крестовникова).
Фиг. 4. Эхинодерматовый известняк. Состоит главным образом из ока-
танных обломков игл морских ежей и других обломков иглокожих, являясь
органогенным песком. На снимке внизу видно сечение створки брахиоподы.
X15. Карбон. Подмосковье, сел. Мячково.
Фиг. 5. Эхинодерматовый известняк, состоящий из остатков иглокожих.
В центре — крупный членик морской лилии типа Pentacrinus с широким
округлым внутренним каналом. Черные участки — окремнелые полости.
Скелет иглокожих выступает в виде светлой сетки; х20. Нижний аален.
СеверныйКавказ, плато Бычисын — междуречье Малки и Кубани (материал
3. В. Тимофеевой, № 227р—2).
Фиг. 6. Поперечное сечение через мелкий придаток (иглу?) морского ежа.
Темные внутренние полости оруденели; Х43. Возраст и местонахождение
те же, что и па фиг. 4.
Фиг. 7, 8. Морской еж в тангенциальном к пластинкам сечении. Фиг. 7 —
в центре находится одна пластинка с двумя крупными отверстиями, а с бо-
ков к ней примыкают две соседние пластинки, захваченные снимком не
полностью. Между пластинками находятся шовные участки, уже перекри-
сталлизованные и потерявшие первоначальную структуру. Отчетливо
видны две различные структуры ячеистой сетки пластины: более крупная
наверху снимка, более тонкая внизу; Х20. Фиг. 8 показывает переход
от мелкой сетки с более правильным характером ячей к крупной с беспоря-
дочно ориентированными ячеями в том же образце и мелкие пятна пере-
кристаллизации с разрушением крупноячеистой сетки; ХбО. Мел. Крым.
Таблица 39
192
Известковые животные организмы
Таблица 40
Иглокожие. Морские ежи
Echinoderniata. Echinoidea
Фиг. 1. Пластинка иглокожего среди шлама различных организмом.
Ячеистая сетка ориентирована в ряды, периферическая часть вторично
изменена; Х40. Силур. Р. Волхов.
Фиг. 2. Оруденелая табличка морского ежа; X 30. Карбон. Подмосковье
(коллекция В. Н. Крестовпикова).
Фиг. 3. Косое сечение иглы морского ежа, сохранились радиальные сплош-
ные септы по периферии иглы. Центральная часть нацело оруденела; Х60.
Девон. Р. Дон.
Фиг. 4, 9. Морской еж Echinocorys sp. (из мела Крыма). Тангенциальные
сечения. Фиг. 4 — видна структура ячей шовной части па краях пластин,
где ячеи ориентированы перпендикулярно краю; X 60. Фиг. 9 — пластинка
с крупными округлыми и продолговатыми порами; х 20.
Фиг. 5. Тангенциальное сечение через шовный участок пластин морского
ежа. Видно рядовое расположение ячей у шва и беспорядочное — более
крупной сетки в удалении от шва; X 50. Мел. Северный Кавказ.
Фиг. 6. Поперечное сечение через иглу морского ежа. Радиальные эле-
менты доминируют во всей игле, кроме осевой зовы. Отчетливо заметно,
что радиальные «септы» имеют расширения, сливающиеся в концентриче-
ские линии и создающие правильную радиально-концентрическую ре-
шетку с увеличением окошек решетки к периферии. Игла находится среди
детритусового известняка с фораминиферами; Х40. Пермь, артинский ярус
Южный Урал, р. Зиган у дер. Гумеровой (коллекция В. Н. Крестовпикова,
№ 271).
Фиг. 7. Придаток морского ежа (игла?, педициллярия?) в поперечном
сечении. Отчетливо видна радиально-концентрическая ориентировка
ячей; X 50. Мел. Крым.
Фиг. 8. Обломок иглокожего, изображенного иа табл. 39, фиг. 4 (в пра-
вом углу) при большом увеличении (Х43). Отчетливо видна радиальная
ориентировка ячей.
Таблица 40
«02 13
194
Известковые животные организмы
Таблица 41
Морские звезды и офиуры
Asteroidea and Ophiuroidea
Фиг. 1. Современная морская звезда в шлифе. Черное — известковые
элементы, серое — остатки органических тканей (мускулы и т. п.); Хб.
Баренцево море.
Фиг. 2. Микроструктура пластинки морской звезды; X 30 (Сауеих,
1916). Преобладающие ячеи шестигранные, расположение рядовое, ре-
шетчатое.
Фиг. 3. Микроструктура современной офиуры Ophiura sarsi Liitken.
«Струйчатая» микроструктура с продолговатыми ячеями, с поперечными
перегородками. Светлые пятна и столбики — известковые выделения;
X100. Баренцево море.
Фиг. 4. Пластинка современной офиуры в скрещенных николях. Микро-
структура однослойная, неправильно-ячеистая, беспорядочная, с сильно
изменчивыми размерами ячей; Х65. Баренцево море.
Фиг. 5, 6. Мелкие пластинки современной офиуры из тонких тяжей,
соединяющие две крупные пластинки (фиг. 5—Х65; фиг. 6—Х40). Ба-
ренцево море.
Фиг. 7—9, И. Микроструктуры известковых элементов современной
офиуры из Баренцева моря: фиг. 7 — налево вверху типичная форма
«позвонка», направо — три элемента со струйчатой микроструктурой,
ХЗО; фиг. 8, И — несколько пластинок вместе со струйчатой микрострук-
турой, округлыми (и слившимися в удлиненные) ячеями, ХЗО; фиг. 9 —
то же, с различной микроструктурой на разных участках: вверху —
струйчатая, внизу — неправильно расположенная структура из непра-
вильных ячей, X 65.
Фиг. 10. Струйчатая микроструктура маргинальной пластинки морской
звезды; Х65. Верхняя юра. Юго-Восточный Крым, р-н сел. Планерское
(коллекция А. Н. Соловьева).
Гиблина 41
196
Известковые животные организмы
Таблица 42
Голотурии и кембрийские иглокожие
Holoturoidea and Cambrian Echinodennata
Фиг. 1 — И, 15. Склериты голотурий (по Deflandre-Rigaut, 1962): фиг. 1 —
Cucumarites dentigerus Defl. —Rig. в скрещенных николях (увелич. около
425). Нижний Оксфорд. Франция, Кальвадос; фиг. 2 — Calclarnnella cla-
vata (Defl. — Rig.) в скрещенных николях, заметно неправильное волни-
стое затухание (увелич. около 425). Олигоцен. ФРГ, Голштсйн; фиг. 3 —
Calclarnnella elliptica (Defl. — Rig.) в скрещенных никелях. Видно волни-
стое угасание (увелич. около 425). Олигоцен. ФРГ, Голштсйн; фиг. 4 —
Calcligula (?) picta Defl. — Rig., в скрещенных николях (увелич. около 425).
Местонахождение то же; фиг. 5 — Paracucumarites porosus (Defl. — Rig.)
в простом свете (увелич. около 425). Оксфорд. Франция, Вийе-сюр-Мер;
фиг. 6 — Cucumarites angulatus Defl. — Rig. в простом свете (увелич.
около 425). Нижний Оксфорд. Франция, Кальвадос; фиг. 7 — Priscopeda-
tus plenus Defl. — Rig. в простом свете (увелич. около 425). Олигоцен.
ФРГ, Голштсйн; фиг. 8 — Calclamnoidea collaris (Defl. — Rig.) в простом
свете (увелич. около 425). Олигоцен. ФРГ, Голштсйн; фиг. 9 — Cucuma-
rites undatus Defl.—Rig. в простом свете (х450). Кимеридж. Юрские горы,
Орбансу; фиг. 10 — Protocaudina mortenseni Defl. — Rig. (диаметр 62 —
80 р). Оксфорд. Франция, Вийе-сюр-Мер; фиг. И — Cucumarites microporus
Defl. — Rig. в скрещенных николях (увелич. около 425). Оксфорд. Фран-
ция, Вийе-сюр-Мер; фиг. 15 — Ornaticanula bonhourei Defl. — Rig. в скре-
щенных николях; заметно неправильное затухание (увелич. около 425).
Оксфорд. Франция, Вийе-сюр-Мер.
Фиг. 12—14. Среднекембрийские иглокожие (ближе неопределимые)
в простом свете. Неправильно-ячеистая микроструктура на фиг. 13 пере-
ходит в струйчатую. На фиг. 12 и 14 интересны зубчатые очертания одной
стороны пластинки; х80. Якутия, р. Батома (материал Н. В. Покров-
ской, № 553).
Таблица 42
198
Известковые животные организмы
Таблица 43
Черви
Vermes
Фиг. 1. Известковые трубки серпулид первого типа (двухслойные, с одним
темным и одним светлым слоем); сечения, приближающиеся к поперечным.
Шлиф; X 70, без анализатора. Визе, серпулово-водорослевый известняк.
Кузнецкий бассейн, р. Кондома (коллекция С. В. Максимовой).
Фиг. 2. Известковые трубки серпулид первого типа в серпулово-водоро-
слевом известняке. В центре снимка и в нижней его половине слева видны
бобовидные и караваеобразные сечения трубок серпулид (спирорбисов).
В правой стороне снимка водорослевые желвачки с нитями различных
диаметров. Наверху сечение в форме восьмерки представляет трубку с одно-
слойной стенкой поперечно-призматического строения, возможно принад-
лежащую примитивным представителям семейства вермитида (гастроподы).
Шлиф; Х26, без анализатора. Кузнецкий бассейн (коллекция С. В. Макси-
мовой).
Фиг. 3. Продольный разрез обломка подковообразно изогнутой трубки
первого типа (в центре снимка). В правой стороне снимка внизу — попе-
речное сечение спирально свернутой трубки, вверху — водорослевый
желвачок, срезан ныи так, что видны только поперечные сечения каналов
(белые кружочки).Шлиф, Х26; тот же образец, что на фиг. 2, без анали-
затора (коллекция С. В. Максимовой).
Фиг. 4. Массовое скопление иловых трубок полихет в микрозернистом
известняке (круги и овалы с толстыми черными стенками). Преобладают
сечения, приближающиеся к поперечным. Шлиф, X 12, без анализатора.
Визе. Кузнецкий бассейн, р. Кондома (коллекция С. В. Максимовой).
Фиг. 5. Известняк, в значительной своей части сложенный сросшимися
трубками серпул. Хорошо видна пузырчатая ткань срастания; х 10, без
анализатора. Эоцен, риштанский ярус. Южная Фергана, Тор-Су (коллек-
ция И. М. Архангельской).
Фиг. 6. Различные сечения трубок Serpula sp. в тонкозернистом извест
няке. Шлиф; X 15, без анализатора. Мэотис. Керченский полуостров, Ста
рый Карантин (коллекция В. II. Маслова).
Таблица 43
200
Известковые животные организмы
Таблица 44
Черви
Vermes
Фиг. 1. Сросток известковых трубок серпулид второго типа. Хорошо
видна сетчатая или пузырчатая ткань срастания, образующаяся между
трубками. Шлиф; Х15, без анализатора. Карбонатная толща верхней
юры. Бухаро-Хивинская область (коллекция Т. И. Бурцевой).
Фиг. 2. Обломки трубок серпулид и ткани срастания (треугольный обло-
мок вверху налево). В обломке, расположенном посредине, в широкой его
части, срезанной более или менее тангенциально, видна поперечная штри-
ховатость, отражающая косую волокнистость наружного слоя трубки.
Тот же шлиф, что на фиг. 1; х46, без анализатора.
Фиг. 3. Стенки двух трубок второго типа в месте соприкосновения (де-
таль при большом увеличении). У нижней стенки хорошо видны изгибаю-
щиеся крутыми параболами волокна наружного темного слоя, внизу пере-
ходящие в слабо различимые, почти горизонтальные фибры (тонкие штрихи
на светлом фоне). У верхней стенки волокна параболического слоя рас-
щеплены, разделены пустыми промежутками и неправильно изогнуты,
т. е. превращены в ткань срастания, что особенно ясно видно в левой сто-
роне снимка. Шлиф; X 42, без анализатора. Карбонатная толща. Верхняя
юра. Бухаро-Хивинская область, Кугитанг-тау (коллекция Т. И. Бурце-
вой).
Фиг. 4. Сросток трубок второго типа. На разрезах видны два слоя, обра-
зующих стенки каждой трубки (внутренний, светло-серый, и наружный,
темный, почти черный), а также ткань срастания (белые пятна и каемки,
облекающие снаружи темный слой). Внизу слева ясно видно сетчатое,
ажурное строение ткани срастания; Х13, без анализатора. Тот же шлиф,
что на фиг. 3.
Фиг. 5. Скошенные поперечное и продольное сечения иловых трубок.
Шлиф; Х43, без анализатора. Возраст и местонахождение те же, что на
фиг. 4.
Фиг. 6. Продольное сечение трубки спирорбис, наросшей на сетчатую
мшанку. Шлиф; X 12, без анализатора. Турне. Кузнецкий бассейн, р. Чу-
мыш (коллекция С. В. Максимовой).
Фиг. 7. Обломки мелких трубок при небольшом увеличении (Х20).
Слои, слагающие стенки, различаются только по цвету. Тот же шлиф,
что на фиг. 3, 4, без анализатора.
Т а б л и ц а 44
202
Известковые животные организмы
Таблица 45
Брахиоподы
В rachiopoda
Фиг. 1. Поперечный разрез раковины современной лингулы. Чередую-
щиеся слои хитина и фосфорнокислого кальция. Фосфатные слои секутся
темными канальчиками. Шлиф; Х110 (по Сауеих).
Фиг. 2. Обломок раковины брахиоподы с волокнистым строением. Шлиф;
Х45. Верхняя пермь, чандалазская свита. Приморье, г. Лысая (коллек-
ция Т. Л. Грунт).
Фиг. 3. Заполнение макушки раковины спирифериды вторичным слоем
волокнистого строения. Шлиф; Х18, 5, николи скрещены. Турне. Кузнец-
кий бассейн (коллекция Г. А. Афанасьевой).
Фиг. 4. Тангенциальный разрез иглистой раковины. Видны многочислен-
ные срезы оснований ребрышек, от которых отходят иглы (темные овалы).
Волокнистость выражена в виде тонких струек. Шлиф; х 18, 5. Турне.
Казахстан (коллекция С. В. Максимовой).
Фиг. 5, G. Разные сечения крупных цилиндрических игл нродуктид:
фиг. 5 — поперечное сечение; фиг. 6 — косое сечение, захватывающее
кусочки створки, от которой отходит игла. Шлиф; Х35. Турне. Казахстан
(коллекция С. В. Максимовой).
Фиг. 7. Скошенный поперечный разрез обломка пористой раковины.
Шлиф; Х45. Турне. Казахстан (коллекция С. В. Максимовой).
Фиг. 8. Тангенциальный разрез пористой раковины. Видны многочислен-
ные черные отверстия пор и обтекающие их волокна раковины. Шлиф;
Х77, николи скрещены. Турне. Казахстан (коллекция С. В. Максимовой).
Фиг. 9. Скошенный тангенциальный разрез раковины Rhipidomella (от-
ряд ортида), у которой пористым является только первичный (наружный)
c.ioii. Черные точки — отверстия пор. Там, где разрез благодаря складча-
тости раковины захватывает пористый вторичный слой, видна только во-
локнистая структура. Шлиф; Х72, николи скрещены. Турне. Казахстан
(коллекция С. В. Максимовой).
Таблица 45
204
Известковые животные организмы
Таблица 46
Брахиоподы
Brachiopoda
Фиг. 1. Обломок двухслойной раковины спирифериды со вторичным слоем
призматического строения. Шлиф; Х45, николи скрещены. Нижний девон.
Кузнецкий бассейн (коллекция С. В. Максимовой).
Фиг. 2. Разрез через спиральный ручной аппарат Cyrtospirijer. Парал-
лельные палочки соответствуют виткам спирали. Шлиф; X 18,5. Нижний
карбон, абышевский горизонт. Кузнецкий бассейн, р. Камышная (кол-
лекция Г. А. Афанасьевой).
Фиг. 3, 4. Поперечные разрезы двуствольных игл Kitakamithyris (се-
мейство ретикулярида). Шлифы, х45. Впзейский ярус (коллекция
Е. Е. Павловой).
Фиг. 5. Скошенный поперечный разрез обломка спирифериды. Видны
оба слоя — волокнистый и призматический, а также бугорки на поверх-
ности раковины (круглые). Шлиф; X 72, николи скрещены. Турнейский
ярус. Казахстан (коллекция С. В. Максимовой).
Фиг. 6. Скошенные продольные сечения раковины рихтгофениид. Видна
ячеистая ткань и поперечные пластины. Шлиф; Х3,5. Верхняя пермь,
чандалазская свита, Приморье, г. Лысая (коллекция Т. А. Грунт).
Фиг. 7. Скошенное поперечное сечение раковины рихтгофенииды, ча-
стично прошедшее через спинную створку. Прямоугольный выступ в верх-
ней части снимка — замок спинной створки, контур которой обрисован
тонкой черной линией, окаймляющей перекристаллизованный участок
в центре. Вокруг спинной створки виден толстый ячеистый слой брюшной
створки и ее тонкий наружный слой (черный). Шлиф; X 3,5. Верхняя пермь,
чандалазская свита. Приморье, г. Лысая (коллекция Т. А. Грунт).
Фиг. 8. Плотный наружный слой раковины рихтгофенииды при большом
увеличении. Видна типичная для брахиопод волокнистость и свойствен-
ные ложнопористым раковинам тонкие стерженьки, пронизывающие во-
локнистую ткань. Шлиф; X100. Верхняя пермь, чандалазская свита.
Приморье, г. Лысая (коллекция Т. А. Грунт).
Фиг. 9. Обломки брахиопод причудливой формы. Шлиф: X 9. Турнейский
ярус. Кузнецкий бассейн.
Фиг. 10. Сечения через целые мелкпе раковинки брахиопод с двумя створ-
ками. Разрез с резко волнистыми линиями характерен для складчатых
ринхонеллид. Шлиф; X 10. Турнейский ярус. Кузнецкий бассейн.
Т а б л н ц а 46
206
Известковые животные организмы
Таблица 47
Мшанки
Bryozoa
Фиг. 1. Мшанки вместе с брахиоподами и иглокожими в биогермном из-
вестняке: древовидные и стержпевидные колонии Trepostomata \Homotry-
pella cribrosa Bassler и Dittopora. colliculata (Eichw)J, двухслойно-симмет-
ричные сетчатые колонии Cryptostomata [Proavella proavus (Eichw.)]
и пластинчатые обрастающие колонии Cystoporata (Favositella sp.); нат.
вел. Средний ордовик, вазалеммский горизонт. Эстонская ССР.
Фиг. 2. Ветвистые колонии мшанок Trepostomata (Batostoma polare
Astr.) в темно-серых тонкокристаллических известняках; нат. вел. Верх-
ний ордовик. Новая Земля, мыс Тимирева.
Фиг. 3. Колонии мшанки Trepostomata (Nicholsonella vaupeliformis Modz.),
нарастающая на членике морской лилии в темно-серых слоистых известня-
ках. Видны трубчатые ячейки с редкими диафрагмами и мезопоры с ча-
стыми диафрагмами, частично закрытые с поверхности известковыми от-
ложениями; Х20. Средний ордовик. Остров Вайгач, Карские ворота.
Фиг. 4. Двухслойно-симметричные лентовидные колонии мшанок Crypto-
stomata (Graptodictya sp. и Rhinidictya sp.) в косых тангенциальных, про-
дольных и поперечных разрезах в темно-сером мшанковом известняке
с обильным карбонатно-глинистым цементом. Видны короткие изогнутые
ячейки, овальные устья, разделяющие их продольные гребни, срединная
пластина, пронизанная капиллярами, и продольные сечения капиллярных
трубочек в стенках ячеек (у Rhinidictya.):, Х20. Средний ордовик. Остров
Вайгач, губа Дыроватая.
Таблица 47
208
Известковые животные организмы
Таблица 48
Мшанки
Bryozoa
Фиг. 1. Нарастающие пластинчатые колонии мшанок Trepostoinata (Ог-
bignyella sp.) в продольном и тангенциальном сечениях и двухслойно-
симметричные лентовидные колонии Gryptostomata в поперечных сечениях
в темно-сером мшанковом известняке с тонкозернистым карбонатно-гли-
нистым цементом; X 20. Средний ордовик. Остров Вайгач, губа Дыроватая.
Фиг. 2. Частично окатанные, переотложенные обломки ветвистых коло-
ний мшанок Trepostoinata (Hallopora sp.) и двухслойно-симметричных
колоний Cryptostomata (Pachydictya sp.) в разных сечениях в красноцвет-
ном обломочном известняке с примесью угловатых зерен кварца. Цемен-
тирующий карбонат с бурыми окислами железа; X 20. Нижний силур,
венлокский ярус. Тува, бассейн р. Сухой.
Фиг. 3. Мшанки отряда Trepostoinata: слоистые колонии Lioclema glo-
meratum Astr., нарастающие на ветвистые колонии Erldotrypa aedilis
Astr. в продольном и поперечном разрезах в черных тонкокристаллических
известняках; X 5. Верхний силур, лудловский ярус. Остров Вайгач,
Карские ворота.
Фиг. 4. Мшанки Cystoporata (Fistulipora sp.) и Cryptostomata (Helopora
sp.) в поперечном и продольном разрезах в обломочном известняке; Х20.
Средний девон. Забайкалье, Нерчинско-Заводский район.
Фиг. 5. Мшанки Trepostoinata (Anomalotoechus sp.) и Cryptostomata
(Helopora sp.) в тангенциальном и поперечных разрезах; X 20. Средний
девон. Забайкалье, Нерчинско-Заводский район.
Фиг. 6. Косое и поперечное сечения двухслойно-симметричной мшанки
Cryptostomata в зернистом известняке. Видна пластинчатая структура
утолщенных стенок и участок двухслойной срединной пластины; X 50.
Средний девон. Кузбасс.
Фиг. 7. Обломки сетчатой колонии Cryptostomata (Semicoscinium sp.)
в тангенциальном косом сечении и ветвистой колонии Trepostoinata
(Lioclema sp.) в обломочном известняке с пиритизированным карбонатом.
Видны сечения ячеек на прутьях сетчатой колонии, разделяющий их киль,
выходы капилляров и струйчатая структура неячеистой стороны колонии?;
X 20. Средний девон. Забайкалье, Нерчинско-Заводский район.
Таблица 48
И 02 14
210
Известковые животные организмы
Таблица 49
Мшанки
Bryozoa
Фиг. 1. Различные сечения колоний мшанок Cryptostomata: ветвистых
(Nicklesopora sp.) и сетчатых (Fenestella sp.) в мшанковых тонкозернистых
известняках; Х12. Нижний карбон. Кузбасс, р. Томь.
Фиг. 2. Обломки сетчатых колоний мшанок Cryptostomata (Fenestella.
sp. и Polypora sp.) в поперечных и косых тангенциальных сечениях в ри-
фовом мшанковом известняке. Видна микроструктура стенок ячеек и ка-
пилляры. Между мшанками вторичный кристаллический кальцит; X 10.
Верхний карбон? Южный Урал, Пещерный лог.
Фиг. 3. Косое тангенциальное сечение участка сетчатой колонии мшанки
Cryptostomata (Feneslella. sp.). Видны угловатые сечения ячеек, бугорки и
струйчатая структура неячеистой стороны колонии; х65. Нижний карбон.
Южный Урал.
Фиг. 4. Поперечное сечение двухслойно-симметричной с вторичными
слоями нарастания мшанки Cystoporata (Hexgonella, sp.). Видны трубчатые
ячейки, почкующиеся в верхнем слое, и многочисленные цистопоры с ча-
стыми выгнутыми перегородками; X 10. Верхний карбон. Южный Урал,
р. Юрюзань.
Фиг. 5. Участок поперечного сечения ветвистой мшанки Trepostomata
с ячейками, заполненными гидроокисью железа. Видны многоугольные
сечения трубчатых ячеек и пластинчатая структура их стенок у периферии
колонии; X 20. Нижняя пермь, артинский ярус. Южный Урал, р. Усолка.
Фиг. 6. Участок тангенциального сечения обрастающей колонии мшанки
отряда Cystoporata (Fistulipora sp.). Видны устья ячеек с лунариями и
многочисленные цистопоры; х 30. Нижняя пермь, артинский ярус. Южный
Урал, р. Зиган.
Фиг. 7. Участки поперечного и тангенциального сечений ветвистой
мшанки Trepostomata (Rhombothypella sp.) в обломочном песчанистом из-
вестняке. Видны овальные устья ячеек и их ромбическое поперечное сече-
ние, мелкие акантопоры и пластинчатая структура утолщенных близ пери-
ферии колоний стенок ячеек; X15. Нижняя пермь, артинский ярус.
Южный Урал.
Таблица 49
212
Известковые животные организмы
Таблица 50
Мшанки
Bryozoa
Фиг. 1. Мшанки отрядов Gyclostomata и Gheilostomata в мелоподобном
известняке; пат. вел. Верхний мел, маастрихтский ярус. Голландия.
Фиг. 2. Поперечное сечение слоистой колонии мшанки Cheilostomata;
Х10. Неоген, миоцен. Закавказье, Шемахинский район.
Фиг. 3. Продольный разрез инкрустирующих колоний мшанок Cheilo-
stomata Nitscheina (Membranipora) в пористом мшанковом известняке
с крустификационной структурой из вторичного кальцита; Х8. Неоген,
миоцен, сарматский ярус. Керченский полуостров, мыс Казантип.
Фиг. 4. Мшанковый пористый известняк с мшанками Cheilostomata.
Ячейки мшанок частично заполнены вторичным кальцитом. В промежут-
ках между колониями — мелкозернистый кальцит; Х15. Неоген, миоцен,
мэотический ярус. Закавказье, Шемахинский район.
Фиг. 5. Косые поперечные сечения ветвистых мшанок Cyclostomata
в мшанковом известняке. Промежутки между колониями заполнены мел-
кими сгустками тонкозернистого кальцита, полости ячеек — вторичным
кальцитом; Х15. Неоген, миоцен, сарматский ярус. Керченский полу-
остров, п-ов Тарханкут.
Таблица 50
214
Известковые животные организмы
Таблица 51
Двустворчатые моллюски, или пластинчатожаберные*
Bivalvia
Фиг. 1. Внешний слой Sauvagerina. Сечение перпендикулярно призмам.
Видны границы призм и мозаичное внутреннее строение их. Простой
свет; X15.
Фиг. 2. Тангенциальное скошенное сечение пластинчатого слоя Chant а
sp., показывающее пучки пластин (псевдопризмы), пересеченные поперек.
Простой свет; X 50.
Фиг. 3. Пластинчатый слой Cardita planicosta в сечении, перпендикуляр-
ном слоям. Простой свет; ХбО.
Фиг. 4. Перпендикулярное к слоям сечение раковины Mytilus edulis.
Простой свет; х40. Темный — пластинчатый, светлый (налево) — призма-
тический слои.
Фиг. 5. То же, что и на фиг. 4, — сечение так называемого призматиче-
ского слоя. Простой свет; х200. Под лупой в этой фотографии можно ви-
деть сложное строение отдельных призм (расщепление).
Фиг. 6, 7. Поперечное сечение раковины Cardium edule: направо — пла-
стинчатый слой (внутренний), налево — так называемый призматический
внешний слой с ветвящимися, сливающимися и выклинивающимися
псевдопризмами или пучками пластин. Простой свет (фиг. 6 — Х100;
фиг. 7 —Х75). По вертикали видны слои нарастания.
Из работы Кайе (Сауеих, 1916).
Таблица 51
216
Известковые животные организмы
Таблица 52
Двустворчатые моллюски, или пластинчатожаберные
Bivalvia
Фиг. 1. Поперечное сечение призматического слоя раковины иноцерама
в скрещенных николях. Видны правильные, четкие, различно затухающие
призмы и слои нарастания; X 20. Верхний мел. Курская область.
Фиг. 2—4. Сечение через так называемый призматический слой устрицы
Ostrea gigantea из среднего эоцена Крыма. Фиг. 2 — поперечный срез в скре-
щенных николях. Видны слои нарастания и неправильные, беспорядочно
затухающие кристаллы без признаков призматического строения (Х20).
Фиг. 3 — та же устрица в тангенциальном сечении призматического слоя
в простом свете. Отчетливо видна «ячеистость» в виде многогранников,
но внутри ячей видны более мелкие, чем ячеи, неправильно ориентирован-
ные кристаллы карбопата (Х20). Фиг. 4 — то же, что и на фиг. 3 (Х40).
Фиг. 5. Поперечное скошенное сечение «призматического» слоя грифеи.
Видны крупные ячеи, заполненные сравнительно мелкокристаллическим
кальцитом. Простой свет; Х40. Мел. Крым.
Фиг. 6. Косой срез раковины Alectrionia acrodonta в поляризованном
свете. Отдельные слои сложены изогнутыми пластинками—кристаллами.
В середине раковины сечения пластинок плоскостные; Х20. Мел. Ма-
астрихт. Крым, Бахчисарай.
Фиг. 7. Косое сечение пластинчатого слоя раковины Exogyra sp. в поля-
ризованном свете. Равномерно затухающие пластинки создают черепитча-
товидную картину; Х95. Мел, Маастрихт. Крым, Бахчисарай.
Фиг. 8. Косое сечение пластинчатого слоя Ostrea gigantea в скрещенных
николях; Х40. Средний эоцен. Крым.
Фиг. 9, 11. Поперечные сечения через раковину Gryphea vesicularis в скре-
щенных николях, прошедшие главным образом через пластинчатый слой.
Белые пятна -г- халцедон. На фиг. 11 в центре видны сферолитоподобные
фигуры затухания, характерные для устриц в плоскостном сечении; х20.
Мел. Крым, Карасубазар.
Фиг. 10. Поперечное сечение современной раковины Cardita в простом
свете. Видны ветвящиеся псевдопризмы; X 60 (Сауенх, 1916).
Таблица 52
218
Известковые животные организмы
Таблица 53
Гастроподы и аммониты
Gastropoda and Aininonoidea
Фиг. 1, 2. Гастроподовый песок. Шлиф через обломки и целые раковины
гастропод. Характерные формы сечений: фиг. 1 — ..<47; фиг. 2— < 12, раз-
рез вдоль оси навивания. Современные. Побережье Черного моря.
Фиг. 3. Поперечное сечение стенки раковины Viriparus sp. в разрезе,
перпендикулярном столбику, в скрещенных николях. Видны ветвящиеся
призмы— пучки пластин; X 35. Неоген. Кавказ, Абхазия.
Фиг. 4. Сечение мелкой гастроподы перпендикулярно столбику; Х15.
Третичные отложения. Керченский полуостров.
Фиг. 5. Продольное сечение (близ оси) через оруденелую гастроподу
Остатки кальцита — светлые. Простой свет; х 30. Третичные отложения
*1*1*1', Рейн.
Фиг. 6. Поперечное к столбику сечение раковины Vipiparus sp., изобра-
женной на фиг. 3. Раковина многослойная, видны чередующиеся слои:
Призматические, пластинчатые и перламутровые (на фото - черные).
Отчетливы два оборота, удваивающие толщину раковины вверху; X 10.
Неоген. Кавказ, Абхазия.
Фиг. 7. Сечение через многослойную перекристаллизованную раковину
гастроподы. Справа — шламовый материал, слева — наросты онколито-
вого характера; ХЗО. Юра. Р. Кунгир, близ Владивостока.
Фиг. Я. 9. Поперечные к столбику сечения раковины Melanopsis spiiiinger
Senin. Видна многослойная структура стенки с чередованием призматиче-
ских слоев (с призмами, ориентированными нормально к поверхности)
и пластинчатых слоев (с ветвящимися пластинками, расположенными косо
к слоям или параллельно им). Перламутровые слои непрозрачны. Простой
свет; фиг. 8 X 50, фиг. 9 — Х20. Неоген. Кавказ, Абхазия.
Фиг. 10, 11. Сечение через раковину Cadoctiiis clalmac Nik. Фиг. 10 —
николи скрещены и видно волнисюе угасание трех слоен. Фиг. 11 - - ни-
коли параллельны; Х40. Нижний келловей. Окрестности г. Елатьмы
на р. Оке.
Таблица 53
220
Известковые животные организмы
Таблица 54
Гаспроподы *
Gastropoda
Фиг. 1, 2. Поперечное сечение среднего слоя листоватых пластин рако-
вины Turritella terebratula: фиг. 1 — простой свет, фиг. 2 — поляризован-
ный свет; X 100. Грубый известняк. Эоцен. Окрестности Парижа.
Фиг. 3. Поперечное сечение стенки раковины ископаемого Conus: внизу —
внутренний листоватый слой, в середине — «плетенчатый» слой, вверху —
внешний пластинчатый слой. Простой свет; х45.
/
Фиг. 4. Поперечное сечение раковины Turritella terebratula: вверху —
внешний слой, внизу — средний пластинчатый в косом сечении. Пластинки
расщепляются и состоят из тонких листочков. Простой свет; х45. Грубый
известняк. Эоцен. Окрестности Парижа.
Фиг. 5. Внешний слой Cyprea bouci. Простой свет; Хб5.
Фиг. 6. Сечение через плетенчатый и внешний листоватопластинчатый
слой раковины Murex sp. Простой свет; х45.
Фиг. 7. Поперечное сечение через раковину Porecellana sp.: внизу —
внутренний, вверху — внешний слой, между ними — плетенчатый сред-
ний слой. Простой свет; х 45,
Фиг. 8. Поперечное сечение через внешний пластинчатый слой Fusus
bulbiformis] Х45.
Из работы Кайё (Сауеих, 1916).
Таблица 54
222
Известковые животные организмы
Таблица 55
Аммониты и белемниты
Ammonoidea and Belemnoidea
Фиг. 1—3. Раковины аммонитов в случайных сечениях в простом свете.
Фиг. 1 — сечение через перегородки в темном пелитоморфном карбонате.
Стенки перекристаллизованы; Х20. Триас, Тироль. Фиг. 2.— обломки
раковины аммонита с сохранившейся структурой стенки. Видны слои
нарастания раковины и трещины вторичного происхождения между тон-
кими пластинками, расположенными перпендикулярно стенкам раковины;
X 50. Мел. Майкопский район. Фиг. 3 — сечение перегородок аммонита
частично перекристаллизованных; Х20. Триас. Тироль.
Фиг. 4—8. Сечения через ростры белемнитов в скрещенных николях.
Фиг. 4, 6, 8 — Hybotites sp.: фиг. 4 — поперечный разрез; фиг. 6, 8 —
продольные сечения. На фиг. 8 светлая горизонтальная полоска — след
сверлильщика, черная вертикальная полоска — трещина в шлифе; X 15.
Апт. Старый Крым. Фиг. 5 — косое поперечное сечение части ростра
Duvalia sp. Балки креста затухания несколько изогнуты; X 15. Нижний
мел. Крым, Феодосия. Фиг. 7 — поперечное сечение половины ростра
Belemnitella americana. Радиальные конусы, вернее, клинья, относительно
правильной формы со спайностью, расположенной косо к границам кри-
сталлов. Отчетливо видно сферолитоподобное затухание клиньев. В центре
структура нарушена пестрыми пятнами халцедона; X 15. Датский ярус.
Крым, Карасубазар.
Т а б .1 н u a 55
224
Известковые животные организмы
Таблица 56
Остракоды
Ostracoda
Фиг. 1—3. Leperditia sp. в сечениях. Фиг. 1 — фрагмент раковины,
стрелка указывает поровый канальчик (?); Х515. Фиг. 2 — фрагмент
раковины, стрелка указывает поровый канальчик (?); Х915. Фиг. 3 —
поперечный раерез вентральной части панциря; Х115.
Фиг. 4. Поперечный разрез через панцирь Т ubulibairdia с lava (Kegel).
Стрелка указывает дорзальный край; Х35.
Фиг. 5. Поперечный разрез через панцирь Bairdtocypris sp.; Х36.
Фиг. 6. Известняк с характерными сечениями остракод; Х40 (Сауеих,
1916).
Фиг. 7. Известняк с многочисленными циприсами; Х35 (Сауеих, 1916).
Фиг. 8. Многослойная (многопанцирная) остракода, не сбрасывающая
старый панцирь; Х40. Девон. Подмосковье (материал В. П. Маслова).
Таблица 56
1102 15
226
Известковые животные организмы
Таблица 57
Трилобиты
Trilobita
Фиг. 1, 2. Сечения через край панциря трилобита в шламовом известняке
в простом свете. По краям в панцирь внедряются кристаллики доломита.
Панцирь имеет однородную «губчатую» структуру или, может быть,
«шагреневую поверхность». На фиг. 2 панцирь изъеден сверлящими орга-
низмами. На фиг. 1 слева в углу — обломки иглокожих; Х25—30. Карбон.
Подмосковье, сел. Мячково.
Фиг. 3, 5. Поперечное сечение панциря с однородной «губчатой» струк-
турой и редкими поперечными каналами. Один и тот же объект: фиг. 3 —
простой свет, фиг. 5 — в скрещенных николях. Затухание волной. Внизу —
поперечное сечение шипа трилобита; Х40. Карбон. Донбасс.
Фиг. 4, 6. Поперечное сечение через различные участки панциря Лха-
phus sp. в поляризованном свете. На фиг. 4 отчетливо видны ромбики вто-
ричного доломита, вкрапленные в панцирь. Заметно также волнистое за-
тухание, особенно на фиг. 6 (фиг. 4—Х20; фиг. 6 — Х20). Нижний силур.
Р. Волхов.
Фиг. 7, 10. Сечения через панцирь Trinucleus sp. (в песчанике); Х50.
Фиг. 7 — тангенциальное сечение с поперечным сечением каналов (столби-
ков, по Кайё). Фиг. 10 — поперечное сечение панциря с продольными се-
чениями каналов (столбиков, по Кайё). Видна светлая оторочка каналов
и поверхности панциря (Сауеих, 1931).
Фиг. 8. Вертикальное сечение панциря Trinucleus goldfussi с поперечными
темными каналами (в песчанике); Х55 (Сауеих, 1931).
Фиг. 9. Поперечное сечение через спинной панцирь трилобита в простом
свете. Характерный контур только для трилобитов. На карях виден заво-
рот или кайма. В панцирь вкраплен пирит в виде мелких точек; X 10.
Карбон. Донбасс.
Фиг. 11. Поперечное сечение Trinucleus goldfussi. Заметна слоистость и
«губчатая» основная структура с тонкими канальцами; Х70 (Сауеих, 1931).
Таблица 57
228
Известковые животные’ организмы
Таблица 58
Трилобиты
Trilobita
Фиг. 1. Представитель группы протоленид. Сечение поперек панциря,
видна кайма с двух сторон в виде «крючков», загнутых вниз. Панцирь
перекристаллизован; Х45. Нижний кембрий. Правый берег р. Лены выше
с. Ботомайское (коллекция Н. В. Покровской, № 111 —16).
Фиг. 2, 3. Известняки, переполненные представителями группы агностид.
Панцири перекристаллизованы; X 10. Средний кембрий. Якутия, р. Бо-
том (коллекция Н. В. Покровской: фиг. 2—№ 67, фиг. 3 — № 553).
Фиг. 4. Сечение трилобита с панцирем хорошей сохранности в скрещен-
ных николях. Участок панциря включает краевую кайму. Видно волновое
затухание кальцита; Х60. Известняк Кв. Карбон. Донбасс.
Фиг. 5, 6. Трилобитовые известняки из представителей группы прото-
ленид. Некоторые сечения дают трехлопастное очертание верхнего спин-
ного панциря. Брюшной панцирь более пологий. Между ними обычно
находится вторичный карбонат, если панцирь не распался на части.
Расположение остатков беспорядочное; X 10. Нижний кембрий. Якутия,
р. Лена (коллекция Н. В. Покровской, № 16).
Т а б л и и а 58
230
Известковые животные организмы
Таблица 59
Позвоночные
Vertebrata
Фиг. 1. Продольное сечение зуба акулы. Свет поляризованный, николи
скрещены. На продольных сечениях остеонов видно волнистое погасание
минерального агрегата, составляющего ткань зуба. На отдельных попе-
речных сечениях остеонов погасание проявляется в виде наложенных тем-
ных крестов; Х80. Палеоген. УССР, окрестности г. Кременчуга Полтав-
ской области.
Фиг. 2. Шлиф чешуи современного карпа. Свет поляризованный, николи
скрещены. Видно волнистое погасание; X 80.
Фиг. 3. Поперечное сечение верхнего панциря черепахи. Свет поляризо-
ванный, николи скрещены. Видны многочисленные поперечные сечения
концентрических остеонов с наложенными темными крестами погасания;
Х80. УССР, сел. Андреевка Николаевской области.
Фиг. 4. Продольное сечение ластовой кости плезиозавра. Свет поляризо*
ванный, николи скрещены. Почти вся костная ткань метосоматически
замещена вторичным минералом, по-видимому, главным образом кварцем.
Видны остатки костной ткани неправильно изогнутой формы с волнистым
погасанием; Х80. Юра. РСФСР, р. Съепсал (приток р. Самары).
Фиг. 5. Продольное сечение левой цевки страуса. Свет поляризованный,
николи скрещены. Видно продольное сечение остеонов и гаверсовых кана-
лов. В костных пластинках видны многочисленные остеоциты. Погасание
волнистое; Х80. Средний плиоцен. УССР, Одесские катакомбы.
Фиг. 6. Продольное сечение трубчатой кости хилотерия. Свет обыкновен-
ный проходящий. Кость минерализована, видны гаверсовы каналы и много-
численные лакуны остеоцитов, заполненные непрозрачными гидроокислами
железа; Х80. Миоцен. МНР, Северное Гоби.
Фиг. 7. То же. Свет Поляризованный, николи скрещены. Видно волнистое
погасание скрытокристаллического агрегата карбонатапатита, слагающего
ископаемую кость'.
Таблица 59
232
Известковые животные организмы
Таблица 60
Позвоночные
Vertebrate
Фиг. 1. Косое сечение трубчатой кости копытного. Свет поляризованный,
николи скрещены. Видны срезанные наискось остеоны, в центре которых
находятся гаверсовы каналы, а в костных пластинках — лакуны остео-
цитов, заполненные непрозрачными гидроокислами железа. Погасание
волнистое; Х80. Мэотис. УССР, сел. Гребенки Одесской области.
Фиг. 2. Продольное сечение трубчатой кости копытного. Свет поляризо-
ванный, николи скрещены. Видно волнистое погасание продольных срезов
остеонов; Х90. Мэотис. УССР, сел. Кубанка Одесской области.
Фиг. 3. Поперечное сечение трубчатой кости цетатерия. Свет поляризо-
ванный, николи скрещены. Видны многочисленные поперечные срезы
остеонов с наложенными темными крестами погасания; Х80. Средний
плиоцен. УССР, Крымская область, Камыш-Бурун.
Фиг. 4. Срез губчатой ткани позвонка крупного копытного. Свет поляри-
зованный, николи скрещены. Кость минерализована. Видны окружающие
крупные поры костные волокна с волнистым погасанием. Поры заполнены
карбонатом (в шлифе — светлые пятна) и непрозрачными гидроокислами
железа; Х80. Поздний плиоцен. УССР, Горный Крым, карстовая шахта.
Фиг. 5. Поперечное сечение свежей трубчатой кости современного быка.
Свет поляризованный, николи скрещены. Видны многочисленные попереч-
ные срезы «коломорфных» остеонов с наложенными темными крестами
погасания; X 80.
Фиг. 6. Сечение зуба Dactylodus sp. в поперечном сечении. Черные по-
лоски — гаверсовы каналы. Внизу — кость, вверху — дентин; X 10.
Карбон. Подмосковье, сел. Мячково (коллекция В. П. Маслова).
Фиг. 7. Плоскостное сечение через пластинку Coccosteus sp. в поляризо-
ванном свете. Видны срезы остеонов с крестами угасания; Х20. Девон.
Р. Сясь (коллекция В. П. Маслова).
Фиг. 8, 9. Зуб акулы Lamna sp. Фиг. 8 — продольное сечение. Крупные
черные и белые пятна — гаверсовы каналы, от которых отходят извилистые
черные канальчики; X 90. Фиг. 9 — поперечное сечение через корень зуба
акулы. В основной темно-бурой массе расположены гаверсовы каналы —
на снимке белые и черные пятна, окруженные светлым ободком из желтой
массы. Этот ободок пересечен тонкими канальчиками, расположенными
перпендикулярно к стенке крупного канала; X 15. Третичные отложения.
Кавказ, Майкопский район (коллекция В. П. Маслова).
Т а б л и ц a 60
234
Известковые животные и спорные организмы
Таблица 61
Спорные организмы и гидроидные полипы
*
Controversial Organisms and Hydrozoa
Фиг. 1—4. Lithochrysites calcareus Masi. Фиг. 1, 4 — простой свет,
фиг. 2, 3 — скрещенные николи. Затемненные места отвечают пятнам
органического вещества, загрязняющего известковые остатки. В скрещен-
ных николях видно, что затухание отдельных клеток часто не зависит от
соседних. Шлифы из агрегатов, выделенных из породы (фиг. 1—3 — X 40;
фиг. 4— Х80). Лёсс. Четвертичные отложения. Украина.
Фиг. 5—9. Distychoplax biserialis (Dietrich) Pia в продольных и косых
сечениях. На всех снимках наблюдаются открытые ячеи, при некоторых
сечениях как бы зазубренные внешние края. Фиг. 5 — по Андрусову (And-
rusov, 1963), X 15; фиг. 6—9 — по Лемуан (Lemoine, 1960): фиг. 6 — X 100;
фиг. 7 — Х60; фиг. 8, 9 — X 150.
Фиг. 10, 11. Uralotimania=Paleoplisina (гидроидный полип). Сечение пер-
пендикулярно пластине. Черные неправильно извитые и ветвящиеся по-
лости — зооидные трубки; светлые промежутки между ними — перекри-
сталлизованные известковые выделения животного. Цементом служит тем-
ный пелитоморфный карбонат. Гидроид ассоциируется с мшанками в био-
гермных известняках от среднекаменноугольного до раннепермского воз-
раста; Х20. Южный Урал, Уфимское плато, р. Юрюзань (фиг. 10 —
внешняя поверхность — направо; фиг. И — внешняя поверхность —
налево).
Таблица 61
236
Известковые спорные органы s.мы
Таблица 62
Спорные организмы
Controversial Organisms
Фиг. 1—7. Pseudovermiporella в разных сечениях: фиг. 1 — .< 30; фиг. 2 —
X 20; фиг. 3—5 — X 46; фиг. 6 — X 10; фиг. 7 — X 30. Пермь, Армения
(материал Р. Л. Аракеляна).
Фиг. 8, 9. Coscinoconus в продольном сечении. Фиг. 8 — С. conicus Masi.;
Х25. Фиг. 9. — С. oblongus Masi.; ХЗО; Верхняя юра. Крым, Яйла.
Фиг. 10, И. Microtubus в продольных и косых сечениях: фиг. 10 — ра-
псе описан под названием Epiphyton (?) jakutii Masi.; Х65. Нижний кем-
брий. Р. Лена (№45/35). Фиг. 11 — Microtubus', X 45. Каменноугольные
отложения. Урал.
Фиг. 12. Lombardia в разных сечениях; ХЗО. Юра. Закарпатье, сел. Пере-
чни (из коллекции Л. В. Липецкой).
Таблица 62
238
Известковые животные и спорные организмы
Таблица 63
Конодонты и сколекодонты Эстонии и контортопоридии
Conodonta and Scolecodonta of Esthonia and Contortoporidia
Фиг. 1. Простые клыкоподобные конодонты в нерастворимом остатке
глауконитового известняка. Видны базальная полость и ось роста; Х60.
Нижний ордовик.
Фиг. 2. Разные остатки конодонтов в глауконитовом песке, темные круги —
зерна глауконита; ХбО. Нижний ордовик.
Фиг. 3. Остаток конодонта в глауконитовом песчанике. Сечение через
базальную часть. Зерна кварца и глауконита покрыты гидроокислом же-
леза с примесью фосфата. Цемент песчаника карбонатный; ХбО. Нижний
ордовик.
Фиг. 4. Остаток конодонта в глауконитовом известняке. Более темные
зерна — глауконит; ХбО. Нижний ордовик.
Фиг. 5. Остаток конодонта в доломите; ХбО. Нижний ордовик.
Фиг. 6. Остаток главного зубца конодонта в глауконитовом песчанике.
Более светлые — хорошо окатанные зерна кварца, темные — зерна глау-
конита. В нижнем левом углу виден обломок кварцевого алевролита.
Цемент песчаника карбонатный; ХбО. Нижний ордовик.
Фиг. 7. То же; кроме остатков конодонтов видны мелкие полосатые об-
ломки брахиопод (оболид?); Х40. Фото В. П. Маслова.
Фиг. 8. Остаток главного зубца в глауконитовом известняке. Видны мел-
кие зерна глауконита. В верхнем левом углу — хорошо окатанное зерно
кварца; ХбО. Нижний ордовик.
Фиг. 9. То же, в доломитовом известняке; ХбО.
Фиг. 10. Пересечение базальной части и главного зубца конодонта.
Шлиф изготовлен из нерастворимого остатка глауконитового известняка;
ХбО. Нижний ордовик.
Фиг. И. Сечение конодонта с хорошо выраженной осью роста в базальной
части; ХбО. Глауконитовый известняк. Нижний ордовик.
Фиг. 12. Главный зубец конодонта с небольшой базальной полостью,
видна ось роста; .<30. Глауконитовый песчаник. Нижний ордовик.
Фиг. 13. То же, при скрещенных николях. Ось роста остается погашен-
ной; Х50.
Фиг. 14. Разные остатки сколекодонтов из нерастворимого остатка карбо-
натного мергеля. Зарисовка по микроскопическим препаратам. Верхний
силур.
Фиг. 15. Остаток сколекодонта из нерастворимого остатка глинистого
доломитистого известняка; Х50. Верхний силур.
Фиг. 16, 17. Contortoporidium в продольных, косых и поперечных сече-
ниях. Фиг. 16 — через всю фигуру проходит продольное сечение Contorto-
poridium в виде вильчатой ленты с внутренними полостями. Светлые не-
правильные пятна — его же поперечные сечения. Около развилки —
сечения через Cribroporidium\ Х35. Фиг. 17 — косые и внизу поперечные
сечения Controtoporidium. В центре снимка — косое сечение Cribropori-
dium\ Х20. Карбон. Донбасс.
Фиг. 18. Проблематический домик червя, ранее описанный как водоросль
Paleonites jakutii Masi.; Х25. Кембрий. Якутия, р. Лена.
Таблица 63
240
Известковые животные организмы
Таблица 64
Хиолиты
llyolitha
Фиг. 1. Поперечное сечение стенки раковины хиолита; X 100. Нижний
кембрий. Сибирская платформа.
Фиг. 2. Поперечное (а) и косое (б) сечения раковины Lenatheca groenlan-
dica (Poulsen) (сем. Orthotheidae); X 10.
Фиг. 3. Поперечное сечение раковины представителя сем. Aliathecidae;
внутри раковины сечения брахиопод (а) и губок Chancelloria (б); X 10;
Фиг. 4. Косопоперечное сечение хиолита, принадлежащего к сем. Hyoli-
thidae; X 20.
Фиг. 5. Участок шлифа с сечениями хиолитов: а — представитель сем. Sul-
cavithidae; б — косое сечение экземпляра, относящегося к сем. Circothe-
cidae, н — поперечное сечение Turcutheca ап пае (Syss.) (сем. Circotheci-
dae); х ю.
Фиг. 6. Поперечное сечение раковины Lenatheca groenlandica (Poulsen);
Х10.
Фиг. 7. a — сечение крышечки хиолита из сем. Orthothecidae; б — по-
перечное сечение раковины сем. Circothecidae; X 10.
Фиг. 8. Косопродольное сечение трубки червя из рода Coleolella">\ Х10.
Фиг. 9. Карман на поверхности размытого пласта, запопиенный облом-
ками раковин хиолитов (а) и брахиопод (б); X 10.
Фиг. 10. Продольные и тангенциальные срезы раковин представителен
сем. Circothecidae. Раковины вставлены одна в другую. У этого семейства
хорошо развиты перегородки; X10.
Фиг. 11. Поперечное сечение раковины хиолита Burithes elongatus (сем. Sul-
cavitidae). Хорошо видны «мускульные впечатления» на внутренней по-
верхности брюшной плоскости по одному с каждого края (а); X 3.
Фиг. 12. а - - обломок раковины рода Allatheca с тремя сохранившимися
перегородками; б — поперечное сечение раковины Circothecidae; раковины
вставлены одна в другую, очевидно, в результате движения воды; в —
поперечное сечение раковины сем. Circothecidae; раковины вставлены одна
в другую. Видна поперечная к стенкам раковины ориентация призматиче-
ских по форме кристаллов; X 10.
Фиг. 13. Поперечное сечение раковины сем. Orthothecidae (род Trapezo-
theca)', X10.
Фиг. 14. Участок шлифа с поперечными сечениями раковин, относящихся
к семействам Orthothecidae (а) и Circothecidae (б); X 10. Нижний кембрий.
Фиг. 15. Поперечное сечение раковины рода Eonovitatus (сем. Allatheci-
dae); X 10.
Фиг. 16. а — поперечное сечение раковины рода Exilitheca (сем. Allathe-
cidae); б — тангенциальный срез раковпны того же вида, что на фиг. 8; X 10.
Фиг. 17. Поперечное сечение трубки червя (?) Hyolithellus tenuis Miss.; X 50.
Таблица 64
1102 %f6
242
Известковые животные организмы. Кремневые водоросли
Таблица 65
Спликофлагеляты и тентакулиты
Silicoflagellata, Tentaculitoidea
Фиг. 1. Vallacerta hortonii Паппа. Сантон —кампан, славгородская свита.
Восточный склон Урала.
Фиг. 2. Dyctyocha triacantha var. inermis Lemm. f. inermis (скелет сверху).
Нижний палеоцен, нижняя часть талицкой свиты. Восточный склон Урала.
Фиг. 3. Dyctyocha triacentha var. hastata Lemm. (скелет снизу). Нижний
палеоцен, нижняя часть талицкой свиты. Восточный склон Урала.
Фиг. 4, 5. Distephanus antiquus Gies. (фиг. 4 — скелет снизу, фиг. 5 —
сверху и сбоку). Нижняя половина верхнего эоцена, верхняя часть верхне-
тасаранской подсвиты. Тургайский прогиб.
Фиг. 6. Distephanus speculum var. pentagonus Lemm. (скелет снизу).
Верхнеэоценовые отложения. Восточный склон Мугоджар.
Фиг. 7. Naviculopsis robusta Defl. (скелет сбоку, виден уплощенный уча-
сток базального кольца). Нижний эоцен, серовская свита. Восточный склон
Урала.
Фиг. 8. Nariculopsis biapiculata (Lemm.) Freng. var. biapiculata (скелет
сверху). Нижняя половина верхнего эоцена, верхнетасаранская подсвита.
Тургайский прогиб.
Фиг. 9, 10. Naviculopsis biapiculata var. minor (Schulz) Gies. (фиг. 9 —
скелет сверху, видна апикальная пластинка; фиг. 10 — скелет сбоку,
видна уплощенная боковая сторона базального кольца). Нижняя поло-
вина верхнего эоцена, верхнетасаранская подсвита. Тургайский прогиб.
Фиг. И, 12. Mesocena elliptica Ehr. Нижняя половина верхнего эоцена,
верхняя часть верхнетасаранской подсвиты. Тургайский прогиб.
Фиг. 13. Mesocena circulus Ehr. Нижняя половина верхнего эоцена,
верхнетасаранская подсвита. Тургайский прогиб. (Фиг. 1 — 13 — X 1000).
Фиг. 14, 15. Косые продольные сечения раковин тептакулитов. Видны
поперечные сечения канальцев в" виде точек, кольцевые ребра и темные
диски. Фиг. 14 — X 20. Девон. Район г. Новгорода. Фиг. 15 — Х46. Де-
вон.
Фиг. 16, Осевое сечение Tentaculites nikifororae G. Ljasch. на конце ра-
ковины с камерами; Х46. Р. Днестр, дер. Кулы (коллекция Г. П. Ляшенко,
№ 825).
Фиг. 17. Осевое сечение в средней части раковины Heteroctenus tichomirovi
G. Ljasch. Видны темные диски в срединной части кольцевых ребер; Х46.
Девон (коллекция Г. П. Ляшенко, № 677).
Таблица 65
Н02 Ш
244
Кремневые животные организмы
Таблица 66
Древние радиолярии*
Old Radiolaria
Фиг. 1. Радиолярит. Многие раковинки радиолярий утратили струк-
туру; Х35. Нижняя пермь. Южный Урал (коллекция И. В. Хворовой).
Фиг. 2. Яшма с обильными радиоляриями. Раковинки утратили струк-
туру, превратившись в халцедоновые шарики. Внизу направо виден тон-
кий шип, отходящий от такого шарика; ХЗО. Карбон. Урал.
Фиг. 3. Силицит с обильными радиоляриями; Х26. Нижний карбон, за-
волжский горизонт. Волго-Уральская область, Дмитриевка, скв. 55
(коллекция А. А. Троховой).
Фиг. 4. Тангенциальное сечение раковинки с хорошо сохранившейся
сетчатой структурой; Х200. Верхний силур. Р. Курейка (коллекция
О. Н. Липиной).
Фиг. 5. Тангенциальное сечение кальцитизированной раковинки с до-
вольно хорошо сохранившейся сетчатой структурой; X 160. Верхний девон,
фаменский ярус. Дмитриевка, скв. 56 (коллекция А. А. Троховой).
Фиг. 6а—в — халцедоновые скелеты радиолярий в фосфорите, сложенном
аморфным фосфатом; Х90. Мезозой. Дальний Восток.
Фиг. 7. Пиритизированная раковинка в перекристаллизованном извест-
няке. Поперечный разрез. Видны контуры двух шаров, вложенных друг
в друга, одна из игл, соединяющих шары и шипы; Х160. Верхний девон,
доманиковый горизонт. Коми АССР (коллекция С. В. Максимовой).
Фиг. 8. Пиритизированная раковинка в тангенциальном сечении. Тот же
шлиф, что на фиг. 7; X 160.
Фиг. 9. Кальцитизированная раковинка радиолярии. Поперечный раз-
рез. Внутренний шар намечен пунктирной окружностью, отражающей
его сетчатое строение; Х160. Верхний девон, доманиковый горизонт.
Коми АССР (коллекция С. В. Максимовой).
Фиг. 10. Поперечный разрез пиритизированной раковинки. Хорошо
видны контуры двух шаров, вложенных друг в друга, и соединяющие их
радиальные иглы, выступающие за внешнюю сферу в виде шипов; X 160.
Верхний девон, доманиковый горизонт. Коми АССР (коллекция С. В. Мак-
симовой).
Фиг. 11. Раковинка радиолярии, утратившая структуру, но сохранившая
шипы. Распространенный тип сохранности; х 160. Нижний карбон, за-
волжский горизонт. Дмитриевка, скв. 55 (коллекция А. А. Троховой).
Фиг. 12. Радиоляриевый известняк из кальцитизированных скелетов
радиолярий. Нижняя пермь, сакмарский ярус. Актюбинская область
(коллекция И. В. Хворовой).
Фиг. 13. Колоколовидная раковинка радиолярии; Х200. Верхняя юра.
Вятские желваковые фосфориты (коллекция Кузнецова).
• Все снимки сделаны без анализатора.
Таблица 66
246
Кремневые животные организмы
Таблица 67
Кремневые губки
Siliceous Spongia
Фиг. 1. Спикуловый известняк с цементом из микрозернистого кальцита.
Видны продольные (палочки) и поперечные (кружки и овалы) сечения
одноосных спикул. Спикулы кальцитизированы, каналы выполнены более
тонкозернистым и темным кальцитом, чем стенки спикул. Шлиф; Х40,
без анализатора. Турнейский ярус. Кузбасс (коллекция С. В. Максимо-
вой).
Фиг. 2. Спикуловый известняк, сложенный густо переплетающимися
мелкими спикулами. Шлиф; Х20, без анализатора. Средний карбон.
Известняк. Донбасс, Белокалитвинский район (коллекция Г. Д. Кире-
евой).
Фиг. 3. Известковистый спонголит. Вверху справа — участок сплошного
окремнения (белое пятно). Мелкие черные точки в этом участке отвечают
поперечным сечениям каналов отдельных спикул, выделяющимся благо-
даря окрашиванию органическим веществом. Шлиф; X10, без анализа-
тора. Турне, лихвинский подъярус. Северный Урал, р. Вижай (коллек-
ция С. В. Максимовой).
Фиг. 4. Перекристаллизованная четырехлучевая спикула в микрозерни-
стом силиците. Верхний девон, франский ярус. Латвийская ССР, скв.
Стамиерепа (В. С. Сорокин, В. Л. Гравптис, 1964).
Фиг. 5. Известковистый спонголит. Видны поперечные сечения спикул
с центральным каналом, различающимся то более, то менее четко. Шлиф;
Х40, без анализатора. Турне, лихвинский подъярус. Северный Урал,
р. Вижай (коллекция С. В. Максимовой).
Фиг. 6, 8. Спикуловый известняк. Два поля зрения одного шлифа. Па
фиг. 6 преобладают продольные сечения спикул (палочки), на фиг.
8 — поперечные (кружочки); Х40. Турнейский ярус. Кузбасс (коллекция
С. В. Максимовой).
Фиг. 7. Обломок шестилучевой (трехосной) спикулы. Шлиф; ХбО, без
анализатора. Мел. Поволжье («Основы палеонтологии», т. 2, табл. III).
Т а б л и ц а 67
248
Кремневые животные организмы
Таблица 68
Кремневые губки
Siliceous Spongia
Фиг. 1, 2. Разновидность одноосных спикул (ризоклоны). Спикулы вы-
делены из породы; Х20. Нижний мел. Крым (коллекция П. Н. Шемя-
кина).
Фиг. 3. Трехосные спикулы на выветрелой поверхности породы; X 5.
Силур. Туркестан («Основы палеонтологии», т. 2, табл. II).
Фиг. 4. Массовое скопление четырехлучевых спикул. Видны поперечные
сечения (кружки) и сечения, приближающиеся к продольным («рогульки»,
«стрелки»), Шлиф; Х35, без анализатора. Верхний карбон. Южный Урал
(коллекция И. В. Хворовой).
Фиг. 5. Остаток «каменистой губки», замещенный пиритом. Шлиф; Х45,
без анализатора. Кампанский ярус. Донбасс (коллекция Г. И. Бушин-
ского).
Фиг. 6. Выделенная при помощи химической препарировки решетка из
спаянных шестилучевых спикул; Х22. Мел.
Фиг. 7. Часть такой же решетки, как на фиг. 6. Шлиф; Х40. Мел. По-
волжье («Основы палеонтологии, т. 2, табл. III).
Фиг. 8. Современный кремнево-губковый осадок, состоящий из беспоря
дочного нагромождения спикул («стеклянная» или «губковая вата»);
Х20. Дальневосточные моря СССР (коллекция В. П. Петелина).
Фиг. 9. Разновидность четырехлучевых спикул (тетраклоны). Спикулы
выделены из породы; Х20. Нижний мел. Крым (коллекция Н. Н. Шемя-
кина).
Фиг. 10. Детритусовый известняк. Видны спикулы губок с сохранивши-
мися каналами. Шлиф; X 20, без анализатора. Пермь. Уфимское плато
(коллекция В. П. Маслова).
89 в п н ir g в £
250
Следы жизнедеятельности организмов
Таблица 69
Стратиферы и онколиты
Stratifera and Oncolithi
Фиг. 1. Строматолитоподобные образования (стратифера) без видимых
ограничений, похожие па слои, смятые давлением или оползанием, по
образующие куполки, а нс складки. Такие образования легко смешать
с механическими или комплексными образованиями. Ордовик, усть-кут-
ская свита. Верховья р. Ангары.
Фиг. 2. Онколит, образовавшийся вокруг обломка строматолита с зерни-
стой и сетчатой микроструктурой. Пришлифовка; ;;3. Ордовик, усть-
кутская свита. Низовья р. Ангары, Кочановская шивера.
Фиг. 3, 4. Онколиты со сгустковой структурой и неравномерным распре
делением сгустков (темное), чередующихся со светлыми слойками и участ-
ками. Онколиты образовались вокруг раковины (фиг. 3) и их обломков
(фиг. 4). Нришлпфовки; <2. Альб, абширская свита. Киргизия, Кувасай
(коллекция 3. 11. Поярковой).
Фиг. 5. Онколитовый оруденелый известняк с зачаточными строматоли-
тами; нат. вел. Кембрий, Малый Каратау (коллекция И. II. Крылова)
Фиг. (>. Онколитовый известняк с желвачками гирванелл. Некоторые онко"
литы вначале росли как строматолиты и затем в последнюю стадию своего
роста переворачивались движением воды; нат. вел. Пришлифовка. Опи-
саны под названием Ottonosia stromatoli.ti.ca Masi. (Маслов, 1937). Нижний
ордовик. I*. Шаманка, приток р. Апгары.
Таблица 69
252
Следы жизнедеятельности организмов
Таблица 70
Микроструктуры строматолитов
Microstructures of Stromatoiithi
Фиг. 1. Стержневая структура на краю колонковидной коллении сгустко-
вой (в основном) структуры. Белые пятна — кластический кварц, ром-
бики — доломит; X 10. Ордовик. 1’. Видим — приток р. Ангары в верхнем
течении.
Фиг. 2. Стержневая структура ио отдельным слоям строматолита. Оруде-
нение слоя рельефно выделяет отдельные конусовидные слои; X 10. Ордо-
вик. 1*. Подкаменная Тунгуска.
Фиг. 3. Стержневая структура (резко выраженная и видимая макроскопи-
чески). Белое — кварцевые зерна и слюда, попавшие между стержнями,
что доказывает рост стержней на дне водоема. Эта структура входит как
основная в строматолит, названный В. 11. Масловым Collenia stylostromica
recta', X 20. Ордовик. Бассейн Ангары, р. Черпая — приток р. Илим.
Фиг. 4, 5. Наросты на слоях строматолита, трактуемые как «подушки»,
образованные пзвестьотлагающей водорослью Pleurocapsites angaricus
Masi.; X 10. Ордовик, низы усть-кутской свиты. Р. Ангара (верхнее тече-
ние, Верхне-Каменный остров).
Фиг. 6—8. Каналы различной формы в строматолите, описанном под назва-
нием Collenia uralica Masi. Фиг. 6 — х20, каналы расположены верти-
кально; фиг. 7 — Х40, каналы извиваются; фиг. 8 — Х40, две системы
каналов: внизу — различно направленные каналы с меняющимся диамет-
ром, вверху — стелющиеся более тонкие каналы. Все эти формы каналов
встречены в одном и том же образце строматолита. Верхняя пермь. Южный
Урал (см. Маслов, 1936).
Фиг. 9. Вертикальное сечение организма, предположительно отнесенного
к губкам Phytospongia cylindrica Masi, со стенками, пронизанными кана-
лами; X 20. Усть-кутская свита. Низовья р. Ангары.
Таблица 70
1102 17
254
Следы жизнедеятельности организмов
Таблица 71
Микроструктуры строматолитов
Microstructures of Stromatolithi
Фиг. 1. Обычная и наиболее распространенная текстура строматолитов
с микрозернистой микроструктурой. Отчетливо видно чередование темных
плотных и более светлых и пористых слойков (сезонных?). Выпуклости
слоев совпадают; Х2. Ордовик. Р. Подкаменная Тунгуска.
Фиг. 2. Другая более редкая текстура со слоями, в которых выпуклости
не совпадают или совпадают на короткий промежуток. Микроструктура
или зернистая или сетчатая; Х4. Верхняя часть ордовика. Низовья
р. Ангары, Гороховская шивера.
Фиг. 3, 4. Почковидная микроструктура строматолита Conophyton. litus
Masi, в сечении, поперечном к оси строматолита (фиг. 3 — Х4; фиг. 4 —
Х15). Докембрий. Р. Нижняя Тунгуска.
Фиг. 5. Доломитизация (эпигенетическая) отдельных слойков стромато-
лита; X 10. Нижний ордовик. Верховья р. Ангары (Каменный ос?ров).
Фиг. 6. Сгустково-зернистая микроструктура в строматолите Conocolle-
nia; X 10. Ордовик, усть-кутская свита. Низовья р. Нюи (приток р. Лены).
Таблица 71
256
Следы жизнедеятельности организмов
Таблица 72
Микроструктуры строматолитов и онколитов
Microstructures of Stroinatolithi and Oncolithi
Фиг. 1. Радиально-лучистый онколит, в котором заметны светлые тонкие
канальцы. Лучистость явно вторичная, видны светлые пятна перекри-
сталлизации и жилки карбоната; X 20. Верхний палеозой. Тянь-Шань,
северный склон хр. Нарын-тау, р. Нарын (коллекция Эктовой, № 14).
Фиг. 2. Микроструктура слоев онколита типа оттонозия в виде «цветной
капусты», описанной под названием Ottonosia amurica Masi.; X 10. Юрские
отложения. Р-н Владивостока, р. Капгауз.
Фиг. 3. Радиальная микроструктура строматолита, внешней формой на-
поминающего морфологический тип Stylophycus Johnson, описанный в ниж-
нем карбоне Колорадо (Johnson, 1960, 1961, табл. 105, стр. 212). Светлые
радиальные полоски — вторичный карбонат. В темных участках (справа
внизу) заметны тонкие светлые канальчики — возможные следы водорос-
лей; X10. Нижний палеозой. Киргизская ССР, левый берег р. Туюк.
Фиг. 4. Обычная самая распространенная микроструктура строматоли-
тов — сгустковая. Она описана Гюрихом под названием Malacostroma
(Giirich, 1906) в карбоне Бельгии. Различная густота сгустков обусло-
влена, вероятно, сезонным развитием растительности; ХЗО. Ордовик.
Низовья р. Апгары.
Фиг. 5. Сгустково-сетчатая или сгустково-нитчатая микроструктура стро-
матолитов с многочисленными порами разного происхождения; X10.
Ордовик. Р. Ангара.
Фиг. 6. Чередование слоев сгустковой и зернистой микроструктур в на-
слоениях строматолита с различной пористостью. Внизу светлые пятна —
кластический кварц. Вообще слои обогащены терригенными частицами;
Х10. Ордовик. Р. Ангара.
Таблица 72
258
Следы жизнедеятельности организмов
Таблица 73
Микроструктуры строматолитов
Microstructures of Stromatolithi
Фиг. 1. Сгустковая структура (по Гюриху — Nalacostroma concentricum
Giirich). Вертикальное сечение наслоения. Видны разной плотности слои
из сгустков. Полости между сгустками принимались Гюрихом за каналы
в ценостеумо животного; Х20. Визе. Бельгия (Giirich, 1906, табл. 17).
Фиг. 2. Комковато-копролитовая структура (по Гюриху — Chondrostroma
problematicum Giirich). Слои состоят главным образом из «стеркомов»
(stercomes), которые Гюрих считал фекалиями тех же животных, которые
образовывали известковый скелет. По-видимому, здесь мы имеем дело
просто с копролитовыми известняками; X10. Визе. Бельгия (Giirich,
1906).
Фиг. 3. Spongiostroma bacilliferum Giirich. Структура состоит из крупных
и мелких копролитов и сгустков, плотно прижатых внизу и разрозненных
наверху, где можно видеть цепочку крупных копролитов (stercomes
Гюриха) по ходу илояда. Структура не относится к строматолитам; Х10.
Визе. Бельгия (Giirich, 1906, табл. 12, фиг. 2).
Фиг. 4. Сетчато-ячеистая структура (по Гюриху — Chondrostroma vermi-
culiferum Giirich); X 10. Визе. Бельгия (Giirich, 1906, табл. 14, фиг. 3).
Фиг. 5. Spongiostroma baccilliferum Giirich состоит из ряда слоев различного
происхождения: внизу сгустково-копролитового, вверху — сгусткового
происхождения. Верхние слои, возможно, строматолиты; Х10. Визе.
Бельгия (Giirich, 1906, табл. 3, фиг. 1).
Таблица 73
260
Следы жизнедеятельности организмов
Таблица 74*
Микрокопролиты
Microcop rol its
Фиг. 1. Копролиты с кишкообразными извилинами внутри Coprolithus
vermicularis Masi.; Х20. Средний карбон. Куйбышевская область, скв.
110/12, глуб. 1220—1222 м (материалы И. И. Далматской).
Фиг. 2. Coprolithus salevensis Parejas (==Favreina Broun). Продольное и
поперечное сечения среди копролитов другого типа. Видны песчанки
внутри темного материала копролита; Х20. Алайский ярус. Таджикская
депрессия.
Фиг. 3. Мелкие оолиты или онколиты, переработанные илоядами. Некото-
рые из них сохранили внутреннюю структуру, другие приобрели внешнюю
форму и внутреннее строение, несравнимые с первоначальными. Внизу
снимка заметен ход илояда, по которому располагаются вытянутые в одном
направлении извилистые копролиты; Х10. Нижний силур. Р. Подкамен-
ная Тунгуска.
Фиг. 4. Возможные копролиты, принятые ранее за онколиты; Х20. Прот-
винский горизонт. Подмосковье.
Фиг. 5. Копролиты, описанные ранее под названием Osagia sibirica Masi.,
из докембрия Прибайкалья. Характерные для копролитов заостренные
концы и включения песчаного материала при отсутствии концентров на-
слоения заставляют отрицать онколитовое происхождение данного об-
разца; Х17. Докембрий, ушаковская свита. Оз. Байкал, падь М. Ка-
дильная.
Фиг. 6. Возможные онколиты—копролиты или переработанные илоядом
онколиты, приобретшие линзовидную форму; Х20. Докембрий, ушаков-
ская свита. Оз. Байкал, падь М. Кадильная.
Фиг. 7, 8. «Хвостатые» копролиты в ордовике р. Подкаменной Тунгуски
(низовья). Такие формы иногда описываются как проблематические водо-
росли или проблематики. На фиг. 8 копролиты ориентированы длинной
осью по горизонтали, но на левой стороне снимка видны вертикально
поставленные копролиты, вероятно, благодаря изменившемуся направле-
нию хода червя; X10.
* Составил В. П. Маслов.
Т а б л и ц а 74
262
Следы жизнедеятельности организмов
Таблица 75*
Микрокопролиты
Microcop rolits
Фиг. 1. Копролиты типа Agregaella Elliott, по форме близкие к копро-
литам некоторых современных Annulata, моллюсков, членистоногих,
голотурий и т. п.; Х15. Третичные отложения. Мангышлак (коллекция
А. Н. Андрусова).
Фиг. 2. Скопление изогнутых и извивающихся копролитов наподобие
копролитов современных червей Lumbrinereis; Х20. Верхний силур —
нижний девон. Северный Урал, р. Вагран, 1 км выше р. Сарайной (мате-
риал Т. В. Прониной).
Фиг. 3. Копролиты, близкие к копролитам мелких рачков типа Artemia
salina; х26. Китай, Цайдам (материал А. И. Левенко).
Фиг. 4. Субсферические копролиты (Coprolithus sphaericus Masi. sp. п.)
с просветлениями центральной части. По форме близки к копролитам
современных моллюсков Масота yoldiformis; X 20. Верхний силур — ниж-
ний девон. Р. Вагран (материал Т. В. Прониной, № 1070).
Фиг. 5, 6. Копролит (Coprolithus spiraliformis Masi. sp. n.) в виде спирально
свернутого цилиндра: фиг. 5 — сечение, параллельное оси спирали, Х20
(№ 37); фиг. 6 — сечение, близкое к перпендикулярному оси спирали,
Х45 (№ 68). При Этом увеличении видно посветление внутри цилиндра
и более уплотненные «стенки» копролита. Девон, Эйфель. Левый берег
р. Вагран, 0,5 км восточнее г. Североуральска (материал Т. В. Прониной).
Фиг. 7. «Хвостатые» копролиты (по Кароции — «псевдоолитовый извест-
няк со смятыми псевдоолитами»); Х21. Средний триас. Тюрингия, Эйх-
родт (Carozzi, 1961, р. 262—274).
Фиг. 8. Обычные широко распространенные эллипсоидальные копролиты
{Coprolithus communis Masi. sp. n.), сцементированные микрозернистым
кальцитом; х40. Карбон, турне. Кузбасс, р. Томь (материал С. В. Макси-
мовой).
Фиг. 9. Копролиты (С. communis Masi.), расположенные по ходу илояда
в детритусово-шламовом известняке; х20. Средний карбон. Донбасс,
известняк М, скв. 127 (материал Г. Д. Киреевой).
Составили В. П. Маслов и С. В. Максимова
SL в n и ir о й i
264
Вторичные минералы в остатках организмов
Т а б л и ц а 76
Вторичные минералы в остатках организмов
Secondary Minerals in Remains of Organisms
Фиг. 1. Окремнение соленопоры. Халцедон (белое) замещает известковые
стенки трубок соленопоры и, расширяясь в стороны, нарушает органиче-
скую структуру. Николи параллельны; Х45. Силур? Узбекистан, 50 км
к северу от Султануиздага (материал А. Е. Шлезингера, скв. 27 п,
глуб. 290 м).
Фиг. 2. Кварц (белые пятнышки), который выкристаллизовался в корочке
литотамния (черное), обволакивающего обломок организма. Николи
параллельны; х45. Датский ярус. Закарпатье, Кобылецкая Поляна,
Раховский район (из Кривина и Маслова, 1962).
Фиг. 3, 4, 6. Окремнение обломков Pseudovermiporella: фиг. 3 — халцедон
(белое) частично заполняет полости и замещает стенки организма; фиг. 4 —
халцедон замещает только стенки, не нарушая органическую структуру;
фиг. 6 — халцедон заполняет только полости в организме, стенки остаются
темными. Николи параллельны, Х46. Пермь. Армения (материалы
Р. А. Аракеляна, обр. № 30).
Фиг. 5. Кальцитизированная раковинка радиолярии с сохранившейся
структурой внутренней сферы; Х40, без анализатора. Верхний девон,
доманнковый горизонт. Коми АССР.
Фиг. 7. Окремнение полостей в обломках организмов. В центре находится
длинный обломок неопределимой раковины, весь источенный сверлиль-
щиками, полости от которых заполнены халцедоном (белое). Внизу и
направо — обломки иглокожих с мелкими ячеями, заполненными основ-
ной илистой массой (черное). Здесь халцедон ведет себя иначе: он выпа-
дает сплошным пятном. Николи параллельны; Х43. Нижний аален,
Северный Кавказ, плато Кубенет-Яйла, правый берег р. Малки (материалы
3. В. Тимофеевой, № 356-2).
Таблица 76
СОДЕРЖАНИЕ
CONTENTS
ПРЕДИСЛОВИЕ 5
ВВЕДЕНИЕ. В. П. Маслов 6
Ч А С Т Ь I. ИЗВЕСТКОВЫЕ ОРГАНИЗМЫ 7
Общие вопросы. В. П. Маслов 7
Водоросли 9
Общий очерк. В. П. М а с л о в. 9
Синезеленые водоросли (Cyanophyta) В. П. Маслов 9
Золотистые водоросли (Chrysophyta) В. П. М а с л о в 14
Кокколитофориды (Cocco lithophoridae) и близкие к ним 15
организмы. С. И. Шумейко
Зеленые водоросли (Chlorophyta) В. П. Маслов 17
Кодиевые сифонеи (Codiaceae) В. П. Маслов 17
Мутовчатые сифонеи (Dasycladaceae) В. П. М а с л о в 20
Багряные водоросли (Rhodophyta) В. П. М а с л о в 22
Харофиты, или харовые водоросли (Charophyta) В. П. М а с- 27
лов, Б. В. Поярков
Животные организмы 32
Фораминиферы (Foraminifera) Д. М. Раузер-Черно- 32
усова
Тинтинниды (Tintinnoidea) Л. В. Линецкая 40
Известковые губки (Calcarea) И. Т. Журавлева 42
Археоциаты (Archaeocyathi) А. Ю. Р о з а н о в 43
Табуляты (Tabulata) И. И. Чудинова 44
Четырех- и шестилучевые кораллы (Tetracoralla, Hexacoralla) 47
Т. Г. И л ь и н а
Гидроидные полипы, ближе неопределимые (Hydrozoa) 53
В. П. М а с л о в
Строматопоры (Stromatoporoidea) В. И. Я в о р с к и й 54
Иглокожие (Echinodermata) 57
Общий очерк. В. П. М а с л о в 57
Морские лилии, или криноидеи (Crinoidea) В. П. М а с л о в 59
Морские ежи (Echinoidea). В. П. М а с л о в 60
Морские звезды (Asteroidea) и офиуры (Ophiuroidea) 61
В. Покорны
Голотурии, или морские огурцы (Holothuroidea) 64
В. Покорны
Черви (Vermes) С. В. Максимова 65
Брахиоподы (Brachiopoda) С. В. М а к с и м о в а, Г. А. А ф а- 68
н а с ь е в а
Мшанки (Bryozoa) Г. Г. Астрова 71
Моллюски (Mollusca) 75
Двустворчатые, или пластинчатожаберные, моллюски 75
(Bivalvia) В. П. М а с л о в
Гастроподы, или брюхоногие моллюски (Gastropoda) 76
В. П. М а с л о в
Аммониты (Ammonoidea) В. П. М а с л о в 77
Белемниты (Belemnoidea) В. П. М а с л о в 78
Ракообразные (Crustacea) 78
Общие замечания В. П. М а с л о в 78
Остракоды (Ostracoda) Ф. Ю. Адамчак 79
Трилобиты (Trilobita) В. П. М а с л о в 80
FOREWORD
INTRODUCTION. V. Р. Maslov
PART I. CALCAREOUS ORGANISMS.
General problems. V. P. M a s I о v
Algae
General sketch. V. P. Maslov
Gyanophyta. V. P. Maslov
Chrysophyta. V. P. Maslov
Coccolithophoridae and related organisms. S. I. Shumenko
Chlorophyta. V. P. Maslov
Codiaceae V. P. Maslov
Dasycladaceae. V. P. Maslov
Rhodophyta. V. P. Maslov
Charophyta. V. P. M a s 1 о v, В. V. P о j а г k о v
Animal organisms
Foraminifera. D. M. Ra user - Chernous.so va
Tintinnoidea. L. V. Linetzkaia
Calcarea. I. T. Zhuravleva
Archaeocyathi. A. Yu. R о z a n
Tabulata. I. I. Chudinova
Tetracoralla, Hexacoralla. T. G.
Hydrozoa. V. P. Maslov
Stromatoporoidea. V. I. Y a v о
Echinodermata.
General sketch. V. P. Mas
Crinoidea. V. P. Maslov
Echinoidea. V. P. Maslov
Asteroidea and Ophiuroidea
Holothuroidea. V. P о k о г n
Vermes. S. V. Maximova
Brachiopoda. S. V. Maximo
Bryozoa. G. G. Astrova
Mollusca
Bivalvia. V. P. Maslov
Gastropoda. V. P. Maslov
о v
Il ’ i n a
г s k у, V. P. M a s 1 о v
1 о v
. V. Pokorny
У
va, G. A. Afanassieva
Ammonoidea. V. P. Maslov
Belemnoidea. V. P. Maslov
Crustacea
General remarks. V. P. Maslov
Ostracoda. F. Yu. Adamchak
Trilobita. V. P. Maslov
Кости позвоночных (Vertebrata) В. П. М а с л о в, Г. А. Б а- 80 ч и н с к и й Спорные и проблематические известковые микроорганизмы 82 В. П. М а с л о в Конодонты (Conodonta) и сколекодонты (Scolecodonta). 84 Э. А. Ю р т е н с о н, В. Я. В и й р а Хиолиты (Hyolitha) В. В. Миссаржевский 86 Тентакулиты (Tentaculitoidea). В. П. М а с л о в 87 Ч А С Т Ь II. КРЕМНЕВЫЕ ОРГАНИЗМЫ 89 Общие замечания. В. П. М а с л о в 89 Диатомовые водоросли (Bacillariophyta) А. П. Ж у з е 89 Кремневые жгутиковые водоросли (Silicoflagellata) 3. И. Г л е- 92 з е р Радиолярии (Radiolaria) 94 Современные радиолярии. С. Б. Кругликова 94 Древние радиолярии. С. В. Максимова 96 Кремневые губки (Silidspongia). С. В. Максимова 97 Ч А С Т Ь Ш. СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ ОРГА- 100 НИЗМОВ Микроструктура строматолитов. В. П. Маслов 100 Желваки органического происхождения В. П. Маслов 101 Следы сверлящих или перфорирующих организмов. 102 В. П. М а с л о в Микрокопролиты. С. В. М а к с и м о в а, В. П. М а с л о в 103 ЛИТЕРАТУРА 107 ТАБЛИЦЫ 1—76 113 Bones of vertebrates. V. Р. Maslov, G. А. Bachinsky Controversial and problematic calcareous microorganisms. V. P. M a s 1 ov Conodonta and Scolecodonta. E. A .Y u r g e n s о n, V. Ya. V i i г a Hyolitha. V. V. Missarzhevsky Tentaculitoidea. V. P. Maslov PART II. SILICEOUSORGANISMS General remarks. V. P. Maslov Bacillariophyta. A. P. Z h u s e Silicoflagellata. Z. I. G 1 e z e r Radiolaria Recent Radiolaria. S. B. Kruglikova Old Radiolaria. S. V. Maximova Silicispongia. S. V. Maximova PART III. TRACES OF ORGANIC LIFE Microstructure of Stromatolithi. V. P. Maslov Concretions of organic origin. V. P. Maslov Traces of boring or perforating organisms. V. P. Maslov Microcoprolites. S. V. M a x i m о v a, V. P. M a s 1 о v BIBLIOGRAPHY PLATES 1-76
АТЛАС ПОРОДООБРАЗУЮЩИХ
ОРГАНИЗМОВ
(известковых и кремневых)
Утверждено к печати
Комиссией
по осадочным породам
при Отделении наук о Земле
и Ордена Трудового
Красного Знамени
Геологическим институтом
Редактор
М. А. Пергамент
Редактор издательства
И. М. Ерофеева
Художник
С. А. Литвак
Художественный редактор
В. Н. Тикунов
Технический редактор
Л. М. Сурикова
Подписано к печати 12/Ш 1973 г.
Формат 60x90%
Усл. печ. л. 33.5
Уч.-изд. л. 32.27
Бумага офсетная № 1
Т-02773. Тираж 1600 экз.
Цена 3 р. 73 к. Тип. зак. 1102
Книга издана офсетным способом
Издательство «Наука», 103717 ГСП,
Москва, К-62, Подсосенский пер., 21
1-я типография издательства «Наука»,
Ленинград, 199034, 9-я линия, 12
Исправления и опечатки
Страница Колонка Строка Напечатано Должно быть
1.7 правая 27 сн. табл. 13, фиг. 1-3 табл. 13, фиг. 1,2,4,7; табл. 14, фиг. 1-3
19 правая 29 св. фиг. 1-3 фиг. 1,5,6
20 левая 13 сн. табл. 13,фиг.4-10 табл. 13,фиг.3,5,6,8-10
113 9 сн. 25-61 25-61 и 63-65
113 8 сн. 61-63 и 63-65 61-63
Атлас породообразующих организмов