/





























































































































































































































































































































































































































































































































































Author: Федоров А.А.
Tags: водоросли живые организмы лишайники водная среда размеры окраска особенности строения
Year: 1977




Text
ВВЕДЕНИЕ ЛКТИНОМИЦЕТЫ ГРИБЫ Главный редактор член-корреспондент АН СССР профессор Ал. А. ФЕДОРОВ ПЛАУНЫ хвощи ПАПОРОТНИКИ OAOCEMEHHbIE РАСТЕНИЯ ЦВЕТКОВЫ Е РАСТЕНИЯ ЦВЕТКОВЫ Е РАСТЕНИЯ П
Ж71 РЕДАКЦИОННАЯ КОЛЛЕГИЯ: член-корреспондент АН СССР Ал. А. ФЕДОРОВ (гл. редактор) академик АН СССР А. Л. КУРСАНОВ академик АН СССР А. Л. ТАХТАДЖЯН академик АН СССР Н. В. ЦИЦИН академик ВАСХНИЛ П. М. ЖУКОВСКИЙ члены-корреспонденты АН СССР М. В. ГОРЛЕНКО Н. А. КРАСИЛЬНИКОВ профессора: М. М. ГОЛЛЕРБАХ А. А. УРАНОВ А. А. ЯЦЕНКО-ХМЕЛЕВСКИЙ кандидат биологических наук С. Г. ЖИЛИН 6050i — 382 ~103(03) 77 Подписное ф Издательство «Просвещение», 1977 г.
ПОД РЕДАКЦИЕИ профессора М. М. ГОЛЛЕРБАХА МОСКВА ((ПРОСВЕЩЕНИЕ» 1977
АВТОРЫ НАСТОЯЩЕГО ТОМА: Доктора биологических наук М. М. ГОЛЛЕРБАХ, А. М. МАТВИЕНКО, И. И. НИКОЛАЕВ, Ю. Е. ПЕТРОВ, В. Е. СЕМЕНЕНКО, Х. Х. ТРАСС, Э. А. ШТИНА. Кандидаты биологических наук К. Л. ВИНОГРАДОВА, М. Г. ВЛАДИМИРОВА, 3. И. ГЛЕ- ЗЕР, Н. С. ГОЛУБКОВА, Т. В. ДОГАДИНА, Э. Г. КУКК, И. В. МАКАРОВА, Г. М. МОРДВИНЦЕВА, Л. А. РУНДИНА, Т. А. САФОНОВА, Т. В. СЕДОВА, И. В. ТАБАЧНИКОВА. Титульное редактирование провел М. М. ГОЛЛЕРБАХ. В томе использованы фотографии Ю. Ф. Астпафьева, Л. И. Волошко, С. И. Генкала, 3. И. Глезер, Н. И. Караевой, Ю. Б. Королева, Г. В. Кузина, И. В. Макаровой, В. И. Михайлова, А. И. Прошкиной-Лавренко, А. И. Рогова, И. Ш. Стирельниковой, В. А. Федоровой.
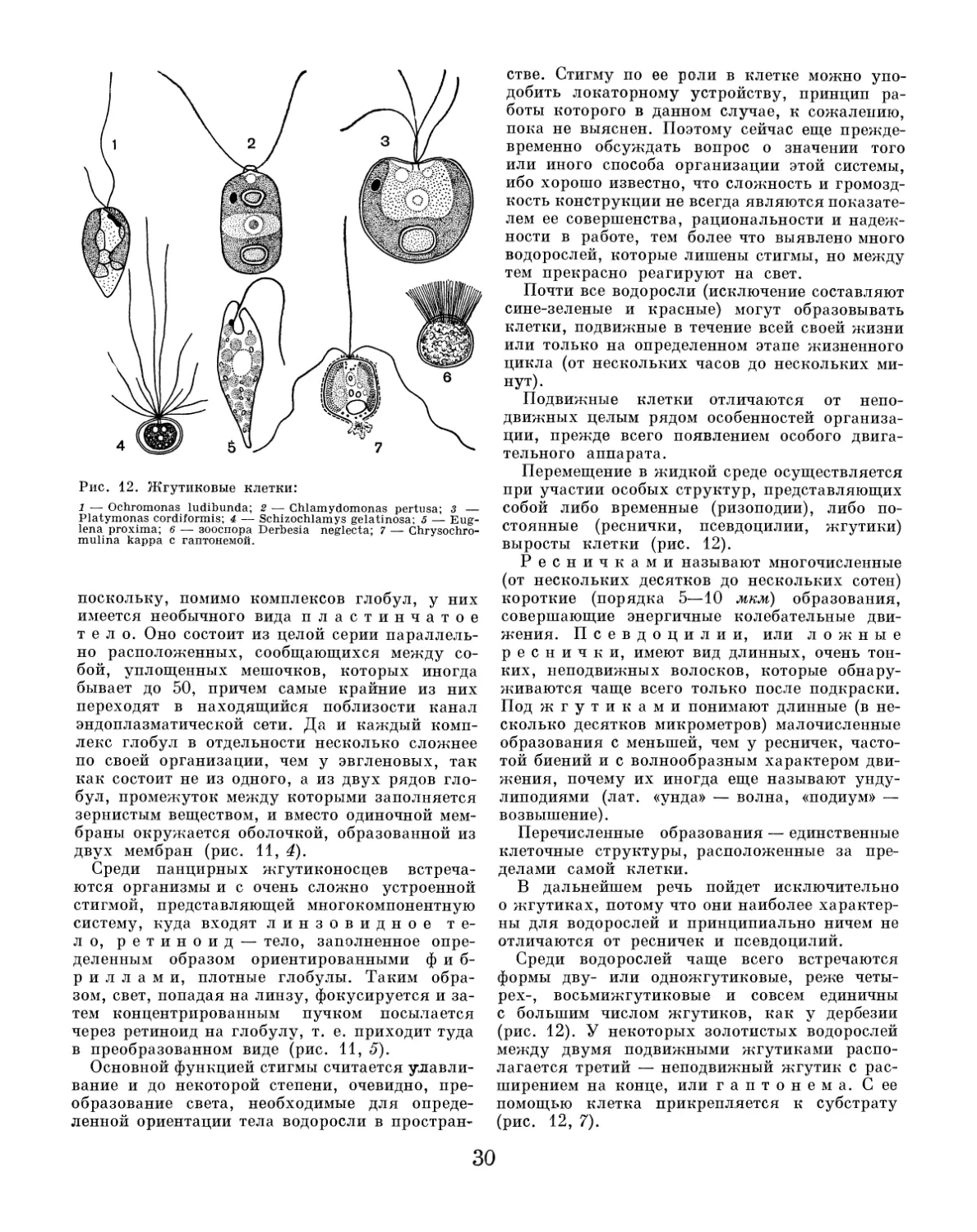
ОБЩЛЯ ХАРАКТЕРИСТИКА БОДОГОСЛЕИ ВОДОРОСЛИ И ИХ ОТЛИЧИЕ ОТ ДРУГИХ РАСТЕНИЙ Мир водорослей огромен. Он занимает в ра- стительном царстве совершенно особое, исклю- чительное по своему значению место как в ис- торическом аспекте, так и по той роли, которая принадлежит ему в общем круговороте веществ в природе. Вместе с тем само понятие «водорос- ли» в научном отношении страдает большой не- определенностью. Это заставляет специально рассмотреть отличие относимых сюда расти- тельных организмов от других представите- лей растительного царства. Действительно, слово «водоросли» означает лишь то, что это — растения, живущие в воде. Однако в ботанике этот термин применяется в более узком смысле, и не все растения, на- блюдаемые нами в водоемах, с научной точки зрения можно называть водорослями. С другой стороны, именно водоросли мы часто попросту не замечаем в водоемах, так как очень многие из них нелегко распознать невооруженным глазом. Приглядываясь к различным водоемам, осо- бенно к озерам и прудам, мы прежде всего за- мечаем благодаря величине и обилию семенные, или цветковые, растения. В каком-нибудь ста- ром, запущенном пруду мы обязательно встре- тим тростник, камыш, рогоз, которые растут в воде у берегов, прикрепляясь корнями к дну пруда, а большую часть стебля с листьями и цветками выставляя над водой. Несколько да- лее от берега можно найти растения с плаваю- щими на поверхности воды листьями и только слегка выступающими над водой цветками, на- пример белые кувшинки или желтые кубышки. Нередко вся поверхность прудов бывает сплошь затянута плавающими мелкими зелеными пла- стинками ряски. Наконец, весьма многочислен- ны растения, целиком погруженные в воду. Некоторые из них прикреплены ко дну, как, например, большинство рдестов, водяная чума и уруть; другие с дном не связаны — таковы пузырчатка и роголистник. Все эти растения, как бы они ни отличались друг от друга, яв- ляются семенными, или цветковыми, хотя не- которые из них почти никогда не цветут и не образуют семян, размножаясь преимуществен- но вегетативно. К этим растениям научный тер- мин «водоросли» неприменим, их называют водяными растениями. Кроме семенных водяных растений, в водое- мах можно встретить и представителей высших споровых растений — мхов и папоротнико- образных. Б ольшинство мхов — влаголюбивые растения, но типично водяных, растущих по- груженными в воду, среди них не так много. Один из наиболее известных — мох фонтиналис, образующий в воде прудов и рек обширные тем- но-зеленые дерновины из стеблей, покрытых крупными (до 8 мм) листьями. Из водяных па- поротникообразных можно отметить растущий вблизи берегов водяной хвощ с характерными мутовками боковых веточек и плавающий на поверхности воды водяной папоротник сальви- нию, встречающуюся у нас местами в средней полосе СССР, преимущественно в речных заво- дях. Сальвиния — маленькое изящное расте- ние, оно имеет горизонтальный стебель с двумя рядами овальных плавающих листьев и отходя- щими книзу видоизмененными листьями, рас- сеченными на отростки, похожие на корни. Наконец, продолжая наш обзор какого- нибудь водоема, мы можем заметить и настоя-
щие водоросли. К ним относятся, например, крупные зеленые скопления так называемой тины, плавающей в летнее время вблизи от поверхности воды в прудах и затишливых мес- тах рек и озер. Разнообразные зеленые и сине- зеленые пленки и войлочные или ватообразные наросты на камнях, бревнах и сваях также образованы водорослями. В летнее время вода в прудах часто бывает окрашена в зеленоватый цвет, и если зачерпнуть ее стаканом, то на про- свет можно заметить в ней мельчайшие водо- росли в форме плавающих точек, хлопьев или шариков. Здесь же нередко встречаются и более крупные водоросли, состоящие из хорошо за- метных на глаз простых или ветвящихся нитей, или совсем крупные харовые водоросли, внеш- не похожие на хвощ, с характерными мутов- ками боковых побегов. С другой стороны, значительное количество микроскопических водорослей, таких же, как в водоемах, произрастает и на суше: на по- верхности почвы и в самой ее толще, на деревь- ях, камнях и т. п. Правда, жизнь этих водорос- лей тоже тесно связана с водой, однако они могут довольствоваться только атмосферной или грунтовой влагой, росой, брызгами водо- падов или фонтанов и т. п. В отличие от «вод- ных» водорослей эти «сухопутные» водоросли легко переносят высыхание и очень быстро оживают при малейшем увлажнении. Чем же все эти растения, объединяемые по- нятием «водоросли», отличаются от других растений? Очевидно, научная характеристика водорослей должна учитывать не только среду их обитания, но и основные, общие для них морфологические и физиологические признаки. В морфологическом отношении для водорос- лей наиболее существенным и действительно всеобъемлющим признаком является отсутст- вие, даже при весьма сложной внешней расчле- ненности тела, настоящих стеблей, листьев и корней, типичных для высших растений. Их тело обозначается как слоевище, слоевцо, или таллом. Иными словами, в царстве растений водоросли относятся к обширному подцарству низших, или слоевцовых, растений, куда входят также бактерии, актиномицеты, слизевики, грибы и лишайники. Как и все низшие расте- ния, водоросли размножаются или вегетативно, или с помощью спор, т. е. относятся к споро- вым растениям (в отличие от семенных, или цветковых, растений). Однако в физиологиче- ском отношении водоросли резко отличаются от остальных низших растений наличием хло- рофилла, благодаря которому они способны ассимилировать на свету углекислый газ, т. е. питаться фототрофно. Правда, такой же спо- собностью обладают и некоторые бактерии, имеющие зеленую окраску. Однако содержащий- ся в них пигмент хоть и близок к хлорофиллу, но не тождествен ему. С другой стороны, име- ются водоросли, вполне типичные по строению, но бесцветные, вторично утратившие хлорофилл и тогда уже полностью питающиеся гетеротроф- но. Кроме того, многим водорослям, обладаю- щим хорошо развитым хлорофиллом, помимо фототрофного, могут быть свойственны и дру- гие типы питания. Тем не менее, несмотря на все эти исключения, сочетание талл омного (слоевцового) строения с наличием хлорофилла достаточно полно характеризует растительные организмы, объединяемые как водоросли. Таким образом, исходя из сказанного, легко вывести точное научное определение водорос- лей. Водоросли — это низшие, т. е. слоевцовые (лишенные расчленения на стебель и листья), споровые растения, содержащие в своих клет- ках хлорофилл и живущие преимущественно в воде. Такое определение, однако, не дает пред- ставления о том огромном разнообразии в строении тела, которое свойственно водорослям. Здесь мы встречаемся и с микроскопическими организмами — одноклеточными, колониальны- ми и многоклеточными, и с крупными форма- ми различного строения. Большого многообра- зия достигают здесь также способы размноже- ния и строение органов размножения. Даже по окраске водоросли неодинаковы, так как одни содержат только хлорофилл, другие- еще ряд дополнительных пигментов, окраши- вающих их в различные цвета. Точный учет всех особенностей в строении, размножении и развитии водорослей привел современную науку к убеждению, что ныне живущие водоросли не представляют собой мо- нолитной группы организмов, объединенных единством строения и происхождения. В настоя- щее время общепризнано, что водоросли пред- ставляют собой совокупность нескольких обособленных отделов (типов) растений, само- стоятельных по своему происхождению и эволю- ции. Каждый из них в систематическом отноше- нии равноценен таким отделам низших расте- ний, как, например, бактерии или грибы. Таким образом, термин «водоросли», столь обычный еще в науке и в общежитии, имеет исключительно биологический смысл, как объе- динение низших хлорофиллоносных растений, живущих преимущественно в воде. Разделение водорослей на систематические группы (таксоны) высшего ранга — отделы (Divisio, Phylum) — в основном совпадает характером их окраски, связанной, конечно, с особенностями строения. Однако относитель- но количества и объема этих отделов в научной литературе нет единства взглядов. В настоящем издании мы принимаем классификацию, наи- более широко распространенную в советской
научной литературе, а именно деление водорос- лей на iO отделов. Приводим их перечень с соблюдением общепринятой русской и латин- ской научной номенклатуры: 1) сине-зеленые водоросли — Cyanophyta; 2) пирофитовые водоросли — Pyrrophyta; 3) золотистые водоросли — Chrysophyta; 4) диатомовые водоросли — Bacillariophyta; 5) желто-зеленые водоросли — Xanthophyta; 6) бурые водоросли — Phaeophyta; 7) красные водоросли — Rhodophyta; 8) эвгленовые водоросли — Euglenophyta; 9) зеленые водоросли — Chlorophyta; 10) харовые водоросли — Charophyta. В научной литературе до сих пор продолжа- ются споры о положении в общей системе, с одной стороны, сине-зеленых водорослей и, с другой стороны, всех тех водорослей, которые представлены одноклеточными подвижными фор- мами, снабженными органами движения— жгутиками (это почти все эвгленовые водоросли, большая часть пирофитовых и золотистых во- дорослей и отдельные классы желто-зеленых и зеленых водорослей). Действительно, сине-зеленые водоросли резко отличаются от других водорослей простотой внутренней организации клеток. Клетки их лишены оформленного ядра, что сближает их с бактериями. Вместе с бактериями сине- зеленые водоросли составляют раздел орга- низмов, обозначаемый как п р о к а р и о т ы (Prokaryota), т. е. «доядерные», в отличие от всех остальных растений и животных, обладаю- щих оформленным клеточным ядром и обозна- чаемых как э у к а р и о т ы (Eukaryota), т. е. «истинно ядерные». Некоторые авторы прида- ют этому признаку основное таксономическое значение и объединяют сине-зеленые водоросли с бактериями, т. е. вообще исключают их из числа собственно водорослей. Что же касается жгутиковых форм водорос- лей, то здесь вопрос осложняется тем, что они во многих случаях близки к подобным же бес- цветным формам, что дало повод для объеди- нения всех их в общую систематическую груп- пу «жгутиковых организмов» и включения в систему животного мира. Такая тенденция сохранилась и до сих пор. В современной 30- ологической систематике жгутиковые рассмат- риваются как один из классов — жгутиконосцы (Mastigophora, или Flagellata) типа одноклеточ- ных животных — простейших (Protozoa), а в пределах этого класса окрашенные жгутиковые объединяются в подкласс растительные жгути- коносцы (Phytomastigina), бесцветные — в под- класс животные жгутиконосцы (Zoomastigina). Не вдаваясь здесь в разбор как первого, так и второго вопроса (это уместнее сделать после описания всех водорослей), отметим только, что мы не можем принять указанные крайние точки зрения. Дело в том, что в процессе эво- люции возникновение хлорофилла у первичных бесцветных гетеротрофных организмов было настолько существенным переломным моментом, что только его и можно считать началом той богатейшей эволюции, которая привела к со- зданию биосферы на Земле в ее современном вы- ражении. И трудно допустить, что такое слож- ное и уникальное органическое соединение, как хлорофилл, давшее организмам возможность фототрофного питания, т. е. совершенно новой энергетики жизненных процессов, основанной на утилизации солнечной энергии, могло воз- никать многократно на разных этапах эволюции. С этих позиций мир водорослей как пер- вичных фототрофных организмов един и цело- стен. Морфологическое многообразие его раз- личных ветвей есть следствие эволюционного взрыва, вызванного появлением фотосинтеза, который обеспечил хлорофиллоносным орга- низмам успешное развитие в чисто абиотиче- ской среде. Учитывая особенности строения клеток сине-зеленых водорослей, следует ду- мать, что возникновение хлорофилла произошло еще на прокариотическом уровне, а наличие в настоящее время сходных хлорофиллоносных и бесцветных эукариотических жгутиковых форм обусловлено морфологическим паралле- лизмом эволюционного развития в разных вет- вях организмов. Во всяком случае, у водорослей подобное явление морфологического парал- лелизма распространено очень широко (напри- мер, строение в пределах Chlorophyta, Xan- thophyta и Chrysophyta). Такая точка зрения хорошо подтверждается еще и тем, что в преде- лах большинства вышеперечисленных отделов водорослей жгутиковые формы тесно связаны переходами с другими, типично «водорослевыми» структурами — неподвижными клетками, ко- лониями и нитями. С другой стороны, в преде- лах некоторых отделов имеются и безусловно вторично обесцветившиеся формы. Таким образом, у нас нет оснований отказы- ваться от рассмотрения водорослей как морфо- физиологической целостности, от выяснения их многообразия в целом, происхождения и взаимных филогенетических связей. Точно так же с этих позиций целостности хорошо выявляются место и роль водорослей в природе: в историче- ском плане они представляют собой первый этап в развитии всего зеленого ствола растительного мира, а в общем круговороте веществ в приро- де играют огромную роль как первичное звено всех пищевых связей в водной среде и гигант- ский поставщик кислорода в атмосферу. Изучение всех этих вопросов составляет предмет особой науки — альгологии (от латин- ского названия водорослей — algae).
спосоБы питАния водорослЕЙ и других хлороФиллоносных РАСТЕНИЙ Несмотря на удивительное многообразие жиз- ненных форм растений, подавляющее большин- ство из них объединяет уникальная особен- ность, которая определяется способом их пита- ния. В отличие от животных организмов и многих бактерий, использующих для своей жизнедеятельности готовые органические сое- динения, у растений выработалась в ходе эво- люции способность использовать для питания такие полностью окисленные вещества, как углекислота и вода, и создавать на их основе органические соединения. Процесс этот осуще- ствляется в природе за счет энергии солнечного света и сопровождается выделением кислорода. Использование световой энергии для биологи- ческих синтезов стало возможно благодаря по- явлению у растений комплекса поглощающих свет пигментов, главнейшим из которых явля- ется хлорофилл. Процесс светового и углерод- ного питания растений получил название фото- синтеза и в общем виде может быть записан следующим суммарным уравнением: 6СО, + 12H,О — С,~„О, + 6Н,0 + хлорофилл + 60, + 2 815 680 Дж. Из уравнения видно, что на каждые 6 грамм- молекул углекислоты и воды синтезируется грамм-молекула глюкозы (С,Н„О,), выделя- ется 6 грамм-молекул кислорода и накапливает- ся 2815 680 Длс энергии. Таким образом, функ- ция фотосинтеза растений является, по сущест- ву, биохимическим процессом преобразования световой энергии в химическую. Водоросли, уже простейшие из них — сине- зеленые, являются первыми организмами, у которых появилась в процессе эволюции спо- собность осуществлять фотосинтез с использо- ванием воды в качестве источника (донора) водорода и выделением свободного кислорода, т. е. процесс, свойственный всем другим водо- рослям, а за ними и высшим растениям. Осуществляемый растениями в грандиозных масштабах процесс преобразования энергии света в химическую энергию продуктов фото- синтеза является практически единственным «руслом», через которое «вливается» в биологи- чески приемлемой форме энергия, необходи- мая для поддержания жизни и круговорота веществ в биосфере нашей планеты. Именно поэтому выдающийся русский естествоиспыта- тель К. А. Т и м и р я з е в говорил о «кос- мической роли зеленых растений». О размерах фотосинтетической деятельности растений в планетарном масштабе можно судить по тому, что весь кислород атмосферы Земли имеет, как сейчас доказано, фотосинтетическое про- исхождение. Залежи каменного угля представ- ляют собой своеобразный «запас» некогда пре- образованной в результате фотосинтеза расте- ний солнечной энергии, складированный в оп- ределенные геологические эпохи. Второй особенностью питания водорослей и других фотосинтезирующих растений, не менее важной, хотя и не такой специфичной, как фотосинтез, является их способность усва- ивать азот, серу, фосфор, калий и другие ми- неральные элементы в виде ионов минеральных солей (ХО,, 804, РО4, К и др.) и исполь- зовать их для синтеза таких важнейших компо- нентов живой клетки, как аминокислоты, бел- ки, нуклеиновые кислоты, макроэргические соединения, вещества вторичного обмена (ал- калоиды, терпены, фекальные соединения, раз- личные витамины, фитогормоны и др.). Среди сине-зеленых водорослей имеются формы, спо- собные осуществлять процесс фиксации свобод- ного азота атмосферы и превращать его в ор- ганические азотистые вещества своего тела. ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ФОТОСИНТЕЗА Каким же образом возник процесс фотосин- теза? Что ему предшествовало и к каким по- следствиям привело появление этого процесса на Земле? Согласно общепризнанной в настоящее время эволюционной теории происхождения и раз- вития жизни, которая более 50 лет назад была сформулирована А. И. О п а р и н ы м, пер- вичные, способные к самовоспроизводству жи- вые образования возникли в результате абио- генной химической эволюции. Будучи окружен- ными близкими по составу, но еще неживыми органическими соединениями, эти первичные существа могли осуществлять в бескислородной среде анаэробный гетеротрофный тип питания с помощью небольшого набора ферментов. Постепенное истощение и деградация органи- ческих веществ, синтезированных а био генным путем, сопровождались накоплением все более окисленных соединений, вплоть до появления наиболее бедного энергией соединения угле- рода — углекислоты. Это влекло за собой не- обходимость все большего и большего совер- шенствования и усложнения ферментативного аппарата, необходимого для ассимиляции все более окисленных веществ. В этих условиях, которые все еще характеризовались отсутствием
в среде кислорода, вполне вероятно возникно- вение первичных автотрофных организмов, ко- торые осуществляли восстановление углеки- слоты за счет химической энергии, полученной из минеральных веществ. Такой тип питания получил название х е м о р е д у к ц и и. Сре- ди современных организмов известна группа сульфатредуцирующих микроорганизмов, ко- торые восстанавливают сульфаты до сероводо- рода, используя для этой цели молекулярный водород. Появление в этот период, который характе- ризовался сильно восстановительными усло- виями среды, светпоглощающих пигментов- фотосенсибилизаторов привело, очевидно, к замене химической энергии в процессах хемо- редукции на световую. Возник простейший тип фотоавтотрофного питания, который получил название фото редукции и бакте- риального фотосинтеза. Такой тип питания осуществляют современные фото- трофные бактерии — пурпурные серобактерии (Thiorhodaceae) и зеленые серобактерии (Chlo- robacteriaceae), у которых роль пигмента-фото- сенсибилизатора выполняет бактериохлоро- филл и которые являются строгими анаэробами. Пурпурные и зеленые серобактерии восстанав- ливают углекислоту за счет энергии света, используя в качестве Н-донора сероводород (Н $): 6СО +12Н S -С Н О бактериохлорофилл + 6Н,О+ 12$ Представленное итоговое уравнение бакте- риального фотосинтеза (фоторедукции) очень напоминает, как мы видим, приведенное выше суммарное уравнение фотосинтеза хлорофил- лоносных растений. В результате сравнитель- ного анализа Ван-Ниль показал, что оба эти процесса могут быть записаны в общем виде одним итоговым уравнением: СО, + 2Н,А — (СН,О) + Н,О + 2А, пигмент где Н,А — донор водорода, в качестве которо- го фотосинтезирующие бактерии используют сероводород, а остальные растения — воду. Вода является более окисленным соединением по сравнению с сероводородом. Использование ее в качестве донора водорода связано с необходи- мостью дополнительной затраты энергии и стало возможно благодаря дальнейшему совершен- ствованию фотохимического аппарата, кото- рое состояло в появлении у растений (начиная с сине-зеленых водорослей) хлорофилла (вме- сто бактериохлорофилла) и дополнительной фотохимической системы, так называемой «фо- тосистемы П». Использование воды в качестве доноре во- дорода привело к тому, что в процессе фотосин- теза стал выделяться кислород, что, в свою очередь, ознаменовало переход от анаэробной к аэробной жизни на нашей планете. На эволюционную связь фоторедукции и фотосинтеза может указывать способность ряда сине-зеленых, зеленых, красных и бурых во- дорослей обратимо переходить к фоторедукции при переводе их в анаэробные условия в ат- мосферу водорода. Таким образом, фотоавтотрофный тип пита- ния и фотосинтез возникли в процессе эволю- ции как «надстройка» над первичным гетеро- трофным типом питания. Появление на Земле фотосинтеза было обусловлено всем ходом пред- шествовавшей биологической эволюции и яви- лось поворотным пунктом в переходе от анаэроб- ного к аэробному типу обмена веществ. Рассмотренная схема дает представление лишь об общих чертах эволюции фотосинтеза и явля- ется в значительной степени гипотетической. Многие этапы эволюции фотосинтеза и тем бо- лее ее детали остаются неясными, ряд моментов по-разному интерпретируется учеными. Неясным, например, остается вопрос о про- исхождении хлоропластов высших растений. Существует точка зрения об эндосимбиотиче- ском их происхождении в результате «захвата» первичных фотосинтезирующих организмов, типа современных сине-зеленых водорослей, гетеротрофным организмом. На такую возмож- ность указывает определенная генетическая автономность хлоропластов, d также подобие их ДНК, ряда важнейших ферментов, свойств рибосом и ряда РНК таковым у прокариотиче- ских организмов, в частности сине-зеленых во- дорослей. Вместе с тем имеется и определенная генетическая подчиненность хлоропласта ядерно- му геному, что может указывать на «прямую» эволюцию фотосинтетического аппарата совре- менных растений от первичных фотосинтези- рующих организмов. Все эти вопросы требуют дальнейшего детального изучения механизмов, молекулярной организации, генетического кон- троля и физиологических свойств фотосинтеза и его аппарата. СТРОЕНИЕ И МОЛЕКУЛЯРНАЯ ОРГАНИЗАЦИЯ ФОТОСИНТЕТИЧЕСКОГО АППАРАТА Фотосинтетический процесс преобразования световой энергии протекает в клетках растений в специализированных мембранных структурах. У высших растений и эукариотических водорос- лей эти мембранные структуры локализованы в особых органеллах — хлоропластах. Ъ' про-
Таблица Распространение в растительном мире хлорофиллов и билипротеинов Билипротеины г Бактериохлорофилл Хлорофилл Подразделения фнко- эритрин фикоц11- анин Бактерии Зеленые серобактерии (Chloro- bacteriaceae) Серные пурпурные бактерии (Thiorhodaceae) Несерные пурпурные бактерии (Athiorh odaceae) Водоросли Сине-зеленые (Cyanophyta) Эвгленовые (Euglenophyta) Зеленые (Chlorophyta) Красные (Rhodophyta) Пирофитовые (Pyrrophyta) Золотистые (Chrysophyta) Желто-зеленые (Xanthophyta) Бурые (Phaeophyta) Диатомовые (Bacillariophyta) Харовые (Charophyta) Высшие растения кариотических сине-зеленых водорослей они располагаются непосредственно в цитоплазме. Мембраны фотосинтетического аппарата име- ют общий с другими биологическими мембрана- ми принцип строения и состоят из липидов и белков. Отличительной особенностью этих мем- бран является то, что в них «встроены» моле- кулы хлорофилла и других сопровождающих пигментов, которые поглощают свет и обеспе- чивают передачу энергии электронного воз- буждения «активному центру» и в фотосинте- тическую цепь переноса электрона. Изучение хлоропластов с помощью электрон- ного микроскопа показало, что мембранная система имеет здесь дифференцированную, очень сложную, но четко упорядоченную структуру, которая различается у разных растений харак- тером упаковки и степенью ее выраженности. Мембранная система погружена в строму (или матрикс) хлоропласта, в которой локализованы ферменты, связанные с восстановлением угле- кислоты и синтезом углеводов. Существенным обстоятельством является то, что в хлороплас- тах, кроме систем, непосредственно участвующих в осуществлении процесса фотосинтеза, имеется также собственная (отличная от ядерной) ДНК, рибосомы и другие компоненты белоксинтези- рующих систем. Для хлоропластов многих водо- рослей характерно также наличие специфическо- го образования — пиреноида, функциональная роль которого остается неясной. Хлоропласты способны к делению и имеют чрезвычайно раз- нообразную форму у разных водорослей. ПИГМЕНТЫ ФОТОСИНТЕТИЧЕСКОГО АППАРАТА Способность растений осуществлять фото- синтез связана с наличием у них пигментов. Главнейшим из них является магнийсодержа- щий порфириновый пигмент — хлорофилл. В природе встречается пять разных типов хлорофилла, которые незначительно различают- ся по своей молекулярной структуре. Хлоро- филл а присутствует у всех водорослей и выс- ших растений; хлорофилл b — у зеленых, харо- вых и эвгленовых и у высших растений; хлоро- филл с — у бурых водорослей, золотистых, диа- томей и динофлагеллат; хлорофилл d — у крас- ных водорослей; хлорофилл е обнаружен лишь однажды, по-видимому, это хлорофилл с; нако- нец, различные виды бактериохлорофилла- у фотосинтезирующих бактерий. Для сине- зеленых и красных водорослей характерно наличие билипротеинов: фикоцианина и фико- эритрина. Наиболее хорошо изучен хлорофилл а. Молекула его состоит из четырех пиррольных колец, с азотом которых связан атом магния, а к одному из колец присоединен одноатомный ненасыщенный спирт фитол. Молекула хлорофилла встроена в мембрану— погружена гидро фобной фитольной цепью в ее липидную часть. Чистый раствор хлорофил- ла а имеет максимум поглощения при 663 нм. В интактной, неповрежденной, нормально функ- ционирующей клетке хлорофилл характеризу- ется еще максимумами поглощения при 672 и
683 нм. Высокая эффективность поглощения света хлорофиллами обусловлена наличием в их молекуле большого числа сопряженных двойных связей. Большинство выделяющих кислород фото- синтезирующих клеток содержат два разных хлорофилла, одним из которых всегда являет- ся х.лорофилл а; другим же, как видно из таб- лицы, у разных растений являются разные хло- рофиллы (b, с, d); в некоторых случаях вместо второго хлорофилла в клетке содержатся били- протеины. Дополнительными рецепторами све- товой энергии являются также входящие в состав фотосинтетических мембран желтые пиг- менты — каротиноиды. Они отличаются от хло- рофилла по положению максимумов поглощения и используют непоглощаемую хлорофиллом часть видимого спектра. Предполагают также, что каротиноиды выполняют защитную функ- цию, предотвращая распад хлорофилла под действием молекулярного кислорода. С помощью тонких методов спектроскопии в составе фотосинтезирующих клеток обнару- жен находящийся в очень небольшом количе- стве пигмент с максимумом поглощения при 700 нм. Этот пигмент получил название пиг- мента Р7ооs Он отличается тем, что легко обес- цвечивается под влиянием света и сильных оки- слителей. Полагают, что пигмент Р„, локали- зован в «активных центрах», представляет собой специализированную форму хлорофилла и выполняет роль «ловушки» квантов энергии, поступающих от основной массы молекул хло- рофилла, функционирующих как «антенна» или своеобразный «сборщик» квантов света. МЕХАНИЗМ ФОТОСИНТЕЗА Преобразование энергии света в химиче- скую энергию продуктов фотосинтеза представ- ляет сложную цепь взаимосвязанных реакций, в которых участвуют многие соединения и структурные компоненты хлоропласта. Цепь реакций, составляющих процесс фото- синтеза, можно разбить на три основные ста- дии: 1. Фотофизическая стадия поглощения кван- тов света пигментами, превращение энергии света в пигментных структурах и передача поглощенной световой энергии «акч'ивному цен- тру». 2. Первичные фотохимические реакции, пе- ренос электрона в электрон-транспортной це- пи фотосинтеза и сопряженные с ним процес- сы образования «восстановительной силы». 3. Использование «восстановительной силы» для восстановления углекислоты и синтеза углеводов. Пигменты фотосинтетического аппарата орга- низованы в хлоропластах растений в два функ- циональных ансамбля, каждый из которых свя- зан, в свою очередь, с определенной цепью переноса электронов. Эти функциональные си- стемы получили название пигментных систем I и II (цв. табл. 1). Как подчеркивалось выше, специфическим свойством фотоавтотрофного питания зеленых растений является то, что для восстановления углекислоты в этом процессе в качестве донора водорода (электрона) используется вода и про- цесс идет с выделением кислорода. Это значит, что перепад окислительно-восстановительных потенциалов (ЛЕ,'), в пределах которого дол- жен быть осуществлен перенос электрона, ле- жит между потенциалом кислородного электро- да (Е ' для системы 0 /Н 0 при рН 7 составля- ет — 0,8 В) и потенциалом, который близок к водородному электроду и при котором идет вос- становление пиридиннуклеотида при участии ферредоксина (E,' для системы Н,/Н' при рН 7 составляет +0,42 В). Таким образом, фотосинтез в интактных растениях идет в диапазоне окис- лительно-восстановительных потенциалов от — 0,8 до +0,42 В, т. е. около 1,2 В, что соответ- ствует увеличению свободной энергии системы (в расчете на 1 грамм-моль) на 504 000 Дж. Каким же образом осуществляется за счет энергии света перенос электрона в таком диапа- зоне окислительно-восстановительных потенци- алов? Принципиальная схема этого процесса представлена на цветной таблице 1. Процесс фотосинтеза начинается с поглоще- ния света хлорофиллом. Молекула пигмента, поглотившая квант света, переходит в возбу- жденное состояние, которое длится около 10 ' сек, после чего молекула возвращается к исходному уровню. При этом энергия возбужде- ния может быть потеряна в виде тепла или флу- оресценции, или передана другим, «соседним» молекулам пигмента при достаточно близком их расположении. В результате такой миграции в ансамбле мо- лекул хлорофилла, расположенных в районе пигмента Р (реакционного центра пигмент- ной системы I) энергия возбуждения (электрон) может быть захвачена пигментом Р„, и пере- дана им акцептору — ферредоксину. При этом потерянный электрон (протон) восполняет- ся у Р„, пластоцианином и снова может быть «переброшен» за счет энергии света ферре- доксину. Аналогичная картина происходит и в пигмент- ной системе II, у которой роль акцептора элек- трона выполняет пластохинон, передавая по- лученный электрон по электрон-транспортной цепи цитохрому b, цитохрому j и, наконец, пла- стоцианину. Таким образом, благодаря окис-
С,н„О H,a' Фн 12 п АЛФ + 12 Ф„ 32НАДФ Н + )2 H+ 'f2 АДФ ~2 АТФ Рис. i. Схема восстановления углерода и образования первичных продуктов фотосинтеза в растениях (пенто- зофосфатпый восстановительный цикл углерода Кальвнна). В осуществлении процесса участвуют l. 3 различных ферментов. Цифры, заключенными в квадрат, обозначают число молекул, образовавшихся в предыдущих реакциях. Цифры, заключенные в кружок, обозначают число молекул, используемых в различ- ных процсссах. лительно-восстановительным превращениям про- межуточных переносчиков, осуществляется по- степенное движение электрона, которое индуци- ровано энергией кванта света. Следует сказать, что донором электрона для пигментной системы I I является в конечном итоге система фотоокисления воды, так же как и донором протона, хотя промежуточные про- дукты фотоокисления воды до сих пор не уда- лось найти и природа промежуточного донора электрона для пигментной системы I I остается неизвестной. Совершенно неясным остается так- же механизм выделения молекулярного ки- слорода. Важным обстоятельством является то, что в процессе передачи электрона в окислительно- восстановительных реакциях электрон-тран- спортной цепи, а именно на этапе передачи электрона от цитохрома b к цитохрому j, про- исходит синтез молекул аденозинтрифосфорной кислоты (АТФ) из аденозиндифосфорной ки- слоты (АДФ) и неорганического фосфата (Ф„) в результате реакции так называемого цикличе- ского фотофосфорилирования: пАДФ + пФ„-пАТФ + пН,О Кроме того, благодаря участию ферредоксина происходит образование АТФ и восстановленно- го никотинамидадениндинуклеотида (НАДФ Н) З реакциях нециклического фотофосфорилиио- вания: 4Фд,„„„+ 2АДФ + Ф„+ + 4Н,О -4Фд„„, +2АТФ+ + О, + 2Н.,О + 4Н+ Возникающие в результате световых реакций АТФ и НАДФ. Н получили название восстано- вительной или ассимиляционной силы и ис- пользуются в дальнейшем в темновых реакциях фотосинтеза для восстановления углекислоты и синтеза углеводов в пентозофосфатном цикле восстановления углерода (цикле Кальвина). Этот цикл, как видно на рисунке 1, представля- ет собой достаточно сложную систему фермен-
тативных реакций, в результате которых про- исходят фиксация и восстановление СО, и циклическое воспроизводство первичного ак- цептора углекислоты — рибулозо-1,5-дифосфата. Последующее преобразование продуктов фо- тосинтеза в реакциях обмена веществ, общих для гетеротрофных и автотрофных организмов, приводит к образованию аминокислот, белков, липидов, нуклеиновых кислот и других важней- ших соединений живой клетки. ФОТОТРОФНЫЙ, ГЕТЕРОТРОФНЫЙ И МИКСОТРОФНЫЙ ТИПЫ ПИТАНИЯ ВОДОРОСЛЕЙ Фотоавтотрофный способ питания с помощью фотосинтеза по масштабам и значимости стал, как мы видели, одним из основных способов питания водорослей и других зеленых растений. В разных отделах водорослей есть виды, кото- рые являются строгими (облигатными) фото- синтетиками. К их числу относятся, например, из сине-зеленых водорослей АпаЬаепа cylin- drica, А. variabilis, Anacystis nidulans, неко- торые штаммы Nostoc muscorum и др.; из зеле- ных — ряд видов Chlorococcum, некоторые виды Chlamydomonas, такие виды Xanthophyta, как Polyedriella helvetica и Monodus subterranea; ряд штаммов диатомей. Однако многие водоросли обладают способ- ностью достаточно легко переключаться в опре- деленных условиях с фотоавтотрофного способа питания на ассимиляцию различных органиче- ских соединений и осуществлять гетеротрофный или фотогетеротрофный тип питания или соче- тать эти способы питания с фотосинтезом. Способность к росту на органических средах в темноте или на свету в отсутствие СО, показа- на для большого числа видов, относящихся к сине-зеленым, зеленым, желто-зеленым, диа- томовым и другим водорослям. Известны также формы водорослей, которые относятся к числу ауксотрофных организмов и нуждаются в экзогенных источниках неко- торых физиологически активных органических веществ, в частности в витаминах В„, В„ в биотине. Таким образом, многие водоросли, обладая способностью осуществлять фотосинтез, не яв- ляются в то же время облигатными фотоавто- трофными организмами и отличаются высокой пластичностью и большим разнообразием типов питания. Способность водорослей усваивать те или иные органические вещества существенным образом зависит не только от химической природы этих веществ, но и от генетических свойств штамма. Так, например, сравнительное изуче- ние способности различных видов и штаммов хлореллы использовать для гетеротрофного роста (в темноте) такие углеводы, как глюкоза, галактоза, манноза, фруктоза, арабиноза, маль- тоза, лактоза и сахароза, показало, что наилуч- шим источником углерода для большинства культур является глюкоза. Вместе с тем среди изученных форм хлореллы были обнаружены и такие штаммы, которые росли на галактозе значительно лучше, чем на глюкозе. Ряд штам- мов эффективно использовали как глюкозу, так и галактозу. Некоторые формы использо- вали фруктозу примерно так же, как глюкозу, но в несколько раз хуже росли на среде с га- ла ктозой. Малопригодными в качестве источ- ника углерода для большинства культур ока- зались манноза, фруктоза, лактоза и сахароза, хотя некоторые штаммы хлореллы могли ис- пользовать и эти сахара. Такое же разнообра- зие свойств штаммов в отношении их способ- ности использовать различные сахара найдено и для других зеленых, а также сине-зеленых и желто-зеленых и некоторых других водорос- лей. В качестве органического источника углеро- да для гетеротрофного роста водорослей при- годны также ацетат, пируват и некоторые дру- гие органические кислоты, хотя степень и «а- рактер их использования, так же как и в слу- чае ассимиляции сахаров, зависит от свойств культур. Гетеротрофный рост водорослей в темноте идет значительно медленнее, чем автотрофный рост на свету. При освещении водорослей, культивируемых на средах, например с глю- козой, но без СО„наблюдается повышение скорости их роста и усвоения глюкозы. При этом культуры переходят к фотогетеротрофному типу питания, когда АТФ, возникающая в реакциях фотосинтетического фосфорилирова- ния, используется для ассимиляции глюкозы. На примере ряда штаммов хлореллы показано, что в случае обеспечения углекислотой куль- тур, выращиваемых на свету на среде с глюко- зой, клетки водорослей могут переходить к миксотрофному типу питания. Скорость роста и продуктивность таких культур превышают скорость их роста при автотрофном питании и равна примерно сумме их продуктивности при фотогетеротрофном и автотрофном пита- нии. Вместе с тем это, очевидно, не является общей закономерностью, и соотношение раз- личных типов питания в клетках водорослей может существенно меняться в зависимости от физиолого-биохимических свойств культу- ры и особенностей организации внутриклеточ- ных регулято рных процессов. В результате изучения путей биохимических превращений органических веществ, поглощаемых клетками разных водорослей в условиях фотогетеротроф-
ного и миксотрофного питания, а также изме- нения активности хлоропластных и цитоплаз- матических ферментов при переходе от авто- трофного к указанным типам питания, сейчас начинают проясняться некоторые внутренние механизмы перестройки метаболизма водорос- лей при смене типов питания. В ряде случаев переход к фотогетеротрофному питанию сопря- жен с индукцией дополнительных ферментных систем, участвующих в преобразовании погло- щаемых клеткой органических веществ. Так, например, показана индукция изоцитратлиазы у Euglena gracilis под влиянием ацетата. У не- которых форм хлореллы найдена под влиянием сахарозы индукция инвертазы, которая расще- пляет молекулу этого сахара на глюкозу и фрук- тозу. Активность некоторых ферментов фото- синтетического аппарата, в частности фермен- тов цикла Кальвина, может несколько снижать- ся при переходе культур водорослей к исполь- зованию органических источников углерода. Поглощаемые клетками органические вещества иногда окисляются до СО„которая в дальней- шем может использоваться в реакциях фото- синтеза. С другой стороны, может иметь место и прямое использование органических источ- ников углерода: ацетата — через цикл глиок- силовой кислоты, глюкозы — через гексокиназ- ную реакцию. Возможность прямой (не опосре- дованной через фотосинтез) ассимиляции угле- рода из органических источников показана, например, для Chlamydomonas mundata n Chlamydobotrys stellata, когда усвоение аце- тата этими водорослями практически не изме- нялось при экспериментальном выключении фотосинтетического аппарата с помощью спе- цифических ингибиторов фотосинтеза. Анало- гичную картину наблюдали на Chlamydomonas reinhardii: фотосинтетический аппарат инги- клЕткА водорослЕЙ Клетка — основная структурная единица те- ла водорослей, представленных либо одно- клеточными, либо многоклеточными формами. (.овершенно уникальную группу составляют си- фоновые водоросли: у них талломы не поделены на клетки, однако в цикле развития имеются одноклеточные стадии. Вполне очевидно, что клетка и здесь сохраняет свое значение как основной элемент, развитие и дифференциация которого приводят к формированию необычно- го слоевища. Особенность одноклеточных форм определя- ется тем, что здесь организм состоит всего из одной клетки, поэтому в ее строении и физио- догии сочетаются клеточные и организменные бировали с помощью рифампицина — специфи- ческого ингибитора синтеза хлоропластных РНК, а рост водорослей поддерживали с по- мощью глюкозы. Помимо использования органических соеди- нений в качестве источника углерода, водорос- ли способны переключаться с ассимиляции неорганического нитратного азота на усвоение органических источников азота — мочевины, амидов и некоторых аминокислот. Гетеротроф- ная ассимиляция азота — усвоение азота из органических соединений — неоднократно до- казана для водорослей как факультативная форма азотного питания. При этом, так же как и в случае усвоения углеводов и органических кислот, использова- ние водорослями аминокислот существенным образом зависит от физиолого-биохимических свойств штаммов. Показано, что такие амино- кислоты, как аргинин, гликокол, орнитин, могут использовать все штаммы хлореллы, в то время как алания, аспарагин, серин, ци- стеин — только отдельные культуры. Лизин, валин, гистидин, метионин и триптофан при- водили к подавлению роста некоторых форм водорослей. Одной из характерных особенностей неко- торых представителей сине-зеленых водорослей является их способность обходиться вообще без связанных форм азота и осуществлять фик- сацию свободного азота атмосферы аналогично азотфиксирующим микроорганизмам. Многообразие и пластичность способов пита- ция водорослей позволяют им иметь широкие ~реалы и занимать разнообразные экологиче- ские ниши. Исследование способов питания водорослей позволило ввести многие из них в промышлен- ную (массовую) кул1ьтуру. черты. Это наложило отпечаток на характер изменчивости одноклеточных форм, наследова- ние признаков, образование популяций и т. д. Представляя собой автономную систему, обла- дающую способностью к росту и самовоспроиз- ведению, мелкая, не видимая простым глазом одноклеточная водоросль выполняет роль свое- образной фабрики, которая добывает сырье (поглощает из окружающей среды растворы ми- неральных солей и углекислоты), его перераба- тывает и производит такие ценные соединения, как белки, углеводы и жиры. Кроме того, важ- ным продуктом ее деятельности считается ки- слород. Таким образом, она активно участвует в круговороте веществ в природе. Одноклеточ-
2 Рис. 2. Схематическое изображение клеточной организации одноклеточных зеленых подвижных водорослей при разных методах исследований: 1 — в прижизненном состоянии (световой микроскоп); 2 — после фиксации и окрашивания (световой микроскоп); 8 — по данным электронной микроскопии: а — жгутики, б — пульсирующие вакуоли, в — стигма, г — лопастной постенный хлоропласт, д- пиреноид, ж — ядро с ядрышком в центре, 3 — базальные тела, и — центриоль, к — ризопласты, л — диктиосома, л~ — мито- хондрии, и — крахмальные зерна, о — эндоплазматическая сеть, и — вакуоля, р — плазмалемма, с — оболочка, т — слизис- тая капсула. Несмотря на более чем 300-летнюю историю изучения, клетка до сих пор не раскрыла всех своих тайн, так как «сцена», на которой здесь разыгрываются события, имеет микроскопич- ескиее размеры, а действующие «герои» изме- ряются микрометрами и ангстремами. Этот ные водоросли иногда образуют временные или постоянные скопления в виде ценобиев и коло- НИИ. Многоклеточные формы возникли после того, как клетка проделала длительный и сложный путь развития в качестве самостоятельного ор- ганизма. В современных растениях сохранились следы этой истории. Переход от одноклеточного к многоклеточному состоянию сопровождался потерей индивидуальности и связанными с этим изменениями в структуре и функциях клетки. Внутри талломов многоклеточных водорослей складываются качественно иные отношения, чем между клетками одноклеточных водорос- лей. С возникновением многоклеточности свя- заны дифференцировка и специализация клеток в талломе, что следует рассматривать как пер- вый шаг на пути становления тканей (г и с т о- генез) и органов (органогенез). В за- висимости от расположения клеток в талломе многоклеточные водоросли могут быть пред- ставлены нитчатыми или пластинчатыми фор- мами. миниатюрный, труднодоступный мир сказочно красив и удивительно гармоничен. Прижизнен- ные наблюдения дают самые общие представ- ления о клетке и ее строении. Для выявления деталей приходится прибегать к специальной обработке материала (фиксации, окрашиванию, удалению воды) и только после этого присту- пать к изучению в световом или электронном микроскопе (рис. 2). Даже при беглом знакомстве с водорослями бросается в глаза чрезвычайное разнообразие как формы (шаровидные, грушевидные, яйце- видные, веретеновидные, спиралевидные, дис- к овидные, бочонковидные, цилиндрические и сложные, не поддающиеся краткому описанию), так и размеров (всего от долей микрометра, как у сине-зеленых, до нескольких десятков сантиметров, как у харовых) их клеток. Наи- большая пестрота картин обнаруживается у свободноживущих одноклеточных водорослей. У водорослей, в отличие от высших расте- ний, встречаются клетки, содержимое которых (п р о т о п л а с т) окружено лишь тонкой, по- о рядка 100 А, пограничной мембраной (п л а з- м а л е м м о й). Такие клетки обычно назы- вают голыми. Они не способны сохранять свою форму и постоянно находятся в метаболирую- щем, амебоидном состоянии. Подобного рода клетки встречаются как среди одноклеточных,
так и многоклеточных водорослей, чаще всего на стадии гамет и зооспор. Клетки некоторых водорослей (эвгленовых, перидиней, желто-зеленых), помимо плазма- леммы, окружены кожистым, эластичным сло- ем, толщина которого колеблется от 80 до о 250 А. Этот слой получил название п е л л и- кулы, или перипласта. Он состоит из фибриллярного вещества, в основном полиса- харидной природы (главным образом из пек- тоз) и имеет сложную, многослойную органи- зацию. Клетки с тонкой пелликулой обычно очень изменчивы по форме. Только толстая, похожая на панцирь пелликула может прочно ее зафиксировать. На поверхности пелликулы иногда возникают складки, выросты в виде зубцов или утолщения, называемые чешуйками. Эти структуры в различных сочетаниях обра- зуют самые причудливые узоры, придавая ор- ганизму неповторимый облик. Но главная их функция — повышение прочности клеточного покрова. У представителей своеобразной группы кок- колитофорид, входящих в состав золотистых водорослей, пелликула снаружи окружена до- полнительным слоем округлых известковых телец — кокколитов. Они лежат в слизи, вы- деляемой протопластом. Иногда в пелликуле обнаруживаются осо- бые структуры в виде блях или шестиуголь- ников, получившие название т р и х о ц и с т. Обладая способностью скручиваться и раскру- чиваться, нити трихоцист при определенных условиях могут мгновенно распрямляться, «выстреливая» наружу. Предполагают, что три- хоцисты выполняют в клетке защитную функ- цию и, может быть, даже участвуют в ее пере- движении. Еще более прочным, чем пелликула, оказался клеточный покров в виде т е к и. У некоторых вольвоксовых и хризомонад тека напоминает причудливой формы домик, в котором находится клетка. Она лежит там свободно, занимая только часть пространства (рис. 69, 8, 4; 195, 196). В структурном отношении тека — сложный мем- бранный комплекс, а по химическому составу представляет собой углеводное соединение, в ос- новном состоящее из галактозы, галактуроновой кислоты и арабинозы. Часто, например, у пе- ридиниевых водорослей, на наружном слое теки появляются шестиугольной формы пластинки, покрытие множеством мелких шипиков, боро- давочек и других выростов. Пластинки распо- лагаются в строгом порядке, срастаются сво- ими краями и минерализуются, образуя соору- жение, напоминающее панцирь. В месте сраста- ния пластинок возникают швы — единствен- ные участки поверхности, сохранившие способ- ность к росту. В теке имеются п о р ы, которые служат для циркуляции веществ между клеткой и внешней средой, для выхода наружу жгути- ков и трихоцист. В делении клетки тека не участвует. Обычно возникающие внутри ее дочерние клетки (или только одна из них) выходят наружу и обра- зуют новую теку. Важным событием, во многом предопределив- шим судьбу всего последующего развития рас- тительной клетки, следует признать появлен- ие наа ее поверхности покрова в виде о б о л о ч- ки — сначала пектино в ой, а затем и ц е л л ю л о з н о й природы. Преимущество этого образования состоит в том, что оно удач- но сочетает защитную и опорную функции с возможностью ростовых процессов и прони- цаемо стью. Клеточные оболочки водорослей весьма раз- нообразны как по своему строению, так и по химическому составу. Толщина оболочки варь- ирует не только от вида к виду, но даже в пре- делах одного вида в зависимости от возраста клетки, ее функционального состояния и усло- вий окружающей среды. Как правило, поко- ящиеся клетки имеют более толстые оболочки по сравнению с репродуктивными или моло- дыми вегетативными клетками (у зигнемы, например, они соответственно равны 1,6 и 0,8 мкм). Оболочки значительно утолщаются при дефиците влаги. У подавляющего большинства водорослей оболочки оказываются сплошными, хотя, как у желто-зеленых, десмидиевых и диатомовых, встречаются и составные, слагающиеся из двух и более частей. По времени заложения и особенностям роста различают п е р в и ч н ы е и в т о р и ч н ы е о б о л о ч к и (рис. 3, 2, 8). В активно деля- щихся клетках обычно образуется только пер- вичная оболочка. Ее рост идет в двух направ- лениях: увеличивается преимущественно по- верхность и в меньшей мере толщина. Переход клеток к покоящемуся состоянию сопровожда- ется заложением снаружи от плазмалеммы вто- ричной оболочки, которая растет главным обра- зом в толщину, отодвигает первичную оболочку от протопласта и вскоре становится преоблада- ющей (рис. 3, 8). Она придает клетке прочность и окончательную форму. При возобновлении в клетке ростовых процессов вторичная обо- лочка подвергается гидратации, становится эластичной и получает возможность растяги- ваться. На долю оболочки в среднем приходится 13 — 15о/о сухой массы клетки. У гидродиктиона, например, этот показатель еще выше и равен приблизительно 39 ~~, при этом жиры состав- ляют 7,2~~, а остальное — белки и углеводы.
Таким образом, белково-углеводный комплекс является основой оболочки, причем у водорос- лей соотношение этих компонентов колеблется в очень широких пределах (у сине-зеленых водорослей они представлены в одинаковой пропорции, а у хламидомонад отмечается сдвиг в сторону увеличения белков). Помимо этого, обращает на себя внимание разнообразный сос- тав полисахаридов. В частности, у хламидомо- нады они образованы галактозами, арабино- зами, в меньшей мере — ксилозами и рамно- зами; у ацетабулярии — маннозами, глюкоза- ми, галактозами и рамнозами; у ооцистиса- глюкозами, а галактозы, арабинозы и другие находятся в виде следов', у валонии — исклю- чительно глюкозами. Создается впечатление, что природа решила поставить на водорослях грандиозный эксперимент по выявлению из множества полисахаридов того, который лучше всего подходил бы в качестве материала кле- точных оболочек. Даже при беглом взгляде бросается в глаза неоднородность, слоистость оболочки водорос- левой клетки. Слои отличаются друг от друга по толщине, плотности и даже по химическому составу. Как правило, внутренние слои оказы- ваются целлюлозными, а наружный — пекти- новым. Он надежно защищает клетку от губи- тельного влияния кислот и других столь же сильных реагентов (конечно, если это воздей- ствие не будет слишком долгим). Изучение оболочки водорослевых клеток в электронном микроскопе расширило наши представления относительно ее организации. Оболочка у подавляющего большинства видов оказалась двухкомпонентной системой, состоя- щей из аморфного основного вещества (с т р а- м ы) и погруженных в него ф и б р и л л цел- о люлозной природы с сечением 200 — 300 А, которые выполняют роль опорного элемента и повышают прочность всей конструкции (рис. 4,1,2,8). Общее количество фибрилл и характер их ориентации варьируют не только в зависимости от формы клетки, но и от слоя к слою в преде- лах одной и той же клетки. Все попытки найти этому объяснение, исходя из теории механи- ческих напряжений, окончились неудачно, по- скольку не учитывались ((TBo рческие способ- ности» самой клетки, живущей по биологичес- ким законам. Изучение оболочек на ультраструктурном уровне позволяет произвести и более точное расчленение оболочек на слои. В большинстве случаев их не больше двух-трех, но бывают и отклонения. У сине-зеленых водорослей обо- лочка, как правило, четырехслойная, причем у осциллатории все слои одинаковой толщины Рис. 3. Последовательные этапы формирования кле- точной оболочки: 1 — молодая вегетативная клетка, лишенная оболочки; 2 — за- ложение первичной оболочки; 8 — появление под ней вторичной оболочки. о (100 А), а у анабены существуют различия, о о и весьма значительные (I — 100 А; II — 140 А; III — 150 — 300 А; IV — 140 А). Многослойные оболочки обнаружены у десмидиевых водорос- лей. Однако не у всех водорослей слоистость оболочки выявляется четко, что, вероятно, служит причиной появления порой весьма про- тиворечивых данных. Так, одни исследователи сообщают о существовании у ценобиальной зе- леной водоросли сценедесмус всего двух слоей- а другие — трех. Отсутствие целлюлозы делает оболочки гомо- генными, что нередко наблюдается у вольвощ- совых (гематококк, вольвокс), улотриксовых (спонгоморфа, акросифония), сифоновых (ко- диум). Иногда там, где фибриллярный компонент отсутствует или развит очень слабо, оболочки интенсивно пропитываются (и н к р у с т и- р у ю т с я) находящимся в среде раствором солей железа (у вольвоксовых), кальция (у ха- ровых) или кремния (у диатомовых), которые укрепляют матрикс, создавая нередко струк- туру, похожую на панцирь. Соли, как известно, в отличие от целлюлозы, упорядочиваются не в линейные, а в трехмерные структуры, что делает оболочку необычайно красиво орнамен- тированной. Нередко оболочки пропитываются органическими соединениями, из которых са-
Рис. 4. Организация клеточной оболочки по данным электронной микроскопии: 1 — многослойная оболочка Glaueocystis nostoehinearum; 2— расположение целлюлозных фибрилл в оболочке Pyroeystis lu- nula; 3 — модель строения оболочки: а — внешняя часть наруж-, ного слоя, б, в, г — внутренний слой оболочки с разным харак- тером упорядоченности целлюлозных фибрилл. мыми распространенными являются лигнин и кутин, секретируемые протопластом клетки. Причем кутин выполняет, помимо опорной, еще и защитную функцию, поскольку задержи- вает губительные ультрафиолетовые лучи и предохраняет клетку от излишней потери воды, тем самым улучшая условия транспирации. От инкрустации следует отличать процесс а д к р у с т а ц и и, под которым понимают скопление вещества на поверхности клетки. Так, кутин, выделяясь наружу, затвердевает и образует к у т и к у л у. Подобного рода обра- зование возникает на вегетативных клетках у порфиры, на оогониях у эдогониума, на эпи- дермальных клетках умногих бурых водорослей. К адк русти рующим веществам относятся и всевозможные слизи. У некоторых сине-зеле- ных водорослей слизистый слой, называемый здесь в л а г а л и щ е м, бывает довольно массивным, порядка 0,5 — 1,0 лкл. Мощная слизистая к а п с у л а появляется у многих одноклеточных зеленых водорослей (пальмел- ла, диктиосфериум, факотус), десмидиевых и в других группах. Она, очевидно, способствует созданию вокруг клетки своеобразного микро- климата. Слизь представляет собой продукт жизнедеятельности оболочки и по своей при- роде может быть либо пектиновой, либо целлю- лозной. От постоянных слизистых форм сле- дует отличать организмы с временно ослиз- няющимися, как у хламидомонад, оболочками. О таких водорослях обычно говорят, что они находятся в п а л ь м е л л е в и д н о м с о с- т о я н и и, которое, по всей вероятности, по- могает им переживать всевозможные неблаго- приятные условия. У красных водорослей на поверхности обо- лочки откладываются в большом количестве сульфированные полисахариды, образующие вокруг клетки мощную капсулу. Поскольку эти полисахариды водорастворимы, то их отложе- ние идет непрерывно. Из подобного рода соеди- нений получают ценный продукт агар-агар, ко- торый находит широкое применение, в част- ности, в кондитерской промышленности. Выделения протопласта поступают на поверх- ность клетки через особые отверстия в оболочке- п о р ы. Их диаметр у десмидиевых равен при- о о близительно 120 — 200 А, у фукуса — 370 А, о у осциллатории — 300 — 600 А, иногда и до о 900 А. В среднем на 1 лкл~ поверхности при- ходится до 20 — 30 пор. Они размещаются рав- номерно или группами. Если у одноклеточных водорослей благодаря порам устанавливается прямая связь протопласта с внешним миром, то у многоклеточных через поры на внутренних стенках — и с соседними клетками талл ома. Иногда через эти поры беспрепятственно про-
ходят цитоплазматические тяжи (п л а з м о- д е с м ы) (рис. 5, 1). Они, по сути, связывают протопласты всех клеток таллома в единое целое и обеспечивают однозначную реакцию многоклеточного организма на изменения в ок- ружающей среде. Не случайно именно у тех водорослей, у которых существуют подобного рода межклеточные коммуникации, можно иногда наблюдать почти фантастическую кар- тину, как, точно по мановению дирижерской палочки, все клетки нити одновременно при- ступают к образованию то акинет, то зооспор. Сравнительное изучение особенностей зало- жения, развития и строения пор показало, что они различаются по происхождению: одни воз- никают в ходе клеточного деления, как резуль- тат неполного срастания перегородки, другие образуются энзиматически, путем разрушения оболочки. Впоследствии поры могут закупори- ваться пробкой, как это нередко наблюдается у сине-зеленых (в особых клетках — гетероци- стах, о которых рассказано ниже) и красных водорослей (рис. 5, 2). Помимо пор, оболочки многих водорослей снабжаются различного рода в ы р о с т а м и в виде щетино к, ш и п и к о в и чешуек. Их роль для клетки неоднозначна: в одних случаях они выполняют, очевидно, защитную функцию, а в других обеспечивают оптималь- ные условия жизни, например поддерживают организмы в состоянии парения. В протопласте любой клетки различают два тесно связанных друг с другом компонента: я д р о (карион, или нуклеус, что в переводе соответственно с греческого и латинского озна- чает «ядро») и ц и т о п л а з м у, причем от степени их развития зависит уровень органи- зации клетки в целом. Водоросли — единст- венная группа растений, где представлены все известные в настоящее время типы клеточной организации: п р о к а р и о т и ч е с к а я- у сине-зеленых водорослей, м е з о к а р и о т и- ч е с к а я — у панцирных жгутиконосцев из группы динофлагеллат, э у к а р и о т и ч е с- к а я — у водорослей остальных отделов. У прокариот отсутствует морфологически оформленное ядро, а его функции выполняет состоящий из микрофибрилл ДНК н у к л е о- и д, который, несмотря на примитивность своей организации, не только содержит, но и точно передает из поколения в поколение всю генети- ческую информацию о клетке. Более того, про- кариоты лишены и многих типичных плазмати- ческих органелл (хлоропластов, митохондрий, диктиосом), но зато снабжены многочисленны- ми и разнообразными как по форме, так и по составу включениями. Мезокариоты уже обладают четко дифферен- цированным ядром, однако в его строении и по- Рис. 5. Строение плазмодесм и гетероцист.' 1 — плазмодесмы в оболочках зеленой водоросли Coleochaete (показаны стрелкой); 2 — продольный срез через гетероцисту сине-зеленой водоросли Anabaena azollae: а — пробка, б — по- лярное утолщение оболочки гетероцисты. ведении сохранились некоторые черты прими- тивности, присущие нуклеоиду. Подобная двойственность проявляется и в других чертах организации клетки, что позволяет говорить о промежуточном положении этих организмов между примитивными прокариотами и достиг- шими в целом наиболее высокого уровня раз- вития эукариотами. У большинства водорослей в клетке присут- ствует всего одно ядро, но известны случаи, когда их бывает два-три и больше. Клетки с не- сколькими десятками, как у кладофоры, или сотнями, как у водяной сеточки (гидродиктион), ядер называют ц е н о ц и т н ы м и. Примеча- тельно, что эти водоросли возвращаются к одно- ядерному состоянию при образовании специа- лизированных клеток бесполого (апланоспоры, зо оси оры) и полового (гаметы) размножения. Форма, размеры и местоположение ядра в клетке сильно варьируют у разных водорос- лей, а также в зависимости от фазы развития, на которой находится тот или иной организм. Очень мелкие ядра (диаметром 1 — 3 лкл) ха- рактерны для подавляющего большинства одно- клеточных водорослей, хотя и здесь встреча- ются такие «рекордсмены», как гематококк (10 лкл), гимнодиниум (12 — 16 лкл), вольвокс
е ° еееиюЬФФчв«е аеу ° ° % ° еъs ° е ° ° ° ъ е ° ° 1 ° ° с ф ф ° ° ° ° ° ъ ° %~в~ ° «лб ° ° ° ° ° ЭЭ р.: ° - ..:. «ф» .. - .- =-.~:, - .Ж е ° 4 — .- -::.а =. -,".-.-~~: ---=-'=-.=' -:.-,- ° . Фэ ф с ... - ~.:Ф"..-,-....—.." ~".Щ;- ==-:-.-.-.' ."Яц~ . -, ~ --...". -с Ф ' Р Рис. 6. Ультратонкое строение клетки.' 1 — продольный срез через клетку одноклеточной зеленой водоросли Pedinomonas minor (электронный микроскоп): а — плазма- лемма, б — пульсирующая вакуоля, в — статическая вакуоля, г — диктиосома, д — митохондрия, е — ядрышко, ж — окру- женное двухмембранной оболочкой ядро с ядрышком, з — пиреноид с крахмальной обкладкой, и — хлоропласт с пучком ламелл; 3 — эндоплазматическая сеть в цитоплазме клетки спирогиры; з — микротрубочки в цитоплазме клетки спирогиры.
(13 — 17 лкл), эремосфера (25 — 30 льл). У много- клеточных водорослей наряду с мелкими ядра- ми (1,5 — 2,5 лкл у улотрикса, 2,5 — 3,5 лкл у стигеоклониума, 2,0 — 4,0 у уронемы) попа- даются крупные (35 — 57 лил у некоторых ви- дов спирогиры) и даже такие гигантские, как у ацетабулярии (1,7лл). Размеры ядер даже в пределах одного тал- лома могут существенно колебаться. Так, у од- ного вида бриопсиса (Bryopsis halymeniae) из сифоновых ядра спорангия имеют диаметр, равный 30 — 45 лил, а ядра вегетативных клеток таллома — 3,0 — 4,5 лил. У другого вида бри- опсиса (Bryopsis hypnoides) ядра в проростке зиготы оказываются крупнее ядра зиготы почти в f700 раз. У водяной сеточки в молодых клет- ках ядра не превышают в диаметре 2,8 лил, а в старых иногда достигают 16 лил. Интерес- но, что перед делением диаметр ядер обычно увеличивается более чем в 2 раза. В этот же период происходит и его перемещение, как пра- вило, в центр (эдогониум) или в переднюю часть клетки (эвглена). В ядре у водорослей выявлены те же самые структуры, что и в ядрах других эукариотов: оболочка, ядерный сок (нуклеоплазма, карио- плазма, кариолимфа), ядрышко (нуклеолус), включения хроматина. Оболочка ядра состоит из двух мемб- ран (внутренней и наружной) и имеет поры (рис. 6, 1 ж). Толщина оболочки, например, у хлореллы (Chlorella pyrenoidosa) равна 300— о 450 А, у клостериума (Closterium ehrenbergii)— о 200 — 300 А, у ацетабулярии (Acetabularia me- o diterranea) — 200 А. Диаметр пор у клостериума о Эренберга составляет 700 А, у ацетабулярии- о 800 А. Наличие пор обусловливает высокую проницаемость оболочки ядра и способствует свободному прохождению через нее крупных макромолекул. Однако оболочка обладает очень высокой избирательностью и пропускает через себя далеко не все из них. Поэтому не следует представлять ее в виде решета, которое чисто механически отсеивает все то, что не соответ- ствует размеру его отверстий. В силу этого о проницаемости оболочки приходится гово- рить как о биологическом процессе, проходя- щем при непосредственном участии самой струк- туры. Сильные преобразования претерпевает обо- лочка ядра во время ядерного деления, которое часто, хотя и не всегда, сопровождается сначала ее фрагментацией, а затем и полным распаде- нием. Все внутреннее пространство ядра заполнено (рис. 7,1 — 6) я д е р н ы м с о к о м, в котором, помимо ядрышка, удается различить х р о м а- ° Р Р.' Ф' Фу ° » Ъ ° ° ° Ю » »° Юе .. ° ° ° ° ф ° » ° » рО ° Д ° в,. ° ° Ф ° % \ ° ° ° » ° » » / 'л»~ М уд ° ° Ъ г ° ° Ф' .,w ° \ ° ° ° O ° » ° ° Ъ 4:..;-- ......-,--:. 8 В ° ~ ° ° ° Ъ ,» 3 pt ° » 4 ° Ъ Ф ° ° Рис. 7. Интерфазные ядра клеток у разных водорос- лей: 1 — Microspora; 2 — Chlamydomonas; 8 — Hydrodictyon; 4— Spirogyra; 5 — Cladophora; 6 — Colacium; 7 — Glenodinium. тиновые структуры в виде скручен- ных нитей и глыб о к; самые крупные глыбкп иногда называют х р ом оцент р а ми, ка- риосомами или л ожными яд рыш- к а м и по чисто морфологическому сходству с последними. Теперь считают, что хроматино- вые нити не что иное, как форма существо- вания хромосом в покоящемся ядре, а хро- моцентры — их более плотные участки. Обычно хроматин скапливается вдоль оболочки; ос- тальная же часть ядра выглядит почти гомо- генной, поскольку там лишь иногда удается обнаружить структуру в виде нежной сеточки с гранулами в ее узлах. Только у очень немно- гих водорослей (эвглена, кладофора) хромати- новые образования распределяются по всему ядру более или менее равномерно, маскируя его содержимое (рис. 7, 5', 6). Переход ядер к делению сопровождается усилением спирализации, укор ачиванием, а затем и формированием морфологически четко выраженных х р о м о с о м. Исключение сос- тавляют мезокариотные водоросли: у них хро= мосомы находятся постоянно в конденсирован- ном состоянии и поэтому всегда хорошо видны (рис. 7, 7). Хромосомы ядра в совокупности составляют хромосомный набор. Каждый вид организмов имеет свой набор строго определен- ных по форме, размерам и количеству хромосом
40 е �e ° ф~~ g<r ° ю с4 r J~рв а~ Ф~~ ю ФГ 'ае 6 Рис. 8. Хромосомные наборы у разных водорослей: 1 — улотрикс (и = 10); 2 — микроспора ( и= 16); 8 — гидро- диктион (и = 19); 4 — эвдорина (n = 7); 5 — ульва (и = 10); 6— трентеполия (и = 24); 7 — вольвулина (и = 7). (к а р и о т и п). Он служит своего рода паспор- том, куда занесены в зашифрованном виде все основные исходные данные об организме. Их расшифровка сопряжена с большими труднос- тями, и здесь предстоит еще очень большая работа. У водорослей в подавляющем боль- шинстве случаев хромосомы очень мелкие, без перетяжек, и имеют вид точек или палочек. В пределах одного набора они, как правило, выглядят все «на одно лицо», поэтому изу-чать их бывает очень трудно (рис. 8, 1 — 7). Каждое ядро обычно содержит одно, но иногда и большее количество я д р ы ш е к. Обычно они бывают диаметром 1 — 2 лкл, хотя встреча- ются и более крупные. Например, у спирогиры (Spirogyra neglecta) они достигают 11 — '14 лила. Ядрышки состоят из очень плотного вещества и лишены мембранного барьера. В них, в от- личие от хроматина, скапливается не ДНК, а РНК. Во время ядерного деления ядрышки претерпевают циклические преобразования, связанные с разрыхлением, потерей красящей способности и диспергированием (рассеивани- ем) во время первой стадии ядерного деления (профазы) и конденсацией, восстановлением кра- сящей способности с полной реконструкцией своего облика к концу этого процесса (в тело- фазе). Однако среди водорослей встречаются отдель- ные группы, где имеются отклонения от подоб- ной схемы, которая признается классической, поскольку присуща подавляющему большин- ству организмов не только растительного про- исхождения. Здесь в первую очередь следует назвать эвгленовые водоросли, где ядрышко делится без предварительного разрыхления пу- тем гантелевидной перетяжки на два дочерних (рис. 199). Несколько иначе ведут себя ядрышки у спирогир: они в той или иной степени раз- рыхляются, слегка диспергируются, но не те- ряют при этом своей красящей способности. В результате ядро оказывается заполненным так называемым ядрышковым веществом, ко- торое очень затрудняет наблюдение за хо- дом ядерного деления, часто маскируя его де- тали. Совокупность всех преобразований, связан- ных с делением ядра, составляет и и т о т и- ч е с к и й ц и к л, или мит о з. У водорослей, как и у других растений, реорганизация ядра следует в строгой последовательности в несколь- ко этапов (фаз). Всего различают 5 фаз: и н т е р- ф а з а, или состояние покоя (самая продолжи- тельная по времени), профаза, метафа- з а, а н а ф а з а и т е л о ф а з а. Из телофазы ядро снова переходит в интерфазу. Второй компонент клетки — ц и т о п л а з- м а — состоит из гомогенного основного вещест- ва (с т р о м ы) и погруженных в него телец (о р г а н е л л) различного размера и формы: митохондрий, диктиосом, эндоплазматической сети и т. д. Этот основной набор органелл при- сущ клеткам не только растений, но и живот- ных. Митохондрии. (рис. 6, 1 д) обычно имеют овальную форму; они покрыты двух- мембранной оболочкой, причем внутренняя мем- брана образует выросты — к р и с т ы. Мито- хондрии располагаются преимущественно в тех местах клетки, где проходят процессы, связан- ные с затратой энергии (около ядра, у основа- ния жгутиков, вокруг стигмы), и выполняют роль дыхательных центров. Число, размеры митохондрий и степень развития крист взаимо- связаны друг с другом и с функциональным состоянием клетки. Д и к т и о с о м ы (рис. 6, 1 г) состоят из собранных в стопки уплощенных мешочков, или цистерн, число которых иногда достигает нескольких десятков. От мешочков по краям отп очко вываются разного размера пузырьки. Диктиосомы на поперечных срезах имеют вид характерных коротких, компактных пучков. Число диктиосом в клетке очень варьирует, и всю их совокупность там называют а п п а- ратом Гольджи, в честь ученого, впер- вые наблюдавшего эти структуры еще в прошлом веке. Диктиосомы осуществляют синтез по- лисахаридов, которые затем переходят в пу- зырьки и транспортируются в разные концы клетки.
ФУ Вся цитоплазма пронизана в той или инои степени развитой сетью каналов. Местами они образуют расширения — цистерны, иногда до- вольно значительные — пузырьки. Вся эта сис- тема получила название э н д о п л а з м а т и- чес кой сети или эндоплазмати- 7 .Эн о- ческого ретикулума (рис.6,2). ндо- плазматическая сеть осуществляет синтез глав- ным образом белков и транспортирует их пре- имущественно в те места, где развертываются интенсивные формо образовательные процессы. Отличительной особенностью клеток водорос- лей следует признать слабое развитие эндо- плазматической сети. У желто-зеленых, золо- тистых, бурых, диатомовых и красных водорос- лей она представлена единственным каналом, огибающим хлоропласт и затем переходящим в наружную мембрану ядерной о олочки, за счет чего устанавливается тесная взаимосвяз ь между ядром и хлоропластом (рис. 9). Совсем недавно в цитоплазме клеток различ- ных организмов, в том числе и водорослей, обнаружены короткие (по сравнению с кана- лами эндоплазматической сети) образования с жесткими гладкими контурами, получившие название микротрубочек (рис. 6, 8). В сечении они имеют вид цилиндров с диамет- о ром п ом просвета 200 — 350 А. Микротрубочки ока- зались крайне динамичными структурами: они могут то появляться, то исчезать, перемещаться из одного района клетки в другой, увеличивать- ся или уменьшаться в числе. Они сосредоточи- ваются преимущественно вдоль плазмалеммы (самый наружный слой цитоплазмы), а в пе- риод клеточного деления перемещаются в об- ласть формирования перегородки. Их скопле- ния обнаруживаются также вокруг ядра, вдоль хлоропласта, около стигмы. Последующие ис- следования показали, что эти структуры при- сутствуют не только в цитоплазме, но и в ядре, хлоропласте, жгутиках. Функции микротрубочек до конца не выяс- нены. Возможно, они участвуют в транспорте веществ по клетке, а характер локализации побуждает рассматривать их как цитоскелет- ные образования, призванные поддерживать форму как отдельных органелл, так и клетки в целом. В клетках водорослей, как впрочем и дру- гих растительных организмов, присутствуют также л и з о с о м ы, где отмечается скопле- ние большого набора ферментов из числа гид- р 7 олаз способных расщеплять важнейшие клас- сы химических соединений, включая белки, углеводы, нуклеиновые кислоты и т. д. Имеются п е р о к с и с о м ы, в составе которых обна- ружены каталаза и целый ряд других фермен- тов пероксидазного комплекса; с ф е р о с о- Рис. 9. Схема структурной связи между ядром, эн- доплазматической сетью и хлоропластом у бурых во- дорослей: а — хлоропласт с ламеллами, собранными в пучки по 3; б — пи- реноид; в — о кл — б адка пиреноида; г — ядро; д — ядрышко; е- я е ная оболочка, состоящая из двух мембран, ж — д — иктиосо- — оля' и — митохондрия; к — фрагмент канала эндо- плазматической сети; л — оболочка хлоропл, р оп аста об азованная вумя мембранами; м — эндоплазматическая мчъ, окру~~~щая двумя хлоропласт и свя занная с наружной мембраной ядерной о о- лочки. м ы, содержащие гидролитические ферменты, липиды и ароматические аминокислоты. Из всего этого видно, что цитоплазма клеток водо- рос ослей достигает очень высокой степени струк- турной организации. В интенсивно растущих клетках, помимо перечисленных органелл, можно наблюдать хорошо развитую систему пузырьков, или в а- к у о л е й. Они обладают способностью сли- ваться друг с другом, что иногда приводит к возникновению одной большой вакуоли, рас- полагающейся обычно в центре клетки. Ее появление компенсирует тот недостаток цито- плазмы, который возникает при резком увели- чении объема клетки в период ее интенсивного роста. В свете данных электронной микроскопии по-новому решается вопрос о природе вакуо- лей. Как теперь выясняется, в их образовании принимает участие не только аппарат Гольджи и эндоплазматическая сеть, но даже плазма- лемма. Среди многочисленных функций, кото- рые выполняет вакуолярная система в клетке, немаловажное место занимает функция осмо- регуляции. К особому типу вакуолей в клетках некото- рых водорослей следует отнести п у л ь с и- рующие вакуоли (рис. 2,1О;6,1б). Они, как правило, появляются в подвижных клет- ках, снабженных жгутиками, но иногда (у не- которых зеленых и диатомовых) образуются и в неподвижных клетках. Эти вакуоли обла-
дают способностью ритмично пульсировать, т. е. попеременно сокращаться и расширяться. В простейших случаях (у многих золотистых и зеленых водорослей) имеется одна или не- сколько простых вакуолей, при своем сокра- щении выбрасывающих содержащуюся в них жидкость в цитоплазму или через специальные выводные канальцы за пределы клетки. В дру- гих (например, у эвгленовых) — налицо целая система вакуолей, состоящая из главной ва- куоли, в которую открывается одна или не- сколько вакуолей второго порядка, часто, в свою очередь, окруженных вакуолями треть- его порядка. При сокращении последних жид- кость поступает в вакуоли второго порядка, которые, сокращаясь, наполняют главную ва- куолю. Выведение жидкости из главной ваку- оли осуществляется через выводной канал. У некоторых перидиней обнаружено еще более сложное устройство системы сократительных вакуолей. Функция пульсирующих вакуолей — уда- ление из клетки излишка воды. Наряду с этим пульсирующие вакуоли, по-видимому, выпол- няют и экскреторную функцию, выбрасывая из клетки ненужные продукты обмена веществ. И все же следует признать, что пульсирующие вакуоли еще во многом остаются загадочны- ми образованиями: они могут то появляться, то исчезать; непонятны и причины их сокра- щения. Своеобразную группу вакуолей составляют так называемые г а з о в ы е в а в у о л и, при- сутствующие в клетках сине-зеленых водорос- лей. Они имеют вид полостей, ограниченных тонкой мембраной и заполненных газом. На- личие газовых вакуолей, несомненно, умень- шает удельный вес организма, и он поднимается в верхние слои воды. Именно в этом многие усматривают биологическое значение газовых вакуолей. Такое объяснение было бы полностью приемлемо, если бы не случаи обнаружения подобных образований и у глубоководных во- дорослей. В связи с этим было высказано пред- положение, что в условиях глубинной жизни при дефиците кислорода начинается внутри- клеточное брожение, приводящее к образова- нию газовых вакуолей. С этой точки зрения «парение» представляется явлением вторич- ным. Газовые в акуоли начинают усиленно фор- мироваться при повышении интенсивности све- та. Очевидно, в этом случае они выполняют защитную функцию, отражая часть света и тем самым предохраняя клетку от его избытка. В отличие от животных (г е т е р о т р о ф- н ы х) организмов растения принадлежат к а в т о т р о ф а м, т. е. обладают способ- ностью образовывать органические вещества из неорганических в процессе фотосинтеза. Правда, некоторые водоросли из числа эвгле- новых, одноклеточных зеленых, желто-зеле- ных и пирофитовых водорослей утрачивают эту способность и переходят к гетеротрофному способу питания, но это свойство у них уже вто- ричного происхождения. У водорослей фото синтетический аппарат представлен разными уровнями организации: от морфологически неоформленного (у сине- зеленых) до типичных хлоропластов (у зеле- ных водорослей). Х л о р о п л а с т ы, первоначально на- званные хроматофорами (и сейчас еще этот термин сохраняется в научной литературе), представляют собой специфические органеллы растительной клетки, несущие зеленый пиг- мент х л о р о ф и л л, существующий в не- скольких формах (а, Ь, с, d, е). Помимо него, в хлоропластах у красных, желто-зеленых, бу- рых и других водорослей присутствуют и та- кие пигменты, как ф и к о ц и а н и н, ф и к о- эритрин, фикоксантин, ркаро- тин, ксантофилл и т. д.,которые часто маскируют основную зеленую окраску крас- ной, желто-зеленой, золотистой, бурой и т. д. В противоположность высшим растениям хло- ропласты водорослей отличаются большим раз- нообразием по форме, числу, местоположению в клетке и, как теперь выясняется, даже по внут- ренней организации. Хлоропласты у водорос- лей могут быть чашевидными, лентовидными, спиралевидными, пластинчатыми, звездчаты- ми и т. д. Как правило, в подвижных клетках у зеленых водорослей присутствует всего один хлоропласт; у водорослей из других отделов их бывает два и больше; у эвгленовых и динофла- геллат в молодых клетках — от 50 до 80, а в ста- рых 200 — 300. Хлоропласты занимают в клетке либо центральное, либо постенное положе- ние. Хлоропласты у водорослей, как и у других растений, окружены оболочкой и состоят из основного вещества (с т р о м ы) и погружен- ных в него пл а стинчатых, или ламел- л я р н ы х, с т р у к т у р, а также различного рода включений, отличающихся по разме- рам, форме и составу своего содержимого (рис. 6, 1и). Оболочка хлор опласта толщиной порядка о 100 — 150 А образована двумя одиночными мем- бранами. Иногда у бурых, золотистых, желто- зеленых, диатомовых водорослей этот барьер усложняется за счет примыкающего к хлоро- пласту канала эндоплазматической сети, в ре- зультате чего вокруг него возникает четырех- мембранная система, а у панцирных жгутико- носцев из группы динофлагеллат и эвгленовых
водорослей мембрана канала эндоплазматиче- ской сети настолько тесно прилегает к наруж- ной мембране оболочки хлоропласта, что гра- ница между ними становится неразличимой, создавая полное впечатление существования здесь трехмембранной системы. Расположенные в строме пластинчатые струк- туры (л а м е л л ы) по виду напоминают мини- атюрные уплощенные мешочки, которые назы- вают т и л а к о и д а м и (греч. «тилакоидес»— мешковидный) или д и с к а м и. Они придают хлоропласту характерную слоистость, что очень хорошо видно на поперечных срезах. Ширина дисков у водорослей разных видов колеблется о от 70 до СОО А, а иногда и более. В отличие от высших растений в хлоропла- стах у водорослей выявлено большое разнооб- разие в расположении дисков (рис. 10). Они могут быть одиночными, как у красных водо- рослей, ы опоясывать весь хлоропласт, ориен- тируясь параллельно его оболочке (поэтому на поперечных срезах выявляется система в виде длинных обособленных пластинок). У остальных водорослей, за исключением зе- леных, диски собираются в пачки по 2 — 4 и могут лежать там порознь или тесно прилегать друг к другу. У харовых и зеленых водорослей число дисков в пачке достигает нескольких де- сятков (у нителлы — от 4 до 20, у хлорококка- от 2 до 40). Причем эти колебания в значитель- ной степени зависят от возраста органеллы (по мере ее старения число дисков в пачке увеличивается), интенсивности освещения (при низкой освещенности число дисков в пачке больше, чем при высокой) и даже от качества света. Увеличение числа дисков в пачке, как прави- ло, сопровождается их укорочением. В хлоропла- стах некоторых зеленых водорослей можно на- блюдать, как короткие многодисковые пачки, чередуясь с единичными длинными дисками, упорядочиваются в штабеля или стопки— структуры, получившие название граноподоб- ных образований, так как по внешнему виду они очень напоминают компоновку дисков в гранах, свойственных хлоропластам высших растений. Диски являются теми структурами, к кото- рым прикрепляются пигменты и в которых осуществляется фотосинтез. Не исключено, что тот или иной способ группировки дисков зави- сит в значительной степени от особенностей качественного состава пигментов, который, как уже говорилось, у водорослей отличается боль- шим разнообразием. В самое последнее время в хлоропластах были обнаружены своеобразные трубчатые элемен- ты — м и к р о т р у б о ч к и сечением около Рис. 10. Схемы расположения дисков в хлоропласта~с различных водорослей: 1 — хлоропласты красных водорослей; 2 — хлоропласты крип- томонад; 3 — хлоропласты бурых водорослей; 4 — хлоропласты золотистых водорослей; 5 — хлоропласты зеленых водорослей~ а — диски, б — оболочка хлоропласта, в = канал эндоплаама тической сети, огибающий хлоропласт.
о 260 А, которые обычно располагаются вдоль оболочки органеллы группами по 5 — 6 штук в каждой и, очевидно, выполняют роль каркаса. В строме хлоропласта, помимо пластинчатых структур, выявлены более плотные зоны, пред- ставляющие скопления ДНК, многочисленные, о рассеянные, мелкие (в среднем порядка 140 А) частицы — р и б о с о м ы, глобулы различно- го размера, формы и состава, а также зерна крахмала, которые образуются в ходе фото- синтеза. Обладая всем необходимым для осуществле- ния белкового синтеза, хлоропласты относятся к числу самовоспроизводящихся органелл. Они размножаются путем перетяжки надвое и, в очень редких случаях, почкованием. Эти про- цессы приурочены к моменту клеточного деле- нпя и идут столь же упорядоченно, как и де- ление ядра, т. е. события следуют здесь в стро- гой последовательности одно за другим: ста- дия роста сменяется периодом дифференциации, за которым наступает состояние зрелости, или готовности к делению. Хлоропласты являются единственным мес- том отложения крахмала в клетке, причем у во- дорослей часть его, и весьма значительная, кон- центрируется вокруг специфических образова- ний, получивших название п и р е н о и д о в (греч. «пирен» — косточка и «эйдос» — вид, т. е. имеющий вид косточки). Пиреноиды са- мым тесным образом, как структурно, так и функционально, связаны с хлоропластом (рис. 9). Среди водорослей только сине-зеленые це- ликом состоят из беспиреноидных форм. Пире- ноиды, как правило, отсутствуют и в клетках высших растений. Исключение составляет лишь небольшая группа мхов из порядка антоцеро- та лес. Пиреноид представляет собой плотное обра- зование белковой природы, окруженное снару- жи обкладкой в виде сплошного кольца или отдельных пластинок в числе от двух и более, обычно крахмальной природы. Однако у бурых водорослей она образована водорастворимым полисахаридом — ламинарином,' у красных— особой формой крахмала, так называемым баг- рянковым крахмалом; у эвгленовых — пара- милоном; у хризомонад — липидами. Изредка, как у желто-зеленых, пиреноиды лишены об- кладки, и поэтому их называют голыми. Об- кладка с ее высоким коэффициентом преломле- ния света делает пиреноиды очень заметной структурой клетки, и они первыми бросаются в глаза при просмотре под микроскопом живого материала. Световой микроскоп не давал возможности проникнуть в детали строения пир еноида, и лишь благодаря электронному микроскопу удалось внести ясность в этот вопрос. Оказа- лось, строма пиреноида плотнее стромы хло- ропласта, 'она пронизана либо одиночными, либо собранными в пачки дисками, которые представляют собой продолжение дисков хлоро- пласта, и не отделена мембранным барьером (рис. 2, 8; 9). По сути, границей между хлоро- пластом и пиреноидом условно служит его обкладка. В голых же пиреноидах этот вопрос трудно решить даже на ультраструктурном уровне, поскольку единственными критериями оста- ются слабоуловимые различия в плотности ст ромы и поведении ламеллярной системы, которая при переходе в пиреноид претерпевает некоторую редукцию в числе дисков, сопровож- дающуюся увеличением их ширины. Обычно пиреноиды располагаются в хлоро- пласте (погруженные пиреноиды). Реже они лежат вне его (выступающие пиреноиды), что особенно характерно для красных водорослей, некоторых родов эвгленовых, пир офитовых, желто-зеленых и бурых водорослей. Чаще всего хлоропласт содержит только один пиреноид, хотя их может быть два и бо- лее, а у отдельных водорослей (спирогира, кладофора) их число даже доходит до несколь- ких десятков. Форма пиреноидов отличается крайним раз- нообразием: от круглой и многоугольной до палочковидной. Размеры пиреноидов колеблются в широких пределах: мелкие не превышают в диаметре 3 лкл, а крупные достигают 10 — 15 лкл, при- чем этот показатель варьирует от вида к виду, у особей внутри одного вида в зависимости от химического состава среды, интенсивности све- та, температуры и т. д. и даже в пределах одной клетки (наряду с одним крупным может су- ществовать несколько мелких), на что, в част- ности, влияет возраст пиреноидов (молодые пи- реноиды всегда мельче закончивших свой рост старых). Новые пиреноиды могут возникать тремя различными способами: делением надвое, фраг- ментацией крупного родительского пиреноида на большое число мелких дочерних и в резуль- тате новообразования. Причем в первом и во втором случаях пиреноид не всегда распадается на равновеликие части, что является еще одной причиной наличия в клетке пиреноидов раз- ного размера. Часто в пределах одного и того же орга- низма можно наблюдать одновременно все спо- собы образования новых пиреноидов. Сейчас накопилось уже много фактов, сви- детельствующих о том, что пиреноид является не просто местом скопления крахмала или иных
резервных веществ, а той зоной хлоропласта, в которой или при участии которой наиболее активно осуществляется их синтез. Возникнув первоначально как центр образования запас- ных веществ и в первую очередь как центр крахмалообразования в хлоропласте, пиреноид как бы остановился на полпути в своем посту- пательном движении, не успев получить того структурного воплощения, которое присуще ти- пичным органеллам. В дальнейшем развитие клетки пошло не по линии совершенствования этого образова- ния, а в направлении создания качественно иной, менее громоздкой структуры с аналогич- ными функциями, о чем свидетельствует от- сутствие пиреноидов в хлоропластах растений, занимающих более высокую ступень развития по сравнению с водорослями. У подвижных или утративших подвижность клеток на переднем конце тела часто можно на- блюдать небольшое, но очень заметное, бла- годаря интенсивно-красной или ярко-оранже- вой окраске, пятно — с т и г м у, или г л а- з о к (рис. 2, 1в). Такая окраска объясняется скоплением здесь большого количества каро- тина. В одних случаях (у зеленых, золотистых, бурых, некоторой части желто-зеленых водо- рослей) стигма располагается в хлоропласте (рис. 11, 1, 2), а в других (у эвгленовых, пан- цирных жгутиконосцев) — за его пределами, в непосредственной близости от двигательного аппарата клетки (рис. 11, 8, 4, о). Вот, собственно, и все сведения, которые удалось собрать о стигме за долгую историю ее изучения с помощью светового микроскопа. Электронный микроскоп намного расширил границы наших знаний в этой области. С его помощью не только были раскрыты тайны строения, но и обнаружены такие особенности тонкой организации, которые убедительно сви- детельствуют о существовании стигм разной степени сложности. Основу стигмы составляют плотные, разного о диаметра (от 750 до 6600 А) п и г м е н т о н е- сущие глобулы в числе от 5 — 6 до нескольких десятков, которые располагаются плотными рядами (их иногда насчитывается до 9). Такой тип строения стигмы, хотя он наи- более распространен, считается самым прими- тивным (рис. 11, 1). Явным признаком усложнения стигмы сле- пует признать группировку глобул по 3 — 5 в комплексы, отграниченные друг от друга тонкой мембраной, что наблюдается у эвгле- новых водорослей (рис. 11, 8). Еще более сложной считается организация стигмы у некоторых панцирных жгутиконосцев, Рис. 11. Различные типы стигм, обнаруженные в клет- ках водорослей: 2 — Chlamydomonas; 2 — Dinobryon; 8 — Ещ;1епа; 4 — Gleno- dinium; б — Nematodium: а — хлоропласт, б — стигма, в- пластинчатое тело, г — линзовидное тело, д — полость, соеди- ненная каналом с цитоплазмой клетки, ж — ретиноид, 3 — пиг- ментные тела.
Рис. 12. Жгутиковые клетки.' 1 — Ochromonas ludibunda; 2 — Chlamydomonas pertusa; 8 Platymonas cordiformis; 4 — Schizochlamys gelatinosa; й — Ещ;- lena proxima; 6 — зооспора Derbesia neglecta; 7 — Chrysochro- mulina kappa с гаптонсмой. поскольку, помимо комплексов глобул, у них имеется необычного вида п л а с т и н ч а т о е т е л о. Оно состоит из целой серии параллель- но расположенных, сообщающихся между со- бой, уплощенных мешочков, которых иногда бывает до 50, причем самые крайние из них переходят в находящийся поблизости канал эндоплазматической сети. Да и каждый комп- лекс глобул в отдельности несколько сложнее по своей организации, чем у эвгленовых, так как состоит не из одного, а из двух рядов гло- бул, промежуток между которыми заполняется зернистым веществом, и вместо одиночной мем- браны окружается оболочкой, образованной из двух мембран (рис. 11, 4). Среди панцирных жгутиконосцев встреча- ются организмы и с очень сложно устроенной стигмой, представляющей многокомпонентную систему, куда входят линзовидное те- л о, р е т и н о и д — тело, заполненное опре- деленным образом ориентированными ф и б- р и л л а м и, плотные глобулы. Таким обра- зом, свет, попадая на линзу, фокусируется и за- тем концентрированным пучком посылается через ретиноид на глобулу, т. е. приходит туда в преобразованном виде (рис. 11, о). Основной функцией стигмы считается улавли- вание и до некоторой степени, очевидно, пре- образование света, необходимые для опреде- ленной ориентации тела водоросли в простран- стве. Стигму по ее роли в клетке можно упо- добить локаторному устройству, принцип ра- боты которого в данном случае, к сожалению, пока не выяснен. Поэтому сейчас еще прежде- временно обсуждать вопрос о значении того или иного способа организации этой системы, ибо хорошо известно, что сложность и громозд- кость конструкции не всегда являются показате- лем ее совершенства, рациональности и надеж- ности в работе, тем более что выявлено много водорослей, которые лишены стигмы, но между тем прекрасно реагируют на свет. Почти все водоросли (исключение составляют сине-зеленые и красные) могут образовывать клетки, подвижные в течение всей своей жизни или только на определенном этапе жизненного цикла (от нескольких часов до нескольких ми- нут). Подвижные клетки отличаются от непо- движных целым рядом особенностей организа- ции, прежде всего появлением особого двига- тельного аппарата. Перемещение в жидкой среде осуществляется при участии особых структур, представляющих собой либо временные (ризоподии), либо по- стоянные (реснички, псевдоцилии, жгутики) выросты клетки (рис. 12). Р е с н и ч к а м и называют многочисленные (от нескольких десятков до нескольких сотен) короткие (порядка 5 — 10 лкл~) образования, совершающие энергичные кол ебательные дви- жения. П с е в д о ц и л и и, или л о ж н ы е р е с н и ч к и, имеют вид длинных, очень тон- ких, неподвижных волосков, которые обнару- живаются чаще всего только после подкраски. Под ж г у т и к а м и понимают длинные (в не- сколько десятков микрометров) малочисленные образования с меньшей, чем у ресничек, часто- той биений и с волнообразным характером дви- жения, почему их иногда еще называют унду- липодиями (лат. «унда» — волна, «подиум»— возвышение). Перечисленные образования — единственные клеточные структуры, расположенные за пре- делами самой клетки. В дальнейшем речь пойдет исключительно о жгутиках, потому что они наиболее характер- ны для водорослей и принципиально ничем не отличаются от ресничек и псевдоцилий. Среди водорослей чаще всего встречаются формы дву- или одножгутиковые, реже четы- рех-, восьмижгутиковые и совсем единичны с большим числом жгутиков, как у дербезии (рис. 12). У некоторых золотистых водорослей между двумя подвижными жгутиками распо- лагается третий — неподвижный жгутик с рас- ширением на конце, или г а п т о н е м а. С ее помощью клетка прикрепл.яется к субстрату (рис. 12, 7).
По соотношению длины жгутиков у клет- ки различаются водоросли р а в н о ж г у т и- ковые и разножгутиковые, или г е т е р о к о н т ы (рис. 12, 1, 2). Иногда, на- пример у эвглен, второй жгутик настолько ре- дуцируется, что удается обнаружить лишь его короткий пенек. Жгутики могут быть направ- лены вперед по ходу движения клетки, что свой- ственно большинству равножгутиковых форм. У разножгутиковых нередко наблюдается раз- ница в частоте биений (г е т е р о д и н а- м и з м) и в положении по отношению к клетке: обычно длинный (д в и г а т е л ь н ы й) жгу- тик направлен вперед и работает энергичнее, чем короткий (р у л е в о й), который обращен в сторону или назад по ходу движения ор- ганизма. Считается, что двигательный жгу- тик обеспечивает поступательное движение клетки, а рулевой — корректирует его на- правление. Как правило, вся жизнь жгутиконосцев про- ходит в непрерывном движении. Некоторые из них развивают скорость до 500 лил/сек и пре- одолевают по 5 — 10 л в сутки — расстояния для столь крошечных существ поистине ма- рафонские. Жгутики бывают г л а д к и м и или о п у- ш е н н ы м и. Последние несут на своей по- верхности выросты в виде тонких, диаметром о не более 50 А, волосков (м а с т и г о н е м) и неминерализованных, разных по форме и раз- мерам ч е ш у е к (рис. 13,1, 2). Опушенным обычно оказывается двигательный жгутик. При- чем у некоторых водорослей (празинокладус, колеохете) жгутики одновременно несут и мас- тигонемы и чешуйки, но чаще имеется какой-то один тип образований. В отличие от чешуек мастигонемы располагаются очень упорядо- ченно, параллельными рядами. И те, и другие закладываются в цитоплазме, в пузырьках с аморфным содержимым и в них переносятся на поверхность жгутика. Несмотря на простоту внешнего облика, строение жгутика оказалось довольно сложным, что особенно хорошо видно при изучении его в электронном микроскопе. Сопоставление кар- тин продольных и поперечных срезов позволи- ло воссоздать в основных чертах принципы внутреннего устройства жгутика (рис. 14). Цилиндрическое, несколько сужающееся к вер- шине тело жгутика окружено снаружи тонкой, о порядка 90 А, мембраной, представляющей про- должение плазмалеммы, и заполнено вещест- вом, которое по виду напоминает цитоплазму и в которое погружена система правильно ориентированных м и к р о т р у б о ч е к. Они располагаются по схеме 9 + 2, что расшифро- вывается следующим образом: по периферии Рис. 13. Поверхностные структуры жгутиков: 2 — мастигонемы на жгутике сперматозоидов бурой водоросли Himanthalia lorea; 2 — чешуйки на жгутиках сперматозоида аровой водоросли Chara eorallina. цилиндра параллельно его поверхности разме- щается 9 пар тесно прилегающих друг к другу микротрубочек, а в центре его лежат две оди- ночные микротрубочки. Причем те и другие связаны между собой целой серией переходных структур, в расположении которых усматри- ваются свои закономерности, что, надо думать, имеет прямое отношение к специфической функ- ции жгутика — осуществлять движение по оп- ределенной траектории. Выявленная в жгути- ках у водорослей система расположения мик- Рис. 14. Схема организации фибриллярных ультра- структур стержня реснички.
основныЕ типы морФологичЕской структуры тЕлА водорослЕЙ рот рубочек присуща подавляющему боль- шинству организмов не только растительного, но и животного происхождения. Отклонения в сторону упрощения или усложнения отме- чан~тся крайне редко и обычно связаны, соот- ветственно, с ослаблением или повышением ча- стоты биения жгутика и с особенностями его движения. При основании каждого жгутика лежит б а- зальное тело (рис. 2,2з). Оно имеет вид короткого, длиной около 2 лкм цилиндра, в периферической части которого располага- ется 9 триплетов микротрубочек, представля~о- щих единую систему с периферическими парами иикротрубочек жгутика. Обычно базальное тело одного жгутика лежит под углом 90 или 180' к базальному телу другого. Иногда ба- зальные тела соединяются перемычкой, что обеспечивает максимальную согласованность в их действиях. От базального тела внутрь цитоплазмы до- вольно часто, но не всегда отходит один или несколько так называемых ж г у т и к о в ы х корешков или ризопластов (рис. 2, И). Ризопласт представляет собой сложную систему, состоящую из двух компонентов: из одиночных или собранных в пучок (по 2 — 9) микротрубочек, которые направляются к плаз- малемме, выполняя там роль цитоскелетных образований, и из поперечно-исчерченной, фиб- риллярной по своей природе структуры, на- правляющейся к ядру и, как у хризомонад, там закрепляющейся. Последнее указывает на связь жгутиков с этой клеточной органеллой. Таким образом, двигательный аппарат клетки представляет сложную систему, состоящую из жгутиков, базальных тел и ризопластов. У подвижных или потерявших подвижность водорослей нередко обнаруживаются структу- ры, по своей внутренней организации очень напоминающие ба зал ьные тела. Они обычно В отличие от высших растений, целиком и полностью характеризующихся одним листосте- бельным типом строения (другая структура у них вызвана вторичным упрощением), водо- росли в пределах слоевцового типа строения обнаруживают исключительное морфологиче- ское разнообразие. Тело водорослей, как уже упоминалось, может быть всех четырех сте- пеней сложности, вообще известных для орга- низмов, — одноклеточным, колониальным, мно- гоклеточным и неклеточным. Размеры их в пре- делах каждой пз этих форм отличаются огром- располагаются около ядра (рис. 2, 2 и), а во время его деления расходятся в противополож- ные стороны, образуя своеобразные полюсы, или центры, деления, за что и получили назва- ние ц е н т р и о л е й. Центриоли — непре- менные структуры клеток животных организ- мов, а среди растений встречаются только у во- дорослей. Как показывают ультраструктурные исследования, центриоли, с одной стороны, связаны с ядром, а с другой — с двигательным аппаратом клетки, объединяя их в своеобраз- ный комплекс. Интересно, что если между двигательным аппаратом клетки и ядром в отдельных слу- чаях все же можно наблюдать отчетливую структурную связь, то между двигательным аппаратом и стигмой, вопреки ожиданиям, та- ковой не обнаружено, хотя взаимодействие, пусть и опосредованное, безусловно должно су- ществовать. В процессе филогенетического развития во- дорослей жгутиковый аппарат, как это случи- лось и с пиреноидом, постепенно редуциро- вался. Это сопровождалось сначала частичной, а затем и полной утратой подвижности, распа- дением жгутиково-ядерного комплекса и исчез- новением центриолей в клетке. В настоящее время не вызывает сомнения, что водоросли дали начало наземным расте- ниям, проложив им дорогу на сушу. Этот акт подготавливался всем ходом развития водо- рослей, в котором решающая роль, по-видимо- му, принадлежала клетке. Большое разнооб- разие строения, состава и свойств отдельных клеточных компонентов свидетельствует о том, что здесь шел интенсивный формообразова- тельный процесс. На его основе методом проб и ошибок отбирались наиболее перспективные структуры и создавался такой тип клеточной организации, который позволил растениям пе- рейти к наземному образу жизни. ным диапазоном — от микроскопических до очень крупных. Так, некоторые виды однокле- точной сине-зеленой водоросли синехоцистис (Synechocystis) едва достигают 1 лил, одно- клеточные зеленые водоросли из рода хлорелла (Chlorella) могут быть в 2 лил, а длина клеток, образующих междоузлия в стеблевидных тал- ломах харовых водорослей, часто составляет 15 — 20 сл. Однако самыми крупными размерами отли- чаются много клеточные морские бурые во- доросли, слоевища которых у отдельных
Рис. 15. Амебоидная ст кт а рослей: я структура у золотистых водо- 2 — одиночные клетки Chrysamoeba' 2 о ино ое а; 2 — рядовое объединение zochrysis. — групповое объединение клеток ВЫ- Рис. тура у зеленых водорослей: онадная структ а rys об 1 — одиночная клет етка Chlamydomonas 2— разованная с астан образованная слизью 1 видов, например у макроцистиса 'Ма pyrifera), мог т о тиса ~ аcI'Ocystis и тонкими ут достигать в длину 30 — 45 л. ими, и тогда их называют и з о ~~~~ы~~ ~~~~~~~~ ~~ж~ б б В ругое название этой струк- месте с TOM Bce это МсЕ чительное многообра исклю- сколько клет а я). Иногда не- разие имеет в своей ток подо ного ст оени несколько хорошо обо б основе ся своими из р ния соединяют- о о особленных типов мо и риз оподиями или аже логической структу ы явл мор~~о- в пл а змодии. даже сливаются -"~~~~~д~~~ строение м~~у м глав мор ологической обретать некоторые о н эволюции. В д~~~~ю~ у (монадная с т б льшей, то мень шеи степени повто м с расывания иуда .ил ются в разных отделах во ря- вания внутрь. Амеб, ус .или втяги- тель ствует об известном ах водорослей, что сви е- д р р ме оидная стр кт а ционного развития в п ом параллелизме эвол лю- тистых и желто-з дставителей пи о ито вых, золо- А б ос типов морфологичес кои стр>кт ы т основных шим типом с строения о нокл является наипростей- рослей. Из н 4 ур тела водо- ма и ес д еточного организ- них относятся к о но ть все основания счита формам, 1 — к дноклеточным эволю ии итать, что в процессе — к неклеточным, осталь ции она ыла самым пе к многоклеточн ьные 4 — ' возникн ервым этапом при чным колониальные ~о мы кновении клеточного овня дучи существенным м этапом на п~ти +îpìû, бу- не исключает т, однако, того чт уровня жизни. Это ния организации водорослей, все же у современных во о о лишь разнови дностью о но же являются быть сле дствием вто дорослеи она может ния) д оклеточного строе- о — ния монадных фо м. ричного упрощени я строе- 1. Амебои оидная структура ппе став 2. ~онадная структ. а сво о — орм. клеточными о га редставлена одно- точным о . а своиственна однокле- организмами, лишенным ~м организмам с, тве о клеточной обол ными твердой лочкой рдой клеточной обо- (рис. 15) Такие о организмы, выпуская п с "изуется наличием т очным перипластом доподии, способ ны совершать полз а я псев- одного дв х или нескольких жг т у таких клеток движения, подобно но простейшим жив зающие мощью котор о ых они активно жгутиков, с по- амебам. Псев опо животным — (рис. 16 . Н о двигаются в воде вдоподии часто бывают д линными но лишены екоторые монадные ормы вторич- ны жгутиков, и тогда как правило У
° ° ° + ° ° ° ° ° ° ° ° ° ° ° ° ° е ' ° е ° е ° е е е ° е ° ° 3 ° е ° е I ° ° е ° е е ° ° ееее ° ° еееее Рис. 17. Коккоидная структура у зеленых водорослей: 1 — одиночная клетка Chlorococcum; 2 — одиночная клетка Cha- racium; 8 — колония Sphaerocystis,,образованная слизью; 4— колония Coelastrum, образованная срастанием клеток. Рис. 18. Пальмеллоидная структура и пальмеллевид- ное состояние: 1 — пальмеллоидная структура у золотистой водоросли Hydru- rus (часть таллома); 2 — пальмеллевидное состояние у зеленой водоросли Chlamydomonas. их тело способно менять свою форму (м е т а- болия). Монадная структура очень широко распро- странена в мире водорослей — она свойственна многим представителям в отделах пирофитовых, золотистых, эвгленовых, желто-зеленых и зе- леных водорослей, причем у первых трех она является преобладающей. Среди водорослей этого уровня организации часто встречаются колониальные формы, пред- ставляющие собой собрание отдельных клеток монадного строения, объединенных слизью в одно целое. Форма таких колоний и строение слизи у разных водорослей могут существенно различаться; в некоторых случаях клетки бы- вают связаны друг с другом п л а з м о д е с- мами. У большинства колониальных форм монад- ного строения клетки внутри колоний не обнаруживают различий, только у немногих из них наблюдается разделение на в е г е- т а т и в н ы е, отличающиеся меньшими раз- мерами, и на более крупные, служащие для размножения (например, у вольвокса и плевдо- рины из отдела зеленых водорослей). Наконец, у многих водорослей во всех отде- лах, за исключением сине-зеленых и красных, независимо от структуры талломов в вегетатив- ном состоянии, монадным строением обладают подвижные воспроизводительные клеткд, слу- жащие для бесполого размножения (з о о с п о- р ы), а при наличии полового процесса — и г а- м е т ы (мужские, а в некоторых случаях и женские). Монадная структура многими альгологами принимается за первичную и исходную в фило- генезе водорослей, однако имеется достаточно веских оснований считать ее вторичной, обра- зовавшейся из амебоидной. Во всяком случае, несомненно, что монадная структура в процессе эволюции водорослей оказалась весьма про- грессивной и сохранилась у них на стадиях размножения вплоть до наиболее сложно орга- низованных форм. 3. Коккоидная структура характеризуется отдельными клетками, снабженными твердой оболочкой и в вегетативном состоянии постоян- но лишенными жгутиков или псевдоподиев (рис. 17). Вследствие такого устройства водо- росли коккоидной структуры не способны к ак- тивному движению (исключение составляют только десмидиевые и диатомовые водоросли, но аппарат движения у них совершенно дру- гой — выделение слизи), они или свободно жи- вут и могут лишь пассивно переноситься водой, или прикрепляются к субстрату. Форма таких клеток необычайно разнообразна — от простой шаровидной до самой причудливой; оболочка у них гладкая или с различными выростами,
цельная или пористая и т. д. Многим водорос- лям коккоидной структуры свойственно обра- зование колоний различной формы, со слизью или без нее. Коккоидная структура чрезвычайно широко распространена у водорослей и в большей или меньшей степени наблюдается во всех отделах, где имеются одноклеточные формы. В отделе диатомовых водорослей, очень многочисленных и широко распространенных, она является единственной. В эволюционном отношении коккоидная структура интересна как такая ступень морфо- логической дифференциации водорослей, на основе которой стало возможно возникновение многоклеточных талломов. 4. Пальмеллоидная структура представляет собой усложненный вариант коккоидной струк- туры, выражающийся в образовании водорос- лями достаточно крупных, преимущественно прикрепленных к субстрату слизистых тел определенной формы, содержащих внутри мно- жество коккоидных клеток (рис. 18). Клетки объединяются в этой слизи чисто механически и плазматических связей не имеют. Такое строение широко распространено среди водо- рослей, но о пальмеллоидной структуре го- ворят лишь тогда, когда оно является постоян- ной формой их вегетативного роста; если же только временной стадией, то тогда его назы- вают пальмеллевидным состоя- н и е м. В пальмеллевидное состояние могут перехо- дить многие одноклеточные подвижные и непод- вижные водоросли (преимущественно при на- ступлении неблагоприятных условий); в мень- шей степени оно наблюдается у водорослей иной структуры. Образующиеся при этом слизистые тела не достигают крупных размеров и не имеют определенной формы. 5. Нитчатая структура, как следует из на- звания, представлена талл омами, состоящими из нескольких, нередко очень многих клеток, расположенных в форме нити. Нити могут быть простыми и разнообразно ветвящимися, сво- бодноживущими, прикрепленными и объеди- ненными в слизистые колонии (рис. 19). Нитчатая структура в мире водорослей яв- ляется простейшей формой много клеточного слоевища и свойственна огромному количеству представителей из разных отделов. Клетки в нитчатых слоевищах тесно связаны друг с другом, во многих случаях здесь доказано наличие пор и плазмодесм, проходящих через поперечные клеточные перегородки. Вместе с тем распадение нитей на участки и даже на отдельные клетки является обычным способом вегетативного размножения многих нитчатых водорослей. ° ° ° ° ° ° \ ':; съ. ° В ° ° tO ° ° ° ° ° ,Ф yl ° ° ° ° а» ° ° % ° ° ° ° ° ° ° ° ° ° ° ° ° » ° Ю ° ° ; ° ° + а~ . ° сс ° ° ° ° ° '1 ° ° ЭФФИ ° е ° Ф ° ° ° » ° ° с Ъ l 4 Рис. i9. Нитчатая структура у сине-зеленых водо- рослей: 1 — простейшее строение нити с диффузным ростом у Oscillato- ria; 2 — нить с дифференцированным основанием у Endonema; 8 — верхушка нити у Rivularia, вытянутая в волосок; 4 — интер- калярный рост у Gloeotrichia; 5 — апикальный рост на концах ветвей у Scytonema; 6 — базальный рост у Calothrix. Рис. 20. Разнонитчатая структура у сине-зеленой водоросли Fischerella.
Рис. 21. Пластинчатая структура у зеленой водоросли Prasiola: однорядная нить, разрастающаяся в одно- слойную пластинку. В простейших случаях талл омы нитчатой структуры слагаются из одного ряда клеток, вполне подобных друг другу. Наряду с этим многочисленны водоросли, у которых нити изменяются к концам — утончаются, или рас- ширяются, или имеют конечные клетки, отли- чающиеся по форме от остальных. Различия в строении концов нитей бывают наиболее от- четливыми при прикреплеаном образе жизни, определяющем полярность нити. При этом нередко нижняя клетка превращается в р и- з о и д или с т о п у, а к вершине нити меня- ется форма клеток. Рост нитчатых талломов происходит по-раз- ному. Е сли способностью делиться обладают все клетки нити, то рост называют д и ф ф у з- н ы м. В других случаях делятся не все клетки, а только часть их, и тогда рост нити происхо- дит в определенных ее участках, называемых зоной роста или меристемаль- н о й з о н о й. В зависимости от положения меристемальной зоны на нити различают, кроме диффузного, еще три типа роста: и н т е р к а- л я р н ы й, если зона роста расположена в средней части нити (обычно ближе к ее верши- не), а п и к а л ь н ы Й рост, осуществляемый делением преимущественно конечных клеток, и б а з а л ь н ы й рост, происходящий за счет деления клеток, расположенных у основания нити. Нитчатая структура в эволюционном отно- шении интересна как исходная ступень для образования ряда более сложных многоклеточ- ных структур, присущих многим водорослям, которые могут быть правильно поняты только как производное нити. 6. Разнонитчатая структура (иногда ее назы- вают г е т е р о т р и х а л ь н о й). Талломы данной структуры, будучи нитчатыми в своей основе, подразделяются на две части: стелю- щуюся по субстрату, горизонтальную, и прямо стоящую, вертикальную (рис. 20). В исходном варианте горизонтальная часть состоит из вет- вящихся, обычно тесно расположенных нитей, которые или отчетливо различимы, или пол- ностью смыкаются в сплошную псевдопаренхи- матическую клеточную пластинку, нарастаю- щую по периферии и уже не имеющую следов нитчатого строения. Вертикальную часть обра- зует одна или многие, часто ветвящиеся нити, на которых обычно развиваются органы раз- множения. В результате вторичных изменений — редук- ции или чрезмерного развития одной из этих частей — такое строение может резко нару- шаться, что в некоторых случаях приводит к полной утрате характерных черт разнонит- чатого строения. Так, при тесном соединении горизонтально расположенных нитей и более или менее полном исчезновении вертикальных нитей (иногда они сохраняются только в форме волосков) слоевище приобретает форму простого однослойного диска, целиком прикрепленного к субстрату. В других случаях недоразвивается или пол- ностью выпадает горизонтальная часть слоеви- ща, тогда как вертикальные нити оказываются сильно развитыми и хорошо дифференциро- ванными. Разнонитчатая структура в том или ином выражении представлена в отделах сине-зеле- ных, зеленых, золотистых, бурых и красных водорослей, где она является или постоянной формой существования, или временным состоя- нием. 7. Пластинчатая структура характеризуется многоклеточными слоевищами в форме пласти- нок, состоящих из одного, двух или нескольких слоев клеток (рис. 21). Образование их всегда начинается с нити и происходит в результате продольного деления клеток, составляющих нить. Если возникающие продольные перего- родки располагаются строго в одном направле- нии, то пластинка сохраняется однослойной; при возникновении перегородок, параллельных первоначальной пластинке, образуется два и более клеточных слоев. Двухслойные пластин- чатые слоевища у некоторых водорослей еще на начальных стадиях их формирования превра-
Рис. 22. Сифональная ст кт а водо осли Са 1 р аи1егра (часть таллома). я структура у морской зеленой Рис. 23. Харофитная структ а: Chara. уктура: молодой экземпляр щаются в т бк ру у или мешок вследствие рас- з ются в связи хождения счоев в сере р охранении их си идающие всем сифону механическую прочность. У ~~ф~~~~~~~~ с~ру~~ура с ло иными. л. б ки становятся одно- в от елах хорошо выражена и. Трубчатые слоевища по п мере аз- д х зеленых и желто-зеленых х — водорослеи. Ю.В растания могут ветвитьс Я. п р — Предположения о путя бр п р — П х ее о разования Пластинчатые б б сих по е е р я до р спростерты по с бс ленными икнемуво ном ру-ррдр д нитчатои с~руктуры леи коккои ной ° результате усложнения ~~>~<>~>~ ур красных водорослей ых и леных, и п чного л, у рощения многоклеточного строения. ру ур~~ своиственна толь и ональная ст арофитная ст к Ю.В некоторым во о осляь только характериз ется к р уа, неклеточны р лям и нередко называе ляг мый слоеви Ф.У крупными многоклеточн 'I й п|. Отличительной чер той его явля- щами линеино-членистого с ьм ется отсутствие внутри стоящимииз гл а вн о г троени'я-" со- 7 и слоевищ клеточных в я м и и го по eга с вет- пе егородок при наличии большог го количест- и си я их в я м и и отходящими снизу р и ~зоадами д р. акие слоевища иногд дящих на нем мутовками членис довольно да достигают в ы х нистых о к о- но крупных размеров, а внешнее х по" егов, у некоторых о м 22). м~~м ыть ~ж~~ с,л~ясны~ (р с. вок на главном побеге называют з л П есново р дные водоросли сифонал части гл авного побега между узл т узлами, туры имеют ви льнои струк- д о з л и я м и ~еют вид или слабоветвящихся нитей л и я м и. Междоузлия и членики боко- различимых простым гл я нитеи, вых побегов всег а об азом, или шаровидных сильно д разованы только одной телец размером с булавочную го сильно вытянутой клеткой но женных а ую головку, снаб- ода ха а , но у многих видов р р (~~ ) р нальные п ружи о раста- представители морских п х водо ослей щ одним слоем линейно распол отличаются большим ра б полнительных клеток о н оложенных о- ол д- об разноо разием внешнего слагающих так назы- о лика и подчас очень сложным о ваемую кору. слоеви сложным расчленением Ха о щ, принимающих у нек ото р ~~итная структура настолько подобие ел оторых видов на что п т ько своео раз- деления на стебли листь У таких сложных фо м тья и ко ни. пути ее возникновения до с ть р и. ются неясными а в эв о сих по оста- о с р ста- * .. ~~орм внутри обычно обра- она является а в эволюционном отношении несомненно, тупиковой.
РАЗМНОЖЕНИЕ И ЦИКЛЫ РАЗВИТИЯ ВОДОРОСЛЕЙ Воспроизведение себе подобных у водорослей совершается посредством вегетативного, бес- полого и полового размножения. Вегетативное размножение одноклеточных во- дорослей заключается в делении особей надвое. У многоклеточных водорослей оно происходит несколькими способами, в том числе при меха- ническом разрушении слоевища на части (вол- нами, течением, в результате погрызов живот- ными) или вследствие процессов, сопровождаю- щихся распадением нитей на многоклеточные или одноклеточные части. Например, делению нитей сине-зеленых водорослей на части неред- ко предшествует отмирание отдельных клеток. Иногда для вегетативного размножения служат специальные образования. На слоевищах сфа- целярии (из бурых водорослей) вырастают поч- ки, которые опадают и прорастают в новые слоевища. Харовые водоросли образуют одно- клеточные или многоклеточные клубеньки, пе- резимовывающие и дающие новые растения. У ряда нитчатых водорослей (например, у улот- рикса из зеленых) отдельные клетки округля- ются, накапливают большое количество запас- ных питательных веществ и пигментов, одно- временно происходит утолщение их оболочки. Такие клетки называют а к и н е т а м и. Они способны переживать неблагоприятные усло- вия, когда обычные вегетативные клетки поги- бают, что ведет к разрушению нити. Подоб- ного типа акинеты есть у нитчатых сине-зеле- ных водорослей, но их подчас называют спора- ми. Некоторые красные, бурые, зеленые и харо- вые водоросли имеют стелющиеся побеги, на которых вырастают новые слоевища. Размножение частями слоевищ не всегда при- водит к возобновлению нормальных растений. Морские водоросли, растущие исключительно на твердых грунтах (камнях и скалах), неред- ко частично или полностью разрушаются под действием волн. Оторванные части или целые слоевища не способны снова закрепиться на твердых грунтах, так как этому мешает посто- янное движение воды. Кроме того, органы при- крепления вновь не образуются. Течения отно- сят такие слоевища в наиболее спокойные ме- ста, обычно с илистым или песчаным дном, где они продолжают расти, лежа на грунте. Со вре- менем более старые части отмирают и отходя- щие от них ветви превращаются в самостоятель- ные слоевища. Таким способом происходит их постоянное вегетативное размножение. При этом вследствие произрастания в спокойных местах такие водоросли сильно видоизменя- ются: ветви их становятся тоньше, уже и слабее разветвляются. В подобных случаях говорят о неприкрепленных или свободноживущих фор- мах соответствующих видов. Иногда они обра- зуют крупные скопления, например неприкреп- ленные формы красных водорослей: филлофора в Черном море, фурцеллярия в Балтийском море, анфельция в дальневосточных морях. Неприкрепленные формы донных водорослей никогда не образуют органов полового и беспо- лого размножения. Органы размножения мож- но наблюдать у них крайне редко — на тех обрывках или слоевищах, которые были отор- ваны после закладки указанных органов. В этих случаях их развитие и созревание завершаются нормально, но в последующем органы размно- жения уже не развиваются вновь. Вегетативное размножение в сущности пред- ставляет собой форму бесполого размножения, осуществляемого вегетативными частями. Бесполое размножение настоящее сопровож- дается, во-первых, делением протопласта клет- ки на части и, во-вторых, выходом продуктов деления из оболочки материнской клетки. При этом перед делением протопласта в нем проис- ходят какие-то не вполне изученные процессы физиологической перестройки, ведущие к его омоложению. Выход продуктов деления из обо- лочки материнской клетки — наиболее суще- ственное отличие настоящего бесполого размно- жения от вегетативного. Бывает, что в клетках образуется по одной споре, но, в отличие от аки- нет, они покидают оболочку материнской клетки. Бесполое размножение водорослей происхо- дит посредством спор или зооспор (спор со жгу- тиками). Они образуются или в клетках, не отличающихся по форме от других клеток, или в особых клетках, называемых с п о р а н г и я- м и, которые нередко имеют другие размеры и форму, чем вегетативные. Главное отличие спорангиев от других клеток заключается в том, что они возникают как выросты обычных кле- ток и выполняют только функцию образования спор. Отличительная особенность спор и зоо- спор — упрощенная по сравнению с обычными клетками форма и мелкие размеры. Они бы- вают шаровидными, эллипсоидными или яйце- видными, покрытыми оболочкой или без нее. Сине-зеленые водоросли, являющиеся прока- риотами, имеют два рода спор — эндоспоры и экзоспоры. Э н д о с п о р ы образуются по не- скольку в клетках в результате дробления со- держимого. Э к з о с п о р ы возникают как вы- рост протопласта на вершине клетки (только у одноклеточных представителей порядка ха- месифоновых); по мере его роста в длину появ- ляются перетяжки, отчленяющие шаровидные споры.
Образованию спор и зооспор у эукариотиче- ских водорослей как в спорангиях, так и в ве- гетативных клетках предшествует деление яд- ра. При этом в зависимости от особенностей цикла развития может происходить редукция числа хромосом (мейоз). Дочерние ядра равно- мерно распределяются в цитоплазме. Одновре- менно делятся хлоропласты и другие органел- лы, после группировки их вокруг отдельных ядер происходит деление цитоплазмы и оконча- тельное формирование спор или зооспор. У не- которых динофитовых зооспоры образуются поч- кованием на поверхности материнской клетки. У большинства эукариотических водорослей бесполое размножение осуществляется посред- ством зооспор. В одной клетке или спорангии их может быть от одной (эдогониум из зеленых) до нескольких сотен (кладофора из зеленых). Зооспоры могут иметь различное строение, что в известной мере отражает различия в строении одноклеточных водорослей, бывших предками соответствующих групп. Зооспоры бывают с од- ним, двумя, четырьмя или множеством жгути- ков; в последцем случае они располагаются венчиком на конце. У водорослей можно встретить несколько типов спор. Многие хлорококковые из зеленых и желто-зеленых имеют споры, одевающиеся оболочкой внутри материнской клетки. Такие споры называют а п л а н о с п о р а м и. При образовании особо утолщенной оболочки их именуют гипноспо рами, так как они способны длительный срок находиться в состоя- нии покоя. Гипноспоры формируются по одной в клетке, но, в отличие от акинет, оболочка материнской клетки не участвует в образова- нии их оболочки. Иногда апланоспоры сразу в материнской клетке приобретают форму, подобную ей. В таких случаях говорят об а в т о с п о р а х. Существуют еще споры, в на- звании которых отражается число их в споран- гии: т е т р а с п о р ы — образуются по 4 (мно- гие красные и диктиотовые из бурых), б и- с п о р ы — две споры в спорангии (некоторые кораллиновые из красных), м о н о с п о р ы- одна спора в спорангии (некоторые красные). Споры и зооспоры обычно выходят в воду через отверстие в стенке спорангия целой группой, окруженные слизистой оболочкой, которая вскоре расплывается. Зооспоры по выходе из спорангия, находясь еще в общей обо- лочке, начинают активно двигаться и после разрыва оболочки моментально расплываются в разные стороны. Подобным образом выходят и гаметы при половом размножении. Половое размножение заключается в слия- нии двух клеток (гамет), в результате чего об- разуется з и г о т а, вырастающая в новую особь или дающая зооспоры. У водорослей Шогамиг emepeemwg бигамия Хоньюааиия Рис. 24. Формы полового процесса у водорослей. половое размножение бывает нескольких ти- пов. В простейшем виде оно представляет сое- динение содержимого двух вегетативных кле- ток. У одноклеточных жгутиковых водорослей (некоторые вольвоксовые) половой процесс сво- дится к слиянию двух особей и называют его г о л о г а м и е й. При слиянии содержимого двух безжгутиковых вегетативных клеток по- ловой процесс называют к о н ъ ю г а ц и е й (рис. 24). Это единственная форма полового размножения в классе конъюгат из зеленых водорослей. Гораздо чаще половое размноже- ние у водорослей, в том числе у одноклеточных жгутиковых, связано с дроблением содержимо- го клеток и образованием внутри их специали- зированных половых клеток — г а м е т. У всех водорослей, кроме конъюгат и красных водорос- лей, по крайней мере мужские гаметы имеют жгутики, а у гамет противоположного пола они имеются не всегда. Образуются гаметы так же, как споры и зооспоры. Особые вместилища га- мет называют г а м е т а н г и я м и. Число га- мет в клетке или гаметангии может колебаться от одной до нескольких сотен. У примитивных водорослей гаметы формируются в вегетатив- ных клетках. В зависимости от относительных размеров гамет, участвующих в слиянии, различают сле- дующие типы полового процесса (рис. 24): 1) и з о г а м и я — гаметы одинаковой ве- личины и формы; 2) г е т е р о г а м и я, или анизогамия,— од- на гамета (женская) крупнее другой (мужской), но сходна с ней;
3) о о г а м и я — женская гамета, называе- мая яйцеклеткой, лишена жгутиков, неподвиж- на и значительно крупнее мужской, которую называют сперматозоидом или антерозоидом, она может быть бесцветной; гаметангии с яйце- клетками именуются оогониями, а гаметангии с мужскими гаметами — сперматангиями или анте ридиями; 4) а в т о г а м и я — особый тип полового процесса, распространенный у части диатомо- вых. Заключается он в том, что ядро клетки предварительно делится с мейозом на 4 ядра, два из них разрушаются, и оставшиеся два ядра сливаются, образуя вновь диплоидное ядро. Автогамия не сопровождается увеличением числа особей, а лишь их омоложением. При гетеро- и оогамии мужские и женские гаметы могут развиваться на одной особи или колонии (обоеполые, или однодом- ные, виды) или на разных (раздельно- полые, или двудомные, виды). Среди водорослей, для которых характерна изога- мия, существуют r о м о т а л л и ч н ы е виды (у них сливаются гаметы из одного слоевища или колонии) и г е т е р о т а л л и ч н ы е (слияние возможно только между гаметами из разных особей), которые ввиду отсутствия мор- фологических различий обозначаются знака- ми + и —, соответственно различают +гаме- ты и — гаметы. В результате слияния гамет образуется ша- ровидная зигота, при этом жгутики отпадают и появляется оболочка. Зиготы некоторых водорослей какое-то время сохраняют жгутики, тогда получается п л а н о з и r о т а, которая способна плавать от нескольких дней до трех недель. В зиготе происходит слияние двух ядер гамет, и она становится диплоидной. В дальней- шем зиготы разных водорослей ведут себя раз- лично. Одни зиготы вырабатывают толстую оболочку (гипнозиготы) и впадают в период покоя, длящийся до нескольких месяцев. Дру- гие зиготы прорастают без периода покоя. В одних случаях из зигот непосредственно вы- растают новые слоевища. В других зиготы де- лятся с мейозом и образованием зооспор; такие зиготы предварительно растут, и из них в зави- симости от размеров выходят 4 — 32 зооспоры. Среди водорослей наблюдаются случаи п а р- теногенетического (без оплодотво- рения) развития женских гамет. Внешне они подобны обычным зиготам, и их называют азиготами или партеноспорами. У одного и того же вида водорослей в зави- симости от времени года и внешних условий наблюдаются разные формы размножения (бес- полое и половое), при этом происходит смена ядерных фаз (гаплоидной и диплоидной). Иск- лючение составляют виды, лишенные полового процесса. Изменения, претерпеваемые особями вида между одноименными стадиями (момента- ми жизни), составляют его цикл развития. У одних видов органы бесполого и полового размножения развиваются на разных особях; тогда растения, образующие споры, называют с п о р о ф и т а м и, а растения, производя- щие гаметы,— г а м е т о ф и т а м и. У других водорослей споры и гаметы образуются на од- них и тех же растениях; одновременно у таких видов могут существовать и особи, дающие только споры, т. е. спорофиты (порфира). Сейчас растения, способные производить и споры (зоо- споры) и гаметы, как правило, называют гамето- фитами. Однако во избежание путаницы с на- стоящими гаметофитами, производящими толь- ко гаметы, их лучше называть г а м е т о с п о- р о ф и т а м и. Развитие органов размножения того или ино- го типа у гаметоспорофитов определяется тем- пературой. Например, пластинчатые слоевища одного из видов порфиры (Porphyra tenera) при температуре ниже +15, +17 'С произво- дят органы полового размножения, а при более высокой температуре — органы бесполого раз- множения. И у других водорослей гаметы обыч- но появляются при более низкой температуре, чем споры. При промежуточной температуре развитие тех или иных органов размножения на гаметоспорофитах определяется другими факторами — интенсивностью света, длиной дня, сезонными изменениями химического со- става воды или солености (для морских водо- рослей). Гаметоспорофиты существуют у уло- триксовых, ульвовых и кладофоровых из зеле- ных водорослей, у эктокарповых, хордарие- вых, сфацеляриевых и пунктариевых из бурых водорослей, бангиевых и некоторых немалие- вых из красных. Спорофиты и гаметофиты (гаметоспорофиты) бывают одинакового строения или разного, и соответственно существуют понятия изоморф- ной (сходной) и гетероморфной (разной) смены форм развития (чередования поколений). В от- ношении большинства водорослей неправильно говорить о чередовании поколений спорофитов и гаметофитов (гаметоспорофитов), так как они нередко существуют одновременно. Иногда они могут расти в несколько различных экологиче- ских условиях. Например, спорофит порфиры имеет вид ветвящихся нитей из одного ряда клеток, которые внедряются в известковый субстрат (раковины моллюсков, известковые скалы) и предпочитают слабое освещение, про- никая на большую глубину. Гаметоспорофит порфиры пластинчатый и растет вблизи уреза воды, в том числе в приливно-отливной зоне. Разница в строении спорофитов и гаметофи- тов (гаметоспорофитов) при гетероморфной сме-
не форм развития может быть очень значитель- ной. Гаметоспорофит или гаметофит может быть многоклеточным, высотой в несколько сантимет- ров, а спорофит — микроскопическим, однокле- точным (акросифония из зеленых). Возможна и обратная картина, когда гаметофит микроско- пический и даже одноклеточный, а спорофит до- стигает в длину 12 м (ламинария японская из бурых). Гаметофиты и спорофиты подавляюще- го большинства водорослей — самостоятельные растения. У ряда водорослей спорофиты растут на гаметофитах (филлофора Броди из красных) или гаметофиты развиваются внутри слоевищ спорофитов (сиклоспоровые из бурых). Поскольку при половом процессе в результа- те слияния гамет и их ядер происходит удвое- ние набора хромосом в ядре, то в песледующем в какой-то момент цикла развития наступает редукСионное деление ядра (мейоз), в резуль- тате которого дочерние ядра получают одинар- ный набор хромосом. Спорофиты многих водо- рослей диплоидные, и мейоз в цикле их разви- тия совпадает с моментом образования спор, из которых развиваются гаплоидные гаметоспо- рофиты или гаметофиты. Такой мейоз называют спорической редукцией (рис. 251). В спорофитах примитивных водорослей (кла- дофора, эктокарпус и многие другие) наряду с гаплоидными спорами могут образовываться диплоидные споры, которые снова развиваются в спорофиты. Споры, появляющиеся на гамето- спорофитах, служат для воспроизведения мате- ринских растений. Спорофиты и гаметофиты водорослей, стоящих на верхних ступенях эво- люции, строго чередуются без самовозобновле- ния (ламинариевые из бурых, многие флоридеи из красных). Ряд водорослей имеют мейоз в зиготе, т. е. зиготическую редукцию (рис. 25, 2). Она характерна для конъюгат из зеленых водорослей. ' Зиготы части пресноводных зеленых водорос- лей, таких, как вольвоксовые, улотриксовые и др., представляют собой одноклеточные спо- рофиты. Они производят до 32 зооспор, что по массе во много раз больше, чем пара родитель- ских гамет. Таким образом, у этих водорослей, по существу, наблюдается спорическая редук- ция. Некоторые группы водорослей имеют г а- м е т и ч е с к у ю р е д у к ц и ю, которая ха- рактерна для царства животных. Мейоз про- исходит при образовании гамет, остальные клетки всегда диплоидные (рис. 25, 3). Такая смена ядерных фаз присуща диатомовым и Сик- лоспоровым водорослям, а также одному из видов кладофоры (Cladophora glomerata). Инте- ресно отметить, что диатомовые преобладают по количеству видов над другими водорослями и ~%плоидные слое5ища илиих vucmu, заослары, заметь~ ~иаковы + ЯИЛЯОиднмв сл08Ащ60лцих чбсяи, 30я~~~рцдщ~~~дц, щ~щщ М. Hecmo пейоза Рис. 25. Смена ядерных фаз у водорослей: 1 — спорическая редукция (ульва); 2 — зиготическая редукция (сфероплея, лишние ядра редуцируются); 8 — гаметическая ре- дукци я (кладофора собранная); 4 — соматическая редукция (праэиола стебельчатая). встречаются во всех местообитаниях, где толь- ко способны расти водоросли. В свою очередь, циклоспоровые относятся к наиболее массо- вым морским водорослям. Видимо, цикл раз- вития с гаметической редукцией дает этим водо- рослям какие-то преимущества. У зеленой водоросли празиолы (Prasiola stipitata) обнаружена с о м а т и ч е с к а я р е д у к ц и я — мейоз совершается в вегета- тивны х клетках верхней части диплоидного гаметофита, при этом появляются участки га- плоидных клеток, в которых вслед за тем обра- зуются гаплоидные гаметы (рис. 25, 4). В цикле развития водорослей, лишенных по- лового размножения (сине-зеленые, криптофи- товые и эвгленовые) или имеющих его в редких случаях (золотистые, желто-зеленые и динофи- товые), наблюдаются только изменения в строе- нии тела. Поэтому применительно к таким во- дорослям принято говорить о ц и к л о м о р- ф о з е. Он может охватывать несколько поко- лений или ограничиваться периодом роста и развития одной особи. В наиболее резкой форме цикломорфоз выражен у гиеллы дернистой (Hyella caespitosa) из сине-зеленых водорослей и у гленодиниума Борге (Glenodinium borgei) из динофитовых. Как циклы развития, так и Сикломорфозы у водорослей отличаются большой пластич- ностью. Их прохождение во многом определя- ется экологическими условиями. Поэтому они далеко не всегда сопровождаются строго по-
) 1 ГС 4 Рис. 26. Циклы развития водорослеи: 1 — гетероморфный цикл с нерегулярной сменой форм развития; 8 — гетероморфный цикл с регулярной сменой форм развития; 8 — изоморфный цикл с регулярной сменой форм развития. С — слорофиты, Г — гаметофиты, ГС — гаметоспорофиты. следов ательным проявлением всех стадий. В зависимости от условий произрастания от- дельные стадии и формы развития могут вы- падать полностью (например, спорофит или га- метоспорофит и гаметофит) или, наоборот, су- ществовать на протяжении нескольких поко- лений, с тем чтобы на период жизни одного поколения уступить место другой форме раз- вития. Строго упорядоченные циклы развития существуют у водорослей, стоящих на верхних ступенях эволюции (рис. 26). Зачатки водорослей в виде спор, гамет и зигот не совсем стихийно разносятся водой, Они обладают различного рода таксисами, определяющими направление их движения в за- висимости от внешних раздражителей: света (фототаксис), температуры (термо- т а к с, и с), химических веществ, содержащих- ся в воде (х е м о т а к с и с). Способностью двигаться обладают не только зооспоры, но и споры, лишенные жгутиков. У них наблю- дается амебоидное движение, при котором сна- чала образуется выступ, а затем в него переме- щается содержимое всей споры. Каждый род таксиса может быть положитель- ным или отрицательным. При положительном таксисе зачатки водорослей двигаются в сто- рону усиления действующего фактора; при отрицательном таксисе — в обратном направле- нии. Характер таксиса определяется интенсив- ностью фактора и физиологическим состоянием двигающихся клеток. Слишком сильное осве- щение вызывает изменение положительного фототаксиса на отрицательный. Фототаксис зоо- спор бентосных (донных) водорослей, перво- начально положительный, со временем сменя- ется на отрицательный, что обеспечивает осе- дание их на дно. Отрицательный фототаксис наблюдается также у зигот донных водорослей. Мужские гаметы обладают хемотаксисом, позво- ляющим им двигаться в сторону неоплодотво- ренных женских гамет, выделяющих особые химические вещества. Обнаружено, что споры некоторых бентосных морских водорослей, ви- димо, путем изменения объема тела и, таким образом, удельного веса концентрируются в слоях воды с определенной температурой и соленостью. В зависимости от направления те- чения в этих слоях споры выносятся к опреде- ленным участкам побережья, где и происходит развитие слоевищ. Течения служат основным средством пере- носа зачатков на большие расстояния. Зооспо- ры сохраняют жизнеспособность в течение не- скольких дней. Более длительное перемещение зачатков водорослей происходит плодонося- щими слоевищами или их частями, которые сохраняются в живом состоянии до конца периода вегетации. Наличие жгутиков у зооспор и некоторых гамет обеспечивает их перемещение в пределах лишь нескольких метров или десятков метров. Скорость движения зооспор и гамет можно сравнить со скоростью одноклеточных орга- низмов со жгутиками — она не превышает 250 мкмlсек, или 0,9 м)ч. Такая небольшая скорость имеет значение для выбора наиболее подходящих слоев воды и места для непосред- ственного прикрепления на дне. На закрепле- ние спор бентосных водорослей оказывает влия- ние присутствие других организмов и особей или спор данного вида и их число на единицу площади. Для прорастания спор и зигот водорослей требуется комплекс условий, включающий оп- ределенные значения температуры, освещен- ности, содержания биогенных и биологически активных веществ. В противном случае они не прорастают. При этом зиготы некоторых водо- рослей, например фукусовых, не принадлежа- щие к гипнозиготам, сохраняют жизнеспособ- ность в течение трех-четырех месяцев. Воспро- изведению и сохранению некоторых водорос- лей в неблагоприятных условиях способствует образование цист. Они известны у золотистых, желто-зеленых, диатомовых и динофитовых во- дорослей. В каждой клетке формируется одна циста. Содержимое клетки округляется, и во- круг него вырабатывается твердая оболочка, содержащая кремнезем. При прорастании цист образуется одна особь, реже несколько.
ВНЕШНИЕ УСЛОВИЯ ЖИЗНИ И ЭКОЛОГИЧЕСКИЕ ГРУППИРОВКИ ВОДОРОСЛЕЙ ВНИПНИЕ УСЛОВИЯ ЖИЗНИ ВОДОРОСЛЕЙ Для водорослей как фототрофных организ- мов первым условием существования является наличие света, источников углерода и мине- ральных веществ, а основной средой жизни для них служит свободная вода. Однако это справедливо не для всех, а только для боль- шинства водорослей и не исчерпывает всего многообразия наблюдающихся у них способов питания и мест поселения. Кроме того, на жизнь водорослей оказывают большое влияние и другие химические и физи- ческие факторы внешней среды — химический состав субстрата, температура и т. д., весьма различные в разных местах. Физиологическая пластичность водорослей и их приспособлен- ность к разным экологическим условиям пои- стине огромны. В силу широкой приспособленности к внеш- ним условиям водоросли в своем расселении по земному шару распределяются в разнооб- разные экологические группи- р о в к и, характеризующиеся более или менее определенным составом слагающих их водо- рослей, приспособленных к определенной амп- литуде экологических факторов. Одним из важнейших факторов в распределе- нии водорослей является общая соленость во- ды. Основную массу природных бассейнов со- ставляют бассейны морские со средней соле- ностью 35 г солей на 1 л воды и бассейны прес- новодные, в которых содержится солей пример- но 0,01 — 0,5 г на 1 л воды. Это определяет разде- ление водорослей на морские и пресноводные, и только очень немногие из них могут жить как в одной, так и в другой воде. Так, напри- мер, почти все бурые и красные водоросли оби- тают в морях, около половины диатомовых во- дорослей живет в морской, другая половина— в пресной воде и т. д. В пределах каждого бассейна — континен- тального и морского — водоросли могут насе- лять толщу воды, свободно плавать в ней, со- ставляя группировку, называемую планкто- ном, или поселяться на дне, образуя бентос. Однако водоросли могут >HBTb HO ToJIbKo B водоемах. При наличии хотя бы периодического увлажнения многие из них успешно развива- ются на различных наземных предметах — ска- лах, коре деревьев и т. д. Вполне благоприятной средой для развития водорослей служит также толща почвенного слоя. Наконец, известны ц такие группировки, которые характеризуются резко отличными экологическими особенностя- ми, выходящими за рамки обычных условий, нормально свойственных всюду распростра- ненным водным и вневодным местообитаниям. Некоторые из них представляют большой инте- рес. Таким образом, здесь рассматриваются сле- дующие экологические группировки, или с о- о б щ е с т в а (ц е н о з ы), водорослей: 1) планктонные водоросли, 2) бентосные водоросли, 3) наземные водоросли, 4) почвенные водоросли, 5) водоросли горячих источников, 6) водоросли снега и льда, 7) водоросли соленых водоемов, 8) водоросли в известковом субстрата.
Рис. 27. Внешние приспособления к планктонному образу жизни у водорослей из разных систематиче- ских групп: 1 — 4 — шиповатые формы (1 — Mallomonas, одноклеточный жгутиконосец из золотистых водорослей с окремнелыми чешуйка- ми на оболочке, снабженными отростками; 2 — колониальная зеленая водоросль педиаструм (Pediastrum) с шипами на краевых клетках; 8 — одноклеточная зеленая водоросль голенкиния (Golenkinia) с шипами, усеивающими оболочку; 4 — одноклеточ- ная диатомея коретрон (Corethron) с тремя венчиками отростков на панцире); б — 6 — парашютные формы (5 — звездчатая колония диатомеи астерионеллы (Asterionella) со слизистыми тяжами между клетками, образующими парашют; 6 — одноклеточная диато- мея планктониелла (Planktoniella) с плоской формой панциря). плАнктонныЕ водоросли Термин п л а н к т о н (греч. «планктон»— блуждающее) впервые был введен в науку Г е и- з е н о м в 1887 г. и по первоначальному пред- ставлению означал совокупность организмов, парящих в воде. Несколько позднее в составе планктона стали различать ф и т о п л а н к- т о н (растительный планктон) и з о о- п л а н к т о н (животный планктон). Следова- тельно, фитопланктоном называют совокуп- ность свободноплавающих (в толще воды) мел- ких, преимущественно микроскопических, рас- тений, основную массу которых составляют во- доросли. Соответственно каждый отдельный организм из состава фитопланктона называют ф и т о п л а н к т е р о м. Экологи считают, что фитопланктон в жизни больших водоемов выполняет ту же роль, что и растения на суше, т. е. производит первичное органическое вещество, за счет которого прямо или косвенно (через цепь питания) существует весь остальной живой мир на суше и в воде. Это справедливо. Однако следует помнить, что в состав фитопланктона, так же как и в состав наземных растительных сообществ, входят гри- бы и бактерии, которые, за редким исключе- нием, не способны сами создавать органическое вещество. Они относятся к той же экологиче- ской группе гетеротрофных организмов, питаю- щихся за счет готового органического вещества, к которой принадлежит весь животный мир. Грибы и бактерии участвуют в разрушении мертвого органического вещества, тем самым выполняя хотя и очень важную роль в кругово- роте веществ, но принципиально иную, чем зеленые растения. Несмотря на это, основной функцией фитопланктона в целом следует все же признать создание органического вещества водорослями. Поэтому далее мы будем говорить здесь лишь о микроскопических водорослях, входящих в состав фитопланктона. Это тем бо- лее оправдано, что состав грибов в сообществе фитопланктона изучен еще очень слабо, а планктонные бактерии (бактериопланктон) в экологии водоемов рассматриваются обычно отдельно. Существование планктонных организмов во взвешенном состоянии в воде обеспечивается некоторыми специальными приспособлениями. У одних видов образуются разного рода вы- росты и придатки тела — шипы, щетинки, ро- говидные отростки, перепонки и т. п. (рис. 27); у других видов происходит накопление в теле веществ с удельным весом меньше единицы, на- пример капель жира, газовых вакуолей (у не- которых сине-зеленых водорослей, рис. 28) и т. д. Облегчается масса клетки также путем уменьшения ее размеров: размеры клеток у планктонных видов, как правило, заметно меньше, чем у близкородственных донных во- дорослей. В планктоне широко распростране-
.'0 '0 IQ В у PI~c. 28. Планктонные сине-зеленые водоросли с газовыми вакуоля и в клетках вызывающие ((цветение» воды 1 — две колонии микроцистиса (Microcystis aeruginosa), образованные бесструктурной слизью; 2 — колония воронихинии (Woroni- chinia naegeliana) с штриховатой наружной слизью; 8, 4 — афанизоменон (Aphanizomenon f los-aquae) (8 — чешуйки из нитей в натуральную величину, 4 — участки нитей при большом увеличении); 5 — собранные в клубочек нити анабены (Anabaena lemmer- mannii); 6 — плавающие отдельные нити анабены (Anabaena scheremetievii); 7, 8 — колония и отдельная нить глеотрихии (Gloeo- trichia echinulata) при разных увеличениях. Газовые вакуоли под микроскопом кажутся черными. ны и мельчайшие организмы, размером в не- сколько микрометров, образующие так назы- ваемый наннопланктон. Состав и экология отдельных представителей водорослевого фитопланктона в разных водое- мах чрезвычайно разнообразны. Фитопланктон существует в водоемах самой разной природы и самых разных размеров — от океана до ма- ленькой лужи. Его нет лишь в водоемах с рез- ко аномальным режимом, в том числе в тер- мальных (при температуре воды выше + 70, +80 'С), заморных (зараженных сероводоро- дом), чистых приледниковых водах, не содер- жащих минеральных питательных веществ. От- сутствует живой фитопланктон также в пещер- ных озерах и на больших глубинах водоемов, где недостает солнечной энергии для фотосин- теза. Общее число видов фитопланктона во всех морских и внутренних водоемах дости- гает 3000. В разных водоемах и даже в одном и том же водоеме, но в разные сезоны года число и соот- ношение видов отдельных таксономических групп весьма различны. Рассмотрим основные его комплексы по главным экологическим ка- тегориям водоемов. Морской фитопланктон состоит в основном ~из диатомовых и перидиниевых водорослей. Применение методов центрифугирования и осаждения помогло обнаружить в планктоне существенное число мел ко размерных видов, ранее неизвестных. Из диатомовых водорослей в морском фитопланктоне особенно многочис- ленны представители класса центрических диа- томей (Centrophyceae), в частности роды хето- церос (Chaetoceros), ризосоления (R hizosole- nia), талассиозира (Thalassiosira), коретрон (Corethron), планктониелла (Planktoniella) и некоторые другие (рис. 29, 1 — 6), совсем от- сутствующие в пресноводном планктоне или представленные в нем лишь небольшим числом видов. Очень разнообразен в морском фитопланк- тоне состав жтутиковых форм пирофитовых водорослей, особенно из класса перидиней (рис. 29, 7 — 10). Эта группа и в пресноводном фитопланктоне довольно разнообразна, но все же насчитывает меньшее число видов, чем в мор- ском, а некоторые роды представлены только в морях: динофизис (Dinophysis), гониаулакс (Goniaulax) и некоторые другие. Весьма много- численны в морском фитопланктоне также известковые жтутиковые — кокколитофориды, представленные в пресных водах лишь несколь- кими видами, и встречающиеся исключительно в морском планктоне кремнежгутиковые, или силикофлагеллаты (табл. 9). Характернейшей морфологической особен- ностью представителей морского фитопланкто- на является образование у них разного рода выростов: щетинок и острых шипов у диатомо- вых, воротничков, лопастей и парашютов у це
Рис. 29. Морские планктонные диатомеи (1 — 6) и перидинеи (7 — 10): 1 — хетоцерос (Chaetoceros); 2 — ризосоления (Rhizosolenia); 3 — талассиозира (Thalassiosira); 4 — дитилиум (Ditylium); 5 — эвкампия (Eucampia); 6 — биддульфия (Biddulphia); 7— динофизис (Р1порйу818); 8 — гониаулакс (Goniaulax); 9 — амфи- соления (Amphisolenia); 10 — подолампас (Podolampas). ридиней. Похожие образования встречаются и у пресноводных видов, но там они выражены значительно слабее. Например, у морских видов церациума (Ceratium) роговидные отростки не только значительно длиннее, чем у пре- сноводных, но у многих видов они еще и загну- ты. Предполагается, что подобные выросты способствуют парению соответствующих орга- низмов. По другим представлениям выросты типа шипов и роговидных образований сфор- мировались как защитное приспособление от выедания фитопланктеров рачками и другими представителями зоопланктона. Хотя морская среда на значительных про- странствах относительно однородна, монотон- ного распределения фитопланктона не наблю- дается. Неоднородность видового состава и различия в численности нередко выражены даже на сравнительно небольших акваториях морских вод, но особенно они рельефны в круп- номасштабном географическом распределении. Здесь проявляется экологическое действие ос- новных факторов среды: солености воды, темпе- ратуры, условий освещенности и содержания питательных веществ. Для морского тропического фитопланктона характерны наибольшее видовое разнообразие, в целом наименьшая продуктивность (за исклю- чением районов апвеллинга, о которых речь будет далее) и наиболее ярко выраженные мор- фологические особенности морского фитопланк- тона (упомянутые выше разного рода выросты). Здесь чрезвычайно разнообразны перидинеи, среди которых встречаются не только отдель- ные виды, но и целые роды, распространенные исключительно или преимущественно в тропиче- ских водах. Тропическая зона — оптимальный биотоп (место существования) и для известко- вых флагеллат — кокколитофорид. Здесь они наиболее разнообразны и местами развиваются в такой массе, что их известковые скелеты образуют особые донные отложения. Тропиче- ские воды по сравнению с холодными водами северных и арктических морей значительно беднее диатомовыми водорослями. Сине-зеле- ные, как и в других морских районах, представ- лены очень небольшим числом видов, и только один из них, относящийся к роду осцил- латория (Oscillatoria erythraea), в некоторых районах тропиков развивается в таком количе- стве, что вызывает «цветение» воды. В отличие от тропиков в полярных и припо- лярных морских водах в фитопланктоне господ- ствуют диатомовые. Это они создают ту огром- ную массу первичной растительной продукции, на базе которой образуются мощные скопления зоопланктона, в свою очередь служащего пищей самым крупным стадам китов в Антарктике, сельдей и китов в приполярных водах Арктики.
Перидинеи в арктических водах представле- ны значительно беднее, чем в морях умерен- ных широт и тем более тропических. Редки здесь и кокколитофориды, зато силикофлагел- латы разнообразны и местами многочисленны. Морские сине-зеленые отсутствуют, в то же время некоторые виды зеленых водорослей раз- виваются в значительном количестве. Не менее значительны различия в составе и продуктивности водорослей и в двух других больших биотопах морей, разграниченных в ши- ротном направлении — океанической и нери- товой областях, особенно если к последней от- нести все внутренние моря. Особые черты океа- нического планктона перечислены выше. Хотя они и различны в тропических и приполярных водах, но в целом отражают характерные осо- бенности морского фитопланктона. Океаниче- ский планктон, и только он, состоит исключи- тельно из таких видов, которые весь свой жиз- ненный цикл совершают в толще воды — в пела- гической зоне водоема, без связи с грунтом. В неритовом планктоне таких видов уже зна- чительно меньше, а в планктоне континенталь- ных вод они могут встречаться лишь как иск- лючение. Неритовая, или шельфовая, зона — это об- ласть моря, простирающаяся от берега до кон- ца материковой отмели, что обычно соответст- вует глубине около 200 м. В некоторых местах она узкая, в других простирается на многие сотни и даже тысячи километров. Главные экологические особенности этой зоны опреде- ляются более выраженной связью с берегом и дном. Здесь наблюдаются значительные от- клонения от океанических условий в солености воды (чаще в сторону понижения); пониженная прозрачность вследствие минеральной и орга- нической взвеси (нередко за счет более высокой продуктивности планктона); отклонения в тем- пературном режиме; более выраженное турбу- лентное перемешивание вод и, что особенно важно для растительного планктона, повышена концентрация питательных веществ. Эти особенности определяют следующие ха- рактерные черты в составе и продуктивности фитопланктона неритовой зоны: 1) многие океа- нические виды выпадают из состава этого сооб- щества, другие представлены в той или иной степени измененными формами (разновидностя- ми); 2) появляется немало специфических мор- ских видов, не встречающихся в океаническом планктоне; 3) складывается комплекс солонова- товодных видов, полностью отсутствующих в океаническом планктоне, а в сильно опрес- ненных водах некоторых внутренних морей, при солености воды ниже 10 — 12",«('/,„, про- милле — тысячная доля какого-либо числа, де- сятая доля процента), значительного разнооб- разия достигают пресноводные виды, которые при опреснении воды до 2 — Зо/„ становятся преобладающими; 4) близость дна и берегов способствует обогащению неритов ого фито- планктона временнопланктонными (меропланк- тонными) видами. Благодаря разнообразию биотопов нерито- вый фитопланктон в целом значительно богаче по видовому составу, чем океанический. В фи- топланктоне неритовой зоны умеренных ши- рот господствуют диатомовые и перидинеи, но среди них немало видов солон оватоводных, в массе развивающихся в опресненных водах внутренних морей (Балтийского, Черного, Азовского и др.). В жизненном цикле многих видов неритового планктона хорошо выражена донная фаза (стадия покоя), что в условиях умеренных широт определяет более четкую сезонную сменность (сукцессию) фитопланкто- на. В целом неритовый фитопланктон в не- сколько раз продуктивнее океанического. Фитопланктон опресненных внутренних мо- рей по составу и продуктивности существенно отличается не только от планктона океаниче- ского, но и от типичного неритового. Примером может служить фитопланктон Балтийского мо- ря. Соленость воды в верхнем слое центральной части Балтики 7 — 8/„, что примерно в 4,5— 5 раз меньше солености океана, но в 20 — 40 раз больше солености пресных вод. В заливах Риж- ском, Финском и Ботническом соленость пони- жается до 5 — 6О/„, у берегов — до 3 — 4О/„, а в устьях рек и в некоторых бухтах-лиманах (Невская губа, Куршский залив и др.) вода совсем пресная. Хотя в фитопланктоне Центральной Балтики и даже в открытой части Рижского, Финского и Ботнического заливов преобладает морской комплекс видов, в строгом смысле морским его можно называть лишь по происхождению. Ти- пичные океанические виды здесь отсутствуют полностью. Даже морской неритовый планктон здесь крайне обеднен и представлен лишь эври- галинными видами — способными переносить широкие колебания солености, хотя и предпо- читающими низкие значения ее. В этом, мор- ском по происхождению, но солоноватоводном по экологии комплексе балтийского фитопланк- тона преобладают виды диатомовых: хетоцерос талассиозира, сцелетонема (Sceletonema), акти- ноциклус (Actinocyclus). Регулярно встреча- ются, но не достигают большой численности перидинеи: гониаулакс, динофизис (Dinophysis baltica) и несколько видов силикофлагеллат. В фитопланктоне Центральной Балтики и особенно ее заливов важную роль играет комп- лекс видов пресноводного происхождения, пре- имущественно из сине-зеленых: анабена (Ana- baena), афанизоменон (Aphanizomenon), ноду-
Рис. 30. Пресноводные планктонные протококковые водоросли.' 1 — тетраэпрон (Tetraedron); 2 — шодателла (Chodatella); 8— актинаструм (Actinastrum); 4 — ооцистис (Qocystis); й — сора- струм (Sorastrum); 6 — селенаструм (Selenastrum). Рис. 31. Пресноводные планктонные диатомеи: 1 — мелозира (Melosira); 2 — стефанопискус (Stephanodiscus); 8 — табеллария (Tabellaria); 4 — фрагилария (Fragilaria). лария (Nodularia), микроцистис (Microcystis), которые летом в устойчивую солнечную погоду развиваются в такой массе, что даже в цент- ральной части моря образуют «цветение» воды (в основном за счет развития афанизоменона и нодуларии, а в южной части моря также мик- роцистиса). В пресноводном комплексе нередки и зеленые водоросли: ооцистис (Oocystis) (по всему морю), виды сценедесмуса (Scenedesmus) и педиаструма (Pe diastrum), более многочисленные в за- ливах. Пресноводный фитопланктон отличается от типично морского огромным разнообразием зе- леных и сине-зеленых водорослей. Особенно многочисленны среди зеленых одноклеточные и колониальные вольвоксовые и протококковые: виды хламидомонад (Chlamydomonas), гониума (Gonium), вольвокса (Volvox), педиаструма, сценедесмуса, ооцистиса, сфероцистиса (Sphae- rocystis) и др. (рис. 30). Среди сине-зеленых многочисленны виды анабены, микроцистиса, афанизоменона, глеотрихии (Gloeotrichia) и др. Видовое разнообразие диатомовых здесь меньше, чем в морях (если не принимать в рас- чет большое разнообразие временнопланктон- ных видов) (рис. 31); по продуктивности же на единицу поверхности воды роль диатомо- вых в пресных и морских водах в среднем со- поставима. Характернейший в морском фитопланктоне род хетоцерос (Chaetoceros) в озерах и прудах совсем отсутствует, а многочисленные в морях ризосолении (Rhizosolenia) в пресных водах представлены лишь несколькими видами. В пресноводном фитопланктоне значительно беднее качественно и количественно представ- лены перидинеи. Обычны среди них виды цера- циума (Ceratium) и перидиниума (Peridinium, рис. 64). В пресных водах отсутствуют кремне- жгутиковые и очень редки кокколитофориды, зато некоторые другие жгутиковые представ- лены здесь разнообразно и нередко в большом количестве. В основном это хризомонады- виды динобриона (Dinebryon), малломонаса (Mallomonas), уроглены (Uroglena) и др. (рис. 68, 69), а также эвгленовые — эвглена (Euglena), трахеломонас (Trachelomonas) и фа- кус (Phacus) (рис. 195, 201, 202); первые преи- мущественно в холодных водах, а вторые— в теплых. Одной из существенных особенностей прес- новодного фитопланктона является обилие в нем временнопланктонных водорослей. Ряд видов, которых принято считать типично планк- тонными, в прудах и озерах имеют донную или перифитонную (прикрепление к какому-либо предмету) фазу в своем жизненном цикле. Та- ким образом, разнообразие экологических усло- вий во внутренних водоемах определяет и зна-
чительно большее разнообразие по сравнению с морями экологических комплексов и видового состава пресноводного планктона. В крупных глубоких озерах выражены менее ярко черты отличия пресноводного фитопланк- тона от морского. В таких озерах-гигантах, как Байкал, Великие озера, Ладожское, Онежское, в фитопланктоне почти круглогодично преоб- ладают диатомовые. Здесь они, как и в морях, создают основную продукцию. Видовой состав диатомов ого озерного планктона отличен от морского, но в их экологии имеется немало общего. Например, мелозира исландская (Me- losira islandica) — массовый вид фитопланк- тона Ладожского и Онежского озер, а также мелозира байкальская (Ме1оя1га baicalensis) из Байкала в фазу покоя после весенней вспышки не опускаются на дно (или опускаются лишь частично), как это наблюдается у других прес- новодных видов в водоемах меньшего размера, а удерживаются в толще воды, образуя на не- которой глубине характерные межсезонные скопления. В больших озерах, как и в морях, велики различия в продуктивности фитопланк- тона: в центральной части водоема продуктив- ность очень низка, а у берегов, особенно в мел- ководных заливах и против устьев рек, она резко повышается. Еще больше сходства с морским в фитопланк- тоне двух самых крупных озер мира с соленой водой — Каспии и Арале. Хотя соленость во- ды в них значительно ниже морской (в Каспии 12 — 13'/„, в Арале 11 — 12",„), но в составе фитопланктона здесь преобладают водоросли морского происхождения, особенно среди диа- томовых: виды хетоцероса (Chaetoceros), ризо- солении (Rhizosolenia) и др. Из жгутиковых характерны солоноватоводные виды эксувиеллы (Exuviella) и др. В опресненных зонах этих озер господствуют пресноводные виды, однако при солености воды даже 3 — 5'/„еще весьма раз- нообразен солоноватоводный фитопланктон морского происхождения. В наиболее типичном виде пресноводный фи- топланктон как по составу и экологии, так и по продукционным свойствам представлен в озе- рах среднего размера умеренной зоны, напри- мер в озерах Балтийского бассейна. Здесь в за- висимости от типа озера и сезона года в фито- планктоне преобладают диатомовые, сине-зеле- ные или зеленые водоросли. Из диатомовых характерны виды мелозиры (Melosira), астерио- неллы (Asterionella), табеллярии (Tabellaria), фрагилярии (Fragilaria), циклотеллы (Cyclo- tella); среди сине-зеленых — виды микро- цистиса (Microcystis), анабены (Anabaena), афа- низоменона (Aphanizomenon), глеотрихии (Gloeo- trichia). Основными представителями зеленых водорослей в озерном планктоне являются пе- речисленные выше Gpo ToII;oII;KoBbIe, а в водах с очень мягкой водой, находящихся под влия- нием болот, многочисленны десмидиевые: виды космариума (Cosmarium), стаураструма (Stau- rastrum), клостериума (Closterium), эуаструма (Euastrum) и др. В мелководных озерах и пру- дах из зеленых водорослей нередко преобла- дают вольвоксовые: вольвокс (Volvox), хлами- домонада (Chlamydomonas), пандорина (Pando- rina), эвдорина (Eudorina). В фитопланктоне озер тундры и северной тайги весьма разнооб- разны х риз омонады: виды дино бриона (Dino- bryon), синуры (Synura), урогленопсиса (Uro- glenopsis), малломонаса (Mallomonas). Харак- тернейшая для морского фитопланктона группа перидиней в пресных водах представлена по- всюду (во всех водоемах), но относительно не- большим числом видов, которые везде, за ред- ким исключением, достигают невысокой чис- ленности. В самых малых водоемах — в неболь- ших озерах и прудах — весьма разнообразны и нередко многочисленны эвгленовые, особенно виды трахеломонаса (Trachelomonas), а в теп- лых водоемах тропиков и субтропиков также эвглены (Euglena), лепоцинклис (Lepocynclis), факус (Phacus) и др. В каждом отдельном водоеме в зависимости от физических и химических особенностей ре- жима и от сезона года преобладает одна или другая из перечисленных групп водорослей, а в периоды очень интенсивного развития гос- подствует нередко всего один вид. В мелких временных водоемах — лужах, выкопанных ямах — весьма обычны мелкие вольвоксовые из рода хламидомонас (Chlamy- domonas), от массового развития которых вада нередко окрашивается в зеленый цвет. В литературе часто выделяют в особую кате- горию пресноводного планктона речной фито- планктон. В больших реках с очень медленным течением, конечно, водоросли успевают размно- жаться в пределах ограниченного участка реки при относительно однородных условиях. Сле- довательно, здесь может сформироваться до некоторой степени особый для данных условий состав фитопланктона. Однако даже в этом случае исходным «материалом» для данного речного сообщества являются организмы, зане- сенные течением из выше расположенного уча- стка реки или из боковых притоков. Чаще же всего в реке состав фитопланктона формируется как смесь фитопланктона притоков, в той или иной степени преобразованная под влиянием условий реки. Преобразующая роль условий реки в форми- ровании ее фитопланктона хорошо проявля- ется, когда большая равнинная река протекает через город или мимо крупного завода, которые загрязняют воду бытовыми и промышленными
стоками. В этом случае состав фитопланктона s реке выше города характеризует чистую воду, а в черте города и сразу за его окраиной под влиянием органических загрязнений фито- планктон сильно обедняется и в нем преобла- дают так называемые сапробные виды — инди- каторы сапробных, т. е. загрязненных, вод. Однако ниже, отчасти в связи с осаждением взвешенных органических веществ, отчасти же в связи с распадом их в результате микробио- логических процессов, вода становится вновь чистой, а фитопланктон приобретает примерно тот же вид, что и выше города. На состав и распределение фитопланктона по отдельным водоемам, на его изменение в пре- делах одного водоема влияет большой комп- лекс факторов. Первостепенное значение из физических факторов имеют световой режим, температура воды, а для глубоких водоемов— вертикальная устойчивость водных масс. Из химических факторов основное значение имеют соленость воды и содержание в ней питатель- ных веществ, в первую очередь солей фосфора, азота, а для некоторых видов также железа и кремния. Рассмотрим некоторые из перечис- ленных факторов. Влияние освещенности как экологического фактора наглядно проявляется в вертикальном и сезонном распределении фитопланктона. В мо- рях и озерах фитопланктон существует лишь в верхнем слое воды. Нижняя граница его в мор- ских, более прозрачных водах находится на глубине 40 — 70 м и лишь в немногих местах достигает 100 — 120 м (Средиземное море, тро- пические воды Мирового океана). В озерных, значительно менее прозрачных водах фито- планктон существует обычно в верхних слоях, на глубине 10 — 15 м, а в водах с очень малой прозрачностью встречается на глубине до 2 — 3 м. Лишь в высокогорных и некоторых крупных озерах (например, Байкале) с проз- рачной водой фитопланктон распространен до глубины 20 — 30 м. Прозрачность воды в дан- ном случае влияет на водоросли не прямо, а косвенно, поскольку она определяет интен- сивность проникновения в водную толщу сол- нечной радиации, без которой невозможен фо- тосинтез. Это хорошо подтверждает сезонный ход развития фитопланктона в водоемах уме- ренных и высоких широт, замерзающих в зим- ний период. Зимой, когда водоем покрыт льдом, часто еще со слоем снега, несмотря на самую высокую в году прозрачность воды, фитопланк- тон почти отсутствует — встречаются лишь весьма редкие физиологически неактивные клетки некоторых видов, а у отдельных водо- рослей — споры или клетки в стадии покоя. При общей большой зависимости фитопланк- тона от освещенностй оптимальные значения последней у отдельных видов варьируют в до- вольно широких пределах. Особенно требова- тельны к этому фактору зеленые водоросли и большинство видов сине-зеленых, в значитель- ном количестве развивающихся в летний сезон. Некоторые виды сине-зеленых в массе разви- ваются только у самой поверхности воды: ос- циллатория (Oscillatoria) — в тропических мо- рях, многие виды микроцистиса (Microcystis), анабены (Anabaena) и др. — в мелких внутрен- них водоемах. Менее требовательны к условиям освещен- ности — диатомовые. Большинство из них из- бегает ярко освещенного приповерхностного слоя воды и более интенсивно развивается лишь на глубине 2 — 3 м в малопрозрачных водах озер и на глубине 10 — 15 м в прозрачных водах морей. Температура воды — важнейший фактор общего географического распределения фито- планктона и сезонных циклов его, но действует этот фактор во многих случаях не прямо, а косвенно. Многие водоросли способны пере- носить большой диапазон колебаний темпера- туры (эвритермные виды) и встречаются в планктоне разных географических широт и в разные сезоны года. Однако зона температур- ного оптимума, в пределах которого наблюда- ется наибольшая продуктивность, для каждого вида обычно ограничена небольшими отклоне- ниями температуры. Например, широко рас- пространенная в озерном планктоне умеренной зоны и субарктики диатомея мелозира исланд- ская (Ме1оз11а islandica) обычно присутствует в планктоне (например, в Онежском и Ладож-. ском озерах, в Неве) при температуре от + 1 до +13 С, максимальное же размножение ее на- блюдается при температуре от +6 до +8 'С. Температурный оптимум у разных видов не совпадает, чем и определяется смена видового состава по сезонам, так называемая сезонная сукцессия видов. Общая схема годового цикла фитопланктона в озерах умеренных широт имеет следующий вид. Зимой подо льдом (осо- бенно когда лед покрыт снегом) фитопланктон почти отсутствует в связи с недостатком сол- нечной радиации. Вегетационный цикл фито- планктона как сообщества начинается в мар- те — апреле, когда солнечной радиации доста- точно для фотосинтеза водорослей даже подо льдом. В это время бывают довольно мно- гочисленными мелкие жгутик овые — крипто- монас (Cryptomonas), хромулина (Chromulina), хризококкус (Chrysococcus) — и начинается по- вышение численности холодноводных видов диатомовых — мелозиры (Melosira), диатомы (Diatoma) и др. Во вторую фазу весны — с момента вскры- тия льда на озере до установления температур-
ной стратификации, что обычно бывает при прогреве верхнего слоя воды до +10, +12 'С, наблюдается бурное развитие холодноводного комплекса диатомовых. В первую фазу летнего сезона, при температуре воды от +10 до +15 'С, холодноводный комплекс диатомовых прекращает вегетацию, в планктоне в это вре- мя еще многочисленны диатомовые, но уже дру- гие виды — умеренно теплов одные: астерио- нелла (Asterionella), табеллария (Tabellaria). Одновременно повышается продуктивность зе- леных и сине-зеленых водорослей, а также хри- зомонад, часть видов которых достигает значи- тельного развития уже во вторую фазу весны. Во вторую фазу лета, при температуре воды выше — 15 'С, наблюдается максимум продук- тивности сине-зеленых и зеленых водорослей. В зависимости от трофического и лимнологиче- ского типа водоема в это время может наблю- даться «цветение» воды, вызванное видами сине- зеленых (Anabaena, Aphanizomenon, Micro- cystis, Gloeotrichia, Oscillatoria) и зеленых водорослей (Scenedesmus, Pediastrum, Oocystis). Диатомовые летом, как правило, занимают подчиненное положение и представлены тепло- водными видами: фрагиларией (Fragilaria) и мелозирой (Melosira granulata). Осенью, с по- нижением температуры воды до +10, +12 'С и ниже, снова наблюдается подъем продуктив- ности холодноводных видов диатомовых. Одна- ко, в отличие от весеннего сезона, в это время заметно большую роль играют сине-зеленые водоросли. В морских водах умеренных широт весенняя фаза в фитопланктоне также выделяется вспыш- кой диатомовых водорослей; летняя же — по- вышением видового разнообразия и обилия пе- ридиней при депрессии продуктивности фито- планктона в целом. Из химических факторов, влияющих на рас- пределение фитопланктона, на первое место следует поставить солевой состав воды. При этом общая концентрация солей является важ- ным фактором качественного (видового) рас- пределения по типам водоемов, а концентра- ция питательных солей, прежде всего солей азота и фосфора, — количественного распре- деления, т. е. продуктивности. Общая концентрация солей нормальных (в экологическом смысле) природных вод варь- ирует в очень широких пределах: примерно от 5 — io до 36 000 — 38 000 мг/л (от 0,005 — 0,01 до 36 — 38'/„). В этом диапазоне солености вы- деляются два основных класса водоемов: мор- ские с соленостью 36 — 38О/«, т. е. 36 000— 38 000 мг/л, и пресные с соленостью от 5 — 10 до 400 — 500 и даже до 1000 мг/л. Промежуточ- ное положение по концентрации солей занимают солоноватые воды. Этим классам вод, как было показано выше, соответствуют и основные груп- пы фитопланктона по видовому составу. Экологическое значение концентрации био- генных веществ проявляется в количественном распределении фитопланктона в целом и со- ставляющих его видов. Продуктивность, или «урожайность», микро- скопических водорослей фитопланктона, как и урожайность крупной растительности, при прочих нормальных условиях в очень большой степени зависит от концентрации питательных веществ в окружающей среде. Из минеральных питательных веществ для водорослей, как и для наземной растительности, в первую очередь необходимы соли азота и фосфора. Средняя концентрация этих веществ в большинстве естественных водоемов очень мала, и поэтому высокая продуктивность фитопланкт она, как устойчивое явление, возможна лишь при усло- вии постоянного поступления минеральных ве- ществ в верхний слой воды — в зону фотосин- теза. Правда, некоторые сине-зеленые водоросли способны еще усваивать элементарный азот из растворенного в воде воздуха, однако таких видов немного и их роль в обогащении азотом бывает существенной лишь для очень мелких водоемов, в частности на рисовых полях. Внутренние водоемы удобряются азотом и фосфором с берега, за счет приноса питатель- ных веществ речной водой с водосборной пло- щади всей речной системы. Поэтому наблюда- ется четкая зависимость продуктивности озер и мелководных внутренних морей от плодоро- дия почв и некоторых других факторов, дейст- вующих в пределах водосборной площади их бассейнов (речных систем). Наименее продук- тивен фитопланктон приледниковых озер, а так- же водоемов, расположенных на кристалличе- ских породах и в районах с большим количе- ством болот в пределах водосборной площади. Примером последних могут служить озера Се- верной Карелии, Кольского полуострова, Се- верной Финляндии, Швеции и Норвегии. На- оборот, водоемы, расположенные в пределах вы- сокоплодо родных почв, отличаются высоким уровнем продуктивности фитопланктона и дру» гих сообществ (Азовское море, нижневолжские водохранилища, Цимлянское водохранилище). Продуктивность фитопланктона зависит и от динамики воды, динамического режима вод. Влияние может быть прямым и косвенным, что, однако, не всегда легко различить. Турбу- лентное перемешивание, если оно не слишком интенсивно, при прочих благоприятных усло- виях прямо способствует повышению продук- тивности диатомовых водорослей, так как мно- гие виды этого отдела, обладая относительно тя- желой оболочкой из кремния, в спокойной'
воде опускаются на дно. Поэтому ряд массо- вых пресноводных видов, в частности из рода мелозира, интенсивно развиваются в планктоне озер умеренных широт лишь весной и осенью, в периоды активного вертикального перемеши- вания воды. При прекращении такого переме- шивания, наступающем при прогреве верхнего слоя до +10, +12 'С и образовании при этом во многих озерах температурного расслоения водной толщи, эти виды из планктона выпадают. Другие водоросли, прежде всего сине-зеле- ные, наоборот, не выносят даже относительно слабого турбулентного перемешивания воды. В противоположность диатомовым многие ви- ды сине-зеленых наиболее интенсивно разви- ваются в предельно спокойной воде. Причины высокой чувствительности их к динамике вод не вполне установлены. Однако в тех случаях, когда вертикальное перемешивание вод распространяется на боль- шую глубину, оно подавляет развитие даже относительно теневыносливых диатомовых. Свя- зано это с тем, что при глубоком перемешива- нии водоросли периодически выносятся токами воды за пределы освещенной зоны — зоны фотосинтеза. Косвенное влияние динамического фактора на продуктивность фитопланктона состоит в том, что при вертикальном перемешивании воды питательные вещества поднимаются из придонных слоев воды, где они не могут быть использованы водорослями вследствие недо- статка света. Здесь проявляется взаимодействие нескольких экологических факторов — свето- вого и динамического режимов и обеспеченности питательными веществами. Такая взаимосвязь характерна для природных процессов. Уже в начале нашего века гидробиологи открыли особое значение фитопланктона в жиз- ни водоемов как основного, а на обширных океанических просторах и единственного про- изводителя первичного органического вещества, на базе которого создается все остальное много- образие водной жизни. Это определило повы- шенный интерес к изучению не только качест- венного состава фитопланктона, но и количест- венного распределения его, а также факторов, регулирующих это распределение. Элементарный метод количественной оценки фитопланктона, который на протяжении не- скольких десятилетий был основным, да и те- перь еще не полностью отвергнут,— метод от- цеживания его из воды с помощью планктон- ных сеток. В сконцентрированной таким пу- тем пробе просчитывают количество клеток и колоний по видам и определяют общую числен- ность их на единицу поверхности водоема. Этот простой и доступный метод имеет, однако, существенный недостаток — он не полностью учитывает даже относительно крупные водо- росли, а самые мелкие (наннопланктон), ко- торые во многих водоемах значительно преоб- ладают, планктонные сетки не улавливают. В настоящее время пробы фитопланктона бе- рут в основном батометром или планктобатомет- ром, позволяющим «вырезать» монолит воды с заданной глубины. Сгущение пробы произ- водится методом осаждения в цилиндрах или фильтрацией через микрофильтры: то и другое гарантирует учет водорослей всех размеров. Когда определились огромные различия в размерах водорослей, составляющих фито- планктон (от нескольких до i000 лкл и более), стало ясно, что для сравнительной оценки про- дуктивности фитопланктона по водоемам вели- чинами численности пользоваться нельзя. Более реальным показателем для этой цели является общая биомасса фитопланктона на единицу площади водоема. Однако в дальнейшем и этот метод был забракован по двум основным при- чинам: во-первых, расчеты биомассы клеток, имеющих у разных видов разную конфигура- цию, очень трудоемки; во-вторых, вклад мел- ких, но быстро размножающихся водорослей в общую продукцию сообщества за единицу времени может быть значительно большим, чем крупных, но медленно размножающихся. Истинным показателем продуктивности фито- планктона является скорость образования им вещества за единицу времени. Для определения этой величины пользуются физиологическим методом. В процессе фотосинтеза, происходя- щем только на свету, поглощается углекислота и выделяется кислород. Наряду с фотосинте- зом происходит и дыхание водорослей. Послед- ний процесс, связанный с поглощением кисло- рода и выделением углекислоты, превалирует в темноте, когда фотосинтез прекращается. Ме- тод оценки продуктивности фитопланктона ос- нован на количественном сопоставлении ре- зультатов фотосинтеза (процесса продукции) и дыхания (процесса деструкции) сообщества по балансу кислорода в водоеме. Для этой цели используются пробы воды в светлых и темных склянках, экспонируемых в водоеме обычно на сутки на разных глубинах. Для повышения чувствительности кислород- ного метода, непригодного для малопродуктив- ных вод, стали применять изотопную (радиоуг- леродную) разновидность его. Однако впослед- ствии выявились недостатки кислородного ме- тода в целом, и в настоящее время широко при- меняют хлорофилльный метод, основанный на определении содержания хлорофилла в коли- чественной пробе фитопланктона. В настоящее время уровень продуктивности фитопланкт она многих внутренних водоемов определяется не столько природными условия-
ми, сколько общественно-экономическими, т. е. плотностью населения и характером хозяйст- венной деятельности в пределах водосборной площади водоема. Эта категория факторов, име- нуемая в экологии антропогенными, т. е. про- исходящими от деятельности человека, приводит к обеднению фитопланктона в одних водоемах, а в других, наоборот, к значительному повышению его продуктивности. Первое происходит в ре- зультате сброса в водоем токсических веществ, содержащихся в сточных водах промышленного производства, а второе — при обогащении во- доема биогенными веществами (особенно сое- динениями фосфора) в минеральной или орга- нической форме, содержащимися в больших концентрациях в водах, стекающих с сельско- хозяйственных территорий, из городов и мел- ких селений (бытовые стоки). Биогены содер- жатся и в сточных водах многих промышлен- ных производств. Второй вид антропогенного влияния — обо- гащение водоема биогенными веществами — по- вышает продуктивность не только фитопланкто- на, но и других водных сообществ, до рыб вклю- чительно, и его следовало бы рассматривать как благоприятный с экономической точки зре- ния процесс. Однако во многих случаях сти- хийное антропогенное обогащение водоемов первичными питательными веществами проис- ходит в таких масштабах, что водоем как эко- логическая система оказывается перегружен- ным биогенами. Следствием этого является чрезмерно бурное развитие фитопланктона («цветение» воды), при разложении которого выделяется сероводород или другие токсиче- ские вещества. Это приводит к гибели живот- ного населения водоема и делает воду непри- годной для питья. Нередки случаи и прижизненного выделения водорослями токсических веществ. В пресно- водных водоемах чаще всего это наблюдается при массовом развитии сине-зеленых водорос- лей, в частности видов рода микроцистис (Mic- rocystis). В морских водах отравление воды нередко вызывается массовым развитием мел- ких жгутиковых. В таких случаях вода иногда окрашивается в красный цвет, отсюда и назва- ние этого явления — «красный прилив». Понижение качества воды в результате ант- ропогенной перегрузки водоема био генными веществами, вызывающей чрезмерное развитие фитопланктона, принято называть явлением антропогенной эвтрофикации водоема. Это одно из печальных проявлений загрязнения окру- жающей среды человеком. О масштабах этого процесса можно судить по тому, что загрязне- ние интенсивно развивается в таких огромных пресных водоемах, как озеро Эри, и даже в не- которых морях. Естественное плодородие морских поверх- ностных вод определяется разными факторами. Пополнение питательными веществами мелко- водных внутренних морей, например Балтий- ского, Азовского, происходит в основном за счет приноса их речными водами. Поверхностные воды океанов обогащаются питательными веществами в районах выхода глубинных вод на поверхность. Явление это вошло в литературу под названием апвеллинга. Очень интенсивен апвеллинг у перуанского по- бережья. На базе высокой продукции фито- планктона здесь чрезвычайно высока продук- ция беспозвоночных, а за счет этого растет численность рыб. Небольшая страна, Перу в 60-х годах по уловам рыбы вышла на первое место в мире. Мощная продуктивность фитопланктона в хо- лодных водах арктических морей и особенно в водах Антарктики определяется также подъе- мом глубинных вод, обогащенных биогенными веществами. Подобное явление наблюдается и в некоторых других районах океана. Проти- воположное явление, т. е. обеднение поверхно- стных вод питательными веществами., тормо- зящее развитие фитопланктона, наблюдается в районах с устойчивой изоляцией поверхност- ных вод от глубинных. Таковы основные особенности типичного фи- то планктона. Среди сообществ мелких растений и живот- ных, населяющих толщу воды, существует комплекс организмов, обитающих только у са- мой поверхности воды — в зоне поверхностной пленки. Этому не столь значительному по ви- довому составу, но очень своеобразному сооб- ществу в 1917 г. Н а у м а н дал особое назва- ние — н е й с т о н (греч. «неин» — плавать), хотя, очевидно, оно является лишь составной частью планктона. Жизнь нейстонных организмов связана с по- верхностной пленкой воды, причем одни из них находятся над пленкой (эпинейстон), другие- под пленкой (гипонейстон). Помимо микроско- пических водорослей и бактерий здесь обитают и мелкие животные — беспозвоночные и даже личинки некоторых рыб. Большие концентрации нейстонных организ- мов вначале были обнаружены в мелких водое- мах — в прудах, выкопанных ямах, в неболь- ших заливах озер — в тихую погоду при спо- койной поверхности воды. Позднее разнообраз- ные нейстонные организмы, в основном мелкие животные, были найдены и в крупных водое- мах, в том числе в морях. В состав пресноводных водорослей нейстона входят виды разных систематических групп. Здесь обнаружен ряд представителей золоти- стых водорослей — хромулина (Chromulina),
ъ Ф Ф Ю.„.ЛЮ Рис. 32. Водоросли нейстона: 1 — чешуйка кремастохризиса (Kremastochrysis) из нескольких «парашютов» с висящими под ними клетками, плавающая на поверхности воды; 2 — конические «парашюты» кремастохлори- са (Кгемаз1осЫО~.1~) на поверхности воды с подвешенными к ним клетками. кромастохризис (Kremastochrysis); из эвгле- новых — эвглена (Euglena), трахеломонас (Tra- chelomonas), а также некоторые зеленые- хламидомонада (Chlamydomonas), кремасто- хлорис (Kremastochloris) — и мелкие прото- кокковые, отдельные виды желто-зеленых и диа- томовых водорослей. У некоторых видов нейстонных водорослей существуют характерные приспособления к су- ществованию у поверхности воды. Например, у видов наутококкуса (Xiautococcus) имеются слизистые парашюты, удерживающие их на поверхностной пленке. У кремастохризиса (рис. 32, 1) для этого служит чешуйчатый па- рашют; у одного вида из зеленых водорослей такой микроскопический парашютик выступает над пленкой поверхностного натяжения в виде конусовидного колпачка (рис. 32, 2). Преимущества существования нейстонных ор- ганизмов на границе водной и воздушной сред неясны, тем не менее в отдельных случаях они развиваются в таком количестве, что покрывают воду сплошной пленкой. Нередко и планктон- ные водоросли (особенно из сине-зеленых) в пе- риод массового развития всплывают к самой поверхности воды, образуя огромные скопле- ния. Обнаружены резко повышенные концент- рации и водных бактерий. В сообществе ней- стона довольно разнообразны и микроскопиче- ские животные, которые даже в морях, в усло- виях почти постоянно волнующейся поверх- ности, временами образуют значительные скоп- ления у нижнего края поверхности воды. БЕНТОСНЫЕ ВОДОРОСЛИ К числу бентосных (донных) водо- рослей относятся водоросли, приспособленные к существованию в прикрепленном состоянии на дне водоемов и на разнообразных предметах, живых и мертвых организмах, находящихся в воде. В зависимости от места произрастания среди бентосных водорослей различаются: 1) э п и- л и т ы, которые растут на поверхности твер- дого грунта (скалы, камни и т. д.); 2) э п и- п е л и т ы, населяющие поверхность рыхлых грунтов (песок, ил); 3) э п и ф и т ы, живущие на поверхности других водорослей; 4) э н д о- л и т ы, или сверлящие водоросли, внедряю- щиеся в известковый субстрат (скалы, ракови- ны моллюсков, панцири ракообразных); 5) э н- д о ф и т ы, которые поселяются в слоевищах других водорослей, но, в отличие от паразити- ческих видов, содержат нормальные хлоропла- сты в клетках; 6) п а р а з и т ы, живущие в слоевищах других водорослей, хлоропласты в клетках не выражены. Иногда водоросли, растущие на предметах, введенных в воду человеком (суда, плоты, буи), относят к п е р и ф и т о н у. Выделе- ние этой группы обосновывают тем, что вхо- дящие в ее состав организмы (водоросли и жи- вотные) живут на предметах, большей частью находящихся в движении или обтекаемых во- дой. Кроме того, эти организмы удалены от дна и, следовательно, находятся в условиях иного светового и температурного режимов, в других условиях поступления био генных веществ, источником которых служат донные отложения. Иногда выделение перифитона обо- сновывают еще и практическими соображе- ниями: это обрастания, которые могут причи- нять практический ущерб — уменьшать ско- рость судов, засорять водозаборные отверстия и трубопроводы., Между эпилитами, эпипелитами и эпифи- тами часто нет резкой грани, особенно это от- носится к микроскопическим бентосным водо- рослям. Правда, существуют виды, которые живут только на других водорослях, и притом только на определенном виде. Например, поли- сифония шерстистая (Polysiphonia lanosa) рас- тет исключительно на аскофиллуме узловатом (Ascophyllum nod osum) . Обычно зачатки водорослей (споры, гаме- ты, зиготы) с водой заносятся на самые разнообразные субстраты. Многие эпилиты про-
израстают в качестве эпифитов. Возможность роста их до зрелого состояния определяется размерами их слоевищ и слоевища хозяина. Когда в качестве эпифита растет крупная во- доросль, слоевище хозяина рвется под дейст- вием волн и течения. Существует определенная связь между разме- рами водорослей, размером частиц грунта, к которым они прикрепляются, и интенсив- ностью движения воды. На песке и иле способ- но расти относительно небольшое число макро- скопических водорослей. В условиях движе- ния воды в морях к этому хорошо приспособ- лены, например, некоторые виды рода халиме- да: их тонкие ризоиды цементируют песок при основании слоевища в плотную массу. Как пра- вило, чем крупнее взрослое слоевище водоросли и чем сильнее движение воды, тем больших раз- меров (большей массы) должны быть камни, на которых они растут. Иначе волны или тече- ния сносят их на большие глубины или выбра- сывают на берег. Зачатки водорослей способны закрепляться на частицах грунта любого размера. По мере их роста происходит отсев и гибель тех слое- вищ, которые растут на недостаточно крупных камнях. При росте водорослей на рыхлых грунтах, например на песке, существует опас- ность перемещения грунта придонными тече- ниями, в результате чего растения или зано- сятся грунтом, или перетираются им. Но даже в таких местах песок не остается совершенно лишенным водорослей. В неровностях песчи- нок живут микроскопические одноклеточные и колониальные виды. Обычно считается, что для роста водорослей особенно важен свет. Однако, выйдя на берег реки, озера или моря, можно заметить, что во- доросли растут далеко не всюду, где имеется достаточно прозрачная вода. Часто бывает нао- борот, бентосные водоросли неплохо растут в местах с мутной водой, но зато находящейся в движении. Как правило, места с интенсивным движе- нием воды отличаются пышным развитием бен- тосных водорослей. В морях это проливы с те- чениями, районы мысов и другие прибрежные участки, где наблюдается постоянный прибой. В реках — это камни на перекатах, где вода течет с большой скоростью. Движение воды оказывает многообразное действие на различные стороны жизни бентос- ных водорослей. Во-первых, усиливается поглощение водорослями биогенных (питатель- ных) веществ за счет того, что в местах с по- стоянным течением или волнами вода непосред- ственно около водорослей непрерывно обновля- ется с большой скоростью (от нескольких сан- тиметров до нескольких метров в секунду). Одновременно фотосинтез уси.ливается в два раза, а дыхание — на 30 — 50о~о. Иными слова- ми, бентосные водоросли, растущие в условиях движения воды, получают преимущества по сравнению с водорослями в тихих местах: один и тот же уровень фотосинтеза может быть достигнут у них при меньшей освещенности, что в итоге способствует росту более крупных слоевищ с большим содержанием белка и угле- водов. Движение воды предотвращает оседание на скалы и камни илистых частиц, которые мешают закреплению зачатков водорослей. Кроме того, движение воды благоприятствует росту бентосных водорослей тем, что с поверх- ности грунта удаляются животные, питающиеся водорослями. Наконец, хотя при сильном те- чении или сильном прибое происходит повреж- дение слоевищ водорослей или отрыв их от грунта, движение воды все же не препятствует поселению микроскопических водорослей и мик- роскопических стадий крупных водорослей. Дело в том, что из-за трения о поверхность грунта скорость движения воды по мере при- ближения к нему уменьшается и на самой по- верхности грунта существует так называемый пограничный слой воды, где течение отсутст- вует. Толщина этого слоя прямо пропорцио- нальна протяженности обтекаемого предмета (например, камня или выступа скалы) и обратно пропорциональна скорости течения. Обычно толщина пограничного слоя бывает не меньше 10 — 100 л~км, что соответствует размерам спор, гамет и зигот большинства бентосных водорос- лей. В пресных водах движение воды имеет ощу- тимое значение в реках, ручьях и горных по- токах. В этих водоемах выделяется группа бен- тосных реофильных организмов, предпочитаю- щих места с постоянным течением. В озерах не бывает сильных течений, и здесь для произ- растания бентосных водорослей приобретают значение волны. В морях волны также оказывают значитель- ное влияние на жизнь бентосных водорослей. По мере приближения к берегу глубина умень- шается, это приводит к возрастанию высоты волн, но до определенного предела. Когда глу- бина становится равной примерно 1,3 высоты волн, происходит их разрушение. Существуют три типа разрушения волн: 1) медленное раз- рушение или рассыпание (скользящий бурун) на малых уклонах дна, где на расстоянии 100 л от берега глубина составляет 1 — 5 л; 2) опро- кидывание гребня (ныряющий бурун) на боль- ших уклонах (глубина на расстоянии 100 м колеблется от 5 до 10 л); 3) опрокидывание всего переднего склона волны при таких укло- нах, где глубина на расстоянии 100 л от берега превышает 10 л. Больше всего благоприятст-
вует росту водорослей разрушение волн по пер- вому типу, которое сопровождается появлением длинных прибойных потоков. В таких усло- виях, так же как в местах с постоянными тече- ниями, водоросли достигают наиболее крупных размеров. Разрушение волн по второму и осо- бенно по третьему типу сопровождается силь- ными ударами больших масс воды о дно, что приводит к повреждению слоевищ водорослей, и в таких местах они становятся короче. Действие волн может сказываться на верти- кальном распределении водорослей. У откры- тых побережий морей и океанов, к которым подходят крупные волны с больших водных пространств, на глубине от 0 до 15 м, в зависи- мости от рельефа дна бывают зоны, лишенные макроскопических (крупных) водорослей. Ис- ключение составляют корковые известковые красные водоросли, способные противостоять сильным ударам волн. Заросли прочих водорос- лей располагаются или на малых глубинах, ку- да доходят ослабленные волны, образующиеся после разрушения более крупных, или на боль- шой глубине, если там достаточно света, где не сказывается разрушительное действие круп- ных волн. Интенсивному развитию бентосных водорос- лей способствует также умеренное (не до за- грязнения) содержание в воде биогенных веществ. В пресных водах это характерно для не- глубоких прудов и берегов озер, заводей по бе- регам рек; в морях — для мелких заливов. Источниками биогенных веществ в воде служат береговые стоки и донные отложения, особенно велика роль последних как аккумуляторов органических остатков. В донных отложениях в результате жизнедеятельности бактерий и грибов происходит минера лиза ция органиче- ских остатков; сложные органические вещества переходят в простые неорганические соедине- ния, доступные для использования фотосинте- зирующими растениями. Наиболее благоприят- ны для развития бентосных водорослей места с отложением органических остатков на не- больших глубинах с достаточным освещением, твердыми грунтами для поселения водорослей и слабым движением воды. Здесь продукты минерализации быстро вовлекаются в процес- сы фотосинтеза, а слабое движение воды содей- ствует их переносу от донных отложений в за- росли водорослей и способствует обогащению придонной воды кислородом, что препятствует появлению в ней сероводорода, губительного для живых организмов. Влияние изменений в содержании биогенных веществ в воде на рост макроскопических бен- тосных водорослей особенно заметно в тропиче- ских морях. В экваториальном поясе воды океа- нов бедны биогенными элементами, соответст- венно по открытым берегам островов такие водоросли встречаются нечасто и имеют мелкие слоевища (высотой до 10 см). В лагунах и зали- вах при наличии илистого песка на малых глу- бинах количество водорослей резко возрастает, а те виды, что встречались в открытых местах, увеличиваются в размерах в 2 — 3 раза. В тро- пическом поясе встречаются большие прост- ранства прибрежных вод, например запад- ное побережье Индии, отличающиеся повышен- ньп| содержанием биогенных веществ из-за апвеллинга — подъема глубинных вод. В та- ких районах тропических морей заросли круп- ных водорослей наблюдаются и на открытых берегах. В местах, где отсутствует движение воды и она не обогащена био генными веществами, бентосные водоросли растут плохо. Это бывает на скалистых берегах бухт с большим уклоном дна и значительными глубинами в центре, из-за чего вещества от донных отложений не выно- сятся в верхние горизонты. Макроскопические морские водоросли могут отсутствовать в от- крытых местах на отвесных стенах скал, когда глубина около них превышает полдлины волны. При таких условиях волны отражаются без разрушения, а значит, отсутствует забуруни- вание и интенсивное движение воды у поверх- ности скал. Зато на тех же скалах макроскопи- ческие водоросли образуют сплошной покров на наклонной поверхности волноприбойной ни- ши, выбитой волнами у уреза воды. Кроме света, движения воды и содержания биогенных веществ, произрастание бентосных водорослей зависит от присутствия растительно- ядных водных животных (морских ежей, брю- хоногих моллюсков, ракообразных, рыб). Это особенно заметно по зарослям ламинариевых водорослей, отличающихся среди морских водо- рослей крупными размерами. Нижняя грани- ца их произрастания нередко определяется при- сутствием морских ежей. Если уничтожать ежей и следить, чтобы они не наползали снова, то ламинариевые начинают расти глубже. У ат- лантических берегов Канады установлена зави- симость между выловом омаров и величиной площади под зарослями ламинариевых водорос- лей. Когда вылавливали много омаров, зарос- ли водорослей сокращались. Достаточно было уменьшить промысел омаров — площадь под зарослями увеличивалась. Оказывается, омары питаются морскими ежами и вылов омаров способствовал увеличению их численности. Замечено, что в тропических морях в некото- рых местах рыбы полностью выедают виды зе- леных, бурых и красных водорослей с мягкими слоевищами, в результате остаются несъедоб- ные для рыб и других животных сине-зеленые водоросли и представители других групп водо-
рослей, имеющих жесткие слоевища. Брюхо- ногие моллюски, ползая по дну, поедают мик- роскопические водоросли и мелкие проростки макроскопических видов. На произрастание определенных видов водо- рослей влияют не только свет, грунт, движение воды, ее химический состав, растительноядные животные, но и присутствие других видов водо- рослей. Лучше всего это заметно на морских видах фукусов. Обычно они растут в зоне, обна- жаемой во время отливов, глубже грунт занят другими водорослями. В Арктике, где число ви- дов водорослей уменьшается, Fucus distichus растет и глубже. То же самое происходит с Fu- cus vesiculosus и в сильно опресненном Балтий- ском море. В настоящее время в связи с усиливающимся загрязнением прибрежных вод наблюдается исчезновение крупных морских водорослей в некоторых бухтах. Но если из чистых мест туда перенести и закрепить крупные слоеви- ща водорослей, то они будут расти. Как уста- новлено, в загрязненных местах усиленно раз- виваются микроскопические бентосные водо- росли, которые быстро обрастают проростки более крупных водорослей и губят их. Немало- важную роль при этом играет то обстоятель- ство, что зачатки крупных водорослей по разме- рам часто не превосходят даже одноклеточных водорослей. Влияние бентосных водорослей друг на дру- га начинается с момента оседания и прораста- ния спор. Например, во время эксперимента зооспоры ламинариевых не прорастали, когда в сосуде находились кусочки слоевища аско- филлума. В холодных морях на бентосные водоросли оказывает влияние лед. В зависимости от его толщины, движения и торошения заросли во- дорослей могут быть уничтожены (стерты) до глубины в несколько метров. Поэтому, напри- мер, в Арктике многолетние водоросли (фу- кус, ламинария) легче всего найти у берега среди валунов и выступов скал. Многообразное влияние на жизнь бентосных водорослей оказывает температура. Наряду с другими факторами она определяет скорость роста, темп и направление развития бентосных водорослей, момент закладки у них органов размножения. Так, во многом именно темпера- тура определяет, в какой момент слоевища не- которых бурых водорослей (например, пункта- рии) из микроскопических и нитевидных разви- ваются в макроскопические и паренхиматиче- ские. При этом имеет значение температура воды в строго определенные моменты жизни водорослей. Так, органы размножения на гаме- тофитах ламинарии японской (Laminaria japo- nica) закладываются только при температуре ниже +-10 'С, причем достаточно, чтобы она продержалась в течение всего лишь одной ночи. До этого момента и после температура может быть выше (+15... +20'С). Температура воды влияет и на глубину про- израстания бентосных водорослей. При умень- шении температуры интенсивность дыхания ослабевает быстрее, чем интенсивность фотосин- теза. Это приводит к тому, что компенсацион- ная точка (момент, когда процессы фотосинте- за и дыхания уравновешивают друг друга) устана вливается при меньшей оцвещенности, т. е. на большей глубине. Поэтому бывают слу- чаи, когда в северном полушарии вид на севере ареала растет на большей глубине, чем на юге. Температура оказывает влияние на поширот- ное географическое распространение водорос- лей. В первую очередь ее действие проявляется косвенным путем — в ускорении или замедле- нии темпов роста и развития отдельных видов, что ведет к вытеснению одних водорослей дру- гими. Лишь иногда на границе распростране- ния некоторых видов наблюдаются признаки их морфологического недоразвития, связанного с непосредственным влиянием температуры. Обычно это происходит в Арктике, где общее число видов сокращается и соответственно осла- бевает конкуренция между оставшимися ви- дами бентосных водорослей. Из всего сказанного можно сделать вывод: свет — основной источник энергии для бептос- ных водорослей как фотосинтезирующих расте- ний, но степень его использования зависит от обеспеченности всех сторон обмена веществ, т. е. от температуры, содержания биогенных и биологически активных веществ, кислорода и неорганических источников углерода, а глав- ное — от темпов поступления этих веществ в слоевище, что находится в зависимости от концентрации веществ и от скорости движения воды. В значительной степени возможность произрастания бентосных водорослей в кон- кретном месте определяется биотическими фак- торами — конкуренцией со стороны других во- дорослей и численностью растительноядных животных. Все это приводит к тому, что отдель- ные виды бентосных водорослей растут далеко не на всех глубинах, где позволяет им свет, и не во всех водоемах с подходящими световым и гидрохимическим режимами. В континентальных водоемах в зависимости от приспособленности видов бентосных водо- рослей к определенным гидрохимическим, све- товым и гидрологическим условиям, от их спо- собности переносить высыхание сложились сообщества мелких стоячих водоемов (луж, прудов), крупных водоемов (озер, водохрани-
Сизигия Солнечный прилив Г Г / Луна -~Солнце Квадратура Луна Лунный прил~~ 1Лунный прил'ив ! Солнечный — -«Солнце прилив Рис. 33. Объяснение сизи~ийных и квадратурных при- ливов. лищ} и текучих вод (ручьев, рек). В глубоких водоемах, где свет не проникает до дна, водо- росли населяют прибрежную зону, называемую иногда литоралью (в морской гидробиологии этот термин имеет иной смысл). Нижняя грани- ца прибрежной зоны в континентальных водое- мах соответствует границе проникновения све- та. Рис. 34. Колебания уровня моря при приливах и рас- членение прибрежной части на зоны. Сообщества бентосных водорослей в море вы- деляются по биотопам (местообитаниям), ха- рактеризующимся иными факторами, чем в кон- тинентальных водоемах. В большинстве морей наблюдаются явления прилива и отлива, выражающиеся в периодиче- ском повышении и понижении уровня воды вследствие влияния сил притяжения Луны и Солнца. Приливы случаются одновременно на противоположных сторонах Земли, лежащих вблизи линии, направленной на Луну и Солн- це. «Горб», обращенный к небесным телам, появляется в результате прямого притяже- ния, а «горб» на противоположной стороне Зем- ли вызван наименьшим притяжением. Из-за вращения Земли в каждом пункте на берегу моря в течение суток должно наблюдаться два прилива и два отлива. Приливы не бывают всю- ду одинаковыми, так как перемещение водной массы (приливной волны) и уровень подъема воды зависят от очертаний суши, широты места, атмосферного давления, ветра и других фак- торов. Поэтому в разных местах Земли бывают приливы: полусуточные (2 прилива в сутки), су- точные (1 прилив в сутки), смешанные с преоб- ладанием по высоте полусуточных приливов, смешанные с преобладанием по высоте суточ- ного прилива. У побережья Баренцева моря, на Кольском полуострове, высота приливов достигает 4 м, в Гижигинском заливе — до 11 м. В то же время во внутренних морях, от- деленных от океана узкими проливами, прилив не заметен. Например, в Таллине он меньше 3 см. Прибрежную часть приливных морей, насе- ленную бентосными водорослями, делят на три зоны: супралиторальная, литоральная, субли- торальная. Л и т о р а л ь н а я з о н а (л и- т о р а л ь) лежит между крайними уровнями прилива и отлива. Выше расположена с у п- ралиторальная зона (супра- л и т о р а л ь). Ниже литоральной зоны нахо- дится сублиторальная (сублито- р а л ь). В бесприливных морях (Черное, Бал- тийское, Каспийское и др.) тоже выделяют три зоны, но литоралью здесь именуют прибойную зону, которую иногда более правильно назы- вают псевдолиторалью. Супралитораль бедна водорослями, так как поверхность грунта увлажняется нерегулярно, только брызгами прибоя при сильном волнении. Значительное влияние на развитие водорослей здесь оказывают атмосферные осадки и сток с суши. Поэтому в супралиторали живут в ос- новном микроскопические водоросли, приспо- собленные к большим колебаниям влажности и солености. По условиям существования бентосных водо- рослей и животных литораль делится на три
горизонта. Это связано с тем, что уровень моря во время приливов и отливов не постоянен. Два раза в месяц, в новолуние и полнолуние, Земля, Луна и Солнце оказываются примерно на одной линии; при этом происходят наиболь- шие приливы и отливы, называемые сизигий- ными. Ими определяются верхняя и нижняя границы литоральной зоны. В фазе первой и последней четвертей Луны, когда силы притя- жения Луны и Солнца направлены под прямым углом друг к другу, амплитуда приливов и от- ливов наименьшая (квадратурные приливы и отливы) (рис. 33). В соответствии с этим раз- личают: I, верхний горизонт — от верхнего уровня сизигийного прилива до верхнего уров- ня квадратурного прилива; I I, средний гори- зонт — от верхнего уровня квадратурного при- лива до нижнего уровня квадратурного отлива; нижний горизонт — от нижнего уровня квадратурного отлива до нижнего уровня си- зигийного отлива (рис. 34). Как видим, время обсыхания водорослей сокращается от верхнего горизонта к нижнему. Если верхний горизонт заливается водой полностью только в сиди- гийный прилив, то нижний горизонт, наоборот, полностью обнажается только в сизигийный от- лив. В зависимости от уклона дна и высоты прили- вов ширина литоральной зоны бывает от не- скольких метров до нескольких километров. В среднем и нижнем горизонтах литорали рас- тут уже заросли макроскопических бентосных водорослей, среди которых в морях СССР есть промысловые растения: виды фукуса, аскофил- лум узловатый (Ascophyllum nodosum). Для литоральных водорослей характерна способ- ность переносить высыхание, сильные колеба- ния солености и температуры. Сублитораль простирается от нижнего уров- ня сизигийного отлива до нижней границы произрастания водорослей. В северных морях ее нижняя граница проходит на глубине 40— 50 л; в тропических и субтропических морях при высокой прозрачности воды она может про- легать на глубине до 200 м. Сублитораль делится на два горизонта: верх- нюю сублитораль и нижнюю сублитораль. Для верхней сублиторали характерны густые за- росли макроскопических водорослей на твер- дых грунтах. Это связано с достаточным осве- щением и постоянным движением воды, вызван- ным разрушением волн у берега. Нижняя граница верхней сублиторали в морях СССР проходит на глубине 6 — 10 л, реже 25 м. Ниж- няя сублитораль характеризуется большим по- стоянством факторов среды и незначительным освещением. Здесь преобладают красные водо- росли, лучше приспособленные к фотосинтезу на больших глубинах, когда остаются в основ- ном зеленые и синие лучи; зеленых и бурых во- дорослей встречается меньше. В тропиках, однако, на больших глубинах (50 — 70 м) пре- обладают сифоновые зеленые водоросли, на этих же глубинах встречаются бурые водорос- ли, в частности виды рода ламинария. Наход- ки Еашюаг1а rodriguezii в Адриатическом море известны с глубины около 200 м. В сообществах бентосных водорослей, в ос- новном морских, когда в их состав входят ви- ды с крупными слоевищами, хорошо выражена ярусность. Иногда можно насчитать до 5 яру- сов, нижний из которых образован видами с корковидными слоевищами. Почти на всех макроскопических бентосных водорослях всегда присутствуют эпифиты. Это могут быть тоже макроскопические виды (вы- сотой 10 — 20 см), но еще больше полумакро- скопических (0,5 — 1 см) и микроскопических водорослей; последние растут как на растении- хозяине, так и на других более крупных эпифи- тах. Эпифитные водоросли могут настолько плотно обрастать слоевище хозяина, что про- исходит его отмирание и разрушение. Особенно много эпифитов развивается на водорослях с кустистыми слоевищами, растущими в местах с ослабленным движением воды и повышенной концентрацией биогенных веществ. Из-за широко распространенного у бентос- ных водорослей явления эпифитизма их за- росли можно сравнить с тропическими леса- ми. Наиболее крупные морские водоросли рас- тут в умеренных и холодных водах. Заросли их бывают настолько густыми, что при погру- жении в них аквалангисты испытывают чувство неуверенности и страха. Это настоящие под- водные джунгли. Только на суше джунгли на- ходятся в экваториальном поясе, а в морях они встречаются в субарктическом, субантарк- тическом и умеренных поясах. НАЗЕМНЫЕ ВОДОРОСЛИ Наземные, или, как их еще называют, воз- душные, водоросли окружают нас повсюду, но мы обычно не замечаем их. И это понятно, так как неспециалисту трудно представить себе, что водоросли, само название которых пред- полагает жизнь в воде, могут обильно разрас- таться вне водоемов. Однако это так, и стоит только внимательно приглядеться к окружаю- щей нас природе, как мы начнем легко узна- вать их. Позеленевшие тротуары под водосточ- ными трубами, зеленые слизистые налеты на срубах колодцев, порошковатые зеленые налеты при основании деревьев в парках — вот неко- торые самые ближайшие примеры этих водорос- лей.
Таким образом, экологическую группиров- ку наземных водорослей составляют все те фор- мы, которые обитают вне водоемов на поверх- ности различных твердых субстратов, благо- даря чему они в течение всей жизни окруже- ны воздухом (отсюда и происходит второе назва- пле этой группировки — воздушные водоросли). Эти водоросли поселяются на стволах деревьев и кустарников, на мхах, валунах и скалах, во влажном тропическом климате — на листьях деревьев и кустарников и надземных частях травянистых растений, а также на заборах, стенах домов, крышах и т. п. Условия существования водорослей на подоб- ного рода местах очень своеобразны и харак- теризуются в первую очередь частой и резкой сменой основных факторов — влажности и тем- пературы. Увлажняются эти водоросли в боль- шинстве случаев только дождем или ежеднев- пой росой, а в засушливые периоды высыхают настолько, что могут быть растерты в порошок; днем они сильно прогреваются, ночью охлаж- даются, а зимой промерзают. Правда, не все указанные местообитания в этом отношении одинаковы и, например на стенах оранжерей или на скалах, орошаемых брызгами от водо- пада, условия жизни довольно постоянны, но все же в целом наземные местообитания суще- ственно отличаются от водных. Естественно, эти группировки дают непосредственные пере- ходы, с одной стороны, к почвенным водорос- лям (постоянные поселения водорослей на по- верхности почвы), с другой стороны, к водным группировкам (поселения водорослей в лужах среди скал, в заболоченных местах и т. п.). К неблагоприятным условиям наземных ме- стообитаний приспособились сравнительно не- многие водоросли, но все же общее количество водорослей, способных вести наземный образ жизни, превышает несколько сотен. Наземные местообитания заселяют микроскопические од- ноклеточные, колониальные и нитчатые водо- росли, как правило, способные развиваться в массовом количестве в виде порошкообразных или слизистых налетов, войлокообразных масс, мягких или твердых пленок и корочек. Боль- шинство из них относится к сине-зеленым (от- дел Cyanophyta) и зеленым (отдел Chlorophyta) водорослям, значительно меньшим числом ви- дов представлены диатомовые водоросли (отдел Bacillariophyta). На коре деревьев самыми обычными поселен- цами являются такие повсеместно распростра- ненные зеленые водоросли, как одноклеточный плеврококк и нитчатая трентеполия (табл. 2, 1, 2). Первая водоросль образует порошковатые ярко-зеленые налеты преимущественно у осно- вания деревьев и пней, а вторая занимает под- час все протяжение ствола доверху, хорошо бро- саясь в глаза кирпично-красным цветом своих налетов, обусловленным скапливающимся в ее клетках оранжевым маслом. Обе они всегда занимают северную (теневую) сторону стволов. Особенно эффектны налеты трентеполии на бе- лом фоне коры берез. Эти водоросли, раз посе- лившись, уже не исчезают, а сохраняются круглый год. В неблагоприятные периоды, когда водоросли зимой промерзают, а летом временами совершенно высыхают, они, конеч- но, не живут, а находятся в состоянии анабио- за, но стоит только наступить благоприятным условиям, как водоросли вновь начинают пыш- ную вегетацию. В этих же условиях могут встретиться виды широко известной хлореллы, хлорококка (табл. 2, 8, 4), некоторых других протококковых и мелких нитчатых зеленых во- дорослей, но они редко образуют такие чистые поросли, как плеврококк и трентеполия. Менее заметны в лесу разрастания водорос- лей среди мхов — на их листьях и стеблях. Здесь широко распространена особая группи- ровка так называемых бриофильных (т. е. мо- холюбивых) диатомовых водорослей, специ- фичны также некоторые формы зеленых и жел- то-зеленых водорослей, поселяющиеся среди торфяных мхов, легко обнаруживаются темно- окрашенные, чернильно-синие поднимающиеся кверху заостренные пучки, образованные сине- зеленой водорослью шизотриксом (Schizothrix friesii, табл. 2, 6). В еще более суровых условиях находятся водоросли, поселяющиеся на поверхности об- наженных скал, но систематический состав их уже другой. Наряду с диатомовыми и некото- рыми, преимущественно одноклеточными, зе- леными здесь наиболее обычны сине-зеленые водоросли, образующие разнообразные налеты и корочки. В сухом состоянии они почти чер- ные и легко крошатся, а при увлажнении ста- новятся слизистыми и значительно светлеют. Из них наиболее часто встречаются на скалах виды колониальной глеокапсы (табл. 2, 5), отличающиеся толстыми слизистыми оболоч- ками, бесцветными или окрашенными в жел- тый, красный или фиолетовый цвет, а также виды стигонемы (табл. 2, 7), толстые ветвящиеся нити которой состоят из нескольких рядов клеток и окружены коричневыми слизистыми об вертками. Этим двум водорослям нередко сопутствуют виды толипотрикса и близкой к нему сцитонемы (табл. 2, 8, 9), некоторые ностоки (рис. 54) и ряд других. В отличие от обнаженной скальной поверх- ности значительно более благоприятные для водорослей условия создаются в мелких и мель- чайших расщелинах и трещинах скал. Многие водоросли способны заселять эти места. Здесь встречаются главным образом сине-зеленые во-
Рис. 35. Схематическое изображение поперечного среза скалы с поселением хазмолитических водорослей по трещинам породы. Рис. 36. Схематическое изображение поперечного среза скалы с поселением эндолитических водорослей в воздушных пространствах между частицами породы, закрытых сверху сплошной коркой. доросли, хотя были обнаружены и одноклеточ- ные зеленые. В расщелинах и трещинах, как бы они ни были малы, водоросли находят защиту и от резкого прямого света, и от быстрого вы- сыхания. А на скалах, подвергающихся вывет- риванию, подобного рода экологические ниши образуются в несметном количестве. Отличи- тельная особенность этих водорослевых разрас- таний заключается в том, что, как бы глубоко они ни заходили в скальную породу, они все же остаются связанными с поверхностью скалы, так как трещины имеют выход наружу (рис. 35). Поэтому и по видовому составу поверхностные наскальные водоросли и водоросли из трещин оказываются сходными, т. е. водо рослевая флора скальных трещин образуется путем про- никновения в трещины водорослей снаружи. Не менее широко распространена на скалах еще одна группировка водорослей — они рас- тут в скальных породах на некоторой глубине от поверхности и без всякой связи с нею. Одна- ко такие поселения возможны лишь на скалах, образованных известняками и песчаниками, ко- торые отличаются светлой окраской и по- ристостью. Водоросли поселяются здесь в мельчайших пространствах между частицами скальной породы. Эти пространства заполнены воздухом и отделены от поверхности скалы сплошной коркой, поры которой забиты мине- ральным веществом (рис. 36). Такая корка тол- щиной не более 3 лш пропускает воздух и воду, но непроходима для клеток водорослей; она служит им хорошей защитой от чрезмерного солнечного света. В большинстве случаев в этих условиях поселяются только виды коло- ниальной сине-зеленой водоросли глеокапсы. На поперечных разломах скальной породы по- селения глеокапсы кажутся невооруженному глазу сплошной тонкой сине-зеленой полоской шириной до 3 лл, расположенной под коркой. При рассматривании таких участков в элект- ронном сканирующем микроскопе хорошо вид- но, как колонии этой водоросли распределя- ются в воздушных камерах породы, прикреп- ляясь группами к кварцевым песчаникам или кристаллам основного породообразующего ве- щества (рис. 37). Характерная особенность этой группировки скальных водорослей — от- сутствие прямого контакта с поверхностью скалы, вследствие чего ее видовой состав от- личается от состава поверхностных разраста- ний. По-видимому, попав в глубь скальной породы, эти водоросли в дальнейшем самостоя- тельно разрастаются только по горизонтали в подкорковом слое скал. Все три типа поселения наземных водорослей на скалах получили специальные названия: на поверхности скал — э п и л и т и ч е с к и е водоросли, в трещинах скал — х а з м о л и- т и ч е с к и е водоросли, в толще скальных пород — э н д о л и т и ч е с к и е водоросли (греч. «эпи» — на, «хазмо» — трещина, «эн-
(~ф' ,л [г ,~р . ~Э~ ФЖ« « .Ф г.„' « ... 1- ё~ Ф у.,: яф. г ..ф ~ 1 4 М р». Я,. Ф к Р ф ' ф Ь~« 3'~ Ф ~д 1 ~4' Ф; « ф аd+-ú=..У= ,.ъ. ~', 3 , В. t~,«, '. .-). 4'-'--:-;-':У '» -Э .~. --.. ) « .ч«ф ) ' ' -'.Э Г 1 » !~r ° у .а4 .,=:. --.:.,-...-.:......:Г,-..-:.=:,, -:,:-.,: -.:-:: -::.-::-: .: —;-. ~г Pl ..« '~~~ Ъ' . ° , '~,~.~ ° - ' ю, ' ~ а Э.' .. ° <г ° .а ( -"-.'-"-::М. -с . «~Ъ» Рис. 37. Микрофотография в электронном сканирую- щем микроскопе поселения эндолитической водоросли глеокапсы (светлые округлые тела) в пространствах между частицами породы при увеличении в 3500 раз. до» — в, внутри, «литос» — камень). Послед- ние два местообитания создают для некоторых водорослей хоть и узкоспециализированные, но все же настолько благоприятные микроусло- вия, что хазмолитические и эндолитические во- доросли получили особенно широкое распро- странение в крайне суровых климатических условиях — в жарких каменистых пустынях обоих полушарий и в холодных пустынях высокого рий. Наконец, весьма своеобразную группу на- земных водорослевых разрастаний составляют виды, поселяющиеся в пещерах — на их стен- ках, сводах, сталактитах и сталагмитах. По мере изучения пещер, которым занимается само- стоятельная наука спел еология, таких «пе- щерных» водорослей находят все больше и боль- ше, и в настоящее время список видов, обнару- женных в этих условиях, насчитывает уже бо- лее 100 названий, относящихся почти ко всем отделам водорослей. В некоторых отношениях пещеры предоставляют водорослям весьма бла- гоприятные условия, отличающиеся к тому же постоянством,— таковы температура и степень увлажнения воздуха (во многих исследованных пещерах относительная влажность достигает 100%). Полное отсутствие света или едва за- метные следы его, казалось бы, делают пещеры крайне неподходящими для жизни водорослей, но тем не менее они там существуют. Как и -«ъ ок~~щм~~ксяух~ . ~ р - с«хаев-,уущ. у~»».зал ~рж«»«лч~». «~р«~~ ф '" у.~ "'ж'~ J Ф Жх ..' ' Р .4 ' ф М.~ "... У'.- ,ф а 'ф Ъ ' .- 'ф",. . yv 4< А ь � Ф;. 'Ф а. .4 гг О г p: ' ъ М '4 C " ф 4 "Ф ' ..а.г ,Ф,3 t. ~'". 1 '.1 C. 4 '' ~::, р'л';,( << 'f .) ~[ ~ ' ~фа «~ (ч У . " ~л"~' 4".м .4 ';,-» ~Ф ~ъ ~. а' Ф. S Ъ фр фр'е .4," .с. А ж.' ' ' - .йл'=Ил '4йс жй Кх -'ай -й;.Й 3'й.Аж . ' А» Рис. 38. Пленчатые разрастания сине-зеленых водорос- лей формидиум (Phormidium) и микроколеус (Micro- coleus) на поверхности глинистых пустынных почв; в ц е н т р е — подушка остролодочника (Oxytropis); для масштаба положен нож. в других наземных местообитаниях, здесь так- же преобладают сине-зеленые водоросли. Таким образом, можно видеть, что наземные поселения водорослей крайне разнообразны и возникают как при вполне благоприятных, так и при крайне суровых климатических ус- ловиях. Внешние и внутренние приспособле- ния наземных водорослей к их образу жизни разнообразны и сходны с тем, что обнаружи- вается у почвенных водорослей, особенно тех, которые обильно разрастаются на поверхности почвы. ПОЧВЕННЫЕ ВОДОРОСЛИ Существование водорослей в почве, на пер- вый взгляд несовместимое с основными особен- ностями этих организмов, в действительности столь же обычно, как обитание их в водоемах. На поверхности почвы нередко можно ви- деть невооруженным глазом различные разрас- тания — кожистые или войлокообразные плен- ки (рис. 38) или слизистые слоевища сине-зе- леных водорослей. Часто наблюдается также общее позеленение почвы, обусловленное мас- совым развитием микроскопических форм, рас- сеянных среди почвенных частиц. Водоросли, населяющие толщу почвы, мож- но обнаружить лишь под микроскопом. Осо-
2О Рис. 39. Наиболее обычные водоросли, обитающие в почве: и из колонии с гетероцистами и спорами; 2 — Cylindrospermum 1 — 8 — сине-зеленые водоросли (1 — Морос microscopicum, нити из кол ни о истой и спо ой 8 — Phormidium autumnale, верхние участки двух нитей, о~на из них с — — (4 — Chlamydomonas atactogama отдельная клетка в хорошо выраженным слизистым влагалище ); м). 4 — 8 — зеленые водоросли — ату тка об азование автоспор и выход их из материнсиой клетки; 6— вегетативном состоянии; б — Chlorella vulgaris, отдельная клетка, образование автоспор и вых $ИсЬососсив ЬасШаг18, короткие нити, распадающиеся на отдельные клетки); 9 — 11 — желто-зеленые ~одоросл~ magna, отдельные клетки разных размеров; 10 — Monodus acuminata, отдельная клетка и образование автоспор; ехi1~8 участок нити) 12 — 14 — диатомовые водоросли (12 — i%avicula mutica; 1 3 — Pinnularia orea is; рЫохув). бенно хорошо заметны водоросли при просмот- ре пробы почвы или почвенной суспензии в лю- минесцентном микроскопе, где хлорофиллонос- ные клетки их выделяются красным свечением (табл. 2, 10). Кроме того, их можно обнаружить с помощью культур, помещая почву в благо- приятную для роста водорослей среду и таким образом способствуя быстрому увеличению чис- ленности имеющихся в почве зачатков водорос- лей. Для выявления почвенных водорослей применяют два вида культур: водные, когда небольшое количество почвы вносят в колбу со стерильной питательной средой, и почвен- ные, когда почву помещают в чашки Петри, увлажняют и на ее поверхность кладут стериль- ные покровные стекла, на которых водоросли хорошо разрастаются. Культуры выдерживают на свету при комнатной температуре и по мере роста водорослей просматривают и определяют их состав. Общее количество обнаруженных в почве видов водорослей приближается уже к 2000. В почвах СССР к настоящему времени найдено около 1400 видов, разновидностей и форм, от- носящихся главным образом к сине-зеленым (438), зеленым (473), желто-зеленым (146) и диатомовым (324) водорослям (рис. 39). Почва как среда обитания характеризуется целым рядом экологических особенностей. Она сходна как с водными, так и с воздушными мес- тообитаниями: в почве есть воздух, но насы- щенный водяными парами, что обеспечивает дыхание атмосферным воздухом без угрозы высыхания. Как промежуточную среду почву широко используют многие организмы при пе- реходе от водного образа жизни к наземному. Для почвы характерно также относительное постоянство ее свойств. Вместе с тем почвен- ная среда крайне неоднородна в горизонталь- ном и вертикальном направлениях. В ней одно- временно содержатся твердые, жидкие и газо- образные вещества, а также разнообразные живые существа — бактерии, грибы, актиноми- цеты, представители микро- и мезофауны, вслед- ствие чего образуются микрозоны. В почве значительнее и резче колебания температуры в сравнении с водной средой, а для поверхности ее характерны неустойчивая влажность и силь- ная инсоляция (освещение солнечными луча- ми). Следовательно, для существования на по- верхности почвы водоросли должны обладать способностью переносить засуху, колебания температуры и яркий свет.
Биологические особенности почвенных водо- рослей. Жизнеспособные водоросли находят на глубине до 2 м в целинных почвах и до 2,7 м в пахотных. Почвенные водоросли отличаются изменчивостью способа питания. На неболь- ших глубинах, в пределах проникновения све- та, они, как и высшие растения, являются ти- пичными фототрофами, используя для фото- синтеза свет слабой интенсивности (,0,04 — 0,1о/о от полного света). Поэтому основную массу водорослей, как правило, обнаруживают в са- мых верхних слоях почвы: при достаточной влажности в слое от 0 до 1 и даже до 0,2 см. С глубиной как численность, так и видовое разнообразие водорослей резко падает. В це- линных почвах, не подвергающихся обработке, уже на глубине 10 — 20 сл количество водорос- лей ничтожно. По-видимому, в более глубокие горизонты водоросли заносятся с поверхности путем вмывания, а также почвенными живот- ными и корнями растений. Однако и в полной темноте они могут оставаться живыми, а в ряде случаев даже размножаться. При невозмож- ности фотосинтеза водоросли переходят на пи- тание готовыми органическими веществами. Правда, их гетеротрофный рост в темноте идет значительно медленнее, чем автотрофный рост на свету. Многие водоросли, несмотря на спо- собность к усвоению органических веществ, нуждаются в свете и в почве сохраняются лишь в покоящемся состоянии. Поэтому в глубоких слоях почвы обнаруживается сравнительно не- большое число видов, преимущественно одно- клеточные зеленые и желто-зеленые водоросли. Были предположения о существовании спе- цифических способов автотрофного питания в темноте, но пока они ничем не доказаны. Разнообразны приспособления почвенных во- дорослей к существованию при низкой влаж- ности почвы и к сохранению жизнеспособности при высыхании. В почве жизнь водорослей свя- зана с водными пленками на поверхности поч- венных частиц. В связи с этим почвенные водо- росли имеют относительно мелкие размеры в сравнении с соответствующими водными фор- мами тех же видов. С уменьшением размеров клеток возрастают их водоудерживающая спо- собность и устойчивость против засухи. У некоторых почвенных водорослей важным приспособлением к защите от засухи является обильное образование слизи — слизистых коло- ний, чехлов и обверток, состоящих из гидро- фильных полисах аридов, способных быстро поглощать и удерживать большие количества во- ды, которые в 8 — 15 раз превышают сухую мас- су водорослей. Кроме того, клеточные оболоч- ки большинства почвенных водорослей тоже способны к ослизнению и накоплению воды. Та- ким путем водоросли не только запасают воду, замедляя высыхание, но и быстро поглощают ее при увлажнении. Особенность почвенных водорослей — «эфе- мерность» их вегетации, т. е. способность бы- стро переходить из состояния покоя к актив- ной жизни и наоборот. Корочки водорослей на почве, высыхающие в сухие периоды, начина- ют расти уже через несколько часов после ув- лажнения. Есть много примеров длительного сохранения жизнеспособности водорослей. Из почв, хра- нившихся в сухом состоянии десятки лет, были выделены многие виды водорослей. Удалось оживить гер барный экземпляр сине-зеленой водоросли носток (Nostoc commune) после 107 лет хранения. Точно так же почвенные водоросли способны переносить разные колебания температуры поч- вы. В опытах многие из них оставались живы- ми при очень высокой (до +100 С) и очень низкой (до — 195 'С) температуре. Холодостой- кость водорослей подтверждается широким рас- пространением их в местообитаниях с посто- янной или длительной низкой температурой. Так, исследования показали, что наземные во- доросли — важнейшая составная часть рас- тительности Антарктиды. Эти водоросли имеют почти черный цвет, благодаря чему темпера- тура их тела оказывается выше по сравнению с температурой окружающей среды. Во многих случаях у водорослей, обитающих на поверхности почвы, появляются приспособ- ления для защиты от избыточного освещения— темные слизистые чехлы вокруг клеток. Особен- но устойчивы против ультрафиолетового облу- чения сине-зеленые водоросли. Почвенные водоросли обладают устойчиво- стью против радиоактивных излучений. Пер- выми растениями, которые появились на грун- те, разрушенном ядерным взрывом при испы- таниях в штате Невада (США), были сине-зе- леные водоросли. Благодаря перечисленным приспособлениям почвенные водоросли способны существовать даже при крайне неблагоприятных условиях среды. Этим объясняются широкое распрост- ранение почвенных и наземных водорослей и быстрота их разрастания даже при кратковре- менном появлении необходимых факторов. Поч- венные водоросли представляют большой обще- биологический интерес как организмы необык- новенной выносливости и устойчивости к край- ним условиям существования. Распространение водорослей в почвах. Ко- личество водорослей в почве очень различно в зависимости от условий среды, в особенности от водного и солевого режимов почвы, от на- земной растительности, а в окультуренных почвах — от агротехники. Так, в целинных
почвах численность всдорослей в разное время года колеблется от 0,5 до 300 тыс. клеток в 1 г почвы в слое от 0 до 10 сл и до 8 млн. клеток в 1 г в слое от 0 до 2 см. В пахотных почвах насчитывается до 1 млн. клеток в 1 г почвы в слое от О до 10 сл. Таким образом, числен- ность водорослей в почвах можно сравнивать с численностью почвенных грибов, актиноми- цетов и простейших. Характер распространения водорослей ме- няется при переходе от одной почвенно-расти- тельной зоны к другой. На участках с разре- женным растительным покровом водоросли за- нимают свободную поверхность почвы, где быстро и интенсивно разрастаются в периоды временного увлажнения и благоприятной тем- пературы. В арктической пустыне и в тундре такие пленки образованы зелеными, желто- зелеными и сине-зелеными водорослями. В тол- ще тундровых почв водоросли (главным обра- зом одноклеточные зеленые) развиваются лишь в самых верхних слоях. Под сомкнутой травянистой растительностью в хорошо сформированных почвах дерново-под- золистой и черноземной зон развиваются диф- фузные группировки почвенных водорослей, распространенные по всему корнеобитаемому слою. При наличии свободных пространств многие из этих видов выходят и на поверхность, образуя временные пленки. К луговым ассоциациям дерново-подзолистой зоны приурочены наиболее сложные группи- ровки водорослей, характеризующиеся боль- шим разнообразием видов и жизненных форм. Биомасса их достигает 300 кгlга. В луговых и ковыльных степях с густым травостоем раз- витие водорослей идет менее интенсивно, по- видимому, из-за сухости верхних слоев почвы и накопления массы степного войлока. Под лесной растительностью в подзолистых и серых лесных почвах водоросли развиваются преимущественно в верхнем слое почвы, а также в опаде. Водорослевые группировки лесных почв однообразны по всей зоне. В них преобла- дают зеленые и желто-зеленые водоросли, ко- личество которых достигает 30 — 85 тыс. клеток в 1 г, а биомасса не превышает 20 кгlга. На южных почвах с разреженным травостоем (сухие степи, полупустыни и пустыни) усили- вается развитие эфемерных поверхностных водорослевых пленок, которые сформированы преимущественно видами сине-зеленых. В од- них случаях это кожистые пленки, образован- ные нитчатыми водорослями (Microcoleus, Phor- midium, Schiz othrix), в других — слизистые, хорошо оформленные слоевища (Nostoc сот- типе с сопутствующими видами), а иногда встречаются войлокообразные скопления раз- ных видов. При окультуривании почвы группировки водорослей сильно изменяются и становятся более однородными. На пахотных почвах не- редко наблюдается интенсивное позелен ение поверхности, так называемое «цветение», обус- ловленное массовым развитием микроскопиче- ских водорослей (до 20 млн. клеток на 1 сл'). Оно происходит при сочетании благоприятных условий (тепла и влажности почвы) с наличием легкодоступной пищи. Народная примета гово- рит, что «цветение» почвы обещает богатый урожай. Рост водорослей в пахотных почвах особенно стимулируется внесением минеральных удоб- рений и известкованием, а также улучшением водного режима. Роль почвенных водорослей в почвообразо- вании и в жизни биогеоценоза. Почвенные во- доросли оказывают разнообразное влияние на жизнь биогеоценоза: на почву, населяющие ее организмы и непосредственно на высшие расте- ния. На первых этапах почвообразования водо- росли участвуют в процессе выветривания гор- ных пород и в создании первичного гумуса на чисто минеральных субстратах. За счет органического вещества, созданного водорос- лями, развиваются низшие гетеротрофные орга- низмы. Такова же роль водорослей в форми- ровании примитивных почв, лишенных расти- тельности. Например, на пустынных почвах такырах водоросли служат главным источни- ком органического вещества, образуя до 500 и даже до 1400 кг сухой массы на гектар. Раз- личные грунты, разрушенные деятельностью человека, например промышленные отвалы, тоже в первую очередь зарастают водоросля- ми. В сформированных почвах, покрытых рас- тительностью, роль водорослей еще более раз- нообразна. Основными процессами, осущест- вляемыми водорослями в почве, являются на- копление органического вещества, фиксация азота атмосферы, закрепление минеральных удобрений, действие на физические свойства почвы. Накопление органического вещества водо- рослями особенно заметно в случае массовых разрастаний их на поверхности почвы. Био- масса водорослей (выраженная в сырой массе) достигает 600 кг/га в слое от 0 до 10 сл и 1,5 т/га в поверхностных разрастаниях. Одна- ко в течение вегетационного периода эта био- масса неоднократно обновляется. Так, в дерно- во-подзолистой почве, поддерживаемой в со- стоянии чистого пара (без растений), и в почве суходольного луга при изменяющейся влаж- ности почвы продукция водорослей за месяц втрое превысила максимальную биомассу, т. е.
жение влияния сине- Рии. 40. Схематическое изо раже ак епление почвенных частиц: зеленых водорослей на закрепле "локообразных разрастаний нитчатых 1 — начальная стадия войлокоо бст ата; 2 — молодая колония ностока Ф р пО рхноСти у р рез) 8 — раз ас ание мик о о- в песок и склеив ниальнои ~~~~~~ (поперечныи леуса (М1сгосо1епв) в форме жгута, развет более мелкие жгуты и отдельные нити, скр ву (поперечный срез). ВОДОРОСЛИ ГОРЯЧИХ ИСТОЧНИКОВ Го ячие источники имеются р в азных местах оряч ито ии СССР их также немало, например на Кавказе, в и ири, л Камчатке. Связанные с под- в С едней зии, на ам температуру воды, доходящую иногда до точки ки кипения. Оказывается, и в таких П ижизненными выделениями свойства почвы. риж ° ° водорослеи, а такж е их отмирающими или ОС- и питаются многие бакте- лабленными клетками / ии и гри ы. лиз о . С истые чехлы и обвертки во- о ослей о ильно заселены различными бакте- ° p б ми и даже водорослями ,.иями, иногда гри ами . Т м образом, клетки или тал- ::,с'~ -"~ дру гих видов. аким почве являются центрами «и ломы водорослеи в почв Г~ более или менее у стойчивых микробных цено- зов — элементарных экосистем. 2 3 ная часть органического вещества 3 начительная в пи ей азличных поч- водорослей становится пищ р венных животных: простейших, щ кле ей, нема- евых че вей, энхитреид, кивсяков, дожд тод, зн ак о на энхи- личинок некоторых насекомых. а, д 320 тыс. клеток хло- т еида съедала за сутки до тр еток ностока. По-видимо- '~'~~Я~'"Э~~ о реллы или 100 тыс. клето "Ю м, почвенные жив вотные являются одним из у у . определяющих динамику важных акторов, оп численности водорослей. г~ ~Ъ.. п еж е всего е о в том что в процессе 3 является р оста водорослей происходит ио. ро поглощение легкора створимых минеральных со- кото ые постепенно освобождаются и ус- й. Поверхностные ваива ются корнями растений. о ослей могут иметь противоэрози- ный ежим поч- значение и влиять на водный режи онное зна .и опле- вы. итчатые в одо росли механически к епляя их, и склеивают тают частицы почвы, закр обильной слизью рис. п о есса говорят такие ци ры. ра Ф.Ю сь по к айней мере проц почв общая длина ма, сса водорослеи обновила об ее коли- на поверхности песчаных ° ° Schizot rex, ° ° аза за месяц. Следовательно, щ й водо,.ослей (Ногт161пт, РЬыпп1й~~) ~ж~~~~~~~ жако~ об азуемого нитеи водо лько десятков водорослями, значи 1 ' (22 — 65 л/сл') при толщине ры, которы ~ у е пол чаются при учете. ыясни- енные взаимо- лось что быстро — а несколько 7 ослей п оисходит почве ми и ко нями расте- удв воение количества водор дей ствия между водоросля ышении влажности почвы. %.Э Н е ко в прикорневои зон ру. е обна жи- при повышен нно частв уют нии. ередк во о ослей, ис- р у одоро . Многие сине-зеле- вается по ко невые выделе- в обогащении почвы азотом. н о осли являются фиксаторами атмос- поль той стороны, известны факты стиму- СССР обнаружено ния. С другои сторон е ного азота. В почвах ли ю его влияния водорослей на ля кото ых экспери- лирующ их экологических груп- � видов водорослеи для оказана азотофиксация. В целин- почвенные водоросли выделяют в ок- ментально д ах ме енной полосы и акопление пировок, поч с е немало иологич ески актив- 17 24 кг~га а ружающую р ду воп ос пока мало азота а водорослями достигает КОИ ЗОНЫ — ДО НЫХ В веществ. Однако этот вопрос п на поливных полях р т опической мов ., изучен. 90 ка~эа. Методом меченых атомов д оказано, ми азот может что фиксированный водорослями о ослями, грибами д усваиваться другими водоро высшими растениями. h оме того, водоросли стиму ру ли ют актив- ность некоторых азотфик ру щ си ю их бактерий, в частности азотобактера . у и кл беньковых ак- тер ий. во о ослей оказы- О ганическое вещество вод р рг как на почвенную микро- г,ает бо б лысое влияние ка так и на физико-химические 6 флору и фауну, так и на и
условиях могут посел они разрастаются з е б яться водо осли. р . Нередко сине-зеленого или здесь ольшими е н д рновинами или коричневого вета ° 4 щими на поверхн ц а, плаваю- ности Bo~ы или а 4 дно и сте нки водоемов. П е ель н д и выстилающими туру воды при которой е е редельную темпера- зывают разную. л осли, разные иссле ов д ватели ука- ю. ля Одних иСточник ° е жизни определяется темпе ат ° ° +75 , + 0 и даже до +84 'С. ше — до К роме высокой темп мп р ур, вода горячих мперат ы о ычно отличается так содержанием с т олеи т кже высоким ° ° ° ° лу так называем , т. е. они относят ся к чис- Водоросли об мых мине альнь р ьных источников. Ф и о ычно поселяются такж загрязненных также в сильно риками, заво горячих во ах д, спускаемых фаб- заводами и электпостан и Самыми типи чными обитателям и горячих вод (~~б~~~~~ ~ж~ их еще называют, те м ° ° ° Ф ° ° Ф зеленые. К пло в одоп ОСЛЯми ями, являются сине- ним относится наи" СтвО ви Ов аибольшее количе- дов, живущих в этих слови сящих наиболее вы условиях и выно- е высокую темпе ат окладус (Маай1аос1 Й 1 мовые, но они обычно ютятся в лодных местах по ятся в "олее хо- меньше всег по окраинам во ое оустонского национального ру о еще боль- ат а источниках амчатки, темпе- было обна ~ем вы да водорослей, из кото ых ем выше темпе ат а 17— ура источников, тем н — лены 17 — д омовым всего ви б ст видов может их н — ee с~ец~~фж~~ Есл — леным. аибол видов ыло вст стречено при +35... +45 'С рячеи воды оказались опять-та сли разбить все ro ячие и зеленые, из 28 в п ь-таки сине- ратурн р ие источники на темпе- встречены и в о г о видов кото~ых тол в д0'~гих во ько 8 были Гре ные r уппы, то реции и объединив даннь е шая часть У ые по ая часть диатомовых и зел оль- следующие ци ы' парку, получим амч зеленых о итала ы: при О атке как в теплы х, так и в холо н дах. при +45 50' ри ... 40 ' сине-зеленых во днако зв~~тны щ т астные цифры. Так, в го олее вает здесь ках Венгрии при тем в, теи и орм. Характерно, обнаружено 29 видов, едставители ~еж~~~ II n о — одноклеточные. В свою ую ри всей обширности сп из нитчатых сине- -зеленых во о осл вою очередь, обна И ти списков водоросле чих источниках п р чих источниках, все х подавляющее большинст е же дов (67аа) относитс льшинство ви- рактерн сильных водорослей ха- вых (Oscillatorial у ц ллаторие- способ оторых считают н— ущ не- р дставителеи ниже + 30 'С ванию при температ е в 7 вых ~ osto(ales~ п им , не так много. уре раз меньше (10'/) К римерно ется что б ередко оказыва- с о ~. оличество ви о сльшая часть во еляющих тот или дов, на- ния гор дорослевого населе- инои иСточник кол рячих источников сос ~~в~~ ж~~~~~е~~~~ — 15 состоит из водорослей дающего, массовог — от до83,ноп реобла- сокой те ЫХ ВОД, ЛИШЬ П ИСП р особившихся к вь- го развития в каж ом емпературе. К числ н достигает всего 2 — 5 дом из них и ши око лу наи олее типичных — видов. Схо ные око известных те мо р илов, распростра- ятся две сине-зеленые нит итчатые
водоросли, способные давать массовые раз- растания, — мастигокладус (Mastigocladus la- minosus, рис. 41) и формидиум (Phormidium laminosum, рис. 42). Наилучшего развития они достигают при температуре от +45 до +.55'С. Условия жизни термофильных водорослей отличаются рядом особенностей. Помимо того что температура воды в таких источниках вы- сока, она не подвержена здесь резким колеба- ниям и даже в зимние месяцы остается выше О С, поэтому водоросли вегетируют круглый год. Следовательно, единственное, к чему они должны быть хорошо приспособлены, это к пе- ренесению высокой температуры, что достига- ется исключительно внутренними физиологи- ческими изменениями в клетках, так как ни- какими внешними особенностями термофильные водоросли не отличаются. Имеется лишь одно Рис. 42. Сине-зеленые водоросли горячих источников.' формидиум (Phormidium laminosum), нити разнооб- разного строения и размеров. отличие от водорослей холодных вод, которое бросается в глаза, — это сравнительно малые размеры их клеток. Наконец, интересной особенностью многих термофильных водорослей является их способ- ность к выделению из воды известковых и крем- нистых отложений, о чем будет рассказано от- дельно. ВОДОРОСЛИ СНЕГА И ЛЬДА П рямую противоположность теплолюбивым (термофильным) водорослям составляет груп- пировка развивающихся на поверхности снега и льда холодолюбивых, или криофильных, во- дорослей. В этих, казалось бы, крайне неблаго- приятных условиях могут жить многие водо- росли, причем они размножаются здесь столь интенсивно, что своей массой явственно окра- шивают поверхность снега и льда. Наибольшую известность с давних пор получило явление так называемого «красного снега». Красный снег встречается во многих местах земного шара, преимущественно в высокогор- ных областях по склонам гор. В пределах СССР он представляет довольно обычное явление на Кавказе, Северном Урале, широко распростра- нен в зоне вечных снегов Камчатки, его нахо- дили также в Сибири (на Алданском хребте) и в Арктике у подошвы снеговых гор в юго-запад- ной части Новой Земли, во многих местах на Земле Франца-Иосифа, в Карском море и т. д. В зависимости от степени развития водорослей, вызывающих эту окраску, интенсивность крас- ного цвета снега может меняться от бледно- розового до кровяно-красного и темно-мали- нового. Нередко красный снег покрывает Ог- ромные пространства. Так, например, в горах Карачая, на перевале Басса на высоте около ЗООО л над уровнем моря, в 1929 г. было найде- но на снегу красное поле площадью в несколько квадратных километров. Толщина окрашен- ного слоя снега измерялась несколькими сан- тиметрами. Главным организмом, вызывающим окрас- ку снега, является один из видов хламидо- монады, названный хламидомонадой снежной (Chlamydomonas nivalis). Большую часть вре- мени эта водоросль находится в состоянии не- подвижных шаровидных клеточек, густоза- полненных красным пигментом гематохромом, но при оттаивании верхних слоев снега она начинает очень быстро размножаться, образуя неподвижные мелкие клетки и типичные под- вижные хламидоманады (рис. 43, 1 — 9). Известно много других случаев, когда водо- росли вызывают «цветение» снега. Окраска снега при этом может быть зеленой, желтои, голубой, бурой и даже черной — в зависимости
от преобладания в нем тех или иных видов снежных водорослей и других организмов. Так, например, на Кавказе в окрашенном снеге было обнаружено 55 видов водорослей, из ко- торых 18 видов относились к зеленым, 1О— к сине-зеленым, 26 — к диатомовым, а 1 вид даже к багрянкам. Все же чаще других встре- чается зеленое «цветение» снега, вызываемое различными видами зеленых водорослей. В частности, в Иеллоустонском парке в США основным организмом зеленого снега была также хламидомонада, но, в отличие от выше- описанной, лишенная красного пигмента гема- тохрома. В Карском море наблюдали зеленый снег, вызванный развитием нитчатой водорос- ли рафидонемы (Rhaphidonema nivale, рис. 43, 10). Нередко такую окраску определяют и десмидиевые водоросли. Коричневую окраску снега вызывают обычно диатомовые, но в Грен- ландии известен случай, когда она была обус- ловлена массовым развитием десмидиевой во- доросли анцилонемы (Ancylonema nordenski- oldii, рис. 43, 11), и т. д. В общей сложности в настоящее время обнаружено уже свыше 100 видов таких «снежных» водорослей. Не менее интенсивное развитие водорослей наблюдается и во льдах арктических и антарк- тических бассейнов. Это — подлинная стихия диатомей, размножающихся здесь в огромных количествах и окрашивающих лед в грязно- бурый или желто-коричневый цвет на таких больших пространствах, что в некоторых мес- тах в летнее время лишь изредка удается встре- тить чисто-белую поверхность ледяных полей. Однако такое «цветение» льда, как показали исследования, в отличие от «цветения» снега, происходит главным образом за счет массового развития водорослей не на поверхности льда, а на нижних частях его — в углублениях и на выступах, погруженных в морскую воду. В этих условиях слизистые скопления диато- мей достигали таких размеров, что с льдины размером 1О — 15 м~ снимали до 1О — 15 жг сы- рой биомассы их. Интенсивное развитие диатомей продолжа- ется в Арктике в течение всего светлого пери- ода, а с наступлением зимы, когда лед снизу начинает нарастать, водоросли, естественно, вмерзают в его толщу. Далее, по мере летнего стаивания льда с поверхности вмерзшие диа- томеи вместе с детритом выходят на поверхность льдов, где и дают те бурые пленки, которые так часто можно наблюдать на льдах полярных бассейнов. Однако здесь, в лужах опресненной воды, водоросли уже не могут размножаться и постепенно отмирают. И все же эти темные пленки имеют важное значение: они, как и все темные предметы, поглощают больше тепловых лучей, чем окружающая их белая поверхность, 10 Рис. 43. Водоросли, вызывающие «цветение» снега.' 1 — 9 — хламидомонада снежная (Chlamydomonas nivalis). вы- зывающая красную окраску снега; 10 — рафидонема снежная (Rhaphidonema nivale), вызывающая зеленую окраску снега; 11 — анцилонема Норденшельда (Ancylonema nord|..nskioeldii), вызывающая коричневую окраску снега и льда. лед под ними тает быстрее, и в итоге образуются глубокие ямки с толстым слоем диатомей на дне. Ямки могут протаивать и до конца, пре- вращаясь в каналы, насквозь пронизывающие лед. «Ледовых» диатомей в арктических и антарк- тических морях обнаружено уже около 8О видов. Все эти водоросли приспособлены к жизни в крайне неблагоприятных условиях низких температур. Находясь в поверхностных слоях снега и льда, они подвергаются очень сильному охлаждению в зимние стужи, когда темпера- тура воздуха опускается на несколько десятков градусов ниже нуля, а в летнее время живут и размножаются в талой воде, т. е. при темпера- туре около О 'С. И если снежная хламидомонада имеет стадию покоя в виде округлых толсто- стенных клеток, то многие другие водоросли, в том числе и диатомеи, лишены каких-либо специальных приспособлений для перенесения столь низких температур. Водоросли снега и льда составляют подавля- ющее большинство организмов, поселяющихся на замерзших субстратах. Такие субстраты получили общее название криобиотопов (греч. «криос» — холод, «топос» — место), их поселен- цы — криобионтов, а отрасль науки, занимаю- щаяся изучением этих организмов, — криобио- логии. По современным данным, общее число водорослей, обнаруженных на криобиотопах во всем мире, достигает 350, но истинных крио-
бионтов, могущих активно вегетировать только в этих условиях, значительно меньше — не- многим более 100 видов. Из них подавляющее большинство относится к зеленым водорослям (около 100 видов); несколькими видами пред- ставлены сине-зеленые, желто-зеленые, золо- тистые, пирофитовые и диатомовые водоросли. Все же остальные найденные здесь водоросли расцениваются либо как просто холодолюбивые формы (среди них наиболее многочисленны диатомовые и сине-зеленые), либо как слу- чайные спутники, попавшие на снег и лед из соседних биотонов. ВОДОРОСЛИ СОЛЕНЫХ ВОДОЕМОВ К числу факторов, создающих особые усло- вия для жизни водорослей, относится также повышенное содержание в воде солей, свойст- венное некоторым связанным с морем и конти- нентальным водоемам. По составу населяющих их водорослей особенно интересны последние. Соленые континентальные водоемы, в част- ности озера, весьма многочисленны на террито- рии СССР. Только в пределах Астраханской области насчитывалось до 700 соленых озер. Происхождение их вполне понятно. Любое озеро, если только оно не проточное, замкнутое, т. е. не имеет стока, рано или поздно должно стать соленым благодаря испарению воды, особенно если оно расположено в засушливых районах и на сильно соленосных породах. Степень соле- ности таких водоемов может быть весьма зна- чительной, вплоть до выпадения солей из на- сыщенного раствора. Так, например, в озерах с преобладанием поваренной соли содержание солей доходит до 285 г на литр воды, а при преобладании глауберовой соли — до 347 г на литр воды. И все-таки даже в таких условиях живут водоросли. Правда, количество видов водорослей по мере увеличения солености убы- вает, и очень высокую соленость переносят только немногие из них, но в целом солевыно- сливых форм немало. Из зеленых водорослей в водоемах с большой концентрацией солей (до 285 г на литр) широко распространена и чрезвычайно характерна дю- налиелла, получившая соответствующее ви- довое название «солевая» (Dunaliella salina, табл. 29, 1 — о). Это — микроскопическая одно- клеточная подвижная водоросль из порядка вольвоксовых. По строению дюналиелла близ- ка к хламидомонадам, но отличается от них большой примитивностью. Тело дюналиеллы грушевидной или яйцевидной формы, заост- ренное на переднем конце, где располагаются два жгута. Явственной, отделимой от прото- пласта оболочки нет — только наружная уп- лотненная пленка. Содержимое клетки такое же, как у хламидомонад', кроме того, имеется еще красный пигмент гематохром, маскирующий зеленый цвет хлоропласта. При массовом раз- множении, когда клетки дюналиеллы отми- рают, ее пигменты сообщают солевому рас- твору (ране) и выпадающей из него соли в пере- солоненных водоемах характерную окраску- от розовой до красной. Эта своеобразная окрас- ка рапы и соли в соленых озерах давно обраща- ла на себя внимание исследователей. Опреде- лить ее причину долго не могли, но в настоя- щее время можно считать твердо установлен- ным, что ее вызывает дюналиелла. Интересно, что эта водоросль придает рапе также харак- терный запах фиалок. Из сине-зеленых водорослей большой инте- рес представляет хлор оглея сарциноидная (Chlorogloea sarcinoides, рис. 44), в огромном количестве развивающаяся в некоторых соле- ных озерах с высокой концентрацией солей, в частности в Мойнакском озере у Евпатории. Здесь она разрастается на подводной гряде известняков, образуя сплошной слой толщиной до 2 см. По мере нарастания сверху отдельные участки этого слоя отрываются волнами и пере- гоняются ветром по всему озеру. При этом они продолжают расти, а затем волны выбрасывают их на берег, образуя мощные подводные и береговые валы синевато-зеленого цвета. Эти залежи, состоящие из массы слизистых крупи- нок разной величины, местное население назы- вает «кашкой». При рассматривании под микро- скопом крупинки хлороглеи оказываются ко- лониями своеобразного строения, состоящими как бы из множества слизистых пакетов (сар- цин), содержащих многочисленные правильно расположенные клетки. Приспособившись к столь необычным усло- виям существования, эти водоросли играют очень большую роль в жизни соленых водое- мов. Сочетание органической массы, образуе- мой водорослями, и большого количества раст- воренных в воде солей обусловливает ряд свое- образных биохимических процессов, свойствен- ных этим водоемам. В частности, хлороглея и ряд других водорослей, тоже размножающихся в массовом количестве, участвуют в некоторых озерах (например, в Мойнакском) в процессе образования лечебных грязей. СВЕРЛЯЩИЕ И ТУФООБРАЗУЮЩИЕ ВОДОРОСЛИ Отдельного рассмотрения заслуживают чрез- вычайно интересные и своеобразные водоросли, обладающие способностью внедряться в суб- страт или отлагать его вокруг себя. И в том и в другом случае жизнь этих водорослей свя-
зана с известью. Они встречаются как в суб- стратах, погруженных в воду, т. е. относятся собственно к бентосу, так и вне воды, вклю- чаясь тем самым в группировку наземных водо- рослей, но в обоих случаях отличаются свое- образным (<активным» отношением к субст- рату. Водоросли, внедряющиеся внутрь известко- вого субстрата, получили название (<сверля- щих». Сверлящие водоросли по количеству ви- дов немногочисленны. Так, например, в прес- ных и соленых водоемах СССР их обнаружено всего 18, из них 12 зеленых и 6 сине-зеленых (рис. 45, 1, 2). Однако распространены они чрезвычайно широко, начиная с многочислен- ных на земном шаре известковых скал и кончая камнями, известковыми раковинами многочис- ленных животных, кораллами, пропитанными известью крупными водорослями и т. д., в прес- ных и морских водах, у поверхности воды и на глубине свыше 2О м, от холодных морей се- вера до вечно теплых морей тропиков. Все сверлящие водоросли — микроскопиче- ские организмы. Основная их особенность состоит в том, что, поселившись сначала на поверхности известкового субстрата, они по- степенно внедряются в глубь его, где и разрас- таются. Глубина проникновения их может быть весьма значительной, до 10 лш и более. Что же позволяет им туда проникать? В процессе жизнедеятельности сверлящие во- доросли выделяют органические кислоты, кото- рые растворяют находящуюся под ними известь. Сначала получается небольшая ямка, которая постепенно все больше и больше углубляется, до тех пор пока водоросль целиком не погружается в субстрат. Однако процесс на этом не прекра- щается, и водоросль проникает все дальше в глубь субстрата. В результате некоторый слой известковой породы (а тонкие раковины живот- ных часто и насквозь) оказывается пронизан- ным многочисленными каналами. Иными сло- вами, сверлящие водоросли разрушают извест- ковый субстрат, в котором они поселяются. Прямо противоположный процесс — процесс созидания известковых пород — осуществля- ют водоросли, способные выделять известь. Они встречаются в воде и в наземных местооби- таниях, в морях и пресных водоемах, в холод- ных и горячих водах (рис. 45, 8, 4). Количество выделяемой водорослями извести различно. Некоторые формы выделяют очень небольшое количество углекислого кальция, который в виде мелких кристаллов располага- ется между особями или образует футляры вокруг клеток и нитей. Другие водоросли вы- деляют известь настолько обильно, что оказы- ваются совершенно погруженными, как бы замкнутыми в нее, и тогда они отмирают, сохра- оо оо оо 0 ор oo Оо~о e +o ~(ОФ фо ОО О Р~~ Оф ООODQD р ро о о4� )+4P4 +DDD~O� ОО О О ° ° go 4 оь оо o Doo~oy оооо + 4 о D~oo ооооО о о ео о о 44 0 Ю 0Р{РО� О О еО ОO ь ро о ° 4 О0 0 ОООЬ0~4 о оо о 0� оор ОО ф 0� Яфьь оь о ооо. «s~о о Оь 0 о 0 oooîî 004 0 О 00 О О 44 00 р оо 0РО 00 +0О о 000 0 О yd 0О О 00 40 Рис. 44. Сине-зеленая водоросль соленых водоемов- хлороглея сарциноидная (Chlorogloea sarcinoides): 1 — общий вид колонии при небольшом увеличении; 2 — учас- ток колонии с характерным расположением клеток. Рис. 45. Сверлящие (1, 2) и туфообразующие (8, 4) сине-зеленые водоросли: 1 — гиелла (Hyell.à caespitosa), отдельная нить; 2 — мастигоко- леус (Mastigocoleus testarum), две нити; 8, 4 — ривулярия (Rivu- laria haematites) (8 — поперечный разрез при небольшом увели- чении через молодую известковую колонию, образовавщую три C',JIP H, N3 HOTOPbIx с RcHBbIMN HHTHMN To JIh80 верхний; 4 — HMx& из верхнего слоя колонии).
Са(НСО ), = СО, + Н,О+ САСО, нарбонат кальция (нерастворимый) бикарбонат кальция (растворимый) СОЖИТЕЛЬСТВО ВОДОРОСЛЕЙ С ДРУГИМИ ОРГАНИЗМАМИ няясь живыми дишь в самых поверхностных слоях тех подчас очень мощных отложений, ко- торые они образуют. Так, например, происхо- дит образование известковых туфов в горячих источниках. Источником извести для водорослей может быть только окружающая их вода. В пресной воде наряду с другими солями всегда имеется некоторое количество растворимого бикарбо- ната кальция, особенно много его в так называе- мых жестких водах. Водоросли используют би- карбонат кальция в качестве источника угле- рода, отщепляя от него углекислоту, в резуль- тате чего выпадает нерастворимый карб онат (известь) и выделяется вода согласно формуле В морской воде процесс идет несколько слож- нее, и основным источником кальция для из- вестковых водорослей здесь служит серно- кислый кальций, который они превращают в углекальциевую соль. С и м б и о з, или сожительство двух орга- низмов, — одно из интереснейших и до сих пор еще во многом загадочных явлений в био- логии, хотя изучение этого вопроса имеет уже почти столетнюю историю. Явление симбиоза впервые было обнаружено швейцарским уче- ным Швенденер ом в 1877 г. при изучении лишайников, которые, как выяснилось, пред- ставляют собой комплексные организмы, сос- тоящие из водоросли и гриба. Термин «симбиоз» появился в научной литературе позднее. Он был предложен в 1879 г. Д е Б а р и. В ряду симбиозов не последнее место зани- мают симбиозы с участием водорослей. Водо- росли способны вступать в симбиотические от- ношения не только друг с другом, но и с пред- ставителями различных систематических групп организмов как животного, так и растительного царства (бактериями, одноклеточными и много- клеточными животными, грибами, мхами, па- поротниками, голосеменными и покрытосемен- ными растениями). Однако список таких водо- рослей весьма ограничен. Из обширнейшей группы сине-зеленых водорослей симбиоз с гри- бами (лишайниковый симбиоз) способны уста- навливать представители не более 5 — 7 родов, из которых чаще других встречаются носток (Nostoc), глеокапса (Gloeocapsa), сцитонема (Scytonema) и стигонема (Stigonema). Пребывание водорослей внутри известкового субстрата не всегда благоприятно отражается на их жизнеспособности. У мощных туфообра- зователей, как уже было сказано, окончательно погрузившиеся в известь особи обычно отми- рают, так как оказываются совершенно изоли- рованными от окружающей среды. Однако при достаточно интенсивном размножении эти во- доросли сохраняются живыми в поверхностных слоях отложений, где еще возможен обмен ве- ществ. Другое дело — сверлящие водоросли. Проникая внутрь субстрата, они сохраняют связь с наружной средой через те каналы, ко- торые они образовали. В этом сверлящие водо- росли уподобляются хазмолитическим водо- рослям, населяющим трещины скал. Поэтому здесь погружение в субстрат можно рассматри- вать как такое приспособление водоросли, ко- торое дает ей преимущества в борьбе за сущест- вование. Занимая столь необычайное место- обитание, сверлящие водоросли избавлены от конкуренции из-за места с другими, так ска- зать, нормальными формами; кроме того, внут- ри субстрата они менее подвержены влиянию неблагоприятных внешних воздействий. Анализ различных симбиозов вскрыл чрез- вычайно многообразный характер взаимоотно- шений между партнерами, разную степень их влияния друг на друга. Одним из простейших случаев является поселение одних организмов на поверхности других. Как известно, растения, обитающие на дру- гих организмах, но питающиеся самостоятель- но, называют э п и ф и т а м и. К эпифитам от- носится и большая группа водорослей. Особенно часто водоросли эпифитируют на подводных растениях и водоплавающих животных, иногда покрывая их плотным налетом (рис. 46). При эпифитировании между участниками устанав- ливаются очень непрочные и кратковременные взаимосвязи, которые, однако, уже можно рас- сматривать как симбиотические. Поскольку эпифитирующая водоросль и хозяин оказывают друг на друга довольно слабое влияние, эпифи- тизм у водорослей принято считать наиболее примитивной формой симбиоза. Его относят даже к разряду «безразличных». С подобным утверждением полностью согласиться трудно. Зпифиты действительно не причиняют прямого вреда организму, к которому прикрепляются, но косвенный ущерб при этом все же наносится. Хорошо известно, например, что обрастающие водорослями ножки водоплавающих клещиков, паучков и жучков становятся менее подвиж-
ными, а растения сильно затеняются расселив- ся на них эпифитами и попадают в усло- н неблагоприятные для фотосинтеза. С яв- лением обрастания нередко приходится стал- киваться при разведении аквариумных расте- ний, которые могут сильно угнетаться обитаю- щими на них водорослями. К сожалению, явление эпифитизма с биологи- ческой точки зрения изучено крайне слабо. Не исключено, что между эпифитом и его хо- зяином устанавливаются взаимоотношения го- раздо более сложные, чем мы обычно себе пред- ставляем. Помимо поверхностного прикрепления, во- дор оросли могут жить в тканях других организ- мов — как внеклеточно (в слизи, межклетниках, редко в оболочках мертвых клеток — р — ис. 47~ так и внутриклеточно (в содержимом живых неповрежденных клеток — рис. 48). Такие во- д,оро оросли по способу обитания относят к группе растений эндофитов. Внеклеточные и особенно внутриклеточные эндофиты из числа водорослей по сравнению с эпифитами образуют более сложные симбио- зы — э нд о с им б и о з ы. Для них характер- но наличие более или менее тесных, постоянных и прочных связей между партнерами. Эндо- симбиозы можно выявить только с помощью специальных цитологических исследований. Наиболее многочисленную группу состав- ляют эндосимбиозы одноклеточных зеленых и желто-зеленых водорослей с одноклеточными животными (рис. 48, 1). Эти водоросли но- сят названия соответственно зоохлорелл и зооксантелл. Из многоклеточных животных зеленые и желто-зеленые водоросли образуют эндосимбиозы с пресноводными губками, гидра- ми и др. (рис. 48, 2). Сине-зеленые водоросли образуют с протозоа и некоторыми другими о ганизмами своеобразную группу эндосимбио- зов, получивших название с и н ц и а н о з о в; возникающий при этом морфологический комп- лекс из двух организмов называют ц и а н о- м о м, а сине-зеленые водоросли в нем — ц и а- н е л л а м и (рис. 48, 8). Сопоставление между собой различных эндо- симбиозов позволяет наметить последовательные ступени усложнения морфологического и функ- ционального соподчинения партнеров. Так, не- которые эндосимбиозы существуют очень не- продолжительное время, а затем распадаются, что является свидетельством их примитивности. Примером этого может служить слизистая ко- лониальная сине-зеленая водоросль ворони- хиния (Woronichinia naegeliana). Почти в 50е4 случаев в слизи, окружающей шаровидные ко- лонии этой водоросли, живут другие сине- зеленые водоросли (Lyngbya endophytica и Sy- nechocysti s end ob i oti ca — рис. 47, 1) . Они интен- Рис. 46. Эпифитизм сине-зеленой водоросли Sokolovia neumaniae на ножках водяного клещика Neuman.ia triangularis: 1 — клещик с конечностями, густо покрытыми эпифитирующей водорослью в виде т н . * онки~ волосков 2 — окончание щетинки на ножке клещика с нитями водоросли. сивно размножаются там, хотя имеют чрезвы- чайно бледную, едва заметную окраску. Это, вероятно, обусловлено появлением у них спо- собности утилизировать уже готовые органи- ческие соединения, которые в изобилии обра- зуются при распаде слизи. G рослей в слизи воронихинии приводит сначала к подавлению клеток, а затем к дезорганиза- ции и гибели всей колонии, а следовательно, и симбиоза в целом. Возникает вопрос: как проникают водоросли в ткани и клетки других организмов? У неко- торых организмов имеются для этого специаль- ные приспособления. Так, у мелкого, плаваю- щего в воде папоротника азоллы (Azolla) на нижней стороне листьев располагаются осо- бые полости с узкими выводными отверстиями, через которые выделяется наружу слизь. В этих полостях, независимо от того, в какой геогра- поселяются колонии строго определенного вида сине-зеленой водоросли — анабены (Anabaena azollae). Со временем полости закрываются и наступает полная изоляция попавших туда водорослей. Попытки заражения азоллы пред- ставителями других родов и даже видов сине- зеленых водорослеи успеха не имел %.Э имели. это сви- детельствует о том, что в процессе возникнове-
Рис. 47. Внеклеточный симбиоз: 1 — поперечный разрез шаровидной кОлонии сине-зеленой водо- росли воронихинии (ее крупные клетки располагаются перифе- рическим слоем), в слизи которой поселяются другие сине-зеле- ные водоросли — синехоцистис (Synechocystis endobiotica— мелкие клетки) и лингбия (Lyngbya endophytica — короткие нити); 2 — ткань ряски, в межклетниках которой поселяется зеленая водоросль хлорохитриум (Chlorochytrium lemnae — круп- ные овальные клетки с толстыми оболочками); 8 — плазмодий желто-зеленой водоросли миксохлорис (Myxochloris sphagni- cola) в мертвой водоносной клетке сфагнума. Рис. 48. Внутриклеточный симбиоз: 1 — амеба с клеточками зеленой водоросли зоохлореллы внутри, вверху — отдельная клетка зоохлореллы при большом увеличе- нии; 2 — продольный разрез через конец щупальца пресновод- ной зеленой гидры (Hydra viridis) с клетками зоохлореллы (тем- ные тельца) в клетках внутреннего слоя гидры; 8 — часть тал- лома обесцветившейся зеленой водоросли геосифон (Geosiphon); разветвяенные нити заканчиваются крупными пузырями, в про- топлазме которых живет сине-зеленая водоросль носток. ния данного симбиоза между участниками уста- навливается довольно специфическая физиоло- гическая взаимозависимость. Этот вывод под- тверждается еще и тем, что вырабатываемые азоллой азотистые соединения полностью усваи- ваются эндосимбиотирующими здесь экземпля- рами анабены, вследствие чего у них отпадает свойственная свободноживущим представите- лям этой сине-зеленой водоросли функция фик- сации атмосферного азота. В свою очередь, анабена дополнительно снабжает ткани хозяи- на кислородом и другими продуктами своей жизнедеятельности. Несмотря на существующую у этих симбион- тов специализацию физиологических процес- сов, ни один из них не претерпевает сколько- нибудь существенных изменений в своей орга- низации. Однако так обстоит дело далеко не у всех эндосимбиозов подобного типа. Эндосимбиоти- ческий образ жизни водорослей чаще всего приводит к частичной или полной редукции их клеточных оболочек. Например, у живущих в тканях морской губки аплизиллы (Aplysilla) особей сине-зеленой водоросли из рода афа- нокапса (Aphanocapsa) редукция клеточной оболочки выражается в уменьшении ее толщи- ны. За счет этого снижаются защитные свойст- ва оболочки, но повышается ее проницаемость. Последнее качество, несомненно, улучшает ус- ловия транспорта веществ между клетками губки и эндосимбиотирующей там водоросли. Эндосимбиозы, относящиеся к разряду в н е- к л е т о ч н ы х, образуют уже довольно ус- тойчивые функциональные и морфологические комплексы. Эта тенденция еще более усили- вается у внутриклеточных эндо- с и м б и о з о в. Механизм проникновения во- дорослей внутрь клеток других организмов без их повреждения и нарушения нормальной жизнедеятельности остается пока нераскрытым. Отчасти предпосылки для возникновения внут- риклеточных эндосимбиозов могут быть зало- жены в сохранении у клеток некоторых Орга- низмов голозойного типа питания. Из всех известных типов питания голозойный тип счи- тается одним из наиболее древних. У организмов с голозойным типом питания захватываемая добыча, в числе которой оказы- ваются и водоросли, поступает непосредственно внутрь клетки и там переваривается. Однако отдельным захваченным особям, вероятно, в силу стечения благоприятных обстоятельств иногда удается не только сохраниться внутри клеток хозяина в неповрежденном виде, но и выработать приспособления к новым, необыч- ным условиям жизни и начать там размножать- ся. В результате между организмами устанав- ливаются отношения нового типа — симбиоти-
ческие. Вероятно, именно так проникают эк- земпляры подвижной одноклеточной водоросли эвглены (Euglena gracilis) в эпителиальные клетки задней кишки личинок некоторых ви- дов стрекоз. Клетки эвглены остаются там зелеными на протяжении всего периода сов- местной жизни. Они, правда, теряют подвиж- ность, но при этом никогда не инцистируются. Очевидно, таким же способом особи однокле- точной зеленой водоросли картерии (Carteria) поселяются в эпидермальных клетках реснич- ного червя конволюта (Convoluta roscoHensis). Как выяснилось, клетки картерии под влия- нием симбиотического образа жизни хотя и претерпевают весьма существенные изменения (полностью редуцируется оболочка, и клетки оказываются окруженными только тонкой плаз- матической мембраной — плазмалеммой, ис- чезает стигма, упрощается внутренняя органи- зация жгутиков), но не прекращают фотосин- тезировать. В свою очередь, червь приобретает способность питаться за счет продуктов жизне- деятельности водоросли, которые вырабаты- ваются в процессе фотосинтеза. В частности, он может жить в течение 4 — 5 недель, не полу- чая никакой пищи извне. Однако, когда про- цесс фотосинтеза прекращается (например, ес- ли опыт проводить в темноте), гибнут и водо- росль, и червь. Более того, личинки червя, лишенные клеток водоросли, не в состоянии вести самостоятельное существование. Искус- ственное их заражение водорослями не удается. Внут риклеточные эндосимбиозы, несомнен- но, легче устанавливаются с теми организма- ми, клетки которых не имеют жесткой оболоч- ки на протяжении всего жизненного цикла или по крайней мере на одной из его стадий. Про- никновение симбионта внутрь клеток с жест- кими оболочками возможно только при усло- вии их частичного или полного разрушения. Последнее может наступить под действием спе- цифических энзимов, вырабатываемых организ- мом, вступающим в симбиотические отношения. Наблюдаемая в ряде случаев строгая специали- зация вступающих в симбиоз организмов, ве- роятно, объясняется именно этим обстоятель- ством. К сожалению, все попытки обнаружить хотя бы следы подобного рода энзимов пока успехом не увенчались. Одни внут риклеточные эндосимбиозы, как это происходит у личинок стрекоз, периодиче- ски распадаются и вновь возобновляются; дру- гие — непрерывно поддерживаются из поколе- ния в поколение, так как в этих случаях между участниками устанавливаются прочные и про- должительные связи. Последняя группа эндо- симбиозов, очевидно, могла возникнуть вслед- ствие утраты той фазы в жизненном цикле организма-хозяина, которая была благоприят- на для проникновения симбионта в его клетки. C этого момента, по-видимому, и начинается тесная совместная жизнь двух организмов. В таких случаях переход к симбиотическому способу существования неизбежно сопровож- дается рядом адаптационных изменений у обо- их организмов. Иногда эти изменения морфо- логически незначительны и симбионт можно узнать (например, носток у геосифона, рис.48,8), а иногда они настолько существенны, что сим- биотирующие водоросли невозможно иденти- фицировать ни с одной из свободноживущих водорослей. Так, в вакуолях одного из видов ресничной инфузории парамеции (Paramecium bursaria) неизменно присутствует одноклеточная зеле- ная водоросль. По морфологии и особенностям поведения ее можно лишь условно отнести к протококковой водоросли из рода хлорелла (Chlorella). Установлено, что клетки водоросли делятся независимо от деления парамеции. Каждая из вновь образующихся дочерних кле- ток (автоспор) водоросли немедленно заклю- чается в особую вакуолю и в таком виде в дальнейшем распределяется между дочерними особями инфузории. В ряде случаев между симбионтами склады- ваются настолько тесные взаимозависимые от- ношения, что вне симбиоза они жить уже не могут. Очевидно, они необратимо утрачивают способность самостоятельно вырабатывать це- лый ряд веществ, которые в готовом виде по- ступают от симбиотирующих с ними водорос- лей. Реальность подобного предположения полностью подтвердилась в опытах с гидрой, которая, оказывается, в нужном количестве получает мальтозу из клетки симбиотирующей там зеленой водоросли, систематическую при- надлежность которой точно установить так и не удалось. Иногда нераспадающиеся эндосимбиозы при- водят к образованию такого комплекса, симбио- тическая природа которого выявляется с боль- шим трудом. Так случилось с двумя водоросля- ми — цианофорой и глаукоцистисом. В 1924 г. была описана новая для науки водоросль, названная цианофорой парадоксаль- ной (Cyanophora paradoxa, табл. 5, 7). Позднее детальное изучение этого организма показало, что цианофора представляет собой симбиоз бесцветной одноклеточной водоросли крипто- монады (отдел Pyrrophyta) и поселяющейся в ней внут риклеточно сине-зеленой водоросли (цианеллы) из рода хроококкус (Chsoococcus, отдел Cyanophyta). Клетки последней под влия- нием симбиотического образа жизни настолько сильно видоизменяются, что теряют свой ти- пичный облик. Это выражается главным обра- зом в сильной редукции клеточной оболочки.
Она уменьшается не только по толщине, но и по числу входящих в ее состав слоев: вместо четырехслойной, обычно характерной для сво- бодноживущих сине-зеленых водорослей, она становится двухслойной. Еще большим преобразованиям подвергают- ся цианеллы, входящие в состав глаукоцистиса (Glaucocystis nostochinearum) — очень свое- образной одноклеточной водоросли, описанной в конце прошлого века. Ее систематическое положение долгое время не удавалось точно определить. На основании сине-зеленой окра- ски ее сначала отнесли к отделу Gyanophyta. В дальнейшем выявление целого ряда призна- ков, абсолютно несвойственных сине-зеленым водорослям (наличие морфологически оформ- ленного ядра, окрашенных телец, размножение посредством автоспор), позволили отнести этот организм к зеленым водорослям (отдел Chlorophyta). Только в 30-е годы текущего столетия было наконец установлено, что глау- коцистис представляет собой крайне своеобраз- ную форму эндосимбиоза обесцветившейся од- ноклеточной водоросли, близкой к роду ооцис- тис (Oocystis), и палочковидной сине-зеленой водоросли, которая претерпела здесь такие сильные преобразования, что установить точно ее систематическую принадлежность не пред- ставляется возможным. В равной степени это может быть любой модифицированный предста- витель из ряда родов одноклеточных палочко- видных сине-зеленых водорослей. В симбиозах подобного рода глаукоцистис является пока единственным примером установления столь тесных взаимоотношений между партнерами. Сине-зеленые водоросли (цианеллы) располага- ются в клетках глаукоцистиса либо упорядо- ченно в виде двух групп, либо беспорядочно, случайно. Цианеллы и свободноживущие сине-зеленые водоросли по своей тонкой организации ничем не отличаются друг от друга. Примечательно, что в цианеллах отсутствуют включения за- пасных питательных веществ, представленных различными метаболическими гранулами. По всей видимости, надобность в этом отпадает, поскольку цианеллы получают необходимые им вещества прямо из клетки хозяина. В то же время цианеллы поставляют в клетки хозяина некоторые продукты, которые вырабатываются ими в процессе фотосинтеза. Об этом свидетель- ствует присутствие в цитоплазме бесцветных клеток организма-хозяина крахмальных зерен. Явление это весьма необычное, поскольку у всех хлорофиллоносных зеленых растений един- ственным местом локализации крахмальных зерен является пластида (хлоропласт). В ус- ловиях симбиоза его участники достигают, вероятно, максимальной специализации, в силу которой симбиотирующие сине-зеленые водо- росли принимают на себя функции хлоропла- стов, но ими не становятся. В пользу послед- него свидетельствует существенная разница в организации цианелл и пластид. У клеток бесцветного симбионта глаукоцистиса утрачи- вается способность к самостоятельному обра- зованию крахмала, который образуется там, очевидно, при непосредственном участии циа- нелл. Изучение с помощью электронного микро- скопа цианелл, входящих в состав глауко- цистиса, выявило у них сильную степень ре- дукции клеточной оболочки. Она сохраняется здесь в виде едва заметного контура, который можно обнаружить лишь при условии высокого качества фиксации и обработки материала. Более тщательное исследование цианелл по- казало, что их окружает лишь тонкая (100 ~ о + 10А) мембрана, называемая плазмалеммой. Такая степень редукции клеточного покрова— уникальное явление среди вступающих в сим- биоз сине-зеленых водорослей. Из приведенной характеристики цианелл вид- но, что они представляют собой не что иное, как клетки сине-зеленых водорослей, лишен- ных запасных веществ и клеточных оболочек. Деление цианелл, как и клеток свободножи- вущих сине-зеленых водорослей, осуществляет- ся путем перетяжки пополам. Оно автономно и не приурочено к периоду размножения клет- ки-хозяина. В каждую его дочернюю клетку обычно попадает по нескольку цианелл. Таким образом обеспечивается непрерывность сим- биоза. В отличие от органелл распределение цианелл между дочерними клетками хозяина носит случайный характер, поэтому их число там сильно варьирует. Ле вызывает никакого сомнения, что само деление и характер рас- хождения цианелл по дочерним клеткам регу- лируется не хозяином, что было бы вполне естественно, если бы они превратились в орга- неллы, а самими цианеллами, сохранившими все свойства клеток. Однако даже в условиях такого высокоразвитого симбиоза, примером которого является глаукоцистис, оба партнера все же сохраняют свои индивидуальные черты и автономность. Об этом свидетельствует их способность к раздельному существованию вне клеток хозяина. В специально подобранной питательной среде изолированные симбионты ведут себя как самостоятельные организмы. Они там не только успешно растут и развивают- ся, но и размножаются. Среди симбиозов, образованных с участием водорослей, наибольший интерес представляет симбиоз водорослей с грибами, известный под названием л и ш а й н и к о в о г о с и м б и о-
з а. В результате этого симбиоза возникла своеобразная группа растительных организ- мов, получивших название лишайников. Под- робнее с ними можно познакомиться в соот- ветствующем разделе данного тома. Не вдаваясь здесь в детальное описание ли- шайникового симбиоза, следует, однако, от- метить его своеобразие. В этом симбиозе возни- кает такое биологическое единство двух орга- низмов, которое приводит к появлению прин- ципиально отличного от них третьего. Вместе с тем каждый партнер сохраняет черты той группы организмов, к которой он относится, и ни у одного из них не появляется тенден- ции к преобразованию в составную часть дру- гого. Пока лишайники являют собой единствен- ный строго доказанный случай возникновения одного совершенно нового организма из двух. Этот факт послужил толчком для поиска синте- тических форм в самых различных систематиче- ских группах растений и животных. Однако все усилия, приложенные в данном направле- нии, оказались пока безрезультатными. Тем не менее предположение о возможности сущест- вования синтетических форм организмов ока- залось настолько заманчивым, что в биологии появляется новое течение. В отличие от обыч- ных, прочно утвердившихся взглядов биологов на происхождение организмов как на переход от простого к сложному путем дифференциации, зарождается новое представление о возникно- вении сложного организма из более простых путем синтеза. Некоторые биологи стали рас- сматривать растительную клетку не как про- дукт постепенной дифференциации протопла- ста, а как симбиотический комплекс, синтези- рованный из нескольких простых организмов. Эти идеи зародились и получили наиболее пол- ное развитие в трудах наших отечественных ученых. Впервые предположение о важной роли ф о р- мативного симбиоза (т. е. симбио- за, приводящего к образованию новых форм) в эволюции организмов было высказано акаде- миком А. С. Фаминциным в 19О7 г. Развивая дальше эти мысли, К. С. М е р е ж- к о в с к и й в 1909 г. сформулировал гипотезу симбиогенного происхождения организмов и назвал ее «теорией симбиогенезиса». В даль- нейшем она получила широкую известность среди биологов. В 2О-е годы ее поддержал и развил дальше известный советский ботаник Б. М. КозоПолянский. В наши дни эти идеи, уже на новом уровне развития биоло- гии, были возрождены американской исследо- вательницей Саган-Маргулис в ее ги- потезе происхождения эукариотических кле- ток. В соответствии с этой гипотезой такие клеточные органеллы, как митохондрии, ба- зальные тела жгутиков и пластиды эукариоти- ческих клеток, возникли из симбиотирующих в них прокариотических клеток сине-зеленых водорослей и бактерий. В качестве основного довода приводятся некоторые черты сходства в составе, строении и поведении перечисленных органелл и прокариотов. Бесспорно, эти факты заслуживают самого пристального внимания. Однако они недостаточны для обоснования гипотезы симбиогенеза, поскольку черты сход- ства, как известно, могут появляться у разных по происхождению структур или организмов и вследствие параллелизма в эволюции. Так, система стигма — жгутик у золотистых, желто- зеленых и бурых водорослей по внешнему виду и функциям очень напоминает палочки сетчатки глаза животных, хотя весь процесс их заложения и ход онтогенетического разви- тия говорят о том, что об общности происхож- дения этих образований не может быть и речи. Исследование симбиотических организмов в электронном микроскопе показывает, что даже у такого высокоразвитого симбиоза, как глаукоцистис, партнеры сохраняют свои инди- видуальные черты и автономность. Анализ сим- биозов водорослей с различными организмами выявляет определенную направленность в раз- витии отношений между партнерами, главным образом по линии максимальной специализа- ции функций и вызванных этим обстоятельст- вом структурных перестроек при сохранении их как самостоятельных организмов; это идет вразрез с положениями гипотезы симбиогене- за. Все это свидетельствует о том, что в настоя- щее время гипотеза симбиогенеза находится на той стадии разработки, когда логические по- строения явно преобладают над фактами. Безусловно, симбиоз может привести к соз- данию новых организмов, что подтверждается появлением такой своеобразной растительной группы, как лишайники. Отрицать роль сим- биоза в эволюции нельзя. И все же очевидно, что это не единственный и не основной путь образования новых форм жизни. С одной стороны, к такому выводу подводит факт су- ществования именно лишайников, поскольку они образуют крайне специализированную и обособленную группу организмов, представ- ляющих слепую ветвь филогенетического раз- вития. С другой стороны, сейчас накапливается большой фактический материал по тонкой орга- низации клетки. Он дает возможность воссоз- дать картину вероятного обособления и услож- нения организации некоторых клеточных орга- нелл у водорослей. Кстати, именно отсутствие подобного рода фактов в свое время как раз и стимулировало зарождение гипотезы симбио- генеза.
СИСТЕМАТИЧЕСКИЙ ОБЗОГ БО~ОГОСЛЕИ ОТДЕЛ СИНЕ-ЗЕЛЕНЫЕ ВОДОРОСЛИ (CYANOPHYTA) Сине-зеленые водоросли (Cyanophyta), дро- бянки, точнее, фикохромовые дробянки (Schi- zophyceae), слизевые водоросли (Myxophyceae)— сколько различных названий получила от ис- следователей эта группа древнейших автотроф- ных растений! Страсти не утихли и до сих пор. Немало таких ученых, которые готовы исклю- чить сине-зеленых из числа водорослей, а не- которые — вообще из царства растений. И не так, «с легкой руки», а с полной уверенностью, что они делают это на серьезной научной ос- нове. «Виноваты» в такой судьбе сине-зеленые водоросли сами. Крайне своеобразное строе- ние клеток, колоний и нитей, интересная биология, большой филогенетический возраст- все эти признаки отдельно и вместе взятые дают основу для множества трактовок систе- млтики этой группы организмов. Нет сомнений в том, что сине-зеленые водо- росли — старейшая группа среди автотроф- ных организмов и среди организмов вообще. Остатки подобных им организмов найдены среди строматолитов (известковые образования с бугорчатой поверхностью и концентрически слоистым внутренним строением из докембрий- ских отложений), возраст которых составлял около трех миллиардов лет. Химический анализ обнаружил в этих остатках продукты разложе- ния хлорофилла. Второе серьезное доказатель- ство древности сине-зеленых водорослей— строение их клеток. Вместе с бактериями они объединены в одну группу под названием до- ядерных организмов (Procaryota). Разные си- стематики по-разному оценивают ранг этой группы — от класса до самостоятельного цар- ства организмов, в зависимости от того, какое значение они придают отдельным признакам или уровню клеточного строения. В система- тике сине-зеленых водорослей еще много не- ясного, большие разногласия возникают на каждом уровне их исследования. Сине-зеленые водоросли встречаются во все- возможных и почти невозможных для сущест- вования местообитаниях, по всем континентам и водоемам Земли. Строение клеток. По форме вегетативных клеток сине-зеленые водоросли можно разде- лить на две основные группы: 1) виды с более или менее шаровидными клетками (шаровид- ные, широкоэллипсоидные, груше- и яйцевид- ные); 2) виды с клетками, сильно вытянутыми (или сжатыми) в одном направлении (удлинен- но-эллипсоидные, веретен овидные, цилиндри- ческие — от короткоцилиндрических и бочон- ковидных до удлиненно-цилиндрических). Клетки живут отдельно, а иногда соединяются в колонии или образуют нити (последние также могут жить отдельно или образовывать дер- новинки или студенистые колонии). Клетки имеют довольно толстые стенки. В сущности, протопласт окружен здесь четырь- мя оболочковыми слоями: двухслойная кле- точная оболочка покрыта сверху внешней вол- нистой мембраной, а между протопластом и оболочкой находится еще и внутренняя клеточ- ная мембрана. В образовании поперечной пере- городки между клетками в нитях участвуют только внутренний слой оболочки и внутренняя мембрана; внешняя мембрана и внешний слой оболочки туда не заходят. Строение клеточной стенки и другие микро- структуры клеток сине-зеленых водорослей изучали с помощью электронного микроскопа (рис. 49).
В клеточной оболочке хотя и содержится целлюлоза, но основную роль играют пекти- новые вещества и слизевые полиса хариды. У одних видов клеточные оболочки хорошо ослизняются и содержат даже пигменты; у дру- гих вокруг клеток образуется специальный слизистый чехол, иногда самостоятельный во- круг каждой клетки, но чаще сливающийся в общий чехол, окружающий группу или весь ряд клеток, называемый у нитчатых форм спе- циальным термином — т р и х о м. У многих сине-зеленых водорослей трихомы окружены настоящими чехлами — влагалищами. Как клеточные, так и настоящие чехлы состоят из тонких переплетающихся волокон. Они могут быть гомогенными или слоистыми: слоистость у нитей с обособленными основаниями и вер- хушкой бывает параллельной или косой, иног- да даже воронкообразной. Настоящие чехлы растут путем наложения новых слоев слизи друг на друга или внедрения новых слоев меж- ду старыми. У некоторых ностоковых (Nostoc, Anabaena) клеточные чехлы образуются путем выделения слизи через поры в оболочках. Протопласт сине-зеленых водорослей лишен оформленного ядра и ранее считался диффуз- ным, разделенным лишь на окрашенную пери- ферическую часть — хроматоплазму — и ли- шенную окраски центральную часть — центро- плазму. Однако различными методами микро- скопии и цитохимии, а также ультрацентри- фугированием было доказано, что такое раз- деление может быть только условным. Клетки сине-зеленых водорослей содержат хорошо вы- раженные структурные элементы, и разное их расположение обусловливает различия между центро- и хроматоплазмой. Некоторые авторы выделяют теперь в протопласте сине-зеленых водорослей три составные части: i) нуклеоплаз- му; 2) фотосинтетические пластины (ламеллы); 3) рибосомы и другие цитоплазматические гра- нулы. Но так как нуклеоплазма занимает район центроплазмы, а ламеллы и другие со- ставные части расположены в районе хромато- плазмы, содержащей пигменты, то нельзя считать ошибкой и старое, классическое разграничение (рибосомы встречаются в обеих частях прото- пласта). Пигменты, сосредоточенные в периферической части протопласта, локализованы в пластинча- тых образованиях — ламеллах, которые рас- полагаются в хроматоплазме по-разному. хао- тично, бывают упакованы в гранулы или ори- ентированы радиально. Подобные системы ла- мелл теперь нередко называют парахроматофо- рами. В хроматоплазме, кроме ламелл и рибосом, встречаются еще эктопласты (цианофициновые зерна, состоящие из липопротеидов) и различ- ного рода кристаллы. В зависимости от физио- логического состояния и возраста клеток все эти структурные элементы могут сильно из- меняться вплоть до полного исчезновения. Центроплазма клеток сине-зеленых водорос- лей состоит из гиалоплазмы и разнообразных палочек, фибрилл и гранул. Последние пред- ставляют собой хроматиновые элементы, кото- рые окрашиваются ядерными красителями. Гиалоплазму и хроматиновые элементы вообще можно считать аналогом ядра, поскольку в этих элементах содержится ДНК; они при делении клеток делятся продольно, и половинки поров- ну распределяются по дочерним клеткам. Но, в отличие от типичного ядра, в клетках си- не-зеленых водорослей вокруг хроматиновых элементов никогда не удается обнаружить ядерной оболочки и ядрышек. Это — ядропо- добное образование в клетке, и называют его нуклеоидом. В нем встречаются и рибосомы, содержащие РНК, вакуоли и полифосфатные гранулы. Установлено, что у нитчатых форм между клетками имеются плазмодесмы. Иногда сое- динены между собой также системы ламелл соседних клеток. Поперечные перегородки в трихоме ни в коем случае нельзя считать ку- сками мертвого вещества. Это живая составная часть клетки, которая постоянно участвует в ее жизненных процессах подобно перипласту жгу- тиковых организмов. Протоплазма сине-зеленых водорослей более густая, чем у других групп растений; она неподвижна и очень редко содержит вакуоли, наполненные клеточным соком. Вакуоли по- являются только в старых клетках, и возник- новение их всегда приводит к гибели клетки. Зато в клетках сине-зеленых водорослей часто встречаются газовые вакуоли (псевдовакуоли). Это полости в протоплазме, наполненные азо- том и придающие клетке в проходящем свете микроскопа черно-бурый или почти черный цвет. Встречаются они у некоторых видов почти постоянно, но есть и такие виды, у ко- торых их не обнаруживают. Присутствие или отсутствие их часто считается таксономически важным признаком, но, конечно, еще далеко не все о газовых вакуолях нам известно. Чаще всего встречаются они в клетках у таких видов, которые ведут планктонный образ жизни (пред- ставители родов Anabaena, Aphanizom enon, Rivularia, Microcystis и др., рис. 50, 58, 1). Нет сомнения в том, что газовые вакуоли у этих водорослей служат своеобразным приспособле- нием к уменьшению удельного веса, т. е. к улучшению «парения» в толще воды. И все же их наличие совсем необязательно, и даже у таких типичных планктеров, как Microcystis aeruginosa и М. f los-aquae, можно наблюдать
К Рис. 49. Схематическое изображение строения клетки сине-зеленых водорослей под электронным микроскопом: а — двухслойная клеточная оболочка с наружной мембраной и внутренней клеточной мембраной (плазмалеммой); б — слизис- тый чехол; в — промежуточная пластина клеточной стенки; a — одна из пор клеточной стенки; д — основная малоструктур- ная клеточная плазма; е — фотосинтетические ламеллы, распо- ложенные пакетом, концентрически и одиночно; лс — рибосомы и полисомы; з — хроматин; и — вакуоля; к — зерна запасных питательных веществ; л — полифосфатные тельца; л — крис- талл; и — цианофициновые зерна. Рис. 50. Газовые вакуоли в клетках нитчатых водо- рослей: 1, 2 — участки нитей анабены Шереметьевой (Anabaena schere- metievii); 8, 4 — участки нитей ностока сливовидного (Nostoc pruniforme). (особенно осенью) почти полное исчезновение газовых вакуолей. У некоторых видов они появляются и исчезают внезапно, часто по неизвестным причинам. У ноппока сливовидного (Nostoc pruniforme, табл. 3, 9), крупные коло- нии которого всегда живут на дне водоемов, они появляются в природных условиях весной, вскоре после таяния льда. Обычно зеленовато- коричневые колонии приобретают тогда серо- ватый, иногда даже молочный оттенок и в течение нескольких дней полностью расплы- ваются. Микроскопирование водоросли в этой стадии показывает, что все клетки ностока набиты газовыми вакуолями (рис. 50) и стали черновато-коричневыми, похожими на клетки планктонных анабен. В зависимости от усло- вий газовые вакуоли сохраняются до десяти дней, но в конце концов исчезают; начинается образование слизистого чехла вокруг клеток и их интенсивное деление. Каждая нить или даже кусок нити дает начало новому организму (колонии). Подобную картину можно наблю- дать и при прорастании спор эпифитных или планктонных видов глеотрихии. Иногда газо- вые вакуоли появляются только в некоторых клетках трихома, например в меристемальной зоне, где происходит интенсивное деление кле- ток и могут возникать гормогонии, выходу которых газовые вакуоли каким-то образом помогают. Газовые вакуоли образуются на границе хромато- и центроплазмы и по очертаниям совсем неправильны. У некоторых видов, жи- вущих в верхних слоях придонного ила (в сап- ропеле), в частности у видов осииллатории, крупные газовые вакуоли располагаются в клетках по сторонам поперечных перегородок. Экспериментально установлено, что появление таких вакуолей бывает вызвано уменьшением в среде количества растворенного кислорода, с прибавлением в среду продуктов сероводо- родного брожения. Можно предполагать, что такие вакуоли возникают в качестве хранилищ или мест отложения газов, которые выделяются при ферментативных процессах, происходя- щих в клетке. Состав пигментного аппарата у сине-зеленых водорослей очень пестрый, у них найдено около 30 различных внутриклеточных пигмен- тов. Они относятся к четырем группам — к хло- рофиллам, каротинам, ксантофиллам и били- протеинам. Из хлорофиллов пока достоверно доказано наличие хлорофилла и; из каротино- идов — u-, P- и е-каротинов; из ксантофил- лов — эхинеона, зеаксантина, криптоксанти- на, миксоксантофилла и др., а из билипротеи- нов — с-фикоцианина, с-фикоэритрина и алло- фикоцианина. Весьма характерно для сине- зеленых водорослей наличие последней группы
Гпюноза Перенос электронов Циклическое фотофосфорилирование — 0,4 + Триозо-+ — 0,3 Ф.Н+Н — 02 Дифосфо глицерат со — 0,1 0 с Щ X О G) о о cU :Е cU о о G5 Ш ! G5 т 0 с cU Цитохром b6 3- фосфоглицерат Пластохинон (D О,1 ? с (D 0,2 о о Я 0,3 о 0 с 0,4 с о X 05 2О Рибулозо-1 5-ди- ф Цит ~иоулозо-.') - Ф Пл 2е , О- 0.6 20Н Н20 Н20 0,7 2 + гОН 2Н О 0,8 Та б л и ц а 1. Схема переноса электронов и взаимосвязи световых и темновых реакций в фотосинтезе. В состав пигментной системы 1 входят хлорофилл а и каротиноиды. В состав пигментной системы II входят хлорофилл а и хлорофилл b.
T а бл и ц а 2. Наземные (1 — 9) и почвенные (10) водоросли: 1 — отдельная клетка и группы клоток плеврококка (Pleurococcus vulgaris); 2 — простые и ветвящиеся нити трентеполии (Trentepohlia piceana) с оранжевым маслом в клетках; 3 — отдельная клетка и размножение хлореллы (Chlorella vulgaris); 4 — группа молодых клеток и взрослая клетка хлорококка (Chlorococcum humicola); 5 — колонии глеокапсы с различной окраской слизистых оболочек: желто-коричневой (Gloeocapsa rupestris), красно-коричневой (Gl. magma) и сине-фиолетовой (Gl. alpina); б — участок таллома шизотрикса (Schizothrix friesii) с двумя трихомами в общем слизистом чехле; 7 — часть нити стигонемы (Stigonema minutum); 8 — нити толипотрикса (Tolypothrix elenkinii); 9 — часть нити сцитонемы (Scytonema mirabile); 10 — общий вид пробы почвы в люминесцентном микроскопе: красным светятся живые клетки водорослей, содержащие хлорофилл.
Та бл и ц а 3. Сине-зеленые водоросли: 1 — S>nechococcus aeruginosus; 2 — Dactylococcopsis rhaphidioides; 3 — Merismopedia glauca; 4 — Microcystis aeruginosa; 5 — Gloeocapsa turgida; б — Gomphosphaeria арошпа; 7 — Chamaesiphon curvatus; 8 — Stigonema ocellatum; 9 — Nostoc pruniforme; 10 — Anabaena hassalii; 11 — Aphanizomenon f los-aquae; 12 — Tolypothrix tenuis; 13 — Calothrix gypsophila; 14 — Oscillatoria chalybea; 15 — Lyngbya confervoi Pes.
Та б л и ц а 4. Сине-зеленые планктонные водоросли: 1 — Gloeocapsa limnetica, скопление клеток ( Х 1600); 2 — Microcystis viridis, скопление колоний ( х 2000); 3 — Nostoc linckia, нити в колонии ( X 400); 4 — Coelosphaerium dubium, скопление колоний ( X 500); 5 — Anabaena scheremetievii, часть Йити (X 800); б — Anabaena planctonica (X 1500); 7 — Gloeothrichia echinulata, скопление колоний (X 15). Фото Г. В. Кузьмина.
~ итовые водоросли'. 1 а. епсапа; 4 — Cryptochrysis corn commutata; 5 Та б л и ц а 5. Крипто итовы — — СЬ1 — .8 — С 1 — о — . 7 — С hora paradoxa; — С. рlatyuris; — у С t monascurvata; б — . р ryp о
Оо D о- о .О О О О .О Р О О. О -.О Ii 1 О .О O ( ~, о.. 0 о С5. О д °, 0 ~~ Cj - ~~ о ( ~3 7 '--Oo -0-. "; О О (-,'. О- (-, О.- .о Т О (~Qo ~:-O -О Р .О '-O ~~ Оъ.- о n' o О.. ' ' '-a 'о»~ о .'~-~ ' 1'С - - '3 р ~. О' оо о о '~- -Q 0 Г' QQ,:- O; 0 QO- 0 Т а б л и и а б. Основные типы структуры тела золотистых водорослей: 1 — амебоидная (Brehmiella chrysohydra); 2 — амебоидная (Chrysarachnion insidians); 3 — монадная (Synochromonas pallida); 4 — коккоидная (Chrysobotrys parvula); 5 — пальмеллоидная (Hydrurus foetidus); б — разнонитчатая (Phaeothamnion borzianum); 7 — пластинчатая (Phaeodermatium rivulare).
Та бл и ц а 7. Окремнелые чешуйки и иглы панциря хризомонад, сфотографированные с помощью электронного микроскопа: 1 — Paraphysomonas vestita ( X 14000); 2 — $упига spinosa ( Х 10000); 3 — $упига sphagnicola ( X 20000); 4 — Chrysosphae- геПа multispina (X 14000); 5 — $упига petersenii (X 10000); б — Mallomonas caudata ( Х 8000); 7 — Mallomonas acrocomos ( Х 12000); 8 — Mallomonas caudata, шип ( Х 4700). Фото И. М. Баланова.
Тп б л и ц а 8. Известковые скелеты ископаемых кокколитофорид из палеогеновых отложений: 1, 2 — Discoaster multiradiatus (1 — вид в сканирующем электронном микроскопе, 2 — вид в световом микроскопе); 3— Coccolithus umbilicus; 4 — Discolithina distincta; 5 — Braarudosphaera bigelovii; б — Discoaster saipanensis; 7— Cyclococcolithus reticulatus. (Увеличение: 1 — х 11400; 2 — х 2000; 3 — 7 — х 3400).
пигментов (встречающейся еще у багрянок и некоторых криптомонад) и отсутствие хлоро- филла b. Последнее еще раз свидетельствует о том, что сине-зеленые водоросли являются древней группой, отделившейся и пошедшей по самостоятельному пути развития еще до возникновения в ходе эволюции хлорофилла Ь, участие которого в фотохимических реакциях фотосинтеза дает наиболее высокий коэффи- циент полезного действия. Разнообразием и своеобразным составом фо- тоассимилирующих пигментных систем объяс- няется устойчивость сине-зеленых водорослей к воздействию продолжительного затемнения и анаэробиоза. Этим же частично объясняется и существование их в крайних условиях обита- ния — в пещерах, богатых сероводородом слоях придонного ила, в минеральных источниках. Продуктом фотосинтеза в клетках сине-зеле- ных водорослей является гликопротеид, кото- рый возникает в хроматоплазме и там же отла- гается. Гликопротеид похож на гликоген — от раствора иода в иодистом калии он приобре- тает коричневый цвет. Между фотосинтетиче- скими ламеллами обнаружены полисахаридные зернышки. Цианофициновые зерна во внешнем слое хроматоплазмы состоят из липодротеидов. Волютиновые зерна в центроплазме представ- ляют собой запасные вещества белкового про- исхождения. В плазме обитателей серных во- поемов появляются зернышки серы. Пестротой пигментного состава можно объяс- нить и разнообразие цвета клеток и трихомов сине-зеленых водорослей. Окраска их варьи- рует от чисто-сине-зеленой до фиолетовой или красноватой, иногда до пурпурной или коричне- вато-красной, от желтой до бледно-голубой или почти черной. Цвет протопласта зависит от систематического положения вида, а также от возраста клеток и условий существования. Очень часто он маскируется цветом слизистых влагалищ или колониальной слизи. Пигменты встречаются и в слизи и придают нитям или колониям желтый, коричневый, красноватый, фиолетовый или синий оттенок. Цвет слизи, в свою очередь, зависит от экологических усло- вий — от света, химизма и рН среды, от коли- чества влаги в воздухе (у аэрофитов). Строение нитей. Немногие сине-зеленые во- доросли растут в виде отдельных клеток, боль- шинству свойственно образование колоний или многоклеточных нитей. В свою очередь, нити могут или образовывать ложнопаренхимные колонии, в которых они тесно сомкнуты, а клетки сохраняют физиологическую самостоя- тельность, или иметь гормогониальное строе- ние, при котором клетки соединены в ряд, слагая так называемый трихом. В трихоме протопласты соседних клеток соединены плазмо- десмами. Трихом, окруженный слизистым вла- галищем, получил название нити. Нитчатые формы могут быть простые и раз- ветвленные. Ветвление у сине-зеленых водо- рослей бывает двояким — настоящим и лож- ным (рис. 51). Настоящим называют такое ветв- ление, когда боковая ветвь возникает в резуль- тате деления одной клетки перпендикулярно к основной нити (порядок Stigonematales). Лож- ным ветвлением называют образование боковой ветви путем разрыва трихома и прорыва его через влагалище в сторону одним или обоими концами. В первом случае говорят об одиноч- ном, во втором — о двойном (или парном) ложном ветвлении. Ложным ветвлением можно считать и петлеобразное ветвление, характер- ное для семейства Scytonemataceae, и редко встречающееся V-образное ветвление — резуль- тат повторного деления и роста двух соседних клеток трихома в двух взаимно противополож- ных направлениях по отношению к длинной оси нити. У очень многих нитчатых сине-зеленых водо- рослей имеются своеобразные клетки, получив- шие название гетероцист. У них хорошо выра- жена двухслойная оболочка, а содержимое всегда лишено ассимиляционных пигментов (оно бесцветное, голубоватое или желтоватое), газовых вакуолей и зерен запасных веществ. Они образуются из вегетативных клеток в раз- ных местах трихома, в зависимости от система- тического положения водоросли: на одном (Rivularia, Calothrix, Gloeotrichia) и обоих (Anabaenopsis, Cylindrospermum) концах три- хома — базально и терминально; в трихоме между вегетативными клетками, т. е. интерка- лярно (Nostoc, Anabaena, Nodularia) или сбоку трихома — латерально (у некоторых Stigone- matales). Гетероцисты встречаются поодиночке или по нескольку (2 — 10) в ряд. В зависимости от расположения в каждой гетероцисте возни- кают одна (у терминальных и латеральных гетероцист) или две, изредка даже три (у интер- калярных) пробки, которые с внутренней сто- роны закупоривают поры между гетероцистой и соседними вегетативными клетками (рис. 5, 2). Гетероцисты называют ботанической загад- кой. В световом микроскопе они выглядят как будто пустыми, но иногда, к большому удивлению исследователей, они вдруг прораста- ли, давая начало новым трихомам. При ложном ветвлении и во время разделения нитей три- хомы чаще всего разрываются возле гетеро- цист, как будто ими ограничивается рост три- хомов. Благодаря этому их раньше называли пограничными клетками. Нити с базальными и терминальными гете роцистами прикрепляются к субстрату при помощи гетероцист. У неко- торых видов с гетероцистами связано образоаа-
Pi«. 51. Схематическое изображение типов ветвления у сине-зеленых водорослей: 1 —; 2 — 5 — ложное ветвление (2 — оди- ночное, 8 — двойное, 4 — петлеобразное, 5 — V-образное). ние покоящихся клеток — спор.' они распола- гаются рядом с гетероцистой по одну (у Cy- lindrospermum, Gloeotrichia, Anabaenopsis ra- ciborskii) или по обе ее стороны (у некоторых Anabaena). Не исключено, что гетероцисты являются хранилищами каких-то запасных ве- ществ или энзимов. Любопытно отметить, что все виды сине-зеленых водорослей, способные фиксировать атмосферный азот, имеют гетеро- ц исты. Размножение. Самым обычным типом раз- множения у сине-зеленых водорослей является деление клеток надвое. Для одноклеточных форм этот способ единственный; в колониях и нитях он приводит к росту нити или колонии. Трихом образуется тогда, когда делящиеся в одном направлении клетки не отходят друг от друга. При нарушении линейного располо- жения возникает колония с беспорядочно рас- положенными клетками. При делении в двух перпендикулярных направлениях в одной пло- скости образуется пластинчатая колония с правильным расположением клеток в виде тетрад (Merismopedia). Объемные скопления в виде пакетов возникают в том случае, когда клетки делятся в трех плоскостях (Eucapsis). Представителям некоторых родов (Gloeocapsa,, Microcystis) свойственно также быстрое деле- ние с образованием в материнской клетке мно- жества мелких клеток — н а н н о ц и т о в. Сине-зеленые водоросли размножаются и другими способами — образованием спор (по. коящихся клеток), экзо- и 'эндоспор, гормого- ниев, гормоспор, гонидиев, кокков и плано- кокков. Одним из самых распространенных видов размножения нитчатых форм является образование r о р м о г о н и е в. Этот способ размножения столь характерен для части сине- зеленых водорослей, что послужил названием целому классу гормогониевых (Hormogoniophy- ceae). Гормогониями принято называть фраг- менты трихома, на которые последний распа- дается. Образование гормогониев — не просто механическое отделение группы из двух, трех или большего числа клеток. Гормогонии обо- собляются благодаря отмиранию некоторых некроидальных клеток, затем с помощью выде- ления слизи они выскальзывают из влагалища (если оно имеется) и, совершая колебательные движения, перемещаются в воде или по суб- страту. Каждый гормогоний может дать начало новой особи. Если группа клеток, похожая на гормогоний, одета толстой оболочкой, ее называют гормоспорой (гормоцис- т о й), которая одновременно выполняет функ- ции и размножения, и перенесения неблаго- приятных условий. У некоторых видов от таллома отделяются одноклеточные фрагменты, которые названы
гонидиями, кокками или плано- к о к к а м и. Гонидии сохраняют слизистую оболочку', кокки лишены ясно выраженных оболочек; планококки тоже голые, но, подобно гормогониям, обладают способностью к актив- ному движению. Причины движения гормогониев, планокок- ков и целых трихомов (у Oscillatoriaceae) дале- ко еще не выяснены. Они скользят вдоль про- дольной оси, колеблясь из стороны в сторону, или вращаются вокруг нее. Движущей силой считают выделение слизи, сокращение трихо- мов по направлению продольной оси, сокраще- ния внешней волнистой мембраны, а также электрокинетические явления. Довольно распространенными органами раз- множения являются споры, особенно у водо- рослей из порядка Nostocales. Они одноклеточ- ные, обычно крупнее вегетативных клеток и возникают из них, чаще из одной. Однако у представителей некоторых родов (Gloeotrichia, Anabaena) они образуются в результате слия- ния нескольких вегетативных клеток, и длина таких спор может достигать 0,5 лл. Не исклю- чено, что в процессе такого слияния происходит и рекомбинация, но пока точных данных об этом нет. Споры покрыты толстой, двухслойной обо- лочкой, внутренний слой которой называют эндоспорием, а наружный — экзоспорием. Обо- лочки гладкие или усеяны сосочками, бес- цветные, желтые или коричневатые. Благодаря толстым оболочкам и физиологическим измене- ниям в протопласте (накопление запасных веществ, исчезновение ассимиляционных пиг- ментов, иногда увеличение количества циано- фициновых зерен) споры могут длительное время сохранять жизнеспособность в неблаго- приятных условиях и при разнообразных сильных воздействиях (при низких и высоких температурах, при высыхании и сильном облу- чении). В благоприятных условиях спора про- растает, ее содержимое делится на клетки— образуются с п о р о г о р м о r о н и и, обо- лочка ослизняется, разрывается или открывает- ся крышкой и гормогоний выходит. Эндо- и экзоспоры встречаются главным об- разом у представителей класса хамесифоновых (Chamaesiphonophyceae). Эндоспоры образуют- ся в увеличенных материнских клетках в боль- шом количестве (свыше ста). Образование их происходит с у к ц е д а н н о (в результате ряда последовательных делений протопласта материнской клетки) или с и м у л ь т а н н о (путем одновременного распадения материн- ской клетки на многие мелкие клетки). Экзо- споры по мере своего образования отчленяются от протопласта материнской клетки и выходят наружу. Иногда они не отделяются от материн- ской клетки, а образуют на ней цепочки (напри- мер, у некоторых видов Chamaesiphon). Половое размножение у сине-зеленых водо- рослей полностью отсутствует. Способы питания и экология. Известно, что большинство сине-зеленых водорослей способ- но синтезировать все вещества своей клетки за счет энергии света. Фотосинтетические про- цессы, происходящие в клетках сине-зеленых водорослей, в своей принципиальной схеме близки процессам, которые совершаются в дру- гих хлорофиллсодержащих организмах. Фотоавтотрофный тип питания является для них основным, но не единственным. Кроме настоящего фотосинтеза, сине-зеленые водо- росли способны к фоторедукции, фотогетеро- трофии, автогетеротрофии, гетероавтотрофии и даже полной гетеротрофии. При наличии в сре- де органических веществ они используют и их в качестве дополнительных источников энер- гии. Благодаря способности к смешанному (миксотрофному) питанию они могут быть ак- тивными и в крайних для фотоавтотрофной жизни условиях. В подобных местообитаниях почти полностью отсутствует конкуренция, и сине-зеленые водоросли занимают доминирую- щее место. В условиях плохой освещенности (в пеще- рах, в глубинных горизонтах водоемов) в клет- ках сине-зеленых водорослей изменяется пиг- ментный состав. Это явление, получившее на- звание хроматической адаптации, представляет собой приспособительное изменение окраски водорослей под влиянием изменения спектраль- ного состава света за счет увеличения количества пигментов, имеющих окраску, дополнитель- ную к цвету падающих лучей. Изменения окраски клеток (хлорозы) происходят и в случае недостатка в среде некоторых компонен- тов, в присутствии токсических веществ, а также при переходе к гетеротрофному типу питания. Есть среди сине-зеленых водорослей и такая группа видов, подобной которой среди других организмов вообще мало. Эти водоросли способ- ны фиксировать атмосферный азот, и это свой- ство сочетается у них с фотосинтезом. Сейчас известно уже около ста таких видов. Как уже указывалось, эта способность свойственна толь- ко водорослям, имеющим rетероцисты, да и им не всем. Большинство сине-зеленых водорослей-азот- фиксаторов приурочено к наземным местооби- таниям. Не исключено, что именно их относи- тельная пищевая независимость как фиксато- ров атмосферного азота позволяет им заселять необитаемые, без малейших следов почвы, ска- лы, как это наблюдалось на острове Кракатау в 1883 r.: через три года после извержения
вулкана на пепле и туфах были найдены слизи- стые скопления, состоящие из представителей родов Anabaena, Gloeocapsa, Nostoc, Caloth- rix, Phormidium и др. Первыми поселенцами острова Сурцей, возникшего в результате из- вержения подводного вулкана в 1963 г. около южного берега Исландии, были тоже азотфик- саторы. Среди них оказались некоторые широко распространенные планктонные виды, вызываю- щие «цветение» воды (Anabaena circinalis, А. cylindrica, А. f los-aquae, Л. lemmermannii, А. scheremetievii, А. spiroides, Anabaenopsis cir- cularis, Gloeotrichia echinulata). Максимальной температурой для существо- вания живой и ассимилирующей клетки счи- тают +65'С, но это не предел для сине-зеленых водорослей (см. очерк о водорослях горячих источников). Такую высокую температуру тер- мофильные сине-зеленые водоросли переносят благодаря своеобразному коллоидному состоя- нию протоплазмы, которая при высокой темпе- ратуре очень медленно коагулирует. Самыми распространенными термофилами являются космополиты Mastigocladus laminosus, Phor- midium laminosum. Сине-зеленые водоросли способны выдерживать и низкую температуру. Некоторые виды без повреждения хранились в течение недели при температуре жидкого воздуха ( — 190'С). В природе такой темпера- туры нет, но в Антарктиде при температуре — 83'С были в большом количестве найдены сине-зеленые водоросли (ностоки). В Антарктиде и в высокогорьях, кроме низ- кой температуры, на водоросли влияет еще и высокая солнечная радиация. Для снижения вредного влияния коротковолнового радиаци- онного излучения сине-зеленые водоросли в хо- де эволюции приобрели ряд приспособлений. Важнейшим из них является выделение слизи вокруг клеток. Слизь колоний и слизистые влагалища нитчатых форм являются хо- рошей защитной обверткой, предохраняющей клетки от высыхания и одновременно действую- щей как фильтр, устраняющий вредное влияние радиации. В зависимости от интенсивности све- та в слизи отлагается больше или меньше пиг- мента, и она окрашивается по всей толще или по слоям. Способность слизи быстро поглощать и дли- тельно удерживать воду позволяет сине-зеле- ньп| водорослям нормально вегетировать и в пустынных районах. Слизь поглощает макси- мальное количество ночной или утренней влаги, колонии набухают, и в клетках начинается ассимиляция. К полудню студенистые колонии или скопления клеток высыхают и превраща- ются в черные хрустящие корочки. В таком состоянии они держатся до следующей ночи, когда снова начинается поглощение влаги. Для активной жизни им вполне достаточно парообразной воды. Сине-зеленые водоросли весьма обычны в поч- ве и в напочвенных сообществах, встреча- ются они и в сырых местообитаниях, а также на коре деревьев, на камнях и т. п. Все эти местообитания часто не постоянно обеспечены влагой и неравномерно освещены (подробнее см. в очерках о наземных и почвенных водо- рослях). Сине-зеленые водоросли встречаются также в криофильных сообществах — на льдах и на снегу. Фотосинтез возможен, конечно, только в том случае, когда клетки окружены прослой- кой жидкой воды, что и происходит здесь при ярком солнечном освещении снега и льда. Солнечная радиация на ледниках и снеж- никах очень интенсивна, значительную часть ее составляет коротковолновое излучение, что вызывает у водорослей защитные приспо- собления. В группу криобионтов входит ряд видов сине-зеленых водорослей, но все же в це- лом представители этого отдела предпочитают местообитания с повышенной температурой (подробнее см. в очерке о водорослях снега и льда). Сине-зеленые водоросли преобладают в планк- тоне эвтрофных (богатых питательными вещест- вами) водоемов, где их массовое развитие часто вызывает «цветение» воды. Планктонному об- разу жизни этих водорослей способствуют га- зовые вакуоли в клетках, хотя они имеются и не у всех возбудителей «цветения» (табл. 4). Прижизненные выделения и продукты посмерт- ного разложения у некоторых из этих сине- зеленых водорослей ядовиты. Массовое разви- тие большинства планктонных сине-зеленых водорослей начинается при высокой темпера- туре, т. е. во второй половине весны, летом и в начале осени. Установлено, что для большин- ства пресноводных сине-зеленых водорослей температурный оптимум находится около+30'С. Есть и исключения. Некоторые виды осциллато- рии вызывают «цветение» воды подо льдом, т. е. при температуре около 0'С. Бесцветные и сероводородолюбивые виды развиваются в мас- совом количестве в глубинных слоях озер. Не- которые возбудители «цветения» явно выходят за границы своего ареала благодаря человече- ской деятельности. Так, виды рода Anabaenopsis за пределами тропических и субтропических областей долгое время совсем не встречались, но потом были найдены в южных районах умерен- ного пояса, а несколько лет назад разви- лись уже в Хельсинкской бухте. Подходящая температура и повышенная эвтрофизация (ор- ганическое загрязнение) позволили этому ор- ганизму развиваться в больших количествах и севернее 60-й параллели.
«Цветение» воды вообще, а вызванное сине- зелеными водорослями особенно считается сти- хийным бедствием, так как вода становится почти ни к чему уже не пригодной. При этом значительно увеличиваются вторичное загряз- нение и заиление водоема, так как биомасса водорослей в «цветущем» водоеме достигает значительных величин (средняя биомасса- до 200 г/м', максимальная — до 450 — 500 г/м'), а среди сине-зеленых очень мало таких видов, которые употреблялись бы другими организ- мами в пищу. Многосторонни отношения между сине-зеле- ными водорослями и другими организмами. Виды из родов Gloeocapsa, Nostoc, Scytonema, Stigonema, Rivularia и Calothrix являются фикобионтами в лишайниках. Некоторые сине- зеленые водоросли живут в других организ- мах в качестве ассимиляторов. В воздушных камерах мхов Anthoceros, Blasia живут виды Anabaena и Nostoc. В листьях водяного папо- ротника Azolla americana обитает Anabaena ахо11ае, в межклетниках Cycas и Zamia — Nostoc punctiforme (подробнее см. в очерке о симбиозе водорослей с другими организмами). Таким образом, сине-зеленые водоросли встре- чаются на всех континентах и во всевозможных местообитаниях — в воде и на суше, в пресных и соленых водах, везде и всюду. Многие авторы придерживаются мнения, что все сине-зеленые водоросли убиквисты и космо- политы, но это далеко не так. Выше уже гово- рилось о географическом распространении рода Anabaenopsis. Подробными исследованиями до- казано, что даже такой распространенный вид, как Nostoc pruniforme, не космополит. Неко- торые роды (например, Nostochopsis, Campty- lonemopsis, Raphidiopsis) целиком приурочены к поясам жаркого или теплого климата, Nostoc 5lagellif ormå — к аридным районам, многие виды рода Chamaesiphon — к холодным и чисто- водным рекам и ручьям горных стран. Отдел сине-зеленых водорослей считают древ- нейшей группой автотрофных растений на Зем- ле. Примитивное строение клетки, отсутствие полового размножения и жгутиковых стадий- все это серьезные доказательства их древности. По цитологии сине-зеленые сходны с бакте- риями, а некоторые их пигменты (билипро- теины) встречаются и у красных водорослей. Однако, учитывая весь комплекс характерных для отдела признаков, можно предполагать, что сине-зеленые водоросли являются самосто- ятельной ветвью эволюции. Свыше трех мил- лиардов лет назад они отошли от основного ствола растительной эволюции и образовали тупиковую ветвь. Говоря о хозяйственном значении сине-зеле- ных, на первое место нужно поставить их роль в качестве возбудителей «цветения» воды. Это, к сожалению, отрицательная роль. Положи- тельное значение их заключается прежде всего в способности усваивать свободный азот. В вос- точных странах сине-зеленые водоросли исполь- зуют даже в пищу, а в последние годы некото- рые из них нашли дорогу в бассейны массовых культур для индустриального производства органического вещества. Систематика сине-зеленых водорослей еще далека от совершенства. Сравнительная про- стота морфологии, относительно малое коли- чество ценных с точки зрения систематики признаков и широкая изменчивость некоторых из них, а также разная трактовка одних и тех же признаков привели к тому, что почти все имею- щиеся системы в той или иной мере субъективны и далеки от естественной. Нет хорошего, обо- снованного разграничения вида как целого и объем вида в разных системах понимается по-разному. Общее количество видов в отделе определяется в 1500 — 2000. По принятой нами системе отдел сине-зеленых водорослей делится на 3 класса, несколько порядков и много се- мейств. КЛАСС ХРООКОККОВЫЕ (CHROOCOCCOPH YCEAE) Сюда входят одноклеточные и колониальные организмы. У большинства колонии образуются за счет выделения значительных масс слизи, реже путем слипания плотно сомкнутых клеток. Располагаются клетки в колониях беспорядочно или правильно, очень редко нитевидно. Клетки в большинстве без дифференцировки на основа- ние и вершину. Размножаются хроококковые делением клеток, реже нанноцитами, плано- кокками и спорами. Эндо- и экзоспоры, а также гетер оцисты отсутствуют. Класс охватывает 35 родов, неравномерно распределяющихся на 2 порядка. ПОРЯДОК ХРООКОККОВЫЕ (CHROOCOCCA LES) Порядок включает широко распространенные одноклеточные и колониальные формы, не об- разующие слоевища, свободноживущие или си- дящие на субстрате. Размножаются эти водорос- ли почти без исключения делением клеток. Основных семейств в порядке 7. Семейство коккобактрейных (Coccobactrea- ceae) содержит около 10 родов, объединяю- щих водоросли с шаровидными, эллипсоидными, цилиндрическими или веретеновидными клетка- ми. Клетки одиночные или соединенные в не- большие колонии с плохо заметной слизью. Наиболее известны 2 рода.
» .Ф ° ° ° ° ° ' ° ° ° 1.".. ° Ф » ..'.. ".» , ° »' ° "..'," ° ° », °...,' ° ° ° ° » ° ° ° ° Рис. 52. Хроококковые: 1 — Aphanothece stagnina; р — Gloeothece palea; 8 — Eucap- sis alpina; 4 — Coelosphaerium kuetzingianum. Род синехоиистис (Synechocystis) охватывает 12 в основном пресноводных или эндофитных (живущих в слизи других водорослей) видов. Клетки у этих водорослей шаровидные, оди- ночные или после деления соединенные попарно. В СССР найдены 9 видов, самый распростра- ненный из них Synechocystis aquatilis. Род сикехожоккус (Synechococcus) содержит 15 видов с эллипсоидными или цилиндрическими клетками. Это обитатели стоячих вод, реже источников или наземных местообитаний, два вида морских. Самые известные виды — Syne- chococcus major и S. aeruginosus (табл. 3, 1). Род дактилококкопсис (Dactylococcopsis) со- держит около 15 видов, отличающихся своеоб- разной формой удлиненных, заостренных на концах клеток, собранных небольшими пучка- ми. Обитают преимущественно в планктоне. Наиболее известный вид — Dactylococcopsis rhaphidioides (табл. 3, 2). Семейство мерисмопедиевых (Merismopediaceae) включает водоросли с пластинчатыми колония- ми, шаровидными или эллипсоидными правиль- но расположенными клетками. Содержит толь- ко 2 рода. Род мерисмопедия (Merismopedia) объединя- ет 13 видов, обитающих преимущественно в пресных водах. Они предпочитают прибреж- ную зону, живут эпифитно или в планктоне. Распространенные виды Merismopedia elegans и М. glauca образуют крупные колонии (табл. 3, 8). Семейство микроиистиевых (Microcystida- ceae) содержит 4 рода со слизистыми шаровид- ными или бесформенными колониями; клетки в них шаровидные, эллипсоидные до па- лочковидно-цилиндрических, располагающиеся в колониях беспорядочно; слизь гомогенная. Род микроиистис (Microcystis) охватывает 20 — 25 трудноопределяемых видов, обитающих в самых разнообразных условиях (около 10 морских). Клетки у них шаровидные, у многих видов с газовыми вакуолями, беспорядочно расположенные в шаровидных, эллипсоидных или продырявленных колониях. Это частые возбудители «цветения» воды почти во всех кли- матических зонах, некоторые содержат (или выделяют) ядовитые вещества. У М. aeruginosa очертания колониальной слизи четкие, диаметр колонии достигает 1 мм (табл. 3, 4, рис. 28, 1). У М. f los-aquae очертания расплывающиеся. M. pulverea встречается в планктоне, бенто- се и даже в наземных местообитаниях и в почве. Род афанотеие (Aphanothece) объединяет око- ло 20 видов, 5 из них обитают в соленых водах. Представители рода отличаются от рода микро- цистис эллипсоидными или цилиндрическими клетками. Колонии иногда макроскопические, величиной до голубиного яйца, встречаются в водных и наземных местообитаниях. Самые известные виды — Aphanothece stagnina и А. elabens (рис. 52, 1). Крупные студенистые коло- нии первой живут на дне водоемов или всплы- вают на поверхность, а колонии второй сначала прикрепляются к растениям и подводным пред- метам, потом плавают в толще воды, вызывая «цветение». Семейство глеокапсовых (Gloeocapsaceae) вклю- чает водоросли, у которых шаровидные или эллипсоидные клетки собраны в микроско- пические колонии. Колонии слизистые, более или менее шаровидные, иногда расплывающие- ся. В колониях клетки в большинстве случаев окружены последовательно включенными друг в друга слизистыми пузырями, реже слизь гомогенная. Как исключение образуются ку- бические колонии. В семействе 3 рода. Род глеокапса (Gloeocapsa) содержит свыше 60 трудноопределяемых видов. Клетки у них шаровидные. Пузыри колониальной слизи хо- рошо выражены, реже слизь однородная. Встре- чаются эти водоросли как в воде, так и в назем- ных местообитаниях; в первом случае у боль- шинства колониальная слизь бесцветная, во втором окрашенная. В колониях со слоистой слизью обычно по 2 — 8 клеток, реже больше; в колониях с гомогенной слизью обыкновенно клеток много. Самые распространенные виды— Gloeocapsa limnetica, Gl. turgida, Gl. minuta, И. magma, Gl. alpina, Gl. rupestris. Первые
три с бесцветной слизью — довольно обычные обитатели планктона, а Gl. turgida (табл. 3, Б) встречается даже в соленой воде, в илу, в боло- тах, в наземных местообитаниях и в горячих источниках. И. magma с розовато-красными до красно-коричневых оболочками, И. rupestris с желтыми до коричневых оболочками и И. alpina с сине-фиолетовыми до черно-фиолетовых оболочками встречаются на влажных скалах, камнях, стенах, реже в воде. Водоросли из рода глеотеае (Gloeothece) по обособленным слизистым оболочкам вокруг клеток напоминают представителей Gloeocapsa, но хорошо отличаются от них удлиненной фор- мой клеток, которые делятся перпендикулярно длинной оси. Видов в роде около 15, большинство из них обитает на влажных скалах, на мхах, на поч- ве, по краям источников (даже теплых), реже в воде. Обычный вид — Gloeothece palea (рис. 52, 2). Представители рода эукапсис (Eucapsis) от- личаются клетками, расположенными в плот- ных кубиках, слизь у них четкая; клетки шаро- видные. Самый известный вид — Eucapsis а1- р1па (рис. 52, 8), обитающий в основном в водо- емах на севере и в горах, реже в торфяных бо- лотах. Семейство целосфериевых (Coelosphaeriaceae) объединяет сине-зеленые водоросли с шаро- видными, реже чуть эллипсоидными или много- угольными клетками, всегда образующими ко- лонии, где они располагаются в один перифе- рический слой. В семействе несколько родов. Род аелосфериум (Coelosphaerium) включает формы со слизистыми, шаровидными или бес- форменными колониями. Содержит около 10 ви- дов, все обитают только в воде, самый обыкно- венный из них — Coelosphaerium kuetzingi- апнт (рис. 52, Ф). Семейство гомфосфериевых (С omphosphaeri- aceae) объединяет водоросли с эллипсоидными или обратнояйцевидными клетками, всегда со- бранными в слизистые колонии, где они си- дят по периферии на ножках, радиально рас- ходящихся от более плотного центрального тела. Род гомфосферия (С omphosphaeria) содер- жит около 10 видов, в СССР встречаются 2: Gomphosphaeria aponina (табл. 3, 6) и С. la- custris, обитающие в планктоне стоячих вод и среди других водорослей. Семейство воронихиниевых (Woronichiniaceae) включает водоросли с удлиненными клетками, расположенными в слизистых колониях в один слой, наружная колониальная слизь радиально- трубчатая. Род воронихиния (Woronichinia) — единствен- ный род с одним видом W. naegeliana (рис. 28, 2), который обитает в планктоне, возбуждая «цве- тение». ПОРЯДОК ЭНТОФИЗАЛИЕВЫЕ (ENTOPHYSALIDALES) Представители этого порядка образуют более или менее полушаровидные или корковидные, плотно прикрепленные к субстрату слизистые слоевища. Последние слагаются из клеток и колоний, которые похожи на колонии глеокап- сы. Колонии срастаются рядами друг с другом. В порядке 2 семейства, объединяющих около 10 родов, но все они довольно редкие, трудно- определимые, некоторые роды морские. КЛАСС ХАМЕСИФОНОВЫЕ (CHAMAESIP HONOPH YCEAE) В этот класс входят одноклеточные предста- вители с дифференцированными на основание и вершину слоевищами, колониальные формы, нитевидные или образующие простые слоевища срастанием нитей боковыми сторонами. Почти все живут прикрепленно, встречаются доволь- но редко. Размножаются хамесифоновые эндо- и экзоспорами. Их развитие на отдельных ста- диях сходно с развитием водорослей из классов. хроококковых и гормогониевых. Хамесифоно- вые растут эпифитно или на камнях, предпо- читая чистоводные и бедные питательными ве- ществами горные ручьи, где их слоевища обра- зуют цветные корочки или пятна на камнях, раковинах и растениях. Некоторые виды оби- тают в морях. В классе 4 порядка, из которых легче дру- гих определяется порядок Dermocarpales, куда входят одноклеточные формы, прикрепляющие- ся одним концом к субстрату поодиночке или группах. Клетки этих водорослей вегетатив- но не делятся, размножение осуществляется только эндо- илг экзоспорами. В порядке 2 семейства. Семейство хамесифоновых (Chamaesiphona- ceae) включает единственный род хамесифок (Chamaesiphon), примерно с 30 пресноводными видами. Клетки у них эллипсоидные, груше- видные или цилиндрические, вокруг нижней части клетки имеется чашка или влагалище. Размножаются хамесифоновые экзоспорами, ко- торые образуются у некоторых видов в боль- шом количестве цепочками, у других понем- ногу, у одних рассеиваются, у других прорас- тают на материнских клетках. Виды Chamae- siphon довольно обыкновенны в чистоводных и холодных горных ручьях и реках. Приме- ром может служить Chamaesi phon cur vatus (табл. 3, 7).
Рис. 5 . 53. Стигонемовые: 1 — 8 — tigonema informe rme; 2 — Hapalosiphon fontinalis КЛАСС ГОРМОГОНИЕВЫЕ (HORMOGONIOPH Y(".EAE) В этот обширный класс вхо ят м ные водоросли кл десм об I друг с др~том пос разуя трихомы. Т их р. осредством плазмо- покрытые сл рихомы голые или ~е слизистыми влагали ам мах имеются щами, в трихо- Размножен или отс тств у уют гете роцисты. ние осуществляется го реже спорами. ся гормогониями, Разные авторы насчитывают чество порядков в ют разное коли- дков в классе — от 12 до 31. ПОРЯДОК СТИГОНЕМОВЫЕ (STIGON EMATA LES) Представители порядка характе из стоящим ветвлени а характеризуются на- лением, хотя здесь вст и ложное ветвление. Т встречается или многорядные ение. рихомы в нит ях одно- или интеркалярные. Н гете роцисты лат еральные видные, подушко об ные. ити соединены в воилоко- новинки. В оо разные или или накипные де- и. порядке около 35 о ов ' 00 видов, распре родов, около предел яющихся в 5 с д ржащих каждое ольших, соде жа у родов с немногими е из тропиков ° .У редкими видами Семейство ст , мореи и гопячих источников. во стигонемовых (Sti~onema во ст 'g nemataceae) са- единяет 14 енное и асп о родов (1 из них мо р ространенное, объ- пических), 70 ви ов морской, 3 тро- ются роды Stigon n видов. аиболее часто еаза и апа1оя встреча- р нно на суше, второй — в воде. Водоросли из рода стиг их окап~ 30 — ж н)т коло видов — характе из ют ронним боковым ризуются всесто- вым ветвлением. Часто зуют темно-ко и то они обра- корковидные дерно . Т ичневые подушкооб а разные или новинки. Т ихомы у НННх обы~ рядные бок или много я оковые ветви тоньше осн рядные, реже одно- аопета 1п1огт ( 53 е рис., 1 вст е х, на мхах и пнях, в т н е St. ~ы11~(~~ ( б 3 8 чащ всего болотах рах и реках. 1 Ро д хапалосифон (Hapalosi hon обычно одноряд и всег рядными главными и всег рядными боковыми и всегда одно- теркалярные. ыми нитями гетероцисты ин- В роде 12 видов. Наиболее Н)1 .111 1 ~ 1 on dna is (рис. 53 2 о чается в торфянь . он встре- яных олотах е источниках. р же в горячих ПОРЯДОК МАСТИГОКЛАДОВЫЕ (MASTIGOCLADALES) Представители порядка ха V б -о разным ветвлен жит родов, 3 из них морские; е с одним видом. Известн мый расп ост тнеиший и са- т раненный вид — Masti aminosus, обитающ — ая gocladus 30 итающий в горячих ис от -1-50 до -1-55' С М от до 60'С с рф~~жически эо вид менчив, выделены различнь ~е стадии
Рис. 54. Ностоковые: нити. 3 — N. unctiforme, колонии разного строения. я — Nostoc commune, колония в натуральную величину и отдельные нити; — . р его развития. Считается, что это филогенети- чески древний вид, реликт термальных место- обитаний. ПОРЯДОК НОСТОКОВЫЕ (NOSTOCALES) У ностоковых трихомы всегда однорядные, всегда с гетероцистами и часто со спорами, не ветвящиеся или ветвящиеся ложно. Они бывают как с влагалищами, так и без них, чаще по одному трихому в каждом влагалище. В поря- док входит 9 семейств. Семейство ностоковых (Nostocaceae) охваты- вает 2 рода — Nostoc (около 50 видов) и Wollea (3 вида). Водоросли из рода Nostoc чрезвычайно ши- роко распространены как в воде, так и на суше. Колонии их имеют разные размеры, форму и консистенцию — от микроскопических до ле- пешек диаметром 30 см, студенистые, мягкие или с крепкой поверхностью, маленькие— часто шаровидные, большие — распростертые. Nostoc commune (рис. 54, 1) встречается на почве и на камнях как в арктических и субарк- тических районах, так и в степях и пустынях. Колонии его в сухом состоянии черные, во влажном черно-оливково-зеленые. На почве они образуют значительные скопления. У N. pruniforme (табл. 3, 9) колонии большей частью шаровидные или эллипсоидные, реже при- плюснутые, диаметром 1 — 8 см, внутри более жидкие, сверху кожистые, распространены пре- имущественно в воде в зоне умеренного климата: на дне водоемов на 1 м' может образовываться до 500 колоний величиной с вишню и более. В умеренной зоне одним из самых распростра- ненных является N. punctiforme (рис. 54, 2), обладающий плотными колониями с трудно- различимыми трихомами. Этот вид успешно растет в стоячих водах, на почве и в почве, а также в лишайниках и в некоторых высших растениях в качестве симбионта. Многие но- стоки живут вначале в прикрепленном состоя- нии, потом колонии освобождаются и живут на дне или в планктоне. Семейство анабеновых (Anabaenaceae) вклю- чает формы, которые по строению нитей похо- жи на представителей семейства ностоковых, но у них никогда не образуются студенистые колонии. Нити у этих водорослей с влагали- щем или без него, соединенные в бесформенные дерновины или одиночные. В семействе 4 рода. Род анабена (Anabaena) широко распростра- нен, он содержит около 100 видов, которые встречаются в воде и в наземных местообита- ниях, среди них много возбудителей «цветения» воды (Anabaena f los-aquae, А. hassalii, А. schere- metievii, А. circinalis, А. lemmermannii и др., табл. 3, 10, рис. 28, 5', 6). Характерные признаки водорослей рода — инте ркалярное расположе- ние гете роцист, образование спор. Форма спор, гетероцист и вегетативных клеток колеб- лется в широких пределах. Наличие газовых вакуолей у некоторых постоянное, но в конце вегетационного периода они могут и исчез- нуть. Отдельные виды, например А. variabilis (рис. 55, 1), часты на почве и в наземных место-
Рис. 55. Ностоковые.' 1 — Anabaena variabilis; 2 — Cylindrospermum muscicola; 8— АпаЬаепоряя raciborskii. обитаниях. Многие анабеновые связывают ат- мосферный азот. Роды анабенопсис (Anabaenopsis) и цилиндро- спермум (Cylindrospermum) имеют одинаковое расположение гете роцист — по обоим концам трихомов, но у Cylindrospermum споры воз- никают рядом с гетероцистой, а у Anabaenopsis без связи с нею. Представители рода Су11пйго- spermum, например С. muscicola (рис. 55, 2), характерны для наземных местообитаний или живут эпифитно на подводных частях цветко- вых растений, а также в верхних слоях при- донного ила, тогда как род Anabaenopsis ти- пично планктонный, распространенный пре- имущественно в тропических и субтропических странах. Примером может быть А. racibor- skii (рис. 55, 8). Семейство афанизоменоновых (Aphanizome- nonaceae) представлено одним родом афанизо- менон (Aphanizomenon), который содержит 10 видов. Трихомы у водорослей из этого рода прямые или изогнутые, одиночные или соеди- ненные в свободноплавающие пучки, дости- гающие значительных размеров. Клетки в сред- ней части трих омов коротко-цилиндрические, с газовыми вакуолями, на концах удлиненные, суживающиеся и часто бесцветные. Гетеро- цисты и споры интеркалярные, разнообразной формы. Самый распространенный вид — Apha- nizomenon f los-aquae (табл. 3, 11; рис. 28, 8, 4) с цилиндрическими спорами, обычный возбудитель «цветения» воды в эвтрофных во- доемах. Семейство суитонемовых (Scytonemataceae) содержит 4 рода, из них наиболее обширны 2. Морфологически они довольно близки. Род сцитонема (Scytonema) характеризуется двойным ложным ветвлением и нередко косой слоистостью влагалищ. Е сли представители этого рода имеют одиночное ветвление, то вла- Рис. 56. Сцитонемовые.' 1 — $cytonema crustaceum; 2 — $с. mirabile; 8, 4 — $с. alatum (8 — нить при малом увеличении, увеличении). 4 — конец нити при ббльшем
Рис. 57. Ривуляриевые: 1 — Calothrix Гияса; 2 — С. braunii; 8 — С. thermalis. Рис. 58. Ривуляриевые: 1 — Rivularia planctonica, колония (при малом увеличении) и части отдельных нитей с газовыми вакуолями в клетках; 2— R. coadunata, верхняя часть кустика из полушаровидной коло- нии; 8 — Gloeotrichia pisum, колонии в натуральную величину и отдельная нить со зрелой спорой. галища у них очень широкие и с воронковид- ными слоями. Трихомы однорядные, дерновинки различной формы и величины. Встречаются обычно на орошаемых скалах и на сырой поч- ве. Очень изменчивый вид Sc. crustaceum (рис. 56, 1) предпочитает известковые субстра- ты, являясь мощным туфообразователем. Са- мый распространенный в СССР вид — Sc. mi- rabile (рис. 56, 2), живущий на скалах, на мхах, в стоячих водах, озерах и гейзерах, по краям грифонов и в стоках. Sc. alatum (рис. 56, 8) образует скопления на орошаемых скалах, в небольших водоемах и в бухтах озер. Для него характерны широкие, с неровными края- ми влагалища и сильно косорасходящаяся слои- стость. Род толипотрикс (Tolypothrix) отличается от предыдущего одиночным ложным ветвле- нием. Встречается в пресной и соленой воде, на орошаемых скалах, в стоячих водоемах и в источниках. Распространенный вид Т. tenuis (табл. 3, 12) и некоторые другие, встречающиеся в воде, ведут сначала прикрепленный образ жизни, потом плавают свободно; обитающие на скалах нередко имеют окрашенную слизь (табл. 2, 8). Семейство ривуляриевых (Bivulariaceae) объе- диняет формы с характерными ассиметричными трихомами и нитями, утончающимися от осно- вания к вершине и часто заканчивающимися бесцветным волоском. Гетероцисты у них ба- 10 Рис. 59. Осциллаториевые: 1 — Oscillatoria sancta; 2 — О. limosa; 8 — О. princeps; 4— О. brevis; 5 — О. formosa; 6 — Spirulina jenneri; 7 — S. major; 8 — Phormidium foveolarum; 9 — Ph. molle; 10 — Ph. autum- nale; 11 — Lyngbya aestuarii. На всех рисунках изображены концы трихомов и нитей.
г ° Ф l / ° 5 ° ° Рис. 6 . 60. Осциллаториевые: 1 — Schizothrix lateritia, слоеви е и 1, слоевище и отдельный трихом; 2 — Я . с1 льшом увеличении 4— при малом и бо — ch. са1с1 — М~сгосо1еп8 чафпаЫз час малом б увеличени ц отдельного трихома; Gloeotrichia na atans, Gl. pisum (рис. 58, 8) жи- жизни вы . ес inulata ве ет пл д анктонный образ ни, вызывая «цветение» во ы' вида всег а с г ды; клетки у этого хда с газовыми вакуолями (табл. 4 7 9 зальные, редко инте каля ное В р алярные. Ветвление лож- ное. семействе 4 рода. од калотрикс (Calothrix) содержит около когда налеты или дерновинки, но ни- не слагаются в ст е уд нистые сферические лонии. части видов ветвление ск но отсутствует вообще. Э ще. кологический диапазон видов довольно ши ок вс ви рок, встречаются эпифит- О Д ОСЦИЛЛАТОРИЕВЫЕ (ОССИ КАТО В1А?.КЯ) рихомы осциллаториевых не со е ж щий в — а о1Ьлх fusca, жив — я всегда лишены спор, они о но- в в — а о1Ь ', у- рядные, с влагалищами и б, нь С 1Ь опоры, w. braunii— или ез них, по вижнь ermalis — в се тывают от 5 о 11 — р~~~~ еко орые 3 ~~~ ним л, ) С Ш~-МЫ1' " 2 ным — очень изменчивый ви ветвлением и ши о ИД С О ИЛЬ- и б — Семейство ос иллат най йденныи на почве камн рокими влагалищами б о ъединяет о ы п и аториевых (Oscillatoriac еае в в воде. амнях и скалах, а также амн, шены влагалищ ил п д 1, представители кото ых х ли- Водоросли о а ному т ихом или заключают толь ько по од- р да ривулярия (Bivularia~ 6- р му в каждом влагалище. Вет разуют студенистые пол ша 1а~ о — отсутствует. етвл ение ви ные полушаровидные или ша о- пол ша р — Род осииллатория (Oscillatoria х нити располаха- доросли кото ьно. екоторые виды инк , у которых трихомы лишены Н нк русти подвижные ны влагалищ, роде олее 100 видов, жив щих В зоне. ivularia в пресной во е ,' о итает в планктоне ек ~м~, В. odt (рис. 58 2'— водныи и солоновато водный его ,' — вид пресно- вызы свая «цветение» во ы в ород, в планктоне, иногда сверху вор онковидно-расши яю и , и т. д. а дне, по бе ег влагалища. р щиеся желтые чаются О. регам и в илу встре- Ро г од глеотрихия (Gloeotrichia~ внешн е о итают также в горячих но отличается наличием м спор. некоторые виды об аз та л. виды о разуют налеты на сыром
ОТДЕЛ ПИРОФИТОВЫЕ ВОДОРОСЛИ (PYRROPHYTA) песке или на почве, например О. brevis, О. formosa (рис. 59, 4, о), а также встречаются в сточных водах. Род спирулина (Spirulina) довольно близок с Oscillatoria, но отличается правильно или неправильно спиральными трихомами. У мел- ких форм поперечные перегородки бывают сов- сем незаметными. Широко распространены Spi- rulina jenneri и S. major (рис. 59, 6, 7). Тропи- ческий вид S. platensis в настоящее время вы- ращивают как объект массовой культуры. Род фарлидиул (Phormidium) объединяет около 100 видов, единственным отличием ко- торых от Oscillatoria является мягкое расплы- вающееся влагалище у нитей, склеивающее их в своеобразные пленчато-кожистые дерновинки. Живут в различных водоемах, в горячих источ- никах, на почве (в том числе на загрязненной) и в грязной воде. Примерами могут служить Phormidium foveolarum, Ph. molle, Ph. autum- nale (рис. 59, 8 — 10). Типичный термофил— Ph. laminosum (рис. 42). Род лингбия (Lyngbya) объединяет свыше 100 видов, которые отличаются от представи- телей вышеприведенных родов наличием креп- ких слизистых влагалищ у всех нитей. Эти К отделу пирофитовых относятся весьма своеобразные, преимущественно одноклеточ- ные, интересные в теоретическом и важные в практическом отношении водоросли. Они представляют собой систематически неоднород- ную группу, объединяющую три довольно обо- собленных подотдела: хлоромонадофитовые, криптофитовые и динофитовые. Наиболее существенным, общим для всех пирофитовых водорослей признаком является спинка-брюшное (дорсовентральное) строение их клеток. У громадного большинства предста- вителей этого отдела в строении клеток отчет- ливо выражены спинная, брюшная и боковые стороны, а также хорошо заметна разница между передним и задним концами (рис. 61 и 62). Наличие у клеток бороздок является вторым важным признаком для всех пирофитовых. Бо- роздок может быть две: продольная и попереч- ная — или одна продольная. Располагается продольная бороздка всегда на брюшной сторо- не клетки. У представителей с двумя бороздка- ми место их пересечения также приурочено к брюшной стороне клетки (рис. 64 и 65). Два разных по длине, строению и даже функ- циям жгутика — третий общий признак для всех подвижных одноклеточных (монадных) форм пирофитовых. Эти жгутики получили водоросли обитают в одинаковых биотопах с ви- дами вышеназванных групп. В горячих источ- никах, в пресной и соленой воде широко рас- пространен очень изменчивый вид L. aestuarii (рис. 59, 11), только в соленых водоемах— ? . confervoides (табл. 3, 1о). Семейство шизотриксовых (Schizothrichaceae) отличается от семейства Oscillatoriaceae нали- чием у его представителей многих трихомов в одном влагалище. Семейство охватывает 6 ро- дов. Род шизотрикс (Schizothrix) наиболее обши- рен. Он включает около 70 видов, которые живут или в наземных местообитаниях, или в воде, предпочитая текучие воды или район прибоя. Некоторые виды характерны для го- рячих источников. Наиболее распространены Schizothrix lateritia, Sch. calcicola, Sch. lar- dacea (рис. 60, 1 — 8), среди влажных мхов обычен Sch. friesii (табл. 2, 6). Род ликроколеус (Microcoleus) характерен для водной среды, лишь М. vaginatus (рис. 60, 4) ведет наземный образ жизни. Почти все виды имеют канатооб разно-переплетенные трихомы. В пресных водах наиболее распространен М. chthonoplastes (рис. 60, о). разные названия: плавательный и рулевой- у хлоромонадофитовых, передний и задний- у криптофитовых, поперечный и продольный- у динофитовых (рис. 62 и 64). Другой очень важный отличительной чер- той является присутствие у громадного боль- шинства одноклеточных пирофитовых так на- зываемой глотки, обычно имеющей вид мешка, трубы, внутреннего кармана или треугольного резервуара, а также наличие особых, сильно преломляющих свет телец — трихоцист. По- следние имеют вид одиночных или многочислен- ных булавовидных или игловидных образова- ний, располагающихся или в периферическом слое цитоплазмы (у хлоромонадофитовых), или продольными рядами по внутренней поверх- ности глотки (у криптофитовых), или внутри протопласта (у динофитовых) (рис. 6i и 62). Особо следует сказать о хлоропластах. По разнообразию их окраски пирофитовым при- надлежит первое место среди водорослей. Обыч- но хлоропласты пирофитовых окрашены в олив- ковый, бурый или коричневый цвет, нередко наблюдаются желтый, золотистый, красный, реже голубой и синий, а также всевозможные оттенки указанных цветов. Лишь чисто зеле- ная окраска хлоропластов у них полностью отсутствует. Как и в составе других отделов,
рас. я. Хлоромонадофвтовыа: 1 — 2 — Gonyostomum semen (1 — вид клетки со спинной сто- роны, 2 — вид сбоку); 8 — Merotrichia capitata; 4 — 5 — Vacuo- laria viridis в продольном и поперечном разрезе: а — перипласт, 6 — глотка, в — жгутик плавательный, г — жгутик рулевой, д — продольная бороздка, е — хлоропласты, лс — ядро, 3— трихоцисты, и — пульсирующая вакуоля, к — слизистые тельца. Рис. 62. Три положения клетки криптомонады: 2 — клетка с брюшной стороны; 2 —; 8 — клетка сверху: а — глотка, б — жгутики, в — продольная бороздка, г — хлоропласты, д — пиреноид, е — ядро, лс — пульсирую- щая вакуоля 3 — тельца Мопа, и — трихоцисты, к — спинная Ф сторона, л — брюшная сторона. здесь имеются также и неокрашенные формы (табл. 5, 9). Громадное большинство пирофитовых водо- рослей характеризуется жгутиковой (монад- ной) структурой тела. Именно вследствие этого пирофитовые водоросли зоологи относят к жи- вотным — к классу жгутиконосцев типа прос- тейших (Protozoa). Однако наличие у них еще и других, в том числе типично растительных структур в организации тела (пальмеллоидной, коккоидной, нитчатой, а также наиболее при- митивной — амебоидной) показывает полное своеобразие этих организмов и необходимость считать их самостоятельным отделом среди водорослей, Таллом пирофитовых (за исключением рода Dinothrix с нитчатой структурой тела, рис. 63, 8) является одноклеточным. К олониальные и ценобиальные формы здесь пока не обнаружены. Строение протопласта в общем простое, но панцирь, имеющийся у многих форм, может быть очень сложным (рис. 64 и 65). Питание автотрофное, реже сап рофитное, иногда сме- шанное. Продуктом ассимиляции является крахмал или масло, изредка лейкозин и волю- тин. Размножение в основном вегетативное, путем продольного деления клетки в подвиж- ном состоянии, реже наблюдается бесполое раз- множение зооспорами и автоспорами. Половой процесс достоверно неизвестен.
рис. 6З. Основные типы структуры тела у Тинофито- ВЫХ '. 1 — амебоидная (Dinamoebidium varians); 2 — монадная (He- midinium nasutum); 8 — 4 — пальмеллоидная (Gloeodinium mon- tanum); 5 — 7 — коккоидная (Cystodinium steinii, Stylodinium sphaera, Tetradinium intermedium); 8 — нитчатая (Dinothrix paradoxa). Рис. 64. Пресноводные динофитовые: 1 — Gymnodinium fuscum; 2 — G. paradoxum; 8 — Amphidinium geitleri; 4 — Katodinium planum; 5 — 6 — Peridinium cinctum; У — Ceratium hirundinella. Пирофитовые водоросли широко распростра- нены в водоемах нашей планеты и обитают как в пресных и солоноватых водах, так и в морях, причем представители каждого из подотделов занимают свои экологические ниши. Хлоро- монадофитовые предпочитают естественные водоемы с кислой реакцией среды и встречаются обычно в сфагновых болотах; криптофитовые живут преимущественно в искусственных во- доемах — отстойниках, различных прудах (био- логических, технических, рыбоводных), обыч- но с загрязненной водой, реже в водохрани- лищах и озерах; динофитовые распространены главным образом в чистых пресных водах или в морях. При сборах и изучении пирофитовых водо- рослей необходимо помнить, что они очень чувствительны к колебаниям внешних условий. При воздействии на них любым фиксатором клетки многих видов изменяются до неузнавае- мости и становятся непригодными для изуче- ния. Поэтому изучать строение клеток многих видов можно лишь на живых объектах. Практическое значение пирофитовых водо- рослей заключается прежде всего в их актив- ном участии в круговороте веществ в водоемах. Они способны давать большую биомассу и слу- жат поэтому важным звеном в цепи питания рыб и других гидробионтов, а также играют существенную роль в отложении сапропелей.
nasutum; 8— 5 — С. tripos. удучи весьма чувствительными к органи е ким загрязнениям в воде или, наоборот, тре- уя для своего развития определенного содер- жания органического вещества в ок в окружающеи среде, многие виды пир офитовых водорослей используются в качестве показательных форм сапробности при биологическом анализе воды, т. е. являются так называемыми биоиндикато- рами при санитарно-биологической оценке вод. Многие пи оф р ~итовые„обитающие в загрязнен- ных и сточных водах выполняю 7 т ~ункцию ак- тивных санитаров этих вод, так как принимают пирофитовых известны и ядовитые формы. На- конец, панцирные формы благодаря окремне- нию и обызвествлению панциря хорошо сохра- няются в осадочных образованиях и, подобно диатомовым, широко используются в геологии и практике поисковых работ для определения Рис. 65. Морские динофитовые: 1 — Amphi di nium extensum; 2 — G yrodi п1шп Peri 1шшп divergens; 4 — Ceratium arcticum; возраста осадочных пород (перидинеи из под- отдела динофитовых). Основываясь на гипотезе амебои оидного про- исхождения растительного ми а можно срав- нительно легко вывести пирофитовые от каких- то первичных примитивных амебоидных фо м, давших начало также золотистым водорослям. Состав пигментов, продуктов ассимиляции и строение цист указывают на родственные свя- зи между этими двумя отделами. Наличие т и- хальных форм и своеобразное строение зооспор орое родство позволяют предположить некото ое о между пирофитовыми и бурыми водорослям Сост ями. став и объем пирофитовых водорослей раз- ные ученые понимают по-разному. Большин- ство разделяет точку зрения А. П а ш е р а, объединившего в 1914 г. криптофитовые и дино- фитовые (перидинеи) в отдел (тогда фил ~ Р r- ro h ta. р у . Некоторые выделяют криптофитовых 7 в самостоятельный отдел сох аня аняя название yrrophyta лишь за динофитовыми. Др гие, признавая динофитовые самостоятельным клас- сом, криптофитовых и хлоромонадофитовых от- носят к жгутиковым невыясненного системати- ческого положения. Мы полагаем, что наиболее удачной является точка зрения, согласно ко- торой под названием Pyrrophyta объединяются в ранге подотделов хлоромонадофитовые, крип- тофитовые и динофитовые. Пе ей ем р дем к последовательному знакомству с этими подотделами. ПОДОТДЕЛ ХЛОРОМОНАДОФИТОВЫЕ (CHLOROMONADOPH YTINA) К этому подотделу относятся представители с одноклеточным талломом монадной й структу- ры (рис. 61). Клетки их уплощены в спинно- брюшном направлении. Продольная бороздка всегда тянется вдоль брюшной стороны, в уг- лу лении на переднем конце клетки она пере- ходит в треугольную глотку. Из глотки выхо- дят два неодинаковых жгутика. Трихоцисты (рис. 61, 1з, 8з) или слизистые тельца (рис. 61, 4к), расположены в поверхностном слое цитоплазмы. Хлоропласты многочисленные, мелкие диско- 7 видные, светло-зеленые, между ними — капли масла. Пигментов меньше, чем в других группах, о наружены только хлорофилл а, 3-каротин и несколько ксантофиллов. Есть и бесцветные формы. Вакуольный аппарат сложный. Ядро крупное. Запасное вещество — капли жира. Хлоромонадофитовые размножаются исключи- тельно делением. Распространены преимущест-- венно в сфагновых болотах, реже в озерах и других стоячих водоемах. Типичных представителей подотдела можно обнаружить в свежепринесенной пробе фито-
планктона из сфагнового болота. Гониоапомум семяподобный (Gonyostomum semen) характе- ризуется сравнительно крупными, длиной до 100 мкм, клетками обратнояйцевидной формы, с хорошо различающимися спинной и брюшной сторонами (рис. 61, 1 — 2). При раосматривании медленно движущейся клетки г они остомума под микроскопом внимание привлекает свет- лый треугольник, расположенный спереди. Это и есть глотка, из которой выходят два до- вольно сильных, неодинаковой длины жгути- ка. Один из них, именуемый плавательным, направлен вперед. Вращаясь вокруг продоль- ной оси клетки, как бы ввинчиваясь в воду, он сообщает клетке поступательно-вращатель- ное движение. Второй жгутик назван рулевым. Выйдя из глотки, он загибается вниз под брюш- ную сторону и тянется вдоль продольной борозд- ки, нередко выступая сзади за пределы клетки. Двигаясь влево и вправо, он и «ведет» клет- ку, как кораблик по заданному курсу. Сбоку, возле треугольной глотки, видна крупная пульсирующая вакуоля, изливаю- щая свой секрет в глотку и снова наполняю- щаяся благодаря деятельности окружающих ее многочисленных мелких пульсирующих ва- куолек. Вращая микрометрическим винтом микроскопа, можно легко увидеть эллипсоид- ное, очень крупное, диаметром до 20 мкм ядро, расположенное почти в центре клетки. Клетка покрыта очень тонким и прозрачным перинластом, сквозь который хорошо видны капельки масла и многочисленные мелкие дисковидные светло-зеленые хлоропласты. Гус- то располагаясь в эктоплазме в один сплошной слой, они становятся шестиугольными или многоугольными. Здесь же обнаруживаются мно- гочисленные, тонко-булавовидные, преломля- ющие свет трихоцисты, расположенные между хлоропластами и торчащие во все стороны. Соприкасаясь во время движения с каким-либо предметом — песчинкой, клеткой или нитью водоросли, трихоцисты выбрасываются в виде длинных слизистых нитей, вызывая этим скач- кообразное перемещение клетки в противопо- ложном направлении. Клетки меротрихии головчатой (Merotri- chia capitata) характеризуются наличием го- ловчатого переднего конца, усаженного целым пучком радиально расходящихся игл овидных трихоцист (рис. 61, 8). Не имеющая трихоцист вакуолярия зеленая (Vacuolaria viridis) имеет вполне «миролюбивый» вид, так как ее клетки способны выделять лишь слизь (рис. 61, 4, б). Из краткого описания трех наиболее типич- ных представителей данной группы видно, что хлоромонадофитовые действительно яв- ляются весьма своеобразной и четко ограни- ченной от других группой водорослей. ПОДОТДЕЛ КРИПТОФИТОВЫЕ (CR YPTOPH YTINA) Подавляющее большинство представителей этого подотдела характеризуются типичной мо- надной структурой тела. Таллом у них всегда одноклеточный, и лишь у некоторых видов при размножении могут возникать скопления неподвижных клеток, соединенных слизью. Значительно реже встречаются здесь пальмел- лоидная и коккоидная структуры. Первая из них представлена родом тетрадиниум (Tet- radinium), а вторая — двумя родами — фео- коккус (Phaeococcus) и феоплакс (Phaeoplax). Клетки криптофитовых (рис. 62) дорсовент- ральные, с выпуклой спинной и плоской или слегка вогнутой брюшной стороной, спереди они обычно косо срезаны, имеют носовидный апикальный вырост над углублением, пере- ходящим внутри клетки в мешковидную глот- ку, выстланную трихоцистами. Из глотки вы- ходят два неодинаковой длины жгутика, на- правленные обычно вперед. Б ороздка одна, продольная, на брюшной стороне клетки. Хло- ропласты обычно крупные, пластинчатые, по- стенные, по 1 — 2 в клетке. Они бывают ок- рашены в самые различные оттенки оливкового, коричневого, бурого, желтого, голубого, си- него и красного цветов (табл. 5). Из пигмен- тов здесь обнаружены хлорофилл а и хлорофилл с, я-каротин, я-каротин, несколько ксанто- филлов (а также, возможно, фикоцианин и фикоэритрин). У некоторых видов имеются пи- реноиды. Выделительная система простая. Яд- ро одно, сферическое, небольших размеров. Отличительной особенностью криптофитовых является наличие в клетке одного-двух осо- бых эллипсоидных зерен, так называемых те- лец Мопа. Криптофитовые водоросли распространены преимущественно в мелких искусственных и естественных стоячих водоемах, обычно с заг- рязненной водой. Они нередко встречаются в массовом количестве, вызывая иногда «цвете- ние» воды. Один вид криптофитовых обнаружен в окрашенном снеге. Известны неокрашенные формы, у некоторых из них наблюдаются циа- неллы (табл. 5, 7, 8). Криптофитовые насчитывают 5 семейств, объединяющих 14 родов, самым крупным из них является род криптомонас (Cryptomonas), содержащий свыше 50 видов. Познакомимся ближе с некоторыми предста- вителями криптофитовых. Поздней осенью, когда с понижением тем- пературы в естественных водоемах активная жизнь водорослей угасает, в искусственных водоемах очистных сооружений сахарных за-
водов и городской канализации она продол- жает бурлить. Рассмотришь под микроскопом каплю живого фитопланктона и не поверишь своим глазам: на дворе ноябрь или декабрь, а активная жизнедеятельность водорослей в разгаре, особенно видов криптомонад. Здесь, в массе стремительно перемещающихся, прыгающих в разных направлениях крипто- монад, обычно хорошо выделяются крупные, длиной до 60 мкм клетки криптомонаса искрив- ленного (Cryptomonas curvata). При подсыхании препарата движение клеток прекращается, и можно увидеть все детали строения клеток этого вида (табл. 5,о). Из криптофитовых, обитающих в чистых во- дах, интересен криптомонас плоскохвостый (Cryptomonas platyuris). Живет он в озерах и других стоячих водоемах, но встречается также в бассейнах оранжерей ботанических садов, где нередко развивается в большом количестве. Там его и можно обнаружить в те- чение всего года. Он отличается от других ви- дов рода уплощенным задним концом, кото- рый, как рулевое весло, качаясь из стороны в сторону, способствует передвижению клетки в толще воды (табл. 5, 6). Не имея существенных отличий в строении клетки, представители других родов крипто- фитовых очень хорошо различаются окраской хлоропластов. Так, виды рода родомонас (Bhodomonas) характеризуются темно-красным или ярко-красным цветом хлоропластов, рода хроомонас (Chroomonas) — синей, рода ииано- монас (Cyanomonas) — сине-зеленой, а рода критпохризис (Cryptochrysis) — бурой окра- ской хлоропластов (табл. 5, 1 — 4). Из неокрашенных форм, обладающих цианел- лами, заслуживает внимания род иианофора (Cyanophora), включающий два вида: С. pa- radoxa с одной и С. tetracyana с четырьмя цианеллами (табл. 5, 7 и 8). Представителем неокрашенных форм, не имеющих цианелл, может служить хиломонас (Chilomonas para- maecium), повсеместно распространенный в за- грязненных водах, нередко в навозной жиже (табл. 5, 9). ПОДОТДЕЛ ДИНОФИТОВЫЕ (DINOPH YTINA) Это наиболее крупный и очень полиморфный подотдел пирофитовых водорослей. Он объе- диняет представителей 5 типов морфологи- ческой структуры: амебоидной, монадной, паль- меллоидной, коккоидной и нитчатой (рис. 63). Коккоидная структура представлена доволь- но богато; амебоидная, пальмеллоидная и нит- чатая одним монотипным родом каждая,' мо- надная структура, как и в двух рассмотрен- ных ранее подотделах, здесь является домини- рующей. Многие монадные формы (их часто называют динофлагеллатами или перидинеями) снабжены панцирем. Панцирь у динофитовых всегда дорсовент- ральный, с двумя бороздками, из них попе- речная делит его на две равные (рис. 64, 1 — 2) или неравные (рис. 64, 8 — 4) части — верх- нюю (эпивальву) и нижнюю (гипо- в а л ь в у), состоящие из пластинок, соеди- ненных швами (рис. 64, 5' — 6). У неподвижных форм имеется настоящая двухслойная целлю- лозная оболочка (рис. 63, 8); у амебоидных форм — перипласт, формирующий псевдопо- дии (рис. 63, 1). У монадных форм всегда два неравных жгутика: поперечный в виде волнистой упло- щенной ленты, который лежит в поперечной бо- роздке и сообщает клетке вращательное дви- жение вокруг продольной оси, и нитевидный продольный жгутик, который тянется вдоль продольной бороздки и определяет поступа- тельное движение клетки вперед (рис. 64, 1,2,4). Хлоропласты у динофитовых обычно много- численные, мелкие, дисковидные, реже эллип- соидные или лентовидные (рис. 64, 1 — 4), окрашенные в темно- или светло-бурый, зо- лотистый, желтый, реже зеленоватый и сине- зеленый цвет. Из пигментов здесь обнаружены хлорофилл а и хлорофилл с, Р-каротин и пять ксантофиллов. Ядро одно, обычно крупных размеров. Основным продуктом фотосинтеза является масло. У некоторых динофитовых водорослей обнаружены трихоцисты. Монадные формы размножаются делением панциря на две более или менее равные части, каждая из которых позже вырабатывает не- достающую половину. У коккоидных, паль- меллоидных и трихальных форм, кроме деле- ния клеток, наблюдается образование зооспор, напоминающих подвижные клетки гимнодини- ума с системой бороздок и глазком; реже обра- зуются автоспоры. Указания на половой про- цесс малодостоверны и требуют дополнитель- ного подтверждения. У динофитовых нередко наблюдаются толстостенные долговечные (до 16 лет) цисты. Динофитовые водоросли обитают преиму- щественно в чистых пресных или соленых водах, встречаются обычно в холодную пору года; известны криофилы, обитающие в снегу и при- дающие ему органогенную окраску. Разви- ваясь в массовых количествах, некоторые виды вызывают «цветение» воды. Перейдем к знакомству с некоторыми пред- ставителями пресноводных динофитовых.
Ранней весной, после таяния снега, повыша- ется уровень воды в многочисленных небольших и незаметных летом стоячих водоемах, в кана- вах, пойменных озерах и сфагновых болотах. Вода в них еще мутная от взвесей и кажется совсем лишенной жизни. Но это не так. Если рассмотреть под микроскопом капель- ку воды, взятой из сфагнового болотца и про- пущенной через центрифугу, то окажется, что она далеко не безжизненна. В этой сгущен- ной капельке фитопланктона вы увидите окра- шенных представителей холодолюбивых форм водорослей. Здесь будут и представители золо- тистых, как светлячки, отражающие лучи све- та, и быстро проносящиеся оливковые, синие или голубые клетки криптофитовых, и темно- коричневые клетки суетящихся динофитовых водорослей. Когда препарат постепенно про- греется и подсохнет, кипучая жизнь в нем замедляется. Теперь можно подробнее позна- комиться с обитателями капельки. Привле- кают внимание желто-коричневые клеточки, у которых верхняя половинка панциря — эпи- вальва — широко округлая, а нижняя поло- винка — гиповальва — обратно-остро-кони- ческая (рис. 64, 1). Это гимнодиниум темный (Gymnodinium fuscum). При вращении его клет- ки вокруг продольной оси тела заметны попе- речная и продольная бороздки, а в месте их пересечения — выход двух жгутиков. Один из них тянется вдоль продольной бороздки и, ка- чаясь из стороны в сторону, как мышиный хвостик, продвигает клетку вперед. Второй жгутик расположен в поперечной бороздке и, спирально извиваясь, сообщает клетке враща- тельное движение вокруг продольной оси. Бла- годаря усилиям этих двух жгутиков клетка гимнодиниума очень быстро передвигается в толще воды. В спокойно лежащей клетке хорошо видны многочисленные мелкие, коричневатые хлоро- пласты, расположенные обычно сплошным слоем по периферии протопласта (рис. 64, 1). Вращая микрометрический винт, можно увидеть в цен- тре клетки ядро и капельки масла. В пробе фитопланктона встречаются и дру- гие представители этого большого рода, на- пример гимнодиниум парадоксальный (G. para- doxum) с хорошо заметным крупным красным глазком (рис. 64, 2). Нередко встречаются также представители других родов — амфи- диниум и катодиниум (рис. 64, 8, 4). Из крупных панцирных форм в зимних и ранневесенних пробах фитопланктона нередко попадается перидиниум опоясанный (Peridinium cinctum). Его аккуратный, шаровидный корич- невый панцирь состоит из постоянного числа Бластинок, определенным образом соединен- ных широкими поперечнополосатыми швами (рис. 64, 5' — 6). Перед жарким летом или силь- ными морозами, когда начинается цистообра- зование, перидиниум опоясанный сбрасывает жгутики, его протопласт внутри панциря сжи- мается, покрывается несколькими толстыми слоистыми оболочками, и он опускается на дно водоема. После периода покоя его содержимое делится и дает молодые подвижные клеточки. Вначале они бледно-желтые, почти голые, с тонким и прозрачным панцирем, но вскоре их протопласт увеличивается в объеме, швы пан- циря разрастаются, пластинки становятся скульптурированными, окрашиваются солями железа в коричневый цвет — и панцирь взро- слой особи готов. Теплолюбивых панцирных форм динофито- вых водорослей сравнительно мало. Типичным представителем их является церациум ласточ- ковый (Ceratium hirundinella). Панцирь его характеризуется наличием выростов, или рогов: одного длинного переднего, или апикального (на эпивальве), и двух-трех коротких задних, антапикальных (на гиповальве). При движе- нии клетки панцирь ее с вытянутым вперед апикальным и более или менее раздвинутыми антапикальными рогами очень напоминает те- ло летящей ласточки, что и послужило поводом для видового названия водоросли (рис. 64, 7). Часто развиваясь в массовом количестве, этот вид вызывает «цветение» воды, придавая ей буровато-белесую окраску. Это явление мож- но наблюдать летом в заводях и пойменных озерах рек Украины. Цветущие там в это время белые кувшинки на фоне буровато-белесой водной глади выглядят необычайно эффектно. Особенно большого морфологического раз- нообразия достигают панцири морских видов динофитовых водорослей монадной струк- туры (рис. 65). Пальмеллоидную структуру тела имеет гле- одиниум горный (Gloeodinium montanum), встречающийся в озерах и сфагновых болотах. Клетки его неподвижны и находятся внутри слизистой слоистой оболочки (рис. 63, 8, 4). Размножаясь простым делением, они могут давать значительные слизистые скопления. В коккоидной группе динофптовых наиболее богато представлен род цистодиниум (Cysto- dinium). В весенней пробе фитопланктона из сфагнового болота мы можем найти одиночные коричневатые клетки полулунной формы — это цистодиниум Штейна (Cystodinium steinii, рис. 63, б). В отличие от мечущихся в препа- рате монадных форм динофитовых это непо- движно лежащие клетки. Когда наступает время размножения, содер- жимое каждой клетки делится; в результате образуются две крохотные, аккуратные зоо- споры. Они покидают материнскую оболочку,
отдкл золотистык водоросли (cHRYs0pH YTA) ~оо некоторое время плавают, а затем сбрасывают жгутики и, разрастаясь в длину, снова развива- ются в полулунную клетку. Перед наступле- нием зимы последние в серии летних поколе- ний клетки формируют толстую, многослой- ную оболочку и опускаются на дно. После зимнего покоя, навстречу весенним солнеч- ным лучам выплывают развившиеся внутри цист зооспоры — родоначальницы новых лет- них поколений. И все повторяется сначала. Сходный цикл развития имеют и прикреп- ленные формы коккоидной структуры из ро- К отделу золотистых относятся водоросли, преимущественно микроскопические, хлоропла- сты которых окрашены в золотисто-желтый цвет. Из пигментов здесь обнаружены хлоро- филл а, однажды — хлорофилл е и много каротиноидов, в том числе каротин и не- сколько ксантофиллов, больше всего золоти- стого фукоксантина. В зависимости от соот- ношения этих пигментов окраска золотистых водорослей может приобретать различные от- тенки: от чисто-золотисто-желтой до зелено- вато-желтой и зеленовато-бурой. Необычайно красочную картину представ- ляют живые клетки золотистых водорослей при рассматривании их в ярко освещенном поле зрения микроскопа, — как будто каждая из них отражает солнечный лучик. Однако при отмирании клеток золотистая окраска хромато- форов исчезает и они становятся зелеными в связи с растворением в воде желтых пигмен- тов и демаскировкой хлорофилла. В процессе фотосинтеза в клетках золотистых водорослей вместо крахмала вырабатывается особый углевод — лейкозин. Распространены золотистые водоросли по все- му земному шару, но чаще встречаются в уме- ренных широтах. Живут преимущественно в чистых пресных водах и особенно характер- ны для кислых вод сфагновых болот. Небольшое число их обитает в морях и соленых озерах, значительно реже встречаются они в загряз- ненных водах. В почвах представлены еди- ничными видами. Развиваются золотистые водоросли обычно в холодную пору года: ранней весной, позд- ней осенью и зимой. Большинство золотистых водорослей — типичные представители планк- тона, в основном это активные планктеры (рис. 68, 4 — 8, 69, 1,4). Некоторые живут в поверхност- ной пленке натяжения воды, будучи предста- вителями нейстона (рис. 68, 1 — 8). Среди золотистых водорослей много эпифитных форм дов стилодиниум (Stylodinium, рис. 63, 6) и тетрадиниум (Tetradinium, рис. 63, 7). Наконец, интересен динотрикс парадок- сальный (Dinothrix paradoxa) с нитчатой струк- турой тела (рис. 63, 8, 9). Это типичный пред- ставитель многоклеточных динофитовых, тал- лом которых разрастается, как и у нитчатых форм других отделов, в результате деления кле- ток. При бесполом размножении этой водорос- ли возникают зооспоры, напоминающие под- вижные вегетативные клетки представителей рода гимнодиниум. (рис. 74), реже встречаются собственно бен- тосные формы (рис. 73). Золотистые водоросли характеризуются боль- шим морфологическим разнообразием. В преде- лах этой группы представлены почти все ос- новные типы структуры тела водорослей: аме- боидная, монадная, пальмеллоидная, кокко- идная, нитчатая, разнонитчатая и даже пла- стинчатая (табл. 6). Золотистые водоросли бывают одноклеточ- ными (рис. 66 и 68), колониальными (рис. 67 и 69) и многоклеточными (рис. 75). Кроме того, среди них имеется один очень своеоб- разный представитель с многоядерным тал- ломом в виде голого плазмодия (рис. 67, 8 — о). Независимо от общего уровня организации тела внутреннее строение клетки золотистых водорослей весьма однотипно (рис. 68,4). В про- топласте размещается обычно один или два постенных корытовидных хлоропласта, на ко- торых у некоторых видов можно обнаружить красный глазок или трудно различимый голый пиреноид. Ядро одно, обычно небольших раз- меров, заметное только после обработки клет- ки специальными красителями. У некоторых видов, обычно в морфологически передней ча- сти клетки, имеется одна или две пульсирую- щие вакуоли. В отличие от внутреннего содержимого по- кровные части клетки у золотистых водоро- слей обнаруживают исключительное разно- образие. У простейших представителей отдела клетка покрыта нежным перипластом, позво- ляющим ей производить выпячивания, полу- чившие в зависимости от их формы названия ризоподий, псевдоподий или аксоподий (рис. 66). У большинства золотистых водорослей пери- пласт более плотный и клетка имеет постоян- ную форму. Многие виды снабжены жгутика- ми, строение, число, размеры и расположение которых имеет важное систематическое значе- ние. У высоко организованных представи-
телей отдела клетки покрыты настоящей цел- люлозной, обычно двухконтурной оболочкой. У многих видов протопласт клетки находится в «домике» (рис. 68, 6 — 8) или покрыт панцирем, состоящим из чешуек, несущих на себе шипы или иглы (рис. 68, б; табл. 7). Размножаются золотистые водоросли про- стым делением клетки (рис. 66, 4), а также пу- тем распада кочоний или многоклеточного таллома на отдельные части. Наблюдается так- же бесполое размножение при помощи одно- жгути тиковых или двужгутиковых зооспор (рис. 68, 8; 73, 2; 75, 1, 2), реже амебоидов (рис. 67, б) или автоспор. Известен также по- ловои проце сс в виде типичной изогамии, хо- логамии или ав втогамии. В результате полового процесса о р об азуются очень разнообразные по характеру скульптурированности о олочки зндогенные кремнистые цисты (рис. 73, 8), которые помогают золотистым водо- ослям переживать неблагоприятные условия. Значение золотистых водорослей как фото- т офных организмов заключается прежде все- го в создании первичной продукции в водое- мах и в участии их в цепи питания гидробионтов, в том числе и рыб. Большую роль они играют также в улучшении газового режима водое- мовивор б азовании отложений сапропеля. а отопая, Яу- Некоторые виды из родов Ма11отоп аз, у- nura, Глпо гуоп D b могут вызывать «цветение» во ы придавая ей запах рыбьего жира и ухуд- шая ее питьевые и технические качес тва. Осо- ыбных хо- бенно тяжелые последствия для ры зяйств имеет «цветение» рыбоводных прудов, вызываемое массовым развитие ием токсического вида примнезиума маленького (Prymnesium par- vum). В соответствии с новейшими данными отдел золотистых водорослей (Chrysophyta) делят на х изомонадовые (Chrysomonadophyceae), хризо- ка апсовые (Chrysocapsophyceae), хризос еровые (Chrysosphaerophyceae) и хризотриховые ( у- вые Ghr- sotrichophyceae). КЛАСС ХРИЗОПОДОВЫЕ (CHRYSOPODOPH YCEAE) К классу хризоподовых относя т наиболее просто устроенных представителей золотис- тых водорослеи. Характерной особенностью ор ганизации их тела является наличие пе и- пласта — тонкой и и нежной оболочки, позволя- ющей клетке изменять свою + р у ~~о м и о разо- вывать выпячивания. Это типичные предста- вители амебоидной (ризоподиальной) струк- т ы. Несмотря на примитивность такой орга- низации тела, хризоподовые р~ в мо ~ологическом Ф ФФ Ф ° ° ° ° ° ° ° еФ ° ° ФФOOO° ° °, ° Ф ° Ф ФФ r Ф ° ФФ ° ° ° ° r Ф ° Ф Рис. 66. Одноклеточные хризоподовые: 1 — 4 — Chrysamoeba radians (1 — ризоподиальное состояние, — с зачаточным жгутиком, 8 — монадная стадия, — tod 8 — Derepyxis Eleutheropyxis arachne; 7 — Х адуп1о p cys о inii; anomala. отношении очень разнообразны. Это проявля- ется главным образом в характере выпячива- ласс ний, строении «домиков» и колоний. ла хризоподовых включает свыше О родов. Хризоподовые представлены одноклеточны- ми, колониальными и плазмодиальными фор- мами. Все они микроскопических размеров, свободно плавают или прикрепляются к су стра- т . Обитают почти исключительно в пресных водах, где встречаются в планктоне, обрастани- ях а также в нейстоне и бентосе, на частичках 7 детрита. Класс хризоподовых подразделяют о ычно на три порядка и шесть семейств. з них наиболее просто устроены хризалебовые. К рассмотрению одного из го из наиболее типичных представителей этого семейства мы и перейдем. Примером мо- жет служить хризамеба лучистая (Chrysamoeha radians), одиночные клетки которой можно обнаружить в планктоне рек, озер, стариц и болот, а также в заболоченных и сильно за- росших вь ысшей водной растительностью кана- вах, прудах и в в других небольших стоячих водоемах.
ъ ° ° ° е ° ° + ° ° е е»в ° ° ° ° ) ° ° . ° с- ° ввв- Ф ° е ° ° ° ° е ° е Ю ° в r~ ~в' ФЧ ФВ л ,е»Ф ° ав ° а ° + ае «Se» ° Ф ° е ° е ° е Ф ° в ° е Ъ Ф Ф ° ФФФФФ ° ° Ф ° åå~ ~~Фее ФФ \ ° ° е Ф ° ° ° ~ ° ° Ъ ° ° » ° аО Ф I ° ° Ф Ф б ве Ф ° ° °, б ° ° ° ° еееее ° ° ° е ° ° . е ° ° ° ° ° ° Ф ° ° ° ° Фе ° ° ° ° ° ° ° ° е- ° ее ° ° 'i ч б еее ее ебе ° äðð е ° б ° Ф в ° ° ° в) Ф1е Ф б ° ~...,„."..Q е ° ° в е ° o~ в ° ° ° е Фе ° е ° ° ° Ô Ô~ ° Ф Ъ е Ф ° ФФ ° Ф ° е ° е ° ее ° ч \ 'Ф б . ,с еб O Ф ° ° ° ° ° Ф ° ° å В ° ° ° ° g<>i, фс~ "е 2 ° е ° ° Ф Фе ° ° беа °, r, °,,' аве, ° аб ° Ф ее ° ° :I б вГ, ° вб ° ° б ° ° е ° O б ° Ф в е в ,вв I с Ф Рис. 67. Колониальные хризоподовые.' 1 — Chrysidiastrum catenatum; 2 — Heliapsis mutabillis, ризо- подии выходят через поры в стенке «домиков»; 8 — 5 — Мухо- с~гу818 paradoxa (8 — молодой, 4 — взрослый плазмодий, 5— образование амебоидов). Клетки хризамебы в очертаниях округлые, диаметром до 17 мкм. Рассматривая живую хри- замебу под микроскопом (рис. 66, 1 — 4), мож- но легко увидеть по периферии клетки нераз- ветвленные, утолщенные у основания ризопо- дии, по длине равные примерно диаметру клетки. Эти ризоподии на глазах у наблюда- теля меняют свою форму: удлиняются и стано- вятся тонкими или, наоборот, утолщаются за счет сокращения длины, они могут исчезать совсем или возникать вновь. За счет этих из- менений происходит перемещение клетки по субстрату. В цитоплазме клетки видны два постенных корытов идных бледно окрашенных в светло- желтый цвет хлоропласта. Между ними, в центральной части клетки, лежит обычно одна крупная блестящая глыб к а лей ко зина. Осве- щенная в препарате, она, подобно призме, преломляет свет и воспринимается наблюдате- лем как миниатюрный голубой фонарик, осве- щающий изнутри хлоропласты. В периферическом слое цитоплазмы в одном- двух, реже трех местах обнаруживаются не- большие светлые пузырьки, которые появляют- ся, постепенно увеличиваются до определенных размеров, исчезают и снова появляются. Это пульсирующие вакуоли, выполняющие в клетке функцию выделительной системы и осуществля- ющие осморегуляцию. Хриз амеба относится к одноядерным орга- низмам, но в живой клетке мы ядра не увидим. Для того чтобы его увидеть, необходимо при- менить специальные красители. Ядро хриза- мебы имеет небольшие размеры и расположено в углублении одного из хлоропластов. Живая клетка хризамебы имеет зачаточный жгутик, причем основание его приурочено имен- но к тому месту в клетке, где расположено ядро. При движении хризамебы по субстрату зачаточный жгутик обычно находится на са- мой возвышенной, обращенной к наблюдате- лю части клетки, т. е. направлен вверх, и, на- ходясь в движении, малозаметен среди много- численных ризоподий. В процессе своей жизнедеятельности хриза- меба лучистая может временно переходить в монадную стадию. Перед этим клетка пре- кращает движение, постепенно втягивает внутрь ризоподии, увеличивается в размерах и становится яйцевидной или шаровидной. Не- втянутым оказывается лишь зачаточный жгу- тик, расположенный на более узком светлом конце клетки (рис. 66, 8). Слегка удлиняясь и продолжая вращаться, жгутик набирает силу и становится на время единственным аппара- том движения. Клетка хризамебы отрывается от субстрата и отправляется в путешествие в толще воды. Быть может, в естественных усло- виях эта прогулка и продолжительна, но в препарате под микроскопом она весьма крат- ковременна, исчисляется минутами. Здесь клет- ка быстро замедляет движение, теряет округ- лую форму, на ее поверхности появляются вы- пячивания, она оседает на предметное стекло и расплывается, как капля жидко заварен- ного крахмала. И снова начинается ее медлен- ное передвижение по субстрату в ту сторону, куда направлена большая часть возникающих ризоподий. Размножается хриз амеба лучистая вегета- тивно, путем простого деления клетки надвое (рис. 66, 4). Деление происходит во время дви- жения клетки и протекает довольно быстро, примерно в течение 5 — 15 мин, при этом на короткое время могут возникать скопления, включающие от 2 до 20 хризамеб. Последние, не будучи соединены плазмодесмами, вскоре расползаются в разные стороны и живут как еамостоятельные одноклеточные организмы. Ин- тересно, что в процессе деления могут появ-
ляться и бесцветные хризамебы, без лородда- стов. Объясняется это тем, что темпы деле- ния клеток иногда обгоняют темпы размноже- ния хлоропластов в пределах клетки, в силу чего одна из дочерних клеток, появившихся в процессе деления, оказывается лишенной хлоропластов. Такой же результат может по- лучиться, если плоскость деления материн- ской клетки окажется смещенной и не разъе- динит хлоропласты. Это один из частных путей возникновения бесцветных форм, которые встречаются почти во всех отделах водорослей. Кроме рода хризамеба, насчитывающего в настоящее время около двух десятков видов, в пределах класса хризоподовых известны другие роды, включающие одноклеточные свободно- плавающие и прикрепленные (рис. 66, о) фор- мы. Имеются виды, клетки которых сидят в «домике» (рис. 66, 6). Последний может быть сидячим (рис. 66, 7), на коротком (рис. 66, 8) или длинном стебельке. У колониальных форм хризоподовых при вегетативном размножении клетки не расхо- дятся, а остаются соединенными плазмодес- мами в колонии в виде цепочки (рис. 67, 1) или сеточки (табл. 6, 2). Клетки такой колонии мо- гут иметь «домики» (рис. 67, 2). Плазмодиальные формы среди хризоподовых представлены одним родом с единственным видом миксохризис парадоксальный (Myxo- chrysis paradoxa), вегетативное тело которого— голая протоплазма с многочисленными ядрами и хлоропластами в ней (рис. 67, 8 — 5). Такие плазмодии образуются путем слияния многих отдельных амеб в одну общую массу. КЛАСС ХРИЗОМОНАДОВЫЕ (CHR YSOMONADO P H Y CEAE) К классу хризомонадовых относят предста- вителей с монадной (жгутиковой) структурой тела. Характерной их особенностью является наличие жгутиков, позволяющих им активно пе- редвигаться в толще воды и находиться во взве- шенном состоянии. Б ольшинство из них- типичные активные планктеры и лишь некото- рые ведут прикрепленный образ жизни. Клетки хризомонадовых (рис. 68) обычно снабжены одним или двумя жгутиками. В по- следнем случае жгутики неравной длины, лишь у немногих представителей оба жгутика одинаковые. Имеются виды с тремя разными жгутиками, или жгутиков может быть четыре в виде двух неодинаковых пар. В этом классе наблюдается очень большое морфологическое разнообразие, заставившее по- делить его на 3 самостоятельных подкласса: Рис. 68. Одноклеточные хризомонадовые: 1 — 8 — Chromulina rosanoffii (1 — метаболия клетки, в н и з у- глыбка лейкозина, в в е р х у — хлоропласт, 2 — циста, 8 — зоос- пора, вышедшая из цисты); 4 — 5 — Mallomonas denticulata (4— клетка, в в е р х у — крупное ядро, по б о к а м — хлоропла- сты, в н и з у — пульсирующие вакуоли, 5 — чешуйки и иглы панциря); 6 — Chrysococcus radiatus; У — Pseudokephyrion acu- tum; 8 — P. elegans. собственно хризомонадовые, или хризомонады (Chrysomonadophycidae), кокколитофориды (Coccolithophoridophycidae) и силикофлагелла- ты (Silicoflagellatophycidae). Однако следует заметить, что, несмотря на это многообразие, преобладание жгутиковых форм в классе хри- зомонадовых дало зоологам повод относить зо- лотистые водоросли к животному царству и помещать их в составе общего класса жгутико- носцев в пределы типа простейших (Protozoa). Как уже указывалось (ср. отдел «Пирофитовые водоросли»), согласиться с этим нельзя. ПОДКЛАСС ХРИЗОМОНАДЫ (СН В ТЯОМОХАРОРНТС1РАЕ} Рассмотрим более подробно одного из наибо- лее известных и хорошо изученных представи- телей современных хризомонад, относящегося к роду хромулина.
~~гот o~ulina rosanoffii (рио б~ 1 — 8 озерах, нру а ~ ~ — ) обитает в х, сфагновых болотах ста рек, цементированн б , старицах танических с ых ассейнах о ранжереи бо- адов, в канавах и водоемах. Ее мо других стоячих е можно найти в отк ыть ослеи, причем част количестве и на п асто в массовом тве и на протяжении всего го типичный предста года. Это тавитель планктона но хорошо представлен но очень нныи и в нейстоне. К 10 мкм в длин х омулины, не п ев р вышающие цикл своего ину, спосо ны ос е ущ ствлять весь о развития в пове хно натяжения во ь Р рхностной пленке воды. При массовом аз ток хромулины развитии кле- ны вода в водоеме ок чневыи цвет. П а мм такой пленки может 7 около 23 000 к ожет приходиться клеток хромулины. Х ромулина очень хорошо вы е б ато ные слов ия, может сп У ся в нак ытых ст ками кристаллизато ах нап стеклянными крыш- ет~ водои из водоема и пост е верное окно комнаты. и поставленных на се- Препараты для изучения х сделать нал у ия хромулины легко ь, наложив зажатое пин ето ное стеклышко цетом покров- сталлизаторе: к о на поверхность р воды в кри- клетки хром лины к стек лышк к прилипают которым и зак ывают воды на предметн У По тном сакле. т каплю од микроскопом при малом вел можно увидеть мн и малом увеличении При большом множество вига д ющихся точек. ом увеличении же п и стро передви вигать препа ат гон у приходится бы- клеткой. гоняясь за одной Рис. 69. К олониальные х изо . изомонадовые: 1 — шаровидная колония Uro 1 rogenopsis americana; колония Hyalobryon ramosum t — на слизистых ножках Спг 2 — колония н — кустистая свободноплаваю ающая колония Dinobr — кустистая гуоп vergens.
Детали строения тела хромулины необхо- димо изучать при иммерсии (иммерсия- введение между объективом микроскопа и рас- сматриваемым предметом жидкости для уве- личения разрешающей способности микроско- па) на временно приостановившейся или при- клеившейся к предметному стеклу клетке. Под микроскопом видно, что клетка хрому- лины не расплывается, как это наблюдалось у хризамебы, не возникают у нее и ризоподии. Здесь протопласт покрыт более плотным пе- рипластом, придающим клетке более или менее определенную форму. Клетка способна лишь в небольших пределах изменять свою форму (метаболировать) от шаровидной до яйцевидной (рис. 68, 1). Внутри клетки хромулины имеются уже из- вестные нам органеллы: хлоропласт, пульси- рующие вакуоли, а после окраски становится заметным и ядро. На морфологически перед- нем конце здесь всегда один жгутик. Размножается хромулина путем продоль- ного деления, чаще всего в пальмеллевидном состоянии. В конце вегетации могут возникать кремнистые, тонкостенные цисты с гладкой по- верхностью и длинной, слегка расширенной у отверстия, шейкой (рис. 68, 2). После периода покоя циста прорастает, из нее выходят 1— 2 — 4 зооспоры (рис. 68, о). Выйдя из оболочки цисты, зооспора становится самостоятельной вегетативной клеткой. В течение лета деление и образование зооспор может повторяться мно- гократно. А так как хромулина является пре- красным кормом для ракообразных, то увели- чение ее численности способствует улучшению кормовой базы для многих гидробионтов во- доема. У многих одноклеточных представителей класса хризомонадовых клетки способны вы- рабатывать или (<домик», отличающийся боль- шим морфологическим разнообразием (рис. 68, 6 — 8), или панцирь, состоящий из кремнистых чешуек, иголочек и шипов (например, у Mallo- топая denticulata, рис. 68, 4 — Б). Изучение кремнистых чешуек хризомонад под электрон- ным микроскопом позволило выявить их чрез- вычайно сложный, иногда ажурный рисунок, оказавшийся постоянным и очень важным си- стематическим признаком (табл. 7). У колониальных форм класса хризомона- довых клетки соединяются при помощи общей слизи (рис. 69, 1), слизистых нитей, или тяжей (рис. 69, 2), или непосредственно своими зад- ними концами (табл. 6, о). У некоторых видов формируются кустистые колонии, состоящие из удлиненных (<домиков» с одиночными клет- ками в верхней части каждого из них, на- пример у гиалобриона (Hyalobryon ramosum, рис. 69, 8). Особого упоминания в этом отно- Рис. 70. Скопление колоний динобриона (Dinobryon divergens, X 300). Микрофотография Г. В. Кузьмина. шении заслуживает род динобрион (Dinobryon, рис. 69, 4, 70), свободноплавающие кустистые ко- лонии которого напоминают канделябры, не- сущие на своих разветвлениях множество зо- лотистых клеток, сидящих у выхода каждого (<домика». Позже — у одних видов постепенно, у других почти одновременно — эти клетки превращаются в золотисто-оранжевые шаровид- ные цисты, закрывающие устье всех «домиков», и тогда колонии становятся похожими на гро- здья спелой рябины. ПОДКЛАСС КОККОЛИТОФОРИДЫ (COCCOLITHOPHORIDOPH YCIDAE) В состав этого подкласса входят как современ- ные, так и ископаемые формы. Современные кокколитофориды — это свое- образные микропланктонные одноклеточные ор- ганизмы, сходные с хризомонадами. Своеобра- зие их заключается в том, что на оболочке они несут различные по форме и размерам известко- вые образования, называемые к о к к о л и т а- м и. Размеры клеток современных кокколито- форид чрезвычайно малы, обычно не более 30 мкм, а кокколиты имеют размеры от 0,25 до 30 мкм. Известковые кокколиты образуются внутри живой массы протоплазмы кокколито- форид, а затем выталкиваются ею на поверх- ность, где распределяются в один слой. Способ облицовки клетки кокколитами самый разно- образный. У некоторых видов кокколиты рас-
10 Рис. 7i. Строение кокколитофорид: 1 — Pontosphaera huxleyi; 2 — Coccolitus pelagicus; 8 — Discosphaera thomsonii; 4 — Rhab dosphaera clavi ger; 5 — 11 — типы кокколитов (0<2000). сеяны по поверхности клетки со значительными промежутками, у других примыкают друг к другу, у третьих перекрываются (рис. 71, 1 — 4). Значение этих известковых образований на теле клетки до сих пор точно не выяснено. Не- которые ученые считают их плавательным уст- ройством. Протопласт кокколитофорид состоит из про- топлазмы, ядра и хлоропластов (в большинстве случаев двух). Строение мембраны, в которую за- ключена протоплазма, изучено слабо. Окраска кокколитофорид зеленоватая или желтовато- коричневая. Кокколитофориды снабжены двумя жгути- ками. У большинства видов жгутики длинные, очень тонкие, хрупкие, нитеобразные, но у не- которых они бывают и короткие. Современные кокколитофориды населяют в морях и океанах толщу воды от поверхности до глубины 150 м. Большинство их распростра- нено в морских бассейнах экваториальных и умеренных широт. В холодных водах обитает лишь несколько видов. Солоноватоводных и пресноводных кокколитофорид известно очень мало. Для их жизнедеятельности важно не только количество растворенных в воде извест- ковистых веществ, но также освещенность и прозрачность воды. Обилие кокколитофорид характерно для открытых вод морей и океа- нов, но иногда их бывает много и в прибреж- ных водах. Систематика кокколитофорид основана как на форме всего панциря или всей клетки в целом, так и на форме отдельных кокколитов. Кокколиты делят на две группы: дисколи- ты — сплошные и тремалиты — с осевым ка- налом в центре (рис. 71, 5' 11; табл. 8, 8, 4). Для отдельных кокколитов, которые, как пра- вило, встречаются в ископаемом состоянии в разобщенном виде, применяют разные клас- сификации, основанные на различиях в их внут- ренней структуре. Обычно их делят на две круп- ные группы: гелиолиты (Heliolithae) — со сфе- рической структурой и ортолиты (Ortholithae)— с кристаллической структурой. Если рассма- тривать гелиолиты под микроскопом в скрещен- ных николях (николь — призма из исландского шпата для поляризации света), то в центре вид- на фигура черного креста с прямыми или изо- гнутыми балками (табл. 8, 7). У ортолитов это- го нет. Ввиду чрезвычайно малых размеров кокколи- тов в последнее время для их изучения стали применять электронные микроскопы с увели- чением в 5000 — 40 000 раз. На полученных снимках видны такие детали строения кокко- литов, которые до этого не были известны (табл. 8,1,2). Применение столь больших уве- личений позволило разработать гораздо более детальные классификации для ископаемых кок- колитов. Если взять кусочек обычного мела и по- скрести его ножом, осыпая порошок в виде пуд- ры в каплю воды на предметном стекле, то, рас- смотрев эту эмульсию под микроскопом (прав- да, при увеличении не менее 1500 раз), мы уви- дим большое количество разнообразных кок- колитов и их обломков в виде звездочек, кру- жочков и других фигур. Известковые панцири кокколитофорид в ископаемом состоянии сла-
гают мощные пласты известняков и мергелей, образуют современные океанические отложения, а также многие породы на материках. Мощность таких пластов может достигать нескольких сотен метров (например, на юге СССР). Некото- рые известняки состоят на 50 — 75оо из кокко- литов. Так, например, в 1 ммз породы третич- ного периода было установлено 5 500 000 эк- земпляров кокколитов. С середины ХХ в. кокколиты стали приме- нять для определения геологического возраста пород, которые их содержат. Оказалось, что очень многие виды кокколитофорид существо- вали по времени очень недолго, но имели ши- рокое, почти глобальное географическое рас- пространение. Одни виды сменяли другие, от- ражая изменение обстановки (климата, соле- ности, прозрачности воды) в различных райо- нах земного шара и эволюционное развитие группы этих организмов в целом. Поэтому, рассматривая под микроскопом соответственно обработанные образцы пород, мы можем уз- нать, к какому периоду геологической истории Земли относятся отложения, представленные этим образцом. В ископаемом состоянии кокколиты извест- ны с кембрия, но описаны и изучены только с юры. Наиболее хорошо изучены меловые и особенно третичные к о к колиты. В третичном периоде особенно широкое развитие получили дискоастериды — одно из семейств кокколито- форид, представители которого имеют вид раз- нообразных звездочек (табл. 8, 5', 6). ПОДКЛАСС СИЛИКОФЛАГЕЛЛАТЫ (SILICOFLAGELLATOPHYCIDAE) Силикофлагеллаты, или кремневые жгутико- вые, как и хризомонады, относятся к водоро- слям с монадной (жгутиковой) структурой тела. От хризомонад они резко отличаются наличием внутри клетки кремневого скелета, состоящего из трубчатых перекладин. К этому подклассу относятся и современные и ископа- емые формы (табл. 9). Впервые скелеты силикофлагеллат были об- наружены около 140 лет назад в меловых по- родах. Позднее аналогичные скелеты нашли в планктоне всех современных морей. Однако, несмотря на многочисленные материалы, си- стематическое положение этих организмов дол- гое время оставалось неясным. Лишь после того, как с большим трудом удалось выловить целые клетки, не разрушив их нежного со- держимого, была установлена принадлежность силикофлагеллат к золотистым водорослям. Строение живой клетки силикофлагеллат хо- рошо изучено у современного морского пред- ставителя рода диктиоха (Dictyocha fibula, табл. 9, 1). Размеры водорослей этого вида ко- леблются от 10 до 30 мкм. Поэтому только при больших увеличениях (в 1000 раз и более) вну- три клетки видны ядро, многочисленные жел- товато-коричневые или зеленовато-коричне- вые зерна хлоропластов и мелкие блестящие капельки запасного питательного вещества- лейкозина. Особой оболочки у клеток нет, поэтому их форма целиком соответствует форме внутреннего скелета. Скелет у этого вида со- стоит из квадратного основания — базального кольца, от углов которого отходят заостренные выросты — радиальные рога; над базальныщ кольцом приподнимается апикальное, обращен- ное кверху образование. Вблизи одного из радиальных рогов отходит жгутик, около ро- гов видны также длинные тонкие нити — псев- доподии. Из строения клетки диктиохи видно, что она очень хорошо приспособлена к планктонному образу жизни. Перекладины скелета полые, трубковидные, очень легкие и вместе с тем прочные. Радиальные рога также увеличива- ют плавучесть клетки, способствуя уменьше- нию ее удельного веса. Благодаря вращатель- ному движению жгутика, клетка может пере- мещаться в толще воды. Роль псевдоподий не совсем ясна. Можно думать, что при их помо- щи клетка опускается и поднимается в толще воды. Возможно также, что в период размноже- ния псевдоподии участвуют в построении ске- лета. Размножаются силикофлагеллаты про- стым делением. Силикофлагеллаты населяют исключительно моря и океаны, развиваясь в значительном ко- личестве пРи солености не менее 20о/оо. Как и все фототрофные организмы, силикофлагелла- ты нуждаются для нормального развития в определенной силе света. В северных и южных широтах, где сила света небольшая, макси- мальное количество силикофлагеллат обита- ет в поверхностном слое воды, на глубине до 1 м; в экваториальной зоне Атлантического оке- ана они встречаются главным образом на глуби- не 15 — 70 м. Наиболее широко распространены в настоя- щее время виды двух родов — диктиоха (Dictyocha) и дистефанус (Distephanus). Рас- смотренная нами Dictyocha ibula — тепло- водный вид, оптимальная температура воды для нее от +18 до +20 'С; массовое развитие этой водоросли отмечалось в экваториальных водах. Для холодноводного вида Distephanus speculum оптимальная температура воды око- ло 0 'С. Известное максимальное число клеток этого вида наблюдалось в фьорде Осло— 50 400 в 1 л воды; в Черном море он развива- ется в значительно меньшем количестве— 2800 клеток в 1 л воды. 107
Рис. 72. Схема эволюционного развития основных родов силикофлагеллат: 1 — Lyramula; 2 — Cornua; 8 — Naviculopsis; 4 — Corbisema; 5 — Dictyocha; 6 — Deflandryocha; 7 — Distephanus; 8 — Can- nopilus. Как уже указывалось, впервые силикофла- геллаты были найдены в виде скелетов в иско- паемом состоянии. В настоящее время во всем мире накоплен обильный палеонтологиче- ский материал, позволивший проследить их эволюцию на протяжении почти 90 млн. лет (рис. 72). Наиболее примитивные из известных нам силикофлагеллат обитали в морях позднеме- лового времени; их скелеты представляли со- бой простые изогнутые трубочки. Описаны они под родовым названием лирамула (Lyramula, табл. 9, 8). В это же время появились силико- флагеллаты рода корнуа (Согпиа, табл. 9, 4), характеризующиеся уже более сложным ске- летом, в котором можно различить зачатки ра- диальных рогов, апикальное образование и другие части. Род корнуа стал родоначальником рода кор- бизема (Corbisema, табл. 9,5), у представите- лей которого имелось уже замкнутое базаль- ное кольцо, но апикальное образование было еще довольно простое, состоящее из трех пере- кладин. В начале палеогенового периода наиболее примитивные роды лирамула и корнуа вымерли, а род корбизема начал завоевывать господствую- щее положение. В палеогене от этого послед- него рода произошли роды диктиоха (Dicty- ocha, табл. 9,1,2) путем дальнейшего усло- жнения апикального образования и нави- кулопсис (Naviculopsis, табл. 9, 6) в результа- те редукции отдельных частей скелета. В морях неогенов ого периода продолжали существовать виды диктиохи и навикулопси- са, но более типичными для этого времени ока- зались виды родов дистефанус (Distephanus, табл. 9, 7) и канноггилус (Cannopilus, табл. 9,8). Оба рода возникли еще в палеогеновое время: первый — непосредственно от диктиохи; вто- рой, имеющий очень сложное апикальное образование,— от дистефануса. В неогене же род дистефанус дал начало еще одному очень своеобразному роду — деф.гандриоха (Def- landryocha, табл. 9, 9). В четвертичное время (в плейстоцене) не появилось ни одного нового рода. В современ- ных морях (в голоцене) сохранились предста- вители только двух родов — диктиохи идисте- фануса. Интересно отметить, что приуроченность ро- да диктиоха к теплым водам является отголо- ском далекого прошлого, когда виды этого рода были господствующими элементами тро- пической и субтропической флоры микроско- пических водорослей. Поскольку эволюция всех организмов тес- нейшим образом связана с историей развития Земли, знание особенностей развития силико- флагеллат дает возможность реконструировать отдельные этапы истории нашей планеты. Так, например, находка в породе крохотных скеле- тов силикофлагеллат позволяет довольно уве- ренно говорить, что данная порода образова- лась в морском бассейне, существовавшем в определенный отрезок геологического времени. КЛАСС ХРИЗОКАПСОВЫЕ (CHRYSOCAPSOPHYCEAE) Представители этого класса золотистых водо- рослей характеризуются дальнейшим услож- нением таллома, представленного здесь паль- меллоидной (слизистой) структурой. В отличие от первых двух классов хризокапсовые вклю- чают неподвижные, прикрепленные или пас- сивно плавающие колониальные формы. Их клетки не имеют ни поверхностных выпячиваний, ни жгутиков. Они объединяются общей слизью колоний, обычно располагаясь в перифери- ческих ее слоях, реже в центральной части. Рост таллома хризокапсовых осуществляет- ся за счет деления клеток. Делиться могут или одновременно все клетки таллома, или только группа верхушечных клеток, или даже одна верхушечная клетка. Класс хризокапсовых небольшой, насчиты- вает около 20 большей частью монотипных
Рис. 73. Хризокапсовые: 1 — 8 — Hydrurus foetidus (1 — конечный участок таллома, 2— зооспора, 8 — циста); 4 — колония Chrysocapsa sordida. Рис. 74. Хризосферовые: 1 — агрегат клеток Epichrysis paludosa на нитчатой зеленой во- доросли (с в е р х у ); 2 — агрегат клеток Е. nitellae на харо- вой водоросли нителла (с б о к Z). родов. Наиболее высокоорганизованным, хо- рошо изученным и широко распространенным является род гидрурус (Hydrurus), с которым мы и познакомимся более подробно. Гидрурус имеет вид слизистых рыжевато- желтых кустиков (табл. 6, б; рис. 73). Это типичный представитель бентоса. Живет он в быстро текущих или низвергающихся холод- ных родниковых или талых чистых водах. Встречается обычно в высокогорных или арк- тических условиях, где вегетирует на протяже- нии всего года. В более теплых районах его также можно изредка встретить, но лишь вес- ной или осенью. Поселяется гидрурус обычно на доломитовых скалах, каменистых ложах рек, ручьев и водопадов, прикрепляясь к субстрату основанием стебелька. В горных местностях нередко можно увидеть рыжева- тые «хвосты» и «космы», извивающиеся в быст- рых потоках воды. Обычно это и есть кустис- тые слизистые талломы гидруруса. Колонии гидруруса состоят из перистых тя- жей и могут быть длиной от 1 до 30 см. Клетки округло-эллипсоидные или веретеновидные; они плотно расположены по периферии и рых- ло — в центральной части слизи таллома. В кон- чике каждого ответвления имеется только од- на верхушечная клетка. За счет деления этих клеток осуществляется верхушечный рост глав- ной оси и каждого ее ответвления (рис. 73, 1). При большом увеличении под микроскопом в клетках виден один крупный корытовидный постенный хлоропласт с одним голым пирено- идом. В живых клетках можно увидеть еще глыбку лейкозина, несколько (до 6) пульси- рующих вакуолей, а после окраски и цент- ральное ядро. Размножается гидрурус одножгутиковыми зоо- спорами тетраэдрической формы (рис. 73, 2). Цисты у этого вида шаровидные, с экватори- альным утолщением оболочки в виде полу- кольца (рис. 73, 8). Они возникают обычно в конечных разветвлениях таллома. Из других представителей класса хризокап- совых следует назвать хризокапсу (Chryso- capsa sordida), неправильно шаровидные сли- зистые колонии которой широко распростра- нены в водоемах различных типов по всему Советскому Союзу (рис. 73, 4). КЛАСС ХРИЗОСФЕРОВЫЕ (CHRYSOSPHAEROPHТСЕАЕ) К классу хризосферовых относятся предста- вители золотистых водорослей с коккоидной структурой тела. Их клетки покрыты плот- ной целлюлозной оболочкой, а ризоподии и жгуты отсутствуют полностью. Это неподвиж- ные одноклеточные, реже колониальные (табл. 6, 4) формы. Последние имеют вид ско- плений клеток, не погруженных в слизь и слабо связанных друг с другом. При раз- множении они не образуют нитей или пла- стинок. Класс хризосферовых насчитывает около 10 родов, преимущественно монотипных. Рас- смотрим одного из наиболее хорошо изучен- ных представителей рода эпихризис. Эпихризис болотный (Epichrysis paludosa, рис. 74, 1), как видно из его названия, обита-
Рис. 75. Хризотриховые: 1 — 4 — нитчатый таллом Nematochr sis 2 — прорастание о з оспоры, 8 — средний часто rysis sessilis (1 — зооспора t нижний участок таллома с ризои ально ризоидальной клеткой); 5 — кустис- аео amnion confervicola. ет преимущественно в болотах хотя его вст е я его можно Живет OH II H р тить также в прудах кана вах и озерах. гл ивет он прикрепляясь к нитям водорос лавным образом эдогониум рослеи, а и микроспоры. Скопления (агрегаты) клеток эпихризиса часто можно наблюдать весной и реже летом. П ине- сенные в лабо ато р рию водоросли этого вида том. рине- могут продолжительное время сохраняться в кристаллизаторе, поставленном ом на окно, об а- щенное на север. ом , ор- Клетки эпих изиса р са болотного шаровидные диаметром о 10 д 0 мкм, одиночные или, чаще, 7 собранные в агрегаты по 16 клеток. Хло о- пласт в клетке о ин д, пластинчатыи, на верхней стороне клетки, обычно он интенсивно но окрашен лейкозина, капли волютина и жира. Живая клетка со е держит две пульсирующие вакуоли, расположенные в нижней об а ной к с бст ат у р ту стороне клетки. Окрасив к о ращен- к спе и ив клет- ожно выявить у ц альными красителями мож в ней одно ядро. Размножение эпихризиса б олотного, как и других представителей хризос~~е с~~еровых, изуче- , что он может но еще недостаточно. Есть данные что размножаться как зооспорам ми, .гак и автоспо- рами. Наблюдалось образование и типичны~ кремнистых цист. Укажем еще один вид этог о рода — апихри- зис нитвлловый (E. nitellae, рис. 74 2) б ° .Ф щ в бассеинах оранжерей на талломах ха о- вой водоросли нителлы. КЛАСС XPII3OTPMXOBbIE (CHRYSOTRICHOPHYgE gE) ~~~~у хризотриховых относятся предста- вители золотистых водорослей с нитчатои, раз- Ф.З Это и нонитчатой и пластинчатой ст к "- турами тела. то исключительно многоклеточнь е прикре- пленные, типично бентосные ормы. аллом здесь имеет вид простых или разветвленных, однорядных или многорядных нитей к ков или па й, кусти- или пар енхиматозных дисков идных пла- стиночек, не погруженных в об Раз а щую слизь. азрастание таллома происходит за счет де- ления клеток в о одной, двух или трех плоско- стях. Бесполое размножение осуществляется при помощи одножгутиковых или двужг- тиковых зооспо жгу- Клас оспор, имеются типичные цис ты. с хризотриховых объединяет в своем со- ставе п есново р дные, реже солоноватоводные и морские формы. Это наиболее высоко органи- зованная группа золотистых водорослей, пред- м виду сходны с ставители которой по внешнем ви схо водорослеи улотриксовыми из отдела зеленых во и гетер отриксовыми из отдела желто-зеле- ных водорослей. Некоторые из них похожи на р устроенных представителей наиболее п осто бурых водорослей. Из з нитчатых неразветвленных о м класса наибо ~орм этого наи олее изученным является немато- 75, 1— хризис сидячий (NeIIIatochrysis 'Г, sessi is, рис. — 4). Рассматривая нематохризис в мик- можно видеть, что его длинные одно- оскоп рядные неразветвленные нити с и состоят из ци- линдрических клеток с довольно толстой в х- контурной целлюлозной оболочкой. Интерес- но, что в талломе этого вида уже наблюдается дифференциация клеток на одну ризоидальн ю и остальные вегетативные. Эти различия не только морфологические, но и функциональ- ные: ризоидальная клетка выполняет функцию прик епления к с р к субстра ту, а вегетативные клетки путем деления обеспечивают рост нити в длину и продуцируют зооспоры. Хлоропла- стов в ка ждой клетке два, они пластинчатые, слегка лопастные, постенные' , кроме того, име- ются лейкозин и капли жи а. В каж ой ира. д такой клетке может возникать по одной крупной двужгутиковой зоосп оре (р ., 1). Выидя за пределы клетки, зооспо- ра некоторое время плавает в воде, а позже при- клеивается передним концом б р к су страт , втя- гивая при этом жгутики. Эта закрепившаяся н я на в коре делится у рате клетка вытягивается и вско е слегка наискось поперечной перегородкой на две неодинаковые клетки. Верхняя пол с е и- ческая клетк тка, именуемая вегетативной, полу- чает почти все со е д ржимое материнской клетки на вскоре делится на две клетки, ис. 75 2~. О каждая из которых в свою оче 7 редь, делится еще
ОТДЕЛ ДИАТОМОВЫЕ ВОДОРОСЛИ (ВАСПД АРДОРНТТА) на две и так далее, в результате чего выраста- ет длинная нить таллома (рис. 75, 8). Нижняя клетка, возникшая после первого деления за- крепившейся на субстрате зооспоры, дальше не делится. Оболочка ее утолщается, разраста- ясь в нижней части в виде подошвы, пропиты- вается солями гидроокиси железа и становится коричневой. Эта клетка, подобно корням выс- ших растений, выполняет механическую функ- цию, и поэтому ее назвали ризоидальной, т. е. подобной корню (рис. 75, 4). Из кустистых форм хризотриховых заслу- живает упоминания род феотамнион (Phaeo- thamnion), виды которого характеризуются еще большей дифференциацией клеток в пре- делах таллома (табл. 6, 6; рис. 75, о). Здесь выражена разнонитчатая структура тела. Вегетативные клетки в таком талломе двоя- кого рода: более крупные, бочонковидные, об- разующие главный стволик, и меньших раз- меров, цилиндрические, слагающие боковые веточки. Последние в основном и принимают участие в размножении, клетки же стволика являются несущими. У основания главной оси (стволика) имеется ризоидальная клетка, вы- полняющая функцию прикрепления таллома к субстрату. Диатомовые водоросли — это совершенно осо- бая группа одноклеточных организмов, резко отличающаяся от остальных водорослей: клет- ка диатомовых снаружи окружена твердой кремнеземной оболочкой, называемой панци- рем. Форма этого панциря настолько разно- образна, затейлива и причудлива, а структу- ра его настолько тонка, изящна и красива, что его можно принять за творение искус- ного художника. Некоторые диатомеи красотой панциря могут поспорить с ювелирными изде- лиями изобретательного мастера. Впервые о существовании диатомовых водо- рослей стало известно в начале XVIII в., когда в практику исследований были введены мик- роскопы Левенгука и сильно увеличивающая лупа. Эта группа водорослей имеет несколько на- учных названий: диатомовые (Diatomeae), кремнеземные (Kieselalgae) и бациллариевые (Bacillariophyta). Первое название они полу- чили благодаря своему размножению делением панциря на две половинки, второе связано с наличием у клетки кремнеземной оболочки и, наконец, последнее название было дано по первому роду, научно описанному в 1788 г.,— бациллария, что обозначает «палочковидный». Виды этого рода — типичные эпифиты. Их кустики, достигающие 250 мкм в длину, встре- чаются обычно в холодную пору года (весной, осенью и зимой) в прибрежной зоне озер, пру- дов, болот, стариц и речек. В качестве субстра- та они используют нитчатые водоросли (кла- дофору, в ашер ию, трибонему, микр оспа ру и др.), мхи и высшие водные растения. Пластинчатый таллом у хризотриховых име- ет феодермаииум речной (Phaeodermatium ri- vulare, табл. 6, 7). Поселяется этот вид в холодных быстрых водах, особенно часто на известковом скалистом ложе горных рек и ру- чьев. В Крыму он встречается в верховьях речек Альмы, Качи и Улу-Узеня, где образует рыжевато-коричневый налет на каменистом ло- же реки. Золотистые водоросли — очень древняя груп- па водорослей, возникшая от каких-то первич- ных амебоидных организмов. По набору пиг- ментов, составу запасных веществ и наличию кремния в оболочках вегетативных клеток и цист золотистые водоросли обнаруживают сход- ство с диатомовыми, желто-зелеными и отчасти бурыми водорослями. Есть основание полагать, что именно золотистые водоросли в свое время дали начало диатомовым водорослям. В русской литературе утвердилось название «диатомовые водоросли» или «диатомеи», наибо- лее современным научным латинским названием является Bacillariophyta. Благодаря прозрачному панцирю и бурой окраске хлоропластов диатомовые легко раз- личить под микроскопом среди остальных од- ноклеточных низших водорослей. Диатомовые водоросли — одноклеточные ми- кроскопические организмы, одиночно живу- щие или объединенные в колонии различного типа: цепочки, нити, ленты, звездочки, кустики или слизистые пленки. Колонии обычно микро- скопические, но иногда макроскопические и тогда видны невооруженным глазом. Размеры клеток — от 4 до 1000 мкм, а у некоторых пред- ставителей — до 2000 мкм. Исследованием клетки диатомей стали зани- маться в начале прошлого столетия. При помо- щи светового микроскопа уже тогда удалось получить довольно полное и правильное пред- ставление о строении клетки и структуре пан- циря. Введение в практику альгологических исследований электронного микроскопа- трансмиссионного и сканирующего — за пос- ледние 10 — 15 лет позволило существенно до- полнить наши знания о строении орга нелл
Рис. 76. Панцири Mastogloia braunii, объемное изоб- ражение при помощи сканирующего электронного микроскопа()( 630). Электронная микрофотография Н. И. Караевой. Рис. 77. Очертания створок: 1, 2 — эллиптическая; 8 — широковллиптическая; 4 — re ТароВпр- ная; 5 — линейная; 6 — булавовидная; 7 —; 8,9— ланиетная; 10 — ромбическая; 11, 18 — полуланцетная; 12— nozzygyroo6paauaa; 14 — си~мои~ная; 15 — прямоу~ол~ная; 16— почковидная. клетки и тонкой структуре панциря, многие детали которой были неизвестны. СТРОЕНИЕ КЛЕТКИ ДИАТОМОВЫХ ВОДОРОСЛЕЙ Клетка диатомовых водорослей состоит из протопласта, окруженного кремнеземной оболоч- кой, называемой панцирем. Протопласт своим наружным уплотненным слоем (плазмалеммой) тесно примыкает к панцирю и заполняет его внут- ренние полости. Целлюлозной оболочки, имею- щейся у большинства водорослей, здесь нет. Хи- мический анализ панциря показал, что он состоит из аморфной формы кремнезема, напоминаю- щей по составу опал, с плотностью 2,07. Тол- щина стенок панциря зависит от концентрации кремния в среде и колеблется в значительных пределах: у тонкостенных форм — от сотых до десятых долей микрометра, а у толстостен- ных достигает 1 — 3 лкл. Стенки панциря про- низаны мельчайшими отверстиями, обеспечи- вающими обмен веществ между протопластом и окружающей средой. Они снабжены также раз- личными форменными элементами, которые со- ставляют структуру панциря и служат основ- ными таксономическими признаками при по- строении системы диатомей. Панцирь и его структура различимы уже при небольших уве- личениях микроскопа. По форме панциря все диатомеи делятся на две группы: центричес- кие — с радиально-симметричным панцирем и пеннатные — с двусторонне-симметричным пан- цирем. Протопласт. Цитоплазма в клетках диатомей располагается постенным слоем или ска- пливается в центре клетки или у ее полюсов. Остальные участки клетки заполнены мно- жеством вакуолей с клеточным соком, которые иногда сливаются в одну крупную вакуолю. Ядро обычно шаровидное и располагается чаще всего близ центра клетки в цитоплазма- тическом мостике или в периферическом слое цитоплазмы. У некоторых диатомей оно имеет Н-образную форму. В ядре различают от 1 до 8 ядрышек. Хлоропласты у диатомей довольно разно- образны по форме, величине и количеству в клетке. У большинства центрических диатомей они мелкие, многочисленные, в форме зерен или дисков, лишенные пиреноида. Реже они более крупные, по одному или нескольку в клетке и имеют форму пластинки с ровными или изрезанными лопастными краями, с одним или несколькими пиреноидами. У пеннатных диатомей хлор опласты обычно крупные, пла- стинчатые, часто с лопастными краями или с перфорациями, они немногочисленные (один или два в клетке), занимают почти всю ее по-
лость, как правило, с пиреноидами. Количество, величина и положение их различно даже среди представителей одного рода (табл. 10). Окраска хлор опластов у диатомовых водо- рослей имеет различные оттенки желто-бурого цвета в зависимости от набора пигментов, среди которых преобладают бурые — каро- тин, ксантофилл и диатомин, маскирующие в живой клетке хлорофиллы и и с. После гибели клетки бурые пигменты растворяются в воде и зеленый хлорофилл становится ясно замет- ным. Интенсивность окраски хлоропластов и их величина различны и зависят от образа жиз- ни водорослей: у планктонных видов они зо- лотисто-желтые, мелкие, дисковидные, а у дон- ных и прикрепленных к субстрату — крупные пластинчатые, темно-бурые, поэтому большие скопления диатомей приобретают хорошо вы- раженную бурую или темно-бурую окраску. В процессе фотосинтеза у диатомовых водо- рослей образуется масло в виде капелек раз- личной величины, иногда в значительном ко- личестве. Оно служит запасным питательным веществом, особенно в период, когда пре- кращается или задерживается деление клетки. Масло, экстрагированное из клеток диатомей, имеет запах рыбьего жира. Кроме масла, для некоторых видов характерно еще присутст- вие в клетках капель волютина, имеющих тусклый голубоватый блеск. Волютин нераст- ворим в эфире и при окраске живой клетки метиленовой синькой приобретает красновато- Рис. 78. Концы створок. 1 — острый; 2 — оттянутый; 8 — клиновидный; 4 — R JIIoBQBHg- ный; 5 — головчатый; 6 — широкозакругленный; 7 — тупой. фиолетовый оттенок. Маленькие капли волю- тина распределяются по всей цитоплазме, а крупные (тельца Бючли) занимают определен- ное положение на концах клетки (виды семей- ства Nitzschiaceae) или по обеим сторонам центрального цитоплазматического мостика (роды семейства Naviculaceae). В качестве Рис. 79. Формы вставочных ободков: 1 — кольцевидные (Lauderia); 2 — воротничковидные (Stephanodiscus); 8 — трапециевидно-чешуевидные; 4 — полуцол~рщщр» ные; б — чешуевидные (8 — 5 — ВЫхояо1еп1а).
~п ~по, ~~ ) ~ПО~ 2! с- ОП с 1 Ц~у ЦП сп 3)~ <v I l С2 1 ге — г -аьу )A в~ зс], нщ mmccl ~.'> т" --у kÅ j 3C2 m C2 в1 г Рис. 80. Составные части панциря (схема): 2 — центрического типа: вверху — вид со створки, внизу — вид с пояска; ДД — диаметр панциря, ВВ — центральная ось пан- циря; 2 — пеннатного типа: вверху слева — вид со створки, вверху справа — вид с пояска, внизу — сечение панциря в по- перечной плоскости; АА — продольная ось, ББ — поперечная ось, ВВ — вертикальная (первальварная) ось панциря. а- эпитека, г — гипотека, с,— створка эпитеки, c2 — створка гипоте- ки, зс, — загиб створки эпитеки, зс, — загиб створки гипотеки, по, — поясковый ободок эпитеки, по, — поясковый ободок гипо- теки, р — ребра (ряды ареол), цу — центральный узелок, цп- центральная пора, ш — шов, ку — конечный (полярный) узелок, кш — конечная (полярная) щель узелка, e — воронка, оп— осевое поле, cn — среднее поле, вщ — внутренняя щель шва, ищ — наружная щель шва. Рис. 81. Расположение осей и плоскостей симметрии панциря (схема). Оси симметрии: АА — продольная, ББ — поперечная, ВВ— центральная. Плоскости симметрии: а — продольная, б — по- перечная, в — створковая (вальварная). питательного вещества в клетках диатомей встречается еще лейк озин. Панцирь и его строение. Панцирь диатомо- вых водорослей вырабатывается самой клеткой в процессе ее жизнедеятельности. Он состоит из двух почти равных частей и по конструкции на- поминает коробку, закрытую крышкой. Наруж- ная, большая часть панциря — э п и т е к а, подобно крышке, находит своими краями на внутреннюю половину — r и п о т е к у, со- ответствующую коробке (рис. 76). Эпитека и гипотека состоят из створки и поясков ого ободка. Створку, принадлежащую эпитеке, на- зывают э пив а л ьв ой, а гипотеке — ги- п о в а л ь в о й. Створка имеет лицевую по- верхность, плоскую или слегка выпуклую, и краевую загнутую часть, называемую загибом створки, иногда отличающуюся структурой. Загиб створки у одних диатомей низкий и вы- ражен довольно слабо, у других он достаточ- но высокий и составляет значительную часть боковой поверхности панциря. Створки бывают самых различных очер- таний: круглые, эллиптические, яйцевидные, ромбические, ланцетные, треугольные, четы- рехугольные, булавовидные, серповидные, ги- таровидные, клиновидные и т. д. Изменчивы и разнообразны концы створок: клюв овидные, головчатые, оттянутые, тупые, острые и пр. (рис. 77, 78). К каждой створке примыкает поясковый (соединительный) ободок, представляющий со- бой широкое или узкое кольцо, окаймляющее загиб створки, но не срастающееся с ним. Поясковый ободок эпитеки своим свободным краем надвигается на поясковый ободок ги- потеки и плотно его охватывает, с ним, однако, не срастаясь. Правда, у представителей неко- TopbIx родов поясковые ободки образуются только во время деления клетки, а эпивальва и гиповальва плотно соединяются друг с другом непосредственно краями загиба створки. Кро- ме того, у многих диатомей между загибом створки и поясковым ободком образуются еще вставочные ободки, от одного до многих. Ка- ждый новый, более молодой вставочный ободок возникает всегда между загибом створки и предыдущим ободком. Они представляют собой дополнительные образования, отличающиеся не только формой, но и структурой. Форма вста- вочных ободков — один из характерных при- знаков рода. Они бывают воротничковидные, кольцевидные, полукольцевидные или состоят из отдельных сегментов, имеющих форму тра- пеции, ромба или чешуйки (рис. 79). Наличие вставочных ободков в панцире имеет большое биологическое значение, так как они способствуют увеличению объема клетки и ее росту.
Часть панциря между эпивальвой и гипо- вальвой, т. е. поясковый ободок гипотеки и на- ходящий на него поясковый ободок эпитеки, а если есть, то и вставочные ободки, называют пояском панциря (рис. 80). Форма панциря зависит от очертаний створки. Он бывает шаровидный, палочковидный, сед- ловидный, в виде апельсиновой дольки, низко- го или высокого цилиндра, параллелепипеда или другой геометрической фигуры. Со сторо- ны пояска он имеет обычно прямоугольную форму. Характерной особенностью панциря яв- ляется геометрическая правильность его строения, в связи с чем для представления о его форме очень важно учитывать соотношение осей и плоскостей симметрии (табл. 11, 12). При двусторонней симметрии у пеннатных ди- атомей определяют несколько осей и плоско- стей симметрии, которые делят панцирь на сим- метричные половины. Известны три основные взаимно перпендикулярные оси симметрии: продольная, поперечная и центральная, про- ходящая через центр двух створок панци- ря. Длина продольной оси определяет длину створки или панциря, длина поперечной оси- ее ширину, а длина центральной оси — высоту панциря. Кроме осей симметрии, различают три плоскости симметрии: продольную, прохо- дящую вдоль панциря перпендикулярно к створкам и делящую его на две равные полови- ны; поперечную, проходящую поперек панциря перпендикулярно к продольной плоскости и к створкам и не всегда делящую панцирь на две равные половины (если панцирь гетеро- польный, а не изопольный, т. е. если концы створок неодинаковы); створковую, перпен- дикулярную к двум предыдущим, но парал- лельную створкам, т. е. проходящую через поясок панциря (рис. 81). У центрических диатомей, обладающих ради- альной симметрией, панцирь имеет только две оси и две плоскости симметрии. Одна ось- это диаметр створки, другая — центральная. Плоскость симметрии, проходящая через центр створки в любом направлении, всегда делит панцирь на две равные части; вторая плоскость симметрии — створковая, как и у пеннатных форм, идущая перпендикулярно к первой. Форма створок и панциря в целом, а также соотношение осей и плоскостей симметрии име- ют важное значение в систематике диатомо- вых водорослей. Однако главным признаком при построении их системы является структу- ра кремнеземного панциря, которая представ- ляет наибольшую трудность при изучении. Под структурой панциря, видимой в световой и электронный микроскопы, подразумевают 'H а ее а ° ° ~файф ЬЩа l Лв Ip~~ggfss~~ ~ ~~рр е ~еф~~;:.' ',,~�и~,ф�;.." "~.4Вф.„i% "4 i .".~~@. 49O'~ .~фр ".:- ДЪд ю а е ~а д ефУ ЧД е е ФФ~Ю ° + eeo~y9y~е l �ÞþÔÞÔÞ Р ВЯ ° В ° % I наружный и внутренний его рисунки, специфи- ческие для разных таксонов. Структурные элементы на створках центрических диатомей имеют радиальное и тангенциальное расположе- ние, а у пеннатных — двустороннее, или попе- речное, расположение, т. е. их структура сим- метрична по отношению к продольной и по- перечной осям. Реже у представителей неко- торых родов панцири бывают асимметричными и не имеют ни одной плоскости симметрии, а Рис. 83. Шов у Navicula radiosa. Створка с проходящей вдоль нее щелью шва, которая в централь- ном узелке соединяется с другой щелью шва, а на конце створки заканчивается конечным узелком (~6000). Электронная микро- фотография Н. И. Караевой. Рис. 82. Строение ареол (продольный разрез через стенку п анциря): 1 — ареола с велумом на внутренней стороне панциря; 2— ареола с велумом на наружной стороне панциря. (По Н. И. Хенди) .
Рис. 84. Схема строения шва навикулоидного типа: 1 — створка со щелевидным швом, 2 — Л — поперечный разрез щели шва, 6 — продольный разрез центрального pçåëêà. Рис. 85. Шов у Nitzschia acuta (часть створки): 4 — п ом 1 — канал-шов 2 — щель каналовидного шва. 8 — ф б — р ежутки или отверстия (><5500). Электронная микрофо- — фи улы; тография В. И. Михайлова. >Baré~ асимметричность выражается толь, в структуре створок. Главная особенность стенок панциря состоит в том, что они пронизаны регулярно повторяю- щимися мельчайшими отверстиями — а р е о л а- м и, обычно затянутыми снаружи или внутри тонкой перфорированной пленкой, получившей латинское название «велум» (рис. 82). Отверстия в стенке панциря необходимы для сообщения протопласта клетки с окружающей средой. При изучении диатомовых водорослей в све- товом микроскопе казалось, что у некоторых видов бесструктурный панцирь, и только вне- дрение в практику альгологических исследо- ваний электронного микроскопа показало, что и эти панцири также имеют чрезвычайно тон- кую пористую стенку. Отверстия, пронизыва- ющие створку, занимают обычно 10 — 75% ее площади, а характер расположения этих от- верстий и их количество специфичны для разных родов и видов. Но на створках есть и участки, лишенные отверстий, например центральное, осевое и боковые поля у шва (см. ниже) и некоторые скульптурные детали структуры. У центрических диатомей ареолы располагаются радиально и тангенциально; у пеннатных — поперечными рядами, либо па- раллельными друг другу, либо к краям створ- ки слегка расходящимися (радиальные ряды) или, наоборот, сходящимися (конвергентные ряды). Иногда ареолы располагаются так, что, кроме поперечных, образуются еще и продоль- ные или взаимно перекрещивающиеся косые ряды (табл. 11). Примечательная особенность пеннатных ди- атомовых водорослей — наличие осевого поля Ъ представляющего собой бесструктурную узкую или широкую полосу по продольной оси створ- ки. У некоторых диатомей осевое поле рас- ширяется на середине створки, образуя сред- нее поле, которое бывает круглым, ромбичес- ким, четырехугольным, иногда доходящим до краев створки. Большинство диатомей пеннатного типа ха- рактеризуется еще одним признаком — присут- ствием шва, представляющего собой короткую или длинную щель или две щели (ветви шва), прорезывающие стенку створки и идущие вдоль створки от ее концов к середине. Строе- ние шва весьма различно — от простого щеле- видного до так называемого каналовидного. Примитивный щелевидный шов представлен двумя короткими изолированными щелями, не доходящими до середины створки. У предста- вителей некоторых родов такой шов находится всего на одной створке, иногда только у од- ного из ее концов, у других — на обеих створ- ках. Хорошо развитый щелевидный шов, ха-
рактерный для водорослей из семейства навику- ловых (Naviculaceae), представлен двумя длин- ными щелями, или ветвями, шва, проходящими по обеим створкам и соединяющимися на середине каждой створки центральным узел- ком, а у концов створки заканчивающими- ся конечными, или полярными, узелками (рис. 83). Щели шва в толще створки коленча- то-изогнуты, так что в поперечном разрезе они имеют вид лежащей буквы V (рис. 84). Щель шва, открывающуюся внутрь клетки, называют внутренней, а открывающуюся на- ружу — наружной. В центральном узелке обе ветви шва соединяются друг с другом, закан- чиваясь здесь центральной порой, а у кон- цов створки — конечной порой. Центральный узелок представляет собой внутреннее утолщение стенки створки, выпук- лость на ее внутренней поверхности, а конеч- ные узелки — внутреннее и наружное утол- щения стенки створки. Наиболее сложное устройство имеет так называемый каналовидный шов — канал, рас- положенный в складке стенки створки. С внешней средой он сообщается узкой щелью, а в полость клетки открывается рядом отвер- стий с кремнеземными перегородками — фи- булами. Каналовидный шов присущ водорослям \ Ю «е в Рис. 86. Схема строения каналовидного шва. ~ — створка с каналовидным швом, 2 — каналовидный шов (средняя часть), а — канал-шов, б — щель каналовидного шва, e — фибулы, г — промежутки, д — центральный узелок. Рис. 87. Расположение каналовидного шва на створке: 2 — шов расположен по спинному краю створки (Rhopalodia); 2 — шов расположен вдоль одного края створки (Nitzschia); 8, 4 — шов расположен на границе створки и ее загиба (8 — Surirejla, 4 — Campylodiscus) (1 — )(3000, 2 — 0<2400, ~, 4— X1000). Электронные микрофотографии: 1 — Н. И. Караевой; 8 — В. И. Михайлова; микрофотографии 8, 4 — А. И. Прошкиной- Лавренко.
'Ф ° ° р ° ее~ ° ~ ° ° Ф +å е ° ее ~,~~ее ° Ю ° ° ° ° ° ° ° Фе , ° ' ° ° и' .Фч: ° ~ю ~му ' ° г, ° ° ° ° ° ° ° ° 4 б ° . Рис. 88. Форма и расположение септ.' 1 — 8 — вставочный ободок с септой (1 — у центрической диатомеи ThalassiosiI'à antlgua var. Вер1а1а, 2, 8 — у пеннатных диато- мей: 2 — у Grammatophora marina, 8 — у Gr. Serpentina); 4 — септа; 5 — полярная септа (Licmophora ehrenbergii); 6 — септа, занимающая половину панциря (Tabellaria fenestrata); 7 — септа с тремя отверстиями (Rhabdonema adriaticum); 8 — септа с многочисленными отверстиями (Climacosphenia moniligera). семейств эпитемиееых (Epithemiaceae), нити- шиееых (Nitzschiaceae) и суриреллоеых (Su- rirellaceae) (рис. 85, 86). Он также имеет цент- ральный узелок, но его положение на створке различно у представителей разных родов. У водорослей из рода эпитемия (Epithemia) вет- ви каналовидного шва соединяются под уг- лом и приближены к брюшному краю; у видов рода ропалодия (ВЬора1ойа) шов тянется по спинному краю; у представителей рода нитгриия (Nitzschia) находится в киле, расположенном вдоль одного из краев створки (рис. 87, 1, 2), а у видов рода дентикула (Denticula) прохо- дит более или менее эксцентрично к продоль- ной оси створки. У водорослей из родов су- рирелла (Surirella) и кампилодискус (Campylo- discus) каналовидный шов лежит на краю крыла створки, находящегося на границе с загибоы створки и опоясывающего ее. Поэтому при рассмотрении панциря со стороны створ- ки он не виден (рис. 87, 8, 4). И только у пред- ставителей рода иилиндротека (Cylindrothe- ca) шов спирально окружает панцирь. Биологическое значение шва в жизни диато- мей очень велико: помимо сообщения протопла- ста клетки с внешней средой, с помощью шва клетки довольно быстро передвигаются по суб- страту и в толще воды. В филогенетическом отношении возникновение шва является про- грессивным признаком, он присущ более моло- дым видам, которые в современных морях и океанах составляют свыше 70% от общего чис- ла диатомей. Кроме названных структур, у большинства диатомовых водорослей на внешней и внутрен- ней поверхности створок имеются различные образования в виде полых или сплошных вы- ростов, выпуклостей, рогов, щетинок, шипов, шипиков, бороздок, камер, ребер и пр., которые выполняют определенные функции: выделяют слизь, объединяют клетки в колонии, увеличи- вают поверхность панциря у планктонных ви- дов, обеспечивая плавучесть клетки в воде и т. д. У некоторых диатомей, имеющих вытянутые створки, на внутренней поверхности вставоч- ных ободков образуются кремнеземные пере- городки, или с е п т ы, вдающиеся в полость панциря параллельно плоскости створок. Сен- ты возникают или по всей внутренней поверх- ности вставочного ободка, или только в одном из его концов. Они обычно хорошо видны со стороны пояска, различны по положению, фор- ме и размерам и имеют одно или несколько отверстий. У диатомей с гетеропольным панци- рем септы чаще всего возникают только в ши- роком его конце (род Licmophora), с изополь- ным панцирем — в любом или в обоих его кон- цах (роды Tetracyclus, Tabellaria). Сенты мо- гут быть узкими, или они глубоко вдаются в полость панциря, вплоть до его середины (рис. 88).
У небольшого числа диатомей образуется еще один тип перегородок, так называемые п с е в д о с е п т ы, развивающиеся на внут- ренней стороне самой створки и вдающиеся в полость панциря в виде короткой и довольно грубой перегородки, видимой со створки и с пояска. В отличие от септ псевдосепты всегда перпендикулярны к створке и возникают одно- временно с ней. Представителям рода масто- глойя (Mastogloia) свойственны особые образо- вания — камеры, представляющие многоуголь- ные, реже удлиненные пустоты в стенке панци- ря, открытые внутрь клетки или наружу круг- лыми отверстиями (рис. 89). Некоторые детали структуры бывают не вид- ны в световом микроскопе, а обнаруживаются только при больших увеличениях с помощью электронного микроскопа. Все перечисленные структуры обладают четкой, правильной фор- мой и определенным количеством элементов на единицу поверхности. Большинство из них вы- полняет определенную функцию, обеспечивая приспособляемость диатомей к условиям суще- ствования. СПОСОБЫ ПИТАНИЯ ДИАТОМОВЫХ ВОДОРОСЛЕЙ Диатомовые водоросли преимущественно фо- тоавтотрофные организмы, которые в процессе фотосинтеза образуют органическое вещество. В хлоропластах диатомей обнаружено девять пигментов: хлорофиллы а и с, р- и а-каротины и пять ксантофиллов — фукоксантин, диаток- сантин, неофукоксантины А и В и диадиноксан- тин. Состав и количество пигментов непостоян- но и зависит от интенсивности света, его каче- ства, содержания в воде биогенов, а также от возраста клетки и особенностей ее жизнедея- тельности. Количество хлорофилла уменьша- ется в старых клетках, а недостаток азота и фосфора резко снижает содержание хлорофил- ла и. Недостаток питательных веществ в воде даже при высокой интенсивности света ведет к уменьшению количества пигментов, а обилие питательных веществ даже при слабой осве- щенности способствует их образованию. Ко- нечным продуктом фотосинтеза являются жиры, а не углеводы. Интенсивность фотосинтеза на единицу био- массы у планктонных и бе нтосных диатомей неодинаковая. У бентосных форм она значи- тельно выше, так как их хлоропласты крупнее и имеют более интенсивную окраску. Кроме того, у подвижных форм фотосинтез протекает актив- нее, чем у неподвижных, и значительно усили- вается в период деления клеток. Условия для фотосинтеза у поверхности воды довольно близки к условиям воздушной среды, но резко меняются с погружением водорослей на глу- бину. Планктонные диатомовые водоросли, оби- тающие в пелагиали морей, могут существовать на глубине 100 м и более при большой прозрач- ности воды. Однако с глубиной меняется не только интенсивность освещения, но и качество света вследствие различного поглощения лучей солнечного спектра разной длины волны, что по-разному отражается на разных видах. Среди планктонных и бентосных диатомей существуют светолюбивые и тенелюбивые виды, отличающиеся интенсивностью ф~утосинтеза и коэффициентом использования солнечной энер- гии при одинаковой радиации. У светолюби- вых видов максимум фотосинтеза приходится на полдень, а у тенелюбивых — на утренние и послеполуденные часы. Изучение диатомовых водорослей в культу- рах позволило выявить большую пластичность диатомей в усвоении как минеральных, так и органических веществ. Особую роль в жизни диатомовых водорослей играет кремний, который им необходим для по- строения панциря. Усвоение его происходит в соответствии с ритмом деления клеток и за- висит от химических и физических свойств окружающей среды. Деление клеток диатомей происходит нормально, если в воде находится Рис. 89. Раздвинутый панцирь у Mastogloia pusilla, видны камеры ()(6000). Электронная микрофотография Н. И. Караевой
Рис. 90. Образование и прораста астание микроспор: та ии образования микроспор у Сояс1- — азличные стадии — 4 — последовательные стадии о раз яс1 а- — 9 — последовательные ст- спор у Сйае1осегоя сп s curvisetus; 6— дии прорастания мин роспор у не менее 5 лг/л кремния, р а п и содержании его О 5 лг/л деление прекращается. около , лгл У ся кремний диатомовы д ми во о ос- Усваивается о ганиче- е к емниевой кислоты и ор лями в виде р я. Пот ебность в крем- ских соединений кремния. отре но ая и зависит от место- нии у диатомей различная ия и изиологического состояния кле- обитания m $mam е виды имеющие ток. Так, напр р, име ентосные в ФР ь н ж аютсяв большем толстостенныи панцир, у д по с авнению с планктон- количестве кремния по р бла ающими тонкостенным ными формами, о лад пе ио обильного размножения, й а неко- кото ое происходит обычно веснои, а у которое прои диатомеи испытывают то ых видов и осенью, ди р ю пот ебность в кремнии: недоста- наибольшую потре но точное содержание его в воде вызыв ление темпов деления и при од во ит к меньше- нию толщины панциря. Помимо неорганических веществ, диатомовым и азвития также нужны водорослям для роста и р о ие количества органических веществ. Очень хорошее воздействие на н В . Из чение органического питания витамин иатомеи показало, что что они н жда д таминах группы В больше, чем другие водо- росли. ли вообще мо- Некото ые диатомовые водоросли е которы тот о ного питания к гете- гут переходить от автотро с бес вет- офном . Известны даже формы с есцвет- все без них — эти хло опластами или вов ными хл р же облигатными гетеро- водоросли являются уже о ли трофами. РАЗМНОЖЕНИЕ ДИАТОМОВЫХ ВОДОРОСЛЕЙ д еление. аще в Ч сего диатомеи размножаются елением клетки на две полови- Вегетативным делением ны; этот от процесс обычно происходит н Темпы деления различн у р ы азных на рассвете. же о ного вида в за- ов и могут меняться даже у одног \У - -- у- --"ру--щ- висимости от сезона или б ается ср д е ы. еснои и в ь " в начале лета на люда е азвитие диатомовых в результа- развити интенсивного деления. аличи тв ет елению и рос- биогенных веществ спосо ству д ту диатомей. что в культуральной Опыты показали, что в к мог т то ые планктонные виды могу среде некоторые п о 3 — 8 раз в сутки. Бентосные делиться до тся го аздо реже— — один раз в виды делятся е более едкого де- 4 дня. И Известны случаи еще олее ней. Но эти сведения ления- ния — один раз за дней. еления могут меняться не абсолютн, еле ы и темпы деле сположения в зависимости от широ отного а ко-химического режима и, водоема, его изико-. конечно, от особенно нностей вида. иатомей своеоб- П оцесс деления клетки у диатомеи сво роц чия тве ого панциря. начала разен из-за наличия твердо капельки начинают скапливаться кап в протопласте на величива- сам п отопласт значительно ув масла, а сам прот эпитека и гипо- д всле ствие чего э и я асходятся, оставаясь тека панциря ра сковых ободков. ными только р к аями своих поясков ве авные части, а П отопласт делится на две равны рот
Рис. 91. Половой процесс у диатомей: 1 — 8 — анизогамный половой процесс у Pinnularia sp. 4 — 5 — оогамный половой процесс у Biddulphia mobiliensis. вместе с ним и хлоропласты. Если хлоропласт один, то он делится пополам; если их много, то в дочерние клетки сначала попадает полови- на их, а затем они делятся, в результате в до- черних клетках образуется исходное количест- во хлор опластов. Ядро делится митотически, часто с хорошо заметными хромосомами и цент- росомоЗ на каждом из образовавшихся полю- сов. После окончательного разделения клетки на две каждая из дочерних клеток, получившая лишь половину материнского панциря, сразу же «достраивает» недостающую половину, но обязательно внутреннюю, т. е. гипотеку. До- полнительные образования панциря — вставоч- ные ободки, септы и другие структурные эле- менты — возникают вскоре после сформирова- ния новой гипотеки. Таким образом, возник- шие в результате деления две дочерние клетки оказываются по размерам несходными: одна клетка, получившая эпитеку, сохраняет размеры материнской клетки, а другая, получившая ма- теринскую гипотеку, ставшую в новой клетке эпитекоЗ, приобретает меньшие размеры. В ре- зультате после многократных делений происхо- дит постепенное уменьшение размеров клеток у половины каждой данной популяции: у цент- рических диатомей уменьшается диаметр кле- ток, а у пеннатных — длина и отчасти ширина клеток. Установлено, что у некоторых видов в процессе делений размеры клеток уменьшают- ся почти в 3 раза по сравнению с первоначаль- ными. Микроспоры. У многих планктонных диа- томеЗ обнаружены так называемые микроспо- ры — мелкие тельца, возникающие в клетках в количестве от 8 до 16 и более, а у некоторых видов их бывает и более 100. Наблюдались микроспоры со жгутиками и без них, с хлоро- пластами и бесцветные. Наиболее часто микро- споры развиваются у видов рода хетпоцерос (Chaetoceros), и наблюдалось даже их прора- стание (рис. 90). Процесс образования микроспор цитологичес- ки не изучен, и природа их точно не установ- лена. Половой процесс и образование ауксоспор. Ауксоспорами, т. е. «растущими спорами», на- зывают такие, которые при своем образовании сильно разрастаются и затем прорастают в клет- ки, резко отличающиеся по размерам от исход- ных. Способность образовывать ауксоспоры свойственна только диатомовым водорослям, но до сих пор не удалось еще полностью объяс- нить этот процесс и порождающие его причины. Образование ауксоспор, по всей вероятности„ вызывается различными причинами. Согласно наиболее распространенному мнению, оно на- ступает вследствие многократных делений, при- водящих, как это описано выше, к мельчанию клеток. Достигнув минимальных размеров, клетки развивают ауксоспоры, что приводит к восстановлению их размеров. Однако другие исследователи считают, что ауксоспорообразо- вание связано просто со старением клеток, так как его нередко удавалось наблюдать и тогда, когда клетки еще не достигли своих минималь- ных размеров. С этих позиций ауксоспорооб- разование рассматривается как процесс «омо- ложения» клетки. Кроме того, есть наблюде- ния, свидетельствующие о развитии ауксоспор
Рис. 92 А - -QH'.<0<>op< 1 — Melosira monilif ormis, цепочка с конечной ауксоспорой; 2, 8 — Rhizosolenia alata (2 — начало образования ауксоспоры, 8 — зрелая конечная ауксоспора); 4— Thalassiosira excentrica, интеркалярная ауксоспора. Рис. 93. Образование покоящихся спор: 1 — 4 — последовательные стадии развития споры у Chaetoceros heterovalvatus. при изменении условий окружающей среды, например при резком понижении температуры воздуха или воды. Каковы бы ни были причины, способствую- щие возникновению ауксоспор, установлено главное: ауксоспорооб разов ание всегда свя- зано с половым процессом. У диатомовых водо- рослей встречаются все три типа полового процесса, вообще известные у водорослей,— изогамный, анизогамный и оогамный, а также некоторые формы редуцированного полового процесса (рис. 91). У пеннатных диатомей по- ловой процесс во всех случаях состоит в сбли- жении двух клеток, в каждой из которых створ- ки раздвигаются и происходит редукционное деление ядра, после чего гаплоидные ядра попарно сливаются и образуется одна или две ауксоспоры. У центрических диатомей попар- ное сближение клеток отсутствует и ауксоспо- ра образуется из одной клетки, в которой сна- чала происходит деление материнского диплоид- ного ядра на четыре гаплоидных ядра, два из них затем редуцируются, а два сливаются в одно диплоидное ядро и образуется ауксо- спора. Все диатомовые водоросли — диплоидные ор- ганизмы, а гаплоидная фаза у них бывает только перед слиянием ядер в ауксоспоре. Как в первом, так и во втором случае после слияния ядер образуется зигота, которая сразу, без стадии покоя, резко увеличивается в разме- рах и развивает ауксоспору. По положению и связи с материнской клеткой ауксоспоры бы- вают разных типов. 'свободная ауксоспора, ко- нечная, боковая, интеркалярная и полуинтер- калярная (рис. 92). После созревания ауксоспоры в ней разви- вается новая клетка, у которой сначала обра- зуется эпитека, а затем гипотека. Первую клет- ку, возникшую из ауксоспоры, называют ини- циальной. По размерам она значительно превы- шает исходную. Покоящиеся споры. Образованию покоящих- ся спор обычно предшествует или обильная ве- гетация вида, или наступление неблагоприят- ных условий. Протопласт клетки сжимается, округляется, и на его поверхности появляется сначала первичная створка споры, а затем вторичная, обе плотно соединяются краями (поясок у них отсутствует). Створки часто от- личаются структурными элементами, они по- крыты шипиками, выростами и некоторыми дру- гими образованиями (рис. 93, 94). Обычно у диатомовых водорослей в клетке развивается только одна спора. Спустя определенное вре- мя покоящаяся спора, подобно ауксоспоре, увеличивается в объеме и дает начало новой клетке, вдвое большей по сравнению с ис- ходной. Покоящиеся споры обычно образуют многие морские неритовые диатомеи, а также некото- рые пресноводные виды. У представителей мно- гих родов они возникают периодически как обычное явление в жизненном цикле.
Рис. 94. Различные споры у видов рода Chaetoceros: 1 — Chaetoceros abnormis, цепочка с молодыми спорами; 2 — Ch. seiracanthus; 8 — Ch. subsecundus; 4 — Ch. lorenzianus; 5 — 6 — Ch. paulsenii; 7 — Ch. compressus; 8 — Ch. rigidus. БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ДИАТОМОВЫХ ВОДОРОСЛЕЙ Образование колоний. Колонии у диатомо- вых водорослей бывают различных размеров и формы, это зависит как от количества общей слизи, так и от способа соединения клеток друг с другом (рис. 95). Образуются колонии всегда из одной клетки в результате последовательных и многократных делений ее самой и всех ос- тальных возникающих при этом клеток. Все клетки остаются самостоятельными, и распад колонии не приводит к их гибели. Соединяются клетки при помощи слизи, выростов, шипиков, щетинок, рогов и пр. Колониальные формы обитают как в планктоне, так и в бентосе на различных субстратах — на растениях и живот- ных, на каменистых, песчаных и илистых грун- тах, на технических сооружениях и других предметах, введенных в воду человеком. В слизистых колониях клетки целиком по- гружены в выделяемую ими самими слизь. Эти колонии часто представлены бесформенными комочками или пленками однородной слизи, в которой клетки расположены беспорядочно. Клетки здесь могут быть подвижными, под мик- роскопом тто создает впечатление движения всей колонии. Есть также вполне оформленные слизистые колонии, имеющие вид простых вет- вистых трубок или листовидных пластинок, которые обычно прикрепляются к субстрату, реже свободно плавают. Они могут достигать макроскопических размеров. Слизь в подоб- ного рода колониях дифференцирована на на- ружный плотный слой и внутренний слой жид- кой консистенции, в котором живут и дви- гаются клетки. Форма колоний, не имеющих общей слизи, зависит от способа соединения клеток и очер- таний створок. Нитевидные колонии образу- ются в том случае, если смежные клетки ци- линдрические, а створки их круглые, причем клетки соединены поверхностью створок (род Leptocylindrus, рис. 95, 1), мелкими шипиками (род Melosira) или специальными трубковидны- ми выростами (роды Detonula, Stephanodiscus, Cyclotella). В лентовидных колониях клетки уплощенные, со створками линейной формы; сое- диняются поверхностью створок посредством слизи или мелких шипиков, расположенных по краю створок (роды Fragilaria, Rhabdonema, Achnanthes, Cymatosira, Dimerogramma). Осо- бенно интересны колонии бациллярии парадок- сальной (Bacillaria paradoxa): благодаря сколь зящему движению отдельных клеток по про- дольной оси створки в обе стороны, ее колония меняет форму от широкой ленты до длинной ступенчатой нити. Если клетки клиновидные и имеют б улав овидные створки, то колония приобретает вееровидную форму (роды Licmo- phora, Meridion, Opephora, табл. 10, 8). Особенно распространены цепочковидные ко- лонии разнообразной формы. В такие колонии клетки объединяются при помощи слизистых
О О О 0 Оо Ih в в в в (y 0 ее о О Рис. 95. Типы колоний: z — нитевидная колония (Leptocylindrus danicus); 2 — цепоч- ковидная колония (Rhizosolenia fragilissima); 8 — цепочковид- ная колония (Sceletonema costatum); 4 — гибкая цепочка (Tha- lassiosira excentrica); 5 — гибкая цепочка (Thalassiosira poly- chorda); 6 — прямая цепочка (Porosira glacialis); 7 — прямая цепочка, образованная при помощи выростов (Biddulphia mo- biliensis); 8 — зигзаговидная цепочка (Thalassionema nitzschioi- des); 9 — зигзаговидная цепочка (Diatoma elongatum); 10— нитевидная колония (Nitzschia seriata). подушечек или тяжей, коротких или длинных трубковидных выростов, рогов, щетинок и ши- пиков (табл. 10, 7; рис. 95, 2 — 7). Клетки, соединенные одним или несколькими довольно длинными слизистыми тяжами, образуют гиб- кие и непрочные цепочки (род Thalassiosira, не- которые виды Cyclotella). Прямые, часто очень длинные и прочные цепочки образуются путем соединения клеток посредством трубковидных выростов — длинных (род Sceletonema) или коротких (роды Bacterosira, Lauderia), а также щетинок (род Chaetoceros), длинных шипов (род Stephanopyxis), мелких шипиков (Rhizo- solenia fragilissima) или рогов (роды Hemiaulus, Biddulphia). В таких цепочках соседние клет- ки обычно расположены на некотором расстоя- нии друг от друга, что повышает плавучесть колонии. При свойственном пеннатным диато- меям соединении клеток в колонии попеременно разными углами прямоугольного панциря обра- зуются зигзаговидные цепочки (роды Tabella- ria, Grammatophora, Thalassionema, Diatoma и др., рис. 95, 8, 9), а если соединение проис- ходит с помощью двух углов одного конца створки, то получается звездчатая колония (роды Asterionella, Thalassiothrix). Клетки в подобных колониях скрепляются слизью, выделяемой специальными слизевыми порами, расположенными на полюсах створки. Прикрепленные формы часто образуют свое- образные пучковидные или кустиковидные ко- лонии. При этом исходная клетка сначала прочно прикрепляется одним концом к суб- страту при помощи слизи, которая выделяется слизевой порой на базальном конце створки и образует слизистую подошву. В процессе по- следующих делений клеток общая слизистая подошва разрастается и разветвляется, бла- годаря чему возникает кустиковидная колония с клетками, сидящими на концах студенистых «веточек» (роды Licmophora, Rhoicosphenia, С omphonema, Cymbella, Didymosphenia, табл. 10, 8). Если такого разрастания слизи не происходит, то клетки сидят пучками непо- средственно на слизистой подошве (роды Sy- nedra, Licmosphenia). Иногда зти колонии до- стигают макроскопических размеров. Совершенно уникальна колония Coenobiodis- cus тигпогт1 я, состоящая из одного слоя в 200 — 500 клеток, соединенных в поясковой зоне перегородками органической природы. Эти колонии воспроизводятся без прохождения од- ноклеточной стадии. Приспособления к планктонному образу жизни. Планктонные диатомеи — это пассив- но плавающие организмы, не связанные в своем жизненном цикле с каким-либо субстратом. Основной особенностью их являются различ- ного рода приспособления для уменьшения
массы клеток и увеличения поверхности панци- ря, способствующие свободному парению в тол- ще воды. Небольшая плотность клеток дости- гается здесь благодаря малому объему цито- плазмы, мелким хлоропластам и наличию в про- топласте значительного количества капелек масла. Интересно, что содержание масла в клет- ках диатомей во много раз выше, чем в клетках других планктонных водорослей. С другой сто- роны, панцирь планктонных диатомей, как пра- вило, тоньше и легче, чем у бентосных форм. Он пронизан густо расположенными мельчайшими отверстиями, а форма его обеспечивает большую поверхность на единицу массы. Так, у многих видов клетки приобрели форму тонких и длин- ных палочек (роды Rhizosolenia, Attheya, Tha- lassiothrix); у других — форму низкого цилинд- ра большого диаметра, часто с выростами и ши- пиками по краю створки (роды Thalassiosira, Bacterosira, Lauderia, Planktoniella). Многие планктонные виды, особенно морские пела- гические, несут на своем панцире тонкие и длинные щетинки (роды Chaetoceros, Bacteri- astrum, Corethron), способствующие повыше- нию плавучести клетки (табл. 13). Наиболее эффективное приспособление к па- рению — это увеличение поверхности путем соединения клеток в колонию. Чем больше по- верхность колонии, тем выше ее плавучесть. Кроме того, слизь, выделяемая клетками планк- тонных видов, легче воды; поэтому она, обво- лакивая клетки, также препятствует погруже- нию отдельных клеток и колонии в целом. Кроме морфологических приспособлений к планктонному образу жизни, диатомеи обла- дают еще физиологической адаптацией, свя- занной с процессами ассимиляции. Мертвые клетки и покоящиеся споры, когда ассимиляция отсутствует, теряют плавучесть, вследствие че- го опускаются на дно. Способы передвижения. Многие пеннатные диатомовые водоросли способны к активному движению. Они передвигаются по-разному: толчками вперед, назад или слегка в сторону, поворотами с боку на бок и медленным враще- нием вокруг своей оси. Особый тип движения наблюдается у бациллярии парадоксальной (Ba- cillaria paradoxa), палочковидные клетки кото- рой, образующие лентовидную колонию, спо- собны перемещаться скользящими движениями одна относительно другой в направлении про- дольной оси панциря. Большинство диатомовых водорослей пере- двигается по субстрату, хотя некоторые дви- жутся и в толще воды. Подвижные диатомеи, как правило, снабжены швом, причем чем слож- нее устройство шва, тем совершеннее их дви- жение. Но все же механизм движения до на- стоящего времени окончательно не объяснен. Существует мнение, что клетки диатомовых водорослей скользят по субстрату благодаря плазматическому потоку в щелевидном шве панциря. Однако движение наблюдалось и у ви- дов, имеющих очень короткий шов на конце створок без центрального узелка, а также в тех случаях, когда клетки были обращены к субстрату поясковой стороной или не сопри- касались с опорой, как, например, при враща- тельном движении. Одна из причин, вызывающих движение диа- томей,— фототаксис. Некоторые виды обладают положительным фототаксисом, другие — отри- цательным. Бентосные диатомеи, обитающие на дне, при сильном освещении погружаются в иловую пленку, а при благоприятном для них освещении перемещаются на ее поверх- ность. Некоторые из них по-разному реаги- руют на различные цвета спектра. Так, на- пример, Navicula radiosa относится положи- тельно к красным лучам спектра и отрицатель- но к голубым. Кроме того, на движение диато- мовых водорослей отрицательно действуют раз- личные вещества, способные растворять слизь. ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ И ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ ДИАТОМОВЫХ ВОДОРОСЛЕЙ Диатомовые водоросли живут повсюду. Мно- гие из них предпочитают водоемы определен- ного типа, с одинаковым физико-химическим режимом; другие обитают в очень разнообраз- ных водоемах. Диатомеи поселяются в верхо- вых болотах и моховых подушках, на камнях и скалах, в почвах и на их поверхности, на снегу и льду. Водные и вневодные местообита- ния неодинаковы как по видовому составу диа- томей, так и по их количеству. Число видов, населяющих вневодные биотопы, невелико, и все они относятся к наиболее широко распро- страненным представителям отдела. Только почвенные сообщества более богаты в видовом отношении. На снегу и льду диатомеи могут развиваться в массе, и тогда они окрашивают их в бурый цвет. Водная среда — основное и первичное место- обитание диатомей; здесь они возникли и про- шли длительный путь эволюции. Они завоевали все типы современных водоемов и принимают участие в образовании различных фитоценозов, преобладая качественно и количественно над другими микроскопическими водорослями. Они живут в океанах, морях, солоноватых, пересо- лоненных и различного типа пресных водое- мах: стоячих — озерах, прудах, болотах, ри- совых полях и т. д. — и текучих — реках, ру- чьях, оросительных каналах и т. д., вплоть 125
до горячих источников с температурой выше +50 'С. В водоемах диатомовые водоросли входят в различные группировки, основные из них — планктон и бентос. Морской планктон подразделяется на при- брежный — неритический, обитающий в при- брежной полосе на глубине примерно до 200 л, и удаленный от берегов — пелагический, насе- ляющий открытую часть моря. Неритический планктон обилен и разнообразен в видовом отношении. Пелагический (или океанический) планктон беднее и по составу и по количеству. Многие неритические виды обитают в пелагиа- ли, а океанические виды лишь иногда встре- чаются в неритическом планктоне: они, как правило, нежны и не могут долго существо- вать в прибрежной полосе из-за разрушающего воздействия прибоя. Морские планктонные виды принадлежат в основном к группе центрических диатомей, хотя к ним примешиваются и некоторые пен- натные формы. В планктоне пресноводных водоемов, наоборот, преобладают пеннатные диатомеи. В неритичес ком планктоне часто встречаются бентосные виды, поднятые водой со дна, часть из них обычно быстро снова опу- скается на дно, тогда как другие могут оста- ваться в толще воды длительное время (табл. 13). Бентос в широком смысле включает диато- мовые водоросли, обитающие непосредственно на дне и обрастающие различные возвышающие- ся над дном субстраты, в том числе и подвижные (буи, корабли, животных и пр.). Жизнь этих диатомей обязательно связана с субстратом- они или прикрепляются к нему, или движутся по его поверхности. Бентосные диатомеи обычно обитают на глубине не более 50 л. В морских и пресных водоемах они очень обильны и раз- нообразны в систематическом отношении (табл. 14). Наиболее разнообразны по видовому соста- ву и количеству диатомей 'ценозы обрастаний. Они состоят из колониальных и одиночно жи- вущих форм. В морях обычны представители родов Licmophora, Сrammatophora, Achnanthes, Mastogloia, Cocconeis, Synedra; в пресных водое- мах — С omphonema, Cymbella, Tabellaria, Diatoma, Rhopalodia, Cocconeis и пр. Особенно значительны и разнообразны обрастания рас- тений. Обрастания животных исследованы еще недостаточно. В частности, весьма интересен случай массового обрастания диатомей Cocco- neis ceticola кожи антарктических китов. Изве- стны диатомовые, живущие на циклопах, тин- тинидах и некоторых других животных. Количество диатомей, обитающих на дне водоемов, зависит от характера грунта и сте- пени его освещенности. На хорошо освещенном илистом грунте они многочисленны, а на пес- чаном или подвижном грунте их значительно меньше. Как правило, донные диатомеи — это одиночно живущие подвижные формы, способ- ные передвигаться к свету и таким образом выбираться на поверхность при заиливании. В морях это виды родов Diploneis, Amphora, Nitzschia, Surirella, Campylodiscus; в пресных водах также еще Pinnularia, Navicula, Cyro- sigma. Видовой состав диатомовых водорослей в во- доемах определяется комплексом физико-хи- мических факторов, из которых большое зна- чение имеет в первую очередь соленость воды. По отношению к солености все диатомеи разде- ляются на морские, солоноватоводные и прес- новодные. Особенно четко проявляется их реак- ция на содержание в воде поваренной соли NaC1, что позволяет различать у них три груп- пы видов. Первую составляют эвгалобы, для развития которых наличие хлоридов обязатель- но. Сюда относятся типично морские обитате- ли (полигалобы) и представители солонова- тых вод (мезогалобы), живущие во внутрен- них морях и опресненных морских бухтах. Во вторую группу входят олигогалобы — обита- тели пресных вод с соленостью не более 5'/„. Среди них различают галофилов, на которых не- значительное повышение содержания в воде NaC1 оказывает стимулирующее действие (Cyc- lotella meneghiniana, Synedra pulchella, Ba- cillaria paradoxa и др.), и индифферентов- типичных представителей пресных водоемов, но способных переносить незначительное при- сутствие в воде NaC1, хотя их развитие при этом и подавляется (Asterionella gracillima, Fragilaria pinnata и многие виды родов Cyclo- tella, С omphonema, Cymatopleura, Surirella). Третья группа — это настоящие пресноводные виды, на которых даже незначительное присут- ствие в воде NaC1 действует губительно (виды родов Eunotia, Pinnularia, Cymbella, Frustulia). Их называют галофобами. Таких индикаторов солености, приуроченных к определенным ее величинам, среди диато- мовых водорослей довольно много, и их список все время пополняется. Многие диатомовые во- доросли настолько чувствительны к содержа- нию в воде NaC1, что не выдерживают даже незначительного изменения солености,— это так называемые стеногалинные (узкосолевые) виды, к которым принадлежат типично мор- ские обитатели. Однако есть виды, степень чув- ствительности которых по отношению к NaC1 не так высока, и они способны существовать в широких пределах изменения солености во- ды, от почти пресной до морской,— это эврига- линные (широкосолевые) виды; они обитают в водоемах, где содержание NaC1 значительно колеблется.
Не менее важным экологическим фактором в развитии диатомей является температура. В общем эти водоросли вегетируют в широких температурных пределах — от 0 до +50 'С, но все же они чутко реагируют на изменения тем- пературы, — это находит свое выражение в се- зонной динамике и пиках развития. Правда, в этом отношении не все диатомеи одинаковы. Существуют эвритермные виды, способные пе- реносить значительные температурные колеба- ния, и стенотермные виды, живущие в узких пределах температурного режима. Для разви- тия большинства диатомей оптимальная темпе- ратура от +10 до +20 'С, но, кроме них, име- ются теплов одные виды, оптимум развития которых приходится на высокую температуру, и холодноводные виды, предпочитающие низ- кую температуру. Промежуточное положение занимают умеренно холодноводные и умеренно тепловодные виды. Степень освещенности и качество света также оказывают существенное влияние на развитие диатомовых водорослей в водоемах и опреде- ляют закономерности их распределения по глубинам. В свою очередь, освещенность за- висит от прозрачности воды, а прозрачность в океанах всегда более высока, чем в пресных водоемах. Диатомовые водоросли, населяющие как во- доемы, так и вневодные биотопы, приурочены к определенным географическим зонам, т. е. имеют определенный ареал. Многие морские виды отличаются строгой зональностью, в то время как другие распространены широко и да- же повсеместно. Особенно часто встречаются космополиты среди диатомей, обитающих в пресных континентальных водоемах. Наобо- рот, известны и эндемичные виды диатомовых, живущие только в каком-нибудь одном или нескольких водоемах одного района. Некото- рые водоемы, например озера Байкал и Тан- ганьика, очень богаты эндемиками, значитель- ное количество их обнаружено и в южных мо- рях СССР. Ограниченные ареалы имеют также реликтовые виды, живущие ныне в некоторых древних пресных водоемах — Байкале, Хуб- сугуле, Эльгыгытгыне, озерах Кольского полу- острова, африканских озерах и др. Известны реликты в Черном, Азовском и Каспийском морях, сохранившиеся от верхнетретичных мо- рей Черноморского бассейна. Закономерности географического распрост- ранения диатомей отчетливее всего проявля- ются в водах Мирового океана. Если принять деление Мирового океана на географические зоны по температурному режиму поверхност- ных слоев воды, то, как показывает анализ, в двух полярных зонах (арктической и антарк- тической), где преобладает низкая температура с незначительными годовыми колебаниями (2— 3'), обитают холодолюбивые стенотермные виды диатомей. Умеренные зоны обоих полушарий— северного (бореальная) и южного (нотальная)— характеризуются температурным режимом ши- рокого диапазона, здесь годовые колебания доходят до 15 — 20 'С. Этим зонам свойственны преимущественно эвритермные, а также уме- ренно холодноводные и умеренно тепловодные виды диатомей, достигающие массового разви- тия в тот или иной сезон. В тропической зоне, где температура поверхностных вод не опуска- ется ниже +15 'С, а годовые температурные колебания незначительны (в среднем около 2'), обитают теплолюбивые стен отермные виды. Некоторые виды диатомей могут обитать в двух смежных зонах — это арктическо-бореальные и бореально-тропические виды, приспособившие- ся к широкому температурному диапазону. Наиболее богата по видовому составу и ко- личеству диатомей бореальная зона, отличаю- щаяся оптимальной для их развития темпера- турой (от +10 до +20 'С). Здесь они вегети- руют почти круглый год, но особенно обильно развиваются весной и осенью. В арктической и тропической зонах вегетация диатомей крат- ковременная: в арктических морях она при- урочена к короткому летнему периоду, так как осенний и весенний -расцветы диатомей здесь по времени сближаются, в тропических — к бо- лее холодному зимнему периоду. Географические закономерности распределе- ния диатомей в континентальных водоемах выражены значительно менее отчетливо из-за их крайнего типологического разнообразия. Влия- ние местных экологических условии на водо- росли здесь настолько велико, что в большой мере нивелирует облик флоры, отвечающий гео- графическому положению каждого данного во- доема. Поэтому различия флористического со- става диатомей часто отчетливо проявляются, например, в двух соседних, но типологически неоднородных озерах, в то время как в различ- ных географических зонах, но в водоемах с одинаковыми экологическими условиями фло- ры диатомей могут оказаться весьма близкими. ЭВОЛЮЦИЯ И ФИЛОГЕНИЯ ДИАТОМОВЫХ ВОДОРОСЛЕЙ Диатомовые водоросли — сравнительно мо- лодая группа, но ее эволюция изучена полнее многих других, так как кремнеземные панцири или створки диатомей способны сохраняться в ископаемом состоянии очень длительное вре- мя. Сейчас известны ископаемые диатомовые водоросли от раннего мела (мезозойская эра- см. рис. на стр. 356) до современности без пе- 127
рерывов в отложениях. Это,дает возможность проследить их эволюцию в целом и установить филогенетические связи между семействами, родами и отдельными видами. Наиболее пол- ное представление получено о видовом составе бентосных ископаемых видов и неритического планктона, в то время как многие пелагические планктонные виды, имеющие тонкий панцирь, который растворяется в толще воды после отмирания клетки, отсутствуют даже в совре- менных донных осадках. Первые обнаруженные представители диато- мовых водорослей были морскими по происхож- дению и принадлежали к центрическому типу. С достоверностью известны панцири диатомей из раннемеловых отложений. В позднем мелу флора диатомей была уже богатой и разнооб- разной по видовому составу; она состояла поч- ти из 300 видов и разновидностей, относящихся к 59 родам. Но это были почти исключительно центрические формы, имеющие довольно при- митивный панцирь из двух створок без пояска. Створки были круглыми, треугольными, реже многоугольными, со структурой из крупных ареол, часто с крупными выростами, шипами и выпуклостями, способствующими объединению клеток в колонию. Б ольшинство древних ро- дов были монотипными или включали по 2— 3 вида. Диатомеи этого периода, по-видимому, не обладали широкой экологической пластич- ностью, а были стеногалинными и стенотерм- ными, что способствовало их быстрому выми- ранию. Некоторые роды вымерли уже к концу мелового периода (табл. 15, 1 — 6). Для меловой флоры диатомей характерно гос- подство видов из родов Hemiaulus и Triceratium, .а также появление первых трех видов пеннат- ного типа. По морфологической структуре пан- циря позднемеловые диатомеи сходны с совре- менными. Наиболее примитивными среди них следует считать представителей родов Stepha- nopyxu и Сladius. Большое видовое разнооб- разие диатомей в меловой период и достаточно сложная структура их панциря заставляют предположить, что к тому времени эта группа водорослей уже прошла длительный эволюци- онный путь развития. Естественно думать, что возникновение диатомей произошло в более раннюю геологическую эпоху. В палеогене (третичный период кайнозой- ской эры) продолжалось развитие центриче- ских диатомей, появлялись новые роды и виды. Для пале огеновой флоры, обнаруженной на территории СССР, известно более 500 видов и разновидностей. Господствующее положение по разнообразию и обилию видов здесь по-прежне- му занимают центрические диатомеи, хотя и пеннатные уже представлены несколькими ро- дами из семейства Fragilariaceae: Grunowiella, Sceptroneis и Rhaphoneis. У водорослей из этих родов панцирь имел еще примитивное строение. Таким образом, эволюция панциря выразилась в удлинении и появлении билате- ральной структуры, однако на этой ступени развития он еще лишен шва и клетки непод- вижны. К концу палеогена появились уже первые представители диатомей со щелевидным швом, развитым только на одной, а затем и на обеих створках (роды Eunotia, Actinella). Позднее возникли отдельные виды с более вы- сокоорганизованным швом из семейства нави- куловых (роды Navicula, Mastogloia, Diploneis, Amphora и др.) и, наконец, каналошовные диа- томеи — единичные виды одного рода. Палео- геновая флора унаследовала от меловой лишь 30оо видов, остальные 70 о о составили новые формы. Однако многие специфические роды, свойственные пал еогену, например Centropo- rus, Porodiscus, Brightwellia, Corona, Kittonia, Jousea, Grunowiella, Kentrodiscus и мно- гие другие, вымерли в последующие эпохи (табл. 12, 1, 2; табл. 15, 8). В неогене (вторая часть третичного периода) видовой состав диатомей еще более увеличил- ся — здесь известно свыше 860 видов и разно- видностей из 102 родов. Преобладали уже пен- натные формы (60оо), хотя в зависимости отме- няющихся экологических условий и палеогео- графической обстановки в различные периоды неогена по численности превалировали то цент- рические, то пеннатные диатомеи. Флора нео- гена унаследовала много родов от палеогена, а некоторые сохранились в ней еще с мела (Melosira, Stephanopyxis, Coscinodiscus, Acti- noptychus), но с иным видовым составом. Многие роды (Thalassiosira, Actinocyclus, Chae- toceros, Synedra, С rammatophora, Cocconeis, Mastogloia, Navicula, Amphora и Nitzschia) переживали в неогене бурный расцвет — для них отмечено интенсивное видообразование и широкое распространение. Все эти роды, и да- же многие неогеновые виды этих родов, дожили до настоящего времени и обитают в морях бо- реальной зоны. Кроме того, для неогена ха- рактерно последовательное возникновение но- вых родов, филогенетически связанных между собой в прямой ряд, показывающий постепен- ные усложнения в строении шва (Rouxia« -«АтрЫр1еига-+Frustulia-~Еат1си1а). В дальнейшем эволюция диатомей шла по пути совершенствования приспособлений кле- ток к условиям среды и освоения новых биото- пов. Появление клеток с дорсовентральным строением панциря и швом, смещенным к брюш- ной стороне створки или находящимся в киле (Amphora — «АтрЫргога — «Denticula — «Rhopa- lodia«Bacillaria«Mediaria«Surirella — «Campy- lodiscus), явилось наиболее прогрессивной сту-
1 а о л п ц а 9. Основные представители современных и ископаемых силикофлагеллат: 1 — Dictyocha fibula, строение клетки: а — базальное кольцо, б — радиальный рог, в — апикальное образование, г- жгутик, д — псевдоподии, е — хлоропласты; 2 — D. fibula, скелет сверху (палеоген, Западно-Сибирская низменность); 3 — Lyramula simplex, скелет (поздний мел, восточный склон Урала); 4 — Cornua poretzkajae, скелет снизу (поздний мел, восточный склон Урала); 5 — Corbisema apiculata, скелет снизу (палеоген, восточный склон Урала); 6 — Naviculopsis biapiculata, скелет сверху (палеоген, восточный склон Урала); 7 — Distephanus speculum, скелет сверху (неоген, Сахалин); 8 — Cannopilus sphaericus, скелет сбоку (неоген, Венгрия); 9 — Deflandryocha naviculoidea, скелет сверху (неоген, Югославия) ( х 1000).
Табл и а уп q О. Клетки диатомовых во о Ю дорослеи с хлоропластами азличнои ормы: v 1 — Pleurosigma sp. 2 — Biddulphia aurita. 3— р а аип~а; — Ме1ояга топ~Иогть; — Н аl — ае1осего~члЬ~~1Ь аг аЬпогт ~ 8 — 1 mis; — icmophora sp. — оси а sp.;
Та б л и ц а 11. Форма и структура панцирей современных видов диатомей: 1 — Melosira nummuloides; 2 — Achnanthes brevipes var. parvula; 3 — Navicula brachium; 4 — Diploneis didyma; 5— Mastogloia braunii; 6 — Rhopalodia musculus var. succincta. Электронные микрофотографии Н. И. Караевой.
/'r«~/.1 0 ц а /2. Многообразие формы и структуры панцирей вымершими видов диатомей: 1 — Kittonia claborata; 2 — Porodiscus sp. 3 — Actinoptychus ~елап~ь(миоцен); 4 — Fntogonia ip. (эоцен); 5 — Triccrайит fins (эоцен/; б — Stranguloncma barbadensis (эоцен). Электронные микрофотографии: /, 2, 3 — В. Уорнардта. 4, 6,— Х. С Кальверат5 — Р. Росса и П. Симс.
Та б.г rr ц а l3. Диатомовые водоросли планктона: I — Stephanodiscus astraea; 2 — Coscinodiscus perforatus var. celulosus; 3 — Asteromphalus robustus; 4 — Thalassiosira pseudonana; 5, 6 — Thalassiosira nordenskioldii (5 — клетка, б — створка); 7 — Sceletonema costatum; 8 — Actinocyclus chrcnbcrgii: 9 — Biddulphia mobiliensis. Микрофотографии: 1 — С.. И. Генкала, 2, 3, 9 — А. И. Прошкиной-Лавренко, 4 — 8 — И. В. Макаровой.
Та б л и ц а 14.,Яиатомовые водоросли бентоса: 1 — Cirammatophora serpentina; 2 — Caloneis liber; 3 — Nitzschia kuetzingiana; 4 — Navicula lyra var. atlantica; 5— Glyphodesmis distans; 6 — Nitzschia vidovichii; 7 — Diploneis bombus; 8 — Соссопе maxima; 9 — Actinoptychus undulatus; 10 — Mastogloia binota; 11 — Podocystis adriaticus. Микрофотографии: 1, 2, 4 — 11 — А. И. Прошкиной-Лавренко, 3— В.И. Михайлова.
Та б л и ц а 15. Ископаемые диатомовые водоросли: 1, 8 — Pyxilla speciosa f. speciosa; 2 — Coscinodiscus dissonus; 3 — Stephanopyxis ornata; 4 — Hemiaulus tschestnovii; 5— Cymatosira saytschenkoi; б — Triceratium cellulosum; 7 — Corona retinervis; 9 — Monopsia mammosa. Микрофотографии: 1 — 3, б, 8, 9 — Н. И. Стрельниковой (мел); 4, 7 — 3. И. Глезер (эоцен); 5 — А. И. Прошкиной-Лавренко (миоцен).
1 Та бл и ц и 16. Бурые водоросли: 1 — макроцистис грушевидный (Macrocystis pyrifera); 2 — нереоцистис Лютке (Nereocystis luetkeana); 3 — дурвиллея антарктическая (Durvillea antarctica); 4 — талассиофиллум решстчатый (ТЬа! <ssiophylluns clathrus); 5 — агарум проды- рявленный (Agarum cribrosum).
пенью. Совершенствование шва в эволюции диатомей занимает главное место, так как имен- но это обеспечивало быстрое развитие пеннат- ных форм, изменившее весь облик флоры водо- рослей в целом, особенно в бентосе морей и океанов. Некоторые роды диатомей, возникнув в нео- гене, просуществовали недолго и вымерли к концу этого периода (Ploiaria, Kisseleviella, Semseya, Mediaria). Наконец, весьма примечательным для неоге- на является сильное развитие диатомей в прес- новодных водоемах. Хотя первые пресноводные диатомеи были зарегистрированы из поздне- палеогеновых отложений озерного происхож- дения, в полную меру они развились только в неогене. Правда, вначале пресноводная флора неоге- на была довольно однообразной и массового развития достигли лишь немногие виды цент- рических (Melosira и Coscinodiscus) и пеннат- ных (Tetracyclus, Fragilaria, Eunotia) диато- мей. Однако уже для поздненеогеновой флоры характерно значительное разнообразие и оби- лие видов диатомовых водорослей. В плейстоцене (начало и середина четвертич- ного периода) происходит дальнейшее развитие и усовершенствование структуры панциря диа- томей, завершающееся в низших слоях голо- цена. Флора диатомей плейстоцена по составу родов уже значительно приближается к со- временной. Филогенетические изменения, кото- рые происходили в это время, выражались глав- ным образом в вымирании одних и возникнове- нии других видов и разновидностей в пределах этих установившихся родов. Большинство ви- дов просуществовало на протяжении всего плей- стоцена, другие — недолгое время. Современ- ные материалы позволяют проследить моменты их появления, расцвета и вымирания. Все основные этапы в развитии плейстоценовых диатомей хорошо представлены комплексами морских, солоноватоводных и пресноводных ви- дов, смена которых происходила под влиянием меняющихся условий среды обитания. Флора современных морей и континенталь- ных водоемов представляет последний этап эво- люционного развития диатомовых водорослей. В ней еще сохранились многие древние роды и некоторые виды. Наибольшая преемствен- ность у современной флоры устанавливается с флорой морей H континентальных водоемов плейстоцена, а наиболее характерная ее чер- та — господство пеннатных форм. В совре- менных морях пеннатные формы составляют 90 о~о бентосных диатомей, а в пресноводном бентосе представители центрических диатомей отсутствуют вовсе. Можно думать, что и в даль- нейшем эволюция диатомовых водорослей бу- дет направлена на процветание пеннатных подвижных форм. Диатомовые водоросли как отдел не имеют прямых родственных связей с другими отделами водорослей. По некоторым отдельным призна- кам, таким, как общность пигментов, сходство продуктов ассимиляции, наличие кремнеземной оболочки и покоящихся спор, обнаруживается отдаленное родство с отделами золотистых (Chrysophyta) и желто-зеленых (Xanthophyta) водорослей. Некоторые альгологи и сейчас объединяют их в качестве классов в общий от- дел Chrysophyta Однако правильнее считать диатомовые во- доросли самостоятельным монолитным и вы- сокоспециализированным отделом, очень рано отошедшим от общего корня с золотистыми и желто-зелеными водорослями. РОЛЬ В ПРИРОДЕ И ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ ДИАТОМОВЫХ ВОДОРОСЛЕЙ Диатомеи занимают совершенно исключитель- ное по своему значению место в общем кругово- роте веществ в природе. Будучи мощным и не- иссякаемым источником органического веще- ства, они служат постоянной кормовой базой и первоначальным звеном в пищевых цепях для многих организмов. Отмирая, они дают массу детрита и растворимых органических веществ, идущих на питание бактерий и простейших. Са- ми по себе они также являются прекрасной пищей для многих беспозвоночных животных. Кроме того, ими питаются некоторые рыбы— сельдь, хамса, сардины и другие, а также мо- лодь многих рыб. Установлено, что питательная ценность планктонных диатомей велика и не уступает ценности пищевых растений, а в некоторых случаях даже превосходит ее. В частности, со- держание белков и жиров у них выше, чем в картофеле и хлебных злаках. Однако диато- меи фитобентоса сублиторали, если учесть про- тяженность береговой линии, имеют еще боль- шее значение в питании различных водных жи- вотных, особенно молоди рыб в прибрежных районах. К тому же H неритический фитопланк- тон увеличивает свою биомассу за счет значи- тельного количества (более 40',о) бентосных диатомей, поднимающихся со дна в толщу воды и способных длительное время существовать в этих условиях. Установлена прямая количест- венная зависимость между количеством и рас- пределением фито- и зоопланктона, фито- и зообентоса и рыбной производительностью во- доемов. В различных географических зонах в опреде- ленные сезоны года численность диатомей мо-
жет достигать нескольких десятков миллионов клеток на 1 л, а биомасса — до нескольких граммов в 1 м'. Даже в сравнительно бедном фитопланктоном Баренцевом море на площади в 1 кл,' насчитывается до 5000 т фитопланкто- на, основную часть которого составляют диа- томеи. По продуктивности их сравнивают с на- земными травами и называют «пастбищем мо- рей». В морских бухтах, загрязненных органиче- скими веществами, планктонные и б енто сные диатомеи деятельно участвуют в процессах есте- ственного очищения воды. Некоторые виды служат хорошими индика- торами загрязнения морской воды различными стоками и нефтепродуктами, их используют при оценке санитарного состояния прибрежных морских вод. Диатомовые водоросли играют первостепен- ную роль в осадконакоплении. Непрерывно опускаясь из поверхностных вод и отмирая в бентосе, панцири диатомей скапливаются на дне в огромном количестве, образуя в океанах и пресных водоемах диатомовые илы. Мощные залежи илов известны в отложениях третичного и четвертичного периодов; их образование про- исходит и в настоящее время, особенно в озе- рах олиготрофного типа: Байкале, Тел ецком, Севане, многих озерах Кольского полуострова, Эстонской CCP и Латвийской CCP и др. Сап- ропели также в значительной мере образованы остатками диатомей, но они отличаются от диа- томовых илов меньшим содержанием кремневой кислоты (20 — 30 о', ) и большим содержанием органики. Сапропелевые илы скапливаются в большом количестве в озерах Среднего и Юж- ного Урала и Казахстана. Известны породы под названием «диатомит», на 50 — 80 о~ состоящлле из панцирей диатомовых водорослей. Диатомит представляет собой очень легкую, часто рыхлую и пористую породу бе- лого или светло-серого цвета с большим со- держанием аморфной кремнекислоты (50— 80 о~о). Благодаря своей пористости и адсорб- ционной способности диатомиты используются в пищевой, химической и медицинской промыш- ленности и в строительстве. При изучении осадочных пород диатомовые водоросли широко используются в стратигра- фических и палеогеографических целях. Видо- вая и экологическая характеристика комплек- сов найденных в осадках створок диатомей позволяет судить о процессе образования осад- ков и условиях среды в период осадконакопле- ния. Количественное распределение диатомей позволяет определять температурные условия, существовавшие во время отложения осадков, и восстанавливать климатическую зональность прошедших эпох. КЛАССИФИКАЦИЯ ДИАТОМОВЫХ ВОДОРОСЛЕЙ Изучением диатомовых водорослей занимают- ся уже почти 150 лет, за этот период была пред- принята не одна попытка их систематизации. Все системы диатомей можно разделить на три группы в зависимости от того, какие признаки были положены в их основу: 1) строение кле- ток и тип колоний; 2) положение, число и фор- ма хлоропластов; 3) форма и детальная струк- тура панциря. Системы первых двух групп оказались неудачными, так как форма колоний и хлоропластов в значительной мере зависит от условий среды H может часто меняться. Да- же в пределах одного рода виды имеют различ- ную форму хлоропластов и различное строение колоний в зависимости от того, где они оби- тают — в планктоне или бентосе. И наоборот, генетически не связанные между собой роды при наличии внешнего сходства по этим при- знакам включались в одну и ту же системати- ческую группу. Системы третьей группы бази- руются на более постоянных признаках — на строении панциря и деталях его структуры, позволяющих устанавливать родственные свя- зи между таксонами. Кроме того, подобные си- стемы имеют еще одно преимущество перед остальными: в них можно охватить не только современные, но и ископаемые формы, а это ва~кно при создании действительно филогене- тической системы диатомей. Первая система, в основу которой была по- ложена морфология панциря, появилась в 40-х годах прошлого столетия. В дальнейшем по мере развития и усовершенствования светового микроскопа эта система упорядочивалась и до- полнялась. Наиболее естественной и отвечаю- щей уровню знаний того времени была система Ф. Ш ю тт а, опубликованная в 1896 г., в ко- торой все диатомовые водоросли впервые были разделены на центрические с радиальной струк- турой и пеннатные с билатеральной (перистой) структурой. Окончательно современная систе- ма диатомей была сформирована в 30-х годах нынешнего столетия Ф. Х у с т е д т о м- крупнейшим диатомологом нашего времени. Однако и в настоящее время система диатомо- вых водорослей продолжает претерпевать изме- нения благодаря новым данным по морфологии, экологии и географии современных и ископае- мых форм. В области морфологии большую роль сыграло введение в практику альгологических исследований сначала трансмиссионного элек- тронного микроскопа (начало 40-х годов), а позднее (60-е годы) сканирующего электрон- ного микроскопа, дающего объемное изображе- ние створки или панциря. Особенно ценным оказался трансмиссионный электронный микро- скоп при изучении панцирей, имеющих очень
малые размеры или очень нежную структуру, неразличимую в световом микроскопе,— такие панцири считали «бесструктурными». Новые данные о тонкой структуре диатомей, полу- ченные за последние 35 лет с помощью электрон- ного микроскопа и сопоставленные с харак- теристиками, полученными в световом микро- скопе, позволяют составить объективное и де- тальное представление о структуре панциря того или иного вида, и эти данные сейчас с успе- хом применяются в таксономии диатомовых водорослей. В настоящее время известно более i0 000 ви- дов современных и ископаемых диатомей, отно- сящихся примерно к 300 родам. В системе, принятой нами для данной книги, все диатомо- вые водоросли рассматриваются как совершенно самостоятельный обособленный отдел Bacilla- riophyta, подразделяющийся на два класса— класс центрических диатомей (Centrophyceae) и класс пеннатных диатомей (Pennatophyceae). В первом классе содержится 5 порядков, во втором — 4 порядка. ризуются преимущественно низкоцилиндриче- ским панцирем с круглыми створками различ- ного строения; у представителей четвертого по- рядка (Soleniales) панцирь высокоцилиндриче- ский или палочкообразный, а створки круглые или эллиптические; диатомеи пятого порядка (Biddulphiales) имеют панцирь призматический или цилиндрический, со створками разных очертаний, часто с выростами или выпуклостя- ми на них. ПОРЯДОК КОСЦИНОДИСКОВЫЕ (COSCINODISCALES) У представителей порядка клетки одиночные или соединены в нитевидные и цепочковидные колонии. Панцирь линзовидный, эллипсоидный, шаровидный и цилиндрический. Створки в очер- тании круглые. Структура стенки створок пред- ставлена ареолами и ребрами, имеются также различного рода выросты. Порядок включает 4 семейства H много родов. Семейство мелозировые (Melosiraceae) КЛАСС ЦЕНТРИЧЕСКИЕ ДИАТОМЕИ (CENTBOPH YCEAE) У центрических диатомей клетки одиночные или соединенные в нитевидные или цепочковид- ные колонии. Форма панциря может быть различной: цилиндрической, дисковидной, лин- зовидной, шаровидной, эллипсоидной, бочонко- видной, реже призматической, со вставочными ободками различной формы. Створки в очер- таниях круглые, эллиптические, треугольные и многоугольные; их поверхность плоская или выпуклая, иногда вогнутая, реже волнистая. Ареолы на створках расположены беспорядочно или радиальными, спиральными и тангенци- альными рядами; по краю створки размещаются различного рода крупные и мелкие выросты или шипы, при помощи которых клетки могут сое- диняться в колонии, кроме того, имеются еще рога, щетинки и пр. Хлоропласты обычно в ви- де многочисленных мелких зерен или одной или нескольких пластинок. Цент рические диатомеи — преимущественно морские формы, обитающие в неритическом и океаническом планктоне; в пресных водоемах их мало. Это очень древняя группа, представи- тели ее известны с раннего мела; к настоящему времени многие роды целиком вымерли, из других родов только некоторые виды. Пять порядков, на которые в настоящее вре- мя делят этот класс, различают в основном по форме панциря и очертаниям створок. Из них водоросли первых трех порядков (Coscinodisca- les, Actinodiscales и Aulacodiscales) характе- Это семейство — одно из основных в поряд- ке. Клетки здесь обычно соединены в колонии, реже одиночные. Панцирь от линзовидного до цилиндрического. Створки плоские, иногда сильно выпуклые. Известны ауксоспоры и по- коящиеся споры. Включает современные и вы- мершие роды. Наиболее развиты и распростра- нены 3 рода. Рис. 96. Створка Hyalodiscus sphaerophorus (X 1000). Микрофотография И. В. Макаровой.
Рис. 97. Различная структура створок у Thalassiosira. 2 — Т. ezcentrica; 2 — Т. pacifica (0<2000). Электронные микрофотографии И. В. Макаровой. Род мелозира (Melosira, табл. 10, 8; 11, 1) широко представлен в планктоне и бентосе во- доемов всех типов — пресноводных, солонова- товодных и морских. Клетки соединены в плот- ные нитевидные колонии. Панцирь цилиндриче- ский, реже эллипсоидный или почти шаровид- ный. Створки плоские. Известны ауксоспоры и покоящиеся споры. Род гиалодискус (Hyalodiscus, табл. 10, 4; рис. 96) объединяет виды с одиночными клет- Рис. 98. Створка с двумя типами трубковидных вы- ростов у Sceletonema costatum (x14 000). По электрон- ной микрофотографии И. В. Макаровой. ками или соединенными слизистыми прослой- ками по 2 — 6. Панцирь у них линзовидный до сферического. Створки выпуклые, но середина их плоская или немного вогнутая, утолщенная, резко отграниченная от остальной части. Из- вестно около 15 современных и ископаемых мор- ских видов. Современные виды встречаются среди обрастаний в литоральной и сублитораль- ной зонах. Род стефанопиксис (Stephanopyxis, табл. 15, 8) отличается клетками, соединенными в цепочки при помощи длинных шипов. Панцирь цилинд- рический или шаровидный. Створки выпук- лые, в форме колпачка или чаши. Известно более 40 видов, преимущественно вымерших. Современные виды обитают в морском нерити- ческом планктоне. Семейство талассиозировые (Thalassiosiraceae) Клетки талассиозировых объединены в ко- лонии при помощи трубковидных выростов, сцепляющихся друг с другом, или студенистых тяжей, выходящих из этих выростов; одиноч- ными клетки бывают редко. Панцирь с пояска четырехугольный, створки круглые. У предста- вителей некоторых родов известны покоящиеся споры. Семейство объединяет 11 родов, обитаю- щих главным образом в морском планктоне. Приводим 3 из них. Род тилассиозира (Thai assiosira, табл. 13, 4 — 6; рис. 97) — обширная группа (около 80 современных и ископаемых видов), широко
Рис. 99. Структура створки у Stephanodiscus tenuis Рис. 100. Структура створкн у Coscinodiscus gigas ()(6400). Микрофотография С. И. Генкала. (>< 1000). Микрофотография И. В. Макаровои. представленная в планктоне морей и солоно- ватых водоемов всех географических зон. Клет- ки у этих водорослей соединены в рыхлые це- почки с помощью одного или нескольких сту- денистых тяжей. Панцирь имеет вид низкого барабана. Створки плоские или слегка вогну- тые или выпуклые посредине. Известны споры и ауксоспоры. Род сцелетонема (Seel etonema, табл. 13, 7; рис. 98) включает виды, у которых клетки сое- динены в плотные цепочки длинными трубко- видными выростами, расположенными по краю диска створки. Форма панциря варьирует от линзовидной до цилиндрической. Створки пло- ские или выпуклые. Видов в состав рода вхо- дит немного. Это морские и солоноватоводные водоросли, исключительно планктонные, эври- галинные и эвритермные. Род стефанодискус (Stephanodiscus, табл. 13,1; рис. 99) характеризуется клетками одиночны- ми или плотно соединенными короткими труб- ковидными выростами в нитевидные колонии. Панцирь низкоцилиндрический. Створки круг- лые, плоские или концентрически-волнистые. Представители рода широко распространены в планктоне пресных и солоноватых водоемов. Известны и ископаемые виды. Семейство косцииодисковые (Coscinodiscaceae) Клетки у косцинодисковых обычно одиноч- ные. Панцирь низкоцилиндрический, эллипсо- идный или бочонковидный. Створки круглые, плоские, выпуклые, вогнутые или концентри- чески-волнистые, с разнообразными выроста- ми. Включает около 10 современных и вымер- ших родов. Род косцинодискус (Coscinodiscus, табл. 13, 2; рис. 100) — основной род семейства. богатый видами и разновидностями (их более 300), преимущественно морскими, пела гическими и неритическими. Многие виды известны и в ис- копаемом состоянии, некоторые вымерли. Клет- ки у этих водорослей всегда одиночные. Пан- цирь низкоцилиндрический, редко клиновид- ный. Створки плоские, выпуклые, изредка кон- центрически- или тангенциально-волнистые. По краям створок часто имеется кольцо мелких щелевидных выростов, а иногда 1 — 2 крупных выроста. ПОРЯДОК АКТИНОДИСКОВЫЕ (ACTINODISCALES) В состав этой группы входят диатомовые во- доросли с одиночными или соединенными в ко- роткие цепочки клетками; панцирь у них низко- цилиндрический или эллипсоидный; створки в очертаниях круглые, эллиптические, реже треугольные или многоугольные. Стенки ство- рок пронизаны ареолами, часто поверхность створок разделена гиалиновыми ребрами на ра- диальные секторы. По краю створок нередко развиты шипы, а на полюсах — выросты. Порядок включает 4 семейства, из которых наиболее широко представлены в современ- ной и ископаемой флоре 2.
К семейству актикоптиховых (Actinoptycha- ceae) относят несколько родов, характеризую- щихся одиночными клетками с низкоцилинд- рическим панцирем и круглыми створками. Поверхность створок слегка вогнутая или ра- диально-волнистая, разделенная ребрами на б или более секторов, имеющих довольно слож- ную структуру. Род актикоптихус (Actinoptychus, табл. 12, 8; табл. 14, 9) основной в семействе. Это древ- няя и обширная группа, включающая около 100 современных и ископаемых видов. Семейство астеролампровых (Asterolampra- ceae) включает одноклеточные диатомеи с низ- коцилиндрическим панцирем. Створки у них круглые или овальные, слегка выпуклые или со слабой радиальной волнисто стью. Центр створки бесструктурный, периферическая часть ареолированная и разделенная гиалиновыми участками на клиновидные секторы. Включает 2 рода. Род астеромфалус (Asteromphalus, табл. 13, 8) — основной в семействе, виды его обитают в южных морях и океанах и известны также в ископаемом состоянии. ПОРЯДОК АУЛАКОДИСКОВЫЕ (AULACODISCALES) У аул акодисковых клетки одиночные, пан- цирь низкоцилиндрический, реже призматиче- ский. Створки в очертаниях круглые, редко трех-четырехугольные, плоские или выпуклые, редко волнистые. Стенки створок пронизаны ареолами, расположенными радиальными ря- дами, часто образующими пучки, реже танген- циальными рядами. На створках имеются так- же выросты, от одного до многих, иногда еще и ребра. Порядок включает 3 семейства, преимущест- венно с вымершими родами. Семейство актикоиикловых (Actinocyclaceae)— основное семейство порядка, содержащее только один род актикоииклус (Actinocyc- lus, табл. 13, 8). Он включает около 10 мор- ских современных и ископаемых видов, оби- тающих в прибрежных, часто опресненных участках морей. ПОРЯДОК СОЛЕНИЕВЫЕ (SOLENIALES) Представители этого порядка имеют клетки одиночные или соединенные в нитевидные ко- лонии. Панцирь у них удлиненно-цилиндриче- ский, часто с многочисленными вставочными ободками различной формы, поэтому почти всегда виден сбоку. Створки в очертаниях круглые или эллиптические, часто очень вы- пуклые, нередко в виде заостренного колпачка с шппом на вершине. Структура стенки ство- рок очень нежная, из мелких ареол, обычно невидимых в световой микроскоп. Порядок объединяет два семейства, которые включают главным образом современные мор- ские планктонные виды. Семейство ризосолениевых (Rhizosoleniaceae)— центральное семейство порядка. В его составе несколько родов. Род ризосоления (Rhizosole- nia, рис. 79, 8 — о; 95, 2) — самый обширный, он объединяет главным образом планктонные морские виды. Клетки у них одиночные, реже соединены в короткие колонии при помощи центрального шипа. Панцирь часто имеет вид длинной палочки, изредка слегка изогнутый, с многочисленными вставочными ободками. Створки обычно в виде эксцентрического кол- пачка с вытянутой верхушкой, заканчиваю- щейся шипом или длинной щетинкой, реже створки слегка выпуклые с шипиком в центре или у края. ПОРЯДОК БИДДУЛЬФИЕВЫЕ (BIDDULPHIALES) У представителей порядка клетки одиночные или соединены в цепочковидные колонии при помощи щетин о к, рогов и шипов; панцирь цилиндрический или призматический, реже эл- липсоидный; створки в очертаниях круглые, эллиптические, трех- или многоугольные, на полюсах с выростами в виде рогов или полых шипов или с длинными щетинками. Структура стенки створок с очень нежными или крупными ареолами. Известны ауксоспоры, микроспоры и покоящиеся споры. Биддульфиевые — обширный порядок, вклю- чающий 5 семейств преимущественно с морски- ми видами. Из них 4 семейства с большим коли- чеством представителей. Семейство хетоиеровых (Chaetoceraceae)— русское название «щетинкорогие» — наряду с вымершими родами содержит большой род хетоиерос (Chaetoceros), хорошо представлен- ный в современной флоре (табл. 10, 7; рис. 101). Он объединяет виды, у которых клетки соеди- нены в цепочки, прямые, изогнутые или скру- ченные вокруг своей оси. Панцирь у них низ- ко- или высокоцилиндрический. Створки круг- лые или эллиптические, плоские, выпуклые или вогнутые, с длинными щетинками, распо- ложенными одна против другой у края створки„ Одноименные щетинки двух соседних створо~ близ своего основания изгибаются, перекрещи- ваются и соприкасаются, а иногда и срастаются, что придает прочность всей колонии. Конеч- ные щетинки в цепочке часто отличаются от остальных щетинок длиной, толщиной и на- правлением. Известны ауксоспоры, микроспо- ры H покоящиеся споры различной формы и структуры (рис. 94). Ископаемые виды пред- 234
ставлены только спорами. Известно более 100 видов этого рода, обитающих в неритическом и океаническом планктоне морей и океанов, а также в солоноватых и пресных (два вида) водоемах. Семейство трииератовых (Triceratiaceae) включает морские роды с большим количест- вом вымерших видов. Род трииератиум (Tricera- tium, табл. 12, б) — обширная группа, предста- вители которой имеют панцирь в виде трех-, че- тырех-, иногда пятигранных призм с грубой структурой. Створки в очертании трех-, четы- рех- или пятиугольные, изредка эллиптические. Семейство биддульфиевых (Biddulphiaceae) ха- рактеризуется клетками, живущими одиночно или соединенными в цепочки, с панцирем в виде коробки, эллипсоида, цилиндра или трех- и многогранной призмы. Створки круглые, эл- липтические, ромбические или трех- и много- угольные, плоские или выпуклые с длинными выростами или низкими выпуклинами на полю- сах. Содержит 5 родов, из которых наиболее интересны 2. Род биддульфия (Biddulphia, табл. 10, 2; табл. 13, 9) имеет клетки, часто соединяющиеся в прямые или зигзаговидные цепочки. Пан- цирь цилиндрический, створки круглые или эллиптические (рис. 102). На полюсах створок по одному или по нескольку выростов различ- ной высоты и формы, на середине — длин- ные щетинки и мелкие шипики. Известно око- ло 30 морских видов, из них многие вымерли. Эти водоросли населяют планктон и бентос литорали. Род киттокия (Kittonia, табл. 12, 1) вклю- чает виды, у которых панцирь эллипсоидный или в виде трехгранной призмы. Створки эл- липтические или треугольные. Поверхность створки неровная, в средней части выпуклая, с кратерообразным углублением в центре. На створках по 2 — 3 длинных полых выроста, рас- положенных под углом к поверхности и закан- чивающихся большой круглой пластинкой в виде воронки. Известно только 3 морских вымерших вида. Семейство хемиауловых (Hemiaulaceae) вклю- чает три морских рода. Род хемиаулус (He- miaulus) — самый обширный и интересный (табл. 15, 4). У его представителей панцирь эллипс оидный или цилиндрический, прямой или слегка изогнутый. Створки эллиптические. По продольной оси створки на ее полюсах имеются длинные или короткие выросты — ро- га различной формы, заканчивающиеся на кон- цах одним — четырьмя шипами. Поверхность створок гладкая или бугристая благодаря бо- роздам, от которых внутрь панциря отходят перегородки (псевдосепты). По краю створки проходит гиалиновый кант, иногда он подни- Рис. 101. Две половинки панциря со скрещенными щетинками соседних створок у Chaetoceros sp. ( ~ 1000). Электронная микрофотография И. В. Макаровой. Рис. 102. Панцирь Biddulphia aurita со стороны пояска (><1000). Микрофотография А. И. Прошкиной-Лавренко. мается до вершины выростов. Обнаружены покоящиеся споры. Известно около 50 морских видов, в большинстве вымерших. КЛАСС ПЕННАТНЫЕ ДИАТОМЕИ (PENNATOPH'1'СЕАЕ) У пеннатных диатомей клетки одиночные или соединены в колонии различного типа. Панцирь симметричный по продольной оси, изопольный, реже гетер опольный, иногда дорсовентраль- ный, редко асимметричный, с пояска линейный, таблитчатый, клиновидный, прямой, иногда Я-об- разно-изогнутый, часто с вставочными обод- ками и септами. Створки в очертаниях бывают
Рпс. 103. Форма п структура панциря у Fragilaria Рис. 105. Synedra baculus, часть створки (>< 1000). pinnata ( x 8800). электронная микрофотография Микрофотография А. И. Прошкиной-Лавренко. Н. И. Караевой. Рис. 104. Asterionella formosa: с л е в а — колония; с п р а в а — конец створки (I — 0<1000, 2 — ><9400). Микрофотографии Л. Н. Волошко. линейные, ланцетные, эллиптические, реже бу- лавовидные, симметричные по отношению к про- дольной и поперечной плоскостям, реже по от- ношению только к одной из плоскостей. Струк- тура створок симметричная, из мелких или крупных ареол, расположенных обычно попе- речными параллельными рядами, которые у по- люсов створки несколько сходятся или ради- ально расходятся к ее краям. Иногда имеются еще поперечные ребра, слизевые поры и оди- ночные щелевидные выросты или мелкие много- численные шипики. По продольной оси створ- ки проходит бесструктурная узкая полсса— осевое поле, прерывающее ряды ареол или штрихи и ребра; в середине створки осевое поле часто расширяется, образуя среднее поле. У большинства видов вдоль середины осевого поля расположен щелевидный шов, иногда шов сдвинут к краю створки или находится в киле (каналовидный шов). Хлоропласты в виде одной или нескольких крупных пластинок, реже мно- гочисленных мелких пластинок. Пеннатные диатомеи — пресноводные и мор- ские формы, обитающие в бентосе на различ- ных субстрата х, и только единичные виды планктонные. Класс пеннатных диатомей мо-
ложе центрических, окончательно он сформи- ровался лишь в конце миоцена. Четыре порядка, на которые в настоящее вре- мя делят этот класс, различаются в основном степенью развития шва, по ним можно просле- дить эволюцию этого признака, начиная со створок без шва (порядок Araphales), затем со щелевидным швом на одной створке (порядок Monoraphales) и на двух створках (порядок Diraphales) и кончая наличием на обеих створ- ках наиболее сложного каналовидного шва (порядок Aulonoraphales). ПОРЯДОК БЕСШОВНЫЕ (ARAPHALES) У представителей порядка клетки одиночные или соединены в пучки или звездчатые и зиг- заговидные колонии. Панцирь прямой, иногда со вставочными ободками и септами. Створки от эллиптических до линейных, иногда булаво- видные. Стенки створок пронизаны ареолами, распо- ложенными поперечными рядами, иногда чере- дующимися с поперечными грубыми ребрами. Осевое поле от нитевидного до широколиней- ного. Щелевидный шов отсутствует. Порядок включает 2 семейства. Семейство фрагилариевые (Fragilariaceae) Клетки фрагилариевых одиночные или соб- раны в колонии разной формы. Панцирь с по- яска линейный, таблитчатый или клиновидный. Створки от линейных до эллиптических, реже ромбические, булавовидные или слегка изо- гнутые. Известно более 20 родов в современных во- доемах и в ископаемом состоянии. Приводим наиболее обширные и интересные. Род фрагилария (Fragilaria, рис. 103) содер- жит около 30 современных и ископаемых ви- дов, обитающих преимущественно в пресных водоемах, в бентосе, реже в планктоне. Клетки собраны в лентовидные или зигзагообразные колонии. Панцирь с пояска узкий, линейный. Створки в очертании от узколинейных до лан- цетных, часто расширенные посередине, иногда волнистые, редко треугольные. Род астерионелла (Asterionella, рис. 104) включает 2 пресноводных и несколько морских видов, все планктонные. Клетки у них образуют звездчатые колонии. Панцирь с пояска линейный, гетеропольный. Створки узколинейные, с головчатыми конца- ми, разной ширины. В пресных водоемах эти диатомеи встречаются очень широко, их легко узнать по характерной форме колоний. Род синедра (Synedra, рис. 105) — обширная и широко распространенная группа. Клетки Рис. 106. Tabellaria fenestrata: 2 — форма и структура створки; я — вид со стороны пояска. этих водорослей живут одиноч- но или соединены в пучковид- ные колонии. Панцирь с пояска палочковидный. Створки пря- мые, линейные, к концам сужен- ные. Часто на одном конце ство- рок бывает щелевидный вырост. В морских, солоноватых и прес- ных водоемах известно более 100 видов, населяющих бентос и обрастания, редко планктон. Род глифодесмис (Glyphodes- mis, табл. 14, о) содержит не- сколько морских сублитораль- ных видов. Клетки соединены в лентовидные ко- лонии. Панцирь с пояска линейный. Створки широколинейные до ланцетных, на середине слегка выпуклые. Род подо пистис (Podocystis, табл. 14, 11) включает виды с одиночными клетками. Пан- цирь у них гетеропольный, створки овально- грушевидные. На месте осевого поля проходит продольное ребро. Видов немного, все они мор- ские, сублиторальные. Семейс гво табелляриевые (Tabellariaceae) Клетки табелляриевых соединены в лентовид- ные или зигзаговидные цепочки, реже в вееро- видные пучки. Панцирь с пояска широколи- нейный до таблитчато го, реже клино видный. Рис. 107. Панцирь Grammatophora marina (X6000). электронная микрофотография Н. И. Караевой.
Створки линейные, ланцетные, удлиненно-эл- липтические, реже булавовидные. Включает около 10 родов, из которых 3 яв- ляются наиболее распространенными в морских и пресных водах. Род табеллярия (Tabellaria, рис. 106, 1, 2) объ- единяет несколько широко распространенных пресноводных видов, встречающихся как в бен- тосе, так H в планктоне. Клетки соединены в лентовидные или зигзаговидные цепочки. Пан- цирь с пояска прямоугольный. Створки изо- польные, линейные или овальные, на концах или на середине расширенные. Род грамматофора (С rammatophora, табл. 14, 1; рис. 107) содержит около 30 мор- ских сублиторальных видов, современных и ис- копаемых. Клетки образуют зигзаговидные це- почки, прикрепленные к субстрату. Панцирь с пояска удлиненно-четырехугольный. Створки .линейные до линейно-эллиптических. Род ликмофора (Licmophora, табл. 10, 8) включает около 30 современных и несколько ископаемых видов. Все виды морские, субли- торальные. Клетки соединены в пучковидные и веер овидные колонии, часто с ветвистыми студенистыми ножками, прикрепленные к суб- страту. Панцирь с пояска клиновидный. Створ- ки гетеропольные, булавовидные. ПОРЯДОК ОДНОШОВНЫЕ (MONORAPHALES) У представителей этого порядка клетки обыч- но одиночные, прикрепляющиеся к субстрату нижней створкой или студенистыми ножками, реже они собраны в лентовидные колонии, так- же прикрепляющиеся к субстрату. Панцирь прямой или изогнутый в продольном или по- перечном направлении. Створки линейные до широкоэллиптических. Структура стенки ство- рок различная: нижняя створка имеет щелевид- ный шов, расположенный по продольной оси створки, верхняя створка без шва, но с про- дольным гладким осевым полем; обе створки с поперечными ребрами, чередующимися с по- перечными рядами ареол. Порядок содержит одно семейство ахнанто- вых (Achnanthaceae). Эта группа включает 5 родов, как ископаемых, так и современных, широко распространенных в морских и прес- ных водах. Род коккокеис (Cocconeis, табл. 14, 8; рис. 108, 1, 2) содержит около 100 видов, ши- роко распространенных в морских, солонова- тых и пресных водах. Эти водоросли встреча- ются в сублиторали, обычно в обрастаниях. Часть видов ископаемые. Клетки одиночные. Панцирь изопольный, слегка изогнутый по поперечной оси. Створки эллиптические, ниж- няя вогнутая, верхняя слегка выпуклая. Род ахнантес (Achnanthes, рис. 109, 1, 2) ха- рактеризуется одиночными клетками или соб- ранными в лентовидные или кустиковидные ко- лонии. Панцирь с пояска прямоугольный, изо- гнутый по продольной оси. Створки линейные до ланцетных, реже эллиптические. Известно более 100 видов, широко распространенных Рис. 108. Створки двух видов Cocconeis: с л е в а — структура наружной поверхности нижней створки у Cocconeis placentula (X 7500); с п р а в а — структура внутренней поверхности верхней створки у Cocconeis pediculus (0<11 500). Электронные микрофотографии Н. И. Караевой.
в морских, солоноватых и пресных водах. В мо- рях они населяют сублитораль, в континенталь- ных водоемах встречаются в бентосе. ПОРЯДОК ДВУХШОВНЫЕ (DIRAPHALES) Здесь клетки большей частью одиночные, подвижные, реже собраны в лентовидные или кустиковидные колонии. Панцирь изопольный, реже гетеропольный, иногда с камерами вдоль краев ободка. Створки в очертаниях линейные до эллиптических, изредка S-образно-изогнутые. Обе створки со щелевидным простым или слож- ным швом. Шов прямой или изогнутый, реже сигмоидный. Структура стенки обеих створок одинаковая, представленная штрихами и ребра- ми или ареолами, расположенными поперечны- ми рядами. Этот обширный порядок разделен на 3 се- мейства, из которых здесь приведены 2. Семейство навикуловые (Naviculaceae) Это одно из центральных и наиболее обшир- ных семейств диатомей. Клетки у навикуловых одиночные или образуют кустиковидные коло- нии. Панцирь с пояска удлиненно-четырех- угольный. Створки линейные, ланцетные, эл- липтические, прямые, сигмоидные или дорсо- вентральные, с разнообразной формой концов. Шов проходит по продольной оси створок. Семейство очень богато родами и видами, оби- тающими в морях, пресных и солоноватых водоемах, обычно в сублиторали. Большое ко- личество видов обнаружено в ископаемом со- стоянии. Ниже приведены 4 наиболее обширных и ин- тересных рода. Род кавикула (Navicula, табл. 14, 4; рис. 110) — самый крупный род диатомовых водорослей, насчитывающий несколько сотен пресноводных, солоноватоводных и морских видов; некоторые виды известны в ископаемом состоянии. Клетки у них одиночные, изредка собранные в лентовидные или кустиковидные колонии. Панцирь с пояска прямоугольный. Створки линейные, ланцетные, реже эллипти- ческие, с острыми, клювовидными, закруглен- ными или головчатыми концами. Шов простой, реже сложный. Род пиккулария (Pinnularia, рис. 111) отно- сится к числу наиболее известных среди диато- мей, так как многие его виды отличаются круп- ными размерами и четкой структурой, благода- ря чему весьма удобны для изучения. Клетки одиночные. Панцирь с пояска прямоугольный. Створки изопольные, линейные, линейно-эл- липтические и ланцетные, иногда с волнистыми краями. Концы створок бывают тупые, за- Рис. 109. Achnanthes bre прея var. parvula. 7 — панцирь; 2 — верхняя створка ()(3000). Электронные мик- рофотографий Н. И. Караевой. кругленные, оттянутые или головчатые. Шов нитевидный или двухконтурный, простой или сложный; центральный и конечные узелки хо- рошо развиты. Известно более 150 видов, на- селяющих преимущественно пресные водоемы, реже моря; часть видов ископаемые. Рис. 110. Navicula intricata, часть створки (><8000) Электронная микрофотография Н. И. Караевой.
PIIs. 114. Ашр~о~а soffeaeformis: с л е в а — панцирь со спинной стороны; с п р а в а — панцирь со стороны створки (xQ4pp). Электронные микрофотографии Н. И. Караевой. Рис. 111. Pinnularia viridis, створка (><1000). Микро- фотография В. А. Федоровой. Рис. 115. Epithemia sorex, панцирь (x 5000). Электрон- ная микрофотография Н. И. Караевой. Рис. 112. P leurosigma elongatum, половина створки (x1000). Микрофотография Н. И. Караевой. Рис. 113. С omphonema lanceolatum, створка (>< 1000). Микрофотография В. А. Федоровой. Рис. 116. Rhopalodia musculus var. succienta, панцирь ()<5700). Электронная микрофотография Н. И. Караевой,
Род мастоглоя (Mastogloia, табл. 14, 10; рис. 76) включает более 100 видов, как вымер- ших, так и coBpeMeHHbIx, встречающихся в бен- тосе морей, co JIoHoBB TIIx и, реже, пресных водоемов. Клетки одиночные или иногда заключен- ные в общую слизь. Панцирь с пояска линей- ный, вдоль краев ободка находятся камеры, открытые в полость панциря. Створки эллип- тические, ланцетные, реже линейно-ланцет- ные. Шов у этих водорослей прямой или слег- ка изогнутый. Род плевросигла (Pleurosigma, рис. 112) имеет одиночные клетки. Панцирь с пояска линей- ный, суженный к концам. Створки ланцет- ные, линейные, S-образно-изогнутые, с тупо- или острозакругленными, иногда оттянутыми концами. Шов S-образно-изогнутый, осевой или эксцентрический. Известно около 50 морских и солоноватоводных видов, современных и ис- копаемых. ПОРЯДОК КАНАЛОШОВНЫЕ (AULONORAPHALES) У каналошовных диатомей клетки обычно одиночные, подвижные, реже сидячие, непод- вижные, очень редко они бывают соединены в подвижные лентовидные и нитевидные коло- нии. Панцирь продольно-, поперечно- или диа- гонально-симметричный. Створки разнообраз- ной формы, линейные, эллиптические, круглые или полулунные. Обе створки с каналовидным швом, расположенным в киле или крыловидном выросте створки. Структура стенки обеих ство- рок одинаковая, представленная ареолами, штрихами, ребрами и другими элементами, рас- положенными поперечными рядами. Это обшир- ный порядок, разделенный на 3 семейства. Семейство эпитемиевые (Epithemiaceae) Это небольшое семейство, интересное строени- ем входящих в него видов. Клетки у них дорсо- вентральные, одиночные, прикрепленные к суб- Семейство гомфоцимбеловые (Gomphocymbel- 1асеае} У гомфоцимбеловых клетки одиночные или образуют кустиковидные колонии, прикреп- ленные к субстрату студенистыми ножками. Панцирь с пояска линейный, клино видный или эллиптический. Створки линейно-ланцет- ные, ладьевидные или полулунные, слегка асимметричные по продольной или попереч- ной оси. Шов обычно слегка изогнутый. Семейство включает 6 родов, из которых наи- более обширны 2. Род голфокема (Gomphonema, рис. 113) со- держит более 100 видов, преимущественно прес- новодных. Часть видов ископаемые. Клетки одиночные, прикрепленные к субстрату студе- нистыми ножками, реже слагаются в ветвистые колонии. Панцирь с пояска клиновидный. Створки асимметричные по отношению к попе- речной оси, булав овидные, ланцетные, часто перешнурованные; концы створок закруглен- ные, головчатые, иногда заостренные. Шов нитевидный или широкий. Род амфора (Amphora, рис. 114, 1, 2) вклю- чает виды с одиночными клетками, подвижными или прикрепленными. Панцирь с пояска от удлиненно-эллиптического до прямоугольного. Створки ладьевидные, с более или менее пря- мым брюшным краем и выпуклым спинным. Шов прямой или изогнутый, сдвинутый к брюш- ному краю. Известно более 100 видов, большинство из них современные. Преобладают морские ви- ды, затем следуют солоноватоводные, а прес- новодных очень мало. ?+ ,, ФФ с+ +' Рис. 117. Структура створок у нитцший: 2 — Nitzschia obtusa var. scalpelliformis; внутренняя поверх- ность створки; 2 — Nitzschia tryblionella var. levidensis, створка ()< 3000). Электронные микрофотографии Н. И. Караевой.
°— '.;Щ~ 1, ~~в~ м~ДФ. Мбб. ЮМФ40Й г 1 Рис. 418 Hacillaria paradoxa: 2 — колония; 2 — створка. = б= ° б 11 — страту брюшной стороной пояс- 1 ковой зоны. Панцирь с пояска ! ! прямоугольный или эллиптиче- ! ! ский. Створки в очертании по- ! 1 лулунные, серповидные или ско- ! бовидные, редко ланцетные. Ка- I ! наловид.ный шов расположен в киле, находящем@я в плоскости створки по продольной оси или на одном из ее краев. = ° Известно 3 рода. 2 ' Род эпителия (Epithemia, рис. 115) включает около 20 со- лоноватоводных и пресно дных видов, часть из них ископаемые. Створки полулунные или бобовидные. Шов коленчато-изогнутый, ветви его у кон- цов створки находятся около брюшного края, на середине створки они изгибаются к спинно- му краю, где обе ветви соединяются, образуя острый угол. Род ропалодия (Rhopalodia, табл. 11, 6; рис. 116) содержит более 10 пресноводных, солоноватоводных и морских (их меньше все- го) видов. Створки полулунные или скобовид- ные. Шов расположен в хорошо заметном киле, проходящем по спинному краю створки. Рис. 119. Surirella fastuosa, створка (X 1000). Микро фотография А. И. Прошкиной-Лавренко. Семейство нитцшиевые (Nitzschiaceae) Это большое семейство, содержащее широко распространенные виды. Клетки у нитцшиевых одиночные, подвижные, реже соединенные в ко- лонии. Панцирь с пояска палочковидный, ве- ретеновидный, иногда S-образно-изогнутый. Створки линейные до эллиптических, с более или менее оттянутыми, клюв овидными, реже головчатыми концами. Каналовидный шов расположен в киле, проходящем по продоль- ной оси створки, или смещен к одному из ее краев. В семействе известно 7 родов, встречающихся как в пресных водоемах, так и в морях. При- водим два из них. Род нитщшия (Nitzschia, табл. 14,8; рис. 117, 1, 2) — обширнейшая группа, содер- жащая несколько сотен повсеместно распро- страненных видов. Клетки у них одиночные, реже соединенные в нитевидные колонии. Створки линейные, реже ланцетные или эллип- тические. Шов находится в киле, расположен- ном по краю створок. Это в основном бен- тосные, реже планктонные виды, встречаю- щиеся в морях, солоноватых и пресных во- доемах. Род бауиллярия (Bacillaria, рис. 118) объеди- няет очень интересные виды со своеобразным движением клеток. У этих водорослей клетки соединены створками в лентовидные колонии, в которых они движутся одна относительно другой, изменяя тем самым форму колонии. Рис. 120. Campylodiscus daemelianus, створка (X1000). Микрофотография И. В. Макаровой.
ОТДЕЛ БУРЫЕ ВОДОРОСЛИ (PHAEOPHYTA) 143 Створки линейные. Шов находится в киле, рас- положенном по продольной оси створок. Известно 4 морских, солоноватоводно-пресно- водных вида, встречающихся и в бентосе, и в планктоне. Семейство сурирелловые (Surirellaceae) Семейство сурирелловых резко отличается от других своеобразной формой и структурой клеток. Клетки здесь одиночные, подвижные. Панцирь в виде низкой коробки, иногда скру- ченный вокруг продольной оси, плоский, седло- видно- или S-образно-изогнутый, с пояска пря- моугольный или трапециевидный, изопольный или гетеропольный. Створки в очертании широ- к олинейные, эллиптические, яйцевидные или почти круглые, с широкозакругленными, реже клиновидными концами. Диск створки на месте перехода в загиб оттянут в узкое или широкое крыло, окружающее весь диск. По краю этого крыла проходит каналовидный шов. По про- К отделу бурых водорослей относятся много- численные, преимущественно макроскопические водоросли, общим внешним признаком которых служит желтовато-бурая окраска их слоевищ, вызванная наличием у них большого количест- ва желтых и бурых пигментов. Хлоропласты клеток бурых водорослей содержат хлорофилл а, хлорофилл с, P- H в-каротин H несколько ксантофиллов — фукоксантин, виолаксантин, антераксантин, зеаксантин. Пиреноиды у бу- рых водорослей мелкие, часто заметные только с помощью электронного микроскопа. Они име- ют грушевидную форму и выступают с поверх- ности хлор опласта в цитоплазму. Подобного типа пиреноиды существуют еще у криптофито- вых и эвгленовых водорослей. Деление ядра у бурых водорослей происходит посредством обычных митоза H мейоза. Запасные питатель- ные вещества — в основном ламинарин и ман- нит (сахароспирт) и в небольших количествах жир. Кроме обычных органелл, в клетках бу- рых водорослей присутствуют физоды, имею- щие вид бесцветных пузырьков в молодых клет- ках и желтых или бурых пузырей в старых. Они содержат танины (дубильные вещества) в виде флороглюцина и других полифенолов. Подвижные клетки бурых водорослей (зооспо- ры и гаметы) грушевидной формы с двумя жгу- '.тиками, прикрепленными сбоку; один жгутик даправлен вперед, перистый; задний жгутик гладкий. У представителей порядка диктиото- зых задний жгутик не развит. дольной оси створки расположено узкое или широкое осевое поле, иногда с одной или не- сколькими поперечными складками. В семей- стве 4 рода. Род сурирелла (Surirella, рис. 1199 содер- жит около 200 морских, солоноватоводных и пресноводных видов. Некоторые виды иско- паемые. Панцирь с пояска линейный или широко- клиновидный. Створки линейные, эллиптиче- ские, овальные, иногда суженные посредине. Поверхность створки попер ечно-волнообраз- ная за счет складок и ребер. Род кампилодискус (Campylodiscus, рис. 1209 объединяет виды с низким, широким, седло- видным панцирем. Створки в очертании почти круглые, повернутые одна относительно дру- гой на 90', так что продольные их оси пере- секаются под прямым углом. Описано много видов, населяющих преимущественно моря и солоноватые водоемы. Часть видов ископа- емые. Бурые водоросли исключительно многокле- точные растения. Оболочка клеток у них со- стоит из внутреннего целлюлозного слоя и на- ружного пектинового слоя, состоящего в основ- ном из альгиновой кислоты и ее солей и соеди- нений с белковыми веществами. Целлюлоза бурых водорослей по своим свойствам отлича- ется от целлюлозы высших растений, поэтому ее иногда называют альгулезой. Альгиновая кислота известна только у бу- рых водорослей. Она представляет линейный гетер ополисахарид, состоящий из связанных остатков D-маннуроновой и ? -гулуроновой кислот. В отличие от других многоклеточных водорос- лей у бурых водорослей, наряду с обычными одногнездными спорангиями (рис. 121, 29, име- ются многогнездные спорангии и гаметангии, неправильно называемые много клеточными (рис. 128, 1 а). Перед образованием зооспор или гамет содержимое многогнездных вмести- лищ делится тонкими перегородками на каме- ры, в которых оказываются заключенными по одному ядру с участком цитоплазмы. В каж- дой камере развивается по одной, реже по две зооспоры или гаметы. На поверхности слоеви- ща многих бурых водорослей развиваются осо- бые многоклеточные волоски, имеющие вид нити из одного ряда клеток с зоной роста при основании; клетки зоны роста делятся чаще других и поэтому имеют мелкие размеры (рис. 121, 1 б).
сразу поднимается пучок однорядных нитей, и одноосевой тип строения, при котором от основания вверх идет одна нить, а от нее ответ- вляются другие нити, идущие рядом с ней. Многие бурые водоросли имеют так называе- мое паренхиматическое строение, когда слоеви- ще состоит из нескольких рядов клеток плот- 7 но соединенных между собой по всей длине. Для бурых водорослей термины «паренхимати- ческий» и «паренхима» имеют относительное значение. У наземных растений паренхимой называют ткань, состоящую из клеток, размеры которых одинаковы по всем направлениям. В слоевищах бурых водорослей это наблюдается редко, в основном там, где идет активное деле- ние клеток. В остальных частях слоевища клет- ки вытянуты вдоль его продольной оси или в другом направлении. Например, «паренхима- тическое» слоевище десларестиевью имеет одно- осевое строение с плотным расположением кле- точных нитей, отходящих от оси. Рис. 121. Строение спорангиев: 1 — стреблонема (Streb lonema): а — многогнез нь нои роста; 2 — мирионема (Myrionema): в — опногнезпные спо- СТРОЕНИЕ СЛОЕВИЩА БУРЫХ ВОДОРОСЛЕЙ Слоевища бурых водорослей бывают от мик- роскопических, размером в несколько десятков микрометров, до гигантских, длиной 50 м (макроцистис, пелагофикус). По форме слое- вища могут быть самыми разнообразными: ни- тевидными или корковидными, шаровидными или мешковидными, пластинчатыми целыми или с разрывами, выростами и многочисленны- ми отве стиями р ями, гладкими или с продольными складками и ребрами, кустообразными. У наи- олее высокоорганизованных бурь урых водорос- лей (саргассул) слоевища кустистые с листо- видными пластинками, снабженными реб ом Слоевища некоторых крупных представителей имеют воздушные пузыри, удерживающие вет- ви в воде в вертикальном положении (табл. 16). Все бурые водоросли растут прикреплен- ными к грунту или к другим водорослям. Пла- вающие или свободно лежащие на грунте слое- вища не способны образовывать органы размно- жения. Для прикрепления к грунту служат всевозможные длинные вырост ты — риз оиды или дисковидное разрастание в основании слое- вища — базальный диск. Слоевища бурых водорослей могут быть об- разованы ветвящимися нитями из одного я а клеток эк одного ряда нити эктокарповые). У ряда представите лей и соединены в пучки, заключенные в слизь (некоторые хордариевые). При этом различают многоосевой тип строения, когда от основания СТРОЕНИЕ КЛЕТКИ БУРЫХ ВОДОРОСЛЕЙ Клетки летки всех бурых водорослей содержат по одному ядру и большей частью по нескольку мелких дисковидных хлоропластов. Реже хло- ропласты бывают лентовидными и пластинча- тыми. Фо рму хлоропластов иногда используют в качестве признака для разграничения родов. В то же время у бурых наблюдается изменение формы хлоропластов по мере старения клеток. Например, для эктокарпуса конфервообразного (Ectocarpus confervoides) обычны узкие лен- товидно-изогнутые хлоропласты, но по мере ста- рения слоевища в клетках появляется значи- тельное количество дисковидных хлоропластов. р д имеются или в хлоропластах веге- тативных клеток (эктокарповые, кутлериевые, хордариевые, пунктариевые), или только в хло- ропластах гамет (сфацеляриевые, фукусовые); у ряда бурых пиреноиды вообще отсутствуют (тилоптеридовые, диктиотовые, спорохновые, десмарестиевые) или встречаются крайне ред- ко (ламинариевые). Содержимое соседних клеток у бурых водо- рослей сообщается посредством плазмодесм. У клеток с толстыми оболочками обычно 7 присущих большим слоевищам, хорошо выра- жены крупные поры. В многорядных слоевищах бурых водорослей наблюдается специализация клеток с образова- нием тканей. В простейшем случае можно аз- личать кору из интенсивно окрашенных клеток, содержащих большое количество хлоропластов и физод, и сердцевину, состоящую из бесцвет- ных часто б 7 олее крупных клеток одинаковой формы. У более сложно организованных б— У- рых водорослей — ламинариевых и фукусо-
вых — коровой слой достигает большой толщи- ны и состоит из сильно окрашенных клеток разного размера и формы. Поверхностные 1 — 4 слоя образованы мелкими клетками, вы- тянутыми по направлению к поверхности,' они способны активно делиться и производить воло- ски и органы размножения. Эти верхние слои называют меристодермой — делящейся покров- ной тканью. Глубже лежит кора из более круп- ных окрашенных клеток, округлых на попереч- ном срезе и слегка удлиненных на продоль- ном. В центральной бесцветной части слоевища таких водорослей можно различить две груп- пы клеток. В центре находятся рыхло или плотно расположенные нити с сильно вытяну- тыми клетками. Это сердцевина. Между серд- цевиной и корой лежат крупные бесцветные клетки — так называемый промежуточный слой. Сердцевина у бурых водорослей не только слу- жит для транспортировки продуктов фотосин- теза (мто было доказано в опытах с радиоактив- ными веществами), но и выполняет механиме- скую функцию: в ней часто имеются тонкие ни- ти с толстыми продольными оболочками. Наи- более сложным анатомическим строением сре- ди бурых водорослей отличаются представите- ли порядка ламинариевых. У них развиваются слизистые каналы с особыми секреторными клетками, своеобразные клеточные нити для транспортировки продуктов фотосинтеза (си- товидные трубки, трубчатые нити). РАЗМНОЖЕНИЕ И ЦИКЛ РАЗВИТИЯ БУРЫХ ВОДОРОСЛЕЙ У бурых водорослей встречаются все формы размножения: вегетативное, бесполое и поло- вое. Вегетативное размножение происходит при случайном отделении ветвей от слоевища, и только у видов рода сфацелярия для этого су- ществуют специальные почки (рис. 131, 2 — 4). Слоевища бурых водорослей, оторванные от грунта, или их части не могут прикрепиться к твердому грунту: у них не образуются орга- ны прикрепления. Такие слоевища сносятся течениями в спокойные места с песчаным или илистым дном, и там они растут в неприкреп- ленном состоянии. Органы бесполого и поло- вого размножения образуются на них лишь в том случае, если их развитие к моменту отрыва слоевища от грунта уже началось. В резуль- тате такие слоевища размножаются только вегетативным путем. Старые нижние части от- мирают и разрушаются, а более молодые ветви становятся самостоятельными растениями. Бесполое размножение бурых водорослей осу- ществляется зооспорами, только у диктиото- вых имеются неподвижные тетраспоры, а у ти- лоптеридовых существуют моноспоры. Половой процесс у бурых водорослей пред- ставлен изогамией, гетерогамией и оогамией, гетерогамия встречается реже двух других форм полового размножения. Зооспоры и подвижные гаметы бурых водо- рослей содержат по одному хлоропласту, ко- торый в мужских гаметах при оогамии может быть бесцветным. Глазок у подвижных клеток бурых обнаруживается не всегда. Образование зооспор и гамет у бурых водо- рослей происходит во вместилищах двух основ- ных типов: одногнездных (рис. 121, 2) и много- гнездных (рис. 128, 1 а). Многогнездные вме- стилища могут функционировать как споран- гии и как гаметангии. Внешне они при этом не отличаются так же, как зооспоры и гаметы. Одногнездные вместилища чаще бывают спо- рангиями. Мейоз у бурых водорослей происхо- дит при образовании спор в одногнездных спо- рангиях, лишь у циклоспоровых он приходится на момент образования гамет. Одно гнездные и многогнездные вместилища часто неправильно называют соответственно од- ноклеточными и многоклеточными. И те и дру- гие могут развиваться из одной и из многих клеток. Когда в однорядных слоевищах группа клеток превращается в одно гнездные споран- гии, то говорят о цепочке одногнездных споран- гиев. Каждый из них при созревании открывает- ся самостоятельным отверстием. В случае мно- гогнездных вместилищ цепочка исходных кле- ток, каждая из которых увеличивается в разме- рах, дает единое многоклеточное многогнезд- ное вместилище (рис. 122, 1, 2). После образо- вания в нем камер можно заметить, что неко- торые поперечные перегородки толще других- это перегородки материнских клеток. При со- зревании содержимое такого вместилища вы- ходит через одно отверстие на вершине. У не- которых бурых водорослей одноклеточные мно- гогнездные вместилища узкие и камеры рас- положены в них в один ряд. Такие образования называют однорядными многогнездными спо- рангиями (гаметангиями, рис. 123). Многогнезд- ные вместилища с гнездами, расположенными в несколько рядов, считаются многорядными. Большинство бурых водорослей встречается в виде двух самостоятельных форм развития- спорофита и гаметофита или гаметоспорофита, которые могут быть как сходного размера и строения, так и разного. Другими словами, у бурых водорослей существуют изоморфная и гетероморфная смены форм развития. При ге- тероморфной смене форм развития одна из них бывает микроскопической, чаще всего это га- метофит. Развитию макроскопимеских слоевищ бурых водорослей нередко предшествует об- разование стелющихся нитей или дисков, на которых потом начинает расти вертикальное 145
Рис. 1 с нг~жоз~~ы1и~ сн 22. илаиелла прибрежная ~Р 1а1е11а коватый (Ectoca ми спорангиями Ш. эктока спорангиями (2) rpus sj. 1сц osus~ с много гнездными хлоропластами 8 . ; клетка эктока п са с л р у лентовидными одо ные образо- макроскопическое слоевище. По б вания называют п р о т о н е м о й. стелю ееся мои. сли такое щееся образование несет органы аз- множения и со в е р менем производит вертикаль- ганы ра�- зпл�етиз- ное слоевище, его называют талл юсом. У наиболее п вые, хо а и римитивных бурых (пункта рие- е ован рд риевые, эктокарповые) ст о т огое че- р д ание форм развития отсутствует. Из спо производимых спо о и твует. з спор, ро~итом, могут развиваться етофитов или гаметоспоро- как слоеви а гамет итон, так и слоевища спорофитов. р д рослей с половым процессом в виде изогамии подвижные клетки (зоои ы которые об аз ютс р у я на гаметофитах, функцио- зооиды нируют или как гаметы, сливаясь попа но и воспроизво я спо д розиты, или как зооспоры, ь попарно и бе п оизводя снова слоевища гамето ит . Т итов. акое есполое размножение гамето итов п вителей выш п то~итов у предста- ышеперечисленных порядков б ых водорослей асп р ространено крайне ши око. в урых ногда в лабо ато р рных культурах наблюдается подряд несколько поколений «г ме не приступаю их к «гамето итов» 7 аким образом, они проявляют себя как спо- и их лучше назы- ро иты и как гаметофиты и вать гаметоспо о р ~итами, чтобы не путать с другими растениями, гаметофиты кото ых не способны к бесполом иты которых не ряда других бурых водорослей спо о ит и гамето~ит ст строго чередуются, особенно у ес- споро ит марестиевых ламин инар иевых и циклоспоровых. у дес- иготы десмарестиевых и ламинариевых аз- виваются в и вых раз- п ик слоев ища спор офитов ост р репленными к оболочкам таваясь ам оогониев. то исключает напрасную гиб ель женских гамет воды и поис- во время нахождения их в толще ка места для прикрепления. ля класса и- клоспоровых характерно развитие гамето итов в слоеви ах спо щ поро~~итов. Некоторые циклоспо- ровые, нап име с р р аргассовые, размножают- ся проростками спорофитов, которые образу- ются в результате оплодотворения и прораста- ния женских гамет н ет на материнском растении. РАСПРОСТРАНЕНИЕ И ЭКОЛОГИЯ БУРЫХ ВОДОРОСЛЕЙ Буюые во о д росли почти исключительно мо- ские юастения ви иды только трех родов аст т н мор- в пресных во ах. О д . Однако некоторые морские тут виды могут обитать в сильно on еснен сненных уча- ах мореи с соленостью меньше 5' Б во о е „. урые всех морях зем- д росли можно встретить во вс ного шара. х заросли известны даже в ур ых местах, как прибрежные воды Аята к- таких тиды и севе ных р островов Канадского арктиче- воды нтарк- ского архипелага. наиболее крупные, длиной в несколько мет ов р , бурые водоросли (ламина- риевые, некоторые фукусовые) распростране-
ны главным образом в морях умеренных и при- полярных зон. Здесь для них благоприятны умеренная и низкая температура воды и повы- шенное по сравнению с водами тропических морей содержание биогенных веществ. Послед- нее обстоятельство обусловлено зимним пере- мешиванием воды: охлажденные поверхност- ные воды, как более тяжелые, опускаются вниз, а на их место поднимаются глубинные воды с повышенным содержанием биогенных веществ. Бурые водоросли населяют по вертикали са- мые различные горизонты. Их заросли встре- чаются от литоральной зоны, где они во время отлива часами находятся вне воды, до глуби- ны 40 — 100 м. Известны даже случаи произрас- тания бурых водорослей на глубине около 200 м. Так, у Гавайских островов на глубине 180 м обнаружены виды рода саргассум (Sar- gassum), в Адриатическом море ламинария Родригеса (Laminaria rodriguezii) была най- дена на глубине 200 м. И все же наиболее гус- тые заросли бурых водорослей располагаются в верхней сублиторали до глубины 6 — 15 м: здесь лучше условия освещения и благодаря прибою и поверхностным течениям существует постоянное движение воды, которое обеспечи- вает интенсивное поступление биогенных ве- ществ к слоевищам и ограничивает поселение растительноядных животных. Обычно бурые водоросли растут на скалах и камнях различ- ной величины (в зависимости от интенсивности движения воды и размеров слоевищ), и только в спокойных местах у берега и на большой глу- бине они могут удерживаться на створках ра- ковин моллюсков и на гравии. Оторванные от грунта слоевища сносятся течением в спо- койные места с илистым или песчаным дном, где при условии достаточной освещенности они продолжают существовать. Такие водоросли встречаются значительно глубже (иногда в 2 ра- за), чем прикрепленные. Виды с воздушными пузырями на слоевище при отрыве от грунта всплывают к поверхности, образуя иногда большие плавающие скопления, особенно в районах со стабильным круговым течением, как, например, в Саргассовом море. Среди бурых водорослей есть виды с эфемер- ными, однолетними и многолетними слоевища- ми. При гетероморфной смене форм развития одна из них может быть многолетней, а дру- гая — эфемерной (соответственно спорофит и гаметофит ламинариевых). На продолжитель- ность существования слоевища большое влия- ние оказывают внешние условия. Наибольшего возраста слоевища достигают при низкой тем- пературе воды. В районах с высоким летним прогревом воды слоевища водорослей, много- летних в холодных водах, функционируют как однолетние. При неблагоприятной осве- Рис. 423. Энтонема эцидиевидная (Entone;na aeci- dioides) на поперечном срезе через пластину ламинарии: а — вегетативные клетки энтонемы; б — опнорядные миогогнезп- ные спорангии; в — волоски с базальной зоной роста. щенности срок созревания, а значит исущест- вования эфемерных слоевищ может растяги- ваться на несколько месяцев вместо двух-трех недель (гаметофиты ламинариевых). Какие-ли- бо особые стадии покоя для переживания не- благоприятных условий у бурых водорослей отсутствуют. В то же время известны случаи, когда, например, зиготы фукусовых остава- лись живыми после пребывания в темноте в те- чение 120 дней, а гаметофиты ламинариевых сохраняли жизнеспособность после семи недель выдер жив ания в темноте. Многолетние слоевища бурых водорослей бы- вают нескольких типов. У одних все слоевище многолетнее, каждый год отмирают только побеги, на которых развивались органы раз- множения (Fucus). Примером другого типа служат ламина риевые — у них многолетние органы прикрепления и ствол. Пластинчатая часть, на которую нередко приходится боль- шая доля длины и массы слоевища, однолет- няя. У некоторых тропических видов саргас- совых многолетним бывает только диск, служа- щий для прикрепления слоевища. Во всех морях земного шара в произрастании бурых водорослей наблюдается сезонность. В морях умеренных и приполярных поясов наибольшего развития они достигают в летние месяцы, но бурный рост их слоевищ начинает- ся уже в конце зимы — ранней весной, когда температура воды еще остается около 0'С. В тропических морях массовое развитие бурых водорослей приурочено к зимним месяцам, ког- да температура воды несколько понижается.
ЗНАЧЕНИЕ БУРЫХ ВОДОРОСЛЕЙ Бурые водоросли — один из основных источ- ников органического вещества в прибрежной зоне, особенно в морях умеренных и приполяр- ных поясов, где их биомасса может достигать десятков килограммов на квадратный метр. Заросли бурых водорослей служат укрытием, местом размножения и питания многих при- брежных животных, кроме того, они создают условия для поселения других микроскопиче- ских и макроскопических водорослей. Роль бурых водорослей в жизни прибрежных вод видна на примере маьроиистииса (Macrocystis), о зарослях которого у берегов Южной Америки писал Ч. Дарвин: «Эти огромные подводные леса южного полушария я могу сравнить толь- ко с наземными лесами тропических областей. И все-таки, если бы в какой-нибудь стране уничтожить лес, то не думаю, чтобы при этом погибло хотя бы приблизительно такое коли- чество видов животных, как с уничтожением этой водоросли». Велика роль бурых водорослей и в хозяйст- венной деятельности человека. Вместе с дру- гими организмами они участвуют в обрастании морских судов и буев, ухудшая их эксплуата- ционные показатели. Но гораздо большее зна- чение имеют бурые водоросли как сырье для получения различного рода веществ. Во-первых, бурые водоросли — это единст- венный источник получения альгинатов — со- лей альгиновой кислоты. В зависимости от того, какие металлы участвуют в образовании альгинатов, они могут быть растворимыми в воде (соли одновалентных металлов) или не- растворимыми (соли поливалентных металлов, кроме магния). Наибольшее применение нахо- дит альгинат натрия, обладающий всеми свой- ствами водорастворимых альгинатов. Он спо- собен поглощать до 300 весовых единиц воды с образованием вязких растворов. Поэтому его широко используют для стабилизации разнооб- разных растворов и суспензий. Добавление не- большого количества альгината натрия повы- шает качество пищевых продуктов (консервов, мороженого, фруктовых соков и т. д.), разно- образных красящих и клеющих веществ. Раст- воры с добавлением альгинатов не теряют своих качеств при замораживании и размораживании. Применение альгинатов повышает качество пе- чатания книг, делает натуральные ткани не- выцветающими и непромокаемыми. Альгинаты используют при производстве пластмасс, син- тетических волокон и пластификаторов, для получения стойких к атмосферным воздейст- виям лакокрасочных покрытий и строитель- ных материалов. С их помощью изготовляют ~ысокекачественные смазочные материалы для машин, растворимые хирургические нити, ма- зи и пасты в фармацевтической и парфюмерной промышленности. В литейном производстве альгинаты улучшают качество формовочной земли. Альгинаты находят применение при брикетировании топлива, в производстве элек- тродов для электросварки, позволяющих полу- чать более высококачественные сварные швы. Трудно назвать отрасль народного хозяйства, где бы не использовались альгинаты. Другое важное вещество, получаемое из бу- рых водорослей, — шестиатомный спирт ман- нит. Он находит применение в фармацевтиче- ской промышленности для изготовления таб- леток, в приготовлении диабетических продук- тов питания, в производстве синтетических смол, красок, бумаги, взрывчатых веществ, при выделке кож. Все больше маннит исполь- зуют при проведении хирургических операций. Бурые водоросли содержат большое количе- ство иода и других микроэлементов. Поэтому они идут на приготовление кормовой муки, используемой как добавка в корм сельскохо- зяйственным животным. Благодаря этому со- кращается падеж скота, повышается его нро- дуктивность, в ряде сельскохозяйственных про- дуктов (яйца, молоко) увеличивается содержа- ние иода, что имеет важное значение для райо- нов, где население страдает от его недостатка. Когда-то бурые водоросли в больших коли- чествах перерабатывались для получения иода, но сейчас с этой целью используют лишь отхо- ды водоросл евой промышленности: в связи с появлением других, более рентабельных ис- точников получения иода, стало выгоднее пе- рерабатывать бурые водоросли на другие веще- ства. Бурые водоросли в свежем и переработан- ном виде применяют в качестве удобрений. Издавна бурые водоросли используют в ме- дицине. Сейчас выявляются все новые направ- ления их применения, например, для изготов- ления заменителей крови, для получения препаратов, предотвращающих свертывание кро- ви, и веществ, способствующих выведению ра- диоактивных веществ из организма. С давних времен бурые водоросли употреб- ляют в пищу, особенно народы Юго-Восточной Азии. Наибольшее значение в этом отношении имеют представители порядка ламинариевых, из них приготавливают наибольшее число са- мых разнообразных блюд. ПРОИСХОЖДЕНИЕ И КЛАССИФИКАЦИЯ БУРЫХ ВОДОРОСЛЕЙ Сходство пигментов и запасных питательных веществ позволяет предположить происхожде- ние бурых водорослей от предков, общих с зо- 148
лотистыми водорослями. Однако современные представители этих групп, кроме перечислен- ных признаков, имеют мало общих черт. К бурым водорослям с неполной достовер- ностью относят растительные остатки, сохра- нившиеся в палеозойских отложениях. В иско- паемом состоянии сохраняется в основном внеш- няя форма слоевища в виде отпечатка, а для достоверного определения принадлежности к от- делу бурых водорослей этого недостаточно, так как слоевища одинаковой формы встреча- ются в разных отделах водорослей. Бурые во- доросли отличаются от других отделов микро- скопическими и биохимическими признаками, которые плохо сохраняются в ископаемом со- стоянии. Полностью достоверные остатки бу- рых водорослей обнаружены в палеогеновых отложениях, имеющих возраст около 50— 60 млн. лет. В них уже появляются представи- тели современных родов иистозейра (Cystosei- ra), фикис (Fucus), аскофиллум (Ascophyllum). В настоящее время известно около 240 ро- дов и 1500 видов бурых водорослей. Из-за боль- шого разнообразия в строении слоевищ и ор- ганов размножения и особенностей циклов раз- вития бурые водоросли делят на несколько порядков (10 — 14), которые относят к одному или нескольким классам. Сначала (в середине XIX в.) бурые водоросли делили на два класса: фэозооспоровые (Phaeozoosporophyceae) и иикло- споровые (Cyclosporophyceae). К циклоспоровым отнесли бурые водоросли, у которых органы размножения развиваются в углублениях (кон- цептакулах) и имеют крупные размеры, позво- ляющие видеть их в препаратах невооруженным глазом. Отсюда и возникло название класса, переводимое на русский язык как «круглоспоро- вые». К фэозооспоровым причислили все осталь- ные бурые водоросли, многие из которых раз- множаются зооспорами, кроме диктиотовых. В Х Х в., когда началось интенсивное изу- чение циклов развития бурых водорослей, швед- ский ботаник Кюлин предложил разделить бурые водоросли на три класса на основе особен- ностей циклов развития. По-прежнему был со- хранен класс циклоспоровых и введено два новых класса: изогенератные (Izogeneratae) с изоморфным циклом развития и гетерогенерат- ные (Heterogeneratae) с гетероморфным циклом развития. Такая классификация бурых водо- рослей на классы получила широкое распрост- ранение. Однако деление на изогенератные и гетерогенератные довольно условно, поскольку в обоих классах в отдельных порядках сущест- вуют представители с противоположным типом смены форм развития. Например, кутлерия (Cutleria), относящаяся к изогенератным, имеет четкую гетероморфную смену форм развития. Мы делим бурые водоросли на два класса. Принятая ниже в тексте схема расположения порядков не претендует па отражение родст- венных связей между ними, она скорее дань традиционному изложению материала. На са- мом деле пути эволюции бурых водорослей нуждаются в коренном пересмотре. Сейчас примитивными считаются водоросли, имеющие слоевище из одного ряда клеток. Но развитие такого слоевища требует более упорядоченного образования перегородок при делении клеток (строго в одном положении без всяких откло- нений), чем, например, при формировании пластинчатого слоевища с расположением кле- ток без особого порядка. Это же подтверждается развитием спорая гиев. У бурых с многоряд- ными или пластинчатыми слоевищами они часто образуются из обычных вегетативных клеток, в то время как у многих представителей с одно- рядными слоевищами они появляются как осо- бые выросты. КЛАСС ФЭОЗООСПОРОВЫЕ (PHAEOZOOSPOROPH YCEAE) У фэозооспоровых спорофиты и гаметофиты существуют как самостоятельные формы раз- вития. При прорастании слоевищ одной формы развития на другой протоплазматическая связь между ними отсутствует. ПОРЯДОК ЭКТОКАРПОВЫЕ (ECTOCARPALES) К эктокарповым относятся бурые водоросли, слоевища которых построены из одно рядных нитей. Эти нити могут ветвиться и образовы- вать кусты высотой до 30, реже до 60 сл, или развиваться в виде микроскопического налета на скалах и других водорослях, или расти в те- ле крупных водорослей и только в период раз- множения выходить на поверхность и образо- вывать органы размножения. Клетки вегета- тивных нитей эктокарповых имеют окрашенные хлоропласты, содержащие пиреноиды. Органами размножения у этих водорослей служат одно- гнездные спорангии и многогнездные вмести- лища, последние могут быть спорангиями или гаметангиями. Половой процесс изо- или гете- рогамный. Гетерогамия наблюдается у видов рода гиффордия (Giffordia); у них известно три типа многогнездных вместилищ, различаю- щихся размерами гнезд (микро-, мезо- и мега- вместилища). Вместилища с самыми мелкими и с самыми крупными гнездами служат гаметан- гиями, с гнездами промежуточного размера- спорангиями. У эктокарповых наблюдается ве- гетативное размножение обрывками нитей. Эктокарповые встречаются во всех морях мира, но особенно много их в холодных морях
и в субтропических морях в холодное время года. Эктокарповые обитают в литорали и в сублиторали, на твердых и мягких грунтах и на более крупных водорослях. Они чаще дру- гих бурых водорослей участвуют в обрастании морских судов. В порядок входит только одно семейство эктиокарповых (Ectocarpaceae), объ- единяющее несколько родов. Род эктпокарпус (Ectocarpus) имеет кустистые слоевища высотой О, 1 — 30 см, крайне редко до 60 см, образованные тонкими нитями тол- щиной 50 — 150 мкм. Основанием служат сте- лющиеся ризоиды; у некоторых видов, расту- щих на водорослях, они могут проникать внутрь их. Крупные вертикальные побеги в основании бывают покрыты корой из ризоидов. Ветви слоевища сужаются к концам и на вершине оканчиваются длинными бесцветными клетка- ми. Волоски со вставочной зоной роста отсутст- вуют. Вертикальные побеги растут за счет деления большого числа клеток, особых зон роста нет. У нитей ризоидов существует верху- шечный рост. Клетки эктокарпуса снабжены пластинчатыми хлоропластами, в том числе лентовидными (рис. 122, 8). Спорангии и гаме- тангии располагаются как боковые выросты ветвей, многогнездные вместилища иногда имеют волосовидную стерильную вершину. Цикл развития у видов рода эктокарпус мо- жет быть изоморфным или гетероморфным. При этом виды, считающиеся ныне самостоятельны- ми, иногда оказываются формами развития од- ного вида. Половоь процесс всегда изогамный. Спорофиты эктокарпуса пучковатиого (Е. fas- ciculatus) в Атлантическом океане у северных берегов Франции при +13 'С несут только одногнездные спорангии, при +20 'С образу- ются многогнездные спорангии и в небольшом количестве одногнездные. Споры из много- гнездных спорангиев снова дают спорофиты. В одногнездных спорангиях происходит мейоз, и из спор вырастают раздельнополые гамето- фиты, напоминающие эктпокарпус драпарналь- диевидный (Е. draparnaldioides). Оплодотворе- ние происходит после прикрепления женских гамет. Из зигот вырастают спорофиты. При партеногенетическом прорастании гамет (без оплодотворения) появляются растения, подоб- ные спорофитам или гаметофитам. Образование разных органов размножения и изменение цикла развития в зависимости от района произрастания наблюдаются у эктокар- пуса аиручковатого (Е. siliculosus, рис. 122, 2). В Белом море и у берегов Норвегии встреча- ются почти исключительно растения с много- гнездными спорангиями — это спорофиты. Око- ло Неаполя зимой растут диплоидные растения с одногнездными спорангиями, производящими в результате мейоза гаплоидные зооспоры. Вес- ной существуют гаплоидные растения с много- гнездными вместилищами; образующиеся в них зооиды функционируют как гаметы или как зооспоры. Летом эти растения сменяются дип- лоидными растениями с одногнездными и многогнездными спорангиями. Из зооспор многогнездных спорангиев вырастают слоевища, растущие зимой и несущие одногнездные споран- гии. Из зооспор одногнездных спорангиев вы- растают гаметофиты, которых сменяют расту- щие зимой диплоидные спорофиты. У эктокарпу- са стручковатого из Неаполя при культивиро- вании в лаборатории наблюдались гаплоидные и диплоидные неветвящиеся спорофиты, кроме того, были обнаружены диплоидные гаметофиты и тетраплоидные спорофиты. У берегов Новой Англии (Атлантическое побережье США) экто- карпус стручковатый весной представлен ди- плоидными растениями с одногнездными и много- гнездными спора нгиями, которые сменяются летом гаметофитами, а осенью и зимой растут диплоидные спорофиты с многогнездными спо- рангиями. Зооспоры эктокарпуса стручковатого из одногнездных и многогнездных спорангиев различаются строением, однако глазок и жгу- тики у них одинаковые. Зооспоры из одногнезд- ных спорангиев крупнее, плавают медленнее и реже меняют направление движения. Род пилайелла (Pylaiella) напоминает экто- карпус (рис. 122, 1). Клетки с мелкими диско- видными хлоропластами. Ветвление односто- роннее и супротивное. Одногнездные спорангии образуются цепочками из вегетативных кле- ток ветвей. В пределах одной цепочки споран- гии созревают одновременно; содержимое их выходит последовательно, начиная с верхнего спорангия, через боковые отверстия, откры- вающиеся на одну сторону, в каждом споран- гии по одному отверстию. Многогнездные спо- рангии многоклеточные, тоже вставочные или в виде боковых веточек. У этих водорослей из- редка происходят продольные деления вегета- тивных клеток, поэтому пилайеллу некоторые исследователи относят к порядку диктиосифо- новых. Цикл развития у пилайеллы изоморфный. Наиболее широко известна пилайелла при- брежная (Р. litoralis). У этого вида растения с одногнездными вместилищами диплоидные. В результате мейоза образуются гаплоидные зооиды, кото",)ые прорастают в гаплоидные растения с многогнездными вместилищами, крайне редко часть зооидов сливаются попарно и снова производят диплоидные растения с одно- гнездными вместилищами. 3ооиды, образуе- мые растениями с многогнездными вместили- щами, функционируют как гаметы или без слияния воспроизводят материнские растения. Иногда, например у растений Тихоокеанского побережья США, смены форм развития не про- 150
с лоеви исходит. природе же и в культуре растут ища только с одногнездными cIIO "ангиями. од фельдманния (Feldmannia) имеет слое- вища высотой до 3 см которые о рые отличаются от йЩ Я -с ~ "у."" ф~, 3 СЛ эктокарпуса наличием в основании в @МЗДА 'Ф'~ '%„„~ ных по вертикаль- 4ИИВ :.:: 'Ф: бегов и волосков вставочных зо н роста, ф Д~ ~~r Ф" состоящих из ряда коротких клеток, а также ,„.ф.,» тем, что клетки со содержат дисковидные хдоо- '~~ � дну.'., у% ' '%. пласты. Боль льшинство видов рода фельдманнин,„,":;,-ф: растут на водорослях. ~ф Род сорокарпус (Sorocarpus) включает ви ы, Ф которые характеризуются многогнездными с о- рангиями, расположенными на на ветвях пучка- 3 ми; одногнездные спорангии неизвестны. од ахелотия (Bachelotia) объединяет водо- щие пилаиед- у. Слоевище у них в высоту достигает 3 . оропласты в клетках вертикал альных побегов;.;::;,': ') '"'" Аф.. удлиненные и расположены одно — ~ п- пами, в кото ых дной-двумя г п- рых ориентированы по радиусам. дно — у руп- клетках ризоидов хлоропласты дисковидные. У наиболее хорошо изученного вида — бахе- 2 "а~~ лотии антильской (В. antillarum) um половои про- у нее зооспоры цесс неизвестен в то же время " нее < из одногнездных спорангиев гоаз о 1 — тилоптерис (Tilopteris) спо анг 15 — 25 х 25 — 35 здО крупнее спорами z, a — гаплоспо 35 мкм) зооспор из многогнезд — ~.й'гнездное ый ных спорангиев (диаметра 4 6,5 р м — 6,5 мкм). Н Зд е вместилище на продольном разрезе, 3 — че- ра. Род стреблонема (Streblonema~ — п мик роскопических эктокарповых аст та~ — пример по о ной или д по нескольку в каждом междо з- пове рхн ости других водорослей , растущих на лии. Ветви н и лишенных со вставоч нередко оканчиваются вол оско уз- м вертикальных вегетативных ветвей, а если о ной зонои роста в основании. все-так сте и есть, то короткие и не отл твен, а если они Своеоб азны ичаются от множения. р у тилоптеридовых органы раз- И Ро э ~ы~~~Ы îî5 ~ж~~ ( ж 121 1' (р ) н р д повые а , растущие в слоевищах других во о о- ает эктокар- наблюдали как в 1 моноспорангии перед выхо- лей ~ ис 123 (р . 123). Их нити растут между клетк у х водорос- дом моноспо ы я р ядро делится с образова- хозяин ду клетками нием четы ех и на. рганы размножения и во б- лоски с а- ис. 124 8 р большего (до 12) числа ядер (р ., ). Míîãîãíåçäíûå вместилища тило аются группами и овых ст лопте- на нитях, расположенных под д наружным слоем до ослей. ридовых устроены иначе, чем у других б ь ых во- клеток или под кутикулой о в доросли-хозяина; полые вн т и и дорослей. Они расположены в основании ветвеи, ~~ы~ y p одним о гнезд о перифе- р --- -- О-" ываю ные и мн ногогнездные вместилища об а жу. дногнезд- ется особым отве рстием. До сих пор не совсем на аз р зных слоевищах водоросл- разуются ясны особенно и-хозяина. домых. По анн сти цикла развития тилопте и- о данным одних исследователей, моно- ПОРЯДОК ТИЛОПТЕРИДОВЫЕ (TILOPTERID рТЕВПЭА~,Е~) у растений одновременно. При этом моноспо- Это слабо изученный порядок. Он объе ры представляют собой оогонии, а многогнез- небольшое е число родов, составляю их к. н о ъединяет ные вместили а- щ — антеридии. Существует мне- огнезд- семей ство, и ограниченное число видов. щих одно ние, что гапл лоспора шаровидная (Haplospora У ~~~~гг~мгг~жй ы~и~йгг~ Ф " тва тилоптеридо- спо а к , аска о- 3 вых (Tilopteridaceae) высота а слоевищ достигает то ит о ра красивая (Scaphospora speciosa— яа' — гаме- см; они кустистые, с нитеви т видными ветвями, ния гамет эти ит одного и того же вида. Однако с н ко слия- у armx других тилоптеридовых р - д с ор б юд лос Дру е сслед- б ви . д. ост слоевищ происходи т членистый т е в основ дит посредством Моноспо ы вном слоевища с моноспоранг нгиями. вставочных зон роста, располо женных часто снова моноспо ы поры давали растения, производ щ я ие моноспоры, и лишь редкие слоевища ,151
ПОРЯДОК ХОРДАРИЕВЫЕ (CHORDARIALES) К ционные нити по ПОЯВЛЯЮТСЯ Г П вместе с органами а руппами только миля ми размножения; ляционные нити в , у других асси- щих вершинах ( и выражены тол олько на расту- у сплахкидиула). лоевища хорда риевых пост щим образом. В построены следую- основании им ная или много меется однослой- слоиная пластин ленные стелющи иеся нити. От ос нка или разветв- отходит большо основания вверх ое число о но точных верти д рядных многокле- кальных нитей п строении) или о дна нить п и о при многоосевом ении). Вертикал ( р одноосевом стро- альные нити аю ковые выросты д т короткие бо- ы, которые многок сивно ветвятся об кратно и интен- щих централ , о разуя слои кл ьныи пучок ве ти леток, окружаю- или единственн ертикальных нитей ную центральн ю в строении слоеви у нить. Различия ныл| числом ве ти вищ хорда иевы х связаны с раз- ертикальных нитей и а также с неоди наковой плотя и их высотой 1 Е Р~ .. дол ьный а ис. 125. Продольный а нити не pазвива- u вертикальные н ьный разрез слоевища мезоглойи разованное стелю ими ли слое- б образовывали ва хордариевы~ нес ли однов ел ьнои зоной ос оста. неко у олоски стилища. р ~енно многогнез Тилоптеридовые обла а Но ью к вегетативном оже однорядные, но го а овые слоевищ а могут об у размножению. ки ассимиляционны ( 2 5 разовываться ас х до см ельных много не толь- также содержат т много хло оп , и их клет- оклеточных часте р пластов. астеи ризо- в тия хорда риевых зак ~х клеток У тилоите заключается боковых ветв п са i Opteris mer nmepu- po ита ьшеи частью к твеи о паз ют омках У ического гамет стающие tensii) a б ру го спо- в новые сло ты, выра- м имеется плети тоспорофита. ~оевища. Тилоп в сублиторали птеридовые м микроскопичес кии спорофит ко тизмоталлюс- глубине в м~~~~~~езд~ые который несет в умеренных мо ачительной оста порангии и может в Атлантическ кого океан д части макроскопич т в виде вы- Амери ки. б~~ы~~ Е н в ропы и з зооспо еское слоевище слоевища пле тизмоталлюса. Мак снова вырастаю т сяоевища спо акроскопические о итов у хо а р p~~û~ роиз- здные или мног гогнездные спо ан- хордариевым отн те и другие однов Р Н- ременно Меиоз вод росли, тосп ческие и макр роскопиче копи- ся ?Х СОСТОИТ И иях. Гаме- видные, п з еские слоевища: я разветвленных нит их стелющих- п узы ревидные, шн а: корко- ются мн ~х нитей, на кото ых а тые (высотой многогнездные вме р х развива- й до 70 с.и1. ~" кусти с- есс- местилища. По прорастает в растеньи аком для Зигота аине редко — гете о — клеток) одно я клеточных нее но гаметоспо о ит д ро иту, а позд- х клетки, со е твленных спо о акроскопическое орофита л оста лоропластов и- шое коли- л тается в ви е кие нит азываю асс 3 б полового п о е они имеют б --"-.-.-.---"- Н ' """-' "-"'"' o цесса снова дают ш~. е~юд1с~ димых поро итам. Из спо п их верхушеч орму за счет тог ~х мак роскопическ р, произво- чные клетки к го,что г т вы кими спо о ита миляционные у врастать как гамето а ми, мо- ные нити бывают х. сси- ро иты. метоспорофиты. так и се это приводит к том 125 127 1) иевых отсутств редование форм телеи, например торых пред- И— развития. р у стилофопы а , ассимиля- вищ з-за большого а разнообразия в ст ищ хорлариевые де строении слое- делят на нес колько семейств. 152
Семейство мирионемовые (Муг1опета(асеае) Слоевище у мирионемовых мелкое, микроско- пическое, состоящее из одно- или двухслойной стелющейся пластинки с короткими вертикаль- ными нитямп (рис. 121, 2). Представители этого семейства часто встречаются на крупных водо- рослях, где их можно обнаружить только с по- мощью микроскопа, а также на камнях. Мири- онемовые произрастают в литоральной и суб- лито ральной зонах всех морей. Возможно, что часть водорослей, относящихся к этому се- мейству, представляют собой микроскопические стадии развития других бурых водорослей. Особенно часто встречаются виды трех родов. Род мирионема (Myrionema) включает виды с дисковидным или полусферическим слоеви- щем, достигающим в диаметре 0,5 — 4 мм. Осно- вание однослойное, из радиально расходящихся плотно сомкнутых нитей. Высота вертикальных нитей достигает 0,2 мм. Спорофит и гаметоспо- рофит могут иметь вертикальные нити разного облика и размера. Гаметоспорофиты с много- гнездными многорядными вместилищами, cGo- рофиты с одногнездными эллипсоидными или грушевидными спорангиями (рис. 121, 2). Род аскониклус (Ascocyclus) отличается от рода мирионема тем, что у его представителей на слоевищах развиваются одноклеточные па- рафизы. Спорангии известны только одноряд- ные многогнездные. Род микроспонгиум (Microspongium) объеди- няет водоросли, у которых слоевища образуют выпуклые слизистые подушечки. В отличие от вышеперечисленных родов вместилища об- разуются не на поверхности горизонтальной части, а по бокам вертикальных нитей. Семейство ральфсиевые (Ralfsiaceae) У ральфсиевых слоевище корковидное, гру- бое, большей частью многолетнее. Оно состоит из горизонтального слоя клеток, образован- ного радиально расходящимися нитями, и из вертикальных плотно сомкнутых рядов клеток (рис. 127, 2). Одногнездные спорангии и мно- гогнездные вместилища образуются как вы- росты поверхностных клеток. Одновременно с органами размножения развиваются много- клеточные парафизы. Ральфсиевые растут в морях умеренных и приполярных поясов. Наиболее широко известна ральфсия бородав- чатиая (Ralfsia verrucosa). Ее темно-коричневые корки достигают в диаметре 10 см и в толщину 2 мм, на них часто выражены концентрические полосы, соответствующие, видимо, периодам интенсивного роста. Ральфсия бородавчатая растет в литоральной зоне в «ваннах» (углуб- ления в скалах, заполненные водой) и на отк- рытых местах, а также в верхней сублиторали. В бесприливных морях она покрывает камни, омываемые прибоем. Органы размножения наб- людаются летом и осенью. Семейство литодермовые (Lithodermataceae) Литодермовые подобны ральфсиевым, но отличаются от них отсутствием парафиз и осооенностями развития органов размножения. Одногнездные спор ангии образуются из по- верхностных клеток, а многогнездные вмести- лища появляются или как боковые выр осты вертикальных нитей, которые вверху обычно неплотно сомкнуты, или путем преобразования их верхних клеток. Среди литодермовых на- ряду с морскими видами имеются представи- тели, растущие в сильно опресненных местах. Виды рода херибаудиелла (Heribaudiella) встре- чаются в реках в местах с сильным течением. Семейство элахистовые (Elachistaceae) Слоевище элахистовых имеет вид маленького пушистого или плотного шарика, несущего густой пучок нитей. Внутри слоевище состоит из вил ьчато-ветвящихся коротких нитей, ко- торые у поверхности переходят в неразветвлен- ные короткие ассимиляционные и длинные пе- риферические нити; последние при сильном развитии придают слоевищу вид пучка нитей. У некоторых представителей ассимиляционные нити отсутствуют. Органы размножения раз- виваются на основных нитях рядом с ассими- ляционными или на периферических нитях. Большинство элахистовых растет в качестве эпифитов на крупных водорослях, некоторые имеют снизу бесцветные нити, внедряющиеся в ткань хозяина. Распространены элахисто- вые в холодных и умеренных морях. Род элахиста (Elachistea) и его вид элахистиа фукусовая (Е. fucicola) наиболее широко извест- ны. Для этих водорослей характерны плотная полушаровидная часть в основании и длинные периферические нити, достигающие в длину 1,5 — 2 см. Периферические нити появляются летом, а осенью опадают, поэтому зимой и весной слоевище может иметь вид плотных шариков. Органами размножения являются одногнездные и многогнездные (однорядные и многорядные) спорангии. Зооспоры из много- гнездных спорангиев служат для воспроизведе- ния материнских растений. В одно гнездных спор ангиях происходит мейоз; образующиеся зооспоры дают микроскопические гаметоспоро- фиты, имеющие вид стелющихся разветвлен- ных нитей; на них развиваются многогнездные гаметангии. Переход от гаметоспорофитов к спо- рофитам осуществляется в результате полового
54 процесса и образования зигот или путем разви- тия слоевища спорофита из выроста на гамето- спорофите. Элахиста фукусовая встречается на бурых водорослях — фукусе и аскофиллуме, в защи- щенных и открытых местах в литорали и в верх- ней сублиторали. Особенно в больших коли- чествах она развивается на видах рода фукус, благодаря чему и получила такое видовое наз- вание. Элахиста фукусовая поселяется и на слое- вищах красных водорослей — халосакционе и родимении, растущих в тех же местах. Распро- странена она в северной части Атлантическо- го океана и в арктических морях. Рис. 426, Хордария плетевидная (Chordaria flagelh- formis). емейство коринофлеевые (Corynophlaeaceae) У коринофлеевых слоевище шаровидное, внутренняя часть его образована дихотомиче- ски-ветвящимися нитями, построенными из крупных бесцветных клеток. Снаружи распола- гаются ассимиляционные нити, а перифери- ческие нити, в отличие от элахистовых, отсут- ствуют. Наиболее крупным слоев ищем обла- дает ллатлзия неоднородная (Leathesia difformis, рис. 129, 1). Она может достигать в диаметре 5 сл. В крупных слоевищах внутренние нити разрушаются, и слоевища превращаются в по- лые шары. На одном слоевище могут одновре- менно развиваться одногнездные и однорядные многогнездные спорангии. Растет леатезия на камнях и на водорослях в литорали и в верх- ней сублиторали морей умеренного пояса. Семейство хордариевые (Chordariaceae) Ф.У Слоевище у хордариевых крупное, высотои до 70 см, кустистое, с цилиндрическими ветвя- ми. В центральной части побегов располагает- ся пучок продольных нитей или одна нить. Поверхность слоевища образована ассимиля- ционными нитями. В отдельных районах морей СССР широко распространены виды пяти родов. Ро эудлсмл (Eudesme) характеризуется неж- од ным слоевищем с побегами в виде сла о р б аз- ветвленных слизистых шнуров рыхлого строе- ния (в препарате при надавливании на покров- ное стекло нити легко расходятся). Ассимиля- ционные нити располагаются пучками. Цен- тральная часть ветвей сложена пучком нитей, слабо соединенных между собой. Органы раз- множения — одногнездные спор ангии. Из видов этого рода особенно известен эудес- мл элллноватый (Е. virescens). В морях СССР его макроскопические слоевища, являющиеся споро~ритами, ми растут в наиболее теплые летние месяцы в защищенных местах на других водо- рослях и камнях. Зимой он существует в виде микроскопических гаметоспорофитов, на кото- рых образуются одногнездные спорангии. Род млэоглойя (Mesogloia) отличается тем, что у его представителей в центре ветвей рас- положена одна нить, а не пучок нитей (рис. 125). Эти водоросли также имеют рыхлое стро- ение. У спорофитов некоторых видов, кроме одногнездных спорангиев, встречаются много- гнездные. Образующиеся в них зооспоры снова дают материнские растения. Род хордария (Chordaria) включает виды с кустистым слоевищем плотного строения, клеточные нити внутри его (кроме ассимиля- ционных) плотно соединены между собои (рис. 126, 127, 1). Центральная часть побегов обра- зована пучком нитей обычного для хордарие-
О о ° .У ез слоевища р»ьФ 1 . вертикальныи разре „дарии плетевиднои аз ез слоевища хо рис. ~;?7. Поперечныи разр ии бородавчатой (~). в более спокойных местах, ены з- этому она растет в ол ними расположены уз- еви а менее грубые. вых диаметра; между лстыми обо- и ее слоевищ обно рассмотреть линных клеток с толсты Ро аналипус (Ana ipus удо кие нити из дли они повышают ют прочность од е е наи олее изве б естного вида аналипу- лочками — гифы, О ганы размнож ) О.....,..„.... слоевища на разрыв. р са японского А. 'а onicus . океана, в морях уме- одногнездные спор ангии. и ной части Тихого океана, в асп остранени ие имеет в север са и о недавнего времени больщв Особенно ир'к" р р . "- О ая С. flagelli огш.1я . стен как гетерохор ария да ия еловая (Не- хордария плетевидная астет в северном по у р ол ша ии во всех в нередко terochordaria abietina). ~х айонах Япон- раст и ме енного поясов, не ое астение в некоторых ра субарктического и ум р частки ка- обычное ра К рильских островов ее заросли покр к ывают большие уча р щищенных ского м р, у мо я Сахалина, ури Камчатки. Аналипус в открытых и защ чного побережья ам менистого грунта в боя слоеви- и восточно ое слоевище высо- мости от силы при оя кий имеет вертикальное с местах. В зависимо ви ной могут ыть б ь то сильно японский см с ент альным п побегом, от которо- щ а хордарии плетевид в более спокойных м естах), тои до сл ц ят ко откие неветвящи иеся веточки. ( r e npmoo5 асс имиляционными твленными ~там, гд й слой образован а то почти нера зветв нгии у споро~ритов у у в имеются Коровой Многогнездные вме стилища разви- сильный). Споранги б я. Гаметоспоро ить ~~ ты микро- нитями. п тем преобразования к клеток ассимиля- с апреля до ноя ря. стилищами. ваются пут х шечной клетки. ногогнездными вмести ых нитей, кроме верх скопические с мног . к льт.р из у зооспор, ционных еви е прикр епляется к грун- В . виях лабораторны" у. услови т спо офиты, снова выра- с- Вертикальное сло щ вой. Иногда оно отходит от ст елющих- которые производят споро и ы а также гаметоспо рофиты. ту подошвой. ногда зветвленных многорядных побегов с плот- азмножать- ся разветвл последние культур р е п одолжали р вым слоем. мест б азуют небольшое тем посредством зоос тикальные по еги о ра ся бесполым пут гаметоспо- боем верт бегов. Могут вы- ы азмножения на га о отких боковых по мечено, что органы р р зкой тем- число кор б и совершенно без ются при более низк ве тикальные по еги рофптах развиваю соответст- растать р п ибойных местах а спо орбитах, что со - ветвей. В еще олее п стелющиеся побеги или вует их пр оизрастанию в природе р азвива ются только стел ющи ок как у раль сии. и слоевища в виде корок, в от илия (Sphaerotrichia) отчасти матохновые (Spermatochnaceae) сп тать по внешнем о лику, Семейство сперматохновые и е сперматохновых кустистое, с од- (S. divari Слоевыце у сперм стое c og- u четы ьмя-пятью осевь са а'„ ная, отсутствуют ги ы. о- хордария плетевидная, о Ы5
образуются только при развитии органов раз- множения и выполняют функцию парафиз — бес- плодных клеточных нитей. Из этого семейства наиболее широко известны род сперльатохнус (Spermatochnus) с одной осевой нитью и род стилофора (Stilophora) с четырьмя-пятью осе- выми нитями. В СССР виды этих родов произ- растают в сублиторали Черного моря. Семейство сплахнидиевые (Splachnidiaceae) К этому семейству относится всего один род сплахнидиум (Splachnidium) с единственным ви- дом. Сплахнидиум морщинистый (Splachnidium rugosum) растет в южном полушарии у мыса Доброй Надежды и по южному берегу Австра- лии. Слоевище у этого растения студенистое, с полостью внутри; оно состоит из центрального толстого вертикального побега с немногочис- ленными слабо разветвленными толстыми бо- ковыми ветвями. В основании слоевища имеет- ся диск для прикрепления к грунту. Органы размножения — одногнездные спорангии- у сплахнидиума морщинистого развиваются в особых углублениях — концептакулах. Кон- цептакулы образуются в некоторых точках поверхности слоевища, вблизи вершин побегов. Здесь начинаются интенсивное деление и рост клеток, при этом возникают впячивания внутрь слоевища. Образованию концептакулов спо- собствует то, что слоевище имеет рыхлое нит- чатое строение и снабжено полостями, запол- ненными слизью. В прошлом сплахнидиум морщинистый относили к фукусовым на том основании, что у него существуют концепта- кулы и на вершинах побегов расположены осо- бые крупные клетки. Однако со временем вы- яснилось, что концептакулы у них образуются совершенно иначе, чем у фукусовых, а особые клетки оказались одноклеточной эндофитной зеленой водорослью кодиолум (Codiolum), ко- торая постоянно присутствует в коровом слое молодых вершин побегов. Ассимиляционные нити у сплахнидиума морщинистого развивают- ся только на молодых частях, потом они опа- дают, и поверхность оказывается образован- ной плотной корой из мелких клеток. У сплах- нидиума имеются многоклеточные волоски с ин- тер калярной зоной роста, которые растут на внутренней поверхности концептакулов и вы- совываются через их отверстия наружу. ПОРЯДОК ДИКТИОСИФОНОВЫЕ (DI|:T YOSIPHONA LES) Высота слоевища диктиосифоновых колеб- лется от нескольких миллиметров до 1 м. Оно целиком или частично паренхиматического строения. Слоевища диктиосифоновых могут быть пластинчатыми или кустистыми с цилин- дрическими ветвями, многорядными или в не- которых местах однорядными. У всех диктио- сифоновых с многорядными слоевищами плот- ный коровой слой, состоящий из тесно сомк- нутых клеток. Макроскопические слоевища мо- гут быть спорофитами или гаметоспорофитами. Органы размножения — одногнездные или мно- гогнездные вместилища — развиваются на по- верхности слоевища; при этом могут появлять- ся парафизы, или же в органы размножения превращаются поверхностные клетки или клет- ки, лежащие под корой. Макроскопические слоевища диктиосифоно- вых образуют одногнездные или многогнезд- ные вместилища или оба типа органов размно- жения одновременно. Мейоз обычно происхо- дит в одногнездных спорангиях. Цикл разви- тия диктиосифоновых подобен циклу развития хордариевых. Он характеризуется, во-первых, нерегулярной сменой форм развития, вызван- ной тем, что они способны к самовозобновлению посредством спор или партеногенетического развития гамет; во-вторых, наличием микро- скопических форм развития, которые могут быть гаметоспорофитами или плетизмоталлю- сами, способными производить многогнездные спорангии и слоевища спорофитов в качестве выростов. Половой процесс у диктиосифоновых в основном изогамный, реже гетерогамный. Диктиосифоновые распространены в морях всех океанов, но особенно много их в морях умеренных и приполярных поясов, где они об- разуют заросли. Встречаются они в самых раз- личных биотопах — в литорали и в сублитора- ли, на камнях и других водорослях. В силу большого разнообразия диктиосифоновые де- лятся на несколько семейств. Семейство пунктариевые (Punctariaceae) Слоевище пунктариевых может быть пластин- чатым, цилиндрическим или нитевидным, не- разветвленным или ветвящимся в самом осно- вании. Одногнездные и многогнездные вмести- лища у пластинчатых слоевищ образуются путем преобразования поверхностных клеток, у нитевидных слоевищ они могут располагать- ся снаружи. Род пунктария (Punctaria) наиболее широко распространен, особенно пукктария широко- листная (Р. latifolia). Ее слоевище имеет вид удлиненной (до 65 см) пластины, сужающейся к основанию. Органом прикрепления к грунту служит маленький диск. Пластина состоит из четырех — семи слоев клеток (рис. 128, 1). Наружные слои образованы более мелкими и интенсивно окрашенными клетками. У рас- тений, растущих в местах, где отсутствует ин-
тенсивное движение в евища плотно сомк ч п воды, клетки вн т у ри сло- прямымц стенками. омкн'етые с п ри оиных местах углы клеток вн т слоев становятся за ток внутренних ся межклеточнь я закругленными ве , у личивают- ныепространства; из е ка лежащие в центре, редка клетки, идущие по нтре, дают нитеви евидные выросты, по межклеточным II Этим достигается б пространствам. чность слоев и тся ольшая эластичн ость и про- р д >~ макроскопиче- ищ. еюедование споро ита и мик оскопи р м.'.,е...,....е,. ита на людается не всег а ча из одно гнездн всегда; часто зооиды дных и многогнез н развиваются д ых вместилищ H в плетизмоталлюс. Эт пространен в морях умепенных с. Этот вид рас- полушарий. В у р ых поясов обоих рий. холодных районах его сл встречаются летом в б ляются зимой и п ом, в олее теплых они появ- и после спороношения к дине лета отми"аю . я, к сере- ная растет в ни рия широколист- в сублиторали т в нижнем горизонте литорали и на камнях и на во о с грубыми слоевищам водорослях с очень сильным дв ищами, отсутств ет в у местах движением воды. Семейство о Никтиосифоновые (D t 1с yosiphonaceae Ф Н т~осиФоновых различ орма слоевищ икти , (~ k л( трубчатые прос ые, нитевидные или И ростые или сильно плотные или с разветвленные о и с полостью. Рост ве х Ф ( p у б ':~ О О:.'::: ..":1.-;-"'-'- . -"" я увеличению собств ) меров и делен ю енных разя- н ю окружающих оказываются пог женнь клеток они П '" И::г,'';., ' (::,Э ««Q,,ð,, ' f:. В макроскопических слоеви ах евища. ~ Оа~ ~1,~-~~ --~= '063 go~ ° 0 РоН диктииосифон (Dict ( tyosiph ) р р морях северного ме А~ж~~~~~ж~~ Т ком и Тихом океанах. Сл у его представителе лоевище тонкие толщи 1— лей разветвле Г б ной — мм, реже о 4 / массовых и ши око ненных видов о относится дикт р о распростра- / о иктиосифон укро- oeniculaceus>. Он п Ъ L 'I название из-за сильно 1 щ с тонкими веточ . Э а — а сильно разветвленного с слоеви- 1 точками. Эта во cTH1'BBT B BbIcoT '1 Р ff у м. Растет в ниж с жнем горжю~ в верхней с блит )~ / ~)~ ', / литорали на ~р~- ослях, реже на камнях. го развития дос стигает летом. . Наибольше- 1 РоН койлодесме (С 1 d 1 в oi о esme~ в rr ми~ной mac~a А 3 анов. Слоевище и тлантического и "' Тихого оке- ще у его поет~ставит или лентови ное с ителеи овальное дутое. Виды этого о ся ° ого о тречают- ого рода в основном вс рослях, некоторые считаются ечные разрезы слоевищ бурых водо- нашло от ажен енными к определенным М; ЭTО — пунк ария широколис' 1— р ение в названии н роко тная (Рц~~~~г~~ ~~~~~~~~а: например д од~ол~е."- дны~ коилодесме истоз toseirae~ одногнездные спорангии б- молодой ВОЛОСОК; 8— ии, — пар афизь ~ с хло оплас aus:а— СЦИТОСИфОН КОЛ р дные многогнездные г а — одно я енчатый (Se to пара изы, в — волоски дные гаметангии, б — бес- 157
Семейство стиктиосифоиовые (Stictyosiphona- ceae) Слоевище у стиктиосифоновых разветвлен- ное, с нитевидными цилиндрическими ветвями, многорядными или местами однорядными. Мно- горядные участки членистые, т. е. клетки в них располагаются ровными поперечными рядами. Спор ангии погружены в поверхностный слой клеток или находятся на поверхности слоеви- ща. Парафизы отсутствуют. Род истмоплея (Isthmoplea) характеризует- ся слоевищем супротивно, мутовчато или по- переменно разветвленным, прикрепляющимся к субстрату многочисленными разветвленными клеточными нитями. На слоевище развиваются одногнездные спорангии, которые располагают- ся супротивно попарно по бокам ветвей, иногда отдельные спорангии размещаются напротив ветви. Спор ангии шаровидные, поэтому один широко распространенный вид получил наз- вание истмоплея шароносная (I. sphaerophora). По внешнему виду ее легко спутать с эктокар- ловыми, с которыми она растет вместе на кам- нях и водорослях в литоральной и сублито- ральной зонах морей северной части Атланти- ческого океана. Род стиктиосифон (Stictyosiphon) встреча- ется во всех морях, кроме тропических. Рас- тения имеют вид пучков прямых грубых нитей с мутовчато, супротивно или попеременно рас- положенными короткими ветвями. Слоевище на большом протяжении многорядное, с бес- цветными волосками, иногда с полостью внут- ри. Прикрепляется к субстрату нитевидными ризоидами, которые могут образовывать диск. Одногнездные или многогнездные спор ангии, развивающиеся на слоевище, погружены в по- верхностный слой или выступают над поверх- ностью. На микроскопической форме развития тоже возникают одногнездные и многогнездные вместилища. В умеренных и арктических морях северного полушария часто встречается стиктиосифон скрученный (S. tortilis). Местами в субаркти- ческих и арктических морях он является одним из наиболее обычных видов. Стиктиосифон скрученный предпочитает расти в нижнем гори- зонте литорали и в верхней сублиторали в за- щищенных местах, в том числе в районах с силь- ным опреснением. Семейство асперококковые (Asperococcaceae) К этому семейству относится один род ас- перококкус (Asperococcus), виды которого встре- чаются в морях умеренных поясов обоих по- лушарий. У них цилиндрическое или мешко- видное полое слоевище (рис. 128, 2). Одногнезд- ные и многогнездные спорангии развиваются на разных индивидах или на одних и тех же. Органы размножения образуются группами на поверхности слоевища, одновременно с ни- ми вырастают многоклеточные парафизы. Мик- роскопическая форма развития имеет много- гнездные вместилища. Виды этого рода растут в качестве эпифитов на других водорослях. Семейство стриариевые (Striariaceae) У стриариевых органы размножения обра- зуются вместе с одноклеточными мешковид- ными парафизами группами на поверхности слоевища. К этому семейству относят один род. Род стриария (Striaria) можно рассмотреть на примере стриарии оттянутой (S. attenu- ata), имеющей кустистое слоевище высотой до 50 см с супротивно, мутовчато или поочередно расположенными ветвями, сужающимися к ос- нованию и к вершине. Макроскопическое слое- вище является спорофитом и несет шаровидные или яйцевидные одногнездные спорангии. Гаме- тофит микроскопический, состоит из стелющих- ся нитей, несущих многогнездные гаметангии. Половой процесс — изогамия. Из зиготы снача- ла вырастают стелющиеся нити — протонема, на которой позднее появляется макроскопическое слоевище спорофита. Полный цикл у стриарии оттянутой осуществляется не всюду: он из- вестен для этих водорослей у берегов Франции; у индивидов же, растущих в Северном море у острова Гельголанд, в культурах наблюда- лось чередование только бесполых слоевищ с одногнездными спорангиями. ПОРЯДОК СЦИТОСИФОНОВЫЕ (SCYTOSIPHONALES) Сцитосифоновые обладают макроскопичес- кими слоевищами, высотой от 1 до 50 см. Форма их разнообразна: они могут иметь вид удлинен- но-вертикальных пластинок, трубок с пере- тяжками, полых шаров, сетчатых пластинок и т. д. Объединяет их то, что они образованы плотно сомкнутыми клетками, мелкими и ин- тенсивно окрашенными в поверхностных сло- ях, а глубже постепенно увеличивающимися в размерах и становящимися бесцветными. Клетки имеют по одному хлоропласту с пире- ноидом. На поверхности слоевища вырастают однорядные или двухрядные многогнездные вместилища. У некоторых родов (сцитосифон, колпомения) среди вместилищ развиваются единичные парафизы, состоящие из одной бес- цветной клетки, более широкой и высокой, чем спорангии. На поверхности слоевища встреча- ются группами и одиночные многоклеточные волоски с базальной зоной роста (рис. 128, о). 158
'4' ° Ф. '4'))' ф" 4 1 ! 1. 1 4 ', i' 4 1, 44 14 .; )л'. ,:Ю« ф 1 " )4. ° ',! .')4, ~ .-«. I i. ф) 41) 1 '. ° . fg .' 4+ ° Е,'. ' лв, 4! л )4 л л I л 4 °:;1 e'. 1 ,,'Ф, @ ' ',,;)~ ':~3 ё -) =:+ -:. ."'4»-. ~-"г.-', . : p" 3 .,)4 I '.Il :Ё::..:, :-':и ,/ :,;, л',. : ':-;:.,с.';. Р.' 4 Ф:4 ~ЛВ'.В :44).~4'В, 'Я '::Лф).' В :Ь '4, ' Ф"' '. / ' 1)) 1 1. ! ti г 44 4%. '444 ! tli) IIII 4 ,1' «414 "-.!4)) t. / ' 4) 'c -44 "В ВВ !I' l; ))Щ 4 , 'v .« :: .::М.В. .~/4 1 ~фЪ ° ) ° ° '« 0;. ..««4.') .. "4= '. ф tl'„ ..-4, +'4 '")ВВ ! ".'4. ° 1 ° 43 ' 4 t. 4' ":,' ~~ВЛ, it" '~ ° ! .ф Вв' 44» фф с :ф.(. 1И~~„ ' 114' .. ВВ \ ., " '.11 4 Э ...';."Ъ," '-.с. . ':~" '~,":с.'',, I 'Л),~ 'ЦВ ':.':.')44.: ° 4j; ': ' P 1.. ' ':":Р' ! ', ~:.'., У,ф .'ф'! k:;. ' ":),),-,В", 4 \ ю; ф. 4 ::В )' 4 ,:"-..г.. , с .«r ;:„-. 'у-., 'В4,:: ",'1 ', фф! 4: )) ..'% ! .4. :):л у". 7 В ',: -;т,ф: .. ~6 ' ф:~. 'Щ )V 1р~ Ю «.:: ~В I 4 ;В «;В«4ф« ~А. В 'В. ВФЯфЪ ~;~ Рис. 129. Бурые водоросли. l — леатезия неоднородная (Leathesia Й1 formis) на красной водоросли фурцеллярии; 2 — сцитосифон коленчатый (Scytosiphon lomentaria); 8 — гаметофит кутлерии (Cutleria); 4 — спорофит кутлерии (Aglaozonia); б — диктиота дихотомическая (Dictyota dichotoma); 6 — падина павлинья (Padina pavonia). . 1" Й 1 4 „фВФ В 1!.4. ")« 4/ ". -Fi. ',~!.: .,-,а В:-..'Э.:, ;;. 4:,;.')~,,) °; I "- ' ~'"~~@~;- УЭ-, е).:: .'-' :.:,. У;;:.-. ° В~'::.:,;"..- .:,': .. 4 - ~З~'-.- ('. -. -.: -..Ъ...," »"... ) / ) c / i .',4 ' ° . '4«$ °,I, ,~ gr Ъ41
Такие макроскопические слоевища сцитосифо- новых считаются гаметофитами, но правильнее их называть гаметоспорофитами. Зооиды, образующиеся во вместилищах без полового процесса или после него, вначале выра- стают в короткую, стелющуюся нить из удли- ненных клеток. В последующем в зависимости от сочетания внешних условий развиваются следующие микроскопические или полумик- роскопические образования, считающиеся спо- рофптами: а) стелющийся диск с короткими вертикальными нитями и одногнездными спо- рангиями; б) корочки, по строению подобные ральфсии и несущие одногнездные спорангии; в~ однорядные нити, которые местами дают полуша ровидные паренхиматические образо- вания, служащие местом развития одногнезд- ных спорангиев; г) длинные стелющиеся нити с многогнездными или одногнездными споран- гиями. Наряду с органами размножения на всех перечисленных растеньицах в культураль- ных условиях наблюдалось развитие пророст- ков макроскопических слоевищ. Развитию вертикальных макроскопических слоевищ гаметоспорофитов из зооспор, которые выходят из одногнездных спорангиев микроско- пических форм развития, тоже предшествует образование стелющихся нитей или многослой- ных дисков. Сцитосифоновые широко распространены в морях, но массового развития они достигают лишь в отдельных районах и в определенные сезоны. В порядок входит только одно семейство сиитосифоновых (Scytosiphonaceae), объединяю- щее несколько родов. Род сиитосифон (Scytosiphon) можно рас- смотреть на примере наиболее широко распро- страненного сиитосифона коленчатиого (S. lomen- taria). Он имеет макроскопическое трубчатое неразиетвленное слоевище с несколькими не- глубокими перетяжками, в высоту оно дости- гает 50 — 70 см, а в ширину — 0,5 — 1 см, реже до 2 см (рис. 129, 2). В северных морях, в част- ности в Белом, его макроскопические слоевища растут летом. Южнее, например под Владиво- стоком, их вегетирование приурочено к весне- начаду лета, после чего слоевища разрушаются. В этот период сцитосифон в заливе Петра Вели- кого в полузащищенных местах покрывает сплошным ковром камни и скалы на больших площадях от уреза воды до глубины 1,5 м. После его отмирания здесь почти не остается макроскопических водорослей. Для сцитосифона характерна небольшая ге- теро гамия. Если женские гаметы, например, длиной 6,5 — 8 мкм, то мужские гаметы— 6,1 — 7,6 мкм. Женские гаметы двигаются мень- ший период времени, чем мужские. Вскоре по- сле выхода из га мета нгиев женские гаметы становятся вялыми и прикрепляются к субст- рату. В то же время мужские гаметы энергично плавают в течение нескольких часов, собираясь в условиях культуры в наиболее темной части сосуда. В конце концов они тоже становятся вялыми и прикрепляются к субстрату. Мужские и женские гаметы у сцитосифона образуются на разных слоевищах. Зиготы начинают про- растать через 1 — 2 дня после образования. Род петалокия (Petalonia) характеризуется слоевищем в виде удлиненной пластинки без полости, лишь иногда на поперечном срезе можно заметить ее образование. Наиболее широко известны петалония лентовидная (Р. f asci a) и петиалония зостиер олистная (Р. zosterifolia). Высота их слоевищ достигает 45 см, у первого вида они широкие, у второ- го — лентовидные. Вполне возможно, что это один вид. Оба вида растут в северном полуша- рии от Арктики до северной границы тропи- ческих вод. Род кольпомения (Colpomenia) отличается шаровидным полым слоевищем диаметром 1 — 4 см, иногда до 10 см. Виды этого рода рас- тут на камнях и на водорослях с кустистыми слоевищами. Они встречаются в теплых морях умеренного пояса и в тех районах тропических морей, где в силу подъема глубинных вод тем- пература воды опускается зимой до +18, +19'С. Слоевища кольпомении глубоковыемчатпой (C. sinuosa) встречаются, например, на юге При- морского края в первых числах апреля при температуре воды около O'С и позднее, в начале лета, а также у северного побережья Индии в Аравийском море в октябре — ноябре при температуре воды выше +20'C. Род хидроклатпрус (Hydroclathrus) обычен в морях субтропического и тропического поя- сов. Его представители имеют волнистую сте- лющуюся по грунту пластинку с многочислен- ными отверстиями. Род иенгария (Iyengaria) распространен там же, где хидроклатрус. У этих водорослей слое- вище тоже пластинчатое и стелющееся по грун- ту, но без отверстий и с вертикальными конусо- видными выростами высотой 1 — 2 см. ПОРЯДОК КУТЛЕРИЕВЫЕ (CUTLERIALES) Для представителей этого порядка характерен трихоталлический рост (греч. «трихос» — воло- сок и «таллос» — отпрыск, молодая ветвь): в растущих частях, по краям пластинчатого слоевища или на вершинах ветвей кустистого слоевища расположены многоклеточные волоски с базальной зоной роста, которая отчленяет клетки наружу — это способствует росту во- лосков по мере отмирания или разрушения 160
1; ) в с поверхности (1); кутлерия (Cutie~'a), р $30 Qpafl слоевища аанардинии (»»r~»») и и (Q) и мужскими (8) гаметангиями. верхних клеток, а также в сторону слоевища (рис. 130, 1). На некотором удалении от зон роста основания волосков сливаются. Благо- даря этому образуется слоевище паренхимати- ческого и плотного строения. В бол р юлее старых частях слоевища можно различить кору из мел- ких окрашенных клеток и сердцевину из круп- ных бесцветны~ клеток. Слоевища у кутлери- евых бывают пластинчатыми: стелющимися корковидными или прямостоячими, сильно разв т ветв ленными. Внешне кутлериевые отли- чаются от других водорослей по наличию бах- ромы из нежных волосков на вершине или по краю слоевища. Цикл развития кутлериевых заключается в чередовании диплоидных спорофитов и гап- лоидных гаметофитов, которые бывают оди- накового или разного строения. Половой Мно- процесс у кутлериевых — гетерогамия. Ъ гогнездные гаметангии развиваются на раз- ветвленных или неразветвленных однорядных выростах (рис. 130, 2, 8). Женские гаметангии с более крупными камерами, чем мужские. Женские гаметы соответственно крупнее муж- ских, содержат несколько хлоропластов и дви- гаются менее продолжительное врем мя. Гаметы у кутлериевых высвобождаются через отвер- стие в каждой камере. Слияние их происходит после того, как женская гамета прикрепилась к грунту или другим водорослям. Неопло- дот отворенные женские гаметы способны к парте- ногенетическому развитию. На спорофитах известны только одногнезд- ные спорангии, которые образуются прямо на поверхности слоевища в виде сплошного слоя. В порядок входит только одно семейство кутлериевых (Cutleriaceae), объединяющее 3 рода. Род микрозония (М1сгохоп1а) изучен оче а ° нь слабо, пока известны только диплоидные спо- р офиты. в с б- Род занардиния (Zanardinia) обитает в су- литорали у южных берегов Европы — в Ат- лантическом океане, в Средиземном и Чер- ном морях. Слоевище имеет вид кож исто го диска диаметром до 20 сл, прикрепленного нижней поверхностью с ризоидами к грунту. Спорофит и гаметофит сходного строения. Га- метангии развиваются на неразветвленных вы- ростах. В спорангиях занардинии образуется только по 4 зооспоры. Род ьутлерия (Cutleria) пользуется наиболь- шей известностью (рис. 129, 8, 4). Он служит классическим примером гетероморфной смены форм развития. У гаметофита слоевище од- нолетнее, вертикальное, вееровидное или кус- тистое высотой до 15 см. Гаметангии развивают- 1 т ся на разветвленных или простых однорядных выростах. Слоевище спорофита однолетнее или многолетнее, корковидное, диаметром до 10 см. В спорангиях образуется от 8 до 32 зоо- спо . Раньше, до того как был изучен цикл развития кутлерии, ее спорофит считали особым родом.
Рис. 431. Сфацелярия (Sphacelaria): 1 — вершина побега; 2 — 4 — вегетативные почки разных видов сфацелярии. Развитие спорофитов и гаметофитов у видов рода кутлерия различно в разных частях аре- ала. У южных берегов Великобритании га- метофиты лучше всего растут летом, а в Сре- диземном море около Неаполя встречаются в основном с декабря по апрель. В прибрежье Скандинавии и Северной Великобритании спо- рофиты встречаются часто, в то время как гаметофиты вовсе не известны. В морях СССР единственный вид кутлерия усыпанная (С. ad- spersa) представлен в Черном море только спорофитами. Этот же вид в Средиземном море существует в основном в виде гаметофитов, спорофиты встречаются редко; предполагается, что гаметофиты воспроизводятся посредством партеногенеза. В лабораторных культурах у разных видов наблюдались случаи развития из зооспор спорофитов вместо гаметофитов. При прорастании неоплодотворенных женских гамет бывают случаи развития из них споро- фитов и гаметофитов. В то же время на обра- зующихся при этом слоевищах гаплоидных спорофитов в качестве выростов развивались ростки гаметофитов. Имеющиеся данные свидетельствуют о том, что гетероморфный цикл развития кутлерии возник из изоморфного. Слоевища спорофитов кутлерии часто имеют в центре короткий вер- тикальный вырост, который можно рассматри- вать как остаток неразвивающегося вертикаль- ного слоевища. Одновременно нынешнее слое- в ище спор офита представляет собой видоиз- мененное и сильно разросшееся основание не- когда вертикального слоевища. Интересно от- метить, что у спорофитов кутлерии отсутствует трихоталлический рост, его заменяет краевой рост посредством крупных клеток, подобно тому как растет диск, которым прикрепляется слое вище гаметофита. ПОРЯДОК СФАЦЕЛЯРИЕВЫЕ (SPHACELARIALES) К этому порядку относятся водоросли с же- сткими кустистыми слоевищами высотой от нескольких миллиметров до 30 сл, ветви у них цилиндрические. В отличие от других бурых водорослей у сфацеляриевых каждая ветвь оканчивается крупной клеткой (рис. l31,1). При росте ветви верхушечная клетка удлиня- ется и делится поперечной перегородкой. Ниж- няя клетка еще раз делится поперечной пере- городкой на две клетки, которые, в свою оче- редь, делятся продольными перегородками на длинные клетки. В дальнейшем образовавшиеся клетки могут делиться поперечными перего- родками и перегородками, расположенными параллельно поверхности ветви. Наиболее активно клетки делятся ближе к поверхности. Диаметр ветви при этом нередко остается таким же, как у верхушечной клетки, а более толстые первичные поперечные перегородки хорошо прослеживаются, поэтому у ветвей заметна членистость. Сформировавшиеся участки вет- вей сложены внутри крупными бесцветными клетками. Снаружи расположен один или не- сколько слоев мелких интенсивно окрашенных клеток, содержащих много хлоропластов и физод. В толщину слоевища сфацеляриевых растут двумя способами: 1) путем активного роста и деления коровых клеток — так образуется вторичная кора; 2) отдельные клетки коры дают тонкие нити (ризоиды), которые, разра- стаясь, оплетают родительскую ветвь толстым слоем, при этом образуется ризоидальная кора. У сфацеляриевых встречаются волоски, ко- торые развиваются по одному из чечевицеоб- разной клетки, отделяющейся на конце вер- хушечной клетки. При дальнейшем росте вер- хушечной клетки волосок смещается на ее боковую поверхность. Для слоевища сфацеляриевых характерно основание в виде корковидной пластинки из нескольких слоев клеток. Наряду с этим могут существовать с т о л о н ы — нити из несколь- ких рядов клеток, стелющиеся по грунту. Столоны производят новые корковидные пла- стинки, на которых вырастают новые верти- кальные побеги. Сфацеляриевые размножаются вегетативным, бесполым и половым способами. Вегетативное
размножение происходит посредством столо- нов, но при этом водоросль продвигается на короткое расстояние. Для распространения на большое расстояние у видов рода сфацелярия (Sphacelaria) имеются специальные вегетатив- ные почки, отделяющиеся от ветвей (рис. 131, 2 — 4). У сфацеляриевых существует изоморфная смена форм развития. Диплоидные спорофиты могут нести многогнездные и одногнездные спорангии. 3ооспоры из многогнездных сп�- оп�анги возобновляют диплоидные растения. Мейоз происходит в одногнездных спорангиях, и гаплоидные зооспоры дают гаметоспорофиты, на которых развиваются только многогнездные вместилища. Образующиеся в них зооиды могут прорастать в растения материнского типа или функционировать как гаметы. У сфацелярие- вых гаметы, как правило, одинаковые по раз- мерам, но имеют физиологические различия. Мужские гаметы дольие сохраняют подвиж- ность. Слияние гамет наступает после того, как женская гамета становится неподвижной. В тех случаях, когда сливаются подвижные гаметы, зигота может сохранять подвижность в течение нескольких часов. Значительно реже, чем изогамия, у сфацеляриевых наблюдается гетерогамия; при этом образуются многогнезд- ные гаметангии, различающиеся размерами камер. У сфацеляриевых известно несколько случаев оогамии. Сфацеляриевые поселяются на камнях и других водорослях. Иногда они сплошным покровом обрастают стволы многолетних во- доросле~~, особенно если у них шероховатая поверхность. Среди сфацеляриевых имеются эндофитные виды, у которых от корковидного основания отходят выросты, внедряющиеся далеко внутрь слоевища водоросли-хозяина. Некоторые эндофитные виды имеют редуциро- ванные (недоразвитые) вертикальные побеги, иногда они совсем не развиваются. Большинство с4ацеляриевых, по-видимому, однолетние, но многослойные корковидные ос- нования могут быть многолетними и каждый год производить новые вертикальные побеги. Порядок содержит одно семейство сфа~еля- риевых (Sphacelariaceae). Наиболее обычны в морях виды четырех родов. Род сфацелярия (Sphacelaria) встречается во всех морях. Слоевище у его представителей небольшое, высотой до 4 сл, неправильно раз- ветвленное; членистость выражена на больиом протяжении ветвей, иногда в их основании развиваются вторичная кора и ризоидальная кора. При вегетативном размножении обра- зуются вегетативные почки, имеющие вид ко- роткой ветви с тремя длинными или короткими лучами на вершине (рис. 131, 2 — 4). Род хетоптерис (Chaetopteris) ооъединяет виды со слоевищем высотой 3 — 10 сл и более; их ветви напоминают птичье перо благодаря коротким веточкам, расположенным в одной плоскости. Все слоевище, за исключением мо- лодых участков ветвей, покрыто вторичной корой, поэтому членистость не выражена. Из видов этого рода наиболее известен хето- nmepuc перистый (С. plumosa), который растет в морях Атлантического и Тихого океанов к се- веру от 50' с. ш., а также в Арктике. Род стипокаулон (Stypocaulon) встречается в нижней части литорали и в сублиторали в умеренных и субтропических морях северного и южного полушарий. Высота слоевища у этих водорослей достигает 15 сл, оно имеет вид крупных пучков грубых разветвленных нитей. Нижние части ветвей покрыты ризоидальной корой. В морях СССР растет стипокаулон метиловидный (S. sco pari um). Род кладостефус (Cladostephus) характери- зуется слоевищами высотой до 20 сл, с ветвя- ми, покрытыми мутовками коротких, серпо- видно-изогнутых веточек длиной 1 — 2 лл. Наиболее широко распространен кладостефус лутовчатый (С. verticillatus). Он встречается в Черном и Средиземном морях, у атланти- ческих берегов Европы и США, около Азор- ских островов, у берегов Австралии и Новой Зеландии. ПОРЯДОК ДИКТИОТОВЫЕ (DICTYOTALES) Представители этого порядка обладают круп- ными, высотой 5 — 50 сл, пластинчатыми, це- лыми, или рассеченными на лопасти, или виль- чато-разветвленными в одной плоскости слое- вищами. Под микроскопом с поверхности вид- ны продольные ряды клеток. На продольных и поперечных срезах тоже прослеживаются ря- ды клеток, идущие параллельно и перпенди- кулярно поверхности. Такое правильное рас- положение клеток связано с тем, что рост диктиотовых происходит за счет деления в оп- ределенных плоскостях специальных крупных клеток, находящихся или по одной на вершине ветви, как, например, у диктиоты (рис. 132, 1, 2), или в виде одного ряда по верхнему краю, если слоевище пластинчатое. Органы размножения диктиотовых устрое- ны иначе, чем у других бурых водорослей. Для бесполого размножения служат тетрас- порангии, в которых образуется по 4 крупных споры (тетраспоры), только у зонарии их всегда бывает 8. Тетраспорангии диктиотовых подобны одно гнездным с по ранги ям других бурых водорослей, и в них образование спор сопровождается мейозом. Они располагаются на поверхности слоевища группами (с о р у-
по я входит только »<muomosbtx (д " о одно семейство Ес1уо[асеае, объе свыше двадцати о ов б ъединяющее растет в тропических и с т родов, ольшинство из к которых диктиотовые вст ечают море (виды о Е родов диктиота, дило с Е оре — в заливе Пет а ор — етра Великого тиоптерис . Род диктиота (Dictyota) ха акте вильчато-разветвл тв ленным слоеви еи скими, обычно ра щ и с пло- расположенными в о скости ветвями без п о 129 о). О рганом прик епления дрически кий ризом — стелющийся р . ния служит цилин- ершина каждой ветви ока днои крупной к леткои рис. 132 1 2 . я один слой к -п 2 окруженный сна жи Ок ружи корои из Рис. 132. В е клеток. мелких интенсив ме нсивно окрашенных ершина ветви диктиоты ( ictyota dichotoma): иоты диуотомической 1— широко распрост анен — вид с поверхности, 2 — п о дихотомическая на диктиота — продольный разрез. ая. е слоевища об аз шие дерновинки и высотой до 20 сл разуют боль- ветвей достигает 4 — 8 сами~ т олько у зонарии т — мл. Диктиота дежд тет ческая интересна на четкими п ДИХОТОДИ- гияди развиваются па распоран- вании и со ериодами в образо- те- я пар афизы, состоя и и созревании гаиетангЕ У икт (рис. 133, 1). гаметангии начинают иев. У берегов диктиотовых у слоеви о в период квад ат инают развиваться вида нер ищ одного и того же атурных приливов а ок едко на людаютс я сл чаи ное созревание и выхо кончатель- содержимого тетр распо ран гиев не на у деления в тече ение нескольких п ил д гамет происходи части, а на две или о на четыре после с приливов, след~.ющих оно совсем не елит амого высокого сиз Е зультате образуются б д ится; в ре- При этом г изигииного прилива. Иногда неподелх тся олее кв ~пные тои гаметы соз евают к . " Евшиеся тетраспо а споры. Такая я периодичность в аждые две неделЕ И Ф растают в новые рангии про- его размножении и св ые слоевища прямо на с лунным ритмом соx вязь скод растении. Ког а на материн- живании слоев охранялась при выдер- огда у проростков об аз- нии слоевищ в лаборатории в те риз оиды, они отделяются от ского растения. л~нлс~ к мн~~ин- Северн А Газ ются от рнои мерики созревание г а мета нгии — оогони в дит с интер гамет происхо- тиотовых тоже об ии и антеви ии- — у дик- начинаетс валом в один дес яц. Развитие гамет о разуются на пове ается за день до сизи слоев ища плотными рхности в полнол зигииного прилива В о ми группами (рис. 133 . нолуние или на сле ю ~~ы~~~~ раы~~аетс~ дующии день, гаметы Н р В--'- ( тся по одной яй ек т или дней сп устя. В заливе Пет- р ть партеногенетиче астат яицеклетки способны п ликого понское до е ФВ чески, однако о ро- встречаются я растения с те р обычно чаще разую HoTopb e проросток вско е по иногда все п ев а тетра спорангиями 9 Антеридии делятс в я говизонта ре погибает. на иа е теринских ращаются в проростки кальными р льными и ве ти- е растениях, и ветви по р,дка и н, 1501-0 Р в каждой из кото ь — 'э 0 камер, прежде матыми, так как про ростки орых появляется по чем отделиться вы 9 антерозоиду. Строе ние антерозоидов из о одному в неско лько миллимет ов. вырастают высотой 6u~mu ombre д одической ~Dict о я растет в ве хней иктиота дихото- о ( yota dicho- тах с 300CIIO г ь9 что, в отличие о от гамет и движением воды. р дру их бурых водорослей он Род дилофус (Dilo hus cx по одоиу ж Ку. леи он .о у его п е ор us) сходен с диктиотой Слоеви 9 ща гаметофитов и спо о ито имеется не один а два и и он . р дставителей вн т и тиотовых имеют одинаковое ст и спорофитов у дик- ных клеток. н, а два и более слоев бесцвет- слоевища оп е строение. Пол Р ределяется во время мей од диктиоптерис Мс1 делении ядер в тет я меиоза при диктиоте ictyopteris), подобно тетраспорангиях об те, характериз ется равное число тетра разуется ленным у я вильчато-разветв- аспор, дающих д . Еи слоев ищем с пло женские слоевища мужские и ветви б оскими ветвями но t м . ~ олее широкие (до 2 сл и более и тельно с продольным еб ьным ре ром, идущим от вер- f64
шины до основания. Из-за широких ветвей и продольного ребра слоевища диктиоптери- са напоминают фукус, но у диктиоптериса ветви менее толстые и имеют иное строение. Вершина каждой ветви у диктиоптериса, в от- личие от диктиоты, несет несколько крупных клеток, расположенных в ряд. В СССР в заливе Петра Великого (Японское море) встречается диктиоптерис растопырен- ный (D. divaricata). Он произрастает с июня по ноябрь на камнях на глубине 0,3 — 2 л. Род падина (Padina) включает виды с пло- ским, вееровидным слоевищем, целым или с несколькими вертикальными разрывами. На поверхности хорошо выражены концентриче- ские полосы, соответствующие рядам волосков. Внутри слоевище образовано тремя-четырьмя слоями бесцветных клеток, окруженных снару- жи одним рядом мелких интенсивно окрашен- ных клеток. Верхний край слоевища закручен внутрь и несет один ряд активно делящихся клеток. Одна сторона слоевища выглядит белесой из-за отложения тонкого слоя извести. Наиболее широко известна падина павли- нья (Р. pavonia). Она распространена у бере- гов Южной Европы и у атлантических берегов Центральной Америки. В СССР этот вид рас- тет в Черном море (Крым, Кавказ), где встре- чается от уреза воды до глубины 5 л (табл. 17, вверху; рис. 129, 6). ПОРЯДОК СПОРОХНОВЫЕ (SP0R0CHNALES) Представителям этого порядка свойственна регулярная гетероморфная смена форм разви- тия. Спорофиты у них крупные, высотой до 0,5 м, разветвленные, с цилиндрическими или уплощенными ветвями, на вершинах их рас- положены пучки волосков (рис. 134). Рост слоевища и отдельных ветвей осуществляется посредством слоя клеток, находящихся на вершине в основании группы многоклеточных волосков с интеркалярной зоной роста (рис. 135, 1). В отличие от кутлериевых волоски у спо- рохновых не принимают участия в росте слое- вища. У сильно разветвл енных представи- телей пучки волосков играют существенную роль в фотосинтезировании органических ве- ществ. Поэтому у видов родов карпомитра и спор охнус спорангии располагаются вблизи вершин ветвей. Крайне своеобразно у спорохновых проис- ходит развитие слоевища спорофита. Новое растение начинает расти на гаметофите в виде вертикальной однорядной нити, снизу у нее появляются ризоиды для прикрепления к грун- ту. В дальнейшем клетки нити видоизменяются, Рис. 133. Продольный разрез слоевища диктиоты (Dictyota): 1 — с тетраспорангиями; 2 — с женскими гаметангиями,. 8— с мужскими гаметангиями.
Рис. 134. Спорохнус (Sporochnus). образуя из верхней части волосок с зоной роста и плоскую меристематическую (способную к активным делениям) клетку. Меристемати- ческая клетка делится в продольном направле- нии в различных плоскостях и образует мери- стематическую пластинку. Клетки ее дают вы- росты вверх, которые преобразуются в мно- гоклеточные волоски с зоной роста в основании, Рис. 135. Нерейя нитевидная (Nereia filiformis): про- дольные разрезы вершины (1) и коры со спорангиями Р) и выросты вниз, превращающиеся в многокле- точные нити, обрастающие первичную нить проростка (рис. 136). Нити, идущие от мери- стематической зоны вниз, срастаясь, слагают слоев ище спорофита. Рост ветвей слоев ища в толщину идет за счет расширения меристе- матической пластинки и продольного деления клеток нитей. Клетки центральной части слое- вища крупные и бесцветные, в наружной части они мельче и интенсивно окрашены из-за боль- шого количества дисковидных хлоропластов и физод. У большинства спорохновых коровой слой плотный, у нерейи он состоит из двул:кле- точных нитей. Ветвление слоевища споровновых про ис- водит посредством прекращения деления кле- ток в меристематической пластинке на опре- деленном участке; при этом возникают две меристематические пластинки, ведущие к росту двух ветвей. На слоевищах спорофитов развиваются толь- ко одногнездные спор ангии, которые образу- ются на простых или разветвленных одноряд- ныв веточках или в основании двух-трехкле- точных парафиз (рис. 135, 2). (порангии рас- полагаются группами по поверхности всего слоевища или в определенныв местах, особенно у вершин ветвей, в таких случаях у последних образуются вздутия. Зооспоры имеют глазок и один влоропласт. Гаметофиты у cIIopoxHQBbIx микроскопичес- кие, образованные однорядными разветвлен- ными стелющимися нитями. Ив клетки содер- жат по одному постенному хлоропласту с ло- пастями, в отличие от нескольких дисковидныв хлоропластов, имеющихся в клетках спорофита. Гаметофиты обоеполые или раздельнополые. Одноклеточные или двухклеточные антеридии имеют вид мелких клеток, появляющихся на поверхности вегетативныв. В каждой клетке антеридия образуется по одному антерозоиду, выходящему через отдельное отверстие. Жен- ские гаметангии представляют собой крупные расширенные клетки на концах боковыв ни- тей. Эти клетки развиваются в спорофиты, не отделяясь от гаметофита; при этом они делят- ся поперечными перегородками и вытяги- ваются в вертикальную нить, единственный влоро пласт дробится на множество диско- видных. Цикл развития спорохновых не вполне ясен, так как процесса оплодотворения у них не наблюдали. В порядок входит только одно семейство спорохновых (SporochIIaceae), объединяющее несколько родов. (порохновые растут главным ооразом в сублиторали и на значительной глу- бине. Большинство их представителей распро- странено в южном полушарии. Предполагают,
что часть видов попала в северное полушарие, участвуя в обрастании судов и перемещаясь с ними. Род нерейя (Nereia) представляет семейство спорохновых в морях СССР. У нас встречается только один вид этого рода — нерейя нитевид- ная (N. filiformis), слоевища спорофитов кото- рой летом растут у берегов Крыма и Кавказа на глубине 7 — 18 м. Они достигают в высоту 25 сл, беспорядочно или перисто разветвлены, ветви у них цилиндрические толщиной 1 — 2 лл с пучками окрашенных волосков длиной 3— 15 мм на вершинах. Помимо Черного моря, нерейя нитевидная распространена в Среди- земном море и у Канарских островов в Атлан- тическом океане. Род карпомитра (Carpomitra) заметно отли- чается от большинства спорохновых, обладаю- щих цилиндрическими ветвями. С. costata, которая растет у западных берегов Европы и у Тихоокеанского побережья Северной Амери- ки, имеет неправильно дихотомически-разветв- ленные слоевища и плоские ветви с продоль- ным ребром. ПОРЯДОК ДЕСМАРЕСТИЕВЫЕ (DESMARESTIALES) К десмарестиевым относятся бурые водоросли с регулярной гетероморфной сменой форм раз- вития. Спорофиты — крупные растения высо- той 0,5 — 4 м, разветвленные, с плоскими или почти цилиндрическими ветвями или пластин- чатые (рис. 137). У плоских ветвей выражено продольное ребро, а у пластинчатых слоевищ наблюдаются ветвящиеся жилки. По краям ветвей у десмарестиевых попеременно распо- ложены короткие шипики, направленные под углом вверх, весной они несут пучок желтых или бурых волосков. Главная особенность десмарестиевых за- ключается в анатомическом строении слоевища спорофита и его развитии. На поперечном срезе среди других клеток в центре слоевища заметна крупная клетка (рис. 138, 2). На продольном срезе видно, что она относится к продольному ряду крупных клеток, именуемому осевой нитью. Поперечные перегородки осевой нити имеют по 4 — 5 групп мелких пор. Присутствие осевой нити в центре слоевища связано с особенностями его развития. Слоевище спорофита десмарестиевых начи- нает развиваться из зиготы как однорядная вертикальная нить. В верхней части ее могут появляться супротивные однорядные ветви. Со временем на некотором удалении от вершины возникает зона роста, которая отчленяет клетки вверх для компенсации разрушения и отмира- ния конечных клеток верхушечного волоска и вниз. За счет клеток, отчленяемых книзу, Рис. 136. Проросток слоевища спорофита нерейи ните- видной (Nereia filiformia). происходит рост осевой нити слоевища. Перио- дически некоторые клетки осевой нити дают однорядные выросты, направленные под углом вверх. Эти выросты превращаются в боковые ветви ограниченного роста. Клетки, входящие в их осевую нить, дают выросты, превращаю- щиеся в клеточные нити, которые обрастают осевую нить слоевища (рис. 138, 1). Клетки таких нитей делятся в разных плоскостях. В ре- зультате получается слоевище псевдопаренхи- матического строения с осевой нитью в центре. Боковые ветви ограниченного роста сохраня- ются в виде упомянутых выше шипиков, за- метных по краям слоевища. На слоевище спорофита развиваются одно- гнездные спорангии, в которых при образова- нии зооспор происходит редукционное деление. Зоос поры десмарестиевых имеют по одному хлоропласту и глазок. У разных представите- лей десмарестиевых спорангии возникают раз- лично. Гаметофиты у десмарестиевых микроскопи- ческие, состоящие из однорядных стелющихся нитей. Половой процесс — оогамия. Десмарестиевые широко распространены во внетропических морях северного и южного полушарий. Иногда вместе с ламинариевыми или фукусовыми они преобладают в сублито- рали над другими водорослями. Развиваясь в массе, десмарестиевые становятся сильными конкурентами промысловых видов ламинарие- вых водорослей.
Рис. 137. Сл Слоевпща десмарестии шиповатой (Desma- restia aculeata) 1 — летнее; 2 — зимнее. звестно три рода десмарестиевых, из них один — фзурус (Phaeurus) растет только в во- дах Антарктики. Остальные два рода расп ю- странены более широко. Из-за больших раз- личий в об а ра зов ании органов размножения Рис. 138. есма leata): рестия шиповатая (Desmarestia ас- асц- 1 — растущая вершина: а — осевая нить, б — ве х аз~ез слоевща: — осевая ни ь на спорофитах их относят к разным семей- ствам. К еэ семеиству десмарестиевых (Desmarestia- ceae) относится только один род десмарестия ( tia). Он имеет то же распространение, что и порядок, но отсутствует в Черном море и крайне редок в Средиземном. У этих водорос- лей слоев ище кустистое или пластинчатое, высотой до 2 — 4 м. Ра концах шипиков вес- ной торчат пучки волосков, опадающие летом (рис. ' 37, 1, 2). Спорангии образуются из по- верхностных клеток слоевища и внешне мало отличаются от вегетативных клеток. Гамето- фиты раздельнополые. Мужские гаметофиты сильнее разветвлены и имеют больше клеток, чем женские. В антеридиях образуется по од- ному антерозоиду. Оогонии содержат по одной яйцеклетке, которая при созревании выходит наружу и остается прикрепленной к краям от- верстия. В т аком положении обычно происхо- дит оплодотворение и последующее развитие спорофита. В морях СССР наиболее часто можно встретить десмарестию шиповатую, десмаре- стим зеленеющую и десмарестию язычковую. Десмарестия шиповатая (D. aculeata) имеет темно-бурое многолетнее слоевище высотой до 2 л с жесткими прутовидными слегка уплощен- ными ветвями шириной 1 — 2 мм. Растет в ос- новном в сублиторали, иногда мелкие слоевища встречаются в лужах нижнего горизонта ли- торали. Северная граница распространения дес- марестии колючей проходит в Карском море и у а надск о го арктического архипелага, юж- К ная граница — у 30 — 40' с. ш. в Атлантичес- ком и Тихом океанах. Десмарестия зеленеющая (D. viridis) отли- чается желто-бурым слоев ищем, ветвящимся супротивно, и цилиндрическими ветвями. Если достать слоевище десмарестии зеленеющей из моря и оставить на воздухе или поместить в со- суд с ограниченным объемом воды, то оно на- чинает зеленеть и распадаться на части водо- 7 росли, лежащие при этом рядом с ней, сначала зеленеют, затем бледнеют и разрушаются. По- этому при коллекционировании морских водо- рослей десмарестию зеленеющую необходимо держать отдельно от других водорослей. Вы- сушивать образцы десмарестии зеленеющей в расправленном состоянии необходимо как мо;кно быстрее (особенно при высокой темпера- туре воздуха летом), используя для обезвожи- вания слоевищ бумагу. Предполагают, что по- зеленение и разрушение слоевищ десмарестии зеленеющей происходит в результате высвобож- дения серной кислоты и разрушения ею снача- ла пигментов, а затем и оболочек клеток. Десмарестия зеленеющая растет в сублито- рали и в «ваннах» нижнего горизонта литорали умеренных и холодных морей северного полу- 168
Рис. 139. Продольный разрез молодого слоевища ламинарии (Laminaria): 1 —; 2 — при большом увеличении: a — трубчатые нити. шария. В Белом море она встречается все лето, в заливе Петра Великого (Японское море) ее слоевища разрушаются в конце июня — в июле. В наибольших количествах десмарестия зеле- неющая растет в местах с ослабленным движе- нием воды. Подобно десмарестии зеленеющей, способна изменять окраску на воздухе и разрушаться десмарестия язычковая (D. ligulata), имеющая плоские ветви шириной 5 — 6 мм с ребром. Она распространена в Атлантическом океане у бе- регов Средней и Южной Европы и в Тихом океане у берегов Северной Японии, южных Курильских островов и южных берегов Са- халина. К семейству артрокладиевых (Arthrocladia- ceae) относится один монотипичный род артро- кладия (Arthrocladia). Единственный вид рода- артрокладия мохнатая (А. villosa) — растет в Черном море и в других субтропических и умеренных морях северной половины Атланти- ческого океана. Слоевище у артрокладии дости- гает в высоту 40 см, с цилиндрическими вет- вями, несущими мутовки простых или разветв- ленных однорядных нитей. Спорангии разви- ваются в виде однорядных цепочек на одноряд- ных клеточных нитях. 3ооспоры прорастают в микроскопические стелющиеся однорядные нити, на которых встречаются образования, напоминающие оо гонии и антеридии; тем не менее половое размножение до сих пор неиз- вестно. В лабораторных условиях наблюдалось не- посредственное развитие макроскопического слоевища из стелющихся нитей. ПОРЯДОК ЛАМИНАРИЕВЫЕ (LAMINARIALES) Ламинариевые имеют гетероморфный цикл развития с обязательным чередованием споро- фитов и гаметофитов. Зрелые спорофиты — крупные растения вы- сотой 0,5 — 6 м, для отдельных видов обычны размеры в 20 м. Самые крупные ламинариевые достигают 50 м. Слоевище ламинариевых имеет одну или несколько пластин, расположенных на простом или разветвленном стволе. Слоеви- ще прикрепляется к грунту диском или разветв- ленными ризоидами. Внутри пластины и ствола можно различить меристодерму и кору, состоя- щие из окрашенных клеток, промежуточный слой из крупных бесцветных клеток и сердце- вину. У одних ламинариевых (подпорядок фил- лариевые) сердцевина состоит из плотно сомк- нутых рядов цилиндрических нешироких кле- ток с оболочками обычной толщины и рядов широких клеток с толстыми боковыми стенка- ми. Сердцевина других ламинариевых (подпо- рядок ламинариевые) образована неплотно рас- положенными клеточными нитями: одни из них идут поперек слоевища от клеток промежуточ- ного слоя; другие ориентированы по длинной оси слоевища, причем клетки последних очень длинные и узкие и только у поперечных перего- родок они воронкообразно расширяются (рис. 139, 1, 2). Эти нити называют т р у б ч а т ы м и н и т я и и, а их поперечные перегородки- ситов идными пластинами, так как в них находятся поры. В краевой части сердце- вины крупных ламинариевых, таких, как макро- циси,ис и нереоцистис, среди тонких нитей рас-
+9~~ß~~ K3C3C3C7( Рис. 140. Слизистые каналы ламинарии северной (La- Pmc. 141. Споранги (а) ~~ па ф (б) minaria hyperborea): е — секреторные клетки. и полагаются с и т о в и д н ы е трубки. Они выделяются клетками большого диаметра по всей длине, их поперечными перегородками слу- жат ситовидные пластины. В коре или проме- жуточном слое многих ламинариевых распо- ложены слизистые каналы, образующие в слое- вище сеть, петли которой вытянуты по его продольной оси. В некоторых участках слизи- стых каналов, особенно в местах их ветвления, располагаются группы секреторных клеток (рис. 140), содержащих большое количество диктиосом. Роль секреторных клеток и слизи- стых каналов ламинариевых пока не ясна; ви- димо, они выполняют какую-то физиологиче- скую функцию. Количество их в слоевищах одного вида изменяется в зависимости от усло- вий произрастания, и иногда они могут совер- шенно не развиваться. Для представителей семейства алариевых ха- рактерно присутствие в поверхностном слое пластин крупных клеток, называемых г л а я- д у л я р н ы м и; они наполнены фукозаном. В молодых частях слоевища эти клетки желтые, в старых частях они темно-бурые или черные. Рост слоевищ спорофитов осуществляется посредством интеркалярной зоны роста. Она занимает основание пластины и вершину ствола. Если ствол разветвленный и имеется несколько пластинок, то интеркалярная зона роста на- ходится~в основании каждой пластины. Интер- калярная зона роста не отграничена резко от остальной части слоевища. Деление клеток и соответственно рост наблюдается и за ее пре- делами. Наряду с процессами роста в пластинах спо- рофита ламинариевых происходят процессы разрушения верхнего конца. Нанося на плас- тину через определенные расстояния отверстия и измеряя, как эти расстояния увеличиваются со временем, установили, что за время роста пластина обновляется по длине 1 — 5 раз, а по массе до 20 раз. Слоевища ламинариевых обла- дают наибольшей скоростью роста, достигаю- щей более 10 сл в сутки. У нереоуистиса Лютке наблюдалось удлинение ствола на 13,5 сл в сут- ки, не считая роста пластин. Особенно быстро слоевища ламинариевых растут в конце зимы и весной, когда температура воды нередко дер- жится около 0'С. В период бурного роста пластина содержит много воды и поэтому, если смять ее рукой, легко трескается. С повышением температуры воды скорость роста замедляется и усиливаются процессы синтеза и накопления органических веществ. Пластины становятся толще, темнее и перестают трескаться при сминании. Наиболее интенсивно накопление органических веществ происходит в основании пластины. В то же время наибольшая скорость фотосинтеза отмечается в верхней ее части. На- личие сердцевины с хорошо развитыми прово- дящими элементами обеспечивает транспорти- ровку веществ в пределах слоевища. Большинство ламинариевых — многолетние растения. Ежегодно после завершения выхода
спор пластинчата я часть слоев ища целиком разрушается. Этот процесс начинается с вер- шины. Тогда же или несколько позднее в интер- калярной зоне роста начинается интенсивное деление клеток и происходит закладка новой пластины. Она появляется между стволом и старой пластиной. У одних ламинариевых меж- ду двумя пластинами хорошо выражен пережим, у других он отсутствует. Обычно новая пласти- на шире старой. По мере роста новой пластины старая отодвигается все дальше вверх, иногда ее остатки могут долго сохраняться на вершине. При низкой температуре воды разрушение старой пластины происходит медленно. В Арк- тике у ламинарии копытной (Laminaria soli- dungula) на вершине новой пластины можно наблюдать цепочку из трех-четырех старых пластин. Как свидетельство многолетнего роста некоторых ламинариевых, в их стволах за- метны кольца. Лучше всего они выражены в са- мом низу — в наиболее старой части ствола. Спорофиты всех ламинариевых образуют органы бесполого размножения одного типа— одноклеточные одногнездные спорангии, кото- рые развиваются на поверхности слоевища среди одноклеточных булавовидных парафиз, содер- жащих в верхнем, расширенном конце хлоро- пласты и физоды (рис. 141). Лишь когда пара- физы достигнут почти полного развития, на- чинают расти спорангии. У всех ламинариевых они имеют сходную форму — эллиптическую или цилиндрическую. Число зооспор в споран- гиях колеблется в зависимости от вида и отчасти от внешних условий от 16 до 128. Образование зооспор сопровождается мейозом. При разви- тии спорангиев на пластинчатых частях слое- вища появляются большие по площади пятна (или одно пятно), слегка (на доли миллиметра) поднимающиеся над поверхностью и незначи- тельно выделяющиеся окраской. После созре- вания и выхода зооспор происходит разрушение спороносных участков пластин, на их месте образуются отверстия, а вскоре разрушается и вся пластина. Массовое развитие спорангиев у ламина риевых начинается в конце лета— начале осени. В холодных морях они сохраня- ются на слоевищах до начала следующего лета. На единичных экземплярах спор ангии могут появляться в начале лета, а также в иные вре- мена года. Зооспоры ламинариевых имеют типичное для подвижных клеток бурых водорослей строение. Они грушевидные, длиной 5 — 10 мкм, с двумя жгутиками, отходящими сбоку. Передний жгу- тик перистый, задний — бичевидный. Отчет- ливо видимая оболочка у зооспор отсутствует. Внутри, как правило, содержится по одному ' хлоропласту. Относительно существования глазка у зооспор ламинариевых имеются IIpo- тиворечивые сведения. При наиболее подходя- щих температурных условиях зооспоры спо- собны к активному движению в течение двух суток. При высокой температуре (для ламина- риевых морей СССР это -т-20'С и выше) они проявляют активность всего несколько минут или вообще не способны двигаться. Если за время активного движения зооспора не найдет место для прикрепления, она погибает. На- правление движения зооспор определяется ин- тенсивностью света. При высокой освещен- ности зооспоры проявляют отрицательный фо- тотаксис. Встретив твердую поверхность, зоо- спора прикрепляется передним жгутиком, ко- торый сокращается и подтягивает тело зооспоры к субстрату. После этого зооспора теряет жгу- тики и округляется, у нее появляется ясно ви- димая оболочка, и зооспора превращается в эмбриосп ор у, которая прорастает без пе- риода покоя. В течение 1 — 2 суток эмбриоспора дает вы- рост — проростковую трубку, вкоторую переходит все содержимое. Направление роста проростковой трубки определяется светом и расстоянием от других эмбриоспор. При не- большом расстоянии между эмбриоспорами их проростковые трубки направлены в разные стороны. Протоплазма, переместившаяся в про- ростковую трубку, отделяется от опустевшей эмбриоспоры поперечной перегородкой (рис. 142). Так образуется первая клетка гаме- тофита. Оболочка эмбриоспоры долго сохраня- ется у гаметофита, часто даже тогда, когда на нем появляется спорофит. Гаметофиты большин- ства л амина риевых, за редким исключением, раздельнополые и различаются морфологически. Различия между мужскими и женскими гаме- тофитами заметны уже на стадии первой клет- ки. У женского гаметофита она короче и шире. В каждом сиорангии ламинариевых образуется равное количество спор, дающих мужские и женские гаметофиты. Повышенная температура воды в первую очередь вызывает гибель жен- ских спор. Гаметофиты ламинариевых многоклеточные, в виде однорядных разветвленных стелющихся нитей; женские гаметофиты могут быть одно- клеточными. Мужские гаметофиты сильнее раз- ветвлены и мельче женских, с более мелкими бледно окрашенными клетками. Обычно гаме- тофиты существуют не более одного — четырех месяцев до созревания гаметангиев. Женские многоклеточные гаметофиты не превышают в диаметре ЗОО лкм. При температуре, слишком высокой для образования гаметангиев, гамето- фиты могут жить больше года и достигать в диа- метре 1 сл~ и более. В экспериментальных ус- ловиях гаметофиты ламинариевых способны к вегетативному размножению. Если разре-
Рис. 142. Цикл развития ламииариевых водорослей: 1 — слоевище со спорами: а — пятно спорангиев; 2 — плавающие зооспоры; 8 — зооспора прикрепилась передним жгутиком к грунту; 4 — эмбриоспоры; 5 — эмбриоспоры с проростковой трубкой; 6 — образование первых клеток гаметофитов (слева- ~венский, справа — мужской); 7 — мужской гаметофит: б — антеридий, в — антерозоиды; 8 — многоклеточный женский гаме- тофит; 9 — одноклеточный женский гаметофит после выхода яйцеклетки из оогония: г — оболочка эмбриоспоры; 10 — 11— начало развития счоевища спорофита: д — первичный ризоид. зать многоклеточный гаметофит на части, из них вырастут новые гаметофиты. Это исполь- зуется при всякого рода исследованиях, когда необходим генетически однородный материал. В природных условиях такое размножение едва ли возможно. Половой процесс у ламина риевых — оога- мия. Антеридии имеют вид мелких бесцветных клеток, располагающихся группами или по одной на поверхности вегетативных клеток. В антеридиях образуется по одному антерозо- иду. После выхода антерозоидов мужские га- метофиты погибают. У многоклеточных жен- ских гаметофитов в оогонии превращаются боль- шей частью конечные клетки ветвей, иногда и предыдущая клетка. Таким образом, один га- метофит несет несколько оогониев. В случае одноклеточных женских гаметофитов единствен- ная клетка со временем превращается в един- ственный оогоний. Образование оогониев сопро- вождается увеличением размеров клеток, со- держимое их окрашивается в темно-бурый цвет. В каждом оогонии формируется по одной яйце- клетке. При созревании она выходит через от- верстие на вершине оогония, но не отделяется, а остается прикрепленной к краям отверстия. В таком положении происходит ее оплодотворе- ние и рост спорофита. После выхода всех яйце- клеток гаметофит отмирает. Однако оболочки его клеток еще долго сохраняются а основании спорофитов. На месте одного многоклеточного женского гаметофита развивается несколько спорофитов, которые располагаются по его пе- риферии. Женские гаметофиты не предостав- ляют никаких питательных веществ развиваю- щимся спорофитам, они обеспечивают им только место для прикрепления на грунте. Это имеет важное значение для существования водорос- лей, поскольку твердые грунты занимают в мо- ре небольшую площадь, и в довершении всего их поверхность в значительной мере занята другими водорослями, а также прикрепленными животными. Основная масса зачатков морских донных водорослей (спор и гамет) погибает из-за того, что они не попадают на подходящую поверхность. Оплодотворенная яйцеклетка, оставаясь при- крепленной к гаметофиту, вырабатывает обо- лочку и без периода покоя трогается в рост. Вначале образуется однорядная нить примерно из семи клеток, затем наряду с поперечными перегородками появляются продольные и обра- зуется однослойная пластинка. Из нижних кле- ток пластинки вырастают тонкие ризоиды. По мере роста пластинки в ней увеличивается число слоев клеток, происходит их специализация, появляются ствол и многорядные ризоиды. Если яйцеклетка почему-либо оказывается не прикрепленной к оболочке оогония, то оплодо- творение происходит, но развитие спорофита
идет ненормально. Образуется шарообразное скопление клеток, которое существует около месяца и погибает. В лабораторных условиях удавалось выра- щивать гаметофиты апоспорически, минуя зооспоры. При содержании стерильных споро- фитов в аквариуме в течение трех-четырех ме- сяцев у них отмирали клетки. Несколько кле- ток оставалось живыми, и они развивались в однорядные разветвленные нити, на которых образовывались оогонии и антеридии. Посколь- ку клетки спорофитов диплоидные, то и гаме- тофиты получались диплоидными. Яйцеклетки таких гаметофитов после оплодотворения раз- вивалнсь в тетраплоидные спорофиты. Таким же путем удавалось выращивать гаметофиты из клеток гаплоидных спорофитов, полученных посредством партеногенеза (из неоплодотворен- ных яйцеклеток) и апогамии (из вегетативных клеток гаметофитов). Партеногенетическому развитию спор офитов предшествует массовая дегенерация яйцеклеток после выхода их из оогониев в условиях отсутствия мужских гаме- тофитов. В нормальные спорофиты при этом партеногенетически развивались немногие яйце- клетки. Апогамные спорофиты вырастают из конечных клеток зрелых ветвей мужских га- метофитов. Ламинариевые обычны для холодных вод северного и южного полушарий, где даже в са- мые теплые месяцы температура воды держится выше +20'С всего лишь в течение двух-трех недель. Причем в северном полушарии растет большее число родов и видов. В последнее вре- мя ламинариевые (виды рода ламинария) обна- ружены в тропических водах около Бразилии и в Филиппинском море, но растут они там на глубине 70 — 85 л, где температура воды ниже, чем у поверхности. Ламинариевые встречаются по вертикали от поверхности моря до глубины 200 л (Laminaria rodriguezii в Средиземном море). В литоральной зоне они растут в основном в нижнем горизонте литорали в тех местах, где во время отлива под- вергаются действию брызг, или там, где посто- янно преобладают пасмурная погода и высокая влажность воздуха. Так, ламинариевые зани- мают большие площади на литорали Коман- дорских островов, отличающихся как раз та- кими погодными условиями. Таким крупным водорослям, как ламинарие- вые, нужны условия, при которых обеспечива- лось бы интенсивное поступление питательных веществ к слоевищам. Этому соответствуют места с постоянным сильным течением или при- боем. Там, где нет достаточных по силе течений или прибоя, ламинариевые растут в водах, обогащенных питательными веществами (но не до степени загрязнения) за счет разного рода стоков, содержащих много органики, или за счет неглубокого залегания илистых грунтов. Поэтому заросли ламинариевых встречаются у открытых берегов, особенно умысов, далеко вдающихся в море. Заросли же видов, способ- ных расти в спокойных местах, располагаются по соседству с илистыми грунтами. Это приво- дит к тому, что ламина риевые можно найти на глубине, в условиях пониженной освещен- ности, но при наличии ила среди камней, а в том же месте у берега, на меньшей глубине, их нет,— если там отсутствует постоянный прибой, а дно имеет большой уклон, из-за чего органические остатки опускаются глубже. Точно так же за- росли ламинариевых легче встретить в местах с мутной водой, подверженной интенсивному движению, чем в местах с прозрачной водой, находящейся в менее активном движении. Подпорядок фиь~гариевые (Phyllarineae) Сердцевина у филлариевых плотного строе- ния; она состоит из нитей большого диаметра с толстостенными клетками и из нитей мень- шего диаметра с тонкостенными клетками; труб- чатые нити, ситовидные трубки и тифы у них отсутствуют. Слизистые каналы неизвестны. К подпорядку относится одно семейство фил- лариевых (Phyllariaceae) с двумя родами:филла- рия (Phyllaria) и саккориза (Saccorhiza), рас- пространенными в северной части Атланти- ческого океана. Слоевище филлариевых имеет одну целую или рассеченную на вертикальные лопасти пластину, переходящую в ствол с ор- ганами прикрепления при основании. У фил- ларии органами прикрепления служат ризоиды. Молодые слоевища саккоризы прикрепляются к субстрату диском; позднее л основании ствола на некоторой высоте над грунтом образуется диск (саккориза кожистая — S. dermatodea) или луковицеобразное расширение (саккориза многополосная — S. polyschides), с нижней сто- роны которых вырастают неразветвленные ри- зоиды. В морях СССР растет один вид — саккориза кожистая. Внешне она напоминает ламинарию. Поверхность пластины выглядит бархатистой, так как она покрыта волосками. Слоевище у с аккоризы кожистой однолетнее, достигает в длину 1 — 2 л. Спорангии развиваются в ок- тябре — декабре в основании пластины и на стволе. Растет этот вид в верхней сублиторали и изредка в нижней литорали в открытых местах с пологим уклоном дна, а также в полузащи- щенных местах. Саккориза кожистая встре- чается у берегов Северной Атлантики от южной части Карского моря до севера Норвегии и у берегов Америки от острова Элсмир до севе- ра штата Массачусетс (США).
Саккориза многополосная растет у берегов Западной Европы. Она интересна тем, что перед развитием спорангиев у нее по бокам ствола вырастает складчатая бахрома, на кото- рой образуются спорангии. Кроме этого, спо- рангии могут располагаться на луковицеоб- разном расширении ствола и в нижней части пластины. Подпорядок лам инириевьсе (Laminarineae) Сердцевина неплотного строения с гифами, трубчатыми нитями или с ситовидными труб- ками. Слизистые каналы имеются или отсут- ствуют. Семейство хордовые (Chordaceae) Слоевище у хордовых однолетнее, шнуровид- ное, нера зветвленное, с полостью, слизистые каналы отсутствуют. К субстрату прикрепля- ется диском. Зона роста расположена под вер- шиной. Семейство включает один род хорда (Chorda), содержащий всего 2 вида. Хорда нитевидная (Chorda filum) имеет олив- ково-желтое слоевище, слабо покрытое волос- ками, часто бесцв.етными. Нижняя клетка во- лосков почти в два раза уже нижележащей клет- ки слоевища. Растет в лужах нижнего гори- зонта литорали и в сублиторали в полуоткры- тых и защищенных местах на скалах и заилен- ном песке на глубине до 15 — 20 м. Хорда ните- видная встречается в северной половине Атлан- тического и Тихого океанов, южная граница ее распространения совпадает с зимней изо- термой воды -+12 'С. Слоевище хорды пушистой (С. tomentosa) обычно оливково-зеленого цвета, густо покрыто интенсивно окрашенными волосками. Нижняя клетка волосков слабо отличается по ширине от нижележащей клетки слоевища. Хорда пу- шистая растет в лужах нижнего горизонта ли- торали и в верхней сублиторали до глубины 5 л в полуоткрытых и защищенных местах. Это растение весенней флоры, южная граница его распространения проходит значительно се- вернее, чем у хорды нитевидной. Семейство ламинариевые (Laminariaceae) У представителей этого семейства на вершине ствола расположено одна пластина, реже не- сколько, на боковых его поверхностях особые пластинки отсутствуют. У многолетних видов пластинка следующего года возникает интер- калярно. Продольные разрывы пластин никог- да не возникают интеркалярно на стыке плас- тины и ствола, разрывы идут от верхнего края ини 3. Род лальинария (Еа~л1паг1а) насчитывает 30 видов, причем только 3 из них растут в юж- ном полушарии: 2 вида у берегов Бразилии на большой глубине (70 — 80 м), 1 вид у южной оконечности Африки. Остальные виды распро- странены в северном полушарии; больше всего видов ламинарий в Тихом океане. Пластина у этих водорослей ровная или морщинистая, без волосков, целая или рассеченная, без от- верстий, иногда с двумя продольными рядами вмятин и выпуклостей или с двумя продоль- ными складками. Органами прикрепления слу- жат диск, ризоиды или ризом (стелющийся побег с ризоидами). Наибольшее промысловое значение имеют ламинария сахаристая, лами- нария пальчатая, ламинария северная, лами- нария японская, ламинария узкая. У ламинарии сахаристой (Ь. saccharina) нерассеченная удлиненная пластина, ровная или морщинистая, с темной продольной поло- сой, иногда с двумя продольными рядами вмя- тин и выпуклостей. Слоевище прикрепляется к субстрату ризоидами. Слизистые каналы у слоевищ, растущих в морях СССР, известны только в пластине, а у растений атлантическо- го прибрежья Северной Америки они бывают и в пластине и в стволе. Спорангии начинают развиваться одновременно на обеих поверх- ностях пластины, при этом очертания пятен спорангиев в точности совпадают. Зрелые спо- рангии наблюдаются с конца августа по ян- варь, а иногда до июня. Слоевище у этой водо- росли многолетнее, максимальная его длина достигает 7 л. Ламинария сахаристая растет в сублиторали на глубине до 20 л, изредка встречается в ниж- нем горизонте литорали, особенно в литораль- ных «ваннах». Это один из наиболее массовых видов рода ламинария в северной части Атлан- тического океана. Ламинария сахаристая рас- пространена от западной части моря Лаптевых и Баффиновой Земли на севере до севера Пор- тугалии и штата Массачусетс (США) на юге. В восточном секторе Арктики и в северной части Тихого океана (до северных островов Японии на юге) широко распространена лами- нария Гурьяновой (Ь. gurjanovae), которую прежде тоже считали ламинарией сахаристой. В дальневосточных морях ламина рией саха- ристой по ошибке еще называют растения ла- минарии курчавой (Ь. cichorioides), слоевища которой в зависимости от скорости движения воды могут иметь различный облик. В спокой- ных местах пластина с сильно курчавыми кра- ями и двумя рядами крупных выпуклостей и вмятин. В местах же с постоянным движением воды пластины гладкие и с ровными краями. Такие слоевища и считают ошибочно ламина- рией сахаристой. Ламинария курчавая отли- 174
чается о.г ламинарии сахаристой и ламинарии Гурьяновой тем, что спорангии у нее начинают развиваться сперва на одной поверхности плас- тины и лишь с большим запозданием на второй. Поэтому очертания пятен спор ангиев на них не совпадают. Имеются и другие отличительные признаки. Распространена ламинария курча- вая в северной половине Японского моря, вок- руг острова Хоккайдо, около острова Куна- шир и островов Малой Курильской гряды, вдоль южного и восточного берегов острова Сахалин до залива Терпения, крайне редко она встречается у острова Итуруп и в северо-запад- ной части Охотского моря. У ламинарии японской (Ь. japonica) пластина нерассеченная, удлиненная (длина ее 2 — 6 м, иногда до 12 м, а ширина iO — 35 см), располо- женная на стволе несколько косо. Орган при- крепления — ризоиды (табл. 39, 1). По продоль- ной оси пластины проходит широкая и толстая срединная полоса, занимающая '/,— '/, ее ши- рины и ограниченная по краям двумя про- дольными складками. Молодые слоевища часто имеют два ряда выпуклостей и вмятин на месте продольных складок. Слизистые каналы в пла- стине расположены между сердцевиной и ко- рой, часто в два ряда, особенно в срединной полосе. Сердцевина в пластине уже, чем у дру- гих видов. У молодых слоевищ ствол цилиндри- ческий, у немолодых — уплощенный или желоб- чатый. Пластина плавно переходит в ствол. Слизистые каналы в стволе всегда имеются. Слоевища ламинарии японской растут два года. На южной границе распространения, где температура воды в море летом бывает выше +20 'С, они однолетние. Спорангии у этого вида образуются на одной или двух поверхнос- тях пластины; в последнем случае они сначала развиваются на нижней, слегка вогнутой по- верхности, и очертания групп спорангиев на двух сторонах пластины не совпадают. В усло- виях естественных зарослей спорангии начи- нают появляться с конца июля, крайне редко спороносящие экземпляры встречаются в июне. Л амина рия японская считается самым цен- ным промысловым видом среди бурых водо- рослей. Она обладает наилучшими вкусовыми качествами. Ее пластина толще и менее жесткая, чем у других видов рода ламинария. Наиболь- шую пищевую ценность имеет утолщенная сре- динная полоса слоевищ второго года. Перво- годние слоевища считаются непромысловыми, хотя они и достигают крупных размеров: плас- тина у них узкая, тонкая и содержит меньше питательных веществ. Ламинарию японскую используют для приготовления первых, вто- рых и третьих блюд, а также разнообразных консервов и кондитерских изделий. Слоевища или их части, непригодные для пищевого ис- пользования, идут в химическую переработку для получения альгинатов, маннита, иода и т. д. Ламинария японская растет в местах с посто- янным движением воды, она встречается от по- верхности до глубины 25 м, ре>ке 35 м. Промыс- ловые заросли находятся в основном на глу- бине 4 — 10 л. Этот вид распространен в север- ной половине Японского моря — от побережья КНДР и северной части острова Хонсю до 50 ' с. ш., а также у южного и юго-восточного берега Сахалина и у южных Курильских остро- вов. Заросли у некоторых островов Малой Курильской гряды дают наибольшую биомассу— 140 кгlм ' В 20-х годах нашего столетия зачатки ла- минарии японской были случайно завезены в Желтое море к побере>кьям Ляодунского и Шаньдунского полуостровов. В КНР были разработаны методы ее искусственного разве- дения, и сейчас ее культивируют вдоль побе- режья КНР вплоть до Кантона. В естественных условиях запасы ламинарии японской подвержены сильным колебаниям. Это отражается на объеме заготовок. Большой урон зарослям наносят сильные штормы и то- рошение льда у берега в холодные зимы; осво- бождаемые во время промысла участки дна нередко заселяются непромысловыми водорос- лями. Поэтому люди издавна пытались искусст- венно увеличить площади под ламинарией япон- ской. Было найдено несколько способов, с ус- пехом применяемых в Японии, КНР и КНДР. Эти способы используются и для выращивания других видов рода ламинария и прочих водо- рослей. Самый простой из них — это чистка твердого грунта от других водорослей и живот- ных в период, когда ожидается появление в во- де спор ламинарии японской, и последующий уход за зарослями, т. е. удаление сорных рас- тений и растительноядных животных. Для рас- ширения площади под зарослями в море с ло- док сбрасывают камни или там, где возможно, взрывают скалы на берегу, обрушивая их в во- ду. На подготовленный таким образом новый грунт или на очищенный старый для ускорения появления зарослей сбрасывают зрелые слое- вища, собранные в других местах. Остальные способы выращивания ламинарии японской свя- заны с погружением в море субстратов, искусст- венно засеянных спорами. В качестве таких субстратов используют камни и бетонные блоки, сбрасываемые на дно, или веревки, которые подвешивают к канатам, поддерживаемым у поверхности поплавками и закрепленным на месте якорями. Посев спор ламинарии японской и других водорослей на субстраты делают следующим образом. Чтобы выход снор был дружным, предварительно производят стимулирование 175
слоевищ. Для этого их выкладывают на воздух в затененном месте на несколько часов или на ночь. Для контроля за ходом стимулирования периодически на поверхность слоевища нано- сят каплю морской воды, через несколько ми- нут эту каплю переносят на предметное стекло под микроскоп. Когда в поле зрения (при увели- чении объектива 8, окуляра — 10~) будет 8 — 10 спор, слоевище считается стимулирован- ным. Его кладут в бассейн, куда предварительно поместили субстраты для последующего куль- тивирования ламинарии. Засеянные спорами камни сбрасывают в море в подходящих местах. Хри культивировании ламинарии на веревках иногда предварительно выращивают рассаду, которую позднее рассаживают с нужными про- межутками на другие веревки. Выращивание ламинарии на веревках дает наибольший уро- жай с единицы площади, и сейчас оно стало наиболее распространенным на плантациях. Для получения максимальных урожаев план- тации ламинарии японской размещают вблизи городских стоков или вносят в воду минераль- ные удобрения. Урожай ламинарии японской при таком культивировании достигает 50— 65 т сухой массы с гектара. Ламинария узкая (Ь. angustata) имеет нерас- сеченную длинную узкую пластину, по продоль- ной оси которой проходит узкая темная полоса, прогнутая в виде мелкого желоба. Вся пласти- на, кроме основания, тоже имеет вид желоба. Орган прикрепления — ризоиды. Слоевища ла- минарии узкой достигают наибольшей длины по сравнению с другими видами рода ламина- рия, известны экземпляры длиной до 22 л. Продолжительность mmmm этой водоросли 3— 4 года. Спорангии появляются в конце июля и располагаются на одной или двух поверхностях пластины, причем очертания групп спорангиев на них не совпадают. Ламинария узкая растет в нижнем горизонте литорали и в сублиторали до глубины 8 — 12 м в местах с постоянным прибоем или течением. Распространение этого вида ограничено островом Кунашир, островами Малой Курильской гряды, островом Хоккайдо и северо-восточной частью Хонсю. Небольшие группы мелких слоевищ встречаются на мысах в Приморском и Хабаровском краях. Ламина- рия узкая наряду с ламинарией японской счи- тается ценным съедобным видом. У ряда видов ламинарий пластина молодых слоевищ целая, но с увеличением размеров под действием движения воды она рвется на про- дольные лопасти. Такова, например, ламинария пальчаторассеченная (Ь. digitata). Она имеет широкоовальную или клиновидную пластину, рассеченную, а в спокойных местах целую— в таком случае она бывает плоской или полу- сферически-изогнутой. Орган прикрепления— ризоиды. Слизистые каналы имеются только в пластине. Спорангии наблюдаются зимой и весной в виде округлых или овальных пятен, очертания которых на обеих поверхностях плас- тины не совпадают. Слоевище у ламинарии пальчаторассеченной многолетнее, длина его достигает в отдельных случаях 5 л, а ширин@ пластины — 1 л. Растет в сублиторали, иногда в прибойных местах встречается в нижнем го- ризонте литорали. Этот вид распространен в Северной Атлантике от Карского моря и Гуд- зонова залива на севере до полуострова Бре- тань и Нью-Иорка на юге. У берегов Европы растет лалинария северная (Ь. hyperborea). Она напоминает ламинарию пальчаторассеченную, но отличается от нее на- личием слизистых каналов в стволе и сильным сужением между старой и новой пластинами, а также другими признаками. Она встречается от Шпицбергена и Кольского полуострова до Средиземного моря (Балеарские острова и Мис- синский пролив). У берегов Северной и Сред- ней Европы она растет от поверхности до глу- бины 15 — 25 л, а в Миссинском проливе ее заросли находятся на глубине 50 — 100 м в местах с постоянным течением. Ламинария северная служит одним из основных источ- ников получения альгинатов в странах Запад- ной Европы и является поэтому объектом все- стороннего изучения. На ней выясняли, как растительноядные водные животные влияют на вертикальное распределение водорослей. Оказалось, что если регулярно уничтожать морских ежей глубже нижней границы ее про- израстания, она начинает расти еще глубже; в Миссинском проливе ламина рия северная не может расти выше 50 л из-за растительно- ядных животных. По берегам северной части Охотского моря в выбросах часто встречаются слоевища двух видов ламинарий с рассеченной пластиной и стволом, лишенным слизистых каналов. Один вид — ламикария прижаторизоидная (Ь. ap- pressirhiza) — характеризуется образованием спорангиев на одной поверхности пластины; при их созревании лопасти свертываются в труб- ки. У другого вида — ламинарии наклонно- ризоидной (Ь. inclinatorhiza) — имеются длин- ные слабоветвящиеся ризоиды, образующие высокий и широкий конус, что позволяет во- доросли расти на грунте, занесенном илом. У нее спорангии развиваются одновременно на обеих поверхностях пластины, очертания пятен спорангиев на них совпадают. Из видов, похожих на ламинарию пальчато- рассеченную, в северной части Тихого океана широко распространена лалинария Бонгарда (Ь. bongardiana). Слизистые каналы у нее раз- виты и в пластине и в стволе. В отличие от лру- 176
гих видов рода ламинария, имеющих рассе- ченную пластину (кроме иногда ламинарии прижаторизоидной), щель каждого разрыва у ламинарии Бонгарда наклонена к поверх- ности пластины, и концы щели на разных по- верхностях смещены друг относительно друга. Спорангии образуются на одной или двух по- верхностях пластины, причем очертания групп спорангиев на них не совпадают. Облик слое- вищ ламинарии Бонгарда крайне изменчив в зависимости от условий произрастания. В нижнем горизонте литорали с пологим укло- ном дна и ровной поверхностью грунта, а также в сублиторали при сильных течениях растут слоевища с коротким стволом и узкой длинной пластиной, часто нерассеченной. Если поверх- ность в нижнем горизонте литорали неровная (нагромо>кдения камней, выступы скал) илп в сублиторали отсутствуют сильные течения, то слоевища имеют широкую и короткую плас- тину и длинный ствол. В мелких литоральных лужах произрастают некрупные слоевища с ши- рокой пластиной и коротким (i — 3 см) стволом. Нередко у слоевищ с широкой пластиной имеет- ся один разрыв; который проходит через всю пластину и затрагивает верхнюю часть ствола. Течением каждая из половинок пластины за- кручивается вокруг своей части ствола. Полу- чается слоевище с вильчато-разветвленным ство- лом и двумя пластинами. Заросли ламинарии Бонгарда встречаются в СССР от острова Иту- руп (Курильские острова), вдоль восточного берега Камчатки до Берингова пролива. Вдоль берегов Северной Америки она распространена до 42 с. ш. Ламггнарггя копытная (Ь. solidungula) — вид, широко распространенный в Арктике, в том числе там, где не встречаются другие ламина- риевые. Пластина ламинарии копытной удли- ненно-яйцевидной формы, ствол расширяется снизу к верхней части и снова сужается при переходе к пластине. Слоевище прикрепляется к грунту диском. В суровых районах Арктики слоевища мелкие, с пластиной длиной 10— 20 сл. Благодаря тому что сохраняются пласти- ны прошлых лет (иногда до трех-четырех плас- тин), общая длина слоевища достигает 60— 70 сл. У самой южной границы распростране- ния, около полуострова Лабрадор, ламинария копытная вырастает до 2,5 л. Такое хорошее развитие вида именно у южной границы рас- пространения позволяет предполагать, что он не встречается южнее не из-за неблагоприят- ных физико-химических факторов, а из-за кон- куренции со стороны других видов, в том числе ламина риевьтх. Ламинария йезоензис (L. yezoensis) растет у дальневосточных берегов СССР и у Северной Америки. У нее рассеченная на лопасти плас- тина, прикрепляющаяся к грунту диском; сли- зистые каналы в стволе отсутствуют, поэтому ее прежде путали с ламинарией пальчаторас- сеченной. Наконец, у трех видов ламинарий слоевище имеет стелющийся по грунту побег с ризоидами (ризом) и вертикальными стволами, несущими по о,",ной пластине. Из этих видов наиболее ши- роко распространена ламинаргся длинноапволь- ная (Ь. longipes). Ее заросли в зоне разрушения волн встречаются вдоль берега Тихого океана от Курильских до Алеутских островов и до 55 ' с. ш. у берегов Северной Америки. При изучении в культуре у ламинарии длинноствольной обнаружены гаиэтофиты од- ного типа, без разделения на мужские и жен- ские, которые были способны к длительному вегетативному росту и очень редко произво- дили спорофиты. Ризом и ствол у спорофита многолетние. Род фггллариелла (Phyllariella) включает все- го один вид — фгглларггеллу охотоморскую (Ph. ochotensis), встречающуюся в северной поло- вине Охотского моря. Пластина у нее гладкая, с пучками волосков, слизистые каналы отсут- ствуют, орган прикрепления — диск. Род челлманиелла (Kjellmaniella) характери- зуется удлиненной, нерассеченной пластиной с вмятинами и выпуклостями в виде сложного орнамента, который расположен поперечными рядами, отходящими от срединной гладкой полосы. Орган прикрепления — ризоиды. К этому роду относятся 2 вида. Челлманиелла тонколистная (K. gyrata) об- ладает тонкой пластиной с узкой неутолщенной срединной полосой. Распространена у южных Курильских островов и у острова Хоккайдо. Челлманггелла толстолггстная (К. crassifolia) имеет толстую пластину с широкой утолщенной срединной полосой. Встречается в Японском море по западному берегу Татарского пролива, у южной оконечности Сахалина и у острова Хоккайдо. Род ггггматэре (Cymathere) включает 3 вида. У них удлиненная пластина, целая, с двумя— четырьмя продольными складками, простыми или утолщенными (складки-ребра). Цггматэре японская (С. japonica), растущая у ю>кных Курильских островов, имеет 4 про- дольные складки. Иногда краевые складки вы- ражены слабо, и тогда этот вид легко спутать с ламинарией японской. Различия между ними заключаются в расположении спорангиев. У ци- матэре японской спорангии развиваются только на одной стороне пластины в ее основании, оставляя свободной срединную полосу, ограни- ченную складками. Две складки-ребра имеет ггггматэре двусклад- чатая (С. fibrosa), которая встречается у сред- 177
них и северных Курильских островов. цила- тэре трехскладчатая (С. triplicata) отличается от остальных видов рода тем, что ее слоевище прикрепляется к субстрату диском и на одной поверхности пластины проходят 3 складки-реб- ра, а на другой поверхности — 4. Циматэре трехскладчатая растет вдоль Тихоокеанского побережья от острова Уруп (Курильские остро- ва) до Командорских и Алеутских островов и у берегов Северной Америки на юг до 48 ' с. ш. Род костария (Costaria) включает всего один вид — костарию ребристую (С. costata), плас- тина которой отличается пятью грубыми про- дольными ребрами, из них 3 находятся на одной поверхности пластины, а 2 расположены между ними на другой поверхности (табл. 17). Кроме ребер, пластина имеет отверстия и пучки во- лосков, поверхность ее изборождена выпук- лостями и вмятинами, число которых, так же как и число отверстий, возрастает в местах с ос- лабленным движением воды. На поверхности ствола заметны мелкие ребрышки. В основании ствола развиты ризоиды. Спорангии начинают появляться с июня в виде многочисленных пя- тен, расположенных в углублениях пластины; со временем пятна спорангиев разрастаются и сливаются в более крупные, выходя за пределы углублений. Слоевища однолетние, они появ- ляются во второй половине зимы. В заливе Петра Великого их разрушение завершается к концу июля — началу августа; на севере Татарского пролива и у Курильских островов слоевища сохраняются до сентября — октября. Костария ребристая растет в большом ко- личестве как в защищенных, так и в прибойных местах, в последнем случае она встречается вместе с другими ламинариевыми, лучше про- тивостоящими сильному движению воды. Ко- старию можно встретить в нижнем горизонте литорали и в сублиторали до глубины 22 л. Распространена она в Японском море от Татар- ского пролива на севере до Корейского полу- острова и северной части Хонсю на юге, в юго-западной части Охотского моря, в при- брежье Северной Америки от островов Шума- гина до Сан-Педро (Калифорния). У рода агарул (Agarum) пластина с одним продольным ребром и многочисленными от- верстиями (табл. 16, 5), пучки волосков и сли- зистые каналы отсутствуют. Прикрепляется слоевище ризоидами. Наиболее широко из- вестен агарум продырявленный (А. cribrosum). Отверстия в его пластине бывают от мелких до крупных, диаметром 2 сл, изменчива и ширина ребра. В спокойных местах растут слоевища с более мелкими и многочисленными отверсти- ями и с более узким ребром. Агарум продыряв- ленный встречается на глубине 5 — 36 л на ска- листых и илистых грунтах в открытых и полу- защищенных местах, за исключением зоны раз- рушения волн. Заросли его наблюдаются в ниж- ней сублиторали, тогда как большинство дру- гих ламинариевых обычно преобладает в верх- ней сублиторали. Такое произрастание агару- ма продырявленного по вертикали связано с тем, что в верхней сублиторали он не выдер- живает конкуренции со стороны других лами- нариевых, а глубже морские ежи и другие 2ас- тительноядные животные предпочитают вы- едать другие ламинариевые, не трогая агарума, обладающего жестким слоевищем. Агарум про- дырявленный распространен в северной части Тихого океана от Берингова пролива до севе- ра Японии, у побережья КНДР, омываемого Японским морем, и до штата Вашингтон (США). Он встречается также у атлантического берега Северной Америки от острова Элсмир до штата Массачусетс (США). Род талассиофиллул (Thalassiophyllum) включает один вид — талассиофиллум peusem- чатый (Т. clathrus), имеющий на стволе одну или несколько дырчатых пластин, свернутых в виде конуса (табл. 16, 4). Отверстия у пла- стины фестончатые, вывернутые на внутреннюю сторону. Молодое слоевище имеет плоскую ши- рокую целую пластину с отверстиями. Позднее по боковым краям пластины образуются две зоны роста, при этом каждый край сворачи- вается в трубку. Одновременно пластина разры- вается вдоль на две половины, каждая из ко- торых растет в дальнейшем за счет своей зоны роста, находящейся внутри конуса. По мере роста каждой пластины увеличивается число витков в конусах. Нижний конец конуса, уплот- няясь, превращается в ветвь. У старых слое- вищ появляются дополнительные пластины, которые возникают как выросты в основании старой пластины. Поэтому старые слоевища приобретают кустистый вид. Талассиофиллум решетчатый растет в откры- тых местах в сублиторали и в лужах нижне- го горизонта литорали у средних и северных Курильских островов, вдоль Тихоокеанско- го побережья Камчатки, у Командорских и Алеутских островов и у южного побережья Аляски. Семейство артротамновые (Arthrothamnaceae) У этих водорослей многолетнее слоевище с разветвленным стволом и несколькими ленто- видными пластинами. Каждая пластина с двумя ушками (полукруглыми выростами) по углам основания. Новые пластины появляются на смену старым как выросты по одной из центра ушек. Таким образом, одна старая пластина дает две новые. Старая пластина разрушается, за исключением узких участков, которые идут
от ствола к новым пластинам: они утолщаются и превращаются в новые ветви ствола. Семейст- во артротамновых включает один род с двумя видами. Артротамнус курильский (Arthrothamnus kurilensis) обладает прямостоячими стволом и ветвями. Растет в открытых местах в нижнем горизонте литорали и в сублиторали до глуби- ны 30 л у юго-западного побережья Сахалина и у Курильских островов (Итуруп, Уруп, Симушир). У артротаинуса раздвоенного (А. bifidus) ствол стелется по грунту и состоит из толстых треугольных пластин с ризоидами по бокам, от двух наружных углов каждой пла- стины отходят по одной такой же пластине или по узкой лентовидной пластине. Этот вид растет у открытых берегов в нижнем горизонте лито- рали и в сублиторали до глубины 25 м в райо- не, включающем остров Хоккайдо, Курильские острова, восточный берег Камчатки до Кроноц- кого залива. Семейство лессониевые (Lessoniaceae) У лессониевых ствол разветвленный, на кон- цах ветвей по одной пластине. Ветвление ствола и увеличение числа пластин происходит за счет продольного разрыва, который возникает ин- теркалярно на стыке основания пластины и вер- шины ствола; удлиняясь, разрыв достигает верхнего края пластины. Спорангии образуют- ся на поверхности пластин (рис. 143, 1). Род лессония (Lessonia) имеет ветви с длин- ными междоузлиями, воздушные пузыри от- сутствуют. Пластины делятся разрывом на две равные части. Известно 5 видов этого рода. Из них 4 вида растут в южном полушарии, где часто образуют настоящие подводные леса. Наиболее крупные слоевища имеет лессония чернеющая (Ь. nigrescens), растущая у берегов Перу, Чили и Фолклендских островов. Ее стволы достигают в толщину 10 сл, а в высоту 4 м. Один вид — леееония лалинариевидная (Ь. laminariaeoides) встречается в северном полушарии, в северной половине Охотского моря, образуя вместе с ламинарией Гурьяновой заросли в нижнем горизонте литорали и верх- ней сублиторали. Слоевище ее достигает в дли- ну 1 — 2 м, первое ветвление ствола расположе- но у самых ризоидов, выше междоузлия до- стигают в длину 30 сл, пластины вытянутые, длиной 20 — 90 сл. Спорангии встречаются с сентября до весны следующего года, они рас- полагаются на обеих поверхностях пластин сплошным пятном. Род макро иистис (Macrocystis) имеет ствол многократно разветвленный. Первые 2 — 3 вет- вления снизу дихотомические, остальные одно- сторонние. Верхушечная пластинка отчленяет новые пластинки с одной своей стороны по- средством косых разрывов. Перед каждой пла- стинкой ветвь обычно расширена в грушевидно- веретеновидный воздушный пузырь. Благода- ря воздушным пузырям и длинным ветвям основная часть слоевища макроцистиса пла- вает у поверхности моря. Это позволяет меха- низировать его добычу. Виды рода макроцисгис перерабатывают для получения альгинатов и других химических продуктов. Макроцистис грушевидный (M. pyrifera, табл. 16,1) прикрепляется к субстрату ризои- дами, слоевище его достигает размеров 30— 50 м, длина пластин 1 — 1,5 л. Это многолетнее растение, отдельные экземпляры живут до 8 — 10 лет. Впервые спорангии появляются на шестимесячных экземплярах. Спорангии обра- зуются на нижних пластинах, сидящих на вильчато-разветвленных коротких побегах в ос- новании слоевища. Эти пластины иногда назы- вают с п о р о ф и л л а м и. Спорангии могут появляться также на верхних пластинах, пла- вающих у поверхности. Созревание спорангиев происходит сначала на верхней, более старой, части пластины. Развитие отдельного спорангия завершается за две недели. Макроцистис грушевидный встречается у от- крытых берегов на глубине 6 — 40 л, иногда его обнаруживали на глубине до 60 м. Распростра- нен в южном полушарии у Южной Америки от мыса Горн до Перу вдоль западного берега и до 50' ю. ш. у восточного берега, около Тасма- нии и Новой Зеландии, у ю>кной оконечности Африки; в северном полушарии растет у Тихо- океанского побережья Северной Америки от Ситки до полуострова Калифорния. В середине прошлого века макроцистис грушевидный на- ходили в Охотском море, у берегов Камчатки и острова Уналашка, сейчас он здесь не встре- чается. Макроцистис грушевидный имеет большое практическое значение, поэтому в настоящее время его экологию интенсивно изучают, осо- бенно у берегов Калифорнии, где находятся наиболее крупные предприятия по его перера- ботке. Обнаружено, что большой урон макро- цистису могут причинять морские ежи. Они способны полностью уничтожать целые зарос- ли этой водоросли. Массовая гибель крупных слоевищ происходит в годы с необычно силь- ным летним прогревом воды. При эхом гибнут побеги, поднимающиеся до перегретого слоя воды. На молодых слоевищах, находящихся глубже, прогрев поверхностных слоев воды не отражается. Эффективный радиус рассеива- ния спор, при котором они дают новые расте- ния, составляет у отдельно растущих слоевищ макроцистиса грушевидного 5 м. В зарослях возобновление слоевищ происходит на удале- 179
° 4 4, n.. j.: .'4 Л 4 .4 .4, ° ' А. 4;.л'К n.-. :.n 'n:-':;. ::;P:4 n..: :0:. :-4 Л :Ф"-" 4'.. 1»ЛГ а . 4 14 4:;С 14.4. :4.- t » ' ° Ъ .а с .Г. ,4.' а '.Ф ° '. r ° 4.. 4 г- '4 ъ 4 а'. а ° ' Са ° d а Ф.л '. ..': )~ 4' :ь- Л:. .'-.4 °" .4,4,.' .n: 'л а,'. - а. у а. а„ ° l r '4, ' °, Ф 'с r с а 4 1 4 I у ю ° Л. 44- 44~ Э.~4" ..41 . -1„„4' .1."Г-,:/ Ф ° аа 1:.;э> '.~ л»4' ° М.- '. \ Ф" 4 а g,t. '.ч 'с»-; ° Рис. 143. Бурые водоросли: 1 — лессония (Lessania); 2 — егрегия (Egregia); 8 — вершина ветви хормозиры Баокса (Hormosira banksii); 4, $ — химанталия удлиненная (Himanthalia elongata): 4 — молодое растение, 5 — растение с рецептакулами (a); 6 — вершина ветви цистозейры бородатой (Cysto;,åirà barbata); 7 —.вершина ветви цистозейры косматой (Cystoseira crinita); 8 — вершина ветви саргассума (Sargassum); 9 — вершина ветви турбинарии (Turbinaria). л .,' г.'.:: f» ,4.' .я r r. ,: ° 4' 4. :., ' 'n., ~С. <а ч 4л 4 4 а' ,. 4 4 а ,с 4 ".. ф ° °, .„' Л ..4." i n '.,'. е. '. ° ° °, i:-.. >', 4 ~с. Ю " Я' с ". ааа у 1:.'-,;Ъ r 4 4 4 Ф рф.-" "Ф '...',. 44.У::.'~ а""ггпу i:,.':,y:,'. „,.".": Г@- g Э; '" .:.::,.:: ь -'..'::.;4 ':::'::-:. --=.:::::«М »р [ф..'...» r .~44 Г У\ -ф ."ч r )4 г-.g Ф ",'ёь а ...':;д; ф ':::,"ф с @'У, )ln ф 6 . '::;1' ~А::� ':.Ъё=':4:."Ф.'."~ c .', .::4 ф .,";; '.':'::, с..;.-Ф у. "Oj 4Р~ 1"'."' ~"':.'"::.::::::: "':~4!! ~'':."4~~."::.:». 4~::::.":::4::'~::::::.':.:;-'. $.:Ú:::~~:.,;::::::, ~~Г- '.~:
нии до 90 л. Расселение на большие расстоя- ния осуществляется посредством оторванных пластин со спорангиями. ЛХакро~истис цельнолистный (M. integrifo- lia) имеет стелющееся основание с ризоидами (ризом). Слоевище многолетнее, высотой до 5 — 8 м, длина пластин 25 — 60 см. Растет до глубины 7 л~ у Тихоокеанского побережья Аме- рики от острова Ванкувер до залива Камель в Калифорнии, у побережья Перу и северной части Чили. В Ботаническом институте АН СССР в Ленинграде хранятся целые экземпля- ры макроцистиса цельнолистного длиной до 0,5 м, собранные в 1870 — 1871 гг. у Сахалина вблизи Александровска-Сахалинского и в за- ливе Анива. Позднее макроцистис у берегов Азии не встречался. Род нереоцистис (Nereocystis) включает один вид — нереоцистис Люпте (N. luetkeana, табл. 19, 2). У него дл. нный ствол, достигаю- щий 15 — 25 л, которыми постепенно расширяет- ся кверху и несет на конце пузырь диаметром 12 — 20 сл. На верхней части пузыря растут две короткие дихотомически-ветвящиеся ве- точки длиной в несколько сантиметров с длин- ными узкими пластинами на концах. Длина пластин достигает 3 — 9 м. Всего таких пластин на слоевище бывает 24 — 40. Спорангии обра- зуются на пластинах. Слоевище прикрепляется к грунту ризоидами, которые образуют конус диаметром до 30 см. Несмотря на гигантские размеры, слоевища у нереоцистиса однолетние. Они появляются в феврале — марте, спороно- сят в ноябре и исчезают (отрываются от грун- та) в декабре — январе; крайне редко единич- ные спороносящие экземпляры встречаются до апреля. Растет нереоцистис на глубине до 20 л, при этом верхняя часть ствола с пузы- рем плавает у поверхности. Часто он образует настолько густые заросли, что через них невозможно пробраться на лодке. Нереоцистис распространен у Тихоокеанского побережья Северной Америки от острова Уналашка до Калифорнии. Свободноплавающие слоевища встречаются у Алеутских островов. В СССР эту водоросль можно найти в выбросах на бе- регах Командорских островов, обычно это стволы с пузырем, но без пластин. Крайне редко плавающие стволы встречаются в Берин- говом проливе. Род пелагофикус (Pelagophycus) включает один вид — пелагофикус пора (Р. porra). В строении его слоевища много общего с нерео- цистисом. Отличие заключается в длинной ветви (до 1,6 м), отходящей от пузыря. Первое ее ветвление дихотомичес кое, следующие— односторонние, как у макроцистиса. Слоевище несет около 10 пластин длиной по 4 — 5,5 м и шириной 29 — 45 см. Общая длина слоевища достигает 50 л~. Растет пелагофикус вместе с макроцистисом у берегов Ю>кной Калифор- нии. Семейство алариевые (Alariaceae) У этих водорослей на вершине ствола рас- положена пластина с продольным ребром или лишенная его. Края пластины ровные или с длинными боковыми лопастями. По бокам сильно или слабо уплощенного ствола у мно- гих представителей имеются пластинки на тон- ких стебельках (спорофиллы) или развивается сплошная складчатая кайма. Спорангии обра- зуются на спорофиллах, кайме или на лопаст- ных выростах центральной пластины. В по- верхностном слое клеток пластинчатых обра- зований часто наблюдаются гландулярные клет- ки, наполненные физодами. Они желтые в мо- лодых частях слоевища и темно-бурые, почти черные в старых. Род алария (Alaria) характеризуется пласти- ной с продольным ребром, идущим. от ствола до вершины. Спорофиллы расположены на тонких стебельках по бокам ствола (рис. 144). Все виды рода алария распространены в се- верном полушарии и предпочитают места с по- стоянным движением воды. В Атлантическом океане наиболее широко распространена алария съедобная (А. esculenta). Ее слоевища достигают в длину 2 — 3 м. Столь же широко распространена в Тихом океане алария окаймленная (А. rnarginata), имеющая такие же размеры. Наиболее крупное слоевище у аларии полой (А. fistulosa). Для нее характерно наличие полого ребра в пластине. Местами полость в ребре пересекается поперечными перегород- ками, в результате получаются как бы длинные камеры, наполненные газом. Благодаря этому слоевище имеет положительную плавучесть, и она сохраняется даже при повреждениях ребра, например при разрушении вершины. Плавающие обрывки пластин ал арии полой встречаются далеко за пределами границ ее произрастания. Обычно алария полая растет так, что вершины ее пластин достигают по- верхности воды и стелются под ней. Макси- мальная глубина, на которой наблюдали этот вид, составляет 35 м. По некоторым данным, у средних Курильских островов встречаются заросли аларии полой, состоящие из растений длиной 41 м. Ал ария полая распространена от острова Итуруп (Курильские острова) до южной части Берингова моря, а также у Командорских и Алеутских островов и вдоль Тихоокеанского по- бережья Аляски. Небольшие заросли ее встре- чаются у Камня Опасности в проливе Лаперуза
(ме>кду островами Сахалин и Хоккайдо). На- роды, живущие по берегам тех мест, где она растет, называют аларию полую бобровой ка- пустой. Это связано с тем, что часто вблизи мест ее произрастания водится калан (его erne называют камчатским бобром). Виды рода ала- рия в холодных водах северного полушария столь же обычны, как виды рода ламинария. Их и используют сходным образом. Некото- рые народы, например населяющие Алеутские и Командорские острова и побережье Чукот- ского полуострова, употребляют виды рода ала- рия в пищу. Особо ценятся спорофиллы ала- рии узкой (А. angusta), отличающиеся большой толщиной и мягкостью. На Чукотке пластинки аларии мочат в пресной воде до тех пор, пока ребро не станет мягким. Представители рода ундария (Undaria) растут в тепловодных местах Японского моря, в Жел- том море и в северной части Восточно-Китай- ского мор.я. Особенно широко распространена ундария перистая (U. pinnatifida). Она имеет цельнокрайнюю или лопастную пластину с пуч- ками волосков. Для прикрепления к грунту у нее служат ризоиды. Слоевища всегда одно- летние. В начале лета по краям ствола начи- нает расти складчатая кайма, на которой обра- зуются спорангии. В августе спорангии созре- вают, и слоевища начинают разрушатьея. Ун- дария перистая обладает нежной пластиной, ее используют в пищу. В Японии ее культиви- руют на специальных морских плантациях. В СССР ундария перистая растет в заливе Петра Великого. Рис. 144. Алария (Alaria). Род эклония (Ecklonia) встречается у берегов Японии, Австралии, Новой Зеландии и южной оконечности Африки. Для эклонии характерно развитие спор ангиев на лопастных выростах по бокам центральной пластинки. У самого крупного вида — эклонии трубовидной (E. buccinalis), растущей у южной оконечности Африки, лопасти во много раз крупнее цент- ральной пластинки. Слоевища эклонии трубо- видной достигают в длину 8 л. По внешнему облику они напоминают нереоцистис. У них длинный (до 5 л) ствол, расширяющийся квер- ху и полыи. Для представителей рода егрегия (Egregia) характерно наибольшее расчленение слоевища среди алариевых и других ламинариевых. Эти водоросли растут у Тихоокеанского побережья Северной Америки (рис. 143, 2). КЛАСС ЦИКЛОСПОРОВЫЕ (C YCLOSPOROPH YCEAE) Гаметофить развиваются в слоевище споро- фита в виде слоя, выстилающего углубления (ко н цент а к ул ы). В отличие от фэозооспоровых все циклоспо- ровые имеют крупные слоевища. Как и у ла- минариевых, слоевища циклоспоровых диффе- ренцированы на ткани: меристодерму, кору, промежуточной слой и сердцевину. Но у цикло- споровых нет характерных для ламинариевых трубчатых нитей, ситовидных трубок, слизистых каналов и гландулярных клеток. Сердцевина у циклоспоровых образована рыхло- или плот- норасположенными клеточными нитями. Основные отличительные особенности цикло- споровых связаны с их циклом развития и раз- множения. Это, во-первых, отсутствие двух самостоятельно растущих форм развития. Во- вторых, гаметангии развиваются в углуолениях на слое клеток, который возникает из одной клетки, именуемой инициальной клеткой кон- цептакула, но мы ее будем называть п р о- с п о р о й. В-третьих, у всех до сих пор изу- ченных циклоспоровых мейоз происходит при образовании гамет. Существует несколько теорий, пытающихся по аналогии с другими водорослями объяснить, как возник такой цикл развития циклоспоро- вых. Напомним, что у большинства растений спорофит диплоидный, а гаметофит гаплоид- ный, циклы с иной сменой ядерных фаз были установлены сравнительно недавно. Поэтому нельзя было просто так принять диплоидные слоев ища цикл осп оров ых за гаметофиты. Своеобразие цикла развития циклоспоровых издавна привлекало внимание ученых: получа- лось, что он подобен циклу животных — у них
тоже гаплоидны только гаметы. Интерес к цик- лу развития циклоспоровых связан и с прак- тическими соображениями. Циклоспоровые произрастают во всех морях от Арктики до Антарктики, включая тропики; они имеют крупные слоев ища длиной 1 — 12 м и часто образуют обширные заросли. Все циклоспоро- вые — промысловые водоросли. Сбор их иног- да осуществляется в больших масштабах, что приводит к уничтожению естественных зарос- лей. В связи с этим необходимо изучение всех особенностей их цикла развития для рацио- нального использования и приумножения имею- щихся ресурсов. У всех циклоспоровых, кроме аскозейры, на- блюдается оогамия. В антеридиях образуется по 64 антерозоида, в оогониях развивается 1 — 8 яйцеклеток. Мейоз происходит при пер- вых двух делениях ядра. В оогониях, несущих даже одну яйцеклетку, всегда образуется 8 ядер. Гаметангии цикл оспоровых выглядят одногнездными, однако у некоторых предста- вителей в антеридиях на стадии 32 ядер на- блюдали нежные перегородки. В оогониях не- которых циклоспоровых яйцеклетки разделе- ны нежными перегородками (хормозира, кси- фофора) или грубыми (пельвеция желобчатая). Эти факты свидетельствуют о том, что гаме- тангии циклоспоровых, видимо, все-таки про- изошли от многогнездных вместилищ других бурых водорослей. В начале нашего века известный ботаник Э. С т р а с б у р г е р утверждал, что оогонии и антеридии циклоспоровых подобны тетраспо- рангиям диктиотовых водорослей, так как они развиваются на диплоидных слоевищах и пер- вое деление ядра сопровождается мейозом. Но сами яйцеклетки и антерозоиды Страсбургер сравнивал с гаметами других водорослей, счи- тая, что тетраспоры сразу, минуя стадию гаме- тофита, делятся и дают гаметы. Крупнейший шведский альголог Г. К ю л и н вначале (в 1916 — 1917 гг.) полагал, что цикл развития цик- лоспоровых возник в результате полного недо- развития гаметофита, а зооспоры (споры) стали сразу функционировать как гаметы. Позднее, в 193( г., Кюлин выдвинул теорию, согласно которой циклоспоровые возникли независимо от остальных бурых водорослей и у них никогда не было гаметофита, а их предки были всегда диплоидными. Автор наиболее полного руко- водства по морфологии и размножению водо- рослей Ф. Ф р и т ч предполагал, что цикл раз- вития циклоспоровых возник в ходе эволюции в результате тенденции неполовых зооспор вес- ти себя как гаметы. Из-за отсутствия убеди- тельных данных, неоспоримо свидетельствую- щих в пользу какой-либо из этих теорий, ни одна из них не стала общепризнанной. Наряду с этим существует другой вопрос. Почему только у циклоспоровых слой, высти- лающий углубления (концептакулы), на кото- ром образуются гаметангии, развивается из одной клетки? Концептакулы с органами раз- мно>кения известны у других бурых (сплахни- диум) и красных (кораллиновые) водорослей. Но здесь они образуются иньп|и способами: а) вследствие впячивания поверхностного слоя клеток слоев ища за счет активного деления и роста клеток в отдельных точках его поверх- ности; б) в результате развития органов раз- множения на каком-то участке и обрастания его окружающими вегатативньп|и тканями; в) путем появления органов размножения на клетках слоевища, выстилающих полость, ко- торая перед этим образовалась вследствие раз- рушения клеток (некоторые известковые крас- ные водоросли). Поскольку до недавнего времени все наши сведения о развитии выстилающего слоя кон- цептакулов циклоспоровых основывались на изучении представителей порядка фукусовых, подавляющее большинство которых имеет апи- кальный рост посредством одной особой клетки на вершине, то И. Рейнке и Ф. Фритч посчита- ли выстилающий слой концептакула ветвью, растущей в обратном направлении. Эта теория благодаря авторитету Фритча нашла многих сто- ронников, к тому же до недавнего времени лишь она учитывала факт развития выстилающего слоя каждого концептакула из одной клетки. В то же время эта теория находится в явном противоречии с тем, что у ряда представителей циклоспоровых рост слоевища происходит по- средством деления нескольких (3 — 8) верху- шечных клеток (хормозира, нотейя) или путем верхушечного роста, но без особых верхушеч- ных клеток (дурвиллея). В довершение всего было обнаружено, что у аскозейры развитие концептакула и его выстилающего слоя начи- нается с деления одной клетки, лежащей на поверхности слоевища, при этом образуется узкая трубка, которая подобно паразитиче- скому организму врастает внутрь, проходит через плотную кору и промежуточный слой и дорастает до сердцевины. Здесь среди рыхло- расположенных клеточных нитей конец труб- ки разрастается в полый шар — концептакул. 8 свете этих данных для объяснения проис- хождения цикла развития циклоспоровых, их гаметангиев и несущих их вместилищ, логичнее всего признать, что инициальнан клетка кон- цептакула является спорой (проспорой), а раз- вивающийся из нее выстилающий слой концеп- такула — гаметофитом. То, что гаметофит у циклоспоровых получается диплоидным, не находится в противоречии с явлениями, наблю- даемыми у других водорослей.
Рис. f45. Начальные этапы развития слоевища фукуса (Fucus!: 1 — 3 —; 4 — продольный разрез через вершину проростка: а — верхушечная клетка. По способу развития гаметофита на спорофи- те класс циклоспоровых делится на несколько таксонов меньшего ранга. ПОДКЛАСС АСКОЗЕЙРОВЫЕ (ASCOSEIROPHТС1ЭАЕ} Подкласс включает один порядок Ascoseira- les с одним семейством Ascoseiraceae. В нем один род с одним видом — аскозейра удивитель- ная (Ascoseira mirabilis). Этот вид растет у Ан- тарктического полуострова, К)жных Шетланд- ских и Южных Сандвичевых островов и у ост- рова Южная Георгия. Здесь он относится к числу наиболее массовых видов. Слоевище аскозейры достигает в длину 1 — 2 м. Оно со- стоит из диска для прикрепления к субстрату, ствола и клиновидной пластины, расширяю- щейся кверху и заканчивающейся множеством лопастей. Ствол может быть вильчаторазвет- вленным с пластинками на конце. Рост слоеви- ща аскозейры осуществляется, по-видимому, как у ламинариевых, посредством интеркаляр- ной зоны роста. Для аскозейры характерны в сердцевине, кроме обычных тонких нитей, толстые прямые нити (проводящие трубки) из длинных клеток большого диаметра, набитых темно-бурым зернистым содержимым. Такие об- разования у других водорослей неизвестны. Концепта кулы у аскозейры расположены внутри слоевища на границе промежуточного слоя и сердцевины. С поверхностью слоевища цолости концептакулов сообщаются узким и длинным каналом. Образуются концептаку- лы следующим образом. Вблизи зоны роста по- верхностные клетки делятся на большое число мелки~ клеток (до 4 — 9). Некоторые из ни~ ста- новятся проспорами. Они удлиняются в сторо- ну, противоположную поверхности, и начинают делиться поперечными и продольными перего- родками. Образуется узкий многорядный тяж клеток, который врастает внутрь слоевища; при достижении более рыхлых тканей его конец разрастается в полый шар, одновременно тяж клеток увеличивается в диаметре, образуя ка- нал. Под микроскопом врастание гаметофита в слоевище аскозейры и образование концепта- кула напоминает внедрение чужеродного орга- низма, паразита, в тело хозяина. Гаметофиты аскозейры производят длинные однорядные ни- ти, верхние клетки которых превращаются в гаметангии. Они содержат по 8 гамет. разде- ленных нежными перегородками. Кроме этих нитей, встречаются единичные волоски с ба- зальной зоной роста, но, в отличие от подоб- ных волосков других циклоспоровых и других бурых, интеркалярная зона у них очень IHIIpo- кая и выражена менее резко. Пробовали наблю- дать за гаметами аскозейры в лаборатории, но они не сливались попарно и совершенно не про- растали. ПОДКЛАСС Ф У К У С О В Ы Е (FUCOPHYCIDAE) У слоевищ фукусовых преобладает верхушеч- ный рост (рис. 145) и гаметофиты начинают раз-
2 Рис. 146. Зрелые концептакулы фукуса пузырчатого (Fucus vesicu]osus): 2 — ЖеКСКИй; 2 — МУЖСКОЙ. виваться в верхних частях ветвей. Концепта- кулы возникают в результате обрастания га- метофитов тканями спорофита (рис. 146). По- ловой процесс — оогамия. ПОРЯДОК ДУРВИЛЛЕЕВЫЕ (DURVILLEALES) Слоевище пластинчатое с лопастями в виде выростов; верхушечный рост, возможно, соче- тается с интеркалярньп|, как уламинариевых. Верхушки лопастей узкие, тонкие; клетки в них расположены плотно; меристодерма состоит из одного ряда клеток. По мере удаления от вер- шины клетки меристодермы растут и делятся горизонтальными перегородками. В результате меристодерма оказывается состоящей из кле- точных нитей, которые непрерывно растут и ветвятся. Проспоры, дающие гаметофиты, за- кладываются там, где нити меристодермы со- стоят по меньшей мере из 5 — 6 клеток. Они образуются в основании нитей из верхушечных клеток боковых выростов. От соседних клеток проспоры отличаются продолговатой формой и более темным содержимым. Окончательно сформировавшаяся проспора окружена тол- стой слизистой оболочкой. После дробления проспоры образуется однослойная горизон- тальная пластинка, состоящая из четырех кле- ток. После образования проспоры нити мери- стодермы, расположенные рядом, дают допол- нительные боковые выросты, которые, разра- стаясь и переплетаясь, образуют вокруг про- споры и в дальнейшем вокруг молодого гаме- тофита защитную ткань. Разрастаясь, гамето- фит занимает все большую площадь, при этом нити меристодермы раздвигаются в стороны, и возникает канал между полостью концепта- кула и внешней средой. Порядок дурвиллеевых включает одно се- мейство Durvilleaeeae с одним родом Durvillea, представители которого растут только в юж- ном полушарии. Наиболее широко распрост- ранена дурвиллея антарктическая (D. ап1агс- tica, табл. 16, 8). Она растет у западного побе- режья Южной Америки от мыса Горн до Валь- параисо, а по восточному берегу — до 50' ю. ш., у Новой Зеландии и субантарктических островов (Южная Георгия, Кергелен, Чатам, Окленд, Кэмпбелл), встречается в выбросах у Южных Шетландских островов. Слоевище этой водоросли внешне напоминает ламинарию, длина его может достигать 10 м, к субстрату оно прикрепляется диском диаметром до 0,5 л. (твол переходит в пластину с длинными лопа- с гями, образующимися как выросты. Толщина пластины бывает около трех сантиметров, внут- ри ее находится полость, разделенная попереч- ными перегородками на камеры. Слоевище дур- виллеи имеет толстую меристодерму, образован- ную длинными вертикальными нитями, насчи- тывающими по 12 — 15 клеток; плазматическая связь между клетками соседних нитей отсут- ствует. В молодых частях, кроме меристодер- мы, различаются кора, промежуточный слой и сердцевина. В немолодых частях строение этих тканей замаскировано обильным разви-
рис. 147. Начальные стадии развития гаметофита у цистозейры (Cystoseira)~ тием ги ф — узких нитей с толстой оболочкой; создается ложное впечатление, что слоев ище состоит из гиф. Оогонии, так же как и антери- дии, развиваются на разветвленных однорядных веточках, поднимающихся от поверхности га- метофитов. В оогониях образуется по 4 яйце- клетки. Дурвиллея антарктическая растет в субли- торали и в нижнем горизонте литорали в местах с сильным прибоем, где ее слоевища постоянно заплескиваются волнами. Заросли у берега бы- вают настолько плотными, что мешают лодкам пристать к берегу. Из-за крупных размеров слоевищ дурвиллея антарктическая представ- ляет ценное сырье для получения альгинатов. В Чили ее употребляют в пищу. В Новой Зе- ландии из дурвиллеи получают ростовые ве- щества и удобрения. ПОРЯДОК ФУКУСОВЫЕ (FUCALES) Слоевище у фукусовых кустистое, обладает верхушечным ростом посредством деления 1 — 8 особых верхуыечных клеток. Образующиеся при их делении новые клетки растут и снова делятся. Клетки, составляющие осевую часть ветви, делятся слабо и вытягиваются в длину, образуя сердцевину. Проспоры, дающие гамето- фиты, развиваются из клеток, отчленяемых верхушечной клеткой (или клетками) в сторо- ну поверхностного ряда клеток слоевища. Обычно такие клетки вытягиваются в высоту и делятся поперечными Берегородками. Клет- ка, превращающаяся в проспору, не удлиня- ется, наоборот, она округляется, приобретает яйцевидную или грушевидную форму, содер- жимое ее становится мелкозернистым и одно- родным. Проспора при развитии гаметофита де- лится вертикальной и горизонтальной перего- родками (пельвеция равновершинная) или на- клонными перегородками (аскофиллум узло- ватый). У проспор остальных фукусовых пер- вая перегородка проходит горизонтально (рис. 147). Верхняя (я з ы ч к о в а я) клетка получается меньше нижней. Язычковая клет- ка делится поперечными перегородками и раз- вивается в волосок с базальной зоной роста. У одних фукусовых он развивается полностью (~истогейра), у других недоразвивается (сар- гассум). Интересно отметить, что с развития волоска начинается развитие проростков у ряда дру- гих бурых водорослей. Нижняя клетка, про- дукт деления проспоры, делится сначала вер- тикальной перегородкой. В последующем в но- вых клетках перегородки закладываются в вер- тикальной и горизонтальной плоскостях. Одно- временно растут и делятся клетки окружающих тканей спорофита, в результате гаметофит ока- зывается погруженным в углубление. Гамето- фиты фукусовых образуют BppTHEBJIbHbIe раз- ветвленные и простые однорядные многокле- точные нити, часть из них несет гаметангии, а остальные служат парафизами. Ближе к кра- ям у гаметофитов могут расти волоски с ба- зальной зоной роста. На слоевищах ряда фукусовых встречаются криптостомы и цекостомы. К р и п т о с т о-
м ы — это ямки, из которых растут многокле- точные волоски с базальной зоной роста. Ц е- к о с т о м ы — микроскопические полости под поверхностью слоевища. Слой клеток, высти- ддющих полости криптостомов и цекостомов, развивается из таких же проспор, как и гаме- тофиты. Это стерильные гаметофиты, выполняю- щие физиологические функции. Криптостомы с волосками, видимо, служат для лучшего об- мена с внешней средой, в частности для погло- щения питательных солей. Криптостомы и це- костомы возникают тогда, когда не заклады- ваются концептакулы, и поэтому они занимают разные участки на ветвях. На границе перехо- да одних образований в другие встречаются углубления, в которых развиваются гаметофи- ты промежуточного типа с недоразвитыми гаме- тангиями и большим количеством волосков на- ряду с парафизами. Гаметофиты у фукусовых бывают либо рассеяны на больших участках, ничем не отличающихся от стерильных, либо сосредоточены на вершинах побегов, становя- щихся из-за этого утолщенными и более широ- кими. Такие части слоевища называют р е- цептакулам и. Нормальное воспроизведение фукусовых воз- можно только половым путем, лишь немногие представители имеют стелющиеся ризомы или ризоиды, дающие на конце новые слоевища (например, саргассул ЛХиябе). Антеридии об- разуются на гаметофитах на концах разветв- ленных веточек, и в их оболочке различимы два слоя. Антерозоиды выходят наружу в виде пакета, окруженного внутренней оболочкой. Субмикроскопическое строение антерозоидов не у всех фукусовых водорослей одинаково. Антерозоиды фукуса, аскофиллума и пель- веции имеют на переднем конце хоботок (рис. 148). Передний жгутик антерозоида химанта- лии снабжен на конце шипом, направленным вбок. Оогонии у фукусовых сидят прямо на по- верхности гаметофита или располагаются на одной поддерживающей клетке. Оогонии фуку- совых снабжены трехслойной оболочкой, наруж- ный слой называют э к з о х и т о н о м, про- межуточный — м е з о х и т о н о м, внутрен- ний — э н д о х и т о н о м. Содержимое вы- ходит в воду, окруженное двумя внутренними слоями. На этой стадии антерозоиды могут проникать в пакет яйцеклеток, но оплодотворе- ния не происходит — этому препятствует по- верхность яйцеклеток. Когда они полностью освобождаются от оболочек, происходит опло- дотворение. Вслед за этим оплодотворенные яйцеклетки вырабатывают собственную тол- стую оболочку. У некоторых фукусовых на- блюдается оплодотворение и прорастание яйце- клеток на материнском растении. Это бывает Рис. 148. Фукус пузырчатый (Fucus vesiculosus): 1 — оогоний; 2 —; 8 — схема строения антерозоида: а — передний жгутик, б — задний жгутик, в — глазок, г — яд- ро, д — хлоропласт, е — хоботок. у тех представителей, у которых образуется по одной яйцеклетке в оогонии. В простейшем случае яйцеклетки прикрепляются слизью око- ло отверстия концептакула (цистозейра), у дру- гих (Sargassaceae, Fucaceae) они прикрепляют- ся слизистым стебельком, тянущимся от оого- ния и представляющим собой вытянутый ме- зохитон. Яйцеклетки остаются прикрепленны- ми к материнскому растению 7 — 16 дней. За это время происходит их оплодотворение и дробление до стадии образования коротких первичных ризоидов, после чего проростки уносятся водой. Дробление яйцеклеток и развитие пророст- ков у всех фукусовых протекает почти одина- ково. После оплодотворения и появления обо- лочки яйцеклетка становится шаровидной, за- тем яйцевидной, ориентированной узким кон- цом вниз. Первая перегородка проходит попе- рек и делит яйцеклетку на две примерно рав- ные части (рис. 145, 1 — 8). В нижней части образуется вторая поперечная перегородка, отделяющая вниз небольшую клетку, называе- мую р и з о и д а л ь н о й, которая дает со временем первичные ризоиды. Последующие пе- регородки, с третьей по шестую, проходят в вер- тикаль.ных плоскостях в верхней и средней клетках. Из ризоидальной клетки вырастает от 1 до 32 ризоидов, в зависимости от рода и ви- да; предварительно эта клетка делится верти- кальными перегородками по числу будущих ризоидов. Положение первой перегородкф
в проростке и направление роста ризоидов определяется, кроме света, соседством других яйцеклеток того же вида. При малом расстоя- нии между яйцеклетками направление их роста целиком определяется этим фактором, вызы- вающим рост вершин в противоположные сто- роны. Установлено, что взаимовлияние яйце- кльток определяется химическими вещества- ми, вь;деляемыми ими в воду. Представители порядка фукусовых отлича- ются большим разнообразием. Они растут во всех морях от Арктики до Антарктики, кроме Каспийского и Аральского морей. Семейство хормозировые (Hormosiraceae) Единственный представитель этого семейст- ва — хормозира Банкса (Hormosira banksiig— имеет дихотомически-разветвленное слоевище, состоящее из шаровидных пузырей, соеди- ненных короткими и узкими перетяжками (рис. 143, 8). Органом прикрепления служит диск. Рост у слоевища верхушечный, осущест- вляется посредством 4 — 8 клеток, которые рас- полагаются в небольшом углублении на верши- не каждой ветви. Концептакулы образуются в поверхностном слое пузырей. Гаметофиты обоеполые, в оогониях созревает по 4 яйце- клетки. Распространена хормозира у берегов Новой Зеландии, острова Тасмания и в не- тропических водах Австралии, растет в лито- ральной зоне. В Новой Зеландии хормозиру используют в качестве корма, особенно для овец. Семейство нотейевые (Notheiaceae) Это семейство включает один род с одним видом. Нотейя аномальная (Notheia anomala) вырастает из концептакулов австралийских фу- кусовых хормозиры и ксифофоры, а также из концептакулов своих же особей. Высота слое- вища достигает 12 сл, побеги прямые, цилинд- рические; боковые ветви вырастают со дна концептакулов. Рост верхушечный, посредст- вом трех клеток. В концептакулах развиваются почти исключительно оогонии с 8 гаметами. Анте ридии с 64 антерозоидами встречаются крайне редко. Одно время ставили под сомне- ние сам факт их существования. В 1960 г. Н и з а м у д д и н и У о м е р с л и, изучая нотейю в культуре, обнаружили, что из оого- ниев, называемых ими макроспорангиями, вы- ходят макроспоры с двумя жгутиками. Мак- роспоры прорастали в разветвленные нити с волосками, наподобие гаметофитов сперма- тохнуса (порядок хордариевые). На этих ни- тях развивались однорядные многогнездные спорангии, снова производящие макроспоры. Судьба этих спор осталась невыясненной. Зоои- ды, выходящие из антеридиев, не сливались с макроспорами и не прорастали. Низамуддин и Уомерсли перенесли нотейю в порядок хор- дариевых в качестве самостоятельного семей- ства. Семейство химанталиевые (Himanthaliaceae) К этому семейству относится один род с од- ним видом. Химанталия удлиненная (Himan- thalia elongatag известна в странах Западной :Европы как «ремневидная водоросль» или «во- доросль-пуговица» (рис. 143, 4, о). Ее слоевп- ще состоит из воронковидного или дисковид- ного образования, прикрепленного эластич- ным стебельком с подошвой ко дну. На второи или третий год жизни из углубления в центре вырастает 1 — 4 дихотомически-разветвленных рецептакула длиной до 2 — 3 л и шириной 1 — 2 сл. Рецептакулы обладают верхушечным ростом, который осуществляется посредством одной верхушечной клетки. Химанталия интересна тем, что в оогониях развивается по одной очень крупной яйцеклетке (длиной до 0,5 лл), и тем, что после выхода га- мет растение обычно отмирает целиком. В от- личие от других фукусовых яйцеклетка химан- талии при развитии в слоевище спорофита уплощается, становится вогнутой с нижней сто- роны и прикрепляется к грунту краями. Позд- нее с нижней стороны вырастают нитевидные ризоиды. В последующем развитие идет, как у других фукусовых. Химанталия растет на камнях и скалах в нижнем горизонте литорали в умеренно при- бойных местах в Атлантическом океане от бе- регов Южной Норвегии до северного побере- жья Марокко; наиболее обычна в Норве- гии, Ирландии, Великобритании и Северной Франции. Семейство фукусовые (Fucaceae) Слоевище у фукусовых кустистое, прикреп- ляется к грунту диском. Рост ветвей осуществ- ляется посредством одной верхушечной клет- ки, имеющей вид четырехгранной усеченной пирамиды. Сердцевина ветвей состоит из не- плотно сомкнутых тонкостенных нитей и тол- стостенных гиф. Оогонии с 1 — 8 яйцеклетками. Ветви плоские или цилиндрические, без «ли- стьев». У водорослей рода пельвеция (Pelvetiag слое- вище ветвится правильно или неправильно дихо- томически. Ветви плоские или цилиндрические, без ребра. Воздушные пузыри одиночные. Криптостомы и цекостомы отсутствуют, изред- ка встречаются ямки без волосков. Рецептаку- 188
лы расположены на вершинах ветвей. Оогонии содержат по две яйцеклетки. Пельвеиия желобчатая (Р. canaliculata) имеет слоевище в виде кустов высотой до 15 сл с ду- гообразными ветвями, согнутыми по продоль- ной оси в виде желоба. Гаметангии созревают с конца июня по сентябрь. Растет пельвеция же- лобчатая над поясом фукусов в среднем и верхнем горизонтах литорали. Распространена она от Белого и Баренцева морей до севера Португалии. Пельвеиия Райта (Р. wrightii) имеет слоевища длиной 10 — 90 сл с прямыми ровными ветвями, несущими иногда одиночные пузыри. Выход гамет происходит в июле — сентябре. Произ- растает в среднем и нижнем горизонтах лито- рали и в сублиторали до глубины 1 л в север- ной половине Японского моря, включая Хок- кайдо и северную часть Хонсю, а также в за- ливе Анива и у Курильских островов до ост- рова Уруп. У представителей рода фукус (Fucus) слоеви- ще дихотомически-разветвленное с плоскими ветвями, имеющими продольное ребро, а также криптостомы и цекостомы. Рецептакулы распо- лагаются на вершинах ветвей. В оогониях обра- зуется по 8 яйцеклеток. Виды рода фукус рас- пространены в холодных и умеренных морях северного полушария. Часто они образуют большие заросли на литорали, это облегчает их сбор и использование. Виды рода фукус при- меняют в качестве удобрений, как корм для скота, для производства кормовой муки, аль- гинатов и других химических веществ. В мо- рях СССР встречается 5 видов этого рода (табл. 18, 19). Фукус зубчатый (F. serratus) выделяется среди остальных видов ветвями с зубчатыми краями. Слоевища его раздельнополые, в дли- ну достигают 1 л. В условиях Баренцева моря гаметы созревают в августе — сентябре, еди- ничные рецептакулы со зрелыми гаметангия- ми встречаются на отдельных растениях почти круглый год, крайне редко их можно найти весной. Фукус зубчатый растет в нижнем и иногда в среднем горизонте литорали в верх- ней сублиторали. Встречается у берегов Евро- пы от Новой Земли до севера Португалии и у берегов Северной Америки в районе залива Святого Лаврентия. Слоевища фукуса пузырчатого (F. vesiculo- sus) часто имеют воздушные пузыри, обычно располагающиеся попарно по бокам ребра. В прибойных местах пузыри не развиваются. Ребро у ветвей резко выражено и прослеживает- ся до вершины. Растения раздельнополые. Мужские экземпляры по сравнению с рядом растущими женскими растениями нередко име- ют слоевища меньшего размера, с более узкими ветвями и более мелкими рецептакулами. Про- споры и гаметофиты появляются на спорофитах с сентября до конца января или до конца Фев- раля, в сильно опресненных местах их заклад- ка прекращается в ноябре. В январе на гамето- фитах появляются первые гаметангии. В нача- ле июня в гаметангиях образуются гаметы; выход их в воду продолжается до конца авгу- ста. Фукус пузырчатый растет в разнообразных условиях — от сильно открытых до защищен- ных мест в среднем и нижнем горизонтах лито- рали и в верхней сублиторали, последнее чаще всего наблюдается в опресненных морях (Ве- лое и Балтийское). Дерновинки мелких слое- вищ, никогда не образующих проспор и гамето- фитов, встречаются на илистых грунтах в верх- нем горизонте литорали и в супралиторали вместе с моховидными и другими наземными растениями. Неприкрепленные слоевища фу- куса пузырчатого встречаются в Белом море до глубины 22 л~. Наиболее пышные заросли фукус пузырчатый образует на литорали в по- лузащищенных местах над поясом аскофиллу- ма, но возможно и обратное их расположение. Фукус пузырчатый распространен у побережья от Карского моря до северо-запада Испании и у берегов Северной Америки от острова Элсмир до штата Северная Каролина (США). В Балтий- ском море он произрастает до восточного берега Выборгского залива. Слоевища фукуса спирального (F. Spiralis) внешне сходны со слоевищами фукуса пузыр- чатого, лишенными пузырей. Отличается этот вид тем, что у него слоевища обоеполые и ре- цептакулы имеют узкую стерильную кайму по краям. Растет фукус спиральный на литорали выше пояса фукуса пузырчатого. Распростра- нен он у берегов Европы от Кольского полуост- рова до Канарских островов и у берегов Север- ной Америки от острова Ньюфаундленд до штата Нью-Йорк (США). Два других вида — фукус двусторонний (F. distichus) и фукус исчезающий (F. evanes- cens) — отличаются от вышерассмотренных ви- дов наличием цекостомов и менее четко выра- женным ребром. Слоевища у них обоеполые. Фукус двусторонний распространен у берегов Азии и Европы от Карского моря до севера Великобритании и у берегов Америки от ост- рова Элсмир до Нью-Йорка. Растет фукус двусторонний в местах с более сильным дви- жением воды и глубже, чем фукус пузырчатый. Развитие органов размножения и созревание гамет происходят у фукуса двустороннего на две-три недели раньше. Фукус исчезающий произрастает в Тихом океане от Приморского края, северной части Хонсю и севера Калифорнии до Чукотского и 189
Восточно-Сибирского морей. Растет он в ниж- нем и среднем горизонтах литорали, в Аркти- ке — в верхней сублиторали. Гаметы созре- вают летом. Род аскофиллул (Ascophyllum) включает один вид — аскофиллум узловатый (А. пойо- sum). Слоевище его, достигающее 1 — 1,5 л в длину, имеет длинные редкодихотомически- ветвящиеся ветви, на которых попеременно или супротивно расположены более корот- кие ветви. Рецептакулы развиваются как ко- роткие булавовидные веточки по бокам круп- ных, при созревании гамет они раздуваются на вершине и становятся похожими на крупные ягоды. Слоевища у аскофиллума раздельнопо- лые. Мужские экземпляры более темной ок- раски, с менее округлыми ветвями и более мелкими рецептакулами. Веточки-рецептаку- лы появляются на слоевищах с середины июля до конца октября, тогда же в них развиваются проспоры и гаметофиты. Первые гаметангии возникают в конце декабря, гаметы созревают в июне, и в течение месяца происходит их вы- ход. Аскофиллум растет в нижнем и отчасти в среднем горизонте литорали и в верхней суб- литорали (в Белом море). Распространен аско- филлум у берегов Азии и Европы от Карского моря до Португалии и у берегов Америки от Баффиновой Земли до штата Нью-Джерси (США). Семейство цистозейровые (Cystoseiraceae) Слоевище цистозейровых кустистое, прикреп- ляется к грунту диском или реже ризоидами. Рост ветвей идет посредством верхушечной клетки, имеющей вид трехгранной усеченной пирамиды. Сердцевина образована плотно сомк- нутыми рядами клеток. В оогониях развива- ется по одной яйцеклетке. Ветви, в отличие от саргассовых, никогда не развиваются из пазух «листьев». «Листья» на ветвях имеются или от- сутствуют. У водорослей из рода иистозейра (Cyctoseira) воздушные пузыри однокамерные, без попереч- ных перегородок. Ветви цилиндрические или многогранные, с «листьями» или без них. В мо- рях СССР виды рода цистозейра произрастают в Черном, Японском и Охотском морях и у Ку- рильских островов. В Черном море растут иистозейра косматая (С. crinita, рис. 143, 7) и иистозейра бородатая (С. barbata, рис. 143, 6). Цистозейра косматая выделяется среди ци- стозейр, растущих в морях СССР, тем, что от диска, которым слоевище прикрепляется к грунту, отходит несколько вертикальных стволов (до 20). Длина стволов составляет '/, длины слоевища, достигающей 0,5 — 1,2 л. Пузыри одиночные, некоторые из них вильча- то-рассеченные или с боковыми выростами. Рецептакулы обоеполые с выступающими боро- давчатыми концептакулами и короткими шипи- ками по бокам. Развиваются рецептакулы на слоевище дважды в год — веснои и осенью. Ветви, несущие рецептакулы, после выхода гамет разрушаются. Растет цистозейра косма- тая в открытых и полузащищенных местах на глубине до 10 — 15 л. Она распространена в Сре- диземном и Черном морях, кроме мелководной северо-западной части последнего. У цистозейры бородатой от диска отходит, как правило, один высокий ствол, который у старых экземпляров становится шерохова- тым. Слоевище достигает в длину 1,7 л. Воз- душные пузыри одиночные или собраны цепоч- ками, располагаются они на конечных веточ- ках или на верхних междоузлиях. Растения обоеполые; рецептакулы у них без шипиков, с гладкой, слегка волнистой поверхностью, раз- виваются дважды в год — весной и осенью. После выхода гамет ветви с рецепта кулами разрушаются. Цистозейра бородатая растет в Атлантическом океане у южного берега Пи- ренейского полуострова, в Средиземном и Чер- ном морях, иногда встречается в Азовском море у южного берега. Цистозейра толстоногая (С. crassipes), про- израстающая в дальневосточных морях СССР, имеет слоевища иного облика, чем предыдущие виды. Ствол у нее короткий и несет на верхнем конце короткие и толстые, иногда булавовид- ные, многолетние ветви, на которых вырастают длинные ветви неограниченного роста с корот- кими боковыми веточками. Слоевище в длину достигает 2 — 6 л. В нижней части слоевища бы- вают узкие «листья» без ребер. Воздушные пу- зыри одиночные или цепочками располагаются на конечных ветвях. Слоевища цистозейры тол- стоногой раздел ьнополые. Гаметангии созре- вают на юге ареала в июне, на севере — в ав- густе. Цистозейра толстоногая растет на камнях и скалах в нижнем горизонте литорали и в суб- литорали на глубине до 25 л в открытых и полу- защищенных местах. Распространена она у бе- регов Азии от северо-восточного побережья Корейского полуострова до материкового по- бережья Охотского моря и от северо-восточной части острова Хонсю до острова Уруп (Ку- рильские острова). У побережья Северной Аме- рики цистозейра толстоногая произрастает от южной части Берингова моря до штата Вашинг- тон (США). Род хорлофига (Hormophysa) включает един- ственный вид хормофига трехгранная (Н. tri- quetra), широко распространенный в тропиче- ских морях. Его нередко относят к роду цисто- зейра. Отличие заключается в том, что у хормо-
физы ветви с широкими пластинчатыми краями, расходящимися по радиусам на три стороны. Род халидрис (Halidrys) отличается от рода цистозейра стручковатыми воздушными пузы- рями, поделенными внутри поперечными пе- регородками на камеры, а также слегка упло- щенными ветвями, расположенными большей частью в одной плоскости. У атлантических берегов Европы от севера Норвегии до Порту- галии распространен халидрис стручковатый (Н. siliquosa). Семейство саргассовые (Sargassaceae) У этих водорослей слоевище кустистое, при- крепляется к субстрату диском, реже ризоида- ми. Все выросты — ветви, воздушные пузыри и рецептакулы — растут из пазух «листьев», которые могут быть широкими с ребром или без него или шиловидными. Когда «листья» разрушаются, то в основании ветвей или дру- гих выростов остается короткий шип. Воздуш- ные пузыри всегда одиночные и располагаются на конечных веточках. Сердцевина ветвей об- разована плотно сомкнутыми рядами клеток, только в старых побегах она разрушается и образуется полость с рыхлопереплетенными ни- тями. Ветви растут посредством верхушечной клетки, имеющей вид трехгранной усеченной пирамиды. Оогонии содержат по одной яйце- клетке. При созревании они выходят из кон- цептакула, но остаются прикрепленными к оо- гонию слизистым стебельком. В таком положе- нии происходит оплодотворение, дробление зи- готы и развитие проростка до начала образова- ния ризоидов, после чего они отделяются. У водорослей из рода саргассул (Sargassum) «листья» пластинчатые с продольным ребром или без него или шиловидные (рис. 143, 8). Ствол короткий, от верхней его части отходят длинные ветви, несущие рецептакулы, эти вет- ви ежегодно отмирают. В тропических морях есть виды, у которых все слоевище однолетнее. Представители рода саргассум распространены в основном в тропических и субтропических морях, отдельные виды заходят в теплые воды морей умеренных поясов. Саргассум исполь- зуют для получения альгинатов, отдельные ви- ды, имеющие нежесткие ветви, употребляют в пищу. В СССР виды рода саргассум растут в дальневосточных морях. Наиболее часто встречаются несколько видов. Саргассул бледный (S. pallidum) прикрепля- ется к грунту конусовидным диском, ствол у него грубый, с широко расставленными вет- вями. Широкие «листья» с ребром; в основании слоевища они бывают овальными или удлинен- ными (длиной до 15 сл). Выше по слоевищу «ли- стья» становятся мельче и уже, вплоть до шило- видных. В условиях сильного движения воды крупные листья не развиваются. Воздушные пузыри шаровидные. Растения у этого вида раз- дельнополые. Выход гамет в заливе Петра Ве- ликого происходит с конца июня до начала ав- густа, на севере Татарского пролива он про- должается до сентября. Длина слоевищ дости- гает 2 — 4,5 л. Этот вид растет в сублиторали на глубине до 20 л, иногда встречается в ниж- нем горизонте литорали. Саргассум бледный распространен в Желтом и Японском морях, у Тихоокеанского побе- режья Японии, вдоль юго-восточного берега Сахалина и у южных Курильских островов до острова Итуруп. Слоевище саргассула Миябе (S. miyabei) при- крепляется к грунту слегка расширенным осно- ванием ствола и ризоидами, которые по всей своей длине прилегают к грунту. При неакку- ратном сборе растений ризоиды легко обламы- ваются и остаются на грунте; при этом кажется, что слоевища прикрепляются диском, как у других видов. Саргассум Миябе отличается еще коротким стволом и мелкими (длиной до 13 лл) клиновидными «листочками», густо по- крывающими молодые побеги. Воздушные пу- зыри у саргассума Миябе эллипсоидные, с ост- pbIM кончиком. Растения раздельнополые, ре- цептакулы мужских растений длиннее женских. Выход гамет у саргассума Миябе, растущего в заливе Петра Великого, происходит в июне— июле, на севере Татарского пролива он продол- жается до октября. Растет саргассум Миябе в открытых и полузащищенных местах в ниж- нем горизонте литорали и в сублиторали. Этот вид встречается в северной половине Японского моря, включая остров Хоккайдо, у юго-восточ- ного побережья Сахалина и южных Куриль- ских островов до острова Итуруп. Своеобразные виды рода саргассум живут в Саргассовом море. В основном там преобла- дают саргассул плавающий (S. fluitans) и сар- гассул погруженный (S. natans). Для обоих видов характерно отсутствие органов прикреп- ления, рецептакулов и какой-либо главной оси. Слоевища этих и других видов рода саргассум образуют спутанную массу, плавающую у по- верхности. Род тпурбинария (Turbinaria, рис. '143, 9) довольно обычен в тропических морях. Для его представителей характерны мясистые листья, расширяющиеся кнаружи в виде конуса. К грунту слоевище турбинарии прикрепляется маленьким диском и жесткими слабо развет- вленными ризоидами. В прибойных местах мо- лодые слоевища представлены слабо разветвлен- ными цилиндрическими побегами и внешне не имеют ничего общего с более крупными слое- вищами.
ОТДЕЛ КРАСНЫЕ ВОДОРОСЛИ (RHODOPHYTA) У берегов Японии и южной половины Япон- ского моря, включая залив Петра Великого, у берега на малой глубине часто встречается представитель рода коккофора (Coccophora)— коккофора Лангсдорфа (С. langsdorfii). Она вы- деляется среди других водорослей прутовид- ными побегами высотой до 0,5 л, отходящими от короткого сильно разветвленного ствола с широким диском при основании. Весной и в начале лета побеги коккофоры покрыты крохотными чешуевидными треугольными «ли- стьями». На вершинах ветвей на коротких веточках расположены грушевидные пузыри, которые одновременно служат рецептакулами. Растения Красные водоросли, или багрянки,— самая обширная среди донных морских водорослей и чрезвычайно своеобразная группа. Насчиты- вают более 600 родов и около 4000 видов баг- рянок. Своеобразие их заключается прежде всего в наборе пигментов. Наряду с двумя хлорофиллами (а и b), дву- мя каротинами (ж и P) и несколькими ксанто- филлами они содержат специфические пигменты билипротеины — красный фикоэритрин и синий фикоцианин, обнаруженные еще лишь у сине- зеленых и, возможно, пирофитовых водорослей. Различное сочетание этих пигментов опреде- ляет окраску багрянок, которая меняется от ярко-красной до голубовато-зеленой и желтой. Продуктом ассимиляции у них служит так на- зываемый багрянковый крахмал, отличаю- щийся от крахмала цветковых растений и близ- кий к амилопектину и гликогену. Представители отдела в подавляющем боль- шинстве многоклеточные организмы сложного морфологического и анатомического строения, и только очень немногие, наиболее примитив- ные, имеют слоевище одноклеточное или коло- ниальное. Большинство багрянок крупные рас- тения, достигающие в длину от нескольких сан- тиметров до метра, но среди них немало и мик- роскопических форм. Багрянки обладают сложным, не встречаю- щимся у других водорослей циклом развития, своеобразным строением женского органа раз- множения — оогония и сложными процессами развития зиготы. Подвижные стадии в цикле развития полностью отсутствуют, их споры и гаметы лишены жгутиков. Подавляющее большинство красных водо- рослей обитает в морях, и только очень немно- гие встречаются в пресных водах. раздельнополые. Гаметы выходят в конце мая и в июне, после этого рецептакулы и несущие их веточки разрушаются, и слоевище преоб- разуется. Ребра крохотных чешуевидных «листьев» вырастают в нитевидные слабоди- хотомически-ветвящиеся веточки длиной до 4 — 15 сл, отходящие перпендикулярно к вет- вям. В связи с этим в конце лета и зимой длинные побеги коккофоры выглядят пушис- тыми. По мере развития новых рецептакулов ните- видные веточки опадают. Яйцеклетки кокко- форы используют в научных исследованиях для выяснения механизмов, определяющих разви- тие растений. Небольшая группа примитивных форм объеди- нена в класс бангиевых, остальные составляют класс флоридеевых. ВНЕШНЯЯ ФОРМА ТЕЛА КРАСНЫХ ВОДОРОСЛЕЙ Внешне слоевища красных водорослей весьма разнообразны, часто красивы и причудливы (табл. 20 — 23). Здесь можно встретить формы нитевидные и пластинчатые, цилиндрические и корковидные, пузыревидные и кораллоподоб- ные, в разной мере рассеченные и разветвлен- ные. Нитевидные слоевища могут быть очень тон- кими, волосовидными, состоящими из одноряд- ных клеточных нитей, как, например, у некото- рых немалиевых и иерамиевых. У многих багря- нок нити более грубые. Они имеют более слож- ное строение и состоят из нескольких рядов клеток. Многим багрянкам со сложным анато- мическим строением свойственны цилиндриче- ские слоевища. Они бывают плотные или с по- лостью внутри, слизистые, хрящеватые или ко- ж истые, в виде шнуров или обильно развет- вленных кустиков. Большого разнообразия до- стигают пластинчатые формы. Встречаются пластины цельные и сложно рассеченные, с до- полнительными выростами по краю и по поверх- ности. Нередко на специальных выростах раз- виваются органы размножения. Мелкие боро- давчатые выросты обычно называют п а п и л л а- ми, более крупные, напоминающие по форме основные ветви, — п р о л и ф и к а ц и я м и. Неж- ные пластины делессериевых имеют листообраз- ную форму и пронизаны «нервами» и «ребрами», напоминающими жилкование листьев цветковых растений. Водоросли, слоевище которых построено в виде корок или пленок, покрывают камни
Та бл и ц а 17. Бурые водоросли: в в е р х у — группа слоевищ надины павлиньей (Padina pavonia); в н и з у — часть слоевища костарии ребристой (Costaria costata), изъеденного животными. Фото Ю. Астафьева и А. Рогова.
Та б л и ц а 18. Бурые водоросли: в в е р х у с л е в а — фукус пузырчатый (Fucus vesiculosus); в в е р х у с п р а в а — фукус двусторонний (F. distichus); в н и з у с л е в а — бурая нитчатая водоросль хордария плетевидная (Chordaria flagelliformis) на фукусе пузырчатом; в н и з у с п р а в а — бурая нитчатая водоросль пилайелла литоральная (Ру)aiella liroralis) на фукусе пузырчатом. Фото Ю. Астафьева.
~ й 6 л и ц а 19. Заросли бурых водорослей в начале отлива на литорали Белого моря. Фото В. Кузина.
Г а б л и ц и 20. Красные водоросли: 1 — порфира (Porphyra variegata); 2 — дюмонтия (Dumontia incrassata); 3 — эутора (Euthora cristata); 4 — хондрус (Chondrus yendoi).
Та б л и ц а 21. Красные водоросли: 1 — ломентария (Lomentaria articulata); 2 — хондрус (Chondrus pinnulatus); 3 — хризимения (Chryzymenia wrightii); 4— родимения (Rhodimenia pertusa).
Та б л и ц и 22. Красные воцоросли: 1 — мсмбраиоптера (Memhranoptera alata); 2 — родимеиия (Rhodimenia stenogona); 3 — фикодрис (Phycodris sin uosa); 4— птилота (Ptilota asplcniotdes).
Та б л и ц а 23. Красные водоросли: 1 — одонталия (Odonthalia ochotensis); 2 — токидодендрон (Tokidodendron bullata); 3 — родомела (Rhodomela lycopodioides); 4 — дазия (Dasia baillouviana).
T cr б л и ц и 24. Красные известковые водоросли: в в е р х у — членистые кораллиновые водоросли — кораллина целебная (Corallina officinalis); в н и з у — корковые кораллиновые водоросли в биоценозе морской травы филлоспадикса. Фото Ю. Астафьева .
или другие водоросли, плотно прирастая к ним. А семейство кораллиновых (Corallinaceae) ха- рактеризуется тем, что их ткани частично или полностью пропитаны известью. Форма этих водорослей отличается чрезвычайным разнооб- разием. У кустистых кораллиновых слоевище со- стоит из пропитанных известью члеников, соединенных между собой короткими некаль- цинированными участками. Встречается здесь большое число кораллоподобных форм и про- сто корок, плотно обволакивающих субстрат. Многообразие внешней формы красных во- дорослей во многом определяется ветвлением— количеством ветвей, их формой, размерами и характером расположения. Основной тип вет- вления, встречающийся у большинства баг- рянок, — моноподиальный. При этой системе ветвления главная ось не прекращает роста, а образует ниже своей точки роста боковые ветви. Реже, но все-таки встречаются здесь и другие типы ветвления — симподиальное и ди- хотомическое. При симподиальном ветвлении главная ось прекращает свой рост или сдви- гается вбок, а ее место занимает боковая ветвь, растущая в направлении главной оси. При ди- хотомическом точка роста разделяется на две новые, дающие одинаково развитые ветви. Не- редко у дихотомически-разветвленных расте- ний в большом количестве развиваются допол- нительные боковые ветви. К таким вторичным боковым ветвям относятся и пролификации, как, например, у хондруса (табл. 21, 2) и мно- гих других. Ветви красных водорослей делятся на две категории. Одни — это основные длинные вет- ви, которые растут в длину в течение всего периода роста растения, называют их ветвями неограниченного роста. Другие растут только до определенного предела и всегда остаются более или менее короткими — это ветви огра- ниченного роста. Расположение ветвей может быть самым разнообразным — беспорядочным, по спирали, мутовчатым. Очень часто в раз- ных группах багрянок можно встретить распо- ложение ветвей в одной плоскости. Поочередное или супротивное отхождение веточек только с двух сторон главных ветвей придает слоевищу перистый вид (табл. 23). Свойственно красным водорослям и дорсо- вентральное строение. При таком строении ос- новные ветви стелются по субстрату. На одной стороне стелющихся ветвей развиваются ри- зоиды, служащие для прикрепления к субстра- ту, на противоположной стороне — вертикаль- ные побеги. Кроме обычных ветвей, у красных водорослей имеются разные специализированные веточки. Так, в порядке церамиевых очень распрост- ранены так называемые т р и х о б л а с т ы, ко- торые состоят из одно рядных разветвленных или неразветвленных клеточных нитей, обычно слабо окрашенных или совсем бесцветных. Располагаются такие веточки в верхней части слоевища и относятся к числу временных обра- зований. Флоридеевые из разных порядков обладают своеобразными закрученными или серповидно- изогнутыми веточками, которыми они цепляются за другие растения или ветви того же растения. Называют такие веточки усиками, так как они, подобно усикам высших растений, делают во- круг ветви, с которой соприкасаются, несколь- ко спиральных витков. Усики отличаются от соседних ветвей и анатомическим строением. Наружные ткани их более сильно развиваются на выпуклой стороне, благодаря чему происхо- дит закручивание. Поверхностные клетки усика в точках соприкосновения с другими ветвями вытягиваются в ризоиды, которые помогают прикреплению к последним. Соединение или срастание ветвей как одного, так и разных растений происходит и у тех багрянок, которым несвойственно образование усиков. Оно обеспечивается развитием ризои- дов, которые растут навстречу друг другу от обеих вступающих в соприкосновение поверх- ностей. СТРОЕНИЕ СЛОЕВИЩА КРАСНЫХ ВОДОРОСЛЕЙ При всем многообразии внешней формы крас- ные водоросли отличаются единым планом строения слоевища — в основе его у всех много- клеточных багрянок лежит клеточная развет- вленная нить. Паренхимный тип организации здесь фактически отсутствует. Единственный пример паренхимного слоевища мы находим в классе бангиевых — у порфиры и бангии. Слоевище бангиевых. Наиболее просто уст- роенным слоев ищем обладают представители класса бангиевых. Только здесь встречаются од- ноклеточные формы. Они растут в виде одиноч- ных клеток или колониями. Самые простые ко- лонии — это бесформенные скопления клеток, соединенные общей слизью, подобные тем, ка- кие образует порфиридиум (Porphyridium, рис. 149, 1). Постепенно у колониальных форм намечается тенденция к нитевидному росту: клетки, заключенные в общую слизистую труб- ку, располагаются в один или несколько рядов. Нитчатые колонии acmepoqumuca(Asterocytis), зониотрихума (Goniotrichum) бывают нераз- ветвленными или ветвятся по типу ложного ветвления (рис. 149, 8, 4). Ложным его назы- вают потому, что возникновение ветвей не связано с боковым ростом клеток нитей.
Рис. 149. Бангиевые.' 1 — Porphyridium; 2 — Chroothece; 8 — 4 — однорядные и мно- горядные нитчатые колонии Goniotrichum. Рис. 150. Строение бангиевых.' 1, 2 — Bangia (1 — основание растения с ризоидами, 2 — часть многорядной нити); 8 — Erythrocladia. Слоевище нитчатых бангиевых может быть вертикальным или стелющимся. Стелющиеся нити обычно ветвятся и образуют на субстрате рыхлую или более или менее плотную пластину неправильных очертаний, как у эритрокла- дии (рис. 150, 8). Если нити расположены плот- но, то такая пластина напоминает паренхим- ную. Большинство бангиевых относится к прямо- стоячим организмам. Усложнение в строении вертикальной нити заключается в том, что клет- ки приобретают способность делиться в не- скольких плоскостях, в результате чего одно- рядная нить преобразуется в многорядную, как, например, у бангии (рис. 150, 1, 2). Кроме нитчатых форм, среди бангиевых име- ется группа водорослей пластинчатого строе- ния. Пластинчатые формы могли образоваться в результате усложнения нитчатых. Приобре- тение клетками способности делиться в двух плоскостях — поперечной и продольной— привело к преобразованию однорядной нити в однослойную пластину паренхимного строе- ния. Если деление происходит в трех плоско- стях, образуется двухслойная пластина. Такое строение мы находим у порфиры, наиболее высокоорганизованной водоросли в классе бангиевых (табл. 20, 1). Но наряду с этим пла- стинчатое вертикальное слоев ище могло воз- никнуть и из стелющейся структуры, подобной эритрокладии: вначале пластина становится многослойной, затем верхний клетчатый слой в ней отделяется с образованием мешковидной структуры, которая, разрываясь, растет дальше как однослойная пластина. Так формируется слоевище порфиропсиса. Вместе с усложнением самого слоевища у вер- тикальных форм усложняется и строение органа прикрепления. Вначале базальная клетка поч- ти не меняет формы и не отличается от осталь- ных клеток, затем у нее появляются выросты, которые создают уже специальный орган при- крепления. У наиболее высокоорганизованных форм ри- зоидные выросты отходят не только от бузаль- ной клетки, но и от соседних клеток слоевища (рис. 150, 1). Нередко их образуется так мно- го, что вместе они складываются в массивную подошву, как, например, у порфиры. Рост у бангиевых диффузный, т. е. проис- ходит в результате деления всех клеток слое- вища. Слоевище флоридеевых. В отличие от бан- гиевых, все представители класса флоридее- вых — многоклеточные организмы. Тело их представляет собой более или менее сложную систему разветвленных нитей. Пуренхимный тип организации у них отсутствует. В наиболее простом случае слоевище состоит из свобод- ных однорядных клеточных нитей. Эта струк- тура свойственна немногим флоридеевым. Ею обладают примитивные представители порядка немалиевых (рис. 151, 1) и некоторые просто организованные аерамиевые (рис. 182, 1 — 3),
о Ооо~ gO Ое Рис. 151. Эволюция ст кт и через слоеви руктуры флоридеевых: 1 — курв!а r— — — czroeam~a Batrachos — ризоиды, в — гонимоблас, т, г — цент альны sp rmum; 8 — продольный с р е клетки, д — перицентральная клетка 8 ; рез 95 EOTOPMIB П ОИЗО р р ошли, вероятно, непосредствен- но от первых. Во р . доросли, слоев ище которых построено из свободных однорядных нитей,— на г преимущественно мелкие организмы ж других водорослях и животных. Многим из них свойственно разнонитчатое строение ' те- по ст лющиеся по су страту нити обычно отлича аются свобо но роению от вертикальных и имеют вид сво одно расположенных нитей или соб аны в плотные пластины У ны. У некоторых из нитчатых со раны форм стелющаяся часть не развивается, и они прикрепляются к грунту одной базальной клет- В процессе эволюции свободно-нитчатое строение претерпело ряд изменений, в ез ль- тате которых сформировалась сложная анато- мическая структура большинства багрянок. Первым шагом в м в усложнении однорядного нитчатого слоевища было возникновение боль- шого числа ко о коротких и обильно разветвленных боковых веточек п , подобно тому, как это проис- ходит у батрахоспермума. Нередко эти боко- вые веточки отхо ят дят супротивно или мутовчато по нескольк у от каждои клетки. В отличие от ° У Ф основных ветвей твей они растут в длину только до определенного предела и поэтому являются ветвями ограниченного роста. У своей вершины веточки or аниченн р енного роста разветвляются более густо чем в н у нижней части, в месте своего отхождения (рис. 151, 2). У многих багрянок с таким ст оени р ем слоевище сильно пропиты- вается студенистым веществом, которое скреп- ляет все многочисленные веточки в те чего ~ о ми , в результа- ~oðMIIð57eTñs компактное слоевище с до- вольно ыхлым Ст роением, имеющее вид слизи- стых шн ов. В ур . дальнеишем по мере усложне- ° У ния строения веточки ограниченного силенно ветв го роста ЗОМ ЧТО ветвятся и разрастаются та б , что смыкаются друг с другом, образуя вок- тный пери ериче- руг центральной части плотный ский слой. и р этом студенистое вещество те- ряет значение скрепляюще го ветви материа- ис. 15i ла и развивается в незначительн ьнои степени (рис., 8). Таким образом формируется ко а баг~янок. П р . ри плотном строении только клет- ки, расположенные у поверхности слоеви а способны во воспринимать свет и участвовать в фотосинтезе и ассимиляции. В отличие от вн т- ренних клеток сл лоевища, они интенсивно окрашены и имеют более мелкие так как п е ст мелкие размеры, как представляют собой конечные аз азвет- роста. тот асси- ения ветвеи ограниченного оста. Эт рой, тогда как глубже расположенные ч веточек ог части ограниченного роста называют внут- ренним коровым слоем. Кора всех баг рянок имеет одно происхожде- ние, но в процессе эволюции у разных форм она приобрела свои морфологические особенности, которые складываются из разме ов слагаю и щих ветви ограниченного роста, степени их разветвленности плотного и р или рыхлого расположения ветвей. олее высокооргани- зованных ~о и к р коровые нити укорачиваются
Рис. 152. Строение Gelidium (1, 2) и Furcellaria (8, 4): 1 — продольный срез через вершину веточки Gelidium; 2— поперечный срез слоевища Gelidium; 8 — продольный срез через вершину Furcellaria; 4 — поперечный срез слоевища Furcellaria: а — внешняя кора, б — внутренняя кора, в — пучок централь- ных нитей, г — ризоиды. и плотнее прилегают друг к другу, так что об- щее строение корового слоя упрощается и ста- новится похожим на паренхимное. Вследствие этого первоначальная нитчатая структура раз- личается с большим трудом. И только у верши- ны слоевища, вблизи точки роста, можно уви- деть, как центральные осевые клетки отчленя- ют от себя боковые клеточные нити, не сообщаю- щиеся с соседними, а только близко примыкаю- щие к ним (рис. 152, 1, 2). Сложная анатомическая структура многих красных водорослей создается в значительной степени за счет особых ризоидных нитей. Уже у форм со свободным нитчатым строением можно наблюдать, как от нижних клеток вет- вей ограниченного роста при их возникнове- нии отходят ризоидные нити, или гифы, имею- щие вид узких клеток с толстой оболочкой, ко- торые растут не в бок, а тянутся вдоль осевых нитей по их поверхности (рис. 151, 2, 8). В дальнейшем по мере усложнения организа- ции слоевища степень развития ризоидов уве- личивается. Это можно проследить у багрянок и с рыхлым и с плотным строением. Нередко они развиваются настолько, что заполняют всю внутреннюю часть слоев ища вокруг основ- ных осевых нитей и клеток внутренней коры (рис. 152, 2; 153, 2). В анатомическом строении флоридеевых раз- личают два основных типа — одноосевой и мно- гоосевой. При одноосевом типе, или, как его еще называют, центр альнонитчатом, в основе строения слоевища лежит единственная одно- рядная клеточная нить с боковыми веточками ограниченного роста (рис. 151; 152, 1, 2). При многоосевом типе центральная ось слоевища состоит из пучка параллельно идущих нитей. Их расположение напоминает струи воды в фон- тане, поэтому этот тип организации называют также фонтанным (рис. 152, 8, 4). Нити пучка могут располагаться плотно в центре слоевища или кольцом, и тогда в центре остается полость, как, например, у родимениевьт. Каждая из нитей центрального пучка растет посредством апикальной клетки и отчленяет в бок веточ- ки ограниченного роста. Тот или иной тип организации характерен для определенных групп водорослей. В одних случаях одинаковое строение имеют нредста- вители целых порядков. Все родимениевые многоосевые, тогда как порядки церамиевых и гелидиевыт построены по одноосевому типу. В остальных порядках представлены оба типа строения, и здесь характер строения высту- пает уже как признак семейства. Иногда, как показали наблюдения за развитием некоторых водорослей, растения одного и того же вида формируются как по одноосевому, так и по многоосевому типу.
В основе строения многих флоридеевых лежит разнонитчатая организация. Более всего она выражена у немалиевых и криптонемиевыт. При прорастании спор у водорослей этих по- рядков вначале образуется стелющаяся струк- тура, от которой позднее возникают более или менее многочисленные вертикальные нити. Позднее из отдельных вертикальных нитей при одноосевом строении и из группы нитей при многоосевом формируется слоевище этих баг- рянок. Разнонитчатую организацию можно разли- чить, хотя и в несколько измененной форме, также в порядках гигартиновыт и родимение- вых. Только у церамиевых стелющаяся часть полностью отсутствует. По-видимому, разно- нитчатая структура представляет собой самый примитивный тип организации, и в процессе эволюции наибольшее развитие получает систе- ма вертикальных нитей, тогда как стелющаяся часть постепенно редуцируется. Такое же на- правление морфологической эволюции показы- вают и бурые водоросли. В отличие от бангиевых, у которых, как уже говорилось, в росте в длину участвуют все клетки, флоридеевые характеризуются апи- кальным ростом. Осевые нити, из которых сла- гается слоевище флоридеевых, растут в длину в результате деятельности верхушечной, или а п и к а л ь н о й, клетки. У многих багрянок верхушечную клетку можно отличить от осталь- ных по куполообразной форме и более крупным размерам, тогда как у некоторых других она не отличается по внешнему виду от нижеле- жащих. У багрянок, построенных по одноосевому типу, имеется одна апикальная клетка, при фонтан- ном типе организации их несколько. Апикаль- ные клетки делятся поперечными перегородка- ми и отчленяют вниз один ряд клеток, которые по своему положению в слоевище являются центральными. Апикальные клетки некоторых высокоорганизованных багрянок имеют две или три отчленяющиеся поверхности. В этом слу- чае при делении клеточные перегородки распо- лагаются косо по отношению к продольной оси, и от одной апикальной клетки отчленяется уже не один, а два-три ряда сегментов. Образо- вавшиеся центральные сегменты отделяют вбок две или четыре клетки, которые располагаются вокруг них или двусторонне. 3то так называемые п е р и ц е н т р а л ь н ы е клетки, которые у боль- шинства флоридеевых функционируют как апи- кальные клетки боковых ветвей ограниченного и неограниченного роста. Отчленив от себя пе- рицентральные клетки, центральные в даль- нейшем уже не подвергаются делению ни по- перечными, ни продольными перегородками и их рост осуществляется только за счет вытя- гивания в длину (рис. 152, 1). Исключение из общего правила составляют некоторые корал- линовые и делессериевые, у которых встречают- ся интеркалярные клеточные деления. В ре- зультате такого способа роста внутренние клет- ки слоевища большинства багрянок, даже с плотным псевдопаренхимным строением, обыч- но длинные и крупные, в то время как клетки, расположенные ближе к периферии, становятся короче и мельче. Часто бывает весьма легко определить, по какому типу — одноосевому или многоосево- му — построено слоевище тех или иных багря- нок. Но иногда это сделать почти невозможно, так как в построении слоевища принимает участие особая ткань — м е р и с т е и а. Иными словами, кроме апикальных, в росте слоевища участвуют многие клетки. У форм с плотным слоевищем в качестве меристемы функциониру- ет обычно поверхностный слой внешней коры, образованный апикальными клетками веточек ограниченного роста. У видов с цилиндриче- ским слоевищем действуех верхушечная мери- стема, у уплощенных фори — краевая. Участие в росте меристемы, по-видимому, следует счи- тать вторичным явлением, возникшим на осно- ве одноосевого и многоосевого типов строения. Кроме того, первоначальная структура может сильно затемняться благодаря развитию внут- ри слоев ища многочисленных ризоидных ни- тей (рис. 153, 1, 2). Процесс формирования слоевища — порядок отчленения клеток, количество перицентраль- ных клеток и т. д. — отличается большой слож- ностью, но у разных групп багрянок харак- теризуется постоянством и специфичностью, поэтому принцип организации слоевища в каж- дом порядке целесообразно рассмотреть от- дельно. Говоря о строении органов прикрепления флоридеевых, следует различать их первичную и вторичную структуру. Первичный орган при- крепления формируется на начальных этапах прорастания, тогда как вторичный характери- зует уже взрослое, сформированное растение. В его создании участвуют многие клетки ниж- них частей слоевища, и он может иметь совсем другое строение по сравнению с первичным. У форм разнонитчатого строения роль органа прикрепления играет стелющаяся структура, состоящая, как известно, из рыхлорасположен- ных нитей или из плотной клеточной пластины. Когда стелющаяся часть не развивается, ба- зальная клетка проростка образует вниз вы- росты, или первичные ризоиды. Из них разви- ваются многообразные структуры, служащие для прикрепления взрослых растений. Многие фло- ридеевые, особенно среди церамиевых, прикреп- ляются одноклеточными, короткими или длин-
00ОО000000000ООО р pg0 P OOО0 ~~ ооОООо О~ООООО000000000000~ООО00000 PP мооро� «( )С ) ~с~«~-~~ оооооoooo~ooo~о~ооooQogyо о оЯор +9~ð(-)oоояоОс >aoс70~( )~ оаооoo~и«оо0о< орррррррррррОРР O+O(7 р рр O~o o O ОO QPPgg ooooo ОО ~О. 000000000000000000000ООООООрОО0000000000000000 000000 00 o o o o o o ODp ~ООООООоаа~ Рис. 153. Строение Callophyllis.' 1 — вершина веточкф; 2 — поперечный срез слоевища. ными, ризоидами, расширяющимися при сопри- косновении с субстратом в присоску. У других базальная часть состоит из многоклеточных побегов. Очень часто орган прикрепления пред- ставляет собой диск овидную подошву, имею- щую клеточное или ризоидное строение. Весь- ма распространено у флоридеевых образование стелющихся побегов, напоминающих столоны высших растений, с многочисленными выро- стами-присосками, которыми растение прикреп- ляется к субстрату. Дополнительные ризоиды могут развиваться и на вертикальных ветвях, особенно в усло- виях, где требуется усиленное прикрепление, например на прибойных скалах. СТРОЕНИЕ КЛЕТКИ КРАСНЫХ ВОДОРОСЛЕЙ Своеобразие внутриклеточного строения крас- ных водорослей складывается как из особенно- стей обычных клеточных компонентов, так и из наличия специфических внутриклеточных вклю- чений. Клеточные оболочки. В клеточных оболоч- ках красных водорослей хорошо различаются два слоя: внутренний, состоящий из целлюлозы, и наружный, построенный пектиновыми сое- динениями. У форм с грубым жестким слоеви- щем снаружи клеточных стенок развивается тонкий слой кутикулы. По составу и строению кутикула багрянок отличается от кутикулы высших растений. Так, например, у порфиры она образуется в результате сгущения моле- кул моносахарида маннозы. У мягких слизи- стых форм кутикула отсутствует. Пектиновые вещества красных водорослей представляют собой соли кальция и магния особых пектиновых кислот. Они обладают спо- собностью растворяться в кипящей воде с об- разованием слизистых растворов. К группе пектиновых веществ относятся также особые коллоидные вещества, которые содержатся в клеточных оболочках и межклетниках мно- гих багрянок. Они представляют собой слож- ную смесь содержащих серу полисахаридов и носят общее название фикоколлоидов. Фико- коллоиды не растворяются в холодной воде, но хорошо растворяются в кипящей с образова- нием коллоидных систем. В настоящее время фикоколлоиды красных водорослей, так же как и бурых, являются самыми важными продукта- ми, получаемыми из морских водорослей. По- скольку эти вещества широко применяются в хозяйственной жизни человека, их химиче- ский состав достаточно хорошо изучен. Фико- коллоиды получены из многих видов, в резуль- тате установлено несколько их разновидностей. Более всего известны агар, каррагинин, нори, агароиды. Эти вещества отличаются друг от друга по составу и свойствам, но обладают об- щей желирующей способностью. Присутствие коллоидных веществ обусловли- вает способность клеточных оболочек к силь- ному набуханию после отмирания, благодаря чему они с трудом поддаются окраске при ис- следовании. Среди красных водорослей встречаются фор- мы, оболочки которых обызвествлены. Сначала известь откладывается в срединной пластинке, между внешним и внутренним слоем оболочки, проникая постепенно в целлюлозный слой и более или менее сильно пропитывая его. Но даже при сильном обызвествлении внутри всегда остается тонкая мембрана, лишенная из- вести и отделяющая плазму от известкового слоя. Состав известковых соединений неодинаков. У кораллиновых откладывается кальцит, у не- которых немалиевых — арагонит. Кроме того, встречаются карбонаты кальция и магния, а также железо. Рост оболочек происходит следующим обра- зом. Новые слои ее закладываются на вершине протопласта апикальной клетки, в то время как наружные слои по мере роста клетки по- степенно разрываются. В результате вся обо- лочка имеет слоистое строение и, так как новые слои откладываются под определенным углом, по виду напоминает воронку. Цитоплазма красных водорослей характери- зуется большой вязкостью и плотно прилегает
к клеточным стенкам. Она очень легко подвер- гается плазмолизу, и поэтому красные водорос- ли весьма чувствительны к изменению условий внешней среды, в частности к опреснению. Ядро. Среди красных водорослей имеется большое число форм с одноядерными клетками. Как правило, это наиболее просто организо- ванные формы. У высокоорганизованных баг- рянок клетки обычно многоядерные, за исклю- чением молодых клеток слоевища — апикаль- ных, коровых и т. п. Одноядерные клетки вет- вей ограниченного роста при определенных со- стояниях становятся много ядерными. Однако можно найти и такие водоросли, у которых старые клетки центральных нитей одноядерные, а окружающие их более молодые клетки, на- оборот, содержат несколько ядер. Репродук- тивные клетки — спермации, карпоспоры, тет- распоры — всегда с одним ядром, но яйцеклет- ки часто окружены многоядерными клетками, даже у форм, все слоевище которых состоит из одноядерных клеток. Ядро у красных водо- рослей мелкое, имеет четкую ядерную оболоч- ку и ядрышко. Хлоропласты. В клетках красных водорос- лей находится один или несколько хлоропластов. У растений класса бангиевых и у примитивных представителей наиболее низкоорганизован- ного порядка немалиевых имеется единствен- ный звездчатый хлоропласт с одним пиренои- дом. Он обычно занимает осевое положение в клетке и тогда состоит из центрального тела и отходящих от него во всех направлениях от- ростков (рис. 149, 2). Пиреноид при осевом хлоропласте находится в его центре. Отростки, возникающие из центрального хлор опласта, могут расширяться на периферии и смыкаться между собой, образуя пристенный хлоропласт неправильной или лентовидной формы. Вероят- но, пристенный хлоропласт большинства багря- нок происходит от осевого в результате утраты центральной части. Роль пиреноида у красных водорослей не очень ясна. В одних случаях его присутствие связано с отложением крахмальных зерен; в других же пиреноид встречается в клетках, не участвующих в процессах ассимиляции. У более высокоорганизованных форм пиреноид исче- зает; этот процесс можно проследить уже в по- рядке немалиевых. У красных водорослей, лишенных пиренои- дов, хлоропласты бывают двух основных типов- лентовидные и линзовидные (или дисковидные) с многочисленными переходами между ними (рис. 154). Виды, стоящие на более низкой ступени эволюции, обладают обычно лентовид- ным хлоропластом; для высокоорганизованных форм, наоборот, более характерны линзовид- ные хлоропласты. То же самое можно сказать 'б. ::1 Рис. 154. Форма хлоропластов багрянок: 1 — Audouinella; 2 — Ceramium; 8 — 4 — Kylinia; 5 — Cystoclo- nium: а — пиреноид. и о количестве хлоропластов — число их с ус- ложнением организации увеличивается. Форма хлоропласта не есть нечто постоянное, она мо- жет меняться с возрастом, с условиями осве- щения, с изменением размеров клеток, хотя нередко крупные группы багрянок характери- зуются хлоропластом определенной формы. У церамиума в крупных клетках междоузлий хлоропласты вытянутые, лентовидные, а в ко- ротких коровых клетках на узлах — это ко- роткие, неправильно лопастные пластинки. Размножение хлор опластов происходит путем простого пер ешнуровывания, как у BI IcIIIIIx растений. По своему тонкому строению, видимому толь- ко под электронным микроскопом, хлоропласты красных водорослей почти не отличаются от хлоропластов других водорослей. Пигменты. Б агрянки отличаются сложным набором пигментов. Кроме обычных для зеле- ных растений, растворимых в спирту хлорофил- ла, каротина и ксантофилла, хлоропласты крас- ных водорослей содержат дополнительные вод- но-растворимые пигменты билипротеины. Это фикоэритрин и фикоцианин. Зеленые растения, как водоросли, так и наземные, содержат две модификации хлорофилла — сине-зеленый хло- рофилл а и желто-зеленый хлорофилл Ь. У крас- ных водорослей найден только хлорофилл а— универсальный пигмент, характерный для всех растений. Кроме того, у некоторых багрянок обнаружен хлорофилл d, природа которого,
однако, остается до сих пор не выясненной. Зеленых пигментов у багрянок по сравнению с высшими растениями содержится немного, и обычно их маскируют дополнительные били- протеины. Замечена некоторая закономерность в изменении количества хлорофилла в зависи- мости от количества света. Водоросли, приспо- собившиеся к жизни при малой освещенности в полярных морях, обычно богаче хлорофил- лом, чем водоросли южных морей. Точно так же глубоководные водоросли богаче хлорофиллом, чем растущие у поверхности воды. Каротиноиды красных водорослей представ- лены а- и р-каротином и ксантофиллами лютеи- ном, зеаксантином и, вероятно, тараксанти- ном. Билипротеинами багрянок являются крас- ный фикоэритрин и голубой фикоцианин. Они близки к пигментам сине-зеленых водорослей, но не идентичны им, так как отличаются по химическому составу. Как показано на много- численных опытах, количество пигментов у баг- рянок возрастает с глубиной; при этом количе- ство фикоэритрина возрастает в большей мере, чем количество хлорофилла. Каждый, кто со- бирал эти водоросли в природе, знает, что ок- рашенные в красный цвет багрянки растут на глубине и что на мелководье они меняют ок- раску. С увеличением количества света они становятся бледно-красными, затем желто-зе- леными, соломенными и наконец полностью обесцвечиваются. Существует теория так называемой хромати- ческой адаптации, по которой проникновение водорослей на те или иные глубины связано с качеством света, проходящего через толщу воды. Как известно, глубже всего проникают лучи из зеленой и синей частей спектра. Крас- ные пигменты багрянок позволяют им фотосин- тезировать в синих лучах, и поэтому, согласно этой теории, они проникают на глубины, не- доступные для других водорослей. Однако на практике эта закономерность наблюдается дале- ко не всегда. Какова же роль билипротеинов в фотосинтезе красных водорослей? В опытах было установлено, что при слабом освещении они участвуют в усиленном поглощении света. Поэтому их можно считать оптическими сенси- билизаторами. Таким образом, проникновение красных водорослей на значительные глубины правильнее объяснить их способностью ус- ваивать малые количества света. В целом баг- рянки — теневыносливые организмы: слабый свет они способны использовать лучше, чем другие водоросли. Если красная окраска водо- рослей при слабом свете получает преимущест- ва, то при более сильном, наоборот, интенсив- ность фотосинтеза багрянок ниже, чем у дру- гих водорослей, как раз благодаря наличию красных пигментов. Для защиты от сильного света у багрянок, живущих на небольших глу- бинах, особенно в тропических и субтропиче- ских морях, служат особые и р ид и р ую щи е т е л ь ц а. Эти мутно-желтые неправильной фор- мы тельца образуются в вакуолях поверхност- ных клеток слоевища и состоят из мелких зер- нышек протеинов ой природы. Они обладают способностью рассеивать и отражать падающие на них солнечные лучи. При очень силь- ном освещении иридирующие тельца рас- полагаются под внешней стенкой клетки, в то время как хлор опласт — на внутренней или боковой, и служат своеобразным занавесом для хлоропласта. При попадании растения в ус- ловия рассеянного света происходит взаимное перемещение и хлоропласт оказывается у внеш- ней стенки. Водоросли, обладающие иридирующими тельцами, имеют обычно в падающем свете го- лубовато-стальной блеск. У некоторых видов в клетках возникают крупные линзообразные тела, которые с понижением освещенности исчезают. Запасные вещества. В качестве продукта ас- симиляции у красных водорослей откладывает- ся полиса харид, называемый багрянковым крахмалом. По химической природе он ближе всего к амилопектину и гликогену и, по-ви- димому, занимает промежуточное положение между обычным крахмалом и гликогеном. От- кладывается багрянковый крахмал в виде мел- ких полутвердых телец различной формы и окраски. Эти тельца могут иметь форму кону- сов или плоских овальных пластинок с углуб- лением на широкой поверхности. Часто на них можно видеть концентрические зоны. Зерна багрянкового крахмала образуются частично в цитоплазме, частично на поверхности хлоро- пластов, но они никогда не образуются внутри пластид, в отличие от обычного крахмала зе- леных растений. У форм, имеющих пиреноид, последний в какой-то мере участвует в синтезе крахмала. Кроме багрянкового крахмала, в качестве запасных веществ у красных водорослей откла- дываются сахара трегалоза, флоридозид, са- хароза и др. У некоторых форм в изобилии встречаются многоатомные спирты. Из жиров известны холестерол, силостерол, фукостерол. Содержание жиров меняется в зависимости от условий среды. Железистые клетки. Особенностью красных водорослей является наличие у некоторых представителей класса флоридеевых особых кле- ток с бесцветным содержимым, сильно прелом- ляющим свет (рис. 155). В литературе они из- вестны как пузырчатые, или железистые, клет- ки. Содержимое этих клеток у разных водорос- лей имеет разную природу; они заполнены ио- 200
дистыми, реже бромистыми, соединениями. Ча- о ще всего железистые клетки встречаются в по- рядке церамиевых. У нитчатой разветвленной Q U водоросли антиталнион они сидят на верхней стороне боковых ветвей (рис. 155, 1). При их 1 развитии сначала отчленяется маленькая линзо- ',::;11~~', образная клетка, содержащая небольшое коли- чество плазмы и мелкие красные хлоропласты. Ядро можно проследить лишь на самой ран- ней стадии развития. Вскоре на дне этой клет- о ки образуется бесцветный светопреломляющий о пузырь. Он растет, и вместе с ним увеличивает- 2 ся в размерах вся клетка. В сформировавшейся Q клетке ббльшая часть занята пузырем, и только в верхней части остается узкий слой плазмы с мелкими хлоропластами (рис. 155, 2 — о). Роль пузырчатых клеток не выяснена хотя на этот 7 3 СФ 1ф счет имеется множество самых различных пред- „, у~ тт д~ положении. у~х считают недоразвитыми спо- рангиями, хранилищем запасных веществ, «пла- вательными пузырями» и т. п. а Желееистые клетки характеркы Лли ллреке- ленных групп водорослей и поэтому служат важным таксономическим признаком. Волоски. Образование волосков — широко распространенное явление в классе флоридее- вых. Настоящие волоски багрянок следует отличать от волосовидных ветвей или ложных волосков. У ви ов с о но слоеви ем можно ви тв д д рядным нитчатым Многоклеточные волоски б б о ычно олее или щ деть, как конечные клет- менее сильно разветв О ки боковых вет р твлены. ни встречаются точек удлиняются и обесцве- только у некоторы \J чиваются, приобретая волосовидное строение ами х водорослеи из по я ка е- д ( ис. 15i 1). Это рамиевых. ак ыло показано в опытах, основ- ~рис., ~. то и есть ложные волоски. На- ная роль волосков закл ков заключается в том, что они и красных водорослеи делятся способствуют поглощению питательных ве- на два типа: одноклеточные и многоклеточные. ществ из окр из окружающеи среды. \J Одноклеточные волоски никогда не ветвятся. П О Уоно ян~ оры. дна из наиболее интересных особен- однорядных нитчатых форм они образуются костей красных во p p красных водорослеи состоит в том, что х шечных клеток ветвей много я- у р д- клетки, слагающие слоевище, соединяются б его воло ных — из поверхностных клеток коры. Клетка между с б ду со ои с помощью специальных образо- удущего волоска отделяется от материнской ваний называ \J , называемых порами. ежду доче ними . М клетки поперечнои перегородкой и сильно вы- к р клетками, т. е. клетками, происходя~ими от тягивается, достигая в длину не е ко милли- метра и более рис. 156 . Она со е жит я р д — однои материнской, соединение осуществляет- (P . ). д ржит ядро и ся посредством первичных пор (рис. 157, 1 — 8). не ольшое количество цитоплазмы. Хлоропласт Они ни формируются в результате неполной пе- по мере роста волоска исчезает, и волоски стано- регородки между д бп вятся бесцветными. Обычно материнская клетка с . П жд двумя вновь о паз ю ими- р у щ летка ся клетками. ервичные поры расположены волоска ничем не отличается отсоседнихвегета- в се едине пе его о к тивных клеток но н л ток, но иногда она намного крупнее рую можно провести линию, соединяю ю и остается хорошо заметной после отпадения во- я ти линию, соединяющую лоска. У ко алл о- ядра дочерних клеток, и представляют собой лоска. кораллиновых волоски не являются тонкую пластинку Ч самостоятельными клетками, а п е ставляют тинк . ерез эту пластинк п о- тинк у р- собо р д вляют ходят тяжи, которые соединяют цитоплаз- со ои только выросты специальных клеток, от му соседних клеток. 1о сам кото ых они не от еля \J клеток. ~о самого последнего клетки намного к пнее ост д яются перегородкои. Эти времени считалось что пе в рвичные поры свои- \J по назв г крупнее остальных и известны ственны только ~ло" и"еев тм ~~л ридеевым и отсутствуют д азванием трихоцитов или гетероцист в классе банг ангиевых — это был один из прин- и,, сли в классе флоридеевых ципиальных признаков, на основании которого о а эти класса. Но недавно первич- одноклеточные волоски встречаются довольно различали б ыли открыты и у представителей часто, то у ангиевых они отсутствуют пол- ные поры был класса бангиевых. Рис. 155. Железистые клетки красных водорослей: 1 — веточка АпШЬапппоп с железистыми клетками; 2 — 5— стадии развития железистой клетки Antithamnion; 6 — попереч- ный срез Opuntiella с железистыми клетками.
Рис. i56. Стадии развития волоска у Cystoclonium. Рис. i57. Схема образования первичных (Z — g~ и в ричных (4 — 8) пор у красных водорослей. Кроме соединения дочерних клеток, у боль- шинства флор идеевых происходит вторичное соединение соседних клеток. Это могут быть клетки одной и той же нити, лежащие рядом клетки соседних нитей или — у паразитиче- ских багрянок — клетки паразита и хозяина. Способностью к вторичному соединению обла- дают более высокоорганизованные представи- тели класса. Она отсутствует в порядке немали- евых и у примитивных водорослей других порядков. Соединение соседних клеток, между которыми первоначально никакой связи не было, осуществляется вторичными порами (рис. 157, 4 — 8). Образование вторичных пор- процесс довольно сложный. Начинается он с того, что ядро в клетке делится и одно из до- черних ядер перемещается к периферии клетки. В том месте, куда попадает ядро, образуется небольшой вырост, который затем отчленяется, как маленькая клетка, соединенная с первой посредством первичной поры. Вытягиваясь, эта дополнительная клетка достигает соседней и сливается с ней. Теперь первичная пора, соединяющая дополнительную клетку с первой, становится вторичной. Иногда две соседние клетки соединены несколькими вторичными порами. Роль этих соединений до конца не вы- яснена, вероятно, они способствуют перено- су питательных веществ от одной клетки к другой. Хотя способ образования первичных и вторичных пор различен, строение их сход- но. И все же не у всех багрянок поры устрое- ны одинаково. В одних случаях между клет- ками имеется непосредственная цитоплазмати- ческая связь, в других — она гораздо менее очевидна. РАЗМНОЖЕНИЕ КРАСНЫХ ВОДОРОСЛЕЙ Размножение красных водорослей чрезвы- чайно сложный и многообразный процесс, от- личающий их от других групп водорослей. Различные формы бесполого размножения, сложное строение половой системы, особенно женского репродуктивного органа, особенно- сти развития зиготы, а также многообразие циклов развития — все эти признаки не только придают своеобразие отделу Rhodophyta, но являются основой, на которой строится клас- сификация багрянок, их разделение на поряд- ки, семейства, роды. Поэтому, чтобы составить целостное представление о красных водорослях, нельзя не остановиться на особенностях их размножения. Собственно говоря, сложным размножением характеризуются представители класса фло- ридеевых; у бангиевых строение репродуктив- ной системы намного проще. Вегетативное размножение флоридеевых про- исходит посредством образования дополнитель- ных побегов, которые берут начало от подош- вы или от стелющихся ветвей. Вертикальное слоевище в определенный период отмирает и остается только базальная часть растения, которая через какое-то время прорастает, об- разуя новые побеги. Самые примитивные баг-
рянки из класса бангиевых размножаются толь- ко вегетативно. У одноклеточных и колониаль- ных форм вегетативное размножение состоит в делении клетки на две или более дочерние клетки. Бесполое размножение. Для бесполого раз- множения служат различного рода споры. У низ- коорганизованных багрянок бесполое размно- жение осуществляется моноспорами. Они ха- рактерны для всех представителей класса бан- гиевых, тогда как среди флоридеевых известны за очень редкими исключениями только в по- рядке немалиевых. Моноспорами называют спо- ры, которые образуются по одной в клетке в результате превращения всего протопласта. Моноспоры не имеют жгутика и оболочки, по- сле выхода из материнской клетки они способ- ны к амебоидному движению. У бангиевых мо- носпоры образуются в любой клетке слоевища и до своего выхода не отличаются от вегетатив- ных клеток. В классе флоридеевых (там, где они встречаются) моноспорангии образуются на веточках ограниченного роста. У водорос- лей, построенных по типу свободных нитей, в моноспорангии превращаются конечные клет- ки коротких боковых веточек. Иногда они об- разуются группами по два-три на одной и той же клетке-ножке (рис. 151, 1). У водоро- слей, имеющих более или менее плотное строе- ние и дифференцированный коровой слой, в мо- носпорангии превращаются внешние клетки ассимиляционных нитей. Моноспоры отличают- ся от вегетативных клеток яйцевидной или шаровидной формой, богатым содержимым и интенсивной окраской. Созревшие моноспоран- гии открываются на вершине; голые моноспоры выходят в окружающую среду, через некото- рое время одеваются оболочкой и начинают прорастать. Для большинства флоридеевых наиболее обычны тетраспоры. Они образуются по четы- ре в клетке, которую называют тетраспоран- гием. По способу деления различают три типа тетраспорангиев: крестообразно, зонально и тетраэдрически разделенные. В крестообразно разделенных тетраспорангиях вначале вся клетка делится пополам поперечной перего- родкой, после чего каждая из двух образовав- шихся клеток делится продольной перегород- кой, при этом обе перегородки проходят в одной плоскости или под прямым углом друг к другу (рис. 158, 5', 7). В последнем случае, если смот- реть на спорангий сбоку, можно различить только три клетки. При зональном делении все три перегородки проходят поперечно и парал- лельно друг другу, в результате чего тет- распоры располагаются в ряд (рис. 158, 4, 6). В тетраэдрически разделенных тетраспорангиях клеточные перегородки ориентированы не под прямым углом, как при крестообразном делении, а косо (рис. 158, 1). Способ деления тетраспорангиев — постоян- ный признак, который закрепился в эволю- ции за той или иной группой. В порядках не- малиевых и гелидиевых имеются только крес- тообразно разделенные спор ангии, у крипто- немиевых и гигартиновых — крестообразно и зонально разделенные. В порядке родимение- вых встречаются как крестообразно, так и тетраэдрически разделенные спорангии. У боль- шинства церамиевых спорангии делятся тетра- эдрически. Как исключение, спорангии делят- ся только на две части, и тогда мы говорим об образовании биспор. Наиболее характерно об- разование биспор для кораллиновых. Место образования тетраспор у разных групп багрянок также постоянный признак, который используется в систематике. Как правило, тетраспоры занимают положение конечных кле- ток веточек ограниченного роста, будь то сво- бодные боковые веточки у просто организован- ных немалиевых или коровые нити у осталь- ных багрянок. Интер калярное возникновение тетраспорангиев, т. е. в середине клеточных нитей, встречается исключительно редко. Од- нако в процессе роста тетраспорангиев — а они обычно бывают намного крупнее соседних вегетативных клеток — они оказываются по- груженными в коровой слой, и их положение конечной клетки в зрелом состоянии не всегда очевидно (рис. 158, 7). Располагаются тетра- спорангии поодиночке или группами. Собрания тетраспорангиев часто имеют вид цепочек. Интеркалярные тетраспорангии образуются из клеток внутренней части коры, как, например, у дилсеи, или в середине вертикальных асси- миляционных нитей у корковых форм. У кор- ковой водоросли круории тетраспорангии пред- ставляют собой боковые выросты вертикальных нитей (рис. 158, 4). Из клеток центральной части слоевища формируются тетраспорангии у таких гигартиновых водорослей, как хондрус и иридея (рис. 158, 2). На слоевище тетраспорангии располагаются по-разному. Очень часто они рассеяны по всему слоевищу или приурочены к какой-то определенной его части, например к верхним ветвям. Но нередки случаи, когда образование тетраспорангиев связано со спе- циальными веточками или выростами. Эти вы- росты располагаются по краю слоевища или на его поверхности у пластинчатых форм и вокруг основных ветвей — у цилиндрических (рис. 158, 8). У некоторых видов тетраспорангии разви- ваются в так называемых н е м а т е ц и я х. 3то специальные образования на поверхности сло- евища. Они состоят из вертикальных клеточ- ных нитей, берущих начало от поверхностных
Рис. 158. Тетраспорангии красных водорослей: g — Callithamnion; 2 — Iridaea; 8 — веточка Gelidium с тетра- спорангиями; 4 — Cruoria; б — Peyssonnelia; 6 — концептакул с тетраспорангиями Corallina; 7 — Gracilaria. коровых клеток, и имеют вид небольших по- душечек. Тетраспорангии развиваются среди нитей нематеция, тесно окруженные ими. Они отчленяются от поверхностных клеток корового слоя, как, например, у сквамариевых (порядок криптонемиевых, рис. 158, о) или от самых нижних веток нитей нематеция, как, например, у эндокладии (порядок криптоне- миевых). У филлофоры и некоторых других гигартиновых все клетки нитей нематеция, за исключением самых поверхностных, превра- щаются в тетраспорангии. Нематеции этих водорослей имеют шаровидную форму, их дол- гое время считали паразитами и относили к дру- гому роду (рис. 177, 1). У кораллиновых тет- распорангии образуются в специальных вме- стилищах — концептакулах (рис. 158, 6). У некоторых церамиевых и других багрянок известны случаи деления одного спорангия на много частей с образованием полиспор. Иног- да встречается до 28 полиспор в одном споран- гии. Они соответствуют тетраспорам и ничем, кроме количества, от них не отличаются. Все бесполые споры багрянок лишены обо- лочек, у некоторых замечена способность к аме- боидному движению. После выхода из споран- гия они через некоторое время одеваются обо- лочкой, оседают и прорастают, как правило, в гаметофиты, которые несут органы полового размножения. Половое размножение присуще всем водо- рослям из класса флоридеевых, тогда как в клас- се бангиевых оно свойственно только наиболее высокоорганизованным формам. У одноклеточ- ных и колониальных бангиевых способность к половому размножению еще не выработалась. Для некоторых флоридеевых известно только бесполое размножение, но отсутствие полового размножения у них всегда результат вторич- ной утраты. Половой процесс у всех красных водорослей оогамный. Мужские и женские гаметы лишены жгутиков. При оплодотворении женские га- меты не выходят в окружающую среду, а ос- таются на растении — гаметофите; мужские гаметы выбрасываются наружу и пассивно переносятся токами воды. В связи с этим жен- ский орган размножения отличается весьма сложным строением. Мужские гаметы, называемые у багрянок с п е р м а ц и я м и, представляют собой шаровид- ные или яйцевидные одноядерные клетки. Спер- мации бангиевых — голые клетки. Что касается флоридеевых, то здесь вопрос о наличии или отсутствии оболочки у спермациев до конца не выяснен. Известно только, что в момент оседа,ния на женское растение они уже окруже- ны тонкой оболочкой. Во всяком случае, спо- собность спермациев к амебоидному движению
позволяет думать, что если не у всех, то у части флоридеевых спермации на первых порах ли- шены оболочек. Спермации флор идеевых, как правило, не имеют хлоропластов и бесцветны. У низших представителей класса остатки хлор опласта в мужских гаметах еще присутствуют, но они уже не играют никакой роли при оплодотво- рении. Постепенную редукцию хлор опласта в спермациях можно проследить уже в классе бангиевых. У простоорганизованных бангие- вых, обладающих половым процессом, в спер- мациях развивается отчетливо видимый хло- ропласт, но у высших представителей класса, например у порфиры, он уже сильно редуциро- ван. Спермации образуются в клетках, которые функционируют как мужские органы размно- жения, называют их сперматангиями. В каж- дом сперматангии образуется по одному спер- мацию. У бангиевых сперматангии возникают в результате повторных делений многочислен- ных клеток нитчатого или пластинчатого слое- вища. Вегетативную клетку, дающую начало сперматангиям, называют материнской. У бан- гиевых любая клетка слоевища, за исключением ризоидных, может стать материнской клеткой сперматангиев, хотя в природе плодоношение обычно охватывает только верхнюю часть рас- тения. Из одной материнской клетки образует- ся 32 — 128 мелких клеток, каждая из которых дает один спермаций. У всех флоридеевых ма- теринские клетки сперматангиев представляют собой конечные клетки веточек ограниченного роста. В соответствии с этим у флоридеевых, об- ладающих свободным нитчатым строением, они сидят на вершине боковых веточек (рис. 159, 1), у багрянок с плотным строением в сперма- тангии превращаются внешние клетки корово- го слоя. Каждая материнская клетка отчленяет от себя один или несколько спер матангиев (рис. 159, 2, 8). После созревания и отпадения первых сперматангиев материнская клетка может отчленить еще несколько вторичных спер- матангиев. По мере усложнения организации флоридеевых материнские клетки сперматан- гиев также постепенно видоизменяются. Их уже можно отличить от соседних вегетативных клеток по размерам и форме, а также по от- сутствию окраски, так как в них происходит редукция хлоропласта, но зато увеличивается количество цитоплазмы. Только у небольшого числа красных водо- рослей сперматангии располагаются по слое- вищу рассеянно. У большинства же они соб- раны более или менее обширными группами. Собрания сперматангиев называют с о р у с а- м и. У нитевидных форм сорусы имеют вид гус- тых пучков, нередко шаровидной формы, распо- ложенных у вершины боковых ветвей. У водо- рослей с более плотным строением сорусы рас- полагаются на поверхности слоевища в виде подушковидных образований. Иногда они ока- зываются погруженными в небольшие углуб- ления на слоевище, а у пропитанных известью кораллиновых приурочены к специальным вместилищам — концептакулам (рис. 159, б). Нередко спермации образуются в специальных нематециях. В этом случае наружные коровые клетки дают начало однорядным параллель- ным нитям, которые выступают над поверх- ностью слоевища. Затем клетки этих нитей образуют по нескольку перицентральных кле- ток, которые служат материнскими клетками и отчленяют каждая по нескольку сперматан- гиев (рис. 159, 4). У высокоорганизованных це- рамиевых сперматангии образуются на спе- циальных коротких веточках — с т и х и д и я х или на трихобластах, окружая эти веточки на- подобие муфты (рис. 159, 6). Женский репродуктивный орган у красных водорослей — оогоний называют к а р п о г о- н о м. Это особая клетка, густо заполненная ци- топлазмой и лишенная хлоропласта. Лишь у при- митивных представителей флоридеевых и у бан- гиевых карпогон содержит окрашенный хлоро- пласт. Карпогон отличается весьма характер- ной, одинаковой для всех флоридеевых формой (рис. 160, 1). Нижняя, или брюшная, часть клетки имеет конусовидную форму, верхняя вытягивается, образуя более или менее длин- ный трубчатый вырост. Вырост служит для улавливания спермациев, называют его т р и- х о г и н о й. Благодаря трихогине карпогон легко отличить о~ остальных клеток слоевища. Ядро карпогона, как правило, находится в брюшной части. Трихогина может быть корот- кой и кеглевидной или длинной, узкой и спи- рально закрученной. У бангиевых собственно трихогина еще отсутствует, а иногда карпого- ны имеют лишь короткие сосочкообразные вы- росты — прообраз будущей трихогины. Поэто- му кар погон бангиевых до оплодотворения трудно отличить от обычных вегетативных клеток, тем более что им может стать любая клетка слоевища, точно так же как это проис- ходит при образовании спермациев и моноспор (рис. 161, 1). Далеко не у всех флоридеевых карпогон раз- вивается как отдельная клетка. Гораздо чаще образуется короткая клеточная нить, состоя- щая из бесцветных клеток, заполненных, как и карпогон, гомогенной цитоплазмой, а уже соб- ственно карпогон развивается как конечная клетка этой нити, или к а р II о г о н н о й в е т в и (рис. 160, 1). Клетку, от которой отходит кар- погонная ветвь, называют несущей или базаль- ной; в состав карпогонной ветви она не входит. 205
EJ рис f59. Сперматаигии (а) <р~~~~х во"оро со спе матангиями Fosliella; 6 — Polysiphonia. G Гd' 4 — Peyssonnelia; 5 — концептакул со сперматангиями Foslie а; 6— 1 — Nemalion; 2, 3 — Ge i ium; Карпогонные ветви флоридеевых играют важ- нимают активное участие в развитии карпогона после оплодотворения. Число клеток в карпо- гонной ветви всегда небольшое, е но непостоян- ное у разных групп багрянок. Если проследить эти изменения от наиболее примитивных к вы- сокоорганизованным представителям класса флоридеевых, то можно заметить, что у низко- организованных немалиевых и гелидиевых кар- погонная ветвь еще отсутствует и имеется толь- ко одна клетка кар а карпогона. У криптонемиевых карпогонная ветвь, будучи еще структу- рой малоспециализированной, характеризуется, с одной стороны, сравнительной многоклеточ- ностью, а с д , а с другой — непостоянным числом клеток. При этом у менее специализированных форм из порядков криптонемиевых и гигарти- новых встречаются даже разветвленные кар- погонные ветви. В порядках родимениевых и церамиевых число клеток в карпогонной ветви меньшается до 3 — 4 и становится уже более постоянным даже для крупных таксонов. Почти у всех флоридеевых карпогонная ветвь об азуется как специальная боковая веточка о разу во внутренней части корового слоя. о низкоорганизованных немалиевых она соот- ветствует обычной боковой веточке слоевища. У багряно к с плотным слоевищем карпогон оказывается погруженным, и тол только т, ихо- гина достигает поверхности слоевища и вы- ступает над ней. У багрянок со свободно нит- чатым строени нием карпогон занимает открытое положение. Строение карпогонной ветви и характер ее образования сильно варьируют азных групп водорослей и служат одним из главных отличительных призна ков в сис- тематике багрянок. Когда принесенные водой спермации сопри- касаются с в вершиной трихогины, оболочки их в месте соприкосновения растворяются и я о спермация перетекает в полость трихоги- ны. Передвигаясь по неи, оно попадае р ную часть сть ка,,погона и там сливается с его После оплодотворения карпогон от- ядром. о еляется от трихогины специальнои про и вскоре трихогина отмирает. Оплодотворен- ная яйцеклетка остается заключенной в обо- лочку карпого огона и собственной оболочки не об аз ет. Этот факт свидетельствует о вероят- ном отсутствии в эволюции багряно д к по виж- ных женских половых клеток. Зигота, не проходя периода покоя, вскоре начинает прорастать. Обычно у водорослей азвивается непосредственно в новые рас- тения, принадлежащие уже к бесполому поко- лению, т. е. , т. е. в спорофиты. У красных водоро- слеи, в отличие от всех остальных, зигота, прежде чем дать начало спорофиту, претерпе- образуются особые споры, дающие начало спо- рофиту. Называют эти споры к арпоспора- м и. ни пред . О п е ставляют собой голые, лишенные жгутиков клетки, неподвижные или изредка
Рис. 160. Карпогонная ветвь (1) и развитие гонимо- бласта (2, 8) у Helminthora: а — карпогон; б — трихогина; в — карпогонная ветвь; г- нити гонимобласта; д — карпоспоры. Рис. 161. Поперечный срез пластины Porphyra: 1 — с карпогонами; 2 — с сперматангиями; 8 — с карпоспорами. обнаруживающие амебоидные движения. Перед началом прорастания они одеваются оболоч- кой. Клетки, в которых образуются карпоспо- ры, называют к а р п о с п о р а н г и я и и. В каж- дом карпоспорангии формируется по одной кар- поспоре. У бангиевых процесс превращения зиготы в карпоспоры весьма прост: после оплодотво- рения карпогон делится с образованием 4— 32 карпоспор (рис. 161, 8). У флоридеевых развитие зиготы — сложный и многообразный процесс, в котором, кроме карпогона, участвуют многие специальные клетки. Зтот процесс протекает неодинаково у разных багрянок, в каждой группе со свои- ми особенностями и многочисленными деталя- ми. Поэтому здесь целесообразно остановиться только на основных чертах развития зиготы. После оплодотворения карпогона развивают- ся специальные нити, которые несут карпо- спорангии. Зти нити состоят из небольшого числа клеток, называют их нитями г о н и м о- 6 л а с т а. В наиболее простом случае — у водо- рослей из порядка немалиевых и гелидиевых- оплодотворенный к ар погон отчленяет от себя несколько боковых клеток, которые, делясь, дают пучок нитей гонимобласта, конечные клет- ки этих нитей превращаются в карпоспоран- гии. Иными словами, применительно к этим водорослям можно говорить о непосредствен- ном развитии нитей гонимобласта из карпого- на (рис. 160, 2, 8). У подавляющего большинства флоридеевых оплодотворенный карпогон вначале сливается с особыми клетками, богатыми питательными веществами, и только после этого развивается гонимобласт. Иногда после оплодотворения кар- погон делится, и уже отчленившаяся от него клетка участвует в процессе слияния. Слияние может быть полным или частичным. Клетки, с которыми сливается карпогон, в литературе часто называют а ук силл яр ными. Но это не совсем верно. По мере того как изучались процессы развития зиготы, было установлено, что клетки, с которыми сливается карпо гон, выполняют разную роль и занимают разное положение. Среди них следует различать два типа клеток: просто питающие клетки, кото- рые образуются до или после оплодотворения и служат только для питания развивающегося гонимобласта, и настоящие ауксиллярные клет- ки. Последние также функционируют как питающие клетки, но их главная и чрезвычай- но важная роль состоит в том, что они пред- ставляют собой исходную клетку, от которой начинается развитие гонимобласта. Иными сло- вами, у тех багрянок, которые имеют ауксил- лярные клетки, гонимобласт развивается не из карпогона, а из ауксиллярной клетки. Зтому процессу обязательно предшествует слияние к ар погона с ауксилляр ной клеткой. Слияние осуществляется при помощи специаль- ных нитей или выростов, которые отходят от карпогона и тянутся, пока не соединятся
с ауксиллярной клеткой. Эти нити называют со- единительными. Так как далеко не всегда аук- силлярные клетки располагаются вблизи кар погона, соединительные нити могут до- стигать значительной длины и состоять из большого числа клеток (рис. 176, 4). По этим нитям ядро оплодотворенного карпогона пере- двигается и переходит в ауксиллярную клет- ку; слияния ядер при этом не происходит. В этом состоит основное назначение соедини- тельных нитей. Ауксиллярные клетки отсутствуют в поряд- ках нем алиевых и гелидиевых, но имеются в остальных порядках класса флоридеевых. Ауксиллярные клетки характеризуются бога- тым содержимым и крупными размерами. Мес- то и время их возникновения различны у раз- ных багрянок. Они могут развиваться на нитях корового слоя и от внутренних клеток слое- вища, вблизи или в удалении от карпогона, до оплодотворения или после него. Все эти особенности имеют большое значе- ние для систематики флоридеевых. Модно предположить, что развитие системы ауксилляр- ных клеток и карпогонной ветви шло неза- висимо в каждом из порядков, но везде связь ауксиллярной системы с карпогоном станови- лась все более тесной. У багрянок, уже обла- дающих ауксиллярными клетками, но стоящих еще на низкой ступени организации, они раз- виваются независимо от кар погонной ветви и бывают удалены от нее на значительное рас- стояние; образование их не приурочено к про- цессу оплодотворения: они возникают до него. У этих водорослей соединительные нити раз- виты очень сильно. По мере дальнейшего раз- вития ауксиллярные клетки начинают возни- кать в непосредственной близости от карпогон- ной ветви и образуют с ней уже единую струк- туру. Соединительные нити заметно укорачи- ваются, редуцируются и в конечном итоге пред- ставляют собой лишь короткий неклеточный отросток карпогона. В тех случаях, когда ауксиллярные клетки развиваются вместе с карпогонной ветвью, образуя единую систе- му нитей, и оплодотворенный карпогон сли- вается со своей ауксиллярной клеткой, мы гово- рим о п р о к а р п е. На ранних этапах разви- тия прокарпа карпогон необязательно сливает- ся с рядом расположенной ауксиллярной клет- кой; соединительные нити могут простираться дальше и соединяться с ауксиллярными клет- ками соседних прокарпов. На следующей ступени ауксиллярные клет- ки развиваются от той же клетки, что и карпогонные ветви, и только после оплодотво- рения карпогона. Таким образом, можно выделить четыре типа настоящих ауксиллярных клеток: 1. Ауксиллярные клетки развиваются перед оплодотворением как отдельные клетки на специальных клеточных нитях — это тип крип- тонемиевых водорослей. 2. Ауксиллярными клетками служат обычные интер калярные клетки слоевища, как прави- ло, клетки внутренней коры, и образуются они перед оплодотворением, это тип гигарти- новых. 3. Ауксиллярные клетки отчленяются перед оплодотворением от дочерней клетки несу- щей клетки к ар погонной ветви; этот тип характерен для родимениевых. 4. Ауксиллярные клетки отчленяются от несущей клетки кар погонной ветви только после оплодотворения. Просто питающие клетки поставляют пита- тельные вещества из материнского растения благодаря полному или частичному слиянию с соединительными нитями или нитями разви- вающегося гонимоб ласта. Когда питающие клетки собраны в большом количестве и их собрания хорошо очерчены, можно говорить о питающей ткани (рис. 167, олс). Питающие клет- ки встречаются у всех флоридеевых, тогда как типичные ауксиллярные клетки характерны только для четырех порядков из шести. Итак, у наиболее примитивных представи- телей класса флоридеевых развитие гонимоб- ласта связано только с карпогонной ветвью. Затем возникают специальные питающие клет- ки, которые частично сливаются с развиваю- щимися нитями гонимобласта и обеспечивают их питательными веществами. Позднее обра- зуется ауксиллярная система, которая сущест- вует наряду с питающими клетками. Постепен- но роль проводника питательных веществ пол- ностью берет на себя ауксиллярная клетка и просто питающая ткань редуцируется. Аук- силлярная система также становится менее громоздкой и более тесно связанной по месту и времени возникновения с оплодотворенным ка рпогоном. Развитие гонимобласта заключается в обра- зовании системы клеточных нитей. Созревая, клетки нитей превращаются в карпоспоры. Различаются багрянки, у которых в карпо- споры превращаются конечные клетки нитей гонимобласта, и багрянки, у которых карпо- споры располагаются цепочкой, так как они образованы рядом клеток. У некоторых фло- ридеевых все клетки гонимобласта преобра- зуются в карпоспоры. Зрелый гонимобласт называют ц и с т о к а р п о м. Строго говоря, под цистокарпом следует понимать не только сам гонимобласт, но и окружающую его вегетатив- ную ткань, когда она имеет определенное мор- фологическое и анатомическое строение и образует обертку гонимобласта. Обертка соз- 208
дается клетками коровых нитей или клетками, расположенными рядом с карпогонными вет- вями. Место возникновения цистокарпов, их форма, характер оболочки, способ выхода карпоспор- все эти признаки имеют важное значение для разграничения родов. Обычно цистокарпы рас- полагаются в молодых частях растений — на ветвях последних порядков или участках близ вершины слоевища. У форм со свободно нит- чатым строением они располагаются на поверх- ности ветвей в виде шаровидных скоплений клеток. Формы с плотным строением характери- зуются погруженными или полупогруженными цистокарпами. Они имеют бугорчатую, шаро- видную или кувшинообразную форму, и вы- ступая над поверхностью слоевища, часто хо- рошо заметны даже невооруженным глазом. Для выхода карпоспор в обертке цистокарпа образуются специальные отверстия — одна или несколько пор. Однако пор может и не быть, тогда карпоспоры выходят после разрушения внешней оболочки цистокарпа. Циклы развития. Как мы видели, красные водоросли размножаются половым и бесполым путем. Оба способа взаимосвязаны и обычно сменяют друг друга в ходе жизненного цикла багрянок. В подавляющем большинстве случа- ев органы полового и бесполого размножения расположены на разных растениях. Споры, служащие для бесполого размножения (моно- споры, тетраспоры, биспоры и т. п.), образуют- ся на растениях-спор офитах, оогонии и сперматангии — на гаметофитах; при этом у большинства багрянок существуют мужские и женские гаметофиты. Иными словами, поло- вая и бесполая формы развития представлены обычно самостоятельными сноб одноживущими поколениями. Однако по мере изучения циклов развития водорослей различных видов все чаще отмечаются случаи, когда одна форма разви- тия возникает на другой, и тогда на одном и том же индивидууме можно найти и тетраспоры, и органы полового размножения. Механизмы этого явления пока остаются невыясненными. В литературе о циклах развития водорослей, особенно в иностранной, гаметофит и спорофит часто называют поколениями. Это было бы верно, если бы половое поколение обязатель- но сменялось бесполым. Но такая строгая смена происходит далеко не всегда. Сплошь и рядом у разных водорослей и гаметофит, и спорофит могут воспроизводить себя в течение нескольких поколений. Поэтому, касаясь вопроса о циклах развития красных водорос- лей, мы будем говорить о смене форм разви- тия, а не о смене поколений. Вопрос о циклах развития красных водорос- лей — один из самых интересных и трудных в альгологии. Ученых интересует не только характер циклов развития сам по себе, его осо- бенности у разных видов, но, главное, происхо- ждение и эволюция этого явления у водорослей отдела Rhodophyta. Говоря о цикле развития, обычно имеют в виду два аспекта этого явле- ния — морфологическую смену форм развития и цитологическую. Особенности цитологиче- ского цикла определяются местом редукционно- го деления и соотношением диплоидной и гап- лоидной фаз. Часто у водорослей морфологи- ческие изменения происходят независимо от цитологических, но у красных водорослей эти явления, по-видимому, взаимосвязаны. У багряно к существует несколько типов циклов развития. В подавляющем большинст- ве случаев происходит смена трех форм разви- тия: тетраспорофита, гаметофита и карпоспо- рофита. При этом карпоспорофитом называют гонимобласт с карпоспорами, который принято считать самостоятельным поколением, ведущим паразитический образ жизни на женском га- метофите и потому морфологически редуциро- ванным. В этом цикле имеются две свободно- живущие формы развития — тетраспорофит и гаметофиты. Редукционное деление происходит при образовании тетраспор. Следовательно, тетраспоры и возникающие из них гаметофиты представляют собой гаплоидную фазу, тогда как зигота, карпоспорофит и тетраспорофит относятся к диплоидной фазе. Такой цикл развития представлен у багрянок двумя разновидностями. Гаметофиты и тетра- спорофит могут иметь совершенно одинаковое морфологическое строение. Этот тип был впер- вые открыт у полисифонии, поэтому его так называют — тип Polysiphonia. У других баг- рянок слоевище тетраспорофита редуцировано по сравнению с гаметофитом и сильно отличает- ся от него по строению. Часто в таких случаях спорофит представлен стелющимися или кор- ковидными растениями. Ярким примером по- добного цикла с так называемой гетероморфной сменой форм развития служит цикл развития порфиры. У этой водоросли пластинчатые крупные гаметофиты, т. е. то, что мы обычно называем порфирой, сменяются микроскопи- ческими стелющимися спорофитами, построен- ными из одно рядных нитей. Среди филлофоровых (порядок гигартиновых) наряду с видами, цикл развития которых про- ходит по типу полисифонии, есть виды с очень своеобразным укороченным циклом. На круп- ных свободноживущих растениях — гаметофи- тах после оплодотворения развиваются не- матеции с тетраспорами, а карпоспорофит от- сутствует. Этот цикл можно объяснить морфо- логической редукцией не только тетраспорофи- та, но и карпоспорофита. Последний редуци- 209
руется до зиготы, которая непосредственно превращается в карпоспору. Та прорастает как паразит тут же, не отделяясь от гамето- фита, в тетраспорофит, тоже морфологически редуцированный. Он напоминает нити немате- ция, собранные в виде подушек и шариков на поверхности слоевища. До тех пор пока не был изучен цикл развития этих водорослей, ре- дуцированные тетраспорофиты принимали за самостоятельные организмы. Редукция гаметофитов в цикле развития красных водорослей встречается гораздо реже. Обычно она выражается в недо развитии га- метофита и прорастании его на тетраспорофите. До последнего времени считали, что у крас- ных водорослей имеются два принципиально отличных типа циклов развития. Кроме уже описанного, в котором участвуют три формы развития, а редукционное деление происходит при образовании тетраспор, существует другой, при котором тетраспорофит отсутствует, а име- ется только одна свободноживущая форма развития — гаметофит с паразитирующим на нем карпоспорофитом. Полагали, что редукци- онное деление в этом цикле происходит сразу после оплодотворения, в самом начале про- растания зиготы, и, следовательно, весь цикл проходит в гаплоидной фазе. Однако недавние исследования показали, что единственное место, где происходит редукционное деление, — тет- распорангии, и поэтому в цикле развития всег- да представлены две цитологические фазы- гаплоидный гаметофит и диплоидные карпо- спорофит и тетраспорофит. Кроме того, оказа- лось, что у большинства видов, которые, как считалось, в природе были представлены толь- ко гаметофитом и карпоспорофитом, имеется и тетраспорофит. Эти тетраспорофиты — очень мелкие, сильно редуцированные растеньица с весьма отличающейся от гаметофита и карпо- спорофита структурой, поэтому исследователи либо не замечали их, либо считали другими организмами. Таким образом, в цикле развития всех фло- ридеевых происходит смена трех форм разви- тия, представленных как в гаплоидной. так и в диплоидной фазе. Но, как мы видели, не всег- да все эти формы развиты одинаково и явля- ются свободноживущими организмами. Какой же тип цикла развития следует считать наибо- лее примитивным и исходным для всего су- ществующего многообразия циклов развития багрянок? Большинство исследователей схо- дится на том, что первоначальный цикл раз- вития заключался в смене трех морфологи- чески одинаковых форм развития, которые существовали независимо одна от другой. Зигота покидала материнское растение и про- растала в свободноживущий карпоспорофит. Следует сказать, что такого рода цикла разви- тия, принимаемого учеными за исходный, в при- роде не существует. Это — гипотетический цикл, выведенный на основе логических умо- заключений. Принимая за исходное состояние свободное существование морфологически раз- витого карпоспорофита, ученые считают, что позднее в процессе эволюции карпоспорофит перешел к паразитическому образу жизни на гаметофите, в результате чего возник цикл раз- вития типа полисифонии. И наконец, дальней- шая эволюция привела к образованию гетеро- морфных циклов развития, когда гаметофит и тетраспорофит заметно различаются по стро- ению и по степени морфологического развития. Можно ли безоговорочно согласиться с этими представлениями об эволюции циклов разви- тия красных водорослей? Отсутствие гипоте- тического исходного цикла развития в природе даже у самых примитивных современных во- дорослей из класса флоридеевых заставляет усомниться в действительном его существова- нии в истории багрянок. Кроме того, ничего подобного нет в других отделах водорослей, где, как правило, зигота развивается в споро- фит, и цикл развития состоит из смены двух форм — гаметофита и спорофита. Зигота баг- рянок не образует собственной оболочки и развивается в стенках карпогона. Не говорит ли это о том, что зигота никогда не покидала гаметофит1 Таким образом, гипотезу об эволюции циклов развития красных водорослей, принятую в на- стоящее время большинством специалистов, нельзя считать до конца обоснованной. Не исключена вероятность того, что карпоспоро- фит багряно к — более позднее эволюционное образование, и первоначальный цикл состоял из двух форм развития. ПРОИСХОЖДЕНИЕ КРАСНЫХ ВОДОРОСЛЕЙ В ископаемом состоянии сохранились лишь немногие багрянки и преимущественно среди обызвествленных форм. Корковидные коралли- новые известны начиная с мела, причем многие из них с трудом можно отличить от современных. Членистые кораллиновые представлены в кай- нозое вымершими видами существующих в на- стоящее время родов. Найдено значительное число ископаемых форм, относимых к отдельному семейству со- ленопоровых, которые отмечаются от ордови- ка до триаса. Однако ни строение этих водо- рослей, ни их систематическое положение окончательно не выяснены. На основании этих немногочисленных палеон- тологических свидетельств невозможно от- ветить на вопрос о происхождении багрянок.
Очевидно лишь то, что это очень древняя груп- па. Многие ученые полагают, что она возникла еще во время археозойской эры. Сравнение с другими водорослями также не проясняет вопроса об их происхождении и родственных связях, так как эта своеобразная группа за- нимает весьма обособленное место среди водо- рослей. Некоторое сходство можно обнаружить только с сине-зелеными водорослями. Харак- терные для красных водорослей пигменты фикоэритрин и фикоцианин найдены, кроме них, только у сине-зеленых. Багрянковый крах- мал близок к крахмалу сине-зеленых. В обеих группах отсутствуют подвижные стадии. Сей- час принято считать, что это сходство не слу- чайно, и многие специалисты выводят самые примитивные багрянки, у которых еще нет полового процесса (класс бангиевых), из си- не-зеленых. Класс флоридеевых выводят из более высокоорганизованных бангиевых, уже обладающих половым процессом. Однако эту гипотезу нельзя считать достаточно обосно- ванной. Дело в том, что пигменты в той и другой группах отличаются по строению. Кроме того, у багрянок имеется специфический хлорофилл d, отсутствующий у сине-зеленых. Поэтому нельзя исключать и другую гипотезу, соглас- но которой оба отдела имеют самостоятельное происхождение и не связаны непосредствен- ным родством. Возможно, что сходство пигмен- тов объясняется лишь тем, что обе группы водорослей возникли в одну и ту же геологи- ческую эпоху в сходных условиях освещения, которое отличалось от современного по качест- ву и было, по-видимому, гораздо более слабым: ведь билипротеины способствуют восприятию малых количеств света. РАСПРОСТРАНЕНИЕ КРАСНЫХ ВОДОРОСЛЕЙ Красные водоросли — типичные морские ра- стения. В морях Мирового океана они распро- странены более широко и представлены более разнообразно, чем бурые и зеленые водоросли. Но, в отличие от бурых водорослей, багрянки иногда встречаются и в пресных водах. Они живут в прудах и реках, лужах и озерах, од- нако предпочитают холодные быстротекущие воды. Самые известные пресноводные багрян- ки — это многочисленные виды батратоспер- мула. Интересно, что в континентальных во- доемах растут представители примитивных, наименее специализированных групп, тогда как высокоорганизованные красные водоросли сосредоточены в море. К пресноводным баг- рянкам относится также небольшое число наземных обитателей. Колонии порфиридиума можно встретить в виде красноватых слизис- тых налетов на различных влажных поверх- ностях — на стенках оранжерей, на почве, по краям садовых луж. Пресноводные красные водоросли редко бывают красного цвета, обычно они зеленые, голубые или даже буровато- черные. В море красные водоросли встречаются пов- семестно, в самых разных условиях. Как и другие крупные донные водоросли, они посе- ляются только на твердых неподвижных грун- тах, поскольку рыхлые — песок, галька, мел- кий щебень — вследствие своей легкой подвиж- ности непригодны для существования макро- скопических форм. Субстратом для красных водорослей служат обычно скалы, рифы, валуны, каменистые россыпи, а также разнообразные искусственные сооружения и другие водоросли. У флоридеевых широко распространены яв- ления эпифитизма и паразитизма. Многочис- ленные багрянки — эпифиты используют дру- гие водоросли (в том числе и красные) только как субстрат, т. е. прикрепляются к их поверх- ности. Но у некоторых форм взаимоотношения становятся более тесными, это проявляется в приуроченности эпифита к определенному хозяину и в проникновении его базальной части внутрь ткани хозяина. Таким образом, наме- чается переход от эпифитного к эндофитному образу жизни. Эндофитизм может быть частич- ным или полным. В последнем случае все слое- вище водоросли растет внутри слоевища дру- гой водоросли и только нити с органами раз- множения выходят на ее поверхность. Об эндофитах мы говорим в том случае, когда водоросль погружена в другое растение, но продолжает питаться автотрофно, за счет фото- синтеза. Однако есть немало багрянок, кото- рые ведут паразитический и полупаразитиче- ский образ жизни; они не только погружены в ткани хозяина, но и питаются за их счет. Как эндофиты, так и паразиты отличаются редуцированным слоевищем, выступающим над поверхностью хозяина в виде шариков или лопастных образований. Весьма интересно, что большинство паразитических форм среди крас- ных водорослей по систематическому положе- нию очень близки к водоросли-хозяину. Они относятся не только к тому же порядку, но даже к одному и тому же семейству. Это тем более поразительно, что красные водоросли растут в богатых видами сообществах, где имеют- ся широкие возможности для выбора хозяина. Причины этого интересного явления остаются пока невыясненными. Диапазон глубин, на которых обитают баг- рянки, заметно шире, чем у бурых водорослей. Дополнительные красные пигменты помогают усваивать им небольшое количество света, и благодаря этому багрянки могут расти на зна-
чительных глубинах. Там, где имеются под- ходящие грунты и хорошая прозрачность воды, они достигают глубины 100 — 200 м. И все же гораздо чаще заросли красных водорослей, так же как и всех макрофитов, кончаются на глу- бине 20 — 40 л. Это связано не столько со светом, сколько с отсутствием на больших глубинах твердых грунтов. Красные водоросли обильно развиваются и в верхних горизонтах моря, в том числе и на литорали. Здесь они подвергаются сильному освещению, а во время отлива — и действию прямой солнечной радиации. В условиях силь- ного освещения цвет багрянок сильно меняет- ся. В их окраске появляются бурые, желтые, зеленые тона. Это обусловлено изменением в составе пигментов и увеличением роли хло- рофилла. Изменение окраски в зависимости от света — процесс обратимый. Даже сухие об- разцы, пролежавшие некоторое время в гер- барии, при отсутствии света приобретали более интенсивную окраску. В тропиках, где инсо- ляция настолько сильна, что порой оказывается губительной, многие багрянки уже не способ- ны расти на литорали и спускаются в субли- тораль. Не только свет обусловливает вертикальное распределение багрянок. С глубиной меняются температура, динамическое действие воды, количество питательных веществ. На литора- ли происходит периодическая смена водной среды на воздушную, в связи с чем здесь резко меняются все условия, в первую очередь тем- пература и соленость. Многие багрянки хоро- шо приспособились к жизни в этой своеобраз- ной зоне моря. Зимой они промерзают, омы- ваются пресной дождевой водой, во время отлива высыхают, становясь хрупкими и ломки- ми, но когда вновь наступает прилив, оказы- ваются вполне жизнеспособными. Особенно рез- кие колебания среды характерны для верхних горизонтов литорали и расположенной выше зоны заплесков и брызг. Здесь живут лишь очень немногие виды и среди них несколько багрянок. На открытых скалистых мысах, где постоянно действует прибой, выше других водорослей поднимаются бангия, порфира, глойопелтис и некоторые другие. Таким обра- зом, и верхняя граница распространения баг- рянок выше, чем бурых водорослей. Красные водоросли характеризуются чрез- вычайно широким географическим распрост- ранением. Они растут во всех морях Мирового океана от тропиков до полюсов. Наиболее раз- нообразно они представлены в тропиках и по числу видов заметно превосходят здесь бурые и зеленые водоросли. В умеренных морях роль их также остается значительной, хотя доля во всей флоре несколько снижается вследствие более обильного по сравнению с тропиками раз- вития здесь бурых водорослей. По направле- нию к высоким широтам общее число видов багрянок постепенно уменьшается и в Аркти- ческом бассейне резко падает. Тропические багрянки, как правило, мелкие организмы, тог- да как в умеренных широтах они могут дости- гать значительных размеров, хотя и уступают в этом более крупным бурым водорослям. Красные водоросли играют заметную роль в жизни моря. Они являются важными компо- нентами биоценозов, нередко доминируя в раз- личных сообществах и определяя характер ра- стительности. Наряду с другими красные водоросли — важный источник органического вещества в море и пища для морских животных. Клетки размножения — тетраспоры, карпо- споры и т. п., образующиеся в большом коли- честве, формируют часть фитопланктона при- брежной части моря. Установлено, что одно растение родимении продырявленной (Rhody- menia pertusa, табл. 21,4), которое может дости- гать в длину 1 м, продуцирует 12 000 000 кар- поспор, а один тетраспорофит этого же вида производит 100 000 000 тетраспор. Известковые кораллиновые водоросли играют важную роль в создании коралловых рифов. Они оказывают цементирующее действие и способствуют под- держанию рифовой структуры, без чего риф не мог бы существовать как единое образование. ИСПОЛЬЗОВАНИЕ КРАСНЫХ ВОДОРОСЛЕЙ Красные водоросли широко используются человеком в хозяйстве и быту. Многие багрян- ки не только съедобны, но и очень полезны. Из них готовят салаты, приправы, гарниры к мясу и рыбе, варят супы. Нередко едят суше- ными или засахаренными. Первое место среди съедобных багрянок занимают родимения и порфира, распространенные во многих наших морях. К сожалению, в отличие от других приморских стран, в СССР употребление водо- рослей в пищу почти не принято, хотя известно, что водоросли — весьма полезные растения, содержащие многие жизненно важные вещест- ва, и что запасы водорослей в наших морях огромны. Охотнее других водоросли употреб- ляют в пищу жители побережья Тихого океана. В Японии даже развито промышленное куль- тивирование порфиры в естественных услови- ях — своего рода морское сельское хозяйство. В прибрежной полосе на мелководье для по- селения порфиры готовят дополнительные пло- щади, укладывая на них камни или пучки ветвей, или растягивают специальные сети из толстых веревок, закрепляя их на бамбуковых шестах так, чтобы они держались у поверхно- сти воды. Эти сети сплошь обрастают порфирой. 212
Промышленное использование красных водо- рослей основывается на присутствии в их обо- лочках фикоколлоидов — слизистых веществ из группы полисахаридов. Самый ценный продукт, получаемый из красных водорослей и имеющий очень широкое применение, — агар. В Японии он известен с 1760 г. До второй мировой войны агар добывали почти исключительно из гели- диума. В настоящее время его производят из большого числа видов; только в Японии ис- пользуют около 30 видов агарофитов. В СССР сырьем для агара служит анфельция. Произ- водство его организовано на Дальнем Востоке и на Белом море. По своим физическим свой- ствам агар похож на животный желатин, но имеет совсем другое химическое строение. Пре- имущество его перед желатином в том, что он остается твердым при более высокой темпе- ратуре. Агар используют в микробиологии для приготовления сред при культивировании микроорганизмов. Его применяют как лекар- ство при расстройствах кишечника, добавля- ют вместо крахмала в хлеб для больных ди- абетом; на агаре делают капсулы и таблетки с антибиотиками, витаминами, сульфопрепа- ратами, особенно когда требуется их медленное рассасывание. Самое широкое применение на- ходит агар в пищевой промышленности. На нем готовят желе, мармелад, мягкие конфеты, варенье, так как он предохраняет их от заса- харивания; его используют также при изготов- лении мясных и рыбных консервов в желе, для очистки вин. Из других фикоколлоидов багрянок извест- ны каррагинин и агароид. Каррагинин полу- чают главным образом из хондруса на побе- режье Атлантического океана. Первыми хон- друс начали использовать жители Северной Ирландии, поэтому он известен под названием «ирландский мох». Агароид добывают в СССР из черноморской водоросли филлофоры, по- лучают его и в ряде других стран. Кроме того, красные водоросли вместе с дру- гими водорослями используют для производ- ства водорослевой муки, которая идет на корм скоту и как удобрение. КЛАССИФИКАЦИЯ КРАСНЫХ ВОДОРОСЛЕЙ Современная система красных водорослей основывается на исследованиях крупного швед- ского альголога К ю л и н а. В целом ее прини- мают все альгологи, противоречия возникают только в отношении систематического поло- жения некоторых мелких таксонов. Принцип, положенный в основу классификации,— строе- ние женских репродуктивных органов и про- цесс развития гонимобласта. По этой системе все красные водоросли делятся на 2 класса— класс бангиевых и класс флоридеевых. Каждый из них содержит по 6 порядков. Основная масса багрянок относится к классу флоридее- вых — в нем 49 семейств. КЛАСС БАНГИЕВЫЕ (BANGIOPHYCEAE) Класс бангиевых объединяет одноклеточ- ные, колониальные и много клеточные формы паренхимного строения. Рост у них диффуз- ный, в результате деления всех клеток слое- вища. Клетки бангиевых одноядерные с одним звездчатым хлоропластом и одним центральным пиреноидом. У многих представителей клас- са хлоропласт имеет осевое положение, кото- рое считается более примитивным по сравнению с пристенным. Половое размножение извест- но только у высокоорганизованных предста- вителей класса. Половые клетки формируются в результате преобразования вегетативной клет- ки. Зигота подвергается делениям и вся прев- ращается в карпоспоры. Бесполое размноже- ние осуществляется моноспо рами, реже аки- нетами. Моноспоры и карпоспоры голые и способны к амебоидным движениям. Большинство представителей класса, осо- бенно среди примитивных, — пресноводные и наземные обитатели. Морские формы распро- странены в прибрежной полосе всех морей, но в умеренных широтах представлены богаче, чем в тропиках. Класс бангиевых невелик по объему. В нем насчитывают около 20 родов и 70 видов. Си- стематики делят его на 6 порядков, наиболее важным из которых является порядок бангие- вых (Bangiales). Всех представителей класса можно разде- лить на две группы, которые, по-видимому, представляют собой две разные ветви эволю- ции. Первая включает многоклеточные нит- чатые и пластинчатые формы, распространен- ные преимущественно в море. Вторую группу со- ставляют одноклеточные и колониальные водо- росли, чаще встречающиеся в пресноводных и наземных местообитаниях. Если первая груп- па принадлежит к типичным багрянкам, то вторая по некоторым признакам сильно тяго- теет к сине-зеленым водорослям. Рассмотрим некоторых представителей класса бангиевых. Наиболее известная одноклеточная водо- росль из порядка порфиридиевых (Porphyridia- les) — порфиридиум (Porphyridium, рис. 149, 1). Округлые клетки этой водоросли обычно со- браны в слизистые колонии, которые в виде кроваво-красных пленок покрывают почву и влажные стены. Среди пигментов порфиридиума
находили модификации фикоэритрина, напо- минающие фикоэритрин сине-зеленых водо- рослей, поэтому считали, что порфиридиум- сине-зеленая водоросль. Запасным веществом у порфиридиума служит багрянковый крах- мал, зерна которого откладываются в непо- средственной близости с хлоропластом. Эти признаки, так же как наличие клеточных ор- ганелл, ясно показывают, что порфиридиум правильнее всего помещать среди красных водорослей в классе бангиевых. Чаще всего порфиридиум размножается ве- гетативным делением пополам. Иногда можно наблюдать образование моноспор, при этом протопласт округляется п покидает материн- скую клетку. Пока неясно, является ли порфи- ридиум примитивным организмом или вторич- но упрощенным. Поскольку эта водоросль при- урочена к специфическим условиям обитания, вполне правдоподобна вторая точка зрения. У другой одноклеточной водоросли — хро- оте~е (Chroothece), также растущей на почве, очень сильного развития достигает слизистая оболочка. Она имеет вид слоистой ножки, пре- выша~ощей длину клетки более чем в 50 раз. Слизь выделяется через многочисленные по- ры, пронизывающие оболочку клетки. 3та во- доросль интересна тем, что после деления до- черние клетки расходятся не сразу, а на ка- кое-то время остаются в материнской оболочке, что приводит к образованию коротких времен- ных нитей. Это явление помогает понять пе- реход от одноклеточного состояния к нитча- тому (рис. 149, 2). У гониотрихума (Goniotrichum), так же как у астероцитиса (Asterocytis), нитчатое слоевище является уже постоянным признаком. У большинства видов клетки собраны в одно- рядные нитчатые колонии, и только у одного Goniotrichum cornu cervi клетки делятся в раз- ных направлениях, так что образующиеся колонии имеют вид многорядных нитей (рис. 149, 4). Для этих водорослей, как и для многих сине-зеленых, характерно ложное вет- вление. В отличие от порфиридиума, гониотри- ховые размножаются преимущественно моно- спорами. Половое размножение у этих водоро- слей полностью отсутствует. Водоросли из порядка бангиевых встречают- ся главным образом в море. Наиболее просто устроенные характеризуются однорядным нит- чатым многоклеточным слоевищем. Типичные представители порядка — водоросли бангия (Bangia) и порфира (Porphyra, табл. 20, 1). На ранних стадиях развития неразветвленные нити бангии сохраняют однорядное строение, но по мере роста слоевища клетки подверга- ются продольным делениям и нити становятся многорядными. Взрослые растения прикреп- ляются ризоидами, которые, достигая субст- рата, образуют небольшую подошву (рис. 150, 1). Морские виды рода бангия очень характер- ны для верхних горизонтов литорали и для зоны брызг. Пресноводные виды обитают обыч- но в эстуариях, образуя бурые и фиолетовые скопления нитей. Формирование пластинчатого слоевища пор- фиры также начинается с однорядной нити, но вскоре клетки начинают делиться в разных направлениях, в результате чего образуется однослойная или двухслойная пластина. Ра- стения порфиры средних размеров, но неред- ко достигают в длину метра и более. В нижней части пластина сужается в небольшой стебе- лек, который переходит в подошву, образо- ванную ризоидами. Когда порфира растет на водорослях, ризоиды могут довольно глубоко проникать в ткани хозяина. У некоторых ви- дов подошва и самые нижние части пластины многолетние, тогда как пластина нарастает заново каждый год. Бесполое размножение осуществляется мо- носпорами, которые могут образовываться в большом количестве — до 10 000 спор на 1 см' слоевища. В отличие от остальных представи- телей класса для водорослей из порядка бан- гиевых характерно половое размножение. Спер- матангии у порфиры и бангии образуются в результате повторных делений вегетативных клеток, так что в одной клетке развивается от 32 до 128 сперматангиев, каждый из которых содержит по одному спермацию (рис. 161, 2). В оогонии, или карпогоны, превращаются обыч- ные вегетативные клетки, каждая клетка — в один карпогон. Трихогина у бангиевых отсут- ствует (рис. 161, 1). Часто карпогон не имеет вообще никаких выр остов, но у некоторых клеток образуются короткие трихогинообраз- ные выросты. У одних видов порфир мужские и женские органы размножения возникают на одном и том же растении, у других — на раз- ных. После оплодотворения в зиготе происходит несколько клеточных делений и образуется 4 — 64 карпоспоры (рис. 161,8). Судьба прора- стающих карпоспор различна. При опти- мальных условиях они снова дают пластин- чатые растения, которые размножаются поло- выми клетками. Однако при изменении усло- вий, например при смене сезонов года, форми- рование пластинчатого слоевища приостанав- ливается и многоклеточные растения остаются карликовыми. В их клетках образуются моно- споры. Очень часто карпоспоры порфиры про- растают в одно рядно-нитчатые разветвленные растения. В клетках этих нитей содержатся лентовидные хлоропласты. Созревая, некоторые клетки слегка раздуваются и функционируют как моноспорангии. В них образуются моно- 214
споры, которые при прорастании дают пла- стинчатые слоевища. Раньше эту нитчатую стадию считали самостоятельной водорослью и называли конхоцелис (Conchocelis). Стадия конхоцелис обитает на раковинах моллюсков, частично проникая внутрь их. Таким образом, у порфиры уже имеется чередование гамето- фита и спорофита — гетер оморфная смена форм развития. Класс бангиевых внутри отдела Rhodophyta составляет весьма обособленную группу, об- ладающую рядом примитивных черт. Диффуз- ный рост, образование органов размножения в любой клетке слоевища и отсутствие специали- зации в их строении, непосредственное прев- ращение зиготы в карпоспоры, отсутствие ти- пичной трихогины — все это говорит об отли- чии бангиевых от флоридеевых. Некоторые уче- ные считали эти отличия столь принципиаль- ными, что выделяли бангиевые из отдела крас- ных водорослей и пытались сближать их с неко- торыми сине-зелеными и даже зелеными водо- рослями. Но, с другой стороны, нельзя не учитывать и важных черт сходства обоих клас- сов. У них одинаковы набор пигмен- тов, запасные вещества, характер спермациев и карпоспор. Интересно отметить, что одним из важных отличительных признаков между ними всегда считалось отсутствие у бангиевых первичных и вторичных пор в месте соедине- ния клеток. Однако при более пристальном изучении у многих бангиевых обнаруживаются типичные для красных водорослей поры. 3то, несомненно, свидетельствует о тесном фило- генетическом родстве всех багрянок. Кроме того, у примитивных флоридеевых можно найти еще много сходства с бангиевыми. Они имеют пиреноиды и звездчатый хлоропласт, одно ядро, споры их также способны к амебо- идному движению. Поэтому логично предпо- ложить, что все красные водоросли едины в своем происхождении. КЛАСС ФЛОРИДЕЕВЫЕ (FLORIDEOPHТСЕАЕ) Класс флоридеевых объединяет формы много- клеточные, преимущественно сложного анато- мического строения. Слоевище флоридеевых представляет собой систему разветвленных ни- тей, паренхимный тип организации у них отсут- ствует. Рост апикальный, в результате деятель- ности верхушечной клетки. Клетки флори- деевых одноядерные, реже — многоядерные с пристенными хлоропластами. Звездчатые хло- ропласты с пиреноидами характерны толь- ко для низкоорганизованных представителей класса. Бесполое размножение осуществля- ется тетраспорами, биспорами, полиспора- ми, только у некоторых — моноспорами. По- ловое размножение свойственно всем флори- деевым, а если оно отсутствует, то в результа- те вторичной утраты. Мужские и женские органы размножения представляют собой спе- циальные образования. Зигота развивается в гонимобласт, на котором формируются кар- поспоры. В развитии гонимобласта важную роль играют специальные питающие и аук- силлярные клетки. Имеется чередование га- метофита и спорофита, сходных или, реже, несходных между собой по морфологическому строению. Подавляющее большинство представителей класса — типичные морские обитатели, распро- страненные во всех морях земного шара. Класс флоридеевых делят на шесть поряд- ков на основании особенностей развития зиго- ты и строения ауксиллярной системы. Каж- дый порядок включает весьма разнообраз- ные по морф ологическому и анатомическому строению водоросли. ПОРЯДОК НЕМАЛИЕВЫЕ (NEMALIALES) Порядок немалиевых объединяет организмы, у которых отсутствуют ауксиллярные клетки. Нити гонимобласта развиваются непосредст- венно от оплодотворенного карпогона или от нижележащей клетки после того, как в нее пе- рейдет ядро карпогона. Специальных питаю- щих клеток у немалиевых тоже нет. У сложно организованных форм во время оплодотво- рения или после него происходит слияние кле- ток карпогонной ветви и несущей клетки в самых разных сочетаниях. Порядок немалиевых — наиболее примитив- ная группа красных водорослей. Он объе- диняет 8 семейств, которые можно разделить на две группы. В первой группе слоевище име- ет одноосевое строение, во второй — оно по- строено по многоосевому типу. Почти все фло- ридеевые, которые обитают в пресных водах, относятся к немалиевым водорослям. Семейство акрохетиевые (Acrochaetiaceae) 3ти водоросли устроены наиболее просто. Слоевище состоит из свободно разветвленных однорядных нитей. Они образуют кустики ми- кроскопических размеров (как правило, вы- сотой до 5 мм), и только как исключение встре- чаются более крупные растения, высотой до 25 мм. Ветвление в этих кустиках попере- менное, неправильное (рис. 151, 1; 162). Часто короткие боковые веточки на вершине заканчиваются волоском, однако имеется не- мало видов, у которых волоски никогда не образуются. 215
Бесполое размножение у акрохетиевых осу- ественно моноспо рами,, В ществляется преимущес о е ко. Спо- U тет аспоры отмечаются довольно редко. по- Ф'. где располагаются одиночно или по 2 — вме- сте рис, етр я клетка, об азно. Карпогон — единственная о разно. к от основных ветвей Ii располагается она с оку или на коротких отких боковых веточках. Изредка ( 1 встречается двухклеточна р я ка погонная ветвь. их ок глых клеток ине боковых веточек Ф -.';-: 7 р асполагаются на вершин о, ° (ри.. 162, 8). 1 "~0..- После оплодотворения и отпадения трихо- Я.': <о ся поперечными перего- "-.1.:.: Ъ гины карпогон делит колько клеток, из которых вы- родками на несколько ста i ис. 162, 4). ет пучок нитей гонимобласта рис. .:У. растает п к охетиевым своиственн но половое ::ф .Ф' е всем ак толь- ие некоторые размножаются о размножение; н с е и них — многие ко бесполым способом, среди них- ы одохортона (Rhodochorton). 4 виды ро офорт В природе акрохетиевые р р р асп ост анены 2 и око. В большинстве своем— чрезвычайно широко. это морские организмы, известные во всех Рис. 162. Строение акрохетиевых.' чки мо ях Мирового океана. Пресноводных видов ния Acrochaetium; 2 — часть веточки МОрНХ Ировог они п инадлежат — с ка погоном и сперма- ~~ c ~~~~~областом. а — нарпого~, — спер- Из морских о итателей о самой характерной и распространеннои вод р о ослью семейства яв- В лите- осли а- ляется акрохетиум (Acrochaetium). ,Лишь немногие акрохетиевые ~одоросли ра- ляется прож . лит стут на грунте — на камнях, х скалах и т. п., боль- ратуре описано ок но в ействительности их, по- д -ви имому, гораз- шинство же предп едпочитает поселяться а дру- о меньше. огда эти ивотных. Особенно часто д о азмножение и циклы гих водорослях и жи х во о ос- культуре, осо енно их р их можно встр стретить на ламинариевых водорос- и азвития, то о на б а~ ~ жилось что многие пре- мшанках, моллюсках. При р иное как стадии или лях, на гидроидах, м к пове х- жние виды — не что ин этом они не т е только прикрепляются к поверх- ж р ткани формы развития в жизненном цикле других но и проникают в тк ности организмов, н видов. о осль семейства — ро- хозяина. тиевых ме- Другая морская водоросл Ст оение базальной части акрохетиевых ме- — отличается более грубым и круп- тся в зависимости от характер у а с бстрата. дохортон — от евищем растущим, как пра авило на няется в е б ь то камни или во- ным слоевищ п ибойной литорали На твердом субстрате, удь то ка аменистом грунте. На при оиной оросли с плотным строением, щ ча е встречаются кам аших северных мореи родохор тон часто о ра- д ы на ыхлом — свобод- наших Ф.У пок ывая зате- псевдопаренхимные подошвы, р т самостоятельныи пояс, р ные стелющиеся нити. В целом д р ля ак охетиевых зует н ю поверхность валунов и и скал темно- акте на азнонитчатая структура. ненную п Наи олее Н б отчетливо она проявляется у во нии так к как при илогенетич отя его и п по)е- разнонитчатом стр роении вначале обычно раз- рохети т а из клеток щают как наи олее п б просто организованную вива ет ется стелющаяся структура, из клеток побеги группу в н начале всей системы лоридеевых, кото рои в озникают вертикальные до сих пор неизв естно говорит ли эта пр Ф.У (рис. 162, 1). та о примитивности его предста вителеи или Внутриклеточное строени р д е п е ставителеи се- та 3 есь мо- она р ено что меиства отличается разнообразием. д езультат вторичного упр щ и п истенным последн В ее время было обнаруж икле аз- жно вс стретить формы с осевым и при ами и без них. некоторые багрянки имеют в своем цикле очно такого же хл р оропла сто), с пиреноидами и ошим п изна- вития с тадии или поколения точно так блю ение Фо ма хлоропластов служит хорош р т оения как акрохетиевые. Это наб д ком для разграничения р д орма хл о ов. У одних они строения, как семейство пред- ые спи аль- позв оляет предположить, что — ставляет собой искусственную группу, к р кото ая но закрученные, у третьих — д с ов. — в ви е дисков.
Рис. 163. Строение Вatrachospermum: ия — го слоевища; 4 — 6 — стадии развития ~т 8 — начало образования взрослого л~жищ~; 1 — внешнии вид ч части растения; 2 — ювенильная стадия; б — т ихогина, в — оплодотворенныи карп погон г — клетки гонимобласта, — р гонимобласта. а — моноспорангии, б — трихогин, Ф.У т ебует более подробного изучения. В этои связи интересно подчеркнуть, что у ряда во- дорослей семейства неизвестно половое раз- множение и что они заметно различаются вну- триклеточным строением. Семейство батрахоспермовые (Batrachosperma- ceae) 3то семейство интересно тем, что в нем мо- жно проследить первые шаги в усложнении о ганизации слоевища багрянок при переходе от свободно-нитчатого к плотному строению. Характерные черты семейства лучше всего проследить, п , познакомившись с одной из самых известных пресноводных багрянок — б р ок — бат, а- хосперлулол (Batrachospermum). Около 50 ви- дов этого рода распространено во всех частях света. Предпочитают они хорошо а эрируемые воды медленно текущих рек и ручьев, но их можно встретить и в прибрежной части озер, там, где отсутствуют сильные колебания тем- ературы. Некоторые проникают даже в тор- пер близи фяные болота, но чаще встречаются вблиз ключей. Мягкое слизистое слоевище батрахосперму- ма состоит из центральной однорядной оси, азветвленной моноподиально или псевдоди- хотомически. Основные ветви густо покрыты ками коротких веточек ограниченного ро- 1 2 ста, расположенных мутовками (рис. 15, 163). Строение батрахоспермума легко пред- ставить как результат усложнения и упорядо- чения строения акрохетиевых, и это подтверж- дается индивидуальным развитием д хоспермума. Вначале образуется стелющаяся по субстрату нить, она ветвится с образованием псевдопаренхимнои подошвы, из Ф.У из клеток ко- то ой возникает множество вертикальных од- но ядных побегов, морфологически почти неот- личимых от акрохетиума. та стад ия кото ю так и называют стадией акрохетиума, может существовать достаточно долгое время и даже размножаться моноспорами (рис. 163 2 . Не- мудрено, что ее часто принимают за самостоя- тельные виды акрохетиума. азвит звитие этой ста- дии в большой степени определяется условиями среды. р . В целом ее можно назвать теневой фор- мой роста, тогда как стадия батрахоспермума- световая форма. В связи с этим в природе они приурочены к разным сезонам. обеги бат а- х оспермума развиваются в лю ом месте ста- дии акрохетиума. Их строение относится к центр альноосевому типу, который в литера- т е часто называют типом батрахоспермума. Ве хушечная клетка центральной о туре ча оси отчле- няет новые сегменты, которые быстро растут в длину и толщину. На каждом из этих сег- ментов образуются мутовки из 4 — 6 мелких клеток, которые постепенно вырастают в пу- чки ветвей ограниченного роста. Конечные клетки боковых веточек нередко преобразуют- ся в волоски. Ветви в пучках связаны большим количеством слизи. Из нижних клеток ветвей
Ст оение Lemanea.' Рис. 164. Стр — о азов ание спарматангиев; — О 23BHTNH ГOHNMOGJIB Т — карпогонная ветвь; вь 4 — начало р б — сперматангий, в — во- 8 — к клетка сперматангия, материнская клетк д — карпогон. JIOCOK, Г— — несущая клетка, ограниченног р о оста в озникают обычно кле- кающиеся вниз ные нити, спуска ризоид и обволакивающие 51 2~. От этих ните ось (рис. 15, ). ветвей ограни- в аться дополни нительные пучки е выми служат ченного роста, к оторые вместе с перв ыми нитями. ассимиляционным — МОНО СПО- лого а�змножени- яр�га беспол р астениях батрахосперм рангии — на ра нь едко, гораздо д, чаще они ча ются очень р астениях, по- на неполовозрелых ра П Вое азмножение акр у у. ети м . олово ото ыми осо енност отличается неко р ля красных во дорослей ке- гина необычнои д илиндрическои глевиднои ил др погон образу ется на вершине вет ветвей ограни- х от нижних клеток mc. 163 4). Шаровндные спер- боко~ы~ матангии развив ива ются на верш 1 — 3 на клетках, кот торые ничем Веточек, по 1— Зрелые гонимо- от вегетативных. не отличаются о р образования, отграниченные бласты — четко имиляцио нных в слизь среди ассим погруженные в с им нитей и плотно окруженные и 4 — 6; 151, 2). вые (Lemaneaceae) Семейство леманеевые о осль леманея (Lemanea)— Пресноводная водоросл ства лемане- е ставитель семеиств единственныи пред етв ленное или п простое трубчатое евых. Рааветвле 64 g) построено слоев ище леман р еманеи атр б т ахоспермум. тому же принц но ядная ось не сет MyToвки центральная о~нор но, в отли- азветвленных ветвеи, н боковых разв ни располагаются чие от батрахос р твятся так, спе м ма, они 3 астаются и вет не свободно, а раз ными ответвле- что смь аются ютвокруг централь- тем самым о разуют киями и т асстоянии от нее, Ф.У на некотором ра овой слои, кот сплошнои коро от т н ю нкцию. ассимиляционну О Ошо ВИДНО, еви е леманеи хо резе через слой образуется из п ченного роста, но эти в веи Ограниче~ р и их клетки ди бразованные густо и ко ы, о ра Внешние клетки, ра и веточками последн разветвленными мелкие и наи олее богатые пиг- ков, самые мелки оженные клетки а гл бже располож ментами, а г окрашенные или менее интенсивно крупнее, м К у батрахоспермума, 6оо етные. ак и у от нижних клеток у леманеи от н которые прости- азуются изоидные нити, ь н ического слоевищ др также имеется стад У леманеи тад ней никогда не о разу ума, но на ней т аспо ы. о сих п ор у этого рода споры, ни тетра ожение. Сперма- о половое размнож погон находятся на тангии и карп р тся на внешних е вые развиваютс же растении. р р ~ и соб аны в виде от тдельных пя- клетках коры и на тех участках ошного кольца на т . ° Я тен или спл овки ветвеи or- г е асположены мут слоевища, где р ская клетка сперма- б оста. Материнска та в длину и н ноклеточный волосок, к вершине однок тчленяется, сдви- . спе матангий от сле того как совсем отпадает н а потом и со гается в сторону, ветвь возникает ( 32 7клето 2 . Карпогонная вет лоем. Она состоит и б самая ниж няя клетка нередко не кото ой также м может развивать- 164 8) Пмж о- ся карпогонная ветвь (рис. ло клеточнои Ф.У ка погон дает нача лодотворения кар й вн т ь слоевища и разветвля- нити, растущей вну пучков нитей азованием густых п 6oo~~bbmmaaH . Не только конечные, ок нитей гонимо ла часть клеток ~ходят после раз- ся в карпоспоры, ь р ото ые вых рыва стенки слоевища. виях они могут При неблаг р гоп иятных услови ое в емя оставаться ся внутри слое- б является своео разнь в ища. Это я няю имся усло овиям среды. ак, лен ием к меня пе иод, когда в м елких водое- су р д, мах на лю б дается недостаток воды, в поспор не происходит.
219 Большой интерес ученых вызывает цикл раз- вития леманеи. Вначале он казался очень про- стым. Были известны только гаметофиты, и поэтому предполагалось, что весь цикл проходит в гаплоидной фазе, диплоидная же ограничена зиготой. Но теперь мы уже знаем, что за внешней п остотой этого цикла скрываются сложные процессы. Когда было найдено действительное место редукционного деления, выяснилось, что первоначально в цикле развития присутство- вал тетраспорофит, но постепенно он претер- пел редукцию и потерял способность образо- вывать тетраспоры. Карпоспоры леманеи про- растают в растения, которые соответствуют тетраспорофиту. Начинается прорастание с об- разования стадии акрохетиума, из нее позднее возникают растения леманеи. В определен- ный период этого роста в клетках леманеи происходит редукционное деление, после чего образуется уже гаплоидная ткань, которая соответствует гаметофиту. У леманеи очень сильно развита способность к вегетативному размножению и регенерации. Она может выдерживать длительное высыха- ние оставаясь жизнеспособной, и, попав вновь ние в условия увлажнения, давать множество но- вых побегов. Растет леманея в континенталь- ных водоемах всего мира, предпочитая мел- кие холодные быстротекущие воды, где водит- ся форель. Семейство боннемезониевые (Воппетаезоп1асеае) Это семейство занимает обособленное место среди немалиевых, построенных по одноосе- вому типу. Боннемезония (Bonnemaesonia) водоросль с довольно ограниченным распро- странением, но она чрезвычайно интересна благодаря особенностям строения и размноже- ния. Слоевище боннемезонии вальковатое или слегка уплощенное, густо покрыто веточками ограниченного роста, которые располагаются в одной плоскости, супротивно, придавая слое- вищу перистый вид (рис. 165, 1). Как же растет Ф.У и формируется слоевище этои водоросли. Апи- кальная клетка делится косой перегородкой, поэтому возникающие сегменты располагаются под углом друг к другу (рис. 165, 2). От этих центральных клеток отделяются по две друг против друга расположенные перицентральные клетки. Они образуются неодновременно, и та, что отчленяется первой, быстро вырастает в очень короткую неразветвленную ветвь огра- ниченного роста, которая при созревании слое- вища всегда остается стерильной. Другая пери- центральная клетка развивается в длинную ветвь неограниченного роста или, оставаясь короткой, несет на себе органы размножения. В ходе дальнейшего развития короткие боковые Рис. 165. Строение боннемезониевых. 1 — внешнии вид веточки Bonnemaesonia; 2 — вершина веточки; 4 — часть про- 8 — троение коры с поверхности слоевища; 4— — СТ . Б 6 — азвитие ци- до льного среза слоевища со сперматангиями;, — ра — железистые клетки, б — материнская кле к — ж, т 8 сток арпа. а — ж Ф.. ьная ось д — пе- сперматангия, в — сперматангии, г — централ риц ентральная клетка, е — несущая клетка, ж — клетки кар- — — пе вая погоннои " ветви з — карпогон, и — питающие нити, к — р клетка гонимобласта, л — клетка слияния, м — р р 1 — ка поспо ы.
веточки становятся многорядными. Это про- исходит следующим образом. Первичные клетки веточек делятся на одну центральную и три расположенные вокруг нее перицентральные клетки. После этого каждая из трех перицентральных клеток образует по четыре клетки, которые составляют первичную кору боннемезониевых. По мере того как цент- ральные клетки растут в длину, коровые обра- зуют вокруг них плотный слой довольно круп- ных клеток. Постепенно снаружи от первич- ной коры образуется мелкоклеточный внешний коровой слой. Внешняя кора не образует сплош- ного покрова, клетки ее имеют сетчатое распо- ложение, и, если смотреть на слоевище с по- верхности, можно заметить, что они сосредото- чены у клеточных стенок первичной коры. В коре разбросано много железистых клеток (рис. 165, 8). Формирование ветвей неограничен- ного роста происходит подобным, но более сложным способом. На поперечном срезе через слоевище можно различить ограниченную коро- вым слоем полость, в центре которой прохо- дит основная ось. Ризоиды, развивающиеся у других немалиевых вокруг центральной оси, здесь отсутствуют. Плодоносные веточки боннемезонии гомоло- гичны ветвям неограниченного роста, поэтому они располагаются напротив коротких веточек и обычно чередуются с ними на оси. Мужские веточки, за исключением вершины и основания, как муфтой, покрыты сперматангиями. При образовании последних клетки внешней коры располагаются сплошным слоем и все превра- щаются в материнские клетки сперматангиев. Одна клетка дает начало двум или трем сперма- тангиям (рис. 165, 4). В каждой женской плодоносной веточке об- разуется, как правило, по одной карпогон- ной ветви; при этом несущей клеткой служит одна из трех перицентральных клеток. Карпо- гонная ветвь трехклеточная с длинной, спи- рально закрученной трихогиной. Интересно, что у боннемезонии оболочка цистокарпа на- чинает формироваться еще до оплодотворения. От всех трех перицентральных клеток, в том числе и от несущей клетки карпогонной ветви, образуются клеточные нити, которые участвуют в построении стенок цистокарпа. Также до оплодотворения подкарпогонная клетка де- лится, и от нее образуется пучок мелкокле- точных разветвленных нитей, которые можно рассматривать как питающие. Уже на первых порах развития оплодотворенного карпогона питающие клетки сливаются друг с другом и с подкарпогонной клеткой, обеспечивая пи- танием развивающийся гонимобласт. Первая клетка гонимобласта отчленяется от карпого- на так, что направлена внутрь молодого цисто- карпа, ближе к его дну, на котором располо- жена группа крупных, богатых содержимым клеток. Первая клетка быстро делится, в ре- зультате образуется клеточный диск из не- скольких слоев клеток. По мере образования этого первичного гонимобласта питательные клетки, соединенные с подкарпогонной, умень- шаются в размерах и теряют свое содержимое. В то же самое время происходит слияние кле- ток диска между собой, а также с несущей клеткой карпогонной ветви, с богатыми содер- жимым клетками на дне цистокарпа и неко- торыми другими. В результате этого процесса образуется крупная многоядерная клетка сли- яния. ат нее, теперь уже по направлению к на- ружной стенке цистокарпа, развиваются корот- кие нити, конечные клетки которых превра- щаются в карпоспоры. Выходят карпоспоры через специальное отверстие на вершине цисто- карпа (рис. 165, о, 6). Долгое время тетраспоры у боннемезонии были неизвестны, и поэтому полагали, что беспо- лая форма развития у нее совсем отсутствует. Но при изучении биологии этой водоросли в культуре оказалось, что карпоспора боннемезо- нии прорастает в растение, построенное из однорядных разветвленных нитей, отчасти сте- лющихся, отчасти прямостоячих, обра зую- щих дернины высотой около 1 — 3 см. Это расте- ние-эпифит раньше было известно как водо- росль с неясным систематическим положением под названием трайлиелла (Trailiella). Раннее развитие при прорастании карпоспор сопро- вождается образованием плотного базального диска. Из этого подробного описания внимательный читатель может заключить, что принцип орга- низации слоевища и особенности размножения боннемезонии заметно отличаются от описан- ных ранее у немалиевых. Эти отличия не слу- чайны, и теперь уже накопилось много данных, которые позволяют усомниться в правильности систематического положения семейства бонне- мезониевых в порядке немалиевых. Некоторые специалисты полагают, что боннемезониевые- самостоятельный порядок, который гораздо бли- же к церамиевым, чем к немалиевым, по таким признакам, как способ прорастания и фор- мирования слоевища, характер карпогонной ветви и сложные процессы слияния при раз- витии гонимобласта, наличие железистых кле- ток и т. п. Рассмотренные семейства немалиевых харак- теризовались одноосевым типом строения. Другую эволюционную линию в порядке пред- ставляют водоросли с фонтанным типом строе- ния. Эта линия представлена главным образом морскими водорослями — гельминтокладиевыми и хетангиевыми. 220
~óюФ16 ° ° ° ° ° ° В Ю ° ° ° ~ ° р ° е ° ~ ° ееефэеее ° ° у °,и ° Ф и~" ° еВ ~Э ° Ъ r ° ° те ° ° ю ° ° г ~ ° ° ° ° Ô ° ° ~ ° б ~ ° ° 3 ° 4 ° ° ° ° ° 1 ° ° ° ° В Р 166. Строение и размножение Nemalion. а а — клетки карпогонной ветви, Рис.. р — р; — 5 — стадии развития гонимобласта. а— 2 — карпогонная ветвь; 8 — — ст 1 — поперечный срез слоевища; ия в — клетки гон имобласта г — карпоспоры. б — карпогон после оплодотворен 7 Семейство гель минтокладиевые (Hel mint пппйос1а- diaceae) П екрасным примером строения ния гельминто- кладиевых служит немалион ~Nemalion> — на- Ф.У широко распространенныи род семейства. се гельминт нтокладиевые — тепловодные организ- из них ог- мы. Распространение большинства из них раничено тропиче скими и субтропическими мо- рями. Виды немалиона заходят в умеренные широты и потому л лучше других гельминтокла- диевых известны в морях С СССР. Слизистое червеобразное слоевище немалиона состоит II3 п у пучка клеточных нитей, каждая из Ф.У кото ых растет п р осредством апикальнои клетки. От пучка радиально расходя ятся многочислен- ные вильчато-разветвленные ветви ограничен- ного роста, о р та образующие рыхлый ассимиляцион- ныи слои, по погруженный в слизь. т нижних ассимиляционных ветвей отходят тон- клеток а кие ризоидные нити, служащие для уьр ния центрального пучка ~рис. ственной немалиону тропической водоросли лиагоры ~Liagora ~ ~слоевище местами пропитано Растения немалиона, несущие по ие половые о га- известью. р ны, бывают однодомными и двудомными. Myæ- ские растения р обычно меньше женских. При формировании сперматанги иев на ве шине ас- р симиляционных веточек вырастают короткие в х-, четырехклеточные нити. Кажд аж ая клет- двух-, ч ка этой нити отчленяет вокру г своей вершины 2 — 4 сперматангия (рис. 159, 1). Карпогонные ветви у немалиона состоят из различного чи- сла клеток, от 4 до 7. Они образуются на месте обычных боковых веточек ассимиляционных нитей рис. ( ис. 166, 2). После оплодотворения кар- кой. Из погон д елится поперечной перегородкой. ве хняя двух о ра б зов авшихся клеток только вер участвует в о раз б овании нитей гонимобласта. етка сливается с остальными клет- ижняя клетка ками ка п в одну крупную, богатую содержимым клет- ку слияния, сл ~ жащую источником пи- ля азвивающегося гонимобласта. тания для раз она — это четко 3 елые гонимобласты немалиона — это ч отг аниченные компактны р е об азования, по- груженные в сли зь среди ассимиляционных Ф-Э нитей (рис. с. 166 8 — о). У других водорослеи семеиства нити нити гонимобласта стелются между ассимиляционными нитями, р егус- о аз я негус- тые пучки веточек с карпоспорами на конце. Ка поспоры у немалиона конечные. ар До последнего времени у пр д п е ставителей семеиства было известно только половое раз- множение, и считалось, что тетраспорофиты тепе ь наши зна- у них отсутствуют. Однако теперь ния о цикле раз вития этих водорослей намного У немал иона червеобразного расширились. гих видов об- (Nemalion vermiculare) и у других видов о- на ружены r спорофиты — карликовые растения,
внешне весьма похожие на растения акрохе- тиума и родохортона, несущие тетраспоры или моноспоры. Прорастание карпоспор, так же как и тетраспор, происходит с образованием вначале простых, а затем и разветвленных однорядных нитей, которые очень напоминают растения акрохетиума. Семейство хетангиевые (Chaetangiaceae) Представители этого семейства, близкого к гельминтокладиевым, построены также по много- осевому принципу. Главные их отличия заклю- чаются в том, что поверхностные клетки асси- миляционных нитей слагаются в сплошной ко- ровой слой, а нити гонимобласта окружены оберткой из тонких перепутанных нитей, в ре- зультате чего формируется мешковидный кон- цептакул с отверстием на вершине для выхода карпоспор. Кроме того, семейство характери- зуется наличием обызвествленных форм. Сре- ди них широко известна галаксаура (С-а1ахаи- га) — одна из самых обычных водорослей в тро- пических морях Мирового океана. В большей или меньшей степени обызвествленное и виль- чато-разветвленное слоевище галаксауры имеет весьма характерный облик. Любопытная осо- бенность видов этого рода (а их описано около 70) заключается в том, что гаметофит и споро- фит, будучи оба макроскопическими растения- ми, заметно различаются по анатомическому строению. Это служило поводом для описания их как отдельных видов, и к настоящему вре- мени систематика этого рода изрядно запутана. ПОРЯДОК ГЕЛИДИЕВЫЕ (GELIDIALES) Гелидиевые — небольшая, хорошо очерчен- ная группа красных водорослей. В ней выде- ляют всего одно семейство и 8 родов. Ауксил- лярные клетки, как и у немалиевых, отсутст- вуют, но еще до оплодотворения рядом с кар- погонной ветвью образуются многочисленные мелкоклеточные нити. Это особая питающая ткань, в которой развиваются нити гонимобла- ста. Все гелидиевые — исключительно морские обитатели, ограниченные в своем распростра- нении тропическими морями и теплыми морями умеренных широт. Во многих странах они из- вестны не только ученым. Среди багрянок, дающих желирующие вещества, гелидиевые за- нимают одно из заметных мест, являясь сырьем для добычи самого ценного и высококачествен- ного фикоколлоида — агара. В нашей стране гелидиевые в качестве источника агара не ис- пользуют, поскольку они встречаются только в южных морях СССР и запасы их здесь неве- лики. Центральный и наиболее обширный (около 40 видов) род этой небольшой единообразной группы водорослей — гелидиум. Познакомив- шись с ним, можно составить представление обо всем порядке в целом. Виды гелидиума — не- крупные хрящеватые растения длиной от не- скольких сантиметров до нескольких десятков сантиметров. Первоначально цилиндрическое слоев ище гелидиума имеет заметно выражен- ную в эволюции тенденцию уплощаться, по- этому у многих видов конечные веточки или даже целые слоевища становятся сдавленно-цилинд- рическими и совсем плоскими, приобретая фор- му ланцетовидных листочков. Одной из харак- терных черт рода, как и всего порядка, являет- ся двустороннее и перистое ветвление, хотя у ряда просто устроенных видов преобладает еще неправильное расположение ветвей вокруг все- го слоевища (рис. 167, 1). Кроме того, прими- тивным видам свойственно ползучее слоевище, которое нередко приобретает дорсовентральное строение. Главные ветви при таком строении стелются по субстрату, цепляясь за него допол- нительными ризоидами — присосками. Склон- ность к образованию стелющихся ветвей про- является не только у пр осто организованных видов. Нередко это связано со специфически- ми условиями обитания. Гелидиевые весьма характерны для прибойной скалистой лито- рали, где более других способны удержать- ся стелющиеся, плотно прикрепленные к кам- ням формы водорослей. Все гелидиевые построены по одноосевому типу. Верхушечная клетка главной оси в ре- зультате деления поперечной перегородкой от- деляет вниз клетки. Каждая из этих клеток, в свою очередь, отчленяет вбок четыре перицент- ральные клетки — вначале по одной справа и слева, позднее еще две, при этом все четыре располагаются крестообразно (рис. 152, 1). Из пер ицентр альных клеток возникают развет- вленные ветви ограниченного роста, клетки которых, будучи очень тесно расположенными, образуют псевдопаренхимную ткань. Конеч- ные ответвления этих веточек, плотно смы- каясь, формируют внешнюю кору. В процес- се формирования слоевища клетки централь- ной оси вытягиваются в длину и становятся очень тонкими, вследствие этого различить центральную ось в относительно старых частях слоевища почти невозможно. От клеток вну- тренней части слоевища развивается множе- ство тонких ризоидов. У гелидиума основная масса ризоидов сосредоточена по периферии внутренней части слоевища ближе к коровому слою. Таким образом, особенностью строения гелидиума является образование псевдопарен- химной структуры в результате плотного рас- положения клеточных нитей, слагающих слое- 222
Рис. 167. Строение и размножение Gelidium: 7 — внешний вид растения; 2 — 5 — развитие карпогонной ветви и гонимобласта; 6 — веточка с цистокарпами. а — инициаль- ная клетка карпогонной ветви, б — несущая клетка, в — карпогон, г — клетка слияния, д — клетки нитей гонимобласта, е- карпоспоры, ж — питающая ткань. 22 вище, и сильное развитие толстостенных ризо- идов, выполняющих, по-видимому, механичес- кую роль. Эти особенности хорошо заметны на поперечном срезе через слоевище (рис. 152, 2). Наличие или отсутствие ризоидов и место, где они образуются, служат хорошим таксономи- ческим признаком для разграничения родов ге- лидиевых. Клетки гелидиевых многоядерные, содержат множество пластинчатых хлор опластов и ли- шены пиреноидов. Размножается гелидиум половым и бесполым способом. При этом отличить гаметофиты от спорофитов можно только по органам размно- жения. Тетраспорангии собраны группами на конечных уплощенных веточках вблизи их вер- шин (рис. 158, 8). Здесь тетраспорангии распо- лагаются беспорядочно или правильными па- раллельными рядами. Материнской клеткой тетраспорангия становится поверхностная коро- вая клетка, которая в процессе созревания спо- рангия погружается в глубь коры. Тетраспо- рангии делятся крестообразно с образованием одноядерных тетраспор, или, изредка, двух- ядерных б испо р. Мужские и женские органы размножения раз- виваются на разных растениях. Сперматангии образуются на вершине молодых веточек и на основных ветвях. Поверхностные коровые клет- ки вытягиваются, теряют окраску и превраща- ются в материнскую клетку сперматангия, ко- торая отчленяет от себя один сперматангий. После его выхода та же клетка может образо- вать еще один сперматангий. Образование спер- матангиев ограничено отдельными участками слоевища, которые имеют форму неправильных пятен. На этих участках все коровые клетки становятся материнскими клетками сперматан- гиев. Карпогоны и затем цистокарпы возникают на специальных коротких веточках, которые в зрелом состоянии легко отличить от вегета- тивных по внешнему строению. Обычно они уплощенные, эллиптической или овальной фор- мы. Клетка, дающая начало карпогону, распо- лагается во внутренней коре, ближе к центру слоевища (рис. 167, 2 — 5). Эта инициальная клетка сильно вытягивается и достигает поверх- ности слоевища. Позднее она делится поперек, отчленяя карпогон с длинной трихогиной. При этом нижняя клетка остается в качестве несу- щей и к карпогонной ветви непосредственно не относится. Таким образом, карпогонная ветвь гелидиума одноклеточная. По мере раз- вития карпо гона, еще до оплодотворения, в непосредственной близости от него возникают дополнительные мелкоклеточные нити, обра- зующие рыхлую питающую ткань. После опло- дотворения карпогон увеличивается в разме- рах и постепенно сливается с несущей клеткой.
В результате этого слияния, а в ряде случаев и слияния несущей клетки также с соседними вегетативными клетками образуется крупная клетка слияния. Она продолжает увеличивать- ся в размерах и окружается вегетативными клетками, но нового слияния между ними здесь уже не происходит. Клетка слияния выпу- скает несколько лопастных отростков. На их вершине образуются клеточные перегород- ки, отделяющие первичные клетки гонимо- бласта, из которых после повторных делений развиваются нити гонимобласта. По мере раз- вития они стелются между клетками централь- ной части слоевища, образуя спутанные мас- сы, а не пучки, как у многих флоридеевых. Процесс оплодотворения стимулирует обра- зование питающих клеток. После него они воз- никают в еще большем количестве и сливаются с нитями гонимобласта. По мере созревания го- нимобласта некоторые клетки его нитей уве- личиваются в размерах, вытягиваются и от- членяют в сторону поверхности слоевища одну или несколько крупных карпоспор. В месте раз- вития гонимобласта кора приподнимается и образует крышу цистокарпа. На ранних стадиях прорастания спор гели- диума (как тетраспор, так и карпоспор) форми- руется плотный узелок клеток, от которого вниз отходит один или несколько первичных ризоидов. Из этого узелка очень быстро вы- растают вертикальные побеги. Характер про- растания гелидиевых весьма своеобразен и не похож на таковой остальных багрянок. Если сравнить гелидиевые с остальными крас- ными водорослями, то сразу бросается в глаза, что это не только четко очерченная, но и до- статочно сильно обособленная группа. До по- следнего времени считали, что провести какие- либо родственные связи между порядком гели- диевых и другими багрянками невозможно. Это объясняется тем, что не были изучены детали развития женской генеративной систе- мы и гонимобласта гелидиевых. Однако на осно- ве современных знаний все-таки можно гово- рить о связи гелидиевых с немалиевыми, с од- ной стороны, и с криптонемиевыми — с другой. Так же как у некоторых немалиевых, карпогон гелидиевых после оплодотворения сливается с клеткой карпогонной ветви или с несущей клеткой последней. Гонимобласт в обеих груп- пах состоит из разветвленных нитей диффузного роста. Слоевище здесь и там имеет одноосевое строение. В обеих группах имеются специаль- ные питающие нити, которые тесно связаны с карпогонной ветвью. Нити гонимобласта раз- виваются из карпогона. С другой стороны, существуют филогенетические связи гелидиевых с водорослями семейства дюмонтиевых (порядок криптонемиевых). У последних уже есть аук- силлярные ветви, но морфологически они на- столько подобны питающим нитям гелидиевых, что это сходство позволяет считать их гомоло- гичными структурами, т. е. имеющими одина- ковое происхождение. ПОРЯДОК КРИПТОНЕМИЕВЫЕ (CRYPTONEMIALES) У криптонемиевых уже имеются специаль- ные ауксиллярные клетки. Они закладываются до оплодотворения на особых дополнительных ветвях в отдалении от карпогонных ветвей или в непосредственной близости к ним, образуя прокарп. Нити гонимобласта развиваются из ауксиллярных клеток после слияния их с кар- погоном. Слоевище криптонемиевых имеет различную форму (от цилиндрической до пластинчатой и корковидной) и различное анатомическое строе- ение (как одноосевого, так и многоосевого ти- па). Целая группа водорослей обладает обыз- вествленным слоевищем. Клетки одноядерные и многоядерные, с одним или несколькими хлоро- пластами пластинчатой или чечевицеобразной формы, лишенными пиреноидов. Рост осуществ- ляется одной или несколькими апик альными клетками, в формировании слоевища ряда во- дорослей участвует особая меристема. Для се- мейства кораллиновых весьма характерен ин- теркалярный рост. У криптонемиевых имеется чередование га- метофита и тетраспорофита, сходных по внеш- нему и внутреннему строению. Тетраспоран- гии, крестообразно или зонально разделенные, развиваются по-разному: одиночно, в сорусах, нематециях, концептакулах. Сперматангии раз- виваются так же, как тетраспорангии. Кар- погонные ветви простые или разветвленные, обычно многоклеточные, они возникают отдель- но друг от друга или по нескольку вместе в со- русах, нематециях или концептакулах. После церамиевых криптонемиевые — самый крупный порядок красных водорослей. Он включает 13 семейств, iiO родов и более 900 видов, распространенных по всему Миро- вому океану. Семейство дюмонтиевые (Dumontiaceae) Наиболее примитивное семейство дюмонти- евых включает множество родов, встречающих- ся преимущественно в морях умеренных зон. Они растут в северных частях Атлантического и Тихого океанов, а также у берегов Южной Австралии; гораздо реже встречаются дюмон- тиевые в тропической зоне. Более других в оте- чественных морях известна водоросль дюмон- тия (Dumontia, табл. 20, 2). Ее можно встре- тить на мелководье, часто в литоральных лу- 224
Рис. 168. Строение и размножение Dumontia! 1 — продольный срез у вершины веточки; 2 — карпогонная ветвь; 8 — эта же ветвь п го~~~обл~с~а з усы~~ярой v~e~vm a — карпо б чщ жах во внутренних частях заливов и бухт. Особенно хорошо развивается дюмонтия в биоценозе двустворчатого моллюска мидии. В скоплениях мидий богатая органика и от- сутствие прямого удара волн сочетаются с хо- рошей аэрацией. Мягкие слизистые растения дюмонтии могут быть длиной 20 — 60 сл. Ее слоев ище построено по одноосевому типу (рис. 168, 1). Как и у других водорослей с та- ким строением, верхушечная клетка попереч- ной или косой перегородкой отделяет вниз сег- менты, от каждого из которых образуется по 4 пер ицентральных клетки, дающих начало радиально расходящимся пучкам разветвлен- ных клеточных нитей. В верхних частях клет- ки этих нитей смыкаются и образуется сплош- ной псевдопаренхимный коровой слой. Единст- венная центральная ось с возрастом теряется, и слоев ище становится полым. Вертикальное слоевище возникает из базальной корки кото- Ф рая состоит из стелющейся системы одноряд- ных нитей и плотно расположенных вертикаль- ных нитей, как правило, коротких и мало раз- ветвленных. При сильном развитии новые кор- ки могут налегать на старые. В отличие от вертикального слоевища базальная часть мно- голетняя и служит для переживания неблаго- приятных условий. При прорастании спор первые деления при- водят к образованию шаровидных клеточных структур, которые при дальнейшем развитии превращаются в к орко видную подошву. От- дельные нити этой подошвы, разрастаясь, да- ют вертикальное слоевище дюмонтии. По сложности организации дюмонтия занимает срединное положение в семействе: в нем можно выделить и роды, построенные более просто, чем дюмонтия, и более специализированные. У низ- коорганизованных дюмонтиевых центральная ось покрыта мутовками разветвленных ассими- ляционных нитей, которые располагаются сво- бодно и соединены лишь сильно развитой слизью. Эти формы очень напоминают растения батрахоспермума. У более высокоорганизован- ных видов эти нити смыкаются, образуя сплош- ной коровой слой. У еще более сложно устроен- ных дюмонтиевых, как, например, у аркти- ческого вида дилсеи съедобной (Dilsea edulis), на ранних стадиях рост пластинчатого слоеви- ща осуществляется верхушечной клеткой, но очень быстро деятельность ее прекращается 7 а из конечных клеток ассимиляционных ни- тей, лежащих ближе всего к верхушечной клетке, формируется краевая поверхностная меристема, за счет которой происходит рост растений. При половом размножении дюмонтии спер- матангии образуются на мужских растениях из поверхностных коровых клеток. Материн- ская клетка сперматангия последовательно от- членяет до трех сперматангиев. При сильном их развитии почти вся поверхность муж-
ских растений бывает покрыта сперматангиями. Обычно мужские растения дюмонтии мельче, чем женские. Особенности развития женской репродук- тивной системы и карпоспорофита дюмонтие- вых заключаются в том, что карпогонная и аук- силлярная ветви развиваются по всему сло- евищу, во внутренней его части отдельно друг от друга (причем ауксиллярных ветвей больше, чем карпогонных). После оплодотворения карпогон сливается со средней клеткой карпо- гонной ветви, иногда с несколькими. Из обра- зовавшейся клетки слияния вырастают две или три длинные соединительные нити, которые достигают ауксиллярных ветвей (рис. 168, 2, 8). Собственно ауксиллярной клеткой служит одна из срединных клеток ауксиллярной ветви. После слияния с соединительными нитями из ауксил- лярной клетки развивается несколько корот- ких разветвленных нитей гонимобласта. При созревании все клетки этих нитей превраща- ются в карпоспоры. Зрелые гонимобласты ли- шены специальной обертки и погружены в ко- ровую ткань растения (рис. 168, 4, о). Если проанализировать строение карпогонной и аук- силлярной ветвей у всех дюмонтиевых, становит- ся очевидным, что это структуры гомологичные, т. е. имеющие одинаковое происхождение. Тетраспорангии образуются из поверхност- ных клеток коры и разбросаны по слоевищу. В месте образования тетраспорангиев кора толще, чем в стерильной части слоевища. У дю- монтии они крестообразно разделенные, но у других членов семейства встречаются и зо- нально разделенные тетраспорангии. Органы полового и бесполого размножения у дюмонтии образуются на сходных по строению макроскопических растениях, т. е. эта водо- росль обладает изоморфной сменой форм раз- вития. Это нельзя сказать обо всех членах се- мейства. У акросимфитона (Acrosymphyton) обнаружен тетраспорофит, который заметно от- личается от половых растений и очень напоми- нает тетраспорофиты некоторых видов бон- немезонии. Дюмонтиевые занимают несколько обособлен- ное положение в порядке криптонемиевых. Они обладают однокарпогонной системой — каждая несущая клетка образует только одну карпогон- ную ветвь. Все остальные семейства порядка (так полагают в настоящее время) связаныбо- лее тесным родством и имеют общего предка, который характеризуется многокарпогонной женской системой, когда на одной несущей клетке развивается несколько карпогонных вет- вей. В ходе эволюции эта многокарпогонная система претерпевала изменения и у многих родов вторично преобразовалась в однокарпо- гонную. Семейство каллимениевые (Kallymeniaceae) Семейство каллимениевых является централь- ным семейством порядка, обладающим много- карпогонным строением женской половой систе- мы. Из шести относимых к семейству родов только три занимают в нем достаточно твердое положение. Это каллиления (Kallymenia), пу- гетпия (Pugetia) и каллофиллис (Callophyllis). Лучше всего изучены виды первого рода, поэто- му на них удобнее всего рассмотреть особенно- сти семейства в целом. Пластинчатые листовидные растения калли- иении почковидной (Kallymenia reniformis) мож- но встретить в северной части Атлантического океана и в Средиземном море. Растет она в суб- литорали на значительных глубинах, но дохо- дит и до нижней границы литорали. Молодые растения каллимении состоят обычно из не- большой цельной пластины, сидящей на корот- кой ножке и прикрепленной к грунту диско- видной подошвой. К концу первого сезона рос- та на пластине развиваются органы размноже- ния. После периода плодоношения большая часть пластины отпадает. В начале следующего сезона по краю оставшейся части пластины воз- никают пролификации, сама пластина разры- вается на лопасти и все слоевище становится многораздельным. Конструкция пластины мно- гоосевая, это можно видеть на поперечном сре- зе через край пластины (рис. 169, 1). Клетки центральных нитей длинные и узкие. Развет- вляясь к поверхности, они образуют внутрен- нюю кору из одного или двух слоев округлых клеток и внешнюю, также из одного-двух слоев, но уже мелких плотно расположенных клеток. Отдельные клетки центральных ни- тей очень сильно увеличиваются в размерах и выпускают множество отростков, которые соединяются вторичными порами с обычными клетками и с отростками других таких же кле- ток. В результате образуется сплошная система звездчатых клеток. Роль этой «ткани», по-ви- димому, механическая: она помогает довольно тонким пластинам противостоять действию волн. Интересно, что по сравнению с другими пластинчатыми багрянками каллимения чаще сохраняет целостность пластины. Возможно, причина этого кроется как раз в наличии свое- образной механической ткани в центре слоеви- ща. В росте каллимении важную роль играет краевая меристема. Особый интерес представляет развитие жен- ской половой системы и гонимобласта у калли- мении. У нее, как и у многих других водорослей этого порядка, несущая клетка карпогонной ветви возникает от клетки внутренней коры и, увеличиваясь в размерах, ориентируется внутрь слоевища. Каждая несущая клетка отчленяет 226
несколько первичных клеток карпогонных вет- вей. Их может быть и шесть, и шестнадцать. Эти клетки, в свою очередь, делятся, образуя трехклеточные карпогонные ветви, и только некоторые из них так и остаются в одноклеточ- ном состоянии, не подвергаясь дальнейшему развитию (рис. 169, 2). Трихогины всех карпо- гонов одной несущей клетки вытягиваются одновременно. Часто это длинные, спирально закрученные образования. Сразу после оплодо- творения происходит кратковременное соедине- ние карпогона с нижней клеткой карпогонной ветви. Вскоре после этого нижние клетки кар- погонных ветвей образуют короткие выросты, которые вытягиваются и сливаются с несущей клеткой. Одновременно с этим трихогина, кар- погон и подкарпогонная клетка уменьшаются в размерах и постепенно дегенерируют. После соединения нижних клеток всех карпогонных ветвей с несущей образуется очень крупная клетка слияния, которая более чем в 2 раза превышает диаметр несущей клетки. От клетки слияния образуется множество лопастных и трубчатых выростов. Раньше считали, что кал- лимении свойствен прокарп, так как клетку слияния принимали за ауксиллярную, а лопаст- ные выросты, идущие от клетки слияния,— за первичные нити гонимобласта. Но потом было установлено, что это соединительные нити, растущие в направлении ауксиллярной клетки и сливающиеся с ней. Ауксиллярная система располагается отдельно, независимо от карпо- гонной. В общей сложности от одной клетки слияния может отходить более десятка соедини- тельных нитей. Они простираются среди нитей центральной части слоевища и могут быть при- няты за вегетативные нити. Соединительные нити толще вегетативных, при значительной длине они не разделяются клеточными перего- родками, весьма редко ветвятся и ие соединя- ются вторичными порами с вегетативными клет- ками. Ауксиллярные клетки также отделяются от клеток внутренней коры. Они очень напоми- нают несущие клетки карпогонных ветвей. Подобно им, ауксиллярные клетки также отчле- няют несколько клеток, которые, однако, даль- нейшим делениям не подвергаются. Нет сом- нения в том, что ауксиллярная и карпогон- ная системы — гомологичные образования. Ве- роятно, первая сформировалась из второй в ре- зультате утраты функции оплодотворения и приобретения питающей функции. После слия- ния с ауксиллярной клеткой соединительная нить вблизи места слияния заметно утолщается и делится, отчленяя от себя первичные клетки гонимобласта. Таким образом, у каллимении гонимобласт берет начало не от ауксиллярной клетки, а от соединительной нити, но только 2 Рис. 169. Kallymenia: 1 — поперечный срез слоевища; 2 — карпогонная система; 8— поперечный срез через зрелый цистокарп. а — несущая клетка, б — карпогонная ветвь. после слияния ее с ауксиллярной клеткой. В зрелом гонимобласте трудно различить нит- чатое строение, скорее он имеет вид скопления клеток. По мере созревания гонимобласта ок- ружающая вегетативная ткань раздувается и образуется тонкий перикарп. Клетки гонимо- бласта превращаются в карпоспоры, крупная ауксиллярная клетка дегенерирует, и остается скопление карпоспор, разбросанных почти по всему слоевищу (рис. 169, 8). Крестообразно разделенные тетраспорангии развиваются на бесполых растениях и разбро- саны среди клеток внешней коры, хотя разви- тие их начинается во внутренней коре. Если сравнить между собой все водоросли семейства, то становится очевидным, что наи- более примитивные члены семейства — это те, у которых внутренняя часть слоевища построе- на узкими ризоидными нитями, на одной несу- щей клетке образуется несколько карпогонных ветвей, а карпогонная и ауксиллярная ветви удалены друг от друга. Наоборот, высокоор- ганизованные представители характеризуются сердцевиной паренхимного вида, одной кар- погонной ветвью на несущей клетке и наличием прокарпа. В соответствии с этим каллимеиию следует считать примитивным представителем семейства, тогда как наиболее специализиро- ванным родом оказывается каллофиллис. Виды каллофиллиса — глубоководные орга- низмы. В нашей стране наибольшего разнооб- 27
'р А' о О 05 Рис. 170. Ст о р ение и размножение Grataeloupia.' 1 — поперечный срез слоевища 2— 3 — а уксиллярна я система; 4 — опло о щ, — карпогонная система — п TBOpeHHbIfI ар о OH 7 карп. а — карпогон б— 7 в — ауксиллярная клетка г- — несущая клетка, поспоры. , г — соединительные нити, д — кар- разия они достигают в морях Дальнего Восток однако общее количество живущих здесь ви- дов назвать весьма затруднительно: обитая в сублиторали труднодоступных районов, этот интересный род остается пока поч ным. В пе ево ти неизучен- ным. переводе на русский язык название этой водоросли звучит как «красиволи т истныи». ло иллиса лоские пластинчатые слоевища калло и атно раз- отличаются от каллимении многок ат ветвленной и разделенной пластиной и вслед- ствие этого очень разнообразной ~о мой. Р слоеви а ща, как у каллимении, происходит в е- зультате деятельности краевой меристемы. На поперечном срезе видно, что внутренняя часть слоевища построена из крупных округлых кле- ток, между которыми простираются нередко в б 7 разные ольшом количестве, тонкие гидо об клеточные нити (рис. 153, 2). Развитие карпогонной системы и первые ста- дии после оплодотворения здесь принципиально не отличаются от этих процессов у каллиме- нии. о в целом между двумя родами имеются серьезные различия, заключающиеся в том, что несущая клетка у большинства видов дает только одну, а не несколько карпогонных вет- вей и гонимобласт начинает развиваться из первой клетки слияния, которая образуется из соединения нижней клетки карпогонной ветви, несущей клетки и дополнительных клеток от- 1 членившихся ранее от несущей. При таком ходе развития гонимобласта несущая клетка карпогонной ветви является одновременно и ауксиллярной. Иными словами, у каллофилли- са можно говорить о наличии прокарпа. М ужские растения каллофиллиса мельче жен- ских, так же как и тетраспоровых. Сперматан- гии образуются по 2 — 3 от поверхностных кле- ток внешней коры. к рестообразно разделенные тетраспорангии разбросаны почти по всему слоевищу. Образу- ются они от клеток внутренней коры как одно- клеточная ветвь. мейство криптонемиевые (Cryptonemiaceae) Водоросли семейства криптонемиевых имеют однокарпогонную половую систему, которая, по-видимому, сформировалась в результате уменьшения числа карпогонных ветвей в пер- воначально многокарпогонной структуре. Семейство криптон емиевых — одно из об- п~ирных в порядке. В нем известно 19 родов, распространенных в теплых морях. В южные моря Советского Союза заходят лишь очень немногие роды с наиболее широкими ареалами. первую очередь из них следует упомянуть род грателупия (Grataeloupia). Внешняя фор- ма видов грателупии весьма разнообразна и служит важным критерием для их разграниче-
ния. Слоевище кожистое или хрящеватое, крупное, длиной от 5 до 75 см, уплощенное или пластинчатое, сильно и неправильно развет- вленное или простое. Построено оно по много- осевому плану. На поперечном срезе через слоевище видно, что внешняя кора состоит из трех-четырех слоев плотно расположенных изо- диаметрических клеток. Внутренняя кора бо- лее рыхлая, так же как и центральная часть. Клетки центральных осевых нитей имеют длин- ные отростки своеобразной звездчатой формы. Среди них развиваются многочисленные ризо- иды, придающие центральной ткани сетчатое строение (рис. 170, 1). Женская репродуктивная система распола- гается на клетках внутренней коры. Несущая клетка двухклеточной карпогонной ветви об- разует, кроме нее, еще несколько дополнитель- ных клеточных нитей, собранных в виде пучка (рис. 170, 2). Точно так же и в ауксиллярной системе одна базальная клетка несет на себе пучок клеточных нитей, при этом более гус- той, чем в карпогонной системе. Эта базальная клетка и служит ауксиллярной (рис. 170, 8). Карпогонная и ауксиллярная ветви отделены друг от друга. Прокарп у изученных предста- вителей семейства до сих пор не обнаружен. Хотя отсутствие про карпа следует считать признаком, свидетельствующим о незначитель- ной специализации, характер карпогонной и ауксиллярной ветвей позволяет думать, что обе происходят из многокарпогонной струк- туры, которую мы встречаем у каллимениевых. Это предположение вызвано наличием допол- нительных клеточных нитей, которые отходят наряду с карпогонной и ауксиллярной ветвями от их несущих клеток. От оплодотворенного карпогона тянутся соединительные нити, ко- торые сливаются с ауксиллярной клеткой (рис. 170, 4). После слияния из нее развиваются разветвленные нити гонимобласта, собранные густым пучком. Созревая, многие клетки го- нимобласта превращаются в карпоспоры. Спе- циальной обертки вокруг гонимобласта не об- разуется, но по мере того, как нити гонимобла- ста растут к поверхности слоевища, коровая ткань приподнимается и в ней образуется от- верстие для выхода карпоспор (рис. 170, о). Бесполое размножение осуществляется тет- распорами, которые возникают в крестообразно разделенных спорангиях и разбросаны по все- му слоевищу. Иногда в месте образования тет- распор наружная кора утолщается, приобретая нематециевидную форму. Семейство хореоколаксовые (Choreocolaceae) Небольшое семейство хореоколаксовых вклю- чает два хорошо известных паразитических ро- да — хореоколакс (Choreocolax) и гарвейелла (Harveiella). Оба рода паразитируют на родо- меловых из порядка церамиевых, образуя на их поверхности бесцветные или слабоокрашен- ные подушечки диаметром до 1 мм. Часть слоевища этих водорослей погружена в ткани хозяина. Она представлена разветвленными нитями, которые соединяются с клетками хо- зяина вторичными порами или специальными присосками. От эндофитных нитей разраста- ются на поверхности подушковидные или боро- давчатые образования, устроенные очень про- сто и состоящие из вильчато-разветвленных нитей, расходящихся веером. К вершине клет- ки нитей уменьшаются в размерах и, смыка- ясь, образуют мелкоклеточную кору. Систематическое положение семейства хорео- колаксовых нельзя считать строго установлен- ным. При развитии женской половой системы несущая клетка, кроме карпогонной ветви, дает начало дополнительным нитям, как у крипто- немиевых. Но, с другой стороны, ауксилляр- ная клетка отделяется от несущей, и только после оплодотворения, а это свойственно це- рамиевым водорослям. Возможно, это семейст- во следует исключить из порядка криптонемие- вых и поместить в порядок церамиевых, тем более что у красных водорослей паразитические виды всегда оказываются родственными орга- низмам-хозяевам. До тех пор пока вопрос этот не решен, семейство Choreocolaceae по тради- ции остается в порядке криптонемиевых. Семейство сквамариевые (Squamariaceae) Водоросли четырех семейств порядка обла- дают одним весьма характерным признаком— органы размножения у них развиваются в не- матециях, т. е. среди специальных нитчатых образований на поверхности слоевища. Наи- более обширными и своеобразными среди этих семейств являются сквамариевые и кораллино- вые. Семейство сквамариевых характеризуется сте- лющимся корковидным слоевищем, более или менее плотно прикрепленным к субстрату нижней поверхностью. Корки сквамариевых мягкие, и только очень немногие представите- ли семейства имеют слегка обызвествленное слоевище. У пейссоннелии (Peyssonnelia) вна- чале образуется однослойная пластина, от кле- ток которой поднимаются простые или раз- ветвленные вертикальные нити, собранные в плотную псевдопаренхимную ткань. На ниж- ней поверхности слоевища развиваются ри- зоиды (рис. 171, 1). Когда сквамариевые, в частности пейссонне- лия, созревают, на поверхности слоевища мож- но видеть образование цематециев. Группы 229
поверхностных клеток прорастают с образова- нием многоклеточных вертикальных нитей. При образовании сперматангиев клетки нитей нема- теция отчленяют вбок несколько перицент- ральных, которые служат материнскими клет- ками сперматангиев (рис. 159, 4). Нити женских нематециев у пейссоннелии состоят из 6 — 8 клеток. Самая нижняя клетка такой нити производит карпогонную или аук- силлярную ветвь. Обе ветви, как правило, че- тырехклеточные (рис. 171, 2). Оплодотворенный карпогон соединяется вначале с двумя клетка- ми карпогонной ветви. Затем эта клетка слия- ния выпускает соединительные нити, которые соединяются с ауксиллярной клеткой — вто- рой снизу на ауксиллярной ветви (рис. 171, 8). В месте слияния начинается развитие гонимо- бласта. Он состоит из 8 — 12 клеток, превра- щающихся в конечном итоге в карпоспоры (рис. 171, 4). Крестообразно разделенные тетраспорангии развиваются также среди нитей нематеция от поверхностных клеток слоевища (рис. 158, б). Из всех многочисленных водорослей семей- ства половое размножение известно только у пейссоннелии. Остальные водоросли размно- жаются тетраспорами. Этот факт позволяет предположить, что сквамариевые, размно- жающиеся только бесполым путем, явля- ются формами развития других водорослей, а все семейство — искусственной группой. И действительно, в последнее время у некото- рых багрянок, в частности у гигартиновых, найдены корковые стадии в циклах развития. Рис. 171. Строение и размножение P eyssonnelia: 1 — вертикальный срез через корку; 2 — карпогонная (а) и ау- ксиллярная (б) ветви; 8 — слияние соединительной нити с ауксил- аярной клеткой; 4 — гонимобласты. в — карпоспоры. Семейство кораллиновые (Corallinaceae) Это семейство занимает в порядке особое ме- сто. Оно характеризуется огромным многооб- разием форм. От остальных водорослей поряд- ка, как и вообще от всех багрянок, коралли- новые отличить легко: слоевище их настолько пропитано известью, что их скорее можно при- нять за кораллы или камни, чем за водоросли. Кроме обызвествления, эти водоросли облада- ют еще одной своеобразной чертой: органы как полового, так и бесполого размножения у них образуются в особых вместилищах — концеп- такулах, которые сообщаются с внешней сре- дой одним или несколькими отверстиями. Систематики делят семейство на два подсе- мейства, различающиеся морфологией слоеви- ща. Корковые и кораллоподобные водоросли объединяют в подсемейство мелобезиевых (Me- lobesioideae), а водоросли с членистым строе- нием слоевища, растущие в форме кустиков, входят в подсемейство кораллиновых (Corallinoi- deae) (табл. 24). Такое деление водорослей се- мейства весьма удобно, потому что признак,
на котором оно основывается, — внешнее строе- ние — легко бросается в глаза, но вряд ли его можно считать естественным. Различаясь строе- нием взрослого слоевища, водоросли обоих под- семейств обнаруживают сходство по некоторым другим довольно важным признакам. Вероят- но, эволюция в семействе шла по нескольким направлениям, каждое из которых включало как корковые, так и членистые формы. Данных об этом сейчас накапливается все больше, но их еще недостаточно, поэтому пересмотр класси- фикации семейства — дело будущего. Как мелобезиевые, так и кораллиновые встречаются во всех морях земного шара от Арктики до тропиков, но более разнообразно представлены в морях теплых широт. Подсемейство мелобезиевые (Melobesioideae) Род мелобезия (Melobesia) отличается наибо- лее простым строением. Виды этого рода растут в виде тонких корочек — чешуек на морских травах, водорослях, раковинах, плотно приле- гая к ним всей нижней поверхностью. В начале развития слоевища на субстрате образуется небольшой клеточный диск. Этот диск растет в ширину за счет того, что от его краевых кле- ток радиально развиваются стелющиеся нити. Они могут располагаться рыхло, но чаще соб- раны в плотную псевдопаренхимную пластину (рис. 172, 1). Таким образом, на первых порах корка мелобезии состоит из одного слоя клеток. Затем каждая клетка этой однослойной пласти- ны отчленяет наверх по одной мелкой клетке. Эти клетки, если смотреть на пластину сверху, покрывают только небольшой передний уча- сток диска и при этом никогда не обызвествля- ются. Называют эти клетки покровными. Позд- нее в клетках диска начинаются деления гори- зонтальными перегородками и образуются короткие плотно сомкнутые вертикальные нити. Корка становится многослойной (рис. 172, 2). Вертикальные нити растут посредством апи- кальных клеток и интеркалярно. Мелобезиевые представляют собой один из редких примеров интеркалярного роста у красных водорослей. Таким образом, в слоевище мелобезии, про- следив за его развитием, можно различить два слоя — первичный базальный и вторичный вер- тикальный. Первый называют — r и п о т а л- л и е м, второй — п е р и т а л л и е м. У остальных мелобезиевых принцип строения остается тот же: все они имеют эти два слоя. Не у всех водорослей подсемейства гипоталлий однослойный — коли- чество клеточных слоев в базальной части раз- лично у разных родов и служит важным крите- рием для их разделения. Когда слоевище сфор- мировано, отличить гипоталлий от периталлия на вертикальном срезе через слоевище можно по форме и цвету клеток, а также по характеру их расположения. Характерной особенностью мелобезиевых яв- ляется образование особых клеток, несущих во- лоски,— трихоцитов. Это крупные, бедные со- держимым клетки, лишенные покровных. Они образуются на поверхности слоевища, чаще по краю пластины, но по мере дальнейшего ее ро- ста в ширину обрастают соседними нитями и оказываются во внутренних частях пластины. Волосок не является самостоятельной клеткой, он представляет собой прозрачный более или менее длинный вырост трихоцита (рис. 172, 8). Через некоторое время волосок отпадает, и толь- ко перед моментом отпадения в его основании возникает поперечная перегородка. После от- падения волоска трихоциты нетрудно разли- чить по крупным по сравнению с соседними клетками размерам и бедному содержимому. Клетки гипоталлия, так же как и соседних нитей периталлия, соединяются между собой, но соединение это происходит не посредством вторичных пор, а в результате частичного раст- ворения оболочек и прямого слияния соседних клеток. С процессом размножения у мелобезиевых связано образование концептакулов. На месте будущего бесполого концептакула из ниж- них клеток периталлия образуются, чередуясь с обычными вегетативными, фертильные (плодо- носные) нити. Каждая такая нить (у мелобезии она может быть очень короткой) оканчивается молодым тетраспорангием, над которым еще в самом начале его образования формируется ма- ленькая клетка с густым содержимым. Вегетатив- ные нити, окружающие спорангии, продолжают расти, но по мере того, как спорангий вытя- гивается в длину, они дегенерируют и исчеза- ют и таким образом образуется полость концеп- та кула. Нетронутыми остаются только верх- ние части стерильных нитей, расположенные выше тетраспорангиев. Эта ткань формирует крышу концептакула. Мелкие клетки над тет- распорангиями постепенно теряют цитоплазму и ядро и, оказываясь расположенными среди нитей крыши концептакула, приобретают вид слизистых пробок, закрывающих многочислен- ные поры, через которые выходят зонально поделенные спорангии, выдавливая при этом пробки (рис. 172, 4). Половые концептакулы мелобезии откры- ваются одним отверстием. У мелобезии Л~ежоли (Melobesia lejolisii) в некоторых местах слое- вища клетки, расположенные ниже покровных, начинают делиться с образованием коротких фертильных нитей (рис. 173, 1). В этих нитях от каждой несущей клетки отчленяется наружу по две, очень редко по три мелкие клетки. Одна из них развивается в двухклеточную карпогон-
Рис. l72. Строение Me|obesia: 1 — корка с поверхности; 2 — вертикальный срез через корку; 3 —; 4 — концептакул с тетраспорангиями. а — волоски, б — покровные клетки. ную ветвь, другая или другие так и остаются Ее- поделенными. Их считают недоразвитыми кар- погонными ветвями. Только изредка можно видеть, как от одной несущей клетки отходит более чем одна карпогонная ветвь. Обычно развитие карпогoHHIIx ветвей происходит ин- тенсивнее в центре плодоносного участка (или концептакула), чем на периферии, где обычно карпогонные ветви вообще не развиваются. По мере развития органов размножения окру- жающие клетки вытягиваются и разрастаются над плодоносным участком в виде клеточной крыши, оставляя лишь одно отверстие в центре. Так формируются женские концептакулы (рис. 173, 2). После оплодотворения карпогон сливается с несущей клеткой, которая таким образом служит ауксиллярной (рис. 173, 8). Затем все ауксиллярные клетки сливаются между собой в одну огромную клетку слияния. Нити гонимобласта вырастают по краю клетки слияния. На конце этих нитей образуются крупные карпоспоры (рис. 173, 4). Мужские концепта кулы образуются на тех же пластинах, что и женские, в непосредствен- ной близости от них. Из клеток, расположенных ниже покровных, развиваются клеточные нити. Самая нижняя клетка функционирует как клет- ка-ножка, ряды выше расположенных клеток являются материнскими клетками сперматан- гиев. Около 25 видов мелобезии обитает в разных морях земного шар~.. Род литотамнион (Lithothamnion) близок к мелобезии. Это название употребляют весьма часто, но не всегда точно. Часто под ним пони- мают разные известковые водоросли, имеющие вид массивных корок. По сравнению с мелобе- зией корки литотамниона значительно толще, часто они бородавчатые или снабжены более или менее длинными кораллоподобными выро- стами. Гипоталлий и периталлий многослой- ные, покровные клетки крупнее, чем у мело- безии, и покрывают поверхность слоевища це- ликом. Соединение клеток происходит в резуль- тате их слияния. Концептакулы располагаются на поверхности слоевища и устроены по тому же принципу, что у мелобезии. Виды литотамниона — а их известно около 100 — описаны из различных морей, но многие из этих описаний сомнительны и требуют про- верки. Род литофиллум (Lithophyllum) по внешнему виду не всегда легко отличить от литотамнио- на. Толщина гипоталлия и периталлия в его многослойном слоевище варьирует таким об- разом, что отделить этот род от других при отсутствии концептакулов практически невоз- можно. Несмотря на большое сходство с лито- тамнионом, между этими родами имеются прин- ципиальные различия. У литофиллума иначе происходит прорастание спор; клетки слоеви- ща соединяются посредством вторичных пор; как бесполые, так и половые концептакулы сообщаются с внешней средой одной порой.
Рис. 173. Развитие гонимобласта Melobesia: 1 — начало образования фертильных нитей; 2 — концептакул со зрелыми карпогонами; 3 — часть концептакула после оплопо- творения. 4 — концептакул со зрелым гонимобластом. а — карпогон с трихогиной, б — несущая клетка, в — ауксилляр- 7 ная клетка, г — соединительная нить, д — клетка слияния, е — карпоспоры. Внутреннее строение концептакулов известно не во всех подробностях. В тетраспоровых кон- цептакулах спорангии образуются только по периферии дна концептакула. Карпоспоран- гии также формируются по краю клетки слияния. В отличие от литотамниона водоросли из группы литофиллума имеют мелкие споры. Подсемейство кораллиновые (Corallinoideae) Членистые кораллиновые характеризуются вертикальным разветвленным слоевищем, в ко- тором обызвествленные членики чередуются с лишенными извести сочленениями. В морях СССР чаще других можно найти кораллину и амфироа. Это наиболее важные представители подсемейства. Каждый из этих родов соединя- ет в себе признаки целой группы родов и отра- жает отдельную линию эволюции. Род кораллина (Corallina) включает невысокие растения, очень красивые благодаря перистому и вильчатому ветвлению (табл. 24; рис. 174,1). Вертикальное слоевище образуется на базаль- ной части, которая здесь развита в меньшей степени, чем у корковых, и намного проще уст- роена. При прорастании образуется многослой- ная корка, в которой можно различить гипо- таллий и периталлий. Вертикальные побеги построены по фонтанному типу. Членики всегда пропитаны известью, хотя обызвествление обыч- но не затрагивает внутренних частей и ограни- чено корой. Нити внутренней части слоевища состоят из клеток одинакового размера, рас- положенных поперечными рядами (на продоль- ном срезе. Несколько слоев коровых клеток заканчивается у поверхности одним слоем по- кровных клеток. Последние отсутствуют толь- ко над апикальными клетками, осуществляю- щими рост в длину. Центральные и коровые клетки часто сливаются друг с другом. Сочле- нение никогда не обызвествляется и состоит из одного ряда параллельно расположенных длинных толстостенных клеток. В начале обра- зования сочленения оно бывает покрыто корой, которая впоследствии разрушается (рис. 174, 2, 8, 4). У кораллины концептакулы образуются на вершине веточек. При образовании бесполых концептакулов вершины конечных веточек уплощаются, апикаль-ные клетки перестают расти и отчленяют начальные клетки тетраспо- рангиев. Одновременно с развитием последних происходит разрастание окружающих клеток, в результате которого образуется крыша концептакула. Спорангии делятся зонально (рис. 158, 6). Не все материнские клетки тетра- спорангиев достигают зрелости. Часть из них остаются стерильными. Половые растения кораллины более непра- вильно разветвлены и гуще покрыты концеп- такулами. Развитие женского концептакула на- чинается с того, что из апик альных клеток центрального пучка нитей образуются плодо-
о в м~0 о0 00 оf 4 ао О Рис. 174. Строение кораллиновых: 1 — внешний вид веточки Corallina; 2 — 5 — продольные срезы (2 — через вершину веточки Corallina, 8 — через членик, 4— через сочленение, Б — через вершину веточки Amphiroa — видны зоны коротких и длинных клеток и двухрядное сочленение). а — апикальные клетки, б — покровные клетки. носные нити. Их строение и развитие подобны таковым у мелобезии. В зрелом концептакуле различается крупная клетка слияния с нитя- ми гонимобласта по краю. Мужские концепта- кулы кораллины имеют на вершине довольно длинный клювик, в середине которого прохо- дит длинный выводной канал. Сперматангии образуются на дне и боковых стенках концеп- такула и имеют весьма характерную форму. Они расширены (наподобие головки) на перед- нем конце и вытянуты в длинный узкий хвост на заднем. Род амфироа (Amphiroa) отличается меньшим числом видов и более ограниченным геогра- фическим распространением. Вильчато-развет- вленные растения амфироа также состоят из отчетливых члеников, однако в их образовании прослеживаются еще некоторые примитивные черты, сближающие этот род с нечленистыми кораллиновыми. Клетки центральных нитей расположены упорядоченно, дугообразными поперечными зонами, причем зона длинных клеток чередуется с зоной коротких. Снаружи центральный пучок ограничен мелкоклеточной корой. Но если у кораллины первое сочлене- ние формируется очень рано, одновременно с возникновением вертикального слоевища, то у амфироа сочленения возникают, когда ткани проростков уже достаточно сильно дифферен- цировались. Сочленения состоят из несколь- ких рядов клеток. Имеются вторичные поры (рис. 174, б). Концептакулы располагаются на боковой поверхности ветвей и формируются за счет ко- ровых клеток. На начало развития любого кон- цептакула указывает секреция поверхностны- ми клетками слизистых веществ, которые обра- зуют своего рода шапку в месте будущего кон- цептакула. В то же самое время начинает формироваться свод концептакула. Группа ко- ровых клеток, лежащих под поверхностными, вытягивается перпендикулярно поверхности слоевища. Это клетки, за счет которых позднее образуется полость концептакула. Над ними продолжается образование мелких коровых клеток, формирующих свод концептакула. Дальнейшее развитие бесполых и половых кон- цептакулов идет разными путями. Гетраспо- рангии берут начало от коровых клеток, ле- жащих на дне концептакула, и располагаются кольцом по периферии дна концептакула. По мере созревания тетраспорангиев длинные клет- ки над ними разрушаются с образованием по- лости. В процессе развития мужских концепта- кулов с момента возникновения материнских клеток сперматангиев начинается разрушение будущей полости концептакула, так же как и клеток первичного свода, так что крыша сформированного концептакула образуется вто- рично в результате разрастания окружаю-
щих тканей. Сперматангии — мелкие овальные клетки — развиваются на дне концептакула. Женские концептакулы формируются так же, как мужские. В зрелом концептакуле можно видеть крупную клетку слияния с кольцом нитей гонимобласта, отходящих по ее краю. Стараясь понять филогенез криптонемиевых, ученые исходят из того, что различные направ- ления развития в порядке (впрочем, как и во всем классе флоридеевых) находят отражение прежде всего в характере строения женской половой системы и в особенностях развития гонимобласта. Основными критериями служат наличие или отсутствие прокарпа, число кле- ток в карпогонной и ауксиллярной ветвях, место образования ауксиллярной клетки. Раньше все семейства порядка делили на две группы — имеющие прокарп и не име- ющие такового. Но чем глубже изучают эти водоросли, тем очевиднее становится, что специализация ауксиллярной системы мо- жет идти у разных водорослей независимо. И действительно, во многих семействах наряду с родами, у которых карпогонная и ауксилляр- ная ветви образуются отдельно, можно найти роды, уже имеющие в той или иной мере разви- тый прокарп. По современным воззрениям (за- метим, что их также нельзя считать непогре- шимыми), одним из важных признаков, на ко- торый опираются при построении классифика- ции криптонемиевых, является одно- или мно- гокарпогонный характер женской половой си- стемы. Считается, что многокарпогонный аппа- рат, свойственный многим криптон емиевым, возник очень давно из однокарпогонного, но позднее у некоторых родов однокарпогонная система могла возникнуть вновь в результате редукции многокарпогонной. Наиболее прими- тивным семейством порядка считают дюмон- тиевые. Они имеют ауксиллярные ветви, изо- лированные от карпогонных, и оба типа ветвей состоят из большого числа клеток. Однокарпо- гонная система здесь — первичное образова- ние. Наиболее отчетливо многокарпогонная структура проявляется в семействе каллиме- ниевых; в других семействах с одной карпо- гонной ветвью на несущей клетке наличие на ней же нескольких стерильных клеточных нитей свидетельствует о том, что, по крайней мере в некоторых случаях, эта структура обра- зовалась в результате редукции числа карпо- гонных ветвей. Таким образом, эволюция в пределах поряд- ка криптонемиевых шла в направлении обра- зования, во-первых, прокарпа, во-вторых, мно- гокарпогонной структуры, а затем вторичного ее упрощения и, в-третьих, усложнения анато- мического строения. Как известно, эволюция различных признаков происходит независимо, и в порядке криптонемиевых сплошь и рядом можно встретить водоросли, которые по одним признакам следует считать высокоорганизован- ными, тогда как по другим — примитивными. В результате оказывается, что понять эволю- цию группы в целом и истинные связи семейств друг с другом трудно. Несомненно, современ- ную классификацию порядка нельзя признать до конца разработанной. Связь порядка с другими багрянками также остается неясной, хотя имеются свидетельства сходства отдельных криптонемиевых с водорос- лями порядка гигартиновых. ПОРЯДОК ГИГАРТИНОВЫЕ (GIGARTINALES) Гигартиновые — чрезвычайно разнообраз- ная группа с нечеткими границами. Это раз- нообразие проявляется во внешнем и внутрен- нем строении, в строении органов размноже- ния, в особенностях развития гонимобласта. Уже само количество входящих в порядок се- мейств — а их насчитывается 21 — говорит о большой его неоднородности. Единственным критерием для выделения порядка служит то, что ауксиллярной клеткой у этих водорослей является обычная клетка слоев ища. Однако есть гигартиновые, у которых ауксиллярной клеткой служит несущая, что можно наблю- дать и у криптонемиевых. По мере изучения этих водорослей границы между упомянутыми порядками становятся все менее ясными и опре- деление этих границ часто доступно лишь опыт- ным специалистам. Идет постоянный процесс перемещения родов из порядка в порядок. Не- редки случаи, когда детали развития оплодотво- ренного карпогона очень трудно разгадать и объяснить и разные исследователи на одном и том же микроскопическом препарате могут видеть разные вещи. По-видимому, справедливо утверждение о том, что стабильность сущест- вующей классификации красных водорослей иллюзорна и что из всех порядков только один — церамиевых — достаточно хорошо от- граничен. Семейство круориевые (Cruoriaceae) Это семейство представляет как раз пример того, о чем было только что сказано. До недав- него времени эту небольшую группу корковых водорослей помещали в порядок криптонемие- вых. По облику круориевые очень напоминают водоросли из семейства сквамариевых, но отли- чаются структурой. Вертикальные нити, под- нимающиеся от гипоталлия, соединены до- вольно рыхло и легко отделяются друг от дру- га (рис. 175, 1). Наиболее известны в семействе круория (Cruoria) и петроиелис (Petrocelis). 235
0 2 Рис. 175. Строение и размножение Cruoria: 1 — вертикальный срез через корку; 2 — карпогонная ветвь; 8 — веточка со сперматангиями. а — базальный слой, б — вер- тикальные нити, в — тетраспорангии. Размножение круориевых изучено до сих пор довольно плохо. У круории двух —,трехкле- точная карпогонная ветвь образуется как ве- точка ассимиляционной вертикальной нити (рис. 175, 2). После оплодотворения из карпо- гона вырастают соединительные нити, которые сливаются с обычными вегетативными клетка- ми соседних нитей. Гонимобласт развивается от соединительных нитей. Клетки в верхних частях ассимиляционных нитей отчленяют по одной боковой клетке, которые дают по 2 — 3 сперматангия (рис. 175, 8). Тетраспорангии, зонально или крестообразно разделенные, об- разуются из клеток вертикальных нитей интер- калярно или как одноклеточные боковые веточ- ки этих нитей. Для многих круориевых извест- но только бесполое размножение. У водорослей остальных 20 семейств гигар- тиновых гонимобласт начинает развиваться из ауксиллярной клетки, а не от соединительных нитей, как у круориевых. Все они делятся на две группы: в первой гонимобласт растет внутрь слоевища, во второй — наружу. Семейство немастомовые (Nemastomaceae) Семейство немастомовых включает 4 тепло- водных рода водорослей различной формы- от плоской широкопластинчатой до цилиндри- ческой вильчато-разветвленной. Анатомическое строение лучше всего изучено у рода платола (Platoma). Слоевище платомы состоит из упло- щенных вильчато- или неправильно разветвлен- ных ветвей. При прорастании спор вначале формируется базальная подушка из плотно соединенных нитей. Из нее вырастают верти- кальные побеги. Группы из 4 — 6 нитей растут вместе, одним пучком, скрепленные слизью (рис. 176, 1). Так образуется центральная ось слоевища фонтанного строения. Пучки боко- вых ветвей располагаются рыхло и соединяются посредством слизи. От нижних клеток боковых ветвей отходят риз оиды, спускающиеся по слоев ищу. Таким образом, строение немастомовых весь- ма напоминает строение примитивных предста- вителей порядков немалиевых и криптонемие- вых (рис. 176, 2). По мере усложнения организации происходит более плотное соединение коровых клеток, так что, например, у шизилении (Schizymenia) нитчатую структуру коры можно различить с большим трудом. Центральная часть слое- вища шизимении построена из тонких рыхлых нитей, что хорошо видно на поперечном срезе. Широкопластинчатое слоевище ее растет в ре- зультате деятельности краевой мер истемы (рис. 176, 8). Каковы же особенности размножения нема- стомовых? У платомы карпогонная ветвь воз- никает на одной из нижних клеток ветвей укороченного роста. Соединительные нити вы- растают прямо из карпогона и сильно вытяги- ваются в длину, пока не соединятся с ауксил- лярной клеткой, которой служит одна из кле- ток соседних ветвей ограниченного роста. Пер- вая клетка гонимобласта отчленяется к по- верхности слоевища и дает густой пучок нитей, большинство клеток которых превращается в карпоспоры. Гонимобласты не имеют обертки и лежат между ассимиляционными нитями (рис. 176, 4). Зонально разделенные тетраспо- рангии разбросаны по слоевищу. Они обра- зуются как одноклеточные боковые веточки коровых нитей. Семейство филлофоровые (Phyl lophoraceae) Семейство филлофоровых интересно с раз- ных точек зрения. С одной стороны, в него входят такие полезные водоросли, как филло- фора и анфельция, промысел которых ведется у нас в Черном, Белом и дальневосточных мо- рях. С другой — некоторым филлофоровым свойственны отклонения от нормального цик- ла развития, изучение которых важно для ре- шения проблемы циклов развития у красных водорослей. Семейство невелико по объему, оно объединяет 8 родов, из которых заслужи- вают упоминания гимногонгрус (С ymnogong-
тпа), филлофора (Phyllophora, рис. 177, 1 — 8) и анфельция (Ahnfeltia, рис. 177, 4). Филлофора распространена почти во всех морях СССР. В Черном море заросли ее, изве- стные под названием «филлофорное поле Зер- нова», достигают промысловых размеров. Слое- вище филлофоры небольшое, пластинчатое, ли- нейной, клиновидной или овальной формы, раз- ветвленное дихотомически, пальчато или не- правильно. Внутренняя часть слоевища, сло- женная крупными бесцветными клетками, огра- ничена тонким коровым слоем. Кора состоит из мелких плотно соединенных клеток, распо- ложенных вертикальными рядами, по 1 — 4 в ряд (рис. 177, 2). Органы размножения обычно располагаются на специальных листочках. Ряд видов филло- форы характеризуется полным циклом разви- тия, в котором представлены все формы- гаметофиты, карпоспорофит и свободноживу- щий тетраспорофит. У Филлофоры перепончато- листпной (Phyllophora membranifolia) половые растения двудомные. При образовании сперма- тангиев из поверхностных клеток возникают ве- точки, на которых, в свою очередь, образуются сперматангии. При этом более длинные плодо- носные веточки чередуются с более корот- кими, и вследствие этого на поверхности слое- вища, под внешней оболочкой, образуются свое- го рода углубления, дно которых выложено сперматангиями. Благодаря такому способу образования сперматангиев поверхность муж- ских листочков становится мелкоячеистой (рис. 177, 8). Трехклеточная карпогонная ветвь сидит на клетке, которая является и ауксиллярной (рис. 177, 2). Нити гонимобласта начинают расти внутрь слоевища среди клеток особой питающей ткани, которая возникает еще до оплодотворения. Большинство клеток гонимо- б ласта превращается в карпоспоры, выходя- щие наружу в результате разрушения стенки цисток арпа, образованной толстой многоряд- ной корой. Сорусы тетраспорангиев располо- жены в нематециях, имеющих форму подуше- чек, на свободноживущих спорофитах. Но есть виды филлофоры, у которых цикл развития проходит иначе. На женских гамето- фитах после оплодотворения карпогона разви- ваются шаровидные нематеции, в которых об- разуются тетраспоры. Долгое время эти нема- теции принимали за другую, паразитическую водоросль. По новейшим данным, это явление можно объяснить следующим образом. По-ви- димому, здесь карпоспорофит редуцируется до зиготы, которая непосредственно превращается в карпоспору. Карпоспора дает начало тетра- спорофиту, который также ~иорфологически ре- дуцирован и развивается на гаметофите, как Рис. 176. Строение и размножение немастомовых.' 1 — ювенильная стадия Platoma; 2 — ч~сть молодого вертикаль- ного побега; 8 — поперечный срез слоезища Schizymenia со зрелым гонимобластом; 4 — развитие гонимобласта y Platoma. а — тетраспорангии, б — железистые клетки, в — карпогонная ветвь, г — соединительная нить, д — ауксилд дуная клетка, е — гонимобласт.
а ° ~е ° ° ° ° ° ~ ~ ° Т ° ° ° % °,1 ° ° ° ° ° ° е ° ° ° ° ° ° ° ~ма ° ъ ф ~~~ Ч ~/~~~~~~'~~ ~ ~И~ ~l ~,',.~~/ ~ gо о а -3 р ис. 177. ~троение и размножение филлофоровмх: Z — астение РЬу11орЬога с нематециями; 2 — поперечный срез слоевища РЬу11орЬога; 3 — сперматангии на поперечном сре е; 4 — растение Ahnfeltia. а — ауксиллярная клетка, б — карпогонная ветвь. Семейство гигартиновые (Gigartinaceae) Небольшое семейство гигартиновых включает пять родов, из них в наших морях обитают четыре — гигартина (С igartina), хондрус (Chon- drus), иридея (Iridea) и родоглоссум (Rhodoglos- sum). Все они обладают сходным строением, но различаются особенностями органов раз- паразит. Сходные явления наблюдаются в цик- ле развития близкого к филлофоре рода гим- ногонгрус. По сравнению с филлофорой этот род относится к более тепловодным организмам и поэтому распространен южнее. Только три вида гимногонгруса (из 40 известных в настоя- щее время) заходят в самые южные моря СССР — Черное и Японское. Род анфельция (табл. 39, рис. 177, 4) распро- странен в северных частях Атлантического и Тихого океанов и заходит в Арктику. Анфель- ция — главное сырье агаровой промышленно- сти в СССР. Ее добывают в Белом море и у побе- режья Сахалина. Для анфельции очень харак- терна неприкрепленная форма роста, которая как раз и образует промысловые скопления. В таких скоплениях анфельция размножается только вегетативным путем. У прикрепленных видов, кроме вегетативного, известно размноже- ние моноспорами, которые развиваются в не- матециях. Способность к половому размноже- нию утеряна у анфельции полностью. множения. Слоевище гигартиновых может быть цилиндрическим, сдавленным или пластин- чатым, разветвленным или рассеченным (табл. 20, 4; 21, 2). Характерной формой ветвле- ния является вильчатое; нередко все ветви покрыты пролификациями. В отличие от фил- лофоровых структура слоевища гигартиновых отчетливо нитчатая. Это хорошо видно на попе- речном срезе через слоевище любой водоросли семейства. В центральной части и во внутрен- ней коре тонкие нити рыхло переплетаются наподобие сети. Внешняя кора построена более плотно, но и в ней нетрудно проследить мел- коклеточные дихотомически-разветвленные нити (рис. 178, 2). Крестообразно разделенные тетраспорангии развиваются группами. У хондруса и иридеи они возникают на клетках центральной части слоевища как короткие разветвленные нити, все клетки которых при созревании превраща- ются в тетраспоры (рис. 158, 2). У гигартины и родоглоссума тетраспорангии образуются из клеток внутренней части коры. В этом случае они располагаются цепочками, так как в тетра- спорангии превращаются несколько клеток каждой коровой нити. Таким образом, если у последних двух родов тетраспорангии соот- ветствуют обычным вегетативным клеткам, то у первых — это дополнительные образования. Характер развития тетраспорангиев — важ- ный, часто единственный признак, помогающий
040.': ° ' а ~..; О Оба'4". о''. Qo L (O g р.', ° ° р 80o00-.:", o o-r'! ~о < ь>о-,: ° @р о .:::,'- .) .:-:::: О ~е +a':, Рис. 178. Строение и размножение гигартиновых.' 1 — продольный срез слоевища Chondrus; 2 — карпогонная ветвь; 8 — начало образования нитей гонимобласта; 4 — зрелыф гонимобласт на поперечном срезе. а — несущая клетка, б — карпогон, в — клетка слияния, г — нити гонимобласта, д- карпоспоры. Семейство грациляриевые (Gracilariaceae) Семейство грациляриевых включает не- сколько небольших родов и один весьма круп- ный — гращилярия (Gracilaria), более 100 ви- дов которого широко распространены в уме- отличить друг от друга роды, во всем осталь- ном, включая анатомическое строение и внеш- нюю форму, чрезвычайно сходные, например ро- доглоссум от иридеи. Строение женской половой системы и разви- тие цистокарпов сходно у всех членов семей- ства. Крупная несущая клетка располагается во внутренней коре (рис. 178, 2). После опло- дотворения она сливается с карпогоном и дает начало гонимобласту (рис. 178, 8). Зрелые го- нимобласты — шаровидные образования, по- груженные в слоевище или выступающие над его поверхностью (рис. 178, 4). Обычно они разбросаны по всему телу водоросли, только у гигартины их образование приурочено к спе- циальным бородавчатым выростам на поверх- ности. По этому признаку род гигартин а хоро- шо отличается от остальных водорослей семей- ства. У хондруса гонимобласт не имеет спе- циальной оболочки. У остальных родов обра- зование гонимобласта связано с особой питаю- щей тканью. От нее вырастают длинные узкие нити, которые окружают зрелый гонимобласт, образуя его обертку. ренных и тропических морях земного шара. Виды грацилярии, как, впрочем, вообще все водоросли порядка, имеют тенденцию упло- щаться, поэтому, кроме цилиндрических, есть немало форм с плоским, вплоть до листовид- ного, слоевищем. Виды грацилярии содержат значительные количества фикоколлоидов и в некоторых странах служат сырьем для агаро- вой промышленности. Благодаря большому со- держанию фикоколлоидов растения грациля- рии жесткие, хрящеватые. Взрослое слоевище имеет «клеточную» структуру, какых-либо сле- дов нитчатого строения проследить не удается. Центральная часть состоит из крупных клеток, кнаружи постепенно уменьшающихся в разме- рах, кора построена из нескольких слоев мел- ких клеток (рис. 179, 1). Рост слоевища про- исходит за счет меристемы, которая сформиро- валась на основе одноосевого строения. Прав- да, центральную ось не удаетс~ проследить даже у проростков, видна только одна апикаль- ная клетка, отчленяющая вниз сегменты от трех своих поверхностей. Особенности размно- жения состоят в том, что ауксиллярная клетка до оплодотворения неразличима. Двухклеточ- ная карпогонная ветвь образуется на клетках внешней коры (рис. 179, 8). После оплодотво- рения карпогон сливается с несколькими сосед- ними клетками, образуя массивную клетку слияния, из которой вырастают нити гонимо- бласта, направленные к поверхности слоеви-
4Ф@ёЩрФе'вве�eg @р 4 ""� ""' убе e�' Уваееёа �Ú:: Ф~-ееа 'ее д ОО Og QQg ц ° Ра ° В 44 ° ° ° o ° аарв ее ° ааааа ° 444 Р р ° 4 ° ° ера Ррр OO ° р 44 ° ° ° еаарре О O 44 044 ° а ° раааа р аврор Оа о о o ° е оа р ° o oo0 0 ° ooo ° рроооО Ор о е 0°, а ре 00 00 ° oo в 4o4 о ооооо оооо 44 евр рер 00 оо оаоав ° РРР 0 Oop O 000 ОЬО о О ° Рарр О 400 ° 0 О ° 4 рррроо oo oo р ор о р о оьр 400 о о 0 'b00000 OO ~00 O О О О О' оо о о ~04 Ооао Орр еа о о о ° F000 0 04 о о рo 0'ое р' Wqe о 00 oeо o ° Ъ|у oор 0 Oo ° oooo 0 0 04 ° «0 О 00 00 Оеаа ° )PO 00 О ° 4 ° 0 ° O ° О ° 00 ° фа 0ОО 0 04 4 О а 4 ееОа ~ О 00 000000 е' ° ° 00 оао g oea Ь ре рае ое ае 44« а 44 О о ее ° а ° оо O ЬФ раааа оа во ° о о ° оо о ооо еа еее оо о ° 6 о о оа о ееа ° О 4 ое ° О О аО ° О ° о о ее РО 04 ° °- оо о о ОО о ае' 0 оа 40 ее. ~~Й. ~ gо ее ° о а ° ° 44 ° р ° еooO ° р ва 0 4 000о вва 4/ ° аве Ooр оаево ° в ео 0 рвов еа 00 Ф 00ао 4 0 ~О ааааа ва ooра ь ° ° 0D DDDDD OO ° е 0 0 ° 444' е ее OD ~еаа ° o Oo ар 04 ~00 ~ва~ер~р ° аа Oooава о 004444~~<<о о~о 444 -t ° 4 ° 00 ра 0~ее ев ° DD4 о о ° е~Ф ° 4040000 аеОР ео ° oo орре А - .: "" " 4- ° 4 ее @~О Рис. 179. Строение и размножение Gracilaria: 1 — продольный срез слоевища; 2 — часть среза со сперматан- гиями; 8 — карпогонная ветвь; 4 — начало развития гонимо- бласта; 5 — зрелый цистокарп. а — несущая клетка, б — клет- ка слияния, в — клетки гонимобласта. ща. После оплодотворения в месте образова- ния прокарпа поверхностные клетки усиленно делятся, образуя толстую мелкоклеточную ко- ру, из которой позднее формируется крыша цистокарпа с отверстием посередине. Цисто- карпы, разбросанные по слоевищу, выступают над его поверхностью в виде мелких шариков. Питание развивающегося гонимобласта осуще- ствляется клеткой слияния и отчасти близлежа- щими клетками слоевища (рис. 179, 4, Б). Сперматангии у грацилярии образуются в специальных углублениях коры, напоминаю- щих концептакулы, с отверстием на вершине (рис. 179, 2). Крестообразно разделенные тетраспорангии разбросаны в коровом слое по всему слоевищу. ПОРЯДОК РОДИМЕНИЕВЫЕ (RHODYMEiVIALES) Родимениевые — небольшой, достаточно хо- рошо отграниченный порядок, занимающий обо- собленное систематическое положение в классе флоридеевых. Основное отличительное свойст- во родимениевых состоит в том, что ауксилляр- ная клетка, которая формируется незадолго до оплодотворения, является производной несу- щей клетки карпогонной ветви. Все родимение- вые имеют прокарп — ауксиллярная и карпо- гонная ветви представляют собой единое об- разование. От несущей клетки развиваются карпогонная ветвь, всегда трех-, четырехклеточ- ная, и двухклеточная ауксиллярная ветвь, верхняя клетка которой служит ауксиллярной. До оплодотворения ауксиллярная клетка пло- хо различима, но после него сильно увеличи- вается и наполняется густым плазматическим содержимым. Все водоросли порядка построены по многоосевому типу. Рост осуществляется в результате деятель- ности апикальных клеток, а также верхушеч- ной или краевой меристем. Порядок делится на два семейства, различаю- щиеся особенностями анатомического строе- ния. Семейство шампиевых (Champiaceae) ха- рактеризуется полым слоевищем, при этом полость ограничена длинными узкими клеточ- ными нитями. Тетраспорангии делятся тетра- эдрически, некоторые роды имеют полиспоры. В семействе родимениевых (Rhodymeniaceae) представители многих родов лишены полости,' у тех же, что имеют ее, длинные узкие нити вокруг полости отсутствуют. Тетраспорангии делятся крестообразно. Несмотря на небольшой объем, порядок от- личается чрезвычайно широким ареалом. Роди- мениевые распространены по всем морям от тропиков до Северного Ледовитого океана, но все-таки наиболее богато они представлены в теплых морях.
Семейство шампиевые (Champiaceae) Строение водорослей семейства шампиевых удобнее всего разобрать на примере хилокла- дии чашевидной (Chylocladia kaliformis) — ви- да, растущего у Атлантического побережья Ев- ропы и в Средиземном море. Трубчатое, полое, с перетяжками слоевище хилокладии имеет членистый вид. В местах перетяжек возникают мутовки веточек. Полость, заполненная сли- зью, прерывается только на перетяжках, так как здесь образуется однослойная клеточная диафрагма. В старых частях слоевища коровой слой состоит преимущественно из одного ряда радиально вытянутых клеток. С внутренней стороны к нему примыкают длинные узкие кле- точные нити, спускающиеся вдоль слоевища и расположенные на одинаковом расстоянии друг от друга. Начинаются эти нити у вершины, где расположены апикальные клетки. Каждая апи- кальная клетка отчленяет вниз сегменты, кото- рые делятся вертикальной перегородкой, обра- зуя две клетки: внешняя формирует коровой слой, внутренняя удлиняется и дает начало продольной нити. Клетки продольных нитей в дальнейшем не подвергаются поперечным делениям, а лишь вытягиваются в длину. Со стороны полости, сбоку от них, отчленяются шаровидные клетки с густым протопластом. Это железистые клетки. Они хорошо заметны в полости слоев ища на поперечном срезе не только у хилокладии, но и у других шампие- вых. В местах ветвления слоевища железистые клетки делятся, образуя в горизонтальной плоскости клеточные ряды, которые, смыка- ясь, складываются в однослойную диафрагму (рис. 180, 1). Другие представители семейства несколько отличаются по внутреннему строению, хотя внешняя перетянутость слоевища в той или иной мере проявляется у всех из них. У шам- пии (Champia) диафрагма, так же как и кора, может быть многослойной. У видов ломента- рии (Lomentaria, табл. 21, 1) диафрагма осо- бого типа. Возникает она за счет того, что вновь образующиеся клетки продольных ни- тей, вместо того чтобы отклоняться друг от друга в процессе роста, тесно соединяются многочисленными вторичными порами. Размножение у всех водорослей порядка родимениевых весьма сходно. В семействе шампиевых у ломентар ии мельобулавовидной (Ь. clavellosa) карпогонная ветвь развивается от коровых клеток, которые соединены с про- дольными нитями первичными порами. От од- ной и той же несущей клетки отчленяется трехклеточная карпогонная и двухклеточная ауксиллярная ветви. Верхняя клетка пос- ледней является собственно ауксиллярной Рис. 180. Строение и размножение шампиевых: 1 — вершина слоевища Chylocladia; 2 — прокарп Lomentaria. а — апикальные клетки, б — клетки коры, в — клетки продоль- ных нитей, г — железистые клетки, д — диафрагма, е — несу- щая клетка, ж — карпогонная ветвь, з — ауксиллярная ветвь. (рис. 180, 2). Развитие гонимобласта происхо- дит как у родимении. Тетраэдрически разде- ленные тетраспорангии собраны сорусами в мо- лодых частях растения. Вокруг места их обра- зования коровой слой разрастается, так что группы спорангиев оказываются в небольших углублениях. Начиная прорастать, споры делятся на четы- ре клетки. Последующие деления приводят к образованию двухслойного диска. Четыре верхние клетки диска являются первичными апикальными клетками, которые, делясь в го- ризонтальной плоскости, формируют верти- кальный проросток. Семейство родимениевые (Rhodymeniaceae) Семейство родимениевых по сравнению с шам- пиевыми гораздо более многочисленная группа. Ядро семейства составляют тропические и суб- тропические виды. В морях СССР оно пред- ставлено немногими родами, но среди них та- кие широко распространенные, как родимения и галосакцион. Род родимения (Rhodymenia) включает около 50 видов, в наших морях их всего четыре. Два близких вида — родимения пальчатая и роди- мения узкоугольная (табл. 22, 2) — обычные обитатели литорали умеренных морей. Первый вид растет в наших северных европейских мо- рях, второй — на Дальнем Востоке. Другие
GOO о роо Оороо ооосо ООО о g pyo<g <ool5 р Рис. 181. Строение и размножение родимениевых.' 1 — поперечный срез Rhodymenia; 2 — часть среза Chrysymenia; 3 — прокарп Rhodymenia до оплодотворения; 4 — он же после оплодотворения. а — тетраспоры, о — железистые клетки, в— несущая клетка, г — ауксиллярная клетка, д — карпогонная ветвь после слияния ее клеток. два вида встречаются в азиатских морях. Родимения четкообразная не выходит за пре- делы Охотского моря, тогда как родимения продырявленная (табл. 21, 4) распространена более широко. Это сублиторальный, глубоко- водный вид. Пластинчатое слоев ище родимении имеет псе вдопаренхимное строение. Центральная часть слоевища сложена из нескольких слоев крупных прозрачных клеток, коровой слой плот- ный, состоит из одного-двух слоев мелких окра- шенных клеток. Между корой и сердцевиной имеется небольшая зона клеток промежуточ- ных размеров (рис. 181, 1). У многих более просто устроенных родов семейства коровой слой более рыхлый и состоит из отчетливо раз- личимых клеточных нитей. Род родимения относится к той группе во- дорослей семейства, у которых полость внутри слоевища отсутствует, хотя в старых частях слоевища клетки центральной части легко рас- ходятся. Род галоспкцион (Halosaccion) близок к роди- мении, но если у родимении расхождение кле- точных слоев — явление эпизодическое, то у га- лосакциона наличие полости — постоянный признак, отсутствует она лишь в самых моло- дых частях слоевища. Однако принцип образо- вания полого слоевища у галосакциона тот же- в результате расхождения клеток в первона- чально плотном слоевище. Виды галосакцио- на — обычные представители литоральной фло- ры наших морей и часто растут вместе с роди- менией. Кроме родов, построенных по типу родиме- нии, в семействе имеется целая группа водорос- лей с первоначально полым слоевищем. Их строение во многом напоминает строение шам- пиевых и отличается от них только отсутствием продольных нитей вокруг полости. У некоторых представителей слоевище может быть просто мешковидно-трубчатым, как, напри- мер, у хризимении (табл. 21, 8), у других оно пе- решнуровано и имеет членистый вид. У ботрио- кладии полость прослеживается в молодом со- стоянии и с возрастом заполняется. Полость ограничена слоем крупных клеток, которые пе- реходят в коровой слой, состоящий из несколь- ких слоев клеток. Вполне возможно, что внут- ренний крупно клеточный слой формируется в результате того, что клетки продольных ни- тей, столь характерных для шампиевых, очень сильно увеличиваются и соединяются друг с другом. Это предположение подтверждается тем, что некоторые из внутренних клеток несут железистые клетки (рис. 181, 2). У родимении несущей клеткой карпогонной ветви служит одна из клеток внутреннего ко- роаого слоя, которую можно отличить от со-
седних по более крупным размерам и богатому содержимому. Карпогонная ветвь трехклеточ- ная. Ауксиллярная клетка формируется до оплодотворения, но только после него стано- вится хорошо различимой, и тогда видно, что это конечная клетка двухклеточной ауксил- лярной ветви, сидящей на той же несущей клетке, что и карпогонная ветвь (рис. 181, 8). Сразу после оплодотворения в прокарпе про- исходит ряд изменений. Клетки карпогонной ветви сливаются, затем происходит слияние с ауксиллярной клеткой (рис. 181, 4). Окру- жающая прокарп ассимиляционная ткань ви- доизменяется и разрастается, образуя стенки цистокарпа. Большинство клеток гонимобласта при созревании превращаются в карпоспоры, только нижние клетки всегда остаются стериль- ными. В процессе развития гонимобластастен- ки цистокарпа приподнимаются, зрелые цисто- карпы имеют полусферическую форму и высту- пают над поверхностью слоевища. Крестообраз- ные тетраспорангии у родимении разбросаны по слоевищу или собраны в сорусы или немате- циевидные образования. Они образуются среди клеток внешней коры и соответствуют однокле- точным боковым веточкам коровых нитей. Приведенное краткое описание обоих се- мейств позволяет говорить о большой их близо- сти и рассматривать весь порядок как единую хорошо очерченную группу. Единственное раз- личие между ними заключается в наличии у шампиевых продольных нитей внутри слое- в ища и в способе деления тетраспорангиев. Судя по тому, что структуру родимениевых можно представить как производную от про- дольных нитей шампиевых, родимениевые пред- ставляют собой более поздний этап эволюции по сравнению с шампиевыми. Однако до конца вопрос о связи обоих семейств, так же как и всего порядка с другими багрянками, остается невыясненным. ПОРЯДОК ЦЕРАМИЕВЫЕ (CERAMIALES) По объему церамиевые намного превосходят все другие порядки в классе флоридеевых. До- статочно сказать, что они объединяют более 250 родов и около 1500 видов. Но, несмотря на столь большой объем, порядок — хорошо очерченная группа, которая делится всего на 4 четко ограниченных семейства. Единооб- разие порядка проявляется прежде всего в структуре и в развитии органов размножения. Всем церамиевым свойствен прокарп. Ауксил- лярная клетка отчленяется непосредственно от несущей клетки четырехклеточной карпогон- ной ветви. Формируется ауксиллярная клетка только после оплодотворения. Цикл разви- тия, как правило, трехфазный со сменой гаме- тофита, карпоспорофита и тетраспорофита; при этом первая и последняя формы развития имеют одинаковое строение. В то время как репродук- тивная система церамиевых построена по еди- ному плану, вегетативная организация их от- личается разнообразием. На этом и основана классификация порядка. Все церамиевые по- строены по одноосевому типу. Из четырех семейств на самой низкой сту- пени развития стоит семейство церамиевых. Остальные три, по-видимому, представляют параллельные линии эволюции, и сейчас труд- но сказать, какое из них стоит на более высокой ступени организации. Распространены церамиевые по всему Миро- вому океану, но особенно богато представлены в тропиках. Семейство церамиевые (Ceramiaceae) Обширное семейство церамиевых включает в себя четвертую часть всех родов и видов по- рядка. В большинстве своем водоросли семей- ства — мелкие, нежные организмы нитчатой или цилиндрической формы, лишенные коры или покрытые ею. В отличие от других у пред- ставителей этого семейства структура никогда не бывает полисифонной. Кроме того, церамие- вые имеют голые гонимобласты. Если обертка и формируется, то она состоит из коротких ве- точек, развивающихся отдельно от гонимобла- ста. У остальных водорослей порядка обертка плотного паренхимного строения. Простейшую организацию мы находим у ро- дов круания и антитамнион. Род круания (Crouania), включающий 10 ви- дов, наиболее богато представлен у берегов Австралии, хотя типовой вид рода широко рас- пространен у Атлантического побережья Ев- ропы и в Средиземном море. Нежные слизистые растения состоят из однорядной оси, покрытой мутовками коротких, многократно разветвлен- ных изогнутых веточек. Каждая мутовка со- стоит из 3 — 4 веточек. Развитие четвертой вет- ви может подавляться или из нее вырастает ветвь неограниченного роста (возникновение основных ветвей вместо укороченных — прими- тивная черта). Формирование ветвей в отдель- ной мутовке не отличается от такового у дру- гих церамиевых. Вторая всегда закладывается напротив первой, третья — между ними, чет- вертая — напротив третьей. У всех церамиевых первые ветви мутовок вдоль оси слоевища рас- полагаются по спирали. Место их возникнове- ния и, следовательно, интервалы, через кото- рые они следуют друг за другом, строго зафик- сированы для каждого рода. У круании они закладываются таким образом, что ветви со- седних мутовок смещены на оси по отношенинз 243
Рис. 182. Строение церамиевых.' ~ — часть веточки Crouania; 2 — веточка Callithamnion; 8 — веточка Antithamnion; 4 — часть слоевища Ceramium; б — про- дольный срез через узел Ceramium. а — тетраспоры, б — центральная клетка, в — перицентральная клетка, г — вторичные коровые клетки. друг к другу на 45' (полный оборот спира- ли составляет 360'). Боковые веточки резко отличаются от основных ветвей толщиной и ветвлением. Первые ветвятся дихотомически и политомически, вторые — моноподиально (рис. 182, 1). По особенностям строения женской половой системы и карпоспорофита круания стоит на низкой ступени развития. В зрелом слоевище одна из перицентральных клеток формируется как несущая и вместо обычной дает начало кар- погонной ветви. Последняя полностью заме- щает вегетативную, никаких стерильных кле- ток рядом с ней не остается (рис. 183, 1). Спер- матангии образуются на вершине обычных вето- чек ограниченного роста. Тетраэдрически раз- деленные тетраспорангии сидят на нижних клетках боковых ветвей и разбросаны по всему слоевищу. Род каллитамнион (Callithamnion) близок к круании, но уже гораздо более специализи- рован. Известно более 50 видов каллитамниона, распространенных в различных морях. Не- сколько видов обитает в Черном море, среди них наиболее часто встречается каллиталнион щитковидный. У каллитамниона на каждом сегменте образуется по одной ветви, никаких следов мутовчатого ветвления в вегетативных ветвях обнаружить не удается (рис. 182, 2). Оно проявляется только в фертильных веточ- ках. Объяснить характер ветвления каллитам- ниона и его происхождение в настоящее время весьма затруднительно. Некоторые виды имеют кору, которая состоит из риз оидных нитей, спускающихся по стенкам основных ветвей слоев ища. Карпогонные ветви образуются попарно из двух перицентральных клеток, возникающих вокруг основной оси. Обе фертильные пери- центральные клетки представляют собой ос- татки ветвей ограниченного роста и вместе с вегетативной ветвью являются единственным проявлением мутовчатого ветвления у калли- тамниона. Род антитамнион (Antithamnion) также характеризуется простой организацией. Как и круания, эти водоросли построены однорядны- ми лишенными коры и свободно разветвлен- ными нитями. Ветви ограниченного роста отхо- дят по две супротивно или по три-четыре му- товками. Вдоль основной оси все ветви огра- ниченного роста располагаются строго друг под другом, а не смещены, как у круании. По внешнему виду они почти не отличаются от основных ветвей. И те и другие ветвятся моно- подиально; в боковых ветвях также прослежи- ваются основная ось и боковые ветви. Харак- терным признаком многих видов служит нали- чие железистых клеток на коротких веточках (рис. 182, 8).
ак у круании, карпогонная ветвь у анти- тамниона берет начало от перицент ральной клетки, но здесь она не полностью замещает вегетативную, а занимает боковое положение на обычной веточке ограниченного роста. Зрелый гонимобласт окружен многочислен- ными соседними вегетативными веточками (рис. 183, 2, 8). Гаметофиты двудомные. Тетра- спорофиты встречаются в природе чаще, чем гаметофиты. На боковых веточках они несут крестообразно поделенные тетра спорангии. Род г~ералиул (Ceramium) относится к той же группе родов, что и антитамнион. Около 60 видов церамиума распространено от тропи- ков до арктических морей. Наибольшего видо- вого разнообразия род достигает в теплых мо- рях. Церамиум интересен как водоросль, имею- щая кору. У некоторых видов кора покрывает сплошь все слоевище; другие же имеют разви- тый коровой слой лишь на отдельных участках, которые повторяются через определенные ин- тервалы, придавая растению членистый вид (рис. 182, 4). Покрытые корой окрашенные «уз- лы» чередуются с голыми прозрачными «междо- узлиями». Эту членистость часто можно заме- тить невооруженным глазом и по ней легко от- личить церамиум от других багрянок. Харак- терной чертой в облике церамиума является также дихотомическое ветвление и вильчато- разделенные вершинки (табл. 25). Как же построено слоевище церамиума. Р Центральная ось состоит из одного ряда круп- ных бесцветных клеток. Делясь косыми пере- городками, они отчленяют вбок перицентраль- ные, которые являются первичными коровыми клетками. Они располагаются кольцом из 6 — 10 довольно крупных клеток вокруг цент- ральной оси. Из первичных клеток коры разви- ваются вверх и вниз по слоевищу мелкокле- точные нити, которые образуют внешний коро- вой слой. Клетки центральной оси при этом вытягиваются (рис. 182, 5). Если коровые ни- ти, развивающиеся от двух соседних осевых клеток, смыкаются, образуется сплошной ко- ровой слой, но чаще между ними остается сво- бодный участок сегмента. Таким образом, ко- ровые нити церамиума соответствуют веточкам ограниченного роста антитамниона, а первич- ные коровые клетки — нижним клеткам этих веточек. Сходство структур подтверждается и тем, что прокарп у церамиума возникает на одной из первичных коровых клеток. Церами- ум — один из высокоорганизованных родов семейства. По-видимому, он происходит от ка- ких-то антитамниевых в результате увеличения числа веточек ограниченного роста в мутовке п более густого их расположения. Гаметофиты церамиума в природе встреча- ются гораздо реже, чем тетраспорофиты, по- Рис. 183. Размножение церамиевых.' 1 — прокарп Crouania; 2 — прокарп Antithamnion; 8 — зрелый цистокарп Antithamnion; 4 — сперматангии Ceramium; 5— моло~ой цистокарп Ceramium; 6 — BbIC TyrIaIoIIIme тетраспо раН- гии Ceramium. а — карпогонная ветвь, б — ауксиллярная клет- ка.
Рис. 184. Строение дазиевых. 1 — вершина веточки Heterosiphonia; — образование коры у Dasya; 8 — стихидий Dasya с тетраспорангиями. этому признаки половых растений меньше используются в систематике рода, чем призна- ки спорофитов. Сперматангии развиваются из поверхностных коровых клеток и, когда их много, как муфтой одевают «узлы» (рис. 183, 4). Возникновение прока рпов приурочено к вер- шинам длинных ветвей слоевища. Карпогон- ная ветвь располагается сбоку первичной ко- ровой клетки. От нее же верхушечно отходит группа стерильных клеток — результат редук- ции обычной веточки ограниченного роста. Зрелые гонимобласты окру~кены короткими ве- гетативными веточками, расположенными ро- зеткой и образующими обертку цистокарпа (рис. 183, о). Тетраздрически разделенные спо- рангии развиваются из коровых клеток и рас- полагаются по-разному: одиночно или кольце- образно на сочленениях, выступая из коры или обрастая ею (рис. 183, 6). У видов со сплошной корой тетраспорангии разбросаны по всему слоевищу. Все эти признаки важны для разгра- ничения видов. Семейство дазиевые (Dasyaceae) Это небольшая группа, насчитывающая 12 ро- дов, приуроченных к тропической и субтропиче- ской зонам. В моря умеренной зоны заходят лишь немногие виды. Среди наиболее обшир- ных и широко распространенных родов следует назвать дазию (Dasya) и гетеросифонию (Hete- rosiphonia). Они me представляют интерес для понимания особенностей строения и размноже- ния всего семейства. Главное отличие дазиевых от остальных водорослей порядка заключается в симподиальной организации. Напомним, что симподиальное ветвление характеризуется тем, что в месте отхождения ветви главная ось сме- щается в сторону, а ее место занимает вновь образовавшаяся ветвь, т. е. постоянной апи- кальной клетки здесь нет, а ее роль принимают на себя апикальные клетки последовательно возникающих ветвей (рис. 184, 1). Слоевище дазии имеет радиальную симметрию. Главные ветви покрыты густо разветвленными одноряд- ными веточками ограниченного роста, что при- дает растениям опушенный вид (табл. 23, 4). Отходят эти веточки по одной от каждого сег- мента и располагаются по спирали. Апикальная клетка отчленяет от себя осе= вую клетку, последняя быстро растет, смещая апикальную клетку в сторону, и сама занимает ее место. Вот эти-то отошедшие в сторону апи- кальные клетки и дают начало однорядным боковым веточкам. По внешнему виду они на- поминают трихоб ласты родомеловых, но по способу возникновения не имеют с ними ничего общего. Сегменты основных ветвей очень скоро становятся многорядными, так как отчленяют пять перицентральных клеток, или сифонов. Их образование оказывается совершенно иным, чем у родомеловых. На клетках центральной оси, в месте возникновения перицент ральной клетки, образуется почковидный выступ, кото-
рый позднее отделяется перегородкой. Пер- воначально перицентральные клетки ко- роче центральных, у родомеловых они того же размера. Порядок, в котором происходит отчленение перицентральных клеток, также отличен от родомеловых. Из нижних частей пе- рицентральных клеток вырастают мелкокле- точные разветвленные ризоидные нити, кото- рые, смыкаясь, формируют многорядную плот- ную кору (рис. 184, 2). В образовании коры так- же принимают участие ризоиды, берущие на- чало от основания коротких боковых веточек. Между собой клетки коры соединяются вто- ричными порами. Строение слоевища дазиевых на ранних стадиях прорастания совсем иное, чем у родомеловых. Проростки дазиевых одно- рядные, нитчатые, свободно разветвленные, тогда как у родомеловых — многорядные и не- разветвленные. Прокарп у дазии закладывается близ верши- ны основных ветвей или на дополнительных бо- ковых веточках. Фертильным обычно становит- ся третий из последовательно возникающих пе- рицентральных сегментов. Его можно отличить уже в самом начале — он крупнее остальных и более богат протоплазматическим содержи- мым. Эта клетка служит несущей и дает начало карпогонной ветви и двум стерильным клеткам. До оплодотворения обе стерильные клетки поч- ти не отличаются от вегетативных и не подвер- гаются делению. После оплодотворения несу- щая клетка дает начало ауксиллярной, кото- рая сливается с карпогоном и образует нити гонимобласта. В карпоспоры превращаются несколько верхних клеток этих нитей. Зрелый гонимобласт находится в полости, окруженной оберткой паренхимного строения. Так же как у делессериевых, формирование обертки на- чинается только после оплодотворения и происходит за счет разрастания соседних с фертильной перицентральных клеток. У родоме- ловых обертка закладывается еще до оплодо- творения. Тетраэдрически-разделенные споран- гии развиваются в специальных коротких ве- точках — стихидиях, располагаясь поперечны- ми рядами среди их клеток (рис. 184, 8). Семейство делессериевые (Delesseriaceae) Представители семейства делессериевых- одни из самых красивых водорослей, и не толь- ко среди багрянок, но, пожалуй, и среди всех водорослей-макрофитов. Растущие на глубине, они обычно окрашены в яркие красные тона. Нежные полупрозрачные пластины многих де- лессериевых пронизаны нервами и жилками и этим очень напоминают листья наземных рас- тений. Сходство с листьями особенно велико у водорослей с изрезанным или зубчатым краем пластины. Когда по мере роста пластины раз- рушалотся, остающиеся нервы можно принять за веточки, на которых сидят молодые листоч- ки (табл. 22, 1, 8; 23, 2). Делессериевые краси- вы как в живом состоянии, только что взятые из моря, так и на бумаге, в расправленном и засу- шенном виде. Особенно красивы зрелые расте- ния — с развитием тетраспор и цистокарпов пластины становятся бахромчатыми и покры- ваются темными пятнышками. Семейство вклю- чает большое число сублиторальных форм, которые распространены по всему Мировому океану, но наибольшего развития достигают в морях южного полушария. Слоевище делессериевых построено по одно- осевому типу. Ветви ограниченного роста, от- ходящие от центральной оси, развиваются в одной плоскости, что приводит к образованию пластинчатого слоевища, в отличие от других водорослей порядка. Рост осуществляется апи- кальной клеткой или краевой меристемой. Для многих характерны интеркалярные клеточные деления. 70 родов дел ессериевых объединяют в два подсемейства. Подсемейство делессериевые (Delesserioideae) Род делессерия (Delesseria) — типичный пред- ставитель подсемейства. Листовидное одно- слойное слоевище делессерии пронизано сред- ним нервом и многочисленными боковыми жил- ками. Новые пластинчатые ветви возникают от среднего нерва, а не от края пластины, как это происходит у некоторых других родов. Как же построено слоевище делессерии? Если посмотреть на молодую пластину с поверх- ности (рис. 185,1), то в ней можно хорошо раз- личить апикальную клетку, которая отчленяет вниз сегменты центральной оси. От централь- ных сегментов берут начало четыре перицент- ральные клетки. Две из них, боковые, располо- женные супротивно, развиваются в клеточные разветвленные нити, за счет которых строится пластина. Среди них можно различить ответвле- ния первого, второго и более высоких поряд- ков. Ветви первого порядка всегда достигают края пластины, тогда как ветви последующих порядков могут кончаться внутри слоевища. Как растут боковые нити, какое количество по- рядков они образуют, какие из них достигают края пластины — все это важные родовые при- знаки. Клетки центральной оси, раз обра- зовавшись, уже не подвергаются поперечным клеточным делениям, а лишь вытягиваются в длину. Клетки боковых нитей растут так же, как и центральная, но, в отличие от нее, в них встречаются интеркалярные деления. В резуль- тате такого роста у делессерии четко просле- 247
Рис. 185. Строение делессериевых: 1 — вершина слоевища Delesseria; 2 — вершина слоевища Phycodrys; 8 — поперечный срез через ребро Phycodrys. живается центральная ось, но расположение клеток в боковых нитях оказывается менее упорядоченным. Отсутствие интеркалярных делений в центральной оси — признак, от- личающий все роды подсемейства. У многих из них интеркалярные деления отсутствуют и в бо- ковых нитях. Поэтому представителей подсе- мейства всегда легко отличить по упорядочен- ному расположению клеток и отчетливой апи- кальной клетке. Две другие перицентральные клетки не получают такого развития. Они фор- мируют кору над центральной осью. За счет этой разрастающейся коры и многочисленных ризоидных нитей, берущих начало от клеток коры, образуется утолщенный срединный нерв, пронизывающий слоевище делессерии. На зрелых растениях можно увидеть малень- кие листочки — в них вдоль среднего нерва образуются прокарпы. Несущими клетками служат перицентральные клетки главной оси плодоносного листочка. Кроме карпогонной ветви, на несущей клетке образуются две груп- пы стерильных клеток. После оплодотворения от нее же отделяется ауксиллярная клетка. Стерильные клетки делятся и наполняются густым содержимым, однако в определенный период оболочки их набухают и ослизняются, а сами клетки постепенно исчезают. В процессе развития гонимобласта из центральной, несу- щей и ауксиллярной клеток образуется клетка слияния, которая служит для питания разви- вающегося гонимобласта. Стенки цистокарпов формируются в резуль- тате разрастания клеток коры. Большинство клеток гонимобласта превращается в карпоспо- ры. Образование сперматангиев и тетраспор также приурочено к особым мелким листочкам. Подсемейство нитофилловые (Nitophylloideae) Род фикодрис (Phycodrys) включает виды, внешне очень напоминающие делессерию. Так же как у делессерии, листовидное слоевище фикодриса пронизано средним нервом и боко- выми жилками, но новые пластины вырастают по краю старой пластины в результате разрас- тания ее лопастей или роста боковых жи- лок. Основные различия между двумя родами и соответственно между двумя подсемействами проявляются в анатомическом строении и размножении, Слоевище фикодриса строится по общему для всех делессериевых принципу, но у него клетки центральной оси легко подвергаются интеркалярным клеточным деле- ниям, в результате чего порядок в расположе- нии клеток нарушается и уже в молодых плас- тинах проследить центральную ось и боковые нити, в отличие от подсемейства делессериевых, не удается (рис. 185, 2). За счет разрастания коровых клеток вдоль центральной оси слоеви- ща у фикодриса образуется ребро, жилки приурочены к боковым ветвям первого порядка. Ризоиды здесь отсутствуют и участия в форми- ровании ребра не принимают (рис. 185, 8).
Рис. 186. Схема строения верхушки слоевища Polysiphonia: в в е р х у — деление апикальной клетки при образовании разветвленных и неразветвленных сегментов; в н и з у — образование перицентральных клеток (поперечный срез). а — центральный сифон, 1 — 5 — перицентральные клетки. У некоторых нитофилловых апикальный рост заменяется ростом за счет краевой меристемы. Для таких форм характерно вовсе беспорядоч- ное расположение клеток и отсутствие апикаль- ной клетки, как, например, у нитофиллума (ХйорЬу11ит). Прокарпы в обоих подсемей- ствах построены по единому плану, но у фико- дриса они разбросаны по всей пластине. Точно так же сперматангии и тетраспорангии, соб- ранные в сорусы, расположены на пластинча- той части, но не на ребрах и жилках. Семейство родомеловые (Rhodomelaceae) Это семейство, включающее более 100 ро- дов,— самое крупное во всем отделе красных водорослей. Оно характеризуется огромным разнообразием вегетативных форм и в то же время поразительным однообразием в строении органов размножения. В существующей клас- сификации родомеловые занимают место наи- более высокоорганизованной группы отдела. Род полисифония (Polysiphonia) — прекрас- ный пример структуры и размножения родо- меловых. В тонконитчатом разветвленном слое- вище полисифонии клетки располагаются пра- вильными ярусами, или члениками. Каждый членик состоит из центральной клетки и рас- положенных вокруг нее нескольких пери- центральных, одинаковой формы и длины (рис. 187,1). Такую структуру называют по- лисифонной. Число перицентральных клеток постоянно для каждого вида, но в пределах рода может заметно меняться. Рост у полисифонии апикальный. Если об- разуется неветвящаяся ветвь, апикальная клет- ка делится поперечной перегородкой, отчленяя центральные сегменты. Образовавшиеся сегмен- ты подвергаются продольным делениям, кото- рые происходят в строгой последовательно- сти — направо и налево от первой перицентраль- ной клетки. В результате образуется полиси- фонный членик. Когда образуется разветвлен- ная ветвь, апика льна я клетка делится косо и образовавшийся клиновидный сегмент дает вырост, который является началом боковой ветви. Только после этого центральный сегмент делится продольно с образованием перицен- тральных клеток (рис. 186). Возникающая ветвь может быть ветвью неограниченного рос- та, и тогда она проходит то же развитие, что и главная ось, или короткой ветвью ограничен- ного роста, оставаясь при этом частично или полностью однорядной. Короткие однорядные веточки — трихобласты — располагаются на слоевище по спирали в строгой последователь- ности и через определенные интервалы, при этом в каждом членике образуется только один трихобласт. Обычно трихобласты вскоре после образования опадают, оставляя лишь маленькую базальную клетку (рис. 187, 1). Длинные ветви неограниченного роста могут возникать в пазухе базальных клеток трихо- бластов или непосредственно от них. Слоевищв
Рис. 187. Строение и размножение Polysiphoøà: 1 — вершина веточки; 2 — 5 — схема развития сперматангиев на трихобласте; 6 — 10 — схема развития гонимобласта. a = карпогон, б — несущая клетка, в — ауксиллярная клетка, г — первая клетка гонимобласта, д — цистокарп. многих видов полисифонии, как и других родов семейства, покрыто корой. Она развива- ется в результате деления перицентральных клеток или образуется возникающими от них же риз оидами. Органы полового размножения располага- ются на плодоносных трихобластах. Прокарп всегда образуется на втором от основания сегменте трихобласта, который становится по- лисифонным в связи с образованием перицен- тральных клеток. Одна из перицентр альных клеток, которая образуется последней, делит- ся, давая несущую клетку и первую клетку карпогонной ветви. Несущая клетка отчленя- ет еще три стерильные клетки. После оплодо- творения карпогон увеличивается в размерах, стерильные клетки делятся, образуя клеточные нити. Несущая клетка отделяет ауксилляр- ную, которая располагается вблизи от карпого- на и сливается с ним посредством маленькой соединительной клетки. В процессе развития гонимобласта несущая, ауксиллярная, цент- ральная и стерильные клетки слоевища слива- ются в одну крупную клетку. Карпоспоры конечные. Зрелый гонимобласт окружен кле- точной оберткой с отверстием на вершине и имеет шаровидную или грушевидную форму (рис. 187, 6 — 10). В мужских трихобластах только один базальный и несколько верхушеч- ных сегментов остаются однорядными. Осталь- ные сегменты скружаются перицентральными клетками, которые подвергаются поперечным делениям с образованием трех или более мате- ринских клеток сперма тангиев. Те, в свою очередь, делятся косыми перегородками и дают по нескольку сперматангиев (рис. 187, 2 — 5). Тетраспорангии возникают на обычных по- лисифонных ветвях. В каждом сегменте обра- зуется по одному тетраспорангию. Перицен- тральная клетка отчленяет от своей внешней поверхности три стерильные покровные клетки. Оставшаяся часть этой клетки делится на крупный тетраспорангий и маленькую клетку- ножку. Цикл развития полисифонии состоит в правильной смене гаметофитов и тетраспоро- фита одинакового морфологического строения. Роды родомела (Rhodomela) и одонтпалия (Odonthalia) тоже наиболее обычные родоме- ловые из флоры наших морей (табл. 23, 1, о). Они относятся к группе родов, у которых пери- центральные клетки располагаются в несколь- ко слоев. Кроме того, органы полового раз- множения образуются у них на специальных коротких полисифонных веточках — стихиди- ях. Тетраспорангии также возникают в спе- циальных веточках. Стерильные трихобласты (если имеются) пигментированы и не опадают. Как уже упоминалось, родомеловые берут начало от каких-то примитивных представи- телей семейства церамиевых и представляют собой самостоятельную, независимую от осталь- ных семейств линию эволюции.
ОТДЕЛ ЖЕЛТО-ЗЕЛЕНЫЕ ВОДОРОСЛИ (XANTHOPHYTA) 25i К отделу желто-зеленых относятся водорос- ли, хлоропласты которых окрашены в светло- или темно-желтый цвет, очень редко зеленый и лишь иногда голубой. Окраска эта определя- ется наличием в хлоропластах основных пигмен- тов — хлорофилла, каротинов и ксантофиллов. Однако в хлоропластах желто-зеленых водорос- лей всегда преобладают каротины, что и обус- ловливает своеобразие их окраски. Кроме того, у них в клетках крахмал отсутствует, а в качестве основного продукта ассимиляции на- капливаются капли масла и лишь у некото- рых, кроме того, глыбки лейкозина и волю- тина. Желто-зеленые водоросли характеризуются большим морфологическим разнообразием. Сре- ди их многочисленных представителей обнару- живаются почти все основные типы структуры тела водорослей: амебоидная, монадная, паль- меллоидная, коккоидная, нитчатая, разнонит- чатая, пластинчатая и сифональная. Распространены желто-зеленые водоросли по всему земному шару. Встречаются они главным образом в чистых пресноводных водо- емах, реже в морях и солоноватых водах, обычны они также в почве; могут обитать как в кислых, так и в щелочных водах; предпочи- тая умеренную температуру, чаще развивают- ся весной и осенью, хотя имеются виды, встре- чающиеся на протяжении всех периодов года, в том числе и зимой. Желто-зеленые водоросли в основном явля- ются представителями планктона, главным образом пассивными планктерами, реже они встречаются в перифитоне и бентосе. Чаще всего их можно найти в скоплениях нитчаток и среди зарослей высших водных растений в прибрежной зоне рек, прудов, озер и водо- хранилищ, реже на чистоводье. Среди желто-зеленых водорослей имеются представители с талломом одноклеточным (рис. 188, 1, 2, Б; 190, 191), колониальным (рис. 189), многоклеточным (рис. 192, 1, 2) и некле- точного строения (рис. 192, 8). Кроме того, здесь известны очень своеобразные водоросли с многоядерным талломом в виде голого плаз- модия (рис. 188, 8). Независимо от внешней структуры, внутрен- нее строение клетки желто-зеленых водорослей довольно однотипно. В протопласте наблю- дается обычно несколько желто-зеленых хлоро- пластов, имеющих дисковидную, корытовидную, пластинчатую, реже лентовидную, звездчатую или чашевидную форму с цельными или лопаст- ными краями. У подвижных форм на переднем конце хлоропласта обычно расположен крас- ный глазок. У немногих видов имеется обычно двус корлупчатый пир еноид. Ядро в клетке одно, обычно небольших размеров, заметно только после обработки клетки специальными красителями, но имеются виды и с многоядерны- ми клетками. У небольшого числа видов в морфологически передней части клетки имеется одна или две пульсирующие вакуоли. Отличительной особенностью желто-зеленых водорослей является наличие у вегетативных клеток монадной структуры и зооспор двух неравных жгутиков. Именно этот признак в свое время послужил основанием для того, чтобы назвать эту группу водорослей разножгу- тиковыми, или гетероконтами (Heterocontae). Помимо различий в длине, жгутики здесь различаются и морфологически: главный жгут состоит из оси и перисто расположенных на ней мерцательных волосков, боковой жгут бичевидный. В отличие от внутреннего содержимого клет- ки ее оболочка у желто-зеленых водорос- лей обнаруживает значительное многообразие. У простейших представителей клетка окружена только тонким и нежным перипластом, позво- ляющим ей производить выпячивания в виде псевдо- и ризоподий (рис. 188,1 — 4). Но у большинства видов клетка покрыта настоящей плотной оболочкой, обусловливающей постоян- ную форму тела. Эта оболочка может быть цельной или двустворчатой, с равными или неравными по величине створками. У большин- ства представителей створки обычно трудно- различимы, они становятся хорошо видными лишь под действием 60',4-ного раствора едкого кали или при окрашивании. Обычно оболочка бесцветная, прозрачная, реже окрашена в бурый или желтоватый цвет. У многих представителей она имеет различные скульптурные украшения, может быть инкру- стирована известью, кремнеземом или солями железа. Размножаются желто-зеленые водоросли про- стым делением клетки или распадом колоний и многоклеточных талломов на отдельные части. Наблюдается также бесполое размножение при помощи двужгутиковых зооспор или автоспор, реже амеб оидов. Половой процесс известен у немногих видов и представлен изо- и оогамией. У некоторых видов в цикле развития известны экзо- и эндогенные цисты с двустворчатой, часто окремневшей оболочкой (рис. 189, 8). Значение желто-зеленых водорослей как фо- тотрофных организмов заключается прежде всего в создании первичной продукции в водо-
««ее««« «« ° е ° « ° е ве~, ,,«« ° Ю С фЬфо r 4-у:-Е:) ~ ё:.'.: Ь «е е ° ° ° ~ о'@ л ° «««ее ° ° ° « ° « ° ° ° « ° ° «' ф ° 4 Ъ е е ° ° ° ° ° ° ° Ф е' 252 ° е ° » с е «« е ° \ ее « « ° ° с\' ««е«С» ф' с ° ° -з- в «« с- ~С«С «е»' «:,фею«рай%»„. диве~. е«ССС а~с~В С ее« ее «« ее» ° «ее ее«ее« ° ° « ~ъ ° е ° ° « ° ° ° ° ««« ° е« ° ° «« ° ««« \е ° ° '«««е«еа със С е ° е ° ° ее « «« ««е ° ее ° ° « «« ° е ««« ° е« ° « '««ее«ее«ее«ее«««е«« ° «««« ф~,.„.:~::ф~фФ"";-'" $:. \ е« е ° ее ° е «е ° ° « «ее«а\« ° «е«ее ««\ ° ° ° ««е« °- ° «««ее ° «««ее ° -« ° ° \е ° «ее««еее С е Ю ° ° ° ° ° « е « «е Ъ, е»- r е « ее Ф «е ° е « ° ° е ° ° ° С ев сй- «« ° ° ° Т«««««ее« « ее ° ее « е Уе ° е еа « r~ ее сс .е ° ° ° е« ,ъ Ю 4-. « 6~~ Ю Се « Ю ~е ~е е O «У Ф» «с Ь « ° е « а « ° \ « ° «е ° « « а ° '« ° « С ° « \ Ю Ъ ° е ° е Ф Ю е«е««~ Ю ° е е Юе ° е Ю ° ° Ъ е в C аа Ю Рнс. 188. Ксантоподовые (1 — 4) и ксантомонадовые (о): 1 — Rhizochloris stigmatica: а — перипласт, б — ризоподии, в — хлоропласт, г — глазок, д — пульсирующие вакуоли; 2 — Stipitococcus гав; 8 — МухосЫог18 sphagnicola, плазмодий в водоносных клетках листьев сфагнума; 4 — Chlorarachnion rep:ans, в разветвленных ризоподиях видны захваченные комоч- ки пищи; 5 — Chlorocardion pleurochloron. емах и в участии их в цепи питания гидробион- тов. Желто-зеленые водоросли наравне со мно- гими другими образуют сапропель (ил). Обитая в водоемах, богатых органическими остатками, они могут служить показательными формами при определении степени загрязнения воды. В почвах они активно участвуют в процессах накопления органических веществ, способствуя повышению плодородия. Желто-зеленые водоросли еще недостаточно изученная группа. Происхождение ее достовер- но не выяснено. В настоящее время все же преобладает мнение, что они являются самосто- ятельным отделом, так как показывают четко выраженный параллелизм форм с золотистыми и зелеными водорослями, выделение которых в самостоятельные отделы ни у кого не вызы- вает сомнений. Несомненно, желто-зеленые во- доросли имеют родственные связи с золотис- тыми и диатомовыми водорослями. В соответствии с новейшими данными отдел желто-зеленых водорослей (Xanthophyta) можно разделить на шесть классов: ксантоподовые (X anthopodophyceae), ксантомонадовые (Xan- thomonadophyceae), ксантокапсовые (Xanthocap- sophyceae), ксантококковые (Xanthococcophyceae), ксантотриховые (Xanthotrichophyceae) и ксан- тосифоновые (Xanthosiphonophyceae). КЛАСС КСАНТОПОДОВЫЕ (XANTHOPODOPHЪ СЕАЕ) К классу ксантоподовых относятся наиболее просто устроенные желто-зеленые водоросли. Это типичные представители амебоидной (ризо- подиальной) структуры. Характерной особен- ностью организации их тела является наличие у клеток только перипласта, позволяющего клетке изменять свою форму и вырабатывать выпячивания в виде более или менее широких псевдоподий или длинных ризоподий. Ксантоподовые представлены в большин- стве своем одноклеточными формами. Все они микроскопических размеров, некоторые заклю- чены в «домик». Живут свободно или прикрепля- ются к нитчатым водорослям. Обитают главным образом в пресных водоемах. Класс ксантоподовых включает один порядок, который подразделяют на два семейства с не- большим числом видов. К наиболее типичным представителям ксантоподовых относится ри- зохлорис глазковый (Rhizochloris stigmatica, рис. 188, 1) — одноклеточный организм, обитаю- щий в придонных слоях воды небольших оли- готрофных (бедных питательными веществами) прудов и глубоких ям. Клетки его с тонким нежным перипластом, в очертаниях округлые, достигают в диаметре 12 — 14 лкл. Они образуют немногочисленные ризоподии, благодаря которым осуществляется движение. В клетке имеются 2 — 3 дисковидных, окра- шенных в светло-желто-зеленый цвет хлоро- пласта, на верхушке одного из них обычно находится маленький темно-красный глазок. Между хлор опластами в центральной части клетки располагаются капли масла и жира и светопреломляющие блестящие кристаллы (возможно, лейкозин); имеются в клетках и пульсирующие вакуоли. Ризохлорис относится к одноядерным орга- низмам, но в живой клетке ядро увидеть не удается. Для того чтобы его разглядеть,
необходимо применить специальные краси~ели. размножается ризохлорис вегетативно, простым делением клетки надвое. Питается он фото- трофно и голозойно, путем захвата ризоподиями бактерий, наннопланктонных водорослей и час- тичек детрита (детрит — продукт распада орга- низмов). Помимо рода ризохлорис, в пределах класса ксантоподовых известны и другие роды, у пред- ставителей которых клетки живут в (<домике» (рис. 188, 2), а также колониальные формы. Среди ксантоподовых имеются плазмодиаль- ные формы, представленные двумя монотипными родами. Особенно интересен миксохлорис сфаг- новый (Myxochloris sphagnicola, рис. 188, 8), плазмодии которого встречаются большими скоплениями в водоносных клетках листьев сфагнума. Плазмодии миксохлориса крупные, содержат большое число дисковидных светло-зеленых хлоропластов, много ядер и пульсирующих вакуолей. Наличие плазмодиальных форм у желто- зеленых водорослей в известной мере подтвер- ждает родственные связи этого отдела с золо- тистыми водорослями, ибо только в этих двух отделах имеются представители с подобной структурой тела (ср. Myxochrysis paradoxa из отдела Chrysophyta). КЛАСС КСАНТОМОНАДОВЫЕ (XANTHOMONADOPH YCEAE) К классу ксантомонадовых относятся пред- ставители желто-зеленых водорослей с монад- ной структурой тела. Характерной их особен- ностью является наличие двух неравных жгу- тов, позволяющих им передвигаться в толще воды. Это типичные активные планктеры. Как и ксантоподовые, ксантомонадовые вклю- чают небольшое число в основном монотипных родов. Один из типичных представителей класса- хлорокардион зеленобокий (Chlorocardion pleuro- chloron, рис. 188, б). Его клетки покрыты уплот- ненным перипластом, неметаболичные, слегка уплощенные. Из небольшого углубления на пе- реднем конце тела выходят жгутики, причем главный обычно в 1,5 раза длиннее самой клет- ки. В клетке хлорокардиона можно видеть от 2 до 4 корытовидных стенкоположных хло- ропластов и одну сократительную в акуолю в переднем конце тела. В центре клетки меж- ду хлоропластами имеются блестящие комоч- ки лейкозина, капли масла и жира. Здесь же после специальной окраски обнаруживает- ся одно ядро. »Ф»»»»ее@»е»е»~ »»Ф»» ° »е» Ф» »Ф» »» ° »Ф ФФ»» Ф » Ъ ФФ г' а ° б с ° Ф Ф с »ФФ Ъ Ф ФФ г Ф r »~»»Ф г а t 3 I 1 I I Ъ \ а I ФФФ» бб а ! Ф» Ф ФФ % с Рис. 189. Есантокапсовые: 1 — Helminthogloea ramosa; 2,8 — Gloeochloris planctonica (2 — часть колонии при большом увеличении, 3 — цисты); 4,5 — Chlorosaccus ulvaceus (4 — общий вид колонии, 5 — часть колонии при большом увеличении). Размножается хлорокардион только веге- тативным путем, простым делением клетки. Встречается он обычно отдельными особями в прибрежной зоне прудов среди зарослей высших растений. КЛАСС КСАНТОКАПСОВЫЕ (XANTHOCAPSOPH YCEAE) Водоросли этого класса характеризуются дальнейшим усложнением талл ома, представ- ленного здесь пальмеллоидной структурой. В отличие от первых двух классов, ксантокап- совые включают неподвижные прикрепленные r r а а а ° ° ° ° ° ° с ° ° ° ° ° а б »» ~»е» с ° Ф % Ф» »» »'4Ь » Ъ » Ъ ЮФФ»» »аа r % а ,ФГ;Ф ° ' '4 ° г Ф ° ° r� ". а а», ° а I I ФФУ Ю а а ~~:,'.": г а ,б 1 %»»» 1 % Ъ г съеФ»г'
или пассивно плавающие колониальные, реже одноклеточные формы. Их клетки постоянной формы, не способны к поверхностным выпячи- ваниям протопласта и лишены жгутиков. Они полностью окружены слизью или выраба- тывают односторонние слизистые образования. Клетки, погруженные в общую слизь колоний, расположены в ней беспорядочно или разме- щаются по периферии в определенном по- рядке. Класс ксантокапсовых небольшой, насчиты- вает всего несколько родов с небольшим числом видов, населяющих как морские, так и пресные и солоноватые воды. Наиболее типичны следую- щие. Гельминтоглея ветвистая (Helminthogloea ramosa, рис. 189, 1), обитающая в засоленных водоемах, отличается слизистым вытянутым разветвленным талломом, прикрепляющимся к суб страту расширенным основанием. Клетки, каждая в слизистой оболочке, распределяются в колониальной слизи беспорядочно. Глеохлорис планктонный (Gloeochloris р1апс- tonica, рис. 189, 2, 8) характеризуется шаровид- ными или эллипсоидными колониями, диаметр которых достигает 100 лкл. Этот вид холодолю- бив и обычно встречается ранней весной в неболь- ших ямах с холодной, талой водой. Под микро- скопом в стекловидной плотной слизи колоний можно увидеть расположенные по периферии эллипсоидные клетки, ориентированные ради- ально своими длинными осями. Клетки имеют от одного до четырех (редко больше) пластинча- тых стенкоположных хлоропластов, одну-две пульсирующие вакуоли и почти черный глазок, а также капли масла и глыбки лейкозина. Размножается зооспорами, известны также ци- сты с гладкой оболочкой, состоящей из двух частей. Третий представитель — хлоросаккус ульвовый (Chlorosaccus ulvaceus, рис. 189, 4, о) — обитает в ручьях и реках на высших водных растениях и на поверхности ила. Он имеет мешковидные, неправильной формы слизистые колонии диа- метром до 100 лкл, на поверхности которых заметны выступы в виде невысоких бугорков. Эти выступы образованы выпячиванием слизи за счет близко расположенных к поверхности клеток зллипсоидной формы. Внутри каждой клеточки имеются по два корытовидных стенко- положных хлоропласта, а между ними, в цен- тре клетки видны капли жира и масла. В про- цессе размножения каждая из клеточек может превратиться в двужгутиковую зооспору, ко- торая выходит из слизи старой колонии, выра- батывает вокруг себя новую слизь и формирует молодую колонию. Интересным представителем класса ксанто- капсовых является также мишококкус конфер- вный (Mischococcus confervicola, табл. 26, 4). Его нежные, дихотомически- или тетрахотоми- чески-ветвящиеся, почти совсем прозрачные, сравнительно крупные колонии довольно час- то встречаются в различных водоемах нашей страны. Особенно часто их можно наблюдать на нитчатых водорослях в пробах перифитона из сфагновых болот, обычно на нитях трибо- немы или эдогония. Обследовав их под микро- скопом, мы обязательно обнаружим колонии мишококкуса. Вначале заметны лишь шаро- видные светло-желто-зеленые клеточки, как бы свободно парящие над нитями водорослей, но затем, внимательно присмотревшись, мы увидим целую систему слизистых тяжей, на верхушках которых «восседают» клеточки. При- менив специальные красители — метиленовую синьку, разбавленные чернила или тушь, мы сможем легко разобраться в скоплении слизис- тых тяжей. Образование кустистых колоний у мишокок- куса связано с тем, что каждая клеточка выде- ляет слизь только в одну сторону, а сама при этом остается на верхушке слизистого тяжа. Разделившись пополам, каждая новая клеточ- ка, в свою очередь, начинает выделять свой собственный слизистый тяж, и таким образом формируются две веточки от старого материн- ского тяжа. Подсчитав «этажи» всей колонии, можно установить число поколений и последо- вательность образования отдельных клеточек колонии. КЛАСС КСАНТОКОККОВЫЕ (XANTHOCOCCOPH YCEAE) К классу ксантококковых относятся желто- зеленые водоросли с коккоидной структурой тела. Их клетки имеют настоящую плотную оболочку, состоящую из двух частей, или сплошную, нередко скул ьптурированную или инкрустированную. Это одноклеточные, реже колониальные формы, причем последние имеют вид скоплений клеток, не погруженных в слизь и слабо связанных между собой. При вегета- тивном размножении такие колонии не обра- зуют нитей и пластинок. Среди ксантококковых имеются как свободноплавающие, так и прик- репленные формы. Класс ксантококковых наиболее богат пред- ставителями и занимает центральное место в отделе желто-зеленых водорослей. В нем насчи- тывают более 50 родов, в большинстве своем достаточно хорошо изученных. Рассмотрим наи- более типичных и часто встречающихся пред- ставителей этого класса. Ботридиопсис эрийский (Botrydiopsis eriensis, рис. 190, 1 — 8) — вид, широко распространен- 254
ный по всему земному шару, живущий в самых различных местообитаниях, но главным обра- зом в почве, на дне прибрежной зоны водоемов различного типа, среди зарослей, в биологичес- кой пленке биофильтров очистных канализацион- ных станций. Встречается он обычно одиночны- ми клетками и в небольшом количестве, но иногда может давать и массовое развитие, придавая в этом случае воде или покрываемой им поверх- ности субстрата нежную желто-зеленую ок- раску. Размножается ботридиопсис преиму- щественно автоспорами, реже зооспорами. Ще- потки любой почвы, собранной во время экс- курсии в лес, на луг, на реку, достаточно для того, чтобы методом культур (см. очерк «Почвенные водоросли») обнаружить этот вид. При первом же просмотре культуры под мик- роскопом мы заметим клетки ботридиопсиса- от крохотных, диаметром около 6 мкл, с двумя хлоропластами, только что вышедших из обо- лочки материнской клетки, до крупных, диа- метром свыше 18 лкл, с явно двухконтурной, иногда желтоватой оболочкой и со множест- вом хлоропластов. Иногда клетки уже напол- иены автоспорами и покрыты разбухшей и ослизненной оболочкой. В планктоне прудов, рек, водохранилищ и дру- гих пресноводных водоемов часто можно встре- тить ксантококковых с разнообразной формой клетки и различными выростами и украшениями оболочки. Таковы, например, тетраэдриелла гигантская (Tetraedriella gigas, рис. 190, 4), псевдостаураструм копьевидный (Pseudostaura- strum hastatum, рис. 190, Б), гониохлорис ту- поконечный (Goniochloris mutica, рис. 190, б) и некоторые другие. Их нередко путают с од- ноклеточными зелеными водорослями, но при внимательном просмотре они довольно четко различаются. Типичным представителем почвенных форм ксантококковых является бумиллериопсис nov- венный (Bumilleriopsis terricola, рис. 190, 7, 8), встречающийся довольно часто и повсеместно. Молодые клетки бумиллериопсиса яйцевидные, взрослые продолговато-эллиптические, с тупо закругленными концами. У всех клеток одна боковая сторона всегда более выпуклая, дру- гая менее выпуклая или почти плоская. Обо- лочка клеток бесцветная и состоит из двух неравных половинок, меньшая из которых явно утолщена и покрывает суженную часть клетки, а при образовании автоспор отделяет- ся в виде колпачка. В клетках наблюдаются немногочисленные пристенные хлоропласты желто-зеленого цве- та, обычно корытовидной или спирально-лен- товидной формы. Автоспоры и молодые клетки одноядерные, взрослые — многоядерные; ядра Рис. 190. Ксантококковые.' 1 — 8 — Botrydiopsis eriensis, клетки различного возраста; обра- зование автоспор; 4 — Tetraedriella gigas; 5 — Pseudostaurastrum hastatum; 6 — Goniochloris mutiea; 7,8 — Bumilleriopsis terrieo- 1а, клетки различного возраста. небольших размеров, видны только после окраски. В различных водоемах на нитчатых водорос- лях, подводных частях высших водных расте- ний, частицах детрита, а также среди обраста- ний на погруженных в воду предметах можно обнаружить изящные, самой разнообразной фор- мы клетки на более или менее длинных ножках. Это — виды рода хараниопсис (Characiopsis, рис. 191, 1, 2). Их насчитывают более 50, и все они ведут прикрепленный (эпифитный) образ жизни. Прикрепление клеток к субстрату осуще- ствляется у них с помощью ножки, размеры и строение которой у разных видов разные. Обыч- но клетки харациопсиса одиночные и разбросаны по поверхности субстрата, но иногда можно обнаружить и их скопления. Внутреннее строение самих клеток здесь очень простое: обычно два хлоропласта да ка- пелька масла, тогда как ядра без специальной окраски не видно. Весьма похожи на харациопсис, но хорошо отличаются от него двустворчатой оболочкой представители рода хлоротениул (Chlorothecium, табл. 26, о). Строение оболочки становится у них особенно отчетливо различимо, когда из клетки выходят зооспоры.
О о 4 Рис. 191. Ксантококковые: 1 — Characiopsis subulata; 2 — СЬ. acuta; 8 — Centritractus belo- nophorus; 4 — OphiocytiulII cochleare. Из других эпифитных форм выделяются своим изящным обликом представители рода перониелла (например, Peroniella curvipes, табл. 26, 6), характеризующиеся клетками, сидящими на длинных тонких ножках. В планктоне самых различных водоемов очень широко распространен типичный пред- ставитель ксантококковых — центр итрактус копьеносный (Centritractus belonophorus puc. 4Ý 1 191, о), у которого оба конца клетки несут длинные выросты оболочки, действительно на- поминающие копья. Виды интересного рода офиоиитиум (Ophio- cytium, рис. 191, 4) чаще всего встречаются в сфагновых болотах. Здесь их клетки могут попадаться с ранней весны до поздней осени, причем, как правило, встречаются одиночными экземплярами. Лишь иногда среди детрита можно наблюдать их в виде рыхлых скопле- ний, напоминающих перепутавшиеся короткие завитки толстого гладкого шнура. Зимой не- редко можно видеть пустые оболочки клеток офиоцитиумов. При рассматривании под микроскопом про- топласта живой клетки офиоцитиума (например, Ophiocytium cochleare, рис. 191, 4) бросается в глаза его необычайная прозрачность. Клетки, имеющие вид более или менее длинных трубо- чек, снаружи покрыты двухконтур ной, глад- кой, бесцветной, реже буроватой, совершенно прозрачной оболочкой. Последняя может иметь лишь полярные утолщения в виде одного-двух более или менее длинных шипиков. Плотно прижимаясь к оболочке, в постенном слое цитоплазмы лежит несколько дисковидных желто-зеленых хлор опластов. Ядра в живой клетке без специальной окраски не видно. Совершенно прозрачная стекловидная цито- плазма придает клетке офиоцитиума своеоб- разный характер «упорядоченности», несвой- ственной обычно другим водорослям. КЛАСС КСАНТОТРИХОВЫЕ (XANTHOTRICHOPH YCEAE) К этому классу относятся исключительно много клеточные формы желто-зеленых водо- рослей, характеризующиеся нитчатой, раз- нонитчатой и пластинчатой структурами тела. Обычно они ведут прикрепленный образ жиз- ни. Таллом здесь имеет вид простых или раз- ветвленных, однорядных или многорядных нитей и кустиков или однослойных и много- слойных паренхиматозных пластинок, клетки которых не погружены в общую слизь. Разрастание таллома ксантотриховых идет за счет деления клеток в одной, двух или трех плоскостях. Бесполое размножение осу- ществляется главным образом при помощи зооспор и автоспор, реже наблюдается обра- зование акинет и цист. Класс ксантотриховых объединяет в своем составе наиболее высоко организованных пред- ставителей желто-зеленых водорослей. По внеш- нему виду они сходны с улотриксовыми из отдела зеленых и хризотриховыми из отдела золотистых водорослей. Из нитчатых неразветвленных форм наибо- лее типичным и распространенным видом яв- ляется трибонема зеленая (Tribonema viride, рис. 192, 1). Нити ее, собранные в ватооб- разные дерновинки, образуют в прибрежной полосе различных водоемов желто-зеленые скопления, мягкие, но не ослизненные на ощупь. Молодые нити трибонемы долго остаются при- крепленными к субстрату и лишь позже, отры- ваясь, образуют скопления, плавающие на поверхности воды или опускающиеся на дно водоема. Под микроскопом нити трибонемы легко отличить от других нитчатых водорослей по характерному очертанию их концов. Это свя- зано с тем, что оболочка клеток у трибонемы крепкая, двустворчатая, разрыв нитей проис- ходит по середине клеток и, когда их содержи- мое выходит наружу в виде зооспор, остав- шаяся оболочка приобретает вид двух рожек на каждой краевой клетке. Хлоропластов в каждой клетке трибонемы обычно несколько, они дисковидные, равномерно расположенные по периферии всей клетки. При наступлении неблагоприятных условий в нитях трибонемы образуются апланоспоры с толстой двуствор-
Х а б л и ц а 25. Красные и бурые нитчатые водоросли: в в е р х у — десмарестия зеленеющая (Desmarestia viridis); в н и з у — церамиум красный (Ceramium гиЬгит) и другие мелкие багрянки на мидиях. Фото Ю. Астафьева .
Та б л и ц а 2б. Желто-зеленые водоросли: 1 — 3 — Botrydium granuiatum (I — поросль на почве при малом увеличении, 2 — отдельная особь при большом увеличе- нии, 3 — образование ризоцист); 4 — Mischococcus confervicola; 5 — Chlorothecium crassiapex, клетка и выход из нее зоо спор; о — Peroniella curvipes.
Та б л и ц а 27. Бесцветиые эвгленовые водоросли: 1 — Cyclidiopsis acus; 2 — Petalomonas praegnans; 3 — Menoidium pellucidum; 4 — Anisonema prosgeobium; 5 — Urceolus cyclostomus; б — Urceolus platyrhynchus; 7 — Peranema pleururum; 8 — Menoidium tortuosum; 9, 10 — Heteronema «cu~, в двух состояниях клетки.
Та б л и ц а 28. Окрашенные эвгленовые водоросли: 1 — Trachelomonas bituricensis; 2 — Strombomonas ensifera; 3 — Euglena sanguinea, заполненная гематохромом; 4 — phacus longicauda; 5 — Euglena ehrenbergii.
Тa б л и ц а 29. Вольвоксовые и протококковые водоросли: 1 — 7 — каротиноносные виды вольвоксовых (1 — 5 — Dunaliella salina, вегетативные клетки с каплями гематохрома (1, 2) и стадии образования цист (3 — 5); б, 7 — Haematococcus pluvialis, вегетативная клетка и апланоспоры); 8 — 11 — прото- кокковые (8 — Chlorangiopsis epizootica, молодая клетка и зооспорангий; 9 — Apiocystis brauniana; 10 — Korschikoviella gracilipes; 11 — Protosiphon botryoides).
Та б л и ц а 30. Улохриксовые водоросли: 1 — стигеоклониум (Stigeoclonium tenue); 2 — улотрикс (Ulothrix zonata); 3 — колеохете (Coleochete scutata); 4 — драпар- нальдия (Draparnaldia glomerata); 5 — эдогониум (Oedogonium stellata); б — прингсхеймиелла (Pringsheimiella scutata).
Та б л и ц а 31. Улохриксовые и сифоновые водоросли: 1 — протомонострома (Protomonostroma undulatum); 2 — энтероморфа (Enteromorpha linza); 3 — акросифония (Acrosiphonia incurva); 4 — кодиум (Codium ritteri); 5 — капсосифон (Capsosiphon groenlandicus); б — ульва (Ulva fene- strata); 7 — монострома (Monostroma grevillei).
Та б л и ц а 32. Улотриксовые и сифоновые водоросли: 1 — удотея (Udotea flabellata); 2 — ацетабулярия (Acetabularia peniculus); 3 — ульвария (Ulvaria obscura); 4 (Гhaetomorpha melagonium); 5 — пеиициллус (Penicillus capitatus); б — кодиум (Codium fragile). хетоморфа
чатой оболочкой, заполненные каплями масла и жира. Из желто-зеленых водорослей, способных образовывать паренхиматозные пластинки, заслуживает упоминания гетеропедия (Hetero- pedia polychloris, рис. 192, 2). Ползучие ветвя- щиеся нити гетеропедии после деления остают- ся соединенными друг с другом и образуют плотное паренхимато зное слоевище. Вегета- тивные клетки в таком слоевище двух типов: внутренние, неправильно многоугольные в очер- таниях, и краевые, несколько более крупные и округлые. В каждой клетке гетер опедии имеется несколько дисковидных хлоропластов (это отражено в видовом названии). Размноже- ние осуществляется двужгутиковыми зооспо- рами, которые образуются главным образом из срединных клеток. Кроме того, могут обра- зовываться и автоспоры. Встречается гетеро- педия преимущественно на влажной почве. КЛАСС КСАНТОСИФОНОВЫЕ (XANTH0SIPHONOPH YCEAE) К этому классу относятся желто-зеленые водоросли с сифональной структурой, т. е. неклеточным строением таллома. Ксантосифо- новые внешне могут иметь сложную форму, но по строению протопласта все они пред- ставляют собой одну гигантскую многоядер- ную клетку, чаще всего м акр оскопических размеров, видимую невооруженным глазом. Как правило, таллом ксантосифоновых назем- ный, прикрепленный, дифференцированный на окрашенную надземную и бесцветную под- земную часть. Одним из типичнейших и часто встречающих- ся представителей этого класса является ботридиум зернистый (Botrydium granulatum, табл. 26, 1 — 3). Обычно он образует целые заросли на глинисто-илистых отложениях у берегов различных водоемов, на дне подсыха- ющих прудов, на влажных богатых питатель- ными веществами почвах. Характерная особенность ботридиума — тре- бовательность к повышенному содержанию извести в почве. Массовое развитие ботридиума можно, на- пример, наблюдать в карповых рыб оводных хозяйствах Украины, когда зимовальные пру- ды переводят на «летование»: в это время кар- повых производителей помещают в нерести- лища, воду из прудов спускают и обнажается слой жирного ила. Буквально через несколько дней подсохшая прибрежная часть, а вскоре и вся поверхность ила покрывается мелкими желто-зелеными шариками, как будто кто-то рассыпал по дну пруда пшено с зеленоватым оттенком. Шарики — это надземная часть тал- ломов ботридиума с типично сифональной струк- турой тела, без внутренних перегородок. При попытке снять эти шарики с субстрата оказы- вается, что это не так легко сделать. Дихото- мически-разветвленная, мощная система бес- цветных ризоидов прочно удерживает голов- чатую окрашенную часть таллома. Чтобы рас- смотреть ботридиум, нужно осторожно взять целый участок заселенного шариками ила, поместить его в кристаллизатор с чистой водой и только после тщательной промывки в совер- шенно чистой воде можно наконец увидеть его отмытые талломы. Для дальнейших ис- следований часть из них можно зафиксировать, перенеся в склянку с водой, разбавленной формалином. Живые талломы ботридиума надо осторожно перенести в чашки Петри или на часовое стекло и при малом увеличении микроскопа или в сильную лупу сначала рассмотреть общее стро- ение надземной и подземной частей. Детали каждой из них можно разглядеть при большом увеличении, приготовив обычный препарат с использованием покровного стекла. В постенном слое цитоплазмы надземной части лежат хлор опласты, пластинчатые или дисковидные. Ниже, под хлоропластами, рас- Рис. 192. Ксантотриховые и ксантосифоновые: 1 — Tribonema viride, часть нити; 2 — Heteropedia polychlo- ris; 8 — Vaucheria sessilis, часть нити, с п р а в а в н и з у- оогоний и антеридий.
положены ядра, еще глубже можно наблюдать множество капель масла и другие включения. В ризоидальной части вышеуказанные орга- неллы и включения отсутствуют, она заполнена лишь цитоплазмой, клеточным соком, а у мо- лодых особей иногда имеются и ядра. Обычно различают главный ризоид и боковые дихото- мически-разветвленные веточки. Оболочка взрослой особи ботридиума слоис- та и состоит из пектина с примесью целлюлозы. У молодых клеток оболочка тонкая, нежная, позже она грубеет и у надземной части покры- вается известковыми гранулами (это послужи- ло поводом для видового названия). Размножается ботридиум главным образом зооспорами, образующимися путем распада про- топласта надземной части таллома на одноядер- ные участки. Полностью сформировавшиеся зооспоры выходят наружу через разрыв оболоч- ки на верхушке. Иногда вместо зооспор могут образовываться автоспоры и апланоспоры. При наступлении неблагоприятных условий (высыхание, понижение температуры) у ботридиу- ма образуются особые цисты — р и з о ц и с т ы. При этом все содержимое надземной окрашенной час- ти разбивается на отдельные участки и переходит в ризоиды (табл. 26,8). Надземная часть после этого отделяется благодаря утолщению оболоч- ки главного ризоида и отмирает. При наступле- нии благоприятных условий ризоцисты либо прорастают непосредственно в новые талломы, либо образуют зооспоры, которые затем также дают новые талломы. К классу ксантосифоновых в настоящее вре- мя относят и представителей рода вошерия (Vaucheria, рис. 192, 3), которых ранее вклю- чали в отдел зеленых водорослей. Основанием для этого служит общность пигментов и про- дуктов ассимиляции. По внешнему виду таллом вошерии представляет собой неправильно и редко ветвящиеся нити нежного светло-зеле- ного цвета, с бесцветными ветвящимися ризо- идами. Внутри, так же как и у ботридиума, перегородок нет — это одна гигантская много- ядерная клетка. Нити вошерии можно встретить и на дне водоемов с быстротекущей водой, и в стоячих водоемах у самого берега, и в виде свободно- плавающих скоплений нитей на поверхности воды, а также на сильно увлажненной почве, где она образует зеленые бархатистые дерно- винки. При сборе материала необходимо лопа- точкой или широким ножом осторожно снять верхний слой почвы с зеленым налетом. Если этот налет вместе с почвой поместить во влажную камеру, то вошерия будет жить долго, представляя интересный объект для наблюдения. Нити вошерии исключительно нежные, по- этому для получения препарата с неповрежден- ным талломом необходимо зеленый налет брать пинцетом очень осторожно, расправляя нити препаровальной иглой в капле воды на пред- метном стекле. Если препарат приготовлен удачно, под микроскопом видна длинная нить, внутри которой можно легко различить мелкие многочисленные желто-зеленые дисковидные хлор опласты и желтовато-оранжевые капли масла. Вошерия — многоядерный организм, но ядра у нее мелкие. Они расположены в пос- тенном слое цитоплазмы под хлоропластами и видны лишь после окрашивания специальны- ми красителями. Всю центральную часть тал- лома вошерии занимает большая вакуоля с клеточным соком. Нередко на конце одного или нескольких разветвлений нити можно заметить более гус- тое и темное содержимое, отделенное перего- родкой. Это зооспорангий, в котором формиру- ется одна зооспора. У вошерии она, в отличие от зооспор многих других водорослей, крупная и несет многочисленные пары жгутиков. В ци- топлазме под каждой такой парой жгутиков располагается ядро, а в более глубоких слоях лежат хлоропласты. Меняя условия культивирования (освещен- ность и влажность), в условиях лаборатории можно вызвать у вошерии образование зооспор и даже увидеть (обычно ночью) их выход из зооспорангия. Имеется у вошерии и половой процесс в ви- де типичной оогамии. При формировании оого- ниев и антеридиев, как и при образовании зооспорангиев, возникают перегородки, отде- ляющие половые органы от остальной части таллома. По строению и расположению на талломе половые органы вошерии разнообраз- ны и служат основными систематическими приз- наками. Поэтому стерильные нити вошерии почти невозможно определить до вида. Желто-зеленые водоросли — самостоятельная в систематическом отношении группа, возник- шая от каких-то первичных амебоидных форм. По набору пигментов, составу запасных веществ и строению оболочки вегетативных клеток и цист желто-зеленые водоросли обнаруживают сходство с золотистыми, диатомовыми и отчасти бурыми водорослями.
ОТДЕЛ ЭВГЛЕНОВЫЕ ВОДОРОСЛИ (EUGLENOPHYTA) Рис. 193. Эвглена зелен (Euglena viridis): а — жгут; б — глотка; в — глаз г — хлоропласт с парамилоновь зернами в центре; д — ядро с рышком. Рис. 194. Изменение клет Euglena mutabilis с помощ метаболичных движений. ° Ф ° ° ° ° ° ° ° ° ° ~е ° ° ° ° ° ° ° ° ° % ° ° ° ° ° ° ° б ° ° ЭФ \~ '1 59 Эвгленовые водоросли — обычные обитатели небольших пресных стоячих водоемов. В летние месяцы можно наблюдать, как в небольшом пруду или луже внезапно зеленеет вода. При- чиной этого позеленения («цветения») может быть массовое развитие эвгленовых водорослей. Под микроскопом в капле воды, взятой из такой лужи, видны многочисленные зеленые веретеновидные клетки. Легко изгибаясь, ме- няя форму тела, они быстро движутся во всех направлениях. Обычно это эвглены (Euglena)— центральный род, по имени которого назван весь отдел. Эвгленовые водоросли — это один из немно- гих отделов водорослей, организация пред- ставителей которого ограничена в основном монадной структурой. Сюда относятся микро- скопические (длиной 4 — 500 мкм) одноклеточ- ные организмы, снабженные одним или двумя жгутами и активно двигающиеся. Движение— характерное свойство эвгленовых водорослей и лишь немногие представители этого отдела, имеющие пальмеллоидную структуру, утрати- ли его. Форма тела эвгленовых водорослей хорошо приспособлена к движению в воде. У большин- ства видов она удлиненная, овальная, эллипсо- идная и веретеновидная. Последняя среди эвгленовых водорослей встречается особенно часто, и поэтому иногда ее называют эвгленооб- разной (рис. 193). Имеются виды и с шаровидны- ми клетками, но они сравнительно немногочис- ленны. Клетки эвгленовых водорослей на попереч- ном сечении обычно округлые. В этом случае их можно было бы считать построенными по радиаль- Рис. 195. «Домики» некоторых видов рода Trachelp- monas.' g — Trachelomonas чо1гос1па; 2 — Tr. globularis; 8 — Tr. arma- ta; 4 — Tr. hystriх; й — Тг, planctonica; б — Tr, curta.
Рис. 196. «Домики» различных видов рода Strombo- monas: 2 — Strombomonas subcurvata; 2 — 81г. acuminata; 8 — 81г. fluviatilis; 4 — 81г. volgensis. Рпс. 197. Схема строения переднего конца клетки эвгленовых водорослей: а — жгут; б — глотка; в — глазок (стигма); г — резервуар; д- фоторецептор; е — вакуоля перец ее опорожнением в резервуар. ной симметрии, но последнюю нарушает спираль- ное расположение штрихов и других элементов орнаментации, а также внутреннее строение клетки. Поэтому более правильно считать клет- ки эвглен овых водорослей асимметричными. Наиболее ярко асимметрия выражена у вы- сокоорганизованного рода факус (Phacus). Клетки факусов сплющены в виде листообраз- ной пластинки, снабжены килевидными вырос- тами и хвостовым отростком, большей частью изогнутым. Некоторые винтообразно-скручен- ные клетки факусов при взгляде сверху напо- минают гребной винт (рис. 202, б). Эвгленовые водоросли не имеют настоящей оболочки, подобной таковой у высших растений. Защитную роль в ыполняет наружный слой эктоплазмы — перипласт. Те виды, у которых перипласт мягкий, эластичный, обладают cпо- собностью менять форму тела. Такие клетки при своем движении то изгибаются, то разду- ваются на одном конце и утончаются на другом, как бы переливая свое содержимое из одного конца клетки в другой (рис. 194). Эту способ- ность эвгленовых водорослей называют мета- болией. Виды с более плотным перипластом сохра- няют постоянную форму тела и не способны к метаболии. Перипласт эвгленовых водорослей редко бы- вает гладким, обычно он покрыт штрихами или рядами точек, бугорков, проходящих спи- рально (реже продольно) от переднего конца к заднему. Под перипластом расположены так называемые слизистые тельца, которые выделяют слизь через поры перипласта. У трахеломонасов (Trachelomonas) и стром- бомонасов (Strombomonas) поверх перипласта образуется плотный «домик» — вначале он тон- кий и бесцветный, но с возрастом пропитыва- ется солями железа и марганца и от этого окра- шивается в различные оттенки бурого, золо- тистого и коричневого цветов. «Домики» стром- бомонасов обычно слегка шероховатые или мор- щинистые, а у трахеломонасов более богато орнаментированы: покрыты точками, бугорка- ми, мелкими щетинками, крупными шипами различного строения. Для выхода жгута в передней части «доми- ка» имеется отверстие, часто окруженное вырос- том стенки «домика» — горльппком (табл. 28, 1, 2; рис. 195, 196). На переднем конце клетки эвгленовых водо- рослей находится углубление, продолжающе- еся внутрь в виде трубки, — «глотка», которая оканчивается расширением — резервуаром (рис. 197). Глотка — органоид выделительной систе- мы. Сократительные вакуоли, в которых скап- ливается жидкость с растворенными продукта- ми обмена веществ, изливают свое содержимое в этот резервуар, откуда через глотку оно вы- водится наружу. Процесс излияния происходит ритмично че- рез 20 — 30 сек. Движение эвгленовых водорослей соверша- ется с помощью жгута. Жгут прикреплен своим основанием к базальному телу на дне резерву- ара. Через глотку он выходит наружу. У боль- шинства видов жгут один; некоторые эвглено- вые снабжены двумя жгутами равной или неравной длины. Но типичным для эвгленовых следует все же считать наличие двух жгутов, так как даже у одножгутиковых форм имеется
тонкий и короткий остаток второго жгута, который не выходит из глотки, а соединяется в ее пределах с длинным жгутом (рис. 197). Движение большинства эвгленовых — это активное плавание. Работая жгутом, клетка одновременно вращается вокруг своей оси и как бы ввинчивается в воду. При этом важную роль играют килевые выросты и конечный отросток — стабилизаторы движения. У неко- торых бесцветных представителей эвгленовых, живущих на дне водоемов среди ила, наряду с плаванием (перанема, гетеронема) наблю- дается и ползание по дну или другому субст- рату (петаломонас, анизонема). Клетка ползает, как бы ощупывая дорогу впереди своим жгу- том. У анизонемы клетка снабжена двумя жгута- ми. Один из них направлен вперед и выполняет роль двигателя; второй волочится сзади, играя, вероятно, роль руля (табл. 27, 4). Среди эвглен существует группа так называ- емых «безжгутиковых» форм. Жгут у них от- сутствует в наружной своей части или значи- тельно укорочен. Движение у этих форм пол- зающее, оно осуществляется за счет метабо- личных изгибов тела. Зеленый цвет эвгленовых водорослей объясня- ется наличием хлорофилла, который содержится у них в виде двух форм — а и Ь. Это те же са- мые формы хлорофилла, которые имеются и у высших растений. Хлорофилл — преобладаю- щий пигмент у эвгленовых водорослей, но, кроме него, присутствуют и другие: сс-, P- и е-ка- ротины и несколько ксантофиллов. Все эти пигменты сосредоточены в хлоропластах. Кроме пигментов, участвующих в фотосин- тезе, у некоторых эвглен (в частности, у Euglena sanguinea) имеется в большом коли- честве пигмент красного цвета — астаксантин. Раствор его в масле (гематохром) выполняет роль световой ширмы: регулирует количество света, попадающего на хлоропласты. В усло- виях интенсивного освещения пигмент скапли- вается в периферической части клетки и зате- няет собой хлоропласты. Клетка при этом окрашивается в различные оттенки красного цвета. При ослаблении освещенности пигмент перемещается к центру клетки, и водоросли становятся ярко-зелеными. Хлоропласты эвгленовых имеют разнообразное строение, особенно у представителей рода эвг- лена. Они могут быть звездчатыми (рис. 198,3), лентовидными (рис. 198, б), крупнопластинча- тыми (рис. 198, 1, 2). Звездчатый хлоропласт Рис. 198. Форма хлоропластов, пиреноидов и парами- лоновых зерен эвгленовых водорослей: 1 — пластинчатый хлоропласт с двускорлупчатым пиреноидом (с б о к у); 2 — пластинчатый хлоропласт с пиреноидом в центре (с в е р х у); 8 — звездчатый хлоропласт с парамилоновыми зернами в центре; 4 — хлоропласт с внутренним пиреноидом, покрытым скорлупкой парамилона; 5 — лентовидные хлоро- пласты; 6 — дисковидные хлоропласты; 7 — 22 — парамилоно- вые зерна. ° ю ° ° Ф ° 3 ° ° ° ° ° ' ~~ ° Э ° ° ~ ° Рис. 199. Схема деления клетки эвглены: 2 —; 2 —; 8 — анафаза; 4 начало продольного разделения клетки.
Рис. 200. Деление эвгленовых водорослей в состоянии пальмеллы: 1 — Euglena geniculata; 2 — Е. pisciformis. е1 ° ° ° ° ° ° Ф' ° ф е ° ° е ° е ° ° ° ° ° ° ° ° ° Фе" ° ° Р. ° е ° ° ° ° ° ° ° ° ° ~а ° ° ° ф1 ° И~ ° ° ° ° ° ° ° Ф ° ° ° ° ° ~ ° е~ ° ° ° 1 ° ° ° ° ° Ие е ° ° Э ° ~ ° ° 1.„1' 'Ве ° м ° 9. м ф~1 ° ~ее е ° ° ° ~ C ° Ф ° ° ° 1 ° ° е ° %:. ° ° ° ° Ф ° ° У ° ° ° ° ° ° ° ° ° ° э~~в ° ° ~ ° ° 'эф ° ° е~~ Ъ Ф, ° ° ° ° ° ° ° ф ° е~~ ° е ° ° ° ° ° ° еЪ е~~е ~ ° ° ° ° ° О~~ ° ~ММе» в' ° « ° ° ° ° у ~ Ф ° ° Ае ' ° . ° ° ~ ° 9~1 Ю ° е ° е ° ~à~ Ф е ° ~ ° Ф Ф ° Ф ° ° ° ° » ° ° ° ° ° ° ° е ICу ~е ° ° ° ° ° ~ÔÝ ° ~еэе ° ° ° ° ° ° O ° ф ф Э ° ° ~Э ° ° ° ° е- ° ° ° е ° ° ° ° O ° ° ° ° е ° ° ° Л ° ° еФ ° ° Фе ° ° ° ° а ° ° ° å ° ° ° ° ° Ъ ° ° ° ° ° ° +% ° вф Ф ° ° ° ФФ ° ° е ° ° ° ° е'е~ ° ° ° ° ~ еа ~е ° ° ° ° ° ° ~~qy ° бабеф ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ~ð ° ° ° ° ° ° ° " ° ° Ъ- ° ° ° ~ ° е~ ° ° ° ° ° ° ° ° ° ° ° ° ° ° ~ ° ° ° ° ре~ ° ° ° ° ° е ° ° l<+ ° ° ° ° ° ° ° ° ° å ° ~~в ° ° ° ° ° ° ° ° ° ° ° ° ° i+ ° ° а~у 2 ° ° ° ° ° ° р ° ° ° ° ° е ° ° "° ° ° ° в Ч~ ° ° y I ° ~е ° ° ååôå %. ° ° +y ° ~ее + see ° ° ° фФ ° ° Ф ° sa е ° е ~ ° е, ° 4 е ° ~ ° ' ° ° ° В ° е -i% ° ° ~ ° ° 1 ° ~ ° ° основной своей частью лежит в центре клет- ки, откуда к периферии расходятся его лучи. Но более распространены у эвгленовых мелкие дисковидные хлор опласты, расположенные в клетках постенно, т. е. такие же, что и у высших растений (рис. 198, 6). В хлоропластах у некоторых видов эвглено- вых водорослей хорошо заметны более плотные образования — пиреноиды (рис. 198, 1, 2), цен- тры образования запасного питательного веще- ства парамилона, производного глюкозы. Пи- реноиды особенно хорошо заметны благодаря покрывающим их скорлупкам пар амилона. Пиреноиды могут откладывать две или одну скорлупку парамилона в форме часовых стекол. У некоторых тра хеломонад и стромбомонад известен так называемый внутренний пирено- ид: он вытянут к центру клетки и покрыт тон- кой наперстковидной скорлупкой парамилона (рис. 198, 4). У ряда эвглен встречается «голый» пиреноид, не образующий парамилона. У форм с мелкими дисковидными хлоропластами пи- реноиды обычно отсутствуют. У видов, не имеющих пиреноидов, парамилон образуется непосредственно в цитоплазме. В во- оемах с большим количеством органических доемах веществ клетки эвгленовых обычно переполн- е- ны массой мелких округлых или удлиненных зерен парамилона. В более «голодных» условиях клетки накапливают меньше парамилона. Кро- ме неопределенного количества мелких зерен, в клетках эвгленовых нередко имеются одно- два более крупных парамилоновых зерна, на- зываемых парамилиями. Парамилии свойст- венны обычно малометаболирующим видам. Форма парамилий, их размеры и положение в клетке постоянны для определенных видов (рис. 198, 7 — 12). По-видимому, парамилии по- вышают остойчивость организмов при движении. У живых эвгленовых водорослей хорошо за- метно красное пятно в передней части клетки— стигма (табл. 28). Стигма прилегает к спинной стороне резервуара; она состоит из мелких оранжево-красных липоидных телец, окрашен- ных каротином и астаксантином. Тельца плотно прилегают друг к другу, образуя пластинку. Вместе с фоторецептором — утолщенным узел- ком на жгуте — стигма выполняет роль свето- чувствительного органа. При движении орга- ° е еае ° ° в ~ е~е 'I~ ° «р ~е ° ° ° ° ° ° ° ° ° ° ° I ° » ~Ц~ с Ъ ° ° ~,~е ig "л' еф ° л ец ее~ Рис. 201. Зеленые ээгленоэые водоросли.' cilis 2 — Е. acus; 8 — Е. convoluta; 4 = Е oxguris 5 — Eutreptia pyrenoidifera; 6 — Lepocinclis mars- sonii.
низка стигма помогает регулировать положе- ние клетки таким образом, чтобы луч света падал на хлоропласт под определенным углом. Зеленые эвгленовые водоросли обладают поло- жительным фототаксисом, но при очень силь- ном освещении они удаляются в более затенен- ные места, т. е. фототаксис становится отрица- тельным. Клеточное ядро эвгленовых водорослей круп- ное, шаровидное или эллипсоидное. Оно распо- ложено в центре клетки или сдвинуто к задне- му концу (рис. 193). У сильно вытянутых кле- ток ядро также вытянуто. Ядро состоит из нуклеоплазмы, содержащей хроматин, и кари- осомы. В отличие от ядрышка высших расте- ний кар иосома эвгленовых не исчезает при митозе, а делится самостоятельно. Эвгленовым водорослям свойственны все три основных типа питания живых организмов: фототрофный, сапрофитный и голозойный (анимальный). Фототрофный тип питания, т. е. усвоение углекислоты с помощью солнечной энергии, характерен для большей части пред- ставителей порядка Euglenales: эвглен, трахе- ломонасов, стромбомонасов, лепоиинклисов и других зеленых форм. Сапрофитный тип пита- ния, т. е. осмотическое потребление растворен- ных в окружающей среде органических веществ, преобладает у бесцветных представителей этого же порядка: астазий, дистигм, мено идиумов и других форм, не имеющих хлоропластов. Наконец, для родов пер анема, гетеронема, энтосифон и других, объединяемых в порядок перанемовых, характерен голозойный (анималь- ный) тип питания: заглатывание оформленных частиц органического вещества или других более мелких организмов. Голозойное питание встречается иногда и у астазиевых, а некоторые перанемовые могут питаться целиком сапро- фитно. Многие зеленые эвгленовые водоросли, усва- ивая углекислоту с помощью солнечной энер- гии, обладают способностью одновременно по- глощать и готовые органические вещества, имеющиеся в растворенном состоянии в окру- жающей среде, т. е. склонны к миксотрофному (смешанному) питанию. Отмечена способность эвгленовых положительно реагировать на име- ющийся в растворе витамин В1 и другие рос- товые вещества. Процесс размножения эвгленовых водорос- лей начинается обычно вечером или рано ут- ром. Заключается он в продольном делении особи надвое (рис. 199). Перед делением эвгле- новые сбрасывают наружную часть жгута. Происходит деление ядра, жгутикового аппа- рата, стигмы и хлоропластов. На переднем конце клетки появляется разделительная щель, которая постепенно увеличивается. Образую- Рис. 202. Зеленые эвгленовые водоросли.' 1 — Phacus monilatus; 2 — Ph. orbicularis; 8 — Ph. longicaudat 4 — Ph. arnoldii; 5 — та же клетка сверху; 6 — Colacium arbus- cula. щиеся при этом дочерние клетки совершают активные метаболичные движения. К концу деления особи связаны лишь своими задними концами. Затем и эта связь прерывается, клет- ки полностью освобождаются друг от друга, вырабатывают жгуты и начинают свое актив- ное движение.
° ' ° .4 ° ° ~фа ++Oe ° ~~ ° ° ° ° 4 ' ' '4''i' в ° ° 4 ° Ф ° ~ + ° °,' ° ° ° Э~у ° а~ 4,4„ ° ° ° И" 4~е ° Ъ ~ ° 8 Рис. 203. Бесцветные эвгленовые водоросли: — i ma roteus; 2 — Astasia inflata; 8 — Gyropaigne cos- costata 5 — Petalomonas 81епш; 6— Entosiphon sulcatum; 7 — Peranema tric ор orum; ronema acus. У п е ставителей рода колааиум (Со1асшт), пред ведущих прикрепленныи образ жизни, р д- Ф.У аз е- лительная щель появляется не на переднем конце, которым они прикрепляются к субстра- ту а на свободном морфологически заднем Tyf конце клетки. Деление клетки может происходить и в так называемом пал ьмеллевидном состоянии ( ис. 200). В этом случае оно сопровождается выделением большого количества слизи. Клет- ки не образуя жгутов, вновь и вновь продол- ~~1 жают делиться, и в результате получается паль- мелла — большое скопление неподвижных кле- ток, погруженных в слизь. Процесс деления может продолжаться от 2— 4 ч ( эвглен) до двух суток (у факусов). В своем цикле развития большинство эвгле- новых водорослей сохраняет подвижный образ ЖИЗНИ. и. Лишь немногие виды способны времен- но пе еходить в пальмеллевидное состояние. Преимущественно неподвижный образ жизни ведут колациум и эвгленпкапса (Euglenocapsa). У эвгленовых водорослей известны и покоя- щиеся стадии. У метаболичных видов это ок- руглые клетки. У видов с постоянной формой тела покоящиеся стадии сохраняют эту же лишь сбрасывают жгут и утолщают пе- уорму, рипласт. Эвгленовые водоросли широко распростра- нены по всей территории земного шара. Нет такой области, нет такого водоема, где они не были бы найдены в том или ином количестве. Ф.У различны к условиям внешнеи среды. Эвгле- новые водо одо росли — это обитатели континен- тальных внутренних водоемов. В морях и оке- анах находки их единичны и случайны. В не- значительном количестве встречаются эвглено- вые в текучих водоемах и в планктоне цент- ральной части крупных озер. Небогат состав этой группы в водоемах Арктики и пустын- ных о ластей. . Излюбленными местообитаниями эвгленовых являются пресные, стоячие, мелко- водные, хорошо прогреваемые, заросшие вод- ными растен тениями и богатые органическими ве- ществами в одоемы лесной и лесостепной зон: заболоченные лесные лужи, канавы, пруды, в особенности удобряемые рыбоводные и биоло- гические, сл ж е, служащие для очистки сточных вод, прибрежные мелководные участки озер. по- добных водоемах эвгленовые водоросли играют большую роль, нередко выступая в качестве доминирующих форм и вызывая «цветение» этих водоемов. П ктическое значение эвгленовых водорос- ракт леи связано прежде всего с их физиологически- ми свойствами. Способность большинства видов питанию позволяет им активно участвовать в самоочищении водоемов, загрязненных орга- ническими веществами. Среди э и эвгленовых во- дорослей существует ряд видов ов Eu lena is- ciformis, Е. viridis, Lepocinclis ovum и др.), которые являются хорошими индикаторами сте- пени загрязнения водоема. Большое содержание эвгленовых водорослей в водоеме всегда указы- вает на повышенную эвтрофность водоема. Неко- то ые эвгленовые водоросли, в частнос па gracilis, служат объектом экспериментального изучения. способность Неприхотливость этого вида, спосо ность интенсивно размножаться в условиях культу- в качестве ры позволили использовать его в эталона для выяснения действии на живой р- ганизм физиологических, химических и дру- гих факторов: различной температуры, анти- биотиков, гербицидов, витаминов, ростовых веществ и др. С помощью эвглен, положительно реагирую- можно Оп е е- На наличие витамина В 2~ ~~жн лить его содержание в различных рас р аство ах.
Отдел эвгленовых водорослей состоит из двух порядков: эвгленовых (Еиа1епа1ез) и перане- мовых (Peranematales). Первый порядок объе- диняет три семейства: эвгленовых (Euglenaceae), колациевых (Colaciaceae) и аппазиевых (Asta- siaceae) . Семейство эвгленовых наиболее богато пред- ставителями. Это зеленые свободноживущие формы. Сюда относится прежде всего централь- ный род — эвглена (Euglena, табл. 28, рис. 201, 1 — 4). Этот род наиболее известен, так как ряд видов эвглен обладает способностью разви- ваться в массе, давая упомянутое выше «цве- тение» воды. Близка к эвгленам эвтрепция (Eutreptia), от- личающаяся от них наличием двух жгутов равной длины и более сильной метаболией. Как исключение среди эвгленовых водорослей, эвтрепция предпочитает морские или солоно- ватые водоемы. Удивительно красивы и своеобразны при рассматривании под микроскопом коричнева- тые «домики» трахеломонасов (Trachelomonas, табл. 28, 1), стенки которых пропитаны гидро- окислами железа. Строение «домика» служит характерным признаком вида. Трахеломонас хорошо развивается в водоемах с темно-бу- рой гумифицированной водой, богатой соеди- нениями железа, тогда как на многие другие водоросли большое количество соединений же- леза производит угнетающее действие. Пред- ставители рода тра хеломонас встречаются в СССР в водоемах Украинского Полесья и в южной тайге Западной Сибири, где они селят- ся в основном в малых лесных заболоченных водоемах. Род лепоцинклис (Lepocinclis, рис. 201, 6) объединяет одноклеточные подвижные одно- жгутиковые организмы, совершенно неспособ- ные к метаболии. По бокам клеток лепоцинк- лиса под микроскопом обычно очень хорошо заметны два сильно преломляющих свет пятна, благодаря которым этот род отличается от некоторых близких видов эвглен. Это парами- лоновые зерна, которые у лепоцинклиса коль- чатые и постенно облегают клетку изнутри. Лепоцинклис распространен южнее, чем тра- хеломонас, и часто встречается в прудах и дру- гих водоемах лесостепи и степи, хотя некоторые виды (например, Lepocinclis ovum) имеют по- всеместное распространение. Род факус (Phacus, табл. 28, 4; рис. 202, 1 — б) морфологически наиболее сложен. Его асим- метричные клетки всегда скручены и снабже- ны килевидными выростами на спинной сто- роне и длинным рулевым отростком. Факусы обычно встречаются в небольшом числе в зарос- лях высших водных растений, случаи массового развития их редки. Семейство колациевых содержит только один род колациум (Colacium, рис. 202, 6). В цикле развития этих водорослей, в отличие от осталь- ных эвгленовых, преобладает неподвижная ста- дия. Колациум — эпипланктонный организм, поселяющийся на мелких планктонных живот- ных: циклопах, дафниях, коловратках. К телу животных колациум прикрепляется с помощью слизистой ножки или подушечки. К семейству аппазиевых относятся бесхлоро- филльные организмы, питающиеся сапрофитно органическими веществами, растворенными в окружающей воде. Питательные вещества осмотически поступают в клетку. Сюда отно- сятся сильно метаболирующие дистигма (Distig- ma, рис. 203, 1) и астазия (Astasia, рис. 203, 2), а также роды меноидиум (Menoidium, табл. 27, 8, 8), гиропаигне (Gyropaigne, рис. 203, 3), ра6- домонас (Rhabdomonas, рис. 203, 4) и цикли- диопсис (Cyclidiopsis, табл. 27, 1), имеющие по- стоянную форму тела. Все эти водоросли часто встречаются в поверхностных слоях ила в раз- личных малых водоемах. Порядок перанемовых включает виды, у ко- торых процесс питания протекает по анималь- ному типу. Относящиеся сюда петаломонас (Petalomonas, рис. 203, б; табл. 27, 2), энтоси- фон (Entosiphon, рис. 203, 6), перанема (Pera- nema, табл. 27, 7; рис. 203, 7), гетеронема (Heteronema, табл. 27, 9, 10; рис. 203, 8), анизо- нема (Anisonema, табл. 27, 4) и урцеолус (Urceo- lus, табл. 27, 5', 6) — хищники. Они заглатывают мелкие оформленные частицы органических ве- ществ и мелкие клетки других водорослей. Од- нако они могут питаться и сапрофитно. В настоящее время известно около 1000 ви- дов эвгленовых водорослей, из них для водо- емов Советского Союза отмечено около поло- вины.
ОТДЕЛ ЗЕЛЕНЫЕ ВОДОРОСЛИ (CHLOROPHYTA) КЛАСС ВОЛЬВОКСОВЫЕ (VOLVOCOP H ТСЕАЕ) 266 Зеленые водоросли — самый обширный от- пел из всех известных в настоящее время от- делов водорослей. По приблизительным под- счетам сюда входит от 13 000 до 20 000 видов. Все они отличаются прежде всего чисто-зеле- ным цветом своих слоевищ, сходным с окраской высших растений и вызванным преобладанием хлорофилла над другими пигментами. Из асси- миляционных пигментов у них обнаружены хлорофиллы а и Ь, и- и р-каротины и около 10 различных ксантофиллов. У некоторых ви- дов и стадий развития зеленая окраска клеток может маскироваться красным пигментом гема- то хромом, накапливающимся в запасных пи- тательных веществах. Клетки зеленых водо- рослей одноядерные и много ядерные, редко голые, в большинстве с целлюлозной и пекти- новой оболочкой. Запасные вещества — крах- мал, реже масло. В морфологичес ком отношении зеленые во- доросли также отличаются наибольшим много- образием по сравнению с другими отделами. Они могут быть одноклеточными, ценобиаль- ными, колониальными и многоклеточными. У них представлены все известные ступени мор- фологической дифференциации тела водорос- лей, кроме амебоидной и тканевой: монадная, коккоидная, пальмеллоидная, нитчатая со всеми ее разновидностями, пластинчатая и неклеточ- ная (сифональная и сифонокладальная). Чрез- вычайно велик и диапазон их размеров от мель- чайших одиночных клеточек диаметром 1— 2 мкм до макроскопических растений, изме- ряемых в длину десятками сантиметров. Здесь встречаются также все основные типы бесполого и полового размножения и все основные типы смены форм развития. Большинство предста- вителей в вегетативном состоянии гаплоидны, некоторые диплоидны. Распространены зеленые водоросли по всему свету. Главным образом они населяют пресные воды, где, по-видимому, возникли и прошли основные этапы своей эволюции, но среди них немало солоноводных и морских форм, а также представителей вневодных наземных и почвен- ных группировок; встречаются эпифиты, па- разиты и симбионты. Классифицируются зеленые водоросли до сих пор весьма по-разному и одной устоявшейся системы их до сих пор нет. Наиболее определен- ными единицами классификации здесь являются порядки, но в классы их группируют в разных системах различно. Наибольшие разногласия до сих пор вызывает положение в системе двух наиболее своеобразных групп, издавна отно- симых к зеленым водорослям: харовых, отли- лающихся лленисто-мутовлатым строением ве- гетативных частей таллома и совершенно не- обычным строением генеративных органов, и конъюгат, объединяемых полным отсутствием подвижных стадий и своеобразным типом поло- вого процесса — конъюгацией. В современных классификациях обе эти группы то включают в отдел Chlorophyta, то выделяют в самостоя- тельные отделы Charophyta и Conjugatophyta (Zygnematophyta, Zygophyta). В первом случае их оценивают то рангом порядка, то как класс, а в отношении конъюгат даже и как подотдел. В настоящей книге харовые рассматриваются как самостоятельный отдел, но конъюгаты включены в отдел зеленых водорослей в ранге самостоятельного класса, характеризующегося более всего специфическим способом полового размножения. Остальные порядки зеленых во- дорослей нам представляется более естествен- ным группировать в классы по тем основным ступеням морфологической дифференциации те- ла, которой они достигли, начиная от наипро- стейшего выражения в рамках каждой ступени и кончая наиболее сложными вариантами, даю- щими переходы к следующему уровню. Таким образом в отделе зеленых водорослей мы принимаем деление на 5 классов: i) вольвок- совые, основная форма вегетативного тела— подвижные жгутиковые клетки и колонии та- ких клеток; 2) протококковые, основная форма вегетативного тела — неподвижные клетки с плотной оболочкой и колонии таких клеток; 3) улотриксовые — многоклеточные нитчатые и пластинчатые формы различной степени усложненности; 4) сифоновые — неклеточные одноядерные и многоядерные формы с различ- ной степенью внешнего расчленения слоевищ и нитчатые формы, образованные многоядер- ными сегментами; 5) конъюгаты — однокле- точные симметричного строения и нитчатые формы с половым размножением типа конъю- гации. К классу вольвоксовых относятся наиболее примитивные представители отдела зеленых водорослей (Chlorophyta), имеющие монадную структуру тела. В вегетативном состоянии они подвижны в течение всей жизни. Громадное большинство вольвоксовых — од- ноклеточные организмы, но есть также цено- биальные формы, а у видов рода вольвокс (Vol- vox) таллом представляет собой колонию. Не- зависимо от строения тела слагающие его клет-
ки всегда несут на переднем конце два, реже четыре одинаковых жгутика; лишь у неболь- шого числа примитивных форм могут наблюдать- ся два или три неравных жгутика; очень редко бывает только один жгутик. Оболочка клеток пектиновая или целлюлозная, обычно доста- точно твердая; она плотно прилегает к прото- пласту или отстает от него, иногда снаружи об- разует выросты разнообразной формы или ос- лизняется, реже у клеток имеется только тон- кий перипласт. Вольвоксовые — одноядерные формы. Ядро у них шаровидное, обычно с хорошо выражен- ным ядрышком, и располагается, как правило, в центре клетки, реже в передней или задней ее части. Хлоропласт, за очень редким исклю- чением, один, стенкоположный, обычно чаше- видный, с одним крупным пиреноидом в утол- щенной части, реже пиреноидов два или не- сколько, иногда они вовсе отсутствуют. На хлоропласте, обычно в передней его части, на- ходится красное пятнышко — глазок. Имеются пульсирующие вакуоли, чаще всего две, рас- положены они спереди, у основания жгутиков, реже их несколько, и они разбросаны по всему протопласту в вырезках хлоропласта. Громадное большинство вольвоксовых — фо- тотрофы, хотя известны случаи гетеротрофного и миксотрофного питания. В процессе фото- синтеза наряду с крахмалом могут вырабаты- ваться масло и волютин. Размножение вол ьвоксовых осуществляется вегетативным, бесполым и половым путем. Осо- бенно большого разнообразия достигают здесь формы полового процесса, в результате кото- рого возникает зигота, прорастающая после периода покоя. При прорастании зиготы обыч- но образуются четыре, реже большее число зооспор, дающих начало новым особям. У не- которых видов зигота вначале подвижна, и тогда ее называют планозиготой, в отличие от неподвижной гипнозиготы. Вольвоксовые — типичные активные планк- теры. Обитают они преимущественно в мелких, стоячих, нередко быстро пересыхающих водо- емах — в лужах, канавках, болотах, отстой- никах, на полях фильтрации, в биологических прудах очистных сооружений и в других водо- емах с органическими загрязнениями воды. Реже они встречаются в водохранилищах, озе- рах, старицах и реках. Известны формы, оби- тающие в водоемах с высокой степенью соле- ности и в почве. Происхождение воль воксовых легко можно вывести из первичных примитивных амебоид- ных форм. Эволюция в пределах класса шла, по-видимому, в трех направлениях: i) по линии усложнения полового процесса — от наиболее примитивного гологамного к наиболее сложно- му оогамному; 2) в направлении усложнения од- ноклеточного таллома через ценобиальные фор- мы к колониальным с хорошо выраженной диф- ференциацией клеток на вегетативные и репро- дуктивные; 3) в направяении утраты подвиж- ности в вегетативном состоянии — одного из наиболее примитивных признаков раститель- ной клетки. Эта линия эволюции привела к фор- мированию протококковых водорослей, сос- тавляющих самостоятельный класс зеленых водорослей. Значение вольвоксовых в природе и в жизни человека велико. Прежде всего это активные санитары загрязненных и сточных вод. Разви- ваясь в массе в многочисленных мелких и силь- но загрязненных водоемах, вольвоксовые при- нимают самое активное участие в процессах самоочищения загрязненных вод. Благодаря способности вольво ксовых выдерживать раз- личную степень загрязнения среды обитания их используют в качестве показательных форм при биологическом анализе загрязненных вод. Вольвоксовые принимают также активное учас- тие в отложении сапропелей, являются одним из звеньев в цепи питания гидробионтов. Некото- рые из них способны вызывать зеленое и крас- ное «цветение» воды в крупных водоемах, где соз- даются оптимальные условия для их массового развития. Из некоторых видов, вызывающих красное «цветение», можно получать каротин, препараты которого широко используются в практике. Вольвоксовые водоросли представляют собой большую и четко очерченную систематическую группу зеленых водорослей, которую естествен- нее всего принимать в ранге класса. Класс воль- воксовых подразделяют на два весьма неоди- наковых по объему подкласса: протохлорино- вые (Protochlorinophycidae) и собственно воль- воксовые (Volvocophycidae). Первый из них, объединяющий в своем составе очень неболь- шое число родов и видов с примитивным строе- нием тела, изучен еще совершенно недостаточ- но, и здесь мы рассматривать его не будем. На- оборот, собственно воль воксовые объединяют очень большое количество видов, и мы рассмот- рим их более подробно. Громадное большинство их имеет хорошо выраженную плотную обо- лочку и обычно два, реже четыре равных жгу- тика. Наиболее просто устроенные представи- тели этого подкласса, с оболочкой в виде тон- кого и нежного перипласта, выделены в поря- док гголиблефаридовых (Polyblepharidales); одно- клеточные формы с хорошо выраженной плот- ной оболочкой выделяют в наиболее обширный порядок хламидомонадовых (Chlamydomona- dales); ценобиальные и колониальные формы объединены в сравнительно небольшой порядок вольвоксовых (Volvocales). 267
Рис. 204. Полиблефаридовые: 1 — Dunaliella salina; 2 — 5 — Phyllocardium complanatum (2— вегетативная клетка, 8 — гологамия, 4 — планозигота, 5— гипнозигота); 6 — Spermatozopsis exsultans; 7 — Pyramimonas reticulata. ПОРЯДОК ПОЛИБЛЕФАРИДОВЫЕ (POLYBLEPHARIDALES) Этот порядок объединяет около 10 неболь- ших родов, состоящих преимущественно из одноклеточных форм. Наиболее изученными, имеющими практическое значение, являются виды рода дюкалиелла (Dunaliella). Это самый крупный род порядка, насчитывающий 38 ви- довых и внутривидовых таксонов. Самый из- вестный и интересный из них — дюналиелла солевая (Dunaliella salina), вызывающая крас- ное «цветение» воды в пересыщенных солью водоемах. Интересна история его открытия. Трудно себе представить красное «цветение» воды, но в природе такое явление уже давно было известно и, конечно, не могло пройти мимо внимания ученых. К выяснению причин красного «цветения» первыми приступили фран- цузские ученые. В 1836 г. Пари~кской Акаде- мией наук такое задание было поручено ака- демику Пайену, который после проведенных исследований пытался объяснить это явление массовым развитием мелкого ракообразного ар- темии. Однако это оказалось ошибочным. Ис- следованиями другого французского ученого- Дюналя — в 1838 г. было показано, что красное «цветение» в сильно засоленных (гипергалин- ных) водоемах вызывается массовым развитием одноклеточной водоросли, которая в 1905 г. была подробно описана румынским ученым Теодореско и названа в честь первооткрывате- тя дюналиеллой. В степях Крыма и Присивашья часто можно увидеть водоем с кирпично-красной водой. Это, как правило, бывает связано с массовым развитием дюналиеллы солевой. Сделав обыч- ный препарат из такой красной воды и рассмат- ривая его при малом увеличении, можно заме- тить лишь крохотные подвижные розовые то- чечки. Переведя микроскоп на большое увели- чение, можно рассмотреть остановившуюся или приклеившуюся к предметному стеклу жгути- ками отдельную клетку. Клетка дюналиеллы солевой (рис. 204, 1; табл. 29, 1 — о) покрыта только тонким и неж- ным перипластом, что позволяет ей изменять форму, но все же чаще всего она удлиненно-яйце- видная. Длина клеток колеблется от 5 до 29 лкл, ширина от 3,8 до 20,3лкл. Сзади клетка широко округлая, спереди суженная и несет два рав- ных жгутика. Внутри клетки имеется чаше- видный хлоропласт, обычно доходящий почти до основания жгутиков. В передней части хло- ропласта расположен бледно-розовый, нечетко выраженный глазок, а внизу, в утолщенном его дне, лежит крупный пиреноид, окруженный крахмальной сферой, состоящей из несколь- ких крупных крахмальных зерен. Ядро одно, шаровидное, оно расположено в центре клетки над пиреноидом, в вырезке хлоропласта, и обычно его трудно заметить. В процессе жизне- деятельности клетки в ней накапливаются ка- ротиноиды, придающие клетке розово-желтую или кирпично-красную окраску. Живые клетки дюналиеллы плавают быстро, но плавно. Это типичные активные планктеры. Размножение у дюналиеллы в основном веге- тативное, путем продольного деления клетки в подвижном состоянии. Известен также поло- вой процесс в виде типичной гологамии. Возник- шая зигота обычно окрашена в зеленовато- красный цвет. Прорастая, она дает начало ве- гетативным клеткам. В цикле развития дюна- лиеллы известны пальмеллы и цисты (табл. 29, 8 — 5). Способность этого и других видов дюнати- елты отлагать в клетке каротиноиды используют в практике — из них добывают пигмент каро- тин, широко применяемый в ряде отраслей про- мыштенности. К порядку полиблефаридовых относятся так- же следующие хорошо изученные и широко распространенные виды: двуясгутиковый фил- локардиум уплощеккый (Phyllocardium com- planatum) и четырехжгутиковые сперлатозопсис прыгающий (Spermatozopsis exsultans) и пира- 268
иимонас сетчатый (Pyramimona рис. 204 3 — 7 s re icu ata, ПОРЯДОК ХЛАМИДОМОНАДОВЫЕ (CHLAMYDOMO- NADA LES) К этому порядку относятся одноклеточные формы с хорошо выраженной оболочкой дв- м я или четырьмя равными жгутиками. Хлоро- пласты у них всегда зеленые, лишь иногда за- маскированные гематохромом в красный цвет, реже клетки бесцветные. Последние выделены в особое семейство политомовых (Polytomaceae). В зависимости от строения оболочки окрашен- ные представители порядка хламидомонадовых подразделяются на три семейства. Основное и наиболее крупное из них — семейство хлали- домонадовых (Chlamydomonadaceae), предста- вители которого характеризуются наличием достаточно мягкой оболочки которая торая при раз- множении клеток обычно растягивается, не распадаясь на отдельные куски. Центральным родом этого семейства является род хлалидоло- 500 ви ов нас (Chlamydomonas), насчитывающий с ий свыше видов. Почти все они — обитатели мелких, хорошо прогреваемых и сильно загрязненных водоемов. Это активные санитары загрязнен- ных вод, в которых они очень быстро размно- жаются и вызывают зеленое «цветение». Наря- ду с автотрофным способом питания их клетки способны всасывать через оболочку растворен- ные в воде органические вещества, что способ- ствует активизации процессов самоочищения загрязненных вод. В сказанном очень легко убедиться, если побывать на очистных сооружен ниях городской канализации и понаблюдать за деятельностью хламидомонадовых в отстойника х или картах полей фильтрации. Здесь хозяйственные и бы- товые стоки могут «зацветать» буквально в те- чение нескольких дней, приобретая ярко-зеле- ную окраску. Под микроскопом можно увидеть, что в капельке такой густой мутной ной зеленои воды жизнь бьет ключом: сотни ярко- ярко-зеленых кле- ток проносятся в поле зрения с неве б роятнои быст- ротой, причем и форма, и размеры, и даже манера плыть не у всех клеток одни и те " . Н же. о иногда в такой зеленой капельке почти все клетки бы- вают совершенно одинаковыми и ведут себя однотипно. Следовательно зеле зеленое «цветение» здесь может быть вызвано нескольки ькими видами или только одним. Но такая активность в по- ведении хламидомонадовых весьма к атк менна. Тот Тот же водоем через несколько дней может оказаться с чистой прозр розрачнои до дна Ф.У и неокрашенной водой. Тех видов — самых активных санитаров сточных в не будет. Их зиготы, цисты или пальмеллы, по- глотив основн ую массу органических веществ, опгстились на на дно и там, в илу, «отдыхают». При благоприятных для них условиях — с по- явлением новых сильных органических загряз- нений в воде, со сбросом стоков — их жизнеде- ятельность опять активизируется. Более подробно с семейством хламидомона- довых можно познакомиться на примере одного из видов рода хламидомонас. Хламидомонас хоботковый (Chlamydomonas pro oscigera), в отличие от дюналиеллы и других полиблефаридовых, имеет плотную двухкон- турную оболочку, спереди образующую замет- ное выпячивание — носик Разме зме ы и орма носика в пределах рода сильно варьируют, но являются постоянными для отдельных видов и используются при их описании как важные систематические признаки. От основания но- ]~ис. 205. Х ламидомонадовые: 1 — 2 — Chlamydomonas proboscigera вз ос 2 — 2 — ~, рОсла клетка и копу- пигаеа cochlearis); 4 — Lobomon ет; — . annuraeae на пан и нцире коловратки (All- ошопаз Ме11аСа; 5 — Ригс111а 51- — arteria crucifera. 269
Рис. 206. Коккомонадовые и факотовые: 2 — 2 — Pedinopera robusta, строение протопласта внутри обо- лочки и оболочка снаружи; 8 — 4 — Pedinoperopsis gracilis, строе- ние протопласта внутри оболочки и оболочка снаружи; 5 — 6— Pteromonas aculeata, клетка с широкой и узкой сто оны. 7— Phacotus coccifer. сика отходят два крепких равных по длине жгу- тика. Всю внутреннюю часть клетки занимает чашевидный хлор опласт, в сильно утолщен- ном дне которого расположен шаровидный пи- реноид с крахмальной сферой. В верхней части ва хлоропласте находится крупный, хорошо заметный красный глазок, а внутри вырезки хлоропласта расположено видимое без спе- циальной окраски крупное шаровидное ядро с отчетливо различимым ядрышком. В перед- ней части клетки, у основания жгутиков, в вы- резке хлоропласта легко заметить две пульси- рующие вакуоли. Кроме бесполого размножения зооспорами, у этого вида наблюдается и половой процесс в виде типичной изогамии. Интересны здесь гаметы. Они имеют копуляционные хоботки У выступающие через оболочку между основани- ями жгутиков (рис. 205, 1, 2), что и послужило основой для видового названия. Хламидомонас хоботковый распространен очень широко. Он часто встречается в неболь- ших стоячих водоемах, лужах, прудах, кана- вах, арыках. Это типичный свободноживущий активный нланктер. Однако среди видов хламидомонас известны и такие, клетки которых прикрепляются к телу водных животных, преимущественно коловра- ток, как, например, хламидомонас аннуреи (Chlamydomonas annuraeae, рис. 205, 8) и не- которые другие. Они потеряли собственную подвижность, но стали передвигаться еще быст- рее, сидя на движущихся коловратках. К этому же семейству относится род гелато- коккус, виды которого (например, Haemato- coccus pluvialis, табл. 29, 6, 7) способны накап- ливать в клетках и особенно в апланоспорах большое количество красного гематохрома, что отражено в названии рода. Интересны также роды лобомонас (Lobomonas, рис. 205, 4) и фурцилла (Furcilla, рис. 205, б), виды которых отличаются разнообразными выростами клеточ- ной оболочки, и род картерия (Carteria), на- считывающий свыше 60 видов, характеризую- щихся наличием у клеток не двух, а четырех одинаковых жгутиков (рис. 205, 6). Кроме рассмотренного большого семейства, в порядке хламидомонадовых имеются еще два небольших, но интересных семейства. Так У представители семейства коккомонадовых (Coc- comon adinaceae) отличаются плотной, пропи- танной известью или содержащей другие вклю- чения сплошной оболочкой, как, например, в роде пединопера (Pedinopera robusta, рис. 206, 1, 2). Представители третьего семейства — факото- вых (Phacotaceae) — характеризуются двуствор- чатой, инкрустированной углекислым каль- цием, кремнеземом или солями железа оболоч- кой, образующей иногда крыловидные выросты. Сюда относятся, например, роды пединопероп- сис (Pedinoperopsis), тперомонас (Pteromonas) и факотус (Phacotus), виды которых часто встре- чаются в ямах, содержащих большое количест- во органических загрязнений (рис. 206, 8 — 7). ПОРЯДОК ВОЛЬВОКСОВЫЕ (VOLVOCALES) Порядок вольвоксовых объединяет наиболее высокоорганизованных представителей клас- са. Сюда относятся исключительно ценобиаль- ные и колониальные формы. Но и здесь отдель- ные клетки, слагающие цен обии и колонии 1 построены по типу хламидоманады. Однако, в отличие от одноклеточных форм, при размно- жении клетки здесь не расходятся, а сраста- ются своими оболочками или остаются соеди- ненными общей слизью. Форма общей слизистой оболочки, количество клеток и способ их раз- мещения в ней, а также способ срастания кле- ток в комплексы являются постоянными при- знаками и широко используются в систематике 270
Обычно порядок вольвоксовых подразделяют на три семейства. Один из наибо- лее типичных представителей семейства спцн- диломоровых (Spondylomoraceae) — пироботрис стройный (Pyrobotrys gracilis, р ., ). Он характеризуется способом срастания кле- ток, расположением их ярусами и отсутствием общей слизистой оболочки вокруг гроздевид- ного ценобия. Этот вид обычно хорошо разви- вается, например, в первой половине мая в от- стойниках очистных сооружений сахарных за- водов Украины в придонных слоях воды. Если взять пипеткой каплю образовавшегося на дне этих водоемов темно-зеленого осадка и приго- товить препар арат то под микроскопом можно видеть почти чистую культуру пироботриса- его четырехъярусные, обычно 16-клеточные це- нобии с клетками различного возраста: от кро- хотных, стройных, грушевидных и еще слабо- х об атно- окрашенных до крупных, толстых, р яиц евидных Ф.У клеток с интенсивно окрашенным темно-зеленым хлоропластом. Часть крупных клеток таких ценобиев вместо протопласта может содержать уже образовавшиеся зооспоры, которые, двиг игаясь в оболочке материнской \J клетки, там же срастаются в новый крохотныи статочно подрастут, они разрывают оболочку материнской клетки и новый молодой ценобий, оказавшийся в воде, начинает жить как само- стоятельный ценобиальный органи изм. В конце вегетационного периода в клетка ц ах енобиев пи робот риса вместо зооспор возникают изо- гаметы. Они попарно копулируют и дают зи- готу, при помощи которой вид переживает не- благоприятные условия до следующего года. В чистых водах рек и озер можно встретить четырехжгутиковую форму, относящуюся к это- м семейству, например спондиломорум чет- веркой (Spondylomorum quaternarium, ри . ис. 207 2), сходный с предыдущим видом. Весьма свое- о разный п б о строению вид — хлор корона о- с. 207 8 гелская (Chlorcorona bohemica, рис., ), в ценобиях которой клетки соединены слизис- тыми тяпками в два 4-клеточных р омбических яруса. е ~Stе- Второе семейство — стефакосферовые ( е- р hanosphaeraceae) — содержит единственный род и вид стефакосферу дождевую (Stephano- sphaera pluvialis, рис. 207, 4). Это типичный холодолюбивый вид (криофил), встречающийся обычно на поверхности снегового покрова в се- верных районах. Его можно найти, например, в Мурманской области, где он вегетирует на протяжении всего лета на поверхности тающего снега и льда в овражках и расщелинах скал каменистой тундры. В пробе воды из такого растаявшего снега при малом увеличении мы увидим шаровидные или эллипсоидные цено- Рис. 207. Спондиломоровые и стефаносферовые: raci lis 2 — Spondylomorum qua ternarium; — —.te hanosphaera pluvialis 8 — Chlorcorona bohemica; 4 — ер бии, внутри которых обычно расположено 8 кле- ток более или менее веретеновидной формы. Клетки покрыты лишь тонким и нежным пери- пластом, позволяющим им вырабатывать раз- ветвленные протоплазматические выросты, ко- торыми они упираются в стенку ценобия. обия клетки расположены поясом внутри цен экватору а их жгутики выходят наружу. тся по В хлоропласте каждой клетки содержится 2 — 5 пиреноидов и округло-треугольный гла- зок, расположенный на наружной его стороне. П льсирующих вакуолей несколько, и они раз- б осаны по всему протопласту. В клетках не- которых ценобиев можно увидеть дробление протопласта на 8, реже 4 или 16 участков. Это начало образования зооспор, которые, сформи- овавшись, не выходят из оболочки материн- ров ской клетки, а размещаются в ней ей по ее эква- тору и слагают новый ценобий. Кроме зооспор, в клетках ценобия может образоваться от 27
Рис. 208. Вольвоксовые: 2 — Gonium pectora е; 2 — u 1; — E dorina elegans; 3 — Pandorina morum. 4 до 32 веретеновидных изогамет, которые, попарно копулируя внутри ценобия, дают одну зиготу. Последняя накапливает гематохром, приобретает красную окраску и переходит в со- стояние покоя. Через некоторое время она дает Последнее семейство — собстпвенно вольвоксо- вие (Volvocaceae) — представлено ценобиальными и колониальными формами. Из ценобиальных форм наиболее обычны представители родов гокиул (Сonium) и эвдорина (Eudorina). У пер- вого из них клетки ценобия расположены в один слой, пакетиком, окруженным общей слизистой обверткой. Яркое впечатление производит под микро- скопом гониум пекторалькый (С onium pec- torale, рис. 208, 1) во время движения, когда его 16-клеточный ценобий парит в толще воды, напоминая сказочный ковер-самолет: клетки ценобия, дружно двигая жгутиками, свисаю- щими вниз бахромой, несут над собой о щую слизистую обвертку. У широко распространенных видов эвдорины ценобии обычно эллипсоидные, состоят из 32 клеток, расположенных не в одной плоскости, а по периферии в поверхностном слое общей слизи в 5 — 8 рядов. Благодаря согласованному движению жгутиков всех 32 клеток ценобий Д овольно быстро передвигается в толще воды. Обычно в ценобии эвдорины все клетки одина- ковые (Eudorina elegans, рис. 208, 2) и все спо- собны, как и у гониума, к образованию беспо- лым путем новых дочерних ценобиев. У пандорикы (Pandorina тогит, рис. 208, 8) в ценобиях обычно 16 клеток, и расположены они не рыхло, как у эвдорины, а компактно, соприкасаясь боковыми сторонами, в связи с чем обычно приобретают многогранную форму. Указанные ценобиальные формы вольвоксо- вых встречаются довольно часто, обычно в не- больших стоячих водоемах, где летом нередко вызывают зеленое «цветение» воды. Размножа- ются они в основном бесполым способом, путем об азования в каждой клетке молодых цено- биев. К концу вегетации у них осуществляе о раз твляется и половой процесс — в виде изогамии у гониу- мов, гетерогамии у пандорин и оогамии у эвдо- рин. Во всех случаях образовавшаяся зигота переходит в состояние покоя. Колониальные формы вольвоксовых представ- лены лишь видами рода вольвокс (Volvox). Это наиболее высокоорганизованные представите- ли класса. Их колонии (рис. 209) имеют вид слизистых, диаметром до 2 лл шаров, в перифе- р ическом слое которых расположено множество, до 20, а у некоторых и до 50тыс. хламидомонадо- подобных клеток, сросшихся своими боковыми ослизненными стенками друг с другом и соеди- ненных одна с другой протоплазменными тяжа- ми (плазмодесмами). Здесь у~ке наблюдается дифференциация или специализация клеток в пределах колонии. Громадное большинство составляют вегетативные клетки, слагающие периферическую часть колонии и не принимаю- щие участие в размножении. Между вегетатив- ными клетками разбросаны более крупные ре- продуктивные клетки. Около десятка из них- клетки бесполого размножения, так называе- мые п а р т е н о г о н и д и и, которые в результате многократных делений дают начало молодым, очерним колониям внутри материнской. Поло- вое размножение представлено типичной оога- мией. Оогонии и антеридии возникают такнсе из репродуктивных клеток. В грушевидных, с темно-зеленым содержимым оогониях возни- кает одна яйцеклетка. В антеридиях образует-
ся по 64 желтоватой окраски антерозоида, сло- женных в виде пакетика. После оплодотворения яйцеклетка превращается в ооспору с много- слойной, шиповатой снаружи оболочкой и бо- гатым запасными веществами внутренним со- держимым. После периода покоя ооспора дает начало новой колонии вольвокса. Наиболее широко распространенными явля- ются два вида этого рода — вольвокс шаровид- ный (Volvox globator) с однодомными колониями и вольвокс золотистый (Volvox aureus), колонии которого двудомны. В центральных районах европейской части СССР оба эти вида часто встречаются в старицах рек и прудах, нередко вызывая дансе «цветение» воды (рис. 209). Из вышесказанного видно, что класс воль- воксовых (Volvocophyceae) — это сравнительно однородная группа отдела зеленых водорослей (Chlorophyta). Они характеризуются только од- ной монадной структурой тела. Таллом их пре- имущественно одноклеточный, у некоторых ти- пично ценобиальный, у немногих колониаль- ный. Строение клетки в общем однородное, почти у всех видов хламидомонадоподобное. Вольвоксовые не являются тупиковой ветвью в эволюции зеленых водорослей. Потеряв под- вижность, представители вольвоксовых на ка- ком-то этапе исторического развития передали эстафету протококковым водорослям. КЛАСС ПРОТОКОККОВЫЕ (PROTOCOCCOPH YCEAE) В отличие от вольвоксовых протококковые водоросли характеризуются в вегетативном со- стоянии полным отсутствием монадной структу- ры тела. У громадного большинства видов тал- лом имеет коккоидную структуру. Реже наблю- дается пальмеллоидная структура, у одного ви- да (Protosiphon botryoides) известна примитив- но сифональная и у нескольких родов прими- тивно нитчатая и примитивно пластинчатая структуры. В основном это одноклеточные и ценобиальные, ре~ке колониальные формы, и лишь у видов с примитивно нитчатой и пластин- чатой структурами можно говорить о много- клеточности таллома. Несмотря на кажущуюся однородность, про- тококковые водоросли представляют собой очень обширную и многообразную группу, ин- тересную в теоретическом и весьма важную в практическом отношении. В теоретическом отношении протококковые водоросли интересны прежде всего тем, что в процессе эволюции отдела зеленых водорослей на этом этапе впервые возникла, утвердилась и приобрела широкое развитие коккоидная, типично растительная структура тела. Именно этот класс стал узловым пунктом в эволюцион- Рис. 209. Вольвоксовые: Volvox aureus. ном развитии зеленых водорослей, именно в его пределах зародились новые ступени морфоло- гической дифференциации тела, представлен- ные ныне сифоновыми, сифонокладовыми, сцеп- лянками и улотриксовыми водорослями. По строению протопласта протококковые на- поминают вольвоксовых. Наиболее примитив- ные из них еще сохраняют пульсирующие ва- куоли, глазок и даже жгутики, хотя послед- ние здесь неподвижны и именуются п с е в д о- ц и л и я м и. Наличие этих рудиментарных органелл позволяет выводить происхождение протококковых непосредственно из вольвоксо- вых. С другой стороны, наличие у некоторых видов протококковых водорослей тенденции да- вать более или менее долговечный нитчатый или пластинчатый таллом, возникающий в результате деления клеток, указывает на пря- мой эволюционный переход от протококковых к улотриксовым водорослям. Громадное большинство протококковых- микроскопические формы, лишь в пределах не- скольких родов таллом может достигать круп- ных размеров. Форма клеток здесь чрезвычайно разнообразна, но преобладающей является ша- ровидная, эллипсоидная и яйцевидная. Встре- чаются также веретеновидные, полулунные, серповидные, почковидные и иной формы клет- ки. Размеры их колеблются примерно от 2 до 30 лил, но у некоторых видов они могут быть крупнее, длиной до 200 — 400 мил, и только у водяной сеточки (Hydrodictyon reticulatum, см. ни~ке) длиной до 1,5 сл при общих размерах всего таллома (ценобия) до 1 л. Оболочка клеток протококковых водорослей всегда сплошная, из целлюлозы, реже с при- месью пектиновых веществ, а у низших форм
чисто пектиновая. У многих видов она снаружи снабжена щетинками, иглами, шипиками, бо- родавками или окружена толстым слоем сли- зи, что помогает клеткам пассивно парить в толще воды. Хлоропласт в клетках обычно один, стенко- положный, чашевидный, реже пластинчатый или сетчатый, изредка центральный, лишь у немногих видов имеется несколько дисковид- ных хлоропластов. Пиреноид в клетке тоже обычно один, ба за льный, реже центральный или боковой, покрытый общей крахмальной сферой или скорлупками крахмала. Лишь у немногих видов пиреноидов в хлоропласте несколько. У громадного большинства прото- кокковых водорослей клетки одноядерные; ядро обычно крупных размеров, с хорошо выражен- ным ядрышком. Сравнительно редко встреча- ются многоядерные клетки. У протококковых водорослей преобладает бесполое размножение, реже наблюдается по- ловой процесс, иногда они размножаются путем простого деления клеток. Бесполое размножение осуществляется при помощи гемизооспор, зооспор и автоспор. Гемизооспоры, или амебо- и д ы, — наиболее примитивные специализирован- ные клетки бесполого размножения. От зооспор они отличаются отсутствием жгутиков, а от авто- спор, кроме того, и отсутствием настоящей обо- лочки, в связи с чем способны производить аме- боидные движения и передвигаться в том или ином направлении. Зооспоры — дальнейший этап в усложнении бесполого размножения протококковых водорослей; они характеризуют- ся наличием жгутиков, но еще лишены настоя- щей оболочки и, как правило, кратковременны. Наконец, автоспоры представляют высший этап в усложнении бесполого размножения прото- кокковых водорослей, они лишены жгутиков и всегда покрыты оболочкой. В громадном большинстве случаев автоспора является ми- ниатюрной копией вегетативной клетки, реже она дооформляется после выхода из оболочки материнской клетки. Половой процесс у протококковых водорос- лей наблюдается сравнительно редко. Обычно это изогамия, реже гетерогамия. Оогамный половой процесс известен только у очень немно- гих родов. Наиболее обычными стадиями покоя у протококковых водорослей являются акинеты и цисты. В экологическом отношении протококковые водоросли исключительно многообразны. В во- доемах они принимают активное участие в сло- жении планктона, бентоса и нейстона, нередко встречаются в перифитоне. Они обычны также в наземных местообитаниях и в почве. Известны среди них эндофитные формы, симбионты и дансе паразиты. Основную массу видов протококко- BbIx составляют все же планктонные формы, размножающиеся автоспорами. Собственно бен- тосных форм сравнительно немного. В сложении перифитона протококковые играют весьма су- щественную роль. Здесь в зависимости от суб- страта, на котором они живут, протококковые четко подразделяются на эпифитные и эпизой- ные формы. Последние поселяются обычно на панцирях циклопов и дафний, ренсе встречают- ся на их личинках, на олигохетах, коловрат- ках, инфузориях, личинках комаров и других гидробионтах, причем иногда могут развиваться в массовых количествах, придавая животному ярко-зеленую окраску и вызывая «цветение» воды. В водоемах протококковые водоросли распро- странены чрезвычайно широко, но наиболее богато они представлены в прудах, в частности в рыбоводных прудах, отстойниках сахарных заводов, биологических прудах и картах полей фильтрации городских очистных сооружений. Самыми распространенными являются виды родов сценедесмус, анкистродесмус, ооцистис, педиаструм, круцигения, хлорелла, но «цвете- ние» воды могут вызывать лишь некоторые виды. Почти все протококковые водоросли относятся к эвригалинным и эвритермным формам, сте- ногалинных и стенотермных видов среди них немного. С другой стороны, многие из прото- кокковых являются типичными сфагнофилами. Известны также представители альпийско-арк- тической флоры, вызывающие образование «зе- леного» и «красного» снега. Физиологические особенности протококко- вых водорослей недостаточно изучены. Исклю- чение составляют только виды родов Ghlorella, отчасти Scenedesmus, Ankistrodesmus и Lagerhei- mia. На этих объектах, ставших классическими моделями растительной клетки, ученые разных стран изучали процесс фотосинтеза и минераль- ного питания зеленых растений, обмен веществ и влияние на растительный организм различ- ных ядов, пестицидов, радиоактивных изото- пов и т. п. Велись также исследования по выяс- нению влияния различных химических, свето- вых и температурных режимов на развитие кор- мовых протококковых водорослей для рыбного хозяйства и животноводства. Полученные дан- ные используются в опытах по интенсификации процессов очистки воды от загрязнений биоген- ного происхождения и по массовому культиви- рованию водорослей на сточной жидкости с целью производства белкового и витаминозного корма. Некоторые виды протококковых водо- рослей специально изучаются для использова- ния в качестве регенераторов воздуха, воды и пищевой биомассы в условиях замкнутых био- логических систем при дальних космических полетах. 274
По количеству видов в водоемах и интенсив- ности развития протококковые водоросли в целом уступают, и то не всегда, лишь диатомо- вым водорослям. Поэтому так велико их зна- чение в общем круговороте веществ и как про- дуцентов кислорода в водоемах, а значит, велика и их роль в жизни человека и планеты в целом. Объем протококковых водорослей как систе- матической группы и номенклатуру их система- тических подразделений понимают по-разному. Некоторые систематики рассматривают их как порядок в пределах класса собственно зеленых водорослей (Chlorophyceae), именуя то протокок- ковыми (Protococcales), то хлорококковыми (Chlo- rococcales); в группу то включают так называе- мые тетраспоровые (Tetrasporales), то не вклю- чают. Другие считают их подклассом или клас- сом. Мы рассматриваем протококковые водоросли как класс и сохраняем за ними латинское наз- вание Protococcophyceae. Кроме двух ранее из- вестных порядков — вакуольных (Vàñèolàlås) и хлорококковых (Chlorococcales), мы устанавли- ваем в пределах этого класса еще и третий по- рядок — прототриховых (Prototrichales), вклю- чая в него представителей, обладающих спо- собностью образовывать примитивные, иногда лишь кратковременно существующие талломы в виде нитей или паренхиматозных пластинок, т. е. представителей с примитивными нитчатой, разнонитчатой и пластинчатой структурами тела. В то время как порядок Vacuolales яв- ляется естественным переходом от класса Vol- vocophyceae к классу Protococcophyceae, поря- док Prototrichales служит связующим звеном между Protococcophyceae и Ulotrichophyceae. Центральное место в пределах класса Protococ- cophyceae принадлежит порядку Chlorococcales. Перейдем теперь к последовательному зна- комству с главнейшими представителями ука- занных трех порядков. ПОРЯДОК ВАКУОЛЬНЫЕ (VACUOLALES) К этому порядку относятся наиболее прими- тивные представители класса, вегетативны е клетки которых всегда неподвижны, но на всех стадиях развития сохраняют еще пульсирую- щие вакуоли (рис. 210). У некоторых из них сохраняются также глазок и редуцированные, ставшие неподвижными жгутики, называемые здесь псевдоцилиями (табл. 29, 9). Размно~кение осуществляется главным образом зооспо рами, реже гемизооспорами. У колониальных форм, кроме того, наблюдается деление клеток, за счет которого происходит разрастание таллома. Половой процесс в виде изогамии или гетеро- гамии известен у немногих видов. По строению протопласта вегетативных кле- ток вакуольные очень близки к представителям семейства хламидомонадовых класса вольвоксо- вых. Сюда входят 8 семейств, объединяющих в своем составе около 30 родов, в том числе все водоросли, по другим системам составляющие порядок тетраспоровых (Tetrasporales), предста- вители которого характеризуются пальмелло- идной структурой тела. Рассмотрим более подробно один из видов рода гипномонас (Hypnomonas chlorococcoides, рис. 210, 1). Знакомиться с этим объектом можно только на живом материале. Получить живую культуру гипномонаса очень легко. Для этого с поверхности почвы в саду, на огороде или просто во дворе берут небольшую почвенную пробу, измельчают ее, освобождают от мусора и высыпают в колбочку с питательным раство- ром или почвенной BbITHiHEofl (см. очерк «Поч- венные водоросли»). При содержании на север- ном окне комнаты через 3 — 5 дней в колбочке по урезу воды появится зеленый налет, особен- но хорошо заметный на ее освещенной стороне. В большинстве случаев этот налет оказывается образованным клетками гипномонаса. Взяв пи- петкой часть налета, готовят висячую каплю или заклеенный препарат. Протопласт крупной зе- леной шаровидной клетки под микроскопом очень напоминает хламидомонаду (рис. 210, 1). Здесь имеется такой же чашевидный хлоро- пласт с вырезкой вверху и крупным пиреноидом в утолщенном дне внизу. В вырезке хлоропласта располагается крупное ядро, с хорошо замет- ным ядрышком, а выше — две пульсирующие вакуоли. Однако, в отличие от хламидомонад, здесь нет жгутиков и даже носика, по бокам которого они обычно выходят из клетки. Кле- точная оболочка здесь толстая, двухконтурная. У громадного большинства вакуольных, как и у гипномонаса, две пульсирующие вакуоли, и находятся они в морфологически переднем конце клетки, в вырезке чашевидного хлоро- пласта. Реже пульсирующих вакуолей несколько и даже много, и тогда они располагаются по периферии протопласта, в вырезках централь- ного звездчатого хлоропласта (Radiosphaera sphaerica, рис. 210, 2) или в прорезях по- стенного, рассеченного на отдельные ло- пасти хлоропласта (Characiochloris characioi- des, рис 210, 8). Большинство вакуольных — прикрепленные формы. Как правило, они встречаются редко, в виде одиночных клеток, поэтому не привле- кают внимания и остаются слабо изученными. В качестве субстрата им обычно служат нитча- тые водоросли и листья водных растений. Из- вестны также виды, поселяющиеся исключи- тельно на мелких ракообразных. Типичной, широко распространенной эпизойной формой 275
276 Рис. 210. Вакуольные: 1 — Hypnomonas chlorococcoides, клетка в оптическом разрезе: а — оболочка, б — хлоропласт, в — пиреноид, г — ядро с яд- рышком, д — пульсирующие вакуоли; 2 — Hadiosphaera sphae- rica, клетка в оптическом разрезе; 8 — Characiochloris characioi- des; 4 — Chlorangiella pygmaea. является, например, хлорангиелла (Chlorangiella pygmaea, рис. 210, 4). Интересно, что клетки этой водоросли еще сохранили сильно укоро- ченные, слегка подвижные рудименты жгути- ков и глазок. Колонии ее, состоящие из клеток с длинными ножками, могут сплошь покрывать тела ракообразных, придавая им зеленую ок- раску и вызывая даже «цветение» воды. Такую картину можно наблюдать обычно ранней вес- ной в сфагновых болотах, когда поверхность водоема по утрам покрыта еще тонким стекло- видным ледком, сквозь который и просвечивает зеленоватая вода болота. Сняв осторожно с поверхности воды лед и процедив через планк- тонную сетку несколько литров воды, можно получить хороший живой материал хлоранги- еллы. Часто поселяются на ракообразных и представители рода хлорангиопсис (например, Chlorangiopsis epizootica, табл. 29, 8). Примером крупных колониальных форм в порядке вакуольных может служить глеофиллул (Gloeophyllum fimbriatum). Его колонии имеют вид нежной тонкой морщинистой пластиночки до 4 с~ в поперечнике, прикрепленной к субст рату в виде раскрытого парашютика. Эта пла- стинка представляет собой два слоя слизи, внут- ри которой попарно или тетрадами (по четыре) лежат клетки, по строению протопласта напо- минающие гипномонас. Делясь в одном или двух взаимно перпендикулярных направлениях, клетки размножаются и таллом разрастается. Из таких же клеток могут возникать зооспоры и гаметы. Эта водоросль — типичный предста- витель перифитона, обитатель чистых пресных вод. Она живет в прибрежной зоне рек, стариц, канав, прикрепляясь обычно к отмершим, пог- руженным в воду листьям рогоза. Весьма интересно строение микроскопических колоний апиоцистиса (Apiocystis brauniana, табл. 29, 9), лишь иногда достигающих высоты до 1,5 лл. Они имеют вид слизистых мешков грушевидной формы, прикрепляющихся к раз- личным растениям или мертвому суб страту. Внутри этих мешков за толстым слоем слизи располагаются многочисленные шаровидные клетки, от каждой из которых, выступая далеко наружу, отходят две неподвижные псевдоци- лии, покрытые толстыми слизистыми чехлами. ПОРЯДОК ХЛОРОКОККОВЫЕ (CHLOROCOCCALES) К этому порядку относится огромное боль- шинство представителей класса протококковых водорослей. Их обычно называют собственно протококковыми водорослями. Они характери- зуются наиболее полно выраженной коккоидной структурой тела, т. е. плотными оболочками и постоянным отсутствием у вегетативных кле- ток пульсирующих вакуолей. Нет у них ни глазка, ни псевдоцилий, не наблюдается и тен- денции к образованию многоклеточного талло- ма. Преимущественно это одноклеточные формы, хотя здесь хорошо представлены колонии и особенно ценобии, отличающиеся столь боль- шим разнообразием сложения, какого нет ни в одном другом отделе водорослей. Они могут быть двухклеточными, четырех клеточными и восьмиклеточными, а иногда состоят и из боль- шего числа, вплоть до множества клеток одной и той же генерации (рис. 213,5). Нередко цено- бии образуют здесь сложные так называемые синценобии, форма которых у хлорококковых такнсе весьма разнообразна (рис. 214). В синце- нобиях клетки обычно остаются соединенными слизью или остатками оболочек старых материн- ских клеток. Форма клеток в пределах порядка хлорокок- ковых чрезвычайно разнообразна, причем осо- бенно большого разнообразия достигают здесь выросты оболочки (рис. 212). Клетки обычно одноядерные, реже ядер в клетке несколько или множество.
Размножаются хлорококковые почти исклю- чительно бесполым путем — зооспорами и ав- тоспорами, реже могут возникать гемизооспо- ры. Половое размножение известно у немно- гих представителей. Хлорококковые наиболее широко представле- ны в фитопланктоне, в большинстве это типич- ные пассивные планктеры. Многие из них встре- чаются часто; развиваясь в большом количестве, они могут вызывать «цветение» воды (главным образом виды родов Ankistrodesmus, Scenedes- mus, Chlorella, Dictyosphaerium, Oocystis, Coe- lastum и Pediastrum, рис. 211, 213). Распрост- ранены они повсюду, в водоемах всех типов, хотя и предпочитают стоячие загрязненные воды. В систематическом отношении порядок хло- рококковых подразделяют на значительное (свы- ше двух десятков) число семейств и подсемейств. Мы познакомимся лишь с некоторыми предста- вителями главнейших семейств. Хлорелла (Chlorella, рис. 211, б — 7) широко распространена и хорошо известна. Ее вегета- тивные клетки устроены очень просто. Это зе- леные шарики, обычно не превышающие в диа- метре 15 лкл, протопласт которых имеет один чашевидный, вернее, колоколообразный хло- ропласт с одним пиреноидом в утолщенной ча- сти. Ядро одно, но в живой клетке без специаль- ной обработки оно невидимо. Покрыт прото- пласт хорошо выраженной двухконтурной оболочкой. Размножается хлорелла исключи- тельно автоспорами, возникающими обычно по 4 — 8 в одной клетке. Хлорелла нетребовательна к условиям обитания и способна весьма интен- сивно размножаться, поэтому очень широко распространена и встречается почти повсемест- но. В водоемах — это типичный планктер, но встречается она и в бентосе, перифитоне и нейсто- не, а также на наземных субстратах и в почве. Она входит в состав лишайников, вступает в симбиоз с разными гидробионтами, образуя так называемые зоохлореллы. Клетка хлореллы ока- залась удобным объектом для разносторонних исследований. По количеству посвященных ей работ, в том числе и научно-популярных, хлорелле принадлежит первое место среди всех водорослей нашей планеты. Она стала основным объектом массового культивирования водорос- лей для практического использования в самых различных направлениях. В таких же местообитаниях, как и хлорелла, но особенно часто в почве встречаются виды рода хлорококкул (Chlorococcum, рис. 211, 1 — 4). Клетки их на первый взгляд кажутся по- хожими на клетки хлореллы, но на самом деле хорошо от них отличаются. По размерам они крупнее хлореллы и имеют глубокочашевидный, лишь с небольшой вырезкой толстостенный « «0 « Рис. 211. Хлорококковые, одноклеточные формы с гладкой оболочкой: 1 — 4 — Chlorococcum sp. (1 — взрослая клетка, 2 — образова- ние зооспор, 8 — зооспора, 4 — молодые клетки, образовавшие- ся из зооспор); 5 — 7 — Chlorella sp. (5 — взрослая клетка, 6— образование автоспор, 7 — выход автоспор из материнской клет- ки); 8 — 9 — Oocystis pseudocoronata, отдельная клетка и обра- зование автоспор; 10 — Selenastrum bibraianum; 11 — Ankist- rodesmus acicularis. хлоропласт с одним пиреноидом. Самое сущест- венное их отличие от хлореллы — размножение только зооспорами. Хлорококк является типичным представите- лем особого семейства хлорококковых (Chlorococ- caceae), куда входит еще шиповатая трейбария (Treubaria, рис. 212, 1) и некоторые другие; хлорелла относится к семейству ооцистовых (Oocystaceae), объединяющему свыше 20 родов, среди которых имеются формы не только с глад- кой оболочкой, как, например, род ооц.истис (Oocystis, рис. 211, 8,9), но и различные шипо- ватые формы, например роды лагертеймия (La- gerheimia, рис. 212, 2), диакантос (Diacanthos, рис. 212, 8), тетраедрон (Tetraedron, рис. 212, 4) и ряд других интересных родов. Из семейства анкистродесмовых (Ankistrodes- шасеае), представители которого характери- зуются узкими заостренными клетками, заслу- живает внимания род анкистродесмус (Ankist- rodesmus, рис. 211, 11). Виды этого рода очень быстро размножаются автоспорами. Наряду с хлореллой они повышают продуктивность рыбоводных прудов и интенсифицируют очист- ку сточных вод в биологических прудах и кар- 77
Рис. 212. Хлорококковые, одноклеточные формы с ши- пами и выростами на оболочке: 1 — Treubaria triappendieulata; 2 — Lagerheimia eitriformis; 8 — Diaeanthos belenophorus; 4 — Tetraedron eaudatum; 5— Golenkinia radiata; 6 — Polyedriopsys spinulosa; 7 — Charaeium aeuminatum. тах полей фильтрации городских очистных сооружений. К этому семейству относится также род селе- наструм (Selenastrum, рис. 211, 10), представи- тели которого отличаются сил ьноизогнутыми клетками, похожими на молодой месяц, и дру- гие интересные роды. К семейству хараггиевых (Characiaceae) отно- сятся водоросли с одиночными, прикрепленны- ми к субстрату клетками. Здесь весьма типичны роды хараиггум (Characium, рис. 212, 7) и кор- шиковиелла (Korschikoviella, табл. 29, 10). Особенно интересны эпизойные виды этих ро- дов, клетки которых способны прикрепляться к телу планктонных ракообразных настолько прочно, что им не опасно быстрое движение животных в толще воды. Возникающие у них зооспоры, несмотря на движение, успевают прикрепиться к хозяину (рачку), прорасти и вскоре превратиться в зооспорангий, чтобы снова дать очередную партию зооспор. Все эти водоросли имеют клетки либо с глад- кими оболочками, либо с немногочисленными выростами. Однако среди хлорококковых очень много видов, отличающихся богатым и разно- образным «оперением» из тонких шипов и ще- тинок, усеивающих оболочку клеток. Очевидно, такое «оперение» помогает им легко парить в толще воды. Таково, например, семейство микрактиниевых (Micractiniaceae), включаю- щее как одноклеточные, так и цен обиальные формы. Примером первых могут служить роды еоленкиния (С olenkinia, рис. 212, о) и иолиэдри- опсис (Polyedriopsis, рис. 212, 6), примером вторых — род микрактиниум (Micractinium, рис. 214, 2). Клетки видов этого рода обычно соединены в очень сложные крупные красивые синценобии, а так как оболочка каждой клетки несет по нескольку щетинок, то эти синцено- бии имеют весьма своеобразный вид. Наиболее изученным и широко распространенным видом этого рода является ликрактиниул маленький (Micractinium pusillum, рис. 214, 2). В отстой- никах на сахарных заводах ранней весной, после сезона сахароварения, отстаиваются сточные, так называемые жомовокислые воды. В это время года они приобретают зеле- ный оттенок, «цветут». В отобранной здесь пробе фитопланктона под микроскопом можно увидеть множество ярко-зеленых синценобиев, от клеток которых во все стороны в виде длин- ных лучей направлены прозрачные, утончаю- щиеся к концу щетинки. Микрактиниум размно- жается автоспорами, возникающими по 4 — 8 в каждой клетке и сразу слагающимися в син- ценобий. Неделей-двумя позже в пробе планк- тона из того же водоема и с поверхности ила можно найти шаровидные шиповатые ооспоры, возникшие в результате оогамного полового процесса. Очень широко распространены в самых раз- личных водоемах виды семейства сценедесмовых (Scenedesmaceae), представляющие собой цено- биальные формы своеобразного строения, раз- мно~кающиеся только автоспорами. Особенно многочисленны виды рода сггенедесмус (Scene- desmus, рис. 213, 2), образующие преимущест- венно четырехклеточные цен обии с редкими шипами на оболочке (у некоторых видов шипы отсутствуют). Сценедесмус наряду с хлореллой стал объектом массовых культур, выращивае- мых в практических целях. Близки к нему, но обладают иной формой клеток и ценобиев роды тетраструм (Tetrastrum, рис. 213, 1) и акти- наструм (Actinastrum, рис. 213, 8). Сложные ценобии (синценобии), состоящие из клеток
двух и более генераций, образуют представи- тели рода круцигения (CruciIrenia, рис. 214, 8). В каждой клетке таких ценобиев образуются по 4 автоспоры, которые тут же срастаются друг с другом характерным для вида образом и выходят из материнской оболочки в виде гото- вого ценобия. Подобным же образом, но только с помощью зооспор, а не автоспор происходит образование ценобиев в семействе гидродикциевых (Hydro- dictyaceae). По этому признаку здесь объедине- ны такие на первый взгляд резко контрастные водоросли, как микроскопический педиасщрущ (Pediastrum, рис. 213, 4), плоские красивые ценобии которого обычны в пресноводном планк- тоне, и гигант среди протококковых водорослей гидродикцион (Hydrodictyon, рис. 213, 5). Пос- ледняя водоросль обычна в тихих заводях рек, прудах, ямах. Она издавна получила русское название «водяная сеточка». Ценобии ее имеют вид сетчатого мешка, размеры которого дости- гают 1 л. Каждая стенка петли — это отдельная клетка длиной 1,5 сл, образовавшаяся из однои зооспоры. Зооспоры могут возникнуть в любои клетке ценобия. У водяной сеточки их образует- ся несколько тысяч в одной материнской клетке, внутри которой они слагаются в новый дочер- ний ценобий. Ценобии у большинства протококковых во- дорослей образуются путем смыкания или сра- стания клеток друг с другом, реже клетки объе- диняются остатками материнских оболочек, обычно расплывающимися в слизь. Примером таких слизистых ценобиев может служить род диктиосфериум (Dictyosphaerium, рис. 214, 1), выделенный вместе с немногими другими родами в особое семейство диктиосфериевых (Dictyo- sphaeriaceae). Некоторые виды диктиосфериума распрост- ранены очень широко. Из краткой характеристики порядка хлоро- кокковых видно, что это большая и разнообраз- ная группа. Общим признаком для всех ее пред- ставителей является наличие коккоидной или пальмеллоидной структуры тела. Но нет правил без исключения. Таким исключением среди хлорококковых является род протосифон (Proto- siphon), содержащий только один вид (P. botry- oides, табл. 29, ll) и образующий самостоятель- ное семейство (Protosiphonaceae). Этот вид обладает сифональной структурой тела и многоядерным протопластом. Подобные ему древние его предки, по-видимому, дали начало современной ветви сифоновых водорослей. Тал- лом протосифона состоит из надземной зеле- ной шаровидной и подземной бесцветной ризои- дальной частей. Обитает этот вид на поверхно- сти почвы, посылая в глубь субстрата длинный узкий ризоид. Рис. 213. Хлорококковые, простые ценобиальные формы: 1 — Tetrastrum triacanthum; 2 — scenedesmus qua- drieauda; 8 — Actinastrum hantzschii; 4 — Pediastrum simplex; 5 — 7 — Hydrodietyon reticulatum (5 — часть ценобия при малом увеличении, 6 — молодой ценобий внутри материнской клетки, 7 — часть молодого ценобия при большем увеличении). ПОРЯДОК ПРОТОТРИХОВЫЕ (PROTOTRICHALES) К этому порядку относятся те представители протококковых водорослей, которые способны образовывать примитивные, иногда лишь крат- ковременно существующие талломы в виде коротких нитей или небольших паренхиматоз- ных пластинок. Благодаря делению клеток в одном или в двух взаимно перпендикулярных направлениях здесь впервые возникает много- клеточный таллом, впервые появляются нит- чатая, разнонитчатая и пластинчатая струк- туры тела. Они здесь еще весьма прими- тивны, имеют зачаточный, порой временный характер, но это то новое в развитии зеленых водорослей, что породила группа протококко- вых. Очевидно, в эволюционном отношении 27
z~»OH««y> ф ф Р t~ r t Фф O Ю Ф г е Og ° ° »»o ° е ° OOyy( ° » ° ° ~ ° е ° е ° е ° е ее 1~ ее ее Фе Ф Ю Ф Ф Ю е е е Ю Ю еее ° е ее ее ° е ° е ° е ° е е е е е е е е Ю е е е Ю е Ф е Ю Ф O Ф е е е е е е е ю Ю е е ° е ее Ф е е е е е е е е \ ° е е В е е е е % е Ъ е ъ Ю % е е е е % Ю \ е е е е е е ее е е е % е е е O е е е O е е е е е е е е е Ю е е е ее Ю еъ ° ее е\ Рис. 214. Хлорококковые, сложные ценобиальные формы: 1 — Dictyosphaerium pulchellum; 2 — Micractinium pusi llum 8 — Crucigenia tetrapedia. порядок прототриховых служит естественным переходом от одноклеточных и колониальных зеленых водорослей к нитчатым, составляющим класс улотриксовых. Рассмотрим несколько представителей этой интересной группы. Виды своеобразного рода злакатотрикс (Ela- katothrix) нередко попадаются в пробах пресно- водного фитопланктона. Особенно отчетливо нитчатое строение проявляется у элакатотрикса студенистого (E. gelatinosa, рис. 215, 1). Он имеет вид слизистого тяжа, внутри которого попарно в один ряд расположены клетки. Пос- ледние, дорастая до предельных размеров, сно- ва делятся поперек, за счет чего удлиняется таллом, сохраняющий характер примитивной НИТИ. Нитчатое строение таллома более типично выражено у видов рода рафидонема (Raphido- nema), например у встречающейся в пресновод- ных водоемах рафидонемы вечнозеленой (R. sem- Рис. 215. Прототриховые. 1 — E lakatothrix gelatinosa; 2 — R aphidonema sempervi rens; 8 — H. nivale; 4,5 — Protococcus viridis. pervirens, рис. 215, 2) или у вызывающей зеле- ное «цветение» снега в горах рафидонемы снежной (R. nivale, рис. 215, 8). Наконец, у повсеместно распространенной водоросли протококкуса зеленого (Protococcus viridis, рис. 215, о), образующего вместе с дру- гими видами зеленый налет на коре и пнях де- ревьев, хорошо выражена способность клеток делиться в двух взаимно перпендикулярных направлениях. Благодаря этому он может обра- зовывать паренхиматозные многоклеточные пластиночки или разнонитчатые талломы. Систематическое положение этого вида рассмат- ривается по-разному, и некоторые альгологи относят его (под названием Pleurococcus vulga- ris) к улотриксовым водорослям. Мы же считаем эту водоросль вершиной того усложнения орга- низации, которое было достигнуто именно в пределах протококковых как самостоятельного класса.
Таким образом, в теоретическом отношении протококковые водоросли особенно интересны тем, что они представляют собой узловой этап эволюции: в пределах этого класса закладыва- лись и формировались качественно новые струк- туры морфологической дифференциации тела водорослей, такие, как примитивно сифоновые, нитчатые, разнонитчатые и пластинчатые, дав- шие начало современным самостоятельным клас- сам. В практическом отношении класс прото- кокковых тоже важен: в его пределах имеются представители с высокими показателями корм- ности и фотосинтетической деятельности, исполь- зуемые человеком в народном хозяйстве. КЛАСС УЛОТРИКСОВЫЕ (ULOTHRICHOPHYCEAE) Класс улотриксовых объединяет очень боль- шое количество крайне разнообразных зеленых водорослей, общим признаком которых явля- ется нитчатое или пластинчатое строение слое- в ища, состоящего у подавляющего большин- ства видов из нескольких или многих одноядер- ных клеток (исключение — многоядерная сфе- роплея). Однако форма слоевища здесь может быть очень разнообразной. В простейшем слу- чае это нить, образованная одним рядом кле- ток. Именно такая форма слу~кит основой и начальной стадией всех последующих более сложных форм. Нити у одних видов свободно- живущие; у других — прикрепленные, и тогда дифференцированные на основание и вершину. Они могут быть ветвящимися и неветвящимися, равномерно цилиндрическими и сужающимися к вершине, распростертыми по субстрату и прямостоящими, иногда и теми и другими в пре- делах одного слоевища (разнонитчатое, или гетеротрихальное, строение), одиночно расту- щими, образующими скопления или дернови- ны и соединенными в слизистые колонии. Плас- тинчатые формы бывают однослойными и двух- слойными или местами многослойными, рас- простертыми по субстрату и прикрепленными только в одном месте, плоскопластинчатыми и мешковидными. Размеры их также разнообраз- ны — от микроскопических нитей, шириной в несколько микрометров до пластинчатых макрофитов с площадью пластин в сотни квад- ратных сантиметров. В их клетках находится по одному, реже по нескольку хлоропластов. Хлоропласты различной формы, преимущест- венно пластинчатые, пристенные, реже осевые. Половое размножение улотриксовых в основе однотипно, но строение гаметангиев весьма разнообразно. Все это многообразие заставляет делить уло- триксовые водоросли на несколько порядков, представляющих частично обособленные, час- тично взаимосвязанные между собой линии эволюционного развития нитчатых форм зеле- ных водорослей. В нашем обзоре мы делим класс улотриксовых на 7 порядков: улотрик- совые (Ulothrichales), ульвовые (Ulvales), хе- тофоровые (Chaetophorales), эдогониевые (Oedo- goniales), сфероплеевые (Sphaeropleales), ци- линдрокапсовые (Cylindrocapsales) и схизого- ниевые (Schizogoniales). ПОРЯДОК УЛОТРИКСОВЫЕ (ULOTHRICHALES) Слоевище улотриксовых построено по типу однорядной неразветвленной нити. Оно сла- гается клетками, подобными друг другу по строению и функции (табл. 30, 2). Потенциально все клетки способны делиться и участвовать в росте растения, точно так же все клетки мо- гут образовывать споры и гаметы. Отличается от остальных лишь клетка в основании нити: с ее помощью осуществляется прикрепление слоевища к субстрату (у прикрепленных форм). Клетки улотриксовых обладают значительной автономностью. С этим свойством связана спо- собность к регенерации и вегетативному раз- множению — отдельные клетки или участки нитей легко отрываются от нитей и переходят к самостоятельному росту. Порядок включает более 16 родов. Несмотря на то что все их представители построены как простая однорядная нить, в их организации можно обнаружить важные различия, на осно- вании которых весь порядок делят на три груп- пы. У водорослей первой группы нить пред- ставляет собой ряд клеток, рыхлорасположен- ных в толстом слизистом чехле. Таковы, напри- мер, водоросли рода геминелла (Geminella, рис. 216, 1). Интересно, что все улотриксовые с по- добным строением — планктонные организмы. Ко второй группе относятся те нитчатые во- доросли, которые вегетируют как одиночные клетки или как короткие цепочки из 2 — 4 кле- ток, очень неплотно соединенных друг с другом. Нити у них образуются редко и на короткое время. Примером подобного строения может слуясить род стихококкус (Stichococeus, рис. 216, 2). Водоросли, входящие в эту группу, ведут наземный образ жизни. Центральной группой порядка является третья группа, включающая водоросли, постро- енные как типичная многоклеточная нить, в ко- торой клетки плотно соединены друг с другом без помощи слизистого футляра. Водоросли, относящиеся к этой группе, в подавляющем большинстве прикрепленные организмы, по крайней мере в молодом состоянии. Их нити— более постоянные образования, они уже не так легко распадаются, и в них можно различить
Рис. 216. Строение улотриксовых.' 2 — часть нити Geminella interrupta; 2 — Stichococcus bacilla- ris; 8 — часть нити Ulothrix subflaccida; 4 — макрозооспора Ulothrix zonata; 5 — микрозооспора того же вида; 6 — прорас- тание зооспор Ulothrix; 7 — одноклеточный спорофит Ulothrix с зооспорами. базальную и верхушечную части. Сюда отно- сится несколько родов, в том числе и централь- ный род порядка — улотрикс (Ulothrix). Виды улотрикса (в настоящее время их из- вестно более 25) обитают преимущественно в пресных водоемах и лишь очень немногие заходят в солоноватые и морские воды. Эти водо- росли могут поселяться и на влажных поверх- ностях, периодически смачиваемых брызгами прибоя или водопадов. Один из самых широко распространенных и хорошо изученных видов — улотрикс опоясан- ный (Ulothrix zonata). Слоевище улотрикса состоит из неразветвлен- ных нитей неопределенной длины, которые в на- чале роста прикрепляются к субстрату базаль- ной клеткой. Клетки нитей цилиндрические или слегка бочонковидные, часто короткие. Клеточные оболочки обычно тонкие, но нередко они утолщаются и могут становиться слоисты- ми. Клетки улотрикса, так же как клетки всех водорослей этого порядка, содержат единствен- ный пристенный хлоропласт с одним или не- сколькими пиреноидами и одно ядро, располо- женное по продольной оси клетки. Хлоропласт имеет форму пояска, который опоясывает весь протопласт или только часть его (рис. 216, 8). Вегетативное размножение улотрикса осу- ществляется фрагментацией: нити распадаются на короткие сегменты и каждый сегмент раз- вивается в новую нить. Однако этим способом улотрикс размножается не так часто, как дру- гие водоросли порядка, имеющие рыхлое стро- ение нитей. Для бесполого размножения служат зооспоры, которые образуются во всех клетках нитей, кроме базальной. Развитие зооспор, так же как и гамет, начинается на вершине нити и постепенно захватывает нижележащие клетки. Зооспоры — яйцевидные клетки с четырьмя жгутиками на переднем конце. Они содержат стигму, несколько сократительных вакуолей и пристенный хлоропласт. У улотрикса опоясан- ного имеются два типа зооспор — макрозоо- споры и микрозооспоры. Крупные макрозоо- споры обладают широкояйцевидной формой, часто с заостренным задним концом, и стиг- мой, расположенной на переднем конце (рис. 216, 4). Микрозооспоры отличаются более мелкими размерами, округлым задним концом и расположением стигмы в середине споры (рис. 216, 5). Природа микрозооспор остается пока не совсем ясной. По-видимому, они пред- ставляют собой переходный тип между макро- зооспорами и гаметами. Выходят зооспоры через отверстия в боковой стенке клетки. Они заключены в общую сли- зистую оболочну, которая через несколько се- кунд после выхода разрывается. Через короткое время зооспоры оседают передним концом на субстрат, покрываются тонкой целлюлозной оболочкой и прорастают. Прорастая, зооспора вытягивается вертикально и дифференциру- ется на две части. Нижняя часть, лишенная хлоропласта, развивается в клетку прикрепле- ния; верхняя — делится с образованием веге- тативных клеток (рис. 216, 6). У улотрикса опоясанного, однако, зооспоры оседают задним концом и начинают расти вбок, а не вертикально. Довольно часто зооспоры не покидают спо- рангий, а выделяют тонкую оболочку и пре- вращаются в апланоспоры. Последние освобож- даются в результате разрушения нити, но иног- да они могут начать прорастать, находясь в спорангии. При половом размножении в нитях точно таким же способом, как зооспоры, образуются гаметы. Как правило, они развиваются в тех же нитях, что и зооспоры, или в подобных им. Чаще всего переход к половому размножению связан с концом активного роста и наступле- нием неблагоприятных условий. В отличие от зооспор, гаметы несут два жгутика. Половой процесс изогамный. Слияние происходит между гаметами одной и той же или разных нитей. Зигота остается подвижной в течение короткого времени, затем оседает, теряет жгутики, оде- вается толстой оболочкой и превращается в од- ноклеточный спорофит. Он впадает в период
покоя, в течение которого происходит накоп- ление запасных веществ. Форма спорофита раз- нообразна, обычно он сферический с гладкой оболочкой, у некоторых морских видов стано- вится яйцевидным и сидит на слизистой нож- ке (рис. 216, 7). После короткого илидлительного периода по- коя зигота начинает прорастать. Первое деление ядра в ней редукционное. Протопласт зиготы делится на 4 — 16 частей, каждая из которых развивается в зооспору или апланоспору. Та- ким образом, для пресноводных видов улот- рикса характерна гетероморфная смена поло- вой и бесполой форм развития: нитчатый многоклеточный гаметофит сменяется однокле- точным спорофитом. У морских видов, например у улотрикса повислого (U. flacca), зигота теряет свое значение покоящейся стадии и может про- растать непосредственно в нить, на которой образуются зооспоры; цикл развития таким образом становится изоморфным. Ьлизкое к улотриксовым положение зани- мает водоросль микроспора (Microspora). Около 20 видов микроспоры широко распространены в пресных водах земного шара. Неразветвлен- ные нити этой водоросли состоят из цилиндри- ческих клеток с тонкой или утолщенной обо- лочкой (рис. 217, 1). Интересной особенностью микроспоры является строение ее клеточных стенок. Они состоят из двух половинок — верх- ней и нижней (рис. 217, 2), которые удер~кива- ются вместе тонким целлюлозным слоем. Если нить специально обработать, оболочки распа- даются на участки, имеющие форму буквы «н». Каждый такой участок состоит из поперечной перегородки и половинок боковых стенок двух соседних клеток (рис. 217, 8). Пристенный хло- ропласт микроспоры, в отличие от хлоропласта улотрикса, выстилает все стенки клетки и часто имеет вид сетчатой пластинки (рис. 217, 1); у некоторых видов он состоит из узких лент, соединенных между собой. Пиреноиды отсутст- вуют. Размножается микроспора бесполым спо- собом двух- или четырехжгутиковыми зооспо- рами или апланоспорами. Половое размноже- ние пока неизвестно. Систематическое положение микроспоры не- ясно. Особенности строения оболочек и хлоро- пласта заставляют одних ученых рассматривать этот род как самостоятельный порядок; другие отводят ему место семейства в порядке Ulo- thrichales; наконец, третьи считают, что это все- го лишь род в семействе улотриксовых. Вообще зеленые нитчатки изучены еще не- достаточно, и, для того чтобы ответить на во- прос, насколько естественна их современная система, потребуются новые исследования. По- рядок же улотриксовых заслуживает особого Рис. 217. Строение микроспоры (Microspora): 1 — клетки нити с поверхности; 2 — часть нити на продольном срезе; 8 — Н-образный участок оболочки. внимания как важная в эволюционном отно- шении группа, давшая начало многим много- клеточным зеленым водорослям. ПОРЯДОК УЛЬВОВЫЕ (ULVALES) Ульвовые — макроскопические водоросли, имеющие, в отличие от нитчатых улотриксовых, паренхимное строение. Возникновение у клет- ки способности делиться в двух и более плоско- стях привело к образованию пластинчатой, мешков идной и трубчатой структуры весьма разнообразной формы (табл. 31, 2, 6, 7). Так же как у улотриксовых, слоевище ульвовых еще слабо дифференцировано. Заметно отличаются от остальных лишь крупные клетки в основа- нии растения, снабженные ризоидными отрост- ками, с помощью которых осуществляется при- крепление растений. Так же как у улотрикса, клетки ульвовых содержат единственный при- стенный хлоропласт с одним или несколькими пиреноидами и одно ядро, расположенное по продольной оси клетки. Наиболее распространенный способ вегета- тивного размножения — прорастание клеток по- дошвы с образованием дополнительных растений. Для бесполого размножения служат четырех- жгутиковые зооспоры или, реже, апланоспоры. Гаметы более мелкие, двужгутиковые. Половой процесс изогамный или анизогамный. Специ- альные органы размножения отсутствуют. Га- меты и споры образуются в обычных вегетатив-
Рис. 218. Строение ульвовых. 1 — 7 — Enteromorpha (1 — поперечный срез через моЛОдое слоевище; 2 — часть поперечного среза с зрелыми спорангиями; 8 — 7 — последовательные стадии прорастания зооспор); 8 — 9 — Ulva (8 — поперечный срез слоевища в ризоидной части, 9 — вид клеток с поверхности слоевища). ных клетках, которые до момента плодоноше- ния ничем не отличаются от остальных. В каж- дой клетке образуется 16 — 64 гаметы и 4 — 16 зооспор. Зооспоры и зиготы способны, попа- дая в неблагоприятные условия, сохранять жизнеспособность в течение долгого времени, но в обычных условиях они вскоре прорастают. По сравнению с улотриксовыми ульвовые характеризуются более полным обособлением гаметофита и спорофита и приобретением изо- морфной смены половой и бесполой форм раз- вития. Только у наименее специализированных форм порядка сохраняется одноклеточныйспо- рофит, который сменяется многоклеточным га- метофитом. Ульвовые, за очень редким исключением, обитают в морских водоемах. Но вряд ли их можно назвать истинно морскими организмами. Подавляющее большинство ульвовых — соло- новатоводные или эвригалинные формы. Очень многие ульвовые ограничены в своем распрост- ранениии лиманами, эсту ар иями (пр иустьевыми участками рек), заболоченными морскими по- бережьями. Ульвовые часто проявляют повы- шенные требования к содержанию в воде орга- нических веществ и лучше других морских во- дорослей выносят загрязнение. Они растут преимущественно в бухтах, причем в бухтах с населенными пунктами по побережью или в их участках, где имеются ручьи или реки. Все представители порядка распространены на мелководье и только в редких случаях дости- гают глубины 20 — 30 м. Ульвовые можно встретить в морях всех кли- матических зон от Арктики до тропиков. Од- нако подавляющее большинство представителей порядка обитает в морях умеренных широт. Здесь их численность особенно высока. До недавнего времени ульвовые считались довольно однородной группой. Однако при бо- лее внимательном изучении оказалось, что структура порядка гораздо сложнее. Сейчас в нем выделено 4 семейства. Центральным и наиболее обширным является семейство ульво- вых (Ulvaceae). Эти водоросли ведут свое про- исхождение непосредственно от улотриксовых предков, они развивались в направлении образо- вания разветвленно-трубчатой и пластинчатой форм слоевища, а тактике приобретения и закре- пления изоморфной смены форм развития. Первый этап усложнения организации вы- разился в превращении однорядной нити в двух- рядную. Такое строение свойственно наиболее примитивному представителю семейства- перкурсарии (Percursaria). Длинные перепутан- ные нити этой водоросли можно встретить в оп- ресненных участках моря — в лагунах, лима- нах, эстуариях. Дальнейшее усложнение строения привело к образованию трубчатой структуры, которая характерна для рода энтероморфа (Enteromor- pha, табл. 31, 2). Трубчатое слоевище эн-
тероморфы может иметь самую разнообраз- ную форму. Здесь и очень узкие, густо раз- ветвленные растения, которые невооружен- ным глазом не всегда отличишь от кладофоры, и более широкие, редко разветвленные, и, наконец, совсем неразветвленные, кишкообраз- ные или пластинчатые, весьма похожие на уль- ву. Образование разветвленно-трубчатой фор- мы слоевища, создавшей наиболее благоприят- ные условия для обмена веществ, позволило роду энтероморфа достичь большого видового разнообразия и более широкого распростране- ния по сравнению с другими родами порядка. Большинство видов энтероморфы обитает в мо- рях, но есть среди них и небольшая группа пресноводных видов. Род характеризуется изоморфной сменой форм развития и отличить половые растения от бес- полых по внешнему виду невозмонсно. Зрелые гаметы и зооспоры выходят в окружающую сре- ду через одно округлое отверстие — пору, об- разующуюся на вершине сосочкообразного из- гиба во внешней стенке клетки (рис. 218, 2). При прорастании зооспор и зигот энтероморфа, как и все водоросли семейства ульвовых, про- ходит стадию однорядной нити (рис. 218, 8 — 7). Клетка, от которой начинается развитие, делит- ся на две — апикальную и базальную. В ре- зультате поперечных делений первой образу- ется вертикальная нить, вторая превращается в первичный ризоид. Позднее нить преобразу- ется в трубчатое слоевище. При изменении ус- ловий среды первые деления могут вести к об- разованию стелющихся нитей, расположенных в форме диска, из клеток которого уже позднее вырастает одна или несколько вертикальных однорядных нитей. Из других водорослей семейства заслуживает упоминания род ульва (Ulva, табл. 31, 6). Виды ульвы хорошо известны под названием «морской салат», так как население многих приморских стран унотребляет их в пищу. На мелководье наших теплых морей, таких, как Черное и Япон- ское, ульва — одна из массовых водорослей. Узнать ее нетрудно по широкому пластинчатому слоевищу ярко-зеленого цвета. Двухслойное пластинчатое слоевище — главный признак этого рода, отличающий его от других водорослей семейства (рис. 218, 8). В остальном строением, размножением и развитием ульва напоминает энтероморфу. По-видимому, пластинчатая структура сформировалась позднее трубчатой и на ее основе. Так же как у энтероморфы, на начальных стадиях развития ульвы образуется однорядная нить, которая переходит в трубча- тую стадию. Затем стенки трубки смыкаются, и дальше она растет как двухслойная пластина. Ульву можно встретить в морях всех клима- тических зон, и все-таки это тепловодная во- доросль, поскольку предпочитает она теплые моря. Здесь род представлен наибольшим ко- личеством видов, отличающихся разнообраз- ным внешним строением. Кроме простых плас- тинчатых видов, характерных для наших морей, здесь растут формы со сложно рассеченной и разветвленной пластиной. Род монострома (Monostroma) — ва~кнейший представитель семейства моностромовых (Mo- nostromataceae). Общее число видов этого рода невелико. В прибрежной полосе наших умерен- ных и холодных морей, особенно в зоне прилива и отлива, можно встретить нежные пластинча- тые или мешковидные растения моностромы Гревиля (Monostroma grevillei, табл. 31, 7). Обычно это небольшие растения длиной от 2 до 15 см, но в тихих бухтах, где они не разруша- ются течениями и прибоем, длина их дости- гает 1 м. Взрослое слоевище моностромы обычно имеет форму однослойной пластины (отсюда и название этого рода). До недавнего времени все ульвовые с однослойным пластинчатым слоевищем объединяли в один род, который и называли «монострома». Но теперь он разделен на несколько, так как выяснилось, что сходные однослойные пластины могут иметь водоросли, резко отличающиеся характером индивидуаль- ного развития, размножением и другими важ- ными признаками. Для моностромы в узком смысле характерна мешковидная структура, которая только на поздних стадиях развития разрывается и становится однослойной, одна- ко нередко она сохраняется и в течение всей жизни водоросли. Одной из характерных черт моностромы, так же как и всего семейства, является способ прорастания и формирования слоевища. Зоо- споры, начиная прорастать, делятся так, что образуются стелющиеся по субстрату разветвлен- ные нити. При повторных делениях эти нити располагаются в форме компактного однослой- ного диска. Постепенно диск становится много- слойным в центральной части, верхний слой клеток здесь отслаивается и разрастается с об- разованием мешковидной структуры (рис. 219, 1 — 6). Макроскопическое слоевище моностромы Гре- виля — гаметофит. Для выхода гамет служит отверстие с неровными краями во внешней стенке клетки (рис. 219,7). Зигота развивается в одноклеточный спорофит. При этом она силь- но увеличивается в размерах и покрывается толстой оболочкой (рис. 219, 8). Через более или менее длительное время содержимое этого одноклеточного растения делится с образова- нием зооспор. Таким образом, здесь смена форм развития гетероморфная. Наличие одноклеточного спорофита свиде- тельствует о тесной связи моностромовых с уло- 285
Рис. 219. Строение моностромы (Monostrorna): 1 — б — последовательные стадии формирования слоевища; 7— часть слоевища с пустыми гаметангиями (вид с поверхности); 8 — зрелый одноклеточный спорофит. триксовыми предками. В системе всего поряд- ка ульвовых семейство моностромовых пред- ставляет боковую ветвь эволюции, отделившую- ся от общего ствола сравнительно рано. ПОРЯДОК ХЕТОФОРОВЫЕ (CHAETOPHORALES) Хетофоровые по разнообразию представите- лей занимают одно из первых мест среди зеле- ных водорослей. Слоевище их, как и у улотрик- совых, слагается из однорядных нитей, но нити эти ветвятся, и все слоевище дифференцирова- но на систему стелющихся и вертикальных разветвленных нитей. Раз нонитчатая структура составляет основ- ное отличие хетофоровых от улотриксовых. Однако в пределах порядка стелющаяся и вертикальная структуры не всегда одинаково хорошо выражены, часто та или другая реду- цируется или недоразвивается. Это создает боль- шое морфологическое разнообразие. Многие хе- тофоровые обнаруживают черты высокой спе- циализации в строении вегетативной и репро- дуктивной систем. Подавляющее большинство водорослей по- рядка имеет на слоевище так называемые во- лоски или щетинки (благодаря этим образова- ниям они и получили свое название). Волоски у хетофоровых бывают по крайней мере двух типов. У одних они представляют конечные клетки веточек, сильно вытянутые и лишенные содержимого. У других — это выросты оболоч- ки. Несмотря на то что волоски присущи почти всем хетофоровым, их, по-видимому, все же нельзя считать принципиальным признаком порядка. Интересно, что бурые и красные во- доросли разнонитчатого строения также несут волоски. Подавляющее большинство хетофоровых- пресноводные организмы, и только небольшое число вторично-упрощенных форм проникает в море. Из пяти семейств порядка наибольший инте- рес представляют три: хетофоровых (Chaeto- phoraceae), трентеполиевых (Trentepohliaceae) и колеохетовых (Coleochaetaceae). В наиболее типичной форме разнонитчатое строение проявляется у рода стигеоклониуль (Stigeoclonium) в семействе хетофоровых. Боль- шинство видов этого рода имеет хорошо раз- витые стелющуюся и вертикальную системы нитей (табл. 30, 1). Неправильно разветвленные вертик альные нити заметно сужаются у вер- шин и заканчиваются длинным волоском, со- стоящим из нескольких бесцветных клеток. Клетки слоевища имеют один пристенный хло- ропласт, расположенный пояском, с одним или несколькими пиреноидами. Строение стелю- щейся части проследить труднее, так как оно
во многом зависит от неровностей субстрата. Она может быть представлена рыхлоразвет- в ленными нитями, состоящими нередко всего из нескольких клеток (рис. 220, 1), или густо разветвленными и плотно расположенными ко- роткими нитями, складывающимися в псевдо- паренхимный однослойный диск (рис. 220, 2). В какой-то мере степени развития вертикальной и стелющейся частей находятся в обратной за- висимости. При сильном развитии стелющейся части вертикальная может редуцироваться до нескольких веточек или быть представленной только волосками. При незначительном раз- витии стелющихся нитей в основании растений развиваются ризоиды (рис. 220, 8). В отличие от прямостоячих нитей, для которых характе- рен диффузный рост, у стелющихся нитей рост апикальный и более короткие клетки. Обычно у стигеоклониума вертикальные ветви не от- личаются по толщине, но у некоторых видов можно выделить главную ось, клетки которой толще, длиннее и беднее содержимым, чем клет- ки веточек. Такое строение можно рассматривать как переходное между стигеоклониумом и драпар- кальдией (Draparnaldia) — другим родом се- мейства (табл. 30, 4). У драпарнальдии стелю- щаяся структура практически отсутствует, тог- да как вертикальная развита значительно и достигает высокой специализации. Она диф- ференцируется на длинные и короткие ветви. Длинные состоят из крупных клеток с зубча- тым хлоропластом, занимающим положение пояска по экватору клетки (рис. 220, 4). Корот- кие веточки густо ветвятся и располагаются мутовками вокруг главной оси. Слагающие их клетки содержат цельный хлоропласт, занимаю- щий большую часть видимой поверхности клет- ки. Вершины этих веточек заканчиваются длин- ным волоском. Очевидно, короткие веточки несут основную ассимиляционную функцию, тогда как главные ветви играют прежде всего механическую, опорную роль. Прикрепляется драпарнальдия редуцированной стелющейся структурой и дополнительными многоклеточ- ными ризоидами (рис. 220, 5). У еще более специализированного рода дра- парнальдиопсис (Draparnaldiopsis) ризоиды раз- виваются по всему слоевищу и настолько силь- но, что образуют плотную обвертку вокруг глав- ной оси (рис. 220, 6). Описанные водоросли характеризуются тактике тем, что их слоевище покрыто тонким слоем сли- зи. В отличие от них у метафоры (Chaetophora) все нити погружены в плотную слизистую об- вертку, которая имеет шаровидную или листо- видную форму (рис. 220, 7, 8). Базальная часть хетофоры состоит из отдельных шаровидных клеток, от которых радиально расходятся вер- Рис. 220. Строение хетофоровых: 1 — 8 — Stigeoclonium (1,2 — строение стелющейся части слое- вища, 8 — ризоиды); 4 — 5 — Draparnaldia (4 — клетка главной оси, 5 — ризоиды); 6 — часть слоевища Draparnaldiopsis; 7 — 9— Chaetophora (7,8 — внешний вид растений, 9 — часть слоевища).
Рис. 22k. Строение хетофоровых: Fritschiella tuberosa. а — стелющиеся нити; б — изои ы в— вертикальные нити. тикальные нити, густо разветвленные у вершины (рис. 220, 9). К этому же семейству относится интересная водоросль фритчиелла (Fritschiella), единствен- ный вид которой приспособился к существова- нию в почве (рис. 221). Под поверхностью почвы простираются стелющиеся нити, от ко- торых вниз отходят ризоиды, а вверх — вер- тикальные нити. Последние, достигая поверх- ности почвы, густо ветвятся, образуя пучки ветвей. Тенденция к редукции вертикальной системы приводит к образованию большого числа стелющихся форм (табл. 30, 6). Все стелющиеся представители семейства, как правило, эпи- фиты или эндофиты, растущие на водных цвет- ковых растениях и водорослях, некоторые из них поселяются на гидроидах и мшанках. Слое- вище этих водорослей устроено по-разному. У одних форм на стелющихся нитях развива- ются очень короткие вертикальные нити, окан- чивающиеся волоском (рис. 222, 1); у других волоски образуются непосредственно на клет- ках стелющихся нитей, как, например, у водо- росли афанохете (Aphanochaete, рис. 222, 2). У близкого рода боабокоаеон (Bolbocoleon) во- лоски берут начало от специальных более мел- ких клеток (рис. 222, 8). Полная потеря верти- кальных нитей и волосков наблюдается у эндо- дерлы (Endoderma), растущей внутри оболочек морских водорослей и мшанок (рис. 222, 4). У морских хетофоровых стелющиеся нити часто соединяются в компактную дисковидную плас- тину. Она остается однослойной или становится многослойной в центре в результате того, что здесь образуются короткие вертикальные нити, плотно соединенные между собой. У водоросли прингсхейм,иелла (Pringsheimiella, табл. 30, 6) ве- гетативные клетки однослойного диска уплоще- ны и радиально вытянуты, тогда как зрелые спо- рангии в центре диска вытягиваются вверх и принимают клиновидную форму. Волоски у ди- сковидных форм обычно отсутствуют. Размножение водорослей семейства хетофо- ровых во многом напоминает размножение уло- трикса. Нередко, кроме четырехжгутиковых макрозооспор, служащих для бесполого раз- множения, можно встретить микрозооспоры. У большинства изученных хетофоровых поло- вой процесс изогамный, а смена форм развития, особенно у морских форм, изоморфная. Только некоторым из них свойственна оогамия. Нали- чие оогамных форм среди стелющихся форм хетофоровых наводит на мысль о том, что в какой-то мере это искусственная группа. Семейство трентеполиевых (Trentepohlia- ceae) объединяет более специализированные по сравнению с хетофоровыми водоросли. Большин- ство родов семейства имеет достаточно хорошо развитую как вертикальную, так и стелющуюся часть и лишено волосков. Спорангии тренте- полиевых — дифференцированные клетки, за- метно отличающиеся по форме от вегетативных клеток. Наиболее интересными особенностями обла- дает род трентеполия (Trentepohlia). Эта ши- роко распространенная водоросль образует по- рошковатые скопления оранжевого или бурого цвета на камнях, пнях, стволах деревьев, де- ревянных постройках. Характерная окраска трентеполии обусловлена присутствием в ее клетках гематох рома — пигмента из группы каротиноидов. Многие виды трентеполии являются водорос- левым компонентом лишайников. Особенно обильно эта водоросль растет в условиях влаж- ного тропического климата. Как правило, слоевище трентеполии имеет одинаково развитые вертикальную и стелющую- ся части, однако нередко преимущественное развитие получает только одна из них. Все нити обильно ветвятся, причем место отхождения ветвей непостоянно, они возникают на вершине материнской клетки или в разных местах ее боковой стенки (рис. 223, 1). Рост нитей апи- кальный в результате деятельности верхушеч- ной клетки. Цилиндрические или бочонковид- ны е клетки одеты толстой целлюлозной обо- лочкой, которая нередко имеет слоистое строе- ние. Поперечные перегородки между клетками всегда имеют посередине пору, через которую
Рис. 222. Строение 1 — Pilinia maritima; б — волосок. стелющихся хетофоровых: 2 — Aphanochaete polychaete' е е; 8 — Bolbocoleon ilif n pi iferum; 4 — Endoderma wit а wi trockii. а — спорангий, 28 проходит цитоплазматический ст оен р ием обладают оболочки апика тяж. йсобым Из огаметы развиваются в о ток. У ряда Э ки образ ется ви оВНа е е р апикальной клет- занимаю и ровидные образования, — 4 разуется слоистый колпачок ' . 22 — ). Клетки трентеполии в отл 'рис. 3, могут быть верхушечными занимаю и на нитях: он йо овых ными, оковыми, инте- ~р~ы~~ Развив аютс они еим ДИСКОВИДНОЙ ИЛИ Л и лентовидной ~о мы о хлоропластов на стелющихс ихся нитях. Эти си ячи они преимущественно aompoa (pic. 223 б- с., — 8). В ста ых уются в результате вели . Э дячие «спорангии» гда множе ство ядер. — р х клетках все- клеток и, в отличие от спо а у личения размеров Больш ую роль в жизни этой не от тделяются от нити пр спорангиев на ножке 7 р вегетативное размнож ает й водоросли иг- ение. летки зм Не всегда развивающ и созревании гамет иесявних в ж легко отделяются ся друг от га нитеи клетки азм змнож ения ве т д у гутиковые ветром. В ви дру и распыляются нере ко о ду себя как гаметы, виде отдельных богатых редко они прорастают без сли мом клеток в водоросль переживает неб гематохро- апланос поры. ияния или дают ятный с хой у период. Во влажное в емя не лагопри- Т енте р полиевые — высокоспе иал происходит беспо полое и половое е время года ная группа в в порядке haeto оспециализирован- трентеполии. размножение бенно б о а р Orales. Это осо- ии. ву- и четырехжг ти росается в глаза п и ее споры образ у иковые зоо- мейством Ch р сравнении с се- ножках (рис. 223 9). П осле ние ранги ях на гом спе циализация т ентеп евидно, во мно- вертикал ьных нитях как о д возникают на тат с пеци ических назем олиевых — резуль- т бчатых дин или несколько СеМ мных условий обитания. д ющихся от нее попе ечной п еля у чатых выростов верхуше чнои клетки, от- емеиство колеохетовых С 1 ставлено единственным о о ем о еос aetaceae п е- ) рд- Эти вы рость ж~ег~р)днюй. 1нюЬаИ ' 14 осты имеют характе н ю ае е'. видов этого о а изогнутую ~~ форму (рис. 223, 10 11~. В рную коленчато- водах к как эпи'иты, образ я по рода растут в пресных уя подушковидные ~~щу~жю~ и, ~~~~~~~~ж Слоев~ в спорангии, отделяется перего о к лоевища колеохетовых име ки-ножки. р р дкои от клет- выраженн ю а ют в разной мере Об р к енную разнонитчатую структу . У п е- ычно зрелые спора ангии от ыв ставителей с развить тыми ве тика у. пред- нити и разно осятся вет ом. Осво" а р аются от ми они об о ычно со аны в ть р льными нитя- р "ождение и про ч у ду у у вии достаточного увлажне ния. усло- гих видов, однако разв усло-,, ивается только базаль часть, которая часто об альная о о разована плотно распо-
рис. f23. Строение трентеиолии (Trentepohlia): — — — б азование колпачка на верхушечной клетке; 5 — 8 — клетки с хлоропластами; — — (c б 0 H ' a — crea~rqaaca часть б — aep~m~aa~~aa 9 — нить со спорангиями на ножках; 10,11 — образование спорангия ~с о к у,. а— нить, в — спорангий на ножке. ложенными нитями и имеет вид компактного округлого диска. Рост слоевища всегда апи- кальный. Одноядерные клетки заполнены круп- ным пристенным хлоропластом неправильной формы с одним или двумя четкими пиреноида- ми. Обычно клетки несут характерные заклю- ченные в футляр щетинки (рис. 224, 8). Бесполое размножение колеохетовых проис- ходит весной и ранним летом двужгутиковыми крупными зооспо рами. Они образуются по одной в обычных клетках и выходят через ок- руглое отверстие на вершине короткого боро- давчатого выроста. Зооспоры отличаются от- сутствием стигмы и тем, что единственный хло- ропласт занимает в них боковое положение. При прорастании зооспоры образуются две лежащие друг над другом клетки, из которых верхняя формирует щетинку, а нижняя делится с образованием клеток диска (рис. 224, 1 — 8). Половое размножение колеохете представ- ляет собой высокоспециализированную оогамию. У колеохете подушковидного (Coleochaete pulvi- nata) оогонии образуются в верхней части вертикальных нитей. Они имеют характерную бутылковидную форму: нижняя, расширенная часть содержит хлоропласт, а верхняя, бесцвет- ная вытянута наподобие трихогины красных водорослей. При созревании вершина оогония разрушается, в результате чего теряется неко- торое количество цитоплазмы. В то же самое время протопласт нижней части округляется, формируя единственную яйцеклетку (рис. 244, 4 — 6). Антеридии развиваются по нескольку вместе на вершине вертикальных нитей, часто тех же самых, которые несут оогонии (рис. 224, 6). Каждый антеридий образует единственный антерозоид шаровидной формы с двумя длин- ными жгутиками на переднем конце. После оплодотворения верхняя, бесцветная часть оогония отделяется перегородкой, тогда как базальная постепенно растет. Одновременно оогоний начинает обрастать разветвленными нитями, возникающими из соседних клеток, которые, смыкаясь, в конце концов образуют плотную обвертку красного или бурого цвета (рис. 224, 7, 8). Эта обвертка позднее разруша- ется, и ооспора переживает период покоя в своей собственной толстой оболочке, которая у нее постепенно развивается. Весной содержимое ооспоры вновь приобретает зеленую окраску и делится с образованием 16 — 32 клиновидных клеток. Эти клетки, превращаясь в зооспоры, выходят наружу в результате разрыва оболочки ооспоры (рис. 224, 9). Подобно многим оогамным зеленым водорос- лям, колеохете занимает обособленное положе- ние в порядке, и связи его с другими хетофо- ровыми, так же как и со всеми зелеными водо- рослями, не совсем ясны. В настоящее время система всех нитчатых зеленых водорослей строится главным обра- зом на морфологическом критерии. Однако
291 нетрудно заметить, что как раз в морфологии водорослей более чем где бы то ни было прояв- ляется параллельное и конвергентное разви- тие, т. е. многие заведомо далекие группы водо- рослей проходят одни и те же этапы морфоло- гической эволюции. Этот факт показывает, что для построения системы водорослей, которая отражала бы эволюцию организмов и их родст- венные связи, одного морфологического кри- терия недостаточно. Требуется всестороннее изучение водорослей, в том числе их размноже- ния и развития. ПОРЯДОК ЭДОГОНИЕВЫЕ (OEDOGONIALES) Порядок эдогониевых — одна из наиболее своеобразных групп зеленых водорослей, отли- чающаяся прежде всего особым типом клеточ- ного деления, строением оболочек и уникаль- ным способом размножения. Порядок включает одно семейство эдогониевых (Oedogoniaceae), представленное тремя родами. Первое место среди них занимает род эдогониум (Oedogonium), насчитывающий 380 видов, рас,— пространенных по всему свету и играющих важ- ную роль в жизни небольших пресных водоемов. Слоевище эдогониума построено по типу длин- ных однорядных неразветвленных нитей прик- 1 репляющихся к субстрату с помощью базальной клетки. Однако взрослые нити, особенно в стоя- чих водоемах, обычно встречаются в свободно- плавающем состоянии. Нити состоят из цилинд- рических клеток с гладкими или слегка волни- стыми оболочками. Верхушечные клетки ча- сто заостряются и заканчиваются щетинкой. ядро летки содержат единственное крупное я о и пристенный сетчатый хлоропласт, который состоит из узких продольных лент, соеди- ненных между собой, и несет множество пире- ноидов (табл. 30, б; рис. 225, 1). Эдогониум отличается своеобразным способом деления и роста клеток. Незадолго до деления в верхней части клетки, ближе к ее вершине, на внутренней стороне боковой стенки возни- кает утолщение в форме валика, опоясывающего клетку. В продольном сечении клетки это утол- щение имеет вид двух незамкнутых колец по обеим сторонам клеточной стенки. Они состоят из внутренней части и более грубого наружного слоя, пограничного с протопластом. Этот пог а- нич ный слой валика своими незамкнутыми в погра- кольцо краями соединен с боковой клеточной стенкой и переходит в нее вверх и вниз от вали- ка. Создается впечатление, что это валикообраз- ное утолщение образовалось в результате втя- гивания внутреннего слоя боковой клеточной стенки (рис. 225, 2). Вслед за этим происходит ядерное деление, которое сопровождается обра- зованием в середине клетки тонкой поперечной Рис. 224. Строение колеохете (Coleochaete): оогония и а 1 — 3 — ранние стадии прорастания зооспоры 4 б 6— нтерипия (4); 7 — оогоний после оплодотворения; — развитие 8 — образование обвертки вокруг оплодотворенного оогония; 9 — прорастающая ооспора. а — щетинка, б — оогоний, в- антеридйй, г — оболочка ооспоры.
Рис. 225. Строение эдогониевых.' 1 — клетка Oedogonium; 2 — кольцевое утолщение на продоль- ном срезе через клетку; 8 — 5 — последовательные стадии кле- точного деления; 6 —,; 7 — проросток; 8 — выход андроспор из андроспорангиев; 9 — веточка Bulbo- chaete. перегородки. Эта перегородка соединяется с боковой стенкой материнской клетки не сразу. Перед этим внешняя клеточная стенка вдоль всего валика снаружи разрывается, а сам валик разглаживается, стенки его вытягиваются, в ре- зультате чего образуется новый участок клеточ- ной стенки, расположенный между двумя стары- ми (рис. 225, 8 — о). Оставшуюся над местом раз- рыва меныяую часть клетки называют колпач- ком, расположенную под ним — футляром. Одно- временно с вытягиванием стенок валика моло- дая поперечная клеточная перегородка смещает- ся с середины материнской клетки вверх до тех пор, пока не займет постоянного места близ нижнего края разрыва клеточной стенки. Здесь она отделяет от футляра образовавшуюся на месте валика новую клетку. Последняя посте- пенно вырастает до размеров обычной вегетатив- ной клетки. При повторных делениях новые кольцевые утолщения образуются всегда под краем колпачка и поэтому новые колпачки ока- зываются непосредственно под старыми. Коли- чество колпачков свидетельствует о количестве делений, которым подвергалась клетка. Нали- чие их — надежный критерий для распознава- ния вегетативных нитей эдогониума среди дру- гих зеленых нитчаток. Вегетативное размножение, кроме обычной фрагментации, осуществляется с помощью аки- нет — наполненных маслом и крахмалом кле- ток красного цвета, образующихся на нити по 2, 4 и более в ряд. Для бесполого размножения служат зооспо- ры, особенностью которых является венец ко- ротких жгутиков на переднем, лишенном хло- ропласта конце. Они возникают по одной в каж- дой клетке и выходят наружу в результате раз- рыва оболочки в верхней части клетки (рис. 225, 6). Проплавав короткое время, зооспоры оседают и прорастают в нить, образуя в основа- нии проростка первичный ризоид (рис. 225, 7). Половой процесс — оогамия, имеются одно- домные и двудомные виды. У ряда видов анте- ридии, так же как и оогонии, образуются на нитях обычного строения, однако большинство двудомных видов характеризуется интересным половым диморфизмом. У таких видов антери- дии образуются на особых карликовых растень- ицах. Если антеридии возникают на обычных нитях, то происходит ряд повторных поперечных де- лений вегетативных клеток, без их разраста- ния, так что антеридии имеют вид расположен- ных в ряд сильно укороченных клеток. В каж- дом антеридии, в свою очередь, образуется по 2 антерозоида. У видов с половым диморфизмом на вегета- тивных нитях образуются не антеридии, а осо- бые спорангии, которые называют а н д р о-
в ° ° Ъ' 8 Рис. 226. Строение сфероилеи (Sphaeroplea): z — 8 — строение хлоропластов; 4 — 6 — разные типы поперечных перегородок', 7 — образование антерозоидов; 8 — яйцеклет- ки в момент оплодотворения и зиготы. а — пиреноид, б — яйцеклетка, в — антерозоид, г — зигота. ломы которого состоят из разветвленных нитей с многочисленными волосками-щетинками. Существует несколько гипотез, пытающихся объяснить происхождение карликовых мужских растений эдогониевых, однако ни одну из них нельзя признать убедительной. Но даже если отвлечься от способа размножения и клеточного деления, связь порядка эдогониевых с другими зелеными водорослями остается далеко не ясной. Наиболее часто их сближают с хетофоровыми. ПОРЯДОК СФЕРОПЛЕЕВЫЕ (SPHAEROPLEALES) с п о р а н г и я м и (рис. 225, 8). Способ их образования не отличается от вышеописанного способа образования антеридиев, но из каждого андроспорангия выходит по одной андроспоре, которая после короткого периода плавания оседает на оогоний или соседнюю с ним клетку и прорастает в карликовое мужское растение (табл. 30, б). Это растение состоит из одной- двух, в лучшем случае из нескольких клеток. Среди них выделяется более крупная ризоидно вытянутая базальная клетка. Очень скоро на мужском растении возникают антеридии. Они отделяются простыми перегородками или по- средством кольцевых утолщений. В этом слу- чае антеридий покрыт колпачком. Антерозоиды выходят в результате разрыва клеточных сте- нок. Это мелкие клетки с венцом жгутиков на переднем конце, как у зооспор. Оогонии- крупные, почти сферические клетки, дающие единственную яйцеклетку. Антерозоид прони- кает в оогоний через специальное отверстие в оболочке или после ее разрыва. Зигота оде- вается толстой, трехслойной, часто скульптур- ной оболочкой, приобретает красно-бурую ок- раску и превращается в ооспору, которая слу- жит покоящейся стадией (табл. 30, 5). При прорастании ооспоры образуется 4 зоо- споры, которые вырастают в новые нитчатые растения. Кроме эдогониума, следует упомянуть еще род бульбохете (Bulbochaete, рис. 225, 9), тал- Водоросли, относящиеся к этому порядку, не часто встречаются в природе, но упомянуть о них необходимо, так как они обладают уни- кальным сочетанием признаков. В порядке один род сфероплея (Sphaeroplea), в котором известно всего 6 видов. Самый известный из них — сфероплея кольцевидная (S. annulina). Однорядные неразветвленные нити сферо- плеи лишены органов прикрепления и всегда встречаются в свободноплавающем состоянии. Клетки нитей весьма варьируют по величине- их длина может превышать ширину только в 2 — 3 раза, а может быть больше ее в 70 — 90 раз. Каждая клетка содержит многочисленные при- стенные хлоропласты, имеющие форму узких замкнутых колец, которые располагаются друг за другом, а между ними находятся крупные вакуоли (рис. 226, 1 — 8). Ядра, которых в клет-
ках сфероплеи множество, находятся в цито- плазме, причем в пределах каждого хлоропласта их чаще всего бывает по два. Вряд ли правильно называть клетками сег- менты, слагающие нити сфероплеи, и не только потому, что они обладают множеством ядер и хлоропластов (и, следовательно, являются явно вторичными образованиями), но и потому, что отделяющие их поперечные перегородки не похожи на клеточные стенки других многокле- точных зеленых водорослей. Они сильно варьи- руют по форме, а также по способу и месту обра- зования (рис. 226, 4 — 6). Часто поперечные пе- регородки имеют вид кольцевых внутренних утолщений на стенках клетки, которые не смы- каются в центре, так что остается отверстие, через которое проходит цитоплазматический тяж (рис. 226, 4). В других случаях вместо пере- городок образуются особые пробки. И, наконец, в любом месте нити могут возникать группы радиально сходящихся тяжей, напоминающих скелетные тяжи каулерпы и играющих механи- ческую роль. Обычно сфероплея размножается половым оогамным способом. Дифференцированные ор- ганы полового размножения здесь отсутству- ют — яйцеклетки и антерозоиды образуются в любом из сегментов (рис. 226, 7,8). После опло- дотворения яйцеклетки образуется ооспора, одетая системой оболочек. Вначале образуется гладкая стекловидная оболочка. Постепенно в ее пределах формируется толстая скульптур- ная стенка. Строение ооспоры специфично для отдельных видов и служит хорошим диагности- ческим признаком. Выделяют два основных типа ооспор: шаровидные с равномерно расположен- ными шипами и вытянутые по однои из осей с крыловидными выростами. Ооспоры покидают нить в результате ее разрушения и остаются в покоящемся состоянии в течение нескольких лет. При прорастании ооспоры образуется 4 зо- оспоры. Зооспоры развиваются, не оседая на субстрат. Они вытягиваются в нить, которая, достигнув в длину 3 — 4 мм, приступает к попе- речным делениям. Вегетативные нити сфероплеи, кроме гамет, изредка дают зооспоры, служащие для бесполо- го размножения. ( истематическое положение и родственные связи этих своеобразных водорослей сейчас определить довольно трудно. По одним призна- кам они напоминают сифоновые, по другим- улотриксовые. С уверенностью можно сказать одно — характер сегментов у сфероплеи сфор- мировался вторично в результате утраты спо- собности образовывать клеточные стенки. Спо- соб размножения сфероплеи, по-видимому, име- ет ту же природу, что и способ размножения других оогамных форм, берущих начало от оогамных форм хламидомонадовых в порядке вольв оксовых. ПОРЯДОК ЦИЛИНДРОКАПСОВЫЕ (CYLINDROCAPSALES) Небольшая группа цилиндрокапсовых вклю- чает всего два рода с очень сходным строением- цилиндрокапса (Cylindrocapsa) и цилиндрокап- сопсис (Cylindrocapsopsis). Их слоевище пост- роено по типу однорядной нера зветвленной нити, целиком заключенной в плотную, часто слоистую слизистую оболочку. В некоторых случаях клетки могут делиться более чем в од- ной плоскости, и тогда на нити образуются мно- горядные псевдопаренхимные участки (рис. 227, 1, 2). Прикрепление нитей к субстрату, особен- но на ранней стадии, осуществляется посредст- вом утолщения и слизистой секреции оболочек, тогда как первичные ризоиды отсутствуют (рис. 227, 8). Клетки содержат одно ядро и один массивный занимающий осевое положение хло- ропласт неправильной или звездчатой формы с одним пиреноидом (рис. 227, 8). Группы клеток или отдельные клетки легко отделяются от нитей и прорастают в новые рас- тения. Так осуществляется вегетативное раз- множение цилиндрокапсовых. Для бесполого размножения служат дву- или четырехжгути- ковые зооспоры, изредка апланоспоры. Они образуются по одной или по две — четыре в каж- дой клетке. Прорастая, зооспоры дают начало новым нитям. Половой процесс оогамный. У цилиндрокап- сы вегетативные клетки перед преобразованием в антеридии подвергаются делению. В резуль- тате этого образуется несколько клеток, содер- жимое которых окрашено в красный цвет. В каждом антеридии формируется, в свою очередь, по одному или несколько антерозоидов (рис. 227, 4, о). Антерозоиды обладают двумя жгутиками и отличаются от зооспор только буровато-крас- ным цветом. Они освобождаются в результате разрыва или растворения стенки антеридия. При образовании оогония клетка увеличивается в размерах, оболочка ее набухает. В каждом оогонии развивается единственная яйцеклетка. Перед оплодотворением в стенке оогония обра- зуется пора, через которую проникают антеро- зоиды. Зигота, или ооспора, заключена в тол- стую оболочку и в зрелом состоянии имеет ярко- красную окраску. У цилиндрокапсопсиса цикл развития более сложный, так как здесь сначала образуются одноклеточные половые растения. Процесс раз- множения начинается с того, что отдельные клетки покидают нить в результате желатини- зации внешней клеточной стенки. В каждой такой клетке возникает по одной четырехжгу- 294
Строение цилиндрокапсы (Су11в (тосарза) 1 — однорядный участок нити; 2 — многорядный участок той же нити' 3 — в х й IHI ) ' 4— а — антеридий, б — оогоний. антеридиями и оогонием; б — зрелый оогоний с яйцеклеткой и антерозоидами. начало непосредственно от одноклеточных под- вижных форм, обладающих осевым хлоропла- стом и оогамным половым процессом, которые мы находим среди хламидомонадовых в порядке вольв оксовых. ПОРЯДОК СХИЗОГОНИЕВЫЕ (SCHIZOGONIALES) 2 тиковой зооспоре. Через некоторое время зоо- споры оседают на то же слоевище или на другие водоросли и преобразуются в одноклеточные женские и мужские половые растения. В даль- нейшем женские растения превращаются в оого- нии с одной яйцеклеткой. В мужских растениях содержимое делится с образованием дву-, четы- рехжгутиковых антерозоидов. Род цилиндрокапса включает всего 4 или 5 видов, род цилиндрокапсопсис — один. Обычно их можно встретить в небольших пресных водо- емах — прудах, болотах, медленно текущих ручьях, осушительных каналах. Некоторые виды растут на лугах в заболоченных местах' 1 на возделываемых землях. Они никогда не об- разуют массовых скоплений и чистых зарослей и предпочитают расти среди других водорослей и цветковых растений. Отмечены цилиндро- капсовые по всему свету — в Северной Америке 1 Европе, Азии. Многие виды встречены в Ин- дии и здесь лучше всего изучены. Порядок цилиндрокапсовых был выделен сравнительно недавно, и многие альгологи до сих пор продолжают считать его семейством в порядке улотриксовых. Главным и единствен- ным основанием для этого служит такое же, как у улотриксовых, нитчатое строение. Однако свойственные цилиндрокапсовым черты внутри- клеточного строения, строения оболочек, раз- множения и развития у других улотриксовых не встречаются. Поэтому выделение их в само- стоятельный порядок справедливо. Что касается происхождения порядка, то он, вероятно, берет Схизогониевые, или празиолевые, — немного- численная, но весьма своеобразная группа одно рядно-нитчатых неразветвленных и пла- стинчатых зеленых водорослей, клетки которых содержат по одному осевому звездообразном хло оропласту с одним пиреноидом и по одному -У ядру. Однорядное нитчатое строение может сохраняться в течение всей жизни водоросли, а может на каком-то этапе преобразовываться. Когда в однорядной нити начинается деление клеток в двух направлениях, то образуются лен- товидные однослойные пластины. Такое строе- ние имеют представители рода схизогониум (Schizogonium, рис. 228, 1). При дальнейшем росте в ширину образуются широкопластин- чатые слоевища, характерные для рода празиола (Prasiola, рис. 228, 2). В нити деление может происходить и в трех направлениях; в этом случае образуются многослойные цилиндричес- кие формы, как, например, у представителей рода розенвингиелла (Rosenvingiella рис. 228 8'). В 7 ' 7 Водоросли, обладающие только той или иной формой строения, можно встретить в при- роде как самостоятельно существующие, но, с другой стороны, много раз приходилось наблю-
Рис. 228. Строение схизогониевых.' 1 — часть нити Schizogonium; 3 — слоевище Prasiola; 8 — часть слоевища Rosenvingiella. 296 дать, что одна структура на определенной ста- дии развития переходит в другую — одноряд- ная в лентовидную или плотноцилиндрическую и затем в пластинчатую. Вполне вероятно, что все эти водоросли представляют собой лишь стадии в индивидуальном развитии или формы роста одной водоросли. Род празиола — наиболее широко распрост- раненный и важный в порядке. Известно около 20 видов празиолы, которые обитают в самых разно- образных условиях. Пресноводные виды обычно растут в холодных водах. Их можно встретить в полярных областях и высоко в горах в неболь- ших речках и источниках, где температура воды держится около 0 С. Пресноводные виды не образуют густых зарослей и обычно не влияют на характер растительности. Исключение пред- ставляет празиола японская, которая растет в массовых количествах. Жители Японии исполь- зуют ее в пищу. Наземные виды растут преиму- щественно там, где имеются в изобилии нитра- ты, поэтому они часто приурочены к местам скоплений морских птиц и млекопитающих, например к колониям пингвинов в Антарктиде. Эти виды образуют нередко огромные пласты. Они могут быть эпифитами, поселяясь на ство- лах деревьев в сообществе с другими водоросля- ми. Так называемые морские виды празиолы по условиям жизни очень сходны с наземными. Они приурочены к зоне заплесков и брызг, особенно в местах птичьих базаров. На ска- листых мысах они поднимаются на высоту 15 — 30 м над уровнем моря и выносят длитель- ное и сильное высыхание. Географическое рас- пространение празиол обычно характеризуется как космополитическое. Они обычны для хо- лодных и умеренных широт, в тропической зоне отмечены лишь в высокогорных областях Америки. Слоевище празиолы построено из мелких клеток и заключено в сильноразвитую общую обвертку, которая не разрушается даже в период размножения. Вегетативное размножение осу- ществляется фрагментами слоевища и акинета- ми. Многоклеточное слоевище празиолевых сла- бо дифференцировано, поэтому оно легко отде- ляет фрагменты, которые, разрастаясь, пре- вращаются в новые растения. При образовании акинет клетки одеваются более толстой оболоч- кой, в них накапливаются запасные вещества. В таком виде клетки способны переносить небла- гоприятные условия неопределенно долгое вре- мя. Когда акинета начинает прорастать, ее де- ление ведет к образованию или непосредственно нити нового растения, или апланоспор, которые уже дают начало нитчатым проросткам. Для бесполого размножения служат лишен- ные жгутиков апланоспоры, которые образуют- ся в любой вегетативной клетке слоевища. В каждой клетке формируется 1 или 2 — 4 аплано- споры. Они располагаются правильными груп- пами, при этом однослойная пластина становит- с я двух-, четырехслойной. Споры, так же как и гаметы, освобождаясь в результате разруше-
297 ния индивидуальных клеточных оболочек, не выходят в окружающую среду, а остаются не- которое время заключенными в общей для всего слоевища оболочке. При одновременном созре- вании спор вся верхняя часть слоевища празио- лы принимает вид пузыря, наполненного спора- ми. Апланоспоры обычно прорастают без перио- да покоя, хотя способны переживать неблаго- приятные условия. Половой процесс празиолевых — оогамия. Женские гаметы крупные, неподвижные; муж- ские — более мелкие, двужгутиковые. При переходе к половому размножению часть клеток в верхней половине слоевища подвергается последовательному делению ц дает начало много- слойной половой ткани. В пределах этой ткани хорошо различаются темные и светлые участки, располагающиеся попарно и соответствующие скоплениям женских и мужских клеток. В каж- дой клетке образуется в конечном итоге по одной гамете. Зигота, как и апланоспоры, прорастает в пластинчатое слоевище. Гаметофит, вплоть до образования половых клеток, морфологиче- ски подобен спорофиту, поэтому цикл развития празиолы можно считать изоморфным. Своеоб- разие его заключается в необычном строении половой ткани. О родственных связях и происхождении порядка известно очень немного. Внешнее сход- ство с пластинчатыми ульвовыми и с нитчатыми улотриксовыми заставляло многих специалистов объединять празиолевые с порядком Ulotricha- les или Ulvales. Однако особенности их внутри- клеточного строения и размножения с очевид- ностью свидетельствуют об обособленности по- рядка от остальных пластинчатых зеленых водорослей. По организации слоевища, внутриклеточному строению, строению оболочек празиолевые об- наруживают много общего с цилиндрокапсо- выми. Очевидно, формы с осевым хлоропла- стом образуют самостоятельную эволюцион- ную ветвь в пределах пресноводных зеленых водорослей, которую можно проследить от одно- клеточных форм через нитчатые цилиндрокапсо- вые к пластинчатым празиолевым. КЛАСС СИФОНОВЫЕ (SIPHONOPHYCEAE) К этому классу относятся те зеленые водо- росли, которые, в отличие от других предста- вителей отдела, не имеют клеточного строения. Их крупное, сложно устроенное, часто даже причудливое по форме слоевище представляет собой одну гигантскую «клетку» или, реже, разделено на участки (сегменты) своеобразными перегородками и содержит одно или множество ядер (у представителей, имеющих перегородки, отдельные сегменты всегда многоядерные). Не- клеточное строение этих водорослей называют сифонным. Несмотря на отсутствие клеточных стенок, они, собственно, уже не одноклеточные организмы, скорее их слоевище — это комплекс не вполне разделившихся клеток. Особенно справедливо это для многоядерных представи- телей класса, так как умножение ядер можно рассматривать как результат незаконченного процесса клеточного деления, когда ядро де- лится, а цитоплазма остается неразделенной. Внутреннее строение сифоновых характери- зуется наличием центральной вакуоли и при- стенного слоя цитоплазмы. В цитоплазме, кроме одного или нескольких ядер, содержится также один или несколько хлоропластов. Многочис- ленные хлоропласты обладают дисковидной или веретеновидной формой; когда хлоропласт один, он имеет сетчатое строение. Принципиальных различий между тем и другим типом хлоропла- стов нет, поскольку сетчатый хлоропласт обра- зовался, по-видимому, из многих мелких хло- ропластов в результате их соединения друг с другом. В хлоропластах, кроме обычных для зеле- ных водорослей пигментов, содержатся еще два специфических из групп каротиноидов- сифонеин и сифоноксантин. Сифоновые — группа чрезвычайно древняя, возникшая ранее многих других зеленых водо- рослей. Общее количество живущих в настоящее время видов сравнительно невелико, не более 400 — 500. Свыше 90< всех современных сифо- новых — морские организмы. Лишь некоторые представители высокоорганизованных семейств проникли в пресные воды и расселились в них. Сифоновые приурочены к тропическим морям, и только некоторые виды проникают в моря умеренной зоны. Класс состоит из трех порядков. Каждый из них представляет самостоятельную, доста- точно хорошо очерченную эволюционную ли- нию, но все они имеют общее происхождение от морских одноклеточных форм, в настоящее время вымерших, и принципиально отличаются от остальных зеленых водорослей. ПОРЯДОК СИФОНОВЫЕ (SIPHONALES) Слоевище водорослей этого порядка лишено перегородок, только в основании ветвей и из- редка гаметангиев возникают своеобразные пробки. В пристенном слое цитоплазмы содер- жатся многочисленные ядра и хлоропласты с пиреноидами или без них. Чтобы познакомиться со строением и биоло- гией водорослей этого порядка, рассмотрим несколько наиболее характерных представите- лей трех различных семейств.
Семейство каулерповые (Саи1еграсеае) Рис. 229. Строение каулерповых.' 1 — б — Caulerpa (1 — 4 — различные типы строения вертикаль- ных ветвей, 5 — «внутренний скелет» ризома на поперечном срезе); 6 — продольный срез слоевища Halimeda; 7 — 11 — ран- ние стадии развития Udotea. Многим известна водоросль каулерпа (Cau- lerpa) — типичный представитель семейства кау- лерповых (Саи1еграсеае). Этот обширный род (около 60 видов) весьма характерен для флоры тропических морей. Лишь немногие виды захо- дят в субтропические широты, и среди них распространенная в Средиземном море каулерпа прораппающая (С. prolifera). Крупные стелю- щиеся растения каулерпы часто покрывают большие пространства морского дна. Поселяясь обычно на различных твердых грунтах — кам- нях, скалистых рифах, мертвых кораллах, кау- лерпа благодаря своему стелющемуся слоевищу и наличию многих ризоидов неплохо чувст- вует себя также на песчаных и илистых грун- тах. Излюбленным местообитанием видов .кау- лерпы является мелководье с более или менее спокойной водой, в удалении от постоянного прибоя, например лагуны, защищенные от действия прибоя коралловыми рифами. Все каулерпы построены по одному плану. Ползучие, распростертые на грунте части слое- вища, или р и з о м ы, имеют вид цилиндриче- ских сифонов. Ризомы весьма напоминают сто- лоны некоторых наземных растений. Они тя- нутся в длину на десятки сантиметров, развет- вляются и отчленяют через определенные ин- тервалы вниз ризоиды и вверх вертикальные побеги. Ризоидов много, на конце они сильно и многократно ветвятся и тем самым обеспечи- вают плотное прикрепление растения к любому субстрату. Вертикальные побеги выполняют роль ассимиляторов. В них сосредоточены хло- ропласты и происходит процесс фотосинтеза. Если ризомы и ризоиды у разных видов почти не отличаются по морфологии, то строение ас- симиляционных ветвей достигает большого раз- нообразия и сложности и служит хорошим приз- наком для разграничения видов (рис. 229, 1— 4). Они могут быть цилиндрическими, подобно ризомам, или уплощенными до листовидных. У большинства видов они снабжены многочис- ленными боковыми выростами разнообразной формы — серповидными, щитковидными, кег- левидными, метельчатыми и т. п. Специфическим свойством каулерповых яв- ляется образование внутреннего скелета. Во всех частях растения через полость сифона про- ходят особые переплетающиеся тяжи, которые ориентированы радиально или перпендикуляр- но поверхности (рис. 229, б). Это отчетливо видно на поперечных срезах через слоевище. Они образуются в цитоплазме близ вершины ветвей. Первоначально они могут быть свобод- ными на обоих концах, но нередко уже в момент возникновения связаны со стенками слоевища. Очень тонкие вначале, они быстро утолщаются
и становятся слоистыми подобно общей оболоч- ке растений. Функция внутреннего скелета остается неясной. Не исключено, что он играет какую-то роль в минеральном питании каулер- пы. Стенки слоевища состоят из каллозы, кроме того, в них можно обнаружить пектиновые ве- щества. Отсутствие целлюлозы отличает кау- лерпу, как и все семейство каулерповых, от остальных водорослей порядка. Кроме зеленых хлоропластов, для каулерпо- вых характерно наличие бесцветных пластид, или лейкопластов, которые участвуют в процессе накопления крахмала. У всех других водорос- лей порядка найдены только хлоропласты. Вегетативное размножение занимает замет- ное место в жизни этих водорослей: отломившие- ся части растений, попадая в новые места, легко прикрепляются к субстрату и разрастаются. Бесполое размножение у каулерповых отсутст- вует, по-видимому, оно было утрачено у совре- менных форм в процессе эволюции. Половое размножение — анизогамия — характеризуется рядом особенностей, отличающих каулерповых от других семейств порядка. Во-первых, у кау- лерпы нет специальных органов размножения- гаметангиев. Гаметы образуются непосредст- венно в ассимиляционных нитях в любой их части. На отдельных участках цитоплазма сгущается, приобретает темно-зеленую окраску, затем сетчатое строение и наконец делится с образованием одноядерных гамет. Какие-либо перегородки, отделяющие место образования гамет, отсутствуют. Такое размножение назы- вают г о л о к а р п и е й. Для выхода гамет на поверхность слоевища образуются довольно длинные выросты — папиллы; гаметы освобож- даются в результате разрыва оболочки на вер- шинах папилл. Долгое время считали, что семейство состоит всего из одного рода каулерпа. Но теперь, когда подробнее изучено строение и размножение дру- гих сифоновых, установлено, что к нему при- надлежит еще несколько родов, которые рань- ше относили к семейству кодиевых. Из них следует упомянуть роды удотея (Udotea) и ха- лиледа (Halimeda). В отличие от каулерпы вертикальные ветви у них тесно переплетаются (рис. 229, 6), в ре- зультате чего формируется псевдопаренхимное слоевище с определенной и довольно сложной морфологией. Виды удотеи характеризуются вееровидным слоевищем различной формы (табл. 32, 1) с ризомами в основании. Представители тро- пических видов этого рода более или менее силь- но пропитаны известью. Интересно, что в преде- лах рода удотея можно проследить переход от форм со свободными ветвями к формам псевдо- паренхимного строения. Наиболее сложным строением характеризу- ется халимеда, у которой слоевище состоит из множества вееровидных сегментов, сильно про- питанных известью (табл. 33, 2). Около 25 видов этого рода известны из тропических и субтро- пических морей. Многие из них играют большую роль в формировании коралловых рифов. Если рассмотреть ранние стадии развития халимеды и удотеи (рис. 229, 7 — 11), то броса- ется в глаза родство всех сифоновых водорослей. Вначале зигота растет, оставаясь какое-то вре- мя одноядерной. Затем, на более поздней ста- дии, она претерпевает ряд цитологических и морфологических изменений. Одно крупное ядро делится, некоторые хлоропласты теряют свой крахмал и превращаются в лейкопласты, вся структура вытягивается в вертикальный сифон с первичным ризоидом в основании. Следующая стадия имеет разнонитчатое строение. И только на самых поздних стадиях развития происхо- дит переплетение и смыкание вертикальных нитей. Семейство бриопсидовые (Bryopsidaceae) Это семейство охватывает четыре рода, тесно связанные между собой. Род дербезия (Derbesia) включает нитчатые водоросли, распространенные не только в тро- пических, но и в умеренных морях. Слоевище их состоит из стелющихся и вертикальных раз- ветвленных нитей. В месте отхождения ветвей, так же как и в основании спорангиев, образуют- ся своеобразные перегородки. Они могут быть простыми или двойными, и тогда между ними остается короткий сегмент с хлоропластами и ядрами. Вегетативное размножение, кроме фрагментации, заключается в образовании на вершине ветвей своеобразных выводковых почек, которые, отделяясь, превращаются в новые растения. Для бесполого размножения служат зооспоры (изредка апланоспоры). Это крупные шаровидные клетки с венцом жгутиков на перед- нем конце (рис. 230, 2 — 8), содержащие несколь- ко ядер и хлоропластов. При их образовании обычные вегетативные веточки меняют свою форму на шаровидную или грушевидную, и, отделяясь от слоевища перегородкой, превра- щаются в спорангии. Споры выходят из спорангиев через отверстие в их стенке и без периода покоя прорастают. Полового размножения нет. Род халицистис (Halicystis) представлен не- сколькими видами, которые всегда растут на известковых красных водорослях типа лито- тамнион. Когда халицистис начинает про- растать, вначале образуются разветвленные нити, стелющиеся на поверхности субстрата. Позднее они проникают внутрь известковых
Рис. 230. Строение бриопсидовых: 1 — 3 — Derbesia (1 — внешний вид слоевища, 2 — зооспора, 3 — зооспорангий); 4 — 6 — Halicystis (4 — первичные нити, б — молодые вертикальные нити, 6 —; 7 — 10 — Bryopsis (7 — веточка, 8 — ризоипы в основании веточек, 9— часть слоевища с гаметангиями, 10 — перегородка в основании гаметангия в продольном сечении); 11 — часть слоевища Pseudo- briopsis с гаметангиями. водорослей, образуя короткие вертикальные побеги, на концах которых формируются пузы- ри диаметром до 3 см (рис. 230, 4 — 6). Таким образом, слоевище халицистиса не просто пузырь. В основе его строения лежит та же сис- тема разветвленных нитей, которая свойственна дербезии, так что принципиальной разницы между этими родами нет. Пузыри же есть не что иное, как видоизмененные вертикальные ветви. Их образование, вероятно, связано со «сверлящим» образом жизни этих водорослей. Вегетативное размножение халицистисов про- исходит путем образования дополнительных пузырей на одном ризоме или нескольких новых пузырей на одном старом. Пузыри легко отделяются от слоевища, сохраняя при этом тургор и способность к размножению. Кроме вегетативного, известно и половое размножение — анизогамия. Гаметы образуют- ся в верхней части пузырей в результате преобразования и деления участков цитоплаз- мы, которые отделяются от остальной части их лишь тонким цитоплазматическим слоем. Гаме- ты выходят под давлением через несколько от- верстий. Один и тот же пузырь плодоносит обыч- но несколько раз. Бесполого размножения нет. В настоящее время установлено, что хали- цистис и дербезия представляют собой разные формы развития одного жизненного цикла, причем размножающаяся зооспорами дербезия является спорофитом, а половые растения ха- лицистиса — гаметофитами. Однако правиль- ная смена гаметофита и спорофита у этих водо- рослей необязательна и каждая форма развития способна к длительному самостоятельному вос- произведению. В природе известно намного больше видов дербезии, чем халицистиса; при этом первых можно встретить в тех районах, где вторые никогда не встречаются. Род бриопсис (Bryopsis) также имеет споро- фитную стадию типа дербезии. Основную часть слоевища бриопсиса составляет система верти- кальных ветвей, тогда как стелющаяся часть развита в меньшей мере и присутствует в виде ризомов или всего лишь развитых ризоидов. В вертикальной части различается главная ось, покрытая ветвями первого и второго порядков. У одних видов слоевища перисто-разветвлен- ные (рис. 230, 7), т. е. ветви у них располагаются супротивно и в одной плоскости; при этом верх- ние короче нижних. Все это делает растение похожим на перо птицы. У других ветви на слоевище располагаются вокруг всей оси ра- диально и тогда имеют метельчатое строение. В основании ветвей могут образовываться до- полнительные ризоиды, которые в начальной стадии своего развития выглядят как корот- кие отростки, при сильном же развитии почти сплошь покрывают главную ось (рис. 230, 8}.
В месте образования ветвей наблюдаются пе- ретяжки, ветви здесь сужаются и внутренняя часть оболочек утолщается. С поверхности эти утолщения можно принять за перегородки, на самом же деле они не отделяют ветвей, и все растение бриопсиса содержит одну центральную вакуолю, ограниченную пристенным слоем ци- топлазмы. Стелющиеся нити 6риопсиса, или ризомы, служат для переживания неблагоприятных ус- ловий и могут быть многолетними, тогда как вертикальная часть существует только в опре- деленный сезон. В основании вертикальных вет- вей постоянно возникают новые ризомы, кото- рые, в свою очередь, опять дают прямостоячие ветви. Благодаря этому бриопсис растет дер- ниной. Обычный способ вегетативного размно- жения заключается в том, что веточки отделя- ются от растений и укореняются при помощи ризоидов, образующихся в их основании. Половой процесс — анизогамия, как и у других сифоновых. Гаметы образуются в конеч- ных веточках, которые, перед тем как стать гаметангиями, не меняя формы, отделяются перегородкой от остального слоевища. Перего- родки образуются как внутренние кольцеобраз- ные утолщения боковой стенки в основании веточек. Эти утолщения, разбухая, оставляют в середине лишь узкий тяж протоплазмы, но постепенно и этот тяж прерывается, так как цитоплазма веточки и главной оси одевается собственной оболочкой, а первоначальное утол- щение остается как студневидная пробка между этими оболочками. Выход гамет осуществляется в результате ослизнения и растворения оболоч- ки на вершине веточки или через боковое отвер- стие в боковой стенке. Таким образом, у бриопсиса мы видим уже специализированные органы полового размно- жения, но, поскольку они сохраняют ту же морфологию, что и вегетативные веточки, мож- но считать, что дифференциация гамет анги ев здесь еще не закончена. Зигота прорастает без периода покоя. Во многих случаях при прорастании вновь форми- руются растения бриопсиса, размножающиеся половым путем, но у некоторых видов обнару- жена смена форм развития. У них зигота про- растает в нитчатое слоевище, ничем не отличаю- щееся от дербезии и размножающееся сходными зооспорами. Зооспоры, в свою очередь, разви- ваются в половые растения бриопсиса. Иными словами, в этой группе водорослей наблюдается интересное явление. Разные роды — халицис- тис и бриопсис — имеют одну и ту же споро- фитную стадию дербезии, с той лишь разницей, что у халицистиса смена гаметофита и спорофита происходит более регулярно и часто, тогда как у бриопсиса это случается реже и большинство видов этого рода размножается только половым путем. Род псевдобриопсис (Pseudobryopsis) обладает наиболее высокой организацией в семействе. По строению и внешнему виду он очень сходен с бриопсисом, но отличается наличием перего- родок в основании вегетативных веточек и более дифференцированными гаметангиями. Веточки, в которых образуются гаметы, видоизменяются, приобретая урновидную или грушевидную фор- му. Освобождаются гаметы через верхушечную пору (рис. 230, 11). Семейство кодиевые (Codiaceae) Семейство кодиевых содержит только один род кодиум (Codium), насчитывающий в настоя- щее время около 50 видов (табл. 31,4; 32, 6; 33, 1). Будучи характерными для флоры субтро- пических морей, они встречаются и в умеренных широтах, проникая в холодные воды северного и южного полушарий. Кодиум хрупкий (С. fra- gile) найден в Норвегии; другой вид — кодиум Рипипера (С. ritteri) — обильно развивается на Командорских островах. В южном полушарии кодиумы доходят до субантарктических райо- нов. Чаще всего их можно встретить на глубине 50 — 230 м и глубже, но некоторые виды выхо- дят на литораль. Они предпочитают чистые воды и поселяются на различных твердых грунтах. Слоевище кодиума обычно небольших разме- ров, длиной 5 — 30 см, но есть крупные виды, как, например, кодиум большой (С. magnum), длиной до 8 м. Темно-зеленое губчатое слоеви- ще имеет псевдопаренхимное строение. Оно сос- тоит из множества более или менее плотно пере- плетенных неклеточных нитей. По форме слое- вищ можно выделить две группы видов. Одни имеют распростертое слоевище, шаровидное или подушко видное; другие — вертикальное, цилиндрическое или сдавленное, обильно раз- ветвленное. Для прикрепления растений слу- жат ризоиды. Принцип анатомического строения кодиума легко понять, глядя на продольный срез через слоевище (рис. 231, 1). В нем можно различить две зоны. Внутренняя образована пучком отно- сительно тонких, бесцветных, более или менее густо разветвленных нитей. От нитей, располо- женных по периферии пучка, отчленяются ши- рокие и короткие пузыри, или у т р и к у л ы. Они располагаются таким образом, что окру- жают внутренний пучок бесцветных нитей, образуя поверхностный, или коровой, слой слоевища. Пузыри окрашены и выполняют асси- миляционную роль. Если внутренние нити у всех видов весьма сходны, то строение пузырей (их форма, размеры, очертания оболочек на вер- шине и т. п.) различно и служит одним из ос-
Рис. 231. Строение кодиума (Codium): 1 — часть продольного среза через слоевище; 2 — пузырь с до- полнительным пузырьком; 8 — пузырь с гаметангиями. \J ионных видовых признаков. 33 верхнеи части пузырей отходят бесцветные, длинные, легко отпадающие волоски, значение которых неясно. Если волоски развиваются обильно, все расте- ние приобретает опушенный вид. Рост слоевища в длину обусловлен апикаль- ным вытягиванием центральных нитей. Обра- зование новых пузырей среди уже существую- их ие только ведет к более плотному их щ расположению, но и представляет собой + р у инте ркалярного роста слоевища. У стелю- щихся подушковидных форм этот способ роста преобладает. Многочисленные мелкие хлоропласты ли- шены пиреноидов. Лейкопласты отсутствуют. Оболочки кодиума состоят из каллозо-пекти- новых веществ, но, в отличие от каулерповых, в них обнаружено небольшое количество цел- люлозы. Известно, что кодиум размножается веге- тативным и половым .путем. Вегетативное раз- множение широко распространено и происхо- дит посредством фрагментации или образования нескольких слоевищ от одного основания. Кроме того, для кодиума характерно образование специальных дополнительных пузырьков с ри- зоидом в основании, которые, отделяясь и уко- реняясь, дают начало новым растениям. Беспо- лое размножение неизвестно; вероятно, у этого род а оно полностью утеряно в процессе эволю- ции. Гаметы у кодиума двужгутиковые, груше- видные. Женские гаметы содержат многочис- ленные хлоропласты с пиреноидами; мужские— всего 1 — 3 хлоропласта, лишенные пиреноидов. Половой процесс, как правило, анизогамия. Дифференцированные гаметангии образуются как боковые выросты грушевидной формы на пузырях и отделяются от них специальными слизистыми перегородками, как у бриопсиса. Когда гаметы созревают, оболочка на вершине гаметангия разрывается, и гаметы выходят в окружающую среду. Зигота прорастает обычно без периода покоя. Новое поколение опять дает гаметы, и цикл развития завершается. Изучение рода кодиум показывает, что семей- ство кодиевых, будучи очень близким к бриоп- сидовым, представляет по сравнению с ними более высокоорганизованную и специализиро- ванную группу. Б олее высокая организация п оявляется в псевдопаренхимном строении, пр в наличии дифференцированных органов р аз- множения и в полной утрате бесполого размно- жения. В настоящее время бриопсидовые и ко- диевые рассматриваются как отдельные семей- ства, но их отличия говорят лишь о большей примитивности первых и эволюционной про- двинутости вторых, принципиальных же раз- личий между ними нет. Оба семейства представ- ляют собой единую в пределах порядка ветвь эволюции, отличную от каулерповых и, по- видимому, более молодую по сравнению с ними. ПОРЯДОК ДАЗИКЛАДОВЫЕ (DASYCLADALES) Дазикладовые образуют хорошо очерченную естественную группу морских водорослей, на- считывающую около 10 родов. Их слоевище, представляющее собой одну гигантскую клетку, имеет весьма сложную морфологию. В основе. их организации лежит радиальная симметрия. У всех дазикладовых слоевище состоит из не- разветвленной центральной оси, покрытой му- товками коротких ветвей и прикрепляющейся к субстрату системой разветвленных ризоидов. За редкими исключениями, для этих водорос- лей характерно более или менее полное обыз- вествление стенок слоевища. В отличие от дру- гих водорослей класса сифоновых слоевище у азикладовых одноядерное. В вегетативном сос- да тоянии ядро располагается в ризоидах. р д гаметообразованием первичное ядро подверга- ется многочисленным делениям и дочерние ядра мигрируют в развивающиеся гаметангии. Хло- ропласты многочисленные, дисковидные или эллипсоидные; пиреноид есть или отсутствует. Среди современных форм наименее специали- зированной структурой обладают представители рода дазикладус (Dasycladus, рис. 23, Центральная ось у этих водорослей, имеющая вид
Рис. 10 ис. 232. Строение дазикладовых.' 1 — 3 — Dasycladus (1 — aH — — ( — внешний вип слоевища, 2 — част вн, — часть поперечного среза че ез сл — п р ину веточки Neomeris' б — 10 — Асе а нтик, 10 — выход гамет из зрелой цисты . а — — танги тангии, в — стерильные побеги. узкого пузыря длиной до 5 см жении, кроме самого основа см, на всем протя- клас нования, густо пок ы- иссле сическим объектом самых б ~х разноо разных та ~~ы~~~~~ из iO — 15 ж~жй. Каж ая чередь, разветвляется дважды ющей зонтик. стерильном состоя- тижы. В р д . еточки второго и третьего или нии астения а ас го порядков цент аль цетабулярии построены п р полагаются также м товкой р ного цилиндра с одной или не о типу т вкой на вершине ми м товками или нескольки- предыдущего сегмента и постепенно ис. 232 нно утончаются ментов. 3 ело утовками неоднократно разветвленн нных сег- наблю ает месте отхождения ка аждои ветви ка, л чи кото релое слоевище имеет форму з у зонти- людается заметное сжатие стенок слоевища, метанги п|и rа- однако перегородок никог а б у и котороро образованы крупньп гда не о разуется и асполо иями, сросшимися друг с с другом или между сегментами остается связь в виде узкого Раны расположенными свободно (табл 32 2). ° ~ ° протоплазматического тяжа. П анние стадии развития всех аз в месте отпа дения ветвей и в основании г у система разветвленных из гиев. Н б аиболее характерной чертой в ст и гаметан- дов и ве тик р альная неразветвленная ось. На ве- изои- дазикла дуса является то, что конечны б рт й в строении шине этой оси и постепенно образуется пе ва а вер- веточки чные оковые товка боковых вая му- и слоевища не срастаются а остаются ве т ых сегментов. К концу сезона свободными. р икальная часть слоевища ых . на роста ща отмирает и на не- ~~щ~~~~ж~жй рода ~ю~~рж (N иная б рис ~ comer>sj летняя и ь много- , более сложная структура л ) р зоидная система. На второй го терно более р, для них харак- зоидов вы астает с год из ри- олее сильное обызвествление. Г сто ильнь расположенные мутовки об аз разованы здесь вет- п оцесс может повто ых ветвеи. тот повторяться в х по вер- пока не наступит время плодоношения. Тог асаются образ ются е огда псев о ру ~~~ж~, М~ж~~~ ~~~~бж днослоиной кото ые за допаренхимной коры (р с. 232 4) К о, аждая Фертильные побеги очень специ т один неразветвленный на о ци"ичны, поэтому волосок. т о на особенностях их строения И наконе т систематика о ов и ния и основывается ретий TMG cTpoeHMÿ ре став е моло щ~~~б~л„зо~ (А~И~ЬЫмж). Э ИЗ ШИ OH 0 р о известных водорослей то одна кладовых повто тх дази- ставшая ковых форм. торяют признаки древних пр пред-
Рис. 233. Строение дазикладовых. Внешний вид (1) и срез через слоевище (2) Bornetella: У современных представителей порядка из- вестно только половое размножение. Половой процесс — изогамия. Гаметы образуются в спе- циальных гаметангиях. У большинства дази- кладовых гаметангии возникают как шаровид- ные выросты на вершине или сбоку ветвей пер- вого порядка и соответствуют видоизмененным ветвям второго порядка (рис. 232, 8, 4). У аце- табулярии гаметангии образуются на коротких сегментах первого порядка как специальные выросты (рис. 232, 9). Внутри гаметангия обра- зуются толстостенные многоядерные цисты со специальной крышечкой (рис. 232, 10). Зрелые цисты содержат множество гамет. При разруше- нии стенок гаметангия цисты выходят во внеш- нюю среду и уже здесь из них высвобождаются гаметы. Цисты, а не зиготы могут служить по- коящимися стадиями. Существует мнение, что образование цист связано с обызвествлением слоевища. У некоторых лишенных извести ро- дов их нет и гаметы образуются непосредственно в гаметангиях. Благодаря способности дазикладовых к бо- лее или менее полному обызвествлению оболо- чек сохранилось достаточно много ископаемых остатков этих водорослей, по которым сейчас в какой-то мере можно проследить их эволюцию (табл. 34). Оказывается, это одна из самых древ- них групп водорослей, пережившая расцвет в отдаленные геологические эпохи и сохранив- шаяся в настоящее время в остаточной форме. Известно более 60 вымерших родов. Самый древ- ний представитель известен в раннем палеозое. Расцвет группы происходил в позднем палео- зое и мезозое; к позднему мезозою их развитие уже идет на спад. Примитивное строение палео- зойских форм характеризуется тем, что боковые выросты еще не разветвлены и располагаются на оси по спирали. К концу палеозоя — началу мезозоя появляется мутовчатое расположение боковых ветвей, при этом они приобретают спо- собность к неоднократному ветвлению. Эти признаки сохраняются до настоящего времени. В эволюции вегетативной структуры можно проследить три линии, которые заканчиваются разными типами строения современных форм. В первой линии боковые ветви располагаются свободно и так называемая кора отсутствует (тип дазикладуса). Вторая линия характери- зуется образованием «коры». У древних форм из этой линии развития кора образуется смыка- нием раздутых вершин неразветвленных боко- вых выростов; у современных — в результате слияния концов ветвей второго порядка (тип неомериса, рис. 233). Третья линия, к которой принадлежит группа ацетабулярии, представ- ляет собой сравнительно недавнюю модифика- цию, которая заключается в том, что в зрелом состоянии система ветвей редуцируется, оста- ется только диск из гаметангиев. У ранних ископаемых форм цисты образо- вывались непосредственно в главной оси слое- вища, специальные гаметангии еще отсутство- вали. Следующая ступень развития — образо- вание цист в боковых ветвях с неизмененной морфологией. Этот тип встречается уже у палео-
зойских форм, но наиболее широко представ- лен в мезозое. У более поздних форм можно встретить специальные гаметангии, которые представляют собой видоизмененные ветви вто- рого, редко третьего порядка. Дазикладовые — типичные морские обитате- ли. Их можно встретить на коралловых рифах, на песчано-каменистом грунте, вблизи мангро- вых зарослей. Они обычны для литорали и верх- них горизонтов сублиторали, но растут и на большой глубине — до 50 — 70 м. Ископаемые формы найдены на территории, которая сейчас находится в широтах с умеренным климатом. Здесь открыты крупные отложения дазикладо- вых, такие, как, например, доломиты в Европе. ПОРЯДОК СИФОНОКЛАДОВЫЕ ($~РНОХОСЬАВАЬЕ$} У сифонокладовых, в отличие от других водорослей класса, сифонное неклеточное слое- вище рано или поздно делится на многоядер- ные участки, или сегменты, и приобретает вид многоклеточного растения. Однако, несмотря на это, сифонокладовые проявляют гораздо больше сходства с водорослями сифонового комп- лекса, чем с остальными зелеными водорослями. Во-первых, образование перегородок наступает у них на более или менее поздней стадии раз- вития, и они достаточно долго вегетируют в фор- ме неклеточного пузыря. Во-вторых, сегменты, образовавшиеся в результате деления, — это не собственно клетки, а вторичные многоядер- ные образования. В-третьих, и это самое глав- ное, образование перегородок не имеет ничего общего с клеточным делением других водорос- лей. Оно известно как сегрегативное д е л е н и е. Протоплазма, сжимаясь, распа- дается на отдельные многоядерные участки раз- личной формы и размера, которые округляются и одеваются собственной оболочкой. По мере роста сформированных участков они приходят в соприкосновение, оболочки их смыкаются и процесс деления заканчивается (рис. 234, о — 8). Самой характерной чертой этого типа деления является полная независимость обра- зования перегородок от деления ядер, тогда как у других водорослей обычное клеточное деление начинается с деления ядра. Внутреннее строение слоевища сифонокладо- вых как на стадии первичного пузыря, так и у взрослых растений весьма сходно с сифоновыми. Под оболочкой находится тонкий слой цито- плазмы с сетчатым хлоропластом, под ним- слой цитоплазмы с множеством мелких ядер, в центре располагается вакуоля. Отличие заклю- чается в строении хлоропласта. Сетчатый хло- ропласт состоит из множества многоугольных или дисковидных участков, соединенных тон- кими тяжами. Из двух своеобразных пигментов, характерных для класса сифоновых, у сифоно- кладовых присутствует один — сифоноксантин. Размножение — там, где оно изучено,— половое и бесполое с.изоморфной сменой форм развития. Специальные органы размножения отсутствуют. Гаметы и споры образуются в любом из сегмен- тов и выходят через одно или несколько отвер- стий. Сифон окладовые — морские обитатели, за исключением кладофоровых, которые проникли из моря в пресные воды и широко в них рассе- лились. Наиболее примитивные члены порядка приурочены к тропикам, тогда как более вы- сокоорганизованные постепенно выходят за пре- делы тропической зоны в умеренные моря, и, наконец, самое высокоорганизованное се- мейство — кладофоровых — распространено вплоть до Арктики. Порядок включает несколько семейств, раз- личающихся строением слоевища. Семейство валониевых (Valoniaceae) включает роды валония (Valonia) и диктиосферия (Dic- tyosphaeria), встречающиеся только в тропиках. У наиболее просто организованного вида- валонии вздутой (V. ventricosa) — слоевище сос- тоит из единственного пузыря сифонной струк- туры, размеры которого могут достигать кури- ного яйца. На нижней части пузыря имеется множество мелких чечевицеобразных клеток, из которых вырастают ризоиды. У видов вало- нии с многопузырчатым слоевищем эти клетки, увеличиваясь в размерах, дают начало новым пузырям, похожим на материнский (рис. 234, 1, 2). Если у валонии новые пузыри образуются снаружи, то у представителей близкого рода диктиосферия образовавшиеся в результате сег- регативного деления сегменты остаются внутри материнского пузыря, создавая эффект компакт- ной паренхимной ткани. Семейство сифонокладовых (Siphonocladaceae), являющееся центральным семейством порядка, характеризуется более сложной структурой. Род сифонокладус (Siphonocladus) имеет раз- ветвленное нитчатое слоевище. В начале его развития образуется булавовидный прямостоя- чий пузырь сифонного строения с ризоидами в основании. Позднее он делится на большое число неправильно расположенных разновели- ких сегментов. По мере роста они смыкаются друг с другом и с оболочкой первичного пузыря. Многие из образовавшихся сегментов дают выросты наружу такой же формы, что и первич- ный пузырь. В боковых выростах происходят повторные сегрегативные деления и образуются вторичные «ветви» (рис. 234, 4 — 8). О размно- жении сифонокладуса известно очень немно- гое. Удалось наблюдать образование и выход клеток размножения через несколько пор в боковой стенке сегментов, но природа этих 305
Рис. 234. Строение сифонокладовых: 1 — Valonia гепСпсова; 2 — Valonia utriculosa; 3 — Dictyo- sphaeria favulosa; 4 — 8 — Siphonocladus (4 — внешний вид слое- вища, б — 8 — стадии развития слоевища); 9 — часть слоевища (;ladophoropsis. а — сегменты, образовавшиеся в результате сегрегативного деления. клеток не выяснена. Из двух видов этого рода сифонокладус тропический (Siphonocladus tro- picus) широко распространен в тропиках, тогда как сифонокладус крохотный (S. pusillus) растет только в Средиземном море. Второй важный род семейства — кладофо- ропсис (Cladophoropsis) своим однорядным нит- чатым слоевищем очень напоминает некоторые виды кладофоры. Однако в организации слоеви- ща у него больше общего с сифонокладусом. Ветви образуются как боковые выросты, а пере- городок в основании ветвей, столь характерных для кладофоры, здесь нет. Виды кладофоропсиса растут на мелководье, образуя, особенно в при- бойных местах, плотные войлокообразные дер- нины. Род представляет связующее звено между двумя семействами — сифонокладовых и будле- евых. Семейство будлеевых (Boodleaceae) включает два пантропических рода — будлея (Boodlea) и струвея (Struvea). Однорядное нитчатое слоеви- ще у представителей этих родов приобретает вид сетчатой пластины или губчатой массы бла- годаря тому, что сегменты различных ветвей соединяются друг с другом специальными корот- кими клетками — присосками, т. е. сильно редуцированными сегментами, расширенными и зубчато разделенными на вершине (рис. 235, 1, 2). В обоих родах рост слоевища с самого начала правильный, симметричный (рис. 235, 8). На каждом сегменте возникает пара боковых ветвей, которые развиваются синхронно и по- вторяют тот же способ роста. Семейство анадиоменовых (Anadyomenaceae) включает два рода — микродиктион (Microdic- tyon) и анадиомене (Anadyomene), представите- ли которых имеют такое же листовидно-сетча- тое строение во взрослом состоянии. Однако в этом семействе первичный пузырь растет не так симметрично, как у будлеевых. Последователь- ность, в которой сегменты образуют боковые выросты, и судьба этих выростов весьма варьи- руют (рис. 235, 4). Кроме того, у анадиоменовых отсутствуют специальные клетки — присоски. Соединение осуществляется всей боковой по- верхностью ветвей или с помощью кольцеобраз- ных зубчатых утолщений оболочки на вершине сегментов. Семейство кладофоровых (Cladophoraceae)— наиболее широко распространенная и хорошо изученная группа в этом порядке. До послед- него времени их рассматривали как самостоя- тельный порядок, очень близкий к улотриксо- вым. Однако пристальное изучение этих водо- рослей показывает, что они гораздо более сход- ны с остальными сифонокладовыми и находятся с ними в тесном родстве. Типичная водоросль семейства — род кладофора (Cladophora). В ли- тературе известно около 150 видов кладофор,
хотя в действительности их, по-видимому, на- много меньше. В европейских водах после кри- тического изучения сейчас насчитывают 27 мор- ских и 11 пресноводных видов. Кустистое слоевище кладофоры построено разветвленными нитями, состоящими из распо- ложенных в один ряд сегментов. На мелководье в прибойных местах, например на скалистых морских берегах, кустики кладофоры неболь- шие, четко оформленные, торчащие. Но очень часто, особенно в местах со стоячей водой и илистым грунтом — в лагунах, озерах, прудах и т. п.,— слоевища кладофоры сильно разраста- ются, образуя огромные спутанные массы раз- ветвленных нитей, или тину. Есть группы видов, у которых отдельные слоевища собираются в плотные шаровидные скопления. Они образуют- ся в местах с твердым грунтом, где постоянное несильное движение воды перекатывает непри- крепленные скопления. Кладофора прикрепляется к субстрату ри- зоидами, возникающими в результате разраста- ния нижней части базальной клетки. Ветви от- ходят от верхней части сегментов, это создает впечатление дихотомического ветвления (или политомического в случае отхождения от одной клетки нескольких ветвей) и очень напоминает ветвление других сифонокладовых (рис. 236, 1). Изредка у кладофоры можно встретить клетки- прицепки и соединение обычно свободных вет- вей друг с другом. При прорастании зооспоры или зиготы сна- чала образуется сифонная булавовидная струк- тура, ориентированная вертикально вверх. Прежде чем в ней начнется образование перего- родок, она достигает заметных размеров, диффе- ренцируясь на базальную (ризоидную) и апи- кальную части. Сегменты всех кладофоровых многоядерные. Хлоропласт имеет сетчатое строе- ние (рис. 236, 2), но, в отличие от остальных си- фонокладовых, у кладофоры тенденция к соеди- нению отдельных хлор опластов выражена в большей мере, поэтому часто хлоропласт вы- глядит как цельная, хотя и продырявленная пластинка, и увидеть, что он состоит из отдель- ных частей, можно лишь с помощью специаль- ной цитологической техники. До последнего времени считалось, что деле- ние у кладофоры принципиально отличается от сегрегативного. Но при внимательном изуче- нии между ними можно найти много общего. Здесь также образуются многоядерные сегменты неравного размера и, главное, деление прото- пласта тоже не связано с делением ядер. Кроме того, недавно у кладофоры было обнаружено и типичное сегрегативное деление. Размножается кладофора половым и беспо- лым способами. Четырехжгутиковые зооспоры и двужгутиковые гаметы образуются в любом Рис. 235. Строение сифонокладовых: 1 — 2 — struvea (1— молодое слоевище, 2 — соединение ветвей); 8 — вершина ве- точки Boodlea; 4 — часть слоевища Microdictyon. Рис. 236. Строение сифонокладовых: 1 — 2 — Cladophora (1 — веточка, 2 — вегетативная клетка m зрелый гаметангий); 8 — часть нити Rhizoclonium с веточкой- ризоидом; 4 — основание слоевища Chaetomorpha.
из сегментов и выходят после созревания через одну пору (рис. 236, 2). Спорофит сменяется гаметофитом того же строения. Входящие в семейство кладофоровых два других рода — ризоклониум (Rizoclonium) и хепхоморфа (Chaetomorpha) — отличаются от кладофоры лишь строением слоевища. У ризо- клониума способность к ветвлению развита уже очень слабо (рис. 236, 8). Настоящие ветви встречаются у него крайне редко, гораздо чаще на нитях образуются короткие боковые выросты из одного или нескольких сегментов, которые больше похожи на ризоиды. Сегменты ризокло- ниума содержат от двух до восьми ядер— меньше, чем у кладофоры и хетоморфы, поэтому нити этой вод()росли обычно тонкие и не дости- гают размеров, характерных для представителей двух других родов. Виды ризоклониума в при- роде почти никогда не встречаются прикреп- ленными к субстрату. Обычно они растут, обра- зуя скопления, запутывающиеся среди ветвей других водорослей. У хетоморфы способность образовывать ветви иотеряна полностью. Одно рядные нити этой водоросли прикрепляются к субстрату базаль- ной клеткой, основание которой расширяется и принимает форму диска с более или менее длинными лопастными выростами (рис. 236, 4). В отличие от других семейств порядка, кла- дофоровые широко распространены в морях всех климатических зон и, кроме того, населяют пресные воды. Однако центр их распростране- ния находится, по-видимому, также в тропиче- ских морях. Здесь сосредоточены виды с наибо- лее древними признаками, по которым кладо- форовые обнаруживают родство с другими тропическими семействами порядка. Кроме того, в тропических водах кладофоровые наиболее разнообразны по видовому составу и морфоло- гии. В умеренных морях среди большого числа видов много близких, с трудом различимых. По-видимому, очаги видообразования возникли здесь сравнительно недавно. От низких широт к высоким число видов кладофоровых уменьша- ется. Систематическое положение всех сифонокла- довых вызывает много споров. Часто происхож- дение порядка связывают с улотриксовыми, однако различия, которые существуют между этими группами, заставляют отказаться от такой точки зрения. Принципиально различны строение хлоропласта (у улотриксовых это цель- ная пластинка), строение и химизм клеточных оболочек, способ клеточного деления. Кроме того, наличие у сифонокладовых на ранних ста- диях развития сифонной структуры — важный признак, не только отличающий их от улотрик- совых, но и свидетельствующий о разном про- исхождении тех и других. Вероятно, сифонокла- довые отделились от какого-то общего с сифо- новыми предка, уже обладающего сифонной многоядерной структурой. У валонии сифонная структура может сохраняться в течение всей жизни, поэтому этот род следует считать наибо- лее примитивным в порядке. Наоборот, наибо- лее высокоорганизованным семейством в этой группе является семейство кладофоровых. Спо- соб образования перегородок достигает здесь высокой специализации. Логично предположить, что при сегрегативном делении, когда образо- вание перегородок не сопровождается делением ядер, наиболее примитивным будет беспоря- дочное деление с образованием фрагментов разной величины и формы. Это можно наблю- дать у валонии и диктиосферии. Более упоря- доченная форма слоевища, свойственная анадио- меновым и кладофоровым, является вторичной. В этом случае однорядная нить хетоморфы представляет наиболее специализированную в пределах порядка морфологическую структуру. КЛАСС КОН ЪЮГАТЫ, ИЛИ СЦЕПЛ ЯНКИ {СОХ J UGATOPH YCEAE) Конъюгаты, или сцеплянки, представляют собой особую, хорошо отграниченную группу преимущественно микроскопических зеленых водорослей. Количество их огромно: в четырех порядках (мезотениевые, гонатозиговые, зиг- немовые, десмидиевые) насчитывается около 4700 видов. Львиная доля (около 4000 видов) принадлежит десмидиевым водорослям. Большинство относящихся к конъюгатам во- дорослей являются одноклеточными (реже ко- лониальными), симметрично построенными ор- ганизмами, клетки которых обычно перешну- рованы посередине и состоят из двух совершен- но одинаковых полуклеток (десмидиевые). Мно- гие формы имеют типично нитевидное строение. При этом клетки, образующие нити, не пере- шнурованы посередине (зигнемовые). Некото- рые из конъюгат образуют очень хрупкие нити, легко распадающиеся на отдельные клетки (гонатозиговые). Освободившиеся после рас- пада нити клетки обычно продолжают жить вполне самостоятельно. И наконец, сюда же относятся очень просто организованные одно- клеточные водоросли, оболочки которых всегда состоят из одного неразрывного куска, без де- ления на особые полуклетки (мезотениевые), чем они резко отличаются от десмидиевых во- дорослей. По простоте строения и ряду других призна- ков среди конъюгат наиболее примитивны мезо- тениевые водоросли. Двумя ветвями в филоге- нетическом древе от них расходятся десмидиевые и зигнемовые. Порядок гонатозиговых занимает
в этой системе крайне условное положение. По сходству признаков его представителей с дру- гими конъюгатами систематики помещают го- натозиговые то между мезотениевыми и дес- мидиевыми, то между мезотениевыми и зигне- мовыми, подчас даже не выделяя их в особый порядок. Самая яркая отличительная черта конъюгат, позволяющая объединять их вместе в особый класс, несмотря на разнообразие внешнего вида, — это полное отсутствие подвижных стадий размножения, т. е. зооспор и подвижных гамет, и наличие своеобразного полового процесса, по- лучившего специальное название — к о н ъ ю- г а ц и я (лат. conjugatio — слияние, соеди- нение). Во время конъюгации происходит сли- яние протопластов двух клеток, которые вы- полняют здесь функцию гамет. В результате такого слияния, происходящего через специ- ально образующийся канал, получается одно- клеточная, покрытая толстой оболочкой зигота. Внешне две сливающиеся (конъюгирующие) клетки совершенно неразличимы; физиологи- ческая дифференциация на мужскую и женскую клетки хотя и определенна, но узнать ее можно лишь косвенно и спустя некоторое время — по положению зиготы. В том случае, когда ско- рости передвижения (вернее, перетекания) сжавшихся протопластов-гамет одинаковы, зи- гота образуется в конъюгационном канале, иногда разрастаясь вплоть до стенок конъюги- рующих клеток-гаметангиев. В этом случае разделение клеток на мужскую и женскую крайне условно, а сам процесс слияния равно- ценных по морфологии и скорости перетекания гамет принято называть и з о г а м и е й (греч. «изос» — равный, «гамос» — брак). Эта форма полового процесса характерна для большинства водорослей-конъюгат. Но у более развитых в эволюционном отношении зигнемовых водо- рослей наряду с изогамией существует и физи- ологическая анизогамия, или гетеро- г а м и я (греч. «анизос» — неравный, «гетерос»— другой, различный, «гамос» — брак) (рис. 24). И здесь протопласты-гаметы обеих конъюгирую- щих клеток одинаковы по размерам, но одна из них (мужская) активно перетекает через конъюгационный канал из отдающей клетки в воспринимающую, где и сливается со сравни- тельно неподвижной гаметой (женской). В этом случае зигота формируется только в восприни- мающей (женской) клетке. После периода покоя зигота прорастает, да- вая начало одному (у зигнемовых и гонатози- говых), двум (у десмидиевых) или четырем (у большинства мезотениевых) новым организ- мам — проросткам. Существует мнение, что уменьшение числа проростков имеет прогрес- сивное эволюционное значение: оно приводит к образованию более жизнеспособного поко- ления. Для нормального протекания конъюгации необходимы благоприятные условия — в пер- вую очередь теплая вода (от +15 до +25 'C) и интенсивная освещенность. Конъюгация длит- ся не более 7 — 14 дней, начиная от образова- ния конъюгационных каналов и кончая созре- ванием зигот. Необходимо отметить, что образование зи- гот у конъюгат — явление довольно редкое, а у многих видов десмидиевых и мезотениевых водорослей зиготы вообще еще не найдены. В виде исключительно редкого явления или как аномалия наблюдается также образование апланоспор, партеноспор или хламидоспор (у десмидиевых). В большинстве случаев одно- клеточные конъюгаты размножаются обычным вегетативным делением в поперечной плоскости, а нитчатые — распадением нитей на отдельные клетки. Конъюгаты характеризуются также чрезвы- чайно разнообразно построенными, иногда очень сложными, осевыми или, реже, постенными хлоропластами. Пиреноиды — носители за- пасного вещества крахмала — в хлоропластах присутствуют всегда. Исключение здесь сос- тавляет только монотипный (с одним видом) род лужоциопсис (Моидеойоря1я) из зигнемо- вых, у которого постоянно отсутствуют пире- ноиды, а также, быть может, некоторые виды рода спиротпения (Spirotaenia) из мезотениевых. Строение клеточных оболочек у конъюгат тоже очень разнообразно. Они могут быть цель- ными, состоящими из одного куска, или сегмен- тированными, состоящими из двух и более частей. Внешний слой оболочки может быть лишен порового аппарата, а может иметь ясно выступающие поры, бывает гладким или с очень богатой орнаментацией, не встречающейся у дру- гих представителей отдела зеленых водорослей. Геометрическая причудливость очертаний клеток в сочетании с изощренностью скульп- турных украшений и изумрудной «подсветкой» хлоропластов — все это давно принесло конъю- гатам славу красивейших водорослей и при- влекло к ним восхищенное внимание исследо- вателей. По существу, группа конъюгат космополитич- на. Их можно встретить на всех континентах, вплоть до Антарктиды. Но их широкое распро- странение не означает «всеядности»: они требу- ют для своей жизни вполне определенных эко- логических условий. Почти все конъюгаты жи- вут в пресных водах, и только некоторые встре- чаются в солоноватых водоемах или в эстуари- ях рек при небольшой солености, но типично морских видов среди них нет. Они ведут и на- земный образ ~кизни, поселяясь на влажных
Рис. 237. Мезотениевые. 1 — Mesotaenium macrococcum, вегетативные клетки, окружен- ные слизью; 2 — Spirotaenia condensata, образование слизистой ножки; 8 — Netrium digitus, поперечный разрез вегетативной клетки; 4 — Mesotaenium chlamydosporum, выход молодых кле- ток из зиготы. мхах, скалах, сырой земле и даже на ледниках в высокогорьях. Класс конъюгат подразделяют на 4 порядка: мезотениевые, гонатозиговые, зигнемовые и десмидиевые. ПОРЯДОК МЕЗОТЕНИЕВЫЕ (MESOTAENIALES) Мезотениевые водоросли специалисты счи- тают самыми примитивными среди конъюгат. В основе такого суждения лежит чрезвычай- ная простота формы клеток и строения оболочки этих водорослей. Их немного по сравнению с огромной группой десмидиевых: в 6 родах насчитывают 47 видов, а в СССР встречено и того меньше — всего 28 видов. И по «красоте» мезотениевые значительно уступают десмидие- вым. Клетки у них прямые или слегка согнутые, без перетяжки посредине, цилиндрические, эллипсоидные или веретеновидные. Живут они поодиночке или объединяются слизью в бес- форменные колонии, и только водоросль ании- лонема (Ancylonema) составляет исключение: ее клетки рыхло соединены в короткие нити (табл. 35, 1). Размеры клеток мезотениевых водорослей весьма различны. Так, некоторые представители рода спиротения (Spirotaenia) выглядят просто «карликами» (ширина клетки 3 — 4 мкм, длина 15 — 27 мкм) по сравнению с «гигантами» из рода нетриум (Netrium), ши- рина клетки которых достигает 83 мкм, а дли- на — 575 мкм (табл. 35, 2, 6). Каждая клетка мезотениевых имеет одно- слойную и сплошную, как бы из одного куска, гладкую оболочку. Двухслойные, иногда рас- слаивающиеся и разрывающиеся оболочки мож- но наблюдать только при делении старых кле- ток. В этом случае внутренний слой очень тон- кий, слабо окрашивающийся от хлор — цинк— иода; наружный слой не дает такой реакции. Известен факт и тонкого обызвествления обо- лочки клеток мезотениума (Mesotaenium) в куль- турах. Для водорослей, обитающих на влаж- ных скалах, мхах, лишайниках и на сырой поч- ве, характерно обильное выделение слизи, ко- торая со всех сторон охватывает клетку и, в от- личие от слизи десмидиевых, всегда бесструк- турна. Лишь иногда слизь состоит из концентри- чески входящих друг в друга слоев, что осо- бенно хорошо выражено у видов рода мезоте- ниум (рис. 237, 1). У одной из самых распрост- раненных мезотениевых водорослей — спиро- тении сжатой (Spirotaenia condensata) — слизь выделяется в форме длинной «ножки», как у десмидиевых. Чем менее подходящи для всех этих водо- рослей условия обитания, тем сильнее они вы- деляют слизь, которая, видимо, предохраняет клетки от неблагоприятных внешних условий, прежде всего от высыхания. Едва ли не самое интересное у мезотениевых- это многообразие хлоропластов: почти каждый род представлен особой формой. И через ряд постепенных усложнений, усовершенствований формы хлоропласта четко выявляется общая морфологическая эволюция родов мезотение- вых водорослей. Наиболее примитивны лезо- тениум и анцилонема. У них осевой пластин- чатый хлоропласт с зазубренными или цель- ными, иногда завернутыми внутрь краями (табл. 35, 1; рис. 238, 1). У ройи (Roya) хлоро- пласт несколько усложняется: появляются продольные ребра-складки и характерный для этого рода срединный вырез у вогнутой сторо- ны клетки (табл. 35, 8). Дальнейшее усложне- ние хлоропласта выражается в увеличении его поверхности за счет развития коротких ради- альных пластинчатых вы ростов у цидиндро- иистиса (табл. 35, 4) или длинных ребер, тоже радиальных, с лопастевидными, глубоко из- резанными или цельными краями у нетриума (табл. 35, 6; рис. 237, 8). У этих двух родов хло-
Рис. 238. Мезотениевые.' 3 — Mesotaenium chlamydosporum, последовательные стадии деления вегетативной клетки; 2,8 — Spirotaenia obscura, веге- тативная клетка — ее внешний вид и поперечный разрез. Рис. 239. Мезотениевые. 1 — С ylindrocystis sculpta, образование зиготы; 2 — 8 — Spiro- taenia condensata (2 — последовательные стадии конъюгации, 8 — зигота). ропласт разделен на две части. Звездчатый хлоропласт цилиндроцистиса очень напоминает таковой зигнемы из нитчатых зигнемовых водо- рослей, а хлоропласт нетриума схож по строе- нию с хлоропластами одноклеточных десмидие- вых. Одиночные клетки спиротении (по внеш- нему виду) очень похожи на клетки однолен- точных видов спирогиры — здесь тот же спи- рально закручивающийся постенный хлоропласт (рис. 237, 2; табл. 35, 2) или спирально закру- чивающиеся ребра вокруг осевой части хлоро- пласта (рис. 238, 2, 8). Пиреноиды у мезотениевых в числе одного, двух и более обычно располагаются в централь- ной части хлоропласта, и только у видов спи- ротении с лентовидно-постенными хлороплас- тами они беспорядочно разбросаны по всему хлоропласту. Форма пиреноидов очень разно- образна, причем у нетприума пальуевидного (Netriurn digitus) и щиликдрощистиса Бребис- сона (Cylindrocystis brebissonii) они достигают иногда очень крупных размеров. Клеточное ядро лежит обычно в центре клет- ки (мезотениум, анцилонема) и относительно тесно связано с хлоропластом, но в тех случаях, когда хлоропласт заполняет всю клетку (ройя, спиротения), оно занимает боковое положение. У видов нетриума и цилиндроцистиса ядро рас- полагается между двумя частями хлоропласта и обычно хорошо заметно. Размеры ядер очень разнообразны и обычно зависят от размеров клеток. Посредине ядра лежит крупное шаро- видное или неправильных очертаний ядрышко. У небольшого числа мезотениевых, как и у десмидиевых, на самых концах клеток образу- ются вакуоли с кристалликами гипса внутри, совершающими броуновское движение. У трех видов спиротении на концах взрослой клетки находятся по два каротиновых шарика, соединенных между собой слабо окрашенным мостиком. Эти шарики коричнево-красного или оранжевого цвета придают водорослям весьма своеобразный вид. У молодых клеток они четко видны только на старых (материнских) концах, полученных от только что поделившейся клет- ки (табл. 35, 2). Деление клеток у мезотениевых может про- исходить различными путями. Так, например, у рода мезотениум сначала наблюдается в хло- ропластах разросшихся, а потому и более круп- ных клеток деление пиреноидов с удвоением их числа, затем наступает деление самого хло- ропласта на два новых, и только после этого образуется поперечная клеточная перегородка, делящая старую клетку на две новые, молодые клетки (рис. 238, 1). У родов нетриум и цилинд- роцистис развитие новых хлоропластов сопро- вождается одновременным образованием кле- точной перегородки, так что иногда еще до окончательного развития хлоропластов старая клетка уже оказывается поделенной на две но- вые, молодые клетки. Для клеточного деления
некоторых видов спиротении характерно обра- зование косой разделительной перегородки. Такие пары клеток, соприкасающихся скошен- ными концами (табл. 35, 2), не обнаружены ни у какого другого рода мезотениевых водорослей. Образование зигот в процессе конъюгации может происходить у мезотениевых тремя пу- тями. При первом, наиболее простом способе их возникновения у двух рядом лежащих кле- ток, различных по половому знаку, но по внеш- нему виду совершенно одинаковых, в любых местах (особенно там, где расстояние между клетками наименьшее) появляются конъюга- ционные выросты, которые затем соприкасаются друг с другом (табл. 35, 4). Клеточные оболочки в месте соприкосновения растворяются или разрываются, и клеточное содержимое из обеих клеток выходит через конъюгационный канал. В результате этого зигота образуется между двумя конъюгирующими клетками (рис. 239,1). Второй способ очень сходен с первым. Конъю- гационные вы росты здесь также возникают в любом месте клетки, но зигота оказывается или целиком заключенной внутри обеих кле- ток, или только частично вклинивается в них (табл. 35, о). Третий, наиболее сложный случай наблюда- ется только у двух видов спиротении. Содержи- мое каждой из двух готовых к конъюгации клеток разделяется на две части, из которых формируются две округлые гаметы. После это- го оболочки клеток расплываются, делаясь совершенно незаметными, и гаметы одной клет- ки сливаются с гаметами другой (рис. 239, 2). В результате всегда получаются две зиготы очень своеобразного и сложного строения. Они шаровидны и окружены ячеистой, как бы сото- образной, окрашенной в бурый цвет оболочкой (рис. 239, 8). Вокруг нее обычно располагается своеобразный слизистый футляр. Форма зигот у мезотениевых очень разнооб- разна. Они могут быть шаровидными или слег- ка овальными, трех-, четырех-, шести- и даже восьмиугольными, с гладкими или, гораздо реже, покрытыми коническими выростами обо- лочками. При прорастании зигот обычно образуются 4 или, значительно реже, только 2 новые клет- ки — проростка (рис. 237, 4). Мезотениевые водоросли встречаются на всех континентах. Они предпочитают торфяные во- доемы: болота, берега озер, мелкие канавы, ямы, лужи. Многие представители этого поряд- ка ведут наземный образ жизни, поселяясь на влажных скалах, сырых стенах, мокрой земле и на влажных мхах. 3То особенно характерно для видов рода мезотениум, нередко образую- щих мощные слизистые скопления. Известны также немногочисленные мезотениевые, живу- щие на снегах и льдах высоко в горах либо на Крайнем Севере. Они интересны еще и тем, что в процессе приспособления к низкой темпера- туре и сильному ультрафиолетовому облуче- нию их клеточный сок окрашивается в фиоле- товый, красно-лиловый и коричневато-пурпур- ный цвет. Содержимое их зигот бывает интен- сивного гранатового цвета. Такие яркие пятна «цветного» снега в нашей стране можно встре- тить на высокогорьях Кавказа и на Земле Франца-Иосифа. Здесь «виновником» цветения снега может быть представитель рода анцило- нема (Ancylonema nordenskjoldii). В горах За- падной Европы, в Эквадоре, в Северной Грен- ландии и Антарктике к нему присоединяются также виды мезотениума (Mesotaenium berggre- nii и М. nivalis). В пределах СССР наиболее широко распрост- ранены и богато представлены виды родов нет- риум и цилиндроцистис. Мезотениевые водоросли надо собирать в ти- хую солнечную погоду, когда они, как и другие одноклеточные конъюгаты, поднимаются на поверхность воды п их изумрудные скопления (вспучивания) хорошо заметны невооруженным глазом. Если среди вегетативных клеток вы встретите конъюгирующие клетки или зиготы- считайте, что вам повезло. ПОРЯДОК ГОНАТОЗИГОВЫЕ (GONATOZYGALES) Гонатозиговые — группа загадочная и спорная по своему положению среди конъюгат. Маленькая по объему (всего 2 рода с 9 видами), причудливо сочетающая признаки десмидие- вых, мезотениевых и зигнемовых водорослей, она долго «терзала» систематиков, прежде чем завоевала свое нынешнее положение. С тех пор как роды гонатпозигон (Gonatozygon) и генику- лярия (Genicularia) в середине прошлого века были описаны Д е Б а р и, их относили либо к десмидиевым, либо к мезотениевым, а по неко- торым данным эти два рода можно было при- числить и к зигнемовым. Сейчас гонатозиговые водоросли принято считать самостоятельным порядком. За ними закреплено положение про- межуточной филогенетической группы между примитивными мезотениевыми и более эволю- ционно продвинутыми зигнемовыми водорос- лями. Каковы же особенности их строения? Клетки гонатозиговых водорослей имеют очень простую цилиндрическую или цилиндри- чески-веретеновидную форму и никогда не бывают перешнурованными посредине (в от- личие от десмидиевых). Ширина клеток варь- ирует от 4,5 — 10 до 25 — 32 лкл, длина их в 10— 20, реже в 35 — 40 раз превосходит ширину, достигая 620 — 760 лкл — почти миллиметра.
Клетки объединены в хрупкие нити, короткие или достигающие очень значительной длины. При малейшем прикосновении они легко рас- падаются на отдельные клетки, так как связь между ними очень слаба и обусловлена лишь незначительным смыканием слегка вздутых концов. После распада нити клетки продолжа- ют жить вполне самостоятельно. Известны слу- чаи, когда некоторых представителей рода го- натозигон наблюдали только в виде одиночных, изолированных, но вполне нормальных клеток. Клеточные оболочки у гонатозиговых, как правило, сплошные. Поэтому у них нет заранее заложенного, определенного места клеточного деления, что сближает гонатозиговых с мезо- тениевыми и зигнемовыми. Так выглядят взрос- лые, вполне сформировавшиеся клетки. У кле- ток, недавно поделившихся и еще не вполне зрелых или находящихся в стадии деления, можно наблюдать различные по своему строе- нию участки оболочки, иногда отделяющиеся друг от друга ясно заметной чертой (рис. 240, 8). Такие участки напоминают пояски (сегменты) некоторых видов рода пениул (Penium) из де- смидиевых. Подобного рода сегментирован- ность наблюдается только у клеток с не вполне развитым наружным слоем оболочки. При росте клетки сегменты смыкаются и распознавание пояска становится совершенно невозможным. Внутренний слой оболочки всегда цельный. Он тонкий, бесструктурный, целлюлозный, ок- рашивающийся от иода и серной кислоты в си- ний цвет. Наружный слой оболочки толстый, пектиновый, при действии тех же реактивов остается бесцветным. Он более толстый на бо- ковых поверхностях клетки и заметно утон- чается — уступом — на самых ее концах. Са- мое интересное и характерное заключается в том, что наружный слой обычно бывает ук- рашен маленькими нежными зернышками или заостренными шипиками, достигающими в дли- ну иногда 9,5 и даже 18 лкл (рис. 240, 4). Однако эти шипики никогда не развиваются на самых концах клетки. Значительно реже оболочки бывают гладкими (рис. 240, 1). Клеточные оболочки у гонатозиговых обыч- но окружены слизистым слоем пектозы, кото- рый при действии метиленовой синьки и индиго обнаруживает явно волокнистую структуру. Волокна располагаются перпендикулярно по- верхности клеточных стенок~ что указывает на присутствие пор в оболочке, через которые и выделяется слизь. Наличием на оболочке пор и скульптурных украшений, и двухслойным строением клеточ- ных оболочек гонатозиговые водоросли хо- рошо отличаются от мезотениевых и зигнемовых, что дало повод некоторым исследователям при- числить их к десмидиевьщ. Рис. 240. Гонатозиговые.' 1 — Gonatozygon kinahanii, вегетативная клетка; 2 — G. bre- bissonii, образование зиготы; 8 — G. monotaenium, сегментиро- ванность оболочки; 4 — G. aculeatum, шиповатость оболочки. Клетки некоторых видов рода гонатози гон обладают способностью отлагать железо в кле- точных оболочках. Особенно много его на кон- цах клетки, причем, чем старше клетка, тем большее количество железа содержится в ее оболочке. Иногда наблюдается развитие конечных, так называемых терминальных вакуолей (рис. 240, 1), которые содержат обычно довольно многочисленные кристаллики гипса. Они, одна- ко, не всегда бывают хорошо заметны. Хлоропласты у гонатозиговых двух типов. У гонатозигона они осевые, извилистые, реже почти прямые, в виде удлиненной пластинки либо бруска, лежат по два или только по од- ному в каждой клетке (табл. 35, 7; рис. 240, 1). Круглые пиреноиды располагаются обычно в виде довольно правильного осевого ряда, приблизительно на одинаковом расстоянии друг от друга. Число их варьирует от 4 до 16 и более. У геникулярии хлоропласты ленто- видно-постенные, спирально извитые или, ре- же, неправильно изогнутые, развивающиеся по два или три в каждой клетке (табл. 35, 9). Пиреноиды у представителей этого рода много- численные и, в отличие от гонатозигона, всегда беспорядочно разбросанные. Клеточное ядро шаровидное, занимает цент- ральное или, гораздо реже, боковое положение, причем последнее наблюдается только у видов гонатозигона с одним хлоропластом.
По внешнему виду — характерным удли- ненным нитям, пластинчатым или спирально извитым хлоропластам — гонатозиговые можно легко спутать с зигнемовыми водорослями: гонатозигон — с муж оцией, а геникулярию- со спирогирой. Однако зернистая поверхность оболочки у геникулярии и шиповатость у го- натозигона хорошо отличают их от зигнемовых, имеющих гладкие оболочки. Вегетативное деление у гонатозиговых про- исходит так же, как и у мезотениевых и зигне- мовых, — закладыванием поперечной клеточ- ной перегородки, делящей старую клетку на две новые, молодые клетки. Место деления не фиксировано, и оно может происходить в различ- ных частях клетки. При этом в росте дочерних клеток до нормальной величины принимает участие вся боковая поверхность клеточной оболочки. Возникающая у молодых клеток гонатозигона сегментированность оболочки вполне естественна: наружный слой ее, отяго- щенный у этих видов украшениями и солями железа, более «консервативен», чем внутренний слой, эластичный и тонкий. При повзросле- нии клетки такие сегменты разрастаются и сливаются. Своеобразие конъюгации у гонатозиговых заключается в том, что она происходит между двумя свободными, одиночными клетками, т. е. после распадения нити. Место разрыва клеточ- ной оболочки для выхода содержимого здесь точно не фиксировано, но обычно оно возникает посередине клетки. Конъюгационный канал очень короткий, к тому же быстро разбухает, и образовавшаяся зигота кажется прилепившейся к обеим конъюгирующим клеткам — гаметан- гиям (табл. 35, 8; рис. 240,2), которые довольно часто принимают коленчато-согнутую форму, что особенно характерно для видов геникуля- рии. Зиготы имеют правильно шаровидную форму и покрыты толстой, гладкой, бесцвет- ной или нередко окрашенной в желто-коричне- вый цвет оболочкой. Слияние ядер в зиготах происходит во время их созревания. В большинстве случаев имеются два последовательных деления ядра; первое со- провождается редукцией (уменьшением) числа хромосом, и из четырех образовавшихся та- ким путем ядер развивается только одно, в результате чего формируется всего один про- росток. Гонатозиговые водоросли живут в планктоне торфяных водоемов: озер, рек, прудов и луж, но нередко обитают также среди обрастаний. Они довольно обычны для сфагновых болот и иногда поселяются в болотах с пушицей и в береговой кайме сфагнов по краям торфяных озер. Наиболее распространены в наших пре- делах три вида гонатозигона (Gonatozygon kinahanii, С. monotaenium, С. brebissonii) и один вид геникулярии (Genicularia spiro- taenia). Лучше всего изучены эти водоросли на севере европейской части нашей страны, где в наиболее удаленных северных районах наряду с вполне типичными по своим измере- ниям экземплярами встречаются особо круп- ные образцы этих видов, намного превышающие стандарт. Но существуют виды, которые пред- почитают низкой температуре горячие источ- ники, или рисовые поля, или богатые известью воды. Сравнительно редкие и немногочисленные, гонатозиговые водоросли заслуживают боль- шого внимания и тщательного изучения. ПОРЯДОК ЗИГНЕМОВЫЕ (ZYGNEMATALES) Нитчатые неветвящиеся ярко-зеленые во- доросли этого порядка чрезвычайно широко распространены в пресных водоемах всех кон- тинентов. Даже в холодных ручьях Антаркти- ды они, хотя и недолго (короткое летнее время), радуют глаз своей изумрудной зеленью. В на- роде их обычно именуют тиной: небольшими рыхлыми подушками плавают они летом у по- верхности воды в прудах, озерах и тихих заво- дях рек. Каждая из таких дерновин-подушек, наполненная пузырьками выделяемого водо- рослями кислорода, состоит из множества нитей, переплетающихся между собой (табл. 36, 1). Слизистые на ощупь нити могут также свободно лежать на дне стоячего водоема или устилать на протяжении многих метров дно рек и ручьев, прилепляясь к песчинкам и кам- ням. Каждая нить сложена из ряда одноядер- ных цилиндрической формы клеток, ширина которых у разных видов колеблется от 3 до 150 — 200 лкл. Клетки довольно плотно при- мыкают друг к другу. Распад нитей на отдель- ные клетки происходит, да и то не у всех пред- ставителей, только при неблагоприятных усло- виях. В отличие от других конъюгат, зигнемовые очень трудно определять в вегетативном со- стоянии даже до рода. Точное систематическое описание их возможно лишь при наличии про- цесса конъюгации и зрелых зигот, которые при сборе материала в природе встречаются дале- ко не часто. В этом и состоит одна из главных трудностей научного определения зигнемовых. К тому же в разных географических областях сроки образования зигот различны, а на сам процесс конъюгации влияют не только темпера- турные и световые условия сезона, но и особен- ности гидрологического и гидро химического режима каждого водоема. В настоящее время известно около 700 видов зигнемовых водорослей, объединенных в 17
родов. Правда, в различных определителях, сводных работах и монографиях количество ро- дов и видов зигнемовых сильно варьирует,, но все же намечается определенная тенденция к их увеличению. Так, за последнее десятиле- тие к 13 известным прежде родам было прибав- дено 4 новых. Выделение новых родов было проведено на основании некоторых отклонений в строении конъюгирующих клеток и особен- ностей самого процесса конъюгации, которым в современной разработке систематики зиг- немовых водорослей уделяется все большее внимание. Важнейшими систематическими признаками при выделении родов у зигнемовых водорослей являются форма и положение хлоропласта. Известны три основные формы хлоропласта: осевой пластинчатый, осевой звездчатый, по- стенный спирально-лентовидный (рис. 241). Исходной формой надо считать осевой хлоро- пласт, хотя нередко и он весьма сложно устро- ен и достигает значительных размеров. По- стенный хлоропласт имеет вторичное проис- хождение — он возник из осевого путем редук- ции массивной центральной части и развития периферических лопастей. Усложнение хло- ропласта сопровождалось значительным уве- личением его поверхности и числа пиреноидов, перемещающихся на периферические части хло- ропласта. Тенденция перехода от осевого хло- ропласта к постенному, по-видимому, харак- терна для конъюгат вообще и для зигнемовых водорослей в частности. Кроме того, строение хлоропластов у зигне- мовых может варьировать в той или иной сте- пени как от вида к виду, так и в зависимости от условий обитания. Форма хлоропласта послужила основой для подразделения порядка Zygnematales на 3 се- мейства. Предполагается, что все они, несмотря на различия в строении хлоропласта, произош- ли от общего предка — мезотениевых водоро- слей и развивались параллельно. В каждое из этих семейств входит по нескольку родов. Три из них — спирогира, мужоция и зигне- ма — самые типичные из зигнемовых, по ним эти семейства и названы.' Spirogyraceae, Mou- geotiaceae, Zygnemamaceae. Перейдем теперь к описанию этих главней- ших представителей и тем самым к обобщаю- щей характеристике порядка зигнемовых во- дорослей. Спирогира (Spirogyra) часто встречается в пресных водах всего земного шара. Эта нит- чатка — один из самых широко известных, до- ступных и наглядных представителей мира водорослей и удобный объект для эксперимен- тального изучения растительной клетки. Спи- рогира — ~-самый обширный род среди зигне- Рис. 241. Участки нитей трех родов зигнемовых водо- рослей (cxeMa): 1 — мужоция; 2 — зигнема; 8 — спирогира. В в е р х у — по- перечные разрезы клеток. мовых. Во всем мире найдено около 340 пред- ставителей этого рода, у нас — свыше 120. Впервые спирогиру (или ее родственников) изобразил в 1741 г. немецкий ботаник Я. Д и л- л е н и у с в виде густо спутанных тонких ни- тей (рис. 242). Она именовалась тогда как «Conf åã÷ а palusmris sericea, crassior em varie exmensa», что в переводе с латинского значит «Конферва болотная шелковистая, толстая и разнообразно распростертая». Первый четкий и понятный рисунок спирогиры дал О. М ю л- л е р в 1782 г., а подробное описание — Я. В о- ш е в 1803 г. В XVIII и начале XIX в. зигнемовые водоросли называли конфервами или конъюгатами (сцеплянками), в большинстве случаев виды у них тогда не различали, а соб- ственно название подменяли пространным опи- санием. В 1820 г. немецкий ботаник Х. Л и н к ввел родовое название «спирогира» (греч. «спи- ра» — спираль, изгиб и «гирос» — извилистый, изогнутый). Родовой признак спирогиры был обозначен как «Interanea in spiras torta», т. е. «содержимое, свитое в спираль». С тех пор прошло 155 лет. Родовой признак спирогиры остался неизменным, но как сильно изменилось представление о строении этой во- доросли! C наивно примитивными рисунками прошлых веков теперь резко контрастируют крупномасштабные современные фотографии зигнемовых водорослей, изученные на субкле- точном уровне под электронным микроскопом
при увеличениях в тысячи и десятки тысяч раз. Однако чтобы хорошо рассмотреть спирогиру, необязательно пользоваться электронным ми- KpocKQIIQM. Даже при небольшом увеличении светового микроскопа можно легко разгля- деть ее строение. Нити у спирогиры, как и у всех зигнемовых, неветвящиеся и состоят из одного ряда одина- ковых, вытянутых, цилиндрических клеток (рис. 241, 8). Рост нити в длину осуществляется по- перечным делением клеток. Нити не длинные, от нескольких миллиметров до 8 — 10 сл, но, ок- руженные общей слизью и переплетенные меж- ду со бой они могут на большом протяжении устилать дно рек и ручьев. У нитей спирогиры, живущих в проточной воде, развиваются ризоиды — вы росты клет- ки отличающиеся утолщенной оболочкой и 7 отсутствием хлоропласта. Обычно они обра- зуются на нижней клетке нити, реже на осталь- ных клетках. Крайне разнообразного вида, от слегка изогнутых неразветвленных до сильно разветвленных или розетковидных и подуш- кообразных, они способствуют прикреплению нитей к суб страту и выживанию водоросли в условиях сильного течения. Прежде иссле- дователи, находившие ризоиды в редких слу- чаях и у немногих видов, рассматривали это явление как исключительное. Теперь выясне- но, что ризоиды характерны для бол ьшого числа видов спирогиры, зигнемы и других зигнемовых, но их образование сильно зависит от внешних условий (рис. 243). Ширина вегетативных клеток спирогиры ко- леблется в довольно большом диапазоне, от 4 до 200 мьм; длина может быть как в половину меньше ширины, так и превышать ее более чем в 20 раз. Оболочки клеток у спирогиры, как и всех зигнемовых, в период деления тонки и двухслойны. Внутренний слой состоит из цел- люлозы; наружный — из пектиновых веществ, дающих с хлор — цинк — иодом желтое окра- шивание. Только у представителей рода сиро- гониум (Sirogonium) пектиновый слой отсут- ствует, что отличает их от спирогиры. Рис. 242. Первый рисунок спирогиры, сделанный Диллениусом в 1741 г, Внешний студенистый покров, который как бы укутывает всю нить и достигает порой зна- чительной толщины, обусловливает ту скольз- кость и шелковистость, которые так присущи зигнемовым водорослям. Толщина этого покро- ва зависит от вида, а у отдельных видов- от внешних условий. Поперечные перегородки клеток, иначе на- зываемые с е п т а м и, у большинства видов спирогиры гладкие, но все же многие виды име- ют складчатые септы, а у двух видов септы в ви- де шайбовидного кольца. Самое яркое и бросающееся в глаза у спиро- гиры — это спирально закрученные зеленые ленты хлоропластов. У разных видов их может быть от одной до шестнадцати в каждой клет- ке (рис. 241, 8). Расположены они по внутрен- ней стороне оболочки, т. е. постенно. Лента хлоропласта у спирогиры представляется же- лобком с продольным срединным гребнем, бла- годаря чему на поперечном срезе она имеет вид двузубой вилки: обоими краями («зубья вил- ки») этот желобок обращен к продольной стен- ке клетки, а гребнем («ручка вилки») направ- лен в полость клетки. Гребень хлоропласта на фоне неправильно изрезанных или волнисто- фестончатых краев выглядит как темно-зеленая полоса. В зависимости от условий среды у спирогиры варьирует как плотность витков, так и поло- жение хлоропластов. При неблагоприятных условиях жизни рост хлоропластов отстает от роста клетки и количество оборотов резко снижается. Нормальный вид они сохраняют в период интенсивного деления клеток. р д У о а сирогониум, близкого к спирогире, ленты хло- ропластов очень узкие и почти параллельные продольным стенкам клетки (табл. 36, 6). По средней линии хлоропласта у спирогиры расположены многочисленные пиреноиды, име- ющие вид небольших округлых бесцветных телец. В процессе фотосинтеза в хлоропластах образуется крахмал, который откладывается вокруг пиреноидов в форме мелких зернистых глыбок. При усиленном фотосинтезе крахмал может откладываться также и в толще хлоро- пласта. Кроме крахмала, в клетках зигнемовых вст ечаются также капли масла, д~ бильного вещества и кристаллы щавелевокислого каль- ция. Центральная клеточная вакуоля запол- нена бесцветным клеточным соком, содержа- щим сахар п другие растворимые вещества, которые не только имеют значение питатель- ных веществ, но и обусловливают тургор клет- ки. Я о спирогиры очень крупное, гомоген- ное, с ясно заметным ядрышком. но р дро у жено в центральной части клетки, окружено
Рис. 243. Ризоиды у зигнемовых водорослей: ы 2 — на срединных клетках спирогиры; 8 — то же у зигнемы. 1 — на нижней клетке спирогиры; — н слоем цитоплазмы и подвешено о на плазмати- ческих тяжах, ах соединяющихся с постенным слоем цитоплазмы. У одних видов ядро имеет шаровидную форму, а у других — линзовид- н ю и тогда ориентировано поперек оси клетки. ную ит Такое разнообразие формы ядра а сочетается с некоторыми ыми другими морфологическими и физиологическими признаками, в первую оче- едь с ширин риной клетки. Деление клетки, про- текающее при благоприятных условиях в ве- чернее время в течение 1— — I 5 ч начинается прежде всего с деления ядра. Для изучения тонкого строения клетки при- меняют соответствующую окр слабым раствором иода в иодистом калии б тро убивает клетки спирогиры и окраши- ыстр р " в синий вает сод р ержащийся в них крахмал Ф.У я о и особенно ядрышко — в желтыи. цвет, а ядро и ос ~ли после него Е одновременно с иодом или после н сли эозина об аботать клетку водным раствором эо о ра отать к получается двойная окраска: ядр я о и пире- то п й вет а кпахмал ноиды окрасятся в розовыи цвет, а вокруг пир еноидов — в синий. Размножение спирогиры, как дру и гих зиг- немовых, мож ет быть вегетативным и половым. В етативное размножение происходит при слу- ег ом азрыве нитей на участки или пр р и п и неблаго- падении их на отдельные клетки пр п иятных условиях. Из кажд р oro отрезка нити и каждой неповрежденной клетки при нормаль- ных условиях для роста и деления образуется новая нить. Известны покоящиеся состоян ия вегетатив- ных клеток, также служащие для размножения. акинеты. Это апланоспоры, партеноспоры, ы об~ аз „ются из вегетативных кле- лпланоспоры о ток путем сокращения их содержимого и раз- вития вторичнои оболочки, р Ф.У кото ая может быть сходной с оболочкой зигот. В роде спироги- ра пять видов р азмножается только апланоспо- рами. Возникновение партеноспор связано с я из аномалиями в конъюгации — они образуютс неслившихся гамет, но имеют форму и строение т. П и об азовании акинет в содержимом ит накопление вегетативных клеток происходит на к о снаб- крахмала и масла а оболочка их, о ычн женная линией разрыва, значительно утол- и акинет — это способ перенесения неблаго- приятных условий. Половое размножение у всех зигнемовых- ха актерная конъюгация. Различают два ос- характе н HQBHbIx типа конъюгации: лестн у вую. В процессе конъюгации нити водоросли довольно сильно изгибаются. При лестничной конъюгации нити распола- гаются параллельно друг дру у r или попарно спирально сворачиваются по д лине. П и этом в двух сближенных нитях у р п отивополож- ных клеток о оболочка начинает образовывать выпуклости, клости направленные навстречу друг г . Эти выпуклости, разрастаясь, превра- другу. щаются в цилиндрические выр ы осты смыкаю- щиеся своими концами. Разделяющая их пере-
Рис. 244. Типы конъюгации у зигнемовых водорослей: 1 — лестничная конъюгация у зигнемы; 2,8 — то же у мужоции; 4 — боковая конъюгация у спирогиры. городка растворяется, и в результате получает- ся сквозной конъюгационный канал, соединяю- щий обе конъюгирующие клетки. Так как обычно этот процесс протекает во многих клет- ках сразу, то образуется подобие миниатюр- ной лестницы (табл. 36, 7; рис. 244, 1). Извест- ны случаи лестничной конъюгации нескольких (до пяти) нитей между собой. Боковая конъюгация протекает в одной и той же нити. В этом случае конъюгируют две соседние клетки с частичным растворением септы и образованием конъюгационного кана- ла. Он обычно не такой длинный, как при лест- ничной конъюгации, и выглядит наподобие изогнутого рукава (рис. 244, 4). Одновременно с образованием конъюгацион- ного канала внутри обеих конъюгирующих клеток происходят значительные изменения. Протопласт клетки сжимается, отстает от обо- лочки, хлор опласты теряют свою обычную форму и отчетливые контуры. У спирогиры од- на из конъюгирующих клеток становится от- дающей, другая — воспринимающей. Сжав- шееся содержимое отдающей клетки в виде аме- бовидной гаметы перетекает через конъюга- ционный канал в другую, воспринимающую клетку, где оба протопласта сливаются. Про- дукт слияния, или зигота, округляется и по- крывается толстой оболочкой. К моменту соз- ревания зигота заполняется запасными пита- тельными веществами, в частности большим количеством капель масла, а ее оболочка корич- невеет. По форме зиготы у зигнемовых большей частью шаровидные или эллипсоидные, неред- ко сплющенные, линзовидные (рис. 244, 245). После периода покоя перед прорастанием зи- готы в ее ядре, образовавшемся путем слияния двух ядер, происходят значительные измене- ния: оно дважды делится пополам и из четырех ядер три небольших дегенерируют, а одно круп- ное остается. Прорастает зигота у всех зиг- немовых одной нитью (рис. 245, 4). Вегетативные клетки в конъюгирующих ни- тях, оставшиеся без конъюгационной пары, обычно не изменяются, но у некоторых видов они могут сильно вздуваться. Так, например, известен один вид спирогиры (Spirogyra bul- lata), у которого стерильные клетки даже при- нимают шаровидную форму и по ширине ста- новятся в 3 — 4 раза больше клеток в вегета- тивной нити. Оболочка зиготы у всех зигнемовых состоит в большинстве случаев из трех слоев, но неред- ко только из двух, а иногда и из большего ко- личества слоев, отличающихся друг от друга толщиной, окраской и скульптурой. Среднюю оболочку, толстую, окрашенную и большей частью снабженную скульптурой, принято на- зывать м е з о с п о р и е м. Другие оболочки называют э н д о с п о р и е м (внутренняя) и э к з о с п о р и е м (наружная). По некото- рым данным, в мезоспории содержится хитин. Скульптура мезоспория при рассматривании с поверхности, особенно при малых увеличе-
нои, однако это не т Н ° .У ниях микроскопа может т казаться о днообраз- р готе неред не так. а зоелой зи .:Ф ть продольн ю г р д у линию разрыва: нее разрывается мезоспо ий растании зиготы. Щ )Ю ы. Ногообвазие ти ~Ю туры мезоспо ия п бр ипов скульп- Д Я орма зигот, строение их о"о Qg тер и сте пень вздутия клеток п и ко х оболочек, харак- Я ( ции — все это при ~9 признаки, кпайне при, р нео" ходимые определения ви ов водорослей. У становление видовой ости собран ых, „"" Рин'д О '.,-',:::;:.':: р р д зигнемовых водо 3 только по вегетативным п и :-О в частности по шир м при накам, р не клетки и числ пластов совершенно ож нно невозмож у хлоро- ожно. Более того тильнои стадии невозмо" 1 не только ви ы зможно различить О о виды, но даже некото ые о например, у рода рые роды. Так, '::::О:,,' да сирогонистм клет "..-.. о;.. -'" хожие в вегет у ки, очень по- , "ЮИ~э~ тативном состоянии на в отличие о на спирогиру . 'Е'':::.'- -'"" ''.. 'Ф.'..:: " ' последней, пе е 1 ' 1. делятся на две р д онъюгацией неравные клетки конъюгационной со" ственно й клеткой стан по размеру, котопая п 4 к своему протоп ел пласту во впемя ел Рис. 245. все содержимое р деления почти иготы зигнемовых во е сестринской сте илья " растание: ых водорослей и их п о- про- ма, кроме того е е 1 — 8 — зиготы аз отсутствует с р ~й канал, взамен кото аз; — прорастающая зигота нов ения коленч нитей сраз п нчато-изогнутых зу происходит раство ение во со спирогирой е е ной оболочки (табл. 36 6' Мжо ия ° ~ ]е ласт у мужоции делится в о у оиия (Mougeotia) была описа ским ботаником К А ются две спи ал но, тогда внутри клетки р льно закрученные лент оказы- ~Ъ~ сть ранцузского врача А. игнема (греч. «Зиг ougeot). Это второй ~ уж о нить «дзигон» — пара, «нема>— не рой по численности о зиг- Q момента мир. H. - . 121 1 встречено свыше 3~ вит елей. ссс Ч 5 — в г' найдено около 20 дов, из них ° ужоция ш гнему бе т ~м~~ ж 3 до 63 руда можно узнать по льл характе по двум . астам звездчатой фо мы широкои осевой пластинки азб ос нем находятся неско лько пи енои р дов, хлоропластам ноид. ент е ложенных хлоропласту или аспо- У р по- как и мужоции, богаты несколько сбоку к хло сле Н~~~ ти зигнемы, имеющие ши ин о пел ек. со енность хлоропласта одеты слизист Ужоции подвижность. П~и — го развитым чем тым чехлом е ще олее мощно 7 хло опла - р р ри ярком осве ен , чем у спироги ы. П ри неблагоприят- ей оси и т я на 90 вокр r сво- иях нити зигнемы ос глаз тогда выглядит т не п ивычно р нои для падаются на от с остальны полоской п стинкои, а узкой зеленой ются на отдельные клетки. проходящей но се е и й '"онъюгация м от одного ее ко конца до редине клетки ни е и яичная и око у ужоции и зиг н остью о другого. С несомнен- вая. Но если спи немы лест- д казано наличие у пирогиры зиготы плазме кле в постенной цито- я только в клетках ст кт клеток чужоции б осо ой спи аль зигнемы они мог ах, то у видов р ут образовываться (продолжите льность осве е пределенных условиях 4), аз аста конъюгационном канале ( б интенси тал. 36 8 ивность) приводит к сп щения и его высо кая образовавших их аясь иногда о . к д наружных стенок похо ю и хлоропласта, становя хожим на хлоропл ласт спироги ы. ящегося образ ю ются, и тог а зи ационные каналы не Сходст- межд к д зиготы лежат свобо ду клетками (рис. 244 2 Т акую конъю- 31
:-О О '~О О О ' j OOOO((1 ° ° ° ° ве ' ° ° ' ° ° ° О '-. ':. (~, о О O оФ (;г.. ""-:3 ° ° В' ° ъ ъ ° ее е ~~Ъ :-<0 :.о О ~ ф ope..' о ""О'1 ~ 4 ~'О""0 "."....."""0" о о о о о О О О О '-) О О О О О О С о о о о -.О О О О О "2 ° ° ° Ъ О~~-~ '='. Q~Ook QQ0QQog 'aC)~p~....: 8 . '~" ~O о о о о о. OOO ,/с~ ю О () �OQ Q Q (g (- v' '"".. f) o C 0 о О О е О о оо О О О ос,' Ъ О О 'Оо ~ 0 л о .-"a "о "б" Q, ''~7~ Q V~~ о > ° ° 4~ ° r т ° ° (e Ф е е О,. Рис. 246. Разнообразные типы скульптуры у зигот зигнемовых водорослей (схематизировано): в в е р х у- поперечный разрез, в н и з у — вид с поверхности. гацию называют экстрацеллюляр- н о й (внеклеточной), в противовес обычной интрацеллюлярной (внутриклеточ- ной) конъюгации. Особенность конъюгации у мужоции заклю- чается в том, что зигота еще до образования оболочки отделяется от образовавших ее кле- ток двумя, тремя или четырьмя перегород- ками. Замечено также, что у мужоции при формировании зиготы в клетках сохраняется остаточная цитоплазма в виде разбросанных мелких глыбок, а у близкого рода тпемногаметпум (Temnogametum) формирование гаметы про- исходит как у сирогониума, т. е. не только из всего содержимого собственно конъюга- ционной клетки, но и большей части сестрин- ской стерильной клетки (табл. 36, о). Форма зигот у видов мужоции и зигнемы очень разнообразна: шаровидная, эллипсоид- ная, кубическая, полиэдрическая, цилиндри- ческая и т. д., но чаще зиготы сплющены и по- этому имеют разный вид с широкой и узкой стороны. Мезоспорий часто и разнообразно скульптурирован, а у некоторых видов окра- шен не в обычный желтовато-коричневый, а в голубой и синевато-зеленый цвета (табл. 36, 4). Зигнемовые водоросли — по преимуществу обитатели пресных вод. Их массовое развитие чаще можно наблюдать в стоячих или слабо- проточных водах, сравнительно чистых, xopo- mo прогреваемых и освещенных: прудах, озер- цах, природниковых лужах, ручьях и при- брежье рек. В больших озерах зигнемовые раз- виваются в литоральной зоне. Здесь часто по- падаются прикрепленные формы. Морских ви- дов среди зигнемовых нет, но все же имеется несколько представителей, выдерживающих широкую амплитуду солености воды — от сильно опресненной до 8,5 — 9,7%,. Замечено также, что виды мужоции предпочитают из- весткованную воду, а широко распространен- ный представитель близкого к зигнеме рода зигогониум (Zygogonium ericetorum) населяет главным образом кислые воды. Зигнемовые водоросли можно встретить и на увлажненной почве, и на орошаемых скалах, и вблизи горя- чих и минерализованных источников. В гор- ных районах отмечены своеобразные «рекорды высотые, достигнутые зигнемовыми: Spirogyra varians была найдена на высоте 3400 м над уровнем моря в Заилийском Алатау, а Spirogy- ra borgeana — на высоте свыше 5600 м над уровнем моря в Тибете. Три рода из зигнемовых водорослей- спирогира, мужоция и зигнема — имеют наи- большее количество видов, которые в боль- шинстве относятся к повсеместно распростра- ненным. Другие роды характеризуются срав- нительно малым количеством видов и ограни- ченным распространением. Так, виды рода темногаметум найдены только в тропических и субтропических областях, а виды рода сиро- кладиул известны только на территории Индии. У родов зигогониум и сирогониум соответ-
Рис. 249. Разнообразие формы клеток у десмидиевых водорослей: 1 — Micrasterias rotata; 2 — Euastrum divaricatum; 8 — Cos- ипапшп protuberans, в н и з у — вид сверху. Рис. 250. Разнообразие формы клеток у десмидиевых водорослеи: 1 — Staurastrum pelagicum; 2 — Staurastrum cyclacanthum, с п р а в а — вид сверху. Он может быть узким и длинным, закрытыщ или открытым, с краями, расходящимися на- ружу от острой или закругленной верхушки. Внешний вид клеток десмидиевых в большин- стве случаев зависит от того, с какой стороны их рассматривают под микроскопом. Различа- ют вид спереди (с широкой, лицевой стороны), вид сбоку и вид сверху (или снизу, рис. 247). Таким образом, клетки имеют три плоскости симметрии. Разнообразие очертаний клетки с различных сторон имеет большое значение в систематике десмидиевых. Несколько видов могут иметь очень сходные очертания при рассматривании их спереди, но при этом резко отличаться друг от друга при рассматривании сбоку или сверху. Однако клетки некоторых видов из родов доцидиум (Docidium) и аитино- тениум (Actinotaenium) сверху округлые и, следовательно, имеют одинаковые очертания при любых поворотах вокруг длинней оси (рис. 248, 1, 2). Своеобразно строение клетки у видов клостериума (Closterium, рис. 248, 8). Они обычно сужены к концам, не перентнуро- ваны и имеют более или менее сериовидную форму, в результате образуются неодинаковые спинной и брюшной края. Спинной край бы- вает более или менее выпуклым, а брюшной— вогнутым или почти прямым. Однако ясная симметрия полуклеток сохраняется. Перешнурованные виды десмидиевых, к которым относится большинство этих водо- рослей, характеризуются огромным разнооб- разием формы клеток (рис. 249, 250). У одних клетки сильно вытянутые, цилиндрические, в по- перечном сечении округленные или звездчатые; у других — сильно сжатые и плоские. У неко- торых представителей развиваются особые отростки и шипы на углах полуклеток. У рых- ло-колониальных и нитчатых десмидиевых клетки соединены особыми образованиями (бу- горками, гранул ами, выступами) или слизью (рис. 248, 4, 5). Одним из наиболее характерных признаков десмидиевых является чрезвычайное многооб- разие очертаний клеток. Края их часто разде- лены на дольки, или сегменты, или сильно изрезаны. Самые сложные очертания — у кле- ток из родов миьрастериас (Micrasterias), ауаструл (Euastrum) и стаураструл (Stau- rastrum) (табл. 37, 4, 5; рис. 249, 1, 2; 250). Форма и очертания клеток имеют большое значение при определении отдельных видов. Оболочка клеток. Клетки десмидиевых об- ладают слоистой оболочкой. Новейшие иссле- дования показали, что она состоит из трех слоев. Самый внешний слой, который до недав- них пор не был известен исследователям, ка- жется под электронным микроскопом то петле- образным, то гладким или имеет вид пчелиных
сот. Химическая природа этого слоя еще неиз- вестна. Предварительные исследования пока- зывают, что он содержит пектинооб разное вещество. У молодых клеток внешний слой оболочки тонкий, у старых достигает значи- тельной толщины. За внешним слоем следует первичная оболочка, затем вто- ричная оболочка — самый внутрен- ний слой. Первичная и вторичная оболочки при рассматривании клетки в световом микро- скопе не отличаются друг от друга и пред- ставляются как один целлюлоз осодержащий слой. Однако с помощью электронного микро- скопа можно хорошо различить их строение. Первичная оболочка состоит из тончайших волоконец — м и к р о ф и б р и л л, имеющих вид пучков соломы, которые перекрещиваются друг с другом. Вторичная оболочка состоит из более широких волоконец — ф и б р и л- л я р н ы х л е н т, расположенных парал- лельно микрофибриллам первичной оболочки. Такая оболочка служит хорошей защитой и опорой для содержимого клетки. Сложность строения оболочки у десмидиевых водорослей усугубляется также наличием в ней особых круглых или овальных отверстий- пор. Они имеют разное значение. Прежде всего поры служат для выделения экскретор- ной слизи, которая постоянно окружает клетки десмидиевых. Через поры удаляются из клетки ненужные продукты жизнедеятельности. Поры служат и для поступления питательных ве- ществ из окружающей среды в клетку, так как внешний слой оболочки сам по себе не- проницаем. Расположение пор в оболочке у разных видов десмидиевых бывает очень разнообразным. У некоторых представителей поры очень мелкие и многочисленные. Они сосредоточиваются в определенных местах или равномерно рас- пределяются по всей оболочке. У других ви- дов поры бывают крупными и немногочислен- ными. Они также располагаются в определенных местах, чаще на концах клеток. У третьих ви- дов имеются и мелкие и крупные поры и рас- положение их также бывает различным. Поры у десмидиевых представляют собой не просто отверстия в оболочке, а имеют часто сложное строение. Исследования под электрон- ным микроскопом показали, что у этих водо- рослей имеются два типа строения порового аппарата — тип клостериума и тип жослариула. У представителей рода клостериум поровый аппарат наиболее примитивен. Он состоит из парового канала обратноворонкообразной фор- мы (рис. 25i, 1), узкая часть которого, являю- щаяся собственно порой, расположена во внешнем слое оболочки, а более широкая часть проходит через первичную и вторичную оболоч- У Ф ° ° Ъ ° ° ° ФЪ Ф ° S~il ° l ФЧ 'a@ ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ~ i ° ФЬ~Ф ° ° Ф ° Ф г ° \ å ~ ° 1 ° i ---I ° ° ° Ф ° ° ° ° 4 1 I ° ° ° ° ° ° ° ° Ф ° ° ~ 1 с; ° ° Ф / Г ° Ф с ° Ф O( Ъ I Ф Рис. 251. Строение порового аппарата клеток у дес- мидиевых водорослей: 1,2 — общие схемы основных типов порового аппарата в разре- зе (в в е р х у ) и с поверхности (в н и з у ) (1 — тип клостериу- ма, 2 — тип космариума); 8 — схематическое изображение электронно-микроскопической картины порового аппарата у Mic- rasterias rotata (поперечный разрез): а — поровые нити, б- конечные запонки, в — пор овая луковица, г — слизистые призмы. ки. Фибриллы этих оболочек пронизывают широкую часть канала, закрывая его своеоб- разным ситом, через которое может проходить клеточное вещество и выбрасываться наружу через узкую часть. Поровый аппарат типа кос- мариума характеризуется наличием свобод- ного сквозного канала (рис. 251, 2), обычно снабженного различными приспособлениями. Самое сложное строение порового аппарата встречается, например, в роде микрастериас (рис. 251, 8). Здесь настоящий поровый канал снабжен особыми п о р о вы ми нитями. Снизу он начинается на внутренней границе вторичной оболочки п о р о в о й л у к о- в и ч к о й, проходит через первичную оболоч- ку и внешний слой и заканчивается над внеш- ним слоем конечными з а п о н к а м и. Поро- вые нити — трубоподобные образования, ко- торые расширяются наружу в бокаловидные конечные запонки. Поровые луковички именует вид коротеньких гвоздиков с широкой шляп- кой, которые погружены своим острием в по- ровый канал: они сидят в поровом канале, как затвор умывальника в отверстии для стока. Это приспособление в паровом аппарате имеет большое значение. С его помощью поры по ме- ре надобности могут то открываться, то закры- ваться. В данном случае поровый механизм служит не только для выделения отбросов и
Рис. 252. Отложение железа на клетках (1 — 8) и сли- зистые покровы (4,5) у десмидиевых водорослей: 2 — Cosmarium quadrum; 2 — Xanthidium aculeatum; 8 — Stau- rastrum gladiosum; 4 — Staurastrum sonthalianum; 5 — Hyalothe- ca mucosa. поступления питательных веществ, но и пре- дохраняет клетку от нежелательной потери клеточного вещества, а также проникновения в клетку вредных веществ снаружи. Кроме пор, структура внешней оболочки десмидиевых может быть усложнена наличием особых скульптурных украшений: гранул, ямок, бородавок и шипов, большинство кото- рых расположены в определенном порядке. Они отдельно или в сочетании с особыми вы- пуклостями, буграми и отростками оболочки создают удивительные узоры, которые придают особую красоту и разнообразие десмидиевым. Эти скульптурные украшения являются важ- ными систематическими признаками при раз- личении таксономических единиц порядка. Внешний слой оболочки у многих десмиди- евых бывает пропитан соединениями железа, отложения которых располагаются у разных видов по-разному, в зависимости от их принад- лежности к тому или другому роду. Железо часто откладывается в местах скульптурных украшений и особенно часто в районе пере- шейка (рис. 252, 1 — 8). У отдельных видов железа откладывается так много, что оболочка их клеток приобретает желтый, ржавый или темно-коричневый цвет. Способность клеток десмидиевых к накоплению железа считает- ся хорошим систематическим признаком, но все же у некоторых родов оно до сих пор не обна- ружено. Внешний слой оболочки благодаря присут- ствию пектина и наличию пор часто образует толстый слизистый покров, полностью окуты- вающий особь или — у нитчатых форм — всю нить (рис. 252, 4, 5). С помощью этого слизи- стого покрова многие десмидиевые приклеи- ваются к высшим водным растениям или другим водорослям; другие же увеличивают свою пла- вучесть для парения в воде. У некоторых представителей клетки объеди- нены в нити посредством слизистых тяжей, проходящих между прилегающими верхушка- ми смежных клеток. При этом связь клеток оказывается настолько крепкой, что нити разрываются намного легче в районе перешей- ка клетки, чем в точках верхушечного при- крепления. Такое склеивание посредством сли- зи, выделяющейся между прилегающими вер- хушками клеток, иногда дополняется разви- тием верхушечных отростков оболочки клеток, как у видов родов сферозосла (Sphaerozosma) и тейликгия (Teilingia) (рис. 248, 4, 5). У колониального рода кослокладиул (Cos- mocladium) клетки удерживаются вместе с по- мощью слизистых стебельков, образуя неболь- шую, разветвленную, свободноплавающую ко- лонию (рис. 260, 1, 2). У рода оокардиул (Oocar- dium) колония имеет вид полушария, достигаю- щего в диаметре 1 — 2 лл и прикрепляющегося к известковым скалам. Она состоит из более или менее параллельных, расходящихся лу- чами слизистых нитей значительной толщины. Каждая нить расширяется по направлению к поверхности колонии и иногда разветвляется. На свободном конце каждой слизистой нити расположена одна клетка. Она размещена так, что ее продольная ось лежит перпендикулярно к оси слизистой нити (рис. 260, 8). У некоторых видов образуется очень плот- ная слизь, которая окружает клетку в виде футляра и служит защитой при неблагоприят- ных условиях: она предохраняет клетку от недостатка влаги и защищает ее от высыхания. Движение клеток. Десмидиевые водоросли обладают способностью двигаться с помощью особой слизи, которая выделяется через спе- циальные крупные поры, размещенные на концах клеток. Движение свойственно всем представителям десмидиевых, но особенно хо- рошо оно изучено у видов рода клостериум. Движение у них осуществляется таким обра- зом. Один конец клетки клостериума временно прикрепляется к субстрату, а другой конец выполняет ритмические колебательные движе- ния под углом в 180'. Через некоторое время свободный конец клетки прикрепляется к суб- страту, а колебательные движения начинает первый конец. Так, прикрепляясь то одним, то другим концом, клетка клостериума передви-
гается по субстрату. При температуре воды ~ ЗЗ'( смена позиции клетки происходит через — мин. у других родов также отмечено не- правильное, порой спазматическое движение, но иногда оно имеет плавный характер. Не- которые виды образуют слизистые «ножки», приподнимающие клетку над субстратом. усе десмидиевые положительно фототаксич- ны. В ясные солнечные дни они всегд.а соби- раются у поверхности воды. Если склянку с живыми десмидиевыми водорослями поставить на окно, то через некоторое время клетки скон- центрируются на освещенной стороне. ~~ротопласт. У десмидиевых протопласт за- полняет всю полость клетки. В цитоплазме находятся хлор опласты, ядро, вакуоли, кри- сталлики гипса и слизистые тельца. Симметрия очертаний клеток и украшений оболочки у десмидиевых отражается на сим- метрии протопласта. Симметрично располо- жены и хлоропласты, что является отличитель- Ф.У ным признаком десмидиевых водорослеи. Хлоропласты. У десмидиевых водорослей хлоропласты бывают двух типов: осевые (или центральные) и постенные. У большинства видов встречается осевой хлор опласт, причем почти всегда по одному в каждой полуклетке. Однако у нескольких очень мелких форм, при- надлежащих к роду космариум, и у некоторых других в центре каждой клетки имеется только один простой хлоропласт с одним пиреноидом. Хлор опласты имеют обычно густо-зеленый цвет и бывают очень разнообразной формы. Они могут иметь вид звездообразных тел (пениум), осевых конусов с радиальными пла- стинками (клостериум), стенкоположных лент (плеуротениум), осевых тел с радиальными лучами (космариум) и т. д. (рис. 253). Постенные хлор опласты встречаются реже, чем осевые, и преимущественно у более круп- ных форм. У некоторых представителей, осо- бенно из родов эуаструм и микрастериас, имею- щих осевые хлоропласты, наблюдается тенден- ция к развитию и постенных хлоропластов. Центральная часть хлор опласта становится у них более тонкой и исчезает совсем, тогда как периферическая часть развивается очень хорошо. Иногда цельные пластинки хлоро- пластов в периферической части клеток разде- ляются на дольки или многочисленные корот- кие отростки, направленные вперед к оболочке клеток. Благодаря этому площадь хлоропласта значительно увеличивается, что улучшает вос- приятие света клеткой. Величина и характер расчленения хлоропласта могут быть различ- ными у разных особей одного и того же вида в зависимости от условий их существования. При хорошем питании и освещении клеток хлоропласт хорошо развит и клетки окрашены Рис. 253. Хлоропласты десмидиевых водорослеи.' 1,2 — Penium spirostriolatum; 8,4 — Closterium ehrenbergii; 5 — Pleurotaenium trabecula; 6,7 — Cosmarium depressuni (2, 4, 7 — вид на поперечном разрезе). в темно-зеленый цвет. В неблагоприятных ус- ловиях (при недостаточном количестве пита- тельных веществ, при отсутствии или, наобо- рот, большом избытке какого-нибудь одного вещества) хлоропласты развиваются слабо, преимущественно у стенок клетки. В таком случае клетки имеют бледно-зеленый или жел- товатый цвет. На внешний вид хлоропласта сильно влияет также количество крахмала, содержащегося в нем в виде свободных крахмальных зерен. Их иногда хорошо видно в живых клетках, но еще легче можно обнаружить, если окрасить клетку раствором иода, который вызывает посинение крахмала. Крахмальные зерна об- разуются и располагаются в основном вокруг пиреноидов. Они могут равномерно покрывать поверхность пиреноида или же располагаться на некотором расстоянии друг от друга. Иног- да зерна крахмала возникают в строме хлоро- пласта. Если крахмала очень мало, то хлоро- пласт имеет тонкую и плотную структуру и кажется гладким. Иногда крахмала в хлоро- пласте бывает очень много, и в этих случаях хлоропласт становится массивнее, приобре- тает более грубую структуру и гранулирован- ный внешний вид. При некоторых условиях крахмальные зерна выбрасываются из пирено- идов в больших количествах и распространяют- ся по хлоропласту. Тогда хлоропласт в резуль- тате разбухания крахмальных зерен увеличи- вается в несколько раз, его структура нарушает-
Рис. 254. Схема вегетативного деления клетки десми- диевых водорослей: 1 — 5 — последовательные стадии деления. ся и он становится бесформенной массой. Это явление иногда вызывает замечательное раз- нообразие формы хлоропласта, особенно у ви- дов клостериума, у которых относительная величина центральных осевых и радиальных ребер хлоропласта в большой степени зависит от количества присутствующего крахмала. Ко- личество и положение хлоропластов имеют большое значение для определения некоторых видов и родов десмидиевых. Количество и положение пиреноидов сильно варьируют у разных представителей десмиди- евых и зависят от формы хлоропласта и усло- вий существования клетки. У представителей большинства видов, имеющих крупные постен- ные хлоропласты, пиреноиды разбросаны без определенного порядка, порой малозаметны. Если виды имеют осевой хлоропласт, пиренои- ды обычно находятся в тех частях клетки, в которых хлоропласт наиболее массивен. Ко- личество пиреноидов, внедренных в ось осево- го хлоропласта, зависит от его величины. У не- которых видов микрастериаса их может быть более ста, а у мелких видов обычно имеется только один пиреноид. В зависимости от массы накопленных питательных веществ количество пиреноидов изменяется. Ядро. Десмидиевые водоросли — одноядер- ные организмы. Только иногда при нарушени- ях процессов деления клетки образуются урод- ливые формы, которые обладают двумя и боль- шим количеством ядер. Ядро у большинства видов десмидиевых за- нимает центральное положение, располагаясь посередине между хлоропластами двух полу- клеток у неперешнурованных видов или в пере- шейке у пер ешнурованных видов. Величина ядра может колебаться от нескольких до 50 лки в поперечнике. Ядро может иметь шаровидно- эллипсоидную, квадратную или прямоугольную форму. Ядра содержат одно или несколько яд- рышек шаровидной формы. У разных видов число ядрышек и их величина зависят от вели- чины клетки. Вакуоли. Для протопласта десмидиевых ха- рактерны вакуоли. Обычно они содержат крис- таллики гипса или слизистые тельца пекти- новой природы. У тех форм, которые имеют постенные хлоропласты, эти вакуоли изредка размещены в цитоплазме, но у видов клостери- ума они могут оказаться в углублениях между продольными радиальными ребрами хлоро- пласта. Кроме того, здесь имеются также хо- рошо заметные вакуоли на концах клеток, содержащие, как правило, один или несколько кристалликов гипса. У видов космариума ва- куоли размещены между хлоропластами обеих полуклеток в области перешейка. При нормальных условиях число и располо- жение вакуолей у одного и того же вида бывают очень постоянными. В большинстве случаев в каждой полуклетке десмидиевых имеются одна крупная (основная) вакуоля и несколько более мелких вакуолей. Величина одних ва- куолей может увеличиваться за счет слияния их с другими. При некоторых ненормальных условиях жизни протоплазма клеток десми- диевых может образовывать многочисленные вакуоли, иногда очень значительных размеров. Вакуольная система десмидиевых послужила предметом многих исследований, но ее полное значение и возможная роль еще до сих пор не совсем ясны. Особенно разнообразные мнения были высказаны по поводу природы и назначе- ния кристаллов гипса в вакуолях на концах кле- ток. Эти кристаллы находятся в постоянном броу- новском движении. Число их в вакуолях зави- сит от возраста клеток и может изменяться под влиянием различных факторов окружающей среды. Иногда их бывает 1, 2, но не больше 10; в других случаях насчитывали более 40 кри- сталлов в каждой вакуоле. Во время деления клетки кристаллы гипса могут полностью ис- чезать. В вакуолях многих десмидиевых встречают- ся еще особые слизистые тельца, имеющие вид маленьких подвижных зерен. Эти тельца со- стоят из органического вещества, так как при нагревании до +64 'С, а также в кислотах и щелочах они растворяются. По-видимому, они представляют собой запасные питательные ве-
щества. связанные каким-то образом с выделе- нием слизи. Размножение десмидиевых осуществляется вегетативным и половым путем. Вегетативное размножение. Деление десми- диевых можно хорошо проследить на примере рода космариум. Сначала ядро, лежащее в рай- оне перешейка, делится, образуя два ядра, которые расходятся к центрам обеих полукле- ток. После того как ядра передвинулись в по- луклетки, в середине перешейка образуется перегородка, тесно примыкающая своими края- ми к внутреннему слою оболочки (рис. 254, 1). Затем область перешейка вытягивается, в результате чего две старые полуклетки от- деляются одна от другой (рис. 254, 2). Верх- няя и нижняя части перешейка, лежащие по обеим сторонам поперечной перегородки, на- чинают раздуваться в виде пузырька, увели- чиваться в размерах, после чего наступает рас- щепление самой перегородки (рис. 254, 8). Образовавшиеся из раздувшихся частей пере- шейка молодые полуклетки начинают быстро расти и достигают нормального облика, оста- ваясь соединенными друг с другом своими вершинами (рис. 254, 4, 5). Когда клетки до- стигают зрелого возраста, их оболочка, являю- щая собой временное образование, отбрасывает- ся и отпадает, наподобие линяющей кожи. Тогда клетки окончательно разъединяются и начинают самостоятельное существование. Таким образом, как уже упоминалось, в каж- дой клетке оказываются более старая и более молодая полуклетки. На самых ранних ста- диях развития содержимое молодых полукле- ток бывает совершенно прозрачным, поэтому клеточные ядра, лежащие в области перешей- ка, бывают хорошо видны даже без специально- го окрашивания. Потом протоплазма становит- ся густозернистой, заполняется вакуолями. Наконец, хлоропласты старых полуклеток удлиняются, делятся, и один из них переходит в молодую полуклетку. Иногда клетки после окончательного деления и завершения роста не распадаются, а образуют ложные нити, ко- торые, однако, легко разрываются. В некото- рых случаях ложные нити образуются путем сцепления верхушечных отростков и лопастей. У неперешнурованных форм, как, например, у представителей родов клостериум или пени- ум, клеточное деление происходит еще более сложным образом. Половое размножение. Как и у других пред- ставителей класса, конъюгация десмидиевых совершается безжгутиковыми апланогаметами, которые выполняют функцию половых клеток. Гаметы двух клеток сливаются, образуя зиго- ту. Зиготы известны для многих представителей десмидиевых, но подробности полового про- ° 4444 ° 4 4 4 е 4 е ъф4 444 ° 4 ° ° ° 44 4 4+ 4 4 % Ф Ю Ю O Ю O 4 Фр Ф ° 4 44 egь r Ф 3 ° ° ° ° ° ° ° ° ° ° ° ее ° ° ° ° ° Рис. 255. Различные стадии конъюгации у Cosmarium botrytis: 1 — ранняя стадия (одна клетка лежит боком, другая — в по- перечной плоскости, вид сверху); 2 — выработка копуляционного канала и начало копуляции; 8,4 — слияние содержимого обеих клеток; 5, 6 — формирование зиготы. цесса были зарегистрированы только у несколь- ких видов. Это связано с тем, что половой про- цесс у этих водорослей довольно редко встре- чается в природе, а культивирование их до сих пор было малоуспешным. У одного из видов космариума половой про- цесс начинается так: пары клеток с различным половым знаком располагаются рядом перпен- дикулярно одна к другой и выделяют слизь, которая их полностью окутывает (рис. 255, 1). Некоторое время спаренные клетки лежат рядом, но к началу слияния гамет отодвигают- ся друг от друга. Первым симптомом наступаю- щего слияния гамет является выработка копу- лирующими клетками цитоплазматических бу- горков. Бугорки увеличиваются, растут на- встречу друг другу и наконец соприкасаются, образуя копуляционный канал (рис. 255, 2). Весь процесс длится в течение 1 ч. Как только бугорки соприкоснулись, протопласты обеих клеток начинают отделяться от клеточных стенок и двигаться к общему центру в копуля- ционный канал (рис. 255, 8, 4). К этому времени в каждой клетке в районе перешейка образуется отверстие, которое расширяется при выходе гаметы. Отделение протопласта от стенок обычно продолжается 13 — 25 лик. Через 4 — 7 лик, после того как протопласты сопри- коснулись, слияние заканчивается и между пустыми оболочками образуется зигота не- 7
правильной формы (рис. 255, 5', 6). Иногда га- меты не отделяются полностью от оболочек клеток, и в таких случаях образовавшаяся зигота бывает прочно прикреплена к пустым оболочкам клеток. В первые минуты образования зиготы в ней появляются многочисленные сокращающиеся вакуоли. Маленькие вакуоли сливаются в более крупные. Каждая вакуоля пульсирует в тече- ние 4 — 7 мин, затем сжимается, и ее содержимое выходит наружу. Действие пульсирующих вакуолей длится от 4 до 32 мин, до тех пор пока прозрачные места зиготы перестают быть видимыми. За этот период (от 1 до 3,5 ч) зигота значительно сокращается. Внутри ее хлоро- пласты и пиреноиды располагаются в перифе- рическом слое. Затем начинают появляться первые признаки орнаментации зиготы — кону- совидные бугорки, разбросанные по ее поверх- ности. Постепенно они вытягиваются и приоб- ретают вид шипов. За время разрастания ши- пов, которое продолжается 2 — 3 ч, зигота опять увеличивается в размерах и становится такой же, как при слиянии гамет. До окончания роста шипов зигота сохра- няет цитоплазматическую оболочку. Когда шипы полностью сформируются, происходит быстрое изменение в их преломляющем ин- дексе, что говорит о том, что в это время обра- зуется новый слой оболочки. В зрелой зиготе оболочка состоит из трех слоев. Э к з о с п о р и й — это самый наружный, целлюлозный слой оболочки зиготы. Во многих случаях он бывает орнаментирован различ- ными скульптурными украшениями. Оконча- тельное формирование экзоспория происхо- дит за несколько дней. Вначале он имеет вид тонкой кожицы, затем на нем образуются вы- росты различной длины и формы, заполненные протоплазмой. В дальнейшем выросты увели- чиваются в размерах, уплотняются и превра- щаются в крепкие зубцы, шипы или развет- вленные отростки. Иногда экзоспорий бывает бородавчатым, ячеистым или пунктированным. У представителей некоторых родов, особенно стаураструм и микрастериас, экзоспорий бы- вает очень сложно и определенно орнаменти- рован, что во многих случаях является хоро- шим признаком при определении вида. Средний слой оболочки зиготы — м е з о с п о р и й- также состоит из целлюлозы, пропитанной пробковидным веществом. Он бывает окрашен в коричневый, красный или оливковый цвет. Самый внутренний слой оболочки зиготы на- зывают э н д о с п о р и е м. Он состоит из целлюлозы и обладает способностью сжимать- ся. Формирование мезоспория требует более длительного времени, чем формирование эк- зоспория. У представителей разных родов десмидиевых существует много мелких, но интересных раз- личий в процессе конъюгации и формирования зигот. Во многих случаях между копулирую- щими клетками возникает копуляционный канал. Это приводит к значительному разно- образию зигот по форме, строению и положению в клетках. Так, кроме зигот, сразу освобож- дающихся от оболочек образовавших их кле- ток (рис. 256, 1, 2), у некоторых десмидиевых возникшие зиготы захватывают часть пустых полуклеток, а их углы оказываются снабжен- ными роговидными выростами (рис. 256, 3). У части представителей нитчатых форм перед началом копуляции происходит распад нити на отдельные клетки. При этом у рода гиа- лотека (Hyalotheca) копуляционный канал со- храняется до окончательного сформирования зиготы. Он значительно расширяется от об- разовавшейся в нем зиготы, и на нем остаются прикрепленными четыре пустые оболочки по- луклеток (рис. 256, 4). У других нитчатых десмидиевых зиготы располагаются или между двумя рядами пустых полуклеток (рис. 256, б), или внутри одной из копулирующих клеток (рис. 256, 6). При этом наблюдается половая дифференциация. Одна из двух гамет — <о|уж- ская клетка» — постепенно переходит в копу- ляционный канал, а затем в другую — «жен- скую клетку». После слияния обеих гамет про- исходит развитие зиготы в «женской клетке». К ней прикреплены копуляционный канал и пустая оболочка «мужской клетки». У десмидиевых может происходить так>ке образование двойных зигот. Они развиваются при копуляции четырех клеток, возникших путем последовательного деления одной особи (рис. 256, 7). У видов клостериума способ об- разования двойных зигот иной: в каждой клет- ке сначала развиваются две особые гаметы, которые, сливаясь, дают двойные зиготы. Очень редко в образовании зигот десмидиевых при- нимают участие три или четыре особи. Конъюгация наиболее часто встречается у форм, имеющих наибольшие возможности для встречи физиологически разных клеток. Такая возможность чаще всего возникает у нитчатых и колониальных водорослей. Поэтому, напри- мер, представители гиалотеки и десмидиума часто встречаются в состоянии конъюгации. Однако у колониального космокладиума, где близость клеток, казалось бы, достаточно обес- печена общей колониальной слизью, конъюга- ция встречается редко. Вероятно, колонии ча- сто представляют здесь чистые клоны (клон— потомство одной клетки, образовавшееся путем вегетативного размножения), и тогда конъюга- ция становится невозможной из-за отсутствия физиологически разных клеток. 328
Рис. 256. Зиготы десмидиевых водорослей: 1 — Cosmarium portianum; 2 — Staurastrum hirsutum; 8 — Closterium swartzii; 6 — Bambusina brebissonii; 7 — Penium didymocarpum. kuetzingii; 4 — Hyalotheca dissiliens; 5 — Desmidium После слияния гамет и образования зиготы наступает длительный период покоя, который может продолжаться всю зиму и больше. В зиготе два из четырех или один из двух хло- ропластов разрушаются (в зависимости от того, сколько их в клетке). Ядра гамет лежат рядом во время периода покоя, а затем сливаются как раз перед прорастанием зиготы. Процесс про- растания зиготы удавалось наблюдать очень редко, и поэтому он еще слабо изучен. Прорастание зигот у десмидиевых происхо- дит весьма своеобразно. В отличие от других зеленых водорослей оболочка зиготы у них разрывается в самом начале прорастания, вы- свобождая в окружающую среду еще не разде- ленное содержимое. Освободившийся прото- пласт окружен только тонкой гиалиновой обо- лочкой, за которой следует тонкий слой, обра- зованный липидными телами. Большую часть протопласта занимают светлая вакуоля и два неслившихся хлоропласта. Далее в течение 1,5 — 2 ч оба хлоропласта постепенно продви- гаются к противоположным сторонам прото- пласта, а его общий объем уменьшается в свя- зи с постепенной потерей воды из вакуоли. Затем происходит деление диплоидного ядра. В результате двух последовательных делений, из которых первое является редукционным, образуются четыре гаплоидных дочерних ядра. Перед этим происходит разделение протопласта на две части. Обе части содержат по одному из первоначальных хлоропластов и по два сест- ринских ядра. Одно из ядер в каждом вновь сформированном протопласте отмирает, а дру- гое увеличивается в размерах. В результате образуются два проростка. В дальнейшем по- середине каждого проростка образуется пере- тяжка, типичная для большинства десмидие- вых водорослей. Оболочка проростка всегда гладкая; он лишен типичных видовых призна- ков. При первом же вегетативном делении про- росток приобретает типовые черты вида, обра- зовавшего зиготу. Два проростка, образован- ные одной зиготой, имеют, как правило, раз- личные половые знаки. Это говорит о том, что разделение полов у десмидиевых происходит при первом делении ядра. Однако у некоторых представителей число проростков достигает че- тырех, а иногда развивается всего один про- росток. У десмидиевых довольно часто встречаются межвидовые половые гибриды, особенно у ро- дов микрастериас и эуаструм, и в этом случае две зрелые полуклетки имеют признаки двух различных видов. В некоторых случаях при определенных усло- виях начавшийся процесс копуляции преры- вается. Тогда содержимое готовой к копуляции клетки округляется и окружается плотной обо- лочкой. Образуются так называемые п а р т е- н о с п о р ы, которые по своей форме и строе- нию очень напоминают зиготы. Однако, в отли-
чие от последних, они бывают значительно меньших размеров и имеют только одно ядро. Изредка встречается еще один вид спор- а п л а н о с п о р ы. Они, как и партеноспо- ры, образуются из всего содержимого клетки и покрываются особой плотной оболочкой, но возникают без всякого участия полового про- цесса, под воздействием резких колебаний тем- пературы или при внезапном и почти одновре- менном старении оболочки обеих полуклеток. У десмидиевых имеются еще так называемые х л а м и д о с п о р ы. Они представляют со- бой бесполые стадии, которые образуются при неблагоприятных для жизни водоросли усло- виях. При их образовании содержимое более старой полуклетки передвигается к границе между двумя пол уклетками, выделяет новую оболочку и отделяется от оболочки старой полу- клетки. Развивающаяся хламидоспора покры- вается толстой бесцветной оболочкой, состоя- щей из двух половинок. В благоприятных усло- виях хламидоспора делится и прорастает, обра- зуя вполне нормальные особи. В некоторых случаях при прорастании зиго- ты, а также при вегетативном делении клеток наблюдаются сильные отклонения формы кле- ток от нормального типа. В результате полу- чаются различные уродливые (т е р а т о л о- г и ч е с к и е) формы. Наблюдения тератоло- гических форм показало, что они могут возни- кать от различных причин. Так, при неполном клеточном делении происходит только деление ядра, а разделительная поперечная перегород- ка между полуклетками не образуется, в ре- зультате чего возникают уродливые клетки, состоящие из трех частей. Крайние части пред- ставляют собой нормальные полуклетки, а по- середине между ними находится уродливая вздутая часть различной формы. Неполное деле- ние встречается у большинства десмидиевых, особенно часто у родов космариум и клостериум. Особенностью некоторых видов является об- разование аномальных форм с неодинаковыми очертаниями вполне развившихся полуклеток и совершенно нормальной оболочкой. У рода клостериум, например, часто наблюдаются сиг- моидные формы, у которых одна полуклетка повернута к другой на 180'. Особенно большое разнообразие аномальных форм однажды удалось наблюдать у одного вида эуаструма (Euastrum didelta), собранного в вы- сокогорном болоте в Украинских Карпатах. Здесь встречались клетки с двойными и трой- ными полярными лопастями, а также с резкими различиями в морфологии материнской и до- черней полуклеток, которые заключались в не- одинаковом числе боковых извилин или в пол- ном отсутствии извилин и более упрощенной полярной лопасти. Иногда лопасти совершенно отсутствовали и одна из полуклеток имела округленные очертания, напоминающие полу- клетку космариума. Некоторые клетки вдоба- вок срастались между собой попарно в одном участке полуклетки, образуя что-то вроде «си- амских близнецов». Аномальные формы воз- никают, по-видимому, при неблагоприятных условиях существования клеток и особенно часто встречаются осенью, однако их наблю- дали и в летнее время — в июле и августе. Воз- можно, прямой причиной являются здесь рез- кие колебания температуры в течение суток, так как аномальные формы особенно обычны в полярных и высокогорных холодных обла- стях. Местообитание и распространение. Десмиди- евые — типично пресноводные организмы, морских видов среди этих водорослей нет. Прав- да, некоторые из них были найдены в слабосо- лоноватых водоемах морских побережий или в более соленых континентальных водоемах, однако существует мнение, что десмидиевые попадают туда случайно и через некоторое время погибают. Десмидиевые предпочитают открытую мест- ность, их редко можно увидеть в тенистых лесах и глубоких канавах. Встречаются они в неболь- ших водоемах и часто обильно населяют пруды, спокойные горные озера со скалистым дном и скалистыми берегами. В быстро текущей воде их, как правило, не обнаруживают. Живут они также в речках среди обрастаний, на влажных скалах, в гипновых и сфагновых болотах. Сфаг- новые болота — постоянные места обитания десмидиевых, однако здесь численность их за- висит от того, к какому типу относится болото. Огромное разнообразие форм десмидиевых встречается в сфагновых низинных болотах (эвтрофных и мезотрофных). Здесь они обра- зуют или слизистые скопления, свободно плавающие в понижениях между кочками, или бурые и зеленые пленки на листьях и стеблях мхов или на отмерших стеблях высших расте- ний. Иногда их можно обнаружить лишь на ощупь — по скользким, влажным стеблям и листьям сфагнума. Лучший способ сбора дес- мидиевых в таких случаях — это выжимание воды из мхов через планктонную сетку или без нее. Олиготрофные сфагновые болота характе- ризуются бедным качественным и количествен- ным составом десмидиевых водорослей. Растут десмидиевые главным образом в мяг- ких водах с низким рН и при наличии гумино- вых кислот (в сфагновых низинных болотах), но некоторые виды клостериума и стаурастру- ма обитают в нейтральных и даже щелочных водах. Большинство видов десмидиевых пред- почитают чистые воды, но ряд представителей родов клостериум, космариум и стаураструм
встречаются и в сильно загрязненных местах, в сточных водах, развиваясь в некоторых слу- чаях в массовом количестве. Некоторые из дес- мидиевых ведут аэрофитный образ жизни, посе- ляясь на влажных скалах, на почве среди мхов и нитчаток, в подушках сфагновых мхов на бо- лотах. Имеются сведения о нахождении десми- диевых в почвах. Большинство десмидиевых — бентосные ор- ганизмы, однако некоторые виды родов стау- раструм, ксантидиум и микрастериас хорошо приспособлены к планктонному существованию благодаря хорошей плавучести за счет развития радиальных полых отростков и шипов и слизи- стых покровов. Десмидиевые встречаются на всех глубинах, где есть подходящий химиче- ский состав воды и достаточное освещение. Распространены десмидиевые не только на равнинах, но и в горах. Их можно встретить на высоте до 3100 м над уровнем моря. В север- ных районах СССР десмидиевые распростране- ны на равнинных местах, богатых торфяными болотами и озерами. В степной полосе количест- во десмидиевых сильно уменьшается, а в пу- стынях их очень мало. Десмидиевые весьма чувствительны к окру- жающим условиям. Главным фактором, опреде- ляющим их распространение и количество, является геологическая природа местности. Об- ласти распространения гранитов, как правило, отличаются большим разнообразием форм этих водорослей; области распространения извест- няков, напротив, очень бедны ими, хотя от- дельные виды десмидиевых селятся в водоемах с большим количеством извести (например, некоторые виды рода оокардиум). Особенно бо- гаты десмидиевыми те области, где подпочва образована палеозойскими и докембрийскими отложениями. Состав почвы отражается на зна- чении рН среды; наибольшее количество видов встречается при рН ниже 7. Осадки также играют немаловажную роль в распространении этих водорослей. Обильная флора десмидиевых развивается там, где BbI- падает большое количество дождей или грун- товые воды подходят близко к поверхности почвы. Десмидневые встречаются по всему миру. Существует мнение, что примерно две трети видов этих водорослей являются космополита- ми или широко распространенными видами. И все же многие виды десмидиевых населяют лишь определенные географические районы и могут быть оценены как эндемики. В тропиках встречаются ряд видов и даже родов, которые до сих пор не обнаружены в других частях земного шара. Собирать десмидиевые лучше всего в ясную солнечную погоду. В небольших неглубоких водоемах их собирают с поверхности воды при помощи небольшого сачка из плотной шелко- вой ткани (лучше всего густое мельничное сито, употребляемое для планктонных сеток). Слизь, которая остается на сите, затем осто- рожно смывают в склянку. Можно просто за- черпывать склянкой или баночкой воду в по- нижениях между кочками или подушками сфагнума. Иногда десмидиевые развиваются на погруженных в воду растениях в таком ко- личестве, что буроватую слизь, в которую они внедрены, можно собрать просто руками. Если слизь очень нежная, то ее можно извлечь из воды, осторожно опустив в нее руку ладонью кверху так, чтобы листья или стебли растения могли проходить между пальцами и как можно ближе к ладони. Сомкнув пальцы и держа ладонь согнутой, поднимают осторожно руку к поверхности, тогда слизь со стеблей скапли- вается на ладони, откуда ее нужно немедленно перелить в склянку. Можно также выжимать эти растения над широким горлышком склянки или соскабливать с них слизь с помощью лож- ки или перочинного ножа. Для сбора планк- тонного материала из больших озер нужны конусообразные планктонные сетки из мель- ничного газа, через которые любым способом фильтруют большое количество воды. Со дна водоемов десмидиевых лучше всего собирать специальным стаканчиком или склянкой. Та- кой материал обычно загрязнен песком и илом, поэтому его полезно помещать в плоскую по- суду и оставлять при. хорошем освещении. Ie- рез некоторое время десмидиевые появятся на поверхности в виде слизистой массы, кото- рую легко удалить с помощью небольшой пипетки. Систематическое положение десмидиевых сре- ди других водорослей сейчас прочно установ- лено, хотя по этому поводу существовали раз- личные, иногда противоречивые мнения. Сна- чала десмидиевых относили к простейшим жи- вотным. Позднее, когда они были причислены к растениям, возникли разные мнения по по- воду их места в общей системе водорослей. Не- которые исследователи объединяли десмидие- вых вместе с диатомовыми, основываясь на сходстве их полового процесса и строения обо- лочки. Однако это объединение было построено на очень поверхностном суждении, так как структура оболочек и их скульптура, поровый аппарат, хлоропласты и пигменты совершенно различны в обеих группах. Иногда десмидие- вых вместе с зигнемовыми считали изолирован- ной группой среди зеленых водорослей и отде- ляли от них совершенно. Сейчас большинство альгологов относят десмидиевых к конъюга- там, которые являются признанной составной частью зеленых водорослей, так как пигменты
Рис. 257. Виды пениума и клостериума: Я — Penium polymorphum; 2 — P. spirostriolatum; 8 — Closteri- um lunula; 4 — С. acerosum; 5 — С. leibleinii. и продукты метаболизма у них в основном те же самые. Десмидиевые входят в состав класса конъю- гат как один из порядков. Положение их внут- ри класса определяется родственными связями с другими его порядками. Существуют две точки зрения на происхож- дение десмидиевых. Согласно одной из них, одноклеточные формы произошли от нитчатых предков в результате их распада на отдельные клетки. Если принять это предположение, то, во-первых, следует признать близкие родст- венные связи десмидиевых с зигнемовыми, а во-вторых, в этом случае приходится счи- тать их более примитивной и вырождающейся группой среди конъюгат, имеющей первона- чальную ретрогрессию от нитчатых форм. Одна- ко против сближения десмидиевых с зигнемо- выми говорят существенные различия в органи- зации их клеток. Нельзя признать и примитив- ность десмидиевых, так как структура их кле- точной оболочки и строение порового аппарата свидетельствуют об их сложности и высокой специализации. Другая точка зрения заключается в том, что нитчатые формы произошли от одноклеточных форм, имеющих простую организацию клетки. Это мнение разделяют большинство альголо- гов. Согласно такому взгляду, наиболее при- митивными среди конъюгат являются мезоте- ниевые водоросли. От них берут начало две линии развития. Одна линия привела к гонато- зиговым и далее к зигнемовым, другая — к вы- сокоразвитым десмидиевым. Самыми примитив- ными среди десмидиевых являются те предста- вители, которые характеризуются цилиндриче- скими, радиально-симметричными клетками с усеченными или округленными верхушками и отсутствием срединной перетяжки. Такие клетки встречаются у представителей рода пе- ниум, отчасти клостериум и плеуротениум. Некоторые виды рода пениум очень близки по своему строению к представителям мезоте- ниевых. Дальнейшая эволюция десмидиевых шла в направлении усложнения формы клеток и структуры их оболочки, а также усложнения и увеличения разнообразия их хлоропластов. До настоящего времени систематика десми- диевых основывается главным образом на мор- фологических и в некоторой мере на цитологи- ческих признаках. Для разграничения родов десмидиевых широко используют форму клет- ки, ее симметрию и орнаментацию. Отдельные роды десмидиевых легко отличаются один от другого, но все же границы многих родов очень неопределенны и расплывчаты. Ни в одной другой группе водорослей нет такого большого количества переходных форм, как у десмидие- вых. В связи с этим систематика десмидиевых
очень запутана и несовершенна. До сих пор в пределах этого класса существуют искусст- венные пол иморфные роды, как, например, космариум (около 2000 видов) и стаураструм (около 1500 видов). Основываясь на форме и структуре клеток, структуре клеточной оболочки и строении по- рового аппарата с учетом данных электронно- микроскопического изучения, мы разделяем порядок десмидиевых на три семейства: пение- вых (Peniaceae), клостериевых (Closteriaceae) и космариевых (Cosmariaceae). Семейство пениевые (Peniaceae) Семейство пениевых содержит только один род пениум (Penium). Клетки у этих водорослей одиночные, прямые, цилиндрические или эл- липсоидные, с округленной или срезанной вер- хушкой, слабо перетянутые или без перетяжки, сверху круглые. Оболочка бывает гладкая, пунктированная, продольно или спирально по- штрихованная, иногда гранулированная, бес- цветная или окрашенная, без пор. Хлоропла- сты осевые, с радиальными пластинками, по од- ному-два в каждой полуклетке. Пирея оиды одиночные, иногда по два в клетке. У некото- рых видов имеются терминальные ваку-оли с движущимися кристаллами гипса. Зиготы шаровидные, эллипсоидные, четырехугольные или неправильных очертаний, образуются меж- ду двумя копулирующими клетками. Для СССР известно 10 видов. Наиболее широко распрост- ранены, например, Penium polymorphum, Р. spi- rostriolatum (рис. 257, 1, 2). Семейство клостериевые (Closteriaceae) Эта группа также содержит только один род клостериум (Closterium). Клетки у его пред- ставителей одиночные, редко склеивающиеся в пучки, обычно серповидно-согнутые, реже прямые, сильно вытянутые и заметно суживаю- щиеся на концах, без срединной перетяжки, на поперечном сечении круглые. Деление всег- да происходит в средней части клетки. Ряд ви- дов образует пояски. Оболочка гладкая или поштрихованная. Поры всегда присутствуют; они бывают разнообразной формы — круглые, овальные или продолговатые. Поровый аппарат состоит из порового канала обратноворонкооб- разной формы, с ситовидным основанием, рас- положенным в фибриллярном слое оболочки, и порового отверстия во внешнем слое оболочки. Хлоропласты осевые, с разным количеством продольных ребер, по одному, реже по два в каждой полуклетке. Пиреноиды шаровидные, эллипсоидные, по 1 — 2 — 20 в клетке; они рас- положены в один осевой ряд или разбросаны беспорядочно по всему хлоропласту. Терми- нальные вакуоли постоянно присутствуют, по одной, реже по две-три на каждом конце клет- ки, с одним-двумя или несколькими движущи- мися кристалликами гипса. Зиготы различной величины и формы — шаровидные, эллипсоид- ные, почти треугольные или четырехугольные, с гладкой или окрашенной оболочкой. В СССР встречается 77 видов. Среди самых распростра- ненных могут быть названы Closterium acero- sum, Cl. leibleinii, Cl. 1ипи1а (рис. 257, 8 — о). Семейство космариевые (Cosmariaceae) Эта группа включает много родов, имеющих клетки очень разнообразной формы и величины, глубоко или, реже, слабо перетянутые, одиноч- ные или соединенные в колонии или нити. Сверху они также имеют очень разнообразную форму и могут быть эллиптическими, ромбовид- ными, круглыми, многоугольными, двух- или многорадиальными. Деление клеток происхо- дит в районе перешейка. Оболочка всегда про- низана порами. Поровый аппарат нередко имеет сложное строение и состоит из порового канала, поровых нитей, поровой луковицы и поровых запонок. Из рода дог~идиул (Docidium) в СССР извест- но несколько видов. Клетки у представителей этого рода вытянутые, прямые, почти цилинд- рические, слабо перетянутые посередине, свер- ху круглые, с волнистым краем. Полуклетки с ровными или волнистыми боковыми сторо- нами, у основания несколько вздутые и снаб- женные венчиком из продольных складочек оболочки, имеющих вид бугорков. Верхушки полуклеток срезанные, без скульптурных укра- шений. Оболочка гладкая, пунктированная или слегка поштрихованная. Хлоропласты осевые, с несколькими неправильными продольными ребрами. Пиреноиды располагаются вдоль оси, число их варьирует, иногда их бывает более 15. Терминальные вакуоли отсутствуют. Зиготы неизвестны. Наиболее распространен Docidium baculum, обычен и D. undulatum (рис. 248, 2). У водорослей из рода плеуротениум (Pleuro- taenium) клетки прямые, удлиненно-цилиндри- ческие, слабо перетянутые посередине, в райо- не перетяжки, у основания полуклетки, они имеют кол ьцеоб разный шов, сверху бывают круглые или звездчато-лопастные. Полуклетки с хорошо заметным базальным вздутием, кото- рое, в отличие от предыдущего рода, никогда не бывает складчатым, и ровными, прямыми или волнистыми боками. Верхушки полуклеток срезанные или притупленно-округленные, глад- кие или снабженные венчиком из гранул или шипов. Оболочка клеток гладкая, пунктиро- ззз
Рис. 258. Десмидиевые: 2 — Pleurotaenium minutum; 3 — Triploceras gracile, часть клет- ки; 8 — Tetmemorus brebissonii; 4 — Т. granulatus. ванная, ямчатая, гранулированная. В каждой полуклетке по одному хлоропласту. Они ленто- видно-постенные, с разбросанными пиреноида- ми. Хорошо заметные терминальные вакуоли содержат кристаллики гипса. Зиготы шаровид- ные, широкоэллипсоидные, с гладкой или по- крытой коническими шипами оболочкой. В СССР известно около 10 видов. Одним из наиболее распространенных является Pleuro- taenium minutum (рис. 258, 1). У представителей рода триплоуерас (Triplo- ceras) клетки вытянутые, умеренно перетяну- тые, прямые, изредка слегка согнутые, с вол- нистыми или гладкими боковыми сторонами, на концах переходящие в различной длины отростки, которые заканчиваются заостренны- ми шипиками. Оболочка гладкая или несет многочисленные кольца шипов или выростов. Хлоропласты осевые, по одному в каждой полу- клетке, с несколькими продольными ребрами и пиреноидами, расположенными вдоль оси. 3иготы неизвестны. В нашей стране известно всего два вида, из них более распространен Triploceras gracile (рис. 258, 2). У рода тетмеморус (Tetmemorus) клетки бывают прямые, удлиненные, почти цилиндри- ческие, веретеновидные или эллипсоидные, сла- бо перетянутые, сверху круглые или широко- эллипсоидные. Полуклетки постепенно сужи- ваются от основания к верхушке, которая обычно округленная и всегда с глубоким узким срединным вырезом. Оболочка пунктированная, поштрихованная или гранулированная, на вер- хушках утолщенная. Хлоропласты осевые, со- стоящие из узкой центральной части и боко- вых продольных ребер или пластинок, по одно- му в каждой полуклетке. Пиреноиды распола- гаются вдоль оси или по одному в центре хло- ропласта. Зиготы шаровидные или четырех- угольные, развиваются между копулирующими клетками. В нашей стране встречаются три вида, из них обычен Tetmemorus brebissonii (рис. 258, 8, 4). У водорослей из рода актинотениум (Actino- taenium) клетки прямые, цилиндрические, ве- ретеновидные или гитаровидные, слабо пере- тянутые, сверху всегда круглые. Полуклетки с более или менее широкоокругленными, вы- пуклыми или срезанными верхушками. Обо- лочка гладкая или ямчатая, иногда утолщен- ная на верхушке с внутренней стороны или неж- но гранулированная на верхушках полуклеток, пронизанная неправильно рассеянными или рас- положенными в определенном порядке крупными или мелкими порами. Хлоропласты звездообраз- ные, пластинчато-звездообразные или ленточно- постенные, с одним-двумя центральными пире- ноидами, реже со многими пиреноидами в каждой ленте постенного хлоропласта. Зиготы шаро- видные, эллипсоидные, угловатые, квадратные, реже двойные, с гладкими или бородавчатыми стенками. В СССР известно около 30 видов. Одним из наиболее распространенных является Actino- taenium cucurbitinum (рис. 248, 1). У рода эуаструм (Euastrum) клетки относи- тельно короткие, обычно заметно сжатые, раз- нообразной формы и величины, глубоко пере- тянутые, с уз колинейным закрытым синусом, слегка раскрывающимся в наружной части, эллиптические или узкоэллиптические. Полу- клетки трехлопастные, имеющие одну поляр- ную лопасть с более или менее узким средин- ным вырезом на верхушке и две боковые лопа- сти, которые бывают цельными или делятся на лопасти второго порядка. В центре каждой полуклетки, а иногда и в углах лопастей име- ются симметрично расположенные вздутия раз- личной величины, часто орнаментированные гранулами или бородавками. Сверху клетки узкоэллиптические, со вздутиями на боковых сторонах или полюсах. Оболочка гладкая, пунктированная, гранулированная или мелко- бородавчатая. Хлоропласты осевые, по одному, редко по два в каждой полуклетке, с одним или мно- гочисленными пиреноидами. Зиготы шаро- видные, редко эллипсоидные, покрыты креп- кими, конусовидными, иногда изогнутыми от- ростками, простыми или разветвленными, ту- пыми или заостренными на концах.
В СССР известно 57 видов. Примерами могут служить Euastrum verrucosum (табл. 37, о) и Е. divaricatum (рис. 249, 2). К роду микрастериас (Micrasterias) относятся наиболее высоко дифференцированные и кра- сивые виды десмидиевых. Клетки у них часто большие, плоские, в некоторых случаях почти дисковидные, круглые или широкоэллиптиче- ские, глубоко перетянутые. Синус узколиней- ный по всей длине или более или менее сильно открывающийся наружу. Полуклетки разде- лены глубокими вырезами на три лопасти: по- лярную и две боковые. Полярная лопасть бы- вает очень разнообразной формы и величины; обычно она возвышается над боковыми, отде- ляясь от них широко открытыми вырезами, и имеет узко- или широковыемчатую верхушку, часто снабженную шипами. Боковые лопасти бывают цельные или разделенные глубокими вырезами на лопасти второго, третьего, четвер- того, пятого и шестого порядков; конечные ло- пасти обычно заканчиваются шипиками. Сверху клетки вытянуто-эллиптические, узкоромбо- видные, часто со слабыми вздутиями на боко- вых сторонах. Оболочка пунктированная, гранулированная или шиповатая. Хлоропласты осевые, по одно- му в полуклетке, с изрезанными краями и мно- гочисленными пиреноидами. Зиготы шаровид- ные, эллипсоидные, покрытые крепкими ши- пами. В нашей стране встречается 21 вид. Хоро- шими примерами могут служить Micrasterias conferva (табл. 37, 4), М. rotata (рис. 249, 1). У представителей рода космариум (Cosmari- ит) клетки относительно короткие, округлен- ных очертаний или правильно угловатые, удли- ненные или укороченные, более или менее глу- боко перетянутые. Синус различной глубины и формы. Полуклетки не разделены на лопасти и не имеют верхушечного выреза, очень разно- образных очертаний: сверху эллиптические, ромбические или круглые, часто с одним-двумя срединными вздутиями. Оболочка гладкая или пунктированная, ямчатая, гранулированная, бородавчатая или покрытая сосочками. Хлоро- пласты обычно осевые, по одному, редко по два в каждой полуклетке, с одним пиреноидом; реже хлоропласты бывают постенные, состоя- щие из узких лентовидных полос, в каждой из которых находится по одному пиреноиду. Зиготы шаровидные, зллипсоидные, кубиче- ские, покрытые ямками, сосочками, простыми или разветвленными шипами. В СССР встречается около 500 видов, многие из них широко распространены, например Cos- marium subtumidum, С. reniforme, С. quinarium (табл. 37, 1 — д), С. caelatum, С. protuberans (рис. 249, 8). У рода ксантидиум (Xanthidium) клетки удлиненные, всегда сжатые, глубоко перетяну- тые. Полуклетки разнообразных очертаний: эллиптические, эллиптическ и-шестиугольные, трапециевидные или многоугольные, всегда с плоской верхушкой; в центре полуклетки имеется утолщение, часто ямчатое, окрашенное в желтый или коричневый цвет, или выступаю- щий бугор, покрытый гранулами, шипами или зубцами. Сверху клетки эллиптические, с буг- рами посередине каждого бока. Оболочка глад- кая или пунктированная, снабженная на углах полуклеток парными или одиночными шипами, иногда шипы имеются по всей оболочке или в центре полуклеток. Хлоропласты постенные, по четыре в полуклетке, с одним или несколь- кими пиреноидами; иногда они бывают звездо- образные, с одним пиреноидом. Зиготы шаро- видные, обычно с простыми или вильчатыми шипами. В СССР известно более 20 видов. Наиболее широко распространен Xanthidium antilopaeum (рис. 259, 1). У представителей рода стауродесмус (Stauro- desmus) клетки относительно короткие, четы- рехугольных очертаний, большей частью глу- боко перетянутые. Синус выемчатый или тре- угольный, с острой или тупой верхушкой, от- крытый. Перешеек часто удлиненный. Полу- нлетки широкоэллиптические, круглые, вере- тенообразные, эллипсоидные, но чаще всего имеют вид срезанной пирамиды; верхние или боковые углы их снабжены одиночными длин- ными или короткими шипами, которые могут быть параллельными, расходящимися или схо- дящимися, иногда шипы в виде маленьких со- сочков на верхних углах. Сверху клетки эллип- тические или 3 — 5-угольные. Оболочка у них гладкая, почти лишенная орнаментации (кроме простых радиальных шипов), пронизанная очень мелкими порами или выступающими по- рами-органеллами, иногда оболочка бывает ям- чатая. Хлоропласты осевые, по одному в полу- клетке, каждый с одним пиреноидом. Зиготы шаровидные, гладкие или снабжены шипами. В нашей стране известно около 50 видов. Наиболее распространенными являются Stau- rodesmus incus, S. convergens (рис. 259, 2, 8) и некоторые другие. У водорослей из рода стаураструм (Staurast- rum) клетки очень разнообразной формы, со сложной структурой, обычно глубоко пере- тянутые. Синус чаще широко открытый, с ост- рой верхушкой. Полуклетки полукруглые, эл- липтические, продолговатые, чашевидные, тре- угольные, углы округленные, острые или вытя- нутые в более или менее длинные отростки или шипы. Сверху клетки трех-, многоугольные, ред- 335
Рис. 259. Десмидиевые: 1 — Xanthidium antilopaeum; 3 — Staurodesmus incus; 8 — S. con- vergens. ко веретенообразные. Оболочка гладкая или шиповатая. Хлоропласты осевые, по одному в каждой полуклетке, с одним центральным пиреноидом, иногда постенные. Зиготы шаро- видные или угловатые, снабженные простыми или вильчатыми шипами. В СССР известно более 200 видов. Многие широко распространены, например: Staurast- rum pelagicum, S. cyclacanthum (рис. 250, 1, 2). У представителей рода космокладиум (Cos- mocladium) клетки соединены в колонии по- средством простых или двойных слизистых не- правильно ветвящихся нитей. Обычно они сво- бодно плавают, но иногда прикрепляются к дру- гим водорослям. Клетки космариевидные, мел- кие, обычно сжатые, симметричные в трех на- правлениях, глубоко перетянутые. Полуклетки почти пирамидальные, эллиптические или поч- ти почковидные, с широким перешейком и уз- ким синусом, сверху эллиптические. О болочка у них гладкая. Хлоропласты осевые, обычно со- держатся по одному в полуклетке. Зиготы более или менее шарообразные, но иногда угловатые, снабженные короткими толстыми и тупыми ши- пами. У нас известно 5 видов. Наиболее часто встре- чаются Cosmocladium saxonicum и С. pusillum (рис. 260, 1, 2). У рода оокардиум (Oocardium) клетки собра- ны в шарообразные колонии диаметром 1— 2 лл. Колонии состоят из радиальных слизи- стых тяжей, инкрустированных известью. Каж- дая нить расширяется к поверхности колонии и иногда разветвляется. На свободном конце каждой нити расположена одна клеткаперпен- дикулярно к оси нити. Клетки космариевидные, мелкие, сжатые, значительно укороченные, сла- бо перетянутые. Оболочка гладкая. Хлоропла- сты осевые, по одному в полуклетке, каждый хлоропласт содержит один пиреноид. Зиготы угловатые, с несколькими сосочковидными гор- биками. Известен всего один вид — Оосагйит stra- tum (рис. 260, 8). У водорослей из рода сферозосла (Sphaero- zosma) клетки собраны в однорядные, иногда скрученные слизистые нити. Обычно они очень мелкие, эллиптические или четырехугольные, часто укороченные, более или менее сжатые, всегда перетянутые посередине, с хорошо обо- значенным перешейком, с боковыми шипами или без них. Синус различной глубины, закры- тый или открытый. На верхушках полуклеток имеется по два небольших, веретенообразных, головчатых отростка, расположенных диаго- нально и плотно крестообразно прилегающих к соответствующим отросткам соседней клетки. Сверху клетки эллиптические. Оболочка у них гладкая, иногда крупнопористая. Хлоропласты осевые, по одному в полуклетке, с одним цент- ральным пир еноидом. Зиготы круглые, четырехугольные или про- долговатые, гладкие или украшенные просты- ми шипами. В СССР известно 5 видов, например Sphaero- zosma filiformis (рис. 248, 4). Род тейлингия (Teilingia) имеет клетки, со- бранные в однорядные, ровные или слегка скру- ченные слизистые нити. Часто клетки удлинен- ные или одинаковые в длину и ширину, более или менее сжатые, эллиптические или четырех- угольные. Синус довольно глубокий, выемча- тый, открытый, с округленной верхушкой. Вер- хушка каждой полуклетки снабжена четырьмя гранулами. Сверху клетки эллиптические. Обо- лочка гладкая или покрыта немногочисленными гранулами. Хлоропласты осевые, с одним цент- ральным пиреноидом. Зиготы шаровидные, эл- липтические, кубические, гладкие или шипова- тые. У нас известно 4 вида, в том числе Teilingia granulata (рис. 248, Б). У водорослей из рода спондилозиум (Spondi- losium) клетки соединены верхушками в длин- ные нити посредством слизистых подушечек. Обычно они небольших размеров, сжатые, всег- да более или менее глубоко перетянутые, с узким, закрытым или открытым синусом раз- нообразной формы. Полуклетки эллиптические, аирамидальные или четырехугольные, часто
Та б л и ц а 33. Сифоновые водоросли: 1 — кодиум (Codjum edule); 2 — халимеда (Haljmeda opuntja); 3 — неомерис (Neomerjs annulata).
Ъ Та б л и ц а 34. Ископаемые представители дазикладовых водорослей (реконструкция): — фо мы с п остыми или разветвленными боковыми выростами, расиоложенными спирально; 2, 3 — форма на попе- стами и с истами асположенными в главной речном и пр родольном срезах с мутовчатыми разветвленными боковыми выро Ц р е вого по я ка на оси, 4, 5 — формы с мутовчато р т расположенными разветвленными выростами и с цистами в выростах п р р д ими по обие ко ы. продольном срезе;,, — р л ; 7, 8, 9 — аз ичные формы с простыми и разветвленными выростами, образующими подо ие коры.
° в в в у в э ° Та б л и ц а 35. Мезотеииевые и гонатозиговые водоросли: 1 — Ancylonema nordenskioeldii, вегетативные клетки; 2 — Spirotaenia erithrocephala, последовательные стадии деления вегетативной клетки; 3 — Roya cambrica, вегетативная клетка; 4 — Cylindrocystis crassa, кон ьюгация; 5 — С. brebissonii, зигота; о — Netrium digitus, вегетативная клетка; 7, 8 — Gonatozygon monotaenium, часть нити и зигота; 9 — Genicularia spirotaenia, часть нити.
Та б л и ц а 36. Зигнемовые водоросли: ° ° ° ° 1 — общии вид живых нитеи в натуральную величину; 2 — вегетативная нить м жоции 3 — 7 — конъюг зигот у Mougeotia gelatinosa (3), Z nemo. s nadel hum ~4) Те п у оции; — — конъюгация и образование ygnema, syna e р ит ( ), Temnogametum mayyanadense (5), Sirogonium megasporum (б),
Та б л и ц а 37. Яесмидиевые водоросли: 1 — Cosmarium subtumidum ( X 1350); 2 — С. reniforme ( X 900); 3 — С. quinarium ( X 900); 4 — Micrasterias conferta ( Х 300); 5 — Euastrum verrucosum.
Та б л и ц а 38. Харовые водоросли, части талломов и ооспоры: 1 — Nitella mucronata; 2 — Tolypella prolifera; 3 — Nitellopsis obtusa; 4 — Chara vulgaris.
Та б л и ц а 39. Промысловые бурые и красные водоросли: 1 — ламинария японская (Laminaria japonica); 2 — алария съедобная (Alaria esculenta); 3 — ундария перистонадрезная (Undaria pinnatifida); 4 — гелидиум тонкий (Gelidium tenue); 5 — анфельция складчатая (Ahnfeltia plicata); g — филло- фора жилковатая (Phyllophora nervosa).
7 ц б л и ц ц 4О. Комплекс установок для массового полупроизводственного культивирования одноклеточных водорослей (Япония).
Рис. 260. Десмидиевые: 2 — Cosmocladium saxonicum; 2 — С. pusillum; 8 — Oocardium stratum. Рис. 261. Десмидиевые: 2 — Spondilosium pulchellum; 2 — Hyalotheca dissiliens; 8— Desmidium swartzii; 4 — Bambuzina brebissonii. с широко срезанной или вогнутой верхушкой, сверху эллиптические, треугольные или трех- лопастные. Оболочка гладкая или гранулиро- ванная. Хлоропласты осевые, по одному в полу- клетке, с одним центральным пиреноидом или несколькими пиреноидами, расположенными в один ряд. Зиготы обычно шаровидные, глад- кие или шиповатые. В СССР известно более 10 видов, чаще дру- гих встречается Spondilosium pulchellum (рис. 261, 1). У рода гиалотека (Hyalotheca) клетки со- единены в однорядные, иногда скрученные нити, заключенные в толстую слизистую обвертку, исчерченную короткими поперечными бороздка- ми. Клетки цилиндрические, очень слабо пере- тянутые или без перетяжки. Полуклетки тра- пециевидные, почти квадратные или продол- говатые, с прямыми или слаб овыпуклыми боками. Сверху клетки круглые или почти круглые. Оболочка гладкая или гранулиро- ванная. Хлоропласты осевые, звездообразные, с одним центральным пиреноидом. Зиготы шаровидные, гладкие, иногда заключенные в особые образования неправильных очертаний, состоящие из гаметангия, соединенного с ши- рокой конъюгационной трубкой. В СССР известно более 6 видов. Наиболее широко распространена Hyalotheca dissiliens (рис. 261, 2). У видов рода десмидиум (Desmidium) клетки соединены в удлиненно-скрученные нити по- средством греб невидных утолщений или усе- ченных отростков, иногда они бывают заклю- чены в толстую слизистую обвертку. Клетки четырехугольные в очертаниях, обычно укоро- ченные, часто сильно сжатые, умеренно перетя- нутые, с более или менее хорошо заметным си- нусом. Сверху клетки эллиптические, треуголь- ные или четырехугольные. Оболочка гладкая; в каждой полуклетке под перешейком имеется один поперечный ряд ямок. Хлоропласты осе- вые, звездообразные, по одному в полуклетке, с массивными, радиально отходящими от цент- ра лопастями и с одним пиреноидом в центре или в каждой лопасти. Зиготы шаровидные или эллипсоидные, глад- кие или с короткими плоскими коническими сосочками. В нашей стране известно около 10 видов. Наиболее широко распространен Desmidium swartzii (рис. 261, 8). У водорослей из рода бамбузина (Bambuzina) клетки, соединенные в слабо скрученные нити, имеют цилиндрическую или бочонкообразную форму, они очень слабо перетянутые, часто вздутые и волнистые в основании каждой полу- клетки над синусом, сверху круглые, часто с двумя противоположными сосочками. Хлоро- пласты осевые, с шестью радиальными пластин- ками, по одному в каждой полуклетке. Зиготы почти шаровидные, овальные, гладкие. В СССР известно 3 вида. Широко распростра- нена Bambuzina brebissonii (рис. 261, 4).
ОТДЕЛ ХАРОВЫЕ ВОДОРОСЛИ (CHAROPHYTA) 338 Харовые водоросли, или, как их еще назы- вают, харофиты, или лучицы, представляют собой совершенно своеобразные крупные расте- ния, резко отличающиеся от всех остальных водорослей. При беглом взгляде они скорее похожи на некоторые высшие растения: одни из них — более всего на хвощ, растущий в ле- сах по тенистым и сырым местам; другие — на водяное растение роголистник. Но это сходство, конечно, чисто внешнее, так как тело харовых водорослей состоит не из стеблей, листьев и корней, а представляет собой настоящее много- клеточное слоевище (таллом), характерное для низших растений, хотя и очень сложно и свое- образно устроенное. Они широко распростра- нены в пресноводных прудах и озерах, особен- но с жесткой известковой водой, а некоторые из них встречаются и в морских заливах, и в солоноватых континентальных водоемах. Как правило, харовые растут не поодиночке, а об- разуют заросли, нередко очень обширные, по- крывающие сплошным ковром дно водоемов. И в этих местообитаниях харовые являются наиболее крупными представителями мира во- дорослей — высота их слоевищ обычно состав- ляет 20 — 30 см, но может достигать 1 и даже 2 м. Все части их тела, включая органы раз- множения, хорошо различимы невооруженным глазом. Самый характерный и легко бросающийся в глаза отличительный признак всех харофи- тов — это внешний облик их слоевищ. Они имеют вид кустисто ветвящихся нитевидных или стеблевидных зеленых побегов членисто- мутовчатого строения, укореняющихся на дне водоемов с помощью многочисленных тонких бесцветных ризоидов (рис. 23; табл. 38). Чле- нисто-мутовчатое строение выражается в том, что на основных побегах, на некотором рассто- янии друг от друга, располагаются мутовки ко- ротких равновеликих боковых побегов, также членистого строения. Обе эти вегетативные части харофитов внешне настолько похожи на органы высших растений, что и в научной литературе их условно называют «стеблями» (основные ветвя- щиеся побеги) и «листьями» (расположенные мутовками боковые побеги). Рост стеблей вер- хушечный, неограниченный; листья обладают предельным ростом. Места расположения му- товок называют узлами, а участки стебля меж- ду ними — междоузлиями. Листья могут быть осевыми, с члениками, расположенными в одну линию, и с «листочками» в узлах, и вильчато- разделенными, когда на вершине первого чле- ника образуются 2 — 4 членика второго порядка, и так до трех раз. Своеобразие харофитов заключается также в существенном различии между клетками уз- лов и междоузлий. Каждое междоузлие — это одна многоядерная гигантская, длиной до не- скольких сантиметров, вытянутая клетка, не- способная к делению (у некоторых харофитов она покрыта еще корой), тогда как каждый узел состоит из собранных в диск нескольких мелких одноядерных клеток, дифференцирую- щихся в процессе деления и образующих как боковые ветви стебля, так и мутовку листьев. Наконец, наибольшего своеобразия достига- ет у харофитов строение органов полового раз- множения, образующихся на листьях на вершине большинства члеников, т. е. в их узлах. Жен- ский орган — оогоний и мужской орган — ан- теридий многоклеточны и развиваются у боль- шинства видов на одном растении, но известны и двудомные виды. Оогонии овальные, длиной до 1мм, состоят из яйцеклетки и наружного ее покрова, стенки которого образованы пятью узкими клетками. Снизу оогоний снабжен одно- клеточной ножкой, а сверху коронкой из пяти или десяти коротких клеточек. Антеридии ша- ровидные, диаметром до 0,5 мм, образованные восемью плоскими, скрепленными краями клет- ками с отходящими внутрь отростками, на ко- торых сложным путем возникает множество мужских половых клеток. Подобное строение в других группах расте- ний не встречается. В настоящее время во всем мире описано около 300 видов харовых водорослей, распре- деляющихся очень неравномерно на шесть ро- дов: нителла (Nitella), толипелла (То1уре11а), нителлопсис (Nitellopsis), лампротамниум (Lam- prothamnium), лихнотамнус (Lychnothamnus) и хара (Chara). По некоторым особенностям строения вегетативных органов и оогониев они отчетливо распадаются на две группы, которые принято называть нителловыми (первые два рода) и собственно харовыми (остальные роды). Самый большой по количеству видов, объеди- няющий наиболее сложно устроенных пред- ставителей — род хара. Рассмотрим более детально основные особен- ности этой своеобразной группы. СТРОЕНИЕ И РАЗМНОЖЕНИЕ ХАРОФИТОВ Строение клеток. Все клетки харовых водо- рослей в начальных стадиях дифференцировки отдельных частей таллома одноядерны (рис. 262, 1). При размножении этих клеток их ядро де- лится митотически (рис. 262, 2). Количество выявляющихся при этом хромосом у разных
Рис. 262. Строение клеток харофитов: 1 — op~a H3 ~zre~o~ талломного ya~a c ядром а ~о~оящемся состоянии (после специальной окраски, выявляющей его строени ) ><; 2 — молодая клетка из растущей части таллома в стадии митотического деления ядра (пос (>< 00);,4 — число (и), размеры и форма хромосом у разных представителей харофитов: 8 — и=12 4 — и=28 ( 1ЧОО; ле такои же окраски) стадии фо ми ования и после ю го ф л дующего амитотического деления ядра из клетки, образующей талломное междоузлие (><1800) — и=, — и= (Х . ); 5 — 7— 8 — концевая часть листа нителлы: левый членик — вид с поверхности правый чле и з н к — вид в продольном разрезе. )э видов различное, от 6 до 70, так же как различ- ны их размеры и форма (рис. 262, 8, 4). Только при последующей дифференцировке, когда оп- ределенные клетки вытягиваются и становятся междоузлиями или другими удлиненными час- тями таллома, в них происходят многократные амитотические деления ядра без образования клеточных перегородок, в результате клетки становятся многоядерными, а ядра — вытяну- тыми, неправильной формы (рис. 262, Б — 7). Хлоропласты в клетках многочисленны и имеют форму мелких дисковидных телец, по- добных хлорофилльным зернам высших расте- ний. Набор ассимиляционных пигментов схо- ден с таковым у зеленых водорослей (хлоро- филлы а и b и почти полностью те же каро- тиноиды), а в качестве запасного питательно- го вещества также вырабатывается крахмал. Вытянутые сформировавшиеся клетки ха- рофитов имеют очень характерное строение (рис. 262, 8). Их оболочка плотная, довольно толстая. Внутренний ее слой образован целлю- лозой; наружный слой состоит из каллозы, в которой может отлагаться углекислая из- весть. Всю центральную часть клетки зани- мает вакуоля клеточного сока, а протоплазма распределена по периферии у внутренней по- верхности оболочки. В тонком, непосредствен- но примыкающем к оболочке слое протоплазмы сосредоточены хлоропласты; в более толстом внутреннем слое, граничащем с вакуолью, рас- положены многочисленные ядра (рис. 262, 8, справа). Оба слоя протоплазмы, если рассмат- ривать клетку под микроскопом, показывают ряд интересных особенностей. При установке фокуса микроскопа на поверхность клетки (рис. 262, 8, слева) прежде всего бросается в гла- за, что хлоропласты распределяются равномер- но, окрашивая всю клетку в сплошной ярко- зеленый цвет. Исключение составляет только узкая бесцветная полоса, идущая вдоль всей клетки чуть наискось — она лишена хлоропла- стов, но определяет их положение в остальных частях клетки: хлоропласты расположены ров- ными рядами параллельно этой бесцветной поло- се. Если сфокусировать микроскоп на внутрен- ний слой протоплазматического мешка, то при достаточно большом увеличении в живых клет- ках открывается удивительная картина стре- мительного движения протоплазмы по всей клетке, параллельно той же продольной бес- цветной линии: по одну ее сторону в одном на- правлении, по другую сторону в прямо проти- воположном направлении. Движение прото- плазмы в клетках, как известно, свойственно всем растениям, но у харовых водорослей оно отличается наибольшей скоростью, равной 1,5— 2 лл в минуту, что при сильных увеличениях
Рис. . 263. Строение верхушки стебля харо итов в п о о 1 — 6 — с харофитов в продольном сечении (схематизировано): — — схема начальных стадий ве х ш ч формирования стеб р ушечного роста - (1 — дв хклеточная теблевого узла и междоузлия 4— на я стадия, 2 — четырехклеточная р >eá . я, — поперечный разрез сформиро~ж~~ж ст щейся ветвью) 7 — ч — асть сте ля ха~ы на п отя пери ерических клеток стеблевого узла 6 — б р р яжении верхних четырех муто — сте левой узел с азвиты рки, г — базальный листовой з вок. а — стеблевой узел, б — м — б . вой узел, д — начальный бугорок боковой в — оковои ветви стебля, e — кле к образ — антеридии, и — оогонии. микроскопа производит впечатление значитель- Строение верхушки основных побегов. Слож- ность строения таллома харовых водорослей легче всего понять, если проследить дифферен- циацию клеток в процессе верхушечного роста это выражено у тех харофитов, талломы кото- рых не покрыты особым слоем клеток, име- нуемым корой, например у видов нителлы. Верхушечная клетка стебля имеет характер- ную плоско-выпуклую форму, т. е. нижняя пе- регородка, отделяющая ее от нижележащей клет- ки, плоская а в , а верхняя свободная стенка пол- сферическая (рис. 263, 1). В течение вегетаци- к длинной оси стебля) перегородкой на две клет- ки, из которых верхняя остается ве-х-ше р ушечнои рической. При следующем делении этих дв х клеток такими ж же горизонтальными перего- нии этих двух родками получается комплекс из четырех кле- является исходным для всего последующего процесса и~~~ ди~~~еренциации и роста таллома. овое деление верхушечной клетки повторяет описанный процесс возникновения цилиндри- ческой клетки а а прежняя цилиндрическая клетка делится на д на верхнюю двояковогнутую клетку и нижнюю двояковыпуклую клетк ку (р ., ). Дальнеишая судьба этих послед- пис. 263 2'. EJ них двух клеток резко различна: верхняя клет- ка делится и становится исходной для форми- рования многоклеточного узла (рис. 263, 8а Ф тогда как нижняя оказывается лишенной спо- собности делиться и дальше только растет в дли- ну, образуя междоузлие (рис. 263, 8 б). Деление исходной узловой клетки происхо- дит с~же не го ризонтальными, а вертикальными (параллельными оси стебля) перегородками, которые располагаются таким образом, что после ряда делений образуются две клетки в центре узла и несколько клеток по его пе и- через узел (рис. 263, 4). В дальнейшем цент- ральные клетки более уже не делятся, а пе и- ферические продолжают делиться и дают начало листьям и боковым ветвям стеб ля а у некото- рых представителей еще так называемым при- листникам и коре). Из каждой периферической клетки стебле- вого узла возникает по одному листу. Листья закладываются первоначально как б как угорки в~, которые лишь постепенно вытя- гиваются и развиваются в листья. При этом периферические узловые клетки стебля выпол- няют роль верхушечных ростовых клеток, по- добных верхушечным клеткам стебля. Однако, в отличие от последних, они делятся не бесп е- д, а отчленяют от себя только оп еделен- ельно а ное количество сегментов. Эти сегменты раз- 1
растаясь и дифференцируясь, образуют чле- нистые листья, состоящие из длинных неделя- щихся клеток и чередующихся с ними узлов, сложенных несколькими мелкими клетками, способными к дальнейшим делениям. Один или несколько конечных членик о в листа лишены узлов. Подобное строение делает листья прин- ципиально сходными со стеблем, но функция узлов у них уже совершенно другая. Из пери- ферических клеток большинства листовых уз- лов образуются органы полового размножения и окружающие их придаточные клетки. Только самый нижний, так называемый базальный узел, находящийся в основании каждого листа (рис. 263, 6 г), имеет другую функцию. Отдель- ные его клетки, расположенные в пазухе листь- ев, дают начало боковым ветвям стебля (рис. 263, бд), другие служат в некоторых родах ха- рофитов для образования так называемых при- листников и коры (рис. 263, 6e). Прилистники. У всех видов хары и у пред- ставителей некоторых других родов харофитов на стеблях при основании листовых мутовок имеется венчик более или менее вытянутых и заостренных, а иногда и очень длинных или, наоборот, почти шаровидных клеток, получив- ших условное название прилистников. Они об- разуются из двух боковых клеток базального узла каждого листа независимо от того, имеется ли у данного вида стеблевая кора или нет. В за- висимости от количества делений каждой та- кой начальной клетки и степени разрастания получающихся от этого клеток может возник- нуть несколько вариантов строения прилист- ников, характерных для разных видов. Если делению подвергается только одна из этих двух клеток, и это деление однократно, а деление второй клетки подавлено, то на каждый лист возникает по одному прилистнику, располагаю- щемуся между листьями (иногда они сдвигают- ся под лист). Такой венчик можно назвать одно- кратным (редкий случай). Если так развиваются обе клетки по обе стороны листа, то венчик ста- новится двукратным (частый случай). В обоих случаях возникает лишь один ряд прилистни- ков, т. е. венчик прилистников может быть или однократным однорядным, или двукратным однорядным. Однако чаще всего встречается третий случай, когда каждая начальная клет- ка делится дважды и развивается еще второй нижний ряд прилистников, т. е. венчик оказы- вается двукратным двухрядным (рис. 264). Строение коры. Междоузлия на стеблях мно- гих харофитов не остаются голыми клетками, как это характерно, например, для рода ни- телла, а обрастают слоем специальных клеток, получившим название коры. Стеблевая кора лучше всего выражена в роде хара, большинст- во видов которого снабжены ею. Стеблевая ко- Рис. 264. Основания листовых мутовок разных видов хары с различно развитым венчиком прилистников.' 1 — однократный однорядный венчик Chara braunii; 2 — дву- кратный однорядный венчик Ch. gymnopitys; 3 — 5 — пвукрат- ные двухрядные венчики (8 — Ch. fragilis, 4 — Ch. vulgaris, 5 — Ch. hispida).
ра харофнтов всегда построена из полос кле- ток, расположенных параллельными рядами вдоль междоузлия слегка наискосок. Эти по- лосы бывают двух сортов — первичные, сос- тавляющие основу коры, и вторичные, услож- няющие ее структуру, но эти последние возни- кают не у всех видов. Количество первичных полос соответствует количеству листьев в му- товке, что позволяет понять их происхождение. От основания каждого листа отходят две первичные коровые полосы: одна — вниз rzo стеблю, другая — вверх. Их образуют две пе- риферические клетки базального листового уз- ла, расположенные посередине: одна — под листом, другая — над ним. Каждая из этих кле- ток выполняет функцию верхушечной росто- вой клетки, она делится и развивается так же, как это было описано для роста стебля. В итоге каждая первичная коровая полоса оказывается состоящей из верхушечной клетки и чередую- щихся узлов и междоузлий. Узловые клетки небольшие, округлые, а клетки междоузлий крупные, вытянутые, более или менее цилинд- рические. По мере роста первичные коровые полосы, идущие вниз по стеблю от листьев од- ной мутовки, примерно в середине стеблевого междоузлия встречаются с другими первичными коровыми полосами, идущими вверх от ниже- лежащей мутовки. Таким образом, количество первичных коровых полос всегда равно коли- честву образовавших их листьев в соответствую- щей мутовке. Когда встречные клетки смыка- ются, рост первичных коровых полос закан- чивается. Волнистая линия смыкания коровых полос бывает хорошо заметна посередине стеб- левых междоузлий. Кора, составленная толь- ко из одних первичных полос, получила назва- ние однополосной (рис. 265, 1). Она встречается у определенной, сравнительно небольшой груп- пы видов рода хара. Строение такой коры усложняется в силу того, что узловые клетки первичных коровых полос способны делиться в двух направлениях. Если плоскости деления располагаются фрон- тально (параллельно поверхности стебля), то это приводит к образованию так называемых шипов — обычно вытянутых и заостренных клеток, одиночных или по 2 — 5 в пучках, на- правленных более или менее перпендикулярно к стеблю. Такие шипы в широкой мере свойст- венны однополосной коре (рис. 266, 1). Второе, боковое направление делений узло- вой клетки коры имеет совершенно другие по- следствия — появление по обе ее стороны двух клеток, разрастающихся по стеблю в длину в промежутках между первичными коровыми по- лосами вверх и вниз каждая. В результате из этих двух клеток получается фигура в форме бук- вы Н с узловой клеткой в середине. Эти боковые клетки, образованные от всех узлов первичной коры, в процессе разрастания по стеблю смы- каются друг с другом и также слагаются в по- лосы, получившие название вторичных коро- вых полос. Однако здесь возможны два случая. У одних видов хары вторичные коровые клет- ки сравнительно короткие, равные примерно половине длины клеток коровых междоузлий, и тогда они могут образовать лишь одну вто- Рис. 265. Схемы строения основных типов стеблевой коры в роде хара (вид с поверхности): 1 — опнополосная кора; 2 — двухполосная кора; 3 — трехполосная кора'. а — первичные коровые полосы, б — их узлы, е — их межпоузлия, г — вторичные коровые полосы.
дове хност@ и ко ои шипами (»+ азлично разви ™ р стебля разных в д и ов хары с ра ко ой 2 — п ые полосы крупнее Рис. 266. Участки с р ко ой: 2 — первичные коровые и в попер ечном разрезе .' ой корои ' 2 — 4 — виды с двухполоснои р осной корой (Chara canescens); — — ко ои. 2 п h. acueo а ~е (Ch contra ) шипы рудиментарнь . г1а 8 — то же, (СЬ й ) 5 мельче вторичных е малочислен- в мя первич- больше, чем с о однополоснои. Ещ е лишенные коры. ричную коро овую полосу между д т ечи концами и противополож- нее хары, воо ще а листьях хар кора о разу б ется сходным ния ко о- ыми путем встр б азованных — п тем встр — ечного нарастани ых (правых с левым ) овых по- образом — у злов вверх и вни, з и ных ~пр них первичных коровых полос из листовых узло тони д у д к аспо- только самый нижнии чл т о %Э в мя первичными кор пе,.вичную нисходящи так как базальныи листо- двум чная на одну пе ого зла вниз, так аз- лага ется одна втори называют д дв „хпо- листово у хо я их коров овых полос не обр у- приходится одна вторичная, вой узел вос д щ р . ение коровых пол лос очень упро- лосной корой. р овые клетки е . т. О нако строен о овая полоса на всем про- Наконец, если вторичные ко ткам междоуз лий первич- ще но — каждая кор и листового м еждоузлия состои т только лине равны кле ит встре- тяжени илиндрических кле ток по д лос то тогда происходи в х одинаковых цил вст етив- ных коровых полос, б,.азованных из дву оседних узлов и встр ных клеток, о их из двух со ча ко жй oporto выросш и той же первичнои хся в середине листов ого междоузлия. у жет быть только в том слу- Л стова на ко аз- полосы по о е ее акие две вто- и рыт корой. днако ва ианты левых с левыми). у бразует каж- чае, ечаются разные вар ав ю и левую о ви ов хары встр х при покрытом р ко ой ричн первичная коров р вая полоса, то по о го междо- раз вити я ко ы. одних е оставаться голы ыми дая п ль всего сте левого е листья могут все ж се чле- вы м сторонам вдоль р кание двух па ,,аллельно сте ле ли гих с ко ой бывают или все ч происходит смы их от без корь, у дру сколько первых ч. ле- узлия п ос п оисходящих или только неско 265 8). ~~~~ ~ж~~, голых концах, образован- идущих в II H длннных д вух соседних пер ия когда м ежду двумя нико ом п остых клеток. Бывает, ч Ко такого строен а. ору ко овыми полосами на т пе вич- ниев. Антерндин н оогон из осо ричные коровые поло ~ы~ отсутствием м ок .глых у . е ическими кл икать или вместе на одном талломе . 266, 3 — о). Видов хары с двух- однодо ли азд в значительно талломах в левой корой и трехполоснои стебл 343
Рис. 267. Строение антеридия харофитов: 1 — 4 — последовательные стадии образования антеридия, вид с поверхности (1) и в продольном разрезе (2 — 4): 1 — начальная восьмиклеточная стадия, 2,8 — последующие стадии обособления наружных щитовых клеток и внутренней части, 4— поздняя стадия с дифференцированной внутренней частью; 5 — зрелый антеридий, вид с поверхности; 6 — два щитка с рукоят- ками, вид изнутри; 7 — рукоятка отдельно с хорошо развитыми головками и антеридиальными нитями; 8 — участок антериди- альной нити с антерозоидами в клетках и снаружи; 9 — антерозоид при большом увеличении. а — исходная клетка основания, б — внутренняя базальная («бутылковидная») клетка, в — наружная базальная клетка, г — клетки листового узла, д- стенка, е — рукоятка, ж — головки, з — антеридиальные нити. Рис. 268. Строение оогония харофитов: 1 — 4 — последовательные стадии образования оогония (1 — начальная стадия, 2 — 4 — последующие стадии формирования яйцеклетки и обволакивающих спиральных клеток); 5 — зрелый оогоний нителлы (Nitella); 6 — зрелый оогоний хары (Chara). а — клетка листового узла, б — яйцеклетка, в — поворотные клетки, г — обволакивающие клетки, д — клетки коронки.
69. Ооспо ы и и и их прорастание: нешний вид з елых релых ооспор нител д ательные стадии п о р растания ~~~>~ð я хары (Chara canescens). Формирование антеридия легко у нителлы. Начал льная клетка ия легко проследить узловой, делится , отчлененная от служит для обра ся на две, из кото орых наружная разования собствен и быстро делится в нно антеридия лярных плоскост я в трех взаимно щихся в шар а ях на восемь о перпендику- клеток, слагаю- р, а прилегающая к ли является исходной к листовому узлу нои клеткой, о ми нование антериди ' 2 р еток в ве хней части происхо и дит прежде всего р й шаровидной разделяющихся е р дольными ере- в итоге чего б оказывается сост удущий антеридий остоящим из восьм восьми средни х и восьми вн т е ми наружных 1 плотно соприкас асающихся д г с утренних клеток 267, 8). Одновре руг с другом (рис. 1 временно исхо ная вания делится д я клетка осно- я горизонтальной на наружную ба зальную клетк о перегородкой далее без особых у, остающуюся ых изменений и базальную та к называем ю б и внутреннюю клетку, сильно о разрастающ юся у утылковидную у ся впоследстви ц с дифференциа ванию восьми пл ции приво ит д к формиро- угольной или че лоских пок овны р ~х клеток тре- етырехугольной стыми утол ен смыкающих ся своими заз б е у щ ниями оболочк и, Эти клетки называемые щитка у ренными краями наружную сте нку антеридия. щитками, образуют к ним изнутри и клетки среднего р д . Примыкающие но вытягиваются го ряда постепен- тся и прев а а рукоятки, нес ру ущие р зо образуют обычно п нтеридия эти основ ные головки дая, и уже на ни о по втопичны х головок каж- ридиальные нити них развиваются я длинные анте- ти, обычно по 4 состоящие приме мерно из 200 на головке 1 в один ряд клеток. В расположенных по одному длин каждой клет ному спиралеви етке образуется антерозоиду с двумя жг тами р идно изогнутому у a~a Ha ~ер~д~ем дый антеридий дает 40 релые антериди т до 0 000 анте роз оидов. вид шарика на дии у всех ха рофитов имеют на короткой ножке ~ред~ем о~~~~ 0,5 ~~, ха характерно пересечен крест зигзагооб а смыкания щитк ра знои линией ков, в каждом и шо заметны по краям а иал д из которых хо оболочки и в ент диальные утолщения реннеи рукоятки ~ . 26 в центре округлое осно ~рис. 267 Р. нование внут- одержащихся Рис. 2 кирпично-красными стов, затем о они становятся ~,г — в ы э 8 — 5— ров ания
Рис. 270. Нижняя часть стебля, ризоиды и клубеньки харофитов: 1 — 8 — многоклеточные (1,2) и одноклеточные (8) корневые клубеньки хары, образовавшиеся на ризоипах (1 — Chara baltica, 2 — Ch. fragifera, 8 — Ch. aspera); 4 — 7 — клубеньки, образовавшиеся из стеблевых узлов на нижних погруженных в ил побе- гах: 4 — у Chara baltica (при слабом увеличении), б — 7 — у Nitellopsis obtusa (при слабом и сильном увеличении). Формирование оогониев происходит значи- тельно проще. Начало этому процессу кладет одна из клеток листового узла, составляющая в дальнейшем ножку оогония. Вверх она от- членяет одну центральную клетку, вбок вок- руг себя — пять будущих обволакивающих клеток (рис. 268, 1). Центральная клетка де- лится один или два раза, в результате возника- ют не равновеликие клетки: крупная клетка, развивающаяся далее в яйцеклетку, и одна- три мелкие стерильные клетки, получившие название «поворотных», так как они, возникнув сбоку или сверху от яйцеклетки, постепенно перемещаются к ее основанию. Их считают сильно редуцированным остатком стенки соб- ственно гаметангия. Одновременно с этим пять боковых клеток разрастаются вокруг яйце- клетки вверх по спирали справа налево и пол- ностью обволакивают ее. На самой вершине каждая из них отчленяет одну или две неболь- шие клеточки, слагающие так называемую ко- р он к у (рис. 268, 2 — 4) . Зрелые оогонии имеют вид эллипсоидных или яйцевидных образований длиной до 1 мм, спи ралевидно-исчерченных обволакивающими клетками и увенчанных небольшой или доволь- но крупной коронкой. Коронка из десяти мел- ких клеток, расположенных в два яруса, ха- рактерна для нителловых (рис. 268, 5); одно- ярусная коронка из пяти более крупных кле- ток — для собственно харовых (рис. 268, 6). Ооспоры и их прорастание. Ооспоры, раз- вивающиеся внутри оогониев в результате опло- дотворения яйцеклетки, имеют у разных видов различную форму и размеры, от почти шаро- видных до узкоэллипсоидных, от очень мелких, длиной около 0,3 мм, до крупных, почти в 1 мм. По мере созревания наружная стенка ооспор затвердевает, пропитывается суберином и крем- неземом и окрашивается. Зрелые ооспоры бы- вают коричневато-желтыми, коричневыми или темно-коричневыми до черных, снаружи они всегда слегка блестящие. Стенки их образова- ны четырьмя оболочками — двумя окрашен- ными и двумя бесцветными. Наиболее прочной и наиболее интенсивно окрашенной является самая наружная оболочка, которая образуется из плотно прилегающих к ооспоре внутренних частей обволакивающих клеток оогония, пре- терпевающих соответствующие изменения. Бла- годаря такому происхождению этой оболочки ооспоры на поверхности несут спирально из- витые ребра, представляющие собою слепки швов соприкосновения спиралевидных обво- лакивающих клеток оогония. Соответственно это- му ребра идут от углов пятигранного основа- ния ооспоры вверх справа налево. Особенно силь- но развиты ребра на ооспорах у представителей семейства нителловых, где они имеют вид ши- рокой бахромы, слагающей наверху ооспоры подобие высокого воротничка (рис. 269, 1). В семействе собственно харовых ребра на ооспо-
рах обычно невысокие, часто едва выдающиеся над поверхностью; наверху ооспоры они часто собираются в небольшой хохолок, а внизу пере- ходят от каждого угла пятигранного основания в длинные коготки (рис. 269, 2). Зрелые ооспоры у многих видов покрываются еще слоем извести; внутри они заполняются запасными питательными веществами, главным образом зернами крахмала и каплями жира. После периода покоя ооспоры прорастают. Прорастание начинается с того, что в верхней части ооспоры грубые крахмальные зерна и капли жира постепенно исчезают, заменяясь зернистой протоплазмой, а находящееся здесь ядро, ставшее после оплодотворения яйцеклет- ки диплоидным, претерпевает сперва одно деле- ние, сопровождаемое мейозом, затем второе, в итоге чего получаются четыре гаплоидных ядра. Далее верхний участок протоплазмы с од- ним из этих ядер отделяется поперечной пере- городкой, образуя линзообразную центральную клетку, от которой берет начало новое расте- ние. Остальная часть ооспоры выполняет функ- ции склада запасного питательного материала. Одновременно с этими внутренними измене- ниями наружная твердая стенка ооспоры рас- щепляется на ее вершине на пять зубцов, да- вая выход проростку, возникающему из цент- ральной клетки (рис. 269, 8). Первое деление центральной клетки происходит поперечной перегородкой, перпендикулярной к ее длин- ной оси, и приводит к образованию двух функ- ционально различных клеток. Из одной, более крупной клетки в дальнейшем образуется стеб- левой побег, который на начальной стадии развития называют предростком, из другой, меньшей клетки — первый ризоид. Оба они растут путем поперечных клеточных делений. Предросток растет вверх и довольно быстро зеленеет, заполняясь хлоропластами, первый ризоид направляется вниз и остается бесцвет- ным (рис. 269, 4). После ряда клеточных деле- ний, сообщающих им строение одно рядных нитей, происходит их дифференцировка на узлы и междоузлия, и дальнейший их верху- шечный рост протекает уже так, как было опи- сано выше для стебля. Из узлов предростка возникают вторичные предростки, мутовки листьев и боковые ветви стебля, из узлов пер- вого ризоида — вторичные ризоиды и их му- товчатые волоски. Таким путем и формируется таллом, состоящий из нескольких стеб левых побегов в верхней части и нескольких сложных ризоидов в нижней части (рис. 269, 5). Нижняя часть стебля, ризоиды и клубеньки. Внешние различия между стеблевой и ризоид- ной частью таллома харофитов отчетливы толь- ко в том случае, если с самого начала стеблевая часть развивалась над грунтом, а ризоидная часть была целиком погружена в донный ил. При нарушении этого полное принципиальное сходство обеих частей становится очевидным. Так, участки стебля, погруженные в грунт, быстро укореняются, образуя из своих узлов ризоиды и проростки, а узлы на участках ри- зоидов, вышедших на поверхность грунта, быстро дают стеблевые побеги. Иными словами, из узловых клеток обеих частей таллома при взаимной перемене условий могут возникать вегетативным путем новые целые растения. Кроме того, у харофитов имеются специальные органы вегетативного размножения, так назы- ваемые корневые и стеблевые клубеньки. Образование клубеньков более всего харак- терно для представителей рода хара. Клубень- ки могут развиваться как на ризоидах, так и на нижних частях стебля, погруженных в грунт. Риз оидные клубеньки бывают многоклеточ- ными (рис. 270, 1, 2) и одноклеточными (рис. 270, 8). В обоих случаях они являются специальными образованиями, заполненными крахмальными зернами и несущими на себе узловые клетки, способные к делению и последующему форми- рованию вегетативных органов. Стеблевые клу- беньки всегда многоклеточны и представляют собой стеблевые узлы с сильно редуцирован- ными и видоизмененными листьями. У видов хары они неправильные, разные по величине и форме (рис. 270, 4), а у нителлопсиса отли- чаются исключительной правильностью и изящ- ной формой звездочек, лучи которых соответст- вуют листьям мутовки (рис. 270, 5 — 7). Эти клубеньки также богаты крахмалом и очень легко прорастают в новые растения. ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ И ГЕОГРАФИЧЕ- СКОЕ РАСПРОСТРАНЕНИЕ ХАРОФИТОВ Харовые водоросли населяют преимущест- венно пресные водоемы разного типа, особенно часто пруды и озера, но они встречаются и во вре- менных небольших водоемах — ямах и канавах, а также, хотя и значительно реже, в ручьях и реках. Некоторые виды в равной мере могут населять и солоноватые воды, есть и такие, которые живут только в солоноватой воде, соленость которой, однако, составляет не более чем две трети солености морской воды. В типич- но морских условиях харовые не встречаются. Чаще всего харофиты поселяются на мягких илистых грунтах, реже в водоемах с песчаным дном, обычно на небольшой — в 1 — 5 м — глу- бине, хотя в озерах и лагунах их можно встре- тить и в очень глубоких местах, на глубине до 30 и даже до 40 м. В большинстве случаев они образуют обширные заросли. Все харофиты предпочитают чистую спокой- ную воду. Для многих из них необходимо по-
вышенное содержание в воде известгл. Степень кислотности воды может быть различной, от среднекислой до сильно щелочной. но все же они чаще встречаются в нейтральных и слабо щелочных водах. Пределы активной реакции среды, установленные в водоемах с харовыми водорослями, составляют рН от 5,2 до 9,8. Наибольшее число видов харовых найдено в водоемах с умеренной температурой воды, од- нако некоторые обнаружены и при резко конт- растных температурных условиях. Например, хара ломкая (Chara fragilis) встречается в ледяной воде на севере Норвегии и в горячих источниках Исландии и Йеллоустонского парка (США). Харовые водоросли, приспособленные в це- лом к умеренным условиям, сильно реагируют на крайнее выражение отдельных экологи- ческих факторов, существенно меняя свой внеш- ний облик. В условиях интенсивного освещения обычно вырастают невысокие растения с много- численными, тесно скученными побегами; при рассеянном свете — вытянутые немногочислен- ные стебли. Образование компактных дерновин характерно и для мест с беспокойной водой, испытывающей частые волнения. При сильном обеднении воды питательными веществами в строении талломов могут возникать различные ненормальные отклонения, например, редукция коры у видов хары, обычно имеющих ее в хоро- шо развитом состоянии, появление раздельно- полых растений у однодомных видов и другие. Географическое распространение харовых во- дорослей отличается значительным своеобрази- ем. Они распространены по всему земному ша- ру, в водоемах всех континентов и на множестве островов, кроме Антарктиды. Общими граница- ми являются 69' с. ш. в Северной Норвегии и 49' ю. ш. на острове Кергелен. Однако из 300из- вестных видов харовых только 6 были найдены на всех континентах, большинство же видов встре- чается только на каком-нибудь одном из них. Будучи преимущественно равнинными рас- тениями, харовые водоросли нередко весьма обильны в водоемах предгорий и даже в горах. Предельная высота, на которой они были най- дены в Гималаях,— 4766 м над уровнем моря. В пределах СССР обнаружено свыше 50 ви- дов харовых водорослей. Наиболее богаты ими водоемы европейской части СССР и Средней Азии; значительно меньше было найдено их на Кавказе, в Западной и Восточной Сибири; менее всего — на Дальнем Востоке и в аркти- ческих районах. РОЛЬ В ПРИРОДЕ И ПРАКТИЧЕСКОЕ ЗНАЧЕЕ1ИЕ ХАРОФИТОВ Место, занимаемое харовыми водорослями в природе, сравнительно невелико, что опреде- ляется их обитанием в основном в водоемах озерного и прудового характера, да и то дале- ко не во всех. Однако там, где они поселяются, влияние их на гидрологический режим и био- логические особенности водоемов весьма зна- чительно, так как они обычно разрастаются в массе, нередко покрывая сплошной зарослью все дно водоема. Имеющиеся наблюдения и специальные исследования показывают, что при массовых разрастаниях xApoBIIx водорос- лей режим водоема становится значительно более устойчивым и в нем создается особый биоценоз. На талломах харофитов почти всегда развивается множество эпифитов — микроско- пических водорослей и сопровождающих их бактерий, которые служат прекрасной кормо- вой базой для многих растительноядных личи- нок насекомых и других представителей бес- позвоночных, в свою очередь поедаемых рыба- ми. В густых зарослях жестких хар находят приют и защиту молодь рыб и другие мелкие животные. С другой стороны, давно было замечено, что во многих случаях в водоемах при обильном развитии харовых водорослей отсутствуют или очень слабо развиты личинки комаров. Эту осо- бенность хотели даже использовать для борь- бы с малярийным комаром, специально разводя харовые в водоемах. Предполагалось, что такое токсическое влияние на личинок комара ока- зывают особые вещества, выделяемые многими харофитами, свидетельством чего служит свой- ственный им острый и неприятный запах. Хотя прямыми опытами такое «антикомариное» вли- яние харофитов и не удалось подтвердить, все же выделение ими каких-то антибиоти- ческих веществ в определенные периоды их развития достаточно доказывается многочис- ленными наблюдениями за ними в лаборатор- ных условиях. Весьма значительна роль харовых водорос- лей в природе как источника превосходной пищи для водоплавающей птицы, особенно для их многотысячных стай на путях осенних перелетов. Замечено, что именно на озерах, богатых харовыми, они охотно останавливаются для отдыха и кормежки. В наибольшей мере птицы используют ооспоры, заполненные крах- малом и каплями жира. Так, например, у уток, подстреленных на кормовых озерах во время перелетов, желудки нередко оказывались сплошь набитыми ооспорами. Прямое использование харовых водорослей человеком невелико и полностью определяется местными обычаями и привычками. В Швей- царии в ряде мест осадки, образовавшиеся на дне прудов и озер, заросших харофитами, вы- гребают на берег, высушивают в кучах и затем используют как удобрение, особенно полезное
~» кислы извести. П х и тяже ри боль лых почв б остатков д др оббиллииюю лаго а в харовых в твенных ско так назь даваемого « аллювиал ьных отл оплениях гут сл-ж «харового ожениях ужить для об мергеля» о зей или испо р р спользоватьс ия лечебных ате риал л екрасный б Р "'Р ую из ительны. ие харовы и очень ~~ ВОДОРОС езвычайно ли пол чи широкое зиологич еских и био о удобног го о ъекта у или в ка- Огромные разме ио изическ доузлия ры клеток их исс следований изучать ь такие яв яют с наиб и ольшей цитопла зматическ~ явления легкостью движен OHHHIl;8803AIIOOCTT ь тенциаль Ы~ побегов р в этом ущую вер у я ее еще не исп изучения ении, можно используют р леточных о я внут ик рекомен о к летка чет неза, так ка к здесь к Х ОСНОВ II роцессов тко ма к каждая о тдельная ослеживает в своем пов ведении и таллома. х этапах ди ди åðåí- В ПРОИС ХОЖДЕН НИЕИ К АЦИЯ ХАРОВЫХ Харовые во ВОДОРОСЛИ , что хорош о по — очень д расте- ревн ность сос ходки их в т нах х в ис- от ема в спи ложению клетках оогония и иральных х о волак извести харофитам издревле б кивающих л егко фосс М, И ЭТО CII особствова ыла сво йственна силизипoB ись в гео а лись и у, что они ологически хорошо такие обр т разования- их напла стованиях. И сохраня- нии и ооспор ры ха рофитов ирова рогониты» получивши сеичас при б ие назва- зем и урении и ественно емных не и других находят д х изучени киваются на П на районы копа динично вст растений. аемых ха о~~ тречающиеся ис ур охранив- б н . а л. кастр. 35 ои ни р ~итов эт . О акого ти иск ос- гически тальный с не могут. е ьно-морфол- о енно ге строение о- водорослей неративных вегетатив- еи не имеет ых органов льных г руппах. аналоги й в харовых И „„РУ'"' Р ко сходство в наб на оре Рис. 271. Пл 8 Д 9 ЩИ~ Л~ЫТ~~ харофитов, хара — ЛИСТ ОДНОДОМНО оцно о го ви ха о, àðàêòåð- дв лы (Nitella елл ре la prolife ); — овой узел м ас вои з л ~ужского рас рари1ов~~ шп; 7 — лист о ыи норы ~ СЬ nophyllà ' о хорошо вы о выраженной
основных пигментов (наличие хлорофиллов а и b) и мутовчатое строение побегов позволяют предполагать, что харовые водоросли произош- ли от каких-то высокоорганизованных мутов- чатых зеленых водорослей, похожих, например, на современные хетофоровые водоросли из родов драпарнальдия (DraparIIaldia) и особенно драпа рнальдиопсис (DraparIIaldiopsis). Ископаемые харофиты при общем сходстве их друг с другом и с современными представи- телями настолько многообразны, что отдел Charophyta в целом мы считаем наиболее пра- вильно рассматривать состоящим из несколь- ких классов. Все современные формы входят в один из этих классов — класс Charophyceae, в пределах которого можно усмотреть лишь один порядок Charales, подразделяемый нами на два семейства — семейство нителловых (Nitellaceae) и семейство собственно харовых (Characeae). Эти семейства представляют собой две самостоятельные линии развития, по-ви- димому, рано разошедшиеся от общего предка, так как нет данных, позволяющих считать какую-нибудь из них эволюционно более про- двинутой. Так, в семействе нителловых сам таллом представляется более примитивным, чем у собственно харовых (отсутствие прилистни- ков и стеблевой коры, вильчатое строение листь- ев), тогда как оогонии устроены сложнее (ко- ронка из 10 клеток). Наоборот, в семействе харовых таллом устроен сложнее (имеются прилистники и у многих представителей стеб- левая и даже листовая кора), а оогонии проще (коронка из 5 клеток). Семейство нителловые (Nitellaceae) Таллом всегда без прилистников и без коры. Листья членисто-вильчатого строения, иногда неправильного, и тогда с более или менее обо- собленной основной осью члеников и многокле- точными листочками в ее узлах, реже простые, без листочков. Оогонии в поперечном сечении округлые или эллиптические, с коронкой из 10 клеток, расположенных в два яруса. Содер- жит 2 рода. Представители рода нителла (Nitella) имеют листья правильно однократно или многократно вильчатые, реже простые. Антеридии образуют- ся на вершине члеников в развилках листа, оогонии — сбоку, у однодомных видов под антеридиями. Ооспоры в поперечном сечении эллиптические. Это обширный род, в СССР известно 10 видов (табл. 38, 1; рис. 271, 1). Род толипелла (То1уре11а) отличается листья- ми неправильно вильчатыми до осевых, с мно- гоклеточньиии листочками в узлах оси, реже простыми. Анте ридии и оогонии образуются сбоку, при развилках листа. Ооспоры в по- перечном сечении округлые. Небольшой род, в СССР известно 4 вида (табл. 38, 3; рис. 271,2). Семейство харовые (Characea e) Таллом с прилистниками и у большинства с корой, иногда кора только стеблевая или от- сутствует. Листья членисто-линейного строе- ния, с основной осью члеников и пучками одно- клеточных листочков в ее узлах. Оогонии в по- перечном сечении округлые, с коронкой из 5 клеток, расположенных в один ярус. Содер- жит 4 реда. Из рода нителлопсис (Nitellopsis) хорошо из- вестен и широко распространен только один двудомный вид — N. Obtusa, встречающийся m в СССР, преимущественно в глубоких прес- ных водоемах. Прилистники рудиментарные, в однорядном венчике. Кора отсутствует. Ли- стья сравнительно простые, дву-, пятичлени- стые, с 1 — 3 длинными листочками в узлах. В нижних частях стеблей всегда образуются клубеньки характерной звездчатой формы. (Табл. 38, 8; рис. 270, 5 — 7; 271, 8 — 5.) Род лампротамниум (LamprothamIIium) не- большой, насчитывающий всего несколько ви- дов. Хорошо известен и широко распространен только один однодомный вид — L. papulosum, обычный у нас в опресненной морской воде бухт, заливов и лагун, реже в солоноватых континентальных водоемах. Прилистники хо- рошо развитые, длинные, в однорядном венчи- ке. Кора отсутствует. Листья из 4 — 7 члени- ков. Листочки IIO 4 и более в каждом листовом узле. Оогонии располагаются ниже антеридиев (рис. 271, 6). Род лихнотамнус (LychIIothamIIus) включает только один однодомный вид — L. barbatus, населяющий пресные водоемы. В пределах СССР он обнаружен лишь в Прибалтике. При- листники хорошо развитые, длинные, в одно- рядном венчике. Стебли с недоразвитой корой или без коры. Листья полностью без коры, из 3 — 5 члеников. Листочки по 4 и более в каждом листовом узле. Оогонии располагаются пооди- ночке между двумя антеридиями (рис. 271, 7). Род хара (Chara) самый обширный, в СССР известно около 40 видов, населяющих как пресные, так и солоноватые воды. Прилистни- ки хорошо развитые, длинные или укорочен- ные, в однорядном или двухрядном венчике. Стебли и листья с хорошо развитой корой, реже без коры, иногда кора развивается только на стеблях. Листья обычно состоят из большого числа члеников. Листочки расположены по 4 и более в каждом листовом узле. Растения одно- домные и двудомные. У однодомных видов оого- нии располагаются выше антеридиев. (Табл. 38, 4; рис. 271, 8, 9.) 350
ПРОИСХОЖУ,ЕНИЕ, ГОЛСТВЕННЫЕ СВЯЗИ И ЭВОЛЮЦИЯ ВОЛОГОСЛЕИ Вопрос о происхождении и эволюции водо- рослей очень сложен из-за разнообразия этих растений, особенно их субмикроскопического строения и биохимических признаков. Решение этой проблемы затруднено, кроме того, плохой сохранностью большинства водорослей в иско- паемом состоянии и отсутствием связующих звеньев между современными отделами расте- ний в виде организмов промежуточного строе- ния. Проще всего решается вопрос о происхождении прокариотических (доядерных) водорослей— сине-зеленых, имеющих много общих призна- ков с фотосинтезирующими бактериями. Веро- ятнее всего, сине-зеленые водоросли произошли от организмов, близких к пурпурным бакте- риям и содержащих хлорофилл а. Относительно происхождения эукариотиче- ских (ядерных) водорослей в настоящее время нет единой точки зрения. Существуют две груп- пы теорий, исходящих или из симбиотического, или из несимбиотического их происхождения. По теории симбиогенеза хлоропласты и ми- тохондрии клеток эукариотических организмов некогда были самостоятельными организмами: хлоропласты — прока риотическими водоросля- ми, митохондрии — аэробными бактериями. Как предполагают, существовало несколько групп прокариотических водорослей, различающихся набором пигментов. В результате захвата аме- боидными гетеротрофными эукариотическими организмами аэробных бактерий и прокариоти- ческих водорослей возникли предки современ- ных групп эукариотических водорослей. Неко- торые исследователи приписывают симбиотиче- ское происхождение хромосомам и жгутикам. Теория симбиотического происхождения эу- кариотических водорослей и др. организмов получила особенно широкое распространение за последнее время. Она основана на давно из- вестных фактах самовоспроизведения упомя- нутых органоидов клеток и на новых данных об автономности хлоропластов и митохондрий и их биохимическом сходстве с прокариотами. Это сходство выражается в следующем: 1. Хлоропласты и митохондрии, подобно про- кариотам, содержат ДНЕ и РНК; в них нахо- дятся рибосомы того же типа, что и у прокарио- тов. 2. Антибиотики, подавляющие рост бакте- рий, тормозят образование и размножение хло- ропластов и митохондрий эукариотических орга- низмов, но не действуют на рост самих клеток и тканей. 3. Механизмы фотосинтеза у сине-зеленых и эукариотических водорослей в значительной ме- ре сходны. Однако наряду с фактами, свидетельствующи- ми в пользу симбиогенеза эукариотических орга- низмов, имеются данные, говорящие об обратном. Например, система переноса электронов в хло- ропластах и набор ферментов, необходимых для образования пигментов, участвующих в фо- тосинтезе, регулируются генами ядра. Синтез ряда ферментов митохондрий зависит от ядра и цитоплазмы. Если учесть эти и другие факты, хлоропласты и митохондрии оказываются не «автономными», а «полуавтономными». Что ка- сается самовоспроизведения, то оно обнару- жено даже у поверхностного (кортикального) слоя инфузорий, признаки которого насле- дуются в ряду поколений независимо от ядра
и цитоплазмы. Это было установлено путем пе- ресадок ядра и цитоплазмы других особей, об- ладающих кортикальным слоем с иными при- знаками. Конечно, никак нельзя представить, что кортикальный слой инфузорий некогда был самостоятельным организмом. На водорослях такие эксперименты не проводились. Автономность хлоропластов и митохондрий в равной мере может свидетельствовать как об их некогда самостоятельном существовании, так и о далеко зашедшем процессе специализа- ции и автономности органоидов клетки. Точно так же и данные о неполной автономности хло- ропластов и митохондрий можно с равным успе- хом принять за доказательство их несимбиоти- ческого происхождения или рассматривать как пример утраты симбионтами полной автоном- ности в ходе эволюции. Нам представляется, что частичная или полная автономность орга- ноидов клетки от ядра является необходимым условием нормальной жизнедеятельности эука- риотической клетки и своевременного реагиро- вания ее органоидов на изменения окружаю- щей среды. Эукариотические клетки в среднем в 10' — 10' раз крупнее прокариотических клеток. При полной зависимости органоидов от ядра ма- лейшее нарушение его нормального функциони- рования, хотя бы на короткий отрезок времени, например при митозе, означало бы нарушение обмена веществ всей клетки. Не в пользу симбиотического происхождения клеток эукариотических водорослей свидетельст- вует также отсутствие достаточного сходства в строении хлоропластов и клеток сине-зеленых водорослей — единственных современных про- кариотических водорослей. Сейчас существует целый ряд бесцветных одноклеточных организ- мов, в которых живут в качестве симбионтов явные сине-зеленые водоросли, выполняющие функции хлоропластов и называемые цианелла- ми. Цианеллы отличаются от свободноживу- щих сине-зеленых водорослей. Интересно, на- пример, что у симбиотического глаукоцистиса (C~laucocystis nostochinearum), изученного по- средством электронного микроскопа, цианел- лы, в отличие от свободноживущих сине-зеле- ных водорослей, не имеют оболочки, т. е. сим- биоз зашел здесь очень далеко. Тем не менее даже в таком «упростившемся» виде цианеллы все же остаются сине-зелеными водорослями и по своей организации и поведению в клетке хозяина решительно не сходны с хлоропластами (подробнее см. раздел «Сожительство водорос- лей с другими организмами»). Таким образом, существующие ныне данные не позволяют принять теорию симбиогенеза в качестве основы для пересмотра путей воз- никновения и эволюции эукариотических водо- рослей. Если исходить из несимбиотического проис- хождения эукариотических водорослей, то при- ходится допустить, что они возникли от предка, общего с сине-зелеными водорослями, имеюще- го хлорофилл а и фотосинтез с выделением кис- лорода (рис. 272). Единственным возражением против этого может быть разница в составе кле- точной стенки: у сине-зеленых водорослей, так же как и у бактерий, в клеточной стенке имеет- ся муреин. В целом по составу и строению кле- точной стенки, а также по реакциям, благода- ря которым идет синтез ее веществ, прокариоты существенно отличаются и от животных, и от остальных растений. В случае принятия такого возражения пришлось бы выводить эукариоти- ческие водоросли от других организмов. Это значило бы признать, что фотосинтез с участием хлорофилла а и выделением кислорода возникал в ходе эволюции жизни на Земле два раза. Однако это, учитывая множество реакций, осуществляемых в процессе фотосинтеза с уча- стием многих ферментов, представляется менее вероятным, чем смена в ходе эволюции веществ клеточной стенки. У эукариотических водорос- лей, очевидно, не сразу появилась твердая жест- кая клеточная стенка из целлюлозы или других веществ. Наиболее примитивной у эукариотиче- ских водорослей, очевидно, следует считать аме- боидную форму строения, а клеточная стенка всех современных прокариот имеет жесткую ос- нову. Следовательно, современные фотосинтези- рующие прокариоты, т. е. сине-зеленые водорос- ли, — это боковая, тупиковая ветвь раститель- ной эволюции. Эукариотические водоросли име- ли с ней лишь общего прокариотического пред- ка, лишенного твердой оболочки. Первым шагом на пути возникновения эука- риотических водорослей было формирование яд- ра и хлоропластов. На этом этапе эволюции появились красные водоросли, для которых ха- рактерно отсутствие жгутиковых стадий, при- митивность строения хлоропластов и появление второй разновидности хлорофилла — хлоро- филла d. Несмотря на сложность цикла разви- тия и анатомического строения, достигнутую некоторыми представителями, красные водорос- ли оказались боковой и слепой ветвью эволю- ции. Важнейшим моментом в становлении органи- ческого мира на Земле явилось развитие у од- ноклеточных эукариотов двигательного аппара- та — жгутиков, построенных весьма своеобраз- но: внутри их по периферии располагаются 9 пар фибрилл и 2 фибриллы находятся в цент- ре. С их появлением зародилась центральная прогрессивная группа — фотосинтезирующие эукариотические жгутиковые. Начиная с этого этапа эволюция эукариотов пошла в нескольких направлениях: у одних организмов в дополне-
Наземные растения с'3И Эвгленов 8одорос Сине-зеленые водоросли Прокариоты, имеющие хлорофилл а и фотосинтезирующие с выделением кислорода. Прокариоты, фотосинтезирующие без выделения кислорода Рис. 272. Схема происхождения и эволюционного развития водорослей. Фотосинтезирующие безжгутиковые эукариоты ние к хлорофиллу а появились хлорофилл b, с или е и ряд новых дополнительных пигментов, а другие организмы утратили фотосинтези- рующие пигменты и полностью перешли к гетеротрофному питанию. Одновременно про- исходила эволюция, сопровождающаяся видоиз- менением первоначального строения жгутиково- го аппарата. Например, у диатомовых исчезла центральная пара фибрилл, у некоторых золо- тистых водорослей появилась г а п т о н е м а (уплощенный «жгутик») с 5 — 8 одиночными фиб- риллами по периферии и тремя мембранами в оболочке. Правда, некоторые исследователи до- пускают, что эукариотические водоросли могли возникнуть от гете рот рофных безжгутиковых эукариотических организмов. Но это было бы возможно, если допустить, что фотосинтез с вы- делением кислорода и жгутики, имеющие одно и то же исходное строение у всех водорослей и гетеротрофных организмов, появлялись дважды. Существуют также предположения, что сине-зеленые водоросли непосредственно дали начало красным водорослям, а водоросли, обладающие жгутиками, произошли от бесцвет- ных жгутиковых. Однако это означало бы, что дважды в ходе эволюции формировалось ядро, имеющее одно и то же строение и делящееся митотически, и дважды создавались одинако- вые фотосинтезирующие системы с хлорофил- лом а и хлоропластами. Все это крайне мало- вероятно. Согласно изложенным представлениям о пу- тях возникновения эукариотических водорос- лей, гетеротрофные жгутиковые приходится выводить из фотосинтезирующих самостоятель- но в нескольких ветвях. Проще допустить неод- нократную утрату хлоропластов, которая, к то- му же, действительно наблюдается у представи- телей многих групп одноклеточных водорослей, чем предположить симбиотическое происхожде- ние эукариот или многократное возникновение фотосинтеза и единообразно устроенных у всех эукариот жгутиков и ядер. Утрата пластид не- которыми жгутиковыми тем более допустима, что фототрофные организмы, как прокариоты, так и эукариоты, никогда не теряли полностью способности к гетеротрофному питанию, унас- ледованному от первичных гетеротрофов, поя- вившихся при зарождении жизни на Земле. Вместе с тем всем организмам, живущим на
Земле, свойственно биохимическое единство независимо от способа питания (автотрофного или гетеротрофного). Дж. Бернал, например, писал: «...при рассмотрении биохимии в целом, включая процессы, протекающие у всех видов животных и растений, а также бактерий и виру- сов, обнаруживалось необычайное единство и экономичность. Все снова и снова мы встреча- емся с одними и теми же химическими реакция- ми и структурами — вплоть до деталей атомной структуры. И даже там, где наблюдаются ва- риации, это вариации на одну и ту же тему. Так, например, порфирины используются в ды- хательных ферментах, при фотосинтезе и при переносе кислорода у высших животных». Первичные фотосинтезирующие жгутиковые эволюционировали по двум главным направле- ниям. У одних жгутиковых преобладающее значение получили бурые пигменты, и у ряда представителей появился второй хлорофилл- с или е, а в числе запасных питательных ве- ществ у многих представителей стали образовы- ваться разные модификации специфического полисахарида ламинарина. Второе направление эволюции жгутиковых характеризуется преоб- ладанием зеленой окраски и наличием хлоро- филла Ь наряду с хлорофиллом а. Промежуточ- ное положение между этими двумя основными направлениями заняли жгутиковые, которые, эволюционируя, привели к появлению разно- родной группы желто-зеленых водорослей, имеющих зеленую окраску. Хлоропласты жел- то-зеленых водорослей не содержат хлорофилла Ь и имеют такое же строение, как у большинства водорослей с бурыми пигментами. В качестве за- пасного вещества может образовываться хри- золаминарин,' крахмал и парамилон отсутст- вуют. В пределах каждого из этих направлений эволюция пошла по нескольким линиям, неред- ко па раллельно, сопровождаясь возникнове- нием сходных форм строения тела. Жгутиковые с преобладанием бурых пигмен- тов эволюционировали несколькими путями, которые привели к возникновению водорослей: золотистых, диатомовых, бурых и пирофитовых. Последние фактически представляют два само- стоятельных отдела, из которых один, объеди- няющий динофитовые водоросли, выделяется строением ядерного аппарата не только среди водорослей с бурыми пигментами, но и среди всех эукариотических организмов. Из числа во- дорослей, окрашенных в бурый цвет, наиболь- шего расцвета достигли диатомовые и бурые. Диатомовые заняли доминирующее положение среди микроскопических водорослей морей и континентальных водоемов. Бурые водоросли оказались наиболее приспособленными к жизни в прибрежной зоне моря. У них возник ряд приспособлений, позднее появившихся на иной основе у наземных растений. Здесь имеется в виду наличие у бурых водорослей ситовидных трубок, подобных по субмикроскопическому строению проводящим элементам флоэмы выс- ших растений, а также развитие спорофита на гаметофите или гаметофита на спорофите и раз- множение некоторых видов посредством сфор- мированных проростков спорофитов. Бурые водоросли захватили в море наиболее благо- приятные для роста донных водорослей места, где существует твердый грунт и почти постоян- ное движение воды. Среди морских донных во- дорослей бурые водоросли заняли такое же по- ложение, как покрытосеменные растения на суше. В планктоне морей значительную долю растительных организмов, кроме диатомовых, составляют еще только кок колитофориды из золотистых водорослей и динофитовые водо- росли. Эволюция жгутиковых с зелеными пигмента- ми шла в основном в одном направлении, охва- тывающем отдел зеленых водорослей. От него очень рано отделились эвгленовые водоросли, которые не эволюционировали дальше однокле- точного состояния. От много клеточных зеле- ных водорослей в качестве боковой ветви эво- люции отделились харовые водоросли. У них наиболее сложно устроенные органы полового размножения среди водорослей с зелеными пиг- ментами, и тем не менее это тупиковая группа, представленная сейчас ограниченным числом родов и видов довольно однотипного строения. В прошлые геологические эпохи пышного раз- вития достигали из зеленых водорослей дази- кладовые и сифоновые, последние и сейчас играют существенную роль в жизни тропиче- ских морей. Эволюция зеленых водорослей в водной среде не привела к появлению сложно устроенных и крупных слоевищ тканевого строения, сравнимых с бурыми и красными во- дорослями. Продолжением эволюции зеленых водорослей явилось возникновение фотосинте- зирующих высших наземных растений. Происхождение наземных растений иногда пытаются связывать с бурыми водорослями из-за существования у них многогнездных спо- раыгиев и гаметангиев, которые ошибочно при- нимают за многоклеточные органы размноже- ния. Вместилища бурых водорослей делятся на камеры в последний момент при формирова- нии в них зооидов, чего не наблюдается у на- земных растений. В то же время бурые водорос- ли сильно отличаются от наземных растений набором пигментов, запасными питательными веществами, химическим составом оболочки.
ИСТОРИЧЕСКАЯ РОЛЬ ВОДОРОСЛЕЙ. ВОДОРОСЛИ КАК ГЕОЛОГИЧЕСКИЙ ФАКТОР 3HAЧЕНИЕ В ОУРО СЛЕИ В ПРИРОДЕ И ЖИЗНИ ЧЕЛОВЕКА Водоросли — одни из древнейших организ- мов, населяющих нашу планету. Пожалуй, только бактерии могут поспорить с ними в древ- ности происхождения и длительности сущест- вования. В прошлые геологические эпохи, как и в настоящее время, водоросли населяли океа- ны, реки, озера и другие водоемы. Обогатив атмосферу кислородом, они вызвали к жизни разнообразный мир животных и способствова- ли развитию аэробных бактерий; они явились родоначальниками растений, заселивших су- шу, и, как это неудивительно, создали мощные толщи горных пород. Для того чтобы лучше представить геологи- ческое значение водорослей, необходимо в об- щих чертах ознакомиться с основными этапами развития Земли, которые отражены в геохро- нологической таблице. Эта таблица — резуль- тат кропотливого труда многих поколений гео- логов и палеонтологов; она составлена с учетом новейших достижений геохимии. Читать ее следует снизу вверх, от наиболее древних пе- риодов к нашему времени (рис. 273). Первые страницы летописи о жизни древ- нейших организмов Земли дошли до нас в виде отдельных обрывков, содержание которых под- час трудно поддается прочтению из-за плохой сохранности остатков вымерших обитателей нашей планеты. По-видимому, остатки многих бесскелетных организмов вообще не дошли до нас. Поэтому геологи назвали первый этап в развитии жизни на нашей планете крипто- зойским, т. е. этапом «скрытой жизни». Крипто- зой (или, как часто говорят, докембрий) объеди- няет две эры — археозойскую и протерозой- скую. Следующий, фанерозойский этап (этап «явной и'.пзни») охватывает только около '/ времени существования жизни на нашей пла- нете, но изучен он гораздо подлее. Обильные остатки организмов, сохранившиеся в слоях фанерозоя, позволили с большей полнотой проследить развитие животных и растений на этом этапе. Фанерозойский этап делят на три эры — па- леозойскую (эра «древней жизни»), мезозой- скую (эра «средней жизни») и кайнозойскую (эра «новой жизни»). Эрой водорослей и бактерий называют про- терозой, однако появились они еще раньше. Уже давно высказывались предположения, что архейские графиты и мраморы — результат жизнедеятельности каких-то древних организ- мов, а в последнее десятилетие удалось найти остатки живых существ, населявших древней- шие моря нашей планеты: в Южной Африке в архейских породах были обнаружены самые древние из известных в настоящее время орга- низмов, жившие на Земле 3 млрд. 200 млн. лет назад. Они представляют собой шаровидные микроскопические тельца (диаметром от 5 до 30 мкм), по своему строению очень похожие на сине-зеленые водоросли.
Этапы Эры Периоды Кайнозойская Четвертичный 2 млн. лет (кайнозой) 67 млн.лет Меловой (мел) 70 млн. лет Юрский (юра) 58 млн. лет Триасовый (триас) 45 млн. л ет Пермский (пермь) 45 млн. лет Палеозой сная (палеозой) 330 млн. лет Девонский (девон) 55 млн. лет Силурийский (силур) 30 млн. лет Рис. 273. Геохронологическая таблица. )- О С X С X ю5 О С9 О CL О Сб -е- )5 2С О )~ О 0Ý О CL О Сб z g)~ 9 Q О Л О О Z ~р~ О K с ~О ~Ьо Мезозойская (мезозой) l73 млн. лет Протерозойская. (протерозой,) 500 млн. лет Археозойская (археозой) 1000 млн.лет Третичный (палеоген,, неоген) 65 млн, лет Каменноугольный (карбон) 75 млн. лет Ордовикский (ордовик) 60 млн. лет Кембрийский (кембрий) 65 млн. лет В Южной Африке было сделано еще одно открытие. Оказалось, что скопление графита в архейских доломитах Трансвааля — это не что иное, как остатки колоний водорослей, живших 2 млрд. 600 млн. лет назад. По своим особенностям они наиболее близки к сине- зеленым. Здесь же были обнаружены и остатки древнейших колоний водорослей в виде мелких куполовидных известковых образований. Гораздо больше следов жизнедеятельности оставили после себя водоросли протерозойской эры. Однако находки самих водорослей — явле- ние чрезвычайно редкое. Огромный интерес для науки представило открытие, сделанное при исследовании под мик- роскопом тончайших срезов (шлифов) горных пород в Канаде. Здесь были обнаружены как бы законсервированные кремнеземом нити во- дорослей, живших в протерозойскую эру. Де- тальное изучение строения этих водорослей дозволило заключить, что еще 2 млрд. лет назад сине-зеленые водоросли так же, как и современные, обладали специальными приспосо- блениями для перенесения неблагоприятных условий и для вегетативного размножения. Что же собой представляют многочисленные памятники жизнедеятельности протерозойских водорослей, дошедшие до наших дней? Во мно- гих районах земного шара распространены толщи своеобразных известняков, получивших названия «плойчатые известняки», «скорлупо- ватые породы», «породы со структурой часовых стекол». Чаще всего их называют «стромато- литы», что в переводе значит «ковровый ка- мень». Внешне эти известняки напоминают мно- гочисленные стопки часовых стекол, плотно прижатые друг к другу (рис. 274). Долгое вре- мя природа этих образований была загадкой, и лишь кропотливый труд многих исследователей позволил выяснить, что строматолиты — это постройки колоний сине-зеленых водорослей. Таким образом, можно утверждать, что на дне прибрежных мелководных участков докем- брийских морей существовали сине-зеленые во- доросли. Они извлекали из воды растворенные соли кальция и откладывали в слизистых чех- лах своих нитей зернышки кальцита. Наиболее интенсивный рост водорослей и накопление кар- боната кальция происходили весной и летом. Нижняя часть колоний водорослей постепенно отмирала, оставляя после себя корочку извест- кового вещества. Осенью и зимой колонии во- дорослей засыпались глинистыми частицами, оседающими из морской воды. Затем с наступ- лением благоприятных условий происходило образование следующего слоя известкового вещества. За многие тысячелетия водоросли образовали слоистые скорлуповатые толщи из- вестняков мощностью до 200 м, простирающиеся
иногда на несколько километров. Особенно мощные, высотой до 1100 м, водорослевые по- стройки описаны в Австралии. Известны они в европейской части СССР, на Урале, в Восточ- ной Сибири, на хребте Ала-Тау, Чукотке, в За- падной Европе, Китае, Маньчжурии, Эква- ториальной Африке, Канаде, США. Изучение строматолитов имеет большое прак- тическое значение, ведь они являются почти единственными следами жизни, дошедшими до нас со времен докембрия. Ероме того, для поисков полезных ископаемых, связанных с докембрийскими породами (железных, мар- ганцевых, кобальтовых и урановых руд, зо- лота, меди, фосфоритов и др.), геологам очень важно знать об условиях, существовавших в то далекое время на Земле. Сейчас по на- ходкам строматолитов определяют береговые линии давно исчезнувших морей и океанов. Массовое развитие водорослей в докембрий- ских морях не только оставило после себя па- мятники в виде толщ горных пород, но и яви- лось подлинной революцией в истории всей нашей планеты. Ведь сине-зеленые докембрий- ские водоросли были первыми на Земле авто- трофными фотосинтезирующими организмами, способными при помощи солнечной энергии усваивать углерод и выделять свободный кис- лород. Считается, что к началу палеозоя атмо- сфера, до этого состоявшая из метана, аммиака и углекислого газа, обогатилась свободным кислородом и ее состав стал близким к совре- менному. В атмосфере появился озон, ставший экраном, предохраняющим Землю от губитель- ного потока радиоактивного излучения. Нали- чие свободного кислорода привело к появлению животных и аэробных бактерий. Водоросли явились мощными создателями не только кис- лорода, но и органического вещества, ставшего пищей животных и бактерий. С палеозоя началось бурное развитие живот- ного мира на нашей планете. Эра безраздель- ного господства бактерий и водорослей закон- чилась. Но водоросли продолжали играть не- малую роль и в более поздние эпохи. Наиболее грандиозные геологические обра- зования, созданные живыми организмами,— это подводные скалы, или рифы. Главными ри- фостроителями в настоящее время являются бес- позвоночные животные кораллы, а из расте- ний — красные водоросли литотампии (Litho- thamnion). Последних называют иногда камен- ными водорослями, так как в оболочках их клеток откладывается такое большое количест- во карбоната кальция и отчасти магния, что они буквально приобретают прочность камня, замуровывая отмирающие части колоний. Особенно широко рифы распространены в тропической части Тихого океана, но извест- Рис. 274. Строматолиты — постройки колоний вы- мерших протер озойских сине-зеленых водорослей. Зарисовка продольного разреза строматолитового из- вестняка. ны они также в Индийском и Атлантическом океанах и в Красном море. О грандиозности сооружений, создаваемых живыми организма- ми, можно судить хотя бы по Большому Барь- ерному рифу, который тянется почти на 2 тыс. км вдоль северо-восточного берега Австралии. В настоящее время рифы — это острова и даже целые архипелаги, на которых живут люди; это надежные гавани, но нередко и грозная опасность для морских судов; в рифах находят пищу и убежище миллиарды живых существ. Образование рифов происходит не только в современную эпоху, этот процесс продолжал- ся в течение всего фанерозоя. Строение рифов и их распространение тесно связаны с распре- делением суши и моря в прошлые геологические эпохи, с развитием морских бассейнов и с эво- люцией организмов-рифостроителей. Древние рифовые постройки — это ценнейшие докумен- ты, позволяющие судить о физико-географиче- ских условиях среды и о развитии жизн и в давно прошедшие времена. Более двухсот лет люди изучают рифы, но до сих пор открыты еще не все тайны этих удивительных сооружений природы. Во вся- ком случае, несомненно, что с древнейших вре- мен постоянными рифостроителями являются различные группы водорослей, способные кон- центрировать в своем теле карбонат кальция; преимущественно это красные, зеленые (сифо- неи) и сине-зеленые водоросли. В некоторых рифах водоросли образуют ос- новную массу, преобладая над кораллами. Так, например, в рифах островов Фиджи в Ти- хом океане водорослей почти в три раза больше чем кораллов.
v..л лi Рис. 275. Отпечаток недоросли эпифитон (Epiphyton parapusillum), найденный в нижнекембрийских по- родах Кузнецкого Алатау. Часто водоросли образуют гребни рифов. Если кораллы, как правило, растут на глу- бине 10 — 20 м, то ближе к поверхности обычно поселяются красные кораллиновые водорос- ли — обитатели подвижных вод. Их гибкие ветки при движении водных масс легко гнутся, не ломаясь. В тропических водах кораллиновые водоросли растут чрезвычайно быстро, удли- няясь на 24 — 26,5 см в год. Имеются наблюде- ния, что судоходные каналы глубиной 2 м, проложенные через рифы, полностью зараста- ли красными водорослями в течение 10 лет. Ветвистые низкие кустики, шаровидные и сте- лющиеся корковидные колонии этих водорос- лей, окрашенные в розовые и красные тона, пе- реплетаясь вместе, образуют по краю корал- лового рифа плотную известковую массу в виде гребня. Активное участие в постройке рифов прини- мают зеленые сифоновые водоросли. В их обо- лочках также концентрируется карбонат каль- ция. В лагунах атолла Фунафути и атоллов Марианских островов в Тихом океане были вскрыты пласты, переполненные остатками си- фонеи халпмеды. Растет халимеда чрезвычайно быстро. На атолле Фунафути (о-ва Эллис) уста- новили, что за два с половиной месяца прирост халимеды составил в высоту 55 см и в диаметре 8 см. Членики халимеды, покрытые ситовидно- продырявленным известковым чехлом, бук- вально оплетают коралловые рифы. Почти во всех районах нашей планеты изве- стны древние рифы, в образовании которых принимали участие водоросли. В кембрии наи- более распространенной водорослью-рифострои- телем был апифитон (Epiphyton, рис. 275). До сих пор существуют разные мнения о систе- матическом положении эпифитона. Одни ис- следователи считают его сине-зеленой водо- рослью, другие относят к красным. На не- сколько километров тянулись рифы, образо- ванные эпифитоном и древними вымершими жи- вотными — археоциатами, в теплом морском бассейне на месте современной Тувы (Восточ- ный Саян). По мере опускания морского дна рифы нарастали все выше и выше, достигнув высоты 500 — 800 м. Рифовые кембрийские из- вестняки, переполненные остатками эпифито- на, известны в Восточной Сибири, Кузнецком Алатау, на Южном Урале, в Антарктике, Ита- лии, Франции, Австралии. Основную массу известняков хребта Кара- Чатыр в Южной Фергане в каменноугольное время создали рифообразующие сифоновые во- доросли. В Крыму по реке Каче обнаружены рифы пермского периода, почти полностью образованные сифоновой водорослью миццией. Такие же мицциевые рифовые известняки опи- саны в Югославии, Греции, Северном Ираке, Японии, США (штаты Оклахома и Колорадо). Мощность толщи известняков, представляющей собой водо рослево-коралловые рифы юрского периода, достигает в Альпах 1000 м. В юрском периоде возникли и рифовые постройки крас- ных водорослей в Крыму, образовав живопис- ные вершины Яйлы, Ай-Петри, Никитскую, Чатырдаг и др. Вдоль Восточных Карпат от города Броды до Каменец-Подольского в районе Днестра тянется гряда высоких (до 70 м) холмов, по- росших лесом, — Медобор. По сторонам гряды располагаются скалистые холмы, известные под названием Толтры. Это древний барьерный риф, построенный в основном красными водо- рослями литотамниями в прибрежной полосе 358
тепловодного неогенового морского бассейна. И таких примеров много. Изучение древних рифов начато сравнитель- но недавно, но оно имеет огромное практиче- ское значение. Рифовые известняки — это цен- ный строительный материал, так как карбонат кальция рифовых построек часто отличается высокой чистотой и его используют на произ- водство самых высоких марок цемента. В Вос- точном Саяне и на восточном склоне Урала к древним рифам приурочены залежи алюми- ниевых бокситовых руд. С рифовыми построй- ками тесно связаны месторождения нефти Баш- кирии: погребенные под мощной толщей более молодых пород, древние рифы, созданные во- дорослями и мшанками в каменноугольное вре- мя, оказались коллекторами огромных зада- сов черного золота. Месторождения нефти в Ираке (Мосул) связаны с мощной грядой во- дорослевых и коралловых рифов конца мело- вого — начала третичного времени. Вмести- лищем нефтяных залежей являются также по- гребенные девонские массивы в США. Не иск- лючено, что масса органического вещества, соз- даваемого водорослями, населявшими древние морские бассейны, явилась одним из источни- ков образования самой нефти. Однако этими грандиозными постройками не ограничивается участие водорослей в обра- зовании карбонатных дород. Хорошо извест- ны, например, так называемые харациты — из- вестняки, сложенные почти исключительно обызвествленными плодовыми телами — оого- ниями — харовых водорослей. Они обнаруже- ны в третичных отложениях Киргизии и Тувы. Двенадцатиметровый пласт раннедевонского ха- рацита, простирающийся на 240 м, найден в штате Колорадо в парке Пери. Какие же мощ- ные заросли харовых должны были развиваться в древних водоемах, чтобы их оогонии, диаметр которых обычно не превышает 1 мм, могли образовать целые пласты известняка! Удивительно и происхождение знакомого всем писчего мела. При рассматривании мела под микроскопом при небольших увеличениях обычно бросаются в глаза главным образом из- вестковые раковины фораминифер, но при уве- личениях более чем в 1000 раз там обнаруживает- ся огромное количество прозрачных пластинок другого происхождения, величина которых не превышает 10 мам. Это так называемые кокко- литы — частицы известкового панциря золо- тистых водорослей кокколитофорид (рис. 71). Электронный микроскоп дозволил установить, что меловые породы почти на 95% состоят из кок колитов и их обломков. Новейшие исследования показали, что и в настоящее время в теплых водах морей и оке- анов происходит образование кокколитовых илов. Развиваясь в огромных количествах (мак- симальное число кокколитофорид в 1 м' воды в тропической части Тихого океана примерно 1 685 000 экземпляров), кокколитофориды из- влекают растворенные в воде соли кальция и используют их для построения своего панциря. Отмирая, клетки опускаются на дно, и их из- вестковая оболочка распадается на отдельные кокколиты, образующие в течение многих ты- сячелетий известковый осадок. Водоросли играют немалую роль не только в круговороте кальция в природе, но и в круго- вороте кремния — одного из самых распрост- раненных элементов. Начиная с позднего ме- ла господствующее положение почти во всех водоемах земного шара завоевали диатомовые водоросли, обладающие кремнеземным панци- рем. Кремний их панциря составляет до 75% от сухой массы всей клетки. Для построения панциря диатомеи извлекают из воды раство- ренную в ней кремниевую кислоту. При благо- приятных условиях они обладают колоссаль- ной энергией размножения. Наблюдения по- казали, что удвоение числа их клеток может происходить в природе каждые 4 ч, в условиях опыта число клеток за 8 ч увеличивалось в 10 раз. Весной во время массового развития, называемого (<цветением» воды, в поверхност- ном слое насчитывалось до 12 млн. клеток диато- мей в 1 л. Расчеты доказывают, что в Мировом океане эти микроскопические водоросли еже- годно извлекают из воды и используют на построение своих панцирей от 72 — 109 до 154 10' т кремнезема. Отмирая, клетки диато- мей опускаются на дно водоемов, где постепен- но накапливается осадок, состоящий из их панцирей. Предполагают, что за 1000 лет может отложиться слой осадков мощностью от 7,5 до 30 см. Эти породы, состоящие из массового скопления панцирей диатомей, так называемые диатомиты, достигают местами мощности в не- сколько сотен метров. Диатомиты широко используются в народ- ном хозяйстве. К 1969 г. добыча диатомитов в капиталистических странах достигла 1,75 млн. т в год. Крупнейшими экспортерами являются США, Франция, ФРГ, Алжир. В СССР место- рождения диатомитов промышленного значения имеются в Поволжье, Приморском крае, на вос- точном склоне Урала, Кавказе, на Сахалине. В настоящее время диатомиты используют почти в 50 отраслях промышленности. Их цен- ные качества — высокая пористость и низкий удельный вес. Большое количество диатоми- тов идет на приготовление легких кирпичей; в частности, плитками из диатомита облицован купол собора святой Софии в Стамбуле. При- меняют диатомиты и в качестве добавки к раз- личным сортам цемента. Но наиболее широкое
РАСПРОСТРАНЕННОСТЬ ВОДОРОСЛЕЙ В СОВРЕМЕННЫХ ВОДОЕМАХ, ИХ БИОМАССА И ПРОДУКЦИЯ Рис. 276. Колонии сине-зеленой водоросли глеокап- соморфы (Gloeocapsomorpha prisca), принимавшей уча- стие в образовании горючих сланцев Эстонии (сильно увеличено) . применение диатомит нашел в качестве фильт- рующего материала при производстве различ- ных масел, жиров, в сахарной и химической промышленности. Была даже осуществлена успешная попытка лечения диатомитом тро- фических язв конечностей. Наконец, в определенных условиях водорос- ли явились исходным веществом для образова- Приспособленность водорослей к жизни при самых разнообразных внешних условиях обус- ловливает повсеместную их распространен- ность. В воде и на суше, в снегу, льдах и горя- чих источниках, по всему земному шару — от просторов Северного Ледовитого океана и его островов до тропиков и от тропиков до снегов и скал Антарктиды, от морских глубин до вы- соких гор — всюду мы находим водоросли. Микроскопические размеры и исключительная жизнестойкость их зачатков способствуют пе- реносу их на большие расстояния. Морские течения разносят их по морям и океанам, ре- ки — с одного участка континента на другой. Такую же роль выполняют и рыбы, особенно проходные, совершающие длинные пути для нереста: обнаружено, что некоторые планктон- ные водоросли нередко застревают у них в жаб- рах. Выброшенные из воды на берег и высох- шие, подхваченные вместе с илом и пылью с по- верхности скал и почвы, водоросли разносятся по воздуху ветром и птицами, звери перено- ния жидких и твердых нефтеподобных соеди- нений. Так, например, горючие сланцы Ле- нинградской области и Эстонии обязаны своим происхождением одноклеточной сине-зеленой водоросли глеокапсоморфе (Gloeocapsomorpha, рис. 276), которая в огромных количествах развивалась в водоемах среднего ордовика. Вещества, созданные другой сине-зеленой во- дорослью — глеокапситосол (Gloeocapsithos), бы- ли исходными при образовании горючих слан- цев Минусинской котловины. В Подмосковье от Тулы до Новомосковска открыты залежи особых разновидностей угля, так называемых гумусово-сапропелевых углей или богхедов. Они образовались за счет органи- ческого вещества сине-зеленых водорослей, в массе развивавшихся в континентальных во- доемах в карбоновое время. Такие же угли известны на Южном Урале, в Шотландии и Пенсильвании. Богхеды являются ценным сы- рьем для получения дегтя. Древние водоросли, условия их существова- ния, их влияние на формирование и развитие атмосферы, животного и растительного мира нашей планеты, участие в образовании горных пород — все эти и многие другие вопросы на- чали изучаться сравнительно недавно. Даль- нейшие исследования, несомненно, помогут лучше понять роль водорослей в истории Зем- ли и полнее использовать богатства, созданные этими интереснейшими организмами. сят их с одного места водопоя на другое и т. д. Пути и способы распространения водорослей исключительно многообразны и полностью обе- спечивают повсеместное их занесение. Кроме приуроченности разных видов водо- рослей к тем или иным экологическим нишам, существуют определенные закономерности и в географическом их размещении по земному шару, связанные с общим комплексом внеш- них условий, меняющихся от географического положения. Эти закономерности особенно от- четливо прослеживаются на морских водорос- лях: планктонных — по всему водному прост- ранству Мирового океана, бентосных — по по- бережью всех шести континентов и бесчислен- ных островов. Наиболее общим выражением их является распределение водорослей по широт- ным зонам: в теплых тропических морях, где условия более благоприятны, мы находим и большее количество видов; в холодных аркти- ческих морях флора водорослей по видовому составу значительно беднее. Существует так-
же ясная приуроченность отдельных видов и целых систематических групп водорос- лей к определенным географическим районам, позволившая разделить Мировой океан на флористические области и подобласти (см. цветную карту 3 в первом томе настоящего издания). Установлено, например, что в отно- сительно холодных морях по количеству видов больше бурых водорослей (40 — 43% от общего состава флоры) и меньше зеленых (12 — 13%), а в теплых — наоборот (18 и 24%), тогда как количество видов красных водорослей изме- няется в малых пределах (для холодных мо- рей — 46%, для теплых — 58%). Вообще мно- гие виды водорослей, особенно среди морских макрофитов, имеют вполне определенные облас- ти распространения, не встречаясь в других местах. Так, например, фукусы характерны для литорали северных морей, а крупный макро- цистис свойствен побережью Тихого океана и т. п. Наряду с этим среди водорослей, особен- но пресноводных, имеется много видов, встре- чающихся повсеместно. Распространенные по всему земному шару, водоросли, несомненно, должны играть значи- тельную роль в жизни природы. Однако, чтобы оценить долю их участия в жизненных процес- сах, протекающих на Земле, необходимо сна- чала установить, насколько велико их количе- ство, т. е. насколько велика та живая органиче- ская масса, обычно называемая биомассой, которую они в совокупности составляют. Ведь на первый взгляд водоросли малозаметны и роль их кажется незначительной, и только в исключительных случаях, как, например, в густых зарослях морских макрофитов или при «цветении» воды, вызываемом планктон- ными водорослями, они поражают своим изо- билием. Произведенные подсчеты показывают, что в масштабах всей Земли роль водорослей в общем балансе живого вещества оказывается поистине огромной. Вот некоторые цифры. Вполне понятно, что количество водорослей, а следовательно, и составляемая ими биомасса значительно колеблются в течение года и в раз- ных водоемах, особенно в планктоне, более из- менчивом в этом отношении, чем бентос. Если в 1 см' воды содержится только 3 особи, что соответствует бедному фитопланктону, то и тогда в 1 мз мы получим уже 3 млн. особей, так что трудно даже представить себе количе- ство планктонных водорослей в сколько-ни- будь значительном объеме воды. Достаточно сказать, что даже в арктических морях, несмотря на, казалось бы, неблагоприят- ные температурные условия, весной в 1 мз воды верхних слоев моря насчитывается от 20 до 30 млн. особей планктонных водорослей. Естественно, в небольших пресноводных хоро- Рис. 277. Представитель протококковых водорослей Botryococcus braunii, нередко вызывающий «цветение» воды. шо прогреваемых водоемах развитие планктон- ных водорослей может быть еще более интен- сивным, и мы уже указывали, что «цветение» воды в них в летнее время представляет собой обычное явление (рис. 277). Так, например, по наблюдениям на водохранилищах, при лет- нем «цветении» воды сине-зелеными водорос- лями в 1 смз ее нередко содержится 1 млн. клеток, а зто значит, что в 1 м' уже 1000 млрд. клеток. В случаях особо сильного «цветения», как это наблюдалось, например, в каскаде днепровских водохранилищ, в прибрежной и мелководной зонах водоемов могут возникать скопления водорослей, подобные густой каше. «Цвести» могут не только пруды, озера и дру- гие пресные водоемы, но и моря, преимущест- венно в бухтах и заливах, подчас на огромных пространствах. В разгар лета в Азовском и Бал- тийском морях вода «цветет» сине-зелеными водорослями, благодаря чему в штиль море производит впечатление темно-зеленого луга, а когда осенью в Азовском море вода «зацветает» диатомеями, море кажется тихим болотом с темно-коричневой водой и характерным болот- ным запахом. О способности водорослей создавать органи- ческую массу можно судить по биомассе — ко- личеству органического вещества в живых ор- ганизмах на единицу поверхности или объема жизненной среды. Определения биомассы рас- тительного планктона в различных водоемах показали, что в арктических морях она может
достигать 6 — 14 г на 1 м' воды, в Каспийском море в среднем равна 1 — 3 г на 1 лз, а в пред- устьевой зоне достигает 100 и даже 140 г на 1 м' и, наконец, в Азовском море в летние ме- сяцы может давать необычайную вспышку- до 270 г на 1 м'. Для донных водорослей при определении на единицу площади было, на- пример, найдено, что в Баренцевом море у Мур- манского прибрежья заросли ламинарий дают от 4,5 до 15 кг сырой массы на 1 м', а в отдель- ных случаях — до 25 — 30 кг на 1 м', тогда как средняя биомасса фукусовых обычно составля- ет 8 — 10 кг на 1 м'. Сходные цифры показывают заросли ламинарий в Белом и Японском мо- рях, в Черном море биом:.сса бурой водоросли цистозейры только местами достигала 6 — 7 кг, в редких случаях — до 13 кг, в среднем же для разных участков зарослей составляла только около 3 кг на 1 м'. В пресных водоемах биомас- са донных водорослей обычно меньше и только в случаях массовых разрастаний кладофоры сухая масса может составлять около 300 г (приблизительно 3 кг сырой массы) на 1 м'. Не меньший интерес представляет и величи- на продукции водорослей за год, т. е. как бы урожай их, или количество органического ве- щества, производимого ими в водоемах в тече- ние года. Ясно, что цифры биомассы не могут этого показать, так как водоросли все время нарождаются и отмирают, их поедают живот- ные и т. п., а биомасса отражает лишь опреде- ленный момент. Годовую продукцию удобнее всего пересчитывать, по аналогии с урожаями наземных растений, на 1 га водной поверх- ности. Годовая продукция планктонных водорослей в морях и пресных водоемах оказалась доволь- но близкой, тогда как продукция донных водо- рослей резко различной. Так, например, в бо- гатом жизнью Баренцевом море годовая про- дукция растительного планктона (в сырой мас- се) определяется в 30 — 50 т на 1 га, а донных бурых водорослей (фукусовых) — в 120 — 231 т (в среднем 192 т) на 1 га. В Черном море про- дукция донных водорослей несколько ниже: в защищенных местах — 100 — 170 т, в откры- том море — 77 m на 1 га. В одном из продук- тивных пресных озер годовая продукция рас- тительного планктона (в сырой массе) состав- ляла 26,5 т, а донных растений (среди которых были, конечно, не только водоросли, но и цвет- ковые) — 4,3 т на 1 га. Подобных примеров можно было бы привести еще немало, но уже из сказанного ясно, что водная толща дает бога- тый урожай водорослей, не уступающий, а под- час и превосходящий продуктивность суши. Далее, чтобы представить себе масштабы участия водорослей в жизни природы, интерес- но посмотреть, какую биомассу может дать тот или иной водоем в целом или значительная его часть. В Черном море, в средней части тре- угольника между Севастополем, Одессой и усть- ем Дуная, на глубине 18 — 60 м располагается огромное «поле» не прикрепленной ко дну крас- ной водоросли филлофоры, занимающее пло- щадь около 11 тыс. км '. Биомасса, образован- ная здесь только одним видом водоросли, со- ставляет около 5,5 млн. m. Это скоплениебыло обнаружено академиком С. А. 3 е р н о в ы м и получило название «Филлофорное поле Зерно- ва». Другой пример — Саргассово море, кото- рое представляет собой огромный овальный участок спокойной воды в северной половине Атлантического океана, ограниченный морски- ми течениями (от 23 до 35' с. ш. и от 30 до 68' з. д.). Здесь массами размножаются пла- вающие крупные водоросли, главным образом бурые из рода саргассум (Sargassum). В этом «море» площадью около 4,5 млн. км' масса во- дорослей, образованная в основном тремя ви- дами, равна примерно 12 — 15 млн. m. В отли- чие от «Филлофорного поля Зернова», где море мелководно и водоросли находятся главным образом на дне, в Саргассовом море глубина огромна — от двух и более километров. При- чем преобладает глубина 4 — 6 км, так что во- доросли здесь плавают у поверхности. Однако эти два примера — исключение; во всех ос- тальных морях донные водоросли сосредото- чены на сравнительно неширокой прибрежной полосе, которая по площади, по имеющимся расчетам, ни в коем случае не превышает '/, всей площади Мирового океана. Поэтому ясно, что в целом наибольшее количество орга- нической массы в воде должны образовывать планктонные водоросли, особенно если учесть, что быстрота их размножения позволяет им давать в течение года продукцию, значительно превосходящую их биомассу в каждый данный момент. Все это хорошо иллюстрируется сле- дующими цифрами, показывающими биомассу и продукцию водорослей по отдельным морям. Биомасса растительного планктона Каспий- ского моря, считая на сырую массу, равна 3,5 млн. m, а продукция его за год составляет уже 1 млрд. m; биомасса донных водорослей равна приблизительно 3 млн. m при такой же величине их годовой продукции. В Черном море общая биомасса планктона (вместе с зоо- планктоном, который, однако, зависит от фи- топланктона и всегда значительно меньше его) измеряется в 12 — 18 млн. т при годовой про- дукции около 225 млн. m, а биомасса донных водорослей только у советских берегов состав- ляет около 12 млн. m. Наконец, в Баренцевом море биомасса растительного планктона оце- нивается примерно в 120 млн. m, а годовая продукция его достигает 5,6 млрд. m, тогда как
биомасса и годовая продукция донных водорос- лей составляет всего несколько миллионов тонн. Что же касается вневодных местообитаний, то биомасса водорослей на них, несомненно, значительно меньше, чем в воде, хотя у поч- венных водорослей она все же достаточно ощу- тима (см. «Почвенные водоросли»). Таким образом, как показывают приведен- ные примеры, общая масса водорослей в при- роде должна быть необычайно велика. Это ста- новится особенно ясно, если представить себе все необозримое пространство воды на земном шаре, содержащее планктон. Известно, что суша занимает на Земле только 29, 2% ее поверхности, а остальные 70,8% прихо- дятся на Мировой океан, что составляет 361 млн. км'. К этому надо добавить еще озера, реки пдругие внутренние водое,мы, ,общая по- верхность которых сейчас определяется при- мерно в 2 млн. кл2, а годовая продукция фито- массы — в 1 млрд. т (правда, здесь сущест- венная доля приходится на высшие растения). Какова же роль этой массы водорослей? Основное значение водорослей в жизни природы вытекает из их физиологических особенностей как зеленых растений: подобно высшим зеленым растениям на суше, водоросли в воде являются основными созидателями органического вещест- ва. Правда, в воде имеются еще и другие авто- трофы — хемосинтезирующие бактерии и выс- шие растения, но удельный вес их по сравне- нию с водорослями весьма незначителен. Та- ким образом, можно без преувеличения ска- зать, что весь остальной мир современных жи- вых существ воды в той или иной мере обязан своим существованием водорослям, так как водоросли, благодаря содержанию в них хло- рофилла, способны созидать органические ве- щества своего тела из неорганических веществ окружающей их воды. Следовательно, они яв- ляются производителями (продуцентами) в во- де той первопищи, которой в дальнейшем поль- зуются все остальные лишенные хлорофилла водные организмы (консументы), или непосред- ственно заглатывая водоросли и образовав- шийся из них детрит, или впитывая органиче- ские вещества, получившиеся от их разложе- ния, или, наконец, поглощая животных, питаю- щихся водорослями. Однако соотношение продуцентов и консу- ментов в разных районах земного шара дале- ко не одинаково, и в этом отношении водная оболочка Земли существенно отличается от суши. По недавним подсчетам В. Г. Б о r о- р о в а, биомасса (в сырой массе) фитопланкто- на в Мировом океане равна 1,5 млрд. т, а его продукция за год — 550 млрд. т, т. е. продук- ция превышает зд сь биомассу в 366 раз. Это означает, что масса планктонных водорослей, отличающихся способностью к интенсивному размножению, возрождается примерно каждые сутки. Иное наблюдается для фитобентоса Ми- рового океана, биомасса и продукция кото- рого определены одинаковыми величинами в 0,2 млрд. т. При сопоставлении этих данных с соответствующими цифрами биомассы и про- дукции консументов, т. е. зоопланктона, зоо- бентоса и крупных водных животных, полу- чается следующая суммарная картина для Ми- рового океана: Таблица II Продукция за год (млрд. т) Биомасса (млрд. т) Группы организмов 550,2 56,2 Водоросли Животные 1,7 32,5 Таким образом, в океане биомасса животных превышает растительную биомассу почти в 20 раз, что обеспечивается огромной продук- тивностью фитопланктона, годовая продукция которого больше годовой продукции животных в 10 раз. Огромное значение имеет также то обстоя- тельство, что водоросли в процессе фотосинте- за выделяют свободный кислород, необходимый для дыхания водных организмов, как живот- ных, так и растительных. В настоящее время считают, что весь свободный кислород Земли является продуктом деятельности зеленых хло- рофиллоносных растений. А в воде, содержа- щей кислород, быстрее завершаются процессы мин ерализации органических веществ. Было установлено, например, что в Черном, Каспий- ском и Азовском морях в летнее время, при наличии аэробных условий, полный бактериаль- ный распад планктонных организмов (расти- тельных и животных) заканчивается примерно в 20 дней, тогда как в северных морях это про- исходит в 3 — 4 раза медленнее. Так осущест- вляется круговорот веществ в воде, и без уча- стия водорослей он был бы немыслим. Но и в масштабах всего земного шара доля водорослей в создании живого вещества также весьма значительна (не говоря об их огромной исторической роли, о чем сказано выше). Правда, до недавнего времени их значение не- сколько преувеличивали. Так, считали, что водорослям и другим растениям, населяющим водную оболочку Земли, принадлежит до 80% всех органических веществ, ежегодно создаю- щихся на всей нашей планете. Однако после многочисленных исследований растительной биомассы, образующейся на суше, и обстоя-
ПРАКТИЧЕСКОЕ ИСПОЛЬЗОВАНИЕ ВОДОРОСЛЕЙ 364 тельных подсчетов выяснилась следующая кар- тина. Если все данные брать в сухой массе, то суммарные запасы растительной биомассы суши следует определять в 2402,5 млрд. т, а в Мировом океане — в 0,17 млрд. т. Иными словами, сухая масса всех водорослей в Миро- вом океане в каждый данный момент в 15 000 раз меньше суммарного запаса сухой фитомассы суши. Однако темпы воспроизводства фито- массы на суше и в воде сглаживают указанную разницу: продукция фитомассы на суше состав- ляет около 7% от ее запаса и суммарно выра- жается в 171,54 млрд. т сухого вещества в год, а в Мировом океане она составляет более Повсеместное распространение водорослей в природе и обильное, а подчас и массовое раз- витие их в водоемах разного типа, на наземных субстратах и в почве определяют огромное значение этих растений как в повседневной жизни человека, так и в его хозяйственной деятельности. И все же имеющиеся возмож- ности практического использования водорослей далеко еще не исчерпаны, а методы управления их жизнью только намечаются. Взаимоотношения «водоросли — человек» очень многогранны и подчас совершенно не- ожиданны. Казалось бы, что может быть об- щего между водорослями и текстильной про- мышленностью, кондитерским делом и изготов- лением бумаги? Однако некоторые вещества, получаемые из морских водорослей, имеют здесь прямое приложение. Разве можно было предполагать хоть какую-нибудь связь между пресноводными водорослями и космическими полетами? Однако выясняется, что дальние рейсы космических кораблей будут, по-види- мому, невозможны без участия этих мельчай- ших зеленых аккумуляторов солнечной энер- гии. И многое, многое другое. Сложность этих взаимоотношений усугубляется еще и тем, что одни водоросли могут приносить человеку поль- зу, другие — вред, а нередко одни и те же проявления их жизнедеятельности в одних условиях оказываются полезными, в других— вредными. Кроме того, влияние водорослей проявляется не однозначно, а несколькими, в основном двумя, путями — косвенным и пря- мым, которые, впрочем, нередко переходят друг в друга. Косвенное влияние выражается в том, что на хозяйстве отражаются те естественные процессы, которые вызывают водоросли в при- роде; человек, учитывая эти процессы, пытает- ся активным вмешательством в природу увели- чить их положительное или уменьшить их отрицательное значение. Прямой путь — это 30 000% от ее запаса и равна 47 — 72 млрд. т„ т. е. всего лишь в 3 раза меньше. Эти материалы позволили оценить годовую первичную продукцию, создаваемую зелеными растениями на всем земном шаре в целом, в 232,54 млрд. т сухого органического вещест- ва, из которых на долю наземных растительных сообществ приходится около 74%. Таким об- разом, водоросли в Мировом океане ежегодно создают чуть более ", всех органических ве- ществ планеты. «Урожайность» водорослей в Мировом океане оценивается, по новейшим дан- ным, в 1 3 — 2 О т сухого вещества на 1 га поверхности воды за год. путь непосредственного использования водо- рослей как растительных продуктов или как сырья для получения различных содержащихся в них веществ, ценных для человека. Приме- нительно к водорослям континентальных водое- мов преобладает косвенный путь, морские во- доросли предоставляют широкие возможности прямого использования. Практическое значение водорослей косвен- ного характера в наибольшей мере проявляется в рыбном, сельском и коммунальном хозяйст- вах, а также при эксплуатации водного транс- порта и гидротехнических сооружений, отча- сти в медицине, тогда как прямое использова- ние их наиболее весомо в качестве пищевого продукта и сырья для ряда отраслей промыш- ленности. Первое из этих направлений отра- жено при описании разных групп водорослей, об основных вехах второго направления речь пойдет здесь. С древнейшего времени, уходящего своими корнями, вероятно, к первым поселениям че- ловека на морском побережье, морские водо- росли используют в пищу и как корм для домашних животных. По берегам морей и океа- нов их употребляют почти всюду, но особенно широко в Японии, где они являются настоящим национальным кушаньем. В европейских стра- нах преобладает кормовое использование мор- ских водорослей. У нас в СССР морские водо- росли употребляет в пищу население главным образом дальневосточного побережья и Кам- чатки, в меньшей степени — жители берегов Мурмана и Белого моря и совсем не едят их на Черном море. Употреблять в пищу можно, по-видимому, почти все водоросли, так как ядовитых форм среди них нет, не годятся лишь слишком гру- бые или невкусные. Так, например, на Сандви- чевых островах из 115 имеющихся видов мест- ное население употребляет в пищу около 60.
Конечно, многое зависит от привычки, способа приготовления и индивидуальных склонностей, но даже и на вкус очень придирчивого человека при соответствующей обработке съедобны весь- ма многие водоросли. Они служат или непосред- ственно как продукт питания, или для пригото- вления различных острых приправ и гарниров. Наибольшей известностью пользуется у нас так называемая морская капуста. Это глав- ным образом ламинария и близкие к ней виды бурых водорослей, например алария и унда- рия (табл. 39, 1 — 8). Продукты, получаемые из ламинариевых, известны в Японии под на- званием «комбу»; для их приготовления при- меняют не менее 12 способов. Собранную мор- скую капусту высушивают на берегу, затем режут на ленты и складывают в пачки. Из морской капусты и комбу приготовляют самые разнообразные кушанья, чаще всего употреб- ляя вместо обыкновенной капусты в супах, с мясом, рыбой, рисом и т. п. Употребляют ее и в кондитерских изделиях — засахаренную, в пастиле, конфетах и т. д. Широко известны и высоко ценятся за неж- ность и вкус некоторые красные водоросли из рода порфира (табл. 20, 1), известные у нас под названием красного морского салата, а в Японии в зависимости от сорта их называют «аманори», «азакузанори», «хошинори» и т. д. Эти водоросли или едят в сыром виде, приго- товляя различные салаты, или варят с мясом, рисом и другими продуктами. Аналогичное применение, особенно в сыром виде в качестве салата, имеет и зеленая водоросль ульва (табл. 31, 6), называемая зеленым морским салатом, а также красная родимения (табл. 21, 4). Многие из водорослей перерабатывают сна- чала в муку, которую затем употребляют для разнообразных печений и других блюд. Из продуктов, выпускаемых нашей пищевой про- мышленностью, наибольшее распространение по всей стране получили консервы из морской капусты с овощами и пастила «Зефир» с при- месью этой же водоросли. Химический анализ наиболее известных представителей пищевых водорослей показы- вает, что в них больше всего углеводов, тогда как белков много только у красного морского салата, а содержание жиров всюду крайне не- значительно. Что же касается усвоения всех этих веществ человеком при питании водорос- лями, то имеются данные, что жиры и угле- воды усваиваются в пределах обычной нормы, а усвоение белков колеблется в пределах от 31,7 до 80%, равняясь в среднем 61,1%. Та- ким образом, морские водоросли вполне пита- тельный продукт. Считают, что усвоение, на- пример, морской капусты не уступает усвоению обычной капусты, перед которой она имеет ряд преимуществ. Однако особую ценность во- дорослевой пище придает то, что, кроме общей питательности, она богата витаминами и обла- дает рядом диетических и лечебных свойств. Так, в морских водорослях точно доказано наличие витаминов А, В1, „„, С и D. Осо- бенно богата витаминами красная водоросль порфира, высокоценимая как пищевой продукт. Витамина В, отсутствие которого в пище вы- зывает у человека болезнь бери-бери, в порфи- ре оказалось лишь вдвое меньше, чем в высоко- витаминозных пивных дрожжах, фактора рос- та — витамина В, — обнаружено в ней вдвое больше, чем в капусте и моркови, а антицингот- ного витамина С в ней столько же, сколько в ли- моне. Большое значение имеет также содер-. жание в морских водорослях иода, брома, мышьяка и некоторых других веществ. Наи- большей известностью как лечебное и профи- лактическое средство пользуется морская ка- пуста, применяемая против некоторых желу- дочно-кишечных расстройств, явлений склеро- за, зоба, рахита и ряда других заболеваний. Ее продают в аптеках в виде сухой крошки, получаемой путем измельчения слоевищ водо- росли. В отличие от морских пресноводные и назем- ные водоросли мало употребляют в пищу. Оче- видно, в данном случае пищевое значение мо- гут иметь лишь те водоросли, которые образу- ют крупные колонии или скапливаются боль- шими компактными массами. Так, считаются съедобными три вида сине-зеленых водорослей из рода носток, образующие слизистые коло- нии: носток сливообразный (Nostoc pruniforme, табл. 3, 9), шаровидные или зллипсоидные хрящевато-слизистые колонии которого разме- ром до куриного яйца нередко скапливаются в массе на дне небольших водоемов; затем по- всеместно распространенный носток обыкновен- ный (N. commune, рис. 54, 1), образующий крупные пластинчатые слизистые сиоевища на влажной почве, и носток войлочный (N.flagelli- forme), широко распространенный в Северном Китае, где он обитает на бесплодных почвах и в сухом состоянии имеет вид войлочных дер- новин из тонких темных нитей, становящихся слизистыми при увлажнении. Для Японии был описан случай образования на склонах неко- торых вулканов мощных пластов студенистой органической массы, состоящей преимущест- венно из колонии одноклеточных сине-зеленых водорослей; местное население называет эти скопления «ячменный хлеб тенгу» (тенгу, по поверью, добрый горный дух) и с давних вре- мен употребляет их в пищу. Советские бота- ники, посетившие в конце 50-х годов районы Южного Китая, обнаружили продававшиеся на
рынке сухие волокнистые бледно-зеленые ле- IIeIIIKH, состоящие из нитчатых зеленых (зиг- немовых) водорослей: как выяснилось, их едят поджаренными на растительном масле. Нако- нец, в самые последние годы в научной литера- туре получил широкую известность один вид сине-зеленой нитчатой водоросли спирулины (Spirulina platensis), образующий скопления в озере Чад (Южная Африка); местное населе- ние издавна употребляет его в пищу. В настоя- щее время эта водоросль стала одним из попу- лярных объектов массового культивирования в различных, в том числе и пищевых, целях. Промышленное использование морских водо- рослей по сравнению с пресноводными также имеет значительно большие масштабы и более давнюю историю. В настоящее время под тер- мином (<водорослевая промышленность» разу- меют промышленную переработку именно мор- ских водорослей. Из них получают как различ- ные органические соединения, так и минераль- ные вещества. Одним из наиболее известных промышленных продуктов водорослевого происхождения явля- ется агар, или кантен, представляющий собой слизистое вещество, извлекаемое из морских водорослей путем их выварки. Все морские во- доросли в той или иной мере отличаются сли- зистостью слоевищ. Если их подвергнуть вывар- ке, то слизь переходит в раствор, застывающий при охлаждении в плотный студень белого или желтоватого цвета (в зависимости от очистки). Его высушивают и выпускают в продажу в виде лент, плиток, кубиков и порошка. Су- хой агар в кипящей воде снова легко перехо- дит в раствор, и даже 20 г его на литр воды дают довольно плотный студень. Агар представляет собой не одно определен- ное вещество — это смесь веществ, более всего углеводов, состав которой зависит от того, из каких водорослей и как ее получают. Наилуч- шие сорта агара дают некоторые красные водо- росли — гелидиум (табл. 39, 4), грацилярия и некоторые другие. В СССР агар получают из красной водоросли анфельции (Ahnfeltia pli- cata, табл. 39, 6), в больших количествах произрастающей в Белом море и в дальневос- точных морях, и близкий к нему агароид, получаемый из красной водоросли филлофоры (Phyllophora nervosa, табл. 39, 6), запасы кото- рой в Черном море неисчерпаемы. Применение агара разнообразно. В больших количествах его используют в пищевой промыш- ленности при изготовлении мармелада, пасти- лы, мороженого, сыра и других, преимущест- венно кондитерских, изделий. Агар добавляют в хлеб, это повышает его качество, и он не так быстро черствеет. В бумажной промышленности его применяют для придания бумаге плотности и глянца, с той же целью его использую~ в текстильной промышленности при аппрету- ре, т. е. отделке тканей, и в ряде других отрас- лей. Наконец, в громадных количествах агар используют в научно-исследовательской работе в качестве твердой среды (после пропитыва- ния питательными солями) для культивирова- ния микроорганизмов. При сходной, но более грубой обработке из водорослей получают клей, который находит применение как в текстиль- ном, так и строительном деле. Прибавление клея к цементу, штукатурке и прочим строитель- ным материалам увеличивает их крепость и во- донепроницаемость. Однако наибольшую ценность из всех орга- нических веществ, получаемых из морских водо- рослей, представляют так называемый альгин, или альгиновая кислота, и ее соли — альги- наты. В чистом виде они обладают большой клеющей силой, в 37 раз превосходящей гумми- арабик и в 14 раз крахмал. Это делает альгин особо ценным для аппретуры тканей и бумаги и для целого ряда других производств, где требуется сильный клей. Морские водоросли долгое время служили сырьем для получения одного из ценных не- органических веществ — иода (позднее были найдены более дешевые способы извлечения его из воды некоторых минеральных источников и соляных зале~кей). Сравнительно недавно их стали применять для получения и других эле- ментов, главным образом калия и натрия. Все эти вещества входят в состав золы, количество которой в морских водорослях весьма велико- от 15 до 45, а у некоторых форм и до 53% от абсолютно сухого веса слоевищ. Доказано, что содержание солей калия в золе водорослей доходит до 35 % . Очевидно, при одностороннем использова- нии морских водорослей происходят большие потери этого ценного сырья. При сжигании на золу пропадают все органические вещества, при выработке только органических веществ пропа- дают все зольные элементы. Поэтому уже давно стремятся выработать методы комплексной пе- реработки водорослей, которые позволили бы получить из одной и той же порции водорослей максимальное количество продуктов. Имею- щийся в этом направлении опыт показывает, что, кроме ранее перечисленных веществ, из морских водорослей действительно можно полу- чить ряд высокоценных продуктов — спирт, уксусную, молочную и другие органические кислоты, маннит, ацетон, эфиры и т. д. Возможности промышленного использования водорослей континентальных водоемов по срав- нению с морскими водорослями значительно более ограничены, и имеющиеся в этом направ- лении попытки еще не вышли за рамки лабора-
МАССОВОЕ КУЛЬТИВИРОВАНИЕ МИКРОСКОПИЧЕСКИХ ВОДОРОСЛЕЙ Одним из реальных методов использования микроскопических водорослей в производствен- ной деятельности человека является их массо- вая культура. Начало этому положили одно- клеточные зеленые водоросли, которые привлек- ли к себе внимание как объекты для промыш- ленного культивирования в силу высокой сте- пени изученности и благодаря ряду ценных биологических и физиологических свойств. По- сле накопления большого опыта стали культи- вировать и другие водоросли. 367 торных изысканий или отдельных производствен- ных задач, не получивших широкого развития. Наибольшее внимание как высококачественное сырье для производства бумажной массы из- давна привлекали огромные запасы нитчатых водорослей кладофоры и ризоклониума в мно- гочисленных озерах Западной Сибири и север- ных районов Казахстана. Оболочка клеток этих водорослей представляет собой почти чис- тую клетчатку. В одних только озерах Барабин- ской и Кулундинской степей общей площадью в несколько десятков тысяч квадратных кило- метров ежегодный урожай этих нитчаток состав- ляет не менее 100 000 т, что может дать товар- ный сбор в пределах 50 000 т. Производствен- ные опыты оказались весьма удачными: из этих водорослей были получены различные сорта беловой бумаги, в смеси с макулатурой— оберточной и обойной бумаги, а при специаль- ной обработке также картон и строительные материалы разного типа. Однако добиться рен- табельности этого производства пока не удалось. В последнее время на Украине начались исследования по практическому применению массы микроскопических планктонных сине- зеленых водорослей, накапливающейся в водо- хранилищах в периоды летнего «цветения» во- Человек издавна ищет в природе и вводит в хозяйственную сферу новые виды растений и животных. И если сейчас, с одной стороны, выведены новые формы и сорта растений и воз- никло понятие «культурные растения», а с дру- гой — развита целая отрасль промышленно- сти — техническая микробиология, использую- щая для получения ценных веществ деятель- ность бактерий и микроскопических грибов, то громадный мир микроскопических водорослей, особенно одноклеточных, до недавнего време- ни оставался вне сферы практической деятель- ности людей. Несмотря на широкую распространенность микроскопических водорослей в природе и гро- мадное значение их в общем балансе фотосин- тетической продукции на Земле, несмотря так- же на то, что первый опыт по выращиванию этих организмов в искусственной среде отно- сится к 1871 г. (А. С. Ф а м и н ц и н), а полу- чение первых бактериологически чистых куль- тур их к 1890 г. (Б е й е р и н к), исследователи лишь в 40-е годы нашего столетия обрати- лись к вопросам искусственного культивиро- вания фотоавтотрофных микроскопических во- дорослей в связи с различными аспектами прак- тического их использования. ды — явления, ставшего в годы жаркого лета подлинным бичом наших южных искусственных водоемов. Исследования показали принципи- альную возможность использования этой мас- сы как в сельском хозяйстве в качестве органи- ческого удобрения под некоторые культуры и кормовой добавки в пищевом рационе домаш- них животных, так и в промышленности для получения некоторых ценных химических ве- ществ. Однако серьезным препятствием на этом пути служит сезонность «цветения» воды, непостоянство этого явления и его зависимость от погодных условий каждого отдельного лета. Поэтому действительно широкий размах полу- чает сейчас другое направление в практиче- ском использовании микроскопических водорос- лей, основанное на возможности управлять про- цессом,— искусственное культивирование их в производственных целях. Исходным объектом такого культивирования послужила однокле- точная водоросль хлорелла, но к настоящему времени в круг подобных объектов вовлечены не только другие зеленые, но и сине-зеленые и диатомовые водоросли. Это направление, по- степенно становящееся особой отраслью ми- кробиологической промышленности, чрезвычай- но интересно и значительно. ОСОБЕННОСТИ МИКРОСКОПИЧЕСКИХ ВОДОРОСЛЕИ КАК ОБЪЕКТОВ МАССОВОГО КУЛЬТИВИРОВАНИЯ Микроскопические водоросли давно стали объектом разносторонних исследований. Бла- годаря малым размерам, относительной про- стоте морфологической организации и спосо- бов размножения эти водоросли сравнитель- но легко поддерживать в лаборатории обычны- ми микробиологическими методами в виде культур на искусственных минеральных или органических, жидких или твердых (агаризо- ванных) питательных средах, на свету или
Q Q~ Ai4 АФ 2 Рис. 278. Схема клеточного цикла и рост синхронной культуры хлореллы: 2 — жизненный цикл клетки, D, L — различные стадии развития клетки; 2 — молодые клетки хлореллы через 2 — 3 часа после выхода автоспор; 8 — делящиеся клетки со сформировавшимися автоспорами. спективны, по мнению ряда исследователей, при массовом выращивании водорослей. Как видно из характера деления клеток, при отсутствии ограничивающих факторов рост чис- ла клеток идет в геометрической прогрессии, в соответствии с уравнением kN = dN dt З6 в темноте. Твердые среды используют преиму- щественно для сохранения водорослей (кол- лекции), а изучение их физиологических и био- химических свойств проводят главным образом на жидких питательных средах. Из микроскопических водорослей в первую очередь стали применять при массовом культи- вировании одноклеточные протококковые водо- росли (отдел Chlorophyta), в частности хлорел- лу и сценедесмус. Размножение этих форм упрощено до предела. У них полностью отсут- ствует половое размножение и рост культуры идет за счет формирования в материнской клет- ке дочерних автоспор, которых может быть в норме 2, 4, 8, 16, 32 (известно образование до 64 автоспор) в зависимости от штамма и усло- вий культивирования. После окончания деле- ния автоспоры выходят из клетки путем разры- ва оболочки; молодые клетки, интенсивно фо- тосинтезируя, растут до стадии созревания, и весь цикл повторяется сначала (рис. 278, 1). В определенные периоды развития по циклу клет- ки нуждаются в свете, а в другие развитие мо- жет проходить в темноте. Эта особенность об- легчает получение синхронных культур водорос- лей, т. е. культур, у которых все клетки нахо- дятся на одной и той же стадии развития (рис. 278, 2, 8). Синхронные культуры представляют интерес для исследования онтогенеза фотосинтезирую- щих клеток. Такие культуры могут быть пер- где Й — удельный коэффициент размножения, N — исходное число клеток, dN — прирост числа клеток за время dt. Однако в обычной накопительной культуре по мере увеличения числа клеток и изменения условий (истощение питательной среды, само- затенение клеток, накопление прижизненных выделений) происходит замедление темпа де- ления и накопления биомассы. В таких куль- турах рост описывается, как и у бактериаль- ных культур, S-образной кривой и может быть разделен на несколько стадий: 1) лаг-фазу, 2) фазу логарифмического, или экспоненци- ального, роста, 3) фазу линейного роста, 4) плато. Длительность лаг-фазы зависит от предыстории культуры, период экспоненци- ального роста у культур водорослей короткий, и чем быстрее происходит нарастание числа клеток, тем быстрее наступает самозатенение культуры, тем короче период экспоненциаль- ного роста. Линейная стадия роста культуры у водорослей наиболее длительная.
Поскольку фотос инте зирующие организмы, в отличие от гетеротрофных, нуждаются в лу- чистой энергии света, проникновение и рас- пределение его в суспензии является сущест- венным фактором обеспечения интенсивного фотосинтеза. В связи с этим немаловажным свойством культур микроскопических водорос- лей является гомогенность суспензии. Разница в величине дочерних и материнских клеток благодаря микроскопическим размерам не оказывает существенного влияния на светорас- пределение в суспензии, и с некоторым прибли- жением суспензии микроскопических водорос- лей можно рассматривать как окрашенный раствор, поглощение и распределение света в котором подчиняются известному закону Бугера — Ламберта — Вера, т. е. находятся в ло- гарифмической зависимости от концентрации клеток. Последнее облегчает расчеты световых полей внутри популяций микроводорослей, ис- ходя из оптических свойств суспензий и особен- ностей световых кривых фотосинтеза клеток различных культур водорослей. Как все зеленые растения, водоросли син- тезируют из СО, и воды в процессе фотосинтеза ценные органические вещества, такие, как бел- ки, жиры, углеводы, витамины и другие физио- логически активные соединения. Протококковые водоросли первоначально привлекли к себе внимание благодаря высо- кому содержанию белка — 50 — 60% от сухой биомассы. В белке хлореллы имеются все не- заменимые аминокислоты, по качеству его мож- но сравнить с белком пивных дрожжей, соевой и арахисовой муки. Его признают равноценным белку сухого молока. В среднем в сухой биомассе хлореллы содер- жится 10 — 20% углеводов. Значительную часть их составляет крахмал, хотя у некоторых пред- ставителей, даже в пределах рода СЫогеИа, углеводы могут быть представлены преимущест- венно гемицеллюлозами. Липиды составляют 20 — ЗОО/о и отличаются значительным содержа- нием ненасыщенных жирных кислот. Соотно- шение жирных кислот у хлореллы сходно с со- отношением, которое характерно для боль- шинства растительных масел. Биомасса протококковых водорослей (хло- реллы, сценедесмуса) содержит богатый набор витаминов, количество которых здесь выше, чем у большинства растительных культур. При- влекает внимание также высокое содержание каротина в культурах хлореллы и сценедес- муса, однако его особенно много у дюналиеллы, относящейся к вольвоксовым. Химический состав интенсивно размножаю- щихся культур хлореллы достаточно стабилен. Вместе с тем обращает на себя внимание чрез- вычайно высокая пластичность метаболизма хлореллы и ее способность радикально менять направленность биосинтеза в зависимости от условий культивирования и при различных воз- действиях. Давно было показано для некоторых культур хлореллы, что в условиях азотного голодания в клетках может накапливаться до 85О/о липидов. На этом основании считали, что хлорелла имеет белково-липидную направлен- ность метаболизма. Однако за последнее деся- тилетие при обследовании различных штаммов хлореллы было показано, что многие культуры обладают в тех же условиях, напротив, белково- углеводной направленностью метаболизма и могут синтезировать до 60% углеводов. У одних культур синтезируется преимущественно крах- мал (до 45О/о от суммы углеводов), у других- гемицеллюлозы. Таким образом, клетки хлореллы в зависи- мости от их генетических свойств и применяе- мых воздействий могут быть превращены в си- стемы, направленно синтезирующие белки, уг- леводы или жиры, что открывает принципи- альные возможности управления не только интенсивностью, но и качественной стороной биосинтеза у микроводорослей. Первые попытки массового культивирова- ния одноклеточных водорослей относятся к 40-и годам. Первоначально проводили исследова- ния по использованию клеток культивируемых водорослей в качестве корма для молоди рыб; затем очень быстро эти водоросли стали рас- сматривать как возможный источник различ- ных веществ, в том числе и пищевых. В настоящее время одноклеточные и некото- рые другие микроскопические фотоавтотроф- ные водоросли широко изучаются не только в СССР, но и в США, Японии, ФРГ, Франции, Италии, Швеции, Бельгии, Чехословакии, Бол- гарии и других странах в связи со следующими аспектами их применения: 1) сельское хозяй- ство — дополнительный источник белка, вита- минов; 2) очистка сточных вод и биосферы; 3) проблема освоения космоса — как звено в замкнутых экологических системах, способ- ное обеспечить биологическую регенерацию воздуха и воспроизводство пищи; 4) получение ценных метаболитов и веществ для медицинской и микробиологической промышленности, а так- же для получения меченых препаратов; 5) био- логическая фиксация атмосферного азота; 6) использование в научно-исследовательских работах в связи с изучением фундаментальных вопросов биологического саморегулирования и биосинтез а фотосинтезирующих организмов. Для решения этих проблем разрабатываются способы культивирования водорослей двух ти- пов — массовая культура под открытым небом и интенсивное культивирование в замкнутых аппаратах в полностью контролируемых усло- 369
3«à фотосинтеза Зона фотосинтеза Активно фото зирующая 3 46 О 1. О 'О Ю m О Х Ф 1. Х Фотосинтез клеток Толщина слоя суспензии Рис. 279. Оптическая п функциональная структура культуры водорослей как фотосинтезирующей систе- мы: справа — распределение света, слева — интенсивность фото- синтеза отдельной клетки в популяции микроводорослей. виях по типу современных микробиологических производств. В обоих случаях задача заклю- чается в получении максимальных выходов биомассы, что требует изучения путей опти- мизации роста и фотосинтетической продуктив- ности культур микроводорослей. ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ КУЛЬТУР МИКРОСКОПИЧЕСКИХ ВОДОРОСЛЕЙ КАК ФОТОСИНТКЗИРУЮЩИХ СИСТЕМ Культуры микроскопических водорослей яв- ляются целостными фотосинтезирующими си- стемами. Для получения высокой фотосинтетиче- ской продуктивности необходимо знание фи- зиологических особенностей этих систем, а не только физиологии отдельной клетки. Как бы ни различались современные методы культивирования водорослей, все они основа- ны на снабжении клеток достаточным количе- ством света, углекислоты и других питатель- ных веществ. Исследование функциональной зависимости продуктивности культур микроводорослей от напряженности и сочетания значимых факто- ров — необходимая предпосылка для разра- ботки физиологически обоснованных режимов их культивирования и принципов интенсифика- ции их продуктивности. Как следует из приведенного на стр. 368 урав- нения, при выращивании в условиях экспонен- циального роста, но при низкой плотности суспензии можно получить высокий коэффи- циент размножения культуры, однако это еще не значит, что будет получен высокий суммар- ный урожай. Такая культура хороша для изу- чения потенциальных свойств клетки, но недо- статочна для использования при массовоя культивировании. В реальных условиях куль- тивирования суммарная фотосинтетическая про- дуктивность водорослей тем выше, чем при бо- лее высокой плотности суспензии сохраняется высокая скорость размножения. И далеко не всегда результаты, полученные при исследо- вании разбавленных суспензий (что может быть приравнено к изучению отдельной клетки), могут быть перенесены на культуры высокой плотности. Например, впервые полученная в 1953 г. термофильная форма хлореллы (С а- р о к и н и М а й е р с) прославилась еще и тем, что для нее в определенных условиях света и температуры в суспензии с низкой плотностью клеток было найдено тысячекрат- ное увеличение числа клеток в сутки (10 удвое- ний). Следовательно, если создать этой культу- ре оптимальные условия экспоненциального роста при плотности хотя бы 50 млн. клеток в 1 мл, то 1 л такой культуры мог бы обеспечить урожай в 500 г/л в сутки и это была бы высоко- интенсивная культура водорослей, размно- жающаяся с максимальной скоростью и даю- щая высокие урожаи за максимально короткий срок. Однако организация такого рода культи- вирования практически трудноосуществима, так как уже при четвертом удвоении плотность культуры поднимается до 800 млн. в 1 лл и, естественно, при этом резко меняются все усло- вия культивирования. Для правильного осве- щения суспензии этот литр культуры требова- лось бы распределить очень тонким слоем по большой поверхности, в результате чего конст- рукция установки стала бы чрезвычайно слож- ной и громоздкой. Таким образом, для правиль- ной организации культивирования водорослей с высокой фотосинтетической продуктивностью необходимо изучение действия хотя бы основ- ных факторов (свет, температура, концентра- ция СО, и т. д.) на фотосинтезирующие системы ВЫСОКОИ ПЛОТНОСТИ. В отличие от популяций гетеротрофных орга- низмов, где каждому из факторов, определяю- щих продуктивность культуры, обеспечен оди- наковый доступ ко всем индивидам в любой рассматриваемый момент времени,, в популя- ции автотрофных клеток исключение составляет световой фактор, который в силу оптических свойств культуры как сильно поглощающей свет среды достаточно быстро становится огра-
Ъ О 3 с О X :Г J) I- О О I- Х , X О 0 О О о I- Б; 600 50 +~ ф 0 0,5 2,0 232 1,5 0 58 116 Нонцентрация Cp, % Интенсивность света Рис. 280. Зависимость интенсивности фотосинтеза (А) и скорости роста (Б) культуры хлореллы от интенсив- ности света, концентрации углекислоты, температуры и от сопряженного их влияния (интенсивность света- в тыс. арг,'см' в сек, интенсивность фотосинтеза — в л Ог с 1 л суспензии в сутки, число клеток — в млн. на 1 лл суспензия) . 37 ничивающим. Кратковременность экспонен- циальной фазы роста в интенсивно растущих культурах и выход на линейную стадию ро- ста объясняется быстрым формированием пол- ностью поглощающего свет слоя, численность клеток в котором при заданной интенсивно- сти света остается величиной постоянной, не- смотря на увеличение плотности по мере роста всей популяции. Формирование такого слоя определяет достаточно резкий переход от процесса возрастания интенсивности фотосин- теза по мере роста культуры к его стацио- нарному состоянию (и постоянству прироста числа клеток и биомассы в единицу времени), несмотря на продолжающееся суммарное уве- личение числа клеток. При этом плотность культуры, при которой происходит указанный переход, различна в реакторах с разной толщи- ной слоя суспензии, однако поверхностная концентрация клеток в момент этого перехода остается величиной постоянной. В плотной, вышедшей на линейную фазу роста культуре суммарная фотосинтетическая продуктивность суспензии определяется в каж- дый элементарный отрезок времени работой только тех клеток, которые находятся в зоне облучения (рис. 279). При этом часть клеток в зоне облучения осуществляет фотосинтез на линейном участке, а часть — на плато свето- вой кривой фотосинтеза отдельной клетки. Остальные клетки (вне «зоны фотосинтеза») в принципе являются балластными и не дают вклад в общий фотосинтез популяции, посколь- ку длительность световых реакций фотосинте- за порядка 10 ' сек существенно короче пе- риода миграции клеток между просвечиваемыми и непросвечиваемыми зонами суспензии, что достигается за счет перемешивания культуры. Описанные закономерности положены в ос- нову ряда разработанных в настоящее время математических моделей фотосинтетической продуктивности микроводорослей и использу- ются при конструкторской разработке техноло- гии, реакторов и управляющей аппаратуры для высокоинтенсивного культивирования этих ор- ганизмов. Углекислота может вводиться в автотрофно выращиваемые культуры водо рослеи различ- ными методами. Наиболее распространенный способ — это подача углекислоты в виде газо- воздушной смеси, осуществляющей, помимо снабжения водорослей источником углерода, функцию переметпивания культуры. Найден- ные применительно к условиям высокоинтен- сивного культивирования хлореллы насыщаю- щие и полунасыщающие концентрации СО, (рис. 280) оказались более высокими, чем опи- саны в литературе для разбавленных суспен- зий, но не превышающими значений 1,6— 1,7% СО2 в газовой фазе (35 — 40 10 ' моля растворенной СО, на 1 л) при насыщающих ин- тенсивностях света и плотности суспензии
Схема взаимосвязи факторов, влияющих на продук- тивность культуры. Интенсивность света Гемпература Угл ени слота ~ б r" г Г г Г г Г/ Г Концентрация элементов финераль н ого литания 600 млн. клеток в 1 мл. На рисунке 280,А в ви- де трехмерного графика представлены семейства углекислотных и световых кривых фотосин- теза хлореллы и зависимость фотосинтетиче- ской продуктивности культуры от взаимосвя- зи этих характеристик. Так, при наибольшей, насыщающей интенсивности света продуктив- ность культуры в 4 раза выше при концентра- .С % Рис. 281. Лабораторная установка для выращивания в условиях интенсивного размножения и фотосинтеза различных культур одноклеточных микроскопических водорослей. ции 1% СО, чем при 0,25% СО, и обратно: при одной и той me концентрации СО, (напри- мер, 1% СО,) продуктивность тем выше, чем интенсивность Водоросли различаются по световым и тем- пературным оптимумам для роста и фото- синтеза. В пределах рода Chlorella есть светолюбивые и теневыносливые культуры с меньшим или большим содержанием хлорофилла, с большей или меньшей активностью единицы хлорофил- ла, есть термофильные, мезофильные и пси- хрофильные (хорошо растущие при температуре +5... +10 'С) штаммы. Каждый штамм в той или иной мере обладает адаптационными спо- собностями. Но у каждой культуры свой гене- тически закрепленный диапазон требований к освещенности и температуре. На основании известных экспериментальных данных можно охарактеризовать в целом груп- пу мезофильных культур как более тенелюби- вую, а термофильных культур как более свето- любивую. Так, мезофильные формы обладают более низким и более узким плато светового насыщения для роста по сравнению с термо- фильными культурами. Применение культур высоких плотностей имеет особенно большой смысл при высо- ких освещенностях, позволяющих максимально приблизиться к потенциально возможным ско- ростям роста культуры. На рисунке 280, Б при- ведены световые и температурные кривые зави- симости роста термофильной культуры. Трех- мерный график хорошо показывает тесную функциональную взаимообусловленность ха- рактера температурных и световых кривых роста и необходимость тщательного подбора оптимальных сочетаний этих параметров для высокопродуктивного культивирования. Для каждой интенсивности света существует своя температурная кривая роста и продуктивности культуры, так же как для каждой температуры имеется своя световая кривая. По мере повы- шения освещенности температурный оптимум смещается в сторону более высоких значений температуры, так же как и при повышении тем- пературы для получения максимальной продук- тивности необходимо соответствующее изме- нение освещенности. Важно, что в культурах высоких плотностей, как и в разбавленных сус- пензиях, четко обнаруживается эффект фото- ингибирования роста (угнетение ростовых про- цессов под действием света) культуры при тем- пературе, не сочетающейся с примененной ин- тенсивностью света. Если учесть, что температурные условия яв- ляются диффузно (или равномерно) действую- щим фактором и не зависят от плотности куль- туры, а облученность отдельных клеток в попу-
ляции, наоборот, сильно зависит от плотности и оптических свойств суспензии, то становится ясно: чтобы найти оптимальные сочетания об- лученности и температуры для каждой культу- ры, необходимо снять световые и температурные характеристики не только для клетки, но и для популяции. Как показано на примерах влияния света, температуры, концентрации СО„для получе- ния высокой продуктивности культуры сущест- венное значение имеет не столько зависимость роста и фотосинтеза водорослей от каждого из этих факторов в отдельности, сколько их сопря- женное действие. Обнаруженные взаимосвязи могут быть показаны в виде схемы на стр. 372. Поскольку в культурах высоких плотностей для обеспечения интенсивного роста применя- ют высокие освещенности, высокие температуры и высокие концентрации солей, т. е. создается напряженность всех факторов среды, «отстава- ние» какого-либо одного фактора, нарушая сбалансированное взаимодействие всех факто- ров, может повлечь за собой не только сниже- ние продуктивности до уровня действия этого минимального фактора, но во многих случаях и гибель культуры. При создании систем культивирования водо- рослей высокой фотосинтетической произво- дительности одним из первых встает вопрос о применении культур, генотипические свой- ства которых отвечали бы заданным условиям. Например, водоросли в интенсивно растущих плотных популяциях должны переносить крат- ковременную переоблученность, т. е. культура должна иметь высокое и продолженное плато светового насыщения для роста и фотосинтеза. Этим свойством в большей степени обладают термофильные штаммы. При массовом культивировании под откры- тым небом более эффективными в определенные периоды могут быть мезофильные культуры. Японские исследователи провели выращивание Рис. 282. Принципиальная схема высокоинтенснвного управляемого проточного культивирования микроводо- рослей в контролируемых условиях. 1 — реактор', 2 — рубашка водяного охлаждения; 8 — ультратермостат; 4 — блок термодатчиков; 5 — схема записи темпера- туры; 6 — прибор, регистрирующий температуру; 7 — 9 — схема контроля и регистрации рН; 10 — датчик плотности; 11— 14 — система контроля, регистрации и регулировки плотности; 15 — дозирующее устройство; 16 — основная и корректирующие питательные среды; 17 — исполнительные электромагнитные клапаны, управляемые системой регулировки рН; 18 — сбор- ник урожая; 19 — циркуляционная помпа; 20 — командное электропневматическое устройство; 21 — газоанализатор СО; 22 — прибор, регистрирующий газовый состав; Л — источник света.
1 Рис. 283. Установки для высокоинтенсивного проточного культивирования водорослей: в в е р х у — реактор с системой световодов, в н и з у — плоскопараллельный реактор барботажно-камерного типа.
смешанной культуры водорослей, состоящей из мезофильного и термофильного штаммов. В жаркие месяцы преимущественно развивалась те рмофильная форма, в остальное время- мезофильная. Отбором в природе и селекционно-генетиче- скими методами получено значительное число мезофильных и те рмофильных высокопродукти- вныхых форм хлореллы. Сравнительное изуче- ние различных штаммов водорослей показы- вает широкое их разнообразие в отношении к температуре, освещенности, способности роста в плотных популяциях. Это открывает возмож- ность направленного подбора форм водорослей применительно к конкретным условиям и це- лям их культивирования. Отбор и поддержание в коллекции перспек- тивных форм является важной задачей. Развитие исследований по массовой культуре водорослей дало толчок к совершенствованию методов лабораторного их выращивания, бла- годаря чему возник метод лабораторного ин- тенсивного культивирования микроводорослей. На рисунке 281 представлен один из способов современного лабораторного культивирования одноклеточных водорослей, обеспечивающий интенсивный рост культур, относящихся к раз- личным систематическим группам (зеленые- СЫогеИа, Scenedesmus, Ankistrodesmus, Duna- liella, Chlamydomonas; сине-зеленые — Anacys- tis, Spirulina, Anabaena, Synechocystis, Apha- nizornenon, Nostoc, Tolypothrix и др.). МЕТОДЫ МАССОВОГО КУЛЬТИВИРОВАНИЯ МИКРОСКОПИЧЕСКИХ ВОДОРОСЛЕЙ Массовая культура под открытым небом. Различного типа установки под открытым не- бом были испытаны и описаны уже в начале 50-х годов Институтом Карнеги (США), в Япо- нии и некоторых других странах. В настоящее время существует много разных конструкций установок для выращивания водорослей под открытым небом (табл. 40, 41). Принципиальная схема такого культивиро- вания состоит в выращивании водорослей в жид- ких питательных средах в бассейнах, лотках и других емкостях с различными способами перемешивания, подачи углекислоты и исполь- зованием солнечного света. Представляет интерес установка для массо- вого культивирования сценедесмуса в Тжебоне (Чехословакия), где суспензия водорослей сте- кает по плоской, специальным образом обору- дованной поверхности, которая является кры- шей оранжереи. В другом варианте эта же установка может размещаться на поверхности грунта и используется, в частности, в Болгарии. В Рупите (Болгария) при строительстве уста- новок массового культивирования применены местные природные ресурсы, обеспечивающие наряду с хорошей инсоляцией использование природного источника СО„воды источника, богатого минеральными солями, и регулирова- ние температуры за счет воды горячих источни- ков. В Советском Союзе Биологическим инсти- тутом ЛГУ разработаны типовые установки лотков ого типа с открытой циркуляционной системой. В Узбекистане используют преиму- щественно круглые бассейны по типу японских установок. Размеры установок варьируют от небольших до 1000 — 2000 лР. В качестве объектов культивирования широ- ко используются зеленые одноклеточные водо- росли, особенно из протококковых (хлорелла и сценедесмус) и из вольвоксовых (дюналиел- ла); ведутся также работы с сине-зеленой нитчатой водорослью спирулиной и некоторы- ми другими. Производительность установок составляет в среднем 15 — 20 г/м' сухой биомассы в сутки, достигая иногда 25 — 30, а в отдельных случа- ях 35 — 40 г/м'. Урожай с единицы объема суспензии в условиях массовой культуры под открытым небом составляет в среднем 0,2 г/л в сутки, поднимаясь в отдельных случаях до 1,2 г/л. Плотность культуры, при которой ве- дется выращивание водорослей, варьирует в зависимости от типа установок и составляет 25 — 400 млн. клеток на 1 мл. Культуры под открытым небом характери- зуются низкими коэффициентами размножения, что свидетельствует, по существу, об экстенсив- ном ведении культуры, при котором получение больших количеств биомассы обеспечивается использованием больших площадей. Постоянно меняющиеся погодные условия являются причиной того, что в установках открытого типа невозможно длительное, ста- бильное снятие устойчивого урожая, что при- ближает этот принцип культивирования водо- рослей к способам возделывания высших рас- тений, когда процесс накопления урожая су- щественным образом зависит от погодных усло- вий. Продуктивность установок под открытым не- бом удается повысить путем организации сис- тем обогрева и охлаждения и досвечивания культуры искусственными источниками осве- щения. Такое культивирование, по существу, занимает промежуточное положение между экс- тенсивной культурой водорослей и интенсив- ным высокоорганизованным культивировани- ем по типу микробиологических производств. Интенсивная культура. В 60-х годах в раз- личных лабораториях был разработан ряд установок и аппаратов высокоинтенсивного 375
управляемого культивирования фотосинтезиру- ющих микроводорослей в полностью контроли- руемых условиях с автоматической стабилиза- цией оптимальных условий и непрерывной автоматической регистрацией таких важных физиологических функций культуры, как ско- рость роста, интенсивность фотосинтеза, мине- ральное питание. Наиболее совершенным из таких методов культивирования является проточное выращи- вание водорослей, при котором по сигналам, получаемым от самой культуры, осуществля- ются автоматический отбор прирастающих кле- ток (урожая), подача свежей питательной среды и стабилизация оптической плотности культуры. Одна из принципиальных схем та- кого типа культивирования представлена на рисунке 282. Главным преимуществом этих методов куль- тивирования является возможность вести дли- тельное непрерывное выращивание водорослей с поддержанием постоянной плотности суспен- зии на оптимальных значениях, при которых на- блюдается максимальная продуктивность куль- туры. С этой целью разработаны специаль- ные реакторы, в которых используются мощные источники света и специальные системы свето- распределения и световодов, что обеспечивает равномерное освещение клеток в достаточно плотных культурах. Примеры таких установок даны на таблице 40 и рисунке 283. Урожаи, в частности, хлореллы, которые получают в таких установках, составляют примерно 30 — 40 г сухой биомассы с 1 л сус- пензии в сутки или 80 — 100 г с 1 м' освеща- емой поверхности. Таким образом, в настоящее время можно считать достаточно детально разработанными физиологические основы культивирования мик- роскопических фотосинтезирующих водорос- лей и некоторые принципы технологии их выращивания как в установках под открытым небом, так и в закрытых аппаратах. Рентабельность массового культивирования водорослей существенным образом зависит от аспектов применения получаемой биомассы и должна оцениваться, очевидно, в каждом конкретном случае индивидуально. Так, мето- ды высокоинтенсивного культивирования водо- рослей, помимо исследовательской работы, направленной на выяснение потенциальной про- дуктивности фотосинтетического аппарата рас- тений и других проблем физиологии, биохимии и генетики фотосинтезирующих клеток, нахо- дят применение для биосинтеза соединений, меченых различными изотопами углерода (С'4, С'з), дейтерированных соединений, а так- же в космической биологии для создания зам- кнутых экологических систем жизнеобеспече- ния. Высокая продуктивность и стабильность работы систем интенсивного проточного куль- тивирования хлореллы позволили осуществить многомесячные эксперименты с испытателями по биологической регенерации воздуха с по- мощью фотосинтеза. Эффективность применения биомассы водо- рослей в сельском хозяйстве остается еще неяс- ной. Биомасса водорослей, получаемая при культивировании под открытым небом, исполь- зуется для изучения их кормовых достоинств как источника белка и физиологически актив- ных соединений. Результаты противоречивы, что свидетельствует о необходимости проведе- ния дальнейшей исследовательской работы. Вместе с тем в более широком плане разра- ботка способов промышленного культивиро- вания одноклеточных фотосинтезирующих мик- роводорослей является одним из путей введения процесса фотосинтеза в промышленное произ- водство. Последствия такого явления своеоб- разной индустриализации фотосинтеза трудно переоценить.
ОБЩЛЯ XAPAKTEPÈCTÈКЛ ANIJJAAH VIKOB ЛИШАЙНИКИ И ИХ ОТЛИЧИЕ ОТ ДРУГИХ РАСТЕНИЙ 379 Лишайники представляют своеобразную груп- пу комплексных организмов, тело которых всег- да состоит из двух компонентов — гриба и водо- росли. Сейчас каждый школьник знает, что в основе биологии лишайников лежит явление симбиоза — сожительства двух различных ор- ганизмов. Но еще немногим более ста лет назад лишайники были для ученых великой загадкой, и открытие Симоном Швенде- н е р о ы в 1867 г. их сущности оценивалось как одно из наиболее удивительных открытий того времени. Однако как организмы лишайники были из- вестны ученым и в народе задолго до открытия пх сущности. Еще великий Т е о ф р а с т (371 — 286 до н. з.), «отец ботаники», дал описа- ние двух лишайников — уснеи (Usnea) и ро- челлы (Воссе11а). Последнюю yme тогда исполь- зовали для получения красящих веществ. По- степенно количество известных видов лишай- ников возрастало. Правда, в те времена их на- зывали нередко то мхами, то водорослями, то даже «хаосом природы» и «убогой нищетой расти- тельности». Сейчас известно более 20 000 лишайников. И каждый год ученые обнаруживают и описыва- ют десятки и сотни новых неизвестных видов. В настоящее время лихенология (лат. lic- hen — лишайник) — наука о лишайниках— изучает сложный комплекс проблем, связанных с возникновением, филогенией, строением, сис- тематикой, биохимией, физиологией, распрост- ранением и экологией лишайников. Ниже эти вопросы будут освещены. Чем же лишайники отличаются от других растений? Во-первых, специфичный признак лишайни- ков — симбиотическое сожительство двух разны~ организмов — гетеротрофного гриба (микобионт) и автотрофной водоросли (фикобионт). Не всякое сожительство гриба и водоросли образует ли- шайник. Лишайниковое сожительство должно быть постоянным и исторически выработавшим- ся, а не случайным, кратковременным. В приро- де бывают случаи, когда гриб и водоросль об- разуют временное смешанное скопление, но это еще не лишайник. В настоящем лишайнике гриб и водоросль вступают в тесные взаимоот- ношения, грибной компонент окружает водо- росли и может даже проникать в их клетки. Во-вторых, лишайники образуют особые морфологические типы, жизненные формы, не встречающиеся отдельно у слагающих лишай- никовое слоевище грибов и водорослей, т. е. лишайники прошли исторический, длительный формообразующий процесс на основе симбиоза, приведший к формированию специфичных мор- фологических форм внешнего и внутреннего строения. В-третьих, для лишайников в целом и каж- дого из их компонентов в отдельности харак- терен особый тип метаболизма. Физиология гриба и водоросли в слоевище лишайника во многом отличается от физиологии свободно- живущих грибов и водорослей. В-четвертых, весьма специфична биохимия лишайников, образование в них вторичных продуктов обмена — лишайниковых веществ, не встречающихся в других группах орга- низмов. Наконец, лишайники существенно отлича- ются от других групп организмов, в том числе
КОМПОНЕНТЫ ЛИШАЙНИКОВ И ИХ ВЗАИМООТНОШЕНИЯ 380 и от свободноживущих грибов и водорослей, особой биологией: способами размножения, мед- ленным ростом, отношением к экологическим условиям и др. На основе этих специфических свойств ли- шайникам можно дать следующее определение: лишайниками являются организмы, тело (слое- вище) которых постоянно состоит из двух компо- нентов — автотрофного фикобионта и гетеро- трофного микобионта, образующих единое сим- биотическое сожительство, отличающееся осо- быми морфологическими типами и особыми фи- зиолого-биохимическими процессами. Однако следует сразу же заметить, что все растения, относящиеся к лишайникам, делятсн на три неравноценные группы, из которых вполне соответствует данному определению лишь первая из них, включающая, правда, подавляющее большинство известных видов. Эту группу видов выделяют как класс сумчатых лишайников, так как грибы, образовавшие их, нвлнютсн сумчатыми грибами. Вторую очень небольшую группу выделяют как класс базиди- альных лишайников в соответствии с тем, что они образованы базидиальными грибами. Ба- ГРИБНОЙ КОМПОНЕНТ (МИКОБИОНТ) Представители трех классов грибов — аско- мицетов, базидиомицетов и фикомицетов, всту- пив в контакт с водорослями, дали начало обра- зованию лишайников. О близости с грибами говорит строение вегетативного тела лишайни- ков и их плодовых тел. Вегетативное тело ли- шайников, называемое, как у всех низших растений, талломом или слоевищем, целиком состоит из переплетения грибных гиф. Водо- росли же либо разбросаны в беспорядке среди грибных гиф по всей толще слоевища, либо расположены отдельным дифференцированным слоем несколько ниже его верхней поверхности. Как известно, вегетативное тело обычных грибов, состоящее из грибных гиф и называе- мое грибницей или мицелием, в большинстве случаев развиваетсн целиком внутри субстрата, а на поверхности мы видим лишь плодовые тела гриба. В отличие от этого вегетативное тело лишайников, также образованное грибными гифами, целиком ~кивет в воздушной среде (исключение составляет лишь небольшая груп- па лишайников, слоевище которых развивается внутри камня или древесного субстрата). Ли- шайники иногда даже называют воздушными грибами. Возникает вопрос: появились ли у лишайникового гриба какие-либо особые при- зидиальные лишайники — менее устойчива н группа и соответствует не всем указанным приз- накам. Наконец, среди лишайников имеется третья группа видов, у которых плодовые тела со спорами не обнаружены, поэтому их место в системе лишайников остаетсн неизвестным. Их объединяют в искусственную группу «не- совершенных лишайников» (Lichenes imperfecti). Со временем у некоторых из них находят аско- карпы и тогда их переносят в соответствующее место в системе класса сумчатых лишайников. В других случаях на действительное место в системе могут указать другие данные- тонкан структура слоевища или химизм. Из несовершенных лишайников самыми распро- страненными нвлнютсн так называемые лепра- рии (Lepraria). Они образуют порошковатые налеты на разных субстратах — камнях, ска- лах, стволах деревьев, мха), обычно беловато- серые (Lepraria aeruginosa), иногда зеленовато- желтые (Lepraria chlorina) или интенсивно- желтые (Lepraria candelaria). В настоящей книге рассматриваютсн только первые два класса, на несовершенных лишайни- ках мы далее не останавливаемсн. способленин в связи с переходом к обитанию в воздушной среде и контакту с водорослями? Отличаются ли гифы, образующие лишайнико- вое слоевище, от гиф грибов? Сначала рассмотрим строение гиф лишайника (рис. 284). Гифы, образующие слоевище ли- шайника, представляют собой простые или раз- ветвленные тонкие (шириной 3 — 10 мкм) нити, растущие вершиной. Гифа покрыта двухслой- ной оболочкой, под которой находится прото- плазма. Обычно гифы бывают поделены попереч- ными перегородками на клетки. У большинства лишайников в центральной части поперечных перегородок имеетсн одно маленькое отверстие, называемое п е р ф о р а ц и е й. Через перфо- рацию протнгиваетсн тончайший цитоплазмати- ческий тнж, который носит название п л а з м о- д е с м ы. Млазмодесма соединяет протопласты соседних клеток. У некоторых лишайников поперечнан перегородка имеет не одну, а три перфорации, расположенные на равных расстоя- ниях друг от друга. Кроме того, перфорации образуютсн не только на поперечных перегород- ках гиф, но и на их продольных стенках, в результате плазмодесмами соединяются прото- пласты двух параллельно расположенных со- седних гиф. Протоплазма лишайников слегка зерниста. В вегетативных клетках почти всегда имеется
одно ядро, но у некоторых видов бывает по два или по многу ядер в одной клетке. Кариосисте- матических исследований лишайников почти не проводилось, поэтому сведения об их хромо- сомах очень скудны. Известно, что они мелкие, трудно различимые и число их невелико. Так, например, у одного из накипных лишайников рода лецидея (Lecidea) 2 хромосомы, а у листо- ватого лишайника из рода дерматокарпон (Der- matocarpon) их 6 или 8. В протоплазме имеются вакуоли шаровидной или вытянутой формы. Теперь вернемся к вопросу: каковы различия в строении гиф лишайников и гиф грибов? Ока- залось, что переход вегетативных гиф лишай- никового слоевища к воздушному образу жизни привел к утолщению их оболочки. По сравне- нию с обычными грибными гифами гифы лишай- ников имеют гораздо более толстые оболочки и более узкий просвет, заполненный протоплаз- мой. Это особенно характерно для высокораз- витых кустистых и листоватых лишайников. Лишайнику, имеющему форму кустика или крупной листовидной пластинки, приходится выдерживать и воздействие сильных ветров, и удары дождевых капель, снега, кусочков льда, и другие влияния окружающей среды. В таких слоевищах грибные гифы с сильно утолщенными стенками выполняют роль механической, укреп- ляющей «ткани». Особенно толстые стенки име- ют гифы, расположенные во внешних, коровых слоях. Например, в коровом слое лишайника цетрарии исландской (Cetraria islandica, табл. 48, б), обитающего в суровых условиях север- ных тундр, оболочки гиф настолько толстые и желатинообразные, а просветы такие узкие и маленькие, что структура этого слоя с трудом просматривается под микроскопом, а отдельные гифы и вовсе невозможно рассмотреть. Далее, в гифах лишайников по сравнению с обычными грибными гифами наблюдается силь- ное утолщение поперечных перегородок. Ка- залось бы, это должно затруднять обмен веще- ствами между клетками, но на самом деле этого не происходит: в местах утолщения поперечных перегородок гифы сами значительно расширяют- ся (рис. 284, 2). Это ведет, в свою очередь, к уве- личению диаметра перфораций в перегородках и, соответственно, к утолщению плазматического тяжа, обеспечивающего обмен веществами меж- ду соседними клетками. Столь большие расши- рения гиф на местах расположения поперечных перегородок также неизвестны у грибов. У целого ряда лишайников оболочки гиф могут сильно разбухать и ослизняться. Это происходит за счет содержания во внешних слоях оболочек пектиновых веществ, легко растворимых в воде углеводов. Особенно сильно ослизняются и набухают при увлажнении гифы так называемых слизистых лишайников, в слое- Рис. 284. Гифы лишайников: 1 — строение гиф; 2 — расширения гиф на месте расположения поперечной перегородки. а — ядро, б — вакуоли, в — резерв- ные вещества, г — плазмодесма. вище которых водорослевым партнером обычно бывает сине-зеленая водоросль. Гифы обычных грибов, как правило, не обладают этой способ- ностью, исключение составляют лишь дрожал- ковые грибы. Интересные, специфичные для лишайнико- вых гиф образования, которые у свободножи- вущих грибов в природных условиях не встре- чаютсл,— так называемые ж и р о в ы е к л е т- ки илиясировые гифы. Ониразвивают- ся обычно в нижней части слоевища, в местах прикрепления к субстрату. Жировые клетки имеют зеленоватую окраску, обусловленную содержанием жира. Они, как правило, крупнее обычных клеток и имеют вздутую мешковидную, иногда почти шаровидную форму (рис. 285). Причины образования жировых клеток и их роль в жизни лишайников до сих пор не выясне- ны. Однако было замечено, что чаще всего жи- ровые клетки встречаются у лишайников, расту- щих на известковом субстрате, особенно в слое- вищах, глубоко погруженных в известняк. В слоевищах лишайников, которые произрастают на гранитных породах и на древесном субстрате, жировые клетки образуются очень редко. Более того, если лишайник растет на горной породе смешанного состава, то жировые клетки можно заметить только в гифах, проходящих через участки карбонатсодерясащих пород, а в гифах того же слоевища на бескарбонатных участках жировые клетки отсутствуют. Это позволило сделать вывод, что образование жира обуслов- ливает углекислота, освобождающаяся при раз- лоясении карбонатов гифами лишайников. Одна- ко это пока лишь одна из гипотез, пытающихся объяснить возникновение этих специфичных для лишайников образований. Тесный контакт с водорослями обусловил появление у микобионта лишайников некото-
Рис. 285. Гифы лишайников с жировыми клетками. Рис. 286. Двигающие гифы в слоевище лишайника: 2 — двигающие гифы, соединенные в пучок, обращенный по направлению к периферии слоевища; 2 — двигающие гифы, про- талкивающие клетку водоросли в маленькую клиновидную по- лость перед ней. а — клетка водоросли, б — двигающие гифы, 8 — полость. рых специальных типов гиф, отсутствующих у грибов. Прежде всего это так называемые ищущие и охватывающие гифы. Как видно из названий, предназначение их- разыскивать и охватывать клетки водорослей. Эти гифы особенно хорошо заметны в начале развития слоевища лишайников из прорастаю- щей споры. В таком слоевище либо вовсе еще нет водорослей, либо их еще очень мало. Ищу- щие гифы являются боковыми ответвлениями обычных гиф. Они имеют вид длинных TQHIiIIx нитей с перегородками лишь у самого основа- ния. После того как ищущие гифы обнаружат водоросль, у них понвлнютсн боковые ответвле- ния, плотно захватывающие и тесно оплетаю- щие клетки или нити водорослей. Эти боковые ответвления в виде тонких повторно ветвящихся гиф, тесно окружающие водоросль, носят наз- вание охватывающих гиф. Позднее охватываю- щие гифы делятся поперечными перегородками на короткие клетки, обычно несколько вздутые, и вступают в контакт с клетками водорослей. Еще один тип специфичных для лишайников гиф, отсутствующий у грибов,— это так назы- ваемые двигающие гифы. Они возни- кают в зоне водорослей и служат длн переноса их клеток в растущий край слоевища, который обычно бывает образован одними только гифамп микобионта и не содержит водорослей. Вначале двигающие гифы окружают отдельную клетку водоросли, затем они становнтсн более круп- ными, богатыми протоплазмой, и соединнютсн в пучок, обращенный к периферии слоевища (рис. 286, 1). Сама водоросль, сдавленная гифа- ми, из шаровидной становится эллипсоидной или грушевидной. Пучок двигающих гиф с за- ключенными внутри их водорослями разраста- ется по направлению к периферии и оказывает определенное давление на окружающие гифы. Гифы, находящиеся впереди пучка, раздвигают- сн, и между ними образуется маленькая клино- видная полость, куда и проталкивается захва- ченная водоросль (рис. 286,2). Так постепенно двигающие гифы переносят водоросль из водо- рослевой зоны в растущий край слоевища. В результате этого процесса, происходящего не- прерывно, в периферической части слоевища постепенно формируется зона водорослей, ко- торан затем соединнетсн в одно целое с зоной водорослей всего остального слоевища. В лишайниковых гифах образуется гораздо большее количество разных химических ве- ществ, чем в грибных гифах. Подробнее об этом рассказано в разделе «Химический состав лишайников». Благодаря воздушному образу жизни и сим- биозу с водорослями у гриба появилась тенден- ция к развитию сложной вегетативной струк- туры. Как правило, микобионт лишайников
образует сложно устроенные талломы, с хорошо дифференцированными анатомическими слоями, с особыми органами прикрепления, которые встречаются лишь у лишайников (см. раздел «Внешнее и внутреннее строение лишайников»). Ученые предполагают, что лишайник овые микобионты не встречаютсн в природе а свобод- ноживущем состоянии. На эту мысль наводит тот факт, что в культуре на искусственных средах изолированные микобионты растут очень медленно и никогда не образуют плодовых тел. Колонии лишайниковых грибов в культуре соВ- сем не похожи по внешнему облику на слоевище лишайника, из которого они были изолированы. В жидкой среде гриб образует в основном сли- зеподобную мицелиальную массу или отдельные комочки, а на агар е — компактные колонии. Обычно месячный прирост изолированных ми- кобионтов в культуре не превышает 1 — 2 мл, а саман быстрорастущая колония лишайнико- вого гриба достигала на агаре за месяц а диа- метре 1 — 2 см. В жидкой среде, например в культуральных сосудах, сухая масса одного из лишайниковых микобионтов составила 100 г, в то время как свободноживущие грибы аспер- гиллы и пенициллы давали в тех же условиях массу, в 4 раза большую. ВОДОРОСЛЕВЫЙ КОМПОНЕНТ (ФикоБионт) Водоросли, встречающиеся в слоевище ли- шайников, называют фикобионтом лишайников. По своему систематическому положению они относнтсн к различным отделам: к сине-зеле- ным (Cyanophyta), зеленым (Chlorophyta), жел- то-зеленым (Xanthophyta) и бурым (Phaeophyta) водорослям. Долгое время считали, что каждому виду лишайника соответствует определенный вид водоросли. Однако, как показали дальней- шие исследования, сравнительно небольшое количество водорослей оказались способными существовать в симбиозе с грибом. Только самые неприхотливые из них, обладающие высокой устойчивостью по отношению к факторам внеш- ней среды, приспособились к жизни в окруже- нии грибных гиф. Экспериментальные исследо- вания показали, что лишайниковый гриб при образовании слоевища, как правило, не про- являет избирательной способности по отноше- нию к водорослям. Если взять чистую культуру гриба, выделенного из слоевища лишайника, и поместить а нее какую-либо свободноживущую водоросль (несвойственную данному лишайнику, а другого типа), то грибные гифы начнут разра- статьсн, давая сложные переплетения, как в на- чале образования слоевища. Однако слоевище а таком случае все-таки не развиваетсн. Б ольшин- ство водорослей, введенных в культуру лишай- Рис. 287. Фикобионты лишайников. 1 — стигонема; 2 — гиелла; 8 — хроококкус; 4 — глеокапса; 5 — коккомикса; 6 — трентеполия; 7 — носток; 8 — требук- сия; 9 — фикопельтис: а — свободноживущий экземпляр, б- та же водоросль в слоевище лишайника. никового гриба, рано или поздно погибает: мно- гие не могут перенести воздействия гиф гриба, причиной гибели других нвлнетсн недостаток света. Водоросли, окруженные со всех стороы грибной тканью, получают гораздо меньше све- та, чем это необходимо длн их нормальной жиз- недеятельности. И если водоросль не может приспособиться к осуществлению процесса фо- тосинтеза при минимальной световой интенсив- ности, она неизбежно погибает. Значит, при образовании слоев ища лишайников не гриб выбирает себе подходящего партнера из числа водорослей. Все зависит от того, сможет ли водоросль существовать в тесном контакте с грибом и при этом нормально жить и разви- ватьсн. Сейчас стало известно, что наиболее широко распространенным фикобионтом лишайников является водоросль требуксия (Trebouxia, рис. 287, 8). Предполагают, что около половины всех известных видов лишайников (7 — 10 тыс. видов) имеют своим фикобионтом именно эту одноклеточную зеленую водоросль. Как пока- зали наблюдения за жизнью этой водоросли в лаборатории, требуксин очень неприхотли-
Рис. 288. Фикобионты лишайников: а — часть колонии свободноживущей водоросли, 1 — носток: а — часть ь йника Pannaria; 2 — ривулярия: — бодноживущей водоросли, б — водор — во о осль в слоевище лишаин — осль а — часть колонии сво одн в слоевище лишайника Lichina, ,Ы.), д..-ри- (Dlchothrlx) р у-р"" (В'- стигонема (Stigonema). Из зеленых водорос- Ф леи а слоевищ евище лишайников встречаютсн пред- '@ ставители хлорококковых и улотриксовых. Из Р ~~' хло ококковых водорослей, как уже указыва- Ф � $ ф. хлорококк лось, самым распространенным ико и является требуксия (Trebouxia). ь.роме того, К о встр ечаетсн также мирмецин � в� ~� (М ' ) псевдохлорелла (Pseudochlorella), Ф � И (мугш.ес1а~, псевд а (Chlo- ,:. o �е э хлорококкум (Chlorococcum), хлорелла о- а: rella), коккомикса (Coccomyxa, рис. 287, ), глео- )! цистис (Gloeocystis) и трохисция (Trochiscia). о..:. (Cephaleuros), хлоросарцина (Chlorosarcina), la "коботрис (Coccobotrys), лептозира Lepto- sira), фикопельтис (Phycopeltis, рис. 28, ), физолинум (Physolinum), плеврококкус (Pleu- cus) псевдоплеврококкус (Pseudopleuro- ОQ съ гососспяу, п енте- О. 2а coccus~, ) стихококкус (Stichococcus) и тр полня (Trentepohlia, рис. 287, 6). Желто-зеленые водоросли очень редко встречаютсн в лишайни- ковых слоевищах. Лишь у двух видов лишай- ников представителей рода веррукария (Ver- ~ В о~аЯ д ~дд о у~, ОО 0 Оё 7 О g rucar>a~, ц ик "обионтом нвлнетсн желто-зеленан Ва во о осль гетерококкус (Heterococcus). Так ясе редки а слоевищах лишайнико ур р "аибыево- Э о осли. В слоевище одного из видов того ясе о р~~-~(:~ %.У ан во о осль .@ l рода веррукарин была наидена буран вод р Мб петродерма (Petroderma). Некоторые роды лишайников очень одно- значны по отношению к водорослн м. Так на- пример, все виды рода пармелия (Parmelia), ши око распространенных листоватых лишай- ников, и кустистые лишайники рода кладонин (Cladonia) имеют фикобионтом требуксию, а х чистой к льту ы слизистые лишаиники семейства ФЛ коллемовых- вый организм. В условинх чистои культур Н г а о ного и того же вида ли- ись ме ленным ростом носток. г~о иногда у одно эти водоросли отличали ь д б обнаружены разные виды п и минимальном шайников ывают и способностью развиватьсн р Н п име у лишайника артопи- ошо пе еносили резкие водорослеи. ~например, освещении, они хорошо пер (Artho renia kelpii) фикобионтом могут перепады температур д т ы и лительные периоды ренин r opyreni сли гиелла глеокапса, псевдоплев- без воды. Клетки требу б ксии могли развиватьсн ыть водоросли ги р " " " стоватого лишайника солорина ком сл чае они пере- рококкус; у листов ФЛ полнои ы~~~~~~, но ~~~~~ у р от о ном питанию. По-види- (Solorina sacca a> в сло ходили к гетер отрофному пи рода коккомикса. мому, все эти свойства, в т. е. высокан устоичи- три вида род жаю ейс е ы, уществует взаи аимосвязь между географиче- вость к влиянию факторов окру.к щ р д м лишайников и преоб- так хо ошо п испособить- ским распространением ли позволили требуксии так хорошо при х слоевищах определенных водорос- сн к жизни внутри сло щ еви а лишайников. ладанием в их сло нной зоне земного шара около йниковых ~ико- лей. ак, в умеренно Как показало изучение лишайниьо меют фикобионтом сине-зеле- е ние го ы в слое- 8% лишайников имеют ико ио бионтов, проведенное а посл д д 9 о/ лишайников — нитчатые тсн п е ставители ные водоросли, о вище лишайников встречаютсн р д еных во о ос- или пластинчатые зелены ые водоросли типа трен- 28 одов водорослей. Из сине-зеленых в д р а ог' ков — хлорококковые р " б нтом в слоевище теполии и 8а gp лишаиников- лей наиболее обычным фикобионтом в щ В т опических и субтро- осли о а носток зеленые водоросли. троп лишайников нвлнютсн водоросли р д х величиваетсн количество 288 1) и глеокапса (Иое- пических странах ув (Nostoc, рис. 287, 7; 28, ) и г ФЛ слоевище которых встречаетсн ocapsa, рис. 287, 4). Встречаются дру и гие лишаиников, в сл нтеполин. Так, в тропиках от кокк с (Chroococcus) и гиел- водоросль трентеполия. а сине-зеленые: хроококкус ~ ro Calo- 5 о 10% лишайников содержат в слоевищах ла (Нуе11а, рис. 287, 2, 8), калотрикс (Calo- до ц ли
Та б л и ц а 41. Комбинированная установка для массового культивирования водорослей, размещенная на крыше оран- жереи (Чехословакия).
Та б л и ц а 42. Накииные (1,2) и листоватые (3 — 7) лишайники: 1 — Rhizocarpon geographicum, ареолированное слоевище с темным подслоевищем; 2 — Haematomma ventosum, ареолиро- ванное слоевище; 3 — Peltigera aphthosa; 4 — Solorina сгосеа; 5 — Umbilicaria muehlenbergii; б — Hypogyrnnia physodes, слоевище сверху и снизу; 7 — Xanthoria parietina.
Та б л и ц а 43. Наскальные лишайники с ареолированными слоевищами: в в е р х у — Lecanora rupicola, внешний вид слоевища с апотециями; в н и з у — Lecanora frustulosa, внешний вид слоевища с апотециями.
Т а б л и ц а 44. Розетковые накипные слоевища: в в е р х у — арктоальпийский наскальный лишайник Placolecanora peltata, внешний вид слоевища с апотециями; в н и 3 у — арктический наскальный лишайник Acarospora molybdina.
Та б л и ц а 45. Чешуйчатый напочвенный пустынный лишайник Acarospora reagens: в в е р х у — отдельная чешуйка с тяжами ризоидов, прикрепляющих ее к почве; в н и з у — внешний вид слоевища.
Ти б л и ц а 4б. Лиетоватые (вверху) и чешуйчатые (внизу) лишайники: в в е р х у с л е в а — $ticta aurata, по краям лопастей кайма ярко-желтых соралей, на нижней поверхности пятнистые ярко-желтые цифеллы; в в е р х у в с е р е д и н е — Parmelia scortea, в центральной части темно-серые изидии; в в е р х у с п р а в а — Xanthoria parietina; в н и з у с л е в а — Psora scalaris; в н и з у с п р а в а — Psora decipiens.
Та б л и ц а 47. Листоватые лишайники: 1 — Lobaria pulmonaria; 2 — Cetraria richardsonii; 3 — Parmelia caperata; 4 — Sticta wrightii; 5 — Cetraria pinastri; 6— Leptogium saturnin um.
Т а б л и ц а 48. Напочвенные кустистые лишайники тундр и сосновых лесов: ~ — С~~~~п~ сиси11~~~; 2 — Cladonia floerkeana; 3 — Thamnolia vermicularis; 4 — Cladonia deformis; 5 — Cetraria islandica; б — Cladonia alpestris.
сине-зеленые водоросли. 45 — 48 о/о лишайни- ков — нитчатые зеленые водоросли типа трен- теполии и такое же количество лишайников имеют фикобионтом остальные зеленые водо- росли. Большинство фикобионтов лишайника встре- чаБтся в свободноживущем состоянии. Но не- которые известны лишь а слоевищах лишайни- ков и до сих пор не были обнаружены в природе. К их числу, например, относятся требуксия, коккомикса, лобококкус и некоторые другие. Водоросли в слоевище лишайника очень сильно изменяют свой внешний облик. Особен- но это касается нитчатых водорослей, которые под влиянием гриба распадаются на отдельные клетки и часто деформируются до неузнавае- мости (рис. 288). Размножаются водоросли внутри слоевища обычно делением и аплано- спорами. Подвижных клеток размножения, характерных для свободноживущих водорос- лей, они не образуют. Но в лаборатории, в усло- виях культуры, фикобионты лишайников, как правило, принимают свой обычный, присущий им в свободноживущем состоянии облик и обра- зуют подвижные зооспоры и гаметы. Сине-зеленые водоросли как в слоевище ли- шайников, так и в лабораторных культурах образуют гетероцисты и гормогонии. Но число гетероцист в слоевище обычно бывает очень небольшим, гораздо меньшим, чем у свободно- живущей водоросли. Все вышеперечисленные особенности фико- бионтов принципиально не отличают их от свободноживущих водорослей, при культиви- ровании на искусственных средах имеющиеся отклонения быстро исчезают, и водоросли вос- станавливают свой обычный внешний вид. По всей вероятности, эти изменения не закреплены генетически и возникают лишь под влиянием жизни водорослей а симбиозе. Однако целый ряд физиологических особен- ностБй, характерных для водорослей, живу- щих в слоевище лишайников, сохраняется у них и при культивировании на искусственных сре- дах. Так, фикобионты лишайников отличаются замедленным ростом в слоевищах, эта же осо- бенность сохраняется у них и при культивиро- вании в лабораторных условиях. Причем если добавлять в культуры изолированных фико- бионтов экстракты, полученные из микобионта лишайника, то рост водоросли усиливается. Интересно, что при таком медленном росте фотосинтетическая способность у лишайнико- вых водорослей ничуть не меньше, чем у сво- бодноживущих. В процессе своей жизнедеятель- ности они способны вырабатывать такое же количество углеводов, как и свободноживущиБ водоросли. Однако эти углеводы фикобионты используют не только для своего развития, зна- чительное количество питательных веществ они отдают грибу (это и приводит к замедлению роста лишайниковых водорослей). Так, опытом было показано, что водоросль носток, выделен- ная из слоБвища одного из видов коллемы и вы- ращенная в культуре, выделяла в среду 5оо азо- та, фиксированного из воздуха. Одновременно в среду поступали полисахариды и витамины (тиамин, биотин, рибофлавин, никотиновая и пантотеновая кислоты). Было установлено, что водоросли а слоевище лишайников полностью или в значительной степени лишены своих обычных избыточных ассимилированных ими веществ, крахмала и запасных веществ, липоидов, зерен цианофицина, гликогБна и некоторых других. Лишайниковые водоросли очень устойчивы к воздействию высокой температуры. Так, фико- бионт, только что выделБнный из слоевища одного из видов кладонии, мог переносить на- грев до +90 'С. После роста этой водоросли в культуре в течение 9 недель она так «изнежи- валась», что погибала уже при температуре +70 'С. Водоросли а слоевище лишайников способны переносить длительное высушивание. Опыты показали, например, что водоросль коккомикса а слоевище одного из лишайников выдерживала высушивание в течение 23 недель, а после нор- мализации условий восстанавливала все про- цессы своей жизнедеятельности. В культуре ~å этот фикобионт погибал без воды уясе через 5 недель. Все эти данные показывают, что в связи с особенностями жизни в контакте с грибом ли- шайниковые водоросли претерпевают не только морфологические, но и физиологические изме- нения. ВЗАИМООТНОШЕНИЯ ГРИБА И ВОДОРОСЛИ В ТЕЛЕ ЛИШАЙНИКА Вопрос взаимоотношения гриба и водоросли в слоевище лишайника занимал умы ученых еще в конце прошлого столетия, да и а наше время продолжает волновать лихенологов. Со дня открытия С. Швенденера прошло более 100 лет. За этот период появилось не менее де- сятка теорий, пытающихся объяснить отноше- ния между грибом и водорослью, однако среди них нет ни одной общепризнанной и оконча- тельно доказанной. С. Ш в е н д е н е р, обна- ружив, что лишайник состоит из гриба и водо- росли, предположил, что гриб а слоевищБ па- разитирует на водоросли. Однако он ошибоч- но отвел грибу роль хозяина, а водоросли— раба. Но уже в те времена некоторые ученые выд- винули мысль о двустороннем паразитизме
компонентов лишайника — гриба на водоросли и водоросли на грибе. При этом было высказа- но предположение, что гриб и водоросль в слое- вище лишайника находятся в полном морфо- физиологическом единстве и связаны между собой так же, как корни и листья цветковых растений. Такое сравнение, безусловно, было совсем необоснованным. Наибольшее распространение среди ученых того времени получила теория мутуалистиче- ского симбиоза. Сторонники этой теории счита- ли, что в слоевище лишайника гриб и водоросль находятся во взаимовыгодном симбиозе: водо- росль «снабжает» гриб органическими вещест- вами, а гриб «защищает» водоросль от чрезмер- ного нагревания и освещения и «обеспечивает» ее водой и неорганическими солями. Однако в 1873 г. этой идеалистической теории был на- несен удар. Известный французский исследо- ватель Е. Б о р н е, изучая анатомическое строе- ние слоевища лишайников, обнаружил внутри водорослевых клеток грибные отростки — гау- стории, всасывающие органы гриба. Это поз- воляло думать, что гриб использует содержи- мое клеток водорослей, т. е. ведет себя как па- разит. За прошедшие со времен Борне 100 лет в слоевище лишайников было открыто и описано много различных форм абсорбционных, или всасывающих, гиф гриба. Эти гифы плотно прижимаются к клетке водоросли или прони- кают в нее и служат, как предполагают, для передачи веществ, которые образуют водоросли в результате своей жизнедеятельности, гриб- ному компоненту. О том, что в слоевище лишайника происхо- дит обмен веществами между грибом и водо- рослью, ученые стали говорить сразу после от- крытия двойственной природы лишайников. Однако некоторые экспериментальные под- тверждения этим предположениям были полу- чены лишь за последние три десятилетия. При- менение новейших методов физиологических исследований с использованием меченых атомов углерода и азота, особых красящих веществ и некоторых других позволило установить, что гриб получает вещества, ассимилируемые водо- рослью, и ведет себя в слоевище лишайника как паразитический организм. Однако для существования как самого гриба, так и лишай- ника в целом необходимо, чтобы водоросль, окруженная со всех сторон грибными гифами, все-таки могла жить и более или менее нормаль- но развиваться. Если гриб начнет проявлять себя слишком активно, поражать все без исклю- чения водоросли и, использовав их содержимое, уничтожать их, это в конце концов может при- вести к гибели всех водорослей слоевища. Но тогда, уничтожив весь свой запас питания, по- гибнет и сам гриб, а значит, перестанет сущест- вовать и лишайник. Гриб должен использовать лишь часть водо- рослей, оставляя резерв — здоровые и нормаль- ные водоросли, содержимым которых он мог бы питаться. Учеными были замечены любопытные защит- ные реакции со стороны лишайниковых водо- рослей. Например, одновременно с проникно- вением гаустория в клетку водоросли эта клетка делилась. При этом плоскость деления, как правило, проходила как раз через участок, занятый гаусторием, а образовавшиеся в ре- зультате этого процесса дочерние клетки были свободны от гаусториев. Было замечено также, что обычно гриб поражает водоросли, уже достигшие определенной стадии зрелости. В мо- лодых растущих водорослях происходит энер- гичное отложение веществ в оболочке клетки и быстрое ее утолщение. Эта толстая оболочка клетки фикобионта препятствует проникнове- нию абсорбционных органов гриба. Однако большей частью защитная реакция водорослей против активности грибного компонента очень слаба. Способность водорослей нормально разви- ваться и даже размножаться в слоевище ли- шайника сохраняется скорее благодаря уме- ренности паразитизма самого гриба. Ученые отмечают, что степень паразитизма гриба на водоросли различна не только у раз- ных видов лишайников, но даже в одном и том же слоевище. Резкий паразитизм обнаружен лишь у примитивных лишайников. Гаустории, проникающие глубоко внутрь протопласта во- доросли, пока что были найдены лишь у наибо- лее просто организованных форм, в слоевище которых еще нельзя различить оформленных дифференцированных слоев. В слоевищах более высокоорганизованных лишайников часть кле- ток водорослей поражена грибными гифами, а остальные продолжают нормально жить и раз- виваться. Обычно у высокоорганизованных форм лишайников паразитизм гриба на водо- росли носит весьма умеренный характер: преж- де чем гриб убьет пораженные им клетки, успе- вает вырасти одно или несколько поколений водорослей. Отношения между мико- п фикобионтом в слоевище лишайника не сводятся только к па- разитизму гриба на водоросли. Ученые пред- полагают, что эти отношения гораздо сложнее. Еще в начале нашего века крупнейший русский лихенолог А. А. Е л е н к и н, изучая анатоми- ческое строение лишайников, обнаружил в их слоевище некральные зоны водорослей — скоп- ления отмерших, потерявших зеленую окраску клеток, расположенные несколько ниже зоны живых водорослей. К этим бесцветным мертвым 386
клеткам водорослей тоже тянулись грибные гифы. Это привело А. А. Еленкина к мысли, что гриб в слоевище лишайника вначале прояв- ляет себя как паразитический организм, пора- жая живые клетки водоросли и используя их содержимое. Затем, убив водоросль, гриб пере- ходит к сапрофитному способу питания, погло- щая и ее мертвые остатки. Таким образом, гриб в слоевище лишайника ведет себя и как паразит, п как сапрофит. И отношения между грибом и водорослью в слоевище лишайника А. А. Елен- кин назвал эндопаразитосапрофи- тизмом. Интересную мысль о взаимоотношении ком- понентов в слоевище лишайника высказал в 60-х годах нашего столетия крупнейший совет- ский лихенолог А. Н. О к с н е р. Он пришел к выводу, что водоросль в слоевище лишайника, полностью изолированная от внешней среды грибной тканью, обязательно должна забирать у грибного компонента все необходимые для своего существования вещества, за исключе- нием тех органических соединений, которые она сама вырабатывает на свету в процессе асси- миляции углекислоты. К этим жизненно необ- ходимым для водоросли веществам относится прежде всего вода, а также минеральные соли, азотистые и некоторые другие неорганические соединения. Следовательно, и водоросль в слое- вище лишайника проявляет себя как паразит. Причем это вовсе не противоречит общему характеру ее питания. Как показало изучение лишайниковых водорослей в чистых культурах, многие из них, будучи большей частью автотроф- IIIIMH организмами, способны и к миксотроф- ному питанию. Таким образом, ученые считают, что водо- рослевый и грибной компоненты лишайника находятся в очень сложных взаимоотношениях. Микобионт ведет себя как паразит и сапро- фит на теле водоросли, а фикобионт, в свою оче- редь, паразитирует на лишайниковом грибе. При этом паразитизм фикобионта всегда носит более умеренный характер, чем паразитизм гриба. Однако все высказанные по этому поводу точки зрения до сих пор остаются лишь догад- ками и большей частью не подтверждены экспе- риментально: лишайники оказались очень труд- ным объектом для физиологических исследова- ний. Ученые пока не научились выращивать и поддерживать в живом состоянии слоевище лишайников в искусственных условиях. Тот контакт между грибом и водорослью, который с такой легкостью достигается в природе (доста- точно вспомнить многообразие лишайников!), никак не удается воспроизвести в лаборатор- ных условиях. Наоборот, при переносе лишай- ников в лабораторию этот контакт легко нару- шается и растение просто погибает. Время от времени появляются сообщения об удачных опытах выращивания лишайника в условиях лаборатории, но пока эти сообщения единичны и не всегда достоверны. Одной из причин неудач подобных попыток можно считать чрезвычайно медленный рост лишайников. Лишайники — многолетние расте- ния. Обычно возраст взрослых слоевищ, кото- рые можно увидеть где-нибудь в лесу на стволе деревьев или на почве, составляет не менее 20 — 50 лет. В северных тундрах возраст неко- торых кустистых лишайников рода кладония достигает 300 лет. Слоевище лишайников, име- ющих вид корочки, в год дает прирост всего 0,2 — 0,3 мм. Кустистые и листоватые лишайники растут несколько быстрее — в год их слоевище уве- личивается на 2 — 3 лм. Поэтому, чтобы вырас- тить взрослый лишайник в лаборатории, тре- буется не менее 20 лет, а может быть, и вся жизнь исследователя. Трудно проводить столь долгов ременный эксперимент! Вот почему физиологические особенности ли- шайников, в том числе взаимоотношения ком- понентов, как правило, изучают на культурах изолированных мико- и фикобионтов. Этот метод очень перспективен, так как позволяет ставить длительные и воспроизводимые опыты. Но, к сожалению, данные, полученные этим мето- дом, не могут полностью отразить те процессы, которые происходят в целом слоевище лишай- ника. И тем более мы не вправе считать, что s природе, в естественных условиях, в слоеви- щах лишайника эти процессы протекают точно так же, как в культурах изолированных сим- бионтов. Вот почему все теории, пытающиеся объяснить взаимоотношения компонентов ли- шайников, остаются пока лишь догадками. Более успешным оказалось изучение форм контакта между гифами гриба и клетками водо- рослей в слоевищах лишайников. Как показа- ли исследования с применением электронной микроскопии, в слоевище лишайников можно встретить по крайней мере пять типов контак- та между грибными гифами и водорослевыми клетками (рис. 289). Чаще всего отдельная клетка водоросли и клетка грибной гифы находятся в непосредст- венном контакте друг с другом. В таком случае гриб образует специальные абсорбционные, вса- сывающие органы, которые проникают внутрь водорослевой клетки или плотно прижимаются к ее оболочке. В настоящее время среди абсорбционных ор- ганов гриба в слоевище лишайников различа- ют несколько типов: гаустории, импрессории и аппрессории. 387
° у еее~ pt ° е Ф ° ' ° Ф ф ° ° ° ° ° j4'~\' ',; ° Ф ° ° Ю Э ° - .:. ° ° Ф ° ' ° ° ю ° ° е \ ю \ ° ° ° с е i / Ю / ° ° ° ° е~ Ф В е г Ъ Ф' в / \ I \ ф 1 ! г .; ~Л~'.: ~р ~'", с М' 1 \ ~с \ I % ° ° ° ° ° ° ° ° г ° ~' ~g м 1„ л ю g % С~~ ~-~ ° ° ° ° ° е ° В б , в Рис. 289. Формы контакта между гифами гриба и клетками водорослей в слсевин.е лишайников: 1 — контакт между клетками водоросли требуксия и грибными гифами в слоевище кладонии; 2 — гаустории, проникающие в клетку водоросли глеокапса в слоевище слизистого лишайника Synalissa symphorea; 8 — интрацеллюлярный гаусторий, про- никающий в протопласт водоросли; 4 — интрацеллюлярный гаусторий, не прорывающий плазмалеммы протопласта клетки водоросли; 5 — интрамембранный гаусторий; 6 — импрессорий, вдавливающий внутрь оболочку клетки водоросли целиком; 7— импрессорий, вдавливающий внутрь лишь внутренний слой оболочки клетки водоросли; 8 — аппрессорий. а — гифа гриба, б — оболочка клетки водоросли, в — плазмалемма, г — плазма клетки водоросли, д — ядро клетки водоросли. терно для лишайников с примитивным строе- нием слоевища. У более высокоорганизованных лишайни- ков образуются только и н т р а м е м б р а н- н ы е г а у с т о р и и. Они прорывают оболоч- ку клетки водоросли и достигают ее протопла- ста, но не углубляются в него, а остаются в оболочке водорослевой клетки (рис. 289, о). Наибольшее количество пнтрамембранных гау- сториев образуется в слоевище лишайников весной, в начале вегетационного периода. С наступлением осени они далеко отступают от протопласта водоросли. Другой тип всасывающих органов гриба— и м п р е с с о р и и — тоже боковые выросты грибных гиф, но, в отличие от гаусториев, они не прорывают оболочку клетки водоросли, а вдавливают ее внутрь (рис. 289, 6, 7). Импрес- сории отмечены у очень многих лишайников, например у широко распространенной пель- тигеры (Peltigera). Г а у с т о р и и — это боковые выросты гиф гриба, которые прорывают оболочку кле гки водоросли и проникают в ее протопласт (рис. 289, 2). Обычно в клетке водоросли развивает- ся один гаусторий, но иногда их может быть и два. В слоевище лишайника гаустории встре- чаются в большом количестве и существуют продолжительное время. Было замечено, что в оболочках молодых гаусториев нет отложе- ний целлюлозы, которая могла бы затруднять обмен между клеткой водоросли и гифой гриба. Старые гаустории почти всегда одеты доволь- но толстым слоем целлюлозы. Различают ин- трацеллюлярные (внутриклеточные) и интра- мембранные (внутриоболочковые) гаустории. И н т р а ц е л л ю л я р н ы е гаустории полностью прорывают оболочку клетки водо- росли и проникают глубоко внутрь ее прото- пласта (рис.289, 8). Интрацеллюлярные гау- сторип образуются в случае резкого парази- тизма гриба на водоросли. Это особенно харак- \ ° Ф ф 1 ° ° ~ с ) с 1 I I r r г ° ° r а ° ° ° ° ф ее i r :.,:::."с. «3':;.. ':. Ъ Ф ° ю ° ю r Ф ° е ° ° 1 р \ Ъ ю Ъ ю 1' ". ° .. 1 ,ее ° Ф I, ° °
Интересно, что в слоевищах, произрастающих во влажных местообитаниях, импрессории поч- ти не развиваются, у тех же видов в сухих местообитаниях они образуются в большом ко- личестве. При длительной засухе число импрес- сориев также увеличивается. Предполагают, что в засушливые периоды и в сухих местообита- ниях гриб, чтобы удовлетворить потребности B питании, увеличивает свою всасывающую по- верхность за счет увеличения количества и размеров импрессориев. В отличие от гаусториев и импрессориев, образованных боковыми отростками гифы, а п- п р е с с о р и и образуются вершиной грибной гифы. Такая вершина гифы плотно прижимает- ся снаружи к оболочке клетки водоросли, никогда не проникая ни в ее протопласт, ни в ее внутренний слой (рис. 289, 8). Наличие в слоевищах многих лишайников абсорбционных органов гриба хорошо доказы- вает паразитическую сущность отношений ми- кобионта к фикобионту. Но во многих случаях у лишайникового гриба все же не удается обна- ружить особых абсорбционных органов, чаще всего у лишайников, фикобионт которых имеет тонкие оболочки клеток. В таких случаях уже внешний контакт гифы гриба и клетки водо- росли может обеспечить обмен веществами между ними. Так, например, обстоит дело у многих видов рода кладония. Фикобионтом кладонии является одноклеточная зеленая водоросль тре- буксия. У этих лишайников отдельные клетки водорослей окружены со всех сторон тонкими тонкостенными гифами, иногда поделенными на мелкие клеточки. Эти гифы, которые носят название обволакивающих или кон- т а к т н ы х, не проникают в протопласт кле- ток водоросли и не внедряются в их оболочку, а просто окружают клетки со всех сторон, так что каждая из них становится похожей на малень- кий шар, охваченный пальцами рук (рис. 289,1). Иногда гифы полностью оплетают водоросли в виде сплошного покрова и при этом, сливаясь своими стенками, даже образуют клеточную псевдопаренхимную ткань. На первый взгляд кажется, что водоросли не особенно страдают от плотного окружения гифами гриба: они долго сохраняют свою зеленую окраску и продолжа- ют интенсивно делиться. Но в более старых участках слоевища мож- но найти немало отмерших обесцвеченных клеток — гриб рано или поздно все-таки уби- вает водоросли. Такой ясе тип контакта между гифами гриба и клетками водорослей был найден у некото- рых слизистых и базидиальных лишайников. У ряда лишайников, в слоевище которых встречаются нитчатые улотриксовые водоросли, можно наблюдать еще один тип контакта. Как правило, в таком случае нити водорослей бы- вают целиком покрыты грибными гифами. При- чем лишь иногда гифы образуют на поверхности водорослевой нити рыхлую сетку. Чаще же они располагаются очень густо и, срастаясь своими стенками, образуют сплошной чехол. Отдельная лопасть такого лишайника имеет вид тончайшего волоса. Под микроскопом она напоминает полую трубку, стенки которой образованы сросшимися грибными гифами; внутри трубки тянется нить водоросли. У слизистых лишайников семейства колле- мовых (Collemataceae) обычно не наблюдается никакого контакта между грибными гифами и клетками водорослей. Слоевище коллемовых не имеет дифференцированной структуры: нити водоросли носток разбросаны в беспорядке среди грибных гиф по всей толще слоевища (рис. 297, 2). Никаких абсорбционных отрост- ков в клетках водорослей обычно не заметно; гифы гриба и нити сине-зеленой водоросли рас- положены друг около друга, не вступая в види- мый контакт. Предполагают, что в данном слу- чае гриб поглощает органические вещества, ассимилируемые водорослями, прямо из слизи, которая обычно окружает нити ностока. Одна- ко более тщательное изучение этих лишайников показало, что у многих видов коллемы (Collema) в слоевище время от времени образуются спе- циальные абсорбционные гифы, которые тесно прижимаются к одной из клеток водорослевой нити, а через некоторое время можно наблюдать отмирание этой клетки. Описанные выше формы контакта между гифами микобионта и клетками водорослей, по всей видимости, не исчерпывают всего много- образия способов, с помощью которых гриб и водоросль в слоевище лишайников устанавли- вают между собой тесную связь. Исследования в этом направлении только начинаются. Можно думать, что дальнейшее изучение тончайших структур лишайникового слоевища с помощью электронного микроскопа не только даст мно- го нового в описании физических контактов между грибным и водорослевым компонентами лишайников, но и откроет новые горизонты в понимании их взаимоотношений.
ВНЕШНЕЕ И ВНУТРЕННЕЕ СТРОЕНИЕ ЛИШАЙНИКОВ ИАКИПИЫЕ ЛИШАЙНИКИ 390 МОРФОЛОГИЯ СЛОЕВИЩА ЛИШАЙНИКОВ Слоевище лишайников очень разнообразно по окраске, размерам, форме и строению. Лишайники окрашены в самые различные цвета: белый, розовый, ярко-желтый, оранже- вый, оранжево-красный, серый, голубовато- серый, серовато-зеленый, желтовато-зеленый, оливково-коричневый, коричневый, черный и некоторые другие. Окраска слоевища лишайни- ков зависит от наличия пигментов, которые откладываются в оболочках гиф, реже в про- топлазме. Наиболее богаты пигментами гифы корового слон лишайников и различные части их плодовых тел. У лишайников различают пять групп пигментов: зеленые, синие, фиолето- вые, красные, коричневые. Механизм образо- вания их до сих пор не выяснен, но совершенно очевидно, что важнейшим фактором, влияющим на этот процесс, нвлнетсн свет. Иногда цвет слоевища зависит от окраски лишайниковых кислот, которые откладываютсн в виде кристаллов или зернышек на поверхно- сти гиф. Большинство лишайпиковых кислот бесцветны, но некоторые из них окрашены, и иногда очень ярко — в желтый, оранжевый, крас- ный и другие цвета. Окраска кристаллов этих веществ определяет и окраску всего слоевища. И здесь важнейшим фактором, способствующим образованию лишайниковых веществ, является свет. Чем ярче освещение в месте произраста- ния лишайника, тем ярче он окрашен. Примером может служить широко распространенный, часто встречающийся на стволах осин лишайник ксантория (Xanthoria parietina, табл. 42, 7). Слоевище у него яркое, красновато-оранжевое. Этот цвет придает ему особое лишайников ое вещество — париетин, которое в виде оранже- вых кристаллов покрывает гифы корового слоя. Если ксанторин растет на солнце, ее слоевище имеет яркую красновато-оранжевую окраску. В условиях слабого освещения этот лишайник полностью тернет оранжевый цвет и становится серовато-зеленоватым. Как правило, очень ярко окрашены лишайники высокогорий и полярных районов Арктики и Антарктики. Это тоже свя- зано с условиями освещения. Для высокогор- ных и полярных районов земного шара харак- терны большая прозрачность атмосферы и вы- сокая интенсивность прямой солнечной радиа- ции, обеспечивающие здесь значительную яркость освещения. В таких условиях в наруж- HhIx слоях слоевищ концентрируетсн большое количество пигментов и лишайниковых кислот, обусловливая яркую окраску лишайников. Предполагают, что окрашенные наружные слои защищают нижележащие клетки водорослей от чрезмерной интенсивности освещения. Интересно, что слоевища многих антаркти- ческих лишайников окрашены в черный или другой темный цвет. Это тоже обънсннетсн высокой концентрацией зеленых, синих и фио- летовых пигментов в коровом слое и плодовых телах этих лишайников. Хорошо пигментиро- ванные наружные слои антарктических лишай- ников не только защищают клетки водорослей от чрезмерно интенсивного освещения, в дан- ном случае темнан окраска слоевища нвлн- етсн приспособительной и служит длн притяги- вания тепловых лучей. Как известно, условия в Антарктике крайне суровы; здесь растениям приходится переносить постоянное воздействие очень низкой температуры и явить фактически без воды. Среднегодовая температура воздуха ледяного континента — 16'С. Зимой она падает до — 45 'С, а летом, которое наступает в янва- ре — феврале, только днем поднимается выше нуля, ночью же снова опускаетсн до — 10 С. Из-за низкой температуры осадки выпадают в Антарктике только в виде снега. В такой фор- ме они не могут быть использованы растениями. Вот здесь-то темнан окраска лишайников и приходит им на помощь. Темноокрашенные слоевища антарктических лишайников за счет высокой солнечной радиации быстро нагрева- ютсн до положительной температуры даже при отрицательной температуре воздуха. Снег, падающий на эти нагретые слоевища, тает, пре- вращаясь в воду, которую лишайник сразу же впитывает. Таким образом он обеспечивает себя водой, необходимой для осуществления процессов дыхания и фотосинтеза. Насколько разнообразны слоевища лишай- ников по окраске, настолько же разнообразны они и по форме. Слоевище моясет иметь вид корочки, листовидной пластинки или кустика. В зависимости от внешнего облика различают три основных морфологических типа: накип- ные, листоватые и кустистые лишайники. Слоевище накипных лишайников имеет вид корочки, плотно сросшейся с субстратом. Тол- щина корочки очень различна. Она может быть весьма тонкой и иметь вид еле заметной накипи или порошкообразного налета; может быть тол- щиной 1 — 2 лл, а иногда бывает и довольно тол- стой, достигая в толщину половины сантимет- ра. Как правило, накипные слоевища небольших размеров, их диаметр составляет всего несколько
мет ов или сантиметров, но ино д ног а может ,1( 0 В роде нередко мо)кн ть и 20 — 3 сл. при ть как небольшие по а лишаиников, ные слоевищ той поверхности ~г~ об азуют на каменистой другом, о р р упные пятна, дости- скал или ство д р лах еревьев крупн х есятков санти- гающие в ди иаметре нескольких де метров (таб бл. 42,1, 3. слоевища плотно п авило, накипные сл сердцевинными гифа- ся с субстратом с некоторых лишаиник ми. Но ун . ик всего бывае тем- оисходит с пом По слоевище чаще всего ы в ища. одс. б азовано темноокра- а ной окраски и и обычно о ра ибными гифами. толстостенными гри н шенными тол орослей (рис. 290). Оно никогда н д р е со е жит водо ща нередко м ° такого подслоевищ Черную каиму т и слоевищ не- ать по пери~уерии наблюд кипных лишайников и которых на слоевища (табл. б орка чи ассимиляц ионного уго 42, 1). п имитивный тип накипного слое- кооб разного е в виде тонкого порош ис.. че ез часток слоевища слоевище в 29О. Поперечный ра~р~~ че ез налета. О Оно носит название слоевищем (а). Лепрозные слоевищ ви а очень п анции с губчатым под скоплений отдельных строени ю. Они состоят из скоплений отд о ослей, окружен- К айников может ыть ц К айн ельной, р — кл бочков водо о очка этих лишайн и~~ами. акие или иметь не е овную поверхнос ть- грЫными чат ю, с различными ши- отрываются а г е п ик епляются к су — о вы остами и т. д. (та л. ными и в другие места, где п ик в емя разраст р е бывает подел ст ату и спустя некоторое р р Нередко слоевищ ел ые площадочки, одина- в новые лепрознь ые слоевища. ча е всего ы ч б вают жел- трещин ками на отдельные и азме у. ти . Э маленькие площа- Леп озные слоевища ч щ вета и ковыепо форме р л а сами слое- епр зеленовато-беловатого цвета ки носят название а р е о л, а а еолированными пок ывают большие пове я ви а называют а р е ованной . Об о они р~ж~~~ю~с щ или стволов дер евьев. ычн . И можно встре- та л. .42; . и оиз астают только на во влажных тенис истых местах. х м кал в узких и структур ой слоевища прои и никогда не встреча- ости отвесных скал р — истом субстрат е и ни ых тить на поверхн сы, ых гни- камен евьев, растительны ельях, в лесах на на почве, стволах дер ии ствочов деревьев ются на ь ях при основании стволо остатках, гниющей древесин ст атах. ля последних характ р р "е но аз- на разлагающихся растительн ских у стр~ г~~. влажненной почве. акипных лишайников со мхах или слегка у и более слож- витие на или порошкообразной Примитивно у р ст оенным, хотя и о кой бо одавчатои или с леп озным, счита ется также де глад р . Если на них и встр ечаются трещинки, Н ым по сравнению с. в ви е отдельных ,.азбро- корочки. бывают неглу окими, б и неопределенны- акипное слоевище в д 3 есь в они обычно Особенно харак- наь чек или зернышек. д ког а не образуют ареол. ФЛ санных бородавочек кт е уже намеч ается не- ми и никогд еоли ованные слоеви ща для лишаини- натомической структур в такой терны ареолир е хности скал в ан ен иация. Водоросли в п оиз астающих на пове азб осаны по всей ных айонах, пу бородавочке не раз р т в ее нижней части, а в верх- высокогорн земного шара с крайн ими условиями ФЛ о бычно отсутствуют в ти " ' но заметить н скопле- областях зем ия астении. ля таких раио- ти бородавочки мо кн для существования р е атуры в неи части ее коровой слой. ны езкие перепады темпера ое слое- нов обычны резки хности скал они ни изованное накипное ок п ичемна поверхн о Более высокооргани ко очки. течение суто мплитуд — 50 — 60 . сплошной плотнои короч ют колоссальных амплит вище имеет вид сп. о же имеют ди фферен- достигают ая ст ктура слоевища накип- Т е слоевища обычн у аки .кт: на поперечном ср езе Ареолированн ру шайников является приспо соблением к ц ированную структуру: н итькоровой с л о й, ных лишаиников езких коле аний б температуры. здесь можно различи е в и н у. перенесению р орослей и сердцевин слой водо
Попытаемся проследить за жизнью какого- нибудь наскального пустынного лишайника в течение суток. Каждый день поверхность скалы, ца которой растет лишайник, нагревается солн- цем до +60, +70 'С и при этом сильно расши- ряется, а ночью с заходом солнца охлаждается иногда до 0 'С и при этом сильно сжимается. Как же в таких условиях ведет себя лишайник? На рассвете после холодной ночи на сильно охлажденных за ночь скалах нередко выпадает роса. С первыми лучами солнца наш наскальный лишайник быстро нагревается, гораздо быстрее, чем поверхность скалы, впитывает влагу росы и начинает активно ассимилировать углекисло- ту, накапливать органические вещества. Такое влажное, разбухшее слоевище заметно увели- чивается в размере, в то время как сама поверх- ность скалы после ночи остается все еще холод- ной и сильно сжатой. Однако постепенно ка- мень все более и более нагревается и начинает расширяться. Лишайник же с повышением температуры воздуха быстро высыхает, его слоевище резко уменьшается в размере и пере- ходит в характерное для него латентное состоя- ние, когда все процессы в нем замирают. И днем, когда температура скал достигает макси- мальной величины, на раскаленной и сильно расширенной каменистой поверхности малень- кий съежившийся лишайник как бы спит. Ночью температура резко падает, поверхность скалы сжимается — гораздо сильнее, чем слоевпще самого лишайника. А утром снова на этой силь- но сжатой от холода каменистой поверхности происходит расширение слоевища лишайника, увлажненного утренней росой. В результате всех этих изменений, происходящих в течение суток, в слоевище возникают очень сильные напряжения, которые и приводят к появлению на его поверхности многочисленных трещинок. Если бы этого не происходило, резкие измене- ния в лишайниковом слоевище, противополож- ные сжатиям и расширениям каменистой по- верхности, на которой оно растет, могли бы привести к отрыву слоевища от субстрата. Благодаря ареолированной структуре слоеви- ща эти напряжения ослабляются. Все перечисленные типы накипных слоевищ являются однообразнонакипными, ибо они оди- наковы по своему строению как в центральной, так и в краевой части слоевища. Дальнейшее усложнение в структуре накипных лишайни- ков происходит путем образования переходов к листоватым формам. Особенно часто такие переходы можно наблюдать у ареолированных слоевищ. В этих случаях ареолы, расположен- ные по периферии лишайника, сильно вытяги- ваются в радиальном направлении и образуют по краям листовидные лопасти. Такие слоевища имеют вид округлых розеток, в центральной своей части ареольно-потрескавшихся, а по периферии лопастных, и носят название фигур- ных или радиальных (табл. 44). У высокооргани- зованных зернистых, бородавчатых или гладко- корковых накипных лишайников по периферии слоевища иногда образуется белый или цветной зонированный край. Обычно по окраске он отличается от остального слоевища, так как состоит из радиально растущих гиф микобионта, еще не содержащих водорослей. Позднее водо- росли переносятся в этот край из водорослевой зоны двигающими гифами. Переходной формой между накипными и листоватыми лишайниками является ч е ш у й- ч а т о е с л о е в ищ е, очень характерное, например, для видов, растущих на почве в пу- стынных областях земного шара (табл. 45; 46, 4, б). В пустынях на поверхности почвы обычно можно заметить коричневатые, серые, желто- ватые и розоватые пятна, образуемые слоеви- щами чешуйчатых лишайников. Диаметр че- шуек колеблется от 2 — 5 лл до 1 см. Они бывают округлыми, угловатыми, с ровными и волнисты- ми, иногда лопастными краями. Чешуйки могут быть расположены на некотором расстоянии друг от друга или расти так тесно, что края одной накладываются на поверхность другой. В отличие от типичных накипных слоевищ че- шуйки обычно менее плотно срастаются с субст- ратом, и их легко можно от него отделить. Чаще они прикрепляются отдельными тонкими гифа- ми, отходящими от нижней поверхности. Не- редко эти гифы отходят только от какого-нибудь одного края чешуйки, в то время как другой остается свободным. В таких случаях чешуйки приподнимаются и растут не горизонтально, а вертикально. Но иногда они прикрепляются к субстрату только в своей центральной части довольно толстыми тяжами, образованнымц склеенными сердцевинными гифами. Эти тяжи у лишайников, растущих на почве, могут до- стигать в длину 0,5 — 1 см и напоминают малень- кий разветвленный корешок (табл. 45). В зависимости от субстрата, на котором про- израстают накипные лишайники, среди них различают несколько экологических групп: э п и л и т н ы е, развивающиеся на поверх- ности горных пород; эпифлеодные- на коре деревьев и кустарников; э п и г е й- н ы е — на поверхности почвы; э п и к с и л ь- н ы е — на обнаженной гниющей древесине. У подавляющего большинства накипных ли- шайников слоевище развивается на поверхности субстрата. Однако существует еще одна срав- нительно небольшая, но интересная группа лишайников, слоевище которых целиком растет внутри камня или коры дерева. Если такое слоевище развивается внутри камня, его назы- вают э н д о л и т н ы м; если внутри коры дере- 392
ва — эндофлеодным или гипофле- о д н ы м. Эти лишайники можно разделить на две группы. У представителей одной из них слоевище полностью погружено в субстрат и никогда не выступает на его поверхность, из- редка выступают лишь плодовые тела лишай- ника; у лишайников второй группы слоевище на поверхности субстрата развивает коровой слой и зону водорослей, а в субстрате — сердце- вину и зону с прикрепляющими гифами. Эндолитные лишайники чаще всего разви- ваются внутри известковых пород, но могут встречаться и внутри силикатных скал, хотя в таком случае видов, полностью погруженных в субстрат, известно немного. Слоевищные гифы эндолитных лишайников способны проникать внутрь камня на значительную глубину, от 1 до 3 см. Чаще всего гифы и водоросли лишай- ника при своем продвижении в глубь камня используют мелкие трешинки, щели, но они обладают способностью проникать и внутрь горных пород, совершенно не тронутых разру- шением. Ока зыв а ется, гифы эндолитных лиша й- ников выделяют кислоты, растворяющие горные скалы. Благодаря этому они могут разрушить даже такие твердые породы, как гранит, кото- рый в этих случаях довольно быстро превраща- ется в тонкозернистую массу, напоминающую глину. Гифы эндолитных лишайников, проникаю- щие в субстрат, обычно очень тонкие (толщина их всего 1 — 3 мкм), нежные, с длинными клет- ками. Часто они не растут прямо, а сгибаются на концах в сторону в виде крючков, охватыва- ющих кусочки субстрата. Иногда на конце этих гиф образуются клетки-щетинки — длинные, тонко заостренные на конце волосковидные клетки (рис. 291). Проникая в горную породу, гифы обходят твердые, плохо растворимые минералы и быстро распространяются по более рыхлым и легче растворимым участкам. Так, например, они до- вольно быстро разрушают слоистые кристаллы слюды. Гифы разъединяют листочки слюды и проникают между ними. Здесь они разветвля- ются и отодвигают пластинки слюды одну от другой. Постепенно разрастаясь и разветвляясь, гифы образуют между пластиночками грибную плектенхиму. Затем в эту плектенхиму прони- кают и клетки водорослей, которые размножа- ются, обвиваются гифами и все больше раздви- гают отдельные листочки слюды. Было заме- чено, что на многих твердых горных породах гифы эндолитных лишайников проникают внутрь камня именно в участках, занятых пла- стинками слюды, а дальше продвигаются уже благодаря химическому разрушению породы. Разрушая твердые горные породы, превра- щая их в зернистую массу, эндолитные лишай- Рис. 291. Гифы эндолитных лишайников: 1 — пуговипно-согнутые гифы, проникающие в толщу камня 8 — конечные клетки-щетинки этих гиф. э ники тем самым выступают как одни из пионе- ров растительности. Они подготавливают по- верхность скал для поселения других растений: листоватых и кустистых лишайников мхов 1 7 цветковых растений и т. д. Но вместе с тем эти лишайники играют в жизни человека и отри- цательную роль. Особый ущерб они наносят памятникам старины, нередко поселяясь на них и разрушая их. Например, широко известно, какой вред причинили лишайники старым вит- ражам церквей в Западной Европе. Эндофлеодные лишайники чаще всего посе- ляются на древесных породах с тонкой или глад- кой корой. Слоевище их обычно имеет вид оваль- ных пятен. Длинная ось такого овального пятна обычно размещена горизонтально. Предполага- ли, что подобная форма слоевища объясняется ростом ствола в толщину. Но оказалось, что она зависит от формы клеток коры дерева. Если они сильно вытянуты в горизонтальном направлении, то и слоевище вытянуто гори- зонтально. Если клетки коры одинаковы по длине и ширине, то и слоевище эндофлеодных лишайников приобретает округлую форму. Слоевище этих лишайников обычно постепен- но проникает в кору дерева. Первыми по малень- ким трещинкам, которые возникают в резуль- тате роста дерева в толщину, проникают вглубь гифы микобионта. А через некоторое время туда же проталкиваются и клетки водорослей, ко- торые из округлых временно становятся удли- ненными. С появлением водоросли начинается быстрый рост лишайника в ширину и дальней- шее проникновение эндофлеодного слоевища в более глубокие слои коры. Через некоторое время развиваются и плодовые тела, которыс у всех эндофлеодных лишайников расположе- ны на поверхности коры дерева.
Обычно гифы эндофлеодных лишайников растут между мертвыми клетками коры, рас- щепляя их на небольшие участки. Способны ли гифы пробивать оболочку клеток коры дере- ва, пока еще неизвестно. Однако вряд ли можно допустить, чтобы гифы, проникая внутрь только по трещинкам в коре, могли образовывать столь оформленные слоевища. Трудно также объяснить разделение пробки коры на неболь- шие участки только механическим воздействи- ем. Скорее всего гифы лишайника оказывают на коровые клетки дерева и химическое воздей- ствие. К этому выводу приводят некоторые наблюдения. Например, в местах контакта гиф лишайника с клетками коры были обнаружены повреждения оболочек клеток пробки, а в не- которых случаях и вообще деформированные одревеснелые оболочки. К роме того, в этих оболочках очень часто отсутствовала окраска, характерная для лигнина. Поэтому ученые до- пускают мысль, что гифы лишайников, расту- щих на коре деревьев и кустарников, обла- дают целлюлоз олитической способностью и содержат ферменты, расщепляющие клетчатку. Иногда типично эндофлеодные лишайники долго остаются полностью погруженными в субстрат, но с изменением условий освещения становятся поверхностными. Большей частью эти изменения зависят от характера коры. Так, эндофлеодные слоевища лишайников, разви- вающиеся в тонкой коре ясеня, в условиях хоро- шего освещения начинают выступать из более глубоких слоев и становятся почти полностью поверхностными. 3начит, один и тот же ли- шайник может быть энда- и эпифлеодным. Еще одна чрезвычайно интересная группа накипных лишайников — это лишайники с ша- ровидной формой слоевища. Они широко из- вестны под названием кочующих лишайников. Встречаются кочующие лишайники в засушли- вых областях земного шара, в равнинных и горных степях, пустынях и иногда в предгор- ных районах. Слоевище у них комковато-ша- ровидной формы (табл. 50) и не прикреплено к субстрату. Такие комочки свободно лежат на поверхности почвы, и ветер или животные переносят их с места на место как маленькие перекати-поле. Форма комочков может быть самой разнообразной — от округлой до угло- ватой, лепешковидной и неправильной. Их поверхность бывает складчатой, бородавчатой, чешуйчатой или покрыта сосочковидными вы- ростами. Кочующий образ жизни в засушли- вых условиях привел к развитию у этих лишайников толстого и плотного корового слоя. Но на поверхности этого слоя можно заметить небольшие беловатые углубления, называемые псевдоцифеллами. Это органы газо- обмена — разрывы коры, через которые воз- дух проникает внутрь слоевища. Обычно в сердцевинном слое этих лишайников между ги- фами скапливаются кристаллы окисла кальция. Эти лишайники, главным образом предста- вителей рода аспицилия (Aspicilia), иногда на- зывают также «лишайниковой манной». Когда- то в пустынных областях в голодные годы их добавляли в пищу. В наше время алжирские крестьяне нередко используют эти лишайники как корм для овец. ЛИСТОВАТЫЕ ЛИШАЙНИКИ Слоевище листоватых лишайников имеет вид листовидной пластинки, горизонтально распро- стертой на субстрате. Наиболее характерна для него округлая форма, которая обусловлена горизонтально-радиальным ростом гиф. В на- чале образования слоевища гифы листоватых лишайников растут от одного зачатка по радиу- сам окружности. У взрослых растений также наблюдается краевой рост гиф. Обычно молодые слоевища имеют правильную округлую форму, но позднее они начинают неравномерно раз- растаться и форма их делается неправильной. Обычно форма слоевища определяется характе- ром субстрата. Чем ровнее его поверхность, тем более правильную округлую форму имеют слое- вища листоватых лишайников. Наиболее простое слоевище листоватых ли- шайников имеет вид одной крупной округлой листовидной пластинки, достигающей в диа- метре 10 — 20 см. Такая пластинка нередко бывает плотной, кожистой, окрашенной в темно- серый, темно-коричневый или черный цвет (рис. 292). Слоевище, состоящее из одной листо- видной пластинки, носит название м о н о- ф и л ь н о r о. Монофильное пластинчатое слое- вище обычно прикрепляется к субстрату только в своей центральной части с помощью толстой короткой ножки, называемой г о м ф о м (рис. 292, За). Такой формой слоевища обладают представители рода умбиликария (Umbilica- ria) и некоторые виды рода дерматокарпон (Der- matocarpon). Это ксерофитные формы, широко встречающиеся на скалах в Арктике, Антарк- тике и высокогорьях. Толстые плотные кожи- стые слоевища этих лишайников, надеж- но прикрепленные к поверхности субстрата толстой короткой ножкой, легко переносят действие сильных ветров, снежные бури, ура- ганы и могут существовать длительные периоды без воды. Иногда такие слоевища состоят не из одной, а из нескольких листовидных пласти- нок, и тогда их называют п о л и ф и л ь н ы- м и (табл. 42, 8,4,5). Нередко пластинчатое слоевище бывает по краям выемчато вырезано или рассечено на широкие доли. Примером могут служить круп- 394
,ю 1~4. ю, з':;. '%~ \ , °, ° ° ~~1 . ~с %- ю 'P !'!у ° ° i i, 1 ,у \ у4 % Ъ ч~ ~,,~ (~Щ ~с Д ( с ° i Ú, ° ~с ' Ъ, у к, Ъ с )~ ~4 ф' .".. Фк "-' <г~ ° 1) 4 "' pj. ~ч Р 3;. ~% ~ Ъ с ъ» ф 'Ъс,.' г с ~А~ ~ъ с, Ъ "1 ) 5 P@q. 292. Моиофильное пластинчатое слоевище: 1 — вид нолистоватые слоевища видов рода пельтигера (Peltigera), которые нередко встречаются в ле- сах на почве среди мха или на основаниях ство- лов деревьев, поросших мхом (табл. 42, 8). Во влажных старых тенистых лесах на стволах деревьев или мшистых скалах растут причуд- ливо вырезанные крупнолистоватые слоевища лобарнй (Lobaria) п стикт (Sticta, табл. 47, 1,4). Эти лишайники обычно довольно рыхло прикрепляются к суострату всей своей нижней поверхностью, свободными остаются лишь при- поднимающиеся кверху края. Более сложным по строению является листо- ватое слоевище, рассеченное на множество мелких лопастей. Эти лопасти бывают самого разного размера и формы: узкими и широкими, слабо- и сильноветвистыми, плоскими и выпук- лыми. тесно сомкнутыми и разделенными, на- легающими друг на друга своими краями или строго отграниченными (табл. 47). Как правило, они собраны в округлые розетки, но иногда образуют слоевища неопределенных, бесконеч- но разнообразных форм — по внешнему облику они напоминают искусно сплетенные кружева, окутывающие разноцветным чехлом стволы и ветви деревьев (табл. 42, 6). Характерной особенностью листоватого слое- вища лишайников является его дорсовентраль- ное строение, при котором верхняя поверхность отличается по строению и окраске от нижней. Например, у представителей рода пельтигера (Peltigera) верхняя сторона слоевища серова- тая или серовато-коричневатая, гладкая или слегка шероховатая, а нижняя — беловатая, розоватая, в центре нередко черно-серая, вой- лочная, с многочисленными жилками и лохма- тыми тяпками прикнепительных тиф. У видов рода гипогнмння (Hypogyrnnia) верхняя сто- слоевища сверху; 2 — вид слоевища сбоку. а — гомф. рона гладкая, серая, а нижняя — черная. У лишайника солорина скортея (Solorina scor- tea) слоевище сверху окрашено в серовато- коричневый цвет, а снизу — в яркий, красновато- оранжевый (табл. 42, 4). Верхняя поверхность слоевища листоватых лишайников бывает ровной, волнистой, ямча- той, голой, глянцевитой или матовой, нередко шероховатой, неровной, покрытой бугорками, бородавочками. Иногда на ней имеются различ- ной формы выросты, реснички, образующие опушение или войлочный налет. Нижняя поверхность также разнообразна по своему строению, но ее характерной особен- ностью является то, что она почти всегда обра- зует особые органы, с помощью которых листо- ватый лишайник прикрепляется к субстрату. В отличие от накипных лишайников, слоевище которых целиком плотно срастается с субстра- том, листов атые лишайники обычно довольно рыхло с ним связаны и в большинстве случаев могут быть легко от него отделены. Только очень немногие виды, например представители рода гипогимния, прикрепляются к субстрату прямо нижним коровым слоем. Но и в таком случае не происходит плотного срастания суб- страта и всей нижней поверхности лишайника- слоевище прикрепляется отдельными участками нижней поверхности. У огромного большинства листоватых лишайников на нижней стороне слоевища образуются особые органы прикреп- ления — ризоиды, ризины или гомф. Они об- разуются тяжами гиф и отличаются друг от дру- га анатомическим строением. У видов рода ан- ция (Anzia), распространенных в тропических и субтропических областях Азии, Америки, Австралии и Океании, а в СССР встречающихся на Дальнем Востоке, образуется губчатое под-
слоевище, с помощью которого эти лишайники прикрепляются к субстрату (рис. 290, а). Листоватые лишайники по сравнению с на- кппными являются значительно более высоко- организованными формами. В эволюционном отношении оказалось выгодным отделение слое- вища от субстрата. Между ними появился не- большой промежуток, и это дало целый ряд преимуществ. Во-первых, в нем заключена про- слойка воздуха, способствующая лучшему га- зообмену внутренних слоев слоевища. Во-вто- рых, там дольше задерживается влага, благо- даря чему слоевище более длительное время мо- жет находиться во влажном состоянии. В-тре- тьих, в узком пространстве между поверхностью субстрата и слоевищем обычно задерживаются различные органические и неорганические ве- щества, которые могут быть использованы ра- стением. Но, с другой стороны, отделившись от суб- страта, лишайник обрек себя на массу неудобств. Он стал более уязвим по отношению к факторам внешней среды — действию ветра, ударам дождя и снега, нападению животных и т. д. И прежде всего оказалась незащищен- ной нижняя поверхность лишайника. Прогрессивная роль отделения лишайника от поверхности, на которой он рос, состояла в усложнении анатомической структуры слое- вища и развитии особых органов прикрепле- ния. Прежде всего на слоевище появился ниж- ний коровой слой, обычно отсутствующий у накипных форм лишайников. Огромное боль- шинство листоватых лишайников имеет на ниж- ней стороне слоевища хорошо развитый коровой слой. Исключение составляют крупнолисто- ватые слоевища пельтигеры. Нижняя поверх- ность этих лишайников выстлана рыхлораспо- ложенными сердцевинными гифами, которые образуют также длинные пучки ризоидальных тяжей, проникающих в субстрат. Эти лишайни- ки обычно растут на почве, среди мхов. Своеоб- разное строение нижней поверхности, отсутст- вие нижнего корового слоя позволяют этим растениям лучше и быстрее втягивать влагу из влажных дерновинок мхов. Однако отделение слоевища от субстрата привело не только к образованию корового слоя на нижней стороне лишайников, но и к усло'к.— нению анатомической структуры всего слоеви- ща. В отличие от накипных лишайников у лп- стоватых форм в слоевищах существует четкая дифференциация анатомических слоев. Как пра- вило, под микроскопом на поперечных срезах таких слоевищ можно различить 4 хорошо дифференцированных слоя: верхний коровой слой, слой водорослей, сердцевину и нижний коровой слой. Особенно боль|ного разнообра- зия у листоватых лишайников достигает струк- тура коровых слоев, которые выполняют здесь не только защитную, но и укрепляющую роль. Среди листоватых лишайников также встре- чаются неприкрепленные, кочующие формы. В степях юга и юго-востока нашей страны и в Монголии на поверхности почвы обычно можно встретить свободнолежащие зеленовато-черные слоевища пармелии блуждающей (Parmelia va- gans, табл. 50). Они не прикреплены к субст- рату и свободно переносятся ветром с места на место. В горных тундрах Сибири и Чукотки на сухих горных склонах и пологих сопках оби- тает другой очень красивый кочующий лишай- ник цетрария Ричардсона (Cetraria richardsonii, табл. 47, 2). Темно-коричневые слоевища его в сухом состоянии сжимаются и свертываются в комки, которые, как миниатюрные перекати- поле, кочуют с помощью ветра с места на место. КУСТИСТЫЕ ЛИШАЙНИКИ Слоевище кустистых лишайников имеет вид прямостоячего или повисающего кустика, реже неразветвленных прямостоячих выростов. По организационному уровню кустистые ли- шайники представляют высший этап развития слоевища. В отличие от накипных и листоватых форм лишайников, для которых характерен горизон- тальный рост гиф, у кустистых лишайников наблюдается вертикально направленный рост гиф и верхушечный рост слоевищ. Это позволя- ет кустистым лишайникам путем изгибов вето- чек в разные стороны занимать наилучшее положение, при котором водоросли могут мак- симально использовать свет для осуществления фотосинтеза. Эти лишайники обычно прикреп- ляются к субстрату только небольшим участ- ком нижней части слоевища. Прямостоячие напочвенные кустистые лишайники чаще всего прикрепляются к почве тонкими нитевидными ризоидами. Прикрепление повисающих слое- вищ кустистых лишайников к коре дерева или поверхности скал происходит с помощью п с е в- д о г о м ф а. Последний имеет вид короткой ножки, расширенной на конце в небольшую пяточку (рис. 293); он напоминает по внешнему виду гомф листоватых лишайников, но отличается от не- го анатомическим строением. Редкое исключение среди кустистых лишайников составляет антарк- тический лишайник гимантормия траурная (Himantormia ligubris). Слоевище его, имеющее вид черных, как бы мертвых кустиков с разбро- санными белыми пятнами, прикрепляется к скалам с помощью подслоевища (табл. 49, 4). Слоевища кустистых лишайников могут быть разных размеров. Высота самы с маленьких сос- тавляет всего несколько миллиметров, а наибо- лее крупных 30 — 50 слю. Повисающие слоеви-
ща кустистых лишайников иногда могут дости- гать колоссальных размеров. Так, длина одно- го из эпифитных лишайников — уснеи длинной (Usnea longissima), свисающей в виде бороды с ветвей лиственниц и кедров в таежных лесах, составляет 7 — 8 м (табл. 49, 8). Слоевища кустистых лишайников чрезвычай- но разнообразны по форме. Наиболее простые имеют вид отдельных прямостоячих неразвет- вленных вырос тов. Ярким примером может служить слоевище тамнолии (Thamnolia, табл. 48, 8). В тундрах Крайнего Севера или высоко- горного пояса на почве среди дерновинок мхов и лишайников нередко можно встретить в бес- порядке разбросанные молочно-белые или блед- но-розоватые заостренные на конце палочки. Эти простые, вертикально растущие палочки, напоминающие по виду маленькие пики, пред- ставляют собой слоевище арктоальпийск ого лишайника тамнолии червеобразной (Thamno- lia vermicularis, табл. 51). Но чаще кустистые лишайники бывают разветвленными и образу- ют слоевище в виде густых компактных дерно- винок (табл. 48; 49, 1, 2, 4, 6, 7). Такой формой слоевища обладают очень многие лесные и тундровые напочвенные лишайники. В сухих сосновых борах, в северных и высокогорных тундрах нередко можно наблюдать на поверх- ности почвы сплошные разноцветные ковры, образованные дерновинками кустистых лишай- ников. Кустистые слоевища эпифитных лишай- ников обычно имеют вид лохматого кустика. Но иногда их лопасти бывают очень тонкими, сильно вытянутыми, почти нитевидными. Такие слоевища напоминают по внешнему облику длинные седые или черные бороды (табл. 49, 8). Среди кустистых лишайников различают слоевища с плоскими и округлыми лопастями. Более примитивным типом строения является слоевище с плоскими лопастями. Эти слоевища ближе всего стоят к листоватым лишайникам, среди них имеются многочисленные переход- ные формы между листоватыми и кустистыми слоевищами. Как и у листоватых лишайников, у кустистых слоевищ с плосколинейными лопа- стями нередко наблюдается дорсовентральное строение. Примером может служить широко известная цетр ария исландская — характер- ный представитель напочвенных лишайников сосновых лесов, болот и ceBepHIIx тундр (табл. 48, о). Слоевище этого кустистого лишай- ника имеет вид рыхлой дернинки высотой до 10 сл и образовано плоскими, местами желоб- чато свернутыми лопастями. Верхняя поверх- ность этих лопастей коричневая или зеленовато- коричневая, блестящая. Нижняя поверхность окрашена светлее, она светло-коричневая, по- чти до белой, с многочисленными белыми пят- нышками, разрывами коры, которые служат для проникновения воздуха. Но, в отличие от листоватых лишайников, на поперечном срезе этих лопастей можно различить уже не четыре, а пять анатомических слоев. Помимо верхнего и нижнего коровых слоев, слоя во- дорослей и сердцевины, у этого лишайника имеется еще один слой водорослей, располо- женный над нижней корой слоевища. Таким образом, уже у кустистых лишайников с пло- сколинейными лопастями благодаря вертикаль- ному росту слоевища происходит увеличение поверхности, занятой водорослями, что способ- ствует более интенсивному процессу фотосин- теза. Далее в эволюционном развитии кустистых лишайников намечается переход к радиальной структуре слоев ища, как наиболее выгодной для лишайника. В цилиндрических лопастях водоросли располагаются по окружности и тем самым достигаются равномерность их освеще- ния со всех сторон и максимальное увеличение ассимиляционной поверхности слоевища. Пе- реход от плосколинейной структуры лопастей Рис. 293. Псевдогомф: 1 — общий вид псевдогомфа у нейропогона (Neuropogon); 2— анатомическое строение псевдогомфа: а — коровой слой, 6— клетки водорослей, в — сердцевинные гифы.
Рис. 294. Ра азличные типы подециев у лишайников из рода кладоний: 1 — ш — шиловидные подеции Cladonia coniocraea; 2 — роговидные подеции Gladonia subulata 8 — кубковидные по ( ф ) adonia fimbriata; 4 — пролифицирующие сцифы Cladonia ver- kieijlata; 5 — разветвленные подеции Cladonia sylvatica. ~с радиальной происходит прежде всего через свертывание плоских лопастей в трубочки. акого типа лопасти можно наблюдать у напоч- венного кустистого лишайника цетрарии клу- бочковой (Cetraria cucullata), характерного обитателя северных и горных тундр. Ярко- желтое или бледно-зеленовато-желтое слоевище его образовано желобчато свернутыми, почти трубчатыми лопастями (табл. 48, 1). У более продвинутых в эволюционном отношении кус- тистых лишайников лопасти слоев ища стано- вятся угловато-округлыми и теряют свое дор- советральное строение. Верхняя и нижняя поверхности этих лопастей одинаково окраше- ны и аналогичны по своему строению. Но, несмотря на радиальную анатомическую струк- туру, по форме эти лопасти не цилиндрические, а слегка сдавленные, почти до плоских и лишь в некоторых участках округлые. Лопасти такой формы имеет, например, широко распростра- ненный эпифитный лишайник летария (Letha- ria thamnodes), напочвенные лишайники рода алектория (Alectoria ochroleuca) и некоторые другие (табл. 49). Своеобразной переходной формой между листоватыми и кустистыми лишайниками явля- ются слоевища видов рода кладония (Cladonia)— широко распространенных напочвенных кус- тистых лишайников. По своему строению эти слоевища считаются уже радиально-кустистыми. У большинства представителей рода кладония тело образовано двумя типами слоевищ: горизон- тальным — чешуйчатым, реже бородавчатым и вертикальным — различной формы вырос- тами или кустиками, растущими вверх от чешу- ек (табл. 48,2, 4, 6). При образовании слоевища у кладонии сначала вырастают горизонталь- ные чешуйки. Эти чешуйки, достигающие обыч- но в диаметре от 2 до 5 лл, но иногда имеющие вид небольших листиков диаметром до 10 — ~ — 20 лл, сверху зеленоватые, снизу бело- ватые или желтоватые, собраны в небольшие дерновинки. По анатомическому строению они обычно трехслойные. В них можно различить верхний коровой слой, слой водорослей и сердцевину. Спустя некоторое время на поверх- ности или по краям этих чешуек появляются вертикально направленные выросты, которые называют п о д е ц и я м и. Форма подециев чрезвычайно разнообразна. Они могут быть простыми, неразветвленными, иметь вид шило- видных или роговидных выростов (рис. 294 1 2~. Х~ 7 ) acro они на конце расширены в виде бокалов (рис. 294, 3). Такие кубковидные подеции на",û- вают с ц и ф а м и . Иногда из центра такого оокальчика вырастает еще один, а из него, в свою очередь, следующий, и таким образом развиваются как бы многоэтажные подецил, образованные сцифами, растущими один из 398
другого (рис. 294, 4). Нередко подеции бывают сильно разветвленными и имеют вид кустика (рис. 294, о). Такая форма подециев, например, очень характерна для группы кладоний, кото- рую называют ягелем или оленьим мхом. Эти- ми лишайниками на Севере питаются олени. Общее количество лишайников, поедаемых оленями, составляет не менее 50 видов. Но особенно олени предпочитают кладонии. Среди них наиболее «любимыми» являются кладония приальпийская (Cia donia alpestris), кладония оленья (C. rangiferina), кладония лесная (С. sylvatica) и кладония мягкая (C. mitis). У этих видов, в отличие от других кладоний, горизонтальное слоевище не чешуйчатое, а бо- родавчатое и рано исчезает. И слоевище этих лишайников фактически состоит лишь из одних подециев, сильно разветвленных и образующих довольно крупные дернинки. Подеции кладоний округлые в сечении и имеют радиальную анатомическую структуру. Снаружи в них можно различить коровой слой, далее слой водорослей, расположенный по окружности, а внутри подеция — сердцевину. Но нередко эти анатомические слои бывают нечетко выражены или вовсе не развиты. Так, например, у группы ягелей на поверхности подециев никогда Бе развивается коровой слой, их поверхность образована рыхлорасполочен- ными сердцевинными гифами (рис. 295). У мно- гих видов кладоний коровой слой развит на поверхности подециев в виде отдельных островков. Слой водорослей также не является непрерывным, а состоит из отдельных разбро- санных группок водорослей. Сердцевина поде- циев, примыкающая к зоне водорослей, обычно образована рыхл орасположенными гифами, а далее по направлению к центру имеет аморф- ное строение и состоит из плотно склеенных гиф. У большинства видов кладоний в центре подециев образуется внутренняя полость, не заполненная сердцевинной тканью. Из лишайников, лопасти которых имеют типично радиальную структуру, можно назвать виды рода уснея (Usnea) и многие виды рода алектория (Alectoria). Лопасти этих лишайников, как правило, округлые, тонкие и сильно вытя- нутые. Чаще всего они образуют длинные, повисающие слоевища в виде редких бледно-зе- леных, серых и буро-черных «бород» (табл. 49, 7, 8). На поперечном разрезе округлых лопастей этих лишайников можно различить снаружи плотный, хорошо развитый коровой слой, под ним расположен слой водорослей, а центральная часть занята сердцевиной. Причем у алекторий сердцевина однородная, она образована беспо- рядочно рыхло расположенными гифами. Но у усней происходит дальнейшее усложнение анатомической структуры слоевища. Серд- л 1 ~~ ~( ) ° if/ с I б / ' / 1 / / ! ,Ф~ I ! Ъ I ° ° Ф с ° ) с ° ° ° ° ю ° ° ) ° ° 1 ) ° ° ° Ъ ° ° ° ° ) ° ° 1 ° ° . ° ~ ° ° ° 'I ) ° ° / % ° ° ° ° Ъ I 1 / 1 / 1 / 1 / 7 / 1 / 1 / / / / / / 2 1 I 1 / 1 I 1 1 / E / 1 I I I ! ! ! ) Рис. 295. Секторы поперечных разрезов через поде- ции кладоний: 1 — Cladonia furcata; 2 — Cladonia rangiferina. а — коровой слой, б — слой водорослей, в — сердцевина, г — центральная ПОЛОСТЬ. цевина этих лишайников неоднородна, в ней можно различить периферическую часть, со- стоящую из рыхлорасположенных гиф, и цен- тральную часть, образованную продольно идущими, плотно склеенными гифами. Эти склеенные между собой толстостенные гифы, расположенные параллельно оси лопастей, образуют так называемый центральный, или осевой, тяч. Осевой тяч придает прочность тонким и сильно вытянутым лопастям этих лишайников (рис. 296). Интересно, что в Антарктике наиболее ши- роко распространенными кустистыми лишай- никами являются виды рода нейропогон (Neu- ropogon). Этот род очень близок к уснеям, его представители также имеют округлые лопасти с осевым тячем в центральной части. Желтовато-зеленоватое, местами зачернен- ное слоевище этих лишайников имеет вид силь- но разветвленного компактного кустика высотой 2 — 10 сл, плотно прикрепляющегося к субстрату расширенной пяточкой псевдогомфа (табл. 49,6). Благодаря морфологическим особенностям строения слоевищ эти лишайники хорошо при- способились к жизни в условиях холодных пустынь Антарктики. Механический тяж, придающий прочность и устойчивость ветвям, и прочное прикрепление к поверхности скал посредством псевдогомфа позволяют этим пря- мостоячим кустистым слоевищам хорошо про- тивостоять постоянным сильным антаркти- ческим ветрам. Зачерненные местами ветви этих лишайников и их черные плодовые тела хорошо притяги- вают тепловые лучи, что также является важ- ной морфологической особенностью для рас-
Рис. 296. Поперечный (1) и продольный (2) разрезы через лопасть кустистого лишайника уснея (Usnea): а — коровой слой; б — клетки водорослей; в — периферическая часть сердцевины, состоящая из рыхлорасположенных гиф; г- центральная часть сердцевины — осевой тяж. тений, живущих в условиях ледяных пустынь. Виды рода нейропогон играют значительную роль в растительном покрове Антарктики. В прибрежных районах в некоторых местах на скалах они образуют сплошные заросли, иногда к ним присоединяются другие лишай- ники и мхи, и тогда они образуют сообщества растений, сплошь покрывающие субстрат. В Антарктике широко распространена своеоб- разная стелющаяся форма кустистых лишай- ников, представленная в этом районе видами рода алектория. Темно-коричневые, почти черные слоев ища этих лишайников, состоя- щие из округлых, местами деформированных лопастей, горизонтально распростерты на по- верхности скал и мелких камней. Это типично ветроустойчивая форма кустистых лишайни- ков: здесь слоевища не только плотно прижи- маются к субстрату, но каждая веточка такого слоевища на своем протяжении в нескольких местах еще прикрепляется к скалам маленьки- ми пяточками. Лопасти этих стелющихся ли- шайников имеют множество вторичных вето- чек, а все слоевище напоминает по внешнему виду плотную корку, состоящую из клубков перепутанных деформированных округлых ло- пастей. Морфологические особенности этих ли- шайников позволяют им хорошо приспособить- ся к жизни в условиях ледяного континента. Кустистые виды алектории со стелющейся формой слоевища чрезвычайно широко рас- пространены в Антарктике; они встречаются не только в прибрежных, более благоприятных районах, но и заходят в глубину материка, поднимаются высоко в горы, где условия для жизни растений особенно суровы. Эти лишай- ники были найдены в крайних пределах су- ществования растительности на земном шаре, всего в 500 кл от Южного полюса. Иногда у крупных кустистых лишайников в условиях тундр и высокогорий развивают- ся добавочные пр икр епительные органы, с помощью которых они прирастают к листьям осок, злаков, к веточкам карликовых берез и дру- гим кустарничкам. Эти добавочные органы при- крепления, называемые r а птер а ми, могут воз- никнуть из любой части слоевища. Например, у цетрарии исландской они нередко развивают- ся из ресничек, окаймляющих края лопастей (табл. 48, Б). Если слоевище цетрарии исландской растет вблизи веточки какого-нибудь кустарни- ка, листа злака, мха или даже слоевища другого лишайника и одна из его ресничек долгое вре- мя находится в соприкосновении с ними, то такая ресничка начинает вытягиваться и на ее конце возникает плоская пластиночка. С по- мощью этой пластиночки лишайник прикреп- ляется к растущему по соседству растению. Причем развитие гаптер и прикрепление слое- вища к новому субстрату происходят довольно быстро. Обычно для их формирования доста- точно одного вегетационного периода. Таким образом лишайники в суровых условиях север- ных и высокогорных тундр предохраняют се- бя от отрыва сильными ветрами и бурями. АНАТОМИЯ СЛОЕВИЩА ЛИШАЙНИКОВ У лишайников в зависимости от анатомиче- ского строения различают два типа слоевищ: 1) r о м е о м е р н о е слоевище, когда водоросли разбросаны по всей толще слоевища; 2) r е т е- р о м е р н о е слоевище, когда водоросли обра- зуют в слоевище обособленный слой. Более примитивным считается слоевище го- меомерного строения. Если рассмотреть попе-
Рис. 297. Анатомическое строение слоевища лишайников: 1 — гетеромерное слоевище (а — верхний коровой слой, б — слой водорослей, в — сердцевина, г — нижний коровой слой); .2 — гомеомерное слоевище слизистого лишайника коллема (Со11ета Г1ассЫит); 8 — гомеомерное слоевище слизистого ли шайника лептогиум (Leptogium saturninum) (а — коровой слой с верхней и нижней стороны слоевища, б — ризоиды). речный срез такого слоевища под микроскопом, то хорошо видно, что оно образовано беспоря- дочными переплетениями гиф гриба, среди ко- торых разбросаны отдельные клетки или нити водорослей (рис. 297, 2, 8). Такое строение наи- более характерно для тех лишайников, фико- бионтом которых являются сине-зеленые во- доросли — носток, глеокапса и некоторые дру- гие. Эти лишайники образуют группу, изве- стную под названием слизистых лишайников. У лишайников, которые имеют фикобионтом зеленые водоросли, слоев ище гомеомерного строения встречается очень редко, лишь у наиболее примитивных накипных форм. Слоевища слизистых лишайников в сухом состоянии имеют вид темных или даже черных твердых и хрупких корок или пленок. Но они обладают одной интересной особенностью— способностью впитывать огромное количество воды, в 20 — 30 раз превышающее их собствен- ную массу. При увлажнении они сильно раз- бухают, ослизняются, приобретают характер студня или желе и становятся оливково-зеле- ными или грязно-зелеными. Примером может служить влажное слоевище слизистого лишай- ника лепт огиума (Le ptogium saturninum), изображенное на цветной таблице 47. Во внешнем облике этих лишайников имеется много общего с некоторыми свободноживу- щими водорослями. Например, у видов рода коллема (Collema), типичных представи- телей слизистых лишайников, форма и цвет слоев ища очень напоминают черные пленки колоний свободноживущей водоросли носток. Иногда кажется, что в таких слоевищах не грибные гифы образуют переплетения, среди которых разбросаны нити водорослей, а просто в колонии водорослей проникли гифы гриба. Действительно, на формирование внешнего об- лика этих слоевищ оказывают большое влияние морфологические особенности их фикобионта- сине-зеленых водорослей. Для клеток этих во- дорослей чрезвычайно характерны слизистые желатинные образования, которые формируют- ся чаще всего в результате ослизнения толстых оболочек клеток или как продукт выделения их содержимого. До последнего времени да- же считали, что сине-зеленые водоросли пол-
Рис. 298. Поперечный разрез через лопасть кустистого лишайника цетрарии исландской (Cetraria islandica): а — верхний коровой слой, б — верхний слой водорослей, в— сердцевина, г — нижний слой водорослей, д — нижний коровой слой, е — псевдоцифелла. костью определяют форму слоев ища у большинства слизистых лишайников. Одна- ко поляризационно-оптические исследования, проведенные в последние годы, показали, что слизистая масса этих слоевищ образована не только водорослью, но и грибом. Значитель- ная часть желатина этих лишайников проду- цируется грибным компонентом. Гриб опреде- ляет размеры слоевища, формирует лопасти и основные детали общего вида. Среди слизистых лишайников встречаются как накипные, так и листоватые и кустистые формы. Но, в отличие от других лишайников, у слизистых усложнение морфологического строения почти не приводит к анатомическим преобразованиям в слоевище. Как у накипных, так и у более высокоразвитых листоватых и кустистых форм этих лишайников структура слоевища гомеомерная, т. е. водоросли всег- да разбросаны по всей толще слоевища. Лишь у видов рода лептогиум (Leptogium) — высоко- организованных форм слизистых лишайников- с верхней и нижней стороны слоевища име- ется однослойный крупноклеточный коровой слой (рис. 297, 8). У наиболее эволюционно продвинутых представителей этого рода все слоевище целиком состоит из крупноклеточ- ной ткани, но среди этих клеток разбросаны в беспорядке нити ностока. Слизистые лишайники насчитывают неболь- шое число видов, всего 750, что составляет толь- ко Зо~о от оощего числа всех известных видов лишайников. Сравнительно небольшое коли- чество этих лишайников, по-видимому, можно объяснить более поздним вовлечением сине-зе- леных водорослей в процесс образования сло- евища лишайников. Но также возможно, что захват грибом сине-зеленых водорослей был менее успешным по сравнению с зелеными во- дорослями. В силу особенностей строения сине-зеленых водорослей, нередко собранных в колонии и обладающих плотными, богатыми слизью оболочками, эти водоросли оказались более трудным объектом для гриба при уста- новлении с ними контакта и при построении слоевища. Безусловно, при захвате этих водо- рослей и их передвижении гриб испытывает гораздо больше трудностей, чем с одноклеточ- ными и другими формами зеленых водорослей. Может быть, именно этим можно объяснить и примитивное анатомическое строение слизис- тых лишайников. Для остального огромного большинства ли- шайников характерна гете ромерная структу- ра, при которой в слоевищах можно различить дифференцированные слои. При этом чем слож- нее морфологическое строение слоевища лишай- ника, тем сложнее и его анатомическая струк- тура. У этих растений в ходе эволюции услож- нение морфологического строения слоевища шло параллельно усложнению его анатоми- ческой структуры. Так, в слоевище накипных лишайников, наиболее примитивных форм, можно различить только три анатомических слоя: коровой слой, слой водо- рослей и сердцевину. Такое же анатомическое строение имеют и некоторые листоватые лишайники, например представи- тели рода пельтпгера (рис. 299, 1). Но у боль- шинства листоватых лишайников, более эволю- ционно продвинутых форм, в связи с отделе- нием от поверхности субстрата происходит образование еще одного корового слоя — с нижней стороны слоевища. В таких слоевищах можно различить уже четыре слоя: верхний коровой слой, слой водорослей, сердцевину и нижний коровой слой (рис. 297, 1). У кустистых лишайников с плоскими лентовидными лопастя- ми образуется еще один анатомический слой— слой водорослей с нижней стороны слоевища. У этих лишайников в слоевище можно разли- чить уже пять анатомических слоев: два коро- вых слоя на верхней и нижней поверхности слоевища, два слоя водорослей, тоже с верхней и нижней стороны лопастей, и сердцевину
Рис. 299. Строение корового слоя лишайников.' 1 — параплектенхимный коровой слой (Peltigera); 2 — прозоплектенхимный коровой слой ровой слой (Teloschistes). а — коровой слой, б — клетки водоросли, в — сердцевинные гифы. (Parmelia); 8 — волокнистый ко» (рис. 298). И наконец, кустистые лишайники с округлыми лопастями, обладающие наиболее высокоорганизованными слоев ищами, имеют радиальную структуру: снаружи лопасти этих лишайников покрыты коровым слоем, под ним расположен слой водорослей, а центральная часть слоевища заполнена сердцевиной (рис. 295). Каждый из перечисленных анатомических слоев слоевища выполняет в жизни лишайника ту или иную функцию и в зависимости от этого имеет совершенно определенное строение. Коровой слой играет в жизни лишайника очень важную роль. Он выполняет сразу две функции: защитную и укрепляющую. Он за- щищает внутренние слои слоевища от воздей- ствия внешней среды, прежде всего водоросли от чрезмерного освещения. Поэтому коровой слой лишайников обычно бывает плотного строения и окрашен в сероватый, коричне- вый, оливковый, желтый, оранжевый или красноватый цвет. Коровой слой служит и для укрепления слоевища. Чем выше слоев ище поднимается над субстратом, тем более оно нуждается в ук- реплении. Укрепляющие механические функ- ции в таких случаях нередко выполняет тол- стый коровой слой. Гифы корового слоя срас- таются своими стенками и могут образовыват~ очень плотные и сложные плектенхимы- клеточные ткани различного строения. Так, например, если срастаются тонкостенные гифы, поделенные поперечными перегородками на одинаковые по ширине и длине клетки, то образуется ткань настоящей клеточной струк- туры (рис. 299, 1). Такую ткань называют пар аплектенхимой. Иногда ее еще называют псевдопаренхимой, так как она по внешнему виду напоминает парен- уиму (клеточную ткань) цветковых растений. Однако сходство этих тканей только внешнее, ибо процесс их образования различен. Если паренхима цветковых растений образуется пу- тем последовательного деления одной клетки, то псевдопаренхима лишайников — путем срастания грибных гиф. У многих лишайников в коровом слое обо- лочки клеток гиф бывают сильно утолщены, а просветы клеток, заполненные протоплаз- мой, очень узкими. При срастании таких гиф образуется коровой слой, представляю- щий сплошную хрящевидную массу, где гра- ницы клеток незаметны, а их просветы ка- жутся почти точками. При этом гифы на сво- ей поверхности выделяют желатин, плотно цементирующий их между собой. Такого типа ткани у лишайников называют п р о з о- нлектенхиматическими. Так уст- роен коровой слой многих кустистых лишай- ников, например цетрарии исландской, лис- товатых лишайников рода пармелия и мно-
Рис. 300. Расположение гиф в слоевище лишайников: 1 — сердцевинный слой в слоевище анаптихии; 2 — палисадный коровой слой в слоевище тамнолии. а — коровой слой, б- клстки водорослей, в — сердцевинные гифы. Рис. 301. Органы прикрепления слоевища листоватых лишайников.' 1 — нижний параплектенхимный коровой слой с отходящими от него ризоидами (Sticta); 2 — ризины пармелии с приирепитель- ными пластинками на конце; 8 — ризина с капелькой слизи на конце. гих других (рис. 299, 2). Плотный хряще- видный коровой слой этих лишайников не только надежно защищает нижележащие слои водорослей, но и хорошо укрепляет припод- нимающееся над землей слоевище. Реже гифы корового слоя лишайников не срастаются, а лежат свободно параллельно друг другу. При этом они могут образовывать два различных типа корового слоя. Если ги- фы расположены параллельными рядами пер- пендикулярно поверхности слоевища, то образу- ется п а л и с а д н ы й к о р о в о й с л о й (рис. 300, 2) . По внешнему виду он напоминает палисадную клеточную ткань в листьях цветковых растений. Такое строение имеет, например, коровой слой тамнолии (табл. 48, 8). Во втором случае (напри- мер, у уснеи) гифы корового слоя лежат не пер- пендикулярно, а параллельно поверхности слое- вища и имеют вид удлиненных волокон (рис. 299, 8). Коровой слой такого строения носит название волокнистого. В слоевищах лишайников верхний и ниж- ний коровые слои могут быть одинаковыми по окраске и строению, но нередко отлича- ются друг от друга, особенно у лис товатых лишайников, для которых характерно дор- совентральное строение слоевища. На нижнем коровом слое лишайников обычно образуются органы прикрепления. Иногда они имеют вид очень тонких нитей, состоящих из одного ряда клеток. Эти нити называют р и з о- и д а м и. Каждая такая нить берет начало от одной клетки нижнего корового слоя (рис. 301,1). Нередко несколько ризоидов соединяются в толстые ризоидальные тяжи. Их можно видеть, например, на нижней стороне крупнолистоватых слоевищ пельтигер. У листо- ватых лишайников рода пармелия образуются более толстые прикрепительные тяжи, назы- ваемые р и з и н а м и (рис. 301, 2). В образо- вании ризин принимают участие не только ги- фы нижнего корового слоя, но и сердцевины. Снаружи они покрыты коровым слоем, а внутри образованы сердцевинными гифами. На са- мом конце ризины клетки гиф вытягиваются и расходятся в разные стороны, образуя кис- точку. Нередко на конце такой кисточки обра- зуется слизистая капелька, с помощью которой слоевище более плотно прикрепляется к суб- страту (рис. 301, 8). Иногда на конце ризины образуется особая прикрепительная пласти- ночка, которой лишайник, как маленькой по- дошвой, прикрепляется к поверхности коры дерева или камня. Как уже отмечалось, у некоторых листова- тых лишайников, имеющих вид более или менее округлой листовидной пластинки, слоевище прикрепляется лишь в своей центральной части с помощью короткой ножки — r о м ф а (рис. 292). Гомф — это также вырост нижней поверх- ности слоевища, и в его образовании прини- мают участие нижний коровой слой и сердце- вина лишайника. Снаружи эта толстая ножка покрыта параплектенхимным коровым слоем, а внутри ее проходят вытянутые сердцевинные гифы. На конце гомфа, на месте прикрепления к субстрату, эти гифы имеют очень толстые темные оболочки и образуют зубцы, с помощью которых слоевище очень плотно прикрепляется к поверхности скал. Некоторые кустистые ли- шайники, виды родов уснея, нейропогон и др.,
прикрепляются к скалам с помощью п с е в д о- r о и ф а, также имеющего вид короткой рас- ширенной на конце ножки. В отличие от гомфа в образовании псевдогомфа принимает участие только сердцевина лишайников. Псевдогомф образован вытянутыми, склеенными сердцевин- ными гифами, также образующими на конце темные зубцы, с помощью которых лишайник плотно прикрепляется к субстрату. В зоне водорослей осуществляются процессы ассимиляции углекислоты и накопление орга- нических веществ. Как известно, для осуществ- ления процессов фотосинтеза водорослям не- обходим солнечный свет. Поэтому слой водо- рослей обычно размещается вблизи верхней поверхности слоевища, непосредственно под верхним коровым слоем, а у вертикально стоя- щих кустистых лишайников еще и над нижним коровьп| слоем. Слой водорослей чаще всего бывает небольшой толщины, и водоросли раз- мещаются в нем так, что находятся почти в оди- наковых условиях освещения. Водоросли в слое- вище лишайника могут образовывать непре- рывный слой, но иногда гифы микобионта де- лят его на отдельные участки. Для осуществле- ния процессов ассимиляции углекислоты и дыхания водорослям необходим также нормаль- ный газообмен. Поэтому грибные гифы в зоне водорослей не образуют плотных сплетений, а расположены рыхло на некотором расстоянии друг от друга. Лишь у некоторых пустынных лишайников водоросли окружены плотной гриб- ной тканью клеточного строения, которая за- щищает их от жаркого и яркого пустынного солнца. Грибные гифы, окружающие водоросли, обычно являются ответвлениями или верши- нами гиф сердцевины. Но, в отличие от сердце- винных гиф, они меньшей толщины, обладают более тонкими стенками и часто бывают поде- лены поперечными -перегородками на множество клеток. Иногда, срастаясь, такие гифы обра- зуют в зоне водорослей рыхлые сетчатые пере- плетения. Под слоем водорослей расположен сердцевин- ный слой. Обычно сердцевина по толщине зна- чительно превышает коровой слой и зону во- дорослей. Особенно мощно она развита по срав- нению с другими слоями у накипных лишайни- ков. От степени развития сердцевины зависит толщина самого слоевища. Основная функция сердцевинного слоя— проведение воздуха к клеткам водорослей, содержащим хлорофилл. Поэтому для боль- шинства лишайников характерно рыхлое рас- положение гиф в сердцевине (рис. 300, 1). Воз- дух, попадающий в слоевище, легко проникает к водорослям по промежуткам между гифами. Сердцевинные гифы слабоветвисты, с ред- кими поперечными перегородками, с гладкими, слабожелатинооб разными толстыми стенками и довольно узким просветом, заполненным про- тоБлазмой. У большинства лишайников серд- цевина белая, так как гифы сердцевинного слоя бесцветны. Хотя на их поверхности, как пра- вило, откладываются кристаллы лишайниковых веществ, но в преобладающем большинстве они бесцветны и не изменяют окраски сердцевины. Если кристаллы лишайниковых веществ окра- шены в тот или иной цвет, то в зависимости от этого и сердцевинный слой приобретает зо- лотисто-желтую, кроваво-красную, серую или другую окраску. Лишайниковые вещества об- ладают одной важной особенностью: они не- растворимы или очень слабо растворимы в хо- лодной воде. Благодаря этому свойству крис- таллы, покрывая поверхность сердцевинных гиф, препятствуют их смачиванию. Поэтому даже во влажном слоевище лишайника его сердцевинный слой может частично оставаться сухим и содержать воздух, необходимый для клеток водорослей. У некоторых кустистых лишайников, слое- вище которых далеко отстает от субстрата, серд- цевинный слой, помимо проводящей функции, выполняет еще одну — укрепляющую. Как от- мечалось, у большинства лишайников укреп- ление слоевища происходит с помощью корово- го слоя, который у высокоразвитых кустистых и листоватых форм постепенно становится все более толстым и плотным. Однако дальнейшее развитие в этом направлении могло бы привести к ухудшению нормальных ассимиляционных процессов: очень толстый и плотный коровой слой препятствовал бы проникновению в слое- вище воздуха и света. Поэтому в процессе раз- вития кустистых форм происходит постепенное снижение роли корового слоя в укреплении слоевища и образование механической ткани под зоной водорослей, т. е. в сердцевинном слое. В сердцевинном слое высокоразвитых кус- тистых лишайников возникают тяжи, которые расположены вдоль ветвей слоевища и состоят из толстостенных, плотно сросшихся друг с дру- гом гиф. У некоторых лишайников, напрпмер из рода летария (Letharia), эти тяжи имеют не- большую толщину; обычно их образуется сразу несколько в сердцевинном слое. Но постепенно в процессе развития кустистых форм происходит слияние этих тяжей в один толстый и плотный центральный тяж. Особенно хорошо развит такой осевой тяж у видов рода уснея, кустис- тых лишайников, свисающих длинными ред- кими «бородами» со стволов и ветвей деревьев. Округлые веточки их слоевищ очень тонкие и нежные, но при этом достигают значительной длины, до iO — 30 сл, а иногда даже до 8 л. К субстрату эти веточки прикрепляются у са- 405
ФФ Оа Рис. 302. Поперечный разрез через цифеллу (Sticta). мого основания; вся остальная часть остается свободной, повисающей. Если бы внутри таких веточек не было плотного осевого тяжа, ма- лейшее дуновение ветра легко могло бы их порвать. Коровой слой усней тонок и непрочен. Если попытаться осторожно растянуть веточку какой-либо уснеи, то можно увидеть, как при этом наружный коровой слой легко разорвется, а в образовавшейся поперечной трещинке об- нажится плотный белый тяж. Такой тяж, со- стоящий из толстостенных с длинными клетками гиф, плотно склеенных хрящевидным вещест- вом, очень прочен и эластичен. Например, тоненькая нежная веточка уснеи (табл. 49, 8) длиной 8 сл способна выдержать груз до 300 г. Как же воздух проникает в слоевище лишай- ника? У некоторых форм на поверхности слое- вища вовсе не образуется корового слоя, напри- мер у кладоний группы ягелей (рис. 295, 2). Поверхность их слоевищ образована рыхлыми сердцевинными гифами. В таком случае воздух беспрепятственно по промежуткам между рыхло- расположенными гифами проникает к водо- рослям, которые разбросаны отдельными груп- пами вблизи верхней поверхности подециев. У крупнолистоватых лишайников рода пель- тигера на нижней стороне слоевищ также не развивается коровой слой, ее образуют рыхло- расположенные сердцевинные гифы. И у этих лишайников воздух свободно проникает в серд- цевину и далее по промежуткам между гифами к клеткам водорослей. Но такие случаи явля- ются исключением. У большинства лишайников на поверхности слоевища образуется плотный коровой слой и доступ воздуха во внутренние слои слоевища сильно затруднен. Для осуществ- ления нормального газообмена на поверхности слоевища лишайников образуются особые от- верстия — специальные органы, которые слу- жат для аэрации внутренних частей растения. Например, у тундрового кустистого лишай- ника цетрарии исландской слоевище покрыто со всех сторон плотным блестящим коровым сло- ем. Он развивается в результате слияния меж- ду собой толстостенных гиф, образующих сплош- ную хрящевидную массу, в которой лишь кое- где видны просветы клеток неправильной фор- мы (рис. 298, а, д). Проникнуть сквозь такой плотный желатинооб разный слой воздух не имел бы никакой возможности, не будь в этом слое особых отверстий. Если внимательно при- смотреться к этому лишайнику, то на нижней, всегда более бледноокрашенной поверхности его лопастей даже невооруженным глазом мож- но заметить беленькие пятнышки неправильной формы (табл. 48, Б). Это разрывы коры, через которые воздух, как через маленькие форточки, проникает внутрь слоевища. Они являются органами газообмена лишайников и носят на- звание псевдоцифелл (или иногда у этого лишайника их называют еще м а к у- л а м и). На поперечном срезе этого лишайника, изображенном на рисунке 298, видно, что в местах образования псевдоцифеллы плотный коровой слой исчезает, а возникшее при этом отверстие заполняется рыхлорасположенными простыми или ветвистыми удлиненными гифа- ми сердцевины. Через промежутки между эти- ми гифами воздух проходит внутрь слоевища. У другого тундрового лишайника — цетрарии сглаженной (Cetraria laevigata), по внешнему облику очень похожей на цетрарию исланд- скую, псевдоцифеллы имеют иную форму. Верх- няя и нижняя поверхности лопастей цетрарии совершенно гладкие, блестящие, без белых пятнышек. Но внимательно присмотревшись, можно увидеть по самому краю трубчато свер- нутых лопастей узенькую белую кайму. Это также псевдоцифеллы, которые образуются в результате разрыва коры вдоль края лопастей. Помимо псевдоцифелл, у лишайников обра- зуются и другие органы газообмена. Так, на нижней поверхности листов атых лишайников рода стикта можно заметить круглые правиль- ной формы белые углубления (табл. 47, 4). Эти разрывы нижней коры лишайника, по фор- ме напоминающие маленькие чашечки, назы- вают ц и ф е л л а м и. Дно их выстлано рых- лорасположенными, шарообразными клетками сердцевины, через промежутки между которы- ми воздух легко проникает в сердцевину (рис. 302). У менегации продырявленной (Mene- gazzia pertusa) вся верхняя поверхность слое- вища покрыта округлыми или овальными от- верстиями, которые носят название п е р ф о-
р а ц и й (табл. 52, 2). Их назначение — тоже проводить воздух внутрь слосвища. Псрфора- ции возникают в результате отмирания малень- ких участков коров ого слоя лишайников. У представителей гипогимний перфорации раз- мещены на вершинах лопастей. У некоторых лишайников органы аэрации слоевища имеют вид маленьких точковидных пор, расположен- ных в нижней коре лишайников. Иногда такие поры возникают на вершине бородавочек, раз- бросанных на верхней поверхности лишайни- ков. В том случае, когда на поверхности слоевища лишайников не образуются специальные ор- ганы газообмена, на помощь приходят различ- ныс трещинки и разрывы в коровом слое. Эти трещинки могут возникнуть как в результате особенностей роста слоевища, так и в резуль- тате повреждения слоевища ветром, животны- ми, человеком. ОРГАНЫ РАЗМНОЖЕНИЯ ЛИШАЙНИКОВ Лишайники размножаются либо спорами, которые образует микобионт половым или бес- полым путем, либо вегетативно — фрагмента- ми слоевища, соредиями и изидиями. Половое размножение. При половом размно- жении на слоевищах лишайников в результате полового процесса формируются половые спо- роношсния в виде плодовых тсл. Среди плодо- вых тел у лишайников различают апотеции, перитеции и гастеротеции. Б ольшинство ли- шайников, свыше 250 родов, формируют откры- тые плодовые тела в виде а п о т е ц и е в- дисковидных образований (рис. 305, 1, 2; 306). Около 70 родов лишайников имеют плодовые тела в форме п с р и т с ц и я — закрытого пло- дового тела, имеющего вид маленького кувшина с отверстием наверху (рис. 307). Небольшое ко- личество лишайников, в основном представи- тели семейства графидовых, образуют узкие плодовые тела удлиненной формы, которые называют r а с т с р о т с ц и я м и (рис. 305,4). В апотециях, перитециях и гастеротециях споры развиваются внутри с у м о к — осо- бых мсшковидных образований (рис. 308). Ли- шайники, формирующие споры в сумках, объ- единяются в большую группу сумчатых лишай- ников (Ascolichenes). Они произошли от грибов класса аскомицетов и представляют основную эволюционную линию развития лишайников. У небольшой группы лишайников споры об- разуются не внутри сумок, а экзогенно, на вершине удлинснно-булавовидных гиф — б а- з и д и й (рис. 303,2). На верхушке базидий имс- ются четыре отростка — с т е р и r мM Iы~, на кон- цах которых развиваются четыре споры. Ли- шайники с таким образованием спор объединя- ются в группу базидиальных лишайников (Ba- sidiolichenes). Несмотря на то что базидиальные лишайники насчитывают всего 20 видов, это совершенно самостоятельная эволюционная линия разви- тия лишайников, берущая свое начало от грибов класса базидиомицетов. Базидиальные лишай- ники в основном тропические растения. По внешнему облику они напоминают тонкослой- ные трутовые грибы — разрушители древе- сины (рис. 303, 1). На нинснсй стороне их слое- вищ, имеющих вид тонких полукруглых пласти- нок, образуется открытый плодущий слой, ко- торый состоит из базидий и бесплодных удли- нснных гиф — п а р а ф и з (рис. 303, Зб). Сов- сем недавно базидиальные лишайники были об- наружсны и в Европе. Однако плодовые тела базидиальных лишайников, произрастающих в горах Средней Европы и в северных тундрах, иного строения. Они имеют вид пальцсвидных или коралловидных выростов либо напоминают по форме маленький шляпочный гриб (рис. 303, 8,4, Б). Половой процесс и развитие плодовых тел у лишайников, особенно базидиальных, изу- чены недостаточно. Эти процессы имеют много общих черт с аналогичными процессами у сво- бодноживущих грибов, хотя и отличаются це- лым рядом особенностей. Было замечено, что развитие плодового тела в различных семей- ствах лишайников протекает неодинаково и изменяется от семейства к семейству. На осно- ве онтогенеза плодовых тел сумчатые лишайники делят на две группы: асколокулярные и аско- гимениальные. Существуют и промежуточные формы (ссмсйство Arthoniaceae), которые объ- единяют черты этих двух основных типов раз- вития плодовых тел. Развитие плодового тела асколокулярных лишайников начинается с возникновения осо- бой ткани — с т р о м ы, состоящей из перепле- тсния грибных гиф, а ужс затем в стромс за- кладывается а р х и к а р п — женский поло- вой орган лишайника. Позднее в строме возни- кают особые камеры, называемые л о к у л а- м и, внутри которых формируются сумки со спорами. Грибная ткань между локулами час- тично отмирает, и от нее остаются лишь от- дельные вертикально расположенные гифы, срастающиеся своими вершинами, их называют п а р а ф и з о и д а м и. Этот тип онтогенеза плодовых тсл наиболее характерен для свободно живущих аскомицет- ных грибов и среди лишайников встречается довольно редко, лишь у представителей двух семейств: артопирснисвых (Arthopyreniaceae) и микопоровых (Mycoporaceae). Подавляющее большинство сумчатых ли- шайников принадлежит к группе аскогимени- 407
альных. Это филогенетически очень древняя группа, представители которой благодаря их симбиотическому образу жизни рано отдели- лись от грибов и развивались далее самостоя- тельно и независимо от них. У аскогимениаль- ных лишайников развитие плодовых тел начи- нается с закладки архикарпа непосредственно среди вегетативных гиф в верхней части серд- цевинного слоя, на границе с зоной водорослей или, реже, в самой зоне водорослей. В даль- нейшем во взрослом плодовом теле аскогиме- ниальных лишайников всегда формируется на- стоящий гимениальный слой, образованный вертикально стоящими сумками со спорами и развивающимися между ними парафизами- бесплодными вытянутыми гифами, свободными на верхнем конце (рис. 305, Б, 6). Женский половой орган лишайников — ар- хикарп — состоит из двух частей (рис. 304, 8, 4). Нижняя часть носит название а с к о г о н а и представляет собой спирально закрученную гифу, более толстую по сравнению с другими тифами и состоящую из 10 — 12 одно- или много- ядерных клеток. От аскогона вверх отходит т р и х о r и н а — тоненькая вытянутая гифа, которая проходит через зону водорослей и ко- Рис. 303: Базидиальные лишайники: 1 — тропический лишайник Cora pavonia; 2 — 4 — базидиальные лишайники с плодовыми телами в форме шляпочного гриба (2,8— Omphalina luteolilacina, внешний вид и поперечный разрез через плодовое тело; 4 — Omphalina ericetorum): а — базидии, б- парафизы, в — споры, 5 — роговидные плодовые тела базидиального лишайника Clavulinopsis septentrionalis.
ровой слой и выходит на поверхность слоевища, возвышаясь над ней своей липкой верхушкой. Как происходит оплодотворение архика па и вооб ообще имеет ли место этот процесс у лишай- р а ников, до сих пор не выяснено. Правда, неко- торым ученым удалось наблюдать, как к клей- кой верхушке трихогины прилипали малень- кие бесцветные тельца — п и к н о к о н и- д и и. Они быстро теряли свое цитоплазмати- ческое содержимое, и на вершине трихогины оставалась лишь их пустая оболочка. Можно было предположить, что содержимое этих телец переходит в трихогину. Это позволило считать пикноконидии мужскими половыми клетками, оплодотворяющими архикарп. Это бе то же представляют собой пикнокон ? идии. то есцветные, очень маленькие клеточки (длиной 2 — 8 лкл, шириной 0,5 — 1 лкл) раз- личной формы: палочковидные, овальные, игло- видные, прямые и изогнутые. Пикноконидии об- разуются в п и к н и д и я х — осооых замкнутых вместилищах шаровидной или овальной формы с маленьким отверстием наверху (рис. 304, 1, 2). Пикнидии нередко можно наблюдать на верхней поверхности слоевища лишайников или на кон- чиках и по краям лопастей, где они бывают за- Рис. 304. Пикнидии и архикарп у лишайников.' 1 — лопасть листоватого лишайника гипогимнии с — м На ывер~юл~ а виде ерных то ек 2— пик нидиями заметны ы) 8— .Ф а — коровои слой слоевища б — се — попереч- ° ыи сз че ло вище коллемы в цент мно н~нтр — o o o р~~~~р (~ — ~~ o~~~ o — рихо — аскогон и трихогина после копуляции ляции с пикноконидией. — ~х гина, высовы-
метны в виде черных, реже красных точек — это видны лишь выводные отверстия пикнидиев, в то время как сами они целиком погружены в слое- вище. Стенки пикнидия состоят из нескольких слоев клеток, и от них перпендикулярно внутрь отходят к о н и д и е н о с ц ы — тонкие гифы, поделенные на маленькие клетки. На кониди- еносцах образуются многочисленные пикноко- нидии. При увлажнении слоевища маленькие пикноконидии в огромной массе выплывают на поверхность слоевища лишайника и некоторые из них, по-видимому, прилипают к высунувшим- ся клейким верхушкам трихогин. Но действи- тельно ли пикноконидии являются мужскими половыми клетками, оплодотворяющими ар- хикарп, пока с уверенностью сказать нельзя. В настоящее время еще нет цитологических данных, показывающих проникновение содер- жимого пикноконидии в трихогину. Многие ученые решительно отрицают, что пикнокони- дии — это мужские половые клетки и, наобо- рот, считают их органами бесполого размноже- ния лишайников. В некоторых случаях уда- валось наблюдать, как пикноконидии в благо- приятных условиях сами прорастали в мицелий, который давал начало образованию слое- вища лишайников. Считается, что для лишайников характерна редукция, утрата процесса оплодотворения. Возможно, что во многих случаях процесса оплодотворения вовсе не происходит и плодо- вое тело формируется из неоплодотворенного аскогона. Причем лишь у слизистых лишайни- ков семейства коллемовых развитие плодового тела происходит из одного архикарпа, как и у свободноживущих грибов. У всех остальных лишайников плодовое тело обычно формиру- ется из многих архикарпов и окружающих их гиф. Из нижней части архикарпа — аскогона- развиваются аскогенные гифы, из которых в дальнейшем формируются сумки. Их разви- тие начинается с разрастания аскогона и попар- ной группировки ядер в его клетках. В резуль- тате этого процесса возникают клетки, каждая из которых содержит по два ядра. Эти двухь- ядерные клетки — материнские клетки сумок— постепенно начинают разрастаться, и из них формируются сумки. Далее, по мере их созре- вания, внутри сумок происходит слияние двух ядер в одно диплоидное, копуляционное ядро. Через некоторое время наблюдается последо- вательное тройное деление этого ядра, в резуль- тате чего в сумке образуется уже 8 ядер, каждое из которых является ядром будущей споры. Постепенно вокруг каждого ядра обособляется часть цитоплазмы сумки, а затем эта обособлен- ная часть цитоплазмы с заключенным внутри ее ядром покрывается оболочкой. Так в сумке образуется 8 спор. Развивающийся аскогон окружается, как футляром, сплетением мелкоклеточных гиф, из которых формируются отдельные части пло- дового тела. Эти ткани называют генеративны- ми. Гифы генеративных тканей отличаются от гиф остального слоевища тонкими клеточными стенками, формой клеток и их богатым содер- жимым, причем структура генеративных тка- ней у различных лишайников сильно варьирует, так же как и процесс дальнейшего формирова- ния плодовых тел. Развитие и созревание плодового тела у ли- шайников — очень медленный процесс, кото- рый длится 4 — 10 лет. Сформировавшееся пло- довое тело тоже является многолетним, способ- ным в течение ряда лет продуцировать споры. Большинство лишайников образуют плодо- вые тела открытого типа в форме апотециев. Эти округлые дисковидные образования, на- поминающие по виду маленькие блюдца, очень часто можно наблюдать на верхней поверхности слоевища накипных и листоватых лишайников или на концах лопастей кустистых слоевищ (табл. 43, 44, 53). Обычно их диаметр не пре- вышает 1 — 2 лл, но у некоторых листоватых и кустистых лишайников он может достигать 1 — 3 сл, а у накипных — всего 0,1 — 0,5 лл. По форме апотеции чаще всего округлые, реже овальные; когда же несколько апотециев раз- вивается рядом друг с другом, они нередко ста- новятся угловатыми. У семейства артониевых (Arthoniaceae) они имеют форму звезд или не- правильных пятен. Обычно апотеции образу- ются на верхней поверхности слоевища лишай- ников, плотно прирастая к ней своей нижней частью или возвышаясь над ней на тонкой ма- ленькой или довольно длинной ножке (рис. 305, 8). У некоторых лишайников они бывают це- ликом погружены в слоевище, например у со- лорины (табл. 42, 4). Только у двух родов ли- шайников — нефрома (Nephroma) и нефромо- псис (Nephromopsis) — апотеции образуются не на верхней, а на нижней стороне лопастей (табл. 49, 5). В апотециях можно различить центральную часть — плоский диск и периферическую— округлый выпуклый валик, окружающий диск. Плодущей частью апотеция является диск. На его поверхности развивается плодущий слой, который носит название гимениального. Он образован вертикально стоящими сумками со спорами и расположенными между ними бес- плодными гифами — парафизами (рис. 305,2,5,6). Парафизы — тонкие, нитевидные гифы (обыч- но толщиной 1,5 — 2 лкл), простые или раз- ветвленные, со свободными верхними концами. Они служат для защиты сумок со спорами. По высоте они обычно несколько превосходят сум- ки, а их свободный верхний конец бывает була-
Рис. 305. Лекано 1— ровые апотеци — внешний ви ии и г и астеротеции: Л апо циев ~~~~~~ЫЙ СЛОЙ б- ни и, — слоевищныЙ к краи, в — па а к р изы, г — с~'мк ми окрашенн и, к рами, д — эпите и и, КИ СО СПО еции, лс — гипоте и 3 4 гас — теротеции; 5 6— — пара-
..: ~-'-:= ':.'::::::::-::'': Ъ: >3> 3 ° >' .'.:,.::"„:фФ. '> .'::;Ч I 1 ° .';: ~~ I .мф> Ф." gl > ,Ъ >'> (' ° 1-. ' .,'."... ".".', ° '' ~> /g 3" I % t' ';;й' > > ,б /' > ..> '-:-М;.. l I > (('ó ( > ° ~ф / r с ° . '.,',1 .: З..":.,(.' ' 41:;: > > т Ъ ° > /t(r (Is ( ° ~ » \~" ,::, -.ц, .ф~-, .-' Щ,:ь .'~~ь Э~ '.>:..>" ':> Ф с ' '>:Ж > .. >) ~ > Рис. 306. Лецидеевые апотеции: 1 — внешний вид темноокрашенных апотециев на накипном слоевище (Lecidea); 2 — поперечный разрез через апотеций (а- дарафизы, б — сумки со спорами, в — зпитеций, г — собственный край, образованный зксципулом, д — слоевище). вовидно утолщен и окрашен в зеленоватый, синеватый, коричневатый, желтоватый и дру- гие цвета. Эти окрашенные утолщенные вер- шинки, плотно прилегая друг к другу, образу- ют над сумками в верхней части гимениального слоя защитный слой, который называют э п и- т е ц и е м. От окраски эпитеция зависит окрас- ка диска апотеция: он может быть коричневым, черным, желтым, оливковым, красным, оранже- вым, розовым и др. (табл. 42, 47,48, 49). Под ги- мениальным слоем расположен r и п о т е ц и й- слой, состоящий из плотно переплетенных гиф. Тонкое цитологическое исследование позволяет различить в нем основания парафиз и аскоген-
ный аппарат, из которого по мере созревания вырастают сумки. Край апотеция, имеющий вид округлого ва- лика, как защитный футляр, одевает плоду- щий диск с боков и снизу. В зависимости от его строения различают несколько типов апоте- циев: леканоровые, лецидеевые и биаторовые. Диск леканоровых апотециев окру- жен слоевищным краем, который всегда содержит внутри водоросли, а строение и окраска его очень похожи на строение и окраску слоевища. На анатомическом срезе в нем можно разли- чить коровой слой, слой водорослей и слой рых- лой сердцевины (рис. 305, 2). Леканоровые апо- теции легко распознаются и по внешнему об- лику: окраска их края всегда отличается от окраски диска. Если диск может быть корич- невьп|, черным, красным, то край чаще всего бывает более светлым, окрашенным в один цвет со слоевищем, — сероватым, беловатым, серо- вато-зеленоватым, светло-коричневым и т. д. (табл. 47). Второй тип апотециев — л е ц и д е е в ы е а п о т е ц и и — также легко можно распоз- нать по внешнему облику. Обычно они черные и очень твердые, их с трудом можно раздавить на предметном стекле. В отличие от леканоро- вых край этих апотециев имеет ту же окраску, что и диск: он тоже черный и носит название собственного края. Собственный край никогда не содержит водорослей и отличается по строе- нию от слоевища. У лецидеевых апотециев он состоит из темноокрашенных, плотно прилегаю- щих друг к другу гиф, которые образуют с трех сторон вокруг диска валик, называемый э к с ц и п у л о м (рис. 306). Б и а т о р о в ы е а п о т е ц и и по строе- нию похожи на лецидеевые: у них тоже имеется собственный край, образованный эксципулом. Но, в отличие от лецидеевых апотециев, гифы, образующие эксципул, и сами биаторовые апо- теции окрашены в светлые тона — они бывают восковидные, желтовато-коричневатые, красно- вато-коричневые, розовато- или беловато-желтые и никогда не бывают черные. Биаторовые апо- теции отличаются очень мягкой консистенци- ей, их легко можно раздавить на предметном стекле, как теплый, расплавленный воск. Гастеротеции, как и апотеции, явля- ются плодовыми телами открытого типа. В них также можно различить диск и край. Но они отличаются от апотециев сильно вытянутой, линейной формой. Обычно гастеротеции имеют вид штрихов, простых или разветвленных ли- ний. Диск у этих плодовых тел очень узкий, на- поминающий тоненькую полоску, в то время как край, который может быть и собственным и слоевищным, сильно выступает и возвышает- ся над глубоко погруженным диском (рис. 305, 4). Гастеротеции у лишайников встречаются довольно редко. Они известны лишь у пред- ставителей двух родов накипных лишайников- графис (Graphis) и опеграфа (Opegrapha). П е р и т е ц и и — плодовые тела закрытого типа, имеющие вид замкнутого кувшина округ- лой или овальной формы с маленьким отвер- стием в верхней части (рис. 307). Через это отверстие, которое носит название у с т ь и ц а, происходит выбрасывание зрелых спор. Обычно перитеции почти незаметны на поверхности ли- шайника: при внимательном рассмотрении мож- но обнаружить лишь черные точки — устьица перитециев, в то время как само плодовое тело целиком погружено в слоевище. Реже перите- ции выступают своими верхушками (рис. 307, 1) или целиком сидят на слоевище. Стенки этого кувшинообразного плодового тела состоят из нескольких слоев (рис. 307, 2). Внутреннюю по- лость перитеция, где образуются сумки со спо- рами, окружает э к с ц и п у л. Это темно- или светлоокрашенная оболочка, состоящая из не- скольких слоев гиф, поделенных на удли- ненные клетки. Нередко эксципул снаружи покрыт еще одной оболочкой — п о к р ы- в а л ь ц е м. Покрывальце, как правило, бы- вает темноокрашенным или черньп|, может охватывать эксципул со всех сторон или по- крывать его лишь до половины; иногда оно раз- вито лишь у устьица, а нередко и вовсе отсут- ствует. Внутри перитеция развиваются сумки со спорами, которые в нижней и боковой частях плодового тела образуют вместе с парафиэами гимениальный сло й (рис. 307, Зб). Гимениальный слой развивается на r и п о- т е ц и и — узком зернистом слое, прилегаю- щем к внутренней стенке перитеция. Харак- терно, что у некоторых лишайников в перите- циях защитные нити — парафиэы — вовсе не образуются или очень рано расплываются в слизь. Это связано с тем, что сумки со спора- ми надежно защищены самими стенками пери- теция и надобность в парафиэах исчезает. Осо- бые защитные нитевидные гифы в перитециях развиваются около устьица (рис. 307, Зг). Они называются п е р и ф и э а м и и служат для защиты всего внутреннего ядра перитеция от воздействий внешней среды. Сумки лишайников могут быть самой раз- личной формы: цилиндрической, булававид- ной, грушевидной, бутылковидной, бесформен- но-мешковидной и др. (рис. 308). Средние их размеры: в длину 40 — 75 мкм и в ширину 15— 25 мкм, но у некоторых родов длина сумок до- стигает 400 мкм, а ширина — 90 мкм. У каждого вида лишайника в сумке разви- вается постоянное количество спор. Чаще всего их бывает 8, но иногда 12 — 32. У некоторых ро- дов (например, у представителей рода акаро-
Ф ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° у ° ° ° Ф г Ф ° ° ° ° ° е ° ° ° е У ° Э ° ° ейск ° ° ° ° ° е ° ° . <."... '.', Зс~~~;~,- ~,л с, \ с. ° ° У ° ° ° ° ° е ° ° ° ° ° ° ° ° ° ° ° спора) в сумке образуется до 200 спор. Однако встречаются лишайники, в сумке которых всег- да развивается лишь одна очень крупная (дли- ной до 300 лкм) спора. Иногда таких спор бывает 2 или 4. Сумки большинства лишайников имеют одну оболочку и называются у н и т у н и к а т н ы- м и. На вершине образуется особый апикаль- ный (верхушечный) аппарат, с помощью кото- рого споры высвобождаются из сумки. Этот аппарат может иметь форму кольца, крышечки и др. У некоторых родов лишайников встреча- ются сумки с двумя оболочками — б и т у н и- к а т н ы е. У них при выбрасывании спор на Рис. 307. Перитеции: 3 — внешний вид перитециев; 2 — поперечный разрез через слоевище с погруженным в него перитецием (д — эксципул, б — гимениальный слой, образованный сумками со спорами и парафизами, в — гипотеций, г — перифизы). некотором расстоянии от вершины происходит поперечный разрыв внешней оболочки и споры выплывают, окруженные тонкой внутренней оболочкой, которая разрывается уже позднее. Битуникатные сумки особенно характерны для асколокулярных лишайников (рис. 309). Спора лишайников представляет собой окру- женный оболочкой небольшой комочек цито- плазмы с заключенным в ней ядром, ядрышком и иногда несколькими каплями жира. Оболоч- ка споры двухслойная. Внешний слой, толстый и нередко окрашенный, носит название э к з о- с п о р и я, внутренний, более тонкий, назы- вают э н д о с п о р и е м. Споры чаще всего Рис. 308. Форма сумок лишайников.' ви ная, Huellia); 2 — цилиндрическая 1 — удлиненно-булавовидная Аг1Ъоп1а); 4 — бутылко- (S haerophorus); 8 — булавовидная (Art, onia; (Th 1 on) 5 — мешковидная (Pertusaria). видная (The ocarpon;
бывают бесцветными или коричневыми. Обыч- но их размеры: в длину 1 — 50 мкм и в ширину 0,5 — 10 лкл. Самые крупные споры — у на- кипного лишайника бацидии (Bacidia margi- nalis), произрастающей на Филиппинских ост- ровах и острове Суматра на листьях вечно- зеленых растений, они достигают величины 525 мкл. По форме споры могут быть шарообразными, эллипсоидными, яйцевидными, веретеновид- ными, игловидными, нитевидными и прямыми, согнутыми, спирально закрученными и др. (рис. 310). Среди них встречаются как одно- клеточные, так и двух- и многоклеточные. По- мимо поперечных перегородок, в многоклеточ- ных спорах нередко встречаются еще и про- дольные перегородки. В таких случаях разме- щение клеток в спорах напоминает кладку кирпичей в стенах и их называют м у р а л ь- н ы м и (тигиз — стена). У лишайников встре- чается еще один интересный тип спор — б и- п о л я р н ы е споры, которые специфичны для этих растений и не встречаются у свободножи- вущих грибов. Биполярные споры двухклеточны и имеют в центральной части толстую пере- городку, внутри которой проходит тончайший Баналец, соединяющий протопласты обеих кле- ток (рис. 310, 4). Как уже отмечалось, плодовые тела у лишай- ников многолетние, образование сумок, их созревание, выход из них спор — сложный про- цесс, который продолжается не один год. После выхода спор стенки сумки распадаются и вместо пустой сумки снизу, из гипотеция, вырастает новая. Считают, что процесс выхода спор- с п о р у л я ц и я приурочена у лишайников к определенной поре года и в различных об- ластях земного шара происходит в разное вре- мя. Например, было замечено, что в Западной Европе у многих лишайников споруляция осу- ществляется особенно активно осенью, зимой и весной, у других происходит круглый год. Но, как показали исследования, у большинства лишайников в условиях эксперимента после увлажнения апотециев споры начинали быстро выбрасываться из сумок. Это привело к мысли, что, возможно, выход спор из сумок связан не столько со временем года, сколько с погод- ньп|и условиями, прежде всего с влажностью воздуха. Лишь у немногих лишайников (порядок Caliciales) происходит пассивное освобождение спор в результате постепенного разрушения тонкой и нежной оболочки сумки. Споры не- которое время остаются склеенными по 8 в ряд, а затем рассыпаются, оставаясь, однако, на плодовом теле и образуя пылистую массу, ко- торая позднее разносится ветром. У большин- ства лишайников споры с силой выбрасываются из сумки. В результате увеличения осмоти- ческого давления внутри сумки и с помощью верхушечного аппарата споры могут быть от- брошены в сторону на 1 — 5 сл и подброшены вверх на 3 сл. При этом их обычно подхватывают воздушные течения и относят порой на довольно Рис. 309. Сумки лишайников. 1 — унитуникатные сумки с апикальным аппаратом (а) на вер- шине, с помощью которого споры высвобождаются из сумок; 2,8 — зрелая и молодая битуникатные сумки; 4 — выбрасыва- ние спор из битуникатной сумки. Рис. 310. Споры лишайников.' 1 — шарообразные; 2 — эллиптические; 8 — удлиненные; 4— биполярные; 5 — двухклеточные; 6 — двухклеточные с сильно и неравномерно утолщенными оболочками; 7 — четырехклеточ- ные вытянутые; 8 — восьмиклеточные вытянутые; 9 — игло- видные многоклеточные; 10 — муральные. lQ
значительные расстояния от материнского пло- дового тела. Сколько же спор способны продуцировать плодовые тела лишайников? Подсчитано, на- пример, что у лишайника солорина (Solorina saccata) в апотеции диаметром 5 лл образуется 31 тыс. сумок, а в каждой сумке обычно раз- вивается по 4 споры. Следовательно, общее ко- личество спор, продуцируемое одним апоте- цием, равно 124 000. В течение одного дня из такого апотеция выбрасывается от 1200 до 1700 спор. Конечно, не все выброшенные из плодового тела споры прорастают. Многие из них, попав в неблагоприятные условия, погибают. Для прорастания споры необходимы прежде всего достаточная влажность и определенная темпе- ратура. Наблюдения в природе позволили вы- яснить, что в умеренном климате споры прораста- ют лучше всего от марта до июня, однако все ус- ловия, необходимые для прорастания спор ли- шайников, пока неизвестны. Например, ока- залось, что некоторым нужно, чтобы в непосред- ственной близости с ними находилась водоросль. При прорастании одноклеточной споры об- разуется одна, редко до пяти ростковых трубо- чек. У муральных спор каждая из их много- численных клеток может развить одну или не- сколько ростковых трубочек, но обычно про- растают лишь их конечные клетки. Ростковые трубочки растут, разветвляются и образуют первичный мицелий, который еще не содержит водорослей. Дальнейшее развитие этого мице- лия и образование слоевища лишайника проис- ходит лишь в том случае, если его гифы встре- чают водоросль, соответствующую данному виду лишайника. В поисках этих водорослей гифы ми- целия посылают во все стороны длинные тонень- кие ищущие гифы. Если на субстрате, где растет первичный мицелий, нужных водорослей нет, этот мицелий обычно скоро погибает. Встретив свойственную данному лишайнику водоросль, гифы первичного мицелия окружают ее со всех сторон, отделяют от субстрата и через некоторое время, воздействуя на нее, вызывают деление водоросли на дочерние клетки, которые тоже, в свою очередь, оплетаются гифами. Так на- чинается формирование слоевища лишайника. У некоторых лишайников в результате сим- биотической жизни лишайникового гриба с во- дорослью появилось своеобразное приспособ- ление: споры гриба выбрасываются из плодовых тел вместе с водорослями. Это возможно пото- му, что в плодовых телах таких лишайников развиваются мелкие гимениальные водоросли. Например, в перитециях чешуйчатого лишай- ника эндокарЫона (Endocarpon) среди сумок, содержащих крупные муральные споры, можно у видеть мелкие водоросли палочковидной, овальной и шарообразной формы. Когда мураль- ные споры этого лишайника выбрасываются из перитеция, вместе с ними бывают выброшены и прилипшие к ним гимениальные водоросли. В дальнейшем эти водоросли увеличиваются в размерах и приобретают нормальную форму, присущую данному лишайниковому фикобион- ту. В этом случае, следовательно, сразу начи- нается формирование слоевища лишайника, так как имеются налицо оба компонента сим- биоза. Однако в общей массе лишайников такие случаи единичны, гимениальные водоросли из- вестны лишь у трех родов лишайников: эндо- карпона (Endocarpon), стауротеле (Staurothele) и гоногимении (Gonohymenia). Бесполые спороношения. Помимо спор, об- разующихся половым путем, у лишайников известны также бесполые спороношения — к о- нидии, пикноконидии и стило- с п о р ы, возникающие экзогенно на поверх- ности к о н и д и е н о с ц е в (рис. ЗО4, Зг). При этом конидии образуются на конидиенос- цах, развивающихся непосредственно на по- верхности слоевища, а пикноконидии и стило- споры в особых вместилищах — п и к н и д и- я х, строение которых описано выше (рис. 304, 1,2) . Из бесполых спороношений лишайники чаще всего формируют пикнидии с пикноконидиями. Пикнидии нередко встречаются на слоевищах многих кустистых и листоватых лишайников, реже их можно наблюдать у накипных форм. У таких листоватых лишайников, как пармелия и гипогимния, пикнидии иногда обильно развиваются на верхней поверхности слоевища, у некоторых видов цетрарии (напри- мер, у цетрарии исландской) они образуются на верхушках ресничек, окаймляющих лопасти этого лишайника, а у кладоний группы ягелей пикнидии всегда находятся на кончиках их разветвленных подециев. В каждом из таких пикнидиев образуются в огромном количестве маленькие одноклеточ- ные споры — пикноконидии. Роль этих столь широко распространенных спороношений в жиз- ни лишайника до сих пор не выяснена. Одни ученые, называя эти споры спермациями, а пикнидии — спермагониями, считают их муж- скими половыми клетками, хотя до сих пор нет ни экспериментальных, ни цитологических дан- ных, доказывающих, что пикноконидии дейст- вительно участвуют в половом процессе лишай- ников. Сомнение, высказанное по этому поводу, подтверждается тем, что пикнидии встречаются одинаково часто как на слоевищах, формирую- щих плодовые тела, так и на слоевищах, кото- рые их никогда не образуют или образуют очень редко. Так, например, кладонии группы яге- лей почти никогда не развивают плодовых тел, 416
° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° e ° ° ° ° ° ° ° ° ° ° ° O ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° В ° В ° ° ° ° ° ° ° ° ° O Рис. 311. Вегетативное размножение лишайников с помощью соредий: 1 — соредии (а — клетки водорослей, б — гифы гриба); 2 — схема образования соредий в слоевище; 8 — 8 — различная форма соралей лишайников:,3 — пятновидная (Physcia orbicularis), 4 — головчатая (Hypogymnia bitteri), 5 — манжетовидная (Me- negazzia pertusa), 6 — губовидная (Physcia tenella), 7 — в виде каймы (Cetraria pinastri), 8 — щелевидная (Parmelia sulcata). ° ° ° В ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° В ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ВВВ ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° В ° ° ° ° ° ° ° ° В ° В ° ° ° ° ° ° ° ° ° ° ° ° ° ° В ° В ° е ° ° ° ° ° ° ° ° ° ° ° ° ° ° е ° ° Ф ° ° в ° ° ° ° а в Э ° ° ° ° ° ° ° ° ° Э ° ° В ° ° ° ° ° ° ° ° Ф ° s ° ° ° ° ° ° ° ° ° ° ° ° В ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° O ° ° ° ° ° В ° ° ° ° ° а' ° ° ° ° ° Э ° ° ° ° ° ° ° ° ° ° ° ° O ° ° ° ° e ° ° ° ° ° ° ° ° ° ° ° а ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° O ° ° ° ° ° ° ° ° ° ° ° а е ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° В ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° а ° ° ° ° ° ° ° ф~ 'Ъ~~, «~, В Я~,В В. V gp ёЗва,в, ' ='ффВВа-~ ° В В М фл ° ° В~
но всегда на кончиках их лопастей присутству- ют пикнидии. С другой стороны, нет доказа- тельств, что пикнидии являются спорами бес- полого размножения. Пока что удавалось на- блюдать прорастание пикноконидий лишь в ис- кусственных условиях, причем эти экспери- менты с проращиванием пикноконидий в куль- туре долго терпели неудачу. Существует пред- положение, что у одних лишайников пикноко- нидии выполняют роль половых элементов, в то время как у других превратились в споры бесполого размножения. Действительно, труд- но допустить, что эти споры, так часто встре- чающиеся у лишайников, не участвуют в их размножении. Иногда в пикнидиях, кроме обычных малень- ких одноклеточных пикноконидий, развиваются крупные (длиной до 50 — 100 мкм) двух- и много- клеточные споры, называемые с т и л о с п о- р а м и. Обычно (но не всегда) и сами пикнидии, образующие стилоспоры, отличаются более крупными размерами. Бывают случаи, когда в одной и той же пикнидии развиваются и пик- ноконидии и стилоспоры. Стилоспоры у ли- шайников образуются гораздо реже, чем пик- ноконидии, однако они известны у представи- телей родов пельтигера, алектория, артония, пармелия и др. Еще реже у лишайников образуются конидии. До сих пор они были обнаружены на слоевищах лишь двух видов лишайников — у слизистого лишайника лемфолемма (Lempholemma minu- tulum) и у накипного лишайника калоплака (Caloplaca decipiens). Чаще их образуют изо- лированные микобионты в условиях культуры. Так, они были обнаружены в чистых куль- турах микобионтов, выделенных из слоевища некоторых видов родов кладония, буеллия, лецидея и некоторых других. Конидии лишай- ников очень разнообразны по форме — палочко- видные, нитевидные, гантелевидные, грушевид- ные, эллипсоидные до почти шаровидных. Они могут быть одноклеточными и поперечно-много- клеточными. Как известно, конидии очень часто встречаются у свободноживущих грибов, они широко распространены у многих гифомицетов, а также обычны у аскомицетов. Редкое образо- вание к онидий у лишайников, по-видимому, связано с утратой ими своего биологического значения. Если у быстро развивающихся и недолговечных грибов образование конидий- это быстрая форма размножения в течение од- ного вегетационного периода, то у лишайни- ков с их многолетними слоевищами и плодо- выми телами, которые непрерывно, круглый год, в течение ряда лет образуют споры, этот сезонный способ размножения излишен. Как и в случае с пикноконидиями, до сих пор не ясно, являются ли стилоспоры и кони- дии спорами бесполого размножения лишайни- ков. Их прорастание также удавалось наблю- дать лишь в условиях культур. Вероятно, у гри- ба лишайников в связи с особенностями жизни в симбиозе с водорослями происходит утрата способа размножения с помощью спор, образо- ванных бесполым путем, столь характерного для свободноживущих грибов. И хотя мико- бионт образует эти бесполые спороношения, роль их в размножении лишайников, по-види- мому, невелика. Более того, у лишайников, этих симбиотиче- ских организмов, в процессе их эволюции на- блюдается также постепенная редукция и по- лового способа размножения, в котором участ- вует тоже лишь микобионт. У лишайников спора, как бесполого споро- ношения, так и отброшенная из плодового тела, должна пройти очень долгий путь, прежде чем из нее разовьется слоевище. Для развития лишайника необходим не только комплекс бла- гоприятных факторов, которые способствовали бы прорастанию споры: даже если спора про- растает и из нее образуется первичный мицелий, это еще вовсе не означает, что из него разовьется слоевище. Развитие лишайника из споры за- висит от многочисленных внешних факторов, главный среди которых — встреча с водорослью, свойственной данному лишайнику, по современ- ным представлениям, носит чисто случайный характер. Размножение с помощью спор проис- ходит чрезвычайно медленно, ведь только на формирование плодовых тел и их созревание требуется не менее 4 — 10 лет. Поэтому в эво- люционном развитии лишайников прослежива- ется тенденция к увеличению в жизни лишай- ников роли вегетативного размножения, в ко- тором принимают участие сразу и грибной и водорослевый компоненты. Вегетативное размножение. Если накипные лишайники, как правило, образуют плодовые тела, то среди более высокоорганизованных листоватых и кустистых лишайников имеется немало представителей, которые размножаются исключительно вегетативным путем. Во многих случаях вегетативное размножение этих лишай- ников осуществляется просто кусочками слое- вища, которые отрываются ветром или отламы- ваются от чрезвычайно хрупких в сухую погоду слоевищ в результате воздействия животных, человека и других факторов внешней среды. Эти небольшие кусочки слоевища переносятся ветром на новые местообитания и при благоприятных услов-иях разрастаются в новые слоевища ли- шайников. Таким образом, например, размно- жаются очень многие тундровые напочвенные лишайники, представители родов цетрария и кладония, мнотие из которых почти никогда не образуют плодовых тел.
Вегетативное размножение лишайников осу- ществляется также соредиями и изидиями- особыми вегетативными образованиями, воз- никшими в процессе эволюции главным обра- зом у высокоорганизованных форм листоватых и кустистых лишайников. Преимущество раз- множения лишайников соредиями и изидиями перед половым способом состоит в том, что в нем принимают участие оба компонента лишайника— и гриб и водоросль. С о р е д и и имеют вид пылинок — микроско- пических клубочков, состоящих из одной или нескольких клеточек водоросли, окруженных гифами гриба. Миллионы таких клубочков со- бираются в мелкозернистую или порошкообраз- ную беловатую, желтоватую или беловато- зеленоватую массу, которая местами покрывает верхнюю поверхность слоевища лишайника или развивается в виде каймы по краям его лопастей (табл. 46, вверху слева). Соредии образуются в слоевище под верхней корой, в зоне водорослей. У разных лишайников процесс образования со- редий происходит неодинаково, однако в общих чертах он всегда сводится к следующему. На ме- Рис. 312. Вегетативное размножение лишайников с помощью изидий: g — слоевище листоватого лишайника P armelia exasperatula с изидиями (a); 2 — различная форма изидий (б — шаровидная, в — булавовидная, г — цилиндрическая, д — коралловидная); 8 — поперечный разрез через слоевище с изидиями (е — коро- вой слой, ж — гифы гриба, з — водоросли).
ФИЗИОЛОГИЧЕСКИЕ И ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ лишлйников 420 сте образования соредий каждая клетка водо- рослей делится на несколько дочерних клеток. В то же время гифы гриба начинают сильно вет- виться, делятся на множество клеток и окружают каждую из дочерних клеток водоросли. В резуль- тате этого процесса под коровым слоем слое- вища образуется масса водорослей, оплетен- ная коротковетвящимися многоклеточными ги- фами. Спустя некоторое время она распадается на отдельные клубочки, каждый из которых представляет собой соредий. Когда клубочков образуется очень много, их масса начинает давить на коровой слой лишайника, тот не выдерживает, разрывается и соредии выступают наружу, на поверхность слоевища (рис. 311, 2). Здесь они образуют порошистую массу, кото- рая легко сдувается и разносится ветром. По- павший в благоприятные условия соредий на- чинает разрастаться и сразу развивается в новое слоевище лишайника. Обычно на поверхности слоевища соредии ооразуют скопления определенной формы, на- зываемые с о р а л я м и. У каждого вида ли- шайника, образующего сорали, их форма и расположение на слоевище постоянны. Напри- мер, у цетрарии морщинистой (Cetraria pinastri) сорали всегда имеют вид ярко-желтой каймы по краям лопастей (рис. 311,7), у эвернии прунастри (Evernia prunastr' ) и лобарии легочной (Loba- ria pulmonaria) они представляют собой белые округлые пятна (табл. 47, 1). У гипогимнии вздутой (Hypogymnia physodes) сорали обра- зуются на вершине лопастей и имеют губовид- ную форму, а у менегации продырявленной они бывают манжетовидной формы и т. д. (табл. 52, 2; рис. 311, 5). Соредии встречаются приблизительно у 30оо видов лишайников, главным образом у высоко- организованных форм. Как правило, виды, об- разующие соредии, очень редко формируют СПОСОБЫ ПИТАНИЯ ЛИШАЙНИКОВ Лишайники представляют для физиологи- ческих исследований сложный объект, так как состоят из двух физиологически противополож- ных компонентов — гетеротрофного гриба и автотрофной водоросли. Поэтому приходится сначала отдельно изучать жизнедеятельность мико- и фикобионта, что делается с помощью культур, а затем жизнь лишайника как це- лостного организма. Понятно, что такая «трой- ная физиология» — трудный путь исследован- ияя, и не удивительно, что в жизнедеятельности плодовые тела, и, наоборот, саредии почти не встречаются у лишайников, развивающих апо- теции или перитеции. Так, их очень редко мож- но наблюдать на слоевищах накипных и сли- зистых лишайников. Соредиеобразование воз- никло, очевидно, уже в более поздний период существования лишайников. И з и д и и — маленькие разнообразной фор- мы выросты верхней поверхности слоевища, одного цвета с ней, а иногда более интенсивной окраски. Они имеют вид бородавочек, зернышек, цилиндрических, булавовидных или коралло- видных выростов, иногда маленьких листочков (рис. 312). Если сделать анатомический разрез через изидии, то можно видеть, что внутри их находятся клетки фиьобионта лишайника, окруженные грибными гифами (рис. 312, 8). Характерным признаком изидий является то, что они, в отличие от соредий, всегда снаружи покрыты коровым слоем. Если соредии обра- зуются в результате разрыва корового слоя, то изидии — в результате его выпячивания. Форма изидий для каждого вида совершенно определенна и постоянна. Оторванные от слоевища изидии в благопри- ятных условиях разрастаются в новые слое- вища. Однако значение изидий в размножении лишайников не так велико по сравнению с со- редиями. Очевидно, это связано с тем, что обыч- но изидйи не отрываются от слоевища так легко, как соредии, а обламываются лишь животными или человеком. Изидии не только служат для размножения, но и выполняют определенную роль в увеличении асдимиляционной поверх- ности слоевищь лишайника. Изидии встречаются реже, чем соредии. Они известны приблизительно у 15 о/о видов лишайни- ков. Как и соредии, они характерны главным образом для высокоразвитых форм — кустис- тых и листоватых лишайников. лишайников кроется еще много загадочного. Однако общие закономерности их обмена ве- ществ все же выяснены. Довольно много исследований посвящено процессу фотосинтеза у лишайников. Так как лишь небольшая часть их слоевища (5 — 10 о~~ объема) образована водорослью, которая тем не менее является единственным источником снабжения органическими веществами, вста- ет существенный вопрос об интенсивности фо- тосинтеза в лишайниках. Как показали измерения, интенсивность фо- тосинтеза у лишайников намного ниже, чем у
высших автотрофных растений. Так, например, отношение продуктивности ассимиляции у ли- стоватых лишайников к продуктивности асси- миляции у картофеля составляет в среднем 1: 16. Но такая интенсивность все же обеспе- чивает нормальную жизнедеятельность ли- шайников, что легко объясняется, если учесть наличие частых периодов значительной эколо- гической депрессии (высыхание) и большую пластичность всего аппарата метаболизма ли- шайников, позволяющую им переносить эти периоды и быстро возвращаться к жизнедея- тельности даже в условиях низкой температу- ры, незначительного содержания углекислоты и пр., в которых другие растения погибают или прекращают жизнедеятельность. Этим же, ко- нечно, следует объяснить и медленный рост ли- шайников. Процесс фотосинтеза у лишайников зави- сит от многих экологических факторов (осве- щенности, температуры, влажности и пр.). Хлоропласты клеток водоросли в слоевище под коровым слоем из гиф получают несколько меньше света, чем хлоропласты под эпидер- мисом в листьях высших растений. Но эта разница небольшая, зато коровой слой, покры- вающий слоевище, выполняет функцию за- щиты от чрезмерно интенсивной радиации в от- крытых местах. Максимальная интенсивность фотосинтеза наблюдается у лишайников при освещенности в пределах 4000 — 23 000 лк- такие показатели освещенности характерны для большинства их местообитаний в тундрах, лесо- тундрах, светлых хвойных лесах. А там, где освещенность выше, слоевище защищено тем- ными пигментами, содержащимися в коровом слое (например, париетином), и лишайниковыми веществами (например, атранорином). Температурный оптимум фотосинтеза для боль- шинства лишайников находится в пределах от +10 до +25 'С, но они поглощают углекислоту и при более высокой (до +35 'С) и более низ- кой температуре (даже до — 25 'С). Особенно примечательна способность лишайников асси- милировать СО при низкой температуре. Мно- гими опытами подтверждено интенсивное по- глощение углекислоты лишайниками при — 5, — 10 'С и даже при более низкой температуре. В таких условиях у большинства высших расте- ний ассимиляции не происходит, в межклетни- ках накапливается лед, происходит обезвожи- вание и повреждение клеток. По-видимому, в слоевищах лишайников совершенно другой тип водного режима, и вода, находящаяся главным образом между гифами, превращаясь в лед, не служит препятствием для их жизнедеятельно- сти и поглощения углекислоты. В то же время высокая температура (выше +35 'С) останав- ливает у них процесс фотосинтеза, и в этом от- ношении лишайники сильно отличаются от высших растений, у которых фотосинтез про- должается даже при температуре от +30 до +50 'С. Для нормальной фотосинтетическ ой актив- ности слоевище должно содержать определен- ное количество воды, зависящее от анатомо- морфологического типа лишайника. В общем в толстых слоевищах оптимальное содержание воды для активного фотосинтеза ниже, чем в тонких и рыхлых слоевищах. При этом весьма существенно то обстоятельство, что многие виды лишайников, особенно в сухих местооби- таниях, вообще редко или по крайней мере очень нерегулярно снабжаются оптимальным количеством внутрислоевищной воды. Ведь регуляция водного режима у лишайников про- исходит совсем по-иному, чем у высших расте- ний, имеющих специальный аппарат, способ- ный контролировать получение и расходова- ние воды. Лишайники усваивают воду (в виде дождя, снега, тумана, росы и пр.) очень быстро, но пассивно всей поверхностью своего тела и отчасти ризоидами нижней стороны. Такое по- глощение слоевищем воды представляет собой простой физический процесс, как, например, впитывание воды фильтровальной бумагой. Ли- шайники способны впитывать воду в очень больших количествах, обычно до 100 — 300% от сухой массы слоевища, а некоторые слизи- стые лишайники (коллемы, лептогиумы и др.) даже до 800 — 3900%. Минимальное содержание воды в лишайниках в природных условиях составляет примерно 2 — 15% от сухой массы слоевища. Отдача воды слоевищем также происходит до- вольно быстро. Насыщенные водой лишайники на солнце через 30 — 60 льин теряют всю свою воду и делаются хрупкими, т. е. содержание воды в слоевище становится ниже минимально необходимого для активного фотосинтеза. Из этого вытекает своеобразная «аритмичность» фотосинтеза лишайников — его продуктив- ность меняется в течение дня, времени года, ряда годов в зависимости от общих экологи- ческих условий, особенно гидрологических и температурных. Имеются наблюдения, что многие лишайники более активно фотосинтезируют в утренние и вечерние часы и что фотосинтез продолжается у них и зимой, а у напочвенных форм даже под нетолстым снеговым покровом. С фотосинтетическ ой деятельностью непос- редственно связано дыхание лишайников. Пока- зано, что оно в общем ниже, чем у высших рас- тений, и составляет в среднем 0,2 — 2,0 мг вы- деляемой СО, на 1 г сухой массы за 1 ч. Так как у большинства лишайников в слоевище по объему и массе преобладает грибной компонент,
то полагают, что интенсивность дыхания в основном обусловливается жизнедеятельно- стью микобионта. Дыхание, как и фотосинтез, зависит от содержания воды в слоевище и от температуры. В общем увеличение содержания воды до максимального насыщения сопровож- дается последовательным повышением интен- сивности дыхания, и, наоборот, при умень- шении содержания воды дыхание ослабляет- ся, все же сохраняясь при очень незначитель- ных количествах воды в слоевище, даже при воздушно-сухом его состоянии. Амплитуда температуры, при которой возможно дыха- ние лишайников, широка: от — 15 до +30, +50 'С, при этом оптимум дыхания обыкно- венно наблюдается в пределах от +15 до +20 'С. При повышении температуры от 0 до +35 'С интенсивность дыхания возрастает, а при +-35 'С поглощение и освобождение СО, уравновешиваются. В общем процессы дыхания у лишайников подчиняются тем же закономерностям, что и у других автотрофных растений, но имеются и некоторые особенности. Основная из них- низкая интенсивность дыхания. С другой сто- роны, для лишайников характерна высокая устойчивость дыхания к высушиванию и низ- кой температуре. Это можно считать приспо- соблением к жизни в неблагоприятных место- обитаниях — полярно-арктических пустынях, с одной стороны, и настоящих аридных пусты- нях, с другой стороны. Общепризнано, что органические вещества, синтезируемые в лишайниковом слоевище фико- бионтом, используются грибным компонентом лишайника. Но как протекает передача асси- милятов от фикобионта к микобионту и в виде каких соединений, не было известно до послед- него времени. Лишь применение радиоактив- ного углерода С'4 внесло в этот сложный во- прос некоторую ясность. Теперь установлено, что в клетках фикобионта из сине-зеленых водорослей (в основном у ностока) образуется при фотосинтезе глюкоза (по некоторым новей- шим данным глюкозан, который под воздей- ствием какого-то грибного энзима превраща- ется в глюкозу), которая поглощается грибом, превращаясь в маннит. У лишайников с фико- бионтом из зеленых и желто-зеленых водо- рослей подвижными углеводами являются мно- гоатомные спирты: у требуксии (Trebouxia) и мирмеции (Myrmecia) — рибит, у тренте- полии (Trentepohlia) и фикопелтиса (Phyco- peltis) — эритрит, у гетерококкуса (Hetero- coccus), хиалококкуса (Hyalococcus) и трохис- ции (Trochiscia) — сорбит. Интересно, что мно- гоатомные спирты выделяют только лишайни- ковые фикобионты, у свободноживущих водо- рослей они не обнаружены. Это указывает на то, что симбиоз изменяет метаболизм водоросли. Далее очевидно, что микобионт активно влия- ет на водорослевые клетки, стимулируя выде- ление необходимых для своего питания асси- милятов. Но как? Определенного ответа на этот вопрос пока еще нет. Предполагают, что гриб выделяет органические кислоты, кото- рые снижают рН, что, в свою очередь, вызы- вает увеличение проницаемости водоросле- вых клеток. Но это может быть и какой-то эн- зим, гидролизирующий продукты ассимиляции, которые в других условиях пошли бы на по- строение клеточных стенок водорослей. Име- ются наблюдения, что лишайниковые вещест- ва (например, усниновая кислота — подроб- нее см. в разделе «Химический состав лишай- ников») также увеличивают проницаемость во- дорослевых клеток. Наконец, некоторые ученые предполагают, что стимуляция фикобионта со стороны гриб- ных гиф в слоевище происходит просто путем физического контакта. Важным компонентом в питании лишайни- ков является азот. Те лишайники, которые име- ют в качестве фикобионта зеленые водоросли (а их большинство), воспринимают соединения азота из водных растворов, когда их слоевища пропитываются водой. Возможно, что часть азотистых соединений лишайники берут и пря- мо из субстрата — почвы, коры деревьев и т. д. Экологически интересную группу состав- ляют так называемые нитрофильные лишай- ники, растущие в местообитаниях, богатых азотистыми соединениями, — на «птичьих камнях», где много экскрементов птиц, на ство- лах придорожных деревьев и т. д. (виды ксан- тории, фисции, калоплаки и др. ). Лишайники, имеющие в качестве фикобионта сине-зеленые водоросли (особенно ностоки), способны фик- сировать атмосферный азот, так как этой спо- собностью обладают содержащиеся в них водо- росли. В опытах с такими видами (из родов коллема, лептогиум, пельтигера, лобария, стикта и др.) было установлено, что их слоеви- ща быстро и активно поглощают атмосфер- ный азот. Эти лишайники часто селятся на суб- стратах, весьма бедных азотистыми соединения- ми. Большая часть азота, фиксированного во- дорослью, направляется микобионту и лишь незначительная часть используется самим фи- кобионтом. Имеются данные, что микобионт в слоевище лишайника ведет активный контроль над освоением и распределением азотистых сое- динений, фиксированных из атмосферы фико- бионтом. Что касается других компонентов питания, участвующих в обмене веществ лишайников, особенно минеральных элементов, то броса- ется в глаза способность лишайников накап- 422
ливать в своем слоевище те вещества, которые в таких количествах для его нормальной жиз- недеятельности, казалось бы, и не нужны. Так, например, определения показали, что обычный напочвенный лишайник диплошистес (Diploschi- stes scruposus) может содержать в своем слое- вище в 10 раз больше цинка (9,34% сухого ве- щества), чем его имеется в том же объеме поч- вы с данного местообитания. Биологический смысл такого избирательного накапливания от- дельных веществ не установлен. ХИМИЧЕСКИЙ СОСТАВ ЛИШАЙНИКОВ В состав лишайников входят многие элементы и вещества. Все их можно разделить на две большие группы — первичные и вторичные. К первичным относятся те вещества, которые не- посредственно принимают участие в клеточном обмене веществ; из них построено тело лишай- ников. К вторичным относятся конечные про- дукты обмена веществ, располагающиеся обыч- но на стенках гиф. Многие из этих вторичных лишайниковых веществ (в более старой лите- ратуре их называли лишайниковыми кислота- ми) специфичны для лишайников и не встре- чаются в организмах из других систематиче- ских групп. Первичные вещества в лишайниках в общем те;ке, что и в других растениях. Оболочки гиф в лишайниковом слоевище составлены в ос- новном углеводами, Часто обнаруживается в гифах хитин (C» Н, М 0„). Характерной со- ставной частью гиф является полисахарид лихенин (C,H»O,)„, называемый лишайнико- вым крахмалом. Реже встречающийся изомер лихенина — изолихенин — найден, кроме обо- лочек гиф, в протопласте. Из высокомолеку- лярных полисахаридов в лишайниках, в част- ности в оболочках гиф, встречаются гемицел- люлозы, являющиеся, очевидно, резервными углеводами. В довольно большом количестве (3 — 5% от воздушно-сухой массы) встречают- ся низкомолекулярные углеводы — дисаха- риды (сахароза, а-трегалоза, умбилицин), поли- спирты (эритрит, D-маннит, волемит, сифулит). В межклеточных пространствах у некоторых лишайников обнаружены пектиновые веще- ства, которые, впитывая в большом количест- ве воду, набухают и ослизняют слоевище. В ли- шайниках встречаются таьже многие фер- менты — инвертаза, амилаза, каталаза, уре- аза, зимаза, лихеназа, в том числе и внеклеточ- ные. Из азотсодержащих веществ в гифах ли- шайников обнаружены многие аминокислоты- аланин, аспарагиновая кислота, глютаминовая кислота, лизин, валин, тирозин, триптофан и др. Фикобионт продуцирует в лишайниках вита- мины, но почти всегда в малых количествах. Обнаружены аскорбиновая кислота (витамин C), биотин (Н), кобаламин (В„), никотиновая ки- слота (В, или PP) и некоторые другие витамины. Лишайники обладают удивительной способ- ностью извлекать из окружающей среды и на- капливать в своем слоевище различнъ~е эле- менты, в том числе и радиоактивные. В США после испытаний атомного оружия в умбили- карии (Umbilicaria mammulata) был обнару- жен радиоактивный цезий в количестве, губи- тельном для высших растений. В лишайниках накапливается намного больше цинка, кадмия, олова и свинца, чем в мхах и цветковых рас- тениях. Иногда наблюдается явный параллелизм ме- жду минеральным составом лишайников и со- держанием веществ в субстрате. В Югославии были проведены химические анализы коры ду- ба и лишайников (эвернии), населявших стволы дубов, которые показали это с полной очевид- ностью (табл. 111). Таблица Ш Минеральный состав коры дуба и лишайников (по Миновичу и Стефановичу) Содержайие веществ (в %) Вещества в лишайниках в коре дуба 9,15 3,95 4,03 1,12 3,20 66,58 1,10 0,66 3,79 0,39 7,60 1,11 10,35 2,54 28,42 6,37 5,53 25,45 5,04 3,16 9,04 0,36 8,96 1,50 Вода Зола ~1О, Р205 804 СаО MgO Ре,Оз А1,Оз МПЗО4 ~,О Ха,О У некоторых лишайников, особенно расту- щих на камнях и скалах, накапливаются соли железа, придающие слоевищу ржаво-красный цвет (например, у ризокарпона — Rhizocar- pon oederi). У многих видов в слоевище нака- пливается щавелевокислый кальций (CaC,О,), иногда в очень большом количестве. Например, у аспицилии съедобной (Aspicilia esculenta) он составляет до 66% от сухой массы. Биоло- гическое значение этого вещества в слоевище лишайников неизвестно. Фотосинтезирующие пигменты хлорофилл а и хлорофилл Ь встречаются в фикобионте ли- шайника в меньшем количестве, чем у высших растений. В последнее время в некоторых видах обнаружены каротины (Ii-каротин, у-каротин) и ксантофиллы.
Рис. 313. Соотношения оливеторовой (белая часть круга) и физодовой кислот (черная часть круга) в слое- вище Pseudevernia furfuracea в Европе и Северной Африке. Вторичные лишайниковые вещества пред- ставляют большую группу органических сое- динений, относящихся к разным биосинтети- ческим группам. Сейчас их известно уже около 250, и каждый год химики обнаруживают еще 3 — 4 новых. Из этого числа примерно 75 являются спе- цифическими лишайниковыми веществами, т. е. встречаются только в лишайниках, ос- тальные содержатся и в других организмах„ особенно в грибах. Количество вторичных лишайниковых ве- ществ в слоевищах лишайников колеблется в довольно широких пределах, обычно их бывает 0,1 — 2% от воздушно-сухой массы, реже до 2 — 5%. Так, например, атранорин содержится в пределах 1,2 — 3%, фумарпро- тоцетрариевая кислота — 0,5 — 1,5%, гирофо- ровая кислота — 1 — 4%, салациновая кислота 4 — 6%, усниновая кислота — 0,2 — 4,0% и т. д. В некоторых исключительных случаях концентрация лишайниковых веществ в слое- вище может быть очень высокой. Так, напри- мер, в пармелии окрашенной (Parmelia tincto- rum) леканоровой кислоты содер;кится 360~î от сухой массы. Что касается генезиса лишайниковых ве- ществ, то об этом известно еще очень мало. Ясно одно — эти вещества образуются микобион- том лишайника в симбиозе с фикобионтом, т. е. углеводы, синтезированные фикобионтом, превращаются в лишайниковые вещества ми- кобионтом. Сам по себе гриб, выделенный из слоевища лишайника, почти никогда специфи- ческого лишайникового вещества не образует. Несмотря на тщательное, в течение почти ста лет, изучение лишайниковых веществ, даже в настоящее время мы еще очень мало знаем об их биологическом значении. На этот счет име- ется несколько объяснений, но большинство из них носит гипотетический характер. Существует мнение, что вторичные вещества защищают лишайники от поедания животными. В то же время известно, что многие кустистые лишайники, содержащие горькие вещества, хорошо поедают животные — северные олени, карибу, улитки, пауки и др. Но лишайники весьма резистентны в отно- шении бактерий. Против них лишайниковые вещества выполняют, несомненно, защитную функцию. Многие исследователи считают лишайнико- вые вещества резервом дополнительного пита- ния, но экспериментальные данные этого не подтверждают — при ухудшении условий пи- тания количество лишайников ых веществ в слоевище не уменьшается. Иногда лишайниковые вещества рассматри- ваются как отходы обмена веществ, что также является сомнительным. Для этого они слиш- ком многообразны в химическом отношении и появляются в одинаковой химической форме у систематически и экологически очень дале- ких видов. Большинство лишайников очень медленно растет, что резко снижает их способность конкурировать с другими организмами за пространство и другие условия существования. По-видимому, лишайников ые вещества явля- ются одним из видов «оружия» в суровой борь- бе за существование. Установлено, что лишай- никовые вещества подавляют рост грибов и мхов (последние чаще всего являются конку- рентами лишайников) и всхожесть семян цвет- ковых растений. Некоторые авторы считают, что биологиче- ское значение лишайниковых веществ может заключаться в защите гиф от чрезмерного смачивания водой и обеспечении внутренней атмосферы для фотосинтеза. Но и это предполо- жение не нашло фактического подтверждения. Более вероятно, что окрашенные лишайнико- вые вещества (пигменты) действуют как свето- фильтры, защищающие фикобионт от чрезмер- ной радиации. Установлено, что некоторые лишайники в более освещенных местообита- ниях всегда содержат больше пигментов (на- пример, париетинар. Интересные данные получены в последнее время в отношении функционального значе-
ния атранорина — вещества, содержащегося во многих лишайниках. Установлено, что оно чаще и в большем количестве встречается у те- нелюбивых и теневыносливых видов. Атра- норин, генерируя коротковолновой свет, по- вышает эффективность фотосинтеза в фико- бионте. Некоторые экспериментальные данные по- казывают, что лишайниковые вещества спо- собствуют передвижению углеводов, синтези- руемых фикобионтом, в гифы микобионта. В этом может заключаться одна из важных функций лишайниковых веществ. Лишайники часто бывают пионерами за- селения свободных субстратов (поверхность скал и камней, древесины и пр.). Установлено, что лишайниковые вещества играют определен- ную роль и в этом процессе. Они разрушающе действуют на твердые минеральные субстраты и являются тем самым зачинателями почвооб- разовательного процесса. Таким образом, очевидно, что лишайниковые вещества могут выполнять различные функции, но необходимо приложить еще много уси- лий, чтобы окончательно раскрыть их биоло- гическую роль. Лишайниковые вещества имеют большое значение в систематике лишайников: опреде- ленные виды, группы видов или роды содержат определенные вещества. При распознавании видов многих родов, особенно морфологически близких, необходимо аналитически опреде- лять лишайниковые вещества. Для этого раз- работана методика микрокристаллизации ве- ществ: под влиянием определенных реактивов образуются специфические для каждого ве- щества кристаллы, которые затем изучают под микроскопом и определяют по эталонам или фотоснимкам. Широко используются и другие, более сложные методы. В простейшем случае применяют реактивы — парафенилендиамин, КОН, Са(С10)„КОН + Ca(C10), и др., кото- рые, реагируя с различными веществами, дают разную окраску коры или сердцевины слоеви- ща лишайников. Цветные реакции широко используются при определении лишайни- ков. Среди лишайников имеется группа видов, у которых в разных частях ареала разный со- став лишайниковых веществ (рис. 313), поэто- му в последнее время в таксономии лишайни- ков возникло особое направление — хемотак- сономия (химическая таксономия). Некоторые лихенологи считают химические вариации морфологически единого вида само- стоятельными видами, другие же называют их просто химическими расами одного и того же вида. РОСТ ЛИШАЙНИКОВ Лишайники считаются медленно растущими организмами, и это действительно так. Однако у различных видов средний годовой прирост неодинаков, и один и тот же вид в различных географических районах и экологических усло- виях может расти с существенно различной. скоростью. Самые медленно растущие лишайники- накипные, особенно литофильные. Так, у ри- зокарпона географического (Rhizocarpon geo- graphicum) радиальный прирост всегда меньше 1 мм в год, а в высокоарктических условиях он составляет лишь 0,25 — 0,5 мм в год. На- много быстрее растут листоватые и кустистые формы. «Рекорд» принадлежит кустистой рама- лине сетчатой (Ramalina reticulata). У этого. вида в Северной Америке был установлен при- рост 90 мм за 7 наиболее влажных и прохлад- ных месяцев или 36,5 мм за средний по влаж- ности год. Весьма быстро растут пельтигеры- обычные крупнопластинчатые лишайники, по- селяющиеся на почве и мхах. Их прирост не- редко равняется 1 — 3 см в год. Из кустистых лишайников сравнительно быстро растут также кладонии, например виды оленьего ягеля (Cladonia rangiferina, С. sylvatica, С. alpestris и др.), которые во всех зонах — в тундрах, лесах, высокогорьях — дают прирост в длину 2 — 7 мм в год. Но и среди листоватых и кусти- стых лишайников встречаются весьма медлен- норастущие. Так, умбиликария цилиндри- ческая (Umbilicaria cylrndrica), обильно по- крывающая скалы и камни в тундрах и высо- когорьях, в Швейцарских Альпах вырастала не более чем на 0,004 мм в год. Скорость роста слоевища лишайников су- щественно зависит от общих экологических условий — температуры, осадков, влажности воздуха. На крупных территориях со сравни- тельно одинаковым климатом она может оста- ваться более или менее одинаковой. Так, виды диплошистеса (Diploschistes) в континенталь- ных районах США вырастали на 0,5 мм, в Ев- ропе — на 2 — 3 мм в год. Прирост лишайников наиболее интенсивен в начальных стадиях развития, когда синте-- зируемые фикобионтом органические вещества поступают довольно равномерно во все части слоевища. Когда слоевище приобретает более крупные размеры и становится толще, его при- рост замедляется, а у некоторых видов совсем прекращается или слоевище в центре разруша- ется и дальше растет лишь в периферических частях. Так, например, пармелия центрифуж- ная (Parmelia centrifuga) образует в Арктике слоевища диаметром до 1 м, но живым и 4ас- тущим является только их наружный краж 425
шириной 5 — 10 см, тогда как вся внутренняя часть их погибает. В течение года лишайники растут неравно- мерно. Точные измерения показали, например, что прирост пармелии в окрестностях Вашинг- тона (США) в сентябре составлял 1,06, в ог- тябре — 0,40, в ноябре — 0,40, в декабре— 0,70, а в январе и феврале — по 0,16 мм. Замедленный рост многих, особенно накип- ных, лишайников обусловливает и длитель- ность их жизни. Так, например, возраст от- дельных слоевищ ризокарпона (Rhizocarpon geographicum) достигает 4000 лет, аспицилии (Aspicilia cinerea) — 1000 лет, умбиликарии (Um- bilicaria cylindrica) — 200 лет и т. д. В среднем возраст большинства листоватых и кустистых форм не превышает 50 — 100 лет. То, что продолжительность жизни многих видов лишайников велика, а прирост их в пре- делах определенной климатической области более или менее постоянен, привело к идее использовать их для определения возраста населяемых ими субстратов. Соответствующий метод был назван лихенометрией. На субстpare измеряют диаметр самых больших слоевищ лишайников и тогда, зная их средне- годовой прирост в данной местности, вычис- ляют минимальный возраст этого субстрата. Так был определен возраст многих ледниковых морен в Арктической Канаде и Гренландии, установлена динамика движения ледников, скартированы геоморфологические движения и пр. Этот метод весьма успешно применяют гляциологи, геологи и другие исследователи в повседневной работе при датировке ланд- шафтных процессов. Однако для лихенометри- ческой датировки применимы лишь те медленно- растущие лишайники (особенно виды ризокар- пона, аспицилии, гематоммы, лецидеи, лека- норы, калоплаки), для которых точно установ- лен среднегодовой прирост слоевища в данной местности. Использование сведений о при- росте, полученных в других областях, недо- пустимо. Достоверные данные о приросте ка- кого-либо вида в определенных местных усло- вия с получают измерениями контрольных слое- вищ из года в год, измерением слоевищ на субстратах с известной давностью и т. д. Интересна история лихенометрической да- тировки гигантских статуй на острове Пасхи, о которых так увлекательно писал Тур Хей- ердал в своей книге «Аку-Аку». Возраст этих статуй (их свыше 600) был для ученых загад- кой. Точные радиокарбонные датировки их не удавались, так как все статуи вытесаны из вулканического туфа. Тогда немецкий лихено- лог Г. Ф о л л ь м а н н применил лихено- метрический метод. В основу были положены летальные фотоснимки, сделанные с 1914 по 1961 г. На них были хорошо видны розетки трех видов лишайников — диплошистеса (Dip- loschistes anactinus), фисции (Physcia picta) и лецидеи (Lecidea paschalis). Сравнение пло- щадей одних и тех же слоевищ на фотоснимках, сделанных в 1914 и 1961 гг., показало, что годовой прирост этих лишайников равен соот- ветственно 8, 12 и 17 мм в год. Теперь было уже легко установить, что минимальный возраст статуй 430 лет. Позже эти результаты совпали с археологическими данными. ОТНОШЕНИЕ ЛИШАЙНИКОВ К СУБСТРАТУ И ДРУГИМ ФАКТОРАМ ВНЕШНЕЙ СРЕДЫ Субстрат. Медленный рост слоевища не да- ет возможности лишайникам в более или менее благоприятных местообитаниях конку- рировать с быстрорастущими цветковыми рас- тениями или мхами. Поэтому обычно лишай- ники заселяют такие экологические ниши, где условия существования слишком суровы для других растений. Так, они обладают уди- вительной способностью расти на самых раз- нообразных субстрата х: каменистых породах (известняках, гранитах, гнейсах, кварцах и др.), почве, коре деревьев, на хвое, листьях вечнозеленых растений, на мхах, гниющей дре- весине, на гниющих растительных остатках. Лишайники поселяются также на стекле, ко- стях, коже, железе, тряпках и других предме- тах, при этом важно лишь одно — чтобы эти предметы пролежали неподвижно довольно длительное время. Хотя среди лишайников и встречаются та- кие, которые способны расти на самых разных субстрата х, все же большинство видов обла- дают избирательной способностью и поселяют- ся на немногих или даже только на одном оп- ределенном субстрате. По отношению к субстрату и другим условиям местообитания среди лишайников различают несколько основных крупных экологических групп. э п и л и т н ы е лишайники — живущие на поверхности горных пород, э п и ф и т н ы е- растущие на коре деревьев и кустарников, э п и к с и л ь н ы е — обитающие на гнию- щей древесине, э п и r е й н ы е — растущие на поверхности почвы, э п и ф и л л ь н ы е- развивающиеся на хвое и листьях вечнозеле- ных растений, эпибриофитные — оби- тающие на дерновинках мхов, и другие. В различных местообитаниях на скалах, почве, стволах деревьев и т. д. лишайники образуют растительные группировки — с и- н у з и и, которые характеризуются определен- ным видовым составом и определенными мор- фологическими типами. На распределение ли- 426
шайников влияют как физические, так и хи- мические свойства субстрата. Так, среди эпи- литных лишайников можно различить группу кальцефилов, поселяющихся исключительно аа известняках, доломитах и других горных породах, содержащих известь, и группу каль- цефобов, растущих только на поверхностях неизвестковых пород: гранитах, гнейсах, квар- цах и др. Обе эти группы отличаются друг от друга видовым составом. Например, на изве- стняках встречается довольно большое коли- чество лишайников с эндолитными слоевищами, целиком или частично погруженными в суб- страт. Это связано с хорошей растворимостью известняков: гифы лишайников, выделяя ли- шайниковые кислоты, легко растворяют из- вестковые породы и проникают на довольно большую глубину — до 10 лл, а иногда даже более 30 лл. Среди кальцефобов очень мало эндолитных лишайников и почти нет видов со слоевищем, целиком погруженным в субстрат. В то же время для них характерно преоблада- ние видов со слоевищем, имеющим форму арео- лированной корочки. Обычно на поверхности скал очень обильно развиваются накипные лишайники (ризокар- поны, лецидеи, калоплаки, леканоры, гематом- мы и др.), слоевища которых образуют на ска- лах пестрые яркие пятна (табл. 42, 1, 2; 55, внизу справа). Но нередко вместе с ними на скалах растут и некоторые листоватые лишай- ники — пармелии, умбиликарии, ксантории, лептогиумы и др. (табл. 42, 5, 7; 47, 8, 6; 55, вверху справа). В горных и арктических рай- онах одним из очень характерных эпилитных лишайников является, например, листоватый красно-оранжевый лишайник ксантория эле- гантная (Xanthoria elegans). В некоторых гор- ных районах Центральной Азии этот вид до- стигает такого колоссального развития, что отдельные горные массивы принимают cIIJIoIII- ной оранжевый оттенок. Среди кустистых ли- шайников типичных эпилитов сравнительно немного (лишайники рода Neuropogon, некото- рые виды Alectoria, Ramalina и др., табл. 55, вверху справа). 3пигейные лишайники чаще всего поселяют- ся на песчаных, торфянистых и сильно щебни- стых почвах, бедных питательными вещест- вами и непригодных для произрастания дру- гих растений. Многие из этих лишайников являются ацидофильными, т. е. живущими на почве с кислой реакцией. Например, кладонии и цетрарии обычно лучше растут на кислых почвах при концентрации водородных ионов (рН) в пределах 3 — 7,4. Среди эпигейных ли- шайников много кустистых видов — кладонии, цетрарии, алектории, стереокаулона и др. (табл. 56). Из листоватых эпигейных лишай- ников наиболее широко распространены виды пельтигеры, а из накипных на почвах чаще всего встречаются представители семейства лецидеевых (Lecidea, Biatora, Bacidia, To- ninia и др.). В степных и пустынных обла- стях земного шара среди напочвенных ли- шайников преобладают виды с чешуйчатым слоевищем (роды Psora„, Dermatocarpon, En- docarpon и др.). На группировки эпигейных лишайников суб- страт оказывает существенное влияние. Так, по видовому составу лишайник овые синузии песчаных почв отличаются от синузий торфя- нистых почв или почв, богатых известью', в значительной степени это обусловлено реак- цией субстрата (рН). Например, на бедных пес- чаных почвах широко встречается синузия накипных лишайников Lecidea uliginosa и Baeomyces roseus; для торфянистых почв ха- рактерна другая синузия, образованная на- кипными лишайниками Icmadophila ericeto- rum и Lecidea granulosa, а для почв, богатых известью, — синузия Toninia coeruleonigri- cans и Fulgensia bracteata. Но особенно сильно физические и химические свойства субстрата влияют на эпифитные ли- шайники. Для них большое значение имеют структура коры, ее расчленение, жесткость, частота отслаивания и другие особенности. Эпифитная лишайниковая растительность раз- ных древесных пород, как правило, разли- чается по составу. Например, на стволах со- сен обычна синузия, образованная гипогим- нией и псевдэвернией (Hypogymnia physodes и Pseudevernia furfuracea). На дубах, липах и других лиственных породах эти лишайники обычно не встречаются, зато здесь обильно развиваются виды листоватой пармелии (особен- но Parmelia sulcata, P. caperata и некоторые другие) и кустистые рамалины и эвернии (Evernia prunastri, табл. 49, 1). Для стволов осин характерны группировки, образованные видами фисции, ксантории, калоплаки и др. (табл. 55, вверху слева). Видовой состав лишайниковых группировок зависит не только от вида дерева, но и от его возраста. В данном случае особенно сильно проявляется влияние физических свойств ко- ры. Так, например, на молодых деревьях лист- венных пород, имеющих гладкую тонкую кору, обычно развиваются накипные лишайники с эндофлеодным слоевищем (виды опеграфы, артонии, графиса и др.). С возрастом свойства коры меняются: она становится грубее, на ней появляются трещины и шероховатости. Меняется и состав живущих на ней лишайни- ков. На такой коре поселяются уже листо- ватые и кустистые лишайники (виды парме- лии, эвернии, рамалины), а из накипных— 427
многочисленные виды леканор, лецидей и др. (табл. 53, 54). Кроме того, состав эпифитных синузий ли- шайников на одном и том же стволе дерева различен в зависимости от высоты над землей; в этом случае на распределение лишайников оказывают влияние не только физические осо- бенности коры, но и экологические факторы— освещенность, влажность и некоторые другие. Однако, несмотря на тесную зависимость, су- ществующую между лишайниками и субстра- том, на котором они поселяются, до сих пор с достоверностью еще неизвестно, исполь- зуют лишайники субстрат только как место прикрепления или они извлекают из него не- которые питательные вещества, необходимые для их жизнедеятельности. С одной стороны, спо- собность лишайников расти на субстрата х, бедных питательными веществами, дает основа- ние считать, что они используют субстрат лишь как место прикрепления. Однако, с другой сто- роны, избирательная способность, проявляе- мая лишайниками при расселении, строгая приуроченность большинства из них к оп- ределенному субстрату, зависимость видо- вого состава лишайниковой растительности не только от физических, но и от химических свойств субстрата невольно наводят на мысль, что лишайники используют субстрат и как дополнительный источник питания. Это под- тверждается и биохимическими исследования- ми, проведенными в последние годы. Например, выяснилось, что у одного и того же вида ли- шайника, растущего на разных древесных по- родах, состав лишайниковых веществ может быть неодинаковым. Еще более очевидным до- казательством служит открытие у лишайников внеклеточных ферментов, которые выделяются во внешнюю среду. Внеклеточные ферменты, такие, например, как инвертаза, амилаза, цел- люлаза и многие другие, представлены в ли- шайниках довольно широко и обладают доста- точно высокой активностью. Причем, как ока- залось, они наиболее активны в нижней части слоевища, которой лишайник прикреплен к субстрату. Это указывает на возможность активного воздействия слоевища лишайников на субстрат с целью извлечения из него до- полнительных питательных веществ. Помимо свойств субстрата, на расселение лишайников большое влияние оказывают так- же климатические факторы — свет, влаж- ность, температура. Свет для лишайников необходим, как для всех фотосинтезирующих организмов. Отсут- ствие света или недостаточная его интенсив- ность препятствует их развитию. Хотя среди лишайников и встречаются тенелюбивые виды, все же большинство из них относятся к свето- любам. Обычно в очень тенистых лесах, особен- но в густых еловых или буковых, где под кро- нами деревьев царит полумрак, лишайников очень мало. И наоборот, в светлых сосновых лесах лишайники развиваются в изобилии„ образуя на почве сплошные ковры. Во влажных тенистых местообитаниях, куда проникает лишь небольшое количество света, как правило„ развиваются накипные лишайники с порошко- видными, соредиозными (покрытыми множе- ством соредий) слоевищами. Свет оказывает решающее влияние и на жизнеспособность большинства лишайников. Так, некоторые из них образуют плодовые те- ла только в ярко освещенных местах (например, широко распространенный лишайник Hypo- gymnia physodes, а также Evernia prunastri, Pseudevernia furfuracea и др.). Избыток света не препятствует развитию лишайников, а вызывает лишь некоторые мор- фологические изменения. У кустистых лишай- ников при ярком освещении появляются формы с более узкими лопастями и развивается более толстая, темноокрашенная кора с высокой кон- центрацией пигментов и окрашенных лишайни- ковых веществ. Как показали исследования, эти «защитные механизмы» обеспечивают ли- шайникам высокую устойчивость к большим дозам ультрафиолетовой и проникающей ра- диации, порядка 3,6 — 10 эргlсл'. Благодаря этому, несмотря на яркое освещение и повышен- ный радиационный режим, характерные для высокого рий и полярных стран, лишайники здесь развиты в изобилии. И все же у различных видов потребность в свете неодинаковая. Это можно хорошо на- блюдать на одном и том же стволе дерева: ли- шайники, растущие на нижней его части, при- способлены к малой интенсивности освеще- ния — они ассимилируют даже при освещенности 400 лк; выше на стволе дерева развиваются другие виды, у которых нижней границей для фотосинтеза является освещенность 1200 лк. Точно так же на крупных валунах синузии эпилитных лишайников на верхней поверхно- сти отличаются от синузий отвесных поверх- ностей и от синузий, образовавшихся при их основании. Но совершенно очевидно, что в обоих случаях на расселение лишайников оказывают влияние условия не только осве- щенности, но и влажности. Влажность является вторым очень важным фактором распределения лишайников по место- обитаниям. Хотя лишайники относятся к чис- лу наиболее засухоустойчивых растений, спо- собных переносить без воды очень длительные периоды и выдерживать такое низкое содер- жание воды в слоевищах, при котором боль- шинство растений погибает, влага все-таки
необходима для их развития. Это объясняется тем, что процессы фотосинтеза и дыхания про- исходят лишь во влажных слоевищах. Лишай- ники черпают влагу из разных источников- из дождевой воды, утренней и вечерней росы, талого снега, туманов, влажного воздуха. Спо- собность использовать влагу воздуха — очень важная особенность лишайников, в этом они значительно превосходят высшие растения. Они могут абсорбировать водяной пар даже из очень сухого воздуха, с относительной влаж- ностью всего 50~~~. Потребность в воде колеблется у лишайников в широких пределах. Одни из них приспособи- лись к существованию в таких крайних место- обитаниях, как поверхность скал или почвы в условиях жарких пустынь, где дождей неред- ко не бывает в течение многих месяцев. Здесь единственным источником воды может быть утренняя роса, но и такого небольшого коли- чества влаги этим лишайникам бывает доста- точно для нормальной вегетации. С другой стороны, есть лишайники, ведущие подводный образ жизни или обитающие на скалах по бе- регам морей, рек и озер, на камнях и валунах в горных речках. Правда, из них сравнительно немногие живут под водой постоянно. Боль- шинство является растениями-амфибиями: часть времени они ведут подводный образ жиз- ни, а другую часть — воздушный, так как обычно поселяются на скальных поверхно- стях, омываемых только брызгами волн, или на морских скалах, затопляемых только во время приливов, в горных речках, наполненных водой лишь во время таяния ледников, а в остальное время имеющих сухие русла и т. д. Благодаря различной потребности лишай- ников в воде фактор влажности является од- ним из основных, определяющих распределе- ние лишайников но различным местообитаниям. По всей видимости, он играет даже большую роль, чем освещенность. Особенно ярко это проявляется в районах с экстремальными условиями существования растений. Примером могут служить холодные пустыни Антарктики или жаркие пустыни Центральной Азии, где местообитания лишайников по своим усло- виям являются одинаково крайне ксероти- ческими. В таких местах именно влажность выступает фактором, лимитирующим развитие лишайников. Так, в Антарктике, например, лишайники поселяются главным образом в рас- щелинах скал, в понижениях между валунами и скалами, где летом скапливается вода от тающего снега, на скалах, расположенных вблизи тающих снежников, на участках скал, куда непрерывно наносится снег с расположен- ного вблизи ледника, и т. п. В пустыне Гоби, где условия существования настолько суровы, что даже лишайниковая растительность пред- ставлена очень бедно, лишайники, как прави- ло, живут в тех местах, где хотя бы время от времени скапливается или стекает вода. На скалах это могут быть места временных водо- токов, трещинки, небольшие ямки, основания скал. Температура в жизни лишайников не иг- рает той роли, какую играют другие климати- ческие факторы. Лишайники проявляют уди- вительную устойчивость к воздействию как очень высокой, так и низкой температуры и относятся к одним из самых тепло- и холодо- устойчивых растений. Температурный фактор никогда не выступает как лимитирующий в раз- витии лишайников. В условиях пустынь, на- пример, лишайники легко переносят ежеднев- ное нагревание до +50, +60'С, в то время как в полярных районах в течение длинной зимы они прекрасно себя чувствуют при температуре — 40, — 50'С и даже ниже. И все же лишайники не безразличны к влия- нию температурного фактора: они способны фотосинтезиров ать только при определенной температуре окружающей среды. Оптимум фо- тосинтеза у лишайников умеренной зоны на- ходится в пределах от +10 до +25 'С, а темпе- ратурный минимум — между — 7 и — 13'С. В по- лярных пустынях эти растения приспособлены к постоянному воздействию отрицательной тем- пературы, это проявляется в сдвиге темпера- турного оптимума ассимиляции углекислоты. Так, у арктических лишайников, например, наблюдалось поглощение углекислоты при — 20... — 25 'С. Лишайники Антарктики способ- ны поддерживать положительный метаболиче- ский баланс при весьма низкой температуре даже находясь под покровом снега, так как выработали способность усваивать СО, при очень небольшом освещении, всего 50О лк. В арктических и антарктических районах воздействие на лишайники отрицательной тем- пературы сказывается также в сокращении периода вегетации этих растений. В Антаркти- ке, например, лишайники вегетируют всего два летних антарктических месяца — декабрь и январь, да и то в течение только нескольких часов днем, когда температура воздуха ста- новится положительной, ибо и летом каждую ночь в этом районе температура падает ниже нуля. Здесь на помощь лишайникам приходит их способность с первыми лучами солнца и повышением температуры быстро восстанав- ливать фотосинтез после долгого периода за- мерзания. Биотические факторы. Помимо особенностей субстрата и климатических факторов, на ли- шайники в природе оказывают воздействие и так называемые биотические факторы: взаимо-
отношения между самими лишайниками, между лишайниками и другими растениями, между лишайниками и животными. Изучение взаимоотношений между компонен- тами в лишайниковых синузиях показало, что внутри этих группировок существуют явления как межвидовой, так и внутривидовой конку- ренции. На любом из субстратов лишайники, разрастаясь, ведут между собой борьбу за пространство и свет, которая сводится к ме- ханическому давлению друг на друга, нараста- нию одного слоевища на другое, подрыву од- ного слоевища другим, воздействию химиче- скими выделениями. Слоевища лишайников, посилившихся на поверхности какой-нибудь скалы, благодаря своему центробежному ро- сту, должны в конце концов неизбежно встре- титься. Довольно редко слоевища одного и того же вида при встрече сливаются без за- метного ущерба друг для друга. Такие случаи можно наблюдать у накипных, реже у листо- ватых лишайников. Обычно встреча слоевищ одного и того же вида, не говоря уже о встрече слоевищ разных видов лишайников, приводит к заметной борьбе и даже к гибели одного из конкурентов. Причем в борьбе представителей одного и того же вида обычно побеждает более молодой, а в борьбе разных видов одерживает верх лишайник, растущий более быстро. Кон- курентные отношения между видами приводят к сменам внутри лишайниковых синузий, что обеспечивает динамику лишайниковых груп- пировок в растительном покрове фитоценозов. По отношению к другим растениям лишай- ники являются слабоканкурентными организ- мами из-за своего медленного роста. При кон- такте с мохообразными и цветковыми растени- ями лишайники редко одерживают победу в борьбе за пространство, чаще бывает наобо- рот: в местах, ранее занятых лишайниками, особенно в молодых местообитаниях, постепен- но поселяются высшие растения, со временем со- вершенно вытесняя лишайники. Правда, извест- ны случаи, когда слоевища крупных листоватых лишайников, развившиеся на мхах, вызывали гибель этих растений. Но обычно при кон- такте с мхами лишайники редко вытесняют их, более известны факты гибели лишайников под мхами. И все же, как показали недавно проведен- ные исследования, лишайники проявляют не- которые «защитные» реакции по отношению к быстрорастущим высшим растениям. Экспе- риментально было показано, что лишайниковые кислоты обладают свойством оказывать за- держивающее действие на рост высших расте- ний. Оказалось, что и в природных условиях лишайниковые кислоты, хотя и в небольших количествах, вымываются из слоевищ и за- тормаживают прорастание семян и развитие проростков как травянистых растений, та~с и древесных лесных пород. При этом у молодых про ростков сильнее всего страдает корневая система. Под воздействием лишайниковых кислот кончики корней утолщаются и ослиз- няются, исчезают корневые волоски, задержи- вается развитие боковых корней. По-видимо- му, в лесных почвах с хорошо развитым ли- шайниковым покровом эти кислоты находятся в достаточной концентрации, чтобы тормозить развитие других растений. Воздействие животных на лишайники может быть прямым и косвенным. С одной стороны, целый ряд беспозвоночных и позвоночных жи- вотных питаются лишайниками или продукта- ми их разрушения. Объедая слоевища, они повреждают их, содействуя их отмиранию и опаду, а затем принимают участие в процессе их разложения. Обламывая веточки слоевищ, животные способствуют также вегетативному размножению этих растений. Особенно боль- шое значение это имеет в тундровой зоне, где многие эпигейные кустистые лишайники раз- множаются исключительно вегетативным пу- тем — кусочками и обломками слоевища. Не- которые животные, например улитки, объ- едают у лишайников диски апотециев и тем самым способствуют рассеиванию их спор. Животные и косвенно влияют на лишайни- ки, выделяя экскременты, уплотняя почву, повреждая субстраты и т. д. Как известно, большинство лишайников могут существовать в таких условиях, где снабжение нитратами крайне скудно, так как многие из них способ- ны фиксировать атмосферный азот или извле- кать его из растворов, вымытых из субстратов дождевой водой. Но существует особая группа нитрофильных лишайников, тесно связанная с нитрогенными местообитаниями. Такие ли- шайники обычно поселяются на местах пти- чьих базаров, на скалах, покрытых экскремен- тами гнездящихся здесь птиц. Приспособление к нитрогенным местообитаниям связано прежде всего с физиологической адаптацией лишайни- ков, их способностью, например, ассимилиро- вать азот в виде аммония. В нитрогенных усло- виях слоевища накипных лишайников нередко сильно разрастаются и принимают форму ма- леньких кустиков. Таким образом, как мы видим, лишайники отличаются удивительной устойчивостью к вли- янию внешних факторов. Они способны расти в самых разнообразных условиях освещения и влажности, легко переносить длительные периоды без воды, резкие колебания темпера- туры, воздействие как высокой, так и низкой температуры, большие дозы ультрафиолетовой и проникающей радиации. И только воздей- 430
ствие антропогенного фактора оказалось гу- бительным для лишайников. Изменяя природ- ные ландшафты, вырубая леса и возделывая землю под пашни, человек способствовал силь- ному обеднению лишайниковой растительности. Особенно губительным для многих лишайников оказалось загрязнение атмосферы, создание крупных индустриальных центров, воздух кото- рых насыщен дымом, гарью и копотью. ЛИШАЙНИКИ И ЗАГРЯЗНЕННОСТЬ ВОЗДУХА Лишайники по-разному реагируют на за- грязненность воздуха: некоторые из них не вы- носят даже малейшего загрязнения и поги- бают; другие, наоборот, живут только в городах и прочих населенных пунктах, хорошо при- способившись к соответствующим антропоген- ным условиям. Изучив это свойство лишайни- ков, можно использовать их для общей оцен- ки степени загрязненности окружающей среды, особенно атмосферного воздуха. На этой ос- нове стало развиваться особое направление индикационной экологии — л и х е н о и н д и- кация. В самом деле, внимательный человек, гуляя в лесу, непременно заметит лишайники, расту- щие на стволах деревьев, — большие светло- серые пятна листоватых пармелий, мучнистые подпалины накипных лишайников, свисающие с веток редкие «бороды» уснеи, алектории и др. Все они здесь живые и нередко покрывают более половины поверхности ствола. Если пройти через какой-нибудь городской парк, то едва ли удастся обнаружить лишайники, разве только маленькие пятнышки хилых фрагмен- тов слоевищ в трещинах коры. Различия между лишайн иковыми флорами естественных и культурных ландшафтов были замечены уже лихенологами прошлого столе- тия. Не имея тогда точных данных относитель- но экологических условий (климат, состав воз- духа и пр.) городов, они могли только предпо- лагать, что часть лишайников чувствительна к каким-то городским условиям, по всей веро- ятности, к составу воздуха. В дальнейшем было установлено, что различные виды лишайников действительно обладают разной чувствитель- ностью. Одни растут только в естественных, не тронутых культурой ландшафтах, другие переносят умеренное влияние цивилизации, сохраняясь в небольших поселках, селах и пр., а третьи способны расти и в крупных городах, по крайней мере на их окраинах. При изучении лишайников многих городов были обнаружены общие закономерности: 1. Чем больше индустриализирован город, чем сильнее загрязнен его воздух, тем меньше встречается в нем видов лишайников, тем меньшую площадь покрывают они на стволах деревьев и других субстратах и тем ниже их жизнеспособность. 2. При повышении степени загрязненности воздуха первыми исчезают кустистые лишай- ники, за ними — листоватые и последними- накипные. Видовой состав лишайников в разных частях городов (в центре, в индустриальных районах, в парках, на окраинах) оказался настолько различным, что ученые стали в пределах го- родов выделять так называемые «зоны лишай- ников». Впервые они были выделены в Сток- гольме, где стали различать лишайниковую «пустыню» (центр города с сильно загрязнен- ным воздухом и фабричные районы) — здесь лишайники почти совсем отсутствуют, зону «соревнования» (части города со средней за- грязненностью воздуха) — флора лишайников бедна, виды с пониженной жизнеспособностью; «нормальную» зону (периферийные районы го- рода), где встречаются многие виды лишай- ников. Позднее такие зоны были установлены и в других городах. Было также обнаружено, что в некоторых из них площадь лишайниковой «пустыни» за последние десятилетия увеличи- лась. Например, в Мюнхене в 1901 г. она со- ставляла 8 км', а в 1957 г. — уже 58 км'. Долгое время не могли объяснить, какие именно факторы приводят к обеднению и даже исчезновению флоры лишайников в городах. В течение последних десятилетий было показа- но, что из компонентов загрязненного воздуха на лишайники самое отрицательное влияние оказывает двуокись серы. Экспериментально установлено, что это вещество уже в концентра- ции 0,08 — 0,10 мг на 1 м' воздуха начинает вредно действовать на многие лишайники: в хлоропластах водорослевых клеток появля- ются бурые пятна, начинается деградация хло- рофилла, плодовые тела лишайников хиреют. Концентрация ЯО„равная 0,5 мг/м', губитель- на для всех видов лишайников, произрастаю- щих в естественных ландшафтах. Интересные данные были получены также методом пересад- ки некоторых лишайников из природных усло- вий в городские, при этом у разных видов вы- явилась различная реакция на изменение усло- вий: одни быстро вымирали, другие приспосаб- ливались даже к отравленной среде. Конечно, в городах на лишайники пагубн~ влияет не только двуокись серы, но и другие загрязнители — окислы азота, окись углеро- да, соединения фтора и др. Кроме того, в го- родах сильно изменены микроклиматические условия — здесь «суше», чем в естественных ландшафтах (примерно на 5 О~), теплее (в раз- личных городах на 1 — 3 'С), меньше света 431
(в Манчестере, например, всего 1100 солнечных часов из возможных 4500). Лишайники пред- почитают сырые местообитания, и эти условия, конечно, оказывают на них определенное влия- ние. И все же первый их «враг» в городах— загрязненный воздух. Теперь уже человек, который хотя бы немного знает лишайники (15 — 20 видов), прогуливаясь по городу, мо- жет сказать, например, что на этой аллее воз- дух сильно загрязнен, количество двуокиси серы в воздухе превышает 0,3 мг/м' (лишайни- ковая «пустыня»), в этом парке воздух за- »рязнен умеренно, количество SO колеблется между 0,05 — 0,2 лг/л' (это можно установить по произрастанию на стволах некоторых вы- носливых по отношению к загрязнителям лишайников — ксантории, фисции, анапти- хии, леканоры и др.), а на этом кладбище воз- дух довольно чистый — SO, менее 0,05 лг/л' (на это указывают произрастающие на стволах виды естественной флоры — пармелии, алек- тории и др.). РОЛЬ ЛИШАЙНИКОВ В ПРИРОДЕ Лишайники чрезвычайно широко распростра- нены на земном шаре, они встречаются почти во всех наземных и даже некоторых водных .экосистемах. Особенно велика их роль в тунд- ровых, лесотундровых и лесных био»еоценозах, где они составляют заметную часть раститель- ного покрова. Развиваясь на почве, стволах деревьев, валу- .нах и скалах, лишайники образуют в биогеоце- нозах определенные растительные группиров- ки — синузии, которые являются компонента- ми био»еоценозов и играют определенную роль в их жизни, динамике и круговороте веществ. Лишайниковые синузии в силу своеобразия ли- шайников как организмов (медленного роста, особого типа питания и обмена веществ, своеоб- разия продуктов метаболизма) обладают неко- торой автономностью развития и рядом специ- фических черт. Так, их видовой состав беднее по сравнению с группировками цветковых растений; они относительно кратковременны вследствие тесной связи с субстратом, свойства которого с течением времени беспрерывно изме- няются, и т. д. И все же, несмотря на некоторую автоном- ность развития, лишайниковые синузии нахо- дятся в определенных отношениях с другими компонентами биогеоценозов. Прежде всего сле- дует отметить, что с лишайниками связана боль- шая группа животных. В основном это беспоз- воночные, но есть и крупные позвоночные жи- вотные, поедающие лишайники. В «лишайнико- вых зарослях» обитает огромное количество клещей, ногохвосток, сеноедов, гусениц, ли- стоедов, тараканов, пауков, клопов, цикад, жу- желиц и др. Всего было зарегистрировано око- ло 300 — 400 видов беспозвоночных животных, жизнь которых так или иначе связана с лишай- никами. Некоторые из них всего лишь пришель- цы из других биотопов — верхних горизонтов почвы, подстилки, стволов и крон деревьев— и используют слоевища лишайников как вре- менное убежище. Но целый ряд животных— клещи, но»охвостки, сеноеды, гусеницы низших бабочек и др.— связаны с лишайниками гораз- до теснее. Они питаются слоевищами лишайни- ков и продуктами их разрушения. В биогеоце- нозах лишайники вместе с некоторыми насеко- мыми и другими беспозвоночными животными, а также со своей микросредой образуют особые биогеосинузии. Занимая такие экологические ниши, как стволы деревьев, поверхность валу- нов и др., эти биогеосинузии усложняют струк- туру биогеоценозов, влияют на круговорот ве- ществ в них, повышают эффективность исполь- зования солнечной радиации. Используя энергию солнечных лучей, погло- щая воду и минеральные соли для построения своего тела, лишайники образуют определен- ную фитомассу. В биогеоценозах разных типов биомасса лишайников различна. Нередко она невелика, но в некоторых био»еоценозах, осо- бенно в тундровых и лесных, лишайники дают значительную биомассу. Так, например, было подсчитано, что общая биомасса лишайников в некоторых биогеоценозах горных тундр со- ставляет 38,65 ц/га, а в долинных тундрах— 19,08 ц/га. В лесных биогеоценозах она несколь- ко ниже, но в лишайниковом сосняке может достигать 20 ц/га, в сосняке брусничном— 5,6 ц/га, в некоторых биогеоценозах широко- лиственных лесов — 1,8 — 6 ц/га. Наряду с накоплением фитомассы, в биогео- ценозах идет и обратный процесс — отмирание лишайников. Вследствие старения и механиче- ского повреждения некоторые слоевища ли- шайников опадают на поверхность почвы. Ско- рость распада этих слоевищ достаточно высока, причем на первых стадиях большую роль в этом процессе играют беспозвоночные живот- ные. В результате разложения различные ве- щества, заключенные в слоев ищах лишайни- ков, попадают в почву и способствуют накопле- нию ряда химических элементов в верхних ее слоях и образованию почвенного гумуса. Эти вещества оказывают также влияние на почвен- ную микрофлору и другие организмы биогео- ценозов. Одно время полагали, что специфические ли- шайниковые вещества нерастворимы в воде и поэтому в природных условиях их влияние на другие организмы биогеоценозов исключается. Однако исследования, проведенные в последние
Та бл и ц а 49. Кустистые (1, 2, 4, б, 7, 8) и листоватые (3, 5)лишайники: 1 — Evernia prunastri; 2 — Stereocaulon tomentosum; 3 — Peltigera canina; 4 — Himantormia ligubris; 5 — Nephroma arcticum; 6 — Neuropogon fasciatus; 7 — Alectoria ochroleuca; 8 — Usnea longissima.
Г а б л и ц а 50. Кочующие степные лишайники (внешний вид слоевиица): в в е р х у — Parmelia vagans; в н и з у — Aspicilia esculenta.
Та бл и ц и 51. Арктоальпийские лишайники: в в е р х у сл е в а — Xanthoria elegans и Thamnolia vermicularis; в в е р х у с п р а в а — Umbilicaria vellea на скалах; в н и з у с fr е в а — Haematomma ventosum; в н и з у с п р а в а — Rhizocarpon tinei.
Та бл и ц а 52. Эпифитные лишайники: 1 — Hypogymnia physodes, внешний вид листоватого слоевища с губовидными соралями ха концах лопастей; 2— Mcncgazzia pertusa, внешний вид листоватого слоевища с округлыми отверстиями на поверхности; 3 — Mycoblastus sanguinarius, внешний вид накипного слоевища с лецилеевыми апотециями.
Та б л и ц а 53. Эпифитные лишайники: в в е р х у — Lecanora subfuscata, внешний вид слоевища с апотециями; в н и з у — Lecanora glabrata, внешний вид слоевища с апотециями.
Та бл и ц а 54. Эпифитные лишайники: в в е р х у — pertusaria globulifera, внешний вид накипного слоевища с соралями; в н и 3 у — Lecanora pachycheila,6axpoM чатый край слоевища.
Та б л и ц а 55. Эпифитные и наскальные лишайниковые группировки: в в е р х у с л е в а — эпифитная синузия на стволе осины, образованная листоватыми лишайниками Xanthoria parietina и Physcia stellaris; в в е р х у в се р е ц и н е — эпифитный кустистый лишайник Ramalina fraxinea; в в е р х у с п р» а— группировка на стволе дуба из Ramalina fraxinea вместе с листоватыми лишайниками Xanthoria parietina, Parmelia sulcata и Physcia hispida; в н и з у с л е в а — группировка на скалах в поясе кедрового стланика, образованная листоватыми лишайниками Parmelia stenophylla и Xanthoria elegans и накипными лишайниками Lecidea, Rhizocarpon и др.; в н и з у с п р а в а — группировка накипных лишайников Caloplaca, Lecidea и Lecanora на скалах в горах.
Та б л и ц а 5б. Тундровые лишайники: в в е р х у с л е в а — группировка напочвенных кустистых лишацнйков: Cladonia alpestris, Cetraria islandica; в в е p x y в с е р е д и н е — Cladonia pleurota; в в е р х у с п р а в а — Pilophorus robustus; в н и з у с л е в а — дернинка Stereocaulon; в н и 3 у с п р а в а — де~нинка Alectoria ochroleuca.
годы, показали, что лишайник овые кислоты все же в какой-то степени растворяются в воде. Например, растворимость усниновой кислоты составляет в среднем 1 + 0,5 мг%. Было по- казано, что усниновая кислота может вымы- в аться из лишайников и, попадая в почву, влиять на развитие почвенной микрофлоры. Например, в почвах под лишайниками, содер- жащими усниновую кислоту, аммонифицирую- щих бактерий было обнаружено в 100 раз мень- ше, чем на соседних участках. Лишайниковые кислоты оказывают также тормозящее действие на прорастание семян и развитие проростков травянистых и древесных растений. Но, с другой стороны, возможно, что лишай- никам в лесных биогеоценозах принадлежит и роль «защитников» деревьев. Это предположе- ние имеет некоторые основания. Известны фак- .ты, показывающие, что дерево, покрытое ли- шайниками, менее подвержено разрушительной деятельности грибов, повреждающих древесину, чем дерево без лишайников. Изучение антибио- тических свойств лишайниковых веществ пока- зало, что ряд лишайниковых кислот (физодо- вая, усниновая, вульпиновая и др.) действи- тельно подавляют рост грибов — разрушителей древесины. Лишайники принимают участие и в химиче- ском выветривании пород. Им нередко принад- лежит роль пионеров растительности при засе- лении свенсеобнаясенных субстратов (камени- стых поверхностей, щебнистого грунта, песча- ных почв и т. д.) в горах, Арктике, Антарктике и других районах земного шара. Обычно пер- выми обнаженные субстраты заселяют бакте- рии, аэрофильные водоросли, актиномицеты и грибы, подготавливающие субстрат для рас- селения лишайников. Лишайники в молодых местообитаниях образуют инициальные стадии растительной жизни. Время заселения лишай- никами того или иного субстрата бывает раз- личным, оно колеблется в небольших преде- лах — от 5 до 20 (40) лет. Первыми поселен- цами на свежеобнаженных субстратах могут быть различные формы лишайников: накипные, листоватые и кустистые. Причем зарастание субстратов происходит постепенно, в несколько сменяющих друг друга стадий. Эти смены ли- шайниковой растительности вызываются рядом причин: постепенным изменением физических и химических свойств субстрата в результате воздействия на него лишайниковых гиф, взаи- моотношением между собой лишайниковых слое- вищ, их отмиранием, изменениями условий сре- ды и др. Все стадии зарастания каменистых поверхно- стей горных пород можно проследить, напри- мер, на ледниковых моренах, постепенно обна- жающихся в результате отступления ледника. На Полярном Урале первые слоевища лишай- ников появляются на моренах через 10 лет после отступления ледника. Среди пионеров, заселяющих обнаженные каменистые поверхно- сти, имеются как накипные лишайники (Leca- nora polytropa, Rhizocarpon tinei, R. concre- tum), так и листоватые (Umbilicaria cylindrica, U. proboscida и др.), которые образуют первые диффузные синузии. Некоторые из этих видов- пионеров (например, виды рода Rhizocarpon) имеют широкую экологическую амплитуду и присутствуют в лишайниковых синузиях доль- ше других. Но обычно на моренах, насчитываю- щих 50 — 70 лет, на каменистых поверхностях доминируют уясе листоватые лишайники (Umbi- licaria hyperborea, U. proboscidea и др.). На древних моренах обычна синузия с участием видов пармелий (например, синузия Parmelia centrifuga — Haematomma ventosum). На пе- реходных участках древних морен в окружаю- щей тундре можно видеть конечную стадию сук- цес сии — дегенерацию лишайников ого покро- ва и появление высших растений. Но в некото- рых случаях можно встретить резкую останов- ку в ходе сукцессий. Например, на древних моренах, насчитывающих 8000 лет, можно на- блюдать одну из синузий накипных лишайников (Lecanora polytropa — Rhizocarpon concretum). На тех же моренах можно проследить и зара- стание мелкоземисто-щебнистых субстратов, ко- торое также происходит в результате сукцес- сионных смен. Например, при формировании синузии Getraria nivalis — Solorina crocea, раз- вивающейся на хорошо освещенных мелкоземи- сто-щебнистых субстратах, определены три ста- дии постепенного зарастания субстрата, причем уже на второй стадии были обнаружены выс- шие растения и мхи. Формирование другой си- нузии (Stereocaulon alpinum — Cetraria cucul- lata), развивающейся в местах, где чередуются небольшие пятна мелкозема и щебенки, прохо- дит через четыре стадии сукцессий. И только на последней стадии появляется ряд высших растений.
СИСТЕМЛТИЧЕСКИИ OV3OV ЛИШАИ НИКОВ 434 КЛАСС СУМЧАТЫЕ ЛИШАЙНИКИ (ASCOLICHENES) Огромное большинство лишайников — почти все известные 20 — 26 тыс. видов — относятся к классу сумчатых. Их объединяет один общий признак — споры у них развиваются внутри сумок. Сумчатые лишайники — филогенетически очень древняя группа, они произошли от до- вольно примитивных форм сапрофитных аскоми- цетных грибов. Часть аскомицетов в симбиозе с зелеными и сине-зелеными, реже с желто-зе- леными и бурыми водорослями в процессе дли- тельного эволюционного развития образовали многочисленные и чрезвычайно разнообразные слоевища листоватых, накипных и кустистых лишайников. Плодовые тела сумчатых лишай- ников — апотеции, перитеции и гастеротеции, с аскогимениальным и асколокулярным типом онтогенеза. Сумчатые лишайники в зависимости от фор- мы плодового тела делят на две основные боль- шие группы (подклассы): пиренокарповые (Py- renocarpeae) и гимнокарповые (Gymnocarpeae). 1IOgMË СС ПИРБНОКАРИОВЫБ ЛИШАÇI- НИИИ (P YRENOCARPEAE) В этот подкласс объединяют лишайники с пло- довыми телами в форме перитециев. Сюда вхо- дит около 70 родов, для которых характерен как аскогимениальный, так и асколокулярный тип онтогенеза. Слоевище у большинства пире- нокарповых лишайников накипное, причем встречается большое количество эндолитных и эндофлеодных форм, со слоев ищем целиком или частично погруженным в субстрат. Реже слоевище бывает листов атым или кустистым. Фикобионтами являются зеленые, желто-зеле- ные и даже бурые водоросли, но наиболее ча- сто — нитчатые зеленые водоросли, особенно трентеполия. Пиренокарповые лишайники широко рас- пространены по всему земному шару, причем наибольшее их количество встречается в уме- ренных областях Голарктики, а также в тро- пиках и субтропиках. В умеренной зоне нашей планеты большинство этих лишайников оби- тает на камнях, скалах и почве, реже на дре- весном субстрате. В тропиках и субтропиках они обычно встречаются на коре деревьев, но среди них имеется немало представителей, раз- вивающихся на толстых, кожистых листьях вечнозеленых тропических растений. В подклассе только один порядок. ПОРЯДОК ПИРЕНОКАРПОВЫЕ (PYRENOCARPALES) Порядок объединяет 15 семейств, представи- тели которых отличаются друг от друга целым рядом признаков: формой слоевища, характе- ром фикобионта, расположением перитециев на слоевище (они погружены в темную ткань слоевища, называемую стромой, равномерно по одному или группами), типом онтогенеза плодовых тел, наличием или отсутствием в пе- ритециях парафиз и парафизоидов. Споры у этих лишайников разнообразные: бесцветные или коричневые, эллипсоидные или несколько вытянутой формы, одноклеточные или с одной или многими поперечными перегородками,
Семейство включает более вид в ов. Они распространены главным образом в умеренных, арктических и BbIcoKoIopHbIx районах земного шара, лишь , чишь очень немногие встречаются в тро- пических странах. Огромное большинство из них — скальные, накипные, эндолитные и эпи- Рис. 314. Pyrenula nitida: ви а с пе итециями; 2 — поперечный разрез — лоеви е б — стенки перитеция перитеция и слоевища (а сл е ище, в — устьице, г — сумки со спорами, со спорами и парафизы. н огда Móðääüíûå. Сумки у бол большинства видоВ - ~P актерный вид пиренула блеетзйш (Руге никатные. Наиболее nula nitI а, рис. Й 314) имеет слоевище в виде битуникатные, редко унитуникат их слабо блестящих, желтовато- о асп остранены се- тонких, гладких, сла многочисленны и широко р р р ливково-коричневых или почти и ве ка иевых. Предста- оливковых, оли мейства пиренулевых и веру р -коричневых пятен на коре деревь- гих семейств например, красновато-ко вители целого ряда других кзотические - асте- ев иногда оно бывает эндофлеодным и тогда Trypetheliaceae) — редкие экзот ния тропиков. 0,6 — 1,0 мл. Вначале они целиком погружены евище только верхушка выдается наружу, б- Семейство пиренулевые (Pyrenulaceae) позднее выступают до половины. Они шароо- о 450 ви ов. разные или слегка приплюснутые, с темноок- Эта группа насчитывает около видов. р ми в верхней части с ма- обитает в т опических и рашенными стенками, в стьицем расположенным на дне не- ких ст анах, поселяясь обычно на леньким усть ц нула блестящая встречается которых встречаются как эпи~~л д ~ р в го ах до верхней границы лес- им на пове хности на равнине и в горах, со слоевищем, произрастающим р была найдена тактике на Дальнем ~~лео ные со слоеви- ного пояса, ыла коры дерева, так и эндо~л д В Китае Северной Африке (Алжире), щем, развитым внутри кор д р ы е ева в глубоких остоке, в ит еве ной и ент альной Америке, на Антиль- слоях перидерм е мы. Слоевище у пиренулевых евернои и ент ез и е ен- ских островах и в Новои Зеландии. ФЛ примитивного строения, нередко ез ди ерен- ских ст кт ы, лишенное коров ого цированной стру ур Ф.У ка иевые V err ucari aceae) СЛОЯ. ФИКО ИОНТОМ . ч~ б нтом является трентеполия. Пе- Семеиство верукари ( ритеции р округлые или полушаровидные, реже слегка приплюснутые, погруженные в слоеви- ще или поверхностные. Характерно развитие в перитециях простых или разветвленных, иногда сетчато-сросшихся пара~ ~. р ~~из. В верхней части перитеция, у устьица, развиваются также бозаметные одноклеточные тоненькие пери- сла о физы. Сумки битуникатные, в них по 8 р. Споры светлые или коричневые, дву- или мно- гоклеточные, с линз о видными, р д ромбов идными или цилиндр дрическими просветами клеток.,;; ~=,:,;;~~, "....- у, В состав семейства входит 9 родов, виды кото- 1 -� '...,ф';,-~ ых отличаются друг от друга расположение личеством клеток в спорах, ~ +р р тов и некоторыми другими признаками. ]I ~' ~~ «» Род пиренула (Pyrenula) включает около '!', ' видов имеющих коричне д у клеточные споры с линзовидными просветами ', ~;,,'„;~', ~а д кчеток и простые неразветвленные свободные 1 Л пара~из ~изы в перитециях. Из описанных видов В Ю C) 93% тропических. От тропической зоны по на- С7 правлению к полюсам количество видов посте- б пенно сокращается и уже в субтропиках про- израста ает лишь немного эндемичных для этих ~(' ( районов форм. В умеренных областях встреча- I ется всего несколько представителей этого ро- й да, характеризующихся широким распростра- О нением, а также 3 — 4 эндемичных вида. В арк- Ф тических и антарктических районах представи- »»»» тарктике был найден один вид. В умеренных районах Голарктики распространение этих ли- шайников связано главным образом с широко- лиственными лесами и лесным поясом гор, где они произрастают на гладкой коре буков, гра- бов и других широколиственных пород.
литные формы со слоев ищем примитивного строения, иногда с хорошо дифференцирован- ным коровым и водорослевым слоями. Фико- бионтом верукариевых лишайников в боль- шинстве случаев являются зеленые водоросли: коккоботрис, плевр око ккус, псевдоплеврокок- кус, мирмеция и трохисция, а также редко встречающиеся в слоев ище лишайников жел- то-зеленая водоросль гетерококкус (у двух видов) и бурая — петродерма (у одного вида). Погруженные или сидячие перитеции верука- риевых имеют толстые стенки, образованные хорошо развитым эксципулом и нередко одетые снаружи черным покрывальцем. В перитециях парафизы и парафизоиды отсутствуют, но около устьиц всегда развиваются ясно различимые, простые или разветвленные перифизы. Сумки битуникатные, содержат от 1 до 8 спор. Споры бесцветные или коричневые, одноклеточные или двух- — многоклеточные, иногда муральные. Род верукария (Verrucaria) — центральный род семейства, насчитывающий около 350 пред- ставителей. Это накипные, обычно эпилитные, реже эндолитные формы с хорошо развитым слоевищем в виде порошковатой, бугорчатой, сплошной или потрескавшейся корочки, часто с хорошо заметным подслоевищем. Споры одно- клеточные, бесцветные, редко светло-коричне- вые. Наибольшее количество видов встречается в умеренных областях земного шара, особенно в горных районах. В Арктике и Антарктике можно насчитать всего около двух десятков видов. Из тропических районов известно толь- ко полдесятка эндемичных форм. За небольшим исключением верукарии- скальные лишайники и особенно часто разви- ваются на известковых скалах и доломитах. Большинство из них ксерофиты, предпочитаю- щие сухие и открытые местообитания, но не ме- нее 50 видов растут на подводных скалах по бе- регам морей и озер. Некоторые из них часть времени ведут наземный образ жизни, а в остальное время обитают под водой. Так, они нередко поселяются на морских побережь- ях на скалах, которые затапливаются водой только во время прилива или лишь периодиче- ски орошаются брызгами волн. Почти всегда их можно найти также на камнях и валунах в прозрачных горных речках, которые летом в период таяния ледников представляют собой стремительно несущиеся потоки, а в остальное время высыхают. Всего несколько видов оби- тает на почве и древесном субстрате. Типичный пример — верукария скальная (Verrucaria rupestris, рис. 315), развивающая- ся на известняках, доломитах, иногда даже на цементе стен. Слоевище у нее эндолитное, заметное на поверхности субстрата в виде бело- ватых, сероватых или коричневых пятен. Пе- ритеции наполовину погружены в субстрат и выдаются над поверхностью камня в виде бу- горков диаметром 0,2 — 0,3 мм, с устьицем в небольшом углублении. Стенки перитеция образованы шаровидным эксципулом, в верхнеи части буро-черным, в нижней — бесцветным. Снаружи эксципул одет черным покрывальцем. Встречается верукария скальная по всей Ев- ропе от арктических районов до гор Апеннин- ского и Пиренейского полуостровов и в евро- пейской части СССР, а также в Северной Аме- рике, Гренландии и Новой Зеландии. Род стауротеле (Staurothele) насчитывает около 50 видов, которые отличаются от верука- рий крупными муральными спорами и нали- чием в перитециях мелких палочковидных, овальных или шарообразных гимениальных во- дорослей. Споры выбрасываются из плодового тела вместе с прикрепляющимися к ним гиме- ниальными водорослями. Все виды стауро- теле скальные. На известковых породах их слоевища нередко бывают эндолитными и тогда обычно имеют очень примитивное строение: они образованы рыхлым переплетением сильно разветвленных гиф, под верхним слоем кото- рых расположена зона водорослей, собранных в клубочки, а сердцевинные гифы довольно глубоко проникают в субстрат. На силикатных горных породах слоевища обычно развиваются на поверхности скал и имеют вид толстых, реже тонких, цельных, бородавчатых или ареолиро- ванных корочек, как правило, целиком состоя- щих из клеточной ткани (параплектенхимы), обычно в верхней части буроватой. Виды рода распространены почти исключи- тельно в северном полушарии, в основном в Средней Европе и Средиземноморской обла- сти, где много эндемиков. В южном полушарии (в тропической области Южной Америки) из- вестно всего 2 — 3 вида. Некоторые представи- тели из умеренных широт продвинулись в вы- сокие широты Арктики, где известно 15 энде- мичных видов. Типичный пример — стауротеле спрятанная (Staurothele clopima, рис. 316), имеющая хоро- шо развитое слоевище в виде довольно толстой, бородавчатой или ареолиров анной серовато- коричневой или темно-коричневой корочки. В каждую бородавочку или ареолку слоевища погружен 1 перитеций диаметром 0,2 — 0,4 лл, выступающий своей верхушкой. Стенки пери- теция образованы только шаровидным эксци- пулом, бесцветным или коричневым в верхней части около устьица. Вид широко распространен в горах Голарк- тики, здесь его можно встретить в сухих откры- тых местообитаниях, а также на поверхности скал, часто омываемых водой, в местах времен- ных водостоков, в углублениях, где скаплива-
° ° ° ° ° ° ° ° ° ° ° ° ° ° ° 4 ° ° ется вода. Растет на силикатных и известковых горных породах, иногда на песчаниках. Часто встречается в Европе от Шпицбергена и Сканди- навии до Апеннинского полуострова, в запад- ных районах европейской части СССР, известен также на Кавказе, в горах Средней Азии, Аф- рики, Северной и Центральной Америки, Грен- ландии. Семейство дерматокарповые (Dermatocarpaceae) ~~РФ(ю ' " ' .~::.1ф~в~ ° ' $gO ° еф ° е ° ° ° ° ° ° ° е ° в Ф е е 1 Эта группа включает более 120 видов. Слое- вище у дерматокарповых в большинстве случа- ев чешуйчатое, прикрепляющееся к субстра- ту подслоевищем, ризоидами и ризоидальными тяжами, реже листов атое, в виде округлых плотных пластин, прикрепляющихся к скалам с помощью гомфа и ризин, достаточно высоко- организованное, покрытое сверху, а иногда п снизу параплектенхимным коровым слоем, иногда целиком параплектенхимное. Фико- бионт — зеленая водоросль плеврококкус. Ие- ритеции у этих лишайников погруженные в слоевище, редко сидячие. Парафизы в пери- тециях отсутствуют, около устьица развивают- ся довольно длинные перифизы. Сумки битуни- катные, содержат по 2 — 8 спор. Споры бесцвет- ные или коричневые, одноклеточные или муральные, реже двух- — четырехклеточные. Большая часть чешуйчатых видов — обитатели почвы. Наиболее часто они встречаются в степных, полупустынных и пустынных районах земного шара, где играют значительную роль в растительном покрове. Вместе с другими лишайниками они образуют степные и пустын- ные напочвенные лишайниковые сообщества. Листоватые виды дерматокарповых — типичные скальные формы горных районов земного шара. Род дерман~окарпон (Dermatocarpon) объеди- няет формы, имеющие одноклеточные, эллип- соидные или почти округлые споры. Перитеции у них обычно целиком погруженные в слоеви- ще, выступающие лишь своими верхушками; их стенки образованы только бесцветным или темно-коричневым эксципулом. Род насчиты- вает около 80 видов. Они распространены глав- ным образом в Голарктике, в палеотропиках и неотропиках известно лишь 12 — 15 видов. Более 50 видов — чешуйчатые формы, разви- вающиеся на почве, реже на поверхности скал, остальные имеют слоевище в виде серых плот- HBIx кожистых пластин. Листоватые формы— типичные представители высок огорий, где их нередко можно встретить на вертикально под- нимающихся и нависающих скалах. Среди них есть обитатели горных речек и озер, развиваю- щиеся на подводных скалах и валунах, иногда они поселяются на скалах, лишь периодически затопляемых водой, или в местах водостоков. ° ° Ю ° ° в е ° ° ° ° ° 1 ф ° ° Рис. 315. Verrucaria rupestris: 1 — общий вид перитециев; 2 поперечный разрезчерезперите- ций (и — эксципул, б покрывальце, в — устьице, г пери- физы); 8 — сумка со спорами. Рис. 316. Staurothele clopima: 1 — общий вид слоевища с перитециями; 2 — поперечный раз- рез через перитеций (а — эксципул, б — сумки, в — гимени- альные водоросли); 8 — сумка со спорами.
° е ° ° ° ° °, ° в ° r % вв Я~У ° в~в в бв ~Ъ в ° ° з(в вВr ° ° в eri ° м ° O Г, в ° ° ° ° % Qf r ° ° r еув 'Ф:ф ° вю ° ° ъЪ~ в в ° 1 ° ° r ° Þ' r ° ° ° ° ° ~ ° ~е Ф юв, ° в r ° r в ° ° r ° ° '~Ъ,»», гу, е ,б' 1 в в fir g ь gt a+. ~,Фв в ° ii% % ° ф % Ф ° ° \ ° ° эЧ ° ъЭ ф' ° Щ%~ Фе в Ь ° ° ° С в ° . в ° ° ~ъ A в ° ° ° в вФ в г в ° вв юв ~в в ° ~~ ф~ ° 3' ° ° . ~. ° а ~~~с ° 'С в й «Ф Ф в 4 Ю~в У Ф Ъ'в ° ° ° ° .. к:-. rO ° вв Ю юф в t ° ,ву ъ ° ° 'r в вЭ ° ~ в в ° ~Ф Ъ в ° ° с ° в % в 4В 'Ф ° ° ° ° ° ° Ф ° Рис. 317. Слоевище лишайника Dermatocarpon minia- tum. Дерлгатокарггон ггеченочный (Dermatocarpon hepaticum) — характерный представитель рода. Его слоевище имеет вид небольших, диаметром до 2 — 7 мм, скученных или обособленно сидя- щих чешуек. Чешуйки округлые, иногда по краям лопастные, рыжевато- или буровато- коричневые, реже глинисто-желтоватые, при- крепляющиеся к почве с помощью черного под- слоевища. Перитеции многочисленные, глубоко погруженные в слоевище, заметные на поверх- ности чешуек в виде черных точек. Стенки пе- ритеция образованы светлым эксципулом, тем- ноокрашенным лишь около устьица. Дермато карп он печеночный — типичный кальцефил, чаще всего встречающийся в горах, по склонам, где он обитает на карбонатной почве, иногда прямо на известняках и доло- митах. Широко распространен по всей Голарктике, хотя встречается довольно редко. Обнаружен в горных районах Европы и Азии, на Кавказе, в Северной Африке, Северной Америке, в Грен- ландии. В южном полушарии известен только в Новой Зеландии. Дерматокарггон лгиниатум (Dermatocarpon miniatum, рис. 317) имеет листоватое слоеви- ще, в виде округлых твердых, ломких, серых или коричнево-серых пластинок, достигающих. в диаметре 5 — 10 см, прикрепляющихся к суб- страту центральным гомфом. Снизу пластинки более светлые, розовато- или желтовато-корич- невые, гладкие, иногда шероховатые от мел- ких. бородавочек. Перитеции достигают в ди- аметре 0,15 — 0,2 лл, они целиком погружены в слоевище и заметны на его поверхности в ви- де черных точек. Стенки перитеция образованы бесцветным, иногда у устьица темноокрашен- ным эксципулом прозоплектенхимной струк- туры, состоящим из клеток с сильно утолщен- ными стенками. Этот лишайник растет на вертикальных, обычно затененных склонах скал, во влажных местах. Встречается чаще всего в горах, но иногда и на равнинах на выходах горных по- род, по всей Голарктике: в Европе, на Кавказе, в Азии (особенно в горах Средней Азии и Юж- ной Сибири), в Северной Америке, в Гренлан- дии, в Мексике, в Северной Африке. Род эндокарггон (Endocarpon) отличается ха- рактерными многоклеточными муральными спорами, по 2 — 6 в сумке, и наличием в пери- тециях гимениальных водорослей. Перитеции у представителей рода целиком погружены в слоевище, со стенками, образованными лишь черным, коричневым, реже светлым эксципу- лом. Слоевище имеет вид мелких листочков или чешуек, иногда оно почти корковидное, прикрепляется к субстрату гифами подслоеви- ща или ризоидами. Сверху и снизу слоевище покрыто пар аплектенхимным коровым слоем, иногда целиком состоит из параплектенхим- ной ткани. Род насчитывает около 30 видов, рассеянных в различных областях земного шара, из них '/, встречаются в Голарктике, преимущественно в Средиземноморье, остальные обитают в тро- пиках и субтропиках Южной Америки и Афри- ки, а также в Австралии. Большинство эндо- карпонов растет на почве в степных и полу- пустынных районах, на растительных остат- ках, на дерновинках мхов в расщелинах. и на карнизах скал, реже они развиваются не- посредственно на каменистом субстрате, не- многие — на прибрежных, омываемых волнами скалах. Имеются виды, растущие на сильно засоленных почвах по берегам соленых озер. Вместе с чешуйчатыми формами рода дерма- токарпон и другими лишайниками эндокар- поны играют определенную роль в образовании напочве нных лишайниковых группировок в степных и полупустынных районах земного шара. Примером может служить эндокарпон мелкий (Endocarpon pusillium), образующий слоеви- ща в виде обособленно расположенных или собранных в дерновинки чешуек. Чешуйки достигают в диаметре 0,5 — 3,5 мл, они оливко- во-желтоватые или темно-коричневые, плоские, по краям зазубренные или лопастные. Перите- ции диаметром 0,2 — 0,4 мл заметны на поверх- ности слоевища в виде черных точек или ма- леньких бугорков. Растет эндок арпон мелкий на известковой, глинистой, реже песчаной почве, чаще всего в горах у основания скал и валунов, распрост-
. лп~папников. Ф.У двое тело (~— — мдзедий), ~z кал~щиевых л азрез Яерез плодо ез через плОДОвое Пло овые тела и спор ПРОДОЛЬНЫИ Р ~ продольп~1И 'Рис. 318. Плодо оцыx ~~~ H ~~~~~~щ~ — б ий вид плодовых тел — — f ( — о ы; 4 — 6 — Соп1осу ~е ц тело, 6 — споры и опе Азии и Северной ранен в о сновном в Европе, .е. Америк Б'ЛА СС Г ГИЛХНОЛЛ РЛ ОВЫ E .ХАША EI- ~7 Щ~тО~АЯРЯАЯ) .НХШ~ГХ (G Подкласс гимно ока ~повых л ишайников вклю- ?х листов етых 250 родов накипных, ~~о м, чрезвычаино ш и кустистых ~орь, у шару. Эти ли- по всему земному страненных по т плодовые тела открытого ти- шайники имеют ии. Онтогенез ии еже гастеротеци . па — апотеции, р ального типа- л аскогимениал .плодовых тел телах образуется гимениальный в плодовых тел спорами и пара- ИИ ИЗ СУМОК СО слой, состоящи лишайников уни- физ. Сумки у гимнокарповых л тавляет лишь по- исключение состав туникатные, вых у представи у ителей которого рядок артониевь тип онтогенеза, наблюдается п юмежуточныи тип асколокулярного в себе черты и ас сочетающий в звития, а сумки льного типа разв и аскопыениа. вляются зеле- Фикобионтами я б ~туникатные. Ф д р, первых наи- ные и сине-зеленые т ебуксия, из аспр остр анена тре более широко ра BTOPI IX — НОСТОК. одкл асс делят По строению плодо овых тел п на 4 порядка. ПОРЯДОК КАЛ ЛИЦИЕВЫЕ (CALICIALES) собенность ю калициевых ли- ана йни ников является образование на д и я — особой поте иев мазеди ности диска апо ц й массы, состоящ ей из спор, порошкооб разной рано освобод ив шихся из сумок, ~х лишаиников, т всех остальных В отличие от 0 ы с сил ои выо асываются из плодового тела лы материнского кального аппарата и. ПОМОЩЬЮ BBIHIHKB я осмотического дав. евых п оисходит из с мок путем ра бождение спор и... а ых оболочек. сво остаются некотор е ия склеенными ВеР-ХНост аспа даются после созревания, оватую массу, па а изами порошков ю вет ом. лаг легко сдуваему шайников не- пп калициевых ли бенности группу оплодньв|и. Фи- ют также порошко . ко ионт Эволюционн р за но главным айников было связа плодных лишаини никновением ряд а приспособле- образом шего ассеивания а калициевых апо У большинства по шаются над слоев ищем на дли 18 напоминая я темные малень- ножках (рис. а высокоорга- или б лавочки, а у кие гвоздики ил . п е ставителеи этого порядка (се- низованных пред ) ни вообще обра- мейство Sphaer p о horaceae они тых азветвленных зуются на вершине кустистых р
странение. Они интересны также тем, что до- вольно часто встречаются формы, тесно связан- ные со свободноживущими грибами: на ран- них стадиях развития в их слоевищах имеются водоросли, а на более поздних их нет. Пред- полагают, что наличие или отсутствие водо- рослей зависит от условий обитания лишайни- ка. Эта особенность дала основание некоторым исследователям отнести калициевые лишайники к свободноживущим грибам. Порядок калициевых насчитывает 17 родов и более 220 видов. Они широко распространены в северном и южном полушариях, преимущест- венно в умеренных, субтропических и тропи- ческих районах. Чаще всего они растут во влажных тенистых лиственных или хвойных лесах, поселяясь на обнаженных корнях у ос- нования стволов, в трещинах коры дубов, буков, грабов, пихт и елей. Один из излюблен- ных субстратов — мертвая гниющая древе- сина. Имеются также виды, паразитирующие на других лишайниках и на грибах, чаще всего трутовых. Очень небольшое количество видов растет на каменистом субстрате. Кустистые формы — обитатели северных и горных тундр. Р11с. 319. Цифелевые и сферофоровые: 1 — 8 — Cyphelium tigilkare: 1 — слоевище с апотециями, 2 продольный разрез через апотеций, 8 — споры; 4 — 5 — Sphaerop- hor>ls globosus: 4 — отдельная веточка слоевища с плодовыми телами (а), 5 — продольный разрез через плодовое тело (6— мазедий, в — слоевищный зксципул, г — устьице, д — столбик). подециев. Сидячие апотеции встречаются здесь редко. Форма апотециев у примитивных видов калициевых конусовидная или цилиндрическая с плоским диском (рис. 318, 1), а у более вы- сокоорганизованных представителей апотеции имеют вид шаровидны~ головок (рис. 318, 4), что также способствует лучшему рассеиванию спор. Хорошо прослеживается также посте- пенное усложнение и в строении слоевища. У одних форм (семейство Caliciaceae) слоеви- ще слаборазвитое, гипофлеодное или в виде зернистой корочки, без дифференцированной структуры; у других (семейство Cypheliaceae) появляются уже мелкочешуйчатые и листоватые слоевища (рис. 319, 1); у наиболее высокоорга- низованных представителей (семейство Sphae- rophoraceae) слоевище имеет вид разветвленных кустиков (рис. 319, 4). Порядок калициевых считается древней группой, рано отделившейся от основной ли- нии развития гимнокарповых лишайников, о чем свидетельствуют их примитивное строе- ние, отсутствие в сумках специальных приспо- соблений для рассеивания спор, большое ко- личество монотипных (содержащих всего один вид) родов и широкое географическое распро- Семейство калициевые (Cal iciaceae) Эта группа включает виды, развивающиеся на гниющей древесине, коре деревьев и мхах, их слоевище имеет вид тонкой, зернистой или порошковатой, желтоватой, серовато-беловатой или зеленоватой корочки, иногда незаметное, целиком развивающееся внутри субстрата. Апо- теции конусовидные или шаровидные, на длинных простых или разветвленных ножках, лишь у представителей рода сфинктрина, па- разитирующих на слоевищах других лишайни- ков и грибов, апотеции сидячие. Головки пло- довых тел обычно покрыты беловатым, корич- неватым, желтоватым или зеленоватым нале- том. Сумки цилиндрические, содержат по 8 спор. Споры одноклеточные или двух- — восьмикле- точные, бесцветные или коричневые. В составе семейства 6 родов, представи- тели которых отличаются друг от друга строением плодовых тел, формой и окраской спор. Большинство из них широко распростра- нены как в северном, так и в южном полушарии. Род стеноцибе (Stenocybe) встречается лишь в Гол арктике, монотипный род пиргидиул (Pyrgidium) — эндемик тропиков Северной Ин- дии. Род калициум (Calicium) включает наиболь- шее число видов — около 100. Для них харак- терны одноклеточные или двухклеточные, ко- ричневые или темно-серые споры веретеновид- ной или яйцевидной формы. Плодоношения с конусовидными или линзовидными головками„
развивающимися на ноясках высотой до 0,5— 2 лм. Слоевище накипное, в виде тонкой по- рошковатой, зернистой или бугорчатой ко- рочки, иногда гипофлеодное. Виды калициу- ма встречаются в обоих полушариях, но многие из них ограничены тропическими и субтропи- ческими районами. В Голарктике они распро- странены главным образом в лесной зоне и лесном поясе гор. В старых тенистых лесах их можно встретить на коре хвойных пород, дубов, грабов, на тоненьких веточках осины, ольхи, орешника, на гниющих пнях и стволах деревьев. Реже они растут на поверхности скал в тенистых влажных лесах. Характерный представитель рода — кали- циум зеленый (Calicium viride, рис. 318, 1 — 3). Слоевище этого лишайника представляет собой желтую или желто-зеленую зернисто-бородав- чатую корочку. Плодовые тела имеют вид ма- леньких гвоздиков высотой до 2 — 3 мм. Го- ловки конусовидной или линзовидной формы, достигающие в диаметре 0,5 — 0,8 лм, с выпук- лым буро-черным диском, снизу коричневые. Высота ножек 2 ли, толщина — 0,2 мл, в верх- ней части они рыжевато-коричневые, внизу черные. Калициум зеленый чаще всего развивается на коре старых хвойных пород, особенно елей, реже на деревьях лиственных пород. Встреча- ется по всей Европе, на Кавказе, на Алтае, а также в Северной Америке, в умеренных об- ластях Южной Америки и в Австралии. Род комиоцибе (Coniocybe) объединяет виды с одноклеточными круглыми, реже овальными, бесцветными или желтоватыми спорами. Го- ловки плодоношений шарообразные, окрашен- ные в бело-желтый или зеленовато-желтый, сероватый, рыжеватый, иногда красноватый цвет. Они развиваются на очень тонких изо- гнутых в разные стороны ножках, достигающих в длину 3 — 5 лш. Слоевище имеет вид однород- ной зернистой или пор ошковатой корочки, иногда незаметное. Встречаются эти лишайники во влажных лесах на оголенных корнях, реже на стволах поваленных деревьев, особенно у их основания, в трещинах и щелях коры, в гнилых дуплах, нередко на гниющих мхах, растительных ос- татках, прямо на почве по тенистым склонам нависающих обрывов, в небольших пещерах. Род насчитывает 24 вида, распространенных в умеренных, тропических и субтропических областях обоих полушарий. В умеренных районах они обитают главным образом в лесной зоне и горных районах, и лишь один вид за- ходит далеко на север. Типичный вид — комиоцибе зернистый (Conio- cybe furfuracea, рис. 318, 4 — 6). Его слоеви- ще имеет вид серо- или зеленовато-желтой по- рошкообразной или крупнозернистой тонкой корочки. Головки плодоношений шарообраз- ные, достигающие в диаметре 0,2 — 0,7 мл, гус- то покрытые сернисто- или зеленовато-желтым порошкообразным налетом. Головки сидят на длинной изогнутой коричневато-черной ножке длиной 1 — 5 мм и толщиной 0,1 — 0,2 мм. Этот лишайник можно встретить в тенистых и влажных местах, по лесистым склонам, под нависающими обрывами, в небольших пеще- рах. Он широко распространен по всей Евро- пе, встречается в Азии, в Северной Америке, а также в крайних районах Арктики — на Чукотском полуострове, островах Шпицберген и в Гренландии. Семейство цифелевые (Cypheliaceae) Семейство насчитывает немногим более 60 видов, объединенных в 8 родов, 5 из которых монотипны. Все они обычно имеют хорошо развитое слоевище в виде струповидной или крупнобородавчатой корочки, по краям неред- ко с листовидными лопастями, иногда оно чешуйчатое или листовидное с восходящими лопастями. Плодовые тела сидячие или даже погруженные в слоевище. Диск апотециев ок- ружен слоевищным краем, иногда двумя края- ми — собственным и слоевищным. Цифелевые лишайники обитают на гнилой древесине, коре деревьев, реже на каменистом субстрате. Широко распространены лишь представители родов цифелиум (Cyphelium) и псевдоколиух (Pseudocolium), они встречаются в умеренных, тропических и субтропических районах се- верного и южного полушарий; четыре рода (Pyrgillus, Tylophoron, Schistophoron п Ty- lophorella) связаны исключительно с тропиками и субтропиками, остальные встречаются в Ев- ропе. Род цифелиум насчитывает 35 видов, для ко- торых характерны двухклеточные, редко одно- клеточные коричневые споры. Плодовые тела погружены в слоевищные бородавочки, реже сидячие, вначале полукруглые или булавовид- ные, с точковидным закрытым диском, затем диск расширяется и становится плоским. Диск апотециев бывает черным, голым, но нередко покрыт желтоватым или желтовато-зеленым налетом. Слоевище беловатое, сероватое или яркое желто-зеленое до желтого, имеет вид по- рошковатой бородавчатой или бугорчаточешуй- чатой корочки, часто с лопастными выростами по краям. Большинство видов встречается в субтропи- ческих областях, особенно в субтропиках Авст- ралии и Северной Америки, около 10 видов известны в умеренных областях Европы и Азии, главным образом в лесах Средней и Западной 441
Рис. 320. Sphaerophorus globosus: слоевище среди мхов и других растений. Европы. Обычно эти лишайники развиваются на коре хвойных пород, реже на дубах, буках, липах и других лиственных породах. Часто их можно найти и на поверхности гниющей древесины, иногда на слоевищах накипных лишайников, изредка — на каменистых субстратах. Широко распространен цифелиум брусковый (Cyphelium tigillare, рис. 319, 1 — 8), растущий на коре хвойных пород и гниющей древесине. Обычно он хорошо бросается в глаза яркой, лимонно-желтой или зеленовато-желтой окрас- кой слоевища, с которой резко контрастируют черные матовые диски апотециев. Апотеции диаметром 0,3 — 0,7 мм погружены в слоевище и лишь в старости несколько выступают над его поверхностью, они с двойным краем— тонким черным собственным и толстым зелено- вато-желтым слоевищным. Встречается этот лишайник в лесах и горных районах Европы, на Кавказе, в Сибири, на Алтае, в Саянах, Якутии, арктических районах Северной Америки. Семейство сферофоровые (Sphaerophoraceae) Эта группа объединяет виды с кустистым слоевищем, состоящим из сильно разветвлен- ных прямостоячих или прижимающихся к суб- страту подециев (рис. 320). Подеции округ- лые или местами сплюснутые, с высокодиффе- ренци рова нной структурой — ясно различи- мыми коровым слоем, водорослевой зоной и сердцевиной, лишь у тропических представите- лей этого семейства слоевище листоватое. Апо- теции располагаются на концах булавовидно- вздутых веточек и глубоко в них погружены (рис. 319, Б). Характерной особенностью пло- довых тел этих лишайников является развитие в апотециях полукруглого столбика (колю- меллы). Споры одноклеточные, коричневые. Сферофоровые, как и другие семейства кали- циевых, богаты монотипными родами (Calici- dium, Pleurocybe, Thysanophoron, Acroscyphus), представители кото рых встречаются исключи- тельно в субтропических и тропических районах южного полушария. Род сферофорус (Sphaerophorus), насчиты- вающий 8 видов, широко распространен и встречается как в северном, так и в южном полушарии. Эти кустистые лишайники — ти- пичные обитатели северных и горных тундр. Они образуют на почве или, реже, на скалах прямостоячие густые дернинки, высота кото- рых инагда достигает 5 — 10 см. Веточки слое- вища округлые или плоские, покрытые плот- ным, хрящевидным коровым слоем с хорошо развитой зоной водорослей, в центральной своей части заполненные сердцевиной. Плодо- ношения размещены на концах веточек, имею- щих вид округлых головок. Сначала апотеции целиком покрыты слоевищем как покрываль- цем, затем этот покров на вершине головок ло- пается, и тогда становится виден темноокра- шенный более или менее округлый гимениаль- ный слой. Сумки цилиндрические, содержат по 8 темных шарообразных или слегка оваль- ных одноклеточных спор. Наиболее широко известен сферофорус круг- лый (Sphaerophorus globosus, рис. 319, 4, Б), имеющий вид прямостоячих коричневых силь- но разветвленных кустиков высотой 1 — 10 см. Веточки его слоевища очень блестящие, с бе- ловатыми кончиками, ветвящиеся во все сторо- ны. Плодоношения развиваются очень редко. Апотеции бывают конечными, диаметром до 1 — 3 мм. Этот лишайник широко встречается в Аркти- ке, в альпийском и субальпийском поясах гор, где растет на земле среди мхов, на каменистой почве, на мшистых скалах (рис. 320). Распрост- ранен на всем побережье Северного Ледовитого океана, на Камчатке, обитает также в горах Европы и Азии, в Северной Америке, Австра- лии, Новой Зеландии и на островах Антарк- тики. ПОРЯДОК АРТОНИЕВЫЕ (ARTHONIALES) У представителей этого порядка плодовые тела — гастеротеции и апотеции, имеющие вид округлых, удлиненных, звездчатых или не- правильной формы плоских пятен, лишенных как слоевищного, так и собственного края. Это очень примитивный признак, тесно связы- вающий артониевые лишайники со свободно- живущими грибами. В отличие от всех осталь-
ных лишайников у артониевых не образуется эксципул, и гимениальный слой возникает поч- ти непосредственно на поверхности вегетатив- ных гиф слоевища — на тоненьком слое слабо- развитого гипотеция. Сумки, как у некоторых представителей аскомицетных асколокуляр- ных грибов, битуникатные и имеют очень толс- тую внешнюю оболочку — экзоаск. В гиме- HHaJIhHQM слое нет настоящих парафиз, вместо них развиваются сетчато-соединенные, несво- бодные на верхнем конце гифы, называемые парафизоидами, образующие рыхлую или гус- тую сетчатую ткань. Слоевища у артониевых очень разнообраз- ные, как примитивные накипные с гомеомерной и гетеромерной структурой, так и высокоорга- низованные кустистые с дифференцированной структурой, но все же преобладают слабораз- витые, примитивные, целиком развивающиеся внутри субстрата в виде простого переплетения гиф гриба с разбросанными между ними клет- ками водорослей. Фикобионтом являются одноклеточные или нитчатые зеленые водо- росли, среди которых наиболее обычна тренте- полия. Большинство представителей этого порядка встречается в субтропических и тропических областях, а также в умеренной и теплоумерен- ной зонах земного шара. Излюбленный субст- рат — кора дерева, реже их можно встретить на каменистых породах, на мхах и гниющей древесине. Семейство артониевые (Arthoniaceae) Семейство включает виды с накипным, сла- боразвитым слоевищем, нередко целиком раз- вивающицся внутри коры дерева, обычно гомео- мерным, реже с едва намечающейся гетеромер- ной структурой. Апотеции или гастеротеции округлые, неправильные или звездчатые, ко- ричневой, красноватой или черной окраски. Сумки булавовидные, толстостенные, с особен- но толстой оболочкой в верхней части, образуют по 8 спор. Споры бесцветные, вытянутые, по- перечно-многоклеточные, иногда муральные. Насчитывают около 650 видов, которые делят на 4 рода, отличающиеся формой спор, распо- ложением сумок в гимениальном слое и некото- рыми другими признаками. Артониевые распространены главным обра- зом в субтропических и тропических странах, а также в умеренной зоне земного шара. Род артония (Arthonia) — центральный в се- мействе, он включает около 500 видов. У этих лишайников споры веретеновидные или вытя- нутые, поперечно-многоклеточные, а сумки рас- положены в гимениальном слое в виде отдель- ных небольших групп. Слоевища накипные, я~й Зф ФЩ 1% с Рис. 321. Arthonia radiata: общий вид слоевищ и апотециев. нередко гипофлеодные, слабозаметные или развивающиеся в виде расплывчатых белова- тых, сероватых, желтоватых или оливково- черных пятен. Излюбленный субстрат артоний- гладкая кора лиственных пород. Особенно часто они встречаются на стволах и веточках тополей, ольхи, молодых дубков, лип, кленов, буков, ясеней, берез, реже на хвойных породах, но имеются виды, растущие и на твердых крем- нисть-.х горных породах, на известняках, а также на гниющей древесине и мхах. Наибольшее число видов — обитатели тропиков и субтро- пиков. В умеренные районы земного шара продвину- лись преимущественно те виды, которые ра- стут на коре деревьев. Здесь они распростра- нены в основном в широколиственных лесах. Характерный представитель рода — арто- ния лучиапая (Arthonia radiata, рис. 321, 322). Слоевище этого лишайника в начале раз- вития бывает целиком погружено в кору дере- ва, затем выступает на ее поверхность и тогда становится заметным в виде тонких беловатых, сероватых или зеленовато-коричневатых пятен, нередко окаймленных черной линией подслое- вища. Плоские черные апотеции развиваются в большом количестве; они погружены в слое- вище или слегка выступают, округлые, оваль- ные, многоугольные или лучисто-звездчатые, шириной 0,5 — 1,5 мм. Этот лишайник нередко встречается на коре ольхи, рябины, ясеня, бука, граба, реже на хвойных породах. Он широко распространен в лесах и горах Европы, на Кавказе, в Южной Сибири и на Дальнем Востоке, в Китае, в Северной Америке, Северной Африке, Австралии, Новой Зеландии.
° ° фЪ ° ° ~ п / С Р~=~С ~~:- РС::::::><-=~ ~- ~~ -~ ~~><.":: фд:~~~~» ~--» =э~-~~=>с == =~ с-=-э~=~~==~ ° .::==) ~:р Фй:~~ >~ ~ < >~= =~~' ~ ~ ~ — >:рс="э~ъ~.-~ ° Ф ° ъ ° а ° ° ° ° ° ° ° ° ° е В ° Ф ~ ° ° е ° ° ° с ° ° ° ° ° ° е ° ° с ° ° ° ° Ф ° ° ° ° ' ° ° ° ° ° ° ° ° ° ° ° % ° ° 4~ Je %% ° а ° ю ° ° с ° ° ° ~ ° ga е ° ° ° ° ~' ° ° ° ° ° ° ° ° Рис. 322. Arthonia radiata: ° У 1 — отдельный апотеций (сильно увеличено); 2 — поперечныи разрез через группу апотециев; 8 — сумка со спорами; 4— споры. Семейство рочелловые (Roccel laceae) Эта группа включает кустистые, реже накип- ные и листоватые формы, растущие в субтро- пических и тропических районах, главным об- разом на скалах морских побережий, реже на коре деревьев или других субстратах. Доста- точно высокоорганизованное кустистое слое- вище этих лишайников с округлыми, плоскими, иногда нитевидными веточками прикрепляется к субстрату маленькой базальной пластиноч- кой. Веточки покрыты хорошо развитым коро- вым слоем, водорослевый слой более или менее обособлен, внутри имеется рыхлая однородная сердцевина. У некоторых видов на слоевище развиваются сорали. Плодовые тела — округ- лые апотеции или вытянутые гастеротеции. Споры вытянутой формы, поперечно-многокле- точные, бесцветные. Семейство охватывает 10 родов, многие из которых монотипны. Родина рочелловых — К)жная Америка. В субтропи- ческих и умеренных районах Чили и К~жного Перу известно 6 эндемичных родов, здесь же отмечено и наибольшее разнообразие морфоло- гических типов рочелловых. В северном полу- шарии встречаются представители только трех родов. Особенности географического распростра- нения, большое количество монотипных родов и своеобразие морфологического строения ро- челловых говорят о древности их происхож- дения. Род рочелла (Roccella) объединяет лишайники с прямостоячим слоевищем, образующим гус- тые, сильно разветвленные лохматые кустики, плотно прикрепляющиеся к скалам маленькой пяточкой. Лопасти слоевища могут быть плос- кими, лентовидными (рис. 323, 1) или более или менее округлыми, иногда в форме пальце- видных выростов. Очень своеобразна окраска этих лишайников: обычно они беловато-серого или сизого, редко грязно-оливкового цвета и похожи по внешнему облику не на живые рас- тения, а на причудливые известковые окаме- нелости. Развивающаяся иногда по краям ло- пастей густая кайма беловатых или грязно- белых соралей только усугубляет это впечат- ление. Округлые апотеции, образующиеся по краям лопастей, также обычно беловато-серой окраски благодаря густому беловатому- налету. Своеобразен по своему строению и коровой слой: он образован рядами гиф, расположен- HbIx перпендикулярно к поверхности слоевища (рис. 323, 8). Коровой слой такого строения носит название палисадного, он чрезвычайно редко встречается у лишайников. Род насчиты- вает 25 видов, из них 17 обитают в Южной Америке. Несмотря на приуроченность к мор- ским побережьям, представители рода широ- ко распространены по всему земному шару- они встречаются в Африке, Австралии, Европе, Азии, Северной Америке. В северном полуша- рии известно 8 видов. В Европе рочеллы рас- пространены главным образом в Средиземно- морье. Характерный представитель рода — рочелла фукоидкая (Roccella fucoides, рис. 3' 3, 9)— ~ «с) имеет вид прямостоячих светло-серых или си- зых кустиков высотой 2 — 10 сл. Веточки толщиной 1 — 3 лл в верхней части повторно дихотомически-ветвящиеся, с разбросанными выпуклыми белыми с ор алями. Лпотеции диа- метром 0,5 — 1 лл, округлые, без краев. Из не- которых видов рочелл со времен средневековья добывали ценные красители (орсей, красный индиго, лакмус, французский пурпур и др.), которые использовали для окраски шерсти, шкур и шелка. Семейство опеграфовые (opegraphaceae) Семейство опеграфовых занимает промежу- точное положение между порядком артоние- вых и порядком графидовых лишайников. Пло- довые тела их — гастеротеции — вытяну~ой или овальной формы. В отличие от остальных ли- шайников порядка артониевых плодовые тела опеграфовых имеют край, образованный угли- сто-черным эксципулом. В семействе один род оиеграфа (Opegrapha), который объединяет накипные лишайники с чрезвычайно примитивно устроенным cлоеви- щем. На поверхности коры деревьев или на скалах они имеют вид беловатых, сероватых или желтоватых пятен, реже гладкой зерни- стой или ареолированной корочки. Ооычно слоевище не имеет дифференцированной струк- туры и состоит из разбросанных в беспорядке
Рис. 323. Рочелловые: 2 — Воссе11а fuciformis; F — Roccella fucoides 8— е а исо1 es; 8 — палисадный коровой слой ха а характерный для этих лишайников. нитчатых во о д рослей из рода трентеполия, окруженных гифами микоб ион Г та. астероте- б ции на слоевище развиваются как правило, в ольшом количестве. они овальн ьные или широ- коверетеновидные с узким или с 7 и слегка расши- реннь:м диском, с заметным собственным к об ~азов краем, зованным углисто-черным замкнутым экс- ципулом. Сумки булавовидные или удлиненные, с тонкими стенками, в них образуется по 8 спор. поры овальные или веретеновидные, попе- речно-многоклеточные бесцветнь 7 ые, в старости коричневые. Известно до 280 видов опеграф, растущих в различных климатических зонах, как в го ных р районах, так и на равнинах. Большое количество видов (45о) встречается в широколиственных лесах Голарктики. Обычно эти лишайники развиваются на коре деревьев, реже на скалах и еще реже на ФУ сине. гнилои древе- Характерный представитель рода — опеграфа токкокрайкая (Opegrapha diaphora 324, рис. ) — имеет гипофлеодное слоев ище, а- стущее на коре деревьев в виде гладкой или бугорчатой, тонкой, светло-серой или светло- коричневой корочки. Гастеротеции в изобилии раз росаны по всему слоевищу; они черные, ма- товые, овальные или удлиненные, длиной до ротециев узкий, щелевидный, у старых — расши- ренный в средней части, окруженный своеоб- разно загнутым краем. Этот вид широко распространен по всей Европе, на востоке доходит до Урала и Запад- ной и ири, а также встречается в Северной Африке (Алжире), в Северной и Южной АМе- рике. ПОРЯДОК ГРАФИДОВЫЕ (GRAPHIDALES Гастеротеции графидовых лишайников име- ют линейную форму и по виду напоминают чер- точки, простые или разветвленные линии. Пло- дущий диск гастеротециев узкий и углублен- ный, а край, который может быть собственным или слоевищным, сильно выст пающии, возвы- Ю-1 шающийся на д диском. Очень редко плодовые тела графидовых бывают в форме округлых апотециев, но диск этих апотециев все равно узкий и углубленный, окруженный сильно вы- ступающим краем. Тип развития плодовых тел аскогимениальный альный. ' умки унитуникатные, сумках образуется о~ 1 до 8 бесцветных или коричневых поперечно-многоклеточных или м- р р, имеющих линзовидные просветы альных спо клеток. Слоевища графидовых лишайников на- кипные, примитивные прик епл 7 яющиеся к су- страту сердцевинными гифами; они или сов- сем лишены ко ов рового слоя, или, реже, покрыты слаборазвитой корой. Фикобионтом их явля- ется чаще всего нитчатая зеленая водоросль трентеполия.
Рис. 324. Opegrapha diaphora: общий виц слоевища и гастеротециев. Рис. 325. Opegrapha diaphora: 1 — поперечный разрез через слоевище и гастеротеций (а— клетки коры дерева, б — слоевище, в — гастеротеций); 2 — сум- ка со; 3 — споры. Графидовые распространены преимуществен- но в тропических и субтропических странах. Они развиваются на коре деревьев, иногда на камнях, почве и листьях вечнозеленых расте- ний. В умеренных областях встречаются лишь немногие виды. Порядок содержит одно се- мейство. Семейство графидовые (Graphidaceae) Это семейство насчитывает около i000 видов. Распространение и богатство форм этих ли- шайников в тропиках дают основание думать, что это семейство тропического происхожде- ния. Предполагают тактике, что графидовые- очень древние представители растительного ми- ра; об этом свидетельствуют ископаемые наход- ки этих лишайников в слоях мезозойской эры. По-видимому, они были широко распростране- ны на земном шаре еще до оформления суще- ствующих ныне континентов и климатов. Все графидовые имеют накипные слоевища или в виде тонких корочек, или целиком погружен- ные в кору дерева. Хорошо различимыми их делают лишь плодовые тела, причудливо раз- рисовывающие гладкую кору дерева. В состав семейства входит ряд родов, пред- ставители которых отличаются друг от друга окраской и формой спор, количеством перего- родок в них, толщиной и формой эксципула, расположением на слоевище апотециев и други- ми признаками. Род графис (Graphis) — один из центральных; он насчитывает 300 видов, для которых харак- терны бесцветные вытянутые двух- или много- клеточные споры с линзовидными или округ- лыми просветами клеток. Гастеротеции у этих лишайников сильно удлиненные, простые или разветвленные, иногда образующие звездчатые группы или распределяющиеся на слоевище правильными параллельными рядами. Их диск, как правило, щелевидный, окаймленный толстым выступающим гладким или бороздча- тым краем. Почти все виды этого рода встре- чаются в тропических и субтропических стра- нах, в умеренные области заходит всего не- сколько видов. В Советском Союзе известны 5 видов, 3 из которых встречаются только в ле- сах Приморского края. Наиболее широко распространен в умерен- ных районах земного шара графис написан- ный (Graphis scripta, рис. 326, 1) — обычней- ший лишайник горных и равнинных широко- лиственных лесов. На коре деревьев его слое- вища имеют вид гладких беловато-серых или желтовато-сизых пятен, иногда слаборазличи- мых. Гастеротеции развиваются в большом ко- личестве. Длинные и узкие (длиной до 3 лл~ и шириной 0,1 — 0,4 лл), простые, прямые, дуго- видно-изогнутые, иногда волнистые и трехвиль- чато-разветвленные, они напоминают нанесен- ные на кору дерева сложные письмена, чем и объясняется название этого лишайника. Диск гастеротециев, имеющий вид щели или желоб- ка, окаймлен нависающим над ним высоким краем, образованным темно-коричневым экс- ципулом. Сумки широкобулавовидные, содер- жат по 8 спор. Этот лишайник предпочитает поселяться на гладкой коре лиственных деревьев, но иногда встречается и на хвойных породах, особенно на пихтах.
2 Рпс. 326. Graphis scripta: Он широко распространен в тропиках и суб- тропиках, в умеренных широтах северного и южного полушарий, в Голарктике, доходя до северной границы широколиственных лесов. ПОРЯДОК КРУГЛОПЛОДНЫЕ (CYCLOCARPALES) Круглоплодные лишайники образуют очень большой порядок, в состав которого входит 30 семейств. Их объединяет один общий приз- нак: плодовые тела этих лишайников — округ- лые апотеции, диск которых, как правило, ок- ружен хорошо развитым собственным или слое- вищным краем. К этой группе принадлежит огромное большинство хорошо заметных кустис- тых, листоватых и накипных лишайников, ши- роко распространенных по всему земному шару. Их слоевища могут быть гомеомерными и гете- ромерными, но у большинства видов они хо- рошо развитые, с ясно дифференцированной анатомической структурой. Фикобионтами круглоплодных бывают как сине-зеленые, так и зеленые водоросли, но наиболее широко рас- пространена одноклеточная зеленая водоросль требуксия. Для плодовых тел характерен аскогимени- альный тип онтогенеза, в гимениальном слое развиваются настоящие парафизы, и только у некоторых представителей — сетчато-сросшиеся парафизоиды. Сумки унитуникатные, тонко- слойные, в них образуется от 1 до 8 и более, иногда до 200 спор. Споры различного строе- ния, одноклеточные и иногоклеточные, бес- цветные и коричневые. При разграничении семейств принимают во внимание такие признаки, как строение апо- тециев, спор, характер фикобионта, анатоми- ческое и морфологическое строение слоевища и др. Семейство коллемовые (Coll emataceae) Эта группа объединяет слизистые лишайники с гомеомерным строением слоевища, их фико- бионт — нитчатая сине-зеленая водоросль но- сток. Листоватые или кустистые, реже накип- ные слоевища коллемовых в сухом состоянии имеют вид темных, черных, оливково-зеленых, темно-коричневых или голубовато-серых лом- ких корочек, пленок или пластинок, но во влажном состоянии они сильно набухают, ос- лизняются и значительно увеличиваются в раз- мерах. В слоевищах большинства этих лишай- ников нельзя обнару~кить никакой дифферен- цированной структуры; они образованы про- стым переплетением гиф микобионта и нитей водорослей, заключенных в большом количест- ве слизи, продуцируемой водорослями и гри- бом (рис. 297, 7), и только у некоторых на верх- 1 — общий вид слоевища и плодовык тел — гастеротециев; 2— гастеротеции в увеличенном виде; 8 — споры; 4 — поперечный срез через слоевище и гастеротеций (а — гимениальный слой, б — эксципул, в — гипотеций, г — слоевище, д — клетки коры дерева). ней и нижней поверхностях слоевища образу- ется крупноклеточный параплектенхимный ко- ровой слой (рис. 297, 8) или все слоевище со- стоит из параплектенхимной ткани. Апотеции чаще всего леканоров ого, реже биаторового или лецидеевого типа. В сумках образуется по 4 — 8 спор, которые могут быть одноклеточны- ми, поперечно-многоклеточными и муральными. Соредии никогда не образуются, но довольно часто развиваются изидии, которые могут быть бородавчатыми, зернистыми, чешуйчатыми и цилиндрически-кораллов идныи и.
Рис. 327. Collem а 1laccidum: общий вид слоевища. Коллемовые встречаются на скалах, порос- ших мхами, на поверхности известковых скал, на коре деревьев, на дернинках мхов, иногда на карбонатной почве. Они широко распростране- ны по всему земному шару, но наиболее богато представлены в тропической и субтропической областях. В состав семейства входит 7 родов, из кото- рых наиболее богаты видами и широко распро- странены два. Род коллежа (Collema) включает главным образом листоватые, очень редко накипные формы чрезвычайно простого гомеомер ного строения (рис. 297, 2). Слоевища у них темно- окрашенные, оливково-черные, зеленовато-ко- ричневые или грязно-зеленые, в форме круп- ных, диаметром до 10 сл, розеток с широкими ли- стоватыми или узкими, многократно рассечен- ными курчавыми лопастями. Слоевища при- крепляются к субстрату гифами нижней по- верхности, реже ризоидами или ризинами. Иногда лопасти бывают прямостоячими, при- поднимающимися и даже округло-коралловид- ными, и тогда слоевище имеет вид маленьких, оливково-черных или черных дернинок. Неред- ко вся поверхность слоевища покрыта крупно- зернистыми черными изидиями или пузыревид- ными вздутиями. Апотеции леканорового типа, с хорошо развитым слоевищным краем, как правило, окрашенные в красноватый, красно- вато-коричневый или рыжеватый цвет и хоро- шо заметные на слоевище. Споры развиваются по 8, редко по 4 в сумке, они бесцветные, двух- клеточные, поперечно-много клеточные или муральные. Род насчитывает около 80 видов, поселяю- щихся во влажных и сырых местообитаниях, на поросших мхами скалах и основаниях ста- рых деревьев, иногда на глинистых почвах, в местах с нарушенным травяным покровом. Некоторые виды предпочитают хорошо освещен- ные сухие места, развиваются на поверхности известковых скал и на карбонатной почве, не- которые являются типичными представителями напочвенных лишайников степей и полу- пустынь. Коллемы широко распространены по всему земному шару, они неизвестны лишь в субан- тарктических и антарктических районах и очень слабо представлены в Новой Зеландии. Особенно богаты видами тропические и суб- тропические районы, а в Голарктике — рай- он Средиземноморья и приокеанические рай- оны Европы и Северной Америки. Примером может служить коллема вялая (Со11ета flaccidum, рис. 327). Слоевище это- го лишайника крупнолистоватое, оно образует темно-зеленоватые или зеленовато-коричне- вые розетки, достигающие в диаметре 6 см и прикрепляющиеся к субстрату толстыми ко- роткими ризинами. Верхняя поверхность слое- вища неровная, складчато-волнистая, покры- тая пузыревидными вздутиями и черноватыми изидиями. Апотеции диаметром 0,5 — 1,5 лл развиваются довольно редко, с красновато- коричневым плоским диском, окруженным тон- ким зазубренным краем. Этот вид обитает на влажных скалах в тенистых местах и на ство- лах деревьев лиственных пород, как на рав- нинах, так и в горах, поднимаясь до высокогор- ного пояса. Встречается в Арктике, по всей Европе, на Кавказе, в Азии, на Филиппинских островах, в Северной Африке, Северной Америке, Авст- ралии и Новой Зеландии. Род лептогиул (Leptogium) объединяет ли- шайники, у которых на поверхности гомеомер- ного слоевища развивается I;pупноклеточный коровой слой, состоящий из одного или несколь- ких рядов клеток (рис. 297, 3). Иногда слоеви- ще целиком образовано крупно клеточной па- раплектенхимной тканью. Слоевища темно-зе- леные, оливково-коричневые или свинцово-се- рые, преимущественно листоватые, образую- щие крупные разрезаннолопастные розетки, мелколистоватые подушки и дернинки из ма- леньких чешуек или бородавочек, реже имеют вид маленьких кустиков, состоящих из мелких цилиндрических или плоских веточек. Апоте- ции леканорового типа, красно-коричневые, бурые или рыжие, сначала погруженные, позднее сидячие на слоевище. Споры у лепто- гиумов бесцветные, поперечно-много клеточные до муральных. Род насчитывает около 100 видов, широко распространенных по всему земному шару, особенно в тропических и субтропических стра- нах. В умеренных областях наибольшее видо- вое разнооб разие наблюдается в приатланти- ческих районах Европы. Обычно эти ли- шайники поселяются во влажных местооби- 448
таниях, на поросших мхами скалах, между дер- нинками мхов на карнизах и в щелях скал, на почве, богатой карбонатами, коре деревьев. Один из обычных видов — лептогиул (Lepto- gium saturninum, табл. 47, 6). Слоевище у этого лишайника крупнолистоватое, свинцово- серое или зеленовато-черное, верхняя поверх- ность более темная, по краям голая, в цен- тральной части густо покрытая зернистыми изидиями, нижняя поверхность светлая, с ко- роткими беловатыми ризоидами, образующими густой пушистый войлок. Лпотеции довольно редкие, достигающие в диаметре 0,5 — 2 лл, с красновато-коричневым диском, окруженным тонким собственным и толстым слоевищным краями. Они рассеяны по всей верхней поверх- ности слоевища. Споры эллипсоидные, сначала с тремя-че- тырьмя поперечными перегородками, в зрело- сти мур альные. Этот лишайник поселяется главным образом в горах при основании стволов деревьев на коре лиственных и хвойных пород, на порос- ших мхами скалах. Встречается в Лрктике, по всей Европе, на Кавказе, Урале, в Западной и Восточной Си- бири, на Алтае, в Саянах, на Дальнем Востоке и Камчатке, в Монголии, Китае и Индии, а также в Северной Лмерике, Гренландии и Новой Зеландии. Семейство пельтигеровые (Peltigeraceae) Семейство объединяет крупнолистоватые ли- шайники, главным образом напочвенные, ре- же развивающиеся на коре деревьев или поверх- ности скал. Их слоевища образуют крупные, выемчато-лопастные розетки или отдельные поднимающиеся среди мхов листовидные пла- стинки, в некоторых случаях редуцированные до небольших чешуек. Верхняя поверхность слоевища гладкая или войлочная, нижняя обыч- но пушисто-войлочная, нередко с разветв- ленной сетью жилок и хорошо заметными пуч- ками ризоидальных тяжей или ризин. Слоевище гетеромерное, с ясно дифференцированной ана- томической структурой, с верхней, а иногда и с нижней крупноклеточной корой. Фикобион- том могут быть одноклеточные зеленые и нит- чатые сине-зеленые водоросли. Апотеции раз- виваются на концах лопастей с верхней и ниж- ней стороны слоевища или бывают погружен- ными в небольшие ямчатые углубления в цент- ральной части слоевища. Характерно, что в начале своего развития апотеции обычно покры- ты тоненьким покрывальцем, образованным ко- ровым слоем слоев ища, позднее покрывал ьце растрескивается и обнажает коричневый диск. Споры бесцветные или светло-коричневые, эл- липсоидные или игловидные, с одной или не- сколькими поперечными перегородками, об- разуются по 2 — 8 или помногу в сумке. Род пельтигера (Peltigera) объединяет лишай- ники, отличающиеся развитием апотециев на концах лопастей с верхней стороны слоевища. Они часто встречаются под кронами деревьев во влажных тенистых, а также в светлых хвой- ных лесах и на открытых, хорошо освещенных пространствах. Слоевища у них обычно круп- ные, достигающие в диаметре 20 — 30 см, они рассечены на широкие, часто извилистые по краям лопасти, а иногда состоят из отдельных вытянутых пластинок шириной 3 — 5 сл, высту- пающих из мхов или травяного покрова своей верхней частью. У некоторых видов слоевища совсем небольшие, в виде листочков диаметром 1 — 2 см, редко до 5 см. Верхняя поверхность слоевища сероватая, серовато-коричневая или серовато-зеленая, при увлажнении иногда яр- ко-зеленая, гладкая, блестящая или матовая. Нижняя поверхность, как правило, хорошо отличается от верхней: она буроватого или черного цвета, по краям светлая, беловато-ро- зовая, иногда целиком светлая, войлочная, с сетью разветвленных выпуклых жилок, иног- да сливающихся в центральной части слоевища, и густым войлоком перепутанных толстых ри- зоидальных тяжей. Коровой слой на нижней поверхности слоевища не развивается, и на по- перечных разрезах можно различить только три анатомических слоя: крупно клеточный верхний коровой слой, образованный бесцвет- ными клетками, далее хорошо развитый слой водорослей и основную толщу слоевища, за- нятую хорошо развитой сердцевиной, обра- зованной слабо разветвленными вытянутыми бесцветными гифами, в нижней части слоевища нередко пигментированными. Обычно эти пиг- ментированные гифы выстилают нижнюю по- верхность слоевища, образуют жилки и пучки ризоидальных тяжей. Фикобионтом у большинства пельтигер яв- ляется нитчатая сине-зеленая водоросль нос- ток, нити которой в слоевищах лишайника рас- падаются на отдельные клетки. Однако у не- которых видов фикобионтом бывает однокле- точная зеленая водоросль коккомикса, в та- ких случаях на поверхности слоевища развива- ются еще особые образования — ц е ф а л о д и и, имеющие вид маленьких бугорков, пластино- чек или чешуек, внутри которых живут коло- нии сине-зеленой водоросли носток (табл. 42,8; рис. 328). Роль цефалодиев в жизни лишайников долгое время была неясной, их считали болез- ненными наростами на слоевищах. Однако позд- нее было замечено, что сине-зеленые водорос- ли, живущие в цефалодиях, фиксируют атмос- ферный азот, и лишь совсем недавно с помощью 449
Рис. 328. Peltigera aphthosa: 2— 1 — лопасть слоеви а — цефалодии в увеличенно ща с рассеянными на н ней цефалодиями; через цефалодий. енном виде 8— енно; — поперечный разрез меченых атомов азота было экспериментально доказано, что азот ~ик , фиксируемый в цефалодиях затем в основном погл 7 оглощается гифами гриба. довании был сделан а основании этих иссле вывод, что в анн д ом случае мы имеем п име симбиоза лишайникового г в одорослями — зеленой а го гриба сразу с дв мя У й, развивающейся в са- алодиях. щ, ине-зеленои, живущей мом слоевище и с в це- Ро п д пельтигера насчитывает 70 р распространенных по всем око ет видов, ши- ру, некоторые из них обитают всему земному ша- но на итают почти повсемест- чается в т опичес- , наи ольшее число вст-еч ких и субтропических раионах главным об а- зом южного полушария. ра- Один из а р спространенных видов — пелыии- 3; рис. 328 гера пупырчатая (Peltigera aphth б ) — имеет слоевища в виде широколопастных пластинок с п иде к пных и выступающими из мхов краями. Ве х- п и няя поверхность слоев ища а зеленовато-се ая ри увлажнении ярко-зелен р ло еная, усеянная це~а- диями в виде округлых, по к а сеченных б го к ~х, по краям часто рас- ами, по пе m~e и р ~ рии беловато-розовая, в центральной части серовато-б бу ой с то- урая до черно- р „сп ными путанными че- и ризоидальными тяжами. Лп вольно е ми. по теции до- р дкие, расположенные на тян тых у лопастеи, плоские или с а концах вы- лобок их и свернутые в же- Этот , их диаметр достигает 3 — 12 — ММ. тот лишайник встреча ется в Ев попе н Кавказе, в Азии Сев А, енлан- вернои Америке, Г енлан- дии, чаще всего в хвОЙных ых лесах на почве среди л мхов, на гниющих пнях в н лов и х, в нижней части ство- ~7 в и на скалах пове х мх хов. ~гельтигеры собачьей Р л., слоевища имеют вид крупных пла- ~/ по краям ям лопасти. е р кие, извилистые ви а се роняя поверхность сл щ рая или коричневато-се ая мат ое- тонковойлочная — розовато-белая, Я, НИЖНЯЯ вы а олее темная, с густой сетью д ющихся светлых жило ю сильно 1 к и с длинными о сл, светлыми или сле 7 гка уроватыми из- ида льными тяжами. Ап р зо- . и. потеции на концах за- уженных лопастей овальн ьные, красновато-ко- ричневые, шириной О 5 — 1 см 7 Этот вид встречается в Европе А С р зии, евер- е жнои Америке Г е нландии, ике на Канарских островах в А BcTp8лии, главным разом в мезофильных условиях, по к о очве в лиственных и смешанных лесах, на лугах, меж ле, . ду скалами в тенистых и влажных местах. Род еолорика (Solorina) включает ч ает лишаиники, по внешнему виду слоевищ очень на- поминают представителей рода пельтигер отличаются а игера, но р сположением апотециев. Слоеви- ща у них листоватые, гете оме омерные, с сетью жи- лок и ризоидами на нижней стороне. К пно- клеточный коровой слой а не. рупно- слой развивается не толь- ко на верхней сто оне н о, в отличие от пельти- гер, местами под апотециями и на н вища. " потеции образуются не краям лопастей а в я не по й, а в центральной части слоеви- ща, причем обычно они бываю ают погружены в не- большие ямчатые углубления. Спо ы дв хкле- том я является о е, по — в сумке. Фикобион- дноклеточная зеленая во о коккомикса, кото ая об доросль торая о разует под коровым сло- ем этих лишайников толс тыйслои,иног ап е ваемый тяжами тиф. Ни цевине, не ги . иже этого слоя, в серд- о ин сл у которых видов развива ется еще д ой из сине-зеленых во носток, представляю одорослей о р да сильно разросшие щии со ой вн т енни ся це алодии. ур ие Распространенный вид — солор ина ша ан- крупнолистоватые слоевищ б а, о разующие ок- руглые розетки диаметром 5 — 10 ные на широкие, по к аям к см, ассечен- о краям курчавые лопасти ириной — сл. Ве х ирин 1 — 2 . рхняя поверхность серо- вато- или светло-ко зеленею ая' ричневая, при увлажнен ии щ, нижняя поверхность красная, с е кими к кирпично- жилками и б ов с е кими красновато-коричневым ми имеют ви т ур атыми риз о идами. Апо потеции д темно-коричневых или черных ок- руглых пятен. Обитает на почве п почве, по склонам и м, на карнизах и в щелях скал. Встречается по всей Арктике в нах Ев опы е, в горных рай- андии. вропы, зии, Северной Америк Г и и рен-
Род нефрома (Nephroma) объединяет виды, характерной особенностью которых является образование апотециев на нижней стороне слое- вища на концах пальчатовытянутых лопастей. Это тоже листоватые лишайники, но, в отличие от представителей двух предыдущих родов, у них на нижней поверхности слоевища не об- разуется жилок, она гладкая, ворсисто-барха- тистая, с редкими ризоидами. Слоевище гете- ромерное, с Hpóïíîêëåòî÷íûì коровым слоем как с верхней, так и с нижней стороны. Споры бесцветные, веретеновидные, двух- — четырех- клеточные, развиваются по 8 в сумке. Многие представители эпифиты, но немало обитающих и на почве, и на заросших мхами скалах. Род насчитывает около 45 видов, широко распространенных на земном шаре, преиму- щественно в горах умеренной зоны. Пример — нефрома арктическая (Nephroma arcticum, табл. 49, Б). Слоевище этого лишай- ника широколистоватое, с округлыми лопа- стями, шириной до 6 см, сверху желтовато- зеленое или соломенно-желтое, снизу волок- нисто-пушистое, по краям светлое, в цент- ральной части черное. Апотеции крупные, ди- аметром до 2,5 сл, с красновато-коричневым диском. С возрастом лопасти слоевища могут поворачиваться, и тогда диск апотециев оказы- вается обращенным кверху. Нефрома арктическая растет на почве, на торфяниках, среди мхов, реже на мшистых ска- лах. Встречается по всей Арктике и высоко в горах Сибири, в Хабаровском крае, на Камчат- ке и Сахалине, а также в северных и горных районах Европы, Северной Америки, на Алеут- ских островах, в Гренландии. Семейство стиктовые (Stictaceae) Семейство стиктовых объединяет крупноли- стоватые лишайники, с высокодифференциро- ванной анатомической структурой. Их слое- вища, снабженные сверху и снизу крупноклеточ- ным коровым слоем, имеют вид оливковых, светло- или темно-коричневых крупных листо- видных пластин, причудливо вырезанных по краям. Они прикрепляются к суострату ооыч- но всей нижней поверхностью с помощью корот- ких нежных ризоидов и лишь по краям слегка приподнимаются, реже только боковой частью, в то время как остальная часть слоевища оста- ется свободной и свисающей. У одних предста- вителей поверхность слоевищ нередко бывает неровной, с характерным сетчатым рисунком— глубокими впадинами, ограниченными выдаю- щимися ребрами; у других такого сетчатого рисунка не образуется. Лпотеции возникают по краям лопастей или на их поверхности, обычно леканорового типа, с хорошо развитым слое- вищным краем. В сумках образуется по 8 спор, споры вытянутые веретеновидные, с одной или несколькими перегородками, бесцветные или коричневые. Кроме того, на слоевищах нередко развиваются соредии и изидии. Фикобионтом могут быть как одноклеточные зеленые водоросли, так и нитчатые сине-зеле- ные. В первом случае, как и у пельтигеровых, на поверхности или внутри слоевища разви- ваются еще и цефалодии, содержащие сине- зеленые водоросли. В состав семейства входит 3 рода, представи- тели которых растут главным образом на стволах и ветвях деревьев, реже на поверх- ности скал. Распространены стиктовые преиму- щественно в тропических, субтропических и умеренно теплых областях южного полушария, причем особенно много их в океанических районах. Род лобария (Lobaria) насчитывает около 70 видов, отличающихся крупнолистоватыми слоевищами с сетчато-складчатой поверхно- стью и распространенных главным образом п тропических, субтропических и умеренно теп- лых областях северного и южного полушарий. Особенно много видов в субтропических и тропических районах Тихого океана. В Голарк- тике наиболее богата лобариями флора Японии, Китая, советского Дальнего Востока, Тихооке- анского побережья Северной Лмерики, в Ев- ропе — приатлантических районов. Один из наиболее широко известных и рас- пространенных видов — лобария легочная (Lo- baria pulmonaria, табл. 47, 1, рис. 329). Слое- вище этого лишайника имеет вид широкой ли- стовидной пластинки, по краям округло вырезанной и образующей короткие и широкие лопасти. Обычно оно прикрепляется к коре дерева или поверхности скал краем, с помощью бокового гомфа; большая часть слоевища оста- ется свободной и свисает. Сверху слоевище светло- или оливково-коричневое, сетчато- складчатое, с глубокими впадинами, окружен- ными выступающими ребрами. Нижняя поверх- ность неровная, с беловатыми голыми выпук- лостями, соответствующими впадинам верх- ней поверхности, отделенными друг от друга темными участками, покрытыми короткими ри- зоидами. На верхней поверхности вдоль ребер обычно расположены крупные бугорчатые бе- лые сорали и изидии. Апотеции образуются ред- ко. Этот лишайник встречается в лесах по всей :Европе, на Кавказе, в Азии, Северной Америке, Африке, на Канарских островах и в Авст- ралии. Лобария легочная издавна известна в на- родной медицине. Когда-то из-за сходства ее внешнего облика с тканью легкого ее приме- няли при лечении легочных болезней. В настоя- 45i
~М 1 Рис. 329. Lobaria pulmonaria: с л е в а — верхняя поверхность слоевища (а — сорали); с п р а в а — нижняя поверхность слоевища. Семейство лецидеевые (Lecideaceae) щее время лишайник используют только в парфюмерной промышленности. Род стикта (Sticta) включает около 170 ви- дов, которые имеют коричневые листоватые слоевища, лишенные сетчатого рисунка свер- ху. На нижней ровной поверхности, которая бы- вает светло- или темно-коричневой, войлочно- пушистой или густо-реснитчатой, всегда раз- бросаны правильно округлые белые или ярко- желтые гладкие пятнышки — цифеллы, пред- ставляющие собой отверстия, через которые воздух проникает внутрь слоевища. Отверстия ведут в чашевидные углубления, дно которых выстлано округлыми клетками сердцевин- ных гиф (рис. 302). Окраска цифелл зависит от окраски сердцевины, которая у этих лишай- ников может быть бесцветной или лимонно- желтой. Соредии, нередко образующиеся по краям лопастей или по всей их поверхности, тоже могут быть лимонно-желтыми или белы- ми (табл. 46, вверху слева). Наибольшее видовое разнообразие стикт на- блюдается в умеренно теплых окраинах юж- ного полушария, особенно в районах с океа- ническим климатом, например в Новой Зелан- дии, в теплых и влажных районах Австралии, Тасмании и в океанических районах тропичес- кой, субтропической и умеренной Южной и Центральной Америки. В субтропики и уме- ренные области северного полушария заходит не более 25 видов, из них в умеренных обла- стях встречается всего 8 видов, один вид рас- тет в полярных районах. Пример — стикта Pauma (Sticta wrightii, табл. 47, 4). Слоевище этого лишайника имеет вид крупных, по краям выемчато-лопастных пластин. Верхняя поверхность слоевища пе- пельно-серая или светло-коричневая, гладкая, слегка блестящая; нижняя — светло-корич- невая, покрытая густым покровом реснитча- тых ризоидов, с белыми цифеллами между ними. На слоевище обычно образуются апо- теции, а соредии и изидии не развиваются. Апотеции в виде плоских коричневых округ- лых дисков диаметром до 8 лл. Этот лишайник растет на коре деревьев, ре- же на поверхности скал. Встречается доволь- но редко, главным образом в горах, известен в .Европе, Азии и Америке. Это одно из основных и довольно обшир- ных семейств лишайников, представителей его объединяет один общий признак: плодовые тела их — апотеции лецидеевого или биаторового типа. Край таких апотециев, называемый соб- ственным, никогда не содержит водорослей и образован хорошо развитым эксципулом, со- стоящим из продольно идущих, плотно прилега-
ющих друг к другу бесцветных или темноокра- шенных гиф. Это главным образом накипные лишайники, образующие на поверхности суб- страта тонкие или довольно толстые порошкова- тые, зернистые, бородавчатые или растрескав- шиеся ареолированные корочки (рис. 330). Сло- евище хотя и гетеромерное, но все же с прими- тивной анатомической структурой, не покры- тое коровым слоем или, редко, со слабораз- витой корой. Оно прикрепляется к субстрату гифами сердцевины или с помощью темного под- слоевища, нередко хорошо заметного между ареолками слоевища или по его краям в виде темной каймы (табл. 55). Лишь у представи- телей некоторых родов слоевище чешуйчатое или в виде маленьких листочков, иногда пузыревидно-вздутых, и тогда оно имеет более или менее хорошо развитый коровой слой и при- крепляется к субстрату либо сердцевинными гифами, либо довольно длинными корнепо- добными ризоидальными тяжами. Фикобион- том могут быть представители целого ряда ро- дов зеленых водорослей, как одноклеточных, так и нитчатых, — мирмеция, псевдохлорелла, требуксия, хлоросарцина, коккоботрис, плев- рококкус. Споры у лецидеевых разнообразного строе- ния, от эллипсоидных одно-двухклеточных до вытянутых поперечно-многоклеточных, иногда споры муральные, бесцветные и темноокра- шенные. Семейство содержит 12 родов, виды которых отличаются друг от друга главным образом формой и строением спор, строением апотециев и слоевища. Лецидеевые растут на скалах, ва- лунах, мелких камнях, почве, коре деревьев, мхах, растительных остатках, на слое вищах других лишайников. Они чрезвычайно широко распространены по всему земному шару, встре- чаются как на равнинах, так и в горных рай- онах, в полярных и высокогорных пустынях, в лесах, в степных и пустынных областях, в тро- пиках и субтропиках. Род лецидея (Lecidea) — центральный в се- мействе. Он насчитывает 970 видов, для кото- рых характерно накипное слоевище, черные, очень твердые лецидеевые апотеции и однокле- точные бесцветные эллипсоидные споры, раз- вивающиеся по 8 (редко по 16 — 32) в сумке. Слоевища имеют вид беловатых, сероватых, желтоватых или коричневатых тонких, зерни- сто-бугорчатых или довольно толстых ячеисто- потрескавшихся корочек. Разбросанные на них характерные углисто-черные, сидячие или, ре- же, погруженные апотеции позволяют уже по внешнему виду лишайника определить его при- надлежность к лецидеям. Однако в пределах рода эти лишайники очень трудно идентифи- цировать. Многочисленные виды, входящие в его состав, отличаются друг от друга главным образом строением апотециев, размерами и окраской их анатомических слоев, формой и величиной спор, толщиной парафиз и другими подобными признаками. При изучении под микроскопом поперечных срезов видно, что внешне однообразно окра- шенные углисто-черные апотеции имеют чрез- вычайно разнообразную окраску анатомичес- ких слоев. Так, верхняя часть гимениального слоя может быть окрашенной в медно-зеленый, изумрудно-синий, зеленовато-черный, бурый, желто- и черно-бурый цвета. Гипотеций быва- ет бесцветным, розоватым, желтовато-коричне- вым, темно-коричневым, рыжим, красным и др. Разнообразна окраска эксципула: синевато- зеленая, черно-зеленая, красно-коричневая, коричневая, часто с фиолетовым оттенком. Каж- дый вид имеет совершенно определенную окрас- ку этих слоев. В качестве субстрата большинство видов предпочитает гранитные скалы, валуны, из- вестняки, доломиты, но нередко они встреча- ются на коре лиственных и хвойных пород, на гниющей обнаженной древесине, на поверх- ности бесплодных почв, на растительных ос- татках, дерновинках мхов, на слоевищах дру- гих лишайников. Наиболее богаты видами об- ласти Голарктики (74' всех известных видов), особенно арктические и горные районы. Око- ло 9 о видов — аборигены палеотропических областей. Неотропические области заметно бед- нее, здесь число видов не превышает 6,2о. Богата эндемичными видами флора Антарк- тики (5,5 ), здесь они встречаются в край- них условиях — всего в 390 км от Южного полюса. Австралийская флора насчитывает око- ло 20 видов, что составляет 2,3о. Бедна вида-' ми капская флора. Один из наиболее широко распространен- ных видов — лецидея бор одавчатиая (Lecid еа glomerulosa, рис. 330). Слоевище этого лишай- ника в виде беловатой, сероватой или серовато- оливковой бугорчатой корочки обычно можно встретить на коре осины, ольхи, тополя и других лиственных пород. На слоевище всегда развиты многочисленные апотеции, достигаю- щие в диаметре 0,3 — 1 мм, черные, голые, сна- чала плоские, с хорошо заметным собственным краем, в зрелости выпуклые, с исчезнувшим краем. Этот вид встречается почти повсеместно в лесах .Европы, Лзии, Северной Лмерики, а также в Гренландии, Северной Африке, на Канарских островах. Чаще всего развивается на коре лиственных пород, реже на хвойных породах и обнаженной древесине, вблизи че- ловеческого жилья (на заборах, старых брев- нах колодцев, сараев). 453
Рис. 330. Lecidea glomerulosa: внешний ви сл с апотециями. . внешнии вид слоевища Род биатора (Biatora) очень близок к преды- дущему роду и нередко считается его под о- дом, так как его представители отличаются от лецидей только строением апотециев. Апо- теции биаторового типа, не углисто-черные и твердые как р, у лецидей, а светлоокрашенные, мягкой консистенции, во влажном состоянии подобные расплавленному воску. Остальные признаки совпадают: это также накипные ли- шайники и в сумках у них как у ц 7 и у леци ей, образуется по 8 небольших эллипсоидных од- ноклеточных бесцветных спор. видов, широко Род насчитывает более 500 видов распространенных по всему земном Более 300 видов (около 60о/о) ограничены в ному шару. своем распространении Голарктикой. В палео- тропических областях известно 90 дов ~18о~' дов ( ~o). Значительно беднее неотропические области з есь д встречается приблизительно 56 видов этого рода (11о/о). Австралийская фло- ра насчитывает 26 аборигенных видов (5,7о~о). Капская и антарктическая флоры имеют каж- дая по 10 эндемичных видов. Выделяются по числу видов флоры Новой Зеландии и Японии Виды биаторы растут на различных субстра- тах, но предпочитают поселяться на ко е е- р, гнилых пнях и другой гниющей или евьев обгорелой древесине на то ФЯ 1 и гумус о %Л р се же нередко, особенно в горах, они азви- ваются и х, они разви- горных на поверхности силикатных пород, андезитах, кварцитах, гнейсах няках. ах, извест- О ин дин из очень широко встречающихся в ле- сах северного полушария видов — б — УШИО~7Я болотная (Biatora uliginosa). Слоевище этого II оливково- лишайника имеет вид оливково оо разнои урой или серо-коричневой порошкооб ии на слое- или зернистой корочки. Апотеции вище развиваются в большом количестве. Они достигают в диаметре 0 4 — 0 7 м — мм, окрашены в бурый или буро-черный цвет, молодые с плоским диском, окруженным тонким собствен- ным краем, старые выпуклые б ез краев. а- стет в хвойных, реже лиственных лесах, на болотах, на торфяной и гумусовой почве, на слабо закрепленных песках, на гнилых пнях, поваленных стволах деревьев Р аспространен этот вид в равнинных и горных лесах Европы, Северной Азии и Северной Америки. Род псора (Psora) очень близок к двум преды- дущим благодаря аналогичному строению апо- тециев и спо р, но отличается строением слое- вищ. Слоевища состоят здесь из коричневых, розо, ирпично-HpBcHbIx, рыжеватых, пе- озовых ки ек, раз ро- пельно-серых или оливковых чеш ек, б санных на поверхности почвы или аст близко г к и растущих друг к другу и образующих корочку. Иногда они прикрепляются к субстрату боко- вой стороной, а остальная часть чешуек оста- ется приподнятой, и тогда чешуйки налегают друг на друга как черепицы крыши. По анато- мическому строению слоевище псор более слож- Ю-1 отдельные слои, сверху оно покрыто пара- плектенхимным коровым слоем, на нижней сто- роне нередко образуются ризоидальные тяжи, длина которых достигает 0,5 см. Иногда че- шуйки прикрепляются к субстрату гифами сердцевины, преимущественно в центральной части. Род насчитывает более 100 видов, из них око- ло 65 встречается в Голарктике. Многие виды— типичные представители напочвенных ли- шайников, в степях и полупустынях они яв- ляются неотъемлемыми компонентами напоч- венной лишайниковой флоры. Некоторые из них встречаются и в холодных пустынях Арк- тики, и в альпийском поясе гор, и в таежных лесах. Палеотропические и неотропические об- ласти бедны этими лишайниками: для первых известно 13 видов, для вторых — 16. Псора обманчивая (Psora decipiens, табл. 46 1 внизу справа) — характерный представитель рода — имеет вид округлых чешуек диаметром 1 — 8мм а , р стущих на почве. Сверху чешуйки 454
кирпично- или коричневато-красные, иног- да с голубовато-белым налетом, часто с белой каимои по краям. Снизу чешуйки пепельно-се- рые с многочисленными ризоидами. Лпотеции достигают в диаметре 0,8 — 1,8 мм, окрашены в черный или буро-черный цвет, располага- ются по краям чешуек. Этот лишайник обитает обычно на карбонат- ной почве. Он очень характерен для флоры лишайников равнинных и горных степей, но встречается также в Арктике и в высокогор- ном поясе. Распространен в арктических об- ластях и горах Европы, на Кавказе, в Средней Азии, на Алтае, в Прибайкалье, Саянах, Якутии, Монголии, Китае, Северной Америке, Гренландии, Африке, Новой Зеландии. Псора лестничная (Psora scalaris, табл. 46, вни- зу слева) образует пепельно- или зеленовато- серые чешуйки диаметром 0,8 — 2 мм, обычно приподнимающиеся и покрывающие друг друга зазуоренными краями. С нижней стороны че- шуйки светлые, покрытые белыми соредиями. Лпотеции развиваются довольно редко, они достигают в диаметре 0,4 — 2 лл, выпуклые, черные, нередко покрытые беловатым налетом, с тонким, извилистым собственным краем. этот лишайник встречается в светлых сос- новых лесах и на открытых местах на коре со- сен, особенно часто на обуглившихся после пожара местах, а также на старых гнилых и горелых пнях и другой гниющей древесине. Распространение вида связано главным обра- зом с зоной хвойных лесов, изредка он захо- дит и в лесотундру. Распространен по всей Европе, в северной части Азии, в Северной и Центральной Америке. Род ба~идия (Bacidia) включает лишайники, для которых характерны многоклеточные спо- ры веретеновидной, шиловидной или нитевид- ной формы, с одной или несколькими попе- речными перегородками. Слоевище у бацидий накипное, чаще всего в виде зернисто-бородав- чатой или порошковатой беловатой, сероватой, зеленовато-серой или оливково-коричневой ко- рочки, иногда гипофлеодное или эндолитное. Оно имеет примитивное анатомическое строе- ние, лишено корового слоя и прикрепляется к субстрату гифами сердцевины или подслоеви- ща. Лпотеции обычно биаторового строения, чаще всего красновато-коричневые, розовато- желтые, светло- или буровато-рыжие, воско- видные, реже темные, буровато- или зелено- вато-черные. Род насчитывает более 600 видов, причем более половины из них встречается в неотро- пических и палеотропических областях. В Гол- арктике бацидии распространены главным об- разом в умеренно теплых областях; в других районах число видов резко сокращается, а в Рис. 331. Bacidia, muscorum: 1 — общий вид слоевища с апотециями (а), развившегося на веточках мха; 2 — споры. Арктику заходят только 3 вида. В Новой Зе- ландии известно более 40, а в Субантарктике— 12 эндемичных видов, в то же время флоры Австралии, Капской области и Антарктики бедны представителями этого рода. Излюблен- ный субстрат бацидий — кора деревьев, раз- рушающиеся дернинки мхов, растительные ос- татки, а также гниющая древесина и слоевища других лишайников, реже они растут на ка- менистых субстратах, силикатных и известко- вых скалах, на поверхности почвы. Существу- ет также группа эпифилльных видов, развива- ющихся на листьях вечнозеленых растений и на хвое. Характерный пример — бацидия моховая (Bacidia muscorum, рис. 331), имеющая вид беловатой, беловато-зеленоватой или серовато- оливковой бугорчато-бородавчатой или зер- нистой корочки, развивающейся на веточках мхов. Лпотеции разбросаны по всему слоеви- щу, диаметром они 0,3 — 1,5 мм, буро-черные, молодые — с плоским диском, окруженным тонким собственным краем, позднее диск ста- новится сильновыпуклым, а край исчезает. Этот лишайник растет на мхах, на богатой кар- бонатами почве, реже на заросших мхами ска- лах. Встречается в лесной зоне, на юге в горах, нередко в степях и полупустынях, но захо- дит также в Арктику. Вид распространен по всей .Европе, в Северной Азии, Северной Аме- рике, Гренландии, Центральной Америке. Род ризокарпон (Rhizocarpon), объединяет лишайники с довольно крупными, двух- — четы- рехклеточными, чаще мур альными, бесцвет-
1 1 ! t I I l с Рис. 332. Rhizocarpon tinei: 1 — апотеций лецидеевого строения (а), расположенный между бугорками слоевища (6) прямо на черном подслоевище (в); 2— сумка со; 8 — муральная спора с толстой внешней бесцветной оболочкой. ными или коричневыми спорами с очень толс- той бесцветной внешней оболочкой студенистой консистенции. Слоевище у них в виде зелено- вато-желтой, сероватой или серовато-коричне- вой ареолированной корочки, развивающейся на черном подслоевище, хорошо заметном на периферии и ме~кду ареолками (табл. 42, 1). Лпотеции черные, лецидеевого типа, развиваю- щиеся обычно между бугорками слоевища не- посредственно на темном подслоевище. Род насчитывает более 150 широко распрост- раненных видов, из них свыше 120 встреча- ются в северном полушарии. Наиболее обиль- но они представлены в Лрктике и в горных районах, меньшее их количество обитает в рав- нинных условиях. В тропических и субтропи- ческих областях известно всего 18 видов, которые в основном встречаются высоко в го- рах. Довольно богата видами флора Антарк- тики — здесь ~кивет 11 видов. Ризокарпоны растут, как правило, на гранитных горных по- родах и только в виде исключения — на из- вестняках или коре деревьев. Они развиваются на поверхности скал и валунов, на мелких кам- нях, покрывающих каменистые почвы в высо- когорных и полярных пустынях. В Арктике и высокогорьях ризокарпоны являются одной из доминирующих групп наскальных лишай- ников, и в этих районах нередко можно на- блюдать, как поверхность скал приобретает зеленоватую или желтоватую окраску благо- даря обильному развитию на них желтоватых слоевищ лишайников этого рода. Обычно они предпочитают поселяться на хорошо освещен- ных солнцем скалах, открытых ветрам, на ко- торых зимой не образуется постоянного снеж- ного покрова. Типичный представитель рода — ризокарпон окрашенный (Rhizocarpon tinei, табл. 51, внизу справа; рис. 332) на поверхности скал имеет вид ярко-желтых, беловато-желтых или зеленовато- желтых пятен, достигающих в диаметре 1— 12 сл. Слоевище состоит из угловатых, плоских или выпуклых ареолок диаметром 2 — 3 лл~, плотно прижатых друг к другу и тогда образу- ющих непрерывную корочку или разбросан- ных на хорошо развитом черном подслоевище, заметном по краям и между ареолками. Апо- теции углисто-черные, достигающие в диамет- ре 0,3 — 1,5 лл, угловатые или округлые, плос- кие. Вид широко распространен в Лрктике и Лнтарктике, в умеренной зоне северного и юж- ного полушарий и высоко в горах тропических и субтропических областей. В умеренной зоне растет как высоко в горах, так и ниже альпий- ского пояса, а также на равнинах, в лесах бореальной зоны, в средиземноморских рай- онах, в океанической части Европы. Семейство кладониевые (Cladoniaceae) К этому семейству принадлежит большой и широко известный род кладония (Cladonia) и некоторые более мелкие роды — глоссодиум (Glossodium), пикнотелия (Pycnothelia) и др. Род кладокия (Cladonia) характеризуется разделением слоевища на две части — первич- ное и вторичное слоевище. Первичное слое- вище состоит из чешуек разной формы и вели- чины (от 1 до 30 лл), покрывающих субстрат (почву, кору, древесину или др.). Иногда они образуют плотный покров или же по мере ста- рения лишайника редеют или совсем исчезают. Из первичного горизонтального слоевища вы- растает вторичное вертикальное слоевище- подеции. Подеции бывают весьма разнообраз- ной формы — простые палочковидные, простые со сцифообразными расширениями, разрежен- но или густо ветвящиеся и пр. Внутри они имеют полость, а снаружи покрыты коровым слоем из густо сплетенных гиф, иногда коровой слой отсутствует. Стенки подециев образованы сердцевиной и содержат водорослевый слой.
На них часто образуются соредии и изидии. Высота подециев у большинства видов 2 — 8 см, редко до 20 см. Подеции нередко покрыты че- шуйками — ф и л л о к л а д и я м и, которые уве- личивают их ассимиляционную поверхность. На подециях (в концах, по краям сциф, редко на стенках) встречаются плодовые тела — апо- теции. Они лецидеевого типа (без слоевищного края), окрашены в коричневый, светло-бурый или красный цвет. В сумках по 8 одноклеточ- ных (редко четырехклеточных) бесцветных спор. Пикнидии расположены на чешуйках первичного слоевища или на подециях. Фико- бионт обычно зеленая одноклеточная требуксия. В химическом отношении кладонии — один из наиболее сложных и пестрых родов среди лишайников. Они содержат десятки разных лишайниковых веществ, причем морфологи- чески единый вид может в разных частях свое- го ареала содержать различные вещества или их комбинации («химические расы», или «хи- мические виды»). Многие из лишайниковых веществ встречаются в определенных видах весьма постоянно и являются для них сущест- венными систематическими признаками. По- этому в современной систематике кладоний нель- зя обойтись без химического анализа гербар- ного материала или по крайней мере без при- менения реактивов, которые, реагируя с ли- шайниковыми веществами, дают определен- ную окраску коры или сердцевины. слоевища. Кладонии чрезвычайно широко распростра- нены во всех растительно-климатических зо- нах, от полярных пустынь до тропиков. Ви- довое разнообразие их также велико. В настоя- щее время известно более 300 видов кладоний, не говоря о сотнях и сотнях внутривидовых единиц — разновидностях, формах. В СССР известно почти 100 видов кладоний. Больше всего их встречается в тундрах, лесотундрах и в зоне хвойных лесов, но их можно найти и в высокогорьях, и даже в сухих степях и пус- 'тынях. В хозяйстве человека самое важное место занимают виды кладоний, издавна называ- емые в народе «оленьи мхи» и «ягели». Эта группа лишайников объединяет более 40 ви- дов, из них в СССР произрастает 12 видов. У видов этой группы (их выделяют в особый подрод (Cladina) первичное корковидное слое- вище по мере роста лишайника скоро исчезает, а вторичное слоевище (подеции) сильно раз- ветвляется и имеет вид кустиков. Поверхность подециев здесь паутинисто-войлочная, почти совсем без корового слоя, без соредий и филло- кладий. Подеции в нижней части постепенно отмирают, а наверху продолжают расти и до- стигают в отдельных случаях в высоту до 20 сл (обычно 5 — 10 см). Многие виды этой группы весьма широко распространены, особенно в уме- ренных и северных зонах, и растут на различ- ных субстратах, чаще всего на земле, но встре- чаются и на древесине, мхах, скалах и пр. Они дают большую биомассу (нередко 10 — 15 и/га) и служат ценнейшим кормом для северных оленей. Кроме того, они находят применение как сырье для получения антибиотически дей- ствующих веществ. Некоторые из них — кладо- ния альпийская (Cladonia alpestris, табл. 48, 6; 56, вверху слева), лесная (Cl. sylvatica), непри- глаженная (Cl. impexa), мягкая (Cl. mitis) и др. — всегда содержат усниновую кислоту, являющуюся материалом для получения ле- карственных препаратов (<Бинан», (<Усно» и др. Из видов этой группы особенно широко рас- пространены кладонии альпийская, лесная, мягкая и оленья (Cl. rangiferina, табл. 56, вверху слева). Кладония альпийская образует белова- то-зеленые или желтоватые куполовидные кус- тики высотой до 20 см; подеции ветвятся дихото- мически, верхушечные веточки прямостоящие; растет в сосновых борах, тундрах, на кочках на верховых болотах. Кладония лесная образует более разрежен- ные по сравнению с предыдущим видом жел- товато-белые дернинк~ высотой до 10 сл, по- деции дихотомически-разветвленные, верхние веточки поникающие, коричневые на концах; растет в сосновых лесах, в тундрах, на старых крышах и т. д. Кладония мягкая по внешнему виду очень похожа на кладонию лесную, но надежно от- личается от нее реакцией на парафенилендиа- мин: у кладонии лесной под действием этого реактива кора окрашивается в красный цвет и, кроме того, ее слоевище на вкус горькое, а у кладонии мягкой парафенилендиамин реак- ции не дает и слоевище не горькое. Кладония мягкая более светолюбивый вид, растет в от- крытых местах в сосновых лесах, в тундрах, на верховых болотах, на замшелых скалах. Кладония оленья хорошо отличается от дру- гих видов этой группы пепельно-серым цветом слоевища и поникающими в одну сторону ко- ричневыми конечными веточками; подеции у нее высотой до 15 см, образуют раскидистые кусти- ки. Растет в сосновых лесах, в тундрах, на вер- ховых болотах, на старых крышах и пнях. Интересную внутриродовую группу образу- ют виды кладоний, имеющие красные апотеции (секция Cocciferae), окраска которых обуслов- лена присутствием родокладониевой кислоты. Принадлежащая к этой группе кладония бес- форменная (Cl. deformis, табл. 48, 4) имеет простые, высотой до 5 см сернисто-желтые по- деции, кончающиеся сцифообразными расши- рениями. Поверхность ее подециев покрыта налетом из мучнистых соредий. Вид содержит 457
458 Рис. 333. Umbilicaria esculenta. в большом количестве (до 8') усниновую ки- слоту и поэтому имеет большое значение как сырье для изготовления антибиотиков. Этот лишайник растет особенно часто на торфянистой почве, но встречается и в сосновых борах, на пнях, у подножия стволов сосен и т. д. Кладо- ния Флёрке (С1. floerkeana) также несет на ко- ротких (до 3 см) разветвленных подециях кра- сивые красные апотеции. Растет на песчаной почве, на гниющей древесине (табл. 48, 2). Род тамнолия (Thamnolia) в последнее вре- мя тоже включают в семейство кладониевых. Раньше его представители считались «несо- вершенными лишайниками», и их относили к семейству уснеевых. Хотя плодовые тела у тамнолии до сих пор не найдены, точными химическими исследованиями установлено род- ство этой группы с кладониевыми. У тамнолии червеобразной (Thamnolia ver- micularis) слоевище состоит из подециевидных белых палочек длиной до 15 см, лежащих на почве, мхах и других лишайниках. «Палочки» внутри полые, к концам заостренные. Каждый, кто внимательно присматривался к раститель- ности тундр или высоко горий, несомненно, замечал эти странные белые образования, иног- да скопляющиеся в густые куртинки (табл. 48, 8; 51, вверху слева). Семейство стереокауловые (Stereocaulaceae) Слоевище стереокауловых, как и кладоние- вых, состоит из двух частей: первичного слое- вища, имеющего вид зернисто-бугорчатой илд мелкочешуйчатой корочки, и вторичного слоеви- ща — псевдоподециев, которые, однако, не по- лые, как подеции у кладониевых, а заполнены внутри толстостенными гифамп. Подеции про- стые или ветвящиеся, они образуют кустики, покрытые филлокладиями разнообразной фор- мы. Очень часто на подециях встречаются це- фалодии в виде светлых или темных головча- то-бугорчатых выростов, содержащих сине-зеле- ные водоросли. Апотеции коричневые или чер- ные, лецидеевого типа. Сумки содержат 6 — 8 cIIop. Споры от четырех- до многоклеточных. Стереокауловые содержат несколько постоянно встречающихся лишайниковых веществ. Род стереокаулон (Stereocaulon) объединяет 80 видов, из них в СССР обнаружено 20. Одним из наиболее распространенных является стерео- каулон войлочный (Stereocaulon tomentosum, табл. 49, 2). У него первичное слоевище почти всегда незаметно. Псевдоподеции густо развет- вленные, образуют беловато-серые дерновинки высотой до 5 см. Растет на песчаной почве на дюнах, в верещатниках, в сосновых лесах. Семейство умбиликариевые (Umhilicariaceae) Это небольшое, но чрезвычайно интересное семейство, очевидно, очень древнее и не имею- щее близких родственных групп. Оно объеди- няет лишайники, слоев ище которых образует листоватые пластинки, иногда довольно боль- шие (диаметром до 20 см), прикрепляющиеся к субстрату при помощи центрального гомфа. Пластинки покрыты с обеих сторон коровым слоем разной окраски, от серовато-белой до черноватой, но никогда не желтой и не зеленой. Нижняя сторона слоевища голая или снабже- на ризинами, которые, однако, не служат для прикрепления слоевища к суб страту. Апоте- ции построены своеобразно, они лецидеев ого типа, но именуются гирофороидными, так как снабжены особыми орнаментальными выроста- ми разной формы, напоминающими извилины головного мозга — гирусы. Парафизы простые, несколько утолщенные. Споры одноклеточные, реже муральные, бесцветные или буроватые. Фикобионтом является одноклеточная зеленая водоросль требуксия. В семейство входит только один род.
Род умбиликария (Umbilicaria) содержит 50 видов, экологической особенностью которых яв- ляется их кальцефобность — они никогда не растут на известковых субстратах. Большинст- во видов встречается на скалах в тундрах, ле- сотундрах и высокогорьях, иногда они растут на камнях и скалах в лесной зоне. Примером может служить умбиликария Мю- ленберга (Umbilicaria muehlenbergii, табл. 42, б). В Японии на скалах встречается в массе умбиликария съедобная (U. esculenta, рис. 333), которую местное население использует в пищу и считает деликатесом. В аркто альпийской зоне обычна умбиликар ия короткошерстная (U. vellea). Семейство акароспоровые (Acarosporaceae) Это семейство объединяет лишайники, имею- щие накипное или чешуйчатое слоевище, и лишь у немногих видов оно листоватое. Сло- евище прикрепляется к субстрату при помощи гиф сердцевинного слоя, подслоевища или гом- фа (у листоватых). Апотеции леканорового или лецидеевого типа, часто погруженные в слое- вище или сидячие. Парафизы простые или раз- ветвленные. Сумки содержат много мелких, одно- или двухклеточных бесцветных спор. Семейство включает 9 родов, из которых наи- более богат видами один. Род акароспора (Acarospora) объединяет 300 видов, характерным признаком которых явля- ется строение слоевища, состоящего из обособ- ленных или сближенных чешуек, несущих апотеции. Иногда слоевище по периферии с ло- пастями или образует сплошную потрескав- шуюся корочку. К субстрату слоевище прикреп- ляется при помощи гиф сердцевинного слоя и подслоевища. Коровой слой всегда хорошо раз- витый, параплектенхимный. Лпотеции лекано- ров ого типа, одиночные или по нескольку вместе, часто погруженные в слоевище. Пара- физы простые, членистые. Сумки вздутые, со- держат множество небольших бесцветных одно- клеточных спор. Интересно, что а к ар оспоры никогда не образуют ни соредий, ни изидий. Виды этого рода широко распространены в нескольких растительно-климатических зонах, от тундр до пустынь. Они обитают как на сили- катных, так и на известковых скалах, на слан- цах, песчаниках, гипсах, солонцеватых поч- вах, на других лишайниках и очень редко на древесине и мхах. В северных областях и высокогорьях встре- чается интересная красновато- или темно-ко- ричневая Acarospora molybdina (табл. 44). Она образует округлые розетки, состоящие в центре из толстой корочки, а по периферии — из уз- ких лопастей. В степях и пустынях на солон- цеватых почвах растет акароспора реагирующая (А. reagens, табл. 45). Там она образует толстые беловато-серые корочки, состоящие из чешуек, покрытых густым белым порошковатым на- летом. Верхний коровой слой ее слоевища при действии едкого кали окрашивается в ржаво- красный цвет (отсюда и название «реагирую- щая»). Другой обычный обитатель солонцева- тых почв — акароспора Шлейхера (А. schlei- cheri) — образует зеленовато-желтые розетки, состоящие из чешуек. Семейство пертузариевые (Pertusariaceae) 3ТО семейство объединяет лишайники, слое- вище у которых накипное, с верхним коровьи| слоем обычно белого или беловато-серого цве- та, очень часто с соралями и изидиями. Лпоте- ции часто отсутствуют, но если они есть, то бывают погружены в слоевище, развиваются в особых плодовых бородавочках. Су-мки с 1— 8 одно- или двухклеточными спорами. Семейство объединяет 4 рода, из которых только один содержит большое число видов. Род пертузария (Pertusaria) объединяет 250 видов, развивающихся на различных субстра- тах, чаще всего на коре деревьев, а также на древесине, скалах, почве, мхах и т. д. Слоеви- ще накипное, прикрепляющееся к субстрату при помощи гиф сердцевинного слоя или под- слоевища, обычно белое или сероватое, часто с изидиями, которые, обламываясь, образуют скопления соредий — сорали. Лпотеции, если они встречаются, развиваются по одному или помногу в особых плодовых бородавочках или погружены в слоевище. Диск апотеция часто лишь точковидный. Парафизы разветвленные, переплетенные. Сумки большие, с 1 — 8 однокле- точными крупными спорами. Фикобионт — плев- рококкус. Виды этого рода содержат большое количество различных лишайниковых веществ. Пертузарии распространены в Лрктике и умеренных зонах северного полушария, осо- бенно много их в Средней Европе и в тундрах. Один из самых обыкновенных видов — nepmy- зария горькая (Pertusaria amara). Это светло- или темно-серый лишайник с белыми сораля- ми, растущий повсеместно на коре деревьев в лиственных и смешанных лесах. Его легко узнать на вкус — он чрезвычайно горький, благодаря содержанию пикролихениновой кис- лоты. В тундрах и лесотундрах, а также в высокогорьях растет на мхах и почве перту- зария пальчатая (Р. dactylina). У нее очень тонкое белое слоевище, из которого выра- стают многочисленные цилиндрические изи- дии высотой 1 — 4 мм. На лиственных поро- дах очень часто встречается пертузарил шариконосная (Р. globulifera, табл. 54), обра- 459
зующая на, коре деревьев большие (диамет- ром до 25 см), зеленовато-серые пятна, окайм- ленные темными и светлыми концентрически- ми зонами. Семейство леканоровые (Lecanoraceae) Леканоровые — одно из наиболее крупных семейств среди лишайников, содержащее более 1О родов с сотнями видов. Слоевище у лекано- ровых накипное или по краям лопастное, с хо- рошо развитым верхним коровым слоем. Апо- теции обычно округлые, сидячие или погружен- ные в слоевище, окруженные слоевищным кра- ем. Сумки с 8 — 32 одноклеточными или попе- речно двух- — многоклеточными спорами. Фико- бионт — типа протококкус. Род леканора (Lecanora) содержит около 400 видов, которые по анатомо-морфологическо- му строению довольно однообразны. Признаки, служащие для их различения, мелкие, часто малозаметные, поэтому идентификация видов сложна. Слоевище накипное, сплошное или ареолированное, редко чешуйчатое, обыкно- венно с хорошо развитым верхним коровым слоем. Лпотеции сидячие на поверхности. Сум- ки с 8 (реже с 16 — 32) одноклеточными бесцвет- ными спорами. Химически виды разнообразны и содержат многие лишайников ые вещества. Распространены эти лишайники очень широ- ко. В Карпатах, на Кавказе и в Крыму на глад- кой коре буков и грабов часто встречается леканора оголенная (Lecanora glabrata, табл. 53), образующая тонкое голое светло-серое с желто- ватым оттенком слоевище, несущее округлые, темно-бурые апотеции. В сухих местообитани- ях на силикатных скалах от Арктики до высо- когорий на юге встречается леканора обломоч- ная (L. frustulosa, табл. 43) с толстым бугорчато- бородавчатым желтоватым слоевищем, несу- щим крупные (диаметром 1 — 3 мм) блестящие коричневые апотеции. Леканора толстогубая (L. pachycheilea, табл. 54) — азиатский вид, встречающийся у нас на Дальнем Востоке на гладкой коре лиственных пород. У нее морщи- нисто-бородавчатое беловатое слоевище, позд- нее бородавки разрушаются и образуют соре- диозный лепр озный покров, среди которого находятся многочисленные коричнево-крас- пые апотеции. Широко распространенный лито- фильный вид — леканора скальная (L.rupicola, табл. 43), образующая на силикатных гор- ных породах довольно крупные, толстые, тре- щиновато-ареолированные серые слоевища с немного углубленными в них выпуклыми апо- тециями, покрытыми сизым налетом. На коре разных лиственных пород очень часто встре- чается леканора коричневая (L. subfuscata, табл. 53). Слоевище у нее тонкое, серое, огра- ниченное черной линией подслоевища, с рыже- вато-бурыми апотециями. Все эти виды встре- чаются в основном в естественных, не трону- тых человеком местообитаниях, но есть и такие, которые предпочитают культурные ландшафты. Самой интересной из них является леканора пылевая (L. conizaevidcs). В Средней и Запад- ной Европе считают, что она растет только в городских условиях. Род аспинилия (Aspicilia) объединяет виды, которые отличаются от лека нор в основном апотециями, погруженными в слоевище. Среди них очень интересную группу составляют так называемые кочующие лишайники, которые к субстрату (почве) не прикрепляются и пере- носятся ветром с места на место. Такова, на- пример, аспиаилия съедобная (Aspicilia escv- lenta, табл. 50), образующая шаровидно-угло- ватые комочки диаметром 1 — 4 см, свободно лежащие на глинистых и солонцеватых почвах в степях, пустынях и горах засушливых об- ластей. Род гематомма (Haematomma) включает ли- шайники, которые образуют бугорчатые или порошковатые слоевища на различных субстра- тах — aa коре деревьев, скалах и пр. У гематпоммы semposou (Haematomma ven- tosum, табл. 42, 2) слоевище толстое, зелено- вато-желтое, с кроваво-красными апотециями. Это очень обычный вид на горных породах в го- рах и Арктике. Род плаколеканора (Placolecanora) объеди- няет виды с лопастным слоевищем. Лла- колеканора щитовидная (Placolecanora peltata, табл. 44) — обычный вид в горах и в Арктике, где растет на силикатных породах. Ее слоеви- ще чешуйчато-лопастное, желтоватое, обра- зует почти правильные розетки диаметром 3— 4 см, прикрепленные к субстрату гомфом. Семейство пармелиевые (Parmeliaceae) 3ТО семейство включает ряд родов с боль- шим количеством широко распространенных видов. Слоевище у них обычно листоватое, реже кустистое, дорсовентральное, с хорошо развитым верхним коровым слоем. Апотеции образуются часто леканоровые, сидячие или, реже, на ножках. Сумки с 8, реже со многими спорами. Споры одноклеточные. Фикобионт- зеленая водоросль требуксия. Род пармелия (Parmelia) — самый большой в семействе, содержит более 600 видов, из них в СССР встречается 80. Слоевища у этих ли- шайников листоватые, различно окрашенные— беловато-серые, серые, желтовато-зеленые, ко- ричневые, черновато-бурые, прикрепляющиеся к субстрату ризинами. Растут пармелии на де- ревьях, древесине, скалах, реже на почве. Рас- 460
пр остр алены во всех растительыо-климатиче- ских зонах. В тундровой и лесотундровой зонах весьма обычна пармелия центробежная (Parmelia cen- trifuga), зеленовато-желтоватое слоев ище ко- торой в центре отмирает, а в периферических частях растет дальше и образует на скалах и камнях иногда большие (диаметром до 1 м) круги. К этому виду близка пармелия извилистая (Р. incurva), на лопастях которой образуются большие головчатые сорали. К той же группе «желтых» пармелий (Xanthoparmelia) отно- сится пармелия блуждаюцая (Р. vagans, табл. 50), широко распространенная в степной и полупустынной зоне. Это типичный кочующий лишайник, слоевище которого не прикрепля- ется к субстрату (почве), а лежит свободно и переносится ветром с одного места на другое. Лопасти слоевища заворачиваются в трубоч- ки (у других видов они плоские). Этот вид ши- роко использовался в народной медицине для лечения ожогов, ран и пр. В лесной зоне из группы (<желтых» пармелий обыкновенны пар- мелия узколистпная (Р. stenophylla) — без изи- дий и пармелия усыпанная (Р. conspersa)— с изидиями. Обе растут на скалах и камнях. Большую группу образуют так называемые «коричневые» пармелии (Melanoparmelia), из которых весьма широко распространена пар- мелия оливковая (Р. olivacea), образующая на коре деревьев, особенно берез, розетковидные блестящие коричневые слоевища, обычно не- сущие большое число апотециев. На камнях и скалах часто растет пармелия темно-бурая (Р. fuliginosa) с тесным скоплением толстых изидий на поверхности слоевищ. Другая груп- па видов, носящая название Amphigymnia, имеет красивые широкие лопасти, окрашенные в желтый или сероватый цвет разных тонов. Из них весьма широко распространена парле- лия козлиная (Р. caperata), образующая на всевозможных субстратах, особенно на коре деревьев, большие желтовато-зеленые розетки. Широкая экологическая амплитуда характери- зует и два светло-серых вида — парлелию бо- роздчатпую (Р. sulcata) и пармелию скальную (Р. saxatilis). Первый вид растет на коре раз- ных древесных пород и на древесине как в ле- сах, так и в населенных пунктах; второй, от- личающийся от первого наличием изидий, рас- тет на камнях и скалах, но нередко, особенно в Сибири и на Дальнем Востоке, поселяется и на коре деревьев. Род гипогимния (Hypogymnia) очень близок к пармелиям, но все же его представители от- личаются от них целым рядом морфологиче- ских и биохимических признаков, более всего полым внутри слоевищем с темной нижней ко- рой. Сюда относится один из самых обыкно- венных и всюду распространенных эпифитных лишайников — гипогимния вздутпая (Hypo- gymnia physodes, табл. 42, 6), серые, узколо- пастные слоевища которой обильно растут на всевозможных субстратах, особенно на ство- лах хвойных. Этот листоватый лишайник с ха- рактерными губовидными соралями на концах лопастей нередко покрывает стволы сплошным однотонным серым покровом. Вид содержит антибиотические вещества и считается перспек- тивным для получения антибактериальных со- единений. Род пармелиопсис (Parmeliopsis) включает виды с узколопастными, прижатыми к субст- рату слоевищами. На хвойных деревьях, осо- бенно при основании стволов, и на древесине часто растет зеленовато-желтый пармелиопсис сомнительный (Parmeliopsis ambigua). Род менегация (Menegazzia) содержит лишай- ники, которые отличаются от гипогимний округлыми или овальными отверстиями в слое- вище и головчатыми соралями. В глухих ле- сах на коре деревьев местами растет харак- терный представитель этого рода менегаиия про- дырявленная (Menegazzia pertusa, табл. 52, 2). Род цетрария (Cetraria) объединяет формы с кустистыми слоевищами, образованными лен- товидно-рассеченными лопастями, плоскими или свернутыми в трубки. В СССР установлено 44 вида этого рода, из которых самым извест- ным и широко распространенным является цетрария исландская (Cetraria islandica, табл. 48, б) — кустистый коричневый лишайник вы- сотой до 10 см, растущий большими куртинами на земле в сосновых лесах, верещатниках, тунд- рах и лесотундрах. Это лекарственное расте- ние, используемое для приготовления раство- ров, применяемых при лечении болезней ды- хательных путей. Местами дает большую био- массу (в северо-западных районах страны в борах до 6 — 7 ц/га), однако собирать этот лишайник следует осторожно, так как прирост его происходит медленно (по 1 — 2 мм в год) и многие местообитания уже истощены. В горных тундрах растет красивая корич- невая цетрйрия с расходящимися подобно ро- гам оленя долями лопастей — иетрария Ричард- сона (C. richardsonii, табл. 47, 2). Часть кусти- стых цетрарий окрашена в желтоватый цвет. Из них широко распространены, особенно в тунд- рах и лесотундрах, цетпрария снежная (C. niva- lis) и цетрария клубочковая (C. cucullata, табл. 48, 1). Оба эти вида образуют кустистые слое- вища, но у первого лопасти плоские, а у второ- го желобчато свернутые или трубчатые. Неко- торые виды цетрарий содержат ядовитую вуль- пиновую кислоту. К ним относятся бореаль- ная цетрария сосновая (C. pinastri) и аркти- ческая цетрария Тилезия (C. tilesii).
Семейство уснеевые (Usneaeeae) Это большое семейство содержит много ро- дов и около 1000 видов, занимающих сущест- венное место в природе и ценных для челове- ка. Их слоевище гетеромерное, кустистое или в виде бороды, свисающее, со всех сторон по- крытое коровым слоем, прикрепляющееся гом- фом или ризоидами. Апотеции леканорового ти- па. Споры одно-, двух- или многоклеточные, бесцветные или коричневые, по 1 — 8 в сумке. Род уснея (Usnea) насчитывает около 600 ви- дов, которые отличаются присутствием в цент- ре слоевищных веточек прочного осевого ци- линдра, образованного плотным сплетением гиф и имеющего механическое значение (рис. 296). Слоевище желтовато-зеленое, раз- личной длины, нередко до 1 — 2 м. Все виды этого рода — эпифнты на коре и ветвях деревьев, реже встречаются на камнях, заборах, брев- нах. Они широко распространены во многих зонах, но особенно их много в умеренной лес- ной зоне; большинство весьма чувствительно к загрязнению воздуха, поэтому их трудно найти в лесах, окружающих большие города, а некоторые виды вообще стали весьма редки- ми. В них в большом количестве содержится специфическая для лишайников усниновая кис- лота, обладающая сильными антибиотически- ми свойствами, из нее уже изготовлено не- сколько медицинских препаратов. Характерный вид еловых лесов — цсиея длин- нейшая (Usnea longissima, табл. 49, 8). Род алектория (Alectoria) включает лишай- ники, которые по внешнему виду близки к ус- неям — слоевище у них имеет вид низких ку- стиков или бород, свисающих с ветвей, но осе- вой цилиндр в нем отсутствует и заменен в цент- ре рыхлой сердцевиной. В роде около 100 ви- дов, многие из них эпифиты, другие растут на скалах и прочих субстратах. В тундре широко распространена алектория желто-белая (Alectoria ochroleuca, табл. 49, 7). Слоевище у нее имеет вид прямостоячих или распростертых кустиков, конечные веточки всегда синевато-черные. Она содержит в боль- шом количестве усниновую кислоту (до 8о~~), ее особенно охотно поедают северные олени. Род эверния (Evernia) объединяет виды, у которых кустистое слоевище, лопасти ленто- видные, мягкие, дихотомически-ветвящиеся. У эвернии сливовой (Evernia prunastri, табл. 49, 1) лопасти дорсовентральные, снизу бело- ватые, сверху беловато-зеленые, по краям с се- ровато-белыми соралями. Этот вид обильно растет на коре деревьев, особенно лиственных. Он имеет большое народнохозяйственное зна- чение: его используют в парфюмерной промыш- ленности под названием «дубовый мох» при изготовлении многих духов и одеколонов — спир- товый экстракт из этого лишайника, или ре- зиноид, служит фиксатором запаха. Род нейропогон (Neuropogon) интересен гео- графически: он распространен биполярно, т. е. встречается только в Арктике и Антарктике. Один из обычных видов — нейропогон пучко- ватый (Neuropogon fasciatus, табл. 49, 6)— имеет кустистое зеленовато-желтое слоевище с верхушками веточек, окрашенными в черно- ватый цвет. Большие апотеции также черно- ватые. Растет на H.BMHHx в самых суровых по- JIHPHbIX УСЛОВИЯХ. Семейство телошистовые (Teloschistaceae) Это большая группа, содержащая ряд родов с широко распространенными видами. Слоеви- ще у них накипное, листоватое или кустистое, обычно желтое или оранжевое, окрашиваю- щееся от едкого кали в фиолетовый цвет. Апо- теции леканорового или лецидеевого типа. Сумки с 8 спорами, споры бесцветные, бипо- лярные, реже одно- или двухклеточные. Фико- бионт — зеленая водоросль требуксия. Род калоплака (Са1ор1аса) насчитывает око- ло 450 видов, слоевище у которых накипное, в виде зернистой корочки, или в центре кор- ковидное, а по периферии лопастное, пре- имущественно желтовато-оранжевое, яркое, прикрепляющееся к субстрату сердцевинными гифами. Апотеции леканорового типа. В сумке обычно по 8 биполярных или одноклеточных спор. Растут на различных субстратах — ска- лах, деревьях, древесине, мхах. Калоплака оранжевая (Са1ор1аса aurantiaca) обычна на коре деревьев, особенно листвен- ных. У нее лимонно-желтое бугорчатое слое- вище, несущее оранжево-желтые апотеции. Ка- лоплака пустынная (С. desertorum) образует на солонцеватых почвах в степях и полупусты- нях маленькие грязновато-желтые подушечки, на которых видны мелкие (диаметром 0,5— 1 мм) оранжево-желтые апотеции. Калоплака настенная (С. тигогит) — обычный вид на камнях, кирпичных стенах и черепицах крыш. Слоевище у нее розетковидное, по периферии с радиально расположенными лопастями, кон- цы которых часто покрыты беловатым налетом, а в центре с многочисленными апотециями. Род телошистес (Teloschistes) включает виды, которые имеют кустовидно-разветвленное сло- евище с округлыми или сплюснутыми лопастя- ми. Коровой слой образован параллельно рас- положенными гифами, сердцевинный слой рых- лый, отчасти полый. Апотеции леканорового типа, расположены на концах лопастей, край апотециев с ресничками (цилиями). Среди ви- дов этого рода много редких. Телошистес жел- 462
теюгций (Teloschistes flavicans) распространен в океанических областях, телошистес ямчатый (Т. lacunosus) — в пустынях и полупустынях. Род ксантория (Xanthoria) объединяет формы с листоватым оранжево-желтым слоевищем, покрытым со всех сторон коровым слоем. К этому роду принадлежит один из самых обычных лишайников — ксантория настенная (Xanthoria parietina, табл. 42, 7), встречающая- ся на различных субстратах (коре деревьев, древесине, скалах, камнях и пр.) на всех конти- нентах. Ее оранжево-желтые розетки, обычно с большим количеством более темных апотециев, хорошо заметны, они часто покрывают стволы деревьев у дороги, в парках, приморские ска- лы и пр. 3ТО нитрофильный лишайник, весьма выносливый к загрязненности воздуха, поэто- му может встречаться и в городах. Семейство фимцимвым (Physciaceae) Семейство объединяет ряд крупных родов, содержащих в целом более 1000 видов. Слоеви- ща у этих лишайников накипные, листоватые или кустистые. Апотеции леканорового или лецидеевого типа. В сумках по 8 коричневых, обычно двухклеточных тонкостенных спор. Род буеллия (Buellia) — самый крупный в семействе, содержит около 600 видов. Слое- вище у них в виде однородной корочки, че- шуйчатое или по периферии лопастное. Апоте- ции черные, лецидеевые. На гладкой коре лиственных пород обычна буеллия дисковггдная (Buellia disciformis), она имеет светлое, зеленоватое слоевище, окаймлен- ное черным подслоевищем и несущее многочис- ленные черные апотеции. Род ринодина (Rinodina) также крупный, в нем 200 видов, которые отличаются от буел- лии строением апотециев: они леканорового типа. Слоевище имеет вид однородной корочки, многда по периферии лопастное. Апотеции тем- ные, сидячие, изредка погруженные в слоеви- ще. В горах на кварцевых породах часто встреча- ется рггнодина горная (Rinodina oreina); у нее бледное зеленовато-желтое розетковидное, плот- но приросшее к субстрату слоевище, имею- щее по периферии лучисто расположенные уз- кие лопасти, в центре много апотециев, часто черноватых. Род фисция (Physcia) содержит широко рас- пространенные лпстоватые лишайники (170 ви- дов), растущие на стволах деревьев, древесине, мхах, скалах и камнях, реже на земле. Многие из них довольно выносливы к загрязненности воздуха и встречаются в городах (в парках, на кладбищах и пр.). Слоевище у фисций ли- стоватое, нередко образующее правильные округлые розетки, сверху с параплектенхим- ным коровым слоем, снизу с параллельно рас- положенными гифами. Эти лишайники при- крепляются к субстрату густыми ризоидами. Апотеции леканорового типа. Споры темные, двух- — четы рехклеточные. Обычный вид на придорожных деревьях, в парках и т. д.— фисция припудренная (Phys- cia pulverulenta). У нее правильное розетко- видное слоевище диаметром до 15 см, сверху оливково-коричневое, очень часто покрытое сизоватым налетом, снизу темное, с черными ризоидами. Апотеции большие (диаметром до 5 мм), с темно-коричневым диском, который нередко покрыт светлым налетом. К этому ви- ду близка фггсцггя моховая (Ph. muscigena), встречающаяся в тундрах и высокогорьях на мшистых почвах. У нее также коричневое сло- евище с налетом, но апотеции встречаются очень редко; лопасти в центре имеют приподни- мающиеся края и расположены черепитчато. Обычной на камнях от тундр до пустынь яв- ляется фисция голубовато-серая (Ph. caesia). У нее розетковидное голубовато-серое слоеви- ще; лопасти узкие (шириной до 1 мм), много- перисто разветвленные, всегда с головчатыми свинцово-серыми соралями. Род анаптихия (Anaptychia) довольно бли- зок к предыдущему, но его представители имеют хороший отличительный признак — волокнис- тый коровой слой, состоящий из гиф, располо- женных параллельно поверхности слоевища, а не параплектенхимный, как у фисций. Слое- вище в виде листовидных розеток или низких кустиков. Самый обычный вид — анаптихия реснигпча- тая (Anaptychia ciliaris) со слоевищем в форме лежащих или слегка приподнимающихся от субстрата кустиков пепельно- или коричнева- то-серого цвета с линейными (шириной 1 — 5 мм), завернутыми вниз краями лопастей, усажен- ными длинными темными фибриллами. Апоте- ции образуются часто, довольно крупные (диа- метром до 8 мм). Встречается в парках, на придорожных де- ревьях, в светлых лиственных лесах на коре деревьев, реже на скалах и древесине. КЛАСС БАЗИДИАЛЬНЫЕ ЛИШАЙНИКИ (BASIDIOLICHENES) Группа базидиальных лишайников невелика, она содержит всего лишь около 20 видов. Сло- евище у этих лишайников образуется из бази- диальных грибов, относящихся к двум поряд- кам — афггллофоровым (Aphyllophorales) и ага- рггковым (Agaricales) — и из некоторых зеленых и сине-зеленых водорослей. Долгое время бази-
диальные лишайники были известны только sтропиках, ,но не так давно они найдены s уме- ренной зоне и даже в Субарктике. Базидиальные лишайники отличаются от сумчатых целым рядом особенностей. Во-пер- вых, плодовые тела у них кратковременные, часто одногодичные, в то время как у сумчатых они существуют долго — десятки и сотни лет. Во-вторых, симбиоз у базидиальных грибов и водорослей не привел к образованию особых жизненных форм, к морфогенетическому обособлению. Базидиальные лишайники имеют такую же внешнюю форму, как соответствую- щие свободноживущие грибы — афиллофоро- вые или агариковые. Следовательно, предста- вители этого класса являются не настоящими лишайниками, а полулишайниками. В-третьих, в базидиальных лишайниках не обнаружены специфические лишайниковые вещества, столь характерные для многих групп сумчатых ли- шайников. У афиллофоровых базидиальных лишай- ников можно наблюдать несколько ступеней образования первичных лишайниковых слое- вищ. У самых простых, например у одон- ции двухцветной (Odontia bicolor) из семей- ства кортициевых (Corticiaceae), слоевище имеет вид тонкой белой или желтой корочки, между гифами которой разбросаны клетки во- доросли коккомиксы, причем гриб не пара- зитирует на водоросли. У ателии эпифилль- ной (Athelia epiphylla), образующей на стволах деревьев круглые слоевища, гриб уже парази- тирует на водоросли протококкус, т. е. обра- зует гаустории, проникающие в ее клетки. На- конец у наиболее развитых базидиальных ли- шайников водоросли образуют в грибе посто- янные и закономерно расположенные внутри- слоевищные слои, в которые гифы проникают своими гаусториями, хотя и у этих лишайников гриб по форме остается типичным .базидиальным грибом. В тропиках самым распространенным видом базидиальных кортициевых лишайников явля- ется кора павонская (Cora pavonia), которая s системе грибов носит название диктионемы горной (Dictyonema montanum). Она растет в виде довольно больших (до 10 см в диаметре) светло-серых пластинок, несущих на нижней стороне гимениальный слой. В центре слоевища расположен слой водорослей, который, однако, иногда отсутствует, и тогда мы имеем дело с на- стоящим грибом (рис. 303, 1). Среди афиллофоровых базидиальных грибов в семействе рогатиковых (Clavariaceae) также имеется ряд видов, которые, сохраняя свой типичный грибной облик, оказываются содер- жащими водоросли. Сюда относится, например, мультиклавула заплесневелая (Multiclavula muci- da), имеющая низкие булавовидные плодовые тела, вырастающие из прикрепленной к гни- лой древесине грибо-водорослевой пленки, фи- кобионтом которой является зеленая водоросль коккомикса. Из пластинчатых грибов (порядок Agarica- les), у которых в слоевище имеется слой водо- рослей, можно назвать, например, корис~иул зеленый (Coriscium viride), в системе грибов он носит название Omphalina luteolilacina. У него между псевдопаренхиматическими коровыми слоями из гиф находится довольно толстая про- слойка из водоросли коккомикса (рис. 303, 2, 3). Систематика базидиальных лишайников еще слабо разработана. В последнее время иссле- дователи находят все новые грибы, которые постоянно или временами состоят в симбиозе с водорослями. В большинстве случаев эти находки указывают на факультативность и эволюционную молодость таких симбиотиче- ских отношений.
ПРОИСХОЖДЕНИЕ ЛИШАЙНИКОВ И ИХ МЕСТО 13 СИСТЕМЕ РАСТИТЕЛЬНОГО МИГА 465 В науке еще очень мало достоверных факти- ческих данных о том, как и когда возникли лишайники. Многие высказывания по этой про- блеме носят сугубо гипотетический характер. Причина такого положения проста — у нас почтп совсем нет данных по ископаемым лишай- никам. Те немногочисленные находки, которые имеются, не позволяют делать общих выводов о возрасте и путях становления лишайников. Самые древние единичные находки ископа- емых лишайников относятся к мезозою, не- много больше их из кайнозоя. Эти находки свидетельствуют, что к тому времени (более 200 млн. лет назад) лишайники уже были вы- сокоразвитыми организмами, листоватыми и кустистыми формами. Когда появились первые примитивные лишайники, неизвестно. Если мы скажем, что они могли существовать уже в на- чале палеозоя (примерно 500 млн. лет назад), то это будет лишь гипотетическое предположе- ние. Необходимо учесть, что лишайники могли возникнуть лишь тогда, когда в процессе эво- люции появились большие группы водорослей и грибов, и при этом в виде крупных, экологи- чески дифференцированных популяций. Эти популяции образовали примитивные смешан- ные группировки — микробиоценозы, в кото- рых между членами (ценобионтами) были раз- нородные взаимовлияния. Очевидно, в этих микробиоценозах, состоящих главным образом из одноклеточных простых организмов, возник- ли первые рыхлые скопления водорослей и гри- бов, первые примитивные (вначале, можно по- лагать, непостоянные) лишайники. Они не име- ли еще определенной формы, взаимовлияние гриба и водоросли лишь в дальнейшем разви- тии привело к определенному формообразова- нию, возникновению специфических лишай- никовых жизненных форм. Гриб мог вступить в контакт с водорослью в том случае, когда для него не оказалось в субстрате достаточного количества питательных веществ,— он стал па- разитировать на водорослях. Этот парази- тизм вначале был разрушительным, но быстрое и непрерывное деление водорослевых клеток сохраняло их популяцию, в то же время у них вырабатывалось сопротивление грибу, и на- конец они стали получать определенную вы- году от такого сожительства. Простой одно- бокий паразитизм не мог обеспечить дальней- шего развития и усовершенствования водо- рослево-грибного соединения. Должны были возобладать мутуалистические, взаимовыгодн- ыее отношения и такие биотипы водорослей, которые не только приобрели определенную устойчивость и выносливость в отношении пара- зитизма гриба, но и стали извлекать из объеди- ненной жизни определенную пользу. Тогда и возникли симбиотические отношения между фико- и микобионтом лишайникового слоевища. Почему возникли постоянные водорослево- грибные скопления — нерешенный вопрос. Ученые называют ряд возможных причин. Од- ной из них мог быть «голод» гриба. В условиях недостатка питательных веществ грибы (в пер- вую очередь сумчатые) нашли себе в лице водо- рослей гибкого партнера, способного обеспе- чивать их необходимыми органическими ве-
ществами. Соединение гриба и водоросли мог- ла стимулировать и сухость местообитания. Имеются наблюдения, что в сухих и недостаточ- но обеспеченных питательными веществами усло- виях грибные гифы охватывают различные округлые предметы, последними в определенных условиях могли быть и водорослевые клетки. Из аморфных в ода рослево-грибных скопле- ний со временем стали развиваться определен- ные жизненные формы лишайников, начался активный формообразовательный процесс. Из временных рыхлых и примитивных скоплений стали вырабатываться более постоянные типы слоевища с определенной формой — корковые, чешуйчатые, листоватые, кустистые и пр. Спе- циальные эффективные способы вегетативного размножения (при помощи соредий, изидий, фрагментов слоевища и др.) обеспечивали ли- шайникам быстрое расселение и завоевание все новых субстратов и местообитаний. Биологиче- ские преимущества лишайников, обусловленные их двойственным характером, — нетребователь- ность к содержанию питательных веществ в субстрате, большая продолжительность суще- ствования и т. д.— позволяют им расти в таких экстремальных условиях, где другие организмы существовать не могут. Для конкурентослабых лишайников это единственная возможность со- храниться в суровой борьбе за существование. Удивительное комплексное сожительство гри- бов и водорослей в одном организме оказалось весьма эффективным в процессе эволюции. На это указывает бурный видообразовательный про- цесс у лишайников. Как уже было сказано, к настоящему времени описано до 26 000 видов лишайников, а в действительности их число может достигать 40 000. Если учесть, что свобод- ноживущих грибов известно примерно 60 000 видов, а водорослей — 40 000 видов, то стано- вится очевидным, что лишайники образуют большую группу организмов. Какое же место занимают они в системе растительного мира? В большинстве систем растительного мира лишайники считались самостоятельным и еди- ным отделом (Divisio, Phylum) или классом. Ввиду того что по массе в слоевище лишайника преобладает гриб и он же образует плодовые тела, выделение классификационных единиц у лишайников производят главным образом по признакам микобионта, его плодового тела. Это дало начало распространяющемуся в послед- нее время взгляду на лишайники как на грибы, точнее лихенизированные грибы. Согласно этой теории, лишайники возникли полифилетически из различных классов и отрядов грибов, кото- рые от гетеротрофного типа питания перешли к использованию водорослей как автотрофов для обеспечения своих потребностей в питатель- ных веществах. По мнению сторонников этой теории, грибы следует классифицировать в единой системе независимо от того, образуют ли они симбиотические сожительства с какими- нибудь другими организмами или нет. Такое утверждение подкрепляется фактами существо- вания симбиотических отношений между други- ми группами организмов, которые не объеди- няются в особые таксономические единицы. Если принять это, то лишайники должны быть рассредоточены в системе грибов и теряют свою обособленность как самостоятельная группа. Но продолжают развиваться и другие взгля- ды. Часть исследователей подчеркивает, что лишайники обладают целым рядом признаков, свидетельствующих об особом, высокоразви- том типе симбиоза, можно сказать «сверхсим- биозе». Симбиоз у лишайников характеризуется исторической выработаны остью и морфогене- зом, что привело к возникновению специфиче- ских жизненных форм, типов строения, не встречающихся в отдельности ни у грибов, ни у водорослей. Лишайники обладают целым рядом особых биологических свойств, не при- сущих другим группам организмов. Это их спо- собы размножения при помощи соредий и изи- дий, своеобразие метаболизма, образование спе- цифических лишайниковых веществ, в синтезе которых принимают участие оба биокомпонен- та лишайникового слоевища. и т. д. Мало того, до последнего времени считалось, что микоб ионт лишайника высокоспециализи- рован к жизни в соединении с водорослью, а последняя более автономна и способна расти вне лишайников ого слоевища в различных экологических нишах. Теперь выяснилось, что, во всяком случае, самый главный тип фикобион- та — водоросль требуксия (ТгеЬоих1а), входя- щая в 7 — 12 тыс. видов лишайников, вне лишай- ника в свободноживущем состоянии не встре- чается, т. е. является специализированным ком- понентом лишайников. Из этого следует вывод о высоком биологическом единстве больших групп лишайников, содержащих эту водоросль. Не исключено также, что в будущем, особенно с применением электронного и сканирующего микроскопов, будет обнаружено, что и другие роды фикобионта отличаются от соответствую- щих свободноживущих водорослей. Тем самым проблема положения лишайников в системе организмов приобретает некоторые новые чер- ты. Очевидно, следует больше учитывать качест- венные признаки, выработавшиеся в резуль- тате длительного эволюционного процесса, от- личающие лишайники как биологически целост- ные организмы от грибов. На этой основе лишай- ники могут рассматриваться как специальный отдел (Divisio Lichenomycota), как биологиче- ски единая группа растительного мира со свои- ми высшими классификационными единицами. 466
ПРАКТИЧЕСКОЕ ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ 467 Экономическое значение лишайников в жизни человека велико. Во-первых, это важнейшие кормовые растения. Лишайники служат основ- ным кормом для северных оленей — животных, играющих большую роль в жизни народов Крайнего Севера. Основу корма северных оленей составляет так называемый олений мох или ягель. Ягелем обычно называют 3 вида кустистых лишайни- ков: кладонию альпийскую (Cladonia alpestris, табл. 48, 6), кладонию лесную (С. sylvatica) и кладонию оленью (С. rangiferina). Однако олени охотно поедают и многие другие лишайники (другие виды Cladonia, Cetraria islandica, С. cu- cullata, С. nivalis, Alectoria ochroleuca и т. д.). Всего олени используют в пищу до 50 видов лишайников, которые составляют '/ от общего количества корма, потребляемого ими на паст- бищах. Олени одинаково охотно поедают лишай- ники и зимой и летом. Но если летом для них Бе менее важным кормом служат различные травы, листья полярной березы и ивы, а также ягоды и грибы, то зимой лишайники — почти единственный источник пищи для этих живот- ных. Олени выкапывают лишайники из-под снега, а когда снежный покров бывает слишком глубок, скусывают лишайники, растущие на скалах, на стволах и ветвях деревьев, особенно повисающие кустистые лишайники (уснея, алек- тория, эверния и др.). Кормовая ценность ли- шайников определяется высоким содержанием углеводов, которые хорошо перевариваются и усваиваются оленями. Однако малое количе- ство витаминов и недостаток зольных и белко- вых веществ делают лишайниковый корм не- полноценным. Кроме того, олени плохо усваи- вают азотистые соединения, входящие в состав лишайников. Поэтому зимой северные олени, питающиеся в это время года преимущественно лишайниками, обычно сильно худеют, их кости становятся хрупкими, а жировые ткани размяг- чаются. Все это является результатом авита- миноза, а также азотистого и зольного голо- дания. Летом, когда рацион оленей пополняет- ся различными травами и листьями кустарни- ков, они быстро откармливаются и толстеют. Однако даже летом недостаток лишайников на летних пастбищах приводит к развитию у жи- вотных поноса, особенно у молодняка. Лишайники служат кормом не только для домашнего северного оленя, но и для диких копытных животных — марала, кабарги, косу- ли, лося. По многочисленным свидетельствам охотников Алтая, в голодное время конца зимы — начала весны эпифитные лишайники могут быть одним из основных кормов марала. Алтайские охотники даже называют уснеи «маральим сеном». Было замечено, что маралы и лоси нередко зимой тщательно объедают ли- шайники с древесных стволов, с сухих и опав- ших веток. Эпифитные лишайники поедают и другие животные, например белки, полевки и др. В северных странах некоторые лишайники, особенно цетрарию исландскую (Cetraria islan- dica), широко использовали как дополнитель- ный корм для домашнего скота. Этот лишайник употребляли и как дополнительный продукт при выпечке хлеба, особенно в Исландии. Че- ловек использует в пищу другие лишайники. Например, в Японии одним из деликатесов является листоватый лишайник умбиликария съедобная (Umbilicaria esculenta Ä 333).
Другая область практического применения лишайников — медицина. Первые сведения об использовании лишайников как лекарственных растений относятся к глубокой древности. Еще древние египтяне за 2000 лет до н. э. употреб- ляли их для медицинских целей. Немало ле- карств, изготовленных из лишайников, имела в своем арсенале средневековая медицина. Однако в то время приготовление этих лекарст- венных препаратов было основано не на знании химических особенностей лишайников, а ско- рее на мистических представлениях и предрас- судках. Уже с V в. н. э. в мировоззрении людей отчетливо сформировалась идея «символов», согласно которой провидение якобы дало расте- ниям форму, указывающую людям, как нужно эти растения использовать. Лекари тех времен пытались найти связь между внешним видом растения и отдельными органами и частями тела человека: существовало представление, что растение может исцелить заболевания того органа, строение которого оно напоминает своим внешним видом. Так, например, лобарию легоч- ную (Lobaria pulmonaria, табл. 47, 1), внешне чем-то напоминающую строение человеческого легкого, использовали при лечении воспаления легких; успею (табл. 49, 8), бородатое слоевище которой имеет некоторое сходство с волосами, применяли при лечении заболеваний волос; желто-оранжевая ксантория (Xanthoria parieti- na) благодаря своей окраске «излечивала» жел- туху. В средние века лишайнику пельтигера собачья (Peltigera canina, табл. 49, 8) приписы- вали свойство излечивать бешенство, отсюда происходит и его своеобразное название. До наших дней сохранился один из рецептов, ко- торый прописывал известный в Англии в XVIII в. врач Ричард Мид для излечения бе- шенства: «Пустить пациенту на руке крови де- вять унций. Взять растения, называемого по- латыни Lichen cinereus terrestris (Peltigera ca- nina. Прим. asmopa), а по-английски — пепель- но-серый печеночный мох, очищенного, высушен- ного и превращенного в порошок, пол-унции и две драхмы черного перца в порошке, хорошень- ко смешать и разделить порошок на четыре до- зы, которые по одной принимаются каждое ут- ро натощак в течение четырех дней в полпинте теплого коровьего молока. После приема этих четырех доз пациент должен принять холодную ванну и купаться в холодном ручье или реке каждое утро натощак в течение месяца. Он дол- жен быть весь покрыт водой (голова под водой), но оставаться в ней не дольше полминуты, если вода очень холодная». Без сомнения, нередко в подобном лечении с помощью лишайников было много абсурдного. Но во многих случаях лишайники, благодаря особенностям своей химической природы, ока- зывали на больного и положительное действие- как стимуляторы, поднимающие тонус организ- ма, или как антибиотики. Так постепенно на- капливался опыт в применении лишайников как лекарственных растений. Уже в XVIII в. их использовали в медицине на более научной основе, с учетом опытных данных; лишайники были внесены в списки лекарственных расте- ний в официальные фармакопеи многих стран. Так, в 1749 г. известный шведский ботаник К. Линней упоминал о семи лекарственных лишайниках (Lichen saxatilis, L. islandicus, L. pulmonarius, L. aphthosus, L. caninus, L. pli- catus, L. cocciferus). Из первого лишайника (современное его название Parmelia saxatilis) в то время делали тампоны для остановки кро- вотечения из носа, из последнего (современное название Cladonia coccifera) готовили средство против кашля у детей. В XIX в. список лекарственных растений пополнился новыми видами лишайников. В одном из обширных обзоров полезных и ядови- тых растений земного шара, изданном в 1862 г., рекомендуется для применения в медицине 32 вида лишайников. Особенно высоко в то вре- мя ценили цетрарию исландскую (Cetraria islan- dica). Так, в одной из полных сводок о практи- ческом использовании лишайников, опублико- ванной в начале XIX в., о цетрарии исландской сообщалось: «Этот лишайник и теперь находит- ся среди самых выдающихся лекарств. Как питающей силой, так укрепляющей и замеча- тельной антисептической силой он выдается. Опыты это утверждают давно. Своею горечью он укрепляет, слизью же питает, и отвар этого лишайника предлагается при чахотке, при внутренних болезнях и при поносе. Очень реко- мендуется при застарелом катаре, кровохар- кании, чахотке и многих других легочных бо- лезнях». Как показали современные исследо- вания химического состава слоевища этого лишайника, цетрария исландская содержит до 70 — 80О/о углеводов, главным образом «лишайник ового крахмала» — лихенина и изо- лихенина, а также сахара (глюкозу и галакто- зу), 0,5 — 3',4 белков, 1 — 2О/о жиров, 1% воска, около Зо/о камеди, около Зо/о пигментов и от 3 до 5О/о лишайниковых кислот (протолихесте- риновой, лихестериновой, фумарпротоцентра- ровой и некоторых других). Именно кислоты придают лишайнику горький вкус и обуслов- ливают его тонизирующее и антибиотическое свойства. Современными исследованиями было показано, что, например, протолихестерино- вая и лихестериновая кислоты проявляют вы- сокую антимикробную активность в отношении стафилококков, стрептококков и некоторых других микроорганизмов. Благодаря этим осо- бенностям цетрарию исландскую применяют как 468
лечебное средство и в современной медицине. Ее широко используют как испытанное народ- ное средство, например, в Швеции. Из этого лишайника готовят отвары для лечения ката- ров и простуды, студни против поносов, упот- ребляют его также как горечь с терапевтиче- скими целями. Как народное средство цетра- рию исландскую применяют и при лечении туберкулеза. В конце XIX — начале XX в. в связи со зна- чительным развитием научной медицины врачи стали все реже обращаться к народным средст- вам, забвению были преданы многие лекарствен- ные растения, в том числе лишайники. В тот период в перечни лекарственных растений ли- шайники либо вовсе не включали, либо указы- вали лишь одну цетрарию исландскую. Однако в начале ХХ в. интенсивное изучение химиче- ских веществ, продуцируемых лишайниками, заставило ученых вновь обратить внимание на эти растения. Открытие в слоевищах лишай- ников огромного количества специфичных для них химических веществ, так называемых ли- шайниковых кислот, повлекло за собой изу- чение их антибиотических свойств. Этому спо- собствовало также открытие в 40-х годах на- шего столетия антимикробных свойств у неко- торых грибов и водорослей. Вслед за этим нача- лись интенсивные поиски новых источников антибиотиков среди низших растений, в том числе среди лишайников. В 40 — 50-е годы почти одновременно и независимо друг от друга в раз- ных странах — в Швейцарии, Финляндии, США, Японии, Испании, Италии и в Советском Союзе — развернулись исследования по изу- чению антимикробных свойств лишайников. В 1944 г. американскими учеными Б е р к- х о л ь д е р о и, Э в е н с о м и некоторыми другими впервые были испытаны 42 вида ли- шайников на наличие у них антимикробных свойств против бактерий Staphylococcus aureus, Escherichia coli и Bacillus subtilis. С этой целью свежесобранные лишайники тщательно из- мельчали и заливали фосфорно-буферным вод- ным раствором. Оказалось, что эти водные экст- ракты с лишайниками подавляют и задержи- вают рост культур вышеуказанных бактерий. Причем различные виды лишайников действо- вали на культуры бактерий неодинаково. Одни лишайники подавляли рост стафил око кков, другие оказывали бактериостатическое влия- ние как на стафилококки, так и на бактерии ба- циллюс, третьи — только на бациллюс и т. д. Это привело ученых к мысли, что, по всей вероятности, лишайники содержат ряд анти- микробных веществ, обладающих избиратель- ными свойствами в отношении различных мик- роорганизмов, и что исследователи имеют дело не с одним антибиотиком, а с целой группой их. Это побудило ученых перейти к изучению анти- бактериальных свойств отдельных веществ, со- держащихся в лишайниках. Исследования про- водили на представителях рода кладония, и оказалось, что слоевища 35 различных видов этих лишайников, проявляющие антимикроб- ные свойства, содержали различные лишайни- ковые вещества: усниновую, фумарпроцетра- ровую, скваматовую, барбатовую и другие кислоты. В большинстве изученных кладоний была обнаружена усниновая кислота. Испы- тание антибиотических свойств этой кислоты показало, что она весьма активна против Bacil- lus subtilis. Вслед за американскими учеными изучение антибиотической активности лишайников было проведено и в других странах. Из всех лишай- никовых веществ особенно выделялась своими антибиотическими свойствами усниновая кис- лота, которая, как было установлено, образу- ется по крайней мере в 70 лишайниках и в зна- чительной степени обусловливает антимикроб- ные свойства многих из них. И уже в 1947 г. немецкими учеными был получен первый анти- биотический препарат из лишайников под наз- ванием «Эвозин». Этот препарат представляет собой смесь эверниевой и усниновой кислот и некоторых других веществ. Получают его главным образом из лишайника эвернии сливо- вой (Evernia prunastri, табл. 49, 1). Препарат «Эвозин» обладает широким антимикробным спектром, преимущественно против стафило- кокков и стрептококков, его используют при местном лечении таких кожных заболеваний, как сикоз, фурункулез, волчанка, а также при болезнях кожи, вызванных развитием патоген- ного грибка Trichophyta. Кроме того, его при- меняют и при лечении мастита крупного рога- того скота. Позднее, в 1952 г., немецкими уче- ными был получен еще один антибиотический препарат из лишайников — «Эвозин-2», или па- рамицин, который с успехом можно применять для лечения открытой формы туберкулеза лег- ких человека. В состав «Эвозина-2» входят такие лишайника вые вещества, как а тра норин, фи- зодова я, каператовая и усниновая кислоты. Сырьем для его получения являются широко распространенные лишайники гипогимния (Hy- pogymnia physodes, табл. 42, 6) и пармелия (Parmelia caperata, табл. 47, 8). В эти же годы (1948 — 1954) испанские ученые также получи- ли новый лечебный препарат из лишайников- уснимицин. Это комбинированный препарат, состоящий из смеси усниновой кислоты со стрептомицином, его используют при лечении туберкулеза и некоторых кожных заболеваний. Ценность уснимицина в том, что он оказывает антибактериальное действие на штаммы тубер- кулезных палочек, устойчивые к стрептомици-
ну. В 1954 г. в Японии получили антибиотиче- ский препарат из лишайников под названием еУснин», который с успехом можно применять против актиномикоза и других кожных забо- леваний. В Финляндии врачи-дерматологи ис- пользовали усниновую кислоту в форме мазей против волчанки. В нашей стране в конце 40-х годов также было начато изучение антибиотических свойств лишайников. В результате этих исследований в Ботаническом институте АН СССР в Ленин- граде был получен новый медицинский препа- рат — натриевая соль усниновой кислоты, или «Бинан». Основой для получения препарата явилась усниновая кислота. Исходным сырьем для приготовления препарата могут служить различные лишайники, содержащие в слоеви- щах усниновую кислоту, — виды кладоний, ус- ней, алекторий, эверний, пармелий и др. Иссле- дование антимикробных свойств препарата по- казало, что он активен против грамположи- тельной бактериальной флоры — золотистого стафилококка, различных стрептококков, пнев- мококков, анаэробов и туберкулезной палочки. Препарат представляет собой эффективное на- ружное антимикробноо средство для лечения нагноительных процессов в раневых поверх- ностях. В настоящее время это лекарство ши- роко продается в аптеках в нескольких формах: в водно-спиртовых новокаиновых растворах, в касторовом масле с анестезином, в пихтовом бальзаме и в виде порошка. Препарат «Бинан» нашел применение в хирургической практике при лечении свежих посттравматических и послеоперационных раневых поверхностей, при лечении варикозных и трофических язв, острого гнойного воспалительного процесса мягких тка- ней, травматических остеомиелитов, при пласти- ческих операциях, при лечении ожогов I I и I I E степени. Его применяют также в гинекологии. Лишайниковые вещества обладают и дру- гими интересными с медицинской точки зрения свойствами. Например, известно антиопухоле- вое действие полипоровой кислоты и кардио- тоническая активность пульвинового дилакто- на. Кроме того, как показали эксперименталь- ные исследования, лишайниковые вещества мо- гут найти себе применение и в фитопатологии. Так, было обнаружено, что усниновая кислота активна против заболеваний томатов (Coryne- bacterium michiganensis); вульпиновая, физо- довая, салациновая и усниновая кислоты— против грибов, разрушающих древесину, а экстракты из лишайников, содержащие лека- норовую, псо ромовую и усниновую кислоты, проявляют активность против вирусного забо- левания «табачной мозаики». Лишайники широко используют и как сырье для парфюмерной промышленности. Издавна было известно, что некоторые из них (Evernia prunastri, Pseudevernia furfuracea, Lobaria pulmonaria и виды рода Ramalina) содержат ароматические вещества, эфирные масла. В древности в Египте и позднее, в XV — XVIII вв., из сухих лишайников получали порошки, ко- торые употребляли затем для изготовления пудр, в частности пудр для париков. В настоя- щее время в парфюмерии используют экстракты этих лишайников. Наибольшее значение как сырье для парфю- мерной промышленности приобрела эверния сливовая (Evernia prunastri), известная на ми- ровом рынке под названием Mousse dechene— «дубовый мох» (табл. 49, 1). Из этого лишайни- ка получают резиноид — концентрированный спиртовой экстракт, имеющий вид густой жид- кости темного цвета. Резиноид — ароматиче- ское вещество, его используют на парфюмер- ных фабриках в качестве ароматического на- чала для некоторых сортов духов. Кроме того, он обладает свойством фиксатора запаха, и парфюмеры в ряде случаев используют его для придачи стойкости духам. Резиноид входит в состав целого ряда духов и одеколонов. Так, в нашей стране на его основе созданы такие ду- хи, как «Бахчисарайский фонтан», «Кристалл», «Кармен», «Подарочные», «Чайка», «Восток» и др., а также одеколоны «Шипр», «Новый» и некоторые другие. Резиноид используют и в других косметических изделиях — в кремах, пудрах, мылах сухих духах. Химическая природа ароматического начала лишайников еще недостаточно ясна. Многие считают, что важнейшие составные части рези- ноида «дубового Mxa» — эверниевая кислота и ее эфиры — являются носителями запаха. Хи- мические исследования резиноида показывают, что это очень сложное по своему составу ве- щество. Оно включает смолы, пигменты (глав- ным образом хлорофилл), углеводы, лишайни- ковые кислоты (усниновая, атранорин, эверни- евая и эверниновая, а также их эфиры), воска и некоторые другие вещества. Со времен глубокой древности лишайники служили сырьем для получения красителей. Эти красители использовали для окраски шер- сти и шелка. Основной цвет красителей, по- лучаемых из лишайниковых веществ, темно-си- ний. Но добавка уксусной кислоты, квасцов и т. д. дает пурпурные, красные и желтые тона. Существенно, что краски из лишайников обла- дают особенно теплыми и глубокими тонами, хотя они и нестойки по отношению к свету. В настоящее время краски получают главным образом синтетическим путем, но до сих пор в Шотландии в текстильной промышленности не- которые определенные виды твидов окрашивают только красителями, добытыми из лишайников. 470
УЕШАТЕЛЬ ТЕРМИНОВ' Базидия 407 Биспора 39, 203, 209, 215, 223 Велум i i5, 116 " Жирным шрифтом отмечены номера страниц с иллюстрациями~ Автоспора 63, 75, 76, 94, 98, 101, 251, 255, 256, 258, 274, 277, 278, 279 Азигота 40 Акинета 21, 38, 213, 256, 274, 292, 296, 317 Амебоид 101, i02, 251, 274 Андроспора 292, 293 Андроспорангий 292, 293 Антеридий 151, 164, 168, 169, 172, 183, 186, 187, 188, 258, 272, 290, 291, 292, 293, 294, 295, 338, 340, 343, 344, 345, 350 Антерозоид 40, 164, 168, 172, 183, i87, 188, 273, 290, 292, 293, 294, 295, 344, 345 Апла ногамета 327 Апланоспора 21, 39, 256, 258, 270, 282, 283, 289, 294, 296, 297, 299, 317, 330 А потеций 410, 411, 412, 413, 415, 430,434, 439, 440, 441, 442, 443, 445, 452, 457, 458, 460, 461, 462, 463 Аппрессорий 387, 388, 389 Ареола ii5, 116, 131, 133, 134, 136, 139, 391, 392, 436, 453, 456 Архикарп 408, 409, 410 Аскогон 408, 409, 410 Аскокарп 380 Ауксоспора 121, 122, 131, 132, 134 Гамета 21, 34, 39, 144, 146, 156, 163, 172, 182, 183, 184, 204, 205, 270, 281, 282, 283, 294, 297, 299, 301, 304, 307, 309, 312, 318, 327, 328, 329 Гаметангий 39, 144, 145, 158, 160, 16i, 163, 164, 165, 171, 183, 186, 187, 189, 190, 281, 301, 302, 303, 304, 307, 314, 346 Гаметоспорофит 40, 42, 146, 152, 154, 155, 156, 157, 160 Гаметофит 40, 42, 146, 147, 151, 161, 671 1 71 ~ i 72~ 1 73 182~ 183, 184, 185, 186, 187, 204, 209, 210, 215, 219, 223, 224, 237, 243, 284, 285 Гаптера 400 Гаптонема 30, 353 Гастеротеций 407, 4ii, 413, 439, 442, 443, 445, 446, 447 Гаусторий 386, 387, 388, 389, 464 Гемизооспора 274, 275, 277 Гетероциста 63, 81, 82, 83, 85, 88, 89, 90, 201, 385 Гипнозигота 40, 42, 267, 268 Гипноспора 39 Гипноциста 152 Гиповальва 98, 99, 114 Гипоталлий 231, 232, 233, 235 Гипотека 114, 120, 121 Гипотеций 411, 412, 413, 414, 415, 443, 447, 453 Гифа 186, 196, 380, 381, 382, 386, 387, 388, 389, 393, 394, 403, 404, 416, 466 Гомф 394., 395, 404, 405, 438, 451, 458, 459, 462 Гонидия 83 Гонимобласт 207, 208, 215, 220, 22i, 222, 224, 227, 228, 229, 230, 233, 235, 236, 239, 243, 247, 248, 250 Гормогоний 80, 82, 83, 88, 385 Гормоспора 82 Зигота 23, 39, 54, 122, 147, 192, 202, 207, 209, 210, 213, 215, 267, 282, 283, 284, 285, 293, 294, 297, 301, 304, 309, 310, 3ii, 3i2, 314, 318, 3i9, 320, 327, 328, 329, 337 Зооид 146, 150, 152, 157, 160, 163, 188, 354 Зооспора 30, 34, 38, 39, 42, 94, 98, 10i, 109, 110, 145, 146, 163, 166, 167, 171, 172, 183, 251, 256, 258, 267, 274, 275, 277, 282, 283, 284, 289, 290, 291, 292, 294, 295, 299, 300, 301 Зооспорангий 258, 278, 300 Изидии 407, 419, 420, 448, 451, 457, 461, 466 Изогамета 269, 271, 272, 289 Импрессорий 387, 388, 389 Карпогон 206, 207, 208, 214, 215, 216, 217, 220, 223, 233, 235 Карпоспора 199, 206, 207, 208, 209, 210, 212, 213, 214, 215, 216, 217, 220, 233 Карпоспорангий 207, 233 Карпоспорофит 209, 210, 226, 237, 243 Кокколит 18, 105, 106, 359 Конидиеносец 409, 410, 416 Конидия 416, 418 Концептакул 156, 182, 183, 184, 204, 205, 206, 224, 230, 231, 232 Конъюгация 39, 266, 309, 311, 312, 314, 318, 319, 320, 327, 328 Криптостома 186, 187, 188, 189
Утрикула 30i Локула 407 Луковичка поровая 323, 333 Мазедий 4i i 439, 440 Макрозооспора 282 Макроспора 188 Макроспорангий 188 Макула 406 Мастигонема 31 Мезоспорий 318, 319, 320, 328 Мезохитон 187 Меристодерма 145, 169, 182, 185 Микобионт 379, 380, 383, 385, 386, 387, 389, 392, 422, 424, 465, 466 Микрозооспора 282 Микроспора 120, 121, 134 Моноспора 39, 151, 203, 209, 2i3, 215 Моноспорангий 151, 203, 215, 2i6, 217, 218 Нанноцит 82, 85 Нематеций 204, 205, 224, 229, 230, 237, 238 Оогоний 20, 40, 146, 151, 183, f87, 192, 209, 214, 258, 272, 290, 338, 344, 345 Ооспора 273, 278, 290, 293, 294, 345, 346, 347, 349, 350 Пал ьмелла 262, 264, 268, 269 Параплектенхима 403, 436 Парафиза 153, 157, 158, 171, 186, 408, 410, 4ii, 414, 434, 439, 443 Парафизоид 407, 434, 443 Партеногонидия 272 Перикарп 227 Периталлий 231, 232, 233 Перитеций 407, 413, 414, 420, 434 Перифизы 413, 414, 436, 437 Перифитон 54, 251, 254, 274, 276, 277 Пикнидий 409, 410, 416, 418, 457 Пикноконидия 409, 410, 416, 418 Планозигота 40, 267, 268 Планококк 82, 83, 85 Плетизмоталлюс 146, 152, 156, 157 Подеций 398, 399, 416, 440, 442, 457, 458 Полиспора 204, 215, 240 Прокарп 208, 227, 228, 229, 240, 24i, 242, 243, 245, 246, 247, 248, 249, 250 Пролификация 192, 193, 226, 238 Проспора 182, 183, 184, 185, 186, 187, 189, 190 Протонема 146, 158 Псевдогомф 396, 397, 399, 405 Псевдоподеций 458 Псе вдосепта 119, 135 Псевдоцифелла 394, 402, 406 Рецептакул 187, 188, 189, 190, 191, 192 Ризина 395, 404, 437, 448, 449 Ризоид 36, 37, 144, 164, 174, 179, 181, 187, 191, 193, i94, 198, 214, 223, 229, 236, 258, 280, 285, 287, 298, 300, 302, 303, 316, 338, 347, 395, 401, 404, 437, 448 Ризом 164, 177, 187, 298, 299, 300, 301 Ризопласт 17, 32 Ризоподий 30, 33, 100, 102, 251, 252, 253 Риз оциста 258 Септа i i8, 121, 135, 316 Синузия 426, 427, 428, 430 Синценобий 276, 278 Синцианоз 73 Сораль 417, 420, 444, 452, 459, 461, 462, 463 Соредий 407, 417, 418, 451, 452, 457, 459, 466 Сорус 163, 164, 205, 224, 237, 241, 249 Сперматангий 205, 206, 209, 214 Спермаций 199, 204, 205, 206, 214, 215, 416 Спермогоний 416 Спора 8, 38, 39, 83, 85, 88, 91, 122, 146, 156, 176, 183, 192, 203, 281, 296, 297, 306, 408, 410, 4ii, 414, 4i5, 418, 434, 439, 443, 445, 447 Сиорангий 38, 144, 146, 149, 152, 156, 158, 1С1, 171, 354 Спор огормогоний 83 Спорофилл 181 Спорофит 40, 42, 146, 147, 151, 152, 156, 161, 162, 163, 165, 166, 167, 169, 170, 171, 182, 206, 209, 210, 215, 223, 283, 284, 297, 354 С пор уляция 415 Стилоспора 416, 418 Стихидий 205, 246, 247, 250 Сцифа 398, 457 Тельца Бючли 113 Тетраспора 39, 145, 163, 164, 203, 209, 215, 218, 219, 224 Тетраспорангий 163, 164, 165, 183, 203, 204, 210, 223, 224, 226, 236, 237, 238, 240, 250 Тетраспорофит 209, 210, 219, 221, 226, 237, 243, 245, "'~0 Трихобласт 193, 246, 250 Трихогина 206, 207, 214, 215, 217, 227, 408, 409, 410 Трихом 79, 81, 83, 88, 90, 9i Трихоциста 18, 93, 94, 97 Трихоцит 201, 231, 232 Фибула 116, 117 Физод 143, 144, 162, 171, 181 Фикобионт 85, 379, 383, 385, 387, 389, 401, 422, 424, 434, 439, 443, 445, 447, 465, 466 Хламидоспора 309, 330 Цекостома 186, 187, 188, 189 Ценобий 271,272,273,276,278,279 Цефалодий 449, 450, 451, 458 Цианелла 73, 75, 76, 97, 98, 352 Цианом 73 Циста 42, 98, 101, 105, 109, 110, 251, 253, 258, 268, 274 Цистокарп 208, 209, 220, 250 Цифелла 406, 452 Экзоаск 443 Экзоспора 38, 82, 85, 87 Экзоспорий 83, 318, 328, 414 Экзохитон 187 Эксципул 412, 413, 414, 440, 444, 446, 452 Эмбриоспора 171, 172 Эндоспора 38, 82, 85, 87 Эндоспорий 83, 318, 328, 414 Эндохитон 187 Эпивальва 98, 99, 114 Эпитека 114, 121 Эпитеций 4И, 412
УКАЗАТЕЛЬ РУССКИХ НАЗВАНЙИ Галаксаура 222 Галосакцион 242 Гарвейелла 229 Гелидиум 213, 222, 223, 366 Гельминтоглея ветвистая 254 Гематококкус 19, 21, 270 Гематомма 426, 427, 460 — ветровая 460, табл. 42 Геминелла 281 Геникулярия 312, 313, 314 Геосифон 74, 75 Гетер онема 260, 261, 265 Гетеропедия многохроматофорная 257 3 Агарум 178 — продырявленный 178, табл. 16 Акароспора 459 — реагирующая 459, табл. 45 — Шлейхера 459 Акросимфитон 226 Акросифония 19, 41 Акрохетиум 216, 217, 218 Актинаструм 48, 278 Актиноптихус 134, табл. 12, 14 Актинотениум 322, 334 Актиноциклус 47, 134, табл. 13 Алария 181, 365 — окаймленная 181 -- полая 182 — съедобная 181, табл. 39 — узкая 182 Алектория 398, 399, 400, 418, 427, 432, 462, 467, 470 — желто-белая 462, табл. 49 Амфироа 234 Амфора 141 Анабена 19, 45, 47, 48, 49, 50, 73, 80, 89 — Шереметьевой 80 Анабеновые 89 — 90 Анабенопсис 90 Анадиомене 306 Аналипус японский 155 А наптихия 404, 463 — реснитчатая 463 Анизонема 261, 265, табл. 27 Анкистродесмус 274, 277 Антитамнион 201, 244 Анфельция 38, 213, 236, 237, 238, 366, табл. 25 Анцилонема 310, 311, 312, табл. 35 Анция 391, 395 Апиоцистис 276, табл. 29 Артония 418, 427, 443 — лучистая 443 Артрокладия мохнатая 169 Артротамнус курильский 179 — раздвоенный 179 Аскозейра 183, 184 — удивительная 184 Аскофиллум 57, 149, 154, 187, 190 — узловатый 54, 59, 186, 190, Аскоциклус 153 Асперо коккус 158 — шипова тый 157 Аспицилия 426, 460 — съедобная 423, 460, табл. 50 Астазия 263, 265 Астерионелла 44, 49, 137 Астеромфалус 134, табл. 13 Астероцитис 193, 214 Ателия эпифильная 464 Аулакодисковые 134 Афанизоменон 45, 47, 49, 90, табл. 3 Афанохете 288 Афанотеце 86 Афиллофоровые 464 Ахнантес 138 Ахнантовые 138 Л цетабулярия 23, 303, табл. 32 Бамбузина 337 Бангиевые 40, 193, 194, 197, 199, 204, 205, 207, 211, 213 — 215 Бангия 193, 194, 212, 214 Батрахоспермум 195, 217, 218 Бахелотия антильская 151 Бацидия 415, 455 — моховая 455 Бациллария 111, 142 — парадоксальная 123, 125 Бесшовные 137 — 138 Биатора 454 — болотная 454 Биддульфия 46, 135, табл. 10, 13 Болбоколеон 288 Боннемезония 219, 220 Ботридиопсис эрийский 255 Ботридиум зернистый 257, табл. 26 Ботриокладия 242 Бриопсис 23, 300, 301 Будлея 306 Буеллия 418, 463 — дисковидная 463 Бульбохете 293 Бумиллериопсис почвенный 255 Вакуольные 275 — 276 Вакуолярия зеленая 97 Валониевые 305 Валонгля 305, 308 — вздутая 305 Верукария 384, 436 — скальная 436 Водяная сеточка 21, 23, 273, 279 Вольвокс 19, 34, 48, 49, 266, 272 — золотистый 273 — шаровидный 273 Воронихиния 45, 73, 87 Вошерия 111, 258
л. 49 427, 314, Гетеросифония 246 Гетерохордария еловая 155 Гиалобрион 105 Гиалодискус 132 Гиалотека 328, 337 Гигартина 238 Гидродиктион 18, 21, 279 Гидрурус 109 Гиелла 71, 384 — дернистая 41 Гимантормия траурная 396, таб Гимногонгрус 236, 238 Гимнодиниум 21, 99, 100 — парадоксальный 99 — темный 99 Гипномонас 275 Гипогимния 395, 407, 409, 416, 461, 469 — вздутая 420, 461, табл. 42 Гиропаигне 265 Гиффордия 149 Глаукоцистис 75, 76, 352 Гленодиниум Борге 41 Глеодиниум горный 99 Глеокапса 60, 61, 62, 72, 86, 384, 388, табл. 2 Глеокапсоморфа 360 Глеотрихия 45, 48, 49, 80 Глеофиллум 276 Глеохлорис планктонный 254 Глифодесмис 137, табл. 14 Голенкиния 44, 278 Гомфо нем а 141 Гомфосферия 87, табл. 3 Гонатоз иго вые 308, 309, 312— 332 Гонатозигон 312, 313, 314 Гониаулакс 45, 46, 47 Гониостомум семяподобный 97 Гониотрихум 193, 214 Гониохлорис тупоконечный 255 Гониум пекторальный 272 Гоногимения 416 Гормогониевые 82, 87, 88 — 92 Грамматофора 138, табл. 14 Грателупия 228 Графис 413, 427 — написанный 446 Грацилярия 239, 240, 366 Дазикладус 302, 304 Дазия 246, табл. 23 Дактилококкопсис 86 Двушовные 139 Делессерия 247, 248 Дентикула 118 Дербезия 299, 300, 301 Дерматокарпон 381, 394, 437 — миниатум 438 — печеночный 438 Десмарестия зеленеющая 168 — шиповатая 168 — язычковая 169 Десмидиум 328, 337 Дефландриоха 108, табл. 9 Диакантос 277 Диатома 50 Диктиоптерис 164 — растопыренный 165 Диктиосифон укроповидный 157 Диктиосфериум 20, 279, 280 Диктиосферия 305, 308 Диктиота 164, 165 — дихотомическая 159, 164 Диктиоха 107, 108, табл. 9 Дилофус 164 Дилсея съедобная 225 Динобрион 48, 49, 105 Динотрикс парадоксальный 100 Динофитовые 93, 95, 96, 98 — 100 Диплошистес 423, 425, 426 Дистефанус 107, 108, табл. 9 Доцидиум 322 Драпарнальдиопсис 287, 350 Драпарнальдия 287, 350, табл. 30 Дурвиллея 183, 185 — антарктическая 185, 186, табл. i6 Дюмонтия 224, 225, табл. 20 Дюналиелла 268, 375 — солевая 70, 268 383, Е грегия 180, 182 3анардиния i 6i Зигнема 18, 3i5, 3i7, Çi8, 320, табл. 36 Зигогониум 320 Зонария 163, 164 Иенгария 160 Иридея 203, 238, 239 Истмоплея ша роно сная 158 Калициум 440 — зеленый 441 Каллимения почковидная 226 Каллитамнион щитковидный 244 Калоплака 418, 426, 427, 462 — настенная 462 — оранжевая 462 — пустынная 462 Каллофилис 226, 227, 228 Калотрикс 92, 384, табл. 3 Кампилодискус 118, 143 Каналошовные 141 — 143 Каннопилус 108, табл. 9 Карпомитра 165, 167 Картерия 75, 270 Катодиниум 99 Каулерпа 294, 298, 299 — прорастающая 298 Киттония 135, Кладония 384, 385, 387, 388, 389, 398, 399, 416, 418, 427, 456, 470 — бесформенная 457 — лесная 399, 457, 467 — мягкая 399 — неприглаженная 457 оленья 399, 457, 467 Кладония приальпийская 399, 457, 467 — Флер ке 458 Кладостефус мутовчатый 163 Кладофора 21, 23, 28, 39, 41, 111, 306, 307, 367 Кладофоропсис 306 Клостериум 23, 49, 322, 323, 324, 327, 328, 330, 331, 332, 333 Кодиолум 156 Кодиум 19, 301, 302 — большой 301 — Риттера 301, табл. 31 — хрупкий 301, табл. 32 Койлодесме цистозейровый 157 Коккоботрис 384, 436, 453 Кокколитофориды 18, 45, 46, 47, 48, 103, 105 — 107, 354, 359 Коккомикса 383, 384, 449, 464 Коккофора Лангсдорфа 192 Колациум 264, 265 Колеохете 31, 289, 29i, 354, табл. 30 — подушковидный 290 Коллема 389, 401, 409, 422, 448 — вялая 448 Кольпомения глубоковыемчатая 160 Кониоцибе 441 — зернистый 441 Конхоцелис 215 Конъюгаты 39, 41, 266, 308 — 338 Кора павонская 464 Кораллина 233 Корбизема 108, табл. 9 Коретрон 44 — 45 Корисциум зеленый 464 Корнуа 108, табл. 9 Коршиковиелла 278, табл. 29 Космариум 49, 323, 325, 327, 330, 335, табл. 37 Космокладиум 324, 328, 336 Костария ребристая 178, табл. 17 К ремас тохло рис 54 Кремастохризис 54 Криптомонады 27, 75, 81, 94, 98 Криптомонас 50, 97 — искривленный 98, табл. 5 — плоскохвостый 98, табл. 5 Криптохризис 98, табл. 5 Круания 243, 244 Круглоплодные 447 — 463 Круория 235 Круцигения 274, 279 Ксантидиум 331, 335 Ксантокапсовые 252, 253 — 254 Ксантококковые 252, 254 — 256 Ксантомонадовые 252, 253 Ксантоподовые 252 — 253 Ксантория 390, 427, 431, 463, 467 — настенная 463, табл. 42 — элегантная 427 Ксантосифоновые 252, 257 — 259 Ксантотриховые 252, 256 — 257 Кутлерия 149, i59, i6i, 162 — усыпанная 162 Лагерхеймия 277 Ламинария 57, 59, 173, 174, 176, 177, 362, 365 — Бонгарда 176, 177 — Гурьяновой 174 длинноствольная 177
Н Ламинария йезоензис 177 — копытная 171, 177 — курчавая 174 — наклонноризоидная 176 — пальчатая 174, 173 — пальчаторассеченная 176 — прижаторизоидная 176, 177 — Родригеса 59, 147, 173 — сахаристая 174 — северная 170, 174, 176 — узкая 174, 176 — японская 40, 57, 174, 175, 176 Лампротамниум 338, 349, 350 Леатезия неоднородная 154, 159 Леканора 426, 427, 432, 460 — коричневая 460 — обломочная 460 — оголенная 460 — пылевая 460 — скальная 460 — толстогубая 460 Леманея 218, 219 Лемфолемма 418 Лепоцинклис 49, 265 Лепрария 380 Лептогиум 40i, 402, 422, 427, 448, табл. 47 — свинцовый 401, 449 Лептозира 384 Лессония 179, 180 — ламинариевидная 179 — чернеющая 179 Летария 398, 405 Лецидея 381, 418, 426, 427, 453, 454 — бородавчатая 453 Лиагора 221 Ликмофора 138 Лингбия 74, 93 Лирамула 108, табл. 9 Литодермовые 153 Литотамнион 232, 357 Литофиллум 232 Лихнотамнус 338, 350 Лобария 395, 422, 451 — легочная 420, 451, 468, табл. 47 Лобококкус 385 Лобомонас 270 Ломентария мелкобулавовидная 241 Макроцистис 169, 179 — грушевидный 179, табл. 16 — цельно листный 181 Малломонас 48, 49 Мастигокладус 68 Мастоглойа 119, 140, табл. i4 Мезоглойа 154 Мезотениум 310, 311, 312 Мелобезия 231, 232, 234 — Лежоли 231 Мелозира 48, 49, 51, 52, 132, табл. 10, 11 — байкальская 49 — исландская 49, 50 Менегация 461 — продырявленная 406, 420, 461, табл. 52 Меноидиум 263, 265 Мерисмопедия 86 Меротрихия головчатая 97 Микопорасиевые 407 Микрактиниум маленький 278 Микрастериас 322, 328, 329, 331, 335, табл. 37 Микродиктион 306 Микрозония 161 Микрококкус 93 Микроколеус 62 Микроспонгиум 153 Микроспора 283 Микроцистис 45, 48, 49, 50, 53, 86 Миксохлорис 74 — сфагновый 253 Миксохризис парадоксальный 103 Мир ионема 144, 153 Мирмеция 383, 436, 453 Мицция 358 Мишоккоккус конфервный 254, табл. 26 Монострома 285, 286 — Гревиля 285, табл. 3i Мужоциопсис 309 Мужоция 315, 3i8, 319, 320 Мультиклавула заплесневелая 464 Навикула 139, табл. i7 Навикулопсис 108, табл. 9 Наутококкус 54 Нейропогон 397, 399, 400, 404, 462 — пучковатый 462, табл. 49 Немалион 221 — червеобразный 22i Нем астомовые 236 Нематохризис сидячий 110 Неомерис 303 Нерейя нитевидная 166, 167 Нереоцистис 169, 181 — Лютке 170, 181, табл. 16 Нетриум 310, 311, 312, табл. 35 — пальцев идный 311 Нефрома 410, 451 — арктическая 451, табл. 49 Нефромопсис 410 Нителла 27, 110, 339, 340, 344, 345, 349, 350, 351, табл. 38 Нителлопсис 338, 349, 350, 351 Нитофиллум 249 Нитцшия 142, табл. 14 Нодулярия 47 — 48 Носток 60, 64, 66, 72, 75, 84, 383, 384, 385, табл. 3, 4 — войлочный 365 — обыкновенный 365 — сливообразный 80, 365 Нотейя 183, 188 — аномальная 188 Одношовные 138 — 139 Одонталия 250, табл. 23 Одонция двухцветная 464 Одуинелла 216 Оокардиум 324, 331, 336 Ооцистис 19, 48, 76, 274, 277 Опеграфа 413, 427, 444 — тонкокрайная 445 Осциллатория 19, 20, 46, 50, 80, 92, табл. 3 Офиоцитиум 256 Падина 165 — павлинья i59, ,165, табл. 17 Пальмелла 20 Пандорина 49, 272 Пармелиопсис сомнительный 461 Пармелия 384, 404, 418, 426, 427, 431, 460, 478 — блуждающая 396, 461, табл. 50 — бороздчатая 461 — извилистая 461 — козлиная 461, табл. 47 — окрашенная 424 — оливковая 461 — скальная 461 — темно-бурая 461 — узколистная 461 — усыпанная 461 — центрифужная 425, 461 Педиаструм 44, 48, 274, 279 Пединопера 270 Пединоперопсис 270 Пейсоннеллия 229, 230 Пелагофикус 144, 181 — пора 181 Пельвеция 187, 188 — желобчатая 183, 189 — равновершинная 186 — Райта 189 Пельтигера 388, 394, 395, 396, 404, 406, 422, 427, 449 — пупырчатая 450, табл. 42 — собачья 450, 468, табл. 49 Пениум 313, 325, 332, 333 Пер анема 265 Пер идиниум 48 — опоясанный 99 Перкурсария 284 Перониелла 256, табл. 26 Пертузария горькая 459 — пальчатая 459 — шариконосная 460, табл. 54 Петаломонас 261, 265, табл. 27 Петалония зостеролистная 160 — лентовидная 160 Петродерма 384 Пикнотелия 456 Пилайелла 150 — прибрежная 146, 150, табл. 18 Пиннулария 139 Пир амимонас сетчатый 268, 269 Пир гидиум 448 Пиренокарповые 434 Пиренула 435 — блестящая 435 Пироботрис стройный 271 Плаколеканора щитовидная 460, табл. 44 Планктониелла 44, 45 Платома 236 Плевдорина 34 Плеврококкус 384, 436, 437, 453 Плевр осигма 141 Плеуротениум 325, 332, 333 Подолампас 46 Подоцистис 137, табл. 14 Полиблефаридовые 267, 268 — 269 Полисифония 209, 210, 249, 250 — шерстистая 54 Политомовые 269 Полиэдриопсис 278 Порфира 20, 40, 193, 194, 198, 205, 212, 214, 365, табл. 20 475
Порфиридиум 193, 211, 213 Порфиропсис 194 Празинокладус 31 Празиола 41, 295 — японская 296 Примнезиум маленький 101 Црингсхеймиелла 288, табл. 30 Протококкус 464 — зеленый 280 Протоси@он 279, табл. 29 Про'готриховые 275, 279 — 281 Протохлориновые 267 Псевдобриопсис 301 Псевдоколиум 441 Псевдоплеврококкус 384 Псевдостаураструм копьевидный 255 Псевдохлорелла 384, 453 Псевдэверния 427 Псора 454 — лестничная 455, табл. 46 — обманчивая 454, табл. 46 Птеромонас 270 Пугетия 226 Пунктария 57, 156 — широколистная 556, 157 Рабдомонас 265 Ральфсия бородавчатая 153, 155 Рамалина 426 — сетчатая 425 Рафидонема 69, 280 — вечнозеленая 280 — снежная 69, 280 Ривулярия 71, 92, 384 Ризокарпон 423, 426, 427, 455, 456 — географический 425 — окрашенный 456, табл. 51 Ризоклониум 308, 367 Ризосоления 45, 46, 48, 49, 134 Ризохлорис 253 — глазковый 252 Ринодина горная 463 Родимения 212, 241, 242, 243, 365 — пальчатая 241 — продырявленная 212, 242, табл. 21 — узкоугольная 241, табл. 22 — четкообразная 242 Родоглоссум 238, 239 Родомела 250 Родомонас 98 Родохортон 216 Розенвингиелла 295 Ройя 310, 311, табл. 35 Ропалодия 118, 142, табл. i i Рочелла 379, 444 — фукоидная 444 Саккориза кожистая 173 — многополосная 173 Саргассум 144, 147, 180, 186, 191, 362 — бледный 191 — Миябе 187, 191 — плавающий 191 — погруженный 191 Селенаструм 48, 278 Силикофлагеллаты 45, 47, 103 Синедра 137 Синехококкус 86 Синехоцистис 32, 74, 86 Синура 49 Сирогониум 316, 319, табл. 36 Сир окладиум 320 Сифоновые 16, 19, 23, 59, 273, 280, 297 — 308, 354, 358 Сифонокладус 306 — крохотный 306 — тропический 306 Скафоспора красивая 151 Сквамариевые 204, 229 — 230 Солениевые 134 Соленопоровые 210 Солорина 384, 395, 416, 450 — Скортея 395, табл. 42 — шафранная 450, табл. 42 Сораструм 48 Сорокарпус 151 Сперматозопсис прыгающий 268 Сперматохнус 156 Спирогира 23, 24, 28, 315 316 317 318, 319 Спиротения 309, 310, 311 — сжатая 310 Спирулина 93, 366, 375 Сплахнидиум 152, 156, 183 — морщинистый 156 Спондилозиум 336 Спондиломорум четверной 271 Спорохнус 165, 166 Стаураструм 49, табл. 37 Стауродесмус 336 Стауротеле 416, 436 — спрятанная 436 Стереокаулон 427, 458 — войлочный 458, табл. 56 Стефанодискус 48, 132, табл. 13 Стефанопиксис 132, табл. 15 Стефаносфера дождевая 271 Стигеоклониум 23, 286, 287, табл. 30 Стиго нема 60, 72, 88, 383, 384, табл. 2 Стикта 395, 422, 452 — Райта 452, табл. 47 Стиктиосифон 158 — скрученный 158 Стиктовые 451 — 452 Стилодиниум 100 Стилофора 152 Стипокаулон метловидный 163 Стихококкус 281, 384, 439 Стреблонема 144, 155 Стриария оттянутая 158 Стромбомонас 260 Струвея 306 Сурирелла 118, 143 Сфацелярия 38, 162, 163 Сферозосма 324, 337 Сфероплея 41, 293, 294 — кольцевидная 293 Сферотрихия развесистая 155 Сферофорус круглый 442 Сфероцистис 48 Сфинктрина 440 Схизогониевые 281, 295 — 297 Схизогониум 295 Сцелетонема 47, 132, табл. 13 Сценедесмус 19, 48, 274, 278, 368, 369, 375 Сцеплянки 273, 308, 315 Сцитонема 60, 72, 90, 383, табл. 2 Сцитосифон 160 — коленчатый 157, 159, 160 Табеллярия 48, 49, 51, 138 Талассиозира 45, 46, 47, 132 Талассиофиллум решетчатый 178, табл. 16 Тамнолия 397, 404, 458 — червеобразная 397, 458, табл. ~ 9. 51 Тейлингия 324, 336 Телошистес 462 — желтеющий 463 — ямчатый 463 Темногаметум 320, табл. 36 Тетмеморус 334 Тетрадиниум 97, 100 Тетраспоровые 275 Тетраструм 278 Тетраэдриелла гигантская 255 Тетраедрон 48, 277 Тилоптерис Мертенса 152 Толипотрикс 60, 91 Толипелла 338, 350, табл. 33 Трайлиелла 220 Трахеломонас 48, 49, 54, 260, 263, 264 Требуксия 383, 384, 388, 389, 439, 466 Трейбария 277 Трентеполия 60, 288, 290, 384, 434 Трибонема 111, 256 — зеленая 256 Триплоцерас 334 Трицератиум 135, табл. 12 Трохисция 383, 436 Турбинария 180, 191 Удотея 299 Улотрикс 23, 38, 282 Улотрикс опоясанный 282 — повислый 283 Ульва 41, 285, 365, табл. 31 Умбиликария 423, 426, 427, 459 — короткошерстная 459, табл. 51 — Мюленберга 459, табл. 42 — съедобная 459, 467 — цилиндрическая 425 Ундария 182, 365 — перистая 182 Уроглена 48 Урогленопсис 49 Уронема 23 Урцеолус 265, табл. 31 Уснея 379, 399, 400, 404, 405, 406, 431, 462, 467, 470 — длиннейшая 397, 462, табл. 49 Факотус 20, 270 Факус 48, 49, 260, 262, 265 Фельдманния 151 Феодермациум речной 111, табл. 6 Феозооспоровые 149 — 182 Феококкус 97 Феоплакс 97 Феотамнион 111, табл. 6 Физолинум 384 Фикодрис 248
Фикопельтис 384, 422 Филлариелла охотоморская 177 Филлария 173 Филлокардиум уплощенный 268 Филлофора 38, 204, 213, 236, 237, 364, 366 — Броди 41 — перепончатолистная 237 Фисция 426, 427, 432, 463 — голубовато-серая 463 — моховая 463 — припудренная 463 Формидиум 62, 68 Фрагилария 48, 49, 51, 137 Фритчиелла 288 Фукус 20, 57, 59, 148, 165, i84, 188, 189, 361 — двусторонний 189, табл. 18 — зубчатый 189 — исчезающий 189 — пузырчатый 185, 187, 189, табл. i8 — спиральный 189 Фурцеллярия 38, 159 Фурцилла 270 Фэурус 168 Халидрис 191 — стручковатый 191 Халимеда 55, 299, 358 Халицистис 299, 300, 301 Халосакцион 154 Хамесифон 87 Хапалосифон 88 Хара 338, 34i, 342, 343, 344, 345, 346, 347, 348, 349, 350, табл. 38 — ломкая 348 Харациопсис 255 Харациум 278 Харофиты 338 — 350 Хемиаулус 135, табл. 15 Херибаудиелла 153 Хетангиевые 220, 222 Хетоморфа 308 Хетоптерис перистый 163 Хетофора 92, 287 Хетоцерос 45, 46, 47, 48, 49, 121, 134, табл. 10 Хидроклатрус 160 Хилокладия чашевидная 241 Хиломонас 98, табл. 5 Химанталия 187, 188 — удлиненная i80, 188 Хламидомонада 19, 20, 48, 49, 54, 69 — снежная 69 Хлорангиелла 276 Хлорангиопсис 276, табл. 29 Хлорелла 15, 23, 32, 60, 66, 75, 277, 367, 368, 369, 370, 372, 373, 375, 376, 383 Хлоркорона богемская 271 Хлороглея сарциноидная 70, 71 Хлорокардион зеленобокий 253 Хлорококкум 27, 60, 277, 384, табл. 2 Хлоромонадофитовые 93, 94, 95, 96— 97 Хлоросаккус ульвовый 254 Хлоросарцина 384, 453 Хлоротециум 255, табл. 30 Хондрус 193, 203, 213, 238, 239, табл. 20, 2i Хорда нитевидная 174 — пушистая 174 Хордария плетевидная 154, 155 Хореоколакс 229 Хормозира 183, 188 — Банкса 180, 188 Хормофиза трехгранная 190 Хризамеба лучистая 101 — 103 Хризимения 242, табл. 21 Хризокапса 109 Хризококкус 50 Хризомонадовые 101, 103 — 108 Хризоподовые 101 — 103 Хризосферовые 101, 109 — 110 Хризотриховые 101, 110 — 111 Хромулина 50, 53, 104 Хроококкус 76, 383, 384 Хроомонас 98 Хроотеце 214 Целосфериум 87 Центритрактус копьеносный 256 Церамиум 245 Церациум 46, 48 — ласточковый 99 Цетрария 416, 427, 461 Цетрария исландская 381, 397, 400, 402, 406, 416, 461, 467, 468, 469, табл. 48 — клубочковая 398, 461, табл. 48 — морщинистая 420 — Ричардсона 396, 461, табл. 47 — сглаженная 406 — снежная 461 — сосновая 461 — Тилезия 461 Цефалейрос 384 Цианомонас 98 Цианофора 75, 98 — парадоксальная 75 Циклидиопсис 265, табл. 27 Циклоспоровые 40, 41, 147, 148, 149, 182 — 192 Цилиндрокапса 294, 295 Цилиндроспермум 90 Цилиндротека 118 Цилиндроцистис 310, 311, 312, табл. 35 — Бребиссона 311, табл. 35 Циматэре двускладчатая 177 — трехскладчатая 178 — японская 177 Цистодиниум Штейна 99 Цистозейра 148, 186, 187, 190, 362 — бородатая 180, 190 — косматая 180, 190 — толстоногая 190 Цифелиум 441 — брусковый 442 Челлманиелла толстолистная 177 — тонколистная 177 7ампия 241 7изим ения 236 7изотрикс 60, 92, табл. 2 7одателла 48 Эвглена 23, 48, 49, 54, 75, 259, 262, 263, табл. 28 — зеленая 259 Эвгленокапса 264 Эвдорина 49, 272 Эверния 423, 427, 462, 467, 470 — прунастри 420 — сливовая 462, 469, 470, табл. 49 Эвкампия 46 Эвтрепция 265 Эдогониум 20, 23, 39, 291 Эклония трубковидная 182 Эксувиелла 49 Эктокарпус 41, 150 — драпа рнальдиевидный 150 — конфервообразный 145 — пучковатый 150 — стручковатый i 46, 150 Элакатотрикс студенистый 280 Элахиста фукусовая 153 Эндодерма 288 Эндокарпон 416, 438 — мелкий 438 Эндокладия 204 Энтероморфа 284, 285, табл. 31 Энтонема 151 Энтосифон 263, 265 Эпитемиевые 118, 141 — 142 Эпитемия 118, 143 Эпифитон 358 Эпихризис болотный 109 — нителловый 110 Эремосфера 23 Эритрокладия 194 Эуаструм 49, 322, 329, 330, 334, табл. 37 Эудесме 154 — зеленоватый 154 Эукапсис 87
УИАЗАТЕЛЬ ЛАТИНСКИХ НА3ВАНИЙ табл. 44 л. 14 16 табл. 49 85, 89, Acarospora molybdina 459, — reagens 459, табл. 45 — schle icheri 459 Acarosporaceae 459 Acetabularia 303, табл. 32 — mediterranea 23 Achnanthaceae 138 Achnanthes 123, 126, 138 — brevipes айаг. parvula 139 Acrochaetiaceae 215 — 217 Acrochaetium 216 Acrosimphyton 226 Actinastrum 48, 278 — hantzschii 279 Actinocyclaceae 134 Actinocyclus 134, табл. 13 Actinod iscales 133 — 134 Actinoptychaceae 134 Actinoptychus 128, 134, таб Actino taenium 322, 334 — cucurb i t inurn 321, 334 Agarum cribrosum 178, табл Ahnfeltia 237, 238 — pl i ca t a 366, табл. 39 Alaria 181 — angusta 182 — esculenta 181, табл. 39 — istulosa 182 — marginata 181 Alectoria 427, 462 — ochroleuca 398, 462, 467, Amphidinium extensum 96 — ge itleri 95 A mph iroa 234 Amphisolenia 46 Amphora 126, 128, 141 — coffeaeformis 140 Anabaena 79, 80, 82, 83, 84, табл. 3 — az о11ае 73, 85 — circinalis 84, 89 — cylindrica 15, 84 Anabaena f los-aquae 84, 89 — hassalii 89, табл. 3 — lemmermannii 45, 84, 89 — scheremetievii 45, 80, 84, табл. 4 — чаг~аЬ~1~я 15, 89, 90 Anabaenopsis 81, 84 — raciborskii 90 Anadyomene 306 Anal ipus j aponicus 155 Anaptychia 463 — ciliaris 463 Ancylonema 310, табл. 35 — nordenskjoldii 69, 312 Anisonema 265, табл. 27 Ankistrodesmus 277 — acicularis 277 Antithamnion 201, 244, 245 Anzia 395 Aphanizomenon 79, 90, 375 — f los-aquae 45, 90, табл. 3 Aphanochaete polychaete 289 Aphanothece 86 — elabens 86 — stagnina 86 Apiocystis brauniana 276, табл. Araphales 137 — 138 Arthonia 414, 443 — radiata 443, 444 Arthopyrenia kelpй 384 Arthrocladia ч~11ояа 169 Arthrothamnus bifidus 179 — curilensis 179 Ascocyclus 153 Ascolichenes 434 — 463 Ascophyllum 149, 190, табл. 23 — nod osum 54, 59, 190 Ascoseira mirabilis 184 Ascoseirales 184 Ascoseirophycidae 184 Asperococcus 158 — echinatus 157 Aspicilia 426, 460 — esculenta 460, табл. 50 Astasia 265 Astasia inflata 264 Asterionella 44, 124, 137 — formosa 136 89, — gracillima 126 Asterocytis 214 Asteromphalus 134 Athelia epiphylla 464 Audouniella 199, 216 Aul acod iscales 131, 134 Aulonoraphales 137, 141 — 143 B acheloti a 151 — antillarum 151 Bacidia 427, 455 — marginalis 415 — muscorum 455 Bacillaria 128, 142 — paradoxa 123, 125, 126, 142 Bacillariophyta 9, 12, 60, 111 — 143 Bambuzina 337 — brebissonii 329, 337 1angia 214 29 Bangiales 213 Basidiolichenes 463 — 464 Batrachospermum 217 В iatora 427, 454 — uliginosa 454 Biddulphia 46, 124, 135, табл. 10, 13 — aurita 135 — тоЫ1iensis 121, 124 Biddulphiales 131, 134 — 135 Bolbocoleon 288 — piliferum 289 Bonnemaezonia 219 Bonnemaezoniaceae 219 Boodlea 306, 307 Bornetella 304 Botrydiopsis eriensis 255 Botrydium granulatum 257, табл. 26 Botryococcus braunii 361 Bryopsis 300 — halymeniae 23
57, 467, 467 408 128, 3, 105— л. 14 124 467, 468, 30 121, л. 10 . 24 табл. 33 479 Bryopsis hypnoides 23 Buellia 414, 463 — disciformis 463 Bulbochaete 292, 293 Bumilleriopsis terricola 255 Caliciales 415, 439 — 442 Calicium 440 — viride 439, 441 Callithamnion 207, 244 Callophyllis 198, 226 Са1ор1аса 462 — aurantfaca 462 — decipiens 418 — desertorum 462 — murorum 462 Calothrix 35, 81, 91, 384 — braunii 91, 92 — gypsophila 91, табл. 3 — fusca 91, 92 — thermal is 91, 92 Campylod iscus 117, 118, 126, 142 — daemelianus 142 Cannopilus 108, табл. 9 Carpomitra 167 — costata 167 Carteria 75, 270 — crucifera 269 Caulerpa 37, 298 — ргоИега 298 Centritractus belonophorus 256 Centrophyceae 45, 131 — 135 Ceramiales 243 — 251 Ceramium 199, 244, 245. табл Ceratium 46, 48 — arct icum 96 — hirundinella 95, 99 — tripoc 96 Cetraria 461 — cucculata 398, 433, 461, табл. 48 — islandica 381, 402, 461, 467, табл. 48 — laevigata 406 Cetraria nivalis 433, 461, 467 — pinastri 417, 420, 462 — richardsonii 396, 461, табл. — tilesii 462 Chaetangiaceae 222 Chaetoceros 45, 46, 48, 49, 120, 124, 125, 128, 134, 135. таб — abnormis 123 — compressus 123 — curvisetus 120 — danicus 120 — lorenz ianus 123 — paulseni i 123 — rigidus 123 — seiracanthus 123 — subsecundus 123 Chaetomorpha 307, 308, табл. 32 Chaetophora 287 Chaetophorales 281, 286 — 291 Chaetopteris 163 — plumosa 163 Chamaesiphon 83, 85, 87 — curvatus 87, табл. 3 Champ ia 241 Chara 37, 338, 345 — aculeolata 343 Chara aspera 346 — baltica 346, 349 — braunii 341 — canescens 343 — contraria 343 — corallina 31 — fragif era 346 — fragilis 341, 343, 348 — gymnophylla 349 — gymnopitys 341 — hispida 341 — rudis 343 — vulgaris 341, табл. 38 Characiochloris characioides 275, 276 Characeac 350 Characiopsis 255 — acuta 256 — subulata 256 Characium 34, 278 — acuminatum 278 Chilomonas paramaecium 98, табл. 5 Chlamydomonadales 267, 269 — 270 Chlamydomonas 29, 34, 269 Chlamydomonas annuraeae 269, 270 — nivalis 69 — pertusa 30 — proboscigera 269 Chlorangiella pygmaea 276 Chlorangiopsis epizootica 276, табл. 29 Chlorarachnion reptans 252 Chlorcorona bohemica 271 Chlorella 32, 274, 277, 369, 372, 374, 384 — vulgaris 63, табл. 2 — pyrenoidosa 23 Chlorocard ion pleurochloron 252, 253 Chlorochytrium lemnae 74 Chlorococcales 275, 276 — 280 Chlorococcum 15, 34, 277, 384, табл. 2 Chlorogloea sarcinoides 70, 71 Chloromonadophytina 96 — 97 Chlorophyta 9, 12, 266 — 338, 368 Chlorothecium 255, табл. 26 Chondrus 238, 239, табл. 20 Chorda 174 — filum 174 — tom entosa 174 Chord aria 154 — flagelliformis 154, 155, табл. 18 Chordariales 152 — 156 Choreocolax 229 Chromulina 50, 53 — rosanoffé 103, 104 Chroococcales 85 — 87 Chroococcus 75 Chroomonas 98, табл. 5 Chroothece 194, 214 Chrysamoeba 33, 101 — 103 — radians 101, 103 Chrysocapsa sordida 109 Chrysomonadophycidae 103 — 105 Chrysophyta 9, 12, 100 — 111, 129 Chrysopodophyceae 101 — 103 Chrysosphaerophyceae 101, 109 — 110 Chrysotrichophyceae 101, 110 — 111 Chrysymenia 242, табл. 21 Chylocladia 241 — kaliformis 241 Cladonia 398, 456, 457 — alpestris 399, 425, 457, 467, табл. 48, 56 — coccifera 468 — con iocraea 398 С1айоп~а deformis 457, табл. — Гimbriata 398 — florkeana 458, табл. 48 — furcata 399 — impexa 457 — mitis 399, 457 — rangiferina 399, 425, 4 табл. 56 — subulata 398 — sylvatica 398, 425, 457, — verticillata 398 Cladophora 306, 307 — glomerata 41 Clad ophoropsis 306 Clad ostephus 163 — verticillatus 163 Clavariaceae 464 Clavulinopsis septentrionalis Closterium 49, 322, 333 — acerosum 332, 334 — ehrenbergii 23, 325 — leibleinii 332, 334 — lunula 332, 334 — manschuricum 321 Coccobactreaceae 85 — 86 Coccolithophoridophycidae 10 107 Coccomonadinaceae 270 Cocconeis 126, 128, 138, таб — reticola 126 Co ceo phora 192 — langsdorfii 192 Codiolum 156 Codium 301, 302 — fragile 301, табл. 32 — magnum 301 — ritteri 301, табл. 31 Coelastrum 34, 277 Coelosphaeriaceae 87 Coelosphaerium 87 — kuetzingianum 86, 87 Coenobiodiscus muriformis Coilodesme 157 — cystoseirae 157 Colacium 262, 264 — arbuscula 263 Coleochaete 289, 291, табл. — pulvinata 290 Collema 389, 401, 448 — flaccidum 401, 448 Colpomenia 160 — sinuosa 160 Coniocybe 441 — furfuraceae 439, 441 Соп jugatophyceae 308 — 337 Cora pavonia 408, 464 Corallina 204, 233, 234, табл Corbisema 108, табл. И Corethron 44, 45, 125 Coriscium viride 464 Cornua 108, табл. 11 Corticiaceae 464 Coscinodiscales 131 — 133 Coscinodiscus 128, 129, 133, 15 — jonesianus 120 — gigas 133 Cosmarium 49 — botrytis 327 — caelatum 335 — de pressum 325 — portianum 329 — protuberans 322, 335 — quadrum 324
480 Cosmarium quinarium 335, табл. 37 — reniforme 335, табл. 37 — subtumidum 335, табл. 37 Cocmocladium 324, 336 Cosmocladium pusillum 336, 337 — saxonicum 336, 337 Costaria 178 — costata 178, табл. 17 Crouania 243, 244, 245 Crucigenia 280 Cruoria 204, 235, 236 Cryptochrysis 98, табл. 5 Cryptomonas 50, 97 — curvata 98, табл. 5 — platyuris 98, табл. 5 Cryptonemiales 224 — 235 Crypto phytina 97 — 98 Cutleria 149, 159, 161 — adspersa 162 Cutleriales 160 Cyanomonas 98, табл. 5 Cyanophora 98 — paradoxa 76, 98, табл. 5 — tetracyana 98, табл. 5 Cyanophyta 9, 12, 78 — 92 Cyclocarpales 447 — 463 Cyclosporophyceae 148, 182 — 192 Cyclotella 49, 123, 124, 126 — meneghiniana 126 Cylindrocapsa 294, 295 Cylindrocapsales 281, 294 — 295 Cylindrocapsopsis 294 Cylindrocystis brebissonii 311, табл. 35 — sculpta 31 i Cylindrospermum 81, 82 — licheniforme 63 Cylindrotheca 117 Cymathere 177 — fibrosa 177 — japonica 177 — triplicata 178 Cyphelium 441 — tigilare 440, 442 Cystoclonium 199, 202 Cystodinium 99 — steinii 95, 99 Cystoseira 148, 186, 190 — barbata 180, 190 — crassipes 190 — crinita 180, 190 Dactylococopsis 86 — rhaphidioides 86, табл. 3 Dasya 246, табл. 23 Dasycladales 302 — 305 Dasycladus 302, 303 Deflanodryocha 108, табл. 9 Delesseria 247, 248 Denticula 118, 128 Derbesia 299, 300 — neglecta 30 Dermatocarpaceae 437 — 439 Dermatocarpon 394, 427, 437 — hepaticum 438 — miniatum 438 Desmarestia 168 — aculeata 168 — ligulata 169 — viridis 168 Desmarestiales 167 — 169 Desmidium 337 — swartzii 329, 337, 337 Desmidiales 320 — 337 Diatoma 50, 124, 126 — e ion ga turn 124 D iacanthos 278 Diacanthos belenophorus 273 Dictyocha 107, 108 — fibula 107, табл. 9 Dictyopteris 164 — divaricata 165 Dictyosiphon 157 — foeniculaceus 157 D ictyosiphonales 156 — 158 D ictyosphaeria 306 — favulosa 306 Dictyosphaerium 277, 279 Dictyota 164, 165 — dichotoma 159, 164 Dictyotales 163 — 165 Dilophus 164 Dilsea edulis 225 Dinobryon 29, 48, 49, 101, 105 — divergens 104, 105 Dinophysis 45, 46 — baltica 47 Dinophytina 98 — 100 Dinothrix 94 — paradoxa 95, 100 Diploschistes 425 — scruposus 423 D iraphales 136, 139 — 141 Distephanus 107, 108 DЬИдта 265 — proteus 264 Docidium 321, 333 — baculum 333 — undulatum 321 Draparnaldia 287, 350, табл. 30 Draparnaldiopsis 287, 350 Dumontia 224, 225, табл. 20 Dunaliella 268, 375 — salina 70, 268, табл. 29 D urvillea 185 — antarctica 185, табл. 16 Durvilleales 185 Ecklonia 182 — buccinal is 182 Ectocarpales 149 — 151 Ectocarpus 150 — confervoides 144 — draparnald io id es 150 — fasciculatus 150 — siliculosus 146, 149, 150 Egregia 180, 182 Elachistea 153 — fucicola 153 Elakatothrix gelatinosa 280 Endocarpon 416, 427, 438 — pusillum 438 End od erma wit trocki i 289 Enteromorpha 284, табл. 31 Entonema 151 — aecidioides 147 Entophysalidales 87 Entosiphon 264, 265 Epichrysis nitellae 109, 110 — pa iud osa 109, 110 Epyphyton prapusillum 358 Epithemia 118, 142 — sorex 140 Erytrocladia 194 Euastrum 49, 322, 335 — divaricatum 322, 335 — verrucosum 335, табл. 37 Eucapsis 82, 87 — alpina 86, 87 Eudesme 154 — virescens 154 Eudorina 33, 49, 272 — elegans 272 Euglena 29, 48, 49, 54, 259, 263 — acus 262 — convoluta 262 — geniculata 262 — gracilis 16, 75, 262 — mutabilis 259 — oxyurus 262 — pisciformis 262 — proxima 30 — sanguinea 261, табл. 28 — viridis 259, 264 Euglenales 265 Euglenocapsa 264 Euglenophyta 9, 12, 259 — 265 Eutreptia 265 — pyrenoidifera 262 Evernia 462, 470 — prunastri 420, 427, 428, 462, 470, табл. 49 Feldmannia 151 Florideophyceae 215 — 251 Fragilaria 48, 49, 51, 123, 136 — pinnata 136 Fragilariaceae 128, 136 — 137 Fritschiella 288 — tub erosa 288 Fucus 147, 149, 184, 189 — distichus 57, 189, табл. 18 — evanescens 189 — serratus 189 — spiral is 189 — vesiculosus 57, 185, 187, 189, табл. 18 Furcellaria 196 Furcilla 270 — bicaudata 269 Gal axuara 222 Gelidiales 222 — 224 Gelidium 196, 204, 206, 223, табл. 39 Geminella 281 Genicularia 312 — spirotaenia 314, табл. 35 Giffordia 149 Gigartina 238 Gigartinales 235 — 240 Glaucocystis nostochinearum 76, 352 Glenodinium 29 — borgei 41 Gloeocapsa 82, 84, 85, 86, 383 — alpina 86, 87, табл. 2 — limnetica 86 — magma 86, 87, табл. 2
pluviа1Ь 270, Gloeocapsa minuta 86 — rupestris 86, 87, табл. 2 — turgida 87, табл. 3 Gloeocapsithos 360 Gloeocapsomorpha prisca 360 Gloeochloris planctonica 253, 254 Gloeocystis 384 Gloeod inium montanum 99 Gloeophyllum fimbriatum 276 Gloeothece 87 — palea 86, 87 Gloeotrichia 35, 81, 82, 83, 91 — echinulata 45, 84, 91, табл. 4 — natans 92 — pisum 91, 92 Glossod ium 456 Glyphodesmis 137, табл. f 4 Golenkinia 44, 278 — rad iata 278 Gomphocymbellaceae 141 Gomphonema 124, 126, 141 — lanceolatum 140 Gomphosphaeria 87 — aponina 87, табл. 3 — lacustris 87 Gonatozygales 312 — 314, 321 Gonatozygon 312 — aculeatum 313 — brebissonii 313, 314 — kinahanii 313, 314 — monotaenium 313, 314, табл. 35 Goniaulax 45, 46 Goniochloris mutica 255 Goniostomum semen 94, 97 Goniotrichum 194, 214 — cornu cervi 214 Gonium 272 — pectorale 272 Gracilaria 204, 239, 240 Grammatophora 124, 126, 128, 138 — marina 118, 137 — serpentina 118, табл. 14 Graph id ales 445 — 447 Graphis 413, 446 — scripta 446, 447 Grateloupia 228 Gymnocarpeae 434, 439 — 463 Gymnodinium fuscum 95, 99 — paradoxum 95, 99 Gymnogongrus 238 Gyropaigne 265 — cosmos 264 Haematococcus табл. 29 Haematomma 460 — чеп1ояпп 460, табл. 42 Halicystis 299, 300 Halidrys 191 — siliquosa 191 Halimeda 298, 299, табл. 33 Halosaccion 242 Hapalosiphon 88 — fontinalis 88 Haplospora globosa 151 Harveiella 229 Helm inthoclad iaceae 221 — 222 Helminthogloea ramosa 253, 254 Helminthora 207 Hemiaulus 124, 128, 135, табл. Heribaudiella 153 Heterochordaria abietina 155 Heterococcus 384, 422 Heteronema 265 — acus 264, табл. 27 Heteroped ia polychloris 257 Heterosiphonia 246 Himanthalia elongata 180, 188 — lorea 31 Himanthaliaceae 188 Himantormia lugubris 396, табл. 49 Hormogoniophyceae 82, 88 — 92 Hormophysa 190 — triquetra 190 Hormosira banksii 180, 188 Hyalobryon ramosum f 04, 105 Hyalod iscus 132, табл. 10 — sphaerophorus 131 Hyalotheca 328, 337 — mucosa 324 — dissiliens 329, 337 Hydroclatrus 160 Hydrodictyaceae 279 Hydrodictyon reticulatum 273, 279 Hydrurus 34, 109 — foetidus 109, табл. 6 Hyella 384 — caespitosa 41, 71 Hypnomonas chlorococcoides 275, 276 Hypogymnia 395, 461 — bitteri 417 — physodes 420, 427, 428, 461, 569, табл. 42, 52 Iengaria 160 Iridaea 204, 238 Istmoplea 158 — sphaerophora 158 Kallymenia 226, 227 — reniformis 226 Kittonia 128, 135, табл. 12 Kjellmaniella 177 — crassifolia 177 — gyrata 177 Korschikoviella 278, табл. 29 Kremastochloris 54 Kremastochrysis 54 Kylinia 195, 199 Lagerheimia 274 — citriformis 278 Ьaminaria f 69, 174 — angustata 176 — appressirhiza 176 — bongardiana 176 — cichorioides 174 — digitata 176 — gurjanovae 174 — hyperborea 170, 176 — inclinatorh zа 176 — japonica 57, 174, табл. 39 — longipes 177 — rodriguez й 59, 147 — saccharina 174 — solidungula 172, 177 Laminaria yezoensis 177 Lam inari ales 169 — 182 I amprothamnium 338, 350 — papulosum 350 Lauderia 113, 124, 125 Leathesia difformis 154, f59 Lecanora 411, 460 — conizaevides 460 Lecanora frustulosa 460, табл. 43 — glabrata 460, табл. 53 — pachycheilea 460, табл. 54 — polytropa 433 — rupicola 460, табл. 53 — subfuscata 460, табл. 53 Lecidea 412, 427, 453 — crustulosa 381 — glomerulosa 453, 454 — granulosa 427 — paschalis 426 — uliginosa 427 Lemanea 195, 218 Lempholemma minutulium 418 Lepocynclis 49, 265 — marssonii 262 — ovum 264, 265 Lepraria 380 Leptocylindrus 123 — danicus 124 Leptogium 402, 448 — saturninum 40f, 449, табл. 44 Lessonia 179 — laminariaeoides 179 — nigrescens 179 Leth aria 405 — thamnodes 398 Liagora 221 Lichen aphthosum 468 — caninus 468 — cinereus terrestris 468 — cocciferus 468 — islandicus 468 — plicatus 468 — pulmonarius 468 — saxatilis 468 Lichenes imperfecti 380 Licmophora 118, 123, 124, 126, 138, табл. 10 — ehrenbergii 118 Liramula 108, табл. 9 Lithodermataceae 153 Lithophyllum 232 Lithothamnion 232, 357 Lobaria 395, 451 — pulmonaria 420, 451, 452, 468, 470, табл. 47 Lobomonas 270 — stellata 269 Lomentaria 241 — clavellosa 241 Lychnotzmnus 338, 350 — barbatus 349 Lyngbia 91 — aestuarii 91 Lyngbia confervoides 93, табл. 3 — end obiotica 73 — end ophytica 74 M acrocyst i< 148, 179 — integrifolia 181 — pyrifera 32, 179, табл. 16 Mallomonas 44, 48, 49, табл. 7
279 О л. 26 30 conf åãv1ñî1à табл. 6 49 — 182 Mallomonas denticulata 103, 105 Mastigocladales 88 — 89 Mastigocladus laminosus 67, 84, 88 Mastogloia 119, 126, 128, 140, табл.i4 — braunii i i2, табл. i i — pusilla i i 9 Melobesia 231, 232, 233 — lejolisii 231 Мelosira 48, 49, 50, 123, 128, 129, 132, табл. iO, ii — baicalensis 49 — granul at a 51 — islandica 49, 50 Menegazzia 461 — pertusa 406, 417, 461, табл. 52 Menoidium 265, табл. 27 Merismopedja 82, 86 — elegans 86 — glauca 86, табл. 3 Merotrichia capitata 94, 97 Mesogloia i52, 154 Mesotaeniales 310 — 312 Mesotaenium 310 — berggrenii 312 — chlamydosporum 310, 31i — macrococcum 3i0 — nivalis 312 Micractinium 278 — pusillum 278, 280 Micrasterias 322, 335 — conferva 335, табл. 37 — rotata 322, 335 Micrococcus 92 — chthonoplastus 92, 93 — чад~паФпя 92, 93 Microcoleus 62, 65, 66 Microcystis 48, 49, 50, 51, 53, 79, 82, 86 — aeruginosa 45, 79, 86, табл. 3 — f los-aquae 80, 86 — pulverea 79 Microspongium 153 Microspora 283 Microdictyon 306, 307 Microzonia 161 Mischococcus 254, табл. 26 Monoraphales 137, 138 — 139 Monostroma 285, 286 — grevillei 285, табл. 3i Mougeotia 319, табл. 36 Mougeotiopsis 309 Multiclavula mucida 464 Mycoporaceae 407 Myrionema i44, 152 Myxochloris sphagnicola 74, 252, 253 Myxochrysis paradoxa i02, 103, 253 IVautococcus 54 Navicula 126, 128, 140, табл. i4 — intricata i39 — mutica 63 — radiosa i i5, 125 Nemaliales 215 — 222 Nemalion 206, 221 — чегппсц1аге 221 Nemastomaceae 236 Nematochrysis sessilis i10 Neomeris 303, табл. 33 Nephroma 410, 451 — arcticum 451, табл. 49 Nephromopsis 410 Nereia 167 — filiformis i66, 167 Nereocystis 181 — luetkeana 181, табл. 16 Netrium 310 — digitus 3iO, 311 Neuropogon 397, 399, 427, 462 — fasciatus 462, табл. 49 Nitella 338, 344, 350 — mucronata 349, табл. 38 Nitellopsis 339, 350 — obtusa 347, 349, 350, табл. 38 Nitophyllum 249 Nitzschia 1177, 126, 128, 142, табл. i4 — acuta ii6 — obtusa айаг. scalpelliformis i41 — seriata 124 — tryblionella айаг. levidensis i4i Nodularia 48, 81 Nostoc 79, 81, 85, 89, 375, 383 — commune 64, 65, 89 — flagelliforme 85, 365 — microscopicum 63 — muscorum 15 — pruniforme 80, 85, 89, 365, табл. 3 — punctiforme 85, 89 Nostocales 67, 83, 89 — 92 Notheia anomala 188 Ochrolechia 470 Odonthalia 250, табл. 23 Odonthia bicolor 464 Oedogoniales 281, 291 — 293 Oedogonium 291, 292, табл. 30 Oocardium 336 — stratum 324, 337 Oocystis 48, 51, 76, 277 — psuedocoronata 277 Omphalina ericetorum 408 — luteolilacina 408 Opegrapha 413, 444 — diaphora 445, 446 Ophiocytium 256 — cochleare 256 Oscillatoria 35, 50, 51, 91 — brevis 9i, 92 — chalybea 92, табл. 3 — erythraea 46 — formosa 9i, 92 — limosa 9i 92 — princeps 9i, 92 — sancta 9i, 92 Oscillatoriales 67, 91 — 93 Padina 165 — pavonia 159, 165, табл. i7 Pandorina 49 — morum 272 Parmelia 360, 384, 403, 470 — caperata 427, 469, табл. 47 — centrifuga 425, 433, 461 — conspersa 461 — exasp era tula 419 Parmelia fuliginosa 461 — fncurva 461 — olivaceae 461 Parmelia saxatilis 461 — stenophylla 461 — sulcata 4i7, 427, 461 — tinctorum 424 — чадапя 396, 461, табл. 50 Parmeliopsis 461 — ambigua 461 Pediastrum 44, 48, 51, 277, — simplex 279 Pedinopera robusta 270 Pedinoperopsis gracilis 270 Pelagophicus 181 — porra 181 Peltigera 388, 395, 403, 449 — aphthosa 450, табл. 42 — canina 450, табл. 49 Pelvetia 188 — canal iculata 189 — wrightii 189 Penium 333 — didymocarpon 329 — polymorphum 333 — spirostriolatum 325, 333 Pennatophyceae 131, 135 — 14 Peranema 265 — trichophorum 264 Peranematales 263 Percursayia 284 Peridinium 48 — cinctum 99 — divergens 96 Peroniella curviceps 256, таб Pertusaria 414, 459 — amara 459, — d act il ina 459, — globulifera 459, табл. 54 Petalomonas 265, табл. 27 — steinii 264 Petalonia 160 — fascia 160 — zosterifolia 160 Рetrocelis 235 Peyssonella 204, 205, 229, 2 Phacotus 270 — coccifer 270 Phacus 48, 49, 265 — arnold й 263 — longicauda 263, табл. 28 — monilatus 263 Phaeococcus 97 Phaeodermatium rivulare 111, Phaeophyta 9, 143 — 192 Phaeoplax 97 Phaeothamnion 111, табл. 6 — conf ervicola i 10 Phaeozoosporophyceae 148, 1 Phaeurus 168 Phormidium 62, 65, 84, 91 — autumnale 63, 9i — foveolarum 9i — laminosum 68, 84, 92 — molle 9i Phycodris 248, табл. 22 Phylariella 177 — ochotensis 177 Phyllaria 173 Phyllocardium complanatum Phyllophora 237, 238 — membranifolia 237 — nervosa 366, табл. 39 Physcia 463 — caesia 463 — muscigena 463 — orbicularis 417
Physcia picta 426 Physcia pulverulenta 463 Pinnularia 121, 126, 140 — borealis 63 — viridis 140 Placolecanora 460 — peltata 460, табл. 44 Planktoniella 44, 45, 125 Platoma 236, 237 Pleurococcus 384 — vulgaris 280, табл. 2 Pleurocybe 442 Pleurosigma elongatum 140 Pleurotaenium 334 — minutum 334 — trabecula 325 Podocystis 137, табл. 14 Polyblepharidales 267, 268 — 269 P olyedriella helve t i ca 15 Polyedriopsis 278 — spinulosa 278 Polysiphonia 206, 209, 249, 250 — lanosa 54 Polytomaceae 269 Porodiscus 128, табл. 12 Porphyra 207, 214, табл. 20 — tenera 40 Porphyridium 193, 194, 213 Prasiola 36, 295, 296 — stipitata 41 Pringsheimiella 288, табл. 30 Protochlorinophycidae 267 Pro tococca les 275 Protococcus viridis 280 Protosiphon 279 — botryoides 273, 279, табл. 29 Prymnesium parvum 101 Pseudobryopsis 300, 301 Pseudocolium 441 Pseud evernia furfuracea 427, 428, 470 Psora 427, 454 — decipiens 454, табл. 46 — scalaris 455, табл. 46 Рteromonas 270 — aculeata 270 Pugetia 226 Punctaria 156 — latifolia 156, 157 Pycnothelia 456 Pylaiella 150 — litoralis 150, табл. 18 Pyramimonas reticulata 268, 269 Pyrenula nitida 435 Pyrenocarpales 434 — 439 Pyrgidium 440 P yrobotrys 33, 271 — gracilis 271 Pyrrophyta 9, 12, 76, 93 — 100 Rad iosphaera sphaerica 275. 276 1 alfsia чеггцсояа 153 Bamalina 427, 470 — reticulata 425 Bhabdomonas costata 264 Bhabdonema 123 — adriaticum 118 Bhaph dопета 280 — nivale 69, 280 — sempervirens 280 Rhizocarpon 433, 455 — concretum 433 — geograficum 425, 426, табл. 42 — tinei 433, 456, табл. 51 Rhizochloris stigmatica 252 Bhizoclonium 307, 308 Rhizosolenia 45, 46, 48, 49, 113, 125, 134 Rhizosolenia alata 122 — fragilissima 124 Rhod imenia 241, 242, табл. 22 — pertusa 212, табл. 21 Rhod imeniales 240 — 243 Rhodochorton 216 Rhodomela 250, табл. 23 Rhodomonas 98, табл. 5 Rhodophyta 9, 12, 192, 251 Rhopalodia 117, 126, 128, 142 — musculus айаг. succienta 142, табл. 11 Rinodina oreina 467 Rivularia 35, 81, 85, 91 — coadunata 91, 92 — planctonica 91, 92 Boccella 444 — fuciformis 445 — fuco ides 444, 445 Rod oglossum 238 Rosenvingiella 295, 296 Roya 310, табл. 35 Saccorhiza 173 — dermatodea 173 — polyschides 173 Sargassum 147, 180, 191 — fluitans 191 — miyabei 191 — natans 191 — pallidum 191 Scaphospora speciosa 151 Sceletonema 124, 132 — costatum 124, 132 табл. 13 Scenedesmus 48, 51, 274, 277, 278, 375 — quadricauda 279 Schizogoniales 281, 295 — 297 Schizogonium 295, 296 Schizothrix 65, 66, 92 — calcicola 92, 93 — friesii 60, 92, табл. 2 — lardaceae 92, 93 — lateritia 92, 9Ë Schizymenia 236, 237 Scytonema 35, 72, 85, 90, 388 — alatum 90 — crustaceum 90 — mirabile 90, табл. 2 Scytosiphon 160 — lomentaria 157, 159, 160 Scytosiphonales 158 Selenastrum 48, 278 — bibraianum 277 Silicoflagellatophycidae 103, 107— 108 Siphonales 297 — 302 S iphonoclad ales 305 — 308 Siphonocladus 305, 306 — pus llus 306 — tropicus 306 S iphonophyceae 297 — 308 Sirogonium 316, табл. 36 Sirogonium sticticum 321 Soleniales 131, 134 Solorina 450 — crocea 433, 450, табл. 42 — saccata 384, 416 — scortea 395 Sorocarpus 151 Spermatochnus 156 Sphacelaria 162, 163 Sphacelariales 162 — 163 Sphaerophorus 442 — globosus 440, 442 Sphaeroplea 293 — annul ina 293 Sphaeropleales 281, 293 — 294 Sphaerotrichia 155 — divaricata 155 Sphaerozosma 324, 337 — filiformis 321, 337 Spirogyra 315, табл. 36 — borgeana 320 — neglecta 24 — чаг~апя 320 Spirotaenia 309 310 табл. 35 — condensata 310 — obscura 311 Spirulina 91, 375 — jenneri 91, 92 — major 91, 92 — platensis 91, 366 S plachnid ium 156 — rugosum 156 Spondilomoraceae 271 Spondylomorum quaternarium 271 S pond ilosium 336 — pulchellum 337 Sporochnales 165 — 167 Sporochnus 166 Squamariaceae 229 — 230 Staurastrum 49, 322, 336 — cyclacanthum 322, 33о — gladiosum 324 — hirsutum 329 — pelagicum 322, 336 — sonthalianum 324 Staurodesmus 336 — .incus 336 — convergens 336 Staurothele 416, 436 — clopima 436, 437 Stenocybe 440 Stephanod iscus 48, 114, 123, 132, табл. 13 — tenuis 133 Stephanopyxis 124, 128, 132, табл. 15 Stephanosphaera pluv ~а1is 271 tereocaulon 458 — alp inurn 433 — tomentosum 458, табл. 49 Stichococcus 281, 384 — bacillaris 63, 282 Sticta 395, 404, 406, 452, табл. 46 — wrightii 452, табл. 47 Stictyosiphon 158 — tortilis 158 Stigeoclonium 286, 287, табл. 30 Stigonema 62, 85, 88, 383 — informe 88 — ocellatum 88, табл. 3, S t igonematales 81, 88 S tylod inium 100 — sphaera 95 S t il ophora 156 Streblonema 144, 151
Х 132, Striaria 158 — attenuata 158 Strombomonas 260 — acuminata 260 — fluviatilis 260 — subcurvata 260 — volgensis 260 Struvae 306, 307 S typo caulon 163 — scoparium 163 Surirella 118, 126, 128, 143 — fastuosa 142 Synechococcus 86 — aeruginosus 86, табл. 3 — major 86 Synechocystis 32, 86, 375 — aquatilis 86 — endobiotica 73, 74 Synedra 124, 126, 128, 136 — baculus 136 — pulchella 126 Synura 49, 101, табл. 7 Tabellaria 48, 124, 138 — fenestrata 118, 137 Teilingia 324, 337 — granulata 321, 337 Teloschistes 403, 462 — flavicans 463 — lacunosus 463 Temnogametum 320, табл. 36 Tetmemorus 334 — brebissonii 334, 335 — granulatus 334 Tetracyclus 117, 129 Tetrad inium 97, 100 — intermedium 95 Tetraedriella gigas 255 Tetraed ron 48, 277 — caudatum 278 Tetrastrum 278 — triacanthum 279 Thallassionema 124 — nitzschioides 124 Thalassiophylum 178 — clathrus 178, табл. 16 Thalassiosira 45, 46, 124, 125, табл. 13 — excentrica 122, 124, 132 Thalassiosira pacifica 132 — polychorda 124 Tamnolia 397, 458 — vermicularis 397, 458, табл. 48, 51 Tilopteris 151 — mertensii 151 Tolypella 339, 351 — prolifera 349, табл. 38 Tolipothrix 90, 375 — tinuis 90, табл. 3 Toninia 427 — coeruleonigricans 427 Trachelomonas 48, 49, 54, 259, 263 — armata 259 — curta 259 — glo bularis 259 — hystrix 259 — planctonica 259 — volvocina 259 Тга il i ella 220 Trebouxia 383, 422, 466 Trentepohli a 288, 290, 384, 422, табл. 2 Treubaria 277 — triappendiculata 278 Tribonema гиМе 256, 257 Triceratium 135 Triploceras 334 — gracile 334 Turbinaria 180, 191 Udotea 298, 299, табл. 32 U1othrich ales 281 — 283, 297 Ulothrix 298 — flacca 283 — subflaccida 282 — zonata 282, табл. 30 Ulva 284, 285, табл. 31 Ulvales 281, 283 — 286, 297 Umbilicaria 394, 459, 470 — cylindrica 425, 426 — esculenta 458, 459, 467 — hyperborea 433 — mammulata 423 — muehlenbergii 459, табл. 42 Und aria 182 — pinnatifida 182, табл. 39 Urceolus 265, табл. 31 Usnea 399, 400, 462 — longissima 397, 462, табл. 49 Vacuolales 275 — 276 Vacuolaria viridis 94, 97 7 alonia 306 — ventricosa 305, 306 — utriculosa 306 Vaucheria 258 — sessilis 257 Verrucaria 384, 436 — rupestris 436, 437 Volvocales 267, 270 — 273 Volvox 49, 266, 272 — aureus 273 — globator 273 Wallea 89 Woronichinia 87 — naegeliana 45, 73, 87 Xanthidium 335 — antilopeum 336 — aculeatum 324 Xanthococcophyceae 252, 254 — 256 Xanthocapsophyceae 252, 253 Xanthomonadophyceae 252, 253 — 254 Xanthophyta 9, 251 — 259, 282 Xanthopodophyceae 252 — 253 Xanthoria 463 — elegans 427, табл. 51 — perietina 395, 463, 468, табл. 42 Xanthosiphonophyceae 252, 257 — 259 Xanthotrichophyceaea 252, 256 — 257 Zanardinia 161 Zygnema 320, табл. 36 Zygnematales 314 — 320 Zygogonium ericetorum 320
СОДЕРЖАНИЕ 103 105 107 10 16 108 109 32 110 38 ВОДОРОСЛИ И СРЕДА 112 119 120 43 44 54 59 62 125 127 129 130 70 131 132 133 72 134 78 135 85 137 87 138 139 88 89 92 141 f42 143 93 96 97 98 . Е. Петров.. Ю 144 100 1~i 5 146 148 101 103 рослеи водоросли ОБЩАЯ ХАРАКТЕРИСТИКА ВОДОРОСЛЕЙ Водоросли и их отличие от других растений. М. М. Голлербах Способы питания водорослей и других хлорофил- лоносных растений. В. Е. Семененко Клетка водорослей. Т. В. Седова Основные типы морфологической структуры тела водорослей. 3f. 3f. Голлербах Размножение и циклы развития водорослей. Ю. . Е. Петров ° ° ° ° ° ° ° ° ° ° ° ° ° ° Внешние условия жизни и экологические груп- пировки водорослей Внешние условия жизни водорослей. 3f. 3f. Г оллер бах ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Планктонные водоросли. И. И Николаев. Бентосные водоросли. Ю. Е. Петров Наземные водоросли. 3f. 3f. Голлербах. Почвенные водоросли. Э. А. Штина. Водоросли горячих источников. 3f. 3f. Голлер- б ах ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Водоросли снега и льда. 3f. 3f. Голлербах. Водоросли соленых водоемов. 3f. 3f. Голлер- б ах ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Сверлящие и туфообразующи<, водоросли. М. М. Голлербах Сожительство водорослей с другими организмами. Т. В. Седова. СИСТЕМАТИЧЕСКИЙ ОБЗОР ВОДОРОСЛЕЙ Отдел сине-зеленые водоросли (Cyanophyta). Э . Г. Кукк. Класс хроококковые (Chroococcophyceae) Порядок хроококковые (Chroococcales) Порядок энтофизалиевые (Entophysalidales) Класс хамесифоновые (Chamaesiphonophyceae) .. Класс гормогониевые (Hormogoniophyceae) Порядок стигонемовые (Stigonematales) Порядок мастигокладовые (Mastigocladales) Порядок ностоковые (Nostocales) Порядок осциллаториевые (Oscillatoriales) Отдел пирофитовые водоросли (Pyrrophyta). А. 3f. Матвиенко. Подотдел хлоромонадофитовые (Chloromonado- phytina) Подотдел криптофитовые (Cryptophytina) Подотдел динофитовые (Dinophytina) Отдел золотистые водоросли (Chrysophyta). А. 3f. Матвиенко. Класс хризоподовые (Chrysopodophyceae). А. М. Матвиенко . Класс хризомонадовые (Chrysomonadophyceae). А. 3f. Матвиенко . П о д к л а с с хризомонады (Chrysomonado- phycidae). А. M. Матвиенко. П о д к л а с с кокколитофориды (Coccolitho- phoridophycidae). И. П. Табачникова П о д к л а с с силикофлагеллаты (Silicoflagel- latophycidae). 3. И. Глезер Класс хризокапсовые (Chrysocapsophyceae). А. 3f. Матвиенко . Класс хризосферовые (Chrysosphaerophyceae). А. 3f. Матвиенко Класс хризотриховые (Chrysotrichophyceae). А. 3f. Матвиенко Отдел диатомовые водоросли (Bacillario- phyta). И. В. Макарова. Строение клетки диатомовых водорослей Способы питания диатомовых водорослей Размножение диатомовых водорослей Биологические особенности диатомовых водород- ° .Ф л еи ° ° ° ° ° ° Экологические особенности и географическое распространение диатомовых водорослей . Эволюция и филогения диатомовых водорослей Роль в природе и практическое значение диато- мовых водорослей Классификация диатомовых водорослей Класс центрические диатомеи (Centrophyceae) Порядок косцинодисковые (Coscinodiscales), Семейство мелозировые (Melosiraceae) Семейство талассиозировые (Thalassiosiraceae) Семейство косцинодисковые (Coscinodiscaceae) Порядок актинодисковые (Actinodiscales) . Порядок аулакодисковые (Aulacodiscales) Порядок солениевые (Soleniales) . Порядок биддульфиевые (Biddulphiales) Класс пеннатные диатомеи (Pennatophyceae) Порядок бесшовные (Araphales) Семейство фрагилариевые (Fragilariaceae) . Семейство табеллариевые (Tabellariaceae) . Порядок одношовные (Monoraphales) Порядок двухшовные (Diraphales) Семейство навикуловые (Naviculaceae) Семейство гомфоцимбеловые (Gomphocymbel- 1 асеае) Порядок каналошовные (Aulonoraphales) . Семейство эпитемиевые (Epithemiaceae) . Семейство нитцшиевые (Nitzschiaceae) Семейство сурирелловые (Surirellaceae) . Отдел бурые водоросли (Phaeophyta). Строение слоевища бурых водорослей Строение клетки бурых водорослей Размножение и цикл развития бурых водоро- с лей ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Распространение и экология бурых водорослей Значение бурых водорослей Происхождение и классификация бурых водо-
Класс фэозооспоровые (Phaezoosporophyceae)... 149 222 224 151 152 153 226 228 229 230 231 154 233 235 236 155 156 238 239 240 241 157 158 246 247 160 162 163 165 167 169 173 174 248 249 25i Класс Класс Класс Класс Класс Класс 252 253 178 179 181 254 256 257 182 184 185 186 188 M. M. Голлербах.. 190 191 268 269 27О 192 Класс протококковые (Protococcophyceae). А. М. К. Л. Виноградова. Матвиенко 193 198 202 210 2ii 212 213 275 276 279 ° ° ° ° Ь ° 281 283 286 291 293 294 295 215 217 218 219 К. д . ВиRO- ° ° ° ° ° ° ° 297 221 222 486 Порядок эктокарповые (Ectocarpales) Порядок тилоптеридовые (Tilopteridales) . Порядок хордариевые (Cbordariales) Семейство мирионемовые (Myrionemataceae) Семейство ральфсиевые (НаЦа1асеае) Семейство литодермовые (Lithodermataceae) Семейство элахистовые (Elacbistaceae) Семейство коринофлеевые (Corynophlaeaceae) Семейство хордариевые (Chordariaceae) Семейство сперматохновые (Spermatochnaceae) Семейство сплахнидиевые (Splachnidiaceae)... Порядок диктиосифоновые (Dictyosiphonales) . Семейство пунктариевые (Punctariaceae) Семейство диктиосифоновые (Dictyosiphona- C еае) ° ° ° ° ° ° ° Семейство стиктиосифоновые (Stictyosiphona- C еае) Семейство асперококковые (Asperococcaceae) Семейство стриариевые (Striariaceae) Порядок сцитосифоновые (Scytosiphonales) . Порядок кутлериевые (Cutleriales) . Порядок сфацеляриевые (Sphacelariales) Порядок диктиотовые (Dictyotales) . Порядок спорохновые (Sporochnales) Порядок десмарестиевые (Desmarestiales) . Порядок ламинариевые (Laminariales) Подпорядок филлариевые (Phyllarineae) . Подпорядок ламинариевые (Laminarineae) Семейство хордовые (Chordaceae) Семейство ламинариевые (Laminariaceae) Семейство артротамновые (Arthrothamnaceae) Семейство лессониевые (Lessoniaceae) . Семейство алариевые (Alariaceae) . Класс циклоспоровые (Cyclosporophyceae) П о д к л а с с аскозейровые (Ascoseiropbycidae) П о д к л а с с фукусовые (Fucophycidae) Порядок дурвиллеевые (Durvilleales) . Порядок фукусовые (Fucales) Семейство хормозировые (Hormosiraceae) Семейство нотейевые (Notheiaceae) Семейство химанталиевые (Himanthaliaceae) Семейство фукусовые (Fucaceae) Семейство цистозейровые (Cystoseiraceae) . Семейство саргассовые (Sargassaceae) . Отдел красные водоросли (Rhodophyta). Внешняя форма тела красных водорослей . Строение слоевища красных водорослей . Строение клетки красных водорослей Размножение красных водорослей Происхождение красных водорослей . Распространение красных водорослей Использование красных водорослей Классификация красных водорослей Класс бангиевые (Bangiophyceae) Класс флоридеевые (Florideophyceae) . Порядок немалиевые (Nemaliales) Семейство акрохетиевые (Acrochaetiaceae) . Семейство батрахоспермовые (Batrachosper- maceae) Семейство леманеевые (Lemaneaceae) Семейство боннемезониевые (Bonnemaesonia- C еае) ° ° Семейство гельминтокладиевые (Helminthocla- п1асеае) Семейство хетангиевые (Chaetangiaceae) . Порядок гелидиевые (Gelidiales) . Порядок криптонемиевые (Cryptonemiales) . Семейство дюмонтиевые (Dumontiaceae) Семейство каллимениевые (Kallymeniaceae) Семейство криптонемиевые (Cryptonemiaceae) Семейство хореоколаксовые (Choreocolaceae) Семейство сквамариевые (Squamariaceae) Семейство кораллиновые (Corallinaceae) . Подсемейство мелобезиевые (Melobesioideae) Подсемейство кораллиновые (Corallinoideae) Порядок гигартиновые (Gigartinales) . Семейство круориевые (Cruoriaceae) . Семейство немастомовые (Nemastomaceae) . Семейство филлофоровые (Phyllophoraceae) .. Семейство гигартиновые (Gigartinaceae) . Семейство грациляриевые (С racilariaceae) . Порядок родимениевые (Rbodymeniales) Семейство шампиевые (Champiaceae) Семейство родимениевые (Rhodymeniaceae) . Порядок церамиевые (Ceramiales) Семейство церамиевые (Ceramiaceae) . Семейство дазиевые (Dasyaceae) . Семейство делессериевые (Delesseriaceae) . Подсемейство делессериевые (Delesserioideae) Подсемейство нитофилловые (Nitophylloi- deae) . Семейство родомеловые (Rhodomelaceae) Отдел желто-зеленые водоросли (Xantho- phyta). Т. В. Яогадина, А. М. Матвиенко. ксантоподовые (Xanthopodophyceae) ксантомонадовые (Xanthomonadophyceae) ксантокапсовые (Xanthocapsophyceae) ксантококковые (Xanthococcophyceae) ксантотриховые (Xanthotrichophyceae) ксантосифоновые (Xanthosiphonophyceae) Отдел эвгленовые водоросли (Euglenophyta). Т. А . Сафо нова. Отдел зеленые водоросли (Chlorophyta). Класс вольвоксовые (Volvocophyceae). А. М. Мат- виенко . Порядок полиблефаридовые (Polyblepharidales) Порядок хламидомонадовые (Chlamydomonadales) Порядок вольвоксовые (Volvocales) Порядок вакуольные (Vacuolales) Порядок хлорококковые (Chlorococcales) . Порядок прототриховые (Prototrichales) Класс улотриксовые (Ulotrichophyceae). R. Л. Ви- ноградова Порядок улотриксовые (Ulotrichales) . Порядок ульвовые (Ulvales) . Порядок хетофоровые (Chaetophorales) . Порядок эдогониевые (Oedogoniales) . Порядок сфероплеевые (Spbaeropleales) . Порядок цилиндрокапсовые (Cylindrocapsales). Порядок схизогониевые (Schizogoniales) Класс сифоновые (Siphonophyceae). градова Порядок сифоновые (Siphonales) Семейство каулерповые (Caulerpaceae)
299 301 302 305 426 431 432 308 СИСТЕМАТИЧЕСКИЙ ОБЗОР ЛИШАЙНИКОВ 310 312 314 321 333 Отдел харовые водоросли ((",harophyta). M. M. Голлербах.............. 338 347 348 349 350 ПРОИСХОЖДЕНИЕ, РОДСТВЕННЫЕ СВЯЗИ И ЭВОЛЮЦИЯ ВОДОРОСЛЕЙ. Ю. F. Петров... 35i 355 360 364 367 ненко . 379 380 383 385 390 400 407 465 467 471 473 478 420 423 425 487 Семейство бриопсидовые (Bryopsidaceae) Семейство кодиевые (Codiaceae) . Порядок дазикладовые (Dasycladales) Порядок сифонокладовые (Siphonocladales) . Класс конъюгаты, или сцеплянки (Conjugatophy- ceae). Л. А. Рукдика . Порядок мезотениевые (Mesotaeniales). Л. А. Рук- д ина. ° ° Порядок гонатозиговые (Gonatozygales). Л. А . P ундина. ° ° ° ° ° ° ° ° ° ° ° ° ° Порядок зигнемовые (Zygnematales). Л. А. Рук- д и на. Порядок десмидиевые (Desmidiales). Г. М. Морд- виниева. ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Семейство пениевые (Peniaceae) Семейство клостериевые (Closteriaceae) Семейство космариевые (Cosmariaceae) С троение и размножение харофитов. Экологические особенности и географическое распространение харофитов Роль в природе и практическое значение харо- итов ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Происхождение и классификация харовых водо- ° . ° р ослеие ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Семейство нителловые (Nitellaceae) Семейство харовые (Characeae) ЗНАЧЕНИЕ ВОДОРОСЛЕЙ В ПРИРОДЕ И ЖИЗНИ ЧЕЛОВЕКА Историческая роль водорослей. Водоросли как геологический фактор. 3. И. Глезер. Распространенность водорослей в современных водоемах, их биомасса и продукция. M. M. Гол- л ербах. ° ° ° ° ° Практическое использование водорослей. M. M. Голлер бах. Массовое культивирование микроскопических водорослей. M Г. Владимирова, В. Е. Семе- ЛИШАЙНИКИ ОБЩАЯ ХАРАКТЕРИСТИКА ЛИШАЙНИКОВ Лишайники и их отличие от других растений. Х . Х. Трасс. Ьомпоненты лишайников и их взаимоотношения. Н. С. Голубкова. Грибной компонент (микобионт) Водорослевый компонент (фикобионт) Взаимоотношения гриба и водоросли в теле лишайника Внешнее и внутреннее строение лишайников. Н. С. Голубкова. Морфология слоевища лишайников Анатомия слоевища лишайников Органы размножения лишайников Физиологические и экологические особенности лишайников Способы питания лишайников. Х. Х. Трасс. Химический состав лишайников. Х. Х. Трасс Рост лишайников. Х. Х. Трасс Отношение лишайников к субстрату и дру- гим факторам внешней среды. П. С. Голубкова Лишайники и загрязненность воздуха. Х. Х. Т расс ° ° ° ° ° ° ° ° ° ° ° 0 Роль лишайников в природе. Н. С. Голубкова Класс сумчатые лишайники (Ascolichenes). Н. С. Голубкова.............. 434 П о д к л а с с пиренокарповые лишайники (Ру- renocarpeae). Н. С. Голубкова. Порядок пиренокарповые (Pyrenocarpales) Семейство пиренулевые (Ругеппlасеае)... 435 Семейство верукариевые (Verrucariaceae) . Семейство дерматокарповые (Dermatocarpa- ceae) . . . . . . . . . . . . . . . . . . . 437 П о д к л а с с гимнокарповые лишайники (Gym- nocarpeae). Н. С. Голубкова......... 439 Порядок калициевые (Caliciales). Н. С. Голубкова. Семейство калициевые (Caliciaceae)..... 440 Семейство цифелевые (Cypheliaceae) .... 441 Семейство сферофоровые (Sphaerophoraceae) 442 Порядок артониевые (Arthoniales). Н. С. Голуб- к ова. ° ° Семейство артониевые (Arthoniaceae).... 443 Семейство рочелловые (R оссеllасеае).... 444 Семейство опеграфовые (Opegraphaceae) . Порядок графидовые (Graphidales). Н. С. Голуб- к ова. ° ° ° ° ° ° ° ° ° ° ° ° ° ° 445 Семейство графидовые (Graphidaceae)... 446 Порядок круглоплодные (Cyclocarpales). Н. С. Го- л убкова.................. 447 Семейство коллемовые (Collemataceae). Н. С. Голубкова. Семейство пельтигеровые (Peltigeraceae). Н. С. Голубкова.............. 449 Семейство стиктовые (Stictaceae). Н. С. Го- лубкова.................. 451 Семейство лецидеевые (Lecideaceae). И. С. Го- л убкова................... 452 Семейство кладониевые (Cladoniaceae). Х. Х. Трасс................ 456 Семейство стереокауловые (Stereocaulaceae). Х. Х. Трасс................ 458 Семейство умбиликариевые (Umbilicariaceae). Х . Х. Трасс. ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Семейство акароспоровые (Acarosporaceae). Х. Х. Трасс................ 459 Семейство пертузариевые (Pertusariaceae). Х . Х. Трасс. ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Семейство леканоровые (Lecanoraceae). Х. Х. Трасс................ 460 Семейство пармелиевые (Parmeliaceae). Х . Х. Трасс. ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Семейство уснеевые (Usneaceae). Х. Х. Трасс. 462 Семейство телошистовые (Teloschistaceae). Х . Х. Трасс. ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Семейство фисциевые (Physciaceae). Х. Х. Трасс................ 463 Класс базидиальные лишайники (Basidiolichenes). Х . Х. Трасс. ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ПРОИСХОЖДЕНИЕ ЛИШАЙНИКОВ И ИХ МЕСТО В СИСТЕМЕ РАСТИТЕЛЬНОГО МИРА. Х ° Х. Трасс. ° ° ь ° е ° ° ° ° ° ь ° ° 3 ° ° ПРАКТИЧЕСКОЕ ИСПОЛЬЗОВАНИЕ ЛИШАЙ- НИКОВ. Н. С. Голубкова.. Указатель терминов . Указатель русских названий Указатель латинских названий
ИБ 1164 жизнь рАстЕний в шести томах том Цена 4 р. 07 к. Редактор М. В. КУЛИКОВА Технический редактор М. Д. КОЗЛОВСКАЯ Корректоры Е. А. БЛИНОВА, Н. И. НОВИКОВА Составитель указателя В. П. ПРОХОРОВ Ответственная за выпуск Н. Н. ФЕДОРОВА 6050i — 382 Ж 103~031 — 77 "'"" "'" 58 Оформление художника И. С. НОВОХАЦКОЙ Цветные иллюстрации В. С. ЮДИНА, А. В. КОНДРАТЬЕВА художественный редактор В. Г. ЕЖКОВ Сдано в набор 22/VI 1976 г. Подписано к печати 1/II 1977 r. 84X108'/ . Бумага тип. Х 1. Печ. л. 30,5+ вкл. 3,5+ форзац 0,25. Условн. л. 51,24+ вкл. 5,88-+ форзац 0,42. Уч.-изд. л. 57,52 + вкл. 6,36+ форзац 0,80. Тираж 300 тыс. экз. Заказ 300. Ордена Трудового Красного Знамени издательство «Просвещение» Государственного комитета Совета Министров РСФСР по делам издательств, полиграфии и книжной торговли. Москва, 3-й проезд Марьиной рощи, 41. Ордена Трудового Красного Знамени Московская типография Х 2 Союзполиграфпрома при Государственном комитете Совета Мини- стров СССР по делам издательств, полиграфии и книжной торговли. Москва, И-85, проспект Мира, 105. Вклейки, суперобложка, форзац отпечатаны на ордена Трудового Красного Знамени Калининском полиграфическом комбинате Союзполиграфпрома при Государственном Комитете Совета Мини- стров СССР по делам издательств, полиграфии и книжной торгов- ли, г. Калинин, пр. Ленина, 5. Жизнь растений. В 6-ти т. Гл. ред. чл.-кор. Ж71 АН СССР, проф. Ал. А. Федоров. Т. 3. Водоросли. Лишайники. Под ред. проф. М. М. Голлербаха. М., (<Просвещение», 1977. 487 с. с ил.; 28 л. ил. Третий том дает читателю обширнейшие сведения о водорослях и ли- шайниках. Водоросли — одна из самых древних групп растений. Их основным местом обитания является водная среда. Размеры, окраска, особенности строения водорослей чрезвычайно разнообразны — от ми- кроскопических одноклеточных диатомей до гигантских бурых водо- рослей, мощные слоевища которых достигают иногда 60 — 80 метров длины. Значительное количество водорослей может существовать и вне воды: в верхних слоях почвы, на стволах деревьев, на скалах и даже на снегу. Известны водоросли, живущие внутри других живых орга- низмов. Лишайники показывают пример удивительного приспособления живых ор ганизмов к условиям существования. Они состоят из двух комвонентов: грибов и водорослей, которые сплетены между собой так тесно, что превратились в единый организм со своеобразной струк- турой и особым обменом веществ.