/






























































































































































































































































































































Text
Дж. Г. Симпсон
ПРИНЦИПЫ
ТАКСОНОМИИ
животных
Перевод с английского
И.Я. Павлинова
Под редакцией и с предисловием
А.И. Шаталкина
Товарищество научных изданий КМК
Москва 2006
УДК: 591
ББК:(Е) 28.6
С37(6)
Симпсон Джордж Г. Принципы таксономии животных. Пер. с
англ. — М.: Товарищество научных изданий КМ К, 2006. — хх +
293 с., илл. Библ. 187.
Монография по теории систематики животных, ставшая одной из клю-
чевых в формировании школы эволюционной систематики. Рассмотре-
ны основные концепции и понятия систематики, проблемы соотношения
между филогенией и классификаций, иерархического построения сис-
темы таксонов. Подробно изложена концепция эволюционного вида.
На примере ряда макротаксонов млекопитающих рассмотрены некото-
рые ведущие правила построения эволюционных классификаций.
Для биологов-систематиков, всех интересующихся классификационны-
ми проблемами в биологии.
ISBN 5-87317-308-7
© И.Я. Павлинов, А.И. Шаталкин, текст. 2006
© 1 -во научных изданий КМ К, издание, 2006
Предисловие к русскому изданию
Джордж Симпсон (1902-1984) и его классический труд
по основаниям систематики
Симпсон был основателем и ведущим представителем одного
из двух фундаментальных направлений систематики — филогене-
тического и эволюционного. Основные положения филогенетиче-
ского подхода были сформулированы Геннигом, эволюционного —
Симпсоном. Иногда говорят о третьем фундаментальном прибли-
жении в систематике — типологическом. Типология, однако, есть
не более чем инструмент, который легко может быть приспособлен
для нужд как эволюционной систематики, так и филогенетики.
Наконец, существуют и будут возникать эклектические приближе-
ния, связанные с попытками сблизить симпсоновский и геннигов-
ский подходы. Но они в большинстве своем основаны на неверном
прочтении того и другого и, скорее всего, не будут иметь продол-
жения.
Оба направления оперируют, по существу, разными предметны-
ми областями и поэтому могут в определенных пределах развивать-
ся независимо. В филогенетике в центре внимания стоят рецентные
формы, тогда как вымершие, если и включаются в анализ, то на
правах рецентных, т.е. по тем правилам, которые приняты для клас-
сификации рецентных форм. Это означает, что в филогенетике ма-
юриал не стратифицирован и не соотнесён с временной шкалой.
iv
Дж. Г. Симпсон. Принципы таксономии животных
Сам Генниг был того мнения, что для каждой исторической эпохи
лучше строить свои классификации. В эволюционном подходе
предметная область стратифицирована, сообразно исторической
преемственности групп. Дальше мы отметим, какие следствия вы-
текают из этого. Сейчас же выделим главное: оба направления на-
целены на изучение разных предметных областей и, следователь-
но, подходят к описанию разнообразия организмов с принципиаль-
но разных сторон, что и определяет их принципиальную несводи-
мость. В силу этих обстоятельств обоим подходам уготована дол-
гая жизнь.
С учетом сказанного мы должны рассматривать Симпсона как
классика систематики. В предлагаемом читателю переводе класси-
ческого труда по теории систематики Симпсон суммировал основ-
ные положения эволюционного подхода. Выделим в его изложении
наиболее важные моменты.
Прежде всего следует остановиться на симпсоновском опреде-
лении систематики, которое как нельзя лучше характеризует его
подход: «Систематика есть научное изучение организмов с точки
зрения их естественных (родовых) объединений (kinds) и разнооб-
разия, а также всевозможных отношений между ними». Определе-
ние акцентирует внимание на трех ключевых моментах — (1) на
особом типе систематических групп, выраженном в понятии рода
(kind — реальная группа в отличие от произвольно выделенной),
(2) на структуре разнообразия, различающегося, как показывает
Симпсон, по объему, непрерывности, упорядоченности и другим
показателям, (3) на специфических системных отношениях, опре-
деляющих возможность существования таксономических групп в
природе, как самостоятельных эволюционных и экологических еди-
ниц. Эти последние находят выражение в таких понятиях как эво-
люционный вид, горизонтальные и вертикальные таксоны, адаптив-
ная зона, экологическая ниша и ряде других.
Представление об особом статусе систематических групп име-
ет давнюю историю. Аристотель различал общие и родовые (сущ-
ностные) понятия, выделяя среди последних вид в качестве наи-
меньшего рода, объединяющего объекты, сходные по сущности.
Аристотелевский взгляд на мир вещей в целом был принят систе-
матикой, которая на первых порах, в период господства типологи-
/ 1редисловие к русскому изданию
V
ческой доктрины видела свою главную задачу в выявлении «есте-
ственных родов» (natural kinds). Они, как считали, отражают суще-
ственные сходства. Симпсон подробно останавливается на пробле-
ме сущностных свойств и, следуя Кейну, приходит к выводу, что
понятие сущности, как оно рассматривалось типологами, если и
имеет смысл, то только применительно к искусственным вещам и
абстрактным построениям. Их природа (сущность), именно пото-
му, что она нам понятна, может быть проанализирована и выражена
в определении (аристотелевских дефинициях). Это — анализиру-
емые сущности (analyzed entities), как назвал их Кейн, например,
треугольник в качестве замкнутой по контуру геометрической фи-
гуры, образованной тремя отрезками. В отношении треугольника не
возникает серьезных проблем с разделением свойств на существен-
ные (вытекающие из определения и (или) каким-то образом взаи-
мозависимые) и несущественные (случайные). Например, сумма
углов в треугольнике равна 180 градусам, квадрат гипотенузы ра-
вен сумме квадратов катетов, в равностороннем треугольнике (ко-
торый можно рассматривать в качестве одного из видов треуголь-
ника), кроме того, равны углы. Конкретная длина сторон в боль-
шинстве случаев является несущественным признаком. Напротив,
организмы являются неанализируемыми сущностями (unanalyzed
entities) и поэтому для такого деления их свойств на существенные
и несущественные необходимо привлечение внешних критериев.
По Симпсону эти критерии должны быть эволюционными по со-
держанию. Заметим, что с точки зрения Аристотеля анализируемые
сущности, раз это наши собственные построения, не имеют сущ-
ности; сущностью обладают лишь природные тела.
Проблема разнообразия, в частности его упорядоченности, так-
же была предметом изучения систематиков с давних пор. Первые
несовершенные попытки изложения этой проблемы были связаны
с представлением организмов в виде лестницы существ. Во време-
на Симпсона структура разнообразия, рассматриваемая с самых
разных сторон, не только с точки зрения его упорядоченности, по-
лучила эволюционное содержание и этому аспекту уделено значи-
тельное место в книге.
Из системных отношений (связей по смежности), к изучению
которых проявляет интерес систематика в связи с проблемой раз-
vi
Дж. Г. Симпсон. Принципы таксономии животных
нообразия, на первом месте стоят связи по происхождению между
предками и потомками. Но Симпсон подробно останавливается так-
же на связях таксонов со средой (адаптивные аспекты), территори-
ей (зоогеографические аспекты) и временем. Основываясь на этом,
он пытается вычленить наиболее важные характеристики система-
тических групп. Но об этом чуть дальше.
Кратко очертив две главные составляющие симпсоновского
определения систематики, мы уже можем сопоставить защищаемый
им эволюционный подход с другими таксономическими приближе-
ниями. Для типологии главной базовой характеристикой система-
тики является первый аспект, касающийся выделения родов как
отражающих сущностное сходство. Для кладистики, напротив, на
первое место выходит второй аспект и, в частности, анализ фило-
генетической (генеалогической) составляющей разнообразия. Коль
скоро генеалогический паттерн определен и выделен, то он авто-
матически будет задавать классификацию и упорядоченность в ней
«естественных родов», как они трактуются в кладистике. Понятие
естественного рода для кладистики является производным и, сле-
довательно, может не включаться в ее базовое определение систе-
матики. Симпсон подробно освещает основные положения типоло-
гии и, на наш взгляд, убедительно показывает, что у типологии нет
критериев, с помощью которых можно было бы обосновать введе-
ние типов. К сожалению, Симпсон, дав добротный критический
анализ типологии, счел свою задачу выполненной и не пошел даль-
ше в поисках объективного содержания типологических характери-
стик. А оно у них есть и сейчас можно уже говорить об их гене-
тической основе. Отличие человека от лягушки не в генах, если
сопоставлять их по белкам, которые они кодируют, но в том регу-
ляторном аппарате (скрытом геноме — hidden genome), который
определяет, как эти белки будут «уложены», чтобы получить опре-
деленный план строения (архетип). Отметим также, что архетип
может быть правильно вычленен лишь через его соотнесение с
филогенией (эволюцией).
Конкретная специфика и своеобразие симпсоновского эволюци-
онного подхода, дающего столь широкое в сравнении с другими
приближениями определение систематики, связана с третьим кру-
гом задач — изучением таксонов как реальных природных единиц.
11редисловие к русскому изданию
vii
Именно сообразуясь с этими задачами, Симпсон выстраивает
принципиально новое эволюционное понимание систематической
группы.
Прежде всего, Симпсон отмечает, что ни сходство, использу-
емое типологами, ни родство в его узком значении не отражают
всего спектра эволюционных процессов, определяющих основные
структурные параметры разнообразия организмов. Раньше недооце-
нивалась роль вымерших организмов. Но классификации, претен-
дующие на то, чтобы называться эволюционными должны вклю-
чать, насколько это возможно, данные по ископаемым животным,
знание которых даст реальную возможность реконструировать
предковые группы и восстановить точные связи между таксонами.
Поэтому главным изучаемым предметом симпсоновского подхода
является филогенетическое дерево. Принятое Симпсоном понима-
ние проистекало из существовавшего в его время представления об
эволюции как постепенном переходе одного вида в другой в резуль-
тате изменения в популяции частоты одних генов и их замещения
другими. Соответственно высшие таксоны появляются ретроспек-
тивно после становления видов и низших таксонов, но не наоборот.
Напротив, в кладистике главным объектом изучения является
современная фауна. Поэтому ни один таксон не может выступать
в качестве предкового для другого. Они воспринимаются как уже
сложившаяся данность, и, следовательно, их рассмотрение в рет-
роспективе в кладистике полностью лишено смысла. Генниг, таким
образом, оперировал в рамках классификаций традиционного вида.
Именно поэтому он пошел иным путем, нежели Симпсон, решая
вопрос относительно того, какой аспект филогении может быть
выражен в классификациях, описывающих современную фауну и
флору.
Заметим, что Генниг считал желательным независимое постро-
ения горизонтальных классификаций для исторически сменяющих-
ся фаун. Ископаемые фауны не сопоставимы с рецентными по
объему и качеству используемых признаков. Вымершая фауна мо-
жет иметь сходный спектр внешних различий, но радикально от-
личаться по своим глубинным характеристикам, определяющим
физиологию, биологию и экологию ее членов. Именно по этим па-
раметрам они уступали представителям новых фаун, что и опреде-
VIII
Дж. Г. Симпсон. Принципы таксономии животных
ляло в конечном итоге их уход с исторической арены. Однако го-
ризонтальные классификации прошлых эпох никогда не имели
большого распространения. Грезы о них были во многом порожде-
нием несбывшихся старых надежд, что можно реконструировать
филогению, описывая и накладывая друг на друга последователь-
ные фауны прошлых эпох. Вернемся однако к книге.
При массовом притоке ископаемых и накоплении по ним дан-
ных будут, как думал Симпсон, постепенно сужаться гиатусы меж-
ду близкими группами. А это выведет работу систематика на прин-
ципиально новый уровень и поставит перед ним ряд новых проблем,
с которыми ранее ему не приходилось сталкиваться. Главная из них —
проблема определения межтаксонных границ и самих таксонов в
условиях, когда имеется более или менее непрерывный эволюцион-
ный ряд. Симпсон считает, что объективных оснований для деле-
ния филетической линии на отрезки не может быть и поэтому пред-
лагает не говорить об объективных (реальных) границах группы, но
только о том, насколько произвольно (arbitrary) или непроизвольно
(nonarbitrary) эти границы установлены с точки зрения рассматри-
ваемых данных и применяемых методов. Вместо абсолютных меж-
таксонных границ, найти которые стремились и стремятся система-
тики, Симпсон, таким образом, предлагает вводить границы, кото-
рые по одному критерию могут оказаться произвольными, по дру-
гому непроизвольными. Например, если по морфологическим раз-
личиям невозможно фиксировать точку перехода одной вымершей
группы к другой, замещающей первую, то ее можно нащупать по
скорости изменений в случае, если этот переход осуществлялся по
типу квантдвой эволюции. Последнее означает, что замещающие
группы существуют в разные исторические эпохи. Из этих рассуж-
дений возникла идея различать вертикальные и горизонтальные
классификации. На этом различии следует остановиться, поскольку
позже оба термина стали использоваться применительно к характе-
ристике дендрограмм, т.е. в несколько ином смысле.
В эволюционной систематике, как ее формулировал Симпсон,
филогенетическое дерево рассматривается в историческом аспекте.
Поэтому для каждого временного сечения дерева можно строить
свою классификацию. Такие классификации Симпсон назвал гори-
зонтальными, противопоставив их вертикальному делению дерева
Предисловие к русскому изданию
ix
на ветви, которые в свою очередь могут быть разделены вертикаль-
но на подветви и т.д. Симпсоновская (горизонтальная) классифи-
кация рецентных форм, как и любая классификация может быть
представлена в виде классификационного дерева. Будучи двумер-
ной конструкцией, любое дерево допускает разбиение с преимуще-
ственным учетом либо вертикальной, либо горизонтальной состав-
ляющих. Вертикальное разбиение классификационного дерева бу-
дет соответствовать кладистическим системам, горизонтальное —
большому спектру классификаций, учитывающих дивергенцию и
включающих парафилетические группы. Таким образом, геннигов-
ские иерархии из строго монофилетических групп, широко исполь-
зуемые кладистами, соответствуют определенному классу симпсо-
новских горизонтальных классификаций. Другой класс горизон-
тальных классификаций, отвечающих традиционным, основан на
исчислении сходственных отношений и описывается через понятия
дивергенции и парафилии.
Вертикальным симпсоновским классификациям и выделяемым
внутри них вертикальным таксонам нет аналогов в других извест-
ных таксономических системах, и они, следовательно, составляют
главный предмет изучения эволюционного подхода. Симпсон под-
робно проясняет специфику вертикальных классификаций. Не по-
скольку в момент написания книги ревизованное изложение фило-
генетической систематики еще только подготавливалось Геннигом
для англоязычного читателя (рукопись была передана в США для
перевода в 1961 г.), Симпсону не с чем было сравнивать свое по-
нимание систематики. Сейчас это легко можно сделать. Во-первых,
монофилетические (голофилетические) группы соответствуют вре-
менному срезу симпсоновских вертикальных таксонов и в этом
смысле они являются внеисторическими единицами. Напротив,
вертикальные таксоны являются развернутыми во времени, т.е. ис-
торическими единицами, которые, следовательно, должны описы-
ваться по своим связям с другими такими же историческими еди-
ницами, причем с момента своего возникновения до терминации.
Анализ преемственности таксонов и их развертывание во времени
предполагает знание широкого круга проблем, непосредственно
связанных с эволюцией. В этом заключается второе принципиаль-
ное отличие симпсоновского подхода от других, разрабатывающих
X
Дж. Г. Симпсон. Принципы таксономии животных
горизонтальные системы, будь то кладистические или типологичес-
кие. В-третьих, таксоны горизонтальных классификаций рассмат-
риваются Симпсоном совсем иначе, чем это делают другие подхо-
ды. Для тех, как уже было сказано, горизонтальные таксоны соот-
ветствуют временным сечениям филогенетического дерева. Доста-
точно взглянуть на рисунок 9а, чтобы стало ясно, что горизонталь-
ный таксон в симпсоновском понимании не является сечением де-
рева, но представляет собой монофилетическое объединение ког-
да-то существовавших групп. Если верна представленная на рисун-
ке филогения, то Creodonta в период своего существования состав-
ляли монофилетическую группу. В другие временные интервалы
они не существовали (поскольку вымерли) и поэтому к ним не при-
ложимо понятие парафилии, если сопоставлять их в единой клас-
сификации с Fissipeda.
Таким образом, Симпсон вводит принципиально новый тип го-
ризонтальных и вертикальных таксонов. Это накладывает отпеча-
ток на все определяемые им базовые понятия систематики. Коснем-
ся, в качестве примера, его определения монофилии как происхож-
дения группы от предкового таксона того же самого или более низ-
кого ранга. Многими это определение рассматривалось вне поло-
жений эволюционного подхода и поэтому не было понято. Стали
говорить о широкой монофилии (в противовес традиционной, уз-
кой), позволяющей будто бы зачислить в число монофилетических
чуть ли не любую полифилетическую группу, если надлежащим
образом расширить объем эволюционно исходной для нее группы.
Здесь надо понять, что Симпсон имеет дело с филогенетическим
деревом, которое ему надо непротиворечиво разбить на монофиле-
тические отрезки. Поэтому какого-либо произвола в выборе пред-
ковых групп быть не может. Они, а это исключительно вымершие
таксоны, также должны определяться с учетом принципа монофи-
лии, как и производные от них группы. Иными словами, Симпсон
имел в виду реальные исходные группы, жившие в прошлые эпо-
хи, тогда как его критики говорят об абстрактных предковых так-
сонах, отвечающих парафилетическим группам. Но в симсоновской
системе нет понятия парафилии, оно является классификационной
конструкцией филограмм, т.е. дендрограмм, в которых учитывает-
ся длина ветвей.
Предисловие к русскому изданию
XI
Столь же непродуктивной является попытка описать симпсо-
новский подход через понятие дивергенции. Мол, Генниг учитывает
лишь ветвление, а желательно дополнить филогенетический анализ
оценкой второй, дивергентной составляющей эволюционного про-
цесса. Но анагенетическую составляющую можно ввести лишь на
дендрограммах, т.е. когда сравниваются виды одного временного
среза. Определяемые на них таксоны статичны и это позволяет
проводить их сравнение по объему. В симпсоновских деревьях так-
соны развернуты во времени. С точки зрения сегодняшнего дня
крылатые насекомые сильно дивергировали от первичнобескры-
лых. Но ведь эти последние когда-то были более разнообразной
группой, чем Pterygota. Следовательно, уровень дивергенции был
иным и его нельзя фиксировать в системе Симпсона. Поэтому по-
нятие дивергенции для эволюционного подхода на самом деле яв-
ляется излишним. Оно лишь затуманивает главную идею Симпсо-
на — как непротиворечиво подразделить филетическую линию на
последовательные отрезки. Применительно к решению этой зада-
чи Симпсон и сформулировал, причем логически безупречно, оп-
ределение монофилии.
Коснемся и другой, уже упоминавшейся нами проблемы произ-
вольности таксономических границ. Обычно мы думаем, что стро-
им вполне объективные классификации или приближаемся к тако-
вым. И, как правило, не ставим под сомнение наше собственное
решение, даже если оно не совпадает с решением наших коллег.
Симпсон, видимо, был первым, кто поставил решение проблемы
объективности или реальности таксономических построений в
практическую плоскость поиска подходящих научных критериев.
Следует подчеркнуть, что речь у него идет лишь о границах меж-
ду таксонами, и о возможности их произвольного или менее про-
извольного проведения. Таксоны для Симпсона реальны, но грани-
цы между ними могут чертиться произвольно, поскольку четких
границ нет в природе. По Симпсону, важнейшими граничными кри-
териями являются внутригрупповая непрерывность и межгруппо-
вая разрывность. Поскольку критерии жестко увязаны с определен-
ными характеристиками, то статус выделяемых групп является в
любом варианте условным, иными словами, он не имеет абсолют-
ного значения. Поэтому Симпсон предлагает говорить не об объек-
xii
Дж Г. Симпсон. Принципы таксономии животных
тивности или реальности групп, но об их произвольном или непро-
извольном выделении. Даже если и не соглашаться с предложен-
ным им решением, проблема поставлена и требует решения. В про-
блеме произвольности таксономических построений есть еще один
момент, непосредственно связанный с эволюционной систематикой.
Поскольку с точки зрения Симпсона горизонтальные и вертикаль-
ные таксоны равноправны, то перед систематиком встает задача
построения сбалансированной классификации, учитывающей оба
эти типа таксонов. А это выводит работу систематика в сферу де-
ятельности, в которой ему приходится часто полагаться на умение
и чутье и которая в этом смысле сродни искусству. Кроме Симп-
сона, по-видимому, никто с такой ясностью не говорил о реальной
систематике как полезном искусстве (умении) строить классифика-
ции с некоторой долей произвольного элемента. Вывод Симеона
однако справедлив: хотя выделяемые нами группы в каких-то ас-
пектах произвольны, тем не менее они представляют ценность в
качестве рабочего аппарата анализа и описания разнообразия орга-
низмов.
Из всего сказанного следует, что в изучении систематических
групп защищавшийся Симпсоном подход, шел, по крайней мере в
своих намерениях, намного дальше филогенетической системати-
ки Геннига. Симпсоновское понятие таксона предполагало рассмот-
рение широкого круга эволюционных проблем, касающихся реаль-
ных аспектов развития таксона во времени, начиная с его проис-
хождения, «жизни» (диверсификации, адаптации к меняющимся
условиям среды, структурой связей с другими группами организ-
мов) и заканчивая его замещением более молодыми таксонами и
уходом с жизненной арены. Поэтому Симпсон с полным правом
мог назвать разбираемое в книге направление эволюционной сис-
тематикой.
Из этой тесной связи с эволюционным учением проистекают и
слабые стороны симсоновского подхода. Хотя эволюционные тео-
рии много дали для развития собственно таксономических идей, но
в исторической перспективе они отражают достигнутый уровень
знаний и в этом смысле имеют преходящее значение. Их главный
недостаток в том, что в отличие от других естественнонаучных те-
орий, они не могут иметь серьезного экспериментального обосно-
Предисловие к русскому изданию Xlii
вания и по этой причине во многом гипотетичны. Последнее ведет
к догматизации эволюционных доктрин и как результат к нараста-
ющему неприятию этого внешнего для таксономии элемента прак-
тическими систематиками. В недавней истории развития система-
тики эта реакция неприятия вылилась в призыв освободить систе-
матику от исторических нарративов (historical narratives), провозг-
лашенный в конце 1970-х гг. Элдреджем (N. Eldredge), Крэкрефтом
(J. Cracraft) и Гулдом (S.J. Gould) и принятый к руководству пат-
терн-кладистикой (G. Nelson, N.I. Platnick). Для последних эволю-
ционный подход оказался не более, чем неудачной попыткой осно-
вать систематику на идеях, истинность которых еще не очевидна и
поддерживается лишь авторитетом принятой научным сообществом
эволюционной парадигмы. Как бы там ни было, реально это был
призыв предать забвению симпсоновскую систематику, что факти-
чески и произошло.
Но времена меняются. Сейчас эволюционное учение стоит на
пороге радикальных изменений, связанных с впечатляющими до-
стижениями молекулярной биологии, в генетике и биологии разви-
тия. Безусловно, новые эволюционные концепции вновь пробудят
интерес у систематиков к эволюционной проблематике, и, будем
надеяться, вдохнут новую жизнь в эволюционную таксономию и в
симпсоновские идеи. Результатом этого будет пересмотр и ревизия
понятийного аппарата самой систематики. Именно по этой причи-
не книга Симпсона важна как одна из составляющих нашего дви-
жения вперед.
В заключение несколько технических замечаний. Настоящий
перевод снабжён комментариями переводчика и редактора, выне-
сенными в Приложение. Ссылки на них даны в основном тексте в
квадратных скобках. В таких же скобках в самом тексте в некото-
рых местах приведены отдельные уточняющие пояснения и англий-
ские термины, если исходный текст допускает слишком широкие
разночтения.
А.И. Шаталкин
Предисловие
Не удивительно ли,
что начинающие систематики обучаются,
словно дрессированные обезьяны,
почти исключительно посредством подражания
и что только в редчайших случаях
они получают инструкции по таксономической теории?
А. Кэйн
Иные авторитеты утверждают, что о таксономии не следует рас-
суждать — ею надлежит лишь заниматься. Именно так я и посту-
пал на протяжении многих лет, однако, подобно несостоявшемуся
философу, сбитому с правильного пути приливами весёлости, я
обнаружил в себе приливы мысли. В 1941-42 гг. (опубликовано в
1945 г.) я изложил некоторые из этих мыслей в связи с моим об-
ширнейшим таксономическим предприятием — классификацией
Mammalia. Тот опус был предназначен для того, чтобы объяснить
и доказать основания данной конкретной классификации, однако
он, как оказалось, имел и несколько более широкий интерес для
таксономистов. С тех самых пор отделение сих теоретических
изысканий от классификации млекопитающих и их самостоятель-
ное издание в более развёрнутой форме стало одним из тех дел,
которые исполняются «по мере наличия времени».
Предисловие
XV
Одно из достоинств всякого цикла лекций — наибольшее из
достоинств, как я полагаю, — состоит в том, что они подвигают
лектора к нахождению времени для приведения своих мыслей в
порядок и представления их в письменной форме. Лестное пред-
ложение присоединиться к числу Джезэпских лекторов [Jesup
lecturers] Колумбийского университета было принято, отчасти оп-
рометчиво, главным образом в расчёте на то, что это вынудит меня
найти время для некогда отправленного в долгий ящик изложения
принципов таксономии. Разумеется, это обернулось в нечто значи-
тельно более серьёзное, нежели простое стряхивание пыли со ста-
рых записей с переписыванием и расширением оных. На самом
деле настоящая книга, основанная на Джезэпских лекциях, прочи-
танных в 1960 г., имеет мало общего с написанным ранее, за ис-
ключением того, что в обоих случаях речь идёт об одном и том же
предмете и что я по-прежнему согласен со многими (безусловно,
не со всеми) идеями, которые я сформулировал 15 или 20 лет тому
назад.
Здесь я предпринял нечто значительно более амбициозное, не-
жели предыдущее издание. В той мере, насколько это позволили
способности и возможности, я попытался исследовать самые глу-
бинные основания таксономии и воздвигнуть на сих основаниях
всё здание зоологической классификации. Дальнейшее прояснение
предмета рассмотрения изложено в начале главы 1, которая будет
прочтена с большим вероятием, а книга, предложенная вашему
вниманию, сама собою покажет, каковы результаты всего предпри-
ятия. Я могу здесь добавить, что хотя рассмотрение означенного
предмета не претендует на элементарность, я всё же включил мно-
гое из того, с чем профессиональный таксономист уже вполне зна-
ком и что он, со своих позиций, сочтёт излишним (многие из по-
добных пассажей опущены в лекциях). Этот материал подан в на-
дежде на то, что начинающие исследователи смогут приобрести и
некие элементарные сведения по мере того, как они, возможно, с
куда большим усердием будут учиться размышлять о таксономии,
а не просто следовать её приёмам.
Др. Энн Роу прочитала весь текст и помогла мне ограничить,
если не полностью исключить, ошибки в изложении. Она заслужи-
вает больше, чем та рядовая благодарность, обычно выражаемая
XVI Дж. Г. Симпсон. Принципы таксономии животных
жёнам. Мисс Холли Ослер сделала значительно больше, чем к тому
обязывает чувство долга, по превращению моей рукописи в прием-
лемый текст, а также помогла в уточнении библиографии и указа-
теля. Рэймонд Дж. Диксон (издательство Колумбийского универси-
тета) отредактировал текст, а мисс Нэнси Диксон разработала ди-
зайн книги. Миссис Нэнси Гэхэн превратила мои карандашные на-
броски в законченные иллюстрации.
Я признателен, возможно, превыше всего, покойному Алексан-
дру Агасси и Гарвардскому университету. Благодаря счастливому
стечению обстоятельств, которые едва ли кто мог предвидеть, они
предоставили мне свободу действий для доведения моих мыслей
до окончательной формы.
Джордж Гэйлорд Симпсон
Кембридж
Апрель 1960 г.
Содержание
1. Систематика, таксономия, классификация,
номенклатура...........................................1
О порядке в природе и упорядочении в науке...........2
Роль систематики, таксономии и классификации.........7
Формы классификации: иерархии и ключи...............13
Линнеевская иерархия................................19
Замечания о теории множеств и символической логике...25
Основания и критерии классификации..................27
Номенклатура........................................32
2. Развитие современной систематики...................41
Схоластика и Линней.................................42
Некоторые другие додарвиновские подходы.............48
Типология...........................................54
Открытие филогении..................................59
Что такое естественная классификация?...............64
Некоторые формальные аспекты отношений..............68
Новая систематика и после неё.......................75
3. Таксономические доказательства и их
эволюционная интерпретация............................78
Определение и доказательство........................79
Категории доказательств.............................82
Сходство и гомология................................90
Некоторые аспекты оснований и методологии...........96
Некоторые критерии гомологии.......................102
Замечание о сериальной гомологии...................108
Последовательности и некоторые критерии
для примитивных и продвинутых признаков..........109
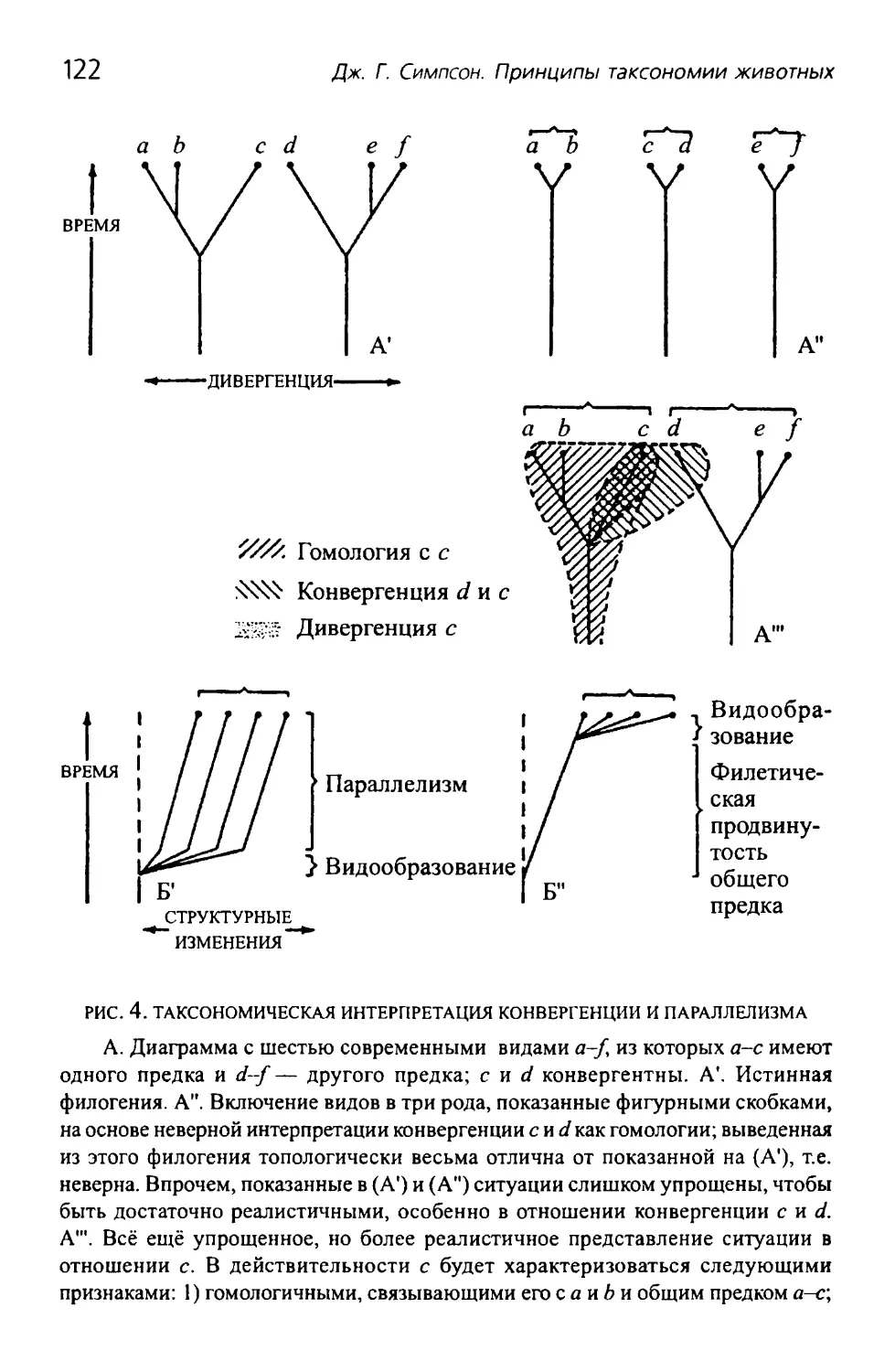
Замечание о параллелизме...........................120
2 — 4338
XVIII
Дж. Г. Симпсон. Принципы таксономии животных
4. От таксономии к классификации....................125
Что делает таксономист?...........................126
Классификация как полезное искусство..............128
Объективность и произвольность....................133
Монофилия и полифилия.............................139
Грады и клады.....................................146
Вертикальные и горизонтальные связи...............150
Дивергенция и разнообразие........................154
Дробительство и объединительство..................158
Относительная древность...........................163
Ранги признаков...................................169
5. Вид и низшие категории...........................171
Генетический вид..................................172
Эволюционный вид..................................177
Некоторые другие категории вида...................181
Вид у однополых организмов........................188
Вид в палеонтологии...............................190
Подвид............................................199
Другие внутривидовые группы.......................205
Надвид............................................209
Типы и гиподигмы..................................213
6. Высшие категории.................................218
Что такое высшая категория?.......................219
Основания для выделения высших таксонов...........223
Определение и характеристики высших категорий.....230
Анализ некоторых филогенетических паттернов.......234
Некоторые примеры из филогении млекопитающих......240
Эволюционная основа таксонов......................256
Цитированные работы.................................265
Приложение..........................................273
Предметный указатель................................279
Указатель латинских названий таксонов...............289
Указатель авторов...................................291
Рисунки
I. Диаграммы отношений.................................69
2. Диаграмма (по Боннэ) непрерывной scala naturae и её
произвольное разделение на таксоны последовательно
возрастающих рангов.................................71
3. Диаграммы в форме деревьев..........................73
4. Таксономическая интерпретация конвергенции и
параллелизма.......................................122
5. Произвольный и непроизвольный аспекты одновременно
существующих таксонов в отношении генетической
непрерывности......................................135
6. Группировка таксонов по разрывам в сходственных
связях.............................................136
7. Три гипотетические филогении.......................139
X. Некоторые схематические представления соотношений
между структурным прогрессом и филогенией..........148
9. Схемы горизонтальной, вертикальной и смешанной
классификаций......................................153
10. Схема, иллюстрирующая дробительский и
объединительский [подходы в классификации]............160
11. Схема роста филогенетического дерева и его отношение
к возникновению высших таксонов........................167
12. Схемы филогении и генетических видов с проходящими
через них филетическими линиями........................192
13. Разделение филетических линий на последовательные
виды..................................................194
14. Альтернативные способы разделения коротких
ветвящихся филетических линий на виды.................196
15. Некоторые проблемы, связанные с выборками
географически смещающихся палеонтологических
видов.................................................197
XX
Дж. Г. Симпсон. Принципы таксономии животных
16. Интерпретация палеонтологических выборок
из различных горизонтов и локалитетов.................198
17. Некоторые соотношения между одновременными и
последовательными подвидами...........................202
18. Анализ концепции надвида..........................211
19. Соотношение между филогенией и высшими и низшими
таксонами.............................................222
20. Критерии разрыва и монофилии для высших категорий..224
21. Влияние заполнения разрыва в группе видов, показанных
на рис. 20 ...........................................228
22. Неприменимость критерия одинаковой древности
происхождения для такоснов одного ранга...............229
23. Некоторые элементарные филогенетические паттерны...236
24. Некоторые элементы паттерна, связанные с экспансией
или радиацией высших [таксонов]........................238
25. Некоторые паттерны полу произвольного подразделения
высших [таксонов]......................................239
26. Радиации, которым в [семействе] Leporidae соответствуют
подсемейства..........................................242
27. Филогения семейств и подотрядов Carnivora.........245
28. Схема филогении и классификация Primates..........247
29. Схема град приматов и их связь с некоторыми
таксонами.............................................250
30. Филогения семейств грызунов.......................252
Систематика, таксономия,
классификация, номенклатура
Всякое обсуждение должно начинаться с ясного пони-
мания того, что подлежит обсуждению. Основная цель настоящей
главы, соответственно, — строго установить, что в данной книге
понимается под таксономией. Сюда относится также рассмотрение
систематики, которая шире таксономии и включает её, наряду с
классификацией и номенклатурой, которые уже таксономии и (в не-
сколько ином качестве) входят в неё. Другой предпосылкой плодо-
творной дискуссии служит обозначение и, по мере необходимости,
создание словаря. Поэтому настоящая глава включает некоторые
определения и объяснения по общей терминологии, которая будет
использоваться в последующих главах. Далее, имеет смысл имен-
но здесь указать определённые общие принципы, которые лежат в
основе всего предмета, некоторые из которых будут проиллюстри-
рованы более предметно и рассмотрены более детально на после-
дующих стадиях изложения. Такого рода предварительное введение
в тему включает также краткое упоминание некоторых проблем,
первичных заключений, процедурных вопросов, а также других
спорных моментов, которые, опять-таки, будут изложены более
подробно и систематически по мере дальнейшего обсуждения. На-
конец, необходимую основу всего последующего составляет рас-
смотрение некоторых формальных схем классификации и правил
номенклатуры.
2
Дж. Г. Симпсон. Принципы таксономии животных
Необходимо особо отметить один момент, прежде чем начинать
обсуждение: название настоящей книги следует понимать букваль-
но. Предметом рассмотрения являются принципы, тогда как отдель-
ные группы организмов или классификации приводятся лишь как
примеры и как основа для индуктивного выведения принципов.
Имеются в виду принципы таксономии, как они будут строго оп-
ределены несколько ниже. Более широкие принципы систематики
в целом привлекаются по мере необходимости, чтобы показать ис-
ходную основу собственно таксономических принципов или про-
яснить их происхождение. При этом имеется в виду именно та так-
сономия, которая применима к животным, под которыми я пони-
маю многоклеточных. Большинство этих принципов применимо
также и к растениям, а некоторые из них, в не столь общей фор-
ме, к простейшим. Впрочем, такие ограничения можно было бы
упомянуть и между строк, они во многом второстепенные. Приме-
ры заимствованы главным образом из млекопитающих, по личным
и практическим причинам. Принципы, полученные при изучении
млекопитающих и применительно к ним, в существенных чертах
являются общими для всех многоклеточных, так что слово «живот-
ные» в названии книги не является избыточным рекламным ходом,
хотя специалист, скажем, по кишечнополостным вполне может
прийти к иным заключениям в свете своего собственного опыта.
Важные различия в точках зрения должны обсуждаться, однако я
не пытался дать извлечения из всей огромной литературы по пред-
мету книги, а ещё менее того — представить некую нейтральную
формулу согласия.
О порядке в природе и упорядочении в науке
«Мир настолько полон бесчисленными вещами» — что это мо-
жет быть в высшей степени, даже безнадёжно обескураживающим.
Если бы каждая из неисчислимых вещей в мире воспринималась
как нечто особенное, отдельное, как вещь сама по себе, не связан-
ная ни с какой другой вещью, восприятие мира было бы низведе-
но к полной бессмыслице. Это было бы так, если бы каждая
вещь — например, каждое дерево — рассматривалась как нечто со-
< истематика, таксономия, классификация, номенклатура
3
всршенно обособленное: разумеется, в таком случае это не было
бы дерево, ибо «дерево» есть обобщающее понятие, не примени-
мое к единичному объекту, рассматриваемому отдельно от всех
прочих. На самом деле (это уже обсуждалось обстоятельно) даже
конкретное дерево нельзя было бы воспринимать как нечто цело-
ciHoe: поскольку всякое чувственное восприятие становится осмыс-
иснным лишь в результате обобщения, без некоторой формы упо-
рядочения и абстракции каждое дерево распадалось бы на бесфор-
менную мозаику «зелёного» (отличного от соответствующего вос-
приятия, идущего от любого другого зеленого дерева), «грубого на
ощупь», «ветвистого» и т.п.
Не было бы связного языка, если бы каждая вещь (или каждое
ощущение) обозначалась бы отдельным словом, и не было бы ра-
«умного мышления, если бы символы не обозначали характеристи-
ки и отношения, общие для некоторых из бесчисленных различных
объектов. Необходимость объединения вещей (или, что является
операциональным эквивалентом, ощущений от них) в классы явля-
ется всеобщим свойством живых существ. Можно было бы усом-
ниться (хотя некоторые с этим не согласны) в возможности приме-
нения таких слов как «осознанность» или «восприятие», например,
к амёбе, однако из реакций амёбы совершенно очевидно, что не-
что в её организации осуществляет акты обобщения. Она не реа-
гирует, скажем, на каждый отдельный кусочек пищи как на некий
уникальный объект, но каким-то образом классифицирует (в неко-
юром смысле этого слова) бесчисленные различные объекты в
группу съедобных вещей. Подобное обобщение, подобная класси-
фикация (в этом значении) является абсолютно необходимым, ми-
нимальным условием приспособления, что, в свою очередь, явля-
с гея абсолютным и минимальным условием существования и про-
должения жизни.
Взаимосвязи между вещами, которые, так или иначе, должны
приниматься в расчёт, весьма и весьма разнообразны и сами по
себе сложным образом взаимозависимы. Среди них следует выде-
лить два типа связей, которые особо значимы. Они приблизитель-
но соответствуют тому, что психологи обычно называют «связью по
смежности [contiguity]» и «связью по сходству [similarity]»: мы мо-
жем заимствовать эти термины применительно к нашим несколь-
4
Дж. Г. Симпсон. Принципы таксономии животных
ко иным целям1. Связь по смежности (для наших целей) — это
структурное или функциональное отношение между вещами, кото-
рое, в терминологии психологов, входит в единый Geschtalt [геш-
тальт]. Связанные данным отношением вещи могут быть весьма
несхожими, или их сходство может не иметь к этому [отношению]
касательства. Таково, например, отношение между растением и
почвой, на которой оно произрастает, между кроликом и лисой,
которая гонится за ним, между отдельными органами, из которых
составлен организм, между всеми деревьями в лесу, или между все-
ми членами данной популяции. Вещи, вступающие друг с другом
в такого рода отношения, структурно и функционально принадле-
жат к тому, что может быть определено в широком, несколько тех-
ническом смысле как единая система.
Связь по сходству — во многом само-объясняющая концепция,
по той единственной причине, что она сама по себе служит осно-
ванием для языковых и иных знаковых систем, которые мы исполь-
зуем при формировании определений. Она означает, разумеется,
просто объединение в группу [класс, множество] многих разных
вещей вследствие того, что все они обладают одним или несколь-
кими совпадающими свойствами. Объекты в таких группах по оп-
ределению являются сходными в некоторой степени, однако какие-
либо структурные или функциональные отношения между ними не
принимаются во внимание, или эти отношения в норме не образу-
ют систему в смысле, принятом в современной теории систем. На-
пример, они не взаимодействуют, или, если взаимодействие проис-
ходит, оно не является частью данного отношения. Они все могут
быть жёлтыми, или гладкими, или с крыльями, или все высотой в
10 футов. Они все могут быть, например, с 4 ножками и спинкой
и будут называться «стульями» — пример того факта, что все сло-
ва языка (за очевидным исключением имён собственных) олицет-
воряют объединения по сходству [1].
Очевидно, в некоторых случаях одно и то же отношение может
рассматриваться, в зависимости от обстоятельств, как связь по 1
1 Эти термины исходно использовались в ныне устаревшем смысле в те-
ории познания. Здесь они применяются непосредственно к ощущениям и не-
явным образом к отношениям, для которых предполагается (или постулирует-
ся) объективное существование в окружающем мире.
Систематика, таксономия, классификация, номенклатура
5
смежности или как сходство. Например, класс всех членов данной
популяции, уже упоминавшейся как пример объединения по смеж-
ности, может также рассматриваться как объединение по предко-
вому сходству [2]. Это не очень строго, поскольку такое сходство
не связывает между собой сами классифицируемые объекты [т.е.
они связаны между собой опосредованно, через предка]. На прак-
тике, как мы все хорошо знаем, в подобной ситуации могут быть
и такие сходства, которые непосредственно соединяют сами объек-
ты, так что связи по сходству сопровождаются или порождаются
связями по смежности, но не идентичны последним. Этот случай
имеет очевидное отношение к таксономии и показывает, что такие
базовые абстрактные рассуждения фундаментальны для нашего
предмета.
Как и в случае многих других характеристик живых существ,
восприятие связей по смежности и по сходству достигло у человека
наибольших высот, не имеющих себе равных ни по глубине, ни по
широте. Мы определённо упорядочиваем наши восприятия внеш-
него мира более полно, более последовательно, более осмысленно,
нежели любые другие организмы, и мы обычно упорядочиваем их
либо каким-то одним, либо сразу обоими способами. Такое упоря-
дочивание наиболее отчётливо в искусстве и в науке — двух, в оп-
ределённом смысле высших формах деятельности человека, прису-
щих только ему. Именно по глубине и совершенству упорядочива-
ния, причём упорядочивания в обеих фундаментальных формах,
науки и искусства едины. И я также полагаю, что именно эту сте-
пень и эти формы упорядочивания надлежит рассматривать как
истинно эстетические (как в искусстве, так и в науке). Уместно
подчеркнуть здесь, что таксономия, которая есть упорядочивание
по самой своей сути, обладает особой эстетической ценностью.
К сожалению, наши изыскания в этом направлении увели бы нас
в сторону, если бы мы углубились в этот изумительный аспект так-
сономии.
Необходимо отметить, что учёные, более чем многие другие
смертные, должны быть терпимы к неопределённости и разочаро-
ваниям (Roe, 1946). Наше столетие сделало всё более очевидным
тот факт, что наука неопределённа по самой своей природе. За ис-
ключением разве что тривиальных и строго наблюдаемых феноме-
6
Дж. Г. Симпсон. Принципы таксономии животных
нов, её результаты редко абсолютны, но чаще устанавливаются с
некоторым уровнем вероятия, или, в более строгих терминах, с не-
которым доверительным интервалом. Учёным следует также ми-
риться с разочарованиями в силу того, что они никогда не могут
наперёд сказать, приведут ли их действия, на которые могут быть
затрачены годы и даже целая жизнь, к результатам с желаемой сте-
пенью доверительности. (Если бы это было возможно, в самих дей-
ствиях отпала бы необходимость.) На самом деле, единственное, в
отношении чего учёные могут быть вполне определённы, — это то,
что полное решение ни одной значимой проблемы недостижимо.
Учёные действительно терпимы к неопределённости и разоча-
рованиям, потому что они должны быть такими. Лишь к одной
вещи они не могут и не должны быть терпимы — к беспорядку.2
Единственная цель теоретической науки — довести хаос до макси-
мально возможной и разумной степени перцепционной редукции,
что началось в низшей и (по всей вероятности) неосознанной фор-
ме одновременно с возникновением жизни. В отдельных случаях
вполне можно задаться вопросом, является ли порядок, таким об-
разом достигаемый, объективной характеристикой феноменов или
это просто артефакт, конструируемый учёным. Такой вопрос воз-
никает время от времени в таксономии животных, и он будет воз-
никать неоднократно в разных обличиях на последующих страни-
цах. Тем не менее, наиболее фундаментальный постулат науки со-
стоит в том, что природа сама по себе упорядочена. В таксономии,
как и в других науках, цель состоит в том, чтобы упорядоченность,
порождаемая наукой, приближалась к порядку природы или отра-
жала его некоторым достойным применения способом. Всякая те-
оретическая наука есть упорядочивание, и если, как мы вскоре бу-
дем обсуждать, систематика равнозначна упорядочиванию, то си-
стематика синонимична теоретической науке. Таксономия, во вся-
ком случае, — это наука, которая наиболее последовательно и явно
занимается упорядочиванием сложных фактов, и в этом отношении
2 Нет явного противоречия в том, что большинство творческих личностей
в науке — часто именно те, которые не только принимают существование бес-
порядка, но к тому же положительно пленимы им. Очевидная причина в том,
что распознавание беспорядка — это и предпосылка, и необходимое условие
творческого акта упорядочивания.
Систематика, таксономия, классификация, номенклатура
7
ей принадлежит особое, в том числе эстетическое (как уже было
сказано) и (как можно было бы сказать) почти наднаучное место
среди наук.
Роль систематики, таксономии и классификации
Представление о том, что всякая наука занимается упорядочи-
ванием и что, таким образом, систематика в общем понимании рав-
нозначна науке, было особо подчёркнуто Геннигом (Hennig, 1950)
в следующем пассаже:
«Для того, чтобы правильно судить о месте систематики в биологии
и о роли, которую она призвана играть в разрешении базовых проблем этой
науки, необходимо прежде всего чётко осознать, что систематика есть не
только в биологии, но что она в значительной степени является объединя-
ющей частью всякой науки. Кажется удивительным и странным, до какой
степени исходное значение этой концепции оказалось забытым в биологии
в результате совершенно неприемлемого, но ныне всеобщего ограничения
представления о систематике как о частном подразделении науки.»3
Такое представление о систематике этимологически оправдано,
оно вполне точно подчёркивает связь между биологической систе-
матикой и упорядочиванием в науке вообще. Оно также указыва-
ет на некорректность отождествления систематики с такой узкой
сферой как, например, классификация. Тем не менее, оно идёт
вразрез с принятым пониманием и к тому же терминологически
неудобно, поскольку мы не нуждаемся в ином термине для теоре-
тической науки в целом, нам необходимо обозначить достаточно
широкие, но всё же специфические и существенные для биологии
аспекты упорядочивания. К несчастью, даже в биологии, по-види-
3 «Um num die Stellung der Systematic im Bahmen der Biologie und die Bolle,
die sie bei der Losung der Grundfragen dieser Wissenschaft zu spielen berufen ist,
richtig beurteilen zu konnen, muss man sich zunachst klarmachen, dass es
‘Systematic nicht nur in der Biologie gibt, dass sie vielmehr einen integrierenden
Bestandteil jeder Wissenschaft iiberhaupt bildet. Es is estraunlich und befremdlich,
zu sechen, wie ser man in der Biologie uber der im Grunde genommen unzulassigen
aber heute allgemein tiblichen Einschrankung des Begriffes der Systematik auf ein
bestimmtes Teilgebiet der Gesamtwissenschaft die Bedeutung vergessen hat, die
diesem Begriff upspranglich zukommt.»
8
Дж. Г. Симпсон. Принципы таксономии животных
мому, все ещё нет единодушия в отношении строгого определения
термина «систематика». Является общепризнанным, что системати-
ка включает формальное, техническое классифицирование организ-
мов. Одна крайность состоит в их отождествлении: систематика
определяется просто как наука классификации. Однако очевидно,
что даже те, кто принимают столь узкое определение, на самом
деле включают в обсуждаемый круг вопросов значительно больше,
чем классификацию в каком бы то ни было частном понимании.
(См., например, Mayr et al., 1953 [3] и особенно Мауг, 1942, где
«систематика» определена как эквивалент таксономии, но при этом
оба термина повсеместно использованы в понимании, значительно
более широком, чем классификация.) Действительно, биологи
склонны давать систематике не слишком ясное расширенное тол-
кование, простирающееся за пределы более чётких, относительно
узких границ классификации. Это стремление компетентно отраже-
но в работе (Blackwelder, Boyden, 1952): «Систематика представля-
ет собой обширную область, имеющую дело с реальными группа-
ми (kind) животных, различением последних, их классификацией
и эволюцией.» В согласии с этой общей тенденцией в современном
использовании термина, ему можно дать ещё более широкое, но не
менее ясное определение:
Систематика есть научное изучение реальных групп [kinds] и
разнообразия организмов, а также всевозможных [any and all]
отношений между ними.
В этом определении слово «отношения» необходимо понимать
не в каком-либо узком, частном смысле (например, в смысле фи-
логенетических связей), а в совершенно общем смысле, включая
все особые случаи связей по смежности и сходству, согласно их
пониманию, изложенному в предыдущем разделе.
Классификация организмов — это деятельность, которая цели-
ком принадлежит систематике, однако эта тесная связь не означа-
ет их тождественности. В отношении к другим биологическим дис-
циплинам систематика держит в фокусе своих интересов получае-
мые ими факты и ищет способы рассмотрения этих фактов. Напри-
мер, молекулярная биология не имеет необходимости и обычно
(если говорить о настоящем времени) не акцентирует внимание на
реальных группах [kind] и разнообразии организмов. Следователь-
Систематика, таксономия, классификация, номенклатура
9
но, в качестве особой биологической дисциплины она не является
частью систематики. Тем не менее, данные молекулярной биоло-
гии, рассматриваемые в сравнительном аспекте на любом уровне
(как внутри, так и между популяциями), весьма решительно вклю-
чаются в сферу интересов систематики: распределение различных
гемоглобинов — только один из бесчисленных более чем уместных
примеров. Систематика, в свою очередь, приложима ко всей моле-
кулярной биологии, показывая ей возможные способы того, как
результаты, получаемые в её области, могут быть объяснены или
осмысленно упорядочены. Можно усмотреть аналогию с эволюци-
онной биологией, предмет рассмотрения которой (возможно, за
исключением филогении) принадлежит исключительно ей, но ко-
торая впитывает данные из всех биологических дисциплин и кото-
рая, в свою очередь, предлагает объяснительные принципы, акту-
альные для всех них. Как я указывал несколько лет назад (Simpson,
1945):
«(Систематика)4 в одно и то же время является и наиболее элементар-
ной, и наиболее объемлющей частью зоологии — наиболее элементарной,
поскольку животные не могут обсуждаться или исследоваться, пока не будет
достигнута некоторая их (систематизация)4, и наиболее объемлющей, по-
скольку (систематика)4 в её различных ипостасях и ответвлениях в конеч-
ном итоге объединяет, использует и обобщает всё, что известно о живот-
ных, будь то данные морфологии, физиологии, психологии или этологии.»
Очевидно, что все сравнительные разделы биологии так или
иначе входят в систематику, поскольку они по необходимости изу-
чают отношения между разнообразными организмами. Это особен-
но верно в отношении сравнительной анатомии, сравнительной
физиологии и сравнительной психологии, в самом широком их по-
нимании, т.е. включая, например, сравнительную цитологию, срав-
нительную биохимию и этологию (один из подходов к сравнитель-
ному изучению поведения). Систематический аспект этих наук зат-
рагивает существенную — впрочем, разумеется, не исчерпываю-
щую — часть объединений по сходству. В некоторых других науках
4 В исходном тексте здесь стоит термин «таксономия», который я в то вре-
мя использовал в смысле, который здесь вкладывается в «систематику». Как
показано ниже, в настоящее время я предпочитаю иное использование. ‘
10
Дж. Г. Симпсон. Принципы таксономии животных
рассматриваются по преимуществу объединения по смежности, в
таком случае их связь с систематикой, хотя не менее реальная и
важная, может быть не столь очевидной. Это в особенности верно
в отношении биогеографии и экологии.
Связь биогеографии с систематикой широко признана, хотя бы
и неявно, и потому заслуживает лишь краткого упоминания. Напри-
мер, в книге «Систематика и происхождение видов» (Мауг, 1942)
повсеместно используются зоогеографические данные, а «Зоогео-
графия...» Дарлингтона (Darlington, 1957) является столь же систе-
матической, сколь и географической [4]. Значение экологии призна-
ётся не так широко, а некоторые систематики его просто отверга-
ют. Так, Blackwelder, Boyden (1952) в уже цитированной статье,
хотя и принимают исключительно широкий взгляд на систематику,
тем не менее исключают экологию, а один из основателей «новой
систематики» Thorpe (1940) утверждал, что:
«(Синэкология) сама по себе оказала небольшое влияние на
исследования по систематике, разве что обязывая её прилагать зна-
чительные усилия для идентификации и отнимая массу времени и
энергии, которые во многих случаях могли бы быть затрачены с
большей пользой.»
Синэкология изучает многовидовые сообщества, в противопо-
ложность аутэкологии, имеющей дело с отношениями между отдель-
ными группами организмов и их средой обитания. Может показать-
ся, что в действительности синэкология не просто близка к систе-
матике, как она здесь очерчена, но является её частью, поскольку
рассматривает некоторые категории отношений (те или иные связи
по смежности) между организмами. Связь аутэкологии с система-
тикой, конечно, не столь прямая, однако в своём сравнительном
разделе она играет ту же роль, как и другие сравнительные биоло-
гические дисциплины, предоставляя данные для систематики, глав-
ным образом в форме связей по сходству. Это, например, существен-
но для изучения конвергенции, которая, как будет показано далее,
составляет одну из ключевых проблем в систематике.
Особо следует указать на тесную связь генетики и системати-
ки, хотя этот факт и так достаточно очевиден. Проблема вида, одна
из центральных в систематике, в своей основе является генетиче-
ской проблемой. Не в меньшей степени и столь же прочно генетика
Систематика, таксономия, классификация, номенклатура
11
вовлечена в широчайшую сферу других проблем систематики, та-
ких как морфогенез (как в онтогении, так и в филогении), измен-
чивость, изоляция, динамика популяций и многие другие, в той или
иной степени затронутые далее в настоящем исследовании.
Активность систематики в изучении отношений (в самом ши-
роком смысле) между разными организмами почти с необходимо-
стью требует создания некой формальной системы, в которой оз-
наченные организмы группируются в классы, и словаря, с помо-
щью которого эти классы обозначаются. Другими словами, как уже
было указано (но не сформулировано в строгих терминах), систе-
матика включает классификацию и номенклатуру, которые в рам-
ках настоящей книги могут быть определены следующим образом:
Зоологическая классификация есть упорядочение животных в
группы (или множества) на основании их отношений, т.е. связей
по смежности, сходству или их обоих.
Зоологическая номенклатура есть применение отличительных
названий для каждой из групп организмов, выделенных в любой зоо-
логической классификации. Номенклатура, таким образом, состав-
ляет существенное дополнение или побочный результат классифи-
кации. (В принципе, «названия» в данном определении не обяза-
тельно должны быть словами, но почти любыми знаками. Однако
хотя другие знаковые формы время от времени предлагаются для
специфических целей, на практике это почти всегда слова, и эта
практика принята в настоящем исследовании.)
Теперь уже должно быть в основных чертах понятно, что хотя
классификация является существенной частью систематики и её
деятельность почти целиком ограничена этой дисциплиной, она
составляет лишь сравнительно узкую её часть, значительно мень-
шую, чем систематика в целом. Однако даже в таком понимании,
более строгом, чем это обыкновенно принимается, слово «класси-
фикация» остаётся несколько двусмысленным. С одной стороны,
как многие аналогичные термины (например, адаптация), оно обыч-
но применяется как к процессу — классифицированию организмов,
так и к результату этого процесса — определённому порядку назва-
ний, присвоенных классам, в которые отдельные организмы были
сгруппированы. Это различие реально и важно, однако в настоящее
время я не вижу оснований для последовательного использования
12
Дж. Г. Симпсон. Принципы таксономии животных
двух разных терминов, поскольку различия в их смысле обычно
очевидны из контекста. Использование одного термина в обоих
смыслах редко приводит к логической путанице.
Имеется, однако, ещё одно различие, которое следует непремен-
но учитывать, чтобы избежать путаницы. Это различие между про-
цессом классификации и методами, приёмами, правилами и прин-
ципами, соответственно которым осуществляется названный про-
цесс. Именно на это различие весомо указал Грегг (Gregg, 1954),
отделив то, что он назвал «собственно таксономией», т.е., по сути,
процесс классификации, от «методологической таксономии». Грегг
пояснил, что «основной сферой приложения исследований в соб-
ственно таксономии является органический мир, тогда как сфера
приложения методологической таксономии — таксономия как та-
ковая». Это различение существенно и должно повсеместно обо-
значаться терминологически, однако только что приведённые тер-
мины мне представляются нескладными, если не сбивающими с
толку.
«Таксономия собственно» — это просто одно из стародавних
значений «классификации», так что я предлагаю продолжать ис-
пользовать этот термин. Таково, например, первое из определений
«классификации» («действие классифицирования») в Оксфордском
словаре английского языка, где оно датируется 1790 г. Возможной
альтернативой для «методологической таксономии» может быть
«метаклассификация», ибо она содержит утверждения о классифи-
кации, так же как, например, метаматематика содержит утвержде-
ния о математике. Однако я не вижу причин, почему для обозна-
чения методологической таксономии не применять более простой
и знакомый термин «таксономия», который во всех вариантах ис-
пользования подразумевал такое толкование.
Таксономия есть теоретическое исследование классификации,
включая её основания, принципы, приёмы [procedures] и правила.
Я бы, в таком случае, перефразировал Грегга и принял, что
предметом классификации являются организмы, а предметом так-
сономии являются классификации. Такое использование этимоло-
гически оправдано, ибо, судя по корням (xa^ta и vopra), слово
«таксономия» можно вольно перевести как «обобщение о клас-
сификации». Большинство прежних определений «таксономии» не
Систематика, таксономия, классификация, номенклатура
13
противоречит данному, будучи (по крайней мере некоторые) просто
более ограниченными. Принятое здесь толкование этого термина
связано с явным разграничением сфер деятельности систематики,
таксономии и классификации5, которое прежде часто отвергалось
или толковалось неправильно.
Формы классификации: иерархии и ключи
Базовый или, можно сказать, начальный шаг в разработке клас-
сификации — это просто объединение в группу индивидуальных
объектов (или понятий) посредством некоторой системы отноше-
ний (или связей) между ними. Эта ментальная операция, дополнен-
ная присвоением каждой группе отличающего её знака, составля-
ет основу языка, и зоологическая классификация на этом началь-
ном уровне уже присутствует в обыденных словарях всех языков.
Слово «кугуар», например, обозначает не отдельный объект (т.е. это
не имя собственное в грамматическом смысле), но целый класс
сходных между собой животных. В данном случае этот класс ока-
зался совпадающим с зоологическим видом (Felis concolor), хотя
чаще всего народные названия шире видов и редко совпадают с
ними.6 Любой, знакомый с этим животным может легко отличить
его от других диких кошек по многим диагностическим признакам
(однотонная окраска, короткий мех на всём туловище, длинный
хвост, округлые уши, крупные размеры и т.п.). Интересно, однако,
отметить, что «нетехническое» распознавание — идентифика-
5 Как и другие, я ранее (например, Simpson, 1945) использовал таксономию
в более широком смысле, включая как систематику, так и таксономию в при-
нятом здесь понимании. Предлагались и другие, действительно вводящие в за-
блуждение определения: например у Блэквельдера и Бойдена (Blackwelder,
Boyden, 1952), которые высказали мнение, что «таксономия может быть разум-
но ограничена описательной, диагностической и номенклатурной стадиями
исследования преимущественно на видовом и более низких уровнях». Я не
знаю других случаев подобного использования «таксономии» и не вижу осно-
ваний для его принятия. В таком понимании это просто классификация на низ-
ших иерархических уровнях.
6 Так называемые обыденные, или народные названия видов, представлен-
ные в полевых определителях и тому подобных справочниках, на самом деле
являются искусственным «переводом» научной номенклатуры.
3 — 4338
14
Дж. Г. Симпсон. Принципы таксономии животных
ция — обычно производится не по отдельным признакам, а по мен-
тальному образу всего животного.
В некоторых диалектах и для некоторых классов объектов лин-
гвистическая классификация не идёт далее этого начального, пред-
варительного этапа. Например, у индейцев камарокото есть обшир-
ный словарь для отдельных разновидностей млекопитающих, но
отсутствует слово, эквивалентное родовому понятию «млекопита-
ющее» или «животное». Однако в обыденной речи, по меньшей
мере, удобно, а в научной систематике абсолютно необходимо
иметь систему [понятий], в которой присутствуют разные уровни
общности, или включённости. На практике это может быть достиг-
нуто двумя различными способами: во-первых, посредством пере-
сечения или объединения неидентичных классов и, во-вторых, со-
подчинением одних классов другим, или включением первых во
вторые. Например, в химии имеется класс соединений натрия и
класс солей соляной кислоты. Ни какой из этих классов не сопод-
чинён другому и не включён в него: они имеют один и тот же ка-
тегориальный ранг (более строго смысл «категории» и «ранга» бу-
дет уточнён позднее). Имеется более узкий класс, именно — хло-
рид натрия, который принадлежит обоим этим классам, т.е. совпа-
дает с областью их пересечения. Сходным образом в зоологии име-
ется класс животных с рогами, класс парнокопытных животных и
класс животных с рогами и парными копытами: последний мень-
ше первых двух и представляет собой область их пересечения.
Классификация посредством соподчинения и включения клас-
сов более обычна в зоологии и в целом более знакома. Так, обы-
денное название «бурый медведь» обозначает класс (соответству-
ющий одному или нескольким видам, в зависимости от мнения зоо-
логов), который находится в отношении соподчинения к большему
классу (в который и включается), обозначенному как «медведь».
Другие классы, такие как «белый медведь», находятся на том же
уровне [соподчинения], что и «бурый медведь», и также включены
в класс «медведь». Такого рода двухсловная номенклатура, где одно
слово обозначает включающий класс или множество, а оба вмес-
те обозначают один из включённых классов или подмножеств, не
обязательна для обыденного языка, хотя и широко распространена.
Она имеет очевидную связь с технической зоологической номенк-
Систематика, таксономия, классификация, номенклатура
15
латурой: «медведь» = Ursus, «бурый медведь» - Ursus arctos, «бе-
лый медведь» = Ursus maritimus. Эта связь не случайна и не про-
сто аналогия. Технический язык возник на основе обыденного, ко-
торый исходно был латынью (во времена оные — обыденный для
научного сообщества), где включающий класс обозначался суще-
ствительным, а включённый класс — чаще всего прилагательным,
ограничивающим или уточняющим слово или фразу. Линней осно-
вал нашу биномиальную систему путём соответственного сокраще-
ния уточняющего обозначения до единственного слова и стремясь
(хотя и не вполне последовательно) отделить номенклатуру от оп-
ределения, сделав второе слово лишь формальным символом, а не
определением (см. Steam, 1959). То, что это только законченная
формализация, соответствующая уже широко распространённой
концептуальной схеме или манере мыслить, замечательно демонст-
рируют некоторые тесные параллели между обыденными названи-
ями и техническими биномиалами. Так, Деннлер (Dennler, 1939)
показал, что в языке южноамериканских индейцев гуарани живот-
ные непременно обозначаются двухсловными названиями, в кото-
рых первое слово является включающим (родовым), а второе уточ-
няющим (видовым), точно так же, как в линнеевских биномиалах:
tatu — броненосец; tatu para, tatu guasii и т.д. — разные виды бро-
неносцев.
Классификация посредством соподчинения, разумеется, не ог-
раничивается единственным шагом между классами двух уровней.
В принципе, эта процедура может охватывать неопределённо боль-
шое число уровней, хотя на практике, по крайней мере, в зоологии,
число шагов редко превосходит 20-25, но может быть и не более 7.
Такое упорядочение [классов] есть иерархия. Упорядочение посред-
ством пересечения классов здесь будет называться ключом, хотя это
не самое обычное определение данного термина или, по меньшей
мере, не вполне обычный взгляд на концепцию, которую он обо-
значает. Формальные определения, соответствующие принятому
подходу, таковы:
Иерархия есть систематическая форма [framework] зоологи-
ческой классификации с такой последовательностью классов (или
множеств) разного уровня, в которой каждый класс, кроме низ-
шего, включает один или более соподчинённых классов.
з*
16
Дж. Г. Симпсон. Принципы таксономии животных
Ключ [определительная таблица] есть систематическая фор-
ма [framework] зоологической классификации (обычно используемая
для идентификации, но не для других целей) с такой последова-
тельностью классов, в которой на каждом уровне более узкий
класс образован пересечением двух или более классов следующего
более высокого уровня.
В принципе, две эти схемы в своих типических чертах совер-
шенно различны, однако, будучи представлены в письменной фор-
ме, они могут быть сходны, так что иногда бывает затруднитель-
но или даже невозможно с уверенностью утверждать, является ли
данная классификация иерархией или ключом. Теоретически, они
резко различны тем, что в иерархии каждый класс принадлежит
единственному классу следующего более высокого уровня, тогда
как в ключе каждый класс принадлежит двум или более классам
следующего более высокого уровня. Таким образом:
Иерархия:
медведи
бурые медведи белые медведи
Ключ: бурые животные медведи белые животные < > ч > бурые медведи белые медведи
Ключи могут быть представлены диаграмматически таким об-
разом, чтобы ясно указать эквивалентность уровня включающих
классов и способ их перекрывания, определяющего класс более
низкого уровня:
Бурые животные
Белые животные
Медведи
бурые медведи
белые медведи
Не медведи
не поименованы
не поименованы
В письменной форме, однако, часто бывает удобнее предста-
вить классы как последовательности, словно между ними есть су-
бординация, даже если логически они относятся к одному уровню,
например:
1. а. Не медведи
(далее не делятся)
аа. Медведи
2. а. Бурые животные
б. Медведи ... бурые медведи
бб. Не медведи ... (далее не делятся)
Систематика, таксономия, классификация, номенклатура
17
б. Бурые ... бурые медведи
бб. Белые ... белые медведи аа. Белые животные
б. Медведи ... белые медведи
бб. Не медведи...
(далее не делятся)
Эти построения, особенно (1), выглядят как иерархии. Тем не
менее, каждое из них предполагает наличие другого, а оба вместе
представляют собой не более чем способы представления одной и
той же информации, как и в предыдущей схеме, где «бурые живот-
ные», «белые животные», «медведи» и «не медведи» являются клас-
сами одного уровня, а «бурые медведи» и «белые медведи» — сле-
дующего более низкого уровня, связанные с классами более высо-
кого уровня в форме ключей, но не иерархии. Наиболее общее и
значительное отличие между ними в том, что в ключе не имеет зна-
чения, какой из классов данного уровня — «медведи» и «не мед-
веди», «бурые животные», «белые животные» — появляется в таб-
лице первым. Мы можем «исключить», т.е. идентифицировать жи-
вотное одинаково успешно, начав с того, что оно белое, или с того,
что это медведь. В самом деле, часто случается, что некто, опо-
знающий (т.е. идентифицирующий) белого медведя, сначала заме-
чает, что он белый, прежде чем обратить внимание на те особен-
ности, которые говорят ему, что это медведь.
В истинной иерархии, в противоположность этому, последова-
тельность уровней и субординация одних классов по отношению к
другим часто является существенным элементом системы. Причём
здесь нет строго формальных и абстрактных логических основа-
ний, почему иерархия
Белые животные Бурые животные
белые медведи песцы «рыжие» лисицы бурые медведи
не должна использоваться вместо иерархии
Медведи Лисицы
белые медведи бурые медведи «рыжие» лисицы песцы
— однако ни один зоолог никогда не отдаст предпочтение первой
вместо второй. Имеются более чем веские причины, настолько оче-
18
Дж. Г. Симпсон. Принципы таксономии животных
видные, что они находят отражение даже в обыденных языках, по-
чему иерархия «медведи или лисицы» предпочтительнее иерархии
«бурые или белые». Причины приоритетности, вообще говоря,
весьма и весьма сложны и потому иногда неопределённы и спор-
ны, однако этот приоритет, по каким бы основаниям он не был за-
дан, с необходимостью встроен в иерархическую классификацию.
Принципы и критерии расстановки приоритетов лежат вне самой
иерархии: они являются таксономическими в широком смысле. Их
распознавание и выбор на самом деле относятся к числу наиболее
фундаментальных проблем таксономии; соответственно, они об-
суждаются в деталях в последующих разделах.
Поскольку ключ не предполагает какого либо принципа при-
оритета [в упорядочении классов] и организован в произвольной,
удобной для пользования последовательности, ключи однозначно
рассматриваются как искусственные. Иерархия же основана на
принципах приоритета, и в той мере, в какой эти принципы выво-
дятся из истинных и естественных отношений между организмами,
иерархические классификации являются естественными. (Выраже-
ние «естественная классификация», впрочем, используется во мно-
гих других, часто взаимоисключающих смыслах, некоторые из ко-
торых будут рассмотрены позже.) Следует признать, что принципы,
используемые при расстановке приоритетов, иной раз сами могут
быть более или менее произвольными, в результате чего получае-
мая классификация не заслуживает в полной мере признания в ка-
честве естественной. Следует также добавить, что хорошие ключи
часто соответствуют последовательности, заданной приоритетами
иерархической организации. В таком случае ключ может считать-
ся столь же естественным, что и иерархия, однако это вытекает из
соответствующей иерархической классификации, а не является соб-
ственным свойством ключа как такового. В действительности то,
что является истинной иерархической классификацией, как она
определена выше, в принципе может быть представлено, как если
бы это был ключ; и наоборот, то, что является по своей сути клю-
чом, может быть (и часто бывает) представлено в форме иерархии.
Систематика, таксономия, классификация, номенклатура
19
Линнеевская иерархия
Разграничение иерархии и ключа и внимание к соотношению
между ними действительно существенно для их понимания. С это-
го момента нас будут заботить почти исключительно классифика-
ции в форме иерархий. Как правило, мы будем использовать толь-
ко одну форму иерархии, которая была принята всеобщим соглаше-
нием для стандартных зоологических классификаций и является
основой для большей части зоологической номенклатуры. Эта
иерархия была развита главным образом в XVII и XVIII веках и
достигла своей почти законченной формы (для зоологов) в 10-м
издании «Системы природы» Линнея в 1758 г., по каковой причи-
не она называется линнеевской иерархией. Её основной чертой яв-
ляется последовательность из семи7 уровней:
Царство
Тип
Класс
Отряд
Семейство
Род
Вид
Вся последовательность, считываемая сверху вниз с отступом
слева, указывают на уменьшающийся объём, или включённость, на
разных уровнях.
Разнообразие организмов, подлежащих классифицированию, в
настоящее время столь велико, что на практике редко бывает дос-
таточно 7 уровней. Недостаток был восполнен главным образом за
счёт добавления уровней, обозначаемых приставками «над-» выше
каждого из основных уровней и, соответственно, «под-» и «инфра-
» ниже них. Неоднократно предлагались дополнительные основные
уровни, однако они не вошли в стандарты и не получили широко-
го применения. Перечислять их полностью нет необходимости,
7 Линней использовал только 5 из них. Тип и семейство были добавлены
позже другими. Линней также использовал ещё один уровень — империю на
вершине иерархии для всего мира феноменов.
20
Дж. Г. Симпсон. Принципы таксономии животных
примеры же по их применению можно найти в работах отдельных
таксономистов. Наиболее популярными, вероятно, стали когорта
между классом и отрядом, а также триба, обычно, но не всегда
помещаемая между семейством и родом. Пример полной иерархии,
использованной в классификации обширной группы животных
(млекопитающие, см. Simpson, 1945), таков:
Царство
Тип
Подтип
Над класс
Класс
Подкласс
Инфракласс
Когорта
Надотряд
Отряд
Подотряд
Инфраотряд
Надсемейство
Семейство
Подсемейство
Триба
Подтриба
Род
Подрод
Вид
Подвид
Это даёт 21 уровень. Использование всех возможных над-, под-
и инфра-уровней между царством и подвидом дало бы 34 уровня,
что, возможно, больше, чем это действительно может пригодиться
в практике. Использование какого-то конкретного числа уровней
является совершенно произвольной договорённостью. Оно не со-
ответствует чему-либо в природе, а выработано потребностями
практического применения иерархии. Однако это не означает, что
группы, распознаваемые на любом данном уровне, всегда в не-
Систематика, таксономия, классификация, номенклатура
21
котором смысле являются искусственными или не соответствуют
неким естественно оформленным сущностям. Они могут или не
могут им соответствовать, и к этому вопросу мы ещё обратимся
далее.
Использование 7 основных линнеевских уровней востребовано
общепринятой практикой, что означает: ни одно животное не мо-
жет считаться удовлетворительно классифицированным, если оно
не отнесено явным или неявным образом к какой-либо группе на
каждом из этих 7 уровней. (На практике, тем не менее, некоторое
количество групп бывает не доведено до завершённой классифика-
ции, поскольку их не всегда удаётся с определённостью отнести к
конкретной группе на некотором уровне.) Использование любого из
других уровней необязательно, зависит от вкусов данного класси-
фикатора и требований, предъявляемых исследуемой им группой
организмов. Всё же имеется одно ограничение свободы действий
в этой сфере: если какой-то подчинённый уровень использован в
некоторой группе, он по мере возможности должен быть исполь-
зован для всех организмов этой группы. Например, если в преде-
лах некоторого семейства использовано подсемейство, то всё это
семейство должно быть разделено на подсемейства и все роды (на-
сколько это позволяют данные) должны быть помещены в то или
иное из этих подсемейств. Однако тот факт, что подсемейства ис-
пользованы в одном из семейств отряда, не означает, что они не-
пременно должны использоваться и в других его семействах.
Здесь необходимо ввести несколько специальных терминов и
понятий для того, чтобы было легче иметь дело с иерархией и
иерархическими классификациями. Первоочередным является воп-
рос: что фактически классифицируется? Он настолько существен и
(как это ни странно) так часто неправильно понимается, что ответ
на него фундаментален для всего, что должно быть сказано о так-
сономии. Кажется очевидным — даже более очевидным, чем это
есть на самом деле (см. главу 5), — что реальной единицей в при-
роде, некой совершенно объективно существующей отдельностью,
не взирая на некоторые исключения, является индивидуальный
организм. Тем не менее, индивид никогда не является и не может
быть предметом классифицирования. Классифицируются только
группы и никакая единица, входящая в классификацию, не может
22
Дж. Г. Симпсон. Принципы таксономии животных
быть индивидом.8 Экземпляр может быть отнесён к данной груп-
пе или включён в неё. Часто это называют «классифицированием»,
однако это неправильное употребление термина. Это соответству-
ет идентификации, что не тождественно классификации. Заключе-
ние о существовании и природе группы могут быть основаны на
экземпляре, но опять-таки это не классификация экземпляра, а ста-
тистический вывод на основе выборки из популяции. То, что клас-
сифицируется, это всегда популяция, понимаемая в самом широком
смысле как любая группа организмов, закономерно взаимосвязан-
ных друг с другом. («Популяция» также иногда используется в си-
стематике в более узком техническом смысле, однако для ясности
мы будем далее обозначать такую популяцию термином «дем».)
Единицей линнеевской иерархии является вид. (Статус подви-
да и других инфравидовых групп разного рода обсуждается в гла-
ве 5 и здесь мы на нём не задерживаемся.) Он, разумеется, состо-
ит из популяции — группы организмов, определённой некоторым
образом, что также будет подробнее обсуждаться позже. Все про-
чие группы, относящиеся к уровням выше видового, состоят из
одного или более видов. Все эти группы, в том числе виды, изве-
стны под общим названием таксон, который может быть опреде-
лён следующим образом:
Таксон есть группа реально существующих организмов, выде-
ленная в качестве формальной единицы на любом уровне иерархи-
ческой классификации.
Porifera (тип), Crustacea (класс), Chelonia (отряд), Didelphidae
(семейство), Bos (род), Homo sapiens (вид) — всё это таксоны.
Мы также нуждаемся в точных терминах для обозначения того,
что было не слишком строго названо иерархическими уровнями и
отношений между ними:
Таксономическая категория или (в данном контексте) просто
категория есть класс, членами которого являются все таксоны,
отнесённые к данному уровню иерархической классификации.
8 Было бы просто трюизмом считать, что последняя особь вымирающей
группы исчерпывает её. Сходным образом, первая особь возникающей группы
(впрочем, реальные группы, происходящие от одного индивида, крайне редки)
не может рассматриваться как составляющая эту группу.
Систематика, таксономия, классификация, номенклатура
23
Ранг категории есть либо её абсолютное положение в данной
иерархической последовательности, либо её положение относитель-
но других категорий.
Ранг таксона есть ранг категории, к которой он относится.
Эти и некоторые другие понятия, представленные в настоящей
главе, недавно специально исследовали Беккнер (Beckner, 1959) и
Грегг (Gregg, 1954), в последнем случае с применением сложного
и строгого аппарата символической логики. (Эти исследования во
многом продолжают более общую работу Вуджера: Woodger, 1937,
1948, 1952.) Излагая их подход в сверхупрощенной форме, таксо-
ном (назван Греггом «таксономической группой») следует считать
множество, члены которого — организмы. Таксономические назва-
ния — суть символы, присваиваемые таксонам, обозначены Грег-
гом как N2. (Собственные имена отдельных организмов, не входя-
щие в язык классификации, обозначаются как №.) Категория (так-
сономическая категория, по Греггу) —это множество множеств,
его членами являются таксоны. Названия категорий (тип, класс,
отряд и т.п.) принадлежат к группе имён N3. Организм является
членом некоторого таксона (или «принадлежит к» таксону) — точ-
нее, целой серии таксонов разного ранга; но он не может быть чле-
ном («принадлежать к») категории. Например, человек не может
логически принадлежать к категории «отряд», он принадлежит к
Primates — таксону, который является членом категории «отряд» и
имеет ранг этой категории.
Беккнер обозначает таксон у-го ранга символом Т, причём J
принимает значения от 1 до п. Т, — обычно вид, но может быть
и подвидом. Более строго, низший таксон является членом низшей
категории, которая выделена в данной системе. Тя — обычно цар-
ство. Каждый Т, кроме Тл, включён в Тч1; Т, — таксон, в который
не включён ни один другой таксон; Тл — таксон, который включает
все прочие таксоны, но сам не включён ни в один другой таксон.
Как было отмечено, в линнеевской классификации число п долж-
но быть равно 7 или выше, оно может варьировать от одной клас-
сификации к другой, будучи, например, равным 21 в работе:
Simpson (1945). Далее, Беккнер оговаривает, что «каждый организм
является членом одного и только одного таксона каждого ранга»,
а также что «имеется общее согласие о том, что число таксонов
24
Дж. Г. Симпсон. Принципы таксономии животных
ранга у+1 должно быть значительно меньше числа таксонов ранга
У». Однако действительная таксономическая практика не столь же-
стка, как эта логическая схема; по моему мнению, она и не долж-
на быть таковой. Я уже отмечал, что п не должна быть постоянной
величиной даже в разных разделах одной и той же классификации.
Например, подсемейства могут выделяться в одном семействе и
отсутствовать в другом. Для такой классификации фиксируется
ранг «подсемейство» с выделением таксонов соответствующего
ранга, однако не обязательно, что все организмы окажутся члена-
ми какого-либо таксона данного ранга.
Ещё одно предложение Беккнера здесь отвергнуто, поскольку
является своего рода аномалией; оно проработано более детально
Греггом. Используя иную символику, Грегг обратил внимание на то,
что в реальных классификациях достаточно обычны монотипичес-
кие таксоны, т.е. Т.+1 нередко включает только один Т. Крайним
примером такого рода является отряд Tubulidentata (трубкозубы),
который включает только один современный вид, так что (исклю-
чая немногие вымершие формы) отряд Tubulidentata, семейство
Orycteropodidae и род Orycteropus все монотипические. Другими
словами, все они имеют членами одни и те же организмы, посколь-
ку включают единственное множество (вид) Orycteropus afer. Вы-
шеприведённое предложение Беккнера означает, что по всеобщему
соглашению такой ситуации не должно быть, однако на деле она
действительно присутствует в огромном числе классификаций,
признанных таксономистами. Грегг принимает этот факт, однако
считает его аномальным или алогичным, поскольку (в принятой
здесь терминологии) он делает таксоны и категории, которые в те-
ории различны, на практике идентичными [5].
Ответ на это возражение таков: реальные классификации затра-
гивают отношения, которые (по крайней мере сейчас) не учитыва-
ются при попытках приложить теорию множеств к иерархической
классификации. Таксон Т р включающий только один таксон Т, тем
не менее отличается от него из-за горизонтальных отношений или
ранговой эквивалентности с другими Т р которые включают более
одного Т. Отряд Tubulidentata отличается от единственного входя-
щего в него современного вида Orycteropus afer самим фактом сво-
его равенства (по уровню) с другими отрядами, а не видами. Соот-
Систематика, таксономия, классификация, номенклатура
25
ветственно, с формальной точки зрения Orycteropus afer отличается
от любого другого вида из других отрядов приблизительно так же,
как отличаются два вида, отнесённые к каким-либо из этих отря-
дов. Такого рода отношение, разбор которого будет дан в последу-
ющем тексте, зависит от зоологических таксономических сообра-
жений, не принятых во внимание Греггом или Беккнером.
Замечания о теории множеств и символической логике
Только что приведённые примеры показывают, что такие при-
ложения теории множеств и символической логики к классифика-
ции не учитывают полностью некоторые отношения, на которых
основаны реальные классификации и которые, по-видимому, явля-
ются вполне логичными. Впрочем, это не означает, что эти отно-
шения не могут быть адекватно включены в такие системы, как
разработанная Греггом, или что этим не стоит заниматься.
Против таких систем (а также против предложенного Вуджером
применения символической логики к другим разделам биологии)
выдвигались возражения, что они тавтологичны и эвристически
несостоятельны. В первом случае имеется в виду, что эти системы
не говорят ничего сверх того, что уже было сказано в более про-
стой форме (хотя и не столь сжато). Во втором случае формальные
системы критикуют за то, что они (пока ещё) не привели к каким-
либо полезным изменениям в таксономической практике или к ка-
ким-либо важным таксономическим открытиям. Оба возражения, с
моей точки зрения, достаточно серьёзны, но не являются решаю-
щими. Отвечая на первое возражение, можно отметить, что тавто-
логичная символическая система всё же может быть полезной: ал-
гебраические символы, например, определённо полезны, хотя и не
говорят ничего такого, что не могло бы быть выражено столь же
ясно обычными словами. Что касается второго возражения, то воз-
можности подобной «символизации» в классификации пока ещё
мало исследованы и нельзя исключить выявления в будущем её
эвристической ценности; пока же демонстрация того, что класси-
фикация может быть связана, по крайней мере в какой-то части, со
строгой абстрактной логикой, сама по себе имеет определённую
ценность.
26
Дж. Г. Симпсон. Принципы таксономии животных
Имеются, однако, и другие возражения против таких систем в
их нынешней форме, которые мне представляются более важными.
Как было показано, они не совершенны и ошибочны в том, что не
принимают в расчёт некоторые логические соображения, суще-
ственные для реальных классификаций. (Имеются и иные упуще-
ния помимо того, которое здесь рассматривается.) Они, как пред-
ставляется, требуют или предполагают необходимость полного раз-
деления множеств или строгой дихотомии А:не-Л (х:х), что не-
применимо при работе с эволюирующими организмами. Судя по
уже имеющимся результатам, они лучше всего или, по крайней
мере в некоторых разделах, исключительно применимы к постоян-
ным множествам, которые определяются совпадением признаков
(characters in common), в то время как члены всех таксонов весь-
ма изменчивы и во многих случаях должны определяться иным
способом. («Политипическая концепция» Беккнера отчасти снимает
это возражение [6]). Они адекватны для описания типологических
и более или менее искусственных классификаций, которые были
обычны для XVIII в., но не для современных, особенно эволюци-
онных классификаций. (Разумеется, Вуджер, Грегг и Беккнер посту-
лируют эволюционное происхождение классифицируемых таксо-
нов, однако, как мне представляется, они не смогли расстаться с
доэволюционными типологическими дефинициями.)
Некоторые из этих и других возражений в равной мере спра-
ведливы и для других подходов формального характера; возмож-
но, что они окажутся снятыми будущим развитием теории мно-
жеств и символической логики. В настоящее время, однако, их
недостатки представляются настолько серьёзными для решения
стоящих передо мной задач, что я воздержусь как от дальнейшего
рассмотрения сего предмета, так и от использования терминов и
символов Вуджера - Грегга - Беккнера при изложении своего
собственного.
Следующий краткий список некоторых наиболее элементарных
символов дан как образец и в целях возможной помощи для тех,
кто проникнется желанием расширить свои знания по данной теме
(что приветствуется). Список также должен продемонстрировать
эквивалентность логических отношений с важными концепциями,
изложенными в данной книге, но в иной терминологии.
Систематика, таксономия, классификация, номенклатура U
е — членство в множестве, х е у означает «х является членом
(элементом) множества у».
с — включение в множество, х с у означает «все члены множе-
ства х являются также членами множества у».
= — равенство, х = у означает «все члены множества х являются
членами у и все члены у являются членами х», что также
может быть записано: х с у и у с х.
Ф — неравенство, х Ф у означает «некоторые члены х не являют-
ся членами у»
х — дополнительность, х является множеством нечленов х [т.е.
всех членов, которые не являются членами х]
п — пересечение, х п у есть множество членов, принадлежащих
одновременно множествам х и у.
и — объединение, х и у есть множество членов, принадлежащих
либо х, либо у, либо обоим множествам.
Основания и критерии классификации
Иерархическая классификация, теперь адекватно охарактеризо-
ванная, по самой своей сути предусматривает определённые опе-
рации и отношения, формальный аспект которых достаточно прост,
хотя их теоретические основания и их практические приложения
чрезвычайно сложны. Первичная операция состоит в (концептуаль-
ном) объединении организмов в таксоны низшего ранга Тг (На
практике при классифицировании, однако, эта операция не всегда
бывает первой, а при идентификации она никогда не бывает тако-
вой.) Вторичные операции состоят просто в последовательном
объединении этих Т( в таксоны более высоких рангов вплоть до Тя,
который в терминах схоластики представляет собой «высший род»,
а в более современной терминологии — множество, не включён-
ное ни в какое другое множество. В этих операциях, независимо
от того, как происходит группирование, принимаются во внимание
два типа отношений или, более точно, отношений между отноше-
ниями. Включение Т в Т.+1 влечет приоритетность [priority] того
отношения, которое объединяет все Т в Т перед отношением,
определяющим группировки в самих Т. В то же время, устанавли-
вается эквивалентность между отношениями, которые задают
28
Дж. Г. Симпсон. Принципы таксономии животных
объединение в конкретное Т и теми отношениями, которые ана-
логичным образом задают прочие Т в данной классификации.
Отношение «по приоритетности» можно проиллюстрировать
отношением между родом и видом [7], а отношение эквивалентно-
сти — отношением между родами в пределах одного семейства
(или подсемейства или трибы, если эти категории выделены). В
одной из схоластических трактовок (не все трактовки одинаковы)
понятия рода и вида были значительно более общими, нежели в
современной зоологии и означали отношение в общем случае, а не
только то, которое установлено между конкретными категориями,
называемыми в настоящее время родом и видом (см. Joseph, 1916;
Cain, 1958а). Видом считалась группа объектов, сходных по сущ-
ности. Родом была та часть сущности, которая объединяет разные
виды, что при экстенсиональном [объемном] выражении означает
группу видов (на любом иерархическом уровне) с некоторыми со-
впадающими атрибутами. Вид и род были двумя предикабилиями
из пяти, которые описывали наиболее общие отношения между ат-
рибутами в операциях логического деления и предикации (отнесе-
ния к классу). Другие предикабилии таковы:
отличие [differentia] — та часть сущности, которая свойствен-
на данному виду и тем самым отличает его от других видов;
свойство [property] — атрибут, общий для всех членов вида, но
не являющийся частью его сущности и не обязательно отличающий
его;
акциденция [accident] — атрибут, имеющийся лишь у некото-
рых, но не у всех членов вида и не являющийся частью его сущ-
ности.
Схоластический подход к логической классификации не толь-
ко полностью неадекватен, но также мало пригоден для современ-
ной таксономии. Он рассматривается здесь и в следующей главе по
двум причинам. Во-первых, он послужил отправным пунктом, с
которого началось развитие зоологической систематики. Её концеп-
ции и терминология были знакомы первым таксономистам, вклю-
чая Линнея, и существенно повлияли на них. Во-вторых, некоторые
сегодняшние таксономисты отстаивают то, что по сути является
возвратом к схоластическим принципам, хотя, по-видимому, не все
из них осознают, какие последствия это может повлечь за собой.
Систематика, таксономия, классификация, номенклатура
29
Они, например, предпочитают определять виды по их «сущности»
(по «существенной природе», «природе» и т.п.), диагностировать их
по «отличиям» и группировать в таксоны более высоких рангов по
«родам» в их исходном схоластическом понимании. «Свойства»
оказываются недиагностическими и потому несущественными, а
просто постоянными признаками видов, при этом «акциденции»
рассматриваются как вариации, не относящиеся к «типу» (несколь-
ко более поздняя концепция) и потому не имеющие таксономичес-
кого значения.
Я надеюсь, что из дальнейшего станет всё более понятно, на-
сколько непрактичны и даже нереальны такого рода системы в так-
сономии. Вместе с тем, схоластическая система дала свой ответ —
при этом ответ, исторически лежащий в основании иерархической
классификации, — на вопрос о том, как должны определяться так-
соны и каким образом между ними должны устанавливаться отно-
шения соподчиненности [priorities] и эквивалентности. Этот вопрос
столь фундаментален, что ему посвящена большая часть настоящей
книги.
Основанием для классификации, разумеется, должны быть от-
ношения между организмами, но эти отношения, понимаемые в
широком смысле, поистине бесчисленны. Из этого следует возмож-
ность существования не только многих разных классификаций, но
также многих разных оснований для классификаций. Гилмур
(прежде всего его работы: Gilmour, 1940, 1951) особо подчёркивал
этот момент; следующие ниже соображения частью основаны на
его доводах, хотя и в значительно изменённой форме.
1. Основное назначение классификации состоит в выделении
классов (нео-линнеевских классификационных таксонов), по пово-
ду которых мы можем делать обобщения. (Гилмур считает именно
это ключевой функцией и указывает обобщения, которые могут
следовать из классификации; однако самой классификации всегда
предшествуют некоторые обобщения, при этом в некоторых случа-
ях никакие дальнейшие обобщения не формулируются.)
2. Классы выделяются в связи с определённой целью, которая
зависит от того, какого рода обобщения считаются важными.
3. Некоторые классификации позволяют делать весьма широкий
круг выводов или более значимые обобщения, чем другие, и в этом
4 — 4338
30
Дж. Г. Симпсон. Принципы таксономии животных
смысле «лучше» или полезнее их. (Гилмур добавляет, что класси-
фикации, обеспечивающие достижение большего числа целей, яв-
ляются естественными, тогда как решающие узкие задачи являются
искусственными. Однако эта трактовка не только не получила рас-
пространения, но вообще неприемлема: понятие «естественная»
применительно к классификации неоднозначно и спорно, как уже
отмечено выше, но едва ли оно когда либо сводилось к большей
полезности. Естественная классификация в определённом смысле
должна быть более осмысленной, но это другой вопрос.) [8]
4. «Не может быть какой-то одной идеальной и абсолютной
схемы классификации для любого конкретного множества объек-
тов, но... всегда должно быть некоторое число классификаций, раз-
личающихся в своей основе, сообразно тем целям, для достижения
которых они разрабатываются» (Gilmour, 1951).
5. Даже классификации, построенные однотипно (как напри-
мер, линнеевская иерархия), для решения одних и тех же задач и
с использованием одних и тех же критериев или принципов, не
универсальны. Одни и те же данные и их интерпретации могут да-
вать одинаково «хорошие», или «валидные» классификации, кото-
рые тем не менее различны у разных специалистов и в зависимо-
сти от времени их разработки. (Этот момент, также обсуждаемый
несколько далее, Гилмуром в указанной статье не отмечен.)
Мы, таким образом, должны признать возможность и даже не-
обходимость не только многих классификаций, но также многих
тынов классификаций, т.е. классификаций, основанных на разных
вариантах отношений и предназначенных для решения разных за-
дач. Например, существуют два общих основания для классифика-
ций, которые очень стары и служат весьма специфическим це-
лям, — экологическое и телеологическое. Экологическая классифи-
кация определяет множества (их не следует называть таксонами)
согласно таким критериям как сообщества, в которых живут орга-
низмы (например, болотные растения, почвенные микроорганизмы,
лесные насекомые), или другие факторы среды (например, альпий-
ские растения, пресноводные рыбы, пещерные саламандры). Чис-
ло таких возможностей, очевидно, очень велико; также очевидна и
полезность таких классификаций для конкретных, весьма частных
задач. Тем не менее, в настоящее время никто не предлагает ис-
Систематика, таксономия, классификация, номенклатура
31
пользовать экологические классификации для более общих це-
лей — например, для идентификации и номенклатуры. В современ-
ной экологии исследуемые организмы идентифицируются и имену-
ются согласно другим классификациям до того, как они рассмат-
риваются в экологической классификации.
Телеологические классификации определяют множества (опять-
таки, не таксоны) соответственно их пользе или отсутствию тако-
вой, обычно для человека. Такими множествами могут быть, напри-
мер: одомашненные животные, которых делят на разводимых ради
мяса, тягловых, домашних любимцев и т.п.; съедобные, несъедоб-
ные и ядовитые рыбы; или растения, классифицируемые согласно
болезням, от которых они могут вылечить. Подобные классифика-
ции прежде были широко распространены, особенно в первых
травниках. Они всё ещё находят применение, хотя не столь универ-
сальное и не в столь систематичной форме, для вполне частных
целей: например, в этноботанике, в поваренных книгах, в сельском
хозяйстве, при изучении природных лекарственных средств. Они не
представляют широкого научного интереса и требуют, чтобы орга-
низмы были прежде рассмотрены в некоторой другой системе.
Хотя Гилмур определённо прав, отвергая как возможность, так
и необходимость всеобщей и абсолютной классификации, тем не
менее истина состоит в том, что одна форма классификации дол-
жна быть применена ко всем организмам (или, в нашем случае, как
минимум к животным), хотя бы в целях номенклатуры, каковы бы
ни были другие формы классификации, вторично разрабатываемые
для более узких групп организмов и целей. Как уже было указано,
по молчаливому согласию такой общей формой является линнеев-
ская иерархия. Далее, также существует общее согласие по пово-
ду того, чтобы наиболее осмысленной и даже совершенно необхо-
димой основой для такой классификации были отношения между
животными как таковыми, рассматриваемыми сами по себе, а не их
отношения к другим вещам, таким как среда (экологическая клас-
сификация) или полезность (телеологическая классификация).
Даже при том, что это общее согласие во многом достигнуто,
в его рамках всё ещё существует множество различных определе-
ний и критериев, которые по-прежнему активно обсуждаются. Под-
ход, наиболее оправданный интуитивно и чаще всего реализуемый
4*
32
Дж Г. Симпсон. Принципы таксономии животных
на практике, основан на объединении по сходству, причём прежде
всего (хотя в настоящее время не исключительно) по морфологи-
ческому сходству. С другой стороны, подход, основанный на одной
из форм объединения по смежности, именно, по эволюционному
происхождению и филогенетическим связям, имеет более широкое
значение, во многом вследствие того, что он принимает во внима-
ние процесс, посредством которого, как сейчас известно, на самом
деле возникли таксоны, тем самым объясняя и позволяя интерпре-
тировать лежащие на поверхности объединения по сходству. Соот-
ветственно, в настоящее время широко признано, что «лучший»
(наиболее осмысленный, наиболее эффективный для многих обоб-
щений) метод классификации — тот, который основан на эволюци-
онных отношениях, а не просто на сходстве организмов. Тем не
менее, некоторые способные и даже выдающиеся таксономисты
отвергают этот вывод. Более того, среди тех, кто согласен с ним в
теории, имеются расхождения относительно того, насколько прак-
тичен этот подход для конкретных классификаций, и даже в боль-
шей степени — как конкретно его применять. Эта проблема здесь
пока что просто обозначена в самых общих словах. Моя собствен-
ная точка зрения, совпадающая с указанной общей позицией, со-
стоит в том, что она действительно лучшая и действительно прак-
тичная, хотя, как и всякая система, она включает множество вто-
ростепенных проблем, требующих дальнейшего решения. Все пос-
ледующие разделы книги представляют собой главным образом
попытку разъяснить эти решения.
Номенклатура
Бэйзер (Bather, 1927), один из тех, кто принял филогенетичес-
кую, или эволюционную9 классификацию, хотя в более поздние
годы и высказавший некоторые глубокие сомнения, отмечал, что:
«Биолог нуждается в системе, которая играет двоякую роль: позволя-
ет ему указать, без опасения ошибиться, те конкретные существа, с кото-
9 Этот подход обычно называют «филогенетическим», однако я полагаю
более подходящим название «эволюционный», причины чего станут ясными
несколько позже.
Систематика, таксономия, классификация, номенклатура
33 8
рыми он имеет дело, и представляет результаты его филогенетическое ис_ -
следования в упорядоченной форме. Если система не может отвечать обо-
им требованиям, она должна отвечать хотя бы одному.»
Представление о том, что классификация должна или ^жетП
решать только задачи указания [объектов], достаточно абсурДНЭ5а
(о чём Бэйзер, конечно, знал), даже если с этим втайне и corj]acHbTci
некоторые зоологи. «Некий крупный зоолог как-то сказал а^Ор^у
этих строк, что поиски естественной классификации не относятсяя
к сфере деятельности музейного куратора. Его работа — иЛенти->'
фикация, а не классификация, и он должен разрабатывать кл^чи
каталожные системы поиска, которые позволяют быстро и легкой
рассортировывать животных по видам» (Caiman, 1940). Этот «Трупп-
ный зоолог» не объяснил, каким образом можно разработать 5ь1СтТ
рый и лёгкий ключ или указатель в отсутствие упорядоченной сК~-‘
стемы, обобщающей прежние исследования, будь то филог^нет1Тч-
ческая или любая другая, равно как он не указал, какие научньхю
задачи можно решать посредством только сортировки видов и не-
чего другого. Конечно же, система, о которой стоит рассу^дат1:ь,
должна решать обе задачи: и представлять в упорядоченно^ ф0^)-
ме результаты исследований животных, и предоставлять ср^дст^а
указания тех групп, которые в ней выделены.
Как уже было отмечено, таким средством указания служц^ при-
своение таксонам в классификации определённых названий в н а-
стоящее время вполне условных. В идеале, к которому с^еду<т
стремиться, но который никогда не будет достигнут, каждь^ та к-
сон должен иметь особое имя и оно должно быть единств^ннь1 ти,
который используют зоологии. Другой идеал, ещё менее до^тийсн-
мый, состоит в том, что это имя, однажды предложенное, ДолЖ^о
быть постоянным. Некоторое непостоянство неизбежно: он<^ зало-
жено в природу зоологического исследования. По мере рос^а зна-
ния о животных неминуемо возникают ситуации, когда д^а или
более вида, ранее полагавшиеся отдельными, объединяются в одцн
вид, или когда нечто, предполагавшееся единым видом, ока^ывает-
ся сборной группой из двух или более видов. Рост знания^ равцо
как и стремление подчеркнуть различия, будет требовать nej^ecTpo-
ек, объединений или дроблений также и таксонов выше в^дового
уровня. Всё это влечёт определённые изменения в номенклатуре
34
Дж. Г. Симпсон. Принципы таксономии животных
Личные предпочтения и пристрастия, нередко влияющие на
перегруппировки надвидовых таксонов, также могут приводить к
изменениям названий, но это не является серьёзной причиной но-
менклатурных изменений: перестройки таксонов без радикального
изменения их определения и состава могут быть достигнуты при
сохранении старых названий. Тот факт, что не существует двух спе-
циалистов, которые признавали бы идентичную систему высших
таксонов, также далеко не всегда приводит к беспорядку в номен-
клатуре, если они применяют одну и ту же форму классификации
и систему номенклатуры, как это чаще всего и бывает.
Наиболее важной причиной нестабильности номенклатуры на
протяжении двух последних столетий был тот факт, что зоологи,
работавшие по всему миру, не так уж редко независимо друг от
друга давали разные названия для обозначения одних и тех же так-
сонов. Предпринимались многочисленные попытки выработки не-
коего кодекса, чтобы решить эти и другие проблемы, и в конце кон-
цов сложился единый свод Международных правил зоологической
номенклатуры, который в момент написания данной книги опять
пересматривается [9]. (Имеются также отдельные кодексы для ра-
стений и для микроорганизмов.) Основным является правило при-
оритета, согласно которому название рода или вида10 — это имя,
которое было первым опубликовано надлежащим образом для его
обозначения после 1758 г. Эта общепринятая точка отсчёта задана
10-м изданием «Системы природы» Линнея (для растений в разных
группах используются иные даты).
На первый взгляд казалось, что принцип приоритета достато-
чен и способен тотчас обеспечить стабильность родовых и видовых
названий, однако на деле задача оказалась и по сию пору остаётся
далекой от решения. Часто случается, даже в самое последнее время,
что некоторые старые названия, опубликованные в малоизвестных
изданиях, оказываются приоритетными относительно тех названий,
которые за долгое время стали общепринятыми среди зоологов. И нет
никакой уверенности в том, что самые старые названия всё ещё не
рассеяны в таких изданиях, так что принцип приоритета грозит
10 Его применение к семействам также крайне необходимо, однако к момен-
ту написания книги это ещё не принято [10].
Систематика, таксономия, классификация, номенклатура
35
нарушениями стабильности номенклатуры почти в той же мере, в
какой он обеспечивает её. Трудоёмким является и определение при-
годности таких изданий, оно само служит предметом рассмотрения
в соответствующих правилах, которые пока ещё разработаны не для
всех случаев. Наконец, имеются многочисленные проблемы, связан-
ные с невозможностью строго определить, каким из ныне признан-
ных таксонов на самом деле соответствуют старые названия.
В последнее время обозначилась чёткая тенденция к решению
этих и других проблем, связанных с номенклатурой, посредством
действий Международной комиссии зоологической номенклатуры,
которой предоставлены полномочия в отдельных случаях толковать
Правила и обозначать nomina conservanda — общеупотребляемые
названия, которые должны использоваться, невзирая на приоритет-
ность. Процедура эта довольно медлительна и громоздка, так что
к настоящему времени лишь незначительная часть зоологических
названий была зафиксирована таким способом. Однако некоторые
зоологи возражают против этой процедуры и по-прежнему наста-
ивают на строгом применении правила приоритета.
Зоологическое содержание таксонов часто и неизбежно изменя-
ется по мере роста знания и появления различий в заключениях и
точках зрениях. Это постоянно порождает неясности в вопросах о
том, является ли ныне выделенный таксон тем же, к которому было
исходно применено некоторое название. Эта проблема решается,
хоть и не полностью, но в целом удовлетворительно, за счёт обо-
значения типов. Типом для названия вида является отдельный эк-
земпляр; общее правило состоит в том, что, независимо от каких-
либо содержательных моментов в определении таксона, название
относится к тому виду, к которому принадлежит соответствующий
типовой экземпляр. Часто случается, что типы двух или более на-
званий помещаются в один вид; именно в таких случаях приори-
тетность и списки nomina conservanda призваны определить, какое
название следует использовать. Типом названия рода является на-
звание вида, которое, таким образом, опосредованно связывает его
с типовым экземпляром. Обычно считается, что типом названия
семейства является название рода, однако пока ещё до конца не
прояснено, как именно они должны быть связаны. Выдвигались
предложения распространить систему типов (и приоритетности) на
36
Дж. Г. Симпсон. Принципы таксономии животных
названия таксонов и более высокого ранга, однако в настоящее вре-
мя это положение не включено в Правило и не вошло в общую
практику [11]. В настоящее время названия таких высших таксонов,
которых значительно меньше, чем родов и видов, определяются
только на основе согласия зоологов и мнения авторитетов: подоб-
ный неформальный подход на этих иерархических уровнях, как
кажется, работает не хуже, чем Правила — на низших уровнях.
К сожалению, отрицательной стороной системы типов являет-
ся то, что она закрепляет старый и в настоящее время совершен-
но неудовлетворительный подход к классификации. Это вызывает
серьёзную путаницу, которая встречается всё ещё слишком широ-
ко. Само слово «тип» закрепляет идею, принятую в XVIII и нача-
ле XIX вв., разработанную схоластами, а до них — в античной Гре-
ции, суть которой в том, что вид определяется посредством неко-
торого фиксированного образа [pattern] (или «сущности»), который,
в других терминах, является его «типом» (также обычно называется
«архетипом»). Типовой экземпляр рассматривается как типичный в
этом смысле и используется, как таковой, для определения вида и
в качестве единственного стандарта сравнения — для идентифика-
ции других экземпляров. Вместе с пониманием того, что вид есть
популяция, каждый член которой в равной степени является пред-
ставителем вида, и что популяции всегда высоко изменчивы, ста-
новится очевидным, что вид просто не может иметь типа ни в об-
щем его понимании, ни в каком-либо узком утилитарном смысле.
Номенклатурный тип — просто нечто, к чему «привязано» назва-
ние посредством чисто договорной системы. Он не должен иметь
ничего общего с процессом определения вида, не относящимся к
номенклатурным задачам, и не должен играть особой роли в иден-
тификации других экземпляров. Современные таксономисты всё с
большим пониманием относятся к различению этих двух аспектов,
однако прежняя путаница по-прежнему присутствует в значитель-
ной части зоологических рассуждений и приёмов. Она, к сожале-
нию, увековечивается Правилами, которые продолжают говорить о
типах видов, родов и т.д., в то время как о них следует говорить
всего лишь как о типах названий. Номиналистическим абсурдом
является смешение множества объектов с названием или символом,
это множество обозначающим.
Систематика, таксономия, классификация, номенклатура
37
В настоящее время, конечно, широко признано, что таксон не
может быть охарактеризован адекватно только по его типу.11 Оче-
видно, для целей номенклатуры название вида должно иметь един-
ственный тип, однако для целей типификации вида или для его
определения необходимы и другие экземпляры (или по крайней мере
их существование должно подразумеваться). Многие таксономисты,
которые признают эти факты, всё же по-прежнему путают тип на-
звания с типификацией или иной формой представления названной
группы. Поэтому они предлагают выделять разные категории «ти-
пов». Это привело к поистине забавной крайности, когда появились
буквально десятки терминов для разных категорий «типов»: топо-
тип, гомотип, метатип т.п. Для целей номенклатуры — единствен-
ных, которые в современной таксономии могут оправдать существо-
вание типов, в действительности нужны только четыре термина:
тип (или голотип) — единственный тип имени;
синтипы — два или более экземпляров, которые были указа-
ны как типы при исходном обозначении имени; эта практика в на-
стоящее время считается неприемлемой, но ранее была широко
распространена;
лектотип — один из двух или более синтипов, в последующем
выбранный в качестве единственного типа;
неотип — замещение типа, который был утрачен или (в ряде
случаев) признан неадекватным в качестве типа. (Неотипы не были
узаконены Правилами, но имеются предложения включить их в
этот свод.)
Ещё один термин следует зафиксировать, поскольку он очень
широко используется в современной таксономической литературе,
хотя (на мой взгляд) он не нужен и вносит путаницу:
napamun — экземпляр, не являющийся типом, но обозначенный
исходным автором названия как использованный при определении
вида [12]. 11
11 В некоторых случаях вид может быть известен по единственному экзем-
пляру, так что носитель имени совпадает с выборкой {гиподигмой, как это оп-
ределено в своём месте), на котором основано определение. Тем не менее, фун-
кции носителя имени, т.е. номенклатурного типа, и таксономической выборки
качественно различны, а определение должно предполагать наличие у суще-
ствующего в природе вида также и других, к тому же отличающихся особей.
38
Дж. Г. Симпсон. Принципы таксономии животных
По договорённости, технические названия животных должны
считаться ново-латинскими. Многие из них действительно являют-
ся латинскими: Homo sapiens, Bos taurus. Многие названия, особен-
но родовые, являются латинской транслитерацией греческих слов:
Synoplotherium, Xenochoerus. Наряду с этим, они могут заимство-
ваться не из классических языков и даже быть вполне вымышлен-
ными: Lama (из языка перуанских индейцев), Dugong (малайское
название), Kogia (от английского разговорного «codger» — скряга),
Tatera (вымышленное).
Латинское название рода состоит из одного слова, начинается
с прописной буквы и считается существительным в именительном
падеже и единственном числе. Название вида состоит из двух слов,
по каковой причине линнеевская номенклатура называется биноми-
альной или бинарной. Первое слово — название рода, к которому
относится вид. Второе слово называется видовым или тривиаль-
ным, начинается со строчной буквы и рассматривается как имя су-
ществительное в форме прилагательного, либо как существитель-
ное (особенно имя собственное) в родительном падеже, либо как
прилагательное, согласованное с родовым названием в числе (един-
ственное) и роде. Названия родов и видов принято писать курси-
вом. Названия подродов имеют ту же форму, что и родовые, и пи-
шутся в скобках после названия рода, к которому они относятся.
Названия подвидов состоят из трёх слов, из которых первых два
обозначают вид, а третье слово — собственно подвидовое. По до-
говорённости, если вид разделён на подвиды, один из них должен
носить тот же тривиальный эпитет, что и вид: один из многочис-
ленных подвидов Rattus rattus называется Rattus rattus rattus.
(Идентичность родового и видового названия, однако, необязатель-
но, в этом отношении приведённый пример нетипичен.)
Названия таксонов рангом выше родового являются однослов-
ными, рассматриваются как латинские существительные во множе-
ственном числе. Они пишутся с прописной буквы, но не курсивят-
ся. Вплоть до надсемейств они образуются путём прибавления оп-
ределённого окончания к корневой основе названия одного из
включённых в данный таксон родов. Род, название которого ис-
пользовано для этого, рассматривается как тип соответствующего
высшего таксона.
Систематика, таксономия, классификация, номенклатура
39
Надсемейство: -oidea (для позвоночных) или -асеае (для беспозвоноч-
ных) [13]. Например'. Hominoidea (от корня homin-, Homo).
Семейство: -idae. Например'. Hominidae.
Подсемейство: -inae. Например'. Homininae.
Названия таксонов рангом выше надсемейства в настоящее вре-
мя образуются вне связи с родовыми названиями, при этом они не
имеют унифицированных окончаний. Выдвигалось предложение
унифицировать окончания для всех таких таксонов, в первую оче-
редь для отрядов, в настоящее время рассматривается включение
этого условия в Правила, однако оно не вошло в общую практику
и, по моему личному мнению, было бы нежелательным для царства
животных [14]. В немногих группах, тем не менее, это стало общей
нормой. Например, названия отрядов птиц составляются из родо-
вого корневого основания и окончания -iformes [15].
В ранних классификациях названия таксонов обычно были
описательными, а не определительными. Например, отряд Carni-
vora был назван так в связи с тем, что, как полагалось, его пред-
ставители являются плотоядными, хотя известно, что не все пло-
тоядные животные относятся к Carnivora. Кроме того, мы знаем,
что некоторые Carnivora на самом деле являются строго раститель-
ноядными. Как уже было упомянуто, видовая номенклатура возник-
ла на основе того, что исходно было краткими латинскими опре-
делениями (или схоластическими отличиями) видов. История по-
степенно, хотя всё ещё не окончательно, привела к пониманию
того, что номенклатура, с одной стороны, и распознавание и опре-
деление поименованных таксонов, с другой стороны, — две совер-
шенно разные вещи. Сведение второй части видовых названий к
единственному слову, грандиозный рост числа описанных видов и
осознание того, что многие высшие таксоны, наподобие Carnivora,
содержат исключения из всякого однословного описательного обо-
значения, сделало очевидным, что практически ни одно название
не может содержать определение или диагноз и что далеко не все
из них могут быть подходящими эпитетами. Заимствованные из
прошлой практики, описательные названия и сейчас нередко пред-
лагаются в таком качестве. Однако в настоящее время понятно, что
техническое зоологическое название служит не характеристикой
таксона, а только его удобной этикеткой. Оно означает сам таксон,
40
Дж. Г. Симпсон. Принципы таксономии животных
а не то, что предполагает его этимологическое происхождение: точ-
но так же, как George Fletcher означает конкретного человека, а не
«фермера, который изготавливает стрелы», Basilosaurus означает не
«короля рептилий», а название рода вымерших морских млекопи-
тающих.
Практикующие классификаторы, разумеется, должны быть зна-
комы со всеми многочисленными деталями зоологической номен-
клатуры. Для этого Майр с соавт. (Mayr, Linsley, Usinger, 1953) под-
готовили великолепное введение, однако необходимо изучать сами
Правила, наряду со многими другими изданиями Международной
комиссии [16]. Представленный здесь краткий обзор с комментари-
ями достаточен для целей книги — он даёт концептуальную основу
и показывает общую связь между номенклатурой и другими аспек-
тами таксономии, которым посвящена основная часть книги.
Зоологическая номенклатура — это просто этикетирование так-
сонов в классификации. У нее нет иных функций в таксономии.
Она разрабатывает словарь для описания и обсуждения животных
и в таком качестве совершенно необходима в зоологии, но сама по
себе она не представляет какого-либо другого зоологического и
вообще научного интереса. Это искусственный инструмент, кото-
рый стал чрезмерно сложной, строго формальной, сугубо юриди-
ческой системой. Некоторые зоологи, по-видимому, получают ис-
тинное удовольствие от этих «крючкотворских», по сути незооло-
гических и по всей видимости бесконечных правил, дискуссий и
актов, но для большинства они — лишь необходимое зло, отнима-
ющее драгоценное время от занятий более важными делами. Со
всеми своими прогрессивными модификациями и таинственными
хитросплетениями, зоологическая номенклатура по-прежнему дале-
ка от решения всех своих проблем, или хотя бы от удовлетворяю-
щего всех решения какой-то одной из них. Тем не менее, присущий
ей беспорядок каким-то образом не мешает ей быть вполне рабо-
тоспособной. И мы не станем более заниматься ею, хотя, несомнен-
но, будем пользоваться её словарём.
2
Развитие современной систематики
У таксономии долгая история, восходящая к античным
грекам и их предшественникам, не столь изощрённым в система-
тизации. Наш интерес здесь сосредоточен на современной таксо-
номии собственно, так что мы опустим многое из её развития
вплоть до середины XVIII в., когда таксономия определённо офор-
милась как самостоятельная ветвь науки. Впрочем, в рамках и этих
двух столетий мы не станем прослеживать исторические этапы.1
Скорее, здесь мы будем иметь дело с немногими наиболее важны-
ми принципами, концепциями и теориями, наиболее активно по-
влиявшими на историческое развитие современной таксономии.
Среди них: теоретические критерии классификации, представления
о естественной классификации, значение открытия эволюции, а
также некоторые формальные аспекты представления структуры от-
ношений между организмами. (Формальная конструкция иерархи-
1 Насколько я знаю, не существует детального исследования или даже при-
стойного обзора истории таксономии или классификации. Есть совсем немного
полезных книг, освещающих этот предмет. Sprague et al. (1950) дали очень
краткую сводку по истории. Циммерманн (Zimmermann, 1953) привёл и про-
комментировал много примеров из додарвиновской таксономии (после-дарви-
новское развитие представлено слабо и неадекватно), а Грегори (Gregory, 1910)
проследил более детально развитие классификации отдельной группы (млеко-
питающие). Разумеется, также имеются многочисленные более краткие иссле-
дования по отдельным моментам, среди которых Cain (1958) — один из пос-
ледних выдающихся примеров.
42
Дж. Г. Симпсон. Принципы таксономии животных
ческой классификации достаточно рассмотрена в первой главе, од-
нако структура отношений — это другой вопрос.) Наконец, будет
предпринята попытка прояснить, что именно отличает «новую си-
стематику» от старой, и охарактеризовать современную таксоно-
мию, которая развивалась уже после «новой систематики» на про-
тяжении последующих 20 лет или около того.
Схоластика и Линней
Как упомянуто в главе 1, среди всевозможных отношений (или
связей), которые могут быть учтены в классификации, наибольший
интерес представляют два: сходства между организмами и связи,
которые обусловлены их эволюцией и происхождением. Их ни в коем
случае нельзя считать независимыми или резко противоположны-
ми. В современной таксономии, которая, по правде сказать, не еди-
на во мнениях, понятия и определения категорий и таксонов фор-
мируются главным образом на основе эволюционных представле-
ний, однако доказательства, которые придают состоятельность этим
понятиям и показывают, что определения значимы, выводятся боль-
шой частью из данных по сходству. Определённо, ни одно из них
нельзя отбросить или, напротив, считать первенствующим. Это
просто разные составляющие общего понимания природного поряд-
ка среди организмов и создания классификаций на этой основе.
Эти отношения не могли быть поняты до тех пор, пока не была
постигнута истина эволюции. Многие догадывались о ней, но Ла-
марк стал первым, кто ввёл эту концепцию в таксономию с доста-
точной определённостью и ясностью. Впрочем, даже для Ламарка
эволюционная идея имела очень незначительное влияние на прак-
тику классификации, и только с Дарвина началось медленное и всё
ещё незавершённое движение в направлении эволюционной таксо-
номии. Однако форма классификации, которой до сих пор пользу-
ются, была разработана до Ламарка, равно как и многие таксоно-
мические принципы, одни из которых до сих пор действуют в пол-
ную силу, а другие, хоть и отвергнутые в их изначальном понима-
нии, оказали неустранимое влияние.
Для того, чтобы разобраться в ситуации времён Линнея, необхо-
димо снова вернуться к схоластической логике, аристотелизму и
Развитие современной систематики
43
томизму.2 Этот корпус знания был поистине основополагающим для
Линнея и пронизывал всё его дело. Согласно данной логической
системе, кратко изложенной в главе 1, целостная «сущность» неко-
торого «вида» (в устаревшем схоластическом смысле) состоит из его
«рода» (в том же смысле) в совокупности с его «отличиями». Одна-
ко Кэйн (Cain, 1958а), чьему внимательному анализу я во многом
обязан при изложении настоящего раздела, указал, что это верно
только в том случае, если между родом и его видами, равно как и
между самими видами, существует некое логическое отношение. Оно
означает применение одного и того же fundamentum divisionis [ос-
нования деления]. Классическим примером является деление рода
«треугольник» на виды «равнобедренный», «равносторонний» и «не-
равносторонний», которые выделены по одному fundamentum
divisionis и, таким образом, полностью исчерпывают логические воз-
можности треугольника. Род «треугольник» можно также разделить
исчерпывающим образом на «прямоугольный», «тупоугольный», «ос-
троугольный» посредством другого fundamentum divisionis. Однако
единая система или классификация не могут логически совмещать
оба эти fundamenta. В примере, более осмысленном биологически,
род «животное с ногами» может иметь виды «двуногое», «четверо-
ногое», «шестиногое» и т.п.; или виды «коротконогое» и «длинно-
ногое»; однако данный род не может одновременно иметь обе кате-
гории видов. (Эти соображения в деталях обсуждены в: Joseph, 1916.)
Строго логическая процедура такого рода даёт таксономию ана-
лизируемых [analyzed] сущностей. Попытки применить её к клас-
сификации организмов показала Линнею, равно как и его предше-
ственникам и современникам, что она неадекватна и частью непри-
годна для этих целей и что в зоологии (и ботанике) на практике
возможна только таксономия неанализируемых [unanalyzed] сущно-
стей. Серьёзной проблемой, которую безуспешно пытаются ре-
шить современные таксономисты, классифицирующие по «суще-
ственной природе», является невозможность в большинстве случаев
2 Если мне будет позволено личное замечание, я не очень склонен брать-
ся за это. Я скорее соглашусь с Роджером Бэконом, что изучение Аристотеля
усугубляет невежество. Тем не менее, сами основатели таксономии были уче-
никами Аристотеля и Аквината (в числе многих других их наследников), так
что данный предмет в определённой мере необходим для моих целей.
44
Дж. Г. Симпсон. Принципы таксономии животных
отличить «сущность» от «свойства» (атрибута, общего для всех чле-
нов вида, но не входящего в род или отличие) или даже от «слу-
чайности» (атрибута, присущего некоторым членам вида). Другая
проблема заключается в том, что в логике один и тот же индивид
в рамках одного рода может принадлежать к любому числу разных
видов соответственно разным принятым fundamenta divisionis, что
совершенно недопустимо в классификации родов и видов в зооло-
гии. Далее, на практике оказалось затруднительным, а в некоторых
группах даже невозможным использовать единый fundamentum для
всех зоологических видов в одном зоологическом роде или сохра-
нять «отличия», логически связанные с родом. Например, автори-
тетная классификация млекопитающих, относящаяся к XVIII в.
(Pennant, 1781), содержит «роды» (в прежнем широком смысле; в
современном понимании соответствующие таксоны имеют более
высокий ранг) «цельнокопытные» и «раздельнопалые»: рассматри-
ваемые как «виды» более высокого «рода», они в своей основе яв-
ляются логическими, поскольку выделены по общему fundamentum.
Однако к «раздельнопалым» в качестве «видов» (в таком же широ-
ком смысле) отнесены «человекоподобные плодоядные», «с увели-
ченными клыками» и «без клыков». Два последних имеют общее
fundamentum divisionis, однако первый выделен по совершенно ино-
му основанию, при этом ни один из этих видов не имеет fundamen-
tum, логически связанный с родом.
В ретроспективной оценке схоластический подход (как основа-
ние зоологической классификации) имеет ряд других недостатков,
столь серьёзных, что это делает его совершенно непригодным для
большинства современных таксономистов. Эти недостатки не мог-
ли и не были в должной мере оценены в дни Линнея и даже теперь
не всеми осознаются. К числу наиболее серьёзных относятся два
следующих. Во-первых, «свойства» и «акциденции» [случайные
атрибуты], в равной мере исключаемые из «сущности» и в силу
этого из определения, в настоящее время являются важной харак-
теристикой таксона, они необходимы для его надлежащего описа-
ния, а также часто и для практического определения. Это почти
всегда верно для «свойств», которые, как правило, имеют генети-
ческую основу. Это часто верно и в отношении «акциденций», т.е.
вариаций в пределах вида, которые всегда столь же важны для по-
Развитие современной систематики
45
нимания вида, а часто столь же существенны и для его определе-
ния и идентификации, как и признаки, постоянные для вида. Во-
вторых, приложение схоластической логической процедуры, хотя
бы и в качестве таксономии анализируемых сущностей, даёт клас-
сификацию признаков, а не организмов. Классификация организмов
также возникает, но параллельно, как побочный продукт той логи-
ческой системы, которая определяет атрибуты организмов, а не
сами организмы. Линней и некоторые другие смутно видели эту
трудность, но не решили вытекающую из неё проблему.
В XVIII в. эта проблема, как она тогда виделась, частью реша-
лась с помощью одновременного использования более чем одного
fundamentum divisionis при установлении «отличий». Знаменитым
классическим примером служит определение человека как «беспё-
рого двуногого». Ни «беспёрость», ни «двуногость» по отдельнос-
ти не могут считаться «родом» или «видом». В нашей терминоло-
гии они оба — объединения по сходству, которые определяют мно-
жества одного ранга, а «человек» является подмножеством, опре-
делённым их пересечением. (Примечательно, что человек на самом
деле является не единственным членом данного подмножества, как
было задумано). Короче говоря, получается некоторым образом
скрытая классификация в форме ключа, как она была определена
ранее. Такое решение почти универсально у Линнея: одним из при-
меров служит определение (логическое «отличие») птицы Fulica
atra: «fronte calva, corpora nigra, digitis lobatus» с тремя очевидно
различными fundamenta divisionis. Тем не менее, использование
единственного fundamentum на разных уровнях обычно считалось
желаемым идеалом, хотя он и не мог быть с очевидностью достиг-
нут на практике ни для одной значительной группы, не говоря уже
обо всём царстве животных. Так, Клейн (Klein, 1751) сумел соста-
вить ряд «четырёхногих», в котором были «когтистые» и «копыт-
ные», а среди последних — «однокопытные», «двукопытные»,
«трёхкопытные», «четырёхкопытные» и «пятикопытные», однако
принятие других fundamenta несомненно разрушило бы эти роды
и виды. Приблизительно в то же время Бриссон (Brisson, 1762) раз-
делил всех копытных (с небольшим числом исключений) по
fundamentum divisionis, связанном с зубами, но привлёк иные
fundamenta на более высоких и низких уровнях. Это, между про-
5 — 4338
46
Дж. Г. Симпсон. Принципы таксономии животных
чим, заставляет поднять вопрос о том, каким образом следует вы-
бирать fundamentunv. почему для Клейна он был непременно свя-
зан с конечностями, а для Бриссона — с зубами, причём прибли-
зительно на одном и том же уровне классификации.
Прежде чем перейти к другим аспектам таксономии XVIII в.,
можно отметить в скобках, что в настоящее время считается жела-
тельным следовать обоим приёмам, указанным в предыдущем аб-
заце, хотя при этом их приходится модифицировать, переосмысли-
вать в иных понятиях и обосновывать исходя из иных причин. Все-
общим правилом стало использование многих признаков на каж-
дом уровне классификации, причём на каждом уровне предпочти-
тельным, насколько это возможно, считается использование одних
и тех же категорий данных (одной и той же совокупности funda-
menta divisionis).
Теперь мы опять обращаемся, как это уже было не раз, к про-
блеме ранжирования или очередности [priority] признаков, исполь-
зуемых в иерархической классификации. Если fundamenta divisionis
«вида» (в схоластическом смысле) логически связаны с таковыми
«рода» (в этом же смысле), проблема решается сама собой, ибо
«отличия» включаются в «род» и исчерпываются его делением.
«Треугольник» очевидным образом задаёт возможность быть «рав-
нобедренным», «равносторонним» и «неравносторонним», а «иметь
ноги» предполагает «двуногость», «четвероногость» и т.д. Однако
если fundamenta меняются от уровня к уровню, ранжирование мо-
жет зависеть от неких соображений, лежащих вне абстрактной ло-
гики. Обычно различают два подхода: априорный, теоретический,
или дедуктивный, с одной стороны, и апостериорный, эмпириче-
ский, или индуктивный, с другой. Первый подразумевает предвари-
тельные рассуждения о том, какие атрибуты следует считать наи-
более важными (наименее несущественными) для классифицируе-
мых организмов, с последующим определением теоретической пос-
ледовательности рангов на этой основе. Второй означает, что сна-
чала исследуется распределение атрибутов на множестве организ-
мов, после чего делаются заключения о том, каким образом следует
расставить приоритеты, при этом ведущим, хотя и не вытекающим
с необходимостью из процедуры, критерием оказывается постоян-
ство атрибута среди членов исследуемой группы.
Развитие современной систематики
47
Различия между этими двумя подходами часто кажутся, по
крайней мере мне, в конкретных случаях неотчётливыми, так что
для меня остаётся открытым вопрос, является ли эта дихотомия
однозначной и исчерпывающей. Несмотря на удивительно деталь-
ное и ясное изложение проблемы Кэйном (особенно в работах:
Cain, 1958, 1959b; см. также Steam, 1959), для меня неясно, какой
именно системе на самом деле стремился следовать Линней в тех
или иных случаях и не является ли его практика на самом деле их
сложной смесью. Очевидно, он считал, что априорный метод, тесно
связанный с его представлением о естественной системе (к ней мы
ещё вернёмся), предпочтительнее с теоретической точки зрения, но
находил неизбежным некоторый элемент эмпиризма. Его афоризм
о том, что не признаки задают род, но род задаёт признаки, по-ви-
димому, подразумевает, что виды сначала объединяются в род, а
затем признаки рода эмпирически выявляются из характеристик
полученного объединения. Впрочем, эти слова могут иметь и иной
подтекст, связанный с существенно иной логикой и философией его
времени.
В любом случае, можно согласиться с Кэйном и другими авто-
рами, что общее движение таксономии от Линнея к Дарвину в од-
ном из своих аспектов было связано с некоторым ослаблением
априоризма и возрастанием роли эмпиризма. Я, впрочем, не впол-
не согласен с Кэйном, который считает, что желательное развитие
эмпиризма было фактически приостановлено под влиянием Дарви-
на, из-за которого одна форма априоризма якобы была просто за-
менена другой. Я бы возразил (достаточно мягко) по следующим
основаниям. Во-первых, я подозреваю, что у эмпиризма XIX в.
было не очень много предложений, когда он был не то чтобы при-
остановлен, а скорее переориентирован Дарвином. Во-вторых, эво-
люционную систему, которая началась с Дарвина, но после него
ушла в своём развитии далеко вперёд, нельзя строго считать ни
априорной, ни эмпирической: скорее, это удачное соединение их
обоих. В третьих, вся ситуация оказалась значительно сложнее,
нежели простое противостояние априоризма и эмпиризма, а в дру-
гих, возможно, более важных отношениях дарвиновская революция
запоздала. Можно также добавить, что школа более или менее чи-
стого эмпиризма ни коим образом не исчезла после Дарвина и се-
5*
48
Дж. Г. Симпсон. Принципы таксономии животных
годня имеет шумных адвокатов, но я не знаю и таких современных
таксономистов, которые были бы строгими априористами.
Некоторые другие додарвиновские подходы
Адансон (1727-1806), молодой современник и соперник Лин-
нея (1707-1778), обычно указывается как основатель эмпиризма в
таксономии (см., среди многих других: Bather, 1927; Cain, 1959а).
Он был ботаником, однако его принципы в равной степени приме-
нимы и в зоологии. Его таксономия (но, надобно сказать, не его
классификация) в некоторых важных аспектах более приемлема
сегодня, чем таковая Линнея, или, скажем, Кювье. Она избежала
тупиков дедуктивных логических систем и тех ошибочных теорий,
которые сегодня отвергнуты. Она была ограничена, насколько по-
зволяли возможности (впрочем, ни тогда, ни сегодня не очень боль-
шие), простой оценкой наблюдаемых данных. С другой стороны,
тот факт, что она в значительной мере была нетеоретической, оз-
начал, что ей недоставало объяснительной силы, осмысленности,
обоснования, которые могут привнести в таксономию и классифи-
кацию некоторую теоретическую оценку данных. Адансонов под-
ход, однако, оказался наилучшим образом приспособленным для
того, чтобы обеспечить предпосылки и эмпирическую основу для
оценок и критериев, которые позже были предложены эволюцион-
ной теорией. Линней, Кювье и Оуэн, а также их последователи,
среди многих других представителей ранних теоретических школ,
внесли в таксономию много ненужного, от чего пришлось со зна-
чительным трудом освобождаться. Адансон же и его последовате-
ли воздвигли довольно осмысленные основания. О современных
адансонианцах (которые себя так не называют) нельзя сказать, что
они неправы, но их работы достаточно поверхностны и далеки от
зрелости.
Ведущий принцип этой школы достаточно прост: исследовать
и регистрировать как можно больше признаков и выделять группы
согласно тому, как указывает большинство совпадающих призна-
ков. Вид состоит из особей с наибольшим числом общих призна-
ков. Род состоит из видов также с наибольшим числом общих при-
знаков. И так далее. В результате шаг за шагом выстраивается не-
Развитие современной систематики
49
обходимая система соподчинения признаков и ранжирования так-
сонов.3 Более того, Адансон (например, Adanson, 1763), Вик д’Азир
(например, Vicq d’Azyr, 1792) и другие представители этой школы
установили, что для членства в группе не требуется совпадения по
всем признакам, определяющим группу, равно как и не требуется
исключения уклонений из её определения или низведения группы
к более низкому рангу. Тем самым они избежали некоторых, хотя
и не всех, ныне критикуемых черт типологии (см. далее) и схола-
стической логики в таксономии. Для формирования таксона доста-
точно, чтобы каждый его член обладал большей частью всех атри-
бутов таксона. На основании этого принципа и используя опреде-
лённую символику (буквы соответствуют атрибутам, цифры — осо-
бям), следующие группировки можно считать валидными видами:
Особи 123 456
Атрибуты
a a b
е е f
f g g
Вид I
Вид II
Такой «вид» можно определить следующим образом:
Вид I есть группа особей с атрибутами a, b, с, d, каждая из ко-
торых обладает большинством (в данном случае тремя) этих атри-
бутов.
Вид II есть группа особей с атрибутами a, b, е, f g, каждая из
которых обладает большинством (в данном случае тремя) этих ат-
рибутов.
Такой «вид» можно диагностировать (т.е. отличить от любо-
го другого, а не ограничить указанием собственных признаков) сле-
дующим образом:
3 Я использую этот и некоторые другие термины для удобства и краткос-
ти, хотя они и не фигурировали в работах, авторы которых никогда не слышали
таких слов. Термин таксон современный, однако классификации Адансона
включали таксоны, так что такая терминология скорее проясняет, а не запуты-
вает ситуацию.
50
Дж. Г. Симпсон. Принципы таксономии животных
В виде I каждый член имеет хотя бы один из атрибутов с, d, ни
один из которых не присутствует в виде II; соответственно, в виде
II каждый член имеет хотя бы один из атрибутов е,/ g, ни один из
которых не присутствует в виде I.
Это есть совершенно чёткий, таксономически обоснованный и
осмысленный приём, который не предусматривает указания всех
совпадающих признаков, несмотря на то, что даже сегодня некото-
рые таксономисты считают совпадение признаков существенным
для концепции таксона.
Определяющие атрибуты не свойственны всем представителям
видов: каждая особь в первом виде обладает только одним атрибу-
том [А], общим для вида в целом; во втором виде — ни один из
трёх атрибутов, свойственных каждой особи, не является общим
для вида; при этом все особи обоих видов обладают атрибутами [а
или А], которые входят в определение противоположного вида. Этот
пример — сверхупрощение, к тому же выходящее за границы до-
пущений, которые, вероятно, принимали ранние таксономисты этой
школы, но он хорошо иллюстрирует общий принцип. Beckner
(1959) более детально прояснил этот принцип (названный им «по-
литипическим»), использовав современную терминологию. Для
дальнейшего изложения полезно отметить, что вид II не имеет типа
(в типологическом смысле), воплощающего весь его морфологичес-
кий образ, и что тип (в этом же смысле) вида I состоит только из
атрибута 6, который, однако, не специфичен для этого вида, но при-
сутствует также в виде II. Отметим также, что вид II не имеет «от-
личия» и что оба вида, хотя для нас они кажутся очевидно близ-
кими, не имеют схоластического, или линнеевского «рода».
В такой форме рассматриваемый принцип по-прежнему лишён
интерпретаций и оценок, разрабатываемых эволюционной таксоно-
мией, однако он вполне соответствует этой таксономии и часто упо-
минается в современных классификациях. С другой стороны, он не
принимался и до сих пор не принимается безоговорочно всеми
эмпиристами, а ещё менее — всеми неэволюционными таксономи-
стами.
Кювье (1769-1832), возможно, оказал большее влияние на так-
сономию, чем кто либо другой от Линнея до Дарвина: его жизнь
охватила как раз период между их днями. Его более всего вспоми-
Развитие современной систематики
51
нают как основателя палеонтологии, что отчасти преувеличено, и
провозвестника принципа органической интеграции, или корреля-
ции. Томпсон (Thompson, 1952) так написал о названном принци-
пе и его отношении к «систематике» (= таксономии, как она здесь
определена):
Кювье определил с галльской чёткостью, что он понимает под основ-
ным философским4 принципом рациональной анатомии и, тем самым, ра-
циональной систематики. «Каждая организованная сущность, — говорил
он, — образует целое, уникальную замкнутую систему, чьи части взаимно
соответствуют друг Другу и каждая вносит свой вклад в единое действие
благодаря взаимодействиям между собой. Ни одна из этих частей не мо-
жет измениться без того, чтобы и прочие также не изменились, и, следо-
вательно, каждая из них, взятая в отдельности, подразумевает все прочие,
указывая на них...» Он был уверен в том, что форма определяется функ-
цией и что связь между формой и функцией управляется законами, столь
же неумолимыми, как и законы физики и математики. Рассматриваемый в
таком ключе, его особенный тип приобретает логичность и необходимость
геометрического определения, из которого мы можем вывести все атрибуты
определяемого объекта.
Этот пассаж достаточно точно суммирует один из аспектов
кювьеровой теоретической системы, однако он ошибочно излага-
ет то, как она связана с таксономий. Тот факт, что организмы пред-
ставляют собой органически интегрированное целое, является дек-
ларированной исходной позицией всей школы современной систе-
матики (см. обсуждение в: Beckner, 1959, и его ссылки), однако для
таксономистов это не было новостью уже задолго до Кювье. Зак-
лючение, ныне выводимое из этого вполне банального факта, со-
стоит в том, что классифицировать следует целые организмы, а не
их части или атрибуты. Однако принцип Кювье не ведёт к этому
заключению и на самом деле даже работает против него. Если вся-
кая часть подразумевает целое и если все атрибуты могут быть
выведены из «типа», то классификация частей и «типов», а не це-
лостного реального организма, и будет не только достаточной, но
и теоретически предпочтительной. Положение дел противоречит
этому, поскольку часть не адекватна целому даже в интегрирован-
4 Я бы сказал «теоретическим». —Д.Г.С.
52
Дж. Г. Симпсон. Принципы таксономии животных
ном организме,5 а отношение «типа» к атрибутам на самом деле
скорее индуктивно, нежели дедуктивно.
Не будучи адекватным даже в своей исходной сфере приложе-
ния, принцип Кювье на самом деле не даёт какого-либо рабочего
инструмента и для систематики. Ни сам Кювье, ни его ближайшие
последователи в XIX в., ни немногие нео-кювьерианцы XX в., та-
кие как Томпсон, не разработали ни одной классификации, соответ-
ствующей этому теоретическому фундаменту. Томпсон раз за разом
отказывается от всех прочих ныне признанных оснований класси-
фикации и в той же статье (1952, с. 5) делает следующее многозна-
чительное заявление:
Хороший систематик способен распознавать [develops] то, что средне-
вековые философы называли habitus [габитус], который есть нечто боль-
шее, чем свойство, и для которого лучшим обозначением был бы термин
secunda natura. Возможно, подобно теннисисту или музыканту, он лучше
всего действует, когда особо не задумывается над тем, как действует.
С этим утверждением согласится любой опытный таксономист.
Он добавил бы, однако (разумеется, Томпсон и сам знает это, чему
иллюстрацией — его собственный опыт), что, используя ту же ана-
логию, музыкант достигает совершенства лишь после того, как изу-
чит правила и технику настолько глубоко, что они станут его вто-
рой природой, и, далее, что сочинение музыки требует изучения и
развития весьма рациональных и строгих теорий и принципов.
Кювьерова таксономия (особенно см. Cuvier, 1835), которую,
как и линнеевскую, недавно компетентно проанализировал Кэйн
(Cain, 1959b), была во многом основана на принципах, отличных
от принципа органической корреляции. Кювье использовал иерар-
хию: в этом состоял его «метод». Он понимал, что для наполнения
этого метода смыслом необходимо располагать неким базисом для
того, чтобы одни атрибуты делать по рангу выше других. В общих
чертах, он пытался установить ранги определяющих атрибутов и,
5 Ложность принципа Кювье была ранее показана на знаменитом приме-
ре халикотерия, чьи конечности соответствуют хищнику, но череп — копыт-
ному [17]. Этот принцип сохранился лишь в преданиях в качестве веры в то,
что палеонтологи могут реконструировать целый скелет по единственной ко-
сточке. Однако сами палеонтологи знают, что это, к сожалению, не так.
Развитие современной систематики
53
тем самым, определяемых ими таксонов на дедуктивной, или ап-
риорной основе. Он утверждал, например, что ощущения и движе-
ния являются атрибутами высшего ранга или наиболее значимы для
животных, поскольку именно они (по его мнению) делают их жи-
вотными. Тем не менее, он столь же часто отстаивал приоритет-
ность [атрибутов] на основе их постоянства, т.е. присутствия в
большом числе групп организмов. Например, кровеносная система
«менее существенна», чем пищеварительная, поскольку не все жи-
вотные с пищеварительной системой также имеют кровеносную
систему. Это уже кажется некой формой скрытого эмпиризма.
Как подчеркнул Кэйн, двойственный подход привёл Кювье к
замкнутому кругу, который сделал всю его систему ложной. Как
можно определить постоянство признаков в группе, а тем самым
и их ранг в иерархии определений, до того, как группа выделена
по какому-то иному основанию? Этот вопрос ни в коей мере не
ограничен кювьеровой таксономией: он неоднократно ставился во
многих теоретических школах, начиная с самых ранних классифи-
каций вплоть до настоящего времени. Очень просто сказать вслед
за Линнеем, что род задаёт признаки. Но что, в таком случае, за-
даёт род? В сущности, Кювье говорил, что это делают признаки,
что замыкает круг, причём нет никакой возможности разорвать его
и начать поступательное движение. Ответ Линнея состоял в том,
что Бог даёт род, однако это не самое практическое средство для
его распознавания. Дедуктивная часть системы Кювье была ещё
одной попыткой ввести внешние критерии в порочный круг, одна-
ко во многих случаях, если не во всех, его дедукции, по-видимо-
му, были просто обратным ходом по кругу, начатому индукциями.
На самом деле, как Кэйн также отмечал в несколько иных выраже-
ниях, многие системы, обнародованные в качестве дедуктивных,
«для себя» строились как индуктивные.
По-видимому, существуют только два осмысленных и практи-
чески значимых способа избежать этого тупика. Один состоит в
том, чтобы придерживаться строго индуктивного, полностью эмпи-
рического подхода; однако, как уже было отмечено, это довольно
поверхностное решение, которое к тому же порой подразумевает
неодолимые сложности с ранжированием, как мы увидим далее.
Другой означает введение критериев ранжирования, особенно эво-
54
Дж. Г. Симпсон. Принципы таксономии животных
люционных дедукций, которые, будучи выведенными индуктивно,
свободны от априоризма Кювье и многих других и являются вне-
шними по отношению к замкнутому кругу.
Ещё один аспект системы Кювье требует особого комментария,
поскольку он представляет значительный раздел додарвиновской (а
также, хоть и в меньше степени, после-дарвиновской) мысли. Ус-
тановив в теории ранги атрибутов, Кювье вводит свой принцип
организменной корреляции, означающий, что все части организма
должны быть совместимы с единой классификацией, которая тем
самым оказывается основанной на общем плане строения. Соглас-
но своим воззрениям, он отбросил очень старую идею о лестнице
природы, или цепи существ, которой во времена Кювье был при-
вержен (с некоторыми модификациями) Ламарк, а также многие до
и немногие после него. Вместо непрерывной последовательности
он признавал иерархию множественных дискретных «планов». Сре-
ди животных он выделил 4 основных «плана», определяющих
4 высших таксона, названных «ответвлениями», под именем Ver-
tebrata, Mollusca, Articulata и Radiata. «Планы» Кювье — это
«типы» таксономической типологии; к этому предмету мы теперь
и обращается.
Типология
Типология берёт начало от Платона и его творений, а в таксо-
номию она проникла вместе с Аристотелем, неоплатониками, схо-
ластами и томистской философией и логикой. Её оформление как
самостоятельной школы таксономии отчётливо связано с натурфи-
лософией и с такими именами, как Гёте (1749-1832), Кювье (1769—
1832), Окен (1779-1851) и Оуэн (1804-1892) (ссылки и обзор воз-
зрений большинства типологов, начиная с античных времён, мож-
но найти в: Zimmermann, 1953). Её базовая концепция, однако,
была значительно более объемлющей, нежели какая-то из частных
систем названных таксономистов, лидирующих среди типологов-
теоретиков. Это не только конкретная таксономическая школа, но
также некий общий подход или концептуальный конструкт, кото-
рый не альтернативен другим школам, возникавшим в разные вре-
мена, но является составной частью почти всех их, даже эмпири-
Развитие современной систематики
55
ческой и эволюционной школ, которые разными способами и в
лице разных таксономистов сделали очень много для того, чтобы
вырваться из тенет типологии.
Основная идея типологии такова: каждая естественная группа
организмов, а тем самым и каждый естественный таксон в класси-
фикации, обладает неким инвариантом — обобщённым или идеа-
лизованным «образом» [pattern], общим для всех членов этой груп-
пы. Образ низшего таксона накладывается на таковой высшего так-
сона, к которому он относится, без существенного изменения выс-
шего образа. Низшие образы являются вариациями на тему высше-
го образа: они воплощают детали, разные для разных таксонов од-
ного уровня, в пределах более генерализованного, менее детализи-
рованного высшего образа. Наиболее детализированный образ ха-
рактеризует вид (вопрос о внутривидовых таксонах здесь не рас-
сматривается). Вариации в пределах вида — «акциденции» схола-
стов — являются досадной помехой, поэтому (или потому что) они
не имеют таксономического значения. Для этих идеализованных
образов было предложено много разных терминов — чаще всего
«тип», также «архетип», «план строения» или «структурный план»,
«морфотип»,6 просто «план» и др.
Теперь мы можем видеть яснее, как возникла выше упомяну-
тая путаница, связанная с использованием термина и концепции
«типа» также и в технической номенклатуре. Согласно типологи-
ческим принципам, каждый экземпляр вида содержит его «тип» и
поэтому адекватен как для определения вида, так и для того, что-
бы служить стандартом для сравнения. С этой точки зрения не воз-
никает проблем с использованием одного и того же экземпляра и
как морфологического, и как номенклатурного типа. Дополнитель-
ные экземпляры, по общему признанию не являющиеся необходи-
мыми для номенклатурных целей, считались таксономически по-
лезными только при необходимости отличить тип от случайных
6 Иногда в «план строения» и «морфотип» вкладывается разный смысл:
первый понимается в более широком смысле абстрактного образа, второй —
как вариант образа одной из нескольких групп, характеризуемых единым пла-
ном (см. Zangerl, 1948). Оба являются типами в общем типологическом смысле,
а их различия, которые во многих случаях недостаточно отчётливы, нет нуж-
ды здесь рассматривать.
56
Дж. Г. Симпсон. Принципы таксономии животных
индивидуальных вариаций. Вместе с тем, даже при допущении со-
стоятельности типологических предпосылок, таксономисты часто
чувствовали, что существуют некие различия в «типе» между, ска-
жем, самцом и самкой или между личинкой и имаго, хотя в прин-
ципе оба имеют один и тот же видовой тип. Отсюда и предложе-
ние о «паратипе» — компромисс, совершенно неоправданный ни
с какой стороны, хотя всё ещё в широком использовании. Номен-
клатура не требует типа и на самом деле может обходиться без
него, за исключением единственного типа, или голотипа. Если ва-
риации в пределах вида не имеют какого-либо таксономического
значения, то, опять-таки, добавления к видовому типу выходят за
пределы разумного. Если же они имеют таксономическое значение,
как полагают практически все современные таксономисты, тогда
следует принимать во внимание все известные вариации, а не толь-
ко те, которые оказались представленными отдельными «паратипа-
ми». Поэтому впредь, чтобы избежать путаницы, я буду использо-
вать оуэновский термин «архетип» для типа в типологическом
смысле и «тип» исключительно для указания номенклатурного
типа.7
Схоластический подход, пример которого дан Линнеем, непос-
редственно ведёт к концепции архетипа, поскольку «сущность»
должна быть единой для всех членов вида. Так, использование не-
скольких fundamenta divisionis неизбежно вело к концепции общ-
ности целого образа, воображаемым представлением которого как
раз служит архетип. Даже осознание того, что в классификации,
вопреки логике архетипа, варианты также входят в целостную кар-
тину, не позволяет избежать этого. Так, Джозеф (Joseph, 1916) рас-
сматривает классификацию не как приложение логических опреде-
лений, а их замещение, однако пишет о классификации, что она
«стремится установить типы: она выбирает некоторые отдельные
характеристики как определяющие тип любого вида... Описание
типа призвано служить для [т.е. вместо] определения.»
7 Было бы несомненно лучше вообще избавиться от ауры «типичного» и
дать другие названия для номенклатурных типов. Некогда я (Simpson, 1940)
предположил, что походящим был бы такой термин как «ономатофор», одна-
ко с тех пор ни я сам, ни кто либо другой не использовали его, просто из-за
того, что мёртвая рука прошлого слишком тяжело давит на всех нас.
Развитие современной систематики
57
Ранний эмпиризм был шагом прочь от типологической теории
и частично реакцией против неё. Ранее в этой главе я показал, что
эмпирические принципы, доведённые до логического завершения,
могут выделять и определять таксоны, для которых нельзя обозна-
чить архетип. Тем не менее, общая тенденция развития эмпириз-
ма оставалась в рамках если не теории, то типологической прак-
тики. Например, два важных недавних предложения для эмпири-
ческого определения «отношения» или «близости» (Sokal, Michener,
1958; Cain, Harrison, 1958) основаны на измерении образа [pattern]
совпадающих признаков и являются количественным представле-
нием типологической по своей сути процедуры, хотя эти авторы в
последующем и интерпретируют результаты в нетипологической
манере.
Эволюционная классификация, хотя ныне и противопоставля-
емая типологической, на деле часто оказывалась пропитанной ти-
пологическими представлениями. Таксономия Дарвина была всё
ещё по преимуществу типологической, а слово «тип», обычное в
его работах, часто можно было прочитать как «архетип» без иска-
жения его мысли. Более наивные эволюционные таксономисты
иногда уравнивали «архетип» с «общим предком», хотя Дарвин
(Darwin, 1859) ясно указал на то, что это не так. По-прежнему ши-
роко признаётся, что число совпадающих признаков прямо пропор-
ционально степени филогенетического родства, что является полу-
правдой или, может быть, на три четверти правдой, но связано с
таким числом оговорок и исключений, что этот принцип нельзя
считать общим для истинно эволюционной классификации. Кирь-
яков (Kiriakoff, 1959) заходит слишком далеко, когда говорит, что
«большинство биологов, которые считают себя филогенетиками, на
самом деле — типологи», однако в этом обвинении есть значитель-
ная доля правды.
Пожалуй, на этом хватит говорить о проникновении типологии
в разные школы систематики, включая и те, которые в теории
обычно противостоят типологии. Признанные типологи (под этим
или иным названием), такие как Гёте или Оуэн, апеллировали к
философско-теоретическим основаниям, которые отвергнуты боль-
шинством современных таксономистов, даже если они в своей
практике и являются типологами. Их вера состояла в том, что ар-
58
Дж. Г. Симпсон. Принципы таксономии животных
хетип в определённом смысле реален и что организмы попросту
являются тенями, отражениями, несовершенными и эфемерными
воплощениями этой трансцендентальной реальности. Архетипы с
философской точки зрения рассматривались как платоновские идеи,
а теологически — как воплощения божественного [плана] творе-
ния. Чтобы ни думали об этих материях, таким метафизическим
верованиям нет места в науке, в них нет эвристической ценности.
Вместе с типологической таксономией, на основе тех же прин-
ципов и как часть той же школы систематики, развивалась идеали-
стическая морфология, задача которой состояла в конструировании
(или распознавании) архетипов как основы для типологических
классификаций. Эта школа не отмерла; напротив, она недавно ис-
пытала возрождение усилиями Нэфа, Келина и других европейских
сравнительных анатомов (обобщение и ссылки см. в: Zangerl,
1948). Утверждается, что архетип (план строения, морфотип), как
минимум — полезный инструмент, как максимум — выражение
анатомических отношений, реально существующих в природе, и
что он не предполагает никаких допущений метафизического или
филогенетического характера. Однако я должен согласиться с Блок-
ком (Block, 1956), что это всё равно не устраняет (частью неосоз-
нанную) засоренность метафизикой.
Изучение и объяснение признаков, совпадающих в группах
организмов, разумеется, должно быть одной из процедур в любой
из систем таксономии. Однако я утверждаю, что сверх этого оче-
видного пункта типологическая теория или строго типологическая
практика не должны присутствовать в современной таксономии.
Возражения должны быть очевидны для любого, кто прочтёт всю
эту книгу, однако некоторые из них, наиболее важные, возможно,
следует здесь хотя бы упомянуть. Типологическая теория запута-
лась в тенетах философского идеализма, который просто по праг-
матическим (если не по каким-то другим!) причинам должен быть
исключён из современной науки. Представление о самостоятельных
статических структурах не может иметь осмысленного применения
к реальным группам организмов, которые являются частью эволю-
ционного континуума и всегда высоко изменчивы. Их изменчи-
вость не второстепенна или «случайна», её нельзя игнорировать ни
на каком из таксономических уровней: она принадлежит самой
Развитие современной систематики
59
природе таксонов и является частью механизмов их возникновения
и длительного существования. Определение таксонов через совпа-
дение признаков является частным случаем, пусть даже вполне
обычным в практических приложениях. Когда окидываешь взгля-
дом весь четырёхмерный аспект царства животных, строго типоло-
гическое определение нередко невозможно, и оно никогда не мо-
жет служить подходящей теоретической основой.
Открытие филогении
После того, как была открыта эволюция, стало очевидно, что
все атрибуты, используемые в классификации, исторически возни-
кают через наследование, сопряженное с изменением [descent with
modification]. Также ясно, что все таксоны до- и неэволюционных
классификаций, а также, разумеется, в эволюционных классифика-
циях, подобным же образом возникают в филогении. Являются ли
они в каждом конкретном случае естественными (в определённом
смысле) филогенетическими единицами — другой вопрос, который
будет рассмотрен далее в настоящей главе. Зоологи, за редчайши-
ми исключениями, согласны с тем, что подходящим теоретическим
базисом таксономии является филогения, которая принципиальна
для понимания и объяснения всех связей, учитываемых в класси-
фикации. Важное расхождение состоит лишь в оценке желаемого
или практически достижимого соотношения между филогенией и
классификацией, особенно в том, может и должна ли филогения
давать критерии для классификации или она может и должна иг-
рать таксономическую роль только в объяснении классификаций,
основанных на иных критериях. Я принимаю первую точку зрения
и буду отстаивать её более детально по мере изложения.
Линней, Бюффон (1707-1788) и некоторые другие доэволюци-
онные таксономисты признавали по крайней мере возможность того,
что виды не полностью статичны, но могли заметно меняться (на-
пример, «дегенерировать») с момента своего творения. Высказыва-
лись даже соображения, что «единицами» творения (Бытие I: 12,21,
24, 25) могли быть роды и даже более высокие таксоны современ-
ности и что виды в пределах каждой единицы могли формироваться
уже после творения. (Эта точка зрения всё ещё всерьёз отстаивает-
60
Дж. Г. Симпсон. Принципы таксономии животных
ся некоторыми фундаменталистами, которые в такой форме пыта-
ются примирить буквальное прочтение Книги бытия с признанием
некоторых фактов эволюции, — например, Marsh, 1944.)
Ламарк (1744-1829) был первым, кто чётко и последовательно
утверждал, что все таксоны возникли эволюционным путём и об-
разуют эволюционный континуум.8 Его идея континуума была го-
раздо более буквальной и широкой, чем у современных филогене-
тиков. Он признавал теоретически, что в природе нет разрывов,
даже между разными филогенетическими линиями, но существует
единый непрерывный ряд, через который проходят все организмы
на всём протяжении истории жизни, и что этот процесс продолжа-
ется и сегодня. Таксоны, в таком случае, — просто условные (и
потому с необходимостью не естественные) подразделения конти-
нуума, причём на основании этого континуума можно заключить,
что ни один таксон не вымер. Конечно, Ламарк признавал, что на
самом деле имеются значительные разрывы между таксонами, ко-
торые реально выделяются в системе и представлены в музеях —
например, между моллюсками и рыбами, и, подобно всем прочим,
использовал эти разрывы для практического разграничения таксо-
нов. Тем не менее, он был уверен, что разрывы будут совершенно
заполнены по мере открытия организмов, пока ещё неизвестных и
не заколлектированных.
Основные выводы для классификации, которые мы сегодня ви-
дим в теории Ламарка, таковы: первое, границы между всеми так-
сонами должны быть условными (и их невозможно исчерпывающе
охарактеризовать архетипами), исключая те случаи, когда они вре-
менно разделены разрывами в знании, а не в природе; и второе,
таксоны на каждом уровне должны составлять естественный ряд от
низших к высшим, от простых к сложным. Сам Ламарк не форму-
лировал первый вывод, да и второй не был им предложен, посколь-
ку тот же ряд классификации был заложен в древнюю неэволюци-
8 В этом состоит наиболее оригинальный и по-прежнему приемлемый
принцип ламаркизма. Ламарк удивился бы, узнай он, что «ламаркизм» теперь
понимается как наследование приобретённых признаков. На самом деле он не
верил, что все приобретённые признаки наследуются, и, кроме того, говоря о
наследовании некоторых таких признаков, он всего лишь повторял то, что все
«знали» со времён античности.
Развитие современной систематики
61
онную концепцию лестницы природы (scala naturae), или цепи су-
ществ. На самом деле классификации Ламарка не отличались в ка-
ких-либо важных чертах от разработанных его современниками-
неэволюционистами. Как и большинство из них, он был озабочен
проблемой ранжирования признаков и в этом отношении его взгля-
ды были существенно такими же, как у его непримиримого против-
ника антиэволюциониста Кювье. «Отношения тем значительнее,
чем существеннее данная часть, на основании которой эти отноше-
ния определены» (Lamarck, 1809 — основной источник по ламар-
ковой теории). У животных наиболее важные отношения выявля-
ются анализом частей, наиболее существенных для поддержания их
жизни, а у растений — наиболее существенных для размножения
(что не слишком отличается от линнеевской системы полов). Ис-
торическим фактом является то, что эволюционизм Ламарка не спо-
собствовал развитию эволюционной таксономии и даже, возмож-
но, затормозил его. Сам он не внёс существенного вклада в это раз-
витие, а его взгляды на сей предмет, насколько они были отличны
от таковых, скажем, Кювье, были почти целиком отвергнуты. На
самом деле они были довольно невразумительны и запутанны, а
многие из них казались абсурдными и в то время, и ещё более
того — сегодня, такие как его вера в то, что все химические соеди-
нения в своём происхождении являются органическими.
Додарвиновские основатели эволюционной таксономии выдви-
гали и иные претензии, однако, по моему мнению, ни одна из них
не заслуживает более серьёзного отношения к себе, чем ламарко-
вы, но и ему я вынуждён отказать в иске (который, впрочем, он и
не предъявлял). Эволюционная таксономия восходит почти исклю-
чительно к Дарвину. Этот предмет постоянно возникает в «Проис-
хождении видов», но особенно в главе «Взаимное родство органи-
ческих существ». (Глава XIII 1-го издания: Darwin, 1859; глава XIV
2-го издания: 1872; для того, чтобы в деталях оценить взгляды Дар-
вина, изложенные в разных изданиях, очень важна книга под ре-
дакцией: Peckam et al., 1959.)
В «Происхождении видов» Дарвина занимает главным образом
выяснение истинной природы эволюции и действенность есте-
ственного отбора как механизма эволюции. Он предложил весьма
богатую концепцию, которую здесь можно было бы проследить, но
6 — 4338
62
Дж. Г. Симпсон. Принципы таксономии животных
в этой своей обобщающей книге он не озаботился её детальным
выстраиванием. Это особенно справедливо в отношении классифи-
кации. Его основная классификационная работа (по рачкам) была
завершена пусть не до формирования, но до публикации его взгля-
дов на эволюцию, и она не до конца эволюционная. Он никогда не
возвращался к детальной классификации (которую на самом деле
невзлюбил) и потому не показал, как используются таксономиче-
ские принципы, подразумеваемые или заявленные в «Происхожде-
нии...». То, что он сделал в действительности, это — взял уже раз-
работанные классификации как данность и показал, во-первых, что
они соответствуют теории об эволюционном происхождении их
таксонов и, во-вторых, что эволюционная филогения может объяс-
нить тот порядок, который уже обнаружен среди организмов. Он
полагал, с полным на то основанием, что такого рода демонстра-
ция является самым сильным из свидетельств истинности эволю-
ции. Именно это, а не перестройка классификаций согласно эволю-
ционным идеям, было его основной целью.
Процесс эволюции порождает группы, распознаваемые в тер-
минах сходства и различия. Поэтому не должно вызывать удивле-
ния, что в своей значительной части филогенетически связанные
группы на самом деле были выявлены до того, как стала понятной
их истинная природа взаимосвязей. Если бы дело ограничилось
только этим, вклад Дарвина в совершенствование таксономии был
бы всё равно выдающимся, однако это в малой степени отразилось
бы на практике классификации. На самом деле Дарвин, почти не-
преднамеренно, сделал больше. Он предложил новые критерии
или, как их назвал Кэйн (Cain, 1959b), априорные правила, которые
в конечном итоге разорвали замкнутый круг кювьерова априориз-
ма. Он также показал на примерах, что эти критерии в некоторых
случаях подтверждают уже разработанные системы организмов, но
в других случаях — нет. Таким образом, они подразумевают изме-
нения не только в теории, но также и в приёмах и результатах клас-
сификации. Сам Дарвин не развивал далее эти идеи, но его после-
дователи сделали это, причём главным образом в рамках разрабо-
танной им схемы.
Сведённые до простейшего уровня, основные принципы соб-
ственно дарвиновской таксономии таковы. Таксономические труп-
Развитие современной систематики
63
пы являются результатом «возникновения путём изменения», или
филогении в более привычной современной терминологии. Каж-
дый валидный таксон имеет общего предка. Наиболее фундамен-
тальным, но не единственным9 критерием ранжирования таксонов
является их родственная близость [propinquity of descent]. Призна-
ки, используемые в определении, следует интерпретировать как
свидетельства филогенетического родства, их надлежит ранжиро-
вать и оценивать согласно их вероятной корреляции с родственной
близостью.
Принципы, обсуждаемые Дарвином, таковы: (1) совпадающие
признаки интерпретируются как унаследованные от общего пред-
ка, (2) цепочки современных таксонов интерпретируются как ре-
зультат происхождения от общего предка с выживанием переход-
ных форм, (3) временная последовательность изменяющихся форм
интерпретируется как истинный ход филогенеза и (4) всё это сви-
детельствует о генетическом единстве. Эти принципы используются
современными систематиками, наряду с некоторыми предложенны-
ми позже (см. главу 3). Что касается (1), Дарвин вполне осознавал,
что совпадающие признаки не обязательно указывают общего
предка и что это было серьезным изъяном большинства неэволю-
ционных классификаций. Поэтому я не могу вслед за Кэйном (Cain,
1959d) считать это основным изъяном дарвиновой таксономии.
Кэйн в целом правильно суммирует идейное наследие Дарвина,
ставя дарвиновской таксономии в вину лишь то, что она будто бы
не даёт аподиктической определённости. Однако представляется
маловероятным, что вообще какая-либо система должна быть апо-
диктически определённой. Всякая определённость, которая может
быть заявлена для такой эмпирической системы как, например,
Кэйна и Харрисона (Cain, Harrison, 1958), мне кажется ненастоя-
щей. Это может быть приемлемо при упорядочении данных, но их
отбор, интерпретация и формализация зависят как от вероятност-
ных суждений, так и от личных вкусов.
Я уже имел возможность (Simpson, 1959с) обсуждать таксоно-
мию Дарвина с большей подробностью, и остальная часть настоя-
9 Выделено курсивом в связи с частым, но неверным утверждением его кри-
тиков, что это является единственным критерием в дарвиновской таксономии.
6*
64
Дж. Г. Симпсон. Принципы таксономии животных
щей книге посвящена в основном результатам развития его осно-
воположений. Поэтому в их дальнейшем анализе здесь нет необ-
ходимости.
Что такое естественная классификация?
Здесь мы, в качестве некой интерлюдии, сделаем паузу, чтобы
рассмотреть постоянно возникающий вопрос, вынесенный в заго-
ловок настоящего раздела. На протяжении двух или трёх столетий
таксономисты настаивали на том, что классификация должна быть
естественной. Большинство из них настаивало на том, что именно
их системы являются естественными, по крайней мере по принци-
пам построения. Другие, хотя и принимая откровенно искусствен-
ный подход, как это сделал Линней, подчёркивали, тем не менее,
теоретическую желательность естественной классификации. Вдоба-
вок ко всему, идеи о том, что на самом деле представляет собой
естественная система, были весьма многообразны и противоречи-
вы, их определения либо отсутствовали, либо были изложены не-
внятно и неудовлетворительно.
Всякая классификация, которая может быть применена к реаль-
ным организмам, несомненно, должна быть как-то связана с харак-
теристиками организмов, присущих их естеству. Использование тер-
мина «естественная» [natural] только в таком смысле было бы со-
вершенно бессмысленным. Естественная классификация должна под-
разумевать, что классификация сама по себе, как таковая, в некото-
ром смысле естественна, а не только что она основана на естествен-
ных характеристиках или критериях. Таксоны естественной класси-
фикации должны иметь некоторое отношение (не обязательно одно-
значное) к группам организмов, реально существующим в природе.
Идеалистически-типологические и креационистские принципы,
которые, как мы видели, часто совпадают и не обязательно явля-
ются альтернативными, постулируют существование в природе уни-
кальной системы архетипов, фиксированных «идей», «типов», «пла-
нов творения». Чтобы создать совершенно естественную классифи-
кацию, необходимо, следовательно, только распознать эти архети-
пы и сгруппировать организмы согласно с ними. Однако слово
«только» маскирует трудность, которая на самом деле непреодоли-
Развитие современной систематики
65
ма: невозможно общее согласие относительно того, как следует
распознавать архетипы, так что на практике постулированная един-
ственная естественная классификация таким способом не может
быть достигнута. На самом деле она едва ли может быть достиг-
нута по той причине, что её просто не существует. Строго архети-
пический подход, не включающий дополнительных критериев на
основе совершенно иных принципов, невозможен даже в теории.
На самом деле, разные архетипы часто выводятся из анализа раз-
личных группировок, полученных до вычленения архетипа и на
иных основаниях, как видно из следующего простого примера:
Виды I II III
а а е
Атрибуты -< b b b
d d
Архетипы ab bd
Действительная ситуация редко бывает столь простой, но эта
двусмысленность постоянно возникает в более сложных формах.
Например, достаточно легко вычленить архетип хищных и архетип
сумчатых после объединения организмов в группы хищных и сум-
чатых. К кому из них следует отнести тасманийского волка (Thyla-
cynus cynicephalus)? Если его собственные атрибуты исследовать
типологически, он в равной степени может принадлежать к любой
из этих групп или к обеим, но в одном случае он будет объединён
с волками (Canis), с которыми он действительно схож по очевид-
ным характеристиками, а в другом — с кенгуру (Macropus), сход-
ство с которым не столь очевидно. Даже если, как это принято,
классификаторы-типологи выберут архетип сумчатых как более
значимый или приоритетный для классифицирования тилацина,
они это будут делать по индукции после того, как сформируют так-
сон Marsupialia (включая Thylacynus) на иных основаниях, а не по
дедукции исходя из принципа архетипа.
Линнеевская, кювьерова и другие системы приписывания при-
оритетов разным категориям атрибутов — всё это попытки решить
неразрешимую по самой своей сути проблему. Уже было показано,
66
Дж. Г. Симпсон. Принципы таксономии животных
что они предусматривают порочный круг, хождение по которому не
позволяет постичь единственную естественную последовательность
архетипов. Многие дискуссии по поводу существования таких ар-
хетипических групп в настоящее время кажутся весьма несерьёз-
ными, поскольку стало достаточно ясно, что ни одна из этих групп
сама по себе не существует в природе. Линней, например, хотя и
с определёнными колебания, как правило, считал, что его роды и,
вероятно, также виды, если правильно диагностированы, соответ-
ствуют истинным единицам творения и, таким образом, совершен-
но естественны, тогда как высшие таксоны в этом смысле менее
естественны или, честнее, неестественны. Правильный диагноз
вида соответствует его природе, которая в свою очередь задаётся
его сущностью, определяемой по роду и видовым отличиям в схо-
ластическом смысле, однако (как уже подчёркивалось) никаких
критериев для распознавания сущности нет и они невозможны
(снова см. статьи Кэйна, особенно Cain, 1958а).
Другой подход, существовавший на протяжении всей истории
классификаций, был основан на идее, что классификация по един-
ственному признаку на каждом уровне, или простая дихотомия, как
в ключах, не является естественной, тогда как классификация, ос-
нованная на одновременном использовании многих атрибутов и,
если нужно, выделении многих подмножеств [на одном уровне],
будет естественной. Хотя и часто повторяемое учёными таксономи-
стами, это банальное утверждение, взятое буквально, граничит с
бессмыслицей. Оно означает, что если вы добавите достаточно ну-
лей (атрибутов, не имеющих естественного обоснования), вы в кон-
це концов получите положительное число (естественное объедине-
ние). Тем не менее, это утверждение не бессмысленно: оно лишь
пренебрегает теми скрытыми постулатами и предпосылками, кото-
рые необходимы для придания ему смысла.
Нужно просто исследовать ряд широко принятых классифика-
ций, чтобы увидеть, что вопрос не столько в том, какое число при-
знаков необходимо принимать во внимание, сколько в том, какие
из них следует выбирать и как интерпретировать. «Животные с во-
лосами», выделенные по одному признаку, характеризуют группу,
которую давно считают естественной все таксономисты, какими бы
ни были их представления о естественности. «Животные с крыль-
Развитие современной систематики
67
ями» такой группой не являются, хотя единственный атрибут, по-
ложенный в её основу, представляется логически эквивалентным по
рангу с атрибутом обволошенности. Действительная причина при-
влечения многих атрибутов не в том, что это само по себе каким-
то образом более естественно, но просто в том, что это повышает
вероятность выделения групп, которые таксономисты в целом бу-
дут воспринимать и принимать как естественные. Одновременно,
что также весьма значимо, это расширяет содержательность и по-
лезность классификации, на что обратил внимание Гилмур (Gil-
mour, 1951), как указано в главе 1. Хотя он кажется мне непосле-
довательным, отождествляя это достоинство с естественностью.
Симпатии и согласия таксономистов имеют очевидное прагма-
тическое значение, но лишены теоретической осмысленности.
В действительности теоретическая дискуссия в истории таксоно-
мии, прикрытая безликим языком и объективистским фасадом, в
своей значительной части была попыткой найти некое теоретиче-
ское обоснование личным и субъективным результатам. Существен-
ную, хотя и не исчерпывающую часть этого обоснования составля-
ет тот простой факт, что легко распознаваемые и определяемые
группы организмов действительно существуют в природе. Несмот-
ря на сомнительные случаи и мириады сложностей, современному
учёному, так же как и доисторическим индейцам гуарани, совер-
шенно очевидно, что естественные виды действительно существу-
ют. Теоретическая проблема, таким образом, заключается в вопро-
се, почему они существуют, а решение этой проблемы должно дать
критерии для определения того, соответствует ли некий таксон ес-
тественной группе. Общее решение сегодня известно, хотя оно не-
сёт с собой множество частных проблем, одни из которых уже ре-
шены, а другие нет. Виды существуют, поскольку они эволюирова-
ли. Такова, в самой сжатой форме, естественная причина существо-
вания видов и, тем самым естественная основа для классификации.
Естественное существование высших таксонов интуитивно менее
очевидно, что и служит причиной всех самокритичных брожений
таксономистов в поисках ответа на вопрос, являются ли такие так-
соны «реальными» или «естественными». Однако эволюция даёт
начало также и высшим таксонам: эта идея служит путеводной ни-
тью при распознавании естественных таксонов на всех уровнях.
68
Дж. Г. Симпсон. Принципы таксономии животных
Вкратце, если нечто, именуемое естественной классификацией,
и может быть достигнуто на разумных основаниях, то это должна
быть эволюционная классификация.
Некоторые формальные аспекты отношений
Разные подходы к классификации связаны с разными способа-
ми формального определения отношений, их восприятия и визуа-
лизации. В настоящем разделе я очень кратко рассмотрю некото-
рые из этих способов и наиболее общие методы преобразования и
представления отношений в форме иерархии.
Отношения вообще и связи по сходству в особенности бывает
полезно разложить на отдельные составляющие. Присутствие ка-
кого-то одного из единичных отношений можно представить как
линию, соединяющую две точки, которые соответствуют либо от-
дельным организмам, либо низшим таксонам. Когда рассматрива-
ется много точек, результатом оказывается нечто вроде сети, свя-
зывающей одни точки и не связывающей другие (рис. 1А). Если
точки соответствуют организмам, множество связанных точек мо-
жет рассматриваться как таксон Т если они соответствуют таксо-
нам Т , получаем таксон Т . Использование многих единичных
отношений позволяет связать все точки: одни — единственной ли-
нией, другие — двумя, тремя, четырьмя и т.д. (рис. 1Б). Множества
точек, связанных большим числом линий, могут считаться низши-
ми таксонами; если точки связаны меньшим числом линий, соот-
ветствующие множества представляют высшие таксоны. Иллюст-
рацией служит рис. 1Б, который показывает, что при определённых
допущения это действительно возможно, но также и то, что иног-
да возможны неожиданные и сомнительные результаты. Например,
точка с наиболее тесно связана с а и Ь, однако между самими а и
Ь связей по рассматриваемым атрибутам нет. Данный подход явля-
ется типологическим: в таких аномалиях (а могут быть и значи-
тельно более сложные), которые действительно возникают при
классификации реальных организмов, проявляется один из дефек-
тов подобной неприемлемой системы. Тем не менее, использова-
ние сети для формализации отношений предложено в таксономии
достаточно давно и по-прежнему служит для систематизации (но
Развитие современной систематики
69
Рис. 1. Диаграммы отношений
Точки, обозначенные буквами, служат представлением индивидов или
низших таксонов.
А. Представление однократного единичного отношения, объединяющего а,
af, е и/в топологический таксон.
Б. Представление многократных единичных отношений; читатель может на
этой основе сам определить таксоны.
В. Диаграмма отношения «группа в группе». Точки представляют особей,
малые окружности — таксоны Тр большие окружности — таксоны Т2,
наибольшие окружности — таксон Тг
не для прямой интерпретации) некоторых данных, таких как ре-
зультат перекрёстной гибридизации генетических популяций.
Другой способ представления такого рода данных гораздо тес-
нее и операциональнее связан с иерархией и по этой, равно как и
по иным причинам имеет значительно более широкое применение.
Он является и более гибким, позволяя представлять более разно-
70
Дж. Г. Симпсон. Принципы таксономии животных
образные отношения. Имеется в виду формализация «группа в
группе» [18], которую легко представить набором кругов или ящи-
ков разного размера и разной степени включённости один в дру-
гой (рис. 1в). Например, окружности одного размера могут отож-
дествляться с таксонами одного ранга, а их включение в окружно-
сти большего размера — с включением в таксоны более высокого
ранга. В терминах типологии, окружности представляют архетипы
возрастающего уровня общности или абстрактности. (Как уже под-
чёркивалось, произвольная оценка приоритетности разных атрибу-
тов [в отношении первоочередности их использования в классифи-
кации], может привести к совершенно разной упорядоченности
одних и тех же данных.) В контексте ранних эволюционных кон-
цепций окружности очерчивают совпадающие признаки, унаследо-
ванные от общего предка: наибольшая окружность соотносится с
признаками, доставшимися от наиболее отдалённого предка и объе-
диняющими всех индивидов, а меньшие окружности соответству-
ют признакам, совпадающим у индивидов в пределах каждой та-
кой окружности и унаследованным от их более близких предков. В
силу аддитивности связей число совпадающих признаков (прибли-
зительно) обратно пропорционально размеру окружностей: инди-
виды в малой окружности обладают не только теми признакам, ко-
торые соотносятся с данной окружностью, но также и теми, кото-
рые соответствуют окружностям, включающим данную. Если пол-
ностью порвать с типологическим мышлением, то окружности бу-
дут указывать происхождение от общего предка, отдалённость ко-
торого приблизительно пропорциональна размеру окружности, од-
нако в данном случае окружности уже не дают прямого указания
на число совпадающих признаков. Остаётся лишь некая статисти-
ческая связь с многими частным исключениями, означающая, что
филогенетически более близкие группы в среднем обычно имеют
больше совпадающих признаков: этот совершенно правильный
вывод часто бывает завуалирован ложными посылками типологи-
ческого подхода.
Третья формализация, заслуживающая упоминания, и проще, и
старше двух предыдущих: это линия, исходно призванная представ-
лять scala naturae (рис. 2). В наиболее явном виде, который отста-
ивал, например, Бонне (Bonnet, 1745), предполагается единствен-
Развитие современной систематики
71
ная линия, неветвящаяся и без разрывов, вдоль которой плотно рас-
пределены организмы: она идёт, согласно системе Бонне, от чело-
века на вершине к «косной материи» (практически совершенно бес-
форменной) у основания. Эта система принципиально нетипологи-
ческая, а её теоретическая связь с иерархией вполне условна, но
столь же ясная и простая, как и типологическая «группа в группе».
Для преобразования в иерархию непрерывная линия произвольно
И без остатка делится на небольшие сегменты, соответствующие
таксонам Тр которые затем последовательно объединяются в не
Рис. 2. Диаграмма (по Боннэ) непрерывной scala naturae и её
произвольное разделение на таксоны последовательно возрастающих
РАНГОВ
72
Дж. Г. Симпсон. Принципы таксономии животных
менее искусственные более крупные сегменты Т2, Т и т.д. (также
показанные на рис. 2). На основе этой scala naturae была вырабо-
тана не вполне эволюционная концепция, которая, подобно типо-
логии, была основана на выборе и неверной интерпретации реаль-
ных явлений, действительно имеющих эволюционную причину
Эта формализация, поэтому, может быть непосредственно преобра-
зована в части истинно эволюционной системы: она достаточно
адекватно представляет единую временную линию популяций, свя-
занных отношением «предок—потомок». Изложение её в терминах
иерархии возможно посредством того же условного приёма наре-
зания сегментов различного объёма.
Этот момент будет рассмотрен далее, однако уже здесь имеет
смысл хотя бы упомянуть, что такое произвольное разбиение не
обязательно приводит к таксонам, которые «нереальны» или «не-
естественны», как иногда утверждают. Простой, но достаточно оче-
видной аналогией в данном случае может служить цепочка бус,
цвет которых плавно меняется, скажем, от синего на одном конце
до зелёного на другом. Разрезание цепочки на два куска — впол-
не произвольный акт, однако получившиеся фрагменты являются
совершенно реальными, естественными частями целого, которое
они до того составляли. Более того, эти два сегмента реально раз-
личаются «средней» окраской и в этом смысле могут быть опреде-
лены естественным образом.
Имеется ещё несколько способов визуализации разных отноше-
ний,10 однако здесь мы ограничимся рассмотрением дерева. Это
широко известная структура со многими ветвящимися в восходя-
щем найравлении линиями. Такой способ визуализации отношений
использовался в работах некоторых доэволюционных авторов, та-
ких как Бюффон [19], но более широко он применяется эволюци-
онистами, поскольку деревья, по-видимому, более всего подходят
для представления эволюционных отношений. Самая ранняя [сре-
ди эволюционистов] древовидная диаграмма была опубликована
Ламарком (Lamarck, 1809), а в «Происхождении видов» Дарвина
она уже единственная (Darwin, 1859). В таком применении дерево
10 Например, перекрывающиеся области для представления ключа или нис-
ходящие ветвящиеся линии для представления родословной.
Развитие современной систематики
73
служит средством представления, по необходимости в схематичной
и незавершённой форме, действительной структуры филогенеза.
Ветви соответствуют филогенетическим линиям, а вертикальная со-
ставляющая — времени. Горизонтальный срез даёт группы, суще-
ствующие в данный момент времени. Таким образом, срез служит
представлением картины, которая может быть более или менее
адекватно показана также диаграммой «группа в группе», а само
Рис. 3. Диаграммы в форме деревьев
А. Гипотетическая филогения. Все сплошные линии составлены
исключительно из видов, каждый из которых имеет протяженность (во времени)
в пределах, обозначенных разделяющими их поперечными штрихами.
Штриховые линии указывают один из нескольких способов объединения видов
в роды.
Б. Дендрограмма без временной размерности. Концевые точки представляют
одновременно существующие виды, также без указания их временной
протяженности. Линии под точками не представляют виды, но формализуют
отношения между видами, представленными точками. Пронумерованные шаги
могут быть использованы при объединении видов в филогенетически
осмысленные высшие таксоны. Хотя эта дендрограмма не является филогенией,
она топологически идентична филогении, показанной на (А), исключая
отсутствие тех вымерших видов, после которых не осталось потомков, поскольку
их существование нельзя указать на основе анализе современных данных.
В. Упорядочение в форме «группа в группе» терминальных видов из (А)
или видов из (Б).
74
Дж. Г. Симпсон. Принципы таксономии животных
дерево даёт эволюционную основу и интерпретацию этой картины.
Эти группы могут быть определены в терминах общего происхож-
дения, показанного на более глубоком уровне дерева, а их иерар-
хические ранги соответствуют (приближённо и с некоторыми бе-
зусловными ограничениями, обсуждаемыми далее) относительной
удалённости общих стволов (рис. ЗА).
Наряду с этим дерево можно использовать для представления
отношений, которые не предполагают филогенетического объясне-
ния, даже для представления строго типологического сходства (см.,
например, Michener, 1957). Такие отношения определяются как
«группа в группе», а затем их подразумеваемые объединения в
стволы, от которых они ответвляются, упорядочиваются таким об-
разом, чтобы отразить иерархические ранги, приписанные таким
образом выделенным группам (рис. ЗБ). В самой простой форме
подобная диаграмма на своих концах представляет таксоны, тогда
как ведущие к ним линии не соответствуют таксонам, но лишь
формализуют выявляемые отношения. Майр с соавт. (Мауг et al.,
1953), кратко, но вполне исчерпывающе обсудившие этот предмет,
предложили для таких деревьев термин дендрограмма." Трактов-
ки дендрограмм варьируют между двумя крайностями: они либо
вообще не имеют отношения к филогении, либо они достаточно
хорошо отражают филогению. Отдельные примеры вполне нагляд-
но воспроизводят каждую из этих точек зрения, однако их ни в
коем случае не следует рассматривать слишком буквально. Денд-
рограмма формализует результаты филогении и едва ли совершен-
но свободна от неё как от причины [этих результатов]. С другой
стороны, дендрограмма, по определению, представляет только вы-
веденную, а не наблюдаемую филогению, и потому вряд ли служит
точным представлением филогении.
11 Они различают дендрограммы и филогенетические деревья или диаграм-
мы, однако отличия не вполне ясны, поскольку их дендрограммы также могут
иметь филогенетический смысл. Кроме того, слово «дендрограмма» этимоло-
гически пригодно для всех древовидных дендрограмм, а различия между теми,
которые предполагают филогенетическую интерпретацию, и теми, которые не
предполагают этого, лучше обозначать как-то по-другому. Однако я не вижу
причины усложнять предмет введением дополнительных терминов [20].
Развитие современной систематики
75
Новая систематика и после неё
Общий интерес к зоологической систематике, который был до-
статочно активным начиная с XVII в. и вплоть до XIX в., стал за-
метно убывать к концу XIX — началу XX вв. В некоторых цент-
рах она стала приравниваться к простой идентификации и этике-
тированию экземпляров, а после первого удивительно плодотвор-
ного соединения таксономии с эволюционными идеями развитие
таксономических принципов дало совсем немного из того, что
можно было бы счесть новым и важным. В самом деле, некоторые
таксономисты, по-видимому, чувствовали, что принципы разрабо-
таны достаточно полно и осталась лишь необъёмная, но скучная
работа по каталогизации. Наметилась тенденция оттока блестящих
молодых умов в новые и, как становилось всё очевиднее, увлека-
тельные области биологии, особенно в генетику и биохимию.
Где-то в 1930-е гг. эта тенденция обратилась вспять. Глубинные
идеи систематики, которые, по счастью, никогда не переставали
занимать таксономистов, стали привлекать всё большее внимание.
Всё более общим становилось понимание того, что систематика
затрагивает и обусловливает новые разделы биологии не в меньшей
степени, чем старые, и что она сама — увлекательный и достой-
ный внимания источник идей и средств в достижении синтеза. Не-
которые систематики чувствовали, что таксономия вовсе не распо-
лагает адекватным устоявшимся набором принципов, но нуждает-
ся в радикальной теоретической ревизии. Другие систематики не
были согласны с этим, но полемика (которая всё ещё не заверше-
на) сама по себе оказывала стимулирующее воздействие.
Одним из поворотных пунктов в этом движении, который под-
вёл итог предшествующим изменениям и обусловил дальнейший
прогресс, стал симпозиум по новой систематике (Huxley, 1940),
который дал название самому движению.12 Правда, только лишь на
основании названной книги едва ли можно составить определённое
представление о том, что нового в этой систематике, или извлечь
формулировку её принципов, однако она ясно показала, что в этой
12 Хотя выражение «новая систематика» было прежде использовано Хаб-
бсом и, возможно, другими.
76
Дж. Г. Симпсон. Принципы таксономии животных
области существует некое брожение умов, и обозначила то общее
направление, в котором надлежит двигаться таксономии. Теперь, два
десятилетия спустя, эпитет «новая» стал весьма относительным, а
систематика в целом и таксономия в частности прошли значитель-
ный путь от состояния, достигнутого к 1940 г. Впрочем, «новое»
всегда относительно, ибо наука всегда меняется, но при этом каж-
дое новшество всегда уходит корнями глубоко в прошлое.
В настоящее время можно составить некий перечень основных
концепций и принципов современной формы новой систематики,
или, быть может, того, что можно назвать «новой новой система-
тикой». Здесь это будет сделан кратко и бегло. Отличия от прежней
систематики (которая, тем не менее, является источником новой) в
основном очевидны из того, что было сказано ранее в этой главе.
Развитие этих принципов и разбор проблем по их применению бу-
дут представлены в последующих разделах книги.
Популяции, а не организмы, являются единицами систематики
и именно они классифицируются. Все популяции изменчивы и эта
изменчивость является существенной частью их природы и их оп-
ределения. У них нет единственного фиксированного образа или
типа. Типы в совершенно ином смысле сохранены только в целях
нормативного регулирования номенклатуры.
Популяции — динамические системы, которые эволюируют как
путём постепенных изменений в собственных границах, так и за
счёт диверсификации и разделения на несколько новых отдельных
систем. Фундаментальной единицей эволюции является вид. Эта
категория- не может быть естественно определена в терминах ста-
тического образа или морфологии, но только в терминах динами-
ческой, эволюционной, генетической концепций, описывающих
отношения между популяциями. Изучение индивидуальной морфо-
логии или других соматических атрибутов важно, но не само по
себе, а для того, чтобы свидетельствовать о том, что данная попу-
ляция соответствует эволюционному определению.
Таксономические исследования всегда статистические по сво-
ей природе. Истинный предмет изучения — природную популя-
цию — редко можно наблюдать непосредственно и во всём объё-
ме. Процедура состоит в умозаключениях, направленных от стати-
стической выборки экземпляров к популяции. В частных случаях
Развитие современной систематики
77
используются статистические методы и параметры, в той мере, в
какой они пригодны и эффективны. Но даже если они не исполь-
зуются, рассмотрение остаётся статистическим по сути.
Таксоны на всех уровнях в принципе определяются не членством,
а связями. Т.е. их нельзя определять исключительно указанием ин-
дивидов, которые к ним относятся или перечислением признаков
этих индивидов, но только указанием явных или подразумеваемых
связей между этими индивидами. Совпадающие признаки и всеоб-
щее сходство не могут играть ведущую роль, но рассматриваются,
о чём было сказано, как свидетельства отношений, которые перво-
степенны. Связи, о которых идёт речь, — эволюционные или, в более
широком смысле (заслуживающем дальнейшего обсуждения), фи-
логенетические. Ранги одновременно сосуществующих подмножеств
Т зависят главным образом, но не исключительно, от удалённости
их общих предков относительно таковых других подмножеств или
тех множеств Т, к которым они принадлежат. Среди других прини-
маемых во внимание суждений, также вполне эволюционных, но
филогенетических только в широком смысле, значимы степень обо-
собленности и разнообразия членов исследуемых таксонов.
Надвидовые таксоны разграничиваются на основании принципа
монофилии: все члены таксона имеют общее филогенетическое
происхождение. Походящий и вполне практический критерий един-
ства происхождения — не возникновение от одного индивида,
пары индивидов, вида или рода, но возникновение от предкового
таксона такого же или более низкого ранга.
Все таксоны обладают временной размерностью, что должно
предусматриваться любым их определением или иным рассмотре-
нием. Для целей формальной классификации непрерывная после-
довательность меняющихся популяций может быть произвольно
поделённой на ранние и более поздние таксоны одного или разных
рангов, однако такие таксоны, хоть и разграничены произвольным
способом, не являются ipso facto неестественными.
Разработка формальных классификаций отдельных групп состав-
ляет существенную часть и является нужным результатом таксоно-
мических изысканий, однако не может считаться единственной или
даже основной их целью. Цель таксономии — понимание биологи-
ческого смысла группировок и отношений между организмами.
7 — 4338
3
Таксономические доказательства
и их эволюционная интерпретация
Современная таксономия является эволюционной, и в
её базис входит филогения, которую невозможно непосредственно
наблюдать; зачастую заключения о ней делаются на основании не-
палеонтологических данных, лишённых существенного временного
измерения. Доказательства того, что таксономические принципы
соблюдены и что классификация филогенетически состоятельна,
таким образом, отличны от [логических] определений (definitio) так-
сономических единиц. Данные, используемые в качестве доказа-
тельств, всё ещё по преимуществу анатомические, но они расши-
ряются за счёт многих других разделов [биологии], наиболее важ-
ные из которых будут указаны и проиллюстрированы в настоящей
главе. Они включают практически всё, что может быть известно о
животном.
Для выделения эволюционных таксонов решающим является
близость по родству, о которой судят главным образом по сходству
между ними. Вместе с тем, сходственные отношения могут быть
разными. Они могут быть связаны с гомологией, параллелизмом,
конвергенцией, мимикрией или быть случайными. Все эти причи-
ны должны быть определены, а затем их следствия должны быть
интерпретированы в терминах эволюционной таксономии. Основ-
ной проблемой является различение между гомологией, которая
отражает родственную близость, и конвергенцией, которая этого не
Таксономические доказательства и их эволюционная интерпретация 79
отражает. Поэтому критерии для распознавания конвергенции зас-
луживают более детального рассмотрения.
В филогенетических и иных таксономических исследованиях
учитываются не только совпадающие признаки, но и последова-
тельности варьирующих признаков в пределах групп. В данном
случае основная проблема состоит в том, чтобы в некоторой пос-
ледовательности уметь различать примитивные и специализирован-
ные признаки и связывать их с [большей или меньшей] близостью
предковых форм.
Наконец, следует установить природу параллелизмов: как будет
выяснено, для интерпретации этого феномена нет нужды в каких-
то действительно важных дополнительных принципах.
Определение и доказательство
Принципы современной таксономии являются эволюционными,
соответственно чему принятый здесь подход к классификации яв-
ляется эволюционным или, в некотором специальном смысле, фи-
логенетическим. Многие эволюционные процессы можно наблю-
дать в действии, как в природе, так и в лаборатории, что также вер-
но и в отношении минимальных сегментов филогении. Эти корот-
кие сегменты имеют большое значение в качестве примеров и для
разработки обоснованных принципов, однако они имеют лишь не-
значительное практическое приложение для классификации выше
некоторого минимального таксономического уровня. Даже протя-
жённые временные серии, известные для многих ископаемых
форм, являются филогенетическими только по предположению:
происходящие в действительности процессы размножения и проис-
хождения не наблюдаемы. (Даже в наиболее детально отслеженных
рядах в высшей степени маловероятно, что предшествующая вы-
борка включает всех или большинство индивидуальных предков
особей последующей выборки.)
Таким образом, истина состоит в том, что эволюционная клас-
сификация в значительной мере полагается на концепции и опре-
деления, для которых исходные данные не могут непосредственно
наблюдаться. Впрочем, это свойственно не только таксономии. Это
в большей или меньшей степени — общая черта всех индуктивных
т
80
Дж. Г. Симпсон. Принципы таксономии животных
наук. Однако по этой причине они не могут считаться менее науч-
ными, а их заключения не обязательно менее определённы, чем в
случае прямых наблюдений. Аналогичным образом, хотя и по
иным причинам, атомная физика имеет дело с объектами, которые
никогда не наблюдались непосредственно, но никто не ставит под
сомнение обоснованность или практичность интерпретаций в тер-
минах частиц и процессов, известных только по умозаключениям.
Сами учёные часто не представляют, какой степени, а подчас
даже какая именно «определённость» (на самом деле это всегда
вероятность) требуется и возможна в науке. Так, Thompson (1952),
способный таксономист, отверг бы саму возможность определения
эволюционной гомологии и, тем самым, существование такой вещи
как эволюционная классификация. «Кто может сказать, что перед-
няя конечность летучей мыши была когда-то чем-то иным, нежели
крыло?» Ответ состоит в следующем: если мы не можем утверж-
дать это с вероятностью столь высокой, что она равна полной оп-
ределённости, то для практических целей мы не сможем обосновать
никакие биологические принципы, представляющие действитель-
ный интерес, и должны отбросить науку, вернувшись к анимизму.
Здесь необходимо ещё раз подчеркнуть различие между опре-
делением и тем доказательством, которое делает это определение
соответствующим [данному случаю]. Мы предлагаем определять
таксономические категории в эволюционных и, более широко, фи-
логенетических терминах, но при этом использовать доказатель-
ства, которые в качестве отдельных наблюдений почти исключи-
тельно, не филогенетические. Несмотря на значительную путаницу
в этом вопросе даже среди некоторых таксономистов, на самом
деле здесь нет особой сложности или таинственности. Пояснени-
ем служит хорошо известный пример однояйцовых («идентичных»)
близнецов, который является больше чем аналогией. Мы определя-
ем таких близнецов как организмы, развившиеся из одной зиготы.
Ни один такой случай в действительности не наблюдался у чело-
века, однако мы распознаём их, если нашему определению соответ-
ствуют доказательства, получаемые по сходству, достаточному для
данного заключения. Эти организмы не потому близнецы, что они
сходны, но наоборот, они сходны, потому что они близнецы. Точ-
но так же, организмы относятся к одному таксону не потому, что
Таксономические доказательства и их эволюционная интерпретация 81
они сходны, но они сходны, потому что относятся к одному таксону
(Линней был совершенно прав, говоря, что род задаёт признаки, а
не наоборот, хотя он и не знал, что именно задаёт род). Это утверж-
дение — центральный элемент эволюционной таксономии, а аль-
тернативная трактовка чётко отграничивает её от неэволюционной
таксономии. Другим способом выражения этой идеи будет утвер-
ждение, что категории определяются в филогенетических терминах,
но что таксоны определяются некими соматическими отношения-
ми, которые возникают в результате филогении и служат доказа-
тельством, что определение категорий состоятельно.
Наиболее резкие возражения против эволюционной таксономии
были связаны с неверным пониманием её принципов и использо-
ванием «соломенного чучела» [т.е. не относящихся к делу фактов
и суждений]. Так, всё тот же Томпсон (Thompson, 1952) полагает,
будто бы эволюционная таксономия утверждает, «что вид — пре-
док организмов, род — предок видов, а семейство — предок ро-
дов.» Это, как намекает Томпсон, бессмыслица, но данная идея
никоим образом не относится к принципам эволюционной таксо-
номии. Ещё одно возражение состоит в том, что филогенетическая
таксономия якобы основана исключительно на отношениях между
предками и потомками, однако филогенетики, конечно же, знают
(в несколько иных терминах), что братья столь же близки генети-
чески, как отцы и сыновья. Другие критики справедливо утверж-
дают, что классификация не может полностью выражать филоге-
нию, однако, если оставить в стороне некоторые вольности языка,
ни один зрелый филогенетик и не думает, что это возможно. Но
лучшим ответом тем, кто критикует эволюционную классификацию
за то, чем она не является, будет объяснение того, что она представ-
ляет собой на самом деле.
Первый шагом должно быть рассмотрение категорий доказа-
тельств, на основании которых в эволюционной таксономии могут
быть эффективно получены таксономические заключения. Наблю-
дение проводятся над экземплярами, но под этим могут и, насколь-
ко это осуществимо, должны подразумеваться целостные индиви-
ды совместно с их естественной средой обитания, как биотической,
так и физической. Что именно здесь имеется в виду, будет рассмот-
рено и проиллюстрировано в следующих разделах.
82
Дж. Г. Симпсон. Принципы таксономии животных
Категории доказательств
Классификация изначально развивалась и по сей день остаётся
в тесной связи с музеями или, по крайней мере, с коллекциями за-
консервированных экземпляров. Лишь в последнее время благодаря
всё более широкому развитию систематики она частью становится
экспериментальной и полевой дисциплиной. Таким образом, дан-
ные, используемые систематикой, всегда в основе своей, хотя и не
исключительно, были связаны с возможностью исследования этих
сохранённых экземпляров. Для мягкотелых беспозвоночных и неко-
торых других групп это в общем случае означает тотально фиксиро-
ванный экземпляр со всей его анатомией, но ничего сверх этого.
В некоторых группах сохраняется и того меньше: для млекопитаю-
щих обычно шкурка и череп, для птиц — только внешние части.
Для ископаемых остатков, за небольшим числом исключений, в до-
ступности были и остаются лишь твёрдые части, причём чаще все-
го (особенно среди наземных позвоночных) только их фрагменты.
По этим чисто практическим причинам данные для классифи-
кации были преимущественно анатомическими или морфологиче-
скими,1 используя обычно лишь ограниченную часть того, что пред-
ставляет анатомия. Распространённым было мнение, что если от-
дельные исследуемые анатомические части достаточно сложны, то
при условии, что они могут быть адекватно описаны, основанные
на них классификации будут в основных чертах такими же, как и
основанные на других анатомических или неанатомических призна-
ках. Например, это было показано для твёрдых частей в противовес
шкуркам при изучении низших таксонов млекопитающих (Simpson,
1943). В определённых группах опытные таксономисты выяснили,
что отдельные части, такие как гениталии у некоторых насекомых,
вполне доступны и адекватны для их целей, тогда как другие части
едва ли нужно исследовать. Такого рода ограниченность данных,
1 Строго говоря, необходимо чётко различать анатомию, связанную с на-
блюдением и описанием структур организма, и морфологию — теоретическую
дисциплину, обобщающую и объясняющую эти наблюдения. Это различие ана-
логично таковому между классификацией и таксономией, принятому в насто-
ящей книге. Однако многие исследователи, в том числе иногда и я сам, чаще
всего используют анатомию и морфологию как взаимозаменяемые понятия.
Таксономические доказательства и их эволюционная интерпретация 83
однако, оправдана лишь для близко родственных групп, таких как
подвиды одного вида или виды одного рода, да и то не всегда. Для
более высоких категорий всегда желательно и даже необходимо
больше данных. Например, виды уток достаточно хорошо разрабо-
таны по внешним признакам, однако рассмотрение других призна-
ков заметно изменило надвидовую систему и значительно улучши-
ло её эволюционную основу (Delacour, Мауг, 1943).
Именно следствием того факта, что классификации подлежат
организмы, а не их признаки, является требование изучение всего
организма, всех его частей и аспектов. Конечно, имеются практи-
ческие ограничения на возможности исследований. Не всё ещё
мало известно даже о наиболее изученном виде — Homo sapiens.
Большинство сведений, которые были бы крайне полезны, всегда
останутся недоступными для вымерших видов. Тем не менее, ак-
сиома современной таксономии состоит в том, что разнообразие
данных должно быть доведено, насколько это возможно, до преде-
лов осуществимости. Объектом классификации должно быть то,
что Генниг (Hennig, 1950)2 назвал голоморфой — совокупностью
всех характеристик индивида на протяжении всей его жизни.
Первый шаг состоит в расширении анатомических данных за
пределы обычных традиционных форм препаровки музейных эк-
земпляров, например, у млекопитающих — кость полового члена
2 Здесь я многим обязан книге Hennig (1950), которая определённо одна из
наиболее значительных, когда либо публиковавшихся по таксономии (как она
здесь определена). Мой подход иной и я развил многие из моих ключевых воз-
зрений (которые я вовсе не хочу приписать исключительно себе) прежде, чем
с явным опозданием прочитал эту книгу. Согласие, конечно, лишь частичное,
но оно весьма существенно. Достойно сожаления, что работа Геннига не ста-
ла доступной англоязычным учёным [21], равно как и то, что в ней нет указа-
теля и словаря сложной терминологии. Наиболее общее возражение, при этом
взаимное, состоит в том, что Генниг, по-видимому, почти не знал огромного
числа исследований в Англии и Америке, которые имеют непосредственное
отношение к его теме. Это проявляется и во мнении ученика Геннига — Ки-
рьякова (Kiriakoff, 1959), что «современная филогенетическая систематика, по-
видимому, совершенно неизвестна в Соединённых Штатах.» Подобное утвер-
ждение возможно лишь вследствие значительного пробела в знаниях. В Соеди-
нённых Штатах, как и в Европе, есть неотипологи, однако ими далеко не ис-
черпываются ни американские таксономисты, ни их воззрения.
84
Дж. Г. Симпсон. Принципы таксономии животных
(например, White, 1953), слуховые косточки (например, Krutzsch,
1954) и, конечно, мягкие ткани (например, Mossman, 1953). Второй
шаг — перенести анатомические наблюдения на более глубокий
уровень цитологии и особенно кариологии (анатомия ядра и в ча-
стности хромосом), значение чего подчёркивал Кэйн (Cain, 1958b),
а примеры можно найти в: Christensen, Nielsen (1955), Matthey
(1952), Patterson, Stone (1952) и многих других.
Другой аспект голоморфы состоит в том, что она имеет времен-
ную размерность. Значение эмбриологии, к чему мы сейчас призы-
ваем систематику, было понято очень давно и, например, доэволю-
ционная работа фон Бэра (Ваег, 1828) до сих пор остаётся осново-
полагающей. Эволюционная интерпретации онтогении, которую
дал Геккель (например, Haeckel, 1875), в своё время казалась бе-
зошибочным путеводителем для реконструкции филогении. К сожа-
лению, она оказалась истинной далеко не во всем и вызвала разо-
чарование (Garstang, 1922). Тем не менее, развивающийся организм
составляет часть голоморфы, а эмбриология остаётся очень важной
частью эволюционных механизмов (см. особенно работу Уоддин-
гтона (Waddington, 1957), хотя и не таксономическую, но показы-
вающую эволюционное значение [эмбриологии]). И ныне эмбрио-
логия продолжает, причем в более зрелой форме, обеспечивать су-
щественными данными эволюционную таксономию. (Это хорошо
показано в: Orton, 1953, 1955 и в ссылках в этих работах).
Физиологическая информация непосредственно недоступна при
изучении музейных экземпляров и потому обычно выпадала из
данных для классификации. Тем не менее, некоторые систематики
прежде использовали физиологию на некой более широкой осно-
ве — например, Кювье и другие, которые пытались установить ап-
риорные критерии важности признаков, о чём шла речь в преды-
дущей главе. Дарвин (Darwin, 1859) сильно сомневался в пригод-
ности физиологии для суждения о родстве, и для этого у него были
достаточные основания, поскольку априорные системы обычно
выпячивают физиологические сходства, которые (в современных
терминах) особенно подвержены конвергенции. Имеется некое
смутное ощущение, что физиологическое сходство следует в целом
рассматривать как аналогичное, а потому (как обсуждается далее)
ненадёжное или вводящее в заблуждение при классификации, и что
Таксономические доказательства и их эволюционная интерпретация 85
только анатомические сходства можно обозначать как гомологич-
ные. На самом деле трудность, как и в случае морфологии, состо-
ит в трактовке немногих простых или генерализованных характе-
ристик, которые часто бывают аналогичными, а не более сложных
и специфичных физиологических черт, которые могут быть таки-
ми же истинными и доказательными гомологиями, как и анатоми-
ческие особенности. Представляется весьма очевидным, что физи-
ологическая функция и анатомическая структура должны быть и
действительно обычно бывают тесно скоррелированы. На этом ос-
новании Блэквельдер (Blackwelder, 1959) заключил, что в таких
частых случаях необходимо исследовать лишь какую-то одну из
этих двух категорий данных. Однако, по-видимому, обе они суще-
ственны и настолько неразделимы, что невозможно понять одну из
них, не анализируя другую.
Более того, имеются многочисленные физиологические данные,
высоко значимые для таксономии, которые могут быть описаны
только в их собственных терминах, а не ссылкой на какую-либо
структуру. Сюда, если понимать «физиологию» в широком смысле,
относятся данные сравнительной биохимии, значение которых в
таксономию всё более возрастает. В этой области наибольшее зна-
чение имеет данные по антигенной реакции на чужеродные белки,
или систематическая серология (например, Boyden, 1953,1958), хотя,
как и во многих других подобных подходах, здесь есть некоторые
всё ещё нерешённые проблемы. Ещё один новый метод получения
сравнительных данных — электрофорез белков и гемоглобина (John-
son, Wicks, 1959, и ссылки в этой работе). Другие современные
биохимические исследования, представляющие специальный инте-
рес для таксономии, связаны, например, с анализом ферментов (Kaplan
et al., 1960) и солей жёлчных кислот (Haslewood, 1959а, Ь)? [22]
3 Указанные исследования выполнены на многоклеточных, где биохимичес-
кие данные дополняют обширный корпус других данных и способствуют луч-
шему пониманию синтетической эволюционной таксономии. Для некоторых
простейших классификация в настоящее время основывается исключительно
на биохимических данных и с такой ограниченной анатомией, что она исполь-
зуется только для дополнительной характеристики. Для них эволюционная
классификация ещё разрабатывается, хотя не исключена по мере роста знаний.
Они в этой книге не рассматриваются.
86
Дж. Г. Симпсон. Принципы таксономии животных
Выше я указал такие данные, которые могут быть получены в
лаборатории при обследовании отдельных организмов. Другой под-
ход, в ограниченной мере и прежде использовавшийся в классифи-
кации, но более свойственный современной таксономии, связан с
«реляционными» данными: наблюдениями над отношениями орга-
низмов между собой и со средой обитания. В минимальном объё-
ме такие дополнительные данные всегда включали сведения о гео-
графическом и геологическом распространении. Поначалу крайне
расплывчатые, они становились всё более и более точными. Их
важность подчёркивается популяционным подходом, поскольку
популяция — это группа организмов, живущих или живших совме-
стно в конкретном пространстве и времени. Эти отношения могут
быть адекватно представлены только на основе полевых данных.
В палеонтологии пионерским исследованием в этой области, у ко-
торого теперь много последователей, была работа Бринкмана
(Brinckman, 1929), проследившего сантиметр за сантиметром изме-
нения в популяциях аммонитов сквозь вертикальную последова-
тельность пород.
Данные по пространственному распространению не только по-
мещают животных в строго локализованные популяции и биотопы,
но также в более широкие зоогеографические рамки. Их значи-
мость для классификации видов и подвидов блестяще показана
Майром (Мауг, 1942), а для высших категорий — Дарлингтоном
(Darlington, 1957). Такие данные широко используются, например,
при распознавании конвергенций (см. далее), которые наиболее
вероятны в группах с широко разобщёнными или изолированными
ареалами. Например, южноамериканские аборигенные грызуны
(«гистрикоморфы» или кавиоморфы) очень сходны с некоторыми
африканскими формами, а некоторые вымершие южноамериканс-
кие сумчатые (боргиениды) похожи на австралийских сумчатых
волков (Thylacynus). Такие сходства рассматривались как показа-
тель особого родства, пока не были получены окончательные до-
казательства глубокой изоляции Южной Америки, решительно ука-
зывающие на конвергентный характер сходства. Позже переиссле-
дования самих этих организмов подтвердили данный вывод (напри-
мер, Wood, 1950 и Haslewood, 1959b — по грызунам; Simpson,
1941 — по сумчатым).
Таксономические доказательства и их эволюционная интерпретация 87
Экологические данные также всегда вовлекались в таксономию
в широких масштабах, а в настоящее время их вклад становится
всё более важным и строгим. Например, сейчас установлено в виде
принципа, что, за немногими ожидаемыми исключениями, виды,
пребывающие в одной пространственно-временной точке (безмер-
ные [nondimensional] виды, как их назвал Майр), экологически раз-
личны. Каждый в общем случае занимает отдельную обособленную
нишу и два и более близких видов обычно довольно чётко разли-
чаются. На этой основе были предложены критерии для разграни-
чения внутривидовых и межвидовых вариаций у животных, как
современных (например, Мауг, 1942), так и ископаемых (например,
Simpson, 1943). Экологические данные дают критерии для распоз-
навания прежде всего низших, но также и высших эволюционных
таксонов. Они также имеют значение и для других таксономиче-
ских проблем, таких как жизненно важная проблема конвергенции,
которая означает экологическое сходство, но филогенетическое раз-
личие между конвергентными группами. (См. историю вопроса: по
современным животным — Allee et al., 1949; по современным и
ископаемым морским животным — Hedgpeth, Ladd, 1957; по иско-
паемым животным — Cloud, 1959).
Среди многих специфических экологических взаимоотноше-
ний, рассматриваемых в современной таксономии, здесь будут ука-
заны только два. Хотя многие животные, особенно насекомые, об-
ладают выраженной пищевой специализацией, она не может харак-
теризовать таксоны разных уровней, несмотря на подверженность
эволюционным изменениям (имеется обзор: Detheir, 1954, с обшир-
ной библиографией). Отношения между хозяевами и их паразита-
ми часто столь резко специфичны, что представляют хорошие так-
сономические данные как для тех, так и для других. Неоднократ-
но утверждалось, что близкие группы паразитов обычно эволюи-
руют совместно с близкими же группами хозяев и вледствие это-
го совпадают с ними. Когда это имеет место, а это с очевидностью
показано в некоторых случаях, одна из групп служит свидетель-
ством о родстве в другой группе. Тем не менее, перенос паразитов
на новых неродственных хозяев, по-видимому, также бывает до-
вольно часто, так что использование данных о паразитах для уста-
новления родства между хозяевами имеет лишь подкрепляющее
88
Дж. Г. Симпсон. Принципы таксономии животных
значение. (Например, Vanzolini, Guimaraes, 1955; также значитель-
ный обзор паразитологических свидетельств о филогении млекопи-
тающих, подготовленный Б. Пэттерсоном и пока не опубликован-
ный).
Наиболее убедительным возможным доказательством принад-
лежности данной особи к данному виду является регистрация того,
что она существует как член данной видовой популяции, особен-
но если наблюдалось её скрещивание с другим членом этой попу-
ляции. Это один из случаев прямых филогенетических данных, ко-
торые можно наблюдать, а не предполагать. Такие данные в про-
шлом были редки, однако в настоящее время они приобретают всё
большее значение для видовых и внутривидовых категорий, по
мере того как таксономисты всё чаще отправляются в поле. Близ-
кий, относительно недавний подход состоит в изучении самого ге-
нетического состава и структуры размножения популяций как в
поле, так и в лаборатории. Детальный анализ возможен пока ещё
для немногих видов и поэтому имеет ограниченное применение в
классификации большинства групп животных. Однако такие иссле-
дования уже приобрели исключительную таксономическую важ-
ность в качестве примеров, а также показывая принципы, которые
могут применяться при генетической оценке популяций, пока ещё
генетически не обследованных. (См. особенно: Dobzhansky, 1951;
о приложении некоторых из этих принципов к вымершим популя-
циям см., например: Simpson, 1953).
На данные по поведению таксономисты нередко взирают с по-
дозрением по той же причине, что была ранее указана для физио-
логических данных. В самом деле, первый уровень, или элементар-
ное поведение, включающее такие акты как позы или геометричес-
ки возможные движения конечностей, является физиологическим
и страдает теми же таксономическими проблемами, что и более
простые, генерализованные физиологические свойства: они особен-
но подвержены конвергенции. Эта проблема, однако, значительно
менее серьёзна для второго уровня, или сложного поведения, вклю-
чающего в той или иной мере усложнённые паттерны или програм-
мы, состоящие из элементарных движений, таких как, например,
брачное поведение у птиц. Майр (Мауг, 1958; несколько глав в Roe,
Simpson, 1958 также имеют отношение к данному предмету) пока-
Таксономические доказательства и их эволюционная интерпретация 89
зал и прекрасно проиллюстрировал таксономическую пригодность
поведения на том уровне, где оно скорее реляционное, чем инди-
видуальное.
Наконец, более чёткое понимание репрезентативности выбор-
ки по отношению к популяции также повлияло на представления
о таксономических данных. Уже со времён античности люди зна-
ли, что многие виды животных включают резко различные формы,
такие, например, как самцы и самки или личинки и имаго. Столь
же стары сведения о том, что многие виды являются полиморфны-
ми в узком смысле, с разными формами, относящимися к одной
жизненной стадии или к одному полу. Необходимость включения
таких различающихся форм достаточно широко, хотя и не слишком
последовательно, признавалась доэволюционными таксономистами.
До сравнительно недавнего времени общее мнение (как обычно, с
исключениями) состояло в том, что достаточно сведений по одно-
му экземпляру для каждой формы, или по одному экземпляру для
вида в целом, если в нём не обнаружены чётко распознаваемые
формы. Другие экземпляры, если они и собирались, считались
«дубликатами» и обычно обменивались с другими музеями или
коллекторами.
Современная таксономическая практика, имеющая дело с попу-
ляциями, а не с типами, использует по возможности серии экзем-
пляров, достаточно обширные для того, чтобы делать выводы об
общей изменчивости популяции, представленной данной выборкой.
Статистические методы предоставляют точные критерии для суж-
дения в каждом случае об адекватности исследуемой выборки в
отношении делаемых на её основании выводов, а также для оцен-
ки размера выборки, необходимого для надёжных выводов при за-
данном уровне вероятности, или доверительности. Конечно, полу-
чение выборки оптимального размера не всегда доступно. Это вер-
но прежде всего в отношении ископаемых форм, даже при том, что
в настоящее время имеются очень большие выборки, особенно по
беспозвоночным.4 Тем не менее, небольшие выборки и единичные
4 Я сомневаюсь, позволят ли когда-нибудь финансы, площади хранилищ и
время, а также наличие в природе, собрать достаточно адекватные выборки по
видам, например, зауроподных динозавров.
90
Дж. Г. Симпсон. Принципы таксономии животных
экземпляры небесполезны. Только они дают недостаточные дове-
рительные интервалы и заниженные оценки популяционной измен-
чивости: важно, чтобы эти недостатки принимались во внимание
(см. Simpson et al., 1960). Хотя неадекватность такого рода со вре-
менем преодолима, неадекватность выборок составляет серьёзную
проблему и в отношении некоторых неонтологических данных.
Например, сравнения семейств и отрядов в серологической систе-
матике всё ещё базируются в значительной мере на анализе еди-
ничных представителей единичных видов, относящихся к сравни-
ваемым высшим таксонам, так что к таксономическим заключени-
ям, сделанным на основании таких данных, следует относиться с
осторожностью.
Сходство и гомология
Как уже упоминалось и будет обсуждаться далее, эволюцион-
ная таксономия не полагается исключительно на объединение по
сходству или на определение (в теории) всех таксонов через совпа-
дение признаков. Регистрация и интерпретация совпадающих при-
знаков, тем не менее, играет существенную роль в эволюционной
таксономии, как и в большинстве систем классификации. Они важ-
ны для решения и других задач, но основное их назначение в том,
что они составляют один — и при этом основной — из несколь-
ких критериев для суждения о родственной близости (здесь умест-
на ссылка на Дарвина). Эта близость, в свою очередь, является од-
ним из важных критериев для присвоения рангов таксонам и при-
писывания признакам организмов соответствующей таксономичес-
кой значимости.
Сходства [23] непосредственно указывают на родственную бли-
зость в том случае, если они уже присутствовали у предкового так-
сона и сохранились во всех или хотя бы некоторых из его таксо-
нов-потомков; иными словами, если они были унаследованы пос-
ледними от общего предка. Как уже было хорошо известно Дарви-
ну и стало ещё яснее в последующем, это не является единствен-
ным способом возникновения сходства в процессе эволюции. Даже
при отсутствии общего предка они могут возникнуть в некоторых
или всех таксонах-потомках в результате параллельного развития,
Таксономические доказательства и их эволюционная интерпретация 91
канализованного предковыми характеристиками, генетическими
или иными. В подобном случае они всё же могут указывать на род-
ственную близость, однако это указание не столь прямое и, возмож-
но, менее определённо. Они могут возникать также в качестве не-
зависимых конвергентных адаптаций к сходному образу жизни у
таксонов с совсем разными предками. В таких нередких случаях
они не имеют отношения к родственной близости и являются ос-
новным потенциальным источником путаницы или ошибок в эво-
люционной таксономии.5 Сходства также могут возникнуть вслед-
ствие мимикрии, в которой сходство как таковое является адапта-
цией, и случайным образом, когда у сходств разные, не связанные
между собой причины. Такие случаи также не имеют отношения к
родственной близости, однако в них сходства столь поверхностны,
что редко вводят в заблуждение.
Одна из действительно фундаментальных проблем эволюцион-
ной таксономии состоит в том, чтобы различать эти категории сход-
ства, особенно те, которые унаследованы и не унаследованы от об-
щего предка. Чтобы их обсуждать, необходим словарь; здесь при-
няты следующие определения.
Гомология есть сходство, унаследованное от общего предка.
Сходные признаки в таком случае являются гомологичными и обо-
значаются именем существительным гомологи.
Гомоплазия есть сходство, не унаследованное от общего пред-
ка. Сходные признаки в данном случае являются гомопластичны-
ми9 для их обозначения подходящего имени существительного пока
нет. Гомоплазия включает параллелизм, конвергенцию, аналогию,
мимикрию и случайное сходство
Параллелизм есть развитие сходных признаков независимо в
двух или более линиях общего предка, обусловленное природой это-
5 Однако неверно считать это проблемой исключительно дарвиновской или
более поздней эволюционной таксономии, как полагают или заявляют Cain
(1959b) и другие. Это даже ещё более серьёзная проблема в типологической
таксономии, поскольку конвергенция также производит «тип», подобно пред-
ковой общности. В типологии нет критериев выбора между альтернативными
или конфликтными типами, так что для этой системы проблема неразрешима.
Эволюционная таксономия, как будет показано, располагает рабочими крите-
риями для решения проблемы, хотя и с разными уровнями вероятия.
92
Дж. Г. Симпсон. Принципы таксономии животных
го предка или процессами канализации развития. Развившиеся та-
ким путем признаки являются параллельными, но опять-таки для их
обозначения подходящего имени существительного нет.
Конвергенция есть развитие сходных признаков независимо в
двух или более линиях вне связи с общим предком, но вследствие
адаптации к одинаковым экологическим условиям. Возникшее та-
ким путем сходство является конвергентным (опять без существи-
тельного).
Аналогия есть функциональное сходство, не связанное с общ-
ностью предка. Признаки, относящиеся к этой категории, являются
аналогичными и обозначаются как аналоги. Конвергентные призна-
ки являются аналогичными в той мере, насколько они могут быть
связаны с функцией, что обычно, а возможно и всегда, имеет место.
Мимикрия есть сходство, приспособительное само по себе и не
связанное с общностью происхождения. Она возникает в пределах
одного сообщества и выражается в том, что одна группа организ-
мов походит на другую, имеющую иное происхождение, вследствие
того, что сходство даёт адаптивное преимущество мимикрирующе-
му организму по тем или иным причинам, выяснение которых нас
здесь не должно заботить. Мимикрирующий организм — это ими-
татор, а тот, которого копируют, — модель. Для обозначения при-
знаков, вовлечённых в мимикрию, общепринятых терминов не су-
ществует.
Случайное сходство есть похожесть по признакам, развив-
шимся в разных таксонах по независимым причинам и вне причин-
ной связи, имеющей отношение к этому сходству как таковому.
Эти термины, как они здесь определены и будут использовать-
ся, подразумевают, что рассматриваемые сходства причинно обус-
ловлены и эволюционно значимы. Они являются познавательными
терминами в области теоретической таксономии; однако, как и в
случае всех состоятельных таксономических концепций, они име-
ют и вполне прикладное значение в классификации. Важно, что это
не просто описательные термины: в самих описаниях и в строгих
типологических схемах на самом деле нет указания ни причин, ни
практических критериев для выделения этих принципиально раз-
личных категорий сходств. Всё, что можно предпринять на уров-
не наблюдения или в полностью эмпирической системе, — это ука-
Таксономические доказательства и их эволюционная интерпретация 93
зать, какие объективные сходства существуют и каковы их степень
и структура.
Сходства, ныне известные как гомологичные, распознавались с
самого начала сравнительной анатомии как науки, начиная с рабо-
ты Белона (Belon, 1555). Ко времени Оуэна (1804-1892), последне-
го великого приверженца гётевской идеалистической морфологии
и кювьеровой классификации, было более или менее отчётливо
установлено, что имеются две категории сходств, различающиеся,
как правило, степенью и не совсем отчётливо осознаваемым каче-
ством. Для их различения Оуэн переопределил старый термин
«аналогия» и ввёл новый термин «гомология». Он (Owen, 1848)
определил аналогию как «часть или орган у одного животного,
имеющие ту же функцию, что и иная часть или иной орган у дру-
гого животного», а гомологию — как «один и тот же орган у раз-
ных животных во всех разнообразных проявлениях формы и фун-
кции». Принимаемые буквально, эти определения совершенно не-
удовлетворительны. Один и тот же орган обычно имеет одну и ту
же функцию у разных животных, и в таких случаях единственное
отличие состоит в том, что гомолог — это «один и тот же орган»,
а аналог — это «другой орган». Оуэн (Owen, 1866) позже усугубил
путаницу, заменив в определении аналогии «другой» [местоимение
another} на «некий» [неопределённый артикль а]. Из чего следует,
что, по определению, большинство [сходных] органов могут быть
как аналогичными, так и гомологичными, поскольку различия отме-
чаются, только если одинаковые органы имеет разные функции, —
и тогда они гомологи, а не аналоги; или если разные органы име-
ют одну функцию — и тогда они аналоги, а не гомологи; при этом
важные уклоняющиеся случаи в определении не уточнены.
Даже если бы различия были указаны более чётко, эти концеп-
ции остаются неудовлетворительными, поскольку они целиком ос-
нованы на представлении о том, что есть и что не есть «одно и то
же». В каком смысле два животных обладают «одним и тем же ор-
ганом»? Органы редко бывают, если вообще бывают, структурно
идентичными в разных таксонах, так что неясно, что имел в виду
Оуэн, когда писал о «разных животных». Имеется широкая града-
ция степеней и категорий сходств и различий между органами, так
что «один и тот же» и «разный» являются не строгой дихотомией,
8 — 4338
94
Дж. Г. Симпсон. Принципы таксономии животных
но нечёткими и целиком субъективными подразделениями сложно-
го континуума. Ни Оуэн, ни кто-либо другой из не-эволюционных
систематиков не выработали чётко сформулированных и работаю-
щих критериев, позволяющих делать различия соответственно их
определениям. Тем не менее, Оуэн и другие опытные систематики
его дней на практике с большим успехом действительно различа-
ли [две категории сходства] на основе скорее интуиции и личного
опыта, чем формальных критериев. В значительном числе случа-
ев они различали гомологии и аналогии точно так же, как мы это
делаем сегодня на эволюционных основаниях. Следует вспомнить
(глава 2), что они также имели значительный успех и в распозна-
вании эволюционных таксонов на неэволюционных основаниях.
Объяснение — одно в обоих случаях. Эволюция производит более
или менее различающиеся группы, которые мы называем таксона-
ми, а также сходства, которые более или менее чётко опознаются
как гомологичные и аналогичные. Доэволюционные таксономисты,
к которым относятся блестящие умы, сделавшие выдающиеся ра-
боты на основе собственных предпосылок, постигли многие (хотя
и далеко не все) из этих результатов эволюции, не осознавая, что
дало им начало или что объективно различает их.
Именно Дарвин дал объяснение и чёткий критерий для гомо-
логий: таковы, как было определено выше, органы или, более ши-
роко, сходства, унаследованные от общего предка. Это совершен-
но ясное и объективное6 определение, призванное заменить невнят-
ное и целиком субъективное «одно и то же». Сам Дарвин не пред-
лагал формального определения, однако оно вскоре было вырабо-
тано эволюционными систематиками и с тех пор принято большин-
ством из них. (Об истории и различных определениях гомологии,
6 Из моего прежнего опыта я знаю, что нужно объяснить данное исполь-
зование «объективного». Филогенетическая история действительно имеет ме-
сто и она действительно предполагает наследование органов и других сходств
голоморфы, или скорее информации и основы для их развития. Это совершен-
но объективный процесс, происходящий в природе. Едва ли стоит говорить,
что наше знание объективного процесса умозрительное и неполное. Аналогия
с атомами ещё раз послужит для прояснения позиции: никто не станет отри-
цать объективное существование атомов, хотя наше знание об атомной струк-
туре умозрительное и неполное.
Таксономические доказательства и их эволюционная интерпретация 95
аналогии и многих связанных с ними терминах см.: Haas, Simpson,
1946; Simpson, 1959с.)
Тем не менее, в современной литературе обширная полемика об
определениях гомологии и аналогии занимает значительное место,
причём некоторая часть способных и опытных систематиков наста-
ивает на возвращении к определениям, в сущности совпадающими
с оуэновскими. Обзор в: Boyden (1947) — один из наиболее ясных
и грамотных, для наших целей он даёт достаточное представление
об этой школе. Бойден и другие близкие к нему по взглядам не от-
рицают наследования сходств от общего предка или того, что оно
является объяснением большинства гомологий по Оуэну. Но и про-
тивоположный лагерь не отрицает того, что оуэновские гомологии
обычно распознаются и без эволюционной терминологии. Если бы
в полемике, как это нередко бывает, вопрос был только в том, ка-
кая концепция должна быть обозначена термином «гомология», в
ней не было бы смысла. Я понимаю, что Бойден использует этот
термин в смысле Оуэна, а он знает, что я использую его в смысле
Дарвина. Я делаю так фактически просто потому, что это соответ-
ствует современному общепринятому пониманию; термин будет
понят именно таким образом, поскольку такова преобладающая
точка зрения. В противном случае я бы использовал другой термин,
такой как «гомогения». Иное использование, по-видимому, также
потребует какого-либо другого термина для гомологии в смысле
Оуэна, а не Дарвина. Подходящим кандидатом является «морфо-
логическое соответствие», которое было определено Вуджером
(Woodger, 1945) с помощью строгой символической логики (одна-
ко, я должен признаться, в такой форме, которая мне представля-
ется трудной или даже невозможной для применения в практичес-
кой таксономии).
На самом деле сердцевину полемики составляет проблема, ко-
торая весьма далека от того, следует ли унаследованность от обще-
го предка называть «гомологией» или как-нибудь ещё. Предметом
спора служит положение концепции гомологии, как она здесь оп-
ределена, в современной таксономии и особенно — составляет ли
она желаемый и практический базис для классификации. Критичес-
ким для этой проблемы является вопрос о том, можно ли в конк-
ретных случаях отличить с достаточной надёжностью таким обра-
8*
96
Дж. Г. Симпсон. Принципы таксономии животных
зом понимаемую гомологию от разных категорий гомоплазии. Как
и многие таксономисты, я полагаю, что можно, и попытаюсь по-
казать это. Однако не следует обещать слишком многого: гомоло-
гию не всегда удаётся распознать; в некоторых случаях её распоз-
навание крайне затруднено, так что при решении этого вопроса мы
всё ещё делаем ошибки. Более того, при существующей практике
в некоторых классификациях (например, для большинства бакте-
рий) это практически неприменимая концепция. Тем не менее, по
крайней мере в классификации многоклеточных она теоретически
необходима и обычно практически реализуема. Возможно, тот факт,
что классификации неооуэнцев всё более и более сближаются с
таковыми неодарвинистов, свидетельствует об этом.
Некоторые аспекты оснований и методологии
Прежде чем обсуждать критерии гомологии, следует вкратце
рассмотреть некоторые общие и немногие частные аспекты подхода
к такого рода проблемам. В первую очередь, эволюционная таксо-
номия должна принять в качестве основания современную эволю-
ционную теорию, значительная часть которой напрямую не связа-
на с таксономией, но которая в целом имеет определённое отноше-
ние к таксономии. Очевидно, почти всё это должно приниматься
здесь без доказательств. Есть несколько книг, которые обобщают
определённый этап развития современной эволюционной теории и,
будучи взятыми вместе, составляют основу этого нового этапа:
Dobzhansky (1937, 2-е изд. — 1941, 3-е изд. — 1951), НеЬегег (1943,
2-е изд. — 1954-1959), Huxley (1942), Lerner (1954), Mayr (1942),
Rensch (1947, 2-е изд. — 1954), Schmalhausen (1949), Simpson (1944
[24], переиздание — 1953), Stebbins (1950). Большая часть после-
дующих идей может быть почерпнута из статей международного
журнала «Evolution», а также из его обзоров и содержащихся там
ссылок.
Во-вторых, необходимо обратить внимание на хорошо извест-
ную, но не всегда различаемую дихотомию в категориях данных,
доступных для эволюционной таксономии. Данные по современ-
ным животным не имеют временной размерности или она слишком
мала, чтобы иметь какое-то реальное значение. Поскольку эволю-
Таксономические доказательства и их эволюционная интерпретация 97
ционная таксономия оперирует временной размерностью, эти дан-
ные неполны в одном очень важном отношении. Они сами по себе
не являются историческими и всё же должны служить источником
для исторических выводов. Это составляет ключевую проблему в
оценке гомологии, а также во многих других аспектах таксономии.
Метод должен быть сравнительным, а выводы должны делаться
так, словно мы имеем дело с последовательностями, в которых со-
хранены только конечные состояния. Одни современные животные
в каких-то свойствах примитивнее других. Поэтому часто оказыва-
ется возможным составлять последовательности из современных
животных, приблизительно соответствующие тем цепочкам собы-
тий, которые разворачивались во времени. Важно помнить, что эти
два ряда — не одно и то же и что вероятность их полного совпа-
дения очень невелика. Но при правильной интерпретации выявлен-
ные последовательности служат существенным дополнением к дан-
ным о совпадающих признаках.
Палеонтологические исследования часто основаны на остатках
одного возраста, и тогда они точно так же являются сравнительны-
ми и внеисторичными, как и данные по современным животным.
Однако будучи основанными на последовательностях в геологиче-
ском времени, а не только связанными с современными фаунами,
палеонтологические исследования действительно вводят истинную
временную размерность: их данные непосредственно исторические.
Несмотря на недостатки в других отношениях (смещённые выбор-
ки, фрагментарная анатомия, отсутствие физиологических данных
и т.п.), ископаемые остатки предоставляют наиболее полный базис
для эволюционной классификации, если, конечно, они сами по себе
достаточно полны. В некоторых обширных группах (например,
млекопитающих) классификации по крайней мере высших таксо-
нов в большей мере основаны на ископаемых, чем на современных
животных. Тем не менее, в царстве животных в целом палеонто-
логические данные недостаточны для лидирующей роли и, вероят-
но, такими и останутся. Надежда на то, что классификация всего
царства животных будет иметь адекватную опору в виде непосред-
ственных исторических данных, более чем несерьёзны. Это особен-
но верно для наиболее многочисленной и в некоторых отношени-
ях наиболее сложной из крупнейших групп — для насекомых.
98
Дж. Г. Симпсон. Принципы таксономии животных
И всё-таки группы, лучше документированные ископаемыми, по-
зволяют разрабатывать принципы сравнительного анализа данных,
применимые в случаях, когда такая документация бедна или её нет.
Неонтологические данные также предоставляют многие принципы,
требуемые для интерпретации и ископаемых остатков. Эволюцион-
ная классификация опирается как на неонтологию, так и на пале-
онтологию, даже если ископаемые остатки отсутствуют или груп-
пы полностью вымерли. (Многие проблемы реконструкции истори-
ческих последовательностей обсуждаются в: Simpson, 1960а.)
Сравнительно простой пример прояснит взаимодействие меж-
ду этими разными подходами к эволюционным интерпретациям
сравнительной анатомии и, тем самым, к филогении. Лошади и
носороги — в какой-то степени очевидные родственники: они, как
минимум, относятся к млекопитающим, которых выделили задол-
го до Линнея. При обследовании почти любой части тела — напри-
мер, конечностей — обнаруживаются и некоторые более детальные
сходства. В обеих группах есть копыта, сходство по которым было
признано важным уже многими доэволюционными таксономиста-
ми, а затем сочтено гомологией в прежнем, ещё не прояснённом
смысле. У них ось конечности проходит через средний палец, ко-
торый у лошадей остаётся единственным функциональным, а у
носорогов — один из нескольких. Это сходство также было подме-
чено некоторыми доэволюционными таксономистами (возможно,
первым был Блэнвиль (Blainville, 1816), но не совсем точно: в ка-
честве общей черты было указано нечётное число пальцев, да и то
как одна из нескольких равнозначных типологических схем,7 между
которыми было мало выбора. Имеются и ещё более тонкие сход-
ства, например, в строении пяточных костей, особенно астрагала
(таранная кость). С другой стороны, лошади отличаются от носо-
рогов достаточно отчётливо наличием только одного пальца и не-
которыми другими свойствами, в числе которых — более тонкая и
обволошенная кожа. По этим и некоторым другим характеристика-
ми носороги схожи с бегемотами больше, чем с лошадьми. На этом
7 Эта неточность была запечатлена в названии Perissodactyla, до сих пор
сохранённом за этой группой. Имеются непарнопалые с чётным числом паль-
цев, равно как и парнопалые (Artiodactyla) с нечётным числом пальцев.
Таксономические доказательства и их эволюционная интерпретация 99
основании можно обозначить несколько архетипов, каждый из ко-
торых и был признан той или иной школой доэволюционных ти-
пологов. Признак «число пальцев» объединяет лошадей в один ар-
хетип, а носорогов и бегемотов — в совершенно другой. Такова
была наиболее распространённая система ранних таксономистов.
Признак «толщина шкуры» даёт такое же сочетание этих трёх
групп: такова была системе Кювье. Другие архетипы дают другие
группировки. Линней, например, поместил носорогов вместе с гры-
зунами в отряд Glires, а лошадей с бегемотами — в отряд Belluae
среди Ungulata (копытные), откуда, однако, были исключены носо-
роги, у которых также есть копыта. (Во всех этих архетипах к при-
знаку, который здесь указан в качестве основного, могут быть до-
бавлены и другие совпадающие признаки, подкрепляющие каждую
из классификаций.)
Только с развитием эволюционной таксономии было прочно
установлено, что указанное сходство лошадей и носорогов является
гомологичным, а другие сходства из числа указанных — нет. В этой
связи можно было бы сказать, что установленный Блэнвилем архе-
тип действительно присутствовал у их общего предка, а другие —
нет.8 Значит, лошади и носороги относятся к одному таксону до-
вольно высокого уровня, из которого должны быть исключены бе-
гемоты: эта система скоро стала всеобще признанной, а прежде
более популярная кювьерова и другие системы были забыты, ис-
ключая разве что историков. Этот вывод основывался на рассмот-
рении многих анатомических частей лошадей и носорогов и на их
сравнении со всеми прочими копытными. Было обнаружено, что
строение конечностей лошадей, носорогов (а также тапиров) обла-
дает сходной сложной функциональной системой и что это сход-
ство сопряжено с другими совпадающими анатомическими черта-
ми. Бегемоты, с другой стороны, имеют столь же сложную, но по-
иному устроенную конечность, в основных чертах сходную с тако-
8 В таких случаях иногда говорят, что типологи предвосхитили эволюци-
онные заключения. Однако это только совпадение. С таким разнообразием
выделенных архетипов было бы очень странно, если бы какой-то один из них,
даже по простой случайности, не совпал с эволюционными фактами без какой-
либо ссылки на эволюцию. Система Блэнвиля в отношении данных животных
просто оказалась удачной.
100
Дж. Г. Симпсон. Принципы таксономии животных
вой у коров, антилоп, свиней и многих других копытных. Как ока-
залось, различия в функционировании конечностей наиболее пол-
но отражены в строении астрагала, нежели в любой другой их ча-
сти. Различия астрагалов непарно- и парнокопытных отмечались
разными типологами, но никто из них не приписал этому призна-
ку наивысшее значение — даже Оуэн, который, по иным и не со-
всем корректным основаниям, выдвинул систему, позднее признан-
ную эволюционной.
В данном случае гомология была установлена без серьёзной
поддержки ископаемыми данными, которые позднее лишь подтвер-
дили её несомненность. Кювье уже знал некоторые вымершие фор-
мы, которые могли бы существенно прояснить проблему, если бы
он счёл их более примитивными родственниками современных
животных, но он этого не сделал. (Его Anoplotherium принадлежит
к тому же надсемейству, что и бегемот, а его Palaeotherium близок
к лошадям и носорогам.) Hyracotherium (валидное техническое на-
звание для «Eohippus») близок к общему предку лошадей и носо-
рогов и действительно обладает признаками, установленными в
качестве гомологов в этих двух группах. Тем не менее, он весьма
отличен от современных лошадей и носорогов и ни в коей мере не
является промежуточным звеном между ними, но это уже другой
вопрос.
Ранние эволюционисты, включая Дарвина, находились под
сильным влиянием типологического стиля мышления. Многие из
них, нО не Дарвин с его более глубокой проницательностью, про-
должали считать, что архетип равен совпадающим признакам и
соответствует предковой организации или хотя бы является её адек-
ватной характеристикой.9 (Они редко используют термин «архе-
тип», но широко применяют соответствующую концепцию.) Про-
стой пример, по-видимому, позволяет обнаружить это заблуждение.
Признаки, общие для лошадей и носорогов, не включают число
пальцев, но их общий предок должен был обладать некоторым оп-
9 Эта наивная точка зрения лежит в основе таких абсурдных обвинений,
вроде упомянутого выше утверждения, что эволюционная таксономия будто бы
считает род прародителем своих видов. Это чисто типологическая, а не эво-
люционно-таксономическая ошибка.
Таксономические доказательства и их эволюционная интерпретация 101
ределённым числом пальцев, не обязательно таким же, как у его
современных потомков. Этот случай указывает на метод определе-
ния родственной близости и характеристик общего предка на ос-
новании сравнительных данных, весьма отличный от основанного
на совпадающих признаках. Не вдаваясь в детали, можно отметить,
что собственная анатомия современных лошадей, в сравнении с
таковой других копытных и вообще млекопитающих, указывает,
что их предки должны были иметь больше одного пальца и что
общий предок лошадей и носорогов должен был иметь не менее
трёх и не более пяти пальцев. Изучение ещё одного современного
непарнокопытного — тапира — делает весьма вероятным, что пре-
док всех непарнокопытных имел четыре пальца на передней и три
пальца на задней ноге, а палеонтологические данные подтвержда-
ют справедливость этого. Носороги (и ещё более — тапиры), та-
ким образом, оказываются структурно более примитивными, или
ближе к предку, чем лошади. Однако это не означает, что носоро-
ги в целом более примитивны: такого рода вопросы требуют де-
тального изучения в каждом отдельном случае.
Несмотря на все комбинации типологов, лошади не настолько
конвергентны с любой другой группой, чтобы современный таксо-
номист ошибся в оценке их гомологий. Однако есть вымершее
южноамериканское копытное Thaotherium, между которым и лоша-
дью отмечается очень тесная конвергенция, особенно в полностью
однопалой конечности. Флорентино Амегино, открывший таотерия,
весьма продвинутый эволюционный таксономист для своего време-
ни (1854-1911), заключил, что это сходство гомологично и что ли-
топтерны (группа, к которой относится Thaotherium) и лошади име-
ют общего предка, который ближе к ним, чем, скажем, общий пре-
док лошадей и носорогов. Это ошибка не была грубой, учитывая,
что, общий предок литоптерн и лошадей не намного древнее; кон-
вергенция между этими двумя группами является одной из наибо-
лее показательных среди известных случаев. Примечательно, что
дальнейшие исследования с использованием и других критериев,
которые ещё будут рассмотрены, убедили, что данное сходство в
строении ног конвергентно, что общий предок литоптерн и лоша-
дей имел пять пальцев и что он был древнее, чем общий предок
лошадей и носорогов.
102
Дж. Г. Симпсон. Принципы таксономии животных
Некоторые критерии гомологии
Установление гомологии никогда не бывает абсолютным, со
стопроцентной определённостью, однако настаивать на этом обсто-
ятельстве как на ущербности эволюционной таксономии — значит,
отрицать ценность любого и всякого научного устремления. Неко-
торые гомологии, например, между плечевой костью человека и
обезьяны, почти столь же определённы, как то, что солнце взойдёт
завтра утром. Другие имеют более низкий уровень вероятности.
Некоторые, казавшиеся почти строгими при слабой изученности,
позже оказывались почти наверняка не гомологиями, а гомоплази-
ями, как в примере с литоптернами и лошадьми. Этот же пример
показывает, что накопление большего объёма данных и примене-
ние более многочисленных и строгих критериев может быть весь-
ма эффективным даже в таких сложных случаях.
Как уже было показано, концепция гомологии выросла из изу-
чения совпадающих признаков. Гомология всегда затрагивает со-
впадающие признаки, однако было убедительно показано и то, что
простое наличие общих признаков или возможность абстрагирова-
ния архетипа, или (в современных терминах) морфотипа, не явля-
ется достаточным критерием гомологии. Имеются также случаи
установленной (т.е. высоко вероятной) гомологии, например, меж-
ду эндостилем и щитовидной железой, когда совпадающих призна-
ков столь мало или они столь тривиальны, что вычлененный архе-
тип может свидетельствовать скорее против гомологии, чем за неё.
При рассмотрении совпадающих признаков вполне очевидны
два критерия: детальность и множественность сходств. В обоих
случаях в сравнение включается больше отдельных единичных
признаков, выделенных таксономистом, и общий принцип состоит
в том, что заключения о родстве (что главным образом означает —
о гомологии) тем весомее, чем по большему числу признаков ус-
тановлено сходство. Вероятности являются кумулятивными (хотя
они не просто суммируются математически), так что много малых
вероятностей, взятых вместе, могут привести к общей высокой ве-
роятности. В этой связи были разработаны полезные, но, к сожа-
лению, порой весьма трудоёмкие методы включения многих при-
знаков в единую операцию сравнения. Примеры такого подхода,
Таксономические доказательства и их эволюционная интерпретация 103
среди многих работ, дают: Stroud (1953), Michener, Sokal (1957),
Cain, Harrison (1958) и Jolicoeur (1959). Однако возникают две про-
блемы, пока ещё не полностью разрешённые. Во-первых, без при-
влечения других критериев гомопластичное сходство может сумми-
роваться вместе с гомологичным. Использование большого числа
признаков, вероятно, позволяет снизить влияние гомоплазии на ре-
зультаты, но в какой-то степени оно всегда возможно, а иногда мо-
жет быть столь велико, что делает несостоятельным результат. Во-
вторых, неверно думать, что при использовании большого числа
признаков вероятность увеличивается в том случае, если признаки
тесно сопряжены генетически или функционально. Признаки, ко-
торые сильно связаны между собой, не могут рассматриваться как
независимые доказательства, они представляют собой одно свиде-
тельство. Одно из предлагаемых средств (Michener, Sokal, 1957)
состоит в использовании очень большого числа разных признаков
в ожидании того, что их взаимные корреляции будут в целом не-
значительными. Это, по-видимому, верно в большинстве случаев,
когда используется действительно очень много признаков, однако
данное исходное допущение не было проверено эмпирически, так
что эта причина ошибок [в установлении гомологий] не может
быть отвергнута. Другой возможностью является вычисление кор-
реляций между всеми признаками, и затем использование тесно
ковариирующих групп в качестве единичных признаков. Подход к
вычленению таких групп разработали Олсон и Миллер (Olson,
Miller, 1958) [25], однако процедура крайне трудоёмка, даже прак-
тически невозможна без применения электронных калькуляторов,
а статистическая значимость результатов, полученных к настояще-
му времени, мне представляются сомнительной.
Сложные адаптивные комплексы едва ли возникают дважды
одним и тем же способом и потому в разных осуществлениях ско-
рее всего конвергентны, а вероятность гомологии тем больше, чем
сложнее адаптации и больше совпадений. С другой стороны, сход-
ная адаптация, различающаяся по признакам, которые не являют-
ся необходимыми для самих адаптаций, служит важным свидетель-
ством конвергентной гомоплазии, а не гомологии. Например, Ин-
гер (Inger, 1958) показал, что у головастиков, приспособленных к
жизни в потоках, хорошая обтекаемость и ротовая присоска воз-
104
Дж. Г. Симпсон. Принципы таксономии животных
никли конвергентно в нескольких филогенетически разных груп-
пах, но что характерные детали ротового диска позволяют распоз-
навать филогенетические группы, в пределах каждой из которых
эти детали гомологичны. Он заключает, что «если рассматривать
только сложные адаптивные структуры, то всякий раз можно рас-
познавать конвергенции», что может быть несколько оптимистич-
но, но по сути верно.
Признаки принципиально разных категорий, как правило, не
бывают генетически или функционально скоррелированы и их со-
впадение может быть решающим при разграничении гомологии и
конвергенции. Например, для уже упоминавшихся южноамерикан-
ских грызунов имеются зоогеографические, анатомические и био-
химические (без явной корреляции с анатомическими) данные.
Каждая из этих совокупностей данных сама по себе ненадёжна и
недостаточна, однако их комбинация делает в высшей степени не-
вероятным, что их сходство с дикобразовыми Старого Света гомо-
логично.
В настоящее время признано, что эмбрионы не представляют
и не повторяют предковые взрослые структуры. Тем не менее, вер-
но то, что процессы индивидуального развития наследуются (на
самом деле именно они, а не взрослые структуры, преимуществен-
но управляются наследственностью) и что они могут представлять
исключительные доказательства гомологии или конвергенции. Эм-
брионы филогенетически родственных форм обычно более сходны,
чем взрослые: этот факт был отмечен фон Бэром ещё до того, как
стало известным его филогенетическое приложение. Тем не менее,
могут быть и противоположные случаи: эмбрионы иногда менее
сходны, чем соответствующие взрослые. Необходимы поиски иных
критериев, чтобы решить, (а) является ли сходство взрослых гомо-
логичным, а эмбриональные различия — результат дивергентного
приспособления к данному периоду жизни, или (б) эмбриональные
различия указывают на различное филогенетическое происхожде-
ние, а взрослое сходство конвергентно. Об этих взаимосвязях меж-
ду эмбрионами и взрослыми см. исчерпывающий обзор в: de Beer
(1951). Что касается отдельных органов, то их сходное эмбриональ-
ное возникновение является важным доказательством гомологии.
Это также было отмечено ещё до того, как стала известна истин-
Таксономические доказательства и их эволюционная интерпретация 105
ная причина гомологии. Оуэн (Owen, 1848) подчёркивал «тесное
общее сходство в способе развития» как критерий гомологии.
Может показаться, что данные по хромосомной и генной иден-
тичности могут быть наиболее решающими из возможных крите-
риев гомологии. Во многих случаях это несомненно так, но имеют-
ся и важные ограничения. В некоторых группах, которые были де-
тально исследованы генетически (например, Drosophila, см. Patter-
son, Stone, 1952), филогенетическая классификация непосредствен-
но на генетических основаниях, если ещё и не полностью достиг-
нута, то во всяком случае несомненно достижима. Однако имеются
показательные частные случаи, где этот метод не имеет широкого
применения. В высшей степени невероятно, что мы когда-либо бу-
дем располагать адекватной генетической информацией, например,
по слонам или глубоководным животным. Даже без прямых данных
по генетическому составу, успешная гибридизация является, по сути,
решающей демонстрацией хромосомной эквивалентности и выте-
кающей из этого гомологичной природы структурных сходств.
Имеется ряд от полной плодовитости между родителями и среди их
потомков через хорошо развитое, но стерильное потомство F} поко-
ления, или с разной степенью ошибок развития в F, до полной роди-
тельской стерильности. Положение в этом ряду, очевидно, соответ-
ствует, в некотором приближении, степени хромосомного сходства.
Эти данные, однако, почти исключительно ограничены внутриви-
довым и видовым уровнями,10 где гомологии обычно можно распоз-
нать по другим, более простым и доступным критериям.
Для таксономии нет явной практической пользы определять
гомологию как сходство, вызванное идентичностью генов, хотя с
теоретической точки зрения возможно и такое определение [26].
Этот критерий редко можно применять, даже когда он необходим.
На самом деле я считаю его нежелательным также и теоретичес-
ки. Немногие варианты структур или других характеристик, наибо-
10 Между животными возможны предположительно межродовые гибриды,
обычно со стерильным потомством. Например, у млекопитающих известны
полученные искусственным путём гибриды Bos х Bison, Equus * * * х Asinus и Ursus
х Thalarctos. По моему мнению, однако, это лучше считать основанием для их
объединения в номинальные роды. Я бы сейчас не присваивал родовой статус
Bison, Asinus и Thalarctos.
106
Дж. Г. Симпсон. Принципы таксономии животных
лее полезных в систематике, особенно на уровне выше видового,
могут быть явственно связаны с единичными или идентифициру-
емыми генами. Большинство из них находится под существенным
влиянием одновременно многих генов и генных взаимодействий.
Крайне маловероятно, что все гены, связанные с гомологичными
признаками, идентичны у некоторого предка и его таксономичес-
ки обособленных потомков. В любом случае такую идентичность,
вероятно, никогда нельзя будет установить операционально. Более
того, в настоящее время известно, что практически идентичные или
очень сходные соматические характеристики могут иметь весьма
разную генетическую основу. Для таксономии, а также для всей
эволюции имеет значение действующий организм как таковой, а не
его гены. Если данная характеристика повсеместно присутствует
как у предка, так и у всех его потомков в пределах филетической
линии, значит, она гомологична у всех них, даже если генетичес-
кий субстрат изменился. Конечно, это не означает отрицание того,
что эволюция всех признаков всегда имеет генетическую основу.
Это только указывает, что генетическая эволюция и соматическая
эволюция не являются идентичными или строго параллельными и
что именно соматическая эволюция имеет более тесное отношение
к таксономии.
Экология в данном случае имеет особое значение, поскольку
имеется тесная связь между адаптацией и гомологией или конвер-
генцией. Хотя мы говорим не совсем строго о полезных атрибутах
организма как об адаптациях, адаптация не является специфиче-
ской характеристикой ни организма, ни среды по отдельности, но
представляет собой результат их взаимодействий. Эти взаимодей-
ствия — экологические по определению. Гомологичные признаки
возникают, обычно лишь с незначительными исключениями, на
основе адаптаций предковой формы и сохраняются у потомков;
соответственно, они становятся гомологами, поскольку сохраняют
у них свою адаптивность или, во всяком случае, не становятся вов-
се инадаптивными. Конвергенция, понимаемая в строгом смысле,
затрагивает приспособление к экологически сходным ситуациям в
двух группах, ведущих своё происхождение от разных предков, по
меньшей мере у одного из которых не было адаптации, общей для
конвергентных потомков.
Таксономические доказательства и их эволюционная интерпретация 107
Таким образом, когда действительные предки непосредственно
неизвестны, конвергенция должна подозреваться для тех групп,
которые сходны между собой только или главным образом по спе-
цифическим приспособлениям к частной разделяемой ими эколо-
гии, а в других отношениях различны. Например, в классическом
случае родов Thylacinus и Canis сходства, хотя многочисленные и
детальные, целиком связаны с особыми хищническими адаптаци-
ями, тогда как по характеристикам, не связанным с такими адап-
тациями, эти животные весьма различны. Следовательно, их сход-
ство явно конвергентно. С другой стороны, собаки (Canidae) и пан-
да (Ailurus) заметно различаются по признакам, связанным с их
экологическими особенностями, но сходны по многим признакам,
не столь узко адаптивным. Эти сходства, таким образом, в боль-
шинстве своём гомологичны. Детальная гомоплазия особенно ве-
роятна в группах, которые существуют (или возникли) в разных
более или менее изолированных регионах, как в случае Thylacinus
и Canis или в других парах экологических викариатов. Подобные
географические соотношения, однако, могут развиться и в группах
общего происхождения и с гомологичными адаптациями, как у ази-
атских и американских тапиров. Обычно, как в данном примере,
гомология вполне очевидна по другим основаниям. Все такие слу-
чаи должны интерпретироваться в свете того, что известно об адап-
тации как общем феномене эволюции, а не только в их таксономи-
ческих аспектах (см., например, Simpson, 1953; Underwood, 1954).
По крайней мере на надвидовом уровне наиболее прямым кри-
терием гомологии является, конечно, открытие общего предка или
ископаемой филетической линии, отчётливо сходящейся назад во
времени к этому предку. Когда такие свидетельства имеются, они
становятся решающими или почти таковыми, что само по себе оче-
видно. Однако уже должно быть ясно, что неправы те неотиполо-
ги, которые настаивают на том, что гомология (или филогения) не
может быть адекватно установлена в отсутствие палеонтологиче-
ских данных. Другие критерии, в том числе уже рассмотренные,
почти всегда достаточны для распознавания гомологии с удовлет-
ворительно высокой степенью вероятия. Яркий пример — порази-
тельная гомология между слуховыми косточками млекопитающих
(молоточек и наковальня) и костями нижней челюсти рептилий (со-
108
Дж. Г. Симпсон. Принципы таксономии животных
членовной и квадратной), которая сначала была выявлена главным
образом на эмбриологических данных по современным организ-
мам, а затем подтверждена и существенно детализирована изуче-
нием ископаемых форм, переходных между двумя классами.
Замечание о сериальной гомологии
Оуэн (Owen, 1866) выделил три категории гомологии: общую,
специальную и сериальную. Общая гомология рассматривалась как
соответствие между структурой конкретного организма и абстрак-
тным архетипом, понятие о котором в настоящее время не имеет
таксономической ценности. Специальная гомология соответствует
более или менее гомологии в современном смысле, но без объяс-
нения ссылкой на общего предка. Сериальная гомология, представ-
ления о которой по-прежнему используются, представляет собой
анатомическое соответствие между повторяющимися или сериаль-
ными структурами в пределах одного организма — например, меж-
ду последовательно расположенными позвонками, перьями или
листьями на дереве. Представляется достаточно очевидным (хотя
это и оспаривалось), что это совершенно особый феномен в срав-
нении с гомологией между разными организмами. Поэтому может
быть лучше называть её, вслед за Бронном, «гомономией» (см.
Remane, 1956 — это ещё одна исключительно ценная книга по так-
сономии в целом).
Гомономная серия или система как целое обычно гомологична
между разными организмами общего происхождения, как волосы
у млекопитающих или листья у деревьев, однако каждый конкрет-
ный волос или каждый конкретный лист не гомологичен таковому
у другого организма. Такие сходства всей системы имеют таксоно-
мическое значение, поскольку они гомологичны в общем смысле,
а не по причине их внутренней гомономии, которая не имеет так-
сономического значения. Однако имеются особые пограничные слу-
чаи. Например, семь шейных позвонков, почти постоянные среди
млекопитающих (с ничтожным числом исключений), в пределах
этого класса могут быть гомологизированы как индивидуально, так
и в качестве серии, однако гомология с шейными позвонками у
амфибий и рептилий, число которых может быть очень различно,
Таксономические доказательства и их эволюционная интерпретация 109
возможна только в отношении всей серии, или метамерного поля,
которое её производит. (Сбивает с толку ещё и то, что гомология
серий у разных животных — не то, что подразумевается под сери-
альной гомологией, и это служит ещё одной причиной предпочесть
термин «гомономия».) Имеются также и сомнительные случаи.
Например, у большеухой лисицы (Otocyon) из Африки есть четы-
ре моляра вместо обычных трёх или менее у плацентарных млеко-
питающих и почти несомненно у их общих предков. Предполага-
ется, что один из моляров был добавлен в конце [зубного ряда] и
что первые три моляра гомологичны типичным предковым. Одна-
ко не менее вероятным представляется, что только молярная серия
гомологична и что в одних случаях её морфогенетическое поле
даёт четыре зуба, в других — три, без чёткой индивидуальной го-
мологии.
Последовательности и некоторые критерии
для примитивных и продвинутых признаков
Выше было упомянуто, что эволюционная интерпретация так-
сономических данных опирается не только на совпадение призна-
ков, что обычно стремились принимать во внимание строгие эмпи-
рики, но и на другие подходы, особенно на интерпретацию после-
довательностей. Что касается последовательностей близких иско-
паемых форм, связанных с реальной временной шкалой, их значе-
ние сравнительно просто и понятно и не требует здесь детального
рассмотрения. Интерпретация последовательностей одновременно
существующих организмов или вообще последовательностей без
заданной временной шкалы более трудна и заслуживает отдельно-
го специального обсуждения. Один аспект данного подхода был
ясен уже Дарвину, который писал (Darwin, 1859):
Есть ракообразные, занимающие противоположные концы ряда, у ко-
торых едва ли можно найти совпадающие признаки; и всё же эти крайние
виды, будучи отчётливо связанные промежуточными формами, можно
опознать как несомненно принадлежащие именно к этому, а никакому дру-
гому классу Articulata.
Следует также помнить (см. главу 2), что даже доэволюцион-
ные таксономисты в принципе признавали, что таксоны могут быть
9 — 4338
110
Дж. Г. Симпсон. Принципы таксономии животных
распознаны и определены на основе равновесного или равномер-
ного (если речь идет о рядах) распределения сходств, независимо
от совпадения признаков и без выделения абстрактных архетипов.
Было также отмечено мимоходом, что Беккнер (Beckner, 1959) ввёл
практически такой же подход в теорию множеств, которая в своей
стандартной форме более типологична. Это он сделал посредством
того, что им было названо «политипической концепцией». Термин
мне кажется неудачным по двум причинам [27]. Во-первых, он свя-
зывает данный подход с типологией, в то время как он может и
должен уводить от типологии в сторону более реалистичной и
прагматичной оценки организмов. Во-вторых, термин «политипи-
ческий» уже широко используется в таксономии, причём в совер-
шенно ином смысле: так обозначается таксон Т , который вклю-
чает более одного Т. (При таком использовании термина, разуме-
ется, невозможно, чтобы таксон Т имел тип или сам был типом.)
Я не собираюсь, однако, заменять термин Беккнера новым, по-
скольку я предлагаю использовать далее эту концепцию в несколько
иной форме и под другим названием. Беккнерово определение «по-
литипического объединения [aggregation]» таково (изложено без
использования специальных символов).
В «политипическом объединении»:
1. Каждый индивид имеет неопределённо большое число
свойств, в совокупности характеризующих объединение как целое;
2. Каждое из этих свойств присуще большому числу этих ин-
дивидов;
3. Ни одно из этих свойств не присуще всем индивидам в объе-
динении.
Последнее условие необходимо для того, чтобы отличить дан-
ное определение объединения, а в конечном итоге таксона в био-
логической систематике, от типологического. Оно не исключает
возможности того, что какой-то таксон может выделяться и по об-
щим (совпадающим) признакам, так что определение таксона (в
данном случае это концептуально иной тип объединения) может
быть выражено и таким образом. На самом деле можно легко со-
ставить объединение, логически подобное «политипическому объе-
динению» Беккнера без применения первых двух из его условий,
как видно из следующего примера:
Таксономические доказательства и их эволюционная интерпретация
111
Т 1 2 3 4 5 6 7
г а b с d е f g
Свойства -< b с d е f g h
1 с d е f g h
Объединение, или Т — и т.д.
Это объединение надёжно очерчивается и легко определяется
на основании того, что каждый индивид Т обладает большинством
свойств, общих со следующим в ряду, однако каждый индивид Т
обладает лишь меньшинством из всего множества свойств и каж-
дое свойство присуще лишь меньшинству индивидов Т. Эта мо-
дель соответствует дарвиновской трактовке Crustacea и постоянно
встречается в биологии, обычно в более сложной форме. Эта мо-
дель соответствует тому, что я впредь буду называть рядом, имея
в виду, что реальные зоологические группы, в которых присутству-
ют ряды, могут иметь и на самом деле обыкновенно имеют совпа-
дающие признаки, но это не слишком влияет на обсуждение рядов
как таковых, а также на то, что одни и те же организмы могут
иметь весьма разные ряды присущих им свойств.
Примем, что «свойства» или признаки, которые соответствуют
соседним членам ряда, достоверно гомологичны. Это означает, что
всякая пара соседних членов имеет общего предка. Однако если,
например, члены пар 1, 2 и 2, 3 имеют общего предка, то и члены
1, 3 также должны иметь общего предка. Это означает, что все [по-
литипическое] объединение должно было иметь общего предка и
может быть представлено как филогенетически валидный таксон,
что соответствует неявному заключению Дарвина в отношении
Crustacea. Таким образом, совершенно обоснованное и, если внут-
ренние гомологии в ряду правильно определены, вполне неизбеж-
ное филогенетическое заключение может быть сделано без (или
независимо от) полного совпадения признаков и без выяснения
того, каким мог быть общий предок всей группы. Тем не менее, мы
хотели бы знать, каким был этот общий предок. Ещё важнее для
классификации знать об этом предке именно то, что позволило бы
нам сделать вывод о филогенетических отношениях между двумя
таксонами, представленными рядами [sequence-taxon]. Оба таксо-
на могли произойти от одного и того же предка, но их эволюция
9*
112
Дж. Г. Симпсон. Принципы таксономии животных
развертывалась по разным совокупностям свойств. Например, пар-
нокопытные образуют ряд, при рассмотрении некоторых их
свойств, таких как число пальцев, или, целое множество частью
совпадающих, частью различных рядов, если рассматривать разные
их свойства, а непарнокопытные имеют другой подобный комплекс
рядов. Наше представление о взаимоотношении между парно- и
непарнокопытными (а также другими высшими таксонами копыт-
ных) будет сильно зависеть от заключения о том, имел ли, напри-
мер, их общий предок один, два, три, четыре или пять пальцев.
(Известный предок, как оказалось, был с пятью пальцами, что вме-
сте с прочими данными указывает на весьма отдалённого предка,
совпадающего с таковым всей когорты копытных.)
Для исследования любого предкового признака, соответствую-
щий ряд, очевидно, должен иметь единый fundamentum, если по-
зволительно использовать схоластический термин; т.е. это должен
быть ряд альтернативных состояний одного и того же признака,
такого как число пальцев или жилкование крыла. Очевидно, у пред-
ка могло быть только одно альтернативное состояние [признака], а
не весь их набор. Из этого следует базовое допущение данного под-
хода к филогенетическому исследованию, а именно: одно из аль-
тернативных состояний в ряду либо совпадает с предковым, либо
наиболее близко к нему. Теперь необходимо иметь критерий для
выбора такого альтернативного состояния. В запутанной и вызыва-
ющей неприятие терминологии Геннига (Hennig, 1950) оно соответ-
ствует «плезиоморфии», тогда как альтернативное состояние, кото-
рое с меньшей вероятностью может быть отнесено к предковому,
будет «апоморфией».
Имеется немного рядов, к сожалению, слишком мало, для ко-
торых очевидное направление эволюции более или менее фикси-
ровано априори. Например, у Drosophila по механическим причи-
нам определённые хромосомные мутации с необходимостью пред-
шествуют другим (Dobzhansky, 1951). Гораздо более многочислен-
ны случаи, для которых информация, внешняя по отношению к
ряду, указывает его вероятное направление. Для тех же хромосом,
известные процессы дупликации и фрагментации делают гораздо
более вероятным происхождение форм с большим числом хромо-
сом от таковых с меньшим, но не наоборот. На основе рецентных
Таксономические доказательства и их эволюционная интерпретация 113
рядов и палеонтологических данных были установлены различные
тренды, столь общие в пределах крупных групп животных, что их
можно применять, хотя бы предварительно, для интерпретации и
других вневременных рядов. Например, у всех без исключения реп-
тилий и млекопитающих число пальцев в норме либо пять, либо
постепенно уменьшается, так что всегда можно предположить с
достаточной вероятностью, что меньшее число пальцев — произ-
водно от большего. Это правило редукции широко применимо к
сериальным структурам, хотя для некоторых из них часты исклю-
чения. Зубная система плацентарных млекопитающих почти всегда
проявляет тенденцию к постепенной редукции примитивной фор-
мулы 3/3 : 1/1 :4/4 : 3/3, однако известны многочисленные исклю-
чения среди китообразных, они также встречаются у неполнозубых
и хищных. Общей тенденцией для позвоночных является редукция
числа позвонков, однако есть и исключения, особенно среди длин-
нотелых рыб и змей, и также изредка в других группах, даже у
млекопитающих. Общая тенденция увеличения размеров тела столь
обычна среди позвоночных, а также известна для некоторых бес-
позвоночных (Newell, 1949), что некоторые таксономисты предпо-
лагали ipso facto исключать крупных животных из числа предков
мелких. Тем не менее, имеется столько исключений, что степень
вероятия более чем умеренна в отсутствие привходящих дополни-
тельных доказательств. Ренш (особенно Rensch, 1954) указал и
проанализировал многие такие правила и закономерности. Он ут-
верждает, что они позволяют не только реконструировать предков,
но и делать предсказания в отношении потомков (также Rensch,
I960).11
Возможно, наиболее общим допущением об эволюционных
рядах является то, что они имеют тенденцию развития от просто-
го к сложному. Этот всеобщий тренд определённо характеризует
прогрессивный характер эволюции как целостного явления. Амё-
ба, какой бы сложной она ни была, очевидно проще, чем страус,
11 Подтверждение подобного «предсказания» может быть получено лишь
благодаря открытию организма, который уже возник в ходе эволюции, а не
событию из действительного будущего, т.е. после времени «предсказания».
Тогда возникает вопрос: является ли это предсказанием в логическом смысле?
Не следует ли его называть «постеказанием» [postdiction]?
114
Дж. Г. Симпсон. Принципы таксономии животных
и несомненно напоминает их общего предка. Однако в пределах
отдельной группы животных этот критерий редко имеет надёжное
применение. Никакой scala naturae на самом деле нет, а при работе
с расходящимися ветвями концепция «более простого» едва ли мо-
жет быть уместной. Проще ли страус, чем кенгуру? Ни один из них
в этом отношении не может считаться более близким к их обще-
му предку, среди современных животных нет строго соответству-
ющих этой концепции рядов, так что весь этот вопрос не представ-
ляется осмысленным. Уровни простоты часто зависят от субъектив-
ного определения и могут отражать предвзятость, основанную со-
вершенно на посторонних мотивах. Абстрактно рассуждая, более
простым может показаться меньшее число костей черепа у млеко-
питающих в сравнении с большим числом костей у их рыбьих
предков, или однопалость Equus в сравнении с многопалостью
Hyracotherium. Опять-таки, концепция простоты может не иметь к
этому отношения или требовать особых дополнительных определе-
ний, чтобы соответствовать фактам, установленным по другим ос-
нованиям. Далее, имеется много случаев, особенно среди парази-
тов, когда объективно более простые организмы происходят от бо-
лее сложных предков.
Серьёзная проблема состоит в том, что почти всякая совокуп-
ность варьирующих объектов по некоторому из своих признаков
может быть организована в монотонную последовательность, одна-
ко такая последовательность не обязательно будет иметь истинную
связь с филогенией. Это обстоятельство породило ныне обычно
отвергаемую (но всё ещё поддерживаемую немногими твёрдолобы-
ми поклонниками) концепцию ортогенеза, которая часто считает-
ся палеонтологической по происхождению, но на самом деле воз-
никла из-за неверной интерпретации искусственных, не имеющих
отношения к филогенезу рядов среди современных млекопитаю-
щих (см. Jepsen, 1949). Простым примером служит число пальцев
у копытных, которое может быть упорядочено в ряд 5-4-3-2-1. Дей-
ствительно, один конец этого ряда (5) отвечает примитивному со-
стоянию, а другой (2 или 1, равноценно) специализированному.
Однако лишь немногие (да и то сомнительно) однопалые копытные
прошли через весь анцестральный ряд 5-4-3-2-1, а двупалые копыт-
ные — через анцестральный ряд 5-4-3-2. В реальных ситуациях
Таксономические доказательства и их эволюционная интерпретация 115
может также оказаться, что ни один из концевых элементов тако-
го ряда не является более примитивным, чем другой. Например,
едва ли у человека какой-то цвет кожи, белый или чёрный, является
примитивным, а для некоторых последовательностей вообще невоз-
можно доказать, что один из элементов примитивнее других.
Во многих затруднительных случаях для распределения призна-
ков могут быть разработаны критерии, сходные с таковыми для
совпадающих признаков,. Например, среди современных копытных
почти все имеют несколько пальцев и только в одном роде Equus
(sensu lato) палец единственный. Среди млекопитающих в целом
пятипалые формы не составляют большинства, однако это число
присутствует у некоторых представителей большинства отрядов и
в пределах каждого отряда ряды предполагают редукцию от пяти-
палого или по меньшей мере от четырехпалого [состояний]. Нет
нужды в подтверждении ископаемыми данными, что 5 — это пред-
ковое число.
Термины «примитивный» и «специализированный», уже рас-
сматривались как существенные для настоящего обсуждения. Их
несогласованное использование таксономистами постоянно вносит
путаницу. Поэтому, прежде чем продолжать, необходимо более
строго определить эти термины.
Понятия примитивности и специализированности относи-
тельны, они бессмысленны, если не связаны определённо с конк-
ретным таксоном, линией или филогенией.
В пределах таксона, характеристики общего предка являются
примитивными, а другие — более или менее специализированными
пропорционально степени их уклонения от предкового состояния.
В пределах единой филетической линии, характеристики,
встречающиеся ранее других, являются более или менее примитив-
ными, а те, которые появляются позже, — более или менее спе-
циализированными пропорционально времени их появления.
В пределах филогении (которая разделена на таксоны и состо-
ит из ветвящихся филетических линий), характеристики любого
общего ствола являются более примитивными, чем различные ха-
рактеристики отходящих от него ветвей; в пределах отдельной
ветви они являются более специализированными в зависимости
более раннего или позднего появления; наконец, при сравнении вет-
116
Дж. Г. Симпсон. Принципы таксономии животных
вей — они более примитивны или более специализированы в зави-
симости от их большего или меньшего отклонения от ствола.
Понятие «генерализованный», которое часто используют вместо
«примитивный», почти всегда двусмысленно, в связи с чем я пред-
почитаю избегать пользоваться им. Иногда оно отсылает к абстрак-
тно совпадающим признакам, которые не могут, или могут только
случайно, сочетаться в реальном организме. В своё время оно при-
менялось в совершенно ином смысле, обозначая такое примитив-
ное состояние, которое не столь глубоко и специфично адаптиро-
вано к некоторой нише в сравнении с более специализированной
формой, возникшей на его основе. Примитивные животные часто
относительно генерализованные в этом смысле, однако, априорное
ожидание не настолько высоко, чтобы считать соответствующее
заключение приемлемым без дополнительного обоснования.
В рядах, отвечающих одновременно существующим организмам,
более примитивными будут те признаки, которые эволюировали
медленнее. Эта зависимость побуждала к поиску таких категорий
признаков, которые обычно эволюируют медленнее и в отношении
которых поэтому можно было предполагать, что они служат надёж-
ной индикацией общих предков. Таких твёрдых оснований не было
найдено, но имеются некоторые критерии для вероятной оценки.
Ранее широко признавалось, что адаптивные признаки менее, а
неадаптивные признаки более надёжны в этом отношении, поскольку
последние предположительно менее изменчивы при действии варьи-
рующих экологических факторов. В настоящее время этот критерий
отброшен. Прежде всего, истинно неадаптивные признаки, по-види-
мому, крайне редки и едва ли могут быть с определённостью иденти-
фицированы. Во-вторых, признаки, которые могут быть неадаптив-
ными или которые не столь отчётливо и строго адаптивны, как пра-
вило, оказываются скорее более лабильными, чем наоборот. Напри-
мер, зуб, который более не окклюдирует или находится в процессе
редукции и утраты важных функций (как зуб мудрости у человека),
исключительно изменчив как по структуре, так и по размерам.
Характеристики более общего значения или те, которые позво-
ляют достичь широкого прогресса в изменчивых экологических
ситуациях, с высокой вероятностью являются более примитивны-
ми. На этом основано понятие «наследственной основы» (heritage),
Таксономические доказательства и их эволюционная интерпретация 117
в противоположность более специфическим адаптациям «габитуса»
[habitus] (Gregory, 1951). У китообразных, например, утрата волос,
обтекаемость тела, плавники и хвостовая лопасть — всё это общие
приспособления к полностью водному образу жизни, относящиеся
к «наследственной основе» [28]. Они с высокой вероятностью яв-
ляются примитивными для данного отряда. Такие признаки как
китовый ус, брюшные борозды, бивни и т.д. являются «габитуса-
ми» в разных низших таксонах Cetacea и с несомненностью воз-
никли позже. Связь с общностью признаков у большего или мень-
шего числа таксонов очевидна. Было бы откровенной ошибкой счи-
тать «наследственную основу» неадаптивной, а «габитус» — адап-
тивным. Китовая наследственная основа по сути даже более адап-
тивна, нежели любой из китовых «габитусов». Следует также по-
мнить, что широко адаптивные признаки наследственной осно-
вы — именно те, которые с наибольшей вероятностью будут кон-
вергентными в разных группах. Уже Дарвин подметил (в других
терминах), что признаки, общие для Cetacea, гомологичны в пре-
делах этой группы, но конвергентны между китообразными и дру-
гими водными позвоночными.
Мы уже отмечали, что сложные, функционально скоординиро-
ванные структуры менее подвержены детальной конвергенции, и
одним из аспектов данного обстоятельства является то, что они
обычно менее лабильны и дольше сохраняют предковые состояния.
Ушной отдел у млекопитающих — именно такая структура, отли-
чающаяся тонкой коадаптацией своих частей. Во всех таких слу-
чаях, вероятно, имеется и соответствующая сложная и тонкая коа-
даптация генетического субстрата. Почти всякое изменение потре-
бует тогда сложных преобразований всего комплекса и будет либо
отвергнуто естественным отбором, либо начнет очень медленно
коадаптироваться. Этот эволюционный принцип объясняет таксо-
номическое наблюдение относительно консервативности таких ком-
плексов, т.е. их низкой изменчивости. Однако изменчивость никог-
да не бывает нулевой. Слуховая структура млекопитающих для
высших категорий является особо «хорошим» таксономическим
признаком, т.е. таким, который с большей вероятностью унаследо-
ван от общих предков, однако классификации, основанные толь-
ко на этой структуре, неудовлетворительны.
118
Дж. Г. Симпсон. Принципы таксономии животных
Серологические реакции, по-видимому, также относительно
малоизменчивы, что определяет повышенный таксономический
интерес к соответствующим данным. Теоретическое обоснование
не вполне удовлетворительно, поскольку большое число отдельных
биохимических веществ, задействованных в них, не было иденти-
фицировано. Поэтому не совсем ясно, почему сочетания этих ве-
ществ должны быть консервативными. Правильность данного зак-
лючения является во многом эмпирической и следует из того фак-
та, что степень серологического сходства обычно хорошо согласу-
ется с суждениями о родстве, ранее сделанными на других осно-
ваниях. Однако есть опасность кругового доказательства при ис-
пользовании такого рода процедур, причём не только в серологии.
Когда они не согласуются, следует ли это рассматривать как недо-
статочную обоснованность выводов или как свидетельство ошиб-
ки в интерпретации данных, используемых для этого обоснования?
Работа с конкретными группами всегда ведёт к эмпирически
обоснованным заключениям относительно того, что некоторые
категории признаков более лабильны, чем другие. Каждый специ-
алист по классификации приобретает «чутьё» на менее и более ла-
бильные признаки в своей группе. Одним из источников является
прямое наблюдение и количественная оценка степени изменчиво-
сти в пределах популяций. Затем, по мере того, как по результатам
согласования всех данных выстраиваются таксоны, становится оче-
видным, что одни свойства действительно характеризуют низшие
таксоны и могут считаться более лабильными, тогда как другие —
высшие таксоны и потому они менее лабильны, являясь с большой
долей вероятия примитивными для всей исследуемой группы. Эти
свойства отличаются от группы к группе и, как правило, должны
рассматриваться отдельно для каждой из них; одни и те же свой-
ства могут характеризоваться весьма разной изменчивостью в раз-
ных группах. Признаки одной категории (или относящиеся к одно-
му fundamenturn) не имеют фиксированного таксономического ран-
га; нет ни одной категории [признаков] с внутренне присущими им
значениями родовых, семейственных или отрядных характеристик,
как иногда склонны думать и на что надеются некоторые таксоно-
мисты. Например, число и форма резцов высоко лабильны и обыч-
но характеризуют роды у эоценовых прозимий, но весьма стабиль-
Систематика, таксономия, классификация, номенклатура
119
ны в пределах семейств у современных прозимий, а также во всём
отряде грызунов. Тем не менее, есть некоторые категории призна-
ков, особенно связанные с окраской, которые почти всегда очень
лабильны, где бы они не исследовались. Мы скорее всего никогда
не узнаем, был ли эогиппус пятнистым, полосатым, однородно ок-
рашенным или каким-либо ещё.
Когда одно состояние признака в некоторой группе или конце-
вое состояние ряда имеет гомолога в другой группе с более отда-
лённым общим предком, то это, возможно, является наиболее ре-
шающим свидетельством примитивности (и тем самым предково-
сти) признака. Среди млекопитающих, например, однопроходные,
сумчатые и плацентарные характеризуются тремя весьма специфич-
ными способами размножения. У однопроходных он в некоторых
отношениях с очевидностью гомологичен таковому большинства
рептилий. Этот способ, таким образом, наиболее примитивен сре-
ди млекопитающих, хотя и наименее обычен среди них. Это обыч-
но используется для предположения, что однопроходные, сумчатые
и плацентарные по способу размножения образуют ряд и что сум-
чатые в этом отношении примитивнее плацентарных. Однако раз-
множение сумчатых не очень сходно с таковым рептилий, так что
весьма вероятно, что оно в некоторых существенных чертах явля-
ется скорее уклоняющейся специализацией, а не примитивным в
сравнении с плацентарными.
Дискуссия касалась главным образом отдельных признаков или
структур, а не животных как целостных организмов. Некоторые
ряды, характеризующие одновременно существующие формы, осо-
бенно линии в пределах одного вида или цепочки близкородствен-
ных видов, в целом могут походить на истинную эволюционную
последовательность (или хроноклину). (Этот и другие аспекты клин
в отношении идентификации примитивных и продвинутых призна-
ков в деталях обсуждал Маслин (Maslin, 1952), причём некоторые
из его выводов также применимы к рядам и на более высоком
уровне.) Тем не менее, обычным правилом является то, что в дан-
ном ряду ни один таксон не будет более примитивным во всех от-
ношениях, чем любой другой. Тапир примитивнее лошади струк-
турой конечности, но нос тапира гораздо более специализирован.
Утконос обычно указывается как наиболее примитивное млекопи-
120
Дж. Г. Симпсон. Принципы таксономии животных
тающее, поскольку он откладывает яйца и имеет сходный с репти-
лиями плечевой пояс, а также некоторые другие примитивные чер-
ты. Однако он высоко специализирован среди млекопитающих по
строению зубов, клюва, амфибийным адаптациям и некоторым дру-
гим особенностям. Такая перекрёстная специализация, когда одна
группа специализирована в одном отношении, а другая родствен-
ная группа — в другом отношении, помогает делать заключения о
родственной близости и характеристиках предков. Предок должен
обладать комбинацией признаков, близких к менее специализиро-
ванным в двух сравниваемых группах, и должен был жить преж-
де возникновения их более продвинутых вариантов.
В некоторых современных группах, таких как Artiodactyla, ни
одна из доживших до современности форм не может рассматри-
ваться как полностью примитивная. Примитивные парнокопытные
были полностью вытеснены и должны были быть во многих от-
ношениях весьма непохожими на их современных потомков, в чём
убеждают ископаемые данные. В других группах большинство сту-
пеней в ряду предков—потомков выжили без особо глубоких изме-
нений. Это особенно явственно в случае приматов. Тупайи — ле-
муры — долгопяты — обезьяны — человекообразные обезьяны —
люди упорядочиваются в данный ряд по многим признакам, так что
имеет смысл рассматривать их как последовательность целых орга-
низмов [не отдельных признаков, с ними связанных], несмотря на
многие уклоняющиеся специализации на низших уровнях в преде-
лах каждого шага ряда. По признакам, которые проходят в виде
рядов через весь отряд Primates, тупайи наиболее примитивны. Эта
последовательность среди современных приматов и её значение для
филогении приматов были детально разработаны в работе: Le Gros
Clark (1959).
Замечание о параллелизме
Как было определено выше, параллелизм — это независимое
присутствие сходных изменений в группах, произошедших от об-
щего предка и вследствие того, что они имеют общего предка. Не-
которые исследователи (например, Хаас в: Haas, Simpson, 1946)
предпочли чисто описательное определение, особенно представлен-
Таксономические доказательства и их эволюционная интерпретация 121
ное в форме геометрической модели параллельных линий, симво-
лизирующих два филетических ствола, которые изменяются таким
образом, что не становятся ни более сходными, ни более различ-
ными. Доводя такое определение до крайности, что логически воз-
можно, но вряд ли будет признано его авторами, можно было бы
говорить о параллелизме между филетическими линиями устриц и
млекопитающих, которые становились больше без других заслужи-
вающих внимание изменений. Эта не очень продуктивная таксоно-
мическая концепция. На деле большинство таксономистов считает,
что термин «параллелизм» следует использовать только в тех слу-
чаях, когда общность предка имеет отношение к данному феномену.
Параллелизм бывает трудно или даже практически невозмож-
но отличить от гомологии, с одной стороны, и от конвергенции, с
другой стороны. Эти три процесса, достаточно обособленные по
своим характерным проявлениям, перекрываются в граничных слу-
чаях. К счастью, полностью параллельное развитие целых организ-
мов, и к тому же с одинаковой скоростью во всех линиях, невоз-
можно. Оно всегда сопровождается гомологиями, а также, если не
во всех случаях, то часто, дивергентной специализацией в каждой
филетической линии. Обычно оно происходит с разными скорос-
тями в разных линиях. Эти линии можно рассортировать по дивер-
гентным признакам, а всякую конвергенцию обычно можно распоз-
нать по критериям, которые уже были рассмотрены. Оставшиеся
сходства, в таком случае, будут или гомологичными, или параллель-
ными. Различие между гомологией и параллелизмом часто может
быть проведено на основе того факта, что гомологичные призна-
ки являются примитивными для данной группы, тогда как парал-
лельные — нет. Если, например, в некоторой совокупности линий,
идущих от общего предка, в одной отмечены зубы с низкой корон-
кой, а в других — с высокой коронкой, и низкокоронковые зубы
примитивны для всей группы, то эволюция высокой коронки в про-
чих линиях, вероятно, шла параллельно. Миченер (Michener, 1949)
привёл более сложный и очень яркий пример выявления эффекта
параллелизма в обширном семействе из разноусых чешуекрылых.
Имеется несколько теоретических принципов, на основании
которых можно понять, а также распознать параллелизм. Структу-
ра предковой группы неизбежно ограничивает направления воз-
122
Дж. Г. Симпсон. Принципы таксономии животных
ВРЕМЯ
-«----ДИВЕРГЕНЦИЯ-
00: Гомология с с
SSS& Конвергенция d и с
Дивергенция с
I
ВРЕМЯ
СТРУКТУРНЫЕ
ИЗМЕНЕНИЯ
* Параллелизм
} Видообразование
. Видообра-
’ зование
Филетиче-
к ская
продвину-
тость
общего
предка
РИС. 4. ТАКСОНОМИЧЕСКАЯ ИНТЕРПРЕТАЦИЯ КОНВЕРГЕНЦИИ И ПАРАЛЛЕЛИЗМА
А. Диаграмма с шестью современными видами a-f из которых а-с имеют
одного предка и d--f — другого предка; с и d конвергентны. А’. Истинная
филогения. А”. Включение видов в три рода, показанные фигурными скобками,
на основе неверной интерпретации конвергенции с и d как гомологии; выведенная
из этого филогения топологически весьма отлична от показанной на (А'), т.е.
неверна. Впрочем, показанные в (А') и (А") ситуации слишком упрощены, чтобы
быть достаточно реалистичными, особенно в отношении конвергенции с и d.
А'". Всё ещё упрощенное, но более реалистичное представление ситуации в
отношении с. В действительности с будет характеризоваться следующими
признаками: 1) гомологичными, связывающими его с а и Ь и общим предком а—с,
Таксономические доказательства и их эволюционная интерпретация 123
можных эволюционных изменений. Этот простой факт значитель-
но увеличивает вероятность того, что среди нескольких близкород-
ственных линий некоторые или даже все будут двигаться в одном
направлении. В географически расширяющейся группе с активным
видообразованием эта вероятность будет к тому же усилена есте-
ственным отбором, если экология в расходящихся линиях остаёт-
ся сходной в отношении адаптаций, вовлечённых в параллелизм.
Степень зависимости от сходной экологии напоминает таковую в
случае конвергенции, однако параллелизм обычно отличается со-
хранением гомологичных признаков от относительно близкого об-
щего предка. Параллельные филетические линии развития (в отли-
чие от конвергентных) начинаются со сходных коадаптированных
генетических систем, и сходные изменения с высокой вероятнос-
тью сохраняют эти коадаптации. Тенденция к генетическому парал-
лелизму также существенно усиливается повторными «гомологич- *
Рис. 4 (продолжение).
2) конвергентными, связывающими его с d, и 3) специфическими для своей линии.
Внимательный анализ этих признаков даёт два рода, показанные на (А"’)
фигурными скобками, их филогенетическая интерпретация корректна. Здесь
показан анализ признаков только для с. Другие виды также характеризуются
специфическими наборами признаков различного таксономического значения.
Б. Диаграмма с четырьмя современными видами, быстро отделившимися
от общего предка и впоследствии эволюировавшими параллельно. Б’. Истинная
филогения. Б”. Интерпретация современных видов без учёта параллелизма ведёт
к ошибочному заключению, что сходство, в действительности возникшее
параллельно, свидетельствует о недавнем видообразовании. Хотя и не столь
ошибочное, как в ситуации в (А), данная картина по-прежнему слишком упрощена
из-за отсутствия многих признаков разной таксономической значимости и
[филогенетической] интерпретации. При рассмотрении альтернативных случаев
(Б’-Б") выбор на основе данных только по терминальным видам может быть
затруднён, однако анализ рядов, вероятно, приведёт опытного таксономиста к
принятию верной схемы (Б’). (Вертикальная штриховая линия означает исходное
состояние для сравнения с современными видами, чтобы сделать более чётким
последовательный ряд.) В любом случае высший таксон, объединяющий эти
четыре вида и обозначенный фигурной скобкой, филогенетически обоснован и
с равным успехом будет выведен как из верной, так и из ошибочной
интерпретации. Более того, структура отношений в (Б") топологически идентична
таковой в (Б1) и неверна лишь в отношении оценки времени расхождения.
124
Дж. Г. Симпсон. Принципы таксономии животных
ными»12 мутациями и сходными относительными темпами мутиро-
вания.
В завершение следует сказать, что параллелизм не всегда мож-
но отличить от гомологии, однако обычно это не имеет особого
значения. Подобно гомологии, параллелизм зависит от общности
предка. В худшем случае он приведёт к преувеличению родствен-
ной близости, но без искажения его природы или структуры род-
ственных связей. Но даже и этого может не быть, так как эволю-
ционные механизмы гарантируют, что в среднем параллелизм
обычно бывает теснее в линиях, близких к общему предку. Сход-
ства, обусловленные параллелизмом, распознаются ли они в каче-
стве таковых или нет, действительно с высокой надёжностью про-
порциональны степени родства. Отличать параллелизм от конвер-
генции жизненно важно, однако обычно это довольно легко сде-
лать, если конвергенция не несет на себе влияния общего предка
(т.е. если она не включает элемента параллелизма). Отличить па-
раллелизм от гомологии труднее, однако по счастью не столь прин-
ципиально. Это не значит, что параллелизм не существенен для
классификации: примеры его важности будут обсуждаться в главе 6.
Корректные и некорректные таксономические интерпретации
одновременно существующих видов с конвергенциями и паралле-
лизмами схематично показаны на рис. 4.
. 12 Это ещё одно использование «гомологии», отличное от обычного в так-
сономии, однако его смысл достаточно очевиден. Такие мутации в параллель-
ных линиях на самом деле также параллельные, а не гомологичные в таксо-
номическом смысле.
4
От таксономии к классификации
Таксономия — это наука, однако её применение в
классификации включает большую долю человеческой изобрета-
тельности и искусности, короче — искусства. В этом искусстве
есть много места для личных вкусов и даже слабостей, однако су-
ществуют также и каноны, которые помогают делать одни класси-
фикации лучше, более осмысленными и пригодными, чем другие.
То, что наилучшая основа для классификации является эволю-
ционной, мы уже решили. Это в первую очередь означает, что клас-
сификация должна быть совместима со всем тем, что может быть
узнано про филогению классифицируемой группы. То, что класси-
фикация может или должна выражать филогению, — очевидная
ошибка. При условии совместимости с соответствующим базисом,
классификация должна быть как можно более стабильной и изме-
няться, только когда новое знание делает её определенно менее
пригодной или выявляет противоречивость.
Дискуссии по поводу «реальности» и «объективности» — или
«нереальности» и «субъективности» — таксонов обыкновенно бы-
вают довольно поверхностными или просто пререканием из-за
слов. Вполне рабочими могут быть концепции «произвольного»
[arbitrary] и «непроизвольного» [nonarbitrary], если их правильно
понимать. Некоторые приёмы классификации должны быть услов-
ными, что не влечёт за собой нарушения принципов, но требует
внимания к самим этим приёмам.
10 — 4338
126
Дж. Г. Симпсон. Принципы таксономии животных
Основной канон совместимости с филогенией подразумевает
монофилию, часто определяемую нестрого или непрактично. Мы
должны найти лучшее определение и оценить его применимость.
Мы должны также на более широкой основе охарактеризовать вза-
имодействие разных категорий отношений в эволюции: отношений
между филетической и остановленной эволюцией, отношений вер-
тикальных в линиях от предков к потомкам и горизонтальных меж-
ду одновременно существующими группами, сравнительную значи-
мость дивергенции [divergence] и разнообразия [diversity]; а также
достоинства и недостатки определения категорий, в общем и в ча-
стности. Мы также должны рассмотреть трудную и во многом за-
путанную проблему значения относительной древности таксонов
для определения их относительных рангов.
Что делает таксономист?
Практическая таксономия обычно означает разработку класси-
фикаций конкретных групп, а также идентификацию экземпляров,
согласно уже имеющейся классификации. За этой практикой стоят
имеющие глубокий смысл теории, принципы, правила и приёмы.
Они, строго говоря, составляют науку таксономию, как она здесь
определена, и являются предметом настоящей книги. Формулиров-
ка и проверка этих теорий, принципов и т.д. — разумеется, самое
важное из того, что делает таксономист. Некоторые из них уже
были обрисованы. Следующий вопрос — на основании каких
таксономических принципов и приёмов должна в действительно-
сти разрабатываться классификация? Когда таксономист приступает
к своей несомненной обязанности — к построению классифика-
ции, которая является практическим приложением его науки, то,
как именно он это делает? В целом, он исполняет следующие дей-
ствия, более или менее в указанной последовательности, но обычно
с заметными перекрываниями шагов, перепрыгиваниями и возвра-
тами [29]:
1. Он отбирает или получает организмы, подлежащие классификации.
Он может исследовать их в поле и оставить их там живыми, но чаще они
коллектируются как музейные экземпляры, предпочтительно, но необяза-
тельно тем таксономистом, который исследовал их прежде. По крайней
От таксономии к классификации
127
мере, для обширных групп он почти всегда вынужден исследовать старые
коллекции, нередко рассредоточенные по разным учреждениям.
2. Он обследует организмы и фиксирует данные о них, а также выбира-
ет из литературы как можно более полные сведения, ранее полученные по
той группе, которую он изучает. Ему нужны данные самых разных категорий,
как отмечено в главе 3. Эта работа перекрывается с указанной в пункте (1),
поскольку одни данные, как минимум о локалитетах и (для ископаемых) го-
ризонтах, в поле могут быть лишь отмечены [observed], а другие (в исключи-
тельных случаях — все) могут быть собраны [obtained] в полевых условиях.
3. Он сортирует организмы по таксономическим единицам, демам (ло-
кальным популяциям, более строго определенным в главе 5), подвидам или
видам соответственно задачам. Затем он анализирует данные по полимор-
фным формам и всем типам изменчивости. Этот анализ в принципе ста-
тистический и в настоящее время обычно проникает глубоко в практику,
позволяя делать выводы о свойствах популяций на основе исследуемой
выборки. Эта работа широко перекрывается с двумя предыдущими, а также
с двумя последующими.
4. Он сравнивает характеристики исследуемых единиц, обращая осо-
бое внимание на типы и степень сходств и различий и на выявленные
ряды. Эти приёмы в основе своей также во многом статистические.
5. Он интерпретирует взаимоотношения, выявленные в результате
Сравнения, в терминах основных таксономических понятий, особенно го-
мологии, параллелизма, конвергенции, примитивности, специализации,
которые рассмотрены в главе 3.
6. На интерпретациях, полученных на предшествующем шаге, он ос-
новывает выводы об эволюционных отношениях между исследуемыми по-
пуляциями. Он может изображать их в виде диаграммы — предпочтитель-
но в виде предполагаемого филогенетического дерева или дендрограммы,
топологически как можно точнее отражающей филогенетическое дерево.
Он может использовать какие-либо другие способы графического представ-
ления результатов или давать целиком их словесное описание. В любом
случае он доводит дело до заключений о родстве соответственно этому де-
реву. Его заключения также будут содержать некоторые рассуждения о сте-
пени дивергенции, которые не могут быть адекватно показаны на древо-
видной диаграмме.
7. Он переводит свои заключения о родстве, дивергенции и т.п. в тер-
мины иерархии, объединяя или разделяя различные группы организмов в
таксоны разных рангов.
8. Согласно правилам и традициям, он выбирает из прежних публика-
ций названия, применимые к выделенным таксонам, а когда подходящих
опубликованных названий нет, он предлагает новые.
10*
128
Дж. Г. Симпсон. Принципы таксономии животных
Одни из этих приёмов уже обсуждались в первых трёх главах.
Другие будут рассмотрены в последующих главах. Некоторые, та-
кие как методы коллектирования экземпляров, хотя и необходимы,
если излагать таксономию [в полном объёме], не столь важны для
нашей темы и потому опущены. Отдельные приёмы, хотя имеют и
к ней более тесное отношение, тем не менее также не рассматри-
ваются по другим основаниям. Последнее особенно верно в отно-
шении технических статистических приёмов, важных на указанных
выше этапах (3) и (4). Их детали не существенны для понимания
таксономических принципов. Их обсуждение, достаточно адекват-
ное для того, чтобы быть действительно полезным, потребовало бы
значительно больше места, чем здесь предоставлено, к тому же
недавно они достаточно полно изложены в другом издании
(Simpson et al., 1960).
В данной главе нас будут заботить, во-первых, некоторые из
практических целей и требований классификации, до этого подроб-
но не обсуждавшихся, и затем некоторые специальные принципы
и приёмы, задействованные главным образом на этапе (7) из пред-
ставленного списка.
Классификация как полезное искусство
Подобно многим другим наукам, таксономия реально представ-
ляет собой соединение науки в строгом смысле и искусства. Её на-
учная сторона преследует своей целью получение приближений —
как хотелось бы надеяться, всё более полных по мере развития
науки, дающих понимание взаимоотношений, существующих в
природе. Одно из словарных определений «искусства» — это «че-
ловеческая изобретательность и искусность»: в этом смысле таксо-
номия во многом оказывается родом искусства, когда речь идёт о
конструировании классификаций. Номенклатура — это в полном
смысле искусство, а не наука, поскольку она относится исключи-
тельно к изобретательности и не соответствует ничему в (лежа-
щей вне человека) природе, хотя и применяется для научного опи-
сания вещей, существующих в природе. Распознавание и упорядо-
чивание таксонов на разных уровнях наполнено научным содержа-
нием, однако и оно смешано с искусством, требуя человеческой
От таксономии к классификации
129
изобретательности и искусности. Уже было упомянуто и будет ста-
новиться всё более очевидным в настоящей и последующих главах,
что даже если бы взаимоотношения в некоторой группе животных
были полностью известны и даже если бы было полное согласие
относительно научных принципов, можно было бы создать бесчис-
ленное множество разных классификаций, совместимых с этими
взаимоотношениями и состоятельными с точки зрения этих прин-
ципов. Выбор из этих альтернатив несомненно является искусст-
вом.
Основополагающий принцип таксономического искусства со-
стоит в том, чтобы его результаты были полезными [useful]. В клас-
сификации это предполагает, среди прочего, три особо важных
вспомогательных принципа:
1. Основанием для классификации должны служить биологи-
чески наиболее значимые отношения между организмами, но вво-
дить их следует не больше, чем это осуществимо на практике.
2. Классификация должна быть совместима с отношениями,
рассматриваемыми в качестве её оснований.
3. Классификация должна быть как можно более стабильной,
если только этим не нарушаются два предыдущих принципа.
Первая позиция уже была рассмотрена в основных чертах. Вто-
рая и третья позиции здесь особенно уместны. Первая и вторая
позиция ограничивают ту область, в которой на практике может
быть достигнута третья. Эволюция, принятая здесь в качестве важ-
нейшего общего базиса классификации, становится всё более изу-
ченной как в деталях, так и в своих общих проявлениях. Лучшее
понимание её теорий и принципов должно влечь за собой некото-
рые изменения в классификациях, с тем чтобы поддерживать их в
наиболее приемлемом соответствии с биологической наукой в це-
лом. Также быстро увеличивается объем информации по отдель-
ным группам организмов, например, рост знаний по их биохимии,
особенно поразительный в наши дни. Даже на сравнительно хоро-
шо возделанном поле анатомии постоянно добавляются новые дан-
ные, но многие всё ещё неизвестны. Такого рода новые данные
делают возможным более надёжные заключения, могущие отрицать
или перевешивать прежние представления, на которых ранее осно-
вывались классификации. По-прежнему открываются новые труп-
130
Дж. Г. Симпсон. Принципы таксономии животных
пы животных. В палеонтологии средняя скорость таких открытий
увеличивается. В некоторых современных группах (например, сре-
ди птиц) она приближается к нулю. В других (например, среди на-
секомых) она по-прежнему высока, но главным образом на низших
уровнях. Тем не менее, увеличивается и число высших современ-
ных таксонов (примерами могут служить Neopilina и Hutchinso-
nielld). Эти новые группы организмов должны не только включать-
ся в классификации: они также обычно требуют более или менее
значительных изменений структуры классификации ранее извест-
ных животных.
Чтобы обеспечивать наибольшую полезность, классификация
должна соответствовать не тому знанию, которое было получено в
прошлом, но по мере возможности — постоянно меняющемуся се-
годняшнему знанию. Поэтому желательно, чтобы классификации не
оставались статичными, но непрерывно изменялись по мере расши-
рения знания. Этот желательный прогресс неизбежно снижает праг-
матическую ценность классификаций. Классификация — абсолют-
но необходимое средство концептуализации, коммуникации и хра-
нения информации о животных. В терминах теории информации,
изменение классификаций (и их номенклатуры) со временем и из-
за личных пристрастий порождает значительный «шум». С этой
точки зрения стабильная классификация, конечно же, наиболее по-
лезна. Хотя в целом для зоологии это не имеет большого значения,
но нельзя забывать и об огромном практическом значении класси-
фикаций для хранителей коллекций: изменения могут ввергать их
в бесполезную во многих отношениях работу по изменению ката-
логов, указателей и всей системы хранения информации.
Должен соблюдаться некий компромисс между полезностью
обновляемых классификаций и полезностью стабильных классифи-
каций.1 Следующий компромисс — определённо наилучший и зас-
луживает того, чтобы на нем было сделано ударение:
1 В действительности многие музейные кураторы принуждены воздержи-
ваться от компромисса и выбирать стабильную, т.е. устаревшую классифика-
цию в своей повседневной работе с коллекциями единственно только из-за не-
достатка времени. Однако так не следует поступать, имея в виду потребности
в исследованиях и коммуникации.
От таксономии к классификации
131
Опубликованную классификацию, находящуюся в научном обо-
роте, следует изменять, если она определённо не соответствует
известным фактам и принятым принципам, но лишь настолько,
насколько это требуется для приведения её в соответствие.
Таксономисты, большинство из которых — выраженные инди-
видуалисты, на практике часто не следуют этому, однако их пренеб-
режение данным правилом причиняет значительное неудобство
другим. Конечно, применение этого правила не означает автомати-
ческого решения проблем в конкретных случаях. В одних и тех же
группах зачастую используются разные классификации. В таких
случаях можно следовать таким рекомендациям:
1. Если какая-либо классификация более соответствует [известным
фактам и принятым принципам], чем другие, принимайте её, если необхо-
димо, с такими изменениями, которые сделают её как можно более адек-
ватной.
2. Если выбор по соответствию не очевиден, принимайте и, если необ-
ходимо, изменяйте ту классификацию, которая используется шире всего.
3. Если выбор по широте использования не очевиден, принимайте и,
если необходимо, изменяйте наиболее авторитетную классификацию, при-
давая должный вес опыту классификатора, осмысленности и множествен-
ности использованных им данных, а также приемлемости его таксономи-
ческих принципов и приёмов.
Концепция соответствия существенна для этих правил, однако
она ещё не была определена здесь. Это сложный предмет, требу-
ющий несколько более пространного обсуждения. Ранее часто ут-
верждалось, а иногда говорится и теперь, что цель эволюционной
классификации — точно выражать филогению. Истина, однако, в
том, что ни одна из ныне выработанных форм классификации,
включая, конечно, и линнеевскую иерархию, не способна реально
выразить филогению, во всяком случае представить её исчерпыва-
юще и непротиворечиво. Это было ясно уже основателю эволюци-
онной таксономии. Имея в виду свою диаграмму филогении, Дар-
вин (Darwin, 1859) писал:
Без использования ветвящейся диаграммы, а только посредством на-
званий групп, написанных в определённой последовательности [как в
иерархической классификации. — Дж.С.], не было бы никакой возможно-
сти представить естественный порядок; и, как общеизвестно, решительно
132
Дж. Г. Симпсон. Принципы таксономии животных
невозможно представить в виде ряда или на плоской поверхности родство,
которое мы раскрываем в природе между существами данной группы.
Эволюционные таксономисты всегда осознавали этот факт, ко-
торый уже Дарвин считал общеизвестным, однако когда они гово-
рили, что классификация выражает филогению, то обычно имели
в виду, что она основана на филогении или является частичным её
выражением. У критиков эволюционной классификации не было
трудностей с демонстрацией того, что она неизбежно является не-
адекватным выражением филогении. Однако, когда они делали зак-
лючение, что эволюционная классификация по этой причине не-
практична и не реализует свои собственные предпосылки, то они
неверно трактовали эти предпосылки и подвергали атаке «соломен-
ное чучело».
Утверждение, что эволюционная классификация основана на
филогении, также служит предметом неверного толкования. Его
воспринимали таким образом, что такая классификация будто бы
строго воспроизводит родословные линии и их ветвление, как это
видно в обычной диаграмме филогенетического дерева. Это (о чём
Дарвин также уже знал) всё ещё не вся правда. Эволюционная
классификация, вообще говоря, требует информации, которая во
многом является филогенетической, но которую практически невоз-
можно включить в древовидную схему. Она также с необходимос-
тью требует некоторых произвольных делений (arbitrary divisions),
которые могут быть применены к дереву как основе, но которые не
вытекают из структуры дерева в его исходном содержании. Эти
факты, хотя они на самом деле не делают классификацию менее
филогенетической по своим основаниям, придают некоторую дву-
смысленность самому термину, вот почему я отдаю предпочтение
обозначению «эволюционная» перед «филогенетической» для той
категории классификаций, о которых я говорю.
Предпочтительнее рассматривать эволюционную классифика-
цию не как выражение филогении и даже не как основанную на
ней (хотя в некотором достаточно широком смысле это верно), но
как совместимую с ней. Совместимая эволюционная классифика-
ция — есть классификация, утверждения которой, выведенные
согласно заявленным критериям, не противоречат воззрениям
классификатора на филогению исследуемой группы. Значит, теперь
От таксономии к классификации
133
мы должны обсудить критерии, которые используются для созда-
ния эволюционной классификации, и посмотреть, каковы её утвер-
ждения. Основные категории критериев, которые надлежит рас-
смотреть, следующие:
1. Критерии, связанные с объективностью, реальностью, услов-
ностью и тому подобное.
2. Критерии, связанные с монофилией, полифилией, кладами и
градами.
3. Критерии, связанные с разными категориями и степенью род-
ства, проявляющегося в филогении.
4. Критерии, связанные с относительной древностью таксонов.
О каждом из них можно столько много сказать, что они будут
рассмотрены один за другим в последующих разделах данной главы.
Объективность и произвольность
Много чернил было пролито, и я думаю, что в основном на-
прасно, по поводу аргументов относительно того, могут ли таксо-
ны быть «реальными», или «объективными». Дискуссия была в
разгаре уже во времена Линнея, который настаивал (в иных терми-
нах) на том, что виды могут быть объективными, роды — обычно
объективны, а более высокие категории — нет. Наиболее частым,
но от этого не менее спорным, было заключение, актуальное и се-
годня, что виды, если они правильно определены, реальны или
объективны, а другие категории — нет (см. ссылки в: Simpson,
1951; см. также обсуждение вида в главе 5 настоящей книги). Вся
дискуссия вместе с этим спорным заключением действительно име-
ет свои основания, однако она обычно облекалась в неверные или,
по меньшей мере, туманные слова. Всякое определение таксона,
если оно таксономически состоятельно,2 ограничивает некоторое
множество реальных, объективных организмов. С другой стороны,
сама концепция таксона — то, что действительно присутствует в
сознании классификатора, что он именует и на что ссылается, —
2 Примеры очевидно несостоятельных определений: «насекомые с 8 нога-
ми» или «животные с одним или более ядрами». Под первое не подпадает ни
одно животное, второму соответствуют все животные.
134
Дж. Г. Симпсон. Принципы таксономии животных
неизменно субъективна, касается ли это вида или любой другой
категории. Таковы же предлагаемые определения и используемые
названия: они не имеют реального отношения к каким-либо объек-
там в природе, если они не выявлены субъективным процессом
классифицирования. Таким образом, всякий таксон в каких-то от-
ношениях или в определённой терминологии реален, или объекти-
вен, а в других в то же самое время — нереален, или субъективен.
Центральный вопрос данной дискуссии, если освободиться от
двусмысленностей и не оставаться в рамках чистой семантики,
можно поставить так: являются ли группы, распознаваемые как
таксоны, непрерывными или дискретными в некотором биологи-
чески значимом, строго очерченном [defined] и определимом
[determinable] смысле. Для того, чтобы прояснить его, я в своё вре-
мя предложил (Simpson, 1951) использовать термины «непроиз-
вольный» [nonarbitrary] и «произвольный» [arbitrary] вместо реаль-
ного и нереального, объективного и субъективного. Группа непро-
извольна по включению [as to inclusion], если все её члены обра-
зуют непрерывный ряд по какому-либо критерию, и непроизволь-
на по исключению [as to exclusion], если она отделена разрывом от
любой другой группы по этому же критерию. Она произвольна по
включению, если внутренне неоднородна, и по исключению, если
не отделена разрывом. Для видов, если следовать обычной практи-
ке их определения у современных млекопитающих (см. главу 5),
подходящим критерием непрерывности является способность к
размножению, реализуемая как вертикально во времени, так и го-
ризонтально в пределах группы. Таким образом, для тех, кто счи-
тает виды объективными, а другие таксоны субъективными, данная
позиция может быть чётко обозначена следующим образом: вид
(определяемый генетически) — единственный таксон, который
обычно бывает непроизвольным как по включению, так и исклю-
чению. Внутривидовые таксоны обычно непроизвольны по вклю-
чению, но произвольны по исключению (см. рис. 5А).
Несмотря на разные сложности и проблемы, с некоторыми из
которых, особо важными, мы будем разбираться в главе 5, данный
критерий обычно пригоден для различения эволюционных внутри-
видовых, видовых и надвидовых таксонов современных животных.
Однако даже в их случае возникает вопрос о том, какие критерии
От таксономии к классификации
135
Рис. 5. Произвольный и непроизвольный аспекты одновременно
СУЩЕСТВУЮЩИХ таксонов в отношении генетической непрерывности
А. Общее представление о произвольности внутри- и надвидовых таксонов.
Особи показаны чёрными точками. Каждый из двух представленных видов,
очерченных штриховыми линиями, формирует сплошную сеть последовательно
скрещивающихся индивидов. Каждый из них является непроизвольным как по
включению, так и исключению. В одном из них точечной линией очерчен подвид,
который смотрится непроизвольным по включению, но произвольным по
исключению. Эти два вида объединены в род, очерченный линией из разомкнутых
кружков: он непроизволен по исключению, но произволен по включению.
Б. Схема генетических связей в пределах вида, разделённого на подвиды,
как показано штриховой линией, между которыми скрещивание менее
эффективно, чем в пределах каждого из них.
Б
использовать для условного разделения видов на внутривидовые
таксоны и для условного объединения видов в надвидовые таксо-
ны. Вопрос определения категорий, к которым относятся эти так-
соны, должен быть отложен до того момента, когда мы будем рас-
полагать несколькими разными критериями. Здесь можно упомя-
нуть два возможных критерия, которые логически связаны с кате-
горией произвольности.
В пределах вида могут существовать границы, обычно просле-
живаемые географически, через которые гибридизация менее успеш-
на, чем в пределах популяций по обе стороны границы. Разделение
вида на внутривидовые таксоны, такие как подвиды, по таким гра-
ницам менее произвольно, чем разрубание достаточно свободно
размножающихся популяций. Подвид в таких случаях, которые да-
леко не универсальны, может быть сравнительно, но не абсолютно,
непроизвольным как по включению, так и исключению (рис. 5Б).
136
Дж. Г. Симпсон. Принципы таксономии животных
Среди видов одни более сходны между собой, чем другие. Сле-
дует признать, что сходства, коль скоро они гомологичные, отража-
ют родственную близость, что позволяет включать другие крите-
рии, обсуждаемые далее. Эти критерии также следует применять в
каждом отдельном случае, но критерий, связанный с условностью,
может основываться на сходстве как таковом. Если существуют
группы видов такие, что все виды одной группы более сходны друг
с другом, чем с какими-либо видами в любой другой группе, то
отождествление этих групп с надвидовыми таксонами является от-
носительно непроизвольным (рис. 6). (Критерий относительной
непрерывности в данном случае означает, разумеется, степень сход-
ства.) Майр и др. (Mayr et al., 1953) определяют категории рода и
семейства главным образом на этой основе, по присутствию «ре-
шающего разрыва» между соответствующими таксонами. Приме-
нение данного критерия не только возможно [теоретически], но и
желательно там, где это возможно [практически]. Однако эта воз-
можность не всегда имеется, а ещё реже реализуется. Поэтому этот
критерий не может быть единственным для определения категорий.
Когда вводится временной параметр, применение критерия про-
извольности усложняется и во многом утрачивает практичность.
Все таксоны животных, будучи прослеженными назад во времени,
Рис. 6. Группировка таксонов по разрывам в сходственных связях
Виды обозначены крестиками, высшие таксоны — штриховыми линиями
Пространственная разобщённость отражает степень сходства.
А. Сравнительно непроизвольная классификация видов в две группы. Вид a
в группе / в действительности дальше от некоторых видов, например, от Ь, в этой
же группе, чем от некоторых видов например, от с, в группе П. Тем не менее,
каждый разрыв в цепочке связей между видами а и b меньше, чем между ним и с
Б. Более чёткая группировка видов в таксоны трёх последовательных рангов
От таксономии к классификации
137
оказываются генетически непрерывными. Однако отдельные ветви
становятся дискретными после обособления, поэтому критерии
произвольности и непроизвольности к ним применимы. Когда рас-
сматривается вся структура во временном аспекте, любые таксоны
могут быть непроизвольными хотя бы по включению: это обеспе-
чивается критерием монофилии, который будем рассмотрен далее.
Однако по исключению ни один таксон (в том числе вид) в этой
общей структуре не может быть строго непроизвольным. Прихо-
дится в каком-то участке непрерывно эволюирующей линии про-
водить совершенно произвольную границу, соответствующую не-
которому моменту времени, и говорить: «здесь заканчивается один
таксон и начинается другой». В пределе это означает, что некое
животное в какое-то мгновение может принадлежать одному виду,
а в следующее — другому. Это очевидно абсурдно, однако не в
большей мере, чем в результате применения других критериев для
обозначения таких по необходимости условных выделений. В дей-
ствительности наши данные всё же не столь полны, чтобы подоб-
ные абсурдные ситуации возникали на практике.
Некоторые [эволюционные] феномены смягчают произволь-
ность такого разделения. Немногие исследователи (например,
Schindewolf, 1950; Goldschmidt, 1940) утверждают, что виды и выс-
шие категории обычно возникают внезапно, одним резким скачком.
Если так, то каждый такой скачок задаёт точку непроизвольного
деления. Такой феномен может иметь место, например, вследствие
полиплоидии, однако общее мнение (с которым я полностью согла-
сен) таково, что это бывает очень редко, особенно у животных, и
что превращение одного высшего таксона в другой не происходят
таким путем. Однако изменения, с которых начинается то, что по-
том становится новым таксоном, особенно на высших уровнях,
происходят обычно, хотя и не универсально, с исключительно вы-
сокими эволюционными скоростями, т.е. посредством того, что я
назвал квантовой эволюцией (Simpson, 1953). В таких случаях пе-
риод быстрого перехода может служить основанием для сравни-
тельно непроизвольного разделения, однако конкретная граничная
точка по-прежнему остаётся произвольной.
На практике эту проблему вынуждены решать только палеон-
тологи, но их данные в этом отношении часто имеют дефекты,
138
Дж. Г. Симпсон. Принципы таксономии животных
которые обращают недостаток в достоинство. Все известные вре-
менные ряды, представленные конкретными ископаемыми форма-
ми, имеют пробелы — временные интервалы, в которых неизвес-
тны остатки исследуемой группы с длительной историей. Хотя это
только перерывы в данных, а не в действительной филогении,
они указывают точки деления, непроизвольные, по крайней, мере
для выборки, которая действительно классифицируется. Все пале-
онтологи используют их в качестве искомых точек деления. Такие
пробелы имеют также то практическое достоинство, что они
обычно обозначены более резко и обнаруживаются чаще в случае
высших, чем низших таксонов. (Этот факт часто приводится как
доказательство действительной прерывистости эволюции, однако
он легко объясним как результат неполной представленности фи-
логений, которые на самом деле непрерывны: см. Simpson, 1953,
1960а).
Необходимость произвольных делений в непрерывных времен-
ных рядах по-прежнему сохраняется и представляет интерес как
для теории таксономии, так и для практики классификации. В на-
стоящее время известно немало коротких или умеренно протяжён-
ных в целом непрерывных рядов видов и родов, которые требуют
произвольного разделения на таксоны. Новые открытия постоянно
заполняют разрывы на всех уровнях. Например, переход от клас-
са Reptilia к классу Mammalia всё более заполняется вновь откры-
ваемыми ископаемыми, так что вопрос о том, где следует провес-
ти произвольную границу между этими классами приобретает всё
большую дискуссионность (см. комментарии Вэн Вэйлена, Рида и
Симпсона в «Evolution», в печати [30]). Очевидно, это больше воп-
рос вкуса или опыта в искусстве построения классификации, чем
теории таксономической науки.
Даже при включении временной шкалы и в предположении,
что филогении детально известны, некоторые группы могут ока-
заться определённо менее условными, чем другие, если использо-
вать «произвольность» в более широком, не столь техническом
смысле, как она определена выше. Этот момент адекватно пред-
ставлен на рис. 7, а его конкретные иллюстрации рассмотрены в
главе 6. В таких случаях вступают в силу другие соображения, ка-
сающиеся монофилии, степени обособленности и разнообразия.
От таксономии к классификации
139
Рис. 7. Три гипотетические филогении
Они аналогичны многим из тех, которые действительно имеют место; их
разделение на таксоны показано штриховой линией. В некотором широком и
нестрого фиксированном смысле филогения в А, очевидно, менее произвольно
разделена, чем в Б, а в Б менее произвольно, чем в В.
Когда имеется выбор, непроизвольная система обычно предпоч-
тительнее произвольной. Однако практически невозможно добиться
полной непроизвольности для всех таксонов, за исключением ви-
дов, хотя часто и для них это невозможно. Нет ничего порочного
в произвольности практики любого искусства, включая искусство
классификации. На самом деле некий элемент произвольности аб-
солютно необходим, а группы, ограниченные произвольно, не ста-
новятся из-за этого менее «реальными». Необъективны границы
между группами, а не сами группы.
Монофилия и полифилия
В общепринятом смысле, приводимом (с вариациями) во мно-
гих словарях, монофилию определяют как эволюционное проис-
хождение от одного предка, а полифилию — как возникновение от
двух или более предков. Это и похожие определения весьма мно-
гозначны, а также неоперациональны, т.е. они настолько нечёткие,
что вырабатываемые на их основе критерии недостаточны для от-
личения одного от другого при анализе данных. Если концепции
предназначены для практических нужд — а они должны быть та-
кими, им следует давать более чёткие определения. Вскоре это бу-
140
Дж. I. Симпсон. Принципы таксономии животных
дет сделано, однако прежде надлежит разобраться с тем фактом,
что в этой области имеются две связанные, но различные концеп-
ции, которые в обширной литературе по данному предмету иног-
да смешиваются (см., например, Remane, 1956 и ссылки у него).
Первый вопрос — о распространённости и природе полифилии в
эволюционном процессе. Второй — может ли вообще и в каком
смысле монофилия использоваться как критерий (канон искусства)
в практике классификации.
Сегодня кажется любопытным, что некоторые из первых эво-
люционных таксономистов, особенно Т. Гексли (Huxley, 1880), счи-
тали, что «законы эволюции» предполагают высокую частоту по-
лифилии, хотя этот вывод не поддерживался Дарвином. Гексли по-
лагал, что каждый отряд млекопитающих независимо проходил ста-
дии «гипотерий», «прототерий», «метатерий» и «эутерий». Совре-
менные однопроходные (отряд Monotremata) не пошли в своей эво-
люции выше стадии прототерий, а современные сумчатые (отряд
Marsupialia) — выше стадии метатерий; при этом названные отря-
ды считались совершенно обособленными от прочих и не имели
отношения, например, к прототериевым и метатериевым предкам
нынешних эутериевых приматов.3 Ещё более крайних воззрений
придерживались некоторые более поздние таксономисты: Кляйнш-
мидт (Kleinschmidt, 1926), например, придерживался точки зрения
раннего пост-дарвинизма, что каждый вид млекопитающих имел
отдельного предка среди простейших} а Роза (Rosa, 1931) утверж-
дал, что общий предок сумчатых и плацентарных должен был быть
беспозвоночным.
Такие воззрения настолько противоречат надёжным доказатель-
ствам, что мы можем лишь улыбнуться по их поводу и пойти да-
лее. Однако это не избавляет нас от необходимости решения дан-
ного вопроса на другом уровне и в связи с более разумным рас-
смотрением доказательств. Параллелизм — широко распространён-
ный феномен эволюции, и не так уж редко обнаруживается, что
3Для Гексли «Hypotheria» были гипотетическими, как видно из названия,
однако данное для них структурное определение очень хорошо соответствует
предполагаемым признакам терапсидных рептилий. Таким образом, точка зре-
ния Гексли означает, что каждый отряд современных млекопитающих имел
отдельного рептилийного предка.
От таксономии к классификации
141
некий общепризнанный таксон возник в результате параллельной
эволюции двумя или более линиями от разных предковых таксонов.
Вот лишь несколько примеров на разных уровнях:
Вымерший род лошадей (Equidae) Mery с hippus в его нынешней
трактовке, как обычно считается, вероятно, произошёл от несколь-
ких видов рода Parahippus (например, Quinn, 1955).
Подотряд приматов Anthropoidea в наиболее поздних классифи-
кациях, очевидно, возник из двух (возможно, даже из трёх) низших
таксонов подотряда Prosimii (например, Le Gros Clark, 1959).
Классический отряд Edentata, признаваемый почти в неизмен-
ной форме, начиная с конца XVIII в. вплоть до XX в., и всё ещё
признаваемый некоторыми нынешними авторитетами, определён-
но имеет три совершенно разных корня, вероятно, относящихся по
меньшей мере к двум разным отрядам (например, Simpson, 1945).
Класс Mammalia, неизменный во всех классификациях начиная
с Линнея, почти наверняка возник от нескольких разных таксонов
в пределах отряда Therapsida, ранее всегда, а в настоящее время
обычно определяемого как часть класса Reptilia (например, Simp-
son, 1959b).
Такие примеры — их можно множить и множить — приводят
ко второму вопросу: как быть с ними в прикладном искусстве так-
сономии. В общем случае имеются четыре возможных подхода:
1. Вернуться к типологической или эмпирической таксономии,
не использующей филогенетические критерии;
2. В эволюционной классификации вместо критерия монофи-
лии или в дополнение к нему принять критерии, связанные с про-
хождением ступеней прогресса, которые в конечном итоге столь же
эволюционны, как и монофилия, и также имеют отношение к фи-
логении в менее узком её понимании;
3. Разработать более строгую, но эволюционно достаточно ос-
мысленную концепцию монофилии, которая свяжет её с рангами
таксонов, что в некоторых, но не во всех случаях сделает таксоны,
возникшие из более чем одной филетической линии, монофилетич-
ными по определению;
4. Принять абсолютное определение монофилии, чётко связан-
ной с каким-то одним таксономическим рангом. В таком случае
возможны два способа соблюдения такого более жёсткого критерия:
11—4338
142
Дж. Г. Симпсон. Принципы таксономии животных
а. Таксон, ранее рассматриваемый в качестве единого, может
быть разделён на два или более таксона одного ранга, каждый из
которых будет монофилетичным согласно принятому определению;
б. Рассматриваемый таксон может быть расширен за счёт вклю-
чения предковых групп, ранее помещаемых в другие таксоны, пока
весь предковый ствол не будет включён в данный таксон, чтобы он
соответствовал принятому определению монофилии.
Каждый из этих подходов применялся в отдельных классифи-
кациях и каждый был поддержан как единственный классифициру-
ющий критерий тем или иным таксономистом. По причинам, ко-
торые должны быть уже совершенно очевидны, я убеждён, что пер-
вый [подход] может быть принят только foute de mieux. Действи-
тельно, есть группы, для которых лучшие решения в настоящее
время недостижимы, особенно среди протистов, но возможно, что
и среди некоторых многоклеточных, о которых у нас просто не хва-
тает достаточно сведений. Однако для большинства многоклеточ-
ных информация, которой мы уже располагаем или которая в прин-
ципе доступна, позволяет прибегнуть к тому или иному из трёх
эволюционных подходов. Эти подходы, перечисленные в пп. (2)-
(4) моего списка, сейчас представляются практически исчерпыва-
ющими. Применение любого двух из них или всех вместе часто
будет давать один и тот же результат, в зависимости главным об-
разом от того, как именно определена монофилия. Все они мне
представляются пригодными в принципе, т.е. совместимыми с при-
нятой теорией эволюционной таксономии. Выбор какого-то одно-
го из них — это вопрос не правильного или ошибочного решения,
но творческого озарения в каждом отдельном случае, когда прихо-
дится принимать во внимание следующее:
желательное совпадение результатов применения двух или всех трех
подходов при решении данной задачи;
практическая возможность делать таксоны легко определяемыми и рас-
познаваемыми, при сохранении их эволюционной состоятельности;
соответствие другим канонам искусства [классификации], таким как
обособленность и разнообразие, обсуждаемым далее в этой главе;
соблюдение условия, что классификацию не следует изменять, если
только это действительно не требуется для соблюдения принципа [моно-
филии].
От таксономии к классификации
143
Часто утверждается, что эволюционной классификации более
всего соответствует подход (4а), очень узко и строго определяющий
монофилию. Нестабильность и неудобство, возникающие во мно-
гих случаях в результате его применения, используются как аргу-
мент против эволюционной классификации. Однако это совсем не
так: эволюционная классификация логически не сводится к подоб-
ному требованию. Подходы (2), (3) и (46) также полностью совме-
стимы с логикой эволюционной таксономии.
На самом деле почти, а то и вовсе невозможно ограничить оп-
ределение монофилии так, чтобы оно было и абсолютным, и прак-
тичным. Если бы «моно-» означало «минимально», соответствую-
щее определение требовало бы происхождения от единственной
особи или семейной пары. Однако с точки зрения такого опреде-
ления монофилии, возможно, лишь немногие таксоны животных
оказались бы монофилетичными. По-видимому, лучшим решени-
ем — именно его обычно имеют в виду, когда определяют этот тер-
мин (большинство таксономистов так и не смогло это сделать), —
было бы [соотносить монофилию] с происхождением от одного
вида. Тем не менее, легко показать, что оно нежелательно в теории
и обычно неприменимо на практике. Чтобы быть действительно
точным, это определение должно подразумевать, что единственный
предковый вид включён в данный таксон, который становится мо-
нофилетичным вследствие его [таксона] анцестрального статуса.4
Но ведь огромное большинство родов имели когда-нибудь два или
более видов, не связанных непосредственно друг с другом отноше-
нием предок—потомок. Значит, если только один предковый вид
поместить в род, которому он дал начало, то либо (а) этот вид ока-
жется включённым в два рода, в одном из которых он возник, а
другому дал начало, что запрещено иерархической системой, либо
(б) он будет объединяться в один и тот же род с видами, с которы-
ми он связан меньшим родством, чем с видами предкового рода (по
причинам, аналогичным тому факту, что по родству человек менее
4 В противном случае данное определение сделает монофилетичным лю-
бое объединение животных — такое, как род, составленный из видов, извест-
ных под названиями Homo sapiens и Drosophila pseudoobscura, поскольку ве-
роятно, что некий единственный вид, предковый для них обоих, действитель-
но входил в какой-либо таксон.
И*
144
Дж. Г. Симпсон. Принципы таксономии животных
близок к своим внукам, чем к своему брату). Последнее возраже-
ние в принципе не столь серьёзно, поскольку подобные аномалии
возникают в любом случае, когда филетические линии разделяют-
ся на временные отрезки, однако на практике это часто может со-
здавать неразрешимые проблемы. Горизонтальные связи между
одновременно существующими видами рода обычно более очевид-
ны, чем между всем родом и предковым для него видом, и они го-
раздо чаще служат тем основанием, на котором распознаётся и дол-
жен распознаваться род. При таком определении монофилии ещё
более ущербным на практике оказывается тот факт, что единствен-
ный предковый вид в действительности редко бывает известен, а
без его идентификации данное определение не имеет практического
значения. С тем большим основанием сходные возражения приме-
нимы ко всякому критерию, согласно которому каждый таксон над-
родового ранга должен происходить от единственного вида, вклю-
чённого в него.
Эти и другие соображения склоняют меня к тому, что опреде-
ление монофилии как происхождения от единственного вида или
от любого другого единственного таксона, ранг которого фиксиро-
ван определением, неприемлемо в классификации и не согласует-
ся с таксономическими принципами. Предпочтительно следующее
определение:
Монофилия есть происхождение таксона одной или нескольки-
ми филетическими линиями (временными последовательностями
популяций, связанных отношением предок—потомок) от одного
ближайшего к нему предкового таксона того же самого или бо-
лее низкого ранга.5 [31].
Согласно этому определению, монофилия может быть разной
степени или уровня, причём этот уровень, как правило, должен
5 Беккнер (Beckner, 1959) следовал той же самой линии мысли в опреде-
лении, которое концептуально почти идентично:
«Будем говорить, что таксон любого ранга является монофилетичным на
уровне таксонов ранга j всякий раз, когда любой член таксона является либо
непосредственным потомком члена данного таксона, либо непосредственным
потомком членов другого таксона ранга].»
Напомним, что в теории множеств члены таксонов являются индивидами,
но не другими таксонами. Члены рода не виды, но организмы.
От таксономии к классификации
145
уточняться для каждого отдельного случая. Уровень монофилии
устанавливается по той категории, к которой относится таксон наи-
меньшего ранга, являющийся единственным ближайшим предком
рассматриваемого таксона. Уровень полифилии устанавливается по
той категории, к которой относятся два или более таксонов наивыс-
шего ранга, являющиеся ближайшими предками рассматриваемо-
го таксона. Таким образом, в выше приведённом примере Merychip-
pus должен считаться монофилетичным на уровне рода, но поли-
филетичным на уровне вида.
Подвиды и виды определяются на основании других критери-
ев, которые перекрывают любой критерий монофилии. Подвид
может произойти от другого подвида, но вряд ли от какого-то фраг-
мента популяции, которому не придан статус таксона в классифи-
кации. Виды обычно минимально монофилетичны, т.е. на видовом
уровне, но они могут быть и полифилетичными, что само по себе
не отрицает современной таксономической теории или практики,
т.е. они могут возникать как частичные или полные межвидовые
гибриды. Для таксонов рангом выше вида я предлагаю следующие
критерии:
Все надвидовые таксоны должны быть минимально монофиле-
тичны, насколько это позволяют имеющиеся данные, т.е. таксон Т
должен быть монофилетичным на уровне С: род — на родовом
уровне, семейство — на уровне семейства и т.д.
При прочих равных условиях (т.е. если другие критерии не пе-
ревешивают явным образом данный) надвидовой таксон Т должен
быть монофилетичным на уровне С , или на более низком, если
позволяют данные и баланс с другими критериями.
Из ранее приведённых примеров, таксоны Anthropoidea и
Mammalia соответствуют второму, более сильному критерию, а род
Merychippus — первому, не столь сильному критерию, но не вто-
рому. Следует ли в данном случае рассматривать другие критерии,
которые перевесили бы более сильный критерий, классификатор
решает в индивидуальном порядке. По моему мнению, сильный
критерий часто перекрывается другими критериями в случае родов
и реже — для таксонов более высокого ранга. Таксон Edentata, как
он был исходно определён, не является монофилетичным ни по
одному из критериев и в настоящее время разделяется на три от-
146
Дж. Г. Симпсон. Принципы таксономии животных
дельных монофилетичных отряда практически всеми таксономис-
тами. Данное определение монофилии и основанные на нём кри-
терии теоретически состоятельны и практически реализуемы.
В своей основе они строго филогенетические. Они гарантируют,
что члены таксона, отвечающего критерию, специфичным образом
взаимосвязаны между собой и что степени родства обычно тесно
скоррелированы с рангом таксона. Информация, имеющаяся или
доступная для большинства многоклеточных, достаточна для опре-
деления с нужной вероятностью, выполняется ли соответствующий
критерий (как обычно, полная определённость невозможна и на ней
нельзя настаивать). Далее, эти концепции монофилии и полифилии
совершенно однозначны, если указаны соответствующие уровни
[иерархии], тогда как любая подобная концепция, не предполагаю-
щая указания уровня, ipso facto неоднозначна.
Грады и клады
Теперь несколько больше внимания будет уделено предмету,
который был затронут мимоходом при обсуждении монофилии:
группам животных, сходным по общему уровню организации, в
противовес группам общего филогенетического происхождения.
Дж. Гексли (Huxley, 1958) недавно обсудил этот вопрос в достаточ-
но ясной и доступной форме и предложил первые группы называть
градами, а вторые — кладами. Самые первые эволюционные так-
сономисты уже делали различия между ними, используя другие
термины, особенно Т. Гексли в связи со своими взглядами на клас-
сификацию млекопитающих, уже упомянутыми выше в настоящей
главе.
В эволюции часто бывает так, что целая группа животных,
представленная многими отдельными филетическими линиями,
развивается со значительным числом параллелизмов, проходя пос-
ледовательный ряд адаптивных зон или всё возрастающих уровней
организации. (О первых см. особенно Simpson, 1953, где приведе-
но много примеров и подробно рассмотрен весь этот феномен; о
вторых см. Huxley, 1958, который подчёркивал этот аспект град.)
В некоторых случаях это довольно устойчивое продвижение, как в
эволюции костистых рыб по направлению к телеостному уровню
От таксономии к классификации
147
на протяжении мезозоя. В других случаях имеет место сравнитель-
но быстрое изменение при переходе от одного уровня к другому с
последующей специализацией и меньшей диверсификацией при
достижении каждого уровня. В таком случае возникают более или
менее чётко выраженные ступени, как в случае листоядных (лес-
ных) и травоядных (пастбищных) лошадей, или между лемурами,
низшими обезьянами, человекообразными обезьянами и людьми.
Концепция град имеет наиболее тесное отношение именно к таким
ступеням.
Существование ступенчатого прогрессивного развития может
не быть очевидным, если филогению рассматривать исключитель-
но как филетическую линию, однако оно может быть чётко прояв-
лено в её структуре (см. рис. 8). В любом случае грады имеют от-
ношение к классификации и порождают определённые проблемы.
Одна из них видна из следующей схемы, заимствованной у Бейзера
(Bather, 1927) и представленной в очень упрощенной форме:
Первая Вторая Третья
a b с Курсив латинский
а b с Обычный латинский
а ₽ Y Греческий
Буквы можно понимать как виды, «Первая», «Вторая» и «Тре-
тья» — как филетические линии, а «Курсив латинский», «Обычный
латинский» и «Греческий» — как грады. Как подчеркнул Бейзер
(см. также Pirie, 1952), для практических целей не имеет никакого
значения, назовём ли мы а «Первым курсивом» или «Курсивом пер-
вым», т.е. имеем ли мы дело с филетической линией «Первая» с
видами а, а и а, или с родом-градой «Курсив» с видами а, b и с.
На самом деле часто применяются оба похода: например, среди
граптолитов род Monograptus исходно обозначал граду и иногда в
этом смысле применяется и сегодня. Однако, как сейчас известно,
этой грады независимо достигло значительное число разных клад,
каждой из которых в большинстве последних работ дано отдель-
ное родовое имя (например, Bulman, 1955).
Я бы сказал, что выбор между филетическими линиями или
градами, выделяемыми в качестве родов, не имеет особого значе-
ния, при условии, что мы можем предположить некую связь, все-
148
Дж. Г. Симпсон. Принципы таксономии животных
Рис. 8. Некоторые схематические представления соотношений между
СТРУКТУРНЫМ прогрессом и филогенией
А. Без чётко выраженных град и таксономически значимых отношений
между градами и кладами: обычный вариант во многих группах животных.
Б. Без чётко выраженных град, но с выраженным параллелизмом между
кладами, которые развиваются в едином направлении и образуют единую
градистическую последовательность; подобная, хотя несомненно более сложная,
картина наблюдается, например, у костистых рыб и грызунов.
В. С градами в форме выраженных ступеней и с кладами, более-менее, но
не строго соответствующих градам; подобная картина отмечается у приматов, а
также в других группах.
гда присутствующую в природе, но отсутствующую в сверхупро-
щенном примере, и далее еще при одном условии, что альтернати-
вы столь же просты и столь же хорошо уравнены, как это предпо-
лагает данный пример, чего на практике почти никогда не бывает.
Мы должны принять, что эти три филетические линии тесно вза-
имосвязаны между собой и что ни одна из них не находится в бо-
лее близкой связи с некоторой другой линией [т.е. с четвертой], чем
с двумя показанными. Если это не так, то упорядочение по градам
совершенно несостоятельно в качестве эволюционной классифика-
ции. На самом деле градистское упорядочение вряд ли можно адап-
тировать даже в предположительно неэволюционную классифика-
цию, поскольку реальные филетические линии демонстрировали
бы характерные несходства и не были бы столь отчётливо парал-
лельными. Таким образом, роды как филетические линии прием-
От таксономии к классификации
149
лемы в любом случае, тогда как грады [в качестве таксонов] при-
емлемы, только если они также удовлетворяют некоему филогене-
тическому критерию, хотя и не столь жёсткому, как видовая моно-
филия. Роды-грады монофи л етичны на родовом уровне, тем самым
соответствуя минимальному из наших критериев. Более того, гра-
ды являются эволюционными, и если минимальное условие моно-
филии выполнено, то их выделение столь же эволюционно, как и
родов-линий.
В реальных филогениях, с их сложным ветвлением и перепле-
тением гомологии, параллелизма, дивергенции и конвергенции (а
не только параллелизма, как в примере Бейзера), грады и клады
редко бывают совершенно уравновешены (я полагаю, что никогда)
и соотносятся друг с другом «под прямым углом» [32]. На самом
деле они обычно идут параллельно друг другу, так что грады и кла-
ды имеют выраженную тенденцию к совпадению, как на рис. 8В.
Концепция грады не имеет особого смысла, если только она не
применена к родственным организмам. И насекомые, и [наземные
позвоночные] [33] имеют (конвергентные, а не гомологичные или
параллельные) глаза, мозг, членистые конечности и т.д., однако
было бы, очевидно, таксономически неоправданно относить их по
этой причине к одной граде. Как минимум, грады должны основы-
ваться на параллелизме, а не на конвергенции, а их применение
никоим образом не должно исключать филогенетическую основу.
По определению, клада монофилетична и возможное совпаде-
ние град и клад в большинстве случаев будет зависеть от приня-
того уровня монофилии. Выше приведённые примеры град у лоша-
дей и приматов удовлетворяют моему критерию для соответству-
ющего уровня монофилии в классификации. То же верно в отно-
шении примеров самого Гексли, а также всех тех, которые мне при-
ходят на ум, хотя возможны и исключения. (Это отчасти артефакт,
потому что в возможных примерах, не отвечающих данному кри-
терию, как в случае отряда Edentata, включающего панголинов,
трубкозуба и броненосцев, вывод о том, что это единая града, не
имеет грамотного обоснования.) В серьёзном исследовании я пред-
почёл бы таксоны-клады, т.е. стал бы искать как можно более низ-
кий уровень монофилии. Однако если какой-нибудь классификатор
выделяет таксоны-грады по причине своих предпочтений в искус-
150
Дж. Г. Симпсон. Принципы таксономии животных
стве или из-за ошибочно принятия параллелизма за гомологию, я
рассматриваю эти таксоны как валидные эволюционные группы:
всегда можно допустить, что имеющиеся доказательства не проти-
воречат их соответствию требованию минимальной монофилии
согласно принятому мной критерию.
Гексли в порядке обсуждения предположил возможность ис-
пользования двух разных классификаций и номенклатур — одну
для град и другую для клад. Концепции и понятия грады и клады
в высшей степени полезны для понимания и обсуждения эволюци-
онных феноменов и классификации. Однако я не вижу какой-либо
необходимости или практичности разрабатывать две системы клас-
сификаций на основе каждой из этих концепций.
Вертикальные и горизонтальные связи
Кому человек более родствен — своему отцу, сыну или брату?
Гены, вовлечённые в родство, могут быть весьма разными, но сте-
пень генетической близости сына к отцу всегда одна и та же — 0.5,
если её выражать долей общих хромосом. Генетические связи бра-
тьев изменчивы — от 1.0 до 0.0 в долях общих хромосом, хотя ве-
роятность этих крайних вариантов исключительно мала, а среднее
значение то же, что и в случае отца и сына. К сожалению, связи
между таксонами не имеют такого фиксированного априорного
ожидания, а также точной оценки. Тем не менее, имеются те же две
категории связей: между последовательными таксонами в филети-
ческой линии предков и потомков, с одной стороны, и между од-
новременно существующими таксонами с более или менее отдалён-
ными общими корнями, с другой стороны. Согласно принятому
способу изображения деревьев, первые связи называются верти-
кальными, вторые — горизонтальными.
Каждый из двух этих типов связей точно так же объективен,
как и другой, и в обоих случаях связи могут быть как близкими,
так и отдалёнными. Более того, представляется очевидным, что оба
[типа связей] в равной степени филогенетические: по аналогии с
человеком, было бы в высшей степени забавным утверждать, что
связи между братьями не имеют никакого отношения к их предкам
или филогенетическому происхождению. Поэтому мы не будем тра-
От таксономии к классификации
151
тить времени на откровенно ошибочную идею, часто высказывае-
мую как сторонниками, так и недоброжелателями эволюционной
классификации, что только вертикальные связи являются филоге-
нетическими. (Осборн, крайний сторонник вертикальной классифи-
кации, обыкновенно повторял, что только вертикальные связи —
эволюционные).
При работе с современными животными или с одновременно
существовавшими ископаемыми фаунами непосредственно доступ-
ны только горизонтальные связи. Однако они интерпретируются в
свете их значения для установления родственной близости, что,
таким образом, вводит в классификации одновременно существу-
ющих животных заключения о вертикальных связях. Во временных
рядах ископаемых форм непосредственно присутствуют вертикаль-
ные связи. В любом случае общие принципы классификации дол-
жны учитывать оба типа связей, которые более или менее адекват-
но отражены на филогенетическом дереве или топологически по-
добной дендрограмме.
Обследование всякого достаточно разветвлённого дерева или
дендрограммы сразу же обнаруживает, что классификация только
по вертикальным или по горизонтальным связям абсолютно невоз-
можна. (Между прочим, это верно и в отношении эмпирической
или типологической классификации, поскольку эти два типа связей
дают разную структуру сходственных отношений или разные архе-
типы, которые рано или поздно вступят во внутренний конфликт и
потребуют выбора какого-то одного из них.) При переводе фило-
гении в таксоны в действие должен вступать компромисс: одни
группировки таксонов будут горизонтальными, другие — верти-
кальными. Выбор какого-то из этих компромиссных решений час-
тью является искусством таксономии, вопросом вкуса и мастерства.
Это является одной из двух причин, почему одна и та же установ-
ленная филогения может быть совместимой со многими разными
классификациями. Другая важная причина — дробительство или
объединительство — будет рассмотрена далее.
По моему мнению, нет хороших прямых критериев для выбо-
ра между вертикальной и горизонтальной классификацией исклю-
чительно на этой основе. Классификаторы разняться свойствами
темперамента, или можно сказать — школами искусства. Одни (на-
152
Дж. Г. Симпсон. Принципы таксономии животных
пример, Осборн), которые вероятнее всего настаивали бы на самом
низком уровне монофилии и являются крайними дробителями, от-
дадут высший приоритет вертикальным связям и согласятся на ком-
промисс с горизонтальными связями только в случае крайней не-
обходимости. Другие (например, Фрешкоп), у которых требования
к монофилии минимальны или их нет вовсе и которые — крайние
объединители, уделяют особое внимание горизонтальным связям и
по возможности игнорируют вертикальные. Обе крайности мне
представляются неоправданными, как и всякие крайности. Однако
в поисках баланса между двумя крайностями имеется достаточно
возможностей для творчества. Это позволяют сделать другие кри-
терии. Абсолютная монофилия чрезмерно подчёркивает вертикаль-
ные связи, но ранее выдвинутые критерии относительной монофи-
лии могут сохранить вертикальные элементы без того, чтобы они
конфликтовали с горизонтальными делениями. Критерий стабиль-
ности благоприятствует горизонтальным элементам, поскольку го-
ризонтальные связи, как правило, легче распознать (особенно среди
современных животных), они часто были раньше обнародованы и
менее подвержены изменениям благодаря новым открытиям, осо-
бенно среди ископаемых.
Необходимость компромисса и его природа могут стать более
очевидными из следующего примера (рис. 9). Отряд млекопитаю-
щих Carnivora появляется в палеоцене и эоцене в составе сразу
нескольких групп, обычно в ранге семейств, которые несомненно
произошли от общего предка, но которые уже резко различны с
первых известных представителей. Одна группа, Miacidae, верти-
кально связана с дожившим до современности подотрядом
Fissipeda (наземные хищные), причём очень близка к его предку
или даже включает его. Однако на уровне палеоцена и эоцена ми-
ациды имеют столь же близкую, но горизонтальную связь с семей-
ствами, не особо связанными вертикально с Fissipeda. Классичес-
кая на сегодняшний день система Мэттью (Matthew, 1909) помеща-
ет все палеоцен-эоценовые группы, включая Miacidae, и их потом-
ков, кроме Fissipeda, в подотряд Creodonta. Это — чётко горизон-
тальная классификация семейств, но с необходимым компромиссом
в отношении вертикальной классификации в пределах самих се-
мейств. Вертикальная классификация семейств помещает Miacidae
От таксономии к классификации
153
Рис. 9. Схемы горизонтальной, вертикальной и смешанной
КЛАССИФИКАЦИЙ
Структура отношений сходна с филогенией семейств наземных Carnivora,
в которой Miacidae занимают правое нижнее положение.
А. Основные группы выделены строго горизонтально, как подотряды
Fissipeda (Fi) и Creodona (Сг) в классификации Мэттью [Matthew].
Б. Основные группы выделены строго вертикально, с тремя подотрядами
(X, Y, Z) в дополнение к Fissipeda.
В. Компромиссный вариант, как в стандартной современной классификации
Carnivora.
в состав Fissipeda, а её логической крайностью является выделение
каждого из палеоцен-эоценовых семейств в отдельный подотряд,
поскольку они, судя по имеющимся данным, друг от друга отлича-
ются так же, как от Miacidae, и приблизительно того же возраста.
Это переносит необходимость горизонтального разделения на уро-
вень предков всех таким образом созданных «подотрядов», что ока-
залось удачным решением на сегодняшний день, поскольку имеется
значительный пробел в знании начальной дивергенции каждого из
постулированных «подотрядов». Тем не менее, насколько мне из-
вестно, никто не предлагал такую систему, очевидно, по той при-
чине, что она даёт подотряды, крайне несбалансированные по сте-
пени обособленности и разнообразия (эти критерии обсуждаются
далее). Компромисс между двумя крайностями, который возвраща-
ет к временам задолго до Мэттью (главным образом, хотя и не бук-
вально, к Соре, 1875), помещает Miacidae «вертикально» в Fissi-
peda, но сохраняет всех остальных паолеоцен-эоценовых хищных
154
Дж. Г. Симпсон. Принципы таксономии животных
в едином «горизонтальном» подотряде Creodonta. Это оказалось
довольно устойчивой системой, признанной и ныне (для дальней-
шего обсуждения см. Simpson, 1945).
Имеется некоторая тенденция к тому, что категории различают-
ся пропорцией горизонтальных и вертикальных связей между от-
носящимися к ним таксонами. К этому мы вернёмся в главах, по-
свящённых низшим и высшим таксонам. Здесь же можно лишь от-
метить, что (имея в виду значительный разброс и некоторую несог-
ласованность данных): роды обычно представляют собой пучки
видов, особенно в неонтологии; подсемейства и семейства могут
иметь преимущественно вертикальные связи, особенно в палеонто-
логии; а в надсемейственных таксонах обычно соблюдается баланс
двух типов связей.
Дивергенция и разнообразие
Достаточно понятно, что концептуально и терминологически
работа над классификацией осуществляется в первую очередь на
том первичном уровне, к которому относится определение степе-
ни сходств и различий. Но как было показано, это необходимая, но
не достаточная основа. Сходства и различия сами могут быть раз-
ных категорий; они часто конфликтуют; они требуют определённых
критериев ранжирования, которые не выработаны в априорных или
эмпирических подходах; но без таких критериев они лишены со-
держательного значения и более широкого научного смысла и по-
лезности. Эти несоответствия восполняются принципами и крите-
риями, специфическими для эволюционной классификации. Это не
значит, что сходства и различия сами по себе не относятся к кри-
териям, применяемым в эволюционной классификации. Совмести-
мость с филогенией — основное требование эволюционной клас-
сификации, однако этому требованию может соответствовать целый
ряд альтернативных классификаций. Тогда вторичными критерия-
ми выбора между этими альтернативами становятся степени сход-
ства и различия по гомологичным и отчасти параллельным призна-
кам, но ни по каким иным.
Критерии, вырабатываемые на основе относительной диверген-
ции [divergence], т.е. суммы различий по всем исследуемым призна-
От таксономии к классификации
155
кам, применяются главным образом для ранжирования уже выде-
ленных таксонов, нежели для их предварительного распознавания.
Они могут применяться для анализа как в пределах таксонов, так
и между ними, в зависимости от обстоятельств. Предлагаемые кри-
терии таковы:
В группе родственных таксонов, относящихся к одной катего-
рии С, желательно, чтобы последовательный ряд различий между
наименее разошедшимися парами Т был приблизительно одинаков.
В группе родственных таксонов одного ранга желательно, что-
бы дивергенция внутри каждого Т была, насколько это практичес-
ки осуществимо, приблизительно равна средней величине для груп-
пы в целом.
Иерархическая организация родственных или соподчинённых
таксонов разных рангов должна быть по возможности такой, что-
бы степень дивергенции как внутри членов С, так и между ними,
была меньше, чем для членов категории С р но больше, чем для
членов категории С. г
Эти рекомендации предлагаются скорее для конкретных обсто-
ятельств, чем в качестве общих правил, поскольку их применение
иногда невозможно, а иногда перекрывается другими критериями.
Первая рекомендация, очевидно, не приложима к Т в случае мо-
нотипических Т р примером служат трубкозубы — единственное
семейство Orycteropodidae в отряде Tubulidentata, в котором нет
различий внутри семейства (Т). Однако именно второе правило
позволяет ранжировать отряд: обособленность трубкозубов от дру-
гих млекопитающих приблизительно соответствует среднему уров-
ню дивергенции между обычно признаваемыми отрядами млекопи-
тающих. Именно подобная ситуация оправдывает и в некоторых
случаях даже требует выделения монотипических таксонов. Отсут-
ствие различий на уровне семейства также делает неприменимой
третью рекомендацию, разграничивающую отряд и семейство.
Возможной четвёртой рекомендацией может быть условие, со-
гласно которому степень дивергенции внутри каждого Т из отно-
сящихся к одному данному Т р была бы меньше, чем между раз-
ными Т [34]. Однако известно немало случаев, в которых общая
сумма различий [divergence] в пределах хорошо определённого
рода, такого как сверхобильный Rattus, превышает дивергенцию
156
Дж. Г. Симпсон. Принципы таксономии животных
между этим родом и близким к нему, таким как Thallomys [35]. Та-
кие случаи столь многочисленны и в принципе могут столь часто
возникать, что данная рекомендация едва ли практична и жела-
тельна.
Приведённые рекомендации просто более точно выражают об-
щий настрой таксономистов, считающих, что таксоны одного ранга
должны быть приблизительно одного «размера» [size] в отношении
морфологических или других отличительных особенностей, тогда
как таксоны более высоких рангов должны быть большего «разме-
ра» в этом же смысле. С этой точки зрения введение подобных ре-
комендаций достаточно понятно и, по-видимому, не требует даль-
нейшего обсуждения. Однако имеется и другой смысл «размера»,
имеющий отношение к классификации: это — число Т. р включён-
ных в Т, т.е. объём разнообразия [diversity] таксона, или уровень
его политипичности.
В данном случае всякая рекомендация должна применяться не
только после ведущих эволюционных критериев, таких как отно-
сительная монофилия, но также и после рекомендаций, относящих-
ся к дивергенции. Имеются огромные различия в разнообразии
даже между таксонами с приблизительно одинаковыми внутренней
и внешней степенями дивергенции. В современной классификации
род Eropeplus включает только один номинальный таксон, тогда как
близкий род Rattus включает более 550 таксонов [36], а есть и бо-
лее разнообразные роды. В таких примерах столь различным по
разнообразию таксонам одинаковый родовой ранг присваивается
главным образом по степени дивергенции. Таксоны, которые про-
тивоположны по разнообразию — либо крайне однородные (осо-
бенно монотипические), либо в высшей степени разнородные
(крайне политипические), тем не менее, должны изучаться в этом
отношении очень внимательно. Монотипический род, такой как
Asinus в некоторых классификациях современных Equidae, может
служить примером необычно и возможно неоправданно низкого
критерия родовой обособленности [divergence]. Если так, то другой
классификатор, возможно, увеличит [пороговую] меру обособлен-
ности, соединив два или более выделенных рода в один (Equus, как
в случае с Asinus), который в этой связи станет соответствовать
более обычному уровню политипии. Однако возможны и несомнен-
От таксономии к классификации
157
ные случаи: ни один современный род не может быть сближен с
Orycteropus настолько, чтобы сделать его политипическим.
С другой стороны, в случае крайней политипии следует преду-
смотреть снижение жёсткости критерия дивергенции, что позволит
разделить таксон на два или более того же ранга с меньшим разнооб-
разием. Это во многом вопрос возможного удобства работы с менее
громоздкими таксонами. Например, большинство наиболее разнооб-
разных групп млекопитающих состоит из так называемых крыс и
мышей и их близких родичей. Традиционно всех их относили к одно-
му семейству Muridae, которое просто необъятно. В настоящее вре-
мя их нередко делят на два семейства — Muridae и Cricetidae, всё
ещё слишком разнообразные, хотя уровень их дивергенции и мень-
ше, чем в среднем для семейств млекопитающих и даже грызунов.6 7
В случаях крайней политипии, в первую очередь следует обра-
щать внимание на снижение громоздкости таксона за счёт исполь-
зования промежуточных таксонов более низкого ранга или под-
групп. Это не может считаться хорошим решением для Muridae и
Cricetidae, поскольку даже при их разделении они требуют введе-
ния подсемейств, чтобы ими можно было манипулировать. Одна-
ко, например, есть немало обильных видами родов, которые, тем
не менее, весьма однообразны (невысокая внутренняя диверген-
ция), поэтому лучше в них выделять подроды, чем делить на само-
стоятельные роды, чему хорошим примером служит RattusJ [37]
Имеется естественная и, в некоторых рамках, оправданная тен-
денция использовать заниженные критерии обособленности в наи-
более разнообразных группах и завышенные — в менее разнооб-
разных. Хищные, например, значительно менее разнообразны, чем
грызуны, так что в большинстве классификаций средняя диверген-
ция между семействами хищных заметно выше, чем между семей-
6 Многие роды, ранее включавшиеся в Muridae, в настоящее время отне-
сены к другим семействам, но главным образом благодаря уточнению их род-
ственных связей или уточнению концепции монофилии.
7 По разным причинам, во многом психологическим, многие классифика-
торы испытывают антипатию к подродам. В широко политипических группах
они, тем не менее, настолько удобны, что заслуживают широкого применения.
К счастью, эта антипатия, по-видимому, не простирается на такие промежуточ-
ные таксоны как подсемейства.
12 — 4338
158
Дж. Г. Симпсон. Принципы таксономии животных
ствами грызунов. Однако при этом следует иметь в виду и другие
соображения. Судя по имеющимся данным, семейства грызунов
должны отличаться меньшей дивергенцией, если применять к ним
более разумные стандарты монофилии.
Дробительство и объедимительство
Со времён Линнея произошло огромное увеличение числа опи-
санных таксонов на всех признанных уровнях. Отчасти это было
прямым следствием новых открытий. Линней имел сведения лишь
по незначительной доле современных животных и почти не знал
вымерших. Возрастание [числа таксонов] по причине открытий
уравновешивалось введением в иерархию новых категорий, кото-
рых в царстве животных теперь достигает 20 и более против 4 у
Линнея [38]. Можно отметить и вторичный эффект, идущий вслед
за тенденцией, отмеченной в предыдущем предложении: по мере
роста знаний о разнообразии снижались критерии дивергенции,
принятые для каждого данного ранга. Это, разумеется, увеличива-
ло число таксонов, которые номинально относились к одному уров-
ню. Однако последнее правильнее было бы рассматривать как [про-
сто] перенос категориальных названий с одного уровня на другой.
Так, часто отмечалось, что практически всем линнеевским родам
животных теперь соответствуют как минимум семейства, а некото-
рым — отряды и даже таксоны более высоких категорий. Число
видов возросло столь сильно, что многие из линнеевских родов,
останься они в прежнем понимании, содержали бы сотни, тысячи
или десятки тысяч видов. Простое включение промежуточных над-
видов и подродов, даже если бы было последовательным (чего на
самом деле нет), не спасло бы ситуацию. Таким образом, всё, что
было сделано на самом деле, было простым перемещением назва-
ний категорий Линнея вверх по шкале, помещая «семейство» там,
где у него был «род», «отряд» — на место его «семейства» и т.д.8
Это добавило необходимые ступени между, скажем, линнеевскими
видом и родом без изменения освящённых временем названий ка-
8 Конечно, это не носило такого упорядоченного характера, поскольку клас-
сификация изменялась и в других отношениях.
От таксономии к классификации
159
тегорий, но изменило их значение, их положение на шкале. Или,
если взглянуть на вопрос по-иному, средняя степень обособленно-
сти [divergence], которую Линней ассоциировал со словом «род»,
стала ассоциироваться со словами «семейство» или «отряд».
Такая перестройка в наибольшей мере сказывается на низших
уровнях иерархии, где на уровне вида отмечен наибольший рост
[числа таксонов]. Среди современных животных Линнею было из-
вестно большинство высших таксонов, так что приведение в соот-
ветствие высших уровней произошло довольно быстро. Только 6 из
118 семейств современных млекопитающих в моей классификации
1945 г. были неизвестны Блэнвилю в 1834 г., хотя конечно его ран-
ги не обязательно были теми же (см. Simpson, 1954). В прошлом
столетии добавление действительно новых категорий было обяза-
но в основном палеонтологии и предполагало, как правило, встав-
ки, а не изменения рангов существующих классификаций. (Пона-
чалу незначительное влияние набирающей силу эволюционной так-
сономии становится более радикальным, но это лежит в стороне от
здесь рассматриваемого вопроса об относительных рангах).
В добавление к тем уточнениям категорийных уровней, которые
позволяют согласовать их число с открытиями множества ранее
неизвестных животных, имели место значительные колебания чис-
ла таксонов, вызванные не новыми открытиями, а изменением кри-
териев, которые применяли разные классификаторы. На уровне
вида типологические подходы и критерии, действующие по «пра-
вилу большого пальца» [39], побуждали некоторых классификато-
ров выделять и именовать в качестве видов бесчисленные морфы
и прочие варианты в пределах видовых популяций. Эта тенденция
сохранилась и позже; она и до сих пор ещё не выправлена даже
среди тех, кто, по меньшей мере, стремится применять эволюци-
онный подход.9 Применение филогенетических критериев заметно
9 Палеонтологи были и во всё возрастающей степени остаются среди наи-
худших грешников в этом отношении. Меня учили в колледже, что всякий ис-
копаемый остаток, размерами на 15% крупнее или мельче типа уже описанного
вида, можно спокойно относить к новому виду. Но и некоторые неонтологи не
лучше. Видный маммолог того же времени говорил мне, что наилучший ви-
довой признак — 100 миль, т.е. экземпляр, пойманный за 100 миль от типа,
всегда должен помещаться в отдельный вид.
12*
160
Дж. Г. Симпсон. Принципы таксономии животных
уменьшило число таким образом сотворённых видов. (Здесь уме-
стно сказать, что эти «виды» сотворены классификаторами, а не
природой.) Современные ревизии давно известных групп по не-
обходимости сопровождаются списками сотен видовых синонимов,
основанных на том, что сейчас считается внутривидовыми вариа-
циями.
Вопреки многим случаям, сомнительным в принципе или недо-
статочно поддержанным необходимыми данными, филогенетичес-
кие критерии (следующий раздел) обычно дают фиксированное
число видов, более или менее не зависящее от личных качеств
классификаторов. На уровнях выше видового, однако, могут возни-
кать классификации с очень разным числом таксонов каждого ран-
га, минимально отвечающие критерию совместимости с известной
филогенией (рис. 10). В данном случае число принимаемых таксо-
нов — во многом вопрос не столько общепринятых научных прин-
Семейства
Роды
Рис. 10. Схема, иллюстрирующая дробительский и объединительский
[подходы в классификации]
Филогения (или дендрограмма) девяти современных видов (чёрные точки),
исходная для обеих классификаций.
А. Дробительская классификация с семью родами и тремя семействами.
Б. Объединительская классификация с тремя родами и одним семейством.
Роды «объединителя» могут быть теми же (только на ином уровне), что и
семейства «дробителя», однако данная схема показывает, что они могут
различаться, даже если их число одинаково. Обе классификации полностью
совместимы с представленной филогенией. Возможны и другие системы, также
совместимые с нею.
От таксономии к классификации
161
ципов таксономии, сколько личных пристрастий к художественным
канонам классификации. Некоторые классификаторы, будучи край-
ними дробителями, предпочитают наиболее жёсткие из возможных
стандартов обособленности и разнообразия и, соответственно, чаще
выделяют на каждом уровне наибольшее число таксонов. Другие,
крайние объединители, предпочитают наиболее широкие из воз-
можных стандартов и выделяют минимальное число таксонов. Дро-
бители предложили не менее 28 родов современных кошачьих
(Felidae), число которых, возможно, даже превышает число видов.
Объединители же помещают всех их в один, в лучшем случае в два
рода — Felis и Acinonyx (последний выделяется для сильно укло-
няющегося гепарда).
Объединительство и дробительство в некоторой степени связа-
ны с целями таксономиста. Тот, кто озабочен, прежде всего, сор-
тировкой своих коллекций на виды и их идентификацией, есте-
ственным образом концентрирует внимание на различиях и диаг-
нозах, разграничивающих таксоны.10 Его подход является аналити-
ческим и он будет вероятнее всего дробителем, по крайней мере,
в рамках своей группы, которую он знает настолько хорошо, что
большое число надвидовых таксонов кажется скорее удобством,
чем помехой. (Многие таксономисты, будучи дробителями в своих
группах, считают, что другие в своих группах должны быть объе-
динителями!) С другой стороны, таксономист, озабоченный пост-
роением иерархической системы для большой группы животных,
должен более применять синтетический подход и сосредотачивать-
ся в значительной мере на сходствах, которые позволяют выделять
сборные таксоны, а также на их определении — на обозначении их
общих границ, а не просто на отличиях от других таксонов. Он
чаще будет объединителем. Вдобавок к этим вполне понятным раз-
личиям в целях, имеются и более смутные персональные различия
между дробителями и объединителями (Roe, 1953): некоторые
люди просто склонны разъединять вещи, а другие — объединять
их.
10 «Диагноз» происходит от греческого 8iccYvyv(D(5K(O, «различать (две)
вещи» [40], а «определение» — от латинского definitio, «заключать в рамки».
Эти понятия существенно различны, однако на практике таксономисты редко
используют их правильно, чаще считая синонимами.
162
Дж. Г. Симпсон. Принципы таксономии животных
Как обычно, имеются аргументы в пользу каждой из крайнос-
тей, равно как и широкие возможности для искусства примирять
их. Следующий непрямой диалог между Дж. Алленом, крайним
дробителем, и мной, откровенным, хотя и, я уверяю, не предель-
ным объединителем, показывают основные точки расхождения.
Alien (1919): Этим группам придан, предварительно и для удобства,
номенклатурный ранг рода. Их оценка будущими исследователями будет
предметом поиска общего согласия с той точки зрения, следует ли всех
кошачьих поместить в род Felis, поскольку они не медведи и не волки, или
можно выделить несколько основных типов, чтобы показать, что домаш-
няя кошечка — и не лев, и совсем не леопард, и не рысь, или лучше по-
казать, что даже среди мелких кошек имеется некоторое разнообразие в
структуре и родстве. Так, последний фаунистический список североамери-
канских Felidae включает 41 форму, из которых 16 помещены в род Lynx,
а 25 — в род Felis. Конечно, самый последний новичок в области есте-
ственной истории с первого взгляда поймёт, что какая-то часть — это
рыси, а остальное — кошки, однако только специалист по номенклатуре
кошачьих сможет распознать, какие из них — большие кошки (как пума и
ягуар) или мелкие кошки, или какие равномерно окрашены, а какие со
сложным рисунком из пятен и полос, или каковы их естественные отно-
шения; исключая случаи подвидов, где ключом служит триномен. После-
дующее использование родовых делений не затемнит того факта, что все
они — кошки, но укажет по крайней мере, что все кошки несхожи между
собой, и возможно воодушевит интерес к выяснению их различий.
Simpson (1945): Этот аргумент почти столь же лицемерный, сколь и
простодушный. В первую очередь, он целиком основан на том, что цель
классификации — в разделении вещей, едва ли принимая во внимание тот
ключевой факт, что осмысленное объединение вещей является более полез-
ной и важной функцией таксономии. Второе, Аллен по-видимому пренеб-
регает существованием каких-либо таксономических градаций между под-
видами и родами. В самом деле, внимательное изучение его классифика-
ции показывает, что он не использует ступень вида каким-либо приемле-
мым способом: в его системе род занимает место вида, а следующей бо-
лее мелкой единицей является «форма» — подвид или локальная раса. Что
касается подродов, он игнорирует также и их. Таким образом, настаивая
на дальнейшем дроблении родов, для того чтобы показать ступени разли-
чий (и сходств, но лишь теории, а не в его практике), он отбрасывает сам
механизм, посредством которого это может быть сделано наилучшим об-
разом. Оказывается практически невозможным показать эти ступени, если
родовой, подродовой и видовой ранги все приведены к родовому уровню.
От таксономии к классификации
163
Возможно, нет необходимости особо подчёркивать черты неискренно-
сти в его аргументации. Аллен несомненно хорошо знал, что обособлен-
ность кошек от медведей и волков показана всеобще признанным отнесе-
нием их к отдельному надсемейству, а не родовой системой. Также невоз-
можно поверить, что Аллен действительно думал, что «новичкам в есте-
ственной истории» больше пользы будет выучить семь родовых названий
вместо одного или что, например, в названии Margay glaucula glaucula есть
нечто, что даст новичку более ясное представление о небольшой серова-
той пятнистой кошке из Мексики, нежели Felis tigrina glaucula. На самом
деле новичок никогда даже не догадается, да и не всякий специалист с ходу
поймёт, что «Margay» — это кошка, поскольку самое название, использу-
емое Алленом, ничего не говорит о её связях, тогда как старое имя сразу
же раскрывает специалисту, а также отчасти и несведущему новчику, всю
последовательность разной степени родства.
Дробители и объединители и дальше будут существовать, а
личностный фактор может быть исключён из классификации не в
большей степени, чем из любого другого искусства. Имеется, од-
нако, разумный канон, который в какой-то мере ограничивает этот
фактор. В самой природе вещей заложено, что некоторое число
монотипических таксонов необходимо в непротиворечивой, хоро-
шо сбалансированной классификации. Однако если классификация
содержит большое число монотипических Т., необходимо подроб-
нее рассмотреть возможность того, что (а) таксоны Т чересчур
раздроблены или (б) таксоны Т чересчур укрупнены. Вообще го-
воря, классификация, хорошо сбалансированная в отношении
дробления/объединения, будет в среднем содержать заметно больше
Т р чем Т., для всех уровней j. Внимание также следует уделять
стабильности (избегать ненужных изменений) и удобству (думай-
те о пользователях, а не о специалистах по данной группе).
Относительная древность
В эволюционной классификации каждый [таксон] Т как ми-
нимум столь же древен, как и любой из включённых в него [так-
сонов] Т , и как правило, с возможными, но несомненно редкими
исключениями, каждый Т f старше, чем средний возраст включён-
ных в него Т. Оба утверждения — необходимые и, как легко по-
казать, очевидные следствия самой природы филогении и органи-
164
Дж. Г. Симпсон. Принципы таксономии животных
зации эволюционных таксонов в иерархической системе. Они важ-
ны для понимания того, о чём они не говорят, не в меньшей сте-
пени, чем то, о чём в них прямо утверждается. Среди прочего, они
не говорят о следующем:
1. Т+| как минимум столь же древние и обычно древнее, чем Т.
(Соотношение соблюдается только для данного Т+1 и включённых в него
Т-)
2. Т.+1 возникают прежде входящих в них Т. (Соотношение соблюда-
ется только для средних, а не индивидуальных возрастов Т.)
3. Т обычно бывают одного возраста. (Ничего из вышеприведённых
утверждений логически не ведёт к данному заключению.)
Эти и сходные заблуждения иногда запутывали классификато-
ров-филогенетиков и время от времени выдвигались в качестве ар-
гументов против практичности филогенетической классификации.
Первый вывод действительно проявляется как тенденция или в
среднем для каждой данной группы, однако он может соблюдать-
ся строго и в общем случае, только если скорости эволюции оди-
наковы, что противоречит известным фактам. Бигелоу (Bigelow,
1958), один из немногих вдумчивых критиков эволюционной клас-
сификации, утверждал, что филогенетическая классификация пред-
полагает одинаковость эволюционных скоростей, однако это не так,
как убедительно показал Бадер (Bader, 1958) в ответ на критику.
Как уже обсуждалось выше в этой главе, эволюционная классифи-
кация способна учитывать и учитывает степень дивергенции, но ни
в коей мере не может из-за этого считаться менее совместимой с
филогенией. Группа, эволюирующая с большей скоростью, будет
дивергировать больше, чем медленно эволюирующая, и по этой
причине ей может быть присвоен более высокий ранг. Быстро эво-
люирующая группа слонов (и мамонтов, которые — просто вымер-
шие слоны) столь сильно дивергировала, что заслуживает ранга
семейства, однако она не столь древняя, как относящийся к тем же
хоботным медленно эволюирующий род Mammut (так называемые
американские мастодонты).
Тем не менее, в пределах каждой отдельной группы (почти
любого объёма и ранга) Т как правило бывают в среднем стар-
ше, чем Т. Все конкретные оценки действительных эволюционных
скоростей (выполненные, например, в: Simpson, 1953) показывают
От таксономии к классификации
165
резко выраженную однопиковую кривую, в которой модальные
(в данном случае, средние) скорости гораздо обычнее, чем замет-
но более быстрые или медленные скорости. Это объясняет тенден-
цию, почему Т +1 бывают старше Т. Однако в разных группах мо-
дальные скорости могут быть очень разными, так что эта тенден-
ция не может быть представлена как общее правило.
То, что высшие таксоны возникают прежде включённых в них
низших таксонов, вменялось в вину филогенетической классифи-
кации в качестве ложной предпосылки, однако на самом деле это
принимали (если вообще принимали) лишь немногие эволюцион-
ные классификаторы, как они понимаются в данной книге. Это
было бы возможно, в некотором смысле, если бы высшие таксоны
происходили в результате единственной мутации, как утверждал
Шиндевольф (Schindewolf, 1950), который был несомненным эво-
люционистом, но чья таксономия была в сущности типологической.
Но даже в его теории таксономии данное утверждение, очевидно,
не может буквально указывать на то, когда возникали высшие так-
соны. Все организмы относятся к видам, не только вследствие при-
нятого определения вида, но по самой природе того явления, кото-
рое мы обозначаем как вид. Каким бы образом ни возникал выс-
ший таксон, его первые члены принадлежат одному или несколь-
ким видам, которые являются ровесниками первого рода, который
является ровесником первого семейства, и это верно для возникно-
вения любого высшего таксона. Исходные низшие таксоны обыч-
но рано или поздно замещаются другими низшими таксонами этого
же высшего таксона, так что в конце концов на некоторый момент
времени не остаётся ни одного низшего таксона, столь же древне-
го, как тот высший таксон, к которому они относятся. Для одновре-
менно существующих животных, таких как входящие в современ-
ную фауну, это может означать, а в действительно обычно и озна-
чает, что ни один известный низший таксон не бывает столь же
древен, как высший таксон, включающий его, — а это, разумеет-
ся, только известные таксоны, представленные в классификации.
Это не отменяет того факта, что высший таксон не может быть
старше включённых в него групп, которые когда-то существовали
и которые, если бы были известны, классифицировались бы как
низшие таксоны.
166
Дж. Г. Симпсон. Принципы таксономии животных
Чтобы далее прояснить этот вопрос, следует взглянуть на так-
соны как на отражения фрагментов филогенетического дерева. Раз
высшие таксоны не возникают за один шаг, что почти все эволю-
ционисты считают в высшей степени невероятным, то они пред-
ставляют собой куски этого дерева, более крупные в сравнении с
соподчинёнными им низшими таксонами, и в целом средние раз-
меры таксонов на соответствующей дендрограмме будут пропорци-
ональны их рангам. В таком случае высший таксон просто не су-
ществует, пока не возникнет соответствующий крупный фрагмент
дерева как результат «роста» низших таксонов (рис. 11). С точки
зрения эволюционного паттерна это будет означать, что высшие
таксоны появляются только ретроспективно после становления вхо-
дящих в них низших таксонов, но не наоборот.
То, что таксоны одного ранга, в пределах одного высшего так-
сона или рассматриваемые вообще, должны быть одного возраста,
абсолютно невозможно для любой осмысленной системы класси-
фикации. Этот критерий мог бы показаться возможным только для
одновременно существующих животных, и попытки таким образом
организовать систему действительно предпринимались. Однако они
оказались совершенно непрактичными. В пределах позвоночных,
последовательность классов (в современном понимании) Osteichthyes,
Amphibia, Reptilia и Mammalia соответствует убыванию возраста
таксонов. Согласно критерию равной древности, у них должен со-
ответственно понижаться ранг: при сохранении Osteichthyes как
класса, Mammalia будут в лучшем случае надотрядом, а то и просто
родом, если не использовать многочисленные промежуточные ран-
ги. Если бы таким образом было построено всё царство животных,
мы обнаружили бы самих себя в таких таксонах как «отряд Lati-
meriae» (в настоящее время это род рыб Latimerid) и «род Primas»
(в настоящее время это отряд Primates). Для того, чтобы сделать та-
кую систему осуществимой, нам потребовалось бы существенно уве-
личить число иерархических рангов, возможно две или три сотни,
если не больше. Всё это можно было бы сделать, но с невероятной
потерей в ясности и удобстве, причём без особой нужды и неоправ-
данно с точки зрения принципов эволюционной классификации.
При любом раскладе, такой критерий можно использовать толь-
ко применительно к одновременно существующим животным. При
От таксономии к классификации
167
Границы между
родами-------
семействами + + + +
Рис. 11. Схема роста филогенетического дерева и его отношение
К ВОЗНИКНОВЕНИЮ ВЫСШИХ ТАКСОНОВ
Горизонтальные линии tp t2 и t3 обозначают три последовательных
временных интервала. Чёрные точки — виды, существовавшие в каждом из
интервалов. (Это обозначение принято для упрощения ситуации применительно
к одновременно существующим видам; в действительности же каждый вид имеет
временную протяжённость и является сегментом филетической линии, а не
точкой на ней.) Штриховые линии очерчивают роды, крестиками очерчены
семейства, выделяемые для каждого из временных интервалов.
А. Два вида на момент tp составляющие (как минимум) один род более
древнего семейства.
Б. Семь видов и два рода на момент t2, составляющие семейство, которое не
существовало на момент tp хотя род, предковый для него и входящий в состав
двух его различных родов, тогда существовал. Два предковых вида теперь поме-
щены в два разных рода, становление которых произошло позже, после времени
существования этих видов. Семейство теперь крупнее, чем каждый из родов.
В. Одиннадцать видов, четыре рода и два семейства на момент t
представляют собой продолжение процесса перехода от (А) к (Б).
исследовании временных последовательностей те биологические
категории, которые среди современных организмов называются ви-
дами, мы должны будем считать типами, по крайней мере в случае
кембрийской фауны, и вся иерархическая система будет разрушена.
168
Дж. Г. Симпсон. Принципы таксономии животных
Таким образом, очевидно, что общий критерий относительной
древности не может иметь практического приложения при ранжи-
ровании эволюционных таксонов. И всё же верно, что в пределах
каждой группы (высшего таксона) одновременно существующих
животных данный критерий обоснован в принципе и применим на
практике. Это особый случай, однако он важен в связи с тем, что
большинство классификаций связано главным образом, если не
исключительно, с одновременно существующими группами. Под-
ходящий для данного случая критерий просто уточняет отношения,
сформулированные в первом предложении данного раздела:
Таксоны одновременно существующих животных должны ран-
жироваться таким образом, чтобы общий предок каждого Т+1 был
не менее древен, чем таковой каждого из включённых в него Т, и
более древен, чем в среднем для всех этих Т.
Имеются также другие способы доказательства относительной
древности (например, уже упоминавшаяся перекрёстная специали-
зация), однако наиболее важно общее сходство по гомологичным
или, с некоторыми оговорками, параллельным характеристикам.
Как правило, чем более сходны по таким характеристикам два (или
более) таксона, тем более недавнего предка они имели, если срав-
нивать их с двумя (или более) другими, причем менее сходными
таксонами, один из которых включает двух (или более) ранее упо-
мянутых. Условие, выделенное курсивом, основано на применении
данного критерия в пределах групп, а не между ними. Опуская это
существенное уточнение, Бигелоу (Bigelow, 1958) решительно воз-
ражал против такого рода доказательства на том основании, что в
принципе нельзя ожидать корреляции между сходством и близос-
тью общего предка.11 Бадер (Bader, 1958) исчерпывающе ответил на
это возражение, опираясь главным образом на характер распреде-
ления скоростей, упомянутое выше. На самом деле, понимание
того, что критерий подразумевает возраст таксонов Т+1 относитель-
но включённых в них Т , а не самих Т р само по себе достаточно
для полного снятия этого возражения.
11 Он также обращается к проблеме конвергенции, которая здесь уже дос-
таточно обсуждалась.
От таксономии к классификации
169
Второе возражение, также выдвинутое Бигелоу и другими, со-
стоит в том, что возраст предков не может быть уверенно опреде-
лён на основании только одновременно существующих животных.
Однако установление абсолютного возраста и не требуется. Тако-
го рода суждения, которые могут быть представлены дендрограм-
мой, не предполагают прямого отображения или реконструкции
предков, однако если они основаны на достаточных данных, интер-
претированных в рамках осмысленных эволюционных принципов,
они с достаточно высокой вероятностью указывают относитель-
ный возраст групп. (Для этих целей действительно бывает полез-
но, но не обязательно, представлять такие заключения в виде ден-
дрограмм.)
Ранги признаков
Искусство классификации в первую очередь связано с выделе-
нием таксонов, а уже затем — с их ранжированием. Было бы при-
ятным упрощением, если бы признаки организмов обладали врож-
дённой связью с разными категориальными рангами. Этот предмет
разбирался в достаточном объёме в предшествующих главах и не
нуждается здесь в более детальном рассмотрении: просто будут
подчёркнуты и подведены некоторые итоги.
Отдельные категории признаков действительно имеют некото-
рую связь с таксономическими рангами, что в известной степени
способствует разграничению таксонов. Однако эта связь сама по
себе не может служить рабочим критерием для ранжирования.
Классифицируются популяции организмов, а не их признаки. Ро-
довой признак действительно характеризует род, однако он не оп-
ределяется заранее. Идентичные признаки в одной группе могут
характеризовать вид (и даже просто внутривидовую вариацию: см.
Simpson, 1937), в другой — род, ещё в одной — семейство.
Достойно сожаления, что нельзя естественным образом группи-
ровать организмы, руководствуясь какими-то простыми отношени-
ями. Скорее, мы должны природу принимать такой, какая она есть.
Одно заключительное замечание. В главе 2 были затрачены зна-
чительные усилия на то, чтобы установить приоритеты между раз-
ными взаимосвязями животных и между характеристиками, кото-
170 Дж. Г. Симпсон. Принципы таксономии животных
рые отражают эти взаимосвязи. Филогенетическая система, с её
степенями близости потомков к общему предку, составляет осно-
ву для определения приоритетов — что буквально означает пред-
шествование во времени, так что эта основа является естественной
в полном смысле этого слова. (Об этом и близких вопросах см.
побуждающую к размышлениям статью Pirie, 1952.)
5
Вид и низшие категории
Определение категории вида долгое время было боль-
шой проблемой в систематике, частью из-за несовместимости ус-
таревших типологических концепций с природой эволюирующего
вида, а частью из-за того, что эволюирующий вид может характе-
ризоваться разными свойствами и его становление в филогении
постепенное. В современной биологической, или более строго —
генетической концепции, вид у двуполых одновременно существу-
ющих организмов рассматривается как группа перекрёстно размно-
жающихся организмов, генетически изолированная от других таких
групп. Это частный случай более широкой эволюционной концеп-
ции вида как филетической линии с особой и единой [unitary] для
его членов эволюционной ролью. Генетический вид приблизитель-
но соответствует одномоментному срезу такой линии в популяции
двуполых организмов. С некоторыми уточнениями практического
свойства эволюционное определение также применимо к однопо-
лым организмам и к таким, которые меняются во времени, таким
образом связывая все их в одну общую концепцию. Надлежит рас-
смотреть и другие концепции, но ни одна из них не подходит для
строго таксономического применения.
Вид во времени, как сегмент филогении, порождает особые
проблемы разделения целостных последовательностей видов и
трактовки ветвящихся филетических линий. Требуемые приёмы по
необходимости произвольны и оставляют место для разных мнений
172
Дж. Г. Симпсон. Принципы таксономии животных
в искусстве классификации, однако можно предложить и ряд прак-
тических критериев.
В эволюционных исследованиях и при описаниях необходимо
принимать во внимание многие формы внутривидовых вариаций,
однако единственным стандартным внутривидовым таксоном явля-
ется подвид. Подвиды имеют широкое, но не универсальное при-
менение, а проблемы их определения связаны с того же рода кри-
териями, что и при разбиении прочих эволюционных континуумов.
Предложенное применение надвида иллюстрирует некоторые
соответствия и различия общего значения между эволюционными
концепциями и таксонами классификации. Наконец, в контексте
этой главы потребуется рассмотреть соотношение между выборка-
ми и популяциями и их значение для классификации.
Генетический вид
Для античных ученых и для схоластиков виды не были ни не-
изменными, ни резко разграниченными. Веру в спонтанное зарож-
дение многих высоко организованных животных (например, пчел,
крыс) сопровождала вера в самые немыслимые мутации, радикаль-
ное вырождение,1 и уродливую гибридизацию между почти любы-
ми животными независимо от их различий (например, между вер-
блюдом и воробьём). Такой порядок среди организмов предпола-
гался, по-видимому, не вследствие, а вопреки изменяемости видов,
представления о чём сохранились ещё в неудавшемся эволюцио-
низме .Ламарка. Более того, наиболее известная доэволюционная
теория порядка, именно scala naturae, предполагала, что все виды
образуют континуум, и поэтому их неотъемлемыми качествами яв-
ляются условность и неразграничимость. Догматическая вера в
фиксированные и резко различные виды является сравнительно
поздним теологическим продуктом, и она долгое время не прини-
малась религиозными натуралистами, пока Линней не сформули-
ровал свой знаменитый афоризм, что видов столько, сколько их
1 Вплоть до почти нынешних времён «вырождение» означало появление
потомков, выпадающих из родительской и предковой норм; оно не обязатель-
но указывало на ухудшение их качеств.
Вид и низшие категории
173
было создано изначально. Это бы потрясло Аквината, который под-
чёркнуто и открыто утверждал, что новые виды возникают посто-
янно (но не посредством эволюции!). (Эта примечательная доэво-
люционная смена концепции вида внимательно рассмотрена в:
Zirkle, 1959).
На протяжении сравнительно короткого спокойного периода
таксономисты верили, что в природе существует строго определён-
ное число чётко разграниченных видов. Прочая иерархия предос-
тавляла много проблем, но распознавание и перечисление видов
казалось в принципе простой задачей. В такой атмосфере доказа-
тельство эволюции сводилось к демонстрации того, что один вид
мог породить другой. Вот почему книга Дарвина, в которой рас-
сматривались все уровни эволюции, а не только одни виды, была
названа «Происхождение видов». Такой образ мыслей сохранялся
как среди антиэволюционных сектантов, так и биологов-эволюци-
онистов, а видообразование продолжало быть столь важным воп-
росом, что нередко считалось почти синонимичным с эволюцией.
Наряду с этим, додарвиновские таксономисты осознавали, что
в природе существуют единицы, которые имеют особый эволюци-
онный статус, в полной мере не присущий таксонам, расположен-
ным в иерархии как выше, так и ниже этих единиц. Эти единицы
часто были столь обособлены, что их можно было различать, не зная
их действительной природы. Многие из них, выявленные задолго
до Дарвина, были названы видами, так что неизбежно этот термин
был перенесён на эволюционные единицы. В большинстве случаев
среди ранних эволюционистов не было понимания того, что этот
термин переносится на новую концепцию, так что существовала
двусмысленная манера размышлять о виде как об эволюционной
единице, но стараться определять его типологически. Некоторые
таксономисты до сих пор не осознают или не принимают это разли-
чие, и продолжающаяся двусмысленность является одной из при-
чин «проблемы вида» — проблемы, на которую в таксономии было
истрачено, вероятно, больше чернил, чем на какую-либо другую, и
которая состоит в том, чтобы разработать всеобще приемлемое и
практическое определение категории, называемой видом.
Существуют ещё три важных причины, почему это — пробле-
ма. Все они возникают из природы самой эволюции и делают
13 — 4338
174
Дж. Г. Симпсон. Принципы таксономии животных
вообще невозможным нахождение общего решения, на что наивно
рассчитывали некоторые таксономисты. Не может быть единого
определения вида, применимого ко всем организмам и однозначно
трактуемого в каждом отдельном случае. Эти три причины
таковы:
1. Члены категории видового уровня не могут быть разделены
на группы по какому-либо абсолютному критерию.
2. Группы, распознаваемые в качестве видов, могут различать-
ся популяционной структурой и формой возникновения, так что их
категории не могут быть однозначно определены на основе единых
эволюционных критериев.
3. Практика использования одной и той же категории как для
временных отрезков филогений, так и для одновременно существу-
ющих организмов, — всеобщая и по-видимому неизбежная, одна-
ко получающиеся таксоны различаются по природе, так что при-
менение единого категориального определения в обоих случаях
весьма затруднительно.
С этого места я приму условие, для которого представлено до-
статочно оснований, что приемлемое современное определение
видовой категории должно быть чётко связано с эволюцией. Зна-
чительная часть настоящей главы посвящена некоторым пробле-
мам, которые возникают вследствие того, что виды действительно
являются эволюционными единицами. Я не буду стараться цитиро-
вать огромную литературу, но уделю особое внимание недавним
симпозиумам (Sylvester-Bradley, 1956; Мауг, 1957), длительной дис-
куссии в журнале Evolution (особенно статьям, рассмотренным и
частью обобщённым в: Simpson, 1951), а также полезной книге
Кэйна (Cain, 1954) [41].
Определения вида, в настоящее время наиболее широко приня-
тые, представляют собой вариации или эквиваленты того, что дал
Майр (Мауг, 1940; также многие последующие статьи и книги
Майра):
Виды — это группы действительно или потенциально скрещи-
вающихся природных популяций, репродуктивно изолированные от
других таких групп.
Это строго генетическая концепция для видов одновременно
существующих двуполых организмов, её часто называют биологи-
Вид и низшие категории
175
ческим понятием или определением вида.2 Значение концепции
вполне понятно, и опытный таксономист может применить её без
особых трудностей, вероятно, по меньшей мере в 90% случаев,
если не больше, в отношении подлежащих классификации живот-
ных. К большинству групп современных животных она приложи-
ма и достаточна, что позволяет её принять и рекомендовать. Тем не
менее, имеются ограничения и трудности, которые, конечно, хоро-
шо известны Майру и другим сторонникам концепции. В первую
очередь, она, даже в принципе, не применима к временным рядам
видов и к видам однополых организмов. Эти особые (но весьма
обыкновенные) случаи, к которым генетическое определение про-
сто не имеет отношения, будут обсуждаться позже. Теперь же бу-
дут рассмотрены основные трудности, частью реальные, а частью
просто мнимые.
Данное определение зависит от критерия скрещиваемости, ко-
торая обычно не наблюдается, а в некоторых случаях и не может
наблюдаться. Это, впрочем, псевдопроблема, как должно быть ясно
из нашего предшествующего обсуждения связи между определени-
ем и доказательством в таксономии. Определение должно опреде-
лять и должно делать это биологически и эволюционно осмыслен-
ным образом. Доказательство, т.е. то, что делает определение со-
ответствующим данному конкретному случаю с достаточным уров-
нем правдоподобия, — это другой вопрос. Доказательство являет-
ся обычно морфологическим, однако заключать на этом основании,
что следует применять и должна применяться морфологическая
концепция категории (не таксона) вида, — либо неверно в принци-
пе, либо означает неоправданный возврат к типологии. О доказа-
тельстве необходимо судить по проверяемым следствиям, вытека-
ющим из заявленной в определении генетической ситуации. Не
единственный, но наиболее важный практический критерий таков:
Если диапазоны популяционной изменчивости (включая поли-
морфизм), выявленные для двух или более выборок, перекрывают-
2 Несколько запутывающее использование термина, поскольку многие до-
полнительные или альтернативные определения в такой же мере биологиче-
ские. На самом деле я не вижу, каким образом любое релевантное определе-
ние категории групп организмов может быть чем либо иным, кроме как био-
логическим.
13*
176
Дж. Г. Симпсон. Принципы таксономии животных
ся по всем исследуемым признакам, то высока вероятность того, что
соответствующие популяции были скрещивающимися на тот момент,
когда экземпляры выборки были живыми, и что по этой причине
эти экземпляры относятся к одному генетическому виду. Если тако-
го значительного перекрывания в выявленных диапазонах нет, то
популяции, скорее всего, были нескрещивающимися в недавнем
прошлом и могут относиться к разным генетическим видам. Одна-
ко если выборки взяты из более или менее удалённых географичес-
ких пунктов, необходимо предусмотреть, насколько это практичес-
ки осуществимо, проверку возможности скрещиваемости исследу-
емых популяций с географически промежуточными популяциями, с
которыми они были объединены в единый генетический вид.
Второе, хоть и реальное, но легко преодолимое препятствие
состоит в неопределённости и трудной проверяемости концепции
потенциальной скрещиваемости. Если, как часто бывает, популяции
разделены территориально, то вероятность того, что происходит
какое-либо скрещивание, крайне низка, а его уровень зависит от
частоты перемещения особей (или гамет, в случае некоторых вод-
ных животных) из одной области в другую, что бывает трудно об-
наружить. Если между ними не возникло заметных генетических
различий, было бы неправильным и в весьма странным настаивать
на том, что каждая такая популяция представляет собой отдельный
генетический вид даже в отсутствие скрещивания. В подобных слу-
чаях можно применять тот же критерий перекрывания, который,
хотя и достаточно условен, но в целом вполне обоснован. Наличие
перекрывания показывает, что скрещивание имело место в недав-
нем прошлом и prima facie сохраняется в потенции. Отсутствие
перекрывания указывает на определённую генетическую диверген-
цию и при отсутствии промежуточных популяций позволяет обо-
снованно сомневаться (хотя это и нельзя опровергнуть) в том, что
потенциал скрещивания сохраняется в достаточной степени, чтобы
соответствовать определению.
Третья трудность, причём единственная по-настоящему серьёз-
ная, [имеющая отношение к оценке того,] насколько определение
вообще уместно [relevant], связана со степенью изоляции. Немно-
гие таксономисты настаивали на абсолютной и постоянной изоля-
ции, т.е. на невозможности получения плодовитых гибридов. Мно-
Вид и низшие категории
177
гие таксономисты согласны, что это условие достаточное, но не
необходимое: популяции, которые не могут давать плодовитых гиб-
ридов (непосредственно или через промежуточные популяции),
являются ipso facto разными видами, однако популяции, которые
могут производить плодовитые гибриды, не являются ipso facto
конспецифичными. Более строгий критерий нежелателен, посколь-
ку популяции, которые во всех других отношениях, несомненно,
являются разными видами, иногда дают плодовитых гибридов.
К тому же это и непрактично, поскольку о возможности получения
плодовитых гибридов редко можно судить с надёжностью на осно-
вании имеющихся данных, исключая виды, которые настолько раз-
личны, что подобный вопрос по отношению к ним вообще не мо-
жет возникнуть.
Виды эволюируют, причём почти всегда постепенно. В случае
эволюционных видов, вероятно, не может быть строгой дихотомии
между свободным скрещиванием и его отсутствием. Имеются все
промежуточные стадии, и нет практически определяемой точки во
времени, когда две внутривидовые популяции внезапно становят-
ся разными видами [42]. К счастью для неонтологов, большинство
современных популяций либо уже прошли эту гипотетическую точ-
ку, либо не слишком близки к ней. Тем не менее, видообразование
активно продолжается сегодня и поэтому многие популяции нахо-
дятся в промежуточном состоянии пониженного скрещивания.
Опять-таки, если имеется отчётливый разрыв между диапазонами
изменчивости признаков, то весьма вероятно, что изоляция доста-
точна для того, чтобы обеспечивать видовую обособленность. Ос-
таются многочисленные сомнительные случаи, когда решение за-
висит от личных предпочтений каждого практикующего искусство
классификации. Настаивать на абсолютном объективном крите-
рии — значит, отрицать факт жизни, прежде всего неумолимый
факт её эволюции.
Эволюционный вид
Именно в результате эволюции генетические виды оказывают-
ся разобщёнными, но она же препятствует их абсолютной, полной
разобщённости. Очевидно поэтому, что генетическое определение
178
Дж. Г. Симпсон. Принципы таксономии животных
вида имеет эволюционный смысл. И всё же странно, что это опре-
деление по-настоящему не предполагает какой-либо эволюционный
критерий и ничего не говорит об эволюции. Его в равной, или даже
в значительно большей степени можно применить к видам, кото-
рые не изменяются. Мауг (1957) указал, что доэволюционные так-
сономисты выдвигали почти такие же определения, например,
Voigt в 1817 г.: «всё, что скрещивается плодовито и размножается,
называется видом».3
Коль скоро генетическое определение вида совместимо с эво-
люцией, то отсутствие в нём какого-либо непосредственного и яв-
ного эволюционного элемента никоим образом не лишает его силы.
Тем не менее, желательно иметь и более широкое теоретическое
определение, которое связывает генетический вид с эволюционным
процессом, благодаря которому он возникает. Я (Simpson, 1951) уже
предлагал такое определение, которое может быть несколько изме-
нено следующим образом:
Эволюционный вид — это филетическая линия (последователь-
ность популяций, связанных отношением предок—потомок), кото-
рая эволюирует независимо от других и обладает своей собствен-
ной единой эволюционной ролью и эволюционными тенденциями.
Это определение не только совместимо с генетическим, но так-
же позволяет прояснить его и исключить некоторые его ограниче-
ния. Эволюционный вид двуполых организмов в каждый отдель-
ный момент времени, т.е. рассматриваемый во временном срезе,
почти полностью совпадает с генетическим видом. Именно поэто-
му генетйческий вид имеет эволюционный смысл и именно поэто-
му, с этой более широкой точки зрения, генетический вид суще-
ствует. Данное определение также позволяет устранить некоторые
из концептуальных трудностей, сохраняющихся в строго генетиче-
ской концепции. Вопрос о том, почему разные популяции следует
помещать в один вид, становится яснее (для меня, по крайней
мере), если представлять, что они сохраняют единую эволюцион-
ную роль, а не просто обладают пониженным потенциалом скре-
щивания. Становится понятнее также и то, что два вида могут скре-
3«Mamm nennt Species... was sich fruchtbar mi einander gettet, fortpflanzt.»
Цитата и ссылка из Майра.
Вид и низшие категории
179
щиваться в некоторой степени без утраты различий в эволюцион-
ных ролях и что это действительно важный момент для эволюци-
онной таксономии. Значит, определение допускает такую степень
скрещивания, которая не вызывает объединения их ролей. Таксо-
номическое значение генетических критериев скрещивания и изо-
ляции заключено не в самих этих характеристиках, а в их свиде-
тельстве способности популяций поддерживать или не поддержи-
вать независимость и единство ролей на протяжении значительного
времени. Скрещивание позволяет сохранить единство роли, а изо-
ляция делает возможным разделение ролей.
Выше приведённое эволюционное определение не включает
критерий скрещиваемости. Скрещивание поддерживает единство
роли и служит свидетельством, что требование этого критерия вы-
полнено. Однако это не единственный способ сохранения единства
[роли], равно как и не единственное свидетельство того, что оно
существует. Эволюционное определение, таким образом, шире, чем
генетическое, в то же время оно включает его как наиболее важ-
ный частный аспект. Однополые популяции также имеют обособ-
ленные и единые роли, так что эволюционное определение может
применяться и к ним, как будет показано далее в этой главе.
Эволюционный вид в явной форме включает элемент времени.
Вид на самом деле имеет значительную временную протяжённость,
поэтому концепция, не рассматривающая этот аспект, если не счи-
тать ее полностью непригодной, то по меньшей мере неполна. Эво-
люционная концепция, таким образом, более чётко связана с пале-
онтологическими последовательностями, что также будет рас-
смотрено далее в этой главе.
Одна значительная трудность в эволюционной концепции ви-
да— определение и распознавание ролей. Она едва ли серьёзна
для полевого натуралиста или эколога, который почти всегда отчёт-
ливо понимает, что разные виды, с которыми он имеет дела, дей-
ствительно имеют отчётливо разные роли. Роли определяемы через
их эквивалентность нишам, если «нишу» понимать как общий об-
раз жизни, или связь популяции животных со средой, а не просто
как микрогеографическое положение.
Здесь снова возникает вопрос о соотношении определения и
доказательства. Роль нельзя непосредственно наблюдать на серии
180
Дж. Г. Симпсон. Принципы таксономии животных
мёртвых музейных экземпляров, современных или ископаемых.
Однако на основании изучения этих экземпляров можно получить
обоснованное и достаточное доказательство обособленности и
единства ролей. Морфологические сходства и различия (популяций,
а не индивидов), если они по своей природе адаптивны, связаны
с ролями. Допущение, что соотношение общего сходства и разли-
чия, хотя бы в среднем, адаптивно, достаточно надёжно обосновы-
вается общей эволюционной теорией. Для чисто таксономических
целей полезно, хоть и не необходимо, иметь возможность уточнить
природу адаптации или роли. Определение требует лишь того, что-
бы роли были обособлены и каждая из них была единой, что с до-
статочной надёжностью правила показывают соматические разли-
чия и сходства между популяциями. На первый взгляд это может
показаться круговой процедурой, но это не так. Хорошая корреля-
ция между ролью и морфологией была детально показана для по-
пуляций, в которых роли были определены независимо на совер-
шенно других основаниях, особенно между средой и поведением.
Другой проверкой и дополнительным средством применения кри-
териев, связанных с ролью, служат адекватные полевые данные,
которые даже для вымерших форм содержат хотя бы некоторые
сведения о среде.
Внутривидовые группы могут в какой-то мере различаться сво-
ими ролями. Однако эти различия, подобно морфологическим раз-
личиям, как правило, образуют плавный ряд через смежные попу-
ляции, так что критерий обособленности ролей не делает такие
группы разными видами. Разумеется, природа вещей такова, что
должны быть сомнительные случаи, ибо формирующиеся виды
приобретают свои самостоятельные роли не вдруг и не момен-
тально.
Определение не утверждает и не подразумевает, что единая
роль какого-либо вида с необходимостью остаётся неизменной.
Эволюция (кроме крайне редких сальтаций) не происходила бы,
если бы роли не менялись в пределах видов. Концепция рассмат-
ривает вид, обладающий единой ролью в каждый данный момент
времени (не обязательно одна и та же роль во все времена), при-
чём его роль в каких-то отношениях (не обязательно одних и тех
же) всегда специфична на протяжении всего его существования.
Вид и низшие категории
181
Некоторые другие категории вида
Используется ещё несколько определений вида, которые нельзя
считать простыми вариантами двух предыдущих: для некоторых из
них предложены особые названия. Я не претендую на их исчерпы-
вающий обзор, но упомяну те из них, которые подразумевают ин-
тересные таксономические концепции или часто упоминаются в
таксономической литературе. Cain (1954) выделяет и называет сле-
дующие категории видов:
Таксономические виды — общее обозначение любого таксона,
который был назван видом и получил видовое название, пригодное
согласно Международным правилам номенклатуры.
Морфовиды — устанавливаются по морфологическому сходству
без учёта иных соображений. Они обсуждаются далее в настоящем
разделе.
Палеовиды4 — развернутые во времени виды внутри единой
филетической линии. Они обсуждаются в отдельном разделе главы.
Биовиды5 — концепция, которую я назвал генетическим видом
и уже рассматривал.
Агамовиды6 — виды у однополых организмов. Также рассмат-
риваются далее в отдельном разделе.
Смысл «морфовидов», включая то, что палеонтологи часто на-
зывают «форм-видами» (в дополнение к «форм-родам» и т.п.) и
«паравидами» (или «паратаксонами» в общем случае), стал весьма
запутанным. Соответствующие термины использовались во многих
разных смыслах, часто плохо определённых или без определения
вообще. Содержание и иногда определение морфовидов и форм-
видов таково, что они «устанавливаются исключительно по морфо-
логическим данным» (Cain, 1954), т.е., с моей точки зрения, совер-
4 Это название не идеально, поскольку огромное большинство палеонто-
логических видов не являются «палеовидами» согласно данному определению.
Однако невозможно сделать этимологию и определение полностью эквивален-
тными, так что если дано чёткое определение, этимология не имеет особого
значения, если только она не вводит в заблуждение.
5 Для «биологического вида»: против этого термина я уже выдвигал воз-
ражения.
6 Также не совсем точная, поскольку дело не в отсутствии гамет, а в од-
нополости.
182
Дж. Г. Симпсон. Принципы таксономии животных
шенно неудовлетворительно. Концепция вида применима к группе
целостных организмов. Если в определении не принимается во
внимание буквально ничего, кроме морфологии, это означает клас-
сификацию признаков, а не организмов. Эта концепция может при-
меняться, хотя и не без оговорок, в некоторых типологических или
строго эмпирических подходах к пониманию вида, но неприемле-
ма в эволюционной таксономии. Наиболее важный момент здесь —
не столько данные, которые используются, сколько то, как они ин-
терпретируются. Генетические и эволюционные виды устанавлива-
ются главным образом или исключительно на основе морфологи-
ческих данных. Однако мне представляется ошибочным подводить
их по этой причине под рубрику «морфовиды» наряду со многими
другими противоречивыми концепциями. Это, конечно, вводило в
заблуждение некоторых таксономистов, которые доказывали, что
поскольку большинство видов, как таксонов, основаны на данных
морфологии, то категория вида не может иметь обоснованного
генетического или эволюционного определения.
Стратиграфы часто устанавливали и называли «форм-виды»
или «морфологические виды» в результате выбора произвольных
«типов» среди организмов, предположительно или несомненно от-
носящихся к единой биологической популяции, или «биовиду». Это
возвращение к чистой типологии пылко защищали, даже в после-
дние годы (например, Eagar, 1956), главным образом на том осно-
вании, что оно полезно при установлении стратиграфических кор-
реляций. Конечно, едва ли кто будет возражать против разработки
стратиграфами каких-либо инструментов, полезных для решения
своих особых задач. На самом же деле есть практикующие стратиг-
рафы, которые устанавливают корреляции по ископаемым остаткам,
не классифицируя, не именуя и даже не рассматривая их как био-
логические объекты, и такая практика не вызывает особых возра-
жений. Однако возникают законные и серьёзные возражения, ког-
да они выделяют несомненно небиологические группы и затем
классифицируют их как биологические, давая им линнеевские на-
звания, с которыми таксономисты-биологи вынуждены иметь дело,
хотя бы в форме синонимов, руководствуясь Международными пра-
вилами. На самом деле такие стратиграфы проигрывают в решении
своих собственных важных задач, поскольку показано, что хорошо
Вид и низшие категории
183
установленные эволюционные виды позволяют устанавливать кор-
реляции более чётко и надёжно, чем «морфовиды» (например,
George, 1956).
Ещё одно понятие форм-таксонов, также наиболее обычное
среди палеонтологов,7 касается полифилетических групп, объеди-
няемых либо по ошибке, либо для удобства по параллельным или
конвергентным морфологическим признакам. Образование родов
(или таксонов более высоких рангов) в таком понимании более
обычно, чем видов, однако концепция в сущности та же. Предше-
ствующая дискуссия о градах и кладах также имеет к этому отно-
шение. Я не вижу причин для намеренного использования форм-
таксонов, когда они по приемлемому критерию монофилии не яв-
ляются монофилетичными. Распознавание и выяснение природы
параллелизмов или конвергенции, равно как и изучение град, пред-
ставляет собой полезный и содержательный результат таксономи-
ческих исследований. Однако это не оправдывает включения таких
групп совершенно иной природы в стандартную систему классифи-
кации, равно как и создания для них параллельной системы клас-
сификации и номенклатуры. Несомненно, ошибочна практика вы-
деления «видов» (и высших таксонов), которые первоначально
предполагаются как генетические или эволюционные, а затем ока-
зываются форм-таксонами. Однако мы вряд ли должны и имеем
право применять определение или концепцию вида для групп, ко-
торые когда-нибудь в будущем будут опознаны не в том качестве,
который мы им приписываем сегодня!
Одна из проблем, наиболее свойственная палеонтологии, —
необходимость каким-то образом классифицировать расчленённые
и изолированные части организмов или их следы. Например, коно-
донты, крайне полезные при вычленении горизонтов, присутству-
ют в целых животных как сложные скопления явно различных не-
больших твёрдых пластинок, игл, гребневидных структур и т.д.
Таких скоплений известно немного, а по большей части классифи-
кации конодонтов приходится основывать на отдельных частях,
7 Называя ущербной практику некоторых палеонтологов, я не имею в виду,
что они представляют палеонтологию в целом. Это не так, ими палеонтоло-
гия не ограничивается.
184
Дж. Г. Симпсон. Принципы таксономии животных
непонятно как связанных между собой. Как явствует из ограничен-
ных данных по естественным скоплениям, неизбежным результа-
том оказывается то, что разные «виды» и «роды» нередко бывают
основаны на частях одного животного (ссылки и примеры см, на-
пример, в: Scott, Collinson, 1959). Ископаемые отпечатки следов лап
представляют собой ту же проблему, поскольку их практически
никогда не удаётся связать с низшими таксонами, известными по
скелетам (например, Lull, 1953).
Одним из решений было бы, конечно, воздержаться от присво-
ения линнеевских названий частям животных, обозначая их каким-
либо существенно иным способом. В отдельных случаях так и пы-
тались делать, однако это не стало приемлемым общим решением,
поскольку отсутствуют надёжные основания для разграничения.
Вся палеонтология основана только на частях животных, и для эво-
люционной биологии стало бы сокрушительным ударом, если бы
стандарты биологической классификации перестали применяться
во всех случаях. Обычной практикой сегодня остаётся — продол-
жать классифицировать несвязанные части, следы и т.д. насколько
это возможно в надежде на будущее: грядущие открытия в конце
концов позволят связать их воедино в целостные организмы. По-
добная ситуация иногда возникает в неонтологии, например, ког-
да разные виды описываются на основании разных стадий жизнен-
ного цикла одного животного. В палеонтологии недавно было пред-
ложено компромиссное решение: группы, основанные на частях
конодонт и подобных разобщённых частях ископаемых организмов,
следует обозначать как паратаксоны и строить систему номенкла-
туры независимо от стандартной (Мооге, 1957). Это предложение
было представлено Международному зоологическому конгрессу в
1958 г., но не получило официального признания [43].
Тот факт, что генетическое определение вида не упоминает
морфологию, предполагает возможность того, что некоторые хоро-
шие генетические виды могут не различаться анатомией. Иногда
это по меньшей мере почти верно, и тогда пары анатомически
очень сходных генетических видов называются криптическими или
видами-двойниками (обширное обсуждение и примеры см. в: Мауг,
1942, 1948). Обнаружено много таких видов, хотя известные слу-
чаи составляют незначительную часть всех видов животных. По-
Вид и низшие категории
185
скольку большинство видов было определено8 только анатомиче-
ски, можно предположить, что виды-двойники на самом деле весь-
ма обычны в природе. Если так, то генетические виды, сохраняя
свой теоретический интерес, оказались бы практически бесполез-
ными в повседневной практике классификации. Этот вывод факти-
чески отстаивал Соннеборн (Sonneborn, 1957), но на данных по
простейшим, которые в этом отношении не сопоставимы с боль-
шинством многоклеточных, однако даже и в отношении простей-
ших вывод более чем сомнителен.9
По крайней мере для многоклеточных есть некоторые основа-
ния сомневаться в том, что эта проблема является столь важной,
как она кажется на первый взгляд. Имеется крайне мало примеров,
в которых виды, называемые двойниками, на самом деле оказалось
невозможно различить в результате более внимательного и одно-
временного изучения всех доступных анатомических признаков. На
деле разграничение так называемых видов-двойников в ряде слу-
чаев было сделано во многом или исключительно на анатомичес-
кой основе. Например, «двойников» каменную и лесную куницы
(Martes foina и М. mattes) путали при попытке определить по не-
диагностическим признакам, однако они без сомнений различимы
при использовании нескольких диагностических признаков (Streuli,
1932; этот случай Майр (Маут, 1942), приводит как один из немно-
гих примеров предположительных видов-двойников среди млеко-
питающих) [44].
Филетические линии, медленно дивергирующие или только
вступающие в видообразование, морфологически различаются не-
значительно, однако во многих случаях эти различия со временем
постепенно увеличиваются. Представляется, что состояние, когда
8 Вид организмов, таксон, может быть и обычно бывает определён анато-
мически (или морфологически), хотя категория, к которой он относится, оп-
ределяется генетически. В этой кажущейся аномалии нет действительного про-
тиворечия, что ещё раз позволяет подчеркнуть различия между определением
и критерием.
’Будет большим, но вряд ли неоправданным сверхупрощением утверждать,
что Соннеборн считает распознавание генетических видов-двойников непрак-
тичным на основании его опыта диагностики многих из них. Весьма вероятно,
что виды-двойники гораздо обычнее среди простейших, чем многоклеточных.
186
Дж. Г. Симпсон. Принципы таксономии животных
различия отсутствуют или минимальны, обычно предшествует
критической точке генетической изоляции или как максимум какое-
то время продолжается после прохождения этой точки. В каждый
момент времени сравнительно немного животных будет находить-
ся именно в этом переходном состоянии установления генетической
изоляции без отчётливых морфологических различий. Вероятно,
что некоторое число видов, которые различаются по обычным при-
знакам, но в очень незначительной степени (как куницы), находятся
как раз в таком состоянии. Применение к ним концепции двойни-
ков имеет сомнительную ценность.
Однако есть несколько проанализированных примеров, в кото-
рых генетическая дивергенция явно значительна, тогда как анато-
мическая дивергенция едва заметна: например, знаменитый случай
Drosophila pseudoobscura и D. persmilus (Dobzhansky, 1951). Он
иллюстрирует предыдущее замечание, показывая, что провозгла-
шаемая связь генетической и соматической эволюции ни в коей
мере не означает их совпадения: в некоторых случаях они могут
лишь незначительно коррелировать. Для того, чтобы быть дей-
ствительно осмысленным, термин «вид-двойник», как я полагаю,
должен быть ограничен такими случаями. При таком более стро-
гом определении известные случаи видов-двойников становятся
крайне редкими. Представляется вероятным, что истинные виды-
двойники действительно редки, но в какой-то степени это чистая
вера, поскольку адекватный генетический анализ выполнен на
очень небольшом числе видов. Пока ещё невозможно сделать та-
кой. анализ рутинной частью общей таксономической процедуры,
а если окажется, что истинные виды-двойники обычны, генети-
ческая концепция вида во многом утратит своё значение для клас-
сификации.
Разные генетические виды, у которых отсутствуют какие-либо
отчётливые анатомические или экологические отличия, согласно
эволюционному определению являются одним видом: у них нет
поддающихся определению отдельных эволюционных ролей. Та-
ким образом определённый сборный эволюционный вид аналоги-
чен однополому виду, в котором единство роли поддерживается
без скрещивания. Применимость к таким случаям — достоинство
эволюционного определения, которое сохраняет свою полезность
Вид и низшие категории
187
для классификации независимо от существования видов-двой-
ников.
Принимая во внимание иные соображения, Рент (Rensch, 1929)
предложил различать Rassenkreis (буквально «круг рас») и Art
(«вид» в строгом понимании). Rassenkreis — это генетический вид,
состоящий из серии интерградирующих, но различающихся локаль-
ных популяций, порой столь различных, что крайние популяции не
могут скрещиваться непосредственно, хотя по-прежнему могут об-
мениваться генами через промежуточные популяции. Art — это ге-
нетический вид без географической дифференциации и, таким об-
разом, более соответствует типологической концепции вида. Мауг
(1942), однако, указывал, что большинство видов представляет со-
бой Rassenkreis и что хотя описание размаха и характера географи-
ческой изменчивости имеет огромное значение, теоретическое раз-
личие между Rassenkreis и Art в эволюционной таксономии не
очень существенно. С этим согласно большинство современных
таксономистов (например, Cain, 1954).
Ещё одним предложением является (Мауг, 1942) разделение
аллопатрических видов, которые распространены в разных облас-
тях, и симпатрических видов, которые распространены, хотя бы
частично, в одной области. Генетическое значение этого состоит в
том, что аллопатрические виды в действительности не имеют воз-
можности скрещиваться. Они могут даже сохранять способность к
скрещиванию, несмотря на то, что реально они не скрещиваются
и подвергаются значительной (но не очень большой) морфологи-
ческой и функциональной дивергенции. Применение к ним генети-
ческого определения может из-за этого оказаться неясным и нео-
пределённым. Применение эволюционного определения в таких
случаях значительно яснее, по крайней мере в теории, хотя может
возникнуть практический вопрос о том, влечёт ли географическое
разделение за собой разделение ролей. Симпатрические виды
в норме остаются таковыми, если скрещиваются только внутри са-
мих себя, но не между собой, и если сохраняют разные роли. К
ним оба определения [генетическое и эволюционное] обычно при-
менимы в равной мере и приводят к выделению одних и тех же
групп в качестве видов.
188
Дж. Г. Симпсон. Принципы таксономии животных
Вид у однополых организмов
Как уже отмечено, обычное генетическое определение вида
применимо только к двуполым организмам. Большинство специа-
листов по таксономии (и генетике) животных склонны игнориро-
вать однополые организмы и не предпринимать попыток опреде-
лить категорию вида с их учётом. Есть некоторое оправдание та-
кой позиции, поскольку длительная облигатная однополость редка
среди многоклеточных, так что практическая проблема возникает
главным образом среди простейших и растений. Даже если двупо-
лость чередуется с однополыми поколениями, то при её достаточ-
ной продолжительности генетическая концепция сохраняет состо-
ятельность. Более того, Понтекорво (Pontecorvo, 1959) указал, что
некоторая форма двуполости, будь то сексуальная или, в его терми-
нологии, парасексу ал ьная,10 присутствует у некоторых членов прак-
тически всех групп организмов, включая простейших и растения,
что означает вероятность того, что все ныне живущие организмы
имели сексуальных или парасексуальных, т.е. двуполых, предков.
Тем не менее, есть облигатно однополые организмы, в том чис-
ле среди животных, и они также должны быть классифицированы.
Также верно, что исследователи этих животных выделяют среди
них группы, которые, по-видимому, относятся к той же категории,
что и виды двуполых организмов, и также считаются видами, в
качестве каковых и именуются. Очевидно, они не могут быть ге-
нетическими видами согласно принятому определению, в связи с
чем Добжанский (Dobzhansky, 1937) настаивал на том, что у одно-
полых организмов не может быть категории вида. Это заблуждение,
каким я его считаю, устраняется категорией эволюционного вида,
как она была определена выше, которая полностью соответствует
генетическому (двуполому) виду, включая его в качестве частного
случая, а также полностью применима к однополым видам в каче-
10 Размножение целиком половое, если оно включает мейоз (редукцию чис-
ла хромосом, предшествующую кариогамии) и кариогамию (объединение на-
боров хромосом от двух организмов в ядре одного организма). Парасексуаль-
ное размножение, выделенное только в последние годы, включает обмен гене-
тическим материалом между отдельными особями несколькими разными спо-
собами без мейоза и кариогамии.
Вид и низшие категории
189
стве ещё одного частного случая.” Meglitsch (1954) в своём исклю-
чительно чётком и проницательном исследовании пришёл к прин-
ципиально сходной эволюционной концепции вида. Он показыва-
ет не только, что эта концепция действительно применима в рав-
ной степени к двуполым и однополым организмам, но также что
генетические последствия для эволюции сопоставимы в обоих слу-
чаях, хотя генетические процессы различны. Его определение та-
ково:
Вид, в случае двуполых и однополых организмов, может быть пред-
ставлен как естественная популяция, эволюирующая как целое в природе
(a unit in actuality) или сохраняющая способность эволюировать как целое
при исчезновении искусственных преград.
(Я понимаю так, что «искусственные» означают преграды, вне-
шние по отношению к самим организмам, а не «творения чело-
века»).
Эволюция в качестве целого, или сохранение единой роли, под-
держивается общей наследственностью, возможностью для генов
распространяться в популяции (которая благодаря этому обладает
генофондом), и запретом на их проникновение в другие популяции.
Все три фактора свойственны как двуполым, так и однополым
организмам. Для общей наследственности это очевидно, но для
двух других факторов не вполне. В обоих случаях поток генов
обеспечивается дифференциальным размножением, частота генов
возрастает, если их обладатели оставляют больше потомства, и сни-
жается или даже сводится к нулю, если потомство меньше. Други-
ми словами, роль определяется и ограничивается естественным
отбором одинаково в обоих случаях. Двуполое размножение при-
водит к рекомбинации генов, возрастанию изменчивости, делая вид
более приспособляемым, однако это не оказывает решающего вли-
яния на поток генов в популяции.
11 Предлагая эволюционное определение (Simpson, 1951), я отметил, что
«подчёркивание единой эволюционной роли может разрешить теоретическую
трудность определения вида в группах с бесполым размножением.» Однако я
неосмотрительно ограничил это применение, включив в определение (в скоб-
ках) слово «скрещивание». В это же время, не зная о моём уточнении, Меглич
прояснил всю ситуацию, так что я надлежащим образом переформулировал
своё определение.
14 — 4338
190
Дж. Г. Симпсон. Принципы таксономии животных
У двуполых организмов поток генов между популяциями вклю-
чает скрещивание, которое несомненно отсутствует у однополых
организмов. Но он включает также перемещение организмов из
одной популяции в другую, которые производят там потомство,
остающееся во второй популяции и размножающееся там — все
эти факторы одинаково актуальны для однополых популяций.
В обоих случаях изоляция поддерживается отсутствием миграций,
невозможностью эффективного размножения в новой обстановке
или интеграции потомства. Двуполость просто добавляет к этому
то, что эффективное размножение может снижаться предпочтени-
ем в выборе партнёров и гибридной стерильностью, а также вне-
шними факторами, которые влияют на обе категории популяций.
Эволюция однополых и двуполых популяций различается во
многих важных отношениях. Но это не отменяет того факта, что в
обоих случаях формируются виды, причём, по принятому опреде-
лению, одной и той же категории.
Вид в палеонтологии
Исходные данные при классификации ископаемых достаточно
специфичны, в отличие от неонтологических, и доставляют много
особого рода проблем. Доступные анатомические данные всегда
неполны, а непосредственные физиологические или поведенческие
данные чаще всего отсутствуют. Экологические данные, хотя и
имеются в достаточном объёме, также гораздо менее полны, чем
для современных животных. Выборки чаще всего невелики, а их
получение бывает связано со значительными затратами. Выборки
могут быть смещёнными по разным, порой весьма своеобычным
причинам. Палеонтологи в общем хорошо осведомлены об этих
проблемах и занимаются активными и в целом успешными поис-
ками способов их решения. Я уже имел возможность обобщить
наиболее важные из них (Simpson, 1960а). Они влияют на решение
скорее частных проблем, чем на общее содержание таксономиче-
ских принципов, поэтому я не буду вдаваться в них в этой книге.
Что касается видов, палеонтологический подход часто принци-
пиально не отличается от неонтологического. Многие выборки ис-
копаемых не имеют существенной временной размерности, т.е. их
Вид и низшие категории
191
протяжённость во времени слишком коротка, чтобы можно было
отметить соматические изменения. В этих многочисленных случа-
ях исследуемые экземпляры могут рассматриваться как одновре-
менно существующие, а определение видовой категории и рас-
познавание видовых таксонов происходит на тех же основаниях,
что и для современных животных. Особая проблема, уникальная
для палеонтологии, возникает в не менее многочисленных случа-
ях временных последовательностей заметно различающихся выбо-
рок из одной или из близко родственных филетических линий.
Выделение видов во временном филогенетическом ряду сходно с
таковым у одновременно существующих организмов, но и ставит
перед необходимостью решения весьма специфичных вопросов.
Эти вопросы обсуждали неоднократно и в деталях, например,
Simpson (1951), Cain (1954), Sylvester-Bradley (1956, введение ре-
дактора), Rhodes (1956), Westoll (1956), Newell (1956), George
(1956), Imbrie (1957) и Grabert (1959), у которых можно также найти
ссылки на обширную литературу по теме.
В ветвящейся филогении (рис. 12А, Б) концепция эволюцион-
ного вида, очевидно, применима к каждой отдельной ветви. Сре-
зы через все ветви превращают каждую из них в вид одновремен-
но существующих животных, в целом соответствующий генетичес-
кому виду. Каждая отдельная выборка, или всякая выборка, кото-
рая не упорядочена во временной ряд, на самом деле может быть
использована для определения эволюционного вида или, если мож-
но предполагать двуполое размножение, в некотором приближе-
нии — генетического вида. Однако прослеживая этот ряд на более
продолжительном отрезке времени, при применении эволюционной
концепции приходится сталкиваться с проблемами двоякого рода.
Эволюционный вид определён как отдельная филетическая ли-
ния (в палеонтологии иногда называется фратрия, gens) с единой
ролью. Если вы начнёте с любой точки в этом ряду и проследите
линию назад во времени, вы не найдёте места, где бы это опреде-
ление не работало. Вы будете всё время оставаться в пределах не-
разрывной, обособленной единой линии и, тем самым, всё время
будете оставаться в пределах вида, от которого вы начали, если
только не будут привлечены какие-то иные критерии выделения
вида (рис. 12В). Если бы ископаемые остатки были полны, вы
14'
192
Дж. Г. Симпсон. Принципы таксономии животных
Рис. 12. Схемы филогении и генетических видов с проходящими
ЧЕРЕЗ НИХ ФИЛЕТИЧЕСКИМИ ЛИНИЯМИ
А. Трёхмерное представление филогении. Горизонтальная размерность в
пределах каждой ветви обозначает диапазон разнообразия, расстояния между
ветвями по горизонтали отражают степень дивергенции отдельных линий и
видов. Дерево представлено так, будто оно разрезано временными плоскостями,
приходящимися на моменты tp t и t3 и каждому пересечению соответствует
совокупность одновременно существующих генетических видов.
Б. Формальное представление той же самой филогении в двух измерениях,
с теми же временными сечениями и генетическими видами (жирные точки).
Каждая линия, ведущая к концевому генетическому виду, — обособленная
филетическая ветвь, очерченная штриховой линией. Проблемы классификации
общих стволов и возможного разделения более длинных ветвей (напр.,/1^)
требуют особого обсуждения.
В. Схема филетической линии, начинающейся (в качестве примера) от
концевого вида b и идущей до начального вида h. Каждый конечный вид имеет
такую ведущую к нему ветвь, и все они в конечном итоге сливаются по мере
движения назад во времени.
смогли бы начать с человека и дойти до простейших, постоянно ос-
таваясь в рамках вида Homo sapiens. Такая классификация явно и
бесполезна, и ошибочна в каких-то своих основах. Разумеется, для
целей классификации филетическая линия должна быть нарублена
на сегменты, и это должно быть сделано произвольно (как опреде-
лено в главе 4), поскольку не существует непроизвольного спосо-
ба разделения непрерывной линии [45]. Это достаточно просто и
полностью оправдано с точки зрения уже рассмотренных таксоно-
мических принципов. На практике, всё, что нужно иметь, — некий
Вид и низшие категории
193
критерий того, насколько большими (и в каком смысле «больши-
ми») должны быть такие сегменты. Следующий критерий, доста-
точно практичный, принимается почти всеми эволюционными
классификаторами ископаемых организмов. (Все палеонтологи,
разумеется, согласны с фактом эволюции, однако не все они — эво-
люционные таксономисты.)
Последовательные (successive) виды должны быть определены
так, чтобы морфологические различия между ними были не мень-
ше, чем различия между одновременно существующими видами
одной группы или близкородственных групп (см. рис. 13А).
Применение этого критерия во многом зависит от личных суж-
дений и снова напоминает нам, что классификация — это искусст-
во. Тем не менее, имеется артистический канон, который ограни-
чивает игру фантазии, создает некоторую основу, которой можно
руководствоваться, и обеспечивает определенное единообразие ре-
шений. Применяя ранее введённый критерий диапазона для одно-
временно существующих видов, данное правило можно сделать
несколько более лёгким в применении и объективным следующим
образом:
Если в филетической линии выявленные диапазоны исследуе-
мых варьирующих признаков для популяций, относящихся к двум
отрезкам времени, не перекрываются, эти популяции могут быть
отнесены к разным последовательным видам, а граница между ви-
дами может быть проведена приблизительно посередине (во вре-
мени) между ними (см. рис. 13Б).
Этот критерий помещает ключевые популяции приблизительно
в середину отрезков, которые соответствуют последовательным
видам. Какие именно выборки будут взяты в качестве представи-
телей популяций, играющих эту роль, во многом зависит от обыч-
ной в практике удачи открытия и от предшествующей истории
классификации данной группы. Если, как часто бывает, один или
два вида были обнаружены и определены прежде, чем филетиче-
ская линия стала более детально изученной, эти виды следует по
мере возможности сохранять. Это означает частичное отступление
от в целом верного утверждения, что тип не влияет в таксономии
ни на что, кроме номенклатуры. Выборка, принятая в качестве да-
тировки средней точки вида, должна включать тип любого видово-
194
Дж. Г. Симпсон. Принципы таксономии животных
Средняя дистанция между видами
Рис. 13. Разделение филетических линий на последовательные виды
А. Критерий, основанный на примерном равенстве горизонтальных и
вертикальных различий между видами в филогении, подобной той, что приведена
на рис. 12. Линии расходятся от узлов приблизительно на равном расстоянии
вдоль ветви, которая соответствует [последовательности] изменяющихся
популяций. Усреднённые различия между видами по горизонтали можно
использовать в качестве шаблона для разделения ветви на последовательные
фрагменты, как показано поперечными штрихами. Последние указывают
приблизительно середину каждого из временных интервалов между ветвлениями,
или «центровые» популяции последовательных видов.
Б. Критерий перекрывания диапазонов в пределах единой филетической
линии. Популяционная изменчивость в каждый момент времени показана нор-
мальной кривой, в сумме образующей единое распределение вдоль линии. Диа-
пазон R2 на момент t2 явно выходит за пределы диапазона Rt на момент tr Соот-
ветствующие популяции отнесены к разным видам, разделение которых произ-
вольно датируется промежуточным моментом, показанным точечной линией.
Вид и низшие категории
195
го названия, ранее предложенного для данного фрагмента филети-
ческой линии. Типы в таком случае помогают определить, где
именно проходят границы между последовательными видами (рис.
13В). (Однако они всё равно не влияют на концепцию вида и не
типифицируют его.) Типы имеют сходный центрирующий эффект
в случае совершенно произвольных подвидов, как одновременно
существующих, так и замещающих один другого во времени (см.
дополнительную дискуссию в конце данной главы).
Как уже было отмечено, виды, представляющие собой отрезки
филетической линии (или фратрии), иногда называются палеовида-
ми. Я предпочитаю называть их последовательными или, вслед за
Imbrie (1957), замещающими [successional]. (Кстати, Имбри назы-
вает виды одновременно существующих организмов, вымерших
или современных, шранзиентпами, transient.) Это иная категория
вида по сравнению с генетическими или иными одновременно су-
ществующими видами, но теперь я думаю, что различие между
ними мной, равно как и другими иногда переоценивалось. Все они
могут рассматриваться как аспекты или проявления эволюционного
вида: в одном случае это сегмент эволюционного вида, приурочен-
ный к определённому отрезку времени, в другом — одномомент-
ный срез через эволюционный вид.
Другая важная проблема, которая возникает при применении
концепции эволюционного вида к ископаемым последовательнос-
тям, связана с ветвлением. Если последовательность представлена
двумя филетическими линиями и их общим стволом, этот ствол,
который не может принадлежать сразу обоим видам-потомкам,
можно отнести либо к одному, либо к другому виду, либо ни к од-
Рис. 13 (продолжение).
В. Влияние типов на разделение филетических линий на последовательные
виды. На В' типы, показанные крестиками, — это экземпляры, взятые из
популяций в моменты tt и t2. Эта ветвь разделена на два вида, граница между
которыми приходится на момент времени, показанный прерывистой линией. На
В” три типа взяты из популяций в моменты tp t2, и tr Три последовательных вида
(или подвида) разделены так, что границы между ними приходятся на моменты
времени, показанные прерывистыми линиями.
196
Дж. Г. Симпсон. Принципы таксономии животных
А
Рис. 14. Альтернативные способы разделения коротких ветвящихся
ФИЛЕТИЧЕСКИХ ЛИНИЙ НА ВИДЫ
А. Три вида разделены в точке ветвления.
Б. Концевая ветвь а не отличается от предкового ствола, с которым поэтому
объединяется, тогда как уклонившаяся ветвь b — самостоятельный вид.
В. Концевая ветвь b мало уклоняется от предкового ствола, с которым
поэтому объединяется, тогда как а уклонилась в большей степени и является
самостоятельным видом.
Г. Разрыв (точки) в летописи позволяет проводить непроизвольную границу
между видами.
ному из них (рис. 14). В принципе, наилучшее решение получает-
ся, когда линии можно разделить на три вида — один предковый
и два потомка, разделённые в точке ветвления. Если, однако, одна
ветвь отклонилась от предка значительно меньше другой, из-за
того, что она более короткая или особенно если она более консер-
вативная (эволюировала с меньшей скоростью), тогда эту ветвь
можно поместить в тот же вид, что и предковый ствол.
Подобные ситуации возникают в палеонтологических класси-
фикациях, однако на самом деле они не столь часты. Существен-
но непрерывные последовательности выборок, содержащие реаль-
ные разветвления филетической линии на две или более, всё ещё
сравнительно редки. Одна из причин состоит в том, что видообра-
зование обычно аллопатрично. Два вида-потомка обычно возника-
ют в разных областях распространения предкового вида, так что
последовательные выборки из любого одного места не могут по-
казать точку ветвления, а немногие локальные выборки из разных
территорий, обычные в палеонтологии, скорее всего, не попадут в
неё. Как уже было разъяснено ранее* такие пробелы позволяют раз-
Вид и низшие категории
197
граничивать виды вполне надежно, насколько это допускают име-
ющиеся фактические данные (рис. 14Г).
Ещё одна проблема, не столько теоретическая, сколько приклад-
ная, возникает в связи с трудностью, а иногда и невозможностью
определения истинной временной последовательности популяций,
представленных палеонтологическими выборками. Стратиграфи-
ческие корреляции всё ещё столь неточны, что часто невозможно
определить относительный возраст выборок приблизительно одно-
го возраста, но из разных локалитетов. Также может случиться, что
некоторый вид, который всегда обнаруживается в более высоких
слоях, чем другой, т.е. более поздний по времени, в каждом дан-
ном локалитете в действительности является современником это-
го второго вида (рис. 15А). Смещение географических областей
распространения родственных популяций или видов порождает
много таких проблем и кажущихся аномалий (другой пример см.
на рис. 15Б).
Две выборки приблизительно одного возраста, но из разных
локалитетов никогда не бывают идентичны. Если различия высо-
Локалитет
----пространство-------
ПРОСТРАНСТВО-------►
Рис. 15. Некоторые проблемы, связанные с выборками географически
смещающихся палеонтологических видов
А. Два вида а и Ь, каждый приуроченный к определённой биоте, замещают
друг друга географически. В каждом локалитете форме b стратиграфически пред-
шествует форма а, т.е. b более поздняя, однако в целом эти два вида — совре-
менники.
Б. Две родственные филетические линии а~е и v~z развиваются в двух смеж-
ных, географически замещающих одна другую биотах. Ни одна из них не пред-
шествует другой ни в одном из локалитетов. Ископаемые остатки в локалитете,
очерченном вертикальными линиями, дают временную последовательность Ь-
х-е, которая не является филетической линией.
198
Дж. Г. Симпсон. Принципы таксономии животных
ПРОСТРАНСТВО
Рис. 16. Интерпретация палеонтологических выборок из различных
горизонтов и локалитетов
А. В данной ситуации (изменчивость популяций показана кривыми) нет
перекрывания диапазонов (разрыв показан прерывистой линией), так что попу-
ляции должны быть отнесены к разным видам.
Б. Интерпретация двух филетических линий (показаны прерывистыми ли-
ниями) с общим корнем. Обнаружение популяции, показанной точечной лини-
ей, усилило бы интерпретацию.
В. Интерпретация последовательных видов одной филетической линии.
Обнаружение популяции, показанной точечной линией, усилило бы интерпре-
тацию. (Ещё одна, третья возможность показана на рис. 17.)
ко значимы в статистическом смысле, это может быть результатом
географической изменчивости, временной эволюции, или их соче-
тания. Промежуточные выборки, если они доступны (в палеонто-
логии они редки) покажут, имеется ли непрерывность между попу-
Вид и низшие категории
199
ляциями, однако не позволят однозначно утверждать, является ли
эта непрерывность географической в пределах генетического вида
или временной в пределах эволюционной линии. В таких очень
частых примерах, с промежуточными выборками или без них, ра-
нее рассмотренные каноны перекрывания или разделения диапазо-
нов могут применяться лишь произвольно. В таком случае класси-
фикатор выделяет виды согласно некоторым приемлемым критери-
ям, но он редко может быть уверен, с видами какой категории име-
ет дело: в частности являются ли они последовательными или от-
носятся к разным одновременно существующим филетическим
линиям (рис. 16).
Подвид
К числу поистине основополагающих и почти универсальных
фактов природы относится изменчивость всех видов. Они варьиру-
ют во времени: циклично — по суткам, сезонам, годам; случай-
но — в форме иррегулярных событий, необычных или уникальных;
долговременно — в форме продолжительных трендов эволюцион-
ной прогрессии. В пространстве (в любой момент времени) они
варьируют: в пределах локальных популяций (или демов, см. ниже)
и географически между неизолированными, частично изолирован-
ными или временно изолированными локальными популяциями.
Большинство доэволюционных таксономистов могли игнорировать
эти факты, столь неудобные для господствующей типологии. Для
временной изменчивости у них практически не было данных по-
мимо коротких (например, годовых) циклов, которые попросту воз-
вращают популяцию к одному и тому же состоянию. Что касается
пространственной изменчивости, то небольшие и, как правило,
широко разобщённые географические выборки, если они заметно
различались, могли считаться (чаще всего так и было) представи-
телями разных видов.
Даже в додарвиновские времена с ростом знаний о географи-
ческом распространении и изменчивости становилось всё очевид-
нее, что популяции, даже полностью интерградирующие и относя-
щиеся к одному виду, понимаемому как реальная дискретная еди-
ница, тем не менее часто могут заметно отличаться в разных час-
200
Дж. Г. Симпсон. Принципы таксономии животных
тях общевидового ареала. Уже в 1844 г. орнитологи стали выделять
в классификации такие географические группы как таксоны и при-
сваивать им трёхсловные названия, добавляя ещё одно, третье на-
звание после линнеевского видового биномена (по-видимому, пер-
вым был Шлегель: см. Sibley, 1954). Эта новация сразу же встре-
тила жёсткую оппозицию, причём возражения против подвида как
отдельного таксона и триномена с тех пор продолжают возникать
и до сих пор.12
Ранние возражения были главным образом типологическими,
хотя обычно не столь откровенными и почти никогда не излагавши-
мися в терминах самой типологии. Если две популяции различают-
ся, они обладают разными архетипами. Воплощением архетипов низ-
шего категориального уровня являются видовые таксоны. Поэтому
подвиды более поздних авторов «на самом деле» являются видами,
которые должны обозначаться так, как если бы они были выделены
в 1758 г, в противном случае это вообще не таксоны, а лишь слу-
чайные вариации в пределах видов. Даже в достаточно поздние годы
таковой была, если не философская, то по крайней мере операцио-
нальная, позиция некоторых немногих таксономистов (например,
Kinsey, 1936; Burma, 1948), которые настаивали на том, что малей-
шие различия между популяциями свидетельствуют об их видовой
обособленности. Такая позиция на протяжении последнего столе-
тия или более (обычно связываемая с «дробительскими» вкусами)
привела к выделению многих тысяч «видов», которые большинство
современных таксономистов рассматривают как невалидные таксо-
ны или в лучшем случае как подвиды.
Огромное большинство современных таксономистов согласно
с тем, что виды следует определять не как минимальные «типы»,
12 Совсем недавняя вспышка этого противостояния последовала после пуб-
ликации работы Уилсона и Брауна (Wilson, Brown, 1953), причём журнал
Systematic Zoology отвёл много места для многочисленных, иногда повторяю-
щихся, иногда несколько непоследовательных дискуссий по этому предмету.
Для моих более общих целей нет необходимости разбирать все эти статьи, рав-
но как и те, которые появились в других журналах: тем, кто особенно инте-
ресуется, можно номер за номером проглядеть подшивку Systematic Zoology.
В качестве примера можно привести статью: Smith, White (1956), в ней содер-
жится один из наиболее полных ответов Уилсону и Брауну.
Вид и низшие категории
201
а в генетических и эволюционных терминах, и что в таком случае
они обычно «политипические» — почти универсально принятый
термин, который тем не менее неудачен, поскольку фиксирует от-
вергнутую [типологическую] концепцию. Сохраняющееся разногла-
сие между истинно эволюционными таксономистами, таким обра-
зом, заключается не в том, имеют ли виды различающиеся геогра-
фические подгруппы (большинство их, очевидно, имеет), а в прак-
тичности и разумности распознавания таких подгрупп как таксонов
и присвоения им названий (триноменов) в обычных формальных
классификациях. Все аргументы можно свести к двум вопросам:
1. Есть ли какой-либо непроизвольный элемент (или «объектив-
ная основа»)в подвиде?
2. Есть ли практический смысл для таксономистов в использо-
вании подвидовой категории (независимо от того, является ли она
полностью произвольной или нет)?
Ответ на оба вопроса таков: «и да, и нет», или «иногда». У не-
которых видов есть почти однородные популяции на довольно об-
ширных территориях, обособленные от соседних отличающихся от
них популяций вдоль линий, соответствующих узким зонам интер-
градации. Эти зоны, как упомянуто в главе 4, действительно раз-
деляют виды на относительно (но не абсолютно) произвольные
подгруппы. Эти подгруппы обычно выделяются как подвиды, ко-
торые «реальны» в том, что существуют как группировки в приро-
де, а не исключительно в сознании таксономиста. У многих дру-
гих видов, однако, нет столь бесспорных непроизвольных или ча-
стично произвольных подгрупп.
Некоторые виды демонстрируют более или менее постепенные
изменения признаков в одном или нескольких географических на-
правлениях — клины. Часто выдвигалось предложение выделять
клины, а не подвиды, однако клины не определяют таксоны, как
будет показано в следующем разделе. Другие виды не разделены на
обширные подгруппы и показывают крайне неупорядоченную гео-
графическую изменчивость, не организованную в клины. (Есть и
такие, редкие, которые показывают столь незначительную геогра-
фическую изменчивость, что в их отношении вопрос о подвидах не
возникает.) Многочисленные примеры всех таких ситуаций пред-
ставлены у Майра (Мауг, 1942, также многие другие его работы;
202
Дж. Г. Симпсон. Принципы таксономии животных
РАССТОЯНИЯ г
И РАЗЛИЧИЯ
В
Рис. 17. Некоторые соотношения между одновременными
и последовательными подвидами
А. Ряд интерградирующих одновременно существующих демов, показан-
ных вариационными кривыми. Различия между двумя заштрихованными кри-
выми, средние значения которых показаны стрелками, удовлетворяют «правилу
75%», соответствующие демы отнесены к разным подвидам, разделёнными гео-
графически как показано прерывистой линией.
Б. Ряд интерградирующих последовательных популяций одной филетичес-
кой линии. При проецировании на единую временную ось или при игнорирова-
нии реального времени, отношения между ними те же, что на (А), так что попу-
Вид и низшие категории
203
см. также его краткие, но содержательные замечания по поводу
полемики о подвиде в: Мауг, 1954).
Когда вид разделён на частично произвольные подгруппы, их
обозначение в качестве подвидов вряд ли вызовет какие-либо воз-
ражения, если данные адекватны, а таксономист считает подвид
полезным для своих целей. Против такого использования не может
служить аргументом то обстоятельство, что подобные подгруппы
можно выделить не во всех видах: подвид не является обязатель-
ной категорией и даже в принципе не обязан применяться повсе-
местно.
Когда географическая изменчивость клинальная или мозаичная,
всякий выделенный подвид будет произвольным. Это, опять-таки,
само по себе не служит аргументом против подвида, поскольку
многие из высших таксонов, которые являются всеобще признан-
ными и необходимыми, выделяются произвольно. Непроизвольные
классификации вообще невозможны. Убедительный ответ на воп-
рос, являются ли они полезными, состоит в том, что большинство
опытных специалистов по многим группам животных (особенно по
высшим позвоночным) считают их таковыми и, тем не менее, по-
стоянно используют. Конечно, [подвидами] можно злоупотреб-
лять — но в равной степени как и любой другой категорией. В од-
ном только виде гофера Thomomys umbrinus на юго-западе США и
севере Мексики в настоящее время выделено 213 подвидов, и те,
Рис. 17 (продолжение).
ляции, помеченные заштрихованными кривыми и стрелками, удовлетворяют
«правилу 75%». Они отнесены к последовательным подвидам, разделённые по
времени как показано горизонтальной линией.
В. Географическая экспансия и морфологическая дивергенция во времени
в единой филетической линии, в результате чего географические и временные
подвиды в сущности эквивалентны. [Временные] подвиды a'-b"-c"' практиче-
ски неотличимы от географических подвидов а*"-Ь"*-с'".
Г. Возможная интерпретация популяций на рис. 16 в качестве подвидов, а
не видов. В пользу этого свидетельствовало бы обнаружение какой-то одной из
заштрихованных популяций или их обеих, указывая на то, что к выявлению ста-
туса этих двух выборок имеют отношение только пространственные, а не
временные различия.
204
Дж. Г. Симпсон. Принципы таксономии животных
которым нравится такая игра, могут продолжать до тех пор, пока
каждая небольшая колония этого гофера не получит своё линнеев-
ское имя. Однако имеются разумные каноны таксономического ре-
месла [art], применение которых обуздывает подобную невоздер-
жанность. Наиболее важные из них — статистические методы, ко-
торые гарантируют, что хотя бы заметная часть особей (сейчас при-
нято, что не менее 75%) соседних подвидов будет вполне иденти-
фицируема (Amadon, 1949). Это обеспечивает высоко значимые
различия в средних значениях признаков между подвидовыми по-
пуляциями (рис. 17А).
Следует исправить два недоразумения, касающиеся подвидов.
Во-первых, подвиды, вопреки мнению типологов, — это не малень-
кие виды, равно как и не зарождающиеся или «почти» виды, как
полагают некоторые эволюционные таксономисты. Они являются
таксонами существенно иного качества, чем виды. Лишь немногие
из них когда-либо станут видами, хотя, по правде сказать, некото-
рые приближаются к этому статусу. Во-вторых, подвиды не служат
выражением географической изменчивости признаков вида и лишь
частично воспроизводят эту изменчивость. Это — формальные так-
сономически обозначенные, обычно произвольные популяционные
единицы, и они могут воспроизводить или полностью описывать
изменчивость этих популяций не больше, чем классификация в це-
лом способна воспроизвести или полностью описать филогению.
Но из всего этого не следует, что они бесполезны при обсуждении
изменчивости.
Подвиды обычно не используются в палеонтологии, и хотя их
применение в палеонтологии, по-видимому, возрастает, здесь нет на
этот счет определённого единодушия. Географические выборки
ископаемых обычно недостаточны для обоснованного выделения
подвидов в точно таком же статусе, как и у современных живот-
ных, хотя работа ведётся, и ведётся всё активнее. Временные под-
виды, т.е. последовательные [successive] подразделения видов, ино-
го качества, чем географические подвиды. Тем не менее, они мо-
гут быть определены на основании сходных приёмов, вроде упомя-
нутого выше «правила 75%», и когда это возможно, они крайне
полезны в стратиграфических исследованиях. Единственной воз-
можной альтернативой является использование неприемлемо дроб-
Вид и низшие категории
205
ных последовательных видов, и на этом основании я полагаю, что
применение последовательных подвидов следует приветствовать.
Некоторые параллели и различия между одновременными и разно-
временными подвидами показаны на рис. 17.
Эта концепция, но не сам термин, в палеонтологии используется
давно, начиная по крайней мере с Ваагена (Waagen, 1869), который
называл последовательные подвиды «мутациями», пока генетики не
захватили это слово. Бэйзер (Bather, 1927) заменил его термином
«транзиент» [transient] (в ином смысле, чем выше упомянутый Им-
бри) (см. Rhodes, 1956). Кастер и Элайес также предложили термин
«ваагенон» (см. Newell, 1947): он наименее двусмысленный, если
желательно отличать внутривидовые группы в палеонтологии от
географических подвидов. Основной аргумент называть ваагеноны
последовательности подвидами состоит в том, что это указывает их
надлежащее место в стандартной иерархии классификации.
Другие внутривидовые группы
Ни вид, ни подвид не является окончательной систематической
популяционной единицей в природе. Наименьшая единица популя-
ции, имеющая эволюционное значение, — локальная группа живот-
ных (одного вида или подвида), которые легко и достаточно часто
контактируют между собой: одновидовые члены некоторого сооб-
щества в наиболее узком смысле. Такая минимальная популяцион-
ная единица называется дем [deme]. У двуполых организмов веро-
ятность того, что половой партнёр будет найдет в такой группе,
значительно выше, чем при спаривании членов из разных групп.13
Как у двуполых, так и у однополых организмов именно внутри
этой группы происходит внутривидовая конкуренция и отбор.
Дем может соответствовать подвиду или даже, в крайне редких
случаях, целому виду. Однако гораздо чаще эта группа существенно
13 По этой причине Dobzhansky (1951) и некоторые другие генетики назы-
вают демы менделевскими популяциями. Я предпочитаю термин «дем», по-
скольку он не ограничивается двуполыми организмами, а также потому, что
термин «менделевская» вызывает некоторые вопросы. Ранее, до введения тер-
мина «дем», таксономисты часто использовали расплывчатый и громоздкий
термин локальная популяция.
15 — 4338
206
Дж. Г. Симпсон. Принципы таксономии животных
меньше, чем каждый из названных таксонов. Некоторые демы обо-
значены чётко: популяции на небольших островах, в отдельной кур-
тине деревьев, в муравьиной куче и тому подобное. Чаще же они
отграничены нечётко или вообще не имеют границ, что зависит от
способа рассмотрения. В пределах равномерно распределённой
видовой популяции дем для особи, входящей в популяцию, охва-
тывает тех других ее представителей, с которыми у данной особи
происходят регулярные контакты.
Хотя демы и являются базовыми популяционными единицами,
они не рассматриваются и не должны рассматриваться в классифи-
кации и получать названия. Причина не в том, что они обычно мо-
гут быть определены лишь произвольно: как мы видели, это вер-
но в отношении большинства таксонов. Демы не классифицируют-
ся, прежде всего, из-за того, что они мимолётны и не обладают эво-
люционной ролью в длительной перспективе. У подвидов также
различия ролей в пределах единого вида невелики и они также
эфемерны, но им свойственна определённая дифференциация и
достаточная устойчивость в масштабах человеческой истории. Со-
седние демы часто не имеют заметных различий, они могут сли-
ваться, разделяться или исчезать на протяжении жизни таксономи-
ста. Такие группы весьма поучительны с эволюционной точки зре-
ния, однако они не предназначены для формальной, даже не слиш-
ком стабильной классификации. Число демов к тому же настоль-
ко велико, что никто не сможет справиться со списком всех их на-
званий, даже в пределах, скажем, одного рода многочисленных
животных. (Ранее упоминавшийся пример с «подвидами» гофера
напоминает классификацию и именование демов, но это плохая
классификация, в которой спутаны концепции подвида и дема, и не
афишируется прямо, что демы как таковые приняты в качестве так-
сонов.)
Одним из наиболее обычных и многозначных терминов в так-
сономии был eapuemem. Согласно типологическим принципам,
каждый вид имеет фиксированный образ. Всё, что не соответствует
этому образу, — «вариетет», концептуальное расширение схоласти-
ческой «акциденция». Как и многое другое из типологического на-
следия, данные термин и понятие были восприняты ранними эво-
люционными таксономистами и продолжали быть бичом и источ-
Вид и низшие категории
207
ником путаницы в науке. Редко бывает понятно, является ли вари-
етет: 1) индивидуальным уклонением, 2) группой таких уклонений,
или морфой, концептуально связанной с собственно изменчивос-
тью, но не с популяцией, или 3) обособленной популяцией в пре-
делах вида, аналогичной или даже идентичной подвиду.’4 Ранние
эволюционисты (и немногие из более поздних) не только не смог-
ли разграничить эти трактовки, но даже не представляли, что они
существуют и жизненно важны для таксономии. Этот недочёт, ве-
роятно, является наиболее серьёзной логической нестрогостью в
обычно скрупулёзных работах Дарвина.
Вариететы, обычно без какого-либо чёткого и определённого
значения, часто вводились в классификации с присвоением трино-
мена, квадриномена или какого-либо ещё символического обозна-
чения. Теперь уже не может быть никаких сомнений, что вариетет
как категория классификации должна быть совершенно отброше-
на (см., например, Newell, 1947). В первых двух значениях он не
имеют отношения к популяциям и поэтому не может относиться к
таксонам, каковые по определению состоят из популяций. В пос-
леднем значении он соотносится с популяциями, признаваемыми в
качестве таксонов, только если совпадает с подвидом.
Такие термины как форма или морфотип часто применялись к
вариететам, понимаемым во втором смысле, т.е. к совокупностям
отдельных вариаций в пределах популяций, но не ко всем варьи-
рующим организмам, составляющим популяцию. Эти термины по-
лезны при описании изменчивости без той неопределённости, ко-
торая присуща вариетету, но они, очевидно, также не могут обозна-
чать категории классификации. Эдвардс (Edwards, 1954) предложил
определять морфы как «диагностируемые симпатричные или син-
хронные популяции единого вида», которые, как он полагает, так-
же могут быть подвидами или демами. На самом деле симпатрич-
ные и синхронные члены единого вида не могут формировать от-
дельные популяции в современном таксономическом смысле: это
просто «вариететы» во втором значении, или формы, или морфо-
14 Сегодня термин «вариетет» более всего известен в значении, перешед-
шем из ботаники и растениеводства. В таком применении часто смешаны трак-
товки (2) и (3). Сам термин и его многозначность вряд ли можно устранить в
растениеводстве, но это не должно беспокоить учёных-таксономистов.
15*
208
Дж. Г. Симпсон. Принципы таксономии животных
типы, короче — морфы полиморфного вида. Это и не подвиды, в
любом их современном понимании, и не демы. Можно только при-
ветствовать предложение Эдвардса не присваивать им технических
названий.
Другого рода концепция, также применяемая к вариациям в
пределах вида, представлена термином «клина», предложенным
Huxley (1938) и в настоящее время получившим широкое призна-
ние. Исходное широкое определение, которое дал Гексли, таково:
«постепенное изменение [gradation] мерного признака». Если не
дано дополнительных уточнений, обычно принимается, что клина
ограничена одним видом и является однонаправленной и плавной,
а не дискретной и ступенчатой. Было выделено несколько разных
вариантов клин, для чего самим Гексли была предложена доволь-
но сложная терминология, дополненная уже после него: геоклина
для географических, экоклина для экологических, хроноклина для
временных клин и многие другие. Чаще других используются гео-
клины, именно их обычно подразумевают, когда говорят просто о
клинах.
Предлагая этот термин, Гексли говорил, что описание с помо-
щью клин предназначено дополнить, а не заменить современные
таксономические методы. Однако почти сразу же он добавил
(Huxley, 1939), что описание с помощью клин в определённых
случаях должно замещать «описание с помощью поименованных
территориальных групп», т.е. подвидов. В какой-то степени это
•предложение встретило понимание, однако в целом клины не
были приняты в классификации как основание для выделения
категорий или присвоения зоологических названий. Концепция
клины оказалась применимой практически во всех группах живот-
ных и крайне полезной для представления и интерпретации их
изменчивости. Она оказалась весьма важной в систематике, одна-
ко это не истинно таксономическая концепция, в строгом понима-
нии таксономии.
Рассматриваемый здесь вопрос хорошо прояснил Майр
(в: Sibley, 1954):
Подвиды и клины — концепции, относящиеся к разным аспектам
[внутривидовой изменчивости]. Популяция может принадлежать только к
одному подвиду, но к нескольким разным клинам. Другими словами, под-
Вид и низшие категории
209
вид есть таксономическая концепция, тогда как клина — эволюционная.15
Мы ничего не приобретаем, присваивая клинам названия.
Клина представляет собой форму упорядочивания признаков, а
не организмов или популяций. Часто бывает, что разные признаки
в пределах одного вида дают совершенно разные клины, идущие
в разных направлениях, даже под прямым углом друг к другу, —
факт, который с самого начала отмечал Гексли и подчеркнул Майр
в только что приведённой цитате. Вкратце, таксономическая клас-
сификация имеет дело с систематизацией популяций, а концепция
клины — нет. Поэтому эту концепцию нельзя использовать для
определения какой-либо таксономической категории. Конечно, как
и любые исследования по внутривидовой изменчивости, она край-
не полезна для систематизации данных классификации, для пони-
мания и определения отдельных таксонов. Тем не менее, в действу-
ющей классификации единственной приемлемой внутривидовой
категорией является подвид.
Надвид
Ближайшая категория выше уровня вида, которая предусмотре-
на Правилами номенклатуры, — широко используемый подрод. Он
обсуждается, наряду с родом и высшими категориями, в главе 6.
Однако иногда используются группировки между видом и подро-
дом: их уместно рассмотреть здесь, поскольку они теснее связаны
с концепцией политипического вида, чем с концепцией рода [46].
Многие крупные (широко политипические) роды содержат
группировки наиболее сходных видов. Авторы монографий для
удобства часто обозначают такие группировки, причём используют
для этого разные термины — например, группы видов. Им не при-
писывается формальный категориальный ранг в иерархии и не при-
сваиваются особые названия: они обозначаются цифрами или дру-
гими символами, чаще же по названию какого-либо из входящих
в них видов, например «группа Drosophila macrospina». В некото-
15 Разумеется, Майр имел в виду не то, что таксономия является неэволю-
ционной, а то, что эта концепция относится к нетаксономическому разделу эво-
люционной систематики, в принятом здесь определении этих терминов.
210
Дж. Г. Симпсон. Принципы таксономии животных
рых случаях такие группировки, по-видимому, более соответству-
ют видам, а включённые в них «виды» соответствуют подвидам.
Обычны также ситуации, когда им можно (а по моему мнению, и
должно) присваивать соответствующее иерархическое обозначение
в качестве подродов: как я уже отмечал, некоторые таксономисты
странным образом не расположены применять подроды.
Категория надвида в иерархии может рассматриваться как до-
полнительная в том же стандартном смысле, что и, например, над-
семейство. Майр (Мауг, 1931, 1942) предложил использовать это
понятие в ситуациях, ранее обозначаемых как Artenkreis, руковод-
ствуясь следующим определением (Мауг et al., 1953):
Hadeud есть монофилетическая группа наиболее близкород-
ственных по преимуществу или целиком аллопатрических видов.
Иными словами, надвиды — это группы популяций, которые по
каким-либо характеристикам (морфология, экология и т.п.) перешли
границу потенциальной скрещиваемости и приобрели самостоя-
тельные эволюционные роли, однако не сделали этого по более
значимым показателям, которые свидетельствовали бы об их обо-
собленности в случае симпатрии. Следует признать, что они нахо-
дятся вблизи критической точки видообразования, точки оконча-
тельной изоляции, и остаётся достаточно неопределённо, перешли
ли они эту точку или чуть-чуть не достигли её. Это становящиеся
виды, которые в случае успешной эволюции составят отдельный
подрод, а может быть и род, но которые пока ещё не достигли та-
кой степени обособленности и разнообразия. Им не присваивают-
ся отдельные названия; во всяком случае, Правила номенклатуры
не оговаривают этого. Обычно они обозначаются по имени какого-
либо из включённых видов, как и в случае групп видов, по край-
ней мере некоторые из которых на самом деле являются надвида-
ми в понимании Майра.
Сильвестр-Брэдли (Sylvester-Bradley, 1951, 1954) предложил
применять термин надвид не только для одновременно существу-
ющих видов, как предполагает определение Майра, но также рас-
ширить его на предковые виды, входящие в хронологический над-
вид в пределах соответствующей филетической линии. Такой сег-
мент филогении, в понимании Сильвестра-Брэдли, в целом будет
надвидом без какого-либо уточнения. Кэйн (Cain, 1953, 1955) ре-
Вид и низшие категории
211
шительно возражает против этого. Их полемика отчасти сводится
к ненужным семантическим софизмам (хотя, конечно, внимание к
терминологическим тонкостям бывает полезно), и ни один из них,
как мне представляется, не достиг ясности в изложении этого важ-
ного вопроса. Кэйн совершенно прав, утверждая, что здесь смеша-
ны три различные концепции, а Сильвестр-Брэдли прав в том, что
всё это — аспекты или части единой, более широкой таксономи-
ческой концепции. Эта проблема заслуживает внимательного рас-
смотрения, поскольку она высвечивает несколько таксономических
принципов.
Подразумеваемая концепцией структура филогенетических свя-
зей схематически показана на рис. 18. В момент времени tj (рис. 18А)
существует монотипический вид, который в момент времени t2 ста-
---Подвиды
*»Виды
0 0 0 Высший таксон
Рис. 18. Анализ концепции надвида
А. Последовательные популяции, связанные отношением «предок—пото-
мок», обозначены вариационными кривыми. На момент tj имеется монотипи-
ческий вид, на момент t2 — вид с тремя подвидами, на момент t3— три близких
вида. Надвид (по Майру) иди географический над вид (по Сильвестру-Брэдли)
соответствует стадии видообразования на момент t3.
Б. Схематичное представление последовательности в (А) в форме филети-
ческих линий. Заштрихованной области под стрелками соответствует стадия, на
которой сохраняется скрещивание и линии не изолированы. Каждая линия (все
они интерградируют в начальной стадии) — хронографический надвид по Силь-
вестру-Брэдли.
В. Разделение представленной схемы филетических линий на пространствен-
но-временные таксоны.
212
Дж. Г. Симпсон. Принципы таксономии животных
новится политипическим с тремя подвидами, а в момент времени t3
это уже надвид в понимании Майра, или географический надвид в
понимании Сильвестра-Брэдли. Таким образом, мы имеем первую
концепцию, а именно — таксономической обособленности, одно из
состояний которой и есть надвид Майра. Вторая концепция (рис. 18Б)
включает последовательное возникновение, показанное на диаграм-
ме тремя линиями, каждая из которых является хронологическим
надвидом Сильвестра-Брэдли. Вид в момент времени Ц и t3 принад-
лежит всем трём. Филетические линии, или «хронологические над-
виды», не могут быть превращены в таксоны без их горизонтально-
го разделения между t2 и t3, что само по себе разбивает «хронологи-
ческие надвиды», которые, следовательно, не могут быть определе-
ны как таксоны. Третья концепция (рис. 18в) является единствен-
ной строго таксономической — именно, иерархическим разделением
филогенетического паттерна, как показано на диаграмме граница-
ми, выделяющими группу-в-группе (group-in-group).
Собственно говоря, предложение Сильвестра-Брэдли состоит в
том, чтобы надвид применялся в стандартной таксономической
форме как таксон, который соответствует фрагменту ветвящейся
филогении, более крупному, чем вид, но более мелкому, чем род
(или, возможно, подрод). Это представляется наиболее последова-
тельным применением, ибо в таком случае надвид естественным
образом включается в последовательность иерархических катего-
рий «подвид—вид—надвид—подрод» и т.д. Однако Сильвестр-
Брэдли использует один и тот же термин для элементов, стадий и
линий, так что в рассмотренном частном случае они не могут од-
нозначно сформировать таксоны в иерархической последовательно-
сти. Эти различия учтены в предложенных Сильвестром-Брэдли
уточняющих эпитетах, однако формальное обозначение всех трёх
концепций в качестве надвида и конфликт с исходным, более по-
пулярным определением Майра всё же запутывает ситуацию.
Если бы была необходима некая таксономическая категория
между видом и подродом, что не очевидно, и если бы она была
чётко определена и указаны условия для образования названий от-
носящихся к ней таксонов, чего не было сделано, надвид мог бы
стать подходящим обозначением этой категории. Как таковой, над-
вид в исходной трактовке Майра представляет собой удачное опи-
Вид и низшие категории
213
сание определённой эволюционной ситуации, однако как определе-
ние таксономической категории он остаётся двусмысленным. По
этой причине я предпочёл бы вернуться к понятию Artenkreis, ко-
торый лучше определён в системе иерархических категорий. (Тот
факт, что он заимствован из немецкого языка и может неправиль-
но произноситься, кажется не большей проблемой, чем неверное
произношение исходно греческого слова «иерархия»).
Типы и гиподигмы
Глава о низших категориях — подходящее место для того, что-
бы подчеркнуть и в какой-то мере развить идею, уже упоминавшу-
юся несколько раз: принципиально важно, что в современной эво-
люционной таксономии таксоны основаны на выборках, а не на
типах. Типы традиционно выполняли три функции: 1) как един-
ственный или ведущий базис для описания и определения таксонов
(прежде всего видов); 2) как стандарт сравнения, соответствие ко-
торому обеспечивает идентификацию экземпляра в качестве пред-
ставителя того же таксона, к которому относится тип; и 3) как не-
кий носитель имени, которое жёстко связано с типом при всех его
перемещениях в связи с разделением или объединением таксонов.
По строго типологическим основаниям нет вообще никакой
причины, почему бы один и тот же единственный экземпляр не мог
служить всем трём целям. В доэволюционной таксономии было
всеобще признанным, что тип может и должен выполнять все три
функции одинаково успешно, поскольку едва ли осознавалось, что
они различны. Эта типологическая идея среди многих прочих была
перенесена почти неосознанно в раннюю эволюционную таксоно-
мию, и она настолько крепко приросла к ней, что её очень трудно
искоренить. На самом деле в истинной и полностью эволюционной
таксономии три указанные функции, приписываемые типу, совер-
шенно несовместимы. Никакой единственный экземпляр не может
соответствовать всем им полностью, а неспособность осознать этот
факт добавила скрытой типологичности к и без того во многом
неоднозначной эволюционной таксономии.
Уже достаточно часто подчёркивалось, что таксоны внутренне
изменчивы и что внимание к их изменчивости существенно при их
214
Дж. Г. Симпсон. Принципы таксономии животных
описании, равно как необходимо при их практическом определе-
нии. Это естественным образом обязывает принимать в расчёт все
доступные экземпляры и предполагает принцип, что никакой един-
ственный экземпляр, отнесённый к данному таксону, для указанных
целей не является ни более важным, ни более типичным, чем лю-
бой другой. Какие-то экземпляры, конечно, ближе к норме [ave-
rage], чем другие, если иметь в виду конкретные признаки с их
средними значениями [mean], хотя это редко случается сразу по
всем признакам организма. Тот простой факт, что выявлена дей-
ствительная норма, означает, что все экземпляры были приняты во
внимание и ни одному из них не был придан особый вес.
Такой подход не означает, что описание единичных экземпля-
ров должно быть отброшено, как было опрометчиво предположе-
но в поразительно типологическом комментарии, приведённом в
одной крупной современной публикации (Shenefelt, 1959). Один из
практических способов рассмотрения изменчивости состоит в де-
тальном описании одного экземпляра, который не является и не
должен считаться номенклатурным типом, и отметить отличия дру-
гих экземпляров в процессе описания. При соответствующей ста-
тистической обработке, однако, все экземпляры рассматриваются
одновременно и на равных основаниях.
При идентификации, разумеется, отдельные экземпляры срав-
ниваются чаще всего, но не обязательно, с номенклатурным типом,
который нередко весьма далёк от нормы, обозначенной соответ-
ствующими диагностическими признаками. Окончательное реше-
ние о принадлежности к одному виду зависит, однако, не от бли-
зости к какому-либо экземпляру, типовому или иному, но от попа-
дания в диапазон изменчивости, полученный для всего таксона.
Этот диапазон выявляется на статистической основе доступными
техническими приёмами при изучении всех экземпляров, ранее от-
несённых к данному таксону. Как при определении, так и при иден-
тификации эти экземпляры рассматриваются в качестве выборки,
на основании которой выявляются характеристики популяций. Если
известен только один экземпляр, то он по необходимости служит
и типом, и единственным основанием для конкретного описания,
однако основанием для сравнения и идентификации остаются ха-
рактеристики популяции, предполагаемые на основании изучения
Вид и низшие категории
215
этого экземпляра, а не он сам по себе. Пригодная выборка, конеч-
но, будет увеличиваться по мере получения новых экземпляров,
причём она будет различной для разных классификаторов, имею-
щих доступ к разным коллекциям. Соответственно, базис для оп-
ределения и сравнения может меняться и, как правило, со време-
нем улучшается. Однако этот базис — не фиксированный тип, ко-
торый раз и навсегда обозначается в первоописании.
Для того, чтобы подчеркнуть значение выборки и определить
его яснее и более сжато, при первом теоретическом рассмотрении
этого вопроса (Simpson, 1940) я предложил термин «гиподигма»,
который может быть определён следующим образом:
Гиподигма, выделенная данным таксономистом в данный момент
времени для данного таксона, состоит из всех экземпляров, извест-
ных ему в это время, рассматриваемых им в качестве несомненных
членов этого таксона и совместно включённых в выборку, на основа-
нии которой он сделал выводы о соответствующей популяции.
Сама концепция (которая, несомненно, предшествует моему
анализу и предложенному термину) уже вошла в общее употребле-
ние, что нельзя сказать о термине, хотя он сейчас также довольно
активно используется, в основном палеонтологами. Выдвигалось
возражение (например, Мауг et al., 1943), что он является излиш-
ним синонимом исследованного материала [47]. Пока концепция
используется, обозначающий её термин не столь важен. Я, тем не
менее, по-прежнему полагаю, что «гиподигму» (или любой аль-
тернативный, должным образом определённый технический тер-
мин) следует использовать вместо «материала» по трём основным
причинам, не считая других: 1) она определена гораздо строже, чем
обыденное слово «материал», даже используемое таксономистами;
2) она концентрирует внимание на концепции и гарантирует, что
процедура действительно основана на исходном принципе; и
3) предлагая особый, технически обозначенный подзаголовок в
формальных описаниях и ревизиях таксонов, она ограничивает все-
гда присутствующий подзаголовок «тип» присущей ему функцией
номенклатурного типа.
Проблемы, связанные с применением концепции гиподигмы, те
же, что в целом присущи составлению выборок. Некоторые из них
подробно обсудил Ньюелл (Newell, 1949b). Наиболее заметная сре-
216
Дж. Г. Симпсон. Принципы таксономии животных
ди них — как указать единственную гомогенную популяцию и га-
рантировать, что вся выборка взята из неё. В идеале каждая выбор-
ка должна представлять отдельный дем в какой-то момент време-
ни, однако это требование редко может быть выполнено, разве что
для минимальных выборок, как для современных, так и для вымер-
ших организмов. К счастью, подобные жёсткие условия на самом
деле не нужны для получения хороших статистических заключений
и могут даже служить помехой. Многие из проблем, связанных с
составлением выборок, обсуждены в: Simpson et al. (1960).
Последующий ревизующий может счесть разумным сужение
или расширение исходной концепции таксона. Для этого он должен
будет точно знать, из каких временных и пространственных границ
таксона были взяты отдельные представители гиподигмы, выделен-
ной автором описания. Он также должен будет знать временную и
пространственную локализацию типа для решения номенклатурных
вопросов, если он делит прежний таксон. Для этого Ньюелл
(Newell, 1949b) предложил сохранить термин monomun для тех эк-
земпляров гиподигмы, «которые произошли [более или менее. —
Дж.С.] строго из того же горизонта, что и голотип [т.е. номенкла-
турный тип или просто тип. — Дж.С.]». Это требование, однако,
может быть выполнено с гораздо большей полнотой и без путаных
концепций «типа» простым указанием горизонта и локалитета для
каждого члена гиподигмы, будь то публикация или запись в коллек-
ции.
Ньюелл полагал также, что следует и дальше использовать па-
ратип, поскольку «его обозначение обеспечивает некий престиж и
служит определённой гарантией того, что гиподигма не будет раз-
делена для целей обмена. Это также подчёркивает документально
опубликованную ценность данных экземпляров.» Однако паратипы
не обладают особой документальной ценностью, а истинная цен-
ность всех экземпляров с документальной значимостью подчёрки-
вается обозначением гиподигмы, отчасти предложенной и по этой
причине. Что касается первого соображения, выдвигалось автори-
тетное предложение выделять паратипы как «дубликаты» типа («го-
лотипа»), чтобы обмениваться ими.
Уже обращалось внимание на тот факт, что типы имеют ещё
одну практическую функцию: обозначают центры полностью про-
Вид и низшие категории 217
извольно очерченных последовательных видов и подвидов, как од-
новременно существующих, так и образующих временные ряды.
Эту их функцию теперь можно пояснить более чётко, хотя она по-
прежнему скорее номенклатурная, чем таксономическая. Произ-
вольно выделенные таксоны вырезаются из непрерывной последо-
вательности популяций, как в пространстве, так и во времени. Обо-
значение центров на самом деле основано не на отдельных особях,
а на популяциях, достаточно чётко ограниченных в рамках одной
последовательности во временном, пространственном или обоих
аспектах. Выбор для этой функции популяции, которая включает
тип, может содействовать стабилизации номенклатуры, и полезен
лишь для этой, но ни для какой другой цели.
6
Высшие категории
Хотя вид является наиболее чётко фиксированной
единицей среди признанных таксономической теорией, даже он в
своей эволюции интерградирует с таксонами выше и ниже его по
рангу. Выражение «высшие категории» имеет разные употребления,
но мы примем, что это надвидовые категории, имея в виду опре-
деление вида, данное в главе 5. Род имеет несколько особый ста-
тус и заслуживает отдельного рассмотрения от прочих высших ка-
тегорий.
Большинство важных принципов, понятий и приёмов, касаю-
щихся выделения высших категорий, уже обсуждалось в кругу об-
щетаксономических принципов в главах 2-4. Одни из них здесь
будут лишь упомянуты, другие будут развиты в данном контексте.
При работе с одновременно существующими животными наиболее
существенны критерии дискретности (разрывов), разнообразия и
дивергенции. Беря ситуацию шире, надлежит рассматривать весь
филогенетический паттерн. Анализ сложных паттернов обычно
обнаруживает в них ряд более простых, присутствующих в них и
так или иначе распознаваемых элементов родства. Это особенно
верно в отношении паттернов диверсификации или радиации, ко-
торые часто, но не всегда обнаруживают существование относи-
тельно непроизвольных (в смысле — «реальных» или «объектив-
ных») высших таксонов и подтаксонов. Имеются многочисленные
частные проблемы — например, то, что группа не может быть еди-
Высшие категории
219
нообразно разделена на таксоны данного уровня. Взаимодействие
разных элементов паттерна должно стать более понятным из кон-
кретных примеров, в данном случае заимствованных в основном из
Mammalia.
Таксономия имеет дело с распознаванием и формализацией от-
ношений между организмами. Все такие отношения в своей осно-
ве и по происхождению являются эволюционными. Признавая и
подчёркивая этот факт, лучше всего завершить данное исследова-
ние по таксономии комментариями о некоторых эволюционных
факторах, которые лежат в основе паттернов таксономических ка-
тегорий.
Что такое высшая категория?
Вероятно, наиболее активно разрабатываемая и, конечно, одна
из наиболее интересных областей частных исследований в сегод-
няшней систематике — эволюционные исследования изменчивос-
ти в пределах видов и групп близкородственных видов. Все такие
исследования имеют значение для таксономистов, но они становят-
ся собственно таксономическими, только если имеют отношение к
выделению и интерпретации таксонов. В настоящее время широ-
ко признанной является только одна внутривидовая категория —
подвид, но даже и он не универсален. Всеобще признанная низшая
категория, обычно обозначаемая как в формальных системах, —
это вид. Он интерградирует с категориями выше и, если использу-
ются подвиды, ниже него, но в целом он уникален в том, что, при
исследовании одновременно существующих животных, он являет-
ся непроизвольным как по включению, так и исключению. Катего-
рии выше вида отличаются тем, что в их состав входит, как пра-
вило, более одной такой непроизвольно выделенной единицы, что
влечёт за собой отсутствие внутренней непрерывности среди одно-
временно существующих групп. Тем самым они получаются про-
извольными [48] по включению, но их определение зависит от
иных критериев, нежели внутренняя непрерывность. Надвидовые
категории, для которых это является общим свойством, в общем
случае и являются тем, что мы называем высшими категориями.
Это отличие не всегда понятно, поэтому предшествующее обсуж-
220
Дж. Г. Симпсон. Принципы таксономии животных
дение низших категорий (глава 5) может быть дополнено тремя
примерами разного понимания этой проблемы.
Меррием (Merriam, 1918) обследовал множество черепов бурых
медведей из Северной Америки и поместил каждую особь, кото-
рую он посчитал достаточно отличной, в отдельный «вид». Эти
основные единицы его системы, на самом деле не являются — и,
возможно, не были даже в теории [in intention] — популяциями в
каком-либо биологически обоснованном смысле1. На самом деле
они, равно как и объединяющие их «роды», — это вообще не так-
соны: они не принадлежат ни к одной из категорий, свойственных
современной классификации.
Кинси (Kinsey, 1936) в одном из наиболее солидных исследо-
ваний, посвящённом, как заявлено, высшим категориям, определил
вид как «низшую из таксономических единиц, выделяемую у рас-
тений и животных», и определил всякую категорию над этим уров-
нем как «высшую». Его «виды» — популяционные единицы, одна-
ко это не виды согласно генетическому или эволюционному опре-
делению. Некоторые, по-видимому, являются демами, тогда как
другие, возможно, — подвидами, т.е. истинными таксонами, но
минимального ранга. «Комплексы», которые Кинси выделяет меж-
ду его «видами» и «подродами», иногда близки к генетическим ви-
дам. Лишь в малой степени его монография о «высших категори-
ях» имеет отношение в высшим категориям, как они здесь пони-
маются.
Великолепная книга Шеффера (Scheffer, 1958) о тюленях и их
родичах (Pinnipedia), среди прочего, касается высших категорий,
удовлетворяющих любому их определению, однако в ней смешаны
уровни рода и вида. В сокращённом и перефразированном изложе-
нии его критерии рода таковы: 1) по меньшей мере один признак
не перекрывается с таковым других родов при сравнении экземп-
ляров одного возраста и пола; 2) образ жизни специфичен, хотя и
не во всём, в сравнении с другими аллопатричными родами; 3) об-
1 Я более не придерживаюсь своего первоначального мнения (Simpson,
1953), что Мерриам имел намерение (или пытался) распознать и назвать демы,
которые хотя и являются единицами популяций, однако, как раз, ниже уровня
практических таксонов в классификации.
Высшие категории
221
ласть размножения ограничена одним или двумя обширными реги-
онами и может быть общей с таковой других родов; 4) разные роды
не скрещиваются в природе; и 5) они обычно распознаются мест-
ными жителями. Это всё характеристики (даже в таком качестве —
не определения) эволюционного вида, а не рода. (Последний из
приведённых критериев необычен, однако, как знают полевые ис-
следователи, он в определённой степени пригоден для видов.)
Шефферовы «роды», таким образом, это настоящие таксоны, но не
высшие.
Другой крайностью является некая тенденция (особенно среди
палеонтологов) рассматривать род как фундаментальный таксон, а
не как добавление к виду. При этом, говоря о высших таксонах,
обычно имеются в виду категории семейства и выше. Понятие
«высший» относительно, и оно неизбежно имеет разный смысл в
разном контексте. Как будет показано, род действительно облада-
ет особым статусом. Тем не менее, наиболее важный разрыв меж-
ду категориями лежит между видом и родом, так что в настоящей
работе все надвидовые группы, противопоставляемые виду, как тот
определен в главе 5, будут считаться высшими таксонами. Высшую
категорию можно определить в терминах филогенетического пат-
терна следующим образом:
Высшая категория такова, что таксон, являющийся ее членом,
включает две или более обособленные (видовые) филетические ли-
нии или сегмент (фратрию) единой линии, объединяющей два или
более последовательных вида.
Вторая часть определения позволяет принять во внимание тот
факт, что высшие таксоны, взятые в некоторый момент времени
(например, современные), часто являются монотипическими, так
что высшие категории не могут быть просто определены как такие,
к которым принадлежат [отношение «е»] таксоны, включающие
[отношение «с»] два или более известных вида.
Здесь вполне уместно окончательно прояснить запутанный мо-
мент, уже упоминавшийся ранее: связь между филогенетическим
деревом и высшими или низшими таксонами. Эта связь часто ил-
люстрируется так (показано на рис. 19А), словно нижняя часть де-
рева представляет высшие таксоны, а ветви — терминальные виды
или низшие таксоны. Эта идея решительно неверна. Корректная
16 — 4338
222
Дж. Г. Симпсон. Принципы таксономии животных
Таксоны
Виды
Роды
Семейство
Рис. 19. Соотношение между филогенией и высшими
И НИЗШИМИ ТАКСОНАМИ
А. Филогенетическое дерево со стволом и ветвями, некорректно переведён-
ное в систему таксонов разного уровня.
Б. То же дерево, корректно разделённое на таксоны.
В. Дендрограмма, на которой представлены только концевые виды (чёрные
точки); ребра под ними соответствуют не филогении или таксонам, а отноше-
ниями между этими видами; таксономические категории показаны скобками
слева, таксоны без временной интерпретации — скобками сверху. В отношении
разделения видов эта система соответствует таковой в (Б) и является коррект-
ным представлением предполагаемой филогении, которая топологически экви-
валентна соотношению между рёбрами в (В). (Диаграмма специально упроще-
на и указывает разбиения, избыточные для такого небольшого числа видов.)
связь пространственно-временных таксонов с филогенией показа-
на на рис. 19Б, а соответствующая ей классификация терминальных
(например, современных) видов показана на рис. 19В.
Высшие категории
223
Основания для выделения высших таксонов
Большинство классификаций на практике основано на одновре-
менно существующих организмах или создано без явного рассмот-
рения временной размерности. Критерии высших категорий обыч-
но отражают это ограничение. В таком контексте наиболее значи-
мы три критерия: степень обособленности (разрывы), масштаб ди-
вергенции и множественность низших таксонов. Они будут рас-
смотрены в первую очередь, причём в связи с некоторыми паттер-
нами филогенеза, которые ответственны за них. Вслед за этим буду
рассмотрены и проиллюстрированы сами филогенетические пат-
терны высших категорий.
Майр и др. (Mayr et al., 1953) определяют все высшие катего-
рии двумя характеристиками: монофилией и разрывом. Их опреде-
ления рода, семейства и более высоких2 категорий через относящи-
еся к ним таксоны могут быть обобщены в иных терминах следу-
ющим образом:
Т t— есть таксон, который включает один или более таксо-
нов Т с предполагаемым общим филогенетическим происхождени-
ем и который отделён от других Т.^ решающим разрывом.
Они, кроме того, рекомендуют, чтобы величина разрыва (т.е.
степень отличия от близких Г.+1) была обратно пропорциональна
объёму данного Т.+1 (т.е. числу входящих в него Г). Общий смысл
и применение этого определения показаны на рис. 20а.
Совместимость этой процедуры с филогенией в основных чер-
тах базируется на двух хорошо обоснованных и ныне признанных
взаимосвязанных принципах:
1. Общность признаков обычно пропорциональна близости об-
щего предка. В таком случае расстояния между низшими таксона-
ми обратно пропорциональны числу их общих признаков.
2. Степень дивергенции обычно пропорциональна отдалённос-
ти общего предка. Величина разрыва (gap) между сохранившими-
ся низшими таксонами, таким образом, обычно прямо пропорци-
онален отдалённости их общего предка.
2 Применение такого определения к категориям выше семейства скорее
подразумевается, нежели вводится явным образом.
16*
224
Дж. Г. Симпсон. Принципы таксономии животных
Семейство
Роды
Виды
Подроды
Роды
Трибы
Подсемейства
Семейство
Рис. 20. Критерии разрыва и монофилии для высших категорий
А. Одновременно существующие виды (чёрные точки), разделённые
разрывами разной величины, которые указывают степень различий между ними.
Скобки сверху указывают распределение видов по высшим таксонам двух
последовательных категорий, снизу — их распределение по таксонам пяти
категорий.
Б -Г. Несколько возможных филогений для видов в (А).
Б. Наиболее вероятная филогения, на которой родственная близость
совпадает со степенью дивергенции, что обеспечивает её совместимость со всеми
таксонами в (А).
Высшие категории
225
Нет сомнений, что таковы общие закономерности и что в отсут-
ствие указаний иного этот критерий действительно обеспечивает (в
качестве некоторой средней тенденции) совместимость с наиболее
вероятной реконструированной филогенией. Если бы число общих
признаков и степень дивергенции, показанные на рис. 20А, стро-
го соответствовали бы этим двум принципам, филогения должна
была бы соответствовать дереву, показанному на рис. 20Б. Миче-
нер (Michener, 1957) в ходе своей более подробной дискуссии, ис-
пользуя несколько иную терминологию, показал, что эти средние
тенденции не всегда воплощаются в деталях и поэтому каждый из
приведённых критериев может давать большие или меньшие откло-
нения от филогении. Филогения, показанная на рис. 20В, суще-
ственно менее вероятна в свете данных на рис. 20А, однако она
может быть возможной причиной этого паттерна. Если бы она была
истинной филогенией, показанные на рис. 20А роды были бы пол-
ностью совместимы с нею, однако ей противоречила бы детальная
схема триб и подсемейств. На рис. 20Г показан другой, ещё менее
вероятный филогенетический паттерн, который также мог бы, но
лишь в виде исключения, дать иерархию, представленную на рис.
20а. Если бы это была истинная филогения, с ней были бы несов-
местимы даже роды, показанные на рис. 20А.
Никакие реконструкции не бывают совершенно определённы-
ми и никакой критерий, или артистический канон, не способен дать
единственную обоснованную классификацию. Кроме того, линей-
ная форма на рис. 20а слишком проста, и если бы представление
было более реалистичным, дополнительный анализ почти наверня-
ка сделал бы возможным более правдоподобный выбор между раз-
ными вероятными филогенетическими паттернами. Одномерный
подход означает редукцию степени сходства между любыми двумя
Рис. 20 (продолжение).
В. Менее вероятная филогения, на которой для видов с- d родственная
близость не совпадает со степенью дивергенции; совместима с «верхними», но
не с «нижними» таксонами в (А).
Г. Ещё менее вероятная филогения, на которой b и с конвергентны; она
несовместима ни с «верхними», ни с «нижними» таксонами в (А).
226
Дж. Г. Симпсон. Принципы таксономии животных
популяциями или видами до единственного измерения, которое,
тем не менее, связано со многими разными признаками организ-
мов. В сущности, таков результат, к которому пришли Сокэл и
Миченер (Sokal, Michener, 1958). Еще более объемлющий, но и еще
более трудоёмкий подход, основанный на дискриминантном анали-
зе, редуцирует многочисленные признаки к двум обобщающим пе-
ременным и показывает вариации и разрывы в двумерном про-
странстве. Это более точное представление всего сравнения, по
сути многомерное, хоть и по-прежнему сверхупрощенное. Эштон
и др. (Ashton et al., 1957) в своём детальном сравнении Ното, ав-
стралопитеков и человекообразных обезьян показали, что выбор
плоскости проекции позволяет отразить не всю, но значительную
относящуюся к делу информацию в двух измерениях.
Результаты Эштона и коллег, кроме того, показывают, что раз-
рывы, выявляемые по разным анатомическим структурам (в данном
случае это зубы) и в разных плоскостях, могут быть весьма различ-
ными по масштабу и положению. В некоторых сравнениях обнару-
жен значимый разрыв между Ното и современными человекооб-
разными обезьянами: он был давно выявлен на качественном уров-
не и служит основанием для выделения семейств. Австралопите-
ки в одних проекциях сближаются с Ното, в других — с челове-
кообразными обезьянами. Таким образом, австралопитеки — это и
не Ното, и не современные понгиды (которые в любом случае со-
храняются), а третья группа, которая некоторым образом, не слиш-
ком очевидным при рассмотрении этих данных, образует связь
между этими двумя группами. Результаты не дают прямого ответа
при решении вопроса о ранге таксона австралопитеков: это может
быть, например, третье семейство, семейство в составе Pongidae
или Hominidae, или род в каком либо из этих семейств [49].
Это и подобные тщательные количественные исследования, по-
видимому, показывают, что масштаб разрывов сам по себе не слу-
жит адекватным критерием для ранжирования высших категорий
и распознавания их взаимоотношений. Сокэл и Миченер идут ещё
дальше в образовании высших категорий на основании сложных
измерений сходства, или «статических отношений». Сокэл (изна-
чально не таксономист), вероятно, считал это адекватным и окон-
чательным результатом таксономического исследования. Миченер
Высшие категории
227
(таксономист) рассматривал эти результаты скорее как хорошо
организованные подготовительные данные, на которых можно за-
тем основывать филогенетическую таксономию в свете иных эво-
люционных соображений (см. также Michener, Sokal, 1957).
В этом вопросе я полностью согласен с Миченером. Измерив
или оценив степень сходства или величину разрыва, таксономист
затем должен привнести в общую картину качественные суждения,
касающиеся гомологии, конвергенции и тому подобное. Например,
если бы схема на рис. 20В была истинной филогенией, то были бы
весьма вероятны гомологии между а и b-с, отсутствующие между
b-с и d, которые свидетельствовали бы о монофилетической груп-
пировке a-b-c несмотря на заметный разрыв между а и Ь. Сход-
ным образом, на рис. 20Г между b и с была бы с высокой вероят-
ностью обнаружена конвергенция, а не гомология, если бы сама
такая возможность предполагалась. Критерий или определение
высших категорий включает монофилию наравне с разрывом (а
также другие критерии). Монофилия обычно связана с разрывом,
но она не может и не должна служить единственным основанием.
Хотя филогенетическая дивергенция почти всегда приводит к
некоторым разрывам, здесь нет необходимой связи, и на самом
деле это бывает без явной закономерности и с исключениями. Сте-
пень дивергенции сама по себе является полезным и действенным
критерием для высших категорий в добавление к критерию моно-
филии, даже при отсутствии чёткого разрыва. Например, рис. 21А
показывает те же виды, что и на рис. 20А, плюс три других, кото-
рые заполняют наиболее крупный разрыв. На основании только
разрывов можно выделить три рода, однако один из них характе-
ризуется общей величиной дивергенции, сопоставимой с диверген-
цией двух других вместе взятых. Соответственно, этот род можно
разделить на два, получив в сумме четыре рода приблизительно
одинакового размера по величине дивергенции. Эти четыре рода,
хотя и различные по своим основаниям, соответствуют показанным
на рис. 20А. Такое упорядочение также было бы совместимо с ве-
роятной филогенией, показанной на рис. 21 Б, или, если добавить
несколько линий, с показанной на рис. 20Б. Принимая окончатель-
ное решение о разделении избыточно обширного рода, разумеется,
необходимо опять обращаться к рассмотрению монофилии.
228
Дж. Г. Симпсон. Принципы таксономии животных
Роды
Виды
Роды
Рис. 21. Влияние заполнения разрыва в группе видов, показанных на рис. 20
А. Виды, показанные на рис. 20, плюс три вида, заполняющие разрыв
(крестики); сверху показано разделение на роды только по критерию разрыва,
снизу — по критериям разрыва и дивергенции.
Б. Сплошные линии указывают вероятную филогению, ведающую к
ситуации в (А) и совместимую с обоими разделениями на роды; штриховые линии
указывают другую возможность, совместимую с «нижним», но не с «верхним»
разделениями на роды в (А).
Соответствующий критерий дивергенции уже был введён ранее,
здесь ему можно дать такую формулировку:
В пределах некоторого высшего таксона Г+2 желательно, что-
бы таксоны следующего более низкого ранга показывали при-
близительно одинаковый уровень дивергенции друг от друга, при
отсутствии противопоказаний таких, как доказательство существен-
ной полифилии таким образом сформированных Г+1.
Критерий разнообразия, второстепенный в отношении уже рас-
смотренных, также был представлен ранее в ином контексте; он
требует лишь незначительной переформулировки:
При отсутствии противопоказаний с точки зрения монофилии,
разрывов и дивергенции, желательно, чтобы таксоны Г+р включён-
ные в некоторый таксон Т 2, включали приблизительно одинаковое
число подтаксонов Т.
Высшие категории
229
Рис. 22. Неприменимость критерия одинаковой древности происхождения
для такоснов одного ранга
А. Простая [филогенетическая схема], разделённая на таксоны двух рангов,
из которой видна невозможность применения критерия одинаковой древности
ко всему филогенетическому паттерну.
Б. Классификация (показана сверху скобками) концевых групп (чёрные
точки) только по одинаковой древности происхождения, показывающая
излишнюю усложнённость и несбалансированность такой классификации.
Прежде уже была подчёркнута мысль, что одинаковая древность
происхождения не является желательным или даже возможным кри-
терием отнесения высших категорий к одному рангу, рис. 22 позво-
ляет напомнить и дополнительно проиллюстрировать эту' позицию.
230
Дж. Г. Симпсон. Принципы таксономии животных
Определение и характеристики высших категорий
Абсолютные определения высших категорий невозможны. Одна
из уникальных особенностей вида состоит в том, что он может
быть определён без ссылки на какую-либо иную категорию. Опре-
деления же высших категорий могут быть даны только относитель-
но таковых других категорий, указывая относительное положение
рангов в иерархии и теоретико-множественные отношения таксо-
нов. Так, например, семейство не может быть определено как та-
ковое, но лишь в его отношениях с категориями следующего более
высокого (обычно надсемейство, подотряд или отряд) или более
низкого (обычно подсемейство или род) рангов. Согласно наиболее
общему определению, любая высшая категория С есть такое мно-
жество таксонов Г, что каждый Т. включает один или чаще не-
сколько подтаксонов Ти и включён (единично или чаще с други-
ми Г.) в единственный надтаксон Т.+|.
Для каждой высшей категории существуют критерии и харак-
теристики, но не строгие определения, которые также являются
сравнительными и связаны с отнесёнными к данной категории так-
сонами. Наиболее общие критерии состоят в том, что каждый так-
сон Т должен быть монофилетичным, по крайней мере в широком
понимании монофилии, и что все таксоны Г, относящиеся к С.,
должны быть сравнимы между собой. Таким образом, на практи-
ке рабочее определение какой-либо категории С, например семей-
ства, состоит в следующем: С есть множество таксонов ранга се-
мейства Г, которые сравнимы между собой и которые отнесены к
категории семейства классификатором, трактующим их именно та-
ким образом. Это определение по необходимости субъективно; до
тех пор, пока сохраняется иерархическое построение системы, ран-
ги приписываются на основании скорее искусства, чем науки. Не
слишком ясное понятие «сравнимый» можно сделать более чётким,
а каноны искусства обеспечиваются специальными критериями,
среди которых наиболее важные — дивергенции, разрыва и разно-
образия. К ним можно добавить ещё два канона:
1. Между видом и некоторой высшей категорией, к которой от-
носятся всеобще признанные и достаточно стабильные таксоны,
промежуточные категории должны вставляться таким образом, что-
Высшие категории
231
бы им соответствовали приблизительно одинаковые шаги в отно-
шении включённости или объёма и различия [distinction] относя-
щихся к ним таксонов. Например, таксон Perissodactyla, ныне рас-
сматриваемый обычно в ранге отряда, оставался почти неизменным
с момента выделения более столетия назад. Современные виды
почти без разногласий признаются теми, кто применяет генетичес-
кое определение. Если, например, лошадей в самом узком понима-
нии (один современный вид) и зебр (три современных вида) поме-
стить в разные семейства, то шаги от вида до семейства будут край-
не малы в сравнении с теми, которые отделяют семейство от отря-
да. По этому канону лошадей и зебр следует в лучшем случае счи-
тать разными родами или даже (по моему мнению) подродами.
2. Когда высший таксон можно достаточно чётко разделить на
«естественные» или полупроизвольные надвидовые группы, припи-
сываемую им категорию следует уравнивать с их уровнем [level],
устанавливаемым в соответствии с предыдущим каноном, а другие
шаги соотносить с этим ранжированием. Для одновременно живу-
щих организмов это обычно согласуется с критерием разрыва, по-
скольку такие группы почти всегда бывают разделены заметным
разрывом на каждом временном отрезке своего существования.
Например, устойчивый таксон Carnivora очевидно и резко разделён
на наземных (фиссипеды) и водных (пиннипеды) на уровне не
ниже под отрядного [50]. Ранги от вида до отряда выровнены по
этому базовому делению. Некоторые из филогенетических паттер-
нов, затрагиваемых такими ситуациями, обсуждаются далее в на-
стоящей главе.
В существующей практике некоторым высшим категориям при-
писываются, хотя и не всегда и зачастую слишком туманно, особые
характеристики, не связанные полностью с рассмотренными кри-
териями и канонами. В психологии таксономистов издавна и глу-
боко укоренилось представление о том, что род — это особая ка-
тегория, отличная от всех категорий более высокого ранга. Такова
была точка зрения Линнея, который заложил её во всю последую-
щую практику, и это даже притом, что его род был значительно
более высокой категорией, чем наш. В сохранившейся поныне лин-
неевской практике родовое название является также частью видо-
вого названия, так что невозможно назвать вид, не называя его род.
232
Дж. Г. Симпсон. Принципы таксономии животных
(Это, конечно же, отдалённое эхо схоластической практики, соглас-
но которой род и вид совместно составляют определение на лю-
бом категориальном уровне.) Родовое название уникально и в том,
что является существительным в единственном числе, тогда как
видовые эпитеты — прилагательные или их производные, а надро-
довые — существительные во множественном числе?
Особый статус рода связан не только с традициями или номен-
клатурным значением. Часто оказывается, что род — более прак-
тичная и надёжная единица классификации, чем вид. Имея дело с
классификациями, не созданными и не ревизованными, согласно
современным и эволюционным стандартам, — ас такими неред-
ко ещё доводится работать — часто приходится ломать голову над
тем, соответствуют ли «виды» этих классификаций нашим опреде-
лениям или это морфы или типологические вариететы, мутанты,
экотипы, демы, подвиды или что-то ещё. «Роды» же с большей ве-
роятностью соответствуют современному определению, приписы-
ваем ли мы им видовой, родовой или какой-либо иной ранг. Даже
в современной практике роды часто могут быть определены и оп-
ределяются более чётко, чем таксоны более низкого или высокого
рангов. Их можно с большим вероятием чётко разграничить на ос-
новании общих диагностических признаков, тогда как более низ-
шие категории, даже виды, разграничены на столь чётко, а высшие
категории имеют меньше общих диагностических признаков, а
иногда их вообще нет (на это обратил внимание также Миченер
(Michener, 1957)).
Сверх того, в некоторых исследованиях по систематике роды
часто бывают более пригодны в качестве рабочих единиц, чем
виды и семейства. Это обычно вытекает из того факта, что они, как
правило, распространены шире, чем виды, особенно в простран-
стве, но также и во времени, и всё же более компактно локализо-
ваны и не столь повсеместны, как многие семейства. Например, в
зоогеографических исследованиях для многих масштабов, исклю-
3 Кэйн (в частности, Cain, 1959) несколько раз предлагал, чтобы род не
имел особого статуса в номенклатуре и что двухсловные видовые названия сле-
дует отбросить. Не считая почти неодолимого давления традиции и жёсткос-
ти Кодекса, проблемы омонимии и мнемоники, по-видимому, сделали бы од-
нословные названия для видов в высшей степени непрактичными.
Высшие категории
233
чая самые узкотерриториальные, виды слишком локальны, а семей-
ства слишком широко распространены для выявления значимых
особенностей, тогда как роды часто оказываются самыми подходя-
щими в этом отношении. В стратиграфической палеонтологии
виды позволяют точные локальные корреляции, однако роды гораз-
до полезнее для широкомасштабных, особенно межконтиненталь-
ных корреляций. Кроме того, в палеонтологических исследовани-
ях по эволюции, роды вообще более удобны, поскольку они оста-
ются достаточно небольшими единицами, но при этом характери-
зуются значительной протяжённостью во времени и более адекват-
но представлены в коллекциях ископаемых остатков (обсуждейие
в: Simpson, 1960а).
Именно при работе с ископаемыми возникает вопрос, напря-
мую и практически связанный с классификацией: речь идет о не-
которой тенденции в использовании подсемейств и даже, хотя и не
в такой степени, семейств как категорий [51] со значительной вре-
менной протяжённостью, т.е. как преимущественно вертикальные
единицы. Здесь, однако, отмечается значительное разнобой [во мне-
ниях], так что это вряд ли можно использовать в качестве прави-
ла или канона. Крайним проявлением служит предложенная Осбор-
ном (Osborn, 1936, 1942) классификация Proboscidea, в которой 21
подсемейство не имеет почти никакой горизонтальной протяжён-
ности: почти каждое соответствует отдельной филетической линии.
Семейство Equidae, как оно сейчас определяется, являет собой бо-
лее сбалансированный пример, с существенной горизонтальной, но
ещё большей вертикальной протяжённостью. Оно также служит
примером [таксона] высшей категории, который, подобно дарви-
новским Crustacea, но по другим причинам, практически не может
быть диагностирован общими признаками. Hyracotherium (или
эогиппус) значительно меньше похож на Equus, чем на какое-либо
животное, отнесённое к другим семействам [непарнокопытных],
так что Hyracotherium и Equus не имеют чётких общих признаков,
которые не присутствовали бы и в других семействах. Тем не ме-
нее, Hyracotherium и Equus явно связаны последовательным сход-
ством через весь временной ряд видов и родов, во многом подоб-
но тому, как последовательными сходствами связаны в простран-
стве между собой Crustacea.
234
Дж. Г. Симпсон. Принципы таксономии животных
Как, наряду с другими, отмечали Майр, Линсли и Юзингер
(Мауг et al., 1953), каждый отдельный надвидовой таксон, прибли-
зительно соответствующий категории семейства, обычно характе-
ризуется однотипным адаптивным обликом, который таксономист
часто распознаёт с первого взгляда. Выше уровня семейства име-
ется «базовый структурный тип», а в виде исключения и очевид-
ный адаптивный облик, как в случае отряда Chiroptera (рукокры-
лые), однако, как правило, «высшие категории не могут безуслов-
но или хотя бы преимущественно выделены по адаптивным при-
знакам.» Этот момент был затронут ранее и ещё раз рассматрива-
ется в конце данной главы. Говоря иными словами, общий адаптив-
ный облик и базовая адаптация окажутся перекрытыми разнообра-
зием по другим признакам или даже утраченными некоторыми чле-
нами группы тем вероятнее, чем выше данная группа поднимется
по шкале категорий.
Анализ некоторых филогенетических паттернов
Выше уже обсуждались многие схемы филогенетического пат-
терна, а некоторые из них были воспроизведены в связи с разны-
ми принципами и приёмами [таксономии]. Теперь разумно рассмот-
реть их более систематическим образом в свете их значения для
выделения высших категорий. Для этого прежде всего будут выч-
ленены более простые филогенетические отношения (хотя здесь и
не так обстоятельно), затем рассмотрены некоторые сочетания этих
элементарных отношений, и наконец приведены примеры реальных
филогений, подтвёрждённых детальными палеонтологическими
данными.
Самых основных категорий эволюционных событий, из кото-
рых строятся реальные филогении, по-видимому, всего четыре:
Прогрессия (progression), или филетическая (т.е. выражаемая в родос-
ловной линии — lineage) эволюция; включает анагенез по Гексли (Huxley,
1957), но нс синонимична ему.
Разветвление (splitting), или диверсификация; кладогенез по Гексли.
Равновесие, персистирование, или остановленная эволюция; стасигенез
по Гексли.
Терминация, или вымирание.
Высшие категории
235
Представляется желательным, чтобы эти аналитические факто-
ры имели техническое обозначения, определённые однозначно, как
это сделал Huxley (1957). Его термины «кладогенез» (заимствован
из работы: Rensch, 1954) и «стасигенез» однозначны. К сожалению,
[термин] «анагенез» оказался не столь определенным. Ренш
(Rensch, 1954) определил его как параллельное повышение органи-
зации или совершенствование функции в разных филетических
линиях некоторой высшей категории. Гексли переопределил анаге-
нез, включив в него «все типы или степени биологического совер-
шенствования». Теперь, когда данный термин используется, возни-
кает вопрос, какой смысл — идущий от Ренша или Гексли —
вкладывается в него.4 В любом случае, ни один из них не соответ-
ствует полностью принятому здесь пониманию, объединяющему
любой тип изменений, последовательно происходящих в единой
филетической линии предков и потомков (без ссылки на какие-либо
ветви, которые линия также может иметь), как показано на рис.
23А. Анагенез в смысле Гексли — частный случай этой более об-
щей концепции. Гексли не предложил особого термина для выми-
рания. Возможно, он и не нужен, однако здесь также имеется дву-
смысленность, поскольку «вымершими» называются и те таксоны,
которые исчезли без последствий, и те, чья эволюция просто при-
вела к появлению того, что следовало бы назвать другими таксо-
нами того же ранга. Терминация, вероятно, с большей очевиднос-
тью соответствует только первому из двух значений. Терминацию
также можно применять к завершению филетической линии в рам-
ках ныне существующей группы, что относится к той же категории
элементов филогенетического паттерна, хотя, конечно, к совер-
шенно иной эволюционной ситуации.
Сочетание прогрессии и разветвления даёт дивергенцию, неко-
торые простейшие паттерны которой показаны на рис. 2 ЗБ-Д. При-
меры, представленные на них, не исчерпывают всех возможностей.
Также очевидно, что эти паттерны не строго альтернативны, но
могут переходить один в другой, равно как и комбинироваться мно-
4 На самом деле этот термин в разных трактовках уже использовался за-
долго до Ренша, но поскольку эти прежние употребления уже практически за-
быты, нет нужды затемнять ими предмет рассмотрения.
236
Дж. Г. Симпсон. Принципы таксономии животных
Рис. 23. Некоторые элементарные филогенетические паттерны
А. Простая прогрессия, штриховая линия указывает предковое состояние,
от которой филетическая линия неуклонно удаляется.
Б. Простое ветвление с последующей дивергенцией.
В-Д. Некоторые из простейших паттернов дивергенции и диверсификации.
В. Неравномерная диверсификация двух расходящихся групп. Г. Диверсификация,
повлекшая несовпадение конечного распределения с последовательностью
ветвления. Д. Диверсификация, повлекшая совпадение конечного распределения
с последовательностью ветвления.
Е-3. Возникновение разрывов в конечном распределении. Е. Разрывы,
возникающие одновременно с дивергенцией. Ж. Разрыв, вызванный вымиранием
промежуточных форм. 3. Разрывы между стабильными нишами или адаптивными
зонами (заштрихованы).
гимн способами. Дивергенция также приводит к разрывам, столь
часто необходимым при выделении и определении высших таксо-
нов, особенно среди одновременно существующих групп (рис. 23Е).
Разрывы также могут получаться в результате терминации проме-
Высшие категории
237
жуточных групп (рис. 23Ж), причём терминация, несомненно, мо-
жет сочетаться с любым паттерном дивергенции, а не только с тем,
который показан в приведённом примере. Разрывы также возника-
ют в результате перехода к другому уровню равновесия, в другую
нишу, адаптивную зону или граду (рис. 233; переход не всегда про-
грессивен, как показано в приведённом примере). Эти три формы
возникновения разрывов не являются взаимно независимыми. Не-
равномерная дивергенция, которая формирует разрывы, может быть
связана исключительно с тем, что разные филетические линии про-
никают в обособленные ниши или адаптивные зоны, возникнове-
ние или поддержание которых может быть связано с вымиранием
линий, занимающих промежуточное положение.
Прогрессия, разветвление и сопровождающая их дивергенция
обычно приводят к возрастанию разнообразия или экспансии (в
эволюционных терминах часто обозначаемых как адаптивная ради-
ация) групп, которые посредством этих процессов становятся выс-
шими таксонами. Элементарной формой экспансии, которая, конеч-
но, никогда не бывает столь простой в природе, была бы непрерыв-
ная дихотомия (рис. 24А). Ветвление может быть изображено бо-
лее реалистично, если оно неравномерно и сопровождается терми-
нацией некоторых линий (рис. 24Б). Сам факт вымирания не обя-
зательно замедляет диверсификацию: группы, показанные на рис.
24А, Б, расширяются с той же самой общей скоростью. Другое
обычное отклонение от этого простого паттерна состоит в том, что
основная диверсификация происходит довольно быстро вблизи ос-
нования высшего таксона с последующей замедленной общей ди-
вергенцией, которая в конце концов затрагивает только низшие так-
сономические уровни (рис. 24В). Этот паттерн является заметным
элементом в эволюции грызунов, ему можно найти огромное мно-
жество примеров среди самых разных групп животных, например,
ископаемых беспозвоночных (например, Nicol et aL, 1959).5 После-
довательные радиации, одна из которых может замещать другую
(рис. 24Г), а также одновременные, но независимые радиации от
общего корня (рис. 24Д) представляют собой две относительно
5 Именно это, как я полагаю, показывают их данные; некоторые из их вы-
водов мне представляются несостоятельными.
17 — 4338
238
Дж. Г. Симпсон. Принципы таксономии животных
Высшие категории
Рис. 24. Некоторые элементы паттерна, связанные с экспансией
или радиацией высших [таксонов] [52]
А. Простая равномерная дихотомия с удвоением филетических линий в
моменты t-t5 через равные промежутки времени.
Б. Равномерная экспансия с той же общей скоростью, как в (А), но с
вымиранием одних филетических линий и политомическим делением других.
В. Быстрая базальная диверсификация с последующей незначительной
диверсификацией на низших таксономических уровнях.
Г. Последовательные радиации общего происхождения, из которых одна,
более поздняя, замещает более раннюю.
Д. Независимые одновременные радиации общего происхождения.
Высшие категории
239
Высшие категории
Рис. 25. Некоторые паттерны полупроизвольного подразделения высших
[таксонов] [53]
Таксоны очерчены штриховыми линиями.
А. Выделение последовательной замещающей радиации.
Б. Выделение последовательной радиации, не замещающей первую.
В. Разделение одновременных радиаций.
Г. Разделение относительно освоенных раздельных зон, как на рис. 233, но
проникновение в зоны одновременное.
Д. Сложный паттерн, отражённый в таксонах со столь различными вариан-
тами возникновения и разнообразия, что они едва ли могут быть сравнимы между
собой или отнесены к одному таксономическому уровню.
17*
240
Дж. Г. Симпсон. Принципы таксономии животных
простые вариации [всё того же паттерна], которые также являют-
ся обычными элементами известных филогений.
Именно дивергенция с разрывами и особенно независимые ра-
диации от общего ствола определяют практику построения многих
из полупроизвольных подразделений высших таксонов, обсуждав-
шихся в главе 4. Четыре наиболее обычных филогенетических
паттерна, которые приводят к такому результату, показаны на рис.
25А-Г. Грады (например, рис. 233) обычно также имеют подобные
таксономические последствия. Многие филогении, несмотря на
сложности, связанные с большим числом филетических линий, и
на разные вероятные нарушения закономерностей, могут быть в
принципе разложены на такие элементы и таксоны, столь же про-
стые, как и рассмотренные паттерны. Тем не менее, имеются груп-
пы, в которых паттерны настолько разнообразны, что выделение
сравнимых таксонов данной категории может оказаться практиче-
ски невозможным, рис. 25Д схематически показывает такую ситу-
ацию: она создаёт проблему в разграничении подотрядов грызунов,
обсуждаемую далее в этой главе. Такие сложные группы могут сво-
дить на нет усилия по равномерному распределению последова-
тельного ряда высших категорий.
Некоторые примеры из филогении млекопитающих
Природа и возникновение надвидовых таксонов были подроб-
но проиллюстрирваны ранее (Simpson, 1959а). Оставшаяся часть
данной главы — в основном краткое изложение, с некоторыми
уточнениями, этой статьи, к которой можно обращаться за деталя-
ми. Примеры из млекопитающих имеют то преимущество, что в
этой группе часто встречаются детально изученные филогении.
Таким образом, млекопитающие предоставляют хорошо обоснован-
ные примеры, которыми можно руководствоваться при интерпре-
тации других групп со слабо или вообще не изученной палеонто-
логией.
Отряд Tubulidentata, семейство Orycteropodidae и современный
род трубкозубов Orycteropus иллюстрируют практически простей-
ший случай иерархического ряда высших таксонов, диагностиру-
емых и ранжируемых на основании дивергенции и разрывов. Име-
Высшие категории
241
ется только один живущий вид, так что все перечисленные таксо-
ны в современной фауне монотипические. Палеонтологические
материалы не обильны, однако по крайней мере начиная с ранне-
го миоцена они указывают на отсутствие диверсификации выше
родового уровня, а в каждом временном отрезке, возможно, — и
выше видового уровня. Тем не менее, эта крайне однородная груп-
па классифицируется как отряд, поскольку её известные предста-
вители отличаются от всех современных, возможно, родственных
отрядов так же, как эти отряды — друг от друга. Отличия от лю-
бого возможно предкового отряда — того же порядка. Эти разры-
вы в настоящее время абсолютны и по своему масштабу считают-
ся отрядными почти всеми таксономистами. Orycteropus ранее по-
мещали в отряд Edentata из-за некоторого сходства, сужающего раз-
рыв с данной группой при чисто эмпирической оценке. Однако эти
сходства полностью конвергентные и должны быть исключены при
рассмотрении сходств и различий в качестве основы эволюционной
классификации.
Базальный разрыв Tubulidentata с их раннетретичными предка-
ми несомненно связан с отсутствием данных. Таким образом, ныне
установленная граница не является произвольной в свете имеющихся
данных, но стала бы почти наверняка произвольной при лучшей
ситуации с данными по группе. Однако даже если между Tubuliden-
tata и предковым отрядом будут найдены переходные формы, ранг
отряда для самих Tubulidentata был бы по-прежнему оправдан, и
нужно было бы также проводить произвольную горизонтальную
границу, отделяющую базальную группу. Терминальный вид труб-
козуба отличается от любого возможного древнего плацентарного
предка как представитель отдельного отряда. Наиболее вероятные
предки, по-видимому, относятся к Condylarthra (см. Colbert, 1941):
это горизонтально ограниченный отряд, который более сходен с его
наиболее вероятными копытными потомками, особенно Perisso-
dactyla и Artiodactyla. [54] Практическая классификация требует
введения нескольких последовательных отрядов и условных границ
между ними, так же как она требует нескольких последовательных
и условно разграниченных семейств, родов и видов.
Семейство Leporidae (зайцы и кролики) — сравнительно про-
стой пример высшего таксона с полупроизвольными подразделени-
242
Дж. Г. Симпсон. Принципы таксономии животных
ями, получившимися в результате более или менее самостоятель-
ных радиаций. Филогения, представленная в общих чертах в:
Dawson (1958) и показанная на рис. 26 в более чем упрощенном и
схематичном виде, позволяет выделить три самостоятельные ради-
ации, частью последовательно-замещающие, частью одновремен-
ные. На основе филетических линий, относящихся к ранне-третич-
ной радиации в Голарктике, возникли две последующие независи-
мые радиации: одна из них явно началась несколько раньше и при-
урочена в основном к Неарктике, а другая исходно палеарктиче-
Рис. 26. Радиации, которым в [семействе] Leporidae соответствуют
ПОДСЕМЕЙСТВА
Схематически показаны филетические линии, отражающие принципиальную
схему радиации, а не конкретные таксоны. Объяснения в тексте. (Упрощенный
вариант схемы, представленной с указанием таксонов в: Dawson, 1958.)
Высшие категории
243
ская. Эти радиации привели к замещению всех линий предшеству-
ющей радиации, которые не были для них предковыми. В обеих
более поздних радиациях имел место последующий взаимный об-
мен между Старым и Новым Светом и исходно палеарктическая
радиация в конечном итоге вытеснила все прочие линии Нового
Света, так же как и Старого. Имеются и другие сложности: напри-
мер, проникновение в Африку и в Южную Азию; однако в целом
эти три радиации хорошо различимы и по морфологическим при-
знакам могут быть определены как подсемейства. Эти подсемей-
ства действительно объективно существуют как единицы филоге-
нии, и они «реальны», т.е. являются полупроизвольными таксона-
ми [55], сравнимыми по своему статусу и объёму.
Как всякий знает, Leporidae крайне многочисленны [по числен-
ности]. Но они не слишком разнообразны: имеется девять совре-
менных родов и приблизительно по столько же было на каждом
этапе эволюции, начиная с олигоцена. Дивергенция в пределах се-
мейства незначительна, отчётливо слабее, чем в среднем для се-
мейств млекопитающих. Все зайцы и кролики очень похожи, и так
было во всей истории группы. Обособленность [divergence] от
единственной близкой группы в составе отряда — пищух (семей-
ство Ochotonidae) — в настоящее время значительнее, чем в пре-
делах Leporidae, однако в среднем меньше, чем между родственны-
ми семействами млекопитающих, а в средне-третичное время эти
две группы были столь сходны, что их крайне трудно диагности-
ровать. По обычным критериям дивергенции, разрыва и разнооб-
разия, если сравнивать с системами других отрядов, было бы со-
вершенно оправданным считать Leporidae и Ochotonidae подсемей-
ствами в составе единого семейства. Однако поскольку это совер-
шенно чётко распознаваемые таксоны внутри отряда и поскольку
удобно считать подгруппы лепорид как семейства, ранг семейства
для лепорид и охотонид в настоящее время всеобще признан.
Таксон Lagomorpha (или Duplicidentata), который включает
Leporidae и Ochotonidae, до сравнительно недавнего времени рас-
сматривался как подотряд в составе Rodentia. Эмпирическое срав-
нение ограниченного набора признаков указывал на небольшой
разрыв между зайцеобразными и собственно грызунами, которым
обычно присваивали ранг отряда. Тем не менее, по мере того, как
244
Дж. Г. Симпсон. Принципы таксономии животных
исследовались другие признаки, разрыв заметно увеличился и ста-
ло очевидно, что многие, если не все специальные сходства конвер-
гентны. При исключении конвергенций данный разрыв несомнен-
но отрядного масштаба. Вывод о том, что Lagomorpha и Rodentia
имеют совершенно разное происхождение и должны возводиться
в ранг самостоятельных отрядов, усиливается данными по ископа-
емым, которые не показывают общих корней на уровне ниже отряд-
ного, хотя их предки на самом деле не были идентифицированы.
Отряд Carnivora даёт пример хорошо определяемых полупро-
извольных высших таксонов, возникших в результате последова-
тельно-замещающей и одновременной радиаций, как и в случае
Leporidae, однако на более высоком категориальном уровне и со
значительной более широкой дивергенцией и диверсификацией.
Филогения, которая в настоящее время представляется наиболее
вероятной, показана на рис. 27. Некоторые из классических се-
мейств (особенно Viverridae, Procyonidae и Mustelidae) характери-
зуются внутренней дивергенцией, превышающей среднюю, поэто-
му для них и для некоторых ископаемых семейств предлагалось
разделение на несколько семейств. Однако достаточно очевидно,
что эти классические семейства — «хорошие» таксоны соответству-
ющего уровня, определяемые по монофилии, и я думаю, что было
бы удобным продолжать считать их семействами. Имеется также
существенный разрыв, как по специализации, так и по известным
ископаемым данным, между наземными (Fissipeda) и водными
(Pinnipedia) хищными, причём иногда этот разрыв считается по
рангу отрядным. Тем не менее, пиннипедии почти наверняка про-
изошли от ранних фиссипед, и я полагаю, что для этого уровня
дивергенции более подходит ранг подотряда [56].
На схеме, представленной на рис. 27, отчётливо видны три ра-
диации, которые легко определимы и в настоящее время обычно
признаются как полу-условные подотряды. Одна филетическая ли-
ния ранней наземной радиации (креодонты), именно семейство
Miacidae, дала начало более поздней наземной радиации (фиссипе-
ды), которая в конечном итоге полностью вытеснила первую. Про-
блема подотрядной принадлежности самих Miacidae уже рассмат-
ривалась (глава 4) как пример горизонтальной и вертикальной клас-
сификаций и компромисса между ними. Третья радиация (пинни-
Высшие категории
245
Рис. 27. Филогения семейств и подотрядов Carnivora
Три подотряда очерчены жирной прерывистой линией. Каждая более или
менее вертикальная непрерывная линия представляет семейство; связи
прерывистыми линиями указывают на наличие надёжных свидетельств, не
подкреплённых известными ископаемыми формами. Arctocyonidae (1) также
близки и к некоторым другим отрядам, их отнесение к Carnivora, хоть и нередкое,
остаётся под вопросом. Обозначения семейств: 1 — Arctocyonidae, 2 —
Mesonychidae, 3 — Oxyaenidae, 4 — Hyaenodontidae, 5 — Miacidae, 6 — Felidae,
7 — Viverridae, 8 — Hyaenidae, 9 — Canidae, 10 — Ursidae, 11 — Procyonidae,
12 — Mustelidae, 13 — Otariidae, 14 — Odobenidae, 15 — Phocidae [57].
246
Дж. Г. Симпсон. Принципы таксономии животных
педии) происходила в основном или целиком одновременно со вто-
рой, но затронула радикальные изменения в экологии, связанные с
переходом из наземной в водную адаптивную зону Уровень дивер-
генции и разнообразие в пределах этой радиации существенно
меньше в сравнении с радиацией фиссипед.
Большинство хищных, относящихся к базальной палеоцен-
эоценовой радиации, характеризовалось хищническими зубами и
(предположительно) хищническим образом жизни, однако в разных
филетических линиях эта общая черта имела свои особенности.
Вопрос недостаточно ясен, но ранние Carnivora, вероятно, не были
хищными ни по морфологии, ни по экологии. Наиболее общий га-
битус, или адаптивный тип отряда, по-видимому, сложился частью
на дивергентной и частью на параллельной основе уже после воз-
никновения отряда как монофилетического таксона. Даже после
того, как этот габитус сложился, он был вовсе утрачен у некоторых
Carnivora (например, панда) или существенно изменён и стал едва
уловим у многих других (например, все ластоногие, а также неко-
торые наземные хищные). Эта смена адаптивных характеристик в
пределах отряда заметно контрастирует, например, со свойствен-
ным Lagomorpha единством адаптивного типа, сложившимся в са-
мом начале эволюции этого отряда и лишь слегка усиленным у
всех поздних представителей.
Отряд Primates также показывает последовательную и одновре-
менную радиации, которые дали начало полу-условным таксонам
на разных категориальных уровнях. Паттерны весьма сложны в
деталях и могут быть интерпретированы таксономически не толь-
ко в терминах радиации, экспансии и сокращения разных групп, но
также в терминах град, которые довольно хорошо соответствуют
таксонам. На рис. 28 очень схематично показан филогенетический
паттерн радиации. Нарисованные линии не представляют какие-то
конкретные филетические линии или временные таксоны, но лишь
дают общее впечатление о паттерне. Впрочем, даже в такой фор-
ме опущены наиболее сложные (и не вполне документированные)
детали в пределах Prosimii.
Базальная радиация, которая породила гораздо большее разно-
образие, чем любая другая более поздняя радиация, происходила в
палеоцене и эоцене. Целый ряд быстро дивергирующих линий
Высшие категории
247
ANTHROPOIDEA
PROSIMH
5 подсемейств
11 семейств
।-----*-----1
Ceboidea
2 семейства
г—I
Cercopithecoidea
1 семейство
Hominoidea
Pongidae
Hominid a e
t
Рис. 28. Схема филогении и классификация Primates
Показанные филетические линии служат схематическим или эскизным
представлением общего паттерна и не указывают конкретные таксоны. Некоторые
инфраотряды, поименованные в верхней части, заключены в штриховые линии.
248
Дж. Г. Симпсон. Принципы таксономии животных
вскоре вымерли, некоторые уже в палеоцене, большинство к началу
олигоцена. Немногие известны по нижним челюстям и зубам, при
этом зубы показывают некоторые замечательные специализации, не
повторяющиеся ни у кого из более поздних приматов. Другие бо-
лее традиционны в сравнении с современными приматами. Сюда
относятся тупайеподобные, лемуроподобные и долгопятоподобные
формы, и именно этот комплекс, понимаемый в самом широком
смысле, несомненно дал начало дожившим до современности ту-
пайям, лемурам, лори и долгопятам, которые не слишком отошли
от своих палеоцен-эоценовых предшественников. Нынешнее таксо-
номическое решение состоит в том, чтобы включать эти сравни-
тельно консервативные формы и их ранних родственников в один
подотряд Prosimii и сделать этот подотряд горизонтальным таксо-
ном за счёт включения в него также и аберрантных, даже не очень
анцестральных ранних форм. Филетические линии менее аберран-
тных членов этого комплекса должны были дать начало также и
высшим приматам — разным представителям подотряда Anthro-
poidea. Однако вопрос не стоит о «заглублении» какого-либо так-
сона в составе Anthropoidea в радиацию базального комплекса, по-
скольку ещё недостаточно чётко установлены связи между этими
двумя группами.
Обезьяны, строго говоря, развивались двумя сходными и одно-
временными радиациями, происходившими (отчасти) параллельно
от вероятно сходных прозимийных предков, но широко разобщён-
ными географически: одна радиация — в американских тропиках,
другая — в Старом Свете. В значительной мере одновременной, но
вероятно начавшейся несколько позже была другая радиация в Ста-
ром Свете, которая дала предков человекообразных обезьян и лю-
дей: она была отделена от радиации обезьян Старого Света боль-
ше экологически, чем географически.6 Эти три радиации вылились
в три сравнимых высших таксона, однако каждая из них значитель-
но менее обособлена и разнообразна, чем радиация прозимий. Пос-
6 В большинстве подобных случаев исходное разделение вероятно бывает
географическим, т.е. предковые группы двух радиаций в основном аллопатрич-
ны. В данном случае, однако, рассматриваемые группы были широко симпат-
ричны на протяжении почти всей истории радиации. Одна не замещала дру-
гую, поскольку они имели разный адаптивный и экологический статус.
Высшие категории
249
ледние классифицируются как подотряд, тогда как три другие ра-
диации объединяются в единый подотряд Anthropoidea и считаются
надсемействами: Ceboidea (обезьяны Нового Света), Cercopithe-
coidea (обезьяны Старого Света) и Hominoidea (человекообразные
обезьяны и люди).
Ни один чёткий диагностический признак или какие-либо при-
знаки наследственной основы [heritage], не отличают отряд Primates
как целое, равно как и примитивных приматов от других прими-
тивных плацентарных млекопитающих. Последующее развитие
разумности, способности пользоваться орудиями, социальности и
т.д. действительно были поздними приобретениями, и они отсут-
ствовали в более примитивных группах, хотя бы и доживших до
нашего времени. Тупайи, пусть и родственные приматам, столь
примитивны и столь близки к общим предкам плацентарных, что
их ранее относили к Insectivora, с чем по-прежнему согласны не-
которые таксономисты [58]. Тем не менее и несмотря на суще-
ственную дивергенцию в пределах грады, обширная группа прози-
мий может быть отделена как от ещё более примитивной анцест-
ральной грады плацентарных, так и от более продвинутой грады
обезьян. Эта последняя града была достигнута независимо и парал-
лельно широконосыми и узконосыми обезьянами. Она характери-
зуется, среди прочего, редукцией и стабилизацией зубной форму-
лы, некоторыми продвинутыми новообразованиями в структуре
мозга и особенностями поведения. Гоминоидная града, вероятно,
возникла от ранних членов грады обезьян, хотя точные связи не
ясны. Её эволюция была сопряжена с дальнейшим развитием моз-
га, утратой хвоста, увеличением размеров, формированием всеяд-
ных зубов, приобретением вертикального положения тела и неко-
торыми другими деталями. Таксон Hominidae, хотя и входит в со-
став таксона Hominoidea, заслуживает (мы, люди, уверены в этом)
отдельной грады, выделяемой по окончательному завершению об-
щей тенденции: полностью вертикальным положением тела, спо-
собностью изготавливать орудя труда, наивысшей разумностью и
речью — вероятно, в такой последовательности.
Каждая из этих град довольно хорошо соответствует, а возмож-
но и идентична, отдельному таксону. Града обезьян не монофиле-
тична в строгом смысле, но почти наверняка едина согласно кри-
ВРЕМЯ
Рис. 29. Схема град приматов и их связь с некоторыми таксонами
Пояснения в тексте.
NJ
СП
О
Высшие категории
251
териям, обсуждавшимся в главе 4, и заслуживает выделения в ка-
честве таксона. Однако не все перечисленные выше грады соответ-
ствуют таксонам одного уровня. По критериям дивергенции, раз-
нообразия и сравнимости гоминидная града характеризует одно
семейство (или даже возникла внутри него); гоминоидная града
обозначает становление надсемейства; града обезьян — общая для
двух надсемейств; наконец, града прозимий эквивалентна подотря-
ду. (В пределах прозимий возможно обозначение град меньшего
масштаба, соответствующих таксонам более низкого ранга.) Эти
отношения представлены схематично на рис. 29.
Последний пример, который здесь будет подробно рассмот-
рен, — отряд Rodentia, крайне запутанный, который демонстриру-
ет все особенности и проблемы, часто присутствующие в крупных
и таксономически сложных группах. Филогения, показанная на рис.
30, основана преимущественно на работах Вуда (Wood, 195 5, 19587,
1959), я лишь отчасти поменял положение таксонов и по-иному
изобразил линии. Вуд представил базальные линии в более древо-
видной форме. Моя схема не означает, что 13 линий возникли спон-
танно или в точности одновременно. Она указывает косвенно на то,
что они сформировались быстро, без известной или предполагае-
мой последовательности, соответствующей отношению «предок—
потомок» между какими-либо из них. За исключением этого момен-
та, линии не являются воображаемыми, но каждая соответствует
конкретному номинальному семейству. Названия отдельных се-
мейств, числом 45, не обязательны для настоящих целей, но их
можно найти в: Wood (1958, 1959); Simpson (1959а; они несколь-
ко по-иному сопряжены с филогенетическим деревом, нежели по-
казано на рис. 30). Здесь для обсуждения можно использовать обо-
значения отдельных базальных стволов, пронумерованных с 1 по
13 на рис. 30.
Несмотря на неизбежные незначительные отличия, главным
образом из-за дробительства и объединительства, семейства грызу-
7 Многие публикации журнала, в котором появилась данная статья, дати-
руются ошибочно: «Wood, 1958» на самом деле была опубликована в 1959,
слишком поздно, чтобы быть рассмотренной в: Simpson, 1959а Изменения,
которые здесь внесены, не меняют основных заключений
Сплошные более или менее вертикальные линии представляют семейства и их известные временные границы.
Филогенетические связи, показанные прерывистыми линиями, недостаточно документированы палеонтологией, основаны
на весомых сравнительных данных. Связи, обозначенные точечными линиями, являются более предположительными. Цифры
внизу обозначают исходные корни, обсуждаемые в тексте. Их последовательность не имеет особого значения, за исключением
того, что (1) обозначает наиболее примитивное семейство, которое, как можно полагать, близко к началу всех прочих.
(В основу положены данные Вуда, особенно из: Wood, 1958,1959, но с существенным изменением формы представления.)
252 Дж. Г. Симпсон. Принципы таксономии животных
Высшие категории
253
нов достаточно хорошо определены и в последние годы не меня-
лись. Отряд в целом также чётко определён. После исключения из
него Lagomorpha он несомненно представляет собой обоснованный
эволюционный таксон, которому неизменно присваивают отрядный
ранг. Такое положение вещей можно считать значительным таксо-
номическим достижением, отражающим более чем удовлетвори-
тельное состояние дел. Тем не менее, было бы удобно, хотя бы для
целей номенклатуры, располагать некими таксонами между един-
ственным отрядом и 45 семействами, и в самом деле специалисты
по грызунам использовали такие промежуточные таксоны на про-
тяжении сотни с лишним лет. В классической системе, формализо-
ванной Брандтом (Brandt, 1855), выделены три подотряда —
Sciuromorpha, Myomorpha и Hystricomorpha — на основе в сущно-
сти единственного комплекса признаков строения жевательной
мышцы. Хотя такой порядок всё ещё широко признан,8 это является
типологической классификацией признаков, но не эволюционной
классификацией животных. Животные же отчётливо показывают,
что каждый из трёх вариантов возникал с теми или иными вариа-
циями независимо в разных предковых группах из более примитив-
ного состояния, неизвестного Брандту. Они несомненно не заслу-
живают выделения в качестве трёх, и только трёх, подотрядов.
Однако приемлемого варианта до сих пор не было предложено.
Обычной альтернативой было умножение числа подотрядов,
пока Wood (1958) не довёл их число до 11, показав логичность та-
кой системы, хотя сам он ей не до конца следовал. Имея в виду
обозначения базальных филетических линий на рис. 30, эта систе-
ма такова:
Protrogomorpha — часть 1 (всё, кроме линии, помеченной «s»).
Caviomorpha — 11.
Myomorpha — 6, 7, 8, 9.
Возможно, новый подотряд или Myomorpha — 10.
Sciuromorpha — часть 1 (линия, помеченная «s»).
Castorimorpha — 5.
8 Я против желания использовал её в 1945 г., хотя и с существенными ого-
ворками. Теперь я думаю, что такие группировки даже при условном приме-
нении совершенно неоправданны.
18 — 4338
254
Дж. Г. Симпсон. Принципы таксономии животных
Theridomorpha — 3.
Новый подотряд (без названия) — 2.
Новый подотряд (без названия) — 4.
Hystricomorpha— 12.
Bathyergomorpha— 13.
В качестве альтернативы Вуд предлагает возможность сохране-
ния только подотрядов, перечисленных выше линии, а прочие се-
мейства (одни в ранге надсемейства, другие нет) просто перечис-
лять как «группы, не отнесённые к подотрядам». Он указывает пре-
имущества и недостатки обеих систем, но не говорит, какую имен-
но предпочитает. Он заключает:
В итоге предполагается, что существующая ситуация, когда имеется
значительное число семейств грызунов, которые кажутся таксономически
изолированными, может быть реальным отражением основополагающего
факта эволюции грызунов и что эти небольшие группы никогда не имели
существенного значения, а их общие предки были в составе примитивно-
го подотряда Protrogomorpha [нижняя часть линии 1 на рис. 30. — Дж.С.],
Я согласен, что именно такова ситуация: другими словами, что
филогенетический паттерн в этом отряде просто не может быть
разделён каким-либо приемлемым способом на небольшое число
сравнимых подотрядов. Очередная отсылка к рис. 30 показывает,
что 11 из 13 базальных линий, которые я выделил, незначительно
или вообще не показывают дивергенции на уровне семейства (две
остались в качестве единственных семейств, семь разделились на
два семейства каждая, а две — на три семейства каждая. И всё-
таки имеются достаточные, хоть и не решающие, свидетельства в
пользу рассмотрения этих базальных линий связанными inter se.9
Они, по крайней мере, сравнимы как таксоны некоторого уровня,
но что касается только этих линий, подходящий для них уровень,
очевидно, не выше надсемейственного. Однако линии 1 и 11 ради-
кально иные. Девять семейств, возникших из 1, эволюировали пу-
тём простого ветвления, экспансии, а затем сужения, так как боль-
шинство этих семейств вымерло. Линия 11 внезапно разветвляет-
9 Вуд всё-таки считает обозначенные мною линии 6, 7, 8 и 9 наиболее близ-
кими, однако доказательства скудны, и он просто склоняется к этому, чтобы
избежать разделения классического подотряда.
Высшие категории
255
ся в самом начале на пять или шесть линий, аналогично тому, как
весь отряд несколько ранее распадается на 13 линий, и вся группа
в конце концов разрастается до 14 семейств.
Филогении, возникшие на основе линий 1 и 11, не полностью
сравнимы, но обе они таковы, что могут считаться подотрядами,
если игнорировать прочих представителей отряда. Кроме того, бо-
лее чем желательно и даже необходимо иметь названия для ссыл-
ки на эти группы несомненно родственных семейств. На самом
деле у них есть названия, данные Вудом: Protrogomorpha (плюс
Sciuridae, которых Вуд исключает по причинам, которые я нахожу
недостаточными) для группы, исходящей от линии 1, и Cavio-
morpha — для группы от линии 11.
Называть группы 1 и 11 подотрядами, а группы 2-10 и 12-13
надсемействами вне подотрядов — значит противоречить примене-
нию и самой логике иерархии. И всё-таки эти две категории групп
несравнимы между собой, и по этой причине называть их одина-
ково либо подотрядами, либо надсемействами нельзя признать
удовлетворительным. Природа в данном случае просто не препод-
несла нам сравнимые таксоны на уровне, промежуточном между
семейством и отрядом, так что если такие таксоны использовать,
единственное решение состоит в том, чтобы согласиться с отнесе-
нием таксонов, которые между собой не сравнимы, к одной кате-
гории. Значит, линии 1-13 на рис. 30 все должны быть или подо-
трядами (с надсемействами в 1 и 11, но не в других), или надсе-
мействами (никакие из них не группируются в подотряды).
Отряд с точки зрения вовлечённых в его становление эволюци-
онных факторов возник благодаря единственной базовой адапта-
ции — приспособления к грызению, которое оказалось столь эф-
фективным, что Rodentia вскоре вытеснили других грызущих и бы-
стро разделились на многие группы в Африке, Евразии и Северной
Америке. Все потомки ранних форм сохранили это приспособле-
ние к грызению. Почти все они проявили тенденцию к её после-
довательному совершенствованию, при этом параллельно несколь-
кими разными способами, что наиболее заметно в перестройке Мас-
сетера. Что касается последнего, возможны три альтернативные и
по-видимому одинаково эффективные специализации, соответству-
ющие типологическим группам Брандта. Та или другая из них воз-
18*
256
Дж. Г. Симпсон. Принципы таксономии животных
никала, по всей видимости почти случайно, в разных ранних ли-
ниях (за исключением единственной группы 1). Раз возникнув, та
или иная специализированная конфигурация в общих чертах сохра-
нялась у всех потомков данной филетической линии, хотя иногда
и со значительными последующими модификациями. Имели мес-
то частые, хотя и не такие всеобщие, параллельные специализации
и по другим параметрам, особенно в развитии гипсодонтных и гре-
бенчатых щёчных зубов. В других отношениях грызуны весьма раз-
нообразны и, в конечном счёте, выработали широкий спектр адап-
тивных типов, практически не имеющих аналогов в других отря-
дах. Эта дивергенция, происходившая независимо в нескольких
обширных регионах, сопровождалась также неоднократной и порой
весьма сложной конвергенцией между грызунами сходной эколо-
гии, но с разным распространением.
Грызуны достигли Южной Америки довольно поздно, прибли-
зительно в конце эоцена. Они там столкнулись с немногочисленны-
ми и большей частью неэффективными конкурентами среди сум-
чатых и мелких копытных, которые вскоре либо вымерли, либо
перешли к иной специализации, где грызуны не составляли конку-
ренции. Таким образом, грызуны получили почти безграничные
возможности на обширном континенте, крайне разнообразном эко-
логически. Они быстро дивергировали и стали весьма разнообраз-
ными, сформировав особый филогенетический паттерн, начиная с
линии 11 на рис. 30.
Эволюционная основа таксонов
Выше постоянно подчёркивалось, что таксоны эволюционной
классификации должны быть совместимы либо с реконструкцией
филогении, либо с филогенетическим процессом как вероятной
причиной возникновения наблюдаемого разнообразия — или с обо-
ими, ибо эти подходы тесно и часто нераздельно взаимосвязаны. В
любом случае таксоны отражают эволюционные факторы, привед-
шие к их возникновению и развитию, и эта преисполненная смысла
связь — одна из наиболее важных среди тех причин, которые по-
буждают настаивать, насколько это применимо на практике, на эво-
люционном подходе к классификации. Подчёркивая эту связь и соот-
Высшие категории
257
нося таксономию со всей сферой систематики и эволюции, необхо-
димо в заключение сказать несколько слов об эволюционном значе-
нии таксонов и их природе, рассматриваемой в ее эволюционном
аспекте. Едва ли имеет смысл здесь вдаваться в наиболее базовые,
элементарные процессы, например, действующие на генетическом
уровне, различные изолирующие механизмы, факторы естественного
отбора. Что действительно следует сделать в завершение этого про-
странного исследования — связать обобщения о таксонах с обоб-
щениями об истории популяций целостных организмов, прежде
всего в терминах адаптации, которая является наиболее важным
эволюционным фактором в интересующем нас аспекте.
Эволюционные виды были определены (глава 5) непосредствен-
но через обособленную и единую природу их эволюционных ролей.
В каждом локальном регионе их роль и адаптивная сущность опре-
деляются их соответствием элементарным экологическим нишам.
Внутривидовые таксономические единицы, являющиеся в подавля-
ющем большинстве случаев аллопатрическими в пределах ареала
единого вида, обычно демонстрируют более тесную адаптацию к
условиям микросреды, на которую влияют также обмен генами,
дрейф генов и мутационный пресс. Адаптивный статус этих внут-
ривидовых популяций, или подвидовых таксонов, не слишком чёт-
ко отличается от соседних популяций или от вида в целом, и такая
[адаптивная] дивергенция, если она и имеет место, то вряд ли смо-
жет поддерживаться длительное время, если только сама внутриви-
довая единица не превратится в вид. Повторение в существенных
чертах адаптаций у разных видов бывает редким и временным, если
только виды не полностью аллопатричны и не разделены географи-
ческими или другими экологическими барьерами.
Таксоны высших категориальных рангов предполагают один
или, за небольшим числом исключений, два типа адаптивных ря-
дов. Во-первых, развертывание от общего предка многих филети-
ческих линий, включающих либо один вид, либо группу видов с
разными адаптивными ролями и способных к обмену генами.10
10 Монофилия в таком самом узком смысле составляет основу рассматри-
ваемых здесь эволюционных процессов. Как уже отмечалось, она не всегда
служит практическим критерием для определения таксонов в формальной клас-
сификации, что является отдельным, хоть и связанным вопросом.
258
Дж. Г. Симпсон. Принципы таксономии животных
Соответствующее различие в ролях в принципе может быть связано
только с распространением, т.е. быть внешним по отношению к
самим организмам, однако в действительности оно почти всегда
включает также соматические различия между организмами. Во-
вторых, любая филетическая линия может адаптивно обособится от
прочих в такой степени, что выделится в самостоятельный таксон.
Такой таксон остаётся политипическим в том смысле, что включает
несколько последовательных видов, каждый из которых по адапта-
циям более или менее отличается от прочих.11 Понятно, что прави-
лом является сочетание этих двух процессов — диверсификации и
дивергенции.
Большинство высших таксонов связано с базовыми адаптаци-
ями, которые возникают либо одновременно с самим таксоном, т.е.
в качестве основы того, что затем станет высшим таксоном, либо
в результате более или менее выраженных параллелизмов в ранних
линиях. Базовая адаптация может соотносится, по существу, с един-
ственной структурой или с ограниченным набором функциональ-
но тесно скоррелированных структур, подобно адаптации грызуще-
го аппарата у грызунов или адаптации к полёту у рукокрылых. С
другой стороны, она может затрагивать целый комплекс многих
разных признаков и структур, подобно приспособлениям к водно-
му образу жизни у китообразных. Очевидно, здесь нет резкого про-
тивопоставления, но есть непрерывный ряд от однопризнаковой до
многопризнаковой адаптации со всеми промежуточными состояни-
ями/Такое возникновение таксонов посредством базовой адапта-
ции обычно связано с переходом из одной адаптивной зоны в дру-
гую, с достаточно чётко выраженной спецификой.
Окончательный ранг таксона, возникающего таким образом,
обычно пропорционален степени отличия (обособленности —
distinction) осваиваемой им зоны и, следовательно, масштабу свя-
занной с этим дивергенции, а также объёму и числу подзон и час-
11 Поэтому в классификации монотипические таксоны таковы лишь из-за
неполноты знания. В принципе, они включают пустые множества — таксоны,
которые присутствуют в природе, но не открыты и не классифицированы. Еди-
ничные исключения могут быть связаны с неким одношаговым процессом, ко-
торый мгновенно производит разрыв на генетическом или более высоком уров-
не, если конечно, это вообще возможно.
Высшие категории
259
тных ниш, т.е. возможности дивергировать в пределах этой зоны.
Среди позвоночных, переход от водного к наземному образу жиз-
ни был связан с масштабной базовой дивергенцией и проникнове-
нием в очень широкую и крайне разнообразную адаптивную зону.
В результате возникло несколько классов, десятки отрядов и мно-
гие тысячи видов. Грызуны, рукокрылые и китообразные, упомяну-
тые в предыдущем абзаце, — примеры появившихся таким путем
отрядов. Окончательно достигнутый ранг, однако, не есть прямая
функция от характера адаптации. Например, адаптация грызущего
аппарата, сходная с таковой у грызунов, возникла в ряде других
отрядов (например, у сумчатых, приматов, различных копытных),
но с меньшей исходной специфичностью и последующей диверси-
фикацией, так что в этих группах она дала начало лишь таксонам
ранга семейства или даже рода.
Адаптации, приводящие к появлению высших таксонов, разли-
чаются как по ширине, т.е. по числу различных экологических си-
туаций, в которых адаптация даёт преимущество (или к которым
открывает доступ), так и по устойчивости, т.е. по тому, в какой
мере адаптация характеризует весь сложившийся высший таксон.
В целом, и это достаточно очевидно, чем шире адаптация, тем ве-
роятнее, что она будет устойчиво сохраняться во всём таксоне, но
здесь нет полной корреляции. Приспособления к грызению у гры-
зунов, к полёту у рукокрылых, водному образу жизни у китообраз-
ных отличаются широтой и сохраняются у всех без исключения
представителей соответствующих отрядов. С другой стороны, на-
земная локомоторная адаптация ранних амфибий была несомнен-
но одной из возможно наиболее широких, и всё-таки она была ут-
рачена многими из их потомков. Эти потомки, однако, не утрати-
ли каких-то других частей комплексной адаптации к наземному
образу жизни, например, воздушного дыхания, даже когда они пе-
рестали быть наземными. Сохранение базовых адаптаций — основ-
ной источник общности признаков. Рассмотрение эволюционных
таксонов с этой точки зрения делает более понятным, почему об-
щность признаков часто полезна при их распознавании, но также
нередко оказывается неадекватной для этих целей.
Широта и устойчивость адаптаций образует непрерывную шка-
лу, которая может быть увязана с [таксономическими] категориями
260
Дж. Г. Симпсон. Принципы таксономии животных
и представлена через них. Таким образом, существуют адаптации
и соответствующие им соматические характеристики, специфичные
для видов, родов, семейств и так далее вплоть до царств, хотя по
мере восхождения по таксономической шкале эта зависимость по
разным причинам подвержена большему числу исключений.
В тех высших таксонах, которые возникают на основе чётко
оформленной единичной адаптации, обычно происходит последу-
ющая более медленная коадаптация других характеристик, так или
иначе связанной с базовой адаптацией. Коадаптация может осуще-
ствляться по-разному в разных филетических линиях, однако она
часто сопряжена с широкими параллелизмами. У грызунов многие
из таких более или менее выраженных параллельных трендов, та-
ких как три типа устройства жевательной мышцы, гипсодонтия и
лофодонтия, могут рассматриваться как коадаптивные по отноше-
нию к базовой адаптации, связанной с грызением.
С другой стороны, когда возникновение высшего таксона свя-
зано с формированием множества адаптаций или нескольких адап-
тивных систем, эти базовые адаптации, вероятно, должны склады-
ваться параллельно в филетических линиях в период начального
становления группы. Разные части комплекса обыкновенно возни-
кают в разное время и с разной скоростью в пределах одной ли-
нии, давая картину, которую ДеБир (de Beer, 1954) назвал «моза-
ичной эволюцией». В качестве примера ДеБир приводит Archaeop-
teryx, у которого одни признаки всё еще полностью рептилийные,
другие уже полностью птичьи, а третьи промежуточные. Тем не
менее, куски такой «мозаики» вставляются или перетасовываются,
формируя новую картину, не случайно. Они также коадаптированы,
пусть даже не жёстко, так что организм в целом остаётся интегри-
рованным. Одни и те же части комплекса также могут формиро-
ваться в разное время и с разной скоростью в разных линиях. В та-
ком случае даже для одной линии нельзя указать какой-то момент
или краткий отрезок времени, в пределах которого появляется весь
адаптивный комплекс становящегося таксона, еще более это про-
блематично сделать для группы линий с параллельной эволюцией
комплекса. Яркий пример такой ситуации — происхождение класса
Mammalia. Их суммарный адаптивный комплекс, общий облик и
повышение организации складывались в процессе параллельной
Высшие категории
261
эволюции в разных филетических линиях на протяжении гигантс-
кого отрезка времени от перми до мела (Simpson, 1959b, 1960b).
Это создало проблемы классификации, которые могут быть реше-
ны только произвольным выбором для целей диагностики какого-
то одного признака, даже незначительного, из всего комплекса,
если он сопряжён с базовой адаптацией и пригоден для практичес-
кого применения.
Одновременно с этими параллельными трендами, пронизыва-
ющими насквозь высший таксон и воплощающимися в его базовых
адаптациях и последующих коадаптациях, происходит дивергенция
соподчинённых таксонов на каждом уровне нисходящего [иерархи-
ческого] ряда вплоть до вида. Так, наряду со становлением адап-
таций, специфичных для класса Mammalia, возникали адаптации,
специфичные для входящих в него отрядов, семейств, родов и ви-
дов.
Медленное формирование сложной базовой адаптации не мо-
жет быть чётко отграничено от случаев, связанных с постепенны-
ми трендами, или совершенствованием в смысле Гексли (Huxley,
1957). В отношении этих случаев не всегда можно утверждать, что
становление таксона на протяжении большей части или всей его
истории было сопряжено именно с трендами, а не с диагностичес-
кой или базовой адаптацией. Оба эти взаимопроникающих процес-
са с высокой вероятностью порождают грады, обычно полифиле-
тические на низших таксономических уровнях, но монофилетичес-
кие близ уровня, которого достигает в конечном итоге становящий-
ся таксон. Общая природа этих феноменов и их связь с таксонами
иллюстрируется примером приматов, обсуждавшимся в предыду-
щем разделе. В этом примере выжили все важнейшие грады и от-
мечено лишь немного случаев замещения низших или более древ-
них град высшими или более поздними. Причина в том, что на са-
мом деле это было не просто улучшение адаптации в пределах од-
ной адаптивной зоны, но также и одновременное развитие других
адаптивных зон, лишь с незначительным их перекрыванием и кон-
куренцией. Однако в случае Carnivora между креодонтами и фис-
сипедами, которые также могут считаться градами, хотя и несколь-
ко иного рода, было заметное перекрывание зон, обусловившее
соревнование и в конце концов вытеснение низшей грады другой,
262
Дж. Г. Симпсон. Принципы таксономии животных
более высоко организованной — т.е. одного таксона другим таксо-
ном того же ранга.
Для высших таксонов характерен баланс между филетической
эволюцией (progression) группы в целом и её дивергенцией от дру-
гих групп того же уровня, с одной стороны, и диверсификацией
группы, связанной с дивергенцией входящих в неё таксонов, с дру-
гой стороны. Lagomorpha демонстрируют некоторую общую про-
грессию, но незначительную внутреннюю дивергенцию. У Carni-
vora была незначительная общая прогрессия, но существенная
внутренняя дивергенция. Rodentia показывают чёткую выражен-
ность обоих феноменов, и почти все высшие таксоны демонстри-
руют в той или иной степени каждый из них. Типичной формой
диверсификации является адаптивная радиация, часто дающая на-
чало группам, которые лишь полупроизвольны и вполне сравнимы,
так что их можно удовлетворительно определять как таксоны од-
ного уровня. Примеры такого рода уже были представлены — та-
кие как подсемейства Leporidae и подотряды Carnivora. В некото-
рых случаях, однако, разовая сравнительно быстрая базальная адап-
тивная радиация порождает группы, резко различающиеся соотно-
шением прогрессии и диверсификации и потому не вполне сравни-
мые: их бывает трудно или невозможно разделить удовлетворитель-
ным образом на таксоны какого-то одного уровня иерархии. Груп-
пы грызунов между семейством и отрядом являют собой прекрас-
ный пример.
•Когда два или более таксона приблизительно или строго одного
категориального уровня возникают от общего предка, это обычно
происходит аллопатрически (в разных регионах, как Ceboidea и
Cercopithecoidea), аллохронно (в разное время, как Prosimii и
Anthropoidea) или обоими путями (как Ceboidea и Hominoidea; дру-
гие примеры всех этих соотношений также рассмотрены в преды-
дущих секциях). В любом случае часто происходит так, что разра-
стающиеся группы после стабилизации оказываются симпатричны-
ми и синхронными. Если они при этом оказываются адаптивно
очень разными, не происходит ничего, кроме обогащения фауны.
Каждая фауна действительно включает множество разных таксо-
нов, которые когда-то имели единого предка. Если, однако, адаптив-
ное сходство всё-таки достаточно для конкуренции, имеет место
Высшие категории
263
взаимодействие двух процессов. С одной стороны, происходит бо-
лее детальная структуризация адаптивных зон с их дроблением на
подзоны, отвечающие различным группам; с другой стороны, имеет
место замещение, или вымирание и замена одной группы другой.
Оба процесса отличались широкими масштабами, примером чему
служит вселение северных грызунов в Южную Америку в позднем
кайнозое, вступивших там в конкуренцию с некоторыми из более
древних местных грызунов.
Конвергенция с наибольшей вероятностью складывается и бы-
вает наиболее глубокой между таксонами аллопатрической радиа-
ции, особенно когда отсутствует или ничтожна экспансия таксонов
одной радиации в «чужую» область. Крайне маловероятно, чтобы
действительно тесная конвергенция возникла между симпатричны-
ми таксонами, равно как и крайне необычно, чтобы конвергентные
таксоны когда-либо стали симпатричными или оставались таковы-
ми сколько-нибудь длительное время. Это одна из причин, почему
зоогеография особенно важна в эволюционной классификации,
позволяя чётко разграничивать конвергенцию и гомологию. Клас-
сические примеры крупномасштабной комплексной конверген-
ции — древние местные фауны млекопитающих Австралии и Юж-
ной Америки. Каждый континент был сценой широкой и сложной
адаптивной радиации, практически изолированной от радиаций на
других континентах. Как в Австралии, так и в Южной Америке
многочисленные группы развили сильные конвергентные сходства
с афро-евразийско-североамериканскими млекопитающими. Когда
по тем или иным причинам конвергентные викариаты разного гео-
графического происхождения оказывались симпатричными, конвер-
гентные австралийские и южноамериканские таксоны в скорости
вымирали — например, сумчатые волки после заселения Австра-
лии собаками, или местные южноамериканские копытные после
вселения на этот континент парно- и непарнокопытных.
Именно с результатами всех такого рода процессов — а их ещё
больше! — таксономия должна иметь дело. Взаимодействие этих
процессов столь сложное, что его результатом оказывается ошелом-
ляющее разнообразие филогенетических паттернов, демонстриру-
емых животными и формируемых из связей по смежности и по
сходству. Результат эволюции — не некая совокупность изящно
264
Дж. Г. Симпсон. Принципы таксономии животных
размещённых ящичков, по которым удобно разложены все её про-
дукты. Всякий таксономический подход, подразумевающий про-
стой естественный порядок, который невозможно обнаружить ни
строго априорными, ни строго эмпирическими средствами, обречён
на неуспех самой природой классифицируемого материала. И всё
же эволюция производит не хаос и не безнадёжный беспорядок.
Это совершенно упорядоченный процесс, но упорядоченный по-
средством многочисленных взаимодействующих, сбалансирован-
ных, а порой даже конфликтующих принципов. Эти принципы мо-
гут быть познаны, но приближение к ним очень постепенное, иног-
да на ощупь. Имеется наука таксономия, которая приспосабливает
принципы эволюции к своей узкой области и добавляет свои соб-
ственные принципы. И эти принципы таксономии применяются к
классификации, которая также является и искусством, основанном
на канонах вкуса, сдержанности и полезности.
Цитированные работы
Adanson М. 1763. Families des plantes. Paris, Vincent.
Allee W. C., Emerson A. E., Park O., Park T., Schmidt К. P. 1949. Principles of
animal ecology. Philadelphia, Saunders.
Allen J. A. 1919. Notes on the synonymy and nomenclature of the smaller spotted
cats of tropical America. Bull. Amer. Mus. Nat. Hist, 41: 341-419.
Amadon D. 1949. The seventy-five per cent rule for subspecies. Condor, 51: 250-
258.
Ashton E. H., M. J. R. Healy, and S. Lipton. 1957. The descriptive use of
discriminant functions in physical anthropology. Proc. Roy. Soc., B, 146: 552-
572.
Bader T. S. 1958. Similarity and recency of common ancestry. Systematic Zoology,
7: 184-187.
Baer К. E. von. 1828. Ueber EntwicMungsgeschichte der Thiere. Beobachtungen
und Reflexion. Konigsberg, Wilhelm Koch.
Bather F. A. 1927. Biological classification: past and future. Quart. Joum. Geol.
Soc., 83: Ixii-civ.
Beckner M. 1959. The biological way of thought. New York, Columbia Univ. Press.
Belon P. 1555. L’histoire de la nature des oiseaux. Paris, Guillaume Cavellat.
Bigelow R. S. 1958. Classification and phylogeny. Systematic Zoology, 7: 49-59.
Blackwelder R. E. 1959. The present status of systematic zoology. Systematic
Zoology, 8: 69-75.
Blackwelder, R. E., Boyden A. 1952. The nature of systematics. Systematic Zoology,
1: 26-33.
Blainville H. M. D. de. 1816. Prodrome d’une nouvelle distribution systematique
du regne animal. Bull. Sci. Soc. Philom. Paris, ser. 3, 3: 105-124.
Block K. 1956. Zur Theorie der naturwissenschaftlichen Systematik unter besonderer
Beriicksichtigung der Biologie. Bibl. Biotheor., 7: 1-138.
Bonnet C. 1745. Traite d’insectologie. Paris, Durand.
Boyden A. 1947. Homology and analogy. Amer. Midland Naturalist, 37: 648-669.
— 1953. Fifty years of systematic serology. Systematic Zoology, 2: 19-30.
— 1958. Comparative serology: aims, methods, and results. Serological and
Biochemical Comparison of Proteins, XIV Annual Protein Conference: 3-24.
Brandt J. F. 1855. Beitrage zur naheren Kenntniss der Saugethiere Russlands. Mem.
Acad. Imp. Sci. St-Petersbourg, ser. 6, 9: 1-375.
Brinkmann R. 1929. Stetistisch-biostratigraphische Untersuchungen an mitteljurassi-
schen Ammoniten iiber Artbegriff und Stammesentwicldung. Abh. Ges. Wiss.
Gottingen, math.-phys. KL, N.F., 8: part 3.
266
Дж. Г. Симпсон. Принципы таксономии животных
Brisson М. J. 1762. Regnum animale in classes IX. 2nd ed. Paris, Cl. Joannem-
Baptistam Bauche.
Bulman, О. M. B. 1955. Graptolithina. Treatise on Invertebrate Paleontology, Part V.
Burma B. J. 1948. Studies in quantitative paleontology: I. Some aspects of the
theory and practice. Jour. Paleont., 22; 725-761.
Cain A. J. 1953. Geography, ecology and coexistence in relation to the biological
definition of the species. Evolution, 7: 76-83.
— 1954. Animal species and their evolution. London, Hutchinson.
— 1955. Not the superspecies. Systematic Zoology, 4: 143-144, 159.
— 1958a. Logic and memory in Linnaeus’s system of taxonomy. Proc. Linnean Soc.
London, 169 session: 144-163.
— 1958b. Chromosomes and taxonomic importance. Proc. Linnean Soc. London,
169 session: 125-128.
— 1959a. The post-Linnaean development of taxonomy. Proc. Linnean Soc.
London, 170: 234-244.
— 1959b. Deductive and inductive methods in post-Linnaean taxonomy. Proc.
Linnean Soc. London, 170: 185-217.
— 1959c. Taxonomic concepts. Ibis, 101: 302-318.
Cain A. J., and Harrison G. A. 1958. An analysis of the taxonomist’s judgement of
affinity. Proc. Zool. Soc. London, 131: 85-98.
Caiman W. T. 1940. A museum zoologist’s view of taxonomy. In Huxley, 1940:
455—459.
Christensen В., С. O. Nielsen. 1955. Studies on Enchytraeidae, 4. Preliminary report
on chromosome numbers of seven Danish genera. Chromosoma, 7: 460-468.
Clark W. E. Le Gros. 1959. The antecedents of man. Chicago, Quadrangle Books.
Cloud P. E., Jr. 1959. Paleoecology — retrospect and prospect. Jour. Paleont, 33:
926-962.
Colbert E. H. 1941. A study of Orycteropus gaudryi from the Island of Samos. Bull.
Amer. Mus. Nat. Hist., 78: 305-351.
Cope E. D. 1875. On the supposed Carnivora of the Eocene of the Rocky
Mountains. Proc. Acad. Nat. Sci. Philadelphia, 27: 444-448.
Cuvier G. 1835. Le regne animal distribue apres son organization. 2nd ed. Paris,
Crochard et Co.
Darlington P. J. 1957. Zoogeography: the geographical distribution of animals. New
York, Wiley.
Darwin C. 1859. On the origin of species by means of natural selection or the
preservation of favoured races in the struggle for life. London, Murray. (For all
subsequent revisions see the variorum edition: Morse Peckham, ed., 1959.
Philadelphia, Univ, of Pennsylvania Press.)
Dawson M. R. 1958. Late Tertiary Leporidae of North America. Univ. Kansas Paleo.
Cont., Vertebrata, art. 6: 1-75.
De Beer G. R. 1951. Embryos and ancestors. Rev. ed. Oxford, Oxford Univ. Press.
— 1954. Archaeopteryx lithographica. A study based upon the British Museum
specimen. London, British Mus. (Nat. Hist.).
Цитированные работы
267
Delacour J., Мауг Е. 1945.The family Anatidae. Wilson Bull., 57: 3-55.
Dennler J. G. 1939. Los nombres indigenas en guarani de los manuferos de la
Argentina у paises limitrofes у su importancia para la sistematica. Physis, 16:
225-244.
Dethier V. G. 1954. Evolution of feeding preferences in phytophagous insects.
Evolution, 8: 32 -54.
Dobzhansky Th. 1937, 1941, 1951. Genetics and the origin of species. 1st ed.
(1937). 2nd ed. (1941). 3rd ed. (1951). New York, Columbia Univ. Press.
Eagar R. M. C 1956. Naming Carboniferous non-marine lamellibranchs. In
Sylvester-Bradley, 1956: 111-116.
Edwards J. G. 1954. A new approach to infraspecific categories. Systematic Zoology,
3: 1-20.
Garstang W. 1922. The theory of recapitulation. Jour. Linnean Soc. London, Zool.,
35: 81-101.
George T. Neville. 1956. Biospecies, chronospecies, and morphospecies. In
Sylvester-Bradley, 1956: 123-137.
Gilmour G. S. L. 1940. Taxonomy and philosophy. In Huxley, 1940: 461 474.
— 1951. The development of taxonomic theory since 1851. Nature, 168: 400-402.
Goldschmidt R. 1940. The material basis of evolution. New Haven, Yale Univ. Press.
Grabert B. 1959. Phylogenetische Untersuchungen an Gaudryina und Spirople-
ctinata (Foram.) besonders aus dem nordwestdeutschen Apt und Alb, Abh.
Senckenberg. Naturforsck Ges., No. 498: 1-71.
Gregg J. R. 1954. The language of taxonomy. New York, Columbia Univ.
Gregory W. К 1910. The orders of mammals. Bull. Amer. Mus. Nat. Hist., 27:
1-524.
— 1951, Evolution emerging. 2 vols. New York, Macmillan.
Haas O., G. G. Simpson. 1946. Analysis of some phylogenetic terms with attempts
at redefinition. Proc. Amer. Phil. Soc, 90: 319-349.
Haeckel E. 1875. Ziel und Wege der heutigen Entwicklungsgeschichte. Jena,
Hermann Dufift.
Haslewood G. A. D. 1959a. Species comparison as an aid in the study of the process
sterols-bile salts. Ciba Symp. on Biosynthesis of Terpines and Sterols: 206-213.
— 1959b. Comparative studies of «bile salts» 12. Application to a problem of rodent
classification: bile salts of the cutting-grass, Thryonomys swinderianus.
Biochem. Jour., 73: 142-144.
Heberer G., ed. 1943, 1954-1959. Die Evolution der Organism. 1st ed. (1943). 2nd
ed. (1954-1959). Stuttgart, Gustav Fischer.
Hedgpeth J. W. (ed. vol. 1), and H. S. Ladd (ed. vol. 2). 1957. Treatise on marine
ecology and paleoecology. 2 vols. Geol. Soc. Amer., Mem. 67.
Hennig W. 1950. Gruncizuge einer Theorie der phylogenetischen Systematik. Berlin,
Deutscher Zentralverlag.
Huxley J. S. 1938. Clines: an auxiliary taxonomic principle. Nature, 142: 219.
— 1939. [No title; discussion of clines.] Proc. Linnean Soc. London, 1938-39:
105-114.
268
Дж. Г. Симпсон. Принципы таксономии животных
—, ed. 1940. The new systematics. Oxford, Oxford Univ. Press.
— 1942. Evolution, the modem synthesis. New York, Harper.
— 1957. The three types of evolutionary process. Nature, 180: 454—455.
— 1958. Evolutionary processes and taxonomy with special reference to grades.
Uppsala Univ. Arssks., 1958: 21-38.
Huxley T. H. 1880. On the application of the laws of evolution to the arrangement
of the Vertebrata and more particularly of the Mammalia. Proc. Zool. Soc.
London, 1880: 649-662.
Imbrie J. 1957. The species problem with fossil animals. In Mayr, 1957: 125-153.
Inger R. F. 1958. Comments on the definition of genera. Evolution, 12: 370-384.
Jepsen G. L. 1949. Selection, «orthogenesis», and the fossil record. Proc. Amer. Phil.
Soc., 93: 479-500.
Johnson M. L., Wicks M. J.. 1959. Serum protein electrophoresis in mammals —
taxonomic implications. Systematic Zoology, 8: 88-95.
Jolicoeur P. 1959. Multivariate geographical variation in the wolf, Canis lupus L.
Evolution, 13: 283-299.
Joseph H. W. B. 1916. An introduction to logic. 2nd ed. Oxford, Oxford Univ. Press.
Kaplan N. О., M. M. Ciotti, M. Hamolosky, and R. E. Bieber. I960. Molecular
heterogeneity and evolution of enzymes. Science, 131: 392-397.
Kinsey A. C. 1936. The origin of higher categories in Cynips. Indiana Univ. Pub.,
Sci. Ser., No. 4: 1-334.
Kiriakofif S. G. 1959. Phylogenetic systematics versus typology. Systematic Zoology,
8: 117-118.
Klein J. T. 1751. Quadrupedum dispositio brevisque historia naturalis. Lipsiae,
Jonam Schmidt.
Kleinschmidt O. 1926. Die Formenkreislehre. Halle-S., Gebauer-Swetschke.
Krutzsch P. H. 1954. North American jumping mice (genus Zapus). Univ. Kansas
Pub., Mus. Nat. Hist., 7: 349-472.
Lamar,ck J.-B. M. de. 1809. Philosophic zoologique. Paris, Dentu.
Lerner I. M. 1954. Genetic homeostasis. New York, Wiley.
Linnaeus C. 1758. Systema naturae. 10th ed. Stockholm, Laurentdi Salvii.
Lull R. S. 1953. Triassic life of the Connecticut Valley. Connecticut State Geol. and
Nat. Hist. Surv., Bull. No. 81.
Marsh F. L. 1944. Evolution, creation and science. Washington, D.C., Review and
Herald Publishing Association.
Maslin T. P. 1952. Morphological criteria of phylogenetic relationships. Systematic
Zoology, 1: 49-70.
Matthew W. D. 1909. The Carnivora and Insectivora of the Bridger basin, middle
Eocene. Mem. Amer. Mus. Nat. Hist, 9: 289-567.
Matthey R. 1952. Chromosomes de Muridae (Microtinae et Cricetinae). Chromo-
soma, 5: 113-138.
Mayr E. 1931. Notes on Halcyon chloris and some of its subspecies. Amer. Mus.
Novitates, No. 469: 1-10.
— 1940. Speciation phenomena in birds. Amer. Naturalist, 74: 249 -278.
Цитированные работы
269
— 1942. Systematics and the origin of species from the viewpoint of a zoologist.
New York, Columbia Univ. Press,
— 1948. The bearing of the new systematics on genetical problems. The nature of
species. Advances in Genetics, 2: 205-237.
— 1954. Notes on nomenclature and classification. Systematic Zoology, 3: 86-89.
—, ed. 1957. The species problem. Amer. Assoc. Adv. Sci., Pub. No. 50.
— 1958. Behavior and systematics. In Roe and Simpson, 1958: 340-362.
Мауг E., Linsley E. G., Usinger R. L. 1953. Methods and principles of systematic
zoology. New York, McGraw-Hill.
Meglitsch P. A. 1954. On the nature of the species. Systematic Zoology, 3: 49-51.
Merriam С. H. 1918. Review of the grizzly and big brown bears of North America
(genus Ursus) with description of a new genus, Vetularctos. N. Amer. Fauna,
41: 1-36.
Michener C. D. 1949. Parallelisms in the evolution of the Satumid moths. Evolution,
3: 129-141.
— 1957. Some bases for higher categories in classification. Systematic Zoology, 6:
160-173.
Michener, CD., and R. S. Sokal. 1957. A quantitative approach to a problem in
classification. Evolution, 11: 130-162.
Moore R. C. 1957. Digest of proposed addition to the «Regies» of provisions
recognizing and regulating the nomenclature of «Parataxa» Jour. Paleont, 31:
1181-1183.
Mossman H. W. 1953. The genital system and fetal membranes as criteria for
mammaliam phylogeny and taxonomy. Jour. Mam., 34: 289-298.
Newell N. D. 1947. Infraspecific categories in invertebrate paleontology. Evolution,
I: 163-171.
— 1949a. Phyletic size increase — an important trend illustrated by fossil
invertebrates. Evolution, 3: 103-124.
— 1949b. Types and hypodigms. Amer. Jour. Sci., 247: 134-142.
— 1956. Fossil populations. In Sylvester-Bradley, 1956: 63-82.
Nicol D., Desborough G. A., Solliday J. R. 1959. Paleontologic record of the
primary differentiation in some major invertebrate groups. Jour. Washington
Acad. Sci., 49: 351-366.
Olson E. C, and R. L. Miller. 1958. Morphological integration. Chicago, Univ, of
Chicago Press.
Orton G. L. 1953. The systematics of vertebrate larvae. Systematic Zoology, 2: 63-
75.
— 1955. The role of ontogeny in systematics and evolution. Evolution, 9: 75-83.
Osborn H. F. 1936, 1942. Proboscidea. 2 vols. New York, Amer. Mus. Nat. Hist.
Owen R. 1848. Report on the archetype and homologies of the vertebrate skeleton.
Rep. 16th Meeting British Assoc. Adv. Sci.: 169-340.
— 1866. On the anatomy of vertebrates. London, Longmans, Green.
Patterson J. T, Stone W. S. 1952. Evolution in the genus Drosophila. New York,
Macmillan.
19
270
Дж. Г. Симпсон. Принципы таксономии животных
Pennant Т. 1781. History of quadrupeds. London, В. White.
Pirie N. W. 1952. Concepts out of context: the pied pipers of science. Brit. Jour.
Phil. Sci., 2: 269-280.
Pontecorvo G. 1959. Present trends in genetic analysis. New York, Columbia Univ.
Press.
Quinn J. H. 1955. Miocene Equidae of the Texas Gulf Coastal Plain. Univ, of Texas
Pub. No. 5516: 1-102.
Remane A. 1956. Die Grundlagen des naturlichen Systems, der vergleichenden
Anatomie und der Phylogenetik. Leipzig, Geest and Portig.
Rensch B. 1929. Das Prinzip geographischer Rassenkreise und das Problem der
Artbildung. Berlin, Gebriider Bomtraeger.
— 1954. Neuere Probleme der Abstammungslehre. Die transspezifische Evolution.
2e Aufl. Stuttgart, Ferdinand Enke.
— I960. The laws of evolution. In The evolution of life, S. Tax, ed., vol. I of
«Evolution after Darwin». Chicago, Univ, of Chicago Press, pp. 95-116.
Rhodes F. H. T 1956. The time factor in taxonomy. In Sylvester-Bradley, 1956: 37-
52.
Roe A. 1946. Artists and their work. Jour. Personality, 15: 1-40.
— 1953. The making of a scientist. New York, Dodd, Mead.
Roe, A., Simpson G. G., eds. 1958. Behavior and evolution. New Haven, Yale Univ.
Press.
Rosa D. 1931. L’Ologenese; nouvelle theorie de revolution et de la distribution
geographique des etres vivants. Paris, Alcan.
Scheffer V. B. 1958. Seals, sea lions, and walruses. Stanford, Stanford Univ. Press.
Schindewolf О. H. 1950. Grundfragen der Palaontologie. Stuttgart, E. Schweizer-
bartische Verlagsbuchhandlung (Erwin Nagele).
Schmalhausen I. I. 1949. Factors of evolution; the theory of stabilizing selection.
Trans, by I. Dordick. Ed. by Th. Dobzhansky. Philadelphia, Blakiston.
Scott A. J., Collinson C. 1959. Intraspecific variability in conodonts: Palmatolepis
glabra Ulrich and Bassler. Jour. Paleont, 33: 550-565.
Shenefelt R. D. 1959. Taxonomic «descriptions» Science, 130: 331.
Sibley C. G. 1954. The contribution of avian taxonomy [to a symposium on
subspecies and clines]. Systematic Zoology, 3: 105-110, 125.
Simpson G. G. 1937. Super-specific variation in nature and in classification from
the view-point of paleontology. Amer. Naturalist, 71: 236-267.
— 1940. Types in modem taxonomy. Amer. Jour. Sci., 238: 413-431.
— 1941. The affinities of the Borhyaenidae. Amer. Mus. Novitates, No. 1118:
1-6.
— 1943. Criteria for genera, species, and subspecies in zoology and paleozoology.
Ann. N.Y. Acad. Sci., 44: 145-178.
— 1944. Tempo and mode in evolution. New York, Columbia Univ. Press.
— 1945. The principles of classification and a classification of mammals. Bull.
Amer. Mus. Nat. Hist., 85: i-xvi, 1-350.
— 1951. The species concept. Evolution, 5: 285-298.
Цитированные работы
271
— 1953. The major features of evolution. New York, Columbia Univ. Press.
— 1954. Tendances actuelles de la systematique des mammiferes. Mammalia, 18:
337-357.
— 1959a. The nature and origin of supraspecific taxa. Cold Spring Harbor Symposia
on Quantitative Biology, 24: 255-271.
— 1959b. Mesozoic mammals and the polyphyletic origin of mammals. Evolution,
13; 405-414.
— 1959c. Anatomy and morphology: classification and evolution: 1859 and 1959.
Proc. Amer. Phil. Soc., 103: 286-306.
— 1960a. The history of life. In The evolution of life, vol. I of «Evolution after
Darwin» Chicago, Univ, of Chicago Press, pp. 117-180.
— 1960b. Diagnoses of the classes Reptilia and Mammalia. Evolution, [in press].
Simpson G. G., Roe A., Lewontin R. C. 1960. Quantitative zoology. Revised ed.
New York, Harcourt, Brace.
Smith H. M„ White F. N. 1956. A case for the trinomen. Systematic Zoology, 5:
183-190.
Sokal R. R., Michener C D. 1958. A statistical method for evaluating systematic
relationships. Univ. Kansas Sci. Bull., 38: 1409-1438.
Sonnebom T. M. 1957. Breeding systems, reproductive methods, and species
problems in Protozoa. In Mayr, 1957: 155-324.
Sprague T. A., Hopwood A. T, Wilmott A. J., Airey Shaw H. K., Smart J. 1950.
Lectures on the development of taxonomy delivered in the rooms of the Linnean
Society during the session 1948-1949. Linnean Soc. London. [A pamphlet of
83 pages, without name of editor or publisher and apparently not generally
distributed, but with the statement that it is «Sold at the Society’s rooms.» I am
indebted to A. T. Hopwood for a copy.]
Steam W. T. 1959. The background of Linnaeus’s contributions to the nomenclature
and methods of systematic biology. Systematic Zoology, 8: 4-22.
Stebbins G. L. 1950. Variation and evolution in plants. New York, Columbia Univ.
Press.
Streuli A. 1932. Zur Frage der Artmerkmale und der Bastardierung von Baum- und
Steinmarder (Martes). Zeitschr. Saugetierk., 7: 58-72.
Stroud С. P. 1953. Factor analysis in termite systematics. Systematic Zoology, 2: 76-
92.
Sylvester-Bradley P. C 1951. The subspecies in paleontology. Geol. Mag., 88: 88-
102.
— 1954. The superspedes. Systematic Zoology, 3: 145-146, 173.
—, ed. 1956. The species concept in paleontology. London, The Systematics
Association.
Thompson W. R. 1952. The philosophical foundation of systematics. Canadian
Entomol., 84: 1-16.
Thorpe W. H. 1940. Ecology and the future of systematics. In Huxley, 1940: 341 —
364.
Underwood G. 1954. Categories of adaptation. Evolution, 8: 365-377.
19*
272
Дж. Г. Симпсон. Принципы таксономии животных
Vanzolini Р. Е., Guimares L. R. 1955. Lice and the history of South American land
mammals. Rev. Brasil. Ent., 3: 13—46.
Vicq d’Azyr F. 1792. Systeme anatomique des quadrupedes. Encyclopedic
methodique, vol. 2. Paris, Vve. Agasse.
Waagen W. 1869. Die Formenkreise des Ammonites subradiatus. Benecke Geol.—
Pal. Beitr., 2: 179-257.
Waddington С. H. 1957. The strategy of the genes. New York, Macmfflan.
Westoll T. S. 1956. The nature of fossil species. In Sylvester-Bradley, 1956: 53-62.
White J. A. 1953. The baculum of the chipmunks of western North America. Univ.
Kansas Pub., Mus. Nat. Hist., 5: 611-631.
Wilson E. O., Brown W. L., Jr. 1953. The subspecies concept. Systematic Zoology,
2: 97-111.
Wood A. E. 1950. Porcupines, paleogeography, and parallelism. Evolution, 4: 87-
98.
— 1955. A revised classification of the rodents. Jour. Mammalogy, 36: 165-187.
— 1958. Are there rodent suborders? Systematic Zoology, 7: 169-173.
— 1959. Eocene radiation and phylogeny of the rodents. Evolution, 13: 354-361.
Woodger J. H. 1937. The axiomatic method of biology. Cambridge, Cambridge Univ.
Press.
— 1945. On biological transformations. In «Essays on growth and form», W. E. Le
Gros Clark and P. B. Medawar, eds.: 95-120.
— 1948. Biological principles. London, Routledge and Kegan Paul.
— 1952. Biology and language. Cambridge, Cambridge Univ. Press.
Zangerl R. 1948. The methods of comparative anatomy and its contribution to the
study of evolution. Evolution, 2: 351-374.
Zimmerman W. 1953. Evolution. Die Ceschichte ihrer Probleme und Erkenntnisse.
Munich, Freiburg.
Zirkle C. 1959. Species before Darwin. Proc. Amer. Phil. Soc., 103: 636-644.
Приложение
Ниже приведены комментарии редактора и переводчика к тек-
сту Дж. Симпсона
[1] На этом примере со стульями можно уяснить соотношение
связей по смежности и сходству. Конкретный стул является объе-
динением по смежности четырех ножек, спинки и т.д. В то же вре-
мя объекты, характеризуемые четырьмя ножками и спинкой, со-
ставляют объединение по сходству, т.е. множество (класс) стульев.
Связи по сходству определяют классы, которые являются абстрак-
тными объектами, тогда как связи по смежности характеризуют
системы в качестве реальных, т.е. существующих в пространстве
и времени, объектов.
[2] Популяцию как систему, объединяемую по смежности, мож-
но рассматривать и как класс, отвлекаясь от ее системных, т.е. про-
странственно-временных характеристик.
[3] Есть русский перевод: Майр Э., Линсли Э., Юзингер R,
1956. Методы и принципы зоологической систематики. М.: Изд-во
иностр, лит.
[4] Есть русский перевод: Дарлингтон Ф., 1966. Зоогеография.
Географическое распространение животных. М.: Прогресс.
[5] Речь здесь не идет о тождестве между таксонами и катего-
риями. С теоретико-множественной точки зрения монотипические
таксоны равны (идентичны) и, следовательно, для них нельзя вве-
сти различающих категорий. Но систематики соотносят их с раз-
ными категориями, и это противоречие составляет содержание так
называемого парадокса Грегга. См.: Шаталкин А.И. 1999. Семан-
тическая структура таксономических имен // Журн. общ. биологии.
Т. 60. № 2. С. 150-163.
[6] В современной литературе предложенная Беккнером «поли-
типическая концепция» обозначается как политетическая концеп-
ция таксона, чтобы «развести» её с политипической концепцией в
традиционном для систематики понимании (т.е. таксон, включаю-
Дж. Г. Симпсон. Принципы таксономии животных
274
щий несколько таксонов следующего более низкого ранга). Поли-
тетическая концепция по смыслу совпадает с концепцией «семей-
ного сходства» Виттгенштейна. См.: Шаталкин А.И. 1988. Биоло-
гическая систематика. М.
[7] Приоритетность отношений можно пояснить следующим
примером двух таксонов Syrphus (Т ) и Syrphidae (Т.+1) из класси-
фикации мух журчалок. Все мухи-журчалки (обозначим их через
а.), являющиеся членами рода Syrphus (символически а. 6 Т), яв-
ляются также членами семейства Syrphidae (а, е Т ;). Но не все
журчалки, входящие в Syrphidae (например, мухи рода Eristalis,
которых вместе с другими мухами, отличными от видов Syrphus,
обозначим через b) являются членами Syrphus. Симпсон говорит,
что отношение a., b. G Т z имеет приоритет над отношением а. 6 Т,.
Из этих двух отношений следует другое, а именно отношение co-
подчиненности Т с Т р означающее, что род Syrphus принадле-
жит, или является частью семейства Syrphidae.
[8] В некоторых концепциях одним из ключевых в понимании
естественной системы является критерий Уэвелла, определяющий
естественность через прогностичность. См.: Розова С.С., 1986.
Классификационная проблема в современной науке. М.
[9] В настоящее время действует Международный кодекс зоо-
логической номенклатуры, принятый в 4-й редакции в 1999 г. См.
русскоязычное издание: «Международный кодекс зоологической
номенклатуры», 2000, СПб.; подробное комментированное изложе-
ние более ранней редакции Кодекса можно найти в книге: Майр Э.,
1971. «Принципы зоологической систематики», М.
[10] В ныне действующей редакции Международного кодекса
(см. Примечание [9]) это предусмотрено; обсуждается также его
применение к таксонам ранга отряда.
[11] См. Примечание 10.
[12] В ныне действующей редакции Международного кодекса
(см. Примечание [9]) паратип лишён номенклатурного статуса.
[13] Это ошибка: окончание -oidea используется для всех жи-
вотных, окончание -асеа — для всех растений (см.: «Международ-
ный кодекс ботанической номенклатуры», 1980, Л.).
[14] В ныне действующей редакции Международного кодекса
(см. Примечание [9]) это предложение по-прежнему не принято.
Приложение
275
[15] Подобная практика принята также для рыбообразных и
активно внедряется в систематику насекомых.
[16] См. Примечание [9].
[17] Данный пример устарел и потому не вполне удачен. В на-
стоящее время выделяется особая жизненная форма крупных рас-
тительноядных наземных млекопитающих, которым когтистые лапы
служат для подтягивания нижних ветвей деревьев. Кроме халикоте-
рия, к нему также относятся вымершие гигантские наземные ленив-
цы {Megatherium, Megalonyx) и лемуры (Megaladap is). См.: О.Л.
Россолимо и др., 2004. Разнообразие млекопитающих, ч. I—III. М.
[18] Такое представление включающей иерархии называют эн-
капсисом, иногда структуру иерархии описывают через такие обра-
зы как «китайский ящик» или «русская матрешка», но это не со-
всем точно.
[19] На самом деле такое представление классификации восхо-
дит к «дереву Порфирия» схоластов, его первое применение в си-
стематике датируется XVII в.
[20] В настоящее время стилизованное филогенетическое дере-
во, на котором отсутствует анагенетическая компонента разнообра-
зия, обозначают термином «кладограмма». См.: Шаталкин А.И.
1988. Биологическая систематика. М.
[21] Первое англоязычное издание книги Геннига появилось в
1966 г. См.: Hennig W., 1966. Phylogenetic Systematics. Urbana.
[22] Исследования по геносистематике, сегодня очень популяр-
ные, в начале 60-х гг. только начинались и, видимо, не были изве-
стны Симпсону.
[23] Очевидно, здесь и далее имеются в виду в основном не
сходства как таковые, а признаки, по которым устанавливаются
сходства. Это следует из того, что сходства не могут «присутство-
вать» у отдельного таксона.
[24] Есть русский перевод: Симпсон Дж.Г, 1948. Темпы и фор-
мы эволюции. М.: Изд-во иностр, лит.
[25] В отечественной литературе он известен как «метод кор-
реляционных плеяд» Тереньтьева. см.: Теренъев П.В., 1959. Метод
корреляционных плеяд // Веста. ЛГУ, № 9: 137-141.
[26] В настоящее время разрабатывается концепция эпигенети-
ческой гомологии, в которой соответствие анатомических структур
276
Дж. Г. Симпсон. Принципы таксономии животных
устанавливается в том числе с учётом их генетических детерминант.
См.: Павлинов И.Я., 2005. Введение в современную филогенетику. М.
[27] См. Примечание [6].
[28] Симпсон дает свое понимание позиции Грегори. По Гре-
гори, наследственная основа — это групповые характеристики, ко-
торые устойчиво сохраняются и не изменяются даже при приспо-
соблении организмов к радикально иной экологической обстанов-
ке, как это имело место при переходе предков китов к водному об-
разу жизни: «Таким образом, наследственная основа китов мам-
мальная, но их габитус рыбий» (Gregory, 1951. Evolution emerging.
New York, Macmillan.V. 1, p. 14). Симпсон под наследственной ос-
новой понимает просто более широкие адаптации в сравнении с
более узкими, которые по отношению к первым будут габитуаль-
ными. Это принципиально иная концепция, поскольку допускает
независимое формирование «наследственной основы», что перечер-
кивает смысл самого понятия: наследственная основа есть то, что
наследуется, а не составляется из разных источников.
[29] Имеется в виду итеративный характер таксономической
процедуры, основанный на методе последовательных приближений.
[30] Эти работы были опубликованы: Evolution, 1960, V. 14, № 3.
Van Valen L. p. 304—314; Reed C.A. p. 314-322; Simpson G.G. p. 388-
392.
[31] Членами рода и подродовых таксонов могут быть и виды,
если понимать их в теоретико-системном значении. Таковы генети-
ческий (биологический) вид Майра и эволюционный вид Симпсо-
на, разбираемые дальше.
[32] Имеются в виду «ортогональные» классификации, т.е. пол-
ностью несовпадающие, исключающие одна другую.
[33] В оригинале здесь указаны рыбы, что выглядит несколько
странно: у них нет по настоящему членистых конечностей, указан-
ных Симпсоном в качестве примера конвергенции с насекомыми.
[34] Скорее всего описка; следует читать: «между разными Т »
[35] Данный пример устарел и потому не вполне удачен: в на-
стоящее время «классический» род Rattus делят на 10-12 родов,
см.: Павлинов ИЛ., 2003. Систематика современных млекопитаю-
щих. М.
[36] См. Примечание [35].
Приложение
ТП
[37] См. Примечание [35].
[38] В современной кладистической систематике число рангов
не фиксировано и поэтому может быть неопределённо большим.
См.: Ereshefsky М., 2001. The poverty of the Linnaean hierarchy: a
philosophical study of biological taxonomy. Cambridge.
[39] Имеются в виду чисто прагматические критерии, направ-
ленные на решения конкретных вопросов.
[40] В более употребимой латыни этому соответствует differentia.
[41] Есть русский перевод: Кэйн А., 1958. Вид и его эволюция.
М.: ИЛ.
[42] Эта проблема отчасти решается концепцией прерывисто-
го равновесия за счёт введения «масштаба времени» в рассмотре-
ние эволюционного процесса. Согласно этому, время перехода от
одного вида к другому на порядки меньше существования самих
видов, что делает их (в соответствующем масштабе времени) ква-
зидискретными единицами. См.: Гулд С.Д. 1986. В защиту концеп-
ции прерывистого равновесия // Катастрофы и история Земли. Но-
вый униформизм. М.
[43] В ныне действующей редакции Международного кодекса
(см. Примечание [7]) признаны паратаксоны, а также ихнотаксо-
ны, основанные на отпечатках следов.
[44] Пример крайне неудачен, а мнение Симпсона частично ус-
тарело; в настоящее время среди мелких млекопитающих открыты
многие десятки комплексов видов-двойников.
[45] См. Примечание [42].
[46] В ныне действующей редакции Международного кодекса
(см. Примечание [9]) официально признана категория «группа ви-
дов», относящаяся к номенклатурной группе вида.
[47] В ныне действующей редакции Международного кодекса
(см. Примечание [9]) этому понятию соответствует типовая серия.
[48] В тексте стоит «непроизвольными (nonarbitrary) по вклю-
чению». На наш взгляд, это описка. Симпсон добавляет, что над-
видовые таксоны выделяются на основании других критериев. По
этим критериям они могут быть произвольными по включению.
[49] По современным данным австралопитеки (Ardipithecus,
Australopithecus, Kenyanthropus и другие) образуют сестринскую
группу относительно рода Ното.
278
Дж. Г. Симпсон. Принципы таксономии животных
[50] В настоящее время эта классификация почти единодушно
отвергнута: признаваемой ныне филогенетической схеме более со-
ответствует базовое деление Carnivora на Feliformia и Caniformia,
к последним отнесены все ластоногие. См.: Павлинов И.Я., 2003.
Систематика современных млекопитающих. М.
[51] Здесь, конечно, правильнее говорить не о категориях, но о
таксонах. Из этого отрывка следует, что Симпсон, как практикую-
щий зоолог, понимал таксоны в теоретико-системном значении,
хотя на протяжении всей книги он рассматривает их как теорети-
ко-множественные конструкции.
[52] В оригинале — «категорий».
[53] См. Примечание [52].
[54] В настоящее время эта концепция пересматривается, осо-
бенно радикально в свете молекулярно-генетических данных. См.:
О.Л. Россолимо и др., 2004. Разнообразие млекопитающих, ч. I-III.
М.
[55] В оригинале — вместо «...таксонами» написано «...выс-
шими категориями», что мы считаем опиской.
[56] Ныне признана существенно иная филогенетическая схе-
ма и соответствующая ей классификация, так что данный пример
представляет в основном дидактический и исторический интерес;
см. также Примечание [50].
[57] Приведённая здесь филогенетическая схема и классифика-
ция в ряде важных деталей устарели. Два семейства, отнесённые
здесь к Creodonta, в настоящее время в наиболее дробных кладис-
тических системах помещаются: Arctocyonidae — в отряд Procreodi,
Mesonychidae — в отряд Acreodi, относящиеся к базальной ради-
ации копытных (в самом широком смысле). Creodonta считается
самостоятельным отрядом. Pinnipedia признаны парафилетической
группой, разные семейства которой произошли от архаичных
Mustelidae (возможно, также Ursidae). См.: McKenna М.С., Bell S.K.,
1997. Classification of mammals above the species level. N.Y.: Colum-
bia Univ. Press; О. Л. Россолимо и др., 2004. Разнообразие млеко-
питающих, ч. I—III. М. См. также Примечание [50].
[58] В настоящее время тупай выделяют в отряд Scandentia,
близкий к приматам.
Предметный указатель
Австралопитеки, 226
Агамовиды, 181
Адаптация базовая, связанная с происхождение высших таксонов, 257-258
Адаптивная зона и разрывы, 236, 258
Адаптивная радиация, 238, 259, 262
Адаптивные признаки, связь с лабильностью, 116
Акциденция, 28, 43, 55, 206
Амёба, 3, 13
Амфибии, 166, 259
Анагенез, 234
Аналогия (по Оуэну), 93; определение, 92
Анатомия, 82; сравнительная, 9; как источник таксономических данных, 82, 129
Антилопы, 100
Апоморфный, 112
Аристотелизм, 42
Архетип(ы), 55-57, 65; у непарнокопытных, 98; связь с общностью признаков,
100
Астрагал, у непарнокопытных, 98
Аутэкология, 9-10
Art. 187
Artenkreis, 210, 213
Бегемоты, 98-100
Белки как источник таксономических данных, 85
Беспозвоночные, 23, 89, 140, 237
Биовид, 181
Биогеография, 10
Биохимия: сравнительная, 9; как источник таксономических данных, 85, 129
Близнецы, аналогия с таксонами, 80
Боргиениды, 86
Броненосец, 15, 149
Ваагенон, 205
Вариетет, 206
Верблюды, 172
Ветвление, связь с видообразованием, 192-193, 236
Ветвящиеся линии, 192, 236
280
Дж. Г. Симпсон. Принципы таксономии животных
Виды: положение в иерархии, 19; как единицы в линнеевской иерархии, 22;
в схоластическом смысле, 28, 43; названия, 37; объективность, 118; генети-
ческие, 171; биологическое или генетическое определение, 174; эволюцион-
ные, 177, 178; категории, 181; таксономические, 181; двойники, 184; алло-
патрические, 187; симпатрические, 187; у однополых организмов, 188; в па-
леонтологии, 190; последовательные, замещающие, 167, 195, 222; и ветвле-
ние, 196; группы, 209; эволюционная основа, 256
Волк сумчатый, тасманийский, 65, 86, 263
Волки, 65, 163
Воробьи, 172
Временная размерность голоморфы, 84
Выборки, 89, 90; в палеонтологии, 170
Вымирание, 234
Высшие категории, см. Категории высшие
Габитус, 52, 117
Гемоглобин (электрофорез), источник таксономических данных, 85
Генезис, 60
Генерализованный (определение), 116
Генетика, 10; популяционная, как источник таксономических данных, 88-89;
и гомология, 105-106
Генофонд, 189
Гены, 189
Геоклина, 208
Гепард, 161
Гибридизация как критерий вида, 175
Гибридизация, 172; и выделение видов, 176-177
Гиподигма, 37, 263; определение, 215
Гипотерии, 140
Гистрикоморфы, 86, 104, 253
Головастики, 103
Голоморфа, 83
Голотип, 37, 56
Гомология, 90; определение, 91; по Оуэну, 93; по Дарвину, 94; критерии 96, 102;
общая, 108; специальная, 108; сериальная, 108; отличие от параллелизмов,
123, 123
Гомономия, 109Гомоплазия, 91, 96
Гомотип, 37
Гоферы, 203, 206
Грады, 176; и разрывы, 237; у приматов, 249; в эволюции высших таксонов, 264
Граптолиты, 147
Грызуны, 157, 237, 240, 251-254, 256, 259; южноамериканские, 104, 256
Дегенерация, 59-172
Предметный указатель
281
Дем, 215
Дендрограмма, 73-74; вертикальные и горизонтальные связи, 150; см. также
Дерево
Дерево филогенетическое, 131; вертикальные и горизонтальные отношения, 153;
и возраст таксонов, 164; рост, 165; определение высших и низших таксонов,
223
Дерево, формализация отношений, 72, диаграммы, 73
Диагноз, 161
Диапазон изменчивости (при разграничении видов), 214
Дивергенция, 154; критерий высших категорий, 223, 228; филогенетический
паттерн, 235; в пределах высших таксонов, 258-260
Диверсификация, 201, 203
Дикобразы, 104
Дискретность таксонов, 134
Доказательство, связь с определением, 78-79, 175; категории доказательств в
таксономии, 82
Долгопяты, 120, 248
Древность таксонов, 163; как критерий ранга, 229
Дробительство, 158, 162, 234
Дюгонь, 38
Зайцы, 241
Зебры, 231
Змеи, 113
Зоогеография: как источник таксономических данных, 86; и становление выс-
ших таксонов, 263
Зубная система, последовательность редукции, 113
Зубы: лабильность в эволюции, 120; параллелизмы, 121
Идеи (по Платону), 58, 64
Идентификация (отличие от классификации), 13, 22-23
Иерархии, 13
Иерархия, подходы, 45
Иерархия: определение, 15; линнеевская, 19
Инфравидовая группа, отличная от подвида, 205
Инфракласс, положение в иерархии, 20
Инфраотряд, положение в иерархии, 20
Ископаемые, см. Фоссилии
Кавиоморфы 86, 253-254
Кариограмма, 188
Кариология как источник таксономических данных, 84
Категории высшие, 218; определение, 219-221; характеристики, 230; эволюци-
онная основа, 256
282
Дж. Г. Симпсон. Принципы таксономии животных
Категория, определение, 22
Квадратная кость, 108
Кенгуру, 65, 114
Киты(ы), 113, 117, 259
Кладогенез, 234-235
Клады, 146
Класс, положение в иерархии, 13
Классификация вертикальная, 150-151
Классификация горизонтальная, 150-151
Классификация компромиссная, 150-151
Классификация: определение, 11; формы, 13; основания и критерии, 27; типы,
30; экологическая, 30; телеологическая, 30-31; эволюционная, 31; естествен-
ная, 64; устойчивость или соответствие, 125-127; как полезное искусство, 125,
128-129; объективность и условность, 125; примеры компромиссов, 153
Клины, 119, 201, 208
Ключи, 13; определение, 16
Коадаптация, 117, 260
Когорта, положение в иерархии, 20
Конвергенция, 106-107; и экология, 84; определение, 92; отличие от паралле-
лизмов, 121-123; в эволюции высших таксонов, 263
Конодонты, 183
Копытные, 99, 256, 259; редукция пальцев, 114
Корреляции принцип (по Кювье), 52
Кошки, 161-163
Креодонты, 152, 244, 261
Кролики, 241
Крысы, 157, 172
Кугуар, 13
Куницы, 185-186
Ламаркизм, 60
Лев, 162
Лектотип, 37
Лемуры, 102, 248
Леопард, 162
Лестница природы, 61, 70, 71, 172
Летучие мыши, 259
Линия эволюционная, 234
Лисицы, 17; большеухие, 103
Литоптерны, 101
Логика схоластическая, 42; символическая, 13, 25
Локальная популяция, 205
Лори, 248
Лошади, 98-101, 119, 141, 231; грады в эволюции, 147
Предметный указатель
283
Мамонты, 164
Мастодонты, 164
Медведи, 17, 163, 220
Международная комиссия зоологической номенклатуры, 35
Международные правила зоологической номенклатуры, 34
Мейоз, 188
Менделевская популяция, 205
Мегатерии, 140
Метатип, 37
Мимикрия, 92
Млекопитающие, 108, 113, 118, 140, 240
Молекулярная биология, 9
Молоточек, 107
Монофилия, 77, 152, 223, 224; определение, 144; связь с разрывами, 227
Морфовид, 181
Морфологическое соответствие (по Вуджеру), 95
Морфология, 82; идеалистическая, 58, 93
Морфотип, 55, 207
Морфы, 207
Музеи, 82
Мутации, 205; гомологичные, 123-124
Мыши, 157
Надвид, 209, 210; хронологичесий, 210; географический, 210
Надкласс, положение в иерархии, 20
Надотряд, положение в иерархии, 20
Надсемейство, положение в иерархии, 20; названия, 38-39
Наковальня (слуховая косточка), 107
Насекомые 97, 130, 149
Наследственность, 116-117
Натурфилософия, 54
Неадаптивные признаки, 116
Нео-кювьерианцы, 52
Неотип, 37
Непарнокопытные, 100, 233, 241, 263
Неполнозубые, 119, 141, 145, 241
Непрерывность таксонов, 134
Непроизвольные (определение), 134
Ниши, 179; отношение к разрывам, 236
Номенклатура, 28; определение, 11; как искусство, 128
Номенклатура бинарная, 38
Номенклатура биномиальная, 38
Носитель имени, 36-37
Носороги, 98-101
284
Дж. Г. Симпсон. Принципы таксономии животных
Noimina cinservanda, 35
Обезьяны человекообразные, 120, 226, 248
Обезьяны, 120, 226, 248
Общность предка, 63, 90
Объединение политипичекское, 110 111
Объединительство, 158, 162
Объективность таксонов, 133
Объём таксона, 156
Однопроходные, 140; размножение, 119
Окраска, эволюционная лабильность, 119
Ономатофор, 56
Определение, 161, 162; и доказательство, 79
Организмы однополоые (при определении вида), 188
Органическая интеграция (по Кювье), 52
Ортогенез, 114
Основание деления, 44, 56
Ответвления (по Кювье), 54
Отличие (схоластический смысл), 28, 66
Отношение «группа-в-группе», 69; определение, 70
Отношение «хозяин—паразит» в таксономии, 87-88
Отношение сетевое, 69
Отношение эквивалентности, 27-28
Отношения вертикальные, 150
Отношения горизонтальные, 150
Отношения, формальные аспекты, 68; горизонтальные, 150
Отряд(ы), положение в иерархии, 19; названия, 39
Палеовид, 181, 195
Палеонтология как источник таксономических данных, 97-98, 130
Панда, 107, 246
Паравид(ы), 181
Паразиты как источник таксономических данных, 87
Параллелизм генетический, 123
Параллелизмы, 120, 121; определение, 91; и полифилия, 141; при становлении
высших таксонов, 260
Парасексуальное отношение, 188
Паратаксон(ы), 181, 184
Паратип(ы), 37, 56, 216
Парнокопытные 100, 112, 120, 263
Паттерны творения, 64; в филогенезе, 234, 235, 239
Персистирование, 234
Пищухи, 243
План структурный, 55, 58
Предметный указатель
285
План: по Кювье, 54; структурный, 55
Плацентарные, 113, 119, 140
Плезиомоорфия, 112
Поведение как источник таксономических данных, 88
Подвид, 199; положение в иерархии, 20; синхронные, 205; последовательные,
204; в палеонтологии, 204; эволюционная основа, 257
Подкласс, положение в иерархии, 20
Подотряд, положение в иерархии, 20
Подразделения полупроизвольные, 244
Подрод, положение в иерархии, 20, 157
Подсемейство, положение в иерархии, 20; названия, 33
Подтип, положение в иерархии, 20
Подтриба, положение в иерархии, 20
Позвонки: гомология, 103; последовательности редукции, 113
Позвоночные, 54, 258
Полиплоидия, 137
Политипии принцип, 50
Политипическая концепция, ПО
Политипия и дивергенция, 156
Полифилия, 139-140, 145
Популяции, 22, 76, 88-90
Порядок в науке, 2-3
Порядок в природе, 2-3
Последовательности, 109; разбиение, 138-139
Правило 75%, 202-204
Предикабилии, 28
Представление отношений: сетевое, 69; линейное, 70-71
Признаки специализированные, 109; примитивные, 109
Признаков общность: связь с архетипом, 86; гомология, 87
Примитивность, 115
Приоритет: в иерархии и ключах, 18; отношение, 27-28; в номенклатуре, 34;
в иерархической классификации, 46, 66, 70
Природа: в смысле сущности, 28, 44; в схоластическом понимании, 66
Прогрессия, 234
Прозимии, 118-119
Произвольность, 125, 134; классификаций, 125.
Произвольный, определение, 115
Простейшие, 140, 185; в эволюционной классификации, 85
Простота и примитивность, 114-115
Прототерий, 140
Психология сравнительная, 9
Птицы, 130
Пумы, 162
Пчёлы, 172
20 — 4338
286
Дж. Г. Симпсон. Принципы таксономии животных
Равновесие, 234; и разрыв, 236
Рвзветвление, 234
Размножение половое, 188
Разнообразие, 154.; критерий высших категорий, 230
Разрывы, 223; как таксономический критерий, 136; по сходству, 136; исполь-
зование для определения высших таксонов, 223.; происхождение, 236
Ранги: категорий, 23; таксонов, 23; признаков, 169; эволюционная основа, 257-
258
Распространение географическое и геологическое, как источник таксономичес-
ких данных, 86
Реальность подразделений, 70-71; таксонов, 133
Род, положение в иерархии, 19; высший, 27; в схоластическом смысле, 26, 41,
5666; названия, 37; особый статус категории, 232
Роды: объективность, 134; грады, 148; линии, 148; форма, 182
Рыбы, 113, 148; грады, 146
Ряды, 11-115
Rassenkreis, 187
Свиньи, 100
Свойство (в схоластическом понимании), 28, 43
Связь по смежности, 3, 4; по сходству 3, 4
Семейство, положение в иерархии, 19; названия, 38
Серология, 118; в систематике, 85, 90
Синтипы, 3\7
Синэкология, 9-10
Систематика: место в науке, 7; определение, 8; новая, 75
Скорость эволюции (в связи с рангами таксонов), 164
Скот домашний, 99
Следы, 184
Слоны, 164
Собаки, 107, 263
Сочленовная кость, 107-108
Специализация, 119
Специализированный, 115
Стасигенез, 234
Статистика в новой систематике, 76; в систематике, 89-90
Страусы, 113-114
Субъективность таксона, 133-134
Сумчатые, 65, 86, 119, 140, 259
Сущность (схоластический смысл понятия), 28, 43, 57, 66
Сходство случайное, 97
Сходство: объединение по, 4, 5; категории, 90
Схоластика: логические связи, 28; у Линнея, 49
Предметный указатель
287
Таксономическая деятельность, 126
Таксономия: определение, 12; анализируемых и неанализируемых сущностей,
43; по Дарвину, 63; задачи в новой систематике, 76; категории доказательств,
81-82, 125-126
Таксоны высшие: основания для выделения, 223; и происхождение адаптаций,
257; и грады в эволюции, 260
Таксоны: определение, 22; в новой систематике, 75-76; политипические, ПО-
111; дискретность и непрерывность, 134; объём, 156; относительная древ-
ность, 163; эволюционная основа, 256
Тапир(ы), 99, 101, 107, 119
Творение, 9
Теория множеств, 25
Терминация, 234
Типы: номенклатурные, 34, 36, 55, 76; в типологии, 56, 65; и последователь-
ные виды, 195, 217; функции, 213
Тип, положение в иерархии, 19
Типология, 54, 91
Томизм, 43
Топотип, 37, 216
Транзиент, 195, 205
Триба, положение в иерархии, 20
Триномен, 200
Трубкозуб, 24, 149, 155, 240
Тупайи, 120, 246, 248
Тюлени, 220
Уместность таксономическая, 133
Уровни линнеевской иерархии, 20; изменения категорий, 159
Утконос, 119
Ушной отдел, 117
Физиология: сравнительная, 9; как источник таксономических данных, 84
Филетическая эволюция, 234, 262
Филогения: реконструкция, 59, 79; представление в виде дерева, 72; связь с
классификацией, 131; выделение высших и низших таксонов, 223-224
Фиссипеды, 152, 231, 246, 261
Форма, 207
Фоссилии: как источник таксономических данных, 82; и гомология, 107; про-
белы в летописи, 138; и проблема классификаций, 190
Халикотерии, 52
Хищные, 65, 113, 152, 231, 244, 246
Хромосомы, 112
Хроноклина, 119, 208
20*
288
Дж. Г. Симпсон. Принципы таксономии животных
Царство (положение в иерархии), 19
Цепь существ, 54, 61; см. также Лестница природы
Цитология: сравнительная, 10; как источник таксономических данных, 84
Человек, 115, 120, 248
Эволюция квантовая, 137; мозаичная, 260; остановленная 234
Экоклина, 208
Экология, 10; и гомология, 106; как источник таксономических данных, 87-88
Экспансия, 235
Эмбриология: как источник таксономических данных, 94; и гомология, 104
Эмпиризм, 47, 53
Эндостиль, 103
Эогиппус, 119, 123
Эстетика, 5
Этология, 10, 88
Ягуары, 162
Указатель латинских названий
таксонов
Ailurus, 10
Amphibia, 166
Anoplotherium, 100
Anthropoidea, 141, 145, 248, 262
Archaeopteryx, 260
Arctocyonidae, 245
Articulata, 54, 109
Artiodactyla, 98, 120, 241
Asinus, 105, 156
Basilosaurus, 40
Bathyergomorpha, 254
Belluae, 99
Bison, 105
Bos, 22, 105
Bos taurus, 38
Canidae, 107, 245
Canis, 65, 107
Carnivora, 39, 55, 97, 152, 231, 244,
246, 261, 262
Castorimorpha, 253
Caviomorpha, 86, 252-253
Ceboidea, 249, 262
Cercopithecoidea, 249, 252
Cetacea, 117
Chelonia, 22
Chiroptera, 234
Condylarthra, 241
Creodonta, 152, 244
Cricetidae, 157
Crustacea, 22, 111, 233
Didelphidae, 22
Drosophila, 105, 112, 186
Drosophila macrospina, 209
Drosophila pseudoobscura, 143, 186
Duplicidentata, 243
Edentata, 141, 145, 149, 241
Eohippus, 100, 233
Equidae, 141, 156, 231
Equus, 105, 114, 156, 231
Eropeplus, 156
Felidae, 161, 162, 245
Felis, 161, 162
Felis concolor, 13
Felis tigrina glaucula, 163
Fissipeda, 152
Fulica atra, 45
Glires, 99
Hominidae, 39, 26, 249
Hominoidea, 249, 262
Homo, 226
Homo sapiens, 22, 38, 83, 143n, 192
Hutchinsoniella, 130
Hyaenidae, 245
Hyaenodontidae, 245
Hyracotherium, 100, 114, 233
Hystricomorpha, 86, 105, 252
Insectivora, 249
Kogia, 38
Lagomorpha, 243, 246, 262
Lama, 38
290
Дж. Г. Симпсон. Принципы таксономии животных
Latimeria, 166
Leporidae, 241-243, 252
Lynx, 162
Macropus, 65
Mammalia, 138, 141, 145, 165, 260, 261
Mammut, 164
Margay glaucula glaucula, 163
Marsupialia, 65, 74, 140, 223
Martes, 185
Merychippus, 141, 145
Mesonychidae, 245
Miacidae, 152, 244, 245
Mollusca, 54
Monograptus, 147
Monotremata, 140
Muridae, 157
Mustelidae, 244, 245
Myomorpha, 253
Neopilina, 130
Ochotonidae, 243
Odobenidae, 245
Orycteropodidae, 24, 155, 240
Orycteropus, 24, 155, 157, 240-241
Osteichthyes, 166
Otariidae, 245
Otocyon, 109
Oxyaenidae, 245
Palaeotherium, 100
Porahippus, 141
Perissodactyla, 98, 231, 241
Phocidae, 245
Pinnipedia, 220, 231, 244
Pongidae, 226
Porifera, 22
Primates, 23, 120, 121, 166, 246, 247
Proboscidea, 231
Procyonidae, 244, 245
Prosimii, 141, 246, 248
Protrogomorpha, 252, 253
Radiata, 54
Rattus, 155-157
Rattus rattus, 38
Reptilia, 138, 141, 166
Rodentia, 243, 244, 251, 255, 262
Sciuridae, 254
Sciuromorpha, 253
Synoplotherium, 38
Tatera, 38
Tatu, 15
Thalarctos, 105
Thallomys, 156
Therapsida, 141
Theridomorpha, 254
Thoatherium, 101
Thylacinus, 65, 86, 107
Thomomys umbrinus, 203
Tubulidentata, 24, 155, 240-241
Ungulata, 99
Ursidae, 245
Ursus, 105
Ursus arctos, 15
Ursus maritimus, 15
Vertebrata, 54
Viverridae, 244, 245
Xenochoerus, 38
Указатель авторов
Адансон (Adanson, М.), 48
Аквинат (Aquinas), 43, 173
Амадон (Amadon, D.), 204
Амегино (Ameghino, Е), 101
Аристотель, 43п, 54
Бейдер (Bader, Т. S.), 164, 158
Бейзер (Bather, F. А.), 32, 48, 147,
149
Беккнер (Beckner, М.), 23-26, 50, 51,
НО
Белон (Belon Р.), 93
Бёрма (Burma, В. J.), 200
Бигелоу (Bigelow, R. S.), 164, 168-169
Блокк (Block, К.), 58
Блэквелдер (Blackwelder, R. Е.), 8, 10,
13, 85
Блэнвиль (Blainville, Н. М. D. de), 98,
99, 159
Бойден (Boyden, А.), 8, 10, 13, 85? 95
Боннэ (Bonnet, С.), 70
Брандт (Brandt, J. Е), 253
Браун (Brown, W. L., Jr.), 20n
Бринкмен (Brinkmann, R.), 86
Бриссо (Brisson, M. J.), 45
Брони (Bronn, H. G.), 108
Булмен (Bulman, О. M. В.), 146
Бэкон (Bacon, Roger), 43n
Бэр (Baer, К. Е. von), 84, 104
Бюффон (Buffon, G. L.), 59, 72
Вааген (Waagen, W.), 205
Ванцолини (Vanzolini, P. E.), 88
Вик д’Азир (Vicq d’Azyr, E), 49
Вуд (Wood, A. E.), 86, 251, 252, 254
Вуджер (Woodger, J. H.), 23, 95
Вэн Вэйлен (Van Valen, L.), 138
Гарстанг (Garstang, W), 84
Геккель (Haeckel, E.), 84
Гексли Дж. (Huxley, J. S.), 75, 96, 146,
208, 234, 261
Гексли T. (Huxley, T. H.), 140, 146
Генниг (Hennig, W.), 7, 83, 112
Гёте (Goethe, J. W. von), 54, 57
Гилмур (Gilmour, G. S. L.), 29, 67
Гимарес (Guimares, L. R.), 88
Гольдшмидт (Goldschmidt, R.), 137
Граберт (Grabert, B.), 191
Грегг (Gregg, J. R.), 12, 23-26
Грегори (Gregory, W. K.), 41, 117
Дарвин (Darwin, C.), 42, 47, 50, 57,
61-63, 79, 84, 90, 95, 100, 109, 111,
117, 131, 140, 173, 207
Дарлингтон (Darlington, R), 10, 86
ДеБир (De Beer, G. R.), 104, 260
Делакур (Delacour, T.), 83
Деннлер (Dennler, V. G.), 15
Джепсен (Jepsen, G. L.), 114
Джозеф (Joseph, H. W. B.), 28, 43, 56
Джонсон (Johnson, M. L.), 85
Джордж (George, T. N.), 183, 191
Дитер (Dethier, V. G.), 87
Добжанский (Dobzhansky, Th.), 88, 96,
112, 188, 205
Доусон (Dawson, M. R ), 242
Erap (Eagar, R. M. C.), 182
Жоликёр (Jolicoeur, R), 103
Зоннеборн (Sonnebom, T. M ), 185
Имбри (Imbrie, J.), 191, 195, 205
292
Дж. Г. Симпсон. Принципы таксономии животных
Ингер (Inger, R. F.), 103
Каплан (Kaplan, N. О ), 83
Кастер (Caster, К. Е.), 205
Кеймен (Caiman, W. Т), 33
Кейн (Cain, А. 1), 28, 41,43, 47,51, 53,
62 -63, 84, 91, 103, 174, 181, 184,
191, 210
Келин (Kalin, J. А.), 58
Кинси (Kinsey, А. С.), 200, 220
Кирьяков (Kiriakoff, S. G.), 57, 83п
Кларк (Clark, W. Е. Le Gros), 120, 141
Клауд (Cloud, Р), 87
Кляйн (Klein, J. T.), 45
Кляйншмидт (Kleinschmidt, О.), 140
Колберт (Colbert, Е. Н.), 241
Коллинсон (Collinson, С.), 184
Коуп (Соре, Е. D.), 153
Кристенсен (Christensen, В.), 84
Крутч (Krutzsch, Р. Н.), 84
Кювье (Cuvier, G.), 48-54, 61, 84, 99,
100
Ламарк (Lamarck, J.-B. М. de), 42, 54,
60-61, 72, 172
Лернер (Lerner, I. М.), 96
Линней (Linnaeus, С.), 15, 19, 28, 34,
42, 47, 59, 81, 99, 133, 141, 158, 173,
231
Линси (Linsley, Е. G.), 40, 234
Лулл (Lull, R. S.), 184
Лэдд (Ladd, Н. S.), 87
Майр (Мауг, Е.), 8, 10, 40, 74, 83, 86-
88, 96, 136, 174, 178, 187, 201, 203,
208, 210, 212, 215, 223, 234
Марш (Marsh, F. L.), 60
Маслин (Maslin, Т. Р.), 119
Маттей (Matthey, R.), 84
Меглитч (Meglitsch, Р. А.), 189
Меррием (Merriam, С. Н.), 220
Миллер (Miller, R. L.), ЮЗ
Миченер (Michener, С. D.), 57, 74, 103,
121, 225-227, 232
Моссмен (Mossman, Н. W.), 84
Мур (Moore, R. С ), 184
Мэттью (Matthew, W. D ), 152-153
Николь (Nicol, D.), 237
Нильсен (Nielsen, С. О.), 84
Ньюелл (Newell, N. D.), 113, 191, 205,
207, 215, 216
Нэф (Naef, А.), 58
Окен (Oken, L.), 54
Олсон (Olson, Е. С.), 103
Ортон (Orton, G. L.), 84
Осборн (Osborn, Н. Е), 129, 158, 233
Оуэн (Owen, R.), 48, 54, 57, 93-95,
100, 105, 108
Пеккэм (Peckham, М.), 61
Пеннант (Pennant, Т.), 44
Пири (Pirie, N. W.), 146, 169
Платон, 54
Понтекорво (Pontecorvo, G.), 188
Пэттерсон Б. (Patterson, В.), 88
Пэттерсон Д. (Patterson, J. Т.), 84,
105
Ремане (Remane, А.), 108, 140
Ренш (Rensch, В.), 96, 113, 187, 235
Рид (Reed, С.), 138
Родс (Rhodes, F. Н. Т), 191, 205
Роза (Rosa, D.), 140
Роу (Roe, А.), 5, 88, 164, 185
Сибли (Sibley, С. G.), 200, 208
Сильвестер-Брэдли (Sylvester-Bradley,
Р. С.), 174, 191, 210, 222
Симпсон (Simpson, G. G.), 9, 13, 20,
23, 56, 63, 82, 86-88, 90, 95-96, 107,
120, 128, 133, 137, 138, 146, 154,
159, 162, 169, 174, 178, 189, 191,
215., 216, 220, 240, 251, 261
Скотт (Scott, A. J.), 184
Смит (Smith, Н. М.), 200п
Сокэл (Sokal, R. S.), 57, 103, 226
Указатель авторов
293
Стеббинс (Stebbins, G. L.), 96
Стим (Steam, W. Т.), 15, 47
Стоун (Stone, W. S.), 84, 105
Страуд (Stroud, С. Р.), 103
Стрюли (Streuli, А.), 185
Томпсон (Thompson, R. W.), 51, 52,
80-81
Торп (Thorpe, W Н.), 10
Уайт Д. (White, J. А.), 84
Уайт Ф. (White, F. N.), 200п
Уестолл (Westoll, Т. S.), 191
Уикс (Wicks, М. I), 85
Уилсон (Wilson, Е. О.), 200п
Ундервуд (Underwood, G.), 107
Уоддингтон (Waddington, С. Н.), 84
Фогг (Voigt, F. S.), 178
Фрешкоп (Frechkop, S.), 152
Хаас (Haas, О.), 95, 120
Хаббс (Hubbs, С. L.), 75п
Харрисон (Harrison, G. А.), 57, 63, 103
Хаслвуд (Haslewood, G. A. D.), 85, 86
Хеберер (Heberer, G.), 96
Хеджпет (Hedgpeth, J. W), 87
Цангерл (Zangerl, R.), 55п, 58
Цёкл (Zirkle, С.), 173
Циммерман (Zimmerman, W.), 41, 54
Шенефельт (Shenefelt, R. D.), 214
Шеффер (Scheffer, V В.), 220
Шиндевольф (Schindewolf, О. Н.),
137, 165
Шлегель (Schlagel, Н.), 200
Шмальгаузен (Schmalhausen, I. I.), 96
Шпраг (Sprague, Т. А.), 41
Эдвардс (Edwards, J. G.), 207
Элайес (Elias, М. К.), 205
Эллен (Allen, J. А.), 162
Элли (Allee, W-С.), 87
Эштон (Ashton, Е. Н.), 226
Юзингер (Usinger, R. L.), 40, 234
Джордж Гейлорд СИМПСОН
ПРИНЦИПЫ ТАКСОНОМИИ животных
Перевод с английского
И.Я. Павлинова
Под редакцией и с предисловием
А.И. Шаталкина
Москва: Товарищество научных изданий КМК. 2006.
хх + 293 с., илл.
Редактор издательства К.Г Михайлов
Оригинал-макет ТА, Горлиной
Для заявок:
123100, Москва, а/я 16, Издательство КМК;
kmk2000@online.ru
см. также:
http:/ /webcenter.ru/~kmk2000
Подписано в печать 25.09.2006. Заказ № 4338
Формат 60x90/16. Объём 20 п.л. Бумага офсетная. Тираж 500 экз.
Отпечатано в ГУП ППП «Типография «Наука» АИЦ РАН,
121099, Москва, Шубинский пер., 6